
Malvaモザイクウイルスおよびウイルス様粒子およびこれらの使用
ポテックスウイルスであるMalvaモザイクウイルス(MaMV)、およびMaMVの外被タンパク質を含むウイルス様粒子(VLP)の、免疫原性特性が開示される。MaMV外被タンパク質から調製されるVLP、VLPを調製するための方法、MaMV外被タンパク質ポリペプチドおよび外被タンパク質をコードするポリヌクレオチドが教示される。さらに、MaMVまたはMaMVの外被タンパク質を含むVLPのみを、またはこれと1もしくは2以上の抗原とを組み合わせて含む免疫原性組成物、および前記組成物の、動物においてワクチン接種する、および/または免疫反応を誘発するための使用もまた教示される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ワクチン製剤およびアジュバントの分野、および特に植物ウイルス粒子に基づくワクチンおよびアジュバントに関する。
【背景技術】
【0002】
ワクチン開発の多数の新しいアプローチの中で、ウイルスヌクレオカプシドから作製されたウイルス様粒子(VLP)は、期待が持てる戦略として登場してきた。現在まで2つのVLPワクチン、B型肝炎ウイルス(HBV)およびヒトパピローマウイルス(HPV)が、ヒトにおいて効率的に機能することが示されている(Fagan et al., 1987, J. Med. Virol, 21:49-56;Harper et al., 2004, Lancet, 364:1757-1765)。例えば、ヒトパピローマウイルス(HPV)主要カプシドタンパク質L1から作製されたVLPは、女性の子宮頸癌の発症に対して100%の防御を与えることが示された(Ault, K.A., 2006, Obstet. Gynecol. Surv. 61 :S26-S31; Harper et al., 2004, Lancet 364: 1757- 1765、また国際特許出願PCT/US01/18701 (WO 02/04007)も参照のこと)。バクテリオファージQβ(Maurer et al., 2005, Eur. J. Immunol. 35:2031-2040)、ウイルスコアタンパク質から作製されたB型肝炎ウイルスVLP(Mihailova et al., 2006, Vaccine 24:4369-4377;Pumpens et al., 2002, Intervirology 45:24-32)、およびパルボウイルスVLP(Antonis et al., 2006, Vaccine 24:5481-5490;Ogasawara et al., 2006, In Vivo 20:319-324)もまた、エピトープを担持し、強い抗体反応を誘発することが示された。同様に、米国特許第6,627,202号には、HBVカプシド結合ペプチドにより架橋された抗原を含むHBVコアタンパク質の、種々のウイルスおよび細菌を標的とした抗原、またはこれらに由来する抗原を含むエピトープ送達システムとしての使用が記載されている。
【0003】
植物ウイルスからのVLPの、エピトープ提示系としての使用は記載されている。植物ウイルスは主に高度に免疫原性であるタンパク質から構成され、複雑な反復性の結晶組織を有する。さらにこれらは、系統発生学的に動物の免疫系からは離れており、そのことが、これらをワクチン開発の良好な候補者にしている。例えば、ササゲ(cowpea)モザイクウイルス(CPMV)、ジョンソングラスモザイクウイルス(JGMV)、タバコモザイクウイルス(TMV)、およびアルファルファモザイクウイルス(AIMV)が、目的のエピトープ提示のために改変された(Canizares, M. C. et al., 2005, Immunol. Cell. Biol. 83:263-270;Brennan et al., 2001, Molec. Biol. 17: 15-26;Saini and Vrati, 2003, J. Virol. 77:3487-3494)。国際特許出願PCT/GB97/01065 (WO 97/39134)には、外被タンパク質および非ウイルス性タンパク質を含み、例えばペプチドエピトープの提示のために用いることができる、キメラウイルス様粒子が記載されている。国際特許出願PCT/USOl/07355 (WO 01/66778)には、リンカーを介してN末端において、病原体微生物のエピトープを含んでよい目的のポリペプチドに融合した、植物ウイルス外被タンパク質、および具体的にはトバモウイルス(tobamovirus)外被タンパク質が記載されている。国際特許出願PCT/USO1/20272 (WO 02/00169)には、外来ペプチドに、および具体的にはニューカッスル病ウイルスまたはヒト免疫不全ウイルス(HIV)エピトープに融合した、ジャガイモウイルスY外被タンパク質または切断型インゲン黄斑モザイクウイルス外被タンパク質のどちらかを含むワクチンが記載されている。また、米国特許第6,042,832号には、例えば病原体エピトープなどのポリペプチドの、アルファルファモザイクウイルスまたはイラルウイルス(ilarvirus)カプシドタンパク質への融合物を動物に投与して、免疫反応を高めるための方法が記載されている。
【0004】
パパイアモザイクウイルス(PapMV)の外被タンパク質由来のVLP、およびそれらの免疫増強剤としての使用が記載されている(国際特許出願PCT/CA03/00985 (WO 2004/004761)および米国特許出願第11/556,678号(US2007/0166322))。大腸菌におけるPapMV外被タンパク質の発現は、反復性かつ結晶様式での数百のCPサブユニットから構成されるVLPの自己集合をもたらす(Tremblay et al., 2006, FEBS J 273: 14)。PapMV CP欠失コンストラクトの発現および精製の研究はさらに、CPサブユニットの自己集合(または多量体形成)が、機能に重要であることを示す(Lecours et al., 2006, Protein Expression and Purification, 47:273-280)。gp100またはインフルエンザウイルスM1タンパク質のどちらかからのエピトープを含むPapMV VLPの能力は、エピトープのMHCクラスI交差提示を誘発し、特異的ヒトT細胞の増幅(expansion)をもたらすことが示された(Leclerc, D., et al, J. Virol, 2007, 81(3):1319-26; Epub. ahead of print November 22, 2006)。さらに、C型肝炎ウイルスE2エンベロープタンパク質由来のエピトープを含むPapMV VLPは、マウスにおいて、PapMV VLPおよびE2ペプチドに対して体液性の反応を誘発することが示された(Denis et al., 2007, Virology, 363(1): 59-68)。
【0005】
plasmodiophora brassicaeの休眠胞子を結合することのできる、親和性ペプチドに融合したパパイアモザイクウイルスVLPの使用も記載されている(Morin et al., 2007, J. Biotechnology, 128: 423-434)。VLPは、P. brassicae胞子を高い親和力で結合できることが示され、P. brassicaeの検出のための抗体の代替として提唱された。
【0006】
HIV、HCV、EBVまたはインフルエンザウイルスからの種々の抗原決定基を担持するジャガイモウイルスX(PVX)由来のVLPが、記載されている(欧州特許出願第1 167 530号)。HIVエピトープを担持するPVX VLPの、マウスにおいて体液性および細胞媒介性経路を介して抗体産生を誘発する能力もまた、記載されている。追加のアジュバントをPVX VLPと併用すると、この効果が増強された。
【0007】
B型肝炎コアタンパク質またはパルボウイルスVLPは、これらが遺伝情報を担持しない場合であってもCTL反応を誘発することが報告されており(Ruedl et al., 2002, Eur. J. Immunol. 32; 818-825;Martinez et al., 2003, Virology, 305; 428-435)、それらが陥入した細胞において活性に複製することができない。リンパ球性脈絡髄膜炎ウイルス(LCMV)または鶏卵アルブミンからのエピトープを担持するVLPの、in vivoでの樹状細胞による交差提示も記載されている(Ruedl et al., 2002、同上;Moron, et al., 2003, J. Immunol. 171 :2242-2250)。LCMVからのエピープを担持するB型肝炎コアタンパク質VLPの、CTL反応をプライムする能力も記載されているが、しかしこのVLPは、それのみで投与された場合にはCTL反応を誘発できず、ウイルスチャレンジに対する効果的な防御を媒介することができなかった。有効なCTL反応は、VLPを抗CD40抗体またはCpGオリゴヌクレオチドと併用した場合にのみ、誘発された(Storni, et al., 2002, J. Immunol. 168:2880-2886)。初期の報告によれば、LCMVからのペプチドを担持する豚パルボウイルス様粒子(PPMV)が、マウスを致死的なLCMVチャレンジに対して保護できたことが示された(Sedlik, et al., 2000, J. Virol. 74:5769-5775)。
【0008】
ポテックスウイルスであるMalva葉脈壊死ポテックスウイルス(MVNV)は、1990年にブラジルにおいてMalva parviflora(ウサギアオイ)に感染するウイルスとして報告され(Brunt et al., (eds.) 1996 onwards. "Plant Viruses Online: Descriptions and Lists from the VIDE Database. Version: 20th August 1996")、Malva parviflora上に局所的な病変部および全体的な葉脈壊死が検出された。
【0009】
この背景情報は、本出願人が本発明に関連する可能性があると信じる既知の情報を明らかにする目的で提供されている。任意の前述の情報が、本発明に対する先行技術を構成するとの認定は必ずしも意図されず、またそのように解釈されるべきではない。
【発明の概要】
【0010】
本発明の目的は、Malvaモザイクウイルスおよびウイルス様粒子(VLP)、ならびにこれらの使用を提供することである。本発明の1つの側面により、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および薬学的に許容し得る担体を含む、免疫原性組成物が提供される。
【0011】
本発明の他の側面により、動物において免疫反応を誘発するための方法であって、前記動物に対して、本発明の免疫原性組成物の有効量を投与することを含む前記方法が提供される。
【0012】
本発明の他の側面により、動物に、疾病、疾患、または感染に対するワクチン接種するための方法であって、該動物対して、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および1または2以上の抗原を含む、免疫原性組成物の有効量を投与することを含む、前記方法が提供される。
【0013】
本発明の他の側面により、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)の、免疫原性組成物の調製における使用が提供される。
【0014】
本発明の他の側面により、Malvaモザイクウイルス(MaMV)外被タンパク質を含むウイルス様粒子(VLP)が提供される。
【0015】
本発明の他の側面により、ウイルス様粒子(VLP)を調製するための方法であって、Malvaモザイクウイルス(MaMV)外被タンパク質を宿主細胞中に発現するステップを含む、前記方法が提供される。
【0016】
本発明の他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質が提供される。
【0017】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質をコードする、単離ポリヌクレオチドが提供される。
【0018】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリポリヌクレオチドにより遺伝子操作された、宿主細胞が提供される。
【0019】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリヌクレオチド、または、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリポリヌクレオチドにより遺伝子操作された宿主細胞の、ウイルス様粒子の調製のための使用が提供される。
【図面の簡単な説明】
【0020】
本発明のこれらおよび他の特徴は、添付の図を参照した以下の詳細な説明において、さらに明らかにされる。
【0021】
【図1A】図1Aは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【図1B】図1Bは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【図1C】図1Cは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【0022】
【図2】図2は、Malvaモザイクウイルスの外被タンパク質のアミノ酸配列を示す(配列番号2)。
【0023】
【図3】図3は、Malvaモザイクウイルスの外被タンパク質をコードするヌクレオチド配列を示す(配列番号3)。
【0024】
【図4】図4は、(A)Malvaモザイクウイルスに感染したMalva種上のモザイク症状を示す写真、(B)Malva neglecta Wallr.に感染した葉からの抽出物を接種した場合の、Chenopodium quinoa(キノア)上のモザイク症状を示す写真、(C)C. quinoaの感染した葉からの抽出物を接種した場合のC. quinoa上に観察されるモザイク病変部を示す写真、および(D)精製したMalvaモザイクウイルスの電子顕微鏡写真(バーは50nmを表わす)である。
【0025】
【図5】Malvaモザイクウイルスの5’末端の解析結果を示す。バーは、配列決定した全てのクローンに見出された各々の配列の頻度を表わす。
【0026】
【図6】図6(A)は、Malvaモザイクウイルス(MaMV)のゲノム構成の模式図であり、(B)は、健康な(レーン1)、または感染した(レーン2)Chenopodium quinoaから、MaMV外被タンパク質に指向するプローブを用いて抽出した全RNAを示す、ノーザンブロットであり、および(C)は、種々のポテックスウイルスのオクタヌクレオチド推定sgプロモーター配列の配列アラインメントを示す(高度に保存された領域は、黒または灰色の四角で強調した)。
【0027】
【図7】図7は、Malvaモザイクウイルス3’非翻訳領域の二次構造の模式図である。
【0028】
【図8】図8は、Malvaモザイクウイルスおよび他のポテックスウイルスのレプリカーゼおよびカプシドタンパク質の系統学的解析結果を示す図である。
【0029】
【図9】図9(A)は、大腸菌から単離された、精製組換えMalvaモザイクウイルス(MaMV)外被タンパク質を表わすSDS−PAGEゲルを、および(B)は、組換え外被タンパク質を含むMaMV VLPの電子顕微鏡写真を示す。バーは50nmである。
【0030】
【図10】図10は、(A)プラスミドpMaMV−CP−6H中に含有されたMaMV外被タンパク質遺伝子のヌクレオチド配列(配列番号23)(開始コドンは下線で、および終止コドンは太字イタリック体で表す);(B)MaMV CP−SMタンパク質をコードするヌクレオチド配列(配列番号62)(イタリック体で下線の配列は、2つの制限酵素認識配列を表す);および(C)MaMV CP gl−SMタンパク質をコードするヌクレオチド配列(配列番号64)(イタリック体で下線の配列は、2つの制限酵素認識配列およびスペーサーをコードする配列を表す)を示す。
【0031】
【図11】図11は、(A)プラスミドpMaMV−CP−6H中に含有されたMaMV外被タンパク質遺伝子によりコードされた外被タンパク質のアミノ酸配列(配列番号24);(B)MaMV CP−SMタンパク質のアミノ酸配列(配列番号63)(イタリック体で下線の配列は、コードされたヌクレオチド配列中の2つの制限酵素認識配列の封入により挿入されたアミノ酸を表す);および(C)MaMV CP gl−SMタンパク質のアミノ酸配列(配列番号65)(イタリック体で下線の配列は、コードされたヌクレオチド配列およびスペーサー配列中の2つの制限酵素認識配列の封入により挿入されたアミノ酸を表す)を示す。それぞれの配列におけるHisタグは太字で示す。
【0032】
【図12】図12は、以下の産生を表すグラフである:(A)、(B)および(C):それぞれ、アジュバント不在のもとで、s.c.によりMaMV VLPを1度注射したBalb/Cマウスにおける、総IgG、IgG1およびIgG2a(ELISAにより測定);および(D)、(E)および(F):それぞれ、同じマウスにおいて、アジュバント不在のもとで、s.c.によりMaMV VLPを2回目(40日目)に注射した場合の、総IgG、IgG1およびIgG2a。ELISAは、最初の免疫化後5日目、10日目、14日目において採取した血液、および2回目の免疫化後4日目に採取した血液で実施した。+の記号は、ELISAのベースラインとして用いた陰性対照(免疫前血清)を表す。
【0033】
【図13】図13は、チフス菌から精製されたポリンであるOmpCおよびOmpFのプロファイルを示すSDS−PAGEゲルの図である。
【0034】
【図14】図14は、gp100(配列番号59)およびインフルエンザM1タンパク質(配列番号60)からのHLA−A*0201エピトープ(太字および下線)を、それぞれそのフランキング配列と共に示した図である。
【0035】
【図15】図15は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV CPを発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV CP;B)組換えMaMV CPを含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV CPが高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0036】
【図16A−C】図16A−Cは、Fluviral(登録商標)に対する(A)総IgG、(B)IgG1および(C)IgG2aの産生を実証するグラフであって、これは、MaMV VLP(3または30μg)またはミョウバンをアジュバントとしたFluviral(登録商標)での免疫化後14日目に、1群当たり5匹のBalb/Cマウスの血清において測定したものである。
【図16D】図16Dは、Fluviral(登録商標)に対するNPタンパク質に対するIgG2aの産生を実証するグラフであって、これは、MaMV VLP(3または30μg)またはミョウバンをアジュバントとしたFluviral(登録商標)での免疫化後14日目に、1群当たり5匹のBalb/Cマウスの血清において測定したものである。
【0037】
【図17】図17は、次をコードするヌクレオチド配列である:(A)MaMV−M1タンパク質(配列番号66)(M1エピトープをコードする配列は下線で示す);(B)MaMV−gp100タンパク質(配列番号68)(gp100エピトープをコードする配列は下線で示す);(C)MaMV g1−F3タンパク質(配列番号70)(F3ペプチドをコードする配列は下線で示す)。
【0038】
【図18】図18は、次のアミノ酸配列である:(A)MaMV−M1タンパク質(配列番号67)(M1エピトープは太字下線で示す);(B)MaMV−gp100タンパク質(配列番号69)(gp100エピトープは太字下線で示す);(C)MaMV g1−F3タンパク質(配列番号71)(F3ペプチドは太字下線で示す)。各配列のHis−tagは太字で示す。
【0039】
【図19】図19は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−SMタンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−SMタンパク質;B)組換えMaMV−SMタンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−SMタンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0040】
【図20】図20は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV g1−SMタンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV g1−SMタンパク質;B)組換えMaMV g1−SMタンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV g1−SMタンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0041】
【図21】図21は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−M1タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−M1タンパク質;B)組換えMaMV−M1タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−M1タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0042】
【図22】図22は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−gp100タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−gp100タンパク質;B)組換えMaMV−gp100タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−gp100タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0043】
【図23】図23は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV g1−F3タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV g1−F3タンパク質;B)組換えMaMV g1−F3タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV g1−F3タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0044】
【図24】図24は、異なるMaMV VLPの平均長(各組換えコンストラクトから100個のVLPを測定した)を示すグラフである。
【0045】
【図25】図25は、CD40活性化Bリンパ球を、蛍光標識パパイアモザイクウイルス(PapMV)VLP、MaMV VLPおよびMaMV−M1 VLPでパルスした、フローサイトメトリーの結果を示す図である。
【0046】
【図26】図26は、(A)T2細胞を、蛍光標識パパイアモザイクウイルス(PapMV)VLPおよびMaMV VLPでパルスしたフローサイトメトリーの結果であり、時間と共にVLPの取り込みを示す;および(B)は、PapMV VLPおよびMaMV VLPのT2細胞による取り込みを、時間に渡り平均蛍光強度(MFI)で測定した結果のグラフ表示である。
【0047】
【図27】図27は、以下を示すグラフである:M1エピトープ(左)もしくはgp100エピトープ(右)を担持するPapMV VLPまたはMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量;(B)M1エピトープ(左)もしくはgp100エピトープ(右)を担持する異なる量のMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量;および(C)4℃にて、M1エピトープ(左)もしくはgp100エピトープ(右)を担持するPapMV VLPまたはMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量。IFN−γの分泌は、ELISAにより評価した。
【0048】
【図28】図28は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#405に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0049】
【図29】図29は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#542に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0050】
【図30】図30は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#614に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0051】
【図31】図31は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#621に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0052】
【図32】図32は、MaMV−SM VLP、MaMV g1−F3 VLP、MaMV g1−SM VLP+F3、MaMV−M1 VLP、またはMaMV−gp100 VLPを注射したマウス(1群当たり10匹)において、(A)ペプチドF3、M1またはgp100、および(B)MaMV CPに対する総IgG応答を示す、ELISAの結果を表す。
【0053】
【図33】図33は、(A)MaMVおよびPapMV CPアミノ酸配列のアラインメントであって、2つのCP間で31.2%の同一性を示した;(B)MaMV CPに指向された抗体は、PapMV VLPを認識できないことを示すELISAの結果;および(C)MaMV CPに指向された抗体は、PapMV VLPを認識できないことを示すELISAの結果である。
【0054】
発明の詳細な説明
本発明は、ポテックスウイルスであるMalvaモザイクウイルス(MaMV)およびMaMVの外被タンパク質を含むウイルス様粒子(VLP)の、本明細書において同定された免疫原性特性に関する。本発明は、MaMVおよびMaMV VLPのこれらの免疫原性特性に基づく、動物における免疫反応の生成に関連する種々の態様を包含する。1つの側面において、例えば、本発明は以下を提供する:MaMVまたはMaMV外被タンパク質を含むVLPの、免疫原性組成物の調製のための使用;MaMVまたはMaMV外被タンパク質を含むウイルス様粒子、および任意に1または2以上の抗原を含む、免疫原性組成物;およびこれら組成物の、動物において免疫反応を誘発するための、および/または動物にワクチン接種するための使用。本発明の他の側面は、MaMV外被タンパク質の、VLPの調製のための使用;MaMV外被タンパク質から調製されたVLP;かかるVLPを調製するための方法;VLPおよびこれをコードするポリヌクレオチドを調製するのに好適な外被タンパク質ポリペプチドを、提供する。
【0055】
いくつかの態様において、MaMVまたはMaMV外被タンパク質を含むVLPのみの投与は、免疫反応を引き起こすのに十分であり、かかる投与は、例えば、抗原の投与の前に、免疫システムを「プライムする」ために用いることができる。MaMVおよびMaMV VLPはまた、アジュバントとして用いることもでき、この状況では、それらの免疫原性特性は、例えば抗原に対する免疫反応を増強するのに有用である。このような状況での抗原は、MaMVまたはVLPの前に、MaMVまたはVLPの後に、またはMaMVもしくはVLPと同時に投与することができる。同時投与の場合、抗原はMaMVまたはVLPに付着させることができ、または別にすることができる。本発明の1つの態様において、抗原は、MaMVまたはMaMV VLPの外被タンパク質に付着させる。MaMVおよびMaMV VLPはまた、ワクチンの調製においても用いることができる。
【0056】
本発明の1つの態様により、MaMVまたはMaMV VLPおよび1または2以上の抗原を含む免疫原性組成物は、動物において体液性および/または細胞性免疫反応を誘発することができる。
【0057】
定義
他の定義がない限り、本明細書で用いる全ての技術的および科学的用語は、本発明が属する分野の業者により一般に理解されるものと同じ意味を有する。
【0058】
本明細書において、用語「約」は、与えられた値からのおよそ+/−10%の変動を意味する。かかる変動は、本明細書に与えられる全ての値において、具体的に言及されるかどうかに関わらず、常に含まれることが理解されるべきである。
【0059】
本明細書において用語「ウイルス様粒子(VLP)」は、ウイルス粒子と類似の外観を有する、自己集合粒子を意味する。VLPは、ウイルス核酸を含んでも含まなくてもよい。VLPは通常、複製不能である。
【0060】
本明細書において用語「偽ウイルス」とは、プラスミド形態の核酸を含むDNAまたはRNAなどの核酸配列を含有するVLPを意味する。偽ウイルスは一般に、複製不能である。
【0061】
本明細書において、「融合タンパク質」は、例えばタンパク質またはタンパク質断片などの2つの別々のポリペプチドをコードする、2種または3種以上のポリヌクレオチドが遺伝的に組み合わされて、2種のポリペプチドの組合せである1つのタンパク質(「融合タンパク質」)をコードする第3のポリヌクレオチドを提供する場合に生成されるタンパク質である。例えば、MaMV外被タンパク質のアミノ酸配列と、抗原のアミノ酸とを含むポリペプチドは、融合タンパク質と考えられる。
【0062】
本明細書において用語「アジュバント」は、動物において免疫反応を増大、刺激、作動、増強、および/または調節する剤を意味する。
【0063】
本明細書において用語「免疫原性」とは、動物において検出可能な免疫反応を誘発する、物質の能力を意味する。
【0064】
本明細書において用語「免疫反応」とは、物質(例えば化合物、分子、材料など)の投与に応答した、動物の免疫系の反応性における変化を意味し、これには、抗体産生、細胞媒介性免疫性の誘発、補体活性化、免疫学的耐性の発達、またはこれらの組合せを含んでよい。
【0065】
本明細書において用語「免疫防御反応」とは、1または2以上の抗原に対する免疫反応であって、これにより、1または2以上の抗原が由来する作用物質によって引き起こされた状態(例えば疾病または疾患)および/または感染に対して防御するための、前記免疫反応を意味する。本発明の目的のために、状態および/または感染に対する免疫防御は、状態または感染の絶対的な防御のみでなく、状態または感染を患う未処置の動物と比べた場合の、処置した動物における、状態または感染の程度または速度における任意の検出可能な低下、または状態の重篤度または作用物質の感染から生じる任意の症状における任意の検出可能な低下をも含む。免疫防御反応は、前に状態を患っていなかった動物において、作用物質に感染していなかった動物において、および/または処置時に状態または感染を有していない動物において、誘発することができる。免疫防御反応はまた、既に状態を患っている動物、または処置時に病原体に感染している動物においても、誘発することができる。免疫防御反応は、体液性および/または細胞性免疫を含む、1または2以上のメカニズムの結果であることができる。
【0066】
本明細書において同義で用いられる用語「免疫刺激(immuno stimulation)」および「免疫刺激(immunostimulation)」とは、MaMV VLPなどの、動物病原体や疾病と関連しない分子の能力であって、免疫系を刺激することによる、および/または前記感染または疾病に応答する免疫系の容量を改善することによる、前記病原体による感染に対する、または前記疾病に対する、防御を提供する前記能力を意味する。免疫刺激は、予防効果、治療効果、またはこれらの組合せを有することができる。
【0067】
本明細書において用語「ワクチン接種」は、免疫防御反応を生成する目的で、対象に対してワクチンを投与することを意味する。ワクチン接種は、予防効果、治療効果、またはこれらの組合せを有することができる。ワクチン接種は、処置する対象に応じて種々の方法を用いて実施でき、これには限定することなく、非経口投与、例えば腹腔内注射(i.p.)、静脈内注射(i.v.)、または筋肉内注射(i.m.);経口投与;鼻腔内投与;皮内投与、経皮投与、および液浸を含む。
【0068】
本明細書において用語「ワクチン」とは、免疫防御反応を生成する目的で、対象に対して投与される組成物を意味する。
【0069】
本明細書において用語「天然の」とは、物に適用する場合、物が自然界で見出せるとの事実を意味する。例えば、生物(ウイルスを含む)、または生物中に存在するポリペプチドまたはポリヌクレオチド配列であって、天然の給源から単離でき、研究室で人間によって意図的に改変されていないものは、天然のものである。
【0070】
本明細書において「ポリペプチド」または「ペプチド」は、ペプチド結合により結合された少なくとも4個のアミノ酸が存在する分子を意味することを意図する。
【0071】
本明細書において表現「ウイルス核酸」とは、ウイルスのゲノムもしくはその一部、または該ゲノムもしくはその一部に対して塩基配列が相補的である核酸分子を意味する。ウイルスRNAに相補的なDNA分子もまた、ウイルスDNAの塩基配列に相補的なRNA分子であるために、ウイルス核酸と考えられる。
【0072】
本明細書において用語「免疫原」および「抗原」とは、1個の分子、複数分子、分子の一部分、分子の複数部分、または分子の組合せであって、全細胞および組織までを含み、それらのみで、またはアジュバントと組み合わせて、対象において免疫反応を誘発することができるものを意味する。免疫原/抗原は、単一のエピトープを含んでよく、または複数エピトープを含んでもよい。例えば、ペプチド、ポリペプチド、タンパク質、糖タンパク質、リポタンパク質、炭水化物、リポ多糖類、核酸、小分子、および種々の微生物の、その全体または一部であって、ウイルス、細菌および寄生生物、および他の感染粒子を含むものはしたがって、免疫反応を誘発することができる限りにおいて、抗原であることができる。ハプテンおよびミモトープ(mimotope)もまた、本明細書において用語「免疫原」および「抗原」に包含されると考えられる。
【0073】
本明細書において、用語「プライムする」およびその文法的な変形は、動物において、抗原投与の前に、免疫反応を刺激すること、および/または作動させることを意味する。
【0074】
本明細書において、用語「処置する(treat)」、「処置される(treated)」、または「処置している(treating)」とは、疾病または疾患などの状態または感染体に関連して用いる場合、対象の、状態または病原体による感染に対する抵抗性を増加させる処置(すなわち、対象が状態になる可能性、または感染体に感染する可能性を低下させる)、および対象が状態になるか感染した後で、状態または感染に対して戦うための処置(例えば、状態もしくは感染またはそれに付随する症状を、低減させ、消滅させ、改善し、または安定化させる)を意味する。
【0075】
本明細書において用語「対象」または「患者」とは、処置の必要な動物を意味する。
【0076】
本明細書において用語「動物」とは、ヒトおよび非ヒト動物の両方を意味する。非ヒト動物は、限定することなく、哺乳類、鳥類、爬虫類および魚類を含み、家畜(ペットを含む)、農場の動物、実験動物、動物園の動物および野生動物を包含し、例えばウシ、ブタ、ウマ、ヤギ、ヒツジ、および他の有蹄動物;イヌ;ネコ;ニワトリ、カモ類おおび他の鳥類;非ヒト霊長類;モルモット;ウサギ;フェレット(クロアシイタチ);ラット;ハムスターおよびマウスである。
【0077】
本明細書において、核酸またはアミノ酸配列との関連で用いる用語「実質的に同一」とは、最適に配列された場合に、例えば以下に記載の方法を用いて、核酸配列またはアミノ酸配列が、規定の第2核酸配列またはアミノ酸配列(または「参照配列」)に対して、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、または少なくとも99%の配列同一性を有することを示す。「実質的な同一性」は、配列の種々の型および長さを指すために用いることができ、例えば全長配列、機能ドメイン、コードおよび/または制限配列、プロモーター、およびゲノム配列などである。2つのアミノ酸配列または核酸配列の間のパーセント同一性は、当分野の業者の技能内である種々の方法により決定することができ、例えば、次のような公開されたコンピューターソフトウェアを用いて決定可能である:Smith Waterman Alignment(Smith, T. F. and M. S. Waterman (1981) JMoI Biol 147: 195-7);「BestFit」(Smith and Waterman, Advances in Applied Mathematics, 482-489 (1981))、これはGeneMatcher Plus(登録商標)Schwarz and Dayhof (1979) Atlas of Protein Sequence and Structure, Dayhof, M. O., Ed pp 353-358に組み込まれている;BLASTプログラム(Basic Local Alignment Search Tool (Altschul, S. F., W. Gish, et al. (1990) J MoI Biol 215: 403-10)、およびこれらの変形であって、BLAST-2、BLAST-P、BLAST-N、BLAST-X、WU-BLAST-2、ALIGN、ALIGN-2、CLUSTALおよびMegalign(DNASTAR)ソフトウェアを含むもの。さらに当業者は、アラインメントを測定するための適切なパラメータを、比較する配列の長さにわたり最大アラインメントを達成するのに必要なアルゴリズムも含んで、決定することができる。一般に、アミノ酸配列に対して、比較配列の長さは少なくも10アミノ酸である。当業者は、アラインメントに用いる実際の長さは、比較する配列全体の長さに依存し、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも200アミノ酸であってよく、またはアミノ酸配列の全長であってもよいことを理解する。核酸については、比較配列の長さは一般に少なくとも25ヌクレオチドであるが、少なくとも50、少なくとも100、少なくとも125、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも500、少なくとも550、または少なくとも600であってよく、または、核酸配列の全長であってもよい。
【0078】
用語「に一致している」または「に一致する」とは、核酸配列が、参照核酸配列の全体または一部と同一であることを示す。これと対照的に、本明細書において用語「に相補的」とは、核酸配列が、参照核酸配列の相補鎖の全体または一部と同一であることを示す。例としては、核酸配列「TATAC」は、参照配列「TATAC」に一致し、参照配列「GTATA」に相補的である。
【0079】
MaMVまたはMaMVウイルス様粒子を含む免疫原性組成物
本発明は、MaMVまたはMaMV VLPおよび任意に1または2以上の抗原を含む免疫原性組成物を提供する。免疫原性組成物はさらに、好適な担体、賦形剤など、および/または前記組成物の安定性、味のよさ、薬物動態、バイオアベイラビリティ等を改善する、医薬組成物の他の標準成分を任意に含んでもよい。
【0080】
Malvaモザイクウイルス
本発明の免疫原性組成物での使用に好適なMaMVの代表的な例は、カナダ・ブリティッシュコロンビアのサマーランド近郊で収穫されたゼニバアオイ(Malva neglecta)から単離され、これは例1に記載されている。本明細書の例1〜4に記載されているように、ウイルスのゲノム配列および物理的特性の分析により、MaMVはポテックスウイルス属のメンバーであり、フレキシウイルス科(Flexiviridae)の可能性が高い。
【0081】
MaMVは、長さが約450nm〜約600nmで幅が約10nm〜約18nmの間の、柔軟な糸状のビリオン(ウイルス粒子:virion)の特徴を有する。図4Dに示す例示のウイルスは、約520〜560nmの長さと、約12〜約16nmの幅である。
【0082】
M. neglectaのMaMVによる感染は、モザイク症状および葉脈の透明化(clearing)をもたらす。MaMVはまた、Chenopodiaceae科のメンバー、例えばChenopodium quinoa上で繁殖するその能力を特徴とする。
【0083】
MaMVのさらなる特徴は、配列番号1に記載の配列と実質的に同一のゲノム配列を有することである(図1およびGenBankアクセッション番号DQ660333も参照)。MaMVゲノムRNAは6858ヌクレオチド(nt)長(ポリ(A)尾を除く)で、GC含有量が45%である。ゲノム構成は他のポテックスウイルスと類似しており、推定RNA依存性RNAポリメラーゼ(RdRp)と、続いてTGBタンパク質をコードする3つのオーバーラップ遺伝子と、最後に外被タンパク質とを含む(図6A参照)。MaMVはさらに、感染の間に3つの主要なウイルスRNAを産生することを特徴とする:1つの大きなゲノムRNA、および、それぞれ2000個および800個のヌクレオチドのRNAとして移動している2つのサブゲノム種である(図6B参照)。
【0084】
MaMVはさらに、配列番号2に記載の配列と実質的に同一のアミノ酸配列を有する外被タンパク質を有することを特徴とする(図2も参照)。
【0085】
当業者に知られているように、ウイルス種はしばしば、同じ特徴を共有する多数の異なる株によって表される。したがって本発明は、例1に記載のMaMVの変異株であって、これと同じ物理的特性および、配列番号1と少なくとも約80%の配列同一性を有するゲノム配列を有する、前記変異株も包含する。本発明の1つの態様において、変異株は、例1に記載のMaMVと同じ物理的特性および、配列番号1と少なくとも約90%の配列同一性を有するゲノム配列を有する。本発明の1つの態様において、変異株は、例1に記載のMaMVと同じ物理的特性および、配列番号1と少なくとも約95%の配列同一性を有するゲノム配列を有する。
【0086】
MaMVウイルス様粒子(VLP)
本発明のMaMV VLPは、MaMV外被タンパク質に由来するのが好ましい。「に由来する」とは、VLPが、野生型MaMV外被タンパク質の配列と実質的に同一のアミノ酸配列を有する外被タンパク質を含み、任意に、以下にさらに詳細に記載するように、前記外被タンパク質に付着した1または2以上の抗原を含んでよいことを意味する。VLPに含まれるMaMV外被タンパク質はしたがって、野生型外被タンパク質またはその組換え型であることができ、野生型タンパク質の配列またはこの配列の改変型であって、多量体化および自己集合可能でVLPを形成することができる前記配列を有してよい。野生型配列および改変配列を有する外被タンパク質の組合せを含むVLPもまた、意図される。
【0087】
MaMV VLPは、多量体化および自己集合する能力を保持したMaMV外被タンパク質から形成される。集合した場合、各々のVLPは、外被タンパク質サブユニットの長いらせん状アレイを含む。VLPが含む外被タンパク質の数は、例えば約40〜約1600まで変化することができる。
【0088】
本発明のVLPは、同一のアミノ酸配列を有する複数の外被タンパク質から、集合した場合の最終的なVLPが、同一の外被タンパク質サブユニットを含むようにして調製することができ、または、VLPは、異なるアミノ酸配列を有する複数の外被タンパク質から、集合した場合の最終的なVLPが、その外被タンパク質サブユニットにおける変化を含むようにして調製することができる。
【0089】
VLPを形成するために用いる外被タンパク質は、全長外被タンパク質または、多量体化してVLPを形成することができる、全長外被タンパク質の一部であってよい。外被タンパク質配列は、天然の(野生型)配列であることができ、または、本明細書に記載のように、改変された外被タンパク質が多量体化しVLP内に集合する能力を保持している限りにおいて、野生型配列の遺伝的な改変型であってよく、例えば、1または2以上のアミノ酸の欠失、挿入、置き換えなどを含む。例1に記載のMaMVの外被タンパク質のアミノ酸配列は、本明細書において配列番号2(図2参照)として提供され、これはまた、GenBankからもアクセス可能である(GenBankアクセッション番号ABG48664)。MaMVの外被タンパク質のヌクレオチド配列もまた、本明細書に配列番号3(図3参照)として提供され、GenBankからもアクセス可能である(GenBankアクセッション番号DQ660333(ヌクレオチド6057〜6788))。
【0090】
上に述べたように、VLPに含まれる外被タンパク質のアミノ酸配列は、配列番号2に記載の野生型配列に正確に一致する必要はなく、すなわち、改変配列であってよい。例えば、外被タンパク質の配列は、1または2以上のアミノ酸残基の置換、挿入または欠失により改変されていてもよく、そのためその変更部位(単数または複数)における残基は、野生型(参照)配列に一致しない。例えば、外被タンパク質をコードするヌクレオチド配列は、酵素切断部位の付加により改変されてよく、その結果、コードされる外被タンパク質の、例えば、1個〜数個のアミノ酸の、C末端、N末端、もしくは両方における、またはこれらの近傍における付加による改変が生じる。当業者はしかし、かかる変異は拡張的(extensive)ではなく、改変外被タンパク質の、多量体化しVLPに集合する能力を妨害しないことを理解する。かかる変異はしかし、MaMV外被タンパク質の自己集合する能力を改善し、および/またはPapMV外被タンパク質について前に示されたような、より長いVLPの形成をもたらす(Laliberte et al., 2008, FEBS Journal, 275: 1474-1484)。MaMV外被タンパク質の改変型の、VLPに集合する能力は、例えば、標準の技法による電子顕微鏡法により評価することができ、例示の方法が本明細書の実施例に提供される。
【0091】
一般に、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約80%同一のアミノ酸配列を含む。1つの態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約85%同一のアミノ酸配列を含む。別の態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約90%同一のアミノ酸配列を含む。他の態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約91%同一の、少なくとも約92%同一の、少なくとも約93%同一の、少なくとも約94%同一の、少なくとも約95%同一の、少なくとも約96%同一の、少なくとも約97%同一の、少なくとも約98%同一の、または少なくとも約99%同一の、アミノ酸配列を含む。
【0092】
多量体化しVLPに集合する能力を保持する野生型タンパク質の断片である外被タンパク質(すなわち、「機能性」断片)もまた、本発明により意図される。例えば、機能性断片は、タンパク質のN末端、C末端または内部からの、1または2以上のアミノ酸の欠失、またはそれらの組合せを含んでよい。一般に、機能性断片は、配列番号2に記載の配列の少なくとも約150の隣接アミノ酸(contiguous amino acid)を含む。本発明の1つの態様において、機能性断片は、配列番号2に記載の配列の少なくとも約180の隣接アミノ酸を含む。別の態様において、機能性断片は、配列番号2に記載の配列の少なくとも約190の隣接アミノ酸を含む。他の態様において、機能性断片は、配列番号2に記載の配列の、少なくとも約200の隣接アミノ酸を、少なくとも約210の隣接アミノ酸を、少なくとも約220の隣接アミノ酸を、少なくとも約225の隣接アミノ酸を、少なくとも約230の隣接アミノ酸を含む。タンパク質のN末端またはC末端での欠失は、タンパク質の多量体化する能力を保持するために、一般に25個以下のアミノ酸を除去すべきである。例えば、1つの態様において、N末端での欠失は、20個以下のアミノ酸を除去する。他の態様において、N末端での欠失は、15個以下のアミノ酸を、12個以下のアミノ酸を、10個以下のアミノ酸を、8個以下のアミノ酸を、および5個以下のアミノ酸を除去する。
【0093】
本発明の1つの態様において、VLPは、MaMV外被タンパク質の組換え型を含む。他の態様において、VLPは、MaMV外被タンパク質の遺伝的改変型を含む。
【0094】
外被タンパク質が、1または2以上のアミノ酸置換を含む改変配列を含む場合、これらは、「保存的」置換または「非保存的」置換であることができる。保存的置換は、1つのアミノ酸残基を、類似の側鎖特性を有する他の残基と置き換えることを含む。当分野で知られているように、20種の天然のアミノ酸は、それらの側鎖の物理化学的特性に従って分類可能である。適切な分類としては、例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニンおよびトリプトファン(疎水性側鎖);グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、およびグルタミン(極性の非荷電側鎖);アスパラギン酸およびグルタミン酸(酸性側鎖)およびリシン、アルギニンおよびヒスチジン(塩基性側鎖)を含む。アミノ酸の他の分類は、フェニルアラニン、トリプトファンおよびチロシン(芳香族側鎖)である。保存的置換は、1つのアミノ酸を、同じ群の別のアミノ酸で置換することが関与する。非保存的置換は、1つのアミノ酸残基を、異なる側鎖特性を有する別の残基で置き換えることが関与し、例えば、酸性残基の中性または塩基性残基による置き換え、中性残基の酸性または塩基性残基による置き換え、疎水性残基の親水性残基による置き換えなどである。
【0095】
外被タンパク質をコードする核酸配列も同様に、野生型配列に正確に一致する必要はないが、遺伝子コードの縮退のために、および/または、上記の改変アミノ酸配列をコードするように、変化してよい。本発明の1つの態様において、したがって、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約70%同一である。他の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約75%同一である。別の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約80%同一である。他の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約85%、または少なくとも約90%同一である。
【0096】
以下にさらに詳細に記載するように、VLP外被タンパク質は任意に、1もしくは2以上の抗原、親和性ペプチドもしくは他の短ペプチド配列、またはこれらの組合せに遺伝子的に融合してよく、1または2以上の抗原の付着を促進する。
【0097】
抗原
免疫原性組成物は任意に、1または2以上の抗原を含んでよく、これらはMaMVまたはMaMV VLPとは別であっても、またはMaMVもしくはMaMV VLPに接合していてもよい。接合および非接合抗原の両方を含む免疫原性組成物もまた、意図される。この後者の場合、非接合抗原は、付加的単離抗原(AIA)として考えることができる。AIAは、接合抗原(単数または複数)と同じであっても、異なっていてもよい。接合は、例えば、外被タンパク質との遺伝子融合、または共有結合、非共有結合もしくは親和性手段を介した結合であることができる。本発明の1つの態様において、1または2以上の抗原は、市販のワクチンの形態で提供されることができる。
【0098】
ワクチン開発に好適な多種多様な抗原が、当分野に知られている。本発明の免疫原性組成物に含めるのに好適な抗原は、例えば、それが指向されるところの疾病または疾患などの免疫原性組成物の所望の最終用途、組成物の様式、組成物が多価ワクチンもしくは単価ワクチンとしての使用を意図されているのか、および/または投与される動物などに基づいて、当業者が容易に選択することができる。
【0099】
例えば、抗原は、動物において、例えば癌、感染症、アレルギー反応、または自己免疫疾患などの疾病または疾患を引き起こすことができる作用物質に由来することができる。またはこれは、薬剤、ホルモンもしくは毒素関連の疾病または疾患に対する免疫反応を誘発するために用いるのに好適な、抗原であることができる。抗原は、当分野に知られている病原体から、例えば細菌、ウイルス、原生動物、真菌、寄生生物、または例えばプリオンなどの感染粒子から由来してよい。または抗原は、腫瘍関連抗原、自己抗原、またはアレルゲンであってよい。あるいは、抗原は、抗原のB細胞エピトープまたはT細胞エピトープを含んでよい。
【0100】
1または2以上の抗原は、免疫原性組成物中に種々の様式で、上記のように、および下にさらに詳細に記載するようにして、含めることができる。免疫原性組成物中に含むための1または2以上の抗原は、したがって、選択した様式に依存してサイズが異なることができる。抗原は、例えば、ペプチド、タンパク質、核酸、多糖類、小分子、またはこれらの組合せであって、病原体全体またはその一部を含み、例えば、生きた、不活性化または減衰型の病原体である。
【0101】
免疫原性組成物が1より多くの抗原を含む場合、組成物中に含めるために選択される抗原は、同一であることができ、または異なっていてもよく、単一の源から、または複数の源から由来してよい。各抗原は、特定の免疫反応を引き起こすことができる1つのエピトープを有するか、または各抗原は、1つより多くのエピトープを有してもよい。
【0102】
抗原は、T細胞、B細胞、NK細胞およびマクロファージ上の表面構造により、またはクラスIもしくはクラスIIのAPC関連細胞表面構造により、またはこれらの組合せにより認識されるエピトープを含んでよい。1つの態様において、本発明は、免疫原性組成物が、小さなおよび/または弱い免疫原性抗原に対して特に有用であることを意図する。
【0103】
本発明の免疫原性組成物に含むための抗原はまた、病原体または他の目的の源から、当分野に知られた方法により選択し、当分野に知られた標準の免疫学的技法を用いて、動物において免疫反応を誘発するそれらの能力についてスクリーニングしてよい。例えば、抗原タンパク質内でのエピトープの予測のための方法は、Nussinov R and Wolfson H J, Comb Chem High Throughput Screen (1999) 2(5):261に記載されており、CTLエピトープの予測のための方法は、Rothbard et al, EMBO J. (1988) 7:93-100およびde Groot M S et al, Vaccine (2001) 19(31):4385-95に記載されている。他の方法は、Rammensee H-G. et al., Immunogenetics (1995) 41: 178-228およびSchirle M et al, Eur J Immunol (2000) 30(18):2216-2225に記載されている。
【0104】
有用なウイルス抗原は、例えば、以下の科のメンバーに由来するものを含む:アデノウイルス科;アレナウイルス科(例えばIppyウイルスおよびラッサウイルス);ビルナウイルス科;ブニヤウイルス科;カリシウイルス科;コロナウイルス科;フィロウイルス科;フラビウイルス科(例えば黄熱病ウイルス、デング熱ウイルスおよびC型肝炎ウイルス);ヘパドナウイルス科(Hepadnaviradae)(例えば、B型肝炎ウイルス);ヘルペスウイルス科(例えばヒト単純ヘルペスウイルス1型);オルトミクソウイルス科(例えば、A型、B型よびC型インフルエンザウイルス);パラミクソウイルス科(例えば流行性耳下腺炎ウイルス、ヒト・メタニューモウイルス、麻疹ウイルス、および呼吸器合胞体ウイルス);ピコルナウイルス科(例えば、ポリオウイルスおよびA型肝炎ウイルス);ポックスウイルス科;レオウイルス科;レトロウイルス科(例えばBLV−HTLVレトロウイルス、HIV−1、HIV−2、ウシ免疫不全ウイルスおよびネコ免疫不全ウイルス);ラブドウイルス科(例えば狂犬病ウイルス)、およびトガウイルス科(例えば風疹ウイルス)。1つの態様において、免疫原性組成物は、以下のような主要なウイルス病原体に由来する1または2以上の抗原を含む:種々の肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、種々のインフルエンザウイルス、ウェストナイルウイルス、呼吸器合胞体ウイルス、狂犬病ウイルス、ヒトパピローマウイルス(HPV)、エプスタインバーウイルス(EBV)、ポリオーマウイルス、またはSARSコロナウイルス。
【0105】
A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、デルタ肝炎ウイルス(HDV)、E型肝炎ウイルス(HEV)、およびG型肝炎ウイルス(HGV)を含む肝炎ウイルス由来のウイルス抗原は、当分野に知られている。例えば、抗原は、HCVコアタンパク質、E1タンパク質、E2タンパク質、NS3タンパク質、NS4タンパク質またはNS5タンパク質から、HVB HbsAg抗原またはHBVコア抗原から、およびHDVデルタ抗原から由来することができる(例えば、米国特許第5,378,814号を参照)。次の米国特許は、例えば、HCVコアタンパク質に基づく種々の抗原について記載している;第6,596,476号;第6,592,871号;第6,183,949号;第6,235,284号;第6,780,967号;第5,981,286号;第5,910,404号;第6,613,530号;第6,709,828号;第6,667,387号;第6,007,982号;第6,165,730号;第6,649,735号、および第6,576,417号。
【0106】
ヘルペスウイルス科からの既知の抗原の非限定的例としては、単純ヘルペスウイルス(HSV)1および2型、例えばHSV−1およびHSV−2糖タンパク質gB、gDおよびgHに由来するものが挙げられる。
【0107】
HIV抗原の非限定的例としては、gp120由来抗原、gp160およびgp41などの種々のエンベロープタンパク質由来の抗原、p24gagおよびp55gagなどのgag抗原、ならびにHIVのpol、env、tat、vif rev、nef vpr、vpuおよびLTR領域に由来するタンパク質が挙げられる。多数のHIV−1およびHIV−2分離株からのgp120配列であって、HIVの種々の遺伝的サブタイプのメンバーを含むものが知られている(例えば、Myers et al., Los Alamos Database, Los Alamos National Laboratory, Los Alamos, N. Mex. (1992)およびModrow et al, J. Virol. (1987) 61:570 578)を参照)。
【0108】
他のウイルス抗原の非限定的例としては、水痘帯状疱疹ウイルス(VZV)、エプスタインバーウイルス(EBV)、およびCMV gBおよびgHを含むサイトメガロウイルス(CMV);および他のヒトヘルペスウイルス、例えばHHV6およびHHV7などからの抗原が挙げられる(例えば、Chee et al. (1990) Cytomegaloviruses (J. K. McDougall, ed., Springer-Verlag, pp. 125 169);McGeoch et al. (1988) J. Gen. Virol. 69:1531 1574;米国特許第5,171,568号;Baer et al. (1984) Nature 310:207 211;およびDavison et al. (1986) J. Gen. Virol. 67:1759 1816を参照)。
【0109】
抗原はまた、インフルエンザウイルスから、例えば赤血球凝集素(HA)、ノイラミニダーゼ(NA)、核タンパク質(NP)、マトリクスタンパク質(M1およびM2)、ポリメラーゼ酸性タンパク質(PA)、およびポリメラーゼ塩基性タンパク質サブユニット(PB1、PB2)から由来することもできる。これらタンパク質の配列は当分野で知られており、全米バイオテクノロジー情報センター(NCBI)により維持されているGenBankデータベースから容易にアクセスできる。HA、NPおよびマトリクスタンパク質の好適な抗原性断片としては、限定はしないが、以下が挙げられる:赤血球凝集素エピトープとして、HA91−108、HA307−319およびHA306−324(Rothbard, Cell, 1988, 52:515-523)、HA458−467(J. Immunol. 1997, 159(10): 4753-61)、HA213−227、HA241−255、HA529−543およびHA535−547(Gao, W. et al, J. Virol, 2006, 80:1959-1964);核タンパク質エピトープとして、NP206−229(Brett, 1991, J Immunol. 147:984-991)、NP335−350およびNP380−393(Dyer and Middleton, 1993, In: Histocompatibility testing, a practical approach (Ed.: Rickwood, D. and Hames, B. D.) IRL Press, Oxford, p. 292; Gulukota and DeLisi, 1996, Genetic Analysis: Biomolecular Engineering, 13:81)、NP305−313(DiBrino, 1993, PNAS 90: 1508-12);NP384−394(Kvist, 1991, Nature. 348:446-448);NP89−101(Cerundolo, 1991, Proc. R. Soc. Lon. 244:169-7);NP91−99(Silver et al, 1993, Nature. 360: 367-369);NP380−388(Suhrbier, 1993, J. Immunology 79: 171- 173);NP44−52およびNP265−273(DiBrino, 1993、同上);およびNP365−380(Townsend, 1986, Cell. 44:959-968);マトリクスタンパク質(M1)エピトープとして、M1 2−22、M1 2−12、M1 3−11、M1 3−12、M1 41−51、M1 50−59、M1 51−59、M1 134−142、M1 145−155、M1 164−172、M1 164−173(全て、Nijman, 1993, Eur. J. Immunol. 23:1215-1219に記載);M1 17−31、M1 55−73、M1 57−68(Carreno, 1992, MoI Immunol 29: 1131-1140);M1 27−35、M1 232−240(DiBrino, 1993、同上)、M1 59−68およびM1 60−68(Eur. J. Immunol. 1994, 24(3): 777-80);およびM1 128−135(Eur. J. Immunol. 1996, 26(2): 335-39);PA 412−422(Heiny AT, et al., 2007, PLoS ONE, 21;2(1l):el190);PB1 15−51(Heiny AT, et al., 2007, PLoS ONE, 21;2(1l):el190)およびPB2 685−696(Heiny AT, et al., 2007, PLoS ONE, 21;2(ll):el190)。
【0110】
その他の例としては、HAタンパク質由来のF3ペプチドであり、これは次の配列を有する:KAYSNCYPYDVPDY(配列番号72)(Lu et al., 2002, Int. Arch. Allergy Immunol, 127: 245-250)、およびM1タンパク質からのCTLエピトープであり、これは次の配列を有する:SPLTKGILGFVFTLTVPSE(配列番号73)、およびGILGFVFTL(配列番号60)。
【0111】
他の関連する抗原性領域およびエピトープも知られている。例えば、インフルエンザイオンチャネルタンパク質(M2)の断片であって、これはM2eペプチド(M2の細胞外ドメイン)を含む。このペプチドの配列は、インフルエンザの異なる株にわたって高度に保存されている。M2eペプチド配列の例は表1に配列番号4として示されている。この配列の変異形が同定されており、そのいくつかも表1に示されている。
表1:M2eペプチドおよびその変異
【表1】
*米国特許出願第2006/0246092号参照
【0112】
M2e配列の全体または一部を用いることができ、例えば、変異体にわたって保存されている部分配列であり、例としてはアミノ酸2〜10により規定される領域内の断片、または保存エピトープEVETPIRN(配列番号9)(M2e配列のアミノ酸6〜13)。6〜13エピトープは、GenBankで利用可能なヒトA型インフルエンザ株の84%において不変であることが見出されている。同定されているこの配列の変異形には以下がある:EVETLTRN(配列番号10)(9.6%)、EVETPIRS(配列番号11)(2.3%)、EVETPTRN(配列番号12)(1.1%)、EVETPTKN(配列番号13)(1.1%)、ならびにEVDTLTRN(配列番号14)、EVETPIRK(配列番号15)およびEVETLTKN(配列番号16)(それぞれ0.6%)(Zou, P., et al., 2005, Int Immunopharmacology, 5:631-635;Liu et al. 2005, Microbes and Infection, 7: 171-177を参照)。
【0113】
他の有用な抗原としては、生ウイルス、弱毒化ウイルス、および不活化ウイルスであって、例えば不活化ポリオウイルス(Jiang et al, J. Biol. Stand., (1986) 14:103-9)、A型肝炎ウイルスの弱毒株(Bradley et al, J. Med. Virol, (1984) 14:373-86)、弱毒麻疹ウイルス(James et al, N. Engl. J. Med, (1995) 332:1262-6)、および百日咳ウイルスのエピトープ(例えば、ACEL-IMUNE(登録商標)無細胞DTP、Wyeth-Lederle Vaccines and Pediatrics)である。
【0114】
抗原はまた、非定型ウイルスまたはウイルス様物質に由来することもでき、例としては、当分野で知られているように、以下のものがある:クールーの原因物質、クロイツフェルトヤコブ病(CJD)、スクラピー、伝染性ミンク脳症、および慢性消耗性疾患、または狂牛病に関連するプリオンなどのタンパク性感染粒子。
【0115】
有用な細菌性抗原としては、例えば、リポ多糖類、莢膜抗原(本質的に、タンパク様または多糖類)などの表在性細菌性抗原成分、または鞭毛成分が挙げられ、これらは次のような疾病に関与する既知の原因物質から得るか、または由来することができる:ジフテリア、百日咳、破傷風、結核、細菌性または真菌性肺炎、コレラ、腸チフス、ペスト、細菌性赤痢またはサルモネラ症、レジオネラ病、ライム病、ハンセン病、マラリア、鉤虫症、オンコセルカ症、住血吸虫症、トリパマソーマ症(trypamasomialsis)、レスマニア症(lesmaniasis)、ジアルジア鞭毛虫症、アメーバ症、フィラリア症、ボレリア、および旋毛虫症。
【0116】
腸内細菌科のグラム陰性菌に由来する抗原の例としては、限定はしないが、腸チフス菌Vi(莢膜多糖類)抗原、大腸菌KおよびCFA(莢膜成分)抗原および、大腸菌線毛付着抗原(K88およびK99)が挙げられる。抗原タンパク質の例としては、ポリン(Secundino et al., 2006, Immunology 117:59)としても知られている外膜タンパク質(Omp);腸チフス菌鉄調節外膜タンパク質などの関連するポリン(IROMP, Sood et al., 2005, MoI Cell Biochem 273:69-78)、および、限定することなく腸チフス菌HSP40(Sagi et al., 2006, Vaccine 24:7135-7141)を含む、熱ショックタンパク質(HSP)が挙げられる。抗原性ポリンの非限定的例としては、OmpCおよびOmpFが挙げられ、これらは多数のサルモネラ種および大腸菌種に見出される。OmpCおよびOmpFのオルソログもまた、他の腸内細菌科に見出されており、これらは本発明の目的のための好適な抗原タンパク質である。さらに、腸内細菌科のポリンタンパク質中に見出される配列の保存領域に基づいて(Diaz-Quinonez et al., 2004, Infect, and Immunity 72:3059- 3062)、Omp1B(Shigella flexneri)、OmpC2(Yersinia pestis)、OmpD(S. enteria)、OmpK36(Klebsiella pneumonie)、OmpN(大腸菌)およびOmpS(S. enterica)も好適であることができる。
【0117】
種々の腸内細菌からの抗原タンパク質の配列は、当分野に知られており、全米バイオテクノロジー情報センター(NCBI)により維持されているGenBankデータベースから容易にアクセスできる。例えば、GenBankアクセッション番号P0A264およびGenBankアクセッション番号NP_804453:OmpC(S. enterica subsp. enterica serovar Typhi Ty2);GenBankアクセッション番号CAD05399:OmpF前駆体タンパク質(S. enterica subsp. enterica serovar Typhi CTl 8);GenBankアクセッション番号16761195:OmpC(S. enterica serovar Typhimurium);GenBankアクセッション番号47797:OmpC(S. enterica serovar Typhi);GenBankアクセッション番号8953564:OmpC(S. enterica serovar Minnesota);GenBankアクセッション番号19743624:OmpC(S. enterica serovar Dublin);GenBankアクセッション番号19743622:OmpC(S. enterica serovar Gallinarum);GenBankアクセッション番号26248604:OmpC(大腸菌);GenBankアクセッション番号24113600:Omp1B(Shigella flexneri);GenBankアクセッション番号16764875:OmpC2(Yersinia pestis);GenBankアクセッション番号16764916:OmpD(S. enterica Serovar Typhimurium);GenBankアクセッション番号151149831:OmpK36(Klebsiella pneumonie);GenBankアクセッション番号3273514:OmpN(大腸菌)、およびGenBankアクセッション番号16760442:OmpS(S. enterica serovar Typhi)。
【0118】
種々の腫瘍関連抗原が当分野に知られている。代表的な例としては、限定はしないが、以下が挙げられる:Her2(乳癌);GD2(神経芽細胞腫);EGF−R(悪性グリア芽細胞腫);CEA(髄様甲状腺癌);CD52(白血病);ヒトメラノーマタンパク質gp100(例えばエピトープ:IMDQVPFSV(配列癌号59)を含む抗原);ヒトメラノーマタンパク質melan−A/MART−1;ヒトDickkopf1(DKK1)タンパク質、ヒトアンギオモチン(Amot)、NA17;NA17−Antタンパク質;p53タンパク質;MAGE1、MAGE2、MAGE3(HLA−A1タンパク質)およびMAGE4を含む種々のMAGE(メラノーマ関連抗原E);種々のチロシナーゼ(HLA−A2ペプチド);変異ras;p97メラノーマ抗原;進行癌に関連するRasぺプチドおよびp53ペプチド;子宮頸癌に関連するHPV16/18およびE6/E7抗原;乳癌関連MUC1−KLH抗原;結腸直腸癌に関連するCEA(癌胎児性抗原)、および前立腺癌関連PSA抗原。
【0119】
有用なアレルゲンとしては、花粉、動物鱗屑、草、カビ、粉塵、抗生物質、刺咬昆虫毒物からのアレルゲン、および種々の環境、薬物および食物アレルゲンが挙げられるが、これらに限定されない。一般的な樹木アレルゲンとしては、ハコヤナギ、ポプラ、セイヨウトリネコ、カバノキ、メープル、オーク、ニレ、ヒッコリー、およびペカンの木からの花粉が挙げられる。一般的な植物アレルゲンとしては、ライ麦、ブタクサ、イギリスオオバコ、スイバ(sorrel dock)およびアカザからのものが挙げられ、植物接触アレルゲンとしては、ウルシ、ツタウルシおよびイラクサからのものが挙げられる。一般的な草のアレルゲンとしては、オオアワガエリ、ジョンソングラス、ギョウギシバ、ウシノケグサ、およびブルーグラスアレルゲンが挙げられる。一般的なアレルゲンは、鱗屑または真菌類から、例えばアルテルナリア属、フザリウム属、ホルモデンドラム属、アスペルギラス属、ミクロポリスポラ属、ケカビ属および好熱性の放線菌からも得ることができる。ペニシリン、スルホンアミドおよびテトラサイクリンは、一般的な抗生物質アレルゲンである。表皮性のアレルゲンは、ハウスダストまたは有機ダスト(典型的には真菌類から)から、イエダニ(dermatphagoides pterosinyssis)などの昆虫から、または羽毛、およびネコおよびイヌの鱗屑などの動物源から、得ることができる。一般的な食物アレルゲンとしては、牛乳およびチーズ(乳製品)、卵、小麦、ナッツ(例えばピーナッツ)、魚介類(例えば、貝)、エンドウ豆、豆、およびグルテンアレルゲンなどが挙げられる。一般的な薬物アレルゲンとしては、局所麻酔薬およびサリチル酸塩アレルゲンが挙げられ、一般的な昆虫アレルゲンとしては、ハチ、スズメバチ、カリバチおよびアリ毒物、およびゴキブリがく(calyx)アレルゲンが挙げられる。
【0120】
特に良好に特徴付けられているアレルゲンとしては、限定はされないが、以下が挙げられる:イエダニアレルゲンDer pIおよびDer pII(Chua, et al., J. Exp. Med., 167:175 182, 1988;およびChua, et al, Int. Arch. Allergy Appl. Immunol, (1990) 91 : 124-129)、Der pIIアレルゲンのT細胞エピトープペプチド(Joost van Neerven, et al, J. Immunol, (1993) 151 :2326-2335を参照)、非常に豊富な抗原E(Amb aI)ブタクサ花粉アレルゲン(Rafnar, et al, J. Biol. Chem., (1991) 266: 1229-1236を参照)、ホスホリパーゼA2(ハチ毒物)アレルゲンおよびその中のT細胞エピトープ(Dhillon, et al, J. Allergy Clin. Immunol, (1992) 42参照)、シラカバ花粉(Betv1)(Breiteneder, et al, EMBO, (1989) 8:1935-1938参照)、Fe1 dI主要な家猫アレルゲン(Rogers, et al, MoI Immunol, (1993) 30:559-568参照)、樹木花粉(Elsayed et al, Scand. J. Clin. Lab. Invest. Suppl, (1991) 204: 17-31参照)、およびマルチエピトープ組換え草(grassアレルゲンrKBG8.3(Cao et al Immunology (1997) 90:46-51)。これらおよび他の好適なアレルゲンは市販されており、および/または既知の技術に従って用意に調製可能である。
【0121】
自己抗原に付随する状態に関連する抗原も、当業者に知られている。かかる抗原の代表的な例としては、限定はしないが、リンホトキシン、リンホトキシン受容体、核因子kBリガンドの受容体アクチベーター(RANKL)、血管内皮成長因子(VEGF)、血管内皮成長因子受容体(VEGF−R)、インターロイキン−5、インターロイキン−17、インターロイキン−13、CCL21、CXCL12、SDF−1、MCP−1、エンドグリン、レジスチン、GHRH、LHRH、TRH、MIF、エオタキシン、ブラジキニン、BLVC、腫瘍壊死因子αおよびアミロイドβペプチド、および免疫学的反応を引き出すために使用可能なこれら各々の断片が挙げられる。
【0122】
有用な毒素は、一般に有毒な植物、動物、および微生物の天然産物であるか、またはこれらの化合物の断片である。かかる化合物は、例えば、アフラトキシン、シガテラ毒素、百日咳毒素およびテトロドトキシンを含む。
【0123】
レクリエーショナルドラッグ中毒に関連して有用な抗原は当分野に知られており、例えば、コデイン、フェンタニル、ヘロイン、モルヒネ、およびアヘンなどの、オピオイドおよびモルヒネ誘導体;アンフェタミン、コカイン、MDMA(メチレンジオキシメタンフェタミン)、メタンフェタミン、メチルフェニデート、およびニコチンなどの興奮薬;LSD、メスカリンおよびプシロシビンなどの幻覚発現物質;ハシシおよびマリファナなどのカンナビノイド(大麻)、その他の常習性薬物または化合物、および派生物、副産物、変異体およびかかる化合物の複合体を含む。
【0124】
本発明の1つの態様において、免疫原性化合物に含まれる抗原(単数または複数)は、タンパク質抗原である。タンパク質抗原は、全長タンパク質、実質的に全長のタンパク質(例えば、約25個未満のアミノ酸のN末端および/またはC末端での欠失を含むタンパク質)、タンパク質の抗原断片、またはこれらの組合せであることができる。全長タンパク質は、適用可能である場合は、タンパク質の前駆体形態またはタンパク質の成熟(処理)形態であってよい。タンパク質は翻訳後修飾されていてもよく、例えば、糖タンパク質またはリポタンパク質であってよい。抗原断片は1または複数のエピトープを含むことができ、したがって数個のアミノ酸(例えば、少なくとも4個のアミノ酸)の1ペプチドから、数百のアミノ酸長のポリペプチドまでの大きさであってよい。本発明の1つの態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約4アミノ酸〜約250アミノ酸の長さである。他の態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約5アミノ酸〜約200アミノ酸の長さである。他の態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約5アミノ酸〜約150アミノ酸の長さ、約5アミノ酸〜約100アミノ酸の長さ、約5アミノ酸〜約75アミノ酸の長さ、約5アミノ酸〜約70アミノ酸の長さ、約5アミノ酸〜約60アミノ酸の長さ、および、約5アミノ酸〜約50アミノ酸の長さである。
【0125】
上記のように、本発明の1つの態様において、免疫原性組成物中に含まれる1または2以上の抗原は、市販のワクチン形態であることができる。種々のヒトワクチンが当分野に知られており、以下に対するワクチンが含まれるが、これらに限定はされない:
炭疽菌(anthrax)、例えばBioThrax(登録商標)(BioPort Corporation);
B型インフルエンザ菌(Hib)、例えばActHIB(登録商標)(Sanofi-aventis)、PedvaxHIB(登録商標)(Merck)およびHibTITER(登録商標)(Wyeth);
A型肝炎、例えばHavrix(登録商標)(GlaxoSmithKline)およびVaqta(登録商標)(Merck);
B型肝炎、例えばEngerix-B(登録商標)(GlaxoSmithKline)およびRecombivax HB(登録商標)(Merck);
帯状疱疹(shingles)、例えばZostavax(登録商標)(Merck);
ヒトパピローマウイルス(HPV)、例えばGardasil(登録商標)(Merck);
インフルエンザ、例えばFluarix(登録商標)およびFluviral(登録商標)(GlaxoSmithKline)、FluLaval(登録商標)(ID Biomedical Corp of Quebec);FluMist(登録商標)(鼻腔内)(Medimmune)、Fluvirin(登録商標)(Chiron)、およびFluzone(登録商標)(Sanofi-aventis);
日本脳炎、例えばJE-Vax(登録商標)(Sanofi-aventis);
麻疹、例えばAttenuvax(登録商標)(Merck);
髄膜炎菌性髄膜炎、例えばMenomune(登録商標)Meningococcal Polysaccharide(Sanofi-aventis);
流行性耳下腺炎、例えばMumpsvax(登録商標)(Merck);
肺炎級菌による疾病、例えばPneumovax 23(登録商標)Pneumococcal Polysaccharide(Sanofi-aventis)およびPrevnar(登録商標)Pneumococcal Conjugate(Wyeth);
ポリオ、例えばIpol(登録商標)(Sanofi-aventis)およびPoliovax(登録商標)(Sanofi-Pasteur);
狂犬病、例えばBioRab(登録商標)(BioPort Corporation)、RabAvert(登録商標)(Chiron)およびImovax(登録商標)Ravies (Sanofi-aventis);
ロタウイルス、例えばRotaTeq(登録商標)(Merck);
風疹、例えばMeruvax II(登録商標)(Merck);
腸チフス菌(腸チフス)、例えばTyphim Vi(登録商標)(Sanofi-aventis)およびVivotif(登録商標)Berna (経口)(Berna);
肺結核(BCG)、例えばTheraCys(登録商標)およびImmuCyst(登録商標)(Sanofi-aventis);TICE(登録商標)BCGおよびOncotice(商標)(Organon Teknika Corporation);Pacis(商標);およびMycobax(登録商標)(Sanofi-Pasteur);
種痘症(天然痘)、例えばDryvax(登録商標)(Wyeth);
水痘(鶏痘)、例えばVarivax(登録商標)(Merck);
黄熱病、例えばYF-Vax(登録商標)(Sanofi-aventis);
A型肝炎/B型肝炎、例えばTwinrix(登録商標)(GlaxoSmithKline);
B型肝炎およびHib、例えばComvax(登録商標)(Merck);
破傷風/Hib、例えばActHIB(登録商標)(Sanofi-Pasteur);
ジフテリア/Hib、例えばHibTITER(登録商標)(Wyeth Pharmaceuticals);
Hib/髄膜炎、例えばPedVaxHIB(登録商標)(Merck & Co);
髄膜炎/ジフテリア、例えばMenactra(登録商標)Meningococcal Conjugate(Sanofi-Pasteur);
破傷風/ジフテリア(Td)、例えばDecavac(登録商標)(Sanofi-aventis);
ジフテリア/破傷風/百日咳(DTaP/DTまたはDTaP)、例えばDaptacel(登録商標)およびTripedia(登録商標)(Sanofi-aventis)およびInfanrix(登録商標)(GlaxoSmithKline);
破傷風/ジフテリア/百日咳(Tdap)、例えばBoostrix(登録商標)(GlaxoSmithKline)およびAdacel(登録商標)(Sanofi-Pasteur);
DTaP/Hib、例えばTriHIBit(登録商標)(Sanofi-aventis);
DTaP/ポリオ/B型肝炎、例えばPediarix(登録商標)(GlaxoSmithKline);
麻疹/流行性耳下腺炎/風疹(MMR)、例えばM-M-R II(Merck)および
麻疹/流行性耳下腺炎/風疹/鶏痘、例えばProQuad(登録商標)(Merck)。
【0126】
獣医による使用の例としては、限定することなく、以下に対するワクチンが挙げられる:
Lawsonia intracellulars(例えば、EnterisolおよびIleitis)、Porphyromonas gulaeおよびP. denticanis (例えば、Priovac)、ストレプトコッカス・エクイ(例えば、Equilis StrepE)、Chlamydophila abortus(例えば、OvilisおよびEnzovax)、Mycoplasma synoviae(例えば、Vaxsafe MS)、Mycoplasma gallisepticum (例えば、Vaxsafe MG)、Bordetella avium(例えば、Art Vax)、Actinobacillus pleuropneumoniae(例えば、PleuroStar APP)、Actinobacillus pleuropneumoniae(例えば、Porcilis APP)、サルモネラ(例えば、Megan VaclおよびMeganEgg)、ウシ流産菌(例えば、RB-51)、アイメリア種(例えば、Coccivac、Immucox、Paracox、Advent、およびNobilis Cox ATM)、アイメリア種(例えば、Inovocox)、E. tenella(例えば、Livacox)、Toxoplasma gondii(例えば、OvilisおよびToxovax)、仮性狂犬病ウイルス(例えば、Suvaxyn Aujeszky)、古典的豚コレラウイルス(例えば、Porcilis PestiおよびBayovac CSF E2)、ウマインフルエンザウイルス(例えば、PROTEQ-FLUおよびRecombitek)、ニューカッスル病ウイルス(例えば、Vectormune FP-ND)、鳥インフルエンザウイルス(例えば、Poulvac FluFend I AI H5N3 RG)、鳥インフルエンザウイルス(例えば、Trovac AI H5)、狂犬病ウイルス(例えば、RaboralおよびPurevax Feline Rabies)、ネコ白血病ウイルス(例えば、EURIFEL FeLV)、イヌパルボウイルス1(例えば、RECOMBITEK Canine Parvo)、イヌコロナウイルス(例えば、RECOMBITEK Corona MLV)、イヌジステンパーウイルス(例えば、RECOMBITEK rDistemperおよびPUREV AXFerret Distemper)、IHNウイルス(例えば、Apex-IHN)。獣医用ワクチンの他の例は、LHRH(例えば、Vaxstrate、Improvac、Equito、イヌゴナドトロピン放出因子免疫治療薬、およびGonaCon)およびアンドロステンジオン(例えば、Fecundin、 AndrovaxおよびOvastim)などの生殖調節ワクチンを含む。
【0127】
本発明の1つの態様において、免疫原性組成物中に含まれる抗原は、既知のインフルエンザワクチンの形態である。ほとんどの市販のインフルエンザワクチンは、表面の糖タンパク質を取り除くためにインフルエンザウイルスを有機溶媒で処理した、スプリット・ウイルスワクチン、サブユニットワクチン、または弱毒化生ウイルスワクチン、またはこれらの組合せである。一般に市販のインフルエンザワクチンは、インフルエンザの3つの株に対する防御を提供することにおいて3価であり、これらは例えば2007〜2008年のシーズンにおいては、A型/ソロモン諸島/3/2006(H1N1)様、A型/ウィスコンシン/67/2005(H3N2)様、およびB型/マレーシア/2506/2004様である。
【0128】
現在市販されているインフルエンザワクチンとしては、限定はしないが、Fluzone(登録商標)およびVaxigrip(登録商標)(Sanofi-aventis)、Fluvirin(登録商標)(Novartis Vaccine)、Fluarix(登録商標)、FluLaval(登録商標)およびFluviral S/F(登録商標)(GlaxoSmithKline)、Afluria(CSL Biotherapies)、FluMist(登録商標)(Medlmmune)およびInfluvac(商標)(Solvay Pharma)が挙げられる。
【0129】
抗原−MaMVおよび抗原 MaMV−VLPの組合せ
上記のように、免疫原性組成物に含まれる1または2以上の抗原は、MaMVまたはMaMV VLPの外被タンパク質に接合することができ、または非接合形態で組成物中に存在してもよく(すなわち、MaMVまたはMaMV VLPと単純に混合されている)、または、接合および非接合形態の両方で存在してもよい。接合は、例えば、外被タンパク質との遺伝子融合、または共有結合、非共有結合もしくは親和性手段を介した結合であることができる。抗原(単数または複数)とMaMVまたはVLPとの組合せは、しかし、抗原の宿主免疫系による認識を、またはMaMVもしくはVLPの免疫反応を増強する能力を妨げてはならない。
【0130】
本発明の1つの態様により、免疫原性組成物に含まれる1または2以上の抗原は、MaMV VLPの外被タンパク質に接合される。VLPが自己集合外被タンパク質の複数のコピーを含むため、抗原の外被タンパク質への付着は、VLPの表面上での複数の抗原の提示を可能とする。
【0131】
VLPの表面上での抗原の提示を可能とし、抗原の免疫認識を強化するために、抗原は、VLPの外表面上に配置された外被タンパク質の領域に付着するのが好ましい。したがって、抗原は、外被タンパク質のアミノ−(N−)またはカルボキシ−(C−)末端の近傍またはその位置に挿入することができ、または、VLPの外表面上に配置された外被タンパク質の内部領域に付着することができる。抗原接合外被タンパク質の、他の融合外被タンパク質、または野生型外被タンパク質と集合して、VLPを形成する能力は保持されるべきであり、この能力は、本明細書に記載のものを含む既知の方法により容易に試験することができる。1つの態様において、抗原は、外被タンパク質のC末端に、またはこの近傍に付着する。
【0132】
本発明の1つの態様により、免疫原性組成物は、1または2以上の抗原に遺伝子的に融合したMaMV外被タンパク質を含む。抗原が、MaMV外被タンパク質−抗原融合体の、VLPに自己集合する能力を妨げる可能性を避けるために、MaMV外被タンパク質への遺伝子融合のために選択される抗原は、一般に、約50個以下のアミノ酸長であり、例えば約45個以下のアミノ酸長である。
【0133】
所望により、スペーサーを、抗原と外被タンパク質の間に含むことができる。この目的のために好適なスペーサーは当分野で知られており、例えば、約3〜約10個のアミノ酸の短いアミノ酸配列を含む。一般に、この状況でのアミノ酸スペーサーは、グリシン、ロイシン、バリンおよびイソロイシンなどの中性アミノ酸から構成される。1つの態様において、抗原がMaMV外被タンパク質に遺伝子的に融合されている場合、融合は、約3〜約10個の中性アミノ酸のペプチドスペーサーを含む。
【0134】
より長い抗原も、MaMVまたはVLPと単純に混合することにより、または化学的架橋または親和性付着により、免疫原性組成物中に組み込むことができ、これは以下にさらに詳細に記載する。
【0135】
抗原(単数または複数)は、例えば、共有結合または非共有結合(例えばイオン性、疎水性、水素結合など)による付着により、化学的に外被タンパク質に架橋することができる。抗原および/または外被タンパク質は、当分野に知られているようにして、かかる架橋を促進するように修飾することができ、これは例えば、官能基または化学部分をタンパク質および/または抗原に、例えばCもしくはN末端または内部の位置において添加することによる。例示の修飾には、S−アセチルメルカプトコハク酸無水物(SAMSA)またはS−アセチルチオアセテート(SATA)などの官能基の添加、または1もしくは2以上のシステイン残基の添加を含む。他の架橋試薬は当分野に知られており、多くは市販されている(例えば、Pierce Chemical Co.およびigma-Aldrichからのカタログ参照)。例としては以下を含むが、これらに限定されない:ジアミン類、例えば1,6−ジアミノヘキサン、1,3−ジアミノプロパンおよび1,3−ジアミノエタン;ジアルデヒド類、例えばグルタールアルデヒド;スクシンイミドエステル類、例えばエチレングリコール−ビス(コハク酸N−ヒドロキシスクシンイミドエステル)、ジスクシンイミジルグルタレート、ジスクシンイミジルスベラート、N−(g−マレイミドブチリルオキシ)スルホスクシンイミドエステルおよびエチレングリコール−ビス(スクシンイミジルスクシネート);ジイソシアネート類、例えばヘキサメチレンジイソシアネート;ビスオキシラン類、例えば1,4−ブタンジイルジグリシジルエーテル;ジカルボン酸類、例えばジサリチル酸サクシニル;3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステル、その他。上述の架橋剤の多くは、親和性部分をVLPから離すためのスペーサーを組み込む。他のスペーサーの使用もまた本発明により意図される。種々のスペーサーが当分野に知られており、限定することなく、6−アミノヘキサン酸;1,3−ジアミノプロパン;1,3−ジアミノエタン;およびポリグリシン配列などの、1〜5アミノ酸の短いアミノ酸配列を含む。
【0136】
1または2以上の抗原の、VLPの外被タンパク質への共有結合的付着を促進するために、外被タンパク質は、VLPの表面に露出され、抗原の化学的付着に対する適切な部位を提供する、短ペプチドまたはアミノ酸リンカーに遺伝子的に融合することができる。例えば、システイン残基を含む短ペプチド、または共有結合を形成可能な側鎖、または当分野で知られているようにして共有結合を形成するように容易に修飾できる側鎖を有するアミノ酸残基(例えば、酸性および塩基性残基)である。アミノ酸リンカーまたはペプチドは、例えば、1〜約20のアミノ酸長であることができる。1つの態様において、外被タンパク質は、1または2以上のレジン残基を含む短ペプチドに融合し、これは例えば抗原のシステイン残基と、上記の好適な架橋剤の使用を通して共有結合することができる。
【0137】
本発明のさらなる態様においては、抗原は、外被タンパク質上に存在する親和性部分を介して付着する。この態様によれば、MaMV VLPは、自己集合の後にVLPの表面上に露出され、抗原に特異的に結合することができる、ペプチドなどの親和性部分を含む。親和性部分は、MaMVまたはVLPに、遺伝子的に融合するか(ペプチドまたはタンパク質断片の場合)、または共有結合もしくは非共有結合的に付着してよい。抗原の、親和性部分への結合は、宿主免疫系による抗原の認識を妨げてはならない。親和性部分は、全タンパク質を結合でき、またはタンパク質断片もしくはペプチドを結合できる。
【0138】
好適な親和性部分の例としては、抗体および抗体断片(例えばFab断片、Fab’断片、Fab’−SH断片、F(ab’)2断片、Fv断片、二機能性抗体(diabody)および単鎖Fv(scFv)分子)、ストレプトアビジン(ビオチン標識抗原を結合する)、抗原に特異的に結合する親和性ペプチドまたはタンパク質断片が挙げられるが、これに限定されない。
【0139】
親和性部分として用いるのに好適なペプチドまたは抗体(抗体断片を含む)は、当分野で知られている技法、例えばファージまたは酵母提示技術により、選択することができる。ペプチドは、天然、組換え、合成またはこれらの組合せであってよい。例えば、ペプチドは天然のタンパク質またはポリペプチドの断片であってよい。ペプチドの用語はまた、ペプチドアナログ、ペプチド誘導体およびペプチド模倣化合物も包含する。かかる化合物は当分野に知られており、天然のペプチドに対して利点を有する可能性があり、これには例えば、より大きな化学的安定性、タンパク質分解に対する抵抗性の増加、および/または抗原性の低下などを含む。
【0140】
親和性部分として用いるのに好適なペプチドは、約3アミノ酸長〜約50アミノ酸長であることができる。本発明の1つの態様により、親和性結合ペプチドは、少なくとも5アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、少なくとも7アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約5〜約50アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約7〜約50アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約5〜約45アミノ酸長、約5〜約40アミノ酸長、約5〜約35アミノ酸長、および約5〜約30アミノ酸長である。本発明の特定の態様により、親和性結合ペプチドは、3、4、5、6、7、8、9、10、11、12、13、14または15アミノ酸長である。当業者により理解されるように、抗原を親和性部分に結合するために選択されるペプチドの長さは、MaMV VLPの自己集合の能力、または宿主の免疫系による、一度結合された抗原の認識の能力を妨げてはならない。
【0141】
MaMVまたはVLPが含む親和性部分は、単一ペプチドまたはペプチドの縦列もしくは複数配置であってよい。所望の場合には、大きな抗原の結合を促進するために、スペーサーを、親和性部分と外被タンパク質の間に含むこともできる。好適なスペーサーは、例えばグリシンなどの中性アミノ酸の短い範囲を含む。例えば、約3〜約10個の中性アミノ酸の範囲である。
【0142】
ファージ提示を用いて、目的の抗原タンパク質に結合する特定のペプチドを選択することができ、ここで標準の技法(例えば、Current Protocols in Immunology, ed. Coligan et al., J. Wiley & Sons, New York, NY参照)、および/または市販のファージ提示キット(例えば、New England Biolabsから入手可能なPh.Dシリーズのキット、およびNovagenより入手可能なT7-Select(登録商標)キット)を用いて行う。ファージ提示によるペプチドの選択の例は、以下の例8にも提供される。
【0143】
MaMVおよびMaMV VLPの調製
本発明は、組換えMaMV外被タンパク質由来のMaMV VLPを、ならびにMaMVまたはMaMV VLPを含む免疫原性組成物を提供する。本発明はさらに、1または2以上の抗原または親和性部分を、外被タンパク質との遺伝子融合で含む、MaMV VLPを提供する。これらの組換え外被タンパク質は、多量体化およびVLPへの集合が可能である。抗原または親和性ペプチドを、抗原へ、また外被たんぱく質への結合のために遺伝子的に融合する方法は当分野に知られており、代表的な例が以下および実施例に記載される。種々の分子をタンパク質に化学的に架橋する方法も当分野でよく知られており、用いることができる。
【0144】
Malvaモザイクウイルス
本明細書の例の節に記載されているように、MaMVはMalva neglecta Wallr(一般のアオイ科植物)から単離することができる。ウイルスは、容易にM. neglectaに、またはアカザ科、例えば例1に記すようにChenopodiaceae quinoaからの植物種に繁殖させることができる。
【0145】
ウイルスは、簡潔に述べると、Chenopodiaceae quinoa(4枚葉ステージ)の健康な葉を、例えば約40μLなどの適切な容積の担体中の約10〜100μgの精製ウイルスでこすることにより、容易に繁殖させることができる。あるいは、乳鉢中で水中の感染した葉(例えば、2mlの水中に1gの感染した葉)をひいて得た液を用いて、健康な植物に接種することができる。カーボランダム(研磨剤)をそっと、ウイルスまたは液でこする前の健康な葉の表面に落とす。感染は、指でそっとこすって微小な損傷部を作ることで開始され、これを通してウイルスは植物細胞中に浸入する。こすった約10分後、接種した葉を水で洗浄して、カーボランダムおよび任意の残留物を取り除く。接種した植物は、感染がモザイクまたは局所病変部の症状を示すまで、さらにおよそ5〜14日間成長させる(光および光周期の環境条件に依存する)。通常植物は、16時間の明期および8時間の暗期で、明期間は22℃で、および暗期間には16℃で成長させる。植物は、製造業者のプロトコルに従い20−20−20の肥料を用いて施肥する。一般に100〜1000gの感染植物の葉が、ウイルス精製のために必要である。
【0146】
ウイルスは、感染した葉から、標準のポテックスウイルス単離技術(例えばAbouHaidar MG, et al. (1998) Methods MoI Cell Biol 81 : 131-143およびTremblay, M.-H., et al., (2006). FEBS J 273: 14-25を参照)により単離することができる。好適な方法のさらなる例は以下である。感染した葉を収穫し、好適な緩衝液中に均質化し、続いてフィルタリングおよび/または遠心分離して砕片を取り除く。得られた懸濁液をブタノールおよび任意に洗浄剤で処理し、次に氷上で撹拌する。溶液を次に遠心分離し、得られたペレットを適切な緩衝液に再懸濁させる。ペレットを均質化し、次に再度遠心分離する。ビリオンを上清から、スクロースクッション上の超遠心分離によりペレット化し、得られたペレットを適切な緩衝液中にホモジナイザーを用いて再懸濁させる。ウイルス溶液は、所望により、0.45μMシリンジフィルターを通す前の追加の遠心分離ステップにより、清浄にすることができる。
【0147】
精製したビリオンは、例えば4℃にて冷蔵保存することができる。
【0148】
MaMV VLP
本発明のVLPの調製に好適な組換えMaMV外被タンパク質は、標準の遺伝子操作技術を用いて、野生型外被タンパク質の配列を提供された当業者により、容易に調製可能である。タンパク質の遺伝子操作の方法は当分野によく知られている(例えば、Ausubel et al. (1994 & updates) Current Protocols in Molecular Biology, John Wiley & Sons, New Yorkを参照)。野生型MaMV外被タンパク質のアミノ酸配列(配列番号2)および野生型タンパク質をコードするヌクレオチド配列(配列番号3)は本明細に提供されており、これらはまた、上記したようにGenBankから公的に入手可能である。
【0149】
野生型タンパク質をコードする核酸配列の単離およびクローニングは、標準の技法を用いて(例えば、Ausubel et al.、同上を参照)、例えばMaMVから標準の技法でRNAを抽出し、次にRNAテンプレートからcDNAを合成する(例えば、RT−PCRにより)ことにより、実施することができる。MaMVは、モザイク症状を示す感染植物の葉から、上記した標準の技法により(本明細書に記載の例1も参照)精製することができる。
【0150】
代替的に、外被タンパク質をコードする遺伝子を、標準の技法を用いて人工的に構成することができる。例えば、いくつか(例えば18〜20個)のオーバーラップしたリン酸化オリゴヌクレオチド(約80ヌクレオチド長)で、遺伝子配列全体を提示するものを、標準技法を用いて合成可能である。各ヌクレオチドは、それらの5’末端または3’末端において、例えば約20個のヌクレオチドがオーバーラップしていなければならず、ただし、3’末端においてのみオーバーラップする最終の5’オリゴと、5’末端においてのみオーバーラップする最終の3’オリゴは除く。オリゴヌクレオチドは適切な緩衝液内(例えば10mMのトリス/HCl、pH8および25mMのNaCl)にプールし、90℃に15分間加熱し、室温にゆっくり冷却して、オリゴヌクレオチド間のアニーリングおよび、全長MaMV CP遺伝子の生成を可能とすることができる。リガーゼ緩衝液へのT4 DNAリガーゼの添加は、オリゴヌクレオチドの集合を完了させる。任意に、MaMV CP遺伝子の5’および3’末端を含むオリゴヌクレオチドはユニークな制限部位を含むことができ、これは、アニールされたDNAを細菌プラスミドなどの適切なベクターにクローニングするのに用いることができる。このベクターは、プラスミド大腸菌などの適切な宿主細胞を形質転換して、プラスミドを増幅し、オリゴヌクレオチドのライゲーションを完了させるために用いることができる。全てのオリゴヌクレオチドのアニーリングは、これらを2つずつ順番にアニーリングし、続いてオリゴヌクレオチドの各ペアを2つずつアニーリングし、これを続けて、全長遺伝子が生成されるまで行うことにより、改善することができる。
【0151】
代替的に、全長MaMV CP遺伝子は、適切なプラスミド(例としてはpET−3D)にクローニングして大腸菌内(例えばBL21(DE3))でタンパク質を発現させる前に、ポリメラーゼ連鎖反応(PCR)により増幅することができる。
【0152】
全長遺伝子配列はまた、合成遺伝子を作製する多くの営利会社の1つによるサービスを通して得ることもできる(例えば、GenScript Corp.(Piscataway, NY)、Geneart AG(Regensberg, Bavaria)およびMolecular Cloning Laboratories(San Francisco, CA))。
【0153】
外被タンパク質をコードする核酸配列は次に、好適な発現ベクターに、直接挿入するか、または1もしくは2以上のサブクローニングステップの後に挿入する。当業者は、用いる正確なベクターは本発明に決定的に重要ではないことを理解する。好適なベクターとしては、プラスミド、ファージミド、コスミド、バクテリオファージ、バキュロウイルス、レトロウイルス、またはDNAウイルスが挙げられるが、これらに限定されない。外被タンパク質は次に、以下にさらに詳細に記載されるようにして、発現および精製することができる。MaMV外被タンパク質を含むベクター(プラスミドpMaMV−CP−6H(MaMV外被タンパク質を含むpET−3D))の例は、例6に記載される。プラスミドpMaMV−CP−6Hに含まれているMaMV外被タンパク質遺伝子のヌクレオチド配列は、図10A(配列番号23)に、およびコードされた外被タンパク質のアミノ酸配列は、図11A(配列番号24)に提供される。
【0154】
任意に、外被タンパク質をコードする核酸配列をさらに操作して、当分野で知られている標準のin vitro部位特異的突然変異誘発技術により、上記のものなどのように1または2以上の変異を導入することができる。突然変異は、コード配列を作っている1または2以上のヌクレオチドの、欠失、挿入、置換、逆位、またはこれらの組合せにより、導入可能である。これは、例えば、1または2以上のヌクレオチドミスマッチ、挿入または欠失を組み込むプライマーが設計されたPCRに基づく技法により、実現することができる。突然変異の存在は、多くの標準技法、例えば、制限解析またはDNA配列決定により確認可能である。
【0155】
上述のように、外被タンパク質はまた、外被タンパク質に融合した1または2以上の抗原、親和性ペプチドおよび/またはスペーサーペプチドを含む、融合タンパク質を産生するように、操作することもできる。融合タンパク質を作製するための方法は、当業者によく知られている。融合タンパク質をコードするDNA配列は、上記のように、好適な発現ベクターに挿入することができる。
【0156】
当業者は、外被タンパク質または融合タンパク質をコードするDNAを、コードされたタンパク質の活性に影響を与えることなく、種々の方法で変更できることを理解する。例えば、DNA配列の変化を用いて、タンパク質を発現するために用いる宿主細胞のコドンの選好を最適化することができ、またはDNA配列の変化は、発現を促進する他の配列の変化を含んでもよい。
【0157】
当業者は、発現ベクターは、外被もしくは融合タンパク質をコードするDNA配列の効率的な転写に必要な調節要素、例えば転写要素などをさらに含んでよいことを理解する。ベクターに組み込むことができる調節要素の例は、プロモーター、エンハンサー、ターミネーター、およびポリアデニル化シグナルを含むが、これに限定しない。したがって本発明は、外被タンパク質または融合タンパク質をコードする核酸配列に動作可能に結合された調節要素を含む、ベクターを提供する。当業者は、好適な調節要素の選択は、タンパク質の発現について選択された宿主細胞に依存し、かかる調節要素は、細菌、真菌、ウイルス、哺乳類または昆虫の遺伝子を含む種々の源に由来し得ることを理解する。
【0158】
必要に応じて、発現ベクターは、当分野に知られているような、発現されたタンパク質の精製を促進する異種(非相同)核酸配列をさらに含んでよい。かかる異種核酸配列の例は、金属親和性タグなどの親和性タグ、ヒスチジンタグ、アビジン/ストレプトアビジンコード配列、グルタチオン−S−トランスフェラーゼ(GST)コード配列およびビオチンコード配列を含むが、これに限定しない。得られた異種アミノ酸配列は、当分野に知られている方法に従って用いる前に、発現されたタンパク質から除去することができる。代替的に、異種アミノ酸配列は、続くVLPへのタンパク質の集合を妨害しない場合には、タンパク質上に保持することもできる。本発明の1つの態様において、外被タンパク質はヒスチジンタグ・タンパク質として発現される。異種アミノ酸配列は、外被タンパク質のカルボキシル末端またはアミノ末端に位置させることができる。
【0159】
発現ベクターは、好適な宿主細胞または組織内に、当分野で知られている種々の方法の1つを用いて、導入することができる。かかる方法は、Ausubel et al.(同上)に一般的に記載されており、例えば、安定または過渡的なトランスフェクション、リポフェクション、エレクトロポレーション、および組換えウイルスベクターによる感染を含む。当業者は、外被タンパク質の発現のための適切な宿主細胞の選択は、選択したベクターに依存することを理解する。宿主細胞の例は、細菌、酵母、昆虫、植物および哺乳類細胞を含むが、これに限定しない。用いる正確な宿主細胞は、本発明に重要ではない。外被タンパク質は、原核生物宿主(例えば、大腸菌、A. salmonicidaまたはB. subtilis)または真核生物宿主(例えば、SaccharomycesまたはPichia;哺乳類細胞、例えばCOS、NIH3T3、CHO、BHK、293、またはHeLa細胞;または昆虫細胞)において産生することができる。本発明の1つの態様において、組換え外被タンパク質は、植物または細菌細胞中に発現させることができる。他の態様において、組換え外被タンパク質は、細菌細胞中に発現させる。他の態様において、組換え外被タンパク質は、大腸菌細胞中に発現させる。
【0160】
必要に応じて、発現されたタンパク質は宿主細胞から、当分野で知られている標準の技法により(例えば、Current Protocols in Protein Science, ed. Coligan, J.E., et al., Wiley & Sons, New York, NYを参照)精製でき、標準のペプチド配列決定技法により、無傷のタンパク質またはそのタンパク質分解断片を用いて配列決定して、タンパク質の同一性を確認することができる。
【0161】
本発明の1つの態様により、組換え外被タンパク質は、多量体化およびVLPへの集合が可能である。一般に、VLP集合は、外被タンパク質を発現する宿主細胞内で起こる。VLPは宿主細胞から、本明細書に記載の実施例の節に記載されているような標準の技法により単離可能である。VLPは、クトマトグラフィなどの標準の技法によりさらに精製して、汚染宿主細胞タンパク質またはLPSなどの他の化合物を除去することができる。
【0162】
本発明の1つの態様において、外被タンパク質は集合して宿主細胞内にウイルスまたは偽ウイルスを提供し、これは核酸およびタンパク質を含む感染ウイルス粒子の産生に用いることができる。これにより、感染ウイルスまたは偽ウイルス粒子による隣接する細胞の感染、およびその中のタンパク質の発現が可能である。この態様において、ウイルスまたは偽ウイルスを複製するために用いる宿主細胞は、ウイルスの複製を可能とする、植物細胞、昆虫細胞、哺乳類細胞または細菌細胞であることができる。これらの細胞は、ウイルスに対する天然の宿主細胞であって、これからウイルス様粒子が導出されるものであってよいが、しかし必ずしもそうである必要はない。宿主細胞は、初めに粒子形態のウイルスまたは偽ウイルス(すなわち、核酸およびタンパク質を含む集合したロッド)により感染されるか、または代替的に、初期感染に用いるウイルス核酸が複製可能であり、全ウイルス粒子の産生をもたらす場合には、核酸形態において(すなわち、ウイルスRNAなどのRNA;cDNAまたはcDNAから調製されるランオフ転写物)感染されることができる。
【0163】
MaMVまたはMaMV VLPのストックの産生
組換えMaMVまたはVLPのストックを、標準技法により調製する。例えば、組換え外被タンパク質を含むMaMVまたは偽ウイルスを、例えばChenopodium quinoaなどの適切な宿主内に、十分なMaMVまたは偽ウイルスが収穫できるように、繁殖させることができる。
【0164】
MaMV VLPのストックは、適切は宿主細胞から、例えば、上述のようにして調製された、VLPを作る組換え外被タンパク質をコードする発現ベクターで形質転換またはトランスフェクトされた大腸菌などから、調製することができる。宿主細胞は次に、当分野で知られているように、コードされたタンパク質の発現に有利な条件下で培養される。発現された外被タンパク質は多量体化して宿主細胞のVLPに集合することができ、これは上述のような標準の技法で、例えば細胞を破壊して、細胞可溶化物を1または2以上のクロマトグラフィによる精製ステップに供することにより、細胞から単離することができる。
【0165】
MaMVおよびMaMV VLPのストックは、例えば4℃にて冷蔵庫内で保存することができる。
【0166】
組換えおよび改変外被タンパク質の特徴
組換え外被タンパク質および、抗原、親和性ペプチドおよび/またはスペーサーペプチドが結合した外被タンパク質を、それらの多量体化およびVLPへ自己集合する能力について、標準の技法により解析することができる。これは例えば、精製タンパク質を電子顕微鏡法により視覚化することによる(例えば、例6を参照)。VLPの形成はまた、超遠心分離法により決定してもよく、円偏光二色性(CD)分光法を用いて、組換えもしくは改変タンパク質とWTウイルスの二次構造を比較することができる(例えば、Tremblay, et al, FEBSJ, 2006, 273: 14-25参照)。
【0167】
VLPおよびMaMVの安定性を、必要に応じて、当分野で知られている技法、例えばSDS−PAGEおよびトリプシン分解分析(例えば、Tremblay, et al, 2006、上記)により、決定することができる。
【0168】
有効性の評価
本発明の免疫原性組成物の、動物において免疫反応を誘発する能力を、当分野に知られた方法、例えば以下および例に記載されたものなどにより、試験することができる。例えば、MaMV、MaMV VLPまたはこれを含む免疫原性組成物を、好適な動物モデルに、例えば皮下注射により、または鼻腔内に投与することができ、抗体の発生を評価する。
【0169】
細胞性免疫反応はまた、当分野で知られている技法により評価することができる。例えば、細胞性免疫反応は、MaMV VLPに発現されたエピトープの、in vitroおよびin vivoでの樹状細胞による特異的Tリンパ球へのプロセシングおよび交差提示(cross presentation)を評価することにより、決定することができる。細胞性免疫(Tリンパ球)の誘発を評価する他の有用な技法としては、T細胞増殖およびIFN−γ分泌放出のモニタリング、例えばELISAによりサイトカインの誘発をモニタリングすること(例えば、Leclerc, D., et al, J. Virol, 2007, 81(3):1319-26参照)などを含む。
【0170】
MaMVまたはMaMV VLPおよび1もしくは2以上の抗原を含む免疫原性組成物の、ワクチンとしての有効性を評価するために、チャレンジ試験を実施することができる。かかる試験は、試験動物(例えばマウス、ラットまたはフェレット)の群に、本発明の免疫原性組成物を標準技法により接種することを含む。非接種動物および/または市販のワクチンを接種した動物を含む対照群、または他の陽性対照は、平行して設定する。ワクチン接種後の適切な時間の後、動物を、天然の抗原含有物質または有機体によりチャレンジする。動物から、接種前および接種後、およびチャレンジ後に採取した血液試料を、ウイルスに対する抗体反応について分析する。抗体反応についての好適な試験は、ウェスタンブロット解析および酵素結合免疫吸着法(ELISA)などを含むがこれに限定されない。動物はまた、抗原含有物質または有機体に関連する状態の症状の発生についてもモニタリングすることができる。
【0171】
同様に、腫瘍関連抗原を含有する免疫原性組成物の、それらの予防効果についての試験は、試験動物に接種し、続いて、癌細胞を動物に例えば皮下的に移植することによるチャレンジ、および動物における腫瘍の発達のモニタリングにより、行うことができる。代替的に、免疫原性組成物の治療効果を、癌細胞の移植および腫瘍の確立後に組成物を試験動物に投与し、腫瘍の増殖および/または転移のモニタリングにより、試験することができる。
【0172】
医薬組成物
上述のように、MaMVまたはMaMV VLPおよび任意に1もしくは2以上の抗原を含む免疫原性組成物は、さらに任意に、好適な担体、賦形剤など、および/または組成物の安定性、味のよさ、薬物動態、バイオアベイラビリティ等を改善する医薬組成物のその他の標準成分を含んでよい。本発明の1つの態様において、免疫原性組成物は、アジュバントとして用いるために製剤化される。他の態様において、免疫原性組成物は、ワクチンとして用いるために製剤化される。
【0173】
組成物は、種々の経路による投与のために製剤化可能である。例えば、組成物は、経口、局所、直腸、鼻腔、または非経口投与用に製剤化でき、または吸入もしくはスプレーによる投与用に製剤化できる。本明細書において用語、非経口的とは、皮下注射、静脈内、筋肉内、髄腔内、胸骨内注射または注入技法を含む。対象への鼻腔内投与は、医薬組成物を、対象の鼻道または鼻腔の粘膜に投与することを含む。本発明の1つの態様において、組成物は局所、直腸もしくは非経口投与用に、または吸入もしくはスプレーによる、例えば鼻腔内経路による投与用に製剤化される。他の態様において、組成物は非経口投与用に製剤化される。
【0174】
組成物は好ましくは、本発明の免疫原性組成物の有効量を含む。本明細書において用語「有効量」とは、検出可能な免疫反応を生成するのに必要な組成物の量をいう。与えられた適用に対しての免疫原性組成物の有効量は最初に、例えば細胞培養アッセイまたは動物モデルにおいて、通常はげっ歯類、ウサギ、イヌ、ブタ、または霊長類において、推定可能である。動物モデルはまた、好適な濃度範囲および投与経路を決定するために用いてもよい。かかる情報は次に、ヒトを含む処置すべき動物における、有用な用量および投与経路の決定に用いることができる。本発明の1つの態様において、単位用量は、約5μg〜約10mgの外被タンパク質を含む。他の態様において、単位用量は、約10μg〜約10mgの外被タンパク質を含む。さらに他の態様において、単位用量は、約5μg〜約5mgの外被タンパク質を含む。他の態様において、単位用量は、約10μg〜約5mgの外被タンパク質を、約10μg〜約2mgの外被タンパク質を、約15μg〜約5mgの外被タンパク質を、および約20μg〜約5mgの外被タンパク質を含む。1また2以上の用量を用いて動物を免疫化することができ、これらは、同じ日に投与することも、または数日もしくは数週間の期間にわたって投与することもできる。
【0175】
上述のように、本発明の免疫原性組成物は複数の抗原(接合および/または非接合形態)を含んでよく、したがって多価ワクチン製剤を提供することができる。複数のVLPを含む多価ワクチン組成物であって、各VLPが異なる抗原に接合しているものもまた、意図される。多価ワクチン製剤は、2価および3価の製剤を、より高い価のワクチンに加えて含む。
【0176】
一定の態様において、複数の(すなわち、2または3以上の)異なる抗原を含む多価ワクチン製剤も、製剤中のより多くの数のエピトープにより、改善された防御を提供することができる。抗原は、接合形態(例えば、各々が異なる抗原に接合した多数の異なるVLPにより)、または非接合形態において、または接合および非接合形態の両方において、含むことができる。
【0177】
経口使用のための組成物は、例えば、錠剤、トローチ剤、ロゼンジ、水性もしくは油性の懸濁液、分散粉末または顆粒、乳剤、硬質もしくは軟質カプセル、またはシロップまたはエリキシル剤として、製剤化可能である。かかる組成物は、当分野で知られている医薬組成物の製造の標準的方法により製造可能であり、甘味料、香味剤、着色剤および保存剤の群から選択される1または2以上の剤を、薬学的に洗練された味のよい調製物を提供するために、含んでよい。錠剤は、免疫原性組成物を、好適な無毒性の薬学的に許容し得る賦形剤、例えば以下に記載の賦形剤と組み合わせて含む:不活性の希釈剤、例えば炭酸カルシウム、炭酸ナトリウム、ラクトース、リン酸カルシウム、またはリン酸ナトリウム;顆粒化または崩壊剤、例えばコーンスターチ、またはアルギン酸;結合剤、例えばスターチ、ゼラチンまたはアカシア、および湿潤剤、例えばステアリン酸マグネシウム、ステアリン酸またはタルク。錠剤は被覆されていなくてもよく、または、消化管での分解および吸収を遅延させ、これにより長期の持続作用を提供するために、既知の技術により被覆されてもよい。例えば、モノステアリン酸グリセリルまたはジステアリン酸グリセリルなどの時間遅延材料を用いてもよい。
【0178】
経口使用のための組成物はまた、免疫原性組成物が不活性の固体希釈剤、例えば炭酸カルシウム、リン酸カルシウムまたはカオリンと混合される、硬質ゼラチンカプセルとして提供することもでき、あるいは、活性成分が水またはピーナツ油、液体パラフィンまたはオリーブ油などの油性媒体と混合される、軟質ゼラチンカプセルとしても提供可能である。
【0179】
鼻腔内投与用の医薬組成物は、例えば、鼻腔用スプレー、鼻腔用のドロップ、懸濁液、溶液、ゲル、軟膏、クリーム、および粉末を含むことができる。組成物は、例えばAccuspray(登録商標)(Becton Dickinson)などの好適な市販の鼻腔用噴霧器を介する投与用に製剤化することができる。鼻腔内投与他の方法は、当分野で知られている。
【0180】
水性懸濁液として製剤化される組成物は、免疫原性組成物を1または2以上の好適な賦形剤と組み合わせて含み、これは例えば、懸濁剤であって例えばカルボキシメチルセルロースナトリウム、メチルセルロース、ヒドロプロピルメチルセルロース、アルギン酸ナトリウム、ポリビニルピロリドン、ヒドロキシプロピル−β−シクロデキストリン、トラガカント・ゴムおよびアカシア・ゴム;分散剤または湿潤剤、例えば天然のホスファチド、例えばレシチン、または酸化アルキレンと脂肪酸との縮合生成物、例えばステアリン酸ポリオキシエチレン、または酸化エチレンと長鎖脂肪族アルコールとの縮合生成物、例えばヘプタ−デカエチレンオキシセタノール、または酸化エチレンと脂肪酸/ヘキシトール由来の部分エステルとの縮合生成物、例えばポリエチレンソルビトールモノオレエート、または酸化エチレンと脂肪酸/ヘキシトール無水物由来の部分エステルとの縮合生成物、例えばポリエチレンソルビタンモノオレエートなどである。水性懸濁液はまた、1もしくは2以上の保存剤、例えばエチル、またはn−プロピルp−ヒドロキシベンゾエート、1もしくは2以上の着色剤、1もしくは2以上の香味剤または1もしくは2以上の甘味料、例えばスクロースもしくはサッカリンを含んでよい。
【0181】
組成物は、油性懸濁液として、免疫原性組成物を例えばラッカセイ油、オリーブ油、ゴマ油またはココナツ油などの植物油中に、または液体パラフィンなどの鉱油中に懸濁させることにより、製剤化することもできる。油性懸濁液は、増粘剤、例えば蜜ろう、硬質パラフィンまたはセチルアルコールなどを含んでよい。上記などの甘味料、および/または香味剤を任意に加えて、口に合う経口製剤を提供することができる。これらの組成物は、アスコルビン酸などの抗酸化剤の添加により、保存することができる。
【0182】
組成物は、分散性の粉末または顆粒として製剤化でき、これは次に、水を添加して水性懸濁液の調製に用いることができる。かかる分散性の粉末または顆粒は、免疫原性組成物を、1または2以上の分散剤または湿潤剤、懸濁剤および/または保存剤と組み合わせて提供する。好適な分散剤または湿潤剤、および懸濁剤は、上記で既に例示されている。追加の賦計剤、例えば、甘味料、香味剤および着色剤もまた、これらの組成物に含むことができる。
【0183】
本発明の組成物はまた、水中油型乳剤としても製剤化可能である。油相は、オリーブ油もしくはラッカセイ油などの植物油、または液体パラフィンなどの鉱油、またはこれらの油の混合物でもよい。これらの組成物に含めるのに好適な乳化剤は、天然のゴム、例えばアカシアゴムもしくはトラガカントゴム;天然のホスファチド、例えば大豆、レシチン;または脂肪酸とヘキシトール由来のエステルもしくは部分エステル、無水物例えばモノオレイン酸ソルビタン、および前記部分エステルと酸化エチレンとの縮合生成物、例えばポリオキシエチレンソルビタンモノオレエートを含む。乳剤はまた、任意に甘味料および香味剤を含むことができる。
【0184】
組成物は、シロップまたはエリキシル剤として、免疫原性組成物を1または2以上の甘味料、例えばグリセロール、プロピレングリコール、ソルビトールまたはスクロースなどと混合することにより製剤化することができる。かかる製剤はまた、1または2以上の粘滑剤、保存剤、香味剤および/または着色剤を任意に含むことができる。
【0185】
組成物は、無菌の注射用水性または油性懸濁液として、当分野で知られている方法により、上述のものなどの、好適な1または2以上の分散剤または湿潤剤および/または懸濁剤を用いて、製剤化することができる。無菌の注射用調製物は、無毒性の非経口的に許容し得る希釈剤または溶媒中の、無菌の注射用溶液または懸濁液であってよく、例えば1,3−ブタンジオール中の溶液である。用いることのできる、許容し得るビヒクルおよび溶媒は、水、リンガー溶液、乳酸化リンガー溶液および等張食塩水を含むが、これに限定されない。他の例には、従来から溶媒または懸濁媒体として用いられている無菌の固定油、および種々の無刺激性の固定油であって、例えば合成モノ−およびジグリセリドなどを含む。オレイン酸などの脂肪酸もまた、注射物の調製に用いることができる。
【0186】
任意に、本発明の組成物は、保存剤例えば抗菌剤、抗酸化剤、キレート剤および不活性ガスなど、および/または安定化剤例えば炭水化物(例えばソルビトール、マンニトール、スターチ、スクロース、グルコース、またはデキストラン)、タンパク質(例えばアルブミンまたはカゼイン)、またはタンパク質含有剤(例えばウシ血清アルブミンまたはスキムミルク)を、好適な緩衝液(例えばリン酸緩衝液)と共に含んでよい。組成物のpHおよび種々の成分の正確な濃度は、周知のパラメータにしたがって調節してよい。
【0187】
さらに、アジュバント活性を有する1または2以上の化合物も、ワクチン組成物に任意に加えてよい。好適なアジュバントは、例えば、水酸化、リン酸化、または酸化アルミニウム;油型乳剤(例えばBayol F(登録商標)またはMarcol52(登録商標));サポニン、またはビタミンE可溶化物を含む。オプソニン化ワクチン組成物も本発明に包含され、例えば、ワクチンにより前に免疫化した動物またはヒトから単離された抗体を含む、ワクチン組成物である。ワクチンにより前に免疫化した動物またはヒトから単離された抗体に基づく組換え抗体も、ワクチン組成物をオプソニン化するために用いることができる。
【0188】
他の医薬組成物および医薬組成物を調製するための方法は、当分野で知られており、例えばRemington: The Science and Practice of Pharmacy”(旧タイトル:“Remingtons Pharmaceutical Sciences”);Gennaro, A., Lippincott, Williams & Wilkins, Philadelphia, PA (2000)に記載されている。
【0189】
さらに、1または2以上の従来のアジュバントも、組成物に任意に加えてよい。好適なアジュバントは、例えば、ミョウバンアジュバント(例えば水酸化、リン酸化、または酸化アルミニウム);油型乳剤(例えばBayol F(登録商標)またはMarcol52(登録商標));サポニン、またはビタミンE可溶化物を含む。ビロソーム(virosome)もアジュバント特性を有するとして知られており(Adjuvant and Antigen Delivery Properties of Virosomes, Glueck, R., et al., 2005, Current Drug Delivery, 2:395-400)、S. typhiポリンタンパク質(例えばOmpC)も同様にこれを有し、本発明の組成物に任意に含むことができる。
【0190】
さらに、本発明は、免疫原性組成物が抗原に融合したMaMV VLPを含む場合、該組成物はまた、組成物中のVLP全体のアジュバント効果を増加するために、非融合タンパク質に由来するMaMV VLPも含んでよいことを意図する。
【0191】
組成物は任意に、オプソニンを、例えば、抗原、MaMV、もしくはMaMV VLPで前に免疫化した動物またはヒトから単離された抗体を、含んでもよい。抗原、MaMV、もしくはMaMV VLPで前に免疫化した動物またはヒトから単離された抗体に基づく組換え抗体もまた、オプソニンとして用いることができる。
【0192】
本発明にさらに包含されるのは、MaMVまたはMaMV VLPを、上記のような市販のワクチンと組み合わせて含む組成物である。
【0193】
免疫原性組成物の使用
本発明は、MaMV、MaMV VLPおよびこれを含む免疫原性組成物の、多くの使用を提供する。非限定的例としては、免疫原性組成物の、アジュバント、免疫促進剤として、またはワクチンとしての使用が挙げられる。1または2以上の抗原に接合したMaMV VLPもまた、抗原(単数または複数)に対する抗体のスクリーニングのために用いることができる。したがって、本発明は、免疫原性組成物を投与することにより、動物に免疫反応を誘発する方法、および免疫原性組成物の、例えばアジュバント、免疫促進剤、ワクチン、および/または医薬組成物などの医薬の調製のための使用も提供する。
【0194】
本発明の免疫原性組成物は、ヒトおよび、家畜用動物と農場用動物を含む非ヒト動物における使用に好適である。免疫原性組成物の投与形態は、任意の他の一般に認められたワクチン接種プログラムと違っている必要はない。例えば、効果的な免疫反応を引き出すのに十分な量での免疫原性組成物の単回投与を用いてよく、または代替的に、免疫原性組成物の初回投与に続いて、抗原のみで、または抗原と免疫原性組成物でのブースティングという他の形態を用いてもよい。同様に、免疫原性組成物または抗原のどちらかによるブースティングは、抗体価が許容レベルを下回った場合には、初回投与の後の時点で行ってもよい。免疫原性組成物の正確な投与方法は、例えば、組成物の成分(例えば、組成物が抗原を含むか、またはアジュバントとして提供されるのか)、処置する動物、および処置の所望の最終効果に依存する。適切な投与方法は、経験ある施術者により容易に決定することができる。
【0195】
免疫原性組成物が1または2以上の非接合抗原を含む場合、MaMVまたはVLP成分は、1または2以上の抗原と同時に投与することができ、または、免疫反応を必要とする対象の必要性に応じて、抗原の投与の前に、またはこれに続いて、投与することができる。
【0196】
免疫原性組成物は、予防的に、例えば、ウイルス、細菌もしくは他の感染粒子による感染を、または疾病もしくは腫瘍の発症を防ぐために用いることができ、または、これは、例えば感染症または癌に関連する疾病もしくは疾患の影響を改善するために治療的に用いてもよい。本発明の1つの態様において、免疫原性組成物は、予防的に用いる。この状況において、免疫原性組成物は、1または2以上の抗原を含むことができ、または免疫原性組成物は、MaMVもしくはMaMV VLPのみを含んでもよく、これは、低レベルの感染などの感染症または疾患に対する抵抗性を誘発するのに十分であり得る。
【0197】
免疫原性組成物は、組成物中の含有について、または組成物との使用について選択された抗原に依存して、種々の疾病または疾患の予防または処置において用いることができる。非限定的な例としては以下が挙げられる:インフルエンザ(種々のインフルエンザウイルスからの抗原を使用)、腸チフス(S. typhiからの抗原を使用)、HCV感染(HCV抗原を使用)、HBV感染(HBV抗原を使用)、HAV感染(HAV抗原を使用)、HIV感染(HIV抗原を使用)、ポリオ(ポリオウイルス抗原を使用)、ジフテリア(ジフテリア毒素由来の抗原を使用)、EBV感染(EBV抗原を使用)、アレルギー反応(種々のアレルゲンを使用)、および癌(種々の癌関連抗原を使用)が挙げられる。他の使用としては、例えば、炎症性疾患(例えば関節炎)、および鳥インフルエンザウイルス、ヒト呼吸器合胞体ウイルス、デング熱ウイルス、麻疹ウイルス、単純ヘルペスウイルス、ヒトパピローマウイルス、偽ウイルス、豚ロタウイルス、豚パルボウイルス、ニューカッスル病ウイルス、口蹄病ウイルス、豚コレラ(hog cholera)ウイルス、豚コレラ(African swine fever)ウイルス、感染性ウシ鼻気管炎ウイルス、感染性咽頭気管炎ウイルス、ラクロスウイルス、新生子牛下痢ウイルス、ウシ呼吸器合胞体ウイルス、ウシウイルス性下痢ウイルス、豚マイコプラズマ肺炎菌、連鎖球菌、淋菌、腸内細菌および寄生生物(例えばリーシュマニアおよびマラリア)が挙げられる。
【0198】
本発明の免疫原性組成物はまた、例えば、該組成物が異なる病原物質からの複数の抗原を含む場合、多価ワクチンとして用いるのに好適である。
【0199】
免疫原性組成物はまた、従来のワクチンと併せて用いて、ワクチンの有効性を改善し、または多価ワクチンを提供することができる。この状況において用いることのできる、市販のワクチンの非限定的例は上に提供されている。市販のワクチンは、ヒトワクチンまたは獣医学的使用を意図されたワクチンであってよい。
【0200】
1つの態様において、本発明はまた、以前にポテックスウイルスベースのワクチンを接種された対象に対するワクチン接種のための、免疫原性組成物の使用も提供する。このアプローチは、第2のワクチンの有効性が、第1のワクチンにおいてポテックスウイルスに対して増殖した抗体との相互作用のために、低下しないことを保証することができる。例えば、PapMVベースのワクチンプラットフォームが記載されている(例えば、国際特許出願PCT/CA03/00985 (WO 2004/004761)および米国特許出願第11/556,678号(US2007/0166322)を参照)。動物またはヒトの、PapMV VLPでアジュバント化したワクチンによる免疫化は、アジュバントの主要成分であるPapMV外被タンパク質に指向された多量の抗体を生成することができる。同じ患者を、同じPapMVプラットフォームで免疫化すると、常在の抗体の新しく投与されたワクチンに対する免疫反応を、PapMVプラットフォームに直接結合することにより制限する可能性がある。したがって、MaMVまたはMaMV VLPに基づく第2のワクチンの使用はこの効果を避けるのに有用であり、より効率的なワクチン接種プログラムの保証を支援する。
【0201】
本発明はまた、1または2以上の抗原に接合したMaMV VLPの、スクリーニング剤としての使用、例えば、1または2以上の抗原に対する抗体をスクリーニングするための使用も提供する。VLPは従来の免疫的技術、例えば酵素結合免疫吸着測定法(ELISA)またはウェスタンブロッティングなどに容易に適合でき、したがって診断および研究の文脈において有用である。
【0202】
キット
本発明はさらに、MaMV、MaMV VLP、または本発明の免疫原性組成物を含むキットの、アジュバントまたはワクチンとしての使用を提供する。MaMV、MaMV VLP、または免疫原性組成物が、市販のワクチンを含む、別の抗原製剤と共に用いることが意図される場合、キットは任意に、該抗原製剤を含むことができる。
【0203】
キットの個々の成分は別々の容器に入れられて、かかる容器に付随して、薬学的または生物学的製品の製造、使用または販売を規制している政府機関による所定の形態における、該機関による製造、使用または販売の承認を反映する注意書きがあってよい。キットは任意に、アジュバントまたはワクチンの使用または投与の方法を概説する使用説明書または指示書を含んでよい。
【0204】
キットの1または2以上の成分が溶液として、例えば水溶液、または無菌の水溶液として提供される場合、容器はそれ自体が、吸入器、シリンジ、ピペット、点眼容器、または他の類似の機器であってよく、これから溶液を対象に投与でき、またはキットの他の成分にこれを適用して混合することができる。
【0205】
キットの成分はまた、乾燥または凍結乾燥形態で提供してもよく、キットは追加して凍結乾燥成分の再構成のための好適な溶媒を含むことができる。容器の数または種類と無関係に、本発明のキットはまた、患者への組成物の投与を支援するための機器も含んでよい。かかる機器は、吸入器、シリンジ、ピペット、鉗子、計量スプーン、点眼容器、または類似の医学的に承認された送達ビヒクルであってよい。
【0206】
抗体検出に用いるための、1または2以上の抗原に接合したMaMV VLPを含むスクリーニングキットも提供される。キットは、診断用キットまたは研究目的のキットであることができる。キットの個別の成分は別々の容器に入れられ、かかる容器に付随して、生物学的製品の製造、使用または販売を規制している政府機関による所定の形態における、該機関による製造、使用または販売の承認を反映する注意書きがあってよい。キットは任意に、免疫原性組成物の使用方法を概説する使用説明書または指示書を含んでよい。
【0207】
本明細書に記載の発明のよりよい理解を得るために、以下の実施例を示す。これらの例は、本発明の例示の態様を記載することを意図し、どのようであれ本発明の範囲の限定を意図するものではないことが理解される。
【0208】
例
例1:Malvaモザイクウイルスの増幅、精製、およびRNA抽出
新しいポテックスウイルスであるMalvaモザイクウイルス(MaMV)を、Malva neglecta Wallr.(一般のアオイ科ゼニアオイ属)から単離した。最初に感染したMalva neglecta植物は、カナダ・ブリティッシュコロンビア州サマーランド周辺の地域で収集した。ウイルス感染によるモザイク症状および葉脈の透明化のみが、Malva neglectaにおいて観察された症状であった(図4A参照)。
【0209】
ウイルスによるChenopodium quinoaの感染は、植物全体を移動する強いモザイクパターンを生成した(図4B参照)。C. quinoaは、したがって、増殖の宿主として用い、これを用いてウイルス精製のための多量の感染葉を収集した。C. quinoaでの増殖のために接種物をC. quinoaの感染葉からとった際に、局所病変が認められた(図4C参照)。局所病変の誘発は、接種に用いる汁液中でのウイルス濃度に関連するようであり(感染したC. quinoaの葉にはより多くのウイルス)、これは、同一宿主上での症状の差を説明する。
【0210】
MaMVを次のようにして増幅および精製した。Malva neglectaの感染した葉を、サマーランドB.C.周辺の地域で収穫し、ケベックにある発明者らの研究室に送った。Malva neglectaの感染した葉を、液体窒素中乳鉢/乳棒で粉砕して、葉の粉末を得た。粉末は水中に再懸濁させて、増殖宿主のChenopodium quinoa上に、葉をカーボランダムでこすることにより接種した。葉は、接種2〜3週間後に感染症状を示した(すなわち、感染した葉の上の、モザイクおよび葉脈透明化)。感染した葉を、10mMのEDTAおよび0.1%の重炭酸ナトリウムを含有する100mMのリン酸ナトリウム緩衝液、pH7.6に均質化した。ホモジェネートを、チーズクロス(粗い綿布)の層を通してフィルタリングし、7,800gで20分間遠心分離した。TritonX-100および2%のブタノールを1滴ずつ上清に加え、氷上で60分間撹拌した。溶液を7,800gで20分間遠心分離した。次に上清を100,000gで90分間超遠心分離し、得られたペレットを100mMのリン酸ナトリウム緩衝液、pH7.6に再懸濁させた。ペレットを、Dyna-Mixホモジナイザー(Fisher Scientific、セッティング2)で均質化し、次に7,800gで5分間、再度遠心分離した。ウイルスは上清から、100,000gで3時間の超遠心分離により、30%のスクロースクッション上にペレット化し、これを再度、10mMのトリス−HCl緩衝液、pH8.0中にDyna-Mixホモジナイザーにより再懸濁させた。ウイルス溶液を、7,800gで5分間の最後の遠心分離により清浄にし、最後に0.45μMシリンジフィルターユニット(Nalgene)を通した。精製ウイルスは、電子顕微鏡法またはウイルスRNA抽出用に用いるまで、4℃で保存した。
【0211】
ウイルスRNAを、ウイルス溶液から、フェノール/クロロホルム抽出とエタノール沈殿により抽出した。プロテイナーゼK処理を用いて、任意の残留タンパク質をRNAから取り除いた。RNAは、必要となるまで−20℃で保存した。
【0212】
ウイルスの電子顕微鏡分析には、精製ウイルス粒子をホルムバール/炭素被覆カッパーグリッド上に載せ、1%タングステン酸メチルアミンで染色した。試料は、JEOL透過電子顕微鏡で分析した。感染したC. quinoaの葉から得た精製ウイルスの電子顕微鏡分析により、ウイルスが約540×13nmの柔軟な繊維状ビリオンであることが示され(図4、バーは50nmを表す)、これはポテックスウイルス科のメンバーに典型的である(Adams et al., 2004. Arch. Virol. 149, 1045-1060)。
【0213】
例2:Malvaモザイクウイルスのクローニングおよび配列決定
MaMVをさらなる試験のために、以下のようにしてクローニングおよび配列決定した。
【0214】
第1のcDNA鎖を、例1に記載のようにして調製したMaMV RNAから、Supercsript(登録商標)のRT−PCR(Invitrogen)用の第1鎖合成システムを製造業者の指示に従って用いて、ランダムヘキサヌクレオチドまたはポリ−dTプライマーで合成した。次に、ターミナルトランスフェラーゼ(New England Biolabs)を製造業者の指示に従って用いて、cDNAの3’末端おいてポリデオキシシチジン尾を添加する前に未使用のプライマーを取り除くため、cDNAをPCR精製キットカラム(QIAGEN)を通した。Asc IまたはPac I制限部位でタグしたポリデオキシグアノシンプライマーをcDNAにハイブリダイズして、製造業者の指示に従って大腸菌DNAポリメラーゼクレノウ断片(New England Biolabs)を用いて、第2鎖を合成した。得られた二本鎖DNAを、フェノール/クロロホルムおよびエタノール沈殿により精製し、次に製造業者の記載に従ってT4 DNAポリメラーゼ(New England Biolabs)で処理して、平滑末端を作製した。平滑末端DNAを適切な制限酵素で消化して、次にpNEB193ベクター(New England Biolabs)中16℃で一晩、T4 DNAリガーゼ(New England Biolabs)にライゲーションした。ライゲーション反応を大腸菌DH5α中に転換し、形質転換細胞をLB寒天プレート上アンピシリン(50μg/ml)を用いて増殖させた。抵抗性コロニーからのプラスミドDNAをQIAprep spin miniprepキット(QIAGEN)を用いて抽出し、次にABI 3730XLシーケンサーで配列決定した。MaMVゲノム配列の3つのオーバーラッピング断片を含有するクローンを得た;1つはCPおよび3つの遺伝子ブロックを含み、1つは複製の残りを含み、いくつかのクローンはMaMVの5’末端を含む(図6A参照)。MaMVゲノムに対応する得られたDNA配列を、図1(配列番号1)に示す。
【0215】
例3:Malvaモザイクウイルスの5’末端の同定
MaMVの5’末端配列は、次のようにして同定した。ゲノムRNA(例1のようにして調製)を最初に、50mMの酢酸ナトリウム、pH6.0、1mMのEDTA、0.1%のβ−メルカプトエタノールおよび0.01%のトリトンX−100中の2Uのタバコ酸性ピロホスファターゼ(Epicentre Biotechnologies)により、37℃で60分間処理して、5’キャップ構造を取り除いた。反応は、フェノール−クロロホルム抽出(0.5vol./0.5vol)により停止させ、RNAを、上記のように逆転写する前に、ただしプライマー0023−11を用いて、エタノール沈殿した;このプライマーは、ゲノムRNAの498〜520位置においてハイブリダイズする(5'-ACATGTAAGCTAAACTAGTGTC-S'、配列番号17)。オリゴヌクレオチドEMSA1(5'-GTGATAAAGTTATGACCATAACCTATGTCGTAGGATATGCATTAACTAAT-3'、配列番号18)を、次にcDNAの3’末端に、20UのRNAリガーゼを用いて製造業者記載の条件下で(New England Biolabs)加えた。第2鎖を次に、キット「Expand High Fidelity PCR System」を製造業者の記載に従って用いて(Roche Diagnostics)、プライマー0023−11およびEMSA2(5'- ATTAGTTAATGCATATCCTACGACATAGGTTATGGTCATAACTTTATCAC-3'、配列番号19)の存在下で、合成して増幅した;これはEMSA1に相補的である。DNAを94℃で2分間変性させ、次に33周期(94℃で45秒間、60℃で45秒間、および72℃で60秒間)のPCRにより増幅した。増幅産物は、TOPO TAクローニングキット(Invitrogen)を用いて、ベクターpCR2.1−TOPO中に直接クローニングした。ライゲーション産物を大腸菌DH5α内に形質転換して増幅し、プラスミドDNAを例2に記載のようにして抽出および配列決定した。いくつかのクローンを正確に配列決定して、MaMVゲノムの5’末端を正確に評価した。
【0216】
MaMVゲノムの5’末端の配列の分析により、以下が決定された。5’非翻訳領域(UTR)は81ヌクレオチドのACに富んだドメインであり、これは、安定性の弱い2つの推定ステムループがそれぞれヌクレオチド26〜36および48〜78に存在することを除いて、ほとんどが非構造化されている。ほとんどのポテックスウイルスと同様に、逆転写の前にタバコ酸性ピロホスファターゼ(TAP)で処理したRNAに観察される優勢な5’末端モチーフは、コンセンサスモチーフGAAAAである(16のうちの10の配列)(図5)。図5は、逆転写反応の前に、タバコ酸性ピロホスファターゼで未処理(−TAP)および前処理(+TAP)されたRNAについて得られた配列を、それぞれ上の図および下の図に示す。バーは、配列決定された全クローンにおいて見出された各配列の頻度を表す。図5でわかるように、タバコ酸性ピロホスファターゼ処理を省略した場合、優勢なモチーフは、ScaVX、CymMVおよびAIsVXにおいて観察されように、GGAAAAであった(17配列中、10)(Wong et al., 1997. Arch. Virol. 142, 383-391;Kim et al., 1998. Mol. Cell. 8, 181-188;Chen et al., 2002. Arch Virol. 147, 683-693;Fuji et al., 2005. Arch Virol. 150, 2377-2385.)。最後のケースの場合、コンセンサスGAAAA配列は2回のみ観察された。この差異は、ほとんどの逆転写酵素がターミナルトランスフェラーゼおよびテンプレートスイッチ活性を有することにより、部分的に説明可能である。Superscript IIは、これがキャップ終端を有するin vitroで転写されたRNAで、製造業者が推奨するバッファー条件を用いて試験された場合に、選好的に1つの追加のシトシン残基をcDNAの3’末端に付加することが示されている(Schmidt et Mueller, 1999. Nucleic Acids Res. 27, e31)。Schmidtはまた、逆転写酵素によるdCMPテーリングが、5’−OH RNAと比べた場合に、キャップされたRNAテンプレートで10倍も効率的であることを観察した。この観察は、本明細書で観察された結果を説明できる可能性があり、その理由は、cDNAの3’末端に付加したシトシン残基は、RNAの5’末端において余分なグアノシンとして読み取られ得るからである。未処理のRNAで観察されたこの追加のG残基は、事実、前に記載した逆転写酵素のターミナルトランスフェラーゼ活性のために人工的(artefactual)であり、そのため、ゲノムRNAの5’末端において見出される正しいモチーフを正確に提示しない。したがって、TAP処理したRNAで得られた結果は、宿主中に見出されるMaMVの天然の集団についてのゲノム配列をより正確に反映すると考えられる。
【0217】
興味深いことには、5’末端のGGAAAAモチーフは、いくつかのポテックスウイルス、例えばAIsVX、LVX、ScaVXなどに見出されている。これらのウイルスが、in vivoでこの特異なGGAAAA配列を示す可能性は完全には排除できないが、上記の結果は、かかるモチーフの発見が用いたプロトコルに結びついており、宿主中に見出されるゲノム集団を正確に表しているわけではなく、コンセンサスGAAAAモチーフは、ポテックスウイルス属に一般に広がっている特長となり得ることを示唆している。
【0218】
例4:Malvaモザイクウイルスの、配列解析、系統発生解析、およびゲノム構成による特徴づけ
MaMVを以下のようにさらに特徴付ける。
【0219】
配列解析
例2に記載のようにして得たMaMV配列をスクリーニングし、全米バイオテクノロジー情報センター(NCBI)のBLASTnデータベースと比較した(Altschul et al., 1997. Nucleic Acids Res. 25, 3389-3402.)。ウイルス配列を、Contig Assembly Program(CAP)ソフトウェア(Huang, 1992. Genomics. 14, 18-25)を用いて解析およびアセンブルした。オープンリーディングフレーム(ORF)を、NCBIのORFファインダーで同定した。同一性/類似性分析をWisconsin(GCG)パッケージversion 10.3からのプログラムGAPにより、アミノ酸比較について8個のギャップ創造(creation)ペナルティー、および2個のギャップ延長(extension)ペナルティーを用いて行った(Anon, 2001. Wisconsin Package version 10.3. Accelrys Inc., San Diego, Ca, USA)。レプリカーゼ、TGB1および種々のポテックスウイルス株からのカプシドタンパク質のアミノ酸配列を決定し、Clustal Wソフトウェア(version 1.83)により生成した複数のアラインメント内に入れて、SeqLabアプリケーション(Wisconsin package version 10.3; Accelrys)の最終目視検査を通して修正した。
【0220】
ポリ(A)尾を除き、MaMVゲノムRNAは、GC含量45%の6858個のヌクレオチド(nt)長である(GenBankアクセッション番号DQ660333)。ゲノム構成は他のポテックスウイルスに類似しており、推定のRNA依存性RNAポリメラーゼ(RdRp)、続いてTGBタンパク質をコードする3つのオーバーラッピング遺伝子、および最後に外被タンパク質を含む(図6A;レプリカーゼは白の四角で表し、3つの遺伝子ブロックタンパク質は薄い灰色の四角で、および外被タンパク質は濃い灰色の四角で表す。黒丸および菱形は、それぞれTGB1およびCPsgプロモーターを示す(略号:MT:メチルトランスフェラーゼ;AlkB:DNA/RNA修復ドメイン;HEL:ヘリカーゼ;POL:RNA依存性RNAポリメラーゼ;CP:外被タンパク質))。レプリカーゼについて示唆されたAUG開始コドンはヌクレオチド81〜83に位置し、この部位からの翻訳により、177.96kDaの計算分子量に対して1571アミノ酸(aa)のタンパク質を産生する。ORF1由来のMaMVレプリカーゼは、少なくとも3つの異なるドメインから構成され(図6A)、これらは、N末端メチルトランスフェラーゼ様ドメイン(aa31〜390)、NTP結合/ヘリカーゼ様ドメイン(aa824〜1059)、およびC末端RdRp2ドメイン(aa1137〜1535)である。これらのドメインは通常良好に保存され、すべてのポテックスウイルスのレプリカーゼ内で提示されている(Rozanov et al., 1992. J. Gen. Virol. 73, 2129-2134;Koonin and Dolja, 1993. Crit. Rev. Biochem. MoI. Biol. 28, 375-430;Longstaff et al., 1993. EMBOJ. 12, 379-386;Davenport and Baulcombe, 1997. J. Gen. Virol. 78, 1247-1251;Batten et al., 2003. Molecular Plant Pathology. 4, 125-131.)。この解析はまた、DNA修復タンパク質AlkBと相同性を共有するドメインの存在を強調する。主としてFlexiviridae科からのいくつかの他の植物RNAウイルスも、これらのレプリカーゼ内に類似のモチーフを有する(Aravind and Koonin, 2001. Genome Biology. 2, research0007.1-0007.8;Bratlie and Drablos, 2005. BMC Genomics. 6, 1-15)。図6Aに示すように、MaMVのアミノ酸配列とゲノム構成は、他のポテックスウイルスで見出されるものと類似している。
【0221】
次の3つのORFは、ポテックスウイルスTGBと類似性を示すオーバーラッピング遺伝子であり、これはウイルス移動に関与する(Beck et al., 1991. Virology. 183, 695-702;Batten et al., 2003.上記;Morozov and Solovyev, 2003. J. Gen. Virol. 84, 1351-1366)。TGB1(ORF2)は、低下した分子量26.3kDaの235aaタンパク質である。これは、非常に高い含量のロイシンと荷電された残基(それぞれ13.2%および21.7%)を有する。これは、全てのポテックスウイルス中最も酸性のTGB1であり、等電点(pI)は4.84である。この配列は、典型的なNTPアーゼ/ヘリカーゼドメインを有し、これのin vitroでのPVXおよび2つのホルデイウイルス(hordeivirus)TGB1タンパク質に対する活性が実証されている(Kalinina, N.O., et al, (2002) Virology 296(2):321-9)。TGB1は、RNAスライシングの阻害において役割を果たし得ることも示唆されている。TGB2タンパク質(ORF3)は、13kDaの計算分子量に対して119aa長であり、その理論的pIは9.42で、一方ORF4またはTGB3は、中性pIの84aaの短いタンパク質(9kDa)である(図7)。PVXにおいて、これら最後の2つのタンパク質は膜および細胞壁と会合し、その機能は主としてTGB1活性を調節することである(Morozov et al., 1991. J. Gen. Virol. 72, 2039-2042;Yang et al., 2000. MoI. Plant-Microbe Interact. 13, 599-605;Morozov and Solovyev, 2003.上記)。MaMV TGBタンパク質の保存された配列および疎水性プロファイルは、他のポテックスウイルスTGBタンパク質のそれらに典型的であり、これらが類似の活性を有し得ることを示唆している。
【0222】
ポテックスウイルスのカプシドまたは外被タンパク質(CP)は、ゲノム保護およびウイルス移動に関与する。MaMV CPは、保存された両親媒性コア配列KYAGFDFFDGVT(配列番号20;nt6545〜6581によりコードされる)を含み、これは、疎水性相互作用を介してポテックスウイルスRNAのCPへの結合に役割を果たすことが提唱されている(Bancroft et al., 1991. J. Gen. Virol. 72, 2173-2181;Dolja et al., 1991. Virology. 184, 79-86;Wong et al., 1997.上記;Cotillon et al., 2002. Arch. Virol. 147, 2231-2238;Thompson and Jelkmann, 2004. Arch. Virol. 149, 1897-1909;Chen et al., 2005. Arch. Virol. 150, 825-832; Fuji et al., 2005. 上記)。MaMV CPは243aaタンパク質であり(図2および配列番号2参照)、推定分子量は26kDaである。
【0223】
MaMVアミノ酸配列を他のポテックスウイルスと比較し、結果を下の表2にまとめて示す。MaMVとより緊密に関連するウイルスにはアスタリスク(*)を付した。MaMVと最大の相同性を有するタンパク質は太字で示した。略号およびGenBankアクセッション番号は以下である:AIsVX:NC_007408;BaMV:NC_001642;CVX:NC_002815;CsCMV:NC_001658;ClYMV:NC_001753;CymMV:NC_001812;FdMV:NC_001483;HdRSV:NC_006943;LVX:NC_007192;MVX;NC_006948;NMV:NC_01441;OVX:NC_006060;PapMV:NC_001748;PepMV:NC_004067;PlAMV:NC_003849;PAMV:NC_03632;PVX:NC_001455;ScaVX:NC_003400;SMYEV:NC_003794;TVX:NC_04322;WClMV:NC_003820;ZVX:NC_06059。
表2:MaMVと他のポテックスウイルスのアミノ酸同一性/相同性
【表2】
aこのタンパク質に対して得られた同一性/相同性の結果は、修正NMV外被タンパク質配列を用いて得られた(下記参照)。
【0224】
この比較から、ORF(レプリカーゼ)はScaVXからの相同タンパク質と最大の同一性を有し、一方他の全てのORF(TGB1、2および3、および外被タンパク質)は、それらのNMV対応物とより関連することが示された。PepMVおよびAlsVXはまた、MaMVと、特にレプリカーゼおよびカプシドタンパク質において良好な局所的相同性を示した。しかし、AlsVXおよびPepMVは全体的にはMaMVから遠く離れており、これは主として、それらのそれぞれのTGBタンパク質間の低い相同性による。上で示したように、相同性解析により、最大のアミノ酸同一性スコアが、MaMVとScaVXのレプリカーゼの間(67.2%)、およびMaMVとNMVのカプシドタンパク質の間(75.6%)で明らかにされた。これらの値は上記のAdams et al. (2004)が確立した種の分画(species demarcation)の分子分類より低いため、MaMVは以前に公開されている全てのポテックスウイルスとは異なる種であると考えることができる。
【0225】
Malva葉脈壊死ポテックスウイルス(MVNV)は、1990年にブラジルにおいてMalva parvifloraに感染するウイルスとして報告されており(Brunt et al., (eds.) 1996 onwards. "Plant Viruses Online: Descriptions and Lists from the VIDE Database. Version: 20th August 1996")、Malva parvifloraには局所病変部と全体の葉脈壊死が検出された。MVNVのヌクレオチド配列は現在利用可能ではない。MVNVが単離された元の宿主は、MaMVについての天然の宿主と類似であるが、この2つのウイルスは次の理由により異なっている:a)電子顕微鏡で観察されたMaMV粒子はMVNVよりも長い、およびb)MVNVはMalva種に局所的病変を誘発することが示されたが、MaMVはモザイク症状を誘発する。
【0226】
系統発生解析
系統発生解析は、MEGA version 3.1(Kumar et al., 2004. Brief Bioinform. 5(2), 150-163)により距離法(distance method)と隣接結合アルゴリズム(neighbour-joining algorithm)を用いて行った。ツリーの位相幾何的精度は、500個のブートストラップ複製を用いて評価した。5’および3’非翻訳領域(UTR)の構造解析は、プログラムmfold version 3.2(Zuker, 2003. Nucleic Acids Res. 31, 3406-3415)を用いて行った。タンパク質ドメインの多重アラインメントおよびプロファイルは、ケンブリッジのWellcome Trust Sanger Instituteのウェブサイトにあるタンパク質ファミリーのPfamデータベース(version 18)により、デフォルトパラメータを用いて行った(Bateman et al., 2004. Nucleic Acids Res. 32, 138-141)。多重アラインメントは、NMV CP(NC_001441)のC末端領域におけるフレームシフトエラーを強調し、これは、このタンパク質の最後の45のaaが他の任意のポテックスウイルスとまったく関連していないことを示す。したがって、修正NMV配列を相同性/系統発生解析に用いた。
【0227】
系統発生解析は、MaMVと他のポテックスウイルスとの間の関係をより正確に試験するために行った。解析は、レプリカーゼおよびカプシドアミノ酸配列のもっとも保存された領域を用いて(図8)、およびTGB1タンパク質の完全な配列を用いて行った。図8は、MaMVと他のポテックスウイルスのレプリカーゼおよびカプシドタンパク質の系統発生解析を示す。レプリカーゼおよび外被タンパク質の系統発生ツリーは、それぞれ右と左の図である。数字は、各枝のブートストラップの値(500複製)を示す。アスタリスクで強調したウイルスは、MaMVの3’末端非翻訳領域において同定されたものと類似の推定偽結節構造を有する。
【0228】
系統発生解析により、ポテックスウイルスに通常検出される保存配列は、MaMV内にも見出されることが示された。系統発生解析は、MaMVが、NMV、ScaVX、AlsVXおよびPepMVを含むポテックスウイルスのサブグループに最も緊密に関連するように見えるとの前の観察を支持し、ここでこれらのウイルスの後者2つは、系統発生学的にわずかに遠い位置にある。解析された全てのツリーにおいて、MaMV、NMVおよびScaVXは常に一緒のグループにあり、この緊密な関係をここでも支持している。しかし、レプリカーゼおよびカプシドタンパク質に対する、このサブグループ内で観察されてブートストラップの低い値(それぞれお66%および39%)は、これら2つのウイルスのどちらに対してMaMVが最も緊密に関係するかについての、明確な結論を引き出すことを許容しなかった。TGB1を用いて行った系統発生解析も類似の結果を示し、MaMVに最も近い親戚についての結論は得られなかった。
【0229】
サブゲノムプロモーターおよび保存RNAプロモーター要素の解析
全RNAを、健康なC. quinoa植物およびMaMV感染のC. quinoa植物から次のようにして抽出した。C. quinoaの感染した葉および非感染の葉を収穫し、液体窒素中で粉砕した。全RNAを、フェノール/クロロホルムを用いて抽出し、RNAスピンカラム(Qiagen)を用いてさらに精製した。全RNAの解析を、ノーザンブロッティングにより次のようにして行った:RNA(20μg)をホルムアルデヒドアガロースゲル(1%)上で分離し、次にナイロン膜(Amersham Biosciences)に移した。不動化したRNAを、PCRによりジゴキシゲニン(Roche Diagnostics)で標識したMaMV外被タンパク質のcDNA断片でプローブした。製造業者のプロトコルに従ってハイブリダイゼーションを行った。ハイブリダイゼーションシグナルを、化学発光基質CDP-Star(Roche Diagnostics)で可視化した。
【0230】
ノーザンブロットにより、3つの主要なウイルスRNAが感染中に産生されることが示された:これらは、1つの大きなゲノムRNAおよび、それぞれ約2000および800ヌクレオチドのRNAとして移動している、2つのサブゲノム種である(図6B)。サブゲノム種は、3重遺伝子ブロックおよびウイルスCP(200nt長のRNA)およびウイルスCPのみ(800ntのRNA)をコードする、ウイルスサブゲノムRNAに対応するはずである。MaMV中のsgプロモーターコンセンサス配列の同定は、TGBとCPが、それらの独自のsgRNAからin vivoで翻訳されることを強く示唆する。しかし、これらが、ゲノムRNAの内部開始により産生可能であるとの可能性も除外できず、その理由は、このメカニズムが少なくともPVX CPに対して前に観察されているからである(Hefferon et al., 1997. J. Gen. Virol. 78, 3051-3059)。
【0231】
オクタヌクレオチドサブゲノムプロモーター配列は、レプリカーゼとTGB1遺伝子の間(nt4851〜4858)およびTGB3コード配列の3’末端のnt5982〜5989の間の遺伝子間領域に見出されている。図6Cは、オクタヌクレオチドの推定sgプロモーター配列の配列アラインメントを示す。右と左の図は、それぞれTGB(黒丸)およびCP(黒の菱形)sgプロモーターコンセンサス配列を示す。コンセンサスオクタヌクレオチド内に高度に保存されたヌクレオチドを、それぞれ黒または灰色の四角で強調する。コンセンサス配列と異なるヌクレオチドは、白字で示す。
【0232】
両方のプロモーターは、ほとんどのポテックスウイルスにおいて取り出される正確なコンセンサス配列GTTAAGTTを有する(Skryabin et al., 1988. FEBS. 240, 33-40;Kim and Hemenway, 1997. Virology. 232, 187-197;Batten et al., 2003.上記)。PVXについて報告されたTGB2/TGB3sgプロモーターコンセンサス配列は、TGB1およびCPのそれらのようには良好に定義されず、その結果、かかるドメインの正確な同定は、より不明確である(Skryabin et al., 1988.上記)。活性なTGB2/TGB3プロモーターに相関する種に対応するsgRNAは、これらの感染植物中に検出されなかった。これらのsgRNA種が存在する場合には、それらはTGB1およびCPのsgRNAのように豊富ではない可能性が高い。
【0233】
3’UTRは70nt長で、他のポテックスウイルスで同定されたように、tRNA様の二次構造内に畳まれていると推定される(Thompson and Jelkmann, 2004.上記)。3’UTRはポリアデニル化シグナルAAUAAAをポリアデニル化部位の14nt上流に含み、また今日まで配列決定された全てのポテックスウイルス中に存在する保存ヘキサマーACUUAAを含む(nt6799〜6804)。両方の配列は、3つのステムループ構造の異なるループ内に局在する(それぞれSL3およびSL1)(図7)。図7は、MaMV3’非翻訳領域の二次構造の模式図である。3つのステムループ構造は、SL1、SL2およびSL3として同定される。全てのポテックスウイルス中で見出されるコンセンサス配列ACUUAAは、SL1内の淡い灰色のヌクレオチドにより強調され、一方、SL3に存在するポリアデニル化シグナルは濃い灰色で示す。プログラムmfoldにより得た二次構造は、挿入図内に示す。SL1とSL2の間の新規な推定偽結節は、点線により示す。クエスチョンマーク(?)は、偽結節が推定のものであり、溶液中のこの構造の畳み込みはまだ決定されていないことを示す。
【0234】
この解析により、SL1とSL2のループに存在するヌクレオチド間の5塩基対の偽結節が明らかにされた。類似の構造が前にPVXについて記載されたが、この場合、相補的領域は、サブゲノムプロモーターからのヌクレオチドとSL1のACUUAA保存配列の間であった(Kim and Hemenway, 1997.上記)。他のポテックスウイルスの3’末端の解析は、類似の偽結節がいくつかの他のウイルス、例えばNMV、CymMV、ScaVX、およびPepMVに検出できることを示した。しかし、後者2つのウイルスにおいて偽結節を形成する相補的配列は、わずか4ヌクレオチド長である。
【0235】
例5:Malvaモザイクウイルス外被タンパク質の産生、精製および大腸菌への自己集合
完全なMaMV外被タンパク質遺伝子を、ゲノムRNAの3’末端全体を包含するシーケンシングプラスミド(CPおよび3重遺伝子ブロックをコードする配列を含有;例2参照)から、PCRにて、フォワードプライマー(5'-GGTACATGTCGAACTCTGGTTCAGCCG-3'、配列番号21)およびリバースプライマー(5'-TACGGATCCTCAATGGTGATGGTGATGGTGGAATTCTGGGGGGGCTTCAA TGG-3'、配列番号22)を用いて増幅した。フォワードプライマーはAf1 III制限部位(下線部)を含み、これはPCR産物をpET−3D発現ベクター内にクローニングするための開始コドンを含み、一方、リバースプライマーは、6X−Hisタグ(下線部)をCP遺伝子の3’末端に付加して、ニッケル親和性カラム上でのさらなるタンパク質の精製を可能とする。PCR反応は、次のように行った: Expand(登録商標)High FidelityPCRシステム(Roche Diagnostics)を用い、製造業者推奨の標準条件化において、94℃で3分間の前インキュベーションの後、94℃で45秒、65℃で45秒、および72℃で60秒を33サイクル。PCR産物はAf1 III/Bam H1制限酵素で消化して、次にpET−3D適合のNco I/Bam H1制限部位にライゲーションした。ライゲーション産物は、大腸菌DH5α中に転換して増幅し、例2に記載のようにして、プラスミドDNAを抽出および配列決定した。
【0236】
CP遺伝子(pMaMV−CP−6H)を含有することが同定されたプラスミドを用いて、大腸菌株BL21(DE3)RIL細胞(Invitrogen)を形質転換した。形質転換細胞を2X YT寒天プレート上に50μg/mlのアンピシリンとともに広げて、37℃で一晩インキュベートした。培養培地(2X YT、50μg/mlのアンピシリン)に、約10個の単離したコロニーの前培養物を播種し、ODが0.6〜0.8となるまで増殖させた。タンパク質の発現を、IPTG(Promega)を最終濃度1mMまで添加して誘発し、培養物を22℃で16時間、225RPMで振とうしつつインキュベートした。次に細胞を遠心分離し、溶解緩衝液中(50mMのNaH2PO4、300mMのNaCl、10mMのイミダゾール、pH8.0)に20μMのPMSF(EM Science)、1Xのプロテアーゼカクテル阻害剤(Roche)および1mg/mlのリソザイム(Sigma)の存在下で再懸濁させ、その後氷上でSonic Dismembrator model 500超音波処理器(Fisher Scientific)で超音波処理した。タンパク質溶液を遠心分離し、上清を、2.5mlのニッケル−NTAアガロースビーズの存在下で4℃で3時間インキュベートし、次に溶出カラム内(Bio Rad)に注いだ。ビーズはそれぞれ25mlの次の緩衝液で洗浄した:洗浄緩衝液1(50mMのNaH2PO4、300mMのNaCl、20mMのイミダゾール、pH8.0)、洗浄緩衝液2(50mMのNaH2PO4、300mMのNaCl、50mMのイミダゾール、pH8.0)、および洗浄緩衝液3(10mMのトリス−HCl、50mMのイミダゾール、pH8.0)。ビーズを次に、溶出緩衝液(10mMのトリス−HCl、1Mのイミダゾール、pH8.0)の存在下で30分間インキュベートして、タンパク質を溶出した。得られた溶出タンパク質を、上記のように10%のSDS−PAGEおよび電子顕微鏡法で解析し、ウイルス様粒子(VLP)の形成をチェックした。
【0237】
タンパク質抽出物のSDS−PAGE解析を用いて、34〜35kDaの優勢な形態を検出した(図9A;レーン1:タンパク質分子量マーカー。分子量マーカーは左側にkDaで示す;レーン2:大腸菌内に産生された精製外被タンパク質;レーン3:感染植物からの精製MaMVから単離された外被タンパク質)。外被タンパク質の、ポリアクリルアミドゲル上での移動性は、他のポテックスウイルス外被タンパク質と整合する(Tremblay et al, 2006. FEBS. 273, 14-25;Hu and Ghabrial 1995, J. Virol. Meth. 55, 367-379;Hammond and Hull, 1981. J. Gen. Virol. 54, 75- 90)。大腸菌内に産生されたMaMV CPの分子量は、野生型タンパク質を比べてわずかに異なるが、これは追加の6X−Hisタグにより説明可能である(図9A)。
【0238】
最後に、精製された組換えタンパク質を電子顕微鏡法で観察し、これにより、MaMV CPの過剰発現が、大腸菌の外被タンパク質の、天然ウイルスと非常によく似たウイルス様粒子への自己集合をもたらすことが確認された(図9B参照、この図は、精製組換えタンパク質MaMV CPから作られたウイルス様粒子の電子顕微鏡写真である(バーは50nmの長さ))。
【0239】
プラスミドpMaMV−CP−6Hに含有される、MaMV外被タンパク質遺伝子のヌクレオチド長を、図10(配列番号23)に示し、コードされた外被タンパク質のアミノ酸配列を図11(配列番号24)に示す。
【0240】
例6:精製組換えMalvaモザイクウイルス外被タンパク質のLC−MS/MS解析による解析
例5からの精製組換え外被タンパク質を、LC−MS/MS解析(Eastern Quebec Proteomics Centre, Centre Hospitalier de l'Universite Laval, Quebec)により解析した。試料は凍結乾燥し、その後還元し、50mMの重炭酸アンモニウム緩衝液中の45mMのジチオトレイトールおよび100mMのヨードアセトアミドでアルキル化した。アセトニトリルに希釈後、トリプシン消化を37℃で一晩、0.2μgの配列決定グレードの修飾トリプシン(Promega)を用いて行った。消化は蟻酸を用いて停止させ、2μLの試料を質量分析計に注入した。ペプチドMS/MSスペクトルを、ナノスプレーインターフェイスを有するLTQ(Thermo-Electron, San Jose, CA, USA)四重極IT質量分析計と連結した、キャピラリー液体クロマトグラフィにより得た。クロマトグラフィ分離は、PicoFritカラムBioBasic C18、10cm×75μm(New Objective, Woburn, MA)上、2〜50%の線形勾配の溶媒B(アセトニトリル、0.1%蟻酸)で30分間、200nL/分にて実施した。ペプチドは、カラムを通り直接LTQ線形イオントラップ質量分析計に溶出したが、この時スプレー電圧は1.8kV、および移送キャピラリー温度は225℃に設定した。質量スペクトルの取得は、Xcalibur 2.0 SR 2ソフトウェア(ThermoElectron Corp)によりデータ依存性取得モードを用いて制御したが、このモードにおいては、各フルスキャン質量スペクトル(400〜2000m/z)に続いて、7つの最も激しいイオンの衝突誘発性解離がある。動的排除機能が可能であり、相対的衝突断片化エネルギーは35%に設定した。
【0241】
得られたMS/MSスペクトルは、MASCOT(Matrix Science, London, UK; version 2.2.0)を用いて解釈し、3つの異なるデータベースを探索した:1つは目的のタンパク質を含むもの、Uniref Humanデータバンク、およびUniref大腸菌データベース(version 8.0、それぞれ94985および40567エントリーを含む)。システインのカルバミドメチル化、およびメチオニンの部分的酸化、2つの欠如した開裂、およびペプチドに対して2.0Daおよび断片に対して0.5Daの誤差耐性を、探索において考慮した。Scaffold(version Scaffold-01_06_18, Proteome Software Inc., Portland, OR)を用いて、MS/MSに基づくペプチドおよびタンパク質の同定を確認した。ペプチドの同定は、それらがPeptide Prophetアルゴリズム(Keller, A et al. 2002; Anal. Chem. 74(20):5383-92)により特定された90.0%を超える確率で確立された場合に、認定された。タンパク質の同定は、それらが90.0%を超える確率で確立され、少なくとも2つの同定されたペプチドを含む場合に、認定された。タンパク質の確率は、Peptide Prophetアルゴリズム(Nesvizhskii, A.I., 2003, Anal Chem. 75(17):4646-58)により割り当てられた。類似のペプチドを含み、MS/MS解析のみに基づいては識別できなかったタンパク質は、節減の原理(principles of parsimony)を満たすためにグループ化した。
【0242】
精製組換えタンパク質のトリプシンによる消化は30個のペプチドを生成し、これはLC−MS/MSにより解析され、全てのペプチドがMaMV CP配列に属することを確認した。MaMV CPは、25,961.2Daの分子量を有する。生成された30個のペプチドは、MaMVタンパク質の予測された断片に100%対応する質量を有した。ペプチドは、MaMV CPのアミノ酸配列の93%をカバーし、精製され大腸菌に過剰発現されたタンパク質の同一性を確認した。表3は、トリプシン消化により生成されたMaMV外被タンパク質ペプチドのLC−MS/MS解析を示す(配列番号25〜53)。
表3:MaMV外被タンパク質ペプチドのLC−MS/MS解析
【表3】
【0243】
例7:MalvaモザイクウイルスVLPの免疫原性効果
MaMV外被タンパク質遺伝子を、大腸菌BL21(pLysS)中に、プラスミドpMaMV−CP−6Hから次のようにして発現させた:pMaMV−CP−6Hを含有する大腸菌BL21(pLysS)の培養物1Lを調製し、コードされた外被タンパク質の発現を、1mMのIPTGにより誘発した。発現は、25℃でO/Nに進むことを可能とした。細胞を収穫し、フレンチプレスを750psi(溶解緩衝液:50mMのNaP緩衝液、pH8、20mMのイミダゾールおよび300mMのNaCl)で用いて細胞溶解した。砕片を10,000gでの遠心分離を2回、それぞれ45分間および30分間行って除去し、上清を保持した。12mlのNi2+ビーズ(Qiagen)を1Lの細菌培養物中に加え、バッチO/Nで4℃にてゆっくり振とうしつつインキュベートした。結合タンパク質を含有するビーズをエコノカラム(econo-column)に入れ、以下により洗浄した:(a)50mlの洗浄緩衝液(10mMのトリス−HCl、pH8+50mMのイミダゾール)、続いて(b)50mlの洗浄緩衝液+0.5%のTriton X-100、(c)50mlの洗浄緩衝液、(d)50mlの洗浄緩衝液+1%のZwittergen、および(e)50mlの洗浄緩衝液。タンパク質を次に1Mのイミダゾールで溶出した。
【0244】
溶出したタンパク質は、3Lの10mMトリス−HCl、pH8に対して1時間透析した。緩衝液を新鮮なものに交換し、透析をさらに1時間続けた。緩衝液を最後に交換し、O/Nで透析を継続した。タンパク質を次に、3LのPBSに対して、20分間隔で緩衝液を3回変えて透析した。次にNaClの濃度を500mMに増加させ、懸濁液を高速(100,000g)で3時間遠心分離して、VLPをペレット化した。
【0245】
VLPペレットを無菌のPBS中に再懸濁させ、0.45ミクロンフィルターを通してろ過した。最後に、タンパク質の濃度を1mg/mlに調節した。VLPの存在を電子顕微鏡を用いて確認し、任意のLPS混入物の存在を、Limulus試験を用いて決定した。
【0246】
MaMV VLPの免疫原性および免疫反応を引き出す能力を、標準のプロトコルに従ってマウスにおいて試験した。簡潔に述べると、5匹のBalb/Cマウスに対し、0日目に100μgのMaMV VLP(アジュバントなし)を注射した。血液試料を免疫化後5、10および14日目に採取した。血清中に存在するVLPに指向されたIgGの総量、IgG1の量、およびIgG2aの量を、標準のELISAにより測定した(図12A〜C参照)。
【0247】
同じマウスを、40日目に、100μgのMaMV VLPでs.c.にてアジュバントなしで2度目に免疫化した。血清中に存在するVLPに指向されたIgGの総量、IgG1の量、およびIgG2aの量を、再度、標準のELISAにより測定した(図12D〜F参照)。
【0248】
結果は、IgGおよびIgG1の高いレベルを示し、これは、MaMV VLPが高度に免疫原性であり、効率的な抗体反応を引き起こし得ることを示す。さらに、IgG2aの存在は、CD4+ T細胞の活性化および抗体クラススイッチの効率的な誘発を示唆する。この結果は、バランスの取れたTH1およびTH2反応が誘発されることを示唆する。
【0249】
例8:腸チフス菌ポリンタンパク質の精製
以下の精製手順を用いて、MaMVまたはMaMV VLPと併用して抗原として用いるのに好適な、腸チフス菌OmpCおよびOmpFタンパク質を精製した。精製手順は、Secundino et al. (2006), Immunology 117:59の記載に基づく。
【0250】
2種のタンパク質を腸チフス菌から共精製した。OmpCおよびOmpFの個別の精製は、OmpC[STYC171(OmpC)]またはOmpF[STYF302(OmpF)]のオープンリーディングフレームのいずれかが分断されている、腸チフス菌のノックアウト変異体を用いて行った。共精製については、細菌のノックアウト変異形態からの個別のタンパク質の精製法に従った。この方法を以下に概説する。
【0251】
細菌株Salmonella typhi 9, 12, Vi:d(ATCC9993)を、酵母抽出物、マグネシウム、およびグルコースを補足した最少培地Aの中で37℃、200rpmで増殖させた。酵母抽出物、マグネシウム、およびグルコースを補足した10Lの最少培地Aの処方は、以下のとおりである:5.0gの無水Na−クエン酸塩(NaC6H5O7:2H2O)、31.0gの一塩基性NaPO4(Na2H2PO4)、70.0gの二塩基性NaPO4(Na2HPO4)、10.0gの(NH4)2SO4、200mLの酵母抽出物溶液5%(300mL中15.0g)。1.434Lの培地を4Lエルレンマイヤーフラスコに分散させた。滅菌を、121℃、1平方インチ当たり15lbの加圧で15分間行った。次に各フラスコに以下を加えた:6.0mLの無菌MgSO4溶液25%、および60.0mLのグルコース溶液12.5%。フラスコに腸チフス菌の一晩培養物を播種し、OD540が1.0に達した時にインキュベーションを停止し、培養物を7,500rpm、4℃で15分間遠心分離した。ペレットを100mLの最終容量のトリス−HCl、pH7.7(6.0gのトリス−塩基/L)に再懸濁させ、バイオマス(生物量)を氷上で90分間超音波処理し、次に7,500rpm、4℃で20分間遠心分離した。10mLの上清の各々に、2.77mLの1MのMgCl2、25mLのRNアーゼA(10,000U/ml)、25mlのDNアーゼA(10,000U/ml)を加えた。混合物を次に120rpm、37℃で30分間遠心分離した。
【0252】
混合物からのポリン抽出は、次のようにして行った:
1.超遠心分離を、45,000rpm、4℃で45分間行い、ペレットを保持した。
2.ペレットを10mLの2%トリス−HCl−SDSに再懸濁させ、均質化した。
3.インキュベーションを、120rpm、32℃で30分間行った。
4.超遠心分離を、40,000rpm、20℃で30分間行い、ペレットを保持した。
5.ペレットを5mLの2%トリス−HCl−SDSに再懸濁させ、均質化した。
6.インキュベーションを、120rpm、32℃で30分間行った。
7.超遠心分離を、40,000rpm、20℃で30分間行い、ペレットを保持した。
8.ペレットを20mLのNikaido緩衝液−1%SDSに再懸濁させ、均質化した。[1LのNikaido緩衝液に対して:6.0gのトリス−塩基、10.0gのSDS、23.4gのNaCl、1.9gのEDTAを水に溶解し、pHを7.7に調整した。次に0.5mLのβ−メルカプトエタノール溶液を加えた]。
9.混合物を120rpm、37℃で120分間インキュベートした。
10.超遠心分離を、40,000rpm、20℃で45分間行った。ポリン抽出物を含む上清を回収した。
【0253】
ポリンを上清から高速タンパク質液体クロマトグラフィ(FPLC)で精製した。β−メルカプトエタノール非含有の0.5XNikaido緩衝液(上記参照)を、精製過程中用いた。タンパク質をSephacryl S-200(FPLC WATERS 650E)を用いて10mL/分のフラックス速度で分離した。カラムに22mLの上清を負荷した。溶出断片を260nmおよび280nmでモニタリングした。精製ポリンを含有する主ピークを保持し、4℃で保管した。精製ポリンは長期間安定であった(1年間以上)。
【0254】
図13は、上記の方法で精製したポリン、OmpCおよびOmpFのSDS−PAGEプロファイルを示す。
【0255】
例9:腸チフス菌ポリンのMalvaモザイクウイルスVLPへの付着に好適な親和性ペプチドの産生
精製OmpCおよびOmpFに対する特異的ペプチドを、Ph.D-7 Phage Display Peptide Library Kit(New England Biolabs, Inc.)を用いて選択した。用いたプロトコルは、「パニング」として知られているin vitro選択法であり、これは製造業者のプロトコルに従って行った。簡潔に述べると、2×1011個のファージを、ELISAプレートのウェルのベースに結合した10μgの精製OmpCまたはOmpFに加え、ウェルの内容物を室温で1時間ゆっくり混合した。非結合ファージを1mlの200mMのグリシン−HCl(pH2.2)で、室温で10分間インキュベートすることにより溶出した。上清を中和するために、およびファージを殺さないために、150μlの1Mのトリス−HCl(pH9.1)を加えた。溶出したファージを次に増幅し、追加の結合/増幅サイクルに供して、結合配列に利するようにプールを豊富化させた。洗浄緩衝液は、パニングの第1ラウンド用に0.1%のTween 20を含有し、これは次のラウンド用に0.5%に増加させた。選択したファージを大腸菌ER2738中、各パニングのラウンドの間増幅させた。このサイクルを3回繰り返して、それぞれのポリンタンパク質に対して最高の親和性のペプチドを選択した。こうして同定したペプチドを表4に示す。
表4:OmpCおよびOmpF親和性ペプチドの配列および出現頻度
【表4】
【0256】
上で同定された親和性ペプチドをMaMV外被タンパク質のC末端に、例えばPCRにより導入することができ、得られた融合タンパク質を大腸菌に発現させて、融合タンパク質からのVLPを産生する。VLPを次に、溶液中でそれらの同族ポリンと、例えば1:1の比率で混合することができ、VLPおよびポリンを含む複合体を提供する。この複合体はそれのみか、または追加のポリンと組み合わせて、続いて腸チフス菌のチャレンジに対してマウスを免疫化するのに用いることができる。
【0257】
例10:市販ワクチン上でのMalvaモザイクウイルスのアジュバント効果
配列番号24に記載の配列(図11A参照)を有する組換えMaMV CPを、大腸菌に22℃で16〜22時間過剰発現させた。細菌をフレンチプレスを用いて溶解し、試料を遠心分離して砕片を取り除き、親和性精製のためにNi2+カラム上に負荷した。親和性クロマトグラフィにより精製した外被タンパク質を図15Aに示す。これらのCPから形成されたVLPは異なる長さを有するが、しかしWTウイルスに類似しており(図15B)、全てのタンパク質が高分子量形態(500kDaを超えるVLP)で見出されるSuperdex 200ゲルろ過からは除外したが、これは、単量体または低い多重形態(500kDa未満)の形態が検出されなかったからである(図15C)。MaMV VLP調製物中のLPSレベルを、24.0EU/mgにおいて評価した。続く免疫化において(下記参照)、3または30μgのVLPを用いたが、これらの量に対してLPS汚染は無視できると考えられる。
【0258】
MaMV VLPのアジュバント特性を測定するために、3価のインフルエンザワクチンFluviral(登録商標)(GlaxoSmithKline)をモデル系として用いた。ヒト用量の5分の1のFluviral(登録商標)を、3μgまたは30μgのどちらかのMaMV VLPで、または従来のアジュバントであるミョウバンで、アジュバント化し、皮下的にBalb/Cマウス(1群あたり5匹)に投与した。免疫化後14日目に出血させ、Fluviral(登録商標)または精製したインフルエンザNP(インフルエンザWSN/33のヌクレオカプシド)タンパク質への免疫反応をELISAにより実施した。Fluviral(登録商標)に対する総IgG(図16A)、IgG1(図16B)、IgG2a(図16C)を測定した。NPに対するIgG2aも測定した(図16D)。
【0259】
図16に示す結果は、30μgのMaMV VLPが、Fluviral(登録商標)に対する免疫反応を大幅に改善できることを示す。総IgG(図16A)、IgG1(図16B)およびIgG2a(図16C)の大幅な増加が、Fluviral(登録商標)タンパク質に対して観察され、一方、ミョウバンは、総IgGおよびIgG2aの量を大幅に改善できなかった。MaMV VLPはまた、TH2免疫反応(IgG1)を誘発し、これについてはミョウバンと類似の有効性を示す(図16B)。さらにMaMV VLPは、NPタンパク質に指向されたIgG2aの産生を誘発することが示された(図16C)。ミョウバンでアジュバント化されたFluviral(登録商標)およびFluviral(登録商標)は、NPタンパク質に対していかなるIgG2aの産生も誘発できなかった。この結果は、インフルエンザウイルス内部に見出され、またFluviral(登録商標)調製物中に存在するNPタンパク質が、MaMV VLPの存在下でのみ、免疫原性となることを示唆する。この結果はまた、クラススイッチが多量のIgG12aアイソタイプの産生をもたらすために、TH1免疫反応が誘発されたことも示唆し、これは、ウイルス感染に対する防御のための理想的な反応であり、MaMV VLPが優れたアジュバント特性を有することを実証する。
【0260】
NPタンパク質は、インフルエンザの全株中もっとも保存されたタンパク質の1つである(92%を超える同一性)。したがって、MaMVベースのアジュバントを用いた、このタンパク質に対する、またはFluviral(登録商標)ワクチンに見出される任意のその他の保存エピトープに対する免疫反応の誘発は、市販のワクチンに見出されるものと無関連であるインフルエンザ株に対する防御を提供することが期待され、すなわち、MaMVベースのアジュバントを市販のインフルエンザワクチンに添加すると、インフルエンザの複数株への防御を提供可能な製剤を提供できる可能性がある。
【0261】
例11:抗原に遺伝子的に融合した外被タンパク質を含むMalvaモザイクルイスVLPの産生および試験
以下の例は、MaMV VLPがワクチンプラットフォームとして使用可能であることを実証する。この例に記載されている全ての外被タンパク質は、前の例に記載されたものと同じCP精製手順を用いて精製した。
【0262】
MaMV外被タンパク質のC末端は、SpeIおよびMluI制限部位を含んで、外被タンパク質のC末端において直接適切な抗原をコードする小さいアニーリングされたオリゴヌクレオチドのクローニングを容易にするように操作された。SpeI/MluI部位を含む配列5' ACTAGTACGCGT 3'(配列番号61)は、図10に示す外被タンパク質遺伝子の最後のアミノ酸(フェニルアラニン:F)の直後で6xHタグの前の位置にクローニングされた。それぞれの酵素の認識配列は以下である:
SpeI=ACTAGT
MluI=ACGCGT
【0263】
得られたMaMV CPは、MaMV CP−SMと名付けられた。このMaMV CP−SMをコードするヌクレオチド配列を図10B(配列番号62)に、およびMaMV CP−SMタンパク質のアミノ酸配列を図11B(配列番号63)に示す。
【0264】
良好に定義された腫瘍抗原gp100(IMQVPFSV;配列番号59)から、およびインフルエンザM1タンパク質(GILGFVFTL;配列番号60)からの9マーHLA−A*201エピトープを、MaMV CPとの融合に選択した。9マーHLA−A*201エピトープを、N−およびC-末端側にそれぞれの天然配列から5残基離して隣接させ(フランクさせ)て、プロテアソームによる自然のプロセシングを有利にした(図14)。インフルエンザまたはgp100エピトープとの融合の際、アミノ酸TSおよびTRは、融合エピトープのN末端およびC末端にそれぞれ融合する。
【0265】
MaMV−SMコンストラクトを用いて、2つの異なる融合物を生成した。1番目(MaMV−M1またはMaMV Flu−M1)は、インフルエンザのM1タンパク質由来のCTLエピトープを含んでいた。MaMV−M1タンパク質をコードするヌクレオチド配列を図17A(配列番号66)に、およびMaMV−M1のアミノ酸配列を図18A(配列番号67)に示す。2番目(MaMV−gp100)は、gp100タンパク質由来のCTLエピトープを含んでいた。MaMV−M1タンパク質をコードするヌクレオチド配列を図17B(配列番号68)に、およびMaMV−M1のアミノ酸配列を図18B(配列番号69)に示す。
【0266】
第3の融合物も、第2MaMV CP(MaMV gl−SM)およびインフルエンザH3株のHAに由来するF3ペプチド(KAYSNCYPYDVPDY;配列番号72)を用いて生成した。MaMV gl−SMはMaMV SMコンストラクトの配列を有するが、しかし、GGGLLLスペーサーを含む。MaMV gl−SMをコードするヌクレオチド配列を図10C(配列番号64)に、およびMaMV gl−SMのアミノ酸配列を図11C(配列番号65)に示す。
【0267】
MaMV gl−SMとF3ペプチドの融合は、コンストラクトMaMV gl−F3をもたらす。MaMV gl−F3をコードするヌクレオチド配列を図17C(配列番号70)に、およびMaMV gl−F3のアミノ酸配列を図18C(配列番号71)に示す。
【0268】
それぞれの外被タンパク質の精製プロファイル、得られたVLPの電子顕微鏡写真、およびVLPのFPLCプロファイルをそれぞれ図19(MaMV−SM)、図20(MaMV gl−SM)、図21(MaMV−M1)、図22(MaMV−gp100)および図23(MaMV gl−F3)に示す。これら組換えタンパク質の各々のVLPの長さを測定したが、それらの間に有意な差は観察されなかった(図24)。VLPの平均長は60nmであるが、200nmを超えるいくつかのVLPも測定された。
【0269】
上記の結果は、MaMV CPが、それらのVLPを形成する能力を失うことなく、いくつかの異なるペプチド配列の融合を耐容できることを実証する。
【0270】
例12:MalvaモザイクウイルスVLPのリンパ球による内在化(インターナリゼーション)
抗原提示細胞(APC)の異なる源の、MaMV VLPを取り込み内在化する能力を、前に記載の技術(Leclerc D, et al. (2007) J Virol 81: 1319-1326)を用いて評価し、パパイアモザイクウイルス(PapMV)の外被タンパク質由来のVLPの取り込みおよび内在化と比較した。
【0271】
簡潔に述べると、PapMV CP、MaMV CPまたはMaMV−M1から形成されたVLP(例11参照)を、蛍光標識(アロフィコシアニンまたはAPC)と接合し、CD40活性化Bリンパ球上で20時間パルスした。CD40活性化Bリンパ球は効率的なAPCであり、樹状細胞(DC)と類似の特性を有する。次に細胞を洗浄し、蛍光をフローサイトメトリーで評価した。
【0272】
結果を図25に示す;PapMV、MaMVおよびMaMV−M1のVLPは類似の効率で内在化されることを実証する。4℃での染色により、全てのVLPが細胞表面に結合可能であることが示唆される。2つのピーク(中程度および高い平均蛍光強度、またはMFI)の存在は、VLPが異なる様式で細胞内に内在化され/細胞と会合するが、しかし、MaMVとMaMV−M1のVLPの間に大きな違いはないことを示唆する。
【0273】
BおよびTリンパ球のヘテロハイブリドーマ細胞株を、次にAPCの源として試験した。この細胞株(T2細胞)は、抗原プロセシング(TAP)と会合するトランスポーターにおいて欠けている。T2細胞による時系列の取り込みアッセイを実施し、ここでT2細胞を図26に示す時点で37℃でパルスし、洗浄し、フローサイトメトリーで分析した。
【0274】
結果を図26Aおよび26Bに示す;PapMV、MaMVおよびMaMV−M1のVLPは時間とともに類似の効率で内在化される。再度、2つのピーク(中程度および高いMFI)が観察され、これは、VLPが異なる様式でT2細胞に内在化され/これと会合することを示唆する。時間に伴うMFIの増加によれば、MaMV VLPはより効率的に取り込まれるように見える。しかし、PapMV VLPはより迅速に分解し、その結果みかけの総MFIがより低くなることも可能である。
【0275】
これらの結果は、MaMV VLPおよびPapMV VLPが同等の有効性でAPCに内在化されることを示し、したがって、これらがワクチンプラットフォームとして同等の有効性を有することを示唆する。
【0276】
例13:MalvaモザイクウイルスVLPの、CTLエピトープのMHCクラスI提示を引き出す能力
PapMVは、プロテアソーム非依存性のメカニズムのもとで、MHCクラスIエピトープ交差提示を媒介する能力を実証した(Leclerc D, et al. (2007) J Virol 81: 1319-1326)。図12に示すように、PapMVおよびMaMVの両方は細胞を結合し、同様に内在化される。したがって、PapMVと同様の様式で、共有結合的に結合したMHCクラスIエピトープの提示を引き出すMaMVの能力を、次に前に記載の技術を用いて検討した(Leclerc et al. (2007)上記)。
【0277】
簡潔に述べると、PapMV外被タンパク質またはMaMV外被タンパク質由来のVLPに、HLA−A*0201インフルエンザM1エピトープをC末端に挿入したもの(PapMV-M1およびMaMV−M1;例11参照)、およびインフルエンザM1エピトープに対応する合成ペプチドを、T2細胞上で20時間パルスした。細胞を洗浄し、関連HLA−A*0201MHCクラスI分子に結合したM1インフルエンザエピトープに特異的なT細胞を、さらに20時間加えた。上清を収穫し、インターフェロン(IFN)−γ分泌をELISAにより評価した。IFN−γは、MHC/ペプチド複合体を認識するT細胞により分泌される。
【0278】
データを図27Aに示す。Gp100は陰性の特異性対照として機能し、これは陰性でなければならない。結果は、PapMVとMaMV両方のVLPが、M1エピトープを交差提示する能力を有することを示す。MaMV VLPについては、10μg/mlにおいて改善された認識が、および50μg/mlでパルスされた場合には認識の顕著な低下が観察された。
【0279】
MaMV−M1を用いて同様の実験を行って、用量反応を評価した(図27B)。Gp100は陰性の特異性対照として機能し、これは陰性でなければならない。
【0280】
プロテアーゼは、上記実験においてAPCを培養するために用いたヒト血清中に存在するため、APCのパルスが、VLPの内在化、プロセシング、およびAPCでの内在化後のMHC負荷からではなく、培養倍地中でのVLPの分解後に達成される可能性がある。この可能性を除外するために、T2細胞をMaMV−M1 VLPで4℃でパルスした低い温度は、VLPの任意の潜在的な分解を阻害するために用いた。ペプチドが外部に放出される場合、これらはMHCクラスIを細胞の外側から負荷しなければならないが、図27Cに示すように、そうではなかった(図27Bに示す実験は同時に実施され、37℃における陽性対照として機能する)。対照として、合成ペプチドを同様にパルスし、これらは4℃または37℃においてパルスした場合に認識された。
【0281】
図27Bおよび図27Cに示す結果は、用量反応がMaMV−M1に対する認識効率と相関することを実証する。しかし、より高い用量でパルスすると(50mg/ml)、認識の顕著な低下をもたらす(図27B)。M1エピトープは、ヒト血清を含有する培養培地中での分解により放出されないことが示された(図27C)。最後に、上記および例12の実験がTAP欠乏したT2細胞中で行われたため、これらの結果により、交差提示メカニズムはプロテアソーム/TAP媒介性ではないことがさらに確認される。
【0282】
例14:MalvaモザイクウイルスVLPによる休止エピトープ特異的Tリンパ球の感作
4人の異なるドナー(#405、#542、#614および#621)からの休止Tリンパ球を用いて、VLPでパルスしたAPCの、インフルエンザM1タンパク質特異的T細胞の増殖(expansion)を誘発する能力を評価した。このアッセイは、「T細胞感作」または「in vitroワクチン接種」と呼ばれる。かかるアッセイにおいて、M1エピトープを担持するVLPが、3週間の培養アッセイにおいて、特異的T細胞を増殖させるのに効率的であるかどうかを評価することができる。
【0283】
各ドナーに対して、自己由来APC(CD40−活性化Bリンパ球)を初めに培養した。これらの細胞を、M1エピトープを担持するPapMV VLPまたはMaMV VLP(例11参照)(25μg/ml)のどちらか、またはM1エピトープを含有する合成ペプチドでパルスした。パルスしたAPCを次に、自己由来血液Tリンパ球で7日間共培養し、つぎに、同じパルスしたAPCで再刺激した。全部で21日の培養後、培養したTリンパ球の、関連するインフルエンザM1エピトープに対する特異性を評価した。
【0284】
産生したTリンパ球の、各ドナーに対する特異性を図28〜31に示す。ドナー#405に対し(図28)、MaMV−M1 VLPは、特異的T細胞の働きの増大をもたらしたが、しかしPapMV−M1 VLPはもたらさなかった。いくつかのペプチド特異株は、MaMV−M1 VLPと比べた場合に、よりアビディティが高いことが観察された。ドナー#542に対し(図29)、PapMV−M1 VLPまたはMaMV−M1 VLPのどちらかを用いて産生されたT細胞株は、MaMV−M1をよりよく認識する。全体としてこのドナーに対して、VLPはペプチドパルスよりも、より高いアビディティのT細胞を生成する傾向があった。ドナー#614に対し(図30)、MaMV−M1 VLPを用いて産生されたT細胞株は、VLP−M1をよりよく認識する。全体として、VLPを用いて生成されたT細胞は、低いペプチド濃度(0.001mM;これは対数スケールであることに注意)でのペプチドパルスに比べて、より高いアビディティのT細胞を産生する。ドナー#621に対し(図31)、PapMV−M1 VLPまたはMaMV−M1 VLPのどちらかを用いて産生されたT細胞株は、VLP−M1を認識する。MaMV−M1T細胞株は、PapMV−M1T細胞株に比べてより高いアビディティを有するようであり、VLPT細胞株は全体としてペプチドパルスより優れていた。
【0285】
結論として、MaMV−M1 VLPおよびPapMV−M1 VLPの両方は、このin vitroのT細胞感作アッセイにおいて抗原特異的Tリンパ球を生成するのに一般に効率的であり、全体として、MaMV VLPおよびPapMV VLPの両方は、ペプチドのみよりも良好に作用した。驚くべきことには、抗プラットフォームTリンパ球はMaMVまたはPapMV VLPの両方について観察されなかった。全体として、MaMV VLPおよびPapMV VLPの両方は、同等であるように見られた。ドナー間での変化は予想された。
【0286】
例12および13は、MaMV−gp100およびMaMV−M1が、これらのCTLエピトープのヒトAPCのMHCクラス1での交差提示を誘発できること、および、CD8+特異的細胞の増殖を誘発できることを実証する。これらの結果は、MaMVプラットフォームが、ヒトまたは動物において、CTL反応を誘発可能であることを、強く示唆する。
【0287】
さらに、例12〜14は、MaMVプラットフォームが、その外被タンパク質の配列がPapMVプラットフォームの外被タンパク質の配列とアミノ酸の69%で非常に異なるにもかかわらず、PapMVプラットフォームと同様の免疫原性特性を有することを実証する。両方のプラットフォームはワクチン開発において大きな可能性を示し、これらが同一のワクチン接種プロトコルにおいて一緒に用いられて、与えられたエピトープへの免疫反応を最適化すること、または同じ患者において交互に用いて、前の処置のプラットフォームに指向された抗体による交差中性化を避けることが、期待される。
【0288】
例15:抗原に融合したMalvaモザイクウイルスVLPの、in vivo免疫反応を引き出す能力
MaMV−gp100、MaMV−M1またはMaMV gl−F3を含むVLP(例11参照)を試験して、融合ペプチドに対する免疫反応が、Balb/Cマウスにおいて検出できるかどうかを確認した。
【0289】
簡潔に述べると、マウスに2回の皮下注射を14日の間隔で表5に示すように施した。各注射の前に血液を採取し(0日目および14日目)、また最後の注射の14日目後にも採取した(28日目)。
表5:マウスの免疫化
【表5】
【0290】
14日目および28日目に採取した血液を用いてELISAを行い、総抗ペプチドおよび総抗MaMV IgG価を定量した。
【0291】
各組換えタンパク質についてLPS含量を評価し、各ケースにおいて15EU/mg未満であった。これは、注射した組成物のLPS含量が基本的に無視できることを示す。
【0292】
驚くべきことには、ほとんどのマウスはMaMV CPに融合したペプチドに対してIgG反応を示さなかった(図32A)。MaMV g1-F3コンストラクトで処置した10匹のうち2匹のマウスは、ペプチドについてセロコンバージョンを示し、MaMV−M1コンストラクトで処置した10匹のうち1匹のマウスは、ペプチドについてセロコンバージョンを示した。MaMV−gp100コンストラクトで処置したマウス、またはF3ペプチドで処置したマウス(F3ペプチドがMaMVプラットフォームに融合しなかった場合)は、いかなる反応も示さなかった。
【0293】
この点について、M1またはgp−100タンパク質由来のCTLエピトープの選択は、マウスにおける体液性の反応を評価するにはおそらく最適ではなかったことが指摘でき、その理由はおそらく、実験に用いたマウスの免疫レパートリーに、これらのペプチドを認識できるB細胞が含まれていないことである。これはまた、F3ペプチドについても同様である。
【0294】
前の例では、MaMVが強力なアジュバントであることを実証した。本例ではしかし、試験動物の免疫レパートリーが抗原を認識できない場合には、強いIgG反応を誘発できないことを確認する。アジュバントは、目的の抗原に特異的なB細胞が利用可能である場合にのみ、B細胞反応を増幅できる。この結論は、MaMV CPに対して全ケースで非常に強いIgG反応が測定できるという事実によって支持され、プラットフォームが高度に免疫原性であることを示す(図32B)。これは、ワクチンプラットフォームとしては重要な性質であり、なぜならば、一般に、HBV VLPまたはPapMV VLPなどのワクチンプラットフォームについて、プラットフォーム自体の本質的な免疫原性が、VLPの表面の融合エピトープまたはペプチドに指向された免疫反応を駆動することが、よく認識されているためである。したがって、MaMV VLPは高度に免疫原性であることが示されたので、より適切なエピトープ(例えば、インフルエンザのM2タンパク質のM2eエピトープ(Denis et al., 2008, Vaccine, 26:3395-3403参照))が、ペプチドに対して常に強い免疫反応を生じさせることが予想される。
【0295】
さらに、将来の実験について、スペーサーを融合ペプチドとMaMVCPの間にクローニングして、ペプチドをVLPの表面に押し出すことが可能であり、これは、この実験では最適化されなかったB細胞との架橋(cross link)を改善することが期待される。
【0296】
例16:パパイアモザイクウイルスVLPおよびMalvaモザイクウイルスVLPに対する抗体の交差反応性
PapMV CPおよびMaMV CPは、31%の配列同一性を共有するのみである(図33A参照)。配列における差はまた、各々の外被タンパク質に特異的な抗体における交差反応性の欠如にも明らかである。例えば図33Bは、MaMV CPに指向された抗体を用いたELISAの結果を示し、これは、PapMV VLPを認識しないことがわかる。PapMV CPに指向された抗体による同様の実験により、PapMV抗体はMaMVプラットフォームに結合しないことも明らかにされた(図33C)。この結果は、2つのVLPの表面は非常に異なっていることを示唆する。2つのプラットフォームはしたがって、ワクチン投与計画において一緒に用いて、より優れた効率的なワクチン接種プログラムを保証することができ、その理由は、第1のプラットフォームに指向された抗体が、第2のプラットフォームを妨害できないからである。
【0297】
例17:Malvaモザイクウイルスの免疫原性効果
以下のプロトコルは、MaMVおよび弱い免疫原を含む組成物の免疫原性効果を評価するために従うことができる、代替的プロトコルである。
【0298】
生得的免疫反応の抗体反応への翻訳は、アジュバントを弱い免疫原性ワクチンと共同投与した場合に観察される。アジュバントは、抗原に対する抗体または細胞の免疫反応を強化または増強可能な物質である。MaMVが他の抗原に対する長期の抗体反応を促進可能なアジュバントであるかどうかを決定するために、BALB/cマウスを弱い免疫原、オボアルブミン(OVA)またはニワトリ卵白リゾチーム(HEL)で、それらのみで、またはMaMVと共に免疫化した。CFAまたはLPSなどの他のアジュバントと、OVAまたはHELとの組合せを陽性対照として用いることができる。次に、OVAまたはHELに特異的なIgG抗体価を、適切な時点でELISAにより測定することができる。
【0299】
MaMV精製
以下の実験で用いるMaMVは、例1に記載のようにして精製することができる。
【0300】
抗原およびアジュバント
LPS非含有OVAグレードVIは、Sigma-Aldrich Chemical Co., St Louis, MOから購入可能であり、ニワトリ卵白リゾチーム(HEL)は、Research Organics Inc. Cleveland, OHから購入可能である。大腸菌O111:B4からのLPSは、Sigma-Aldrich, St Louis, MOから購入可能である。
【0301】
免疫化
6〜8週齢のBALB/cマウスを、医薬品安全性試験実施基準に準拠した条件下で飼育し維持する。試験アジュバントの効果を試験するために、マウスの群を2mgのOVAもしくはHELのみ、または30mgのMaMV、CFA1:1(v/v)、または5mgの大腸菌O111:B4からのLPS(Sigma-Aldrich)で、0日目に腹腔内で免疫化した。対照マウスには、生理食塩水のみを注射した。血液試料は、後方眼窩洞から様々な時点で採取した。個々の血清試料は分析まで−20℃で保存した。各実験に3匹のマウスを用いた。
【0302】
ELISAによる抗体価の決定
高結合96ウェルポリスチレンプレート(Corning(登録商標)、New York, NY)を、0.1Mの炭酸−重炭酸緩衝液(pH9.5)中の1mg/mLのMaMV、100mg/mLのHEL、または150mg/mLのOVAで被覆した。プレートを37℃で1時間、次に4℃で一晩インキュベートした。次の朝、使用前に、プレートを0.05%Tween-20(Sigma-Aldrich)含有のPBS(pH7.2)(PBS−T)で3回洗浄した。非特異的結合を、PBSに希釈した5%の脱脂粉乳(PBS−M)を用いて37℃で1時間ブロックした。洗浄後、マウス血清をPBS−M中1:40に希釈し、2倍連続希釈液をウェルに加えた。プレートを37℃で1時間インキュベートし、次にPBS−Tで4回洗浄した。ペルオキシダーゼ接合ウサギ抗マウスIgM(最適希釈1:1000)IgG、IgG1、IgG2a、IgG2b抗体(Zymed, San Francisco, CA)またはIgG3(最適希釈1:3000)(Rockland, Gilbertsville, PA)を加え、プレートを37℃で1時間インキュベートし、PBS−Tで3回洗浄した。30%過酸化水素含有の0.1Mのクエン酸緩衝液(pH5.6)中のオルトフェニレンジアミン(0.5mg/mL;Sigma-Aldrich)を、酵素基質として用いた。反応を2.5NのH2SO4で停止させ、吸収を、BIOLINX 2.22ソフトウェアと自動ELISAプレートリーダー(Dynex Technologies MRII, Chantilly, VA, USA)を用いて、490nmにて決定した。抗体価は、−log2希釈×40として与える。正の力価は、陰性対照の平均値の上の3SDとして定義した。
【0303】
例18:MalvaモザイクウイルスおよびMalvaモザイクウイルスVLPによる、亜致死量のインフルエンザウイルスに対する抵抗性の誘発
MaMVまたは組換えMaMV VLPの、亜致死量のインフルエンザウイルスに対する抵抗性を、抗原の不在下において誘発する能力を、マウスにおいて試験することができる。簡潔に述べると、マウスを鼻腔内経路で100ugのMaMVで7日間隔で3回処置する。最後の処置の2日後に、動物を亜致死量のインフルエンザウイルスに暴露する(A/WSN/33)(1LD50)。動物中で複製されるウイルスの量を、感染後に2日の間隔で鼻での分泌をサンプリングして測定する。動物の体温と体重を、チャレンジ後7日間毎日モニタリングする。感染による病的状態を、感染後の10日間、スコア化する。
【0304】
本発明を、ある特定の態様を参照して記載したが、これらについての種々の改変は、本発明の精神および範囲から逸脱することなく、当業者には明らかである。当業者に明らかであるかかる改変の全ては、以下の特許請求の範囲に包含されることが意図される。
【技術分野】
【0001】
本発明は、ワクチン製剤およびアジュバントの分野、および特に植物ウイルス粒子に基づくワクチンおよびアジュバントに関する。
【背景技術】
【0002】
ワクチン開発の多数の新しいアプローチの中で、ウイルスヌクレオカプシドから作製されたウイルス様粒子(VLP)は、期待が持てる戦略として登場してきた。現在まで2つのVLPワクチン、B型肝炎ウイルス(HBV)およびヒトパピローマウイルス(HPV)が、ヒトにおいて効率的に機能することが示されている(Fagan et al., 1987, J. Med. Virol, 21:49-56;Harper et al., 2004, Lancet, 364:1757-1765)。例えば、ヒトパピローマウイルス(HPV)主要カプシドタンパク質L1から作製されたVLPは、女性の子宮頸癌の発症に対して100%の防御を与えることが示された(Ault, K.A., 2006, Obstet. Gynecol. Surv. 61 :S26-S31; Harper et al., 2004, Lancet 364: 1757- 1765、また国際特許出願PCT/US01/18701 (WO 02/04007)も参照のこと)。バクテリオファージQβ(Maurer et al., 2005, Eur. J. Immunol. 35:2031-2040)、ウイルスコアタンパク質から作製されたB型肝炎ウイルスVLP(Mihailova et al., 2006, Vaccine 24:4369-4377;Pumpens et al., 2002, Intervirology 45:24-32)、およびパルボウイルスVLP(Antonis et al., 2006, Vaccine 24:5481-5490;Ogasawara et al., 2006, In Vivo 20:319-324)もまた、エピトープを担持し、強い抗体反応を誘発することが示された。同様に、米国特許第6,627,202号には、HBVカプシド結合ペプチドにより架橋された抗原を含むHBVコアタンパク質の、種々のウイルスおよび細菌を標的とした抗原、またはこれらに由来する抗原を含むエピトープ送達システムとしての使用が記載されている。
【0003】
植物ウイルスからのVLPの、エピトープ提示系としての使用は記載されている。植物ウイルスは主に高度に免疫原性であるタンパク質から構成され、複雑な反復性の結晶組織を有する。さらにこれらは、系統発生学的に動物の免疫系からは離れており、そのことが、これらをワクチン開発の良好な候補者にしている。例えば、ササゲ(cowpea)モザイクウイルス(CPMV)、ジョンソングラスモザイクウイルス(JGMV)、タバコモザイクウイルス(TMV)、およびアルファルファモザイクウイルス(AIMV)が、目的のエピトープ提示のために改変された(Canizares, M. C. et al., 2005, Immunol. Cell. Biol. 83:263-270;Brennan et al., 2001, Molec. Biol. 17: 15-26;Saini and Vrati, 2003, J. Virol. 77:3487-3494)。国際特許出願PCT/GB97/01065 (WO 97/39134)には、外被タンパク質および非ウイルス性タンパク質を含み、例えばペプチドエピトープの提示のために用いることができる、キメラウイルス様粒子が記載されている。国際特許出願PCT/USOl/07355 (WO 01/66778)には、リンカーを介してN末端において、病原体微生物のエピトープを含んでよい目的のポリペプチドに融合した、植物ウイルス外被タンパク質、および具体的にはトバモウイルス(tobamovirus)外被タンパク質が記載されている。国際特許出願PCT/USO1/20272 (WO 02/00169)には、外来ペプチドに、および具体的にはニューカッスル病ウイルスまたはヒト免疫不全ウイルス(HIV)エピトープに融合した、ジャガイモウイルスY外被タンパク質または切断型インゲン黄斑モザイクウイルス外被タンパク質のどちらかを含むワクチンが記載されている。また、米国特許第6,042,832号には、例えば病原体エピトープなどのポリペプチドの、アルファルファモザイクウイルスまたはイラルウイルス(ilarvirus)カプシドタンパク質への融合物を動物に投与して、免疫反応を高めるための方法が記載されている。
【0004】
パパイアモザイクウイルス(PapMV)の外被タンパク質由来のVLP、およびそれらの免疫増強剤としての使用が記載されている(国際特許出願PCT/CA03/00985 (WO 2004/004761)および米国特許出願第11/556,678号(US2007/0166322))。大腸菌におけるPapMV外被タンパク質の発現は、反復性かつ結晶様式での数百のCPサブユニットから構成されるVLPの自己集合をもたらす(Tremblay et al., 2006, FEBS J 273: 14)。PapMV CP欠失コンストラクトの発現および精製の研究はさらに、CPサブユニットの自己集合(または多量体形成)が、機能に重要であることを示す(Lecours et al., 2006, Protein Expression and Purification, 47:273-280)。gp100またはインフルエンザウイルスM1タンパク質のどちらかからのエピトープを含むPapMV VLPの能力は、エピトープのMHCクラスI交差提示を誘発し、特異的ヒトT細胞の増幅(expansion)をもたらすことが示された(Leclerc, D., et al, J. Virol, 2007, 81(3):1319-26; Epub. ahead of print November 22, 2006)。さらに、C型肝炎ウイルスE2エンベロープタンパク質由来のエピトープを含むPapMV VLPは、マウスにおいて、PapMV VLPおよびE2ペプチドに対して体液性の反応を誘発することが示された(Denis et al., 2007, Virology, 363(1): 59-68)。
【0005】
plasmodiophora brassicaeの休眠胞子を結合することのできる、親和性ペプチドに融合したパパイアモザイクウイルスVLPの使用も記載されている(Morin et al., 2007, J. Biotechnology, 128: 423-434)。VLPは、P. brassicae胞子を高い親和力で結合できることが示され、P. brassicaeの検出のための抗体の代替として提唱された。
【0006】
HIV、HCV、EBVまたはインフルエンザウイルスからの種々の抗原決定基を担持するジャガイモウイルスX(PVX)由来のVLPが、記載されている(欧州特許出願第1 167 530号)。HIVエピトープを担持するPVX VLPの、マウスにおいて体液性および細胞媒介性経路を介して抗体産生を誘発する能力もまた、記載されている。追加のアジュバントをPVX VLPと併用すると、この効果が増強された。
【0007】
B型肝炎コアタンパク質またはパルボウイルスVLPは、これらが遺伝情報を担持しない場合であってもCTL反応を誘発することが報告されており(Ruedl et al., 2002, Eur. J. Immunol. 32; 818-825;Martinez et al., 2003, Virology, 305; 428-435)、それらが陥入した細胞において活性に複製することができない。リンパ球性脈絡髄膜炎ウイルス(LCMV)または鶏卵アルブミンからのエピトープを担持するVLPの、in vivoでの樹状細胞による交差提示も記載されている(Ruedl et al., 2002、同上;Moron, et al., 2003, J. Immunol. 171 :2242-2250)。LCMVからのエピープを担持するB型肝炎コアタンパク質VLPの、CTL反応をプライムする能力も記載されているが、しかしこのVLPは、それのみで投与された場合にはCTL反応を誘発できず、ウイルスチャレンジに対する効果的な防御を媒介することができなかった。有効なCTL反応は、VLPを抗CD40抗体またはCpGオリゴヌクレオチドと併用した場合にのみ、誘発された(Storni, et al., 2002, J. Immunol. 168:2880-2886)。初期の報告によれば、LCMVからのペプチドを担持する豚パルボウイルス様粒子(PPMV)が、マウスを致死的なLCMVチャレンジに対して保護できたことが示された(Sedlik, et al., 2000, J. Virol. 74:5769-5775)。
【0008】
ポテックスウイルスであるMalva葉脈壊死ポテックスウイルス(MVNV)は、1990年にブラジルにおいてMalva parviflora(ウサギアオイ)に感染するウイルスとして報告され(Brunt et al., (eds.) 1996 onwards. "Plant Viruses Online: Descriptions and Lists from the VIDE Database. Version: 20th August 1996")、Malva parviflora上に局所的な病変部および全体的な葉脈壊死が検出された。
【0009】
この背景情報は、本出願人が本発明に関連する可能性があると信じる既知の情報を明らかにする目的で提供されている。任意の前述の情報が、本発明に対する先行技術を構成するとの認定は必ずしも意図されず、またそのように解釈されるべきではない。
【発明の概要】
【0010】
本発明の目的は、Malvaモザイクウイルスおよびウイルス様粒子(VLP)、ならびにこれらの使用を提供することである。本発明の1つの側面により、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および薬学的に許容し得る担体を含む、免疫原性組成物が提供される。
【0011】
本発明の他の側面により、動物において免疫反応を誘発するための方法であって、前記動物に対して、本発明の免疫原性組成物の有効量を投与することを含む前記方法が提供される。
【0012】
本発明の他の側面により、動物に、疾病、疾患、または感染に対するワクチン接種するための方法であって、該動物対して、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および1または2以上の抗原を含む、免疫原性組成物の有効量を投与することを含む、前記方法が提供される。
【0013】
本発明の他の側面により、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)の、免疫原性組成物の調製における使用が提供される。
【0014】
本発明の他の側面により、Malvaモザイクウイルス(MaMV)外被タンパク質を含むウイルス様粒子(VLP)が提供される。
【0015】
本発明の他の側面により、ウイルス様粒子(VLP)を調製するための方法であって、Malvaモザイクウイルス(MaMV)外被タンパク質を宿主細胞中に発現するステップを含む、前記方法が提供される。
【0016】
本発明の他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質が提供される。
【0017】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質をコードする、単離ポリヌクレオチドが提供される。
【0018】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリポリヌクレオチドにより遺伝子操作された、宿主細胞が提供される。
【0019】
他の側面により、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリヌクレオチド、または、抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む融合タンパク質をコードするポリポリヌクレオチドにより遺伝子操作された宿主細胞の、ウイルス様粒子の調製のための使用が提供される。
【図面の簡単な説明】
【0020】
本発明のこれらおよび他の特徴は、添付の図を参照した以下の詳細な説明において、さらに明らかにされる。
【0021】
【図1A】図1Aは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【図1B】図1Bは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【図1C】図1Cは、Malvaモザイクウイルスのゲノム配列を示す(配列番号1)。配列は、GenBankアクセッション番号DQ660333のもとで入手可能である。
【0022】
【図2】図2は、Malvaモザイクウイルスの外被タンパク質のアミノ酸配列を示す(配列番号2)。
【0023】
【図3】図3は、Malvaモザイクウイルスの外被タンパク質をコードするヌクレオチド配列を示す(配列番号3)。
【0024】
【図4】図4は、(A)Malvaモザイクウイルスに感染したMalva種上のモザイク症状を示す写真、(B)Malva neglecta Wallr.に感染した葉からの抽出物を接種した場合の、Chenopodium quinoa(キノア)上のモザイク症状を示す写真、(C)C. quinoaの感染した葉からの抽出物を接種した場合のC. quinoa上に観察されるモザイク病変部を示す写真、および(D)精製したMalvaモザイクウイルスの電子顕微鏡写真(バーは50nmを表わす)である。
【0025】
【図5】Malvaモザイクウイルスの5’末端の解析結果を示す。バーは、配列決定した全てのクローンに見出された各々の配列の頻度を表わす。
【0026】
【図6】図6(A)は、Malvaモザイクウイルス(MaMV)のゲノム構成の模式図であり、(B)は、健康な(レーン1)、または感染した(レーン2)Chenopodium quinoaから、MaMV外被タンパク質に指向するプローブを用いて抽出した全RNAを示す、ノーザンブロットであり、および(C)は、種々のポテックスウイルスのオクタヌクレオチド推定sgプロモーター配列の配列アラインメントを示す(高度に保存された領域は、黒または灰色の四角で強調した)。
【0027】
【図7】図7は、Malvaモザイクウイルス3’非翻訳領域の二次構造の模式図である。
【0028】
【図8】図8は、Malvaモザイクウイルスおよび他のポテックスウイルスのレプリカーゼおよびカプシドタンパク質の系統学的解析結果を示す図である。
【0029】
【図9】図9(A)は、大腸菌から単離された、精製組換えMalvaモザイクウイルス(MaMV)外被タンパク質を表わすSDS−PAGEゲルを、および(B)は、組換え外被タンパク質を含むMaMV VLPの電子顕微鏡写真を示す。バーは50nmである。
【0030】
【図10】図10は、(A)プラスミドpMaMV−CP−6H中に含有されたMaMV外被タンパク質遺伝子のヌクレオチド配列(配列番号23)(開始コドンは下線で、および終止コドンは太字イタリック体で表す);(B)MaMV CP−SMタンパク質をコードするヌクレオチド配列(配列番号62)(イタリック体で下線の配列は、2つの制限酵素認識配列を表す);および(C)MaMV CP gl−SMタンパク質をコードするヌクレオチド配列(配列番号64)(イタリック体で下線の配列は、2つの制限酵素認識配列およびスペーサーをコードする配列を表す)を示す。
【0031】
【図11】図11は、(A)プラスミドpMaMV−CP−6H中に含有されたMaMV外被タンパク質遺伝子によりコードされた外被タンパク質のアミノ酸配列(配列番号24);(B)MaMV CP−SMタンパク質のアミノ酸配列(配列番号63)(イタリック体で下線の配列は、コードされたヌクレオチド配列中の2つの制限酵素認識配列の封入により挿入されたアミノ酸を表す);および(C)MaMV CP gl−SMタンパク質のアミノ酸配列(配列番号65)(イタリック体で下線の配列は、コードされたヌクレオチド配列およびスペーサー配列中の2つの制限酵素認識配列の封入により挿入されたアミノ酸を表す)を示す。それぞれの配列におけるHisタグは太字で示す。
【0032】
【図12】図12は、以下の産生を表すグラフである:(A)、(B)および(C):それぞれ、アジュバント不在のもとで、s.c.によりMaMV VLPを1度注射したBalb/Cマウスにおける、総IgG、IgG1およびIgG2a(ELISAにより測定);および(D)、(E)および(F):それぞれ、同じマウスにおいて、アジュバント不在のもとで、s.c.によりMaMV VLPを2回目(40日目)に注射した場合の、総IgG、IgG1およびIgG2a。ELISAは、最初の免疫化後5日目、10日目、14日目において採取した血液、および2回目の免疫化後4日目に採取した血液で実施した。+の記号は、ELISAのベースラインとして用いた陰性対照(免疫前血清)を表す。
【0033】
【図13】図13は、チフス菌から精製されたポリンであるOmpCおよびOmpFのプロファイルを示すSDS−PAGEゲルの図である。
【0034】
【図14】図14は、gp100(配列番号59)およびインフルエンザM1タンパク質(配列番号60)からのHLA−A*0201エピトープ(太字および下線)を、それぞれそのフランキング配列と共に示した図である。
【0035】
【図15】図15は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV CPを発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV CP;B)組換えMaMV CPを含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV CPが高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0036】
【図16A−C】図16A−Cは、Fluviral(登録商標)に対する(A)総IgG、(B)IgG1および(C)IgG2aの産生を実証するグラフであって、これは、MaMV VLP(3または30μg)またはミョウバンをアジュバントとしたFluviral(登録商標)での免疫化後14日目に、1群当たり5匹のBalb/Cマウスの血清において測定したものである。
【図16D】図16Dは、Fluviral(登録商標)に対するNPタンパク質に対するIgG2aの産生を実証するグラフであって、これは、MaMV VLP(3または30μg)またはミョウバンをアジュバントとしたFluviral(登録商標)での免疫化後14日目に、1群当たり5匹のBalb/Cマウスの血清において測定したものである。
【0037】
【図17】図17は、次をコードするヌクレオチド配列である:(A)MaMV−M1タンパク質(配列番号66)(M1エピトープをコードする配列は下線で示す);(B)MaMV−gp100タンパク質(配列番号68)(gp100エピトープをコードする配列は下線で示す);(C)MaMV g1−F3タンパク質(配列番号70)(F3ペプチドをコードする配列は下線で示す)。
【0038】
【図18】図18は、次のアミノ酸配列である:(A)MaMV−M1タンパク質(配列番号67)(M1エピトープは太字下線で示す);(B)MaMV−gp100タンパク質(配列番号69)(gp100エピトープは太字下線で示す);(C)MaMV g1−F3タンパク質(配列番号71)(F3ペプチドは太字下線で示す)。各配列のHis−tagは太字で示す。
【0039】
【図19】図19は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−SMタンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−SMタンパク質;B)組換えMaMV−SMタンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−SMタンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0040】
【図20】図20は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV g1−SMタンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV g1−SMタンパク質;B)組換えMaMV g1−SMタンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV g1−SMタンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0041】
【図21】図21は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−M1タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−M1タンパク質;B)組換えMaMV−M1タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−M1タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0042】
【図22】図22は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV−gp100タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV−gp100タンパク質;B)組換えMaMV−gp100タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV−gp100タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0043】
【図23】図23は、A)SDS−PAGEゲルであって、それぞれ以下を示す:第1レーン:分子量マーカー、第2レーン:インサートなしで発現ベクターpET−3Dのみを含有する細菌溶解物、第3レーン:MaMV g1−F3タンパク質を発現する大腸菌細胞の溶解物、および第4レーン:親和性クロマトグラフィにより精製されたMaMV g1−F3タンパク質;B)組換えMaMV g1−F3タンパク質を含むMaMVウイルス様粒子の電子顕微鏡写真;C)ゲルろ過クロマトグラフィの溶出プロファイルであって、精製MaMV g1−F3タンパク質が高分子量であり、カラムから排除されたことを示し、すなわち、全てのタンパク質が、500kDaを超える分子量のオリゴマー化状態にあることを示唆する。
【0044】
【図24】図24は、異なるMaMV VLPの平均長(各組換えコンストラクトから100個のVLPを測定した)を示すグラフである。
【0045】
【図25】図25は、CD40活性化Bリンパ球を、蛍光標識パパイアモザイクウイルス(PapMV)VLP、MaMV VLPおよびMaMV−M1 VLPでパルスした、フローサイトメトリーの結果を示す図である。
【0046】
【図26】図26は、(A)T2細胞を、蛍光標識パパイアモザイクウイルス(PapMV)VLPおよびMaMV VLPでパルスしたフローサイトメトリーの結果であり、時間と共にVLPの取り込みを示す;および(B)は、PapMV VLPおよびMaMV VLPのT2細胞による取り込みを、時間に渡り平均蛍光強度(MFI)で測定した結果のグラフ表示である。
【0047】
【図27】図27は、以下を示すグラフである:M1エピトープ(左)もしくはgp100エピトープ(右)を担持するPapMV VLPまたはMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量;(B)M1エピトープ(左)もしくはgp100エピトープ(右)を担持する異なる量のMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量;および(C)4℃にて、M1エピトープ(左)もしくはgp100エピトープ(右)を担持するPapMV VLPまたはMaMV VLP、または対応するM1もしくはgp100ペプチドによってパルスされたT2細胞に添加された場合の、M1特異的Tリンパ球により分泌されたIFN−γの量。IFN−γの分泌は、ELISAにより評価した。
【0048】
【図28】図28は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#405に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0049】
【図29】図29は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#542に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0050】
【図30】図30は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#614に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0051】
【図31】図31は、インフルエンザM1ペプチドを担持するPapMV VLPまたはMaMV VLPを用いた、ドナー#621に対するin vitroでのT細胞の感作結果を示す:(A)表示のようにパルスしたT2細胞との共培養により評価した、産生されたT細胞の特異性を示し;(B)対照として、特異的T細胞系を、インフルエンザM1ペプチドまたはgp100ペプチドの滴定濃度と共培養した結果を示す。
【0052】
【図32】図32は、MaMV−SM VLP、MaMV g1−F3 VLP、MaMV g1−SM VLP+F3、MaMV−M1 VLP、またはMaMV−gp100 VLPを注射したマウス(1群当たり10匹)において、(A)ペプチドF3、M1またはgp100、および(B)MaMV CPに対する総IgG応答を示す、ELISAの結果を表す。
【0053】
【図33】図33は、(A)MaMVおよびPapMV CPアミノ酸配列のアラインメントであって、2つのCP間で31.2%の同一性を示した;(B)MaMV CPに指向された抗体は、PapMV VLPを認識できないことを示すELISAの結果;および(C)MaMV CPに指向された抗体は、PapMV VLPを認識できないことを示すELISAの結果である。
【0054】
発明の詳細な説明
本発明は、ポテックスウイルスであるMalvaモザイクウイルス(MaMV)およびMaMVの外被タンパク質を含むウイルス様粒子(VLP)の、本明細書において同定された免疫原性特性に関する。本発明は、MaMVおよびMaMV VLPのこれらの免疫原性特性に基づく、動物における免疫反応の生成に関連する種々の態様を包含する。1つの側面において、例えば、本発明は以下を提供する:MaMVまたはMaMV外被タンパク質を含むVLPの、免疫原性組成物の調製のための使用;MaMVまたはMaMV外被タンパク質を含むウイルス様粒子、および任意に1または2以上の抗原を含む、免疫原性組成物;およびこれら組成物の、動物において免疫反応を誘発するための、および/または動物にワクチン接種するための使用。本発明の他の側面は、MaMV外被タンパク質の、VLPの調製のための使用;MaMV外被タンパク質から調製されたVLP;かかるVLPを調製するための方法;VLPおよびこれをコードするポリヌクレオチドを調製するのに好適な外被タンパク質ポリペプチドを、提供する。
【0055】
いくつかの態様において、MaMVまたはMaMV外被タンパク質を含むVLPのみの投与は、免疫反応を引き起こすのに十分であり、かかる投与は、例えば、抗原の投与の前に、免疫システムを「プライムする」ために用いることができる。MaMVおよびMaMV VLPはまた、アジュバントとして用いることもでき、この状況では、それらの免疫原性特性は、例えば抗原に対する免疫反応を増強するのに有用である。このような状況での抗原は、MaMVまたはVLPの前に、MaMVまたはVLPの後に、またはMaMVもしくはVLPと同時に投与することができる。同時投与の場合、抗原はMaMVまたはVLPに付着させることができ、または別にすることができる。本発明の1つの態様において、抗原は、MaMVまたはMaMV VLPの外被タンパク質に付着させる。MaMVおよびMaMV VLPはまた、ワクチンの調製においても用いることができる。
【0056】
本発明の1つの態様により、MaMVまたはMaMV VLPおよび1または2以上の抗原を含む免疫原性組成物は、動物において体液性および/または細胞性免疫反応を誘発することができる。
【0057】
定義
他の定義がない限り、本明細書で用いる全ての技術的および科学的用語は、本発明が属する分野の業者により一般に理解されるものと同じ意味を有する。
【0058】
本明細書において、用語「約」は、与えられた値からのおよそ+/−10%の変動を意味する。かかる変動は、本明細書に与えられる全ての値において、具体的に言及されるかどうかに関わらず、常に含まれることが理解されるべきである。
【0059】
本明細書において用語「ウイルス様粒子(VLP)」は、ウイルス粒子と類似の外観を有する、自己集合粒子を意味する。VLPは、ウイルス核酸を含んでも含まなくてもよい。VLPは通常、複製不能である。
【0060】
本明細書において用語「偽ウイルス」とは、プラスミド形態の核酸を含むDNAまたはRNAなどの核酸配列を含有するVLPを意味する。偽ウイルスは一般に、複製不能である。
【0061】
本明細書において、「融合タンパク質」は、例えばタンパク質またはタンパク質断片などの2つの別々のポリペプチドをコードする、2種または3種以上のポリヌクレオチドが遺伝的に組み合わされて、2種のポリペプチドの組合せである1つのタンパク質(「融合タンパク質」)をコードする第3のポリヌクレオチドを提供する場合に生成されるタンパク質である。例えば、MaMV外被タンパク質のアミノ酸配列と、抗原のアミノ酸とを含むポリペプチドは、融合タンパク質と考えられる。
【0062】
本明細書において用語「アジュバント」は、動物において免疫反応を増大、刺激、作動、増強、および/または調節する剤を意味する。
【0063】
本明細書において用語「免疫原性」とは、動物において検出可能な免疫反応を誘発する、物質の能力を意味する。
【0064】
本明細書において用語「免疫反応」とは、物質(例えば化合物、分子、材料など)の投与に応答した、動物の免疫系の反応性における変化を意味し、これには、抗体産生、細胞媒介性免疫性の誘発、補体活性化、免疫学的耐性の発達、またはこれらの組合せを含んでよい。
【0065】
本明細書において用語「免疫防御反応」とは、1または2以上の抗原に対する免疫反応であって、これにより、1または2以上の抗原が由来する作用物質によって引き起こされた状態(例えば疾病または疾患)および/または感染に対して防御するための、前記免疫反応を意味する。本発明の目的のために、状態および/または感染に対する免疫防御は、状態または感染の絶対的な防御のみでなく、状態または感染を患う未処置の動物と比べた場合の、処置した動物における、状態または感染の程度または速度における任意の検出可能な低下、または状態の重篤度または作用物質の感染から生じる任意の症状における任意の検出可能な低下をも含む。免疫防御反応は、前に状態を患っていなかった動物において、作用物質に感染していなかった動物において、および/または処置時に状態または感染を有していない動物において、誘発することができる。免疫防御反応はまた、既に状態を患っている動物、または処置時に病原体に感染している動物においても、誘発することができる。免疫防御反応は、体液性および/または細胞性免疫を含む、1または2以上のメカニズムの結果であることができる。
【0066】
本明細書において同義で用いられる用語「免疫刺激(immuno stimulation)」および「免疫刺激(immunostimulation)」とは、MaMV VLPなどの、動物病原体や疾病と関連しない分子の能力であって、免疫系を刺激することによる、および/または前記感染または疾病に応答する免疫系の容量を改善することによる、前記病原体による感染に対する、または前記疾病に対する、防御を提供する前記能力を意味する。免疫刺激は、予防効果、治療効果、またはこれらの組合せを有することができる。
【0067】
本明細書において用語「ワクチン接種」は、免疫防御反応を生成する目的で、対象に対してワクチンを投与することを意味する。ワクチン接種は、予防効果、治療効果、またはこれらの組合せを有することができる。ワクチン接種は、処置する対象に応じて種々の方法を用いて実施でき、これには限定することなく、非経口投与、例えば腹腔内注射(i.p.)、静脈内注射(i.v.)、または筋肉内注射(i.m.);経口投与;鼻腔内投与;皮内投与、経皮投与、および液浸を含む。
【0068】
本明細書において用語「ワクチン」とは、免疫防御反応を生成する目的で、対象に対して投与される組成物を意味する。
【0069】
本明細書において用語「天然の」とは、物に適用する場合、物が自然界で見出せるとの事実を意味する。例えば、生物(ウイルスを含む)、または生物中に存在するポリペプチドまたはポリヌクレオチド配列であって、天然の給源から単離でき、研究室で人間によって意図的に改変されていないものは、天然のものである。
【0070】
本明細書において「ポリペプチド」または「ペプチド」は、ペプチド結合により結合された少なくとも4個のアミノ酸が存在する分子を意味することを意図する。
【0071】
本明細書において表現「ウイルス核酸」とは、ウイルスのゲノムもしくはその一部、または該ゲノムもしくはその一部に対して塩基配列が相補的である核酸分子を意味する。ウイルスRNAに相補的なDNA分子もまた、ウイルスDNAの塩基配列に相補的なRNA分子であるために、ウイルス核酸と考えられる。
【0072】
本明細書において用語「免疫原」および「抗原」とは、1個の分子、複数分子、分子の一部分、分子の複数部分、または分子の組合せであって、全細胞および組織までを含み、それらのみで、またはアジュバントと組み合わせて、対象において免疫反応を誘発することができるものを意味する。免疫原/抗原は、単一のエピトープを含んでよく、または複数エピトープを含んでもよい。例えば、ペプチド、ポリペプチド、タンパク質、糖タンパク質、リポタンパク質、炭水化物、リポ多糖類、核酸、小分子、および種々の微生物の、その全体または一部であって、ウイルス、細菌および寄生生物、および他の感染粒子を含むものはしたがって、免疫反応を誘発することができる限りにおいて、抗原であることができる。ハプテンおよびミモトープ(mimotope)もまた、本明細書において用語「免疫原」および「抗原」に包含されると考えられる。
【0073】
本明細書において、用語「プライムする」およびその文法的な変形は、動物において、抗原投与の前に、免疫反応を刺激すること、および/または作動させることを意味する。
【0074】
本明細書において、用語「処置する(treat)」、「処置される(treated)」、または「処置している(treating)」とは、疾病または疾患などの状態または感染体に関連して用いる場合、対象の、状態または病原体による感染に対する抵抗性を増加させる処置(すなわち、対象が状態になる可能性、または感染体に感染する可能性を低下させる)、および対象が状態になるか感染した後で、状態または感染に対して戦うための処置(例えば、状態もしくは感染またはそれに付随する症状を、低減させ、消滅させ、改善し、または安定化させる)を意味する。
【0075】
本明細書において用語「対象」または「患者」とは、処置の必要な動物を意味する。
【0076】
本明細書において用語「動物」とは、ヒトおよび非ヒト動物の両方を意味する。非ヒト動物は、限定することなく、哺乳類、鳥類、爬虫類および魚類を含み、家畜(ペットを含む)、農場の動物、実験動物、動物園の動物および野生動物を包含し、例えばウシ、ブタ、ウマ、ヤギ、ヒツジ、および他の有蹄動物;イヌ;ネコ;ニワトリ、カモ類おおび他の鳥類;非ヒト霊長類;モルモット;ウサギ;フェレット(クロアシイタチ);ラット;ハムスターおよびマウスである。
【0077】
本明細書において、核酸またはアミノ酸配列との関連で用いる用語「実質的に同一」とは、最適に配列された場合に、例えば以下に記載の方法を用いて、核酸配列またはアミノ酸配列が、規定の第2核酸配列またはアミノ酸配列(または「参照配列」)に対して、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも96%、少なくとも97%、少なくとも98%、または少なくとも99%の配列同一性を有することを示す。「実質的な同一性」は、配列の種々の型および長さを指すために用いることができ、例えば全長配列、機能ドメイン、コードおよび/または制限配列、プロモーター、およびゲノム配列などである。2つのアミノ酸配列または核酸配列の間のパーセント同一性は、当分野の業者の技能内である種々の方法により決定することができ、例えば、次のような公開されたコンピューターソフトウェアを用いて決定可能である:Smith Waterman Alignment(Smith, T. F. and M. S. Waterman (1981) JMoI Biol 147: 195-7);「BestFit」(Smith and Waterman, Advances in Applied Mathematics, 482-489 (1981))、これはGeneMatcher Plus(登録商標)Schwarz and Dayhof (1979) Atlas of Protein Sequence and Structure, Dayhof, M. O., Ed pp 353-358に組み込まれている;BLASTプログラム(Basic Local Alignment Search Tool (Altschul, S. F., W. Gish, et al. (1990) J MoI Biol 215: 403-10)、およびこれらの変形であって、BLAST-2、BLAST-P、BLAST-N、BLAST-X、WU-BLAST-2、ALIGN、ALIGN-2、CLUSTALおよびMegalign(DNASTAR)ソフトウェアを含むもの。さらに当業者は、アラインメントを測定するための適切なパラメータを、比較する配列の長さにわたり最大アラインメントを達成するのに必要なアルゴリズムも含んで、決定することができる。一般に、アミノ酸配列に対して、比較配列の長さは少なくも10アミノ酸である。当業者は、アラインメントに用いる実際の長さは、比較する配列全体の長さに依存し、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも200アミノ酸であってよく、またはアミノ酸配列の全長であってもよいことを理解する。核酸については、比較配列の長さは一般に少なくとも25ヌクレオチドであるが、少なくとも50、少なくとも100、少なくとも125、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも350、少なくとも400、少なくとも450、少なくとも500、少なくとも550、または少なくとも600であってよく、または、核酸配列の全長であってもよい。
【0078】
用語「に一致している」または「に一致する」とは、核酸配列が、参照核酸配列の全体または一部と同一であることを示す。これと対照的に、本明細書において用語「に相補的」とは、核酸配列が、参照核酸配列の相補鎖の全体または一部と同一であることを示す。例としては、核酸配列「TATAC」は、参照配列「TATAC」に一致し、参照配列「GTATA」に相補的である。
【0079】
MaMVまたはMaMVウイルス様粒子を含む免疫原性組成物
本発明は、MaMVまたはMaMV VLPおよび任意に1または2以上の抗原を含む免疫原性組成物を提供する。免疫原性組成物はさらに、好適な担体、賦形剤など、および/または前記組成物の安定性、味のよさ、薬物動態、バイオアベイラビリティ等を改善する、医薬組成物の他の標準成分を任意に含んでもよい。
【0080】
Malvaモザイクウイルス
本発明の免疫原性組成物での使用に好適なMaMVの代表的な例は、カナダ・ブリティッシュコロンビアのサマーランド近郊で収穫されたゼニバアオイ(Malva neglecta)から単離され、これは例1に記載されている。本明細書の例1〜4に記載されているように、ウイルスのゲノム配列および物理的特性の分析により、MaMVはポテックスウイルス属のメンバーであり、フレキシウイルス科(Flexiviridae)の可能性が高い。
【0081】
MaMVは、長さが約450nm〜約600nmで幅が約10nm〜約18nmの間の、柔軟な糸状のビリオン(ウイルス粒子:virion)の特徴を有する。図4Dに示す例示のウイルスは、約520〜560nmの長さと、約12〜約16nmの幅である。
【0082】
M. neglectaのMaMVによる感染は、モザイク症状および葉脈の透明化(clearing)をもたらす。MaMVはまた、Chenopodiaceae科のメンバー、例えばChenopodium quinoa上で繁殖するその能力を特徴とする。
【0083】
MaMVのさらなる特徴は、配列番号1に記載の配列と実質的に同一のゲノム配列を有することである(図1およびGenBankアクセッション番号DQ660333も参照)。MaMVゲノムRNAは6858ヌクレオチド(nt)長(ポリ(A)尾を除く)で、GC含有量が45%である。ゲノム構成は他のポテックスウイルスと類似しており、推定RNA依存性RNAポリメラーゼ(RdRp)と、続いてTGBタンパク質をコードする3つのオーバーラップ遺伝子と、最後に外被タンパク質とを含む(図6A参照)。MaMVはさらに、感染の間に3つの主要なウイルスRNAを産生することを特徴とする:1つの大きなゲノムRNA、および、それぞれ2000個および800個のヌクレオチドのRNAとして移動している2つのサブゲノム種である(図6B参照)。
【0084】
MaMVはさらに、配列番号2に記載の配列と実質的に同一のアミノ酸配列を有する外被タンパク質を有することを特徴とする(図2も参照)。
【0085】
当業者に知られているように、ウイルス種はしばしば、同じ特徴を共有する多数の異なる株によって表される。したがって本発明は、例1に記載のMaMVの変異株であって、これと同じ物理的特性および、配列番号1と少なくとも約80%の配列同一性を有するゲノム配列を有する、前記変異株も包含する。本発明の1つの態様において、変異株は、例1に記載のMaMVと同じ物理的特性および、配列番号1と少なくとも約90%の配列同一性を有するゲノム配列を有する。本発明の1つの態様において、変異株は、例1に記載のMaMVと同じ物理的特性および、配列番号1と少なくとも約95%の配列同一性を有するゲノム配列を有する。
【0086】
MaMVウイルス様粒子(VLP)
本発明のMaMV VLPは、MaMV外被タンパク質に由来するのが好ましい。「に由来する」とは、VLPが、野生型MaMV外被タンパク質の配列と実質的に同一のアミノ酸配列を有する外被タンパク質を含み、任意に、以下にさらに詳細に記載するように、前記外被タンパク質に付着した1または2以上の抗原を含んでよいことを意味する。VLPに含まれるMaMV外被タンパク質はしたがって、野生型外被タンパク質またはその組換え型であることができ、野生型タンパク質の配列またはこの配列の改変型であって、多量体化および自己集合可能でVLPを形成することができる前記配列を有してよい。野生型配列および改変配列を有する外被タンパク質の組合せを含むVLPもまた、意図される。
【0087】
MaMV VLPは、多量体化および自己集合する能力を保持したMaMV外被タンパク質から形成される。集合した場合、各々のVLPは、外被タンパク質サブユニットの長いらせん状アレイを含む。VLPが含む外被タンパク質の数は、例えば約40〜約1600まで変化することができる。
【0088】
本発明のVLPは、同一のアミノ酸配列を有する複数の外被タンパク質から、集合した場合の最終的なVLPが、同一の外被タンパク質サブユニットを含むようにして調製することができ、または、VLPは、異なるアミノ酸配列を有する複数の外被タンパク質から、集合した場合の最終的なVLPが、その外被タンパク質サブユニットにおける変化を含むようにして調製することができる。
【0089】
VLPを形成するために用いる外被タンパク質は、全長外被タンパク質または、多量体化してVLPを形成することができる、全長外被タンパク質の一部であってよい。外被タンパク質配列は、天然の(野生型)配列であることができ、または、本明細書に記載のように、改変された外被タンパク質が多量体化しVLP内に集合する能力を保持している限りにおいて、野生型配列の遺伝的な改変型であってよく、例えば、1または2以上のアミノ酸の欠失、挿入、置き換えなどを含む。例1に記載のMaMVの外被タンパク質のアミノ酸配列は、本明細書において配列番号2(図2参照)として提供され、これはまた、GenBankからもアクセス可能である(GenBankアクセッション番号ABG48664)。MaMVの外被タンパク質のヌクレオチド配列もまた、本明細書に配列番号3(図3参照)として提供され、GenBankからもアクセス可能である(GenBankアクセッション番号DQ660333(ヌクレオチド6057〜6788))。
【0090】
上に述べたように、VLPに含まれる外被タンパク質のアミノ酸配列は、配列番号2に記載の野生型配列に正確に一致する必要はなく、すなわち、改変配列であってよい。例えば、外被タンパク質の配列は、1または2以上のアミノ酸残基の置換、挿入または欠失により改変されていてもよく、そのためその変更部位(単数または複数)における残基は、野生型(参照)配列に一致しない。例えば、外被タンパク質をコードするヌクレオチド配列は、酵素切断部位の付加により改変されてよく、その結果、コードされる外被タンパク質の、例えば、1個〜数個のアミノ酸の、C末端、N末端、もしくは両方における、またはこれらの近傍における付加による改変が生じる。当業者はしかし、かかる変異は拡張的(extensive)ではなく、改変外被タンパク質の、多量体化しVLPに集合する能力を妨害しないことを理解する。かかる変異はしかし、MaMV外被タンパク質の自己集合する能力を改善し、および/またはPapMV外被タンパク質について前に示されたような、より長いVLPの形成をもたらす(Laliberte et al., 2008, FEBS Journal, 275: 1474-1484)。MaMV外被タンパク質の改変型の、VLPに集合する能力は、例えば、標準の技法による電子顕微鏡法により評価することができ、例示の方法が本明細書の実施例に提供される。
【0091】
一般に、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約80%同一のアミノ酸配列を含む。1つの態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約85%同一のアミノ酸配列を含む。別の態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約90%同一のアミノ酸配列を含む。他の態様において、好適な改変外被タンパク質は、配列番号2に記載された野生型配列と少なくとも約91%同一の、少なくとも約92%同一の、少なくとも約93%同一の、少なくとも約94%同一の、少なくとも約95%同一の、少なくとも約96%同一の、少なくとも約97%同一の、少なくとも約98%同一の、または少なくとも約99%同一の、アミノ酸配列を含む。
【0092】
多量体化しVLPに集合する能力を保持する野生型タンパク質の断片である外被タンパク質(すなわち、「機能性」断片)もまた、本発明により意図される。例えば、機能性断片は、タンパク質のN末端、C末端または内部からの、1または2以上のアミノ酸の欠失、またはそれらの組合せを含んでよい。一般に、機能性断片は、配列番号2に記載の配列の少なくとも約150の隣接アミノ酸(contiguous amino acid)を含む。本発明の1つの態様において、機能性断片は、配列番号2に記載の配列の少なくとも約180の隣接アミノ酸を含む。別の態様において、機能性断片は、配列番号2に記載の配列の少なくとも約190の隣接アミノ酸を含む。他の態様において、機能性断片は、配列番号2に記載の配列の、少なくとも約200の隣接アミノ酸を、少なくとも約210の隣接アミノ酸を、少なくとも約220の隣接アミノ酸を、少なくとも約225の隣接アミノ酸を、少なくとも約230の隣接アミノ酸を含む。タンパク質のN末端またはC末端での欠失は、タンパク質の多量体化する能力を保持するために、一般に25個以下のアミノ酸を除去すべきである。例えば、1つの態様において、N末端での欠失は、20個以下のアミノ酸を除去する。他の態様において、N末端での欠失は、15個以下のアミノ酸を、12個以下のアミノ酸を、10個以下のアミノ酸を、8個以下のアミノ酸を、および5個以下のアミノ酸を除去する。
【0093】
本発明の1つの態様において、VLPは、MaMV外被タンパク質の組換え型を含む。他の態様において、VLPは、MaMV外被タンパク質の遺伝的改変型を含む。
【0094】
外被タンパク質が、1または2以上のアミノ酸置換を含む改変配列を含む場合、これらは、「保存的」置換または「非保存的」置換であることができる。保存的置換は、1つのアミノ酸残基を、類似の側鎖特性を有する他の残基と置き換えることを含む。当分野で知られているように、20種の天然のアミノ酸は、それらの側鎖の物理化学的特性に従って分類可能である。適切な分類としては、例えば、アラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニン、フェニルアラニンおよびトリプトファン(疎水性側鎖);グリシン、セリン、スレオニン、システイン、チロシン、アスパラギン、およびグルタミン(極性の非荷電側鎖);アスパラギン酸およびグルタミン酸(酸性側鎖)およびリシン、アルギニンおよびヒスチジン(塩基性側鎖)を含む。アミノ酸の他の分類は、フェニルアラニン、トリプトファンおよびチロシン(芳香族側鎖)である。保存的置換は、1つのアミノ酸を、同じ群の別のアミノ酸で置換することが関与する。非保存的置換は、1つのアミノ酸残基を、異なる側鎖特性を有する別の残基で置き換えることが関与し、例えば、酸性残基の中性または塩基性残基による置き換え、中性残基の酸性または塩基性残基による置き換え、疎水性残基の親水性残基による置き換えなどである。
【0095】
外被タンパク質をコードする核酸配列も同様に、野生型配列に正確に一致する必要はないが、遺伝子コードの縮退のために、および/または、上記の改変アミノ酸配列をコードするように、変化してよい。本発明の1つの態様において、したがって、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約70%同一である。他の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約75%同一である。別の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約80%同一である。他の態様において、外被タンパク質をコードする核酸配列は、配列番号3に記載の配列と少なくとも約85%、または少なくとも約90%同一である。
【0096】
以下にさらに詳細に記載するように、VLP外被タンパク質は任意に、1もしくは2以上の抗原、親和性ペプチドもしくは他の短ペプチド配列、またはこれらの組合せに遺伝子的に融合してよく、1または2以上の抗原の付着を促進する。
【0097】
抗原
免疫原性組成物は任意に、1または2以上の抗原を含んでよく、これらはMaMVまたはMaMV VLPとは別であっても、またはMaMVもしくはMaMV VLPに接合していてもよい。接合および非接合抗原の両方を含む免疫原性組成物もまた、意図される。この後者の場合、非接合抗原は、付加的単離抗原(AIA)として考えることができる。AIAは、接合抗原(単数または複数)と同じであっても、異なっていてもよい。接合は、例えば、外被タンパク質との遺伝子融合、または共有結合、非共有結合もしくは親和性手段を介した結合であることができる。本発明の1つの態様において、1または2以上の抗原は、市販のワクチンの形態で提供されることができる。
【0098】
ワクチン開発に好適な多種多様な抗原が、当分野に知られている。本発明の免疫原性組成物に含めるのに好適な抗原は、例えば、それが指向されるところの疾病または疾患などの免疫原性組成物の所望の最終用途、組成物の様式、組成物が多価ワクチンもしくは単価ワクチンとしての使用を意図されているのか、および/または投与される動物などに基づいて、当業者が容易に選択することができる。
【0099】
例えば、抗原は、動物において、例えば癌、感染症、アレルギー反応、または自己免疫疾患などの疾病または疾患を引き起こすことができる作用物質に由来することができる。またはこれは、薬剤、ホルモンもしくは毒素関連の疾病または疾患に対する免疫反応を誘発するために用いるのに好適な、抗原であることができる。抗原は、当分野に知られている病原体から、例えば細菌、ウイルス、原生動物、真菌、寄生生物、または例えばプリオンなどの感染粒子から由来してよい。または抗原は、腫瘍関連抗原、自己抗原、またはアレルゲンであってよい。あるいは、抗原は、抗原のB細胞エピトープまたはT細胞エピトープを含んでよい。
【0100】
1または2以上の抗原は、免疫原性組成物中に種々の様式で、上記のように、および下にさらに詳細に記載するようにして、含めることができる。免疫原性組成物中に含むための1または2以上の抗原は、したがって、選択した様式に依存してサイズが異なることができる。抗原は、例えば、ペプチド、タンパク質、核酸、多糖類、小分子、またはこれらの組合せであって、病原体全体またはその一部を含み、例えば、生きた、不活性化または減衰型の病原体である。
【0101】
免疫原性組成物が1より多くの抗原を含む場合、組成物中に含めるために選択される抗原は、同一であることができ、または異なっていてもよく、単一の源から、または複数の源から由来してよい。各抗原は、特定の免疫反応を引き起こすことができる1つのエピトープを有するか、または各抗原は、1つより多くのエピトープを有してもよい。
【0102】
抗原は、T細胞、B細胞、NK細胞およびマクロファージ上の表面構造により、またはクラスIもしくはクラスIIのAPC関連細胞表面構造により、またはこれらの組合せにより認識されるエピトープを含んでよい。1つの態様において、本発明は、免疫原性組成物が、小さなおよび/または弱い免疫原性抗原に対して特に有用であることを意図する。
【0103】
本発明の免疫原性組成物に含むための抗原はまた、病原体または他の目的の源から、当分野に知られた方法により選択し、当分野に知られた標準の免疫学的技法を用いて、動物において免疫反応を誘発するそれらの能力についてスクリーニングしてよい。例えば、抗原タンパク質内でのエピトープの予測のための方法は、Nussinov R and Wolfson H J, Comb Chem High Throughput Screen (1999) 2(5):261に記載されており、CTLエピトープの予測のための方法は、Rothbard et al, EMBO J. (1988) 7:93-100およびde Groot M S et al, Vaccine (2001) 19(31):4385-95に記載されている。他の方法は、Rammensee H-G. et al., Immunogenetics (1995) 41: 178-228およびSchirle M et al, Eur J Immunol (2000) 30(18):2216-2225に記載されている。
【0104】
有用なウイルス抗原は、例えば、以下の科のメンバーに由来するものを含む:アデノウイルス科;アレナウイルス科(例えばIppyウイルスおよびラッサウイルス);ビルナウイルス科;ブニヤウイルス科;カリシウイルス科;コロナウイルス科;フィロウイルス科;フラビウイルス科(例えば黄熱病ウイルス、デング熱ウイルスおよびC型肝炎ウイルス);ヘパドナウイルス科(Hepadnaviradae)(例えば、B型肝炎ウイルス);ヘルペスウイルス科(例えばヒト単純ヘルペスウイルス1型);オルトミクソウイルス科(例えば、A型、B型よびC型インフルエンザウイルス);パラミクソウイルス科(例えば流行性耳下腺炎ウイルス、ヒト・メタニューモウイルス、麻疹ウイルス、および呼吸器合胞体ウイルス);ピコルナウイルス科(例えば、ポリオウイルスおよびA型肝炎ウイルス);ポックスウイルス科;レオウイルス科;レトロウイルス科(例えばBLV−HTLVレトロウイルス、HIV−1、HIV−2、ウシ免疫不全ウイルスおよびネコ免疫不全ウイルス);ラブドウイルス科(例えば狂犬病ウイルス)、およびトガウイルス科(例えば風疹ウイルス)。1つの態様において、免疫原性組成物は、以下のような主要なウイルス病原体に由来する1または2以上の抗原を含む:種々の肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、種々のインフルエンザウイルス、ウェストナイルウイルス、呼吸器合胞体ウイルス、狂犬病ウイルス、ヒトパピローマウイルス(HPV)、エプスタインバーウイルス(EBV)、ポリオーマウイルス、またはSARSコロナウイルス。
【0105】
A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、デルタ肝炎ウイルス(HDV)、E型肝炎ウイルス(HEV)、およびG型肝炎ウイルス(HGV)を含む肝炎ウイルス由来のウイルス抗原は、当分野に知られている。例えば、抗原は、HCVコアタンパク質、E1タンパク質、E2タンパク質、NS3タンパク質、NS4タンパク質またはNS5タンパク質から、HVB HbsAg抗原またはHBVコア抗原から、およびHDVデルタ抗原から由来することができる(例えば、米国特許第5,378,814号を参照)。次の米国特許は、例えば、HCVコアタンパク質に基づく種々の抗原について記載している;第6,596,476号;第6,592,871号;第6,183,949号;第6,235,284号;第6,780,967号;第5,981,286号;第5,910,404号;第6,613,530号;第6,709,828号;第6,667,387号;第6,007,982号;第6,165,730号;第6,649,735号、および第6,576,417号。
【0106】
ヘルペスウイルス科からの既知の抗原の非限定的例としては、単純ヘルペスウイルス(HSV)1および2型、例えばHSV−1およびHSV−2糖タンパク質gB、gDおよびgHに由来するものが挙げられる。
【0107】
HIV抗原の非限定的例としては、gp120由来抗原、gp160およびgp41などの種々のエンベロープタンパク質由来の抗原、p24gagおよびp55gagなどのgag抗原、ならびにHIVのpol、env、tat、vif rev、nef vpr、vpuおよびLTR領域に由来するタンパク質が挙げられる。多数のHIV−1およびHIV−2分離株からのgp120配列であって、HIVの種々の遺伝的サブタイプのメンバーを含むものが知られている(例えば、Myers et al., Los Alamos Database, Los Alamos National Laboratory, Los Alamos, N. Mex. (1992)およびModrow et al, J. Virol. (1987) 61:570 578)を参照)。
【0108】
他のウイルス抗原の非限定的例としては、水痘帯状疱疹ウイルス(VZV)、エプスタインバーウイルス(EBV)、およびCMV gBおよびgHを含むサイトメガロウイルス(CMV);および他のヒトヘルペスウイルス、例えばHHV6およびHHV7などからの抗原が挙げられる(例えば、Chee et al. (1990) Cytomegaloviruses (J. K. McDougall, ed., Springer-Verlag, pp. 125 169);McGeoch et al. (1988) J. Gen. Virol. 69:1531 1574;米国特許第5,171,568号;Baer et al. (1984) Nature 310:207 211;およびDavison et al. (1986) J. Gen. Virol. 67:1759 1816を参照)。
【0109】
抗原はまた、インフルエンザウイルスから、例えば赤血球凝集素(HA)、ノイラミニダーゼ(NA)、核タンパク質(NP)、マトリクスタンパク質(M1およびM2)、ポリメラーゼ酸性タンパク質(PA)、およびポリメラーゼ塩基性タンパク質サブユニット(PB1、PB2)から由来することもできる。これらタンパク質の配列は当分野で知られており、全米バイオテクノロジー情報センター(NCBI)により維持されているGenBankデータベースから容易にアクセスできる。HA、NPおよびマトリクスタンパク質の好適な抗原性断片としては、限定はしないが、以下が挙げられる:赤血球凝集素エピトープとして、HA91−108、HA307−319およびHA306−324(Rothbard, Cell, 1988, 52:515-523)、HA458−467(J. Immunol. 1997, 159(10): 4753-61)、HA213−227、HA241−255、HA529−543およびHA535−547(Gao, W. et al, J. Virol, 2006, 80:1959-1964);核タンパク質エピトープとして、NP206−229(Brett, 1991, J Immunol. 147:984-991)、NP335−350およびNP380−393(Dyer and Middleton, 1993, In: Histocompatibility testing, a practical approach (Ed.: Rickwood, D. and Hames, B. D.) IRL Press, Oxford, p. 292; Gulukota and DeLisi, 1996, Genetic Analysis: Biomolecular Engineering, 13:81)、NP305−313(DiBrino, 1993, PNAS 90: 1508-12);NP384−394(Kvist, 1991, Nature. 348:446-448);NP89−101(Cerundolo, 1991, Proc. R. Soc. Lon. 244:169-7);NP91−99(Silver et al, 1993, Nature. 360: 367-369);NP380−388(Suhrbier, 1993, J. Immunology 79: 171- 173);NP44−52およびNP265−273(DiBrino, 1993、同上);およびNP365−380(Townsend, 1986, Cell. 44:959-968);マトリクスタンパク質(M1)エピトープとして、M1 2−22、M1 2−12、M1 3−11、M1 3−12、M1 41−51、M1 50−59、M1 51−59、M1 134−142、M1 145−155、M1 164−172、M1 164−173(全て、Nijman, 1993, Eur. J. Immunol. 23:1215-1219に記載);M1 17−31、M1 55−73、M1 57−68(Carreno, 1992, MoI Immunol 29: 1131-1140);M1 27−35、M1 232−240(DiBrino, 1993、同上)、M1 59−68およびM1 60−68(Eur. J. Immunol. 1994, 24(3): 777-80);およびM1 128−135(Eur. J. Immunol. 1996, 26(2): 335-39);PA 412−422(Heiny AT, et al., 2007, PLoS ONE, 21;2(1l):el190);PB1 15−51(Heiny AT, et al., 2007, PLoS ONE, 21;2(1l):el190)およびPB2 685−696(Heiny AT, et al., 2007, PLoS ONE, 21;2(ll):el190)。
【0110】
その他の例としては、HAタンパク質由来のF3ペプチドであり、これは次の配列を有する:KAYSNCYPYDVPDY(配列番号72)(Lu et al., 2002, Int. Arch. Allergy Immunol, 127: 245-250)、およびM1タンパク質からのCTLエピトープであり、これは次の配列を有する:SPLTKGILGFVFTLTVPSE(配列番号73)、およびGILGFVFTL(配列番号60)。
【0111】
他の関連する抗原性領域およびエピトープも知られている。例えば、インフルエンザイオンチャネルタンパク質(M2)の断片であって、これはM2eペプチド(M2の細胞外ドメイン)を含む。このペプチドの配列は、インフルエンザの異なる株にわたって高度に保存されている。M2eペプチド配列の例は表1に配列番号4として示されている。この配列の変異形が同定されており、そのいくつかも表1に示されている。
表1:M2eペプチドおよびその変異
【表1】
*米国特許出願第2006/0246092号参照
【0112】
M2e配列の全体または一部を用いることができ、例えば、変異体にわたって保存されている部分配列であり、例としてはアミノ酸2〜10により規定される領域内の断片、または保存エピトープEVETPIRN(配列番号9)(M2e配列のアミノ酸6〜13)。6〜13エピトープは、GenBankで利用可能なヒトA型インフルエンザ株の84%において不変であることが見出されている。同定されているこの配列の変異形には以下がある:EVETLTRN(配列番号10)(9.6%)、EVETPIRS(配列番号11)(2.3%)、EVETPTRN(配列番号12)(1.1%)、EVETPTKN(配列番号13)(1.1%)、ならびにEVDTLTRN(配列番号14)、EVETPIRK(配列番号15)およびEVETLTKN(配列番号16)(それぞれ0.6%)(Zou, P., et al., 2005, Int Immunopharmacology, 5:631-635;Liu et al. 2005, Microbes and Infection, 7: 171-177を参照)。
【0113】
他の有用な抗原としては、生ウイルス、弱毒化ウイルス、および不活化ウイルスであって、例えば不活化ポリオウイルス(Jiang et al, J. Biol. Stand., (1986) 14:103-9)、A型肝炎ウイルスの弱毒株(Bradley et al, J. Med. Virol, (1984) 14:373-86)、弱毒麻疹ウイルス(James et al, N. Engl. J. Med, (1995) 332:1262-6)、および百日咳ウイルスのエピトープ(例えば、ACEL-IMUNE(登録商標)無細胞DTP、Wyeth-Lederle Vaccines and Pediatrics)である。
【0114】
抗原はまた、非定型ウイルスまたはウイルス様物質に由来することもでき、例としては、当分野で知られているように、以下のものがある:クールーの原因物質、クロイツフェルトヤコブ病(CJD)、スクラピー、伝染性ミンク脳症、および慢性消耗性疾患、または狂牛病に関連するプリオンなどのタンパク性感染粒子。
【0115】
有用な細菌性抗原としては、例えば、リポ多糖類、莢膜抗原(本質的に、タンパク様または多糖類)などの表在性細菌性抗原成分、または鞭毛成分が挙げられ、これらは次のような疾病に関与する既知の原因物質から得るか、または由来することができる:ジフテリア、百日咳、破傷風、結核、細菌性または真菌性肺炎、コレラ、腸チフス、ペスト、細菌性赤痢またはサルモネラ症、レジオネラ病、ライム病、ハンセン病、マラリア、鉤虫症、オンコセルカ症、住血吸虫症、トリパマソーマ症(trypamasomialsis)、レスマニア症(lesmaniasis)、ジアルジア鞭毛虫症、アメーバ症、フィラリア症、ボレリア、および旋毛虫症。
【0116】
腸内細菌科のグラム陰性菌に由来する抗原の例としては、限定はしないが、腸チフス菌Vi(莢膜多糖類)抗原、大腸菌KおよびCFA(莢膜成分)抗原および、大腸菌線毛付着抗原(K88およびK99)が挙げられる。抗原タンパク質の例としては、ポリン(Secundino et al., 2006, Immunology 117:59)としても知られている外膜タンパク質(Omp);腸チフス菌鉄調節外膜タンパク質などの関連するポリン(IROMP, Sood et al., 2005, MoI Cell Biochem 273:69-78)、および、限定することなく腸チフス菌HSP40(Sagi et al., 2006, Vaccine 24:7135-7141)を含む、熱ショックタンパク質(HSP)が挙げられる。抗原性ポリンの非限定的例としては、OmpCおよびOmpFが挙げられ、これらは多数のサルモネラ種および大腸菌種に見出される。OmpCおよびOmpFのオルソログもまた、他の腸内細菌科に見出されており、これらは本発明の目的のための好適な抗原タンパク質である。さらに、腸内細菌科のポリンタンパク質中に見出される配列の保存領域に基づいて(Diaz-Quinonez et al., 2004, Infect, and Immunity 72:3059- 3062)、Omp1B(Shigella flexneri)、OmpC2(Yersinia pestis)、OmpD(S. enteria)、OmpK36(Klebsiella pneumonie)、OmpN(大腸菌)およびOmpS(S. enterica)も好適であることができる。
【0117】
種々の腸内細菌からの抗原タンパク質の配列は、当分野に知られており、全米バイオテクノロジー情報センター(NCBI)により維持されているGenBankデータベースから容易にアクセスできる。例えば、GenBankアクセッション番号P0A264およびGenBankアクセッション番号NP_804453:OmpC(S. enterica subsp. enterica serovar Typhi Ty2);GenBankアクセッション番号CAD05399:OmpF前駆体タンパク質(S. enterica subsp. enterica serovar Typhi CTl 8);GenBankアクセッション番号16761195:OmpC(S. enterica serovar Typhimurium);GenBankアクセッション番号47797:OmpC(S. enterica serovar Typhi);GenBankアクセッション番号8953564:OmpC(S. enterica serovar Minnesota);GenBankアクセッション番号19743624:OmpC(S. enterica serovar Dublin);GenBankアクセッション番号19743622:OmpC(S. enterica serovar Gallinarum);GenBankアクセッション番号26248604:OmpC(大腸菌);GenBankアクセッション番号24113600:Omp1B(Shigella flexneri);GenBankアクセッション番号16764875:OmpC2(Yersinia pestis);GenBankアクセッション番号16764916:OmpD(S. enterica Serovar Typhimurium);GenBankアクセッション番号151149831:OmpK36(Klebsiella pneumonie);GenBankアクセッション番号3273514:OmpN(大腸菌)、およびGenBankアクセッション番号16760442:OmpS(S. enterica serovar Typhi)。
【0118】
種々の腫瘍関連抗原が当分野に知られている。代表的な例としては、限定はしないが、以下が挙げられる:Her2(乳癌);GD2(神経芽細胞腫);EGF−R(悪性グリア芽細胞腫);CEA(髄様甲状腺癌);CD52(白血病);ヒトメラノーマタンパク質gp100(例えばエピトープ:IMDQVPFSV(配列癌号59)を含む抗原);ヒトメラノーマタンパク質melan−A/MART−1;ヒトDickkopf1(DKK1)タンパク質、ヒトアンギオモチン(Amot)、NA17;NA17−Antタンパク質;p53タンパク質;MAGE1、MAGE2、MAGE3(HLA−A1タンパク質)およびMAGE4を含む種々のMAGE(メラノーマ関連抗原E);種々のチロシナーゼ(HLA−A2ペプチド);変異ras;p97メラノーマ抗原;進行癌に関連するRasぺプチドおよびp53ペプチド;子宮頸癌に関連するHPV16/18およびE6/E7抗原;乳癌関連MUC1−KLH抗原;結腸直腸癌に関連するCEA(癌胎児性抗原)、および前立腺癌関連PSA抗原。
【0119】
有用なアレルゲンとしては、花粉、動物鱗屑、草、カビ、粉塵、抗生物質、刺咬昆虫毒物からのアレルゲン、および種々の環境、薬物および食物アレルゲンが挙げられるが、これらに限定されない。一般的な樹木アレルゲンとしては、ハコヤナギ、ポプラ、セイヨウトリネコ、カバノキ、メープル、オーク、ニレ、ヒッコリー、およびペカンの木からの花粉が挙げられる。一般的な植物アレルゲンとしては、ライ麦、ブタクサ、イギリスオオバコ、スイバ(sorrel dock)およびアカザからのものが挙げられ、植物接触アレルゲンとしては、ウルシ、ツタウルシおよびイラクサからのものが挙げられる。一般的な草のアレルゲンとしては、オオアワガエリ、ジョンソングラス、ギョウギシバ、ウシノケグサ、およびブルーグラスアレルゲンが挙げられる。一般的なアレルゲンは、鱗屑または真菌類から、例えばアルテルナリア属、フザリウム属、ホルモデンドラム属、アスペルギラス属、ミクロポリスポラ属、ケカビ属および好熱性の放線菌からも得ることができる。ペニシリン、スルホンアミドおよびテトラサイクリンは、一般的な抗生物質アレルゲンである。表皮性のアレルゲンは、ハウスダストまたは有機ダスト(典型的には真菌類から)から、イエダニ(dermatphagoides pterosinyssis)などの昆虫から、または羽毛、およびネコおよびイヌの鱗屑などの動物源から、得ることができる。一般的な食物アレルゲンとしては、牛乳およびチーズ(乳製品)、卵、小麦、ナッツ(例えばピーナッツ)、魚介類(例えば、貝)、エンドウ豆、豆、およびグルテンアレルゲンなどが挙げられる。一般的な薬物アレルゲンとしては、局所麻酔薬およびサリチル酸塩アレルゲンが挙げられ、一般的な昆虫アレルゲンとしては、ハチ、スズメバチ、カリバチおよびアリ毒物、およびゴキブリがく(calyx)アレルゲンが挙げられる。
【0120】
特に良好に特徴付けられているアレルゲンとしては、限定はされないが、以下が挙げられる:イエダニアレルゲンDer pIおよびDer pII(Chua, et al., J. Exp. Med., 167:175 182, 1988;およびChua, et al, Int. Arch. Allergy Appl. Immunol, (1990) 91 : 124-129)、Der pIIアレルゲンのT細胞エピトープペプチド(Joost van Neerven, et al, J. Immunol, (1993) 151 :2326-2335を参照)、非常に豊富な抗原E(Amb aI)ブタクサ花粉アレルゲン(Rafnar, et al, J. Biol. Chem., (1991) 266: 1229-1236を参照)、ホスホリパーゼA2(ハチ毒物)アレルゲンおよびその中のT細胞エピトープ(Dhillon, et al, J. Allergy Clin. Immunol, (1992) 42参照)、シラカバ花粉(Betv1)(Breiteneder, et al, EMBO, (1989) 8:1935-1938参照)、Fe1 dI主要な家猫アレルゲン(Rogers, et al, MoI Immunol, (1993) 30:559-568参照)、樹木花粉(Elsayed et al, Scand. J. Clin. Lab. Invest. Suppl, (1991) 204: 17-31参照)、およびマルチエピトープ組換え草(grassアレルゲンrKBG8.3(Cao et al Immunology (1997) 90:46-51)。これらおよび他の好適なアレルゲンは市販されており、および/または既知の技術に従って用意に調製可能である。
【0121】
自己抗原に付随する状態に関連する抗原も、当業者に知られている。かかる抗原の代表的な例としては、限定はしないが、リンホトキシン、リンホトキシン受容体、核因子kBリガンドの受容体アクチベーター(RANKL)、血管内皮成長因子(VEGF)、血管内皮成長因子受容体(VEGF−R)、インターロイキン−5、インターロイキン−17、インターロイキン−13、CCL21、CXCL12、SDF−1、MCP−1、エンドグリン、レジスチン、GHRH、LHRH、TRH、MIF、エオタキシン、ブラジキニン、BLVC、腫瘍壊死因子αおよびアミロイドβペプチド、および免疫学的反応を引き出すために使用可能なこれら各々の断片が挙げられる。
【0122】
有用な毒素は、一般に有毒な植物、動物、および微生物の天然産物であるか、またはこれらの化合物の断片である。かかる化合物は、例えば、アフラトキシン、シガテラ毒素、百日咳毒素およびテトロドトキシンを含む。
【0123】
レクリエーショナルドラッグ中毒に関連して有用な抗原は当分野に知られており、例えば、コデイン、フェンタニル、ヘロイン、モルヒネ、およびアヘンなどの、オピオイドおよびモルヒネ誘導体;アンフェタミン、コカイン、MDMA(メチレンジオキシメタンフェタミン)、メタンフェタミン、メチルフェニデート、およびニコチンなどの興奮薬;LSD、メスカリンおよびプシロシビンなどの幻覚発現物質;ハシシおよびマリファナなどのカンナビノイド(大麻)、その他の常習性薬物または化合物、および派生物、副産物、変異体およびかかる化合物の複合体を含む。
【0124】
本発明の1つの態様において、免疫原性化合物に含まれる抗原(単数または複数)は、タンパク質抗原である。タンパク質抗原は、全長タンパク質、実質的に全長のタンパク質(例えば、約25個未満のアミノ酸のN末端および/またはC末端での欠失を含むタンパク質)、タンパク質の抗原断片、またはこれらの組合せであることができる。全長タンパク質は、適用可能である場合は、タンパク質の前駆体形態またはタンパク質の成熟(処理)形態であってよい。タンパク質は翻訳後修飾されていてもよく、例えば、糖タンパク質またはリポタンパク質であってよい。抗原断片は1または複数のエピトープを含むことができ、したがって数個のアミノ酸(例えば、少なくとも4個のアミノ酸)の1ペプチドから、数百のアミノ酸長のポリペプチドまでの大きさであってよい。本発明の1つの態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約4アミノ酸〜約250アミノ酸の長さである。他の態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約5アミノ酸〜約200アミノ酸の長さである。他の態様において、免疫原性組成物中に含まれるのに好適な抗原断片は、約5アミノ酸〜約150アミノ酸の長さ、約5アミノ酸〜約100アミノ酸の長さ、約5アミノ酸〜約75アミノ酸の長さ、約5アミノ酸〜約70アミノ酸の長さ、約5アミノ酸〜約60アミノ酸の長さ、および、約5アミノ酸〜約50アミノ酸の長さである。
【0125】
上記のように、本発明の1つの態様において、免疫原性組成物中に含まれる1または2以上の抗原は、市販のワクチン形態であることができる。種々のヒトワクチンが当分野に知られており、以下に対するワクチンが含まれるが、これらに限定はされない:
炭疽菌(anthrax)、例えばBioThrax(登録商標)(BioPort Corporation);
B型インフルエンザ菌(Hib)、例えばActHIB(登録商標)(Sanofi-aventis)、PedvaxHIB(登録商標)(Merck)およびHibTITER(登録商標)(Wyeth);
A型肝炎、例えばHavrix(登録商標)(GlaxoSmithKline)およびVaqta(登録商標)(Merck);
B型肝炎、例えばEngerix-B(登録商標)(GlaxoSmithKline)およびRecombivax HB(登録商標)(Merck);
帯状疱疹(shingles)、例えばZostavax(登録商標)(Merck);
ヒトパピローマウイルス(HPV)、例えばGardasil(登録商標)(Merck);
インフルエンザ、例えばFluarix(登録商標)およびFluviral(登録商標)(GlaxoSmithKline)、FluLaval(登録商標)(ID Biomedical Corp of Quebec);FluMist(登録商標)(鼻腔内)(Medimmune)、Fluvirin(登録商標)(Chiron)、およびFluzone(登録商標)(Sanofi-aventis);
日本脳炎、例えばJE-Vax(登録商標)(Sanofi-aventis);
麻疹、例えばAttenuvax(登録商標)(Merck);
髄膜炎菌性髄膜炎、例えばMenomune(登録商標)Meningococcal Polysaccharide(Sanofi-aventis);
流行性耳下腺炎、例えばMumpsvax(登録商標)(Merck);
肺炎級菌による疾病、例えばPneumovax 23(登録商標)Pneumococcal Polysaccharide(Sanofi-aventis)およびPrevnar(登録商標)Pneumococcal Conjugate(Wyeth);
ポリオ、例えばIpol(登録商標)(Sanofi-aventis)およびPoliovax(登録商標)(Sanofi-Pasteur);
狂犬病、例えばBioRab(登録商標)(BioPort Corporation)、RabAvert(登録商標)(Chiron)およびImovax(登録商標)Ravies (Sanofi-aventis);
ロタウイルス、例えばRotaTeq(登録商標)(Merck);
風疹、例えばMeruvax II(登録商標)(Merck);
腸チフス菌(腸チフス)、例えばTyphim Vi(登録商標)(Sanofi-aventis)およびVivotif(登録商標)Berna (経口)(Berna);
肺結核(BCG)、例えばTheraCys(登録商標)およびImmuCyst(登録商標)(Sanofi-aventis);TICE(登録商標)BCGおよびOncotice(商標)(Organon Teknika Corporation);Pacis(商標);およびMycobax(登録商標)(Sanofi-Pasteur);
種痘症(天然痘)、例えばDryvax(登録商標)(Wyeth);
水痘(鶏痘)、例えばVarivax(登録商標)(Merck);
黄熱病、例えばYF-Vax(登録商標)(Sanofi-aventis);
A型肝炎/B型肝炎、例えばTwinrix(登録商標)(GlaxoSmithKline);
B型肝炎およびHib、例えばComvax(登録商標)(Merck);
破傷風/Hib、例えばActHIB(登録商標)(Sanofi-Pasteur);
ジフテリア/Hib、例えばHibTITER(登録商標)(Wyeth Pharmaceuticals);
Hib/髄膜炎、例えばPedVaxHIB(登録商標)(Merck & Co);
髄膜炎/ジフテリア、例えばMenactra(登録商標)Meningococcal Conjugate(Sanofi-Pasteur);
破傷風/ジフテリア(Td)、例えばDecavac(登録商標)(Sanofi-aventis);
ジフテリア/破傷風/百日咳(DTaP/DTまたはDTaP)、例えばDaptacel(登録商標)およびTripedia(登録商標)(Sanofi-aventis)およびInfanrix(登録商標)(GlaxoSmithKline);
破傷風/ジフテリア/百日咳(Tdap)、例えばBoostrix(登録商標)(GlaxoSmithKline)およびAdacel(登録商標)(Sanofi-Pasteur);
DTaP/Hib、例えばTriHIBit(登録商標)(Sanofi-aventis);
DTaP/ポリオ/B型肝炎、例えばPediarix(登録商標)(GlaxoSmithKline);
麻疹/流行性耳下腺炎/風疹(MMR)、例えばM-M-R II(Merck)および
麻疹/流行性耳下腺炎/風疹/鶏痘、例えばProQuad(登録商標)(Merck)。
【0126】
獣医による使用の例としては、限定することなく、以下に対するワクチンが挙げられる:
Lawsonia intracellulars(例えば、EnterisolおよびIleitis)、Porphyromonas gulaeおよびP. denticanis (例えば、Priovac)、ストレプトコッカス・エクイ(例えば、Equilis StrepE)、Chlamydophila abortus(例えば、OvilisおよびEnzovax)、Mycoplasma synoviae(例えば、Vaxsafe MS)、Mycoplasma gallisepticum (例えば、Vaxsafe MG)、Bordetella avium(例えば、Art Vax)、Actinobacillus pleuropneumoniae(例えば、PleuroStar APP)、Actinobacillus pleuropneumoniae(例えば、Porcilis APP)、サルモネラ(例えば、Megan VaclおよびMeganEgg)、ウシ流産菌(例えば、RB-51)、アイメリア種(例えば、Coccivac、Immucox、Paracox、Advent、およびNobilis Cox ATM)、アイメリア種(例えば、Inovocox)、E. tenella(例えば、Livacox)、Toxoplasma gondii(例えば、OvilisおよびToxovax)、仮性狂犬病ウイルス(例えば、Suvaxyn Aujeszky)、古典的豚コレラウイルス(例えば、Porcilis PestiおよびBayovac CSF E2)、ウマインフルエンザウイルス(例えば、PROTEQ-FLUおよびRecombitek)、ニューカッスル病ウイルス(例えば、Vectormune FP-ND)、鳥インフルエンザウイルス(例えば、Poulvac FluFend I AI H5N3 RG)、鳥インフルエンザウイルス(例えば、Trovac AI H5)、狂犬病ウイルス(例えば、RaboralおよびPurevax Feline Rabies)、ネコ白血病ウイルス(例えば、EURIFEL FeLV)、イヌパルボウイルス1(例えば、RECOMBITEK Canine Parvo)、イヌコロナウイルス(例えば、RECOMBITEK Corona MLV)、イヌジステンパーウイルス(例えば、RECOMBITEK rDistemperおよびPUREV AXFerret Distemper)、IHNウイルス(例えば、Apex-IHN)。獣医用ワクチンの他の例は、LHRH(例えば、Vaxstrate、Improvac、Equito、イヌゴナドトロピン放出因子免疫治療薬、およびGonaCon)およびアンドロステンジオン(例えば、Fecundin、 AndrovaxおよびOvastim)などの生殖調節ワクチンを含む。
【0127】
本発明の1つの態様において、免疫原性組成物中に含まれる抗原は、既知のインフルエンザワクチンの形態である。ほとんどの市販のインフルエンザワクチンは、表面の糖タンパク質を取り除くためにインフルエンザウイルスを有機溶媒で処理した、スプリット・ウイルスワクチン、サブユニットワクチン、または弱毒化生ウイルスワクチン、またはこれらの組合せである。一般に市販のインフルエンザワクチンは、インフルエンザの3つの株に対する防御を提供することにおいて3価であり、これらは例えば2007〜2008年のシーズンにおいては、A型/ソロモン諸島/3/2006(H1N1)様、A型/ウィスコンシン/67/2005(H3N2)様、およびB型/マレーシア/2506/2004様である。
【0128】
現在市販されているインフルエンザワクチンとしては、限定はしないが、Fluzone(登録商標)およびVaxigrip(登録商標)(Sanofi-aventis)、Fluvirin(登録商標)(Novartis Vaccine)、Fluarix(登録商標)、FluLaval(登録商標)およびFluviral S/F(登録商標)(GlaxoSmithKline)、Afluria(CSL Biotherapies)、FluMist(登録商標)(Medlmmune)およびInfluvac(商標)(Solvay Pharma)が挙げられる。
【0129】
抗原−MaMVおよび抗原 MaMV−VLPの組合せ
上記のように、免疫原性組成物に含まれる1または2以上の抗原は、MaMVまたはMaMV VLPの外被タンパク質に接合することができ、または非接合形態で組成物中に存在してもよく(すなわち、MaMVまたはMaMV VLPと単純に混合されている)、または、接合および非接合形態の両方で存在してもよい。接合は、例えば、外被タンパク質との遺伝子融合、または共有結合、非共有結合もしくは親和性手段を介した結合であることができる。抗原(単数または複数)とMaMVまたはVLPとの組合せは、しかし、抗原の宿主免疫系による認識を、またはMaMVもしくはVLPの免疫反応を増強する能力を妨げてはならない。
【0130】
本発明の1つの態様により、免疫原性組成物に含まれる1または2以上の抗原は、MaMV VLPの外被タンパク質に接合される。VLPが自己集合外被タンパク質の複数のコピーを含むため、抗原の外被タンパク質への付着は、VLPの表面上での複数の抗原の提示を可能とする。
【0131】
VLPの表面上での抗原の提示を可能とし、抗原の免疫認識を強化するために、抗原は、VLPの外表面上に配置された外被タンパク質の領域に付着するのが好ましい。したがって、抗原は、外被タンパク質のアミノ−(N−)またはカルボキシ−(C−)末端の近傍またはその位置に挿入することができ、または、VLPの外表面上に配置された外被タンパク質の内部領域に付着することができる。抗原接合外被タンパク質の、他の融合外被タンパク質、または野生型外被タンパク質と集合して、VLPを形成する能力は保持されるべきであり、この能力は、本明細書に記載のものを含む既知の方法により容易に試験することができる。1つの態様において、抗原は、外被タンパク質のC末端に、またはこの近傍に付着する。
【0132】
本発明の1つの態様により、免疫原性組成物は、1または2以上の抗原に遺伝子的に融合したMaMV外被タンパク質を含む。抗原が、MaMV外被タンパク質−抗原融合体の、VLPに自己集合する能力を妨げる可能性を避けるために、MaMV外被タンパク質への遺伝子融合のために選択される抗原は、一般に、約50個以下のアミノ酸長であり、例えば約45個以下のアミノ酸長である。
【0133】
所望により、スペーサーを、抗原と外被タンパク質の間に含むことができる。この目的のために好適なスペーサーは当分野で知られており、例えば、約3〜約10個のアミノ酸の短いアミノ酸配列を含む。一般に、この状況でのアミノ酸スペーサーは、グリシン、ロイシン、バリンおよびイソロイシンなどの中性アミノ酸から構成される。1つの態様において、抗原がMaMV外被タンパク質に遺伝子的に融合されている場合、融合は、約3〜約10個の中性アミノ酸のペプチドスペーサーを含む。
【0134】
より長い抗原も、MaMVまたはVLPと単純に混合することにより、または化学的架橋または親和性付着により、免疫原性組成物中に組み込むことができ、これは以下にさらに詳細に記載する。
【0135】
抗原(単数または複数)は、例えば、共有結合または非共有結合(例えばイオン性、疎水性、水素結合など)による付着により、化学的に外被タンパク質に架橋することができる。抗原および/または外被タンパク質は、当分野に知られているようにして、かかる架橋を促進するように修飾することができ、これは例えば、官能基または化学部分をタンパク質および/または抗原に、例えばCもしくはN末端または内部の位置において添加することによる。例示の修飾には、S−アセチルメルカプトコハク酸無水物(SAMSA)またはS−アセチルチオアセテート(SATA)などの官能基の添加、または1もしくは2以上のシステイン残基の添加を含む。他の架橋試薬は当分野に知られており、多くは市販されている(例えば、Pierce Chemical Co.およびigma-Aldrichからのカタログ参照)。例としては以下を含むが、これらに限定されない:ジアミン類、例えば1,6−ジアミノヘキサン、1,3−ジアミノプロパンおよび1,3−ジアミノエタン;ジアルデヒド類、例えばグルタールアルデヒド;スクシンイミドエステル類、例えばエチレングリコール−ビス(コハク酸N−ヒドロキシスクシンイミドエステル)、ジスクシンイミジルグルタレート、ジスクシンイミジルスベラート、N−(g−マレイミドブチリルオキシ)スルホスクシンイミドエステルおよびエチレングリコール−ビス(スクシンイミジルスクシネート);ジイソシアネート類、例えばヘキサメチレンジイソシアネート;ビスオキシラン類、例えば1,4−ブタンジイルジグリシジルエーテル;ジカルボン酸類、例えばジサリチル酸サクシニル;3−マレイミドプロピオン酸N−ヒドロキシスクシンイミドエステル、その他。上述の架橋剤の多くは、親和性部分をVLPから離すためのスペーサーを組み込む。他のスペーサーの使用もまた本発明により意図される。種々のスペーサーが当分野に知られており、限定することなく、6−アミノヘキサン酸;1,3−ジアミノプロパン;1,3−ジアミノエタン;およびポリグリシン配列などの、1〜5アミノ酸の短いアミノ酸配列を含む。
【0136】
1または2以上の抗原の、VLPの外被タンパク質への共有結合的付着を促進するために、外被タンパク質は、VLPの表面に露出され、抗原の化学的付着に対する適切な部位を提供する、短ペプチドまたはアミノ酸リンカーに遺伝子的に融合することができる。例えば、システイン残基を含む短ペプチド、または共有結合を形成可能な側鎖、または当分野で知られているようにして共有結合を形成するように容易に修飾できる側鎖を有するアミノ酸残基(例えば、酸性および塩基性残基)である。アミノ酸リンカーまたはペプチドは、例えば、1〜約20のアミノ酸長であることができる。1つの態様において、外被タンパク質は、1または2以上のレジン残基を含む短ペプチドに融合し、これは例えば抗原のシステイン残基と、上記の好適な架橋剤の使用を通して共有結合することができる。
【0137】
本発明のさらなる態様においては、抗原は、外被タンパク質上に存在する親和性部分を介して付着する。この態様によれば、MaMV VLPは、自己集合の後にVLPの表面上に露出され、抗原に特異的に結合することができる、ペプチドなどの親和性部分を含む。親和性部分は、MaMVまたはVLPに、遺伝子的に融合するか(ペプチドまたはタンパク質断片の場合)、または共有結合もしくは非共有結合的に付着してよい。抗原の、親和性部分への結合は、宿主免疫系による抗原の認識を妨げてはならない。親和性部分は、全タンパク質を結合でき、またはタンパク質断片もしくはペプチドを結合できる。
【0138】
好適な親和性部分の例としては、抗体および抗体断片(例えばFab断片、Fab’断片、Fab’−SH断片、F(ab’)2断片、Fv断片、二機能性抗体(diabody)および単鎖Fv(scFv)分子)、ストレプトアビジン(ビオチン標識抗原を結合する)、抗原に特異的に結合する親和性ペプチドまたはタンパク質断片が挙げられるが、これに限定されない。
【0139】
親和性部分として用いるのに好適なペプチドまたは抗体(抗体断片を含む)は、当分野で知られている技法、例えばファージまたは酵母提示技術により、選択することができる。ペプチドは、天然、組換え、合成またはこれらの組合せであってよい。例えば、ペプチドは天然のタンパク質またはポリペプチドの断片であってよい。ペプチドの用語はまた、ペプチドアナログ、ペプチド誘導体およびペプチド模倣化合物も包含する。かかる化合物は当分野に知られており、天然のペプチドに対して利点を有する可能性があり、これには例えば、より大きな化学的安定性、タンパク質分解に対する抵抗性の増加、および/または抗原性の低下などを含む。
【0140】
親和性部分として用いるのに好適なペプチドは、約3アミノ酸長〜約50アミノ酸長であることができる。本発明の1つの態様により、親和性結合ペプチドは、少なくとも5アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、少なくとも7アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約5〜約50アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約7〜約50アミノ酸長である。本発明の他の態様により、親和性結合ペプチドは、約5〜約45アミノ酸長、約5〜約40アミノ酸長、約5〜約35アミノ酸長、および約5〜約30アミノ酸長である。本発明の特定の態様により、親和性結合ペプチドは、3、4、5、6、7、8、9、10、11、12、13、14または15アミノ酸長である。当業者により理解されるように、抗原を親和性部分に結合するために選択されるペプチドの長さは、MaMV VLPの自己集合の能力、または宿主の免疫系による、一度結合された抗原の認識の能力を妨げてはならない。
【0141】
MaMVまたはVLPが含む親和性部分は、単一ペプチドまたはペプチドの縦列もしくは複数配置であってよい。所望の場合には、大きな抗原の結合を促進するために、スペーサーを、親和性部分と外被タンパク質の間に含むこともできる。好適なスペーサーは、例えばグリシンなどの中性アミノ酸の短い範囲を含む。例えば、約3〜約10個の中性アミノ酸の範囲である。
【0142】
ファージ提示を用いて、目的の抗原タンパク質に結合する特定のペプチドを選択することができ、ここで標準の技法(例えば、Current Protocols in Immunology, ed. Coligan et al., J. Wiley & Sons, New York, NY参照)、および/または市販のファージ提示キット(例えば、New England Biolabsから入手可能なPh.Dシリーズのキット、およびNovagenより入手可能なT7-Select(登録商標)キット)を用いて行う。ファージ提示によるペプチドの選択の例は、以下の例8にも提供される。
【0143】
MaMVおよびMaMV VLPの調製
本発明は、組換えMaMV外被タンパク質由来のMaMV VLPを、ならびにMaMVまたはMaMV VLPを含む免疫原性組成物を提供する。本発明はさらに、1または2以上の抗原または親和性部分を、外被タンパク質との遺伝子融合で含む、MaMV VLPを提供する。これらの組換え外被タンパク質は、多量体化およびVLPへの集合が可能である。抗原または親和性ペプチドを、抗原へ、また外被たんぱく質への結合のために遺伝子的に融合する方法は当分野に知られており、代表的な例が以下および実施例に記載される。種々の分子をタンパク質に化学的に架橋する方法も当分野でよく知られており、用いることができる。
【0144】
Malvaモザイクウイルス
本明細書の例の節に記載されているように、MaMVはMalva neglecta Wallr(一般のアオイ科植物)から単離することができる。ウイルスは、容易にM. neglectaに、またはアカザ科、例えば例1に記すようにChenopodiaceae quinoaからの植物種に繁殖させることができる。
【0145】
ウイルスは、簡潔に述べると、Chenopodiaceae quinoa(4枚葉ステージ)の健康な葉を、例えば約40μLなどの適切な容積の担体中の約10〜100μgの精製ウイルスでこすることにより、容易に繁殖させることができる。あるいは、乳鉢中で水中の感染した葉(例えば、2mlの水中に1gの感染した葉)をひいて得た液を用いて、健康な植物に接種することができる。カーボランダム(研磨剤)をそっと、ウイルスまたは液でこする前の健康な葉の表面に落とす。感染は、指でそっとこすって微小な損傷部を作ることで開始され、これを通してウイルスは植物細胞中に浸入する。こすった約10分後、接種した葉を水で洗浄して、カーボランダムおよび任意の残留物を取り除く。接種した植物は、感染がモザイクまたは局所病変部の症状を示すまで、さらにおよそ5〜14日間成長させる(光および光周期の環境条件に依存する)。通常植物は、16時間の明期および8時間の暗期で、明期間は22℃で、および暗期間には16℃で成長させる。植物は、製造業者のプロトコルに従い20−20−20の肥料を用いて施肥する。一般に100〜1000gの感染植物の葉が、ウイルス精製のために必要である。
【0146】
ウイルスは、感染した葉から、標準のポテックスウイルス単離技術(例えばAbouHaidar MG, et al. (1998) Methods MoI Cell Biol 81 : 131-143およびTremblay, M.-H., et al., (2006). FEBS J 273: 14-25を参照)により単離することができる。好適な方法のさらなる例は以下である。感染した葉を収穫し、好適な緩衝液中に均質化し、続いてフィルタリングおよび/または遠心分離して砕片を取り除く。得られた懸濁液をブタノールおよび任意に洗浄剤で処理し、次に氷上で撹拌する。溶液を次に遠心分離し、得られたペレットを適切な緩衝液に再懸濁させる。ペレットを均質化し、次に再度遠心分離する。ビリオンを上清から、スクロースクッション上の超遠心分離によりペレット化し、得られたペレットを適切な緩衝液中にホモジナイザーを用いて再懸濁させる。ウイルス溶液は、所望により、0.45μMシリンジフィルターを通す前の追加の遠心分離ステップにより、清浄にすることができる。
【0147】
精製したビリオンは、例えば4℃にて冷蔵保存することができる。
【0148】
MaMV VLP
本発明のVLPの調製に好適な組換えMaMV外被タンパク質は、標準の遺伝子操作技術を用いて、野生型外被タンパク質の配列を提供された当業者により、容易に調製可能である。タンパク質の遺伝子操作の方法は当分野によく知られている(例えば、Ausubel et al. (1994 & updates) Current Protocols in Molecular Biology, John Wiley & Sons, New Yorkを参照)。野生型MaMV外被タンパク質のアミノ酸配列(配列番号2)および野生型タンパク質をコードするヌクレオチド配列(配列番号3)は本明細に提供されており、これらはまた、上記したようにGenBankから公的に入手可能である。
【0149】
野生型タンパク質をコードする核酸配列の単離およびクローニングは、標準の技法を用いて(例えば、Ausubel et al.、同上を参照)、例えばMaMVから標準の技法でRNAを抽出し、次にRNAテンプレートからcDNAを合成する(例えば、RT−PCRにより)ことにより、実施することができる。MaMVは、モザイク症状を示す感染植物の葉から、上記した標準の技法により(本明細書に記載の例1も参照)精製することができる。
【0150】
代替的に、外被タンパク質をコードする遺伝子を、標準の技法を用いて人工的に構成することができる。例えば、いくつか(例えば18〜20個)のオーバーラップしたリン酸化オリゴヌクレオチド(約80ヌクレオチド長)で、遺伝子配列全体を提示するものを、標準技法を用いて合成可能である。各ヌクレオチドは、それらの5’末端または3’末端において、例えば約20個のヌクレオチドがオーバーラップしていなければならず、ただし、3’末端においてのみオーバーラップする最終の5’オリゴと、5’末端においてのみオーバーラップする最終の3’オリゴは除く。オリゴヌクレオチドは適切な緩衝液内(例えば10mMのトリス/HCl、pH8および25mMのNaCl)にプールし、90℃に15分間加熱し、室温にゆっくり冷却して、オリゴヌクレオチド間のアニーリングおよび、全長MaMV CP遺伝子の生成を可能とすることができる。リガーゼ緩衝液へのT4 DNAリガーゼの添加は、オリゴヌクレオチドの集合を完了させる。任意に、MaMV CP遺伝子の5’および3’末端を含むオリゴヌクレオチドはユニークな制限部位を含むことができ、これは、アニールされたDNAを細菌プラスミドなどの適切なベクターにクローニングするのに用いることができる。このベクターは、プラスミド大腸菌などの適切な宿主細胞を形質転換して、プラスミドを増幅し、オリゴヌクレオチドのライゲーションを完了させるために用いることができる。全てのオリゴヌクレオチドのアニーリングは、これらを2つずつ順番にアニーリングし、続いてオリゴヌクレオチドの各ペアを2つずつアニーリングし、これを続けて、全長遺伝子が生成されるまで行うことにより、改善することができる。
【0151】
代替的に、全長MaMV CP遺伝子は、適切なプラスミド(例としてはpET−3D)にクローニングして大腸菌内(例えばBL21(DE3))でタンパク質を発現させる前に、ポリメラーゼ連鎖反応(PCR)により増幅することができる。
【0152】
全長遺伝子配列はまた、合成遺伝子を作製する多くの営利会社の1つによるサービスを通して得ることもできる(例えば、GenScript Corp.(Piscataway, NY)、Geneart AG(Regensberg, Bavaria)およびMolecular Cloning Laboratories(San Francisco, CA))。
【0153】
外被タンパク質をコードする核酸配列は次に、好適な発現ベクターに、直接挿入するか、または1もしくは2以上のサブクローニングステップの後に挿入する。当業者は、用いる正確なベクターは本発明に決定的に重要ではないことを理解する。好適なベクターとしては、プラスミド、ファージミド、コスミド、バクテリオファージ、バキュロウイルス、レトロウイルス、またはDNAウイルスが挙げられるが、これらに限定されない。外被タンパク質は次に、以下にさらに詳細に記載されるようにして、発現および精製することができる。MaMV外被タンパク質を含むベクター(プラスミドpMaMV−CP−6H(MaMV外被タンパク質を含むpET−3D))の例は、例6に記載される。プラスミドpMaMV−CP−6Hに含まれているMaMV外被タンパク質遺伝子のヌクレオチド配列は、図10A(配列番号23)に、およびコードされた外被タンパク質のアミノ酸配列は、図11A(配列番号24)に提供される。
【0154】
任意に、外被タンパク質をコードする核酸配列をさらに操作して、当分野で知られている標準のin vitro部位特異的突然変異誘発技術により、上記のものなどのように1または2以上の変異を導入することができる。突然変異は、コード配列を作っている1または2以上のヌクレオチドの、欠失、挿入、置換、逆位、またはこれらの組合せにより、導入可能である。これは、例えば、1または2以上のヌクレオチドミスマッチ、挿入または欠失を組み込むプライマーが設計されたPCRに基づく技法により、実現することができる。突然変異の存在は、多くの標準技法、例えば、制限解析またはDNA配列決定により確認可能である。
【0155】
上述のように、外被タンパク質はまた、外被タンパク質に融合した1または2以上の抗原、親和性ペプチドおよび/またはスペーサーペプチドを含む、融合タンパク質を産生するように、操作することもできる。融合タンパク質を作製するための方法は、当業者によく知られている。融合タンパク質をコードするDNA配列は、上記のように、好適な発現ベクターに挿入することができる。
【0156】
当業者は、外被タンパク質または融合タンパク質をコードするDNAを、コードされたタンパク質の活性に影響を与えることなく、種々の方法で変更できることを理解する。例えば、DNA配列の変化を用いて、タンパク質を発現するために用いる宿主細胞のコドンの選好を最適化することができ、またはDNA配列の変化は、発現を促進する他の配列の変化を含んでもよい。
【0157】
当業者は、発現ベクターは、外被もしくは融合タンパク質をコードするDNA配列の効率的な転写に必要な調節要素、例えば転写要素などをさらに含んでよいことを理解する。ベクターに組み込むことができる調節要素の例は、プロモーター、エンハンサー、ターミネーター、およびポリアデニル化シグナルを含むが、これに限定しない。したがって本発明は、外被タンパク質または融合タンパク質をコードする核酸配列に動作可能に結合された調節要素を含む、ベクターを提供する。当業者は、好適な調節要素の選択は、タンパク質の発現について選択された宿主細胞に依存し、かかる調節要素は、細菌、真菌、ウイルス、哺乳類または昆虫の遺伝子を含む種々の源に由来し得ることを理解する。
【0158】
必要に応じて、発現ベクターは、当分野に知られているような、発現されたタンパク質の精製を促進する異種(非相同)核酸配列をさらに含んでよい。かかる異種核酸配列の例は、金属親和性タグなどの親和性タグ、ヒスチジンタグ、アビジン/ストレプトアビジンコード配列、グルタチオン−S−トランスフェラーゼ(GST)コード配列およびビオチンコード配列を含むが、これに限定しない。得られた異種アミノ酸配列は、当分野に知られている方法に従って用いる前に、発現されたタンパク質から除去することができる。代替的に、異種アミノ酸配列は、続くVLPへのタンパク質の集合を妨害しない場合には、タンパク質上に保持することもできる。本発明の1つの態様において、外被タンパク質はヒスチジンタグ・タンパク質として発現される。異種アミノ酸配列は、外被タンパク質のカルボキシル末端またはアミノ末端に位置させることができる。
【0159】
発現ベクターは、好適な宿主細胞または組織内に、当分野で知られている種々の方法の1つを用いて、導入することができる。かかる方法は、Ausubel et al.(同上)に一般的に記載されており、例えば、安定または過渡的なトランスフェクション、リポフェクション、エレクトロポレーション、および組換えウイルスベクターによる感染を含む。当業者は、外被タンパク質の発現のための適切な宿主細胞の選択は、選択したベクターに依存することを理解する。宿主細胞の例は、細菌、酵母、昆虫、植物および哺乳類細胞を含むが、これに限定しない。用いる正確な宿主細胞は、本発明に重要ではない。外被タンパク質は、原核生物宿主(例えば、大腸菌、A. salmonicidaまたはB. subtilis)または真核生物宿主(例えば、SaccharomycesまたはPichia;哺乳類細胞、例えばCOS、NIH3T3、CHO、BHK、293、またはHeLa細胞;または昆虫細胞)において産生することができる。本発明の1つの態様において、組換え外被タンパク質は、植物または細菌細胞中に発現させることができる。他の態様において、組換え外被タンパク質は、細菌細胞中に発現させる。他の態様において、組換え外被タンパク質は、大腸菌細胞中に発現させる。
【0160】
必要に応じて、発現されたタンパク質は宿主細胞から、当分野で知られている標準の技法により(例えば、Current Protocols in Protein Science, ed. Coligan, J.E., et al., Wiley & Sons, New York, NYを参照)精製でき、標準のペプチド配列決定技法により、無傷のタンパク質またはそのタンパク質分解断片を用いて配列決定して、タンパク質の同一性を確認することができる。
【0161】
本発明の1つの態様により、組換え外被タンパク質は、多量体化およびVLPへの集合が可能である。一般に、VLP集合は、外被タンパク質を発現する宿主細胞内で起こる。VLPは宿主細胞から、本明細書に記載の実施例の節に記載されているような標準の技法により単離可能である。VLPは、クトマトグラフィなどの標準の技法によりさらに精製して、汚染宿主細胞タンパク質またはLPSなどの他の化合物を除去することができる。
【0162】
本発明の1つの態様において、外被タンパク質は集合して宿主細胞内にウイルスまたは偽ウイルスを提供し、これは核酸およびタンパク質を含む感染ウイルス粒子の産生に用いることができる。これにより、感染ウイルスまたは偽ウイルス粒子による隣接する細胞の感染、およびその中のタンパク質の発現が可能である。この態様において、ウイルスまたは偽ウイルスを複製するために用いる宿主細胞は、ウイルスの複製を可能とする、植物細胞、昆虫細胞、哺乳類細胞または細菌細胞であることができる。これらの細胞は、ウイルスに対する天然の宿主細胞であって、これからウイルス様粒子が導出されるものであってよいが、しかし必ずしもそうである必要はない。宿主細胞は、初めに粒子形態のウイルスまたは偽ウイルス(すなわち、核酸およびタンパク質を含む集合したロッド)により感染されるか、または代替的に、初期感染に用いるウイルス核酸が複製可能であり、全ウイルス粒子の産生をもたらす場合には、核酸形態において(すなわち、ウイルスRNAなどのRNA;cDNAまたはcDNAから調製されるランオフ転写物)感染されることができる。
【0163】
MaMVまたはMaMV VLPのストックの産生
組換えMaMVまたはVLPのストックを、標準技法により調製する。例えば、組換え外被タンパク質を含むMaMVまたは偽ウイルスを、例えばChenopodium quinoaなどの適切な宿主内に、十分なMaMVまたは偽ウイルスが収穫できるように、繁殖させることができる。
【0164】
MaMV VLPのストックは、適切は宿主細胞から、例えば、上述のようにして調製された、VLPを作る組換え外被タンパク質をコードする発現ベクターで形質転換またはトランスフェクトされた大腸菌などから、調製することができる。宿主細胞は次に、当分野で知られているように、コードされたタンパク質の発現に有利な条件下で培養される。発現された外被タンパク質は多量体化して宿主細胞のVLPに集合することができ、これは上述のような標準の技法で、例えば細胞を破壊して、細胞可溶化物を1または2以上のクロマトグラフィによる精製ステップに供することにより、細胞から単離することができる。
【0165】
MaMVおよびMaMV VLPのストックは、例えば4℃にて冷蔵庫内で保存することができる。
【0166】
組換えおよび改変外被タンパク質の特徴
組換え外被タンパク質および、抗原、親和性ペプチドおよび/またはスペーサーペプチドが結合した外被タンパク質を、それらの多量体化およびVLPへ自己集合する能力について、標準の技法により解析することができる。これは例えば、精製タンパク質を電子顕微鏡法により視覚化することによる(例えば、例6を参照)。VLPの形成はまた、超遠心分離法により決定してもよく、円偏光二色性(CD)分光法を用いて、組換えもしくは改変タンパク質とWTウイルスの二次構造を比較することができる(例えば、Tremblay, et al, FEBSJ, 2006, 273: 14-25参照)。
【0167】
VLPおよびMaMVの安定性を、必要に応じて、当分野で知られている技法、例えばSDS−PAGEおよびトリプシン分解分析(例えば、Tremblay, et al, 2006、上記)により、決定することができる。
【0168】
有効性の評価
本発明の免疫原性組成物の、動物において免疫反応を誘発する能力を、当分野に知られた方法、例えば以下および例に記載されたものなどにより、試験することができる。例えば、MaMV、MaMV VLPまたはこれを含む免疫原性組成物を、好適な動物モデルに、例えば皮下注射により、または鼻腔内に投与することができ、抗体の発生を評価する。
【0169】
細胞性免疫反応はまた、当分野で知られている技法により評価することができる。例えば、細胞性免疫反応は、MaMV VLPに発現されたエピトープの、in vitroおよびin vivoでの樹状細胞による特異的Tリンパ球へのプロセシングおよび交差提示(cross presentation)を評価することにより、決定することができる。細胞性免疫(Tリンパ球)の誘発を評価する他の有用な技法としては、T細胞増殖およびIFN−γ分泌放出のモニタリング、例えばELISAによりサイトカインの誘発をモニタリングすること(例えば、Leclerc, D., et al, J. Virol, 2007, 81(3):1319-26参照)などを含む。
【0170】
MaMVまたはMaMV VLPおよび1もしくは2以上の抗原を含む免疫原性組成物の、ワクチンとしての有効性を評価するために、チャレンジ試験を実施することができる。かかる試験は、試験動物(例えばマウス、ラットまたはフェレット)の群に、本発明の免疫原性組成物を標準技法により接種することを含む。非接種動物および/または市販のワクチンを接種した動物を含む対照群、または他の陽性対照は、平行して設定する。ワクチン接種後の適切な時間の後、動物を、天然の抗原含有物質または有機体によりチャレンジする。動物から、接種前および接種後、およびチャレンジ後に採取した血液試料を、ウイルスに対する抗体反応について分析する。抗体反応についての好適な試験は、ウェスタンブロット解析および酵素結合免疫吸着法(ELISA)などを含むがこれに限定されない。動物はまた、抗原含有物質または有機体に関連する状態の症状の発生についてもモニタリングすることができる。
【0171】
同様に、腫瘍関連抗原を含有する免疫原性組成物の、それらの予防効果についての試験は、試験動物に接種し、続いて、癌細胞を動物に例えば皮下的に移植することによるチャレンジ、および動物における腫瘍の発達のモニタリングにより、行うことができる。代替的に、免疫原性組成物の治療効果を、癌細胞の移植および腫瘍の確立後に組成物を試験動物に投与し、腫瘍の増殖および/または転移のモニタリングにより、試験することができる。
【0172】
医薬組成物
上述のように、MaMVまたはMaMV VLPおよび任意に1もしくは2以上の抗原を含む免疫原性組成物は、さらに任意に、好適な担体、賦形剤など、および/または組成物の安定性、味のよさ、薬物動態、バイオアベイラビリティ等を改善する医薬組成物のその他の標準成分を含んでよい。本発明の1つの態様において、免疫原性組成物は、アジュバントとして用いるために製剤化される。他の態様において、免疫原性組成物は、ワクチンとして用いるために製剤化される。
【0173】
組成物は、種々の経路による投与のために製剤化可能である。例えば、組成物は、経口、局所、直腸、鼻腔、または非経口投与用に製剤化でき、または吸入もしくはスプレーによる投与用に製剤化できる。本明細書において用語、非経口的とは、皮下注射、静脈内、筋肉内、髄腔内、胸骨内注射または注入技法を含む。対象への鼻腔内投与は、医薬組成物を、対象の鼻道または鼻腔の粘膜に投与することを含む。本発明の1つの態様において、組成物は局所、直腸もしくは非経口投与用に、または吸入もしくはスプレーによる、例えば鼻腔内経路による投与用に製剤化される。他の態様において、組成物は非経口投与用に製剤化される。
【0174】
組成物は好ましくは、本発明の免疫原性組成物の有効量を含む。本明細書において用語「有効量」とは、検出可能な免疫反応を生成するのに必要な組成物の量をいう。与えられた適用に対しての免疫原性組成物の有効量は最初に、例えば細胞培養アッセイまたは動物モデルにおいて、通常はげっ歯類、ウサギ、イヌ、ブタ、または霊長類において、推定可能である。動物モデルはまた、好適な濃度範囲および投与経路を決定するために用いてもよい。かかる情報は次に、ヒトを含む処置すべき動物における、有用な用量および投与経路の決定に用いることができる。本発明の1つの態様において、単位用量は、約5μg〜約10mgの外被タンパク質を含む。他の態様において、単位用量は、約10μg〜約10mgの外被タンパク質を含む。さらに他の態様において、単位用量は、約5μg〜約5mgの外被タンパク質を含む。他の態様において、単位用量は、約10μg〜約5mgの外被タンパク質を、約10μg〜約2mgの外被タンパク質を、約15μg〜約5mgの外被タンパク質を、および約20μg〜約5mgの外被タンパク質を含む。1また2以上の用量を用いて動物を免疫化することができ、これらは、同じ日に投与することも、または数日もしくは数週間の期間にわたって投与することもできる。
【0175】
上述のように、本発明の免疫原性組成物は複数の抗原(接合および/または非接合形態)を含んでよく、したがって多価ワクチン製剤を提供することができる。複数のVLPを含む多価ワクチン組成物であって、各VLPが異なる抗原に接合しているものもまた、意図される。多価ワクチン製剤は、2価および3価の製剤を、より高い価のワクチンに加えて含む。
【0176】
一定の態様において、複数の(すなわち、2または3以上の)異なる抗原を含む多価ワクチン製剤も、製剤中のより多くの数のエピトープにより、改善された防御を提供することができる。抗原は、接合形態(例えば、各々が異なる抗原に接合した多数の異なるVLPにより)、または非接合形態において、または接合および非接合形態の両方において、含むことができる。
【0177】
経口使用のための組成物は、例えば、錠剤、トローチ剤、ロゼンジ、水性もしくは油性の懸濁液、分散粉末または顆粒、乳剤、硬質もしくは軟質カプセル、またはシロップまたはエリキシル剤として、製剤化可能である。かかる組成物は、当分野で知られている医薬組成物の製造の標準的方法により製造可能であり、甘味料、香味剤、着色剤および保存剤の群から選択される1または2以上の剤を、薬学的に洗練された味のよい調製物を提供するために、含んでよい。錠剤は、免疫原性組成物を、好適な無毒性の薬学的に許容し得る賦形剤、例えば以下に記載の賦形剤と組み合わせて含む:不活性の希釈剤、例えば炭酸カルシウム、炭酸ナトリウム、ラクトース、リン酸カルシウム、またはリン酸ナトリウム;顆粒化または崩壊剤、例えばコーンスターチ、またはアルギン酸;結合剤、例えばスターチ、ゼラチンまたはアカシア、および湿潤剤、例えばステアリン酸マグネシウム、ステアリン酸またはタルク。錠剤は被覆されていなくてもよく、または、消化管での分解および吸収を遅延させ、これにより長期の持続作用を提供するために、既知の技術により被覆されてもよい。例えば、モノステアリン酸グリセリルまたはジステアリン酸グリセリルなどの時間遅延材料を用いてもよい。
【0178】
経口使用のための組成物はまた、免疫原性組成物が不活性の固体希釈剤、例えば炭酸カルシウム、リン酸カルシウムまたはカオリンと混合される、硬質ゼラチンカプセルとして提供することもでき、あるいは、活性成分が水またはピーナツ油、液体パラフィンまたはオリーブ油などの油性媒体と混合される、軟質ゼラチンカプセルとしても提供可能である。
【0179】
鼻腔内投与用の医薬組成物は、例えば、鼻腔用スプレー、鼻腔用のドロップ、懸濁液、溶液、ゲル、軟膏、クリーム、および粉末を含むことができる。組成物は、例えばAccuspray(登録商標)(Becton Dickinson)などの好適な市販の鼻腔用噴霧器を介する投与用に製剤化することができる。鼻腔内投与他の方法は、当分野で知られている。
【0180】
水性懸濁液として製剤化される組成物は、免疫原性組成物を1または2以上の好適な賦形剤と組み合わせて含み、これは例えば、懸濁剤であって例えばカルボキシメチルセルロースナトリウム、メチルセルロース、ヒドロプロピルメチルセルロース、アルギン酸ナトリウム、ポリビニルピロリドン、ヒドロキシプロピル−β−シクロデキストリン、トラガカント・ゴムおよびアカシア・ゴム;分散剤または湿潤剤、例えば天然のホスファチド、例えばレシチン、または酸化アルキレンと脂肪酸との縮合生成物、例えばステアリン酸ポリオキシエチレン、または酸化エチレンと長鎖脂肪族アルコールとの縮合生成物、例えばヘプタ−デカエチレンオキシセタノール、または酸化エチレンと脂肪酸/ヘキシトール由来の部分エステルとの縮合生成物、例えばポリエチレンソルビトールモノオレエート、または酸化エチレンと脂肪酸/ヘキシトール無水物由来の部分エステルとの縮合生成物、例えばポリエチレンソルビタンモノオレエートなどである。水性懸濁液はまた、1もしくは2以上の保存剤、例えばエチル、またはn−プロピルp−ヒドロキシベンゾエート、1もしくは2以上の着色剤、1もしくは2以上の香味剤または1もしくは2以上の甘味料、例えばスクロースもしくはサッカリンを含んでよい。
【0181】
組成物は、油性懸濁液として、免疫原性組成物を例えばラッカセイ油、オリーブ油、ゴマ油またはココナツ油などの植物油中に、または液体パラフィンなどの鉱油中に懸濁させることにより、製剤化することもできる。油性懸濁液は、増粘剤、例えば蜜ろう、硬質パラフィンまたはセチルアルコールなどを含んでよい。上記などの甘味料、および/または香味剤を任意に加えて、口に合う経口製剤を提供することができる。これらの組成物は、アスコルビン酸などの抗酸化剤の添加により、保存することができる。
【0182】
組成物は、分散性の粉末または顆粒として製剤化でき、これは次に、水を添加して水性懸濁液の調製に用いることができる。かかる分散性の粉末または顆粒は、免疫原性組成物を、1または2以上の分散剤または湿潤剤、懸濁剤および/または保存剤と組み合わせて提供する。好適な分散剤または湿潤剤、および懸濁剤は、上記で既に例示されている。追加の賦計剤、例えば、甘味料、香味剤および着色剤もまた、これらの組成物に含むことができる。
【0183】
本発明の組成物はまた、水中油型乳剤としても製剤化可能である。油相は、オリーブ油もしくはラッカセイ油などの植物油、または液体パラフィンなどの鉱油、またはこれらの油の混合物でもよい。これらの組成物に含めるのに好適な乳化剤は、天然のゴム、例えばアカシアゴムもしくはトラガカントゴム;天然のホスファチド、例えば大豆、レシチン;または脂肪酸とヘキシトール由来のエステルもしくは部分エステル、無水物例えばモノオレイン酸ソルビタン、および前記部分エステルと酸化エチレンとの縮合生成物、例えばポリオキシエチレンソルビタンモノオレエートを含む。乳剤はまた、任意に甘味料および香味剤を含むことができる。
【0184】
組成物は、シロップまたはエリキシル剤として、免疫原性組成物を1または2以上の甘味料、例えばグリセロール、プロピレングリコール、ソルビトールまたはスクロースなどと混合することにより製剤化することができる。かかる製剤はまた、1または2以上の粘滑剤、保存剤、香味剤および/または着色剤を任意に含むことができる。
【0185】
組成物は、無菌の注射用水性または油性懸濁液として、当分野で知られている方法により、上述のものなどの、好適な1または2以上の分散剤または湿潤剤および/または懸濁剤を用いて、製剤化することができる。無菌の注射用調製物は、無毒性の非経口的に許容し得る希釈剤または溶媒中の、無菌の注射用溶液または懸濁液であってよく、例えば1,3−ブタンジオール中の溶液である。用いることのできる、許容し得るビヒクルおよび溶媒は、水、リンガー溶液、乳酸化リンガー溶液および等張食塩水を含むが、これに限定されない。他の例には、従来から溶媒または懸濁媒体として用いられている無菌の固定油、および種々の無刺激性の固定油であって、例えば合成モノ−およびジグリセリドなどを含む。オレイン酸などの脂肪酸もまた、注射物の調製に用いることができる。
【0186】
任意に、本発明の組成物は、保存剤例えば抗菌剤、抗酸化剤、キレート剤および不活性ガスなど、および/または安定化剤例えば炭水化物(例えばソルビトール、マンニトール、スターチ、スクロース、グルコース、またはデキストラン)、タンパク質(例えばアルブミンまたはカゼイン)、またはタンパク質含有剤(例えばウシ血清アルブミンまたはスキムミルク)を、好適な緩衝液(例えばリン酸緩衝液)と共に含んでよい。組成物のpHおよび種々の成分の正確な濃度は、周知のパラメータにしたがって調節してよい。
【0187】
さらに、アジュバント活性を有する1または2以上の化合物も、ワクチン組成物に任意に加えてよい。好適なアジュバントは、例えば、水酸化、リン酸化、または酸化アルミニウム;油型乳剤(例えばBayol F(登録商標)またはMarcol52(登録商標));サポニン、またはビタミンE可溶化物を含む。オプソニン化ワクチン組成物も本発明に包含され、例えば、ワクチンにより前に免疫化した動物またはヒトから単離された抗体を含む、ワクチン組成物である。ワクチンにより前に免疫化した動物またはヒトから単離された抗体に基づく組換え抗体も、ワクチン組成物をオプソニン化するために用いることができる。
【0188】
他の医薬組成物および医薬組成物を調製するための方法は、当分野で知られており、例えばRemington: The Science and Practice of Pharmacy”(旧タイトル:“Remingtons Pharmaceutical Sciences”);Gennaro, A., Lippincott, Williams & Wilkins, Philadelphia, PA (2000)に記載されている。
【0189】
さらに、1または2以上の従来のアジュバントも、組成物に任意に加えてよい。好適なアジュバントは、例えば、ミョウバンアジュバント(例えば水酸化、リン酸化、または酸化アルミニウム);油型乳剤(例えばBayol F(登録商標)またはMarcol52(登録商標));サポニン、またはビタミンE可溶化物を含む。ビロソーム(virosome)もアジュバント特性を有するとして知られており(Adjuvant and Antigen Delivery Properties of Virosomes, Glueck, R., et al., 2005, Current Drug Delivery, 2:395-400)、S. typhiポリンタンパク質(例えばOmpC)も同様にこれを有し、本発明の組成物に任意に含むことができる。
【0190】
さらに、本発明は、免疫原性組成物が抗原に融合したMaMV VLPを含む場合、該組成物はまた、組成物中のVLP全体のアジュバント効果を増加するために、非融合タンパク質に由来するMaMV VLPも含んでよいことを意図する。
【0191】
組成物は任意に、オプソニンを、例えば、抗原、MaMV、もしくはMaMV VLPで前に免疫化した動物またはヒトから単離された抗体を、含んでもよい。抗原、MaMV、もしくはMaMV VLPで前に免疫化した動物またはヒトから単離された抗体に基づく組換え抗体もまた、オプソニンとして用いることができる。
【0192】
本発明にさらに包含されるのは、MaMVまたはMaMV VLPを、上記のような市販のワクチンと組み合わせて含む組成物である。
【0193】
免疫原性組成物の使用
本発明は、MaMV、MaMV VLPおよびこれを含む免疫原性組成物の、多くの使用を提供する。非限定的例としては、免疫原性組成物の、アジュバント、免疫促進剤として、またはワクチンとしての使用が挙げられる。1または2以上の抗原に接合したMaMV VLPもまた、抗原(単数または複数)に対する抗体のスクリーニングのために用いることができる。したがって、本発明は、免疫原性組成物を投与することにより、動物に免疫反応を誘発する方法、および免疫原性組成物の、例えばアジュバント、免疫促進剤、ワクチン、および/または医薬組成物などの医薬の調製のための使用も提供する。
【0194】
本発明の免疫原性組成物は、ヒトおよび、家畜用動物と農場用動物を含む非ヒト動物における使用に好適である。免疫原性組成物の投与形態は、任意の他の一般に認められたワクチン接種プログラムと違っている必要はない。例えば、効果的な免疫反応を引き出すのに十分な量での免疫原性組成物の単回投与を用いてよく、または代替的に、免疫原性組成物の初回投与に続いて、抗原のみで、または抗原と免疫原性組成物でのブースティングという他の形態を用いてもよい。同様に、免疫原性組成物または抗原のどちらかによるブースティングは、抗体価が許容レベルを下回った場合には、初回投与の後の時点で行ってもよい。免疫原性組成物の正確な投与方法は、例えば、組成物の成分(例えば、組成物が抗原を含むか、またはアジュバントとして提供されるのか)、処置する動物、および処置の所望の最終効果に依存する。適切な投与方法は、経験ある施術者により容易に決定することができる。
【0195】
免疫原性組成物が1または2以上の非接合抗原を含む場合、MaMVまたはVLP成分は、1または2以上の抗原と同時に投与することができ、または、免疫反応を必要とする対象の必要性に応じて、抗原の投与の前に、またはこれに続いて、投与することができる。
【0196】
免疫原性組成物は、予防的に、例えば、ウイルス、細菌もしくは他の感染粒子による感染を、または疾病もしくは腫瘍の発症を防ぐために用いることができ、または、これは、例えば感染症または癌に関連する疾病もしくは疾患の影響を改善するために治療的に用いてもよい。本発明の1つの態様において、免疫原性組成物は、予防的に用いる。この状況において、免疫原性組成物は、1または2以上の抗原を含むことができ、または免疫原性組成物は、MaMVもしくはMaMV VLPのみを含んでもよく、これは、低レベルの感染などの感染症または疾患に対する抵抗性を誘発するのに十分であり得る。
【0197】
免疫原性組成物は、組成物中の含有について、または組成物との使用について選択された抗原に依存して、種々の疾病または疾患の予防または処置において用いることができる。非限定的な例としては以下が挙げられる:インフルエンザ(種々のインフルエンザウイルスからの抗原を使用)、腸チフス(S. typhiからの抗原を使用)、HCV感染(HCV抗原を使用)、HBV感染(HBV抗原を使用)、HAV感染(HAV抗原を使用)、HIV感染(HIV抗原を使用)、ポリオ(ポリオウイルス抗原を使用)、ジフテリア(ジフテリア毒素由来の抗原を使用)、EBV感染(EBV抗原を使用)、アレルギー反応(種々のアレルゲンを使用)、および癌(種々の癌関連抗原を使用)が挙げられる。他の使用としては、例えば、炎症性疾患(例えば関節炎)、および鳥インフルエンザウイルス、ヒト呼吸器合胞体ウイルス、デング熱ウイルス、麻疹ウイルス、単純ヘルペスウイルス、ヒトパピローマウイルス、偽ウイルス、豚ロタウイルス、豚パルボウイルス、ニューカッスル病ウイルス、口蹄病ウイルス、豚コレラ(hog cholera)ウイルス、豚コレラ(African swine fever)ウイルス、感染性ウシ鼻気管炎ウイルス、感染性咽頭気管炎ウイルス、ラクロスウイルス、新生子牛下痢ウイルス、ウシ呼吸器合胞体ウイルス、ウシウイルス性下痢ウイルス、豚マイコプラズマ肺炎菌、連鎖球菌、淋菌、腸内細菌および寄生生物(例えばリーシュマニアおよびマラリア)が挙げられる。
【0198】
本発明の免疫原性組成物はまた、例えば、該組成物が異なる病原物質からの複数の抗原を含む場合、多価ワクチンとして用いるのに好適である。
【0199】
免疫原性組成物はまた、従来のワクチンと併せて用いて、ワクチンの有効性を改善し、または多価ワクチンを提供することができる。この状況において用いることのできる、市販のワクチンの非限定的例は上に提供されている。市販のワクチンは、ヒトワクチンまたは獣医学的使用を意図されたワクチンであってよい。
【0200】
1つの態様において、本発明はまた、以前にポテックスウイルスベースのワクチンを接種された対象に対するワクチン接種のための、免疫原性組成物の使用も提供する。このアプローチは、第2のワクチンの有効性が、第1のワクチンにおいてポテックスウイルスに対して増殖した抗体との相互作用のために、低下しないことを保証することができる。例えば、PapMVベースのワクチンプラットフォームが記載されている(例えば、国際特許出願PCT/CA03/00985 (WO 2004/004761)および米国特許出願第11/556,678号(US2007/0166322)を参照)。動物またはヒトの、PapMV VLPでアジュバント化したワクチンによる免疫化は、アジュバントの主要成分であるPapMV外被タンパク質に指向された多量の抗体を生成することができる。同じ患者を、同じPapMVプラットフォームで免疫化すると、常在の抗体の新しく投与されたワクチンに対する免疫反応を、PapMVプラットフォームに直接結合することにより制限する可能性がある。したがって、MaMVまたはMaMV VLPに基づく第2のワクチンの使用はこの効果を避けるのに有用であり、より効率的なワクチン接種プログラムの保証を支援する。
【0201】
本発明はまた、1または2以上の抗原に接合したMaMV VLPの、スクリーニング剤としての使用、例えば、1または2以上の抗原に対する抗体をスクリーニングするための使用も提供する。VLPは従来の免疫的技術、例えば酵素結合免疫吸着測定法(ELISA)またはウェスタンブロッティングなどに容易に適合でき、したがって診断および研究の文脈において有用である。
【0202】
キット
本発明はさらに、MaMV、MaMV VLP、または本発明の免疫原性組成物を含むキットの、アジュバントまたはワクチンとしての使用を提供する。MaMV、MaMV VLP、または免疫原性組成物が、市販のワクチンを含む、別の抗原製剤と共に用いることが意図される場合、キットは任意に、該抗原製剤を含むことができる。
【0203】
キットの個々の成分は別々の容器に入れられて、かかる容器に付随して、薬学的または生物学的製品の製造、使用または販売を規制している政府機関による所定の形態における、該機関による製造、使用または販売の承認を反映する注意書きがあってよい。キットは任意に、アジュバントまたはワクチンの使用または投与の方法を概説する使用説明書または指示書を含んでよい。
【0204】
キットの1または2以上の成分が溶液として、例えば水溶液、または無菌の水溶液として提供される場合、容器はそれ自体が、吸入器、シリンジ、ピペット、点眼容器、または他の類似の機器であってよく、これから溶液を対象に投与でき、またはキットの他の成分にこれを適用して混合することができる。
【0205】
キットの成分はまた、乾燥または凍結乾燥形態で提供してもよく、キットは追加して凍結乾燥成分の再構成のための好適な溶媒を含むことができる。容器の数または種類と無関係に、本発明のキットはまた、患者への組成物の投与を支援するための機器も含んでよい。かかる機器は、吸入器、シリンジ、ピペット、鉗子、計量スプーン、点眼容器、または類似の医学的に承認された送達ビヒクルであってよい。
【0206】
抗体検出に用いるための、1または2以上の抗原に接合したMaMV VLPを含むスクリーニングキットも提供される。キットは、診断用キットまたは研究目的のキットであることができる。キットの個別の成分は別々の容器に入れられ、かかる容器に付随して、生物学的製品の製造、使用または販売を規制している政府機関による所定の形態における、該機関による製造、使用または販売の承認を反映する注意書きがあってよい。キットは任意に、免疫原性組成物の使用方法を概説する使用説明書または指示書を含んでよい。
【0207】
本明細書に記載の発明のよりよい理解を得るために、以下の実施例を示す。これらの例は、本発明の例示の態様を記載することを意図し、どのようであれ本発明の範囲の限定を意図するものではないことが理解される。
【0208】
例
例1:Malvaモザイクウイルスの増幅、精製、およびRNA抽出
新しいポテックスウイルスであるMalvaモザイクウイルス(MaMV)を、Malva neglecta Wallr.(一般のアオイ科ゼニアオイ属)から単離した。最初に感染したMalva neglecta植物は、カナダ・ブリティッシュコロンビア州サマーランド周辺の地域で収集した。ウイルス感染によるモザイク症状および葉脈の透明化のみが、Malva neglectaにおいて観察された症状であった(図4A参照)。
【0209】
ウイルスによるChenopodium quinoaの感染は、植物全体を移動する強いモザイクパターンを生成した(図4B参照)。C. quinoaは、したがって、増殖の宿主として用い、これを用いてウイルス精製のための多量の感染葉を収集した。C. quinoaでの増殖のために接種物をC. quinoaの感染葉からとった際に、局所病変が認められた(図4C参照)。局所病変の誘発は、接種に用いる汁液中でのウイルス濃度に関連するようであり(感染したC. quinoaの葉にはより多くのウイルス)、これは、同一宿主上での症状の差を説明する。
【0210】
MaMVを次のようにして増幅および精製した。Malva neglectaの感染した葉を、サマーランドB.C.周辺の地域で収穫し、ケベックにある発明者らの研究室に送った。Malva neglectaの感染した葉を、液体窒素中乳鉢/乳棒で粉砕して、葉の粉末を得た。粉末は水中に再懸濁させて、増殖宿主のChenopodium quinoa上に、葉をカーボランダムでこすることにより接種した。葉は、接種2〜3週間後に感染症状を示した(すなわち、感染した葉の上の、モザイクおよび葉脈透明化)。感染した葉を、10mMのEDTAおよび0.1%の重炭酸ナトリウムを含有する100mMのリン酸ナトリウム緩衝液、pH7.6に均質化した。ホモジェネートを、チーズクロス(粗い綿布)の層を通してフィルタリングし、7,800gで20分間遠心分離した。TritonX-100および2%のブタノールを1滴ずつ上清に加え、氷上で60分間撹拌した。溶液を7,800gで20分間遠心分離した。次に上清を100,000gで90分間超遠心分離し、得られたペレットを100mMのリン酸ナトリウム緩衝液、pH7.6に再懸濁させた。ペレットを、Dyna-Mixホモジナイザー(Fisher Scientific、セッティング2)で均質化し、次に7,800gで5分間、再度遠心分離した。ウイルスは上清から、100,000gで3時間の超遠心分離により、30%のスクロースクッション上にペレット化し、これを再度、10mMのトリス−HCl緩衝液、pH8.0中にDyna-Mixホモジナイザーにより再懸濁させた。ウイルス溶液を、7,800gで5分間の最後の遠心分離により清浄にし、最後に0.45μMシリンジフィルターユニット(Nalgene)を通した。精製ウイルスは、電子顕微鏡法またはウイルスRNA抽出用に用いるまで、4℃で保存した。
【0211】
ウイルスRNAを、ウイルス溶液から、フェノール/クロロホルム抽出とエタノール沈殿により抽出した。プロテイナーゼK処理を用いて、任意の残留タンパク質をRNAから取り除いた。RNAは、必要となるまで−20℃で保存した。
【0212】
ウイルスの電子顕微鏡分析には、精製ウイルス粒子をホルムバール/炭素被覆カッパーグリッド上に載せ、1%タングステン酸メチルアミンで染色した。試料は、JEOL透過電子顕微鏡で分析した。感染したC. quinoaの葉から得た精製ウイルスの電子顕微鏡分析により、ウイルスが約540×13nmの柔軟な繊維状ビリオンであることが示され(図4、バーは50nmを表す)、これはポテックスウイルス科のメンバーに典型的である(Adams et al., 2004. Arch. Virol. 149, 1045-1060)。
【0213】
例2:Malvaモザイクウイルスのクローニングおよび配列決定
MaMVをさらなる試験のために、以下のようにしてクローニングおよび配列決定した。
【0214】
第1のcDNA鎖を、例1に記載のようにして調製したMaMV RNAから、Supercsript(登録商標)のRT−PCR(Invitrogen)用の第1鎖合成システムを製造業者の指示に従って用いて、ランダムヘキサヌクレオチドまたはポリ−dTプライマーで合成した。次に、ターミナルトランスフェラーゼ(New England Biolabs)を製造業者の指示に従って用いて、cDNAの3’末端おいてポリデオキシシチジン尾を添加する前に未使用のプライマーを取り除くため、cDNAをPCR精製キットカラム(QIAGEN)を通した。Asc IまたはPac I制限部位でタグしたポリデオキシグアノシンプライマーをcDNAにハイブリダイズして、製造業者の指示に従って大腸菌DNAポリメラーゼクレノウ断片(New England Biolabs)を用いて、第2鎖を合成した。得られた二本鎖DNAを、フェノール/クロロホルムおよびエタノール沈殿により精製し、次に製造業者の記載に従ってT4 DNAポリメラーゼ(New England Biolabs)で処理して、平滑末端を作製した。平滑末端DNAを適切な制限酵素で消化して、次にpNEB193ベクター(New England Biolabs)中16℃で一晩、T4 DNAリガーゼ(New England Biolabs)にライゲーションした。ライゲーション反応を大腸菌DH5α中に転換し、形質転換細胞をLB寒天プレート上アンピシリン(50μg/ml)を用いて増殖させた。抵抗性コロニーからのプラスミドDNAをQIAprep spin miniprepキット(QIAGEN)を用いて抽出し、次にABI 3730XLシーケンサーで配列決定した。MaMVゲノム配列の3つのオーバーラッピング断片を含有するクローンを得た;1つはCPおよび3つの遺伝子ブロックを含み、1つは複製の残りを含み、いくつかのクローンはMaMVの5’末端を含む(図6A参照)。MaMVゲノムに対応する得られたDNA配列を、図1(配列番号1)に示す。
【0215】
例3:Malvaモザイクウイルスの5’末端の同定
MaMVの5’末端配列は、次のようにして同定した。ゲノムRNA(例1のようにして調製)を最初に、50mMの酢酸ナトリウム、pH6.0、1mMのEDTA、0.1%のβ−メルカプトエタノールおよび0.01%のトリトンX−100中の2Uのタバコ酸性ピロホスファターゼ(Epicentre Biotechnologies)により、37℃で60分間処理して、5’キャップ構造を取り除いた。反応は、フェノール−クロロホルム抽出(0.5vol./0.5vol)により停止させ、RNAを、上記のように逆転写する前に、ただしプライマー0023−11を用いて、エタノール沈殿した;このプライマーは、ゲノムRNAの498〜520位置においてハイブリダイズする(5'-ACATGTAAGCTAAACTAGTGTC-S'、配列番号17)。オリゴヌクレオチドEMSA1(5'-GTGATAAAGTTATGACCATAACCTATGTCGTAGGATATGCATTAACTAAT-3'、配列番号18)を、次にcDNAの3’末端に、20UのRNAリガーゼを用いて製造業者記載の条件下で(New England Biolabs)加えた。第2鎖を次に、キット「Expand High Fidelity PCR System」を製造業者の記載に従って用いて(Roche Diagnostics)、プライマー0023−11およびEMSA2(5'- ATTAGTTAATGCATATCCTACGACATAGGTTATGGTCATAACTTTATCAC-3'、配列番号19)の存在下で、合成して増幅した;これはEMSA1に相補的である。DNAを94℃で2分間変性させ、次に33周期(94℃で45秒間、60℃で45秒間、および72℃で60秒間)のPCRにより増幅した。増幅産物は、TOPO TAクローニングキット(Invitrogen)を用いて、ベクターpCR2.1−TOPO中に直接クローニングした。ライゲーション産物を大腸菌DH5α内に形質転換して増幅し、プラスミドDNAを例2に記載のようにして抽出および配列決定した。いくつかのクローンを正確に配列決定して、MaMVゲノムの5’末端を正確に評価した。
【0216】
MaMVゲノムの5’末端の配列の分析により、以下が決定された。5’非翻訳領域(UTR)は81ヌクレオチドのACに富んだドメインであり、これは、安定性の弱い2つの推定ステムループがそれぞれヌクレオチド26〜36および48〜78に存在することを除いて、ほとんどが非構造化されている。ほとんどのポテックスウイルスと同様に、逆転写の前にタバコ酸性ピロホスファターゼ(TAP)で処理したRNAに観察される優勢な5’末端モチーフは、コンセンサスモチーフGAAAAである(16のうちの10の配列)(図5)。図5は、逆転写反応の前に、タバコ酸性ピロホスファターゼで未処理(−TAP)および前処理(+TAP)されたRNAについて得られた配列を、それぞれ上の図および下の図に示す。バーは、配列決定された全クローンにおいて見出された各配列の頻度を表す。図5でわかるように、タバコ酸性ピロホスファターゼ処理を省略した場合、優勢なモチーフは、ScaVX、CymMVおよびAIsVXにおいて観察されように、GGAAAAであった(17配列中、10)(Wong et al., 1997. Arch. Virol. 142, 383-391;Kim et al., 1998. Mol. Cell. 8, 181-188;Chen et al., 2002. Arch Virol. 147, 683-693;Fuji et al., 2005. Arch Virol. 150, 2377-2385.)。最後のケースの場合、コンセンサスGAAAA配列は2回のみ観察された。この差異は、ほとんどの逆転写酵素がターミナルトランスフェラーゼおよびテンプレートスイッチ活性を有することにより、部分的に説明可能である。Superscript IIは、これがキャップ終端を有するin vitroで転写されたRNAで、製造業者が推奨するバッファー条件を用いて試験された場合に、選好的に1つの追加のシトシン残基をcDNAの3’末端に付加することが示されている(Schmidt et Mueller, 1999. Nucleic Acids Res. 27, e31)。Schmidtはまた、逆転写酵素によるdCMPテーリングが、5’−OH RNAと比べた場合に、キャップされたRNAテンプレートで10倍も効率的であることを観察した。この観察は、本明細書で観察された結果を説明できる可能性があり、その理由は、cDNAの3’末端に付加したシトシン残基は、RNAの5’末端において余分なグアノシンとして読み取られ得るからである。未処理のRNAで観察されたこの追加のG残基は、事実、前に記載した逆転写酵素のターミナルトランスフェラーゼ活性のために人工的(artefactual)であり、そのため、ゲノムRNAの5’末端において見出される正しいモチーフを正確に提示しない。したがって、TAP処理したRNAで得られた結果は、宿主中に見出されるMaMVの天然の集団についてのゲノム配列をより正確に反映すると考えられる。
【0217】
興味深いことには、5’末端のGGAAAAモチーフは、いくつかのポテックスウイルス、例えばAIsVX、LVX、ScaVXなどに見出されている。これらのウイルスが、in vivoでこの特異なGGAAAA配列を示す可能性は完全には排除できないが、上記の結果は、かかるモチーフの発見が用いたプロトコルに結びついており、宿主中に見出されるゲノム集団を正確に表しているわけではなく、コンセンサスGAAAAモチーフは、ポテックスウイルス属に一般に広がっている特長となり得ることを示唆している。
【0218】
例4:Malvaモザイクウイルスの、配列解析、系統発生解析、およびゲノム構成による特徴づけ
MaMVを以下のようにさらに特徴付ける。
【0219】
配列解析
例2に記載のようにして得たMaMV配列をスクリーニングし、全米バイオテクノロジー情報センター(NCBI)のBLASTnデータベースと比較した(Altschul et al., 1997. Nucleic Acids Res. 25, 3389-3402.)。ウイルス配列を、Contig Assembly Program(CAP)ソフトウェア(Huang, 1992. Genomics. 14, 18-25)を用いて解析およびアセンブルした。オープンリーディングフレーム(ORF)を、NCBIのORFファインダーで同定した。同一性/類似性分析をWisconsin(GCG)パッケージversion 10.3からのプログラムGAPにより、アミノ酸比較について8個のギャップ創造(creation)ペナルティー、および2個のギャップ延長(extension)ペナルティーを用いて行った(Anon, 2001. Wisconsin Package version 10.3. Accelrys Inc., San Diego, Ca, USA)。レプリカーゼ、TGB1および種々のポテックスウイルス株からのカプシドタンパク質のアミノ酸配列を決定し、Clustal Wソフトウェア(version 1.83)により生成した複数のアラインメント内に入れて、SeqLabアプリケーション(Wisconsin package version 10.3; Accelrys)の最終目視検査を通して修正した。
【0220】
ポリ(A)尾を除き、MaMVゲノムRNAは、GC含量45%の6858個のヌクレオチド(nt)長である(GenBankアクセッション番号DQ660333)。ゲノム構成は他のポテックスウイルスに類似しており、推定のRNA依存性RNAポリメラーゼ(RdRp)、続いてTGBタンパク質をコードする3つのオーバーラッピング遺伝子、および最後に外被タンパク質を含む(図6A;レプリカーゼは白の四角で表し、3つの遺伝子ブロックタンパク質は薄い灰色の四角で、および外被タンパク質は濃い灰色の四角で表す。黒丸および菱形は、それぞれTGB1およびCPsgプロモーターを示す(略号:MT:メチルトランスフェラーゼ;AlkB:DNA/RNA修復ドメイン;HEL:ヘリカーゼ;POL:RNA依存性RNAポリメラーゼ;CP:外被タンパク質))。レプリカーゼについて示唆されたAUG開始コドンはヌクレオチド81〜83に位置し、この部位からの翻訳により、177.96kDaの計算分子量に対して1571アミノ酸(aa)のタンパク質を産生する。ORF1由来のMaMVレプリカーゼは、少なくとも3つの異なるドメインから構成され(図6A)、これらは、N末端メチルトランスフェラーゼ様ドメイン(aa31〜390)、NTP結合/ヘリカーゼ様ドメイン(aa824〜1059)、およびC末端RdRp2ドメイン(aa1137〜1535)である。これらのドメインは通常良好に保存され、すべてのポテックスウイルスのレプリカーゼ内で提示されている(Rozanov et al., 1992. J. Gen. Virol. 73, 2129-2134;Koonin and Dolja, 1993. Crit. Rev. Biochem. MoI. Biol. 28, 375-430;Longstaff et al., 1993. EMBOJ. 12, 379-386;Davenport and Baulcombe, 1997. J. Gen. Virol. 78, 1247-1251;Batten et al., 2003. Molecular Plant Pathology. 4, 125-131.)。この解析はまた、DNA修復タンパク質AlkBと相同性を共有するドメインの存在を強調する。主としてFlexiviridae科からのいくつかの他の植物RNAウイルスも、これらのレプリカーゼ内に類似のモチーフを有する(Aravind and Koonin, 2001. Genome Biology. 2, research0007.1-0007.8;Bratlie and Drablos, 2005. BMC Genomics. 6, 1-15)。図6Aに示すように、MaMVのアミノ酸配列とゲノム構成は、他のポテックスウイルスで見出されるものと類似している。
【0221】
次の3つのORFは、ポテックスウイルスTGBと類似性を示すオーバーラッピング遺伝子であり、これはウイルス移動に関与する(Beck et al., 1991. Virology. 183, 695-702;Batten et al., 2003.上記;Morozov and Solovyev, 2003. J. Gen. Virol. 84, 1351-1366)。TGB1(ORF2)は、低下した分子量26.3kDaの235aaタンパク質である。これは、非常に高い含量のロイシンと荷電された残基(それぞれ13.2%および21.7%)を有する。これは、全てのポテックスウイルス中最も酸性のTGB1であり、等電点(pI)は4.84である。この配列は、典型的なNTPアーゼ/ヘリカーゼドメインを有し、これのin vitroでのPVXおよび2つのホルデイウイルス(hordeivirus)TGB1タンパク質に対する活性が実証されている(Kalinina, N.O., et al, (2002) Virology 296(2):321-9)。TGB1は、RNAスライシングの阻害において役割を果たし得ることも示唆されている。TGB2タンパク質(ORF3)は、13kDaの計算分子量に対して119aa長であり、その理論的pIは9.42で、一方ORF4またはTGB3は、中性pIの84aaの短いタンパク質(9kDa)である(図7)。PVXにおいて、これら最後の2つのタンパク質は膜および細胞壁と会合し、その機能は主としてTGB1活性を調節することである(Morozov et al., 1991. J. Gen. Virol. 72, 2039-2042;Yang et al., 2000. MoI. Plant-Microbe Interact. 13, 599-605;Morozov and Solovyev, 2003.上記)。MaMV TGBタンパク質の保存された配列および疎水性プロファイルは、他のポテックスウイルスTGBタンパク質のそれらに典型的であり、これらが類似の活性を有し得ることを示唆している。
【0222】
ポテックスウイルスのカプシドまたは外被タンパク質(CP)は、ゲノム保護およびウイルス移動に関与する。MaMV CPは、保存された両親媒性コア配列KYAGFDFFDGVT(配列番号20;nt6545〜6581によりコードされる)を含み、これは、疎水性相互作用を介してポテックスウイルスRNAのCPへの結合に役割を果たすことが提唱されている(Bancroft et al., 1991. J. Gen. Virol. 72, 2173-2181;Dolja et al., 1991. Virology. 184, 79-86;Wong et al., 1997.上記;Cotillon et al., 2002. Arch. Virol. 147, 2231-2238;Thompson and Jelkmann, 2004. Arch. Virol. 149, 1897-1909;Chen et al., 2005. Arch. Virol. 150, 825-832; Fuji et al., 2005. 上記)。MaMV CPは243aaタンパク質であり(図2および配列番号2参照)、推定分子量は26kDaである。
【0223】
MaMVアミノ酸配列を他のポテックスウイルスと比較し、結果を下の表2にまとめて示す。MaMVとより緊密に関連するウイルスにはアスタリスク(*)を付した。MaMVと最大の相同性を有するタンパク質は太字で示した。略号およびGenBankアクセッション番号は以下である:AIsVX:NC_007408;BaMV:NC_001642;CVX:NC_002815;CsCMV:NC_001658;ClYMV:NC_001753;CymMV:NC_001812;FdMV:NC_001483;HdRSV:NC_006943;LVX:NC_007192;MVX;NC_006948;NMV:NC_01441;OVX:NC_006060;PapMV:NC_001748;PepMV:NC_004067;PlAMV:NC_003849;PAMV:NC_03632;PVX:NC_001455;ScaVX:NC_003400;SMYEV:NC_003794;TVX:NC_04322;WClMV:NC_003820;ZVX:NC_06059。
表2:MaMVと他のポテックスウイルスのアミノ酸同一性/相同性
【表2】
aこのタンパク質に対して得られた同一性/相同性の結果は、修正NMV外被タンパク質配列を用いて得られた(下記参照)。
【0224】
この比較から、ORF(レプリカーゼ)はScaVXからの相同タンパク質と最大の同一性を有し、一方他の全てのORF(TGB1、2および3、および外被タンパク質)は、それらのNMV対応物とより関連することが示された。PepMVおよびAlsVXはまた、MaMVと、特にレプリカーゼおよびカプシドタンパク質において良好な局所的相同性を示した。しかし、AlsVXおよびPepMVは全体的にはMaMVから遠く離れており、これは主として、それらのそれぞれのTGBタンパク質間の低い相同性による。上で示したように、相同性解析により、最大のアミノ酸同一性スコアが、MaMVとScaVXのレプリカーゼの間(67.2%)、およびMaMVとNMVのカプシドタンパク質の間(75.6%)で明らかにされた。これらの値は上記のAdams et al. (2004)が確立した種の分画(species demarcation)の分子分類より低いため、MaMVは以前に公開されている全てのポテックスウイルスとは異なる種であると考えることができる。
【0225】
Malva葉脈壊死ポテックスウイルス(MVNV)は、1990年にブラジルにおいてMalva parvifloraに感染するウイルスとして報告されており(Brunt et al., (eds.) 1996 onwards. "Plant Viruses Online: Descriptions and Lists from the VIDE Database. Version: 20th August 1996")、Malva parvifloraには局所病変部と全体の葉脈壊死が検出された。MVNVのヌクレオチド配列は現在利用可能ではない。MVNVが単離された元の宿主は、MaMVについての天然の宿主と類似であるが、この2つのウイルスは次の理由により異なっている:a)電子顕微鏡で観察されたMaMV粒子はMVNVよりも長い、およびb)MVNVはMalva種に局所的病変を誘発することが示されたが、MaMVはモザイク症状を誘発する。
【0226】
系統発生解析
系統発生解析は、MEGA version 3.1(Kumar et al., 2004. Brief Bioinform. 5(2), 150-163)により距離法(distance method)と隣接結合アルゴリズム(neighbour-joining algorithm)を用いて行った。ツリーの位相幾何的精度は、500個のブートストラップ複製を用いて評価した。5’および3’非翻訳領域(UTR)の構造解析は、プログラムmfold version 3.2(Zuker, 2003. Nucleic Acids Res. 31, 3406-3415)を用いて行った。タンパク質ドメインの多重アラインメントおよびプロファイルは、ケンブリッジのWellcome Trust Sanger Instituteのウェブサイトにあるタンパク質ファミリーのPfamデータベース(version 18)により、デフォルトパラメータを用いて行った(Bateman et al., 2004. Nucleic Acids Res. 32, 138-141)。多重アラインメントは、NMV CP(NC_001441)のC末端領域におけるフレームシフトエラーを強調し、これは、このタンパク質の最後の45のaaが他の任意のポテックスウイルスとまったく関連していないことを示す。したがって、修正NMV配列を相同性/系統発生解析に用いた。
【0227】
系統発生解析は、MaMVと他のポテックスウイルスとの間の関係をより正確に試験するために行った。解析は、レプリカーゼおよびカプシドアミノ酸配列のもっとも保存された領域を用いて(図8)、およびTGB1タンパク質の完全な配列を用いて行った。図8は、MaMVと他のポテックスウイルスのレプリカーゼおよびカプシドタンパク質の系統発生解析を示す。レプリカーゼおよび外被タンパク質の系統発生ツリーは、それぞれ右と左の図である。数字は、各枝のブートストラップの値(500複製)を示す。アスタリスクで強調したウイルスは、MaMVの3’末端非翻訳領域において同定されたものと類似の推定偽結節構造を有する。
【0228】
系統発生解析により、ポテックスウイルスに通常検出される保存配列は、MaMV内にも見出されることが示された。系統発生解析は、MaMVが、NMV、ScaVX、AlsVXおよびPepMVを含むポテックスウイルスのサブグループに最も緊密に関連するように見えるとの前の観察を支持し、ここでこれらのウイルスの後者2つは、系統発生学的にわずかに遠い位置にある。解析された全てのツリーにおいて、MaMV、NMVおよびScaVXは常に一緒のグループにあり、この緊密な関係をここでも支持している。しかし、レプリカーゼおよびカプシドタンパク質に対する、このサブグループ内で観察されてブートストラップの低い値(それぞれお66%および39%)は、これら2つのウイルスのどちらに対してMaMVが最も緊密に関係するかについての、明確な結論を引き出すことを許容しなかった。TGB1を用いて行った系統発生解析も類似の結果を示し、MaMVに最も近い親戚についての結論は得られなかった。
【0229】
サブゲノムプロモーターおよび保存RNAプロモーター要素の解析
全RNAを、健康なC. quinoa植物およびMaMV感染のC. quinoa植物から次のようにして抽出した。C. quinoaの感染した葉および非感染の葉を収穫し、液体窒素中で粉砕した。全RNAを、フェノール/クロロホルムを用いて抽出し、RNAスピンカラム(Qiagen)を用いてさらに精製した。全RNAの解析を、ノーザンブロッティングにより次のようにして行った:RNA(20μg)をホルムアルデヒドアガロースゲル(1%)上で分離し、次にナイロン膜(Amersham Biosciences)に移した。不動化したRNAを、PCRによりジゴキシゲニン(Roche Diagnostics)で標識したMaMV外被タンパク質のcDNA断片でプローブした。製造業者のプロトコルに従ってハイブリダイゼーションを行った。ハイブリダイゼーションシグナルを、化学発光基質CDP-Star(Roche Diagnostics)で可視化した。
【0230】
ノーザンブロットにより、3つの主要なウイルスRNAが感染中に産生されることが示された:これらは、1つの大きなゲノムRNAおよび、それぞれ約2000および800ヌクレオチドのRNAとして移動している、2つのサブゲノム種である(図6B)。サブゲノム種は、3重遺伝子ブロックおよびウイルスCP(200nt長のRNA)およびウイルスCPのみ(800ntのRNA)をコードする、ウイルスサブゲノムRNAに対応するはずである。MaMV中のsgプロモーターコンセンサス配列の同定は、TGBとCPが、それらの独自のsgRNAからin vivoで翻訳されることを強く示唆する。しかし、これらが、ゲノムRNAの内部開始により産生可能であるとの可能性も除外できず、その理由は、このメカニズムが少なくともPVX CPに対して前に観察されているからである(Hefferon et al., 1997. J. Gen. Virol. 78, 3051-3059)。
【0231】
オクタヌクレオチドサブゲノムプロモーター配列は、レプリカーゼとTGB1遺伝子の間(nt4851〜4858)およびTGB3コード配列の3’末端のnt5982〜5989の間の遺伝子間領域に見出されている。図6Cは、オクタヌクレオチドの推定sgプロモーター配列の配列アラインメントを示す。右と左の図は、それぞれTGB(黒丸)およびCP(黒の菱形)sgプロモーターコンセンサス配列を示す。コンセンサスオクタヌクレオチド内に高度に保存されたヌクレオチドを、それぞれ黒または灰色の四角で強調する。コンセンサス配列と異なるヌクレオチドは、白字で示す。
【0232】
両方のプロモーターは、ほとんどのポテックスウイルスにおいて取り出される正確なコンセンサス配列GTTAAGTTを有する(Skryabin et al., 1988. FEBS. 240, 33-40;Kim and Hemenway, 1997. Virology. 232, 187-197;Batten et al., 2003.上記)。PVXについて報告されたTGB2/TGB3sgプロモーターコンセンサス配列は、TGB1およびCPのそれらのようには良好に定義されず、その結果、かかるドメインの正確な同定は、より不明確である(Skryabin et al., 1988.上記)。活性なTGB2/TGB3プロモーターに相関する種に対応するsgRNAは、これらの感染植物中に検出されなかった。これらのsgRNA種が存在する場合には、それらはTGB1およびCPのsgRNAのように豊富ではない可能性が高い。
【0233】
3’UTRは70nt長で、他のポテックスウイルスで同定されたように、tRNA様の二次構造内に畳まれていると推定される(Thompson and Jelkmann, 2004.上記)。3’UTRはポリアデニル化シグナルAAUAAAをポリアデニル化部位の14nt上流に含み、また今日まで配列決定された全てのポテックスウイルス中に存在する保存ヘキサマーACUUAAを含む(nt6799〜6804)。両方の配列は、3つのステムループ構造の異なるループ内に局在する(それぞれSL3およびSL1)(図7)。図7は、MaMV3’非翻訳領域の二次構造の模式図である。3つのステムループ構造は、SL1、SL2およびSL3として同定される。全てのポテックスウイルス中で見出されるコンセンサス配列ACUUAAは、SL1内の淡い灰色のヌクレオチドにより強調され、一方、SL3に存在するポリアデニル化シグナルは濃い灰色で示す。プログラムmfoldにより得た二次構造は、挿入図内に示す。SL1とSL2の間の新規な推定偽結節は、点線により示す。クエスチョンマーク(?)は、偽結節が推定のものであり、溶液中のこの構造の畳み込みはまだ決定されていないことを示す。
【0234】
この解析により、SL1とSL2のループに存在するヌクレオチド間の5塩基対の偽結節が明らかにされた。類似の構造が前にPVXについて記載されたが、この場合、相補的領域は、サブゲノムプロモーターからのヌクレオチドとSL1のACUUAA保存配列の間であった(Kim and Hemenway, 1997.上記)。他のポテックスウイルスの3’末端の解析は、類似の偽結節がいくつかの他のウイルス、例えばNMV、CymMV、ScaVX、およびPepMVに検出できることを示した。しかし、後者2つのウイルスにおいて偽結節を形成する相補的配列は、わずか4ヌクレオチド長である。
【0235】
例5:Malvaモザイクウイルス外被タンパク質の産生、精製および大腸菌への自己集合
完全なMaMV外被タンパク質遺伝子を、ゲノムRNAの3’末端全体を包含するシーケンシングプラスミド(CPおよび3重遺伝子ブロックをコードする配列を含有;例2参照)から、PCRにて、フォワードプライマー(5'-GGTACATGTCGAACTCTGGTTCAGCCG-3'、配列番号21)およびリバースプライマー(5'-TACGGATCCTCAATGGTGATGGTGATGGTGGAATTCTGGGGGGGCTTCAA TGG-3'、配列番号22)を用いて増幅した。フォワードプライマーはAf1 III制限部位(下線部)を含み、これはPCR産物をpET−3D発現ベクター内にクローニングするための開始コドンを含み、一方、リバースプライマーは、6X−Hisタグ(下線部)をCP遺伝子の3’末端に付加して、ニッケル親和性カラム上でのさらなるタンパク質の精製を可能とする。PCR反応は、次のように行った: Expand(登録商標)High FidelityPCRシステム(Roche Diagnostics)を用い、製造業者推奨の標準条件化において、94℃で3分間の前インキュベーションの後、94℃で45秒、65℃で45秒、および72℃で60秒を33サイクル。PCR産物はAf1 III/Bam H1制限酵素で消化して、次にpET−3D適合のNco I/Bam H1制限部位にライゲーションした。ライゲーション産物は、大腸菌DH5α中に転換して増幅し、例2に記載のようにして、プラスミドDNAを抽出および配列決定した。
【0236】
CP遺伝子(pMaMV−CP−6H)を含有することが同定されたプラスミドを用いて、大腸菌株BL21(DE3)RIL細胞(Invitrogen)を形質転換した。形質転換細胞を2X YT寒天プレート上に50μg/mlのアンピシリンとともに広げて、37℃で一晩インキュベートした。培養培地(2X YT、50μg/mlのアンピシリン)に、約10個の単離したコロニーの前培養物を播種し、ODが0.6〜0.8となるまで増殖させた。タンパク質の発現を、IPTG(Promega)を最終濃度1mMまで添加して誘発し、培養物を22℃で16時間、225RPMで振とうしつつインキュベートした。次に細胞を遠心分離し、溶解緩衝液中(50mMのNaH2PO4、300mMのNaCl、10mMのイミダゾール、pH8.0)に20μMのPMSF(EM Science)、1Xのプロテアーゼカクテル阻害剤(Roche)および1mg/mlのリソザイム(Sigma)の存在下で再懸濁させ、その後氷上でSonic Dismembrator model 500超音波処理器(Fisher Scientific)で超音波処理した。タンパク質溶液を遠心分離し、上清を、2.5mlのニッケル−NTAアガロースビーズの存在下で4℃で3時間インキュベートし、次に溶出カラム内(Bio Rad)に注いだ。ビーズはそれぞれ25mlの次の緩衝液で洗浄した:洗浄緩衝液1(50mMのNaH2PO4、300mMのNaCl、20mMのイミダゾール、pH8.0)、洗浄緩衝液2(50mMのNaH2PO4、300mMのNaCl、50mMのイミダゾール、pH8.0)、および洗浄緩衝液3(10mMのトリス−HCl、50mMのイミダゾール、pH8.0)。ビーズを次に、溶出緩衝液(10mMのトリス−HCl、1Mのイミダゾール、pH8.0)の存在下で30分間インキュベートして、タンパク質を溶出した。得られた溶出タンパク質を、上記のように10%のSDS−PAGEおよび電子顕微鏡法で解析し、ウイルス様粒子(VLP)の形成をチェックした。
【0237】
タンパク質抽出物のSDS−PAGE解析を用いて、34〜35kDaの優勢な形態を検出した(図9A;レーン1:タンパク質分子量マーカー。分子量マーカーは左側にkDaで示す;レーン2:大腸菌内に産生された精製外被タンパク質;レーン3:感染植物からの精製MaMVから単離された外被タンパク質)。外被タンパク質の、ポリアクリルアミドゲル上での移動性は、他のポテックスウイルス外被タンパク質と整合する(Tremblay et al, 2006. FEBS. 273, 14-25;Hu and Ghabrial 1995, J. Virol. Meth. 55, 367-379;Hammond and Hull, 1981. J. Gen. Virol. 54, 75- 90)。大腸菌内に産生されたMaMV CPの分子量は、野生型タンパク質を比べてわずかに異なるが、これは追加の6X−Hisタグにより説明可能である(図9A)。
【0238】
最後に、精製された組換えタンパク質を電子顕微鏡法で観察し、これにより、MaMV CPの過剰発現が、大腸菌の外被タンパク質の、天然ウイルスと非常によく似たウイルス様粒子への自己集合をもたらすことが確認された(図9B参照、この図は、精製組換えタンパク質MaMV CPから作られたウイルス様粒子の電子顕微鏡写真である(バーは50nmの長さ))。
【0239】
プラスミドpMaMV−CP−6Hに含有される、MaMV外被タンパク質遺伝子のヌクレオチド長を、図10(配列番号23)に示し、コードされた外被タンパク質のアミノ酸配列を図11(配列番号24)に示す。
【0240】
例6:精製組換えMalvaモザイクウイルス外被タンパク質のLC−MS/MS解析による解析
例5からの精製組換え外被タンパク質を、LC−MS/MS解析(Eastern Quebec Proteomics Centre, Centre Hospitalier de l'Universite Laval, Quebec)により解析した。試料は凍結乾燥し、その後還元し、50mMの重炭酸アンモニウム緩衝液中の45mMのジチオトレイトールおよび100mMのヨードアセトアミドでアルキル化した。アセトニトリルに希釈後、トリプシン消化を37℃で一晩、0.2μgの配列決定グレードの修飾トリプシン(Promega)を用いて行った。消化は蟻酸を用いて停止させ、2μLの試料を質量分析計に注入した。ペプチドMS/MSスペクトルを、ナノスプレーインターフェイスを有するLTQ(Thermo-Electron, San Jose, CA, USA)四重極IT質量分析計と連結した、キャピラリー液体クロマトグラフィにより得た。クロマトグラフィ分離は、PicoFritカラムBioBasic C18、10cm×75μm(New Objective, Woburn, MA)上、2〜50%の線形勾配の溶媒B(アセトニトリル、0.1%蟻酸)で30分間、200nL/分にて実施した。ペプチドは、カラムを通り直接LTQ線形イオントラップ質量分析計に溶出したが、この時スプレー電圧は1.8kV、および移送キャピラリー温度は225℃に設定した。質量スペクトルの取得は、Xcalibur 2.0 SR 2ソフトウェア(ThermoElectron Corp)によりデータ依存性取得モードを用いて制御したが、このモードにおいては、各フルスキャン質量スペクトル(400〜2000m/z)に続いて、7つの最も激しいイオンの衝突誘発性解離がある。動的排除機能が可能であり、相対的衝突断片化エネルギーは35%に設定した。
【0241】
得られたMS/MSスペクトルは、MASCOT(Matrix Science, London, UK; version 2.2.0)を用いて解釈し、3つの異なるデータベースを探索した:1つは目的のタンパク質を含むもの、Uniref Humanデータバンク、およびUniref大腸菌データベース(version 8.0、それぞれ94985および40567エントリーを含む)。システインのカルバミドメチル化、およびメチオニンの部分的酸化、2つの欠如した開裂、およびペプチドに対して2.0Daおよび断片に対して0.5Daの誤差耐性を、探索において考慮した。Scaffold(version Scaffold-01_06_18, Proteome Software Inc., Portland, OR)を用いて、MS/MSに基づくペプチドおよびタンパク質の同定を確認した。ペプチドの同定は、それらがPeptide Prophetアルゴリズム(Keller, A et al. 2002; Anal. Chem. 74(20):5383-92)により特定された90.0%を超える確率で確立された場合に、認定された。タンパク質の同定は、それらが90.0%を超える確率で確立され、少なくとも2つの同定されたペプチドを含む場合に、認定された。タンパク質の確率は、Peptide Prophetアルゴリズム(Nesvizhskii, A.I., 2003, Anal Chem. 75(17):4646-58)により割り当てられた。類似のペプチドを含み、MS/MS解析のみに基づいては識別できなかったタンパク質は、節減の原理(principles of parsimony)を満たすためにグループ化した。
【0242】
精製組換えタンパク質のトリプシンによる消化は30個のペプチドを生成し、これはLC−MS/MSにより解析され、全てのペプチドがMaMV CP配列に属することを確認した。MaMV CPは、25,961.2Daの分子量を有する。生成された30個のペプチドは、MaMVタンパク質の予測された断片に100%対応する質量を有した。ペプチドは、MaMV CPのアミノ酸配列の93%をカバーし、精製され大腸菌に過剰発現されたタンパク質の同一性を確認した。表3は、トリプシン消化により生成されたMaMV外被タンパク質ペプチドのLC−MS/MS解析を示す(配列番号25〜53)。
表3:MaMV外被タンパク質ペプチドのLC−MS/MS解析
【表3】
【0243】
例7:MalvaモザイクウイルスVLPの免疫原性効果
MaMV外被タンパク質遺伝子を、大腸菌BL21(pLysS)中に、プラスミドpMaMV−CP−6Hから次のようにして発現させた:pMaMV−CP−6Hを含有する大腸菌BL21(pLysS)の培養物1Lを調製し、コードされた外被タンパク質の発現を、1mMのIPTGにより誘発した。発現は、25℃でO/Nに進むことを可能とした。細胞を収穫し、フレンチプレスを750psi(溶解緩衝液:50mMのNaP緩衝液、pH8、20mMのイミダゾールおよび300mMのNaCl)で用いて細胞溶解した。砕片を10,000gでの遠心分離を2回、それぞれ45分間および30分間行って除去し、上清を保持した。12mlのNi2+ビーズ(Qiagen)を1Lの細菌培養物中に加え、バッチO/Nで4℃にてゆっくり振とうしつつインキュベートした。結合タンパク質を含有するビーズをエコノカラム(econo-column)に入れ、以下により洗浄した:(a)50mlの洗浄緩衝液(10mMのトリス−HCl、pH8+50mMのイミダゾール)、続いて(b)50mlの洗浄緩衝液+0.5%のTriton X-100、(c)50mlの洗浄緩衝液、(d)50mlの洗浄緩衝液+1%のZwittergen、および(e)50mlの洗浄緩衝液。タンパク質を次に1Mのイミダゾールで溶出した。
【0244】
溶出したタンパク質は、3Lの10mMトリス−HCl、pH8に対して1時間透析した。緩衝液を新鮮なものに交換し、透析をさらに1時間続けた。緩衝液を最後に交換し、O/Nで透析を継続した。タンパク質を次に、3LのPBSに対して、20分間隔で緩衝液を3回変えて透析した。次にNaClの濃度を500mMに増加させ、懸濁液を高速(100,000g)で3時間遠心分離して、VLPをペレット化した。
【0245】
VLPペレットを無菌のPBS中に再懸濁させ、0.45ミクロンフィルターを通してろ過した。最後に、タンパク質の濃度を1mg/mlに調節した。VLPの存在を電子顕微鏡を用いて確認し、任意のLPS混入物の存在を、Limulus試験を用いて決定した。
【0246】
MaMV VLPの免疫原性および免疫反応を引き出す能力を、標準のプロトコルに従ってマウスにおいて試験した。簡潔に述べると、5匹のBalb/Cマウスに対し、0日目に100μgのMaMV VLP(アジュバントなし)を注射した。血液試料を免疫化後5、10および14日目に採取した。血清中に存在するVLPに指向されたIgGの総量、IgG1の量、およびIgG2aの量を、標準のELISAにより測定した(図12A〜C参照)。
【0247】
同じマウスを、40日目に、100μgのMaMV VLPでs.c.にてアジュバントなしで2度目に免疫化した。血清中に存在するVLPに指向されたIgGの総量、IgG1の量、およびIgG2aの量を、再度、標準のELISAにより測定した(図12D〜F参照)。
【0248】
結果は、IgGおよびIgG1の高いレベルを示し、これは、MaMV VLPが高度に免疫原性であり、効率的な抗体反応を引き起こし得ることを示す。さらに、IgG2aの存在は、CD4+ T細胞の活性化および抗体クラススイッチの効率的な誘発を示唆する。この結果は、バランスの取れたTH1およびTH2反応が誘発されることを示唆する。
【0249】
例8:腸チフス菌ポリンタンパク質の精製
以下の精製手順を用いて、MaMVまたはMaMV VLPと併用して抗原として用いるのに好適な、腸チフス菌OmpCおよびOmpFタンパク質を精製した。精製手順は、Secundino et al. (2006), Immunology 117:59の記載に基づく。
【0250】
2種のタンパク質を腸チフス菌から共精製した。OmpCおよびOmpFの個別の精製は、OmpC[STYC171(OmpC)]またはOmpF[STYF302(OmpF)]のオープンリーディングフレームのいずれかが分断されている、腸チフス菌のノックアウト変異体を用いて行った。共精製については、細菌のノックアウト変異形態からの個別のタンパク質の精製法に従った。この方法を以下に概説する。
【0251】
細菌株Salmonella typhi 9, 12, Vi:d(ATCC9993)を、酵母抽出物、マグネシウム、およびグルコースを補足した最少培地Aの中で37℃、200rpmで増殖させた。酵母抽出物、マグネシウム、およびグルコースを補足した10Lの最少培地Aの処方は、以下のとおりである:5.0gの無水Na−クエン酸塩(NaC6H5O7:2H2O)、31.0gの一塩基性NaPO4(Na2H2PO4)、70.0gの二塩基性NaPO4(Na2HPO4)、10.0gの(NH4)2SO4、200mLの酵母抽出物溶液5%(300mL中15.0g)。1.434Lの培地を4Lエルレンマイヤーフラスコに分散させた。滅菌を、121℃、1平方インチ当たり15lbの加圧で15分間行った。次に各フラスコに以下を加えた:6.0mLの無菌MgSO4溶液25%、および60.0mLのグルコース溶液12.5%。フラスコに腸チフス菌の一晩培養物を播種し、OD540が1.0に達した時にインキュベーションを停止し、培養物を7,500rpm、4℃で15分間遠心分離した。ペレットを100mLの最終容量のトリス−HCl、pH7.7(6.0gのトリス−塩基/L)に再懸濁させ、バイオマス(生物量)を氷上で90分間超音波処理し、次に7,500rpm、4℃で20分間遠心分離した。10mLの上清の各々に、2.77mLの1MのMgCl2、25mLのRNアーゼA(10,000U/ml)、25mlのDNアーゼA(10,000U/ml)を加えた。混合物を次に120rpm、37℃で30分間遠心分離した。
【0252】
混合物からのポリン抽出は、次のようにして行った:
1.超遠心分離を、45,000rpm、4℃で45分間行い、ペレットを保持した。
2.ペレットを10mLの2%トリス−HCl−SDSに再懸濁させ、均質化した。
3.インキュベーションを、120rpm、32℃で30分間行った。
4.超遠心分離を、40,000rpm、20℃で30分間行い、ペレットを保持した。
5.ペレットを5mLの2%トリス−HCl−SDSに再懸濁させ、均質化した。
6.インキュベーションを、120rpm、32℃で30分間行った。
7.超遠心分離を、40,000rpm、20℃で30分間行い、ペレットを保持した。
8.ペレットを20mLのNikaido緩衝液−1%SDSに再懸濁させ、均質化した。[1LのNikaido緩衝液に対して:6.0gのトリス−塩基、10.0gのSDS、23.4gのNaCl、1.9gのEDTAを水に溶解し、pHを7.7に調整した。次に0.5mLのβ−メルカプトエタノール溶液を加えた]。
9.混合物を120rpm、37℃で120分間インキュベートした。
10.超遠心分離を、40,000rpm、20℃で45分間行った。ポリン抽出物を含む上清を回収した。
【0253】
ポリンを上清から高速タンパク質液体クロマトグラフィ(FPLC)で精製した。β−メルカプトエタノール非含有の0.5XNikaido緩衝液(上記参照)を、精製過程中用いた。タンパク質をSephacryl S-200(FPLC WATERS 650E)を用いて10mL/分のフラックス速度で分離した。カラムに22mLの上清を負荷した。溶出断片を260nmおよび280nmでモニタリングした。精製ポリンを含有する主ピークを保持し、4℃で保管した。精製ポリンは長期間安定であった(1年間以上)。
【0254】
図13は、上記の方法で精製したポリン、OmpCおよびOmpFのSDS−PAGEプロファイルを示す。
【0255】
例9:腸チフス菌ポリンのMalvaモザイクウイルスVLPへの付着に好適な親和性ペプチドの産生
精製OmpCおよびOmpFに対する特異的ペプチドを、Ph.D-7 Phage Display Peptide Library Kit(New England Biolabs, Inc.)を用いて選択した。用いたプロトコルは、「パニング」として知られているin vitro選択法であり、これは製造業者のプロトコルに従って行った。簡潔に述べると、2×1011個のファージを、ELISAプレートのウェルのベースに結合した10μgの精製OmpCまたはOmpFに加え、ウェルの内容物を室温で1時間ゆっくり混合した。非結合ファージを1mlの200mMのグリシン−HCl(pH2.2)で、室温で10分間インキュベートすることにより溶出した。上清を中和するために、およびファージを殺さないために、150μlの1Mのトリス−HCl(pH9.1)を加えた。溶出したファージを次に増幅し、追加の結合/増幅サイクルに供して、結合配列に利するようにプールを豊富化させた。洗浄緩衝液は、パニングの第1ラウンド用に0.1%のTween 20を含有し、これは次のラウンド用に0.5%に増加させた。選択したファージを大腸菌ER2738中、各パニングのラウンドの間増幅させた。このサイクルを3回繰り返して、それぞれのポリンタンパク質に対して最高の親和性のペプチドを選択した。こうして同定したペプチドを表4に示す。
表4:OmpCおよびOmpF親和性ペプチドの配列および出現頻度
【表4】
【0256】
上で同定された親和性ペプチドをMaMV外被タンパク質のC末端に、例えばPCRにより導入することができ、得られた融合タンパク質を大腸菌に発現させて、融合タンパク質からのVLPを産生する。VLPを次に、溶液中でそれらの同族ポリンと、例えば1:1の比率で混合することができ、VLPおよびポリンを含む複合体を提供する。この複合体はそれのみか、または追加のポリンと組み合わせて、続いて腸チフス菌のチャレンジに対してマウスを免疫化するのに用いることができる。
【0257】
例10:市販ワクチン上でのMalvaモザイクウイルスのアジュバント効果
配列番号24に記載の配列(図11A参照)を有する組換えMaMV CPを、大腸菌に22℃で16〜22時間過剰発現させた。細菌をフレンチプレスを用いて溶解し、試料を遠心分離して砕片を取り除き、親和性精製のためにNi2+カラム上に負荷した。親和性クロマトグラフィにより精製した外被タンパク質を図15Aに示す。これらのCPから形成されたVLPは異なる長さを有するが、しかしWTウイルスに類似しており(図15B)、全てのタンパク質が高分子量形態(500kDaを超えるVLP)で見出されるSuperdex 200ゲルろ過からは除外したが、これは、単量体または低い多重形態(500kDa未満)の形態が検出されなかったからである(図15C)。MaMV VLP調製物中のLPSレベルを、24.0EU/mgにおいて評価した。続く免疫化において(下記参照)、3または30μgのVLPを用いたが、これらの量に対してLPS汚染は無視できると考えられる。
【0258】
MaMV VLPのアジュバント特性を測定するために、3価のインフルエンザワクチンFluviral(登録商標)(GlaxoSmithKline)をモデル系として用いた。ヒト用量の5分の1のFluviral(登録商標)を、3μgまたは30μgのどちらかのMaMV VLPで、または従来のアジュバントであるミョウバンで、アジュバント化し、皮下的にBalb/Cマウス(1群あたり5匹)に投与した。免疫化後14日目に出血させ、Fluviral(登録商標)または精製したインフルエンザNP(インフルエンザWSN/33のヌクレオカプシド)タンパク質への免疫反応をELISAにより実施した。Fluviral(登録商標)に対する総IgG(図16A)、IgG1(図16B)、IgG2a(図16C)を測定した。NPに対するIgG2aも測定した(図16D)。
【0259】
図16に示す結果は、30μgのMaMV VLPが、Fluviral(登録商標)に対する免疫反応を大幅に改善できることを示す。総IgG(図16A)、IgG1(図16B)およびIgG2a(図16C)の大幅な増加が、Fluviral(登録商標)タンパク質に対して観察され、一方、ミョウバンは、総IgGおよびIgG2aの量を大幅に改善できなかった。MaMV VLPはまた、TH2免疫反応(IgG1)を誘発し、これについてはミョウバンと類似の有効性を示す(図16B)。さらにMaMV VLPは、NPタンパク質に指向されたIgG2aの産生を誘発することが示された(図16C)。ミョウバンでアジュバント化されたFluviral(登録商標)およびFluviral(登録商標)は、NPタンパク質に対していかなるIgG2aの産生も誘発できなかった。この結果は、インフルエンザウイルス内部に見出され、またFluviral(登録商標)調製物中に存在するNPタンパク質が、MaMV VLPの存在下でのみ、免疫原性となることを示唆する。この結果はまた、クラススイッチが多量のIgG12aアイソタイプの産生をもたらすために、TH1免疫反応が誘発されたことも示唆し、これは、ウイルス感染に対する防御のための理想的な反応であり、MaMV VLPが優れたアジュバント特性を有することを実証する。
【0260】
NPタンパク質は、インフルエンザの全株中もっとも保存されたタンパク質の1つである(92%を超える同一性)。したがって、MaMVベースのアジュバントを用いた、このタンパク質に対する、またはFluviral(登録商標)ワクチンに見出される任意のその他の保存エピトープに対する免疫反応の誘発は、市販のワクチンに見出されるものと無関連であるインフルエンザ株に対する防御を提供することが期待され、すなわち、MaMVベースのアジュバントを市販のインフルエンザワクチンに添加すると、インフルエンザの複数株への防御を提供可能な製剤を提供できる可能性がある。
【0261】
例11:抗原に遺伝子的に融合した外被タンパク質を含むMalvaモザイクルイスVLPの産生および試験
以下の例は、MaMV VLPがワクチンプラットフォームとして使用可能であることを実証する。この例に記載されている全ての外被タンパク質は、前の例に記載されたものと同じCP精製手順を用いて精製した。
【0262】
MaMV外被タンパク質のC末端は、SpeIおよびMluI制限部位を含んで、外被タンパク質のC末端において直接適切な抗原をコードする小さいアニーリングされたオリゴヌクレオチドのクローニングを容易にするように操作された。SpeI/MluI部位を含む配列5' ACTAGTACGCGT 3'(配列番号61)は、図10に示す外被タンパク質遺伝子の最後のアミノ酸(フェニルアラニン:F)の直後で6xHタグの前の位置にクローニングされた。それぞれの酵素の認識配列は以下である:
SpeI=ACTAGT
MluI=ACGCGT
【0263】
得られたMaMV CPは、MaMV CP−SMと名付けられた。このMaMV CP−SMをコードするヌクレオチド配列を図10B(配列番号62)に、およびMaMV CP−SMタンパク質のアミノ酸配列を図11B(配列番号63)に示す。
【0264】
良好に定義された腫瘍抗原gp100(IMQVPFSV;配列番号59)から、およびインフルエンザM1タンパク質(GILGFVFTL;配列番号60)からの9マーHLA−A*201エピトープを、MaMV CPとの融合に選択した。9マーHLA−A*201エピトープを、N−およびC-末端側にそれぞれの天然配列から5残基離して隣接させ(フランクさせ)て、プロテアソームによる自然のプロセシングを有利にした(図14)。インフルエンザまたはgp100エピトープとの融合の際、アミノ酸TSおよびTRは、融合エピトープのN末端およびC末端にそれぞれ融合する。
【0265】
MaMV−SMコンストラクトを用いて、2つの異なる融合物を生成した。1番目(MaMV−M1またはMaMV Flu−M1)は、インフルエンザのM1タンパク質由来のCTLエピトープを含んでいた。MaMV−M1タンパク質をコードするヌクレオチド配列を図17A(配列番号66)に、およびMaMV−M1のアミノ酸配列を図18A(配列番号67)に示す。2番目(MaMV−gp100)は、gp100タンパク質由来のCTLエピトープを含んでいた。MaMV−M1タンパク質をコードするヌクレオチド配列を図17B(配列番号68)に、およびMaMV−M1のアミノ酸配列を図18B(配列番号69)に示す。
【0266】
第3の融合物も、第2MaMV CP(MaMV gl−SM)およびインフルエンザH3株のHAに由来するF3ペプチド(KAYSNCYPYDVPDY;配列番号72)を用いて生成した。MaMV gl−SMはMaMV SMコンストラクトの配列を有するが、しかし、GGGLLLスペーサーを含む。MaMV gl−SMをコードするヌクレオチド配列を図10C(配列番号64)に、およびMaMV gl−SMのアミノ酸配列を図11C(配列番号65)に示す。
【0267】
MaMV gl−SMとF3ペプチドの融合は、コンストラクトMaMV gl−F3をもたらす。MaMV gl−F3をコードするヌクレオチド配列を図17C(配列番号70)に、およびMaMV gl−F3のアミノ酸配列を図18C(配列番号71)に示す。
【0268】
それぞれの外被タンパク質の精製プロファイル、得られたVLPの電子顕微鏡写真、およびVLPのFPLCプロファイルをそれぞれ図19(MaMV−SM)、図20(MaMV gl−SM)、図21(MaMV−M1)、図22(MaMV−gp100)および図23(MaMV gl−F3)に示す。これら組換えタンパク質の各々のVLPの長さを測定したが、それらの間に有意な差は観察されなかった(図24)。VLPの平均長は60nmであるが、200nmを超えるいくつかのVLPも測定された。
【0269】
上記の結果は、MaMV CPが、それらのVLPを形成する能力を失うことなく、いくつかの異なるペプチド配列の融合を耐容できることを実証する。
【0270】
例12:MalvaモザイクウイルスVLPのリンパ球による内在化(インターナリゼーション)
抗原提示細胞(APC)の異なる源の、MaMV VLPを取り込み内在化する能力を、前に記載の技術(Leclerc D, et al. (2007) J Virol 81: 1319-1326)を用いて評価し、パパイアモザイクウイルス(PapMV)の外被タンパク質由来のVLPの取り込みおよび内在化と比較した。
【0271】
簡潔に述べると、PapMV CP、MaMV CPまたはMaMV−M1から形成されたVLP(例11参照)を、蛍光標識(アロフィコシアニンまたはAPC)と接合し、CD40活性化Bリンパ球上で20時間パルスした。CD40活性化Bリンパ球は効率的なAPCであり、樹状細胞(DC)と類似の特性を有する。次に細胞を洗浄し、蛍光をフローサイトメトリーで評価した。
【0272】
結果を図25に示す;PapMV、MaMVおよびMaMV−M1のVLPは類似の効率で内在化されることを実証する。4℃での染色により、全てのVLPが細胞表面に結合可能であることが示唆される。2つのピーク(中程度および高い平均蛍光強度、またはMFI)の存在は、VLPが異なる様式で細胞内に内在化され/細胞と会合するが、しかし、MaMVとMaMV−M1のVLPの間に大きな違いはないことを示唆する。
【0273】
BおよびTリンパ球のヘテロハイブリドーマ細胞株を、次にAPCの源として試験した。この細胞株(T2細胞)は、抗原プロセシング(TAP)と会合するトランスポーターにおいて欠けている。T2細胞による時系列の取り込みアッセイを実施し、ここでT2細胞を図26に示す時点で37℃でパルスし、洗浄し、フローサイトメトリーで分析した。
【0274】
結果を図26Aおよび26Bに示す;PapMV、MaMVおよびMaMV−M1のVLPは時間とともに類似の効率で内在化される。再度、2つのピーク(中程度および高いMFI)が観察され、これは、VLPが異なる様式でT2細胞に内在化され/これと会合することを示唆する。時間に伴うMFIの増加によれば、MaMV VLPはより効率的に取り込まれるように見える。しかし、PapMV VLPはより迅速に分解し、その結果みかけの総MFIがより低くなることも可能である。
【0275】
これらの結果は、MaMV VLPおよびPapMV VLPが同等の有効性でAPCに内在化されることを示し、したがって、これらがワクチンプラットフォームとして同等の有効性を有することを示唆する。
【0276】
例13:MalvaモザイクウイルスVLPの、CTLエピトープのMHCクラスI提示を引き出す能力
PapMVは、プロテアソーム非依存性のメカニズムのもとで、MHCクラスIエピトープ交差提示を媒介する能力を実証した(Leclerc D, et al. (2007) J Virol 81: 1319-1326)。図12に示すように、PapMVおよびMaMVの両方は細胞を結合し、同様に内在化される。したがって、PapMVと同様の様式で、共有結合的に結合したMHCクラスIエピトープの提示を引き出すMaMVの能力を、次に前に記載の技術を用いて検討した(Leclerc et al. (2007)上記)。
【0277】
簡潔に述べると、PapMV外被タンパク質またはMaMV外被タンパク質由来のVLPに、HLA−A*0201インフルエンザM1エピトープをC末端に挿入したもの(PapMV-M1およびMaMV−M1;例11参照)、およびインフルエンザM1エピトープに対応する合成ペプチドを、T2細胞上で20時間パルスした。細胞を洗浄し、関連HLA−A*0201MHCクラスI分子に結合したM1インフルエンザエピトープに特異的なT細胞を、さらに20時間加えた。上清を収穫し、インターフェロン(IFN)−γ分泌をELISAにより評価した。IFN−γは、MHC/ペプチド複合体を認識するT細胞により分泌される。
【0278】
データを図27Aに示す。Gp100は陰性の特異性対照として機能し、これは陰性でなければならない。結果は、PapMVとMaMV両方のVLPが、M1エピトープを交差提示する能力を有することを示す。MaMV VLPについては、10μg/mlにおいて改善された認識が、および50μg/mlでパルスされた場合には認識の顕著な低下が観察された。
【0279】
MaMV−M1を用いて同様の実験を行って、用量反応を評価した(図27B)。Gp100は陰性の特異性対照として機能し、これは陰性でなければならない。
【0280】
プロテアーゼは、上記実験においてAPCを培養するために用いたヒト血清中に存在するため、APCのパルスが、VLPの内在化、プロセシング、およびAPCでの内在化後のMHC負荷からではなく、培養倍地中でのVLPの分解後に達成される可能性がある。この可能性を除外するために、T2細胞をMaMV−M1 VLPで4℃でパルスした低い温度は、VLPの任意の潜在的な分解を阻害するために用いた。ペプチドが外部に放出される場合、これらはMHCクラスIを細胞の外側から負荷しなければならないが、図27Cに示すように、そうではなかった(図27Bに示す実験は同時に実施され、37℃における陽性対照として機能する)。対照として、合成ペプチドを同様にパルスし、これらは4℃または37℃においてパルスした場合に認識された。
【0281】
図27Bおよび図27Cに示す結果は、用量反応がMaMV−M1に対する認識効率と相関することを実証する。しかし、より高い用量でパルスすると(50mg/ml)、認識の顕著な低下をもたらす(図27B)。M1エピトープは、ヒト血清を含有する培養培地中での分解により放出されないことが示された(図27C)。最後に、上記および例12の実験がTAP欠乏したT2細胞中で行われたため、これらの結果により、交差提示メカニズムはプロテアソーム/TAP媒介性ではないことがさらに確認される。
【0282】
例14:MalvaモザイクウイルスVLPによる休止エピトープ特異的Tリンパ球の感作
4人の異なるドナー(#405、#542、#614および#621)からの休止Tリンパ球を用いて、VLPでパルスしたAPCの、インフルエンザM1タンパク質特異的T細胞の増殖(expansion)を誘発する能力を評価した。このアッセイは、「T細胞感作」または「in vitroワクチン接種」と呼ばれる。かかるアッセイにおいて、M1エピトープを担持するVLPが、3週間の培養アッセイにおいて、特異的T細胞を増殖させるのに効率的であるかどうかを評価することができる。
【0283】
各ドナーに対して、自己由来APC(CD40−活性化Bリンパ球)を初めに培養した。これらの細胞を、M1エピトープを担持するPapMV VLPまたはMaMV VLP(例11参照)(25μg/ml)のどちらか、またはM1エピトープを含有する合成ペプチドでパルスした。パルスしたAPCを次に、自己由来血液Tリンパ球で7日間共培養し、つぎに、同じパルスしたAPCで再刺激した。全部で21日の培養後、培養したTリンパ球の、関連するインフルエンザM1エピトープに対する特異性を評価した。
【0284】
産生したTリンパ球の、各ドナーに対する特異性を図28〜31に示す。ドナー#405に対し(図28)、MaMV−M1 VLPは、特異的T細胞の働きの増大をもたらしたが、しかしPapMV−M1 VLPはもたらさなかった。いくつかのペプチド特異株は、MaMV−M1 VLPと比べた場合に、よりアビディティが高いことが観察された。ドナー#542に対し(図29)、PapMV−M1 VLPまたはMaMV−M1 VLPのどちらかを用いて産生されたT細胞株は、MaMV−M1をよりよく認識する。全体としてこのドナーに対して、VLPはペプチドパルスよりも、より高いアビディティのT細胞を生成する傾向があった。ドナー#614に対し(図30)、MaMV−M1 VLPを用いて産生されたT細胞株は、VLP−M1をよりよく認識する。全体として、VLPを用いて生成されたT細胞は、低いペプチド濃度(0.001mM;これは対数スケールであることに注意)でのペプチドパルスに比べて、より高いアビディティのT細胞を産生する。ドナー#621に対し(図31)、PapMV−M1 VLPまたはMaMV−M1 VLPのどちらかを用いて産生されたT細胞株は、VLP−M1を認識する。MaMV−M1T細胞株は、PapMV−M1T細胞株に比べてより高いアビディティを有するようであり、VLPT細胞株は全体としてペプチドパルスより優れていた。
【0285】
結論として、MaMV−M1 VLPおよびPapMV−M1 VLPの両方は、このin vitroのT細胞感作アッセイにおいて抗原特異的Tリンパ球を生成するのに一般に効率的であり、全体として、MaMV VLPおよびPapMV VLPの両方は、ペプチドのみよりも良好に作用した。驚くべきことには、抗プラットフォームTリンパ球はMaMVまたはPapMV VLPの両方について観察されなかった。全体として、MaMV VLPおよびPapMV VLPの両方は、同等であるように見られた。ドナー間での変化は予想された。
【0286】
例12および13は、MaMV−gp100およびMaMV−M1が、これらのCTLエピトープのヒトAPCのMHCクラス1での交差提示を誘発できること、および、CD8+特異的細胞の増殖を誘発できることを実証する。これらの結果は、MaMVプラットフォームが、ヒトまたは動物において、CTL反応を誘発可能であることを、強く示唆する。
【0287】
さらに、例12〜14は、MaMVプラットフォームが、その外被タンパク質の配列がPapMVプラットフォームの外被タンパク質の配列とアミノ酸の69%で非常に異なるにもかかわらず、PapMVプラットフォームと同様の免疫原性特性を有することを実証する。両方のプラットフォームはワクチン開発において大きな可能性を示し、これらが同一のワクチン接種プロトコルにおいて一緒に用いられて、与えられたエピトープへの免疫反応を最適化すること、または同じ患者において交互に用いて、前の処置のプラットフォームに指向された抗体による交差中性化を避けることが、期待される。
【0288】
例15:抗原に融合したMalvaモザイクウイルスVLPの、in vivo免疫反応を引き出す能力
MaMV−gp100、MaMV−M1またはMaMV gl−F3を含むVLP(例11参照)を試験して、融合ペプチドに対する免疫反応が、Balb/Cマウスにおいて検出できるかどうかを確認した。
【0289】
簡潔に述べると、マウスに2回の皮下注射を14日の間隔で表5に示すように施した。各注射の前に血液を採取し(0日目および14日目)、また最後の注射の14日目後にも採取した(28日目)。
表5:マウスの免疫化
【表5】
【0290】
14日目および28日目に採取した血液を用いてELISAを行い、総抗ペプチドおよび総抗MaMV IgG価を定量した。
【0291】
各組換えタンパク質についてLPS含量を評価し、各ケースにおいて15EU/mg未満であった。これは、注射した組成物のLPS含量が基本的に無視できることを示す。
【0292】
驚くべきことには、ほとんどのマウスはMaMV CPに融合したペプチドに対してIgG反応を示さなかった(図32A)。MaMV g1-F3コンストラクトで処置した10匹のうち2匹のマウスは、ペプチドについてセロコンバージョンを示し、MaMV−M1コンストラクトで処置した10匹のうち1匹のマウスは、ペプチドについてセロコンバージョンを示した。MaMV−gp100コンストラクトで処置したマウス、またはF3ペプチドで処置したマウス(F3ペプチドがMaMVプラットフォームに融合しなかった場合)は、いかなる反応も示さなかった。
【0293】
この点について、M1またはgp−100タンパク質由来のCTLエピトープの選択は、マウスにおける体液性の反応を評価するにはおそらく最適ではなかったことが指摘でき、その理由はおそらく、実験に用いたマウスの免疫レパートリーに、これらのペプチドを認識できるB細胞が含まれていないことである。これはまた、F3ペプチドについても同様である。
【0294】
前の例では、MaMVが強力なアジュバントであることを実証した。本例ではしかし、試験動物の免疫レパートリーが抗原を認識できない場合には、強いIgG反応を誘発できないことを確認する。アジュバントは、目的の抗原に特異的なB細胞が利用可能である場合にのみ、B細胞反応を増幅できる。この結論は、MaMV CPに対して全ケースで非常に強いIgG反応が測定できるという事実によって支持され、プラットフォームが高度に免疫原性であることを示す(図32B)。これは、ワクチンプラットフォームとしては重要な性質であり、なぜならば、一般に、HBV VLPまたはPapMV VLPなどのワクチンプラットフォームについて、プラットフォーム自体の本質的な免疫原性が、VLPの表面の融合エピトープまたはペプチドに指向された免疫反応を駆動することが、よく認識されているためである。したがって、MaMV VLPは高度に免疫原性であることが示されたので、より適切なエピトープ(例えば、インフルエンザのM2タンパク質のM2eエピトープ(Denis et al., 2008, Vaccine, 26:3395-3403参照))が、ペプチドに対して常に強い免疫反応を生じさせることが予想される。
【0295】
さらに、将来の実験について、スペーサーを融合ペプチドとMaMVCPの間にクローニングして、ペプチドをVLPの表面に押し出すことが可能であり、これは、この実験では最適化されなかったB細胞との架橋(cross link)を改善することが期待される。
【0296】
例16:パパイアモザイクウイルスVLPおよびMalvaモザイクウイルスVLPに対する抗体の交差反応性
PapMV CPおよびMaMV CPは、31%の配列同一性を共有するのみである(図33A参照)。配列における差はまた、各々の外被タンパク質に特異的な抗体における交差反応性の欠如にも明らかである。例えば図33Bは、MaMV CPに指向された抗体を用いたELISAの結果を示し、これは、PapMV VLPを認識しないことがわかる。PapMV CPに指向された抗体による同様の実験により、PapMV抗体はMaMVプラットフォームに結合しないことも明らかにされた(図33C)。この結果は、2つのVLPの表面は非常に異なっていることを示唆する。2つのプラットフォームはしたがって、ワクチン投与計画において一緒に用いて、より優れた効率的なワクチン接種プログラムを保証することができ、その理由は、第1のプラットフォームに指向された抗体が、第2のプラットフォームを妨害できないからである。
【0297】
例17:Malvaモザイクウイルスの免疫原性効果
以下のプロトコルは、MaMVおよび弱い免疫原を含む組成物の免疫原性効果を評価するために従うことができる、代替的プロトコルである。
【0298】
生得的免疫反応の抗体反応への翻訳は、アジュバントを弱い免疫原性ワクチンと共同投与した場合に観察される。アジュバントは、抗原に対する抗体または細胞の免疫反応を強化または増強可能な物質である。MaMVが他の抗原に対する長期の抗体反応を促進可能なアジュバントであるかどうかを決定するために、BALB/cマウスを弱い免疫原、オボアルブミン(OVA)またはニワトリ卵白リゾチーム(HEL)で、それらのみで、またはMaMVと共に免疫化した。CFAまたはLPSなどの他のアジュバントと、OVAまたはHELとの組合せを陽性対照として用いることができる。次に、OVAまたはHELに特異的なIgG抗体価を、適切な時点でELISAにより測定することができる。
【0299】
MaMV精製
以下の実験で用いるMaMVは、例1に記載のようにして精製することができる。
【0300】
抗原およびアジュバント
LPS非含有OVAグレードVIは、Sigma-Aldrich Chemical Co., St Louis, MOから購入可能であり、ニワトリ卵白リゾチーム(HEL)は、Research Organics Inc. Cleveland, OHから購入可能である。大腸菌O111:B4からのLPSは、Sigma-Aldrich, St Louis, MOから購入可能である。
【0301】
免疫化
6〜8週齢のBALB/cマウスを、医薬品安全性試験実施基準に準拠した条件下で飼育し維持する。試験アジュバントの効果を試験するために、マウスの群を2mgのOVAもしくはHELのみ、または30mgのMaMV、CFA1:1(v/v)、または5mgの大腸菌O111:B4からのLPS(Sigma-Aldrich)で、0日目に腹腔内で免疫化した。対照マウスには、生理食塩水のみを注射した。血液試料は、後方眼窩洞から様々な時点で採取した。個々の血清試料は分析まで−20℃で保存した。各実験に3匹のマウスを用いた。
【0302】
ELISAによる抗体価の決定
高結合96ウェルポリスチレンプレート(Corning(登録商標)、New York, NY)を、0.1Mの炭酸−重炭酸緩衝液(pH9.5)中の1mg/mLのMaMV、100mg/mLのHEL、または150mg/mLのOVAで被覆した。プレートを37℃で1時間、次に4℃で一晩インキュベートした。次の朝、使用前に、プレートを0.05%Tween-20(Sigma-Aldrich)含有のPBS(pH7.2)(PBS−T)で3回洗浄した。非特異的結合を、PBSに希釈した5%の脱脂粉乳(PBS−M)を用いて37℃で1時間ブロックした。洗浄後、マウス血清をPBS−M中1:40に希釈し、2倍連続希釈液をウェルに加えた。プレートを37℃で1時間インキュベートし、次にPBS−Tで4回洗浄した。ペルオキシダーゼ接合ウサギ抗マウスIgM(最適希釈1:1000)IgG、IgG1、IgG2a、IgG2b抗体(Zymed, San Francisco, CA)またはIgG3(最適希釈1:3000)(Rockland, Gilbertsville, PA)を加え、プレートを37℃で1時間インキュベートし、PBS−Tで3回洗浄した。30%過酸化水素含有の0.1Mのクエン酸緩衝液(pH5.6)中のオルトフェニレンジアミン(0.5mg/mL;Sigma-Aldrich)を、酵素基質として用いた。反応を2.5NのH2SO4で停止させ、吸収を、BIOLINX 2.22ソフトウェアと自動ELISAプレートリーダー(Dynex Technologies MRII, Chantilly, VA, USA)を用いて、490nmにて決定した。抗体価は、−log2希釈×40として与える。正の力価は、陰性対照の平均値の上の3SDとして定義した。
【0303】
例18:MalvaモザイクウイルスおよびMalvaモザイクウイルスVLPによる、亜致死量のインフルエンザウイルスに対する抵抗性の誘発
MaMVまたは組換えMaMV VLPの、亜致死量のインフルエンザウイルスに対する抵抗性を、抗原の不在下において誘発する能力を、マウスにおいて試験することができる。簡潔に述べると、マウスを鼻腔内経路で100ugのMaMVで7日間隔で3回処置する。最後の処置の2日後に、動物を亜致死量のインフルエンザウイルスに暴露する(A/WSN/33)(1LD50)。動物中で複製されるウイルスの量を、感染後に2日の間隔で鼻での分泌をサンプリングして測定する。動物の体温と体重を、チャレンジ後7日間毎日モニタリングする。感染による病的状態を、感染後の10日間、スコア化する。
【0304】
本発明を、ある特定の態様を参照して記載したが、これらについての種々の改変は、本発明の精神および範囲から逸脱することなく、当業者には明らかである。当業者に明らかであるかかる改変の全ては、以下の特許請求の範囲に包含されることが意図される。
【特許請求の範囲】
【請求項1】
Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および薬学的に許容し得る担体を含む、免疫原性組成物。
【請求項2】
1または2以上の抗原をさらに含む、請求項1に記載の免疫原性組成物。
【請求項3】
1または2以上の抗原がMaMVまたはVLPに付着している、請求項2に記載の免疫原性組成物。
【請求項4】
免疫原性組成物がVLPを含み、1または2以上の抗原が前記VLPに含まれる外被タンパク質に遺伝子的に融合している、請求項2に記載の免疫原性組成物。
【請求項5】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項6】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項7】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項8】
動物において免疫反応を誘発するための方法であって、前記動物に対して、請求項1、2、3、4、5、6または7のいずれか一項に記載の免疫原性組成物の有効量を投与することを含む、前記方法。
【請求項9】
免疫反応が、抗体の産生を含む、請求項8に記載の方法。
【請求項10】
免疫反応が、細胞性反応を含む、請求項8に記載の方法。
【請求項11】
免疫原性組成物を注射により投与する、請求項8、9または10のいずれか一項に記載の方法。
【請求項12】
免疫原性組成物を鼻腔内に投与する、請求項8、9または10のいずれか一項に記載の方法。
【請求項13】
動物が哺乳類、鳥類、魚類または爬虫類である、請求項8、9、10、11または12のいずれか一項に記載の方法。
【請求項14】
動物がヒトである、請求項8、9、10、11または12のいずれか一項に記載の方法。
【請求項15】
動物に、疾病、疾患、または感染に対してワクチン接種するための方法であって、前記動物に対して、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および1または2以上の抗原を含む免疫原性組成物の、有効量を投与することを含む、前記方法。
【請求項16】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項15に記載の方法。
【請求項17】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項15に記載の方法。
【請求項18】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項15に記載の方法。
【請求項19】
1または2以上の抗原がMaMVまたはVLPに付着している、請求項15、16、17または18のいずれか一項に記載の方法。
【請求項20】
免疫原性組成物がVLPを含み、1または2以上の抗原が、前記VLPに含まれる外被タンパク質に遺伝子的に融合している、請求項15、16、17または18のいずれか一項に記載の方法。
【請求項21】
免疫原性組成物および1または2以上の抗原を、注射により投与する、請求項15、16、17、18、19または20のいずれか一項に記載の方法。
【請求項22】
動物が哺乳類である、請求項15、16、17、18、19、20または21のいずれか一項に記載の方法。
【請求項23】
動物がヒトである、請求項15、16、17、18、19、20または21のいずれか一項に記載の方法。
【請求項24】
Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)の、免疫原性組成物の調製における使用。
【請求項25】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項24に記載の使用。
【請求項26】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項24に記載の使用。
【請求項27】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項24に記載の使用。
【請求項28】
免疫原性組成物が、MaMVまたはVLPに付着している1または2以上の抗原をさらに含む、請求項24、25、26または27のいずれか一項に記載の使用。
【請求項29】
免疫原性組成物が、VLPを含み、前記VLPに含まれる外被タンパク質に遺伝子的に融合している1または2以上の抗原をさらに含む、請求項24、25、26または27のいずれか一項に記載の使用。
【請求項30】
Malvaモザイクウイルス(MaMV)外被タンパク質を含む、ウイルス様粒子(VLP)。
【請求項31】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項30に記載のVLP。
【請求項32】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項30に記載のVLP。
【請求項33】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項30に記載のVLP。
【請求項34】
VLPに付着している1または2以上の抗原をさらに含む、請求項30、31、32または33のいずれか一項に記載のVLP。
【請求項35】
MaMV外被タンパク質に遺伝子的に融合している1または2以上の抗原をさらに含む、請求項30、31、32または33のいずれか一項に記載のVLP。
【請求項36】
ウイルス様粒子(VLP)を調製するための方法であって、Malvaモザイクウイルス(MaMV)外被タンパク質を宿主細胞中に発現するステップを含む、前記方法。
【請求項37】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項36に記載の方法。
【請求項38】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項36に記載の方法。
【請求項39】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項36に記載の方法。
【請求項40】
MaMV外被タンパク質が、外被タンパク質に遺伝子的に融合した1または2以上の抗原を含む融合タンパク質である、請求項36、37、38または39のいずれか一項に記載の方法。
【請求項41】
宿主細胞が細菌細胞である、請求項36、37、38、39または40のいずれか一項に記載の方法。
【請求項42】
宿主細胞が大腸菌細胞である、請求項36、37、38、39または40のいずれか一項に記載の方法。
【請求項43】
抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質。
【請求項44】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項45】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項46】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項47】
請求項43、44、45または46のいずれか一項に記載の融合タンパク質をコードする、単離ポリヌクレオチド。
【請求項48】
請求項47に記載のポリヌクレオチドにより遺伝子操作された、宿主細胞。
【請求項49】
請求項43、44、45または46のいずれか一項に記載の融合タンパク質、請求項47に記載のポリヌクレオチド、または請求項48に記載の宿主細胞の、ウイルス様粒子を調製するための使用。
【請求項1】
Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および薬学的に許容し得る担体を含む、免疫原性組成物。
【請求項2】
1または2以上の抗原をさらに含む、請求項1に記載の免疫原性組成物。
【請求項3】
1または2以上の抗原がMaMVまたはVLPに付着している、請求項2に記載の免疫原性組成物。
【請求項4】
免疫原性組成物がVLPを含み、1または2以上の抗原が前記VLPに含まれる外被タンパク質に遺伝子的に融合している、請求項2に記載の免疫原性組成物。
【請求項5】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項6】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項7】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項1、2、3または4のいずれか一項に記載の免疫原性組成物。
【請求項8】
動物において免疫反応を誘発するための方法であって、前記動物に対して、請求項1、2、3、4、5、6または7のいずれか一項に記載の免疫原性組成物の有効量を投与することを含む、前記方法。
【請求項9】
免疫反応が、抗体の産生を含む、請求項8に記載の方法。
【請求項10】
免疫反応が、細胞性反応を含む、請求項8に記載の方法。
【請求項11】
免疫原性組成物を注射により投与する、請求項8、9または10のいずれか一項に記載の方法。
【請求項12】
免疫原性組成物を鼻腔内に投与する、請求項8、9または10のいずれか一項に記載の方法。
【請求項13】
動物が哺乳類、鳥類、魚類または爬虫類である、請求項8、9、10、11または12のいずれか一項に記載の方法。
【請求項14】
動物がヒトである、請求項8、9、10、11または12のいずれか一項に記載の方法。
【請求項15】
動物に、疾病、疾患、または感染に対してワクチン接種するための方法であって、前記動物に対して、Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)、および1または2以上の抗原を含む免疫原性組成物の、有効量を投与することを含む、前記方法。
【請求項16】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項15に記載の方法。
【請求項17】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項15に記載の方法。
【請求項18】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項15に記載の方法。
【請求項19】
1または2以上の抗原がMaMVまたはVLPに付着している、請求項15、16、17または18のいずれか一項に記載の方法。
【請求項20】
免疫原性組成物がVLPを含み、1または2以上の抗原が、前記VLPに含まれる外被タンパク質に遺伝子的に融合している、請求項15、16、17または18のいずれか一項に記載の方法。
【請求項21】
免疫原性組成物および1または2以上の抗原を、注射により投与する、請求項15、16、17、18、19または20のいずれか一項に記載の方法。
【請求項22】
動物が哺乳類である、請求項15、16、17、18、19、20または21のいずれか一項に記載の方法。
【請求項23】
動物がヒトである、請求項15、16、17、18、19、20または21のいずれか一項に記載の方法。
【請求項24】
Malvaモザイクウイルス(MaMV)またはMaMV外被タンパク質を含むウイルス様粒子(VLP)の、免疫原性組成物の調製における使用。
【請求項25】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項24に記載の使用。
【請求項26】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項24に記載の使用。
【請求項27】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項24に記載の使用。
【請求項28】
免疫原性組成物が、MaMVまたはVLPに付着している1または2以上の抗原をさらに含む、請求項24、25、26または27のいずれか一項に記載の使用。
【請求項29】
免疫原性組成物が、VLPを含み、前記VLPに含まれる外被タンパク質に遺伝子的に融合している1または2以上の抗原をさらに含む、請求項24、25、26または27のいずれか一項に記載の使用。
【請求項30】
Malvaモザイクウイルス(MaMV)外被タンパク質を含む、ウイルス様粒子(VLP)。
【請求項31】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項30に記載のVLP。
【請求項32】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項30に記載のVLP。
【請求項33】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項30に記載のVLP。
【請求項34】
VLPに付着している1または2以上の抗原をさらに含む、請求項30、31、32または33のいずれか一項に記載のVLP。
【請求項35】
MaMV外被タンパク質に遺伝子的に融合している1または2以上の抗原をさらに含む、請求項30、31、32または33のいずれか一項に記載のVLP。
【請求項36】
ウイルス様粒子(VLP)を調製するための方法であって、Malvaモザイクウイルス(MaMV)外被タンパク質を宿主細胞中に発現するステップを含む、前記方法。
【請求項37】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項36に記載の方法。
【請求項38】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項36に記載の方法。
【請求項39】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項36に記載の方法。
【請求項40】
MaMV外被タンパク質が、外被タンパク質に遺伝子的に融合した1または2以上の抗原を含む融合タンパク質である、請求項36、37、38または39のいずれか一項に記載の方法。
【請求項41】
宿主細胞が細菌細胞である、請求項36、37、38、39または40のいずれか一項に記載の方法。
【請求項42】
宿主細胞が大腸菌細胞である、請求項36、37、38、39または40のいずれか一項に記載の方法。
【請求項43】
抗原に融合したMalvaモザイクウイルス(MaMV)外被タンパク質を含む、融合タンパク質。
【請求項44】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも80%同一であるアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項45】
MaMV外被タンパク質が、配列番号2に記載の配列と少なくとも90%同一であるアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項46】
MaMV外被タンパク質が、配列番号2に記載のアミノ酸配列を含む、請求項43に記載の融合タンパク質。
【請求項47】
請求項43、44、45または46のいずれか一項に記載の融合タンパク質をコードする、単離ポリヌクレオチド。
【請求項48】
請求項47に記載のポリヌクレオチドにより遺伝子操作された、宿主細胞。
【請求項49】
請求項43、44、45または46のいずれか一項に記載の融合タンパク質、請求項47に記載のポリヌクレオチド、または請求項48に記載の宿主細胞の、ウイルス様粒子を調製するための使用。
【図1A】


【図1B】

【図1C】

【図2】

【図3】

【図4】
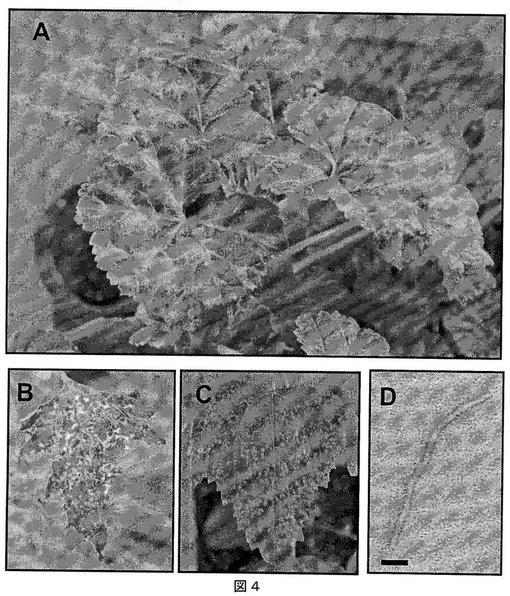
【図5】
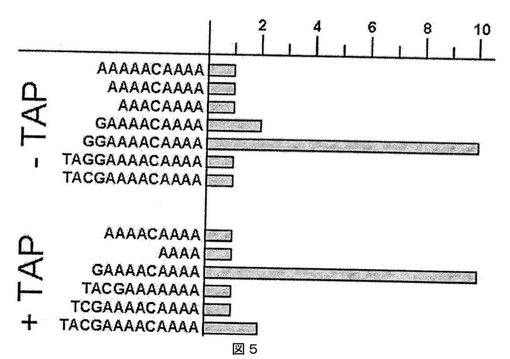
【図6】
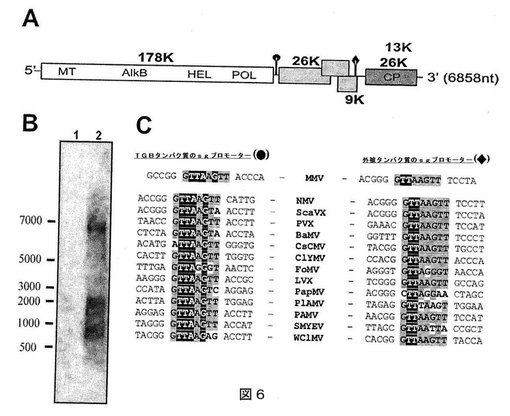
【図7】
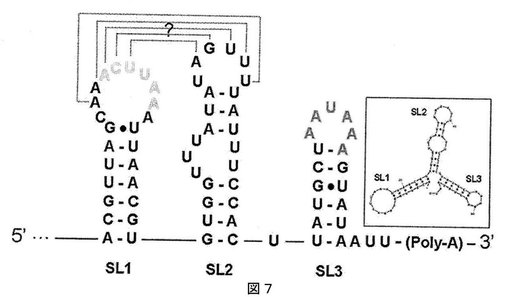
【図8】
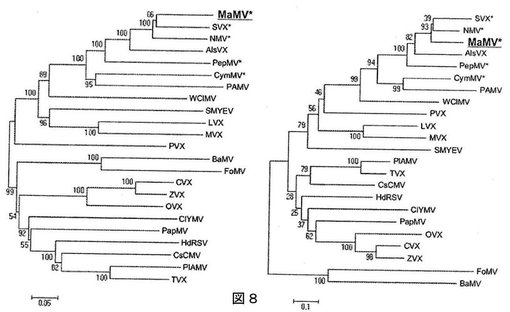
【図9】
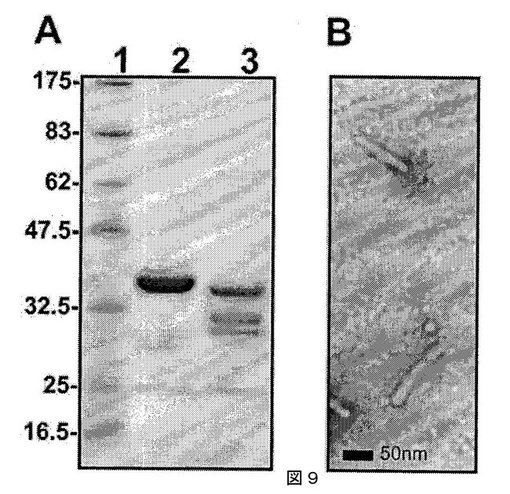
【図10】

【図11】

【図12】
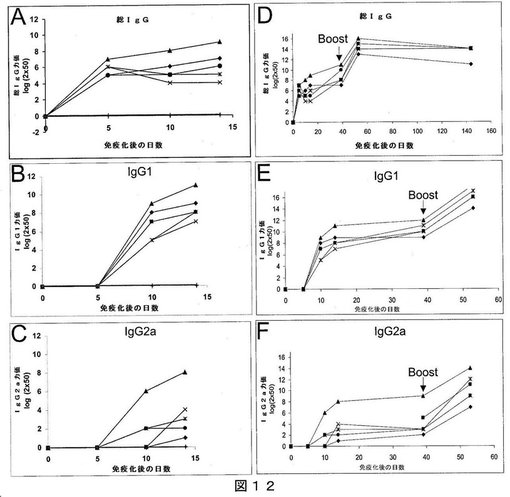
【図13】
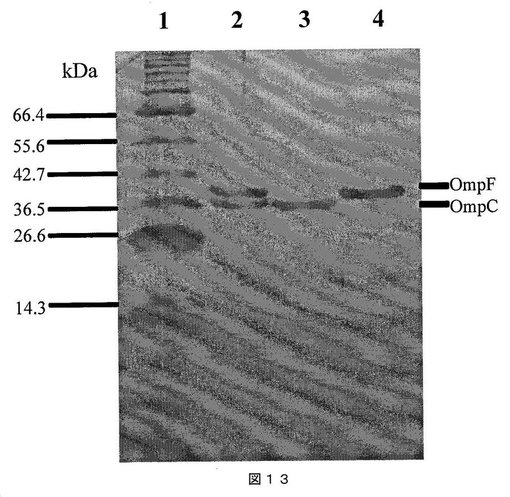
【図14】

【図15】
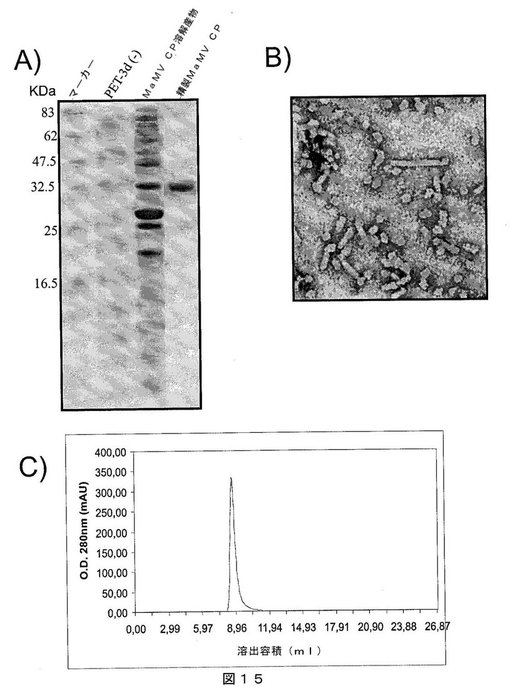
【図16A−C】
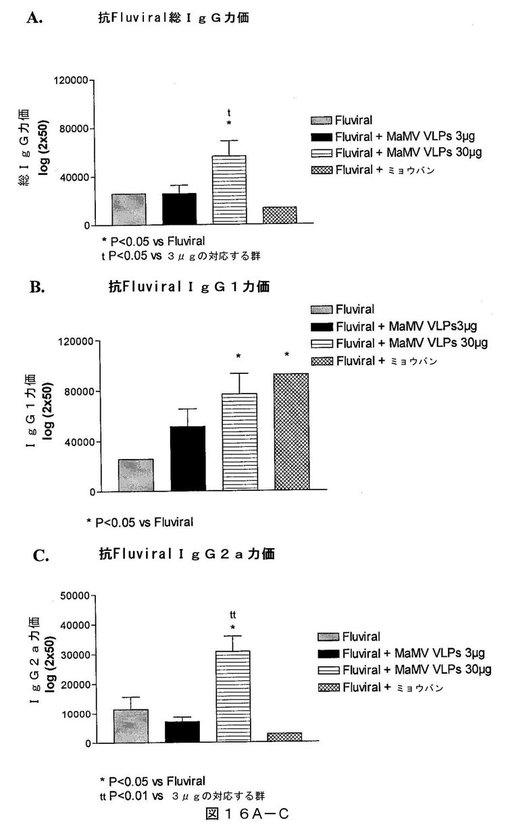
【図16D】
【図17】

【図18】

【図19】
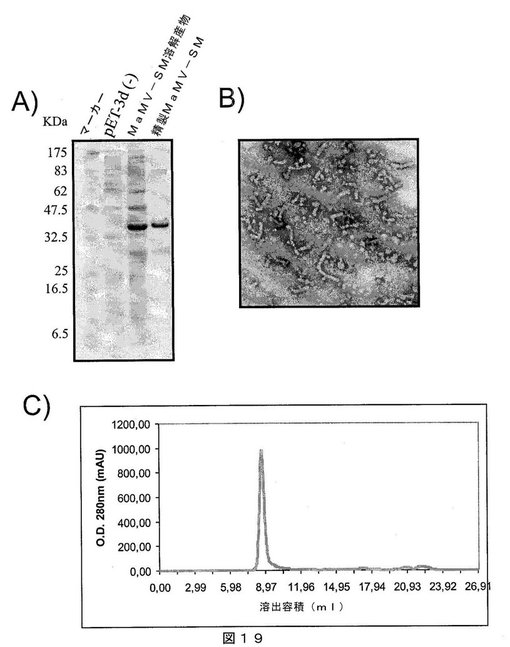
【図20】
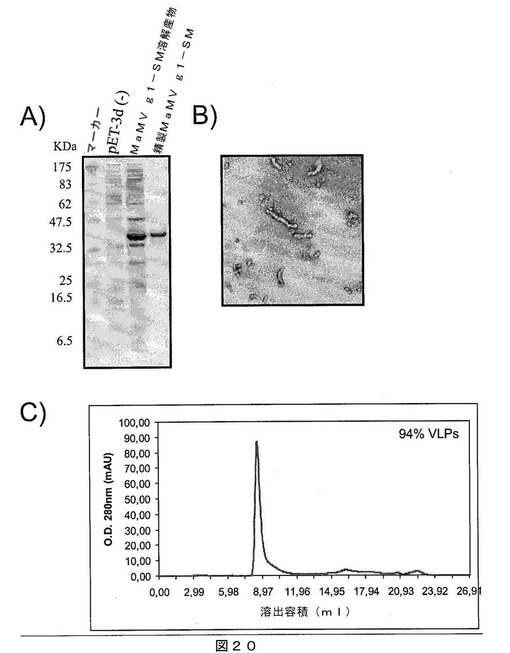
【図21】
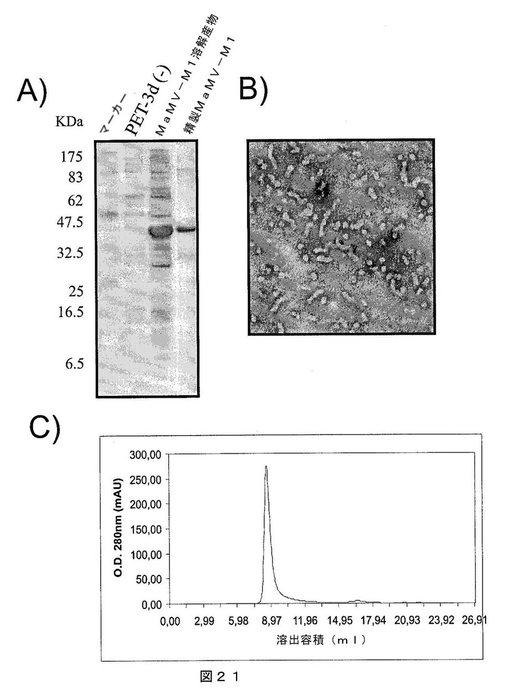
【図22】
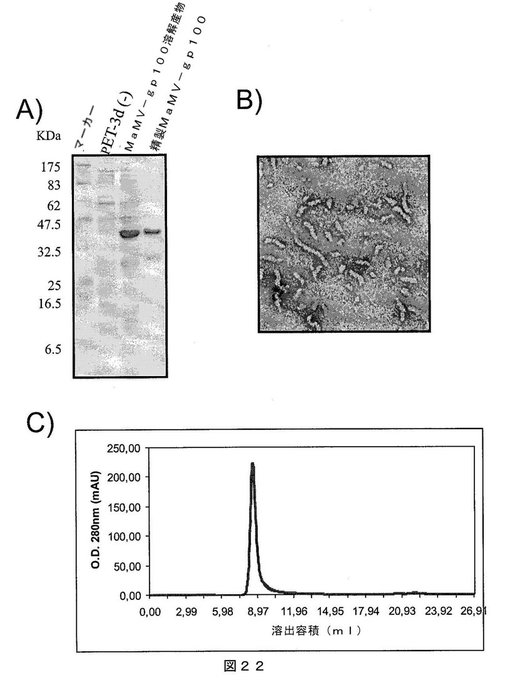
【図23】
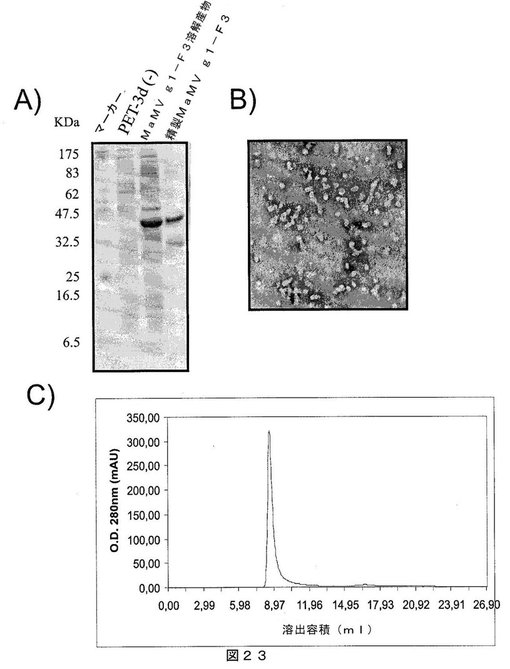
【図24】
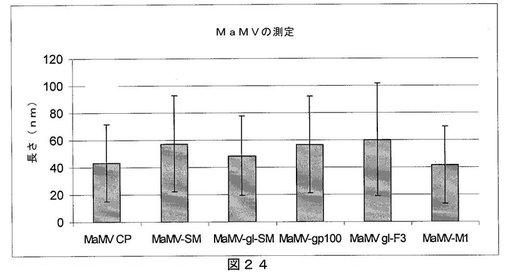
【図25】
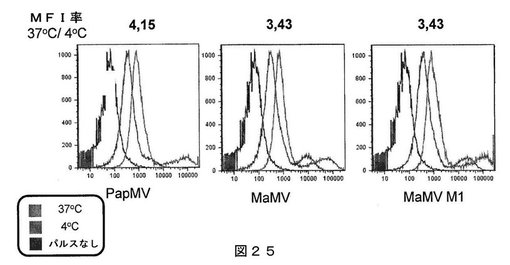
【図26】
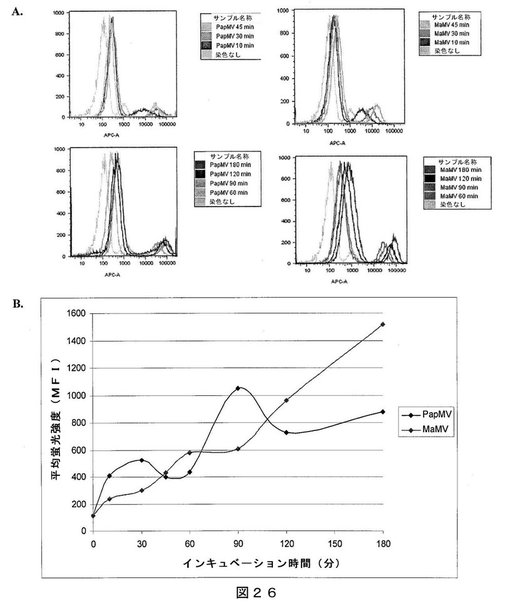
【図27】
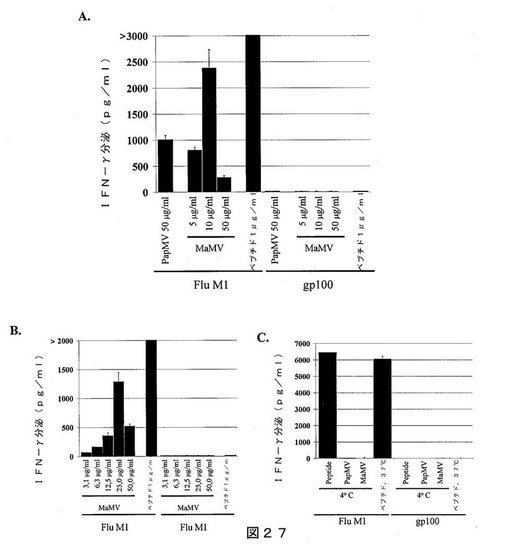
【図28】
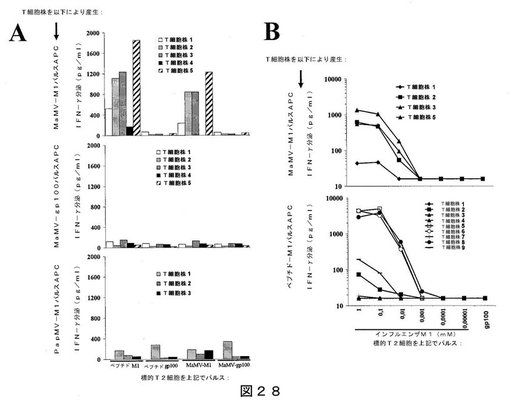
【図29】
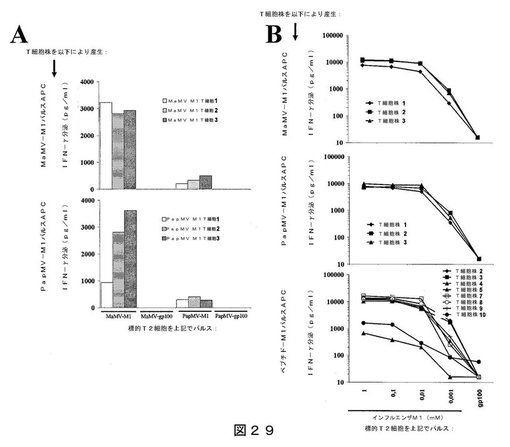
【図30】
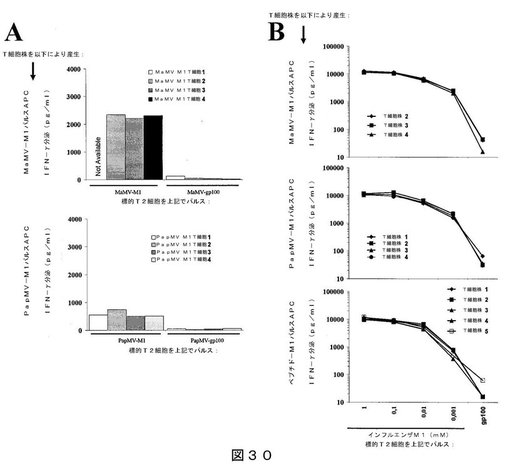
【図31】
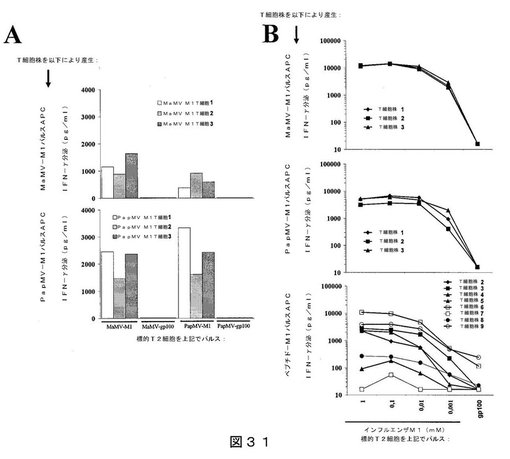
【図32】
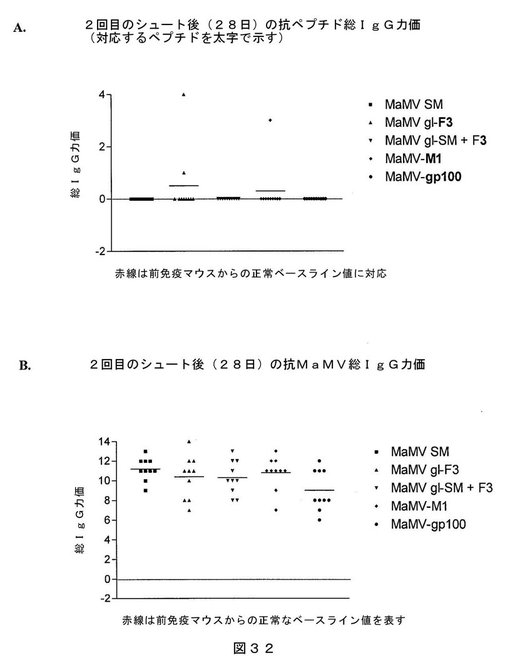
【図33】
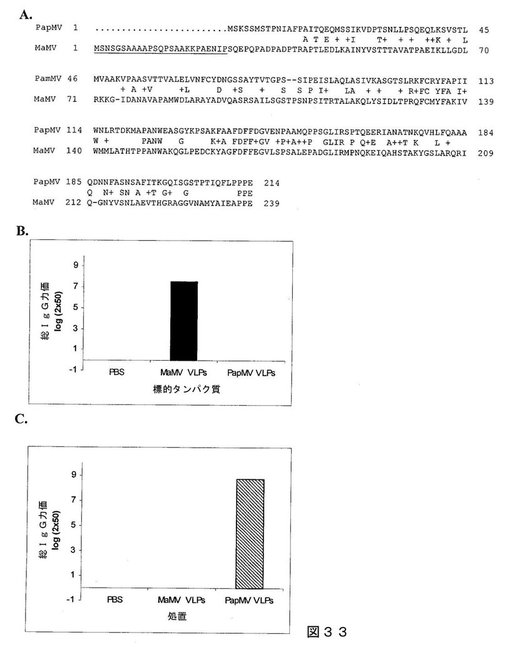
【図1B】
【図1C】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16A−C】
【図16D】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【公表番号】特表2010−538619(P2010−538619A)
【公表日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願番号】特願2010−524318(P2010−524318)
【出願日】平成20年9月11日(2008.9.11)
【国際出願番号】PCT/CA2008/001603
【国際公開番号】WO2009/033276
【国際公開日】平成21年3月19日(2009.3.19)
【出願人】(510066374)ユニベルシテ ラヴァル (1)
【Fターム(参考)】
【公表日】平成22年12月16日(2010.12.16)
【国際特許分類】
【出願日】平成20年9月11日(2008.9.11)
【国際出願番号】PCT/CA2008/001603
【国際公開番号】WO2009/033276
【国際公開日】平成21年3月19日(2009.3.19)
【出願人】(510066374)ユニベルシテ ラヴァル (1)
【Fターム(参考)】
[ Back to top ]