
MN遺伝子およびMN蛋白質
【課題】腫瘍性疾患および/または前腫瘍性疾患を効果的に治療する。
【解決手段】一つもしくはそれ以上の実質的に純粋なMN蛋白質および/またはポリペプチドが免疫原となる量で生理学的に許容される非毒性基剤中に分散されたワクチンによりMN蛋白質またはポリペプチドに対する免疫応答を誘導し、あるいはMN遺伝子から転写されたmRNAに実質的に相補的なアンチセンス核酸配列を投与することによりMN遺伝子の発現を阻害する。
【解決手段】一つもしくはそれ以上の実質的に純粋なMN蛋白質および/またはポリペプチドが免疫原となる量で生理学的に許容される非毒性基剤中に分散されたワクチンによりMN蛋白質またはポリペプチドに対する免疫応答を誘導し、あるいはMN遺伝子から転写されたmRNAに実質的に相補的なアンチセンス核酸配列を投与することによりMN遺伝子の発現を阻害する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は医学遺伝子学の一般的領域に属し、生化学工学および免疫化学の分野に属する。より詳細には、本発明は新規な遺伝子であるMN遺伝子(該遺伝子はMN蛋白質をコードする細胞性遺伝子である)の同定に関する。本発明者らは、MN蛋白質が腫瘍形成に関与していることを発見した。患者の試料中からMN抗原、さらに該抗原に特異的な抗体を同定することは、癌の診断/予後の分析の基礎を提供する。
【背景技術】
【0002】
珍しい特性を有する新規な疑似ウイルス体が、ヒト胸腺癌細胞と共培養したHeLa細胞[ヒト子宮頚管腺癌由来の細胞株]内において、熱不安定性表面G蛋白質を有する水疱性口内炎ウイルス(vesicular surface virus, VSV)変異体を補足する能力により検出された(非特許文献1から5を参照)。該疑似ウイルス体は、おそらくヒト乳腺腫瘍(mammary tumor )から得られたことからMaTuと呼ばれる。
【0003】
MaTuを研究し、特性を明らかにすることに医学的に大きな興味がひかれた。MaTuは生細胞への全く新しい型の寄生分子と思われ、また、ヒト腫瘍細胞由来の可能性があるからである。本明細書には、MN遺伝子およびMNタンパク質を発見するに至った、MaTuの生物学的および分子学的性質を記述している。本発明者らは、MaTuが2つのコンポーネント、すなわち、外因性の輸送コンポーネントMX、および内因性の細胞性コンポーネントMNからなることを発見した。本明細書に記載しているように、MNコンポーネントは細胞性遺伝子として発見され、既知のDNA配列とは相同性(ホモロジー)がほとんどない。MN遺伝子は試験した全ての脊椎動物の染色体DNA内に存在することが見出されており、該遺伝子の発現は腫瘍形成性に強く関与していることが明らかになっている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】ザバダ(Zavada)ら、Nature New Biol., 240 :124(1972)
【非特許文献2】ザバダ(Zavada)ら、J. Gen. Viol., 24 :327(1974);
【非特許文献3】J.ザバダ(Zavada)、 Arch. Viol., 50 :1(1976);
【非特許文献4】J.ザバダ(Zavada)、J. Gen. Viol., 63 :15-24(1982)
【非特許文献5】J.ザバダ(Zavada)、Arch. Viol., 118 :189(1991)
【発明の概要】
【発明が解決しようとする課題】
【0005】
本明細書に記述しているのは、バクテリアベクター内におけるMN遺伝子のクローニングと配列決定、およびMN遺伝子にコードされた蛋白質の産生についてである。このように遺伝子工学的に処理されたMN蛋白質は、他のMN蛋白質/ポリペプチドと同様に、本発明に従い、MN特異的抗体を検出する血清学的分析に使用することができる。さらに、MN抗原と反応するそのようなMN蛋白質/ポリペプチドと抗体は、本発明に従い、MN抗原を検出および/または定量する免疫学的検定(イムノアッセイ)に使用することができる。このような分析は、腫瘍性および/または前腫瘍性の疾患の診断および/または予後に利用される。
【課題を解決するための手段】
【0006】
本明細書は、MaTuの内因性コンポーネントである細胞性遺伝子、すなわち、MN遺伝子について開示している。イントロンを有しないと考えられる該遺伝子の実質的な全cDNA配列は、第1A図−第1B図[配列番号1]に示す。
【0007】
本発明は、該MN遺伝子、その断片およびそれに関係するcDNAに関し、これらは、たとえば次のように有用である。1)生化学工学によりMN蛋白質/ポリペプチドを産生。2)被検材料の細胞内のMN遺伝子の存在を確認するための核酸プローブの調製。3)適切なポリメラーゼ連鎖反応(PCR)プライマーの調製。これは、たとえば、PCRに基づく分析や核酸プローブの産生などに使用する。4)MN蛋白質およびポリペプチド、ならびにそれらと相同あるいはほぼ相同なポリペプチドの同定。5)各種の組織および細胞株に存在するMN遺伝子から転写されたさまざまなmRNAの同定。6)MN遺伝子の突然変異の同定。本発明はさらに、MN遺伝子またはその断片、あるいは関連するcDNAまたはその断片からなる、精製単離されたDNAに関する。
【0008】
さらに、本発明は、これまで未知であった蛋白質MN(MN遺伝子によりコードされている)の発見に関する。MN蛋白質の発現は、高密度培養中において成長細胞により誘導され、そのような発現は腫瘍形成性細胞と関与していることが見出された。
【0009】
MN蛋白質は、インビトロ(in vitro)でいくつかのヒト腫瘍細胞株において産生されることがわかっている。たとえば、HeLa(子宮頚管癌由来)、T24(膀胱癌由来)およびT47D(乳腺癌由来)およびSK−Mel 1477(メラノーマ由来)などの細胞株、腫瘍形成性ハイブリッド細胞など。また、インビボ(in vivo)でいくつかのヒト癌細胞においても産生される。たとえば、子宮頚部細胞、卵巣および子宮内膜癌細胞、さらに乳頭腫などのようないくつかの良性腫瘍細胞など。MN蛋白質は、非腫瘍形成性ハイブリッド細胞や正常組織の細胞においては見出されない。このことから、MN蛋白質は腫瘍特異的であると考えられる。
【0010】
HeLa細胞株および腫瘍形成性HeLa細胞と線維芽細胞とのハイブリッド(H/F/T)細胞株においては、MN蛋白質は「双子(twin)」蛋白質p54/58Nとして表現される。該蛋白質は、グリコシル化され、ジスルフィド結合しているオリゴマーの形をとっている。還元性ゲルを用いた電気泳動によって測定すると、MN蛋白質は約40Kdから約70Kdの範囲の分子量を有しており、好ましくは約45Kdから約65Kdの範囲、より好ましくは約48Kdから約58Kdの範囲の分子量を有しているのがよい。非還元性のゲルにおいては、オリゴマー型のMN蛋白質は約145Kdから約160Kdの範囲の分子量を有しており、好ましくは約150Kdから約155Kdの範囲、より好ましくは約152Kdから約154Kdの範囲の分子量を有しているのがよい。本発明における好ましいMN蛋白質の推定アミノ酸配列は第1A図−第1B図に示す。
【0011】
MN遺伝子とMN蛋白質およびそれらにコードされた実質的に相補性のMN遺伝子とMN蛋白質の発見により、MN蛋白質の発現が腫瘍形成性と関連していることが見いだされた。この発見の結果、癌および前癌状態の診断/予後の方法が確立された。脊椎動物、好ましくはホ乳類、より好ましくはヒトの細胞と組織抽出物を含んだ患者の試料中からMN抗原を検出および/または定量することにより、腫瘍性疾患の発症や存在を同定ための方法や材料が提供される。そのようなMN抗原は体液中からも検出される。
【0012】
MN蛋白質およびMN遺伝子は、癌の診断/予後における腫瘍形成の分子メカニズムの解明の研究に利用されており、また、癌の免疫療法に応用され得る。
【0013】
本発明は、広範な腫瘍性および/または前腫瘍性疾患の検出に有効である。腫瘍性疾患の例としては、乳腺、膀胱、卵巣、子宮、子宮頚管、子宮内膜、偏平細胞および腺偏平癌などの腫瘍;頭および首の癌;神経芽細胞腫および網膜芽腫などの中胚葉性腫瘍;骨肉腫およびユーイング肉腫(Ewing′s sarcoma)などの肉腫;およびメラノーマが挙げられる。特に興味深いものとしては、頭および首の癌、卵巣、子宮頚管、腟、子宮内膜および陰門の癌を含む婦人科の癌;胃、結腸および食道の癌などの胃腸系の癌;膀胱および腎臓の癌などの尿路系の癌;皮膚癌;肝臓癌;前立腺癌;肺癌および乳癌がある。中でも特に興味があるのは、婦人科の癌;乳癌;尿路系の癌、ことに膀胱癌;肺癌;胃、結腸および食道の癌などの胃腸系の癌;および肝臓癌である。さらにとりわけ興味が強いのは、婦人科の癌および乳癌である。婦人科の癌の中で特に関心があるのは子宮頚管、子宮内膜および卵巣の癌であるが、子宮頚管偏平細胞腫瘍、腺偏平細胞腫瘍、腺腫などを含む婦人科の癌と同様に、後形体子宮頚管組織やコンジロームなどの婦人科の前癌状態も非常に興味深い。
【0014】
本発明はさらに、MN遺伝子、その断片あるいは関連するcDNAを生化学的に処理することに関する。たとえば、該遺伝子またはその断片、あるいは関連するcDNAは適切な発現ベクターに組込まれ、宿主細胞がそのような発現ベクターに形質転換され、MN蛋白質/ポリペプチド、好ましくはMN蛋白質がその中で発現する。そのような組換え蛋白質あるいはポリペプチドは、グリコシル化されているかまたはいないかであるが、好ましくはグリコシル化されているのがよく、ほぼ純粋の状態に精製され得る。本発明はさらに、合成的にあるいはその他の生化学的手法で調製されたMN蛋白質/ポリペプチドにも関する。
【0015】
該MN蛋白質/ポリペプチドは、患者の試料中のMN抗原を検出する分析、あるいはMN特異的抗体を調べる血清学的検定に用いることができる。本発明のMN蛋白質/ポリペプチドは、血清学的に活性で、免疫原性があり、および/または抗原性がある。該MN蛋白質/ポリペプチドは、さらに、MN特異的抗体(ポリクローナルおよび/またはモノクローナル)を産生させたり、T細胞免疫応答を起こす免疫原として用いることもできる。
【0016】
さらに本発明は、MN特異的抗体に関するものであり、該抗体は診断/予後に用いられ、また、治療に用いることもできる。MN特異的抗体は次のような用途に用いられる。たとえば、免疫蛍光顕微鏡や免疫組織化学染色による実験室での解析、臨床サンプル中のMN抗原の検出および/または定量のための免疫測定法の構成分の一つとして;MN抗原を検出するイムノブロット法のプローブ;細胞内のMN蛋白質および/またはポリペプチドの局在を示す金コロイドビーズを用いた免疫電子顕微鏡法;およびMN遺伝子、その断片あるいは関連するcDNAをクローニングする遺伝子工学など。そのようなMN特異的抗体は、たとえばインビトロ(in vitro)での組織片を用いた診断および/または予後用のキットの構成材料として用いられる。そのような抗体はまた、たとえば、適切な放射活性同位体を用いて抗体を適切にラベルし、インビボ(in vivo)での抗体の局在の転移をシンチグラフィーで追跡することにより、インビボ(in vivo)での診断/予後に用いることができる。さらに、そのような抗体は、毒性因子および/または細胞増殖抑制因子をそれらに結合させて、あるいはさせずに、インビボ(in vivo)療法として癌患者の加療に使用することもできる。また、そのような抗体は、腫瘍性および/または前腫瘍性疾患の存在を検出するためにインビボ(in vivo)で用いることができる。さらに、そのような抗体はMN蛋白質およびポリペプチドのアフィニティー精製に使用することができる。
【0017】
典型的なMN特異的抗体であるモノクローナル抗体M75を産生するハイブリドーマは、1992年9月17日にATCC番号HB 11128としてATCC(American Type Culture Collection,米国、メリーランド州、ロックヴィル)に寄託された。このM75抗体は、MN蛋白質を発見、同定するために用いられたものであるが、ホルマリン固定された組織サンプルから、ウエスタンブロット、ラジオイムノアッセイおよび免疫組織化学的方法により、MN抗原を簡単に検出するために用いることができる。
【0018】
本発明はさらに、MN蛋白質またはポリペプチドをコードするDNA配列を有する組換えDNA、ならびに、MN蛋白質またはポリペプチドをコードするのみならず非MN蛋白質またはポリペプチドのアミノ酸配列をもコードする組換えDNAに関する。該アミノ酸配列はヒトに免疫原性を有しないこと、およびヒト体液中の抗体に一般的な反応性を有しないことが望ましい。そのようなDNA配列の例としては、β−ガラクトシダーゼのα−ペプチドコード領域、およびグルタチオンS−トランスフェラーゼをコードする配列とその断片などが挙げられる。さらに、実質的に純粋であり、天然には存在しないそのような組換え融合タンパク質/ポリペプチドも本発明に含まれる。本発明の融合タンパク質の例としてはpGEX−3X−MNがある。
【0019】
本発明は、また、腫瘍性疾患および/または前腫瘍性疾患の治療方法に関し、該方法は、MN遺伝子から転写されたmRNAと実質的に相補的であるアンチセンス核酸配列を投与することにより、MN遺伝子の発現を抑制することからなる。アンチセンス核酸配列は、第1A図−第1B図に示すように、MN cDNAの5’末端において実質上相補的なものであることが好ましい。このアンチセンス核酸配列とは、オリゴヌクレオチドであることが好ましい。
【0020】
本発明はまた、実質的に純粋な一つもしくはそれ以上のMN蛋白質および/またはポリペプチドを免疫原量として十分に含むワクチンに関する。MN蛋白質および/またはポリペプチドは生理学的に許容性で非毒性の基剤に分散し、MN蛋白質の発現と関係している腫瘍性疾患に対して、脊椎動物、好ましくはホ乳類、より好ましくはヒトに十分な免疫効果を上げる量を使用する。該蛋白質は、組換え、合成あるいは他の生化学的手法によりつくられる。組換えMN蛋白質としては、pGEX−3X−MNなどのような融合蛋白質が挙げられる。該ワクチンの特徴的な使用法として、再発および/または転移の阻止がある。たとえば、MN関与性腫瘍を外科的に切除した患者に腫瘍の再発を防ぐために該ワクチンを投与することができる。
【0021】
本発明はさらに、MN遺伝子の核酸配列と実質的に相補的な核酸プローブに関する。本発明における好ましい核酸プローブとは、その配列が、第1A図−第1B図に示すように、MN cDNAの配列に実質的に相補的なものである。本発明に従う試験キット(テストキット)は、腫瘍性および/または前腫瘍性疾患の診断/予後に有効なプローブから構成されている。好ましい試験キットは、前記プローブとMN遺伝子またはMN遺伝子のmRNA産物とのハイブリダイゼーションを視覚化などにより検出あるいは測定する手法により構成されている。
【0022】
本発明に従うイムノアッセイは、MN蛋白質/ポリペプチドおよび/またはMN特異的抗体からなる試験キットとして具体化される。そのような試験キットは、固相状態であるが、それに限定されるわけではなく、液相状態であってもよく、非増幅あるいはアビジン/ビオチン法などを用いて増幅した状態で、ELISA法、粒子アッセイ、放射測定あるいは蛍光測定アッセイなどに基づくものである。
【0023】
略語表
本明細書内で使用する略語は以下の通りである:
AA −アミノ酸
ATCC −アメリカン タイプ カルチャー コレクション(American Type Culture Collection)
bp −塩基対
BSA −ウシ血清アルブミン
Ci −キュリー
cm −センチメーター
cpm −カウント/分
C-末端 −カルボキシル末端
℃ −摂氏
DMEM −ダルベッコ変法イーグル培地(Dulbecco modified Eagle medium)
EDTA −エチレンジアミン四酢酸
EIA −酵素免疫測定法、エンザイムイムノアッセイ
ELISA −酵素免疫吸着測定法
F −線維芽細胞
FCS −ウシ胎児血清
FIBR −線維芽細胞
FITC −フルオレセインイソチオシアネート
H −HeLa細胞
HEF −ヒト胚線維芽細胞
HeLa K −標準型HeLa細胞
HeLa S −スタンブリッジ(Stanbridge)変異HeLa細胞 D98/AH.2
H/F−T −ハイブリッドHeLa線維芽細胞(腫瘍原性)、HeLa細胞 D98/AH.2由来
H/F−N −ハイブリッドHeLa線維芽細胞(非腫瘍原性)、HeLa細胞 D98/AH.2由来
HGPRT− −ヒポキサンチングアニンフォスフォリボシルトランスフェラーゼ欠損
HRP −西洋ワサビ(ホースラディッシュ)ペルオキシダーゼ
IPTG −イソプロピル−β−D−チオガラクト−ピラノサイド
kb −キロベース
kd −キロダルトン
M −モル濃度
mA −ミリアンペア
MAb −モノクローナル抗体
ME −メルカプトエタノール
MEM −最少必須培地
mg −ミリグラム
ml −ミリリットル
mM −ミリモル濃度
MTV −乳腺腫瘍ウイルス
N −規定濃度
ng −ナノグラム
N-末端 −アミノ末端
ODN −オリゴデオキシヌクレオチド
PAGE −ポリアクリルアミドゲル電気泳動
PBS −リン酸緩衝生理食塩水
PEST −プロリン(proline)、グルタミン酸(glutamic acid)、生理食塩水(serine)、スレオニン(threonine)の頭文字の組合せ
pI −等電点
RIP −放射性免疫沈降法
RIPA −放射性免疫沈降測定法
SAC −プロテインA(黄色ブドウ球菌(Staphylococcus aureus)由来)
SDS −ドデシル硫酸ナトリウム
SDS−PAGE −ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動
SSPE −NaCl(0.18M)、リン酸ナトリウム(0.01M)、EDTA(0.001M)
TCA −トリクロロ酢酸
TC培地 −組織培養培地
μCi −マイクロキュリー
μg −マイクログラム
μl −マイクロリットル
μM −マイクロモル濃度
VSV −水疱性口内炎ウイルス(vesicular surface virus)
X−MLV −異種マウス白血病ウイルス
【0024】
細胞株(セルライン)
本明細書に記述している実験例において使用した細胞株を以下に示す:
HeLa K −標準型HeLa細胞,異数倍数性、上皮様の細胞株、ヒト子宮頚管腺腫より単離され(ゲイ(Gey)ら、Cancer Res., 12: 264(1952)、ジョーンズ(Jones)ら、Obstet. Gynecol., 38: 945-949(1971)参照)、B.コリシュ教授(Korych)(チャールス大学(Charles University)医学微生物学および免疫学研究所(Insutitute of Medical Microbiology and Immunology)、チェコスロバキア、プラハ)より入手
HeLa D98/AH.2(またはHeLa S) −変異HeLaクローンであり、ヒポキサンチングアニンフォスフォリボシルトランスフェラーゼ欠損(HGPRT−)、エリック J.スタンブリッジ(Eric J. Stanbridge)(カリフォルニア大学医学部微生物学科(Department of Microbiology, College of Medicine, University of California)、米国、カリフォルニア州、アーヴァン)より供与され、スタンブリッジ(Stanbridge)らにより報告されている(Science, 215: 252-259(1982年1月15日号))。ハイブリッド細胞H/F−Nの親株も同じくE.J.スタンブリッジ(Stanbridge)から入手
NIH−3T3 −マウス線維芽細胞株、アーロンソン(Aaronson)により報告されている(Science, 237: 178(1987))
T47D −ヒト乳腺癌由来の細胞株(ケイダー(Keydar)ら、Eur. J. Cancer, 15: 659-670(1979))。J.ケイダー(Keydar)(ハダッサー医科大学(Haddasah Medical School)、イスラエル、エルサレム)より供与
T24 −膀胱癌由来の細胞株(ブベニク(Bubenik)ら、Int. J. Cancer, 11: 765-773(1973))。J.ブベニク(Bubenik)(チェコスロバキア科学アカデミー分子遺伝学研究所(Insutitute of Melecular Genetics, Czechoslovak Academy of Sciences)、チェコスロバキア、プラハ)より供与
HMB2 −メラノーマ由来の細胞株(スヴェク(Svec)ら、Neoplasma, 35: 665-681(1988))
HEF −ヒト胚線維芽細胞(ザバダ(Zavada)ら、Nature New Biology, 240: 124-125(1972))
SIRC −ウサギ角膜由来の細胞株(対照およびX−MLV感染)(ザバダ(Zavada)ら、Virology, 82: 221-231(1977))
ベロ細胞(Vero cells) −アフリカミドリザル由来の細胞株(ザバダ(Zavada)ら、1977年)
ミエローマ細胞株NS−0 −モノクローナル抗体の産生において融合親細胞として用いられるミエローマ細胞株(ガルフレ(Galfre)とミルシュテイン(Milstein)、Methods Enzymol.,73: 3-46(1981) )
SK−Mel 1477 −ヒトメラノーマ細胞株。K.E.ヘルストロン(Hellstrom)(フレッド・ハトキンス癌研究センター腫瘍免疫部門(Division of Tumor Immunology, Fred Hutchins Cancer Research Center)、米国、ワシントン州、シアトル)より供与
XC −ラット横紋筋肉腫由来の細胞であり、ラウス肉腫(Rous sarcoma)ウイルスによって誘導された誘導ラット肉腫(スヴォボダ(Svoboda),J.、国立癌センター研究所モノグラフ第17巻(Natl. Cancer Center Institute Monograph No.17)より「鳥類腫瘍ウイルスに関する国際学会(International Conference on Avian Tumor Viruses)」(J.W.ベアード(Beard)編)、pp.277-298(1964年)。ジャン スヴォボダ(Svoboda)(チェコスロバキア科学アカデミー分子遺伝学研究所(Insutitute of Melecular Genetics, Czechoslovak Academy of Sciences)、チェコスロバキア、プラハ)より供与
Rat2−Tk− −チミジンキナーゼ欠損細胞株。L.クチノヴァ(Kutinova)(血清およびワクチン研究所(Institute of Sera and Vaccines)、チェコスロバキア、プラハ)より供与
CGL1 −H/F−Nハイブリッド細胞(HeLa D98/AH.2起源)
CGL2 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
CGL3 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
CGL4 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
【0025】
ヌクレオチドおよびアミノ酸配列記号
本明細書では、以下の記号をヌクレオチドを表す記号として使用する:
塩 基 記 号
アデニン A
シトシン C
グアニン G
チミン T
ウラシル U
【0026】
主要アミノ酸は20個あり、それらのおのおのは3個の隣接するヌクレオチド(三つ組(トリプレット)コードまたはコドン)の異なる組合せによって特定されており、さらに、特徴的な蛋白質を形成するために特定の順番で結合される。本明細書においては、該アミノ酸を表記するために、たとえば第1A図−第1B図に示すような3文字記号を使用する。それらを以下に示す:
アミノ酸名 記 号
アラニン Ala
アルギニン Arg
アスパラギン Asn
アスパラギン酸 Asp
システイン Cys
グルタミン酸 Glu
グルタミン Gln
グリシン Gly
ヒスチジン His
イソロイシン Ile
ロイシン Leu
リジン Lys
メチオニン Met
フェニルアラニン Phe
プロリン Pro
セリン Ser
スレオニン Thr
トリプトファン Trp
チロシン Tyr
バリン Val
【図面の簡単な説明】
【0027】
【図1A】図1Aは、本明細書に記載しているようにMN cDNAクローンから単離されたヌクレオチド配列およびcDNAによってコードされている推定アミノ酸配列[それぞれ配列番号1および2]を表している。シークエンスデータはEMBLデータライブラリー(EMBL Data Library)(ドイツ、ハイデルベルグ)に送付済みであり、受入番号X66839として利用可能である。
【図1B】図1Bは、図1Aの続きである。
【図2】図2(AおよびB)は、ヒト線維芽細胞(F)、HeLa細胞(H)およびH/F−NとH/F−Tハイブリッド細胞内におけるMN特異的蛋白質およびMX特異的蛋白質の発現をグラフに表したものであり、MX感染細胞およびMX非感染細胞における発現を対比させている。実施例5に操作と結果について詳しく記載している。
【図3】図3(AおよびB)は、125I−pGEX−3X−MN蛋白質およびいろいろな抗体を用いた放射免疫沈降実験の結果をグラフに表したものである(実施例8に記述)。放射活性蛋白質(15×103cpm/チューブ)は、次の腹水あるいは血清およびSACと沈降した。(A)MAb M75を含む腹水;(B)ウサギ抗MaTu血清;(C)正常ウサギ血清;(D)ヒト血清L8;(E)ヒト血清KH;(F)ヒト血清M7。
【図4】図4は、MN抗原に対するラジオイムノアッセイの結果を示す(実施例8に記述)。腹水(沈降が50%の放射活性となるように希釈)は、次のものと2時間反応させた。(A)「コールド(cold)」(ラベルしていない)蛋白質pGEX−3X−MN;あるいは以下の細胞の抽出物(B)HeLa+MX;(C)Rat−2TK−;(D)HeLa;(E)ラットXC;(F)T24および(G)HEF。次に、125IでラベルしたpGEX−3X−MN蛋白質(25×103cpm/チューブ)を加え、さらに2時間インキュベートした。最後に、放射活性MAb M75をSACに吸着させ、測定した。
【発明を実施するための形態】
【0028】
MaTu−−MXおよびMNコンポーネント
本明細書に記述するように、MaTuは2コンポーネントシステムからなっている。コンプレックスの一つの部分である外因性MXは、透過性であり、蛋白質p58Xを発現する。該蛋白質は、ヒトおよびさまざまな動物の天然の血清と反応する細胞質抗原である。もう一つのコンポーネントであるMNは、ヒト細胞に内因するものである。
【0029】
MNは細胞性遺伝子であり、既知のDNA配列とはほとんど相同性がない。MNは保存性であり、多くの脊椎動物の染色体DNA内で単一のコピー遺伝子として存在している。本明細書には、MN cDNAのクローニングおよび配列決定、また融合蛋白質(MN+グルタチオンS−トランスフェラーゼのC末端からなる:アフィニティークロマトグラフィーにより容易に精製できる)の遺伝子工学的処理について記述している。
【0030】
MNはHeLa細胞において双子蛋白質p54/58Nとして表現され、細胞表面および核内に局在している。p54/58Nに反応するモノクローナル抗体(MAb M75)を用いたイムノブロットにおいて54kdと58kdの2本のバンドが確認できる。これらの2本のバンドは、一種類の蛋白質に関して、グリコシル化のパターンあるいはプロセッシングの過程が異なることによるものかもしれない。(p54Nおよびp58Nは、両方ともマンノースを含むオリゴ糖残基によりグリコシル化されているが、p58Nだけはグルコサミンも含有している。)本明細書で使用する「双子蛋白質(twin protein)」とはp54/58Nを指している。
【0031】
MNは、HeLa細胞の急速成長する希薄培養中では出現しないが、細胞を高密度培養中で維持することにより、あるいは、さらに効率的な手段として、細胞にMXを感染させることにより誘導される。p54/58NのみがMaTuに感染したHeLa細胞内で再生された水疱性口内炎ウイルス(vesicular surface virus, VSV)のビリオンに結合する。双子蛋白質p54/58Nはグリコシル化され、ジスルフィド結合で連結されたオリゴマーの形をとっているのに対し、p58Nはグリコシル化されておらず、ジスルフィド結合で連結されたオリゴマー形ではない。
【0032】
VSVは、HeLa細胞内でp54/58Nをビリオン中に集めるが、このことは、該双子蛋白質がVSVのG蛋白質変異型の相補性と、VSV(MaTu)のプソイド型の形成に関与していることを示唆している。エンベロープを有するウイルスのみが、感染性で機能的なプソイド型を形成するための表面糖蛋白質を産生し、ビリオンの細胞への吸着および侵入という特殊な機能を発揮することができる(ザバダ(Zavada), J., J. Gen. Virol.,63: 15-24(1982))。このことはMN遺伝子が疑似ウイルスの配列として行動することを示している。
【0033】
エンベロープを有するウイルスの表面蛋白質は、VSVのプソイド型の形成に関与しており、MN双子蛋白質p54/58Nと同様に、グリコシル化されている。また、MN蛋白質は、オリゴマー(好ましくは三量体または四量体)の形成においてはウイルスの糖蛋白質と類似している。そのようなオリゴマー形成においては、S−S結合(ジスルフィド結合)の関与は必須ではないが、ビリオンの集積には必須である(クレイス(Kreis)とロディッシュ(Lodish), Cell, 46: 929-937(1986))。ジスルフィド結合は2−メルカプトエタノールを用いて還元することにより分裂する。
【0034】
パストレコヴァ(Pastorekova)らにより報告されているように(Virology, 187: 620-626(1992))、メルカプトエタノールを用いて還元した後は、細胞抽出物あるいはVSV由来のp54/58Nはイムノブロットにおいて非常に類似している。還元を行わない場合、細胞抽出物のp54/58Nは150kd付近に複数のバンドとして現れ、このことは細胞が数種の異なるオリゴマー(おそらくp54:p58の比率が異なる)を含んでいることを示唆している。しかし,VSVにおいては選択的にその中の一つ、分子量約153kdに集まっている。該オリゴマーは三量体もしくは四量体であり、54kdおよび58kdの蛋白質からなっている。還元状態でのVSVサンプルの分析において54kdと58kdのバンドの強さがほぼ同等であることから、VSVビリオンにおいてはp54:p58が等モル比であることが示された。
【0035】
MN蛋白質の発現は、腫瘍性疾患の診断/予後にみられる。MN双子蛋白質p54/58Nは、HeLa細胞およびスタンブリッジ(Stanbridge)の腫瘍形成性(H/F−T)ハイブリッド細胞において発現が確認されているが(スタンブリッジ(Stanbridge)ら、Somatic Cell Genet, 7: 699-712(1981)およびスタンブリッジ(Stanbridge)ら、Science, 215: 252-259(1982) )、線維芽細胞あるいは非腫瘍形成性(H/F−N)ハイブリッド細胞においては確認されていない(スタンブリッジ(Stanbridge)ら、同上)。ヒト卵巣、子宮内膜および子宮頚管癌、またいくつかの良性腫瘍(乳頭腫など)を用いて行ったイムノブロットにおいてはMN蛋白質が確認されるが、正常な卵巣、子宮内膜、子宮あるいは胎盤の組織からは確認されない。MXに感染したHeLa細胞内では、微細構造の交替が顕著に行われており、このことは、細胞表面でのおびただしい糸状体の形成およびミトコンドリアの増幅を意味している。免疫金(immunogold)標識法を用いると、p54/58Nは糸状体の表面および核、特に核小体上で観察される。すなわち、MN蛋白質は正常非腫瘍細胞では産生されていないことから、腫瘍特異的であるといえる。
【0036】
本明細書の実施例において、MNおよびMXは二つの異なる存在であり、互いに独立して存在することが示されている。外因性の透過性物質であるMXは、線維芽細胞あるいは非腫瘍形成性(H/F−N)ハイブリッド細胞において増殖するが、これらの細胞はMN関連蛋白質は発現しない(第2A図−第2B図)。そのような細胞内ではMXはMN蛋白質の産生を誘導しないのである。第2A図−第2B図および実施例5と6に示されるように、MXの非存在下でもMN蛋白質はHeLa細胞および他の腫瘍細胞内で産生される。しかしながら、MXはHeLa細胞内におけるMN蛋白質の強力な誘導剤である。非感染細胞内において、MXはMN蛋白質の産生を濃度にして30倍増加させた(下記の実施例5と8、実施例8の表1参照)。
【0037】
MN遺伝子−−クローニングおよび塩基配列決定
第1A図−第1B図は、本項に記載されている方法に従って単離されたMN cDNAクローンの塩基配列を示している。遺伝子コドンの縮重から、一つのコドンが一つ以上のアミノ酸をコードしており(たとえば、TTA、TTG、CTT、CTC、CTAおよびCTGはいずれもロイシン(leu)というアミノ酸をコードしている)、また、たとえば第1A図−第1B図に示すように、一つのコドンが他のコドンと入れ替わるヌクレオチド配列の多様性により、本発明と実質的に同等な蛋白質およびポリペプチドが産生される。MN cDNAのヌクレオチド配列および相補的な核酸配列に関するそのような変形もすべて本発明の範ちゅうに含まれる。
【0038】
さらに、本明細書に記述し、第1A図−第1B図に示しているヌクレオチド配列は、単離され、本明細書で説明しているcDNAヌクレオチド配列のうち、はっきりした構造のみを表したものである。わずかに変更されたヌクレオチド配列が見つかることもあろうし、また、たとえば、同様のエピトープを有する等の、実質的に同等なMN蛋白質およびポリペプチドをコードするように当該分野で知られた技術により変形することも可能である。そしてそのような蛋白質/ポリペプチドは本発明の目的に適合する。MN蛋白質/ポリペプチドと相同あるいはほぼ相同な蛋白質/ポリペプチドをコードする合成核酸配列のように、同等なコドンを有するDNAおよびRNAは本発明の範ちゅうに含まれる。遺伝子コードの縮重がなければ、これらの核酸配列はやはり前記cDNAヌクレオチド配列にハイブリダイズする。本明細書で説明しているように、核酸配列が修飾されたり変形される結果、MN配列およびその断片と実質的に同等の配列が作り出される。
【0039】
MN遺伝子を見つけるために、MX感染HeLa細胞からλgt11によるcDNAライブラリーを調製した。MX感染HeLa細胞からの全RNAはグアニジンチオシアネートCsCl法を用いて抽出し、mRNAはオリゴdTセルロースを用いるアフィニティーにより分離した。cDNAの合成およびgt11へのそのクローニングはアマシャム(Amarsham)社のキットを用いて行ったが、EcoRI−NotIアダプターだけはストラタジーン(Stratagene)社(米国、カリフォルニア州、ラ・ホーラ(La Jolla))のものを使用した。モノクローナル抗体M75とアルカリフォスファターゼを縮合したヤギ抗マウス抗体とを組み合わせたイムノスクリーニングにライブラリーをかけた。このイムノスクリーニング法は、ヤング(Young)とデイヴィス(Davis)により報告されている(PNAS(USA), 80: 1194-1198(1983))。350,000のプラーク(全ライブラリーのおおよそ半分にあたる)をスクリーニングし、1個のポジティブクローンを取り出した。
【0040】
ポジティブクローンをpBluescript KS(ストラタジーン(Stratagene)社)のNotI部位に組み込んでサブクローニングを行い、pBluescript-MNを作った。Erase-a-BaseTMキット(プロメガ(Promega)社、米国、ウィスコンシン州、マディソン)を使用して、方向が反対で重なる2個の欠失を作り、T7シークエンス用キット(ファルマシア(Pharmacia)社、米国、ニュージャージー州、ピスカタウェイ)を用いてジデオキシ法により配列を決定した。配列はcDNAクローンの一部を表しており、インサートの長さは1397bpであった。本配列を第1A図−第1B図に示す(配列番号1)。配列は、大きな1290bpのオープンリーディングフレームおよびポリAシグナル(AATAAA)を含む107bpの3’非翻訳領域からなっている。該配列のもう一つの特徴は、mRNAの不安定性に関与する領域(1389番目のAUUUA)が存在することである。この領域は、ある種の腫瘍遺伝子およびリンホカインのmRNAに特異的なものである(ショウ(Shaw)とカーメン(Kamen), Cell, 46: 659-667(1986))。MNクローンの大きさと対応するmRNAのそれとをノーザンブロットにより比較すると(実施例12)、このcDNAは、その配列の5’末端から約100bpが欠損していることがわかった。 MN cDNAクローンのオープンリーディングフレームは、約48kdの推定蛋白質をコードしている(第1A図−第1B図、配列番号2)。推定翻訳アミノ酸(AA)配列の分析では、既報の蛋白質の配列とは高い相同性を示さなかった。最も近い相同性が見られたのは、MN蛋白質およびいろいろな型の炭酸脱水素酵素のC末端である(170−200AAの重なりにおいて約30−35%)。炭酸脱水素酵素の活性部位は、Zn2+結合ドメインと同様に、MN蛋白質においてもよく保存されている。しかしながら、MN遺伝子は、ヒトゲノム由来の新規な配列であることは明かである。
【0041】
上述したように、MN遺伝子は既知の炭酸脱水素酵素といくらかの相同性を有するが、いくつかの面でそれらとは異なっている。7個の炭酸脱水素酵素が報告されている(ドッジソン(Dodgson)ら(編)、炭酸脱水素酵素(The Carbonic Anhydrases)、(プレナムプレス(Plenum Press)社、ニューヨーク/ロンドン(1991年))。それらのおのおのは7個のイントロンを含んでいるが、MN遺伝子はイントロンを含まないようである。また、既知の炭酸脱水素酵素はすべておよそ30kdの蛋白質であり、これらはMN遺伝子のp54/58N関連の産生物より小さい。さらに、炭酸脱水素酵素はMN関連蛋白質のようなオリゴマーを形成しない。
【0042】
推定アミノ酸配列から、MN遺伝子の産生物は、303−313のアミノ酸位置に一つの活性なN−グリコシル化部位を有する塩基性蛋白質(pI9.08)であることが明かである。これらの事実は、HeLa細胞由来のp54/58N蛋白質が、Endo HおよびEndo Fによる分裂(おのおの約3kdの欠損を起こす)に感受性であることに対応している。親水性プロフィルは、アミノ酸の親水性配列(371−395位)を示しており、これはプラズマ膜にかかる領域を表していると考えられ、また、分裂シグナルも含むと考えられる。該プロフィルは、p54/58N蛋白質が細胞膜に局在していることとよく一致する。MNアミノ酸配列にはPEST領域は存在しないことから、MN遺伝子の産生物は安定で永続性の蛋白質であることが示唆される(ロジャース(Rogers)ら、Science, 234: 364-368(1986))。そのような特性から、発明者らのp54/58Nの代謝ラベルが非効率的であったことを説明できる。推定アミノ酸配列はさらにほかの特徴をも示す。すなわち、10個の活性リン酸化部位と7個のミリスチル化部位および3個の抗原決定因子を有する。
【0043】
p54/58N蛋白質の両方が一つの遺伝子によってコードされているか否かを確認するために、MN遺伝子の発現を特異的に阻害するアンチセンスODNsを用いた。(このようなアンチセンスODNsの使用法については、ステイン(Stein)とコーエン(Cohen)により総説されている(Cancer Res., 48: 2659-2668(1988))。)これらの実験については実施例11に詳述している。実験の結果、ODNsと共に培養したHeLa細胞においてはp54/58Nの合成がかなり阻害されていることがわかり、一方、いろいろなHeLa細胞の蛋白質の産生量はほぼ同程度に保たれていた。さらに、イムノブロットにおいて重要な結果が得られた。すなわち、ODNsによる特異的阻害はp54/58N蛋白質のの両方に影響を与えていた(実施例11)。これらのことから、MN遺伝子はHeLa細胞においてp54/58N蛋白質の両方をコードしていると結論づけられる。
【0044】
クローニングされた遺伝子がp54/58N特異的蛋白質をコードしているか否かを確認するために、該遺伝子をバクテリア発現ベクターpGEX−3X(ファルマシア(Pharmacia)社、スゥエーデン、ウプサラ)にサブクローニングし、グルタチオンS−トランスフェラーゼのC末端を有する融合蛋白質を発現するように構築した。このサブクローニングは、本発明におけるMN関連蛋白質の遺伝子工学的手法の一つを示すものである。以下の記述は例示であり、如何なる意味においても本発明を限定するものではない。
【0045】
融合蛋白質pGEX−3X−MNの産生
上述のpBluescript-MN由来のcDNA挿入体(インサート)は、プラスミドDNAをNotIで消化(切断(digesting))することにより切出した。該cDNA挿入体は、平滑末端を得るためにS1ヌクレアーゼで処理し、pGEX−3X(ファルマシア(Pharmacia)社)の脱リン酸化したSmaI部位にクローニングした。XL1-Blueの形質転換およびIPGTによる誘導の結果、融合蛋白質が得られた。
【0046】
融合蛋白質であるMNグルタチオンS−トランスフェラーゼは、グルタチオンS−セファロース4B(ファルマシア(Pharmacia)社)を用いたアフィニティークロマトグラフィーにより精製した。10%ゲルを用いたSDS−PAGEにより分離された2つの同様なサンプルから、20ミリグラムの精製された組換え蛋白質が得られた。一つのサンプル(A)はクマジーブリリアントブルーで染色し、他方のサンプル(B)はハイボンドCメンブレン(Hybond C membrane)(アマシャム(Amarsham)社、英国、バックス、アリスバリー)にブロットした。このブロットは、125IでラベルしたMAb M75を用いてオートラジオグラフィーにより展開した。
【0047】
SDS−PAGE分析から興味深い結果が得られた。すなわち、異なる分子量を有する一連の蛋白質のバンドが存在することである。本発明に従って産生された別の融合蛋白質である、β−ガラクトシダーゼMN(λ gt11溶原)においても同様なDSD−PAGEパターンが得られた。MN配列内に9個のAGGAGGコドンタンデムが存在することによる翻訳エラーのために、これらのパターンが現れたものと思われる。対応するtRNAが短いため、バクテリア遺伝子内でこれらのコドンを使用することは絶対に避けられている。かくして、外来性mRNAからのAGGAGGタンデムの翻訳の際に、+1のリボソーム上でのフレームシフトが高い頻度(約50%)で起こる(スパンヤード(Spanjaard)ら、Nuc. Acid Res., 18: 5031-5036(1990))。
【0048】
イムノブロッティングにおいて、同様なパターンが得られた。すなわち、染色されたSDS−PAGEゲル上で確認された全てのバンドは、MN特異的MAb M75に反応し、このことは、全ての蛋白質のバンドがMN特異的であることを示している。また、これらの結果から、MAb M75への結合部位は、フレームシフトの影響を受けないMN蛋白質のN末端部分にあることが示された。
【0049】
下記の実施例8に示すように、融合蛋白質pGEX−3X−MNは、MN特異的抗体およびMN抗原のラジオイムノアッセイに使用した。
【0050】
MN蛋白質および/またはポリペプチド
本明細書で用いている「MN蛋白質および/またはポリペプチド」(MN蛋白質/ポリペプチド)とは、MN遺伝子あるいはその断片によりコードされている蛋白質および/またはポリペプチドを意味している。好ましいMN蛋白質の例は、推定アミノ酸配列が第1A図−第1B図に示されているものである(配列番号2)。好ましいMN蛋白質/ポリペプチドは、第1A図−第1B図に示すMN蛋白質と実質的に相同性を有する蛋白質/ポリペプチドである。
【0051】
「ポリペプチド」とは、ペプチド結合によるアミノ酸の共有結合鎖のことであり、本明細書では、50あるいはそれ以下のアミノ酸から構成されるものと考えている。本明細書における「蛋白質」とは、50より多くのアミノ酸から構成されるポリペプチドと定義される。
【0052】
インビボ(in vivo)の腫瘍細胞から産生される蛋白質/ポリペプチドの配列が細胞培養内の腫瘍細胞から産生される蛋白質/ポリペプチドのものと異なることがある。すなわち、MN蛋白質/ポリペプチドが、アミノ酸置換、伸張、欠損、削除およびそれらの組合せ(これらに限定されるわけではないが)のようなアミノ酸配列変化を有していても、それらはすべて本発明の範ちゅうに属する。体液中に残存する蛋白質は蛋白質分解などの分解処理を受けることがある。すなわち、血清などの体液中にはかなりの削除が行われたMN蛋白質およびMNポリペプチドが見いだされる。本明細書で使用している「MN抗原」とは、MN蛋白質/ポリペプチドを包含している。
【0053】
さらに、MN蛋白質およびポリペプチドのアミノ酸配列は、遺伝子工学によって変化させることもできる。1個またはそれ以上のアミノ酸を削除したり置換することができる。そのようなアミノ酸の変化も、生物学的活性に有意の変化をもたらさず、本発明の範囲に含まれる蛋白質やポリペプチドを生じさせることができる。
【0054】
本発明のMN蛋白質およびポリペプチドは、本発明の方法にしたがって、さまざまな手段で調製できる。たとえば、組換え、合成、あるいはその他の生物学的手法、すなわち、長い蛋白質およびポリペプチドを酵素および/または化学的に解裂する等の方法が挙げられる。MN蛋白質を調製する好ましい方法は組換え法である。組換えによるMN蛋白質の産生のために特に好ましい方法は、融合蛋白質pGEX−3X−MNに関して上述した方法である。
【0055】
MN蛋白質およびポリペプチドの組換え産生
第1A図−第1B図に示すMN蛋白質およびその断片を調製する代表的な方法は、適切なMN cDNA断片を上で例示したような適切な発現ベクターに挿入することである。本明細書に記載しているように、単離されたMN DNAのクローニングには、広範な種類の宿主−クローニングベクターの組合せを用いることができる。たとえば、有用なクローニング媒体としては次のようなものを挙げることができる。染色体DNA、非染色体DNA、合成のDNAであって、たとえば、pBR322等の各種の既知のバクテリアプラスミド、その他の大腸菌(E. Coli)プラスミドおよびそれらの誘導体、ならびに広い宿主範囲のプラスミド、たとえば、RP4やファージDNA(たとえば、NB989等のλファージの多数の誘導体)、ファージDNAの発現をコントロールする配列を有する修飾プラスミド等のプラスミドとファージDNAの組合せから作られたベクターなど。プラスミドpGEX−3Xが好ましいクローニング媒体である。
【0056】
宿主として有用な細胞は真核性でも原核性でもよく、次のようなものが例示される。バクテリア宿主、たとえば、大腸菌(E. Coli)とその他のバクテリア株、酵母およびその他の菌類等、動物または植物の培養細胞等の動物あるいは植物宿主、昆虫細胞およびその他の宿主。もちろん、全ての宿主が同じ効果を有するわけではない。本明細書に記載されている原則を考慮しながら本発明の範囲から逸脱しないように、当業者によって宿主−クローニング媒体の組合せの選択がなされる。
【0057】
組換えDNA分子を形成するため、選択されたDNA断片をクローニング媒体へ挿入する特定部位の決定は、さまざまな因子に影響される。これらの因子としては、発現させる蛋白質およびポリペプチドのサイズと構造、所望する蛋白質およびポリペプチドの宿主細胞の成分による内部酵素分解に対する感受性、宿主細胞蛋白質によるコンタミネーション、開始および終了コドンの局在などの発現特性、および当業者により知られているその他の因子などが挙げられる。
【0058】
MN遺伝子、その断片あるいはMN遺伝子由来のcDNAを含む組換え核酸分子は宿主の形質転換に用いられるが、その形質転換により、宿主(形質転換体)にその構造蛋白質および断片を発現させ、かつ、ハイブリッドDNAがコードしている蛋白質およびポリペプチドを産生させる。組換え核酸分子はまた、MN核酸およびその断片の源としてさらに組換え核酸分子を複製生産させるために宿主の形質転換に用いられる。これらのそれぞれの使用に対する適切な宿主の選択は、当該分野で知られている多くの因子により定められている。これらの因子としては、たとえば、選択したベクターとの適合性、共生産物の毒性、所望する蛋白質およびポリペプチドの回収のしやすさ、発現特性、生物学的安全性およびコストが挙げられる。
【0059】
宿主細胞として大腸菌(E. Coli)などの原核細胞を用いる場合は、DNAを取り込むことのできるコンピテントな細胞は、指数増殖期の後に細胞を回収(ハーベスト)し、続いて既知の手法であるCaCl2法で処理することにより調製できる。宿主細胞のプロトプラストを作った後、形質転換を行うことができる。
【0060】
宿主細胞として原核細胞を用いる場合には、リン酸カルシウム沈降法、エレクトロポレーション、マイクロインジェクション等の従来からの機械的手法、赤血球あるいはリポソームに封入したプラスミドの挿入、リゾフォスファチジルコリン等の薬剤を用いた細胞処理、あるいはウイルスベクターの使用などの方法がとられる。
【0061】
蛋白質およびポリペプチドの産生量は3つの大きな因子に左右される。(1)細胞内の遺伝子またはDNA配列のコピー数、(2)該遺伝子および配列が転写、翻訳される効率、および(3)mRNAの安定性。転写、翻訳(これらが共同して発現を司る)の効率は、核酸配列、一般には所望のコード配列の前に位置している核酸配列に依存する。これらの核酸配列、すなわち発現コントロール配列は、とりわけ、転写を開始するためにRNAポリメラーゼが相互作用する位置(プロモーター配列)、および翻訳を開始するためにリボソームがmRNA(転写産物)と結合し、相互作用する位置を定めている。そのような全ての発現コントロール配列が同じ効果を発揮するわけではない。そのため、所望する蛋白質に特異的なコード配列を近傍の核酸配列から切り出し、その代わりに既知の発現コントロールベクターに融合して希望する高レベルの発現を得られるようにするのがよい。上述の操作を行ってから、処理されたこの新しいDNA断片が多数のプラスミドまたはバクテリオファージ誘導体に挿入され、細胞内で遺伝子または配列のコピー数を増し、さらに、発現蛋白質の収量を増加させる。
【0062】
発現コントロール配列として数種のものが用いられる。これらの中には、オペレーター、プロモーター、大腸菌(E. Coli)のラクトースオペロン(lac 系)のリボソーム結合および相互作用配列、大腸菌(E. Coli)のトリプトファン合成系(trp 系)に対応する配列、trpとlacプロモーターの融合(tac 系)、λフ
ァージの主要オペレーターとプロモーター(OLPLとORPR)、およびfdファージのコート蛋白質のコントロール領域などが含まれる。これらの配列を含むDNA断片は、lacあるいはtrpオペロンを有する形質導入ファージのDNA、またはλあるいはfdファージのDNAから制限酵素を用いた開裂により切出せる。つぎに、これらの断片を操作して、必須コントロール配列が、コード配列の開始コドンの非常に近傍または並列に位置しているようになった限定された分子集団を得る。
【0063】
融合体は次に、適切な宿主の形質転換あるいはトランスフェクションのためのクローニング媒体に挿入し、抗原産生量を測定する。そして、最も効率的な発現をする細胞を選択する。別の方法としては、開始コドンに接続されたlac、trpまたはλPLコントロール系を有するクローニング媒体を用い、MN蛋白質およびポリペプチドをコードする配列を含む断片に融合し、それによって遺伝子あるいは配列がクローニング媒体の開始コドンから正しく翻訳されるようにすることもできる。
【0064】
本明細書で使用している「組換え核酸分子」とは、少なくとも2つの核酸配列からなるハイブリッドヌクレオチド配列を指しており、ここで、第一の配列は、通常、自然状態では第二の配列と共存していない。
【0065】
本明細書で使用している「発現コントロール配列」とは、構造遺伝子と結合した場合にその発現を制御または調整するヌクレオチド配列のことである。
【0066】
MN蛋白質およびポリペプチドの合成および生物工学的発現
本発明のMN蛋白質およびポリペプチドは組換え法のみでなく、合成およびその他の生物学的手法によっても調製することができる。蛋白質またはポリペプチドの合成による産生は、当該分野でよく知られた方法に従って所望するアミノ酸鎖を化学的に合成していくことからなる。所望するポリペプチドまたは蛋白質を調製するためのその他の生物学的手法の例としては、所望するアミノ酸配列を含む長いMNポリペプチドまたは蛋白質を選択的に蛋白分解することが挙げられる。たとえば、長いポリペプチドまたは蛋白質を化学試薬または酵素で分解することなどである。
【0067】
ペプチドの化学合成は従来技術であり、たとえば、メリフィールド(Merrifield)固相合成法(メリフィールド(Merrifield),J., Am. Chem. Soc., 85: 2149-2154(1963)、ケント(Kent)ら、「生物学および医学における合成ペプチド(Synthetic Peptides in Biology and Medicine)29ページ」、アリターロ(Alitalo)ら編、エルセヴィール科学出版(Elsevier Science Publishers)社、1985年、およびハーグ(Haug),J.D.「ペプチド合成および保護基戦略(Peptide Synthesis and Protecting Group Strategy)」, American Biotechnology Laboratory, 5(1): 40-47(1987年、1/2月号)などにより実施される。
【0068】
化学的ペプチド合成の方法には、市販の保護アミノ酸を用いる自動ペプチド合成機の使用も含まれる。合成機としては、たとえばバイオサーチ(Biosearch)社(米国、カリフォルニア州、サンラファエル)の9500型および9600型、アプライド バイオシステムズ(Applied Biosystems)社(米国、カリフォルニア州、フォスターシティー)の430型、ミリジェン(Milligen)社(ミリポア(Millipore)社の子会社、米国、マサチューセッツ州、ベッドフォード)の9050型、およびデュポン(DuPont)のRAMP(高速自動複数ペプチド合成機)(デュポンコンパス(Dupont Compass)社、米国、デラウェア州、ウィルミントン)などがある。
【0069】
核酸プローブおよび試験キット
本発明の核酸プローブは、第1A図−第1図Bに示すMN cDNA配列またはMN遺伝子配列と実質的に相補的な配列からなる。本明細書で使用する「実質的に相補的」という語は、当該技術分野で広く理解されている意味と同じであり、それゆえ、通常のハイブリダイゼーションの状態の意味で用いる。ハイブリダイゼーションの程度は、相補正の精度に応じて変わり得る。
【0070】
前記プローブは、MN DNAおよび/またはRNAの検出に使用でき、したがって、患者の細胞内のMN遺伝子の存在や欠如、増殖、変異、あるいは遺伝子再配列の試験に使用できる。たとえば、MN遺伝子の過剰発現は、本発明のプローブを使用したノーザンブロットにより検出できる。増幅、転座、逆位ならびに欠失などの遺伝子変化は本発明のプローブを用いることにより検出でき、このとき、該プローブは、細胞分裂中期の染色体の広がった状態でも間期の核のいずれの状態であっても、患者の細胞由来の染色体とインサイチュー(in situ)ハイブリダイゼーションする。また、本発明のプローブを用いたサザンブロットによってMN遺伝子の増幅あるいは欠失を検出することもできる。該プローブを用いた制限酵素断片長多型(RFLP)分析は、遺伝子変化、変異および欠失の検出として好ましい方法である。該プローブはまた、いろいろな組織由来のMN遺伝子から転写された各種のmRNAとハイブリダイゼーションすることにより、MN蛋白質および/またはポリペプチドならびにそれらと相同またはほぼ相同な蛋白質および/またはポリペプチドを検出するために用いられる。
【0071】
このように、該プローブは診断/予後に有用である。該プローブは試験キットとして具体化でき、好ましくは、該プローブが適切なMN遺伝子またはMN mRNAターゲットとハイブリダイゼーションした時に、視覚化できる適切な手段を伴っているのがよい。そのような試験のサンプルとしては、組織標本、体液ならびに組織および細胞の抽出物が挙げられる。
【0072】
分析
本発明に従う分析は、脊椎動物のサンプル、好ましくはホ乳類のサンプル、より好ましくはヒトのサンプル中のMN抗原またはMN特異的抗体の検出および/または定量である。そのようなサンプルとしては、組織標本、体液、組織抽出物および細胞抽出物が挙げられる。MN抗原は、イムノアッセイ、免疫組織学的染色、免疫電子および走査顕微鏡観察、とりわけ、免疫金沈降(immunogold)を用いる技術により検出できる。
【0073】
MN抗原分析の好ましいサンプルは組織および/または細胞抽出物である(下記の実施例7と8が代表的なものである)。しかし、MN抗原は体液、とりわけ、血液、血清、プラズマ、精液、乳汁、唾液、涙、喀痰、粘液、尿、リンパ液、サイトゾル、腹水、胸水、羊水、膀胱洗浄液、気管支肺胞洗浄液、随液からも検出できる。試験前に大量の体液からMN抗原を濃縮することが望ましい。分析に好ましい体液は、試験する癌の型にもよるが、一般的に好ましい体液は、乳汁、胸水および腹水である。
【0074】
血液、プラズマ、血清、リンパ液、粘液、涙、尿、髄液および唾液などの体液サンプル中の活性MN蛋白質/ポリペプチドにMN特異的抗体は血清学的に結合するが、そのような抗体は血液、プラズマおよび血清、好ましくは血清に普通にみられる。MN特異的抗体の検出のための代表的な分析は、下記の実施例8に示すものであり、その実施例では融合蛋白質pGEX−3X−MNを用いている。MN抗原およびそれらと反応するMN特異的抗体の検出および/または定量試験の結果の相関から、患者の病状の好ましいプロファイルが示される。
【0075】
本発明の分析は、診断および/または予後の両方、すなわち、診断/予後である。本明細書で使用している「診断/予後」とは、臨床的状況に依存して以下の手順のそれぞれ、または、それらの手順のいくつかが重複していることを意味する。疾病の存在の判断、疾病の特性の判断、ある疾病と他の疾病との区別、病状の帰結予想、患者の様子と症状から示される疾病からの回復の見込み、患者の病状のモニタリング、疾病の再発に関する患者のモニタリング、および/または患者に対する好ましい治療方法の決定など。本発明における診断/予後の方法はたとえば以下のような場合に有用である。腫瘍性および/または前腫瘍性疾患の存在に対する集団のスクリーニング、腫瘍性疾患の進展の危険性の判断、腫瘍性および/または前腫瘍性疾患の存在の診断、腫瘍性疾患の患者の病状のモニタリング、および/または腫瘍性疾患の経過に対する予後の判断など。
【0076】
本発明は、広範な種類の腫瘍性疾患の存在のスクリーニングに有用である。たとえば、乳腺、尿路、卵巣、子宮、子宮頚管、子宮内膜、偏平細胞および腺偏平細胞などの腫瘍;頭および首の癌;神経芽細胞腫および網膜芽種などの中胚葉性の腫瘍;骨肉腫およびユーイング肉腫(Ewing′s sarcoma)などの肉腫;あるいはメラノーマなどが含まれる。特に興味があるのは婦人科の癌であり、この中には卵巣、子宮、子宮頚管、膣、陰門、子宮内膜癌が含まれるが、中でもとりわけ興味があるのは卵巣、子宮頚管および子宮内膜の癌である。同様に特に興味があるのは、胸、食道を含む胃、結腸、腎臓、前立線、肝臓、膀胱を含む尿路系、肺、および頭と首の癌である。
【0077】
本発明は、たとえば、宿主から取出した直後の細胞群を使用して、悪性腫瘍あるいは前悪性腫瘍性細胞の存在の可能性を推し量るための方法および構成物を提供する。そのような分析は、腫瘍の発見、それらの増殖の計測および疾病の診断と予後に役立つ。この分析はまた、癌の転移の存在の発見、ならびに、手術、癌の化学療法および/または放射線療法の後、全ての腫瘍組織がなくなっているか除去されているかを確認するのに役立つ。さらに、診断は、癌の化学療法および腫瘍の再発をモニターするのに役立つ。
【0078】
MN抗原または抗体の存在は、多くの既に確立された診断分析を用いて検出および/または定量することができる。当業者であれば、従来から存在する任意のイムノアッセイ法を適用して、MN抗原および/または抗体を検出および/または定量することができる。実施例8に本発明の好ましい診断方法であるラジオイムノアッセイ法について詳しく述べている。もちろん、MN抗原およびMN特異的抗体の検出にはほかの多くの方式も利用可能である。たとえば、ウエスタンブロット、ELISA(酵素結合免疫吸着法)、RIA(ラジオイムノアッセイ)、競合EIA、二重抗体サンドイッチ法、その他診断研究において通常使用される全ての分析法などがある。そのようなイムノアッセイにおける結果の解釈は、抗体あるいは抗体の組合せは、MNに関係のないサンプル中に存在する他の蛋白質および蛋白質の断片とは交差反応しないという仮定に基づいている。
【0079】
MN抗原検出の代表的な一つのELISA試験は、MN蛋白質/ポリペプチドに対して作られた抗体あるいはMN蛋白質を発現する細胞全体に対して作られた抗体をコートしたマイクロタイタープレートに、組織あるいは細胞抽出物のような患者のサンプルを加える方法である。ある抗原が抗体と結合するように一定時間インキュベートした後、プレートを洗浄し、酵素が連結している別の抗MN抗体を加え、反応が生じるようにインキュベートし、プレートを再洗浄する。その後、マイクロタイタープレートに酵素基質を加え、酵素が基質に作用するように一定時間インキュベートし、最終試料の吸光度を測定する。吸光度が大きく変化することは陽性の結果を示している。
【0080】
患者の体液、組織および/または細胞中のMN抗原の存在を検出および/または定量するためにMN蛋白質/ポリペプチドが使用できることもイムノアッセイの分野の当業者には明かである。そのような実施態様の一つとして競合イムノアッセイがあるが、この方法では、MN蛋白質/ポリペプチドはラベルされ、MN蛋白質/ポリペプチドに特異的な抗体への該ラベル化MN蛋白質/ポリペプチドの結合に競合させるために体液を加える。そのような分析は実施例8に記載しているようなMN抗原の検出および/または定量に使用できる。
【0081】
別の実施例としては、MN蛋白質またはポリペプチドに対するラベルされた抗体を用いるイムノメトリック分析がある。そのような分析においては、抗原結合抗体とコンプレックスを形成するラベル化抗体の量は、試料中のMN抗原の量と直接的に比例する。
【0082】
MN特異的抗体を検出する代表的な分析は競合分析であり、この分析では、ラベルされたMN蛋白質/ポリペプチドはサンプル中の抗体、たとえば、MN蛋白質/ポリペプチドを認識するモノクローナル抗体と結合して沈降する。当業者であれば、従来から存在する任意のイムノアッセイ方法を適用してMN抗原の検出および/または定量することができる。該抗体のMN蛋白質/ポリペプチドとの結合の検出は、当業者に知られた多くの方法、たとえば、ヒトにおいては抗ヒトラベルIgGの使用などにより行うことができる。
【0083】
本発明に従い、脊椎動物のサンプルからMN抗原を検出および/または定量するイムノアッセイの方法の例は、以下のステップから構成されている:
a)MN抗原に結合する一組あるいはそれ以上の組の抗体(一つの抗体かあるいは複数の抗体)(そのうち一組はラベルされているかまたはその他の手法で検出可能となっている)と脊椎動物のサンプルをインキュベートする;
b)MN抗原と該抗体からなる免疫コンプレックスが存在するかについて、インキュベートしたサンプルを調べる。
【0084】
本発明に従う別のイムノアッセイ法の例は競合イムノアッセイであり、脊椎動物サンプル中のMN抗原を検出および/または定量するために使用され、以下のステップから構成されている:
a)脊椎動物のサンプルを一組あるいはそれ以上の組の抗体および一定量のラベルされたもしくは他の手段で認識できるようになっているMN蛋白質/ポリペプチドとインキュベートし、このとき、該MN蛋白質/ポリペプチドが抗体との結合において、サンプル中に存在するMN抗原と競合する;
b)インキュベートしたサンプルを調べて、抗体に結合しているラベルした/検出可能なMN蛋白質/ポリペプチドの量を測定する;
c)ステップb)の検査から、前記サンプル中に存在するMN抗原および/または前記サンプル中に存在するMN抗原の量を決定する。
【0085】
望ましい特性を有する抗体(生物学的に活性な抗体の断片を含む)が調製されると、特定の抗体抗原コンプレックスの形成を判定するための広範な免疫学的分析法が可能である。多くの競合、非競合蛋白質結合分析に関して化学文献および特許明細書に記述されており、そのような分析法の多くは購入可能である。血清中の抗原の検出に適したイムノアッセイの例としては、以下の米国特許に記載されているものが含まれる。米国特許第3,791,932号、第3,817,837号、第3,839,153号、第3,850,752号、第3,850,578号、第3,853,987号、第3,867,517号、第3,879,262号、第3,901,654号、第3,935,074号、第3,984,533号、第3,996,345号、第4,034,074号、および第4,098,876号。
【0086】
分析に用いられる抗体は、ラベルされているかまたはラベルされていないものである。ラベルされていない抗体は凝集に用いられ、ラベルされた抗体は、多くの種類のラベルを施すことにより、広範な分析に使用される。
【0087】
適切な検出には、放射核、酵素、補酵素、発蛍光剤、化学発光剤、色素原、酵素基質あるいは補助因子、酵素阻害剤、フリーラジカル、パーティクル、染料などのラベルを用いるものが挙げられる。そのようなラベル試薬は、既知のさまざまな分析、たとえば、ラジオイムノアッセイ、ELISA等の酵素免疫アッセイ、蛍光免疫アッセイなどに用いられる。たとえば、米国特許第3,766,162号、第3,791,932号、第3,817,837号および第4,233,402号を参照のこと。
【0088】
本発明の分析において有用な抗体を調製する方法を以下に示す。以下の例は、本発明に従う代表的な分析について詳述したものである。
【0089】
イムノアッセイ試験キット
今までに概要を示した分析は、MN抗原および/またはMN特異的抗体(生物学的に活性な抗体の断片を含む)を検出および/または定量する試験キットとして具体化される。MN抗原を検出および/または定量する試験キットは、MN蛋白質/ポリペプチドおよび/または、(ポリクローナルおよび/またはモノクローナルの)MN特異的抗体からなる。そのような診断/予後用の試験キットは、一組またはそれ以上の組のポリクローナルおよび/またはモノクローナル抗体から構成されており、サンドイッチ法の場合には、抗体がMN抗原のエピトープを認識し、一組は適切にラベルされているか、その他の手段により検出可能である。 ラベルされた(あるいは他の手段で検出可能な)MN蛋白質/ポリペプチドとサンプル中のMN抗原との間で抗体に対する結合に関して競合がある分析方法の試験キットは、ラベルされた蛋白質/ポリペプチドと抗体の量を組合わせて、最適の感度と精度が得られるように構成されている。
【0090】
MN特異的抗体検出のための試験キットは、ラベルされた/検出可能なMN蛋白質/ポリペプチドから構成されていることが好ましく、必要であれば、たとえば、下記の実施例8に概要を示しているような好ましい分析を行うために、その他の成分を含んでいてもよい。そのような試験キットは、その他の適切な手段を含んで、従来からあるような分析に応用することもできる。
【0091】
MN特異的抗体の調製
本明細書で使用している「抗体」という語は、抗体全体を指すだけでなく、生物学的に活性な抗体の断片、好ましくは抗原結合部位を有する断片をも指している。そのような抗体は、従来からの手法および/または遺伝子工学により調製できる。抗体の断片は、遺伝子操作により、好ましくは超可変領域を含む、短鎖および/または長鎖の可変領域(VHとVL)から調製されるのがよく、さらに好ましくは、VH領域とVL領域の両方から調製されるのがよい。たとえば、本明細書で使用している「抗体」という語は、ポリクローナルおよびモノクローナル抗体ならびにそれらの生物学的に活性な断片をも意味しており、とりわけ、「一価」抗体(グレニー(Glennie)ら、Nature, 295: 712(1982));共有結合性または非共有結合性凝集をするFab’およびF(ab’)2断片を含むFab蛋白質;好ましくは短鎖および長鎖の可変領域(VHとVL領域)を含み、より好ましくは超可変領域(該VHとVL領域の相補性決定領域(CDRs)としても知られている)を含む短鎖または長鎖単独;Fc蛋白質;1以上の抗体と結合可能な「ハイブリッド」抗体;定常−可変領域キメラ;異なる起源由来の長鎖および短鎖からなる「複合」免疫グロブリン;通常の組換え手法により、またはオリゴヌクレオチドの依存突然変異誘発法(ダルバディー−マクファーランド(Dalbadie-McFarland)ら、PNAS (USA), 79: 6409(1982))により調製された、特異性やその他の特性を改良した「改造」抗体などが含まれる。
【0092】
治療および/またはイメージングに使用する抗体は、生物学的に活性な抗体の断片であることが好ましく、より好ましくは遺伝子工学的に処理された断片がよく、さらに好ましくはVHおよび/またはVL領域から遺伝子工学的に処理された断片がよく、さらにより好ましいのは、それらの超可変領域からなる断片である。 ポリクローナルおよびモノクローナル抗体を作出する従来からの技術は、イムノアッセイの分野では周知である。MN特異的抗体を産生するための免疫原には、MN蛋白質および/またはポリペプチド、好ましくは純粋で、MXに感染した腫瘍細胞株、たとえば、MX感染HeLa細胞およびその他の免疫原が含まれる。
【0093】
抗ペプチド抗体は、ヨーロッパ特許出願公開第44,710号(1982年1月27日公開)に記載されているような当該分野の従来法によっても作出される。該方法を要約すると、第1A図−第1B図に示すようなMNアミノ酸配列からペプチドを選択し、化学的に合成し、適切な免疫原蛋白質に結合し、適切な動物、通常はウサギあるいはマウスに注入し、ポリクローナルまたはモノクローナル抗体を作らせることにより、抗ペプチド抗体が調製される。モノクローナル抗体はコーラー−ミルシュテイン(Kohler-Milstein)法に従って作出される。
【0094】
従来のハイブリドーマ法だけでなく、新しい技術も本発明に従う抗体を産生するために用いることができる。たとえば、クローン作成および抗体のV遺伝子の発現にポリメラーゼ連鎖反応(PCR)法を用い、結合活性を有する断片をコードしている抗体遺伝子の選択にファージディスプレイ法を用いることにより、免疫されたマウスおよびヒト由来のPCR増幅したV遺伝子の集団から抗体の断片を単離することができた(マークス(Marks)ら、BioTechnology, 10: 779(1992年7月号)参照のこと。シャン(Chiang)ら、BioTechniques, 7(4): 360(1989);ワード(Ward)ら、Nature, 341: 544(1989年10月12日号);マークス(Marks)ら、J. Mol. Biol., 222: 581(1991);クラクソン(Clackson)ら、Nature, 352: 624-628(1991年8月15日号);ムリナックス(Mullinax)ら、PNAS (USA), 87: 8095(1990年10月号))。
【0095】
組換え技術を用いた抗体(本明細書では生物学的に活性な抗体の断片を含む)の調製法に関する記載は下記に見出される。米国特許第4,816,567号(1989年3月28日付与)、ヨーロッパ特許出願公開番号(EP)第338,745号(1989年10月25日公開)、EP第368,684号(1990年6月16日公開)、EP第239,400号(1987年9月30日公開)、WO第90/14424号(1990年11月29日公開)、WO第90/14430号(1990年5月16日公開)、ヒューセ(Huse)ら、Science, 246: 1275(1989年12月8日号);マークス(Marks)ら、BioTechnology, 10: 779(1992年7月号);ラ・サストリー(La Sastry)ら、PNAS(USA), 86: 5728(1989年8月号);シャン(Chiang)ら、BioTechniques, 7(4): 360(1989);オーランディ(Orlandi)ら、PNAS(USA), 86: 3833(1989年5月号);ワード(Ward)ら、Nature, 341: 544(1989年10月12日号);マークス(Marks)ら、J. Mol. Biol., 222: 581(1991);フーゲンブーム(Hoogenboom)ら、Nucleic Acids Res., 19(15): 4133(1991)。
【0096】
モノクローナル抗体の調製
本発明の分析で使用するモノクローナル抗体は、当該分野において既知の方法により得られる。たとえば、ガルフレ(Galfre)と ミルシュテイン(Milstein)「モノクローナル抗体の調製:戦略および方法(Preparation of Monoclonal Antibodies: Strategies and Procedures)」(酵素学的方法:免疫化学的手法(Methods in Enzymology: Immunochemical Techniques)73巻1−46ページより)(ランゴーン(Langone)とヴァナティス(Vanatis)編、アカデミック・プレス(Academic Press)社(1981年)、および古典的な参考として、ミルシュテイン(Milstein)とコーラー(Kohler)、Nature, 256: 495-497(1975)がある。
【0097】
本発明の代表的なハイブリドーマは、マウス細胞株の融合により調製されるが、ヒト/ヒトハイブリドーマ(オルソン(Olsson)ら、PNAS(USA), 77: 5429(1980))およびヒト/マウスハイブリドーマ(シュローム(Schlom)ら、PNAS(USA), 77: 6841(1980)、シェアマン(Shearman)ら、J. Immunol., 146: 928-935(1991)、ゴーマン(Gorman)ら、PNAS(USA), 88: 4181-4185(1991))からも調製され得る。そのようなヒト型のモノクローナル抗体は、治療およびイメージングに使用するのに好ましいモノクローナル抗体である。
【0098】
本発明に用いるモノクローナル抗体は、適切なホ乳類、好ましくは齧歯類、より好ましくはウサギまたはマウスに、適切な免疫原、たとえば、MaTuに感染したHeLa細胞や、あるいは、必要であればキャリアー蛋白質をつけたMN蛋白質/ポリペプチドを用いて免疫することにより調製される。例として、MAb M75を分泌するハイブリドーマVU−M75の作出について以下に記載している。MAb M75は、多くの実験室での診断試験、たとえば、培養腫瘍細胞、臨床サンプルなどにおけるMN蛋白質/ポリペプチドの同定に役立っている。MAb M75を産生する方法により、MAb M16(IgG2Bのアイソタイプ)およびMAb M67(IgG1のアイソタイプ)も産生された。
【0099】
MAb M75
モノクローナル抗体M75(MAb M75)は、マウスリンパ球ハイブリドーマVU−M75から産生されるが、該ハイブリドーマは、当初スロヴァク科学アカデミーウイルス研究所(Institute of Virology, Slovak Academy of Sciences)のハイブリドーマコレクション(Collection of Hybridomas)(チェコスロバキア、ブラティスラヴァ)に寄託され、また、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(米国、メリーランド州、ロックヴィル)に1992年9月17日にATCC受入番号HB11128として寄託されたものである。
【0100】
ハイブリドーマVU−M75は、ガーハード(Gerhard)、W.によって記載されている方法(「懸濁液中での細胞融合および調整培地中でのハイブリッドの発芽後生育(Fusion of cells in suspension and outgrowth of hybrids in conditioned medium)」モノクローナル抗体・ハイブリドーマ:生物学的分析の新しい方向(Monoclonal Antibodies. Hybridomas: A New Dimension in Biological Analysis)より、370ページ(ケネット(Kennet)ら編、プレナム(Plenum)社、米国、ニューヨーク))に従って産生される。BALB/CマウスをMaTuに感染したHeLa細胞で免疫し、それらの脾細胞をミエローマ細胞株NS−0と融合した。ハイブリドーマの組織培養培地をモノクローナル抗体のスクリーニングにかけた。用いた抗原は、MaTuに感染したHeLa細胞の抽出物とウサギ抗MaTu血清の免疫沈降物であるp58、および黄色ブドウ球菌(Staphylococcus aureus)細胞(SAC)のプロテインAであり(ザバダ(Zabada)とザバドバ(Zavadova)、Arch. Virol., 118: 189-197(1991))、SDS−PAGEゲルにより分離した。モノクローナル抗体は、プロテインA−セファロースを用いたアフィニティークロマトグラフィー(ハーロウ(Harlow)とレイン(Lane)、「抗体:実験室手順書(Antibodies: A Laboratory Manual)」コールドスプリングハーバー(Cold Spring Harbor)社、米国、ニューヨーク州、コールドスプリングハーバー、1988年)により、TC培地から精製された。
【0101】
本発明に従ってMN蛋白質/ポリペプチドを同定するのに有用なモノクローナル抗体は、従来からの任意の方法によってラベルすることができる。たとえば、西洋ワサビペルオキシダーゼ(HRP)のような酵素、蛍光化合物、125Iなどの放射活性同位体などである。本発明に従う好ましいラベルは125Iであり、好ましい抗体ラベル化の方法は、クロラミン−T法(ハンター(Hunter)、W.H.、「ラジオイムノアッセイ(Radioimmunoassay)」実験免疫学の手引(Handbook of Experimental Immunology)より、14.1-14.40(D.W.ウェイアー(Weir)編、ブラックウェル(Blackwell)社、オックスフォード/ロンドン/エディンバラ/メルボルン、1978年)である。
【0102】
MAb H460
モノクローナル抗体H460(MAb H460)は、MAb M75 と同様な方法によって調製された。ただし、マウスをMaTuに感染していないHeLa細胞により免疫し、マウスの脾細胞ではなくリンパ球をミエローマ細胞株NS−0の細胞と融合した点が異なる。MAb H460は、どんなヒト細胞ともほぼ同様に反応する。
【0103】
MN特異的抗体の治療への応用
本発明のMN特異的抗体、モノクローナル抗体および/またはポリクローナル抗体、好ましくはモノクローナル抗体、より好ましくはMAb M75は、腫瘍および/または前腫瘍性疾患の治療に用いることができ、単独あるいは化学療法剤または毒性剤(たとえば、リシンA等)と組み合わせて用いることができる。治療用としてより好ましいのは、本明細書に記載しているような生物学的に活性な抗体の断片である。同様に、治療用として好ましいMN特異的抗体は、ヒト型モノクローナル抗体である。
【0104】
MN特異的抗体は、好ましくは生理学的に許容性の非毒性液体基剤に分散された形で、治療効果を発揮するのに十分な量が投与される。
【0105】
イメージングへの抗体の応用
さらに、本発明のMN特異的抗体は、放射核などのイメージング剤と結合させた場合には、イメージングに利用できる。生物学的に活性な抗体の断片あるいはヒト型モノクローナル抗体がイメージング用としては好ましい。
【0106】
たとえば、腫瘍部位や転移の位置などが患者の腫瘍組織から同定できる。適切にラベルしたあるいはイメージング剤と結合させた抗体を生理学的に許容性のキャリアーと共に患者に投与し、結合した抗体は、ラベルあるいはイメージング剤の検出に適した方法、たとえば、シンチグラフィーなどにより検出される。
【0107】
アンチセンスMN核酸配列
本発明のMN遺伝子は、腫瘍遺伝子と推定され、それによってコードされている蛋白質は、腫瘍性蛋白質であると推定される。MN遺伝子から転写されたmRNAと実質的に相補性のアンチセンス核酸配列は、実施例11のアンチセンスオリゴデオキシヌクレオチド(ODNs)に代表されるものであり、MN遺伝子の発現を減少あるいは抑制するのに用いることができる(ザメクニック(Zamecnick)、P.C.「イントロダクション:遺伝子情報解読のモジュレーターとしてのオリゴヌクレオチド塩基ハイブリダイゼーション(Introduction: Oligonicleotide Base Hybridization as a Modulator of Genetic Message Readout)」1−6ページ、癌およびAIDSへのアンチセンス核酸療法の予測(Prospects for Antisense Nucleic Acid Therapy of Cancer and AIDS )より(ウィレイ−リス(Wiley-Liss)社、米国、ニューヨーク州、ニューヨーク、1991年);ウィックストーム(Wickstorm)、E.「HL−60前骨髄球性白血病細胞に対するアンチセンスDNA療法:末端配列の変化と標的配列への依存性(Antisense DNA Treatment of HL-60 Promyelocytic Leukemia Cells: Terminal Differenciation and Dependence on Target Sequence)」7−24ページ、同上;レザマン(Leserman)ら、「腫瘍遺伝子の発現に干渉するアンチセンスオリゴヌクレオチドの標的化と細胞内分布(Targeting and Intracellular Delivery of Antisense Oligonucleotides Interfering with Oncogene Expression)」25−34ページ、同上;ヨコヤマ(Yokoyama)、K.「アンチセンスRNAによる腫瘍原遺伝子c-mycの転写制御(Transcriptional Regulation of c-myc Proto-oncogene by Antisense RNA)」35−52ページ、同上;ヴァンデンベルク(van den Berg)ら、「染色体の異常世代を抑制するアンチセンスfosオリゴデオキシリボヌクレオチド(Antisense fos Oligodeoxyribonucleotides Suppress the Generation of Chromosomal Aberrations)」63−70ページ、同上;メルコーラ(Mercola)、D.「アンチセンスfosおよびfunRNA(Antisense fos and fun RNA)」83−114ページ、同上;イノウエ(Inouye)、Gene, 72: 25-34(1988)、ミラー(Miller)とティソー(Ts′o)、Ann. Reports Med. Chem., 23: 295-304(1988);ステイン(Stein)とコーエン(Cohen)、Cancer Res., 48: 2659-2668(1988);ステヴェンソン(Stevenson)とインヴァーセン(Inversen)、J. Gen. Virol., 70: 2673-2682(1989);グッドチャイルド(Goodchild)「オリゴヌクレオチドによる遺伝子発現の抑制(Inhibition of Gene Expression by Oligonucleotides)」53−77ページ、オリゴデオキシヌクレオチド:遺伝子発現のアンチセンス抑制剤(Oligodeoxynucleotides: Antisense Inhibitors of Gene Expression)、(コーエン(Cohen)、J.S.編、CRCプレス(CRC Press)社、米国、フロリダ州、ボカ・ラートン、1989年);デルヴァン(Dervan)ら、「三重らせん形成による二重らせんDNAのオリゴヌクレオチドの認識(Oligonucleotide Recognition of Double- helical DNA by Triple-helix Formation)」197−210ページ、同上;ネッカーズ(Neckers)、L.M.「細胞制御研究の道具としてのアンチセンスオリゴデオキシヌクレオチド:取込のメカニズムと腫瘍遺伝子機能の研究への応用(Antisense Oligodeoxynucleotides as a Tool for Studying Cell Regulation: Mechanisms of Uptake and Application to the Study of Oncogene Function)」211−232ページ、同上;レイトナー(Leitner)ら、PNAS(USA), 87: 3430-3434(1990);ベヴィラッカ(Bevilacqua)ら、PNAS(USA), 85: 831-835(1988);ローク(Loke)ら、Curr. Top. Microbiol. Immunol., 141: 282-288(1988);サリン(Sarin)ら、PNAS(USA), 85: 7448-7451(1988);アグラワル(Agrawal)ら、「アンチセンスオリゴヌクレオチド:化学療法およびAIDSへのアプローチの可能性(Antisense Oligonucleotides: A Possible Approach for Chemotherapy and AIDS)」核酸療法に関する生化学会国際会議(International Union of Biochemistry Conference on Nucleic Acid Therapeutics)(1991年1月13−17日、米国、フロリダ州、クリアウォタービーチ);アームストロング(Armstrong)、L.、Ber. Week、88−89ページ(1990年3月5日号);ウエイントラウブ(Weintraub)ら、Trends, 1: 22-25(1985))。そのようなアンチセンス核酸配列、好ましくは、オリゴヌクレオチドはMN mRNA、特にリボソーム結合部位と翻訳開始点の近傍でハイブリダイゼーションすることにより、mRNAの翻訳を阻害する。それゆえ、そのようなアンチセンス核酸配列を使用することは、癌の治療の一つの方法と考えられる。
【0108】
本発明に従う好ましいアンチセンスオリゴヌクレオチドは、遺伝子特異的ODNsあるいはMN mRNAの5’末端に相補的なオリゴヌクレオチドである。特に好ましいのは、下記の実施例11に配列が示されているような、29−mer ODN1および19−mer ODN2である。これらのアンチセンスODNsは、MN遺伝子の発現を抑制する機能を有する多くのアンチセンス核酸配列の中の代表的なものである。当業者であれば、第1A図−第1B図の核酸配列から適切なアンチセンス核酸配列、好ましくはアンチセンスオリゴヌクレオチドを確定することができる。
【0109】
ワクチン
本発明のMN蛋白質およびポリペプチドは、腫瘍性疾患に対して防御免疫を誘起でき、腫瘍形成活性を弱める効果を有するようなワクチンに組込むことができる。MN蛋白質および/またはポリペプチドは、合成あるいは組換えやその他の生物学的手法により調製されて、単量体、または多量体型のMN蛋白質の1またはそれ以上のエピトープに対応する、1またはそれ以上のアミノ酸配列から構成されるようにすることができる。つぎに、これらの蛋白質および/またはポリペプチドは、防御免疫を誘起できるワクチンに組込まれる。そのようなポリペプチドの免疫原性を上げる方法は、多量体構造に組込むこと、キーホールリンペットヘモシアニン(KLH)あるいはジフテリア毒素等の高免疫原性蛋白質キャリアーに結合させること、およびアジュバントあるいはその他の免疫応答強化剤と組み合わせて投与することを含む。
【0110】
本発明に従うワクチンにおいて使用される好ましいMN蛋白質/ポリペプチドは、遺伝子工学的に処理されたMN蛋白質である。好ましい組換えMN蛋白質は、本発明に従って産生された融合蛋白質pGEX−3X−MNである。
【0111】
本発明のそのようなワクチンの好ましい使用例は、MN関与性一次癌が外科的に切除された患者へ投与することである。ワクチンは患者の体内で能動免疫を誘起し、再発あるいは転移を防ぐことができる。
【0112】
さらに、MN蛋白質/ポリペプチドに対する抗体に対する抗イディオタイプ抗体もワクチンとして有用であり、同様に製剤化できる。
【0113】
単量体、または多量体型のMN蛋白質/ポリペプチドのエピトープに対応するアミノ酸配列は、化学的合成、あるいは遺伝的に変更された微生物やそれらの培養培地などの生物源を精製することによっても得られる(ラーナー(Lerner)「合成ワクチン(Synthetic Vaccines)」Sci. Am., 248(2): 66-74(1983)を参照)。蛋白質/ポリペプチドは、他の蛋白質/ポリペプチド(他のタンパク質の断片を含む)と組合わされて一つのアミノ酸を形成することがあり、たとえば、融合蛋白質として合成される場合や、合成または生物由来の抗原性あるいは非抗原性の他のポリペプチドに結合する場合などがある。
【0114】
「MN蛋白質/ポリペプチドのエピトープに対応する」という言葉は、天然に存在する蛋白質またはポリペプチドのアミノ酸配列の変化が抗原性を与えることがあり、腫瘍性疾患に対する防御免疫および/または抗腫瘍形成性効果を付与することがあるという実際的な可能性を含むものとする。配列の変化の可能性としては、アミノ酸の置換、伸長、欠失、削除、挿入およびこれらの組合せが挙げられるが、これらに限定されるわけではない。そのような変形も本発明の範ちゅうに含まれる。ただし、それらを含有する蛋白質またはポリペプチドが免疫原性であり、そのような蛋白質またはポリペプチドによって分泌された抗体は、天然に存在するMN蛋白質またはポリペプチドと交差反応し、その分泌量は、ワクチンとして投与したときに防御免疫および/または抗腫瘍形成性活性を誘起するのに十分な量であるものとする。
【0115】
そのようなワクチンの組成物は、生理学的に許容し得る基剤、たとえば、免疫的に許容される希釈剤およびキャリアー、また、フロイントの完全アジュバント(Freund′s Complete Adjubant)、サポニン、明ばん等の通常用いられるアジュバントなどと組合せられる。投与は、免疫学的に有効な量のMN蛋白質またはポリペプチドで行うが、好ましい投与量ユニットは、被投与体の体重1kgあたり0.01から10.0μgの免疫学的に活性なMN蛋白質および/またはポリペプチドである。防御に有効な総投与量は、抗原量として0.1から約100μgの範囲である。
【0116】
投与経路、抗原量、投与の回数と頻度はすべて最適の条件で行うが、これらは当該分野の通常の技術範囲の範ちゅうに含まれる。
【実施例】
【0117】
以下の実施例は説明をするためのものであり、如何なる意味においても本発明を限定するものではない。
【0118】
材料および方法
次の材料および方法は、以下の実施例で使用するものである。
【0119】
MaTu感染および非感染HeLa細胞
MaTu剤(ザバダ(Zabada)ら、Nature New Biol., 240:124-125(1972)、ザバダ(Zabada)ら、J. Gen. Virol., 24: 327-337(1974))は、「MaTu」細胞由来であり(ウィドメイアー(Widmaier)ら、Arch. Geschwulstforsch, 44: 1-10(1974))、これは、マイトマイシンCで処理したMaTu細胞と共培養することにより、発明者らのHeLa細胞ストックに移したものであり、これにより、対照とMaTu感染細胞を対比できるようなった。MaTu細胞は、5μg/mlのマイトマイシンC(カルバイオケム(Calbiochem)社、米国、カリフォルニア州、ラ・ホーラ(La Jolla))を含む培地で37℃で3時間インキュベートした。混合培養は、培地5mlあたり、2×105個のマイトマイシンC処理細胞および4×105個の新鮮感受性細胞とからなるようにした。3日後に最初の継代を行い、さらに週に1−2回継代した。
【0120】
対照のHeLa細胞は、ザバダ(Zabada)らの記載(Nature New Biol.,240:124-125(1972))と同様に扱った。
【0121】
血清
癌患者、各種の非腫瘍性症状に苦しむ患者および健康な女性からのヒト血清は、大学院医学研究科産婦人科病院(Clinics of Obstetrics and Gynaecology at the Postgraduate Medical School)(チェコスロバキア、ブラチスラヴァ)から入手した。
【0122】
ヒト血清KHは、切除後14カ月の50才の乳腺腫患者から得た。該血清は、401の血清サンプルの中で、ザバダ(Zabada)らにより記載(Nature New Biol., 24
0:124-125(1972))されているVSV(MaTu)のプソイド型に対する中和抗体を含有する2つの血清の内の一つである。血清L8は、パジェット病(Paget′s disease)の患者から得た。血清M7は、健康な献血者から得た。
【0123】
ウサギ抗MaTu血清は、MaTuに感染した生存HeLa細胞10−5×107個を30日の間隔で3回ウサギに免疫することにより調製した。
【0124】
RIPとPAGE
RIPとPAGEは基本的には、ザバダ(Zabada)とザバドヴァ(Zabadova)による記載(Arch. Virol., 118: 189-197(1991))に従って行ったが、本明細書に記載している実験において異なる点は、[35S]メチオニン(NEN)、10μCi/mlのメチオニン非含有MEM培地、2%のFCSを添加した3%の完全MEM培地を使用した点である。細胞の全面ペトリ皿培養を、該培地で終夜インキュベートした。
【0125】
RIPには、SAC法(ケスラー(Kessler)、J., J. Immunol., 115: 1617-1624(1975))を用いた。インキュベーションおよび遠心分離はすべて0−4℃で行った。単層細胞は、RIPA緩衝液(0.14Mの塩化ナトリウム、7.5mMのリン酸緩衝液(pH7.2)、1%のトライトンX−100(Triton X-100)、0.1%のデオキシコール酸ナトリウム、1mMのフッ化フェニルメチルスルフォニルおよびトラシロール(Trasylol)からなる)で抽出した。非特異反応を減少させるため、抗血清をウシ胎児血清(バーバシッド(Barbacid)ら、PNAS(USA), 77: 1617-1621(1980))およびSAC処理した抗原抽出物に前吸着させた。
【0126】
PAGE(還元状態)には、10%のSDSゲルを用いた(レムリ(Laemmli)、Nature, 227: 680-685(1970))。対照マーカー蛋白質としては、シグマキット(Sigma Kit)(製品番号MW-SDS-200)を使用した。フルオログラフィーには、サリチル酸塩を用いた(ヘーガード(Heegaard)ら、Electrophoresis 5: 263-269(1984))。
【0127】
イムノブロット
本明細書に記載しているイムノブロットは、トウビン(Towbin)らの方法(PNAS(USA) 76: 4350-4354(1979))に従って行った。蛋白質は、レムリ(Laemmli)の電気泳動緩衝液を蒸留水で1:10に希釈し、メタノールやSDSを含まない状態でゲルからニトロセルロース(シュレイカー・アンド・シュエル(Schuleicher and Schuell)社、ドイツ、ダッセル、0.45μm多孔性)に移した。移送は1.75mA/cm2で2時間半かけて行った。ブロットは、125IでラベルしたMAbと共に展開し、増感フィルターを使用してX線フィルムを−70℃で感光することによりオートラジオグラフィーを行った。
【0128】
細胞培養の抽出物には少量のMN抗原しか含まれていないため、0.5から1mlの抽出物に50μlの10%SAC懸濁液(MAb M75が含まれている)を添加して濃縮した。本方法は、ヒトIgGを含む臨床サンプルのMN抗原の濃縮にも使用できる。予備対照実験の結果から、そのような方法がSAC吸着M75へのMN抗原の結合に影響しないことが示されている。組織抽出物は、組織をモルタル、乳棒、砂(分析用グレード)と共に粉砕することにより得られた。このホモジネートに、RIPA緩衝液を当初の組織に対して10:1(重量当りの容量)の割合で加えた。抽出物は、エッペンドルフ(Eppendorf)遠心分離に3分間かけ、清澄にした。
【0129】
実施例1
MaTu特異的抗原の免疫蛍光法
対照細胞、および上述の方法で調製されたモノクローナル抗体を含むMaTu感染HeLa細胞について、免疫蛍光実験を行った。該モノクローナル抗体は、MaTu関連抗原に特異的である。モノクローナル抗体の存在確認には、FITC結合抗マウスIgGを用いた。細胞のギムザ(Giemsa)染色から、対照とMaTu感染HeLa細胞にははっきりとした違いはないことが示された。
【0130】
MaTu関連抗原に特異的であることが事前の試験で証明されているMAbは、免疫蛍光において二つの異なる反応性を示した。第一のグループを代表するMAb M67は、MaTu感染HeLa細胞において顆粒状の細胞質蛍光反応を示したが、これはアセトンで固定した細胞のみにみられ、生細胞は蛍光を示さなかった。MAb M16も同様の蛍光を示した。M67およびM16のいずれについても、対照のHeLa細胞においてごく弱い「バックグラウンド」蛍光がみられた。
【0131】
もう一つのMAbであるM75は、生きたMaTu感染HeLa細胞において顆粒状の膜蛍光反応を示し、アセトン固定細胞においては顆粒状の核蛍光反応を示した。しかしながら、M75は、非感染HaLa細胞においても、非常に弱いが類似の蛍光を示すことがあった。増殖の条件に基づく関係が見いだされた。すなわち、MaTuに非感染のHeLa細胞においては、細胞を高密度培養中で数回継代して増殖したもののみがMAb M75による両方の型の蛍光を示し、希薄培養中で増殖したものは示さない。
【0132】
M75反応性の細胞表面抗原の量は、細胞蛍光測定法により測定し、その量は、細胞培養の密度およびMaTuの感染に依存していた。対照および MaTu感染HeLa細胞は、高密度あるいは希薄培養中で12日間増殖させた。細胞は、ヴァーセン(Versene)(EDTA)によって遊離させ、MAb M75と共にあるいはMAbなしでインキュベートし、続いてFITC結合抗マウスIgGと共にインキュベートした。蛍光の強さを測定した。
【0133】
抗原結合MAb M75は、誘導性のものと思われる。すなわち、該抗体は、希薄培養で増殖した対照のHeLa細胞にはなく、高密度培養でのHeLa細胞の増殖あるいはMaTuの感染によって誘起されることがわかった。これら二つの因子は、付加的あるいは共働作用効果を有することがわかった。これらの知見と本明細書に記載したその他の結果により、次のことが示唆される。すなわち、二つの異なる物質があり、その一つはM67に反応する外因性、透過性のMX、もう一つはMAb M75により検出される内因性、誘導性のMNである。
【0134】
実施例2
MAb M75に反応する蛋白質のイムノブロット分析
MAb M75が、非感染およびMaTu感染HeLa細胞の両方において同じ蛋白質に反応するか否かを確かめ、さらに、該蛋白質の分子量を求めるために、それらの細胞の抽出物をPAGEおよびイムノブロッティング(上述の方法による)により分析した。高密度あるいは希薄培養中で12日間増殖させた非感染およびMaTu感染HeLa細胞を5cmのペトリ皿にまき、すべての変異型について細胞数が5×105個となるようにした。2日後に細胞をRIPA緩衝液で抽出し(上述)、200μl/皿の量にした。抽出物は、6%のメルカプトエタノールを含有する2倍濃縮のレムリ(Laemmli)のサンプル緩衝液と混合し、5分間煮沸した。蛋白質は、SDS−PAGEにより分離し、ニトロセルロースにブロットした。ブロットは、125IでラベルしたMAb M75を用いて展開し、オートラジオグラフィーにかけた。
【0135】
MAb M75は、54kdと58kdの2本のMN特異的蛋白質のバンドと反応し、この結果は高密度で増殖した非感染HeLa細胞、ならびにMaTu感染HeLa細胞において同じであったことから、M75は、非感染およびMaTu感染HeLa細胞の両方で同じ蛋白質を認識していることが明らかになった。細胞蛍光測定の結果と併せて考えると、抗原量は、細胞密度とMaTuの感染に依存しており、後者がより強力なp54/58Nの誘導因子である。
【0136】
実施例3
インサイチュー(in situ)でのMaTu特異的抗原のラジオイムノアッセイ
M75での結果とは対照的に、別のMAbである、M67は、外因性で透過性の物質MXに特異的と思われる。M67においては、細胞が、高密度培養で増殖されたかあるいは希薄培養で増殖されたかに関係なく、対照のHeLa細胞では免疫蛍光を示さなかった。この相違は、125IでラベルしたMAbs M67およびM75を用いたラジオイムノアッセイ実験によってはっきり示された。
【0137】
そのような実験においては、非感染およびMaTu感染HeLa細胞を高密度培養あるいは希薄培養で同時培養して増殖させた。培養細胞は、生きたもの(固定しない)か、固定したもの(メタノールで5分間処理後風乾)かである。培養細胞は、125IでラベルしたMAbsを6×104cm/皿加えたペトリ皿で2時間インキュベートした。その後、培養細胞をPBSで4回洗浄し、1ml/皿の2N NaOHで溶解し、γカウンターで放射活性を測定した。
【0138】
本例の簡単な放射活性法は、ペトリ皿の培養細胞に直接行うものである。ラジオイムノアッセイの16の変異型から、MXおよびMNが細胞表面あるいは細胞内に存在しているかが確認でき、これら二つの抗原の発現がMaTuの感染およびペトリ皿にまく前の細胞増殖の時の密度にどのように依存しているかが確認された。生きた固定されていない細胞では、細胞表面の抗原のみがMAbsと結合できる。これらの細胞においては、M67は培養細胞の如何なる変異型とも反応を示さなかったが、一方、M75は上述の実施例1および2の結果に示すように反応した。
【0139】
メタノールで細胞を固定することにより、細胞膜はMAbsを透過させるようになる。M67は、事前の細胞密度に関係なくMaTu感染HeLa細胞と反応するが、対照のHeLa細胞には結合しない。メタノール固定された細胞内のMAb M75により、希薄培養由来の非感染HeLa細胞には対応する抗原が存在しないこと、さらに、該抗原は、高密度培養による増殖あるいはMaTuの感染により誘導されることが確認された。
【0140】
実施例4
動物血清に反応性またはVSVビリオンに関連するMaTu成分の同定
非感染またはMaTu感染HeLa細胞由来のRIPA抽出物、あるいは、対照またはMaTu感染HeLa細胞内で再産生された精製VSV由来のMaTu特異的蛋白質のイムノブロット分析を行うこと、抗原p58Xあるいはp54/58Nのちどの抗原が動物血清と放射免疫沈降するかということ、さらに、VSV変異株の相補性およびプソイド型(pseudotype)のビリオンの形成に関与しているかということを調べた。方法の詳細については、パストレコヴァ(Pastorekova)らによる記載(Virology, 187: 620-626(1992))を参照のこと。
【0141】
MaTu感染HeLa細胞で免疫したウサギの血清は、MAb M67およびMAb M75反応性の蛋白質(p58Xおよびp54/58Nの両方)の両方と免疫沈降したのに対し、正常ウサギ、ヒツジあるいは白血病ウシの「自発的(spontaneously)」血清は、M67反応性蛋白質(p58X)とのみ免疫沈降した。一方、MaTu感染HeLa細胞により産生され、精製されたVSVは、p54/58NのM75反応性のバンドのみが存在した。このことから、MXとMNは、MaTuの独立した構成成分であると考えられ、VSV変異株に相補性であり、プソイド型のビリオンに集積されるのはp54/58Nである。
【0142】
第2A図に示し、下記の実施例5に述べているように、MX抗原は、MaTu感染線維芽細胞内に存在することがわかっている。ザバダ(Zavada)とザバドバ(Zavadova)は(1991年)、MX感染線維芽細胞由来のp58のバンドは、ウサギ抗MaTu血清を用いたRIPでは検出できなかったと報告している。該血清中には、MX抗原よりもMN抗原に対する抗体が多く含まれていた。この矛盾は、感染培養細胞内におけるMXの伝播が極端に遅いことで説明できる。ザバダ(Zavada)とザバドバ(Zavadova)による報告は(1991年)、感染後6週目の線維芽細胞に関するものであり、他方、後の試験は感染4カ月後の線維芽細胞に関してである。イムノブロットの結果から、MXは、まず、感染の4週間後にはH/F−NハイブリッドおよびH/F−Tハイブリッドの両方から検出され、6週後にはHeLa細胞に検出され、線維芽細胞においては感染10週後になってようやく検出されることを発明者らは見出した。
【0143】
実施例5
MN特異的およびMX特異的蛋白質の発現
第2A図−第2B図は、ヒト線維芽細胞、HeLa細胞およびH/F−Nハイブリッド細胞とH/F−Tハイブリッド細胞内におけるMN特異的およびMX特異的蛋白質の発現をグラフとして示したものであり、MX感染および非感染細胞での発現を対比している。細胞は、マイトマイシンCで処理したMX感染HeLa細胞と共培養することにより感染させた。感染および非感染細胞は、高密度培養中で3回継代して増殖させた。感染4カ月後に、感染細胞を非感染細胞と同時にペトリ皿で増殖し、高密度単相を形成させた。
【0144】
全面ペトリ皿(5cm)培養した細胞にラジオイムノアッセイを直接行い、メタノールで固定した(基本的には上記の実施例3と同様)。細胞の単層はメタノールで固定し、125IでラベルしたMAbs M67(外因性のMX抗原特異的)またはM75(内因性MN抗原特異的)を6×104cpm/皿加えて処理した。結合放射活性を測定した。結果は第2A図−第2B図に示す。
【0145】
第2A図は、試験した4つの細胞株すべて、すなわち、ヒト胚線維芽細胞、HeLa細胞、およびH/F−NハイブリッドとH/F−Tハイブリッド細胞、にMXが透過したことを示しており、同時に、対応する4つの非感染細胞株にはMXが存在しないことを示している。第2B図は、MX感染および非感染HeLa細胞ならびにH/F−T細胞にはMN抗原が存在するが、線維芽細胞には存在しないことを示している。対照のH/F−NにはMNは全く検出されず、MaTu感染H/F−NにおいてMN抗原のバックグラウンドを越えるわずかな増加が見られただけであった。このことから、ハイブリッドにおいては、MN抗原の発現が腫瘍形成性と強く関係していることが明らかになった。
【0146】
これらの結果は、イムノブロッティングにより得られた結果と一致する。MN特異的双子蛋白質p54/58Nは、イムノブロッティングによりHeLa細胞株(標準型の両方、すなわちHeLa K、およびスタンブリッジ(Stanbridge)突然変異HeLa細胞、すなわちD98/AH.2あるいはHeLa S)および腫瘍形成性H/F−Tにおいて検出されたが、線維芽細胞あるいは非腫瘍形成性H/F−Nでは、放射活性を検出するために使用したフィルムをゆっくりと長時間感光してもp54/58Nは検出されなかった。MXのHeLa細胞への感染により、p54/58N蛋白質の急激な濃度上昇がみられた。
【0147】
ハイブリッド細胞H/F−NおよびH/F−Tは、エリック J.スタンブリッジ(Eric J. Stanbridge)によって作出された(スタンブリッジ(Stanbridge)ら、Somatic Cell Genetics, 7: 699-712(1981)、スタンブリッジ(Stanbridge)ら、Science, 215: 252-259(1982))。彼の本来のハイブリッドは、HeLa細胞とヒト線維芽細胞との融合によって作られ、ヌードマウスにおいては腫瘍形成性ではなかったが、形質転換細胞内のいくつかの特性、たとえば、軟寒天上での増殖など、を保持していた。
【0148】
染色体11を失ったハイブリッド由来の分離体(セグレガント)が腫瘍形成性であることはまれである。これらの分離体における腫瘍形成性に関する最も妥当な説明は、染色体11が抑制遺伝子(抗腫瘍遺伝子)を有しており、これが未知の腫瘍遺伝子の発現をブロックしているということである。該腫瘍遺伝子によってコードされている腫瘍蛋白質は、ヌードマウス内で腫瘍を形成するH/Fハイブリッドの能力に対して重大な働きをする。p54/58N蛋白質は、H/Fハイブリッドの腫瘍形成性に関連があり、推定腫瘍蛋白質の一つの候補である。
【0149】
実施例6
ヒト腫瘍細胞培養由来およびヒト組織の臨床標本由来のMN抗原のイムノブロット
実施例5に示したように、H/Fハイブリッド細胞内の腫瘍形成性において、MN抗原が関与することから、その他のヒト腫瘍細胞培養および臨床標本内のMN抗原の存在を調べることにした。予備実験の結果から、その他のヒト腫瘍細胞培養の抽出物内のMN抗原の濃度はHeLa細胞よりも低いことが示された。このことから、オートラジオグラフィーにおいては、長時間の感光が必要であることがわかった。これにより、上述の材料および方法:イムノブロッティングに従う方法(ここでは、MAb M75を結合させたSACを用いて沈澱させることにより、MN抗原を濃縮している)により、該方法の感度を増加させた。
【0150】
細胞培養の抽出物内のMN蛋白質のイムノブロットは、次のものから調製した。(A)MX感染HeLa細胞、(B)ヒト線維芽細胞、(C)T24、(D)T47D、(E)SK−Mel 1477、および(F)MX非感染HeLa細胞。蛋白質は、3%のメルカプトエタノールを含むかまたは含まない状態(+MEまたは0MEと表す)でサンプル緩衝液に加えて加熱した後、PAGEにより分離した。つまり、おのおのの抽出物に関し、+MEについて、次に0MEについて一回の泳動を行った。レーン(A)(MX感染HeLa細胞由来の細胞培養抽出物)については直接分析を行ったが(10μl/レーン)、その他の抽出物(レーンB−E)由来の抗原については、500μlの抽出物をMAb M75とSACを用いて沈澱させることによりおのおの濃縮した。イムノブロットの結果、2つの他のヒト癌細胞株がMN関連蛋白質を含有することが示された。すなわち、T24(膀胱癌;レーンC)とT47D(乳腺癌;レーンD)である。これらの細胞は、MAb M75と反応し、還元状態では54kdと56kdの分子量を含有し、非還元状態では約153kdの分子量を有する蛋白質を含有している。これらのバンドの強度は、HeLa細胞由来のp54/58N双子蛋白質のそれの少なくとも10分の1以下である。
【0151】
ヒトメラノーマ細胞由来の抽出物(SK−Mel 1477;レーンE)の還元状態では、おおよそ52kdの位置にごく弱いバンドが観察されたが、ヒト線維芽細胞抽出物(レーンB)では、還元状態でも非還元状態でもそのようなバンドはみられなかった。
【0152】
外科標本を含むヒト組織抽出物のイムノブロットをMX感染HeLa細胞由来の細胞抽出物(レーンA)のそれと比較した。他のレーンの組織抽出物は次のものから調製した。(B)満期の胎盤、(C)子宮体部、(D,M)子宮内膜腺癌、(E,N)卵巣腺癌、(F,G)トロホブラスト、(H)正常卵巣、(I)子宮筋腫、(J)乳頭腫、(K)正常乳腺、(L)高増殖性子宮内膜、(O)子宮頚管癌、および(P)メラノーマ。レーンA(10μl/レーンで直接分析した)を除くその他の全ての抽出物由来のMN関連抗原は、上述のように、まず1mlの抽出物から濃縮した。MN蛋白質は、子宮内膜(レーンDとM)、卵巣(レーンEとN)および子宮頚管(レーンO)の癌について見つかった。これらの抽出物においては、MN関連蛋白質は、約48kdから約58kdの間の分子量を有する3本のバンドとして現れた。乳腺癌の組織抽出物中には別のMN関連蛋白質が存在しており、該蛋白質は、約48kdの一本のバンドとして現れた(レーンJ)。
【0153】
満期の胎盤(レーンB)、正常乳腺(レーンK)、高増殖性子宮内膜(レーンL)、正常卵巣(レーンH)、および子宮筋腫(レーンI)からの抽出物では全く何も現れてこなかった。トロホブラスト(レーンFとG)およびメラノーマ(レーンP)からの抽出物においては、ごく弱いMN関連のバンドが現れただけであった。
【0154】
p54/58Nに関連する抗原は、いくつかの型のヒトの癌の臨床標本内では発現するが、対応する器官の正常組織においては発現しないという所見から、MN抗原の腫瘍形成性との関係がさらにはっきりしてきた。しかしながら、ヒト腫瘍においては、腫瘍は成熟した分化後の細胞から生じるものではなく、分岐および分化能を有するある種の基幹細胞から生じると考えられるので、正常細胞が本当に適切な対照となるわけではないことに注意しなければならない。体内器官では、そのような細胞はきわめて稀である。
【0155】
実施例7
動物細胞株のMN抗原
試験した全ての脊椎動物の染色体DNAにはMN遺伝子が存在していたことから、数種の動物種の正常組織由来および腫瘍由来の細胞株についてもMN関連抗原を調べた。MN関連抗原は、二つのラット細胞株から見つかった。一つは、ルイス肉腫ウイルス(Rous sarcoma virus)により誘起されたラット横紋筋肉腫由来のXC細胞株であり、もう一つは、Rat2−Tk−細胞株である。これら両方のラット細胞株の抽出物において、一本の蛋白質のバンドがブロット上に認められた。還元ゲルおよび非還元ゲルにより得られたブロットの分子量は、それぞれ53.5kdおよび153kdであった。
【0156】
MN蛋白質のイムノブロットは、(A)MX感染HeLa細胞、および(B)Rat2−TK−細胞株(上述のように、3%のメルカプトエタノールを含む(+ME)かまたは3%のメルカプトエタノールを含まない(0ME)状態でサンプル緩衝液に加えて加熱した後、PAGEにより分離した)から調製した。これら二つの細胞株内のMN抗原の濃度は、+MEおよび0MEの泳動においてほぼ同じであった。抽出物は直接分析した(40μl/レーン)。
【0157】
XC細胞のMN関連蛋白質は、還元および非還元状態のいずれにおいてもRat2−Tk−細胞のそれと同様のパターンを示したが、濃度は30分の1であった。二つのラット細胞株(上述のイムノブロットおよび実施例4より)においてMN関連蛋白質p53.5Nが発見されたことにより、モデル系の基礎が作られた。
【0158】
MN抗原を濃縮し、高感度イムノブロット法を使用しても、試験した他の動物細胞株にはいずれも検出できる量のMN抗原は含まれていなかった。MNを含有していなかった細胞は次のものである。ベロ(Vero)細胞(アフリカミドリザル);マウスL細胞;マウスNIH−3T3細胞の正常型、サル白血病ウイルス感染型あるいはハーヴェイ肉腫ウイルス(Harvey sarcoma virus)形質転換型;GR細胞(MTVにより誘起されたマウス乳腺腫細胞);およびNMG細胞(正常マウス乳腺)。
【0159】
実施例8
MN特異的抗体およびMN抗原検出のための組換えMN蛋白質を用いた液相ラジオイムノアッセイ
グルタチオンSトランスフェラーゼと融合させ、上述のように調製、精製した遺伝子組換えMN蛋白質、pGEX−3X−MNを、クロラミンT法(ハンター(Hunter)ら、1978年)により125Iでラベルした。精製した蛋白質により、MN特異的抗体ならびにMN抗原の定量RIAを行うことができた。抗体の希釈および抗原の希釈はすべて、1%のウシ胎児血清(FCS)を添加したRIPA緩衝液(1%のトライトンX−100(TRITON X-100)および0.1%のデオキシコール酸ナトリウムのPBS(リン酸緩衝生理食塩水、pH7.2)溶液で行った。組織および細胞の抽出物は、1mMのフッ化フェニルメチルスルフォニルおよび1mlあたり200トリプシン阻害ユニットのトラシロール(Trasylol)(アプロチニン)を添加し、FCSを含まないRIPA緩衝液で調製した。125IでラベルしたpGEX−3X−MN(2.27μCi/μgのTCA沈降活性を有する蛋白質)は、使用前に1%のFCSを含むRIPAで希釈し、非特異結合放射活性をプロテインA固定黄色ブドウ球菌(Staphylococcus aureus)(SAC)の懸濁液で吸着した。 MN特異的抗体に関するRIAにおいては、MAbを含む腹水あるいは試験血清を125Iでラベルした蛋白質と混合し、総量が1mlとなるようにして室温で2時間反応させた。次に、50μlのSACの10%懸濁液(ケスラー(Kessler)ら、同上)を加え、得られた混合物を30分インキュベートした。最後に、SACを沈澱させ、RIPAで3回洗浄し、結合放射活性をγカウンターで測定した。
【0160】
MN抗原に対する抗体の滴定の結果を第3A図−第3B図に示す。M75ハイブリドーマ細胞を含有するマウスの腹水(A)は、1:1.4×10−6希釈において50%終点を有することを示している。同時に、MX蛋白質(M16およびM67)に特異的なMAbsを含有する腹水は、1:200希釈においても125IでラベルしたpGEX−3X−MNとは沈降しなかった。正常ウサギ血清(C)は、MN抗原とははっきりとは沈降しなかった(結果は示していない)。生きたMX感染HeLa細胞で免疫したウサギから得られたウサギ抗MaTu血清(B)は、1:200に希釈したときに放射活性MN蛋白質と7%の沈降を示した。実施例4(上述)に示すようなイムノブロットにより、ウサギ抗MaTu血清は、MXおよびMNのいずれの蛋白質とも沈降を示した。
【0161】
試験した180のヒト血清(90の対照、および、乳房、卵巣、子宮頚管癌の患者からの90の血清)の中からただ一つだけが、放射活性ラベルされた組換えMN蛋白質と明かに沈降した。その血清L8(D)をイムノブロット(実施例4)で再試験したが、MX感染HeLa細胞由来のp54/58Nのいずれとも沈降しなかった。さらに、KH(E)を含む6つの他のヒト血清もイムノブロットで反応しなかった。つまり、RIAでただ一つ反応したヒト血清L8は、遺伝子組換え産物とのみ反応し、HeLa細胞で発現した、生来のp54/58Nとは反応しなかった。
【0162】
MN抗原に対するRIAにおいては、MAb M75の希釈液(事前の試験で50%の最大沈降放射活性(=1:1.4×10−6希釈)を示した)を細胞抽出物の希釈物と混合し、2時間反応させた。次に、125IでラベルしたpGEX−3X−MN(25×103cpm/チューブ)を加え、さらに2時間反応させた。最後に、MAb M75に結合した放射活性をSACで沈降させ、上述のように洗浄した。100%の沈降(=阻害なし)を、用いた希釈MAbによる最大放射活性結合と見なした。試験に用いた細胞抽出物中のMN抗原の濃度は、「コールド(cold)」のpGEX−3X−MNを標準として使用した阻害曲線から計算した(pGEX−3X−MNを標準として使用した阻害曲線から計算した(第4図のA)。
放射活性ラベルしたpGEX−3X−MN蛋白質とMAb M75との反応から、細胞抽出物中のMN抗原を直接的に定量することができた。第4図は、3ngの「コールド」のpGEX−3X−MN(A)が、「ホット(hot)」のpGEX−3X−MNと50%の沈降阻害を起こしたことを示すものである。同量のMN抗原が、MaTu感染HeLa細胞(B)およびRat2−Tk−細胞(C)から抽出した3×103ngの蛋白質にも存在している。本実験のRIAで測定した細胞抽出物中の蛋白質の濃度を下記の表1に示す。細胞抽出物中のMN抗原は、ある程度の異なる大きさであるということ、また、遺伝子操作されたMN蛋白質は、さまざまな大きさの分子を含有するものであることから、計算値は絶対的なものではないことに注意されたい。
【表1】

【0163】
実施例9
MX抗原のRIP
p58X蛋白質のおよその濃度は、MaTu感染HeLa細胞の抽出物のRIPにより求めることができ、ここで、該抽出物は、[35S]メチオニンあるいは[14C]アミノ酸混合物で代謝的にラベルしたものである。これらの結果を表2に示す。
【表2】
【0164】
表2に示す結果から、p58Xは、細胞抽出物中の蛋白質のおよそ0.8%を占めることがわかった。
【0165】
ラベルしたアミノ酸と共に終夜培養した培養細胞および並行培養(アミノ酸を完全に補充した「コールド」の培地でさらに24時間インキュベートしたもの)の培養細胞からも非常に近似した値が得られた。これらの結果は、得られた放射活性の値が既に平衡状態にあることを反映したものであり、取り込みの速度を表したものではないことを示しており、該値は、抽出物中のp58Xの実際の含量と大差はない。[35S]メチオニンでラベルした細胞の抽出物のp58Xの値は、[14C]アミノ酸混合物でラベルした培養の抽出液内のそれと同程度であった。
【0166】
実施例10
対照およびMX感染HeLa細胞の免疫電子および走査顕微鏡法
上記の実施例1に示したように、MAb M75を用いた直接免疫蛍光法により検出されたMN抗原は、MX感染HeLa細胞の膜表面および核内あるいは高密度培養で増殖させたHaLa細胞内に存在している。MN抗原の所在をさらに明らかにさせるために、免疫電子顕微鏡法を用いた。該方法においては、MN抗原に結合したMAb M75を免疫金(immunogold)ビーズで視覚化した(ヘルゾーグ(Herzog)ら、「細胞表面域判定のためのコロイド金ラベリング法(Colloidal gold labeling for determining cell surface area) 」コロイド金(Colloidal Gold)第3巻(ハヤット(Hayat),M.A.編)より、139−149ページ(アカデミックプレス(Academic Press)社、米国、カリフォルニア州、サンディエゴ)参照)。
【0167】
MX非感染(対照)およびMX感染HeLa細胞の超薄片を、免疫金を含むMAb M75および含まないMAb M75で染色した。(いくつかの細胞は、包埋し、切断する前にM75と免疫金で固定し、処理した。この操作により、免疫金装飾が細胞表面の抗原に対してのみ施される。いくつかの細胞は、包埋し、切断した後にのみM75と免疫金で処理した。こうすると、細胞内の抗原も装飾される。免疫金で処理しなかったいくつかの細胞は、細胞分裂の末期のものである)。
【0168】
染色した細胞の免疫電子および走査顕微鏡像から、細胞内のMN抗原の局在が示され、さらに、対照およびMX感染HeLa細胞とでは微細構造にかなりの違いがあることが見出された。対照のHeLa細胞は、その表面に、金ビーズで視覚化されたMN抗原をほとんど有していないことが示された。細胞表面は比較的滑らかで、2つの小さい突起を有するだけであった。細胞質内にはミトコンドリアは見られなかった。これとは対照的に、MX感染HeLa細胞では、表面にたくさんの密集した糸状突起がみられた。ほとんどのMN抗原はこれらの糸状体の上に局在しており、該抗原は、染色に免疫金を用いた場合には免疫金によって修飾されていた。MX感染HeLa細胞の細胞質には多くのミトコンドリアを含んでいるのが見られた。MN抗原は、MX感染HeLa細胞の核にあった。MN抗原は核質の中にいくらか存在していたが(おそらくクロマチンと結合しているのであろう)、より高濃度のMN抗原は核小体に存在していた。繰り返すと、正常HeLa細胞の細胞表面は比較的滑らかであり、一方、MX感染HeLa細胞の表面には多数の糸状体と「水疱(blebs)」がある。糸状体のいくつかは、近隣の細胞とつながる橋を形成しているように見える。
【0169】
インビトロ(in vitro)で形質転換された細胞とそれらの正常親細胞を比較したいくつかの例において注目すべきことは、相違点の一つが、正常細胞の表面は滑らかであるが、形質転換された細胞の表面には多数の毛髪状の糸状体を有するということである(ダーネル(Darnell)ら、「分子細胞生物学(Molecular Cell Biology)」(第2版)、サイエンスアメリカンブックス(Sci. Am. Books)、W.H.フリーマン(Freeman)株式会社、米国、ニューヨーク州(1990年))。これらの特徴からすると、本実施例の顕微鏡像で明らかにされたように、MX感染HeLa細胞は超形質転換した外観を呈している。
【0170】
さらに、数種の腫瘍では、ミトコンドリアの増幅が報告されている(ベーンハード(Bernhard)、W.,「分子細胞学ハンドブック(Handbook of Molecular Cytology)」、687−715ページ、リマ・デ・ファリア(Lima de Faria)編、ノースホランド(North-Holland)出版株式会社、アムステルダム−ロンドン(1972年))。そのような増幅は、ヤーヌス緑(Janus′ green )で特にミトコンドリアが強く染色されたMX感染HeLa細胞において顕著であり、一方、対照のHeLa細胞では弱くしか染色されなかった。
【0171】
電子顕微鏡の熟練者によっても、腫瘍細胞に特異的な構造的特徴は見出すことができなかったことを付記しておく。
【0172】
実施例11
MN遺伝子発現を阻害するアンチセンスODNs
p54/58N蛋白質の両方が一つの遺伝子によってコードされているか否かを確認するために、アンチセンスODNsを用いて以下の実験を行った。過密培養を作るために、事前に希薄培養していたHeLa細胞をまき、MN mRNAの5’末端に相補的で2つの遺伝子に特異的なODNsの非存在下および存在下で130時間インキュベートした。HeLa細胞は、10%のFCSを含むDMEM1mlあたり8×105個となるように継代した。同時に、次のようにODNsを培地に添加した。(A)29−mer ODN1(5’CGCCCAGTGGGTCATCTTCCCCAGAAGAG3’(配列番号3)、44−72位に相補的)を4μM(最終濃度)、(B)19−mer ODN2(5’GGAATCCTCCTGCATCCGG3’(配列番号4)、12−30位に相補的)を4μM(最終濃度)、(C)CDN1およびODN2をおのおの2μM(最終濃度)、(D)同様に処理したが、ODNsを加えずにインキュベートしたものであり、対照とする。130時間後に細胞から抽出物を調製し、125IでラベルしたMAbs M75を用いたイムノブロットにより分析した。細胞の蛋白質抽出物は、MAbs M75を用いたイムノブロットおよびRIAにより分析した。 ODNsを添加したHeLa細胞の培養の結果、p54/58N合成がかなり阻害されていることがわかった。最終濃度4μMの19−mer ODN2が非常に効果があり、RIAで測定したところ、40%の阻害を起こした。一方、29−mer ODN1(4μM)および2つのODNsの組合せ(おのおの最終濃度2μM)では効果は低くなり、RIAで23−35%のMN関連蛋白質の減少を示した。このとき、特異的MAb H460を用いたRIAで測定した、別のHeLa細胞蛋白質の量は、全ての細胞変異種においてほぼ同程度であった。最も重要なことは、イムノブロットにおけるODNsによる特異的阻害は、p54/58N蛋白質の両方に影響を与えていることである。このことから、発明者らがクローンしたMN遺伝子は、HeLa細胞内においてp54/58N蛋白質の両方をコードしていると結論づけられる。
【0173】
実施例12
腫瘍形成性細胞由来および非腫瘍形成性細胞株由来のMN mRNAのノーザンブロッティング
ヒト細胞株のMN mRNAのノーザンブロッティングを行った。全RNAは、次の細胞株からグアニジンチオシアネートCsCl法により調製した。高密度(A)および希薄(B)培養で増殖したHeLa細胞;(C)CGL1(H/F−N)ハイブリッド細胞;(D)CGL3分離体および(E)CGL4分離体(両者ともH/F−T)、ならびに(F)ヒト胚線維芽細胞。15μgのRNAを1.2%のホルムアルデヒドゲルで分離し、ハイボンドCスーパーメンブレン(Hybond C Super membrane)(アマシャム(Amersham)社)の上にブロットした。MN cDNAのNotIプローブを、ランダムプライミング(マルチプライムDNAラベリングシステム(Multiprime DNA labelling system)、(アマシャム(Amersham)社)を用いてラベルした。50%のホルムアミド存在下42℃でハイブリダイゼーションを行い、0.1%のSSPEと0.1%のSDSを用いて65℃で最終洗浄を行った。RNAラダー(RNA ladder)(0.24−9.5kb)(ベセスダ研究所(BRL)(Bethesda Research Laboratories)、米国、メリーランド州、ベセスダ)をサイズマーカーとして用いた。
【0174】
検出されたのは1.5kbのMN特異的mRNAで、2つの腫瘍形成性分離体クローン、CGL3とCGL4(H/F−T)からのみであり、非腫瘍形成性ハイブリッドクローンCGL1(H/F−N)あるいは正常ヒト線維芽細胞からは検出されなかった。さらに、1.5kbのmRNAは、高密度培養で増殖したHeLa細胞からは検出されたが、希薄培養のものからは検出されなかった。
【0175】
このように、MN関連蛋白質が腫瘍形成性と関連しているということについてノーザンブロッティングの結果は、上述の実施例の結果と一致している。
【0176】
実施例13
各種の脊椎動物種由来のゲノムDNAのサザンブロッティングによるMN遺伝子の検出
いろいろな脊椎動物のゲノムDNA中のMN遺伝子をサザンブロッティングにより検出した。次のものの染色体DNAをSstIで消化した。(A)ニワトリ、(B)ウシ、(C)ネコ、(D)MX感染HeLa細胞、(E)マウスNIH 3T3細胞、(F)ヒト胎盤細胞、(G)HeLa細胞、(H)ヒツジ、(I)ヒトメラノーマ細胞、および(J)サルベロ(Vero)細胞。制限酵素による断片を、0.7%の寒天ゲルで分離し、ハイボンドNメンブレン(Hybond N membrane)(アマシャム(Amersham)社)の上にアルカリブロットした。MN cDNAプローブのラベリングおよびハイブリダイゼーションの操作は、実施例12のノーザンブロット分析と同様に行った。
【0177】
染色体DNAのSstI制限酵素断片を用いたサザンブロット分析は、いずれの種においても、約1.5kbのところに唯一のバンドを示した。さらに、XhoIおよびSalIによる切断によって生じた制限酵素断片を用いたハイブリダイゼーションにおいては、おのおのの染色体DNAサンプルは4.5kbおよび4.7kbにそれぞれ1本のバンドを示した。これらの結果から、脊椎動物のゲノムにおいては、MN遺伝子は単一のコピーとして存在していることが示唆される。さらに、これらの結果は、MN遺伝子がその側面配列と共に保存性であることを示唆している。
【0178】
SstIの開裂部位がMN遺伝子における自然な境界を形成し、MN mRNAのサイズがノーザンブロットにおけるMN遺伝子のサイズと同じであることから(実施例12の結果と比較されたい)、MN遺伝子にはイントロンが存在しないと推論される。この結論は、MN cDNAの制限酵素パターンとMN特異的ゲノムのSstI断片のそれとが同じであることからも支持される。
【0179】
下記に挙げる材料は、アメリカンタイプカルチャーコレクション(American Type Culture Collection)(ATCC)(米国、メリーランド州、ロックヴィル、パークローン通り(Parklawn Drive)12301番地、郵便番号20852)に寄託されている。寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約(Budapest Treaty on the International Recognition of Deposited Microorganisms for the Purposes of Patent Procedure and Regulations thereunder)(ブダペスト条約)に基づいてなされた。生細胞培養の維持は寄託の日から30年間保障されている。微生物は、ブダペスト条約の規定に基づいてATCCから入手可能であり、寄託者とATCCは、関連する米国特許が付与されると制限されることなく入手できることについて合意している。寄託した株が入手可能できるとは言っても、ある政府機関がその国の特許法に基づいて許可する権利に違反して発明を実施する実施権を与えるということではない:
ハイブリドーマ 寄託日 ATCC番号
VU−M75 1992年9月17日 HB 11128
【0180】
本発明に関する以上の具体例の記述は、例示および説明のためのものである。これらの具体例は全てではなく、また、開示された特定のものに発明を限定するものでもない。さらに、以上の教示から多くの修正や変形が可能なことは明かである。以上の具体例は、本発明の要旨を説明するため、および、当業者が意図する特定の使用に適するようにさまざまな修正を加えた各種の態様で本発明を利用できるようにするために、選択して記載したものである。本発明の範囲は、明細書に添付された請求項によって定められる。
【0181】
引用した全ての文献は、参考のために本明細書に取り入れられている。
【技術分野】
【0001】
本発明は医学遺伝子学の一般的領域に属し、生化学工学および免疫化学の分野に属する。より詳細には、本発明は新規な遺伝子であるMN遺伝子(該遺伝子はMN蛋白質をコードする細胞性遺伝子である)の同定に関する。本発明者らは、MN蛋白質が腫瘍形成に関与していることを発見した。患者の試料中からMN抗原、さらに該抗原に特異的な抗体を同定することは、癌の診断/予後の分析の基礎を提供する。
【背景技術】
【0002】
珍しい特性を有する新規な疑似ウイルス体が、ヒト胸腺癌細胞と共培養したHeLa細胞[ヒト子宮頚管腺癌由来の細胞株]内において、熱不安定性表面G蛋白質を有する水疱性口内炎ウイルス(vesicular surface virus, VSV)変異体を補足する能力により検出された(非特許文献1から5を参照)。該疑似ウイルス体は、おそらくヒト乳腺腫瘍(mammary tumor )から得られたことからMaTuと呼ばれる。
【0003】
MaTuを研究し、特性を明らかにすることに医学的に大きな興味がひかれた。MaTuは生細胞への全く新しい型の寄生分子と思われ、また、ヒト腫瘍細胞由来の可能性があるからである。本明細書には、MN遺伝子およびMNタンパク質を発見するに至った、MaTuの生物学的および分子学的性質を記述している。本発明者らは、MaTuが2つのコンポーネント、すなわち、外因性の輸送コンポーネントMX、および内因性の細胞性コンポーネントMNからなることを発見した。本明細書に記載しているように、MNコンポーネントは細胞性遺伝子として発見され、既知のDNA配列とは相同性(ホモロジー)がほとんどない。MN遺伝子は試験した全ての脊椎動物の染色体DNA内に存在することが見出されており、該遺伝子の発現は腫瘍形成性に強く関与していることが明らかになっている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】ザバダ(Zavada)ら、Nature New Biol., 240 :124(1972)
【非特許文献2】ザバダ(Zavada)ら、J. Gen. Viol., 24 :327(1974);
【非特許文献3】J.ザバダ(Zavada)、 Arch. Viol., 50 :1(1976);
【非特許文献4】J.ザバダ(Zavada)、J. Gen. Viol., 63 :15-24(1982)
【非特許文献5】J.ザバダ(Zavada)、Arch. Viol., 118 :189(1991)
【発明の概要】
【発明が解決しようとする課題】
【0005】
本明細書に記述しているのは、バクテリアベクター内におけるMN遺伝子のクローニングと配列決定、およびMN遺伝子にコードされた蛋白質の産生についてである。このように遺伝子工学的に処理されたMN蛋白質は、他のMN蛋白質/ポリペプチドと同様に、本発明に従い、MN特異的抗体を検出する血清学的分析に使用することができる。さらに、MN抗原と反応するそのようなMN蛋白質/ポリペプチドと抗体は、本発明に従い、MN抗原を検出および/または定量する免疫学的検定(イムノアッセイ)に使用することができる。このような分析は、腫瘍性および/または前腫瘍性の疾患の診断および/または予後に利用される。
【課題を解決するための手段】
【0006】
本明細書は、MaTuの内因性コンポーネントである細胞性遺伝子、すなわち、MN遺伝子について開示している。イントロンを有しないと考えられる該遺伝子の実質的な全cDNA配列は、第1A図−第1B図[配列番号1]に示す。
【0007】
本発明は、該MN遺伝子、その断片およびそれに関係するcDNAに関し、これらは、たとえば次のように有用である。1)生化学工学によりMN蛋白質/ポリペプチドを産生。2)被検材料の細胞内のMN遺伝子の存在を確認するための核酸プローブの調製。3)適切なポリメラーゼ連鎖反応(PCR)プライマーの調製。これは、たとえば、PCRに基づく分析や核酸プローブの産生などに使用する。4)MN蛋白質およびポリペプチド、ならびにそれらと相同あるいはほぼ相同なポリペプチドの同定。5)各種の組織および細胞株に存在するMN遺伝子から転写されたさまざまなmRNAの同定。6)MN遺伝子の突然変異の同定。本発明はさらに、MN遺伝子またはその断片、あるいは関連するcDNAまたはその断片からなる、精製単離されたDNAに関する。
【0008】
さらに、本発明は、これまで未知であった蛋白質MN(MN遺伝子によりコードされている)の発見に関する。MN蛋白質の発現は、高密度培養中において成長細胞により誘導され、そのような発現は腫瘍形成性細胞と関与していることが見出された。
【0009】
MN蛋白質は、インビトロ(in vitro)でいくつかのヒト腫瘍細胞株において産生されることがわかっている。たとえば、HeLa(子宮頚管癌由来)、T24(膀胱癌由来)およびT47D(乳腺癌由来)およびSK−Mel 1477(メラノーマ由来)などの細胞株、腫瘍形成性ハイブリッド細胞など。また、インビボ(in vivo)でいくつかのヒト癌細胞においても産生される。たとえば、子宮頚部細胞、卵巣および子宮内膜癌細胞、さらに乳頭腫などのようないくつかの良性腫瘍細胞など。MN蛋白質は、非腫瘍形成性ハイブリッド細胞や正常組織の細胞においては見出されない。このことから、MN蛋白質は腫瘍特異的であると考えられる。
【0010】
HeLa細胞株および腫瘍形成性HeLa細胞と線維芽細胞とのハイブリッド(H/F/T)細胞株においては、MN蛋白質は「双子(twin)」蛋白質p54/58Nとして表現される。該蛋白質は、グリコシル化され、ジスルフィド結合しているオリゴマーの形をとっている。還元性ゲルを用いた電気泳動によって測定すると、MN蛋白質は約40Kdから約70Kdの範囲の分子量を有しており、好ましくは約45Kdから約65Kdの範囲、より好ましくは約48Kdから約58Kdの範囲の分子量を有しているのがよい。非還元性のゲルにおいては、オリゴマー型のMN蛋白質は約145Kdから約160Kdの範囲の分子量を有しており、好ましくは約150Kdから約155Kdの範囲、より好ましくは約152Kdから約154Kdの範囲の分子量を有しているのがよい。本発明における好ましいMN蛋白質の推定アミノ酸配列は第1A図−第1B図に示す。
【0011】
MN遺伝子とMN蛋白質およびそれらにコードされた実質的に相補性のMN遺伝子とMN蛋白質の発見により、MN蛋白質の発現が腫瘍形成性と関連していることが見いだされた。この発見の結果、癌および前癌状態の診断/予後の方法が確立された。脊椎動物、好ましくはホ乳類、より好ましくはヒトの細胞と組織抽出物を含んだ患者の試料中からMN抗原を検出および/または定量することにより、腫瘍性疾患の発症や存在を同定ための方法や材料が提供される。そのようなMN抗原は体液中からも検出される。
【0012】
MN蛋白質およびMN遺伝子は、癌の診断/予後における腫瘍形成の分子メカニズムの解明の研究に利用されており、また、癌の免疫療法に応用され得る。
【0013】
本発明は、広範な腫瘍性および/または前腫瘍性疾患の検出に有効である。腫瘍性疾患の例としては、乳腺、膀胱、卵巣、子宮、子宮頚管、子宮内膜、偏平細胞および腺偏平癌などの腫瘍;頭および首の癌;神経芽細胞腫および網膜芽腫などの中胚葉性腫瘍;骨肉腫およびユーイング肉腫(Ewing′s sarcoma)などの肉腫;およびメラノーマが挙げられる。特に興味深いものとしては、頭および首の癌、卵巣、子宮頚管、腟、子宮内膜および陰門の癌を含む婦人科の癌;胃、結腸および食道の癌などの胃腸系の癌;膀胱および腎臓の癌などの尿路系の癌;皮膚癌;肝臓癌;前立腺癌;肺癌および乳癌がある。中でも特に興味があるのは、婦人科の癌;乳癌;尿路系の癌、ことに膀胱癌;肺癌;胃、結腸および食道の癌などの胃腸系の癌;および肝臓癌である。さらにとりわけ興味が強いのは、婦人科の癌および乳癌である。婦人科の癌の中で特に関心があるのは子宮頚管、子宮内膜および卵巣の癌であるが、子宮頚管偏平細胞腫瘍、腺偏平細胞腫瘍、腺腫などを含む婦人科の癌と同様に、後形体子宮頚管組織やコンジロームなどの婦人科の前癌状態も非常に興味深い。
【0014】
本発明はさらに、MN遺伝子、その断片あるいは関連するcDNAを生化学的に処理することに関する。たとえば、該遺伝子またはその断片、あるいは関連するcDNAは適切な発現ベクターに組込まれ、宿主細胞がそのような発現ベクターに形質転換され、MN蛋白質/ポリペプチド、好ましくはMN蛋白質がその中で発現する。そのような組換え蛋白質あるいはポリペプチドは、グリコシル化されているかまたはいないかであるが、好ましくはグリコシル化されているのがよく、ほぼ純粋の状態に精製され得る。本発明はさらに、合成的にあるいはその他の生化学的手法で調製されたMN蛋白質/ポリペプチドにも関する。
【0015】
該MN蛋白質/ポリペプチドは、患者の試料中のMN抗原を検出する分析、あるいはMN特異的抗体を調べる血清学的検定に用いることができる。本発明のMN蛋白質/ポリペプチドは、血清学的に活性で、免疫原性があり、および/または抗原性がある。該MN蛋白質/ポリペプチドは、さらに、MN特異的抗体(ポリクローナルおよび/またはモノクローナル)を産生させたり、T細胞免疫応答を起こす免疫原として用いることもできる。
【0016】
さらに本発明は、MN特異的抗体に関するものであり、該抗体は診断/予後に用いられ、また、治療に用いることもできる。MN特異的抗体は次のような用途に用いられる。たとえば、免疫蛍光顕微鏡や免疫組織化学染色による実験室での解析、臨床サンプル中のMN抗原の検出および/または定量のための免疫測定法の構成分の一つとして;MN抗原を検出するイムノブロット法のプローブ;細胞内のMN蛋白質および/またはポリペプチドの局在を示す金コロイドビーズを用いた免疫電子顕微鏡法;およびMN遺伝子、その断片あるいは関連するcDNAをクローニングする遺伝子工学など。そのようなMN特異的抗体は、たとえばインビトロ(in vitro)での組織片を用いた診断および/または予後用のキットの構成材料として用いられる。そのような抗体はまた、たとえば、適切な放射活性同位体を用いて抗体を適切にラベルし、インビボ(in vivo)での抗体の局在の転移をシンチグラフィーで追跡することにより、インビボ(in vivo)での診断/予後に用いることができる。さらに、そのような抗体は、毒性因子および/または細胞増殖抑制因子をそれらに結合させて、あるいはさせずに、インビボ(in vivo)療法として癌患者の加療に使用することもできる。また、そのような抗体は、腫瘍性および/または前腫瘍性疾患の存在を検出するためにインビボ(in vivo)で用いることができる。さらに、そのような抗体はMN蛋白質およびポリペプチドのアフィニティー精製に使用することができる。
【0017】
典型的なMN特異的抗体であるモノクローナル抗体M75を産生するハイブリドーマは、1992年9月17日にATCC番号HB 11128としてATCC(American Type Culture Collection,米国、メリーランド州、ロックヴィル)に寄託された。このM75抗体は、MN蛋白質を発見、同定するために用いられたものであるが、ホルマリン固定された組織サンプルから、ウエスタンブロット、ラジオイムノアッセイおよび免疫組織化学的方法により、MN抗原を簡単に検出するために用いることができる。
【0018】
本発明はさらに、MN蛋白質またはポリペプチドをコードするDNA配列を有する組換えDNA、ならびに、MN蛋白質またはポリペプチドをコードするのみならず非MN蛋白質またはポリペプチドのアミノ酸配列をもコードする組換えDNAに関する。該アミノ酸配列はヒトに免疫原性を有しないこと、およびヒト体液中の抗体に一般的な反応性を有しないことが望ましい。そのようなDNA配列の例としては、β−ガラクトシダーゼのα−ペプチドコード領域、およびグルタチオンS−トランスフェラーゼをコードする配列とその断片などが挙げられる。さらに、実質的に純粋であり、天然には存在しないそのような組換え融合タンパク質/ポリペプチドも本発明に含まれる。本発明の融合タンパク質の例としてはpGEX−3X−MNがある。
【0019】
本発明は、また、腫瘍性疾患および/または前腫瘍性疾患の治療方法に関し、該方法は、MN遺伝子から転写されたmRNAと実質的に相補的であるアンチセンス核酸配列を投与することにより、MN遺伝子の発現を抑制することからなる。アンチセンス核酸配列は、第1A図−第1B図に示すように、MN cDNAの5’末端において実質上相補的なものであることが好ましい。このアンチセンス核酸配列とは、オリゴヌクレオチドであることが好ましい。
【0020】
本発明はまた、実質的に純粋な一つもしくはそれ以上のMN蛋白質および/またはポリペプチドを免疫原量として十分に含むワクチンに関する。MN蛋白質および/またはポリペプチドは生理学的に許容性で非毒性の基剤に分散し、MN蛋白質の発現と関係している腫瘍性疾患に対して、脊椎動物、好ましくはホ乳類、より好ましくはヒトに十分な免疫効果を上げる量を使用する。該蛋白質は、組換え、合成あるいは他の生化学的手法によりつくられる。組換えMN蛋白質としては、pGEX−3X−MNなどのような融合蛋白質が挙げられる。該ワクチンの特徴的な使用法として、再発および/または転移の阻止がある。たとえば、MN関与性腫瘍を外科的に切除した患者に腫瘍の再発を防ぐために該ワクチンを投与することができる。
【0021】
本発明はさらに、MN遺伝子の核酸配列と実質的に相補的な核酸プローブに関する。本発明における好ましい核酸プローブとは、その配列が、第1A図−第1B図に示すように、MN cDNAの配列に実質的に相補的なものである。本発明に従う試験キット(テストキット)は、腫瘍性および/または前腫瘍性疾患の診断/予後に有効なプローブから構成されている。好ましい試験キットは、前記プローブとMN遺伝子またはMN遺伝子のmRNA産物とのハイブリダイゼーションを視覚化などにより検出あるいは測定する手法により構成されている。
【0022】
本発明に従うイムノアッセイは、MN蛋白質/ポリペプチドおよび/またはMN特異的抗体からなる試験キットとして具体化される。そのような試験キットは、固相状態であるが、それに限定されるわけではなく、液相状態であってもよく、非増幅あるいはアビジン/ビオチン法などを用いて増幅した状態で、ELISA法、粒子アッセイ、放射測定あるいは蛍光測定アッセイなどに基づくものである。
【0023】
略語表
本明細書内で使用する略語は以下の通りである:
AA −アミノ酸
ATCC −アメリカン タイプ カルチャー コレクション(American Type Culture Collection)
bp −塩基対
BSA −ウシ血清アルブミン
Ci −キュリー
cm −センチメーター
cpm −カウント/分
C-末端 −カルボキシル末端
℃ −摂氏
DMEM −ダルベッコ変法イーグル培地(Dulbecco modified Eagle medium)
EDTA −エチレンジアミン四酢酸
EIA −酵素免疫測定法、エンザイムイムノアッセイ
ELISA −酵素免疫吸着測定法
F −線維芽細胞
FCS −ウシ胎児血清
FIBR −線維芽細胞
FITC −フルオレセインイソチオシアネート
H −HeLa細胞
HEF −ヒト胚線維芽細胞
HeLa K −標準型HeLa細胞
HeLa S −スタンブリッジ(Stanbridge)変異HeLa細胞 D98/AH.2
H/F−T −ハイブリッドHeLa線維芽細胞(腫瘍原性)、HeLa細胞 D98/AH.2由来
H/F−N −ハイブリッドHeLa線維芽細胞(非腫瘍原性)、HeLa細胞 D98/AH.2由来
HGPRT− −ヒポキサンチングアニンフォスフォリボシルトランスフェラーゼ欠損
HRP −西洋ワサビ(ホースラディッシュ)ペルオキシダーゼ
IPTG −イソプロピル−β−D−チオガラクト−ピラノサイド
kb −キロベース
kd −キロダルトン
M −モル濃度
mA −ミリアンペア
MAb −モノクローナル抗体
ME −メルカプトエタノール
MEM −最少必須培地
mg −ミリグラム
ml −ミリリットル
mM −ミリモル濃度
MTV −乳腺腫瘍ウイルス
N −規定濃度
ng −ナノグラム
N-末端 −アミノ末端
ODN −オリゴデオキシヌクレオチド
PAGE −ポリアクリルアミドゲル電気泳動
PBS −リン酸緩衝生理食塩水
PEST −プロリン(proline)、グルタミン酸(glutamic acid)、生理食塩水(serine)、スレオニン(threonine)の頭文字の組合せ
pI −等電点
RIP −放射性免疫沈降法
RIPA −放射性免疫沈降測定法
SAC −プロテインA(黄色ブドウ球菌(Staphylococcus aureus)由来)
SDS −ドデシル硫酸ナトリウム
SDS−PAGE −ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動
SSPE −NaCl(0.18M)、リン酸ナトリウム(0.01M)、EDTA(0.001M)
TCA −トリクロロ酢酸
TC培地 −組織培養培地
μCi −マイクロキュリー
μg −マイクログラム
μl −マイクロリットル
μM −マイクロモル濃度
VSV −水疱性口内炎ウイルス(vesicular surface virus)
X−MLV −異種マウス白血病ウイルス
【0024】
細胞株(セルライン)
本明細書に記述している実験例において使用した細胞株を以下に示す:
HeLa K −標準型HeLa細胞,異数倍数性、上皮様の細胞株、ヒト子宮頚管腺腫より単離され(ゲイ(Gey)ら、Cancer Res., 12: 264(1952)、ジョーンズ(Jones)ら、Obstet. Gynecol., 38: 945-949(1971)参照)、B.コリシュ教授(Korych)(チャールス大学(Charles University)医学微生物学および免疫学研究所(Insutitute of Medical Microbiology and Immunology)、チェコスロバキア、プラハ)より入手
HeLa D98/AH.2(またはHeLa S) −変異HeLaクローンであり、ヒポキサンチングアニンフォスフォリボシルトランスフェラーゼ欠損(HGPRT−)、エリック J.スタンブリッジ(Eric J. Stanbridge)(カリフォルニア大学医学部微生物学科(Department of Microbiology, College of Medicine, University of California)、米国、カリフォルニア州、アーヴァン)より供与され、スタンブリッジ(Stanbridge)らにより報告されている(Science, 215: 252-259(1982年1月15日号))。ハイブリッド細胞H/F−Nの親株も同じくE.J.スタンブリッジ(Stanbridge)から入手
NIH−3T3 −マウス線維芽細胞株、アーロンソン(Aaronson)により報告されている(Science, 237: 178(1987))
T47D −ヒト乳腺癌由来の細胞株(ケイダー(Keydar)ら、Eur. J. Cancer, 15: 659-670(1979))。J.ケイダー(Keydar)(ハダッサー医科大学(Haddasah Medical School)、イスラエル、エルサレム)より供与
T24 −膀胱癌由来の細胞株(ブベニク(Bubenik)ら、Int. J. Cancer, 11: 765-773(1973))。J.ブベニク(Bubenik)(チェコスロバキア科学アカデミー分子遺伝学研究所(Insutitute of Melecular Genetics, Czechoslovak Academy of Sciences)、チェコスロバキア、プラハ)より供与
HMB2 −メラノーマ由来の細胞株(スヴェク(Svec)ら、Neoplasma, 35: 665-681(1988))
HEF −ヒト胚線維芽細胞(ザバダ(Zavada)ら、Nature New Biology, 240: 124-125(1972))
SIRC −ウサギ角膜由来の細胞株(対照およびX−MLV感染)(ザバダ(Zavada)ら、Virology, 82: 221-231(1977))
ベロ細胞(Vero cells) −アフリカミドリザル由来の細胞株(ザバダ(Zavada)ら、1977年)
ミエローマ細胞株NS−0 −モノクローナル抗体の産生において融合親細胞として用いられるミエローマ細胞株(ガルフレ(Galfre)とミルシュテイン(Milstein)、Methods Enzymol.,73: 3-46(1981) )
SK−Mel 1477 −ヒトメラノーマ細胞株。K.E.ヘルストロン(Hellstrom)(フレッド・ハトキンス癌研究センター腫瘍免疫部門(Division of Tumor Immunology, Fred Hutchins Cancer Research Center)、米国、ワシントン州、シアトル)より供与
XC −ラット横紋筋肉腫由来の細胞であり、ラウス肉腫(Rous sarcoma)ウイルスによって誘導された誘導ラット肉腫(スヴォボダ(Svoboda),J.、国立癌センター研究所モノグラフ第17巻(Natl. Cancer Center Institute Monograph No.17)より「鳥類腫瘍ウイルスに関する国際学会(International Conference on Avian Tumor Viruses)」(J.W.ベアード(Beard)編)、pp.277-298(1964年)。ジャン スヴォボダ(Svoboda)(チェコスロバキア科学アカデミー分子遺伝学研究所(Insutitute of Melecular Genetics, Czechoslovak Academy of Sciences)、チェコスロバキア、プラハ)より供与
Rat2−Tk− −チミジンキナーゼ欠損細胞株。L.クチノヴァ(Kutinova)(血清およびワクチン研究所(Institute of Sera and Vaccines)、チェコスロバキア、プラハ)より供与
CGL1 −H/F−Nハイブリッド細胞(HeLa D98/AH.2起源)
CGL2 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
CGL3 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
CGL4 −H/F−Tハイブリッド細胞(HeLa D98/AH.2起源)
【0025】
ヌクレオチドおよびアミノ酸配列記号
本明細書では、以下の記号をヌクレオチドを表す記号として使用する:
塩 基 記 号
アデニン A
シトシン C
グアニン G
チミン T
ウラシル U
【0026】
主要アミノ酸は20個あり、それらのおのおのは3個の隣接するヌクレオチド(三つ組(トリプレット)コードまたはコドン)の異なる組合せによって特定されており、さらに、特徴的な蛋白質を形成するために特定の順番で結合される。本明細書においては、該アミノ酸を表記するために、たとえば第1A図−第1B図に示すような3文字記号を使用する。それらを以下に示す:
アミノ酸名 記 号
アラニン Ala
アルギニン Arg
アスパラギン Asn
アスパラギン酸 Asp
システイン Cys
グルタミン酸 Glu
グルタミン Gln
グリシン Gly
ヒスチジン His
イソロイシン Ile
ロイシン Leu
リジン Lys
メチオニン Met
フェニルアラニン Phe
プロリン Pro
セリン Ser
スレオニン Thr
トリプトファン Trp
チロシン Tyr
バリン Val
【図面の簡単な説明】
【0027】
【図1A】図1Aは、本明細書に記載しているようにMN cDNAクローンから単離されたヌクレオチド配列およびcDNAによってコードされている推定アミノ酸配列[それぞれ配列番号1および2]を表している。シークエンスデータはEMBLデータライブラリー(EMBL Data Library)(ドイツ、ハイデルベルグ)に送付済みであり、受入番号X66839として利用可能である。
【図1B】図1Bは、図1Aの続きである。
【図2】図2(AおよびB)は、ヒト線維芽細胞(F)、HeLa細胞(H)およびH/F−NとH/F−Tハイブリッド細胞内におけるMN特異的蛋白質およびMX特異的蛋白質の発現をグラフに表したものであり、MX感染細胞およびMX非感染細胞における発現を対比させている。実施例5に操作と結果について詳しく記載している。
【図3】図3(AおよびB)は、125I−pGEX−3X−MN蛋白質およびいろいろな抗体を用いた放射免疫沈降実験の結果をグラフに表したものである(実施例8に記述)。放射活性蛋白質(15×103cpm/チューブ)は、次の腹水あるいは血清およびSACと沈降した。(A)MAb M75を含む腹水;(B)ウサギ抗MaTu血清;(C)正常ウサギ血清;(D)ヒト血清L8;(E)ヒト血清KH;(F)ヒト血清M7。
【図4】図4は、MN抗原に対するラジオイムノアッセイの結果を示す(実施例8に記述)。腹水(沈降が50%の放射活性となるように希釈)は、次のものと2時間反応させた。(A)「コールド(cold)」(ラベルしていない)蛋白質pGEX−3X−MN;あるいは以下の細胞の抽出物(B)HeLa+MX;(C)Rat−2TK−;(D)HeLa;(E)ラットXC;(F)T24および(G)HEF。次に、125IでラベルしたpGEX−3X−MN蛋白質(25×103cpm/チューブ)を加え、さらに2時間インキュベートした。最後に、放射活性MAb M75をSACに吸着させ、測定した。
【発明を実施するための形態】
【0028】
MaTu−−MXおよびMNコンポーネント
本明細書に記述するように、MaTuは2コンポーネントシステムからなっている。コンプレックスの一つの部分である外因性MXは、透過性であり、蛋白質p58Xを発現する。該蛋白質は、ヒトおよびさまざまな動物の天然の血清と反応する細胞質抗原である。もう一つのコンポーネントであるMNは、ヒト細胞に内因するものである。
【0029】
MNは細胞性遺伝子であり、既知のDNA配列とはほとんど相同性がない。MNは保存性であり、多くの脊椎動物の染色体DNA内で単一のコピー遺伝子として存在している。本明細書には、MN cDNAのクローニングおよび配列決定、また融合蛋白質(MN+グルタチオンS−トランスフェラーゼのC末端からなる:アフィニティークロマトグラフィーにより容易に精製できる)の遺伝子工学的処理について記述している。
【0030】
MNはHeLa細胞において双子蛋白質p54/58Nとして表現され、細胞表面および核内に局在している。p54/58Nに反応するモノクローナル抗体(MAb M75)を用いたイムノブロットにおいて54kdと58kdの2本のバンドが確認できる。これらの2本のバンドは、一種類の蛋白質に関して、グリコシル化のパターンあるいはプロセッシングの過程が異なることによるものかもしれない。(p54Nおよびp58Nは、両方ともマンノースを含むオリゴ糖残基によりグリコシル化されているが、p58Nだけはグルコサミンも含有している。)本明細書で使用する「双子蛋白質(twin protein)」とはp54/58Nを指している。
【0031】
MNは、HeLa細胞の急速成長する希薄培養中では出現しないが、細胞を高密度培養中で維持することにより、あるいは、さらに効率的な手段として、細胞にMXを感染させることにより誘導される。p54/58NのみがMaTuに感染したHeLa細胞内で再生された水疱性口内炎ウイルス(vesicular surface virus, VSV)のビリオンに結合する。双子蛋白質p54/58Nはグリコシル化され、ジスルフィド結合で連結されたオリゴマーの形をとっているのに対し、p58Nはグリコシル化されておらず、ジスルフィド結合で連結されたオリゴマー形ではない。
【0032】
VSVは、HeLa細胞内でp54/58Nをビリオン中に集めるが、このことは、該双子蛋白質がVSVのG蛋白質変異型の相補性と、VSV(MaTu)のプソイド型の形成に関与していることを示唆している。エンベロープを有するウイルスのみが、感染性で機能的なプソイド型を形成するための表面糖蛋白質を産生し、ビリオンの細胞への吸着および侵入という特殊な機能を発揮することができる(ザバダ(Zavada), J., J. Gen. Virol.,63: 15-24(1982))。このことはMN遺伝子が疑似ウイルスの配列として行動することを示している。
【0033】
エンベロープを有するウイルスの表面蛋白質は、VSVのプソイド型の形成に関与しており、MN双子蛋白質p54/58Nと同様に、グリコシル化されている。また、MN蛋白質は、オリゴマー(好ましくは三量体または四量体)の形成においてはウイルスの糖蛋白質と類似している。そのようなオリゴマー形成においては、S−S結合(ジスルフィド結合)の関与は必須ではないが、ビリオンの集積には必須である(クレイス(Kreis)とロディッシュ(Lodish), Cell, 46: 929-937(1986))。ジスルフィド結合は2−メルカプトエタノールを用いて還元することにより分裂する。
【0034】
パストレコヴァ(Pastorekova)らにより報告されているように(Virology, 187: 620-626(1992))、メルカプトエタノールを用いて還元した後は、細胞抽出物あるいはVSV由来のp54/58Nはイムノブロットにおいて非常に類似している。還元を行わない場合、細胞抽出物のp54/58Nは150kd付近に複数のバンドとして現れ、このことは細胞が数種の異なるオリゴマー(おそらくp54:p58の比率が異なる)を含んでいることを示唆している。しかし,VSVにおいては選択的にその中の一つ、分子量約153kdに集まっている。該オリゴマーは三量体もしくは四量体であり、54kdおよび58kdの蛋白質からなっている。還元状態でのVSVサンプルの分析において54kdと58kdのバンドの強さがほぼ同等であることから、VSVビリオンにおいてはp54:p58が等モル比であることが示された。
【0035】
MN蛋白質の発現は、腫瘍性疾患の診断/予後にみられる。MN双子蛋白質p54/58Nは、HeLa細胞およびスタンブリッジ(Stanbridge)の腫瘍形成性(H/F−T)ハイブリッド細胞において発現が確認されているが(スタンブリッジ(Stanbridge)ら、Somatic Cell Genet, 7: 699-712(1981)およびスタンブリッジ(Stanbridge)ら、Science, 215: 252-259(1982) )、線維芽細胞あるいは非腫瘍形成性(H/F−N)ハイブリッド細胞においては確認されていない(スタンブリッジ(Stanbridge)ら、同上)。ヒト卵巣、子宮内膜および子宮頚管癌、またいくつかの良性腫瘍(乳頭腫など)を用いて行ったイムノブロットにおいてはMN蛋白質が確認されるが、正常な卵巣、子宮内膜、子宮あるいは胎盤の組織からは確認されない。MXに感染したHeLa細胞内では、微細構造の交替が顕著に行われており、このことは、細胞表面でのおびただしい糸状体の形成およびミトコンドリアの増幅を意味している。免疫金(immunogold)標識法を用いると、p54/58Nは糸状体の表面および核、特に核小体上で観察される。すなわち、MN蛋白質は正常非腫瘍細胞では産生されていないことから、腫瘍特異的であるといえる。
【0036】
本明細書の実施例において、MNおよびMXは二つの異なる存在であり、互いに独立して存在することが示されている。外因性の透過性物質であるMXは、線維芽細胞あるいは非腫瘍形成性(H/F−N)ハイブリッド細胞において増殖するが、これらの細胞はMN関連蛋白質は発現しない(第2A図−第2B図)。そのような細胞内ではMXはMN蛋白質の産生を誘導しないのである。第2A図−第2B図および実施例5と6に示されるように、MXの非存在下でもMN蛋白質はHeLa細胞および他の腫瘍細胞内で産生される。しかしながら、MXはHeLa細胞内におけるMN蛋白質の強力な誘導剤である。非感染細胞内において、MXはMN蛋白質の産生を濃度にして30倍増加させた(下記の実施例5と8、実施例8の表1参照)。
【0037】
MN遺伝子−−クローニングおよび塩基配列決定
第1A図−第1B図は、本項に記載されている方法に従って単離されたMN cDNAクローンの塩基配列を示している。遺伝子コドンの縮重から、一つのコドンが一つ以上のアミノ酸をコードしており(たとえば、TTA、TTG、CTT、CTC、CTAおよびCTGはいずれもロイシン(leu)というアミノ酸をコードしている)、また、たとえば第1A図−第1B図に示すように、一つのコドンが他のコドンと入れ替わるヌクレオチド配列の多様性により、本発明と実質的に同等な蛋白質およびポリペプチドが産生される。MN cDNAのヌクレオチド配列および相補的な核酸配列に関するそのような変形もすべて本発明の範ちゅうに含まれる。
【0038】
さらに、本明細書に記述し、第1A図−第1B図に示しているヌクレオチド配列は、単離され、本明細書で説明しているcDNAヌクレオチド配列のうち、はっきりした構造のみを表したものである。わずかに変更されたヌクレオチド配列が見つかることもあろうし、また、たとえば、同様のエピトープを有する等の、実質的に同等なMN蛋白質およびポリペプチドをコードするように当該分野で知られた技術により変形することも可能である。そしてそのような蛋白質/ポリペプチドは本発明の目的に適合する。MN蛋白質/ポリペプチドと相同あるいはほぼ相同な蛋白質/ポリペプチドをコードする合成核酸配列のように、同等なコドンを有するDNAおよびRNAは本発明の範ちゅうに含まれる。遺伝子コードの縮重がなければ、これらの核酸配列はやはり前記cDNAヌクレオチド配列にハイブリダイズする。本明細書で説明しているように、核酸配列が修飾されたり変形される結果、MN配列およびその断片と実質的に同等の配列が作り出される。
【0039】
MN遺伝子を見つけるために、MX感染HeLa細胞からλgt11によるcDNAライブラリーを調製した。MX感染HeLa細胞からの全RNAはグアニジンチオシアネートCsCl法を用いて抽出し、mRNAはオリゴdTセルロースを用いるアフィニティーにより分離した。cDNAの合成およびgt11へのそのクローニングはアマシャム(Amarsham)社のキットを用いて行ったが、EcoRI−NotIアダプターだけはストラタジーン(Stratagene)社(米国、カリフォルニア州、ラ・ホーラ(La Jolla))のものを使用した。モノクローナル抗体M75とアルカリフォスファターゼを縮合したヤギ抗マウス抗体とを組み合わせたイムノスクリーニングにライブラリーをかけた。このイムノスクリーニング法は、ヤング(Young)とデイヴィス(Davis)により報告されている(PNAS(USA), 80: 1194-1198(1983))。350,000のプラーク(全ライブラリーのおおよそ半分にあたる)をスクリーニングし、1個のポジティブクローンを取り出した。
【0040】
ポジティブクローンをpBluescript KS(ストラタジーン(Stratagene)社)のNotI部位に組み込んでサブクローニングを行い、pBluescript-MNを作った。Erase-a-BaseTMキット(プロメガ(Promega)社、米国、ウィスコンシン州、マディソン)を使用して、方向が反対で重なる2個の欠失を作り、T7シークエンス用キット(ファルマシア(Pharmacia)社、米国、ニュージャージー州、ピスカタウェイ)を用いてジデオキシ法により配列を決定した。配列はcDNAクローンの一部を表しており、インサートの長さは1397bpであった。本配列を第1A図−第1B図に示す(配列番号1)。配列は、大きな1290bpのオープンリーディングフレームおよびポリAシグナル(AATAAA)を含む107bpの3’非翻訳領域からなっている。該配列のもう一つの特徴は、mRNAの不安定性に関与する領域(1389番目のAUUUA)が存在することである。この領域は、ある種の腫瘍遺伝子およびリンホカインのmRNAに特異的なものである(ショウ(Shaw)とカーメン(Kamen), Cell, 46: 659-667(1986))。MNクローンの大きさと対応するmRNAのそれとをノーザンブロットにより比較すると(実施例12)、このcDNAは、その配列の5’末端から約100bpが欠損していることがわかった。 MN cDNAクローンのオープンリーディングフレームは、約48kdの推定蛋白質をコードしている(第1A図−第1B図、配列番号2)。推定翻訳アミノ酸(AA)配列の分析では、既報の蛋白質の配列とは高い相同性を示さなかった。最も近い相同性が見られたのは、MN蛋白質およびいろいろな型の炭酸脱水素酵素のC末端である(170−200AAの重なりにおいて約30−35%)。炭酸脱水素酵素の活性部位は、Zn2+結合ドメインと同様に、MN蛋白質においてもよく保存されている。しかしながら、MN遺伝子は、ヒトゲノム由来の新規な配列であることは明かである。
【0041】
上述したように、MN遺伝子は既知の炭酸脱水素酵素といくらかの相同性を有するが、いくつかの面でそれらとは異なっている。7個の炭酸脱水素酵素が報告されている(ドッジソン(Dodgson)ら(編)、炭酸脱水素酵素(The Carbonic Anhydrases)、(プレナムプレス(Plenum Press)社、ニューヨーク/ロンドン(1991年))。それらのおのおのは7個のイントロンを含んでいるが、MN遺伝子はイントロンを含まないようである。また、既知の炭酸脱水素酵素はすべておよそ30kdの蛋白質であり、これらはMN遺伝子のp54/58N関連の産生物より小さい。さらに、炭酸脱水素酵素はMN関連蛋白質のようなオリゴマーを形成しない。
【0042】
推定アミノ酸配列から、MN遺伝子の産生物は、303−313のアミノ酸位置に一つの活性なN−グリコシル化部位を有する塩基性蛋白質(pI9.08)であることが明かである。これらの事実は、HeLa細胞由来のp54/58N蛋白質が、Endo HおよびEndo Fによる分裂(おのおの約3kdの欠損を起こす)に感受性であることに対応している。親水性プロフィルは、アミノ酸の親水性配列(371−395位)を示しており、これはプラズマ膜にかかる領域を表していると考えられ、また、分裂シグナルも含むと考えられる。該プロフィルは、p54/58N蛋白質が細胞膜に局在していることとよく一致する。MNアミノ酸配列にはPEST領域は存在しないことから、MN遺伝子の産生物は安定で永続性の蛋白質であることが示唆される(ロジャース(Rogers)ら、Science, 234: 364-368(1986))。そのような特性から、発明者らのp54/58Nの代謝ラベルが非効率的であったことを説明できる。推定アミノ酸配列はさらにほかの特徴をも示す。すなわち、10個の活性リン酸化部位と7個のミリスチル化部位および3個の抗原決定因子を有する。
【0043】
p54/58N蛋白質の両方が一つの遺伝子によってコードされているか否かを確認するために、MN遺伝子の発現を特異的に阻害するアンチセンスODNsを用いた。(このようなアンチセンスODNsの使用法については、ステイン(Stein)とコーエン(Cohen)により総説されている(Cancer Res., 48: 2659-2668(1988))。)これらの実験については実施例11に詳述している。実験の結果、ODNsと共に培養したHeLa細胞においてはp54/58Nの合成がかなり阻害されていることがわかり、一方、いろいろなHeLa細胞の蛋白質の産生量はほぼ同程度に保たれていた。さらに、イムノブロットにおいて重要な結果が得られた。すなわち、ODNsによる特異的阻害はp54/58N蛋白質のの両方に影響を与えていた(実施例11)。これらのことから、MN遺伝子はHeLa細胞においてp54/58N蛋白質の両方をコードしていると結論づけられる。
【0044】
クローニングされた遺伝子がp54/58N特異的蛋白質をコードしているか否かを確認するために、該遺伝子をバクテリア発現ベクターpGEX−3X(ファルマシア(Pharmacia)社、スゥエーデン、ウプサラ)にサブクローニングし、グルタチオンS−トランスフェラーゼのC末端を有する融合蛋白質を発現するように構築した。このサブクローニングは、本発明におけるMN関連蛋白質の遺伝子工学的手法の一つを示すものである。以下の記述は例示であり、如何なる意味においても本発明を限定するものではない。
【0045】
融合蛋白質pGEX−3X−MNの産生
上述のpBluescript-MN由来のcDNA挿入体(インサート)は、プラスミドDNAをNotIで消化(切断(digesting))することにより切出した。該cDNA挿入体は、平滑末端を得るためにS1ヌクレアーゼで処理し、pGEX−3X(ファルマシア(Pharmacia)社)の脱リン酸化したSmaI部位にクローニングした。XL1-Blueの形質転換およびIPGTによる誘導の結果、融合蛋白質が得られた。
【0046】
融合蛋白質であるMNグルタチオンS−トランスフェラーゼは、グルタチオンS−セファロース4B(ファルマシア(Pharmacia)社)を用いたアフィニティークロマトグラフィーにより精製した。10%ゲルを用いたSDS−PAGEにより分離された2つの同様なサンプルから、20ミリグラムの精製された組換え蛋白質が得られた。一つのサンプル(A)はクマジーブリリアントブルーで染色し、他方のサンプル(B)はハイボンドCメンブレン(Hybond C membrane)(アマシャム(Amarsham)社、英国、バックス、アリスバリー)にブロットした。このブロットは、125IでラベルしたMAb M75を用いてオートラジオグラフィーにより展開した。
【0047】
SDS−PAGE分析から興味深い結果が得られた。すなわち、異なる分子量を有する一連の蛋白質のバンドが存在することである。本発明に従って産生された別の融合蛋白質である、β−ガラクトシダーゼMN(λ gt11溶原)においても同様なDSD−PAGEパターンが得られた。MN配列内に9個のAGGAGGコドンタンデムが存在することによる翻訳エラーのために、これらのパターンが現れたものと思われる。対応するtRNAが短いため、バクテリア遺伝子内でこれらのコドンを使用することは絶対に避けられている。かくして、外来性mRNAからのAGGAGGタンデムの翻訳の際に、+1のリボソーム上でのフレームシフトが高い頻度(約50%)で起こる(スパンヤード(Spanjaard)ら、Nuc. Acid Res., 18: 5031-5036(1990))。
【0048】
イムノブロッティングにおいて、同様なパターンが得られた。すなわち、染色されたSDS−PAGEゲル上で確認された全てのバンドは、MN特異的MAb M75に反応し、このことは、全ての蛋白質のバンドがMN特異的であることを示している。また、これらの結果から、MAb M75への結合部位は、フレームシフトの影響を受けないMN蛋白質のN末端部分にあることが示された。
【0049】
下記の実施例8に示すように、融合蛋白質pGEX−3X−MNは、MN特異的抗体およびMN抗原のラジオイムノアッセイに使用した。
【0050】
MN蛋白質および/またはポリペプチド
本明細書で用いている「MN蛋白質および/またはポリペプチド」(MN蛋白質/ポリペプチド)とは、MN遺伝子あるいはその断片によりコードされている蛋白質および/またはポリペプチドを意味している。好ましいMN蛋白質の例は、推定アミノ酸配列が第1A図−第1B図に示されているものである(配列番号2)。好ましいMN蛋白質/ポリペプチドは、第1A図−第1B図に示すMN蛋白質と実質的に相同性を有する蛋白質/ポリペプチドである。
【0051】
「ポリペプチド」とは、ペプチド結合によるアミノ酸の共有結合鎖のことであり、本明細書では、50あるいはそれ以下のアミノ酸から構成されるものと考えている。本明細書における「蛋白質」とは、50より多くのアミノ酸から構成されるポリペプチドと定義される。
【0052】
インビボ(in vivo)の腫瘍細胞から産生される蛋白質/ポリペプチドの配列が細胞培養内の腫瘍細胞から産生される蛋白質/ポリペプチドのものと異なることがある。すなわち、MN蛋白質/ポリペプチドが、アミノ酸置換、伸張、欠損、削除およびそれらの組合せ(これらに限定されるわけではないが)のようなアミノ酸配列変化を有していても、それらはすべて本発明の範ちゅうに属する。体液中に残存する蛋白質は蛋白質分解などの分解処理を受けることがある。すなわち、血清などの体液中にはかなりの削除が行われたMN蛋白質およびMNポリペプチドが見いだされる。本明細書で使用している「MN抗原」とは、MN蛋白質/ポリペプチドを包含している。
【0053】
さらに、MN蛋白質およびポリペプチドのアミノ酸配列は、遺伝子工学によって変化させることもできる。1個またはそれ以上のアミノ酸を削除したり置換することができる。そのようなアミノ酸の変化も、生物学的活性に有意の変化をもたらさず、本発明の範囲に含まれる蛋白質やポリペプチドを生じさせることができる。
【0054】
本発明のMN蛋白質およびポリペプチドは、本発明の方法にしたがって、さまざまな手段で調製できる。たとえば、組換え、合成、あるいはその他の生物学的手法、すなわち、長い蛋白質およびポリペプチドを酵素および/または化学的に解裂する等の方法が挙げられる。MN蛋白質を調製する好ましい方法は組換え法である。組換えによるMN蛋白質の産生のために特に好ましい方法は、融合蛋白質pGEX−3X−MNに関して上述した方法である。
【0055】
MN蛋白質およびポリペプチドの組換え産生
第1A図−第1B図に示すMN蛋白質およびその断片を調製する代表的な方法は、適切なMN cDNA断片を上で例示したような適切な発現ベクターに挿入することである。本明細書に記載しているように、単離されたMN DNAのクローニングには、広範な種類の宿主−クローニングベクターの組合せを用いることができる。たとえば、有用なクローニング媒体としては次のようなものを挙げることができる。染色体DNA、非染色体DNA、合成のDNAであって、たとえば、pBR322等の各種の既知のバクテリアプラスミド、その他の大腸菌(E. Coli)プラスミドおよびそれらの誘導体、ならびに広い宿主範囲のプラスミド、たとえば、RP4やファージDNA(たとえば、NB989等のλファージの多数の誘導体)、ファージDNAの発現をコントロールする配列を有する修飾プラスミド等のプラスミドとファージDNAの組合せから作られたベクターなど。プラスミドpGEX−3Xが好ましいクローニング媒体である。
【0056】
宿主として有用な細胞は真核性でも原核性でもよく、次のようなものが例示される。バクテリア宿主、たとえば、大腸菌(E. Coli)とその他のバクテリア株、酵母およびその他の菌類等、動物または植物の培養細胞等の動物あるいは植物宿主、昆虫細胞およびその他の宿主。もちろん、全ての宿主が同じ効果を有するわけではない。本明細書に記載されている原則を考慮しながら本発明の範囲から逸脱しないように、当業者によって宿主−クローニング媒体の組合せの選択がなされる。
【0057】
組換えDNA分子を形成するため、選択されたDNA断片をクローニング媒体へ挿入する特定部位の決定は、さまざまな因子に影響される。これらの因子としては、発現させる蛋白質およびポリペプチドのサイズと構造、所望する蛋白質およびポリペプチドの宿主細胞の成分による内部酵素分解に対する感受性、宿主細胞蛋白質によるコンタミネーション、開始および終了コドンの局在などの発現特性、および当業者により知られているその他の因子などが挙げられる。
【0058】
MN遺伝子、その断片あるいはMN遺伝子由来のcDNAを含む組換え核酸分子は宿主の形質転換に用いられるが、その形質転換により、宿主(形質転換体)にその構造蛋白質および断片を発現させ、かつ、ハイブリッドDNAがコードしている蛋白質およびポリペプチドを産生させる。組換え核酸分子はまた、MN核酸およびその断片の源としてさらに組換え核酸分子を複製生産させるために宿主の形質転換に用いられる。これらのそれぞれの使用に対する適切な宿主の選択は、当該分野で知られている多くの因子により定められている。これらの因子としては、たとえば、選択したベクターとの適合性、共生産物の毒性、所望する蛋白質およびポリペプチドの回収のしやすさ、発現特性、生物学的安全性およびコストが挙げられる。
【0059】
宿主細胞として大腸菌(E. Coli)などの原核細胞を用いる場合は、DNAを取り込むことのできるコンピテントな細胞は、指数増殖期の後に細胞を回収(ハーベスト)し、続いて既知の手法であるCaCl2法で処理することにより調製できる。宿主細胞のプロトプラストを作った後、形質転換を行うことができる。
【0060】
宿主細胞として原核細胞を用いる場合には、リン酸カルシウム沈降法、エレクトロポレーション、マイクロインジェクション等の従来からの機械的手法、赤血球あるいはリポソームに封入したプラスミドの挿入、リゾフォスファチジルコリン等の薬剤を用いた細胞処理、あるいはウイルスベクターの使用などの方法がとられる。
【0061】
蛋白質およびポリペプチドの産生量は3つの大きな因子に左右される。(1)細胞内の遺伝子またはDNA配列のコピー数、(2)該遺伝子および配列が転写、翻訳される効率、および(3)mRNAの安定性。転写、翻訳(これらが共同して発現を司る)の効率は、核酸配列、一般には所望のコード配列の前に位置している核酸配列に依存する。これらの核酸配列、すなわち発現コントロール配列は、とりわけ、転写を開始するためにRNAポリメラーゼが相互作用する位置(プロモーター配列)、および翻訳を開始するためにリボソームがmRNA(転写産物)と結合し、相互作用する位置を定めている。そのような全ての発現コントロール配列が同じ効果を発揮するわけではない。そのため、所望する蛋白質に特異的なコード配列を近傍の核酸配列から切り出し、その代わりに既知の発現コントロールベクターに融合して希望する高レベルの発現を得られるようにするのがよい。上述の操作を行ってから、処理されたこの新しいDNA断片が多数のプラスミドまたはバクテリオファージ誘導体に挿入され、細胞内で遺伝子または配列のコピー数を増し、さらに、発現蛋白質の収量を増加させる。
【0062】
発現コントロール配列として数種のものが用いられる。これらの中には、オペレーター、プロモーター、大腸菌(E. Coli)のラクトースオペロン(lac 系)のリボソーム結合および相互作用配列、大腸菌(E. Coli)のトリプトファン合成系(trp 系)に対応する配列、trpとlacプロモーターの融合(tac 系)、λフ
ァージの主要オペレーターとプロモーター(OLPLとORPR)、およびfdファージのコート蛋白質のコントロール領域などが含まれる。これらの配列を含むDNA断片は、lacあるいはtrpオペロンを有する形質導入ファージのDNA、またはλあるいはfdファージのDNAから制限酵素を用いた開裂により切出せる。つぎに、これらの断片を操作して、必須コントロール配列が、コード配列の開始コドンの非常に近傍または並列に位置しているようになった限定された分子集団を得る。
【0063】
融合体は次に、適切な宿主の形質転換あるいはトランスフェクションのためのクローニング媒体に挿入し、抗原産生量を測定する。そして、最も効率的な発現をする細胞を選択する。別の方法としては、開始コドンに接続されたlac、trpまたはλPLコントロール系を有するクローニング媒体を用い、MN蛋白質およびポリペプチドをコードする配列を含む断片に融合し、それによって遺伝子あるいは配列がクローニング媒体の開始コドンから正しく翻訳されるようにすることもできる。
【0064】
本明細書で使用している「組換え核酸分子」とは、少なくとも2つの核酸配列からなるハイブリッドヌクレオチド配列を指しており、ここで、第一の配列は、通常、自然状態では第二の配列と共存していない。
【0065】
本明細書で使用している「発現コントロール配列」とは、構造遺伝子と結合した場合にその発現を制御または調整するヌクレオチド配列のことである。
【0066】
MN蛋白質およびポリペプチドの合成および生物工学的発現
本発明のMN蛋白質およびポリペプチドは組換え法のみでなく、合成およびその他の生物学的手法によっても調製することができる。蛋白質またはポリペプチドの合成による産生は、当該分野でよく知られた方法に従って所望するアミノ酸鎖を化学的に合成していくことからなる。所望するポリペプチドまたは蛋白質を調製するためのその他の生物学的手法の例としては、所望するアミノ酸配列を含む長いMNポリペプチドまたは蛋白質を選択的に蛋白分解することが挙げられる。たとえば、長いポリペプチドまたは蛋白質を化学試薬または酵素で分解することなどである。
【0067】
ペプチドの化学合成は従来技術であり、たとえば、メリフィールド(Merrifield)固相合成法(メリフィールド(Merrifield),J., Am. Chem. Soc., 85: 2149-2154(1963)、ケント(Kent)ら、「生物学および医学における合成ペプチド(Synthetic Peptides in Biology and Medicine)29ページ」、アリターロ(Alitalo)ら編、エルセヴィール科学出版(Elsevier Science Publishers)社、1985年、およびハーグ(Haug),J.D.「ペプチド合成および保護基戦略(Peptide Synthesis and Protecting Group Strategy)」, American Biotechnology Laboratory, 5(1): 40-47(1987年、1/2月号)などにより実施される。
【0068】
化学的ペプチド合成の方法には、市販の保護アミノ酸を用いる自動ペプチド合成機の使用も含まれる。合成機としては、たとえばバイオサーチ(Biosearch)社(米国、カリフォルニア州、サンラファエル)の9500型および9600型、アプライド バイオシステムズ(Applied Biosystems)社(米国、カリフォルニア州、フォスターシティー)の430型、ミリジェン(Milligen)社(ミリポア(Millipore)社の子会社、米国、マサチューセッツ州、ベッドフォード)の9050型、およびデュポン(DuPont)のRAMP(高速自動複数ペプチド合成機)(デュポンコンパス(Dupont Compass)社、米国、デラウェア州、ウィルミントン)などがある。
【0069】
核酸プローブおよび試験キット
本発明の核酸プローブは、第1A図−第1図Bに示すMN cDNA配列またはMN遺伝子配列と実質的に相補的な配列からなる。本明細書で使用する「実質的に相補的」という語は、当該技術分野で広く理解されている意味と同じであり、それゆえ、通常のハイブリダイゼーションの状態の意味で用いる。ハイブリダイゼーションの程度は、相補正の精度に応じて変わり得る。
【0070】
前記プローブは、MN DNAおよび/またはRNAの検出に使用でき、したがって、患者の細胞内のMN遺伝子の存在や欠如、増殖、変異、あるいは遺伝子再配列の試験に使用できる。たとえば、MN遺伝子の過剰発現は、本発明のプローブを使用したノーザンブロットにより検出できる。増幅、転座、逆位ならびに欠失などの遺伝子変化は本発明のプローブを用いることにより検出でき、このとき、該プローブは、細胞分裂中期の染色体の広がった状態でも間期の核のいずれの状態であっても、患者の細胞由来の染色体とインサイチュー(in situ)ハイブリダイゼーションする。また、本発明のプローブを用いたサザンブロットによってMN遺伝子の増幅あるいは欠失を検出することもできる。該プローブを用いた制限酵素断片長多型(RFLP)分析は、遺伝子変化、変異および欠失の検出として好ましい方法である。該プローブはまた、いろいろな組織由来のMN遺伝子から転写された各種のmRNAとハイブリダイゼーションすることにより、MN蛋白質および/またはポリペプチドならびにそれらと相同またはほぼ相同な蛋白質および/またはポリペプチドを検出するために用いられる。
【0071】
このように、該プローブは診断/予後に有用である。該プローブは試験キットとして具体化でき、好ましくは、該プローブが適切なMN遺伝子またはMN mRNAターゲットとハイブリダイゼーションした時に、視覚化できる適切な手段を伴っているのがよい。そのような試験のサンプルとしては、組織標本、体液ならびに組織および細胞の抽出物が挙げられる。
【0072】
分析
本発明に従う分析は、脊椎動物のサンプル、好ましくはホ乳類のサンプル、より好ましくはヒトのサンプル中のMN抗原またはMN特異的抗体の検出および/または定量である。そのようなサンプルとしては、組織標本、体液、組織抽出物および細胞抽出物が挙げられる。MN抗原は、イムノアッセイ、免疫組織学的染色、免疫電子および走査顕微鏡観察、とりわけ、免疫金沈降(immunogold)を用いる技術により検出できる。
【0073】
MN抗原分析の好ましいサンプルは組織および/または細胞抽出物である(下記の実施例7と8が代表的なものである)。しかし、MN抗原は体液、とりわけ、血液、血清、プラズマ、精液、乳汁、唾液、涙、喀痰、粘液、尿、リンパ液、サイトゾル、腹水、胸水、羊水、膀胱洗浄液、気管支肺胞洗浄液、随液からも検出できる。試験前に大量の体液からMN抗原を濃縮することが望ましい。分析に好ましい体液は、試験する癌の型にもよるが、一般的に好ましい体液は、乳汁、胸水および腹水である。
【0074】
血液、プラズマ、血清、リンパ液、粘液、涙、尿、髄液および唾液などの体液サンプル中の活性MN蛋白質/ポリペプチドにMN特異的抗体は血清学的に結合するが、そのような抗体は血液、プラズマおよび血清、好ましくは血清に普通にみられる。MN特異的抗体の検出のための代表的な分析は、下記の実施例8に示すものであり、その実施例では融合蛋白質pGEX−3X−MNを用いている。MN抗原およびそれらと反応するMN特異的抗体の検出および/または定量試験の結果の相関から、患者の病状の好ましいプロファイルが示される。
【0075】
本発明の分析は、診断および/または予後の両方、すなわち、診断/予後である。本明細書で使用している「診断/予後」とは、臨床的状況に依存して以下の手順のそれぞれ、または、それらの手順のいくつかが重複していることを意味する。疾病の存在の判断、疾病の特性の判断、ある疾病と他の疾病との区別、病状の帰結予想、患者の様子と症状から示される疾病からの回復の見込み、患者の病状のモニタリング、疾病の再発に関する患者のモニタリング、および/または患者に対する好ましい治療方法の決定など。本発明における診断/予後の方法はたとえば以下のような場合に有用である。腫瘍性および/または前腫瘍性疾患の存在に対する集団のスクリーニング、腫瘍性疾患の進展の危険性の判断、腫瘍性および/または前腫瘍性疾患の存在の診断、腫瘍性疾患の患者の病状のモニタリング、および/または腫瘍性疾患の経過に対する予後の判断など。
【0076】
本発明は、広範な種類の腫瘍性疾患の存在のスクリーニングに有用である。たとえば、乳腺、尿路、卵巣、子宮、子宮頚管、子宮内膜、偏平細胞および腺偏平細胞などの腫瘍;頭および首の癌;神経芽細胞腫および網膜芽種などの中胚葉性の腫瘍;骨肉腫およびユーイング肉腫(Ewing′s sarcoma)などの肉腫;あるいはメラノーマなどが含まれる。特に興味があるのは婦人科の癌であり、この中には卵巣、子宮、子宮頚管、膣、陰門、子宮内膜癌が含まれるが、中でもとりわけ興味があるのは卵巣、子宮頚管および子宮内膜の癌である。同様に特に興味があるのは、胸、食道を含む胃、結腸、腎臓、前立線、肝臓、膀胱を含む尿路系、肺、および頭と首の癌である。
【0077】
本発明は、たとえば、宿主から取出した直後の細胞群を使用して、悪性腫瘍あるいは前悪性腫瘍性細胞の存在の可能性を推し量るための方法および構成物を提供する。そのような分析は、腫瘍の発見、それらの増殖の計測および疾病の診断と予後に役立つ。この分析はまた、癌の転移の存在の発見、ならびに、手術、癌の化学療法および/または放射線療法の後、全ての腫瘍組織がなくなっているか除去されているかを確認するのに役立つ。さらに、診断は、癌の化学療法および腫瘍の再発をモニターするのに役立つ。
【0078】
MN抗原または抗体の存在は、多くの既に確立された診断分析を用いて検出および/または定量することができる。当業者であれば、従来から存在する任意のイムノアッセイ法を適用して、MN抗原および/または抗体を検出および/または定量することができる。実施例8に本発明の好ましい診断方法であるラジオイムノアッセイ法について詳しく述べている。もちろん、MN抗原およびMN特異的抗体の検出にはほかの多くの方式も利用可能である。たとえば、ウエスタンブロット、ELISA(酵素結合免疫吸着法)、RIA(ラジオイムノアッセイ)、競合EIA、二重抗体サンドイッチ法、その他診断研究において通常使用される全ての分析法などがある。そのようなイムノアッセイにおける結果の解釈は、抗体あるいは抗体の組合せは、MNに関係のないサンプル中に存在する他の蛋白質および蛋白質の断片とは交差反応しないという仮定に基づいている。
【0079】
MN抗原検出の代表的な一つのELISA試験は、MN蛋白質/ポリペプチドに対して作られた抗体あるいはMN蛋白質を発現する細胞全体に対して作られた抗体をコートしたマイクロタイタープレートに、組織あるいは細胞抽出物のような患者のサンプルを加える方法である。ある抗原が抗体と結合するように一定時間インキュベートした後、プレートを洗浄し、酵素が連結している別の抗MN抗体を加え、反応が生じるようにインキュベートし、プレートを再洗浄する。その後、マイクロタイタープレートに酵素基質を加え、酵素が基質に作用するように一定時間インキュベートし、最終試料の吸光度を測定する。吸光度が大きく変化することは陽性の結果を示している。
【0080】
患者の体液、組織および/または細胞中のMN抗原の存在を検出および/または定量するためにMN蛋白質/ポリペプチドが使用できることもイムノアッセイの分野の当業者には明かである。そのような実施態様の一つとして競合イムノアッセイがあるが、この方法では、MN蛋白質/ポリペプチドはラベルされ、MN蛋白質/ポリペプチドに特異的な抗体への該ラベル化MN蛋白質/ポリペプチドの結合に競合させるために体液を加える。そのような分析は実施例8に記載しているようなMN抗原の検出および/または定量に使用できる。
【0081】
別の実施例としては、MN蛋白質またはポリペプチドに対するラベルされた抗体を用いるイムノメトリック分析がある。そのような分析においては、抗原結合抗体とコンプレックスを形成するラベル化抗体の量は、試料中のMN抗原の量と直接的に比例する。
【0082】
MN特異的抗体を検出する代表的な分析は競合分析であり、この分析では、ラベルされたMN蛋白質/ポリペプチドはサンプル中の抗体、たとえば、MN蛋白質/ポリペプチドを認識するモノクローナル抗体と結合して沈降する。当業者であれば、従来から存在する任意のイムノアッセイ方法を適用してMN抗原の検出および/または定量することができる。該抗体のMN蛋白質/ポリペプチドとの結合の検出は、当業者に知られた多くの方法、たとえば、ヒトにおいては抗ヒトラベルIgGの使用などにより行うことができる。
【0083】
本発明に従い、脊椎動物のサンプルからMN抗原を検出および/または定量するイムノアッセイの方法の例は、以下のステップから構成されている:
a)MN抗原に結合する一組あるいはそれ以上の組の抗体(一つの抗体かあるいは複数の抗体)(そのうち一組はラベルされているかまたはその他の手法で検出可能となっている)と脊椎動物のサンプルをインキュベートする;
b)MN抗原と該抗体からなる免疫コンプレックスが存在するかについて、インキュベートしたサンプルを調べる。
【0084】
本発明に従う別のイムノアッセイ法の例は競合イムノアッセイであり、脊椎動物サンプル中のMN抗原を検出および/または定量するために使用され、以下のステップから構成されている:
a)脊椎動物のサンプルを一組あるいはそれ以上の組の抗体および一定量のラベルされたもしくは他の手段で認識できるようになっているMN蛋白質/ポリペプチドとインキュベートし、このとき、該MN蛋白質/ポリペプチドが抗体との結合において、サンプル中に存在するMN抗原と競合する;
b)インキュベートしたサンプルを調べて、抗体に結合しているラベルした/検出可能なMN蛋白質/ポリペプチドの量を測定する;
c)ステップb)の検査から、前記サンプル中に存在するMN抗原および/または前記サンプル中に存在するMN抗原の量を決定する。
【0085】
望ましい特性を有する抗体(生物学的に活性な抗体の断片を含む)が調製されると、特定の抗体抗原コンプレックスの形成を判定するための広範な免疫学的分析法が可能である。多くの競合、非競合蛋白質結合分析に関して化学文献および特許明細書に記述されており、そのような分析法の多くは購入可能である。血清中の抗原の検出に適したイムノアッセイの例としては、以下の米国特許に記載されているものが含まれる。米国特許第3,791,932号、第3,817,837号、第3,839,153号、第3,850,752号、第3,850,578号、第3,853,987号、第3,867,517号、第3,879,262号、第3,901,654号、第3,935,074号、第3,984,533号、第3,996,345号、第4,034,074号、および第4,098,876号。
【0086】
分析に用いられる抗体は、ラベルされているかまたはラベルされていないものである。ラベルされていない抗体は凝集に用いられ、ラベルされた抗体は、多くの種類のラベルを施すことにより、広範な分析に使用される。
【0087】
適切な検出には、放射核、酵素、補酵素、発蛍光剤、化学発光剤、色素原、酵素基質あるいは補助因子、酵素阻害剤、フリーラジカル、パーティクル、染料などのラベルを用いるものが挙げられる。そのようなラベル試薬は、既知のさまざまな分析、たとえば、ラジオイムノアッセイ、ELISA等の酵素免疫アッセイ、蛍光免疫アッセイなどに用いられる。たとえば、米国特許第3,766,162号、第3,791,932号、第3,817,837号および第4,233,402号を参照のこと。
【0088】
本発明の分析において有用な抗体を調製する方法を以下に示す。以下の例は、本発明に従う代表的な分析について詳述したものである。
【0089】
イムノアッセイ試験キット
今までに概要を示した分析は、MN抗原および/またはMN特異的抗体(生物学的に活性な抗体の断片を含む)を検出および/または定量する試験キットとして具体化される。MN抗原を検出および/または定量する試験キットは、MN蛋白質/ポリペプチドおよび/または、(ポリクローナルおよび/またはモノクローナルの)MN特異的抗体からなる。そのような診断/予後用の試験キットは、一組またはそれ以上の組のポリクローナルおよび/またはモノクローナル抗体から構成されており、サンドイッチ法の場合には、抗体がMN抗原のエピトープを認識し、一組は適切にラベルされているか、その他の手段により検出可能である。 ラベルされた(あるいは他の手段で検出可能な)MN蛋白質/ポリペプチドとサンプル中のMN抗原との間で抗体に対する結合に関して競合がある分析方法の試験キットは、ラベルされた蛋白質/ポリペプチドと抗体の量を組合わせて、最適の感度と精度が得られるように構成されている。
【0090】
MN特異的抗体検出のための試験キットは、ラベルされた/検出可能なMN蛋白質/ポリペプチドから構成されていることが好ましく、必要であれば、たとえば、下記の実施例8に概要を示しているような好ましい分析を行うために、その他の成分を含んでいてもよい。そのような試験キットは、その他の適切な手段を含んで、従来からあるような分析に応用することもできる。
【0091】
MN特異的抗体の調製
本明細書で使用している「抗体」という語は、抗体全体を指すだけでなく、生物学的に活性な抗体の断片、好ましくは抗原結合部位を有する断片をも指している。そのような抗体は、従来からの手法および/または遺伝子工学により調製できる。抗体の断片は、遺伝子操作により、好ましくは超可変領域を含む、短鎖および/または長鎖の可変領域(VHとVL)から調製されるのがよく、さらに好ましくは、VH領域とVL領域の両方から調製されるのがよい。たとえば、本明細書で使用している「抗体」という語は、ポリクローナルおよびモノクローナル抗体ならびにそれらの生物学的に活性な断片をも意味しており、とりわけ、「一価」抗体(グレニー(Glennie)ら、Nature, 295: 712(1982));共有結合性または非共有結合性凝集をするFab’およびF(ab’)2断片を含むFab蛋白質;好ましくは短鎖および長鎖の可変領域(VHとVL領域)を含み、より好ましくは超可変領域(該VHとVL領域の相補性決定領域(CDRs)としても知られている)を含む短鎖または長鎖単独;Fc蛋白質;1以上の抗体と結合可能な「ハイブリッド」抗体;定常−可変領域キメラ;異なる起源由来の長鎖および短鎖からなる「複合」免疫グロブリン;通常の組換え手法により、またはオリゴヌクレオチドの依存突然変異誘発法(ダルバディー−マクファーランド(Dalbadie-McFarland)ら、PNAS (USA), 79: 6409(1982))により調製された、特異性やその他の特性を改良した「改造」抗体などが含まれる。
【0092】
治療および/またはイメージングに使用する抗体は、生物学的に活性な抗体の断片であることが好ましく、より好ましくは遺伝子工学的に処理された断片がよく、さらに好ましくはVHおよび/またはVL領域から遺伝子工学的に処理された断片がよく、さらにより好ましいのは、それらの超可変領域からなる断片である。 ポリクローナルおよびモノクローナル抗体を作出する従来からの技術は、イムノアッセイの分野では周知である。MN特異的抗体を産生するための免疫原には、MN蛋白質および/またはポリペプチド、好ましくは純粋で、MXに感染した腫瘍細胞株、たとえば、MX感染HeLa細胞およびその他の免疫原が含まれる。
【0093】
抗ペプチド抗体は、ヨーロッパ特許出願公開第44,710号(1982年1月27日公開)に記載されているような当該分野の従来法によっても作出される。該方法を要約すると、第1A図−第1B図に示すようなMNアミノ酸配列からペプチドを選択し、化学的に合成し、適切な免疫原蛋白質に結合し、適切な動物、通常はウサギあるいはマウスに注入し、ポリクローナルまたはモノクローナル抗体を作らせることにより、抗ペプチド抗体が調製される。モノクローナル抗体はコーラー−ミルシュテイン(Kohler-Milstein)法に従って作出される。
【0094】
従来のハイブリドーマ法だけでなく、新しい技術も本発明に従う抗体を産生するために用いることができる。たとえば、クローン作成および抗体のV遺伝子の発現にポリメラーゼ連鎖反応(PCR)法を用い、結合活性を有する断片をコードしている抗体遺伝子の選択にファージディスプレイ法を用いることにより、免疫されたマウスおよびヒト由来のPCR増幅したV遺伝子の集団から抗体の断片を単離することができた(マークス(Marks)ら、BioTechnology, 10: 779(1992年7月号)参照のこと。シャン(Chiang)ら、BioTechniques, 7(4): 360(1989);ワード(Ward)ら、Nature, 341: 544(1989年10月12日号);マークス(Marks)ら、J. Mol. Biol., 222: 581(1991);クラクソン(Clackson)ら、Nature, 352: 624-628(1991年8月15日号);ムリナックス(Mullinax)ら、PNAS (USA), 87: 8095(1990年10月号))。
【0095】
組換え技術を用いた抗体(本明細書では生物学的に活性な抗体の断片を含む)の調製法に関する記載は下記に見出される。米国特許第4,816,567号(1989年3月28日付与)、ヨーロッパ特許出願公開番号(EP)第338,745号(1989年10月25日公開)、EP第368,684号(1990年6月16日公開)、EP第239,400号(1987年9月30日公開)、WO第90/14424号(1990年11月29日公開)、WO第90/14430号(1990年5月16日公開)、ヒューセ(Huse)ら、Science, 246: 1275(1989年12月8日号);マークス(Marks)ら、BioTechnology, 10: 779(1992年7月号);ラ・サストリー(La Sastry)ら、PNAS(USA), 86: 5728(1989年8月号);シャン(Chiang)ら、BioTechniques, 7(4): 360(1989);オーランディ(Orlandi)ら、PNAS(USA), 86: 3833(1989年5月号);ワード(Ward)ら、Nature, 341: 544(1989年10月12日号);マークス(Marks)ら、J. Mol. Biol., 222: 581(1991);フーゲンブーム(Hoogenboom)ら、Nucleic Acids Res., 19(15): 4133(1991)。
【0096】
モノクローナル抗体の調製
本発明の分析で使用するモノクローナル抗体は、当該分野において既知の方法により得られる。たとえば、ガルフレ(Galfre)と ミルシュテイン(Milstein)「モノクローナル抗体の調製:戦略および方法(Preparation of Monoclonal Antibodies: Strategies and Procedures)」(酵素学的方法:免疫化学的手法(Methods in Enzymology: Immunochemical Techniques)73巻1−46ページより)(ランゴーン(Langone)とヴァナティス(Vanatis)編、アカデミック・プレス(Academic Press)社(1981年)、および古典的な参考として、ミルシュテイン(Milstein)とコーラー(Kohler)、Nature, 256: 495-497(1975)がある。
【0097】
本発明の代表的なハイブリドーマは、マウス細胞株の融合により調製されるが、ヒト/ヒトハイブリドーマ(オルソン(Olsson)ら、PNAS(USA), 77: 5429(1980))およびヒト/マウスハイブリドーマ(シュローム(Schlom)ら、PNAS(USA), 77: 6841(1980)、シェアマン(Shearman)ら、J. Immunol., 146: 928-935(1991)、ゴーマン(Gorman)ら、PNAS(USA), 88: 4181-4185(1991))からも調製され得る。そのようなヒト型のモノクローナル抗体は、治療およびイメージングに使用するのに好ましいモノクローナル抗体である。
【0098】
本発明に用いるモノクローナル抗体は、適切なホ乳類、好ましくは齧歯類、より好ましくはウサギまたはマウスに、適切な免疫原、たとえば、MaTuに感染したHeLa細胞や、あるいは、必要であればキャリアー蛋白質をつけたMN蛋白質/ポリペプチドを用いて免疫することにより調製される。例として、MAb M75を分泌するハイブリドーマVU−M75の作出について以下に記載している。MAb M75は、多くの実験室での診断試験、たとえば、培養腫瘍細胞、臨床サンプルなどにおけるMN蛋白質/ポリペプチドの同定に役立っている。MAb M75を産生する方法により、MAb M16(IgG2Bのアイソタイプ)およびMAb M67(IgG1のアイソタイプ)も産生された。
【0099】
MAb M75
モノクローナル抗体M75(MAb M75)は、マウスリンパ球ハイブリドーマVU−M75から産生されるが、該ハイブリドーマは、当初スロヴァク科学アカデミーウイルス研究所(Institute of Virology, Slovak Academy of Sciences)のハイブリドーマコレクション(Collection of Hybridomas)(チェコスロバキア、ブラティスラヴァ)に寄託され、また、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(米国、メリーランド州、ロックヴィル)に1992年9月17日にATCC受入番号HB11128として寄託されたものである。
【0100】
ハイブリドーマVU−M75は、ガーハード(Gerhard)、W.によって記載されている方法(「懸濁液中での細胞融合および調整培地中でのハイブリッドの発芽後生育(Fusion of cells in suspension and outgrowth of hybrids in conditioned medium)」モノクローナル抗体・ハイブリドーマ:生物学的分析の新しい方向(Monoclonal Antibodies. Hybridomas: A New Dimension in Biological Analysis)より、370ページ(ケネット(Kennet)ら編、プレナム(Plenum)社、米国、ニューヨーク))に従って産生される。BALB/CマウスをMaTuに感染したHeLa細胞で免疫し、それらの脾細胞をミエローマ細胞株NS−0と融合した。ハイブリドーマの組織培養培地をモノクローナル抗体のスクリーニングにかけた。用いた抗原は、MaTuに感染したHeLa細胞の抽出物とウサギ抗MaTu血清の免疫沈降物であるp58、および黄色ブドウ球菌(Staphylococcus aureus)細胞(SAC)のプロテインAであり(ザバダ(Zabada)とザバドバ(Zavadova)、Arch. Virol., 118: 189-197(1991))、SDS−PAGEゲルにより分離した。モノクローナル抗体は、プロテインA−セファロースを用いたアフィニティークロマトグラフィー(ハーロウ(Harlow)とレイン(Lane)、「抗体:実験室手順書(Antibodies: A Laboratory Manual)」コールドスプリングハーバー(Cold Spring Harbor)社、米国、ニューヨーク州、コールドスプリングハーバー、1988年)により、TC培地から精製された。
【0101】
本発明に従ってMN蛋白質/ポリペプチドを同定するのに有用なモノクローナル抗体は、従来からの任意の方法によってラベルすることができる。たとえば、西洋ワサビペルオキシダーゼ(HRP)のような酵素、蛍光化合物、125Iなどの放射活性同位体などである。本発明に従う好ましいラベルは125Iであり、好ましい抗体ラベル化の方法は、クロラミン−T法(ハンター(Hunter)、W.H.、「ラジオイムノアッセイ(Radioimmunoassay)」実験免疫学の手引(Handbook of Experimental Immunology)より、14.1-14.40(D.W.ウェイアー(Weir)編、ブラックウェル(Blackwell)社、オックスフォード/ロンドン/エディンバラ/メルボルン、1978年)である。
【0102】
MAb H460
モノクローナル抗体H460(MAb H460)は、MAb M75 と同様な方法によって調製された。ただし、マウスをMaTuに感染していないHeLa細胞により免疫し、マウスの脾細胞ではなくリンパ球をミエローマ細胞株NS−0の細胞と融合した点が異なる。MAb H460は、どんなヒト細胞ともほぼ同様に反応する。
【0103】
MN特異的抗体の治療への応用
本発明のMN特異的抗体、モノクローナル抗体および/またはポリクローナル抗体、好ましくはモノクローナル抗体、より好ましくはMAb M75は、腫瘍および/または前腫瘍性疾患の治療に用いることができ、単独あるいは化学療法剤または毒性剤(たとえば、リシンA等)と組み合わせて用いることができる。治療用としてより好ましいのは、本明細書に記載しているような生物学的に活性な抗体の断片である。同様に、治療用として好ましいMN特異的抗体は、ヒト型モノクローナル抗体である。
【0104】
MN特異的抗体は、好ましくは生理学的に許容性の非毒性液体基剤に分散された形で、治療効果を発揮するのに十分な量が投与される。
【0105】
イメージングへの抗体の応用
さらに、本発明のMN特異的抗体は、放射核などのイメージング剤と結合させた場合には、イメージングに利用できる。生物学的に活性な抗体の断片あるいはヒト型モノクローナル抗体がイメージング用としては好ましい。
【0106】
たとえば、腫瘍部位や転移の位置などが患者の腫瘍組織から同定できる。適切にラベルしたあるいはイメージング剤と結合させた抗体を生理学的に許容性のキャリアーと共に患者に投与し、結合した抗体は、ラベルあるいはイメージング剤の検出に適した方法、たとえば、シンチグラフィーなどにより検出される。
【0107】
アンチセンスMN核酸配列
本発明のMN遺伝子は、腫瘍遺伝子と推定され、それによってコードされている蛋白質は、腫瘍性蛋白質であると推定される。MN遺伝子から転写されたmRNAと実質的に相補性のアンチセンス核酸配列は、実施例11のアンチセンスオリゴデオキシヌクレオチド(ODNs)に代表されるものであり、MN遺伝子の発現を減少あるいは抑制するのに用いることができる(ザメクニック(Zamecnick)、P.C.「イントロダクション:遺伝子情報解読のモジュレーターとしてのオリゴヌクレオチド塩基ハイブリダイゼーション(Introduction: Oligonicleotide Base Hybridization as a Modulator of Genetic Message Readout)」1−6ページ、癌およびAIDSへのアンチセンス核酸療法の予測(Prospects for Antisense Nucleic Acid Therapy of Cancer and AIDS )より(ウィレイ−リス(Wiley-Liss)社、米国、ニューヨーク州、ニューヨーク、1991年);ウィックストーム(Wickstorm)、E.「HL−60前骨髄球性白血病細胞に対するアンチセンスDNA療法:末端配列の変化と標的配列への依存性(Antisense DNA Treatment of HL-60 Promyelocytic Leukemia Cells: Terminal Differenciation and Dependence on Target Sequence)」7−24ページ、同上;レザマン(Leserman)ら、「腫瘍遺伝子の発現に干渉するアンチセンスオリゴヌクレオチドの標的化と細胞内分布(Targeting and Intracellular Delivery of Antisense Oligonucleotides Interfering with Oncogene Expression)」25−34ページ、同上;ヨコヤマ(Yokoyama)、K.「アンチセンスRNAによる腫瘍原遺伝子c-mycの転写制御(Transcriptional Regulation of c-myc Proto-oncogene by Antisense RNA)」35−52ページ、同上;ヴァンデンベルク(van den Berg)ら、「染色体の異常世代を抑制するアンチセンスfosオリゴデオキシリボヌクレオチド(Antisense fos Oligodeoxyribonucleotides Suppress the Generation of Chromosomal Aberrations)」63−70ページ、同上;メルコーラ(Mercola)、D.「アンチセンスfosおよびfunRNA(Antisense fos and fun RNA)」83−114ページ、同上;イノウエ(Inouye)、Gene, 72: 25-34(1988)、ミラー(Miller)とティソー(Ts′o)、Ann. Reports Med. Chem., 23: 295-304(1988);ステイン(Stein)とコーエン(Cohen)、Cancer Res., 48: 2659-2668(1988);ステヴェンソン(Stevenson)とインヴァーセン(Inversen)、J. Gen. Virol., 70: 2673-2682(1989);グッドチャイルド(Goodchild)「オリゴヌクレオチドによる遺伝子発現の抑制(Inhibition of Gene Expression by Oligonucleotides)」53−77ページ、オリゴデオキシヌクレオチド:遺伝子発現のアンチセンス抑制剤(Oligodeoxynucleotides: Antisense Inhibitors of Gene Expression)、(コーエン(Cohen)、J.S.編、CRCプレス(CRC Press)社、米国、フロリダ州、ボカ・ラートン、1989年);デルヴァン(Dervan)ら、「三重らせん形成による二重らせんDNAのオリゴヌクレオチドの認識(Oligonucleotide Recognition of Double- helical DNA by Triple-helix Formation)」197−210ページ、同上;ネッカーズ(Neckers)、L.M.「細胞制御研究の道具としてのアンチセンスオリゴデオキシヌクレオチド:取込のメカニズムと腫瘍遺伝子機能の研究への応用(Antisense Oligodeoxynucleotides as a Tool for Studying Cell Regulation: Mechanisms of Uptake and Application to the Study of Oncogene Function)」211−232ページ、同上;レイトナー(Leitner)ら、PNAS(USA), 87: 3430-3434(1990);ベヴィラッカ(Bevilacqua)ら、PNAS(USA), 85: 831-835(1988);ローク(Loke)ら、Curr. Top. Microbiol. Immunol., 141: 282-288(1988);サリン(Sarin)ら、PNAS(USA), 85: 7448-7451(1988);アグラワル(Agrawal)ら、「アンチセンスオリゴヌクレオチド:化学療法およびAIDSへのアプローチの可能性(Antisense Oligonucleotides: A Possible Approach for Chemotherapy and AIDS)」核酸療法に関する生化学会国際会議(International Union of Biochemistry Conference on Nucleic Acid Therapeutics)(1991年1月13−17日、米国、フロリダ州、クリアウォタービーチ);アームストロング(Armstrong)、L.、Ber. Week、88−89ページ(1990年3月5日号);ウエイントラウブ(Weintraub)ら、Trends, 1: 22-25(1985))。そのようなアンチセンス核酸配列、好ましくは、オリゴヌクレオチドはMN mRNA、特にリボソーム結合部位と翻訳開始点の近傍でハイブリダイゼーションすることにより、mRNAの翻訳を阻害する。それゆえ、そのようなアンチセンス核酸配列を使用することは、癌の治療の一つの方法と考えられる。
【0108】
本発明に従う好ましいアンチセンスオリゴヌクレオチドは、遺伝子特異的ODNsあるいはMN mRNAの5’末端に相補的なオリゴヌクレオチドである。特に好ましいのは、下記の実施例11に配列が示されているような、29−mer ODN1および19−mer ODN2である。これらのアンチセンスODNsは、MN遺伝子の発現を抑制する機能を有する多くのアンチセンス核酸配列の中の代表的なものである。当業者であれば、第1A図−第1B図の核酸配列から適切なアンチセンス核酸配列、好ましくはアンチセンスオリゴヌクレオチドを確定することができる。
【0109】
ワクチン
本発明のMN蛋白質およびポリペプチドは、腫瘍性疾患に対して防御免疫を誘起でき、腫瘍形成活性を弱める効果を有するようなワクチンに組込むことができる。MN蛋白質および/またはポリペプチドは、合成あるいは組換えやその他の生物学的手法により調製されて、単量体、または多量体型のMN蛋白質の1またはそれ以上のエピトープに対応する、1またはそれ以上のアミノ酸配列から構成されるようにすることができる。つぎに、これらの蛋白質および/またはポリペプチドは、防御免疫を誘起できるワクチンに組込まれる。そのようなポリペプチドの免疫原性を上げる方法は、多量体構造に組込むこと、キーホールリンペットヘモシアニン(KLH)あるいはジフテリア毒素等の高免疫原性蛋白質キャリアーに結合させること、およびアジュバントあるいはその他の免疫応答強化剤と組み合わせて投与することを含む。
【0110】
本発明に従うワクチンにおいて使用される好ましいMN蛋白質/ポリペプチドは、遺伝子工学的に処理されたMN蛋白質である。好ましい組換えMN蛋白質は、本発明に従って産生された融合蛋白質pGEX−3X−MNである。
【0111】
本発明のそのようなワクチンの好ましい使用例は、MN関与性一次癌が外科的に切除された患者へ投与することである。ワクチンは患者の体内で能動免疫を誘起し、再発あるいは転移を防ぐことができる。
【0112】
さらに、MN蛋白質/ポリペプチドに対する抗体に対する抗イディオタイプ抗体もワクチンとして有用であり、同様に製剤化できる。
【0113】
単量体、または多量体型のMN蛋白質/ポリペプチドのエピトープに対応するアミノ酸配列は、化学的合成、あるいは遺伝的に変更された微生物やそれらの培養培地などの生物源を精製することによっても得られる(ラーナー(Lerner)「合成ワクチン(Synthetic Vaccines)」Sci. Am., 248(2): 66-74(1983)を参照)。蛋白質/ポリペプチドは、他の蛋白質/ポリペプチド(他のタンパク質の断片を含む)と組合わされて一つのアミノ酸を形成することがあり、たとえば、融合蛋白質として合成される場合や、合成または生物由来の抗原性あるいは非抗原性の他のポリペプチドに結合する場合などがある。
【0114】
「MN蛋白質/ポリペプチドのエピトープに対応する」という言葉は、天然に存在する蛋白質またはポリペプチドのアミノ酸配列の変化が抗原性を与えることがあり、腫瘍性疾患に対する防御免疫および/または抗腫瘍形成性効果を付与することがあるという実際的な可能性を含むものとする。配列の変化の可能性としては、アミノ酸の置換、伸長、欠失、削除、挿入およびこれらの組合せが挙げられるが、これらに限定されるわけではない。そのような変形も本発明の範ちゅうに含まれる。ただし、それらを含有する蛋白質またはポリペプチドが免疫原性であり、そのような蛋白質またはポリペプチドによって分泌された抗体は、天然に存在するMN蛋白質またはポリペプチドと交差反応し、その分泌量は、ワクチンとして投与したときに防御免疫および/または抗腫瘍形成性活性を誘起するのに十分な量であるものとする。
【0115】
そのようなワクチンの組成物は、生理学的に許容し得る基剤、たとえば、免疫的に許容される希釈剤およびキャリアー、また、フロイントの完全アジュバント(Freund′s Complete Adjubant)、サポニン、明ばん等の通常用いられるアジュバントなどと組合せられる。投与は、免疫学的に有効な量のMN蛋白質またはポリペプチドで行うが、好ましい投与量ユニットは、被投与体の体重1kgあたり0.01から10.0μgの免疫学的に活性なMN蛋白質および/またはポリペプチドである。防御に有効な総投与量は、抗原量として0.1から約100μgの範囲である。
【0116】
投与経路、抗原量、投与の回数と頻度はすべて最適の条件で行うが、これらは当該分野の通常の技術範囲の範ちゅうに含まれる。
【実施例】
【0117】
以下の実施例は説明をするためのものであり、如何なる意味においても本発明を限定するものではない。
【0118】
材料および方法
次の材料および方法は、以下の実施例で使用するものである。
【0119】
MaTu感染および非感染HeLa細胞
MaTu剤(ザバダ(Zabada)ら、Nature New Biol., 240:124-125(1972)、ザバダ(Zabada)ら、J. Gen. Virol., 24: 327-337(1974))は、「MaTu」細胞由来であり(ウィドメイアー(Widmaier)ら、Arch. Geschwulstforsch, 44: 1-10(1974))、これは、マイトマイシンCで処理したMaTu細胞と共培養することにより、発明者らのHeLa細胞ストックに移したものであり、これにより、対照とMaTu感染細胞を対比できるようなった。MaTu細胞は、5μg/mlのマイトマイシンC(カルバイオケム(Calbiochem)社、米国、カリフォルニア州、ラ・ホーラ(La Jolla))を含む培地で37℃で3時間インキュベートした。混合培養は、培地5mlあたり、2×105個のマイトマイシンC処理細胞および4×105個の新鮮感受性細胞とからなるようにした。3日後に最初の継代を行い、さらに週に1−2回継代した。
【0120】
対照のHeLa細胞は、ザバダ(Zabada)らの記載(Nature New Biol.,240:124-125(1972))と同様に扱った。
【0121】
血清
癌患者、各種の非腫瘍性症状に苦しむ患者および健康な女性からのヒト血清は、大学院医学研究科産婦人科病院(Clinics of Obstetrics and Gynaecology at the Postgraduate Medical School)(チェコスロバキア、ブラチスラヴァ)から入手した。
【0122】
ヒト血清KHは、切除後14カ月の50才の乳腺腫患者から得た。該血清は、401の血清サンプルの中で、ザバダ(Zabada)らにより記載(Nature New Biol., 24
0:124-125(1972))されているVSV(MaTu)のプソイド型に対する中和抗体を含有する2つの血清の内の一つである。血清L8は、パジェット病(Paget′s disease)の患者から得た。血清M7は、健康な献血者から得た。
【0123】
ウサギ抗MaTu血清は、MaTuに感染した生存HeLa細胞10−5×107個を30日の間隔で3回ウサギに免疫することにより調製した。
【0124】
RIPとPAGE
RIPとPAGEは基本的には、ザバダ(Zabada)とザバドヴァ(Zabadova)による記載(Arch. Virol., 118: 189-197(1991))に従って行ったが、本明細書に記載している実験において異なる点は、[35S]メチオニン(NEN)、10μCi/mlのメチオニン非含有MEM培地、2%のFCSを添加した3%の完全MEM培地を使用した点である。細胞の全面ペトリ皿培養を、該培地で終夜インキュベートした。
【0125】
RIPには、SAC法(ケスラー(Kessler)、J., J. Immunol., 115: 1617-1624(1975))を用いた。インキュベーションおよび遠心分離はすべて0−4℃で行った。単層細胞は、RIPA緩衝液(0.14Mの塩化ナトリウム、7.5mMのリン酸緩衝液(pH7.2)、1%のトライトンX−100(Triton X-100)、0.1%のデオキシコール酸ナトリウム、1mMのフッ化フェニルメチルスルフォニルおよびトラシロール(Trasylol)からなる)で抽出した。非特異反応を減少させるため、抗血清をウシ胎児血清(バーバシッド(Barbacid)ら、PNAS(USA), 77: 1617-1621(1980))およびSAC処理した抗原抽出物に前吸着させた。
【0126】
PAGE(還元状態)には、10%のSDSゲルを用いた(レムリ(Laemmli)、Nature, 227: 680-685(1970))。対照マーカー蛋白質としては、シグマキット(Sigma Kit)(製品番号MW-SDS-200)を使用した。フルオログラフィーには、サリチル酸塩を用いた(ヘーガード(Heegaard)ら、Electrophoresis 5: 263-269(1984))。
【0127】
イムノブロット
本明細書に記載しているイムノブロットは、トウビン(Towbin)らの方法(PNAS(USA) 76: 4350-4354(1979))に従って行った。蛋白質は、レムリ(Laemmli)の電気泳動緩衝液を蒸留水で1:10に希釈し、メタノールやSDSを含まない状態でゲルからニトロセルロース(シュレイカー・アンド・シュエル(Schuleicher and Schuell)社、ドイツ、ダッセル、0.45μm多孔性)に移した。移送は1.75mA/cm2で2時間半かけて行った。ブロットは、125IでラベルしたMAbと共に展開し、増感フィルターを使用してX線フィルムを−70℃で感光することによりオートラジオグラフィーを行った。
【0128】
細胞培養の抽出物には少量のMN抗原しか含まれていないため、0.5から1mlの抽出物に50μlの10%SAC懸濁液(MAb M75が含まれている)を添加して濃縮した。本方法は、ヒトIgGを含む臨床サンプルのMN抗原の濃縮にも使用できる。予備対照実験の結果から、そのような方法がSAC吸着M75へのMN抗原の結合に影響しないことが示されている。組織抽出物は、組織をモルタル、乳棒、砂(分析用グレード)と共に粉砕することにより得られた。このホモジネートに、RIPA緩衝液を当初の組織に対して10:1(重量当りの容量)の割合で加えた。抽出物は、エッペンドルフ(Eppendorf)遠心分離に3分間かけ、清澄にした。
【0129】
実施例1
MaTu特異的抗原の免疫蛍光法
対照細胞、および上述の方法で調製されたモノクローナル抗体を含むMaTu感染HeLa細胞について、免疫蛍光実験を行った。該モノクローナル抗体は、MaTu関連抗原に特異的である。モノクローナル抗体の存在確認には、FITC結合抗マウスIgGを用いた。細胞のギムザ(Giemsa)染色から、対照とMaTu感染HeLa細胞にははっきりとした違いはないことが示された。
【0130】
MaTu関連抗原に特異的であることが事前の試験で証明されているMAbは、免疫蛍光において二つの異なる反応性を示した。第一のグループを代表するMAb M67は、MaTu感染HeLa細胞において顆粒状の細胞質蛍光反応を示したが、これはアセトンで固定した細胞のみにみられ、生細胞は蛍光を示さなかった。MAb M16も同様の蛍光を示した。M67およびM16のいずれについても、対照のHeLa細胞においてごく弱い「バックグラウンド」蛍光がみられた。
【0131】
もう一つのMAbであるM75は、生きたMaTu感染HeLa細胞において顆粒状の膜蛍光反応を示し、アセトン固定細胞においては顆粒状の核蛍光反応を示した。しかしながら、M75は、非感染HaLa細胞においても、非常に弱いが類似の蛍光を示すことがあった。増殖の条件に基づく関係が見いだされた。すなわち、MaTuに非感染のHeLa細胞においては、細胞を高密度培養中で数回継代して増殖したもののみがMAb M75による両方の型の蛍光を示し、希薄培養中で増殖したものは示さない。
【0132】
M75反応性の細胞表面抗原の量は、細胞蛍光測定法により測定し、その量は、細胞培養の密度およびMaTuの感染に依存していた。対照および MaTu感染HeLa細胞は、高密度あるいは希薄培養中で12日間増殖させた。細胞は、ヴァーセン(Versene)(EDTA)によって遊離させ、MAb M75と共にあるいはMAbなしでインキュベートし、続いてFITC結合抗マウスIgGと共にインキュベートした。蛍光の強さを測定した。
【0133】
抗原結合MAb M75は、誘導性のものと思われる。すなわち、該抗体は、希薄培養で増殖した対照のHeLa細胞にはなく、高密度培養でのHeLa細胞の増殖あるいはMaTuの感染によって誘起されることがわかった。これら二つの因子は、付加的あるいは共働作用効果を有することがわかった。これらの知見と本明細書に記載したその他の結果により、次のことが示唆される。すなわち、二つの異なる物質があり、その一つはM67に反応する外因性、透過性のMX、もう一つはMAb M75により検出される内因性、誘導性のMNである。
【0134】
実施例2
MAb M75に反応する蛋白質のイムノブロット分析
MAb M75が、非感染およびMaTu感染HeLa細胞の両方において同じ蛋白質に反応するか否かを確かめ、さらに、該蛋白質の分子量を求めるために、それらの細胞の抽出物をPAGEおよびイムノブロッティング(上述の方法による)により分析した。高密度あるいは希薄培養中で12日間増殖させた非感染およびMaTu感染HeLa細胞を5cmのペトリ皿にまき、すべての変異型について細胞数が5×105個となるようにした。2日後に細胞をRIPA緩衝液で抽出し(上述)、200μl/皿の量にした。抽出物は、6%のメルカプトエタノールを含有する2倍濃縮のレムリ(Laemmli)のサンプル緩衝液と混合し、5分間煮沸した。蛋白質は、SDS−PAGEにより分離し、ニトロセルロースにブロットした。ブロットは、125IでラベルしたMAb M75を用いて展開し、オートラジオグラフィーにかけた。
【0135】
MAb M75は、54kdと58kdの2本のMN特異的蛋白質のバンドと反応し、この結果は高密度で増殖した非感染HeLa細胞、ならびにMaTu感染HeLa細胞において同じであったことから、M75は、非感染およびMaTu感染HeLa細胞の両方で同じ蛋白質を認識していることが明らかになった。細胞蛍光測定の結果と併せて考えると、抗原量は、細胞密度とMaTuの感染に依存しており、後者がより強力なp54/58Nの誘導因子である。
【0136】
実施例3
インサイチュー(in situ)でのMaTu特異的抗原のラジオイムノアッセイ
M75での結果とは対照的に、別のMAbである、M67は、外因性で透過性の物質MXに特異的と思われる。M67においては、細胞が、高密度培養で増殖されたかあるいは希薄培養で増殖されたかに関係なく、対照のHeLa細胞では免疫蛍光を示さなかった。この相違は、125IでラベルしたMAbs M67およびM75を用いたラジオイムノアッセイ実験によってはっきり示された。
【0137】
そのような実験においては、非感染およびMaTu感染HeLa細胞を高密度培養あるいは希薄培養で同時培養して増殖させた。培養細胞は、生きたもの(固定しない)か、固定したもの(メタノールで5分間処理後風乾)かである。培養細胞は、125IでラベルしたMAbsを6×104cm/皿加えたペトリ皿で2時間インキュベートした。その後、培養細胞をPBSで4回洗浄し、1ml/皿の2N NaOHで溶解し、γカウンターで放射活性を測定した。
【0138】
本例の簡単な放射活性法は、ペトリ皿の培養細胞に直接行うものである。ラジオイムノアッセイの16の変異型から、MXおよびMNが細胞表面あるいは細胞内に存在しているかが確認でき、これら二つの抗原の発現がMaTuの感染およびペトリ皿にまく前の細胞増殖の時の密度にどのように依存しているかが確認された。生きた固定されていない細胞では、細胞表面の抗原のみがMAbsと結合できる。これらの細胞においては、M67は培養細胞の如何なる変異型とも反応を示さなかったが、一方、M75は上述の実施例1および2の結果に示すように反応した。
【0139】
メタノールで細胞を固定することにより、細胞膜はMAbsを透過させるようになる。M67は、事前の細胞密度に関係なくMaTu感染HeLa細胞と反応するが、対照のHeLa細胞には結合しない。メタノール固定された細胞内のMAb M75により、希薄培養由来の非感染HeLa細胞には対応する抗原が存在しないこと、さらに、該抗原は、高密度培養による増殖あるいはMaTuの感染により誘導されることが確認された。
【0140】
実施例4
動物血清に反応性またはVSVビリオンに関連するMaTu成分の同定
非感染またはMaTu感染HeLa細胞由来のRIPA抽出物、あるいは、対照またはMaTu感染HeLa細胞内で再産生された精製VSV由来のMaTu特異的蛋白質のイムノブロット分析を行うこと、抗原p58Xあるいはp54/58Nのちどの抗原が動物血清と放射免疫沈降するかということ、さらに、VSV変異株の相補性およびプソイド型(pseudotype)のビリオンの形成に関与しているかということを調べた。方法の詳細については、パストレコヴァ(Pastorekova)らによる記載(Virology, 187: 620-626(1992))を参照のこと。
【0141】
MaTu感染HeLa細胞で免疫したウサギの血清は、MAb M67およびMAb M75反応性の蛋白質(p58Xおよびp54/58Nの両方)の両方と免疫沈降したのに対し、正常ウサギ、ヒツジあるいは白血病ウシの「自発的(spontaneously)」血清は、M67反応性蛋白質(p58X)とのみ免疫沈降した。一方、MaTu感染HeLa細胞により産生され、精製されたVSVは、p54/58NのM75反応性のバンドのみが存在した。このことから、MXとMNは、MaTuの独立した構成成分であると考えられ、VSV変異株に相補性であり、プソイド型のビリオンに集積されるのはp54/58Nである。
【0142】
第2A図に示し、下記の実施例5に述べているように、MX抗原は、MaTu感染線維芽細胞内に存在することがわかっている。ザバダ(Zavada)とザバドバ(Zavadova)は(1991年)、MX感染線維芽細胞由来のp58のバンドは、ウサギ抗MaTu血清を用いたRIPでは検出できなかったと報告している。該血清中には、MX抗原よりもMN抗原に対する抗体が多く含まれていた。この矛盾は、感染培養細胞内におけるMXの伝播が極端に遅いことで説明できる。ザバダ(Zavada)とザバドバ(Zavadova)による報告は(1991年)、感染後6週目の線維芽細胞に関するものであり、他方、後の試験は感染4カ月後の線維芽細胞に関してである。イムノブロットの結果から、MXは、まず、感染の4週間後にはH/F−NハイブリッドおよびH/F−Tハイブリッドの両方から検出され、6週後にはHeLa細胞に検出され、線維芽細胞においては感染10週後になってようやく検出されることを発明者らは見出した。
【0143】
実施例5
MN特異的およびMX特異的蛋白質の発現
第2A図−第2B図は、ヒト線維芽細胞、HeLa細胞およびH/F−Nハイブリッド細胞とH/F−Tハイブリッド細胞内におけるMN特異的およびMX特異的蛋白質の発現をグラフとして示したものであり、MX感染および非感染細胞での発現を対比している。細胞は、マイトマイシンCで処理したMX感染HeLa細胞と共培養することにより感染させた。感染および非感染細胞は、高密度培養中で3回継代して増殖させた。感染4カ月後に、感染細胞を非感染細胞と同時にペトリ皿で増殖し、高密度単相を形成させた。
【0144】
全面ペトリ皿(5cm)培養した細胞にラジオイムノアッセイを直接行い、メタノールで固定した(基本的には上記の実施例3と同様)。細胞の単層はメタノールで固定し、125IでラベルしたMAbs M67(外因性のMX抗原特異的)またはM75(内因性MN抗原特異的)を6×104cpm/皿加えて処理した。結合放射活性を測定した。結果は第2A図−第2B図に示す。
【0145】
第2A図は、試験した4つの細胞株すべて、すなわち、ヒト胚線維芽細胞、HeLa細胞、およびH/F−NハイブリッドとH/F−Tハイブリッド細胞、にMXが透過したことを示しており、同時に、対応する4つの非感染細胞株にはMXが存在しないことを示している。第2B図は、MX感染および非感染HeLa細胞ならびにH/F−T細胞にはMN抗原が存在するが、線維芽細胞には存在しないことを示している。対照のH/F−NにはMNは全く検出されず、MaTu感染H/F−NにおいてMN抗原のバックグラウンドを越えるわずかな増加が見られただけであった。このことから、ハイブリッドにおいては、MN抗原の発現が腫瘍形成性と強く関係していることが明らかになった。
【0146】
これらの結果は、イムノブロッティングにより得られた結果と一致する。MN特異的双子蛋白質p54/58Nは、イムノブロッティングによりHeLa細胞株(標準型の両方、すなわちHeLa K、およびスタンブリッジ(Stanbridge)突然変異HeLa細胞、すなわちD98/AH.2あるいはHeLa S)および腫瘍形成性H/F−Tにおいて検出されたが、線維芽細胞あるいは非腫瘍形成性H/F−Nでは、放射活性を検出するために使用したフィルムをゆっくりと長時間感光してもp54/58Nは検出されなかった。MXのHeLa細胞への感染により、p54/58N蛋白質の急激な濃度上昇がみられた。
【0147】
ハイブリッド細胞H/F−NおよびH/F−Tは、エリック J.スタンブリッジ(Eric J. Stanbridge)によって作出された(スタンブリッジ(Stanbridge)ら、Somatic Cell Genetics, 7: 699-712(1981)、スタンブリッジ(Stanbridge)ら、Science, 215: 252-259(1982))。彼の本来のハイブリッドは、HeLa細胞とヒト線維芽細胞との融合によって作られ、ヌードマウスにおいては腫瘍形成性ではなかったが、形質転換細胞内のいくつかの特性、たとえば、軟寒天上での増殖など、を保持していた。
【0148】
染色体11を失ったハイブリッド由来の分離体(セグレガント)が腫瘍形成性であることはまれである。これらの分離体における腫瘍形成性に関する最も妥当な説明は、染色体11が抑制遺伝子(抗腫瘍遺伝子)を有しており、これが未知の腫瘍遺伝子の発現をブロックしているということである。該腫瘍遺伝子によってコードされている腫瘍蛋白質は、ヌードマウス内で腫瘍を形成するH/Fハイブリッドの能力に対して重大な働きをする。p54/58N蛋白質は、H/Fハイブリッドの腫瘍形成性に関連があり、推定腫瘍蛋白質の一つの候補である。
【0149】
実施例6
ヒト腫瘍細胞培養由来およびヒト組織の臨床標本由来のMN抗原のイムノブロット
実施例5に示したように、H/Fハイブリッド細胞内の腫瘍形成性において、MN抗原が関与することから、その他のヒト腫瘍細胞培養および臨床標本内のMN抗原の存在を調べることにした。予備実験の結果から、その他のヒト腫瘍細胞培養の抽出物内のMN抗原の濃度はHeLa細胞よりも低いことが示された。このことから、オートラジオグラフィーにおいては、長時間の感光が必要であることがわかった。これにより、上述の材料および方法:イムノブロッティングに従う方法(ここでは、MAb M75を結合させたSACを用いて沈澱させることにより、MN抗原を濃縮している)により、該方法の感度を増加させた。
【0150】
細胞培養の抽出物内のMN蛋白質のイムノブロットは、次のものから調製した。(A)MX感染HeLa細胞、(B)ヒト線維芽細胞、(C)T24、(D)T47D、(E)SK−Mel 1477、および(F)MX非感染HeLa細胞。蛋白質は、3%のメルカプトエタノールを含むかまたは含まない状態(+MEまたは0MEと表す)でサンプル緩衝液に加えて加熱した後、PAGEにより分離した。つまり、おのおのの抽出物に関し、+MEについて、次に0MEについて一回の泳動を行った。レーン(A)(MX感染HeLa細胞由来の細胞培養抽出物)については直接分析を行ったが(10μl/レーン)、その他の抽出物(レーンB−E)由来の抗原については、500μlの抽出物をMAb M75とSACを用いて沈澱させることによりおのおの濃縮した。イムノブロットの結果、2つの他のヒト癌細胞株がMN関連蛋白質を含有することが示された。すなわち、T24(膀胱癌;レーンC)とT47D(乳腺癌;レーンD)である。これらの細胞は、MAb M75と反応し、還元状態では54kdと56kdの分子量を含有し、非還元状態では約153kdの分子量を有する蛋白質を含有している。これらのバンドの強度は、HeLa細胞由来のp54/58N双子蛋白質のそれの少なくとも10分の1以下である。
【0151】
ヒトメラノーマ細胞由来の抽出物(SK−Mel 1477;レーンE)の還元状態では、おおよそ52kdの位置にごく弱いバンドが観察されたが、ヒト線維芽細胞抽出物(レーンB)では、還元状態でも非還元状態でもそのようなバンドはみられなかった。
【0152】
外科標本を含むヒト組織抽出物のイムノブロットをMX感染HeLa細胞由来の細胞抽出物(レーンA)のそれと比較した。他のレーンの組織抽出物は次のものから調製した。(B)満期の胎盤、(C)子宮体部、(D,M)子宮内膜腺癌、(E,N)卵巣腺癌、(F,G)トロホブラスト、(H)正常卵巣、(I)子宮筋腫、(J)乳頭腫、(K)正常乳腺、(L)高増殖性子宮内膜、(O)子宮頚管癌、および(P)メラノーマ。レーンA(10μl/レーンで直接分析した)を除くその他の全ての抽出物由来のMN関連抗原は、上述のように、まず1mlの抽出物から濃縮した。MN蛋白質は、子宮内膜(レーンDとM)、卵巣(レーンEとN)および子宮頚管(レーンO)の癌について見つかった。これらの抽出物においては、MN関連蛋白質は、約48kdから約58kdの間の分子量を有する3本のバンドとして現れた。乳腺癌の組織抽出物中には別のMN関連蛋白質が存在しており、該蛋白質は、約48kdの一本のバンドとして現れた(レーンJ)。
【0153】
満期の胎盤(レーンB)、正常乳腺(レーンK)、高増殖性子宮内膜(レーンL)、正常卵巣(レーンH)、および子宮筋腫(レーンI)からの抽出物では全く何も現れてこなかった。トロホブラスト(レーンFとG)およびメラノーマ(レーンP)からの抽出物においては、ごく弱いMN関連のバンドが現れただけであった。
【0154】
p54/58Nに関連する抗原は、いくつかの型のヒトの癌の臨床標本内では発現するが、対応する器官の正常組織においては発現しないという所見から、MN抗原の腫瘍形成性との関係がさらにはっきりしてきた。しかしながら、ヒト腫瘍においては、腫瘍は成熟した分化後の細胞から生じるものではなく、分岐および分化能を有するある種の基幹細胞から生じると考えられるので、正常細胞が本当に適切な対照となるわけではないことに注意しなければならない。体内器官では、そのような細胞はきわめて稀である。
【0155】
実施例7
動物細胞株のMN抗原
試験した全ての脊椎動物の染色体DNAにはMN遺伝子が存在していたことから、数種の動物種の正常組織由来および腫瘍由来の細胞株についてもMN関連抗原を調べた。MN関連抗原は、二つのラット細胞株から見つかった。一つは、ルイス肉腫ウイルス(Rous sarcoma virus)により誘起されたラット横紋筋肉腫由来のXC細胞株であり、もう一つは、Rat2−Tk−細胞株である。これら両方のラット細胞株の抽出物において、一本の蛋白質のバンドがブロット上に認められた。還元ゲルおよび非還元ゲルにより得られたブロットの分子量は、それぞれ53.5kdおよび153kdであった。
【0156】
MN蛋白質のイムノブロットは、(A)MX感染HeLa細胞、および(B)Rat2−TK−細胞株(上述のように、3%のメルカプトエタノールを含む(+ME)かまたは3%のメルカプトエタノールを含まない(0ME)状態でサンプル緩衝液に加えて加熱した後、PAGEにより分離した)から調製した。これら二つの細胞株内のMN抗原の濃度は、+MEおよび0MEの泳動においてほぼ同じであった。抽出物は直接分析した(40μl/レーン)。
【0157】
XC細胞のMN関連蛋白質は、還元および非還元状態のいずれにおいてもRat2−Tk−細胞のそれと同様のパターンを示したが、濃度は30分の1であった。二つのラット細胞株(上述のイムノブロットおよび実施例4より)においてMN関連蛋白質p53.5Nが発見されたことにより、モデル系の基礎が作られた。
【0158】
MN抗原を濃縮し、高感度イムノブロット法を使用しても、試験した他の動物細胞株にはいずれも検出できる量のMN抗原は含まれていなかった。MNを含有していなかった細胞は次のものである。ベロ(Vero)細胞(アフリカミドリザル);マウスL細胞;マウスNIH−3T3細胞の正常型、サル白血病ウイルス感染型あるいはハーヴェイ肉腫ウイルス(Harvey sarcoma virus)形質転換型;GR細胞(MTVにより誘起されたマウス乳腺腫細胞);およびNMG細胞(正常マウス乳腺)。
【0159】
実施例8
MN特異的抗体およびMN抗原検出のための組換えMN蛋白質を用いた液相ラジオイムノアッセイ
グルタチオンSトランスフェラーゼと融合させ、上述のように調製、精製した遺伝子組換えMN蛋白質、pGEX−3X−MNを、クロラミンT法(ハンター(Hunter)ら、1978年)により125Iでラベルした。精製した蛋白質により、MN特異的抗体ならびにMN抗原の定量RIAを行うことができた。抗体の希釈および抗原の希釈はすべて、1%のウシ胎児血清(FCS)を添加したRIPA緩衝液(1%のトライトンX−100(TRITON X-100)および0.1%のデオキシコール酸ナトリウムのPBS(リン酸緩衝生理食塩水、pH7.2)溶液で行った。組織および細胞の抽出物は、1mMのフッ化フェニルメチルスルフォニルおよび1mlあたり200トリプシン阻害ユニットのトラシロール(Trasylol)(アプロチニン)を添加し、FCSを含まないRIPA緩衝液で調製した。125IでラベルしたpGEX−3X−MN(2.27μCi/μgのTCA沈降活性を有する蛋白質)は、使用前に1%のFCSを含むRIPAで希釈し、非特異結合放射活性をプロテインA固定黄色ブドウ球菌(Staphylococcus aureus)(SAC)の懸濁液で吸着した。 MN特異的抗体に関するRIAにおいては、MAbを含む腹水あるいは試験血清を125Iでラベルした蛋白質と混合し、総量が1mlとなるようにして室温で2時間反応させた。次に、50μlのSACの10%懸濁液(ケスラー(Kessler)ら、同上)を加え、得られた混合物を30分インキュベートした。最後に、SACを沈澱させ、RIPAで3回洗浄し、結合放射活性をγカウンターで測定した。
【0160】
MN抗原に対する抗体の滴定の結果を第3A図−第3B図に示す。M75ハイブリドーマ細胞を含有するマウスの腹水(A)は、1:1.4×10−6希釈において50%終点を有することを示している。同時に、MX蛋白質(M16およびM67)に特異的なMAbsを含有する腹水は、1:200希釈においても125IでラベルしたpGEX−3X−MNとは沈降しなかった。正常ウサギ血清(C)は、MN抗原とははっきりとは沈降しなかった(結果は示していない)。生きたMX感染HeLa細胞で免疫したウサギから得られたウサギ抗MaTu血清(B)は、1:200に希釈したときに放射活性MN蛋白質と7%の沈降を示した。実施例4(上述)に示すようなイムノブロットにより、ウサギ抗MaTu血清は、MXおよびMNのいずれの蛋白質とも沈降を示した。
【0161】
試験した180のヒト血清(90の対照、および、乳房、卵巣、子宮頚管癌の患者からの90の血清)の中からただ一つだけが、放射活性ラベルされた組換えMN蛋白質と明かに沈降した。その血清L8(D)をイムノブロット(実施例4)で再試験したが、MX感染HeLa細胞由来のp54/58Nのいずれとも沈降しなかった。さらに、KH(E)を含む6つの他のヒト血清もイムノブロットで反応しなかった。つまり、RIAでただ一つ反応したヒト血清L8は、遺伝子組換え産物とのみ反応し、HeLa細胞で発現した、生来のp54/58Nとは反応しなかった。
【0162】
MN抗原に対するRIAにおいては、MAb M75の希釈液(事前の試験で50%の最大沈降放射活性(=1:1.4×10−6希釈)を示した)を細胞抽出物の希釈物と混合し、2時間反応させた。次に、125IでラベルしたpGEX−3X−MN(25×103cpm/チューブ)を加え、さらに2時間反応させた。最後に、MAb M75に結合した放射活性をSACで沈降させ、上述のように洗浄した。100%の沈降(=阻害なし)を、用いた希釈MAbによる最大放射活性結合と見なした。試験に用いた細胞抽出物中のMN抗原の濃度は、「コールド(cold)」のpGEX−3X−MNを標準として使用した阻害曲線から計算した(pGEX−3X−MNを標準として使用した阻害曲線から計算した(第4図のA)。
放射活性ラベルしたpGEX−3X−MN蛋白質とMAb M75との反応から、細胞抽出物中のMN抗原を直接的に定量することができた。第4図は、3ngの「コールド」のpGEX−3X−MN(A)が、「ホット(hot)」のpGEX−3X−MNと50%の沈降阻害を起こしたことを示すものである。同量のMN抗原が、MaTu感染HeLa細胞(B)およびRat2−Tk−細胞(C)から抽出した3×103ngの蛋白質にも存在している。本実験のRIAで測定した細胞抽出物中の蛋白質の濃度を下記の表1に示す。細胞抽出物中のMN抗原は、ある程度の異なる大きさであるということ、また、遺伝子操作されたMN蛋白質は、さまざまな大きさの分子を含有するものであることから、計算値は絶対的なものではないことに注意されたい。
【表1】
【0163】
実施例9
MX抗原のRIP
p58X蛋白質のおよその濃度は、MaTu感染HeLa細胞の抽出物のRIPにより求めることができ、ここで、該抽出物は、[35S]メチオニンあるいは[14C]アミノ酸混合物で代謝的にラベルしたものである。これらの結果を表2に示す。
【表2】
【0164】
表2に示す結果から、p58Xは、細胞抽出物中の蛋白質のおよそ0.8%を占めることがわかった。
【0165】
ラベルしたアミノ酸と共に終夜培養した培養細胞および並行培養(アミノ酸を完全に補充した「コールド」の培地でさらに24時間インキュベートしたもの)の培養細胞からも非常に近似した値が得られた。これらの結果は、得られた放射活性の値が既に平衡状態にあることを反映したものであり、取り込みの速度を表したものではないことを示しており、該値は、抽出物中のp58Xの実際の含量と大差はない。[35S]メチオニンでラベルした細胞の抽出物のp58Xの値は、[14C]アミノ酸混合物でラベルした培養の抽出液内のそれと同程度であった。
【0166】
実施例10
対照およびMX感染HeLa細胞の免疫電子および走査顕微鏡法
上記の実施例1に示したように、MAb M75を用いた直接免疫蛍光法により検出されたMN抗原は、MX感染HeLa細胞の膜表面および核内あるいは高密度培養で増殖させたHaLa細胞内に存在している。MN抗原の所在をさらに明らかにさせるために、免疫電子顕微鏡法を用いた。該方法においては、MN抗原に結合したMAb M75を免疫金(immunogold)ビーズで視覚化した(ヘルゾーグ(Herzog)ら、「細胞表面域判定のためのコロイド金ラベリング法(Colloidal gold labeling for determining cell surface area) 」コロイド金(Colloidal Gold)第3巻(ハヤット(Hayat),M.A.編)より、139−149ページ(アカデミックプレス(Academic Press)社、米国、カリフォルニア州、サンディエゴ)参照)。
【0167】
MX非感染(対照)およびMX感染HeLa細胞の超薄片を、免疫金を含むMAb M75および含まないMAb M75で染色した。(いくつかの細胞は、包埋し、切断する前にM75と免疫金で固定し、処理した。この操作により、免疫金装飾が細胞表面の抗原に対してのみ施される。いくつかの細胞は、包埋し、切断した後にのみM75と免疫金で処理した。こうすると、細胞内の抗原も装飾される。免疫金で処理しなかったいくつかの細胞は、細胞分裂の末期のものである)。
【0168】
染色した細胞の免疫電子および走査顕微鏡像から、細胞内のMN抗原の局在が示され、さらに、対照およびMX感染HeLa細胞とでは微細構造にかなりの違いがあることが見出された。対照のHeLa細胞は、その表面に、金ビーズで視覚化されたMN抗原をほとんど有していないことが示された。細胞表面は比較的滑らかで、2つの小さい突起を有するだけであった。細胞質内にはミトコンドリアは見られなかった。これとは対照的に、MX感染HeLa細胞では、表面にたくさんの密集した糸状突起がみられた。ほとんどのMN抗原はこれらの糸状体の上に局在しており、該抗原は、染色に免疫金を用いた場合には免疫金によって修飾されていた。MX感染HeLa細胞の細胞質には多くのミトコンドリアを含んでいるのが見られた。MN抗原は、MX感染HeLa細胞の核にあった。MN抗原は核質の中にいくらか存在していたが(おそらくクロマチンと結合しているのであろう)、より高濃度のMN抗原は核小体に存在していた。繰り返すと、正常HeLa細胞の細胞表面は比較的滑らかであり、一方、MX感染HeLa細胞の表面には多数の糸状体と「水疱(blebs)」がある。糸状体のいくつかは、近隣の細胞とつながる橋を形成しているように見える。
【0169】
インビトロ(in vitro)で形質転換された細胞とそれらの正常親細胞を比較したいくつかの例において注目すべきことは、相違点の一つが、正常細胞の表面は滑らかであるが、形質転換された細胞の表面には多数の毛髪状の糸状体を有するということである(ダーネル(Darnell)ら、「分子細胞生物学(Molecular Cell Biology)」(第2版)、サイエンスアメリカンブックス(Sci. Am. Books)、W.H.フリーマン(Freeman)株式会社、米国、ニューヨーク州(1990年))。これらの特徴からすると、本実施例の顕微鏡像で明らかにされたように、MX感染HeLa細胞は超形質転換した外観を呈している。
【0170】
さらに、数種の腫瘍では、ミトコンドリアの増幅が報告されている(ベーンハード(Bernhard)、W.,「分子細胞学ハンドブック(Handbook of Molecular Cytology)」、687−715ページ、リマ・デ・ファリア(Lima de Faria)編、ノースホランド(North-Holland)出版株式会社、アムステルダム−ロンドン(1972年))。そのような増幅は、ヤーヌス緑(Janus′ green )で特にミトコンドリアが強く染色されたMX感染HeLa細胞において顕著であり、一方、対照のHeLa細胞では弱くしか染色されなかった。
【0171】
電子顕微鏡の熟練者によっても、腫瘍細胞に特異的な構造的特徴は見出すことができなかったことを付記しておく。
【0172】
実施例11
MN遺伝子発現を阻害するアンチセンスODNs
p54/58N蛋白質の両方が一つの遺伝子によってコードされているか否かを確認するために、アンチセンスODNsを用いて以下の実験を行った。過密培養を作るために、事前に希薄培養していたHeLa細胞をまき、MN mRNAの5’末端に相補的で2つの遺伝子に特異的なODNsの非存在下および存在下で130時間インキュベートした。HeLa細胞は、10%のFCSを含むDMEM1mlあたり8×105個となるように継代した。同時に、次のようにODNsを培地に添加した。(A)29−mer ODN1(5’CGCCCAGTGGGTCATCTTCCCCAGAAGAG3’(配列番号3)、44−72位に相補的)を4μM(最終濃度)、(B)19−mer ODN2(5’GGAATCCTCCTGCATCCGG3’(配列番号4)、12−30位に相補的)を4μM(最終濃度)、(C)CDN1およびODN2をおのおの2μM(最終濃度)、(D)同様に処理したが、ODNsを加えずにインキュベートしたものであり、対照とする。130時間後に細胞から抽出物を調製し、125IでラベルしたMAbs M75を用いたイムノブロットにより分析した。細胞の蛋白質抽出物は、MAbs M75を用いたイムノブロットおよびRIAにより分析した。 ODNsを添加したHeLa細胞の培養の結果、p54/58N合成がかなり阻害されていることがわかった。最終濃度4μMの19−mer ODN2が非常に効果があり、RIAで測定したところ、40%の阻害を起こした。一方、29−mer ODN1(4μM)および2つのODNsの組合せ(おのおの最終濃度2μM)では効果は低くなり、RIAで23−35%のMN関連蛋白質の減少を示した。このとき、特異的MAb H460を用いたRIAで測定した、別のHeLa細胞蛋白質の量は、全ての細胞変異種においてほぼ同程度であった。最も重要なことは、イムノブロットにおけるODNsによる特異的阻害は、p54/58N蛋白質の両方に影響を与えていることである。このことから、発明者らがクローンしたMN遺伝子は、HeLa細胞内においてp54/58N蛋白質の両方をコードしていると結論づけられる。
【0173】
実施例12
腫瘍形成性細胞由来および非腫瘍形成性細胞株由来のMN mRNAのノーザンブロッティング
ヒト細胞株のMN mRNAのノーザンブロッティングを行った。全RNAは、次の細胞株からグアニジンチオシアネートCsCl法により調製した。高密度(A)および希薄(B)培養で増殖したHeLa細胞;(C)CGL1(H/F−N)ハイブリッド細胞;(D)CGL3分離体および(E)CGL4分離体(両者ともH/F−T)、ならびに(F)ヒト胚線維芽細胞。15μgのRNAを1.2%のホルムアルデヒドゲルで分離し、ハイボンドCスーパーメンブレン(Hybond C Super membrane)(アマシャム(Amersham)社)の上にブロットした。MN cDNAのNotIプローブを、ランダムプライミング(マルチプライムDNAラベリングシステム(Multiprime DNA labelling system)、(アマシャム(Amersham)社)を用いてラベルした。50%のホルムアミド存在下42℃でハイブリダイゼーションを行い、0.1%のSSPEと0.1%のSDSを用いて65℃で最終洗浄を行った。RNAラダー(RNA ladder)(0.24−9.5kb)(ベセスダ研究所(BRL)(Bethesda Research Laboratories)、米国、メリーランド州、ベセスダ)をサイズマーカーとして用いた。
【0174】
検出されたのは1.5kbのMN特異的mRNAで、2つの腫瘍形成性分離体クローン、CGL3とCGL4(H/F−T)からのみであり、非腫瘍形成性ハイブリッドクローンCGL1(H/F−N)あるいは正常ヒト線維芽細胞からは検出されなかった。さらに、1.5kbのmRNAは、高密度培養で増殖したHeLa細胞からは検出されたが、希薄培養のものからは検出されなかった。
【0175】
このように、MN関連蛋白質が腫瘍形成性と関連しているということについてノーザンブロッティングの結果は、上述の実施例の結果と一致している。
【0176】
実施例13
各種の脊椎動物種由来のゲノムDNAのサザンブロッティングによるMN遺伝子の検出
いろいろな脊椎動物のゲノムDNA中のMN遺伝子をサザンブロッティングにより検出した。次のものの染色体DNAをSstIで消化した。(A)ニワトリ、(B)ウシ、(C)ネコ、(D)MX感染HeLa細胞、(E)マウスNIH 3T3細胞、(F)ヒト胎盤細胞、(G)HeLa細胞、(H)ヒツジ、(I)ヒトメラノーマ細胞、および(J)サルベロ(Vero)細胞。制限酵素による断片を、0.7%の寒天ゲルで分離し、ハイボンドNメンブレン(Hybond N membrane)(アマシャム(Amersham)社)の上にアルカリブロットした。MN cDNAプローブのラベリングおよびハイブリダイゼーションの操作は、実施例12のノーザンブロット分析と同様に行った。
【0177】
染色体DNAのSstI制限酵素断片を用いたサザンブロット分析は、いずれの種においても、約1.5kbのところに唯一のバンドを示した。さらに、XhoIおよびSalIによる切断によって生じた制限酵素断片を用いたハイブリダイゼーションにおいては、おのおのの染色体DNAサンプルは4.5kbおよび4.7kbにそれぞれ1本のバンドを示した。これらの結果から、脊椎動物のゲノムにおいては、MN遺伝子は単一のコピーとして存在していることが示唆される。さらに、これらの結果は、MN遺伝子がその側面配列と共に保存性であることを示唆している。
【0178】
SstIの開裂部位がMN遺伝子における自然な境界を形成し、MN mRNAのサイズがノーザンブロットにおけるMN遺伝子のサイズと同じであることから(実施例12の結果と比較されたい)、MN遺伝子にはイントロンが存在しないと推論される。この結論は、MN cDNAの制限酵素パターンとMN特異的ゲノムのSstI断片のそれとが同じであることからも支持される。
【0179】
下記に挙げる材料は、アメリカンタイプカルチャーコレクション(American Type Culture Collection)(ATCC)(米国、メリーランド州、ロックヴィル、パークローン通り(Parklawn Drive)12301番地、郵便番号20852)に寄託されている。寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約(Budapest Treaty on the International Recognition of Deposited Microorganisms for the Purposes of Patent Procedure and Regulations thereunder)(ブダペスト条約)に基づいてなされた。生細胞培養の維持は寄託の日から30年間保障されている。微生物は、ブダペスト条約の規定に基づいてATCCから入手可能であり、寄託者とATCCは、関連する米国特許が付与されると制限されることなく入手できることについて合意している。寄託した株が入手可能できるとは言っても、ある政府機関がその国の特許法に基づいて許可する権利に違反して発明を実施する実施権を与えるということではない:
ハイブリドーマ 寄託日 ATCC番号
VU−M75 1992年9月17日 HB 11128
【0180】
本発明に関する以上の具体例の記述は、例示および説明のためのものである。これらの具体例は全てではなく、また、開示された特定のものに発明を限定するものでもない。さらに、以上の教示から多くの修正や変形が可能なことは明かである。以上の具体例は、本発明の要旨を説明するため、および、当業者が意図する特定の使用に適するようにさまざまな修正を加えた各種の態様で本発明を利用できるようにするために、選択して記載したものである。本発明の範囲は、明細書に添付された請求項によって定められる。
【0181】
引用した全ての文献は、参考のために本明細書に取り入れられている。
【特許請求の範囲】
【請求項1】
単離されたMN遺伝子および/またはその断片、あるいは該MN遺伝子またはその断片と実質上相補的である、単離された核酸配列を含むことを特徴とする組成物。
【請求項2】
第1A図−第1B図[配列番号1]に示される塩基配列からなるcDNA配列あるいはそのcDNA配列とストリンジェントな条件下でハイブリダイズする塩基配列からなるcDNA配列またはその断片に実質上相補的な核酸配列を含むことを特徴とする請求項1記載の組成物。
【請求項3】
前記MN遺伝子またはその断片、あるいは前記MN遺伝子またはその断片と実質上相補的である核酸配列がベクターに含まれていることを特徴とする請求項1記載の組成物。
【請求項4】
前記核酸配列が、前記ベクター内の発現コントロール配列に機能的に結合していることを特徴とする請求項3記載の組成物。
【請求項5】
請求項4記載の組成物で形質転換またはトランスフェクトした原核細胞または真核細胞である単細胞宿主。
【請求項6】
MN遺伝子および/またはその断片によりコードされている蛋白質および/またはポリペプチドを含むことを特徴とする組成物。
【請求項7】
前記蛋白質および/またはポリペプチドが、第1A図−第1B図[配列番号2]に示されるアミノ酸配列あるいはそのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列またはその一部、あるいは前記アミノ酸配列またはその一部の変異体および/または第1A図−第1B図に示されるアミノ酸配列あるいはそのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列に実質上相補的な蛋白質および/またはポリペプチドからなることを特徴とする請求項6記載の組成物。
【請求項8】
pGEX−3X−MNであることを特徴とする請求項7記載の組成物。
【請求項9】
MN蛋白質/ポリペプチドに特異的に結合する単一の抗体または複数の抗体を含むことを特徴とする組成物。
【請求項10】
前記抗体がモノクローナル抗体であることを特徴とする請求項9記載の組成物。
【請求項11】
前記モノクローナル抗体がM75と称され、ATCCにATCC番号HB 11128として寄託されているハイブリドーマVU−M75から産生されることを特徴とする請求項10記載の組成物。
【請求項12】
前記抗体が、化学療法剤もしくは毒性剤および/またはイメージング剤と結合していることを特徴とする請求項9記載の組成物。
【請求項13】
MN蛋白質/ポリペプチドに対する特異的結合性を有するモノクローナル抗体を産生するハイブリドーマを含むことを特徴とする組成物。
【請求項14】
患者の腫瘍性疾患をイメージングする方法であって、
a.請求項12記載の抗体を適切にラベルして該患者に投与し、次に、
b.該ラベル化した抗体の結合を検出する
各工程を含むことを特徴とする方法。
【請求項15】
癌細胞に化学療法剤または毒性剤を輸送する方法であって、請求項12記載の抗体に該細胞を接触させる工程を含むことを特徴とする方法。
【請求項16】
MN蛋白質またはポリペプチドを組換えにより産生する方法であって、
a.請求項4記載の組成物を用いて単細胞宿主を形質転換し、
b.該単細胞宿主を培養してMN蛋白質および/またはポリペプチドを発現させ、次に、
c.該MN蛋白質および/またはポリペプチドを抽出し単離する、
各工程を含むことを特徴とする方法。
【請求項17】
脊椎動物サンプル中のMN抗原を検出および/または定量する方法であって、
a.該サンプルを請求項9記載の単一または複数の抗体と接触させ、
b.該サンプル中の抗原に対する該抗体の結合を検出および/または定量する、
各工程を含むことを特徴とする方法。
【請求項18】
脊椎動物サンプル中のMN特異的抗体を検出および/または定量する方法であって、
a.脊椎動物のサンプルをMN蛋白質/ポリペプチドと接触させてインキュベートし、次に、
b.該サンプル中の抗体に対する該MN蛋白質/ポリペプチドの結合を検出および/または定量する、
各工程を含むことを特徴とする方法。
【請求項19】
一つもしくはそれ以上の実質的に純粋なMN蛋白質および/またはポリペプチドが免疫原となる量で存在して、生理学的に許容性で非毒性の基剤中に分散されたワクチンであって、該量がMNタンパク質の発現に関連する腫瘍性疾患に対して脊椎動物を免疫するのに有効な量であることを特徴とするワクチン。
【請求項20】
MN遺伝子から転写されたmRNAに実質的に相補的なアンチセンス核酸配列を投与することにより、MN遺伝子の発現を阻害することを含むことを特徴とする、腫瘍性疾患および/または前腫瘍性疾患の治療法。
【請求項1】
単離されたMN遺伝子および/またはその断片、あるいは該MN遺伝子またはその断片と実質上相補的である、単離された核酸配列を含むことを特徴とする組成物。
【請求項2】
第1A図−第1B図[配列番号1]に示される塩基配列からなるcDNA配列あるいはそのcDNA配列とストリンジェントな条件下でハイブリダイズする塩基配列からなるcDNA配列またはその断片に実質上相補的な核酸配列を含むことを特徴とする請求項1記載の組成物。
【請求項3】
前記MN遺伝子またはその断片、あるいは前記MN遺伝子またはその断片と実質上相補的である核酸配列がベクターに含まれていることを特徴とする請求項1記載の組成物。
【請求項4】
前記核酸配列が、前記ベクター内の発現コントロール配列に機能的に結合していることを特徴とする請求項3記載の組成物。
【請求項5】
請求項4記載の組成物で形質転換またはトランスフェクトした原核細胞または真核細胞である単細胞宿主。
【請求項6】
MN遺伝子および/またはその断片によりコードされている蛋白質および/またはポリペプチドを含むことを特徴とする組成物。
【請求項7】
前記蛋白質および/またはポリペプチドが、第1A図−第1B図[配列番号2]に示されるアミノ酸配列あるいはそのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列またはその一部、あるいは前記アミノ酸配列またはその一部の変異体および/または第1A図−第1B図に示されるアミノ酸配列あるいはそのアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列に実質上相補的な蛋白質および/またはポリペプチドからなることを特徴とする請求項6記載の組成物。
【請求項8】
pGEX−3X−MNであることを特徴とする請求項7記載の組成物。
【請求項9】
MN蛋白質/ポリペプチドに特異的に結合する単一の抗体または複数の抗体を含むことを特徴とする組成物。
【請求項10】
前記抗体がモノクローナル抗体であることを特徴とする請求項9記載の組成物。
【請求項11】
前記モノクローナル抗体がM75と称され、ATCCにATCC番号HB 11128として寄託されているハイブリドーマVU−M75から産生されることを特徴とする請求項10記載の組成物。
【請求項12】
前記抗体が、化学療法剤もしくは毒性剤および/またはイメージング剤と結合していることを特徴とする請求項9記載の組成物。
【請求項13】
MN蛋白質/ポリペプチドに対する特異的結合性を有するモノクローナル抗体を産生するハイブリドーマを含むことを特徴とする組成物。
【請求項14】
患者の腫瘍性疾患をイメージングする方法であって、
a.請求項12記載の抗体を適切にラベルして該患者に投与し、次に、
b.該ラベル化した抗体の結合を検出する
各工程を含むことを特徴とする方法。
【請求項15】
癌細胞に化学療法剤または毒性剤を輸送する方法であって、請求項12記載の抗体に該細胞を接触させる工程を含むことを特徴とする方法。
【請求項16】
MN蛋白質またはポリペプチドを組換えにより産生する方法であって、
a.請求項4記載の組成物を用いて単細胞宿主を形質転換し、
b.該単細胞宿主を培養してMN蛋白質および/またはポリペプチドを発現させ、次に、
c.該MN蛋白質および/またはポリペプチドを抽出し単離する、
各工程を含むことを特徴とする方法。
【請求項17】
脊椎動物サンプル中のMN抗原を検出および/または定量する方法であって、
a.該サンプルを請求項9記載の単一または複数の抗体と接触させ、
b.該サンプル中の抗原に対する該抗体の結合を検出および/または定量する、
各工程を含むことを特徴とする方法。
【請求項18】
脊椎動物サンプル中のMN特異的抗体を検出および/または定量する方法であって、
a.脊椎動物のサンプルをMN蛋白質/ポリペプチドと接触させてインキュベートし、次に、
b.該サンプル中の抗体に対する該MN蛋白質/ポリペプチドの結合を検出および/または定量する、
各工程を含むことを特徴とする方法。
【請求項19】
一つもしくはそれ以上の実質的に純粋なMN蛋白質および/またはポリペプチドが免疫原となる量で存在して、生理学的に許容性で非毒性の基剤中に分散されたワクチンであって、該量がMNタンパク質の発現に関連する腫瘍性疾患に対して脊椎動物を免疫するのに有効な量であることを特徴とするワクチン。
【請求項20】
MN遺伝子から転写されたmRNAに実質的に相補的なアンチセンス核酸配列を投与することにより、MN遺伝子の発現を阻害することを含むことを特徴とする、腫瘍性疾患および/または前腫瘍性疾患の治療法。
【図1A】


【図1B】

【図2】
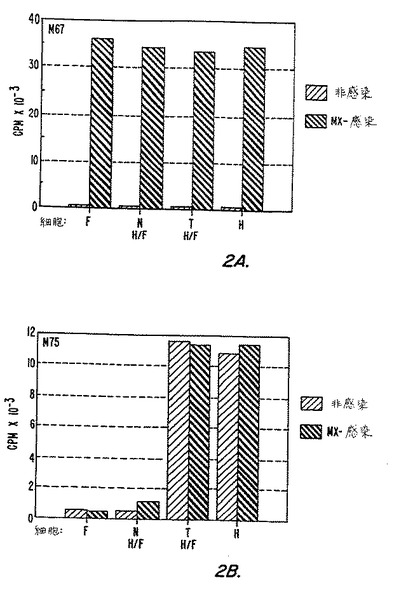
【図3】
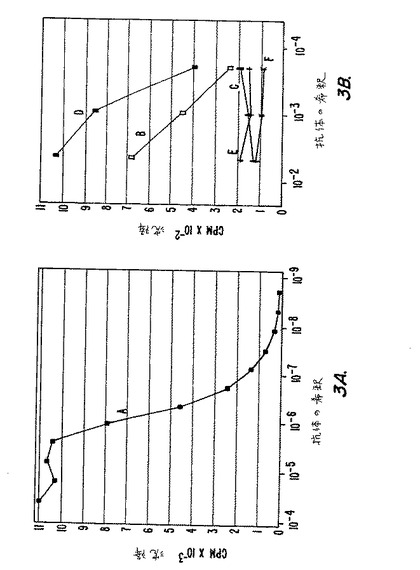
【図4】
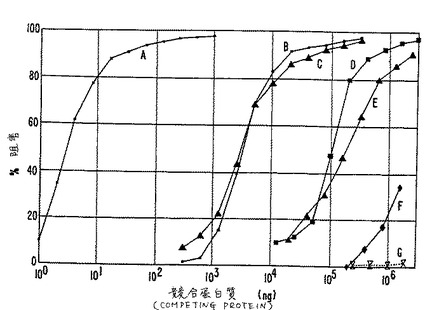
【図1B】
【図2】
【図3】
【図4】
【公開番号】特開2011−36254(P2011−36254A)
【公開日】平成23年2月24日(2011.2.24)
【国際特許分類】
【出願番号】特願2010−200720(P2010−200720)
【出願日】平成22年9月8日(2010.9.8)
【分割の表示】特願2005−297594(P2005−297594)の分割
【原出願日】平成5年3月8日(1993.3.8)
【出願人】(300058891)バイエル コーポレーション (5)
【出願人】(505343985)インスティテュート オブ ヴァイロロジー (2)
【Fターム(参考)】
【公開日】平成23年2月24日(2011.2.24)
【国際特許分類】
【出願日】平成22年9月8日(2010.9.8)
【分割の表示】特願2005−297594(P2005−297594)の分割
【原出願日】平成5年3月8日(1993.3.8)
【出願人】(300058891)バイエル コーポレーション (5)
【出願人】(505343985)インスティテュート オブ ヴァイロロジー (2)
【Fターム(参考)】
[ Back to top ]