
NAD+依存型脱水素酵素を用いたL−トリプトファン定量法およびそれに用いるキット
【課題】試料中のL-トリプトファンを、これまでの方法に比べ安価で簡易的に定量可能である酵素的定量法を提供する。この酵素的定量法を実施する際に利用できる測定用のキット及び酵素センサーを提供する。
【解決手段】検体に、L-トリプトファン脱水素酵素を作用させ、生じたNADHを定量することを含む、前記試料に含有されるL-トリプトファンの測定方法。トリプトファン脱水素酵素を用いることを特徴とする酵素センサー。以下の(1)及び(2)の試薬を含むL-トリプトファンの測定用キット。
(1)L-トリプトファン脱水素酵素
(2)NAD+
【解決手段】検体に、L-トリプトファン脱水素酵素を作用させ、生じたNADHを定量することを含む、前記試料に含有されるL-トリプトファンの測定方法。トリプトファン脱水素酵素を用いることを特徴とする酵素センサー。以下の(1)及び(2)の試薬を含むL-トリプトファンの測定用キット。
(1)L-トリプトファン脱水素酵素
(2)NAD+
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、試料溶液中のL-トリプトファンを迅速かつ簡易的に定量する方法およびそれに用いるキットに関するものである。
【背景技術】
【0002】
L-トリプトファンは、タンパク質構成アミノ酸の一つであり、さまざまな二次代謝産物の生合成原料にもなる。
【0003】
L-トリプトファンの定量は、これまで主に自働アミノ酸分析装置やプレカラム誘導体化逆相高速液体クロマトグラフィーによる分離と誘導体化を伴う機器分析的手法で行われている。また、L-トリプトファンと直接反応する発色試薬を用いる方法もあるが、類似化合物と反応するため生体試料への適用が困難である。酵素を用いたL-トリプトファンの定量方法も知られている。
【0004】
酵素を用いた方法として、ヒトヨタケ属担子菌(Copurinus sp.)由来のL-トリプトファン用アミノ酸オキシダーゼを用いる方法が報告されている(特許文献1および非特許文献1)。L-トリプトシルtRNA合成酵素を用いた方法も知られている(非特許文献2)。また、L-トリプロファン2-モノオキシゲナーゼを用いた酵素センサーも知られている(非特許文献3)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2001-69974号公報
【非特許文献】
【0006】
【非特許文献1】Biosci. Biotechnol. Biochem. 64, 1468-1493 (2000)
【非特許文献2】Anal. Biochem. 154, 618-623 (1986)
【非特許文献3】Biosen.and Bioelect. 12, 363-371 (1997)
【非特許文献4】J. Am. Chem. Soc., 130, 15260-15261 (2008)
【発明の概要】
【発明が解決しようとする課題】
【0007】
上記のように、L-トリプトファンは、主にHPLCを中心とした機器分析的手法により定量が行われてきた。しかし、トリプトファンを含むアミノ酸の定量に用いられる機器は、一般に高価のものが多く特に生体試料に適用するには、高度な分離能が必要となり、高価である。
【0008】
L-トリプトファンと直接作用する発色剤を用いた方法もあるが、生体試料などを測定対象とした場合、特異性に問題が生じる。
【0009】
特許文献1および非特許文献1に記載の方法で用いられるヒトヨタケ由来のアミノ酸オキシダーゼは、L-フェニルアラニンに7%の相対活性を有している。また、オキシダーゼ活性の他にジオキシゲナーゼ活性も持つため、定量的に反応が進行しない。このように、ヒトヨタケ由来のアミノ酸オキシダーゼは、L-フェニルアラニンに対しても若干の活性があり、オキシゲナーゼ活性も有するため定量には適さない。
【0010】
非特許文献2に記載の方法は、放射性同位体元素を使用しているため、汎用的に使用する事は出来ない。
【0011】
非特許文献3に記載の酵素センサーに用いられるL-トリプトファン2−モノオキシゲナーゼは、L-トリプトファン以外にL-フェニルアラニンにも反応してしまう。そのため、L-トリプトファン2-モノオキシゲナーゼを用いた酵素電極では、生体試料の測定においては、十分な精度が得られない。
【0012】
そこで、本発明は、酵素的定量方法であって、試料溶液中のL-トリプトファンを簡便且つ高い精度で実施できる方法を提供することを目的とする。
【0013】
さらに本発明は、上記酵素的定量方法を実施する際に利用できる測定用キット及び酵素センサーを提供することも目的とする。
【課題を解決するための手段】
【0014】
発明者らが種々検討した結果、ある特定のL-トリプトファン脱水素酵素をL-トリプトファンを含有する試料溶液に添加することで、L-トリプトファン脱水素酵素が試料溶液中のL-トリプトファンに特異的に作用して、共存するNAD+がNADHへと変換され、生成するNADHの量を測定することで、試料溶液中のL-トリプトファンを定量することが可能であることが明らかとなった。
【0015】
本発明は、以下に示す通りである。
[1]
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの定量方法であって、
前記L-トリプトファン脱水素酵素が、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する前記方法。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[2]
工程(A)において検体との混合に用いられる前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤の存在下で保存されているものである、[1]に記載の方法。
[3]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[2]に記載の方法。
[4]
前記ラン藻がノストク・パンクチフォルム(Nostoc punctiform )である[1]〜[3]のいずれかに記載の方法。
[5]
前記工程(C)で計測する反応生成物がNADHである[1]〜[4]のいずれかに記載の方法。
[6]
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファン脱水素酵素、および
(K2)NAD+
但し、前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[7]
前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物である、[6]に記載のキット。
[8]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[7]に記載のキット。
[9]
(K3)反応用緩衝液)および/または(K4)NADH検出試薬をさらに含む、[6]〜[8]のいずれかに記載のキット。
[10]
L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物であって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する組成物。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[11]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[10]に記載の組成物。
[12]
L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーであって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する酵素センサー。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[13]
前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置される、[12]に記載の酵素センサー。
[14]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[13]に記載の酵素センサー。
【発明の効果】
【0016】
本発明によれば、特定のL-トリプトファン脱水素酵素を用いることで、この酵素が特異的にL-トリプトファンに作用し、さまざまな化合物を含有する試料中であっても高い選択性でL-トリプトファンを簡便に定量することができる。
【0017】
また、本発明は、さまざまな試料に対し有効であり、NADHの検出法により、紫外部吸収法、発色法、蛍光法、さらには、電極型酵素センサーを提供することもできる。
【図面の簡単な説明】
【0018】
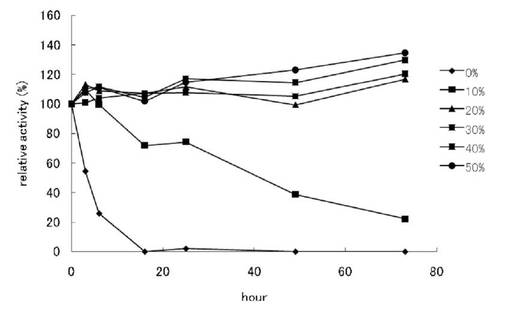
【図1】L-トリプトファン脱水素酵素の安定化試験結果を示す。
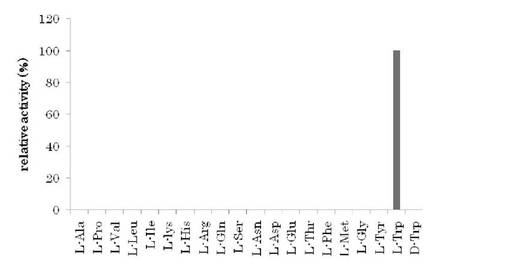
【図2】L-トリプトファン脱水素酵素の基質特異性の試験結果を示す。
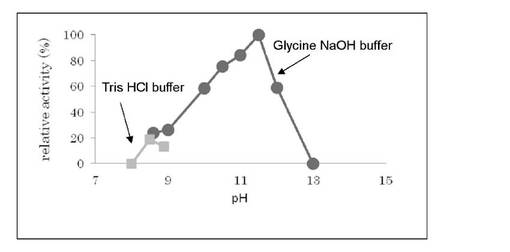
【図3】L-トリプトファン脱水素酵素活性に対するpHの影響を示す。
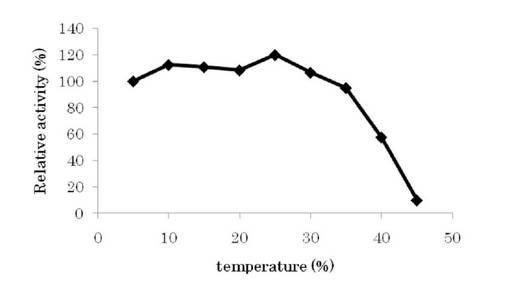
【図4】L-トリプトファン脱水素酵素の温度安定性試験結果を示す。
【図5】L-トリプトファン脱水素酵素のL-トリプトファンに対するミカエリス・メンテンプロットを示す。
【図6】L-トリプトファン脱水素酵素を用いたL-トリプトファン定量の検量線を示す。
【発明を実施するための形態】
【0019】
[L-トリプトファンの定量方法]
本発明のL-トリプトファンの定量方法は、
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含み、前記工程を経ることで、前記検体中のL-トリプトファンを定量することができる。
【0020】
さらに上記方法に用いるL-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【0021】
<L-トリプトファン定量用酵素>
本発明に用いるL-トリプトファン脱水素酵素は、下記式Aに示す反応を触媒して、L-トリプトファンとインドールピルビン酸とアンモニウムイオンを生産する際にNAD+をNADHに変換する酵素である。
【0022】
【化1】

【0023】
本発明で用いるL-トリプトファン脱水素酵素の第1の態様は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【0024】
配列表の配列番号1に記載のアミノ酸配列は、実施例で示すように、ノストク・パンクチフォルム(Nostoc punctiform)ATCC29133のゲノムDNAから調製したL-トリプトファン脱水素酵素遺伝子(NpR1275)を発現させたL-トリプトファン脱水素酵素のアミノ酸配列である。
【0025】
この配列番号1に記載のアミノ酸配列を有するL-トリプトファン脱水素酵素は、図2に結果を示すように、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用しない。さらに、この前記L-トリプトファン脱水素酵素は、NAD+をNADHに変換する脱水素酵素活性を有するものである。NAD+をNADHに変換する脱水素酵素活性を有することは、図6の検量線の結果から明らかである。前記L-トリプトファン脱水素酵素が、NAD+をNADHに変換する脱水素酵素活性を有することは、実施例に示すように、NADHの生成を340nmの吸光度の変化を測定することで確認することができる。尚、タンパク質構成アミノ酸とは、L-アラニン、L-プロリン、L-バリン、L-ロイシン、L-イソロイシン、L-リジン、L-ヒスチジン、L-アルギニン、L-グルタミン、L-セリン、L-アスパラギン酸、L-グルタミン酸、L-スレオニン、L-フェニルアラニン、L-メチオニン、L-グリシン、L-チロシン、L-アスパラギンを意味する。L-システインについては、本酵素が活性を示す中性から塩基性のpHでは、速やかに不溶性のシスチンへ変換されることから、本発明においては、上記タンパク質構成アミノ酸には含まれない。
【0026】
配列番号1に記載のアミノ酸配列を有するL-トリプトファン脱水素酵素についての、上記基質特異性以外の特性は、実施例において具体的に記載する。
【0027】
さらに、このL-トリプトファン脱水素酵素は、NAD+の存在下、L-トリプトファンに特異的に作用してインドールピルビン酸とアンモニアを生成する。この点は、非特許文献4において述べられている本酵素のscytonemin合成経路上の位置づけと実施例で示すL-トリプトファン依存的なNADHの生産から明らかである。 L-トリプトファン脱水素酵素が、NAD+の存在下、L-トリプトファンに特異的に作用することは、(i)L-トリプトファン依存的なNADHの生産、(ii)L-トリプトファン以外のアミノ酸を基質とした場合NADHの生産が無いことで確認する事が出来る。
【0028】
本明細書で言う「1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から数個」の範囲は、欠失等を有するタンパク質が、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである限り、特に限定されない。前記「1から数個」の範囲は、前記脱水素酵素活性を有するタンパク質である割合が高いことから、例えば、1から30個、好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から7個、一層好ましくは1から5個、特に好ましくは1から3個程度であることができる。
【0029】
本明細書で言う「配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列」における相同性は、前記アミノ酸配列の相同性を有するタンパク質が、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである限り、特に限定されない。前記アミノ酸配列の相同性は、90%以上であれば特に限定されないが、好ましくは95%以上、さらに好ましくは96%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0030】
本発明で用いるL-トリプトファン脱水素酵素の第2の態様は、(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
【0031】
実施例で示すように、ノストク・パンクチフォルム(Nostoc punctiform) ATCC29133のゲノムDNAから調製したL-トリプトファン脱水素酵素遺伝子(NpR1275)を発現させたL-トリプトファン脱水素酵素は、(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のアミノ酸には作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。さらに前述のように、このL-トリプトファン脱水素酵素は、配列表の配列番号1に記載のアミノ酸配列を有する。
【0032】
L-トリプトファン脱水素酵素[EC 1.4.1.19]は、一部の植物におけるインドール酢酸生合成経路上に存在する事が古くから指摘されている。しかし、単一のタンパク質として精製されたことはなく、遺伝子も不明である。一方、ラン藻において、紫外線防御物質として生産されるscytoneminの生合成経路の初発反応を行う酵素がL-トリプトファンに作用するNAD+依存型脱水素酵素であることがBalskusらによって報告されている(非特許文献4)。しかし、これまでの報告では、scytonemin生合成遺伝子クラスター上にコードされているNpR1275遺伝子配列が、既知のアミノ酸脱水素酵素と相同性があることと、大腸菌で発現させたNpR1275遺伝子産物がL-トリプトファンに対し活性がありL-チロシンに対する活性がほとんどないことが示されたのみである。これまでに、本酵素の基質特異性等の酵素の性質に関しては明らかにされておらず、ましてや、本酵素を用いたL-トリプトファン定量について検討されたことはない。
【0033】
本発明で用いるL-トリプトファン脱水素酵素の第2の態様は、特定の種より生産されるものに限られるものではなく、ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素またはラン藻由来生物中でscytonemin合成経路を担っていると推定されるホモログの遺伝子としてコードされている酵素から選択され、かつL-トリプトファンを基質として作用するNAD+依存型の脱水素酵素を意味する。L-トリプトファン脱水素酵素としては、例えば、ラン藻ノストク・パンクチフォルム(Nostoc punctiform)のscytonemin生合成遺伝子クラスター上にコードされているNpR1275遺伝子産物及びそのホモログが挙げられる。表1にBLAST上で検索されたNpR1275遺伝子のホモログと推定される遺伝子の例を挙げる。これらホモログは、NpR1275遺伝子同様にscytonemin生合成遺伝子クラスター上に存在しアミノ酸脱水素酵素をコードすると推定される遺伝子であることから、その遺伝子産物は、NpR1275遺伝子産物と同様にL-トリプトファン脱水素酵素としての機能を持つものと推定される。
【0034】
【表1】
【0035】
さらに、本発明に用いるL-トリプトファン脱水素酵素は、同様の活性を有するものであれば、自然界より分離された生物に由来するもの、本酵素をコードする遺伝子を大腸菌や他の生物を宿主として発現させて得られる酵素、更にこれらの遺伝子を適宜改変して作製した酵素も含まれる。
【0036】
また、異種発現による生産法としては、例えば、上記ノストク・パンクチフォルム(Nostoc punctiform)より抽出したゲノムDNAから該当する遺伝子(配列表の配列番号2に示す)をPCRにて増幅しpETもしくはpUCなどに組み込んだプラスミドベクターを構築したのち、BL21、JM109などの宿主菌株に形質転換し、培養する方法が挙げられる。これら以外の公知の方法も適宜用いることができる。
【0037】
本発明で用いるL-トリプトファン脱水素酵素の取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組換え技術により作製した組換えタンパク質でもよい。組換えタンパク質を作製する場合には、後述するように当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、上記L-トリプトファン脱水素酵素を産生することができる。
【0038】
上記L-トリプトファン脱水素酵素は、上記L-トリプトファン脱水素酵素をコードする遺伝子をベクター上に搭載し、このベクターによって宿主細胞を形質転換した後、形質転換させた宿主細胞を培養して培養物中に前記遺伝子がコードするタンパク質を蓄積し、蓄積したタンパク質を収集することを含む、生産方法により調製することができる。
【0039】
上記L-トリプトファン脱水素酵素をコードする遺伝子の取得方法は特に限定されない。本発明のL-トリプトファン脱水素酵素をコードする遺伝子は、例えば、配列番号1に記載のアミノ酸配列および配列番号2に記載した塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0040】
例えば、配列表の配列番号2に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0041】
本明細書中の配列表の配列番号1に記載したアミノ酸配列または配列番号2に示す塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いてノストク・パンクチフォルム(Nostoc punctiform )ATCC29133のゲノムライブラリーより本発明の遺伝子を単離することができる。ゲノムライブラリーは、ノストク・パンクチフォルム(Nostoc punctiform )ATCC29133から常法により作製することができる。
【0042】
PCR法により本発明のL-トリプトファン脱水素酵素をコードする遺伝子を取得することもできる。上記ノストク・パンクチフォルム(Nostoc punctiform )ATCC29133のゲノムDNAを鋳型として使用し、配列番号2に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌(E. coli)等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0043】
上記したプローブ又はプライマーの調製、ゲノムライブラリーの構築、ゲノムライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0044】
上記L-トリプトファン脱水素酵素の遺伝子は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製するベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、ベクターは発現ベクターである。発現ベクターにおいて上記遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモータは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0045】
細菌細胞で作動可能なプロモータとしては、バチルス・ステアロテルモフィルス・マルトジェニック・アミラーゼ遺伝子(Geobacillus stearothermophilus maltogenic amylase gene)、バチルス・リケニホルミスαアミラーゼ遺伝子(Bacillus licheniformis alpha-amylase gene)、バチルス・アミロリケファチエンス・BANアミラーゼ遺伝子(Bacillus amyloliquefaciens BAN amylase gene)、バチルス・サブチリス・アルカリプロテアーゼ遺伝子(Bacillus Subtilis alkaline protease gene)もしくはバチルス・プミルス・キシロシダーゼ遺伝子(Bacillus pumilus xylosldase gene)のプロモータ、またはファージ・ラムダのPR若しくはPLプロモータ、大腸菌(E. coli)のlac、trp若しくはtacプロモータなどが挙げられる。
【0046】
哺乳動物細胞で作動可能なプロモータの例としては、SV40プロモータ、MT−1(メタロチオネイン遺伝子)プロモータ、またはアデノウイルス2主後期プロモータなどがある。昆虫細胞で作動可能なプロモータの例としては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュウロウイルス即時型初期遺伝子1プロモータ、またはバキュウロウイルス39K遅延型初期遺伝子プロモータ等がある。酵母宿主細胞で作動可能なプロモータの例としては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータなどが挙げられる。糸状菌細胞で作動可能なプロモータの例としては、ADH3プロモータまたはtpiAプロモータなどがある。
【0047】
また、上記L-トリプトファン脱水素酵素の遺伝子は必要に応じて、適切なターミネータに機能的に結合されてもよい。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターは更に、ポリアデニレーションシグナル(例えばSV40またはアデノウイルス5E1b領域由来のもの)、転写エンハンサ配列(例えばSV40エンハンサ)などの要素を有していてもよい。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターは更に、該ベクターが宿主細胞内で複製することを可能にするDNA配列を具備してもよく、その一例としてはSV40複製起点(宿主細胞が哺乳類細胞のとき)が挙げられる。
【0048】
L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、ジヒドロ葉酸レダクターゼ(DHFR)またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、または例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン若しくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。L-トリプトファン脱水素酵素の遺伝子、プロモータ、および所望によりターミネータおよび/または分泌シグナル配列をそれぞれ連結し、これらを適切なベクターに挿入する方法は当業者に周知である。
【0049】
L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターを導入される宿主細胞は、L-トリプトファン脱水素酵素の遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌および高等真核細胞等が挙げられる。
【0050】
細菌細胞の例としては、バチルスまたはストレプトマイセス等のグラム陽性菌又は大腸菌(E. coli)等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、または公知の方法でコンピテント細胞を用いることにより行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も公知であり、例えば、エレクトロポーレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0051】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0052】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0053】
昆虫細胞を宿主として用いる場合には、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる(例えば、Baculovirus Expression Vectors, A Laboratory Manua1;及びカレント・プロトコールズ・イン・モレキュラー・バイオロジー、Bio/Technology, 6, 47(1988)等に記載)。
【0054】
バキュロウイルスとしては、例えば、ヨトウガ科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、Spodoptera frugiperdaの卵巣細胞であるSf9、Sf21〔バキュロウイルス・エクスプレッション・ベクターズ、ア・ラボラトリー・マニュアル、ダブリュー・エイチ・フリーマン・アンド・カンパニー(W. H. Freeman and Company)、ニューヨーク(New York)、(1992)〕、Trichoplusia niの卵巣細胞であるHiFive(インビトロジェン社製)等を用いることができる。
組換えウイルスを調製するための、昆虫細胞への組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法又はリポフェクション法等を挙げることができる。
【0055】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明で用いるL-トリプトファン脱水素酵素を単離精製するには、通常のタンパク質の単離、精製法を用いればよい。例えば、本発明で用いるL-トリプトファン脱水素酵素が、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のタンパク質の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、本発明のL-トリプトファン脱水素酵素を精製標品として得ることができる。
【0056】
[L-トリプトファンの定量方法]
本発明のL-トリプトファンの定量方法は、
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む。
【0057】
工程(A)
検体とL-トリプトファン脱水素酵素とNAD+とを混合する。
前記式Aに示すように、酵素反応においては、NAD+を電子受容体としL-トリプトファンのα位の脱アミノ化が進行する。この結果、インドールピルビン酸、アンモニウムイオン、NADHが生じる。従って本発明における定量法では、試料溶液に含まれるL-トリプトファンの他、L-トリプトファン脱水素酵素とNAD+が含まれていれば、上記反応は進行する。
【0058】
本発明の測定対象となる検体は、L-トリプトファンを含む可能性があり、L-トリプトファン脱水素酵素の活性及び生産されるNADHの検出を損なう成分(例えば、340 nm付近に吸収を有する化合物など)を含まない試料であれば、如何なるものでもよい。試料溶液中のL-トリプトファンにL-トリプトファン脱水素酵素を作用させて生じるNADHを340 nmの吸収及びジアホラーゼやNADHオキシダーゼを用いることにより濃度を測定することができる。さらに、NADHを340 nmの吸光度で行う場合、測定対象は、過度のNADH及びL-トリプトファン脱水素酵素を阻害する物質、340 nm付近の吸収をもつ化合物もしくはNADHと容易に反応する化合物が含まれていないことが望ましい。この他、上記の発色・蛍光による定量を行う場合は、用いる酵素や電子伝達体に影響を与える化合物が含まれていないことが望ましい。
【0059】
検体が上記のようなL-トリプトファン脱水素酵素の活性またはNADHの検出を損なう成分等を含む場合には、検体をこれらの成分を除去するように前処理することもできる。
【0060】
上記L-トリプトファン脱水素酵素は、基質特異性及び親和性により、定量用酵素としての利用が制限される。例えば、後述の実施例に示すノストク・パンクチフォルム(Nostoc punctiform)のL-トリプトファン脱水素酵素の場合、トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンに対しては、活性が検出されない。また、L-トリプトファンに対するKm値は、0.0759 mMであり、比較的L-トリプトファン量が少ない(0〜1 mM)生体試料を対象としも使用可能である。このような観点からL-トリプトファン脱水素酵素の混合量は、10〜100 mU/mLの範囲とすることが適当であり、NAD+の混合量は、1〜50 mMの範囲とすることが適当である。
【0061】
さらに、L-トリプトファン脱水素酵素およびNAD+に加えて、好ましくは、L-トリプトファン脱水素酵素の至適pHおよびNADHの安定性を考慮したpHを示す緩衝液を含むことができる。さらに、L-トリプトファン脱水素酵素の安定化剤をさらに含むこともできる。L-トリプトファン脱水素酵素の安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0062】
L-トリプトファン脱水素酵素は、特に、ノストク・パンクチフォルム(Nostoc punctiform)のL-トリプトファン脱水素酵素の場合、安定性に乏しく、従って、上記工程(A)に用いる酵素は、工程(A)に使用される前には、上記安定化剤の存在下で保存されていたものであることが、再現性良く定量できるという観点で好ましい。
【0063】
反応液のpHは、L-トリプトファン脱水素酵素及びNADHの定量に酵素を用いる場合には、使用する酵素及びその他の試薬の安定性、可溶性が許す範囲においていずれのpHでも可能であるが、好ましくはその酵素の至適pH付近で行うことが望ましい。特に上述のノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を用いる場合は、塩基性(pH8.5〜11.5)が望ましい。また、本発明により定量可能なL-トリプトファン濃度は、使用するL-トリプトファン脱水素酵素とNADHの検出法に依存する。例えば後述の実施例に従いノストク・パンクチフォルム(Nostoc punctiform)菌由来のL-トリプトファン脱水素酵素を用い、340 nmの吸収によりNADHを検出し、end-point法で定量する場合、実施例に示す通り本酵素には、基質阻害が確認されるため、試料中のL-トリプトファン濃度としては、例えば、0〜1mMが適当であり、より高濃度の試料については、希釈することにより定量が可能となる。
【0064】
工程(B)
工程(B)では、前記混合により得られた反応液を所定時間放置する。
具体的には、被検体を含む試料溶液に対し、L-トリプトファン脱水素酵素を作用させる反応及び生じたNADHを定量する反応は、一般的に酵素反応が可能な4〜80℃の範囲で、L-トリプトファン脱水素酵素及びNADHの検出反応の至適温度を考慮して適宜決定される。上述のノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を用いる場合、好ましくは、常温付近である15℃〜30℃の範囲の温度で実施することが望ましい。
【0065】
工程(C)
工程(C)では、放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する。
本発明においては、定量には、end-point法を用いることができるが、この他に反応の初速度を用いる初速度法を用いることもできる。
【0066】
本酵素によって触媒される反応でNAD+より変換されて生じるNADHは、340 nmに極大吸収を持つことから、吸光度の測定により容易に検出する事が可能である。この生産量は、化学量論的に反応によって消費されたL-トリプトファンと等量であることから、L-トリプトファンの定量に適している。
【0067】
また、NADHの検出法としては、340 nmの吸光度を測定するほか、発色性の電子受容体を用いることもできる。発色性電子受容体としては、2-(4-indophenyl)-3-(4-nitrophenyl)-5-phenyl-2H-tetrazolium chloride(INT)、レサズリン、フェロシアン化合物等を例として挙げることができる。これらの電子伝達体は、還元されることにより蛍光もしくは発色するため、容易に測定する事が可能である。またこの際、NADHからの電子伝達を触媒するジアホラーゼを共存させることもできる。
【0068】
また、NADHの検出法として、NADHを酸化する別の酵素を検出用試薬としてNADHオキシダーゼを用いることができる。この場合、更に発色・蛍光試薬としてペルオキシダーゼを用いる必要がある。ペルオキシダーゼの例としては、例えば西洋わさびペルオキシダーゼを用いることができる。更に発色試薬としてペルオキシダーゼの基質を加えなければならない。ペルオキシダーゼの基質としては、2-アミノアンチピリン・フェノールやABTSを挙げることができる。
【0069】
さらに、NADHの検出に電極を使用する事もできる。検出には、前述の電子伝達体を単独で使用するか、ジアホラーゼやNADHオキシダーゼと組み合わせることもできる。また、L-トリプトファン脱水素酵素及びジアホラーゼ,NADHオキシダーゼは、遊離の状態で使用する事もできるが、公知の方法により直接あるいは、間接的に電極へ固定化する事も出来る。
【0070】
[L-トリプトファンの定量用キット]
本発明のL-トリプトファンの定量用キットは、以下の試薬を含む。
(K1)L-トリプトファン脱水素酵素および
(K2)NAD+
【0071】
前記(K1)のL-トリプトファン脱水素酵素は、上記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
【0072】
(K1)のL-トリプトファン脱水素酵素は、天然の酵素であっても、遺伝子組換技術により生産された酵素であってもよい。天然の酵素は、例えば、ラン藻など微生物の培養物よりL-トリプトファン脱水素酵素を採取し、必要により精製することで調製できる。遺伝子組換技術により生産された酵素とは、当該活性を有することが確認された酵素の遺伝子、若しくはそのホモログ、またはその改変体に由来する発現タンパク質がL-トリプトファン脱水素酵素活性を有していればよい。これらの遺伝子を導入した形質転換体、例えば、組み換え大腸菌を常法により培養、タンパク質の発現を誘導し、得られた菌体からL-トリプトファン脱水素酵素を採取し、必要により精製することで調製できる。
【0073】
本発明のキットにおいては、前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物であることが好ましい。前記酵素の安定化剤との混合物は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲である。安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である。
【0074】
(K2)のNAD+は、L-トリプトファン脱水素酵素による反応に必要な電子伝達体である。L-トリプトファン脱水素酵素は、NAD+に依存する酵素である。このため、L-トリプトファンの2位の脱アミノ化は、NAD+のNADHへの変換と共役して起こる。従って、変換されて生じるNADHは、消費されたL-トリプトファン量と化学量論的に等量である。
【0075】
本発明のキットは、(K3)反応用緩衝液および/または(K4)NADH検出試薬をさらに含むことができる。
【0076】
(K3)反応用緩衝液は、反応液中を定量反応に適したpHに維持するために用いられる。後述の実施例に示すノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を含め、ほとんどのNAD+依存型脱水素酵素は、塩基性水溶液で高い活性を示す。このため使用する緩衝液は、中性より塩基性側のpHであることが望まれ、より好ましくは、pH 9よりも塩基性の緩衝液が望ましい。
【0077】
(K4)NADH検出試薬は、NADHの検出を発色もしくは蛍光によって行う場合に必要となる。発色で検出を行う場合、例えばINTなどNADHから直接電子をうけると事が出来る発色試薬を用いることができる。また、ジアホラーゼやNADHオキシダーゼを用いることにより間接的にNADHから発色性物質へ電子を伝達し発色反応を行うこともできる。ジアホラーゼと併用する発色もしくは蛍光性基質としては、レサズリンを挙げることができる。NADHオキシダーゼをNADH検出試薬として用いる場合、NADHオキシダーゼにより生じる過酸化水素を検出するため、ペルオキシダーゼ及びペルオキシダーゼに対する発色基質が必要となる。NADHオキシダーゼと併用する発色試薬の組み合わせとしては、西洋わさびペルオキシダーゼと2-アミノアンチピリン・フェノールの組み合わせを挙げることができる。
【0078】
[L-トリプトファンの定量用組成物]
本発明は、L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物を包含する。
この組成物で用いるL-トリプトファン脱水素酵素は、前記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
さらに、前記酵素の安定化剤は、前記と同様に、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。酵素と安定化剤との混合比は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲である。
【0079】
[酵素センサー]
本発明は、L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーを包含する。この酵素センサーで用いるL-トリプトファン脱水素酵素は、前記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
【0080】
本発明の酵素センサーは、酵素センサーを構成する検出用電極とL-トリプトファン脱水素酵素を固定化の有無にかかわらずL-トリプトファンの定量に用いるものである。本発明の酵素センサーは、L-トリプトファン脱水素酵素によりL-トリプトファン依存的に生成するNADHを直接または間接的に定量的に検出できるものである。例えば、L-トリプトファン脱水素酵素に加え、ジアホラーゼなどのNADHを検出用電極で感度良く検出するための酵素を、上記検出用電極と固定化の有無にかかわらず併用するものであることもできる。前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置されることが好ましい。前記安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0081】
加えて、本発明の酵素センサーは、上記酵素以外に、電子伝達物質やペルオキシダーゼなど検出に寄与する他のタンパク質を、上記検出用電極と固定化の有無に関わらず併用したものであることもできる。それ以外の構成は、公知の酵素センサーで採用されている構成をそのまま、または適宜改変して利用することができる。
【0082】
本発明の酵素センサーは、被検体を含有する試験溶液に少なくとも検出用電極部分を浸漬し、L-トリプトファン脱水素酵素によりL-トリプトファン依存的に生産されるNADHを直接あるいは間接的に電気化学的に検出する。
【0083】
本発明における酵素センサーは、公知の酵素電極の基本的な構成にL-トリプトファン脱水素酵素を組み合わせた物であり、直接もしくは間接的にL-トリプトファン脱水素酵素および前述のNADH検出用酵素を固定化した酵素電極を用いる電極型酵素センサーであることができる。さらに、この酵素電極の検出用電極付近に酵素,NADHと電極の間の電子の授受を容易にする電気化学メディエーターを存在させたものであることもできる。
【実施例】
【0084】
以下に実施例を挙げて本発明を具体的に説明するが本発明はこれに限定されるものではない。
【0085】
1.L-トリプトファン脱水素酵素の調製例
ノストク・パンクチフォルム(Nostoc punctiforme)ATCC29133よりゲノムDNAを調製し、データベース上にある同株のゲノムシークエンス(CP001037)を元に設計した下記のプライマー(i)および(ii)を用いてPCRによりL-トリプトファン脱水素酵素遺伝子(NpR1275)を増幅した。増幅産物をpT7Blue vector を用いてJM109を宿主としてサブクローニングを行った。さらに、サブクローニングされたpT7Blue-NpR1275より下記のプライマー(iii)および(ii)を用いSacI 及びXhoIの切断配列を導入したPCR産物を作製し、同制限酵素で処理したのち、同様の制限酵素で処理したpET21(+)とライゲーションし、BL21(DE3)に導入した。このプラスミドは、発現されるタンパク質のC末端に6残基のヒスチジンタグが導入されるため、Bio-Rad Ni-IMACを用い精製が可能である。
【0086】
上記発現株、[BL21(DE3)/pET21(+)NpR1275]をアンピシリンを含むLB培地(500 mL)に植菌後37℃で90分、25℃で60分、15℃で90分培養したのち、終濃度0.4 mMのIPTGを加えて15℃で20時間培養した。培養液を6000xg,5分遠心分離後、上清を取り除き、残渣に対し5倍量の20 mM リン酸カリウムバッファーを加えて懸濁後、超音波ホモジナイザーにて細胞を破砕した。この破砕液を20000xg,30分遠心分離し、上清を無細胞抽出液とした。無細胞抽出液は、直ちにBio-Rad Ni-IMACを用いて精製し、必要に応じて後述の安定化剤を加え、同様に安定化剤を含む20 mMリン酸カリウムバッファーに透析し、SDS-PAGE的に均一な酵素を得た。この酵素の塩基配列を決定し、配列番号2に示す。さらに酵素のアミノ酸配列を配列番号1に示す。
【0087】
NpR1275増幅用プライマー
(i)NpR1275_f1:ggaattccatatgctgctatttgaaactgttag (配列番号3)
(ii)NpR1275_r1:cttgctcgagagctgcgatcgctttagac (配列番号4)
(iii)NpR1275_f2:ctagtgagctcgatgagaggtccatatgctgctattg (配列番号5)
【0088】
2.L-トリプトファン脱水素酵素の性質
・L-トリプトファン脱水素酵素活性の測定法
L-トリプトファン脱水素酵素活性の測定は、分光光度計もしくは、マイクロプレートリーダーを用い340 nmの吸光度を測定して行った。
L-トリプトファン脱水素酵素活性の測定には、以下の終濃度となるように調製した反応液に対し任意の酵素溶液を加えて行った。
【0089】
100 mM グリシン-NaOH バッファー(pH 11.5)
10 mM NAD+
0.4 mM L-トリプトファン
【0090】
分光光度計を使用する場合は、1 mL、マイクロプレートリーダーを使用する場合は、0.2 mLの最終液量で行った。活性は、1分間あたりにNADHを1 mmol生産する酵素量を1 Uと定義した。また、タンパク質量は、Bio-Rad protein assayを用い、BSAで作製した検量線を元に測定した。
【0091】
・L-トリプトファン脱水素酵素の安定化
本酵素は、調製後急速に活性が低下することから、安定化剤としてグリセロールの添加を検討した。Ni-IMACからの溶出直後の酵素液に対し、0, 10, 20, 30, 40, 50 % (W/V)となるようにグリセロールを加え、氷中に保持して活性の経時変化を測定した(図1)。図1に示す通り、グリセロール非添加では、24時間以内に活性が検出限界以下にまで低下したが、20%以上のグリセロール添加により3日以上の間活性に有意な減少は見られなく立った。このため、これ以降の実験では、Ni-IMACからの溶出直後に終濃度20%となるようにグリセロールを加え、同様に20%グリセロールを含む20 mMリン酸緩衝液(pH 7.0)に透析したものを用いることとした。
【0092】
また、スクロース、ソルビトール、トレハロースについても別途検討したところ、表2に示す通り48時間、氷中で保持した場合、それぞれ20%以上の添加により、より優れた安定化効果が認められた。
【0093】
【表2】
*調製直後に測定した値に対する48時間経過後の測定値の割合
【0094】
・基質特異性
上記の活性測定法に基づきL-トリプトファンを含む18種のタンパク質構成アミノ酸及びD-トリプトファン(いずれも0.4 mM)に対する本酵素の活性を調べた結果を図2に示す。本酵素は、L-トリプトファン以外を基質とする活性は、確認できずL-トリプトファン特異的に働くことを示した。
【0095】
・至適pH
上記の活性測定法に基づきpH7から14までの範囲で活性を測定した結果を図3に示す。この結果、pH11.5の緩衝液中で最も高い活性を示した。
【0096】
・温度安定性
酵素溶液(20%グリセロールを含む)を5から45℃の各温度で15分間静置し、直後に氷中で冷却したのち、上記の方法により活性を測定した。図4に示す通り、20%グリセロール存在下で本酵素は、0〜35℃までは、15分間の処理では、活性の大きな低下は見られず、40℃で半減する事が明らかとなった。
【0097】
・反応速度論的解析
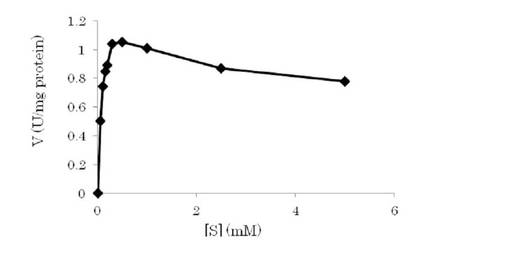
図5に示すミカエリス・メンテンプロットの結果、本酵素は、L-トリプトファンにより、0.5 mM以上の基質濃度で基質阻害を受けることが明らかとなった。基質阻害の影響が少ない終濃度0.3 mMまでの基質濃度で算出したKmは、0.0759 mMであった。
【0098】
L-トリプトファン定量法
L-トリプトファン定量法として、ノストク・パンクチフォルム(Nostoc punctiform)由来L-トリプトファン脱水素酵素を用い340 nm の吸光度によりNADHを検出する方法を検討した。
【0099】
(1)L-トリプトファン定量用試薬の調製例
L-トリプトファン定量反応液(10回分)
【0100】
(2)L-トリプトファンの検量線
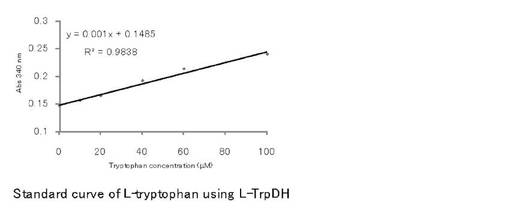
前述のL-トリプトファン定量法に則り0, 0.05, 0.1, 0.2, 0.3, 0.5 mMのL-トリプトファン水溶液を標準試料として用い、エンドポイント法による測定を行った結果を図6に示す。定量反応は、30℃で行い、反応開始30分の値を用いて検量線の作製を行った。図6で明らかなように本定量法では、0〜0.1 mMの範囲で、直線的な吸光度の増加がみられた。得られた直線の関係は、y = 0.001x +0.1485(R2 =0.9838)[ここでのyは340 nmの吸光度、xは、L-トリプトファン濃度、Rは、相関係数を示す]であって正確なL-トリプトファン定量が可能であることが分かった。
【産業上の利用可能性】
【0101】
L-トリプトファンは、必須アミノ酸であり、極端に不足した場合、ナイアシン欠乏症状を示す事が風土病ペラグラとして知られている。また、過剰に摂取した場合、好酸球増加や筋肉痛症候群の原因になるか可能性が指摘されている。このため、L-トリプトファンの定量は、食品分析、医薬品・サプリメントの品質管理、過剰症・欠乏症時の血液検査及び酵素センサーとして利用が考えられる。また、代謝経路上、ナイアシンやNAD+など生体内で重要な機能を持つ物質の原材料となることから、アミノグラムによる疾病診断「アミノインデックス」においても様々な疾病のバイオマーカーを構成する数値の一つとして活用できる。これらの理由から、食品や生体試料などについて、L-トリプトファン量を測定することは、産業的にも医学的にも重要な技術となると考えられるが、現状では、高速液体クロマトフラフィーをはじめとする非常に高価な機器及び試薬が必要となる方法しか実用化されていない。したがって、本発明は、L-トリプトファン定量用キットや酵素センサーなどの形態で商品化が可能であり、安価かつ簡便なL-トリプトファン定量法として事業化できる。
【技術分野】
【0001】
本発明は、試料溶液中のL-トリプトファンを迅速かつ簡易的に定量する方法およびそれに用いるキットに関するものである。
【背景技術】
【0002】
L-トリプトファンは、タンパク質構成アミノ酸の一つであり、さまざまな二次代謝産物の生合成原料にもなる。
【0003】
L-トリプトファンの定量は、これまで主に自働アミノ酸分析装置やプレカラム誘導体化逆相高速液体クロマトグラフィーによる分離と誘導体化を伴う機器分析的手法で行われている。また、L-トリプトファンと直接反応する発色試薬を用いる方法もあるが、類似化合物と反応するため生体試料への適用が困難である。酵素を用いたL-トリプトファンの定量方法も知られている。
【0004】
酵素を用いた方法として、ヒトヨタケ属担子菌(Copurinus sp.)由来のL-トリプトファン用アミノ酸オキシダーゼを用いる方法が報告されている(特許文献1および非特許文献1)。L-トリプトシルtRNA合成酵素を用いた方法も知られている(非特許文献2)。また、L-トリプロファン2-モノオキシゲナーゼを用いた酵素センサーも知られている(非特許文献3)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2001-69974号公報
【非特許文献】
【0006】
【非特許文献1】Biosci. Biotechnol. Biochem. 64, 1468-1493 (2000)
【非特許文献2】Anal. Biochem. 154, 618-623 (1986)
【非特許文献3】Biosen.and Bioelect. 12, 363-371 (1997)
【非特許文献4】J. Am. Chem. Soc., 130, 15260-15261 (2008)
【発明の概要】
【発明が解決しようとする課題】
【0007】
上記のように、L-トリプトファンは、主にHPLCを中心とした機器分析的手法により定量が行われてきた。しかし、トリプトファンを含むアミノ酸の定量に用いられる機器は、一般に高価のものが多く特に生体試料に適用するには、高度な分離能が必要となり、高価である。
【0008】
L-トリプトファンと直接作用する発色剤を用いた方法もあるが、生体試料などを測定対象とした場合、特異性に問題が生じる。
【0009】
特許文献1および非特許文献1に記載の方法で用いられるヒトヨタケ由来のアミノ酸オキシダーゼは、L-フェニルアラニンに7%の相対活性を有している。また、オキシダーゼ活性の他にジオキシゲナーゼ活性も持つため、定量的に反応が進行しない。このように、ヒトヨタケ由来のアミノ酸オキシダーゼは、L-フェニルアラニンに対しても若干の活性があり、オキシゲナーゼ活性も有するため定量には適さない。
【0010】
非特許文献2に記載の方法は、放射性同位体元素を使用しているため、汎用的に使用する事は出来ない。
【0011】
非特許文献3に記載の酵素センサーに用いられるL-トリプトファン2−モノオキシゲナーゼは、L-トリプトファン以外にL-フェニルアラニンにも反応してしまう。そのため、L-トリプトファン2-モノオキシゲナーゼを用いた酵素電極では、生体試料の測定においては、十分な精度が得られない。
【0012】
そこで、本発明は、酵素的定量方法であって、試料溶液中のL-トリプトファンを簡便且つ高い精度で実施できる方法を提供することを目的とする。
【0013】
さらに本発明は、上記酵素的定量方法を実施する際に利用できる測定用キット及び酵素センサーを提供することも目的とする。
【課題を解決するための手段】
【0014】
発明者らが種々検討した結果、ある特定のL-トリプトファン脱水素酵素をL-トリプトファンを含有する試料溶液に添加することで、L-トリプトファン脱水素酵素が試料溶液中のL-トリプトファンに特異的に作用して、共存するNAD+がNADHへと変換され、生成するNADHの量を測定することで、試料溶液中のL-トリプトファンを定量することが可能であることが明らかとなった。
【0015】
本発明は、以下に示す通りである。
[1]
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの定量方法であって、
前記L-トリプトファン脱水素酵素が、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する前記方法。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[2]
工程(A)において検体との混合に用いられる前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤の存在下で保存されているものである、[1]に記載の方法。
[3]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[2]に記載の方法。
[4]
前記ラン藻がノストク・パンクチフォルム(Nostoc punctiform )である[1]〜[3]のいずれかに記載の方法。
[5]
前記工程(C)で計測する反応生成物がNADHである[1]〜[4]のいずれかに記載の方法。
[6]
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファン脱水素酵素、および
(K2)NAD+
但し、前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[7]
前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物である、[6]に記載のキット。
[8]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[7]に記載のキット。
[9]
(K3)反応用緩衝液)および/または(K4)NADH検出試薬をさらに含む、[6]〜[8]のいずれかに記載のキット。
[10]
L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物であって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する組成物。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[11]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[10]に記載の組成物。
[12]
L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーであって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する酵素センサー。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
[13]
前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置される、[12]に記載の酵素センサー。
[14]
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、[13]に記載の酵素センサー。
【発明の効果】
【0016】
本発明によれば、特定のL-トリプトファン脱水素酵素を用いることで、この酵素が特異的にL-トリプトファンに作用し、さまざまな化合物を含有する試料中であっても高い選択性でL-トリプトファンを簡便に定量することができる。
【0017】
また、本発明は、さまざまな試料に対し有効であり、NADHの検出法により、紫外部吸収法、発色法、蛍光法、さらには、電極型酵素センサーを提供することもできる。
【図面の簡単な説明】
【0018】
【図1】L-トリプトファン脱水素酵素の安定化試験結果を示す。
【図2】L-トリプトファン脱水素酵素の基質特異性の試験結果を示す。
【図3】L-トリプトファン脱水素酵素活性に対するpHの影響を示す。
【図4】L-トリプトファン脱水素酵素の温度安定性試験結果を示す。
【図5】L-トリプトファン脱水素酵素のL-トリプトファンに対するミカエリス・メンテンプロットを示す。
【図6】L-トリプトファン脱水素酵素を用いたL-トリプトファン定量の検量線を示す。
【発明を実施するための形態】
【0019】
[L-トリプトファンの定量方法]
本発明のL-トリプトファンの定量方法は、
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含み、前記工程を経ることで、前記検体中のL-トリプトファンを定量することができる。
【0020】
さらに上記方法に用いるL-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【0021】
<L-トリプトファン定量用酵素>
本発明に用いるL-トリプトファン脱水素酵素は、下記式Aに示す反応を触媒して、L-トリプトファンとインドールピルビン酸とアンモニウムイオンを生産する際にNAD+をNADHに変換する酵素である。
【0022】
【化1】
【0023】
本発明で用いるL-トリプトファン脱水素酵素の第1の態様は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有し、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【0024】
配列表の配列番号1に記載のアミノ酸配列は、実施例で示すように、ノストク・パンクチフォルム(Nostoc punctiform)ATCC29133のゲノムDNAから調製したL-トリプトファン脱水素酵素遺伝子(NpR1275)を発現させたL-トリプトファン脱水素酵素のアミノ酸配列である。
【0025】
この配列番号1に記載のアミノ酸配列を有するL-トリプトファン脱水素酵素は、図2に結果を示すように、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用しない。さらに、この前記L-トリプトファン脱水素酵素は、NAD+をNADHに変換する脱水素酵素活性を有するものである。NAD+をNADHに変換する脱水素酵素活性を有することは、図6の検量線の結果から明らかである。前記L-トリプトファン脱水素酵素が、NAD+をNADHに変換する脱水素酵素活性を有することは、実施例に示すように、NADHの生成を340nmの吸光度の変化を測定することで確認することができる。尚、タンパク質構成アミノ酸とは、L-アラニン、L-プロリン、L-バリン、L-ロイシン、L-イソロイシン、L-リジン、L-ヒスチジン、L-アルギニン、L-グルタミン、L-セリン、L-アスパラギン酸、L-グルタミン酸、L-スレオニン、L-フェニルアラニン、L-メチオニン、L-グリシン、L-チロシン、L-アスパラギンを意味する。L-システインについては、本酵素が活性を示す中性から塩基性のpHでは、速やかに不溶性のシスチンへ変換されることから、本発明においては、上記タンパク質構成アミノ酸には含まれない。
【0026】
配列番号1に記載のアミノ酸配列を有するL-トリプトファン脱水素酵素についての、上記基質特異性以外の特性は、実施例において具体的に記載する。
【0027】
さらに、このL-トリプトファン脱水素酵素は、NAD+の存在下、L-トリプトファンに特異的に作用してインドールピルビン酸とアンモニアを生成する。この点は、非特許文献4において述べられている本酵素のscytonemin合成経路上の位置づけと実施例で示すL-トリプトファン依存的なNADHの生産から明らかである。 L-トリプトファン脱水素酵素が、NAD+の存在下、L-トリプトファンに特異的に作用することは、(i)L-トリプトファン依存的なNADHの生産、(ii)L-トリプトファン以外のアミノ酸を基質とした場合NADHの生産が無いことで確認する事が出来る。
【0028】
本明細書で言う「1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列」における「1から数個」の範囲は、欠失等を有するタンパク質が、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである限り、特に限定されない。前記「1から数個」の範囲は、前記脱水素酵素活性を有するタンパク質である割合が高いことから、例えば、1から30個、好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1から7個、一層好ましくは1から5個、特に好ましくは1から3個程度であることができる。
【0029】
本明細書で言う「配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列」における相同性は、前記アミノ酸配列の相同性を有するタンパク質が、L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである限り、特に限定されない。前記アミノ酸配列の相同性は、90%以上であれば特に限定されないが、好ましくは95%以上、さらに好ましくは96%以上、さらに好ましくは97%以上、さらに好ましくは98%、特に好ましくは99%以上である。
【0030】
本発明で用いるL-トリプトファン脱水素酵素の第2の態様は、(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。
【0031】
実施例で示すように、ノストク・パンクチフォルム(Nostoc punctiform) ATCC29133のゲノムDNAから調製したL-トリプトファン脱水素酵素遺伝子(NpR1275)を発現させたL-トリプトファン脱水素酵素は、(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のアミノ酸には作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有するものである。さらに前述のように、このL-トリプトファン脱水素酵素は、配列表の配列番号1に記載のアミノ酸配列を有する。
【0032】
L-トリプトファン脱水素酵素[EC 1.4.1.19]は、一部の植物におけるインドール酢酸生合成経路上に存在する事が古くから指摘されている。しかし、単一のタンパク質として精製されたことはなく、遺伝子も不明である。一方、ラン藻において、紫外線防御物質として生産されるscytoneminの生合成経路の初発反応を行う酵素がL-トリプトファンに作用するNAD+依存型脱水素酵素であることがBalskusらによって報告されている(非特許文献4)。しかし、これまでの報告では、scytonemin生合成遺伝子クラスター上にコードされているNpR1275遺伝子配列が、既知のアミノ酸脱水素酵素と相同性があることと、大腸菌で発現させたNpR1275遺伝子産物がL-トリプトファンに対し活性がありL-チロシンに対する活性がほとんどないことが示されたのみである。これまでに、本酵素の基質特異性等の酵素の性質に関しては明らかにされておらず、ましてや、本酵素を用いたL-トリプトファン定量について検討されたことはない。
【0033】
本発明で用いるL-トリプトファン脱水素酵素の第2の態様は、特定の種より生産されるものに限られるものではなく、ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素またはラン藻由来生物中でscytonemin合成経路を担っていると推定されるホモログの遺伝子としてコードされている酵素から選択され、かつL-トリプトファンを基質として作用するNAD+依存型の脱水素酵素を意味する。L-トリプトファン脱水素酵素としては、例えば、ラン藻ノストク・パンクチフォルム(Nostoc punctiform)のscytonemin生合成遺伝子クラスター上にコードされているNpR1275遺伝子産物及びそのホモログが挙げられる。表1にBLAST上で検索されたNpR1275遺伝子のホモログと推定される遺伝子の例を挙げる。これらホモログは、NpR1275遺伝子同様にscytonemin生合成遺伝子クラスター上に存在しアミノ酸脱水素酵素をコードすると推定される遺伝子であることから、その遺伝子産物は、NpR1275遺伝子産物と同様にL-トリプトファン脱水素酵素としての機能を持つものと推定される。
【0034】
【表1】
【0035】
さらに、本発明に用いるL-トリプトファン脱水素酵素は、同様の活性を有するものであれば、自然界より分離された生物に由来するもの、本酵素をコードする遺伝子を大腸菌や他の生物を宿主として発現させて得られる酵素、更にこれらの遺伝子を適宜改変して作製した酵素も含まれる。
【0036】
また、異種発現による生産法としては、例えば、上記ノストク・パンクチフォルム(Nostoc punctiform)より抽出したゲノムDNAから該当する遺伝子(配列表の配列番号2に示す)をPCRにて増幅しpETもしくはpUCなどに組み込んだプラスミドベクターを構築したのち、BL21、JM109などの宿主菌株に形質転換し、培養する方法が挙げられる。これら以外の公知の方法も適宜用いることができる。
【0037】
本発明で用いるL-トリプトファン脱水素酵素の取得方法は特に制限されず、化学合成により合成したタンパク質でもよいし、遺伝子組換え技術により作製した組換えタンパク質でもよい。組換えタンパク質を作製する場合には、後述するように当該タンパク質をコードする遺伝子(DNA)を取得する。このDNAを適当な発現系に導入することにより、上記L-トリプトファン脱水素酵素を産生することができる。
【0038】
上記L-トリプトファン脱水素酵素は、上記L-トリプトファン脱水素酵素をコードする遺伝子をベクター上に搭載し、このベクターによって宿主細胞を形質転換した後、形質転換させた宿主細胞を培養して培養物中に前記遺伝子がコードするタンパク質を蓄積し、蓄積したタンパク質を収集することを含む、生産方法により調製することができる。
【0039】
上記L-トリプトファン脱水素酵素をコードする遺伝子の取得方法は特に限定されない。本発明のL-トリプトファン脱水素酵素をコードする遺伝子は、例えば、配列番号1に記載のアミノ酸配列および配列番号2に記載した塩基配列の情報に基づいて、化学合成、遺伝子工学的手法又は突然変異誘発などの当業者に既知の任意の方法で作製することができる。
【0040】
例えば、配列表の配列番号2に記載の塩基配列を有するDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的手法等を用いて行うことができる。遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989(以下、モレキュラークローニング第2版と略す)、Current Protocols in Molecular Biology, Supplement 1〜38, John Wiley & Sons (1987-1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)等に記載の方法に準じて行うことができる。
【0041】
本明細書中の配列表の配列番号1に記載したアミノ酸配列または配列番号2に示す塩基配列の情報に基づいて適当なブローブやプライマーを調製し、それらを用いてノストク・パンクチフォルム(Nostoc punctiform )ATCC29133のゲノムライブラリーより本発明の遺伝子を単離することができる。ゲノムライブラリーは、ノストク・パンクチフォルム(Nostoc punctiform )ATCC29133から常法により作製することができる。
【0042】
PCR法により本発明のL-トリプトファン脱水素酵素をコードする遺伝子を取得することもできる。上記ノストク・パンクチフォルム(Nostoc punctiform )ATCC29133のゲノムDNAを鋳型として使用し、配列番号2に記載した塩基配列を増幅できるように設計した1対のプライマーを用いてPCRを行う。PCRの反応条件は適宜設定することができ、例えば、94℃で30秒間(変性)、55℃で30秒〜1分間(アニーリング)、72℃で2分間(伸長)からなる反応工程を1サイクルとして、例えば30サイクル行った後、72℃で7分間反応させる条件などを挙げることができる。次いで、増幅されたDNA断片を、大腸菌(E. coli)等の宿主で増幅可能な適切なベクター中にクローニングすることができる。
【0043】
上記したプローブ又はプライマーの調製、ゲノムライブラリーの構築、ゲノムライブラリーのスクリーニング、並びに目的遺伝子のクローニングなどの操作は当業者に既知であり、例えば、モレキュラークローニング第2版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載の方法に準じて行うことができる。
【0044】
上記L-トリプトファン脱水素酵素の遺伝子は適当なベクター中に挿入して使用することができる。本発明で用いるベクターの種類は特に限定されず、例えば、自立的に複製するベクター(例えばプラスミド等)でもよいし、あるいは、宿主細胞に導入された際に宿主細胞のゲノムに組み込まれ、組み込まれた染色体と共に複製されるものであってもよい。好ましくは、ベクターは発現ベクターである。発現ベクターにおいて上記遺伝子は、転写に必要な要素(例えば、プロモーター等)が機能的に連結されている。プロモータは宿主細胞において転写活性を示すDNA配列であり、宿主の種類に応じて適宜選択することができる。
【0045】
細菌細胞で作動可能なプロモータとしては、バチルス・ステアロテルモフィルス・マルトジェニック・アミラーゼ遺伝子(Geobacillus stearothermophilus maltogenic amylase gene)、バチルス・リケニホルミスαアミラーゼ遺伝子(Bacillus licheniformis alpha-amylase gene)、バチルス・アミロリケファチエンス・BANアミラーゼ遺伝子(Bacillus amyloliquefaciens BAN amylase gene)、バチルス・サブチリス・アルカリプロテアーゼ遺伝子(Bacillus Subtilis alkaline protease gene)もしくはバチルス・プミルス・キシロシダーゼ遺伝子(Bacillus pumilus xylosldase gene)のプロモータ、またはファージ・ラムダのPR若しくはPLプロモータ、大腸菌(E. coli)のlac、trp若しくはtacプロモータなどが挙げられる。
【0046】
哺乳動物細胞で作動可能なプロモータの例としては、SV40プロモータ、MT−1(メタロチオネイン遺伝子)プロモータ、またはアデノウイルス2主後期プロモータなどがある。昆虫細胞で作動可能なプロモータの例としては、ポリヘドリンプロモータ、P10プロモータ、オートグラファ・カリホルニカ・ポリヘドロシス塩基性タンパクプロモータ、バキュウロウイルス即時型初期遺伝子1プロモータ、またはバキュウロウイルス39K遅延型初期遺伝子プロモータ等がある。酵母宿主細胞で作動可能なプロモータの例としては、酵母解糖系遺伝子由来のプロモータ、アルコールデヒドロゲナーゼ遺伝子プロモータ、TPI1プロモータ、ADH2-4cプロモータなどが挙げられる。糸状菌細胞で作動可能なプロモータの例としては、ADH3プロモータまたはtpiAプロモータなどがある。
【0047】
また、上記L-トリプトファン脱水素酵素の遺伝子は必要に応じて、適切なターミネータに機能的に結合されてもよい。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターは更に、ポリアデニレーションシグナル(例えばSV40またはアデノウイルス5E1b領域由来のもの)、転写エンハンサ配列(例えばSV40エンハンサ)などの要素を有していてもよい。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターは更に、該ベクターが宿主細胞内で複製することを可能にするDNA配列を具備してもよく、その一例としてはSV40複製起点(宿主細胞が哺乳類細胞のとき)が挙げられる。
【0048】
L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターはさらに選択マーカーを含有してもよい。選択マーカーとしては、例えば、ジヒドロ葉酸レダクターゼ(DHFR)またはシゾサッカロマイセス・ポンベTPI遺伝子等のようなその補体が宿主細胞に欠けている遺伝子、または例えばアンピシリン、カナマイシン、テトラサイクリン、クロラムフェニコール、ネオマイシン若しくはヒグロマイシンのような薬剤耐性遺伝子を挙げることができる。L-トリプトファン脱水素酵素の遺伝子、プロモータ、および所望によりターミネータおよび/または分泌シグナル配列をそれぞれ連結し、これらを適切なベクターに挿入する方法は当業者に周知である。
【0049】
L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターを適当な宿主に導入することによって形質転換体を作製することができる。L-トリプトファン脱水素酵素の遺伝子を含む組換えベクターを導入される宿主細胞は、L-トリプトファン脱水素酵素の遺伝子を発現できれば任意の細胞でよく、細菌、酵母、真菌および高等真核細胞等が挙げられる。
【0050】
細菌細胞の例としては、バチルスまたはストレプトマイセス等のグラム陽性菌又は大腸菌(E. coli)等のグラム陰性菌が挙げられる。これら細菌の形質転換は、プロトプラスト法、または公知の方法でコンピテント細胞を用いることにより行えばよい。哺乳類細胞の例としては、HEK293細胞、HeLa細胞、COS細胞、BHK細胞、CHL細胞またはCHO細胞等が挙げられる。哺乳類細胞を形質転換し、該細胞に導入されたDNA配列を発現させる方法も公知であり、例えば、エレクトロポーレーション法、リン酸カルシウム法、リポフェクション法等を用いることができる。
【0051】
酵母細胞の例としては、サッカロマイセスまたはシゾサッカロマイセスに属する細胞が挙げられ、例えば、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)またはサッカロマイセス・クルイベリ(Saccharomyces kluyveri)等が挙げられる。酵母宿主への組換えベクターの導入方法としては、例えば、エレクトロポレーション法、スフェロブラスト法、酢酸リチウム法等を挙げることができる。
【0052】
他の真菌細胞の例は、糸状菌、例えばアスペルギルス、ニューロスポラ、フザリウム、またはトリコデルマに属する細胞である。宿主細胞として糸状菌を用いる場合、DNA構築物を宿主染色体に組み込んで組換え宿主細胞を得ることにより形質転換を行うことができる。DNA構築物の宿主染色体への組み込みは、公知の方法に従い、例えば相同組換えまたは異種組換えにより行うことができる。
【0053】
昆虫細胞を宿主として用いる場合には、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を発現させることができる(例えば、Baculovirus Expression Vectors, A Laboratory Manua1;及びカレント・プロトコールズ・イン・モレキュラー・バイオロジー、Bio/Technology, 6, 47(1988)等に記載)。
【0054】
バキュロウイルスとしては、例えば、ヨトウガ科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、Spodoptera frugiperdaの卵巣細胞であるSf9、Sf21〔バキュロウイルス・エクスプレッション・ベクターズ、ア・ラボラトリー・マニュアル、ダブリュー・エイチ・フリーマン・アンド・カンパニー(W. H. Freeman and Company)、ニューヨーク(New York)、(1992)〕、Trichoplusia niの卵巣細胞であるHiFive(インビトロジェン社製)等を用いることができる。
組換えウイルスを調製するための、昆虫細胞への組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法又はリポフェクション法等を挙げることができる。
【0055】
上記の形質転換体は、導入された遺伝子の発現を可能にする条件下で適切な栄養培地中で培養する。形質転換体の培養物から、本発明で用いるL-トリプトファン脱水素酵素を単離精製するには、通常のタンパク質の単離、精製法を用いればよい。例えば、本発明で用いるL-トリプトファン脱水素酵素が、細胞内に溶解状態で発現した場合には、培養終了後、細胞を遠心分離により回収し水系緩衝液に懸濁後、超音波破砕機等により細胞を破砕し、無細胞抽出液を得る。該無細胞抽出液を遠心分離することにより得られた上清から、通常のタンパク質の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)セファロース等のレジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィ一法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、本発明のL-トリプトファン脱水素酵素を精製標品として得ることができる。
【0056】
[L-トリプトファンの定量方法]
本発明のL-トリプトファンの定量方法は、
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む。
【0057】
工程(A)
検体とL-トリプトファン脱水素酵素とNAD+とを混合する。
前記式Aに示すように、酵素反応においては、NAD+を電子受容体としL-トリプトファンのα位の脱アミノ化が進行する。この結果、インドールピルビン酸、アンモニウムイオン、NADHが生じる。従って本発明における定量法では、試料溶液に含まれるL-トリプトファンの他、L-トリプトファン脱水素酵素とNAD+が含まれていれば、上記反応は進行する。
【0058】
本発明の測定対象となる検体は、L-トリプトファンを含む可能性があり、L-トリプトファン脱水素酵素の活性及び生産されるNADHの検出を損なう成分(例えば、340 nm付近に吸収を有する化合物など)を含まない試料であれば、如何なるものでもよい。試料溶液中のL-トリプトファンにL-トリプトファン脱水素酵素を作用させて生じるNADHを340 nmの吸収及びジアホラーゼやNADHオキシダーゼを用いることにより濃度を測定することができる。さらに、NADHを340 nmの吸光度で行う場合、測定対象は、過度のNADH及びL-トリプトファン脱水素酵素を阻害する物質、340 nm付近の吸収をもつ化合物もしくはNADHと容易に反応する化合物が含まれていないことが望ましい。この他、上記の発色・蛍光による定量を行う場合は、用いる酵素や電子伝達体に影響を与える化合物が含まれていないことが望ましい。
【0059】
検体が上記のようなL-トリプトファン脱水素酵素の活性またはNADHの検出を損なう成分等を含む場合には、検体をこれらの成分を除去するように前処理することもできる。
【0060】
上記L-トリプトファン脱水素酵素は、基質特異性及び親和性により、定量用酵素としての利用が制限される。例えば、後述の実施例に示すノストク・パンクチフォルム(Nostoc punctiform)のL-トリプトファン脱水素酵素の場合、トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンに対しては、活性が検出されない。また、L-トリプトファンに対するKm値は、0.0759 mMであり、比較的L-トリプトファン量が少ない(0〜1 mM)生体試料を対象としも使用可能である。このような観点からL-トリプトファン脱水素酵素の混合量は、10〜100 mU/mLの範囲とすることが適当であり、NAD+の混合量は、1〜50 mMの範囲とすることが適当である。
【0061】
さらに、L-トリプトファン脱水素酵素およびNAD+に加えて、好ましくは、L-トリプトファン脱水素酵素の至適pHおよびNADHの安定性を考慮したpHを示す緩衝液を含むことができる。さらに、L-トリプトファン脱水素酵素の安定化剤をさらに含むこともできる。L-トリプトファン脱水素酵素の安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0062】
L-トリプトファン脱水素酵素は、特に、ノストク・パンクチフォルム(Nostoc punctiform)のL-トリプトファン脱水素酵素の場合、安定性に乏しく、従って、上記工程(A)に用いる酵素は、工程(A)に使用される前には、上記安定化剤の存在下で保存されていたものであることが、再現性良く定量できるという観点で好ましい。
【0063】
反応液のpHは、L-トリプトファン脱水素酵素及びNADHの定量に酵素を用いる場合には、使用する酵素及びその他の試薬の安定性、可溶性が許す範囲においていずれのpHでも可能であるが、好ましくはその酵素の至適pH付近で行うことが望ましい。特に上述のノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を用いる場合は、塩基性(pH8.5〜11.5)が望ましい。また、本発明により定量可能なL-トリプトファン濃度は、使用するL-トリプトファン脱水素酵素とNADHの検出法に依存する。例えば後述の実施例に従いノストク・パンクチフォルム(Nostoc punctiform)菌由来のL-トリプトファン脱水素酵素を用い、340 nmの吸収によりNADHを検出し、end-point法で定量する場合、実施例に示す通り本酵素には、基質阻害が確認されるため、試料中のL-トリプトファン濃度としては、例えば、0〜1mMが適当であり、より高濃度の試料については、希釈することにより定量が可能となる。
【0064】
工程(B)
工程(B)では、前記混合により得られた反応液を所定時間放置する。
具体的には、被検体を含む試料溶液に対し、L-トリプトファン脱水素酵素を作用させる反応及び生じたNADHを定量する反応は、一般的に酵素反応が可能な4〜80℃の範囲で、L-トリプトファン脱水素酵素及びNADHの検出反応の至適温度を考慮して適宜決定される。上述のノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を用いる場合、好ましくは、常温付近である15℃〜30℃の範囲の温度で実施することが望ましい。
【0065】
工程(C)
工程(C)では、放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する。
本発明においては、定量には、end-point法を用いることができるが、この他に反応の初速度を用いる初速度法を用いることもできる。
【0066】
本酵素によって触媒される反応でNAD+より変換されて生じるNADHは、340 nmに極大吸収を持つことから、吸光度の測定により容易に検出する事が可能である。この生産量は、化学量論的に反応によって消費されたL-トリプトファンと等量であることから、L-トリプトファンの定量に適している。
【0067】
また、NADHの検出法としては、340 nmの吸光度を測定するほか、発色性の電子受容体を用いることもできる。発色性電子受容体としては、2-(4-indophenyl)-3-(4-nitrophenyl)-5-phenyl-2H-tetrazolium chloride(INT)、レサズリン、フェロシアン化合物等を例として挙げることができる。これらの電子伝達体は、還元されることにより蛍光もしくは発色するため、容易に測定する事が可能である。またこの際、NADHからの電子伝達を触媒するジアホラーゼを共存させることもできる。
【0068】
また、NADHの検出法として、NADHを酸化する別の酵素を検出用試薬としてNADHオキシダーゼを用いることができる。この場合、更に発色・蛍光試薬としてペルオキシダーゼを用いる必要がある。ペルオキシダーゼの例としては、例えば西洋わさびペルオキシダーゼを用いることができる。更に発色試薬としてペルオキシダーゼの基質を加えなければならない。ペルオキシダーゼの基質としては、2-アミノアンチピリン・フェノールやABTSを挙げることができる。
【0069】
さらに、NADHの検出に電極を使用する事もできる。検出には、前述の電子伝達体を単独で使用するか、ジアホラーゼやNADHオキシダーゼと組み合わせることもできる。また、L-トリプトファン脱水素酵素及びジアホラーゼ,NADHオキシダーゼは、遊離の状態で使用する事もできるが、公知の方法により直接あるいは、間接的に電極へ固定化する事も出来る。
【0070】
[L-トリプトファンの定量用キット]
本発明のL-トリプトファンの定量用キットは、以下の試薬を含む。
(K1)L-トリプトファン脱水素酵素および
(K2)NAD+
【0071】
前記(K1)のL-トリプトファン脱水素酵素は、上記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
【0072】
(K1)のL-トリプトファン脱水素酵素は、天然の酵素であっても、遺伝子組換技術により生産された酵素であってもよい。天然の酵素は、例えば、ラン藻など微生物の培養物よりL-トリプトファン脱水素酵素を採取し、必要により精製することで調製できる。遺伝子組換技術により生産された酵素とは、当該活性を有することが確認された酵素の遺伝子、若しくはそのホモログ、またはその改変体に由来する発現タンパク質がL-トリプトファン脱水素酵素活性を有していればよい。これらの遺伝子を導入した形質転換体、例えば、組み換え大腸菌を常法により培養、タンパク質の発現を誘導し、得られた菌体からL-トリプトファン脱水素酵素を採取し、必要により精製することで調製できる。
【0073】
本発明のキットにおいては、前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物であることが好ましい。前記酵素の安定化剤との混合物は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲である。安定化剤は、例えば、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である。
【0074】
(K2)のNAD+は、L-トリプトファン脱水素酵素による反応に必要な電子伝達体である。L-トリプトファン脱水素酵素は、NAD+に依存する酵素である。このため、L-トリプトファンの2位の脱アミノ化は、NAD+のNADHへの変換と共役して起こる。従って、変換されて生じるNADHは、消費されたL-トリプトファン量と化学量論的に等量である。
【0075】
本発明のキットは、(K3)反応用緩衝液および/または(K4)NADH検出試薬をさらに含むことができる。
【0076】
(K3)反応用緩衝液は、反応液中を定量反応に適したpHに維持するために用いられる。後述の実施例に示すノストク・パンクチフォルム(Nostoc punctiform)由来のL-トリプトファン脱水素酵素を含め、ほとんどのNAD+依存型脱水素酵素は、塩基性水溶液で高い活性を示す。このため使用する緩衝液は、中性より塩基性側のpHであることが望まれ、より好ましくは、pH 9よりも塩基性の緩衝液が望ましい。
【0077】
(K4)NADH検出試薬は、NADHの検出を発色もしくは蛍光によって行う場合に必要となる。発色で検出を行う場合、例えばINTなどNADHから直接電子をうけると事が出来る発色試薬を用いることができる。また、ジアホラーゼやNADHオキシダーゼを用いることにより間接的にNADHから発色性物質へ電子を伝達し発色反応を行うこともできる。ジアホラーゼと併用する発色もしくは蛍光性基質としては、レサズリンを挙げることができる。NADHオキシダーゼをNADH検出試薬として用いる場合、NADHオキシダーゼにより生じる過酸化水素を検出するため、ペルオキシダーゼ及びペルオキシダーゼに対する発色基質が必要となる。NADHオキシダーゼと併用する発色試薬の組み合わせとしては、西洋わさびペルオキシダーゼと2-アミノアンチピリン・フェノールの組み合わせを挙げることができる。
【0078】
[L-トリプトファンの定量用組成物]
本発明は、L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物を包含する。
この組成物で用いるL-トリプトファン脱水素酵素は、前記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
さらに、前記酵素の安定化剤は、前記と同様に、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。酵素と安定化剤との混合比は、例えば、酵素100質量部に対して、10〜70質量部の安定化剤を含むことが、酵素を長期間安定に保存できるという観点から好ましい。安定化剤の量は、好ましくは、酵素100質量部に対して、20〜60質量部の範囲である。
【0079】
[酵素センサー]
本発明は、L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーを包含する。この酵素センサーで用いるL-トリプトファン脱水素酵素は、前記L-トリプトファンの定量方法で説明したL-トリプトファン脱水素酵素と同様である。
【0080】
本発明の酵素センサーは、酵素センサーを構成する検出用電極とL-トリプトファン脱水素酵素を固定化の有無にかかわらずL-トリプトファンの定量に用いるものである。本発明の酵素センサーは、L-トリプトファン脱水素酵素によりL-トリプトファン依存的に生成するNADHを直接または間接的に定量的に検出できるものである。例えば、L-トリプトファン脱水素酵素に加え、ジアホラーゼなどのNADHを検出用電極で感度良く検出するための酵素を、上記検出用電極と固定化の有無にかかわらず併用するものであることもできる。前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置されることが好ましい。前記安定化剤は、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種であることができる。
【0081】
加えて、本発明の酵素センサーは、上記酵素以外に、電子伝達物質やペルオキシダーゼなど検出に寄与する他のタンパク質を、上記検出用電極と固定化の有無に関わらず併用したものであることもできる。それ以外の構成は、公知の酵素センサーで採用されている構成をそのまま、または適宜改変して利用することができる。
【0082】
本発明の酵素センサーは、被検体を含有する試験溶液に少なくとも検出用電極部分を浸漬し、L-トリプトファン脱水素酵素によりL-トリプトファン依存的に生産されるNADHを直接あるいは間接的に電気化学的に検出する。
【0083】
本発明における酵素センサーは、公知の酵素電極の基本的な構成にL-トリプトファン脱水素酵素を組み合わせた物であり、直接もしくは間接的にL-トリプトファン脱水素酵素および前述のNADH検出用酵素を固定化した酵素電極を用いる電極型酵素センサーであることができる。さらに、この酵素電極の検出用電極付近に酵素,NADHと電極の間の電子の授受を容易にする電気化学メディエーターを存在させたものであることもできる。
【実施例】
【0084】
以下に実施例を挙げて本発明を具体的に説明するが本発明はこれに限定されるものではない。
【0085】
1.L-トリプトファン脱水素酵素の調製例
ノストク・パンクチフォルム(Nostoc punctiforme)ATCC29133よりゲノムDNAを調製し、データベース上にある同株のゲノムシークエンス(CP001037)を元に設計した下記のプライマー(i)および(ii)を用いてPCRによりL-トリプトファン脱水素酵素遺伝子(NpR1275)を増幅した。増幅産物をpT7Blue vector を用いてJM109を宿主としてサブクローニングを行った。さらに、サブクローニングされたpT7Blue-NpR1275より下記のプライマー(iii)および(ii)を用いSacI 及びXhoIの切断配列を導入したPCR産物を作製し、同制限酵素で処理したのち、同様の制限酵素で処理したpET21(+)とライゲーションし、BL21(DE3)に導入した。このプラスミドは、発現されるタンパク質のC末端に6残基のヒスチジンタグが導入されるため、Bio-Rad Ni-IMACを用い精製が可能である。
【0086】
上記発現株、[BL21(DE3)/pET21(+)NpR1275]をアンピシリンを含むLB培地(500 mL)に植菌後37℃で90分、25℃で60分、15℃で90分培養したのち、終濃度0.4 mMのIPTGを加えて15℃で20時間培養した。培養液を6000xg,5分遠心分離後、上清を取り除き、残渣に対し5倍量の20 mM リン酸カリウムバッファーを加えて懸濁後、超音波ホモジナイザーにて細胞を破砕した。この破砕液を20000xg,30分遠心分離し、上清を無細胞抽出液とした。無細胞抽出液は、直ちにBio-Rad Ni-IMACを用いて精製し、必要に応じて後述の安定化剤を加え、同様に安定化剤を含む20 mMリン酸カリウムバッファーに透析し、SDS-PAGE的に均一な酵素を得た。この酵素の塩基配列を決定し、配列番号2に示す。さらに酵素のアミノ酸配列を配列番号1に示す。
【0087】
NpR1275増幅用プライマー
(i)NpR1275_f1:ggaattccatatgctgctatttgaaactgttag (配列番号3)
(ii)NpR1275_r1:cttgctcgagagctgcgatcgctttagac (配列番号4)
(iii)NpR1275_f2:ctagtgagctcgatgagaggtccatatgctgctattg (配列番号5)
【0088】
2.L-トリプトファン脱水素酵素の性質
・L-トリプトファン脱水素酵素活性の測定法
L-トリプトファン脱水素酵素活性の測定は、分光光度計もしくは、マイクロプレートリーダーを用い340 nmの吸光度を測定して行った。
L-トリプトファン脱水素酵素活性の測定には、以下の終濃度となるように調製した反応液に対し任意の酵素溶液を加えて行った。
【0089】
100 mM グリシン-NaOH バッファー(pH 11.5)
10 mM NAD+
0.4 mM L-トリプトファン
【0090】
分光光度計を使用する場合は、1 mL、マイクロプレートリーダーを使用する場合は、0.2 mLの最終液量で行った。活性は、1分間あたりにNADHを1 mmol生産する酵素量を1 Uと定義した。また、タンパク質量は、Bio-Rad protein assayを用い、BSAで作製した検量線を元に測定した。
【0091】
・L-トリプトファン脱水素酵素の安定化
本酵素は、調製後急速に活性が低下することから、安定化剤としてグリセロールの添加を検討した。Ni-IMACからの溶出直後の酵素液に対し、0, 10, 20, 30, 40, 50 % (W/V)となるようにグリセロールを加え、氷中に保持して活性の経時変化を測定した(図1)。図1に示す通り、グリセロール非添加では、24時間以内に活性が検出限界以下にまで低下したが、20%以上のグリセロール添加により3日以上の間活性に有意な減少は見られなく立った。このため、これ以降の実験では、Ni-IMACからの溶出直後に終濃度20%となるようにグリセロールを加え、同様に20%グリセロールを含む20 mMリン酸緩衝液(pH 7.0)に透析したものを用いることとした。
【0092】
また、スクロース、ソルビトール、トレハロースについても別途検討したところ、表2に示す通り48時間、氷中で保持した場合、それぞれ20%以上の添加により、より優れた安定化効果が認められた。
【0093】
【表2】
*調製直後に測定した値に対する48時間経過後の測定値の割合
【0094】
・基質特異性
上記の活性測定法に基づきL-トリプトファンを含む18種のタンパク質構成アミノ酸及びD-トリプトファン(いずれも0.4 mM)に対する本酵素の活性を調べた結果を図2に示す。本酵素は、L-トリプトファン以外を基質とする活性は、確認できずL-トリプトファン特異的に働くことを示した。
【0095】
・至適pH
上記の活性測定法に基づきpH7から14までの範囲で活性を測定した結果を図3に示す。この結果、pH11.5の緩衝液中で最も高い活性を示した。
【0096】
・温度安定性
酵素溶液(20%グリセロールを含む)を5から45℃の各温度で15分間静置し、直後に氷中で冷却したのち、上記の方法により活性を測定した。図4に示す通り、20%グリセロール存在下で本酵素は、0〜35℃までは、15分間の処理では、活性の大きな低下は見られず、40℃で半減する事が明らかとなった。
【0097】
・反応速度論的解析
図5に示すミカエリス・メンテンプロットの結果、本酵素は、L-トリプトファンにより、0.5 mM以上の基質濃度で基質阻害を受けることが明らかとなった。基質阻害の影響が少ない終濃度0.3 mMまでの基質濃度で算出したKmは、0.0759 mMであった。
【0098】
L-トリプトファン定量法
L-トリプトファン定量法として、ノストク・パンクチフォルム(Nostoc punctiform)由来L-トリプトファン脱水素酵素を用い340 nm の吸光度によりNADHを検出する方法を検討した。
【0099】
(1)L-トリプトファン定量用試薬の調製例
L-トリプトファン定量反応液(10回分)
【0100】
(2)L-トリプトファンの検量線
前述のL-トリプトファン定量法に則り0, 0.05, 0.1, 0.2, 0.3, 0.5 mMのL-トリプトファン水溶液を標準試料として用い、エンドポイント法による測定を行った結果を図6に示す。定量反応は、30℃で行い、反応開始30分の値を用いて検量線の作製を行った。図6で明らかなように本定量法では、0〜0.1 mMの範囲で、直線的な吸光度の増加がみられた。得られた直線の関係は、y = 0.001x +0.1485(R2 =0.9838)[ここでのyは340 nmの吸光度、xは、L-トリプトファン濃度、Rは、相関係数を示す]であって正確なL-トリプトファン定量が可能であることが分かった。
【産業上の利用可能性】
【0101】
L-トリプトファンは、必須アミノ酸であり、極端に不足した場合、ナイアシン欠乏症状を示す事が風土病ペラグラとして知られている。また、過剰に摂取した場合、好酸球増加や筋肉痛症候群の原因になるか可能性が指摘されている。このため、L-トリプトファンの定量は、食品分析、医薬品・サプリメントの品質管理、過剰症・欠乏症時の血液検査及び酵素センサーとして利用が考えられる。また、代謝経路上、ナイアシンやNAD+など生体内で重要な機能を持つ物質の原材料となることから、アミノグラムによる疾病診断「アミノインデックス」においても様々な疾病のバイオマーカーを構成する数値の一つとして活用できる。これらの理由から、食品や生体試料などについて、L-トリプトファン量を測定することは、産業的にも医学的にも重要な技術となると考えられるが、現状では、高速液体クロマトフラフィーをはじめとする非常に高価な機器及び試薬が必要となる方法しか実用化されていない。したがって、本発明は、L-トリプトファン定量用キットや酵素センサーなどの形態で商品化が可能であり、安価かつ簡便なL-トリプトファン定量法として事業化できる。
【特許請求の範囲】
【請求項1】
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの定量方法であって、
前記L-トリプトファン脱水素酵素が、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する前記方法。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項2】
工程(A)において検体との混合に用いられる前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤の存在下で保存されているものである、請求項1に記載の方法。
【請求項3】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項2に記載の方法。
【請求項4】
前記ラン藻がノストク・パンクチフォルム(Nostoc punctiform)である請求項1〜3のいずれかに記載の方法。
【請求項5】
前記工程(C)で計測する反応生成物がNADHである請求項1〜4のいずれかに記載の方法。
【請求項6】
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファン脱水素酵素、および
(K2)NAD+
但し、前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項7】
前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物である、請求項6に記載のキット。
【請求項8】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項7に記載のキット。
【請求項9】
(K3)反応用緩衝液)および/または(K4)NADH検出試薬をさらに含む、請求項6〜8のいずれかに記載のキット。
【請求項10】
L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物であって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する組成物。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項11】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項10に記載の組成物。
【請求項12】
L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーであって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する酵素センサー。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項13】
前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置される、請求項12に記載の酵素センサー。
【請求項14】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項13に記載の酵素センサー。
【請求項1】
検体とL-トリプトファン脱水素酵素とNAD+とを混合する工程(A)、
前記混合により得られた反応液を所定時間放置する工程(B)、
放置後の反応液中に存在する前記酵素の作用による反応生成物の少なくとも一種の量を計測する工程(C)
を含む、前記検体中のL-トリプトファンの定量方法であって、
前記L-トリプトファン脱水素酵素が、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する前記方法。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項2】
工程(A)において検体との混合に用いられる前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤の存在下で保存されているものである、請求項1に記載の方法。
【請求項3】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項2に記載の方法。
【請求項4】
前記ラン藻がノストク・パンクチフォルム(Nostoc punctiform)である請求項1〜3のいずれかに記載の方法。
【請求項5】
前記工程(C)で計測する反応生成物がNADHである請求項1〜4のいずれかに記載の方法。
【請求項6】
以下の試薬を含むL-トリプトファンの定量用キット。
(K1)L-トリプトファン脱水素酵素、および
(K2)NAD+
但し、前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項7】
前記(K1)L-トリプトファン脱水素酵素は、前記酵素の安定化剤との混合物である、請求項6に記載のキット。
【請求項8】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項7に記載のキット。
【請求項9】
(K3)反応用緩衝液)および/または(K4)NADH検出試薬をさらに含む、請求項6〜8のいずれかに記載のキット。
【請求項10】
L-トリプトファン脱水素酵素及び前記酵素の安定化剤を含むL-トリプトファンの定量用組成物であって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する組成物。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項11】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項10に記載の組成物。
【請求項12】
L-トリプトファン脱水素酵素を検出用電極の表面または検出用電極の近傍に配置したL-トリプトファンの検出または定量用酵素センサーであって、
前記L-トリプトファン脱水素酵素は、(a1)下記(1)〜(3)の何れかのアミノ酸配列を有するか、または(a2)ラン藻由来生物中でscytonemin合成経路の一部を担っている酵素から選択され、かつ(a3)L-トリプトファン以外のタンパク質構成アミノ酸及びD-トリプトファンには作用せず、NAD+の存在下、L-トリプトファンに特異的に作用して前記NAD+をNADHに変換する脱水素酵素活性を有する酵素センサー。
(1)配列表の配列番号1に記載のアミノ酸配列;
(2)配列表の配列番号1に記載のアミノ酸配列において1から数個のアミノ酸の欠失、置換及び/又は付加を有するアミノ酸配列;又は
(3)配列表の配列番号1に記載のアミノ酸配列に対して90%以上の相同性を有するアミノ酸配列
【請求項13】
前記L-トリプトファン脱水素酵素は、前記酵素の安定化剤と共に検出用電極の表面または検出用電極の近傍に配置される、請求項12に記載の酵素センサー。
【請求項14】
前記安定化剤が、グリセロール、スクロース、ソルビトール及びトレハロースから成る群から選ばれる少なくとも1種である、請求項13に記載の酵素センサー。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−183018(P2012−183018A)
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2011−48099(P2011−48099)
【出願日】平成23年3月4日(2011.3.4)
【出願人】(000236920)富山県 (197)
【Fターム(参考)】
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成23年3月4日(2011.3.4)
【出願人】(000236920)富山県 (197)
【Fターム(参考)】
[ Back to top ]