
Nアセチルガラクトサミンの製造方法
【課題】Nアセチルガラクトサミンの有効な製造方法の提供。
【解決手段】出発原料であるNアセチルグルコサミンを、酵素を含む反応溶液と、ATPおよびUDP−糖の存在下で作用させることにより、Nアセチルガラクトサミンを製造する方法。
【解決手段】出発原料であるNアセチルグルコサミンを、酵素を含む反応溶液と、ATPおよびUDP−糖の存在下で作用させることにより、Nアセチルガラクトサミンを製造する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Nアセチルグルコサミンを、ATPおよびUDP−糖の存在下にて、酵素と反応させることにより、Nアセチルガラクトサミンを製造する方法に関する。
【背景技術】
【0002】
Nアセチルガラクトサミンは糖鎖中によく見られる単糖の一つであり、糖鎖を利用した機能性物質の原料として期待されている。しかしながら、Nアセチルガラクトサミンを豊富に含んだ天然産の糖質は多量に存在しない。これまでに、Nアセチルガラクトサミンの製造方法としては、鮭軟骨などから抽出されるコンドロイチン硫酸の酸加水分解分解物からガラクトサミンを抽出後N−アセチル化することにより製造する方法が知られている(非特許文献1を参照)。
【0003】
しかしながら、この方法では、出発物質の含有量の低さなどから、産業的に有効な製造法とはいえず、そのためにNアセチルガラクトサミンは非常に高価な物質であった。
【0004】
【非特許文献1】Jeanloz, RZ編、“The Amino Sugars /volume 1A”, Academic Press (1969)
【発明の開示】
【発明が解決しようとする課題】
【0005】
従来法では、安価に効率的にNアセチルガラクトサミンを製造することはできなかった。従って、Nアセチルガラクトサミンの、有効な製造方法の確立が望まれていた。
【0006】
本発明は、Nアセチルグルコサミンを、ATPおよびUDP−糖の存在下にて、酵素と反応させることにより、Nアセチルガラクトサミンを製造する方法を提供する。
【課題を解決するための手段】
【0007】
本発明者らは、上記課題を解決すべく、鋭意検討した結果、出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下、以下の(i)〜(iii)の酵素:(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素、と同時に作用させることにより、Nアセチルガラクトサミンを製造できることを見出し、本発明を完成させるに至った。
【0008】
すなわち、本発明は以下のとおりである。
[1] 出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下で、以下の(i)〜(iii)の酵素と同時に作用させることにより、Nアセチルガラクトサミンを製造する方法:
(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;
(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに
(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素。
【0009】
[2] UDP−糖が、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンまたはそれらの少なくとも2つの混合物からなる群から選択される、[1]の方法。
[3] UDP−糖が、UDP−グルコースまたはUDP−Nアセチルグルコサミンである、[2]の方法。
【0010】
[4] (i)の酵素が、UDP−グルコース4−エピメラーゼである、[1]〜[3]のいずれかの方法。
[5] UDP−グルコース4−エピメラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[4]の方法。
【0011】
[6] (ii)の酵素が、UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼである、[1]〜[3]のいずれかの方法。
[7] UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[6]の方法。
【0012】
[8] (iii)の酵素が、Nアセチルヘキソサミン1キナーゼである、[1]〜[3]のいずれかの方法。
[9] Nアセチルヘキソサミン1キナーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[8]の方法。
【0013】
[10] (i)の酵素としてUDP−グルコース4−エピメラーゼ、(ii)の酵素としてUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ、(iii)の酵素としてNアセチルヘキソサミン1キナーゼを同時に作用させる、[1]〜[3]のいずれかの方法。
[11] 作用させる全ての酵素が、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する[10]の方法。
[12] さらに、製造されたNアセチルガラクトサミンを精製する工程を含む、[1]〜[9]のいずれかの方法。
【発明の効果】
【0014】
本発明では安価に得られるNアセチルグルコサミンを出発原料として、ATPおよびUDP−糖の存在下、(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素、(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素、ならびに(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素を同時に作用させることにより一段階でNアセチルガラクトサミンを製造できる。
【発明を実施するための最良の形態】
【0015】
出発原料であるNアセチルグルコサミンの濃度は、特に限定されるものではないが、好ましくは10mM〜2M、より好ましくは100mM〜1M、さらに好ましくは600mMである。
【0016】
(i)の酵素は、UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する活性を有する限り、特に限定されないが、UDP−グルコース4−エピメラーゼのうち、UDP−Nアセチルグルコサミンにも活性を示す酵素を用いるのが好ましい。このような酵素としては、ヒト由来、Bacillus属細菌由来、ビフィズス菌由来のUDP−グルコース4−エピメラーゼ(Thoden, J. B.ら Structural analysis of the Y299C mutant of Escherichia coli UDP-galactose 4-epimerase. Teaching an old dog new tricks. The Journal of Biological Chemistry. 277:27528-27534 (2002)、再表01/038555、Genbank登録番号AB303839)を用いることができるが、好ましくは、ビフィドバクテリウム・ロンガムに由来するUDP−グルコース4−エピメラーゼを用いる。
【0017】
(ii)の酵素は、UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応を触媒する活性を有する限り、特に限定しないが、ビフィズス菌由来のUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ(西本ら、第58回日本生物工学会大会講演要旨集p. 92 (2006)、Genbank登録番号AB303839)を用いるのが好ましい。特に好ましくは、ビフィドバクテリウム・ロンガムに由来するUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼを用いる。
【0018】
(iii)の酵素は、NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる活性を持つ酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる活性を持つ酵素、またはその両活性を持つ酵素である限り、特に限定されないが、ビフィズス菌由来のNアセチルヘキソサミン1キナーゼ(特開2007−97517、Genbank登録番号AB303839)を用いるのが好ましい。特に好ましくは、ビフィドバクテリウム・ロンガムに由来するNアセチルヘキソサミン1キナーゼを用いる。
【0019】
通常、(iii)に示す様な活性の酵素はキナーゼと呼ばれており、一般的にはその逆反応の利用はほとんど省みられたことはなかった。この逆反応は、本発明者により初めて見出されたものであり、本発明では、この酵素の逆反応をNアセチルガラクトサミンを生じる最終段階で利用している。
【0020】
これらの酵素は、天然の生物または遺伝子組換え生物(もしくは細胞)により産生されたもの、あるいは市販されているものを用いることができる。遺伝子組換え生物(もしくは細胞)は、当業者に公知である分子生物学的手法を用いて、当該酵素をコードする遺伝子を適当なベクターに組み込み、そのベクターを用いて適当な宿主生物(例えば、細菌、酵母、動物細胞、昆虫細胞、植物細胞など)を形質転換することにより得ることができる(Sambrook J.ら、“Molecular Cloning A LBORATORY MANUAL /second edition”, Cold Spring Harbor Laboratory Press (1989)参照)。
【0021】
上記生物により産生された酵素は、通常の方法を用いて、当該生物の培養物より単離精製できる。ここで「培養物」とは、培養上清、培養細胞もしくは培養菌体または細胞もしくは細胞の破砕物を意味する。培養後、目的の酵素が菌体内または細胞内に生産される場合には、細胞を遠心分離により回収し、緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を回収する。あるいは、目的の酵素が菌体外または細胞外に生産される場合には、遠心分離などにより菌体または細胞を除去し、培養液を回収する。この無細胞抽出液を遠心分離し、得られた上清から、または上記培養液から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、陰イオン交換クロマトグラフィー法、陽イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用いて、精製酵素を得ることができる。
【0022】
本発明方法において用いられる酵素は、精製された形態であっても、上記無細胞抽出液または培養液のような粗製物の形態であってもよい。
【0023】
本発明における反応において、上記酵素群は、以下の一連の反応を触媒し、ATPおよびUDP−糖の存在下、Nアセチルグルコサミンより、Nアセチルガラクトサミンを生成する:

図中の略語:
GlcNAc:Nアセチルグルコサミン
GlcNAc1P:Nアセチルグルコサミン1−リン酸
UDP−GalNAc:UDP−Nアセチルガラクトサミン
UDP−GlcNAc:UDP−Nアセチルグルコサミン
GalNAc1P:Nアセチルガラクトサミン1−リン酸
GalNAc:Nアセチルガラクトサミン。
【0024】
反応液中の酵素の濃度は、0.01U〜200U/ml、好ましくは、0.05U〜100U/mlの範囲で適宜設定でき、例えば、Nアセチルヘキソサミンキナーゼであれば1.0U〜10U/ml、UDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼであれば0.05U/ml、UDPグルコース−4−エピメラーゼであれば100U/mlで用いるのが好ましい。
【0025】
反応液中のATPの濃度は、特に限定されるものではないが、好ましくは0.01mM〜100mM、より好ましくは0.1mM〜10mM以下、さらに好ましくは、0.75mM〜2.0mMである。
【0026】
用いるUDP−糖としては、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンのいずれかまたはそれらの少なくとも2つの混合物を用いることができる。好ましくは、UDP−グルコースまたはUDP−Nアセチルグルコサミンを用いる。反応液中のUDP−糖の濃度は、特に限定されるものではないが、好ましくは0.005mM〜50mM、より好ましくは0.05mM〜10mM、さらに好ましくは、0.2mM〜0.6mMである。
【0027】
反応液のpHは好ましくは5〜9、さらに好ましくは、6.5である。反応温度は特に限定されるものではないが好ましくは5℃〜80℃、より好ましくは20℃〜60℃、さらに好ましくは、30℃である。反応時間は、1時間〜30日間、好ましくは24時間〜10日間、さらに好ましくは5日間である。必要に応じて撹拌しながら反応を行っても良い。
【0028】
反応系は特に限定されるものではないが通常水溶液あるいは緩衝液中で行われる。緩衝液としては、酵素反応に一般的に用いられるもの、例えば、トリス塩酸緩衝液、トリス酢酸緩衝液、HEPES緩衝液、MOPS緩衝液、リン酸緩衝液、酢酸ナトリウム緩衝液、クエン酸ナトリウム緩衝液などを使用できるが、MOPS緩衝液を用いるのが好ましい。使用濃度は、10mM〜500mM、好ましくは10mM〜300mM、さらに好ましくは100mMである。
【0029】
反応は、固定化酵素リアクターを用いて行うことも可能である。酵素を固定化することによって、酵素が安定化され、連続反復使用が可能となる点において有効である。酵素の固定化は、担体結合法、架橋法、包括法を用いて行うことができる。担体結合法では、酵素を担体(例えば、セルロース、デキストラン、アガロースなどの多糖類の誘導体、ポリアクリルアミドゲル、ポリスチレン樹脂、多孔性ガラス、金属酸化物など)に、物理的吸着、イオン結合および共有結合等を用いて結合させることができる。架橋法では、2個またはそれ以上の官能基を持つ試薬を用いて、酵素同士を互いに架橋することによって固定化する。架橋試薬としては、Schiff塩基をつくるグルタルアルデヒド、ペプチド結合をするイソシアン酸誘導体、N,N’−エチレンマレイミド、ジアゾカップリングをするビスジアゾベンジン、あるいはアルキル化するN,N’−ポリメチレンビスヨードアセトアミドなどを用いることができる。包括法では、高分子ゲルの細かい格子の中に酵素を取り込む格子型と、半透膜の高分子の皮膜によって酵素を皮膜するマイクロカプセル型を用いる。格子型の方法では、合成高分子物質のポリアクリルアミドゲル、ポリビニルアルコール、光硬化性樹脂および天然高分子物質のデンプン、コンニャク粉、ゼラチン、アルギン酸、カラギーナンなどの高分子化合物を用いることができる。マイクロカプセル型の方法では、ヘキサメチレンジアミン、セバコイルクロリド、ポリスチレン、レシチンなどを用いることができる(福井三郎、千畑一郎、鈴木周一、「酵素工学」東京化学同人発行、1981年)。
【0030】
担体は、3種の酵素を固定化したものを用いてもよいし、それぞれの酵素を固定化した複数の担体を用いてもよい。
【0031】
本発明の方法により、用いたNアセチルグルコサミンの少なくとも20%、好ましくは少なくとも25%がNアセチルガラクトサミンに変換される。
【0032】
生成したNアセチルガラクトサミンは、例えば、イオン交換クロマトグラフィー、吸着クロマトグラフィー、アフィニティクロマトグラフィー、ゲル濾過法など各種のクロマトグラフィー、向流分配、向流抽出など二液相間の分配を利用する方法、濃縮、冷却、有機溶媒添加など溶解度の差を利用する方法、および、塩析などの通常の精製法を単独で、あるいは適宜組み合わせて用いて、出発原料であるNアセチルグルコサミンと分離することが可能である。好ましくは、Nアセチルガラクトサミンは、強イオン性陽イオン交換クロマトグラフィーのような方法により、Nアセチルグルコサミンと分離することができる。
【0033】
得られたNアセチルガラクトサミンは、例えば医薬品、化粧品、食品添加物等の材料として好適に用いることができる。
【実施例】
【0034】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0035】
実施例1:酵素の調製
Nアセチルヘキソサミン1キナーゼの調製
ビフィドバクテリウム・ロンガムのNアセチルヘキソサミン1キナーゼの遺伝子配列(配列番号1)を基に、以下のプライマー作製し:
フォーワードプライマー:aacggaccccatatgaccgaaagcaatgaagttttattc(配列番号2)
リバースプライマー:gctgacctcgagcctggcagcctccatgatgtcggctac(配列番号3)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてNアセチルヘキソサミン1キナーゼのORF全長をPCRにより増幅した。
【0036】
PCR産物を制限酵素NcoI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0037】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質を、Ni−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製して、精製酵素標品を得た。得られた精製酵素の比活性は1.5U/mgであった。
【0038】
UDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼの調製
ビフィドバクテリウム・ロンガムのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼの配列(配列番号4)を基に、以下のプライマーを作製し:
フォーワードプライマー:gatatatacatatgaacgatcagctgaccgaggta(配列番号5)
リバースプライマー:tgacctctcgagcctagcggcaaaaccaaggctttcga(配列番号6)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ遺伝子をPCRにより増幅した。
【0039】
PCR産物を制限酵素NdeI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET30(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0040】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質はNi−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製して、精製酵素標品を得た。得られた精製酵素の比活性は1.1U/mgであった。
【0041】
UDPグルコース−4−エピメラーゼの調製
ビフィドバクテリウム・ロンガムのUDPグルコース−4−エピメラーゼの配列(配列番号7)を基に、以下のプライマーを作製し:
フォーワードプライマー:gatatacatatgactactgttctggttacgggc(配列番号8)
リバースプライマー:ctgctcctcgagctccgcgtcgcggaaaccgttggggttc(配列番号9)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ遺伝子をPCRにより増幅した。
【0042】
PCR産物を制限酵素NdeI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET30(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0043】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質はNi−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製し、精製酵素標品を得た。得られた精製酵素の比活性は158U/mgであった。
以上のようにして得た酵素を以下の反応液に用いた。
【0044】
実施例2:反応液の調製
[反応液1]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.6mMのUDP−Nアセチルグルコサミン、2.0mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、1.0U/mlのNアセチルヘキソサミン1キナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、50U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液1を調製した。
【0045】
[反応液2]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.6mMのUDP−グルコース、0.75mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、10U/mlのNアセチルヘキソサミンキナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、100U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液2を調製した。
【0046】
[反応液3]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.2mMのUDP−グルコース、1.0mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、10U/mlのNアセチルヘキソサミンキナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、100U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液を調製した。
【0047】
実施例3:Nアセチルガラクトサミンの生成
実施例2で調製した反応液1、反応液2、および反応液3をそれぞれ恒温器で30℃、5日間インキュベートし、酵素反応を行った。反応終了後、反応液を適宜希釈して高速液体クロマトグラフィー(島津製作所製)を用いて、NアセチルグルコサミンとNアセチルガラクトサミンの濃度を調べ、モル収率を算出した。
【0048】
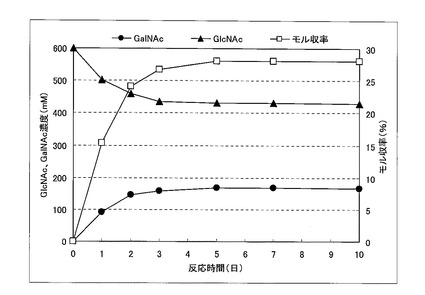
この結果、反応液1、反応液2、および反応液3におけるNアセチルガラクトサミンのモル収率はそれぞれ27.7%、28.0%、24.7%であった。反応液1のタイムコースを図1に示す。
【0049】
実施例4:Nアセチルガラクトサミンの精製
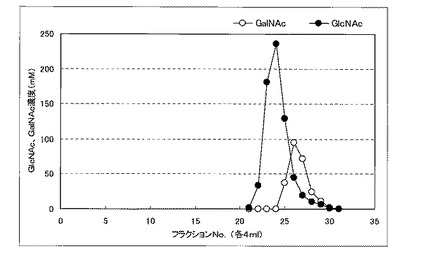
NアセチルグルコサミンとNアセチルガラクトサミンの分離は、カラム剤:イオン交換樹脂(ダウエックス、ザ・ダウ・ケミカル・カンパニー製)、流速:0.5ml/min、移動相:水、の条件下で行うことができる。反応液を電気透析し脱塩した後、サンプル5mlをカラム(2.6cm×40cm、200mlカラム剤)に供し、4mlずつ分画した。各フラクションをHPLCで分析した結果を図2に示す。図2に示すようにNアセチルグルコサミンとNアセチルガラクトサミンを分離し、Nアセチルガラクトサミンを精製することができる。
【0050】
以上の結果より、本願発明に係る方法を用いることによって、出発原料のNアセチルグルコサミンのうち約30%程度をNアセチルガラクトサミンに変換できることが明らかとなった。
【産業上の利用可能性】
【0051】
本発明は、Nアセチルグルコサミンを出発原料として用いて、Nアセチルガラクトサミンを製造することを可能とする。本発明は、Nアセチルガラクトサミンの有効な製造法である上に、Nアセチルガラクトサミンを安価に、かつ一工程で製造できるという利点を有する。製造されたNアセチルガラクトサミンは食品、医薬品あるいはそれらの原料として用いることが可能である。
【図面の簡単な説明】
【0052】
【図1】実施例3における、反応液1のタイムコースを示す図である。
【図2】実施例4における各フラクションをHPLCで分析した結果を示す図である。
【技術分野】
【0001】
本発明は、Nアセチルグルコサミンを、ATPおよびUDP−糖の存在下にて、酵素と反応させることにより、Nアセチルガラクトサミンを製造する方法に関する。
【背景技術】
【0002】
Nアセチルガラクトサミンは糖鎖中によく見られる単糖の一つであり、糖鎖を利用した機能性物質の原料として期待されている。しかしながら、Nアセチルガラクトサミンを豊富に含んだ天然産の糖質は多量に存在しない。これまでに、Nアセチルガラクトサミンの製造方法としては、鮭軟骨などから抽出されるコンドロイチン硫酸の酸加水分解分解物からガラクトサミンを抽出後N−アセチル化することにより製造する方法が知られている(非特許文献1を参照)。
【0003】
しかしながら、この方法では、出発物質の含有量の低さなどから、産業的に有効な製造法とはいえず、そのためにNアセチルガラクトサミンは非常に高価な物質であった。
【0004】
【非特許文献1】Jeanloz, RZ編、“The Amino Sugars /volume 1A”, Academic Press (1969)
【発明の開示】
【発明が解決しようとする課題】
【0005】
従来法では、安価に効率的にNアセチルガラクトサミンを製造することはできなかった。従って、Nアセチルガラクトサミンの、有効な製造方法の確立が望まれていた。
【0006】
本発明は、Nアセチルグルコサミンを、ATPおよびUDP−糖の存在下にて、酵素と反応させることにより、Nアセチルガラクトサミンを製造する方法を提供する。
【課題を解決するための手段】
【0007】
本発明者らは、上記課題を解決すべく、鋭意検討した結果、出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下、以下の(i)〜(iii)の酵素:(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素、と同時に作用させることにより、Nアセチルガラクトサミンを製造できることを見出し、本発明を完成させるに至った。
【0008】
すなわち、本発明は以下のとおりである。
[1] 出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下で、以下の(i)〜(iii)の酵素と同時に作用させることにより、Nアセチルガラクトサミンを製造する方法:
(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;
(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに
(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素。
【0009】
[2] UDP−糖が、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンまたはそれらの少なくとも2つの混合物からなる群から選択される、[1]の方法。
[3] UDP−糖が、UDP−グルコースまたはUDP−Nアセチルグルコサミンである、[2]の方法。
【0010】
[4] (i)の酵素が、UDP−グルコース4−エピメラーゼである、[1]〜[3]のいずれかの方法。
[5] UDP−グルコース4−エピメラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[4]の方法。
【0011】
[6] (ii)の酵素が、UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼである、[1]〜[3]のいずれかの方法。
[7] UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[6]の方法。
【0012】
[8] (iii)の酵素が、Nアセチルヘキソサミン1キナーゼである、[1]〜[3]のいずれかの方法。
[9] Nアセチルヘキソサミン1キナーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、[8]の方法。
【0013】
[10] (i)の酵素としてUDP−グルコース4−エピメラーゼ、(ii)の酵素としてUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ、(iii)の酵素としてNアセチルヘキソサミン1キナーゼを同時に作用させる、[1]〜[3]のいずれかの方法。
[11] 作用させる全ての酵素が、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する[10]の方法。
[12] さらに、製造されたNアセチルガラクトサミンを精製する工程を含む、[1]〜[9]のいずれかの方法。
【発明の効果】
【0014】
本発明では安価に得られるNアセチルグルコサミンを出発原料として、ATPおよびUDP−糖の存在下、(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素、(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素、ならびに(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素を同時に作用させることにより一段階でNアセチルガラクトサミンを製造できる。
【発明を実施するための最良の形態】
【0015】
出発原料であるNアセチルグルコサミンの濃度は、特に限定されるものではないが、好ましくは10mM〜2M、より好ましくは100mM〜1M、さらに好ましくは600mMである。
【0016】
(i)の酵素は、UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する活性を有する限り、特に限定されないが、UDP−グルコース4−エピメラーゼのうち、UDP−Nアセチルグルコサミンにも活性を示す酵素を用いるのが好ましい。このような酵素としては、ヒト由来、Bacillus属細菌由来、ビフィズス菌由来のUDP−グルコース4−エピメラーゼ(Thoden, J. B.ら Structural analysis of the Y299C mutant of Escherichia coli UDP-galactose 4-epimerase. Teaching an old dog new tricks. The Journal of Biological Chemistry. 277:27528-27534 (2002)、再表01/038555、Genbank登録番号AB303839)を用いることができるが、好ましくは、ビフィドバクテリウム・ロンガムに由来するUDP−グルコース4−エピメラーゼを用いる。
【0017】
(ii)の酵素は、UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応を触媒する活性を有する限り、特に限定しないが、ビフィズス菌由来のUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ(西本ら、第58回日本生物工学会大会講演要旨集p. 92 (2006)、Genbank登録番号AB303839)を用いるのが好ましい。特に好ましくは、ビフィドバクテリウム・ロンガムに由来するUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼを用いる。
【0018】
(iii)の酵素は、NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる活性を持つ酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる活性を持つ酵素、またはその両活性を持つ酵素である限り、特に限定されないが、ビフィズス菌由来のNアセチルヘキソサミン1キナーゼ(特開2007−97517、Genbank登録番号AB303839)を用いるのが好ましい。特に好ましくは、ビフィドバクテリウム・ロンガムに由来するNアセチルヘキソサミン1キナーゼを用いる。
【0019】
通常、(iii)に示す様な活性の酵素はキナーゼと呼ばれており、一般的にはその逆反応の利用はほとんど省みられたことはなかった。この逆反応は、本発明者により初めて見出されたものであり、本発明では、この酵素の逆反応をNアセチルガラクトサミンを生じる最終段階で利用している。
【0020】
これらの酵素は、天然の生物または遺伝子組換え生物(もしくは細胞)により産生されたもの、あるいは市販されているものを用いることができる。遺伝子組換え生物(もしくは細胞)は、当業者に公知である分子生物学的手法を用いて、当該酵素をコードする遺伝子を適当なベクターに組み込み、そのベクターを用いて適当な宿主生物(例えば、細菌、酵母、動物細胞、昆虫細胞、植物細胞など)を形質転換することにより得ることができる(Sambrook J.ら、“Molecular Cloning A LBORATORY MANUAL /second edition”, Cold Spring Harbor Laboratory Press (1989)参照)。
【0021】
上記生物により産生された酵素は、通常の方法を用いて、当該生物の培養物より単離精製できる。ここで「培養物」とは、培養上清、培養細胞もしくは培養菌体または細胞もしくは細胞の破砕物を意味する。培養後、目的の酵素が菌体内または細胞内に生産される場合には、細胞を遠心分離により回収し、緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を回収する。あるいは、目的の酵素が菌体外または細胞外に生産される場合には、遠心分離などにより菌体または細胞を除去し、培養液を回収する。この無細胞抽出液を遠心分離し、得られた上清から、または上記培養液から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、陰イオン交換クロマトグラフィー法、陽イオン交換クロマトグラフィー法、疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用いて、精製酵素を得ることができる。
【0022】
本発明方法において用いられる酵素は、精製された形態であっても、上記無細胞抽出液または培養液のような粗製物の形態であってもよい。
【0023】
本発明における反応において、上記酵素群は、以下の一連の反応を触媒し、ATPおよびUDP−糖の存在下、Nアセチルグルコサミンより、Nアセチルガラクトサミンを生成する:
図中の略語:
GlcNAc:Nアセチルグルコサミン
GlcNAc1P:Nアセチルグルコサミン1−リン酸
UDP−GalNAc:UDP−Nアセチルガラクトサミン
UDP−GlcNAc:UDP−Nアセチルグルコサミン
GalNAc1P:Nアセチルガラクトサミン1−リン酸
GalNAc:Nアセチルガラクトサミン。
【0024】
反応液中の酵素の濃度は、0.01U〜200U/ml、好ましくは、0.05U〜100U/mlの範囲で適宜設定でき、例えば、Nアセチルヘキソサミンキナーゼであれば1.0U〜10U/ml、UDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼであれば0.05U/ml、UDPグルコース−4−エピメラーゼであれば100U/mlで用いるのが好ましい。
【0025】
反応液中のATPの濃度は、特に限定されるものではないが、好ましくは0.01mM〜100mM、より好ましくは0.1mM〜10mM以下、さらに好ましくは、0.75mM〜2.0mMである。
【0026】
用いるUDP−糖としては、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンのいずれかまたはそれらの少なくとも2つの混合物を用いることができる。好ましくは、UDP−グルコースまたはUDP−Nアセチルグルコサミンを用いる。反応液中のUDP−糖の濃度は、特に限定されるものではないが、好ましくは0.005mM〜50mM、より好ましくは0.05mM〜10mM、さらに好ましくは、0.2mM〜0.6mMである。
【0027】
反応液のpHは好ましくは5〜9、さらに好ましくは、6.5である。反応温度は特に限定されるものではないが好ましくは5℃〜80℃、より好ましくは20℃〜60℃、さらに好ましくは、30℃である。反応時間は、1時間〜30日間、好ましくは24時間〜10日間、さらに好ましくは5日間である。必要に応じて撹拌しながら反応を行っても良い。
【0028】
反応系は特に限定されるものではないが通常水溶液あるいは緩衝液中で行われる。緩衝液としては、酵素反応に一般的に用いられるもの、例えば、トリス塩酸緩衝液、トリス酢酸緩衝液、HEPES緩衝液、MOPS緩衝液、リン酸緩衝液、酢酸ナトリウム緩衝液、クエン酸ナトリウム緩衝液などを使用できるが、MOPS緩衝液を用いるのが好ましい。使用濃度は、10mM〜500mM、好ましくは10mM〜300mM、さらに好ましくは100mMである。
【0029】
反応は、固定化酵素リアクターを用いて行うことも可能である。酵素を固定化することによって、酵素が安定化され、連続反復使用が可能となる点において有効である。酵素の固定化は、担体結合法、架橋法、包括法を用いて行うことができる。担体結合法では、酵素を担体(例えば、セルロース、デキストラン、アガロースなどの多糖類の誘導体、ポリアクリルアミドゲル、ポリスチレン樹脂、多孔性ガラス、金属酸化物など)に、物理的吸着、イオン結合および共有結合等を用いて結合させることができる。架橋法では、2個またはそれ以上の官能基を持つ試薬を用いて、酵素同士を互いに架橋することによって固定化する。架橋試薬としては、Schiff塩基をつくるグルタルアルデヒド、ペプチド結合をするイソシアン酸誘導体、N,N’−エチレンマレイミド、ジアゾカップリングをするビスジアゾベンジン、あるいはアルキル化するN,N’−ポリメチレンビスヨードアセトアミドなどを用いることができる。包括法では、高分子ゲルの細かい格子の中に酵素を取り込む格子型と、半透膜の高分子の皮膜によって酵素を皮膜するマイクロカプセル型を用いる。格子型の方法では、合成高分子物質のポリアクリルアミドゲル、ポリビニルアルコール、光硬化性樹脂および天然高分子物質のデンプン、コンニャク粉、ゼラチン、アルギン酸、カラギーナンなどの高分子化合物を用いることができる。マイクロカプセル型の方法では、ヘキサメチレンジアミン、セバコイルクロリド、ポリスチレン、レシチンなどを用いることができる(福井三郎、千畑一郎、鈴木周一、「酵素工学」東京化学同人発行、1981年)。
【0030】
担体は、3種の酵素を固定化したものを用いてもよいし、それぞれの酵素を固定化した複数の担体を用いてもよい。
【0031】
本発明の方法により、用いたNアセチルグルコサミンの少なくとも20%、好ましくは少なくとも25%がNアセチルガラクトサミンに変換される。
【0032】
生成したNアセチルガラクトサミンは、例えば、イオン交換クロマトグラフィー、吸着クロマトグラフィー、アフィニティクロマトグラフィー、ゲル濾過法など各種のクロマトグラフィー、向流分配、向流抽出など二液相間の分配を利用する方法、濃縮、冷却、有機溶媒添加など溶解度の差を利用する方法、および、塩析などの通常の精製法を単独で、あるいは適宜組み合わせて用いて、出発原料であるNアセチルグルコサミンと分離することが可能である。好ましくは、Nアセチルガラクトサミンは、強イオン性陽イオン交換クロマトグラフィーのような方法により、Nアセチルグルコサミンと分離することができる。
【0033】
得られたNアセチルガラクトサミンは、例えば医薬品、化粧品、食品添加物等の材料として好適に用いることができる。
【実施例】
【0034】
本発明を以下の実施例によって具体的に説明するが、本発明はこれらの実施例によって限定されるものではない。
【0035】
実施例1:酵素の調製
Nアセチルヘキソサミン1キナーゼの調製
ビフィドバクテリウム・ロンガムのNアセチルヘキソサミン1キナーゼの遺伝子配列(配列番号1)を基に、以下のプライマー作製し:
フォーワードプライマー:aacggaccccatatgaccgaaagcaatgaagttttattc(配列番号2)
リバースプライマー:gctgacctcgagcctggcagcctccatgatgtcggctac(配列番号3)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてNアセチルヘキソサミン1キナーゼのORF全長をPCRにより増幅した。
【0036】
PCR産物を制限酵素NcoI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0037】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質を、Ni−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製して、精製酵素標品を得た。得られた精製酵素の比活性は1.5U/mgであった。
【0038】
UDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼの調製
ビフィドバクテリウム・ロンガムのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼの配列(配列番号4)を基に、以下のプライマーを作製し:
フォーワードプライマー:gatatatacatatgaacgatcagctgaccgaggta(配列番号5)
リバースプライマー:tgacctctcgagcctagcggcaaaaccaaggctttcga(配列番号6)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ遺伝子をPCRにより増幅した。
【0039】
PCR産物を制限酵素NdeI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET30(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0040】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質はNi−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製して、精製酵素標品を得た。得られた精製酵素の比活性は1.1U/mgであった。
【0041】
UDPグルコース−4−エピメラーゼの調製
ビフィドバクテリウム・ロンガムのUDPグルコース−4−エピメラーゼの配列(配列番号7)を基に、以下のプライマーを作製し:
フォーワードプライマー:gatatacatatgactactgttctggttacgggc(配列番号8)
リバースプライマー:ctgctcctcgagctccgcgtcgcggaaaccgttggggttc(配列番号9)、
ビフィドバクテリウム・ロンガムJCM 1217株のゲノムDNAを鋳型としてUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ遺伝子をPCRにより増幅した。
【0042】
PCR産物を制限酵素NdeI及びXhoIで消化後、同様に処理した市販の遺伝子発現用プラスミドpET30(ノバジェン社製)にDNAライゲーションキット(宝酒造株式会社製)を用いて連結した。さらに、これらのプラスミドを用いてSambrook,J.,Fritsch, E. F. and Maniatis, T. "Molecular Cloning, A Laboratory Manual 第2版”1.74章 Vo1. 1 (1989)に記載された方法に従い、大腸菌を形質転換した。
【0043】
以上のようにして得た形質転換体を用い、常法に従って該遺伝子の発現及び組換えタンパク質の生産を行った。得られた組換えタンパク質はNi−NTAアガロース(キアゲン社製)を用いたカラムクロマトグラフィーにより精製し、精製酵素標品を得た。得られた精製酵素の比活性は158U/mgであった。
以上のようにして得た酵素を以下の反応液に用いた。
【0044】
実施例2:反応液の調製
[反応液1]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.6mMのUDP−Nアセチルグルコサミン、2.0mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、1.0U/mlのNアセチルヘキソサミン1キナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、50U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液1を調製した。
【0045】
[反応液2]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.6mMのUDP−グルコース、0.75mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、10U/mlのNアセチルヘキソサミンキナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、100U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液2を調製した。
【0046】
[反応液3]
100mM MOPS緩衝液(pH6.5)中に、最終濃度が600mMのNアセチルグルコサミン、0.2mMのUDP−グルコース、1.0mMのATP、10mMの塩化マグネシウム、0.05%アジ化ナトリウム、10U/mlのNアセチルヘキソサミンキナーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、0.05U/mlのUDPグルコース−ヘキソース−1−リン酸ウリジリルトランスフェラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)、100U/mlのUDPグルコース−4−エピメラーゼ(ビフィドバクテリウム・ロンガム由来精製酵素)となるように、それぞれを加えて反応液を調製した。
【0047】
実施例3:Nアセチルガラクトサミンの生成
実施例2で調製した反応液1、反応液2、および反応液3をそれぞれ恒温器で30℃、5日間インキュベートし、酵素反応を行った。反応終了後、反応液を適宜希釈して高速液体クロマトグラフィー(島津製作所製)を用いて、NアセチルグルコサミンとNアセチルガラクトサミンの濃度を調べ、モル収率を算出した。
【0048】
この結果、反応液1、反応液2、および反応液3におけるNアセチルガラクトサミンのモル収率はそれぞれ27.7%、28.0%、24.7%であった。反応液1のタイムコースを図1に示す。
【0049】
実施例4:Nアセチルガラクトサミンの精製
NアセチルグルコサミンとNアセチルガラクトサミンの分離は、カラム剤:イオン交換樹脂(ダウエックス、ザ・ダウ・ケミカル・カンパニー製)、流速:0.5ml/min、移動相:水、の条件下で行うことができる。反応液を電気透析し脱塩した後、サンプル5mlをカラム(2.6cm×40cm、200mlカラム剤)に供し、4mlずつ分画した。各フラクションをHPLCで分析した結果を図2に示す。図2に示すようにNアセチルグルコサミンとNアセチルガラクトサミンを分離し、Nアセチルガラクトサミンを精製することができる。
【0050】
以上の結果より、本願発明に係る方法を用いることによって、出発原料のNアセチルグルコサミンのうち約30%程度をNアセチルガラクトサミンに変換できることが明らかとなった。
【産業上の利用可能性】
【0051】
本発明は、Nアセチルグルコサミンを出発原料として用いて、Nアセチルガラクトサミンを製造することを可能とする。本発明は、Nアセチルガラクトサミンの有効な製造法である上に、Nアセチルガラクトサミンを安価に、かつ一工程で製造できるという利点を有する。製造されたNアセチルガラクトサミンは食品、医薬品あるいはそれらの原料として用いることが可能である。
【図面の簡単な説明】
【0052】
【図1】実施例3における、反応液1のタイムコースを示す図である。
【図2】実施例4における各フラクションをHPLCで分析した結果を示す図である。
【特許請求の範囲】
【請求項1】
出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下で、以下の(i)〜(iii)の酵素と同時に作用させることにより、Nアセチルガラクトサミンを製造する方法:
(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;
(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに
(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素。
【請求項2】
UDP−糖が、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンおよびそれらの少なくとも2つの混合物からなる群から選択される、請求項1に記載の方法。
【請求項3】
UDP−糖が、UDP−グルコースまたはUDP−Nアセチルグルコサミンである、請求項2に記載の方法。
【請求項4】
(i)の酵素が、UDP−グルコース4−エピメラーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
UDP−グルコース4−エピメラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項4に記載の方法。
【請求項6】
(ii)の酵素が、UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項7】
UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項6に記載の方法。
【請求項8】
(iii)の酵素が、Nアセチルヘキソサミン1キナーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項9】
Nアセチルヘキソサミン1キナーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項8に記載の方法。
【請求項10】
(i)の酵素としてUDP−グルコース4−エピメラーゼ、(ii)の酵素としてUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ、(iii)の酵素としてNアセチルヘキソサミン1キナーゼを同時に作用させる、請求項1〜3のいずれか1項に記載の方法。
【請求項11】
作用させる全ての酵素が、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項10に記載の方法。
【請求項12】
さらに、製造されたNアセチルガラクトサミンを精製する工程を含む、請求項1〜9のいずれか1項に記載の方法。
【請求項1】
出発原料であるNアセチルグルコサミンを、ATPおよびUDP−糖の存在下で、以下の(i)〜(iii)の酵素と同時に作用させることにより、Nアセチルガラクトサミンを製造する方法:
(i) UDP−NアセチルグルコサミンをUDP−Nアセチルガラクトサミンに変換する酵素;
(ii) UDP−NアセチルグルコサミンとNアセチルガラクトサミン1−リン酸間でUMP単位の交換反応をする酵素;ならびに
(iii) NアセチルグルコサミンとATPからNアセチルグルコサミン1−リン酸とADPを生じる酵素およびNアセチルガラクトサミンとATPからNアセチルガラクトサミン1−リン酸とADPを生じる酵素、またはその両活性を持つ酵素。
【請求項2】
UDP−糖が、UDP−グルコース、UDP−ガラクトース、UDP−Nアセチルグルコサミン、UDP−Nアセチルガラクトサミンおよびそれらの少なくとも2つの混合物からなる群から選択される、請求項1に記載の方法。
【請求項3】
UDP−糖が、UDP−グルコースまたはUDP−Nアセチルグルコサミンである、請求項2に記載の方法。
【請求項4】
(i)の酵素が、UDP−グルコース4−エピメラーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
UDP−グルコース4−エピメラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項4に記載の方法。
【請求項6】
(ii)の酵素が、UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項7】
UDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項6に記載の方法。
【請求項8】
(iii)の酵素が、Nアセチルヘキソサミン1キナーゼである、請求項1〜3のいずれか1項に記載の方法。
【請求項9】
Nアセチルヘキソサミン1キナーゼが、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項8に記載の方法。
【請求項10】
(i)の酵素としてUDP−グルコース4−エピメラーゼ、(ii)の酵素としてUDP−グルコース−ヘキソース1リン酸ウリジリルトランスフェラーゼ、(iii)の酵素としてNアセチルヘキソサミン1キナーゼを同時に作用させる、請求項1〜3のいずれか1項に記載の方法。
【請求項11】
作用させる全ての酵素が、ビフィドバクテリウム・ロンガム(Bifidobacterium longum)に由来する、請求項10に記載の方法。
【請求項12】
さらに、製造されたNアセチルガラクトサミンを精製する工程を含む、請求項1〜9のいずれか1項に記載の方法。
【図1】

【図2】
【図2】
【公開番号】特開2009−27971(P2009−27971A)
【公開日】平成21年2月12日(2009.2.12)
【国際特許分類】
【出願番号】特願2007−194867(P2007−194867)
【出願日】平成19年7月26日(2007.7.26)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度独立行政法人農業・食品産業技術総合研究機構生物系特定産業技術研究支援センター委託研究による成果(産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成21年2月12日(2009.2.12)
【国際特許分類】
【出願日】平成19年7月26日(2007.7.26)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成19年度独立行政法人農業・食品産業技術総合研究機構生物系特定産業技術研究支援センター委託研究による成果(産業活力再生特別措置法第30条の適用を受けるもの)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]