
N−グリカンコアβ−ガラクトシルトランスフェラーゼおよびその使用
本発明は、新しいガラクトシルトランスフェラーゼ、それらをコードしている核酸、ならびにそれらに関連する組換えベクター、宿主細胞、抗体、使用および方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新しいガラクトシルトランスフェラーゼ、それらをコードしている核酸、ならびにそれらに関連する組換えベクター、宿主細胞、抗体、使用および方法に関する。
【背景技術】
【0002】
「回虫」または「線虫」は、擬体腔動物の中の最も多様な門、およびすべての動物の中で最も多様なものの1つである。線虫種は、識別するのが困難であり、80,000を超える種が記載されていて、そのうち15,000を超える種が寄生虫である。回虫種の合計数は、500,000より多いと推定されてきた。線虫は、淡水、海洋環境および陸上環境においていたるところに存在する。多くの寄生虫形態は、大部分の植物、動物における、またヒトにおける病原体を含む。
【0003】
カエノラブディティス・エレガンス (Caenorhabditis elegans)は、モデル線虫であり、分節がなく、蠕虫型であり、左右相称であり、クチクラ外皮、4つの主な表皮性神経索および体液で満たされた擬体腔を有する。野生において、カエノラブディティス・エレガンスは、腐食した植物質に発生する細菌を餌にしている。Hannemannら(Glycobiology、16、874、2006)は、カエノラブディティス・エレガンスのN-グリカンコアにおいて、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-D-GlcNAc(Gal-Fuc)エピトープを単離し、構造的に特性決定した。カエノラブディティス・エレガンスのN-グリコシル化パターンは、Paschingerら(Carbohydrate Res.、343、2041、2008)において最近概説された。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】US 5,582,981
【非特許文献】
【0005】
【非特許文献1】Hannemannら(Glycobiology、16、874、2006)
【非特許文献2】Paschingerら(Carbohydrate Res.、343、2041、2008)
【非特許文献3】Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986
【非特許文献4】Altschulら、J. Mol. Biol.、215、403〜410、1990
【非特許文献5】HuangおよびMiller、Adv. Appl. Math.、12、337〜357、1991
【非特許文献6】BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894
【非特許文献7】SambrookおよびRussell、Molecular cloning: A laboratory manual (3 volumes)、2001
【非特許文献8】http://www.-ncbi.nlm.nih.gov/entrez/query.fcgi?db=nucleotide
【非特許文献8】Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニア
【非特許文献9】Methods in Enzymology、350、248、2002
【非特許文献10】Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カースルバッド、カリフォルニア
【非特許文献11】Novagen Insect Cell Expression Manual、Merck Chemicals Ltd.、ノッティンガム、イギリス
【非特許文献12】Applied Microbiology and Biotechnology、72、211、2006
【非特許文献13】LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009
【非特許文献14】KohlerおよびMilstein、Nature、256、495〜497、1975
【非特許文献15】Winterら、Annu. Rev. Immunol. 12、433〜455、1994
【非特許文献16】Schaffitzelら、J. Immunol. Methods、231、119〜135、1999
【非特許文献17】Giovannoniら、Nucleic Acids Res. 29、E27、2001
【非特許文献18】HolligerおよびHudson、Biotechnol.、23(9)、1126〜36、2005
【非特許文献19】Binzら、Nature Biotechnol.、23(10)、1257〜1268、2005
【非特許文献20】Jayasena、Clin. Chem.、45、1628〜1650、1999
【非特許文献21】KlugおよびFamulok、M. Mol. Biol. Rep.、20、97〜107、1994
【非特許文献22】Nolteら、Nat. Biotechnol.、14、1116〜1119、1996
【非特許文献23】Klussmannら、Nat. Biotechnol.、14、1112〜1115、1996
【非特許文献24】Besteら、Proc. Natl. Acad. Sci. USA、96、1898〜1903、1999
【非特許文献25】Skerra、Biochem. Biophys. Acta、1482、337〜350ページ、2000
【非特許文献26】Skerra、J. Mol. Recognition、13、167〜287、2000
【非特許文献27】Hey、Trends in Biotechnology、23、514〜522、2005
【非特許文献28】Zhangら、Glycobiology、7、1153〜1158、1997
【非特許文献29】Takahashiら、Eur. J. Biochem.、270、2627〜2632、2003
【非特許文献30】Wuhrerら、Biochem. J.、378、625〜632、2004
【非特許文献31】Iskratschら、Anal. Biochem.、368、133〜146、2009
【非特許文献32】Tatenoら、Glycobiology、19(5)、527〜536、2009
【非特許文献33】Paschingerら、Glycobiology、15(5)、463〜474、2005
【非特許文献34】Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001
【非特許文献35】Roitingerら、Glycoconj. J.、15(1)、89〜91、1998
【非特許文献36】Brenner, S.(Genetics 77(1)、71〜94、1974
【非特許文献37】Mullerら、J. Biol. Chem. 277(36)、32417〜32420、2002
【非特許文献38】Gutterniggら、J. Biol. Chem. 282(38)、27825〜27840、2007
【発明の概要】
【発明が解決しようとする課題】
【0006】
Gal-Fuc含有(多/オリゴ)糖およびGal-Fuc含有複合糖質の組換えによる作製のための新しい手段を提供することが、本発明の目的である。さらなる目的は、Gal-Fuc含有多糖/オリゴ糖およびGal-Fuc含有複合糖質のための新しい使用を提供することである。
【課題を解決するための手段】
【0007】
第1の態様において、この目的は、
(i)配列番号1、3、5、7および9に記載されている核酸配列からなる群から選択される少なくとも1つの核酸配列、好ましくは配列番号1の核酸配列を含む核酸;
(ii)配列番号1、3、5および7に記載されている核酸配列からなる群から選択される核酸配列、好ましくは配列番号1の核酸配列と、少なくとも60、65、70または75%の同一性、好ましくは、少なくとも80、85または90%の同一性、より好ましくは、少なくとも95%の同一性、最も好ましくは、少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される単離および精製された核酸によって解決される。
【0008】
好ましい一態様において、単離および精製された核酸は、
(i)配列番号1、3、7および9に記載されている核酸配列ならびに配列番号5の最初の1428核酸からなる群から選択される少なくとも1つの核酸配列、好ましくは配列番号1の核酸配列を含む核酸;
(ii)配列番号1、3および7に記載されている核酸配列ならびに配列番号5の最初の1428核酸からなる群から選択される核酸配列、好ましくは配列番号1の核酸配列と、少なくとも60、65、70または75%の同一性、好ましくは、少なくとも80、85または90%の同一性、より好ましくは、少なくとも95%の同一性、最も好ましくは、少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる、核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される。
【0009】
好ましくは、上記の核酸は、本発明のポリペプチド、好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6-(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986による命名)含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、好ましくは酵素ガラクトシルトランスフェラーゼ活性を有するポリペプチド、より好ましくはβ-1,4-ガラクトシルトランスフェラーゼ活性を有するポリペプチドをコードしている。
【0010】
本明細書において使用される場合、ガラクトシルトランスフェラーゼ活性は、活性化されたドナー型(すなわち、ヌクレオチド活性化ガラクトース、好ましくは、UDP-Gal)からアクセプターへの、ガラクトース残基の酵素転移を記述することを意味する。本明細書において使用される場合、β-1,4-ガラクトシルトランスフェラーゼ活性は、ガラクトシルトランスフェラーゼ活性の特異性、すなわち、β1,4-配座でのガラクトースのアクセプター分子への転移を記述することを意味する。本明細書において使用される場合、アクセプター基質としてのL-フコシドに対するβ-1,4-ガラクトシルトランスフェラーゼ活性は、アクセプター基質としてのL-フコシドへのβ- 1,4-結合型転移におけるガラクトシルトランスフェラーゼ活性の特異性を記述することを意味する。本明細書において意味される場合、L-フコシドは、α、最も好ましくは、α-1,6配座において、例えば、MMF6またはGnGnF6(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986)の部分として、末端L-フコースを含有するアクセプター基質としての多糖/オリゴ糖または複合糖質を記述することを意味する。
【0011】
最も好ましい実施形態において、コードされているポリペプチドは、配列番号2、4、6、8および10に記載されているポリペプチド配列からなる群から選択されるポリペプチド配列、好ましくは、配列番号2のポリペプチド配列、またはこれらのうちいずれかの機能的フラグメントまたは機能的誘導体を含む。
【0012】
配列番号1は、配列番号2をコードしている核酸配列である:
(NCBIにおいて、参照配列NM_072144.4として、Wormbaseにおいて、M03F8.4としても記載されており、カエノラブディティス・エレガンスのガラクトシルトランスフェラーゼ[実施例セクションにおいてGalTと称されている]をコードしている)
ATGCCTCGAATCACCGCCAGTAAAATAGTTCTTCTAATTGCATTATCATTTTGTATTACTGTTATTTATCACTTTCCAATAGCAACGAGAAGCAGTAAGGAGTACGATGAATATGGAAATGAATATGAAAACGTTGCATCGATAGAGTCGGATATAAAAAATGTACGTCGATTACTTGACGAGGTACCGGATCCCTCACAAAACCGTCTACAATTCCTGAAACTTGATGAGCATGCTTTTGCATTCTCGGCCTACACAGACGATCGAAATGGAAATATGGGGTACAAATATGTCCGAGTCCTGATGTTTATCACGTCACAAGACAACTTTTCCTGTGAAATAAACGGGAGAAAGTCCACAGATGTATCACTTTACGAGTTCTCGGAAAATCACAAAATGAAGTGGCAAATGTTTATTTTGAATTGTAAACTACCCGATGGTATAGATTTCAATAATGTTAGCTCTGTAAAGGTCATAAGAAGCACAACCAAGCAGTTTGTTGATGTGCCGATTCGGTATAGAATTCAAGATGAGAAAATAATTACGCCAGACGAATATGACTATAAAATGTCAATTTGTGTTCCAGCATTGTTTGGAAATGGATATGATGCAAAGCGAATTGTTGAGTTTATTGAGCTGAATACTTTGCAAGGAATCGAGAAAATATACATTTACACTAATCAAAAAGAGCTTGATGGATCCATGAAGAAAACGTTGAAATACTATTCGGATAATCACAAAATAACATTAATTGATTACACATTACCATTCAGAGAGGATGGTGTTTGGTATCACGGGCAATTGGCAACTGTTACTGATTGTTTACTGAGAAACACTGGAATCACAAAATACACATTTTTCAATGATTTTGATGAGTTCTTCGTCCCCGTTATCAAAAGTCGGACTCTCTTTGAAACAATCAGTGGGCTTTTTGAAGATCCCACTATTGGATCGCAACGAACAGCTTTGAAGTATATAAATGCAAAAATCAAGAGCGCTCCGTATTCACTGAAAAATATTGTTTCCGAAAAACGAATTGAAACAAGATTCACGAAATGTGTAGTTCGACCGGAAATGGTTTTGAACAGGGTATTCATCATACGAGTAGAGTGATTCAAGACAACTATAAAACGGTTTCCCATGGCGGATCCCTTCTACGGGTTTATCATTACAAGGATAAAAAGTATTGTTGCGAAGACGAGAGCCTCTTGAAAAAACGGCATGGAGATCAACTTCGGGAAAAATTCGATTCAGTTGTTGGTCTTTAGACTTGTAG
配列番号2(NCBIにおいて参照配列NP_504545.2とも記載されている)
MPRITASKIVLLIALSFCITVIYHFPIATRSSKEYDEYGNEYENVASIESDIKNVRRLLDEVPDPSQNRLQFLKLDEHAFAFSAYTDDRNGNMGYKYVRVLMFITSQDNFSCEINGRKSTDVSLYEFSENHKMKWQMFILNCKLPDGIDFNNVSSVKVIRSTTKQFVDVPIRYRIQDEKIITPDEYDYKMSICVPALFGNGYDAKRIVEFIELNTLQGIEKIYIYTNQKELDGSMKKTLKYYSDNHKITLIDYTLPFREDGVWYHGQLATVTDCLLRNTGITKYTFFNDFDEFFVPVIKSRTLFETISGLFEDPTIGSQRTALKYINAKIKSAPYSLKNIVSEKRIETRFTKCVVRPEMVFEQGIHHTSRVIQDNYKTVSHGGSLLRVYHYKDKKYCCEDESLLKKRHGDQLREKFDSVVGLLDL
配列番号3は、配列番号4をコードしている核酸配列である:
(NCBIにおいて参照配列XM_001674213.1とも記載されており、カエノラブディティス・ブリグサエ(Caenorhabditis briggsae)のガラクトシルトランスフェラーゼをコードしている)
ATGCCACGAA TAACGGCAAG CAAAATAGTG TTATTATCTG TATTATCCTT ACTAACAGTT TTCTATCTGA ATACATTTTC GTCTATTAAA ATTGAAAACG ATCTCGACGG GACTGATTAC GACTTGGATT ACATAGAATC TGATATCAAA AAGACGCGTC GATTACTCAA TGAAATCCCT GATCCATCTC AAAACCGAGT TCAATTTTTT AAACTCGATG ATAATGGATA TGCATTCTCA GCATATACAG ATAATAGGAA AGGAAATATG GGTCACAAAT ATGTCAGAAT ATTAGTGTTC CTAACTAAAT TTGATGATTT TTCTTGCGAA ATTAACTCGA AGAAATCCTA TGTTGTTACA CTCTACGAGC TATCAGAAAA TCACAATATG AAGTGGAAAA TGTATATTTT GAATTGTTTA CTTCCCGATG GAATCACTTT CAACGATGTG AATTCTGTAA AAATATCTAG AAGTTCTTCA AAACTTTCAG TCCAAATCCC GATCAGATAT AGAATTCAAG ATGAGAAAAT GATGACTCCA GATGAATACG ATTATAAGTT GTCGATTTGT GTTCCTGCAC TTTTTGGAAA CGTTTATTAT CCAAGGAGGA TTATTGAATT TGTGGAACTA AACAGCTTGC AAGACATCGA CAAAATCTAC ATCTACTACA ATCCTTTAGA AATGACAGAT GAGGCCACAG AAAGGACTTT GAAGTTTTAT TCCAATAATG GGAAAATCAA TTTAATAGAA TTCATTCTCC CATTTCTAC TCGAGATGTT TGGTATTATG GGCAATTGGC CACCGTTACA GATTGTCTTC TCCGTAACAC TGGAATAACT CAATACACAT TTTTCAATGA TTTGGATGAA TTTTTCGTGC CAGTACTGGA CAACCAAACT CTCTCTGAAA CTGTGTCAGG ATTATTTGAA AATCGAAAAA TTGCCTCTCA GAGAACGGCC TTGAAATTTA TTAGTACAAA AATCAATCGA TCTCCTGTAA CTCTCAATAA TATTGTGTCT TCTAAAAATT TTGAAACGAG ATTCACAAAA TGCGTCGTAC GGCCGGAAAT GGTTTTTGAG CAGGGCATTC ACCATACGAG TAGAGTAATA CAAGACGACT ACGAAACCCC ATCCCATGAT GGATCACTTT TGCGTGTGTA TCACTACAGA GAACCAAGAT ATTGCTGCGA AAACGAGAAT CTTCTAAAAC AAAGATACGA TAAGAAGCTT CAAGAAGTTT TTGATGCTGT AGTTCTTATA TTGCATGTCA CATTTGATGT ATGGATATAT CACCTGAAAA ACACCCTCTA A
配列番号4(NCBI参照配列XP_001674265.1においても記載されている)
MPRITASKIV LLSVLSLLTV FYLNTFSSIK IENDLDGTDY DLDYIESDIK KTRRLLNEIP DPSQNRVQFF KLDDNGYAFS AYTDNRKGNM GHKYVRILVF LTKFDDFSCE INSKKSYVVT LYELSENHNM KWKMYILNCL LPDGITFNDV NSVKISRSSS KLSVQIPIRY RIQDEKMMTP DEYDYKLSIC VPALFGNVYY PRRIIEFVEL NSLQDIDKIY IYYNPLEMTD EATERTLKFY SNNGKINLIE FILPFSTRDV WYYGQLATVT DCLLRNTGIT QYTFFNDLDE FFVPVLDNQT LSETVSGLFE NRKIASQRTA LKFISTKINR SPVTLNNIVS SKNFETRFTK CVVRPEMVFE QGIHHTSRVI QDDYETPSHD GSLLRVYHYR EPRYCCENEN LLKQRYDKKL QEVFDAVVLI LHVTFDVWIY HLKNTL
配列番号5は、配列番号6、その後に終止コドンおよびさらに68ヌクレオチドをコードしている核酸配列(1428核酸)である:(NCBI参照配列XM_001629141.1においても記載されており、ネマトステラ・ベクテンシス(Nematostella vectensis)のガラクトシルトランスフェラーゼをコードしている)
ATGCGATGCT ATATTTACAA ATTGAGGTTG TCCGTTTGTC TGTTTGTAGT GCTCTTCACA GCACTGCTTT TCATCACCTA TTTAAACCAC TCAGAGCTTG AATCAGCAGA GAAAAGTAGC GGAAAAAGGA AGACGCGACA TCGTAAACGA ACACGTTCAC GCAAACAACA CGAGAGCCAT TTTCAGAAAG CTCGACTACA AGAAAGAGAA CTAGTATTAA GATCTACAGC GCCACCAACA TTACGAAGAG AAGTACAAGC GCATCGATTA GGGCAGATCC GTGGCAAGAA CACGGACCAG GGGATAACTG GAAAGTTCAC AGAGATCGCT AAAGACACGC ATATTTATTC AGCGTTTTAC GACGATGCCA AGTCAAATCC ATTCATTCGT CTTATCATCC TCTCGGGAAA ACACTACCAG CCTGGATTAT CTTGCCAATT TTGCGAACCT TTGTCCGCCA GTTGTAGTTT TGCGGACTCT AAAGCTGAAT ACTACACGAC CAACGAGAAC CATGGGAGAG TATTTGGCGG GTTCATTGCG AGTTGCCTCG TGCCTGATGG ATTCAATGCA GTGCCATTGT TTGTTGACAT AACGGCCGAT GTTAAGGGGG AGAAAAGCAA GGCACGGGTA CCTGTGGTGT CTAATGCACA TCTCTACTAC CCTATTAAAT ACGCAATCTG CGTCCCACCC CTCCGATCAG AGAAACTAAC AGCGAAAAGA CTCATAGAGT TTGTCGAGCT AACCAAACTT TTAGGCGCTA ACCATTTTAC TTTTTATGAC TTCAAAACGG ACCCGGAAGT CAATAACGTT TTAAGATATT ACCAGGAGAC ACAAGTAGCA AATGTTCTGC CATGGAATCT ACCTTCAAAT TTGGTATCCA GGCCGAACGA TATTTGGTAC TTTGGTCAGG TTTTGGCTAT TCTAGATTGC TTGTATCGCT ACAAGAACAG GGCAAAATTT GTAGCCTTCA ATGACGTAGA TGAGTTTATC GTTCCGCTAA GGAACAGCTC GATAGTGGAA ATACTAAACG CGTTTCACCG GCCATACCAC TGTGGACATT GCTTTCAGAG CGTGGTGTTC AGCTCAAACG CGAGATTTCC CAGGCAAAAA AGCGAGTTAG TTTCTCAGCG GTTCTTCCAC AGGACCCAGG AAACCATCCC TCTCCTCTCG AAATGCATTG TGGATCCTTT GAGAGTGTTC GAGATGGGGA TTCACCACAT AAGCAAGGCT ACAGGTCTGC GGTATTCCGT CAACTCAGTA CACGAGAGTG ACGCGGTTAT CTTCCATTAC AGGACTTGCA CTACGTCATT TGGTATACGT CATCAGTGCA TGAACCTAGT GCATGATGGG ACCATGGCCA AATATGGAAA ACGACTTCAG AAAATGTTTA GAAAGGTTGT AAATGATTTA AAACTTTTGG CACCAACGTA GCTATTTCGT AACACTTCAC ACTTTCATTG TTATAACAGA ATACAGAATA AATTAATGAT TGTTGTGCC
配列番号6(NCBI参照配列XP_001629191においても記載されている)
MRCYIYKLRL SVCLFVVLFT ALLFITYLNH SELESAEKSS GKRKTRHRKR TRSRKQHESH FQKARLQERE LVLRSTAPPT LRREVQAHRL GQIRGKNTDQ GITGKFTEIA KDTHIYSAFY DDAKSNPFIR LIILSGKHYQ PGLSCQFCEP LSASCSFADS KAEYYTTNEN HGRVFGGFIA SCLVPDGFNA VPLFVDITAD VKGEKSKARV PVVSNAHLYY PIKYAICVPP LRSEKLTAKR LIEFVELTKL LGANHFTFYD FKTDPEVNNV LRYYQETQVA NVLPWNLPSN LVSRPNDIWY FGQVLAILDC LYRYKNRAKF VAFNDVDEFI VPLRNSSIVE ILNAFHRPYH CGHCFQSVVF SSNARFPRQK SELVSQRFFH RTQETIPLLS KCIVDPLRVF EMGIHHISKA TGLRYSVNSV HESDAVIFHY RTCTTSFGIR HQCMNLVHDG TMAKYGKRLQ KMFRKVVNDL KLLAPT
配列番号7は、配列番号8をコードしている核酸配列である:
(NCBI参照配列XM_002189335においても記載されており、キンカチョウ(Taeniopygia guttata)のガラクトシルトランスフェラーゼをコードしている)
ATGACTGTAA CTTTAATGCT TGTGGTTTCT TATCTGAGAT TACAGAGACT TTCTCATCAG CCAAAAGTAA TTCAAGAAAG TAGAAGATGT AGAGGGAAAA TTGCCCTTAG CACAATAACA GCATTGGAAG GTAACAAAAC TGATATTATA TCCCCATACT TTGATGACAG AGAAAACAAA ATCACTCGTC TGATTGGGAT TGTTCACCAT AAAGATGTAA AACAACTGTT CTGCTGGTTC TGCTGTCAAG CCAATGGAAA GATATATGTA TCAAAAGCAG AAATAGATGT TCACTCGGAT AGATTTGGAT TCCCTTATGG TGCAGCAGAT ATAATTTGTT TGGAACCTGA AAACTGTGAT CCAACACATG TATCAATTCA TCAGTCTCCA TATGGAAATA TTGACCAGCT GCCGAGGTTT GAAATTAAAA ATCGCAGGCC TGAGACCTTT TCTGTTGACT TCACCGTGTG CATTTCTGCC ATGTTTGGAA ACTACAACAA TGTCTTGCAG TTTGTACAGA GTATGGAAAT GTATAAGATT CTTGGAGTAC AGAAAGTGGT GATCTATAAG AACAACTGCA GCCATCTGAT GGAGAAAGTC TTGAAATTTT ATATAGAAGA AGGAACTGTT GAGGTAATTC CCTGGCCAAT AGACTCACAC CTCAGGGTTT CTTCTAAATG GCGCTTCATG GAAGACGGGA CACACATTGG CTACTATGGA CAAATCACAG CTCTAAATGA CTGTATATAC CGCAACATGG AAAGGACCAA GTTTGTGGTC CTTAATGACG CTGATGAAAT AATTCTTCCC CTTAAACACC CAGACTGGAA AACAATGATG AACAGTCTTC AGGAGCAAAA CCCAGGGACT AGTGTTTTCC TTTTTGAGAA CCATATCTTC CCAGAAACTG TATTTTCTCC CATGTTCAAC ATTTCATCTT GGAATACTGT GCCAGGTGTT AACATATTGC AGCATGTGTA CAGAGAGCCT GACAGGAAAC ATGTAATCAA TCCCAGGAAA ATGATAGTTG ATCCACGAAA GGTGATTCAG ACTTCAGTCC ATTCTGTCCT ACGTGCTTAT GGGAAGAGCG TGAATGTTCC CATGGAAGTT GCCCTCATTT ATCACTGTCG GAAGGCCCTT CAAGGAAACC TTCCCAGAGA ATCTCTCATC AGGGATACAA CACTGTGGAG ATATAACTCA TCATTAATCA TGAATGTTAA CAAGGTTCTA TCTCAAACCA TGCTGCAAAC TCAAAATTGA
配列番号8(NCBI参照配列XP_002189371においても記載されている)
MTVTLMLVVS YLRLQRLSHQ PKVIQESRRC RGKIALSTIT ALEGNKTDII SPYFDDRENK ITRLIGIVHH KDVKQLFCWF CCQANGKIYV SKAEIDVHSD RFGFPYGAAD IICLEPENCD PTHVSIHQSP YGNIDQLPRF EIKNRRPETF SVDFTVCISA MFGNYNNVLQ FVQSMEMYKI LGVQKVVIYK NNCSHLMEKV LKFYIEEGTV EVIPWPIDSH LRVSSKWRFM EDGTHIGYYG QITALNDCIY RNMERTKFVV LNDADEIILP LKHPDWKTMM NSLQEQNPGT SVFLFENHIF PETVFSPMFN ISSWNTVPGV NILQHVYREP DRKHVINPRK MIVDPRKVIQ TSVHSVLRAY GKSVNVPMEV ALIYHCRKAL QGNLPRESLI RDTTLWRYNS SLIMNVNKVL SQTMLQTQN
配列番号9は、配列番号10をコードしている核酸配列である:
(NCBI参照配列XM_626032においても記載されており、クリプトスポリジウム・パルバム(Cryptosporidium parvum)のガラクトシルトランスフェラーゼをコードしている)
ATGCAAAGTA AAGTCATTTT TAGGATCTTG GTATTGATCA TTTCGGTGAT TGGATCCTTA TACTCAATAA TTCAATTAAT GCTAAAGGAG CTATCAAGTA ACAAAAATAT TCAAGAGGTT AGTCATTCAA GGAGGCTAAT AAGTGAACCT TACAGTGAAA GTATTAATGA ACAAAATGAT CAAGATTGGA AAGAACTAAA GCTAATAATT CCAAATCATT CTCAAATTAA CCAGCAGGAA AAAAATGGTA ATTTGATTGA GTTTAAAGTT TATATATACT CAGCATATTA TGATTGGAGA ATAGATAGGA TACGAATAAA TTCACTTATC CCATCGAATT TTTATGATCG AATAGAAATG GAATGTGCAA TAATCTTGGA CAAAAATATT TACACAGGAA CTATTAAAAA AGTGATTCAT AAGGAGCACC ATAATAAAGA ATATGTATCA TCGACTTTAC TCTGCGAAAT TGCAAAAAAT GAAATTAAAT TTGAGGATAT TTCAAGGAAA GTTTTGATAA CAATTTTGGA AAATGGAAAC AGCACAAATA AATCAGAAAT ATGGATAACT CTAAAAAAAA TTCCAAAAAA TAGCTCTAAT AATCATGAGC TGACTGTTTG TGTGAGACCT TGGTGGGGAG AGCCAATAAA GAATGGAAAC TTGGGAAATA AACAAAAATT TAACAATTCA GGGTTAATGC TTGAATTTAT TAATTCATAT TTATTCTTAG GAGCAAATAA ATTTTATTTA TATCAAAATT ACTTGGACAT TGACGAAGAT GTAAGAAATA TAATAAATTA TTATTCTAAT ATCAAAAATG TTTTGGAAAT TATTCCATAC TCATTACCAA TAATTCCATT TAAACAAGTT TGGGATTTCG CACAAACAAC AATGATACAG GACTGCCTAC TAAGAAATAT TGGAAAAACA AAATACTTGT TATTCGTAGA TACCGATGAA TTTGTATTTC CAAACTTGAA AAATTATAAC TTAATGGATT TTTTAAATTT ATTAGAAGCC AACAATCCTT ATTATAAAAA CAAAGTCGGG GCAATGTGGA TTCCAATGTA TTTTCATTTT TTAGAGTGGG AATCTGATAA AAATAATTTG AAGAAATATT CAACAATTGA GAAAAAAATT AAGAAAAAGA TGGCAAATAT TGAGTTTGTT CTATATCGTA AAACATGTAG AATGTTAAGT TCTGGAACAA AAAAAAGTGA CAAGACGAGA AGAAAAGTTA TTATTAGACC TGAAAGAGTT TTGTATATGG GTATACATGA AACAGAAGAG ATGCTAAGCA AAAAATTTCA TTTCATTAGA GCTCCTGTAA TTAATGTGGG TGGAGGAAAC GAACTAAGTA TATATTTACA TCATTATAGA AAAGCAAAAG GTATTGTAAA CAATGATCCC AAACAAAGAG AACTTGTGAA TATGTATTTA GAAAATGTTT GTTCAGATAA GCTGTTAGAT TCAGGGGGAG ATTCCATTCA AGATGGAGTA ATTGTCGACA ATACTGTTTG GGAGATATTT GGAACACACT TATACCAGAT AATTTTTGAG CATATTAAAG AAATCCAAGA TATGTACACA AATAAGGAAA TAATTAATGG AAATAAAAAT TTAAGTGTTG AAGAATTACA TAATTAA
配列番号10(NCBI参照配列XP_626032においても記載されている)
MQSKVIFRIL VLIISVIGSL YSIIQLMLKE LSSNKNIQEV SHSRRLISEP YSESINEQND QDWKELKLII PNHSQINQQE KNGNLIEFKV YIYSAYYDWR IDRIRINSLI PSNFYDRIEM ECAIILDKNI YTGTIKKVIH KEHHNKEYVS STLLCEIAKN EIKFEDISRK VLITILENGN STNKSEIWIT LKKIPKNSSN NHELTVCVRP WWGEPIKNGN LGNKQKFNNS GLMLEFINSY LFLGANKFYL YQNYLDIDED VRNIINYYSN IKNVLEIIPY SLPIIPFKQV WDFAQTTMIQ DCLLRNIGKT KYLLFVDTDE FVFPNLKNYN LMDFLNLLEA NNPYYKNKVG AMWIPMYFHF LEWESDKNNL KKYSTIEKKI KKKMANIEFV LYRKTCRMLS SGTKKSDKTR RKVIIRPERV LYMGIHETEE MLSKKFHFIR APVINVGGGN ELSIYLHHYR KAKGIVNNDP KQRELVNMYL ENVCSDKLLD SGGDSIQDGV IVDNTVWEIF GTHLYQIIFE HIKEIQDMYT NKEIINGNKN LSVEELHN
【0013】
本発明との関連において使用される場合、「ポリペプチドをコードしている核酸」という用語は、対立変異および遺伝子コードの冗長性を含むことを意味する。
【0014】
当業者に知られているようにおよび本明細書において使用されるように、「%(パーセント)同一性」という用語は、配列間の合致によって決定される2つ以上の核酸分子間の関連性の程度を示す。「同一性」のパーセンテージは、ギャップおよび別の配列特性を考慮した上での2つ以上の配列における同一な領域のパーセンテージの結果である。
【0015】
関連する核酸分子の同一性は、既知の方法による支援によって決定することができる。一般に、この作業の特定の要求に適応させるために適合させたアルゴリズムを使用する特別なコンピュータプログラムが用いられる。同一性を決定するための好ましい方法は、比較される配列間で最大の程度の同一性の作成から始まる。2つの核酸配列間の同一性を決定するための好ましいコンピュータプログラムは、BLASTN(Altschulら、J. Mol. Biol.、215、403〜410、1990)およびLALIGN(HuangおよびMiller、Adv. Appl. Math.、12、337〜357、1991)を含むがこれらに限定的されない。BLASTプログラムは、米国立生物工学情報センター(NCBI:National Center for Biotechnology Information)から、および別の供給源(BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894)から入手することができる。
【0016】
本発明による核酸分子は、当業者によく知られている方法によって合成により調製してもよいが、また、適したDNAライブラリーおよび別の公的に利用可能な核酸の供給源から単離してもよく、その後、任意選択により、突然変異させてもよい。そのようなライブラリーまたは突然変異の調製は、当業者によく知られている。
【0017】
好ましい一実施形態において、本発明の核酸分子は、二重鎖または単鎖のいずれか(すなわちセンスまたはアンチセンス鎖のいずれか)の、cDNA、ゲノムDNA、合成DNA、RNAまたはPNAである。本発明の範囲内に包含される核酸分子およびそのフラグメントは、例えば、ポリメラーゼ連鎖反応(PCR)によって作製してもよく、または、DNA合成を使用して合成によって、もしくはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムのmRNAを使用して逆転写によって生成してもよい。
【0018】
場合によって、本発明はまた、特に参照されている核酸配列と、好ましくはストリンジェントな条件下で、ハイブリダイズする能力を有することを特徴とする、本発明のポリペプチドをコードしている新規な核酸も提供する。特に参照されている核酸配列とストリンジェントな条件下でハイブリダイズする能力を決定するための、従来技術において一般的なおよび/または標準的なプロトコール(例えば、SambrookおよびRussell、Molecular cloning: A laboratory manual (3 volumes)、2001)の次に、例えば、上記のBLASTN(Altschulら、J. Mol. Biol.、215、403〜410、1990)およびLALIGNアラインメントツールなどのアラインメントツールを用いて遺伝子データベース(例えば、http://www.-ncbi.nlm.nih.gov/entrez/query.fcgi?db=nucleotide)において見出すことができるヌクレオチド配列を比較することによって、特に参照されている核酸配列とストリンジェントな条件下でハイブリダイズする能力を分析および決定することが好ましい。
【0019】
最も好ましくは、本発明の核酸が、核酸、例えば、配列番号1、3、5、7および/または9のいずれかにおいて記載されている核酸とハイブリダイズする能力は、以下の条件下で、サザンブロットアッセイにおいて確認される:45℃での6×塩化ナトリウム/クエン酸ナトリウム(SSC)、その後0.2×SSC、0.1%SDS中で65℃での洗浄。
【0020】
本発明の核酸は、本発明のポリペプチドをインビトロまたはインビボで産生するために適したベクターおよび/または宿主細胞における発現を支配するプロモーターに作動可能に連結されていることが好ましい。
【0021】
単離および精製された核酸への作動可能な連結に適したプロモーターは、当技術分野において知られている。好ましい一実施形態において、本発明の核酸は、ピキア・パストリス(Pichia pastoris)AOX1またはGAPプロモーター(例えば、Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)GAL1、ADH1、ADH2、MET25、GPDまたはTEFプロモーター(例えば、Methods in Enzymology、350、248、2002を参照されたい)、バキュロウイルスポリヘドリンp10またはie1プロモーター(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カースルバッド、カリフォルニア、およびNovagen Insect Cell Expression Manual、Merck Chemicals Ltd.、ノッティンガム、イギリスを参照されたい)、大腸菌(E.coli)T7、araBAD、rhaP BAD、tetA、lac、trc、tacまたはpLプロモーター(Applied Microbiology and Biotechnology、72、211、2006を参照されたい)、植物CaMV35S、ocs、nos、Adh-1、Tetプロモーター(例えば、LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009を参照されたい)またはSambrookおよびRussell(2001)に記載の、哺乳動物細胞用の誘導可能なプロモーターからなる群から選択されるプロモーターに作動可能に連結されているものである。
【0022】
好ましくは、単離および精製された核酸は、エピソームベクターまたはウイルスベクターなどの組換えベクターの形態である。適したベクターおよび発現制御配列の選択ならびにベクター構築は、当業者の範囲内である。好ましくは、ウイルスベクターは、バキュロウイルスベクター(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)である。プロモーターおよび別の発現制御配列とのコード配列の作動可能な連結を含むベクター構築は、当業者の範囲内である。
【0023】
したがって、またさらなる態様において、本発明は、本発明の核酸を含む組換えベクターに関する。
【0024】
本発明のさらなる一態様は、本発明の、好ましくは、本発明のポリペプチドを産生する、核酸および/またはベクターを含む宿主細胞を対象とする。本発明のポリペプチドを産生するための好ましい宿主細胞は、酵母細胞、好ましくは、サッカロマイセス・セレビジエ(例えば、Methods in Enzmology、350、248、2002を参照されたい)、ピキア・パストリス細胞(例えば、Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)、大腸菌細胞(BL21(DE3)、K-12および誘導体)(例えば、Applied Microbiology and Biotechnology、72、211、2006を参照されたい)、植物細胞、好ましくは、タバコ(Nicotiana tabacum)またはヒメツリガネゴケ(Physcomitrella patens)(例えば、LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009を参照されたい)、NIH-3T3哺乳動物細胞(例えば、SambrookおよびRussell、2001を参照されたい)および昆虫細胞、好ましくは、sf9昆虫細胞(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)からなる群から選択される。
【0025】
本発明の別の重要な態様は、
(a)配列番号2、4、6、8および10からなる群から選択されるアミノ酸配列、好ましくは配列番号2のアミノ酸配列を有するポリペプチド、
(b)本発明の核酸によってコードされているポリペプチド、
(c)(a)および/または(b)のポリペプチドと、少なくとも25、30、または40%、好ましくは、少なくとも50または60%、より好ましくは、少なくとも70または80%、最も好ましくは、少なくとも90または95%のアミノ酸配列同一性を有するポリペプチド、
(d)(a)、(b)または(c)のフラグメントおよび/または機能的誘導体
からなる群から選択される、単離および精製されたポリペプチドを対象とする。
【0026】
関連するアミノ酸分子の同一性は、既知の方法による支援によって決定することができる。一般に、この作業の特定の要求に適応させるために適合させたアルゴリズムを使用する特別なコンピュータプログラムが用いられる。同一性を決定するための好ましい方法は、比較すべき配列間で最大の程度の同一性を作成することから始まる。2つのアミノ酸配列間の同一性を決定するための好ましいコンピュータプログラムは、TBLASTN、BLASTP、BLASTXまたはTBLASTX(Altschulら、J. Mol. Biol.、215、403〜410、1990)を含むがこれらに限定的されない。BLASTプログラムは、米国立生物工学情報センター(NCBI:National Center for Biotechnology Information)から、および別の供給源(BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894)から入手することができる。
【0027】
好ましくは、前記ポリペプチドは、本発明の上記の核酸によってコードされている。
【0028】
好ましい一実施形態において、本発明のポリペプチド、フラグメントおよび/または誘導体は、機能的である、すなわち、好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-および最も好ましくは、GnGnF6-(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986による命名)含有多糖/オリゴ糖または複合糖質をアクセプター基質として、酵素ガラクトシルトランスフェラーゼ活性、好ましくは、酵素β-1,4-ガラクトシルトランスフェラーゼ活性、より好ましくは、酵素β-1,4-ガラクトシルトランスフェラーゼ活性を有する。
【0029】
例えば、本発明による、ポリペプチド、そのフラグメントおよび誘導体の機能性、すなわち酵素活性を決定するための好ましいアッセイは、下記の実施例4において示されている。
【0030】
本発明のポリペプチドの「機能的誘導体」という用語は、その誘導体が、測定可能な程度まで、例えば、もとの非修飾ポリペプチドの少なくとも約1〜10%の、上記の酵素活性の少なくとも1つを依然として有する限り、例えば、アミノ酸残基の付加、置換および/または欠失によってそのアミノ酸配列が化学的にまたは遺伝子的に修飾されている、および/または、例えば、付加、欠失、再編成、酸化、還元などによって、その原子および/または化学的官能基の少なくとも1つが化学的に修飾されている、任意のポリペプチドまたはそのフラグメントを含むことを意味する。
【0031】
この状況において、本発明の機能的フラグメントは、本発明のポリペプチドまたは誘導体の一部を形成し、かつ測定可能な程度で、例えば、完全なタンパク質の少なくとも約1〜10%の、上記の酵素活性の少なくとも1つを依然として有するものである。
【0032】
「単離および精製されたポリペプチド」という用語は、本明細書で使用される場合、例えば、カエノラブディティス・エレガンス組織またはその画分中の、天然に存在する対応物がない(例えば、ペプチド模倣物)、または、それに天然に付随する成分から分離または精製されている、ポリペプチドまたはペプチドフラグメントを指す。好ましくは、ポリペプチドは、乾燥調製物の少なくとも60重量%を占める場合、「単離および精製された」、したがって、最も天然に存在するポリペプチドおよび/またはそれに天然に付随する有機分子を含まないとみなされる。好ましくは、本発明のポリペプチドは、乾燥調製物の少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは、少なくとも99重量%を占める。乾燥ポリペプチド調製物の少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは、少なくとも99重量%を占める本発明によるポリペプチドがより好ましい。化学的に合成されたポリペプチドは、上記の状況において本来「単離および精製されて」いる。
【0033】
単離された本発明のポリペプチドは、例えば、天然の供給源、例えば、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムからの抽出によって;宿主、好ましくは、異種宿主における、そのポリペプチドをコードしている組換え核酸の発現によって;または、化学合成によって得ることができる。それが天然に由来する供給源とは異なる細胞系において産生されるポリペプチドは、それに天然に付随する成分から分離されているため、「単離および精製されて」いる。単離および/または純度の程度は、任意の適切な方法、例えば、カラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、HPLC分析、NMR分光法、ガス液体クロマトグラフィー、または質量分析によって測定することができる。
【0034】
さらに、一態様において本発明は、本発明のポリペプチドに特異的に結合する抗体、その機能的フラグメントおよび機能的誘導体に関する。これらは、標的抗原が利用可能となれば、ハイブリドーマ技術(KohlerおよびMilstein、Nature、256、495〜497、1975)、抗体ファージディスプレイ(Winterら、Annu. Rev. Immunol. 12、433〜455、1994)、リボソームディスプレイ(Schaffitzelら、J. Immunol. Methods、231、119〜135、1999)および反復コロニーフィルタースクリーニング(Giovannoniら、Nucleic Acids Res. 29、E27、2001)によって、常法に従って利用可能である。抗体を機能的生成物に断片化するための典型的なプロテアーゼは、よく知られている。結果として生じるフラグメントが、特定の高親和性および、好ましくは、マイクロモルからピコモル範囲の解離定数を有する限り、別の断片化技術も同様に使用することができる。
【0035】
標的化用途のために非常に好都合な抗体フラグメントは、可変重鎖および可変軽鎖ドメインが、ポリペプチドリンカーによって一緒に結合されている単鎖Fvフラグメントである。本発明のポリペプチドを特定するための別の抗体フラグメントは、Fabフラグメント、Fab2フラグメント、ミニ抗体(低分子免疫タンパク質とも呼ばれる)、タンデムscFv-scFv融合体ならびに適したドメインとの(例えば、免疫グロブリンのFc部分との)scFv融合体を含む。特定の抗体フォーマットに関する概説については、HolligerおよびHudson、Biotechnol.、23(9)、1126〜36、2005を参照されたい。
【0036】
本発明において使用するための抗体の「機能的誘導体」という用語は、その誘導体が、そのもとの抗原に関して実質的に同じ結合親和性を有する、好ましくは、マイクロモル、ナノモルまたはピコモルの範囲内の解離定数を有する限り、例えば、アミノ酸残基の付加、置換および/または欠失によって、そのアミノ酸配列が化学的にまたは遺伝子的に修飾されている、および/または、例えば、付加、欠失、再編成、酸化、還元などによって、その原子および/または化学官能基のうちの少なくとも1つが化学的に修飾されている、任意の抗体またはそのフラグメントを含むことを意味する。
【0037】
好ましい一実施形態において、本発明において使用するための抗体、そのフラグメントまたは機能的誘導体は、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒト化抗体、CDR移植抗体、Fvフラグメント、FabフラグメントおよびFab2フラグメントおよび抗体様結合タンパク質、例えば、アフィリン(affilin)、アンチカリン(anticalin)およびアプタマーからなる群から選択されるものである。
【0038】
抗体様結合タンパク質の概説については、非免疫グロブリンドメインからの結合タンパク質のエンジニアリングに関する、Binzら、Nature Biotechnol.、23(10)、1257〜1268、2005を参照されたい。「アプタマー」という用語は、高親和性を有するポリペプチドに結合する核酸を記述する。アプタマーは、様々な単鎖RNA分子の大きなプールから、SELEX(例えば、Jayasena、Clin. Chem.、45、1628〜1650、1999;KlugおよびFamulok、M. Mol. Biol. Rep.、20、97〜107、1994;US 5,582,981を参照されたい)などの選択方法によって単離することができる。アプタマーはまた、合成し、その鏡像体から、例えば、L-リボヌクレオチドとして選択することもできる(Nolteら、Nat. Biotechnol.、14、1116〜1119、1996;Klussmannら、Nat. Biotechnol.、14、1112〜1115、1996)。このように単離された形態は、それらが、天然に存在するリボヌクレアーゼによって分解されず、したがって、より高い安定性を有するという利点を有する。
【0039】
別の抗体様結合タンパク質および伝統的抗体の代替物は、いわゆる「タンパク質足場」、例えば、リポカリンをベースとするアンチカリン(Besteら、Proc. Natl. Acad. Sci. USA、96、1898〜1903、1999)である。リポカリンの、例えば、レチノール結合タンパク質またはビリン結合タンパク質の天然のリガンド結合部位は、例えば、「コンビナトリアルタンパク質設計」手法を用いることによって、それらが選択されたハプテンに結合するように(Skerra、Biochem. Biophys. Acta、1482、337〜350ページ、2000)変更することができる。別のタンパク質足場については、これらは抗体の代替物であることがまた知られている(Skerra、J. Mol. Recognition、13、167〜287、2000;Hey、Trends in Biotechnology、23、514〜522、2005)。
【0040】
要約すると、機能的抗体誘導体という用語は、上記のタンパク質由来の抗体代替物、すなわち、ポリペプチド、そのフラグメントまたは誘導体を特異的に認識する、抗体様結合タンパク質、例えば、アフィリン、アンチカリンおよびアプタマーを含むことを意味する。
【0041】
さらなる態様は、本発明によるモノクローナル抗体を発現するハイブリドーマ細胞系に関する。
【0042】
本発明の核酸、ベクター、宿主細胞、ポリペプチドおよび抗体は、多くの新しい用途を有する。
【0043】
一態様において、本発明は、ガラクトシド含有オリゴ糖/多糖および/または複合糖質、好ましくは、ガラクトシル-フコシド含有オリゴ糖/多糖および複合糖質、より好ましくは、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-GlcNAc含有オリゴ糖/多糖および複合糖質、最も好ましくは、GnGnF6Gal-またはMMF6Gal含有オリゴ糖/多糖および複合糖質を作製するための、ポリペプチド、本発明のポリペプチドを含む細胞抽出物、好ましくは、線虫抽出物、より好ましくは、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムの抽出物、および/または本発明の宿主細胞の使用に関する。
【0044】
本明細書で使用される場合、複合糖質という用語は、非糖成分の性質に関して限定するものではないことが理解される。好ましくは、複合糖質の非糖成分は、ポリペプチド/オリゴペプチドである。
【0045】
ガラクトシル-フコシル特異的オリゴ糖および複合糖質の酵素的合成は、高度に特異的であり、制御されており、環境に優しく、生成物は、それを必要とするヒトまたは別の哺乳動物などの対象における、寄生虫感染、好ましくは線虫およびアピコンプレックス感染の、治療および予防のために、高度に寄生虫特異的な(このエピトープはまた、タコ[Zhangら、Glycobiology、7、1153〜1158、1997]、イカ[Takahashiら、Eur. J. Biochem.、270、2627〜2632、2003]およびカサガイ[Wuhrerら、Biochem. J.、378、625〜632、2004]において存在していることのみが知られている)ワクチン成分として作用することができる。
【0046】
例示的なおよび好ましいガラクトシル-フコシル特異的オリゴ糖および複合糖質は、N-結合グリカン、N-糖タンパク質、糖脂質および脂質結合オリゴ糖(LOS)からなる群から選択される。本明細書において使用される場合、「複合糖質」という用語は、オリゴ糖成分および非糖成分の、任意のタイプのコンジュゲート、必然的ではないが好ましくは共有結合しているもの、例えば、共有結合性リンカーにより結合しているもの、例えば、生理学的に許容でき、さらには、例えば、線虫毒性などを付与する免疫刺激アジュバントとして、所望の生理的機能を有し得る、ポリペプチドまたは任意の別のタイプの有機または無機担体を含むことを意味する。
【0047】
例えば、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムまたは本発明のポリペプチドを産生する組換え昆虫細胞の粗抽出物は、Gal-Fuc含有コンジュゲート、例えば、遊離のGal-Fucグリカン、Gal-Fuc-ペプチド、Gal-Fuc-ポリペプチド、Gal-Fuc-フォールディングしたタンパク質を生成することができる。α-1,6-結合フコシドは、α-1,3-結合フコシドよりも非常に好ましい。
【0048】
本発明の別の態様は、以下の段階:
(i)少なくとも1つの本発明のポリペプチドを提供する段階、
(ii)少なくとも1つのフコシル化されたアクセプター基質を提供する段階、
(iii)好ましくは、マンガン(II)、コバルト(II)および/または鉄(II)イオンから選択され、より好ましくはマンガン(II)である少なくとも1つの適した二価の金属陽イオン補因子と、少なくとも1つの活性化された糖基質、好ましくは、ウリジン二リン酸(UDP)-ガラクトースとの存在下、本発明のポリペプチドの酵素活性に適した条件下で、(i)および(ii)をインキュベートする段階、
(iv)ガラクトシル-フコース誘導体を任意選択により単離する段階
を含む、ガラクトシル-フコシル誘導体を作製するための方法を対象とする。
【0049】
本発明のポリペプチドは、緩衝液、宿主細胞、細胞抽出物またはその酵素活性を持続させ、その基質および活性化された糖基質へのアクセスを可能にする任意の別の系において、乾燥形態または可溶性形態の単離ポリペプチドとして提供されてもよい。フコシル化されたアクセプター基質は、任意選択により、単離された形態の、または本発明のポリペプチドにより酵素的に修飾することができる系の成分としての、任意の種類のフコシル含有基質である。活性化された糖基質は、好ましくは、UDP-ガラクトースであるが、フコシル化されたアクセプター基質へ転移することができる、活性化された、好ましくは、リン酸活性化されたガラクトシル誘導体の任意の別のタイプとすることもできる。本発明の方法好ましくは、ガラクトピラノシル-β-1,4-L-フコピラノシル-誘導体、より好ましくは、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-βGlcNAc(Gal-Fuc)誘導体をもたらす。
【0050】
本発明のポリペプチドは、基質が適したフコシル部分を特徴として有する限り、広い基質特異性を有する。ガラクトシル-トランスフェラーゼ活性が、例えば、フコシル糖、フコシルペプチド、フコシルポリペプチド、さらには複合体およびフォールディングしたフコシルポリペプチドなどの基質について実証された。例えば、ガラクトシル-トランスフェラーゼ活性が、主要なエピトープとしてGnGnF6糖鎖構造を有する糖タンパク質、ヒトIgG1について実証された。これらのIgG1グリカンは、PNGaseFによって消化可能であることが知られている。ヒトIgG1のグリコシル化は、カエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼを含有する未精製のsf9昆虫細胞抽出物を用いて実証された。カエノラブディティス・エレガンスからの酵素抽出物の存在下における、放射性標識されたUDP-GalとのヒトIgG1のインキュベーションは、基質ガラクトシル化をもたらした。加えて、ガラクトシル化は、主要なエピトープとしてGnGnF6糖鎖構造を保有する再構築したヒトトランスフェリンに関して実証された。この目的のために、ヒトアポトランスフェリンを、シアリダーゼ(Iskratschら、Anal. Biochem.、368、133〜146、2009)、アスペルギルス・オリゼ(Aspergillus oryzae)のβ1,4-ガラクトシダーゼおよびピキア・パストリスにおいて発現された組換えハマダラカ属(Anopheles)コアα1,6-FucTで連続的に処理して、主要なエピトープとしてGnGnF6糖鎖構造を有する糖タンパク質が作製された。カエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼを含有する未精製のsf9昆虫細胞抽出物とのインキュベーションは、ガラクトシル化をもたらし、これは、フコース特異的ヒイロチャワンタケ(Aleuria aurantia)レクチンを用いたドットブロット法によって、および、様々なネオグリコフォーム(neoglycoform)のトリプシンペプチドのMALDI-TOF MSによってモニタリングされた。
【0051】
哺乳動物、特に、ヒトにおいて、慢性肝炎および肝硬変などの良性の肝臓疾患は、コアがフコシル化されたAFPをもたらさないことから、コアがフコシル化されたαフェトプロテイン(AFP)の血清含量が肝細胞癌(HCC)に高度に特異的であることが、ごく最近示された(Tatenoら、Glycobiology、19(5)、527〜536、2009を参照されたい)。
【0052】
したがって、さらなる態様において、コアがフコシル化されたAFPは、本発明のポリペプチドのアクセプター基質として選択的に適しているため、本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物は、ガラクトシル化合物を、コアがフコシル化されたαフェトプロテイン(AFP)に共有結合させるために、好ましくは、好ましくはHCC患者の血液からのコアがフコシル化されたαフェトプロテイン(AFP)を選択的に標識することによって、肝細胞癌(HCC)細胞を検出および/または定量するために使用することができる。
【0053】
したがって、本発明は、コアがフコシル化されたAFPを検出するため、すなわち、肝細胞癌(HCC)細胞を検出および/または定量するための診断手段を調製するための本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物に関する。
【0054】
また、本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物は、その出現が別のタイプの癌細胞と相互関係を示す、さらにコアがフコシル化されたマーカー糖タンパク質を検出するための診断手段を調製するために有用である。
【0055】
好ましい一実施形態において、本発明は、以下の段階:
(i)AFPを含む血液またはその画分、好ましくは血清を提供する段階と、
(ii)前記血液または前記その画分を、(a)本発明のポリペプチド、本発明の宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物とともに、および(b)活性化されたガラクトシル誘導体、好ましくは、標識されたガラクトシル誘導体、好ましくは標識されたUDP-ガラクトースとともに、活性化されたガラクトースの、コアがフコシル化されたAFP(AFP-L3)へのガラクトシル転移を可能にする条件下で、インキュベートする段階と、
(iii)ガラクトース標識された、したがって、コアがフコシル化されたAFP(AFP-L3)を検出する段階と
を含む診断方法に関する。
【0056】
上記の方法を実践するための、活性化されたガラクトシル誘導体の標識は、同位体、例えば14C、化学修飾、例えば、ハロゲン置換および別の選択的に検出可能な修飾、例えば、ビオチン、アジドなど、からなる群から選択される。好ましくは、段階(i)から(iii)のすべては、生体外で、すなわち、インビトロで実施される。
【0057】
本発明のさらなる一態様は、対象とする試料、例えば、ヒトまたは哺乳動物の試料において、好ましくは、細胞画分または抽出物試料において、線虫およびアピコンプレックス、好ましくは、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、およびクリプトスポリジウム・パルバムをそれぞれ同定および/または定量するための、本発明のポリペプチド、好ましくは、配列番号2、4、6、8および/または10のいずれかから選択される配列を有するポリペプチドに特異的に結合する抗体の使用を対象とする。典型的な抗体アッセイ、例えば、ELISAの設計および開発は、当業者の範囲内であり、さらに詳しく述べる必要はない。
【0058】
本発明を、好ましい実施形態および典型的な例に重点をおいて説明してきた。しかしながら、好ましい実施形態の変形形態が使用されてもよいこと、および本発明が、本明細書に具体的に記載されているものとは別の方法で実践されてもよいことが意図されることが、当業者には明らかであろう。さらに、前述の例は純粋に例示の目的で含まれているため、これらはいかなる点においても本発明の範囲を限定すると解釈されるべきではない。したがって、本発明は、本明細書に添付されている特許請求の範囲によって定義されている本発明の精神および範囲内に包含されるすべての変更形態を含む。
【図面の簡単な説明】
【0059】
【図1】バキュロウイルス感染sf9全細胞抽出物の抗FLAG免疫ブロット法を示す図である。空ベクター対照(e.v.)、N末端にFLAGをタグ付けしたM03F8.4(FLAG-GalT)およびタグなしM03F8.4(GalT)を含有するバキュロウイルスの様々なクローン。充填容量約150k細胞/スロット、SDS-PAGE12%、α-FLAG(1:2000、SIGMA)、α-マウス-HRP(1:2000、Santa Cruz Biotechnology)、ECL(Pierce、2s曝露)。
【図2】バキュロウイルス感染sf9全細胞抽出物のSDS-PAGE分析の図である。空ベクター対照(e.v.)、N末端にFLAGをタグ付けしたM03F8.4(FLAG-GalT)およびタグなしM03F8.4(GalT)を含有するバキュロウイルスの様々なクローン。充填容量約150k細胞/スロット、SDS-PAGE12%、銀染色による検出。(タンパク質は少量発現され、未精製の抽出物において空ベクター構築体に関して銀染色では検出不能である。)
【図3】Mn2+、Mg2+およびEDTAの存在下において、GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化ターンオーバーを示すカラムチャートであり、金属イオン依存性を示す;MES、pH6、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 2369/(m/z 2207+m/z 2369)]*100)によって決定されたターンオーバー。
【図4】GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化、タグ付きおよびタグなしの構築体の機能性を示すカラムチャートである;MES、pH6、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 2369/(m/z 2207+m/z 2369)]*100)によって決定されたターンオーバー。
【図5】MS分析による、GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化、タグ付きおよびタグなしの構築体の機能性を示す図である(MES pH6、室温、2.5時間)。上段スペクトル:UDP-Galなしの反応、中段スペクトル: UDP-Galあり、下段スペクトル:中段スペクトルの生成物の、アスペルギルス属β-ガラクトシダーゼによる消化産物(クエン酸緩衝液、pH5、室温、2d)。酵素は、ガラクトースをβ-ガラクトシダーゼによって消化することができるこのアクセプター基質に明らかに付加し、したがって、GalTによって組み込まれたβ-結合Gal残基を示す。さらなるGlcNAc除去は、未精製の昆虫細胞抽出物中のヘキソサミニダーゼの存在のために、長い反応時間(>2d)の後に起こる。
【図6】図5の、アクセプター(上段スペクトル)およびガラクトシル化された反応生成物(下段スペクトル)のMS/MSスペクトルの比較を示す図である。MS/MS分析は、ダブシル化されたGENRペプチドに結合されているHex-dHex-HexNAcモチーフに相当する二次イオン1272.61から観察される通り、コアのフコースに結合しているガラクトースを明らかに示す。
【図7】ガラクトシルトランスフェラーゼのドナー特異性の比較分析を示す図である(ダンシル-N[GnGnF6]ST、MES pH6.5、Mn2+、室温、13時間)。酵素は、UDP-Galに対する高い特異性を有するようであり、UDP-Glcに関する残留活性は無視できる。
【図8】アクセプター特異性の分析のカラムチャートである:カエノラブディティス・エレガンスGalTは、α-1,3-結合したフコースよりα-1,6結合したフコースを選択的にガラクトシル化する;ダブシルGEN-[MMF6/3]R、MES pH6.5、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 1963/(m/z 1801+m/z 1963)]*100)によって決定されたターンオーバー。
【図9】UDP-Galについて、タグなしガラクトシルトランスフェラーゼのKm(見かけ)のグラフによる決定を示す図である: Km(見かけ、UDP-Gal)=約40μM。
【図10】本発明のガラクトシルトランスフェラーゼの温度依存性の分析を示す図である(ダンシル-N[GnGnF6]ST、UDP-Gal、MES pH6.5、Mn2+、2.5時間)。
【図11】本発明のポリペプチド、すなわちカエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼによる、ヒトIgG1のグリコシル化(GnGnF6エピトープを有する)を示すカラムチャートである。
【図12】本発明のポリペプチド、すなわちカエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼによる、再構築したヒトトランスフェリン(GnGnF6エピトープを有する)のグリコシル化を示すMALDI-TOF MSスペクトルを示す図である。示されているm/z値は、GnGn(3813)、GnGnF6(3957)およびGnGnF6Gal(4119)をそれぞれ保有するペプチド622〜642に相当する。
【0060】
(実施例)
実験手順
化学製品および供給業者
UDP-Gal(VWR InternationalおよびSigma)、UDP-Glc、UDP-GlcNAc、UDP-GalNAc(すべてSIGMA)、UDP-14C-Gal(GE Healthcare)、GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man、Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc、MMF6、GnGnF6(すべてDextra Laboratories、イギリス)、Fuc-α-1,6-GlcNAc(Carbosynth Ltd.、イギリス)、ダブシル-GEN[GnGnF6]R(Paschingerら、Glycobiology、15(5)、463〜474、2005)、ダブシル-GEN[MMF6]R(Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001)、ダブシル-GEN[MMF3]R(Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001)およびダンシル-N[GnGnF6]ST(Roitingerら、Glycoconj. J.、15(1)、89〜91、1998)は、以前に公開されている方法に従って得た。
【0061】
(実施例1)
カエノラブディティス・エレガンスcDNAの単離およびM03F8.4の発現ベクター中へのクローニング
線虫系統:
カエノラブディティス・エレガンスを培養するための方法は、Brenner, S.(Genetics 77(1)、71〜94、1974)において記載されている。野生型BristolN2系統を、大腸菌OP50を播種した標準的なNGM寒天プレート上で、20℃で増殖させた。
【0062】
カエノラブディティス・エレガンスM03F8.4cDNAの単離:
カエノラブディティス・エレガンス混合培養物を、標準的なNGM寒天プレートから回収し、滅菌したM9緩衝液(22mM KH2PO4、42mM Na2HPO4、85mM NaCl、1mM MgSO4)中で2回洗浄した。NucleoSpin(登録商標)RNA II RNA単離キット(MACHEREY-NAGEL AG)を使用して、全RNAを抽出した。Superscript(商標)III Platinum Two-Step qRT-PCR Kit(Invitrogen AG)の第1鎖cDNA合成ステップを使用して、0.5μgの全RNAを用いてcDNA合成を実施した。
【0063】
sf9昆虫細胞における組換え遺伝子発現のためのpFastBac1ドナープラスミドの構築:
M03F8.4cDNAを、以前に調製されたcDNAライブラリーから、提供されたマニュアルに従ってPhusion High-Fidelity DNAポリメラーゼ(Finnzymes)を使用したPCRによって単離した。タグなし型の構築では、SalIおよびXbaI制限部位にそれぞれ隣接している以下のフォワードおよびリバースプライマーを使用した:5'-TTTGTCGA-CACTTCTGAATGCCTCG-3'(配列番号11)および5'-TTTTCTAGACTACAAGTCTAA-AAGACCAAC-3'(配列番号12)。結果として生じたフラグメントを、適切な制限酵素で消化し、pFastBac1ドナープラスミド(Invitrogen)中にクローニングした。N末端FLAGタグ付き型の構築では、開始コドンを欠くフォワードプライマーを使用した:5'-TTTGTCGACCCTCGAATCACCGCC-3'(配列番号13)。結果として生じたフラグメントを、N末端FLAG配列を含有するpFastBac1ドナープラスミド中にクローニングした(Mullerら、J. Biol. Chem. 277(36)、32417〜32420、2002)(両ベクターは、チューリッヒ大学、Institute of Physiology、Thierry Hennetから好意により提供された)。
【0064】
(実施例2)
組換えタンパク質の発現
カエノラブディティス・エレガンスのコアβ-1,4-GalT候補cDNA(N末端FLAGタグありおよびなし)および空ベクター対照を含有する組換えバキュロウイルスを、製造業者の使用説明書(Invitrogen)に従って作製した。2×106のS.フルギペルダ(S.frugiperda)(sf9)付着性昆虫細胞に組換えバキュロウイルスを感染させ、28℃で72時間インキュベーションした後、細胞を、2体積%Triton-X100およびプロテアーゼ阻害剤カクテル(Roche、complete EDTA-free)を含有する150μLのTris緩衝生理食塩水(pH7.4)において、振とう(4℃、15分)により溶解させた。溶解混合物を遠心分離にかけ(2000×g、5分)、除核後の上清を回収して、すべてのさらなる酵素試験に使用した。
【0065】
(実施例3)
変性ゲル電気泳動分析および免疫ブロット法
感染sf9細胞(2×106細胞、上記参照)を、200μLのLaemmli緩衝液中でボルテックスにかけ、加熱(95℃、5分)によってタンパク質を変性させた。室温まで冷却後、試料を遠心分離にかけ(16krpm、5分)、上清をさらなる分析に使用した。試料を、SDS-PAGEによって分離した(12%アクリルアミド、120V)。結果として生じたゲルを、銀染色またはニトロセルロース膜上へのブロッティングのいずれかによって分析した。膜をブロッキング(PBST中5%BSA)した後、抗FLAG抗体M2(SIGMA、PBST+1%BSA中1:2000希釈)、その後、徹底的に洗浄(PBST)した後に抗マウスHRP(Santa Cruz Biotechnology、PBST+1%BSA中1:10000希釈)とのインキュベーション、ならびにECL(Pierce)および写真用フィルムへの曝露を使用した最終的な検出によって、免疫検出を実施した。
【0066】
(実施例4)
グリコシルトランスフェラーゼアッセイ
塩化マンガン(II)(10μM)、UDP-ガラクトース(1mM)およびアクセプターフコシド(グリカンまたは糖(ポリ)ペプチド、40μM)を含有する2.5μLの最終体積のMES緩衝液(pH6.5、40μM)中で、0.5μlのsf9細胞粗抽出物(空ベクター対照バクミド、推定GalT発現バクミドまたは推定FLAGタグ付きGalT発現バクミドのいずれかを含有する)を使用して、適切な炭水化物または複合糖質に対する酵素活性を評価した。グリコシル化反応は。典型的には、別段記載のない限り、室温で2時間行った。ドナー特異性分析では、UDP-ガラクトースを、等濃度のUDP-Glc、UDP-GlcNAcまたはUDP-GalNAc(Sigma)でそれぞれ置き換えた。補因子特異性分析では、MnCl2を、等濃度の様々な金属塩化物またはNa2EDTAで置き換えた。アクセプターグリカン中へのガラクトースの組込みを定量するために、全UDP-Gal濃度を、10%UDP-14C-Gal(25nCi、GE Healthcare)でドープした。反応混合物(100μLのH2Oでクエンチさせた)を、陰イオン交換樹脂のカラム(AG1-X8、Cl-型、Bio-Rad Laboratories、200mg)に充填すること、および荷電していない生成物(H2O、900μL)の溶出によって、過剰の放射活性(UDP-14C-Gal)を除去した。
【0067】
非放射性UDP-Galが存在せず、UDP-14C-Gal(75nCi)で置き換えられていること以外は上記と同じ緩衝液、塩および酵素条件を使用して、50μLの全体積において、ヒトIgG1(5μlの3g/L、Calbiochem)のグリコシル化を実施した。反応は、室温で終夜実施した。セファロース-プロテインGビーズ(Amersham Biosciences、10μL)のPBS(200μL)中の懸濁液を添加し、振とう(4℃、1時間)によってIgG1のビーズへの結合を行った。ビーズをPBS(5×200μL)で洗浄し、20mMのHCl水溶液(3×100μL)でIgG1を溶出した。ドナー特異性に関する、蛍光標識されたグリコペプチドの直接のMALDI-TOF質量分析、HPLC分析、または放射標識アッセイのシンチレーション計数のいずれかによって、反応生成物の分析(下記参照)を実施した。
【0068】
ヒトアシアロトランスフェリンN-グリカンの段階的な再構築を、以下の通り実施した:
アシアロトランスフェリン(GalGal)は、ヒトアポトランスフェリンのシアリダーゼ処理によって以前に調製された(Iskratschら、Anal. Biochem.、368、133〜146、2009)。
【0069】
アシアロガラクトトランスフェリン(GnGn)を作製するために、β1,4-ガラクトシダーゼ(3U、アスペルギルス・オリゼ)を、約1mgのGalGalに添加し、試料を、48時間37℃でインキュベートした(全体積50μl)。
【0070】
GnGnF6を得るために、試料を0.5μlの1M NaOHで中性pHにした後、50nmolのGDP-フコースおよび15μlのピキア・パストリスにおいて発現された組換えハマダラカ属のコアα1,6-FucT調製物を添加した。調製物を終夜インキュベートした後、さらに50nmolのGDP-フコースおよびさらなる15μlの酵素(FucT)を添加し、終夜、37℃で再度インキュベートした。合計で、約1mgのGnGnF6が得られた。
【0071】
GalFuc-トランスフェリンを調製するために、1μlの組換えカエノラブディティス・エレガンスGalTの調製物、0.2mmolのMnCl2および20nmolのUDP-ガラクトースをGnGnF6(300μg)のアリコートに添加し、終夜30℃でインキュベートした。再度、さらなる基質(UDP-ガラクトース)および酵素(GalT)の添加後、2回目の終夜のインキュベーションにより所望のグリカン構造を増加させた。
【0072】
トランスフェリンの修飾の程度は、フコース特異的ヒイロチャワンタケレクチンを用いたドットブロット法によって、および様々なネオグリコフォームのトリプシンペプチドのMALDI-TOF MSによってモニタリングした。
【0073】
(実施例5)
構造分析
ダブシル-GEN[GnGnF6]Rをガラクトシル化条件に曝露させた後、結果として生じた未精製の混合物を、50mMナトリウムクエン酸およびpH4.5に調整し、アスペルギルス・オリゼβガラクトシダーゼ(27mU)(Gutterniggら、J. Biol. Chem. 282(38)、27825〜27840、2007を参照されたい)で2日間、30℃で消化した。試料を、MALDI-TOF質量分析によって分析した(下記参照)。
【0074】
HPLC分析:
ドナー特異性およびドナー濃度に対する反応速度依存の分析の両方で、逆相Hypersil ODS C18カラム(4×250mm、5μm)においてアイソクラティック溶媒系(0.7mL/分、0.05体積%TFA水溶液中9%MeCN(95体積%))、および蛍光検出(315nmで励起、発光は550nmで検出した)を室温で使用して、ダンシル-N[GnGnF6]STアクセプター基質を、ガラクトシル化された反応生成物から分離した。Shimadzu HPLCシステムは、SCL-10A制御装置、2つのLC10APポンプおよびClass-VPソフトウェア(V6.13SP2)を使用してパーソナルコンピュータにより制御されるRF-10AXL蛍光検出器からなっていた。ダンシル-N[GnGnF6]STは、9.09分の保持時間で、およびガラクトシル化された反応生成物は8.06分で溶出した。
【0075】
質量分析:
BRUKER Ultraflex TOF/TOF装置において、α-シアノ-4-ヒドロキシ桂皮酸マトリックスを使用して、MALDI-TOF質量分析によってグリカンを分析した。外部較正にはペプチド標準混合物(Bruker)を使用した。
【0076】
シンチレーション計数:
陰イオン交換樹脂カラムおよびプロテインGビーズの溶出液を、シンチレーション流体(Irga-Safe Plus、Packard、4mL)と完全に混合し、Perkin Elmer Tri-Carb 2800TRを用いて測定した。
【0077】
炭水化物の省略形:
Fuc-L-フコース、Gal-D-ガラクトース、GalNAc-D-N-アセチルガラクトサミン、Glc-D-グルコース、GlcNAc-D-N-アセチルグルコサミン、Man-D-マンノース
複合グリカンの省略形(Schachter命名法[Biochem Cell Biol 64(3)、163〜181、1986]に従う):
GalGal Gal-β-1,4-GlcNAc-β-1,2-Man-α-1,6-[Gal-β-1,4-GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-GlcNAc
GnGn GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-GlcNAc
GnGnF6 GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc
GnGnF6Gal GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[Gal-β-1,4-Fuc-α-1,6]-GlcNAc
MMF6 Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc
MMF6Gal Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[Gal-β-1,4-Fuc-α-1,6]-GlcNAc
MMF3 Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,3-Fuc]-GlcNAc
【技術分野】
【0001】
本発明は、新しいガラクトシルトランスフェラーゼ、それらをコードしている核酸、ならびにそれらに関連する組換えベクター、宿主細胞、抗体、使用および方法に関する。
【背景技術】
【0002】
「回虫」または「線虫」は、擬体腔動物の中の最も多様な門、およびすべての動物の中で最も多様なものの1つである。線虫種は、識別するのが困難であり、80,000を超える種が記載されていて、そのうち15,000を超える種が寄生虫である。回虫種の合計数は、500,000より多いと推定されてきた。線虫は、淡水、海洋環境および陸上環境においていたるところに存在する。多くの寄生虫形態は、大部分の植物、動物における、またヒトにおける病原体を含む。
【0003】
カエノラブディティス・エレガンス (Caenorhabditis elegans)は、モデル線虫であり、分節がなく、蠕虫型であり、左右相称であり、クチクラ外皮、4つの主な表皮性神経索および体液で満たされた擬体腔を有する。野生において、カエノラブディティス・エレガンスは、腐食した植物質に発生する細菌を餌にしている。Hannemannら(Glycobiology、16、874、2006)は、カエノラブディティス・エレガンスのN-グリカンコアにおいて、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-D-GlcNAc(Gal-Fuc)エピトープを単離し、構造的に特性決定した。カエノラブディティス・エレガンスのN-グリコシル化パターンは、Paschingerら(Carbohydrate Res.、343、2041、2008)において最近概説された。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】US 5,582,981
【非特許文献】
【0005】
【非特許文献1】Hannemannら(Glycobiology、16、874、2006)
【非特許文献2】Paschingerら(Carbohydrate Res.、343、2041、2008)
【非特許文献3】Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986
【非特許文献4】Altschulら、J. Mol. Biol.、215、403〜410、1990
【非特許文献5】HuangおよびMiller、Adv. Appl. Math.、12、337〜357、1991
【非特許文献6】BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894
【非特許文献7】SambrookおよびRussell、Molecular cloning: A laboratory manual (3 volumes)、2001
【非特許文献8】http://www.-ncbi.nlm.nih.gov/entrez/query.fcgi?db=nucleotide
【非特許文献8】Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニア
【非特許文献9】Methods in Enzymology、350、248、2002
【非特許文献10】Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カースルバッド、カリフォルニア
【非特許文献11】Novagen Insect Cell Expression Manual、Merck Chemicals Ltd.、ノッティンガム、イギリス
【非特許文献12】Applied Microbiology and Biotechnology、72、211、2006
【非特許文献13】LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009
【非特許文献14】KohlerおよびMilstein、Nature、256、495〜497、1975
【非特許文献15】Winterら、Annu. Rev. Immunol. 12、433〜455、1994
【非特許文献16】Schaffitzelら、J. Immunol. Methods、231、119〜135、1999
【非特許文献17】Giovannoniら、Nucleic Acids Res. 29、E27、2001
【非特許文献18】HolligerおよびHudson、Biotechnol.、23(9)、1126〜36、2005
【非特許文献19】Binzら、Nature Biotechnol.、23(10)、1257〜1268、2005
【非特許文献20】Jayasena、Clin. Chem.、45、1628〜1650、1999
【非特許文献21】KlugおよびFamulok、M. Mol. Biol. Rep.、20、97〜107、1994
【非特許文献22】Nolteら、Nat. Biotechnol.、14、1116〜1119、1996
【非特許文献23】Klussmannら、Nat. Biotechnol.、14、1112〜1115、1996
【非特許文献24】Besteら、Proc. Natl. Acad. Sci. USA、96、1898〜1903、1999
【非特許文献25】Skerra、Biochem. Biophys. Acta、1482、337〜350ページ、2000
【非特許文献26】Skerra、J. Mol. Recognition、13、167〜287、2000
【非特許文献27】Hey、Trends in Biotechnology、23、514〜522、2005
【非特許文献28】Zhangら、Glycobiology、7、1153〜1158、1997
【非特許文献29】Takahashiら、Eur. J. Biochem.、270、2627〜2632、2003
【非特許文献30】Wuhrerら、Biochem. J.、378、625〜632、2004
【非特許文献31】Iskratschら、Anal. Biochem.、368、133〜146、2009
【非特許文献32】Tatenoら、Glycobiology、19(5)、527〜536、2009
【非特許文献33】Paschingerら、Glycobiology、15(5)、463〜474、2005
【非特許文献34】Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001
【非特許文献35】Roitingerら、Glycoconj. J.、15(1)、89〜91、1998
【非特許文献36】Brenner, S.(Genetics 77(1)、71〜94、1974
【非特許文献37】Mullerら、J. Biol. Chem. 277(36)、32417〜32420、2002
【非特許文献38】Gutterniggら、J. Biol. Chem. 282(38)、27825〜27840、2007
【発明の概要】
【発明が解決しようとする課題】
【0006】
Gal-Fuc含有(多/オリゴ)糖およびGal-Fuc含有複合糖質の組換えによる作製のための新しい手段を提供することが、本発明の目的である。さらなる目的は、Gal-Fuc含有多糖/オリゴ糖およびGal-Fuc含有複合糖質のための新しい使用を提供することである。
【課題を解決するための手段】
【0007】
第1の態様において、この目的は、
(i)配列番号1、3、5、7および9に記載されている核酸配列からなる群から選択される少なくとも1つの核酸配列、好ましくは配列番号1の核酸配列を含む核酸;
(ii)配列番号1、3、5および7に記載されている核酸配列からなる群から選択される核酸配列、好ましくは配列番号1の核酸配列と、少なくとも60、65、70または75%の同一性、好ましくは、少なくとも80、85または90%の同一性、より好ましくは、少なくとも95%の同一性、最も好ましくは、少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される単離および精製された核酸によって解決される。
【0008】
好ましい一態様において、単離および精製された核酸は、
(i)配列番号1、3、7および9に記載されている核酸配列ならびに配列番号5の最初の1428核酸からなる群から選択される少なくとも1つの核酸配列、好ましくは配列番号1の核酸配列を含む核酸;
(ii)配列番号1、3および7に記載されている核酸配列ならびに配列番号5の最初の1428核酸からなる群から選択される核酸配列、好ましくは配列番号1の核酸配列と、少なくとも60、65、70または75%の同一性、好ましくは、少なくとも80、85または90%の同一性、より好ましくは、少なくとも95%の同一性、最も好ましくは、少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる、核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される。
【0009】
好ましくは、上記の核酸は、本発明のポリペプチド、好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6-(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986による命名)含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、好ましくは酵素ガラクトシルトランスフェラーゼ活性を有するポリペプチド、より好ましくはβ-1,4-ガラクトシルトランスフェラーゼ活性を有するポリペプチドをコードしている。
【0010】
本明細書において使用される場合、ガラクトシルトランスフェラーゼ活性は、活性化されたドナー型(すなわち、ヌクレオチド活性化ガラクトース、好ましくは、UDP-Gal)からアクセプターへの、ガラクトース残基の酵素転移を記述することを意味する。本明細書において使用される場合、β-1,4-ガラクトシルトランスフェラーゼ活性は、ガラクトシルトランスフェラーゼ活性の特異性、すなわち、β1,4-配座でのガラクトースのアクセプター分子への転移を記述することを意味する。本明細書において使用される場合、アクセプター基質としてのL-フコシドに対するβ-1,4-ガラクトシルトランスフェラーゼ活性は、アクセプター基質としてのL-フコシドへのβ- 1,4-結合型転移におけるガラクトシルトランスフェラーゼ活性の特異性を記述することを意味する。本明細書において意味される場合、L-フコシドは、α、最も好ましくは、α-1,6配座において、例えば、MMF6またはGnGnF6(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986)の部分として、末端L-フコースを含有するアクセプター基質としての多糖/オリゴ糖または複合糖質を記述することを意味する。
【0011】
最も好ましい実施形態において、コードされているポリペプチドは、配列番号2、4、6、8および10に記載されているポリペプチド配列からなる群から選択されるポリペプチド配列、好ましくは、配列番号2のポリペプチド配列、またはこれらのうちいずれかの機能的フラグメントまたは機能的誘導体を含む。
【0012】
配列番号1は、配列番号2をコードしている核酸配列である:
(NCBIにおいて、参照配列NM_072144.4として、Wormbaseにおいて、M03F8.4としても記載されており、カエノラブディティス・エレガンスのガラクトシルトランスフェラーゼ[実施例セクションにおいてGalTと称されている]をコードしている)
ATGCCTCGAATCACCGCCAGTAAAATAGTTCTTCTAATTGCATTATCATTTTGTATTACTGTTATTTATCACTTTCCAATAGCAACGAGAAGCAGTAAGGAGTACGATGAATATGGAAATGAATATGAAAACGTTGCATCGATAGAGTCGGATATAAAAAATGTACGTCGATTACTTGACGAGGTACCGGATCCCTCACAAAACCGTCTACAATTCCTGAAACTTGATGAGCATGCTTTTGCATTCTCGGCCTACACAGACGATCGAAATGGAAATATGGGGTACAAATATGTCCGAGTCCTGATGTTTATCACGTCACAAGACAACTTTTCCTGTGAAATAAACGGGAGAAAGTCCACAGATGTATCACTTTACGAGTTCTCGGAAAATCACAAAATGAAGTGGCAAATGTTTATTTTGAATTGTAAACTACCCGATGGTATAGATTTCAATAATGTTAGCTCTGTAAAGGTCATAAGAAGCACAACCAAGCAGTTTGTTGATGTGCCGATTCGGTATAGAATTCAAGATGAGAAAATAATTACGCCAGACGAATATGACTATAAAATGTCAATTTGTGTTCCAGCATTGTTTGGAAATGGATATGATGCAAAGCGAATTGTTGAGTTTATTGAGCTGAATACTTTGCAAGGAATCGAGAAAATATACATTTACACTAATCAAAAAGAGCTTGATGGATCCATGAAGAAAACGTTGAAATACTATTCGGATAATCACAAAATAACATTAATTGATTACACATTACCATTCAGAGAGGATGGTGTTTGGTATCACGGGCAATTGGCAACTGTTACTGATTGTTTACTGAGAAACACTGGAATCACAAAATACACATTTTTCAATGATTTTGATGAGTTCTTCGTCCCCGTTATCAAAAGTCGGACTCTCTTTGAAACAATCAGTGGGCTTTTTGAAGATCCCACTATTGGATCGCAACGAACAGCTTTGAAGTATATAAATGCAAAAATCAAGAGCGCTCCGTATTCACTGAAAAATATTGTTTCCGAAAAACGAATTGAAACAAGATTCACGAAATGTGTAGTTCGACCGGAAATGGTTTTGAACAGGGTATTCATCATACGAGTAGAGTGATTCAAGACAACTATAAAACGGTTTCCCATGGCGGATCCCTTCTACGGGTTTATCATTACAAGGATAAAAAGTATTGTTGCGAAGACGAGAGCCTCTTGAAAAAACGGCATGGAGATCAACTTCGGGAAAAATTCGATTCAGTTGTTGGTCTTTAGACTTGTAG
配列番号2(NCBIにおいて参照配列NP_504545.2とも記載されている)
MPRITASKIVLLIALSFCITVIYHFPIATRSSKEYDEYGNEYENVASIESDIKNVRRLLDEVPDPSQNRLQFLKLDEHAFAFSAYTDDRNGNMGYKYVRVLMFITSQDNFSCEINGRKSTDVSLYEFSENHKMKWQMFILNCKLPDGIDFNNVSSVKVIRSTTKQFVDVPIRYRIQDEKIITPDEYDYKMSICVPALFGNGYDAKRIVEFIELNTLQGIEKIYIYTNQKELDGSMKKTLKYYSDNHKITLIDYTLPFREDGVWYHGQLATVTDCLLRNTGITKYTFFNDFDEFFVPVIKSRTLFETISGLFEDPTIGSQRTALKYINAKIKSAPYSLKNIVSEKRIETRFTKCVVRPEMVFEQGIHHTSRVIQDNYKTVSHGGSLLRVYHYKDKKYCCEDESLLKKRHGDQLREKFDSVVGLLDL
配列番号3は、配列番号4をコードしている核酸配列である:
(NCBIにおいて参照配列XM_001674213.1とも記載されており、カエノラブディティス・ブリグサエ(Caenorhabditis briggsae)のガラクトシルトランスフェラーゼをコードしている)
ATGCCACGAA TAACGGCAAG CAAAATAGTG TTATTATCTG TATTATCCTT ACTAACAGTT TTCTATCTGA ATACATTTTC GTCTATTAAA ATTGAAAACG ATCTCGACGG GACTGATTAC GACTTGGATT ACATAGAATC TGATATCAAA AAGACGCGTC GATTACTCAA TGAAATCCCT GATCCATCTC AAAACCGAGT TCAATTTTTT AAACTCGATG ATAATGGATA TGCATTCTCA GCATATACAG ATAATAGGAA AGGAAATATG GGTCACAAAT ATGTCAGAAT ATTAGTGTTC CTAACTAAAT TTGATGATTT TTCTTGCGAA ATTAACTCGA AGAAATCCTA TGTTGTTACA CTCTACGAGC TATCAGAAAA TCACAATATG AAGTGGAAAA TGTATATTTT GAATTGTTTA CTTCCCGATG GAATCACTTT CAACGATGTG AATTCTGTAA AAATATCTAG AAGTTCTTCA AAACTTTCAG TCCAAATCCC GATCAGATAT AGAATTCAAG ATGAGAAAAT GATGACTCCA GATGAATACG ATTATAAGTT GTCGATTTGT GTTCCTGCAC TTTTTGGAAA CGTTTATTAT CCAAGGAGGA TTATTGAATT TGTGGAACTA AACAGCTTGC AAGACATCGA CAAAATCTAC ATCTACTACA ATCCTTTAGA AATGACAGAT GAGGCCACAG AAAGGACTTT GAAGTTTTAT TCCAATAATG GGAAAATCAA TTTAATAGAA TTCATTCTCC CATTTCTAC TCGAGATGTT TGGTATTATG GGCAATTGGC CACCGTTACA GATTGTCTTC TCCGTAACAC TGGAATAACT CAATACACAT TTTTCAATGA TTTGGATGAA TTTTTCGTGC CAGTACTGGA CAACCAAACT CTCTCTGAAA CTGTGTCAGG ATTATTTGAA AATCGAAAAA TTGCCTCTCA GAGAACGGCC TTGAAATTTA TTAGTACAAA AATCAATCGA TCTCCTGTAA CTCTCAATAA TATTGTGTCT TCTAAAAATT TTGAAACGAG ATTCACAAAA TGCGTCGTAC GGCCGGAAAT GGTTTTTGAG CAGGGCATTC ACCATACGAG TAGAGTAATA CAAGACGACT ACGAAACCCC ATCCCATGAT GGATCACTTT TGCGTGTGTA TCACTACAGA GAACCAAGAT ATTGCTGCGA AAACGAGAAT CTTCTAAAAC AAAGATACGA TAAGAAGCTT CAAGAAGTTT TTGATGCTGT AGTTCTTATA TTGCATGTCA CATTTGATGT ATGGATATAT CACCTGAAAA ACACCCTCTA A
配列番号4(NCBI参照配列XP_001674265.1においても記載されている)
MPRITASKIV LLSVLSLLTV FYLNTFSSIK IENDLDGTDY DLDYIESDIK KTRRLLNEIP DPSQNRVQFF KLDDNGYAFS AYTDNRKGNM GHKYVRILVF LTKFDDFSCE INSKKSYVVT LYELSENHNM KWKMYILNCL LPDGITFNDV NSVKISRSSS KLSVQIPIRY RIQDEKMMTP DEYDYKLSIC VPALFGNVYY PRRIIEFVEL NSLQDIDKIY IYYNPLEMTD EATERTLKFY SNNGKINLIE FILPFSTRDV WYYGQLATVT DCLLRNTGIT QYTFFNDLDE FFVPVLDNQT LSETVSGLFE NRKIASQRTA LKFISTKINR SPVTLNNIVS SKNFETRFTK CVVRPEMVFE QGIHHTSRVI QDDYETPSHD GSLLRVYHYR EPRYCCENEN LLKQRYDKKL QEVFDAVVLI LHVTFDVWIY HLKNTL
配列番号5は、配列番号6、その後に終止コドンおよびさらに68ヌクレオチドをコードしている核酸配列(1428核酸)である:(NCBI参照配列XM_001629141.1においても記載されており、ネマトステラ・ベクテンシス(Nematostella vectensis)のガラクトシルトランスフェラーゼをコードしている)
ATGCGATGCT ATATTTACAA ATTGAGGTTG TCCGTTTGTC TGTTTGTAGT GCTCTTCACA GCACTGCTTT TCATCACCTA TTTAAACCAC TCAGAGCTTG AATCAGCAGA GAAAAGTAGC GGAAAAAGGA AGACGCGACA TCGTAAACGA ACACGTTCAC GCAAACAACA CGAGAGCCAT TTTCAGAAAG CTCGACTACA AGAAAGAGAA CTAGTATTAA GATCTACAGC GCCACCAACA TTACGAAGAG AAGTACAAGC GCATCGATTA GGGCAGATCC GTGGCAAGAA CACGGACCAG GGGATAACTG GAAAGTTCAC AGAGATCGCT AAAGACACGC ATATTTATTC AGCGTTTTAC GACGATGCCA AGTCAAATCC ATTCATTCGT CTTATCATCC TCTCGGGAAA ACACTACCAG CCTGGATTAT CTTGCCAATT TTGCGAACCT TTGTCCGCCA GTTGTAGTTT TGCGGACTCT AAAGCTGAAT ACTACACGAC CAACGAGAAC CATGGGAGAG TATTTGGCGG GTTCATTGCG AGTTGCCTCG TGCCTGATGG ATTCAATGCA GTGCCATTGT TTGTTGACAT AACGGCCGAT GTTAAGGGGG AGAAAAGCAA GGCACGGGTA CCTGTGGTGT CTAATGCACA TCTCTACTAC CCTATTAAAT ACGCAATCTG CGTCCCACCC CTCCGATCAG AGAAACTAAC AGCGAAAAGA CTCATAGAGT TTGTCGAGCT AACCAAACTT TTAGGCGCTA ACCATTTTAC TTTTTATGAC TTCAAAACGG ACCCGGAAGT CAATAACGTT TTAAGATATT ACCAGGAGAC ACAAGTAGCA AATGTTCTGC CATGGAATCT ACCTTCAAAT TTGGTATCCA GGCCGAACGA TATTTGGTAC TTTGGTCAGG TTTTGGCTAT TCTAGATTGC TTGTATCGCT ACAAGAACAG GGCAAAATTT GTAGCCTTCA ATGACGTAGA TGAGTTTATC GTTCCGCTAA GGAACAGCTC GATAGTGGAA ATACTAAACG CGTTTCACCG GCCATACCAC TGTGGACATT GCTTTCAGAG CGTGGTGTTC AGCTCAAACG CGAGATTTCC CAGGCAAAAA AGCGAGTTAG TTTCTCAGCG GTTCTTCCAC AGGACCCAGG AAACCATCCC TCTCCTCTCG AAATGCATTG TGGATCCTTT GAGAGTGTTC GAGATGGGGA TTCACCACAT AAGCAAGGCT ACAGGTCTGC GGTATTCCGT CAACTCAGTA CACGAGAGTG ACGCGGTTAT CTTCCATTAC AGGACTTGCA CTACGTCATT TGGTATACGT CATCAGTGCA TGAACCTAGT GCATGATGGG ACCATGGCCA AATATGGAAA ACGACTTCAG AAAATGTTTA GAAAGGTTGT AAATGATTTA AAACTTTTGG CACCAACGTA GCTATTTCGT AACACTTCAC ACTTTCATTG TTATAACAGA ATACAGAATA AATTAATGAT TGTTGTGCC
配列番号6(NCBI参照配列XP_001629191においても記載されている)
MRCYIYKLRL SVCLFVVLFT ALLFITYLNH SELESAEKSS GKRKTRHRKR TRSRKQHESH FQKARLQERE LVLRSTAPPT LRREVQAHRL GQIRGKNTDQ GITGKFTEIA KDTHIYSAFY DDAKSNPFIR LIILSGKHYQ PGLSCQFCEP LSASCSFADS KAEYYTTNEN HGRVFGGFIA SCLVPDGFNA VPLFVDITAD VKGEKSKARV PVVSNAHLYY PIKYAICVPP LRSEKLTAKR LIEFVELTKL LGANHFTFYD FKTDPEVNNV LRYYQETQVA NVLPWNLPSN LVSRPNDIWY FGQVLAILDC LYRYKNRAKF VAFNDVDEFI VPLRNSSIVE ILNAFHRPYH CGHCFQSVVF SSNARFPRQK SELVSQRFFH RTQETIPLLS KCIVDPLRVF EMGIHHISKA TGLRYSVNSV HESDAVIFHY RTCTTSFGIR HQCMNLVHDG TMAKYGKRLQ KMFRKVVNDL KLLAPT
配列番号7は、配列番号8をコードしている核酸配列である:
(NCBI参照配列XM_002189335においても記載されており、キンカチョウ(Taeniopygia guttata)のガラクトシルトランスフェラーゼをコードしている)
ATGACTGTAA CTTTAATGCT TGTGGTTTCT TATCTGAGAT TACAGAGACT TTCTCATCAG CCAAAAGTAA TTCAAGAAAG TAGAAGATGT AGAGGGAAAA TTGCCCTTAG CACAATAACA GCATTGGAAG GTAACAAAAC TGATATTATA TCCCCATACT TTGATGACAG AGAAAACAAA ATCACTCGTC TGATTGGGAT TGTTCACCAT AAAGATGTAA AACAACTGTT CTGCTGGTTC TGCTGTCAAG CCAATGGAAA GATATATGTA TCAAAAGCAG AAATAGATGT TCACTCGGAT AGATTTGGAT TCCCTTATGG TGCAGCAGAT ATAATTTGTT TGGAACCTGA AAACTGTGAT CCAACACATG TATCAATTCA TCAGTCTCCA TATGGAAATA TTGACCAGCT GCCGAGGTTT GAAATTAAAA ATCGCAGGCC TGAGACCTTT TCTGTTGACT TCACCGTGTG CATTTCTGCC ATGTTTGGAA ACTACAACAA TGTCTTGCAG TTTGTACAGA GTATGGAAAT GTATAAGATT CTTGGAGTAC AGAAAGTGGT GATCTATAAG AACAACTGCA GCCATCTGAT GGAGAAAGTC TTGAAATTTT ATATAGAAGA AGGAACTGTT GAGGTAATTC CCTGGCCAAT AGACTCACAC CTCAGGGTTT CTTCTAAATG GCGCTTCATG GAAGACGGGA CACACATTGG CTACTATGGA CAAATCACAG CTCTAAATGA CTGTATATAC CGCAACATGG AAAGGACCAA GTTTGTGGTC CTTAATGACG CTGATGAAAT AATTCTTCCC CTTAAACACC CAGACTGGAA AACAATGATG AACAGTCTTC AGGAGCAAAA CCCAGGGACT AGTGTTTTCC TTTTTGAGAA CCATATCTTC CCAGAAACTG TATTTTCTCC CATGTTCAAC ATTTCATCTT GGAATACTGT GCCAGGTGTT AACATATTGC AGCATGTGTA CAGAGAGCCT GACAGGAAAC ATGTAATCAA TCCCAGGAAA ATGATAGTTG ATCCACGAAA GGTGATTCAG ACTTCAGTCC ATTCTGTCCT ACGTGCTTAT GGGAAGAGCG TGAATGTTCC CATGGAAGTT GCCCTCATTT ATCACTGTCG GAAGGCCCTT CAAGGAAACC TTCCCAGAGA ATCTCTCATC AGGGATACAA CACTGTGGAG ATATAACTCA TCATTAATCA TGAATGTTAA CAAGGTTCTA TCTCAAACCA TGCTGCAAAC TCAAAATTGA
配列番号8(NCBI参照配列XP_002189371においても記載されている)
MTVTLMLVVS YLRLQRLSHQ PKVIQESRRC RGKIALSTIT ALEGNKTDII SPYFDDRENK ITRLIGIVHH KDVKQLFCWF CCQANGKIYV SKAEIDVHSD RFGFPYGAAD IICLEPENCD PTHVSIHQSP YGNIDQLPRF EIKNRRPETF SVDFTVCISA MFGNYNNVLQ FVQSMEMYKI LGVQKVVIYK NNCSHLMEKV LKFYIEEGTV EVIPWPIDSH LRVSSKWRFM EDGTHIGYYG QITALNDCIY RNMERTKFVV LNDADEIILP LKHPDWKTMM NSLQEQNPGT SVFLFENHIF PETVFSPMFN ISSWNTVPGV NILQHVYREP DRKHVINPRK MIVDPRKVIQ TSVHSVLRAY GKSVNVPMEV ALIYHCRKAL QGNLPRESLI RDTTLWRYNS SLIMNVNKVL SQTMLQTQN
配列番号9は、配列番号10をコードしている核酸配列である:
(NCBI参照配列XM_626032においても記載されており、クリプトスポリジウム・パルバム(Cryptosporidium parvum)のガラクトシルトランスフェラーゼをコードしている)
ATGCAAAGTA AAGTCATTTT TAGGATCTTG GTATTGATCA TTTCGGTGAT TGGATCCTTA TACTCAATAA TTCAATTAAT GCTAAAGGAG CTATCAAGTA ACAAAAATAT TCAAGAGGTT AGTCATTCAA GGAGGCTAAT AAGTGAACCT TACAGTGAAA GTATTAATGA ACAAAATGAT CAAGATTGGA AAGAACTAAA GCTAATAATT CCAAATCATT CTCAAATTAA CCAGCAGGAA AAAAATGGTA ATTTGATTGA GTTTAAAGTT TATATATACT CAGCATATTA TGATTGGAGA ATAGATAGGA TACGAATAAA TTCACTTATC CCATCGAATT TTTATGATCG AATAGAAATG GAATGTGCAA TAATCTTGGA CAAAAATATT TACACAGGAA CTATTAAAAA AGTGATTCAT AAGGAGCACC ATAATAAAGA ATATGTATCA TCGACTTTAC TCTGCGAAAT TGCAAAAAAT GAAATTAAAT TTGAGGATAT TTCAAGGAAA GTTTTGATAA CAATTTTGGA AAATGGAAAC AGCACAAATA AATCAGAAAT ATGGATAACT CTAAAAAAAA TTCCAAAAAA TAGCTCTAAT AATCATGAGC TGACTGTTTG TGTGAGACCT TGGTGGGGAG AGCCAATAAA GAATGGAAAC TTGGGAAATA AACAAAAATT TAACAATTCA GGGTTAATGC TTGAATTTAT TAATTCATAT TTATTCTTAG GAGCAAATAA ATTTTATTTA TATCAAAATT ACTTGGACAT TGACGAAGAT GTAAGAAATA TAATAAATTA TTATTCTAAT ATCAAAAATG TTTTGGAAAT TATTCCATAC TCATTACCAA TAATTCCATT TAAACAAGTT TGGGATTTCG CACAAACAAC AATGATACAG GACTGCCTAC TAAGAAATAT TGGAAAAACA AAATACTTGT TATTCGTAGA TACCGATGAA TTTGTATTTC CAAACTTGAA AAATTATAAC TTAATGGATT TTTTAAATTT ATTAGAAGCC AACAATCCTT ATTATAAAAA CAAAGTCGGG GCAATGTGGA TTCCAATGTA TTTTCATTTT TTAGAGTGGG AATCTGATAA AAATAATTTG AAGAAATATT CAACAATTGA GAAAAAAATT AAGAAAAAGA TGGCAAATAT TGAGTTTGTT CTATATCGTA AAACATGTAG AATGTTAAGT TCTGGAACAA AAAAAAGTGA CAAGACGAGA AGAAAAGTTA TTATTAGACC TGAAAGAGTT TTGTATATGG GTATACATGA AACAGAAGAG ATGCTAAGCA AAAAATTTCA TTTCATTAGA GCTCCTGTAA TTAATGTGGG TGGAGGAAAC GAACTAAGTA TATATTTACA TCATTATAGA AAAGCAAAAG GTATTGTAAA CAATGATCCC AAACAAAGAG AACTTGTGAA TATGTATTTA GAAAATGTTT GTTCAGATAA GCTGTTAGAT TCAGGGGGAG ATTCCATTCA AGATGGAGTA ATTGTCGACA ATACTGTTTG GGAGATATTT GGAACACACT TATACCAGAT AATTTTTGAG CATATTAAAG AAATCCAAGA TATGTACACA AATAAGGAAA TAATTAATGG AAATAAAAAT TTAAGTGTTG AAGAATTACA TAATTAA
配列番号10(NCBI参照配列XP_626032においても記載されている)
MQSKVIFRIL VLIISVIGSL YSIIQLMLKE LSSNKNIQEV SHSRRLISEP YSESINEQND QDWKELKLII PNHSQINQQE KNGNLIEFKV YIYSAYYDWR IDRIRINSLI PSNFYDRIEM ECAIILDKNI YTGTIKKVIH KEHHNKEYVS STLLCEIAKN EIKFEDISRK VLITILENGN STNKSEIWIT LKKIPKNSSN NHELTVCVRP WWGEPIKNGN LGNKQKFNNS GLMLEFINSY LFLGANKFYL YQNYLDIDED VRNIINYYSN IKNVLEIIPY SLPIIPFKQV WDFAQTTMIQ DCLLRNIGKT KYLLFVDTDE FVFPNLKNYN LMDFLNLLEA NNPYYKNKVG AMWIPMYFHF LEWESDKNNL KKYSTIEKKI KKKMANIEFV LYRKTCRMLS SGTKKSDKTR RKVIIRPERV LYMGIHETEE MLSKKFHFIR APVINVGGGN ELSIYLHHYR KAKGIVNNDP KQRELVNMYL ENVCSDKLLD SGGDSIQDGV IVDNTVWEIF GTHLYQIIFE HIKEIQDMYT NKEIINGNKN LSVEELHN
【0013】
本発明との関連において使用される場合、「ポリペプチドをコードしている核酸」という用語は、対立変異および遺伝子コードの冗長性を含むことを意味する。
【0014】
当業者に知られているようにおよび本明細書において使用されるように、「%(パーセント)同一性」という用語は、配列間の合致によって決定される2つ以上の核酸分子間の関連性の程度を示す。「同一性」のパーセンテージは、ギャップおよび別の配列特性を考慮した上での2つ以上の配列における同一な領域のパーセンテージの結果である。
【0015】
関連する核酸分子の同一性は、既知の方法による支援によって決定することができる。一般に、この作業の特定の要求に適応させるために適合させたアルゴリズムを使用する特別なコンピュータプログラムが用いられる。同一性を決定するための好ましい方法は、比較される配列間で最大の程度の同一性の作成から始まる。2つの核酸配列間の同一性を決定するための好ましいコンピュータプログラムは、BLASTN(Altschulら、J. Mol. Biol.、215、403〜410、1990)およびLALIGN(HuangおよびMiller、Adv. Appl. Math.、12、337〜357、1991)を含むがこれらに限定的されない。BLASTプログラムは、米国立生物工学情報センター(NCBI:National Center for Biotechnology Information)から、および別の供給源(BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894)から入手することができる。
【0016】
本発明による核酸分子は、当業者によく知られている方法によって合成により調製してもよいが、また、適したDNAライブラリーおよび別の公的に利用可能な核酸の供給源から単離してもよく、その後、任意選択により、突然変異させてもよい。そのようなライブラリーまたは突然変異の調製は、当業者によく知られている。
【0017】
好ましい一実施形態において、本発明の核酸分子は、二重鎖または単鎖のいずれか(すなわちセンスまたはアンチセンス鎖のいずれか)の、cDNA、ゲノムDNA、合成DNA、RNAまたはPNAである。本発明の範囲内に包含される核酸分子およびそのフラグメントは、例えば、ポリメラーゼ連鎖反応(PCR)によって作製してもよく、または、DNA合成を使用して合成によって、もしくはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムのmRNAを使用して逆転写によって生成してもよい。
【0018】
場合によって、本発明はまた、特に参照されている核酸配列と、好ましくはストリンジェントな条件下で、ハイブリダイズする能力を有することを特徴とする、本発明のポリペプチドをコードしている新規な核酸も提供する。特に参照されている核酸配列とストリンジェントな条件下でハイブリダイズする能力を決定するための、従来技術において一般的なおよび/または標準的なプロトコール(例えば、SambrookおよびRussell、Molecular cloning: A laboratory manual (3 volumes)、2001)の次に、例えば、上記のBLASTN(Altschulら、J. Mol. Biol.、215、403〜410、1990)およびLALIGNアラインメントツールなどのアラインメントツールを用いて遺伝子データベース(例えば、http://www.-ncbi.nlm.nih.gov/entrez/query.fcgi?db=nucleotide)において見出すことができるヌクレオチド配列を比較することによって、特に参照されている核酸配列とストリンジェントな条件下でハイブリダイズする能力を分析および決定することが好ましい。
【0019】
最も好ましくは、本発明の核酸が、核酸、例えば、配列番号1、3、5、7および/または9のいずれかにおいて記載されている核酸とハイブリダイズする能力は、以下の条件下で、サザンブロットアッセイにおいて確認される:45℃での6×塩化ナトリウム/クエン酸ナトリウム(SSC)、その後0.2×SSC、0.1%SDS中で65℃での洗浄。
【0020】
本発明の核酸は、本発明のポリペプチドをインビトロまたはインビボで産生するために適したベクターおよび/または宿主細胞における発現を支配するプロモーターに作動可能に連結されていることが好ましい。
【0021】
単離および精製された核酸への作動可能な連結に適したプロモーターは、当技術分野において知られている。好ましい一実施形態において、本発明の核酸は、ピキア・パストリス(Pichia pastoris)AOX1またはGAPプロモーター(例えば、Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)GAL1、ADH1、ADH2、MET25、GPDまたはTEFプロモーター(例えば、Methods in Enzymology、350、248、2002を参照されたい)、バキュロウイルスポリヘドリンp10またはie1プロモーター(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カースルバッド、カリフォルニア、およびNovagen Insect Cell Expression Manual、Merck Chemicals Ltd.、ノッティンガム、イギリスを参照されたい)、大腸菌(E.coli)T7、araBAD、rhaP BAD、tetA、lac、trc、tacまたはpLプロモーター(Applied Microbiology and Biotechnology、72、211、2006を参照されたい)、植物CaMV35S、ocs、nos、Adh-1、Tetプロモーター(例えば、LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009を参照されたい)またはSambrookおよびRussell(2001)に記載の、哺乳動物細胞用の誘導可能なプロモーターからなる群から選択されるプロモーターに作動可能に連結されているものである。
【0022】
好ましくは、単離および精製された核酸は、エピソームベクターまたはウイルスベクターなどの組換えベクターの形態である。適したベクターおよび発現制御配列の選択ならびにベクター構築は、当業者の範囲内である。好ましくは、ウイルスベクターは、バキュロウイルスベクター(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)である。プロモーターおよび別の発現制御配列とのコード配列の作動可能な連結を含むベクター構築は、当業者の範囲内である。
【0023】
したがって、またさらなる態様において、本発明は、本発明の核酸を含む組換えベクターに関する。
【0024】
本発明のさらなる一態様は、本発明の、好ましくは、本発明のポリペプチドを産生する、核酸および/またはベクターを含む宿主細胞を対象とする。本発明のポリペプチドを産生するための好ましい宿主細胞は、酵母細胞、好ましくは、サッカロマイセス・セレビジエ(例えば、Methods in Enzmology、350、248、2002を参照されたい)、ピキア・パストリス細胞(例えば、Pichia Expression Kit Instruction Manual、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)、大腸菌細胞(BL21(DE3)、K-12および誘導体)(例えば、Applied Microbiology and Biotechnology、72、211、2006を参照されたい)、植物細胞、好ましくは、タバコ(Nicotiana tabacum)またはヒメツリガネゴケ(Physcomitrella patens)(例えば、LauおよびSun、Biotechnol Adv. 27、1015〜1022、2009を参照されたい)、NIH-3T3哺乳動物細胞(例えば、SambrookおよびRussell、2001を参照されたい)および昆虫細胞、好ましくは、sf9昆虫細胞(例えば、Bac-to-Bac Expression Kit Handbook、Invitrogen Corporation、カールズバッド、カリフォルニアを参照されたい)からなる群から選択される。
【0025】
本発明の別の重要な態様は、
(a)配列番号2、4、6、8および10からなる群から選択されるアミノ酸配列、好ましくは配列番号2のアミノ酸配列を有するポリペプチド、
(b)本発明の核酸によってコードされているポリペプチド、
(c)(a)および/または(b)のポリペプチドと、少なくとも25、30、または40%、好ましくは、少なくとも50または60%、より好ましくは、少なくとも70または80%、最も好ましくは、少なくとも90または95%のアミノ酸配列同一性を有するポリペプチド、
(d)(a)、(b)または(c)のフラグメントおよび/または機能的誘導体
からなる群から選択される、単離および精製されたポリペプチドを対象とする。
【0026】
関連するアミノ酸分子の同一性は、既知の方法による支援によって決定することができる。一般に、この作業の特定の要求に適応させるために適合させたアルゴリズムを使用する特別なコンピュータプログラムが用いられる。同一性を決定するための好ましい方法は、比較すべき配列間で最大の程度の同一性を作成することから始まる。2つのアミノ酸配列間の同一性を決定するための好ましいコンピュータプログラムは、TBLASTN、BLASTP、BLASTXまたはTBLASTX(Altschulら、J. Mol. Biol.、215、403〜410、1990)を含むがこれらに限定的されない。BLASTプログラムは、米国立生物工学情報センター(NCBI:National Center for Biotechnology Information)から、および別の供給源(BLAST handbook、Altschulら、NCB NLM NIH Bethesda、MD 20894)から入手することができる。
【0027】
好ましくは、前記ポリペプチドは、本発明の上記の核酸によってコードされている。
【0028】
好ましい一実施形態において、本発明のポリペプチド、フラグメントおよび/または誘導体は、機能的である、すなわち、好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-および最も好ましくは、GnGnF6-(Schachter、Biochem. Cell. Biol. 64(3)、163〜181、1986による命名)含有多糖/オリゴ糖または複合糖質をアクセプター基質として、酵素ガラクトシルトランスフェラーゼ活性、好ましくは、酵素β-1,4-ガラクトシルトランスフェラーゼ活性、より好ましくは、酵素β-1,4-ガラクトシルトランスフェラーゼ活性を有する。
【0029】
例えば、本発明による、ポリペプチド、そのフラグメントおよび誘導体の機能性、すなわち酵素活性を決定するための好ましいアッセイは、下記の実施例4において示されている。
【0030】
本発明のポリペプチドの「機能的誘導体」という用語は、その誘導体が、測定可能な程度まで、例えば、もとの非修飾ポリペプチドの少なくとも約1〜10%の、上記の酵素活性の少なくとも1つを依然として有する限り、例えば、アミノ酸残基の付加、置換および/または欠失によってそのアミノ酸配列が化学的にまたは遺伝子的に修飾されている、および/または、例えば、付加、欠失、再編成、酸化、還元などによって、その原子および/または化学的官能基の少なくとも1つが化学的に修飾されている、任意のポリペプチドまたはそのフラグメントを含むことを意味する。
【0031】
この状況において、本発明の機能的フラグメントは、本発明のポリペプチドまたは誘導体の一部を形成し、かつ測定可能な程度で、例えば、完全なタンパク質の少なくとも約1〜10%の、上記の酵素活性の少なくとも1つを依然として有するものである。
【0032】
「単離および精製されたポリペプチド」という用語は、本明細書で使用される場合、例えば、カエノラブディティス・エレガンス組織またはその画分中の、天然に存在する対応物がない(例えば、ペプチド模倣物)、または、それに天然に付随する成分から分離または精製されている、ポリペプチドまたはペプチドフラグメントを指す。好ましくは、ポリペプチドは、乾燥調製物の少なくとも60重量%を占める場合、「単離および精製された」、したがって、最も天然に存在するポリペプチドおよび/またはそれに天然に付随する有機分子を含まないとみなされる。好ましくは、本発明のポリペプチドは、乾燥調製物の少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは、少なくとも99重量%を占める。乾燥ポリペプチド調製物の少なくとも80%、より好ましくは、少なくとも90%、および最も好ましくは、少なくとも99重量%を占める本発明によるポリペプチドがより好ましい。化学的に合成されたポリペプチドは、上記の状況において本来「単離および精製されて」いる。
【0033】
単離された本発明のポリペプチドは、例えば、天然の供給源、例えば、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムからの抽出によって;宿主、好ましくは、異種宿主における、そのポリペプチドをコードしている組換え核酸の発現によって;または、化学合成によって得ることができる。それが天然に由来する供給源とは異なる細胞系において産生されるポリペプチドは、それに天然に付随する成分から分離されているため、「単離および精製されて」いる。単離および/または純度の程度は、任意の適切な方法、例えば、カラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、HPLC分析、NMR分光法、ガス液体クロマトグラフィー、または質量分析によって測定することができる。
【0034】
さらに、一態様において本発明は、本発明のポリペプチドに特異的に結合する抗体、その機能的フラグメントおよび機能的誘導体に関する。これらは、標的抗原が利用可能となれば、ハイブリドーマ技術(KohlerおよびMilstein、Nature、256、495〜497、1975)、抗体ファージディスプレイ(Winterら、Annu. Rev. Immunol. 12、433〜455、1994)、リボソームディスプレイ(Schaffitzelら、J. Immunol. Methods、231、119〜135、1999)および反復コロニーフィルタースクリーニング(Giovannoniら、Nucleic Acids Res. 29、E27、2001)によって、常法に従って利用可能である。抗体を機能的生成物に断片化するための典型的なプロテアーゼは、よく知られている。結果として生じるフラグメントが、特定の高親和性および、好ましくは、マイクロモルからピコモル範囲の解離定数を有する限り、別の断片化技術も同様に使用することができる。
【0035】
標的化用途のために非常に好都合な抗体フラグメントは、可変重鎖および可変軽鎖ドメインが、ポリペプチドリンカーによって一緒に結合されている単鎖Fvフラグメントである。本発明のポリペプチドを特定するための別の抗体フラグメントは、Fabフラグメント、Fab2フラグメント、ミニ抗体(低分子免疫タンパク質とも呼ばれる)、タンデムscFv-scFv融合体ならびに適したドメインとの(例えば、免疫グロブリンのFc部分との)scFv融合体を含む。特定の抗体フォーマットに関する概説については、HolligerおよびHudson、Biotechnol.、23(9)、1126〜36、2005を参照されたい。
【0036】
本発明において使用するための抗体の「機能的誘導体」という用語は、その誘導体が、そのもとの抗原に関して実質的に同じ結合親和性を有する、好ましくは、マイクロモル、ナノモルまたはピコモルの範囲内の解離定数を有する限り、例えば、アミノ酸残基の付加、置換および/または欠失によって、そのアミノ酸配列が化学的にまたは遺伝子的に修飾されている、および/または、例えば、付加、欠失、再編成、酸化、還元などによって、その原子および/または化学官能基のうちの少なくとも1つが化学的に修飾されている、任意の抗体またはそのフラグメントを含むことを意味する。
【0037】
好ましい一実施形態において、本発明において使用するための抗体、そのフラグメントまたは機能的誘導体は、ポリクローナル抗体、モノクローナル抗体、キメラ抗体、ヒト化抗体、CDR移植抗体、Fvフラグメント、FabフラグメントおよびFab2フラグメントおよび抗体様結合タンパク質、例えば、アフィリン(affilin)、アンチカリン(anticalin)およびアプタマーからなる群から選択されるものである。
【0038】
抗体様結合タンパク質の概説については、非免疫グロブリンドメインからの結合タンパク質のエンジニアリングに関する、Binzら、Nature Biotechnol.、23(10)、1257〜1268、2005を参照されたい。「アプタマー」という用語は、高親和性を有するポリペプチドに結合する核酸を記述する。アプタマーは、様々な単鎖RNA分子の大きなプールから、SELEX(例えば、Jayasena、Clin. Chem.、45、1628〜1650、1999;KlugおよびFamulok、M. Mol. Biol. Rep.、20、97〜107、1994;US 5,582,981を参照されたい)などの選択方法によって単離することができる。アプタマーはまた、合成し、その鏡像体から、例えば、L-リボヌクレオチドとして選択することもできる(Nolteら、Nat. Biotechnol.、14、1116〜1119、1996;Klussmannら、Nat. Biotechnol.、14、1112〜1115、1996)。このように単離された形態は、それらが、天然に存在するリボヌクレアーゼによって分解されず、したがって、より高い安定性を有するという利点を有する。
【0039】
別の抗体様結合タンパク質および伝統的抗体の代替物は、いわゆる「タンパク質足場」、例えば、リポカリンをベースとするアンチカリン(Besteら、Proc. Natl. Acad. Sci. USA、96、1898〜1903、1999)である。リポカリンの、例えば、レチノール結合タンパク質またはビリン結合タンパク質の天然のリガンド結合部位は、例えば、「コンビナトリアルタンパク質設計」手法を用いることによって、それらが選択されたハプテンに結合するように(Skerra、Biochem. Biophys. Acta、1482、337〜350ページ、2000)変更することができる。別のタンパク質足場については、これらは抗体の代替物であることがまた知られている(Skerra、J. Mol. Recognition、13、167〜287、2000;Hey、Trends in Biotechnology、23、514〜522、2005)。
【0040】
要約すると、機能的抗体誘導体という用語は、上記のタンパク質由来の抗体代替物、すなわち、ポリペプチド、そのフラグメントまたは誘導体を特異的に認識する、抗体様結合タンパク質、例えば、アフィリン、アンチカリンおよびアプタマーを含むことを意味する。
【0041】
さらなる態様は、本発明によるモノクローナル抗体を発現するハイブリドーマ細胞系に関する。
【0042】
本発明の核酸、ベクター、宿主細胞、ポリペプチドおよび抗体は、多くの新しい用途を有する。
【0043】
一態様において、本発明は、ガラクトシド含有オリゴ糖/多糖および/または複合糖質、好ましくは、ガラクトシル-フコシド含有オリゴ糖/多糖および複合糖質、より好ましくは、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-GlcNAc含有オリゴ糖/多糖および複合糖質、最も好ましくは、GnGnF6Gal-またはMMF6Gal含有オリゴ糖/多糖および複合糖質を作製するための、ポリペプチド、本発明のポリペプチドを含む細胞抽出物、好ましくは、線虫抽出物、より好ましくは、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムの抽出物、および/または本発明の宿主細胞の使用に関する。
【0044】
本明細書で使用される場合、複合糖質という用語は、非糖成分の性質に関して限定するものではないことが理解される。好ましくは、複合糖質の非糖成分は、ポリペプチド/オリゴペプチドである。
【0045】
ガラクトシル-フコシル特異的オリゴ糖および複合糖質の酵素的合成は、高度に特異的であり、制御されており、環境に優しく、生成物は、それを必要とするヒトまたは別の哺乳動物などの対象における、寄生虫感染、好ましくは線虫およびアピコンプレックス感染の、治療および予防のために、高度に寄生虫特異的な(このエピトープはまた、タコ[Zhangら、Glycobiology、7、1153〜1158、1997]、イカ[Takahashiら、Eur. J. Biochem.、270、2627〜2632、2003]およびカサガイ[Wuhrerら、Biochem. J.、378、625〜632、2004]において存在していることのみが知られている)ワクチン成分として作用することができる。
【0046】
例示的なおよび好ましいガラクトシル-フコシル特異的オリゴ糖および複合糖質は、N-結合グリカン、N-糖タンパク質、糖脂質および脂質結合オリゴ糖(LOS)からなる群から選択される。本明細書において使用される場合、「複合糖質」という用語は、オリゴ糖成分および非糖成分の、任意のタイプのコンジュゲート、必然的ではないが好ましくは共有結合しているもの、例えば、共有結合性リンカーにより結合しているもの、例えば、生理学的に許容でき、さらには、例えば、線虫毒性などを付与する免疫刺激アジュバントとして、所望の生理的機能を有し得る、ポリペプチドまたは任意の別のタイプの有機または無機担体を含むことを意味する。
【0047】
例えば、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウまたはクリプトスポリジウム・パルバムまたは本発明のポリペプチドを産生する組換え昆虫細胞の粗抽出物は、Gal-Fuc含有コンジュゲート、例えば、遊離のGal-Fucグリカン、Gal-Fuc-ペプチド、Gal-Fuc-ポリペプチド、Gal-Fuc-フォールディングしたタンパク質を生成することができる。α-1,6-結合フコシドは、α-1,3-結合フコシドよりも非常に好ましい。
【0048】
本発明の別の態様は、以下の段階:
(i)少なくとも1つの本発明のポリペプチドを提供する段階、
(ii)少なくとも1つのフコシル化されたアクセプター基質を提供する段階、
(iii)好ましくは、マンガン(II)、コバルト(II)および/または鉄(II)イオンから選択され、より好ましくはマンガン(II)である少なくとも1つの適した二価の金属陽イオン補因子と、少なくとも1つの活性化された糖基質、好ましくは、ウリジン二リン酸(UDP)-ガラクトースとの存在下、本発明のポリペプチドの酵素活性に適した条件下で、(i)および(ii)をインキュベートする段階、
(iv)ガラクトシル-フコース誘導体を任意選択により単離する段階
を含む、ガラクトシル-フコシル誘導体を作製するための方法を対象とする。
【0049】
本発明のポリペプチドは、緩衝液、宿主細胞、細胞抽出物またはその酵素活性を持続させ、その基質および活性化された糖基質へのアクセスを可能にする任意の別の系において、乾燥形態または可溶性形態の単離ポリペプチドとして提供されてもよい。フコシル化されたアクセプター基質は、任意選択により、単離された形態の、または本発明のポリペプチドにより酵素的に修飾することができる系の成分としての、任意の種類のフコシル含有基質である。活性化された糖基質は、好ましくは、UDP-ガラクトースであるが、フコシル化されたアクセプター基質へ転移することができる、活性化された、好ましくは、リン酸活性化されたガラクトシル誘導体の任意の別のタイプとすることもできる。本発明の方法好ましくは、ガラクトピラノシル-β-1,4-L-フコピラノシル-誘導体、より好ましくは、D-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-βGlcNAc(Gal-Fuc)誘導体をもたらす。
【0050】
本発明のポリペプチドは、基質が適したフコシル部分を特徴として有する限り、広い基質特異性を有する。ガラクトシル-トランスフェラーゼ活性が、例えば、フコシル糖、フコシルペプチド、フコシルポリペプチド、さらには複合体およびフォールディングしたフコシルポリペプチドなどの基質について実証された。例えば、ガラクトシル-トランスフェラーゼ活性が、主要なエピトープとしてGnGnF6糖鎖構造を有する糖タンパク質、ヒトIgG1について実証された。これらのIgG1グリカンは、PNGaseFによって消化可能であることが知られている。ヒトIgG1のグリコシル化は、カエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼを含有する未精製のsf9昆虫細胞抽出物を用いて実証された。カエノラブディティス・エレガンスからの酵素抽出物の存在下における、放射性標識されたUDP-GalとのヒトIgG1のインキュベーションは、基質ガラクトシル化をもたらした。加えて、ガラクトシル化は、主要なエピトープとしてGnGnF6糖鎖構造を保有する再構築したヒトトランスフェリンに関して実証された。この目的のために、ヒトアポトランスフェリンを、シアリダーゼ(Iskratschら、Anal. Biochem.、368、133〜146、2009)、アスペルギルス・オリゼ(Aspergillus oryzae)のβ1,4-ガラクトシダーゼおよびピキア・パストリスにおいて発現された組換えハマダラカ属(Anopheles)コアα1,6-FucTで連続的に処理して、主要なエピトープとしてGnGnF6糖鎖構造を有する糖タンパク質が作製された。カエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼを含有する未精製のsf9昆虫細胞抽出物とのインキュベーションは、ガラクトシル化をもたらし、これは、フコース特異的ヒイロチャワンタケ(Aleuria aurantia)レクチンを用いたドットブロット法によって、および、様々なネオグリコフォーム(neoglycoform)のトリプシンペプチドのMALDI-TOF MSによってモニタリングされた。
【0051】
哺乳動物、特に、ヒトにおいて、慢性肝炎および肝硬変などの良性の肝臓疾患は、コアがフコシル化されたAFPをもたらさないことから、コアがフコシル化されたαフェトプロテイン(AFP)の血清含量が肝細胞癌(HCC)に高度に特異的であることが、ごく最近示された(Tatenoら、Glycobiology、19(5)、527〜536、2009を参照されたい)。
【0052】
したがって、さらなる態様において、コアがフコシル化されたAFPは、本発明のポリペプチドのアクセプター基質として選択的に適しているため、本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物は、ガラクトシル化合物を、コアがフコシル化されたαフェトプロテイン(AFP)に共有結合させるために、好ましくは、好ましくはHCC患者の血液からのコアがフコシル化されたαフェトプロテイン(AFP)を選択的に標識することによって、肝細胞癌(HCC)細胞を検出および/または定量するために使用することができる。
【0053】
したがって、本発明は、コアがフコシル化されたAFPを検出するため、すなわち、肝細胞癌(HCC)細胞を検出および/または定量するための診断手段を調製するための本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物に関する。
【0054】
また、本発明のポリペプチド、本発明のポリペプチドを含む宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物は、その出現が別のタイプの癌細胞と相互関係を示す、さらにコアがフコシル化されたマーカー糖タンパク質を検出するための診断手段を調製するために有用である。
【0055】
好ましい一実施形態において、本発明は、以下の段階:
(i)AFPを含む血液またはその画分、好ましくは血清を提供する段階と、
(ii)前記血液または前記その画分を、(a)本発明のポリペプチド、本発明の宿主細胞ならびに/または、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物とともに、および(b)活性化されたガラクトシル誘導体、好ましくは、標識されたガラクトシル誘導体、好ましくは標識されたUDP-ガラクトースとともに、活性化されたガラクトースの、コアがフコシル化されたAFP(AFP-L3)へのガラクトシル転移を可能にする条件下で、インキュベートする段階と、
(iii)ガラクトース標識された、したがって、コアがフコシル化されたAFP(AFP-L3)を検出する段階と
を含む診断方法に関する。
【0056】
上記の方法を実践するための、活性化されたガラクトシル誘導体の標識は、同位体、例えば14C、化学修飾、例えば、ハロゲン置換および別の選択的に検出可能な修飾、例えば、ビオチン、アジドなど、からなる群から選択される。好ましくは、段階(i)から(iii)のすべては、生体外で、すなわち、インビトロで実施される。
【0057】
本発明のさらなる一態様は、対象とする試料、例えば、ヒトまたは哺乳動物の試料において、好ましくは、細胞画分または抽出物試料において、線虫およびアピコンプレックス、好ましくは、カエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、およびクリプトスポリジウム・パルバムをそれぞれ同定および/または定量するための、本発明のポリペプチド、好ましくは、配列番号2、4、6、8および/または10のいずれかから選択される配列を有するポリペプチドに特異的に結合する抗体の使用を対象とする。典型的な抗体アッセイ、例えば、ELISAの設計および開発は、当業者の範囲内であり、さらに詳しく述べる必要はない。
【0058】
本発明を、好ましい実施形態および典型的な例に重点をおいて説明してきた。しかしながら、好ましい実施形態の変形形態が使用されてもよいこと、および本発明が、本明細書に具体的に記載されているものとは別の方法で実践されてもよいことが意図されることが、当業者には明らかであろう。さらに、前述の例は純粋に例示の目的で含まれているため、これらはいかなる点においても本発明の範囲を限定すると解釈されるべきではない。したがって、本発明は、本明細書に添付されている特許請求の範囲によって定義されている本発明の精神および範囲内に包含されるすべての変更形態を含む。
【図面の簡単な説明】
【0059】
【図1】バキュロウイルス感染sf9全細胞抽出物の抗FLAG免疫ブロット法を示す図である。空ベクター対照(e.v.)、N末端にFLAGをタグ付けしたM03F8.4(FLAG-GalT)およびタグなしM03F8.4(GalT)を含有するバキュロウイルスの様々なクローン。充填容量約150k細胞/スロット、SDS-PAGE12%、α-FLAG(1:2000、SIGMA)、α-マウス-HRP(1:2000、Santa Cruz Biotechnology)、ECL(Pierce、2s曝露)。
【図2】バキュロウイルス感染sf9全細胞抽出物のSDS-PAGE分析の図である。空ベクター対照(e.v.)、N末端にFLAGをタグ付けしたM03F8.4(FLAG-GalT)およびタグなしM03F8.4(GalT)を含有するバキュロウイルスの様々なクローン。充填容量約150k細胞/スロット、SDS-PAGE12%、銀染色による検出。(タンパク質は少量発現され、未精製の抽出物において空ベクター構築体に関して銀染色では検出不能である。)
【図3】Mn2+、Mg2+およびEDTAの存在下において、GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化ターンオーバーを示すカラムチャートであり、金属イオン依存性を示す;MES、pH6、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 2369/(m/z 2207+m/z 2369)]*100)によって決定されたターンオーバー。
【図4】GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化、タグ付きおよびタグなしの構築体の機能性を示すカラムチャートである;MES、pH6、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 2369/(m/z 2207+m/z 2369)]*100)によって決定されたターンオーバー。
【図5】MS分析による、GnGnF6アクセプター基質(ダブシル-GEN[GnGnF6]R)のガラクトシル化、タグ付きおよびタグなしの構築体の機能性を示す図である(MES pH6、室温、2.5時間)。上段スペクトル:UDP-Galなしの反応、中段スペクトル: UDP-Galあり、下段スペクトル:中段スペクトルの生成物の、アスペルギルス属β-ガラクトシダーゼによる消化産物(クエン酸緩衝液、pH5、室温、2d)。酵素は、ガラクトースをβ-ガラクトシダーゼによって消化することができるこのアクセプター基質に明らかに付加し、したがって、GalTによって組み込まれたβ-結合Gal残基を示す。さらなるGlcNAc除去は、未精製の昆虫細胞抽出物中のヘキソサミニダーゼの存在のために、長い反応時間(>2d)の後に起こる。
【図6】図5の、アクセプター(上段スペクトル)およびガラクトシル化された反応生成物(下段スペクトル)のMS/MSスペクトルの比較を示す図である。MS/MS分析は、ダブシル化されたGENRペプチドに結合されているHex-dHex-HexNAcモチーフに相当する二次イオン1272.61から観察される通り、コアのフコースに結合しているガラクトースを明らかに示す。
【図7】ガラクトシルトランスフェラーゼのドナー特異性の比較分析を示す図である(ダンシル-N[GnGnF6]ST、MES pH6.5、Mn2+、室温、13時間)。酵素は、UDP-Galに対する高い特異性を有するようであり、UDP-Glcに関する残留活性は無視できる。
【図8】アクセプター特異性の分析のカラムチャートである:カエノラブディティス・エレガンスGalTは、α-1,3-結合したフコースよりα-1,6結合したフコースを選択的にガラクトシル化する;ダブシルGEN-[MMF6/3]R、MES pH6.5、室温、2.5時間、未精製の反応混合物のMALDI-MSピーク強度の比([m/z 1963/(m/z 1801+m/z 1963)]*100)によって決定されたターンオーバー。
【図9】UDP-Galについて、タグなしガラクトシルトランスフェラーゼのKm(見かけ)のグラフによる決定を示す図である: Km(見かけ、UDP-Gal)=約40μM。
【図10】本発明のガラクトシルトランスフェラーゼの温度依存性の分析を示す図である(ダンシル-N[GnGnF6]ST、UDP-Gal、MES pH6.5、Mn2+、2.5時間)。
【図11】本発明のポリペプチド、すなわちカエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼによる、ヒトIgG1のグリコシル化(GnGnF6エピトープを有する)を示すカラムチャートである。
【図12】本発明のポリペプチド、すなわちカエノラブディティス・エレガンスのコアガラクトシルトランスフェラーゼによる、再構築したヒトトランスフェリン(GnGnF6エピトープを有する)のグリコシル化を示すMALDI-TOF MSスペクトルを示す図である。示されているm/z値は、GnGn(3813)、GnGnF6(3957)およびGnGnF6Gal(4119)をそれぞれ保有するペプチド622〜642に相当する。
【0060】
(実施例)
実験手順
化学製品および供給業者
UDP-Gal(VWR InternationalおよびSigma)、UDP-Glc、UDP-GlcNAc、UDP-GalNAc(すべてSIGMA)、UDP-14C-Gal(GE Healthcare)、GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man、Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc、MMF6、GnGnF6(すべてDextra Laboratories、イギリス)、Fuc-α-1,6-GlcNAc(Carbosynth Ltd.、イギリス)、ダブシル-GEN[GnGnF6]R(Paschingerら、Glycobiology、15(5)、463〜474、2005)、ダブシル-GEN[MMF6]R(Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001)、ダブシル-GEN[MMF3]R(Fabiniら、J. Biol. Chem. 276(30)、28058〜28067、2001)およびダンシル-N[GnGnF6]ST(Roitingerら、Glycoconj. J.、15(1)、89〜91、1998)は、以前に公開されている方法に従って得た。
【0061】
(実施例1)
カエノラブディティス・エレガンスcDNAの単離およびM03F8.4の発現ベクター中へのクローニング
線虫系統:
カエノラブディティス・エレガンスを培養するための方法は、Brenner, S.(Genetics 77(1)、71〜94、1974)において記載されている。野生型BristolN2系統を、大腸菌OP50を播種した標準的なNGM寒天プレート上で、20℃で増殖させた。
【0062】
カエノラブディティス・エレガンスM03F8.4cDNAの単離:
カエノラブディティス・エレガンス混合培養物を、標準的なNGM寒天プレートから回収し、滅菌したM9緩衝液(22mM KH2PO4、42mM Na2HPO4、85mM NaCl、1mM MgSO4)中で2回洗浄した。NucleoSpin(登録商標)RNA II RNA単離キット(MACHEREY-NAGEL AG)を使用して、全RNAを抽出した。Superscript(商標)III Platinum Two-Step qRT-PCR Kit(Invitrogen AG)の第1鎖cDNA合成ステップを使用して、0.5μgの全RNAを用いてcDNA合成を実施した。
【0063】
sf9昆虫細胞における組換え遺伝子発現のためのpFastBac1ドナープラスミドの構築:
M03F8.4cDNAを、以前に調製されたcDNAライブラリーから、提供されたマニュアルに従ってPhusion High-Fidelity DNAポリメラーゼ(Finnzymes)を使用したPCRによって単離した。タグなし型の構築では、SalIおよびXbaI制限部位にそれぞれ隣接している以下のフォワードおよびリバースプライマーを使用した:5'-TTTGTCGA-CACTTCTGAATGCCTCG-3'(配列番号11)および5'-TTTTCTAGACTACAAGTCTAA-AAGACCAAC-3'(配列番号12)。結果として生じたフラグメントを、適切な制限酵素で消化し、pFastBac1ドナープラスミド(Invitrogen)中にクローニングした。N末端FLAGタグ付き型の構築では、開始コドンを欠くフォワードプライマーを使用した:5'-TTTGTCGACCCTCGAATCACCGCC-3'(配列番号13)。結果として生じたフラグメントを、N末端FLAG配列を含有するpFastBac1ドナープラスミド中にクローニングした(Mullerら、J. Biol. Chem. 277(36)、32417〜32420、2002)(両ベクターは、チューリッヒ大学、Institute of Physiology、Thierry Hennetから好意により提供された)。
【0064】
(実施例2)
組換えタンパク質の発現
カエノラブディティス・エレガンスのコアβ-1,4-GalT候補cDNA(N末端FLAGタグありおよびなし)および空ベクター対照を含有する組換えバキュロウイルスを、製造業者の使用説明書(Invitrogen)に従って作製した。2×106のS.フルギペルダ(S.frugiperda)(sf9)付着性昆虫細胞に組換えバキュロウイルスを感染させ、28℃で72時間インキュベーションした後、細胞を、2体積%Triton-X100およびプロテアーゼ阻害剤カクテル(Roche、complete EDTA-free)を含有する150μLのTris緩衝生理食塩水(pH7.4)において、振とう(4℃、15分)により溶解させた。溶解混合物を遠心分離にかけ(2000×g、5分)、除核後の上清を回収して、すべてのさらなる酵素試験に使用した。
【0065】
(実施例3)
変性ゲル電気泳動分析および免疫ブロット法
感染sf9細胞(2×106細胞、上記参照)を、200μLのLaemmli緩衝液中でボルテックスにかけ、加熱(95℃、5分)によってタンパク質を変性させた。室温まで冷却後、試料を遠心分離にかけ(16krpm、5分)、上清をさらなる分析に使用した。試料を、SDS-PAGEによって分離した(12%アクリルアミド、120V)。結果として生じたゲルを、銀染色またはニトロセルロース膜上へのブロッティングのいずれかによって分析した。膜をブロッキング(PBST中5%BSA)した後、抗FLAG抗体M2(SIGMA、PBST+1%BSA中1:2000希釈)、その後、徹底的に洗浄(PBST)した後に抗マウスHRP(Santa Cruz Biotechnology、PBST+1%BSA中1:10000希釈)とのインキュベーション、ならびにECL(Pierce)および写真用フィルムへの曝露を使用した最終的な検出によって、免疫検出を実施した。
【0066】
(実施例4)
グリコシルトランスフェラーゼアッセイ
塩化マンガン(II)(10μM)、UDP-ガラクトース(1mM)およびアクセプターフコシド(グリカンまたは糖(ポリ)ペプチド、40μM)を含有する2.5μLの最終体積のMES緩衝液(pH6.5、40μM)中で、0.5μlのsf9細胞粗抽出物(空ベクター対照バクミド、推定GalT発現バクミドまたは推定FLAGタグ付きGalT発現バクミドのいずれかを含有する)を使用して、適切な炭水化物または複合糖質に対する酵素活性を評価した。グリコシル化反応は。典型的には、別段記載のない限り、室温で2時間行った。ドナー特異性分析では、UDP-ガラクトースを、等濃度のUDP-Glc、UDP-GlcNAcまたはUDP-GalNAc(Sigma)でそれぞれ置き換えた。補因子特異性分析では、MnCl2を、等濃度の様々な金属塩化物またはNa2EDTAで置き換えた。アクセプターグリカン中へのガラクトースの組込みを定量するために、全UDP-Gal濃度を、10%UDP-14C-Gal(25nCi、GE Healthcare)でドープした。反応混合物(100μLのH2Oでクエンチさせた)を、陰イオン交換樹脂のカラム(AG1-X8、Cl-型、Bio-Rad Laboratories、200mg)に充填すること、および荷電していない生成物(H2O、900μL)の溶出によって、過剰の放射活性(UDP-14C-Gal)を除去した。
【0067】
非放射性UDP-Galが存在せず、UDP-14C-Gal(75nCi)で置き換えられていること以外は上記と同じ緩衝液、塩および酵素条件を使用して、50μLの全体積において、ヒトIgG1(5μlの3g/L、Calbiochem)のグリコシル化を実施した。反応は、室温で終夜実施した。セファロース-プロテインGビーズ(Amersham Biosciences、10μL)のPBS(200μL)中の懸濁液を添加し、振とう(4℃、1時間)によってIgG1のビーズへの結合を行った。ビーズをPBS(5×200μL)で洗浄し、20mMのHCl水溶液(3×100μL)でIgG1を溶出した。ドナー特異性に関する、蛍光標識されたグリコペプチドの直接のMALDI-TOF質量分析、HPLC分析、または放射標識アッセイのシンチレーション計数のいずれかによって、反応生成物の分析(下記参照)を実施した。
【0068】
ヒトアシアロトランスフェリンN-グリカンの段階的な再構築を、以下の通り実施した:
アシアロトランスフェリン(GalGal)は、ヒトアポトランスフェリンのシアリダーゼ処理によって以前に調製された(Iskratschら、Anal. Biochem.、368、133〜146、2009)。
【0069】
アシアロガラクトトランスフェリン(GnGn)を作製するために、β1,4-ガラクトシダーゼ(3U、アスペルギルス・オリゼ)を、約1mgのGalGalに添加し、試料を、48時間37℃でインキュベートした(全体積50μl)。
【0070】
GnGnF6を得るために、試料を0.5μlの1M NaOHで中性pHにした後、50nmolのGDP-フコースおよび15μlのピキア・パストリスにおいて発現された組換えハマダラカ属のコアα1,6-FucT調製物を添加した。調製物を終夜インキュベートした後、さらに50nmolのGDP-フコースおよびさらなる15μlの酵素(FucT)を添加し、終夜、37℃で再度インキュベートした。合計で、約1mgのGnGnF6が得られた。
【0071】
GalFuc-トランスフェリンを調製するために、1μlの組換えカエノラブディティス・エレガンスGalTの調製物、0.2mmolのMnCl2および20nmolのUDP-ガラクトースをGnGnF6(300μg)のアリコートに添加し、終夜30℃でインキュベートした。再度、さらなる基質(UDP-ガラクトース)および酵素(GalT)の添加後、2回目の終夜のインキュベーションにより所望のグリカン構造を増加させた。
【0072】
トランスフェリンの修飾の程度は、フコース特異的ヒイロチャワンタケレクチンを用いたドットブロット法によって、および様々なネオグリコフォームのトリプシンペプチドのMALDI-TOF MSによってモニタリングした。
【0073】
(実施例5)
構造分析
ダブシル-GEN[GnGnF6]Rをガラクトシル化条件に曝露させた後、結果として生じた未精製の混合物を、50mMナトリウムクエン酸およびpH4.5に調整し、アスペルギルス・オリゼβガラクトシダーゼ(27mU)(Gutterniggら、J. Biol. Chem. 282(38)、27825〜27840、2007を参照されたい)で2日間、30℃で消化した。試料を、MALDI-TOF質量分析によって分析した(下記参照)。
【0074】
HPLC分析:
ドナー特異性およびドナー濃度に対する反応速度依存の分析の両方で、逆相Hypersil ODS C18カラム(4×250mm、5μm)においてアイソクラティック溶媒系(0.7mL/分、0.05体積%TFA水溶液中9%MeCN(95体積%))、および蛍光検出(315nmで励起、発光は550nmで検出した)を室温で使用して、ダンシル-N[GnGnF6]STアクセプター基質を、ガラクトシル化された反応生成物から分離した。Shimadzu HPLCシステムは、SCL-10A制御装置、2つのLC10APポンプおよびClass-VPソフトウェア(V6.13SP2)を使用してパーソナルコンピュータにより制御されるRF-10AXL蛍光検出器からなっていた。ダンシル-N[GnGnF6]STは、9.09分の保持時間で、およびガラクトシル化された反応生成物は8.06分で溶出した。
【0075】
質量分析:
BRUKER Ultraflex TOF/TOF装置において、α-シアノ-4-ヒドロキシ桂皮酸マトリックスを使用して、MALDI-TOF質量分析によってグリカンを分析した。外部較正にはペプチド標準混合物(Bruker)を使用した。
【0076】
シンチレーション計数:
陰イオン交換樹脂カラムおよびプロテインGビーズの溶出液を、シンチレーション流体(Irga-Safe Plus、Packard、4mL)と完全に混合し、Perkin Elmer Tri-Carb 2800TRを用いて測定した。
【0077】
炭水化物の省略形:
Fuc-L-フコース、Gal-D-ガラクトース、GalNAc-D-N-アセチルガラクトサミン、Glc-D-グルコース、GlcNAc-D-N-アセチルグルコサミン、Man-D-マンノース
複合グリカンの省略形(Schachter命名法[Biochem Cell Biol 64(3)、163〜181、1986]に従う):
GalGal Gal-β-1,4-GlcNAc-β-1,2-Man-α-1,6-[Gal-β-1,4-GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-GlcNAc
GnGn GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-GlcNAc
GnGnF6 GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc
GnGnF6Gal GlcNAc-β-1,2-Man-α-1,6-[GlcNAc-β-1,2-Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[Gal-β-1,4-Fuc-α-1,6]-GlcNAc
MMF6 Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,6-Fuc]-GlcNAc
MMF6Gal Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[Gal-β-1,4-Fuc-α-1,6]-GlcNAc
MMF3 Man-α-1,6-[Man-α-1,3-]-Man-β-1,4-GlcNAc-β-1,4-[α-1,3-Fuc]-GlcNAc
【特許請求の範囲】
【請求項1】
(i)配列番号1、3、5、7および9に記載されている核酸配列からなる群から選択される少なくとも1つの核酸配列を含む核酸;
(ii)配列番号1に記載されている核酸配列と、少なくとも60または70%の同一性、好ましくは、少なくとも80または90%の同一性、より好ましくは少なくとも95%の同一性、最も好ましくは少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される、単離および精製された核酸。
【請求項2】
DNA、RNAまたはPNA、好ましくはDNAまたはPNA、より好ましくはDNAである、請求項1に記載の核酸。
【請求項3】
好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、ガラクトシルトランスフェラーゼ活性、好ましくは、β-1,4-ガラクトシルトランスフェラーゼ活性を有するタンパク質をコードしている、請求項1または2に記載の核酸。
【請求項4】
(a)配列番号2、4、6、8および10からなる群から選択されるアミノ酸配列、好ましくは配列番号2のアミノ酸配列を有するポリペプチド、
(b)請求項1から3のいずれかに記載の核酸によりコードされているポリペプチド、
(c)(a)および/または(b)のポリペプチドと、少なくとも25、30または40%、好ましくは、少なくとも50または60%、より好ましくは、少なくとも70または80%、最も好ましくは、少なくとも90または95%のアミノ酸配列同一性を有するポリペプチド、
(d)(a)、(b)または(c)のフラグメントおよび/または機能的誘導体
からなる群から選択される、単離および精製されたポリペプチド。
【請求項5】
好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、ガラクトシルトランスフェラーゼ活性、好ましくは、β-1,4-ガラクトシルトランスフェラーゼ活性を有する、請求項4に記載のポリペプチド。
【請求項6】
請求項1から3のいずれかに記載の核酸を含む組換えベクター、好ましくは、ウイルスまたはエピソームのベクター、好ましくはバキュロウイルスベクター。
【請求項7】
酵母細胞、好ましくは、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)細胞、大腸菌(E.coli)細胞、植物細胞、好ましくはタバコ(Nicotiana tabacum)またはヒメツリガネゴケ(Physcomitrella patens)細胞、NIH-3T3哺乳動物細胞および昆虫細胞からなる群から選択される、より好ましくは、sf9昆虫細胞である、請求項1から3および6のいずれかに記載の核酸またはベクターを含む宿主細胞。
【請求項8】
請求項4または5に記載のポリペプチドに特異的に結合する抗体。
【請求項9】
モノクローナル抗体である、請求項8に記載の抗体。
【請求項10】
請求項5に記載のポリペプチドに特異的に結合するモノクローナル抗体を発現する、ハイブリドーマ細胞系。
【請求項11】
ガラクトシル含有オリゴ糖/多糖および/または複合糖質、好ましくはガラクトシル-フコシド含有オリゴ糖/多糖および/または複合糖質、より好ましくはD-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-GlcNAc含有オリゴ糖/多糖および/または複合糖質、最も好ましくはGnGnF6Gal-および/またはMMF6Gal含有オリゴ糖および複合糖質を作製するための、請求項4または5に記載のポリペプチド、請求項4または5に記載のポリペプチドを含む細胞抽出物、好ましくは、カエノラブディティス・エレガンス(Caenorhabditis elegans)、カエノラブディティス・ブリグサエ(Caenorhabditis briggsae)、ネマトステラ・ベクテンシス(Nematostella vectensis)、キンカチョウ(Taeniopygia guttata)もしくはクリプトスポリジウム・パルバム(Cryptosporidium parvum)の抽出物、および/または請求項7の宿主細胞の使用。
【請求項12】
以下の段階:
(i)少なくとも1つの本発明のポリペプチドを提供する段階、
(ii)少なくとも1つのフコシル化されたアクセプター基質を提供する段階、
(iii)好ましくはマンガン(II)、コバルト(II)および/または鉄(II)イオンから選択され、より好ましくはマンガン(II)である少なくとも1つの適した二価の金属陽イオン補因子と、少なくとも1つの活性化された糖基質、好ましくはウリジン二リン酸(UDP)-ガラクトースとの存在下、本発明のポリペプチドの酵素活性に適した条件下で、(i)および(ii)をインキュベートする段階、
(iv)ガラクトシル-フコース誘導体を任意選択により単離する段階
を含む、ガラクトシル-フコシル誘導体を作製するための方法。
【請求項13】
ガラクトシル化合物をコアがフコシル化されたαフェトプロテイン(AFP)に共有結合させるための、好ましくは、肝細胞癌(HCC)を検出および/または定量するための、請求項4または5に記載の少なくとも1つのポリペプチド、請求項7に記載の宿主細胞ならびに/またはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物の使用。
【請求項14】
以下の段階:
(i)AFPを含む血液またはその画分、好ましくは血清を提供する段階と、
(ii)前記血液または前記その画分を、(a)請求項4または5に記載のポリペプチド、請求項7に記載の宿主細胞ならびに/またはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物とともに、および(b)活性化されたガラクトシル誘導体、好ましくは標識されたガラクトシル誘導体、好ましくは標識されたUDP-ガラクトースとともに、活性化されたガラクトースの、コアがフコシル化されたAFP(AFP-L3)へのガラクトシル転移を可能にする条件下で、インキュベートする段階と、
(iii)ガラクトース標識された、したがって、コアがフコシル化されたAFP(AFP-L3)を検出する段階と
を含む診断方法。
【請求項15】
対象とする試料、例えば、ヒトまたは哺乳動物の試料において、好ましくは細胞画分または抽出物試料において、線虫または病原体、好ましくはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエおよび/またはクリプトスポリジウム・パルバムを同定および/または定量するための、請求項8または9に記載の抗体の使用。
【請求項1】
(i)配列番号1、3、5、7および9に記載されている核酸配列からなる群から選択される少なくとも1つの核酸配列を含む核酸;
(ii)配列番号1に記載されている核酸配列と、少なくとも60または70%の同一性、好ましくは、少なくとも80または90%の同一性、より好ましくは少なくとも95%の同一性、最も好ましくは少なくとも98%の同一性の配列を有する核酸;
(iii)(i)または(ii)の核酸とハイブリダイズする核酸;
(iv)(i)、(ii)または(iii)の核酸のうちの1つの置換、付加および/または欠失によって誘導できる核酸;
(v)(i)の核酸とハイブリダイズする、(i)から(iv)の核酸のうちのいずれかのフラグメント
からなる群から選択される、単離および精製された核酸。
【請求項2】
DNA、RNAまたはPNA、好ましくはDNAまたはPNA、より好ましくはDNAである、請求項1に記載の核酸。
【請求項3】
好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、ガラクトシルトランスフェラーゼ活性、好ましくは、β-1,4-ガラクトシルトランスフェラーゼ活性を有するタンパク質をコードしている、請求項1または2に記載の核酸。
【請求項4】
(a)配列番号2、4、6、8および10からなる群から選択されるアミノ酸配列、好ましくは配列番号2のアミノ酸配列を有するポリペプチド、
(b)請求項1から3のいずれかに記載の核酸によりコードされているポリペプチド、
(c)(a)および/または(b)のポリペプチドと、少なくとも25、30または40%、好ましくは、少なくとも50または60%、より好ましくは、少なくとも70または80%、最も好ましくは、少なくとも90または95%のアミノ酸配列同一性を有するポリペプチド、
(d)(a)、(b)または(c)のフラグメントおよび/または機能的誘導体
からなる群から選択される、単離および精製されたポリペプチド。
【請求項5】
好ましくはL-フコシド-、より好ましくはα-L-フコシド-、より好ましくはFuc-α-1,6-GlcNAc-、最も好ましくはGnGnF6含有多糖/オリゴ糖または複合糖質をアクセプター基質とする、ガラクトシルトランスフェラーゼ活性、好ましくは、β-1,4-ガラクトシルトランスフェラーゼ活性を有する、請求項4に記載のポリペプチド。
【請求項6】
請求項1から3のいずれかに記載の核酸を含む組換えベクター、好ましくは、ウイルスまたはエピソームのベクター、好ましくはバキュロウイルスベクター。
【請求項7】
酵母細胞、好ましくは、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)、ピキア・パストリス(Pichia pastoris)細胞、大腸菌(E.coli)細胞、植物細胞、好ましくはタバコ(Nicotiana tabacum)またはヒメツリガネゴケ(Physcomitrella patens)細胞、NIH-3T3哺乳動物細胞および昆虫細胞からなる群から選択される、より好ましくは、sf9昆虫細胞である、請求項1から3および6のいずれかに記載の核酸またはベクターを含む宿主細胞。
【請求項8】
請求項4または5に記載のポリペプチドに特異的に結合する抗体。
【請求項9】
モノクローナル抗体である、請求項8に記載の抗体。
【請求項10】
請求項5に記載のポリペプチドに特異的に結合するモノクローナル抗体を発現する、ハイブリドーマ細胞系。
【請求項11】
ガラクトシル含有オリゴ糖/多糖および/または複合糖質、好ましくはガラクトシル-フコシド含有オリゴ糖/多糖および/または複合糖質、より好ましくはD-ガラクトピラノシル-β-1,4-L-フコピラノシル-α-1,6-GlcNAc含有オリゴ糖/多糖および/または複合糖質、最も好ましくはGnGnF6Gal-および/またはMMF6Gal含有オリゴ糖および複合糖質を作製するための、請求項4または5に記載のポリペプチド、請求項4または5に記載のポリペプチドを含む細胞抽出物、好ましくは、カエノラブディティス・エレガンス(Caenorhabditis elegans)、カエノラブディティス・ブリグサエ(Caenorhabditis briggsae)、ネマトステラ・ベクテンシス(Nematostella vectensis)、キンカチョウ(Taeniopygia guttata)もしくはクリプトスポリジウム・パルバム(Cryptosporidium parvum)の抽出物、および/または請求項7の宿主細胞の使用。
【請求項12】
以下の段階:
(i)少なくとも1つの本発明のポリペプチドを提供する段階、
(ii)少なくとも1つのフコシル化されたアクセプター基質を提供する段階、
(iii)好ましくはマンガン(II)、コバルト(II)および/または鉄(II)イオンから選択され、より好ましくはマンガン(II)である少なくとも1つの適した二価の金属陽イオン補因子と、少なくとも1つの活性化された糖基質、好ましくはウリジン二リン酸(UDP)-ガラクトースとの存在下、本発明のポリペプチドの酵素活性に適した条件下で、(i)および(ii)をインキュベートする段階、
(iv)ガラクトシル-フコース誘導体を任意選択により単離する段階
を含む、ガラクトシル-フコシル誘導体を作製するための方法。
【請求項13】
ガラクトシル化合物をコアがフコシル化されたαフェトプロテイン(AFP)に共有結合させるための、好ましくは、肝細胞癌(HCC)を検出および/または定量するための、請求項4または5に記載の少なくとも1つのポリペプチド、請求項7に記載の宿主細胞ならびに/またはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物の使用。
【請求項14】
以下の段階:
(i)AFPを含む血液またはその画分、好ましくは血清を提供する段階と、
(ii)前記血液または前記その画分を、(a)請求項4または5に記載のポリペプチド、請求項7に記載の宿主細胞ならびに/またはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエ、ネマトステラ・ベクテンシス、キンカチョウおよび/もしくはクリプトスポリジウム・パルバムの細胞抽出物とともに、および(b)活性化されたガラクトシル誘導体、好ましくは標識されたガラクトシル誘導体、好ましくは標識されたUDP-ガラクトースとともに、活性化されたガラクトースの、コアがフコシル化されたAFP(AFP-L3)へのガラクトシル転移を可能にする条件下で、インキュベートする段階と、
(iii)ガラクトース標識された、したがって、コアがフコシル化されたAFP(AFP-L3)を検出する段階と
を含む診断方法。
【請求項15】
対象とする試料、例えば、ヒトまたは哺乳動物の試料において、好ましくは細胞画分または抽出物試料において、線虫または病原体、好ましくはカエノラブディティス・エレガンス、カエノラブディティス・ブリグサエおよび/またはクリプトスポリジウム・パルバムを同定および/または定量するための、請求項8または9に記載の抗体の使用。
【図1】


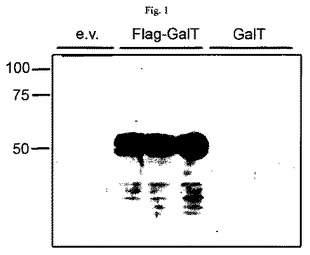
【図2】
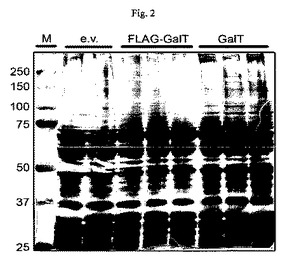
【図9】
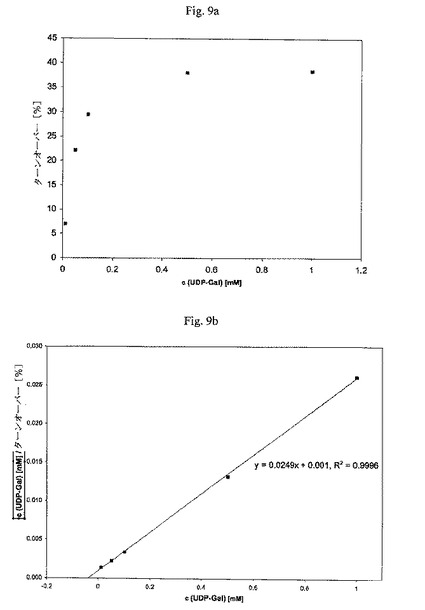
【図10】
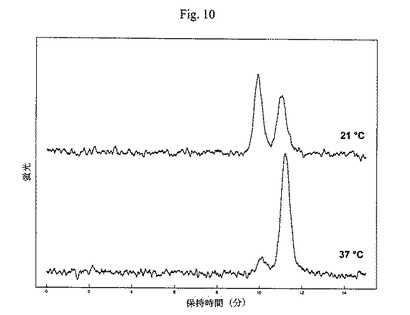
【図11】
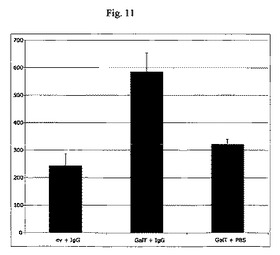
【図3】
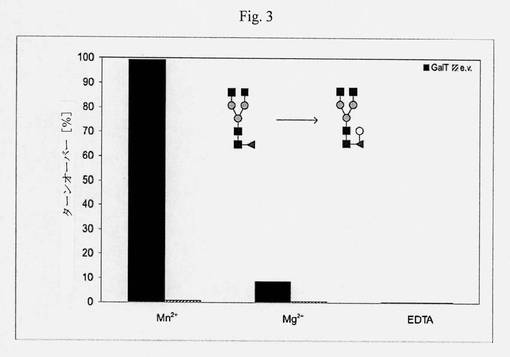
【図4】
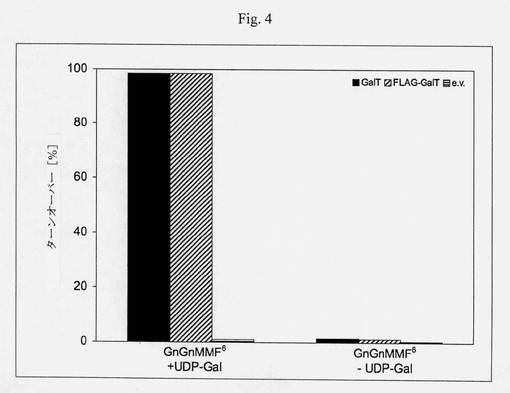
【図5】
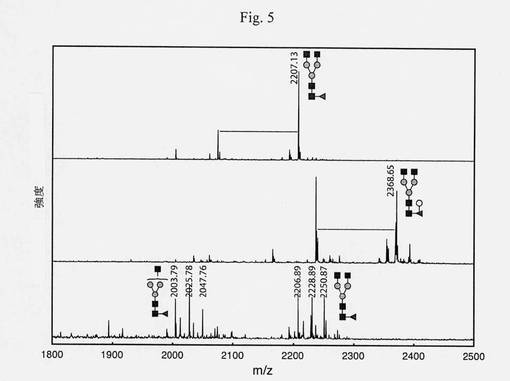
【図6】
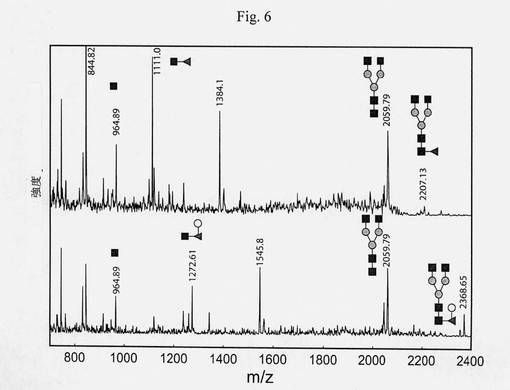
【図7】
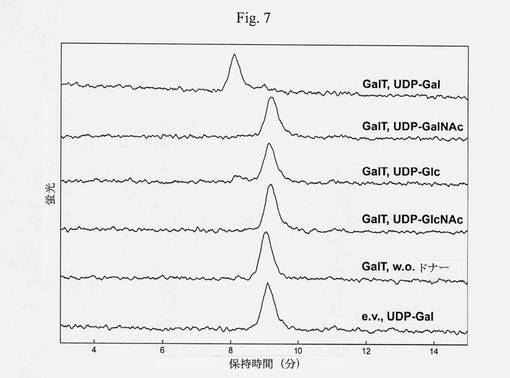
【図8】
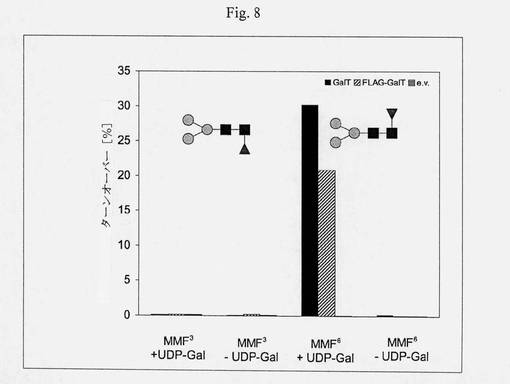
【図12】
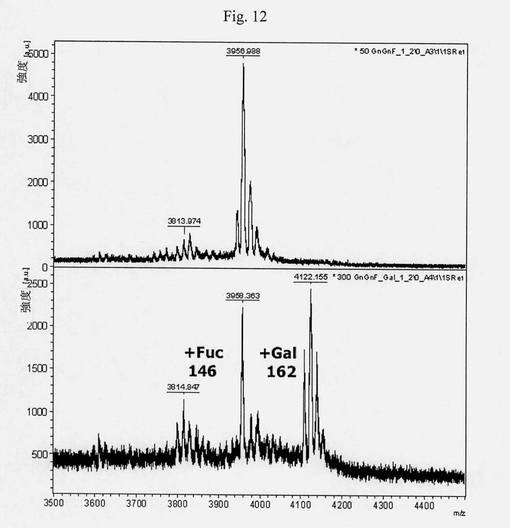
【図2】
【図9】
【図10】
【図11】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図12】
【公表番号】特表2012−527223(P2012−527223A)
【公表日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願番号】特願2012−511203(P2012−511203)
【出願日】平成22年5月28日(2010.5.28)
【国際出願番号】PCT/EP2010/003249
【国際公開番号】WO2010/136209
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(506110634)
【出願人】(509217725)ユニヴェルシタット・チューリッヒ (2)
【出願人】(511282656)ユニヴェルシタット・フュア・ボーデンクルトゥール・ウィーン (1)
【Fターム(参考)】
【公表日】平成24年11月8日(2012.11.8)
【国際特許分類】
【出願日】平成22年5月28日(2010.5.28)
【国際出願番号】PCT/EP2010/003249
【国際公開番号】WO2010/136209
【国際公開日】平成22年12月2日(2010.12.2)
【出願人】(506110634)
【出願人】(509217725)ユニヴェルシタット・チューリッヒ (2)
【出願人】(511282656)ユニヴェルシタット・フュア・ボーデンクルトゥール・ウィーン (1)
【Fターム(参考)】
[ Back to top ]