
MHCクラスII抗原提示系及びCD4+T細胞の活性化方法
【課題】未感作CD4+T細胞を活性化させるとともに、進行中の活性化状態をTh1またはTh2細胞のどちらかの好ましい分化集団に移行させるための合成抗原提示マトリックスを提供する。
【解決手段】本発明は、合成抗原提示マトリックス、それらの製造方法およびそれらの使用方法に関する。そのようなマトリックスの1つは、MHC抗原提示分子を1つ以上のアクセサリー分子とともに産生させるためにトランスフェクトした細胞である。当該マトリックスは、未感作CD4+T細胞を活性化させるとともに、進行中の活性化状態をTh1またはTh2細胞のどちらかの好ましい分化集団に移行させるために用いられる。
【解決手段】本発明は、合成抗原提示マトリックス、それらの製造方法およびそれらの使用方法に関する。そのようなマトリックスの1つは、MHC抗原提示分子を1つ以上のアクセサリー分子とともに産生させるためにトランスフェクトした細胞である。当該マトリックスは、未感作CD4+T細胞を活性化させるとともに、進行中の活性化状態をTh1またはTh2細胞のどちらかの好ましい分化集団に移行させるために用いられる。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
この出願は上記と同じ発明の名称を有する、1996年5月23日に出願された同時継続米国特許仮出願第60/018,175号(これらの開示が参考として本明細書に含まれる)の一部継続出願である。
【0002】
技術分野
本発明は特別な抗原ペプチドに対する特異性を有するCD4+T細胞を活性化する物質及び方法、種々の症状の治療のためのインビボでの活性化T細胞の使用、並びにこれらの使用に適した組成物に関する。
【背景技術】
【0003】
背景
T細胞レパートリーは主として胸腺中のT細胞発生中に成形されるが、成熟CD4+T細胞がまた胸腺外で調節される。活性化の或る条件はアネルギーまたはクローン排除を反映する寛容をもたらすが、その他の条件は観察される応答の型の変化をもたらす。CD4+T細胞について、機能性表現型のこの変化は主として産生されるサイトカインのパターンの変化である。急性活性化を受けるCD4+T細胞は多種サイトカインを産生する能力を維持するが、慢性刺激の条件下で得られたT細胞はサイトカイン産生の更に制限されたパターンを頻繁に示す。例えば、インビトロでの反復刺激により維持されたT細胞クローンは、Th1型細胞及びTh2型細胞と称されるCD4+T細胞の2種の主要な機能性カテゴリーを形成した。Th1型細胞は主としてインターロイキン−2 (IL−2) 、インターフェロン−γ(IFN- γ)及び腫瘍壊死因子(TNF) を産生し、これらの全てが炎症性サイトカインと称される。対照的に、Th2型細胞は典型的にはIL−4、IL−5、及びIL−10を産生し、抗体産生及びTh1型細胞の応答の調節に重要である。
【0004】
サイトカイン産生におけるこのような極度の分離はしばしばインビボT細胞応答中に見られないが、リーシュマニアの如き或る型の感染症からの回復はIL−2/IFN-γの優先産生と関連する。リーシュマニアに対するTh2応答を上昇するマウスは感染を抑制することができず、最終的に死亡する。Th2型応答のサイトカインの不適当な産生はアレルギー型疾患、例えば、喘息及び接触過敏症に頻繁に関連していた。CD4+T細胞の活性化及びアレルギー疾患における役割に関する総説について、Hetzel及びLamb, Clinical Immunol.Immunopath., 73:1−10 (1994)を参照のこと。
【0005】
おそらく、ヒト疾患とサイトカイン産生の非対称のパターンの最も強い関連はTh1応答及びTh1型サイトカインと自己免疫疾患の関連である。実験モデルにおける強力な証拠は、糖尿病を含む自己免疫の多くの型、多発性硬化症、自己免疫甲状腺炎等の実験モデルがTh1型CD4+T細胞により媒介されることを示す。これらのモデル中のIL−4の如きTh2関連サイトカインの発現は自己免疫疾患の発生を阻害する。Th2型サイトカインはTh1型細胞の応答を低下させ、一方、Th1型サイトカインはTh2型応答の発生を拮抗作用する。
【0006】
特別な活性化T細胞サブセットと特別な症状の関連に鑑みて、それ故、疾患の進行を変化するのに極めて有益であるプロセスである所望のT細胞サブセットへのCD4+T細胞の増殖及び活性化を誘導することができるようにとの要望が存する。一つの可能な解決は必要によりアレルギーまたは自己免疫症状を有してもよい被験者から最初に単離されるCD4+T細胞をインビトロで活性化して好ましいサイトカインプロフィールを分泌する細胞を生産することである。次いで得られる活性化T細胞が被験者に再度導入されて疾患の進行を変化し、そしておそらく長期治癒さえも与える。
【0007】
本発明により今解決された、このアプローチにおける挑戦は所望の治療サイトカインプロフィールを生じるCD4+T細胞サブセットを再現して生じる活性化条件を特定することの難しさである。特別なサイトカインの発現は特別な抗原提示細胞(APC) 及びそれらの関連アクセサリー(補助)分子と関連している。T細胞同時刺激に関係するアクセサリー分子として利用できる表面タンパク質の総説について、Mondino 及びJenkins, J.Leukocyte Biol., 55:805-815 (1994) を参照のこと。APCにより産生されたサイトカイン及び協調して発現されたアクセサリー分子の両方それ自体が抗原の型、T細胞レセプター(TCR) −抗原相互作用のアフィニティー、抗原濃度等を含む多くの因子により調節されるので、抗原提示後のT細胞活性化の成果の予想は伝統的に非常に困難である。実際に、付加的なアクセサリー分子がインビボでの活性化プロセスに提案されていたので、多くの多様な分子がT細胞応答の調節に関係し、コンビナトリアル様式で作用してT細胞活性化の成果に影響することが次第に明らかになってきている。
【0008】
本発明の以前には、一種以上の選択されたアクセサリー分子と一緒に選択されたMHCクラスII分子の同時発現は可能ではなかった。今、本発明はT細胞の再現可能な活性化により好ましいT細胞表現型を予測可能に生成してTh1またはTh2型T細胞を生じることの解決を提供する。本発明は特定のアクセサリー分子と組み合わせてMHCクラスII分子を中性バックグラウンドで提供する合成APCの生成を記載する。MHCクラスII分子及び特定のアクセサリー分子は非哺乳類昆虫細胞中で発現され、その分子を示す昆虫細胞を含む合成APCの種々の形態で提示し得る。
【0009】
本発明のMHCクラスII/アクセサリー分子組成物について発現及び提示ビヒクルとして昆虫細胞を使用することの利点は、細胞が調節サイトカインを内因的に産生せず、しかも哺乳類アクセサリー分子を発現しないことである。これはT細胞応答を変化することができる多くの分子を発現する哺乳類APCを使用することの特有の予測不能を解決する。加えて、本発明に記載された昆虫細胞発現系は或る種の制限環境、例えば、温度要件下で生成し得る結合ペプチドを含まないMHCクラスII分子(即ち、“エンプティー”分子)の発現を与える。生理温度で、これらの“エンプティー”分子は通常細胞表面に達することができない。何となれば、結合ペプチドを含まないクラスIIは非常に熱不安定であるからである。本発明は内因的にローディングされたカウンターパートを与える能力とともに選択されたペプチドの外因的ローディングを可能にする“エンプティー”MHCクラスII組成物の能力を利用する。
【0010】
組換えグリコシル−ホスファチジルイノシトール(GPI) 修飾MHCクラスI分子(HLA-A2.1:GPI/β2m) がTykocinskiの国際公開番号WO 96/12009 に記載されたようにして上記昆虫細胞系中で生成されて抗原提示細胞を生産した。その公開において、組換えGPI修飾MHCクラスI分子はその後の細胞膜への再とり込みのためにアフィニティー精製により昆虫細胞から単離される。その他の局面において、その公開はGPI修飾B7.1同時刺激分子と細胞膜に同時固定されたGPI修飾MHCクラスI分子の調製を記載している。その公開はGPI修飾MHCクラスII分子がMHCクラスIのそれらについて記載されたようにして調製し得ることを記載しているが、その公開はこのような調製について詳細を示していない。
【0011】
対照的に、本発明はB7.1の如き一種以上のアクセサリー分子と一緒に選択されたMHCクラスIIハプロタイプを同時発現してCD4+T細胞を活性化して特別なT細胞サブセット、Th1細胞またはTh2細胞(これらは好ましいサイトカインプロフィールに影響する)への分化をもたらすことに基く特異な手段を提供し、記載する。本発明は好ましいT細胞サブセットへインビトロでCD4+T細胞を選択的に活性化し、その後に患者への活性化T細胞の再導入を可能にするという利点を与える。こうして、本発明は個々の提示分子を選択された組み合わせの発現のための特別なアクセサリー分子と化合する能力を与え、これがその他のアプローチでは利用可能ではない所望のT細胞サブセットへのCD4+T細胞の選択的活性化について再現性及び予測可能性を可能にする。
【発明の概要】
【課題を解決するための手段】
【0012】
発明の簡単な要約
同時刺激分子及び付着分子を含む選択されたアクセサリー分子と組み合わせて発現された組換えMHCクラスII分子はMHCクラスII分子ヘテロダイマーがペプチドとの複合体形成のために発現される標的細胞を認識する武装化(armd)エフェクターT細胞になるようにCD4+T細胞を活性化するのに有効であることが今発見された。活性化は特別なサイトカインを分泌するエフェクターT細胞サブセット、Th1及びTh2への増殖及び分化を特徴とする。Th1型T細胞及びTh2型T細胞は夫々炎症細胞及びTヘルパー細胞と称される。
【0013】
こうして、本発明はCD4+T細胞を活性化するために一種以上のアクセサリー分子と組み合わせて哺乳類、好ましくはヒトMHCクラスII分子を生成し、提示するための合成抗原提示系(また、APCと称される)に関する。
【0014】
一実施態様において、その系は担体と担体に機能的に結合され、かつ選択されたペプチドに結合することができるMHCクラスIIヘテロダイマー分子の少なくとも細胞外部分とを有する合成抗原提示マトリックスに関する。また、マトリックスは担体に機能的に結合されたアクセサリー分子を含む。アクセサリー分子はCD4+T細胞に存在する特定のレセプターと相互作用する。MHCクラスII分子及びアクセサリー分子は、ペプチドがMHC分子の細胞外部分に結合される時にMHCクラスII/ ペプチド組み合わせに特異性のCD4+T細胞の集団を活性化するのに充分な数で存在する。
【0015】
アクセサリー分子と一緒に、MHCクラスIIヘテロダイマーまたはそのMHC に特異性のペプチドをローディングされたその少なくとも細胞外部分を有する抗原提示マトリックスはCD4+T細胞を活性化する際に相乗反応を与えることがわかった。アクセサリー分子の例はB7.1及びB7.2を含む同時刺激分子、付着分子、例えば、細胞内付着分子−1 (ICAM−1) 及びリンパ球機能関連抗原−3 (LFA−3)、並びにサバイバル分子(survival molecule)、例えば、Fasリガンド(FasL)及びCD70である。幾つかの実施態様において、このようなアクセサリー分子の細胞外部分がまた本発明に使用し得る。
【0016】
マトリックスに使用される担体は幾つかの異なる形態をとり得る。担体の例として、固体担体、例えば、金属またはプラスチック、多孔質材料、例えば、樹脂または変性セルロースカラム、ミクロビーズ、ミクロタイタプレート、赤血球及びリポソームが挙げられる。
【0017】
担体の別の型は細胞フラグメント、例えば、細胞膜フラグメントである。全細胞がまた担体として意図されている。この実施態様において、マトリックスは実際に少なくとも一種のアクセサリー分子とともにMHCクラスIIα鎖及びβ鎖の発現のための遺伝子を含む一種以上の発現ベクターで形質転換された細胞である。次いで発現されたタンパク質が細胞膜に輸送され、そこでクラスII鎖のトランスメンブランドメインがアンカーを与え、細胞外ドメインが外細胞表面に露出されることを可能にし、それにより合成抗原提示細胞(APC) を生じる。発現ベクターは構成的または誘導性であるプロモーターに機能的に結合された選択された遺伝子(cDNA配列の形態であることが好ましい)を含む。
【0018】
MHC α鎖及びβ鎖は一緒に会合してMHCクラスIIヘテロダイマーを形成し、これがそのヘテロダイマーに特異性のペプチドに結合する。本発明により、ペプチドをMHCクラスIIヘテロダイマーにローディングする二つの方法が意図されている。一実施態様において、ペプチドがペプチドフラグメントへの内在化無傷タンパク質のタンパク質分解プロセシング後に細胞内にローディングされる。次いでペプチドが新たに生成されたMHCクラスII分子にローディングされ、その間それらは依然として細胞内にある。また、MHCクラスII分子は細胞表面でエンプティー分子として発現され、次いで合成または処理ペプチド試薬がMHCクラスIIヘテロダイマーに細胞外でローディングされる。
【0019】
ベクター中のプロモーターに機能的に結合された少なくとも一種のアクセサリー分子遺伝子をコードするためのヌクレオチド配列がまた細胞に導入される。発現後に、アクセサリー分子はMHCクラスII/ ペプチド複合体に特異性のCD4+T細胞リンパ球の集団を活性化するのに充分な数でMHCクラスIIヘテロダイマーと一緒に細胞の表面に協調的に固定される。抗原プロセシング補助分子と称されるその他の分子がまた組換えAPCを生成する際の使用について意図されている。これらの分子はAPCとして使用される細胞により、または上記のような発現ベクター系により外因的に与えられる。このような抗原プロセシング補助分子の例として、不変鎖、リソソーム酵素並びにH2−M分子及びH2−O分子が挙げられる。
【0020】
細胞系は、上記遺伝子の少なくとも一つが細胞系が由来する細胞中に自然に存在しないという点で合成である。変温細胞系を使用することが好ましい。何となれば、MHC分子が熱不安定であるからである。或る範囲の種がこの目的に有益である。例えば、この使用のための多数の種を説明するPetersenらの米国特許第5,314,813 号(その開示が参考として本明細書に含まれる)を参照のこと。真核生物細胞及び好ましくは昆虫細胞がAPCとして使用される。好ましい昆虫細胞として、ドロソフィラ(Drosophila)(ショウジョウバエ)及びスポドプテラ(Spodoptera)(チョウ)が挙げられる。
【0021】
MHCクラスII分子は昆虫細胞、例えば、ドロソフィラ細胞及びスポドプテラ細胞中で発現された。これらの細胞は哺乳類免疫系の全ての成分を有しないので、ペプチドローディング機構に関係する種々のタンパク質がこのような細胞には不在である。哺乳類ペプチドローディング機構の欠如は、細胞が28℃のような熱安定温度制限条件で培養される時に導入された哺乳類MHCクラスII分子が細胞表面でエンプティー分子として発現されることを可能にする。対照的に、37℃で、エンプティークラスI分子は熱不安定であり、崩壊する傾向がある。こうして、MHCクラスII発現ドロソフィラ細胞を固定MHCクラスII分子に特異的に結合するペプチドとともにインキュベートすることにより、実際に夫々のクラスII分子が一つの同じペプチドでローディングされる。更に、本発明はあらゆる既知のMHCクラスIIα鎖及びβ鎖遺伝子を発現ベクターに導入し、それによりいずれか一種の哺乳類中で発現されるMHCクラスII分子の数の固有の制限を解消する手段を提供する。
【0022】
本発明において、CD4+T細胞をサイトカインインターロイキン−2 (IL−2) の増加を特徴とするTh1型応答に誘導するのに特に有効な相乗反応はペプチド、同時刺激分子、及び付着分子で結合されたMHCクラスII分子を有するドロソフィラ抗原提示細胞から生じる。特に、CD4+増殖と対にされたIL−2産生の高度に有効な相乗的発生はB7.2及びICAM−1の組み合わせから生じる。対照的に、ICAM−1ではなく、B7.1またはB7.2では、ペプチドをローディングされたドロソフィラAPC系はIL−4の増加を特徴とするTh2型応答を誘導した。こうして、ICAM−1はTh2型応答を拮抗作用してTh1型表現型をもたらす。
【0023】
増殖応答と対にされたIL−2産生を特徴とするTh1表現型はまたB7.2を用いて、またはそれを用いずにICAM−1と同時に発現されたCD70を有する合成抗原提示細胞から生じた。
【0024】
それ故、本発明のAPC中の発現のための少なくとも一種のアクセサリー分子遺伝子と組み合わせたMHCクラスII遺伝子の選択はCD4+T細胞の増殖及び表現型活性化を行うのに所望される成果に応じて適応し得る。
【0025】
また、本発明は、MHCクラスIIヘテロダイマー及びアクセサリー分子の遺伝子を含む少なくとも一種の発現ベクターが導入される上記合成APC系をつくる方法に関する。
【0026】
インビトロで活性化CD4+T細胞を生産する方法がまた意図されている。一つの好ましい方法はインビトロでCD4+細胞を抗原特異的様式でCD4+T細胞の集団を活性化するのに充分な期間にわたって上記の合成MHCクラスII/アクセサリー分子を有するAPCと接触することを含む。更に、その方法は(1) 活性化CD4+細胞を抗原提示マトリックスから分離し、(2) 活性化CD4+細胞を許されるキャリヤーまたは賦形剤中で懸濁させ、そして(3) その懸濁液を治療を要する個体に投与することを含んでもよい。先に説明したように、抗原は天然または未分解のタンパク質またはポリペプチドを含んでもよく、またはそれらは哺乳類MHCクラスIIヘテロダイマー分子と一緒のインキュベーションの前に少なくとも8アミノ酸残基を含むペプチドフラグメントに開裂または合成された抗原ポリペプチドを含んでもよい。
【0027】
上記の所望のT細胞サブセットへのCD4+T細胞の活性化を誘導することができるという実用性に加えて、あらゆるMHCクラスII分子を発現する能力はその特別なMHCクラスII分子に特異性の未知のCD4+活性化ペプチドを同定する手段を与える。このようなものとして、本発明は特別なMHCクラスIIヘテロダイマーを発現する合成APCによるペプチドライブラリーのスクリーニングによるCD4+T細胞の活性化を意図している。
【0028】
更に別の実施態様において、本明細書に記載された合成APC系は細胞の異種集団からの反応性CD4+T細胞の単離に有益である。このような単離は患者の症状で進行中のCD4+T細胞介在性応答を監視することができることを与える。
【0029】
上記の別の変化において、CD4+T細胞を好ましいサイトカインプロフィールを産生するための特別なT細胞サブセットに選択的に活性化することができることに鑑みて、本発明は望ましくないCD4+応答により媒介される患者の症状の治療方法に関する。Th1型応答またはTh2型応答を特徴とするこのような症状として、自己免疫疾患、アレルギー及び癌が挙げられる。治療目標は好ましいT細胞サブセットに活性化されたCD4+T細胞を導入して進行中のCD4+T細胞応答を拮抗作用することである。こうして、その方法は(1) 患者から静止または未感作(naive)CD4+細胞を含む液体サンプルを得、(2) インビトロでCD4+細胞を抗原特異的様式でCD4+細胞を活性化するのに充分な期間にわたって本発明の選択された合成ペプチドローディングされたAPCと接触させ、そして(3) 活性化CD4+細胞を患者に投与することを含む。
その他の実施態様は当業者に明らかである。
【図面の簡単な説明】
【0030】
【図1A】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図1B】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図1C】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図2】図2は表面発現されたMHCクラスIIヘテロダイマーにローディングされたオボアルブミン(OVA)ペプチドを用いて、またはそれを用いずにドロソフィラ細胞系の存在下で培養した時のDO10T細胞レセプター(TCR) トランスジェニックマウス細胞系(Tg細胞)の増殖応答を示す。増殖は実施例5に記載されるようにして分析される。増殖はX軸の次第に増加する濃度(μM)のOVAペプチドに対しY軸にプロットされたようにカウント/分(cpm)×1000で測定される。T-S(ダイヤモンドでマークした線)は対照脾臓APCによる応答を示す。組換えMHCクラスII単独による増殖応答が閉じた円を有する線で示される。同時刺激分子B7.1またはB7.2と組み合わされたMHCクラスIIによる増殖応答が夫々開いた正方形及び閉じた正方形を有する線で示される。
【図3】図3は組換えMHCクラスII単独(閉じた円を有する線)、MHCクラスII+B7.2( 閉じた正方形を有する線)、MHCクラスII+ICAM−1(開いた三角形を有する線)、及びMHCクラスII+B7.2及びICAM−1(開いたダイヤモンドを有する線)の増殖応答を示す。その他の詳細について図2脚注を参照のこと。
【図4】図4A〜4Dは組換えMHCクラスII単独またはB7.1もしくはB7.2同時刺激分子と組み合わせた組換えMHCクラスIIを有するドロソフィラAPCの存在下で培養した時のCD4+T細胞の活性化に応答して生じたサイトカインプロフィールを示す。脾臓APC(T-Sと標識した)は対照アッセイである。アッセイは実施例5に記載されるようにして行われる。図4A〜4DはY軸にプロットされたようにサイトカインIL−2、IL−4、IFN-γ及びIL−10 (単位ng/ml)を夫々示す。サイトカインプロフィールは培養の3日目〜5日目の3日間にわたって評価された。
【図5】図5A〜5Dは組換えMHCクラスII単独またはB7.2同時刺激分子、ICAM−1並びにB7.2とICAM−1と組み合わせた組換えMHCクラスIIを有するドロソフィラAPCの存在下で培養した時のCD4+T細胞の活性化に応答して生じたサイトカインプロフィールを示す。図5A〜5DはY軸にプロットされたようにサイトカインIL−2、IL−4、IFN-γ及びIL−10 (単位ng/ml)を夫々示す。その他の詳細について図4脚注を参照のこと。
【発明を実施するための形態】
【0031】
発明の詳細な説明
A.定義
アミノ酸残基:ペプチド結合の位置におけるポリペプチドの化学消化(加水分解)後に生成されたアミノ酸。本明細書に記載されたアミノ酸残基は“L”異性体であることが好ましい。しかしながら、“D”異性体の残基は、所望の機能性がポリペプチドにより保持される限り、あらゆるL−アミノ酸残基を置換し得る。NH2 はポリペプチドのアミノ末端に存在する遊離アミノ基を表す。COOHはポリペプチドのカルボキシ末端に存在する遊離カルボキシ基を表す。通常のポリペプチド命名法(J.Biol.Chem., 243: 3552-3559 (1969) に記載され、37 CFR§1.822(b)(2)で採用される)を遵守して、アミノ酸残基の略号が下記の対応表に示される。
【0032】
対応表
記号 アミノ酸
1文字 3文字
Y Tyr チロシン
G Gly グリシン
F Phe フェニルアラニン
M Met メチオニン
A Ala アラニン
S Ser セリン
I Ile イソロイシン
L Leu ロイシン
T Thr スレオニン
V Val バリン
P Pro プロリン
K Lys リシン
H His ヒスチジン
Q Gln グルタミン
E Glu グルタミン酸
Z Glx Glu 及び/またはGln
W Trp トリプトファン
R Arg アルギニン
D Asp アスパラギン酸
N Asn アスパラギン
B Asx Asn 及び/またはAsp
C Cys システイン
X Xaa 未知またはその他
【0033】
式により本明細書に表される全てのアミノ酸残基配列はアミノ末端からカルボキシ末端への通常の方向で左から右への配向を有することが注目されるべきである。加えて、“アミノ酸残基”という用語は対応表にリストされたアミノ酸並びに修飾アミノ酸及び特異アミノ酸、例えば、37 CFR§1.822 (b)(4)にリストされ、また本明細書に参考として含まれるものを含むように広く定義される。更に、アミノ酸残基配列の最初または最後のダッシュは一つ以上のアミノ酸残基の更に別の配列に結合されたペプチドまたはNH2 もしくはアセチルの如きアミノ末端基もしくはCOOHの如きカルボキシ末端基への共有結合を示す。
【0034】
組換えDNA(rDNA)分子:二つのDNAセグメントを機能的に結合することにより生成されたDNA分子。こうして、組換えDNA分子は自然には通常一緒に見られない少なくとも二つのヌクレオチド配列を含むハイブリッドDNA分子である。共通の生物起源を有しない、即ち、進化上異なるrDNAが“異種”と称される。
【0035】
ベクター:細胞中で自律複製することができ、DNAセグメント、例えば、遺伝子またはポリヌクレオチドが付着セグメントの複製をもたらすように機能的に結合し得るrDNA分子。一種以上のポリペプチドをコードする遺伝子の発現を誘導することができるベクターが本明細書中“発現ベクター”と称される。
【0036】
上流:DNA転写の方向と反対の方向で、それ故、非コーディングストランドでは5'から3'に進み、またはmRNAでは3'から5'に進む。
【0037】
下流:配列転写または読み取りの方向にDNA配列に沿って、即ち、DNAの非コーディングストランドに沿って3'から5'の方向またはRNA転写産物に沿って5'から3'の方向に移動している。
【0038】
読み取り枠:遺伝子の構造タンパク質をコードする部分、または構造遺伝子を形成する翻訳に使用される連続のヌクレオチドトリプレット(コドン)の特別な配列。読み取り枠は翻訳開始コドンの位置に依存する。
【0039】
ポリペプチド:連続アミノ酸残基のα−アミノ基とカルボキシ基の間のペプチド結合により互いに連結された線状の一連のアミノ酸残基。
【0040】
タンパク質:ポリペプチドのように互いに連結された線状の一連の50より大きいアミノ酸残基。
【0041】
レセプター:レセプターは別の分子に特異的に(非ランダムに)結合し得る分子、例えば、タンパク質、糖タンパク質等である。
【0042】
実質的に精製され、または単離された:ポリペプチドまたはタンパク質に関して使用される場合、その用語はそれらと自然に随伴する成分から分離された分子を記載する。典型的には、モノマータンパク質は、サンプルの少なくとも約60%〜75%が単一ポリペプチド主鎖を示す時に実質的に純粋である。少量の変異体または化学修飾が典型的には同じポリペプチド配列を共有する。実質的に精製されたタンパク質は典型的にはタンパク質サンプルの約85%〜90%以上、更に通常約95%を構成し、好ましくは約99%以上純粋であろう。タンパク質またはポリペプチドの純度または等質性は当業界で公知の幾つかの手段、例えば、サンプルのポリアクリルアミドゲル電気泳動、続いて染色によるその視覚化により示し得る。或る目的には、高分解能が必要とされ、高速液体クロマトグラフィー(HPLC)または同様の精製手段が利用される。
【0043】
合成ペプチド:天然産タンパク質及びそのフラグメントを含まないペプチド結合により一緒に結合されたアミノ酸残基の化学的に生成された鎖。
【0044】
B.MHCクラスIIヘテロダイマー、アクセサリー分子及び抗原プロセシング補助分子
本発明はCD4+T細胞を活性化するのに使用するための合成抗原提示系に関する。T細胞レパートリーは主として胸腺中のT細胞発生中に形成されるが、成熟CD4+T細胞がまた胸腺外で調節される。活性化の或る条件がアネルギーまたはクローン排除を反映するトレランスをもたらすが、その他の条件は観察される応答の型の変化をもたらす。CD4+T細胞について、機能性表現型のこの変化は主として産生されるサイトカインのパターンの変化である。急性活性化を受けるCD4+T細胞は多種サイトカインを産生する能力を維持するが、慢性刺激の条件下で得られたT細胞はサイトカイン産生の更に制限されたパターンを頻繁に示す。例えば、インビトロの反復刺激により維持されたT細胞クローンはTh1型細胞及びTh2型細胞と称されるCD4+T細胞の二つの主要な機能性カテゴリーを形成した。Th1型細胞は主としてインターロイキン−2 (IL−2) 、インターフェロン−γ(IFN- γ)及び腫瘍壊死因子(TNF) を産生し、これらの全てが炎症性サイトカインと称される。このようなものとして、Th1細胞は時折炎症性T細胞と称され、これらはその後にマクロファージを活性化して、CD4+T細胞に提示された病原タンパク質のプロセシングから生成されたペプチドにより媒介されるようにそれらが宿している小胞内病原体を死滅させる。対照的に、Th2型細胞は典型的にはIL−4、IL−5、及びIL−10 を産生し、抗体産生及びTh1型細胞の応答の調節に重要である。このようなものとして、Th2細胞はヘルパーT細胞と時折称され、これらはB細胞を活性化して、CD4+T細胞細胞に提示される細胞外病原体及び毒素に由来するペプチドに応答して抗体をつくる。
【0045】
CD4+T細胞はCD3複合体のタンパク質と会合したα:βまたはγ:Δヘテロダイマーレセプターの存在と一緒にその細胞表面でCD4+コレセプターを有するT細胞またはTリンパ球と定義される。
【0046】
CD4+T細胞サブセットの活性化は抗原提示細胞により誘発された刺激の型に依存性の上記のサイトカインの選択産生と協調した応答性T細胞集団の増殖を特徴とする。後者は抗原を処理でき、リンパ球活性化に必要とされる分子と一緒に細胞表面でそれらのペプチドフラグメントを露出する高度に特殊化された細胞と定義される。CD4+T細胞活性化の特異性は抗原提示細胞(APC) の表面のMHCクラスIIヘテロダイマーに結合されたペプチド抗原のT細胞抗原レセプター(TCR)認識に基いている。T細胞の主要APCは樹状細胞、マクロファージ及びB細胞である。加えて、APC誘導非抗原性同時刺激シグナルがCD4+T細胞活性化に寄与する役割を果たす。
【0047】
本発明は上記の自然免疫応答メカニズムに基いて合成抗原提示系を使用して、同時刺激分子と広く称される一種以上のアクセサリー分子と一緒に組換えMHC クラスII分子によりCD4+T細胞の活性化を操作する。後者として、特定の同時刺激分子、付着分子及びサバイバル分子等が挙げられる。本発明のその他の局面において、合成抗原提示系はペプチドローディングされたMHCクラスII分子を生成するのに有益である抗原プロセシング補助分子を更に含む。本発明に関して、合成抗原提示系はCD4+T細胞をインビトロ及びインビボで活性化するのに有益である。これらの局面がCD4+細胞応答の変更方法に関する節Eに説明される。
【0048】
こうして、上記のように、本発明の抗原提示系は少なくとも2種の主成分を有する。第一成分はTCR による認識によりCD4+T細胞活性化の特異性を与えるペプチドに結合することができる組換えMHCクラスIIヘテロダイマーの少なくとも細胞外の部分である。第二主成分はCD4+T細胞の活性化において非抗原特異性同時刺激シグナルを与える少なくとも一種のアクセサリー分子の少なくとも細胞外の部分である。別の実施態様において、抗原提示系及び非抗原同時刺激シグナルの全分子が使用される。
【0049】
説明を容易にするために、MHC分子の細胞外部分が本発明の或る局面で使用し得ることを理解して、MHCクラスIIヘテロダイマーが一般に説明される。本発明に必要なMHC分子の部分はCD4+T細胞への提示のために抗原ペプチドに結合する部分である。
【0050】
本発明は組換えMHCクラスIIヘテロダイマーがMHCクラスIIヘテロダイマーと既に複合体形成されたペプチドでベクター形質転換された合成抗原提示細胞により産生されることを可能にする。また、それらと複合体形成されたペプチドを未だ有していないエンプティーMHCクラスIIヘテロダイマーが生成される。この後者の実施態様が特に有益である。何となれば、それは特別なペプチドとの複合体形成またはMHCクラスIIヘテロダイマーが発現された後のペプチドのライブラリーのスクリーンを可能にするからである。
【0051】
1.MHCクラスII遺伝子及びコードされたヘテロダイマー
MHCクラスII分子は二つの鎖、α及びβ(これらの両方が膜をスパンする)の非共有結合複合体からなる細胞表面糖タンパク質である。ペプチド結合クレフトが共同鎖の間に形成される。MHCクラスII分子を結合するペプチドは長さが可変であり、ペプチドの末端から種々の距離にあるアンカー残基を有し、それにより結合ポケットのクレフト中にしっかりと結合されていない末端を有するペプチドをもたらす。Janeway 及びTravers, Immunobiology, Section 4-4, Current Biology LTD,第2編, 1996を参照のこと。抗原提示ペプチドの更に別の局面が節Dに説明される。
【0052】
インビボでは、エンプティーMHCクラスIIヘテロダイマーは不安定化されるようになり、続いて細胞表面から除去され、それによりMHC分子が周囲の細胞外液体からのペプチド(これらはT細胞に特異的に有害に作用するであろう)を獲得することを阻止する。本発明において、合成抗原提示細胞系は不安定化イベントを受けていない細胞表面でエンプティーMHCクラスIIヘテロダイマーの生成を可能にする。その結果として、表面発現された組換えMHCクラスIIヘテロダイマーへの抗原ペプチドのローディングがCD4+T細胞活性化及びサイトカイン産生を操作する際のその後の使用のために促進される。
【0053】
本発明、協調抗原との選択されたMHCクラスII分子の特別な組み合わせはMHCクラスII遺伝子中のコンセンサスヌクレオチド配列の存在により促進される。以下更に記載されるこれらの領域は哺乳類中のMHC複合体中の多種MHCクラスII遺伝子並びに夫々の遺伝子の多種対立遺伝子の読み出し及び使用を可能にする。換言すれば、MHCは幾つかの遺伝子及び夫々の遺伝子の多種対立遺伝子を有する多遺伝子性かつ多形性の両方である。
【0054】
ヒトでは、MHCはHLAと称され、一方、マウスでは、それはH−2と称される。ヒトのMHCクラスIIα鎖及びβ鎖遺伝子の三つの対がHLA−DP、HLA−DQ及びHLA−DRと称された。HLA−DRクラスターは特別のβ鎖遺伝子を含む。このようなものとして、これらの遺伝子の三つの組がMHCクラスII分子の四つの型を生じ得る。MHCクラスII遺伝子及びヒト遺伝子から得られるコードされたα鎖及びβ鎖はヒト起源であると言われる。マウスでは、MHCクラス遺伝子がH2−M、H2−A及びH2−Eと称される。夫々のMHCクラスII分子は異なる範囲のペプチドを結合するので、多遺伝子座の存在は、夫々のクラスの唯一のMHCクラスII分子が細胞表面で発現された場合よりも広範囲の異なるペプチドを提示する能力を個体に与える。
【0055】
多形性MHCクラスII遺伝子は唯一または二三のアミノ酸だけ変化する相当するタンパク質をコードするが、異なる対立遺伝子変異体は20までのアミノ酸を異にする。その結果として、MHCクラスII多様性はT細胞による抗原認識の能力を拡大する。更に、MHC制限により、T細胞は特別なMHC分子に関してペプチドを認識するが、別の分子に提示された時に認識しないことが示された。こうして、T細胞レセプター特異性はペプチド及びそれを結合するMHC分子の両方により付与される。
【0056】
選択されたα鎖及びβ鎖を含む本発明のMHCクラスIIヘテロダイマーはオリゴヌクレオチドプライマーの特定の対によるMHCクラスIIをコードする遺伝子及びその対立遺伝子変異体の増幅により得られる。プライマーのヌクレオチド配列は遺伝子のカテゴリー内の遺伝子中に存在する5'及び3'コンセンサスヌクレオチド配列に基いてMHCクラスII遺伝子及びその対立遺伝子変異体の多様性の増幅を可能にする。ヒトHLA−DP遺伝子、HLA−DQ遺伝子及びHLA−DR遺伝子のα鎖及びβ鎖を増幅するためのプライマー対、並びにマウスIAdをコードするヘテロダイマーを増幅するためのプライマー対の特定のヌクレオチド配列が実施例2Aに示される。
【0057】
MHCクラスIIをコードする遺伝子はB細胞、マクロファージ及び樹状細胞(これらの全てが血液中に存在する)を含む種々の細胞源から増幅可能である。本発明のプライマーで増幅されたMHCクラスIIをコードする遺伝子を得るための増幅条件が節Cに記載される。
【0058】
本発明のα鎖及びβ鎖を含むMHCヘテロダイマーは固定形態または可溶性形態で有益である。固定形態では、組換えMHCヘテロダイマーはそれが発現される合成抗原提示細胞に固定される。また、組換えMHCヘテロダイマーは可溶性形態で分泌された後に本発明に記載された担体を含むマトリックスに固定される。後者は、終止コドンが増幅操作中またはその後にトランスメンブランドメインに先行する特別のMHCクラスIIα鎖及びβ鎖をコードするヌクレオチド配列に操作される時に生成される。
【0059】
2.アクセサリー遺伝子及びコードされた分子
同時刺激分子、付着分子及びサバイバル分子を含む本発明のアクセサリー分子はCD4+T細胞を活性化して標的細胞を認識する武装化エフェクターT細胞になる際にペプチドと複合体形成されたMHCクラスIIヘテロダイマーと協力して有効である。未感作T細胞は、それらがAPCの表面でペプチドローディングされたMHCクラスIIヘテロダイマーにより提示された時のそれらの特異的抗原と出会う時に活性化されて武装化エフェクターT細胞に増殖し、分化する。活性化はMHCクラスIIヘテロダイマーに結合された外来ペプチドフラグメントの認識を必要とするだけでなく、それはAPCにより同時発現された同時刺激シグナルの同時の送出を必要とする。
【0060】
こうして、本発明の合成APCまたはマトリックスは特別なMHCクラスIIヘテロダイマーの存在だけでなく、アクセサリー分子と広く定義される一種以上の同時刺激分子の存在を特徴とする。特定の同時刺激分子、付着分子、及びサバイバル分子を含むアクセサリー分子の少なくとも三つの型が本発明の合成APCまたはマトリックスを調製する際の使用について意図されている。
【0061】
a.同時刺激分子
アクセサリー分子の第一の型はB7.1(B7として既に知られており、またCD80として知られている)及びT細胞のCD28に結合するB7.2(またCD86として知られている)の如き同時刺激分子を含む。B7.1及びB7.2は免疫グロブリンスーパーファミリーのホモダイマー員である構造関連糖タンパク質である。その他の同時刺激分子は抗CD28抗体またはこのような抗体の機能性部分、例えば、CD28に結合するFab部分である。上記分子によるCD28の結合が未感作T細胞の成長を同時刺激することが示された。活性化T細胞では、付加的なレセプターであるCTLA−4がCD28とのアフィニティーより高いアフィニティーでB7分子を結合する。
【0062】
本発明の合成APCまたはマトリックスに使用するための組換えB7同時刺激分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0063】
b.付着分子
本発明のアクセサリー分子の別の主要な型はT細胞活性化にまた機能する付着分子である。アクセサリー付着分子として、種々のICAM分子が挙げられ、これらとして、細胞間付着分子(ICAM) ICAM−1 、ICAM−2、ICAM−3、リンパ球機能関連抗原(LFA) LFA−1 及びLFA−3 が挙げられる。これらの分子の全てが免疫グロブリンスーパーファミリーの員である。全てのICAM関連員がT細胞インテグリン、LFA−1 に結合する。樹状細胞、マクロファージ及びB細胞を含むAPCで発現されることに加えて、ICAM−1及びICAM−2はまた内皮で発現され、それにより細胞付着及び循環白血球と内皮の間のその後の遊出を媒介する。しかしながら、ICAM−3は白血球にのみ発現され、T細胞とAPCの間の付着に重要な役割を果たすと考えられる。
【0064】
ICAM−1、ICAM−2及びICAM−3の間の相互作用は、APC及びT細胞表面で夫々発現されるLFA−3 (CD58)とLFA−2(CD2) の間の第二の付着相互作用と相乗作用する。
【0065】
本発明の合成APCまたはマトリックスに使用するための組換え付着分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0066】
c.サバイバル分子
サバイバル分子は刺激から細胞死滅誘発に至る範囲の代謝応答に役割を果たすアクセサリー分子の別の型である。こうして、サバイバル分子はまた細胞死滅調節分子と称される。サバイバル分子は典型的にはタンパク質であるが、その他の型の巨大分子、例えば、炭水化物、脂質等を含んでもよい。本発明の組成物及び方法に使用するためのサバイバル分子として、Fasリガンド、TNF-レセプター、TNF、CD70、II型トランスメンブランタンパク質(これはTNFレセプターファミリーの員であるCD27に結合するTNFファミリーの員である)が挙げられる。FasリガンドはFasと称されるレセプターに結合し、レセプター占有がFasレセプターを発現する細胞のアポトーシス細胞死滅の誘発をもたらす。CD27は静止T細胞及びB細胞で発現され、一方、CD70は活性化T細胞及びB細胞で発現される。そのレセプター、CD27へのCD70の結合はT細胞同時刺激を誘発し、その相互作用が未感作T細胞プールからのT細胞の動員に重要であるかもしれない。その他の或る条件下で、TNFによるTNFレセプターの活性化が同様の応答を生じる。
【0067】
本発明の合成APCまたはマトリックス中の使用について上記された組換えサバイバル分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0068】
実施例に示されるように、一種以上の上記組換えアクセサリー分子と一緒に使用される組換えMHCクラスII分子に結合されたペプチドの特別な組み合わせはT細胞を武装化エフェクターT細胞(これらはTh1炎症性T細胞及びTh2ヘルパーT細胞に区別できる)に活性化する。
【0069】
3.抗原プロセシング補助遺伝子及びコードされた分子
a.HLA−DM
ヒト中のHLA−DM及びマウス中のH2−MはMHCクラスII様分子であり、これはまたMHCクラスII遺伝子クラスター中でコードされる。HLA−DMは、MHCクラスIIのように、ヘテロダイマーを形成するα鎖遺伝子及びβ鎖遺伝子の両方を含む。しかしながら、MHCクラスIIと違って、ペプチドローディングが分子を安定化するのに必要とされない。HLA−DMは以下に更に記載されるような非変異鎖の除去後に新たに形成されたMHCクラスIIヘテロダイマーへのタンパク質のローディングを促進する。組換えHLA−DMは内部処理されたペプチドのローディングを補助するために本発明の組成物及び方法における使用について意図されている。
【0070】
b.非変異鎖
非変異鎖は新たに形成されたMHCクラスIIヘテロダイマーに結合し、それによりMHCクラスIIヘテロダイマーの夫々のサブユニットとトリマーを形成する特殊化タンパク質である。トリマー化分子は小胞体中に存在する細胞内ペプチドのローディングを阻止するが、それはまたその区画からのその分子の輸送を促進する。その後、非変異鎖は多工程により開裂されてMHCクラスIIヘテロダイマーを生じ、次いでこれが処理ペプチドと複合体形成し得る。
【0071】
こうして、組換え非変異鎖は内部処理されたペプチドのローディングを補助するために本発明の組成物及び方法における使用について意図されている。
【0072】
C.核酸及びポリヌクレオチド
1.MHCクラスII及びアクセサリー分子をコードする遺伝子を得るためのPCR
本発明のMHCクラスII分子、アクセサリー分子及び抗原プロセシング補助分子をコードする核酸配列は、直接合成、クローニング、このような遺伝子を含む細胞からのDNAの精製等を含む当業者に良く知られている幾つかの方法で得られる。本明細書に記載された組成物及び方法に使用される分子をコードするための遺伝子を得るための一つの適切な手段は以下に更に記載されるような選択されたオリゴヌクレオチドプライマー対による選択された核酸鋳型のポリメラーゼ連鎖(PCR)増幅による。
【0073】
既知の部分及び推定のヒト白血球抗原(HLA) 、ヒトMHCの遺伝的名称、アミノ酸配列及びコンセンサス配列を含むヌクレオチド配列が公表されており(例えば、Zemmour 及びParham, Immunogenetics 33: 310-320 (1991) を参照のこと)、HLA 変異体を発現する細胞系が同様に知られており、一般にATCCから多く利用できる。それ故、PCRを使用して、MHCクラスIIをコードするヌクレオチド配列が本発明の発現ベクターに容易に機能的に結合され、次いでこれがその中の発現に適した細胞を形質転換するのに使用される。
【0074】
本発明の組換え分子を生成するのに特に好ましい方法は本明細書に記載されるようにPCR反応産物を生成するためにPCRのプライマーとして前もって選択されたオリゴヌクレオチドの使用に頼る。
【0075】
遺伝子がPCR増幅により得られる場合、一般に、PCRプライマー対を含む2種のプライマーが増幅される核酸の夫々のストランドについて使用される。簡素化のために、例示のMHCクラスIIヘテロダイマーをコードする遺伝子の合成が説明されるが、記載されるPCR増幅方法は完全配列が現在知られていないものを含むMHCクラスII対立遺伝子変異体、アクセサリー分子及び抗原プロセシング補助分子の合成に同等に適用可能であることが明らかに理解されるべきである。
【0076】
一般に、第一プライマーはフォワード(forward) プライマーまたは5'と称される。何となれば、それは鋳型DNAの上部ストランドと同じ配列を有し、こうして下部相補ストランドにハイブリッドを形成するからである。
【0077】
第二プライマーはバックワード(backward)プライマーまたは3'と称される。何となれば、それは下部ストランドと同じ配列を有し、こうして上部ストランドの相補ストランドにハイブリッドを形成するからである。典型的には、換言すれば、一つのプライマーはヌクレオチド配列の負(-) または下部のストランドに相補性であり、別のプライマーは正(+) または上部のストランドに相補性である。
【0078】
好ましい局面において、第一プライマー及び第二プライマーの両方がMHCクラスII遺伝子内の保存領域にハイブリッドを形成するように(即ち、相補性であるように)選ばれる。しかしながら、プライマーはコンセンサス配列ではなく特異な配列にハイブリッドを形成することにより特定のMHCクラスII遺伝子及びその対立遺伝子変異体を増幅するように設計し得る。この局面について、鋳型配列がこのようなプライマー対の設計について知られていることが好ましい。
【0079】
第一プライマー及び第二プライマーの一方または両方はエンドヌクレアーゼ認識部位を形成するヌクレオチド配列を増幅産物に導入するように設計し得る。その部位は増幅されるMHCクラスII遺伝子に異種であってもよく、典型的にはプライマーの5'末端付近に現れる。それはまた4-塩基スペーサー配列を制限部位に近位に配置して酵素による増幅産物の切断の効率を改良するのに有益であり得る。
【0080】
特定のヌクレオチド配列を単離するための本発明のプライマーは相当するヌクレオチド配列を有するかなりの数の核酸の重合の特定の開始を与えるように充分な長さ及び適当な配列のオリゴヌクレオチドを含む。詳しくは、本明細書に使用されるオリゴヌクレオチドプライマーという用語は二つ以上、好ましくは三つより多く、更に好ましくはほぼ20のデオキシリボヌクレオチドまたはリボヌクレオチドを含む配列を表し、その配列はプライマー伸長産物の合成を開始することができる。
【0081】
合成を行う実験条件として、ヌクレオチドトリホスフェートの存在並びに重合及び伸長のための薬剤、例えば、熱安定性ポリメラーゼ、及び好適な緩衝剤、温度及びpHが挙げられる。プライマーは増幅の最大効率のために一本鎖であることが好ましいが、二本鎖であってもよい。二本鎖の場合、プライマーは最初に処理されて二つのストランドを分離し、その後に伸長産物を調製するのに使用される。プライマーはオリゴデオキシリボヌクレオチドであることが好ましい。プライマーはヌクレオチドの重合及び伸長の誘導剤の存在下で伸長産物の合成を開始するのに充分に長く、かつ実質的に相補性である必要がある。プライマーの正確な長さは温度、緩衝剤、及びヌクレオチド組成を含む多くの因子に依存するであろう。オリゴヌクレオチドプライマーは典型的には15〜22 またはそれ以上のヌクレオチドを含むが、それは更に少ないヌクレオチドを含んでもよい。また、当業界で公知であるように、ヌクレオシドトリホスフェートの混合物がバイアスされて突然変異の形成に影響して特異なペプチドをCD4+T細胞に提示するのに使用するための突然変異した組換えMHCクラスIIをコードする分子のライブラリーを得ることができる。
【0082】
本発明のオリゴヌクレオチドプライマーは指数増殖量のヌクレオチド配列を生成する酵素連鎖反応であるPCR増幅方法に使用される。変性核酸へのプライマーのアニール、続いて熱安定性ポリメラーゼ、例えば、テルモフィルス・アクアチクス(Taq)及びピロコッカス・フリオサス(Pfu) (Hoffman La-Roche, Basal,スイス)、及びヌクレオチドによる伸長が新たに合成された(+) 鎖及び(-) 鎖をもたらす。これらの新たに合成された配列はまた鋳型であるので、変性、プライマーアニール、及び伸長の反復サイクルがプライマーにより形成されたDNAフラグメントの指数的生成をもたらす。連鎖反応の産物は使用された特定のプライマーの末端に相当する末端を有する不連続の核酸二重らせんである。当業者は標的核酸のコピー数を増加するのにまた使用し得るその他の増幅方法を知っているであろう。これらとして、例えば、連結反応活性化転写(LAT) 、リガーゼ連鎖反応(LCR)、及びストランド置換活性化(SDA) が挙げられるが、以下にリストされる米国特許に記載されたようなPCRが好ましい方法である。
【0083】
本発明のオリゴヌクレオチドプライマーは、あらゆる好適な方法、例えば、相補オリゴヌクレオチドの合成またはその自動化実施態様について上記された通常のホスホトリエステル方法及びホスホジエステル方法を使用して調製し得る。修飾固体担体上にオリゴヌクレオチドを合成するための一つの方法が米国特許第4,458,066 号明細書に記載されている。
【0084】
MHCクラスII遺伝子、アクセサリー分子遺伝子、及び抗原プロセシング補助分子遺伝子を増幅するのに好ましいプライマーが実施例2に記載される。
【0085】
PCR増幅方法が米国特許第4,683,192 号、同第4,683,202 号、同第4,800,159号、同第4,965,188 号及び同第5,395,750 号(これらの開示が参考として本明細書に含まれる)、並びに“PCR Technology: Principles and Applications for DNA Amplification", H.Erlich編集, Stockton Press, New York (1989);及び“PCR Protocols: A Guide to Methods and Applications", Innisら編集, Academic Press, San Diego, California (1990) を含む少なくとも幾つかの書籍に詳しく記載されている。ここに使用される種々の好ましい方法及びプライマーが以下に記載され、またZemmour ら, Immunogenetics, 33: 310-20 (1991) 、Ausebelら, In Current Protocols in Molecular Biology, Wiley and Sons, New York (1993) 及びSambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, (1989)により記載されている。当業者に公知であるネステッド(nested)PCR 、オーバーラップPCR、逆転写酵素−PCR等を含む特別なPCR方法が本発明の組換え分子を得る際の使用について意図されている。
【0086】
別の実施態様において、PCR方法は種々のMHCクラスIIをコードする分子を生成するのに使用されるだけでなく、高度に多形性のMHC遺伝子座中に観察されるものと匹敵し得る突然変異を誘発し、または単一の親クローンから多様性を生じ、それにより更に大きな異質性を有するMHCクラスIIをコードするDNA“ライブラリー" を得るのに使用される。
【0087】
2.発現ベクター
本発明はMHCクラスIIをコードする遺伝子、アクセサリー分子遺伝子及び抗原プロセシング補助遺伝子の発現を誘導して相当する組換えタンパク質を生成することができる実質的に純粋な形態のプラスミド発現ベクターを意図している。簡素化のために、上記遺伝子がポリペプチドをコードするヌクレオチド配列と本明細書で集約的に称される。それらが機能的に結合される遺伝子の発現を誘導することができるベクターは本明細書で“発現ベクター”または“発現プラスミド”と称され、これらの両方がまた“プラスミド”と称される。
【0088】
本明細書に使用される“ベクター”または“プラスミド”という用語は異なる遺伝子環境の間でそれが機能的に結合された別の核酸を輸送することができる核酸分子を表す。好ましいベクターはそれらが機能的に結合されるDNAセグメント中に存在する構造遺伝子産物の自律複製及び発現の可能なベクターである。それ故、ベクターは宿主選択系と適合性であるレプリコン及び選択可能なマーカーを含むことが好ましい。好ましいベクターの一つの型はエピソーム、即ち、染色体外複製の可能な核酸である。
【0089】
本発明のプラスミドは本発明の翻訳可能なポリペプチドをコードするヌクレオチド配列の転写を活性化することができる要素を有する調節領域を少なくとも含む環状二本鎖プラスミドである。プラスミドは、本発明の所望のコードされたポリペプチドが発現される翻訳可能なヌクレオチド配列を更に含む。こうして、ベクターは相当する発現可能な遺伝子からコードされたような本明細書に記載された組換えポリペプチドの発現を誘導することができると言われる。
【0090】
本発明の使用に好ましいベクターはプラスミドである。それは高コピー数プラスミドであることが更に好ましい。また、特別なベクターが選択された宿主中の発現に最適であることが好ましい。
【0091】
このような発現ベクターは宿主中の挿入された遺伝子配列の有効な転写を促進する調節領域中にプロモーター配列を含む。ベクターは誘導プロモーター配列を含むことが好ましい。何となれば、誘導プロモーターはこのようなベクター(これらは未変性またはキメラのヌクレオチド配列を有するようにしばしば構築される)が導入された細胞に対し選択圧力を制限する傾向があるからである。また、発現ベクターは典型的には複製起点並びに形質転換細胞の表現型選択を可能にする特定の遺伝子を含む。DNAセグメントは調節要素、例えば、プロモーター(例えば、T7プロモーター、メタロチオネインIプロモーター、またはポリヘドリンプロモーター)に機能的に結合されたベクター中に存在し得る。
【0092】
別の実施態様において、プラスミドはまたその発現が宿主細胞に導入または形質転換された時に選択的利点、例えば、薬剤耐性をその細胞に与える遺伝子を含む。典型的な原核生物及び真核生物の薬剤耐性遺伝子は夫々アンピシリンまたはテトラサイクリン及びネオマイシン(G418またはゲネチシン)に対する耐性を与える。その他の薬剤耐性マーカーとして、クロラムフェニコール、カナマイシン、ストレプトマイシン、カルベニシリン、水銀、リファムプシン、リファムピシン、フサル酸等が挙げられる。
【0093】
本発明の調節領域及びポリペプチドをコードするヌクレオチド配列が機能的に結合されるベクターの選択は、当業界で公知であるように、所望される機能的性質、例えば、複製またはタンパク質発現、及び形質転換される宿主細胞に直接依存し、これらは組換えDNA分子を構築する技術に固有の制限である。
【0094】
機能的な結合はDNAの一つのストランド(一本鎖形態または二本鎖形態を問わない)への好ましくは通常のホスホジエステル結合によるヌクレオチド配列の共有結合を表す。更に、ヌクレオチド配列の結合は本明細書に記載されたようなプロモーター及び下流のポリペプチドをコードするヌクレオチド配列との調節領域中の応答要素の如き機能要素の結合をもたらす。
【0095】
インサートを発現プラスミドに機能的に結合するための一つの典型的な方法は方向性連結反応による。これは方向性連結反応に適しているヌクレオチドの配列により行われる。このような配列は(1) 複製のために機能的に結合し、上流及び下流の翻訳可能なDNA配列を輸送し、(2) ベクターへのDNA配列の方向性連結反応のための部位または手段を与えるDNA発現ベクターの領域であるポリリンカーと普通称される。典型的には、方向性ポリリンカーは二つ以上の制限エンドヌクレアーゼ認識配列、または制限部位を形成するヌクレオチドの配列である。制限開裂後に、二つの部位は、翻訳可能なDNA配列がDNA発現ベクターにつながれる付着末端を生じる。二つの制限部位は、制限開裂後に、非相補性であり、それにより、カセットへの翻訳可能なDNA配列の方向性挿入を可能にする付着末端を与えることが好ましい。
【0096】
種々の宿主−発現ベクター系がヌクレオチド配列によりコードされたポリペプチドを発現するのに使用し得る。これらとして、微生物、例えば、ポリペプチドをコードするヌクレオチド配列を含む組換えバクテリオファージDNA、プラスミドDNAまたはコスミドDNA発現ベクターで形質転換されたバクテリア;ポリペプチドをコードするヌクレオチド配列を含む組換え酵母発現ベクターで形質転換された酵母;ポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV; タバコモザイクウイルス、TMV)で感染され、または組換えプラスミド発現ベクター(例えば、Tiプラスミド)で形質転換された植物細胞系;ポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、バキュロウイルス)で感染された昆虫細胞系;またはポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、レトロウイルス、アデノウイルス、ワクシニアウイルス)で感染された動物細胞系、または安定な発現について操作された形質転換された動物細胞系が挙げられるが、これらに限定されない。グリコシル化が重要であり得る場合、翻訳修飾及び後翻訳修飾を与える発現系、例えば、哺乳類発現系、昆虫発現系、酵母発現系または植物発現系が使用し得る。
【0097】
これらの系のいずれもが本発明の方法を実施するのに有益である。上記発現系のいずれかにより、選択された宿主がその後にペプチドのローディングに実際のメカニズムに応じて、即ち、内部または外部に応じて抗原プロセシング補助分子を用いて、または用いずに少なくとも一種のMHCクラスIIヘテロダイマー単独または少なくとも一種のアクセサリー分子と一緒の発現に使用される。
【0098】
使用される宿主/ベクター系に応じて、構成的プロモーター及び誘導プロモーター、転写エンハンサー要素、転写ターミネーター等を含む幾つかの好適な転写要素及び翻訳要素のいずれかが発現ベクター中に使用し得る(例えば、Bitterら,Methods in Enzymology, 153: 516-544, (1987) を参照のこと)。例えば、バクテリア系中のクローニングの場合、誘導プロモーター、例えば、バクテリオファージλのP1、Plac、Ptrp、Ptac(Ptrp−lacハイブリッドプロモーター)等が使用し得る。哺乳類細胞系中のクローニングの場合、哺乳類細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)または哺乳類ウイルスに由来するプロモーター(例えば、レトロウイルス長末端リピート;アデノウイルス後期プロモーター;ワクシニアウイルス7.5Kプロモーター)が使用し得る。組換えDNAまたは合成技術により生成されたプロモーターがまたポリペプチドをコードするヌクレオチド配列の転写を与えるのに使用し得る。
【0099】
バクテリア系では、幾つかの発現ベクターが本発明の方法に従ってポリペプチドをコードするヌクレオチド配列の発現に有利に選択し得る。例えば、多量が生成される場合、容易に精製し得る融合タンパク質産物の高レベルの発現を誘導するベクターが望ましいかもしれない。開裂部位を含んでタンパク質の回収を助けるように操作されるものが好ましい。このようなベクターとして、E.coli発現ベクターpUR278 (Rutherら, EMBO J., 2: 1791, (1983)) (そのポリペプチドをコードするヌクレオチド配列はハイブリッドポリペプチド−LacZタンパク質が生成されるようにLacZコーディング領域とインフレームでベクターにつながれてもよい);pINベクター(Inouye &Inouye, Nuc.Acids Res., 13: 3101-3109,(1985);Van Heeke &Schuster, J.Biol.Chem., 264: 5503-5509, (1989)) 等が挙げられるが、これらに限定されない。
【0100】
一実施態様において、使用されるベクターはバクテリア中のベクターの増殖を促進する原核生物配列、即ち、バクテリア宿主細胞に導入された時に染色体外で組換えDNA分子の自律複製及び維持を誘導する能力を有するDNA配列を含む。このようなレプリコンは当業界で公知である。
【0101】
また、原核生物レプリコンを含むこれらのベクターは典型的には本発明の組換えDNA分子の挿入に都合の良い制限部位を含む。このようなベクタープラスミドの代表はバイオラド・ラボラトリィズ(リッチモンド、CA)からのpUC8、ぷC9、pBR322、及びpBR329及びファーマシア(ピスキャットアウェイ、NJ)からのpPL、並びにストラタゲン(ラジョラ、CA)から入手し得るpブルースクリプト及びpBSである。また、本発明のベクターはMolecular Cloning: A Laboratory Manual,第二編, Maniatisら編集, Cold Spring Harbor, NY (1989) に記載されたλベクターを含むλファージであってもよい。
【0102】
別の好ましい実施態様において、本発明に使用するためのプラスミドベクターはまた真核生物細胞と適合性である。真核細胞発現ベクターは当業界で公知であり、幾つかの商業源から入手し得る。典型的には、このようなベクターは所望の組換えDNA分子の挿入に都合の良い制限部位を与え、更に先に説明したような真核生物細胞中で発現することができるコードされた遺伝子の発現のためのプロモーターを含む。このようなベクターの代表はpSVO及びpKSV−10 (ファーマシア)、及びpPVV−1/PML2d(インターナショナル・バイオテクノロジー社)、並びにpTDT1 (ATCC、No. 31255)である。
【0103】
加えて、真核生物プラスミドでは、真核生物細胞中でのみ発現される一つ以上の転写単位が存在する。真核生物転写単位は非コーディング配列及び選択可能なマーカーをコードする配列からなる。また、本発明の発現ベクターは正確かつ有効なポリアデニル化に必要とされる特異な配列要素を含む。加えて、成熟mRNAを発生するためのスプライシングシグナルがベクター中に含まれる。真核生物プラスミド発現ベクターはウイルスレプリコンを含むことができ、その存在がクローン化遺伝子の発現のレベルの増加を与える。好ましい複製配列がシミアンウイルス40またはSV40パポバウイルスにより与えられる。
【0104】
本発明に使用するための組換え分子を生成するのに好ましい発現系は昆虫系である。一つのこのような系において、オートグラファ・カリフォルニカ核多角体病ウイルス(AcNPV) が外来遺伝子を発現するためのベクターとして使用される。そのウイルスはスポドプテラ・フルギペルダ(Sf9) 細胞中で増殖する。本発明のポリペプチドをコードするヌクレオチド配列はそのウイルスの非必須領域(スポドプテラ・フルギペルダ中、例えば、ポリヘドリン遺伝子)にクローン化され、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置かれてもよい。ポリペプチドをコードするヌクレオチド配列の成功裏の挿入はポリヘドリン遺伝子の不活化及び非閉塞性(non-occluded)組換えウイルス(即ち、ポリヘドリン遺伝子によりコードされたタンパク質外殻を欠いているウイルス)の生産をもたらすであろう。次いでこれらの組換えウイルスは、挿入遺伝子が発現される細胞を感染するのに使用される。Smith ら, J.Biol.Chem., 46: 584, (1983); Smithの米国特許第4,215,051 号を参照のこと。
【0105】
好ましい実施態様において、宿主細胞集団はp25−lacZ (Bello 及びCouble, Nature, 346: 480 (1990) を参照のこと)またはpRmHa−1 、−2、もしくは−3 (Bunchら, Nucl.Acids Res., 16: 1043−1061 (1988) を参照のこと)のようなベクターに機能均等のベクターを含む適合性ベクターを必要とするドロソフィラ細胞培養物である。好ましい実施態様において、ベクターはpRmHa−3 であり、これが図1Cに示される。このベクターはメタロチオネインプロモーターを含み、これはMHC配列が挿入される部位の上流にあることが好ましく、ポリアデニル化部位が前記MHC配列の下流にあることが好ましい。昆虫細胞、特にドロソフィラ細胞が本発明によれば好ましい宿主である。節Dに更に記載されるようなシュナイダー−2(S2)細胞の如きドロソフィラ細胞はプロモーターの活性化に必要とされる必要なトランス作用性因子を有し、こうして更に好ましい。
【0106】
発現ベクターpRmHa−3 はバクテリアプラスミドpRmHa−1 (図1A)をベースとし、その後者はプラスミドpUC18をベースとし、ATCC(ロックビル、MD) に寄託され、受理番号37253 を有する。pRmHa−3 ベクターは図1Cに示されるようにプロモーター、EcoRI 部位及びStu I 部位を除去されたメタロチオネイン遺伝子の5'未翻訳リーダー配列を含む。それはまたポリアデニル化部位を含むドロソフィラADH遺伝子の3'部分を含む。それ故、クローン化DNAはメタロチオネインプロモーターにより転写調節され、ポリアデニル化される。pRmHa−1 プラスミドの構築がBunch ら, Nucl.Acids Res. 16: 1043−1061 (1988)に記載されてい。pRmHa−3 プラスミド及びpRmHa−2 プラスミド(その後者はEcoRI フラグメントとして除去されてもよいメタロチオネインプロモーター配列を有する)の構築が実施例に記載される。pRmHa−3 、本発明による使用に好ましいプラスミドに関して、PstI、SphI及びHindIIIがプロモーターフラグメント中にあり、それ故、特異ではない。XbaIはADHフラグメント(その3'末端から4塩基)にあり、また特異ではない。しかしながら、下記の制限部位がpRmHa−3 中で特異であり、本発明の組換え遺伝子のクローニングを促進する。EcoRI、SacI、KpnI、SmaI、BamHI、SalI、Hinc2、及びAccI。
【0107】
発現を誘導するために組換えウイルスまたはウイルス要素を使用する哺乳類細胞系が操作し得る。例えば、アデノウイルス発現ベクターを使用する場合、ポリペプチドのコーディング配列がアデノウイルス転写/翻訳調節複合体、例えば、後期プロモーター及び三部分リーダー配列に結合されてもよい。このキメラ遺伝子はその後にインビトロまたはインビボ組換えによりアデノウイルスゲノムに挿入されてもよい。ウイルスゲノムの非必須領域(例えば、領域E1またはE3)中の挿入は、生存しており、感染宿主中でポリペプチドを発現することができる組換えウイルスを生じるであろう(例えば、Logan &Shenk, Proc.Natl.Acad.Sci., USA,81: 3655-3659, (1984) を参照のこと)。また、ワクシニアウイルス7.5Kプロモーターが使用されてもよい(例えば、Mackett ら, Proc.Natl.Acad.Sci., USA,79: 7415-7419, (1982); Mackettら, J.Virol., 49: 857-864 (1984); Panicaliら, Proc.Natl.Acad.Sci., USA, 79: 4927-4931, (1982) を参照のこと)。染色体要素として複製する能力を有するウシパピローマウイルスをベースとするベクターが特に重要である(Sarver ら, Mol.Cell.Biol., 1: 486, (1981)) 。マウス細胞へのこのDNAのエントリーの直後に、プラスミドが細胞当たり約100 〜200 のコピーまで複製する。挿入されたcDNAの転写は宿主の染色体へのプラスミドの組込みを必要とせず、それにより高レベルの発現を生じる。これらのベクターは、プラスミド中に選択可能なマーカー、例えば、neo 遺伝子を含むことにより安定な発現に使用し得る。また、レトロウイルスゲノムが宿主細胞中の本発明のポリペプチドをコードするヌクレオチド配列の発現を導入し、誘導することができるベクターとしての使用のために修飾し得る(Cone &Mulligan, Proc.Natl.Acad.Sci., USA, 81: 6349-6353, (1984)) 。また、メタロチオニンIIAプロモーター及び熱ショックプロモーターを含むが、これらに限定されない誘導プロモーターを使用して、高レベルの発現が達成し得る。
【0108】
組換えタンパク質の長期の高収率の生産には、安定な発現が好ましい。ウイルス複製起点を含む発現ベクターを使用するのではなく、宿主細胞が適当な発現調節要素(例えば、プロモーター配列及びエンハンサー配列、転写ターミネーター、ポリアデニル化部位等)により調節されたcDNA、及び選択可能なマーカーで形質転換し得る。上記のように、組換えプラスミド中の選択可能なマーカーは選択に対する耐性を与え、細胞がプラスミドをそれらの染色体に組込み、成長してフォーカスを形成し、これらが順に細胞系にクローン化され、拡大されることを可能にする。
【0109】
例えば、外来DNAの導入後に、操作された細胞が濃縮培地中で1〜2日間増殖され、ついで選択培地に切り換えられる。単純ヘルペスウイルスチミジンキナーゼ(Wigler ら, Cell, 11: 223, (1977))、ヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ(Szybalska&Szybalski, Proc.Natl.Acad.Sci., USA, 48: 2026, (1962))、及びアデニンホスホリボシルトランスフェラーゼ(Lowy ら,Cell, 22: 817, (1980))遺伝子(これらは夫々tk−細胞、hgprT細胞またはaprT細胞中で使用し得る)を含むが、これらに限定されない幾つかの選択系が使用し得る。また、抗代謝産物耐性を与える遺伝子が選択の基礎として使用し得る。例えば、dhfr(これはメトトレキセートに対する耐性を与える(Wigler ら,Proc.Natl.Acad.Sci., USA, 77: 3567, (1980); O'Hareら, Proc.Natl.Acad.Sci.,USA, 78: 1527, (1981));gpt (これはミコフェノール酸に対する耐性を与える)(Mulligan &Berg, Proc.Natl.Acad.Sci., USA, 78: 2072, (1981)); neo ( これはアミノグリコシドG−418に対する耐性を与える)(Colberre-Garapinら, J.Mol.Biol., 150:1, (1981));及びhygro (これはヒグロマイシンに対する耐性を与える)(Santerre ら, Gene, 30: 147, (1984))の遺伝子。最近、付加的な選択可能な遺伝子、即ち、trpB(これは細胞にトリプトファンに代えてインドールを使用させる);hisD(これは細胞にヒスチジンに代えてヒスチノールを使用させる)(Hartman&Mulligan, Proc.Natl.Acad.Sci., USA, 85: 804, (1988)); 及びODC (オルニチンデカルボキシラーゼ)(これはオルニチンデカルボキシラーゼインヒビター、2−(ジフルオロメチル)−DL−オルニチン、DFMOに対する耐性を与える) (McConlogue L., In: Current Communications in Molecular Biology, Cold Spring Harbor Laboratory編集, (1987)が記載されている。
【0110】
原核生物発現ベクター及び真核生物発現ベクターの両方がベクター構築の当業者に良く知られており、Ausebel ら, In Current Protocols in Molecular Biology, Wiley and Sons, New York (1993) 及びSambrookら, Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory, (1989)により記載されている。
【0111】
本発明の組成物及び方法に使用するための組換えMHCクラスIIα鎖及びβ鎖、アクセサリー分子及び抗原プロセシング補助分子を生成するために、夫々のヌクレオチド領域が本明細書に記載された本発明の発現ベクターに機能的に挿入される。節Bに記載されたように、本発明の組換えポリペプチドをコードする核酸は幾つかの方法で得られ、その一つがPCR増幅による。ベクター配列に機能的に結合されるヌクレオチドセグメントの一つはMHCクラスIIα鎖及びβ鎖の少なくとも一部をコードする。完全α鎖及びβ鎖をコードする夫々のヌクレオチド配列がそれからの発現のための発現ベクターに別々に挿入されることが好ましい。しかしながら、また幾つかの非コーディングMHC配列を同様に含むベクターを構築することが実施可能である。アクセサリー分子及び抗原プロセシング補助分子をコードする配列が別の発現ベクターに同様に挿入される。
【0112】
また、本発明は同じベクター中に存在する一つより多いポリペプチドをコードする遺伝子の存在を意図しており、その発現はプロモーターの如き別の調節要素により誘導される。換言すれば、α鎖及びβ鎖の両方をコードする核酸がアクセサリー分子をコードする一種以上の核酸配列を用いて、または用いずに同じ発現ベクターに機能的に結合されてもよい。こうして、本発明の組換えタンパク質を生成するための発現ベクター構築の全ての可能な組み合わせが意図されている。
【0113】
上記の完全なコードする核酸配列に加えて、本発明の発現された組換えポリペプチドの可溶性形態が意図されている。可溶性形態は、それがトランスメンブランドメインまたはその他の機能性配置の前に挿入された“終止" コドンを含んで可溶性非膜固定可能なタンパク質を生じる点で非可溶性形態とは異なる。
【0114】
D.合成抗原提示細胞及びペプチド提示のためのマトリックス
1.合成抗原提示細胞及びマトリックス
本発明によれば、組換えMHCクラスIIヘテロダイマー及び少なくとも一種のアクセサリー分子が担体を含むマトリックスに機能的に結合され、その結果、MHC分子の細胞外部分に複合体形成されたペプチドで提示された時に、MHC クラスII及びアクセサリー分子がCD4+T細胞リンパ球の集団を活性化するのに充分な数で存在する。ペプチドは、それが担体に結合される前または後にMHCクラスIIヘテロダイマーに結合し得る。
【0115】
担体は多くの異なる形態をとり得る。それは固体担体、例えば、プラスチック材料または金属材料であってもよく、それは分離カラムに普通に使用されるような多孔質材料であってもよく、それはリポソームまたは赤血球であってもよく、またはそれは細胞または細胞フラグメントであってもよい。以下に更に詳しく説明されるように、細胞が担体として利用できる場合、MHCクラスII分子及びアクセサリー分子がその細胞上の提示のために、または別の細胞を含み得る別の担体上の提示のために細胞により生成し得る。
【0116】
前者の状況では、MHC分子はその後にMHCクラスIIの可溶性形態中に存在しない細胞質ドメインではないとしても少なくともトランスメンブランドメインにより細胞に結合される。後者の状況では、MHCクラスII分子及びアクセサリー分子の細胞外部分が担体に固定された抗体に反応するエピトープを用意することにより担体に結合し得る。加えて、MHC分子またはアクセサリー分子は担体の部分を形成する際にニッケルと反応する(His)6で生成され、またはそれに結合し得る。MHC分子を担体に固定または結合するためのその他の手段が当業界で公知である。
【0117】
先に説明したように、担体は細胞膜または全細胞であってもよい。このような場合、真核生物細胞系がMHCクラスIIに関するペプチドをT細胞リンパ球に提示するのに使用するための合成抗原提示細胞系になるように修飾される。エンプティーMHC分子は熱不安定性であるので、細胞培養物は変温性であることが好ましく、種々の細胞系が以下に詳しく説明される。
【0118】
本発明の好ましい細胞系は培養中に連続増殖することができ、かつ細胞表面で哺乳類MHCクラスII分子及びアクセサリー分子を発現することができる。バクテリア細胞系、酵母細胞系、昆虫細胞系、及び哺乳類細胞系を含む種々の形質転換された細胞もしくは細胞系または未形質転換細胞もしくは細胞系のいずれもがこの目的に適している(例えば、種々の細胞系、例えば、E.coli及びサルコミセス・セレビジエを培養し、使用するための要約及び操作について、Current Protocols in Molecular Biology, John Wiley&Sons, NY (1991) を参照のこと)。
【0119】
細胞系は真核細胞系であることが好ましい。細胞系は変温性(即ち、哺乳類細胞系よりも温度攻撃に対し感受性ではない)であることが更に好ましい。それは昆虫細胞系であることが更に好ましい。蛾(ATCC CCL 80) 、アワヨトウガ(ATCC CCL 1711) 、蚊の幼虫(ATCC 系列CCL 125 、CCL 126 、CRL 1660、CRL 1591、CRL 6585、CRL 6586) 、カイコ(ATCC CRL 8851) 及びチョウ(スポドプテラ・フルギペルダ(Sf9細胞、ATCC CRL 1711)を含む種々の昆虫細胞系が本発明による使用に利用できる。好ましい実施態様において、細胞系はドロソフィラ細胞系、例えば、シュナイダー細胞系である(Schneider, J.Embryol.Exp.Morph., 27: 353-365 (1972)) 。細胞系はM3培地中の増殖に適したシュナイダー2(S2)細胞系(S2/M3) であることが好ましい(Lindquistら, Drosophila Information Service, 58:163 (1982)を参照のこと)。シュナイダー2(S2)細胞はブタペスト条約要件に従って1992年2月18日にATCC(ロックビル、MD) に寄託され、受理番号CRL 10974を指定された。
【0120】
本発明の合成抗原提示細胞を生成するため、一種以上のアクセサリー分子と一緒に選択されたMHCクラスIIヘテロダイマーの発現を誘導するための一種以上の発現ベクターがレシピエント宿主細胞に導入される。加えて、別の実施態様において、HLA−DM及び非変異鎖を含む抗原プロセシング補助分子を発現するためのベクターがまたレシピエント細胞に導入される。上記のための遺伝子が節B及びCに記載された。
【0121】
こうして、合成抗原提示細胞またはマトリックスを調製するために、本発明の組換えポリペプチドをコードする発現ベクターが選択された宿主細胞にトランスフェクト、即ち、導入される。発現ベクターの選択並びにその構築は以下に繰り返して言われるように先に説明したようなCD4+活性化に所望の成果に依存する。トランスフェクション(また、形質転換と称される)はリン酸カルシウム方法、DEAE−デキストラン方法、安定移入方法、エレクトロポレーションを含む多数の方法により、またはリポソーム媒介方法により行われてもよい。多数の書籍が入手でき、これらは既知のトランスフェクション方法及びヌクレオチドを細胞に導入するためのその他の操作を記載している。例えば、Current Protocols in Molecular Biology, John Wiley &Sons, NY (1991) を参照のこと。一種以上のベクターの導入後に、レシピエント細胞は形質転換されると言われ、その選択は一時的または安定のいずれであってもよい。
【0122】
細胞の培養物が最初に樹立される。細胞系がトランスフェクションについて選択される。何となれば、それは導入される遺伝子の少なくとも一つを欠いているからである。昆虫細胞が有利であることがわかった。何となれば、それらが変温性であるからだけでなく、それらがこれらの遺伝子及びメカニズムを欠いているからであり、そうしないとペプチドに結合されたMHC分子を生成するであろう。これはペプチド結合MHC分子の生成、及びエンプティーMHC分子の生成に対する更に大きな調節を可能にする。
【0123】
次いで選択された細胞が第一プロモーターに機能的に結合された発現可能なMHCクラスIIα鎖遺伝子及び第二プロモーターに機能的に結合された発現可能なMHCクラスIIβ鎖遺伝子を含む発現ベクターの導入により形質転換される。ベクター中の第三プロモーターに機能的に結合された第一の発現可能なアクセサリー分子遺伝子がまた上記細胞に導入される。更に別の実施態様において、ベクター中の第四プロモーターに機能的に結合された発現可能な抗原プロセシング補助遺伝子が上記細胞に導入される。
【0124】
更に好ましい実施態様において、ベクターは節Cに記載されたドロソフィラ発現プラスミドpRmHa−3 を含み、これには上記組換えタンパク質をコードする発現可能なヌクレオチド配列が本明細書に開示された技術を使用して挿入されていた。MHCクラスII鎖をコードするヌクレオチド配列、少なくとも一種のアクセサリー分子をコードするヌクレオチド配列及び抗原プロセシング補助分子をコードするヌクレオチド配列は培養された細胞に個々に同時トランスフェクトされる別々の発現プラスミドに機能的に結合されることが好ましい。また、ヌクレオチド配列は同じ発現プラスミド中の別々のプロモーターに機能的に結合され、その同じプラスミドにより同時トランスフェクトされてもよい。MHCクラスIIα鎖及びβ鎖は異種からのものであることが好ましく、定温動物、例えば、哺乳類、最適にはヒトからのものであることが更に好ましい。
【0125】
遺伝子の少なくとも一種、特に、MHCクラスII鎖遺伝子は誘導プロモーターに結合されることが好ましい。これはMHC分子の生成に調節を与え、その結果、関係するペプチドが内部または外部で利用でき、培養物中で提示されて生成されたMHC分子と反応する時にのみそれらが生成される。これは望ましくないMHC分子/ペプチド複合体を最小にする。
【0126】
こうして、好ましい細胞系は夫々が第一プロモーター及び第二プロモーターに夫々機能的に結合されたMHCクラスIIα鎖及びβ鎖を含む別々のベクターを有する変温性細胞系である。プロモーターはMHCクラスII鎖の発現を調節するように誘導性であることが好ましい。加えて、細胞は第三プロモーターに機能的に結合された第一の発現可能なアクセサリー分子遺伝子を少なくとも含む第三ベクターを含む。更に別の実施態様において、細胞はまた第四プロモーターに機能的に結合された発現可能な抗原プロセシング補助遺伝子を含む第四ベクターを含む。細胞はエンプティーMHC分子を集合し、それらを細胞表面に提示し、その結果、特別なMHCクラスIIハプロタイプまたは変異体対立遺伝子に特異性のペプチドが所望に応じて選択し得ることが好ましい。
【0127】
一種以上の特別なアクセサリー分子をコードする遺伝子と一緒に使用するための適合性MHCクラスIIα鎖遺伝子及びβ鎖遺伝子の選択は所望されるT細胞活性化プロフィールに依存する。例えば、実施例に記載されるように、組換えB7.1またはB7.2は単独で、またはドロソフィラAPCの表面で発現された組換えマウスIAdMHCクラスIIと一緒に、IL−4及びIL−10 の増大された産生のTh2プロフィールを有するCD4+T細胞の増殖をもたらした。対照的に、B7.1またはB7.2が同じMHC分子とともにICAM−1と一緒にドロソフィラAPCの表面で発現された時、CD4+T細胞の活性化は増大されたIL−2産生並びに減少されたIL−4及びIL−10産生を有するTh1プロフィールをもたらした。
【0128】
こうして、本発明は本発明のアクセサリー分子のいずれか一つとのMHCクラスIIハプロタイプヘテロダイマーのあらゆる組み合わせを細胞表面に有する合成APCの生成を意図している。特に好ましい組み合わせはB7.1またはB7.2を含む同時刺激分子、ICAM−1、ICAM−2、ICAM−3またはLFA−3 を含む付着分子、またはFasリガンド(FasL)を含むサバイバル分子とともにMHCクラスIIを含む。上記のように、アクセサリー分子の夫々のカテゴリーの一種より多くが、例えば、B7.1及びB7.2として同時発現し得る。別の好ましい実施態様は、異なるカテゴリーの2種のアクセサリー分子が細胞表面で同時発現される順列を含む。換言すれば、同時刺激分子と一緒の付着分子、及びサバイバル分子と一緒の付着分子、サバイバル分子と一緒の同時刺激分子。更に別の実施態様において、本明細書に記載された異なるカテゴリーの3種のアクセサリー分子がAPC表面で同時発現される。また、これらの実施態様の全てにおいて、特別なカテゴリーの一種より多くの員がその他のカテゴリーの一種より多くの員と一緒に発現し得ることが意図されている。特別な選択された組み合わせは、それらが発現される細胞の表面で有効であり、または本発明のマトリックスの表面に固定された時に有効である。調製される実際の組み合わせは抗原ペプチドと複合体形成された発現されたMHCクラスII分子に鑑みてT細胞活性化の成果に基いて選択される。こうして、MHCクラスII/ペプチドの複合体に応じて、T細胞活性化の成果は同じ発現されたアクセサリー分子を有するにもかかわらず異なり得る。
【0129】
更に別の実施態様において、抗原プロセシング補助遺伝子は上記組み合わせのいずれかで同時トランスフェクトされて増進された内部ペプチドプロセシング及びローディングを与える。こうして、この局面はその後のペプチド複合体形成のための細胞表面におけるエンプティーMHCクラスII分子の生成を伴わない。むしろ、非変異鎖、HLA−DMまたはリソソーム酵素の発現が利用されて細胞内在化後のタンパク質分解ペプチドフラグメントの最適のプロセシング及びローディングを可能にする。こうして、本発明のAPCは細胞内または細胞外でローディングされる組換えMHCクラスIIヘテロダイマーを有するモチーフで機能し得る。
【0130】
成功裏に形質転換された細胞、即ち、本発明のヌクレオチド配列の発現を誘導することができる少なくとも一種の発現ベクターを含む細胞は公知の技術により同定し得る。例えば、本発明のcDNAまたはrDNAの導入により得られる細胞はクローン化されて個々のコロニーを生じ得る。これらのコロニーからの細胞が回収され、溶解され、それらのDNA含量がSouthern, J.Mol.Biol. 98: 503(1975)により記載された方法の如き方法を使用してrDNAの存在について試験し得る。rDNAの存在について直接分析することに加えて、成功裏の形質転換またはトランスフェクションは、rDNAがアクセサリー分子の主題MHC クラスIIタンパク質の発現を誘導することができる時に公知の免疫学的方法により確認し得る。例えば、一種以上の発現ベクターで成功裏に形質転換された細胞は、適当な抗体、例えば、特別なハプロタイプの抗クラスIIを使用して容易に測定される特別な抗原性を示すタンパク質を生産し得る。加えて、成功裏の形質転換/トランスフェクションは上記されたようなマーカー配列、例えば、ネオマイシン耐性を有する付加的なベクターの使用により確かめられてもよい。
【0131】
また、培養物は安定であり、低下した温度で持続成長することができることが好ましい。例えば、培養物はほぼ室温、例えば、約24〜27℃で維持されることが好ましい。別の実施態様において、培養物は、特にCD4+T細胞の活性化のプロセス中に高温で維持される。こうして、本発明の培養物は約30℃〜約37℃の温度攻撃に耐えることができることが好ましい。
本発明の少なくとも一種のアクセサリー分子及び必要により抗原プロセシング補助分子と一緒にエンプティーまたはMHCクラスII分子の発現のための培養物を調製するために、培養物は前もって決められた期間にわたって、例えば、CuSO4誘導による刺激を最初に必要とし得る。好適な誘導期間、例えば、約12〜48時間後に、ペプチドが前もって決められた濃度(例えば、約0.2 μg/ml〜20μg/ml)で添加し得る。内部ローディング及び外部ローディングの両方のためのタンパク質及びペプチドは以下に説明されるように調製される。更なるインキュベーション期間、例えば、27℃で約12時間後に、培養物がCD4+T細胞の活性化における使用に供される。この追加のインキュベーション期間は短縮され、またはおそらく省かれてもよいが、培養物は静止または未感作CD4+T細胞の添加の前に或る時間にわたってインキュベートされる場合に温度攻撃に対し次第に安定になる傾向がある。例えば、ペプチドが添加された本発明の培養物は、37℃で延長された期間にわたってインキュベートされる時でさえもペプチドローディングされたMHCクラスII分子のかなりの量を発現することができる。
【0132】
形質転換された宿主細胞の培養に有益な栄養培地は当業界で公知であり、多数の商業源から入手し得る。宿主細胞が哺乳類である実施態様において、“無血清"培地が使用されることが好ましい。
【0133】
得られる組換え発現されたMHCクラスII分子は特別なペプチドに結合し、APCの表面で少なくとも一種のアクセサリー分子とともに充分な数で存在してMHCクラスII/ペプチド複合体に対しT細胞リンパ球の集団を活性化する。
【0134】
細胞系が一種以上の所望の分子を既に生産する場合、培養物を細胞中に欠けている遺伝子について発現可能な遺伝子でトランスフェクトすることのみが必要である。例えば、細胞がそれらの表面でMHC分子を既に提示する場合、培養物をアクセサリー分子について発現可能な遺伝子を含むベクターでトランスフェクトすることのみが必要である。
【0135】
既に説明したように、タンパク質またはペプチドは、細胞が内部プロセシングのためにMHCクラスII分子を生産している時に細胞培養物に導入し得る。浸透圧ショックの如き方法により、ペプチドが細胞に導入でき、生産されたMHC分子に結合する。また、特に変温性細胞系の場合、MHC分子は細胞表面でエンプティーに提示されるであろう。次いでペプチドが培養物に添加されて、所望されるようにMHC分子に結合し得る。簡素化のために、一種のペプチドが本明細書に記載されるが、本発明の方法は本発明の治療方法に使用するための新規な抗原ペプチドを同定するためのペプチドライブラリーのスクリーニングを意図している。
【0136】
細胞表面にMHCクラスIIヘテロダイマー及び少なくとも一種のアクセサリー分子を有する細胞が生産された後、細胞が凍結乾燥されてCD4+T細胞リンパ球の集団を活性化するのに使用するための細胞フラグメントを生成し得る。
【0137】
また、細胞のトランスフェクトされた培養物がMHCクラスII及びアクセサリー分子の細胞外部分を生成するのに使用される。固体担体の如き担体と一緒の細胞外部分の使用は生産の或る種の利点を有する。生細胞が合成抗原提示細胞を得るのに使用される場合、少なくとも3種の遺伝子、MHCクラスIIヘテロダイマーを生成するための2種とアクセサリー分子のための一種が細胞に導入される必要がある。しばしば、抗生物質耐性のための追加の遺伝子がまたトランスフェクトされる。
【0138】
固体担体系が使用されている場合、一種の細胞系がMHCクラスII分子の細胞外部分を生成するのに使用され、一方、別の細胞系がアクセサリー分子の細胞外部分を生成するのに使用される。次いでMHCクラスII部分及びアクセサリー分子部分がそれらの夫々の培養物から回収される。次いでこれらの分子がT細胞の集団を活性化するのに充分な数で適当な担体に結合される。生産の観点から、2種の異なる培養物が使用し得るが、同じ培養物を使用することが可能であるが、培養物がアクセサリー分子の細胞外部分を発現するための追加の遺伝子でトランスフェクトされることを必要とする。
【0139】
この実施態様の更なる改良は、発現可能な第二のアクセサリー分子遺伝子でトランスフェクトされる細胞の第三培養物を用意することである。例えば、細胞の第二培養物は同時刺激分子の細胞外部分を生産し、一方、細胞の第三培養物は付着分子の細胞外部分を生産する。付着分子部分が回収され、担体に結合される。担体に結合されるMHCクラスIIヘテロダイマーの細胞外部分を調製する際に、可溶性分子が既に説明されたように調製される。これらの分子は一般にMHC分子中にトランスメンブラン及び細胞質ドメインを欠いている。
【0140】
2.ペプチド
潜在的能力をもつMHCクラスIIに特異的なペプチドとして機能する関連ペプチドフラグメントを生成するために、ウイルス抗原の他に実質的に全ての細胞タンパク質を用いることができる。本発明の方法および組成物は、CD4+ 細胞を特異的に活性化させる能力が増強されたMHCクラスII分子を提供する。
【0141】
本発明のペプチドはMHCクラスII分子と結合する。この結合は、インビトロと同様にインビボでも作り出すことができる生物学的条件下で生じる。本発明の実施に当該ペプチドの結合の正確な性質を知ることは必要ではない。
【0142】
MHCクラスII分子と結合するペプチドの長さは様々で、さらにそのアンカー残基の当該ペプチド末端からの距離も様々である。ある実施態様では、MHCクラスII分子に付加するために調製されるペプチドは単一種に由来する。すなわち、MHCに付加される全てのペプチドのサイズおよび配列は同一で、単抗原ペプチド付加MHCクラスII分子を生じる。また別の実施態様では、ペプチドは均質ではなく、ペプチドのランダムライブラリーを含み、先に述べたように所望のT細胞活性化プロフィールを生じる固有のペプチドの選別を可能にすることができる。合成ランダムペプチドライブラリーの生成およびスクリーニングは当業者には周知で、米国特許第5556762 号明細書、同5510240 号明細書、同5498530 号明細書、同5432018 号明細書、同5382513 号明細書、同5338665 号明細書および同5270170 号明細書(これらの文献は参照により本明細書に含まれる)に記載されている。
【0143】
ペプチドは種々の手段によって細胞に与えることができる。好ましくは、ペプチドは、細胞内ペプチドプールに入ることができるような態様で与えられる。例えば、ペプチドは浸透圧付加によって与えることができる。典型的には、ペプチドは培養液に添加される。ペプチドは、完全なポリペプチドまたはタンパク質の形状で培養に添加される。後者は、その後細胞の一連の作用(例えば酵素分解)によって分解される。また別には、完全なポリペプチドまたはタンパク質は、細胞培養に添加する前に例えば化学的消化(例えば臭化シアン)またはプロテアーゼ(例えばキモトリプシン)のような他の手段によって分解できる。他の実施態様では、ペプチドは、特定のMHCクラスIIハプロタイプと結合したより小さなセグメント(これは抗原性アミノ酸配列を含んでいてもいなくてもよい)として提示される。
【0144】
好ましくは、十分量のタンパク質またはペプチドは、細胞培養または合成マトリックスに添加され、MHCクラスII分子が結合して、続いて本発明の合成APCまたはマトリックス上に高密度でペプチド(好ましくはMHCヘテロダイマーの各々と結合した同じ種類のペプチド)を提示することを可能にする。
【0145】
本発明の別の実施態様では、ペプチドは、細胞によって発現されるMHC分子の耐熱性を強化するために、本発明のトランスフェクト細胞に添加される。上記に記したようにペプチドは好ましくは培養液に添加される。MHCクラスII分子に結合する抗原性ペプチドは、MHC分子に耐熱性を付与し、また細胞表面発現を高めるために役立つ。MHC分子に結合するペプチドを添加された培養は、したがって添加ペプチドのない培養よりも温度によるチャレンジに極めて耐性を有する。
【0146】
E.CD4+T細胞反応を変化させる方法
1.Th1およびTh2CD4+T細胞仲介疾患
未感作T細胞を所望の活性化T細胞型に誘導するか、または活性化T細胞のエフェクター機能をTh1型からTh2型にもしくはその逆に偏向させることは、特にCD4+T細胞仲介疾患を処置する治療方法について本発明の目的の1つである。
【0147】
分裂中のCD4+T細胞が炎症性T細胞またはヘルパーT細胞のどちらに分化するかは、感染体によって産生されるサイトカイン、主にIL−12およびIL−4、アクセサリー分子の影響、並びにMHCクラスII/ペプチド複合体の性質によって左右される。先に考察したように、細胞介在性免疫は、最初に細胞内寄生体(例えばマイコバクテリウム(Mycobacterium)、リーシュマニア(Leishmania)、ニューモシスチス(Pneumocystis)などの寄生体を含む)に誘導されたTh1炎症性細胞によって活性化されたマクロファージによる細胞内病原体の破壊を必要とする。対照的に、液性免疫は、最初に細胞外病原体(クロストリジウム(Clostridium)、スタフィロコッカス(Staphylococcus)、ストレプトコッカス(Streptococcus)、ポリオウイルス、ニューモシスチス(Pneumocystis)などを含む)に誘導されたヘルパーT細胞によって活性化されたB細胞による抗体の産生に依存する。
【0148】
例えば、ある種のタイプの感染(例えばリーシュマニア)からの回復は、もっぱらIL−2/IFN−γの産生を必要とする。リーシュマニアに対するTh2応答が上昇したマウスは感染を抑制できず、最終的に死亡する。Th2型応答の不適切なサイトカイン産生は、しばしばアレルギー型疾患(例えば喘息および接触過敏症)と密接に関係する。
【0149】
ヒトの疾患と歪んだサイトカイン産生パターンとの最も強い関わりは、おそらくTh1応答およびTh1型サイトカインの自己免疫疾患との関わりであろう。実験モデルにおける強力な証拠は、多くのタイプの自己免疫疾患(糖尿病、多発性硬化症の実験モデル、自己免疫性甲状腺炎など)はTh1型CD4+T細胞によって仲介されること示唆している。これらのモデルにおけるTh2付随サイトカイン(例えばIL−4)の発現は自己免疫疾患の進行に干渉する。Th2型サイトカインはTh1型細胞の反応を低下させ、一方、Th1型サイトカインはTh2型応答の進行に拮抗する。
【0150】
特定の活性化T細胞サブセットと特定の疾患とのつながりという観点から、したがって所望のT細胞サブセットへCD4+T細胞の分裂および活性化、すなわち疾患の流れを変えるために極めて有益である一連の作用を誘導することができることが必要である。有望な解決の1つは、場合によってアレルギーまたは自己免疫疾患をもつ対象者から最初に単離したCD4+T細胞をインビトロで活性化し、好ましいサイトカインプロフィールを分泌する細胞を産生することである。得られた活性化T細胞を続いて当該対象者に再導入し、疾患の流れを変え、長期的治癒を提供する。
【0151】
また別の実施態様は、Th1またはTh2応答の何れか(その者にとって応用可能ないずれか)の進行に対して潜在的な反応性をもつ個体を" 免疫する”能力を目的とする。換言すれば、特定のT細胞サブセットの選択的誘発は、未感作細胞が望ましくない表現型へと変化することを抑制することによって達成できる。例えば、潜在的にアトピーをもつ個体の場合、有害なTh2応答を防止することはヘツェルとラム(Hetzel & Lamb, Clinical Immunol. Immunopath., 73:1−10(1994))が記載したように有益であろう。
【0152】
さらに別の実施態様では、本発明の組成物および方法は、未感作T細胞を所望の表現型へ発展させるために活発に刺激するために有用である。現時点で存在する治療モデルには、ヘツェルとラム(上掲書)が概説したように、抗サイトカイン抗体と担体タンパク質の使用、外因性サイトカインの作用を長く続かせるためのイディオタイプ/GM−CSF融合タンパク質ワクチンの使用、選択アジュバントの使用、リポソーム被包化アレルゲンの使用、ペプチド類似体の使用などが含まれる。しかしながら、著者らは、インビボでのアレルゲンに対する長期的Th2応答については、Th2応答の所望のダウンレギュレーションを達成する可能性に対する実験的データは殆ど存在しないと述べている。
【0153】
前述の観点から、本発明の組成物および方法は、上記で考察した治療的介入を達成する価値ある手段を提供する。本発明は、所望の治療用サイトカインプロフィールを産生するCD4+T細胞サブセットの生成を再現することができる活性化条件を特定することを可能にする。特定のサイトカインの発現は、特定の抗原提示細胞(APC)およびそれらの付随アクセサリー分子と連携している。APCによって産生されるサイトカインおよび統合的に発現されるアクセサリー分子は両方とも、多数の要素(抗原のタイプ、T細胞レセプター(TCR)−抗原相互作用の親和性、抗原濃度などを含む)によってそれ自体調節されるので、抗原提示の際のT細胞活性化の発現を予測することはこれまでのところ極めて困難である。実際、新たなアクセサリー分子がインビボでの活性化過程について候補に挙げられたので、多くの異なる分子がT細胞反応の調節に必要とされ、連携的態様で作用してT細胞活性化を達成することがますます明らかになってきた。
【0154】
本発明は、好ましくは非哺乳類昆虫細胞で発現する限定されたアクセサリー分子と結合したMHCクラスII分子を中性的バックグラウンドで提示する合成APCの生成を提供する。本発明のMHCクラスII/アクセサリー分子組成物を発現および提示する担体として昆虫細胞を使用する利点は、昆虫細胞は調節性サイトカインを内因的に産生することはなく、さらに哺乳類のアクセサリー分子を発現しないということである。このことによって、T細胞反応を変化させることができる多数の分子を発現させる哺乳類APCを用いる場合の固有の非予測性が克服される。したがって本発明は、他の手段では得ることができない再現性および予測性を可能にする選択された組み合わせでの発現に向けて個々の提示分子およびアクセサリー分子を単離する能力を提供する。
【0155】
2.治療方法
上記で述べたように、本発明は、CD4+T細胞を活性化して武装エフェクターT細胞サブタイプに分化させる方法に関する。本方法は、ペプチドを結合することができる組換えMHCクラスIIヘテロダイマーを外部表面に定着させた合成APCまたはマトリックスを提供することに関する。この組成物はまた、当該細胞表面に提示された少なくとも1つのアクセサリー分子を有する。未感作または活性化CD4+ T細胞は処置される個体から取り出すことによって得ることができる。続いて、T細胞を活性化して所望のT細胞表現型に分裂させ分化させるために十分な時間当該CD4+T細胞を抗原提示細胞と接触させる。
【0156】
活性化CD4+T細胞をこの細胞株から分離し、許容可能な担体中に懸濁して当該個体に投与する。
【0157】
ヒトの遺伝子を用いるのが好ましく、したがってヒト分子類似体が製造される。先の米国特許第5314813号明細書に示されたように、ネズミの系はT細胞活性化の実施をテストするために有用なモデルを提供し、この工程をヒトの系で応用することができることを示した(Sykulevら、Immunity, 1:15-22(1994)もまた参照のこと)。
【0158】
a.休止または活性化CD4+T細胞の単離
MHCクラスIIに関して提示される特異的抗原を標的とするように未だ活性化されていない休止(または未感作)CD4+T細胞を活性化CD4+T細胞と同様に個体から抽出し、本発明の形質転換培養と保温するかまたはこれら培養に暴露する。未感作細胞は、主に細胞表面マーカーCD455RAおよびCD45を基準にプライマー細胞と区別できる。
【0159】
さらにまた、特異的に活性化されるべきCD4+ 細胞の能力と干渉する可能性がある他の処置または療法を開始する前に、個体からCD4+ 細胞を得ることが望ましい。例えば、自己免疫疾患の患者を治療しようとしている場合は、ステロイド治療のような補助的治療の開始前、または当該患者が全く治療を受けていない期間中に細胞および培養サンプルを得ることが好ましい。
【0160】
患者のT細胞仲介免疫反応を変化させるためにCD4+T細胞を活性化させるとき、適切な対抗治療(対抗T細胞表現型およびサイトカインの産生を含む)を開始するために、当該患者を先ず分析してT細胞表現型の症状を調べなければならない。サイトカインプロフィールは、ATCCから入手可能な抗サイトカイン抗体を用い、さらに米国特許第5405751号明細書、同第5322787号明細書および同第5209920号明細書(これらの文献は参照により本明細書に含まれる)に記載された方法によって確定される。好ましいサイトカイン分析には、インターロイキン−2(IL−2)、インターフェロン−γ(IFN−γ)、腫瘍壊死因子(TNF)、インターロイキン−4(IL−4)、インターロイキン−10(IL−10)などが含まれる。先に考察したように、特定のサイトカインプロフィールがT細胞表現型および疾患に付随している。
【0161】
特に、当該症状が自己免疫疾患(多発性硬化症、自己免疫性甲状腺炎、全身性紅斑性狼瘡、重症筋無力症、クローン病および炎症性大腸疾患を含む)の場合、サイトカインプロフィールは、IL−2、IFN−γおよびTNFの増加を特徴とするTh1型応答によって生じる。対照的に、当該症状がアレルギー(例えば喘息および接触過敏症)である場合は、サイトカインプロフィールは、IL−4およびIL−10の増加を特徴とするTh2型応答によって生じる。
【0162】
患者のサイトカインプロフィールおよび症状を分析した後、下記に述べるように本発明の合成APC、細胞フラグメントまたはマトリックスと患者から単離されたCD4+T細胞をインビトロで十分な量で十分な時間接触させ、当該接触細胞を機能的に対抗するサイトカインプロフィールを産生する活性化CD4+T細胞に分裂し分化するように誘発する。すなわち、患者がTh1型応答者としての特徴を有する場合、患者のCD4+T細胞に提示されるべき抗原は、細胞がTh2型に分裂し分化するように誘発するために必要なものであろう。これと反対の治療様式は、患者がTh2型応答を特徴とする場合に実施される。したがって、いったん対抗する活性化細胞が患者に戻された場合、T細胞表現型の反応における変化を達成する治療成果が得られる。
【0163】
リンパ球の抽出方法および培養方法は周知である。例えば、米国特許第4690915号明細書(Rosenberg)には、リンパ球フェレーシスによって大量のリンパ球を得る方法が記載されている。用いられる適切な培養条件は哺乳類細胞についてであり、典型的には37℃で実施される。
【0164】
CD4+ 細胞培養の分離および/または濃縮のために種々の方法がまた用いられる。細胞の分離のためのいくつかの一般的方法には、特異的に被覆した表面に間接的に細胞を結合させるものが含まれる。別の例では、CD4+ 細胞を含むヒトの末梢血リンパ球(PBL)がフィコールハイパーク(Ficoll-Hypaque)勾配遠心沈殿(Pharmacia, Piscataway, ニュージャージ)によって単離される。PBLリンパ芽球はその後直ちに用いてもよいが、また10%DMSO(Sigma Chemical Co., セントルイス、ミズーリ)含有FBS(これは細胞生命活性およびリンパ球機能を保護する)中で凍結させた後液体窒素で保存してもよい。
【0165】
前駆体細胞培養を分離および/または濃縮するまた別の方法には正および負の選別方法が含まれる。正の選別方法については、リンパ球濃縮PBL集団を全血から調製した後、CD4補助レセプター抗原を標的とする親和性(アフィニティー)を基準にした分離技術によってCD4+ リンパ球の亜集団を当該集団から調製する。これらのアフィニティーを基準にした技術には、蛍光活性化細胞分類(FACS)、細胞粘着、磁性ビーズ分離などの方法が含まれる。(例えば以下の成書を参照されたい:Scher & Mage,「基礎免疫学(Fundamental Immunology)」、W.E. Paul 編、pp.767-780, River Press 刊、ニューヨーク(1984))。アフィニティー法は、アフィニティー試薬の供給源として抗CD4補助レセプター抗体を利用する。また別には、CD4レセプターの天然のリガンドまたはリガンド類似体をアフィニティー試薬として用いてもよい。本方法で使用できる種々の抗T細胞抗体および抗CD4単クローン性抗体は、アメリカ菌培養収集所(ロックビル、メリーランド)およびファーミンゲン(Pharmingen, サンディエゴ、カリフォルニア)を含む多様な市販源から入手できる。
【0166】
負の選別方法は、CD4+細胞から非CD4細胞を除去することによって実施される。この技術ではリンパ球フェレーシスを受けた患者のTおよびB細胞集団からCD4+細胞が濃縮される。目的とする抗原にしたがって適切な抗体は異なるであろう。例えば、ヒトTリンパ球、ヒトT細胞サブセットおよび活性化T細胞と反応するものとして、それぞれ単クローン性抗体OKT4(抗CD4、ATCC No.CRL8002)、OKT5(ATCC No.8013および8016)、OKT8(抗CD8、ATCC No.CRL8014)およびOKT9(ATCC No.CRL8021)がATCCの細胞株およびハイブリドーマカタログ(ロックビル、メリーランド)で特定できる。他の種々の抗体もまたT細胞種(前駆体細胞並びに未感作および活性化メモリー成熟末梢T細胞を含む)の特定および単離のために利用できる。
【0167】
b.CD4+ 細胞のインビトロ活性
特定のCD4+T細胞表現型の生成のためのインビトロ条件を最適化するために、抗原提示細胞培養は適切な培養液で維持される。好ましくは、完全な細胞である本発明の担体を用いる場合は、抗原提示細胞はショウジョウバエ(Drosophila、ドロソフィラ)細胞で、この細胞は好ましくは血清非含有培養液(例えばエクセル(Excell)400)で維持される。しかしながらまた別の実施態様では、担体が細胞フラグメントまたは先に述べたように人工的担体マトリックスである場合は、培養液は標的細胞の生命活性を維持するために選択される。
【0168】
本発明の合成APC、細胞フラグメントまたはマトリックスを活性化させるべきT細胞と接触させる前に、ある量の抗原性ペプチドを当該APCまたはマトリックスに十分量で提供し、当該APCまたはマトリックス表面での発現のためにヒトMHCクラスII分子上に付加させる。先に考察したように、ペプチド付加は細胞内または細胞外で生じるであろう。したがって両態様が本明細書で述べる活性化過程に含まれるが、単純化のためにペプチドの付加はMHCクラスIIヘテロダイマーへのペプチドの提示として包括的に述べる。付加については既に考察したとおりである。さらにまた、ペプチドライブラリーと同様に個々のペプチドも、先に考察したように活性化CD4+T細胞の調製での使用が意図されている。本発明にしたがって、十分量のペプチドとは、約200から約500000、好ましくは約200から1000以上のMHCクラスII分子が各合成APCまたはマトリックスの表面で発現されるべきペプチドで付加される量である。好ましくは、上記の組成物は0.2μg/mlから20μg/mlまでのペプチドとともに保温される。
【0169】
続いてCD4+ 細胞を活性化させるために十分な時間、単離CD4+ 細胞は、合成APCまたはマトリックス上に発現された適切なペプチド付加MHCクラスIIヘテロダイマーと培養される。したがってCD4+ 細胞は好ましくは抗原特異的態様で活性化されるであろう。CD4+ 細胞対抗原提示細胞の比は個体間で変動し、さらに、例えば各人のリンパ球の培養条件に対する順応性並びに疾患の性質および重篤度または処方された治療様式で用いられる他の条件のような変数によって左右されるであろう。しかしながら、好ましくはリンパ球:抗原提示細胞またはマトリックス比は約1:1から300:1の範囲であろう。
【0170】
エフェクター/抗原提示培養は、治療的に使用可能数または有効数のCD4+細胞集団を活性化させるか、またはそのような集団を濃縮するために必要な時間維持されるであろう。一般的には最適期間は約1日から5日で、最大特異レベルは一般に3から5日培養後に認められるであろう。本発明のある実施態様では、CD4+細胞のインビトロ活性化は、細胞株にトランスフェクトした後短時間内に検出される。
【0171】
好ましくは、CD4+細胞の活性化は抗原提示細胞に暴露してから1週間以内が最適である。好ましい実施態様ではその後、活性化細胞は、密度勾配、抗体赤血球細胞調製物によるロゼット法、カラムクロマトグラフィーなどを含む単離方法によってさらに精製される。当該精製に続いて、得られたCD4+細胞調製物は109 の活性化CD4+細胞集団を得るためにある期間培養を維持することによってさらに増殖させる。この期間は細胞の増殖時間にしたがって変動させることができるが、一般には14日であろう。
【0172】
c.合成APCまたはマトリックスからのCD4+ 細胞の分離
活性化CD4+ 細胞は、種々の既知の方法の1つを用いて本発明の抗原提示組成物から効果的に分離できる。例えば、MHCクラスIIヘテロダイマー、それに付加されたペプチド、またはCD4+細胞(またはそのセグメント)に対して特異的な単クローン性抗体を用いてそれらの適切な相補的リガンドに結合させることができる。続いて、適切な手段、例えば周知のフローサイトメトリーまたは磁性ビーズ分離法によって抗体標識細胞を刺激体−エフェクター細胞混合物から抽出することができる。密度勾配もまた芽細胞を分離するために使用することができる。
【0173】
d.活性化CD4+細胞による治療
活性化CD4+細胞の有効量は、CD4+集団の活性化に用いた抗原性ペプチドを発現している標的細胞の量および型に関する場合と同様に、インビトロの使用とインビボの使用との間で変動するであろう。この量はまた患者の状態によっても変動し、医師によって適切な全ての要素を考慮して決定されなければならない。しかしながら、マウスで用いられる5×106 から5×107 細胞と比較すると好ましくは約1×106 から約1×1012、より好ましくは約1×108 ら約1×1011、さらに好ましくは約1×109 から約1×1010の活性化CD4+細胞が成人に用いられる。
【0174】
上記で考察したように好ましくは、処置される患者にCDr+細胞を投与する前に、活性化CD4+ 細胞は培養されている合成APCまたはマトリックスから集められる。しかしながら、現在の治療様式または推奨されている治療様式と異なり、本方法の好ましい実施態様ではドロソフィラAPCまたは無細胞マトリックスが用いられていることを特記する必要がある。したがって、細胞および完全な活性化CD4+細胞の分離が達成されない場合、哺乳類の腫瘍促進細胞の投与は極めて危険であるが、一方、少数の合成APCまたはマトリックスの投与に付随する既知の固有の危険性は存在しない。
【0175】
細胞性成分を再導入する方法は当技術分野で既知であり、そのような方法には、米国特許第4844893 号明細書(Honskiら)および米国特許第4690915 号明細書(Rosenberg)に例示されたような方法が含まれる。例えば、静脈内輸液による活性化CD4+細胞の投与が適切である。
【0176】
実施例
以下の実施例は説明を目的とし、本発明を制限するものではない。
【0177】
1.pRmHa−3発現ベクターの調製
本発明で述べたようにドロソフィラシュナイダー(Schneider)2(S2)細胞でMHCタンパク質を発現する際に用いるpRmHa−3発現ベクターを、SphIで直線化したpRmHa−1DNA発現ベクターをsPhI制限酵素で消化したpRmHa−2発現ベクターから得たDNAフラグメントと連結して下記に述べるpRmHa−3発現ベクターを形成することによって構築した。直線化pRmHa−1のpRmHa−2フラグメントによる本態様での連結は、pRmHa−1に存在する2つのEcoRI制限エンドヌクレアーゼクローニング部位のうちの1つを除去するために実施された。したがって、得られたpRmHa−3発現ベクターは、実施例で述べるように種々のMHCクラスIIをコードするDNAフラグメントが挿入されている多クローニング部位(ポリリンカー)内にただ1つのEcoRI制限部位を含んでいた。
【0178】
A.pRmHa−1発現ベクターの調製
メタルチオネインプロモーター、金属反応コンセンサス配列(MTと称する)およびポリアデニル化シグナル(Drosophila melanogasterから単離)含有アルコールデヒドロゲナーゼ(ADH)遺伝子を含むpRmHa−1発現ベクターを文献(Bunchら、Nucl. Acids Res., 16:1043-61(1988))にしたがって構築した。プラスミド発現ベクター、pUC18(ATCCアクセッション番号37253)を本明細書で述べる以降のベクターの由来源として用いた。pUC18プラスミドは、多クローニング部位の5’から3’に以下の制限部位を含む(図1のpUC由来ベクターの模式図にその全てが示されているわけではない):EcoRI;SacI;KpnI;SmaIおよび同じ位置に存在するSmaI;BamHI;XbaI;SalI;AccIおよび同じ位置のHincII;pSTI;SphIおよびHindIII。pUC18ベクターを先ずHindIIIで消化し、直線化pUC18を形成する。続いて、マニアーティスら(「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、 Cold Spring Harbor Laboratory刊、ニューヨーク(1982))の記載にしたがって、DNAポリメラーゼIの大型フラグメントで当該HindIII末端を充填することによって平滑端(blunt end)を作出した。
【0179】
ドロソフィラ・メラノガスター(Drosophila melanogaster)ポリアデニル化シグナルを含むADH遺伝子のHinfIフラグメント(740塩基対(bp))と得られた直線化平滑端pUC18ベクターを連結した。連結するADH対立遺伝子は先ず、ゴールドバーグら(Goldbergら、Proc. Natl. Acad. Sci. USA, 77:5794-5798(1980))が記載したプラスミドpSACIから以下のようにして単離した:HinfIで消化し、続いてクレノウ(Klenow)を用いて平滑端を作り配列番号:1に挙げたヌクレオチド配列を得る。ADH対立遺伝子を含むpSACIベクターは、任意の高分子量(15kb以上)を含むバクテリオファージラムダライブラリーから選別したドロソフィラDNAのEcoRIフラグメント(4.7kb)をpBR322(ATCCアクセッション番号31344)でサブクローニングすることによって構築した。5’HinfI制限部位は、文献(Kreitman, Nature,304:412-417(1983))に記載されているように1770位のADH遺伝子内に天然に存在する。3’HinfI部位は、ADH遺伝子がクローニングされたpUC18ベクターに由来した。この位置はADH遺伝子の2500位にあるXbaI部位から3’側に4塩基離れていた。ADHセグメントは、ADHmRNAの3’側非翻訳部分のポリアデニル化/切断配列の35bp上流からポリアデニル化シグナルの700bp下流に広がっていていた。得られたADH遺伝子フラグメントを含むpUC18由来ベクターは図1Aに示すようにpHa−1と称した。
【0180】
pHa−1に挿入するためのMT遺伝子のEcoRI/StuIフラグメント(421bp)は、ドロソフィラ・メラノガスターのゲノムDNAライブラリーの約15.3kbのDNAを含むクローンから得た。昆虫DNAのMboIによる部分消化で調製したライブラリーをラムダ由来EMBL4でクローニングした。421bpフラグメントが、ドロソフィラのMT遺伝子のMTプロモーターおよび金属反応コンセンサス成分を含んでいた(Maroniら、Genetics, 112:493-504(1986))。プロモーターおよびヌクレオチドの+1の位置の転写開始部位を含むこの領域は、MT遺伝子の−370の位置からヌクレオチドの+54の位置に対応していた(配列番号:2)。続いて、得られたフラグメントを先にEcoRIとSmaIで直線化した上記で調製したpHa−1発現ベクターに連結した。StuI消化で作出したMTの3’側平滑端は、SmaI消化で作出したpHa−1の平滑端と適合した。ドロソフィラの5’MT遺伝子フラグメントおよび3’ADH遺伝子フラグメントを含む得られたpUC18由来ベクターをpRmHa−1と称した。このpRmHa−1発現ベクターは、複製開始点(ori)および図1Aに示すようにpHa−1ベクター上のpUC由来のアンピシリン耐性(Ampr)を付与するベータ・ラクタマーゼ遺伝子を含んでいた。pRmHa−1はまた、5’から3’に向けてMT遺伝子、多クローニング部位およびADH遺伝子フラグメントを含んでいた。pRmHa−1ベクターは、下記で述べるようにpRmHa−3発現ベクターの構築で用いられた。
【0181】
B.pRmHa−2発現ベクターの調製
図1Aに示したpRmHa−2発現ベクターを構築するために、pRmHa−1を構築するために上記で述べた方法に少しの改変を加えながら、pUC18由来ベクターpHa−1に上記で調製したMTフラグメントを挿入した。上記で調製したEcoRI/StuI単離MT遺伝子フラグメントのStuI部位にEcoRIリンカーを添加して、両端にEcoRI制限部位をもつメタロチオネインフラグメントを形成する。続いて得られたフラグメントを、EcoRIで先に直線化したADHフラグメント含有pUC18発現ベクターに連結した。5’ドロソフィラMT遺伝子フラグメントおよび、多クローニング部位に対して5’側に2つのEcoRI制限部位を有する3’ADH遺伝子フラグメントを含む得られたpUC18由来ベクターをpRmHa−2と称した。このpRmHa−2発現ベクターは複製開始点(ori)およびpUC由来のアンピシリン耐性(Ampr )を付与するベータ・ラクタマーセ遺伝子を含んでいた。pRmHa−2の模式図はまた、MT遺伝子フラグメントの5’から3’までの連続した部位、多クローニング部位およびADH遺伝子フラグメントを示している。このpRmHa−2ベクターをpRmHa−1とともに下記で述べるようにpRmHa−3発現ベクターの構築に用いた。
【0182】
C.pRmHa−3発現ベクターの調製
ただ1つのEcoRI制限部位を有するpRmHa−3発現ベクターを調製するために、pRmHa−2由来フラグメントをpRmHa−1に連結した。この構築のために、上記で調製したpRmHa−2を先ずSphIで消化した。得られたSphIフラグメント(MT遺伝子の中央で始まり、多クローニング部位のSphI部位まで広がる)を先ずpRmHa−2ベクターから単離し、続いてpRmHa−1につないだ。この連結の前に、pRmHa−1は、MT遺伝子フラグメントに対して5’側にあるEcoRI制限部位を除去して改変し続いてSphIで直線化した。この過程は図1Bに模式的に示した。pRmHa−1のEcoRI部位を除去するために、このベクターを先ずEcoRIで消化し、直線化ベクターを作製し、続いてマングビーン(Mung Bean)のヌクレアーゼで平滑端を作り再び連結した。
【0183】
pRmHa−3ベクターの模式図は図1Cに示す。pRmHa−3が誘導されたpUC18ベクターから由来した種々の制限部位の相対的な位置は図に示されている。pUC18から誘導されたpRmHa−3ベクターは、pUC18の複製開始点およびアンピシリン耐性を付与するベータ・ラクタマーゼ遺伝子を含む。したがって、本発明で調製しpRmHa−3の多クローニング部位でクローニングしたMHCクラスIIをコードするDNAフラグメントは、MTプロモーターによる転写によって調節され、ADH遺伝子を介してポリアデニル化された。
【0184】
2.発現性MHCクラスII遺伝子の調製と発現
A.MHCクラスII遺伝子の増幅と発現
好ましい哺乳類のMHCクラスIIのいずれかをコードする遺伝子またはcDNAはポリメラーゼ連鎖反応(PCR)を用いてクローニングされる。本明細書に記載するプライマーを適切なクラスIIcDNAを増幅するために別々の反応で用い、続いてこれら増幅cDNAをクローニングして配列を決定する。MHCクラスIIタンパク質を生成するために、完全な長さのネズミIAd α鎖およびβ鎖cDNAを先ず入手し、続いてPCR増幅および改変を実施した。ネズミIAd α鎖cDNAの完全なヌクレオチド配列はベノイストら(Benoistら、Cell, 34:169-177(1983))が報告しており、またジェンバンク(GenBank)のリストにも記載されている(アクセッション番号K01923)。ネズミIAd β鎖cDNAの完全なヌクレオチド配列はマリセンら(Malissenら、Science, 221:750-754(1983))が記載し、ジェンバンクにも登録されている(アクセッション番号K00007およびK00008)。各鎖を増幅するために全RNAの供給源としてマウスの脾細胞を用いた。第一の鎖のcDNAは、オリゴ(dT)およびニワトリ骨髄芽球症ウイルスの逆転写酵素を用いて合成した。得られたcDNAをPCR増幅反応に用いた。増幅反応では、下記に述べる適切なプライマーおよびジーンアンプ(GeneAmp)キットおよび温度循環装置(Perkin-Elmer/Cetus, ノーウォーク、コネチカット)を利用した。反応条件は、好ましくは1μgのcDNA鋳型、および200nMの各オリゴヌクレオチドプライマーを含んでいた。以下のように30サイクル実施した:(a)92℃で1分;(b)60℃で1分;(c)72℃で1分。続いてPCR反応物を99℃で10分加熱し、Taqポリメラーゼを不活化し、さらにDNA端をT4ポリメラーゼ(Stratagene, ラホイヤ、カリフォルニア)で平滑端にした。
【0185】
実施例で示したオリゴヌクレオチドプライマーの全てについて、5’プライマーはまたフォワードプライマーまたはセンスプライマーと称する。なぜならば、これは、相補的な下の鎖とハイブリダイズさせるためのcDNAの上の鎖と同じ配列を有するからである。対照的に、3’プライマーは相補的な上の鎖とハイブリダイズさせるためのcDNAの下の鎖と同じ配列を有するのでバックワードプライマーまたはアンチセンスとも称される。両方のプライマーとも5’から3’方向で記載されている。5'CTTGAATTCCACCATGCCGTGCAGCAGAGCTCTGA3'(配列番号:3)なるヌクレオチド配列を有する5’プライマーを用いて完全な長さのIAd α鎖cDNAを増幅した。またこの5’プライマーをEcoRI制限部位をもつようにデザインして、当該増幅生成物を方向性をもって受容体の発現ベクターに連結することを可能にした。
【0186】
5'TTTGGATCCTCATAAAGGCCCTGGGTGTC3'(配列番号:4)なる3’プライマーもまたBamHI制限部位をもつようにデザインした。
【0187】
5'CTTGAATTCCACCATGGCTCTGCAGATCCCCA3'(配列番号:5)なるヌクレオチド配列を有する5’前進プライマーを用いて完全な長さのIAd β鎖cDNAを増幅した。この5’プライマーもまたEcoRI制限部位をもつようにデザインし、当該増幅生成物を方向性をもって受容体の発現ベクターに連結することを可能にした。
【0188】
5'TTTGGATCCTCACTGCAGGAGCCCTGCT3'(配列番号:6)なる3’プライマーをBamHI制限部位を含むようにデザインした。ネズミIAd α鎖およびIAd β鎖をコードする改変cDNAを別々に先ずメタロチオネイン駆動pRmHa−3ベクターのEcoRIおよびBamHI制限部位のポリリンカーに方向性をもたせてクローニングした。続いて色素ターミネータ技術によってアプライドバイオシステム(Applied Biosystem)373A自動配列決定装置で配列を調べた。
【0189】
pRmHa−3でクローニングしたネズミIAd α鎖およびIAd β鎖の増幅領域の完全なヌクレオチド配列は、それぞれ配列番号:7および8で示されている。
【0190】
続いて、別の場所(Jacksonら、Proc. Natl. Acad. Sci. USA, 89:12117-21(1992))で述べたように、ドロソフィラ・メラノガスターのシュナイダー2細胞(ATCCCRL10974、ロックビル、メリーランド))に別々のプラスミドをトランスフェクトした。2つのクラスII鎖をコードするプラスミドの同じ量を、ネオマイシン耐性遺伝子(プラスミドphshsneo(Bunchら、Nuc. Acids Res., 16:1043-1061(1988))とともに1:30の割合で同時にトランスフェクトして安定な細胞株を作製した。この細胞株は、10%ウシ胎児血清(55℃で1時間加熱処理)、100単位/mlのペニシリン、100μg/mlのストレプトマイシンおよび1mMのグルタミンを補充したシュナイダーのドロソフィラ培養液(Gibco/brl, グランドアイランド、ニューヨーク)での4週間を越えるG418選別によって得られた。具体的には、トランスフェクトした後に上清を注意深く取り除き、500μg/mlのジェネチシン(G418)(Gibco/BRL, グランドアイランド、ニューヨーク)を含む全容量12mlのシュナイダー培養液を入れた75cm2 のフラスコに細胞を移した。4日後、500μg/mlのG418とともに6mlのシュナイダー培養液を含む新しいフラスコに4mlの培養を移した。この操作を安定な細胞集団が出現するまで4〜7日毎に繰り返した。安定な細胞集団は弱くフラスコに付着し、約24時間の分裂時間で増殖した。その後これらの細胞を培養し、上記で述べたように選別培地で継代した。安定的にトランスフェクトされた凍結部分標本は、遠心沈殿によって5−20×106細胞を採集し、1mlの細胞凍結培養液(93%ウシ胎児血清/7%ジメチルスルホキシド)に再懸濁することによって調製された。続いて部分標本を−70℃に1週間置き、その後液体窒素貯蔵庫に移した。安定的にトランスフェクトされたMHCクラスIIのIAd α鎖およびβ鎖の遺伝子の発現は、0.7mMの硫酸第二銅を用いて27℃で24時間誘発された。
【0191】
トランスフェクトしたドロソフィラ細胞の細胞表面でのMHCクラスIIのIAd ヘテロダイマーの発現は、MKD6(IAd 特異的単クローン性抗体(Kapplerら、J. Exp. Med., 153:1198(1981))による染色後フローサイトメトリーで調べた。簡単に記せば、細胞の一部分(5×105 )を氷上の試験管に移し、遠心沈殿(1000×g5分)によって集め、適切な一次抗体(MKD6)を含有する5%ウマ血清を含む0.1mlのドロソフィラ培養液に再懸濁した。室温で20分保温した後、ウマ血清を含む3mlのシュナイダー培養液で2回洗浄し、0.1mlのFITC標識二次抗体(Cappell, Durham,ノースカロライナ)含有培養液に再懸濁した。氷上で20分維持した後、ウマ血清含有シュナイダー培養液で2回洗浄し、この緩衝液に1×106 /mlの濃度で再懸濁した。沃化プロピジウムを添加して死細胞を分析から排除した。続いてサンプルをファクスキャン(FACScan)またはファクソート(FACSort)装置(Becton Dickison)で分析した。1つ以上のアクセサリー分子とともにネズミIAd MHCクラスIIハプロタイプ(本明細書で述べまた当技術分野でも周知である)を発現する本発明の合成抗原提示細胞は下記のように製造された。
DR、DQおよびDPに対する完全な長さのヒトMHCクラスIIα鎖およびβ鎖を末梢血細胞(B細胞、マクロファージおよび樹状突起細胞を含む)から増幅する。各々のハプロタイプを増幅するために用いた5’および3’プライマーは以下の配列を有する:
【0192】
DRα:5’プライマー=5'CCACCATGGCCATTAGTGGAGTC3'(配列番号:9)
3’プライマー=5'TTTGGATCCTTACAGAGGCCCCCTGCGTT3'(配列番号:10);
DRβ:5’プライマー=5'CCACCATGGTGTGTCTGAGGCTCC3'(配列番号:11)
3’プライマー=5'TTTGGATCCTCAGCTCAGGAATCCTCTTG3'(配列番号:12);
DQα:5’プライマー=5'CCACCATGGTCCTAAACAAAGCTCTGAT3'(配列番号:13)
3’プライマー=5'TTTGGATCCTCACAAGGGCCCTTGGTGTCT3'(配列番号:14);
DQβ:5’プライマー=5'CCACCATGGCTTGGAAGAAGGCCTTT3'(配列番号:15)
3’プライマー=5'TTTAGATCTCAGTGCAGAAGCCCTTT3'(配列番号:16);
DPα:5’プライマー=5'CCACCATGGGCCCTGAAGACAGAAT3'(配列番号:17)
3’プライマー=5'TTTGGATCCTCACAGGGTCCCCTGGGC3'(配列番号:18);
DPβ:5’プライマー=5'CCACCATGGTTCTGCAGGTTTCTGCG3'(配列番号:19)
3’プライマー=5'TTTGGATCCTTATGCAGATCCTCGTTGAA3'(配列番号:20);
【0193】
上記ヒトハプロタイプを得るための増幅条件は、ネズミの対応物を得るために上記で述べた条件と同一である。
1つ以上のアクセサリー分子とともにヒトMHCクラスIIハプロタイプ(本明細書で述べまた当技術分野でも周知である)を発現する本発明の合成抗原提示細胞は下記のように製造される。
【0194】
B.不変鎖およびHLA−DM鎖cDNAの増幅
詳細な説明の節Bで述べた本発明の幾つかの態様のために、不変鎖並びにHLA−DMαおよびHLA−DMβを選択したMHCクラスIIヘテロダイマーおよび少なくとも1つの補助因子とともに同時発現させる。本事例では、MHCクラスIIα鎖およびβ鎖、不変鎖並びにHLA−DMαおよびHLA−DMβを、5:5:8:1:1のモル比で受容体の抗原提示細胞にトランスフェクトする。他の遺伝子のモル比に対するアクセサリー分子の比は1:1である。続いて、得られたトランスフェクト細胞を上記のように外因性遺伝子の発現のために誘発して本発明の合成抗原提示細胞株を作製する。
【0195】
1)不変鎖
ネズミの不変鎖cDNAをマウスの脾臓細胞から作製した。それぞれ
【0196】
5'AAGAATTCACTAGAGGCTAGAGCCAT3'(配列番号:21)および、
5'AAGGATCCTCACAGGGTGACTTGACC3'(配列番号:22)
【0197】
のヌクレオチド配列を有する5’および3’プライマーのプライマー対を用いて当該供給源から完全な長さの不変鎖を増幅させる。上記で述べたように、5’および3’プライマーをそれぞれEcoRIおよびBamHI制限部位を含むようにデザインした。増幅不変鎖をEcoRI/BamHI消化pRmHa−3ベクターでクローニングした後、得られたクローンの配列を調べ、配列番号:23に挙げた配列を有することが分かった。
【0198】
本発明で用いるヒト不変鎖cDNAは、γ−インターフェロン誘発HeLa細胞(ATCCアクセッション番号CCL2)由来RNAを用いて作製した。続いて、それぞれ
【0199】
5'AAGAATTCACCATGGATGATCAGCGCGACCTT3'(配列番号:24)および、
5'AAAGGATCCTCACATGGGGACTGGGCCCAGA3'(配列番号:25)
【0200】
のヌクレオチド配列をもつ5’および3’オリゴヌクレオチドプライマーを用いて、得られたcDNAはPCRで鋳型として使用する。得られたPCRフラグメントをEcoRIおよびBamHIで切断し、同様に消化したpRmHa−3に連結する。
【0201】
2)HLA−DM
またγ−インターフェロン誘発HeLa細胞由来mRNAを用いてHLA−DMcDNAのαおよびβ鎖を合成する。続いてこれらをPCRで鋳型として用いる。α鎖は、それぞれ
【0202】
5'AAACCATGGGTCATGAACAGAACCA3'(配列番号:26)および、
5'TTTGTCGACTCAGTCACCTGAGCAAGG3'(配列番号:27)
【0203】
のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅する。β鎖は、それぞれ
【0204】
5'AAACCATGGTCTCATTCCTGCC3'(配列番号:28)および、
5'TTTGTCGACCTAGGAAATGTGCCATCC3'(配列番号:29)
【0205】
のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅する。得られたPCRフラグメントをNcoIおよびSalIで切断し、同様に消化したpRmHa−3に別々に連結する。
【0206】
C.アクセサリー分子をコードする遺伝子の増幅
1)付着分子
a.ICAM−1、ICAM−2およびICAM−3
ネズミICAM−1を単離するために、脾臓細胞をBalb/cマウスから単離した。mRNAの単離前に、脾臓細胞を先ずファストトラック(Fast Track)キット(Invitrogen, サンディエゴ、カリフォルニア)を用い製造元の指示にしたがってconAで刺激した。cDNAは上記のようにmRNAから合成した。続いて、公表されたcDNAヌクレオチド配列(G. Siuら、J. Immunol., 143:3813−3820(1989))を基にデザインしたそれぞれ以下の5’および3’プライマーを用いて得られたcDNAをPCRに付した:
【0207】
5'TTTAGAATTCACCATGGCTTCAACCCGTGCCAAG3'(配列番号:30)および、
5'TTTAGTCGACTCAGGGAGGTGGGGCTTGTCC3'(配列番号:31)。
【0208】
続いて、PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0209】
続いて、上記で述べたリン酸カルシウム法を用い、上記で調製したネズミIAdα鎖およびβ鎖遺伝子とともに上記の発現構築物を、ICAM−1:α鎖:β鎖を1:1:1のモル比でドロソフィラS2細胞に同時にトランスフェクトした。安定的にトランスフェクトされた細胞が上記のように得られた。細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子ICAM−1を発現している合成抗原提示ドロソフィラS2細胞を、続いて先に述べたように発現を誘発することによって製造した。
【0210】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様ではまた、ネズミIAd 分子と一緒にICAM−1、B7.1および/またはB7.2をコードする遺伝子を含む合成抗原提示細胞を製造した。
【0211】
ヒトICAM−1は同様にヒトの細胞株K562から単離したmRNAから増幅する。当該細胞株は、ヒトの慢性骨髄性白血病に由来し(ATCCアクセッション番号CCL-243)、推奨された条件下で培養した(すなわち、10%ウシ胎児血清添加RPMIで5%CO2 下37℃)。ヒトアクセサリー分子を増幅するPCRプライマーは利用可能な既知の配列を基に、適切なベクターでクローニングするために必要な5’および3’クローニング部位を考慮してデザインされる。ヒトICAM−1cDNAのヌクレオチド配列は、ジェンバンクアクセッション番号GB-J03132により入手できる。ICAM−1の5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0212】
5'ACCCTTGAATTCATGGCTCCCAGCAGCCCCCGGCCC3'(配列番号:32)および、
5'ATTACCGGATCCTCAGGGAGGCGTGGCTTGTGTGTTCGG3'(配列番号:33)。
【0213】
増幅ヒトICAM−1を得るために、先で述べたようにこれらプライマーを用いてPCRを実施する。続いて、得られたPCR生成物を先に述べたようにpRmHa−3でクローニングし、その後、これもまた先に述べたように受容細胞にトランスフェクトする。
【0214】
同様に、ヒトICAM−2およびICAM−3(これらのヌクレオチド配列はそれぞれジェンバンクアクセッション番号GB-X15606 およびGB-S50015 により入手できる)は、該当するプライマー対を用いて増幅される。ICAM−2を増幅する5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0215】
5'AAGGTACCCGTGGAGACTGCCAGAGAT3'(配列番号:34)および、
5'TTTGGATCCCTATGGCCGGAAGGCCTG3'(配列番号:35)。
【0216】
ICAM−3を増幅する5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0217】
5'AAGAATTCCTGTCAGAATGGCCACCAT3'(配列番号:36)および、
5'TTTAGATCTTCACTCAGCTCTGGACGGT'(配列番号:37)。
【0218】
続いて、得られた増幅ICAM生成物を別々にpRmHa−3につなぎ、別々にドロソフィラS2細胞にトランスフェクトする。
【0219】
選択したヒトMHCクラスIIハプロタイプとともにアクセサリー分子ICAM−1を発現している合成抗原提示ドロソフィラS2細胞は、続いて先に述べたように発現を誘発することによって製造される。
【0220】
他の実施態様では、実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、ヒトハプロタイプと合わせてB7.1および/またはB7.2と一緒にICAM−1、ICAM−2またはICAM−3をコードする遺伝子を含む合成抗原提示細胞がまた作製される。本発明で使用することを目的とするまた別の変更には、LFA−3および/またはFasリガンド(FasL)(これらはともに下記で述べる他のアクセサリー分子である)と上記の組み合わせが含まれる。
【0221】
b.LFA−3
ヒトLFA−3は血中リンパ球から単離される。ヒトLFA−3のcDNAのヌクレオチド配列は、ジェンバンクアクセッション番号GB-I09083 によって入手できる。したがって、ヒトLFA−3はそれぞれ以下のヌクレオチドを有する5’および3’プライマーを用いて増幅される:
【0222】
5'ACCCTTGAGCTCATGGTTGCTGGGAGCGACGCGGGG3'(配列番号:38)および、
5'ATTACCGGATCCTTAAAGAACATTCATATACAGCACAATACA3'(配列番号:39)。
【0223】
実施例5で述べる活性化アッセイで使用する合成抗原提示細胞を作製するために、先に述べたようにヒトハプロタイプとともにヒトLFA−3のみをコードするか、または本明細書で述べた他のアクセサリー分子と一緒にヒトLFA−3をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0224】
2)同時刺激分子
a.B7.1
ICAM−1について上記で述べたように、マウスから単離したmRNAからcDNAを作製した。続いて、以下のそれぞれ5’および3’オリゴヌクレオチドプライマーを用いて得られたcDNAをPCRに付した(これらプライマーは、公表されたcDNAヌクレオチド配列(Freemanら、J. Exp. Med., 174:625-631(1991))を基にデザインし5’から3’の方向で表示する):
【0225】
5'TTTAGAATTCACCATGGCTTGCAATTGTCAGTTG3'(配列番号:40)および、
5'TTTAGTCGACCTAAAGGAAGACGGTCTGTTC3'(配列番号:41)。
【0226】
PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0227】
続いて、上記で調製したネズミのIAd α鎖およびβ鎖遺伝子とともに上記の発現構築物を、B7.1:α鎖:β鎖のモル比を1:1:1として上記で述べたように燐酸カルシウム法を用いてドロソフィラS2細胞に同時にトランスフェクトした。上記で述べたように安定的にトランスフェクトされた細胞を得た。続いて、細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子B7.1を発現する合成抗原提示細胞ドロソフィラS2細胞を、上記で述べたように誘発することによって作製した。
【0228】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ネズミIAd 分子と一緒にネズミICAM−1、B7.1および/またはB7.2をコードする遺伝子を含む合成抗原提示細胞もまた製造される。
【0229】
ヒトB7.1は同様にK562細胞から単離され、上記で述べたようにpRmHa−3でクローニングされる。ヒトB7.1cDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-M83071 により入手できる。5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0230】
5'ACCCTTGAATCCATGGGCCACACACGGAGGCAG3'(配列番号:42)および、
5'ATTACCGGATCCTTATACAGGGCGTACACTTTCCCTTCT3'(配列番号:43)。
【0231】
続いて、得られたPCR生成物をpRmHa−3に挿入し、続いてこれを増幅させクローニングしたヒトハプロタイプとともにドロソフィラS2細胞に同時にトランスフェクトした。
【0232】
続いて先に述べたように発現を誘発することによって、細胞表面に選択したヒトMHCクラスIIハプロタイプとともにアクセサリー分子B7.1を発現する合成抗原提示細胞ドロソフィラS2細胞を作製する。
【0233】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ヒトハプロタイプとともに本明細書で述べたアクセサリー分子を種々に組み合わせたB7.1をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0234】
b.B7.2
ネズミIC−21細胞(ATCCアクセッション番号TIB186)を10%ウシ胎児血清含有RPMI1640培養液で増殖させた。上記のようにこれらの細胞から単離したmRNAからcDNAを合成した。続いて、得られたcDNAを以下のそれぞれ5’および3’オリゴヌクレオチドプライマーを用いてPCRに付した(これらプライマーは、公表されたcDNAヌクレオチド配列(Freemanら、J. Exp. Med., 178:2185-2192(1993))を基にデザインし5’から3’の方向で示す):
【0235】
5'TTTAGAATTCACCATGGACCCCAGATGCACCATGGG3'(配列番号:44)および、
5'TTTAGTCGACTCACTCTGCATTTGGTTTTGCTGA3'(配列番号:45)。
【0236】
PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0237】
続いて、上記で調製したネズミのIAd α鎖およびβ鎖遺伝子とともに上記の発現構築物を、B7.2:α鎖:β鎖のモル比を1:1:1として上記で述べたように燐酸カルシウム法を用いてドロソフィラS2細胞に同時にトランスフェクトした。上記で述べたように安定的にトランスフェクトされた細胞を得た。続いて、細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子B7.2を発現する合成抗原提示細胞ドロソフィラS2細胞を、上記で述べたように誘発することによって作製した。
【0238】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ICAM−1およびネズミIAd 分子と一緒にネズミB7.2をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0239】
ヒト細胞株HL60(前骨髄球性白血病由来)(ATCCアクセッション番号CCL−240)からヒトB7.2を単離する。ヒトB7.2cDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-M83071 により入手できる。ヒトB7.2を増幅するための5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0240】
5'ACCCTTGAGCTCATGGATCCCCAGTGCACTATG3'(配列番号:46)および、
5'ATTACCCCCGGGTTAAAAACATGTATCACTTTTGTCGCATGA3'(配列番号:47)。
【0241】
選択したヒトハプロタイプのための構築物と一緒にドロソフィラS2細胞にトランスフェクトするために、増幅B7.2生成物を続いて上記で述べたようにpRmHa−3でクローニングする。トランスフェクトされた遺伝子の発現は上記のように誘発される。
【0242】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ヒトハプロタイプとともに本明細書で述べたアクセサリー分子を種々に組み合わせたヒトB7.2をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0243】
3)サバイバル分子
ヒトFasリガンドは活性化ヒトT細胞から単離される。ヒトFasリガンドcDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-U08137 により入手できる。したがって、ヒトFasリガンドは、それぞれ以下のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅される:
【0244】
5'AAAGGATCCACCATGCAGCAGCCCTTCAATT3'(配列番号:48)および、
5'TTTGGATCCTTAGAGCTTATATAAGCCGA3'(配列番号:49)。
【0245】
ヒトCD70(T細胞上に発現されるCD27のためのリガンド)は、それぞれ以下のヌクレオチド配列を有する5’および3’プライマー対を用いて上記のように増幅される:
【0246】
5'AAAGAATTCGGTACCATGCCGGAGGAGGGTTCGG3'(配列番号:50)および、
5'TTTGGATCCTCAGGGGCGCACCCACTGCA3'(配列番号:51)。
【0247】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、上記で述べたように、ヒトハプロタイプとともにヒトFasリガンドのみをコードするか、または本明細書で述べた他のアクセサリー分子を種々に組み合わせたヒトFasリガンドをコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0248】
3.合成担体に定着させたMHCクラスIIの調製
ここで述べる実施例は、大量のMHCクラスII分子および単一種のアクセサリー分子またはそれらの種々の組み合わせを、単クローン性抗体結合およびロゼット形成実験(T細胞レセプター結合)で判定したとき固有の構成で種々の表面(ハエの細胞、赤血球細胞、ラテックスビーズ)に固定させる新規な方法を提供する。本方法は、人工リン脂質膜を含む他の合成表面に範囲を広げることができる。アビジン結合リン脂質とともにホスファチジルエタノールアミンは特に本発明に適切である。これらのリン脂質は、リペックスバイオメンブレン社(Lipex Biomembrane Inc., バンクーバー、ブリティッシュコロンビア、カナダ)から市販ルートで入手できる。
【0249】
A.アビジン被覆赤血球細胞上へのビオチン付加MHCクラスIIの固定
NHS-LCビオチン、ニュートロアビジン(neutravidin)およびビオチン−BMCCはピアース(Pierce, ロックフォード、イリノイ)から購入する。ヒツジ赤血球はコロラドシーラムカンパニー(Colorado Serum Company, デンバー、コロラド)から入手する。MHCクラスIIを発現しているドロソフィラS2細胞は上記のように調製する。ハイブリドーマ細胞培養上清として、単クローン性抗体MKD6およびM5−114(MKD6分泌ハイブリドーマについてはATCCアクセッション番号TIB−120)を用いる。用いたプロトコルは文献に記載されている(Muzykantov & Taylor, Anal. Biochem., 223:142-148(1994))。簡単に記せば、SRBCを4回PBSで洗浄し、NHS-LCビオチンでビオチン付加し、再びPBSで4回洗浄し、3%ウシ胎児血清および0.02%アジ化ナトリウムを含むPBS中で4℃で保存する。組換え発現MHCクラスIIα鎖およびβ鎖はビオチン−BMCCを用いてビオチン付加する。ビオチン−BMCCは、チオール基と反応するマレイミド結合ビオチンである。ビオチン付加は製造元の推奨にしたがって実施する。未反応ビオチンはセントリコン(Centricon)10で除去する。
【0250】
ビオチン付加MHCクラスIIα鎖およびβ鎖は、最終濃度0.2mg/mlでアビジン被覆SRBCとともに30分保温し、続いて10%ウシ胎児血清含有DMEMで洗浄することによって固定される。MHCクラスII結合SRBCは直ちに使用する。
【0251】
アビジン被覆SRBC上へのビオチン付加MHCクラスIIα鎖およびβ鎖の固定は上記のように実施する。結合は、上記の抗体を用いてフローサイトメトリーにより調べる。ロゼット形成アッセイのためには、MHCクラスII発現ドロソフィラS2細胞またはMHCクラスII被覆SRBCのどちらかを、特定のペプチド(実施例4参照)(0.02mM)または無関係ペプチド(0.02mM)とともに30分氷上で保温する。続いて、CD4+T細胞を添加する(割合は、10個のT細胞に対して1個のドロソフィラS2細胞または10個のSRBCに対して1個のT細胞)。その後混合物を沈澱させ、少なくとも30分氷上で保温する。続いて、細胞を注意深く再懸濁させてロゼットを数える。ロゼットは、少なくとも3個のCD4+T細胞と結合したドロソフィラS2細胞、または少なくとも3個のSRBCと結合したCD4+T細胞である。
【0252】
B.アビジン被覆ラテックスビーズ上へのビオチン付加MHCクラスIIの固定
直径が6ミクロンのラテックススルフェートビーズをインターフェイシャルダイナミクス社(Interfacial Dynamics Corporation, ポートランド、オレゴン)から購入し、上記のプロトコルにしたがってビオチン付加する。アビジン被覆ラテックスビーズは、1%のラテックスビーズ懸濁液を1mg/mlのニュートロアビジンを含むPBS中で1時間室温で保温して調製する。等容量の10%ウシ胎児血清含有PBSを続いて添加する。室温で1時間保温した後、ビーズを3回洗浄し、組換えビオチン付加MHCクラスIIの結合のために使用する。
【0253】
組換えビオチン付加MHCクラスIIを最終濃度0.2mg/mlでアビジン被覆ラテックスビーズと30分保温し、続いて10%ウシ胎児血清含有DMEM中で洗浄することによって固定される。結合MHCクラスIIを有するSRBCは直ちに使用する。ロゼット形成実験は先に述べたように実施する。
【0254】
C.種々の固形担体(例えばプラスチックマイクロウェルプレート)に結合したMHCクラスIIの固定と検出
MHCクラスII分子は、以下のようにマイクロタイタープレート(Corning)に直接結合によって固定し検出する:MHCクラスIIを所望の濃度にPBSで希釈し(例えば100ng/ウェルのために1mg/ml)、続いてプラスチック製マイクロタイタープレートの各ウェルに添加する。プレートを1時間室温で保温する。保温後、プレートをPBSで1回洗浄し、さらにPBS+トゥイーン(PBST)中の2%ウシ血清アルブミン200μl(0.05%)を添加し、さらに1時間室温で保温する。プレートを3回PBSTで洗浄し、2%BSA含有PBS中のビオチン付加抗MHCクラスII単クローン性抗体を添加する(1:2500)。プレートをさらに1時間室温で保温し、PBSTで3回洗浄する。アビジン結合HRPを2%BSA含有PBS中で添加する(1:2500)。室温でさらに1時間保温した後、プレートをPBSTで3回洗浄し、H2O2 またはトフェニルジアミン(thophenyldiamine)を添加する。反応はH2SO4 で停止させる。反応生成物は490nmで比色計により検出する。また別には、組換えMHCクラスII分子は、基質とのビオチン−アビジン連携相互反応により結合させることができる。この実施態様では、マイクロウェルプレートは、PBSで0.001mg/mlの濃度に希釈した100μlのアビジンで被覆される。過剰なアビジンはPBSによる洗浄で除去する。MHCクラスII結合の提示検出のための上記の過程がその後に続く。
【0255】
また別には、組換えMHC分子は、基質に結合させたニッケルと相互反応するMHCに添加したポリヒスチジンタッグによる架橋によって固定される。結合および検出のための上記の過程の後は、ニッケルキレート被覆マイクロウェルプレート(Xenopore)およびポリヒスチジンタッグを付加された発現MHC分子組換え体を使用する。
【0256】
4.ペプチド生成
本発明の抗原性ペプチドは、本明細書で述べたものを含む天然に存在する供給源から得るか、または既知の方法を用いて合成してもよい。本明細書で開示する種々の実施例では、全てのペプチドは、アプライド・バイオシステム社の装置ABI−431A(Foster City, カリフォルニア)を基にT−BOC法によって別々に化学的に合成され、C18逆相カラムにより精製される。アミノ酸残基のランダムアソートメントにより生成されるペプチドの単離または合成もまた適切であるかもしれない。特に、CD4細胞をおそらく刺激すると考えられるペプチドを空のMHC分子に付加するために特定のエピトープを突き止めようとしている場合は適切であろう。ランダム化ペプチドの混合物は、プロテアソームを用いることによって(タンパク質またはポリペプチドを分解過程(例えばキモトリプシンによる消化)に付すことにより)または合成によって得られる。
【0257】
本発明の細胞株は、タンパク質またはポリペプチドをヒトMHCクラスII分子に付加できるより小さなペプチドに分解することができるが、ペプチドを直接細胞に導入してより迅速な付加および発現作用を促進することが好ましい。
【0258】
ペプチドがアミノ酸残基を任意に取り込みながら合成される場合は、修飾アミノ酸とともにLまたはD型の全てのアミノ酸変種が、各合成サイクル時に好ましくは取り込まれるであろう。しかしながら、種々のパラメーター(例えばある種のアミノ酸の溶媒不適合)が、ある種のアミノ酸を欠くペプチドを含む混合物を生じるかもしれない。したがって、この過程は、溶媒および反応条件を変更することにより必要とされるように調節して最も多様なペプチドを生成しなければならない。ネズミMHCクラスIIに関して実施例5で述べるようにペプチド付加のために用いられるペプチドは、アミノ酸残基配列ISQAVHAAHAEINEAGR(配列番号:52)を有するオボアルブミン323-339 (OVA)である。ヒトMHCクラスIIハプロタイプに特異的な好ましいペプチドには以下のペプチドが含まれる:1)HLA−DRB1*0101と結合するインフルエンザヘマグルチニン306-318 PKYVKQNTLKLAT(配列番号:53);2)HLA-DRB1*0301と結合する結核菌(M. tuberculosis)由来HSP652-12 KTIATDEEARR(配列番号:54);3)HLA-DRB1*0301と結合するクラミジア・トラコマチス(C. trachomatis)のミトコンドリア外側タンパク質(MOMP) QASLALSYRLNMFTP(配列番号:55);4)HLA-DRB1*0401と結合するHLA−A233-45 FVRFDSDAASQRM(配列番号:56)。ペプチドリガンドがどのように限定されるかについての記載とともに既知のクラスII結合ペプチドの豊富な記載はラメンシーらの報告によって提示されている(Rammenseeら、Immunogenetics, 41:178-228(1995))。本発明の方法を実施するときに使用するペプチドはまたキロンミメトープ(Chiron Mimetope)により入手できる。他の実施態様では、本明細書で述べた合成抗原提示細胞に関する本発明の方法を実施する場合はペプチドライブラリー(当業者は容易に作製できる)の使用もまた意図される。
【0259】
5.ペプチドとのインビトロ保温による空のMHCクラスII分子の付加
下記に提示する結果によって示すように、選択アクセサリー分子とともにドロソフィラ細胞株の表面に発現されたネズミMHCクラスII分子は、用量依存態様で特定のペプチドと結合し、さらに抗原特異的T細胞にこれらペプチドを提示してT細胞活性化をもたらすことができる。(これらのドロソフィラ細胞株は、MHCクラスII分子およびアクセサリー分子をコードする種々の組み合わせのプラスミドをトランスフェクトされている。)
【0260】
下記で実施する分裂および活性化アッセイのために、ドロソフィラ・メラノガスターシュナイダー2(SC2)細胞(ATCCアクセッション番号CRL10974)を、実施例2で述べたH2−Ad のMHCクラスIIα鎖およびβ鎖をコードする遺伝子で形質転換した。選択補助細胞がMHCクラスII分子と一緒に発現される他のアッセイのためには、B7.1、B7.2およびICAM−1のための遺伝子もまた先に述べたMHC遺伝子含有細胞に導入した。
【0261】
アクセサリー分子は下記に規定するような選択濃度および組み合わせで発現させ、再現性をもつ一定のCD4+ 細胞活性化を可能にした。発現(硫酸銅による処置によって誘発される)に続いて、発現分子の濃度をモニターする。所望レベルの分子を発現している細胞の選択はフローサイトメトリーによる細胞株の分類によって実施する。表1は、ネズミIAd MHCクラスII分子のみまたは種々のアクセサリー分子と一緒に発現するドロソフィラ細胞株を示す。この表はまた、下記でさらに述べる分裂効果の結果の要約を提示する(" −”は効果無し、" +”は陽性分裂効果を示す)。合成抗原提示細胞の各タイプによって産生されたサイトカインの種類の要約もまた表示し、合わせて個々のサイトカインの産生がアップレギュレートであるか、ダウンレギュレートであるかをさらに矢印によって示す(これらは下記でもさらに説明する)。
【0262】
表1
アクセサリー分子 分裂反応 産生サイトカイン
無し − −
B7.1 + IL−4 IL−10 IL−2
B7.2 + IL−4 IL−10 IL−2
ICAM−1 − −
B7.1+B7.2 + IL−4 IL−10 IL−2
B7.1+ICAM−1 + ↑IL−2 ↓IL−4
↓IL−10
B7.2+ICAM−1 + ↑IL−2 ↓IL−4
↓IL−10
CD70+ICAM−1 + IL−2
CD70 − −
CD70+ICAM−1 + ↑IL−2
+B7.2
【0263】
抗原提示および最終的T細胞反応の効果を評価するために、抗原特異的T細胞は、T細胞レセプター遺伝子導入マウスから得たCD4+ 細胞であった。本明細書で用いた当該遺伝子導入系統は、オボアルブミンペプチド323−339に特異的なT細胞レセプターを発現するD01であった。D01マウス系統はデニス博士(Dr. Dennis Lo)から入手した。組換え体の表面発現MHCクラスII分子に対するT細胞反応性の評価のために、ホストの全てのAPCを反応集団から除外するようにデザインした技術を組み合わせてCD4+ 細胞を先ずリンパ節から精製した。続いて、これら高度に精製したCD4+ 細胞の少数(5×104 )をovaペプチドの存在下または非存在下でドロソフィラ細胞株とともに培養した。
【0264】
アクセサリー分子(B7.1、B7.2またはB7.2+ICAM−1)とともに、およびアクセサリー分子無しでMHCクラスIIの存在下でペプチド提示の効果を、文献の記載にしたがって先ず分裂アッセイで調べた(Webbら、Cell, 63:1249(1990))。簡単に記せば、5×104 CD4+T細胞を5×104 ドロソフィラ細胞とともに300μlの培養液でマイクロタイターウェルで培養した。採集18時間前に、100μlの上清をサイトカイン分析のために取り出し、1μCiの3 H−チミジンを各ウェルに添加した。表示した期間で(3、4または5日)それぞれのウェルをガラスファイバー上に採集し、液体シンチレーション計測器で計測した。
【0265】
図2で示されるように、MHCクラスII分子のみを発現している細胞株は、放射性チミジンの低い取り込みで示されるとおりT細胞の分裂反応を刺激できなかった。すなわち、より高いカウントが、μMで示した刺激ペプチドの量に対してプロットした分裂の増加を示している。しかしながら、B7.1またはB7.2同時刺激分子のどちらかの同時発現は、MHCクラスII提示ovaペプチドの能力に強力な分裂反応刺激を付与した。比較のために、脾臓APC(T−Sと表示)を用いた結果を表2に示す。
【0266】
図3に示すように、付着分子ICAM−1とともにMHCクラスII分子を発現するドロソフィラ細胞株は刺激されなかった。しかしながら、B7.2と一緒にICAM−1を発現させることによって、低濃度のペプチド抗原で分裂反応を刺激する合成APC細胞が得られた。これらの結果は、ネズミH2−Ad MHCクラスII分子および同時刺激分子(B7.1、B7.2およびB7.2+ICAM−1)を発現しているドロソフィラ細胞株は、分裂の増加によって測定した場合未感作CD4+T細胞を活性化できることを示している。
【0267】
分裂に加えて、特定のサイトカインを産生する再現性のあるCD4+T細胞の活性化が所望される。したがって、測定されるサイトカインに特異的な抗体を用いて標準的なELISAによって上記の培養で産生されたサイトカインもまた調べた。図4A−4Dに示したデータは、APC上の刺激分子に対して産生されたサイトカインの量をng/mlの濃度で表示している。
【0268】
これらのアッセイの結果は、組換えMHCクラスIIを発現するドロソフィラAPC上のアクセサリー分子の種々の組み合わせによって、図4A−4Dおよび図5A−5Dで示すように種々のサイトカインの産生がもたらされることを示している。IL−2産生(Th1型応答)のためには、B7.1またはB7.2と一緒にMHCクラスIIを発現する細胞株は図4Aで示されるように極めて弱く刺激された。顕著なレベルのIL−2を達成するためには、ドロソフィラ細胞株はICAM−1、B7およびクラスIIの同時発現を要求した(図5A)。B7.1およびB7.2を発現するドロソフィラ細胞株によるγ−インターフェロンの産生もまた貧弱に誘発された(図4D)。しかしながら、IL−2に関する結果とは対照的に、ICAM−1の同時発現はγ−IFNの産生に影響を与えなかった(図5C)。脾臓APC(T−Sと表示)は強いγ−IFN産生を誘発し、新たな分子がこのサイトカインの産生を調節していることを示唆した。
【0269】
IL−4産生(Th2型応答)は、一般には未感作CD4+T細胞の一次刺激中には観察されない。これらの観察とともに、脾臓APC(T−S)は、D01細胞およびovaペプチドとの培養後顕著なIL−4を誘発することができなかった(図5B)。しかしながら、B7.1もしくはB7.2のどちらかまたはその両方と一緒にクラスIIMHC分子を同時発現しているドロソフィラ細胞は、培養中に強いIL−4産生を誘発した(図4Bおよび5B)。興味深いことには、ICAM−1の添加はこのサイトカインの産生に対して拮抗的影響を示した(図5B)。同様に、図4Dおよび5Dに示すようにIL−10はIL−4に対して調節された。
【0270】
上記に述べたように、Th1型応答よりはむしろTh2型応答のサイトカインプロフィールに関する結果が、B7およびMHCクラスIIを発現している細胞株について得られた。対照的に、B7およびICAM−1と組み合わせたMHCクラスIIの同時発現はIL−4およびIL−10の産生と拮抗し、一方IL−2は強力に促進し、それによってTh1型応答を誘発し、CD4+T細胞の所望のT細胞サブセットへの活性化を駆動する能力についての本発明を支持する。これらの結果は、種々のパターンのサイトカインを産生する所望のCD4+T細胞集団を得る場合に使用するために、ドロソフィラ細胞株を規定のアクセサリー分子の組み合わせを発現するように仕立てることができることを決定的に示している。したがって、本明細書で述べた組成物および方法は、本発明の組成物および使用方法が有効であることを立証した。
【0271】
前述の記載は本発明を説明することを目的としており、制限を意図したものではない。数多くの変型および変更が本発明の範囲および本質から外れることなく実施可能であろう。
【0272】
【化1】

【0273】
【化2】
【0274】
【化3】
【0275】
【化4】
【0276】
【化5】
【0277】
【化6】
【0278】
【化7】
【0279】
【化8】
【0280】
【化9】
【0281】
【化10】
【0282】
【化11】
【0283】
【化12】
【0284】
【化13】
【0285】
【化14】
【0286】
【化15】
【0287】
【化16】
【0288】
【化17】
【0289】
【化18】
【0290】
【化19】
【0291】
【化20】
【0292】
【化21】
【0293】
【化22】
【0294】
【化23】
【0295】
【化24】
【0296】
【化25】
【0297】
【化26】
【0298】
【化27】
【0299】
【化28】
【0300】
【化29】
【0301】
本発明は以下に関するものである。
1.CD4+T細胞の活性化ための合成抗原提示細胞(APC)であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができるMHCクラスIIα鎖遺伝子、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができるMHCクラスIIβ鎖遺伝子(ここでα鎖およびβ鎖遺伝子が発現したとき、当該α鎖およびβ鎖は、ペプチドをローディングすることができるMHCクラスIIヘテロダイマーを形成する)および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる少なくとも1つのアクセサリー分子遺伝子を含み、クラスII遺伝子およびアクセサリー分子遺伝子の少なくとも1つを欠いていることを特徴とする細胞。
【0302】
2.当該α鎖およびβ鎖遺伝子がヒト起源である前記1のAPC。
【0303】
3.少なくとも1つのプロモーターが誘導性である前記1のAPC。
【0304】
4.当該α鎖、β鎖およびアクセサリー分子遺伝子が同じベクターに存在する前記1のAPC。
【0305】
5.当該α鎖、β鎖およびアクセサリー分子遺伝子の少なくとも1つが別のベクターに存在する前記1のAPC。
【0306】
6.当該APCが昆虫細胞である前記1のAPC。
【0307】
7.当該昆虫細胞がスポドプテラおよびドロソフィラからなる群から選ばれる前記6のAPC。
【0308】
8.さらにベクターに機能的に結合したネオマイシン耐性遺伝子を含む前記1のAPC。
【0309】
9.当該アクセサリー分子遺伝子が同時刺激分子をコードする前記1のAPC。
【0310】
10.当該同時刺激分子がB7.1またはB7.2である前記9のAPC。
【0311】
11.当該アクセサリー分子遺伝子が付着分子をコードする前記1のAPC。
【0312】
12.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記11のAPC。
【0313】
13.当該アクセサリー分子遺伝子がサバイバル分子をコードする前記1のAPC。
【0314】
14.当該サバイバル分子がFasリガンドまたはCD70である前記13のAPC。
【0315】
15.第一のアクセサリー分子のための遺伝子および第二のアクセサリー分子のための遺伝子を有する前記1のAPC。
【0316】
16.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記15のAPC。
【0317】
17.当該同時刺激分子がB7.1またはB7.2で、当該付着分子がICAM−1である前記16のAPC。
【0318】
18.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記15のAPC。
【0319】
19.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記15のAPC。
【0320】
20.当該サバイバル分子がCD70で、当該付着分子がICAM−1である前記19のAPC。
【0321】
21.当該第一および第二のアクセサリー分子が同時刺激分子である前記15のAPC。
【0322】
22.当該同時刺激分子がB7.1およびB7.2である前記21のAPC。
【0323】
23.第一のアクセサリー分子のための遺伝子、第二のアクセサリー分子のための遺伝子および第三のアクセサリー分子のための遺伝子を有する前記1のAPC。
【0324】
24.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記23のAPC。
【0325】
25.当該同時刺激分子がB7.2で、当該付着分子がICAM−1で、当該サバイバル分子がCD70である前記24のAPC。
【0326】
26.当該MHCクラスIIヘテロダイマーおよびアクセサリー分子が、ペプチドが当該ヘテロダイマー上にローディングされるときにCD4+T細胞を活性化させるために十分な量で当該APCの外側表面に存在する前記1のAPC。
【0327】
27.当該ペプチドが細胞外でローディングされる前記26のAPC。
【0328】
28.当該ペプチドが細胞内でローディングされる前記26のAPC。
【0329】
29.ベクター内の第四のプロモーターに機能的に結合した、抗原プロセシング補助分子を発現することができる抗原プロセシング補助遺伝子をさらに含む前記1のAPC。
【0330】
30.CD4+T細胞を活性化するために細胞フラグメント上に機能的に結合したMHCクラスIIヘテロダイマーおよび少なくとも1つのアクセサリー分子を有する前記1のAPC由来の細胞フラグメント。
【0331】
31.当該MHCクラスIIヘテロダイマーがエンプティである前記30の細胞フラグメント。
【0332】
32.ペプチドが当該MHCクラスIIヘテロダイマー上にローディングされる前記30の細胞フラグメント。
【0333】
33.CD4+T細胞を活性化するための合成抗原提示マトリックスであって、
a)担体、
b)担体に機能的に連結され、さらに選択ペプチドをローディングすることができるMHCクラスIIヘテロダイマーの細胞外部分、および、
c)ペプチドが当該ヘテロダイマーの細胞外部分にローディングされたときにCD4+T細胞を活性化させるために十分な数で当該MHCクラスIIヘテロダイマーおよびアクセサリー分子の細胞外部分が当該マトリックス上に存在するように、当該担体に機能的に結合した少なくとも1つのアクセサリー分子の細胞外部分を含むことを特徴とするマトリックス。
【0334】
34.当該担体が細胞フラグメントである前記33のマトリックス。
【0335】
35.当該担体が細胞である前記33のマトリックス。
【0336】
36.当該MHC分子の細胞外部分が、当該MHCクラスIIヘテロダイマーのトランスメンブレンドメインによって当該細胞に結合している前記35のマトリックス。
【0337】
37.当該担体がリポソームである前記33のマトリックス。
【0338】
38.当該担体が固体表面である前記33のマトリックス。
【0339】
39.当該MHCクラスIIヘテロダイマーの細胞外部分が、当該担体に当該部分を連結させるために抗体と反応するエピトープに結合している前記33のマトリックス。
【0340】
40.当該MHCクラスIIヘテロダイマーの細胞外部分が、当該担体に当該部分が連結させるためにニッケルと反応する(His)6 に結合している前記33のマトリックス。
【0341】
41.当該担体が多孔性物質である前記33のマトリックス。
【0342】
42.当該ペプチドがMHCクラスIIヘテロダイマーの細胞外部分にローディングされる前記33のマトリックス。
【0343】
43.当該MHCクラスIIヘテロダイマーの細胞外部分がエンプティである前記33のマトリックス。
【0344】
44.当該アクセサリー分子が同時刺激分子である前記33のマトリックス。
【0345】
45.当該同時刺激分子がB7.1またはB7.2である前記44のマトリックス。
【0346】
46.当該アクセサリー分子が付着分子である前記33のマトリックス。
【0347】
47.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記46のマトリックス。
【0348】
48.当該アクセサリー分子がサバイバル分子である前記33のマトリックス。
【0349】
49.当該サバイバル分子がFasリガンドまたはCD70である前記48のマトリックス。
【0350】
50.第一のアクセサリー分子および第二のアクセサリー分子を有する前記33のマトリックス。
【0351】
51.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記50のマトリックス。
【0352】
52.当該同時刺激分子がB7.1またはB7.2で、当該付着分子がICAM−1である前記51のマトリックス。
【0353】
53.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記50のマトリックス。
【0354】
54.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記50のマトリックス。
【0355】
55.当該サバイバル分子がCD70で、当該付着分子がICAM−1である前記54のマトリックス。
【0356】
56.当該第一および第二のアクセサリー分子が同時刺激分子である前記50のマトリックス。
【0357】
57.当該同時刺激分子がB7.1およびB7.2である前記56のマトリックス。
【0358】
58.さらに第三のアクセサリー分子を含む前記50のマトリックス。
【0359】
59.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記58のマトリックス。
【0360】
60.当該同時刺激分子がB7.2で、当該付着分子がICAM−1で、当該サバイバル分子がCD70である前記59のマトリックス。
【0361】
61.合成抗原提示細胞(APC)を製造する方法であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができる発現可能MHCクラスIIα鎖遺伝子で細胞を形質転換する工程、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができる発現可能MHCクラスIIβ鎖遺伝子で細胞を形質転換する工程、および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる発現可能なアクセサリー分子遺伝子で細胞を形質転換する工程を特徴とする方法。
【0362】
62.当該細胞が、当該α鎖、β鎖およびアクセサリー分子遺伝子の少なくとも1つをコードする遺伝子を欠く前記61の方法。
【0363】
63.さらに、ベクター内の第四のプロモーターに機能的に結合した、抗原プロセシング補助分子を発現することができる発現可能な抗原プロセシング補助遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0364】
64.当該α鎖およびβ鎖遺伝子がヒト起源である前記61の方法。
【0365】
65.少なくとも1つのプロモーターが誘導性である前記61の方法。
【0366】
66.当該α鎖、β鎖およびアクセサリー分子遺伝子が同じベクターに存在する前記61の方法。
【0367】
67.当該α鎖、β鎖およびアクセサリー分子遺伝子が別個のベクターに存在する前記61の方法。
【0368】
68.当該細胞が昆虫細胞である前記61の方法。
【0369】
69.当該昆虫細胞がスポドプテラおよびドロソフィラからなる群から選ばれる前記61の方法。
【0370】
70.さらにベクターに機能的に結合した発現可能なネオマイシン耐性遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0371】
71.当該アクセサリー分子遺伝子が同時刺激分子をコードする前記61の方法。
【0372】
72.当該同時刺激分子がB7.1またはB7.2である前記71の方法。
【0373】
73.当該アクセサリー分子遺伝子が付着分子をコードする前記61の方法。
【0374】
74.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記73の方法。
【0375】
75.当該アクセサリー分子遺伝子がサバイバル分子をコードする前記61の方法。
【0376】
76.当該サバイバル分子がFasリガンドまたはCD70である前記75の方法。
【0377】
77.さらに、第二のアクセサリー分子のための遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0378】
78.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記77の方法。
【0379】
79.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記77の方法。
【0380】
80.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記77の方法。
【0381】
81.さらに、第三のアクセサリー分子のための遺伝子で細胞を形質転換する工程を含む前記77の方法。
【0382】
82.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記81の方法。
【0383】
83.合成抗原提示細胞(APC)を製造する方法であって、
a)MHCクラスIIα鎖、MHCクラスIIβ鎖およびアクセサリー分子の少なくとも1つをコードする遺伝子を欠く細胞を提供する工程、及び、
b)当該細胞に欠けている(a)の遺伝子の各々のために発現可能であって、当該遺伝子を発現する能力を有するベクター内のプロモーターに機能的に結合した遺伝子で当該細胞を形質転換する工程からなることを特徴とする方法。
【0384】
84.合成抗原提示細胞(APC)を製造する方法であって、
a)MHCクラスIIα鎖、MHCクラスIIβ鎖、アクセサリー分子および抗原プロセシング補助分子の少なくとも1つをコードする遺伝子を欠く細胞を提供しする工程、及び、
b)当該細胞に欠けている(a)の遺伝子の各々のために発現可能であって、当該遺伝子を発現する能力を有するベクター内の機能できる第一のプロモーターに結合した遺伝子で当該細胞を形質転換する工程からなることを特徴とする方法。
【0385】
85.合成抗原提示マトリックスを製造する方法であって、
a)組換えMHCクラスIIヘテロダイマーの細胞外部分を提供する工程、
b)少なくとも1つの組換えアクセサリー分子の細胞外部分を提供する工程、及び、
c)MHCクラスIIヘテロダイマーにペプチドがローディングされたときにCD4+T細胞を活性化させるために十分な数で、担体に当該MHCクラスIIヘテロダイマーおよびアクセサリー分子を結合する工程からなることを特徴とする方法。
【0386】
86.当該アクセサリー分子が同時刺激分子である前記85の方法。
【0387】
87.当該同時刺激分子がB7.1またはB7.2である前記86の方法。
【0388】
88.当該アクセサリー分子が付着分子である前記86の方法。
【0389】
89.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記88の方法。
【0390】
90.当該アクセサリー分子がサバイバル分子である前記85の方法。
【0391】
91.当該サバイバル分子がFasリガンドまたはCD70である前記90の方法。
【0392】
92.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記26のAPCを提供する工程、及び
b)工程a)のAPCをCD4+T細胞と接触させ、それによって接触CD4+T細胞を増殖させ、さらに活性化CD4+T細胞に分化させる工程からなることを特徴とする方法。
【0393】
93.さらに、c)当該APCから活性化CD4+T細胞を分離する工程を含む前記92の方法。
【0394】
94.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記93の方法。
【0395】
95.さらに、当該懸濁物を患者に投与する工程を含む前記94の方法。
【0396】
96.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記30の細胞フラグメントを提供する工程、
b)当該MHCクラスIIヘテロダイマーにペプチドをローディングする工程、および、
c)当該ペプチドがローディングした細胞フラグメントをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなるを特徴とする方法。
【0397】
97.さらに、当該細胞フラグメントから活性化CD4+T細胞を分離する工程を含む前記96の方法。
【0398】
98.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記97の方法。
【0399】
99.さらに、当該懸濁物を患者に投与する工程を含む前記98の方法。
【0400】
100.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記33のマトリックスを提供する工程、
b)当該MHCクラスIIヘテロダイマーにペプチドをローディングする工程、および、
c)当該ペプチドをローディングした細胞マトリックスをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなることを特徴とする方法。
【0401】
101.さらに、当該マトリックスから活性化CD4+T細胞を分離する工程を含む前記100の方法。
【0402】
102.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記101の方法。
【0403】
103.さらに、当該懸濁物を患者に投与する工程を含む前記102の方法。
【0404】
104.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記1のAPC、前記30の細胞フラグメントまたは前記33のマトリックスを十分な量でペプチドライブラリーとインビトロで十分な時間接触させ、ペプチドがローディングしたMHCクラスIIヘテロダイマーを作製する工程、及び、
b)工程b)のペプチド積載MHCクラスIIヘテロダイマーをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなることを特徴とする方法。
【0405】
105.CD4+T細胞仲介免疫反応を変化させて患者の症状を治療する方法であって、
a)患者に固有のサイトカインプロフィールについて患者を分析する工程、
b)当該患者からCD4+T細胞を採取する工程、
c)当該CD4+T細胞を前記26のAPCと十分な量で十分な時間インビトロで接触させ、それによって当該接触CD4+T細胞の増殖分裂および、工程a)で得られたプロフィールに機能的に対抗するサイトカインプロフィールを産生する活性化CD4+T細胞への分化を誘発する工程、および、
d)当該活性化CD4+T細胞を当該患者に戻す工程からなることを特徴とする方法。
【0406】
106.当該症状が自己免疫疾患である前記105の方法。
【0407】
107.当該免疫疾患が、糖尿病、多発性硬化症、自己免疫性甲状腺炎、全身性紅斑性狼瘡、重症筋無力症、クローン病および炎症性大腸症候群から成る群から選ばれる前記106の方法。
【0408】
108.当該患者に固有のサイトカインプロフィールがCD4+Th1型応答によって生じる前記106の方法。
【0409】
109.当該患者に固有のサイトカインプロフィールが、インターロイキン−2、インターフェロン−γおよび腫瘍壊死因子からなる群から選ばれるサイトカインを含む前記108の方法。
【0410】
110.当該症状がアレルギーである前記105の方法。
【0411】
111.当該アレルギーが、喘息および接触過敏症からなる群から選ばれる前記110の方法。
【0412】
112.当該患者に固有のサイトカインプロフィールがCD4+Th2型応答によって生じる前記110の方法。
【0413】
113.当該患者に固有のサイトカインプロフィールが、インターロイキン−4およびインターロイキン−10からなる群から選ばれるサイトカインを含む前記112の方法。
【技術分野】
【0001】
関連出願の相互参照
この出願は上記と同じ発明の名称を有する、1996年5月23日に出願された同時継続米国特許仮出願第60/018,175号(これらの開示が参考として本明細書に含まれる)の一部継続出願である。
【0002】
技術分野
本発明は特別な抗原ペプチドに対する特異性を有するCD4+T細胞を活性化する物質及び方法、種々の症状の治療のためのインビボでの活性化T細胞の使用、並びにこれらの使用に適した組成物に関する。
【背景技術】
【0003】
背景
T細胞レパートリーは主として胸腺中のT細胞発生中に成形されるが、成熟CD4+T細胞がまた胸腺外で調節される。活性化の或る条件はアネルギーまたはクローン排除を反映する寛容をもたらすが、その他の条件は観察される応答の型の変化をもたらす。CD4+T細胞について、機能性表現型のこの変化は主として産生されるサイトカインのパターンの変化である。急性活性化を受けるCD4+T細胞は多種サイトカインを産生する能力を維持するが、慢性刺激の条件下で得られたT細胞はサイトカイン産生の更に制限されたパターンを頻繁に示す。例えば、インビトロでの反復刺激により維持されたT細胞クローンは、Th1型細胞及びTh2型細胞と称されるCD4+T細胞の2種の主要な機能性カテゴリーを形成した。Th1型細胞は主としてインターロイキン−2 (IL−2) 、インターフェロン−γ(IFN- γ)及び腫瘍壊死因子(TNF) を産生し、これらの全てが炎症性サイトカインと称される。対照的に、Th2型細胞は典型的にはIL−4、IL−5、及びIL−10を産生し、抗体産生及びTh1型細胞の応答の調節に重要である。
【0004】
サイトカイン産生におけるこのような極度の分離はしばしばインビボT細胞応答中に見られないが、リーシュマニアの如き或る型の感染症からの回復はIL−2/IFN-γの優先産生と関連する。リーシュマニアに対するTh2応答を上昇するマウスは感染を抑制することができず、最終的に死亡する。Th2型応答のサイトカインの不適当な産生はアレルギー型疾患、例えば、喘息及び接触過敏症に頻繁に関連していた。CD4+T細胞の活性化及びアレルギー疾患における役割に関する総説について、Hetzel及びLamb, Clinical Immunol.Immunopath., 73:1−10 (1994)を参照のこと。
【0005】
おそらく、ヒト疾患とサイトカイン産生の非対称のパターンの最も強い関連はTh1応答及びTh1型サイトカインと自己免疫疾患の関連である。実験モデルにおける強力な証拠は、糖尿病を含む自己免疫の多くの型、多発性硬化症、自己免疫甲状腺炎等の実験モデルがTh1型CD4+T細胞により媒介されることを示す。これらのモデル中のIL−4の如きTh2関連サイトカインの発現は自己免疫疾患の発生を阻害する。Th2型サイトカインはTh1型細胞の応答を低下させ、一方、Th1型サイトカインはTh2型応答の発生を拮抗作用する。
【0006】
特別な活性化T細胞サブセットと特別な症状の関連に鑑みて、それ故、疾患の進行を変化するのに極めて有益であるプロセスである所望のT細胞サブセットへのCD4+T細胞の増殖及び活性化を誘導することができるようにとの要望が存する。一つの可能な解決は必要によりアレルギーまたは自己免疫症状を有してもよい被験者から最初に単離されるCD4+T細胞をインビトロで活性化して好ましいサイトカインプロフィールを分泌する細胞を生産することである。次いで得られる活性化T細胞が被験者に再度導入されて疾患の進行を変化し、そしておそらく長期治癒さえも与える。
【0007】
本発明により今解決された、このアプローチにおける挑戦は所望の治療サイトカインプロフィールを生じるCD4+T細胞サブセットを再現して生じる活性化条件を特定することの難しさである。特別なサイトカインの発現は特別な抗原提示細胞(APC) 及びそれらの関連アクセサリー(補助)分子と関連している。T細胞同時刺激に関係するアクセサリー分子として利用できる表面タンパク質の総説について、Mondino 及びJenkins, J.Leukocyte Biol., 55:805-815 (1994) を参照のこと。APCにより産生されたサイトカイン及び協調して発現されたアクセサリー分子の両方それ自体が抗原の型、T細胞レセプター(TCR) −抗原相互作用のアフィニティー、抗原濃度等を含む多くの因子により調節されるので、抗原提示後のT細胞活性化の成果の予想は伝統的に非常に困難である。実際に、付加的なアクセサリー分子がインビボでの活性化プロセスに提案されていたので、多くの多様な分子がT細胞応答の調節に関係し、コンビナトリアル様式で作用してT細胞活性化の成果に影響することが次第に明らかになってきている。
【0008】
本発明の以前には、一種以上の選択されたアクセサリー分子と一緒に選択されたMHCクラスII分子の同時発現は可能ではなかった。今、本発明はT細胞の再現可能な活性化により好ましいT細胞表現型を予測可能に生成してTh1またはTh2型T細胞を生じることの解決を提供する。本発明は特定のアクセサリー分子と組み合わせてMHCクラスII分子を中性バックグラウンドで提供する合成APCの生成を記載する。MHCクラスII分子及び特定のアクセサリー分子は非哺乳類昆虫細胞中で発現され、その分子を示す昆虫細胞を含む合成APCの種々の形態で提示し得る。
【0009】
本発明のMHCクラスII/アクセサリー分子組成物について発現及び提示ビヒクルとして昆虫細胞を使用することの利点は、細胞が調節サイトカインを内因的に産生せず、しかも哺乳類アクセサリー分子を発現しないことである。これはT細胞応答を変化することができる多くの分子を発現する哺乳類APCを使用することの特有の予測不能を解決する。加えて、本発明に記載された昆虫細胞発現系は或る種の制限環境、例えば、温度要件下で生成し得る結合ペプチドを含まないMHCクラスII分子(即ち、“エンプティー”分子)の発現を与える。生理温度で、これらの“エンプティー”分子は通常細胞表面に達することができない。何となれば、結合ペプチドを含まないクラスIIは非常に熱不安定であるからである。本発明は内因的にローディングされたカウンターパートを与える能力とともに選択されたペプチドの外因的ローディングを可能にする“エンプティー”MHCクラスII組成物の能力を利用する。
【0010】
組換えグリコシル−ホスファチジルイノシトール(GPI) 修飾MHCクラスI分子(HLA-A2.1:GPI/β2m) がTykocinskiの国際公開番号WO 96/12009 に記載されたようにして上記昆虫細胞系中で生成されて抗原提示細胞を生産した。その公開において、組換えGPI修飾MHCクラスI分子はその後の細胞膜への再とり込みのためにアフィニティー精製により昆虫細胞から単離される。その他の局面において、その公開はGPI修飾B7.1同時刺激分子と細胞膜に同時固定されたGPI修飾MHCクラスI分子の調製を記載している。その公開はGPI修飾MHCクラスII分子がMHCクラスIのそれらについて記載されたようにして調製し得ることを記載しているが、その公開はこのような調製について詳細を示していない。
【0011】
対照的に、本発明はB7.1の如き一種以上のアクセサリー分子と一緒に選択されたMHCクラスIIハプロタイプを同時発現してCD4+T細胞を活性化して特別なT細胞サブセット、Th1細胞またはTh2細胞(これらは好ましいサイトカインプロフィールに影響する)への分化をもたらすことに基く特異な手段を提供し、記載する。本発明は好ましいT細胞サブセットへインビトロでCD4+T細胞を選択的に活性化し、その後に患者への活性化T細胞の再導入を可能にするという利点を与える。こうして、本発明は個々の提示分子を選択された組み合わせの発現のための特別なアクセサリー分子と化合する能力を与え、これがその他のアプローチでは利用可能ではない所望のT細胞サブセットへのCD4+T細胞の選択的活性化について再現性及び予測可能性を可能にする。
【発明の概要】
【課題を解決するための手段】
【0012】
発明の簡単な要約
同時刺激分子及び付着分子を含む選択されたアクセサリー分子と組み合わせて発現された組換えMHCクラスII分子はMHCクラスII分子ヘテロダイマーがペプチドとの複合体形成のために発現される標的細胞を認識する武装化(armd)エフェクターT細胞になるようにCD4+T細胞を活性化するのに有効であることが今発見された。活性化は特別なサイトカインを分泌するエフェクターT細胞サブセット、Th1及びTh2への増殖及び分化を特徴とする。Th1型T細胞及びTh2型T細胞は夫々炎症細胞及びTヘルパー細胞と称される。
【0013】
こうして、本発明はCD4+T細胞を活性化するために一種以上のアクセサリー分子と組み合わせて哺乳類、好ましくはヒトMHCクラスII分子を生成し、提示するための合成抗原提示系(また、APCと称される)に関する。
【0014】
一実施態様において、その系は担体と担体に機能的に結合され、かつ選択されたペプチドに結合することができるMHCクラスIIヘテロダイマー分子の少なくとも細胞外部分とを有する合成抗原提示マトリックスに関する。また、マトリックスは担体に機能的に結合されたアクセサリー分子を含む。アクセサリー分子はCD4+T細胞に存在する特定のレセプターと相互作用する。MHCクラスII分子及びアクセサリー分子は、ペプチドがMHC分子の細胞外部分に結合される時にMHCクラスII/ ペプチド組み合わせに特異性のCD4+T細胞の集団を活性化するのに充分な数で存在する。
【0015】
アクセサリー分子と一緒に、MHCクラスIIヘテロダイマーまたはそのMHC に特異性のペプチドをローディングされたその少なくとも細胞外部分を有する抗原提示マトリックスはCD4+T細胞を活性化する際に相乗反応を与えることがわかった。アクセサリー分子の例はB7.1及びB7.2を含む同時刺激分子、付着分子、例えば、細胞内付着分子−1 (ICAM−1) 及びリンパ球機能関連抗原−3 (LFA−3)、並びにサバイバル分子(survival molecule)、例えば、Fasリガンド(FasL)及びCD70である。幾つかの実施態様において、このようなアクセサリー分子の細胞外部分がまた本発明に使用し得る。
【0016】
マトリックスに使用される担体は幾つかの異なる形態をとり得る。担体の例として、固体担体、例えば、金属またはプラスチック、多孔質材料、例えば、樹脂または変性セルロースカラム、ミクロビーズ、ミクロタイタプレート、赤血球及びリポソームが挙げられる。
【0017】
担体の別の型は細胞フラグメント、例えば、細胞膜フラグメントである。全細胞がまた担体として意図されている。この実施態様において、マトリックスは実際に少なくとも一種のアクセサリー分子とともにMHCクラスIIα鎖及びβ鎖の発現のための遺伝子を含む一種以上の発現ベクターで形質転換された細胞である。次いで発現されたタンパク質が細胞膜に輸送され、そこでクラスII鎖のトランスメンブランドメインがアンカーを与え、細胞外ドメインが外細胞表面に露出されることを可能にし、それにより合成抗原提示細胞(APC) を生じる。発現ベクターは構成的または誘導性であるプロモーターに機能的に結合された選択された遺伝子(cDNA配列の形態であることが好ましい)を含む。
【0018】
MHC α鎖及びβ鎖は一緒に会合してMHCクラスIIヘテロダイマーを形成し、これがそのヘテロダイマーに特異性のペプチドに結合する。本発明により、ペプチドをMHCクラスIIヘテロダイマーにローディングする二つの方法が意図されている。一実施態様において、ペプチドがペプチドフラグメントへの内在化無傷タンパク質のタンパク質分解プロセシング後に細胞内にローディングされる。次いでペプチドが新たに生成されたMHCクラスII分子にローディングされ、その間それらは依然として細胞内にある。また、MHCクラスII分子は細胞表面でエンプティー分子として発現され、次いで合成または処理ペプチド試薬がMHCクラスIIヘテロダイマーに細胞外でローディングされる。
【0019】
ベクター中のプロモーターに機能的に結合された少なくとも一種のアクセサリー分子遺伝子をコードするためのヌクレオチド配列がまた細胞に導入される。発現後に、アクセサリー分子はMHCクラスII/ ペプチド複合体に特異性のCD4+T細胞リンパ球の集団を活性化するのに充分な数でMHCクラスIIヘテロダイマーと一緒に細胞の表面に協調的に固定される。抗原プロセシング補助分子と称されるその他の分子がまた組換えAPCを生成する際の使用について意図されている。これらの分子はAPCとして使用される細胞により、または上記のような発現ベクター系により外因的に与えられる。このような抗原プロセシング補助分子の例として、不変鎖、リソソーム酵素並びにH2−M分子及びH2−O分子が挙げられる。
【0020】
細胞系は、上記遺伝子の少なくとも一つが細胞系が由来する細胞中に自然に存在しないという点で合成である。変温細胞系を使用することが好ましい。何となれば、MHC分子が熱不安定であるからである。或る範囲の種がこの目的に有益である。例えば、この使用のための多数の種を説明するPetersenらの米国特許第5,314,813 号(その開示が参考として本明細書に含まれる)を参照のこと。真核生物細胞及び好ましくは昆虫細胞がAPCとして使用される。好ましい昆虫細胞として、ドロソフィラ(Drosophila)(ショウジョウバエ)及びスポドプテラ(Spodoptera)(チョウ)が挙げられる。
【0021】
MHCクラスII分子は昆虫細胞、例えば、ドロソフィラ細胞及びスポドプテラ細胞中で発現された。これらの細胞は哺乳類免疫系の全ての成分を有しないので、ペプチドローディング機構に関係する種々のタンパク質がこのような細胞には不在である。哺乳類ペプチドローディング機構の欠如は、細胞が28℃のような熱安定温度制限条件で培養される時に導入された哺乳類MHCクラスII分子が細胞表面でエンプティー分子として発現されることを可能にする。対照的に、37℃で、エンプティークラスI分子は熱不安定であり、崩壊する傾向がある。こうして、MHCクラスII発現ドロソフィラ細胞を固定MHCクラスII分子に特異的に結合するペプチドとともにインキュベートすることにより、実際に夫々のクラスII分子が一つの同じペプチドでローディングされる。更に、本発明はあらゆる既知のMHCクラスIIα鎖及びβ鎖遺伝子を発現ベクターに導入し、それによりいずれか一種の哺乳類中で発現されるMHCクラスII分子の数の固有の制限を解消する手段を提供する。
【0022】
本発明において、CD4+T細胞をサイトカインインターロイキン−2 (IL−2) の増加を特徴とするTh1型応答に誘導するのに特に有効な相乗反応はペプチド、同時刺激分子、及び付着分子で結合されたMHCクラスII分子を有するドロソフィラ抗原提示細胞から生じる。特に、CD4+増殖と対にされたIL−2産生の高度に有効な相乗的発生はB7.2及びICAM−1の組み合わせから生じる。対照的に、ICAM−1ではなく、B7.1またはB7.2では、ペプチドをローディングされたドロソフィラAPC系はIL−4の増加を特徴とするTh2型応答を誘導した。こうして、ICAM−1はTh2型応答を拮抗作用してTh1型表現型をもたらす。
【0023】
増殖応答と対にされたIL−2産生を特徴とするTh1表現型はまたB7.2を用いて、またはそれを用いずにICAM−1と同時に発現されたCD70を有する合成抗原提示細胞から生じた。
【0024】
それ故、本発明のAPC中の発現のための少なくとも一種のアクセサリー分子遺伝子と組み合わせたMHCクラスII遺伝子の選択はCD4+T細胞の増殖及び表現型活性化を行うのに所望される成果に応じて適応し得る。
【0025】
また、本発明は、MHCクラスIIヘテロダイマー及びアクセサリー分子の遺伝子を含む少なくとも一種の発現ベクターが導入される上記合成APC系をつくる方法に関する。
【0026】
インビトロで活性化CD4+T細胞を生産する方法がまた意図されている。一つの好ましい方法はインビトロでCD4+細胞を抗原特異的様式でCD4+T細胞の集団を活性化するのに充分な期間にわたって上記の合成MHCクラスII/アクセサリー分子を有するAPCと接触することを含む。更に、その方法は(1) 活性化CD4+細胞を抗原提示マトリックスから分離し、(2) 活性化CD4+細胞を許されるキャリヤーまたは賦形剤中で懸濁させ、そして(3) その懸濁液を治療を要する個体に投与することを含んでもよい。先に説明したように、抗原は天然または未分解のタンパク質またはポリペプチドを含んでもよく、またはそれらは哺乳類MHCクラスIIヘテロダイマー分子と一緒のインキュベーションの前に少なくとも8アミノ酸残基を含むペプチドフラグメントに開裂または合成された抗原ポリペプチドを含んでもよい。
【0027】
上記の所望のT細胞サブセットへのCD4+T細胞の活性化を誘導することができるという実用性に加えて、あらゆるMHCクラスII分子を発現する能力はその特別なMHCクラスII分子に特異性の未知のCD4+活性化ペプチドを同定する手段を与える。このようなものとして、本発明は特別なMHCクラスIIヘテロダイマーを発現する合成APCによるペプチドライブラリーのスクリーニングによるCD4+T細胞の活性化を意図している。
【0028】
更に別の実施態様において、本明細書に記載された合成APC系は細胞の異種集団からの反応性CD4+T細胞の単離に有益である。このような単離は患者の症状で進行中のCD4+T細胞介在性応答を監視することができることを与える。
【0029】
上記の別の変化において、CD4+T細胞を好ましいサイトカインプロフィールを産生するための特別なT細胞サブセットに選択的に活性化することができることに鑑みて、本発明は望ましくないCD4+応答により媒介される患者の症状の治療方法に関する。Th1型応答またはTh2型応答を特徴とするこのような症状として、自己免疫疾患、アレルギー及び癌が挙げられる。治療目標は好ましいT細胞サブセットに活性化されたCD4+T細胞を導入して進行中のCD4+T細胞応答を拮抗作用することである。こうして、その方法は(1) 患者から静止または未感作(naive)CD4+細胞を含む液体サンプルを得、(2) インビトロでCD4+細胞を抗原特異的様式でCD4+細胞を活性化するのに充分な期間にわたって本発明の選択された合成ペプチドローディングされたAPCと接触させ、そして(3) 活性化CD4+細胞を患者に投与することを含む。
その他の実施態様は当業者に明らかである。
【図面の簡単な説明】
【0030】
【図1A】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図1B】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図1C】図1A〜1C は発現プラスミドpRmHa−2及びpRmHa−3の構築を図示する。図1A中、pRmHa−2構築が示され、図1B中、pRmHa−3構築が示され、また図1C中、pRmHa−3ベクターが示され、制限部位、ポリリンカー、プロモーター、及びポリアデニル化部位、並びにヌクレオチド配列が発現のために挿入し得る部位を示す。
【図2】図2は表面発現されたMHCクラスIIヘテロダイマーにローディングされたオボアルブミン(OVA)ペプチドを用いて、またはそれを用いずにドロソフィラ細胞系の存在下で培養した時のDO10T細胞レセプター(TCR) トランスジェニックマウス細胞系(Tg細胞)の増殖応答を示す。増殖は実施例5に記載されるようにして分析される。増殖はX軸の次第に増加する濃度(μM)のOVAペプチドに対しY軸にプロットされたようにカウント/分(cpm)×1000で測定される。T-S(ダイヤモンドでマークした線)は対照脾臓APCによる応答を示す。組換えMHCクラスII単独による増殖応答が閉じた円を有する線で示される。同時刺激分子B7.1またはB7.2と組み合わされたMHCクラスIIによる増殖応答が夫々開いた正方形及び閉じた正方形を有する線で示される。
【図3】図3は組換えMHCクラスII単独(閉じた円を有する線)、MHCクラスII+B7.2( 閉じた正方形を有する線)、MHCクラスII+ICAM−1(開いた三角形を有する線)、及びMHCクラスII+B7.2及びICAM−1(開いたダイヤモンドを有する線)の増殖応答を示す。その他の詳細について図2脚注を参照のこと。
【図4】図4A〜4Dは組換えMHCクラスII単独またはB7.1もしくはB7.2同時刺激分子と組み合わせた組換えMHCクラスIIを有するドロソフィラAPCの存在下で培養した時のCD4+T細胞の活性化に応答して生じたサイトカインプロフィールを示す。脾臓APC(T-Sと標識した)は対照アッセイである。アッセイは実施例5に記載されるようにして行われる。図4A〜4DはY軸にプロットされたようにサイトカインIL−2、IL−4、IFN-γ及びIL−10 (単位ng/ml)を夫々示す。サイトカインプロフィールは培養の3日目〜5日目の3日間にわたって評価された。
【図5】図5A〜5Dは組換えMHCクラスII単独またはB7.2同時刺激分子、ICAM−1並びにB7.2とICAM−1と組み合わせた組換えMHCクラスIIを有するドロソフィラAPCの存在下で培養した時のCD4+T細胞の活性化に応答して生じたサイトカインプロフィールを示す。図5A〜5DはY軸にプロットされたようにサイトカインIL−2、IL−4、IFN-γ及びIL−10 (単位ng/ml)を夫々示す。その他の詳細について図4脚注を参照のこと。
【発明を実施するための形態】
【0031】
発明の詳細な説明
A.定義
アミノ酸残基:ペプチド結合の位置におけるポリペプチドの化学消化(加水分解)後に生成されたアミノ酸。本明細書に記載されたアミノ酸残基は“L”異性体であることが好ましい。しかしながら、“D”異性体の残基は、所望の機能性がポリペプチドにより保持される限り、あらゆるL−アミノ酸残基を置換し得る。NH2 はポリペプチドのアミノ末端に存在する遊離アミノ基を表す。COOHはポリペプチドのカルボキシ末端に存在する遊離カルボキシ基を表す。通常のポリペプチド命名法(J.Biol.Chem., 243: 3552-3559 (1969) に記載され、37 CFR§1.822(b)(2)で採用される)を遵守して、アミノ酸残基の略号が下記の対応表に示される。
【0032】
対応表
記号 アミノ酸
1文字 3文字
Y Tyr チロシン
G Gly グリシン
F Phe フェニルアラニン
M Met メチオニン
A Ala アラニン
S Ser セリン
I Ile イソロイシン
L Leu ロイシン
T Thr スレオニン
V Val バリン
P Pro プロリン
K Lys リシン
H His ヒスチジン
Q Gln グルタミン
E Glu グルタミン酸
Z Glx Glu 及び/またはGln
W Trp トリプトファン
R Arg アルギニン
D Asp アスパラギン酸
N Asn アスパラギン
B Asx Asn 及び/またはAsp
C Cys システイン
X Xaa 未知またはその他
【0033】
式により本明細書に表される全てのアミノ酸残基配列はアミノ末端からカルボキシ末端への通常の方向で左から右への配向を有することが注目されるべきである。加えて、“アミノ酸残基”という用語は対応表にリストされたアミノ酸並びに修飾アミノ酸及び特異アミノ酸、例えば、37 CFR§1.822 (b)(4)にリストされ、また本明細書に参考として含まれるものを含むように広く定義される。更に、アミノ酸残基配列の最初または最後のダッシュは一つ以上のアミノ酸残基の更に別の配列に結合されたペプチドまたはNH2 もしくはアセチルの如きアミノ末端基もしくはCOOHの如きカルボキシ末端基への共有結合を示す。
【0034】
組換えDNA(rDNA)分子:二つのDNAセグメントを機能的に結合することにより生成されたDNA分子。こうして、組換えDNA分子は自然には通常一緒に見られない少なくとも二つのヌクレオチド配列を含むハイブリッドDNA分子である。共通の生物起源を有しない、即ち、進化上異なるrDNAが“異種”と称される。
【0035】
ベクター:細胞中で自律複製することができ、DNAセグメント、例えば、遺伝子またはポリヌクレオチドが付着セグメントの複製をもたらすように機能的に結合し得るrDNA分子。一種以上のポリペプチドをコードする遺伝子の発現を誘導することができるベクターが本明細書中“発現ベクター”と称される。
【0036】
上流:DNA転写の方向と反対の方向で、それ故、非コーディングストランドでは5'から3'に進み、またはmRNAでは3'から5'に進む。
【0037】
下流:配列転写または読み取りの方向にDNA配列に沿って、即ち、DNAの非コーディングストランドに沿って3'から5'の方向またはRNA転写産物に沿って5'から3'の方向に移動している。
【0038】
読み取り枠:遺伝子の構造タンパク質をコードする部分、または構造遺伝子を形成する翻訳に使用される連続のヌクレオチドトリプレット(コドン)の特別な配列。読み取り枠は翻訳開始コドンの位置に依存する。
【0039】
ポリペプチド:連続アミノ酸残基のα−アミノ基とカルボキシ基の間のペプチド結合により互いに連結された線状の一連のアミノ酸残基。
【0040】
タンパク質:ポリペプチドのように互いに連結された線状の一連の50より大きいアミノ酸残基。
【0041】
レセプター:レセプターは別の分子に特異的に(非ランダムに)結合し得る分子、例えば、タンパク質、糖タンパク質等である。
【0042】
実質的に精製され、または単離された:ポリペプチドまたはタンパク質に関して使用される場合、その用語はそれらと自然に随伴する成分から分離された分子を記載する。典型的には、モノマータンパク質は、サンプルの少なくとも約60%〜75%が単一ポリペプチド主鎖を示す時に実質的に純粋である。少量の変異体または化学修飾が典型的には同じポリペプチド配列を共有する。実質的に精製されたタンパク質は典型的にはタンパク質サンプルの約85%〜90%以上、更に通常約95%を構成し、好ましくは約99%以上純粋であろう。タンパク質またはポリペプチドの純度または等質性は当業界で公知の幾つかの手段、例えば、サンプルのポリアクリルアミドゲル電気泳動、続いて染色によるその視覚化により示し得る。或る目的には、高分解能が必要とされ、高速液体クロマトグラフィー(HPLC)または同様の精製手段が利用される。
【0043】
合成ペプチド:天然産タンパク質及びそのフラグメントを含まないペプチド結合により一緒に結合されたアミノ酸残基の化学的に生成された鎖。
【0044】
B.MHCクラスIIヘテロダイマー、アクセサリー分子及び抗原プロセシング補助分子
本発明はCD4+T細胞を活性化するのに使用するための合成抗原提示系に関する。T細胞レパートリーは主として胸腺中のT細胞発生中に形成されるが、成熟CD4+T細胞がまた胸腺外で調節される。活性化の或る条件がアネルギーまたはクローン排除を反映するトレランスをもたらすが、その他の条件は観察される応答の型の変化をもたらす。CD4+T細胞について、機能性表現型のこの変化は主として産生されるサイトカインのパターンの変化である。急性活性化を受けるCD4+T細胞は多種サイトカインを産生する能力を維持するが、慢性刺激の条件下で得られたT細胞はサイトカイン産生の更に制限されたパターンを頻繁に示す。例えば、インビトロの反復刺激により維持されたT細胞クローンはTh1型細胞及びTh2型細胞と称されるCD4+T細胞の二つの主要な機能性カテゴリーを形成した。Th1型細胞は主としてインターロイキン−2 (IL−2) 、インターフェロン−γ(IFN- γ)及び腫瘍壊死因子(TNF) を産生し、これらの全てが炎症性サイトカインと称される。このようなものとして、Th1細胞は時折炎症性T細胞と称され、これらはその後にマクロファージを活性化して、CD4+T細胞に提示された病原タンパク質のプロセシングから生成されたペプチドにより媒介されるようにそれらが宿している小胞内病原体を死滅させる。対照的に、Th2型細胞は典型的にはIL−4、IL−5、及びIL−10 を産生し、抗体産生及びTh1型細胞の応答の調節に重要である。このようなものとして、Th2細胞はヘルパーT細胞と時折称され、これらはB細胞を活性化して、CD4+T細胞細胞に提示される細胞外病原体及び毒素に由来するペプチドに応答して抗体をつくる。
【0045】
CD4+T細胞はCD3複合体のタンパク質と会合したα:βまたはγ:Δヘテロダイマーレセプターの存在と一緒にその細胞表面でCD4+コレセプターを有するT細胞またはTリンパ球と定義される。
【0046】
CD4+T細胞サブセットの活性化は抗原提示細胞により誘発された刺激の型に依存性の上記のサイトカインの選択産生と協調した応答性T細胞集団の増殖を特徴とする。後者は抗原を処理でき、リンパ球活性化に必要とされる分子と一緒に細胞表面でそれらのペプチドフラグメントを露出する高度に特殊化された細胞と定義される。CD4+T細胞活性化の特異性は抗原提示細胞(APC) の表面のMHCクラスIIヘテロダイマーに結合されたペプチド抗原のT細胞抗原レセプター(TCR)認識に基いている。T細胞の主要APCは樹状細胞、マクロファージ及びB細胞である。加えて、APC誘導非抗原性同時刺激シグナルがCD4+T細胞活性化に寄与する役割を果たす。
【0047】
本発明は上記の自然免疫応答メカニズムに基いて合成抗原提示系を使用して、同時刺激分子と広く称される一種以上のアクセサリー分子と一緒に組換えMHC クラスII分子によりCD4+T細胞の活性化を操作する。後者として、特定の同時刺激分子、付着分子及びサバイバル分子等が挙げられる。本発明のその他の局面において、合成抗原提示系はペプチドローディングされたMHCクラスII分子を生成するのに有益である抗原プロセシング補助分子を更に含む。本発明に関して、合成抗原提示系はCD4+T細胞をインビトロ及びインビボで活性化するのに有益である。これらの局面がCD4+細胞応答の変更方法に関する節Eに説明される。
【0048】
こうして、上記のように、本発明の抗原提示系は少なくとも2種の主成分を有する。第一成分はTCR による認識によりCD4+T細胞活性化の特異性を与えるペプチドに結合することができる組換えMHCクラスIIヘテロダイマーの少なくとも細胞外の部分である。第二主成分はCD4+T細胞の活性化において非抗原特異性同時刺激シグナルを与える少なくとも一種のアクセサリー分子の少なくとも細胞外の部分である。別の実施態様において、抗原提示系及び非抗原同時刺激シグナルの全分子が使用される。
【0049】
説明を容易にするために、MHC分子の細胞外部分が本発明の或る局面で使用し得ることを理解して、MHCクラスIIヘテロダイマーが一般に説明される。本発明に必要なMHC分子の部分はCD4+T細胞への提示のために抗原ペプチドに結合する部分である。
【0050】
本発明は組換えMHCクラスIIヘテロダイマーがMHCクラスIIヘテロダイマーと既に複合体形成されたペプチドでベクター形質転換された合成抗原提示細胞により産生されることを可能にする。また、それらと複合体形成されたペプチドを未だ有していないエンプティーMHCクラスIIヘテロダイマーが生成される。この後者の実施態様が特に有益である。何となれば、それは特別なペプチドとの複合体形成またはMHCクラスIIヘテロダイマーが発現された後のペプチドのライブラリーのスクリーンを可能にするからである。
【0051】
1.MHCクラスII遺伝子及びコードされたヘテロダイマー
MHCクラスII分子は二つの鎖、α及びβ(これらの両方が膜をスパンする)の非共有結合複合体からなる細胞表面糖タンパク質である。ペプチド結合クレフトが共同鎖の間に形成される。MHCクラスII分子を結合するペプチドは長さが可変であり、ペプチドの末端から種々の距離にあるアンカー残基を有し、それにより結合ポケットのクレフト中にしっかりと結合されていない末端を有するペプチドをもたらす。Janeway 及びTravers, Immunobiology, Section 4-4, Current Biology LTD,第2編, 1996を参照のこと。抗原提示ペプチドの更に別の局面が節Dに説明される。
【0052】
インビボでは、エンプティーMHCクラスIIヘテロダイマーは不安定化されるようになり、続いて細胞表面から除去され、それによりMHC分子が周囲の細胞外液体からのペプチド(これらはT細胞に特異的に有害に作用するであろう)を獲得することを阻止する。本発明において、合成抗原提示細胞系は不安定化イベントを受けていない細胞表面でエンプティーMHCクラスIIヘテロダイマーの生成を可能にする。その結果として、表面発現された組換えMHCクラスIIヘテロダイマーへの抗原ペプチドのローディングがCD4+T細胞活性化及びサイトカイン産生を操作する際のその後の使用のために促進される。
【0053】
本発明、協調抗原との選択されたMHCクラスII分子の特別な組み合わせはMHCクラスII遺伝子中のコンセンサスヌクレオチド配列の存在により促進される。以下更に記載されるこれらの領域は哺乳類中のMHC複合体中の多種MHCクラスII遺伝子並びに夫々の遺伝子の多種対立遺伝子の読み出し及び使用を可能にする。換言すれば、MHCは幾つかの遺伝子及び夫々の遺伝子の多種対立遺伝子を有する多遺伝子性かつ多形性の両方である。
【0054】
ヒトでは、MHCはHLAと称され、一方、マウスでは、それはH−2と称される。ヒトのMHCクラスIIα鎖及びβ鎖遺伝子の三つの対がHLA−DP、HLA−DQ及びHLA−DRと称された。HLA−DRクラスターは特別のβ鎖遺伝子を含む。このようなものとして、これらの遺伝子の三つの組がMHCクラスII分子の四つの型を生じ得る。MHCクラスII遺伝子及びヒト遺伝子から得られるコードされたα鎖及びβ鎖はヒト起源であると言われる。マウスでは、MHCクラス遺伝子がH2−M、H2−A及びH2−Eと称される。夫々のMHCクラスII分子は異なる範囲のペプチドを結合するので、多遺伝子座の存在は、夫々のクラスの唯一のMHCクラスII分子が細胞表面で発現された場合よりも広範囲の異なるペプチドを提示する能力を個体に与える。
【0055】
多形性MHCクラスII遺伝子は唯一または二三のアミノ酸だけ変化する相当するタンパク質をコードするが、異なる対立遺伝子変異体は20までのアミノ酸を異にする。その結果として、MHCクラスII多様性はT細胞による抗原認識の能力を拡大する。更に、MHC制限により、T細胞は特別なMHC分子に関してペプチドを認識するが、別の分子に提示された時に認識しないことが示された。こうして、T細胞レセプター特異性はペプチド及びそれを結合するMHC分子の両方により付与される。
【0056】
選択されたα鎖及びβ鎖を含む本発明のMHCクラスIIヘテロダイマーはオリゴヌクレオチドプライマーの特定の対によるMHCクラスIIをコードする遺伝子及びその対立遺伝子変異体の増幅により得られる。プライマーのヌクレオチド配列は遺伝子のカテゴリー内の遺伝子中に存在する5'及び3'コンセンサスヌクレオチド配列に基いてMHCクラスII遺伝子及びその対立遺伝子変異体の多様性の増幅を可能にする。ヒトHLA−DP遺伝子、HLA−DQ遺伝子及びHLA−DR遺伝子のα鎖及びβ鎖を増幅するためのプライマー対、並びにマウスIAdをコードするヘテロダイマーを増幅するためのプライマー対の特定のヌクレオチド配列が実施例2Aに示される。
【0057】
MHCクラスIIをコードする遺伝子はB細胞、マクロファージ及び樹状細胞(これらの全てが血液中に存在する)を含む種々の細胞源から増幅可能である。本発明のプライマーで増幅されたMHCクラスIIをコードする遺伝子を得るための増幅条件が節Cに記載される。
【0058】
本発明のα鎖及びβ鎖を含むMHCヘテロダイマーは固定形態または可溶性形態で有益である。固定形態では、組換えMHCヘテロダイマーはそれが発現される合成抗原提示細胞に固定される。また、組換えMHCヘテロダイマーは可溶性形態で分泌された後に本発明に記載された担体を含むマトリックスに固定される。後者は、終止コドンが増幅操作中またはその後にトランスメンブランドメインに先行する特別のMHCクラスIIα鎖及びβ鎖をコードするヌクレオチド配列に操作される時に生成される。
【0059】
2.アクセサリー遺伝子及びコードされた分子
同時刺激分子、付着分子及びサバイバル分子を含む本発明のアクセサリー分子はCD4+T細胞を活性化して標的細胞を認識する武装化エフェクターT細胞になる際にペプチドと複合体形成されたMHCクラスIIヘテロダイマーと協力して有効である。未感作T細胞は、それらがAPCの表面でペプチドローディングされたMHCクラスIIヘテロダイマーにより提示された時のそれらの特異的抗原と出会う時に活性化されて武装化エフェクターT細胞に増殖し、分化する。活性化はMHCクラスIIヘテロダイマーに結合された外来ペプチドフラグメントの認識を必要とするだけでなく、それはAPCにより同時発現された同時刺激シグナルの同時の送出を必要とする。
【0060】
こうして、本発明の合成APCまたはマトリックスは特別なMHCクラスIIヘテロダイマーの存在だけでなく、アクセサリー分子と広く定義される一種以上の同時刺激分子の存在を特徴とする。特定の同時刺激分子、付着分子、及びサバイバル分子を含むアクセサリー分子の少なくとも三つの型が本発明の合成APCまたはマトリックスを調製する際の使用について意図されている。
【0061】
a.同時刺激分子
アクセサリー分子の第一の型はB7.1(B7として既に知られており、またCD80として知られている)及びT細胞のCD28に結合するB7.2(またCD86として知られている)の如き同時刺激分子を含む。B7.1及びB7.2は免疫グロブリンスーパーファミリーのホモダイマー員である構造関連糖タンパク質である。その他の同時刺激分子は抗CD28抗体またはこのような抗体の機能性部分、例えば、CD28に結合するFab部分である。上記分子によるCD28の結合が未感作T細胞の成長を同時刺激することが示された。活性化T細胞では、付加的なレセプターであるCTLA−4がCD28とのアフィニティーより高いアフィニティーでB7分子を結合する。
【0062】
本発明の合成APCまたはマトリックスに使用するための組換えB7同時刺激分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0063】
b.付着分子
本発明のアクセサリー分子の別の主要な型はT細胞活性化にまた機能する付着分子である。アクセサリー付着分子として、種々のICAM分子が挙げられ、これらとして、細胞間付着分子(ICAM) ICAM−1 、ICAM−2、ICAM−3、リンパ球機能関連抗原(LFA) LFA−1 及びLFA−3 が挙げられる。これらの分子の全てが免疫グロブリンスーパーファミリーの員である。全てのICAM関連員がT細胞インテグリン、LFA−1 に結合する。樹状細胞、マクロファージ及びB細胞を含むAPCで発現されることに加えて、ICAM−1及びICAM−2はまた内皮で発現され、それにより細胞付着及び循環白血球と内皮の間のその後の遊出を媒介する。しかしながら、ICAM−3は白血球にのみ発現され、T細胞とAPCの間の付着に重要な役割を果たすと考えられる。
【0064】
ICAM−1、ICAM−2及びICAM−3の間の相互作用は、APC及びT細胞表面で夫々発現されるLFA−3 (CD58)とLFA−2(CD2) の間の第二の付着相互作用と相乗作用する。
【0065】
本発明の合成APCまたはマトリックスに使用するための組換え付着分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0066】
c.サバイバル分子
サバイバル分子は刺激から細胞死滅誘発に至る範囲の代謝応答に役割を果たすアクセサリー分子の別の型である。こうして、サバイバル分子はまた細胞死滅調節分子と称される。サバイバル分子は典型的にはタンパク質であるが、その他の型の巨大分子、例えば、炭水化物、脂質等を含んでもよい。本発明の組成物及び方法に使用するためのサバイバル分子として、Fasリガンド、TNF-レセプター、TNF、CD70、II型トランスメンブランタンパク質(これはTNFレセプターファミリーの員であるCD27に結合するTNFファミリーの員である)が挙げられる。FasリガンドはFasと称されるレセプターに結合し、レセプター占有がFasレセプターを発現する細胞のアポトーシス細胞死滅の誘発をもたらす。CD27は静止T細胞及びB細胞で発現され、一方、CD70は活性化T細胞及びB細胞で発現される。そのレセプター、CD27へのCD70の結合はT細胞同時刺激を誘発し、その相互作用が未感作T細胞プールからのT細胞の動員に重要であるかもしれない。その他の或る条件下で、TNFによるTNFレセプターの活性化が同様の応答を生じる。
【0067】
本発明の合成APCまたはマトリックス中の使用について上記された組換えサバイバル分子はMHCクラスII分子について記載されたようなPCRにより得られる。好ましいオリゴヌクレオチドプライマー及びそれからの増幅のための細胞源が実施例2Cに記載される。
【0068】
実施例に示されるように、一種以上の上記組換えアクセサリー分子と一緒に使用される組換えMHCクラスII分子に結合されたペプチドの特別な組み合わせはT細胞を武装化エフェクターT細胞(これらはTh1炎症性T細胞及びTh2ヘルパーT細胞に区別できる)に活性化する。
【0069】
3.抗原プロセシング補助遺伝子及びコードされた分子
a.HLA−DM
ヒト中のHLA−DM及びマウス中のH2−MはMHCクラスII様分子であり、これはまたMHCクラスII遺伝子クラスター中でコードされる。HLA−DMは、MHCクラスIIのように、ヘテロダイマーを形成するα鎖遺伝子及びβ鎖遺伝子の両方を含む。しかしながら、MHCクラスIIと違って、ペプチドローディングが分子を安定化するのに必要とされない。HLA−DMは以下に更に記載されるような非変異鎖の除去後に新たに形成されたMHCクラスIIヘテロダイマーへのタンパク質のローディングを促進する。組換えHLA−DMは内部処理されたペプチドのローディングを補助するために本発明の組成物及び方法における使用について意図されている。
【0070】
b.非変異鎖
非変異鎖は新たに形成されたMHCクラスIIヘテロダイマーに結合し、それによりMHCクラスIIヘテロダイマーの夫々のサブユニットとトリマーを形成する特殊化タンパク質である。トリマー化分子は小胞体中に存在する細胞内ペプチドのローディングを阻止するが、それはまたその区画からのその分子の輸送を促進する。その後、非変異鎖は多工程により開裂されてMHCクラスIIヘテロダイマーを生じ、次いでこれが処理ペプチドと複合体形成し得る。
【0071】
こうして、組換え非変異鎖は内部処理されたペプチドのローディングを補助するために本発明の組成物及び方法における使用について意図されている。
【0072】
C.核酸及びポリヌクレオチド
1.MHCクラスII及びアクセサリー分子をコードする遺伝子を得るためのPCR
本発明のMHCクラスII分子、アクセサリー分子及び抗原プロセシング補助分子をコードする核酸配列は、直接合成、クローニング、このような遺伝子を含む細胞からのDNAの精製等を含む当業者に良く知られている幾つかの方法で得られる。本明細書に記載された組成物及び方法に使用される分子をコードするための遺伝子を得るための一つの適切な手段は以下に更に記載されるような選択されたオリゴヌクレオチドプライマー対による選択された核酸鋳型のポリメラーゼ連鎖(PCR)増幅による。
【0073】
既知の部分及び推定のヒト白血球抗原(HLA) 、ヒトMHCの遺伝的名称、アミノ酸配列及びコンセンサス配列を含むヌクレオチド配列が公表されており(例えば、Zemmour 及びParham, Immunogenetics 33: 310-320 (1991) を参照のこと)、HLA 変異体を発現する細胞系が同様に知られており、一般にATCCから多く利用できる。それ故、PCRを使用して、MHCクラスIIをコードするヌクレオチド配列が本発明の発現ベクターに容易に機能的に結合され、次いでこれがその中の発現に適した細胞を形質転換するのに使用される。
【0074】
本発明の組換え分子を生成するのに特に好ましい方法は本明細書に記載されるようにPCR反応産物を生成するためにPCRのプライマーとして前もって選択されたオリゴヌクレオチドの使用に頼る。
【0075】
遺伝子がPCR増幅により得られる場合、一般に、PCRプライマー対を含む2種のプライマーが増幅される核酸の夫々のストランドについて使用される。簡素化のために、例示のMHCクラスIIヘテロダイマーをコードする遺伝子の合成が説明されるが、記載されるPCR増幅方法は完全配列が現在知られていないものを含むMHCクラスII対立遺伝子変異体、アクセサリー分子及び抗原プロセシング補助分子の合成に同等に適用可能であることが明らかに理解されるべきである。
【0076】
一般に、第一プライマーはフォワード(forward) プライマーまたは5'と称される。何となれば、それは鋳型DNAの上部ストランドと同じ配列を有し、こうして下部相補ストランドにハイブリッドを形成するからである。
【0077】
第二プライマーはバックワード(backward)プライマーまたは3'と称される。何となれば、それは下部ストランドと同じ配列を有し、こうして上部ストランドの相補ストランドにハイブリッドを形成するからである。典型的には、換言すれば、一つのプライマーはヌクレオチド配列の負(-) または下部のストランドに相補性であり、別のプライマーは正(+) または上部のストランドに相補性である。
【0078】
好ましい局面において、第一プライマー及び第二プライマーの両方がMHCクラスII遺伝子内の保存領域にハイブリッドを形成するように(即ち、相補性であるように)選ばれる。しかしながら、プライマーはコンセンサス配列ではなく特異な配列にハイブリッドを形成することにより特定のMHCクラスII遺伝子及びその対立遺伝子変異体を増幅するように設計し得る。この局面について、鋳型配列がこのようなプライマー対の設計について知られていることが好ましい。
【0079】
第一プライマー及び第二プライマーの一方または両方はエンドヌクレアーゼ認識部位を形成するヌクレオチド配列を増幅産物に導入するように設計し得る。その部位は増幅されるMHCクラスII遺伝子に異種であってもよく、典型的にはプライマーの5'末端付近に現れる。それはまた4-塩基スペーサー配列を制限部位に近位に配置して酵素による増幅産物の切断の効率を改良するのに有益であり得る。
【0080】
特定のヌクレオチド配列を単離するための本発明のプライマーは相当するヌクレオチド配列を有するかなりの数の核酸の重合の特定の開始を与えるように充分な長さ及び適当な配列のオリゴヌクレオチドを含む。詳しくは、本明細書に使用されるオリゴヌクレオチドプライマーという用語は二つ以上、好ましくは三つより多く、更に好ましくはほぼ20のデオキシリボヌクレオチドまたはリボヌクレオチドを含む配列を表し、その配列はプライマー伸長産物の合成を開始することができる。
【0081】
合成を行う実験条件として、ヌクレオチドトリホスフェートの存在並びに重合及び伸長のための薬剤、例えば、熱安定性ポリメラーゼ、及び好適な緩衝剤、温度及びpHが挙げられる。プライマーは増幅の最大効率のために一本鎖であることが好ましいが、二本鎖であってもよい。二本鎖の場合、プライマーは最初に処理されて二つのストランドを分離し、その後に伸長産物を調製するのに使用される。プライマーはオリゴデオキシリボヌクレオチドであることが好ましい。プライマーはヌクレオチドの重合及び伸長の誘導剤の存在下で伸長産物の合成を開始するのに充分に長く、かつ実質的に相補性である必要がある。プライマーの正確な長さは温度、緩衝剤、及びヌクレオチド組成を含む多くの因子に依存するであろう。オリゴヌクレオチドプライマーは典型的には15〜22 またはそれ以上のヌクレオチドを含むが、それは更に少ないヌクレオチドを含んでもよい。また、当業界で公知であるように、ヌクレオシドトリホスフェートの混合物がバイアスされて突然変異の形成に影響して特異なペプチドをCD4+T細胞に提示するのに使用するための突然変異した組換えMHCクラスIIをコードする分子のライブラリーを得ることができる。
【0082】
本発明のオリゴヌクレオチドプライマーは指数増殖量のヌクレオチド配列を生成する酵素連鎖反応であるPCR増幅方法に使用される。変性核酸へのプライマーのアニール、続いて熱安定性ポリメラーゼ、例えば、テルモフィルス・アクアチクス(Taq)及びピロコッカス・フリオサス(Pfu) (Hoffman La-Roche, Basal,スイス)、及びヌクレオチドによる伸長が新たに合成された(+) 鎖及び(-) 鎖をもたらす。これらの新たに合成された配列はまた鋳型であるので、変性、プライマーアニール、及び伸長の反復サイクルがプライマーにより形成されたDNAフラグメントの指数的生成をもたらす。連鎖反応の産物は使用された特定のプライマーの末端に相当する末端を有する不連続の核酸二重らせんである。当業者は標的核酸のコピー数を増加するのにまた使用し得るその他の増幅方法を知っているであろう。これらとして、例えば、連結反応活性化転写(LAT) 、リガーゼ連鎖反応(LCR)、及びストランド置換活性化(SDA) が挙げられるが、以下にリストされる米国特許に記載されたようなPCRが好ましい方法である。
【0083】
本発明のオリゴヌクレオチドプライマーは、あらゆる好適な方法、例えば、相補オリゴヌクレオチドの合成またはその自動化実施態様について上記された通常のホスホトリエステル方法及びホスホジエステル方法を使用して調製し得る。修飾固体担体上にオリゴヌクレオチドを合成するための一つの方法が米国特許第4,458,066 号明細書に記載されている。
【0084】
MHCクラスII遺伝子、アクセサリー分子遺伝子、及び抗原プロセシング補助分子遺伝子を増幅するのに好ましいプライマーが実施例2に記載される。
【0085】
PCR増幅方法が米国特許第4,683,192 号、同第4,683,202 号、同第4,800,159号、同第4,965,188 号及び同第5,395,750 号(これらの開示が参考として本明細書に含まれる)、並びに“PCR Technology: Principles and Applications for DNA Amplification", H.Erlich編集, Stockton Press, New York (1989);及び“PCR Protocols: A Guide to Methods and Applications", Innisら編集, Academic Press, San Diego, California (1990) を含む少なくとも幾つかの書籍に詳しく記載されている。ここに使用される種々の好ましい方法及びプライマーが以下に記載され、またZemmour ら, Immunogenetics, 33: 310-20 (1991) 、Ausebelら, In Current Protocols in Molecular Biology, Wiley and Sons, New York (1993) 及びSambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, (1989)により記載されている。当業者に公知であるネステッド(nested)PCR 、オーバーラップPCR、逆転写酵素−PCR等を含む特別なPCR方法が本発明の組換え分子を得る際の使用について意図されている。
【0086】
別の実施態様において、PCR方法は種々のMHCクラスIIをコードする分子を生成するのに使用されるだけでなく、高度に多形性のMHC遺伝子座中に観察されるものと匹敵し得る突然変異を誘発し、または単一の親クローンから多様性を生じ、それにより更に大きな異質性を有するMHCクラスIIをコードするDNA“ライブラリー" を得るのに使用される。
【0087】
2.発現ベクター
本発明はMHCクラスIIをコードする遺伝子、アクセサリー分子遺伝子及び抗原プロセシング補助遺伝子の発現を誘導して相当する組換えタンパク質を生成することができる実質的に純粋な形態のプラスミド発現ベクターを意図している。簡素化のために、上記遺伝子がポリペプチドをコードするヌクレオチド配列と本明細書で集約的に称される。それらが機能的に結合される遺伝子の発現を誘導することができるベクターは本明細書で“発現ベクター”または“発現プラスミド”と称され、これらの両方がまた“プラスミド”と称される。
【0088】
本明細書に使用される“ベクター”または“プラスミド”という用語は異なる遺伝子環境の間でそれが機能的に結合された別の核酸を輸送することができる核酸分子を表す。好ましいベクターはそれらが機能的に結合されるDNAセグメント中に存在する構造遺伝子産物の自律複製及び発現の可能なベクターである。それ故、ベクターは宿主選択系と適合性であるレプリコン及び選択可能なマーカーを含むことが好ましい。好ましいベクターの一つの型はエピソーム、即ち、染色体外複製の可能な核酸である。
【0089】
本発明のプラスミドは本発明の翻訳可能なポリペプチドをコードするヌクレオチド配列の転写を活性化することができる要素を有する調節領域を少なくとも含む環状二本鎖プラスミドである。プラスミドは、本発明の所望のコードされたポリペプチドが発現される翻訳可能なヌクレオチド配列を更に含む。こうして、ベクターは相当する発現可能な遺伝子からコードされたような本明細書に記載された組換えポリペプチドの発現を誘導することができると言われる。
【0090】
本発明の使用に好ましいベクターはプラスミドである。それは高コピー数プラスミドであることが更に好ましい。また、特別なベクターが選択された宿主中の発現に最適であることが好ましい。
【0091】
このような発現ベクターは宿主中の挿入された遺伝子配列の有効な転写を促進する調節領域中にプロモーター配列を含む。ベクターは誘導プロモーター配列を含むことが好ましい。何となれば、誘導プロモーターはこのようなベクター(これらは未変性またはキメラのヌクレオチド配列を有するようにしばしば構築される)が導入された細胞に対し選択圧力を制限する傾向があるからである。また、発現ベクターは典型的には複製起点並びに形質転換細胞の表現型選択を可能にする特定の遺伝子を含む。DNAセグメントは調節要素、例えば、プロモーター(例えば、T7プロモーター、メタロチオネインIプロモーター、またはポリヘドリンプロモーター)に機能的に結合されたベクター中に存在し得る。
【0092】
別の実施態様において、プラスミドはまたその発現が宿主細胞に導入または形質転換された時に選択的利点、例えば、薬剤耐性をその細胞に与える遺伝子を含む。典型的な原核生物及び真核生物の薬剤耐性遺伝子は夫々アンピシリンまたはテトラサイクリン及びネオマイシン(G418またはゲネチシン)に対する耐性を与える。その他の薬剤耐性マーカーとして、クロラムフェニコール、カナマイシン、ストレプトマイシン、カルベニシリン、水銀、リファムプシン、リファムピシン、フサル酸等が挙げられる。
【0093】
本発明の調節領域及びポリペプチドをコードするヌクレオチド配列が機能的に結合されるベクターの選択は、当業界で公知であるように、所望される機能的性質、例えば、複製またはタンパク質発現、及び形質転換される宿主細胞に直接依存し、これらは組換えDNA分子を構築する技術に固有の制限である。
【0094】
機能的な結合はDNAの一つのストランド(一本鎖形態または二本鎖形態を問わない)への好ましくは通常のホスホジエステル結合によるヌクレオチド配列の共有結合を表す。更に、ヌクレオチド配列の結合は本明細書に記載されたようなプロモーター及び下流のポリペプチドをコードするヌクレオチド配列との調節領域中の応答要素の如き機能要素の結合をもたらす。
【0095】
インサートを発現プラスミドに機能的に結合するための一つの典型的な方法は方向性連結反応による。これは方向性連結反応に適しているヌクレオチドの配列により行われる。このような配列は(1) 複製のために機能的に結合し、上流及び下流の翻訳可能なDNA配列を輸送し、(2) ベクターへのDNA配列の方向性連結反応のための部位または手段を与えるDNA発現ベクターの領域であるポリリンカーと普通称される。典型的には、方向性ポリリンカーは二つ以上の制限エンドヌクレアーゼ認識配列、または制限部位を形成するヌクレオチドの配列である。制限開裂後に、二つの部位は、翻訳可能なDNA配列がDNA発現ベクターにつながれる付着末端を生じる。二つの制限部位は、制限開裂後に、非相補性であり、それにより、カセットへの翻訳可能なDNA配列の方向性挿入を可能にする付着末端を与えることが好ましい。
【0096】
種々の宿主−発現ベクター系がヌクレオチド配列によりコードされたポリペプチドを発現するのに使用し得る。これらとして、微生物、例えば、ポリペプチドをコードするヌクレオチド配列を含む組換えバクテリオファージDNA、プラスミドDNAまたはコスミドDNA発現ベクターで形質転換されたバクテリア;ポリペプチドをコードするヌクレオチド配列を含む組換え酵母発現ベクターで形質転換された酵母;ポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、カリフラワーモザイクウイルス、CaMV; タバコモザイクウイルス、TMV)で感染され、または組換えプラスミド発現ベクター(例えば、Tiプラスミド)で形質転換された植物細胞系;ポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、バキュロウイルス)で感染された昆虫細胞系;またはポリペプチドをコードするヌクレオチド配列を含む組換えウイルス発現ベクター(例えば、レトロウイルス、アデノウイルス、ワクシニアウイルス)で感染された動物細胞系、または安定な発現について操作された形質転換された動物細胞系が挙げられるが、これらに限定されない。グリコシル化が重要であり得る場合、翻訳修飾及び後翻訳修飾を与える発現系、例えば、哺乳類発現系、昆虫発現系、酵母発現系または植物発現系が使用し得る。
【0097】
これらの系のいずれもが本発明の方法を実施するのに有益である。上記発現系のいずれかにより、選択された宿主がその後にペプチドのローディングに実際のメカニズムに応じて、即ち、内部または外部に応じて抗原プロセシング補助分子を用いて、または用いずに少なくとも一種のMHCクラスIIヘテロダイマー単独または少なくとも一種のアクセサリー分子と一緒の発現に使用される。
【0098】
使用される宿主/ベクター系に応じて、構成的プロモーター及び誘導プロモーター、転写エンハンサー要素、転写ターミネーター等を含む幾つかの好適な転写要素及び翻訳要素のいずれかが発現ベクター中に使用し得る(例えば、Bitterら,Methods in Enzymology, 153: 516-544, (1987) を参照のこと)。例えば、バクテリア系中のクローニングの場合、誘導プロモーター、例えば、バクテリオファージλのP1、Plac、Ptrp、Ptac(Ptrp−lacハイブリッドプロモーター)等が使用し得る。哺乳類細胞系中のクローニングの場合、哺乳類細胞のゲノムに由来するプロモーター(例えば、メタロチオネインプロモーター)または哺乳類ウイルスに由来するプロモーター(例えば、レトロウイルス長末端リピート;アデノウイルス後期プロモーター;ワクシニアウイルス7.5Kプロモーター)が使用し得る。組換えDNAまたは合成技術により生成されたプロモーターがまたポリペプチドをコードするヌクレオチド配列の転写を与えるのに使用し得る。
【0099】
バクテリア系では、幾つかの発現ベクターが本発明の方法に従ってポリペプチドをコードするヌクレオチド配列の発現に有利に選択し得る。例えば、多量が生成される場合、容易に精製し得る融合タンパク質産物の高レベルの発現を誘導するベクターが望ましいかもしれない。開裂部位を含んでタンパク質の回収を助けるように操作されるものが好ましい。このようなベクターとして、E.coli発現ベクターpUR278 (Rutherら, EMBO J., 2: 1791, (1983)) (そのポリペプチドをコードするヌクレオチド配列はハイブリッドポリペプチド−LacZタンパク質が生成されるようにLacZコーディング領域とインフレームでベクターにつながれてもよい);pINベクター(Inouye &Inouye, Nuc.Acids Res., 13: 3101-3109,(1985);Van Heeke &Schuster, J.Biol.Chem., 264: 5503-5509, (1989)) 等が挙げられるが、これらに限定されない。
【0100】
一実施態様において、使用されるベクターはバクテリア中のベクターの増殖を促進する原核生物配列、即ち、バクテリア宿主細胞に導入された時に染色体外で組換えDNA分子の自律複製及び維持を誘導する能力を有するDNA配列を含む。このようなレプリコンは当業界で公知である。
【0101】
また、原核生物レプリコンを含むこれらのベクターは典型的には本発明の組換えDNA分子の挿入に都合の良い制限部位を含む。このようなベクタープラスミドの代表はバイオラド・ラボラトリィズ(リッチモンド、CA)からのpUC8、ぷC9、pBR322、及びpBR329及びファーマシア(ピスキャットアウェイ、NJ)からのpPL、並びにストラタゲン(ラジョラ、CA)から入手し得るpブルースクリプト及びpBSである。また、本発明のベクターはMolecular Cloning: A Laboratory Manual,第二編, Maniatisら編集, Cold Spring Harbor, NY (1989) に記載されたλベクターを含むλファージであってもよい。
【0102】
別の好ましい実施態様において、本発明に使用するためのプラスミドベクターはまた真核生物細胞と適合性である。真核細胞発現ベクターは当業界で公知であり、幾つかの商業源から入手し得る。典型的には、このようなベクターは所望の組換えDNA分子の挿入に都合の良い制限部位を与え、更に先に説明したような真核生物細胞中で発現することができるコードされた遺伝子の発現のためのプロモーターを含む。このようなベクターの代表はpSVO及びpKSV−10 (ファーマシア)、及びpPVV−1/PML2d(インターナショナル・バイオテクノロジー社)、並びにpTDT1 (ATCC、No. 31255)である。
【0103】
加えて、真核生物プラスミドでは、真核生物細胞中でのみ発現される一つ以上の転写単位が存在する。真核生物転写単位は非コーディング配列及び選択可能なマーカーをコードする配列からなる。また、本発明の発現ベクターは正確かつ有効なポリアデニル化に必要とされる特異な配列要素を含む。加えて、成熟mRNAを発生するためのスプライシングシグナルがベクター中に含まれる。真核生物プラスミド発現ベクターはウイルスレプリコンを含むことができ、その存在がクローン化遺伝子の発現のレベルの増加を与える。好ましい複製配列がシミアンウイルス40またはSV40パポバウイルスにより与えられる。
【0104】
本発明に使用するための組換え分子を生成するのに好ましい発現系は昆虫系である。一つのこのような系において、オートグラファ・カリフォルニカ核多角体病ウイルス(AcNPV) が外来遺伝子を発現するためのベクターとして使用される。そのウイルスはスポドプテラ・フルギペルダ(Sf9) 細胞中で増殖する。本発明のポリペプチドをコードするヌクレオチド配列はそのウイルスの非必須領域(スポドプテラ・フルギペルダ中、例えば、ポリヘドリン遺伝子)にクローン化され、AcNPVプロモーター(例えば、ポリヘドリンプロモーター)の制御下に置かれてもよい。ポリペプチドをコードするヌクレオチド配列の成功裏の挿入はポリヘドリン遺伝子の不活化及び非閉塞性(non-occluded)組換えウイルス(即ち、ポリヘドリン遺伝子によりコードされたタンパク質外殻を欠いているウイルス)の生産をもたらすであろう。次いでこれらの組換えウイルスは、挿入遺伝子が発現される細胞を感染するのに使用される。Smith ら, J.Biol.Chem., 46: 584, (1983); Smithの米国特許第4,215,051 号を参照のこと。
【0105】
好ましい実施態様において、宿主細胞集団はp25−lacZ (Bello 及びCouble, Nature, 346: 480 (1990) を参照のこと)またはpRmHa−1 、−2、もしくは−3 (Bunchら, Nucl.Acids Res., 16: 1043−1061 (1988) を参照のこと)のようなベクターに機能均等のベクターを含む適合性ベクターを必要とするドロソフィラ細胞培養物である。好ましい実施態様において、ベクターはpRmHa−3 であり、これが図1Cに示される。このベクターはメタロチオネインプロモーターを含み、これはMHC配列が挿入される部位の上流にあることが好ましく、ポリアデニル化部位が前記MHC配列の下流にあることが好ましい。昆虫細胞、特にドロソフィラ細胞が本発明によれば好ましい宿主である。節Dに更に記載されるようなシュナイダー−2(S2)細胞の如きドロソフィラ細胞はプロモーターの活性化に必要とされる必要なトランス作用性因子を有し、こうして更に好ましい。
【0106】
発現ベクターpRmHa−3 はバクテリアプラスミドpRmHa−1 (図1A)をベースとし、その後者はプラスミドpUC18をベースとし、ATCC(ロックビル、MD) に寄託され、受理番号37253 を有する。pRmHa−3 ベクターは図1Cに示されるようにプロモーター、EcoRI 部位及びStu I 部位を除去されたメタロチオネイン遺伝子の5'未翻訳リーダー配列を含む。それはまたポリアデニル化部位を含むドロソフィラADH遺伝子の3'部分を含む。それ故、クローン化DNAはメタロチオネインプロモーターにより転写調節され、ポリアデニル化される。pRmHa−1 プラスミドの構築がBunch ら, Nucl.Acids Res. 16: 1043−1061 (1988)に記載されてい。pRmHa−3 プラスミド及びpRmHa−2 プラスミド(その後者はEcoRI フラグメントとして除去されてもよいメタロチオネインプロモーター配列を有する)の構築が実施例に記載される。pRmHa−3 、本発明による使用に好ましいプラスミドに関して、PstI、SphI及びHindIIIがプロモーターフラグメント中にあり、それ故、特異ではない。XbaIはADHフラグメント(その3'末端から4塩基)にあり、また特異ではない。しかしながら、下記の制限部位がpRmHa−3 中で特異であり、本発明の組換え遺伝子のクローニングを促進する。EcoRI、SacI、KpnI、SmaI、BamHI、SalI、Hinc2、及びAccI。
【0107】
発現を誘導するために組換えウイルスまたはウイルス要素を使用する哺乳類細胞系が操作し得る。例えば、アデノウイルス発現ベクターを使用する場合、ポリペプチドのコーディング配列がアデノウイルス転写/翻訳調節複合体、例えば、後期プロモーター及び三部分リーダー配列に結合されてもよい。このキメラ遺伝子はその後にインビトロまたはインビボ組換えによりアデノウイルスゲノムに挿入されてもよい。ウイルスゲノムの非必須領域(例えば、領域E1またはE3)中の挿入は、生存しており、感染宿主中でポリペプチドを発現することができる組換えウイルスを生じるであろう(例えば、Logan &Shenk, Proc.Natl.Acad.Sci., USA,81: 3655-3659, (1984) を参照のこと)。また、ワクシニアウイルス7.5Kプロモーターが使用されてもよい(例えば、Mackett ら, Proc.Natl.Acad.Sci., USA,79: 7415-7419, (1982); Mackettら, J.Virol., 49: 857-864 (1984); Panicaliら, Proc.Natl.Acad.Sci., USA, 79: 4927-4931, (1982) を参照のこと)。染色体要素として複製する能力を有するウシパピローマウイルスをベースとするベクターが特に重要である(Sarver ら, Mol.Cell.Biol., 1: 486, (1981)) 。マウス細胞へのこのDNAのエントリーの直後に、プラスミドが細胞当たり約100 〜200 のコピーまで複製する。挿入されたcDNAの転写は宿主の染色体へのプラスミドの組込みを必要とせず、それにより高レベルの発現を生じる。これらのベクターは、プラスミド中に選択可能なマーカー、例えば、neo 遺伝子を含むことにより安定な発現に使用し得る。また、レトロウイルスゲノムが宿主細胞中の本発明のポリペプチドをコードするヌクレオチド配列の発現を導入し、誘導することができるベクターとしての使用のために修飾し得る(Cone &Mulligan, Proc.Natl.Acad.Sci., USA, 81: 6349-6353, (1984)) 。また、メタロチオニンIIAプロモーター及び熱ショックプロモーターを含むが、これらに限定されない誘導プロモーターを使用して、高レベルの発現が達成し得る。
【0108】
組換えタンパク質の長期の高収率の生産には、安定な発現が好ましい。ウイルス複製起点を含む発現ベクターを使用するのではなく、宿主細胞が適当な発現調節要素(例えば、プロモーター配列及びエンハンサー配列、転写ターミネーター、ポリアデニル化部位等)により調節されたcDNA、及び選択可能なマーカーで形質転換し得る。上記のように、組換えプラスミド中の選択可能なマーカーは選択に対する耐性を与え、細胞がプラスミドをそれらの染色体に組込み、成長してフォーカスを形成し、これらが順に細胞系にクローン化され、拡大されることを可能にする。
【0109】
例えば、外来DNAの導入後に、操作された細胞が濃縮培地中で1〜2日間増殖され、ついで選択培地に切り換えられる。単純ヘルペスウイルスチミジンキナーゼ(Wigler ら, Cell, 11: 223, (1977))、ヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ(Szybalska&Szybalski, Proc.Natl.Acad.Sci., USA, 48: 2026, (1962))、及びアデニンホスホリボシルトランスフェラーゼ(Lowy ら,Cell, 22: 817, (1980))遺伝子(これらは夫々tk−細胞、hgprT細胞またはaprT細胞中で使用し得る)を含むが、これらに限定されない幾つかの選択系が使用し得る。また、抗代謝産物耐性を与える遺伝子が選択の基礎として使用し得る。例えば、dhfr(これはメトトレキセートに対する耐性を与える(Wigler ら,Proc.Natl.Acad.Sci., USA, 77: 3567, (1980); O'Hareら, Proc.Natl.Acad.Sci.,USA, 78: 1527, (1981));gpt (これはミコフェノール酸に対する耐性を与える)(Mulligan &Berg, Proc.Natl.Acad.Sci., USA, 78: 2072, (1981)); neo ( これはアミノグリコシドG−418に対する耐性を与える)(Colberre-Garapinら, J.Mol.Biol., 150:1, (1981));及びhygro (これはヒグロマイシンに対する耐性を与える)(Santerre ら, Gene, 30: 147, (1984))の遺伝子。最近、付加的な選択可能な遺伝子、即ち、trpB(これは細胞にトリプトファンに代えてインドールを使用させる);hisD(これは細胞にヒスチジンに代えてヒスチノールを使用させる)(Hartman&Mulligan, Proc.Natl.Acad.Sci., USA, 85: 804, (1988)); 及びODC (オルニチンデカルボキシラーゼ)(これはオルニチンデカルボキシラーゼインヒビター、2−(ジフルオロメチル)−DL−オルニチン、DFMOに対する耐性を与える) (McConlogue L., In: Current Communications in Molecular Biology, Cold Spring Harbor Laboratory編集, (1987)が記載されている。
【0110】
原核生物発現ベクター及び真核生物発現ベクターの両方がベクター構築の当業者に良く知られており、Ausebel ら, In Current Protocols in Molecular Biology, Wiley and Sons, New York (1993) 及びSambrookら, Molecular Cloning:A Laboratory Manual, Cold Spring Harbor Laboratory, (1989)により記載されている。
【0111】
本発明の組成物及び方法に使用するための組換えMHCクラスIIα鎖及びβ鎖、アクセサリー分子及び抗原プロセシング補助分子を生成するために、夫々のヌクレオチド領域が本明細書に記載された本発明の発現ベクターに機能的に挿入される。節Bに記載されたように、本発明の組換えポリペプチドをコードする核酸は幾つかの方法で得られ、その一つがPCR増幅による。ベクター配列に機能的に結合されるヌクレオチドセグメントの一つはMHCクラスIIα鎖及びβ鎖の少なくとも一部をコードする。完全α鎖及びβ鎖をコードする夫々のヌクレオチド配列がそれからの発現のための発現ベクターに別々に挿入されることが好ましい。しかしながら、また幾つかの非コーディングMHC配列を同様に含むベクターを構築することが実施可能である。アクセサリー分子及び抗原プロセシング補助分子をコードする配列が別の発現ベクターに同様に挿入される。
【0112】
また、本発明は同じベクター中に存在する一つより多いポリペプチドをコードする遺伝子の存在を意図しており、その発現はプロモーターの如き別の調節要素により誘導される。換言すれば、α鎖及びβ鎖の両方をコードする核酸がアクセサリー分子をコードする一種以上の核酸配列を用いて、または用いずに同じ発現ベクターに機能的に結合されてもよい。こうして、本発明の組換えタンパク質を生成するための発現ベクター構築の全ての可能な組み合わせが意図されている。
【0113】
上記の完全なコードする核酸配列に加えて、本発明の発現された組換えポリペプチドの可溶性形態が意図されている。可溶性形態は、それがトランスメンブランドメインまたはその他の機能性配置の前に挿入された“終止" コドンを含んで可溶性非膜固定可能なタンパク質を生じる点で非可溶性形態とは異なる。
【0114】
D.合成抗原提示細胞及びペプチド提示のためのマトリックス
1.合成抗原提示細胞及びマトリックス
本発明によれば、組換えMHCクラスIIヘテロダイマー及び少なくとも一種のアクセサリー分子が担体を含むマトリックスに機能的に結合され、その結果、MHC分子の細胞外部分に複合体形成されたペプチドで提示された時に、MHC クラスII及びアクセサリー分子がCD4+T細胞リンパ球の集団を活性化するのに充分な数で存在する。ペプチドは、それが担体に結合される前または後にMHCクラスIIヘテロダイマーに結合し得る。
【0115】
担体は多くの異なる形態をとり得る。それは固体担体、例えば、プラスチック材料または金属材料であってもよく、それは分離カラムに普通に使用されるような多孔質材料であってもよく、それはリポソームまたは赤血球であってもよく、またはそれは細胞または細胞フラグメントであってもよい。以下に更に詳しく説明されるように、細胞が担体として利用できる場合、MHCクラスII分子及びアクセサリー分子がその細胞上の提示のために、または別の細胞を含み得る別の担体上の提示のために細胞により生成し得る。
【0116】
前者の状況では、MHC分子はその後にMHCクラスIIの可溶性形態中に存在しない細胞質ドメインではないとしても少なくともトランスメンブランドメインにより細胞に結合される。後者の状況では、MHCクラスII分子及びアクセサリー分子の細胞外部分が担体に固定された抗体に反応するエピトープを用意することにより担体に結合し得る。加えて、MHC分子またはアクセサリー分子は担体の部分を形成する際にニッケルと反応する(His)6で生成され、またはそれに結合し得る。MHC分子を担体に固定または結合するためのその他の手段が当業界で公知である。
【0117】
先に説明したように、担体は細胞膜または全細胞であってもよい。このような場合、真核生物細胞系がMHCクラスIIに関するペプチドをT細胞リンパ球に提示するのに使用するための合成抗原提示細胞系になるように修飾される。エンプティーMHC分子は熱不安定性であるので、細胞培養物は変温性であることが好ましく、種々の細胞系が以下に詳しく説明される。
【0118】
本発明の好ましい細胞系は培養中に連続増殖することができ、かつ細胞表面で哺乳類MHCクラスII分子及びアクセサリー分子を発現することができる。バクテリア細胞系、酵母細胞系、昆虫細胞系、及び哺乳類細胞系を含む種々の形質転換された細胞もしくは細胞系または未形質転換細胞もしくは細胞系のいずれもがこの目的に適している(例えば、種々の細胞系、例えば、E.coli及びサルコミセス・セレビジエを培養し、使用するための要約及び操作について、Current Protocols in Molecular Biology, John Wiley&Sons, NY (1991) を参照のこと)。
【0119】
細胞系は真核細胞系であることが好ましい。細胞系は変温性(即ち、哺乳類細胞系よりも温度攻撃に対し感受性ではない)であることが更に好ましい。それは昆虫細胞系であることが更に好ましい。蛾(ATCC CCL 80) 、アワヨトウガ(ATCC CCL 1711) 、蚊の幼虫(ATCC 系列CCL 125 、CCL 126 、CRL 1660、CRL 1591、CRL 6585、CRL 6586) 、カイコ(ATCC CRL 8851) 及びチョウ(スポドプテラ・フルギペルダ(Sf9細胞、ATCC CRL 1711)を含む種々の昆虫細胞系が本発明による使用に利用できる。好ましい実施態様において、細胞系はドロソフィラ細胞系、例えば、シュナイダー細胞系である(Schneider, J.Embryol.Exp.Morph., 27: 353-365 (1972)) 。細胞系はM3培地中の増殖に適したシュナイダー2(S2)細胞系(S2/M3) であることが好ましい(Lindquistら, Drosophila Information Service, 58:163 (1982)を参照のこと)。シュナイダー2(S2)細胞はブタペスト条約要件に従って1992年2月18日にATCC(ロックビル、MD) に寄託され、受理番号CRL 10974を指定された。
【0120】
本発明の合成抗原提示細胞を生成するため、一種以上のアクセサリー分子と一緒に選択されたMHCクラスIIヘテロダイマーの発現を誘導するための一種以上の発現ベクターがレシピエント宿主細胞に導入される。加えて、別の実施態様において、HLA−DM及び非変異鎖を含む抗原プロセシング補助分子を発現するためのベクターがまたレシピエント細胞に導入される。上記のための遺伝子が節B及びCに記載された。
【0121】
こうして、合成抗原提示細胞またはマトリックスを調製するために、本発明の組換えポリペプチドをコードする発現ベクターが選択された宿主細胞にトランスフェクト、即ち、導入される。発現ベクターの選択並びにその構築は以下に繰り返して言われるように先に説明したようなCD4+活性化に所望の成果に依存する。トランスフェクション(また、形質転換と称される)はリン酸カルシウム方法、DEAE−デキストラン方法、安定移入方法、エレクトロポレーションを含む多数の方法により、またはリポソーム媒介方法により行われてもよい。多数の書籍が入手でき、これらは既知のトランスフェクション方法及びヌクレオチドを細胞に導入するためのその他の操作を記載している。例えば、Current Protocols in Molecular Biology, John Wiley &Sons, NY (1991) を参照のこと。一種以上のベクターの導入後に、レシピエント細胞は形質転換されると言われ、その選択は一時的または安定のいずれであってもよい。
【0122】
細胞の培養物が最初に樹立される。細胞系がトランスフェクションについて選択される。何となれば、それは導入される遺伝子の少なくとも一つを欠いているからである。昆虫細胞が有利であることがわかった。何となれば、それらが変温性であるからだけでなく、それらがこれらの遺伝子及びメカニズムを欠いているからであり、そうしないとペプチドに結合されたMHC分子を生成するであろう。これはペプチド結合MHC分子の生成、及びエンプティーMHC分子の生成に対する更に大きな調節を可能にする。
【0123】
次いで選択された細胞が第一プロモーターに機能的に結合された発現可能なMHCクラスIIα鎖遺伝子及び第二プロモーターに機能的に結合された発現可能なMHCクラスIIβ鎖遺伝子を含む発現ベクターの導入により形質転換される。ベクター中の第三プロモーターに機能的に結合された第一の発現可能なアクセサリー分子遺伝子がまた上記細胞に導入される。更に別の実施態様において、ベクター中の第四プロモーターに機能的に結合された発現可能な抗原プロセシング補助遺伝子が上記細胞に導入される。
【0124】
更に好ましい実施態様において、ベクターは節Cに記載されたドロソフィラ発現プラスミドpRmHa−3 を含み、これには上記組換えタンパク質をコードする発現可能なヌクレオチド配列が本明細書に開示された技術を使用して挿入されていた。MHCクラスII鎖をコードするヌクレオチド配列、少なくとも一種のアクセサリー分子をコードするヌクレオチド配列及び抗原プロセシング補助分子をコードするヌクレオチド配列は培養された細胞に個々に同時トランスフェクトされる別々の発現プラスミドに機能的に結合されることが好ましい。また、ヌクレオチド配列は同じ発現プラスミド中の別々のプロモーターに機能的に結合され、その同じプラスミドにより同時トランスフェクトされてもよい。MHCクラスIIα鎖及びβ鎖は異種からのものであることが好ましく、定温動物、例えば、哺乳類、最適にはヒトからのものであることが更に好ましい。
【0125】
遺伝子の少なくとも一種、特に、MHCクラスII鎖遺伝子は誘導プロモーターに結合されることが好ましい。これはMHC分子の生成に調節を与え、その結果、関係するペプチドが内部または外部で利用でき、培養物中で提示されて生成されたMHC分子と反応する時にのみそれらが生成される。これは望ましくないMHC分子/ペプチド複合体を最小にする。
【0126】
こうして、好ましい細胞系は夫々が第一プロモーター及び第二プロモーターに夫々機能的に結合されたMHCクラスIIα鎖及びβ鎖を含む別々のベクターを有する変温性細胞系である。プロモーターはMHCクラスII鎖の発現を調節するように誘導性であることが好ましい。加えて、細胞は第三プロモーターに機能的に結合された第一の発現可能なアクセサリー分子遺伝子を少なくとも含む第三ベクターを含む。更に別の実施態様において、細胞はまた第四プロモーターに機能的に結合された発現可能な抗原プロセシング補助遺伝子を含む第四ベクターを含む。細胞はエンプティーMHC分子を集合し、それらを細胞表面に提示し、その結果、特別なMHCクラスIIハプロタイプまたは変異体対立遺伝子に特異性のペプチドが所望に応じて選択し得ることが好ましい。
【0127】
一種以上の特別なアクセサリー分子をコードする遺伝子と一緒に使用するための適合性MHCクラスIIα鎖遺伝子及びβ鎖遺伝子の選択は所望されるT細胞活性化プロフィールに依存する。例えば、実施例に記載されるように、組換えB7.1またはB7.2は単独で、またはドロソフィラAPCの表面で発現された組換えマウスIAdMHCクラスIIと一緒に、IL−4及びIL−10 の増大された産生のTh2プロフィールを有するCD4+T細胞の増殖をもたらした。対照的に、B7.1またはB7.2が同じMHC分子とともにICAM−1と一緒にドロソフィラAPCの表面で発現された時、CD4+T細胞の活性化は増大されたIL−2産生並びに減少されたIL−4及びIL−10産生を有するTh1プロフィールをもたらした。
【0128】
こうして、本発明は本発明のアクセサリー分子のいずれか一つとのMHCクラスIIハプロタイプヘテロダイマーのあらゆる組み合わせを細胞表面に有する合成APCの生成を意図している。特に好ましい組み合わせはB7.1またはB7.2を含む同時刺激分子、ICAM−1、ICAM−2、ICAM−3またはLFA−3 を含む付着分子、またはFasリガンド(FasL)を含むサバイバル分子とともにMHCクラスIIを含む。上記のように、アクセサリー分子の夫々のカテゴリーの一種より多くが、例えば、B7.1及びB7.2として同時発現し得る。別の好ましい実施態様は、異なるカテゴリーの2種のアクセサリー分子が細胞表面で同時発現される順列を含む。換言すれば、同時刺激分子と一緒の付着分子、及びサバイバル分子と一緒の付着分子、サバイバル分子と一緒の同時刺激分子。更に別の実施態様において、本明細書に記載された異なるカテゴリーの3種のアクセサリー分子がAPC表面で同時発現される。また、これらの実施態様の全てにおいて、特別なカテゴリーの一種より多くの員がその他のカテゴリーの一種より多くの員と一緒に発現し得ることが意図されている。特別な選択された組み合わせは、それらが発現される細胞の表面で有効であり、または本発明のマトリックスの表面に固定された時に有効である。調製される実際の組み合わせは抗原ペプチドと複合体形成された発現されたMHCクラスII分子に鑑みてT細胞活性化の成果に基いて選択される。こうして、MHCクラスII/ペプチドの複合体に応じて、T細胞活性化の成果は同じ発現されたアクセサリー分子を有するにもかかわらず異なり得る。
【0129】
更に別の実施態様において、抗原プロセシング補助遺伝子は上記組み合わせのいずれかで同時トランスフェクトされて増進された内部ペプチドプロセシング及びローディングを与える。こうして、この局面はその後のペプチド複合体形成のための細胞表面におけるエンプティーMHCクラスII分子の生成を伴わない。むしろ、非変異鎖、HLA−DMまたはリソソーム酵素の発現が利用されて細胞内在化後のタンパク質分解ペプチドフラグメントの最適のプロセシング及びローディングを可能にする。こうして、本発明のAPCは細胞内または細胞外でローディングされる組換えMHCクラスIIヘテロダイマーを有するモチーフで機能し得る。
【0130】
成功裏に形質転換された細胞、即ち、本発明のヌクレオチド配列の発現を誘導することができる少なくとも一種の発現ベクターを含む細胞は公知の技術により同定し得る。例えば、本発明のcDNAまたはrDNAの導入により得られる細胞はクローン化されて個々のコロニーを生じ得る。これらのコロニーからの細胞が回収され、溶解され、それらのDNA含量がSouthern, J.Mol.Biol. 98: 503(1975)により記載された方法の如き方法を使用してrDNAの存在について試験し得る。rDNAの存在について直接分析することに加えて、成功裏の形質転換またはトランスフェクションは、rDNAがアクセサリー分子の主題MHC クラスIIタンパク質の発現を誘導することができる時に公知の免疫学的方法により確認し得る。例えば、一種以上の発現ベクターで成功裏に形質転換された細胞は、適当な抗体、例えば、特別なハプロタイプの抗クラスIIを使用して容易に測定される特別な抗原性を示すタンパク質を生産し得る。加えて、成功裏の形質転換/トランスフェクションは上記されたようなマーカー配列、例えば、ネオマイシン耐性を有する付加的なベクターの使用により確かめられてもよい。
【0131】
また、培養物は安定であり、低下した温度で持続成長することができることが好ましい。例えば、培養物はほぼ室温、例えば、約24〜27℃で維持されることが好ましい。別の実施態様において、培養物は、特にCD4+T細胞の活性化のプロセス中に高温で維持される。こうして、本発明の培養物は約30℃〜約37℃の温度攻撃に耐えることができることが好ましい。
本発明の少なくとも一種のアクセサリー分子及び必要により抗原プロセシング補助分子と一緒にエンプティーまたはMHCクラスII分子の発現のための培養物を調製するために、培養物は前もって決められた期間にわたって、例えば、CuSO4誘導による刺激を最初に必要とし得る。好適な誘導期間、例えば、約12〜48時間後に、ペプチドが前もって決められた濃度(例えば、約0.2 μg/ml〜20μg/ml)で添加し得る。内部ローディング及び外部ローディングの両方のためのタンパク質及びペプチドは以下に説明されるように調製される。更なるインキュベーション期間、例えば、27℃で約12時間後に、培養物がCD4+T細胞の活性化における使用に供される。この追加のインキュベーション期間は短縮され、またはおそらく省かれてもよいが、培養物は静止または未感作CD4+T細胞の添加の前に或る時間にわたってインキュベートされる場合に温度攻撃に対し次第に安定になる傾向がある。例えば、ペプチドが添加された本発明の培養物は、37℃で延長された期間にわたってインキュベートされる時でさえもペプチドローディングされたMHCクラスII分子のかなりの量を発現することができる。
【0132】
形質転換された宿主細胞の培養に有益な栄養培地は当業界で公知であり、多数の商業源から入手し得る。宿主細胞が哺乳類である実施態様において、“無血清"培地が使用されることが好ましい。
【0133】
得られる組換え発現されたMHCクラスII分子は特別なペプチドに結合し、APCの表面で少なくとも一種のアクセサリー分子とともに充分な数で存在してMHCクラスII/ペプチド複合体に対しT細胞リンパ球の集団を活性化する。
【0134】
細胞系が一種以上の所望の分子を既に生産する場合、培養物を細胞中に欠けている遺伝子について発現可能な遺伝子でトランスフェクトすることのみが必要である。例えば、細胞がそれらの表面でMHC分子を既に提示する場合、培養物をアクセサリー分子について発現可能な遺伝子を含むベクターでトランスフェクトすることのみが必要である。
【0135】
既に説明したように、タンパク質またはペプチドは、細胞が内部プロセシングのためにMHCクラスII分子を生産している時に細胞培養物に導入し得る。浸透圧ショックの如き方法により、ペプチドが細胞に導入でき、生産されたMHC分子に結合する。また、特に変温性細胞系の場合、MHC分子は細胞表面でエンプティーに提示されるであろう。次いでペプチドが培養物に添加されて、所望されるようにMHC分子に結合し得る。簡素化のために、一種のペプチドが本明細書に記載されるが、本発明の方法は本発明の治療方法に使用するための新規な抗原ペプチドを同定するためのペプチドライブラリーのスクリーニングを意図している。
【0136】
細胞表面にMHCクラスIIヘテロダイマー及び少なくとも一種のアクセサリー分子を有する細胞が生産された後、細胞が凍結乾燥されてCD4+T細胞リンパ球の集団を活性化するのに使用するための細胞フラグメントを生成し得る。
【0137】
また、細胞のトランスフェクトされた培養物がMHCクラスII及びアクセサリー分子の細胞外部分を生成するのに使用される。固体担体の如き担体と一緒の細胞外部分の使用は生産の或る種の利点を有する。生細胞が合成抗原提示細胞を得るのに使用される場合、少なくとも3種の遺伝子、MHCクラスIIヘテロダイマーを生成するための2種とアクセサリー分子のための一種が細胞に導入される必要がある。しばしば、抗生物質耐性のための追加の遺伝子がまたトランスフェクトされる。
【0138】
固体担体系が使用されている場合、一種の細胞系がMHCクラスII分子の細胞外部分を生成するのに使用され、一方、別の細胞系がアクセサリー分子の細胞外部分を生成するのに使用される。次いでMHCクラスII部分及びアクセサリー分子部分がそれらの夫々の培養物から回収される。次いでこれらの分子がT細胞の集団を活性化するのに充分な数で適当な担体に結合される。生産の観点から、2種の異なる培養物が使用し得るが、同じ培養物を使用することが可能であるが、培養物がアクセサリー分子の細胞外部分を発現するための追加の遺伝子でトランスフェクトされることを必要とする。
【0139】
この実施態様の更なる改良は、発現可能な第二のアクセサリー分子遺伝子でトランスフェクトされる細胞の第三培養物を用意することである。例えば、細胞の第二培養物は同時刺激分子の細胞外部分を生産し、一方、細胞の第三培養物は付着分子の細胞外部分を生産する。付着分子部分が回収され、担体に結合される。担体に結合されるMHCクラスIIヘテロダイマーの細胞外部分を調製する際に、可溶性分子が既に説明されたように調製される。これらの分子は一般にMHC分子中にトランスメンブラン及び細胞質ドメインを欠いている。
【0140】
2.ペプチド
潜在的能力をもつMHCクラスIIに特異的なペプチドとして機能する関連ペプチドフラグメントを生成するために、ウイルス抗原の他に実質的に全ての細胞タンパク質を用いることができる。本発明の方法および組成物は、CD4+ 細胞を特異的に活性化させる能力が増強されたMHCクラスII分子を提供する。
【0141】
本発明のペプチドはMHCクラスII分子と結合する。この結合は、インビトロと同様にインビボでも作り出すことができる生物学的条件下で生じる。本発明の実施に当該ペプチドの結合の正確な性質を知ることは必要ではない。
【0142】
MHCクラスII分子と結合するペプチドの長さは様々で、さらにそのアンカー残基の当該ペプチド末端からの距離も様々である。ある実施態様では、MHCクラスII分子に付加するために調製されるペプチドは単一種に由来する。すなわち、MHCに付加される全てのペプチドのサイズおよび配列は同一で、単抗原ペプチド付加MHCクラスII分子を生じる。また別の実施態様では、ペプチドは均質ではなく、ペプチドのランダムライブラリーを含み、先に述べたように所望のT細胞活性化プロフィールを生じる固有のペプチドの選別を可能にすることができる。合成ランダムペプチドライブラリーの生成およびスクリーニングは当業者には周知で、米国特許第5556762 号明細書、同5510240 号明細書、同5498530 号明細書、同5432018 号明細書、同5382513 号明細書、同5338665 号明細書および同5270170 号明細書(これらの文献は参照により本明細書に含まれる)に記載されている。
【0143】
ペプチドは種々の手段によって細胞に与えることができる。好ましくは、ペプチドは、細胞内ペプチドプールに入ることができるような態様で与えられる。例えば、ペプチドは浸透圧付加によって与えることができる。典型的には、ペプチドは培養液に添加される。ペプチドは、完全なポリペプチドまたはタンパク質の形状で培養に添加される。後者は、その後細胞の一連の作用(例えば酵素分解)によって分解される。また別には、完全なポリペプチドまたはタンパク質は、細胞培養に添加する前に例えば化学的消化(例えば臭化シアン)またはプロテアーゼ(例えばキモトリプシン)のような他の手段によって分解できる。他の実施態様では、ペプチドは、特定のMHCクラスIIハプロタイプと結合したより小さなセグメント(これは抗原性アミノ酸配列を含んでいてもいなくてもよい)として提示される。
【0144】
好ましくは、十分量のタンパク質またはペプチドは、細胞培養または合成マトリックスに添加され、MHCクラスII分子が結合して、続いて本発明の合成APCまたはマトリックス上に高密度でペプチド(好ましくはMHCヘテロダイマーの各々と結合した同じ種類のペプチド)を提示することを可能にする。
【0145】
本発明の別の実施態様では、ペプチドは、細胞によって発現されるMHC分子の耐熱性を強化するために、本発明のトランスフェクト細胞に添加される。上記に記したようにペプチドは好ましくは培養液に添加される。MHCクラスII分子に結合する抗原性ペプチドは、MHC分子に耐熱性を付与し、また細胞表面発現を高めるために役立つ。MHC分子に結合するペプチドを添加された培養は、したがって添加ペプチドのない培養よりも温度によるチャレンジに極めて耐性を有する。
【0146】
E.CD4+T細胞反応を変化させる方法
1.Th1およびTh2CD4+T細胞仲介疾患
未感作T細胞を所望の活性化T細胞型に誘導するか、または活性化T細胞のエフェクター機能をTh1型からTh2型にもしくはその逆に偏向させることは、特にCD4+T細胞仲介疾患を処置する治療方法について本発明の目的の1つである。
【0147】
分裂中のCD4+T細胞が炎症性T細胞またはヘルパーT細胞のどちらに分化するかは、感染体によって産生されるサイトカイン、主にIL−12およびIL−4、アクセサリー分子の影響、並びにMHCクラスII/ペプチド複合体の性質によって左右される。先に考察したように、細胞介在性免疫は、最初に細胞内寄生体(例えばマイコバクテリウム(Mycobacterium)、リーシュマニア(Leishmania)、ニューモシスチス(Pneumocystis)などの寄生体を含む)に誘導されたTh1炎症性細胞によって活性化されたマクロファージによる細胞内病原体の破壊を必要とする。対照的に、液性免疫は、最初に細胞外病原体(クロストリジウム(Clostridium)、スタフィロコッカス(Staphylococcus)、ストレプトコッカス(Streptococcus)、ポリオウイルス、ニューモシスチス(Pneumocystis)などを含む)に誘導されたヘルパーT細胞によって活性化されたB細胞による抗体の産生に依存する。
【0148】
例えば、ある種のタイプの感染(例えばリーシュマニア)からの回復は、もっぱらIL−2/IFN−γの産生を必要とする。リーシュマニアに対するTh2応答が上昇したマウスは感染を抑制できず、最終的に死亡する。Th2型応答の不適切なサイトカイン産生は、しばしばアレルギー型疾患(例えば喘息および接触過敏症)と密接に関係する。
【0149】
ヒトの疾患と歪んだサイトカイン産生パターンとの最も強い関わりは、おそらくTh1応答およびTh1型サイトカインの自己免疫疾患との関わりであろう。実験モデルにおける強力な証拠は、多くのタイプの自己免疫疾患(糖尿病、多発性硬化症の実験モデル、自己免疫性甲状腺炎など)はTh1型CD4+T細胞によって仲介されること示唆している。これらのモデルにおけるTh2付随サイトカイン(例えばIL−4)の発現は自己免疫疾患の進行に干渉する。Th2型サイトカインはTh1型細胞の反応を低下させ、一方、Th1型サイトカインはTh2型応答の進行に拮抗する。
【0150】
特定の活性化T細胞サブセットと特定の疾患とのつながりという観点から、したがって所望のT細胞サブセットへCD4+T細胞の分裂および活性化、すなわち疾患の流れを変えるために極めて有益である一連の作用を誘導することができることが必要である。有望な解決の1つは、場合によってアレルギーまたは自己免疫疾患をもつ対象者から最初に単離したCD4+T細胞をインビトロで活性化し、好ましいサイトカインプロフィールを分泌する細胞を産生することである。得られた活性化T細胞を続いて当該対象者に再導入し、疾患の流れを変え、長期的治癒を提供する。
【0151】
また別の実施態様は、Th1またはTh2応答の何れか(その者にとって応用可能ないずれか)の進行に対して潜在的な反応性をもつ個体を" 免疫する”能力を目的とする。換言すれば、特定のT細胞サブセットの選択的誘発は、未感作細胞が望ましくない表現型へと変化することを抑制することによって達成できる。例えば、潜在的にアトピーをもつ個体の場合、有害なTh2応答を防止することはヘツェルとラム(Hetzel & Lamb, Clinical Immunol. Immunopath., 73:1−10(1994))が記載したように有益であろう。
【0152】
さらに別の実施態様では、本発明の組成物および方法は、未感作T細胞を所望の表現型へ発展させるために活発に刺激するために有用である。現時点で存在する治療モデルには、ヘツェルとラム(上掲書)が概説したように、抗サイトカイン抗体と担体タンパク質の使用、外因性サイトカインの作用を長く続かせるためのイディオタイプ/GM−CSF融合タンパク質ワクチンの使用、選択アジュバントの使用、リポソーム被包化アレルゲンの使用、ペプチド類似体の使用などが含まれる。しかしながら、著者らは、インビボでのアレルゲンに対する長期的Th2応答については、Th2応答の所望のダウンレギュレーションを達成する可能性に対する実験的データは殆ど存在しないと述べている。
【0153】
前述の観点から、本発明の組成物および方法は、上記で考察した治療的介入を達成する価値ある手段を提供する。本発明は、所望の治療用サイトカインプロフィールを産生するCD4+T細胞サブセットの生成を再現することができる活性化条件を特定することを可能にする。特定のサイトカインの発現は、特定の抗原提示細胞(APC)およびそれらの付随アクセサリー分子と連携している。APCによって産生されるサイトカインおよび統合的に発現されるアクセサリー分子は両方とも、多数の要素(抗原のタイプ、T細胞レセプター(TCR)−抗原相互作用の親和性、抗原濃度などを含む)によってそれ自体調節されるので、抗原提示の際のT細胞活性化の発現を予測することはこれまでのところ極めて困難である。実際、新たなアクセサリー分子がインビボでの活性化過程について候補に挙げられたので、多くの異なる分子がT細胞反応の調節に必要とされ、連携的態様で作用してT細胞活性化を達成することがますます明らかになってきた。
【0154】
本発明は、好ましくは非哺乳類昆虫細胞で発現する限定されたアクセサリー分子と結合したMHCクラスII分子を中性的バックグラウンドで提示する合成APCの生成を提供する。本発明のMHCクラスII/アクセサリー分子組成物を発現および提示する担体として昆虫細胞を使用する利点は、昆虫細胞は調節性サイトカインを内因的に産生することはなく、さらに哺乳類のアクセサリー分子を発現しないということである。このことによって、T細胞反応を変化させることができる多数の分子を発現させる哺乳類APCを用いる場合の固有の非予測性が克服される。したがって本発明は、他の手段では得ることができない再現性および予測性を可能にする選択された組み合わせでの発現に向けて個々の提示分子およびアクセサリー分子を単離する能力を提供する。
【0155】
2.治療方法
上記で述べたように、本発明は、CD4+T細胞を活性化して武装エフェクターT細胞サブタイプに分化させる方法に関する。本方法は、ペプチドを結合することができる組換えMHCクラスIIヘテロダイマーを外部表面に定着させた合成APCまたはマトリックスを提供することに関する。この組成物はまた、当該細胞表面に提示された少なくとも1つのアクセサリー分子を有する。未感作または活性化CD4+ T細胞は処置される個体から取り出すことによって得ることができる。続いて、T細胞を活性化して所望のT細胞表現型に分裂させ分化させるために十分な時間当該CD4+T細胞を抗原提示細胞と接触させる。
【0156】
活性化CD4+T細胞をこの細胞株から分離し、許容可能な担体中に懸濁して当該個体に投与する。
【0157】
ヒトの遺伝子を用いるのが好ましく、したがってヒト分子類似体が製造される。先の米国特許第5314813号明細書に示されたように、ネズミの系はT細胞活性化の実施をテストするために有用なモデルを提供し、この工程をヒトの系で応用することができることを示した(Sykulevら、Immunity, 1:15-22(1994)もまた参照のこと)。
【0158】
a.休止または活性化CD4+T細胞の単離
MHCクラスIIに関して提示される特異的抗原を標的とするように未だ活性化されていない休止(または未感作)CD4+T細胞を活性化CD4+T細胞と同様に個体から抽出し、本発明の形質転換培養と保温するかまたはこれら培養に暴露する。未感作細胞は、主に細胞表面マーカーCD455RAおよびCD45を基準にプライマー細胞と区別できる。
【0159】
さらにまた、特異的に活性化されるべきCD4+ 細胞の能力と干渉する可能性がある他の処置または療法を開始する前に、個体からCD4+ 細胞を得ることが望ましい。例えば、自己免疫疾患の患者を治療しようとしている場合は、ステロイド治療のような補助的治療の開始前、または当該患者が全く治療を受けていない期間中に細胞および培養サンプルを得ることが好ましい。
【0160】
患者のT細胞仲介免疫反応を変化させるためにCD4+T細胞を活性化させるとき、適切な対抗治療(対抗T細胞表現型およびサイトカインの産生を含む)を開始するために、当該患者を先ず分析してT細胞表現型の症状を調べなければならない。サイトカインプロフィールは、ATCCから入手可能な抗サイトカイン抗体を用い、さらに米国特許第5405751号明細書、同第5322787号明細書および同第5209920号明細書(これらの文献は参照により本明細書に含まれる)に記載された方法によって確定される。好ましいサイトカイン分析には、インターロイキン−2(IL−2)、インターフェロン−γ(IFN−γ)、腫瘍壊死因子(TNF)、インターロイキン−4(IL−4)、インターロイキン−10(IL−10)などが含まれる。先に考察したように、特定のサイトカインプロフィールがT細胞表現型および疾患に付随している。
【0161】
特に、当該症状が自己免疫疾患(多発性硬化症、自己免疫性甲状腺炎、全身性紅斑性狼瘡、重症筋無力症、クローン病および炎症性大腸疾患を含む)の場合、サイトカインプロフィールは、IL−2、IFN−γおよびTNFの増加を特徴とするTh1型応答によって生じる。対照的に、当該症状がアレルギー(例えば喘息および接触過敏症)である場合は、サイトカインプロフィールは、IL−4およびIL−10の増加を特徴とするTh2型応答によって生じる。
【0162】
患者のサイトカインプロフィールおよび症状を分析した後、下記に述べるように本発明の合成APC、細胞フラグメントまたはマトリックスと患者から単離されたCD4+T細胞をインビトロで十分な量で十分な時間接触させ、当該接触細胞を機能的に対抗するサイトカインプロフィールを産生する活性化CD4+T細胞に分裂し分化するように誘発する。すなわち、患者がTh1型応答者としての特徴を有する場合、患者のCD4+T細胞に提示されるべき抗原は、細胞がTh2型に分裂し分化するように誘発するために必要なものであろう。これと反対の治療様式は、患者がTh2型応答を特徴とする場合に実施される。したがって、いったん対抗する活性化細胞が患者に戻された場合、T細胞表現型の反応における変化を達成する治療成果が得られる。
【0163】
リンパ球の抽出方法および培養方法は周知である。例えば、米国特許第4690915号明細書(Rosenberg)には、リンパ球フェレーシスによって大量のリンパ球を得る方法が記載されている。用いられる適切な培養条件は哺乳類細胞についてであり、典型的には37℃で実施される。
【0164】
CD4+ 細胞培養の分離および/または濃縮のために種々の方法がまた用いられる。細胞の分離のためのいくつかの一般的方法には、特異的に被覆した表面に間接的に細胞を結合させるものが含まれる。別の例では、CD4+ 細胞を含むヒトの末梢血リンパ球(PBL)がフィコールハイパーク(Ficoll-Hypaque)勾配遠心沈殿(Pharmacia, Piscataway, ニュージャージ)によって単離される。PBLリンパ芽球はその後直ちに用いてもよいが、また10%DMSO(Sigma Chemical Co., セントルイス、ミズーリ)含有FBS(これは細胞生命活性およびリンパ球機能を保護する)中で凍結させた後液体窒素で保存してもよい。
【0165】
前駆体細胞培養を分離および/または濃縮するまた別の方法には正および負の選別方法が含まれる。正の選別方法については、リンパ球濃縮PBL集団を全血から調製した後、CD4補助レセプター抗原を標的とする親和性(アフィニティー)を基準にした分離技術によってCD4+ リンパ球の亜集団を当該集団から調製する。これらのアフィニティーを基準にした技術には、蛍光活性化細胞分類(FACS)、細胞粘着、磁性ビーズ分離などの方法が含まれる。(例えば以下の成書を参照されたい:Scher & Mage,「基礎免疫学(Fundamental Immunology)」、W.E. Paul 編、pp.767-780, River Press 刊、ニューヨーク(1984))。アフィニティー法は、アフィニティー試薬の供給源として抗CD4補助レセプター抗体を利用する。また別には、CD4レセプターの天然のリガンドまたはリガンド類似体をアフィニティー試薬として用いてもよい。本方法で使用できる種々の抗T細胞抗体および抗CD4単クローン性抗体は、アメリカ菌培養収集所(ロックビル、メリーランド)およびファーミンゲン(Pharmingen, サンディエゴ、カリフォルニア)を含む多様な市販源から入手できる。
【0166】
負の選別方法は、CD4+細胞から非CD4細胞を除去することによって実施される。この技術ではリンパ球フェレーシスを受けた患者のTおよびB細胞集団からCD4+細胞が濃縮される。目的とする抗原にしたがって適切な抗体は異なるであろう。例えば、ヒトTリンパ球、ヒトT細胞サブセットおよび活性化T細胞と反応するものとして、それぞれ単クローン性抗体OKT4(抗CD4、ATCC No.CRL8002)、OKT5(ATCC No.8013および8016)、OKT8(抗CD8、ATCC No.CRL8014)およびOKT9(ATCC No.CRL8021)がATCCの細胞株およびハイブリドーマカタログ(ロックビル、メリーランド)で特定できる。他の種々の抗体もまたT細胞種(前駆体細胞並びに未感作および活性化メモリー成熟末梢T細胞を含む)の特定および単離のために利用できる。
【0167】
b.CD4+ 細胞のインビトロ活性
特定のCD4+T細胞表現型の生成のためのインビトロ条件を最適化するために、抗原提示細胞培養は適切な培養液で維持される。好ましくは、完全な細胞である本発明の担体を用いる場合は、抗原提示細胞はショウジョウバエ(Drosophila、ドロソフィラ)細胞で、この細胞は好ましくは血清非含有培養液(例えばエクセル(Excell)400)で維持される。しかしながらまた別の実施態様では、担体が細胞フラグメントまたは先に述べたように人工的担体マトリックスである場合は、培養液は標的細胞の生命活性を維持するために選択される。
【0168】
本発明の合成APC、細胞フラグメントまたはマトリックスを活性化させるべきT細胞と接触させる前に、ある量の抗原性ペプチドを当該APCまたはマトリックスに十分量で提供し、当該APCまたはマトリックス表面での発現のためにヒトMHCクラスII分子上に付加させる。先に考察したように、ペプチド付加は細胞内または細胞外で生じるであろう。したがって両態様が本明細書で述べる活性化過程に含まれるが、単純化のためにペプチドの付加はMHCクラスIIヘテロダイマーへのペプチドの提示として包括的に述べる。付加については既に考察したとおりである。さらにまた、ペプチドライブラリーと同様に個々のペプチドも、先に考察したように活性化CD4+T細胞の調製での使用が意図されている。本発明にしたがって、十分量のペプチドとは、約200から約500000、好ましくは約200から1000以上のMHCクラスII分子が各合成APCまたはマトリックスの表面で発現されるべきペプチドで付加される量である。好ましくは、上記の組成物は0.2μg/mlから20μg/mlまでのペプチドとともに保温される。
【0169】
続いてCD4+ 細胞を活性化させるために十分な時間、単離CD4+ 細胞は、合成APCまたはマトリックス上に発現された適切なペプチド付加MHCクラスIIヘテロダイマーと培養される。したがってCD4+ 細胞は好ましくは抗原特異的態様で活性化されるであろう。CD4+ 細胞対抗原提示細胞の比は個体間で変動し、さらに、例えば各人のリンパ球の培養条件に対する順応性並びに疾患の性質および重篤度または処方された治療様式で用いられる他の条件のような変数によって左右されるであろう。しかしながら、好ましくはリンパ球:抗原提示細胞またはマトリックス比は約1:1から300:1の範囲であろう。
【0170】
エフェクター/抗原提示培養は、治療的に使用可能数または有効数のCD4+細胞集団を活性化させるか、またはそのような集団を濃縮するために必要な時間維持されるであろう。一般的には最適期間は約1日から5日で、最大特異レベルは一般に3から5日培養後に認められるであろう。本発明のある実施態様では、CD4+細胞のインビトロ活性化は、細胞株にトランスフェクトした後短時間内に検出される。
【0171】
好ましくは、CD4+細胞の活性化は抗原提示細胞に暴露してから1週間以内が最適である。好ましい実施態様ではその後、活性化細胞は、密度勾配、抗体赤血球細胞調製物によるロゼット法、カラムクロマトグラフィーなどを含む単離方法によってさらに精製される。当該精製に続いて、得られたCD4+細胞調製物は109 の活性化CD4+細胞集団を得るためにある期間培養を維持することによってさらに増殖させる。この期間は細胞の増殖時間にしたがって変動させることができるが、一般には14日であろう。
【0172】
c.合成APCまたはマトリックスからのCD4+ 細胞の分離
活性化CD4+ 細胞は、種々の既知の方法の1つを用いて本発明の抗原提示組成物から効果的に分離できる。例えば、MHCクラスIIヘテロダイマー、それに付加されたペプチド、またはCD4+細胞(またはそのセグメント)に対して特異的な単クローン性抗体を用いてそれらの適切な相補的リガンドに結合させることができる。続いて、適切な手段、例えば周知のフローサイトメトリーまたは磁性ビーズ分離法によって抗体標識細胞を刺激体−エフェクター細胞混合物から抽出することができる。密度勾配もまた芽細胞を分離するために使用することができる。
【0173】
d.活性化CD4+細胞による治療
活性化CD4+細胞の有効量は、CD4+集団の活性化に用いた抗原性ペプチドを発現している標的細胞の量および型に関する場合と同様に、インビトロの使用とインビボの使用との間で変動するであろう。この量はまた患者の状態によっても変動し、医師によって適切な全ての要素を考慮して決定されなければならない。しかしながら、マウスで用いられる5×106 から5×107 細胞と比較すると好ましくは約1×106 から約1×1012、より好ましくは約1×108 ら約1×1011、さらに好ましくは約1×109 から約1×1010の活性化CD4+細胞が成人に用いられる。
【0174】
上記で考察したように好ましくは、処置される患者にCDr+細胞を投与する前に、活性化CD4+ 細胞は培養されている合成APCまたはマトリックスから集められる。しかしながら、現在の治療様式または推奨されている治療様式と異なり、本方法の好ましい実施態様ではドロソフィラAPCまたは無細胞マトリックスが用いられていることを特記する必要がある。したがって、細胞および完全な活性化CD4+細胞の分離が達成されない場合、哺乳類の腫瘍促進細胞の投与は極めて危険であるが、一方、少数の合成APCまたはマトリックスの投与に付随する既知の固有の危険性は存在しない。
【0175】
細胞性成分を再導入する方法は当技術分野で既知であり、そのような方法には、米国特許第4844893 号明細書(Honskiら)および米国特許第4690915 号明細書(Rosenberg)に例示されたような方法が含まれる。例えば、静脈内輸液による活性化CD4+細胞の投与が適切である。
【0176】
実施例
以下の実施例は説明を目的とし、本発明を制限するものではない。
【0177】
1.pRmHa−3発現ベクターの調製
本発明で述べたようにドロソフィラシュナイダー(Schneider)2(S2)細胞でMHCタンパク質を発現する際に用いるpRmHa−3発現ベクターを、SphIで直線化したpRmHa−1DNA発現ベクターをsPhI制限酵素で消化したpRmHa−2発現ベクターから得たDNAフラグメントと連結して下記に述べるpRmHa−3発現ベクターを形成することによって構築した。直線化pRmHa−1のpRmHa−2フラグメントによる本態様での連結は、pRmHa−1に存在する2つのEcoRI制限エンドヌクレアーゼクローニング部位のうちの1つを除去するために実施された。したがって、得られたpRmHa−3発現ベクターは、実施例で述べるように種々のMHCクラスIIをコードするDNAフラグメントが挿入されている多クローニング部位(ポリリンカー)内にただ1つのEcoRI制限部位を含んでいた。
【0178】
A.pRmHa−1発現ベクターの調製
メタルチオネインプロモーター、金属反応コンセンサス配列(MTと称する)およびポリアデニル化シグナル(Drosophila melanogasterから単離)含有アルコールデヒドロゲナーゼ(ADH)遺伝子を含むpRmHa−1発現ベクターを文献(Bunchら、Nucl. Acids Res., 16:1043-61(1988))にしたがって構築した。プラスミド発現ベクター、pUC18(ATCCアクセッション番号37253)を本明細書で述べる以降のベクターの由来源として用いた。pUC18プラスミドは、多クローニング部位の5’から3’に以下の制限部位を含む(図1のpUC由来ベクターの模式図にその全てが示されているわけではない):EcoRI;SacI;KpnI;SmaIおよび同じ位置に存在するSmaI;BamHI;XbaI;SalI;AccIおよび同じ位置のHincII;pSTI;SphIおよびHindIII。pUC18ベクターを先ずHindIIIで消化し、直線化pUC18を形成する。続いて、マニアーティスら(「分子クローニング:実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、 Cold Spring Harbor Laboratory刊、ニューヨーク(1982))の記載にしたがって、DNAポリメラーゼIの大型フラグメントで当該HindIII末端を充填することによって平滑端(blunt end)を作出した。
【0179】
ドロソフィラ・メラノガスター(Drosophila melanogaster)ポリアデニル化シグナルを含むADH遺伝子のHinfIフラグメント(740塩基対(bp))と得られた直線化平滑端pUC18ベクターを連結した。連結するADH対立遺伝子は先ず、ゴールドバーグら(Goldbergら、Proc. Natl. Acad. Sci. USA, 77:5794-5798(1980))が記載したプラスミドpSACIから以下のようにして単離した:HinfIで消化し、続いてクレノウ(Klenow)を用いて平滑端を作り配列番号:1に挙げたヌクレオチド配列を得る。ADH対立遺伝子を含むpSACIベクターは、任意の高分子量(15kb以上)を含むバクテリオファージラムダライブラリーから選別したドロソフィラDNAのEcoRIフラグメント(4.7kb)をpBR322(ATCCアクセッション番号31344)でサブクローニングすることによって構築した。5’HinfI制限部位は、文献(Kreitman, Nature,304:412-417(1983))に記載されているように1770位のADH遺伝子内に天然に存在する。3’HinfI部位は、ADH遺伝子がクローニングされたpUC18ベクターに由来した。この位置はADH遺伝子の2500位にあるXbaI部位から3’側に4塩基離れていた。ADHセグメントは、ADHmRNAの3’側非翻訳部分のポリアデニル化/切断配列の35bp上流からポリアデニル化シグナルの700bp下流に広がっていていた。得られたADH遺伝子フラグメントを含むpUC18由来ベクターは図1Aに示すようにpHa−1と称した。
【0180】
pHa−1に挿入するためのMT遺伝子のEcoRI/StuIフラグメント(421bp)は、ドロソフィラ・メラノガスターのゲノムDNAライブラリーの約15.3kbのDNAを含むクローンから得た。昆虫DNAのMboIによる部分消化で調製したライブラリーをラムダ由来EMBL4でクローニングした。421bpフラグメントが、ドロソフィラのMT遺伝子のMTプロモーターおよび金属反応コンセンサス成分を含んでいた(Maroniら、Genetics, 112:493-504(1986))。プロモーターおよびヌクレオチドの+1の位置の転写開始部位を含むこの領域は、MT遺伝子の−370の位置からヌクレオチドの+54の位置に対応していた(配列番号:2)。続いて、得られたフラグメントを先にEcoRIとSmaIで直線化した上記で調製したpHa−1発現ベクターに連結した。StuI消化で作出したMTの3’側平滑端は、SmaI消化で作出したpHa−1の平滑端と適合した。ドロソフィラの5’MT遺伝子フラグメントおよび3’ADH遺伝子フラグメントを含む得られたpUC18由来ベクターをpRmHa−1と称した。このpRmHa−1発現ベクターは、複製開始点(ori)および図1Aに示すようにpHa−1ベクター上のpUC由来のアンピシリン耐性(Ampr)を付与するベータ・ラクタマーゼ遺伝子を含んでいた。pRmHa−1はまた、5’から3’に向けてMT遺伝子、多クローニング部位およびADH遺伝子フラグメントを含んでいた。pRmHa−1ベクターは、下記で述べるようにpRmHa−3発現ベクターの構築で用いられた。
【0181】
B.pRmHa−2発現ベクターの調製
図1Aに示したpRmHa−2発現ベクターを構築するために、pRmHa−1を構築するために上記で述べた方法に少しの改変を加えながら、pUC18由来ベクターpHa−1に上記で調製したMTフラグメントを挿入した。上記で調製したEcoRI/StuI単離MT遺伝子フラグメントのStuI部位にEcoRIリンカーを添加して、両端にEcoRI制限部位をもつメタロチオネインフラグメントを形成する。続いて得られたフラグメントを、EcoRIで先に直線化したADHフラグメント含有pUC18発現ベクターに連結した。5’ドロソフィラMT遺伝子フラグメントおよび、多クローニング部位に対して5’側に2つのEcoRI制限部位を有する3’ADH遺伝子フラグメントを含む得られたpUC18由来ベクターをpRmHa−2と称した。このpRmHa−2発現ベクターは複製開始点(ori)およびpUC由来のアンピシリン耐性(Ampr )を付与するベータ・ラクタマーセ遺伝子を含んでいた。pRmHa−2の模式図はまた、MT遺伝子フラグメントの5’から3’までの連続した部位、多クローニング部位およびADH遺伝子フラグメントを示している。このpRmHa−2ベクターをpRmHa−1とともに下記で述べるようにpRmHa−3発現ベクターの構築に用いた。
【0182】
C.pRmHa−3発現ベクターの調製
ただ1つのEcoRI制限部位を有するpRmHa−3発現ベクターを調製するために、pRmHa−2由来フラグメントをpRmHa−1に連結した。この構築のために、上記で調製したpRmHa−2を先ずSphIで消化した。得られたSphIフラグメント(MT遺伝子の中央で始まり、多クローニング部位のSphI部位まで広がる)を先ずpRmHa−2ベクターから単離し、続いてpRmHa−1につないだ。この連結の前に、pRmHa−1は、MT遺伝子フラグメントに対して5’側にあるEcoRI制限部位を除去して改変し続いてSphIで直線化した。この過程は図1Bに模式的に示した。pRmHa−1のEcoRI部位を除去するために、このベクターを先ずEcoRIで消化し、直線化ベクターを作製し、続いてマングビーン(Mung Bean)のヌクレアーゼで平滑端を作り再び連結した。
【0183】
pRmHa−3ベクターの模式図は図1Cに示す。pRmHa−3が誘導されたpUC18ベクターから由来した種々の制限部位の相対的な位置は図に示されている。pUC18から誘導されたpRmHa−3ベクターは、pUC18の複製開始点およびアンピシリン耐性を付与するベータ・ラクタマーゼ遺伝子を含む。したがって、本発明で調製しpRmHa−3の多クローニング部位でクローニングしたMHCクラスIIをコードするDNAフラグメントは、MTプロモーターによる転写によって調節され、ADH遺伝子を介してポリアデニル化された。
【0184】
2.発現性MHCクラスII遺伝子の調製と発現
A.MHCクラスII遺伝子の増幅と発現
好ましい哺乳類のMHCクラスIIのいずれかをコードする遺伝子またはcDNAはポリメラーゼ連鎖反応(PCR)を用いてクローニングされる。本明細書に記載するプライマーを適切なクラスIIcDNAを増幅するために別々の反応で用い、続いてこれら増幅cDNAをクローニングして配列を決定する。MHCクラスIIタンパク質を生成するために、完全な長さのネズミIAd α鎖およびβ鎖cDNAを先ず入手し、続いてPCR増幅および改変を実施した。ネズミIAd α鎖cDNAの完全なヌクレオチド配列はベノイストら(Benoistら、Cell, 34:169-177(1983))が報告しており、またジェンバンク(GenBank)のリストにも記載されている(アクセッション番号K01923)。ネズミIAd β鎖cDNAの完全なヌクレオチド配列はマリセンら(Malissenら、Science, 221:750-754(1983))が記載し、ジェンバンクにも登録されている(アクセッション番号K00007およびK00008)。各鎖を増幅するために全RNAの供給源としてマウスの脾細胞を用いた。第一の鎖のcDNAは、オリゴ(dT)およびニワトリ骨髄芽球症ウイルスの逆転写酵素を用いて合成した。得られたcDNAをPCR増幅反応に用いた。増幅反応では、下記に述べる適切なプライマーおよびジーンアンプ(GeneAmp)キットおよび温度循環装置(Perkin-Elmer/Cetus, ノーウォーク、コネチカット)を利用した。反応条件は、好ましくは1μgのcDNA鋳型、および200nMの各オリゴヌクレオチドプライマーを含んでいた。以下のように30サイクル実施した:(a)92℃で1分;(b)60℃で1分;(c)72℃で1分。続いてPCR反応物を99℃で10分加熱し、Taqポリメラーゼを不活化し、さらにDNA端をT4ポリメラーゼ(Stratagene, ラホイヤ、カリフォルニア)で平滑端にした。
【0185】
実施例で示したオリゴヌクレオチドプライマーの全てについて、5’プライマーはまたフォワードプライマーまたはセンスプライマーと称する。なぜならば、これは、相補的な下の鎖とハイブリダイズさせるためのcDNAの上の鎖と同じ配列を有するからである。対照的に、3’プライマーは相補的な上の鎖とハイブリダイズさせるためのcDNAの下の鎖と同じ配列を有するのでバックワードプライマーまたはアンチセンスとも称される。両方のプライマーとも5’から3’方向で記載されている。5'CTTGAATTCCACCATGCCGTGCAGCAGAGCTCTGA3'(配列番号:3)なるヌクレオチド配列を有する5’プライマーを用いて完全な長さのIAd α鎖cDNAを増幅した。またこの5’プライマーをEcoRI制限部位をもつようにデザインして、当該増幅生成物を方向性をもって受容体の発現ベクターに連結することを可能にした。
【0186】
5'TTTGGATCCTCATAAAGGCCCTGGGTGTC3'(配列番号:4)なる3’プライマーもまたBamHI制限部位をもつようにデザインした。
【0187】
5'CTTGAATTCCACCATGGCTCTGCAGATCCCCA3'(配列番号:5)なるヌクレオチド配列を有する5’前進プライマーを用いて完全な長さのIAd β鎖cDNAを増幅した。この5’プライマーもまたEcoRI制限部位をもつようにデザインし、当該増幅生成物を方向性をもって受容体の発現ベクターに連結することを可能にした。
【0188】
5'TTTGGATCCTCACTGCAGGAGCCCTGCT3'(配列番号:6)なる3’プライマーをBamHI制限部位を含むようにデザインした。ネズミIAd α鎖およびIAd β鎖をコードする改変cDNAを別々に先ずメタロチオネイン駆動pRmHa−3ベクターのEcoRIおよびBamHI制限部位のポリリンカーに方向性をもたせてクローニングした。続いて色素ターミネータ技術によってアプライドバイオシステム(Applied Biosystem)373A自動配列決定装置で配列を調べた。
【0189】
pRmHa−3でクローニングしたネズミIAd α鎖およびIAd β鎖の増幅領域の完全なヌクレオチド配列は、それぞれ配列番号:7および8で示されている。
【0190】
続いて、別の場所(Jacksonら、Proc. Natl. Acad. Sci. USA, 89:12117-21(1992))で述べたように、ドロソフィラ・メラノガスターのシュナイダー2細胞(ATCCCRL10974、ロックビル、メリーランド))に別々のプラスミドをトランスフェクトした。2つのクラスII鎖をコードするプラスミドの同じ量を、ネオマイシン耐性遺伝子(プラスミドphshsneo(Bunchら、Nuc. Acids Res., 16:1043-1061(1988))とともに1:30の割合で同時にトランスフェクトして安定な細胞株を作製した。この細胞株は、10%ウシ胎児血清(55℃で1時間加熱処理)、100単位/mlのペニシリン、100μg/mlのストレプトマイシンおよび1mMのグルタミンを補充したシュナイダーのドロソフィラ培養液(Gibco/brl, グランドアイランド、ニューヨーク)での4週間を越えるG418選別によって得られた。具体的には、トランスフェクトした後に上清を注意深く取り除き、500μg/mlのジェネチシン(G418)(Gibco/BRL, グランドアイランド、ニューヨーク)を含む全容量12mlのシュナイダー培養液を入れた75cm2 のフラスコに細胞を移した。4日後、500μg/mlのG418とともに6mlのシュナイダー培養液を含む新しいフラスコに4mlの培養を移した。この操作を安定な細胞集団が出現するまで4〜7日毎に繰り返した。安定な細胞集団は弱くフラスコに付着し、約24時間の分裂時間で増殖した。その後これらの細胞を培養し、上記で述べたように選別培地で継代した。安定的にトランスフェクトされた凍結部分標本は、遠心沈殿によって5−20×106細胞を採集し、1mlの細胞凍結培養液(93%ウシ胎児血清/7%ジメチルスルホキシド)に再懸濁することによって調製された。続いて部分標本を−70℃に1週間置き、その後液体窒素貯蔵庫に移した。安定的にトランスフェクトされたMHCクラスIIのIAd α鎖およびβ鎖の遺伝子の発現は、0.7mMの硫酸第二銅を用いて27℃で24時間誘発された。
【0191】
トランスフェクトしたドロソフィラ細胞の細胞表面でのMHCクラスIIのIAd ヘテロダイマーの発現は、MKD6(IAd 特異的単クローン性抗体(Kapplerら、J. Exp. Med., 153:1198(1981))による染色後フローサイトメトリーで調べた。簡単に記せば、細胞の一部分(5×105 )を氷上の試験管に移し、遠心沈殿(1000×g5分)によって集め、適切な一次抗体(MKD6)を含有する5%ウマ血清を含む0.1mlのドロソフィラ培養液に再懸濁した。室温で20分保温した後、ウマ血清を含む3mlのシュナイダー培養液で2回洗浄し、0.1mlのFITC標識二次抗体(Cappell, Durham,ノースカロライナ)含有培養液に再懸濁した。氷上で20分維持した後、ウマ血清含有シュナイダー培養液で2回洗浄し、この緩衝液に1×106 /mlの濃度で再懸濁した。沃化プロピジウムを添加して死細胞を分析から排除した。続いてサンプルをファクスキャン(FACScan)またはファクソート(FACSort)装置(Becton Dickison)で分析した。1つ以上のアクセサリー分子とともにネズミIAd MHCクラスIIハプロタイプ(本明細書で述べまた当技術分野でも周知である)を発現する本発明の合成抗原提示細胞は下記のように製造された。
DR、DQおよびDPに対する完全な長さのヒトMHCクラスIIα鎖およびβ鎖を末梢血細胞(B細胞、マクロファージおよび樹状突起細胞を含む)から増幅する。各々のハプロタイプを増幅するために用いた5’および3’プライマーは以下の配列を有する:
【0192】
DRα:5’プライマー=5'CCACCATGGCCATTAGTGGAGTC3'(配列番号:9)
3’プライマー=5'TTTGGATCCTTACAGAGGCCCCCTGCGTT3'(配列番号:10);
DRβ:5’プライマー=5'CCACCATGGTGTGTCTGAGGCTCC3'(配列番号:11)
3’プライマー=5'TTTGGATCCTCAGCTCAGGAATCCTCTTG3'(配列番号:12);
DQα:5’プライマー=5'CCACCATGGTCCTAAACAAAGCTCTGAT3'(配列番号:13)
3’プライマー=5'TTTGGATCCTCACAAGGGCCCTTGGTGTCT3'(配列番号:14);
DQβ:5’プライマー=5'CCACCATGGCTTGGAAGAAGGCCTTT3'(配列番号:15)
3’プライマー=5'TTTAGATCTCAGTGCAGAAGCCCTTT3'(配列番号:16);
DPα:5’プライマー=5'CCACCATGGGCCCTGAAGACAGAAT3'(配列番号:17)
3’プライマー=5'TTTGGATCCTCACAGGGTCCCCTGGGC3'(配列番号:18);
DPβ:5’プライマー=5'CCACCATGGTTCTGCAGGTTTCTGCG3'(配列番号:19)
3’プライマー=5'TTTGGATCCTTATGCAGATCCTCGTTGAA3'(配列番号:20);
【0193】
上記ヒトハプロタイプを得るための増幅条件は、ネズミの対応物を得るために上記で述べた条件と同一である。
1つ以上のアクセサリー分子とともにヒトMHCクラスIIハプロタイプ(本明細書で述べまた当技術分野でも周知である)を発現する本発明の合成抗原提示細胞は下記のように製造される。
【0194】
B.不変鎖およびHLA−DM鎖cDNAの増幅
詳細な説明の節Bで述べた本発明の幾つかの態様のために、不変鎖並びにHLA−DMαおよびHLA−DMβを選択したMHCクラスIIヘテロダイマーおよび少なくとも1つの補助因子とともに同時発現させる。本事例では、MHCクラスIIα鎖およびβ鎖、不変鎖並びにHLA−DMαおよびHLA−DMβを、5:5:8:1:1のモル比で受容体の抗原提示細胞にトランスフェクトする。他の遺伝子のモル比に対するアクセサリー分子の比は1:1である。続いて、得られたトランスフェクト細胞を上記のように外因性遺伝子の発現のために誘発して本発明の合成抗原提示細胞株を作製する。
【0195】
1)不変鎖
ネズミの不変鎖cDNAをマウスの脾臓細胞から作製した。それぞれ
【0196】
5'AAGAATTCACTAGAGGCTAGAGCCAT3'(配列番号:21)および、
5'AAGGATCCTCACAGGGTGACTTGACC3'(配列番号:22)
【0197】
のヌクレオチド配列を有する5’および3’プライマーのプライマー対を用いて当該供給源から完全な長さの不変鎖を増幅させる。上記で述べたように、5’および3’プライマーをそれぞれEcoRIおよびBamHI制限部位を含むようにデザインした。増幅不変鎖をEcoRI/BamHI消化pRmHa−3ベクターでクローニングした後、得られたクローンの配列を調べ、配列番号:23に挙げた配列を有することが分かった。
【0198】
本発明で用いるヒト不変鎖cDNAは、γ−インターフェロン誘発HeLa細胞(ATCCアクセッション番号CCL2)由来RNAを用いて作製した。続いて、それぞれ
【0199】
5'AAGAATTCACCATGGATGATCAGCGCGACCTT3'(配列番号:24)および、
5'AAAGGATCCTCACATGGGGACTGGGCCCAGA3'(配列番号:25)
【0200】
のヌクレオチド配列をもつ5’および3’オリゴヌクレオチドプライマーを用いて、得られたcDNAはPCRで鋳型として使用する。得られたPCRフラグメントをEcoRIおよびBamHIで切断し、同様に消化したpRmHa−3に連結する。
【0201】
2)HLA−DM
またγ−インターフェロン誘発HeLa細胞由来mRNAを用いてHLA−DMcDNAのαおよびβ鎖を合成する。続いてこれらをPCRで鋳型として用いる。α鎖は、それぞれ
【0202】
5'AAACCATGGGTCATGAACAGAACCA3'(配列番号:26)および、
5'TTTGTCGACTCAGTCACCTGAGCAAGG3'(配列番号:27)
【0203】
のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅する。β鎖は、それぞれ
【0204】
5'AAACCATGGTCTCATTCCTGCC3'(配列番号:28)および、
5'TTTGTCGACCTAGGAAATGTGCCATCC3'(配列番号:29)
【0205】
のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅する。得られたPCRフラグメントをNcoIおよびSalIで切断し、同様に消化したpRmHa−3に別々に連結する。
【0206】
C.アクセサリー分子をコードする遺伝子の増幅
1)付着分子
a.ICAM−1、ICAM−2およびICAM−3
ネズミICAM−1を単離するために、脾臓細胞をBalb/cマウスから単離した。mRNAの単離前に、脾臓細胞を先ずファストトラック(Fast Track)キット(Invitrogen, サンディエゴ、カリフォルニア)を用い製造元の指示にしたがってconAで刺激した。cDNAは上記のようにmRNAから合成した。続いて、公表されたcDNAヌクレオチド配列(G. Siuら、J. Immunol., 143:3813−3820(1989))を基にデザインしたそれぞれ以下の5’および3’プライマーを用いて得られたcDNAをPCRに付した:
【0207】
5'TTTAGAATTCACCATGGCTTCAACCCGTGCCAAG3'(配列番号:30)および、
5'TTTAGTCGACTCAGGGAGGTGGGGCTTGTCC3'(配列番号:31)。
【0208】
続いて、PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0209】
続いて、上記で述べたリン酸カルシウム法を用い、上記で調製したネズミIAdα鎖およびβ鎖遺伝子とともに上記の発現構築物を、ICAM−1:α鎖:β鎖を1:1:1のモル比でドロソフィラS2細胞に同時にトランスフェクトした。安定的にトランスフェクトされた細胞が上記のように得られた。細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子ICAM−1を発現している合成抗原提示ドロソフィラS2細胞を、続いて先に述べたように発現を誘発することによって製造した。
【0210】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様ではまた、ネズミIAd 分子と一緒にICAM−1、B7.1および/またはB7.2をコードする遺伝子を含む合成抗原提示細胞を製造した。
【0211】
ヒトICAM−1は同様にヒトの細胞株K562から単離したmRNAから増幅する。当該細胞株は、ヒトの慢性骨髄性白血病に由来し(ATCCアクセッション番号CCL-243)、推奨された条件下で培養した(すなわち、10%ウシ胎児血清添加RPMIで5%CO2 下37℃)。ヒトアクセサリー分子を増幅するPCRプライマーは利用可能な既知の配列を基に、適切なベクターでクローニングするために必要な5’および3’クローニング部位を考慮してデザインされる。ヒトICAM−1cDNAのヌクレオチド配列は、ジェンバンクアクセッション番号GB-J03132により入手できる。ICAM−1の5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0212】
5'ACCCTTGAATTCATGGCTCCCAGCAGCCCCCGGCCC3'(配列番号:32)および、
5'ATTACCGGATCCTCAGGGAGGCGTGGCTTGTGTGTTCGG3'(配列番号:33)。
【0213】
増幅ヒトICAM−1を得るために、先で述べたようにこれらプライマーを用いてPCRを実施する。続いて、得られたPCR生成物を先に述べたようにpRmHa−3でクローニングし、その後、これもまた先に述べたように受容細胞にトランスフェクトする。
【0214】
同様に、ヒトICAM−2およびICAM−3(これらのヌクレオチド配列はそれぞれジェンバンクアクセッション番号GB-X15606 およびGB-S50015 により入手できる)は、該当するプライマー対を用いて増幅される。ICAM−2を増幅する5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0215】
5'AAGGTACCCGTGGAGACTGCCAGAGAT3'(配列番号:34)および、
5'TTTGGATCCCTATGGCCGGAAGGCCTG3'(配列番号:35)。
【0216】
ICAM−3を増幅する5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0217】
5'AAGAATTCCTGTCAGAATGGCCACCAT3'(配列番号:36)および、
5'TTTAGATCTTCACTCAGCTCTGGACGGT'(配列番号:37)。
【0218】
続いて、得られた増幅ICAM生成物を別々にpRmHa−3につなぎ、別々にドロソフィラS2細胞にトランスフェクトする。
【0219】
選択したヒトMHCクラスIIハプロタイプとともにアクセサリー分子ICAM−1を発現している合成抗原提示ドロソフィラS2細胞は、続いて先に述べたように発現を誘発することによって製造される。
【0220】
他の実施態様では、実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、ヒトハプロタイプと合わせてB7.1および/またはB7.2と一緒にICAM−1、ICAM−2またはICAM−3をコードする遺伝子を含む合成抗原提示細胞がまた作製される。本発明で使用することを目的とするまた別の変更には、LFA−3および/またはFasリガンド(FasL)(これらはともに下記で述べる他のアクセサリー分子である)と上記の組み合わせが含まれる。
【0221】
b.LFA−3
ヒトLFA−3は血中リンパ球から単離される。ヒトLFA−3のcDNAのヌクレオチド配列は、ジェンバンクアクセッション番号GB-I09083 によって入手できる。したがって、ヒトLFA−3はそれぞれ以下のヌクレオチドを有する5’および3’プライマーを用いて増幅される:
【0222】
5'ACCCTTGAGCTCATGGTTGCTGGGAGCGACGCGGGG3'(配列番号:38)および、
5'ATTACCGGATCCTTAAAGAACATTCATATACAGCACAATACA3'(配列番号:39)。
【0223】
実施例5で述べる活性化アッセイで使用する合成抗原提示細胞を作製するために、先に述べたようにヒトハプロタイプとともにヒトLFA−3のみをコードするか、または本明細書で述べた他のアクセサリー分子と一緒にヒトLFA−3をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0224】
2)同時刺激分子
a.B7.1
ICAM−1について上記で述べたように、マウスから単離したmRNAからcDNAを作製した。続いて、以下のそれぞれ5’および3’オリゴヌクレオチドプライマーを用いて得られたcDNAをPCRに付した(これらプライマーは、公表されたcDNAヌクレオチド配列(Freemanら、J. Exp. Med., 174:625-631(1991))を基にデザインし5’から3’の方向で表示する):
【0225】
5'TTTAGAATTCACCATGGCTTGCAATTGTCAGTTG3'(配列番号:40)および、
5'TTTAGTCGACCTAAAGGAAGACGGTCTGTTC3'(配列番号:41)。
【0226】
PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0227】
続いて、上記で調製したネズミのIAd α鎖およびβ鎖遺伝子とともに上記の発現構築物を、B7.1:α鎖:β鎖のモル比を1:1:1として上記で述べたように燐酸カルシウム法を用いてドロソフィラS2細胞に同時にトランスフェクトした。上記で述べたように安定的にトランスフェクトされた細胞を得た。続いて、細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子B7.1を発現する合成抗原提示細胞ドロソフィラS2細胞を、上記で述べたように誘発することによって作製した。
【0228】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ネズミIAd 分子と一緒にネズミICAM−1、B7.1および/またはB7.2をコードする遺伝子を含む合成抗原提示細胞もまた製造される。
【0229】
ヒトB7.1は同様にK562細胞から単離され、上記で述べたようにpRmHa−3でクローニングされる。ヒトB7.1cDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-M83071 により入手できる。5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0230】
5'ACCCTTGAATCCATGGGCCACACACGGAGGCAG3'(配列番号:42)および、
5'ATTACCGGATCCTTATACAGGGCGTACACTTTCCCTTCT3'(配列番号:43)。
【0231】
続いて、得られたPCR生成物をpRmHa−3に挿入し、続いてこれを増幅させクローニングしたヒトハプロタイプとともにドロソフィラS2細胞に同時にトランスフェクトした。
【0232】
続いて先に述べたように発現を誘発することによって、細胞表面に選択したヒトMHCクラスIIハプロタイプとともにアクセサリー分子B7.1を発現する合成抗原提示細胞ドロソフィラS2細胞を作製する。
【0233】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ヒトハプロタイプとともに本明細書で述べたアクセサリー分子を種々に組み合わせたB7.1をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0234】
b.B7.2
ネズミIC−21細胞(ATCCアクセッション番号TIB186)を10%ウシ胎児血清含有RPMI1640培養液で増殖させた。上記のようにこれらの細胞から単離したmRNAからcDNAを合成した。続いて、得られたcDNAを以下のそれぞれ5’および3’オリゴヌクレオチドプライマーを用いてPCRに付した(これらプライマーは、公表されたcDNAヌクレオチド配列(Freemanら、J. Exp. Med., 178:2185-2192(1993))を基にデザインし5’から3’の方向で示す):
【0235】
5'TTTAGAATTCACCATGGACCCCAGATGCACCATGGG3'(配列番号:44)および、
5'TTTAGTCGACTCACTCTGCATTTGGTTTTGCTGA3'(配列番号:45)。
【0236】
PCR生成物を制限酵素EcoRIおよびSalIで切断し、同様に消化したpRmHa−3に連結した。
【0237】
続いて、上記で調製したネズミのIAd α鎖およびβ鎖遺伝子とともに上記の発現構築物を、B7.2:α鎖:β鎖のモル比を1:1:1として上記で述べたように燐酸カルシウム法を用いてドロソフィラS2細胞に同時にトランスフェクトした。上記で述べたように安定的にトランスフェクトされた細胞を得た。続いて、細胞表面にIAd α鎖およびβ鎖とともにアクセサリー分子B7.2を発現する合成抗原提示細胞ドロソフィラS2細胞を、上記で述べたように誘発することによって作製した。
【0238】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ICAM−1およびネズミIAd 分子と一緒にネズミB7.2をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0239】
ヒト細胞株HL60(前骨髄球性白血病由来)(ATCCアクセッション番号CCL−240)からヒトB7.2を単離する。ヒトB7.2cDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-M83071 により入手できる。ヒトB7.2を増幅するための5’および3’プライマーはそれぞれ以下のヌクレオチド配列を有する:
【0240】
5'ACCCTTGAGCTCATGGATCCCCAGTGCACTATG3'(配列番号:46)および、
5'ATTACCCCCGGGTTAAAAACATGTATCACTTTTGTCGCATGA3'(配列番号:47)。
【0241】
選択したヒトハプロタイプのための構築物と一緒にドロソフィラS2細胞にトランスフェクトするために、増幅B7.2生成物を続いて上記で述べたようにpRmHa−3でクローニングする。トランスフェクトされた遺伝子の発現は上記のように誘発される。
【0242】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、他の実施態様では、ヒトハプロタイプとともに本明細書で述べたアクセサリー分子を種々に組み合わせたヒトB7.2をコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0243】
3)サバイバル分子
ヒトFasリガンドは活性化ヒトT細胞から単離される。ヒトFasリガンドcDNAのヌクレオチド配列はジェンバンクアクセッション番号GB-U08137 により入手できる。したがって、ヒトFasリガンドは、それぞれ以下のヌクレオチド配列を有する5’および3’プライマー対を用いて増幅される:
【0244】
5'AAAGGATCCACCATGCAGCAGCCCTTCAATT3'(配列番号:48)および、
5'TTTGGATCCTTAGAGCTTATATAAGCCGA3'(配列番号:49)。
【0245】
ヒトCD70(T細胞上に発現されるCD27のためのリガンド)は、それぞれ以下のヌクレオチド配列を有する5’および3’プライマー対を用いて上記のように増幅される:
【0246】
5'AAAGAATTCGGTACCATGCCGGAGGAGGGTTCGG3'(配列番号:50)および、
5'TTTGGATCCTCAGGGGCGCACCCACTGCA3'(配列番号:51)。
【0247】
実施例5で述べる活性化アッセイで用いる合成抗原提示細胞を作製するために、上記で述べたように、ヒトハプロタイプとともにヒトFasリガンドのみをコードするか、または本明細書で述べた他のアクセサリー分子を種々に組み合わせたヒトFasリガンドをコードする遺伝子を含む合成抗原提示細胞もまた作製される。
【0248】
3.合成担体に定着させたMHCクラスIIの調製
ここで述べる実施例は、大量のMHCクラスII分子および単一種のアクセサリー分子またはそれらの種々の組み合わせを、単クローン性抗体結合およびロゼット形成実験(T細胞レセプター結合)で判定したとき固有の構成で種々の表面(ハエの細胞、赤血球細胞、ラテックスビーズ)に固定させる新規な方法を提供する。本方法は、人工リン脂質膜を含む他の合成表面に範囲を広げることができる。アビジン結合リン脂質とともにホスファチジルエタノールアミンは特に本発明に適切である。これらのリン脂質は、リペックスバイオメンブレン社(Lipex Biomembrane Inc., バンクーバー、ブリティッシュコロンビア、カナダ)から市販ルートで入手できる。
【0249】
A.アビジン被覆赤血球細胞上へのビオチン付加MHCクラスIIの固定
NHS-LCビオチン、ニュートロアビジン(neutravidin)およびビオチン−BMCCはピアース(Pierce, ロックフォード、イリノイ)から購入する。ヒツジ赤血球はコロラドシーラムカンパニー(Colorado Serum Company, デンバー、コロラド)から入手する。MHCクラスIIを発現しているドロソフィラS2細胞は上記のように調製する。ハイブリドーマ細胞培養上清として、単クローン性抗体MKD6およびM5−114(MKD6分泌ハイブリドーマについてはATCCアクセッション番号TIB−120)を用いる。用いたプロトコルは文献に記載されている(Muzykantov & Taylor, Anal. Biochem., 223:142-148(1994))。簡単に記せば、SRBCを4回PBSで洗浄し、NHS-LCビオチンでビオチン付加し、再びPBSで4回洗浄し、3%ウシ胎児血清および0.02%アジ化ナトリウムを含むPBS中で4℃で保存する。組換え発現MHCクラスIIα鎖およびβ鎖はビオチン−BMCCを用いてビオチン付加する。ビオチン−BMCCは、チオール基と反応するマレイミド結合ビオチンである。ビオチン付加は製造元の推奨にしたがって実施する。未反応ビオチンはセントリコン(Centricon)10で除去する。
【0250】
ビオチン付加MHCクラスIIα鎖およびβ鎖は、最終濃度0.2mg/mlでアビジン被覆SRBCとともに30分保温し、続いて10%ウシ胎児血清含有DMEMで洗浄することによって固定される。MHCクラスII結合SRBCは直ちに使用する。
【0251】
アビジン被覆SRBC上へのビオチン付加MHCクラスIIα鎖およびβ鎖の固定は上記のように実施する。結合は、上記の抗体を用いてフローサイトメトリーにより調べる。ロゼット形成アッセイのためには、MHCクラスII発現ドロソフィラS2細胞またはMHCクラスII被覆SRBCのどちらかを、特定のペプチド(実施例4参照)(0.02mM)または無関係ペプチド(0.02mM)とともに30分氷上で保温する。続いて、CD4+T細胞を添加する(割合は、10個のT細胞に対して1個のドロソフィラS2細胞または10個のSRBCに対して1個のT細胞)。その後混合物を沈澱させ、少なくとも30分氷上で保温する。続いて、細胞を注意深く再懸濁させてロゼットを数える。ロゼットは、少なくとも3個のCD4+T細胞と結合したドロソフィラS2細胞、または少なくとも3個のSRBCと結合したCD4+T細胞である。
【0252】
B.アビジン被覆ラテックスビーズ上へのビオチン付加MHCクラスIIの固定
直径が6ミクロンのラテックススルフェートビーズをインターフェイシャルダイナミクス社(Interfacial Dynamics Corporation, ポートランド、オレゴン)から購入し、上記のプロトコルにしたがってビオチン付加する。アビジン被覆ラテックスビーズは、1%のラテックスビーズ懸濁液を1mg/mlのニュートロアビジンを含むPBS中で1時間室温で保温して調製する。等容量の10%ウシ胎児血清含有PBSを続いて添加する。室温で1時間保温した後、ビーズを3回洗浄し、組換えビオチン付加MHCクラスIIの結合のために使用する。
【0253】
組換えビオチン付加MHCクラスIIを最終濃度0.2mg/mlでアビジン被覆ラテックスビーズと30分保温し、続いて10%ウシ胎児血清含有DMEM中で洗浄することによって固定される。結合MHCクラスIIを有するSRBCは直ちに使用する。ロゼット形成実験は先に述べたように実施する。
【0254】
C.種々の固形担体(例えばプラスチックマイクロウェルプレート)に結合したMHCクラスIIの固定と検出
MHCクラスII分子は、以下のようにマイクロタイタープレート(Corning)に直接結合によって固定し検出する:MHCクラスIIを所望の濃度にPBSで希釈し(例えば100ng/ウェルのために1mg/ml)、続いてプラスチック製マイクロタイタープレートの各ウェルに添加する。プレートを1時間室温で保温する。保温後、プレートをPBSで1回洗浄し、さらにPBS+トゥイーン(PBST)中の2%ウシ血清アルブミン200μl(0.05%)を添加し、さらに1時間室温で保温する。プレートを3回PBSTで洗浄し、2%BSA含有PBS中のビオチン付加抗MHCクラスII単クローン性抗体を添加する(1:2500)。プレートをさらに1時間室温で保温し、PBSTで3回洗浄する。アビジン結合HRPを2%BSA含有PBS中で添加する(1:2500)。室温でさらに1時間保温した後、プレートをPBSTで3回洗浄し、H2O2 またはトフェニルジアミン(thophenyldiamine)を添加する。反応はH2SO4 で停止させる。反応生成物は490nmで比色計により検出する。また別には、組換えMHCクラスII分子は、基質とのビオチン−アビジン連携相互反応により結合させることができる。この実施態様では、マイクロウェルプレートは、PBSで0.001mg/mlの濃度に希釈した100μlのアビジンで被覆される。過剰なアビジンはPBSによる洗浄で除去する。MHCクラスII結合の提示検出のための上記の過程がその後に続く。
【0255】
また別には、組換えMHC分子は、基質に結合させたニッケルと相互反応するMHCに添加したポリヒスチジンタッグによる架橋によって固定される。結合および検出のための上記の過程の後は、ニッケルキレート被覆マイクロウェルプレート(Xenopore)およびポリヒスチジンタッグを付加された発現MHC分子組換え体を使用する。
【0256】
4.ペプチド生成
本発明の抗原性ペプチドは、本明細書で述べたものを含む天然に存在する供給源から得るか、または既知の方法を用いて合成してもよい。本明細書で開示する種々の実施例では、全てのペプチドは、アプライド・バイオシステム社の装置ABI−431A(Foster City, カリフォルニア)を基にT−BOC法によって別々に化学的に合成され、C18逆相カラムにより精製される。アミノ酸残基のランダムアソートメントにより生成されるペプチドの単離または合成もまた適切であるかもしれない。特に、CD4細胞をおそらく刺激すると考えられるペプチドを空のMHC分子に付加するために特定のエピトープを突き止めようとしている場合は適切であろう。ランダム化ペプチドの混合物は、プロテアソームを用いることによって(タンパク質またはポリペプチドを分解過程(例えばキモトリプシンによる消化)に付すことにより)または合成によって得られる。
【0257】
本発明の細胞株は、タンパク質またはポリペプチドをヒトMHCクラスII分子に付加できるより小さなペプチドに分解することができるが、ペプチドを直接細胞に導入してより迅速な付加および発現作用を促進することが好ましい。
【0258】
ペプチドがアミノ酸残基を任意に取り込みながら合成される場合は、修飾アミノ酸とともにLまたはD型の全てのアミノ酸変種が、各合成サイクル時に好ましくは取り込まれるであろう。しかしながら、種々のパラメーター(例えばある種のアミノ酸の溶媒不適合)が、ある種のアミノ酸を欠くペプチドを含む混合物を生じるかもしれない。したがって、この過程は、溶媒および反応条件を変更することにより必要とされるように調節して最も多様なペプチドを生成しなければならない。ネズミMHCクラスIIに関して実施例5で述べるようにペプチド付加のために用いられるペプチドは、アミノ酸残基配列ISQAVHAAHAEINEAGR(配列番号:52)を有するオボアルブミン323-339 (OVA)である。ヒトMHCクラスIIハプロタイプに特異的な好ましいペプチドには以下のペプチドが含まれる:1)HLA−DRB1*0101と結合するインフルエンザヘマグルチニン306-318 PKYVKQNTLKLAT(配列番号:53);2)HLA-DRB1*0301と結合する結核菌(M. tuberculosis)由来HSP652-12 KTIATDEEARR(配列番号:54);3)HLA-DRB1*0301と結合するクラミジア・トラコマチス(C. trachomatis)のミトコンドリア外側タンパク質(MOMP) QASLALSYRLNMFTP(配列番号:55);4)HLA-DRB1*0401と結合するHLA−A233-45 FVRFDSDAASQRM(配列番号:56)。ペプチドリガンドがどのように限定されるかについての記載とともに既知のクラスII結合ペプチドの豊富な記載はラメンシーらの報告によって提示されている(Rammenseeら、Immunogenetics, 41:178-228(1995))。本発明の方法を実施するときに使用するペプチドはまたキロンミメトープ(Chiron Mimetope)により入手できる。他の実施態様では、本明細書で述べた合成抗原提示細胞に関する本発明の方法を実施する場合はペプチドライブラリー(当業者は容易に作製できる)の使用もまた意図される。
【0259】
5.ペプチドとのインビトロ保温による空のMHCクラスII分子の付加
下記に提示する結果によって示すように、選択アクセサリー分子とともにドロソフィラ細胞株の表面に発現されたネズミMHCクラスII分子は、用量依存態様で特定のペプチドと結合し、さらに抗原特異的T細胞にこれらペプチドを提示してT細胞活性化をもたらすことができる。(これらのドロソフィラ細胞株は、MHCクラスII分子およびアクセサリー分子をコードする種々の組み合わせのプラスミドをトランスフェクトされている。)
【0260】
下記で実施する分裂および活性化アッセイのために、ドロソフィラ・メラノガスターシュナイダー2(SC2)細胞(ATCCアクセッション番号CRL10974)を、実施例2で述べたH2−Ad のMHCクラスIIα鎖およびβ鎖をコードする遺伝子で形質転換した。選択補助細胞がMHCクラスII分子と一緒に発現される他のアッセイのためには、B7.1、B7.2およびICAM−1のための遺伝子もまた先に述べたMHC遺伝子含有細胞に導入した。
【0261】
アクセサリー分子は下記に規定するような選択濃度および組み合わせで発現させ、再現性をもつ一定のCD4+ 細胞活性化を可能にした。発現(硫酸銅による処置によって誘発される)に続いて、発現分子の濃度をモニターする。所望レベルの分子を発現している細胞の選択はフローサイトメトリーによる細胞株の分類によって実施する。表1は、ネズミIAd MHCクラスII分子のみまたは種々のアクセサリー分子と一緒に発現するドロソフィラ細胞株を示す。この表はまた、下記でさらに述べる分裂効果の結果の要約を提示する(" −”は効果無し、" +”は陽性分裂効果を示す)。合成抗原提示細胞の各タイプによって産生されたサイトカインの種類の要約もまた表示し、合わせて個々のサイトカインの産生がアップレギュレートであるか、ダウンレギュレートであるかをさらに矢印によって示す(これらは下記でもさらに説明する)。
【0262】
表1
アクセサリー分子 分裂反応 産生サイトカイン
無し − −
B7.1 + IL−4 IL−10 IL−2
B7.2 + IL−4 IL−10 IL−2
ICAM−1 − −
B7.1+B7.2 + IL−4 IL−10 IL−2
B7.1+ICAM−1 + ↑IL−2 ↓IL−4
↓IL−10
B7.2+ICAM−1 + ↑IL−2 ↓IL−4
↓IL−10
CD70+ICAM−1 + IL−2
CD70 − −
CD70+ICAM−1 + ↑IL−2
+B7.2
【0263】
抗原提示および最終的T細胞反応の効果を評価するために、抗原特異的T細胞は、T細胞レセプター遺伝子導入マウスから得たCD4+ 細胞であった。本明細書で用いた当該遺伝子導入系統は、オボアルブミンペプチド323−339に特異的なT細胞レセプターを発現するD01であった。D01マウス系統はデニス博士(Dr. Dennis Lo)から入手した。組換え体の表面発現MHCクラスII分子に対するT細胞反応性の評価のために、ホストの全てのAPCを反応集団から除外するようにデザインした技術を組み合わせてCD4+ 細胞を先ずリンパ節から精製した。続いて、これら高度に精製したCD4+ 細胞の少数(5×104 )をovaペプチドの存在下または非存在下でドロソフィラ細胞株とともに培養した。
【0264】
アクセサリー分子(B7.1、B7.2またはB7.2+ICAM−1)とともに、およびアクセサリー分子無しでMHCクラスIIの存在下でペプチド提示の効果を、文献の記載にしたがって先ず分裂アッセイで調べた(Webbら、Cell, 63:1249(1990))。簡単に記せば、5×104 CD4+T細胞を5×104 ドロソフィラ細胞とともに300μlの培養液でマイクロタイターウェルで培養した。採集18時間前に、100μlの上清をサイトカイン分析のために取り出し、1μCiの3 H−チミジンを各ウェルに添加した。表示した期間で(3、4または5日)それぞれのウェルをガラスファイバー上に採集し、液体シンチレーション計測器で計測した。
【0265】
図2で示されるように、MHCクラスII分子のみを発現している細胞株は、放射性チミジンの低い取り込みで示されるとおりT細胞の分裂反応を刺激できなかった。すなわち、より高いカウントが、μMで示した刺激ペプチドの量に対してプロットした分裂の増加を示している。しかしながら、B7.1またはB7.2同時刺激分子のどちらかの同時発現は、MHCクラスII提示ovaペプチドの能力に強力な分裂反応刺激を付与した。比較のために、脾臓APC(T−Sと表示)を用いた結果を表2に示す。
【0266】
図3に示すように、付着分子ICAM−1とともにMHCクラスII分子を発現するドロソフィラ細胞株は刺激されなかった。しかしながら、B7.2と一緒にICAM−1を発現させることによって、低濃度のペプチド抗原で分裂反応を刺激する合成APC細胞が得られた。これらの結果は、ネズミH2−Ad MHCクラスII分子および同時刺激分子(B7.1、B7.2およびB7.2+ICAM−1)を発現しているドロソフィラ細胞株は、分裂の増加によって測定した場合未感作CD4+T細胞を活性化できることを示している。
【0267】
分裂に加えて、特定のサイトカインを産生する再現性のあるCD4+T細胞の活性化が所望される。したがって、測定されるサイトカインに特異的な抗体を用いて標準的なELISAによって上記の培養で産生されたサイトカインもまた調べた。図4A−4Dに示したデータは、APC上の刺激分子に対して産生されたサイトカインの量をng/mlの濃度で表示している。
【0268】
これらのアッセイの結果は、組換えMHCクラスIIを発現するドロソフィラAPC上のアクセサリー分子の種々の組み合わせによって、図4A−4Dおよび図5A−5Dで示すように種々のサイトカインの産生がもたらされることを示している。IL−2産生(Th1型応答)のためには、B7.1またはB7.2と一緒にMHCクラスIIを発現する細胞株は図4Aで示されるように極めて弱く刺激された。顕著なレベルのIL−2を達成するためには、ドロソフィラ細胞株はICAM−1、B7およびクラスIIの同時発現を要求した(図5A)。B7.1およびB7.2を発現するドロソフィラ細胞株によるγ−インターフェロンの産生もまた貧弱に誘発された(図4D)。しかしながら、IL−2に関する結果とは対照的に、ICAM−1の同時発現はγ−IFNの産生に影響を与えなかった(図5C)。脾臓APC(T−Sと表示)は強いγ−IFN産生を誘発し、新たな分子がこのサイトカインの産生を調節していることを示唆した。
【0269】
IL−4産生(Th2型応答)は、一般には未感作CD4+T細胞の一次刺激中には観察されない。これらの観察とともに、脾臓APC(T−S)は、D01細胞およびovaペプチドとの培養後顕著なIL−4を誘発することができなかった(図5B)。しかしながら、B7.1もしくはB7.2のどちらかまたはその両方と一緒にクラスIIMHC分子を同時発現しているドロソフィラ細胞は、培養中に強いIL−4産生を誘発した(図4Bおよび5B)。興味深いことには、ICAM−1の添加はこのサイトカインの産生に対して拮抗的影響を示した(図5B)。同様に、図4Dおよび5Dに示すようにIL−10はIL−4に対して調節された。
【0270】
上記に述べたように、Th1型応答よりはむしろTh2型応答のサイトカインプロフィールに関する結果が、B7およびMHCクラスIIを発現している細胞株について得られた。対照的に、B7およびICAM−1と組み合わせたMHCクラスIIの同時発現はIL−4およびIL−10の産生と拮抗し、一方IL−2は強力に促進し、それによってTh1型応答を誘発し、CD4+T細胞の所望のT細胞サブセットへの活性化を駆動する能力についての本発明を支持する。これらの結果は、種々のパターンのサイトカインを産生する所望のCD4+T細胞集団を得る場合に使用するために、ドロソフィラ細胞株を規定のアクセサリー分子の組み合わせを発現するように仕立てることができることを決定的に示している。したがって、本明細書で述べた組成物および方法は、本発明の組成物および使用方法が有効であることを立証した。
【0271】
前述の記載は本発明を説明することを目的としており、制限を意図したものではない。数多くの変型および変更が本発明の範囲および本質から外れることなく実施可能であろう。
【0272】
【化1】
【0273】
【化2】
【0274】
【化3】
【0275】
【化4】
【0276】
【化5】
【0277】
【化6】
【0278】
【化7】
【0279】
【化8】
【0280】
【化9】
【0281】
【化10】
【0282】
【化11】
【0283】
【化12】
【0284】
【化13】
【0285】
【化14】
【0286】
【化15】
【0287】
【化16】
【0288】
【化17】
【0289】
【化18】
【0290】
【化19】
【0291】
【化20】
【0292】
【化21】
【0293】
【化22】
【0294】
【化23】
【0295】
【化24】
【0296】
【化25】
【0297】
【化26】
【0298】
【化27】
【0299】
【化28】
【0300】
【化29】
【0301】
本発明は以下に関するものである。
1.CD4+T細胞の活性化ための合成抗原提示細胞(APC)であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができるMHCクラスIIα鎖遺伝子、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができるMHCクラスIIβ鎖遺伝子(ここでα鎖およびβ鎖遺伝子が発現したとき、当該α鎖およびβ鎖は、ペプチドをローディングすることができるMHCクラスIIヘテロダイマーを形成する)および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる少なくとも1つのアクセサリー分子遺伝子を含み、クラスII遺伝子およびアクセサリー分子遺伝子の少なくとも1つを欠いていることを特徴とする細胞。
【0302】
2.当該α鎖およびβ鎖遺伝子がヒト起源である前記1のAPC。
【0303】
3.少なくとも1つのプロモーターが誘導性である前記1のAPC。
【0304】
4.当該α鎖、β鎖およびアクセサリー分子遺伝子が同じベクターに存在する前記1のAPC。
【0305】
5.当該α鎖、β鎖およびアクセサリー分子遺伝子の少なくとも1つが別のベクターに存在する前記1のAPC。
【0306】
6.当該APCが昆虫細胞である前記1のAPC。
【0307】
7.当該昆虫細胞がスポドプテラおよびドロソフィラからなる群から選ばれる前記6のAPC。
【0308】
8.さらにベクターに機能的に結合したネオマイシン耐性遺伝子を含む前記1のAPC。
【0309】
9.当該アクセサリー分子遺伝子が同時刺激分子をコードする前記1のAPC。
【0310】
10.当該同時刺激分子がB7.1またはB7.2である前記9のAPC。
【0311】
11.当該アクセサリー分子遺伝子が付着分子をコードする前記1のAPC。
【0312】
12.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記11のAPC。
【0313】
13.当該アクセサリー分子遺伝子がサバイバル分子をコードする前記1のAPC。
【0314】
14.当該サバイバル分子がFasリガンドまたはCD70である前記13のAPC。
【0315】
15.第一のアクセサリー分子のための遺伝子および第二のアクセサリー分子のための遺伝子を有する前記1のAPC。
【0316】
16.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記15のAPC。
【0317】
17.当該同時刺激分子がB7.1またはB7.2で、当該付着分子がICAM−1である前記16のAPC。
【0318】
18.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記15のAPC。
【0319】
19.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記15のAPC。
【0320】
20.当該サバイバル分子がCD70で、当該付着分子がICAM−1である前記19のAPC。
【0321】
21.当該第一および第二のアクセサリー分子が同時刺激分子である前記15のAPC。
【0322】
22.当該同時刺激分子がB7.1およびB7.2である前記21のAPC。
【0323】
23.第一のアクセサリー分子のための遺伝子、第二のアクセサリー分子のための遺伝子および第三のアクセサリー分子のための遺伝子を有する前記1のAPC。
【0324】
24.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記23のAPC。
【0325】
25.当該同時刺激分子がB7.2で、当該付着分子がICAM−1で、当該サバイバル分子がCD70である前記24のAPC。
【0326】
26.当該MHCクラスIIヘテロダイマーおよびアクセサリー分子が、ペプチドが当該ヘテロダイマー上にローディングされるときにCD4+T細胞を活性化させるために十分な量で当該APCの外側表面に存在する前記1のAPC。
【0327】
27.当該ペプチドが細胞外でローディングされる前記26のAPC。
【0328】
28.当該ペプチドが細胞内でローディングされる前記26のAPC。
【0329】
29.ベクター内の第四のプロモーターに機能的に結合した、抗原プロセシング補助分子を発現することができる抗原プロセシング補助遺伝子をさらに含む前記1のAPC。
【0330】
30.CD4+T細胞を活性化するために細胞フラグメント上に機能的に結合したMHCクラスIIヘテロダイマーおよび少なくとも1つのアクセサリー分子を有する前記1のAPC由来の細胞フラグメント。
【0331】
31.当該MHCクラスIIヘテロダイマーがエンプティである前記30の細胞フラグメント。
【0332】
32.ペプチドが当該MHCクラスIIヘテロダイマー上にローディングされる前記30の細胞フラグメント。
【0333】
33.CD4+T細胞を活性化するための合成抗原提示マトリックスであって、
a)担体、
b)担体に機能的に連結され、さらに選択ペプチドをローディングすることができるMHCクラスIIヘテロダイマーの細胞外部分、および、
c)ペプチドが当該ヘテロダイマーの細胞外部分にローディングされたときにCD4+T細胞を活性化させるために十分な数で当該MHCクラスIIヘテロダイマーおよびアクセサリー分子の細胞外部分が当該マトリックス上に存在するように、当該担体に機能的に結合した少なくとも1つのアクセサリー分子の細胞外部分を含むことを特徴とするマトリックス。
【0334】
34.当該担体が細胞フラグメントである前記33のマトリックス。
【0335】
35.当該担体が細胞である前記33のマトリックス。
【0336】
36.当該MHC分子の細胞外部分が、当該MHCクラスIIヘテロダイマーのトランスメンブレンドメインによって当該細胞に結合している前記35のマトリックス。
【0337】
37.当該担体がリポソームである前記33のマトリックス。
【0338】
38.当該担体が固体表面である前記33のマトリックス。
【0339】
39.当該MHCクラスIIヘテロダイマーの細胞外部分が、当該担体に当該部分を連結させるために抗体と反応するエピトープに結合している前記33のマトリックス。
【0340】
40.当該MHCクラスIIヘテロダイマーの細胞外部分が、当該担体に当該部分が連結させるためにニッケルと反応する(His)6 に結合している前記33のマトリックス。
【0341】
41.当該担体が多孔性物質である前記33のマトリックス。
【0342】
42.当該ペプチドがMHCクラスIIヘテロダイマーの細胞外部分にローディングされる前記33のマトリックス。
【0343】
43.当該MHCクラスIIヘテロダイマーの細胞外部分がエンプティである前記33のマトリックス。
【0344】
44.当該アクセサリー分子が同時刺激分子である前記33のマトリックス。
【0345】
45.当該同時刺激分子がB7.1またはB7.2である前記44のマトリックス。
【0346】
46.当該アクセサリー分子が付着分子である前記33のマトリックス。
【0347】
47.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記46のマトリックス。
【0348】
48.当該アクセサリー分子がサバイバル分子である前記33のマトリックス。
【0349】
49.当該サバイバル分子がFasリガンドまたはCD70である前記48のマトリックス。
【0350】
50.第一のアクセサリー分子および第二のアクセサリー分子を有する前記33のマトリックス。
【0351】
51.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記50のマトリックス。
【0352】
52.当該同時刺激分子がB7.1またはB7.2で、当該付着分子がICAM−1である前記51のマトリックス。
【0353】
53.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記50のマトリックス。
【0354】
54.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記50のマトリックス。
【0355】
55.当該サバイバル分子がCD70で、当該付着分子がICAM−1である前記54のマトリックス。
【0356】
56.当該第一および第二のアクセサリー分子が同時刺激分子である前記50のマトリックス。
【0357】
57.当該同時刺激分子がB7.1およびB7.2である前記56のマトリックス。
【0358】
58.さらに第三のアクセサリー分子を含む前記50のマトリックス。
【0359】
59.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記58のマトリックス。
【0360】
60.当該同時刺激分子がB7.2で、当該付着分子がICAM−1で、当該サバイバル分子がCD70である前記59のマトリックス。
【0361】
61.合成抗原提示細胞(APC)を製造する方法であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができる発現可能MHCクラスIIα鎖遺伝子で細胞を形質転換する工程、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができる発現可能MHCクラスIIβ鎖遺伝子で細胞を形質転換する工程、および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる発現可能なアクセサリー分子遺伝子で細胞を形質転換する工程を特徴とする方法。
【0362】
62.当該細胞が、当該α鎖、β鎖およびアクセサリー分子遺伝子の少なくとも1つをコードする遺伝子を欠く前記61の方法。
【0363】
63.さらに、ベクター内の第四のプロモーターに機能的に結合した、抗原プロセシング補助分子を発現することができる発現可能な抗原プロセシング補助遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0364】
64.当該α鎖およびβ鎖遺伝子がヒト起源である前記61の方法。
【0365】
65.少なくとも1つのプロモーターが誘導性である前記61の方法。
【0366】
66.当該α鎖、β鎖およびアクセサリー分子遺伝子が同じベクターに存在する前記61の方法。
【0367】
67.当該α鎖、β鎖およびアクセサリー分子遺伝子が別個のベクターに存在する前記61の方法。
【0368】
68.当該細胞が昆虫細胞である前記61の方法。
【0369】
69.当該昆虫細胞がスポドプテラおよびドロソフィラからなる群から選ばれる前記61の方法。
【0370】
70.さらにベクターに機能的に結合した発現可能なネオマイシン耐性遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0371】
71.当該アクセサリー分子遺伝子が同時刺激分子をコードする前記61の方法。
【0372】
72.当該同時刺激分子がB7.1またはB7.2である前記71の方法。
【0373】
73.当該アクセサリー分子遺伝子が付着分子をコードする前記61の方法。
【0374】
74.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記73の方法。
【0375】
75.当該アクセサリー分子遺伝子がサバイバル分子をコードする前記61の方法。
【0376】
76.当該サバイバル分子がFasリガンドまたはCD70である前記75の方法。
【0377】
77.さらに、第二のアクセサリー分子のための遺伝子で細胞を形質転換する工程を含む前記61の方法。
【0378】
78.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子である前記77の方法。
【0379】
79.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子がサバイバル分子である前記77の方法。
【0380】
80.当該第一のアクセサリー分子がサバイバル分子で、当該第二のアクセサリー分子が付着分子である前記77の方法。
【0381】
81.さらに、第三のアクセサリー分子のための遺伝子で細胞を形質転換する工程を含む前記77の方法。
【0382】
82.当該第一のアクセサリー分子が同時刺激分子で、当該第二のアクセサリー分子が付着分子で、当該第三のアクセサリー分子がサバイバル分子である前記81の方法。
【0383】
83.合成抗原提示細胞(APC)を製造する方法であって、
a)MHCクラスIIα鎖、MHCクラスIIβ鎖およびアクセサリー分子の少なくとも1つをコードする遺伝子を欠く細胞を提供する工程、及び、
b)当該細胞に欠けている(a)の遺伝子の各々のために発現可能であって、当該遺伝子を発現する能力を有するベクター内のプロモーターに機能的に結合した遺伝子で当該細胞を形質転換する工程からなることを特徴とする方法。
【0384】
84.合成抗原提示細胞(APC)を製造する方法であって、
a)MHCクラスIIα鎖、MHCクラスIIβ鎖、アクセサリー分子および抗原プロセシング補助分子の少なくとも1つをコードする遺伝子を欠く細胞を提供しする工程、及び、
b)当該細胞に欠けている(a)の遺伝子の各々のために発現可能であって、当該遺伝子を発現する能力を有するベクター内の機能できる第一のプロモーターに結合した遺伝子で当該細胞を形質転換する工程からなることを特徴とする方法。
【0385】
85.合成抗原提示マトリックスを製造する方法であって、
a)組換えMHCクラスIIヘテロダイマーの細胞外部分を提供する工程、
b)少なくとも1つの組換えアクセサリー分子の細胞外部分を提供する工程、及び、
c)MHCクラスIIヘテロダイマーにペプチドがローディングされたときにCD4+T細胞を活性化させるために十分な数で、担体に当該MHCクラスIIヘテロダイマーおよびアクセサリー分子を結合する工程からなることを特徴とする方法。
【0386】
86.当該アクセサリー分子が同時刺激分子である前記85の方法。
【0387】
87.当該同時刺激分子がB7.1またはB7.2である前記86の方法。
【0388】
88.当該アクセサリー分子が付着分子である前記86の方法。
【0389】
89.当該付着分子がICAM−1、ICAM−2、ICAM−3またはLFA−3である前記88の方法。
【0390】
90.当該アクセサリー分子がサバイバル分子である前記85の方法。
【0391】
91.当該サバイバル分子がFasリガンドまたはCD70である前記90の方法。
【0392】
92.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記26のAPCを提供する工程、及び
b)工程a)のAPCをCD4+T細胞と接触させ、それによって接触CD4+T細胞を増殖させ、さらに活性化CD4+T細胞に分化させる工程からなることを特徴とする方法。
【0393】
93.さらに、c)当該APCから活性化CD4+T細胞を分離する工程を含む前記92の方法。
【0394】
94.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記93の方法。
【0395】
95.さらに、当該懸濁物を患者に投与する工程を含む前記94の方法。
【0396】
96.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記30の細胞フラグメントを提供する工程、
b)当該MHCクラスIIヘテロダイマーにペプチドをローディングする工程、および、
c)当該ペプチドがローディングした細胞フラグメントをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなるを特徴とする方法。
【0397】
97.さらに、当該細胞フラグメントから活性化CD4+T細胞を分離する工程を含む前記96の方法。
【0398】
98.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記97の方法。
【0399】
99.さらに、当該懸濁物を患者に投与する工程を含む前記98の方法。
【0400】
100.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記33のマトリックスを提供する工程、
b)当該MHCクラスIIヘテロダイマーにペプチドをローディングする工程、および、
c)当該ペプチドをローディングした細胞マトリックスをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなることを特徴とする方法。
【0401】
101.さらに、当該マトリックスから活性化CD4+T細胞を分離する工程を含む前記100の方法。
【0402】
102.さらに、当該活性化CD4+T細胞を許容可能な担体または賦形剤に添加し懸濁物を生成する工程を含む前記101の方法。
【0403】
103.さらに、当該懸濁物を患者に投与する工程を含む前記102の方法。
【0404】
104.インビトロでCD4+T細胞を活性化させる方法であって、
a)前記1のAPC、前記30の細胞フラグメントまたは前記33のマトリックスを十分な量でペプチドライブラリーとインビトロで十分な時間接触させ、ペプチドがローディングしたMHCクラスIIヘテロダイマーを作製する工程、及び、
b)工程b)のペプチド積載MHCクラスIIヘテロダイマーをCD4+T細胞と接触させ、それによって当該接触CD4+T細胞の増殖および活性化CD4+T細胞への分化を誘発する工程からなることを特徴とする方法。
【0405】
105.CD4+T細胞仲介免疫反応を変化させて患者の症状を治療する方法であって、
a)患者に固有のサイトカインプロフィールについて患者を分析する工程、
b)当該患者からCD4+T細胞を採取する工程、
c)当該CD4+T細胞を前記26のAPCと十分な量で十分な時間インビトロで接触させ、それによって当該接触CD4+T細胞の増殖分裂および、工程a)で得られたプロフィールに機能的に対抗するサイトカインプロフィールを産生する活性化CD4+T細胞への分化を誘発する工程、および、
d)当該活性化CD4+T細胞を当該患者に戻す工程からなることを特徴とする方法。
【0406】
106.当該症状が自己免疫疾患である前記105の方法。
【0407】
107.当該免疫疾患が、糖尿病、多発性硬化症、自己免疫性甲状腺炎、全身性紅斑性狼瘡、重症筋無力症、クローン病および炎症性大腸症候群から成る群から選ばれる前記106の方法。
【0408】
108.当該患者に固有のサイトカインプロフィールがCD4+Th1型応答によって生じる前記106の方法。
【0409】
109.当該患者に固有のサイトカインプロフィールが、インターロイキン−2、インターフェロン−γおよび腫瘍壊死因子からなる群から選ばれるサイトカインを含む前記108の方法。
【0410】
110.当該症状がアレルギーである前記105の方法。
【0411】
111.当該アレルギーが、喘息および接触過敏症からなる群から選ばれる前記110の方法。
【0412】
112.当該患者に固有のサイトカインプロフィールがCD4+Th2型応答によって生じる前記110の方法。
【0413】
113.当該患者に固有のサイトカインプロフィールが、インターロイキン−4およびインターロイキン−10からなる群から選ばれるサイトカインを含む前記112の方法。
【特許請求の範囲】
【請求項1】
CD4+T細胞の活性化ための合成抗原提示細胞(APC)であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができるMHCクラスIIα鎖遺伝子、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができるMHCクラスIIβ鎖遺伝子(ここでα鎖およびβ鎖遺伝子が発現したとき、当該α鎖およびβ鎖は、ペプチドをローディングすることができるMHCクラスIIヘテロダイマーを形成する)および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる少なくとも1つのアクセサリー分子遺伝子を含み、クラスII遺伝子およびアクセサリー分子遺伝子の少なくとも1つを欠いていることを特徴とする細胞。
【請求項1】
CD4+T細胞の活性化ための合成抗原提示細胞(APC)であって、
a)ベクター内の第一のプロモーターに機能的に結合した、MHCクラスIIα鎖を発現することができるMHCクラスIIα鎖遺伝子、
b)ベクター内の第二のプロモーターに機能的に結合した、MHCクラスIIβ鎖を発現することができるMHCクラスIIβ鎖遺伝子(ここでα鎖およびβ鎖遺伝子が発現したとき、当該α鎖およびβ鎖は、ペプチドをローディングすることができるMHCクラスIIヘテロダイマーを形成する)および、
c)ベクター内の第三のプロモーターに機能的に結合した、アクセサリー分子を発現することができる少なくとも1つのアクセサリー分子遺伝子を含み、クラスII遺伝子およびアクセサリー分子遺伝子の少なくとも1つを欠いていることを特徴とする細胞。
【図1A】

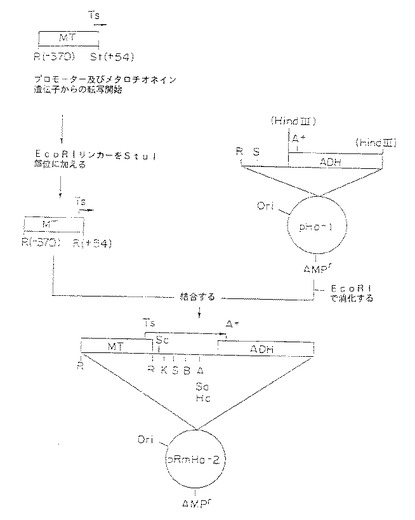
【図1B】
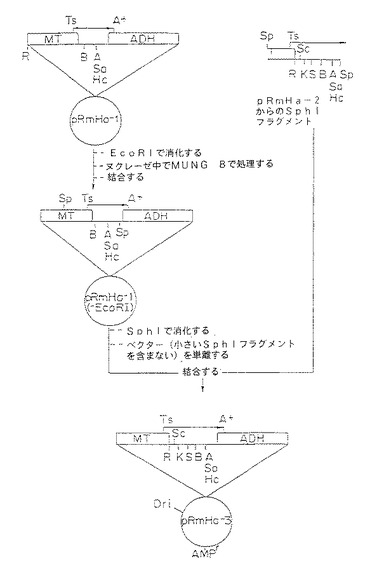
【図1C】
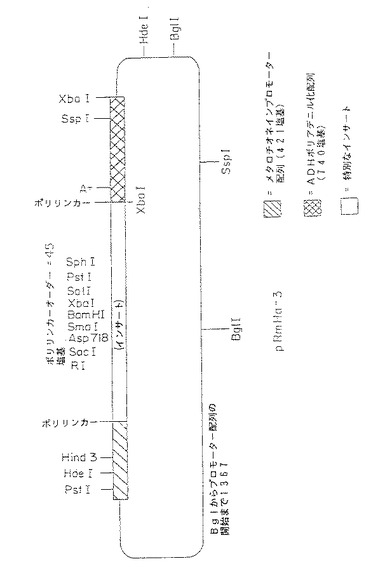
【図2】
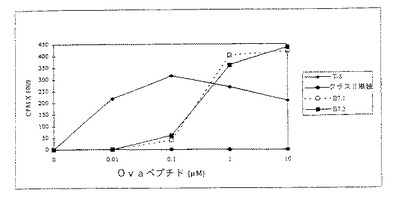
【図3】
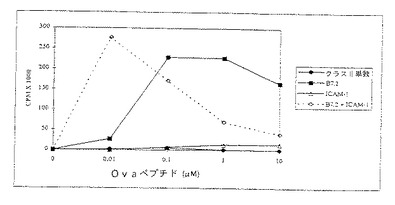
【図4】
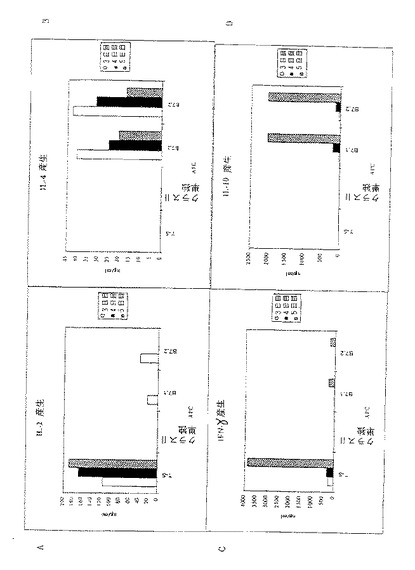
【図5】
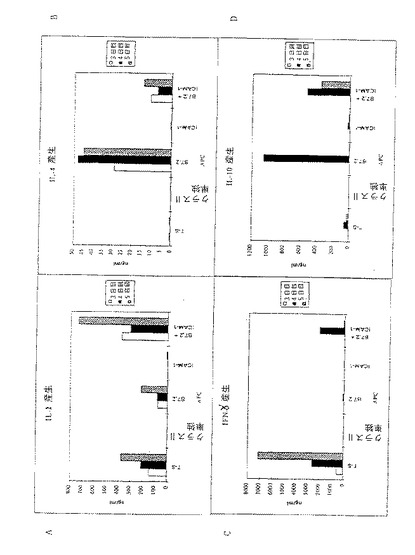
【図1B】
【図1C】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−143245(P2012−143245A)
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【出願番号】特願2012−77703(P2012−77703)
【出願日】平成24年3月29日(2012.3.29)
【分割の表示】特願平10−500616の分割
【原出願日】平成9年5月22日(1997.5.22)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【Fターム(参考)】
【公開日】平成24年8月2日(2012.8.2)
【国際特許分類】
【出願日】平成24年3月29日(2012.3.29)
【分割の表示】特願平10−500616の分割
【原出願日】平成9年5月22日(1997.5.22)
【出願人】(399038620)ザ スクリプス リサーチ インスティチュート (51)
【Fターム(参考)】
[ Back to top ]