
MHCクラスI制限hTERTエピトープをコードするポリヌクレオチド、それらの類似体またはポリエピトープ
【課題】本発明は、抗癌療法の分野に関するものであり、ヒトテロメラーゼ逆転写酵素(hTERT)由来の免疫原性ペプチドの同定に関する。
【解決手段】本発明は、MHCクラスI分子に制限されたhTERTエピトープをコードするポリヌクレオチド、それらの類似体、並びに前記エピトープ及び/または類似体を含有するポリエピトープに関する。また、本発明は、前記ポリヌクレオチドを含むベクター及び細胞を含む。本発明はまた、癌の治療及び/または予防に使用するための、hTERTポリペプチド、対応するポリヌクレオチド、ベクター及び細胞を含む組成物に関する。
【解決手段】本発明は、MHCクラスI分子に制限されたhTERTエピトープをコードするポリヌクレオチド、それらの類似体、並びに前記エピトープ及び/または類似体を含有するポリエピトープに関する。また、本発明は、前記ポリヌクレオチドを含むベクター及び細胞を含む。本発明はまた、癌の治療及び/または予防に使用するための、hTERTポリペプチド、対応するポリヌクレオチド、ベクター及び細胞を含む組成物に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗癌療法の分野に関するものであり、ヒトテロメラーゼ逆転写酵素(hTERT)由来の免疫原性ペプチドの同定に関する。本発明は、MHCクラスI分子に制限されたhTERTエピトープをコードするポリヌクレオチド、それらの類似体、並びに前記エピトープ及び/または類似体を含有するポリエピトープに関する。また、本発明は、前記ポリヌクレオチドを含むベクター及び細胞を含む。本発明はまた、癌の治療及び/または予防に使用するための、hTERTポリペプチド、対応するポリヌクレオチド、ベクター及び細胞を含む組成物に関する。
【背景技術】
【0002】
現在の抗癌療法手法の欠陥という観点から、抗腫瘍療法は、あらためて関心がもたれている。最近の研究では、腫瘍回帰に関連する抗メラノーマ免疫応答についての本発明者らの理解を深めている。これらのデータをまとめると、活性化された腫瘍特異的なCD8 CTLは、効力のある抗腫瘍療法の選択において免疫学的武器であることを示唆されている。異なる由来の腫瘍に対して細胞毒性Tリンパ球を作動させる過剰発現タンパク質の研究もまた進行している。
【0003】
テロメラーゼは、タンパク質成分のTERT、及び染色体の末端上に付加された反復単位(T2AG3)の合成のためのテンプレートを含有するRNA成分(TR)からなるリボ核タンパク質複合体であって、複製の間染色体を安定化し、末端間融合を防ぐ。一定のテロメア長の維持により、細胞を老化から防ぎ、不死性を付与する(Hahnら、Nat Med 1999年;5:1164〜70頁)。ヒト癌の85%超に高いhTERT活性が見られたが、一方、大部分の正常ヒト成人組織は、テロメラーゼ活性を示さないか、またはほとんど示さない(Counterら、Blood 1995年;85:2315〜20頁)。
【0004】
腫瘍におけるテロメラーゼの広範な発現は、hTERTのペプチド断片が、腫瘍特異的抗原として役立ち得ることを示しており、このことはいくつかの報告により確認されている(Vonderheideら、Immunity 1999年;10:673〜9頁)。第一相臨床試験からの最近のデータは、HLA-A2+患者におけるhTERTに対してワクチンの実現可能性を立証し、治療的ワクチン接種のためのhTERTの使用への道を開いた(Vonderheideら、Clin Cancer Res 2004年;10:828〜39頁;Parkhurstら、Clin Cancer Res 2004年;10:4688〜98頁)。それにもかかわらず、今日まで同定されている免疫原性hTERTペプチドは、1つのMHC対立遺伝子HLA-A2.1に制限され、当該集団の44.2%及び40%にそれぞれ示される2つのHLA上位型のHLA-A3及びHLA-A24に関する最初の2つの報告のみである。
【0005】
従って、以下の刊行物は、hTERTペプチドの同定を報告した:
−HLA-A2に制限されているhTERTペプチドILAKFLHWL(Vonderheideら、Immunity 1999年;10:673〜9頁)、
−HLA-A関連(context)内の情報予測によって同定されたhTERTペプチドMPRAPRCRA、RPAEEATSL、RPSFLLSSL、及びAPRCRAVRS(WO00/02581)。しかし、これらのペプチドは、HLA-B7関連または任意のHLA関連のいずれにおいても、効率的なエピトープであることが実験結果によって確認されていない。
−HLA-A3に制限されているhTERTペプチドKLFGVLRLK(K973)(Vonderheideら、Clin Cancer Res 2001年;7:3343〜8頁)、並びに
−HLA-A24に制限されているhTERTペプチドVYAETKHFL(TEL324)及びVYGFVRACL(TEL461)(Araiら、Blood 2001年;9:2903〜7頁)。
【0006】
その結果、これまでに同定されたhTERTペプチドはすべての集団を網羅しておらず、それゆえ、多くの患者を除外している。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 00/02581
【特許文献2】WO 99/55892
【特許文献3】WO 01/27300
【特許文献4】WO 01/27304
【非特許文献】
【0008】
【非特許文献1】Hahnら、Nat Med 1999年;5:1164〜70頁
【非特許文献2】Counterら、Blood 1995年;85:2315〜20頁
【非特許文献3】Vonderheideら、Immunity 1999年;10:673〜9頁
【非特許文献4】Vonderheideら、Clin Cancer Res 2004年;10:828〜39頁
【非特許文献5】Parkhurstら、Clin Cancer Res 2004年;10:4688〜98頁
【非特許文献6】Vonderheideら、Clin Cancer Res 2001年;7:3343〜8頁
【非特許文献7】Araiら、Blood 2001年;9:2903〜7頁
【非特許文献8】Firatら(1999年、Eur. J. of Immunol. 29:3112〜3121頁)
【非特許文献9】Firatら、2002年、The Journal of Gene Medicine 4:38〜45頁
【非特許文献10】Esslingerら、Hum Gene Ther 2002年;13:1091〜100頁
【非特許文献11】Breckpotら、J Gene Med 2003年;5:654〜67頁
【非特許文献12】Esslingerら、J Clin Invest 2003年;111:1673〜81頁
【非特許文献13】Tartourら、2000年、Immunol Lett Sep 15;74(1):1〜3頁
【非特許文献14】Tartourら、1996年、Presse Med. Nov 16;25(25):1717〜22頁
【非特許文献15】Minevら(2000年、PNAS;97(9):4796〜4801頁)
【非特許文献16】Pascoloら、J Exp Med、1997年;185:2043〜51頁
【非特許文献17】Rohrlichら、Int Immunol、2003年;15:765〜72頁
【非特許文献18】Smithら、J Immunol、1996年;156:3755〜64頁
【非特許文献19】Lu及びCelis E Cancer Res、2000年;60:5223〜7頁
【非特許文献20】Counterら、Proc Natl Acad Sci U.S.A.1998年;95:14723〜8頁
【非特許文献21】Zennouら、Cell、2000年;14:101:173頁
【非特許文献22】Miyahiraら、J Immunol Methods、1995年;181:45〜54頁
【非特許文献23】Hernandezら、Proc Natl Acad Sci U.S.A.、2002年;99:12275〜80頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
この欠陥を少なくとも一部において克服し、それによってヒト集団の遺伝子多様性をより良好に網羅するために、本発明は、HLA-A3及びHLA-A24とは異なる特定のHLAに制限されたhTERT由来の新規なエピトープを同定する。多くの異なる対立遺伝子の中で、本出願は、集団の約25%に発現されるHLA-B7上位型、特にヒトHLA-B集団において最も多く発現される第2の対立遺伝子:HLA-*B0702(ヒト集団における個体の15〜20%に存在する対立遺伝子)に関心を有する。
【課題を解決するための手段】
【0010】
テロメラーゼのアイソフォーム-1の遺伝子は、4015の塩基対(bp)長(NCBI登録番号AF015950)であり、1132のアミノ酸のタンパク質(NCBI登録番号AAC51672.1)をコードする(図1)。
【0011】
本発明は、ヒトテロメラーゼ逆転写酵素(hTERT)ペプチドをコードするポリヌクレオチドに関する。本発明の特定の実施形態において、コード化ペプチドは、長さが9個のアミノ酸(9量体)または長さが10個のアミノ酸(10量体)であり、ゆえに、ポリヌクレオチドは、27または30のヌクレオチドを有する。一般に、コード化ペプチドは15未満のアミノ酸であり、ポリヌクレオチドは45未満のヌクレオチドを有する。
【0012】
本発明はまた、MHCクラスI分子に制限されたエピトープ、特にHLA-B7に制限された免疫応答を誘発するのに好適であるエピトープであるhTERTペプチドをコードするポリヌクレオチドに関する。本発明のポリヌクレオチドのヌクレオチド配列は、特定の実施形態において、hTERTペプチドをコードする配列に限定される。このようなペプチドは、MPRAPRCRA(p1;1〜9のアミノ酸残基)、APRCRAVRSL(p4;4〜13のアミノ酸残基)、APSFRQVSCL(p68;68〜77のアミノ酸残基)、RPAEEATSL(p277;277〜285のアミノ酸残基)、RPSFLLSSL(p342;342〜350のアミノ酸残基)、RPSLTGARRL(p351;351〜360のアミノ酸残基)、DPRRLVQLL(p444、444〜452のアミノ酸残基)、FVRACLRRL(p464、464〜472のアミノ酸残基)、AGRNMRRKL(p966、966〜974のアミノ酸残基)、LPGTTLTAL(p1107、1107〜1115のアミノ酸残基)、及びLPSPKFTIL(p1123、1123〜1131のアミノ酸残基)からなる群から選択されうる。これらのポリヌクレオチドすべては、HLA-B7制限免疫応答を誘発するために使用されうる。特定の実施形態において、本発明は特に、RPSLTGARRL(p351)、APSFRQVSCL(p68)、APRCRAVRSL(p4)、DPRRLVQLL(p444)、FVRACLRRL(p464)、AGRNMRRKL(p966)、LPGTTLTAL(p1107)、及びhLPSPKFTIL(p1123)からなる群から選択される、HLA-B7制限hTERTエピトープをコードするポリヌクレオチドに関する。
【0013】
本明細書に定義されているように、「エピトープ」は、抗原決定基、すなわち免疫系の細胞(免疫細胞)によって認識されるペプチド部位、特に免疫応答を誘発するのに必要な部位である。用語エピトープは、それに対する連続アミノ酸(特に、9または10)が免疫細胞によって認識される線状エピトープ、及びそれが適切な立体配置または立体配座をとる範囲で免疫細胞がアミノ酸を認識する立体配置的エピトープの両方を含む。その結果、いくつかのエピトープにおいて、立体配座(三次元構造)は、アミノ酸配列(一次構造)と同様に重要である。
【0014】
表現「MHCクラスI制限」とは、クラスIのMHC(主要組織適合性複合体)分子に対して親和性を有する特定のペプチドまたはエピトープに対する能力を表す。同様に、表現「HLA-B7制限」とは、このタイプのHLA分子に対する親和性を有する特定のペプチドまたはエピトープに対する能力を表す。
【0015】
要するに、MHC遺伝子は、外来性ペプチドに結合するだけでなく、過剰発現したまたは自己でないペプチド、あるいは変異型自己ペプチドにも結合しうる細胞表面多型分子をコードして、適切な免疫細胞、特にT細胞による認識を可能にする細胞の細胞表面にそれらを提示する。マウスにおいてH-2及びヒトにおいてHLA(ヒト白血球抗原)と称される前記MHC分子は、クラスI分子(HLA-A、B、またはCと表される)またはクラスII分子(DP、DQ、またはDRと表される)として分類される。
【0016】
従って、MHCクラスI分子は、細胞毒性Tリンパ球上に発現されるCD8分子(TCD8+とも称される)に特異的に結合するが、一方、MHCクラスII分子は、ヘルパーTリンパ球上に発現されるCD4分子(TCD4+)に特異的に結合する。
【0017】
MHCクラスI分子は、タンパク分解的に分解されたタンパク質、特に細胞によって内因的に合成されたタンパク質に由来するペプチドに結合する。それによって得られた小型ペプチドは小胞体に輸送され、そこでそれらは新生MHCクラスI分子と結合してから、ゴルジ体を介して輸送されれ、細胞毒性Tリンパ球による認識のために細胞表面に提示される。
【0018】
本発明において、前記同定ペプチドは、一方では高いまたは中程度の親和性でMHCクラスI分子に結合し、他方、MHC/エピトープ複合体として細胞の細胞表面に効率的に輸送されることが示されている。好ましい実施形態において、MHCクラスI分子はHLA-B7上位型ファミリーのMHC対立遺伝子であり;hTERTエピトープは前記HLA-B7上位型制限である。前記ファミリーは、対立遺伝子B0702、B0703、B0704、B0705、B1508、B3501、B3502、B3503、B51、B5301、B5401、B5501、B5502、B5601、B5602、B6701、及びB7801を含み、それらのファミリーからHLA-B0702が好ましい(HLA-B0702制限hTERTエピトープ)。
【0019】
MHC安定化アッセイを用いて、参照として本明細書に組み込まれているFiratら(1999年、Eur. J. of Immunol. 29:3112〜3121頁)に記載されているもの等、特定のHLAクラスI分子に対するペプチドの親和性(相対結合活性)を試験することができる。要するに、MHCクラスI分子でトランスフェクションされた細胞を、
−ペプチドの非存在下(ネガティブコントロール)、
−参照ペプチドの存在下(ポジティブコントロール)、及び
−試験されるペプチド(この場合、hTERTペプチド)の存在下で、
100ng/mlのヒトβ2-ミクログロブリンを補充された無血清培地AIM-V(Invitrogen社、Gibco)中、96ウェルプレート中に2×105細胞/ウェルで一晩インキュベーションする。
ペプチドを、0.1〜100μMの範囲の種々の最終濃度で(1μM及び10μMの中間濃度で)インキュベーションする。次いで、トランスフェクションされた細胞を、特定のHLA MHCクラスI分子を認識する抗体の飽和濃度で標識し、次に2回洗浄し、最終的にフローサイトメトリー前に二次抗体で染色する。結果は相対結合活性値として表され、これは、参照ペプチドの値を参照して最大結合の20%に達するのに必要な試験ペプチドの濃度比率である。それゆえ、この値が低くなるほど、結合が強くなる。この方法の後に、RA<1の場合、ペプチドは特定のHLAクラスI分子に対して高い相対親和性を有するといわれる。対照的に、RAが1〜5、好ましくは1〜3である場合、中程度の相対親和性と結論付けられる。
【0020】
hTERT9量体及び10量体、特に前記同定ペプチドの中で、以下のMHCクラスI制限hTERTエピトープ:MPRAPRCRA(p1)、APRCRAVRSL(p4)、及びAPSFRQVSCL(p68)を、MHCクラスI分子に対して高い相対親和性を有するものとして分類することができる。同じ手法を用いて、以下のMHCクラスI制限hTERTエピトープ:RPAEEATSL(p277)、RPSFLLSSL(p342)、及びRPSLTGARRL(p351)を、MHCクラスI分子に対して中程度の相対親和性を有するものとして分類することができる。
【0021】
本発明はまた、MHCクラスI制限エピトープ類似体、すなわち前記のようなクラスI制限hTERTエピトープと比較して少なくとも1つのアミノ酸置換を有するエピトープ、特にHLA-B7制限エピトープ類似体をコードするポリヌクレオチドに関する。
【0022】
本明細書に定義される用語「類似体」とは、その配列が、保存的、半保存的、または非保存的な少なくとも1つのアミノ酸置換により、前記のhTERTペプチドから得られるペプチドに関する。類似体は、そのヌクレオチド及び/またはアミノ酸配列が、いわゆる野生型ペプチドまたはポリヌクレオチドを定義するために、参照の分子として本出願内で考慮される図1に開示されている参照hTERT遺伝子またはタンパク質に見られないという事実により、エピトープと対立するものである。このような類似体は、前記アミノ酸をコードするコドンにおける1つまたはいくつかの核塩基の対応するヌクレオチド配列の置換により生じ得る。従って、ポリヌクレオチド類似体は、置換されるアミノ酸残基をコードするコドンにおいて少なくとも1つの置換、好ましくは1つ、2つ、または3つの置換により、その野生型対応物(ペプチド類似体が由来する参照配列でhTERTペプチドをコードするポリヌクレオチド)とは異なる。一例として、APRRLVQLLペプチド(p444*と称される)は、最初のアミノ酸残基(D->A)の置換によりp444ペプチドから得られる。
【0023】
HLA-B7制限類似体の特定の類似体は、それが由来するエピトープと同じ長さを有するか、またはそれよりも短い。
【0024】
特定の実施形態として、前記のhTERTエピトープまたはその類似体は、それらの一次構造において、2位にプロリン(P)、及び/または最後のC末端位置に以下のアミノ酸:A、L、I、M、V、F、W、またはYのうちの1つを常に保存する。従って、類似体を含むhTERTペプチドは、以下のコンセンサス配列:X-P-X6〜7-[ALIMVFWY]を有し、式中、Xは任意のアミノ酸を表し、X6〜7はアミノ酸数を表し、且つ[ALIMVFWY]はこれらのアミノ酸のうちの1つを表す。
【0025】
従って、エピトープ類似体を提供するために、当該ポリヌクレオチドにおけるアミノ酸置換または対応するコドン置換は、2位(または第2コドン)に位置しない。好ましい実施形態において、たとえ最後のC末端アミノ酸を等価のアミノ酸、すなわちA、L、I、M、V、F、W、またはYで置換できたとしても、C末端位置において置換は行われない。最後に、好ましい実施形態において、置換は1位に位置し、任意のアミノ酸はアラニン(A)で置換される。
【0026】
本発明のさらなる実施形態において、生じた9量体がX-P-X6〜7-[ALIMVFWY]コンセンサス配列を維持するか、または選択するという条件で、10量体の最後のC末端アミノ酸を欠失させて、9量体を得る。同じ方法で、生じた10量体がX-P-X6〜7-[ALIMVFWY]コンセンサス配列を維持するか、または選択するという条件で、9量体のC末端にA、L、I、M、V、F、W、及びYの中から選択するアミノ酸を付加させて、10量体を生じさせる。
【0027】
個々にまたは組合せとして選択され、特に前記に説明されたものを含む、置換、欠失、または付加の場合、ペプチド類似体の三次元立体配座は、野生型対応物の1つに対して同じかまたはわずかに改変されたものでなければならず、それにより、当該類似体の正しい折りたたみ及びMHCクラスI分子に対するその正しい結合を確保する。前記のMHC安定化アッセイを用いて、このような制約が満たされることをチェックすることができる。
【0028】
生じた類似体は、特定のMHCクラスI分子、特にHLA-B7分子に対する親和性に関して、その野生型対応物と少なくとも同じ特徴を有し、且つ細胞表面上のエピトープ/MHC複合体と実質的に同じ輸送される能力を有し、及び/または同じ条件で試験された場合、免疫原性応答を誘発する実質的に同じ能力を有する。
【0029】
好ましい実施形態において、開始ペプチドは、中程度の親和性を有するhTERTエピトープであり、生じた類似体は、その野生型対応物よりも高い親和性を有する。別の実施形態において、類似体は、その野生型対応物よりも高い免疫原性を有する。一例として、前記に示されたp444*ペプチド類似体は、それが由来するp444ペプチドと比較して、MHCクラスI分子に対して増大した親和性を有する。
【0030】
本発明の目的は、免疫応答、特にCTL応答(細胞毒性Tリンパ球)を誘発できるhTERTエピトープまたは類似体を提供することである。しかしながら、Tリンパ球は、抗原提示細胞を介して前記リンパ球のみを刺激するために使用される前記類似体を認識しない。本発明の実施形態において、前記の類似体は、その免疫原性挙動を維持し、且つhTERTエピトープを過剰発現する細胞に対する免疫応答、すなわちCTLはエピトープ類似体により刺激されたとしても、野生型エピトープを認識することを誘発できる。特定の実施形態において、本発明のエピトープ類似体により刺激されたリンパ球は、hTERTエピトープを過剰発現しない細胞に対して反応しない。従って、刺激されたリンパ球は、他のエピトープを過剰発現する細胞と反応(交差反応)しないか、または基底レベルとしてのhTERTエピトープを発現する細胞(健常細胞)と反応しない。特定の実施形態において、前記のすべての特徴は、HLA-B7環境内にあり、好ましくは、HLA-B0702関連にある。
【0031】
参照として本明細書に組み込まれているFiratの刊行物(1999年、Eur. J. of Immunol. 29:3112〜3121頁)にあるように、標準的4〜5時間の51Cr放出アッセイを使用することによって従来の細胞毒性アッセイを実施して、標的細胞に対して反応する刺激されたリンパ球の能力を試験することができる。要するに、CTLを含む細胞懸濁液をin vivoでペプチド(この場合、hTERTペプチドまたは類似体)+自己MHCクラスI分子で活性化する。次いで、あらかじめ51Crとインキュベーションされた標的細胞(対応するhTERTエピトープを発現しているまたは発現しない)を、活性化リンパ球とともにインキュベーションする。活性化CTLによる標的細胞の認識により、標的細胞のアポトーシス及び51Crの放出が生じ、前記放出は死滅した標的細胞数に比例する。標的細胞とコントロールペプチドとのインキュベーションを、51Crの自発的放出を算出するためにネガティブコントロールとして使用する。試験されるペプチドに関して得られた細胞溶解に対してコントロールペプチドで観察された非特異的細胞溶解を差し引くことによって、細胞溶解の具体的なパーセンテージを算出する。そのパーセンテージが高いほど、CTLにより死滅した標的は多い。特異的細胞溶解を、エフェクター(CTL)対標的細胞(E:T)のいくつかの比率で判定する。特定の細胞溶解を、[実験的放出−自発的放出]と[全放出−自発放出]との間の比率として算出する。
【0032】
本発明はまた、ポリエピトープをコードするポリヌクレオチドに関する。ポリエピトープとは、MHCクラスI制限、特に本発明のHLA-B7制限hTERTエピトープ(p1、p4、p68、p277、p342、p351、p444、p464、p966、p1107、及びp1123)及び/またはMHCクラスI制限、特にHLA-B7制限エピトープ類似体の中から選択される少なくとも2つのエピトープを有するポリペプチドとして定義される。本発明のポリヌクレオチドは、前記エピトープまたは類似体をコードする少なくとも2つのポリヌクレオチド単位を含むか、またはそれらからなる。ポリヌクレオチド単位は、本明細書に開示された本発明のエピトープまたは類似体に対するコード配列として定義される。
【0033】
本発明は特に、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)、または前記に定義されるそれらの類似体の中から選択される少なくとも2つのエピトープを含む、ポリエピトープをコードするポリヌクレオチドに関する。ポリエピトープをコードする別のポリヌクレオチドは、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)に対応するHLA-B7制限hTERTエピトープ及び/または前記に定義されるそれらの類似体をコードするポリヌクレオチドの群から選択される少なくとも1つのポリヌクレオチド単位、並びに配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)、あるいは前記に定義されるそれらの類似体をコードするポリヌクレオチドの群から選択される少なくとも1つのポリヌクレオチド単位を含む。
【0034】
本発明のポリエピトープをコードするポリヌクレオチドのいずれも、全長hTERTのコード配列と一致しない。
【0035】
本発明の特定の実施形態において、調製されたポリエピトープにおけるMHCクラスI制限hTERTエピトープ及び/または類似体の数は、30に制限される。別の実施形態において、HLA-B7制限hTERTエピトープ及び/または類似体の数は、およそ30に制限され、好ましくは2、3、4、5、6、7、8、9、10、15、20、25、または30である。別の実施形態において、HLA-B0702制限hTERTエピトープ及び/または類似体の数は、およそ10に制限され、好ましくは2、3、4、5、6、7、8、9、または10である。
【0036】
従って、ポリエピトープをコードするポリヌクレオチドは、30以下のポリヌクレオチド単位、特に2、3、4、5、6、7、8、9、10、15、20、25、または30、あるいはこの範囲内の任意の数を有する。
【0037】
特定の実施形態において、前記ポリヌクレオチドのポリヌクレオチド単位(ゆえに、エピトープ)は連続的である。
【0038】
本発明のこのような特定の実施形態において、連続的hTERTエピトープまたは類似体をコードする核酸配列のサイズは、3000bp未満であり、好ましくは2000bp、1000bp、500bp、400bp、300bp、200bp、または100bp未満である。
【0039】
特定の実施形態において、複数エピトープ(ポリエピトープ)をコードするポリヌクレオチドは、hTERTタンパク質の切断型または変異型をコードする核酸分子からなる。好ましい実施形態において、hTERTタンパク質の切断型または変異型は、その触媒活性を失い、すなわちテロメア長の維持に関与する染色体の末端における反復単位(T2AG3)の合成を方向付けることができない。その触媒活性、すなわち逆転写酵素活性が失われたそのようなhTERTタンパク質は非機能的といわれる。従って、別の特定の実施形態において、hTERTタンパク質の切断型または変異型をコードする核酸分子は、hTERTの触媒活性ドメインを欠失している。特定の実施形態において、hTERTタンパク質の切断型をコードする核酸分子は、少なくとも500の最後のC末端アミノ酸からなるタンパク質をコードする。
【0040】
特定の実施形態において、当該ポリヌクレオチドは、図1(野生型)のヌクレオチド2654〜2662に対応し、欠失部位を表す図9のヌクレオチド2658に隣接するヌクレオチド2657を提供するアミノ酸867〜869(VDD配列)を欠失した、あるいは、図1(野生型)のヌクレオチド2645〜2671に対応し、欠失部位を表す図10のヌクレオチド2649に隣接するヌクレオチド2648を提供するアミノ酸864〜872を欠失したhTERTタンパク質をコードする。特定の実施形態において、コードされたhTERTタンパク質は、少なくともアミノ酸残基867〜869を含む欠失、すなわち3個のアミノ酸残基(VDD配列)よりも大きい欠失を有する。一例として、前記の864〜872欠失、並びにアミノ酸残基857から出発して879までの22のアミノ酸欠失(図1によれば)、あるいはVDD配列(ヌクレオチド2639〜2679に対応する、図1によればアミノ酸862〜アミノ酸874)に対して5個のN末端アミノ酸及び5個のC末端アミノ酸を含む欠失である。特定の実施形態において、本発明は、図9または図10に示された核酸配列を含むか、またはそれからなるポリヌクレオチドに関する。
【0041】
本発明の複数エピトープをコードするポリヌクレオチド単位は連続的に配列され得、すなわち第1のポリヌクレオチド単位の3'末端は、第2のポリヌクレオチド単位(など)の5'末端に直接連結し、連続的エピトープのみを含むペプチド配列をコードするポリヌクレオチドを生じる。このようなポリヌクレオチドは、p1、p4、p68、p277、p342、p351、p444、p464、p966、p1107、及びp1123ペプチドを含むか、またはそれらからなるポリエピトープをコードし得る。特定の実施形態において、前記ポリヌクレオチドは、以下のペプチド配列MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLをコードし、ゆえに6個のMHCクラスI制限、特にHLA-B7制限hTERTエピトープを含む。
【0042】
あるいは、本発明の複数エピトープは、1個のアミノ酸スペーサーまたはペプチドスペーサーによって分離され得る。すなわち、異なるポリヌクレオチド単位は、それぞれ1個またはいくつかのアミノ酸をそれぞれコードする1個またはいくつかのコドンによって分離されることを意味する。複数エピトープのプロセシングを促進するスペーサーとして、C末端位置におけるアルギニン(R)及び他の位置における親水性残基(A、K、D、及び/またはT)を含む4個のアミノ酸ペプチドが好ましい。特にC末端位置における陽電荷残基または酸性残基を有する4個のアミノ酸ペプチドを、他の位置における親水性残基(A、K、D、及び/またはT)に依存して、またはそれらから独立して使用できる。特定の実施形態において、前記スペーサーは、複数エピトープのより良好なプロセシングを可能にし、且つ重複した切断により生じる新たなペプチドのプロセシングを避ける、エンドソームまたはリソソームプロセシング配列等の内在的プロセシング配列である。そのようなスペーサーに依存した分離を用いて、ポリヌクレオチド単位のすべて、またはそれに反して一部、ゆえにエピトープのすべてまたは一部を分離することができる。
【0043】
エピトープが配列される順序は、以下の:いくつかの順序は、ポリヌクレオチドの転写及び/または翻訳を促進し得、特に三次元立体配座がその特性に影響を与える場合、小胞体(ER)においてその生じた発現ポリエピトープの輸送を促進し得、且ついくつかのエピトープまたは類似体におけるポリエピトープのプロセシングを促進して、重複するエピトープのプロセシングを避けることができる;という基準に従って当業者により決定され得る。
【0044】
本発明のポリエピトープは、単一の動物またはヒトにおいて、同時に、ポリエピトープ中に含まれる少なくとも1つ、特にいくつかのエピトープまたは類似体に対するCTL応答の誘発を可能にする。
【0045】
特定の実施形態において、本発明のポリエピトープをコードするポリヌクレオチドは、少なくとも2つのエピトープの最もN末端側のエピトープをコードするポリヌクレオチド単位に作動可能に連結された、標的シグナルをコードするポリヌクレオチドをさらに含む。本明細書に用いられる「作動可能に連結された」とは、標的シグナルを作動可能にでき、すなわちポリエピトープを正しい細胞コンパートメントまたはドメインに標的化できる方法で、標的シグナル(上流配列)をN末端エピトープ(下流配列)に連結させることを意味する。従って、2つの配列間の連結により、各配列が異なる位置及び/または異なる段階でそれ自身の機能を果たす。特定の実施形態において、前記標的シグナルは、小胞体シグナル配列であり、適切なプロセシング及びMHCクラスI分子との結合のために、ポリエピトープをERに方向付けさせる。
【0046】
さらなる実施形態において、翻訳過程が開始コドンを必要とする場合、メチオニン残基をコードするコドンを最もN末端側のエピトープをコードする配列の上流に付加させて、ポリヌクレオチドの正しい翻訳を可能にする。最もN末端側のエピトープがその1位にメチオニン残基を有さない場合にのみ、このようなコドンを付加する。
【0047】
本発明はまた、前記に示した定義に従って、
a. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
b. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
c. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
d. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
e. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、
f. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)、
g. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
h. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
i. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
j. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
k. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
l. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
からなる群から選択される少なくとも2つのポリヌクレオチド単位を含むか、またはそれらからなるポリヌクレオチドに関する。
【0048】
以下のポリヌクレオチド:
−a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位、並びに
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位を含むポリヌクレオチド、
−a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位を含むポリヌクレオチドもまた、本発明の一部を構成する。
【0049】
遺伝子コードの縮重を考慮すると、本発明のエピトープ、類似体、またはポリエピトープをコードするポリヌクレオチドまたはポリヌクレオチド単位もまた本発明に含まれる。従って、hTERTヌクレオチド参照配列からのコドンによって、または前記アミノ酸をコードする任意のコドンによって各アミノ酸をコードすることができる。
【0050】
本発明はまた、前記群から選択されるこれらのポリヌクレオチド単位またはそれらの類似体の少なくとも2つの任意の組合せを含むか、またはそれらからなるポリヌクレオチドに関するものであって、前記ポリヌクレオチド単位または類似体は、MHCクラスI制限、特にHLA-B7制限hTERTエピトープをコードする。
【0051】
エピトープ、類似体、ポリエピトープ、それらの組合せ、スペーサー、標的シグナル配列等に関して前記に示されたすべての特徴は、本発明の任意のポリペプチド、並びに対応するポリヌクレオチド配列に適用可能である。
【0052】
前記に定義された本発明のポリヌクレオチドを含むか、またはそれからなる組換えベクターもまた、本発明の目的の1つである。前記組換えベクターは、真核生物または原核生物発現用のベクター、例えばプラスミド、細菌導入用のファージ、酵母を形質転換するためのYAC、ウイルスベクター、特にレトロウイルスベクター、または任意の発現ベクターであってよい。本明細書に定義される発現ベクターを選択して、in vitroまたはin vivoのいずれかで、前記に定義されたエピトープまたは類似体またはポリエピトープの産生を可能にする。
【0053】
従って、前記ポリヌクレオチドに加えて、本発明のベクターは、転写制御領域(プロモーター、エンハンサー、リボソーム結合部位(RBS)、ポリAシグナル)、終結シグナル、原核生物もしくは真核生物の複製起点、及び/または選択遺伝子をさらに含むことができる。プロモーターの特徴は、必要とされる発現、すなわち構成的、一過性または誘導性、強いまたは弱い、組織特異的及び/または発生段階特異的プロモーターを考慮して、当業者によって容易に決定され得る。従って、組織特異的プロモーターは、このベクターを含有する組成物が投与される、例えば注入される臓器に応じて、及び必要とされる発現強度に応じて選択され得る。特定の実施形態において、プロモーターはCMVプロモーター(ヒトサイトメガロウイルス)である。前記ベクターはまた、条件付発現を可能にする配列、例えばCre/Loxシステムまたは類似システムの配列等を含むこともできる。
【0054】
本発明の発現ベクターは、ウイルスベクター、特にレトロウイルス由来等のウイルス発現ベクター、特にHIV-、FIV-、またはSIV-由来のベクター等のレンチウイルス由来のベクターであってよい。とりわけ、レンチウイルス由来のベクターは、ヒトレンチウイルス由来のベクター、例えばHIV発現ベクター、特にHIV-1またはHIV-2由来のベクター等である。レトロウイルス由来のベクターは、通常、プラスミド等のDNA構築物中に含まれ、ウイルス粒子中に発現されるレトロウイルスベクターゲノムを含み、前記レトロウイルスベクターゲノムは、逆転写に必要な要素、特に部分的欠失、特にU3領域における欠失を含む変異可能なLTRを含む。いずれの場合にも、レトロウイルス由来のベクターは、全長レトロウイルスタンパク質をコードするすべてのヌクレオチド配列含有することはない。場合により、前記タンパク質またはその機能性断片をコードしないという条件で、それは、1つまたはいくつかの前記ヌクレオチド配列の一部を含有する。前記レトロウイルスベクターゲノムを含む前記DNA構築物は、レトロウイルスヌクレオチド配列で組換えられた対象のDNAをさらに含み、前記対象のDNAは、本発明のポリヌクレオチドを含むか、またはそれからなる。
【0055】
好ましい実施形態において、レトロウイルス由来のベクターゲノムは、下記のようなDNAフラップ及び本発明の少なくとも1つのポリヌクレオチドを含む。好ましい実施形態において、レトロウイルス由来のベクターは、下記のようなDNAフラップ及び本発明の少なくとも1つのポリヌクレオチドを含むHIV発現ベクターである。従って、HIVベクターは、2つのHIV LTRの間に含まれる本発明のポリヌクレオチドを含んだ対象の核酸のみを発現し、ゆえに、5〜6kbまでの大きな配列を収容し得る。本発明の特定の実施形態は、HIV発現ベクター、とりわけHIV-1またはHIV-2発現ベクターであり、HIV-1 LTRまたはHIV-2 LTRはそれぞれ、U3ドメインのプロモーター及びエンハンサーを欠失されている(ΔU3)。この特定の欠失は、特にプロモーターと結合される場合、当該ベクター中に含まれる核酸の発現を増加させることが以前に示されている。別の特定の実施形態において、当該ベクターは、U3ドメインのプロモーター及びエンハンサーを欠失したLTR、CMVまたはEF1αプロモーター等のプロモーター、及び本発明のポリヌクレオチドを含む。
【0056】
別の実施形態において、レトロウイルス由来のベクターに導入された本発明のポリヌクレオチドは、発現カセット中に含まれる。
【0057】
DNAフラップ(またはWO99/55892、WO01/27300及びWO01/27304に開示されたトリプレックス)は、2つの必須領域、すなわちcPPT(セントラルポリプリン配列)及びCTS領域(シス作用終結領域)を含むレトロウイルスまたはレトロウイルス様由来のヌクレオチド配列であって、前記cPPT及びCTS領域は、三本鎖DNA構造を誘導する。本発明に適したDNAフラップは、レトロウイルスまたはレトロウイルス様生物、例えば合成的(化学合成)またはポリメラーゼ連鎖反応(PCR)等の任意のレトロウイルス核酸からのDNAフラップの増幅により調製されたレトロトランスポゾン等から得ることができる。DNAフラップを得ることができるレトロウイルスは、特にレトロウイルスまたはレンチウイルス、とりわけヒトレトロウイルスまたはレンチウイルスであり、特にHIVレトロウイルス、CAEV(ヤギ関節炎脳炎ウイルス)ウイルス、EIAV(ウマ感染性貧血ウイルス)ウイルス、VISNAウイルス、SIV(サル免疫不全ウイルス)ウイルス、またはFIV(ネコ免疫不全ウイルス)ウイルスである。より好ましい実施形態において、DNAフラップは、HIVレトロウイルス、例えばHIV-1またはHIV-2ウイルス、あるいはこれら2つのタイプの任意の異なる単離株から得られる。前記DNAフラップは、その天然(ウイルスゲノム)ヌクレオチド関連、すなわちレトロウイルス中に天然に含まれるpol遺伝子から単離されて使用されることが注目に値する。従って、本発明において使用されるDNAフラップは、pol遺伝子の不必要な5'及び3'部分を欠失しており、異なる由来の配列で組換えられている。
【0058】
DNAフラップは、ベクターの核内への移入のシス決定基として作用し、組換え並びに非分裂及び分裂細胞の両方への核酸の組込みにとって非常に興味深い。DNAフラップ配列(TRIPベクター)を含む発現レトロウイルス由来のベクター、特にHIV由来のベクターは、DNAフラップを欠失する他のHIVベクターよりも10倍高い効率で、初代B細胞及びT細胞、マクロファージ、樹状細胞等を形質導入することができる。細胞の80〜90%の形質導入を、日常的に得ることができる。
【0059】
好ましい実施形態において、in vivo発現及びワクチン戦略に適したベクターは、レトロウイルスベクター、特にレンチウイルスベクターである(WO 99/55892、WO 01/27300、及びWO 01/27304を参照)。このようなベクターは、それらのゲノムが改変された場合、特に効率的で安全であることが示されている(Firatら、2002年、The Journal of Gene Medicine 4:38〜45頁)。実際、これらのベクターは、治療遺伝子またはレポーター遺伝子を、多種多様な細胞及び組織、例えば造血幹細胞、脳、肝臓、及び網膜等のに効率的かつ安定的に移行させる能力を有する。さらに、この高い形質導入効率は、標的細胞の増殖状態に無関係である。特に、これらのベクターは、in vivoだけでなくex vivoで高い効率で樹状細胞(DC)等の抗原提示細胞を形質導入する能力により、マウスにおけるin vivo及びヒトにおけるex vivoの両方でCD8+ T細胞応答を効率的に誘発することが示されている(Esslingerら、Hum Gene Ther 2002年;13:1091〜100頁;Breckpotら、J Gene Med 2003年;5:654〜67頁;Esslingerら、J Clin Invest 2003年;111:1673〜81頁)。
【0060】
図7に定義されているベクターは、本発明のエピトープ、類似体、またはポリエピトープのin vitroまたはin vivo発現に使用されている。本発明のベクターの由来となり得るベクターの例は以下のとおりであり、すべてCNCM(Collection Nationale de Culture de Microorgnismes at Institut Pasteur:パスツール研究所にある国立微生物コレクション、パリ、フランス)に寄託されている。
【表1】

【0061】
特許出願WO 01/27304に記載されているベクター、特にTRIPΔU3 Efα1 GFP及びTRIPΔU3 PL CMV GFPもまた、本発明のベクターを得るために使用することができる。
【0062】
本発明の特定のベクターは、2006年7月28日に番号CNCM I-3660の下でCNCM(パスツール研究所(Institut Pasteur)、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターである。このベクターを培養するのに適した増殖培地は、場合によってはヒグロマイシンを補充されたTB培地である。本発明の別の発現ベクターは、2006年7月28日に番号CNCM I-3660の下でCNCMに寄託されたpTRIP-CMV-ΔhTERTベクターであり、欠失したhTERT配列は、本発明の任意のポリヌクレオチドによって置換されている。
【0063】
本発明はまた、本発明のポリヌクレオチドまたはポリヌクレオチド単位を含む細胞に関する。
【0064】
一実施形態において、当該細胞は、当業者に周知の方法によって、すなわち化学的トランスフェクション(リン酸カルシウム、リポフェクタミン)、脂質ベース方法(リポソーム)、エレクトロポレーション、光穿孔、ウイルスベクターの使用等によって、本発明のベクターでトランスフェクションされる。別の実施形態において、相同的細胞配列での組換えまたは細胞ゲノム内への挿入のいずれかによって、細胞ゲノム内へのポリヌクレオチドの組込みを可能にする方法で、細胞を本発明のポリヌクレオチドで形質転換または形質導入する。トランスフェクション、感染、または形質導入は、ex vivo、すなわち生体外の人工的環境で生じる得る。
【0065】
ワクチン戦略で特に関心のある細胞としては、免疫系の細胞、特に抗原提示細胞(APC)がある。特定の実施形態において、これらの細胞は、樹状細胞(DC)のようなMHCクラスI認識、あるいはマクロファージまたはBリンパ球等のMHCクラスII認識のいずれかに関与するAPCである。DCとしては、ex vivoで完全に成熟させたDC、すなわちエピトープまたは類似体によってin vitroで成熟されたDCが好ましい。
【0066】
本明細書に用いられている用語「トランスフェクションされた」、「形質転換された」、または「感染された」とは、本発明のベクターを含む細胞(一過性発現)を表し、一方、用語「遺伝的に形質転換された」とは、そのゲノムが、本発明のポリヌクレオチドによって決定的に改変されている細胞(永続的発現)を表す。
【0067】
前記一過的または安定的に形質転換された細胞は、任意の原核生物(細菌)細胞または真核生物(酵母、哺乳動物、特にヒトを含む動物)細胞であってよい。一実施形態において、細胞は非ヒト細胞である。特定の実施形態において、本発明の細胞は、単離されたヒト細胞であり、「単離された」とは、その天然環境の外部を意味する。
【0068】
特定の宿主は、2006年7月28日に番号CNCM I-3660の下でCNCMに寄託された大腸菌(E. coli)株である。
【0069】
本発明はまた、本発明のポリヌクレオチドを記載する際に前記に定義されたエピトープ、類似体、またはポリエピトープ、特に本発明のポリヌクレオチドまたはポリヌクレオチド単位によってコードされた任意のポリペプチドに関する。特定のポリペプチドは:
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるMHCクラスI制限、特にHLA-B7制限hTERTエピトープである。
【0070】
特定のHLA-B7制限hTERTエピトープは:
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択される。
【0071】
特定の群は、以下のMHCクラスI制限、特にHLA-B7制限hTERTエピトープ:
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、及び
d. RPSLTGARRL(p351)
からなる。
【0072】
別の群は、以下のHLA-B7制限hTERTエピトープ:RPAEEATSL(p277)及びRPSFLLSSL(p342)からなる。これらのエピトープによって標的化される好ましいHLA-B7対立遺伝子は、HLA-B0702である。
【0073】
本発明はまた、前記に開示され、少なくとも1つのアミノ酸置換を有するペプチドの類似体に関する。これらの類似体に関する特徴は、特に前記の頁に記載されている。特定のペプチド類似体は、以下のペプチド配列APRRLVQLLを有するp444*である。
【0074】
最後に、本発明はまた、本明細書に記載されているhTERTエピトープ、類似体、またはポリエピトープをコードする任意のポリヌクレオチドに関する。
【0075】
本発明はまた、前記の少なくとも2つのエピトープ及び/または類似体を含むポリエピトープに関する。本発明のポリエピトープは、全長hTERTタンパク質ではない。前記ポリエピトープのサイズは、15〜1000個、特に50または100〜1000個のアミノ酸であってよく、とりわけ特に約100、200、300、400、500、または1000個のアミノ酸である。このようなエピトープは、2〜30個のエピトープまたは類似体、特に2〜20個または2〜10個のエピトープ及び/または類似体を含むか、またはそれらからなる。特定のポリエピトープは、6個の連続エピトープを含むか、またはそれらからなり、以下の配列:MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLを有する。別の特定のポリエピトープは、前記p1、p4、p68、p277、p342、及びp351エピトープを含むか、またはそれらからなり、前記エピトープは、得られたポリエピトープにおいて互いに連続的であるか、ペプチドスペーサーによってそれらのすべてもしくは一部は分離されている。
【0076】
本発明の別のポリエピトープは、少なくとも2つのエピトープを含み、少なくとも1つが:
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、且つ少なくとも1つが:
j. MPRAPRCRA(p1)、
k. RPAEEATSL(p277)、
l. RPSFLLSSL(p342)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、前記ポリエピトープは、全長hTERTではない。
【0077】
本発明のポリペプチドは、化学的に合成され得るか、あるいは細胞系で対応する核酸配列の発現後にin vitro(無細胞系)またはin vivoで産生され得る。
【0078】
図1に示される全長hTERTタンパク質並びに対応する全長コード配列は、本発明から除外される。RPALLTSRLペプチドも本発明から除外される。これらのペプチドは、HLA-B7認識の関連において特に除外される。
【0079】
本発明はまた、hTERTに対するHLA-B7制限免疫応答を誘発させるか、またはそれを提供することに関与する使用のために、前記に定義されたエピトープ、類似体、ポリエピトープまたはポリヌクレオチド、発現ベクターまたは宿主細胞に関する。
【0080】
本発明はまた、本発明のポリヌクレオチド、ベクター、宿主細胞、及び/またはポリペプチドを含む組成物に関する。特定の実施形態において、前記組成物は、in vivo投与に適しており、すなわち前記組成物は、注入用に調製されるか、またはより一般的には、投与用に生理的に許容され得る溶液または乳濁液とともに調製される。前記組成物は、特に注入によって全身または局所投与のいずれかに使用され得、製薬的に適した賦形剤(水、生理食塩水、デキストロース、グリセロール、エタノール、及びそれらの組合せを含む)、あるいはキャリア及び/またはビヒクルをさらに含むことができる。
【0081】
特定の実施形態において、前記組成物は、エピトープ、類似体をコードするか、または前記のポリエピトープをコードする、本発明のポリヌクレオチドを含む。前記組成物は、HLA-B7のものとは異なるMHCクラスI対立遺伝子に制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをコードする他の核酸分子を含むことができる。種々のHLA上位型または対立遺伝子に制限されたhTERTエピトープの組合せは、単一の上位型または対立遺伝子よりも治療を必要としている患者のより大きな集団を網羅することができる。最後に、HLA-A1、-A2、-A3、及びA24が好ましい。
【0082】
別の実施形態において、当該組成物は、MHCクラスIIに制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをコードする核酸分子を含む。前記組成物は、前記の核酸分子と、本発明の少なくとも1つのHLA-B7制限hTERTエピトープ、類似体、またはポリエピトープとの任意の組合せを含むことができる。核酸分子の組成物において、種々の免疫細胞(クラスIに対するTリンパ球またはNK細胞対クラスIIに対するヘルパーリンパ球)の反応、及び/または種々の免疫応答(液性応答対細胞性応答)の誘発を可能にする、クラスI及びクラスII制限エピトープをコードするポリヌクレオチドの組合せもまた、本発明内に入る。
【0083】
別の実施形態において、当該組成物は、1つの腫瘍特異的抗原(TSA)をコードする少なくとも核酸分子、並びに/あるいは少なくとも1つの腫瘍関連抗原(TAA)、例えば前立腺特異的抗原(PSA)、前立腺特異的膜抗原(PSMA)、または前立腺酸性ホスファターゼ(PAP)等を含む核酸分子を含む(Tartourら、2000年、Immunol Lett Sep 15;74(1):1〜3頁;Tartourら、1996年、Presse Med. Nov 16;25(25):1717〜22頁)。
【0084】
いくつかのタイプの治療用組成物を、本発明のエピトープまたは類似体に対して免疫応答を誘発させるために使用することができる。
【0085】
本発明ポリヌクレオチドを含む組成物は、宿主に投与され、例えば注入され(DNAワクチン接種として知られている)、前記核酸は、本発明の複数エピトープを含むか、またはそれらからなるポリペプチドをin vivoで発現する。このようなDNAワクチン接種は、通常、前記のプラスミドベクターからなる。裸のDNAの送達は、効率に乏しいことが示されており、細胞内へのDNAの送達及び取込みを改善するためにいくつかのキャリアを必要とする。2つのタイプのキャリア:(1)ウイルスキャリア(アデノウイルス、レンチウイルス、麻疹ウイルス)、または(2)非ウイルス性キャリア、例えばポリマー(特にカチオン性ポリマー類)、カプセル化DNA(DNA及びRNA等のポリアニオンとの同時且つ迅速に相互作用するカチオン性脂質を含み、リポソーム/核酸複合体を生じるリポソーム)、または金微粒子に連結されたDNA等が現在までのところ開発されている。さらに、核酸の細胞取込みを補助する薬剤、例えばカルシウムイオン、細菌タンパク質(赤痢菌(Shigella)タンパク質)、ウイルスタンパク質、及び他のトランスフェクション促進剤等を有利に使用することができる。本発明による組成物の別のタイプは、前記のベクターまたはベクターゲノムを含むレンチウイルス擬似粒子を含む組成物である。
【0086】
別のタイプの組成物は、本発明のエピトープ、類似体、またはポリエピトープを含む。このような組成物は免疫原性であり、すなわちそれは、それが投与される宿主に免疫応答を誘発させ得る。しかしながら、本発明のポリペプチドの免疫原性特性を増大させるために、アジュバントを前記ポリペプチドとともに投与して、免疫応答を誘発または改善することができる。アジュバントは、前記アジュバントと混合された抗原の免疫原性を増強する任意の物質として定義される。いくつかのアジュバントは、可溶性抗原を小粒子、例えば水酸化アルミニウムゲル、水中油乳濁液、または免疫刺激複合体(ISCOM)等に変換する。別のクラスのアジュバントは、無菌の細菌成分、例えば細胞壁または多糖、フロイントアジュバント等を含む。最後に、乳化剤またはpH緩衝剤を用いて、前記エピトープまたは類似体の免疫原性挙動を増強することもできる。
【0087】
前記に引用されたすべての組成物を、種々の経路:皮下(s.c.)、皮内(i.d.)、筋内(i.m.)、または静脈内(i.v.)注射、経口投与、及び粘膜投与、特に鼻腔内投与または吸入を介して宿主内に注入することができる。投与される量(投与量)は、患者の病態、個々の免疫系の状態、投与経路、及び宿主のサイズを含む、治療を受ける対象に依存する。好適な投与量は、200μg〜1mgの範囲であり、状況に応じて当業者により変更することができる。
【0088】
本発明の組成物は、hTERTの過剰発現により生じる腫瘍を含む制御できない細胞増殖から生じる患者における悪性状態の予防及び/または治療、並びにこのような悪性状態、例えば癌に伴う有害な結果の治療に有用である。表現「治療」とは、本発明の組成物により達成される治癒効果、及び治療を受けている患者に対する有益な効果を含み、前記効果は、細胞レベルまたは臨床レベルで得られ、その結果として、患者の病態の改善、並びに/あるいは寛解状態または健康状態の回復を含む。特定の実施形態において、本発明の組成物は、腫瘍の予防または治療に有用な付加的な活性化合物、すなわち一般的な化合物または組織特異的な癌において活性があることが証明された化合物をさらに含む。
【0089】
本発明はまた、
a. Tリンパ球を提供する工程;及び
b. 前記Tリンパ球を、本発明の少なくとも1つのエピトープまたはエピトープ類似体、またはポリエピトープとともに、前記リンパ球の活性化を可能にする条件でin vitro培養する工程;
を含む、クラスI制限、特にHLA-B7制限hTERTエピトープに対してTリンパ球を活性化させる方法に関する。
特定の実施形態において、活性化Tリンパ球は、細胞毒性Tリンパ球(CTL)である。Tリンパ球を活性化させる従来の条件は、インターロイキン(IL)2、IL-7、IL-12、及び/またはIL-15を用い、参照として本明細書に組み込まれているMinevら(2000年、PNAS;97(9):4796〜4801頁)に記載されている。
【0090】
本発明はまた、
a. Tリンパ球を、本発明の少なくとも1つのエピトープまたはエピトープ類似体、またはポリエピトープとともに、適切な条件下でin vitro培養することによる、前記Tリンパ球を活性化させる工程;
b.前記活性化リンパ球を、MHCクラスI分子に結合している本発明のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件下でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動を調査する方法に関する。
【0091】
hTERTエピトープの免疫原性挙動を調査するための特定の方法において、個々に、すなわちポリエピトープの形態下ではなく用いられる場合、当該エピトープは、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)の中から選択され、類似体の例は、前記に定義されたp444*である。
【0092】
hTERTペプチドの免疫原性挙動及びHLA-B7制限性を調査する方法は、
a.前記に記載されたTリンパ球を、MPRAPRCRA(p1)、APRCRAVRSL(p4)、APSFRQVSCL(p68)、RPAEEATSL(p277)、RPSFLLSSL(p342)、RPSLTGARRL(p351)、DPRRLVQLL(p444)、FVRACLRRL(p464)、AGRNMRRKL(p966)、LPGTTLTAL(p1107)、及びLPSPKFTIL(p1123)の中から選択されるエピトープまたはAPRRLVQLL(p444*)等のエピトープ類似体、または前記に定義されたポリエピトープで活性化する工程;
b.前記活性化リンパ球を、HLA-B7分子に結合している本発明のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件下でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;を含む。
【0093】
リンパ球の活性化は、抗原提示細胞による前記エピトープまたは類似体の、天然の(活性化されていない)Tリンパ球に対する提示を含む。天然のTリンパ球が本発明のエピトープまたは類似体を認識すると、特定のクラスI HLA分子の関連において、それらは「活性化」といわれ、標的細胞の細胞表面の前記エピトープを認識する状態にある。前記活性化リンパ球(エフェクター細胞)と標的細胞(それによりリンパ球が活性化されたエピトープを発現している)との間の接触により、分子が分泌され、標的細胞が死滅する。リンパ球を活性化させるために用いられたエピトープまたは類似体が、前記活性化リンパ球による前記エピトープを有する標的細胞の効率的な破壊をもたらす場合、前記エピトープは、前記エピトープを発現している細胞に対してin vitroのみならずin vivoでのT細胞反応を可能にするのに十分に免疫原性であるものとして見なされ得る。標的細胞/リンパ球認識に適した条件は、RPMI培地中において37℃で4時間の接触である。
【0094】
本発明はまた、MHCクラスI制限、特にHLA-B7制限hTERTエピトープに対して、細胞、特に抗原提示細胞(APC)、B細胞、T細胞、及び/または樹状細胞をin vitroで成熟させる方法に関する。細胞をin vitroで成熟させるための前記方法は、:
a. 細胞を提供する工程;
b. 少なくとも1つのMHCクラスI制限、特に本発明のHLA-B7制限hTERTエピトープまたはエピトープ類似体、またはポリエピトープを用いて、前記細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟細胞の増殖を促進する工程;
を含む。
【0095】
前記のとおり、リンパ球の活性化は、成熟抗原提示細胞によるエピトープの提示を必要とする。本発明の好ましい実施形態において、前記細胞は、少なくとも1つのHLA-B7対立遺伝子を発現する。それらのうちの1つである樹状細胞(DC)は、MHCクラスIに制限された内因性エピトープのTリンパ球に対する提示において特に有効である。前記細胞の成熟の目的の1つは、成熟させた時点での治療を必要としている患者への投与である。前記成熟DCの投与により、患者のリンパ球の活性化、及び前記エピトープ(DCを形質転換している1つ)を発現している細胞に対する迅速な反応をin vivoで生じるあろう。
【0096】
特定の実施形態において、樹状細胞をin vitroで成熟させる方法は、:
a. 樹状細胞を提供する工程;
b. 本発明の少なくとも1つのMHCクラスI制限hTERTエピトープまたはエピトープ類似体、またはポリエピトープを用いて、前記樹状細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟樹状細胞の増殖を促進する工程;
を含む。
【0097】
特定の実施形態において、前記樹状細胞は、循環血液または骨髄細胞から単離される。他の実施形態において、樹状細胞は、治療を必要としている患者に対する投与後の拒絶反応を避けるために、前記患者またはHLA適合ドナーから単離される。
【0098】
DCの成熟は、本発明のポリヌクレオチドを用いた前記樹状細胞の遺伝的形質転換によって、本発明のベクターを用いた前記樹状細胞のトランスフェクションによって、または本発明の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープとの前記樹状細胞の接触によって達成され得る。遺伝的形質転換は、その効率性、及びDCゲノム内に挿入されたポリヌクレオチドによってコードされたエピトープ、類似体、またはポリエピトープの永続的発現が理由で、好ましい。
【0099】
本発明はまた、癌の予防及び/または治療に使用するための、HLA-B7-制限hTERTエピトープをコードするポリヌクレオチドに関する。特定の実施形態において、癌の予防及び/または治療に使用するための前記ポリヌクレオチドは、HLA-B7-制限hTERTエピトープまたはその類似体、あるいは本出願に記載されている少なくとも1つのHLA-B7-制限hTERTエピトープまたはその類似体を含むポリエピトープをコードする。ポリエピトープをコードするポリヌクレオチドに関する限り、それは全長hTERTのコード配列に一致しない。他方、それは、hTERTの変異型または欠失型に一致し得、前記変異または欠失は、ヒトテロメラーゼの触媒活性を抑制する。特定の実施形態において、前記ポリヌクレオチドは、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)、
からなる群から選択されるHLA-B7-制限hTERTエピトープをコードする少なくとも1つのポリヌクレオチド単位、あるいはAPRRLVQLL(p444*)等の前記HLA-B7-制限hTERTの少なくとも1つの類似体、あるいは前記HLA-B7エピトープ及び/または類似体をコードする少なくとも2つのポリヌクレオチド単位を有するポリヌクレオチドを形成する任意の組合せを含む。
【0100】
本発明は、癌の予防及び/または治療に使用するための、HLA-B7 hTERTエピトープ、特にHLA-B0702制限エピトープに関する。本発明はまた、癌の予防及び/または治療に使用するための、本発明のポリヌクレオチド、ベクター、宿主細胞、またはポリペプチドに関する。
【0101】
本発明はまた、癌の予防及び/または治療のための薬剤の製造のための、HLA-B7 hTERTエピトープ、(または対応するポリヌクレオチド)の使用に関する。特定のHLA-B7 hTERTエピトープ(または対応するポリヌクレオチド)は、前記のもの、並びにそれらを含むか、またはそれらからなるベクター、細胞、または組成物である。特定の実施形態において、癌の予防及び/または治療のための薬剤の製造における、本発明のポリエピトープを含むか、またはそれからなるポリヌクレオチド、ベクター、宿主細胞、またはポリペプチドの使用は、前記に定義した少なくとも1つのHLA-B7対立遺伝子、特に少なくとも1つのHLA-B0702対立遺伝子を有する患者に提供されることを意図されている。
【0102】
本明細書に提供された各定義は、それぞれの及び任意のペプチド(エピトープ、類似体、またはポリエピトープ)、並びに個別に選択された(それ自体が)または群に包含されたそれぞれの及び任意のポリヌクレオチドに適用される。
【図面の簡単な説明】
【0103】
【図1A】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1B】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1C】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1D】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1E】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1F】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図2】hTERT由来ペプチドは、HLA-B0702トランスジェニックマウスにおいてプロセシングされる。HLA-B7 Tgマウス及び1匹の無処理マウス(N)を、hTERTをコードする100μgのDNAで免疫化した。14日目に、各マウスからの脾臓細胞を、種々のhTERT由来ペプチドを用いて別々にin vitro刺激した。6日後、材料及び方法に記載されている関連ペプチド(黒い四角)またはコントロールペプチド(白い四角)を負荷させたRMA-B7標的に対して、エフェクター細胞をアッセイした。60/1比における溶解パーセンテージを示す(2つの独立した実験からの結果)。
【図3】健常な血液ドナーからのPBMCにおける、hTERTに対するCTL応答の誘発。HLA-*B0702+健常ドナーからのTリンパ球細胞を、材料及び方法に記載されている6種のhTERTペプチドパルス化自系PBMCのそれぞれで活性化した。週1回の刺激の4ラウンド後、関連ペプチド(黒い四角)またはコントロールペプチド(白い四角)でパルス化したエフェクター細胞を、51Cr標識T2-B7細胞に対する溶解活性についてアッセイした。20/1のエフェクター対標的比における溶解パーセンテージを示す。10のドナーのうち8のドナーからの結果を示す(d1〜d8)。
【図4】腫瘍細胞に対するCTLp351の細胞毒性に及ぼす抗HLAクラスI mAbの効果。HLA mAbs(抗HLAクラスI mAbまたは抗クラスII mAb(HLA-DR))の非存在または存在下で前処理されたHLA-*B0702+腫瘍細胞系Mamo及びU293Tに対するCTLp351系の細胞毒性を、10/1のエフェクター対標的比において、標準的51Cr放出アッセイにより測定した。
【図5】Lv-hTERT免疫化後のhTERT特異的T細胞応答のex vivo検出。(A)HLA-B7トランスジェニックマウスを、組換えTrip-hTERT粒子またはコントロールTrip-GFP(1500ng)で免疫化した。12日後、各マウスのIFNγを産生するhTERTペプチド特異的T細胞を、新鮮な脾臓細胞中、IFNγ-ELISPOTアッセイによってex vivo検出した。ネガティブコントロール値を減算後、IFNγ SFCの数を算出した。3つの独立した実験からの結果を示す。(B)前記のように、HHDマウスをTrip-hTERTで免疫化した。前記のように、IFNγを産生するhTERTペプチド特異的T細胞を、ELISPOTによってex vivo検出した。2つの独立した実験からの結果を示す。
【図6】Trip-hTERT免疫化後のHLA-*B0702トランスジェニックマウスにおける特異的CD8+T細胞応答の刺激(priming)。HLA-*B0702トランスジェニックマウスを、Trip-hTERT(初めの4本の黒色のバー)またはコントロール(終りの2本の縦縞のバー)のいずれかで免疫化した。12日後、IFN-γELISPOTアッセイによって、各マウスの脾細胞中のIFN-γ産生単細胞をex vivo検出した。個々の免疫化マウスの新鮮な単離脾細胞からのフィコール精製リンパ球を、5μg/mlの各HLA-*B0702制限hTERT由来ペプチドとともに、またはそれなしで、24時間直接培養した。ペプチドなしのコントロールに関して得られた非特異的値を減算後、特異的IFN-γSFCの数を算出し(<15SFC)、当該応答は、SFC≧30に対して陽性と見なされた。
【図7】pTrip-hTERTの略図。このレンチウイルス由来ベクターは、HIV-1ゲノムのpsi配列、cPPT、及びCTS中心シス活性配列(フラップ)、並びに対象の遺伝子の発現を可能にするCMVプロモーターを含む。さらに、3'LTRにおいて、U3ドメインが欠失している(ΔU3)。
【図8】(A)DNA pTRIP-CMV-ΔhTERT免疫化は、HHDマウスにおいて、hTERT特異的CD8+T細胞応答を誘発した。HHDマウス(HLA-A2.1 Tg)を、hTERTの機能的でない形態(pTRIP-CMV-ΔhTERT)をコードするDNAを用いてDNA免疫化した。10日後、IFN-γELISPOTアッセイにより、IFNγ産生ペプチド特異的T細胞をex vivo検出した。個々の免疫化マウスの脾細胞からのフィコール精製リンパ球を、5μg/mlの各HLA-A2.1制限hTERT由来ペプチドとともに、またはそれなしで、24時間直接培養した。特異的IFN-γSFCの数を前記のように算出した。当該応答は、SFC≧30に対して陽性と見なされた。(B)pTRIP-CMV-ΔhTERT免疫化後のHHDマウスにおける、短期CTL応答の誘発。HHDマウスを、hTERTの機能的でない形態(pTRIP-CMV-ΔhTERT)をコードするDNAを用いて、10日間免疫化した。個々のマウスからの脾臓細胞を、HLA-A2.1制限hTERT由来のp540及びpY572ペプチドを用いて、6日間、in vitroで再刺激した。関連ペプチドまたは非関連ペプチドのいずれかを負荷したHHDでトランスフェクションされたEL4細胞に対する51Cr放出アッセイにおいて、エフェクター細胞を試験した。
【図9A】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9B】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9C】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9D】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9E】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9F】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10A】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10B】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10C】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10D】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10E】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10F】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【発明を実施するための形態】
【0104】
[実施例]
I−材料及び方法
<血液ドナー>
末梢血サンプルは、成人の健常血小板ドナー(centre de transfusion sanguine de I'hopital Mondor、Creteil、フランス)から、書面によるインフォームドコンセントに従って得た。末梢血ドナーのHLA分類は、H.Mondor.Hospital Creteil(フランス)のHLA研究室において実施された。前記試験は、フランス血液バンク機関(French Blood Bank Institute)によって承認された。
【0105】
<マウス>
構成的マウスβ2-m分子(HLA-B7mα3)及びキメラHLA-A2.1/H2-Db分子を発現しているHHDトランスジェニックマウスと組み合わせて、HLA-B0702α1α2、H2-Kdα3キメラ構築物を発現しているHLA-*B0702トランスジェニック(Tg)マウスは、以前に記載されているように、それらのH2-Db及びH2-kb遺伝子を欠失していた(Pascoloら、J Exp Med、1997年;185:2043〜51頁;Rohrlichら、Int Immunol、2003年;15:765〜72頁)。これらのマウスはC57BL/6をバックグラウンドとしており、本発明者らの動物施設において、特定病原体未感染の条件下で飼育され、維持された。
【0106】
<腫瘍細胞系>
T-BハイブリッドT1、EBV形質転換B細胞JY、腎臓癌細胞系U293T、及びBurkittリンパ腫細胞Rajiは、アメリカン・タイプ・カルチャー・コレクション(American type Culture Collection:ATCC)から入手した。メラノーマ細胞系(SK23MEL、LB34、及びKUL68)は、P.Coulie(ブリュッセル、ベルギー)からの好意で提供され、EBV形質転換B細胞BBG.1及びBC3は、H.Collandre(R.A.H.P.、グレノーブル、フランス)からの好意で提供された。
【0107】
HLA-*B0702でトランスフェクションされたTAP欠失T2細胞(T2-B7)は、P.Cresswell(Smithら、J Immunol、1996年;156:3755〜64頁)からの好意で提供された。マウスリンパ腫細胞系RMA及びEL4は、ATCCから入手し、これらの細胞もまた、HLA-*B0702遺伝子でトランスフェクションされ、標的細胞として用いられた。
【0108】
<エピトープ選択ペプチド合成>
HLA-*B0702に結合することが予測される9アミノ酸(9量体)または10アミノ酸(10量体)ペプチドの存在についてhTERTのアミノ酸配列を分析するために、予測的アルゴリズム「SYFPEITHI」(Lu及びCelis E、Cancer Res、2000年;60:5223〜7頁)を用いた。標準的HLA-B7結合アンカー、2位にPro、及びカルボキシル末端に疎水性脂肪族(AlaまたはLeu)を含有する候補ペプチドを、それらの最高予測スコアに従って選択した。6種のペプチド、すなわちp1(MPRAPRCRA、残基1〜9)、p277(RPAEEATSL、残基277〜285)、及びp342(RPSFLLSSL、残基342〜350)と称される3種の10アミノ酸ペプチド、並びにp4(APRCRAVRSL、残基4〜13)、p68(APSFRQVSCL、残基68〜77)、及びp351(RPSLTGARRL、残基351〜360)と称される3種の9アミノ酸ペプチド(アンカー位置に下線を引いた)が保持され、合成された。
【0109】
ヒトサイトメガロウイルスpp65に由来するペプチド、RPHERNGFTV(R10TV)、及びヒト免疫不全ウイルス1型IPRRIRQGLを合成し、コントロールペプチドとして用いた。マウスにおけるペプチド免疫化用のヘルパーペプチドとして、B型肝炎ウイルスコア128〜140由来のペプチド(TPPATRPPNAPIL)を用いた。ペプチドは、最低純度80%までPRIMから購買し、2mg/mlの濃度で蒸留水またはDMSO中で再構成した。
【0110】
<HLA-B0702結合/安定化アッセイ>
HLA-B0702でトランスフェクションされたT2(T2-B7)細胞に関する、MHC安定化アッセイを用いることによって、記載された(Rohrlichら、Int Immunol、2003年;15:765〜72頁)参照ペプチド(R10TV)と比較した、HLA-*B0702に対するhTERT由来ペプチドの相対的結合活性を測定した。要するに、種々の最終濃度(100、10、1、及び0.1μM)における、参照ペプチドR10VまたはhTERTペプチドのいずれかの非存在下(ネガティブコントロール)または存在下で、100ng/mlのヒトβ2-ミクログロブリンを添加した無血清培地AIM-V(Invitrogen社、Gibco)中、96ウェルプレートにおいて、T2-B7を2×105細胞/ウェルで一晩インキュベーションした。T2-B7細胞を飽和濃度の抗HLA-B7mAb、ME.1で標識し、次いで2回洗浄し、最後にFITC接合F(ab')2ヤギ抗マウスIgで染色して、フローサイトメトリーを行った。
【0111】
結果を、参照ペプチドの濃度に対する、最高結合度(参照ペプチドに関して得られた)の20%に達するのに必要な試験ペプチドの濃度の比率である相対的結合活性の値として表す。従って、当該値が低いほど、結合性が強い。
【0112】
<CTL誘発用のHLA-*B0702トランスジェニックマウスのペプチド免疫化>
8〜10週齢のメスHLA-*B0702トランスジェニックマウスに、フロイント不完全アジュバント(Difco、デトロイト、ミシガン州)中、共乳化された140μgのヘルパーペプチドを補充した50μgの個々のHLA-B0702制限hTERTペプチドを、尾基部に皮下注射(s.c.)した。10日後、個々のマウスの脾臓細胞を6ウェルのプレート中で、関連ペプチドを用いてin vitroで再活性化した。関連ペプチドまたはネガティブコントロールペプチドでパルス化された、HLA-*B0702でトランスフェクションされたRMA細胞(RMA-B7)を用いて、エフェクターCTL細胞を、標準的な4〜5時間51Cr放出アッセイにおいて試験した。特異的溶解≧10%が観察された場合、マウスを応答動物であると見なす。
【0113】
<HLA-*B0702トランスジェニックマウスにおけるDNA免疫化>
CMVプロモーターの制御下、hTERT遺伝子をコードするLvCMV-hTERTプラスミドベクターを、エンドトキシンのない条件下でのプラスミドGigaキットカラム(Qiagen)上で精製した。麻酔したHLA-*B0702トランスジェニックマウスの再生前脛骨筋内に、前記プラスミドを注入した(各側に50μg)。14日後、個々のマウスの脾臓細胞を、Con Aで活性化したラット脾臓細胞からの10%上清を補充した完全培地中、ペプチドでパルス化された(10μg/ml)同系のγ照射(50Gy)LPS-リンパ芽球を用いてin vitroで再活性化した。記載されたとおり、細胞毒性アッセイを6日間実施した。
【0114】
<レンチウイルスベクター構築物及び製造>
まず、pBABE-hygro-hTERTプラスミド(Counterら、Proc Natl Acad Sci U.S.A.1998年;95:14723〜8頁)由来のEcoRI-SalI hTERT挿入物を、pSP73ベクター(Promega)内にサブクローニングすることによって、pTRIP-デルタU3-CMV-hTERT(TRIPLv-hTERTまたはLv-hTERT、またはpTrip-hTERTと称される)(図7)構築物を作製した。次いで、BamHI及びXhoIで切断されたpTRIP-CMVプラスミド内に、BglII-SalI断片を挿入した。記載されているとおり、293T細胞の一過性の(48時間)トランスフェクションにより、擬似組換えレトロウイルス粒子を作製した(Zennouら、Cell、2000年;14:101:173頁;Firatら、J Gene Med、2002年;4:38〜45頁)。組換えレトロウイルス粒子を、超遠心によって濃縮し、PBS中に再懸濁した。ベクター粒子の量を、市販のELISAアッセイにおけるp24タンパク質のものから推定した(NEN、DUPONT、France Perkin Elmer)。
【0115】
2006年7月28日にCNCM I-3660の番号でCNCM(Institut Pasteur、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターを、前記段落に記載されたように実施した。しかし、hTERTタンパク質は、図1のヌクレオチド2654〜2662(野生型)に相当するアミノ酸867〜869の欠失によって機能的ではなくなっていた。QuickChange XL部位特異的突然変異誘発キット(Stratagene)を用いて、アミノ酸残基867〜869の欠失を作り出すことによって、触媒効力を失ったhTERT RT変異体(ΔhTERT)を作製し、配列決定によって確認した。
【0116】
<MHCクラスIトランスジェニックマウスにおける免疫化及びCTL欠失>
1500ngのTripLv-hTERT懸濁液またはコントロールベクターの単回皮下(尾基部)注入と同様に、TRIPLv-hTERTを用いた免疫化を実施した。
【0117】
500μlのPBS中1500ngのp24抗原に等価の、組換えレンチウイルス粒子のpTRIP-CMV-ΔhTERTまたはコントロールとしてのTrip-GFPの単回腹腔内注入として、HLA.A2トランスジェニックマウスにおける免疫化を実施した。
【0118】
12日後、ELISPOTアッセイにより、脾細胞中のhTERTペプチド特異的なTを検出した(下記を参照)。前記のようにペプチドパルス化によるin vitro刺激後、同一の免疫脾細胞集団に対して、細胞毒性アッセイを実施した。
【0119】
<ex vivoにおけるIFN-γELISPOTアッセイによる、T細胞応答の評価>
以前に記載されているようなIFN-γELISPOTアッセイによって、免疫化マウスからのペプチド特異的T細胞を検出した(Miyahiraら、J Immunol Methods、1995年;181:45〜54頁)。抗マウスIFN-γmAb's(3μg/ml;Pharmigen、Becton Dickinson biosciences)を、96ウェルニトロセルロースマイクロプレート(マルチスクリーン;Millipore社、モルスハイム、フランス)上にコーティングした。赤血球溶解後、個々のマウスの新鮮な単離脾臓リンパ球(5×105、2.5×105、及び1.25×105細胞/ウェル)を、5μgの天然hTERTペプチドとともに、またはなしで、37℃で18時間、直接培養した。洗浄後、当該プレートを、ビオチン化抗マウスIFN-γ(2μg/ml;Pharmigen、Becton Dickinson biosciences)とともに2時間インキュベーションした。最後に、当該プレートを洗浄し、アルカリホスファターゼ接合ストレプトアビジン(Roche molecular biochemicals、マンハイム、ドイツ)とともに37℃で1時間インキュベーションした。ポジティブコントロールは、ホルボール・ミリステート・アセテート(100ng/ml、Sigma)、及びイオノマイシン(1μg/ml)で刺激した細胞を含む。ペルオキシダーゼ基質(BCIP/NBT、Promega社、マジソンW;米国)を加えることによって、IFN-γスポット形成細胞(SFC)を展開し、自動化画像分析システムBioreader 2000(Biosys、カルベン、ドイツ)を用いてカウントした。ネガティブコントロール値(<10SFC)を減算した後、特異的SFCの数を算出した。刺激されたウェル中のSFCの平均が、ネガティブコントロールウェル中のSFCの平均+2S.D.よりも大きく、且つ50SFC/106細胞よりも大きい場合、応答は陽性であった。
【0120】
<細胞溶解アッセイ>
以前に記載されているような標準的な4〜5時間51Cr放出アッセイを用いることによって、細胞毒性アッセイを実施した(Firatら、J Gene Med、2002年;4:38〜45頁)。コントロールペプチド関して観察された非特異的溶解を減算することことによって、%での特異的溶解を算出した。特異的溶解≧10%が観察された場合、マウスを応答動物と見なした。
【0121】
<ヒトにおけるhTERTペプチド特異的CTLの産生>
以前に記載されているような、HLA-B0702制限のhTERTペプチドを用いた、4週間のin vitroでのPBMC再活性化後、ドナーからヒトCTLを得た(Hernandezら、Proc Natl Acad Sci U.S.A.、2002年;99:12275〜80頁)。要するに、フィコール精製ヒトPBMCを解凍し、加熱不活化10%ヒト血清(Institut Jacques Boy、レイムス、フランス)を補充したRPMI 1640、1mMのピルビン酸ナトリウム、100IU/mlのペニシリン、100μg/mlのストレプトマイシン、10mMのHEPES、5×10〜5Mの2-メルカプトエタノール中、24ウェルプレート中でインキュベーションした(4×106/ウェル)。それらを、各hTERTペプチド(10μg/ml)で刺激し、組換えヒトIL-7(20ng/ml;R&D Systems)を加えた。
【0122】
7日目、ペプチドパルス化γ照射自系PBMC(50Gy)を用いて、リンパ球を再活性化した。翌日、20IU/mlのヒトIL-2(Roche、マンハイム、ドイツ)を当該培養物に加えた。CTL系を、4サイクルの間、毎週再活性化した。一部のドナーに対して、CD8マイクロビーズ(Miltenyi Biotec、ベルギッシュグラッドバッハ、ドイツ)をメーカーの推奨に従って用いることによって、3回サイクル後にCD8*T細胞を精製し、機能試験前に1回活性化した。種々の51Cr-標識化標的:試験したhTERTペプチドまたは非関連ペプチドでパルス化したT2-B7、または腫瘍細胞系に対する最後の再活性化の6日後に、細胞溶解アッセイを実施した。
【0123】
いくつかの実験において、最適濃度(10μg/mL)における抗HLAクラスIフレームワークmAb、w6/32(BD Pharmigen)、または抗HLA-*B0702 mAbのME1、または抗HLA-DR mAbBのG46.6(BD Pharmigen)とともに、腫瘍細胞を30分間インキュベーションし、細胞毒性がHLAクラスIに限定されているかどうかを判定した。
【0124】
II−結果
1.HLA-*B0702で予測されたhTERT由来のペプチドの免疫原性
T細胞エピトープ予測プログラムを用いることによって、本発明者らは、hTERTタンパク質配列を分析し、それらの高い予測スコア(表1)により、6種のペプチド(3種の9量体、3種の10量体)を維持した。次に、それらのHLA-B0702分子への結合能を、HLA-B0702遺伝子でトランスフェクションされた抗原−トランスポーター(TAP)欠失T2細胞(T2-B7)を用いることによって試験した。3種のペプチド、p1、p4、及びp68は、それぞれ高い相対的親和性(RA≦1)を示し、他の3種、p277、p342、及びp351は、中程度のRA>1を示した(表1)。これらのデータは、これら6種のペプチドがHLA-B0702に対する優れた結合物質であることを示している。従って、小胞体内部で形成されたこのような複合体が腫瘍細胞の表面に到達し、CTL認識に利用可能であることが予測され得る。
【0125】
これらのペプチドがin vivoで免疫原性であるかどうかを試験するために、本発明者らはHLA-*B0702トランスジェニックマウスを免疫化した。前記6種のペプチドは、差は認めされたものの、CTL応答を誘発し得ることが示された(表1)。HLA-B7分子に対して高い親和性で結合する2種のペプチドp4及びp68は、試験されたすべてのマウスにおいて強いCTL応答を誘発する。対照的に、HLA-B7分子に対して低い親和性を有するペプチド(p277及びp342)は、試験されたマウスの50%のみで中程度の特異的CTLの産生を可能にする。トランスジェニックマウスから産生された、hTERTエピトープに特異的なCTL系は、それぞれ特異的なペプチドでパルス化されたヒトT2-B7を認識し(データは示していない)、複合体MHC/ペプチドに対するそれらのTCRの高い親和性を示した。従って、HLA-B0702の結合/安定化の結果とHLA-B7トランスジェニックマウスにおけるin vivoでのCTL応答との間には全体的な相関性がある。
【0126】
【表2】
【0127】
2.hTERT由来のペプチドはHLA-B0702トランスジェニックマウスにおいてプロセシングされた
同定されたこれら6種のhTERTエピトープに対するHLA-B0702分子との関連で、内在的に合成されたhTERTペプチドの提示を評価するために、HLA-*B0702トランスジェニックマウスを、hTERTをコードするcDNAで免疫化し、40日後、個々のマウスの脾臓細胞内のペプチド特異的CTL応答を評価した。図2に示されるように、p4、p68、p1、p277、及びp351に関して、マウスの50%〜80%の大部分の免疫化マウス(M)において、hTERTペプチド特異的CTLが誘発された。対照的に、p342特異的CTLは、試験されたマウスの約15%で誘発され得た。有意なhTERT特異的CTL応答は、非免疫化無処理マウスからも誘発されなかった。従って、これら6種のhTERTエピトープは、細胞内で効果的にプロセシングされる。さらに、アミノ酸配列または構造の点において合成ペプチドと類似した天然ペプチドは、当該細胞表面上の対応するHLA-*B0702分子によって提示される。
【0128】
さらに、これらのデータは、単一マウスにおいて、いくつかのhTERTエピトープに対して複数のCTL特異性を同時に誘発させることができることを示しており、且つ候補ワクチンを試験するためのそれらの可能性について、本発明者らのHLAクラスIトランスジェニックマウスモデルの有効性を実証している。
【0129】
3.hTERTペプチドによる健常ドナーからの一次CTL応答の誘発
In vitro免疫化プロトコルにおけるHLA-B0702健常ドナーのPBMCを用いることによって、hTERTペプチドがHLA-B7制限CTLの上昇において有効であるかどうかを試験した。10個体のうち8個体(d1〜d8)においてCTL応答が生じ、ペプチド特異的CTL応答は、p342(20%)を除いて、ドナーの少なくとも50%で得られた(図3)。In vitroで生じたCTLによるhTERTエピトープの認識は、それらの遺伝的バックグラウンドに応じてドナー間で変化する(図3)。従って、HLA-B0702健常ドナーのランダム試験によって、これらのhTERTペプチドはヒトにおいて免疫原性であることが明白に確認され、hTERTに対する特異的CTL前駆体は末梢成人レパートリーにおいて欠失していないことが示唆される。従って、健常ドナーから生じたCTL系がHLA適合hTERT+腫瘍細胞を死滅させることができるかどうかを探索した。
【0130】
4.特異的hTERT CTLは種々の由来の腫瘍を溶解できた
種々の由来のヒト腫瘍細胞系を溶解させる能力について、ドナーからのhTERT特異的CTLを試験した。表2に示されている結果は、健常ドナー由来のin vitroで生じたCTL系はHLA-B0702+腫瘍細胞を死滅させるが、他方、HLA-B0702-腫瘍に対する細胞毒性は検出されなかったことを示している。[例えば、KU L268または293-UT標的における、d1、d2、及びd3でのCTLp351(それぞれ、52、25、20%、及び34、41、19%)対T1またはBBG1標的におけるもの(それぞれ、9、2、6%、及び0、0、2%)を参照]。CTL特異性に従って、腫瘍認識に違いが観察されたが、このことは、腫瘍細胞の表面上のhTERTペプチドの差異的な差異によって説明できるであろう。重要なことには、種々のドナーから生じたp351特異的CTL系が、試験された腫瘍細胞系の大多数を認識することである(表2)。対照的に、すべてのp4特異的CTL系が、試験された腫瘍のすべてのタイプを溶解するわけではない。p1及びp68ペプチドに特異的なCTL系は、T1-B7標的のみを認識する。p342特異的CTL系は、メラノーマ細胞(LB 34及びKU 268標的)のみを認識する。最後に、p277特異的CTL系は、腎臓癌(293 UT)を認識するが、メラノーマ細胞もリンパ腫細胞のどちらも認識しない。他方、正常PBMC及びCD40活性化B細胞は、HLAのタイプに関わらず、これらのhTERTペプチド特異的CTL系によって溶解されなかった(表2の最後の2つの系)。
【0131】
図4に示されるように、HLA-B7+腫瘍細胞に対するCTLp351系の細胞毒性活性は、抗クラスI mAb抗HLA-B0702によって阻害されるが、抗HLA-DR mAb(MHC クラスII)によっては阻害されない。同様のデータが、他のペプチド特異的CTL系でも得られており(データは示さず)、CTL系が、HLA-B0702制限様式でhTERT+腫瘍細胞に対して細胞毒性を及ぼすことを示唆している。まとめると、これらの結果は、これら6種のhTERT由来ペプチドが腫瘍細胞表面で等しく天然に発現されているのではないこと、及びhTERTペプチド特異的CTLが、HLA-B0702分子との関連でhTERTペプチドの認識によって、腫瘍細胞と正常細胞を識別できることを示している。
【0132】
5.hTERTをコードするレンチウイルスベクターのワクチン接種により、マウスにおいて有効なペプチド特異的T細胞応答が誘発される
次いで、本発明者らは、HIV由来フラップベクター内に導入された、全長hTERT遺伝子または機能的でないhTERT遺伝子のいずれかを含む候補ワクチンを試験した(図7)。このタイプのレンチウイルスベクターは、in vitro及びin vivoで樹状細胞を標的とし、動物において強い多特異的抗腫瘍CTL応答を誘発することが、以前のデータによって示されている。従って、本発明者らは、組換えLv-hTERTまたはpTRIP-CMV-ΔhTERTを用いて、HLA-*B0702トランスジェニックマウスを免疫化した。12日後、個々のマウスの脾臓細胞を、ex vivo ELISPOTアッセイによって評価した。
【0133】
図5Aに示されるように、Lv-GFPコントロールベクターを投与されたマウスと比較して、HLA-B0702制限hTERTエピトープに対するペプチド特異的CD8+ T細胞応答が得られた。In vitro刺激後のクロム放出アッセイにおいて、当該誘導されたペプチド特異的CD8+細胞の機能分析により、ex vivo ELISPOTデータが裏付けられ(表3)、さらにLv-hTERTの単回注入後のマウスの約50〜70%において及びTRIPLv-hTERTによる増強後のマウスの100%において、これら6種のペプチドに対する有効な特異的CTL応答が生じることが示されている(図6)。このことはまた、すべてのマウスにおける強いCTL応答とも関連していた(表3)。さらに、表3に示されるように、HLA-A2.1に対してトランスジェニックのHHDマウスを同一ベクターで免疫化すると、顕性(p540)及び潜性(p572)として以前に分類された2つのHLA-A2.1.1制限エピトープに対して特異的な強力なCTL応答が誘発された。まとめると、これらの結果は、Lv-hTERTの投与により、マウスにおいて、非常に有効な多特異的T細胞応答の誘発がもたらされること明白に示され、hTERTが癌免疫療法のためのポリエピトープ及びポリ対立遺伝子の(polyallelic)TAAとして作用し得ることを示唆している。
【0134】
図8に示されるように、組換えpTRIP-CMV-ΔhTERTの単回注入後のHLA-A2トランスジェニック(Tg)マウスにおいて、hTERTペプチド特異的CD8+T細胞応答がex vivoで検出された。本発明者らは、p540及びPY572エピトープに特異的なCD8+T細胞が、免疫化マウスの少なくとも50%において誘発されたことを示した(図8)。これらの結果は、前記2つのエピトープが正確に内在的にプロセシングされ、pTRIP-CMV-ΔhTERTによる免疫化後のHLA-A2 Tgマウスにおいて提示されたことを明白に示した。
【0135】
まとめると、これらの結果は、TRIP-hTERTの単回注入により、両方のHLAトランスジェニックマウス群において、強力な多特異的抗hTERT CD8+T細胞応答の誘発がもたらされることを示した。
【0136】
【表3】
【0137】
【表4】
【0138】
III−結論
In vivo免疫原であり、且つH-2クラスIノックアウトHLA-B0702トランスジェニックマウスにおいてプロセシングされる、新規なhTERTエピトープが同定された。さらに、in vitroにおいて、健常ドナーからのHLA-B702+PBLを用いたhTERTペプチド免疫化は、種々の由来のhTERT+腫瘍を認識する特異的CTL応答を誘発し、ヒトT細胞レパートリーにおいてこれらのエピトープに対する欠失がないことを示唆している。さらに、腫瘍の由来に応じて細胞表面上に発現されるペプチドレパートリーは質的に異なっていることが示され、hTERTを癌細胞の周囲の(circumvent)抗原変異性に対するポリエピトープ腫瘍関連抗原として特徴づけするための有用性を明確に示している。最後に、ヒト化HLA-*B0702及びHLA-A2 1トランスジェニックマウスを用いて、新世代のレンチウイルス由来フラップベクター内に挿入された機能的でないテロメラーゼ遺伝子からなる候補ワクチンを試験した。すべての当該HLAトランスジェニックマウスにおいて、強力なhTERT特異的CD8+T細胞応答が観察された。これらのデータは、癌患者における治療的ワクチン接種の使用を支持し、多くの癌患者を網羅するための治療標的としてのhTERTの潜在的適用性を広げている。
【0139】
(参考文献)
【技術分野】
【0001】
本発明は、抗癌療法の分野に関するものであり、ヒトテロメラーゼ逆転写酵素(hTERT)由来の免疫原性ペプチドの同定に関する。本発明は、MHCクラスI分子に制限されたhTERTエピトープをコードするポリヌクレオチド、それらの類似体、並びに前記エピトープ及び/または類似体を含有するポリエピトープに関する。また、本発明は、前記ポリヌクレオチドを含むベクター及び細胞を含む。本発明はまた、癌の治療及び/または予防に使用するための、hTERTポリペプチド、対応するポリヌクレオチド、ベクター及び細胞を含む組成物に関する。
【背景技術】
【0002】
現在の抗癌療法手法の欠陥という観点から、抗腫瘍療法は、あらためて関心がもたれている。最近の研究では、腫瘍回帰に関連する抗メラノーマ免疫応答についての本発明者らの理解を深めている。これらのデータをまとめると、活性化された腫瘍特異的なCD8 CTLは、効力のある抗腫瘍療法の選択において免疫学的武器であることを示唆されている。異なる由来の腫瘍に対して細胞毒性Tリンパ球を作動させる過剰発現タンパク質の研究もまた進行している。
【0003】
テロメラーゼは、タンパク質成分のTERT、及び染色体の末端上に付加された反復単位(T2AG3)の合成のためのテンプレートを含有するRNA成分(TR)からなるリボ核タンパク質複合体であって、複製の間染色体を安定化し、末端間融合を防ぐ。一定のテロメア長の維持により、細胞を老化から防ぎ、不死性を付与する(Hahnら、Nat Med 1999年;5:1164〜70頁)。ヒト癌の85%超に高いhTERT活性が見られたが、一方、大部分の正常ヒト成人組織は、テロメラーゼ活性を示さないか、またはほとんど示さない(Counterら、Blood 1995年;85:2315〜20頁)。
【0004】
腫瘍におけるテロメラーゼの広範な発現は、hTERTのペプチド断片が、腫瘍特異的抗原として役立ち得ることを示しており、このことはいくつかの報告により確認されている(Vonderheideら、Immunity 1999年;10:673〜9頁)。第一相臨床試験からの最近のデータは、HLA-A2+患者におけるhTERTに対してワクチンの実現可能性を立証し、治療的ワクチン接種のためのhTERTの使用への道を開いた(Vonderheideら、Clin Cancer Res 2004年;10:828〜39頁;Parkhurstら、Clin Cancer Res 2004年;10:4688〜98頁)。それにもかかわらず、今日まで同定されている免疫原性hTERTペプチドは、1つのMHC対立遺伝子HLA-A2.1に制限され、当該集団の44.2%及び40%にそれぞれ示される2つのHLA上位型のHLA-A3及びHLA-A24に関する最初の2つの報告のみである。
【0005】
従って、以下の刊行物は、hTERTペプチドの同定を報告した:
−HLA-A2に制限されているhTERTペプチドILAKFLHWL(Vonderheideら、Immunity 1999年;10:673〜9頁)、
−HLA-A関連(context)内の情報予測によって同定されたhTERTペプチドMPRAPRCRA、RPAEEATSL、RPSFLLSSL、及びAPRCRAVRS(WO00/02581)。しかし、これらのペプチドは、HLA-B7関連または任意のHLA関連のいずれにおいても、効率的なエピトープであることが実験結果によって確認されていない。
−HLA-A3に制限されているhTERTペプチドKLFGVLRLK(K973)(Vonderheideら、Clin Cancer Res 2001年;7:3343〜8頁)、並びに
−HLA-A24に制限されているhTERTペプチドVYAETKHFL(TEL324)及びVYGFVRACL(TEL461)(Araiら、Blood 2001年;9:2903〜7頁)。
【0006】
その結果、これまでに同定されたhTERTペプチドはすべての集団を網羅しておらず、それゆえ、多くの患者を除外している。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】WO 00/02581
【特許文献2】WO 99/55892
【特許文献3】WO 01/27300
【特許文献4】WO 01/27304
【非特許文献】
【0008】
【非特許文献1】Hahnら、Nat Med 1999年;5:1164〜70頁
【非特許文献2】Counterら、Blood 1995年;85:2315〜20頁
【非特許文献3】Vonderheideら、Immunity 1999年;10:673〜9頁
【非特許文献4】Vonderheideら、Clin Cancer Res 2004年;10:828〜39頁
【非特許文献5】Parkhurstら、Clin Cancer Res 2004年;10:4688〜98頁
【非特許文献6】Vonderheideら、Clin Cancer Res 2001年;7:3343〜8頁
【非特許文献7】Araiら、Blood 2001年;9:2903〜7頁
【非特許文献8】Firatら(1999年、Eur. J. of Immunol. 29:3112〜3121頁)
【非特許文献9】Firatら、2002年、The Journal of Gene Medicine 4:38〜45頁
【非特許文献10】Esslingerら、Hum Gene Ther 2002年;13:1091〜100頁
【非特許文献11】Breckpotら、J Gene Med 2003年;5:654〜67頁
【非特許文献12】Esslingerら、J Clin Invest 2003年;111:1673〜81頁
【非特許文献13】Tartourら、2000年、Immunol Lett Sep 15;74(1):1〜3頁
【非特許文献14】Tartourら、1996年、Presse Med. Nov 16;25(25):1717〜22頁
【非特許文献15】Minevら(2000年、PNAS;97(9):4796〜4801頁)
【非特許文献16】Pascoloら、J Exp Med、1997年;185:2043〜51頁
【非特許文献17】Rohrlichら、Int Immunol、2003年;15:765〜72頁
【非特許文献18】Smithら、J Immunol、1996年;156:3755〜64頁
【非特許文献19】Lu及びCelis E Cancer Res、2000年;60:5223〜7頁
【非特許文献20】Counterら、Proc Natl Acad Sci U.S.A.1998年;95:14723〜8頁
【非特許文献21】Zennouら、Cell、2000年;14:101:173頁
【非特許文献22】Miyahiraら、J Immunol Methods、1995年;181:45〜54頁
【非特許文献23】Hernandezら、Proc Natl Acad Sci U.S.A.、2002年;99:12275〜80頁
【発明の概要】
【発明が解決しようとする課題】
【0009】
この欠陥を少なくとも一部において克服し、それによってヒト集団の遺伝子多様性をより良好に網羅するために、本発明は、HLA-A3及びHLA-A24とは異なる特定のHLAに制限されたhTERT由来の新規なエピトープを同定する。多くの異なる対立遺伝子の中で、本出願は、集団の約25%に発現されるHLA-B7上位型、特にヒトHLA-B集団において最も多く発現される第2の対立遺伝子:HLA-*B0702(ヒト集団における個体の15〜20%に存在する対立遺伝子)に関心を有する。
【課題を解決するための手段】
【0010】
テロメラーゼのアイソフォーム-1の遺伝子は、4015の塩基対(bp)長(NCBI登録番号AF015950)であり、1132のアミノ酸のタンパク質(NCBI登録番号AAC51672.1)をコードする(図1)。
【0011】
本発明は、ヒトテロメラーゼ逆転写酵素(hTERT)ペプチドをコードするポリヌクレオチドに関する。本発明の特定の実施形態において、コード化ペプチドは、長さが9個のアミノ酸(9量体)または長さが10個のアミノ酸(10量体)であり、ゆえに、ポリヌクレオチドは、27または30のヌクレオチドを有する。一般に、コード化ペプチドは15未満のアミノ酸であり、ポリヌクレオチドは45未満のヌクレオチドを有する。
【0012】
本発明はまた、MHCクラスI分子に制限されたエピトープ、特にHLA-B7に制限された免疫応答を誘発するのに好適であるエピトープであるhTERTペプチドをコードするポリヌクレオチドに関する。本発明のポリヌクレオチドのヌクレオチド配列は、特定の実施形態において、hTERTペプチドをコードする配列に限定される。このようなペプチドは、MPRAPRCRA(p1;1〜9のアミノ酸残基)、APRCRAVRSL(p4;4〜13のアミノ酸残基)、APSFRQVSCL(p68;68〜77のアミノ酸残基)、RPAEEATSL(p277;277〜285のアミノ酸残基)、RPSFLLSSL(p342;342〜350のアミノ酸残基)、RPSLTGARRL(p351;351〜360のアミノ酸残基)、DPRRLVQLL(p444、444〜452のアミノ酸残基)、FVRACLRRL(p464、464〜472のアミノ酸残基)、AGRNMRRKL(p966、966〜974のアミノ酸残基)、LPGTTLTAL(p1107、1107〜1115のアミノ酸残基)、及びLPSPKFTIL(p1123、1123〜1131のアミノ酸残基)からなる群から選択されうる。これらのポリヌクレオチドすべては、HLA-B7制限免疫応答を誘発するために使用されうる。特定の実施形態において、本発明は特に、RPSLTGARRL(p351)、APSFRQVSCL(p68)、APRCRAVRSL(p4)、DPRRLVQLL(p444)、FVRACLRRL(p464)、AGRNMRRKL(p966)、LPGTTLTAL(p1107)、及びhLPSPKFTIL(p1123)からなる群から選択される、HLA-B7制限hTERTエピトープをコードするポリヌクレオチドに関する。
【0013】
本明細書に定義されているように、「エピトープ」は、抗原決定基、すなわち免疫系の細胞(免疫細胞)によって認識されるペプチド部位、特に免疫応答を誘発するのに必要な部位である。用語エピトープは、それに対する連続アミノ酸(特に、9または10)が免疫細胞によって認識される線状エピトープ、及びそれが適切な立体配置または立体配座をとる範囲で免疫細胞がアミノ酸を認識する立体配置的エピトープの両方を含む。その結果、いくつかのエピトープにおいて、立体配座(三次元構造)は、アミノ酸配列(一次構造)と同様に重要である。
【0014】
表現「MHCクラスI制限」とは、クラスIのMHC(主要組織適合性複合体)分子に対して親和性を有する特定のペプチドまたはエピトープに対する能力を表す。同様に、表現「HLA-B7制限」とは、このタイプのHLA分子に対する親和性を有する特定のペプチドまたはエピトープに対する能力を表す。
【0015】
要するに、MHC遺伝子は、外来性ペプチドに結合するだけでなく、過剰発現したまたは自己でないペプチド、あるいは変異型自己ペプチドにも結合しうる細胞表面多型分子をコードして、適切な免疫細胞、特にT細胞による認識を可能にする細胞の細胞表面にそれらを提示する。マウスにおいてH-2及びヒトにおいてHLA(ヒト白血球抗原)と称される前記MHC分子は、クラスI分子(HLA-A、B、またはCと表される)またはクラスII分子(DP、DQ、またはDRと表される)として分類される。
【0016】
従って、MHCクラスI分子は、細胞毒性Tリンパ球上に発現されるCD8分子(TCD8+とも称される)に特異的に結合するが、一方、MHCクラスII分子は、ヘルパーTリンパ球上に発現されるCD4分子(TCD4+)に特異的に結合する。
【0017】
MHCクラスI分子は、タンパク分解的に分解されたタンパク質、特に細胞によって内因的に合成されたタンパク質に由来するペプチドに結合する。それによって得られた小型ペプチドは小胞体に輸送され、そこでそれらは新生MHCクラスI分子と結合してから、ゴルジ体を介して輸送されれ、細胞毒性Tリンパ球による認識のために細胞表面に提示される。
【0018】
本発明において、前記同定ペプチドは、一方では高いまたは中程度の親和性でMHCクラスI分子に結合し、他方、MHC/エピトープ複合体として細胞の細胞表面に効率的に輸送されることが示されている。好ましい実施形態において、MHCクラスI分子はHLA-B7上位型ファミリーのMHC対立遺伝子であり;hTERTエピトープは前記HLA-B7上位型制限である。前記ファミリーは、対立遺伝子B0702、B0703、B0704、B0705、B1508、B3501、B3502、B3503、B51、B5301、B5401、B5501、B5502、B5601、B5602、B6701、及びB7801を含み、それらのファミリーからHLA-B0702が好ましい(HLA-B0702制限hTERTエピトープ)。
【0019】
MHC安定化アッセイを用いて、参照として本明細書に組み込まれているFiratら(1999年、Eur. J. of Immunol. 29:3112〜3121頁)に記載されているもの等、特定のHLAクラスI分子に対するペプチドの親和性(相対結合活性)を試験することができる。要するに、MHCクラスI分子でトランスフェクションされた細胞を、
−ペプチドの非存在下(ネガティブコントロール)、
−参照ペプチドの存在下(ポジティブコントロール)、及び
−試験されるペプチド(この場合、hTERTペプチド)の存在下で、
100ng/mlのヒトβ2-ミクログロブリンを補充された無血清培地AIM-V(Invitrogen社、Gibco)中、96ウェルプレート中に2×105細胞/ウェルで一晩インキュベーションする。
ペプチドを、0.1〜100μMの範囲の種々の最終濃度で(1μM及び10μMの中間濃度で)インキュベーションする。次いで、トランスフェクションされた細胞を、特定のHLA MHCクラスI分子を認識する抗体の飽和濃度で標識し、次に2回洗浄し、最終的にフローサイトメトリー前に二次抗体で染色する。結果は相対結合活性値として表され、これは、参照ペプチドの値を参照して最大結合の20%に達するのに必要な試験ペプチドの濃度比率である。それゆえ、この値が低くなるほど、結合が強くなる。この方法の後に、RA<1の場合、ペプチドは特定のHLAクラスI分子に対して高い相対親和性を有するといわれる。対照的に、RAが1〜5、好ましくは1〜3である場合、中程度の相対親和性と結論付けられる。
【0020】
hTERT9量体及び10量体、特に前記同定ペプチドの中で、以下のMHCクラスI制限hTERTエピトープ:MPRAPRCRA(p1)、APRCRAVRSL(p4)、及びAPSFRQVSCL(p68)を、MHCクラスI分子に対して高い相対親和性を有するものとして分類することができる。同じ手法を用いて、以下のMHCクラスI制限hTERTエピトープ:RPAEEATSL(p277)、RPSFLLSSL(p342)、及びRPSLTGARRL(p351)を、MHCクラスI分子に対して中程度の相対親和性を有するものとして分類することができる。
【0021】
本発明はまた、MHCクラスI制限エピトープ類似体、すなわち前記のようなクラスI制限hTERTエピトープと比較して少なくとも1つのアミノ酸置換を有するエピトープ、特にHLA-B7制限エピトープ類似体をコードするポリヌクレオチドに関する。
【0022】
本明細書に定義される用語「類似体」とは、その配列が、保存的、半保存的、または非保存的な少なくとも1つのアミノ酸置換により、前記のhTERTペプチドから得られるペプチドに関する。類似体は、そのヌクレオチド及び/またはアミノ酸配列が、いわゆる野生型ペプチドまたはポリヌクレオチドを定義するために、参照の分子として本出願内で考慮される図1に開示されている参照hTERT遺伝子またはタンパク質に見られないという事実により、エピトープと対立するものである。このような類似体は、前記アミノ酸をコードするコドンにおける1つまたはいくつかの核塩基の対応するヌクレオチド配列の置換により生じ得る。従って、ポリヌクレオチド類似体は、置換されるアミノ酸残基をコードするコドンにおいて少なくとも1つの置換、好ましくは1つ、2つ、または3つの置換により、その野生型対応物(ペプチド類似体が由来する参照配列でhTERTペプチドをコードするポリヌクレオチド)とは異なる。一例として、APRRLVQLLペプチド(p444*と称される)は、最初のアミノ酸残基(D->A)の置換によりp444ペプチドから得られる。
【0023】
HLA-B7制限類似体の特定の類似体は、それが由来するエピトープと同じ長さを有するか、またはそれよりも短い。
【0024】
特定の実施形態として、前記のhTERTエピトープまたはその類似体は、それらの一次構造において、2位にプロリン(P)、及び/または最後のC末端位置に以下のアミノ酸:A、L、I、M、V、F、W、またはYのうちの1つを常に保存する。従って、類似体を含むhTERTペプチドは、以下のコンセンサス配列:X-P-X6〜7-[ALIMVFWY]を有し、式中、Xは任意のアミノ酸を表し、X6〜7はアミノ酸数を表し、且つ[ALIMVFWY]はこれらのアミノ酸のうちの1つを表す。
【0025】
従って、エピトープ類似体を提供するために、当該ポリヌクレオチドにおけるアミノ酸置換または対応するコドン置換は、2位(または第2コドン)に位置しない。好ましい実施形態において、たとえ最後のC末端アミノ酸を等価のアミノ酸、すなわちA、L、I、M、V、F、W、またはYで置換できたとしても、C末端位置において置換は行われない。最後に、好ましい実施形態において、置換は1位に位置し、任意のアミノ酸はアラニン(A)で置換される。
【0026】
本発明のさらなる実施形態において、生じた9量体がX-P-X6〜7-[ALIMVFWY]コンセンサス配列を維持するか、または選択するという条件で、10量体の最後のC末端アミノ酸を欠失させて、9量体を得る。同じ方法で、生じた10量体がX-P-X6〜7-[ALIMVFWY]コンセンサス配列を維持するか、または選択するという条件で、9量体のC末端にA、L、I、M、V、F、W、及びYの中から選択するアミノ酸を付加させて、10量体を生じさせる。
【0027】
個々にまたは組合せとして選択され、特に前記に説明されたものを含む、置換、欠失、または付加の場合、ペプチド類似体の三次元立体配座は、野生型対応物の1つに対して同じかまたはわずかに改変されたものでなければならず、それにより、当該類似体の正しい折りたたみ及びMHCクラスI分子に対するその正しい結合を確保する。前記のMHC安定化アッセイを用いて、このような制約が満たされることをチェックすることができる。
【0028】
生じた類似体は、特定のMHCクラスI分子、特にHLA-B7分子に対する親和性に関して、その野生型対応物と少なくとも同じ特徴を有し、且つ細胞表面上のエピトープ/MHC複合体と実質的に同じ輸送される能力を有し、及び/または同じ条件で試験された場合、免疫原性応答を誘発する実質的に同じ能力を有する。
【0029】
好ましい実施形態において、開始ペプチドは、中程度の親和性を有するhTERTエピトープであり、生じた類似体は、その野生型対応物よりも高い親和性を有する。別の実施形態において、類似体は、その野生型対応物よりも高い免疫原性を有する。一例として、前記に示されたp444*ペプチド類似体は、それが由来するp444ペプチドと比較して、MHCクラスI分子に対して増大した親和性を有する。
【0030】
本発明の目的は、免疫応答、特にCTL応答(細胞毒性Tリンパ球)を誘発できるhTERTエピトープまたは類似体を提供することである。しかしながら、Tリンパ球は、抗原提示細胞を介して前記リンパ球のみを刺激するために使用される前記類似体を認識しない。本発明の実施形態において、前記の類似体は、その免疫原性挙動を維持し、且つhTERTエピトープを過剰発現する細胞に対する免疫応答、すなわちCTLはエピトープ類似体により刺激されたとしても、野生型エピトープを認識することを誘発できる。特定の実施形態において、本発明のエピトープ類似体により刺激されたリンパ球は、hTERTエピトープを過剰発現しない細胞に対して反応しない。従って、刺激されたリンパ球は、他のエピトープを過剰発現する細胞と反応(交差反応)しないか、または基底レベルとしてのhTERTエピトープを発現する細胞(健常細胞)と反応しない。特定の実施形態において、前記のすべての特徴は、HLA-B7環境内にあり、好ましくは、HLA-B0702関連にある。
【0031】
参照として本明細書に組み込まれているFiratの刊行物(1999年、Eur. J. of Immunol. 29:3112〜3121頁)にあるように、標準的4〜5時間の51Cr放出アッセイを使用することによって従来の細胞毒性アッセイを実施して、標的細胞に対して反応する刺激されたリンパ球の能力を試験することができる。要するに、CTLを含む細胞懸濁液をin vivoでペプチド(この場合、hTERTペプチドまたは類似体)+自己MHCクラスI分子で活性化する。次いで、あらかじめ51Crとインキュベーションされた標的細胞(対応するhTERTエピトープを発現しているまたは発現しない)を、活性化リンパ球とともにインキュベーションする。活性化CTLによる標的細胞の認識により、標的細胞のアポトーシス及び51Crの放出が生じ、前記放出は死滅した標的細胞数に比例する。標的細胞とコントロールペプチドとのインキュベーションを、51Crの自発的放出を算出するためにネガティブコントロールとして使用する。試験されるペプチドに関して得られた細胞溶解に対してコントロールペプチドで観察された非特異的細胞溶解を差し引くことによって、細胞溶解の具体的なパーセンテージを算出する。そのパーセンテージが高いほど、CTLにより死滅した標的は多い。特異的細胞溶解を、エフェクター(CTL)対標的細胞(E:T)のいくつかの比率で判定する。特定の細胞溶解を、[実験的放出−自発的放出]と[全放出−自発放出]との間の比率として算出する。
【0032】
本発明はまた、ポリエピトープをコードするポリヌクレオチドに関する。ポリエピトープとは、MHCクラスI制限、特に本発明のHLA-B7制限hTERTエピトープ(p1、p4、p68、p277、p342、p351、p444、p464、p966、p1107、及びp1123)及び/またはMHCクラスI制限、特にHLA-B7制限エピトープ類似体の中から選択される少なくとも2つのエピトープを有するポリペプチドとして定義される。本発明のポリヌクレオチドは、前記エピトープまたは類似体をコードする少なくとも2つのポリヌクレオチド単位を含むか、またはそれらからなる。ポリヌクレオチド単位は、本明細書に開示された本発明のエピトープまたは類似体に対するコード配列として定義される。
【0033】
本発明は特に、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)、または前記に定義されるそれらの類似体の中から選択される少なくとも2つのエピトープを含む、ポリエピトープをコードするポリヌクレオチドに関する。ポリエピトープをコードする別のポリヌクレオチドは、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)に対応するHLA-B7制限hTERTエピトープ及び/または前記に定義されるそれらの類似体をコードするポリヌクレオチドの群から選択される少なくとも1つのポリヌクレオチド単位、並びに配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)、あるいは前記に定義されるそれらの類似体をコードするポリヌクレオチドの群から選択される少なくとも1つのポリヌクレオチド単位を含む。
【0034】
本発明のポリエピトープをコードするポリヌクレオチドのいずれも、全長hTERTのコード配列と一致しない。
【0035】
本発明の特定の実施形態において、調製されたポリエピトープにおけるMHCクラスI制限hTERTエピトープ及び/または類似体の数は、30に制限される。別の実施形態において、HLA-B7制限hTERTエピトープ及び/または類似体の数は、およそ30に制限され、好ましくは2、3、4、5、6、7、8、9、10、15、20、25、または30である。別の実施形態において、HLA-B0702制限hTERTエピトープ及び/または類似体の数は、およそ10に制限され、好ましくは2、3、4、5、6、7、8、9、または10である。
【0036】
従って、ポリエピトープをコードするポリヌクレオチドは、30以下のポリヌクレオチド単位、特に2、3、4、5、6、7、8、9、10、15、20、25、または30、あるいはこの範囲内の任意の数を有する。
【0037】
特定の実施形態において、前記ポリヌクレオチドのポリヌクレオチド単位(ゆえに、エピトープ)は連続的である。
【0038】
本発明のこのような特定の実施形態において、連続的hTERTエピトープまたは類似体をコードする核酸配列のサイズは、3000bp未満であり、好ましくは2000bp、1000bp、500bp、400bp、300bp、200bp、または100bp未満である。
【0039】
特定の実施形態において、複数エピトープ(ポリエピトープ)をコードするポリヌクレオチドは、hTERTタンパク質の切断型または変異型をコードする核酸分子からなる。好ましい実施形態において、hTERTタンパク質の切断型または変異型は、その触媒活性を失い、すなわちテロメア長の維持に関与する染色体の末端における反復単位(T2AG3)の合成を方向付けることができない。その触媒活性、すなわち逆転写酵素活性が失われたそのようなhTERTタンパク質は非機能的といわれる。従って、別の特定の実施形態において、hTERTタンパク質の切断型または変異型をコードする核酸分子は、hTERTの触媒活性ドメインを欠失している。特定の実施形態において、hTERTタンパク質の切断型をコードする核酸分子は、少なくとも500の最後のC末端アミノ酸からなるタンパク質をコードする。
【0040】
特定の実施形態において、当該ポリヌクレオチドは、図1(野生型)のヌクレオチド2654〜2662に対応し、欠失部位を表す図9のヌクレオチド2658に隣接するヌクレオチド2657を提供するアミノ酸867〜869(VDD配列)を欠失した、あるいは、図1(野生型)のヌクレオチド2645〜2671に対応し、欠失部位を表す図10のヌクレオチド2649に隣接するヌクレオチド2648を提供するアミノ酸864〜872を欠失したhTERTタンパク質をコードする。特定の実施形態において、コードされたhTERTタンパク質は、少なくともアミノ酸残基867〜869を含む欠失、すなわち3個のアミノ酸残基(VDD配列)よりも大きい欠失を有する。一例として、前記の864〜872欠失、並びにアミノ酸残基857から出発して879までの22のアミノ酸欠失(図1によれば)、あるいはVDD配列(ヌクレオチド2639〜2679に対応する、図1によればアミノ酸862〜アミノ酸874)に対して5個のN末端アミノ酸及び5個のC末端アミノ酸を含む欠失である。特定の実施形態において、本発明は、図9または図10に示された核酸配列を含むか、またはそれからなるポリヌクレオチドに関する。
【0041】
本発明の複数エピトープをコードするポリヌクレオチド単位は連続的に配列され得、すなわち第1のポリヌクレオチド単位の3'末端は、第2のポリヌクレオチド単位(など)の5'末端に直接連結し、連続的エピトープのみを含むペプチド配列をコードするポリヌクレオチドを生じる。このようなポリヌクレオチドは、p1、p4、p68、p277、p342、p351、p444、p464、p966、p1107、及びp1123ペプチドを含むか、またはそれらからなるポリエピトープをコードし得る。特定の実施形態において、前記ポリヌクレオチドは、以下のペプチド配列MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLをコードし、ゆえに6個のMHCクラスI制限、特にHLA-B7制限hTERTエピトープを含む。
【0042】
あるいは、本発明の複数エピトープは、1個のアミノ酸スペーサーまたはペプチドスペーサーによって分離され得る。すなわち、異なるポリヌクレオチド単位は、それぞれ1個またはいくつかのアミノ酸をそれぞれコードする1個またはいくつかのコドンによって分離されることを意味する。複数エピトープのプロセシングを促進するスペーサーとして、C末端位置におけるアルギニン(R)及び他の位置における親水性残基(A、K、D、及び/またはT)を含む4個のアミノ酸ペプチドが好ましい。特にC末端位置における陽電荷残基または酸性残基を有する4個のアミノ酸ペプチドを、他の位置における親水性残基(A、K、D、及び/またはT)に依存して、またはそれらから独立して使用できる。特定の実施形態において、前記スペーサーは、複数エピトープのより良好なプロセシングを可能にし、且つ重複した切断により生じる新たなペプチドのプロセシングを避ける、エンドソームまたはリソソームプロセシング配列等の内在的プロセシング配列である。そのようなスペーサーに依存した分離を用いて、ポリヌクレオチド単位のすべて、またはそれに反して一部、ゆえにエピトープのすべてまたは一部を分離することができる。
【0043】
エピトープが配列される順序は、以下の:いくつかの順序は、ポリヌクレオチドの転写及び/または翻訳を促進し得、特に三次元立体配座がその特性に影響を与える場合、小胞体(ER)においてその生じた発現ポリエピトープの輸送を促進し得、且ついくつかのエピトープまたは類似体におけるポリエピトープのプロセシングを促進して、重複するエピトープのプロセシングを避けることができる;という基準に従って当業者により決定され得る。
【0044】
本発明のポリエピトープは、単一の動物またはヒトにおいて、同時に、ポリエピトープ中に含まれる少なくとも1つ、特にいくつかのエピトープまたは類似体に対するCTL応答の誘発を可能にする。
【0045】
特定の実施形態において、本発明のポリエピトープをコードするポリヌクレオチドは、少なくとも2つのエピトープの最もN末端側のエピトープをコードするポリヌクレオチド単位に作動可能に連結された、標的シグナルをコードするポリヌクレオチドをさらに含む。本明細書に用いられる「作動可能に連結された」とは、標的シグナルを作動可能にでき、すなわちポリエピトープを正しい細胞コンパートメントまたはドメインに標的化できる方法で、標的シグナル(上流配列)をN末端エピトープ(下流配列)に連結させることを意味する。従って、2つの配列間の連結により、各配列が異なる位置及び/または異なる段階でそれ自身の機能を果たす。特定の実施形態において、前記標的シグナルは、小胞体シグナル配列であり、適切なプロセシング及びMHCクラスI分子との結合のために、ポリエピトープをERに方向付けさせる。
【0046】
さらなる実施形態において、翻訳過程が開始コドンを必要とする場合、メチオニン残基をコードするコドンを最もN末端側のエピトープをコードする配列の上流に付加させて、ポリヌクレオチドの正しい翻訳を可能にする。最もN末端側のエピトープがその1位にメチオニン残基を有さない場合にのみ、このようなコドンを付加する。
【0047】
本発明はまた、前記に示した定義に従って、
a. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
b. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
c. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
d. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
e. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、
f. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)、
g. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
h. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
i. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
j. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
k. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
l. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
からなる群から選択される少なくとも2つのポリヌクレオチド単位を含むか、またはそれらからなるポリヌクレオチドに関する。
【0048】
以下のポリヌクレオチド:
−a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位、並びに
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位を含むポリヌクレオチド、
−a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、または前記に定義されたそれらの類似体から選択される少なくとも1つのポリヌクレオチド単位を含むポリヌクレオチドもまた、本発明の一部を構成する。
【0049】
遺伝子コードの縮重を考慮すると、本発明のエピトープ、類似体、またはポリエピトープをコードするポリヌクレオチドまたはポリヌクレオチド単位もまた本発明に含まれる。従って、hTERTヌクレオチド参照配列からのコドンによって、または前記アミノ酸をコードする任意のコドンによって各アミノ酸をコードすることができる。
【0050】
本発明はまた、前記群から選択されるこれらのポリヌクレオチド単位またはそれらの類似体の少なくとも2つの任意の組合せを含むか、またはそれらからなるポリヌクレオチドに関するものであって、前記ポリヌクレオチド単位または類似体は、MHCクラスI制限、特にHLA-B7制限hTERTエピトープをコードする。
【0051】
エピトープ、類似体、ポリエピトープ、それらの組合せ、スペーサー、標的シグナル配列等に関して前記に示されたすべての特徴は、本発明の任意のポリペプチド、並びに対応するポリヌクレオチド配列に適用可能である。
【0052】
前記に定義された本発明のポリヌクレオチドを含むか、またはそれからなる組換えベクターもまた、本発明の目的の1つである。前記組換えベクターは、真核生物または原核生物発現用のベクター、例えばプラスミド、細菌導入用のファージ、酵母を形質転換するためのYAC、ウイルスベクター、特にレトロウイルスベクター、または任意の発現ベクターであってよい。本明細書に定義される発現ベクターを選択して、in vitroまたはin vivoのいずれかで、前記に定義されたエピトープまたは類似体またはポリエピトープの産生を可能にする。
【0053】
従って、前記ポリヌクレオチドに加えて、本発明のベクターは、転写制御領域(プロモーター、エンハンサー、リボソーム結合部位(RBS)、ポリAシグナル)、終結シグナル、原核生物もしくは真核生物の複製起点、及び/または選択遺伝子をさらに含むことができる。プロモーターの特徴は、必要とされる発現、すなわち構成的、一過性または誘導性、強いまたは弱い、組織特異的及び/または発生段階特異的プロモーターを考慮して、当業者によって容易に決定され得る。従って、組織特異的プロモーターは、このベクターを含有する組成物が投与される、例えば注入される臓器に応じて、及び必要とされる発現強度に応じて選択され得る。特定の実施形態において、プロモーターはCMVプロモーター(ヒトサイトメガロウイルス)である。前記ベクターはまた、条件付発現を可能にする配列、例えばCre/Loxシステムまたは類似システムの配列等を含むこともできる。
【0054】
本発明の発現ベクターは、ウイルスベクター、特にレトロウイルス由来等のウイルス発現ベクター、特にHIV-、FIV-、またはSIV-由来のベクター等のレンチウイルス由来のベクターであってよい。とりわけ、レンチウイルス由来のベクターは、ヒトレンチウイルス由来のベクター、例えばHIV発現ベクター、特にHIV-1またはHIV-2由来のベクター等である。レトロウイルス由来のベクターは、通常、プラスミド等のDNA構築物中に含まれ、ウイルス粒子中に発現されるレトロウイルスベクターゲノムを含み、前記レトロウイルスベクターゲノムは、逆転写に必要な要素、特に部分的欠失、特にU3領域における欠失を含む変異可能なLTRを含む。いずれの場合にも、レトロウイルス由来のベクターは、全長レトロウイルスタンパク質をコードするすべてのヌクレオチド配列含有することはない。場合により、前記タンパク質またはその機能性断片をコードしないという条件で、それは、1つまたはいくつかの前記ヌクレオチド配列の一部を含有する。前記レトロウイルスベクターゲノムを含む前記DNA構築物は、レトロウイルスヌクレオチド配列で組換えられた対象のDNAをさらに含み、前記対象のDNAは、本発明のポリヌクレオチドを含むか、またはそれからなる。
【0055】
好ましい実施形態において、レトロウイルス由来のベクターゲノムは、下記のようなDNAフラップ及び本発明の少なくとも1つのポリヌクレオチドを含む。好ましい実施形態において、レトロウイルス由来のベクターは、下記のようなDNAフラップ及び本発明の少なくとも1つのポリヌクレオチドを含むHIV発現ベクターである。従って、HIVベクターは、2つのHIV LTRの間に含まれる本発明のポリヌクレオチドを含んだ対象の核酸のみを発現し、ゆえに、5〜6kbまでの大きな配列を収容し得る。本発明の特定の実施形態は、HIV発現ベクター、とりわけHIV-1またはHIV-2発現ベクターであり、HIV-1 LTRまたはHIV-2 LTRはそれぞれ、U3ドメインのプロモーター及びエンハンサーを欠失されている(ΔU3)。この特定の欠失は、特にプロモーターと結合される場合、当該ベクター中に含まれる核酸の発現を増加させることが以前に示されている。別の特定の実施形態において、当該ベクターは、U3ドメインのプロモーター及びエンハンサーを欠失したLTR、CMVまたはEF1αプロモーター等のプロモーター、及び本発明のポリヌクレオチドを含む。
【0056】
別の実施形態において、レトロウイルス由来のベクターに導入された本発明のポリヌクレオチドは、発現カセット中に含まれる。
【0057】
DNAフラップ(またはWO99/55892、WO01/27300及びWO01/27304に開示されたトリプレックス)は、2つの必須領域、すなわちcPPT(セントラルポリプリン配列)及びCTS領域(シス作用終結領域)を含むレトロウイルスまたはレトロウイルス様由来のヌクレオチド配列であって、前記cPPT及びCTS領域は、三本鎖DNA構造を誘導する。本発明に適したDNAフラップは、レトロウイルスまたはレトロウイルス様生物、例えば合成的(化学合成)またはポリメラーゼ連鎖反応(PCR)等の任意のレトロウイルス核酸からのDNAフラップの増幅により調製されたレトロトランスポゾン等から得ることができる。DNAフラップを得ることができるレトロウイルスは、特にレトロウイルスまたはレンチウイルス、とりわけヒトレトロウイルスまたはレンチウイルスであり、特にHIVレトロウイルス、CAEV(ヤギ関節炎脳炎ウイルス)ウイルス、EIAV(ウマ感染性貧血ウイルス)ウイルス、VISNAウイルス、SIV(サル免疫不全ウイルス)ウイルス、またはFIV(ネコ免疫不全ウイルス)ウイルスである。より好ましい実施形態において、DNAフラップは、HIVレトロウイルス、例えばHIV-1またはHIV-2ウイルス、あるいはこれら2つのタイプの任意の異なる単離株から得られる。前記DNAフラップは、その天然(ウイルスゲノム)ヌクレオチド関連、すなわちレトロウイルス中に天然に含まれるpol遺伝子から単離されて使用されることが注目に値する。従って、本発明において使用されるDNAフラップは、pol遺伝子の不必要な5'及び3'部分を欠失しており、異なる由来の配列で組換えられている。
【0058】
DNAフラップは、ベクターの核内への移入のシス決定基として作用し、組換え並びに非分裂及び分裂細胞の両方への核酸の組込みにとって非常に興味深い。DNAフラップ配列(TRIPベクター)を含む発現レトロウイルス由来のベクター、特にHIV由来のベクターは、DNAフラップを欠失する他のHIVベクターよりも10倍高い効率で、初代B細胞及びT細胞、マクロファージ、樹状細胞等を形質導入することができる。細胞の80〜90%の形質導入を、日常的に得ることができる。
【0059】
好ましい実施形態において、in vivo発現及びワクチン戦略に適したベクターは、レトロウイルスベクター、特にレンチウイルスベクターである(WO 99/55892、WO 01/27300、及びWO 01/27304を参照)。このようなベクターは、それらのゲノムが改変された場合、特に効率的で安全であることが示されている(Firatら、2002年、The Journal of Gene Medicine 4:38〜45頁)。実際、これらのベクターは、治療遺伝子またはレポーター遺伝子を、多種多様な細胞及び組織、例えば造血幹細胞、脳、肝臓、及び網膜等のに効率的かつ安定的に移行させる能力を有する。さらに、この高い形質導入効率は、標的細胞の増殖状態に無関係である。特に、これらのベクターは、in vivoだけでなくex vivoで高い効率で樹状細胞(DC)等の抗原提示細胞を形質導入する能力により、マウスにおけるin vivo及びヒトにおけるex vivoの両方でCD8+ T細胞応答を効率的に誘発することが示されている(Esslingerら、Hum Gene Ther 2002年;13:1091〜100頁;Breckpotら、J Gene Med 2003年;5:654〜67頁;Esslingerら、J Clin Invest 2003年;111:1673〜81頁)。
【0060】
図7に定義されているベクターは、本発明のエピトープ、類似体、またはポリエピトープのin vitroまたはin vivo発現に使用されている。本発明のベクターの由来となり得るベクターの例は以下のとおりであり、すべてCNCM(Collection Nationale de Culture de Microorgnismes at Institut Pasteur:パスツール研究所にある国立微生物コレクション、パリ、フランス)に寄託されている。
【表1】
【0061】
特許出願WO 01/27304に記載されているベクター、特にTRIPΔU3 Efα1 GFP及びTRIPΔU3 PL CMV GFPもまた、本発明のベクターを得るために使用することができる。
【0062】
本発明の特定のベクターは、2006年7月28日に番号CNCM I-3660の下でCNCM(パスツール研究所(Institut Pasteur)、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターである。このベクターを培養するのに適した増殖培地は、場合によってはヒグロマイシンを補充されたTB培地である。本発明の別の発現ベクターは、2006年7月28日に番号CNCM I-3660の下でCNCMに寄託されたpTRIP-CMV-ΔhTERTベクターであり、欠失したhTERT配列は、本発明の任意のポリヌクレオチドによって置換されている。
【0063】
本発明はまた、本発明のポリヌクレオチドまたはポリヌクレオチド単位を含む細胞に関する。
【0064】
一実施形態において、当該細胞は、当業者に周知の方法によって、すなわち化学的トランスフェクション(リン酸カルシウム、リポフェクタミン)、脂質ベース方法(リポソーム)、エレクトロポレーション、光穿孔、ウイルスベクターの使用等によって、本発明のベクターでトランスフェクションされる。別の実施形態において、相同的細胞配列での組換えまたは細胞ゲノム内への挿入のいずれかによって、細胞ゲノム内へのポリヌクレオチドの組込みを可能にする方法で、細胞を本発明のポリヌクレオチドで形質転換または形質導入する。トランスフェクション、感染、または形質導入は、ex vivo、すなわち生体外の人工的環境で生じる得る。
【0065】
ワクチン戦略で特に関心のある細胞としては、免疫系の細胞、特に抗原提示細胞(APC)がある。特定の実施形態において、これらの細胞は、樹状細胞(DC)のようなMHCクラスI認識、あるいはマクロファージまたはBリンパ球等のMHCクラスII認識のいずれかに関与するAPCである。DCとしては、ex vivoで完全に成熟させたDC、すなわちエピトープまたは類似体によってin vitroで成熟されたDCが好ましい。
【0066】
本明細書に用いられている用語「トランスフェクションされた」、「形質転換された」、または「感染された」とは、本発明のベクターを含む細胞(一過性発現)を表し、一方、用語「遺伝的に形質転換された」とは、そのゲノムが、本発明のポリヌクレオチドによって決定的に改変されている細胞(永続的発現)を表す。
【0067】
前記一過的または安定的に形質転換された細胞は、任意の原核生物(細菌)細胞または真核生物(酵母、哺乳動物、特にヒトを含む動物)細胞であってよい。一実施形態において、細胞は非ヒト細胞である。特定の実施形態において、本発明の細胞は、単離されたヒト細胞であり、「単離された」とは、その天然環境の外部を意味する。
【0068】
特定の宿主は、2006年7月28日に番号CNCM I-3660の下でCNCMに寄託された大腸菌(E. coli)株である。
【0069】
本発明はまた、本発明のポリヌクレオチドを記載する際に前記に定義されたエピトープ、類似体、またはポリエピトープ、特に本発明のポリヌクレオチドまたはポリヌクレオチド単位によってコードされた任意のポリペプチドに関する。特定のポリペプチドは:
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるMHCクラスI制限、特にHLA-B7制限hTERTエピトープである。
【0070】
特定のHLA-B7制限hTERTエピトープは:
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択される。
【0071】
特定の群は、以下のMHCクラスI制限、特にHLA-B7制限hTERTエピトープ:
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、及び
d. RPSLTGARRL(p351)
からなる。
【0072】
別の群は、以下のHLA-B7制限hTERTエピトープ:RPAEEATSL(p277)及びRPSFLLSSL(p342)からなる。これらのエピトープによって標的化される好ましいHLA-B7対立遺伝子は、HLA-B0702である。
【0073】
本発明はまた、前記に開示され、少なくとも1つのアミノ酸置換を有するペプチドの類似体に関する。これらの類似体に関する特徴は、特に前記の頁に記載されている。特定のペプチド類似体は、以下のペプチド配列APRRLVQLLを有するp444*である。
【0074】
最後に、本発明はまた、本明細書に記載されているhTERTエピトープ、類似体、またはポリエピトープをコードする任意のポリヌクレオチドに関する。
【0075】
本発明はまた、前記の少なくとも2つのエピトープ及び/または類似体を含むポリエピトープに関する。本発明のポリエピトープは、全長hTERTタンパク質ではない。前記ポリエピトープのサイズは、15〜1000個、特に50または100〜1000個のアミノ酸であってよく、とりわけ特に約100、200、300、400、500、または1000個のアミノ酸である。このようなエピトープは、2〜30個のエピトープまたは類似体、特に2〜20個または2〜10個のエピトープ及び/または類似体を含むか、またはそれらからなる。特定のポリエピトープは、6個の連続エピトープを含むか、またはそれらからなり、以下の配列:MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLを有する。別の特定のポリエピトープは、前記p1、p4、p68、p277、p342、及びp351エピトープを含むか、またはそれらからなり、前記エピトープは、得られたポリエピトープにおいて互いに連続的であるか、ペプチドスペーサーによってそれらのすべてもしくは一部は分離されている。
【0076】
本発明の別のポリエピトープは、少なくとも2つのエピトープを含み、少なくとも1つが:
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、且つ少なくとも1つが:
j. MPRAPRCRA(p1)、
k. RPAEEATSL(p277)、
l. RPSFLLSSL(p342)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、前記ポリエピトープは、全長hTERTではない。
【0077】
本発明のポリペプチドは、化学的に合成され得るか、あるいは細胞系で対応する核酸配列の発現後にin vitro(無細胞系)またはin vivoで産生され得る。
【0078】
図1に示される全長hTERTタンパク質並びに対応する全長コード配列は、本発明から除外される。RPALLTSRLペプチドも本発明から除外される。これらのペプチドは、HLA-B7認識の関連において特に除外される。
【0079】
本発明はまた、hTERTに対するHLA-B7制限免疫応答を誘発させるか、またはそれを提供することに関与する使用のために、前記に定義されたエピトープ、類似体、ポリエピトープまたはポリヌクレオチド、発現ベクターまたは宿主細胞に関する。
【0080】
本発明はまた、本発明のポリヌクレオチド、ベクター、宿主細胞、及び/またはポリペプチドを含む組成物に関する。特定の実施形態において、前記組成物は、in vivo投与に適しており、すなわち前記組成物は、注入用に調製されるか、またはより一般的には、投与用に生理的に許容され得る溶液または乳濁液とともに調製される。前記組成物は、特に注入によって全身または局所投与のいずれかに使用され得、製薬的に適した賦形剤(水、生理食塩水、デキストロース、グリセロール、エタノール、及びそれらの組合せを含む)、あるいはキャリア及び/またはビヒクルをさらに含むことができる。
【0081】
特定の実施形態において、前記組成物は、エピトープ、類似体をコードするか、または前記のポリエピトープをコードする、本発明のポリヌクレオチドを含む。前記組成物は、HLA-B7のものとは異なるMHCクラスI対立遺伝子に制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをコードする他の核酸分子を含むことができる。種々のHLA上位型または対立遺伝子に制限されたhTERTエピトープの組合せは、単一の上位型または対立遺伝子よりも治療を必要としている患者のより大きな集団を網羅することができる。最後に、HLA-A1、-A2、-A3、及びA24が好ましい。
【0082】
別の実施形態において、当該組成物は、MHCクラスIIに制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをコードする核酸分子を含む。前記組成物は、前記の核酸分子と、本発明の少なくとも1つのHLA-B7制限hTERTエピトープ、類似体、またはポリエピトープとの任意の組合せを含むことができる。核酸分子の組成物において、種々の免疫細胞(クラスIに対するTリンパ球またはNK細胞対クラスIIに対するヘルパーリンパ球)の反応、及び/または種々の免疫応答(液性応答対細胞性応答)の誘発を可能にする、クラスI及びクラスII制限エピトープをコードするポリヌクレオチドの組合せもまた、本発明内に入る。
【0083】
別の実施形態において、当該組成物は、1つの腫瘍特異的抗原(TSA)をコードする少なくとも核酸分子、並びに/あるいは少なくとも1つの腫瘍関連抗原(TAA)、例えば前立腺特異的抗原(PSA)、前立腺特異的膜抗原(PSMA)、または前立腺酸性ホスファターゼ(PAP)等を含む核酸分子を含む(Tartourら、2000年、Immunol Lett Sep 15;74(1):1〜3頁;Tartourら、1996年、Presse Med. Nov 16;25(25):1717〜22頁)。
【0084】
いくつかのタイプの治療用組成物を、本発明のエピトープまたは類似体に対して免疫応答を誘発させるために使用することができる。
【0085】
本発明ポリヌクレオチドを含む組成物は、宿主に投与され、例えば注入され(DNAワクチン接種として知られている)、前記核酸は、本発明の複数エピトープを含むか、またはそれらからなるポリペプチドをin vivoで発現する。このようなDNAワクチン接種は、通常、前記のプラスミドベクターからなる。裸のDNAの送達は、効率に乏しいことが示されており、細胞内へのDNAの送達及び取込みを改善するためにいくつかのキャリアを必要とする。2つのタイプのキャリア:(1)ウイルスキャリア(アデノウイルス、レンチウイルス、麻疹ウイルス)、または(2)非ウイルス性キャリア、例えばポリマー(特にカチオン性ポリマー類)、カプセル化DNA(DNA及びRNA等のポリアニオンとの同時且つ迅速に相互作用するカチオン性脂質を含み、リポソーム/核酸複合体を生じるリポソーム)、または金微粒子に連結されたDNA等が現在までのところ開発されている。さらに、核酸の細胞取込みを補助する薬剤、例えばカルシウムイオン、細菌タンパク質(赤痢菌(Shigella)タンパク質)、ウイルスタンパク質、及び他のトランスフェクション促進剤等を有利に使用することができる。本発明による組成物の別のタイプは、前記のベクターまたはベクターゲノムを含むレンチウイルス擬似粒子を含む組成物である。
【0086】
別のタイプの組成物は、本発明のエピトープ、類似体、またはポリエピトープを含む。このような組成物は免疫原性であり、すなわちそれは、それが投与される宿主に免疫応答を誘発させ得る。しかしながら、本発明のポリペプチドの免疫原性特性を増大させるために、アジュバントを前記ポリペプチドとともに投与して、免疫応答を誘発または改善することができる。アジュバントは、前記アジュバントと混合された抗原の免疫原性を増強する任意の物質として定義される。いくつかのアジュバントは、可溶性抗原を小粒子、例えば水酸化アルミニウムゲル、水中油乳濁液、または免疫刺激複合体(ISCOM)等に変換する。別のクラスのアジュバントは、無菌の細菌成分、例えば細胞壁または多糖、フロイントアジュバント等を含む。最後に、乳化剤またはpH緩衝剤を用いて、前記エピトープまたは類似体の免疫原性挙動を増強することもできる。
【0087】
前記に引用されたすべての組成物を、種々の経路:皮下(s.c.)、皮内(i.d.)、筋内(i.m.)、または静脈内(i.v.)注射、経口投与、及び粘膜投与、特に鼻腔内投与または吸入を介して宿主内に注入することができる。投与される量(投与量)は、患者の病態、個々の免疫系の状態、投与経路、及び宿主のサイズを含む、治療を受ける対象に依存する。好適な投与量は、200μg〜1mgの範囲であり、状況に応じて当業者により変更することができる。
【0088】
本発明の組成物は、hTERTの過剰発現により生じる腫瘍を含む制御できない細胞増殖から生じる患者における悪性状態の予防及び/または治療、並びにこのような悪性状態、例えば癌に伴う有害な結果の治療に有用である。表現「治療」とは、本発明の組成物により達成される治癒効果、及び治療を受けている患者に対する有益な効果を含み、前記効果は、細胞レベルまたは臨床レベルで得られ、その結果として、患者の病態の改善、並びに/あるいは寛解状態または健康状態の回復を含む。特定の実施形態において、本発明の組成物は、腫瘍の予防または治療に有用な付加的な活性化合物、すなわち一般的な化合物または組織特異的な癌において活性があることが証明された化合物をさらに含む。
【0089】
本発明はまた、
a. Tリンパ球を提供する工程;及び
b. 前記Tリンパ球を、本発明の少なくとも1つのエピトープまたはエピトープ類似体、またはポリエピトープとともに、前記リンパ球の活性化を可能にする条件でin vitro培養する工程;
を含む、クラスI制限、特にHLA-B7制限hTERTエピトープに対してTリンパ球を活性化させる方法に関する。
特定の実施形態において、活性化Tリンパ球は、細胞毒性Tリンパ球(CTL)である。Tリンパ球を活性化させる従来の条件は、インターロイキン(IL)2、IL-7、IL-12、及び/またはIL-15を用い、参照として本明細書に組み込まれているMinevら(2000年、PNAS;97(9):4796〜4801頁)に記載されている。
【0090】
本発明はまた、
a. Tリンパ球を、本発明の少なくとも1つのエピトープまたはエピトープ類似体、またはポリエピトープとともに、適切な条件下でin vitro培養することによる、前記Tリンパ球を活性化させる工程;
b.前記活性化リンパ球を、MHCクラスI分子に結合している本発明のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件下でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動を調査する方法に関する。
【0091】
hTERTエピトープの免疫原性挙動を調査するための特定の方法において、個々に、すなわちポリエピトープの形態下ではなく用いられる場合、当該エピトープは、(a)RPSLTGARRL(p351)、(b)APSFRQVSCL(p68)、(c)APRCRAVRSL(p4)、(d)DPRRLVQLL(p444)、(e)FVRACLRRL(p464)、(f)AGRNMRRKL(p966)、(g)LPGTTLTAL(p1107)、及び(h)LPSPKFTIL(p1123)の中から選択され、類似体の例は、前記に定義されたp444*である。
【0092】
hTERTペプチドの免疫原性挙動及びHLA-B7制限性を調査する方法は、
a.前記に記載されたTリンパ球を、MPRAPRCRA(p1)、APRCRAVRSL(p4)、APSFRQVSCL(p68)、RPAEEATSL(p277)、RPSFLLSSL(p342)、RPSLTGARRL(p351)、DPRRLVQLL(p444)、FVRACLRRL(p464)、AGRNMRRKL(p966)、LPGTTLTAL(p1107)、及びLPSPKFTIL(p1123)の中から選択されるエピトープまたはAPRRLVQLL(p444*)等のエピトープ類似体、または前記に定義されたポリエピトープで活性化する工程;
b.前記活性化リンパ球を、HLA-B7分子に結合している本発明のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件下でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;を含む。
【0093】
リンパ球の活性化は、抗原提示細胞による前記エピトープまたは類似体の、天然の(活性化されていない)Tリンパ球に対する提示を含む。天然のTリンパ球が本発明のエピトープまたは類似体を認識すると、特定のクラスI HLA分子の関連において、それらは「活性化」といわれ、標的細胞の細胞表面の前記エピトープを認識する状態にある。前記活性化リンパ球(エフェクター細胞)と標的細胞(それによりリンパ球が活性化されたエピトープを発現している)との間の接触により、分子が分泌され、標的細胞が死滅する。リンパ球を活性化させるために用いられたエピトープまたは類似体が、前記活性化リンパ球による前記エピトープを有する標的細胞の効率的な破壊をもたらす場合、前記エピトープは、前記エピトープを発現している細胞に対してin vitroのみならずin vivoでのT細胞反応を可能にするのに十分に免疫原性であるものとして見なされ得る。標的細胞/リンパ球認識に適した条件は、RPMI培地中において37℃で4時間の接触である。
【0094】
本発明はまた、MHCクラスI制限、特にHLA-B7制限hTERTエピトープに対して、細胞、特に抗原提示細胞(APC)、B細胞、T細胞、及び/または樹状細胞をin vitroで成熟させる方法に関する。細胞をin vitroで成熟させるための前記方法は、:
a. 細胞を提供する工程;
b. 少なくとも1つのMHCクラスI制限、特に本発明のHLA-B7制限hTERTエピトープまたはエピトープ類似体、またはポリエピトープを用いて、前記細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟細胞の増殖を促進する工程;
を含む。
【0095】
前記のとおり、リンパ球の活性化は、成熟抗原提示細胞によるエピトープの提示を必要とする。本発明の好ましい実施形態において、前記細胞は、少なくとも1つのHLA-B7対立遺伝子を発現する。それらのうちの1つである樹状細胞(DC)は、MHCクラスIに制限された内因性エピトープのTリンパ球に対する提示において特に有効である。前記細胞の成熟の目的の1つは、成熟させた時点での治療を必要としている患者への投与である。前記成熟DCの投与により、患者のリンパ球の活性化、及び前記エピトープ(DCを形質転換している1つ)を発現している細胞に対する迅速な反応をin vivoで生じるあろう。
【0096】
特定の実施形態において、樹状細胞をin vitroで成熟させる方法は、:
a. 樹状細胞を提供する工程;
b. 本発明の少なくとも1つのMHCクラスI制限hTERTエピトープまたはエピトープ類似体、またはポリエピトープを用いて、前記樹状細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟樹状細胞の増殖を促進する工程;
を含む。
【0097】
特定の実施形態において、前記樹状細胞は、循環血液または骨髄細胞から単離される。他の実施形態において、樹状細胞は、治療を必要としている患者に対する投与後の拒絶反応を避けるために、前記患者またはHLA適合ドナーから単離される。
【0098】
DCの成熟は、本発明のポリヌクレオチドを用いた前記樹状細胞の遺伝的形質転換によって、本発明のベクターを用いた前記樹状細胞のトランスフェクションによって、または本発明の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープとの前記樹状細胞の接触によって達成され得る。遺伝的形質転換は、その効率性、及びDCゲノム内に挿入されたポリヌクレオチドによってコードされたエピトープ、類似体、またはポリエピトープの永続的発現が理由で、好ましい。
【0099】
本発明はまた、癌の予防及び/または治療に使用するための、HLA-B7-制限hTERTエピトープをコードするポリヌクレオチドに関する。特定の実施形態において、癌の予防及び/または治療に使用するための前記ポリヌクレオチドは、HLA-B7-制限hTERTエピトープまたはその類似体、あるいは本出願に記載されている少なくとも1つのHLA-B7-制限hTERTエピトープまたはその類似体を含むポリエピトープをコードする。ポリエピトープをコードするポリヌクレオチドに関する限り、それは全長hTERTのコード配列に一致しない。他方、それは、hTERTの変異型または欠失型に一致し得、前記変異または欠失は、ヒトテロメラーゼの触媒活性を抑制する。特定の実施形態において、前記ポリヌクレオチドは、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)、
からなる群から選択されるHLA-B7-制限hTERTエピトープをコードする少なくとも1つのポリヌクレオチド単位、あるいはAPRRLVQLL(p444*)等の前記HLA-B7-制限hTERTの少なくとも1つの類似体、あるいは前記HLA-B7エピトープ及び/または類似体をコードする少なくとも2つのポリヌクレオチド単位を有するポリヌクレオチドを形成する任意の組合せを含む。
【0100】
本発明は、癌の予防及び/または治療に使用するための、HLA-B7 hTERTエピトープ、特にHLA-B0702制限エピトープに関する。本発明はまた、癌の予防及び/または治療に使用するための、本発明のポリヌクレオチド、ベクター、宿主細胞、またはポリペプチドに関する。
【0101】
本発明はまた、癌の予防及び/または治療のための薬剤の製造のための、HLA-B7 hTERTエピトープ、(または対応するポリヌクレオチド)の使用に関する。特定のHLA-B7 hTERTエピトープ(または対応するポリヌクレオチド)は、前記のもの、並びにそれらを含むか、またはそれらからなるベクター、細胞、または組成物である。特定の実施形態において、癌の予防及び/または治療のための薬剤の製造における、本発明のポリエピトープを含むか、またはそれからなるポリヌクレオチド、ベクター、宿主細胞、またはポリペプチドの使用は、前記に定義した少なくとも1つのHLA-B7対立遺伝子、特に少なくとも1つのHLA-B0702対立遺伝子を有する患者に提供されることを意図されている。
【0102】
本明細書に提供された各定義は、それぞれの及び任意のペプチド(エピトープ、類似体、またはポリエピトープ)、並びに個別に選択された(それ自体が)または群に包含されたそれぞれの及び任意のポリヌクレオチドに適用される。
【図面の簡単な説明】
【0103】
【図1A】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1B】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1C】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1D】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1E】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図1F】hTERTタンパク質をコードする遺伝子及び対応するアミノ酸配列。当該コード配列は、ヌクレオチド56〜3454に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図2】hTERT由来ペプチドは、HLA-B0702トランスジェニックマウスにおいてプロセシングされる。HLA-B7 Tgマウス及び1匹の無処理マウス(N)を、hTERTをコードする100μgのDNAで免疫化した。14日目に、各マウスからの脾臓細胞を、種々のhTERT由来ペプチドを用いて別々にin vitro刺激した。6日後、材料及び方法に記載されている関連ペプチド(黒い四角)またはコントロールペプチド(白い四角)を負荷させたRMA-B7標的に対して、エフェクター細胞をアッセイした。60/1比における溶解パーセンテージを示す(2つの独立した実験からの結果)。
【図3】健常な血液ドナーからのPBMCにおける、hTERTに対するCTL応答の誘発。HLA-*B0702+健常ドナーからのTリンパ球細胞を、材料及び方法に記載されている6種のhTERTペプチドパルス化自系PBMCのそれぞれで活性化した。週1回の刺激の4ラウンド後、関連ペプチド(黒い四角)またはコントロールペプチド(白い四角)でパルス化したエフェクター細胞を、51Cr標識T2-B7細胞に対する溶解活性についてアッセイした。20/1のエフェクター対標的比における溶解パーセンテージを示す。10のドナーのうち8のドナーからの結果を示す(d1〜d8)。
【図4】腫瘍細胞に対するCTLp351の細胞毒性に及ぼす抗HLAクラスI mAbの効果。HLA mAbs(抗HLAクラスI mAbまたは抗クラスII mAb(HLA-DR))の非存在または存在下で前処理されたHLA-*B0702+腫瘍細胞系Mamo及びU293Tに対するCTLp351系の細胞毒性を、10/1のエフェクター対標的比において、標準的51Cr放出アッセイにより測定した。
【図5】Lv-hTERT免疫化後のhTERT特異的T細胞応答のex vivo検出。(A)HLA-B7トランスジェニックマウスを、組換えTrip-hTERT粒子またはコントロールTrip-GFP(1500ng)で免疫化した。12日後、各マウスのIFNγを産生するhTERTペプチド特異的T細胞を、新鮮な脾臓細胞中、IFNγ-ELISPOTアッセイによってex vivo検出した。ネガティブコントロール値を減算後、IFNγ SFCの数を算出した。3つの独立した実験からの結果を示す。(B)前記のように、HHDマウスをTrip-hTERTで免疫化した。前記のように、IFNγを産生するhTERTペプチド特異的T細胞を、ELISPOTによってex vivo検出した。2つの独立した実験からの結果を示す。
【図6】Trip-hTERT免疫化後のHLA-*B0702トランスジェニックマウスにおける特異的CD8+T細胞応答の刺激(priming)。HLA-*B0702トランスジェニックマウスを、Trip-hTERT(初めの4本の黒色のバー)またはコントロール(終りの2本の縦縞のバー)のいずれかで免疫化した。12日後、IFN-γELISPOTアッセイによって、各マウスの脾細胞中のIFN-γ産生単細胞をex vivo検出した。個々の免疫化マウスの新鮮な単離脾細胞からのフィコール精製リンパ球を、5μg/mlの各HLA-*B0702制限hTERT由来ペプチドとともに、またはそれなしで、24時間直接培養した。ペプチドなしのコントロールに関して得られた非特異的値を減算後、特異的IFN-γSFCの数を算出し(<15SFC)、当該応答は、SFC≧30に対して陽性と見なされた。
【図7】pTrip-hTERTの略図。このレンチウイルス由来ベクターは、HIV-1ゲノムのpsi配列、cPPT、及びCTS中心シス活性配列(フラップ)、並びに対象の遺伝子の発現を可能にするCMVプロモーターを含む。さらに、3'LTRにおいて、U3ドメインが欠失している(ΔU3)。
【図8】(A)DNA pTRIP-CMV-ΔhTERT免疫化は、HHDマウスにおいて、hTERT特異的CD8+T細胞応答を誘発した。HHDマウス(HLA-A2.1 Tg)を、hTERTの機能的でない形態(pTRIP-CMV-ΔhTERT)をコードするDNAを用いてDNA免疫化した。10日後、IFN-γELISPOTアッセイにより、IFNγ産生ペプチド特異的T細胞をex vivo検出した。個々の免疫化マウスの脾細胞からのフィコール精製リンパ球を、5μg/mlの各HLA-A2.1制限hTERT由来ペプチドとともに、またはそれなしで、24時間直接培養した。特異的IFN-γSFCの数を前記のように算出した。当該応答は、SFC≧30に対して陽性と見なされた。(B)pTRIP-CMV-ΔhTERT免疫化後のHHDマウスにおける、短期CTL応答の誘発。HHDマウスを、hTERTの機能的でない形態(pTRIP-CMV-ΔhTERT)をコードするDNAを用いて、10日間免疫化した。個々のマウスからの脾臓細胞を、HLA-A2.1制限hTERT由来のp540及びpY572ペプチドを用いて、6日間、in vitroで再刺激した。関連ペプチドまたは非関連ペプチドのいずれかを負荷したHHDでトランスフェクションされたEL4細胞に対する51Cr放出アッセイにおいて、エフェクター細胞を試験した。
【図9A】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9B】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9C】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9D】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9E】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図9F】機能的でないhTERTタンパク質配列(アミノ酸867〜869の欠失)。当該コード配列は、ヌクレオチド59〜3348に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10A】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10B】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10C】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10D】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10E】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【図10F】機能的でないhTERTタンパク質配列(アミノ酸864〜872の欠失)。当該コード配列は、ヌクレオチド59〜3430に位置している。開始コドン及び終止コドンには下線が引かれている。1行目はヌクレオチド配列であり、2行目は対応するアミノ酸配列である。3行目は最初のアミノ酸としての開始コドンから出発する、当該hTERTコード配列の番号である。
【発明を実施するための形態】
【0104】
[実施例]
I−材料及び方法
<血液ドナー>
末梢血サンプルは、成人の健常血小板ドナー(centre de transfusion sanguine de I'hopital Mondor、Creteil、フランス)から、書面によるインフォームドコンセントに従って得た。末梢血ドナーのHLA分類は、H.Mondor.Hospital Creteil(フランス)のHLA研究室において実施された。前記試験は、フランス血液バンク機関(French Blood Bank Institute)によって承認された。
【0105】
<マウス>
構成的マウスβ2-m分子(HLA-B7mα3)及びキメラHLA-A2.1/H2-Db分子を発現しているHHDトランスジェニックマウスと組み合わせて、HLA-B0702α1α2、H2-Kdα3キメラ構築物を発現しているHLA-*B0702トランスジェニック(Tg)マウスは、以前に記載されているように、それらのH2-Db及びH2-kb遺伝子を欠失していた(Pascoloら、J Exp Med、1997年;185:2043〜51頁;Rohrlichら、Int Immunol、2003年;15:765〜72頁)。これらのマウスはC57BL/6をバックグラウンドとしており、本発明者らの動物施設において、特定病原体未感染の条件下で飼育され、維持された。
【0106】
<腫瘍細胞系>
T-BハイブリッドT1、EBV形質転換B細胞JY、腎臓癌細胞系U293T、及びBurkittリンパ腫細胞Rajiは、アメリカン・タイプ・カルチャー・コレクション(American type Culture Collection:ATCC)から入手した。メラノーマ細胞系(SK23MEL、LB34、及びKUL68)は、P.Coulie(ブリュッセル、ベルギー)からの好意で提供され、EBV形質転換B細胞BBG.1及びBC3は、H.Collandre(R.A.H.P.、グレノーブル、フランス)からの好意で提供された。
【0107】
HLA-*B0702でトランスフェクションされたTAP欠失T2細胞(T2-B7)は、P.Cresswell(Smithら、J Immunol、1996年;156:3755〜64頁)からの好意で提供された。マウスリンパ腫細胞系RMA及びEL4は、ATCCから入手し、これらの細胞もまた、HLA-*B0702遺伝子でトランスフェクションされ、標的細胞として用いられた。
【0108】
<エピトープ選択ペプチド合成>
HLA-*B0702に結合することが予測される9アミノ酸(9量体)または10アミノ酸(10量体)ペプチドの存在についてhTERTのアミノ酸配列を分析するために、予測的アルゴリズム「SYFPEITHI」(Lu及びCelis E、Cancer Res、2000年;60:5223〜7頁)を用いた。標準的HLA-B7結合アンカー、2位にPro、及びカルボキシル末端に疎水性脂肪族(AlaまたはLeu)を含有する候補ペプチドを、それらの最高予測スコアに従って選択した。6種のペプチド、すなわちp1(MPRAPRCRA、残基1〜9)、p277(RPAEEATSL、残基277〜285)、及びp342(RPSFLLSSL、残基342〜350)と称される3種の10アミノ酸ペプチド、並びにp4(APRCRAVRSL、残基4〜13)、p68(APSFRQVSCL、残基68〜77)、及びp351(RPSLTGARRL、残基351〜360)と称される3種の9アミノ酸ペプチド(アンカー位置に下線を引いた)が保持され、合成された。
【0109】
ヒトサイトメガロウイルスpp65に由来するペプチド、RPHERNGFTV(R10TV)、及びヒト免疫不全ウイルス1型IPRRIRQGLを合成し、コントロールペプチドとして用いた。マウスにおけるペプチド免疫化用のヘルパーペプチドとして、B型肝炎ウイルスコア128〜140由来のペプチド(TPPATRPPNAPIL)を用いた。ペプチドは、最低純度80%までPRIMから購買し、2mg/mlの濃度で蒸留水またはDMSO中で再構成した。
【0110】
<HLA-B0702結合/安定化アッセイ>
HLA-B0702でトランスフェクションされたT2(T2-B7)細胞に関する、MHC安定化アッセイを用いることによって、記載された(Rohrlichら、Int Immunol、2003年;15:765〜72頁)参照ペプチド(R10TV)と比較した、HLA-*B0702に対するhTERT由来ペプチドの相対的結合活性を測定した。要するに、種々の最終濃度(100、10、1、及び0.1μM)における、参照ペプチドR10VまたはhTERTペプチドのいずれかの非存在下(ネガティブコントロール)または存在下で、100ng/mlのヒトβ2-ミクログロブリンを添加した無血清培地AIM-V(Invitrogen社、Gibco)中、96ウェルプレートにおいて、T2-B7を2×105細胞/ウェルで一晩インキュベーションした。T2-B7細胞を飽和濃度の抗HLA-B7mAb、ME.1で標識し、次いで2回洗浄し、最後にFITC接合F(ab')2ヤギ抗マウスIgで染色して、フローサイトメトリーを行った。
【0111】
結果を、参照ペプチドの濃度に対する、最高結合度(参照ペプチドに関して得られた)の20%に達するのに必要な試験ペプチドの濃度の比率である相対的結合活性の値として表す。従って、当該値が低いほど、結合性が強い。
【0112】
<CTL誘発用のHLA-*B0702トランスジェニックマウスのペプチド免疫化>
8〜10週齢のメスHLA-*B0702トランスジェニックマウスに、フロイント不完全アジュバント(Difco、デトロイト、ミシガン州)中、共乳化された140μgのヘルパーペプチドを補充した50μgの個々のHLA-B0702制限hTERTペプチドを、尾基部に皮下注射(s.c.)した。10日後、個々のマウスの脾臓細胞を6ウェルのプレート中で、関連ペプチドを用いてin vitroで再活性化した。関連ペプチドまたはネガティブコントロールペプチドでパルス化された、HLA-*B0702でトランスフェクションされたRMA細胞(RMA-B7)を用いて、エフェクターCTL細胞を、標準的な4〜5時間51Cr放出アッセイにおいて試験した。特異的溶解≧10%が観察された場合、マウスを応答動物であると見なす。
【0113】
<HLA-*B0702トランスジェニックマウスにおけるDNA免疫化>
CMVプロモーターの制御下、hTERT遺伝子をコードするLvCMV-hTERTプラスミドベクターを、エンドトキシンのない条件下でのプラスミドGigaキットカラム(Qiagen)上で精製した。麻酔したHLA-*B0702トランスジェニックマウスの再生前脛骨筋内に、前記プラスミドを注入した(各側に50μg)。14日後、個々のマウスの脾臓細胞を、Con Aで活性化したラット脾臓細胞からの10%上清を補充した完全培地中、ペプチドでパルス化された(10μg/ml)同系のγ照射(50Gy)LPS-リンパ芽球を用いてin vitroで再活性化した。記載されたとおり、細胞毒性アッセイを6日間実施した。
【0114】
<レンチウイルスベクター構築物及び製造>
まず、pBABE-hygro-hTERTプラスミド(Counterら、Proc Natl Acad Sci U.S.A.1998年;95:14723〜8頁)由来のEcoRI-SalI hTERT挿入物を、pSP73ベクター(Promega)内にサブクローニングすることによって、pTRIP-デルタU3-CMV-hTERT(TRIPLv-hTERTまたはLv-hTERT、またはpTrip-hTERTと称される)(図7)構築物を作製した。次いで、BamHI及びXhoIで切断されたpTRIP-CMVプラスミド内に、BglII-SalI断片を挿入した。記載されているとおり、293T細胞の一過性の(48時間)トランスフェクションにより、擬似組換えレトロウイルス粒子を作製した(Zennouら、Cell、2000年;14:101:173頁;Firatら、J Gene Med、2002年;4:38〜45頁)。組換えレトロウイルス粒子を、超遠心によって濃縮し、PBS中に再懸濁した。ベクター粒子の量を、市販のELISAアッセイにおけるp24タンパク質のものから推定した(NEN、DUPONT、France Perkin Elmer)。
【0115】
2006年7月28日にCNCM I-3660の番号でCNCM(Institut Pasteur、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターを、前記段落に記載されたように実施した。しかし、hTERTタンパク質は、図1のヌクレオチド2654〜2662(野生型)に相当するアミノ酸867〜869の欠失によって機能的ではなくなっていた。QuickChange XL部位特異的突然変異誘発キット(Stratagene)を用いて、アミノ酸残基867〜869の欠失を作り出すことによって、触媒効力を失ったhTERT RT変異体(ΔhTERT)を作製し、配列決定によって確認した。
【0116】
<MHCクラスIトランスジェニックマウスにおける免疫化及びCTL欠失>
1500ngのTripLv-hTERT懸濁液またはコントロールベクターの単回皮下(尾基部)注入と同様に、TRIPLv-hTERTを用いた免疫化を実施した。
【0117】
500μlのPBS中1500ngのp24抗原に等価の、組換えレンチウイルス粒子のpTRIP-CMV-ΔhTERTまたはコントロールとしてのTrip-GFPの単回腹腔内注入として、HLA.A2トランスジェニックマウスにおける免疫化を実施した。
【0118】
12日後、ELISPOTアッセイにより、脾細胞中のhTERTペプチド特異的なTを検出した(下記を参照)。前記のようにペプチドパルス化によるin vitro刺激後、同一の免疫脾細胞集団に対して、細胞毒性アッセイを実施した。
【0119】
<ex vivoにおけるIFN-γELISPOTアッセイによる、T細胞応答の評価>
以前に記載されているようなIFN-γELISPOTアッセイによって、免疫化マウスからのペプチド特異的T細胞を検出した(Miyahiraら、J Immunol Methods、1995年;181:45〜54頁)。抗マウスIFN-γmAb's(3μg/ml;Pharmigen、Becton Dickinson biosciences)を、96ウェルニトロセルロースマイクロプレート(マルチスクリーン;Millipore社、モルスハイム、フランス)上にコーティングした。赤血球溶解後、個々のマウスの新鮮な単離脾臓リンパ球(5×105、2.5×105、及び1.25×105細胞/ウェル)を、5μgの天然hTERTペプチドとともに、またはなしで、37℃で18時間、直接培養した。洗浄後、当該プレートを、ビオチン化抗マウスIFN-γ(2μg/ml;Pharmigen、Becton Dickinson biosciences)とともに2時間インキュベーションした。最後に、当該プレートを洗浄し、アルカリホスファターゼ接合ストレプトアビジン(Roche molecular biochemicals、マンハイム、ドイツ)とともに37℃で1時間インキュベーションした。ポジティブコントロールは、ホルボール・ミリステート・アセテート(100ng/ml、Sigma)、及びイオノマイシン(1μg/ml)で刺激した細胞を含む。ペルオキシダーゼ基質(BCIP/NBT、Promega社、マジソンW;米国)を加えることによって、IFN-γスポット形成細胞(SFC)を展開し、自動化画像分析システムBioreader 2000(Biosys、カルベン、ドイツ)を用いてカウントした。ネガティブコントロール値(<10SFC)を減算した後、特異的SFCの数を算出した。刺激されたウェル中のSFCの平均が、ネガティブコントロールウェル中のSFCの平均+2S.D.よりも大きく、且つ50SFC/106細胞よりも大きい場合、応答は陽性であった。
【0120】
<細胞溶解アッセイ>
以前に記載されているような標準的な4〜5時間51Cr放出アッセイを用いることによって、細胞毒性アッセイを実施した(Firatら、J Gene Med、2002年;4:38〜45頁)。コントロールペプチド関して観察された非特異的溶解を減算することことによって、%での特異的溶解を算出した。特異的溶解≧10%が観察された場合、マウスを応答動物と見なした。
【0121】
<ヒトにおけるhTERTペプチド特異的CTLの産生>
以前に記載されているような、HLA-B0702制限のhTERTペプチドを用いた、4週間のin vitroでのPBMC再活性化後、ドナーからヒトCTLを得た(Hernandezら、Proc Natl Acad Sci U.S.A.、2002年;99:12275〜80頁)。要するに、フィコール精製ヒトPBMCを解凍し、加熱不活化10%ヒト血清(Institut Jacques Boy、レイムス、フランス)を補充したRPMI 1640、1mMのピルビン酸ナトリウム、100IU/mlのペニシリン、100μg/mlのストレプトマイシン、10mMのHEPES、5×10〜5Mの2-メルカプトエタノール中、24ウェルプレート中でインキュベーションした(4×106/ウェル)。それらを、各hTERTペプチド(10μg/ml)で刺激し、組換えヒトIL-7(20ng/ml;R&D Systems)を加えた。
【0122】
7日目、ペプチドパルス化γ照射自系PBMC(50Gy)を用いて、リンパ球を再活性化した。翌日、20IU/mlのヒトIL-2(Roche、マンハイム、ドイツ)を当該培養物に加えた。CTL系を、4サイクルの間、毎週再活性化した。一部のドナーに対して、CD8マイクロビーズ(Miltenyi Biotec、ベルギッシュグラッドバッハ、ドイツ)をメーカーの推奨に従って用いることによって、3回サイクル後にCD8*T細胞を精製し、機能試験前に1回活性化した。種々の51Cr-標識化標的:試験したhTERTペプチドまたは非関連ペプチドでパルス化したT2-B7、または腫瘍細胞系に対する最後の再活性化の6日後に、細胞溶解アッセイを実施した。
【0123】
いくつかの実験において、最適濃度(10μg/mL)における抗HLAクラスIフレームワークmAb、w6/32(BD Pharmigen)、または抗HLA-*B0702 mAbのME1、または抗HLA-DR mAbBのG46.6(BD Pharmigen)とともに、腫瘍細胞を30分間インキュベーションし、細胞毒性がHLAクラスIに限定されているかどうかを判定した。
【0124】
II−結果
1.HLA-*B0702で予測されたhTERT由来のペプチドの免疫原性
T細胞エピトープ予測プログラムを用いることによって、本発明者らは、hTERTタンパク質配列を分析し、それらの高い予測スコア(表1)により、6種のペプチド(3種の9量体、3種の10量体)を維持した。次に、それらのHLA-B0702分子への結合能を、HLA-B0702遺伝子でトランスフェクションされた抗原−トランスポーター(TAP)欠失T2細胞(T2-B7)を用いることによって試験した。3種のペプチド、p1、p4、及びp68は、それぞれ高い相対的親和性(RA≦1)を示し、他の3種、p277、p342、及びp351は、中程度のRA>1を示した(表1)。これらのデータは、これら6種のペプチドがHLA-B0702に対する優れた結合物質であることを示している。従って、小胞体内部で形成されたこのような複合体が腫瘍細胞の表面に到達し、CTL認識に利用可能であることが予測され得る。
【0125】
これらのペプチドがin vivoで免疫原性であるかどうかを試験するために、本発明者らはHLA-*B0702トランスジェニックマウスを免疫化した。前記6種のペプチドは、差は認めされたものの、CTL応答を誘発し得ることが示された(表1)。HLA-B7分子に対して高い親和性で結合する2種のペプチドp4及びp68は、試験されたすべてのマウスにおいて強いCTL応答を誘発する。対照的に、HLA-B7分子に対して低い親和性を有するペプチド(p277及びp342)は、試験されたマウスの50%のみで中程度の特異的CTLの産生を可能にする。トランスジェニックマウスから産生された、hTERTエピトープに特異的なCTL系は、それぞれ特異的なペプチドでパルス化されたヒトT2-B7を認識し(データは示していない)、複合体MHC/ペプチドに対するそれらのTCRの高い親和性を示した。従って、HLA-B0702の結合/安定化の結果とHLA-B7トランスジェニックマウスにおけるin vivoでのCTL応答との間には全体的な相関性がある。
【0126】
【表2】
【0127】
2.hTERT由来のペプチドはHLA-B0702トランスジェニックマウスにおいてプロセシングされた
同定されたこれら6種のhTERTエピトープに対するHLA-B0702分子との関連で、内在的に合成されたhTERTペプチドの提示を評価するために、HLA-*B0702トランスジェニックマウスを、hTERTをコードするcDNAで免疫化し、40日後、個々のマウスの脾臓細胞内のペプチド特異的CTL応答を評価した。図2に示されるように、p4、p68、p1、p277、及びp351に関して、マウスの50%〜80%の大部分の免疫化マウス(M)において、hTERTペプチド特異的CTLが誘発された。対照的に、p342特異的CTLは、試験されたマウスの約15%で誘発され得た。有意なhTERT特異的CTL応答は、非免疫化無処理マウスからも誘発されなかった。従って、これら6種のhTERTエピトープは、細胞内で効果的にプロセシングされる。さらに、アミノ酸配列または構造の点において合成ペプチドと類似した天然ペプチドは、当該細胞表面上の対応するHLA-*B0702分子によって提示される。
【0128】
さらに、これらのデータは、単一マウスにおいて、いくつかのhTERTエピトープに対して複数のCTL特異性を同時に誘発させることができることを示しており、且つ候補ワクチンを試験するためのそれらの可能性について、本発明者らのHLAクラスIトランスジェニックマウスモデルの有効性を実証している。
【0129】
3.hTERTペプチドによる健常ドナーからの一次CTL応答の誘発
In vitro免疫化プロトコルにおけるHLA-B0702健常ドナーのPBMCを用いることによって、hTERTペプチドがHLA-B7制限CTLの上昇において有効であるかどうかを試験した。10個体のうち8個体(d1〜d8)においてCTL応答が生じ、ペプチド特異的CTL応答は、p342(20%)を除いて、ドナーの少なくとも50%で得られた(図3)。In vitroで生じたCTLによるhTERTエピトープの認識は、それらの遺伝的バックグラウンドに応じてドナー間で変化する(図3)。従って、HLA-B0702健常ドナーのランダム試験によって、これらのhTERTペプチドはヒトにおいて免疫原性であることが明白に確認され、hTERTに対する特異的CTL前駆体は末梢成人レパートリーにおいて欠失していないことが示唆される。従って、健常ドナーから生じたCTL系がHLA適合hTERT+腫瘍細胞を死滅させることができるかどうかを探索した。
【0130】
4.特異的hTERT CTLは種々の由来の腫瘍を溶解できた
種々の由来のヒト腫瘍細胞系を溶解させる能力について、ドナーからのhTERT特異的CTLを試験した。表2に示されている結果は、健常ドナー由来のin vitroで生じたCTL系はHLA-B0702+腫瘍細胞を死滅させるが、他方、HLA-B0702-腫瘍に対する細胞毒性は検出されなかったことを示している。[例えば、KU L268または293-UT標的における、d1、d2、及びd3でのCTLp351(それぞれ、52、25、20%、及び34、41、19%)対T1またはBBG1標的におけるもの(それぞれ、9、2、6%、及び0、0、2%)を参照]。CTL特異性に従って、腫瘍認識に違いが観察されたが、このことは、腫瘍細胞の表面上のhTERTペプチドの差異的な差異によって説明できるであろう。重要なことには、種々のドナーから生じたp351特異的CTL系が、試験された腫瘍細胞系の大多数を認識することである(表2)。対照的に、すべてのp4特異的CTL系が、試験された腫瘍のすべてのタイプを溶解するわけではない。p1及びp68ペプチドに特異的なCTL系は、T1-B7標的のみを認識する。p342特異的CTL系は、メラノーマ細胞(LB 34及びKU 268標的)のみを認識する。最後に、p277特異的CTL系は、腎臓癌(293 UT)を認識するが、メラノーマ細胞もリンパ腫細胞のどちらも認識しない。他方、正常PBMC及びCD40活性化B細胞は、HLAのタイプに関わらず、これらのhTERTペプチド特異的CTL系によって溶解されなかった(表2の最後の2つの系)。
【0131】
図4に示されるように、HLA-B7+腫瘍細胞に対するCTLp351系の細胞毒性活性は、抗クラスI mAb抗HLA-B0702によって阻害されるが、抗HLA-DR mAb(MHC クラスII)によっては阻害されない。同様のデータが、他のペプチド特異的CTL系でも得られており(データは示さず)、CTL系が、HLA-B0702制限様式でhTERT+腫瘍細胞に対して細胞毒性を及ぼすことを示唆している。まとめると、これらの結果は、これら6種のhTERT由来ペプチドが腫瘍細胞表面で等しく天然に発現されているのではないこと、及びhTERTペプチド特異的CTLが、HLA-B0702分子との関連でhTERTペプチドの認識によって、腫瘍細胞と正常細胞を識別できることを示している。
【0132】
5.hTERTをコードするレンチウイルスベクターのワクチン接種により、マウスにおいて有効なペプチド特異的T細胞応答が誘発される
次いで、本発明者らは、HIV由来フラップベクター内に導入された、全長hTERT遺伝子または機能的でないhTERT遺伝子のいずれかを含む候補ワクチンを試験した(図7)。このタイプのレンチウイルスベクターは、in vitro及びin vivoで樹状細胞を標的とし、動物において強い多特異的抗腫瘍CTL応答を誘発することが、以前のデータによって示されている。従って、本発明者らは、組換えLv-hTERTまたはpTRIP-CMV-ΔhTERTを用いて、HLA-*B0702トランスジェニックマウスを免疫化した。12日後、個々のマウスの脾臓細胞を、ex vivo ELISPOTアッセイによって評価した。
【0133】
図5Aに示されるように、Lv-GFPコントロールベクターを投与されたマウスと比較して、HLA-B0702制限hTERTエピトープに対するペプチド特異的CD8+ T細胞応答が得られた。In vitro刺激後のクロム放出アッセイにおいて、当該誘導されたペプチド特異的CD8+細胞の機能分析により、ex vivo ELISPOTデータが裏付けられ(表3)、さらにLv-hTERTの単回注入後のマウスの約50〜70%において及びTRIPLv-hTERTによる増強後のマウスの100%において、これら6種のペプチドに対する有効な特異的CTL応答が生じることが示されている(図6)。このことはまた、すべてのマウスにおける強いCTL応答とも関連していた(表3)。さらに、表3に示されるように、HLA-A2.1に対してトランスジェニックのHHDマウスを同一ベクターで免疫化すると、顕性(p540)及び潜性(p572)として以前に分類された2つのHLA-A2.1.1制限エピトープに対して特異的な強力なCTL応答が誘発された。まとめると、これらの結果は、Lv-hTERTの投与により、マウスにおいて、非常に有効な多特異的T細胞応答の誘発がもたらされること明白に示され、hTERTが癌免疫療法のためのポリエピトープ及びポリ対立遺伝子の(polyallelic)TAAとして作用し得ることを示唆している。
【0134】
図8に示されるように、組換えpTRIP-CMV-ΔhTERTの単回注入後のHLA-A2トランスジェニック(Tg)マウスにおいて、hTERTペプチド特異的CD8+T細胞応答がex vivoで検出された。本発明者らは、p540及びPY572エピトープに特異的なCD8+T細胞が、免疫化マウスの少なくとも50%において誘発されたことを示した(図8)。これらの結果は、前記2つのエピトープが正確に内在的にプロセシングされ、pTRIP-CMV-ΔhTERTによる免疫化後のHLA-A2 Tgマウスにおいて提示されたことを明白に示した。
【0135】
まとめると、これらの結果は、TRIP-hTERTの単回注入により、両方のHLAトランスジェニックマウス群において、強力な多特異的抗hTERT CD8+T細胞応答の誘発がもたらされることを示した。
【0136】
【表3】
【0137】
【表4】
【0138】
III−結論
In vivo免疫原であり、且つH-2クラスIノックアウトHLA-B0702トランスジェニックマウスにおいてプロセシングされる、新規なhTERTエピトープが同定された。さらに、in vitroにおいて、健常ドナーからのHLA-B702+PBLを用いたhTERTペプチド免疫化は、種々の由来のhTERT+腫瘍を認識する特異的CTL応答を誘発し、ヒトT細胞レパートリーにおいてこれらのエピトープに対する欠失がないことを示唆している。さらに、腫瘍の由来に応じて細胞表面上に発現されるペプチドレパートリーは質的に異なっていることが示され、hTERTを癌細胞の周囲の(circumvent)抗原変異性に対するポリエピトープ腫瘍関連抗原として特徴づけするための有用性を明確に示している。最後に、ヒト化HLA-*B0702及びHLA-A2 1トランスジェニックマウスを用いて、新世代のレンチウイルス由来フラップベクター内に挿入された機能的でないテロメラーゼ遺伝子からなる候補ワクチンを試験した。すべての当該HLAトランスジェニックマウスにおいて、強力なhTERT特異的CD8+T細胞応答が観察された。これらのデータは、癌患者における治療的ワクチン接種の使用を支持し、多くの癌患者を網羅するための治療標的としてのhTERTの潜在的適用性を広げている。
【0139】
(参考文献)
【特許請求の範囲】
【請求項1】
HLA-B7制限免疫応答の誘発に使用するための、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるhTERT(ヒトテロメラーゼ逆転写酵素)エピトープをコードするポリヌクレオチド。
【請求項2】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)
からなる群から選択されるHLA-B7制限hTERTエピトープをコードするポリヌクレオチド。
【請求項3】
少なくとも1つのアミノ酸置換によって改変され、且つHLA-B7制限エピトープ類似体をコードする、請求項1または2に記載のポリヌクレオチド。
【請求項4】
前記少なくとも1つの置換が2位に位置していない、請求項3に記載のポリヌクレオチド。
【請求項5】
前記少なくとも1つの置換が最後のC末端位に位置していない、請求項3または4に記載のポリヌクレオチド。
【請求項6】
前記少なくとも1つの置換が1位に位置する、請求項3から5のいずれか一項に記載のポリヌクレオチド。
【請求項7】
1位に位置したアミノ酸がアラニン(A)によって置換される、請求項3から6のいずれか一項に記載のポリヌクレオチド。
【請求項8】
ペプチド配列APRRLVQLL(p444*)をコードする、請求項3から7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
前記類似体が、その野生型対応物よりもHLA-B7分子に対して高い親和性を有する、請求項3から8のいずれか一項に記載のポリヌクレオチド。
【請求項10】
前記類似体が、その野生型対応物よりも高い免疫原性を有する、請求項3から9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
ポリエピトープをコードし、且つ請求項2に記載のHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/または請求項3から9のいずれか一項に記載のHLA-B7制限エピトープ類似体をコードするポリヌクレオチドの間で選択される少なくとも2つのポリヌクレオチド単位を含むポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項12】
ポリエピトープをコードし、且つ
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/またはそれらの類似体をコードするポリヌクレオチドの間で選択される少なくとも2つのポリヌクレオチド単位を含むポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項13】
−請求項2に記載のHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/または請求項3から9のいずれか一項に記載のHLA-B7制限エピトープ類似体をコードするポリヌクレオチドの間で選択される少なくとも1つのポリヌクレオチド単位;並びに
−配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)、あるいはそれらの類似体の間で選択される少なくとも1つのポリヌクレオチド単位であって、前記ポリヌクレオチドが、全長hTERTのコード配列ではないポリヌクレオチド単位;
を含む、ポリエピトープをコードする請求項12に記載のポリヌクレオチド。
【請求項14】
各ポリヌクレオチド単位がMHCクラスI制限hTERTエピトープをコードする2〜10のポリヌクレオチド単位を含む、請求項11から13のいずれか一項に記載のポリエピトープをコードするポリヌクレオチド。
【請求項15】
hTERTタンパク質の切断型または変異型をコードする核酸分子からなる、請求項11から14のいずれか一項に記載のポリヌクレオチド。
【請求項16】
hTERTタンパク質の前記切断型または変異型がその触媒活性を失っている、請求項15に記載のポリヌクレオチド。
【請求項17】
hTERTタンパク質の前記切断型または変異型が触媒活性ドメインを欠失している、請求項15に記載のポリヌクレオチド。
【請求項18】
前記少なくとも2つのポリヌクレオチド単位が連続的に配列されている、請求項11から13のいずれか一項に記載のポリヌクレオチド。
【請求項19】
前記p1、p4、p68、p277、p342、及びp351エピトープ、またはそれらの類似体をコードするポリヌクレオチド単位を少なくとも含む、請求項11から18のいずれか一項に記載のポリヌクレオチド。
【請求項20】
ポリペプチド配列:
MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLをコードする核酸分子を含むか、またはそれからなる、請求項19に記載のポリヌクレオチド。
【請求項21】
前記少なくとも2つのポリヌクレオチド単位が、1つのアミノ酸スペーサーまたはペプチドスペーサーをコードするポリヌクレオチドによって分離されている、請求項11から17のいずれか一項に記載のポリヌクレオチド。
【請求項22】
前記スペーサーが:
a. C末端位に陽電荷残基または酸性残基を有する4アミノ酸ペプチド;
b. C末端位に陽電荷残基または酸性残基を、且つ他の位置に親水性残基(A、K、D、及び/またはT)を有する4アミノ酸ペプチド;並びに
c. C末端位にアルギニン(R)を、且つ他の位置に親水性残基(A、K、D、及び/またはT)を含む4アミノ酸ペプチド;
の群から選択される、請求項21に記載のポリヌクレオチド。
【請求項23】
少なくとも2つのエピトープのうち最もN末端側のエピトープをコードするポリヌクレオチド単位に作動可能に連結された、標的シグナルをコードするポリヌクレオチドをさらに含む、請求項11から22のいずれか一項に記載のポリヌクレオチド。
【請求項24】
前記標的シグナルが小胞体シグナル配列である、請求項23に記載のポリヌクレオチド。
【請求項25】
前記最もN末端側のエピトープが、1位にメチオニン残基を有さない場合、少なくとも2つのエピトープのうち前記最もN末端側のエピトープをコードするポリヌクレオチド単位の上流に位置し、且つ作動可能に連結されたメチオニン残基をコードするコドンを含む、請求項11から24のいずれか一項に記載のポリヌクレオチド。
【請求項26】
HLA-B7制限免疫応答の誘発に使用するための、請求項11から25のいずれか一項に記載のポリヌクレオチド。
【請求項27】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)、
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
から選択される少なくとも1つのポリヌクレオチド単位を含むか、またはこれらのポリヌクレオチド単位もしくはその類似体のうち少なくとも2つの任意の組合せを含み、前記ポリヌクレオチド単位が、MHCクラスI制限hTERTエピトープをコードする、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項28】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、またはそれらの類似体
から選択される少なくとも1つのポリヌクレオチド単位、並びに
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、またはそれらの類似体
から選択される少なくとも1つのポリヌクレオチド単位を含む、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項29】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、及び
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、またはそれらの類似体
から選択される少なくとも2つのポリヌクレオチド単位を含む、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項30】
図9または図10に示されているヌクレオチド配列を含むか、またはそれからなる、請求項16に記載のポリヌクレオチド。
【請求項31】
請求項1から30のいずれか一項に記載のポリヌクレオチドを含む発現ベクター。
【請求項32】
2006年7月28日に番号CNCM I-3660の下でCNCM(パスツール研究所、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターである、請求項31に記載の発現ベクター。
【請求項33】
2006年7月28日に番号CNCM I-3660の下でCNCMに寄託されたpTRIP-CMV-ΔhTERTベクターであって、欠失したhTERT配列が請求項1から30のいずれか一項に定義されたポリヌクレオチドによって置換されている、請求項31に記載の発現ベクター。
【請求項34】
請求項31から33のいずれか一項に記載のベクターを含む宿主細胞。
【請求項35】
請求項2から30のいずれか一項に記載のポリヌクレオチドで遺伝的に形質転換された宿主細胞。
【請求項36】
抗原提示細胞(APC)である、請求項34または35に記載の宿主細胞。
【請求項37】
樹状細胞(DC)である、請求項36に記載の宿主細胞。
【請求項38】
ex vivoで完全に成熟させたDCである、請求項37に記載の宿主細胞。
【請求項39】
非ヒト哺乳動物細胞である、請求項34から38のいずれか一項に記載の宿主細胞。
【請求項40】
単離ヒト細胞である、請求項34から38のいずれか一項に記載の宿主細胞。
【請求項41】
2006年7月28日に番号CNCM I-3660の下でCNCMに寄託された宿主細胞である、請求項34または35に記載の宿主細胞。
【請求項42】
hTERTに対するHLA-B7制限免疫応答の誘発に使用するための、請求項31から41のいずれか一項に記載の発現ベクターまたは宿主細胞。
【請求項43】
請求項2から29のいずれか一項に記載のポリヌクレオチドによってコードされているポリペプチド。
【請求項44】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択されるMHCクラスI制限hTERTエピトープまたは類似体。
【請求項45】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. MPRAPRCRA(p1)、
d. APRCRAVRSL(p4)、
e. RPAEEATSL(p277)、
f. RPSFLLSSL(p342)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択される少なくとも2つのエピトープを含むポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項46】
少なくとも2つのエピトープを含み、少なくとも1つは、
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、且つ少なくとも1つが、
j. MPRAPRCRA(p1)、
k. RPAEEATSL(p277)、
l. RPSFLLSSL(p342)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択される請求項45に記載のポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項47】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体の群から選択される少なくとも2つのエピトープを含む請求項45に記載のポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項48】
2〜10のhTERTエピトープまたはそれらの類似体を含む、請求項45から47のいずれか一項に記載のポリエピトープ。
【請求項49】
ペプチド配列:
MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLを含むか、またはそれからなる、請求項45から48のいずれか一項に記載のポリエピトープ。
【請求項50】
a. 請求項2から30のいずれか一項に記載のポリヌクレオチド;
b. 請求項31から33のいずれか一項に記載のベクター;
c. 請求項34から41のいずれか一項に記載の宿主細胞;及び
d. 請求項43から49のいずれか一項に記載のポリペプチド;
からなる群から選択される少なくとも1つの成分を含む組成物。
【請求項51】
in vivo投与に適している、請求項50に記載の組成物。
【請求項52】
前記ポリヌクレオチドが、HLA-B7のものとは異なるMHCクラスI対立遺伝子に制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをさらにコードする、請求項50に記載の組成物。
【請求項53】
前記ポリヌクレオチドが、MHCクラスIIに制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをさらにコードする、請求項50から52のいずれか一項に記載の組成物。
【請求項54】
前記ポリヌクレオチドが、少なくとも1つの腫瘍特異的抗原(TSA)をさらにコードする、請求項50から53のいずれか一項に記載の組成物。
【請求項55】
前記ポリヌクレオチドが、少なくとも1つの腫瘍関連抗原(TAA)をさらにコードする、請求項50から53のいずれか一項に記載の組成物。
【請求項56】
アジュバント、ビヒクル、及び/または製薬上許容され得るキャリアをさらに含む、請求項50から55のいずれか一項に記載の治療用組成物。
【請求項57】
細胞による前記ポリヌクレオチドの取込みを促進させる少なくとも1つの成分をさらに含む、請求項56に記載の治療用組成物。
【請求項58】
a. Tリンパ球を提供する工程;及び
b. 前記Tリンパ球を、請求項43から49のいずれか一項に記載の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)の中から選択されるエピトープとともに、前記リンパ球の活性化を可能にする条件でin vitro培養する工程;
を含む、HLA-B7制限hTERTエピトープに対するTリンパ球の活性化方法。
【請求項59】
a. 請求項58に記載のTリンパ球を、請求項43から49のいずれか一項に記載のエピトープまたはエピトープ類似体またはポリエピトープで活性化する工程;
b.前記活性化リンパ球を、MHCクラスI分子に結合している請求項2に記載のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動を調査する方法。
【請求項60】
a. 請求項58に記載のTリンパ球を、請求項43から49のいずれか一項に記載のエピトープまたはエピトープ類似体またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)の中から選択されるエピトープで活性化する工程;
b.前記活性化リンパ球を、HLA-B7分子に結合している請求項2に記載の、あるいは配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)を有するhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動及びHLA-B7制限性を調査する方法。
【請求項61】
ステップc.において、前記活性化リンパ球が前記標的細胞を死滅させる、請求項60に記載の方法。
【請求項62】
活性化を可能にする条件が、RPMI培地中、37℃で4時間の接触である、請求項60または61に記載の方法。
【請求項63】
a. 細胞を提供する工程;
b. 請求項43から49に記載の少なくとも1つのHLA-B7制限hTERTエピトープまたはエピトープ類似体またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、RPSFLLSSL(p342)の中から選択されるエピトープを用いて、前記細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟細胞の増殖を促進する工程;
を含む、HLA-B7制限hTERTエピトープに対する細胞のin vitro成熟方法。
【請求項64】
前記細胞が樹状細胞である、請求項63に記載の方法。
【請求項65】
前記樹状細胞が、循環血液または骨髄細胞のいずれかから単離される、請求項64に記載の方法。
【請求項66】
前記樹状細胞が、治療を必要としている患者からまたはHLA適合ドナーから単離される、請求項64または65に記載の方法。
【請求項67】
前記細胞が少なくとも1つのHLA-B7対立遺伝子を発現している、請求項63から66のいずれか一項に記載の方法。
【請求項68】
ステップb.が、請求項2から29のいずれか一項に記載のポリヌクレオチド、または配列MPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)をコードするポリヌクレオチド、または配列ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、もしくはCGGCCCTCCTTCCTACTCAGCTCTCTG(n342)からなるポリヌクレオチドによる、前記樹状細胞の遺伝的形質転換を含む、請求項64から67のいずれか一項に記載の方法。
【請求項69】
ステップb.が、請求項30に記載のベクターによる前記樹状細胞の形質転換を含む、請求項64から67のいずれか一項に記載の方法。
【請求項70】
ステップb.が、前記樹状細胞と、請求項39から45に記載の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、RPSFLLSSL(p342)の中から選択されるエピトープとの接触を含む、請求項64から67のいずれか一項に記載の方法。
【請求項71】
癌の予防及び/または治療に使用するための、HLA-B7制限hTERTエピトープまたはその類似体、あるいは少なくとも1つのHLA-B7制限hTERTエピトープまたはその類似体を含むポリエピトープをコードするポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項72】
前記HLA-B7制限hTERTエピトープまたは類似体が、
a. APRCRAVRSL(p4)、
b. APSFRQVSCL(p68)、
c. RPSLTGARRL(p351)
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択される、請求項71に記載のポリヌクレオチド。
【請求項73】
癌の予防及び/または治療に使用するための、請求項2から30のいずれか一項に記載のポリヌクレオチド、請求項31から33のいずれか一項に記載のベクター、請求項34から42のいずれか一項に記載の宿主細胞、または請求項43から49のいずれか一項に記載のポリペプチド。
【請求項74】
HLA-B7制限免疫応答の誘発に使用するための、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、
k. LPSPKFTIL(p1123)、及び
l. APRRLVQLL(p444*)
からなる群から選択されるhTERTエピトープまたは類似体。
【請求項1】
HLA-B7制限免疫応答の誘発に使用するための、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるhTERT(ヒトテロメラーゼ逆転写酵素)エピトープをコードするポリヌクレオチド。
【請求項2】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)
からなる群から選択されるHLA-B7制限hTERTエピトープをコードするポリヌクレオチド。
【請求項3】
少なくとも1つのアミノ酸置換によって改変され、且つHLA-B7制限エピトープ類似体をコードする、請求項1または2に記載のポリヌクレオチド。
【請求項4】
前記少なくとも1つの置換が2位に位置していない、請求項3に記載のポリヌクレオチド。
【請求項5】
前記少なくとも1つの置換が最後のC末端位に位置していない、請求項3または4に記載のポリヌクレオチド。
【請求項6】
前記少なくとも1つの置換が1位に位置する、請求項3から5のいずれか一項に記載のポリヌクレオチド。
【請求項7】
1位に位置したアミノ酸がアラニン(A)によって置換される、請求項3から6のいずれか一項に記載のポリヌクレオチド。
【請求項8】
ペプチド配列APRRLVQLL(p444*)をコードする、請求項3から7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
前記類似体が、その野生型対応物よりもHLA-B7分子に対して高い親和性を有する、請求項3から8のいずれか一項に記載のポリヌクレオチド。
【請求項10】
前記類似体が、その野生型対応物よりも高い免疫原性を有する、請求項3から9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
ポリエピトープをコードし、且つ請求項2に記載のHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/または請求項3から9のいずれか一項に記載のHLA-B7制限エピトープ類似体をコードするポリヌクレオチドの間で選択される少なくとも2つのポリヌクレオチド単位を含むポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項12】
ポリエピトープをコードし、且つ
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)
からなる群から選択されるHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/またはそれらの類似体をコードするポリヌクレオチドの間で選択される少なくとも2つのポリヌクレオチド単位を含むポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項13】
−請求項2に記載のHLA-B7制限hTERTエピトープをコードするポリヌクレオチド及び/または請求項3から9のいずれか一項に記載のHLA-B7制限エピトープ類似体をコードするポリヌクレオチドの間で選択される少なくとも1つのポリヌクレオチド単位;並びに
−配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)、あるいはそれらの類似体の間で選択される少なくとも1つのポリヌクレオチド単位であって、前記ポリヌクレオチドが、全長hTERTのコード配列ではないポリヌクレオチド単位;
を含む、ポリエピトープをコードする請求項12に記載のポリヌクレオチド。
【請求項14】
各ポリヌクレオチド単位がMHCクラスI制限hTERTエピトープをコードする2〜10のポリヌクレオチド単位を含む、請求項11から13のいずれか一項に記載のポリエピトープをコードするポリヌクレオチド。
【請求項15】
hTERTタンパク質の切断型または変異型をコードする核酸分子からなる、請求項11から14のいずれか一項に記載のポリヌクレオチド。
【請求項16】
hTERTタンパク質の前記切断型または変異型がその触媒活性を失っている、請求項15に記載のポリヌクレオチド。
【請求項17】
hTERTタンパク質の前記切断型または変異型が触媒活性ドメインを欠失している、請求項15に記載のポリヌクレオチド。
【請求項18】
前記少なくとも2つのポリヌクレオチド単位が連続的に配列されている、請求項11から13のいずれか一項に記載のポリヌクレオチド。
【請求項19】
前記p1、p4、p68、p277、p342、及びp351エピトープ、またはそれらの類似体をコードするポリヌクレオチド単位を少なくとも含む、請求項11から18のいずれか一項に記載のポリヌクレオチド。
【請求項20】
ポリペプチド配列:
MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLをコードする核酸分子を含むか、またはそれからなる、請求項19に記載のポリヌクレオチド。
【請求項21】
前記少なくとも2つのポリヌクレオチド単位が、1つのアミノ酸スペーサーまたはペプチドスペーサーをコードするポリヌクレオチドによって分離されている、請求項11から17のいずれか一項に記載のポリヌクレオチド。
【請求項22】
前記スペーサーが:
a. C末端位に陽電荷残基または酸性残基を有する4アミノ酸ペプチド;
b. C末端位に陽電荷残基または酸性残基を、且つ他の位置に親水性残基(A、K、D、及び/またはT)を有する4アミノ酸ペプチド;並びに
c. C末端位にアルギニン(R)を、且つ他の位置に親水性残基(A、K、D、及び/またはT)を含む4アミノ酸ペプチド;
の群から選択される、請求項21に記載のポリヌクレオチド。
【請求項23】
少なくとも2つのエピトープのうち最もN末端側のエピトープをコードするポリヌクレオチド単位に作動可能に連結された、標的シグナルをコードするポリヌクレオチドをさらに含む、請求項11から22のいずれか一項に記載のポリヌクレオチド。
【請求項24】
前記標的シグナルが小胞体シグナル配列である、請求項23に記載のポリヌクレオチド。
【請求項25】
前記最もN末端側のエピトープが、1位にメチオニン残基を有さない場合、少なくとも2つのエピトープのうち前記最もN末端側のエピトープをコードするポリヌクレオチド単位の上流に位置し、且つ作動可能に連結されたメチオニン残基をコードするコドンを含む、請求項11から24のいずれか一項に記載のポリヌクレオチド。
【請求項26】
HLA-B7制限免疫応答の誘発に使用するための、請求項11から25のいずれか一項に記載のポリヌクレオチド。
【請求項27】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)、
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
から選択される少なくとも1つのポリヌクレオチド単位を含むか、またはこれらのポリヌクレオチド単位もしくはその類似体のうち少なくとも2つの任意の組合せを含み、前記ポリヌクレオチド単位が、MHCクラスI制限hTERTエピトープをコードする、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項28】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、及び
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)、またはそれらの類似体
から選択される少なくとも1つのポリヌクレオチド単位、並びに
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、またはそれらの類似体
から選択される少なくとも1つのポリヌクレオチド単位を含む、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項29】
a. AGGCCCAGCCTGACTGGCGCTCGGAGGCTC(n351)
b. GCCCCCTCCTTCCGCCAGGTGTCCTGCCTG(n68)、
c. GCTCCCCGCTGCCGAGCCGTGCGCTCCCTG(n4)、
d. GACCCCCGTCGCCTGGTGCAGCTGCTC(n444)、
e. TTCGTGCGGGCCTGCCTGCGCCGGCTG(n464)、
f. GCTGGGAGGAACATGCGTCGCAAACTC(n966)、
g. CTCCCGGGGACGACGCTGACTGCCCTG(n1107)、
h. CTGCCCTCAGACTTCAAGACCATCCTG(n1123)、
i. GCCCCCCGTCGCCTGGTGCAGCTGCTC(n444*)
j. ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、
k. AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、及び
l. CGGCCCTCCTTCCTACTCAGCTCTCTG(n342)、またはそれらの類似体
から選択される少なくとも2つのポリヌクレオチド単位を含む、請求項1から26のいずれか一項に記載のポリヌクレオチド。
【請求項30】
図9または図10に示されているヌクレオチド配列を含むか、またはそれからなる、請求項16に記載のポリヌクレオチド。
【請求項31】
請求項1から30のいずれか一項に記載のポリヌクレオチドを含む発現ベクター。
【請求項32】
2006年7月28日に番号CNCM I-3660の下でCNCM(パスツール研究所、パリ、フランス)に寄託されたpTRIP-CMV-ΔhTERTベクターである、請求項31に記載の発現ベクター。
【請求項33】
2006年7月28日に番号CNCM I-3660の下でCNCMに寄託されたpTRIP-CMV-ΔhTERTベクターであって、欠失したhTERT配列が請求項1から30のいずれか一項に定義されたポリヌクレオチドによって置換されている、請求項31に記載の発現ベクター。
【請求項34】
請求項31から33のいずれか一項に記載のベクターを含む宿主細胞。
【請求項35】
請求項2から30のいずれか一項に記載のポリヌクレオチドで遺伝的に形質転換された宿主細胞。
【請求項36】
抗原提示細胞(APC)である、請求項34または35に記載の宿主細胞。
【請求項37】
樹状細胞(DC)である、請求項36に記載の宿主細胞。
【請求項38】
ex vivoで完全に成熟させたDCである、請求項37に記載の宿主細胞。
【請求項39】
非ヒト哺乳動物細胞である、請求項34から38のいずれか一項に記載の宿主細胞。
【請求項40】
単離ヒト細胞である、請求項34から38のいずれか一項に記載の宿主細胞。
【請求項41】
2006年7月28日に番号CNCM I-3660の下でCNCMに寄託された宿主細胞である、請求項34または35に記載の宿主細胞。
【請求項42】
hTERTに対するHLA-B7制限免疫応答の誘発に使用するための、請求項31から41のいずれか一項に記載の発現ベクターまたは宿主細胞。
【請求項43】
請求項2から29のいずれか一項に記載のポリヌクレオチドによってコードされているポリペプチド。
【請求項44】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択されるMHCクラスI制限hTERTエピトープまたは類似体。
【請求項45】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. MPRAPRCRA(p1)、
d. APRCRAVRSL(p4)、
e. RPAEEATSL(p277)、
f. RPSFLLSSL(p342)、
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、及び
k. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択される少なくとも2つのエピトープを含むポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項46】
少なくとも2つのエピトープを含み、少なくとも1つは、
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択され、且つ少なくとも1つが、
j. MPRAPRCRA(p1)、
k. RPAEEATSL(p277)、
l. RPSFLLSSL(p342)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体からなる群から選択される請求項45に記載のポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項47】
a. RPSLTGARRL(p351)、
b. APSFRQVSCL(p68)、
c. APRCRAVRSL(p4)、
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、及び
h. LPSPKFTIL(p1123)、
または少なくとも1つのアミノ酸残基の置換によって得られるそれらの類似体の群から選択される少なくとも2つのエピトープを含む請求項45に記載のポリエピトープであって、全長hTERTではないポリエピトープ。
【請求項48】
2〜10のhTERTエピトープまたはそれらの類似体を含む、請求項45から47のいずれか一項に記載のポリエピトープ。
【請求項49】
ペプチド配列:
MPRAPRCRAAPRCRAVRSLAPSFRQVSCLRPAEEATSLRPSFLLSSLRPSLTGARRLを含むか、またはそれからなる、請求項45から48のいずれか一項に記載のポリエピトープ。
【請求項50】
a. 請求項2から30のいずれか一項に記載のポリヌクレオチド;
b. 請求項31から33のいずれか一項に記載のベクター;
c. 請求項34から41のいずれか一項に記載の宿主細胞;及び
d. 請求項43から49のいずれか一項に記載のポリペプチド;
からなる群から選択される少なくとも1つの成分を含む組成物。
【請求項51】
in vivo投与に適している、請求項50に記載の組成物。
【請求項52】
前記ポリヌクレオチドが、HLA-B7のものとは異なるMHCクラスI対立遺伝子に制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをさらにコードする、請求項50に記載の組成物。
【請求項53】
前記ポリヌクレオチドが、MHCクラスIIに制限された、少なくとも1つのhTERTエピトープまたはその類似体、あるいはポリエピトープをさらにコードする、請求項50から52のいずれか一項に記載の組成物。
【請求項54】
前記ポリヌクレオチドが、少なくとも1つの腫瘍特異的抗原(TSA)をさらにコードする、請求項50から53のいずれか一項に記載の組成物。
【請求項55】
前記ポリヌクレオチドが、少なくとも1つの腫瘍関連抗原(TAA)をさらにコードする、請求項50から53のいずれか一項に記載の組成物。
【請求項56】
アジュバント、ビヒクル、及び/または製薬上許容され得るキャリアをさらに含む、請求項50から55のいずれか一項に記載の治療用組成物。
【請求項57】
細胞による前記ポリヌクレオチドの取込みを促進させる少なくとも1つの成分をさらに含む、請求項56に記載の治療用組成物。
【請求項58】
a. Tリンパ球を提供する工程;及び
b. 前記Tリンパ球を、請求項43から49のいずれか一項に記載の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)の中から選択されるエピトープとともに、前記リンパ球の活性化を可能にする条件でin vitro培養する工程;
を含む、HLA-B7制限hTERTエピトープに対するTリンパ球の活性化方法。
【請求項59】
a. 請求項58に記載のTリンパ球を、請求項43から49のいずれか一項に記載のエピトープまたはエピトープ類似体またはポリエピトープで活性化する工程;
b.前記活性化リンパ球を、MHCクラスI分子に結合している請求項2に記載のhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動を調査する方法。
【請求項60】
a. 請求項58に記載のTリンパ球を、請求項43から49のいずれか一項に記載のエピトープまたはエピトープ類似体またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)の中から選択されるエピトープで活性化する工程;
b.前記活性化リンパ球を、HLA-B7分子に結合している請求項2に記載の、あるいは配列MPRAPRCRA(p1)、RPAEEATSL(p277)、またはRPSFLLSSL(p342)を有するhTERTエピトープを細胞表面で発現している標的細胞とともに、好適な条件でin vitro培養する工程;及び
c. 前記活性化リンパ球が、前記標的細胞に対して反応するかどうかを判定する工程;
を含む、hTERTペプチドの免疫原性挙動及びHLA-B7制限性を調査する方法。
【請求項61】
ステップc.において、前記活性化リンパ球が前記標的細胞を死滅させる、請求項60に記載の方法。
【請求項62】
活性化を可能にする条件が、RPMI培地中、37℃で4時間の接触である、請求項60または61に記載の方法。
【請求項63】
a. 細胞を提供する工程;
b. 請求項43から49に記載の少なくとも1つのHLA-B7制限hTERTエピトープまたはエピトープ類似体またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、RPSFLLSSL(p342)の中から選択されるエピトープを用いて、前記細胞の成熟を可能にする工程;及び
c. 場合によっては、前記成熟細胞の増殖を促進する工程;
を含む、HLA-B7制限hTERTエピトープに対する細胞のin vitro成熟方法。
【請求項64】
前記細胞が樹状細胞である、請求項63に記載の方法。
【請求項65】
前記樹状細胞が、循環血液または骨髄細胞のいずれかから単離される、請求項64に記載の方法。
【請求項66】
前記樹状細胞が、治療を必要としている患者からまたはHLA適合ドナーから単離される、請求項64または65に記載の方法。
【請求項67】
前記細胞が少なくとも1つのHLA-B7対立遺伝子を発現している、請求項63から66のいずれか一項に記載の方法。
【請求項68】
ステップb.が、請求項2から29のいずれか一項に記載のポリヌクレオチド、または配列MPRAPRCRA(p1)、RPAEEATSL(p277)、及びRPSFLLSSL(p342)をコードするポリヌクレオチド、または配列ATGCCGCGCGCTCCCCGCTGCCGAGCC(n1)、AGACCCGCCGAAGAAGCCACCTCTTTG(n277)、もしくはCGGCCCTCCTTCCTACTCAGCTCTCTG(n342)からなるポリヌクレオチドによる、前記樹状細胞の遺伝的形質転換を含む、請求項64から67のいずれか一項に記載の方法。
【請求項69】
ステップb.が、請求項30に記載のベクターによる前記樹状細胞の形質転換を含む、請求項64から67のいずれか一項に記載の方法。
【請求項70】
ステップb.が、前記樹状細胞と、請求項39から45に記載の少なくとも1つのエピトープ、エピトープ類似体、またはポリエピトープ、あるいはMPRAPRCRA(p1)、RPAEEATSL(p277)、RPSFLLSSL(p342)の中から選択されるエピトープとの接触を含む、請求項64から67のいずれか一項に記載の方法。
【請求項71】
癌の予防及び/または治療に使用するための、HLA-B7制限hTERTエピトープまたはその類似体、あるいは少なくとも1つのHLA-B7制限hTERTエピトープまたはその類似体を含むポリエピトープをコードするポリヌクレオチドであって、全長hTERTのコード配列ではないポリヌクレオチド。
【請求項72】
前記HLA-B7制限hTERTエピトープまたは類似体が、
a. APRCRAVRSL(p4)、
b. APSFRQVSCL(p68)、
c. RPSLTGARRL(p351)
d. DPRRLVQLL(p444)、
e. FVRACLRRL(p464)、
f. AGRNMRRKL(p966)、
g. LPGTTLTAL(p1107)、
h. LPSPKFTIL(p1123)、及び
i. APRRLVQLL(p444*)
からなる群から選択される、請求項71に記載のポリヌクレオチド。
【請求項73】
癌の予防及び/または治療に使用するための、請求項2から30のいずれか一項に記載のポリヌクレオチド、請求項31から33のいずれか一項に記載のベクター、請求項34から42のいずれか一項に記載の宿主細胞、または請求項43から49のいずれか一項に記載のポリペプチド。
【請求項74】
HLA-B7制限免疫応答の誘発に使用するための、
a. MPRAPRCRA(p1)、
b. APRCRAVRSL(p4)、
c. APSFRQVSCL(p68)、
d. RPAEEATSL(p277)、
e. RPSFLLSSL(p342)、
f. RPSLTGARRL(p351)
g. DPRRLVQLL(p444)、
h. FVRACLRRL(p464)、
i. AGRNMRRKL(p966)、
j. LPGTTLTAL(p1107)、
k. LPSPKFTIL(p1123)、及び
l. APRRLVQLL(p444*)
からなる群から選択されるhTERTエピトープまたは類似体。
【図1A】


【図1B】

【図1C】

【図1D】

【図1E】

【図1F】

【図2】
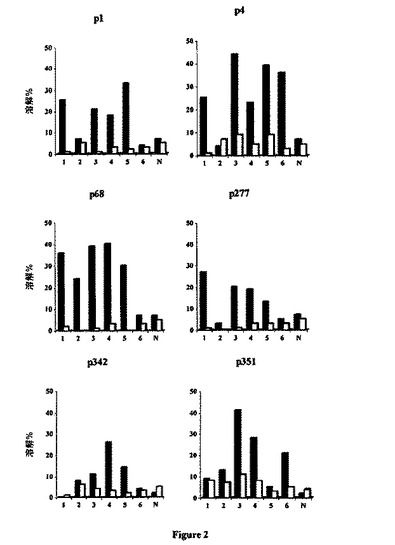
【図3】

【図4】
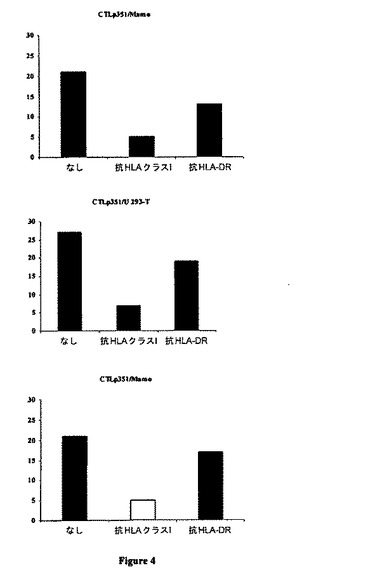
【図7】
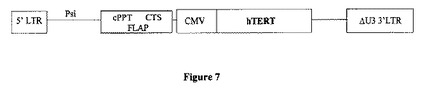
【図8】
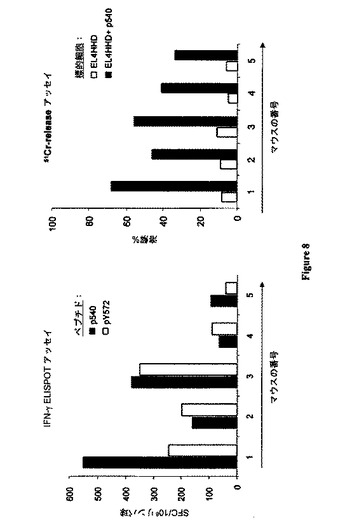
【図9A】

【図9B】

【図9C】

【図9D】

【図9E】

【図9F】

【図10A】

【図10B】

【図10C】

【図10D】

【図10E】

【図10F】

【図5】
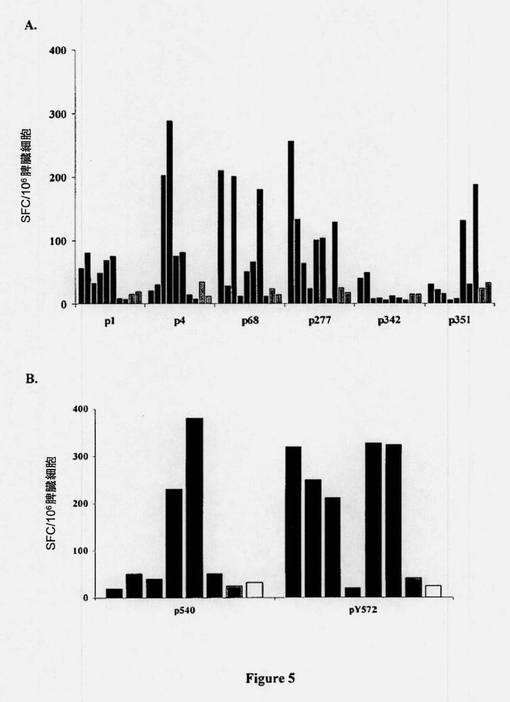
【図6】
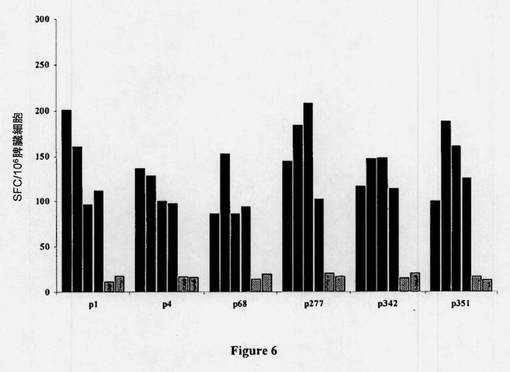
【図1B】
【図1C】
【図1D】
【図1E】
【図1F】
【図2】
【図3】
【図4】
【図7】
【図8】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図10A】
【図10B】
【図10C】
【図10D】
【図10E】
【図10F】
【図5】
【図6】
【公開番号】特開2012−183076(P2012−183076A)
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−133834(P2012−133834)
【出願日】平成24年6月13日(2012.6.13)
【分割の表示】特願2008−523267(P2008−523267)の分割
【原出願日】平成18年7月31日(2006.7.31)
【出願人】(508029653)
【出願人】(507241492)アンスティトゥート・ナシオナル・ドゥ・ラ・サンテ・エ・ドゥ・ラ・ルシャルシュ・メディカル・(インセルム) (9)
【Fターム(参考)】
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−133834(P2012−133834)
【出願日】平成24年6月13日(2012.6.13)
【分割の表示】特願2008−523267(P2008−523267)の分割
【原出願日】平成18年7月31日(2006.7.31)
【出願人】(508029653)
【出願人】(507241492)アンスティトゥート・ナシオナル・ドゥ・ラ・サンテ・エ・ドゥ・ラ・ルシャルシュ・メディカル・(インセルム) (9)
【Fターム(参考)】
[ Back to top ]