
MECP2E1遺伝子
【課題】MECP2E1タンパク質をコードする配列を含む、単離された核酸分子を提供する。
【解決手段】MECP2遺伝子の新規のオープンリーディングフレームを同定した。新規MECP2E1スプライス変異体およびその相当するポリペプチドである。神経精神障害または発達障害の医学的診断および処置における、これらの核酸配列およびタンパク質の使用。
【解決手段】MECP2遺伝子の新規のオープンリーディングフレームを同定した。新規MECP2E1スプライス変異体およびその相当するポリペプチドである。神経精神障害または発達障害の医学的診断および処置における、これらの核酸配列およびタンパク質の使用。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、新規のMECP2E1遺伝子、およびその遺伝子によりコードされるタンパク質、MeCP2E1に関する。遺伝子における突然変異は神経精神障害および発達障害と関連する。
【背景技術】
【0002】
発明の背景
世界保健機関によれば、神経精神障害は、世界中で最も影響が大きい10種の疾患のうち6種を占める。米国経済への負担は1000億ドル―医師を訪れる4人に1人は診断可能な精神障害を有する。
【0003】
レット症候群(RTT)(OMIM #312750)は、女児において、生後一年の後半における神経発達の漸進的な遅滞の発生から4歳までに停滞に向かい、その後、獲得した優れた運動技能およびコミュニケーション技能を退化および喪失することにより特徴付けられる。温存された歩行、異常なコミュニケーション、および常同性手絞り(hand wringing)という病像が初期の自閉症に近い、偽静止期が後に続く。しかしながら、退化は潜行性に進行し続け、最終的に著明な精神遅滞をもたらす(1)。
【0004】
RTTを有する患者の最大80%が、MeCP2転写リプレッサーをコードする4エキソンのMECP2遺伝子(図1a)(4)のエキソン3および4に突然変異を有する(2、3)。患者の残り20%における突然変異は分かりにくいままである。公知の遺伝子転写産物においては、4つのエキソン全てが用いられ、翻訳開始部位はエキソン2にあり、エキソン1およびエキソン2の大部分は、5'非翻訳領域(UTR)を形成する(4)。明確にするため、この転写産物をMECP2E2(以前はMECP2A)、およびそのコードするタンパク質をMeCP2E2(以前はMeCP2A)と名付ける。
【0005】
MeCP2E2定義のエキソン2に特異的な突然変異は、数百人の患者がこのエキソン中の突然変異について分析されたにも関わらず、今日まで見出されていない(31の刊行物;参考文献3に最も良く概説される)。これらの研究は、エキソン1は非コード性と見なされたため、エキソン1を含まない。
【0006】
非不活性化型のMECP2突然変異もまた精神遅滞および自閉症のような、RTTと重複する表現型に関連付けられている。RTTの診断および処置方法を同定できるように、残り20%のRTT患者の原因となるさらなる突然変異の同定が必要である。
【0007】
レット症候群遺伝子、MECP2における突然変異は、自閉症患者(20〜22)、ならびに幼時期発生精神病(23)、アンゲルマン症候群、非症候性精神遅滞、および新生児脳症(encepalopathy)(24)の患者にもまた見出され、MECP2の突然変異にさまざまな表現型の結果が存在し得ることを示している。
【発明の概要】
【0008】
発明の概要
本発明者らは、MECP2E1と呼ばれる、MECP2遺伝子の新規のオープンリーディングフレームを同定した。5'UTRの検査から、エキソン2はATGの上流に多数のインフレームの終結を有するが、エキソン1はATGを含むオープンリーディングフレームをその全長にわたって含むことが明らかとなった。このオープンリーディングフレームは、MECP2遺伝子のエキソン1、3、および4からなる転写産物をコードする。MECP2E1は、エキソン2のスプライシング切り出しに相当する、ヌクレオチド71〜193が非存在であることを除き、MECP2E2(GenBankアクセッション番号NM_004992、図5(a))と類似している。
【0009】
従って、本発明は、MECP2E1タンパク質をコードする配列を含む、単離された核酸分子を提供する。本発明は、相当するポリペプチドMeCP2E1もまた含む。
【0010】
一つの態様において、精製および単離された核酸分子は、以下を含む:
(a)図6(b)(SEQ ID No. 4)に示したタンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【0011】
本発明の特異的な態様において、単離された核酸分子は図6(a)(SEQ ID No. 3)に示した配列、またはその断片もしくは変異体を有するよう提供される。
【0012】
本発明者らは、レット症候群および精神遅滞のような神経精神障害または発達障害を有する患者が、MECP2E1遺伝子のエキソン1に突然変異を有することを見出した。従って、本発明は、MECP2E1配列(図6(a)(SEQ ID No. 3))のエキソン1における突然変異もしくは欠失を検出する段階を含む、神経精神障害または発達障害を検出する方法を提供する。突然変異は、プライマーX1F/X1R:突然変異スクリーニングプライマー(図1)を用いたゲノムDNA由来のPCR産物のシーケンシングを行うことにより検出され得る。挿入または欠失突然変異の検出には、PCR産物を適切なプラスミドベクターにクローニングし、その後大腸菌(E.Coli)へトランスフェクトし、単離されたコロニーからクローンのシーケンシングを行うことが必要であり得る。または、突然変異は、4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対を用いる多重ライゲーション依存性プローブ増幅(multiplex ligation-dependent probe amplification)(MLPA)により検出され得る。変異または欠失は、MECP2E1によりコードされるタンパク質産物についてアッセイすることによってもまた検出され得る。
【0013】
本発明の他の特徴および効果は以下の詳細な説明から明らかになる。しかしながら、詳細な説明および特異的な実施例は、本発明の好ましい態様を示すが、この詳細な説明から、本発明の精神および範囲内におけるさまざまな変化および改変が当業者には明らかであることから、例証としてのみ示される。
【図面の簡単な説明】
【0014】
ここで本発明を以下の図面との関連において記載する:
【図1】MECP2 5'スプライス変異体を示す。a)MECP2遺伝子の構造。番号付けされたボックスはエキソンを示し;アスタリスクはインフレームの終止コドンを示す。伝統的なMECP2E2スプライス変異体では、開始コドンはエキソン2にある。MECP2E1では、エキソン2は存在せず、開始コドンはエキソン1にある。HF/HR1およびMF/MR:パネルcに示すrtPCR実験に用いたヒトおよびマウスのプライマー対。HR2:別のヒト逆方向プライマーで、これによりHR1を用いて得た結果を確認する(データは図示せず)。X1F/X1R:突然変異スクリーニングプライマー(図2を参照されたい)。プライマー配列(5'-3'):HF-ctcggagagagggctgtg(SEQ ID NO. 5)、HR1-cttgaggggtttgtccttga(SEQ ID No. 6)、HR2-cgtttgatcaccatgacctg(SEQ ID No. 7)、MF-aggaggcgaggaggagagac(SEQ ID No. 8)、MR-ctggctctgcagaatggtg(SEQ ID No. 9)、X1F-ccatcacagccaatgacg(SEQ ID No. 19)、X1R-agggggagggtagagaggag(SEQ ID No. 20)。b)MECP2 ESTの例。c)示された成体組織(他に示された場合を除く)および細胞培養由来のcDNAに対する、(a)のプライマー(HF/HR1およびMF/MR)を用いたPCR結果;d.p.c.:性交後日数。d)示された組織または細胞培養由来のcDNAに対する転写産物特異的リアルタイム定量的PCR(SYBR Green検出法)。e)3'mycタグ化したMeCP2E1(およびMeCP2E2)は主に核内および細胞質の未確定の点内に局在する。f)示されたタンパク質のN末端;ダッシュはアミノ酸がないことを示す。
【図2】患者V1における欠失突然変異を示す。a1)X1F/X1Rプライマー(図1a)を用いたゲノムDNA由来のPCR産物の配列。混合された配列であることに留意されたい。a2)およびa3)患者の野生型および突然変異対立遺伝子のクローンそれぞれの配列;赤いボックスは、突然変異された対立遺伝子において欠失された11ヌクレオチドを示す。b)同一のクローニングされた野生型および欠失された対立遺伝子の電気泳動図。c)HF/HR1プライマーを用いた、示されたcDNAに対するPCR(図1a、c)。レーン1および2(2.5%高分解能アガロース上)は、それぞれ対照および患者の全血に由来する。レーン3〜8(6%変性ポリアクリルアミド上)は、対照血(3)、患者血(4)、対照胎児脳(5)、対照成体脳(6)、対照精巣(7)、および対照ゲノムDNA(8)に由来する。11bpのエキソン1欠失を有する患者のMECP2E2転写産物の発現(266bpのバンド)は、非欠失対立遺伝子(277bp)と比較して減少していないことに留意されたい。141および152bpのバンドはそれぞれ、欠失、および非欠失のMECP2E1転写産物である。
【図3】患者V2における欠失突然変異を示す。MECP2の多重ライゲーション依存性プローブ増幅(MLPA)ピークプロファイルを示す。対照座位を最上部に沿って示す。四角で囲われた領域(E1〜E4)はMECP2エキソン1〜4を示す。a)正常対照のMLPAプロファイル。b)患者V2のMLPAプロファイルはヘミ接合のエキソン1欠失(アスタリスク)を示す。結果は一貫して再現性があり、シーケンシングにより、SNPがMLPA反応のライゲーション効率を阻害する可能性が排除された。
【図4】rtPCRR(A)およびIF(B)による脳細胞初代培養の特徴を示す。(A)Map2、Gfap、およびネスチン発現から、B-27培地中の培養物は主に神経細胞からなり、G-5培地中のものはグリア細胞であることが示された。同一の胚由来の繊維芽細胞もまた培養され、負の対照として用いられた。全脳組織(15.5dpc)をMap2およびネスチンの正の対照として用いた。(B)マウス抗MAP2抗体およびウサギ抗GFAP抗体を用いて神経細胞の二重染色を行った。それらはDAPI(青色)でもまた対比染色された。大部分の細胞はMAP2(緑色)について陽性の染色を示す神経細胞であり、およびGFAP(赤色)について陽性の染色を示すグリア細胞によるわずかな割合の混入が検出された。
【図5A−1】5'および3'非翻訳領域を含むMECP2E2核酸配列(SEQ ID. No. 1)を示す。
【図5A−2】5A−1の続きを示す。
【図5A−3】5A−2の続きを示す。
【図5A−4】5A−3の続きを示す。
【図5B】MeCP2E2タンパク質配列(SEQ ID No. 2)を示す。
【図6A】MECP2E1核酸配列(SEQ ID No. 3)を示す。
【図6B】MeCP2E1タンパク質配列(SEQ ID No. 4)を示す。
【図7】女性MR患者において同定された5種のMECP2エキソン1変異体のヌクレオチド配列を示す。全ての配列は、ヘテロ接合PCR産物をpDRIVEベクター(Qiagen)へクローニングした後に、単一のコロニーから得た。可能な場合は、ATG開始コドンを赤色のボックスにより示す。結果的に生じたアミノ酸配列もまた、赤色で示される野生型配列と共に示し、および変化を緑色の活字で示す。
【図8】負の対照(レーン1および2)、3bp挿入(レーン3および4)、9bp挿入(レーン5)、および2bp欠失(レーン6)についてのMECP2エキソン1に対するPCR産物の高解像度アガロースゲル(2.2%)を示す。サイズラダー(M)100bpラダー(MBI Fermentas)は、PCRレーンに隣接している。
【発明を実施するための形態】
【0015】
発明の詳細な説明
本発明者らは、レット症候群および精神遅滞のような神経精神障害を有する患者において突然変異を含み得る、新たなコード配列に寄与するMECP2スプライス変異体を同定した。
【0016】
I. 本発明の核酸分子
本明細書において先に述べたように、本発明は、単離されたMECP2E1核酸分子に関する。用語「単離された」は、組換えDNA技術によって作製した場合、細胞材料もしくは培地が実質的にない核酸、または化学的に合成した場合、化学物質前駆体、もしくは他の化学物質を指す。
【0017】
用語「核酸」は、DNAおよびRNAを含むことが意図され、二本鎖または一本鎖のいずれであってもよい。この用語は、核酸分子ならびに核酸類似体および/もしくはヌクレオチド類似体の混合物か、または完全に核酸類似体および/もしくはヌクレオチド類似体からなる鎖を含むこともまた意図する。
【0018】
大まかに提示して、本発明は、MECP2遺伝子のMECP2E1転写産物をコードする配列を含む、単離された核酸分子を提供する。従って、本発明は図6(b)に示したMECP2E1をコードする配列、またはその断片、変異体、もしくは類似体を含む、単離された核酸分子を提供する。
【0019】
一つの態様において、精製および単離された核酸分子は、以下を含む:
(a)図6(b)(SEQ ID No. 4)に示したMECP2E1タンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【0020】
本発明の特異的な態様において、単離された核酸分子は図6(a)(SEQ ID No. 3)に示した配列、またはその断片もしくは変異体を有する。
【0021】
用語「MECP2E1」は、エキソン1、3、および4を含むがエキソン2を欠く、MECP2遺伝子のアイソフォームを意味する。この遺伝子は以前、MECP2Bと呼ばれたが、現在はMECP2E1と呼ばれ、翻訳開始部位がエキソン1にあることを示している。用語「MECP2E1」は、図6(a)(SEQ ID No. 3)に示した配列、ならびに神経精神障害および発達障害と関連した、その突然変異、変異体、および断片を含む。
【0022】
以下に述べるように、本発明が、本発明のMeCP2E1タンパク質の切り詰め、ならびに本発明のMeCP2E1タンパク質の類似体および相同体、およびそれらの切り詰めをコードする核酸分子を含むことが理解される。
【0023】
さらに、本発明が、本発明の核酸配列およびその断片と実質的配列相同性を有する核酸配列を含む核酸分子を含むことが理解される。用語「実質的配列相同性を有する配列」は、これらの配列から、軽度または瑣末な配列変異を有するこれらの核酸配列、すなわち、実質的に同一の様式で機能し、機能的に同等なタンパク質を産生する配列を意味する。変異は局所的な突然変異または構造的な改変に起因し得る。
【0024】
一般的に、実質的相同性を有する核酸配列は、本発明の核酸配列と、少なくとも70%、好ましくは80〜90%の同一性を有する核酸配列を含む。
【0025】
配列同一性は最も好ましくはBLASTバージョン2.1プログラム・アドバンスド・サーチのアルゴリズムによって評価される(BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト(Gap existence cost)11;残基あたりのギャップ・コスト(gap cost)1;ラムダ比0.85デフォルト))。例えば、ヌクレオチド配列(「配列A」と呼ばれる)が図6(a)のヌクレオチド配列の部分と90%の同一性を有するならば、配列Aが図6(a)のヌクレオチド配列の参照された部分の100ヌクレオチドごとに最大10個の点突然変異、例えば他のヌクレオチドとの置換を含み得ることを除き、配列Aは図6(a)のヌクレオチド配列の参照された部分と同一である。MECP2E1転写産物と機能的に同等なヌクレオチド配列は、以下に記載されるように、さまざまな形態で生じ得る。
【0026】
用語「類似体である核酸配列」は、(a)、(b)、または(c)の配列と比較して、修飾された核酸配列であって、修飾が本明細書に記載されている配列の有用性を変更しない核酸配列を意味する。修飾された配列または類似体は、(a)、(b)、または(c)に示した配列にわたって改善された性質を有していてもよい。類似体を調製する修飾の一つの例は、図6(a)に示した配列の天然に存在する塩基(すなわち、アデニン、グアニン、シトシン、またはチミジン)のうちの1つを、以下のものなどの修飾塩基と置換することである:キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル、2-プロピル、およびその他のアルキルアデニン、5-ハロウラシル、5-ハロシトシン、6-アザウラシル、6-アザシトシン、および6-アザチミン、プソイドウラシル、4-チオウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオールアルキルアデニン、8-ヒドロキシルアデニン、およびその他の8位置換されたアデニン、8-ハログアニン、8アミノグアニン、8-チオールグアニン、8-チオールアルキルグアニン、8-ヒドロキシルグアニン、およびその他の8位置換されたグアニン、その他のアザおよびデアザウラシル、チミジン、シトシン、アデニン、またはグアニン、5-トリフルオロメチルウラシル、ならびに5-トリフルオロシトシン。
【0027】
修飾のもう一つの例は、図6(a)に示した核酸分子において、リン酸骨格における修飾された亜リン酸もしくは酸素へテロ原子、短鎖アルキルもしくはシクロアルキルの糖間結合、または短鎖ヘテロ原子もしくは複素環の糖間結合を含む。例えば、核酸配列は、ホスホロチオアート、リン酸トリエステル、メチルホスホナート、およびホスホロジチオアートを含み得る。
【0028】
本発明の核酸分子の類似体のさらなる例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸骨格が、ペプチドにおいて見られるものと同様のポリアミド骨格で置換されたペプチド核酸(PNA)である(P.E. Nielsen et al., Science 1991, 254, 1497)。PNA類似体により、酵素による分解に耐性であること、ならびにインビボおよびインビトロでの生存が延長されることが示されている。また、PNA鎖とDNA鎖の間には電荷相反がないため、PNAは、相補的なDNA配列に対してより強く結合する。その他の核酸類似体は、重合体骨格、環状骨格、または非環式骨格を含むヌクレオチドを含み得る。例えば、ヌクレオチドは、モルホリノ骨格構造(米国特許第5,034,506号)を有していてもよい。また、類似体は、レポーター基、核酸配列の薬物動態学的または薬力学的な性質を改善するための基などの基を含んでいてもよい。
【0029】
本発明の他の局面は、ハイブリダイゼーション条件下で、本発明の核酸分子とハイブリダイズする核酸分子、および少なくとも15塩基を有するその断片を提供する。そのような核酸分子は、好ましくは、MECP2E1の全体もしくは部分、またはその相補体に、本明細書で定義されるようなストリンジェントな条件下でハイブリダイズする(Sambrook et al.(最新版)Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Ausubel et al.(eds.), 1995, Current Protocols in Molecular Biology,(John Wiley & Sons, NY))。ハイブリダイズする核酸分子の部分は典型的に、少なくとも15(例えば、20、25、30、または50)ヌクレオチド長である。ハイブリダイズする核酸のハイブリダイズする部分は、MeCP2E1ポリペプチドをコードする核酸の部分もしくは全体の配列、またはその相補体と、少なくとも80%、例えば少なくとも95%または少なくとも98%同一である。本明細書において記載される型のハイブリダイズする核酸は、例えば、クローニングプローブ、プライマー(例えば、PCRプライマー)、または診断用プローブとして用いられ得る。核酸試料とオリゴヌクレオチドプローブのハイブリダイゼーションは、典型的には、ストリンジェントな条件下で行われる。核酸二重鎖またはハイブリッドの安定性は、プローブが標的DNAから解離する温度である、融解温度またはTmとして表される。融解温度は必要なストリンジェンシー条件を定義するために用いられる。配列がプローブと関連しており、かつ同一ではないが実質的に同一であることを同定するならば、第一に、特定の塩(例えば、SSCまたはSSPE)濃度における、相同的ハイブリダイゼーションのみが起こる最低温度を確立することが有用である。次いで、1%のミスマッチがTmの1℃の低下を生じると想定し、ハイブリダイゼーション反応における最終洗浄の温度がそれに応じて低下される(例えば、プローブと95%より大きな同一性を有する配列が探索される場合は、最終洗浄温度は5℃低下される)。実際には、Tmの変化は1%ミスマッチにつき0.5℃〜1.5℃の間であり得る。低ストリンジェンシー条件は約:50℃で1×SSC、0.1%SDSでのハイブリダイゼーションを含む。高ストリンジェンシー条件は:65℃で0.1×SSC、0.1%SDSである。中程度のストリンジェンシーは約、60度で1×SSC、0.1%SDSである。塩濃度および温度のパラメーターは、プローブと標的核酸の間の同一性の最適レベルを達成するため変化され得る。
【0030】
遺伝暗号の縮重のために、図6(a)に示した核酸配列とは異なる配列を有する単離および精製された核酸分子もまた本発明の範囲内である。遺伝暗号が縮重しているため、MeCP2E1アミノ酸配列(図6(b))と同一のポリペプチドをコードする他の核酸分子もまた用いられ得る。
【0031】
本発明は、表1に列挙した特異的な突然変異を含む、神経精神障害または発達障害に関連した、突然変異型のMECP2E1もまた含む。具体的には、以下の突然変異はレット症候群に関連している:(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置70〜71における配列TGの欠失。
【0032】
以下の突然変異は発育遅延に関連している:(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数コピーのGCCトリヌクレオチド配列の挿入;(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、ヌクレオチド配列GGAの挿入;(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【0033】
発育遅延群における突然変異(4)および(5)について、これらはSEQ ID No. 1に示したヌクレオチド1の上流である。GenBankアクセッション番号BX538060は上流配列を有する。従って、より明確には、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失からなる突然変異(4)は、配列BX538060のヌクレオチド11〜12に相当し;およびヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失からなる突然変異(5)はBX538060のヌクレオチド30〜31に相当する。
【0034】
MECP2E1由来の核酸分子は、図6(a)に示した核酸配列の全体または一部に基づく標識された核酸プローブを調製すること、およびこの標識された核酸プローブを適切なDNAライブラリー(例えば、cDNAまたはゲノムDNAライブラリー)のスクリーニングに用いることにより単離され得る。cDNAまたはゲノムDNAライブラリーのスクリーニングにより単離された核酸は、標準的な技術によりシーケンシングされ得る。他の方法は、MECP2E1配列を他の配列と比較する段階、例えば、データベースサーチまたはアライメントストラテジーのようなバイオインフォマティックスの技術を用い、MECP2E1核酸配列の存在を検出する段階を含む。
【0035】
また、本発明の核酸分子は、ポリメラーゼ連鎖反応(PCR)法およびcDNAまたはゲノムDNAを使用して、核酸を選択的に増幅することによって単離することができる。PCRに使用するために、図6(a)に示すとおりの核酸分子から合成オリゴヌクレオチドプライマーをデザインすることができる。核酸は、これらのオリゴヌクレオチドプライマーおよび標準的なPCR増幅技術を使用して、cDNAまたはゲノムDNAから増幅することができる。こうして増幅された核酸は、適切なベクターにクローン化することができ、DNA配列解析によって特徴付けることができる。cDNAは、種々の技術によって、例えば、Chirgwin et al., Biochemistry, 18, 5294-5299 (1979)のチオシアン酸グアニジニウム(guanidinium-thiocyanate)抽出法を使用することによって、全細胞mRNAを単離することにより、mRNAから調製してもよいことが認識される。次いで、逆転写酵素(例えば、Gibco/BRL, Bethesda, MDから入手可能なモロニーMLV逆転写酵素、またはSeikagaku America, Inc., St. Petersburg, FLから入手可能なAMV逆転写酵素)を使用して、mRNAからcDNAが合成される。
【0036】
RNAである本発明の単離された核酸分子は、適切なベクターに本発明の新規のタンパク質をコードするcDNAをクローン化することによって単離することができ、これにより、cDNAを転写してMeCP2E1タンパク質をコードするRNA分子を産生することができる。例えば、cDNAは、ベクターのバクテリオファージプロモーター(例えば、T7プロモーター)の下流にクローン化することができ、cDNAをインビトロにおいてT7ポリメラーゼで転写させ、生じるRNAを標準的な技術によって単離することができる。
【0037】
また、本発明の核酸分子は、標準的な技術を使用して化学的に合成され得る。ポリデオキシヌクレオチドを化学的に合成する種々の方法が公知であり、ペプチド合成のように、市販されているDNA合成装置での完全に自動化された固相合成を含む(例えば、Itakuraら、米国特許第4,598,049号;Caruthersら、米国特許第4,458,066号;ならびにItakura米国特許第4,401,796号および同第4,373,071号を参照されたい)。
【0038】
本発明の核酸分子の開始コドンおよび非翻訳配列は、PC/Gene(IntelliGenetics Inc., Calif.)などの本目的のためにデザインされた現在利用できるコンピュータ・ソフトウェアを使用して決定してもよい。調節エレメントは、従来の技術を使用して同定することができる。エレメントの機能は、エレメントに機能的に結合されたリポーター遺伝子を発現するようにこれらのエレメントを使用して確認することができる。これらの構築物は、標準的な手順を使用して培養細胞に導入してもよい。DNA中の調節エレメントを同定することに加えて、このような構築物はまた、当技術分野において公知の技術を使用して、エレメントと相互作用するタンパク質を同定するために使用してもよい。
【0039】
アンチセンス核酸分子を産生するため転写のための通常の提示に対して、本発明の核酸分子の配列を逆にしてもよい。好ましくは、アンチセンス配列は、開始コドンまたは保存されていない領域の前の領域を逆にすることによって構築される。アンチセンス核酸分子を産生するため転写のための通常の提示に対して、特に、本発明の核酸分子もしくはこれらの断片、好ましくは図6(a)に示した核酸配列を逆にしてもよい。
【0040】
本発明のアンチセンス核酸分子もしくはこれらの断片は、天然に存在するヌクレオチドを使用して化学的に合成してもよく、または分子の生物学的安定性を増大するように、もしくはmRNAまたは天然の遺伝子と形成される二重鎖の物理安定度を増大するようにデザインされた多様に修飾されたヌクレオチド、例えばホスホロチオアート誘導体およびアクリジン置換されたヌクレオチドでもよい。アンチセンス配列は、アンチセンス配列が、高効率制御領域(その活性は、ベクターが導入される細胞型によって決定してもよい)の制御下で産生される組換えプラスミド、ファージミド、または弱毒ウイルスの形態で細胞に導入された発現ベクターを使用して生物学的に産生してもよい。
【0041】
また、本発明は、本発明の新規タンパク質、および選択されたタンパク質、または選択可能なマーカータンパク質を含む融合タンパク質をコードする核酸を提供する(下記を参照されたい)。
【0042】
II. 本発明の新規のタンパク質
本発明は、本発明の核酸分子によってコードされる単離されたMeCP2E1タンパク質をさらに含む。本発明の文脈の範囲内で、本発明のタンパク質、生物活性を保持する、種々のタンパク質一次構造形態を含んでいてもよい。
【0043】
大まかに提示して、本発明はMECP2遺伝子のエキソン1、3、および4によりコードされる、単離されたタンパク質を提供する。
【0044】
本発明の好ましい態様において、MeCP2E1タンパク質は図6(b)(SEQ ID No. 4)に示したアミノ酸配列、またはその断片もしくは変異体を有する。
【0045】
本発明は、神経精神障害または発達障害と関連した突然変異型のMeCP2E1タンパク質もまた含む。具体的には、本発明は表1に記載されたMECP2E1の突然変異を含む。
【0046】
全長アミノ酸配列に加え、本発明のタンパク質は、本明細書において記載されるように、タンパク質の切り詰め、ならびにタンパク質の類似体および相同体、ならびにそれらの切り詰めもまた含む。切り詰められたタンパク質は、少なくとも15アミノ酸残基のペプチドを含み得る。
【0047】
本明細書において記載されるように、図6(b)に示したアミノ酸配列を有するタンパク質の類似体もしくは変異体、および/またはそれらの切り詰めは、1つもしくは複数のアミノ酸置換、挿入、および/または欠失を含むアミノ酸配列を含み得るが、これらに限定されない。アミノ酸置換は、保存的または非保存的な性質であってもよい。保存的アミノ酸置換は、同様の電荷、大きさ、および/または疎水性の特徴のアミノ酸で、本発明のタンパク質の1つもしくは複数のアミノ酸を置換することを含む。保存的置換だけが作製されるとき、生じる類似体は、機能的に同等物であるはずである。非保存的置換は、異なる電荷、大きさ、および/または疎水性の特徴を有する1つもしくは複数のアミノ酸でアミノ酸配列の1つもしくは複数のアミノ酸を置換することを含む。
【0048】
1つまたは複数のアミノ酸挿入が、図6(b)に示したアミノ酸配列に導入され得る。アミノ酸挿入は、単一のアミノ酸残基または長さが2〜15アミノ酸の範囲の一連のアミノ酸からなってもよい。例えば、アミノ酸の挿入は、タンパク質がもはや活性ではないように、標的配列を破壊するために使用され得る。この手順は、インビボで本発明のタンパク質の活性を阻害するために使用され得る。
【0049】
欠失は、図6(b)に示したアミノ酸配列からの1つもしくは複数のアミノ酸または離れた部分の除去からなっていてもよい。欠失されるアミノ酸は、隣接していても、または隣接していなくてもよい。欠失突然変異で生じる類似体の下限の長さは、約10アミノ酸、好ましくは100アミノ酸である。
【0050】
本発明のタンパク質の類似体は、タンパク質をコードするヌクレオチド配列に突然変異を導入することによって調製してもよい。本発明のタンパク質の類似体の発現のために構築されるヌクレオチド配列の突然変異は、コード配列のリーディングフレームを保存しなければならない。さらに、突然変異は、好ましくは、受容体mRNAの翻訳に悪影響を与え得る、ハイブリダイズして、ループまたはヘアピンなどの二次mRNA構造を生じさせ得る相補領域を生じない。
【0051】
突然変異は、制限部位に隣接して変異配列を含むオリゴヌクレオチドを合成して、天然の配列の断片に対するライゲーションを可能にすることによって、特定の遺伝子座に導入され得る。ライゲーションに続いて、結果的に生じる再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する類似体をコードする。
【0052】
または、必要とされる置換、欠失、または挿入に従って変更された特定のコドンを有する変更された遺伝子を提供するために、オリゴヌクレオチド部位特異的突然変異法を使用してもよい。本発明のタンパク質の欠失または切り詰めは、所望の欠失に隣接する制限エンドヌクレアーゼ部位を利用することによってもまた構築され得る。制限後に、オーバーハングを埋めて、DNAを再結合してもよい。上述した変更を作製する例示的な方法は、Sambrook et al(Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989)によって開示される。
【0053】
本発明のタンパク質は、本明細書において記載されるように、図6(b)に示したエキソン1領域を有するアミノ酸配列の相同体、および/またはそれらの切り詰めもまた含む。
【0054】
相同タンパク質は、図6(b)に示したアミノ酸配列と、少なくとも70%、好ましくは80〜90%の同一性を有するアミノ酸配列を有するタンパク質を含み、かつMeCP2E1タンパク質に特徴的なエキソン1領域を含む。本発明の核酸分子については、同一性は当技術分野において公知の方法に従って計算される。配列同一性は最も好ましくはBLASTバージョン2.1アドバンスド・サーチのアルゴリズムによって評価される。BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト11;残基あたりのギャップ・コスト1;ラムダ比0.85デフォルト)。
【0055】
本発明は、本発明のタンパク質のアイソフォームもまた意図する。アイソフォームは、本発明のタンパク質と同じアミノ酸の数および種を含むが、そのアイソフォームは異なる分子構造を有する。本発明によって意図されるアイソフォームは、本明細書に記載されているように、本発明のタンパク質と同じ性質を有するものである。
【0056】
本発明は、融合タンパク質を産生するために、選択されたタンパク質、または選択可能なマーカータンパク質(下記を参照されたい)と結合した本発明のタンパク質もまた含む。加えて、本発明のタンパク質の免疫原性部分も本発明の範囲内である。
【0057】
本発明のタンパク質(切り詰め、類似体などを含む)は、組換えDNA法を使用して調製してもよい。従って、本発明のタンパク質をコードする配列を有する本発明の核酸分子を、タンパク質の良好な発現を保証する適切な発現ベクターに公知の様式で組み込んでもよい。可能な発現ベクターは、ベクターが使用される宿主細胞と適合性を持つ限り、コスミド、プラスミド、または改変されたウイルス(例えば、複製欠損のレトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)を含むが、これらに限定されない。「宿主細胞のトランスフォーメーションのために適切な」発現ベクターは、発現ベクターが、本発明の核酸分子、および機能的に核酸分子に結合された、発現のために使用される宿主細胞を基礎として選択された調節配列を含むことを意味する。機能的に結合されるとは、核酸が、核酸を発現できる様式で調節配列に結合されことを意味することが意図される。
【0058】
従って、本発明は、本発明の核酸分子、またはその断片、ならびに挿入されたタンパク質配列の転写および翻訳に必要な調節配列を含む本発明の組換え発現ベクターを意図する。適切な調節配列は、細菌、真菌、またはウイルスの遺伝子を含む種々の供与源に由来してもよい(例えば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)に記載されている調節配列を参照されたい)。適切な調節配列の選択は、選択される宿主細胞に依存的であり、当業者によって容易に達成され得る。このような調節配列の例は、転写プロモーターおよびエンハンサーまたはRNAポリメラーゼ結合配列、翻訳開始シグナルを含むリボソーム結合配列を含む。その上、選択される宿主細胞および使用されるベクターに応じて、複製開始点、さらなるDNA制限部位、エンハンサー、および転写誘導能を与える配列などのその他の配列を発現ベクターに組み込んでもよい。必要な調節配列が、天然のタンパク質および/またはその隣接領域によって供給され得ることもまた認識される。
【0059】
本発明は、アンチセンス方向で発現ベクターにクローン化された本発明のDNA核酸分子を含む組換え発現ベクターをさらに提供する。すなわち、DNA分子は、DNA分子の転写によって、図6(a)に示したヌクレオチドを含むヌクレオチド配列に対してアンチセンスであるRNA分子を発現することができる様式で、調節配列に機能的に結合される。アンチセンスRNA分子を持続的に発現させる、アンチセンス核酸に機能的に結合された調節配列を選択することができる。
【0060】
本発明の組換え発現ベクターは、本発明の組換え分子を形質転換またはトランスフェクトした宿主細胞の選択を容易にする選択可能なマーカー遺伝子もまた含み得る。選択可能なマーカー遺伝子の例は、特定の薬物に対する耐性を与える、G418およびハイグロマイシンなどのタンパク質、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼをコードする遺伝子である。選択可能なマーカー遺伝子の転写は、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼなどの選択可能なマーカータンパク質の濃度の変化によってモニターされる。選択可能なマーカー遺伝子が、ネオマイシン耐性などの抗生物質耐性を与えるタンパク質をコードする場合、形質転換体細胞は、G418によって選択することができる。選択可能なマーカー遺伝子を組み込んだ細胞は、生存すると考えられるが、その他の細胞は死滅する。これにより、本発明の組換え発現ベクターの発現の視覚化およびアッセイを可能にし、特に発現および表現型に対する突然変異の効果を決定することが可能となる。選択可能なマーカーは、関心対象の核酸から分離したベクターに導入することができることが理解される。
【0061】
組換え発現ベクターは、組換えタンパク質の発現の増大;組換えタンパク質の溶解性の増大;およびアフィニティー精製においてリガンドとして作用することによって標的組換えタンパク質の精製の援助をもたらす融合部分をコードする遺伝子もまた含み得る。例えば、タンパク質分解切断部位を標的組換えタンパク質に付加して、融合タンパク質の精製後に融合部分から組換えタンパク質を分離させてもよい。
【0062】
組換え発現ベクターを宿主細胞に導入して、形質転換された宿主細胞を作製することができる。用語「形質転換された宿主細胞」は、本発明の組換え発現ベクターで形質転換またはトランスフェクトされた原核細胞および真核細胞を含むことが意図される。用語「で形質転換され」、「をトランスフェクトされ」「形質転換」、および「トランスフェクション」は、当技術分野において公知の多くの可能な技術のうちの1つによって、細胞に核酸(例えば、ベクター)を導入することを包含することが意図される。例えば、原核細胞は、電気穿孔法または塩化カルシウムを媒介したトランスフォーメーションによって核酸で形質転換することができる。核酸は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE-デキストランを媒介したトランスフェクション、リポフェクチン、電気穿孔法、または微量注入などの従来の技術を介して哺乳類細胞に導入することができる。宿主細胞を形質転換し、トランスフェクトするための適切な方法は、Sambrook et al.(Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press (1989))、およびその他の実験教科書において見出すことができる。
【0063】
適切な宿主細胞は、多種多様な原核生物および真核生物の宿主細胞を含む。例えば、本発明のタンパク質は、大腸菌などの細菌細胞、昆虫細胞(バキュロウイルスを使用する)、酵母細胞、または哺乳類細胞に発現され得る。その他の適切な宿主細胞は、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1991)において見出すことができる。
【0064】
本発明のタンパク質は、固相合成などのタンパク質の化学において周知の技術を使用する化学合成(Merrifield, 1964, J. Am. Chem. Assoc. 85:2149-2154)、または同種溶液における合成(Houbenweyl, 1987, Methods of Organic Chemistry, ed. E. Wansch, Vol.15 I and II, Thieme, Stuttgart)によってもまた調製され得る。
【0065】
III. 適用
A.診断適用
前述したように、本発明者らはMECP2遺伝子の新規スプライス変異体、MECP2E1を単離し、レット症候群もしくは精神遅滞のような神経精神障害または発達障害を有する人々においてエキソン1が欠失または突然変異されていることを示した。その結果、本発明は、MECP2E1核酸またはタンパク質における突然変異または欠失の検出による、神経精神障害または発達障害の検出方法もまた含む。
【0066】
本明細書において使用されるように、用語「神経精神障害」は自閉症/自閉症領域障害、てんかん、アンゲルマン症候群、プラダー-ウィリ症候群、脳症、精神分裂症、双極気分障害、鬱病、強迫性障害、パニック障害、注意欠陥多動性障害、および運動失調を含むが、これらに限定されない。
【0067】
用語「発達障害」は精神遅滞を含むが、これに限定されない。
【0068】
i)核酸配列中の突然変異の検出
一つの態様において、本発明は、動物、好ましくは哺乳動物、より好ましくはヒトから取得した試料中で、MECP2遺伝子のエキソン1における欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法を提供する。
【0069】
実施例および表1は、レット症候群または発育遅延を有する患者において、MECP2E1内に見出された突然変異のいくつかを要約する。(それらはセクションIにもまた記載される)。それぞれの突然変異に対するスクリーニングアッセイも開発され得る。突然変異を検出するために用いられ得る方法の例は、シーケンシング、ポリメラーゼ連鎖反応法、逆転写ポリメラーゼ連鎖反応法、変性HPLC、電気泳動移動度、核酸ハイブリダイゼーション、蛍光インサイチューハイブリダイゼーション、および多重ライゲーション依存性プローブ増幅を含む。用いられ得るスクリーニングアッセイの詳細は、実施例3、4、または5に提供される。
【0070】
レット症候群は、MECP2のエキソン1における欠失により生じることが示された。これらの欠失についてホモ接合の患者は、X1F/X1Rプライマーを用いるPCR増幅ならびにエキソン1および隣接配列のシーケンシングにより検出され得る。結果的に、本発明は、以下の段階を含む方法による、MECP2遺伝子のエキソン1における欠失を決定するための方法を含み:
(a)複製連鎖反応において、プライマーX1F(5'-CCATCACAGCCAATGACG- 3')(SEQ ID No. 19)およびX1R(5'- AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)を用いて試料中の核酸配列を増幅する段階;
(b)同一のプライマーを用いて、対照由来の核酸配列を増幅する段階;
(c)増幅された配列のシーケンシングを行う段階;ならびに
(d)対照配列と試料配列を比較する段階;
ここで、対照配列と比較した試料配列中のヌクレオチドの欠失は、試料がレット症候群を有する動物由来であることを示す。
【0071】
PCR反応により検出できないさらなるエキソン1突然変異は、4つ全てのエキソンにおける多重ライゲーション依存性プローブ増幅(MLPA)を用いて同定され得る。MLPA分析は、参照により本明細書に組み入れられる、参考文献5、およびSchouten、米国特許出願10/218,567(公開番号2003/0108913)に記載される。従って、本発明は4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対を用いるMLPA分析の実施による、MECP2遺伝子のエキソン1における欠失を決定するための方法を含む。
【0072】
当業者は、上記および実施例において論議したものに加え、他の方法をMECP2のエキソン1における突然変異を検出するために使用し得ることを理解する。例えば、試料から核酸を単離するために、本発明の核酸配列からヌクレオチドプローブを調製することができる。加えて、本明細書において記載される核酸プローブ(例えば、図1を参照されたい)もまた使用され得る。ヌクレオチドプローブは、32P、3H、14Cなどの適切なシグナルを提供し、十分な半減期を有する放射性の標識などの、検出可能なマーカーで標識され得る。使用され得る他の検出可能なマーカーは、特異的に標識された抗体によって認識される抗原、蛍光化合物、酵素、標識された抗原に特異的な抗体、および化学発光化合物を含む。適切な標識は、検出されるヌクレオチドに対するプローブのハイブリダイゼーションおよび結合の割合、ならびにハイブリダイゼーションに利用できるヌクレオチドの量を考慮して選択してもよい。
【0073】
従って、本発明は、試料を、核酸分子にハイブリダイズし、かつ検出可能なマーカーで標識された1つまたは複数のヌクレオチドプローブと、ハイブリダイゼーション条件下で接触させる段階、および試料中の核酸分子とヌクレオチドプローブとの間のハイブリダイゼーションの程度を決定する段階を含む、試料中のMECP2遺伝子のエキソン1を含む核酸分子の存在を検出する方法にもまた関する。
【0074】
本発明の方法に使用してもよいハイブリダイゼーション条件は、当技術分野において公知であり、例えばSambrook J, Fritch EF, Maniatis T. In: Molecular Cloning, A Laboratory Manual,1989. (Nolan C, Ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYに記載されている。ハイブリダイゼーション産物は、当技術分野において公知の技術を使用してアッセイしてもよい。ヌクレオチドプローブは、本明細書において記載したように、検出可能なマーカーで標識してもよく、ハイブリダイゼーション産物は、検出可能なマーカーによって生じる検出可能なマーカーまたは検出可能な変化を検出することによってアッセイしてもよい。
【0075】
DNAプローブと試料をハイブリダイズさせる前に、試料中の核酸配列を増幅するために、MECP2遺伝子に隣接するプライマーで試料を処理することができる。使用するプライマーは、本出願に記載されているものであってもよい。例えば、ヒトMECP2に特異的なプライマーは、HF(ctcggagagagggctgtg)(SEQ ID No. 5)、HR1(cttgaggggtttgtccttga)(SEQ ID No. 6)、HR2(cgtttgatcaccatgacctg)(SEQ ID No. 7)を含む。マウスMEC2Pに対するプライマーは、MF(aggaggcgaggaggagagac)(SEQ ID NO. 8)およびMR(ctggctctgcagaatggtg)(SEQ ID No. 9)を含む。加えて、また本明細書において提供されるMECP2遺伝子の配列により、本発明の核酸分子を増幅するためのプライマーとして使用され得る、新たなヌクレオチド配列の同定および単離、または合成が可能となる。プライマーは、その他の種のゲノムDNAを増幅するために使用してもよい。PCR増幅された配列を、様々な種の遺伝子間の関係を決定するために調べることができる。
【0076】
PCRに使用するためのプライマーの長さおよび塩基は、これらが所望の配列の異なる鎖に対してハイブリダイズするように、および配列に沿った相対的位置で、テンプレートから分離するとき1つのプライマーから合成された伸長産物が、定義された長さの核酸へのその他のプライマーの伸長のテンプレートとして役立つことができるように選択される。本発明に使用してもよいプライマーは、オリゴヌクレオチド、すなわち本発明の核酸分子の2つ以上のデオキシリボヌクレオチドを含む分子であり、これらは、精製された制限エンドヌクレアーゼ消化物として天然に存在するか、または例えば、リン酸トリエステルおよびリン酸ジエステル法(Good et al Nucl. Acid Res 4:2157, 1977を参照されたい)または自動技術(例えば、Conolly, B. A. Nucleic Acids Res. 15: 15 (7): 3131, 1987を参照されたい)などの当技術分野において公知の技術を使用して合成的に産生される。プライマーは、本発明のDNA配列に対して相補的であるプライマー伸長産物の合成を可能にする条件下に、すなわちヌクレオチド基質、DNAポリメラーゼなどの重合のための薬剤の存在下において、ならびに適切な温度およびpHに配置されたときに合成の開始位置として作用することができる。好ましくは、プライマーは、プライマーのその他のコピーと塩基対形成することによって二次構造を形成しない配列か、またはヘアピン立体配置を形成する配列である。プライマーは、好ましくは約7〜25個のヌクレオチドを含む。
【0077】
プライマーは、増幅産物の検出を可能にする検出可能なマーカーで標識してもよい。適切な検出可能なマーカーは、32P、35S、125I、および3Hなどの放射性のマーカー、化学発光マーカー、好ましくはルミノール、ならびに蛍光マーカー、好ましくは塩化ダンシル、フルオルセイン-5-イソチオシアナート、および4-フルオロ-7-ニトロベンズ-2-axa-1,3ジアゾールなどの発光のマーカー、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、アセチルコリンエステラーゼ、またはビオチンなどの酵素マーカーである。
【0078】
プライマーは、プライマーの十分な量が、増幅される本発明の核酸分子またはそれらのオリゴヌクレオチド断片に対して相補的な配列を含むことを条件として、非相補的な配列を含み得ることが理解される。制限部位リンカーをプライマーに組み入れて、適切な制限酵素での増幅された産物の消化を可能にして、増幅産物のクローニングおよびシーケンシングを容易にもまたし得る。
【0079】
本発明のある態様において、増幅配列を形成することができる条件下において、ポリメラーゼ連鎖反応法において核酸分子または予め定められたそのオリゴヌクレオチド断片を増幅することができるプライマーで試料を処理して増幅配列を形成する段階、および増幅された配列をアッセイする段階を含む、本発明の核酸分子の存在を決定する方法が提供される。
【0080】
ポリメラーゼ連鎖反応法は、一般的に参照により本明細書に組み入れられる、Innis et al, Academic Press, 1990、Mullis et al., 米国特許第4,863,195号およびMullis, 米国特許第4,683,202号に記載されている、標的核酸配列を増幅するためのプロセスをいう。核酸テンプレートを増幅するための条件は、同様に参照により本明細書に組み入れられるM.A. Innis and D.H. Gelfand, PCR Protocols, A Guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989に記載されている。
【0081】
増幅産物は、当技術分野において公知の技術を使用して、単離され、それらのそれぞれのサイズに基づいて区別され得る。例えば、増幅後に、DNA試料をアガロースゲル上で分離して、臭化エチジウムで染色後に紫外(UV)光下で視覚化することができる。DNAは、所望のレベルに増幅され得、さらなる伸長反応により、放射性標識されたか、またはビオチン標識されたヌクレオシド三リン酸などの検出可能なマーカーを有するヌクレオチド誘導体を組み入れてもよい。プライマーは、上述したように検出可能なマーカーを用いてもまた標識され得る。検出可能なマーカーは、制限および電気泳動分離、または当技術分野において公知のその他の技術によって解析され得る。
【0082】
PCRを使用する本発明の方法において使用され得る条件は、試料中のDNAおよび適切な相補的ハイブリダイゼーション・プライマーの存在下において、ハイブリダイゼーションおよび増幅反応を進行させることができるものである。ポリメラーゼ連鎖反応法のために適した条件は、一般に当技術分野において公知である。例えば、参照により本明細書に組み入れられる、M.A. Innis and D.H. Gelfand, PCR Protocols, A guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989を参照されたい。好ましくは、PCRは、好熱性細菌サーマス・アクアティクス(Thermus aquatics)(Taqポリメラーゼ、GeneAmpキット、Perkin Elmer Cetus)から得られたポリメラーゼを利用し、またはその他の耐熱性ポリメラーゼを使用して、DNA鋳型鎖が増幅され得る。
【0083】
リガーゼ連鎖反応法(LCR)およびNASBAなどのその他の技術を本発明の核酸分子を増幅するために用いてもよいことが理解される(Barney in "PCR Methods and Applications", August 1991, Vol.1(1), page 5、および1989年6月14日に公開された欧州特許出願第0320308号、ならびにMalekの米国特許出願第5,130,238号)。
【0084】
(ii)MECP2E1タンパク質の検出
他の態様において、本発明は、動物由来の試料中のMECP2E1タンパク質における欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法を提供する。
【0085】
本発明のMECP2E1タンパク質は、後述する種々の免疫アッセイ法を使用してMECP2E1に特異的である抗体を使用して、生体試料中で検出してもよい。
【0086】
抗体を調製するために、従来法を使用することができる。例えば、本発明のMECP2E1タンパク質由来のペプチドを使用することにより、ポリクローナル抗血清またはモノクローナル抗体を標準的な方法を使用して作製することができる。哺乳動物(例えば、マウス、ハムスター、またはウサギ)に、哺乳動物の抗体反応を引き出すペプチドの免疫原形態で免疫することができる。ペプチドに免疫原性を与えるための技術は、キャリアとの抱合または当技術分野において周知のその他の技術を含む。例えば、ペプチドをアジュバントの存在下において投与することができる。免疫の過程は、プラズマまたは血清中の抗体力価の検出によってモニターすることができる。抗体のレベルを評価するために、標準的なELISAまたはその他の免疫アッセイ手順では、抗原として免疫原を使用することができる。免疫化に続いて、抗血清を得ることができ、必要であれば、ポリクローナル抗体を血清から単離することができる。
【0087】
モノクローナル抗体を産生するためには、抗体産生細胞(リンパ球)を免疫した動物から収集し、標準的な体細胞融合手順によって骨髄腫細胞と融合させることによって、これらの細胞を不死化し、ハイブリドーマ細胞を産生することができる。このような技術は、当技術分野において周知である(例えば、当初Kohlerおよび Milstein (Nature 256, 495-497 (1975))によって開発されたハイブリドーマ技術、ならびにヒトB-細胞ハイブリドーマ技術(Kozbor et al., Immunol. Today 4, 72 (1983))、ヒトモノクローナル抗体を産生するためのEBVハイブリドーマ技術(Cole et al., Monoclonal Antibodies in Cancer Therapy (1985) Allen R. Bliss, Inc., pages 77-96)、およびコンビナトリアル抗体ライブラリーのスクリーニング(Huse et al., Science 246, 1275 (1989))などのその他の技術)。ペプチドと特異的に反応する抗体の産生について、ハイブリドーマ細胞を免疫化学的にスクリーニングすることができ、モノクローナル抗体を単離することができる。したがって、本発明はまた、本発明のタンパク質に対する特異性を有するモノクローナル抗体を分泌するハイブリドーマ細胞を想定する。
【0088】
本明細書に使用される用語「抗体」は、本発明のタンパク質、またはこれらのペプチドと特異的に反応するこれらの断片も含むことが意図される。上記のものと同じ方法で利用するために、従来技術を使用して抗体を断片化し、断片をスクリーニングすることができる。例えば、F(ab')2断片は、ペプシンで抗体を処理して作製することができる。生じるF(ab')2断片を処理してジスルフィド架橋を還元し、Fab'断片を産生することができる。
【0089】
キメラ抗体誘導体、すなわちヒト以外の動物の可変領域とヒトの定常領域を組み合わせた抗体分子も、本発明の範囲内において想定される。キメラ抗体分子は、例えば、ヒト定常領域を有する、マウス、ラット、またはその他の種の抗体に由来する抗原結合ドメインを含むことができる。CipAタンパク質を認識する免疫グロブリン可変領域を含むキメラ抗体を作製するために、従来法を使用してもよい(例えばMorrison et al., Proc. Natl. Acad. Sci. U.S.A. 81,6851(1985);Takeda et al., Nature 314, 452(1985)、Cabilly et al., 米国特許第4,816,567号;Boss et al.,米国特許第4,816,397号;Tanaguchi et al.,欧州特許第171496号;欧州特許第0173494号、英国特許第GB 2177096B号を参照されたい)。
【0090】
本明細書に記載されている本発明のタンパク質と特異的に反応するモノクローナルまたはキメラ抗体は、可変領域の一部、特に抗原結合ドメインの保存されたフレームワーク領域が、ヒト起源であり、かつ高頻度可変領域だけが非ヒト起源であるヒト定常領域キメラを産生することによって、さらにヒト化することができる。このような免疫グロブリン分子は、当技術分野において公知の技術によって作製され得る(例えばTeng et al., Proc. Natl. Acad. Sci. U.S.A., 80, 7308-7312 (1983); Kozbor et al., Immunology Today, 4, 7279 (1983); Olsson et al., Meth. Enzymol., 92, 3-16 (1982))、および国際公開公報第92/06193号または欧州特許第0239400号)。また、ヒト化抗体は、商業的に産生することができる(Scotgen Limited, 2 Holly Road, Twickenham, Middlesex, Great Britain)。
【0091】
本発明のタンパク質に対して反応性の、特異的な抗体、または抗体断片は、本発明の核酸分子から産生されるペプチドを用いて、細菌で発現した免疫グロブリン遺伝子、またはこれらの部分をコードする発現ライブラリーをスクリーニングすることによって作製され得る。例えば、完全Fab断片、VH領域、およびFV領域は、ファージ発現ライブラリーを使用して、細菌で発現させることができる(Ward et al., Nature 341, 544-546: (1989); Huse et al., Science 246, 1275-1281 (1989);および McCafferty et al., Nature 348, 552-554 (1990)を参照されたい)。
【0092】
また、抗体は、DNA免疫を使用して調製してもよい。例えば、本発明の(上記のとおりの)核酸を含む発現ベクターをマウスなどの適切な動物に注射してもよい。したがって、本発明のタンパク質がインビボで発現されて、抗体が誘導されると考えられる。抗体は、単離することができ、タンパク質免疫化のために上記のとおりに調製することができる。
【0093】
抗体は、種々の酵素、蛍光物質、発光材料、および放射性物質を含む検出可能なマーカーで標識してもよい。適切な酵素の例は、西洋ワサビペルオキシダーゼ、ビオチン、アルカリホスファターゼ、β-ガラクトシダーゼ、またはアセチルコリンエステラーゼを含み;適切な蛍光物質の例は、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド、またはフィコエリトリンを含み;発光材料の例は、ルミノールを含み;および適切な放射性物質の例は、S-35、Cu-64、Ga-67、Zr-89、Ru-97、Tc-99m、Rh-105、Pd-109、In-111、I-123、I-125、I-131、Re-186、Au-198、Au-199、Pb-203、At-211、Pb-212、およびBi-212を含む。また、抗体は、リガンド結合対の一方のパートナーに対して標識するか、または抱合してもよい。代表例は、アビジン-ビオチンおよびリボフラビン-リボフラビン結合タンパク質を含む。上記した代表的標識をもつ、上で論議した抗体を抱合、または標識するための方法は、従来技術を使用して容易に達成し得る。
【0094】
本発明のタンパク質(例えば、酵素抱合体または標識された誘導体)に対する抗体反応性は、種々の試料中における本発明のタンパク質を検出するために使用してもよく、例えば、これらは、本発明のタンパク質の抗原決定基と抗体との間の結合相互作用に依存する任意の公知の免疫アッセイ法に使用してもよい。このようなアッセイ法の例は、放射免疫アッセイ法、酵素免疫測定法(例えば、ELISA)、免疫蛍光、免疫沈降、ラテックス凝集反応法、赤血球凝集法、および組織化学的試験である。したがって、レット症候群の存在を診断するために、試料中の本発明のタンパク質の量を同定、または定量化するため抗体を使用してもよい。
【0095】
本発明の方法では、試料または濃縮された試料の予め定められた量が、抗体または標識抗体と混合される。本方法に使用される抗体の量は、選択される標識剤に依存的である。生じる、抗体または標識された抗体に結合したタンパク質は、従来の単離技術、例えばば塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。
【0096】
試料または抗体は、不溶化してもよく、例えば、試料または抗体は、公知の方法を使用して適切なキャリアと反応させることができる。適切なキャリアの例は、セファロースまたはアガロースビーズである。不溶化された試料または抗体が使用されるときは、抗体または反応していない抗体に結合したタンパク質は、洗浄することによって単離される。例えば、試料がニトロセルロース膜にブロットされるときは、本発明のタンパク質に結合した抗体は、緩衝液、例えばウシ血清アルブミン(BSA)を含むリン酸緩衝食塩水(PBS)で洗浄することにより、反応していない抗体から分離される。
【0097】
標識された抗体が使用されるときは、MECP2E1の存在は、試料中の本発明のタンパク質に結合した標識された抗体の量または反応していない標識された抗体の量を測定することによって決定することができる。標識された物質を測定する適切な方法は、標識剤に依存する。
【0098】
標識されていない抗体を本発明の方法に使用するときは、MECP2E1の存在は、凝集または沈澱を生じさせるために、抗体と特異的に相互作用する物質を使用してタンパク質に結合した抗体の量を測定することによって決定することができる。特に、本発明のタンパク質に特異的な抗体に対する標識された抗体を、反応混合物に添加することができる。本発明のタンパク質の存在は、標識剤のタイプに応じて、すでに記載されている技術の中から適切な方法によって決定することができる。本発明のタンパク質に特異的な抗体に対する抗体は、本明細書に記載した当技術分野において公知の従来法によって調製し、および標識することができる。本発明のタンパク質に特異的な抗体に対する抗体は、種特異的な抗免疫グロブリン抗体またはモノクローナル抗体であってもよく、例えば、ヤギ抗ウサギ抗体を本発明のタンパク質に特異的なウサギ抗体を検出するために使用してもよい。
【0099】
(iii)キット
本発明の方法を実施するために適した試薬は、適切な容器にパックされて、必要な材料を提供する便利なキットにパックされ得る。このようなキットは、本明細書に記載されている方法、および任意に本発明の方法を行う際に有用な適切な支援によって、試料中の本発明の核酸分子またはタンパク質を検出するために必要とされる全ての試薬を含み得る。
【0100】
本発明の一つの態様において、キットは、本発明の核酸分子またはこれらの予め定められたオリゴヌクレオチド断片を増幅することができるプライマー、ポリメラーゼ連鎖反応法において増幅された核酸分子またはこれらの予め定められた断片を産生するために必要とされる全ての試薬、および増幅された配列をアッセイするための手段を含む。また、キットは、PCR産物を消化するための制限酵素も含んでいてもよい。本発明のもう一つの態様において、キットは、本発明の核酸分子とハイブリダイズするヌクレオチド・プローブ、核酸分子とヌクレオチド・プローブのハイブリダイゼーションのために必要とされる試薬、およびその使用説明書を含む。本発明のさらなる態様において、キットは、本発明の抗体、および試料中において本発明のタンパク質に対して抗体を結合するために必要とされる試薬を含む。
【0101】
キットは、神経精神障害および発達障害を検出または処置するために、本発明の核酸分子、タンパク質、または抗体(上述)を、それらの使用のための説明書と共に含み得る。
【0102】
本発明の方法およびキットを用いて、レット症候群および精神遅滞のような神経精神障害ならびに発達障害を検出し得る。試験され得る試料は、血液、尿、血清、涙、唾液、糞便、組織、器官、細胞などの身体材料を含む。ヒト試料に加えて、試料は、非ヒト霊長類などのような哺乳動物から採取され得る。
【0103】
本明細書において記載される方法に従って試料を試験する前に、試料は、遠心分離および濾過のような当技術分野において公知の技術を使用して濃縮され得る。本明細書において記載されるハイブリダイゼーションおよび/またはPCRに基づいた方法のために、核酸は、当技術分野において公知の技術を使用して試験試料の細胞抽出物から抽出され得る。
【0104】
B.治療適用
上述したように、本発明の核酸分子は神経精神障害および発達障害を有する人々において欠失または突然変異されている。従って、本発明は、神経精神障害および発達障害を処置または予防するためにMECP2E1スプライス変異体の十分な部分を含む核酸配列を投与することにより、神経精神障害および発達障害を処置または予防する方法を提供する。本発明は、神経精神障害および発達障害を処置または検出するための、本発明の核酸分子およびタンパク質の使用を含む。
【0105】
核酸分子またはその断片を含む組換え分子は、レトロウイルスベクター、アデノウイルスベクター、およびDNAウイルスベクターのような送達媒体を用いて、インビボで細胞または組織に直接導入され得る。それらはまた、微量注入および電気穿孔法のような物理的技術、または共沈およびリポソームへのDNAの取り込みのような化学的方法を用いても細胞に導入され得る。組換え分子はまた、エアロゾルの形態で、または洗浄によっても送達され得る。
【0106】
核酸配列は、インビボでの投与に適した生物学的に適合性を有する形態で、対象への投与のための薬学的組成物に調剤され得る。「インビボでの投与に適した生物学的に適合性を有する形態」とは、治療的な効果が任意の有毒作用を上回って投与されるような物質の形態を意味する。物質は、ヒト、および動物を含む、生きている生物に投与してもよい。本発明の薬学的組成物の治療的に有効な量の投与は、所望の結果を達成するために必要な用量で、および期間で有効な量として定義される。例えば、物質の治療的に有効な量は、個々の疾病状態、年齢、性、および重量、ならびに個々に所望の反応を引き出すための抗体の能力などの因子によって変更してもよい。投与計画は、最適な治療反応を提供するように調整してもよい。例えば、いくつかの分割された用量を毎日投与してもよく、または用量は、治療状況の緊急性によって示されるように、比例的に減らされ得る。
【0107】
活性物質は、注射(皮下の、静脈内の、その他)、経口投与、吸入、経皮適用、または直腸投与のような、便利な様式で投与され得る。投与の経路に応じて、活性物質は、酵素、酸、および化合物を不活性化し得るその他の天然の条件の作用から化合物を保護する材料中にコーティングされ得る。
【0108】
本明細書において記載されている組成物は、薬学的に許容される組成物の調製のためのそれ自体公知の方法によって調製することができ、これを、有効な量の活性物質が薬学的に許容される媒体との混合物中に組み合わされるように被検者に投与することができる。適切な媒体は、例えばRemington’s Pharmaceutical Sciences (Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pa., USA 1985)に記載されている。これに基づいて、組成物は、限定的ではないものの、1つもしくは複数の薬学的に許容される媒体または希釈液と組み合わされ、適切なpHを有し、かつ生理液と等浸透圧の緩衝液に含まれる、物質の溶液を含む。
【0109】
C.実験モデル
本発明は、MECP2遺伝子およびMeCP2E1タンパク質の機能を研究するための方法、ならびに実験モデルを含む。MECP2E1スプライス変異体を欠いた、もしくはMeCP2E1発現を部分的に欠いた細胞、組織、および非ヒト動物は、MECP2E1遺伝子に特異的な欠失または変異を有する組換え発現ベクターを使用して開発してもよい。組換え発現ベクターを、相同組換えによってMECP2遺伝子を不活性化または変化させるために使用して、これにより、MECP2E1欠損細胞、組織、または動物を作製してもよい。特に、MECP2E2は変化させず、MECP2E1の欠損をまねくよう標的突然変異が設計され得る。これはMECP2遺伝子のエキソン1を標的とすることで達成され得る。
【0110】
ヌル対立遺伝子は、欠失突然変異によって胚幹性細胞などの細胞に作製され得る。また、組換えMECP2遺伝子を、MECP2E1を不活性化する挿入突然変異を含むようにもまた操作し得る。次いで、このような構築物を、トランスフェクション、電気穿孔法、注入等の技術によって、胚幹性細胞などの細胞に導入してもよい。次いで、例えばサザンブロッティング、ノーザンブロッティングによって、または本明細書において記載されている方法を使用してMECP2E1をアッセイすることによって、無傷のMECP2遺伝子を欠いている細胞を同定してもよい。次いで、そのような細胞を胚幹細胞に融合して、MECP2E1が欠損したトランスジェニック非ヒト動物を作製してもよい。突然変異の生殖系列伝達は、例えば、インビトロで、8細胞胚などの初期胚で胚幹細胞を凝集すること;生じる胚盤胞をレシピエント女性へ移すこと、および;生じる凝集キメラの生殖系列伝達を発生させることによって達成してもよい。そのような変異体動物を使用して、通常はMECP2E1発現に依存的な、特異的な細胞群、発達パターン、およびインビボ・プロセスを定義してもよい。本発明は、MECP2E1変異体の組織特異的ノックアウトの調製もまた含む。
【0111】
以下の非限定の実施例は、本発明の例示である:
【実施例】
【0112】
実施例1
MECP2E1スプライス変異体の同定
5'UTRの検査から、エキソン2はATGの上流に多数のインフレームの終結を有するが、エキソン1はATGを含むオープンリーディングフレームをその全長にわたって含むことが明らかとなった。エキソン1、3、および4からなる理論上の構築物を、その周囲のコザックヌクレオチド状況の重要性に基づきATGが開始コドンになる可能性を予測する、ATGprプログラム(http://www.hri.co.jp/atgpr/)で処理すると、MECP2E2の64%と比較して97%の信頼性スコアが返された。ESTデータベースの検索から、本発明者らの理論上の転写産物(MECP2E1と名付けた)を8例同定した(図1b)(対して、MECP2E2は14例)。MECP2E1は、エキソン1により決定された、より長いN末端を代わりに有する、新たな変異体MeCP2E1をコードすると予測される。
【0113】
実施例2
MECP2E1の発現
MECP2E1が実際に発現され、cDNAライブラリー調製物のアーチファクトではないことを確認するため、さまざまな組織由来のcDNAをエキソン1内の5'プライマーおよびエキソン3内の3'プライマーを用いてPCR増幅した(図1a)。サイズおよび配列によりMECP2E2およびMECP2E1に相当する2種類のPCR産物を、胎児および成体の脳を含む全ての組織、ならびに脳の小領域で得た(図1c)。マウスでの結果は類似していた(図1c)。成体ヒト脳における2種類の転写産物の発現レベルを定量化した。MECP2E1の発現はMECP2E2より10倍高かった(図1d)。3'mycタグ化したMECP2E1のCOS-7細胞へのトランスフェクション後の、MeCP2E1の細胞内局在は主に核内であることが見出された(図1e)。
【0114】
MECP2E1は以前の発現研究では検出されなかった。ノーザン分析は、代替のポリアデニル化シグナルの使用に起因して、サイズが異なる1.9、5、および10.1kbの3種類の転写産物を明らかにした(4、6、8)(図1a)。MECP2E1は、124ヌクレオチドのエキソン2を欠く点でMECP2E2と異なる。ゲル上の5および10.1kb位置では、2種類の転写産物は分離不可能である。1.9kbの範囲においては、公表されたノーザンブロットは2種類の転写産物に相当する可能性のある、太いかまたは二重のバンドを示す。同様に、従来型のウェスタンブロット分析も2種類のMeCP2アイソフォームの分解は不可能である(分子量の差<0.9kD;図1f)。
【0115】
実施例3
レット症候群におけるMECP2E1の突然変異
レット症候群において新たなコード領域が突然変異を受けているか否か決定するため、他のエキソンにおいて突然変異が見出されなかった、典型的RTTを有する19人の少女において、エキソン1および隣接配列をPCR増幅し、シーケンシングした。1人の患者(V1)は、エキソン1に11bpの欠失突然変異を有することが見出された(図2)。欠失は、MECP2E1の予測されたエキソン1オープンリーディングフレーム内で起こり、ミスセンスアミノ酸配列、続いてアミノ酸36の後に未成熟な終止コドンを生じるフレームシフトをもたらす。これはMECP2E2のコード配列には影響しない。この配列変化は患者の両親および兄弟を含む200人の対照個体においては見出されなかった。
【0116】
残りの患者において、本発明者らのPCR反応によっては検出できなかったさらなるエキソン1欠失を検索するため、多重ライゲーション依存性プローブ増幅(MLPA)(5)を4つ全てのエキソンについて実施し、1人の患者においてエキソン1のヘミ接合欠失を検出した(患者V2;図3)。最終的に、エキソン1に限定されるMLPA検出の欠失を有するさらに一人の患者について、最近抄録の形で文書化されたが、MECP2E1への影響は認識されていなかった(S. Boulanger et al. Am J Hum Genet 73, 572(2003))。
【0117】
対照的に、数百人の患者がエキソン2における突然変異について分析されたにも関わらず、MeCP2E2定義性のエキソン2に特異的な突然変異は今日まで見出されていない(31の刊行物;参考文献3に最も良く概説される)。これらの研究は、エキソン1を非コード性と見なしたためにエキソン1を含まない。
【0118】
エキソン1の欠失により、MeCP2E1タンパク質の非存在または切断がもたらされる。しかしながら、それらはまたMECP2E2の5'UTRの短縮をもたらし得、その発現に影響する可能性もあり得る。全血に対するRT-PCRにより、患者V1においてこの可能性について試験した。MECP2E2の発現の減少は存在しなかった(図2c)。結論として、突然変異のデータは、MeCP2E1の不活性化がRTTに十分であり、現在のところ、MeCP2E2については同じことを言明できない。
【0119】
材料および方法
PCR、手動シーケンシング、クローニング、rtPCR、ゲルブロット
PCR増幅を、1Mベタイン、50%デアザ(deaza)dGTPを含む200μM dNTPを含む[NH4]2SO4含有PCR緩衝液(MBI Fermentas)を用いて、3分間の95℃変性段階、続いて30秒間の95℃、30秒間の55℃、45秒間の72℃のサイクルを30サイクル繰り返し、次いで72℃7分間の浸漬段階により行った。手動シーケンシングは、1%アガロースゲルからの抽出後、Thermosequenase(商標)キット(USB/Amersham)を用いて、6%変性ポリアクリルアミドゲルに3時間泳動して行った。PCR産物をpDRIVEベクター(Qiagen PCR cloning kit)を用いてクローニングした。全血RNAを、PAXgene Blood RNAキット(Qiagen)を用いて抽出した。逆転写は、ランダム六量体および標準的なSuperscript IIIプロトコル(Invitrogen)を用いて行った。ヒト脳小領域のcDNAをOriGeneより得た。ポリアクリルアミドゲル(図2c)をHybond N+(Amersham)上にブロットし、デオキシヌクレオチド転移酵素(MBI Fermentas)を用いて3'末端が[α32P]-dCTPで標識されたHFプライマーとハイブリダイズさせた。
【0120】
神経およびグリア培養の調製
大脳皮質を性交後15.5日(15.5dpc)のCD-1マウス胚から調製した。Yamasaki et al.(Yamasaki et al. Hum Mol Genet 12: 837-847, 2003)の手順を用いた。簡単には、髄膜を除いた胎児大脳皮質を機械的粉砕により分離し、EDTAとともに0.25%トリプシンにより消化した。ウシ胎児血清(FBS;GIBCO BRL)の添加後、ろ過された細胞を遠心分離により回収した。細胞のペレットを、神経細胞の増殖のためにB-27(GIBCO BRL)か、またはグリア細胞の増殖のためにG-5(GIBCO BRL)を補ったNeurobasal(GIBCO BRL)培地に再懸濁した。細胞をポリエチレンイミンコーティングされたプラスチックディッシュに2×106細胞/mlの密度で平板培養した。神経細胞およびグリア細胞の培養物は、5% CO2、37℃で、それぞれ6日間および12日間維持された。単離された脳細胞をRT-PCR、ならびに神経細胞に対してMAP2(microtubule-associated protein 2)、グリア細胞に対してGFAP(glial fibrillary acidic protein)、および前駆細胞に対してネスチンというマーカーを用いる免疫蛍光法(IF)により特徴付けた。IFには、以下の特異的抗体を用いた:マウスモノクローナル抗MAP2(CHEMICON)、およびウサギポリクローナル抗GFAP(DAKO)。rtPCRに用いたプライマーはYamasaki et al.と同一である。半定量的PCRを得るため、内部対照としてGapdh増幅に従って、至適cDNA濃度およびサイクル数を決定した。図4はrtPCR(A)およびIF(B)による脳細胞初代培養の特徴を示す。
【0121】
定量的rtPCR
異なる組織におけるMECP2転写産物の量を決定するため、本発明者らは、SYBRグリーン検出法(PE Applied Biosystems, ABI PRISM 7900 Sequence Detection System)を用いた転写産物特異的リアルタイム定量的PCRアッセイを開発した。以下のMECP2E2特異的順方向プライマー(25nM)(エキソン2内)を設計した:5'-ctcaccagttcctgctttgatgt-3'(SEQ ID No. 12)。MECP2E1特異的プライマー(25nM)はエキソン1と3の接続部に設置した:5'-aggagagactggaagaaaagtc-3'(SEQ ID No. 10)。両方のアッセイとも、161bp(MECP2E2)および65bp(MECP2E1)の断片を産生するエキソン3内の同一の逆方向プライマー(25nM)を用いた:5'-cttgaggggtttgtccttga-3'(SEQ ID No. 11)。マウスのmecp2転写産物(mecp2e2 167bpおよびmecp2e1 71bp)についての相当する転写産物特異的プライマー(25nM)は、5'-ctcaccagttcctgctttgatgt-3'(SEQ ID No. 12)(MECP2E2);5'-aggagagactggaggaaaagtc-3'(SEQ ID No. 13)(MECP2E1)、および共通の逆方向プライマー5'-cttaaacttcagtggcttgtctctg-3'(SEQ ID No. 14)であった。PCR条件は:50℃2分間、95℃10分間、および95℃15秒間、60℃85秒間を40サイクル、であった。PCR反応は別個のチューブで実施し;かつMECP2E2およびMECP2E1転写産物の絶対的定量は、ヒト成体脳、小脳、繊維芽細胞、およびリンパ芽球(Clontech, Palo Alto, USA)、ならびにマウス神経細胞およびグリア細胞培養物(上述)由来のcDNAから行った。結果を、製造者の使用説明書(PE Applied Biosystems, ABI PRISM 7900 Sequence Detection System)に従って標準曲線法を用いて分析した。標準曲線は、精製した転写産物特異的PCR産物の希釈溶液を用いて作製した。
【0122】
免疫蛍光光学顕微鏡検査
3'mycタグ化したMECP2E2およびMECP2E1構築物(pCDNA3.1A-MECP2E2-mycおよびpcDNA3.1A-MECP2E1-myc)を、BamH1(5')およびXbaI(3')制限部位を付した、それぞれの転写産物の全長cDNAのPCR増幅、ならびにその後のpcDNA3.1 version A(Invitrogen)へのmycとインフレームでのクローニングにより作製した。MECP2E2に対する順方向プライマーはエキソン2内の開始コドンを含み(5'-tatggatccATGgtagctgggat-3')(SEQ ID No. 15)、一方MECP2E1に対する順方向プライマーはエキソン1内の開始コドンを含む(5'-tatggatccggaaaATGgccg-3')(SEQ ID No. 16)(BamHI制限部位を下線で示した、開始コドンは大文字である)。逆方向プライマーは両方の増幅で同一であった(5'-gcgtctagagctaactctct-3')(SEQ ID No. 17)(XbaI制限部位を下線で示した)。PCRに用いたテンプレートは、MECP2E2については小腸cDNAであり、およびMECP2E1については骨格筋cDNAであった。pcDNA3.1A-MECP2E2-mycおよびpcDNA3.1A-MECP2E1-myc(2μg)をCOS-7細胞にlipofectamine(Invitrogen)を用いてトランスフェクトし、脂質DNA複合体をDMEM(GIBCO)中で5時間曝露した。トランスフェクトションから48時間後に、培養物をPBS中でリンスし、アセトン:メタノール(1:1)混合物中で15分間、-20℃で固定し、1時間ブロックして(PBS中10%BSA)、室温で45分間、抗myc(Santa Cruz Biotechnology、ブロッキング緩衝液中1:50)と共にインキュベートした。PBSを用いて洗浄した後、スライドをブロッキング溶液中の二次抗体(FITC標識されたヤギ抗マウス(Jackson Immunoresearch labo)、1:400、緑色フィルターを介して検出可能)と共にインキュベートし、Dako Anti-Fadeを用いて封入し、免疫蛍光光学顕微鏡検査によって分析した。
【0123】
MLPA分析
MLPAはSchouten et al.、前記により記載されたように、およびSchouten、前記により記載されたように行った。MRC-Holland、Amsterdam、Netherlands(http://www.mrc-holland.com)のMECP2試験キットを利用し、キットは4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対からなっていた。簡単には、100〜200ngのゲノムDNAを変性させ、プローブ混合物と一晩、60℃でハイブリダイズさせた。翌朝、熱安定性のLigase-65を用いて、15分間、54℃で対のプローブを結合させた。結合後、各結合産物の末端にハイブリダイズする共通のプライマー対によるPCRを行った。PCRプライマーの一方はFAM標識され、PCR条件は以下の通りであった:95℃30秒間、60℃30秒間、および72℃1分間。結果的に生じた単位複製配列を、ABI 3100キャピラリー電気泳動法装置およびABI Genescanソフトウェアで分析した。データ管理および正常対照との比較は全てExcelソフトウェアを用いて行った。
【0124】
考察
最近、カエル(アフリカツメガエル(Xenopus laevis))における研究から、神経発生の転写調節におけるMeCP2の役割についての重要な洞察が提供された。MeCP2は、Notch-Delta9による発生段階特異的媒介の後に、神経分化に関わる遺伝子の調節に関わるSMRT複合体の構成成分であることが示された。これらの実験においてサイレンシングの標的とされたカエルMecp2転写産物は、MECP2E1のオルソログであった(図1f)。実際、非哺乳類脊椎動物においては、MeCP2E1がMeCP2の唯一の形態であるようである(図1f)。
【0125】
新たなMeCP2 N末端は、ポリアラニンおよびポリグリシン区域を含む、特徴的な21アミノ酸ペプチドである(MAAAAAAAPSGGGGGGEEERL)(SEQ ID No. 18)(図1f)。類似のN末端は、複数のシグナル伝達経路の重要な共通構成成分である、ERK1(MAPK3)、細胞外シグナル調節性キナーゼ(extracellular signal-regulated kinase)にも見られる(図1f)。興味深いことに、神経細胞では、ERK110およびMeCP211の両方が核に加えてシナプス後区画内に存在することが示され、かつ前者は2つの区画間を転位し、シナプス活性と転写調節を結びつけていることが示された10。MeCP2E1も同様にシナプス活性、この場合、神経発生時のシナプス接合ガイダンスと転写調節を結びつけている可能性がある。連続したポリアラニンおよびポリグリシン区域が見出される他の唯一のタンパク質群は、ホメオボックス(HOX)ファミリーのいくつかのメンバー12に含まれる。MeCP2と同様に、これらは発生時の転写調節因子である。
【0126】
最後に、非不活性化型のMECP2突然変異が、精神遅滞および自閉症のようなRTTと重複した表現型と関連付けられている13。本研究において発見されたMeCP2変異体は、これらの障害への関与の候補である。
【0127】
実施例4
精神遅滞におけるMECP2E1の突然変異
本発明者らは、N=401の自閉症発端者、および非特異的精神遅滞を有するN=493の患者において、MECP2E1遺伝子をスクリーニングした。Autism Genetic Resource Exchange(AGRE;N=242;女性100、男性142)からの多発性の家系の発端者のみならず、Hospital for Sick Children in Toronto(N=146;男性114、女性32)を通じ、およびLondon, UK(N=13;男性10、女性3)から募られた自閉症発端者もまたスクリーニングした。各地の施設倫理審議会の認可を得、参加者から書面による同意を得た。Hospital for Sick ChildrenのDepartment of Pediatric Laboratory Medicineで、脆弱性X試験について参照された(しかし試験結果は陰性)、非特異的発育遅延/精神遅滞を有する、女性293人および男性200人の患者について、匿名化されたDNA試料もまた得られた。実施例3に前述したようなPCRプライマーおよび条件を用いて、ポリメラーゼ連鎖反応の後に変性高速液体クロマトグラフィー(DHPLC)を突然変異検出に用いた。配列変異体を有すると疑われる女性個体に由来するPCR産物をpDRIVEベクター(Qiagen)にクローニングし、少なくとも4クローンを、順方向および逆方向で、自動化BigDye(商標)シーケンシング(ABI 3100)を用いてシーケンシングした。男性由来のPCR産物をアガロースゲルから切り出し、カラム精製した後、同様に順方向および逆方向の両方で、自動化BigDye(商標)シーケンシング(ABI 3100)を用いてシーケンシングした。自閉症スクリーニングセットにおいては突然変異は同定されなかったが、配列変異体は8人の女性MR症例に同定され(図7を参照されたい)、うち3人はポリアデニン反復ストレッチ内にアミノ酸の挿入または欠失を生じ、およびうち2人はMECP2E1のN末端部分のポリグリシン反復内にグリシン残基の挿入が生じていた。同定された最初の個体は、推定MECP2E1開始コドンの45〜46bp上流に位置するGpCジヌクレオチドの欠失についてヘテロ接合であった。この欠失は、潜在的なSP1転写因子結合部位(http://www.gene-regulation.com/pub/programs/alibaba2/index.htmlのAliBaba2.1を用いて予測された)を破壊し得、および潜在的にメチル化可能なシトシン残基もまた除去し得る。他の個体は、MECP2E1開始コドンの26bp上流でApGジヌクレオチドの欠失についてヘテロ接合であった。2個体がポリ[GGA]ストレッチ内へのGGAトリヌクレオチド挿入についてヘテロ接合であり、それにより予測されたポリグリシンストレッチ内に追加のグリシン残基がもたらされる。5個体目は、ポリアラニンをコードするトリプレット反復ストレッチ内での、GCCトリヌクレオチド欠失についてヘテロ接合であった。2個体は、GCCトリヌクレオチド反復/ポリアラニン領域においてもまた、9bpの挿入についてヘテロ接合であり、ポリアラニンストレッチの7から10残基への伸長をもたらす。
【0128】
女性の非特異的MR症例の〜2%における、精神遅滞症候群に以前より関連付けられていたタンパク質の新たなアイソフォームについてのアミノ酸配列変異は非常に興味深い。さらに、多数の脊椎動物種間で保存されている、タンパク質の部分内で変異が生じているという事実もまた興味を増大させる(チンパンジー、オランウータン、マカク、ネコ、およびイヌのMECP2E1アミノ酸配列に対し、100%同一性)。変異体を有する個体間で任意の特異的な表現型の特徴が存在するか、症状の重症度はいかほどか、およびレット症候群の表現型との重複部分または差異が存在するかを知ることは特に有用である。しかしながら、DNAが匿名化されていたため、今回は、見出された突然変異を表現型の特徴または重症度と相関させることは不可能である。この問題に取り組む試みにおいて、Greenwood Genetic Center、South Carolinaに由来するMR症例(女性188、男性96)の第二の試料セットをスクリーニング後、シーケンシングした。男性においては変異体は見出されず、女性2人が余分なグリシン残基をコードするGGA挿入を有していた。
【0129】
本研究において、3人の女性MR患者が、MECP2E1のN末端のポリグリシンストレッチ内に余分なグリシン残基をもたらす3bpの挿入を有することを同定された。これまでにグリシン反復内の伸張に関して疾病関連性は報告されていない。植物におけるToc75タンパク質の研究から、このタンパク質内のポリグリシンストレッチがタンパク質の葉緑体外包膜への正確なターゲティングに必須であることが示唆されている(14)にも関わらず、MECP2E1タンパク質の文脈においてまたはより一般的にのいずれにおいても、ポリグリシンストレッチの機能は未知である。類似のタンパク質輸送機能が、ポリグリシンストレッチを有する哺乳類タンパク質、およびMECP2E1についての真相であることもまたあり得る。
【0130】
ポリアラニン区域内の変異体は特に関心対象である。それは、それらがまれにしか多型を示さず、かつそのような区域内の多くの小伸張(または重複)が、鎖骨頭蓋骨異形成症(RUNX2)、眼球咽頭型筋ジストロフィー(PABPN1)、および精神遅滞(ARX;この遺伝子もまたX染色体性で、非常に多岐にわたる表現型を有する-上記を参照されたい)に及ぶ疾患を引き起こすことが報告されているためである(15)。ポリアラニン疾患遺伝子の大部分は転写因子をコードするが、PABPN1遺伝子はポリアデニレート結合タンパク質をコードする。一方では、これらの疾患のうちで、転写因子遺伝子内の最も小さな病原性の反復は一般的に20アラニン長より大きく、従ってMECP2E1によりコードされるほど短いアラニンストレッチが病原性になり得ることはあり得ないと考えられ、1または3つのアラニン残基の変化は、まれな多型であろうと考えられ得る。現在のところ、ARX遺伝子内の1または3つのアラニン残基の小伸張が病原性であり得るか、または無害な変異体であり得るかについてはいくらかの不確定性が存在する(16、17)。他方では、眼球咽頭型筋ジストロフィーはPABPN1遺伝子のGCG区域内の突然変異により引き起こされる。この突然変異は、ポリアラニン区域をちょうど10個のアラニン残基から12〜17個のアラニン残基に伸張させる。さらに、MECP2E1のポリアラニン区域と同じく、PABPN1のポリアラニン区域はちょうど遺伝子のN末端にあり、従って、タンパク質のN末端部分内の反復ストレッチ内の比較的小さな突然変異が、タンパク質の中央部分に位置するより大きな突然変異より有害であり得る可能性がある。
【0131】
最近公表された研究は、エキソン2、3、または4に突然変異を有さない97人のレット患者、および146人の対照においてMECP2エキソン1における突然変異についてスクリーニングした(18)。レット患者のうち一人はポリアラニンをコードする[GCC]ストレッチ内に6bpの挿入を有することが見出されたが、そのような変異は対照には観察されなかった。この変異体は非罹患の母親から遺伝され、従ってこの変異は病因学的に関連していないようであると結論された(18)。しかしながら、最近、マウスでのMecp2の発現における微細な変化でさえ、深刻な神経学的および行動的結果を有し得ることが示された(19)。同一のMECP2突然変異を有する患者が非常に異なる表現型の特徴および重症度を有し得ることは明らかであり、かつX不活性化パターンにおける変異が、この不整合性において役割を果たしているようである。従って、ポリアラニンもしくはポリグリシンストレッチ長の変化をもたらす反復ストレッチ内か、または開始コドンの直上流の領域内のいずれかで、エキソン1内の変異が、機能または発現レベルに影響して神経病理学的な表現型をもたらし得ることは非常に可能性が高い。
【0132】
実施例5
レット症候群におけるMECP2E1のさらなる突然変異
エキソン1、2、3、および4のコード領域全体、ならびにそれらのイントロン隣接配列を分析した。エキソン2〜4をPCRにより、Human Genome Project working draft site(UCSC、www.genome.ucsc.edu)由来のゲノム配列情報およびLasergene Primer select programを用いて設計したプライマー対を用いて増幅した。dHPLC(WAVE Nucleic Acid Fragment Analysis System、Transgenomic, San Jose, CA)による塩基変化分析の前に増幅を確認するため、PCR産物を2%アガロースゲルにロードした。溶媒Aは0.1mol/Lトリエチルアンモニウムアセテート(TEAA)および25%アセトニトリルからなり、溶媒Bは1M TEAA、25%アセトニトリルを含んでいた。dHPLCにおいてクロマトグラフィー上の変異を示したPCR産物を自動シーケンサー(Gene Reader 4200)で直接シーケンシングした。シーケンシングデータをDNA Star software SeqMan(Lasergene)を用いて分析した。最近記載されたように(13)、全患者についてエキソン1をPCR増幅し、かつシーケンシングした。
【0133】
第一のエキソン1突然変異は、エキソン1イントロン1境界での、欠落した2塩基対からなる。この領域における配列の性質のため、本発明者らは欠落した2ヌクレオチドが、イントロン1の最初の2塩基対(GT)であるか、またはエキソン1の最後のヌクレオチド(T)およびイントロンの最初のヌクレオチド(G)であるか解明できなかった。どちらの場合であっても、欠落したヌクレオチドの対は、予測されたコンセンサススプライス部位を破壊し、イントロン1の読み過しをもたらす(データは図示せず)。エキソン1突然変異を有する第二の患者では、1A→T置換(ATG -> TTG)が最初のメチオニンコドンをロイシンに変化させる。予測では、開始コドンの非存在のために、MECP2E1の翻訳は著しくまたは完全に妨げられる。MECP2E2は正常に製造される(および、疾患の表現型をレスキューできないようである)。
【0134】
本発明は、好ましい実施例であると現在みなされるものに関して記載したが、本発明は、開示された実施例に限定されないことが理解される。対照的に、本発明は、添付の特許請求の範囲の精神および範囲内に含まれる種々の改変および同等の処理を包含することが意図される。
【0135】
全ての刊行物、特許、および特許出願は、あたかも個々の刊行物、特許、または特許出願が具体的に、および個々に、その全体が参照により組み入れられることを示したのと同程度まで、これらの全体が参照により本明細書に組み入れられる。
【0136】
(表1)今日までに同定されたMECP2E1突然変異または変異体。「del」は欠失を示し;「ins」は挿入を示す。

【0137】
本明細書において参照された参考文献の完全な引用
【技術分野】
【0001】
発明の分野
本発明は、新規のMECP2E1遺伝子、およびその遺伝子によりコードされるタンパク質、MeCP2E1に関する。遺伝子における突然変異は神経精神障害および発達障害と関連する。
【背景技術】
【0002】
発明の背景
世界保健機関によれば、神経精神障害は、世界中で最も影響が大きい10種の疾患のうち6種を占める。米国経済への負担は1000億ドル―医師を訪れる4人に1人は診断可能な精神障害を有する。
【0003】
レット症候群(RTT)(OMIM #312750)は、女児において、生後一年の後半における神経発達の漸進的な遅滞の発生から4歳までに停滞に向かい、その後、獲得した優れた運動技能およびコミュニケーション技能を退化および喪失することにより特徴付けられる。温存された歩行、異常なコミュニケーション、および常同性手絞り(hand wringing)という病像が初期の自閉症に近い、偽静止期が後に続く。しかしながら、退化は潜行性に進行し続け、最終的に著明な精神遅滞をもたらす(1)。
【0004】
RTTを有する患者の最大80%が、MeCP2転写リプレッサーをコードする4エキソンのMECP2遺伝子(図1a)(4)のエキソン3および4に突然変異を有する(2、3)。患者の残り20%における突然変異は分かりにくいままである。公知の遺伝子転写産物においては、4つのエキソン全てが用いられ、翻訳開始部位はエキソン2にあり、エキソン1およびエキソン2の大部分は、5'非翻訳領域(UTR)を形成する(4)。明確にするため、この転写産物をMECP2E2(以前はMECP2A)、およびそのコードするタンパク質をMeCP2E2(以前はMeCP2A)と名付ける。
【0005】
MeCP2E2定義のエキソン2に特異的な突然変異は、数百人の患者がこのエキソン中の突然変異について分析されたにも関わらず、今日まで見出されていない(31の刊行物;参考文献3に最も良く概説される)。これらの研究は、エキソン1は非コード性と見なされたため、エキソン1を含まない。
【0006】
非不活性化型のMECP2突然変異もまた精神遅滞および自閉症のような、RTTと重複する表現型に関連付けられている。RTTの診断および処置方法を同定できるように、残り20%のRTT患者の原因となるさらなる突然変異の同定が必要である。
【0007】
レット症候群遺伝子、MECP2における突然変異は、自閉症患者(20〜22)、ならびに幼時期発生精神病(23)、アンゲルマン症候群、非症候性精神遅滞、および新生児脳症(encepalopathy)(24)の患者にもまた見出され、MECP2の突然変異にさまざまな表現型の結果が存在し得ることを示している。
【発明の概要】
【0008】
発明の概要
本発明者らは、MECP2E1と呼ばれる、MECP2遺伝子の新規のオープンリーディングフレームを同定した。5'UTRの検査から、エキソン2はATGの上流に多数のインフレームの終結を有するが、エキソン1はATGを含むオープンリーディングフレームをその全長にわたって含むことが明らかとなった。このオープンリーディングフレームは、MECP2遺伝子のエキソン1、3、および4からなる転写産物をコードする。MECP2E1は、エキソン2のスプライシング切り出しに相当する、ヌクレオチド71〜193が非存在であることを除き、MECP2E2(GenBankアクセッション番号NM_004992、図5(a))と類似している。
【0009】
従って、本発明は、MECP2E1タンパク質をコードする配列を含む、単離された核酸分子を提供する。本発明は、相当するポリペプチドMeCP2E1もまた含む。
【0010】
一つの態様において、精製および単離された核酸分子は、以下を含む:
(a)図6(b)(SEQ ID No. 4)に示したタンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【0011】
本発明の特異的な態様において、単離された核酸分子は図6(a)(SEQ ID No. 3)に示した配列、またはその断片もしくは変異体を有するよう提供される。
【0012】
本発明者らは、レット症候群および精神遅滞のような神経精神障害または発達障害を有する患者が、MECP2E1遺伝子のエキソン1に突然変異を有することを見出した。従って、本発明は、MECP2E1配列(図6(a)(SEQ ID No. 3))のエキソン1における突然変異もしくは欠失を検出する段階を含む、神経精神障害または発達障害を検出する方法を提供する。突然変異は、プライマーX1F/X1R:突然変異スクリーニングプライマー(図1)を用いたゲノムDNA由来のPCR産物のシーケンシングを行うことにより検出され得る。挿入または欠失突然変異の検出には、PCR産物を適切なプラスミドベクターにクローニングし、その後大腸菌(E.Coli)へトランスフェクトし、単離されたコロニーからクローンのシーケンシングを行うことが必要であり得る。または、突然変異は、4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対を用いる多重ライゲーション依存性プローブ増幅(multiplex ligation-dependent probe amplification)(MLPA)により検出され得る。変異または欠失は、MECP2E1によりコードされるタンパク質産物についてアッセイすることによってもまた検出され得る。
【0013】
本発明の他の特徴および効果は以下の詳細な説明から明らかになる。しかしながら、詳細な説明および特異的な実施例は、本発明の好ましい態様を示すが、この詳細な説明から、本発明の精神および範囲内におけるさまざまな変化および改変が当業者には明らかであることから、例証としてのみ示される。
【図面の簡単な説明】
【0014】
ここで本発明を以下の図面との関連において記載する:
【図1】MECP2 5'スプライス変異体を示す。a)MECP2遺伝子の構造。番号付けされたボックスはエキソンを示し;アスタリスクはインフレームの終止コドンを示す。伝統的なMECP2E2スプライス変異体では、開始コドンはエキソン2にある。MECP2E1では、エキソン2は存在せず、開始コドンはエキソン1にある。HF/HR1およびMF/MR:パネルcに示すrtPCR実験に用いたヒトおよびマウスのプライマー対。HR2:別のヒト逆方向プライマーで、これによりHR1を用いて得た結果を確認する(データは図示せず)。X1F/X1R:突然変異スクリーニングプライマー(図2を参照されたい)。プライマー配列(5'-3'):HF-ctcggagagagggctgtg(SEQ ID NO. 5)、HR1-cttgaggggtttgtccttga(SEQ ID No. 6)、HR2-cgtttgatcaccatgacctg(SEQ ID No. 7)、MF-aggaggcgaggaggagagac(SEQ ID No. 8)、MR-ctggctctgcagaatggtg(SEQ ID No. 9)、X1F-ccatcacagccaatgacg(SEQ ID No. 19)、X1R-agggggagggtagagaggag(SEQ ID No. 20)。b)MECP2 ESTの例。c)示された成体組織(他に示された場合を除く)および細胞培養由来のcDNAに対する、(a)のプライマー(HF/HR1およびMF/MR)を用いたPCR結果;d.p.c.:性交後日数。d)示された組織または細胞培養由来のcDNAに対する転写産物特異的リアルタイム定量的PCR(SYBR Green検出法)。e)3'mycタグ化したMeCP2E1(およびMeCP2E2)は主に核内および細胞質の未確定の点内に局在する。f)示されたタンパク質のN末端;ダッシュはアミノ酸がないことを示す。
【図2】患者V1における欠失突然変異を示す。a1)X1F/X1Rプライマー(図1a)を用いたゲノムDNA由来のPCR産物の配列。混合された配列であることに留意されたい。a2)およびa3)患者の野生型および突然変異対立遺伝子のクローンそれぞれの配列;赤いボックスは、突然変異された対立遺伝子において欠失された11ヌクレオチドを示す。b)同一のクローニングされた野生型および欠失された対立遺伝子の電気泳動図。c)HF/HR1プライマーを用いた、示されたcDNAに対するPCR(図1a、c)。レーン1および2(2.5%高分解能アガロース上)は、それぞれ対照および患者の全血に由来する。レーン3〜8(6%変性ポリアクリルアミド上)は、対照血(3)、患者血(4)、対照胎児脳(5)、対照成体脳(6)、対照精巣(7)、および対照ゲノムDNA(8)に由来する。11bpのエキソン1欠失を有する患者のMECP2E2転写産物の発現(266bpのバンド)は、非欠失対立遺伝子(277bp)と比較して減少していないことに留意されたい。141および152bpのバンドはそれぞれ、欠失、および非欠失のMECP2E1転写産物である。
【図3】患者V2における欠失突然変異を示す。MECP2の多重ライゲーション依存性プローブ増幅(MLPA)ピークプロファイルを示す。対照座位を最上部に沿って示す。四角で囲われた領域(E1〜E4)はMECP2エキソン1〜4を示す。a)正常対照のMLPAプロファイル。b)患者V2のMLPAプロファイルはヘミ接合のエキソン1欠失(アスタリスク)を示す。結果は一貫して再現性があり、シーケンシングにより、SNPがMLPA反応のライゲーション効率を阻害する可能性が排除された。
【図4】rtPCRR(A)およびIF(B)による脳細胞初代培養の特徴を示す。(A)Map2、Gfap、およびネスチン発現から、B-27培地中の培養物は主に神経細胞からなり、G-5培地中のものはグリア細胞であることが示された。同一の胚由来の繊維芽細胞もまた培養され、負の対照として用いられた。全脳組織(15.5dpc)をMap2およびネスチンの正の対照として用いた。(B)マウス抗MAP2抗体およびウサギ抗GFAP抗体を用いて神経細胞の二重染色を行った。それらはDAPI(青色)でもまた対比染色された。大部分の細胞はMAP2(緑色)について陽性の染色を示す神経細胞であり、およびGFAP(赤色)について陽性の染色を示すグリア細胞によるわずかな割合の混入が検出された。
【図5A−1】5'および3'非翻訳領域を含むMECP2E2核酸配列(SEQ ID. No. 1)を示す。
【図5A−2】5A−1の続きを示す。
【図5A−3】5A−2の続きを示す。
【図5A−4】5A−3の続きを示す。
【図5B】MeCP2E2タンパク質配列(SEQ ID No. 2)を示す。
【図6A】MECP2E1核酸配列(SEQ ID No. 3)を示す。
【図6B】MeCP2E1タンパク質配列(SEQ ID No. 4)を示す。
【図7】女性MR患者において同定された5種のMECP2エキソン1変異体のヌクレオチド配列を示す。全ての配列は、ヘテロ接合PCR産物をpDRIVEベクター(Qiagen)へクローニングした後に、単一のコロニーから得た。可能な場合は、ATG開始コドンを赤色のボックスにより示す。結果的に生じたアミノ酸配列もまた、赤色で示される野生型配列と共に示し、および変化を緑色の活字で示す。
【図8】負の対照(レーン1および2)、3bp挿入(レーン3および4)、9bp挿入(レーン5)、および2bp欠失(レーン6)についてのMECP2エキソン1に対するPCR産物の高解像度アガロースゲル(2.2%)を示す。サイズラダー(M)100bpラダー(MBI Fermentas)は、PCRレーンに隣接している。
【発明を実施するための形態】
【0015】
発明の詳細な説明
本発明者らは、レット症候群および精神遅滞のような神経精神障害を有する患者において突然変異を含み得る、新たなコード配列に寄与するMECP2スプライス変異体を同定した。
【0016】
I. 本発明の核酸分子
本明細書において先に述べたように、本発明は、単離されたMECP2E1核酸分子に関する。用語「単離された」は、組換えDNA技術によって作製した場合、細胞材料もしくは培地が実質的にない核酸、または化学的に合成した場合、化学物質前駆体、もしくは他の化学物質を指す。
【0017】
用語「核酸」は、DNAおよびRNAを含むことが意図され、二本鎖または一本鎖のいずれであってもよい。この用語は、核酸分子ならびに核酸類似体および/もしくはヌクレオチド類似体の混合物か、または完全に核酸類似体および/もしくはヌクレオチド類似体からなる鎖を含むこともまた意図する。
【0018】
大まかに提示して、本発明は、MECP2遺伝子のMECP2E1転写産物をコードする配列を含む、単離された核酸分子を提供する。従って、本発明は図6(b)に示したMECP2E1をコードする配列、またはその断片、変異体、もしくは類似体を含む、単離された核酸分子を提供する。
【0019】
一つの態様において、精製および単離された核酸分子は、以下を含む:
(a)図6(b)(SEQ ID No. 4)に示したMECP2E1タンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【0020】
本発明の特異的な態様において、単離された核酸分子は図6(a)(SEQ ID No. 3)に示した配列、またはその断片もしくは変異体を有する。
【0021】
用語「MECP2E1」は、エキソン1、3、および4を含むがエキソン2を欠く、MECP2遺伝子のアイソフォームを意味する。この遺伝子は以前、MECP2Bと呼ばれたが、現在はMECP2E1と呼ばれ、翻訳開始部位がエキソン1にあることを示している。用語「MECP2E1」は、図6(a)(SEQ ID No. 3)に示した配列、ならびに神経精神障害および発達障害と関連した、その突然変異、変異体、および断片を含む。
【0022】
以下に述べるように、本発明が、本発明のMeCP2E1タンパク質の切り詰め、ならびに本発明のMeCP2E1タンパク質の類似体および相同体、およびそれらの切り詰めをコードする核酸分子を含むことが理解される。
【0023】
さらに、本発明が、本発明の核酸配列およびその断片と実質的配列相同性を有する核酸配列を含む核酸分子を含むことが理解される。用語「実質的配列相同性を有する配列」は、これらの配列から、軽度または瑣末な配列変異を有するこれらの核酸配列、すなわち、実質的に同一の様式で機能し、機能的に同等なタンパク質を産生する配列を意味する。変異は局所的な突然変異または構造的な改変に起因し得る。
【0024】
一般的に、実質的相同性を有する核酸配列は、本発明の核酸配列と、少なくとも70%、好ましくは80〜90%の同一性を有する核酸配列を含む。
【0025】
配列同一性は最も好ましくはBLASTバージョン2.1プログラム・アドバンスド・サーチのアルゴリズムによって評価される(BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト(Gap existence cost)11;残基あたりのギャップ・コスト(gap cost)1;ラムダ比0.85デフォルト))。例えば、ヌクレオチド配列(「配列A」と呼ばれる)が図6(a)のヌクレオチド配列の部分と90%の同一性を有するならば、配列Aが図6(a)のヌクレオチド配列の参照された部分の100ヌクレオチドごとに最大10個の点突然変異、例えば他のヌクレオチドとの置換を含み得ることを除き、配列Aは図6(a)のヌクレオチド配列の参照された部分と同一である。MECP2E1転写産物と機能的に同等なヌクレオチド配列は、以下に記載されるように、さまざまな形態で生じ得る。
【0026】
用語「類似体である核酸配列」は、(a)、(b)、または(c)の配列と比較して、修飾された核酸配列であって、修飾が本明細書に記載されている配列の有用性を変更しない核酸配列を意味する。修飾された配列または類似体は、(a)、(b)、または(c)に示した配列にわたって改善された性質を有していてもよい。類似体を調製する修飾の一つの例は、図6(a)に示した配列の天然に存在する塩基(すなわち、アデニン、グアニン、シトシン、またはチミジン)のうちの1つを、以下のものなどの修飾塩基と置換することである:キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル、2-プロピル、およびその他のアルキルアデニン、5-ハロウラシル、5-ハロシトシン、6-アザウラシル、6-アザシトシン、および6-アザチミン、プソイドウラシル、4-チオウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオールアルキルアデニン、8-ヒドロキシルアデニン、およびその他の8位置換されたアデニン、8-ハログアニン、8アミノグアニン、8-チオールグアニン、8-チオールアルキルグアニン、8-ヒドロキシルグアニン、およびその他の8位置換されたグアニン、その他のアザおよびデアザウラシル、チミジン、シトシン、アデニン、またはグアニン、5-トリフルオロメチルウラシル、ならびに5-トリフルオロシトシン。
【0027】
修飾のもう一つの例は、図6(a)に示した核酸分子において、リン酸骨格における修飾された亜リン酸もしくは酸素へテロ原子、短鎖アルキルもしくはシクロアルキルの糖間結合、または短鎖ヘテロ原子もしくは複素環の糖間結合を含む。例えば、核酸配列は、ホスホロチオアート、リン酸トリエステル、メチルホスホナート、およびホスホロジチオアートを含み得る。
【0028】
本発明の核酸分子の類似体のさらなる例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸骨格が、ペプチドにおいて見られるものと同様のポリアミド骨格で置換されたペプチド核酸(PNA)である(P.E. Nielsen et al., Science 1991, 254, 1497)。PNA類似体により、酵素による分解に耐性であること、ならびにインビボおよびインビトロでの生存が延長されることが示されている。また、PNA鎖とDNA鎖の間には電荷相反がないため、PNAは、相補的なDNA配列に対してより強く結合する。その他の核酸類似体は、重合体骨格、環状骨格、または非環式骨格を含むヌクレオチドを含み得る。例えば、ヌクレオチドは、モルホリノ骨格構造(米国特許第5,034,506号)を有していてもよい。また、類似体は、レポーター基、核酸配列の薬物動態学的または薬力学的な性質を改善するための基などの基を含んでいてもよい。
【0029】
本発明の他の局面は、ハイブリダイゼーション条件下で、本発明の核酸分子とハイブリダイズする核酸分子、および少なくとも15塩基を有するその断片を提供する。そのような核酸分子は、好ましくは、MECP2E1の全体もしくは部分、またはその相補体に、本明細書で定義されるようなストリンジェントな条件下でハイブリダイズする(Sambrook et al.(最新版)Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.; Ausubel et al.(eds.), 1995, Current Protocols in Molecular Biology,(John Wiley & Sons, NY))。ハイブリダイズする核酸分子の部分は典型的に、少なくとも15(例えば、20、25、30、または50)ヌクレオチド長である。ハイブリダイズする核酸のハイブリダイズする部分は、MeCP2E1ポリペプチドをコードする核酸の部分もしくは全体の配列、またはその相補体と、少なくとも80%、例えば少なくとも95%または少なくとも98%同一である。本明細書において記載される型のハイブリダイズする核酸は、例えば、クローニングプローブ、プライマー(例えば、PCRプライマー)、または診断用プローブとして用いられ得る。核酸試料とオリゴヌクレオチドプローブのハイブリダイゼーションは、典型的には、ストリンジェントな条件下で行われる。核酸二重鎖またはハイブリッドの安定性は、プローブが標的DNAから解離する温度である、融解温度またはTmとして表される。融解温度は必要なストリンジェンシー条件を定義するために用いられる。配列がプローブと関連しており、かつ同一ではないが実質的に同一であることを同定するならば、第一に、特定の塩(例えば、SSCまたはSSPE)濃度における、相同的ハイブリダイゼーションのみが起こる最低温度を確立することが有用である。次いで、1%のミスマッチがTmの1℃の低下を生じると想定し、ハイブリダイゼーション反応における最終洗浄の温度がそれに応じて低下される(例えば、プローブと95%より大きな同一性を有する配列が探索される場合は、最終洗浄温度は5℃低下される)。実際には、Tmの変化は1%ミスマッチにつき0.5℃〜1.5℃の間であり得る。低ストリンジェンシー条件は約:50℃で1×SSC、0.1%SDSでのハイブリダイゼーションを含む。高ストリンジェンシー条件は:65℃で0.1×SSC、0.1%SDSである。中程度のストリンジェンシーは約、60度で1×SSC、0.1%SDSである。塩濃度および温度のパラメーターは、プローブと標的核酸の間の同一性の最適レベルを達成するため変化され得る。
【0030】
遺伝暗号の縮重のために、図6(a)に示した核酸配列とは異なる配列を有する単離および精製された核酸分子もまた本発明の範囲内である。遺伝暗号が縮重しているため、MeCP2E1アミノ酸配列(図6(b))と同一のポリペプチドをコードする他の核酸分子もまた用いられ得る。
【0031】
本発明は、表1に列挙した特異的な突然変異を含む、神経精神障害または発達障害に関連した、突然変異型のMECP2E1もまた含む。具体的には、以下の突然変異はレット症候群に関連している:(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置70〜71における配列TGの欠失。
【0032】
以下の突然変異は発育遅延に関連している:(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数コピーのGCCトリヌクレオチド配列の挿入;(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、ヌクレオチド配列GGAの挿入;(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【0033】
発育遅延群における突然変異(4)および(5)について、これらはSEQ ID No. 1に示したヌクレオチド1の上流である。GenBankアクセッション番号BX538060は上流配列を有する。従って、より明確には、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失からなる突然変異(4)は、配列BX538060のヌクレオチド11〜12に相当し;およびヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失からなる突然変異(5)はBX538060のヌクレオチド30〜31に相当する。
【0034】
MECP2E1由来の核酸分子は、図6(a)に示した核酸配列の全体または一部に基づく標識された核酸プローブを調製すること、およびこの標識された核酸プローブを適切なDNAライブラリー(例えば、cDNAまたはゲノムDNAライブラリー)のスクリーニングに用いることにより単離され得る。cDNAまたはゲノムDNAライブラリーのスクリーニングにより単離された核酸は、標準的な技術によりシーケンシングされ得る。他の方法は、MECP2E1配列を他の配列と比較する段階、例えば、データベースサーチまたはアライメントストラテジーのようなバイオインフォマティックスの技術を用い、MECP2E1核酸配列の存在を検出する段階を含む。
【0035】
また、本発明の核酸分子は、ポリメラーゼ連鎖反応(PCR)法およびcDNAまたはゲノムDNAを使用して、核酸を選択的に増幅することによって単離することができる。PCRに使用するために、図6(a)に示すとおりの核酸分子から合成オリゴヌクレオチドプライマーをデザインすることができる。核酸は、これらのオリゴヌクレオチドプライマーおよび標準的なPCR増幅技術を使用して、cDNAまたはゲノムDNAから増幅することができる。こうして増幅された核酸は、適切なベクターにクローン化することができ、DNA配列解析によって特徴付けることができる。cDNAは、種々の技術によって、例えば、Chirgwin et al., Biochemistry, 18, 5294-5299 (1979)のチオシアン酸グアニジニウム(guanidinium-thiocyanate)抽出法を使用することによって、全細胞mRNAを単離することにより、mRNAから調製してもよいことが認識される。次いで、逆転写酵素(例えば、Gibco/BRL, Bethesda, MDから入手可能なモロニーMLV逆転写酵素、またはSeikagaku America, Inc., St. Petersburg, FLから入手可能なAMV逆転写酵素)を使用して、mRNAからcDNAが合成される。
【0036】
RNAである本発明の単離された核酸分子は、適切なベクターに本発明の新規のタンパク質をコードするcDNAをクローン化することによって単離することができ、これにより、cDNAを転写してMeCP2E1タンパク質をコードするRNA分子を産生することができる。例えば、cDNAは、ベクターのバクテリオファージプロモーター(例えば、T7プロモーター)の下流にクローン化することができ、cDNAをインビトロにおいてT7ポリメラーゼで転写させ、生じるRNAを標準的な技術によって単離することができる。
【0037】
また、本発明の核酸分子は、標準的な技術を使用して化学的に合成され得る。ポリデオキシヌクレオチドを化学的に合成する種々の方法が公知であり、ペプチド合成のように、市販されているDNA合成装置での完全に自動化された固相合成を含む(例えば、Itakuraら、米国特許第4,598,049号;Caruthersら、米国特許第4,458,066号;ならびにItakura米国特許第4,401,796号および同第4,373,071号を参照されたい)。
【0038】
本発明の核酸分子の開始コドンおよび非翻訳配列は、PC/Gene(IntelliGenetics Inc., Calif.)などの本目的のためにデザインされた現在利用できるコンピュータ・ソフトウェアを使用して決定してもよい。調節エレメントは、従来の技術を使用して同定することができる。エレメントの機能は、エレメントに機能的に結合されたリポーター遺伝子を発現するようにこれらのエレメントを使用して確認することができる。これらの構築物は、標準的な手順を使用して培養細胞に導入してもよい。DNA中の調節エレメントを同定することに加えて、このような構築物はまた、当技術分野において公知の技術を使用して、エレメントと相互作用するタンパク質を同定するために使用してもよい。
【0039】
アンチセンス核酸分子を産生するため転写のための通常の提示に対して、本発明の核酸分子の配列を逆にしてもよい。好ましくは、アンチセンス配列は、開始コドンまたは保存されていない領域の前の領域を逆にすることによって構築される。アンチセンス核酸分子を産生するため転写のための通常の提示に対して、特に、本発明の核酸分子もしくはこれらの断片、好ましくは図6(a)に示した核酸配列を逆にしてもよい。
【0040】
本発明のアンチセンス核酸分子もしくはこれらの断片は、天然に存在するヌクレオチドを使用して化学的に合成してもよく、または分子の生物学的安定性を増大するように、もしくはmRNAまたは天然の遺伝子と形成される二重鎖の物理安定度を増大するようにデザインされた多様に修飾されたヌクレオチド、例えばホスホロチオアート誘導体およびアクリジン置換されたヌクレオチドでもよい。アンチセンス配列は、アンチセンス配列が、高効率制御領域(その活性は、ベクターが導入される細胞型によって決定してもよい)の制御下で産生される組換えプラスミド、ファージミド、または弱毒ウイルスの形態で細胞に導入された発現ベクターを使用して生物学的に産生してもよい。
【0041】
また、本発明は、本発明の新規タンパク質、および選択されたタンパク質、または選択可能なマーカータンパク質を含む融合タンパク質をコードする核酸を提供する(下記を参照されたい)。
【0042】
II. 本発明の新規のタンパク質
本発明は、本発明の核酸分子によってコードされる単離されたMeCP2E1タンパク質をさらに含む。本発明の文脈の範囲内で、本発明のタンパク質、生物活性を保持する、種々のタンパク質一次構造形態を含んでいてもよい。
【0043】
大まかに提示して、本発明はMECP2遺伝子のエキソン1、3、および4によりコードされる、単離されたタンパク質を提供する。
【0044】
本発明の好ましい態様において、MeCP2E1タンパク質は図6(b)(SEQ ID No. 4)に示したアミノ酸配列、またはその断片もしくは変異体を有する。
【0045】
本発明は、神経精神障害または発達障害と関連した突然変異型のMeCP2E1タンパク質もまた含む。具体的には、本発明は表1に記載されたMECP2E1の突然変異を含む。
【0046】
全長アミノ酸配列に加え、本発明のタンパク質は、本明細書において記載されるように、タンパク質の切り詰め、ならびにタンパク質の類似体および相同体、ならびにそれらの切り詰めもまた含む。切り詰められたタンパク質は、少なくとも15アミノ酸残基のペプチドを含み得る。
【0047】
本明細書において記載されるように、図6(b)に示したアミノ酸配列を有するタンパク質の類似体もしくは変異体、および/またはそれらの切り詰めは、1つもしくは複数のアミノ酸置換、挿入、および/または欠失を含むアミノ酸配列を含み得るが、これらに限定されない。アミノ酸置換は、保存的または非保存的な性質であってもよい。保存的アミノ酸置換は、同様の電荷、大きさ、および/または疎水性の特徴のアミノ酸で、本発明のタンパク質の1つもしくは複数のアミノ酸を置換することを含む。保存的置換だけが作製されるとき、生じる類似体は、機能的に同等物であるはずである。非保存的置換は、異なる電荷、大きさ、および/または疎水性の特徴を有する1つもしくは複数のアミノ酸でアミノ酸配列の1つもしくは複数のアミノ酸を置換することを含む。
【0048】
1つまたは複数のアミノ酸挿入が、図6(b)に示したアミノ酸配列に導入され得る。アミノ酸挿入は、単一のアミノ酸残基または長さが2〜15アミノ酸の範囲の一連のアミノ酸からなってもよい。例えば、アミノ酸の挿入は、タンパク質がもはや活性ではないように、標的配列を破壊するために使用され得る。この手順は、インビボで本発明のタンパク質の活性を阻害するために使用され得る。
【0049】
欠失は、図6(b)に示したアミノ酸配列からの1つもしくは複数のアミノ酸または離れた部分の除去からなっていてもよい。欠失されるアミノ酸は、隣接していても、または隣接していなくてもよい。欠失突然変異で生じる類似体の下限の長さは、約10アミノ酸、好ましくは100アミノ酸である。
【0050】
本発明のタンパク質の類似体は、タンパク質をコードするヌクレオチド配列に突然変異を導入することによって調製してもよい。本発明のタンパク質の類似体の発現のために構築されるヌクレオチド配列の突然変異は、コード配列のリーディングフレームを保存しなければならない。さらに、突然変異は、好ましくは、受容体mRNAの翻訳に悪影響を与え得る、ハイブリダイズして、ループまたはヘアピンなどの二次mRNA構造を生じさせ得る相補領域を生じない。
【0051】
突然変異は、制限部位に隣接して変異配列を含むオリゴヌクレオチドを合成して、天然の配列の断片に対するライゲーションを可能にすることによって、特定の遺伝子座に導入され得る。ライゲーションに続いて、結果的に生じる再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する類似体をコードする。
【0052】
または、必要とされる置換、欠失、または挿入に従って変更された特定のコドンを有する変更された遺伝子を提供するために、オリゴヌクレオチド部位特異的突然変異法を使用してもよい。本発明のタンパク質の欠失または切り詰めは、所望の欠失に隣接する制限エンドヌクレアーゼ部位を利用することによってもまた構築され得る。制限後に、オーバーハングを埋めて、DNAを再結合してもよい。上述した変更を作製する例示的な方法は、Sambrook et al(Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989)によって開示される。
【0053】
本発明のタンパク質は、本明細書において記載されるように、図6(b)に示したエキソン1領域を有するアミノ酸配列の相同体、および/またはそれらの切り詰めもまた含む。
【0054】
相同タンパク質は、図6(b)に示したアミノ酸配列と、少なくとも70%、好ましくは80〜90%の同一性を有するアミノ酸配列を有するタンパク質を含み、かつMeCP2E1タンパク質に特徴的なエキソン1領域を含む。本発明の核酸分子については、同一性は当技術分野において公知の方法に従って計算される。配列同一性は最も好ましくはBLASTバージョン2.1アドバンスド・サーチのアルゴリズムによって評価される。BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト11;残基あたりのギャップ・コスト1;ラムダ比0.85デフォルト)。
【0055】
本発明は、本発明のタンパク質のアイソフォームもまた意図する。アイソフォームは、本発明のタンパク質と同じアミノ酸の数および種を含むが、そのアイソフォームは異なる分子構造を有する。本発明によって意図されるアイソフォームは、本明細書に記載されているように、本発明のタンパク質と同じ性質を有するものである。
【0056】
本発明は、融合タンパク質を産生するために、選択されたタンパク質、または選択可能なマーカータンパク質(下記を参照されたい)と結合した本発明のタンパク質もまた含む。加えて、本発明のタンパク質の免疫原性部分も本発明の範囲内である。
【0057】
本発明のタンパク質(切り詰め、類似体などを含む)は、組換えDNA法を使用して調製してもよい。従って、本発明のタンパク質をコードする配列を有する本発明の核酸分子を、タンパク質の良好な発現を保証する適切な発現ベクターに公知の様式で組み込んでもよい。可能な発現ベクターは、ベクターが使用される宿主細胞と適合性を持つ限り、コスミド、プラスミド、または改変されたウイルス(例えば、複製欠損のレトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)を含むが、これらに限定されない。「宿主細胞のトランスフォーメーションのために適切な」発現ベクターは、発現ベクターが、本発明の核酸分子、および機能的に核酸分子に結合された、発現のために使用される宿主細胞を基礎として選択された調節配列を含むことを意味する。機能的に結合されるとは、核酸が、核酸を発現できる様式で調節配列に結合されことを意味することが意図される。
【0058】
従って、本発明は、本発明の核酸分子、またはその断片、ならびに挿入されたタンパク質配列の転写および翻訳に必要な調節配列を含む本発明の組換え発現ベクターを意図する。適切な調節配列は、細菌、真菌、またはウイルスの遺伝子を含む種々の供与源に由来してもよい(例えば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)に記載されている調節配列を参照されたい)。適切な調節配列の選択は、選択される宿主細胞に依存的であり、当業者によって容易に達成され得る。このような調節配列の例は、転写プロモーターおよびエンハンサーまたはRNAポリメラーゼ結合配列、翻訳開始シグナルを含むリボソーム結合配列を含む。その上、選択される宿主細胞および使用されるベクターに応じて、複製開始点、さらなるDNA制限部位、エンハンサー、および転写誘導能を与える配列などのその他の配列を発現ベクターに組み込んでもよい。必要な調節配列が、天然のタンパク質および/またはその隣接領域によって供給され得ることもまた認識される。
【0059】
本発明は、アンチセンス方向で発現ベクターにクローン化された本発明のDNA核酸分子を含む組換え発現ベクターをさらに提供する。すなわち、DNA分子は、DNA分子の転写によって、図6(a)に示したヌクレオチドを含むヌクレオチド配列に対してアンチセンスであるRNA分子を発現することができる様式で、調節配列に機能的に結合される。アンチセンスRNA分子を持続的に発現させる、アンチセンス核酸に機能的に結合された調節配列を選択することができる。
【0060】
本発明の組換え発現ベクターは、本発明の組換え分子を形質転換またはトランスフェクトした宿主細胞の選択を容易にする選択可能なマーカー遺伝子もまた含み得る。選択可能なマーカー遺伝子の例は、特定の薬物に対する耐性を与える、G418およびハイグロマイシンなどのタンパク質、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼをコードする遺伝子である。選択可能なマーカー遺伝子の転写は、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼなどの選択可能なマーカータンパク質の濃度の変化によってモニターされる。選択可能なマーカー遺伝子が、ネオマイシン耐性などの抗生物質耐性を与えるタンパク質をコードする場合、形質転換体細胞は、G418によって選択することができる。選択可能なマーカー遺伝子を組み込んだ細胞は、生存すると考えられるが、その他の細胞は死滅する。これにより、本発明の組換え発現ベクターの発現の視覚化およびアッセイを可能にし、特に発現および表現型に対する突然変異の効果を決定することが可能となる。選択可能なマーカーは、関心対象の核酸から分離したベクターに導入することができることが理解される。
【0061】
組換え発現ベクターは、組換えタンパク質の発現の増大;組換えタンパク質の溶解性の増大;およびアフィニティー精製においてリガンドとして作用することによって標的組換えタンパク質の精製の援助をもたらす融合部分をコードする遺伝子もまた含み得る。例えば、タンパク質分解切断部位を標的組換えタンパク質に付加して、融合タンパク質の精製後に融合部分から組換えタンパク質を分離させてもよい。
【0062】
組換え発現ベクターを宿主細胞に導入して、形質転換された宿主細胞を作製することができる。用語「形質転換された宿主細胞」は、本発明の組換え発現ベクターで形質転換またはトランスフェクトされた原核細胞および真核細胞を含むことが意図される。用語「で形質転換され」、「をトランスフェクトされ」「形質転換」、および「トランスフェクション」は、当技術分野において公知の多くの可能な技術のうちの1つによって、細胞に核酸(例えば、ベクター)を導入することを包含することが意図される。例えば、原核細胞は、電気穿孔法または塩化カルシウムを媒介したトランスフォーメーションによって核酸で形質転換することができる。核酸は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE-デキストランを媒介したトランスフェクション、リポフェクチン、電気穿孔法、または微量注入などの従来の技術を介して哺乳類細胞に導入することができる。宿主細胞を形質転換し、トランスフェクトするための適切な方法は、Sambrook et al.(Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press (1989))、およびその他の実験教科書において見出すことができる。
【0063】
適切な宿主細胞は、多種多様な原核生物および真核生物の宿主細胞を含む。例えば、本発明のタンパク質は、大腸菌などの細菌細胞、昆虫細胞(バキュロウイルスを使用する)、酵母細胞、または哺乳類細胞に発現され得る。その他の適切な宿主細胞は、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1991)において見出すことができる。
【0064】
本発明のタンパク質は、固相合成などのタンパク質の化学において周知の技術を使用する化学合成(Merrifield, 1964, J. Am. Chem. Assoc. 85:2149-2154)、または同種溶液における合成(Houbenweyl, 1987, Methods of Organic Chemistry, ed. E. Wansch, Vol.15 I and II, Thieme, Stuttgart)によってもまた調製され得る。
【0065】
III. 適用
A.診断適用
前述したように、本発明者らはMECP2遺伝子の新規スプライス変異体、MECP2E1を単離し、レット症候群もしくは精神遅滞のような神経精神障害または発達障害を有する人々においてエキソン1が欠失または突然変異されていることを示した。その結果、本発明は、MECP2E1核酸またはタンパク質における突然変異または欠失の検出による、神経精神障害または発達障害の検出方法もまた含む。
【0066】
本明細書において使用されるように、用語「神経精神障害」は自閉症/自閉症領域障害、てんかん、アンゲルマン症候群、プラダー-ウィリ症候群、脳症、精神分裂症、双極気分障害、鬱病、強迫性障害、パニック障害、注意欠陥多動性障害、および運動失調を含むが、これらに限定されない。
【0067】
用語「発達障害」は精神遅滞を含むが、これに限定されない。
【0068】
i)核酸配列中の突然変異の検出
一つの態様において、本発明は、動物、好ましくは哺乳動物、より好ましくはヒトから取得した試料中で、MECP2遺伝子のエキソン1における欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法を提供する。
【0069】
実施例および表1は、レット症候群または発育遅延を有する患者において、MECP2E1内に見出された突然変異のいくつかを要約する。(それらはセクションIにもまた記載される)。それぞれの突然変異に対するスクリーニングアッセイも開発され得る。突然変異を検出するために用いられ得る方法の例は、シーケンシング、ポリメラーゼ連鎖反応法、逆転写ポリメラーゼ連鎖反応法、変性HPLC、電気泳動移動度、核酸ハイブリダイゼーション、蛍光インサイチューハイブリダイゼーション、および多重ライゲーション依存性プローブ増幅を含む。用いられ得るスクリーニングアッセイの詳細は、実施例3、4、または5に提供される。
【0070】
レット症候群は、MECP2のエキソン1における欠失により生じることが示された。これらの欠失についてホモ接合の患者は、X1F/X1Rプライマーを用いるPCR増幅ならびにエキソン1および隣接配列のシーケンシングにより検出され得る。結果的に、本発明は、以下の段階を含む方法による、MECP2遺伝子のエキソン1における欠失を決定するための方法を含み:
(a)複製連鎖反応において、プライマーX1F(5'-CCATCACAGCCAATGACG- 3')(SEQ ID No. 19)およびX1R(5'- AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)を用いて試料中の核酸配列を増幅する段階;
(b)同一のプライマーを用いて、対照由来の核酸配列を増幅する段階;
(c)増幅された配列のシーケンシングを行う段階;ならびに
(d)対照配列と試料配列を比較する段階;
ここで、対照配列と比較した試料配列中のヌクレオチドの欠失は、試料がレット症候群を有する動物由来であることを示す。
【0071】
PCR反応により検出できないさらなるエキソン1突然変異は、4つ全てのエキソンにおける多重ライゲーション依存性プローブ増幅(MLPA)を用いて同定され得る。MLPA分析は、参照により本明細書に組み入れられる、参考文献5、およびSchouten、米国特許出願10/218,567(公開番号2003/0108913)に記載される。従って、本発明は4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対を用いるMLPA分析の実施による、MECP2遺伝子のエキソン1における欠失を決定するための方法を含む。
【0072】
当業者は、上記および実施例において論議したものに加え、他の方法をMECP2のエキソン1における突然変異を検出するために使用し得ることを理解する。例えば、試料から核酸を単離するために、本発明の核酸配列からヌクレオチドプローブを調製することができる。加えて、本明細書において記載される核酸プローブ(例えば、図1を参照されたい)もまた使用され得る。ヌクレオチドプローブは、32P、3H、14Cなどの適切なシグナルを提供し、十分な半減期を有する放射性の標識などの、検出可能なマーカーで標識され得る。使用され得る他の検出可能なマーカーは、特異的に標識された抗体によって認識される抗原、蛍光化合物、酵素、標識された抗原に特異的な抗体、および化学発光化合物を含む。適切な標識は、検出されるヌクレオチドに対するプローブのハイブリダイゼーションおよび結合の割合、ならびにハイブリダイゼーションに利用できるヌクレオチドの量を考慮して選択してもよい。
【0073】
従って、本発明は、試料を、核酸分子にハイブリダイズし、かつ検出可能なマーカーで標識された1つまたは複数のヌクレオチドプローブと、ハイブリダイゼーション条件下で接触させる段階、および試料中の核酸分子とヌクレオチドプローブとの間のハイブリダイゼーションの程度を決定する段階を含む、試料中のMECP2遺伝子のエキソン1を含む核酸分子の存在を検出する方法にもまた関する。
【0074】
本発明の方法に使用してもよいハイブリダイゼーション条件は、当技術分野において公知であり、例えばSambrook J, Fritch EF, Maniatis T. In: Molecular Cloning, A Laboratory Manual,1989. (Nolan C, Ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYに記載されている。ハイブリダイゼーション産物は、当技術分野において公知の技術を使用してアッセイしてもよい。ヌクレオチドプローブは、本明細書において記載したように、検出可能なマーカーで標識してもよく、ハイブリダイゼーション産物は、検出可能なマーカーによって生じる検出可能なマーカーまたは検出可能な変化を検出することによってアッセイしてもよい。
【0075】
DNAプローブと試料をハイブリダイズさせる前に、試料中の核酸配列を増幅するために、MECP2遺伝子に隣接するプライマーで試料を処理することができる。使用するプライマーは、本出願に記載されているものであってもよい。例えば、ヒトMECP2に特異的なプライマーは、HF(ctcggagagagggctgtg)(SEQ ID No. 5)、HR1(cttgaggggtttgtccttga)(SEQ ID No. 6)、HR2(cgtttgatcaccatgacctg)(SEQ ID No. 7)を含む。マウスMEC2Pに対するプライマーは、MF(aggaggcgaggaggagagac)(SEQ ID NO. 8)およびMR(ctggctctgcagaatggtg)(SEQ ID No. 9)を含む。加えて、また本明細書において提供されるMECP2遺伝子の配列により、本発明の核酸分子を増幅するためのプライマーとして使用され得る、新たなヌクレオチド配列の同定および単離、または合成が可能となる。プライマーは、その他の種のゲノムDNAを増幅するために使用してもよい。PCR増幅された配列を、様々な種の遺伝子間の関係を決定するために調べることができる。
【0076】
PCRに使用するためのプライマーの長さおよび塩基は、これらが所望の配列の異なる鎖に対してハイブリダイズするように、および配列に沿った相対的位置で、テンプレートから分離するとき1つのプライマーから合成された伸長産物が、定義された長さの核酸へのその他のプライマーの伸長のテンプレートとして役立つことができるように選択される。本発明に使用してもよいプライマーは、オリゴヌクレオチド、すなわち本発明の核酸分子の2つ以上のデオキシリボヌクレオチドを含む分子であり、これらは、精製された制限エンドヌクレアーゼ消化物として天然に存在するか、または例えば、リン酸トリエステルおよびリン酸ジエステル法(Good et al Nucl. Acid Res 4:2157, 1977を参照されたい)または自動技術(例えば、Conolly, B. A. Nucleic Acids Res. 15: 15 (7): 3131, 1987を参照されたい)などの当技術分野において公知の技術を使用して合成的に産生される。プライマーは、本発明のDNA配列に対して相補的であるプライマー伸長産物の合成を可能にする条件下に、すなわちヌクレオチド基質、DNAポリメラーゼなどの重合のための薬剤の存在下において、ならびに適切な温度およびpHに配置されたときに合成の開始位置として作用することができる。好ましくは、プライマーは、プライマーのその他のコピーと塩基対形成することによって二次構造を形成しない配列か、またはヘアピン立体配置を形成する配列である。プライマーは、好ましくは約7〜25個のヌクレオチドを含む。
【0077】
プライマーは、増幅産物の検出を可能にする検出可能なマーカーで標識してもよい。適切な検出可能なマーカーは、32P、35S、125I、および3Hなどの放射性のマーカー、化学発光マーカー、好ましくはルミノール、ならびに蛍光マーカー、好ましくは塩化ダンシル、フルオルセイン-5-イソチオシアナート、および4-フルオロ-7-ニトロベンズ-2-axa-1,3ジアゾールなどの発光のマーカー、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、アセチルコリンエステラーゼ、またはビオチンなどの酵素マーカーである。
【0078】
プライマーは、プライマーの十分な量が、増幅される本発明の核酸分子またはそれらのオリゴヌクレオチド断片に対して相補的な配列を含むことを条件として、非相補的な配列を含み得ることが理解される。制限部位リンカーをプライマーに組み入れて、適切な制限酵素での増幅された産物の消化を可能にして、増幅産物のクローニングおよびシーケンシングを容易にもまたし得る。
【0079】
本発明のある態様において、増幅配列を形成することができる条件下において、ポリメラーゼ連鎖反応法において核酸分子または予め定められたそのオリゴヌクレオチド断片を増幅することができるプライマーで試料を処理して増幅配列を形成する段階、および増幅された配列をアッセイする段階を含む、本発明の核酸分子の存在を決定する方法が提供される。
【0080】
ポリメラーゼ連鎖反応法は、一般的に参照により本明細書に組み入れられる、Innis et al, Academic Press, 1990、Mullis et al., 米国特許第4,863,195号およびMullis, 米国特許第4,683,202号に記載されている、標的核酸配列を増幅するためのプロセスをいう。核酸テンプレートを増幅するための条件は、同様に参照により本明細書に組み入れられるM.A. Innis and D.H. Gelfand, PCR Protocols, A Guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989に記載されている。
【0081】
増幅産物は、当技術分野において公知の技術を使用して、単離され、それらのそれぞれのサイズに基づいて区別され得る。例えば、増幅後に、DNA試料をアガロースゲル上で分離して、臭化エチジウムで染色後に紫外(UV)光下で視覚化することができる。DNAは、所望のレベルに増幅され得、さらなる伸長反応により、放射性標識されたか、またはビオチン標識されたヌクレオシド三リン酸などの検出可能なマーカーを有するヌクレオチド誘導体を組み入れてもよい。プライマーは、上述したように検出可能なマーカーを用いてもまた標識され得る。検出可能なマーカーは、制限および電気泳動分離、または当技術分野において公知のその他の技術によって解析され得る。
【0082】
PCRを使用する本発明の方法において使用され得る条件は、試料中のDNAおよび適切な相補的ハイブリダイゼーション・プライマーの存在下において、ハイブリダイゼーションおよび増幅反応を進行させることができるものである。ポリメラーゼ連鎖反応法のために適した条件は、一般に当技術分野において公知である。例えば、参照により本明細書に組み入れられる、M.A. Innis and D.H. Gelfand, PCR Protocols, A guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989を参照されたい。好ましくは、PCRは、好熱性細菌サーマス・アクアティクス(Thermus aquatics)(Taqポリメラーゼ、GeneAmpキット、Perkin Elmer Cetus)から得られたポリメラーゼを利用し、またはその他の耐熱性ポリメラーゼを使用して、DNA鋳型鎖が増幅され得る。
【0083】
リガーゼ連鎖反応法(LCR)およびNASBAなどのその他の技術を本発明の核酸分子を増幅するために用いてもよいことが理解される(Barney in "PCR Methods and Applications", August 1991, Vol.1(1), page 5、および1989年6月14日に公開された欧州特許出願第0320308号、ならびにMalekの米国特許出願第5,130,238号)。
【0084】
(ii)MECP2E1タンパク質の検出
他の態様において、本発明は、動物由来の試料中のMECP2E1タンパク質における欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法を提供する。
【0085】
本発明のMECP2E1タンパク質は、後述する種々の免疫アッセイ法を使用してMECP2E1に特異的である抗体を使用して、生体試料中で検出してもよい。
【0086】
抗体を調製するために、従来法を使用することができる。例えば、本発明のMECP2E1タンパク質由来のペプチドを使用することにより、ポリクローナル抗血清またはモノクローナル抗体を標準的な方法を使用して作製することができる。哺乳動物(例えば、マウス、ハムスター、またはウサギ)に、哺乳動物の抗体反応を引き出すペプチドの免疫原形態で免疫することができる。ペプチドに免疫原性を与えるための技術は、キャリアとの抱合または当技術分野において周知のその他の技術を含む。例えば、ペプチドをアジュバントの存在下において投与することができる。免疫の過程は、プラズマまたは血清中の抗体力価の検出によってモニターすることができる。抗体のレベルを評価するために、標準的なELISAまたはその他の免疫アッセイ手順では、抗原として免疫原を使用することができる。免疫化に続いて、抗血清を得ることができ、必要であれば、ポリクローナル抗体を血清から単離することができる。
【0087】
モノクローナル抗体を産生するためには、抗体産生細胞(リンパ球)を免疫した動物から収集し、標準的な体細胞融合手順によって骨髄腫細胞と融合させることによって、これらの細胞を不死化し、ハイブリドーマ細胞を産生することができる。このような技術は、当技術分野において周知である(例えば、当初Kohlerおよび Milstein (Nature 256, 495-497 (1975))によって開発されたハイブリドーマ技術、ならびにヒトB-細胞ハイブリドーマ技術(Kozbor et al., Immunol. Today 4, 72 (1983))、ヒトモノクローナル抗体を産生するためのEBVハイブリドーマ技術(Cole et al., Monoclonal Antibodies in Cancer Therapy (1985) Allen R. Bliss, Inc., pages 77-96)、およびコンビナトリアル抗体ライブラリーのスクリーニング(Huse et al., Science 246, 1275 (1989))などのその他の技術)。ペプチドと特異的に反応する抗体の産生について、ハイブリドーマ細胞を免疫化学的にスクリーニングすることができ、モノクローナル抗体を単離することができる。したがって、本発明はまた、本発明のタンパク質に対する特異性を有するモノクローナル抗体を分泌するハイブリドーマ細胞を想定する。
【0088】
本明細書に使用される用語「抗体」は、本発明のタンパク質、またはこれらのペプチドと特異的に反応するこれらの断片も含むことが意図される。上記のものと同じ方法で利用するために、従来技術を使用して抗体を断片化し、断片をスクリーニングすることができる。例えば、F(ab')2断片は、ペプシンで抗体を処理して作製することができる。生じるF(ab')2断片を処理してジスルフィド架橋を還元し、Fab'断片を産生することができる。
【0089】
キメラ抗体誘導体、すなわちヒト以外の動物の可変領域とヒトの定常領域を組み合わせた抗体分子も、本発明の範囲内において想定される。キメラ抗体分子は、例えば、ヒト定常領域を有する、マウス、ラット、またはその他の種の抗体に由来する抗原結合ドメインを含むことができる。CipAタンパク質を認識する免疫グロブリン可変領域を含むキメラ抗体を作製するために、従来法を使用してもよい(例えばMorrison et al., Proc. Natl. Acad. Sci. U.S.A. 81,6851(1985);Takeda et al., Nature 314, 452(1985)、Cabilly et al., 米国特許第4,816,567号;Boss et al.,米国特許第4,816,397号;Tanaguchi et al.,欧州特許第171496号;欧州特許第0173494号、英国特許第GB 2177096B号を参照されたい)。
【0090】
本明細書に記載されている本発明のタンパク質と特異的に反応するモノクローナルまたはキメラ抗体は、可変領域の一部、特に抗原結合ドメインの保存されたフレームワーク領域が、ヒト起源であり、かつ高頻度可変領域だけが非ヒト起源であるヒト定常領域キメラを産生することによって、さらにヒト化することができる。このような免疫グロブリン分子は、当技術分野において公知の技術によって作製され得る(例えばTeng et al., Proc. Natl. Acad. Sci. U.S.A., 80, 7308-7312 (1983); Kozbor et al., Immunology Today, 4, 7279 (1983); Olsson et al., Meth. Enzymol., 92, 3-16 (1982))、および国際公開公報第92/06193号または欧州特許第0239400号)。また、ヒト化抗体は、商業的に産生することができる(Scotgen Limited, 2 Holly Road, Twickenham, Middlesex, Great Britain)。
【0091】
本発明のタンパク質に対して反応性の、特異的な抗体、または抗体断片は、本発明の核酸分子から産生されるペプチドを用いて、細菌で発現した免疫グロブリン遺伝子、またはこれらの部分をコードする発現ライブラリーをスクリーニングすることによって作製され得る。例えば、完全Fab断片、VH領域、およびFV領域は、ファージ発現ライブラリーを使用して、細菌で発現させることができる(Ward et al., Nature 341, 544-546: (1989); Huse et al., Science 246, 1275-1281 (1989);および McCafferty et al., Nature 348, 552-554 (1990)を参照されたい)。
【0092】
また、抗体は、DNA免疫を使用して調製してもよい。例えば、本発明の(上記のとおりの)核酸を含む発現ベクターをマウスなどの適切な動物に注射してもよい。したがって、本発明のタンパク質がインビボで発現されて、抗体が誘導されると考えられる。抗体は、単離することができ、タンパク質免疫化のために上記のとおりに調製することができる。
【0093】
抗体は、種々の酵素、蛍光物質、発光材料、および放射性物質を含む検出可能なマーカーで標識してもよい。適切な酵素の例は、西洋ワサビペルオキシダーゼ、ビオチン、アルカリホスファターゼ、β-ガラクトシダーゼ、またはアセチルコリンエステラーゼを含み;適切な蛍光物質の例は、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド、またはフィコエリトリンを含み;発光材料の例は、ルミノールを含み;および適切な放射性物質の例は、S-35、Cu-64、Ga-67、Zr-89、Ru-97、Tc-99m、Rh-105、Pd-109、In-111、I-123、I-125、I-131、Re-186、Au-198、Au-199、Pb-203、At-211、Pb-212、およびBi-212を含む。また、抗体は、リガンド結合対の一方のパートナーに対して標識するか、または抱合してもよい。代表例は、アビジン-ビオチンおよびリボフラビン-リボフラビン結合タンパク質を含む。上記した代表的標識をもつ、上で論議した抗体を抱合、または標識するための方法は、従来技術を使用して容易に達成し得る。
【0094】
本発明のタンパク質(例えば、酵素抱合体または標識された誘導体)に対する抗体反応性は、種々の試料中における本発明のタンパク質を検出するために使用してもよく、例えば、これらは、本発明のタンパク質の抗原決定基と抗体との間の結合相互作用に依存する任意の公知の免疫アッセイ法に使用してもよい。このようなアッセイ法の例は、放射免疫アッセイ法、酵素免疫測定法(例えば、ELISA)、免疫蛍光、免疫沈降、ラテックス凝集反応法、赤血球凝集法、および組織化学的試験である。したがって、レット症候群の存在を診断するために、試料中の本発明のタンパク質の量を同定、または定量化するため抗体を使用してもよい。
【0095】
本発明の方法では、試料または濃縮された試料の予め定められた量が、抗体または標識抗体と混合される。本方法に使用される抗体の量は、選択される標識剤に依存的である。生じる、抗体または標識された抗体に結合したタンパク質は、従来の単離技術、例えばば塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。
【0096】
試料または抗体は、不溶化してもよく、例えば、試料または抗体は、公知の方法を使用して適切なキャリアと反応させることができる。適切なキャリアの例は、セファロースまたはアガロースビーズである。不溶化された試料または抗体が使用されるときは、抗体または反応していない抗体に結合したタンパク質は、洗浄することによって単離される。例えば、試料がニトロセルロース膜にブロットされるときは、本発明のタンパク質に結合した抗体は、緩衝液、例えばウシ血清アルブミン(BSA)を含むリン酸緩衝食塩水(PBS)で洗浄することにより、反応していない抗体から分離される。
【0097】
標識された抗体が使用されるときは、MECP2E1の存在は、試料中の本発明のタンパク質に結合した標識された抗体の量または反応していない標識された抗体の量を測定することによって決定することができる。標識された物質を測定する適切な方法は、標識剤に依存する。
【0098】
標識されていない抗体を本発明の方法に使用するときは、MECP2E1の存在は、凝集または沈澱を生じさせるために、抗体と特異的に相互作用する物質を使用してタンパク質に結合した抗体の量を測定することによって決定することができる。特に、本発明のタンパク質に特異的な抗体に対する標識された抗体を、反応混合物に添加することができる。本発明のタンパク質の存在は、標識剤のタイプに応じて、すでに記載されている技術の中から適切な方法によって決定することができる。本発明のタンパク質に特異的な抗体に対する抗体は、本明細書に記載した当技術分野において公知の従来法によって調製し、および標識することができる。本発明のタンパク質に特異的な抗体に対する抗体は、種特異的な抗免疫グロブリン抗体またはモノクローナル抗体であってもよく、例えば、ヤギ抗ウサギ抗体を本発明のタンパク質に特異的なウサギ抗体を検出するために使用してもよい。
【0099】
(iii)キット
本発明の方法を実施するために適した試薬は、適切な容器にパックされて、必要な材料を提供する便利なキットにパックされ得る。このようなキットは、本明細書に記載されている方法、および任意に本発明の方法を行う際に有用な適切な支援によって、試料中の本発明の核酸分子またはタンパク質を検出するために必要とされる全ての試薬を含み得る。
【0100】
本発明の一つの態様において、キットは、本発明の核酸分子またはこれらの予め定められたオリゴヌクレオチド断片を増幅することができるプライマー、ポリメラーゼ連鎖反応法において増幅された核酸分子またはこれらの予め定められた断片を産生するために必要とされる全ての試薬、および増幅された配列をアッセイするための手段を含む。また、キットは、PCR産物を消化するための制限酵素も含んでいてもよい。本発明のもう一つの態様において、キットは、本発明の核酸分子とハイブリダイズするヌクレオチド・プローブ、核酸分子とヌクレオチド・プローブのハイブリダイゼーションのために必要とされる試薬、およびその使用説明書を含む。本発明のさらなる態様において、キットは、本発明の抗体、および試料中において本発明のタンパク質に対して抗体を結合するために必要とされる試薬を含む。
【0101】
キットは、神経精神障害および発達障害を検出または処置するために、本発明の核酸分子、タンパク質、または抗体(上述)を、それらの使用のための説明書と共に含み得る。
【0102】
本発明の方法およびキットを用いて、レット症候群および精神遅滞のような神経精神障害ならびに発達障害を検出し得る。試験され得る試料は、血液、尿、血清、涙、唾液、糞便、組織、器官、細胞などの身体材料を含む。ヒト試料に加えて、試料は、非ヒト霊長類などのような哺乳動物から採取され得る。
【0103】
本明細書において記載される方法に従って試料を試験する前に、試料は、遠心分離および濾過のような当技術分野において公知の技術を使用して濃縮され得る。本明細書において記載されるハイブリダイゼーションおよび/またはPCRに基づいた方法のために、核酸は、当技術分野において公知の技術を使用して試験試料の細胞抽出物から抽出され得る。
【0104】
B.治療適用
上述したように、本発明の核酸分子は神経精神障害および発達障害を有する人々において欠失または突然変異されている。従って、本発明は、神経精神障害および発達障害を処置または予防するためにMECP2E1スプライス変異体の十分な部分を含む核酸配列を投与することにより、神経精神障害および発達障害を処置または予防する方法を提供する。本発明は、神経精神障害および発達障害を処置または検出するための、本発明の核酸分子およびタンパク質の使用を含む。
【0105】
核酸分子またはその断片を含む組換え分子は、レトロウイルスベクター、アデノウイルスベクター、およびDNAウイルスベクターのような送達媒体を用いて、インビボで細胞または組織に直接導入され得る。それらはまた、微量注入および電気穿孔法のような物理的技術、または共沈およびリポソームへのDNAの取り込みのような化学的方法を用いても細胞に導入され得る。組換え分子はまた、エアロゾルの形態で、または洗浄によっても送達され得る。
【0106】
核酸配列は、インビボでの投与に適した生物学的に適合性を有する形態で、対象への投与のための薬学的組成物に調剤され得る。「インビボでの投与に適した生物学的に適合性を有する形態」とは、治療的な効果が任意の有毒作用を上回って投与されるような物質の形態を意味する。物質は、ヒト、および動物を含む、生きている生物に投与してもよい。本発明の薬学的組成物の治療的に有効な量の投与は、所望の結果を達成するために必要な用量で、および期間で有効な量として定義される。例えば、物質の治療的に有効な量は、個々の疾病状態、年齢、性、および重量、ならびに個々に所望の反応を引き出すための抗体の能力などの因子によって変更してもよい。投与計画は、最適な治療反応を提供するように調整してもよい。例えば、いくつかの分割された用量を毎日投与してもよく、または用量は、治療状況の緊急性によって示されるように、比例的に減らされ得る。
【0107】
活性物質は、注射(皮下の、静脈内の、その他)、経口投与、吸入、経皮適用、または直腸投与のような、便利な様式で投与され得る。投与の経路に応じて、活性物質は、酵素、酸、および化合物を不活性化し得るその他の天然の条件の作用から化合物を保護する材料中にコーティングされ得る。
【0108】
本明細書において記載されている組成物は、薬学的に許容される組成物の調製のためのそれ自体公知の方法によって調製することができ、これを、有効な量の活性物質が薬学的に許容される媒体との混合物中に組み合わされるように被検者に投与することができる。適切な媒体は、例えばRemington’s Pharmaceutical Sciences (Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pa., USA 1985)に記載されている。これに基づいて、組成物は、限定的ではないものの、1つもしくは複数の薬学的に許容される媒体または希釈液と組み合わされ、適切なpHを有し、かつ生理液と等浸透圧の緩衝液に含まれる、物質の溶液を含む。
【0109】
C.実験モデル
本発明は、MECP2遺伝子およびMeCP2E1タンパク質の機能を研究するための方法、ならびに実験モデルを含む。MECP2E1スプライス変異体を欠いた、もしくはMeCP2E1発現を部分的に欠いた細胞、組織、および非ヒト動物は、MECP2E1遺伝子に特異的な欠失または変異を有する組換え発現ベクターを使用して開発してもよい。組換え発現ベクターを、相同組換えによってMECP2遺伝子を不活性化または変化させるために使用して、これにより、MECP2E1欠損細胞、組織、または動物を作製してもよい。特に、MECP2E2は変化させず、MECP2E1の欠損をまねくよう標的突然変異が設計され得る。これはMECP2遺伝子のエキソン1を標的とすることで達成され得る。
【0110】
ヌル対立遺伝子は、欠失突然変異によって胚幹性細胞などの細胞に作製され得る。また、組換えMECP2遺伝子を、MECP2E1を不活性化する挿入突然変異を含むようにもまた操作し得る。次いで、このような構築物を、トランスフェクション、電気穿孔法、注入等の技術によって、胚幹性細胞などの細胞に導入してもよい。次いで、例えばサザンブロッティング、ノーザンブロッティングによって、または本明細書において記載されている方法を使用してMECP2E1をアッセイすることによって、無傷のMECP2遺伝子を欠いている細胞を同定してもよい。次いで、そのような細胞を胚幹細胞に融合して、MECP2E1が欠損したトランスジェニック非ヒト動物を作製してもよい。突然変異の生殖系列伝達は、例えば、インビトロで、8細胞胚などの初期胚で胚幹細胞を凝集すること;生じる胚盤胞をレシピエント女性へ移すこと、および;生じる凝集キメラの生殖系列伝達を発生させることによって達成してもよい。そのような変異体動物を使用して、通常はMECP2E1発現に依存的な、特異的な細胞群、発達パターン、およびインビボ・プロセスを定義してもよい。本発明は、MECP2E1変異体の組織特異的ノックアウトの調製もまた含む。
【0111】
以下の非限定の実施例は、本発明の例示である:
【実施例】
【0112】
実施例1
MECP2E1スプライス変異体の同定
5'UTRの検査から、エキソン2はATGの上流に多数のインフレームの終結を有するが、エキソン1はATGを含むオープンリーディングフレームをその全長にわたって含むことが明らかとなった。エキソン1、3、および4からなる理論上の構築物を、その周囲のコザックヌクレオチド状況の重要性に基づきATGが開始コドンになる可能性を予測する、ATGprプログラム(http://www.hri.co.jp/atgpr/)で処理すると、MECP2E2の64%と比較して97%の信頼性スコアが返された。ESTデータベースの検索から、本発明者らの理論上の転写産物(MECP2E1と名付けた)を8例同定した(図1b)(対して、MECP2E2は14例)。MECP2E1は、エキソン1により決定された、より長いN末端を代わりに有する、新たな変異体MeCP2E1をコードすると予測される。
【0113】
実施例2
MECP2E1の発現
MECP2E1が実際に発現され、cDNAライブラリー調製物のアーチファクトではないことを確認するため、さまざまな組織由来のcDNAをエキソン1内の5'プライマーおよびエキソン3内の3'プライマーを用いてPCR増幅した(図1a)。サイズおよび配列によりMECP2E2およびMECP2E1に相当する2種類のPCR産物を、胎児および成体の脳を含む全ての組織、ならびに脳の小領域で得た(図1c)。マウスでの結果は類似していた(図1c)。成体ヒト脳における2種類の転写産物の発現レベルを定量化した。MECP2E1の発現はMECP2E2より10倍高かった(図1d)。3'mycタグ化したMECP2E1のCOS-7細胞へのトランスフェクション後の、MeCP2E1の細胞内局在は主に核内であることが見出された(図1e)。
【0114】
MECP2E1は以前の発現研究では検出されなかった。ノーザン分析は、代替のポリアデニル化シグナルの使用に起因して、サイズが異なる1.9、5、および10.1kbの3種類の転写産物を明らかにした(4、6、8)(図1a)。MECP2E1は、124ヌクレオチドのエキソン2を欠く点でMECP2E2と異なる。ゲル上の5および10.1kb位置では、2種類の転写産物は分離不可能である。1.9kbの範囲においては、公表されたノーザンブロットは2種類の転写産物に相当する可能性のある、太いかまたは二重のバンドを示す。同様に、従来型のウェスタンブロット分析も2種類のMeCP2アイソフォームの分解は不可能である(分子量の差<0.9kD;図1f)。
【0115】
実施例3
レット症候群におけるMECP2E1の突然変異
レット症候群において新たなコード領域が突然変異を受けているか否か決定するため、他のエキソンにおいて突然変異が見出されなかった、典型的RTTを有する19人の少女において、エキソン1および隣接配列をPCR増幅し、シーケンシングした。1人の患者(V1)は、エキソン1に11bpの欠失突然変異を有することが見出された(図2)。欠失は、MECP2E1の予測されたエキソン1オープンリーディングフレーム内で起こり、ミスセンスアミノ酸配列、続いてアミノ酸36の後に未成熟な終止コドンを生じるフレームシフトをもたらす。これはMECP2E2のコード配列には影響しない。この配列変化は患者の両親および兄弟を含む200人の対照個体においては見出されなかった。
【0116】
残りの患者において、本発明者らのPCR反応によっては検出できなかったさらなるエキソン1欠失を検索するため、多重ライゲーション依存性プローブ増幅(MLPA)(5)を4つ全てのエキソンについて実施し、1人の患者においてエキソン1のヘミ接合欠失を検出した(患者V2;図3)。最終的に、エキソン1に限定されるMLPA検出の欠失を有するさらに一人の患者について、最近抄録の形で文書化されたが、MECP2E1への影響は認識されていなかった(S. Boulanger et al. Am J Hum Genet 73, 572(2003))。
【0117】
対照的に、数百人の患者がエキソン2における突然変異について分析されたにも関わらず、MeCP2E2定義性のエキソン2に特異的な突然変異は今日まで見出されていない(31の刊行物;参考文献3に最も良く概説される)。これらの研究は、エキソン1を非コード性と見なしたためにエキソン1を含まない。
【0118】
エキソン1の欠失により、MeCP2E1タンパク質の非存在または切断がもたらされる。しかしながら、それらはまたMECP2E2の5'UTRの短縮をもたらし得、その発現に影響する可能性もあり得る。全血に対するRT-PCRにより、患者V1においてこの可能性について試験した。MECP2E2の発現の減少は存在しなかった(図2c)。結論として、突然変異のデータは、MeCP2E1の不活性化がRTTに十分であり、現在のところ、MeCP2E2については同じことを言明できない。
【0119】
材料および方法
PCR、手動シーケンシング、クローニング、rtPCR、ゲルブロット
PCR増幅を、1Mベタイン、50%デアザ(deaza)dGTPを含む200μM dNTPを含む[NH4]2SO4含有PCR緩衝液(MBI Fermentas)を用いて、3分間の95℃変性段階、続いて30秒間の95℃、30秒間の55℃、45秒間の72℃のサイクルを30サイクル繰り返し、次いで72℃7分間の浸漬段階により行った。手動シーケンシングは、1%アガロースゲルからの抽出後、Thermosequenase(商標)キット(USB/Amersham)を用いて、6%変性ポリアクリルアミドゲルに3時間泳動して行った。PCR産物をpDRIVEベクター(Qiagen PCR cloning kit)を用いてクローニングした。全血RNAを、PAXgene Blood RNAキット(Qiagen)を用いて抽出した。逆転写は、ランダム六量体および標準的なSuperscript IIIプロトコル(Invitrogen)を用いて行った。ヒト脳小領域のcDNAをOriGeneより得た。ポリアクリルアミドゲル(図2c)をHybond N+(Amersham)上にブロットし、デオキシヌクレオチド転移酵素(MBI Fermentas)を用いて3'末端が[α32P]-dCTPで標識されたHFプライマーとハイブリダイズさせた。
【0120】
神経およびグリア培養の調製
大脳皮質を性交後15.5日(15.5dpc)のCD-1マウス胚から調製した。Yamasaki et al.(Yamasaki et al. Hum Mol Genet 12: 837-847, 2003)の手順を用いた。簡単には、髄膜を除いた胎児大脳皮質を機械的粉砕により分離し、EDTAとともに0.25%トリプシンにより消化した。ウシ胎児血清(FBS;GIBCO BRL)の添加後、ろ過された細胞を遠心分離により回収した。細胞のペレットを、神経細胞の増殖のためにB-27(GIBCO BRL)か、またはグリア細胞の増殖のためにG-5(GIBCO BRL)を補ったNeurobasal(GIBCO BRL)培地に再懸濁した。細胞をポリエチレンイミンコーティングされたプラスチックディッシュに2×106細胞/mlの密度で平板培養した。神経細胞およびグリア細胞の培養物は、5% CO2、37℃で、それぞれ6日間および12日間維持された。単離された脳細胞をRT-PCR、ならびに神経細胞に対してMAP2(microtubule-associated protein 2)、グリア細胞に対してGFAP(glial fibrillary acidic protein)、および前駆細胞に対してネスチンというマーカーを用いる免疫蛍光法(IF)により特徴付けた。IFには、以下の特異的抗体を用いた:マウスモノクローナル抗MAP2(CHEMICON)、およびウサギポリクローナル抗GFAP(DAKO)。rtPCRに用いたプライマーはYamasaki et al.と同一である。半定量的PCRを得るため、内部対照としてGapdh増幅に従って、至適cDNA濃度およびサイクル数を決定した。図4はrtPCR(A)およびIF(B)による脳細胞初代培養の特徴を示す。
【0121】
定量的rtPCR
異なる組織におけるMECP2転写産物の量を決定するため、本発明者らは、SYBRグリーン検出法(PE Applied Biosystems, ABI PRISM 7900 Sequence Detection System)を用いた転写産物特異的リアルタイム定量的PCRアッセイを開発した。以下のMECP2E2特異的順方向プライマー(25nM)(エキソン2内)を設計した:5'-ctcaccagttcctgctttgatgt-3'(SEQ ID No. 12)。MECP2E1特異的プライマー(25nM)はエキソン1と3の接続部に設置した:5'-aggagagactggaagaaaagtc-3'(SEQ ID No. 10)。両方のアッセイとも、161bp(MECP2E2)および65bp(MECP2E1)の断片を産生するエキソン3内の同一の逆方向プライマー(25nM)を用いた:5'-cttgaggggtttgtccttga-3'(SEQ ID No. 11)。マウスのmecp2転写産物(mecp2e2 167bpおよびmecp2e1 71bp)についての相当する転写産物特異的プライマー(25nM)は、5'-ctcaccagttcctgctttgatgt-3'(SEQ ID No. 12)(MECP2E2);5'-aggagagactggaggaaaagtc-3'(SEQ ID No. 13)(MECP2E1)、および共通の逆方向プライマー5'-cttaaacttcagtggcttgtctctg-3'(SEQ ID No. 14)であった。PCR条件は:50℃2分間、95℃10分間、および95℃15秒間、60℃85秒間を40サイクル、であった。PCR反応は別個のチューブで実施し;かつMECP2E2およびMECP2E1転写産物の絶対的定量は、ヒト成体脳、小脳、繊維芽細胞、およびリンパ芽球(Clontech, Palo Alto, USA)、ならびにマウス神経細胞およびグリア細胞培養物(上述)由来のcDNAから行った。結果を、製造者の使用説明書(PE Applied Biosystems, ABI PRISM 7900 Sequence Detection System)に従って標準曲線法を用いて分析した。標準曲線は、精製した転写産物特異的PCR産物の希釈溶液を用いて作製した。
【0122】
免疫蛍光光学顕微鏡検査
3'mycタグ化したMECP2E2およびMECP2E1構築物(pCDNA3.1A-MECP2E2-mycおよびpcDNA3.1A-MECP2E1-myc)を、BamH1(5')およびXbaI(3')制限部位を付した、それぞれの転写産物の全長cDNAのPCR増幅、ならびにその後のpcDNA3.1 version A(Invitrogen)へのmycとインフレームでのクローニングにより作製した。MECP2E2に対する順方向プライマーはエキソン2内の開始コドンを含み(5'-tatggatccATGgtagctgggat-3')(SEQ ID No. 15)、一方MECP2E1に対する順方向プライマーはエキソン1内の開始コドンを含む(5'-tatggatccggaaaATGgccg-3')(SEQ ID No. 16)(BamHI制限部位を下線で示した、開始コドンは大文字である)。逆方向プライマーは両方の増幅で同一であった(5'-gcgtctagagctaactctct-3')(SEQ ID No. 17)(XbaI制限部位を下線で示した)。PCRに用いたテンプレートは、MECP2E2については小腸cDNAであり、およびMECP2E1については骨格筋cDNAであった。pcDNA3.1A-MECP2E2-mycおよびpcDNA3.1A-MECP2E1-myc(2μg)をCOS-7細胞にlipofectamine(Invitrogen)を用いてトランスフェクトし、脂質DNA複合体をDMEM(GIBCO)中で5時間曝露した。トランスフェクトションから48時間後に、培養物をPBS中でリンスし、アセトン:メタノール(1:1)混合物中で15分間、-20℃で固定し、1時間ブロックして(PBS中10%BSA)、室温で45分間、抗myc(Santa Cruz Biotechnology、ブロッキング緩衝液中1:50)と共にインキュベートした。PBSを用いて洗浄した後、スライドをブロッキング溶液中の二次抗体(FITC標識されたヤギ抗マウス(Jackson Immunoresearch labo)、1:400、緑色フィルターを介して検出可能)と共にインキュベートし、Dako Anti-Fadeを用いて封入し、免疫蛍光光学顕微鏡検査によって分析した。
【0123】
MLPA分析
MLPAはSchouten et al.、前記により記載されたように、およびSchouten、前記により記載されたように行った。MRC-Holland、Amsterdam、Netherlands(http://www.mrc-holland.com)のMECP2試験キットを利用し、キットは4つのMECP2エキソン、6つのX連鎖対照領域、および10個の常染色体対照領域を標的にした20のプローブ対からなっていた。簡単には、100〜200ngのゲノムDNAを変性させ、プローブ混合物と一晩、60℃でハイブリダイズさせた。翌朝、熱安定性のLigase-65を用いて、15分間、54℃で対のプローブを結合させた。結合後、各結合産物の末端にハイブリダイズする共通のプライマー対によるPCRを行った。PCRプライマーの一方はFAM標識され、PCR条件は以下の通りであった:95℃30秒間、60℃30秒間、および72℃1分間。結果的に生じた単位複製配列を、ABI 3100キャピラリー電気泳動法装置およびABI Genescanソフトウェアで分析した。データ管理および正常対照との比較は全てExcelソフトウェアを用いて行った。
【0124】
考察
最近、カエル(アフリカツメガエル(Xenopus laevis))における研究から、神経発生の転写調節におけるMeCP2の役割についての重要な洞察が提供された。MeCP2は、Notch-Delta9による発生段階特異的媒介の後に、神経分化に関わる遺伝子の調節に関わるSMRT複合体の構成成分であることが示された。これらの実験においてサイレンシングの標的とされたカエルMecp2転写産物は、MECP2E1のオルソログであった(図1f)。実際、非哺乳類脊椎動物においては、MeCP2E1がMeCP2の唯一の形態であるようである(図1f)。
【0125】
新たなMeCP2 N末端は、ポリアラニンおよびポリグリシン区域を含む、特徴的な21アミノ酸ペプチドである(MAAAAAAAPSGGGGGGEEERL)(SEQ ID No. 18)(図1f)。類似のN末端は、複数のシグナル伝達経路の重要な共通構成成分である、ERK1(MAPK3)、細胞外シグナル調節性キナーゼ(extracellular signal-regulated kinase)にも見られる(図1f)。興味深いことに、神経細胞では、ERK110およびMeCP211の両方が核に加えてシナプス後区画内に存在することが示され、かつ前者は2つの区画間を転位し、シナプス活性と転写調節を結びつけていることが示された10。MeCP2E1も同様にシナプス活性、この場合、神経発生時のシナプス接合ガイダンスと転写調節を結びつけている可能性がある。連続したポリアラニンおよびポリグリシン区域が見出される他の唯一のタンパク質群は、ホメオボックス(HOX)ファミリーのいくつかのメンバー12に含まれる。MeCP2と同様に、これらは発生時の転写調節因子である。
【0126】
最後に、非不活性化型のMECP2突然変異が、精神遅滞および自閉症のようなRTTと重複した表現型と関連付けられている13。本研究において発見されたMeCP2変異体は、これらの障害への関与の候補である。
【0127】
実施例4
精神遅滞におけるMECP2E1の突然変異
本発明者らは、N=401の自閉症発端者、および非特異的精神遅滞を有するN=493の患者において、MECP2E1遺伝子をスクリーニングした。Autism Genetic Resource Exchange(AGRE;N=242;女性100、男性142)からの多発性の家系の発端者のみならず、Hospital for Sick Children in Toronto(N=146;男性114、女性32)を通じ、およびLondon, UK(N=13;男性10、女性3)から募られた自閉症発端者もまたスクリーニングした。各地の施設倫理審議会の認可を得、参加者から書面による同意を得た。Hospital for Sick ChildrenのDepartment of Pediatric Laboratory Medicineで、脆弱性X試験について参照された(しかし試験結果は陰性)、非特異的発育遅延/精神遅滞を有する、女性293人および男性200人の患者について、匿名化されたDNA試料もまた得られた。実施例3に前述したようなPCRプライマーおよび条件を用いて、ポリメラーゼ連鎖反応の後に変性高速液体クロマトグラフィー(DHPLC)を突然変異検出に用いた。配列変異体を有すると疑われる女性個体に由来するPCR産物をpDRIVEベクター(Qiagen)にクローニングし、少なくとも4クローンを、順方向および逆方向で、自動化BigDye(商標)シーケンシング(ABI 3100)を用いてシーケンシングした。男性由来のPCR産物をアガロースゲルから切り出し、カラム精製した後、同様に順方向および逆方向の両方で、自動化BigDye(商標)シーケンシング(ABI 3100)を用いてシーケンシングした。自閉症スクリーニングセットにおいては突然変異は同定されなかったが、配列変異体は8人の女性MR症例に同定され(図7を参照されたい)、うち3人はポリアデニン反復ストレッチ内にアミノ酸の挿入または欠失を生じ、およびうち2人はMECP2E1のN末端部分のポリグリシン反復内にグリシン残基の挿入が生じていた。同定された最初の個体は、推定MECP2E1開始コドンの45〜46bp上流に位置するGpCジヌクレオチドの欠失についてヘテロ接合であった。この欠失は、潜在的なSP1転写因子結合部位(http://www.gene-regulation.com/pub/programs/alibaba2/index.htmlのAliBaba2.1を用いて予測された)を破壊し得、および潜在的にメチル化可能なシトシン残基もまた除去し得る。他の個体は、MECP2E1開始コドンの26bp上流でApGジヌクレオチドの欠失についてヘテロ接合であった。2個体がポリ[GGA]ストレッチ内へのGGAトリヌクレオチド挿入についてヘテロ接合であり、それにより予測されたポリグリシンストレッチ内に追加のグリシン残基がもたらされる。5個体目は、ポリアラニンをコードするトリプレット反復ストレッチ内での、GCCトリヌクレオチド欠失についてヘテロ接合であった。2個体は、GCCトリヌクレオチド反復/ポリアラニン領域においてもまた、9bpの挿入についてヘテロ接合であり、ポリアラニンストレッチの7から10残基への伸長をもたらす。
【0128】
女性の非特異的MR症例の〜2%における、精神遅滞症候群に以前より関連付けられていたタンパク質の新たなアイソフォームについてのアミノ酸配列変異は非常に興味深い。さらに、多数の脊椎動物種間で保存されている、タンパク質の部分内で変異が生じているという事実もまた興味を増大させる(チンパンジー、オランウータン、マカク、ネコ、およびイヌのMECP2E1アミノ酸配列に対し、100%同一性)。変異体を有する個体間で任意の特異的な表現型の特徴が存在するか、症状の重症度はいかほどか、およびレット症候群の表現型との重複部分または差異が存在するかを知ることは特に有用である。しかしながら、DNAが匿名化されていたため、今回は、見出された突然変異を表現型の特徴または重症度と相関させることは不可能である。この問題に取り組む試みにおいて、Greenwood Genetic Center、South Carolinaに由来するMR症例(女性188、男性96)の第二の試料セットをスクリーニング後、シーケンシングした。男性においては変異体は見出されず、女性2人が余分なグリシン残基をコードするGGA挿入を有していた。
【0129】
本研究において、3人の女性MR患者が、MECP2E1のN末端のポリグリシンストレッチ内に余分なグリシン残基をもたらす3bpの挿入を有することを同定された。これまでにグリシン反復内の伸張に関して疾病関連性は報告されていない。植物におけるToc75タンパク質の研究から、このタンパク質内のポリグリシンストレッチがタンパク質の葉緑体外包膜への正確なターゲティングに必須であることが示唆されている(14)にも関わらず、MECP2E1タンパク質の文脈においてまたはより一般的にのいずれにおいても、ポリグリシンストレッチの機能は未知である。類似のタンパク質輸送機能が、ポリグリシンストレッチを有する哺乳類タンパク質、およびMECP2E1についての真相であることもまたあり得る。
【0130】
ポリアラニン区域内の変異体は特に関心対象である。それは、それらがまれにしか多型を示さず、かつそのような区域内の多くの小伸張(または重複)が、鎖骨頭蓋骨異形成症(RUNX2)、眼球咽頭型筋ジストロフィー(PABPN1)、および精神遅滞(ARX;この遺伝子もまたX染色体性で、非常に多岐にわたる表現型を有する-上記を参照されたい)に及ぶ疾患を引き起こすことが報告されているためである(15)。ポリアラニン疾患遺伝子の大部分は転写因子をコードするが、PABPN1遺伝子はポリアデニレート結合タンパク質をコードする。一方では、これらの疾患のうちで、転写因子遺伝子内の最も小さな病原性の反復は一般的に20アラニン長より大きく、従ってMECP2E1によりコードされるほど短いアラニンストレッチが病原性になり得ることはあり得ないと考えられ、1または3つのアラニン残基の変化は、まれな多型であろうと考えられ得る。現在のところ、ARX遺伝子内の1または3つのアラニン残基の小伸張が病原性であり得るか、または無害な変異体であり得るかについてはいくらかの不確定性が存在する(16、17)。他方では、眼球咽頭型筋ジストロフィーはPABPN1遺伝子のGCG区域内の突然変異により引き起こされる。この突然変異は、ポリアラニン区域をちょうど10個のアラニン残基から12〜17個のアラニン残基に伸張させる。さらに、MECP2E1のポリアラニン区域と同じく、PABPN1のポリアラニン区域はちょうど遺伝子のN末端にあり、従って、タンパク質のN末端部分内の反復ストレッチ内の比較的小さな突然変異が、タンパク質の中央部分に位置するより大きな突然変異より有害であり得る可能性がある。
【0131】
最近公表された研究は、エキソン2、3、または4に突然変異を有さない97人のレット患者、および146人の対照においてMECP2エキソン1における突然変異についてスクリーニングした(18)。レット患者のうち一人はポリアラニンをコードする[GCC]ストレッチ内に6bpの挿入を有することが見出されたが、そのような変異は対照には観察されなかった。この変異体は非罹患の母親から遺伝され、従ってこの変異は病因学的に関連していないようであると結論された(18)。しかしながら、最近、マウスでのMecp2の発現における微細な変化でさえ、深刻な神経学的および行動的結果を有し得ることが示された(19)。同一のMECP2突然変異を有する患者が非常に異なる表現型の特徴および重症度を有し得ることは明らかであり、かつX不活性化パターンにおける変異が、この不整合性において役割を果たしているようである。従って、ポリアラニンもしくはポリグリシンストレッチ長の変化をもたらす反復ストレッチ内か、または開始コドンの直上流の領域内のいずれかで、エキソン1内の変異が、機能または発現レベルに影響して神経病理学的な表現型をもたらし得ることは非常に可能性が高い。
【0132】
実施例5
レット症候群におけるMECP2E1のさらなる突然変異
エキソン1、2、3、および4のコード領域全体、ならびにそれらのイントロン隣接配列を分析した。エキソン2〜4をPCRにより、Human Genome Project working draft site(UCSC、www.genome.ucsc.edu)由来のゲノム配列情報およびLasergene Primer select programを用いて設計したプライマー対を用いて増幅した。dHPLC(WAVE Nucleic Acid Fragment Analysis System、Transgenomic, San Jose, CA)による塩基変化分析の前に増幅を確認するため、PCR産物を2%アガロースゲルにロードした。溶媒Aは0.1mol/Lトリエチルアンモニウムアセテート(TEAA)および25%アセトニトリルからなり、溶媒Bは1M TEAA、25%アセトニトリルを含んでいた。dHPLCにおいてクロマトグラフィー上の変異を示したPCR産物を自動シーケンサー(Gene Reader 4200)で直接シーケンシングした。シーケンシングデータをDNA Star software SeqMan(Lasergene)を用いて分析した。最近記載されたように(13)、全患者についてエキソン1をPCR増幅し、かつシーケンシングした。
【0133】
第一のエキソン1突然変異は、エキソン1イントロン1境界での、欠落した2塩基対からなる。この領域における配列の性質のため、本発明者らは欠落した2ヌクレオチドが、イントロン1の最初の2塩基対(GT)であるか、またはエキソン1の最後のヌクレオチド(T)およびイントロンの最初のヌクレオチド(G)であるか解明できなかった。どちらの場合であっても、欠落したヌクレオチドの対は、予測されたコンセンサススプライス部位を破壊し、イントロン1の読み過しをもたらす(データは図示せず)。エキソン1突然変異を有する第二の患者では、1A→T置換(ATG -> TTG)が最初のメチオニンコドンをロイシンに変化させる。予測では、開始コドンの非存在のために、MECP2E1の翻訳は著しくまたは完全に妨げられる。MECP2E2は正常に製造される(および、疾患の表現型をレスキューできないようである)。
【0134】
本発明は、好ましい実施例であると現在みなされるものに関して記載したが、本発明は、開示された実施例に限定されないことが理解される。対照的に、本発明は、添付の特許請求の範囲の精神および範囲内に含まれる種々の改変および同等の処理を包含することが意図される。
【0135】
全ての刊行物、特許、および特許出願は、あたかも個々の刊行物、特許、または特許出願が具体的に、および個々に、その全体が参照により組み入れられることを示したのと同程度まで、これらの全体が参照により本明細書に組み入れられる。
【0136】
(表1)今日までに同定されたMECP2E1突然変異または変異体。「del」は欠失を示し;「ins」は挿入を示す。
【0137】
本明細書において参照された参考文献の完全な引用
【特許請求の範囲】
【請求項1】
MeCP2E1タンパク質をコードする配列を含む、単離された核酸分子。
【請求項2】
以下を含む、請求項1記載の単離された核酸分子:
(a)図6(b)(SEQ ID No. 4)に示したような配列を有する、MeCP2E1タンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【請求項3】
図6(a)(SEQ ID No. 3)に示した配列を有する、請求項1記載の核酸、またはその断片もしくは変異体。
【請求項4】
突然変異が以下からなる群より選択される、図5(a)(SEQ ID No.1)に示した核酸配列中に突然変異を含む単離された核酸分子:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置69〜70における配列TGの欠失。
【請求項5】
突然変異が以下からなる群より選択される、図5(a)(SEQ ID No. 1)に示した核酸配列中に突然変異を含む単離された核酸分子:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数コピーのGCCトリヌクレオチド配列の挿入;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、ヌクレオチド配列GGAの挿入;
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに
(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【請求項6】
請求項1〜5のいずれか一項記載の核酸分子、およびプロモーターが宿主細胞における核酸分子の転写を増強するように機能的に結合されたプロモーター配列を含む、組換え核酸分子。
【請求項7】
請求項1〜5のいずれか一項記載の核酸分子を含むベクター。
【請求項8】
請求項6記載の組換え核酸分子もしくは請求項7記載のベクターを含む宿主細胞、または宿主細胞の子孫。
【請求項9】
哺乳動物細胞、真菌細胞、酵母細胞、細菌細胞、微生物、および植物細胞からなる群より選択される、請求項8記載の宿主細胞。
【請求項10】
単離されたMECP2E1タンパク質。
【請求項11】
図6(a)(SEQ ID No. 3)に示した核酸配列によりコードされる、請求項10記載のタンパク質、または該タンパク質の断片もしくは変異体。
【請求項12】
図6(b)(SEQ ID No. 4)に示したアミノ酸配列を有する請求項10記載の単離されたタンパク質、または該タンパク質の断片もしくは変異体。
【請求項13】
図6(a)(SEQ ID No. 3)に示した核酸配列のエキソン1によりコードされるペプチドに結合する抗体。
【請求項14】
動物由来の試料中の請求項1〜3のいずれか一項記載の核酸配列内に、突然変異または欠失を検出する段階を含む、神経精神障害または発達障害を検出する方法。
【請求項15】
核酸配列のエキソン1内に突然変異または欠失を検出する段階を含む、請求項14記載の方法。
【請求項16】
以下の段階を含む、請求項15記載の方法であって:
(a)ポリメラーゼ連鎖反応において、プライマーX1F(5'-CCATCACAGCCAATGACG-3')(SEQ ID No. 19)およびX1R(5'-AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)を用いて試料中の核酸配列を増幅する段階;
(b)同一のプライマーを用いて、対照由来の核酸配列を増幅する段階;
(c)増幅された配列のシーケンシングを行う段階;ならびに
(d)対照配列と試料配列を比較する段階;
ここで、対照配列と比較した試料配列中のヌクレオチドの欠失は、試料が神経精神障害または発達障害を有する動物由来であることを示す、方法。
【請求項17】
図5(a)(SEQ ID No. 1)に示したMECP2遺伝子配列の4つのエキソン全てにおいて、多重ライゲーション依存性プローブ増幅(multiplex ligation-dependent probe amplification)を行う段階を含む、請求項14記載の方法。
【請求項18】
請求項10〜12のいずれか一項記載のタンパク質中に欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法。
【請求項19】
神経精神障害がレット症候群である、請求項14〜18のいずれか一項記載の方法。
【請求項20】
検出された突然変異または欠失が以下からなる群より選択される、請求項19記載の方法:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置70〜71における配列TGの欠失。
【請求項21】
発達障害が精神遅滞である、請求項14〜18のいずれか一項記載の方法。
【請求項22】
突然変異または欠失が以下からなる群より選択される、請求項21記載の方法:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数のGCCトリヌクレオチド配列の挿入;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、GGAヌクレオチド配列の挿入;
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに
(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【請求項23】
請求項1〜5のいずれか一項記載の核酸配列もしくはその断片を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項24】
請求項1〜5のいずれか一項記載の核酸分子を増幅可能な一つまたは複数のプライマーを、それらの使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項25】
プライマーがX1F(5'-CCATCACAGCCAATGACG-3')(SEQ ID No. 19)およびX1R(5'-AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)である、請求項24記載のキット。
【請求項26】
請求項10〜12のいずれか一項記載のタンパク質またはその断片を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項27】
請求項13記載の抗体を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項28】
障害が、レット症候群または精神遅滞からなる群より選択される、請求項23〜27のいずれか一項記載のキット。
【請求項29】
神経精神障害または発達障害の処置または検出における、請求項1〜5のいずれか一項記載の核酸分子の使用。
【請求項30】
神経精神障害または発達障害の処置または検出における、請求項10〜12のいずれか一項記載のタンパク質の使用。
【請求項1】
MeCP2E1タンパク質をコードする配列を含む、単離された核酸分子。
【請求項2】
以下を含む、請求項1記載の単離された核酸分子:
(a)図6(b)(SEQ ID No. 4)に示したような配列を有する、MeCP2E1タンパク質をコードする核酸配列;
(b)(a)に相補的な核酸配列;
(c)(a)もしくは(b)に対して実質的相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;
(e)少なくとも15塩基、好ましくは20〜30塩基であり、かつストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする、(a)〜(d)の断片;または
(f)遺伝暗号の縮重のために、コドン配列が(a)〜(c)の任意の核酸と異なる核酸分子。
【請求項3】
図6(a)(SEQ ID No. 3)に示した配列を有する、請求項1記載の核酸、またはその断片もしくは変異体。
【請求項4】
突然変異が以下からなる群より選択される、図5(a)(SEQ ID No.1)に示した核酸配列中に突然変異を含む単離された核酸分子:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置69〜70における配列TGの欠失。
【請求項5】
突然変異が以下からなる群より選択される、図5(a)(SEQ ID No. 1)に示した核酸配列中に突然変異を含む単離された核酸分子:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数コピーのGCCトリヌクレオチド配列の挿入;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、ヌクレオチド配列GGAの挿入;
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに
(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【請求項6】
請求項1〜5のいずれか一項記載の核酸分子、およびプロモーターが宿主細胞における核酸分子の転写を増強するように機能的に結合されたプロモーター配列を含む、組換え核酸分子。
【請求項7】
請求項1〜5のいずれか一項記載の核酸分子を含むベクター。
【請求項8】
請求項6記載の組換え核酸分子もしくは請求項7記載のベクターを含む宿主細胞、または宿主細胞の子孫。
【請求項9】
哺乳動物細胞、真菌細胞、酵母細胞、細菌細胞、微生物、および植物細胞からなる群より選択される、請求項8記載の宿主細胞。
【請求項10】
単離されたMECP2E1タンパク質。
【請求項11】
図6(a)(SEQ ID No. 3)に示した核酸配列によりコードされる、請求項10記載のタンパク質、または該タンパク質の断片もしくは変異体。
【請求項12】
図6(b)(SEQ ID No. 4)に示したアミノ酸配列を有する請求項10記載の単離されたタンパク質、または該タンパク質の断片もしくは変異体。
【請求項13】
図6(a)(SEQ ID No. 3)に示した核酸配列のエキソン1によりコードされるペプチドに結合する抗体。
【請求項14】
動物由来の試料中の請求項1〜3のいずれか一項記載の核酸配列内に、突然変異または欠失を検出する段階を含む、神経精神障害または発達障害を検出する方法。
【請求項15】
核酸配列のエキソン1内に突然変異または欠失を検出する段階を含む、請求項14記載の方法。
【請求項16】
以下の段階を含む、請求項15記載の方法であって:
(a)ポリメラーゼ連鎖反応において、プライマーX1F(5'-CCATCACAGCCAATGACG-3')(SEQ ID No. 19)およびX1R(5'-AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)を用いて試料中の核酸配列を増幅する段階;
(b)同一のプライマーを用いて、対照由来の核酸配列を増幅する段階;
(c)増幅された配列のシーケンシングを行う段階;ならびに
(d)対照配列と試料配列を比較する段階;
ここで、対照配列と比較した試料配列中のヌクレオチドの欠失は、試料が神経精神障害または発達障害を有する動物由来であることを示す、方法。
【請求項17】
図5(a)(SEQ ID No. 1)に示したMECP2遺伝子配列の4つのエキソン全てにおいて、多重ライゲーション依存性プローブ増幅(multiplex ligation-dependent probe amplification)を行う段階を含む、請求項14記載の方法。
【請求項18】
請求項10〜12のいずれか一項記載のタンパク質中に欠失または突然変異を検出する段階を含む、神経精神障害または発達障害を検出するための方法。
【請求項19】
神経精神障害がレット症候群である、請求項14〜18のいずれか一項記載の方法。
【請求項20】
検出された突然変異または欠失が以下からなる群より選択される、請求項19記載の方法:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54における11bpの欠失;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド1〜69を含むエキソン1の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置8におけるアデノシンからトレオニンへの変化;および
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド位置70〜71における配列TGの欠失。
【請求項21】
発達障害が精神遅滞である、請求項14〜18のいずれか一項記載の方法。
【請求項22】
突然変異または欠失が以下からなる群より選択される、請求項21記載の方法:
(1)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間への、一つまたは複数のGCCトリヌクレオチド配列の挿入;
(2)図5(a)(SEQ ID No. 1)に示したヌクレオチド11〜29の間からの、一つまたは複数コピーのGCCトリヌクレオチド配列の欠失;
(3)図5(a)(SEQ ID No. 1)に示したヌクレオチド38〜54の間への、GGAヌクレオチド配列の挿入;
(4)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-38および-39におけるヌクレオチド配列GCの欠失;ならびに
(5)図5(a)(SEQ ID No. 1)に示したヌクレオチド1の上流、ヌクレオチド-19および-20におけるヌクレオチド配列AGの欠失。
【請求項23】
請求項1〜5のいずれか一項記載の核酸配列もしくはその断片を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項24】
請求項1〜5のいずれか一項記載の核酸分子を増幅可能な一つまたは複数のプライマーを、それらの使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項25】
プライマーがX1F(5'-CCATCACAGCCAATGACG-3')(SEQ ID No. 19)およびX1R(5'-AGGGGGAGGGTAGAGAGGAG-3')(SEQ ID No. 20)である、請求項24記載のキット。
【請求項26】
請求項10〜12のいずれか一項記載のタンパク質またはその断片を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項27】
請求項13記載の抗体を、その使用のための説明書と共に含む、神経精神障害または発達障害を処置または検出するためのキット。
【請求項28】
障害が、レット症候群または精神遅滞からなる群より選択される、請求項23〜27のいずれか一項記載のキット。
【請求項29】
神経精神障害または発達障害の処置または検出における、請求項1〜5のいずれか一項記載の核酸分子の使用。
【請求項30】
神経精神障害または発達障害の処置または検出における、請求項10〜12のいずれか一項記載のタンパク質の使用。
【図1】

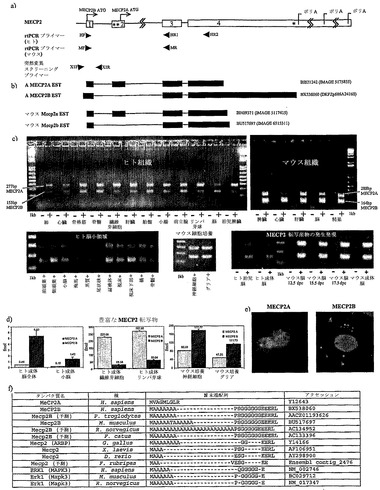
【図2】
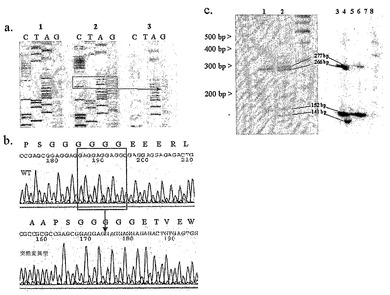
【図3】
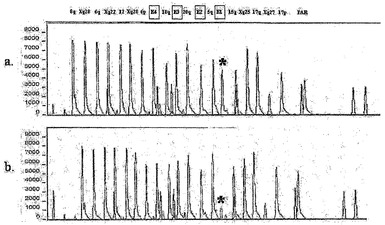
【図4】
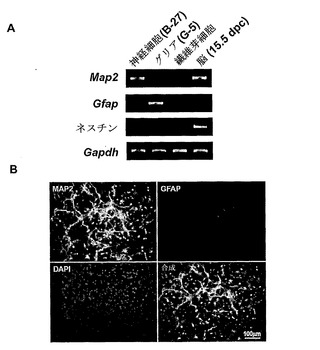
【図5A−1】

【図5A−2】

【図5A−3】

【図5A−4】

【図5B】

【図6A】

【図6B】

【図7】
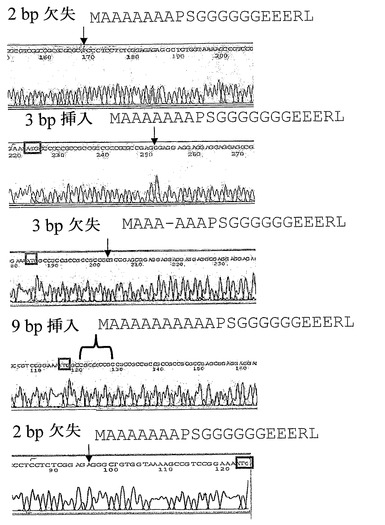
【図8】
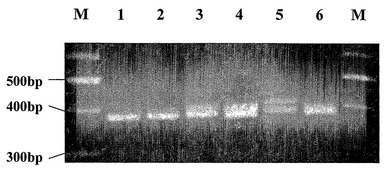
【図2】
【図3】
【図4】
【図5A−1】
【図5A−2】
【図5A−3】
【図5A−4】
【図5B】
【図6A】
【図6B】
【図7】
【図8】
【公開番号】特開2012−10703(P2012−10703A)
【公開日】平成24年1月19日(2012.1.19)
【国際特許分類】
【出願番号】特願2011−167298(P2011−167298)
【出願日】平成23年7月29日(2011.7.29)
【分割の表示】特願2006−553398(P2006−553398)の分割
【原出願日】平成17年2月17日(2005.2.17)
【出願人】(506281886)
【出願人】(506280535)
【Fターム(参考)】
【公開日】平成24年1月19日(2012.1.19)
【国際特許分類】
【出願日】平成23年7月29日(2011.7.29)
【分割の表示】特願2006−553398(P2006−553398)の分割
【原出願日】平成17年2月17日(2005.2.17)
【出願人】(506281886)
【出願人】(506280535)
【Fターム(参考)】
[ Back to top ]