
NADHの製造方法及びギ酸脱水素酵素の製造方法
【課題】従来の知見からは予測できないほどに高い比活性を示すギ酸脱水素酵素を利用し、当該ギ酸脱水素酵素の有用な利用用途を提供する。
【解決手段】アカカビ由来のギ酸脱水素酵素は従来公知のギ酸脱水素酵素のなかでも抜群に高い比活性を示すことを明らかにした。アカカビ由来のギ酸脱水素酵素としては、例えば特定のアミノ酸配列からなるタンパク質を挙げることができる。このアカカビ由来のギ酸脱水素酵素を利用してNADHを製造することができる。
【解決手段】アカカビ由来のギ酸脱水素酵素は従来公知のギ酸脱水素酵素のなかでも抜群に高い比活性を示すことを明らかにした。アカカビ由来のギ酸脱水素酵素としては、例えば特定のアミノ酸配列からなるタンパク質を挙げることができる。このアカカビ由来のギ酸脱水素酵素を利用してNADHを製造することができる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ギ酸脱水素酵素を用いたNADHの製造方法、及びギ酸脱水素酵素の製造方法に関する。
【背景技術】
【0002】
酸化型ニコチンアミドアデニンジヌクレオチド(NAD+)は、ギ酸と水の存在下でギ酸脱水素酵素(EC.1.2.1.2)の触媒反応によって還元型ニコチンアミドアデニンジヌクレオチド(NADH)に還元される。このように、ギ酸脱水素酵素は、NAD+からNADHを再生する系に利用されている。従来、ギ酸脱水素酵素としては、例えば特許文献1に記載されるようなカンジダボイジニイ(Candida boidinii)(ATCC32195)由来のギ酸脱水素酵素、特許文献2に開示されるようなバチルス属細菌由来のNAD+依存性ギ酸脱水素酵素、特許文献3に開示されるようなマイコバクテリウム・バッカエ(Mycobacterium vaccae)由来のギ酸脱水素酵素が知られていた。
【0003】
また、非特許文献1には、これら以外にも種々の微生物や植物由来のギ酸脱水素酵素が開示されている。しかしながら、非特許文献1に記述されているように、ギ酸脱水素酵素は、様々な酵素のなかでも比活性の低いほうである。換言すると、ギ酸脱水素酵素によるNADHへの還元反応を利用したNADHの生産方法は、ギ酸脱水素酵素の比活性の低さに起因して生産性が悪いといえる。
【0004】
【特許文献1】特開2003−180383号公報
【特許文献2】特開2002−233395号公報
【特許文献3】特願平10−023896号公報
【非特許文献1】BIOCHEMISTRY (Moscow) Vol. 69 No. 11 2004 pp. 1252-1267.(Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.の英訳)
【発明の開示】
【発明が解決しようとする課題】
【0005】
そこで、本発明は、上述した実情に鑑み、従来の知見からは予測できないほどに高い比活性を示すギ酸脱水素酵素を利用したNADHの製造方法を提供するとともに、高い比活性を示すギ酸脱水素酵素の製造方法及び当該ギ酸脱水素酵素の有用な利用用途を提供することを目的としている。
【課題を解決するための手段】
【0006】
上述した目的を達成するため本発明者らが鋭意検討した結果、アカカビ由来のギ酸脱水素酵素は従来公知のギ酸脱水素酵素のなかでも抜群に高い比活性を示すことを明らかにした。なお、このアカカビ由来のギ酸脱水素酵素については、そのアミノ酸配列がデータベースにエントリーされているものの、このデータは塩基配列情報からの推定アミノ酸配列であって、実際に単離されておらず遺伝子もクローニングされていなかった。また、本発明者らが、鋭意検討したところ、アカカビ由来のギ酸脱水素酵素を定法に従って発現させても全くと言っていいほど活性型として取得することはできず、所定の条件で発現させることで初めて高い比活性を示す酵素として単離することに成功した。
【0007】
以上の知見に基づく本発明は以下を包含する。
(1)ギ酸及びNAD+を含む反応系にアカカビ由来のギ酸脱水素酵素を作用させる、NADHの製造方法。
本発明に係るNADHの製造方法において、上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることが好ましい。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
また、本発明に係るNADHの製造方法において、上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることが好ましい。
【0008】
(2)アカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導し、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養し、上記ギ酸脱水素酵素を取得することを特徴とする、ギ酸脱水素酵素の製造方法。
【0009】
本発明に係るギ酸脱水素酵素の製造方法において上記アカカビ由来のギ酸脱水素酵素は、上記(a)〜(c)のいずれかのタンパク質であることが好ましい。また、上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることが好ましい。
さらに、本発明に係るギ酸脱水素酵素の製造方法において上記対数増殖を過ぎた時は定常期であることが好ましい。また、本発明に係るギ酸脱水素酵素の製造方法において、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度は15〜37℃とすることができる。
【発明の効果】
【0010】
本発明に係るNADHの製造方法によれば、非常に高い比活性を示すギ酸脱水素酵素を利用するため、非常に高価な物質として知られるNADHを優れた生産性で製造することができる。本発明に係るNADHの製造方法を適用することによって、NADHの工業的製造が可能となる。
【0011】
また、本発明に係るギ酸脱水素酵素の製造方法によれば、アカカビ由来のギ酸脱水素酵素を非常に高い比活性を示す状態で製造することができる。このアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に高い比活性を示すため、NADHの製造方法等に広く利用することができる。
【発明を実施するための最良の形態】
【0012】
以下、本発明を図面を参照して詳細に説明する。
【0013】
本発明においては、アカカビ由来のギ酸脱水素酵素が従来公知のギ酸脱水素酵素と比較して抜群に優れ比活性を示すといった知見に基づいている。先ず、アカカビ由来のギ酸脱水素酵素について説明する。
【0014】
アカカビ由来のギ酸脱水素酵素
本発明において使用可能なギ酸脱水素酵素は、アカカビ由来、すなわちアカカビが本来有しているギ酸脱水素酵素である。ここで、アカカビ由来のギ酸脱水素酵素を取得するためには、アカカビ(学名:Gibberella zeae)として保存されている、従来公知の種々の菌株を使用することができる。例えば、American Type Culture Collection (ATCC)には、ATCC番号10910、20271、20272、20274、24689、28106又は48063として保存されているアカカビを使用することができる。また、ATCCには、登録名がFusarium graminearumとして保存されている場合もあるが、別名としてGibberella zeaeが登録されている場合にはこれらも使用することができる。なお、Gibberella zeaeは、Fusarium graminearumの完全世代(テレオモルフ)のことを示している。また、独立行政法人製品評価技術基盤機構の生物遺伝資源部門(NBRC)には、NBRC番号4474、5269、6608、7160、7520、7772、8850又は9462として保存されているアカカビを使用することができる。
【0015】
また、上述したようなアカカビを野生型として、ランダムに突然変異を導入した変異株を使用して、アカカビ由来のギ酸脱水素酵素を取得してもよい。さらに、ATCCやNBRCといった機関に保存された菌株ではなく、自然界から独自に単離したアカカビを使用してアカカビ由来のギ酸脱水素酵素を取得しても良い。
【0016】
一例として、NBRC番号4474で保存されたアカカビから単離したギ酸脱水素酵素遺伝子の塩基配列及びギ酸脱水素酵素のアミノ酸配列をそれぞれ配列番号1及び2に示す。なお、この塩基配列及びアミノ酸配列は、Genbankのデータベースに登録番号NW_059922として登録されているものと100%一致している。しかし、登録番号NW_059922として登録された塩基配列及びアミノ酸配列は、コンピュータ解析による予測によりギ酸脱水素酵素遺伝子のコーディング領域とされたに過ぎず、実験的にギ酸脱水素酵素遺伝子が単離された結果として同定されたものではない。
【0017】
本発明において使用可能なギ酸脱水素酵素は、配列番号2に示したアミノ酸配列からなるものに限定されず、例えば、配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から30個、好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記遺伝子をコードする塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA社製)を用いて変異が導入される。
【0018】
また、本発明においては、配列番号2に示すアミノ酸配列に対して、例えば85%以上、好ましくは90%以上、より好ましくは95%以上、最も好ましくは98%以上の相同性を有するアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するギ酸脱水素酵素を使用することもできる。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0019】
さらに、本発明においては、配列番号1に示す塩基配列の一部又は全部に対して相補的なポリヌクレオチドに対してストリンジェントな条件下でハイブリダイズするポリヌクレオチドにコードされたタンパク質であってギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質をギ酸脱水素酵素として使用することができる。ここで、ストリンジェントな条件下でハイブリダイズするとは、60℃で2×SSC洗浄条件下で結合を維持することを意味する。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0020】
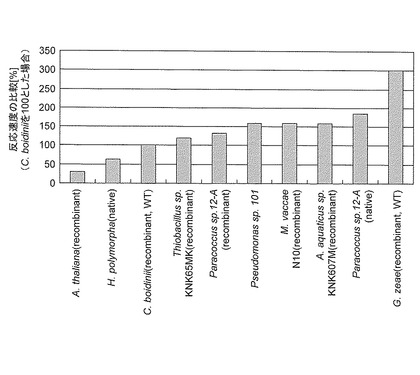
以上で説明したアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に優れた比活性を有している。ここで比較対象となる従来公知のギ酸脱水素酵素としてはNeurospora crassa由来、Saccharomyces cerevisiae由来、Emericella nidulans由来又はCandida boidinii由来のギ酸脱水素酵素を挙げることができる。また、BIOCHEMISTRY (Moscow) Vol. 69 No. 11 2004 pp. 1252-1267.(Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.の英訳)には、従来公知のギ酸脱水素酵素の比活性が掲載されており、これら掲載されたギ酸脱水素酵素の比活性と比較することもできる。いずれの従来公知のギ酸脱水素酵素と比較しても、アカカビ由来のギ酸脱水素酵素の比活性は、2〜10倍以上の値を示している。なお、従来公知のギ酸脱水素酵素のなかでは、Paracoccus sp.12-A由来のギ酸脱水素酵素の比活性が最も高い値を示している。Paracoccus sp.12-A由来のギ酸脱水素酵素の比活性と比較すると、アカカビ由来のギ酸脱水素酵素の比活性は約2倍となっている。
【0021】
アカカビ由来のギ酸脱水素酵素の製造
上述したアカカビ由来のギ酸脱水素酵素は、従来公知のタンパク質の製造方法では取得することができない。本発明に係るギ酸脱水素酵素の製造方法は、上述したアカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を準備する。そして当該宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導する。次に、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養することでギ酸脱水素酵素を宿主内に発現させる。
【0022】
本発明に係るギ酸脱水素酵素の製造方法において、誘導型プロモーターとしては、特に限定されず、従来公知のものを使用することができる。例えば、上記の宿主として大腸菌を使用する場合には、イソプロピル-β-チオガラクトピラノシド(IPTG)の存在下に転写活性を示す誘導型プロモーターを使用することができる。このようなプロモーターの例としては、Trpプロモーター、Lacプロモーター、Trcプロモーター及びTacプロモーターを挙げることができる。また、IPTG以外の誘導物質の存在下に転写活性を示す他のプロモーターや、培地成分及び温度等の培養条件に応じて転写活性を示す他のプロモーターも、誘導型プロモーターとして使用することができる。
【0023】
また、本発明に係るギ酸脱水素酵素の製造方法において、ベクターとしては、上記の宿主内で複製可能なものであれば特に限定されず、如何なるベクターをも使用することができる。例えば、上記宿主として大腸菌を使用する場合には、ベクターとしてはプラスミドベクター、ファージベクターのいずれであっても良い。具体的なベクターとしては、pCDFシリーズ、pRSFシリーズ、pETシリーズ等を例示列挙することができる。
【0024】
さらに宿主としては、発現ベクターに組み込まれたプロモーターから転写可能な宿主であれば特に限定されないが、例えば、発現ベクターがpET(T7プロモーター)系の場合には大腸菌BL21(DE3)を使用することができる。上述したベクターを宿主に導入する手法としては、一般的に形質転換法として知られる各種の手法を適用することができる。具体的な手法としては、例えば、リン酸カルシウム法、エレクトロポレーション法、リポフェクション法等を適用することができる。
【0025】
特に本発明に係るギ酸脱水素酵素の製造方法においては、ベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導する。ギ酸脱水素酵素遺伝子の発現を誘導する前において、宿主の培養条件としては、何ら限定されず、例えば当該宿主の生育至適温度や生育至適pHを勘案して適宜、設定すればよい。しかし、培養を継続しながら宿主の増殖を観察し、いわゆる対数増殖期を過ぎた時点で以下の条件を満足するような培養条件に変更する。すなわち、条件1としてギ酸脱水素酵素遺伝子の発現を誘導すし、条件2として宿主の生育至適温度よりも低く且つ宿主が生存可能な温度で培養する。
【0026】
ここで、対数増殖期を過ぎた時とは、横軸に培養時間及び縦軸に細胞数の対数をとった増殖曲線において、所定の傾きの略直線で表される部分から、接線の傾きが低下し始める時点を意味する。なお、培養曲線は培地中のOD600nmを測定することによって作製することができる。また、ギ酸脱水素酵素遺伝子の発現を誘導する際には、対数増殖期を過ぎて定常期に入ってからが好ましい。ここで定常期とは、上述した増殖曲線の接線の傾きがほぼ0となる期間である。
【0027】
また、宿主の生育至適温度とは、宿主毎に異なる温度範囲として公知である、例えば、大腸菌B株を宿主とした場合、生育至適温度は37℃である。例えば、大腸菌B株を宿主とした場合、生育可能な温度は15〜37℃である。したがって、大腸菌B株を宿主とした場合、宿主の生育至適温度よりも低く且つ宿主が生存可能な温度としては15〜37℃を意味する。特に、大腸菌B株を宿主とした場合、対数増殖期を過ぎた時に約20℃にして培養を継続することが好ましい。
【0028】
宿主の対数増殖期を過ぎたときに上記温度範囲に設定することでギ酸脱水素酵素遺伝子が発現し、宿主内に非常に高い比活性を示すギ酸脱水素酵素が生成することとなる。培養後、目的のギ酸脱水素酵素が宿主内に生産されるため菌体又は細胞を破砕し、粗酵素懸濁液を調製する。この粗酵素懸濁液には、非常に高い比活性を示すギ酸脱水素酵素が含まれる。したがって、得られた粗酵素懸濁液をそのまま利用することができる。なお、得られた粗酵素懸濁液からギ酸脱水素酵素を単離精製することもできる。このとき、蛋白質の単離精製に用いられる一般的な生化学的方法、例えば硫酸アンモニウム沈殿、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等を単独で又は適宜組み合わせて用いることができる。単離精製されたギ酸脱水素酵素は、所定のpHの緩衝液等に懸濁された状態で利用することができる。
【0029】
アカカビ由来のギ酸脱水素酵素の利用形態
以上のようにして製造できるアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に高い比活性を示すため、従来公知のギ酸脱水素酵素が使用されている反応系のいずれにも優れた代替物として利用することができる。すなわち、従来公知のギ酸脱水素酵素が使用されている反応系において、ギ酸脱水素酵素として上述したアカカビ由来のギ酸脱水素酵素を使用することによって、従来の酵素反応と比較して優れた反応効率を達成することができる。
【0030】
例えば、アカカビ由来のギ酸脱水素酵素の利用形態としては、ギ酸脱水素酵素を利用したNADHの再生系を挙げることができる。NADHは種々の酵素反応で利用されNAD+へと変換される。NADHは、例えば、化学工業や製薬工業の分野において光学異性体を生物的に合成する際の補酵素として利用される。NADHの再生系とは、反応系に残存するNAD+を還元してNADHとし、NADHを回収して再び上記酵素反応に利用することを意味する。
【0031】
上述したアカカビ由来のギ酸脱水素酵素をギ酸及びNAD+を含む反応系に作用させることによって、当該NAD+を還元してNADHを合成することができる。
【0032】
以上のようにアカカビ由来のギ酸脱水素酵素をNADHの再生系に利用することによって、反応系に含まれるNAD+からNADHを効率よく製造することができる。特に、アカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して非常に優れた比活性を示すため、従来公知のギ酸脱水素酵素の使用量と比較して小量でNADHを製造することができる。換言すると、アカカビ由来のギ酸脱水素酵素を利用することによって、従来公知のギ酸脱水素酵素の使用した場合と比較してNADHの生産スピードを向上させることができる。
【実施例】
【0033】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0034】
〔実施例1〕
本実施例ではアカカビ由来のギ酸脱水素酵素遺伝子をクローニングした。先ず、アカカビ(Gibberella zeae)として、独立行政法人製品評価技術基盤機構のNBRCに保存されている株を購入し、指定される方法で復元したものを使用した。比較対照としてEmericella nidulansを同様に購入し復元したものを使用した。またこれらの微生物株はPD培地(組成:Potato Dextrose 24g/L、pH7に調整後オートクレーブ)培地を用いて培養した。本実施例で準備した微生物を表1に示す。
【0035】
【表1】

【0036】
次に、上述した微生物株を培養して得た菌体から、RNeasy Plant Mini Kit(QIAGEN社製)を用いてtotal RNAを調製した。次に、RNA PCR Kit(TaKaRa社製)を用いて、得られたTotal RNAを鋳型としてcDNA合成を行った。反応液組成は以下の通りである。また、反応サイクルは50℃(2時間)の後、99℃(5分間)とし、その後4℃とした。
(反応液組成)
最終濃度
5mM MgCl2
1X RT buffer
1mM dNTP mixture
0.5U RNase Inhibitor
0.25U AMV Reverse Transcriptase XL
0.125μM Oligo dT-Adaptor primer
5μg Total RNA
RNase free H2Oを加え、液量が10μlとなるようにした。
【0037】
次に、合成されたcDNAを鋳型とし、データベースに保存された塩基配列に基づいて設計したプライマーを用いてPCRを行った。Gibberella zeae由来のcDNAを用いたPCRには表2に示したプライマーセットを用いた。Emericella nidulans由来のcDNAを用いたPCRには表3に示したプライマーセットを用いた。
【0038】
【表2】
【0039】
【表3】
【0040】
なお、PCRの際にはPyrobest DNA polymerase(TaKaRa社製)を酵素として使用した。反応液50μl中の成分組成は以下の通りである。また、反応サイクルは、95℃で1分間維持した後、95℃で30秒間、60℃で30秒間及び72℃で1分間を1サイクルとして25サイクル行い、最後に72℃で10分間維持し、4℃を維持した。
(反応液組成)
1x Pyrobest buffer
200μM dNTPs mixture
2.5U Pyrobest DNA polymerase
50pmol Primer(forward)
50pmol Primer(reverse)
10μl cDNA溶液
滅菌水を加え、液量が50μlとなるようにした。
【0041】
次に、得られたPCR産物のサイズをアガロースゲル電気泳動により確認後、MiniElute Gel Extraction Kit(QIAGEN製)を用いてアガロースゲルより精製したPCR産物を、pT7 Blue T-vector(Novagen社製)、大腸菌JM109株 competent cellを用いてサブクローニングした。作製できたプラスミドをGzFDH/pT7、EnFDH/pT7とする。ここでGzはGibberella zeae、EnはEmericella nidulans由来であることを意味する。クローン化されたPCR産物の塩基配列はDye Terminator cycle sequence Kit、ABI PRIZM 3100 Genetic Analyzer(アプライドバイオシステムズ社製)および、遺伝子情報処理プログラムGENETYX-WIN(ソフトウェア開発株式会社)を用いて解析した。Gibberella zeae由来のPCR産物の塩基配列解析に使用したプライマーを表4に示し、Emericella nidulans由来のPCR産物の塩基配列解析に使用したプライマーを表5に示す。
【0042】
【表4】
【0043】
【表5】
【0044】
その結果、Gibberella zeae由来ギ酸脱水素酵素遺伝子の塩基配列を配列番号1に示し、Gibberella zeae由来ギ酸脱水素酵素のアミノ酸配列を配列番号2示した。なお、解析の結果として得られた配列番号2に示したアミノ酸配列は、データベース上に公開されている配列(Genbank NW_059922)と100%の一致を示していた。
【0045】
次に、単離されたギ酸脱水素酵素遺伝子を発現ベクターに組み込んだ。具体的には、作製されたプラスミド(GzFDH/pT7、EnFDH/pT7)を制限酵素NdeI及びEcoRIで処理した(37℃、2時間)。反応後の溶液を0.8%アガロースゲル電気泳動に供し、NdeI/EcoRI断片としてベクターから切り出された各ギ酸脱水素酵素遺伝子(約1.2kb)をMini Elute Gel Extraction Kit(QIAGEN製)を用いて精製した。
【0046】
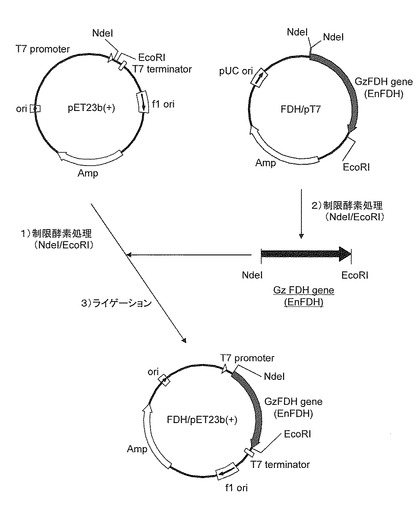
次に、各種FDH/pT7と同様に、制限酵素NdeI及びEcoRIで処理した遺伝子発現用ベクターpET23b(+)(Novagen社製)のNdeI/EcoRIサイトにTaKaRa ligation kit Ver.2.1(TaKaRa製)を用いて上記ギ酸脱水素酵素遺伝子断片を導入後、大腸菌JM109 competent cell(TaKaRa製)によるサブクローニングをおこなった。Fowardプライマー;pET-F(5’-ACT CAC TAT AGG GAG ACC ACA AC-3’)およびReverseプライマー;pET-R(5’-TAT TGC TCA GCG GTG GCA G-3’)を用いたコロニーPCRにより目的の大きさのDNA断片を含むクローンを選抜したあと、Plasmid mini prep kit(QIAGEN製)を用いてプラスミドを精製した。更に、制限酵素マッピングにより計画通りに遺伝子の導入が確認されたプラスミドをGzFDH/pET23b(+)、EnFDH/pET23b(+)とした。なお、上述した発現ベクターの構築の工程を図1に示した。
【0047】
〔実施例2〕
本実施例では、実施例1で作製した発現ベクターを用いて、大腸菌においてギ酸脱水素酵素の発現を検証した。
【0048】
実施例1で作製した発現ベクターを含む大腸菌形質転換体を5mlのLB-アンピシリン培地(組成:Tryptone 10g/L、Yeast Extract 5g/L、NaCl 5g/L及びAmpicillin(SIGMA社製) 50mg/L)で一晩培養(37℃)した後、100mlの同組成培地に1%植菌し、好気的に振盪培養した。OD600の値が0.5〜0.9に達したところで対数増殖期を過ぎたと判断し、最終濃度1mMとなるようにIPTG(イソプロピル-1-チオ-β-D-ガラクトシド)(TaKaRa社製)を添加することでギ酸脱水素酵素遺伝子の発現誘導を開始した。その後、培養温度を20℃として、更に16時間培養した。
【0049】
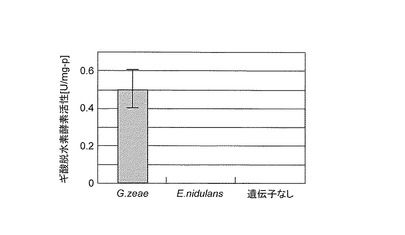
次に、得られた培養液を遠心分離(13krpm,4℃,10min)により集菌後、1ml/0.1g(菌体湿重量)の0.1Mリン酸カリウムバッファーで懸濁し、1g/0.1g(菌体湿重量)の0.1mm径のガラスビーズを加えマルチビーズショッカー(安井器械社製)を用いて菌体を破砕し、粗酵素液を得た。粗酵素液のギ酸脱水素酵素活性の測定は、162mM ギ酸ナトリウム、1.62mM NADを含む0.1Mリン酸カリウムバッファー(pH7.5)中で、37℃でのNADHの生成に伴う340nmの吸光度の増加を測定することによりおこなった。また、タンパク質の定量はBovine serum albumin(BSA)(BioRad社製)を標準タンパク質としてBradford法によりおこなった。その結果を図2に示した。
【0050】
図2に示すように、アカカビ由来のギ酸脱水素酵素遺伝子を含む形質転換体由来の粗酵素液ではNADHの生成に伴う340nmの吸光度の増加が確認された。一方、Emericella nidulans由来のギ酸脱水素酵素遺伝子を含む形質転換体由来の粗酵素液では340nmの吸光度増加は確認されなかった。同様にギ酸脱水素酵素遺伝子を含まないベクターのみによって形質転換された大腸菌粗酵素液も340nmの吸光度の増加が確認されなかった。以上の結果から、本実施例に示した発現誘導方法によれば、アカカビ由来のギ酸脱水素酵素を、大腸菌発現系において生産可能であることが確認された。
【0051】
〔実施例3〕
本実施例では、実施例2で得られたアカカビ由来のギ酸脱水素酵素を精製し、精製されたアカカビ由来のギ酸脱水素酵素の比活性を従来公知のギ酸脱水素酵素との間で比較した。
【0052】
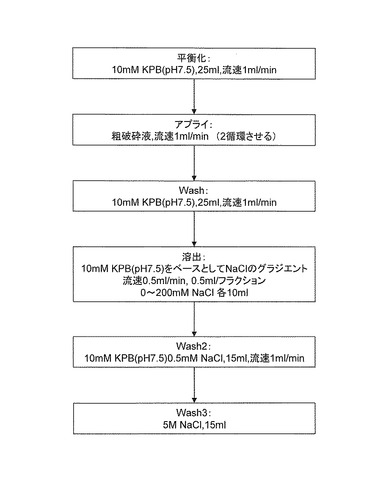
先ず実施例2で得られたアカカビ由来のギ酸脱水素酵素を含む粗破砕液をサンプルとして、HiTrap Q FFカラム 5ml(アマシャムバイオサイエンス社製)を使用し、図3に示す手順でカラム精製を実施した。溶出画分のうちギ酸脱水素酵素を含む10フラクション(全体の1/3)を2次精製のサンプルとした。
【0053】
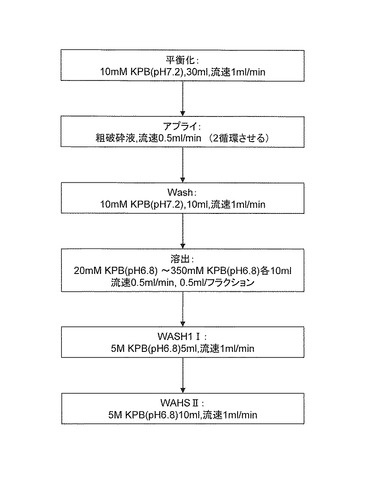
そして、上記で得られたギ酸脱水素酵素を含む画分は、Amicon Ultra-15(30kDaカット)を用いて脱塩後、10mM KPB(pH7.2)で適量(3bed=15ml程度)に希釈した。二次精製要カラムとしてEcono-Pac CHT-IIカラム 5mlを使用し、図4の手順でカラム精製を実施した。二次精製後に精製度80%以上の画分を得た。
【0054】
得られた画分は、アカカビ由来のギ酸脱水素酵素(GzFDH)のギ酸分解反応における活性比較を行うため濃度調整した。対照として性状が公知であるCandida boidinii由来FDH(CbFDH)(シグマ社製)を用いた。CbFDHはリン酸カリウム緩衝液に溶解後、本実施例で上述した方法と同様の方法で精製したものを標品とした。精製したGzFDHとCbFDHのそれぞれについて、BSAを標準タンパク質としたBradford法で定量後、濃度が一定となるよう調製した。Bradford法染色液としてBio-Safe CBB G-250ステイン(BioRad社製)を使用した。
【0055】
精製して濃度調整済みのGzFDH、CbFDHの2種についてギ酸分解反応のエンドポイント法による活性測定を行い、反応速度を比較した。FDH触媒によるギ酸分解反応は次式で示される。
HCOO- + NAD+ → CO2 + NADH
【0056】
ここに電子伝達物質であるMethoxy PMS(mPMS)と、酸化還元発色指示薬であるWST1(共にDOJINDO社製)を加えることで次式のように反応が進むため、波長438nmの吸光度で黄色ホルマザンを測定することで、ギ酸分解量の定量が可能となる。なお黄色ホルマザンの吸光係数はNADHの約6倍であり、NADHの直接測定よりも高感度での定量が可能となる。
NADH + mPMS → NAD+ + mPMS(還元型)
mPMS(還元型) + WST1 → mPMS + 黄色ホルマザン(37000 / M・cm、438 nm)
【0057】
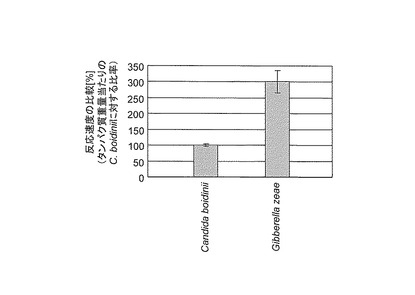
反応液の組成は、162mMのHCOONa、1.62mMのNAD、3.33μg/mlのmPMS、0.33mg/mlのWST1、0.02mg/mlのGzFDH又はCbFDH及び100mMのリン酸カリウム緩衝液(pH7.5)とし、全量を100μlとした。反応液を混和後、25℃で30分間の反応を行った後、プレートリーダー(TECAN社製)によってA430を測定した。反応後の黄色ホルマザン生成量(反応速度)の比較を図5に示す。また従来公知のギ酸脱水素酵素の性状を網羅した論文データ(Biochemistry vol.69, No.11 2004 991252-1267)との比較を図6に示す。これらの結果および公知データとの比較から、今回取得したアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較すると世界最高レベルの比活性を有していることが示された。
【0058】
〔比較例1〕
比較例1では、実施例1で作製した発現ベクターを含む大腸菌形質転換体を用いて、対数増殖期に発現誘導を行う以外は実施例2と同様にしてアカカビ由来のギ酸脱水素酵素の生産を試みる。比較例1では、上記発現有の結果としてタンパク質の発現は確認されるが、ギ酸脱水素酵素活性を検出することができない。
【0059】
〔比較例2〕
比較例2では、実施例1で作製した発現ベクターを含む大腸菌形質転換体を用いて、対数増殖期を過ぎた時点で発現誘導し、その後、37℃で培養を継続する以外は実施例2と同様にしてアカカビ由来のギ酸脱水素酵素の生産を試みる。比較例2では、上記発現有の結果としてタンパク質の発現は確認されるが、ギ酸脱水素酵素活性を検出することができない。
【図面の簡単な説明】
【0060】
【図1】GzFDH/pET23b(+)及びEnFDH/pET23b(+)の構築の工程を示す模式図である。
【図2】実施例2で調製した粗酵素液を用いて酵素反応を行いNADHの生成に伴う340nmの吸光度を検出した結果を示す特性図である。
【図3】実施例3で行ったカラム精製の手順を示すフローチャートである。
【図4】実施例3で行った二次精製の手順を示すフローチャートである。
【図5】精製したGzFDH及びCbFDHについて反応速度を比較した結果を示す特性図である。
【図6】精製したGzFDH及びCbFDHについて、従来公知のギ酸脱水素酵素の性状を網羅した論文データ(Biochemistry vol.69, No.11 2004 991252-1267)と比較した結果を示す特性図である。
【技術分野】
【0001】
本発明は、ギ酸脱水素酵素を用いたNADHの製造方法、及びギ酸脱水素酵素の製造方法に関する。
【背景技術】
【0002】
酸化型ニコチンアミドアデニンジヌクレオチド(NAD+)は、ギ酸と水の存在下でギ酸脱水素酵素(EC.1.2.1.2)の触媒反応によって還元型ニコチンアミドアデニンジヌクレオチド(NADH)に還元される。このように、ギ酸脱水素酵素は、NAD+からNADHを再生する系に利用されている。従来、ギ酸脱水素酵素としては、例えば特許文献1に記載されるようなカンジダボイジニイ(Candida boidinii)(ATCC32195)由来のギ酸脱水素酵素、特許文献2に開示されるようなバチルス属細菌由来のNAD+依存性ギ酸脱水素酵素、特許文献3に開示されるようなマイコバクテリウム・バッカエ(Mycobacterium vaccae)由来のギ酸脱水素酵素が知られていた。
【0003】
また、非特許文献1には、これら以外にも種々の微生物や植物由来のギ酸脱水素酵素が開示されている。しかしながら、非特許文献1に記述されているように、ギ酸脱水素酵素は、様々な酵素のなかでも比活性の低いほうである。換言すると、ギ酸脱水素酵素によるNADHへの還元反応を利用したNADHの生産方法は、ギ酸脱水素酵素の比活性の低さに起因して生産性が悪いといえる。
【0004】
【特許文献1】特開2003−180383号公報
【特許文献2】特開2002−233395号公報
【特許文献3】特願平10−023896号公報
【非特許文献1】BIOCHEMISTRY (Moscow) Vol. 69 No. 11 2004 pp. 1252-1267.(Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.の英訳)
【発明の開示】
【発明が解決しようとする課題】
【0005】
そこで、本発明は、上述した実情に鑑み、従来の知見からは予測できないほどに高い比活性を示すギ酸脱水素酵素を利用したNADHの製造方法を提供するとともに、高い比活性を示すギ酸脱水素酵素の製造方法及び当該ギ酸脱水素酵素の有用な利用用途を提供することを目的としている。
【課題を解決するための手段】
【0006】
上述した目的を達成するため本発明者らが鋭意検討した結果、アカカビ由来のギ酸脱水素酵素は従来公知のギ酸脱水素酵素のなかでも抜群に高い比活性を示すことを明らかにした。なお、このアカカビ由来のギ酸脱水素酵素については、そのアミノ酸配列がデータベースにエントリーされているものの、このデータは塩基配列情報からの推定アミノ酸配列であって、実際に単離されておらず遺伝子もクローニングされていなかった。また、本発明者らが、鋭意検討したところ、アカカビ由来のギ酸脱水素酵素を定法に従って発現させても全くと言っていいほど活性型として取得することはできず、所定の条件で発現させることで初めて高い比活性を示す酵素として単離することに成功した。
【0007】
以上の知見に基づく本発明は以下を包含する。
(1)ギ酸及びNAD+を含む反応系にアカカビ由来のギ酸脱水素酵素を作用させる、NADHの製造方法。
本発明に係るNADHの製造方法において、上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることが好ましい。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
また、本発明に係るNADHの製造方法において、上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることが好ましい。
【0008】
(2)アカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導し、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養し、上記ギ酸脱水素酵素を取得することを特徴とする、ギ酸脱水素酵素の製造方法。
【0009】
本発明に係るギ酸脱水素酵素の製造方法において上記アカカビ由来のギ酸脱水素酵素は、上記(a)〜(c)のいずれかのタンパク質であることが好ましい。また、上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることが好ましい。
さらに、本発明に係るギ酸脱水素酵素の製造方法において上記対数増殖を過ぎた時は定常期であることが好ましい。また、本発明に係るギ酸脱水素酵素の製造方法において、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度は15〜37℃とすることができる。
【発明の効果】
【0010】
本発明に係るNADHの製造方法によれば、非常に高い比活性を示すギ酸脱水素酵素を利用するため、非常に高価な物質として知られるNADHを優れた生産性で製造することができる。本発明に係るNADHの製造方法を適用することによって、NADHの工業的製造が可能となる。
【0011】
また、本発明に係るギ酸脱水素酵素の製造方法によれば、アカカビ由来のギ酸脱水素酵素を非常に高い比活性を示す状態で製造することができる。このアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に高い比活性を示すため、NADHの製造方法等に広く利用することができる。
【発明を実施するための最良の形態】
【0012】
以下、本発明を図面を参照して詳細に説明する。
【0013】
本発明においては、アカカビ由来のギ酸脱水素酵素が従来公知のギ酸脱水素酵素と比較して抜群に優れ比活性を示すといった知見に基づいている。先ず、アカカビ由来のギ酸脱水素酵素について説明する。
【0014】
アカカビ由来のギ酸脱水素酵素
本発明において使用可能なギ酸脱水素酵素は、アカカビ由来、すなわちアカカビが本来有しているギ酸脱水素酵素である。ここで、アカカビ由来のギ酸脱水素酵素を取得するためには、アカカビ(学名:Gibberella zeae)として保存されている、従来公知の種々の菌株を使用することができる。例えば、American Type Culture Collection (ATCC)には、ATCC番号10910、20271、20272、20274、24689、28106又は48063として保存されているアカカビを使用することができる。また、ATCCには、登録名がFusarium graminearumとして保存されている場合もあるが、別名としてGibberella zeaeが登録されている場合にはこれらも使用することができる。なお、Gibberella zeaeは、Fusarium graminearumの完全世代(テレオモルフ)のことを示している。また、独立行政法人製品評価技術基盤機構の生物遺伝資源部門(NBRC)には、NBRC番号4474、5269、6608、7160、7520、7772、8850又は9462として保存されているアカカビを使用することができる。
【0015】
また、上述したようなアカカビを野生型として、ランダムに突然変異を導入した変異株を使用して、アカカビ由来のギ酸脱水素酵素を取得してもよい。さらに、ATCCやNBRCといった機関に保存された菌株ではなく、自然界から独自に単離したアカカビを使用してアカカビ由来のギ酸脱水素酵素を取得しても良い。
【0016】
一例として、NBRC番号4474で保存されたアカカビから単離したギ酸脱水素酵素遺伝子の塩基配列及びギ酸脱水素酵素のアミノ酸配列をそれぞれ配列番号1及び2に示す。なお、この塩基配列及びアミノ酸配列は、Genbankのデータベースに登録番号NW_059922として登録されているものと100%一致している。しかし、登録番号NW_059922として登録された塩基配列及びアミノ酸配列は、コンピュータ解析による予測によりギ酸脱水素酵素遺伝子のコーディング領域とされたに過ぎず、実験的にギ酸脱水素酵素遺伝子が単離された結果として同定されたものではない。
【0017】
本発明において使用可能なギ酸脱水素酵素は、配列番号2に示したアミノ酸配列からなるものに限定されず、例えば、配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するものであっても良い。ここで、複数個のアミノ酸としては、例えば、1から30個、好ましくは1から20個、より好ましくは1から10個、さらに好ましくは1個から5個、特に好ましくは1個から3個を意味する。なお、アミノ酸の欠失、置換若しくは付加は、上記遺伝子をコードする塩基配列を、当該技術分野で公知の手法によって改変することによって行うことができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA社製)を用いて変異が導入される。
【0018】
また、本発明においては、配列番号2に示すアミノ酸配列に対して、例えば85%以上、好ましくは90%以上、より好ましくは95%以上、最も好ましくは98%以上の相同性を有するアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するギ酸脱水素酵素を使用することもできる。ここで、相同性の値は、blastアルゴリズムを実装したコンピュータプログラム及び遺伝子配列情報を格納したデータベースを用いてデフォルトの設定で求められる値を意味する。
【0019】
さらに、本発明においては、配列番号1に示す塩基配列の一部又は全部に対して相補的なポリヌクレオチドに対してストリンジェントな条件下でハイブリダイズするポリヌクレオチドにコードされたタンパク質であってギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質をギ酸脱水素酵素として使用することができる。ここで、ストリンジェントな条件下でハイブリダイズするとは、60℃で2×SSC洗浄条件下で結合を維持することを意味する。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0020】
以上で説明したアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に優れた比活性を有している。ここで比較対象となる従来公知のギ酸脱水素酵素としてはNeurospora crassa由来、Saccharomyces cerevisiae由来、Emericella nidulans由来又はCandida boidinii由来のギ酸脱水素酵素を挙げることができる。また、BIOCHEMISTRY (Moscow) Vol. 69 No. 11 2004 pp. 1252-1267.(Biokhimiya, Vol. 69, No. 11, 2004, pp. 1537-1554.の英訳)には、従来公知のギ酸脱水素酵素の比活性が掲載されており、これら掲載されたギ酸脱水素酵素の比活性と比較することもできる。いずれの従来公知のギ酸脱水素酵素と比較しても、アカカビ由来のギ酸脱水素酵素の比活性は、2〜10倍以上の値を示している。なお、従来公知のギ酸脱水素酵素のなかでは、Paracoccus sp.12-A由来のギ酸脱水素酵素の比活性が最も高い値を示している。Paracoccus sp.12-A由来のギ酸脱水素酵素の比活性と比較すると、アカカビ由来のギ酸脱水素酵素の比活性は約2倍となっている。
【0021】
アカカビ由来のギ酸脱水素酵素の製造
上述したアカカビ由来のギ酸脱水素酵素は、従来公知のタンパク質の製造方法では取得することができない。本発明に係るギ酸脱水素酵素の製造方法は、上述したアカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を準備する。そして当該宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導する。次に、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養することでギ酸脱水素酵素を宿主内に発現させる。
【0022】
本発明に係るギ酸脱水素酵素の製造方法において、誘導型プロモーターとしては、特に限定されず、従来公知のものを使用することができる。例えば、上記の宿主として大腸菌を使用する場合には、イソプロピル-β-チオガラクトピラノシド(IPTG)の存在下に転写活性を示す誘導型プロモーターを使用することができる。このようなプロモーターの例としては、Trpプロモーター、Lacプロモーター、Trcプロモーター及びTacプロモーターを挙げることができる。また、IPTG以外の誘導物質の存在下に転写活性を示す他のプロモーターや、培地成分及び温度等の培養条件に応じて転写活性を示す他のプロモーターも、誘導型プロモーターとして使用することができる。
【0023】
また、本発明に係るギ酸脱水素酵素の製造方法において、ベクターとしては、上記の宿主内で複製可能なものであれば特に限定されず、如何なるベクターをも使用することができる。例えば、上記宿主として大腸菌を使用する場合には、ベクターとしてはプラスミドベクター、ファージベクターのいずれであっても良い。具体的なベクターとしては、pCDFシリーズ、pRSFシリーズ、pETシリーズ等を例示列挙することができる。
【0024】
さらに宿主としては、発現ベクターに組み込まれたプロモーターから転写可能な宿主であれば特に限定されないが、例えば、発現ベクターがpET(T7プロモーター)系の場合には大腸菌BL21(DE3)を使用することができる。上述したベクターを宿主に導入する手法としては、一般的に形質転換法として知られる各種の手法を適用することができる。具体的な手法としては、例えば、リン酸カルシウム法、エレクトロポレーション法、リポフェクション法等を適用することができる。
【0025】
特に本発明に係るギ酸脱水素酵素の製造方法においては、ベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導する。ギ酸脱水素酵素遺伝子の発現を誘導する前において、宿主の培養条件としては、何ら限定されず、例えば当該宿主の生育至適温度や生育至適pHを勘案して適宜、設定すればよい。しかし、培養を継続しながら宿主の増殖を観察し、いわゆる対数増殖期を過ぎた時点で以下の条件を満足するような培養条件に変更する。すなわち、条件1としてギ酸脱水素酵素遺伝子の発現を誘導すし、条件2として宿主の生育至適温度よりも低く且つ宿主が生存可能な温度で培養する。
【0026】
ここで、対数増殖期を過ぎた時とは、横軸に培養時間及び縦軸に細胞数の対数をとった増殖曲線において、所定の傾きの略直線で表される部分から、接線の傾きが低下し始める時点を意味する。なお、培養曲線は培地中のOD600nmを測定することによって作製することができる。また、ギ酸脱水素酵素遺伝子の発現を誘導する際には、対数増殖期を過ぎて定常期に入ってからが好ましい。ここで定常期とは、上述した増殖曲線の接線の傾きがほぼ0となる期間である。
【0027】
また、宿主の生育至適温度とは、宿主毎に異なる温度範囲として公知である、例えば、大腸菌B株を宿主とした場合、生育至適温度は37℃である。例えば、大腸菌B株を宿主とした場合、生育可能な温度は15〜37℃である。したがって、大腸菌B株を宿主とした場合、宿主の生育至適温度よりも低く且つ宿主が生存可能な温度としては15〜37℃を意味する。特に、大腸菌B株を宿主とした場合、対数増殖期を過ぎた時に約20℃にして培養を継続することが好ましい。
【0028】
宿主の対数増殖期を過ぎたときに上記温度範囲に設定することでギ酸脱水素酵素遺伝子が発現し、宿主内に非常に高い比活性を示すギ酸脱水素酵素が生成することとなる。培養後、目的のギ酸脱水素酵素が宿主内に生産されるため菌体又は細胞を破砕し、粗酵素懸濁液を調製する。この粗酵素懸濁液には、非常に高い比活性を示すギ酸脱水素酵素が含まれる。したがって、得られた粗酵素懸濁液をそのまま利用することができる。なお、得られた粗酵素懸濁液からギ酸脱水素酵素を単離精製することもできる。このとき、蛋白質の単離精製に用いられる一般的な生化学的方法、例えば硫酸アンモニウム沈殿、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等を単独で又は適宜組み合わせて用いることができる。単離精製されたギ酸脱水素酵素は、所定のpHの緩衝液等に懸濁された状態で利用することができる。
【0029】
アカカビ由来のギ酸脱水素酵素の利用形態
以上のようにして製造できるアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して抜群に高い比活性を示すため、従来公知のギ酸脱水素酵素が使用されている反応系のいずれにも優れた代替物として利用することができる。すなわち、従来公知のギ酸脱水素酵素が使用されている反応系において、ギ酸脱水素酵素として上述したアカカビ由来のギ酸脱水素酵素を使用することによって、従来の酵素反応と比較して優れた反応効率を達成することができる。
【0030】
例えば、アカカビ由来のギ酸脱水素酵素の利用形態としては、ギ酸脱水素酵素を利用したNADHの再生系を挙げることができる。NADHは種々の酵素反応で利用されNAD+へと変換される。NADHは、例えば、化学工業や製薬工業の分野において光学異性体を生物的に合成する際の補酵素として利用される。NADHの再生系とは、反応系に残存するNAD+を還元してNADHとし、NADHを回収して再び上記酵素反応に利用することを意味する。
【0031】
上述したアカカビ由来のギ酸脱水素酵素をギ酸及びNAD+を含む反応系に作用させることによって、当該NAD+を還元してNADHを合成することができる。
【0032】
以上のようにアカカビ由来のギ酸脱水素酵素をNADHの再生系に利用することによって、反応系に含まれるNAD+からNADHを効率よく製造することができる。特に、アカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較して非常に優れた比活性を示すため、従来公知のギ酸脱水素酵素の使用量と比較して小量でNADHを製造することができる。換言すると、アカカビ由来のギ酸脱水素酵素を利用することによって、従来公知のギ酸脱水素酵素の使用した場合と比較してNADHの生産スピードを向上させることができる。
【実施例】
【0033】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0034】
〔実施例1〕
本実施例ではアカカビ由来のギ酸脱水素酵素遺伝子をクローニングした。先ず、アカカビ(Gibberella zeae)として、独立行政法人製品評価技術基盤機構のNBRCに保存されている株を購入し、指定される方法で復元したものを使用した。比較対照としてEmericella nidulansを同様に購入し復元したものを使用した。またこれらの微生物株はPD培地(組成:Potato Dextrose 24g/L、pH7に調整後オートクレーブ)培地を用いて培養した。本実施例で準備した微生物を表1に示す。
【0035】
【表1】
【0036】
次に、上述した微生物株を培養して得た菌体から、RNeasy Plant Mini Kit(QIAGEN社製)を用いてtotal RNAを調製した。次に、RNA PCR Kit(TaKaRa社製)を用いて、得られたTotal RNAを鋳型としてcDNA合成を行った。反応液組成は以下の通りである。また、反応サイクルは50℃(2時間)の後、99℃(5分間)とし、その後4℃とした。
(反応液組成)
最終濃度
5mM MgCl2
1X RT buffer
1mM dNTP mixture
0.5U RNase Inhibitor
0.25U AMV Reverse Transcriptase XL
0.125μM Oligo dT-Adaptor primer
5μg Total RNA
RNase free H2Oを加え、液量が10μlとなるようにした。
【0037】
次に、合成されたcDNAを鋳型とし、データベースに保存された塩基配列に基づいて設計したプライマーを用いてPCRを行った。Gibberella zeae由来のcDNAを用いたPCRには表2に示したプライマーセットを用いた。Emericella nidulans由来のcDNAを用いたPCRには表3に示したプライマーセットを用いた。
【0038】
【表2】
【0039】
【表3】
【0040】
なお、PCRの際にはPyrobest DNA polymerase(TaKaRa社製)を酵素として使用した。反応液50μl中の成分組成は以下の通りである。また、反応サイクルは、95℃で1分間維持した後、95℃で30秒間、60℃で30秒間及び72℃で1分間を1サイクルとして25サイクル行い、最後に72℃で10分間維持し、4℃を維持した。
(反応液組成)
1x Pyrobest buffer
200μM dNTPs mixture
2.5U Pyrobest DNA polymerase
50pmol Primer(forward)
50pmol Primer(reverse)
10μl cDNA溶液
滅菌水を加え、液量が50μlとなるようにした。
【0041】
次に、得られたPCR産物のサイズをアガロースゲル電気泳動により確認後、MiniElute Gel Extraction Kit(QIAGEN製)を用いてアガロースゲルより精製したPCR産物を、pT7 Blue T-vector(Novagen社製)、大腸菌JM109株 competent cellを用いてサブクローニングした。作製できたプラスミドをGzFDH/pT7、EnFDH/pT7とする。ここでGzはGibberella zeae、EnはEmericella nidulans由来であることを意味する。クローン化されたPCR産物の塩基配列はDye Terminator cycle sequence Kit、ABI PRIZM 3100 Genetic Analyzer(アプライドバイオシステムズ社製)および、遺伝子情報処理プログラムGENETYX-WIN(ソフトウェア開発株式会社)を用いて解析した。Gibberella zeae由来のPCR産物の塩基配列解析に使用したプライマーを表4に示し、Emericella nidulans由来のPCR産物の塩基配列解析に使用したプライマーを表5に示す。
【0042】
【表4】
【0043】
【表5】
【0044】
その結果、Gibberella zeae由来ギ酸脱水素酵素遺伝子の塩基配列を配列番号1に示し、Gibberella zeae由来ギ酸脱水素酵素のアミノ酸配列を配列番号2示した。なお、解析の結果として得られた配列番号2に示したアミノ酸配列は、データベース上に公開されている配列(Genbank NW_059922)と100%の一致を示していた。
【0045】
次に、単離されたギ酸脱水素酵素遺伝子を発現ベクターに組み込んだ。具体的には、作製されたプラスミド(GzFDH/pT7、EnFDH/pT7)を制限酵素NdeI及びEcoRIで処理した(37℃、2時間)。反応後の溶液を0.8%アガロースゲル電気泳動に供し、NdeI/EcoRI断片としてベクターから切り出された各ギ酸脱水素酵素遺伝子(約1.2kb)をMini Elute Gel Extraction Kit(QIAGEN製)を用いて精製した。
【0046】
次に、各種FDH/pT7と同様に、制限酵素NdeI及びEcoRIで処理した遺伝子発現用ベクターpET23b(+)(Novagen社製)のNdeI/EcoRIサイトにTaKaRa ligation kit Ver.2.1(TaKaRa製)を用いて上記ギ酸脱水素酵素遺伝子断片を導入後、大腸菌JM109 competent cell(TaKaRa製)によるサブクローニングをおこなった。Fowardプライマー;pET-F(5’-ACT CAC TAT AGG GAG ACC ACA AC-3’)およびReverseプライマー;pET-R(5’-TAT TGC TCA GCG GTG GCA G-3’)を用いたコロニーPCRにより目的の大きさのDNA断片を含むクローンを選抜したあと、Plasmid mini prep kit(QIAGEN製)を用いてプラスミドを精製した。更に、制限酵素マッピングにより計画通りに遺伝子の導入が確認されたプラスミドをGzFDH/pET23b(+)、EnFDH/pET23b(+)とした。なお、上述した発現ベクターの構築の工程を図1に示した。
【0047】
〔実施例2〕
本実施例では、実施例1で作製した発現ベクターを用いて、大腸菌においてギ酸脱水素酵素の発現を検証した。
【0048】
実施例1で作製した発現ベクターを含む大腸菌形質転換体を5mlのLB-アンピシリン培地(組成:Tryptone 10g/L、Yeast Extract 5g/L、NaCl 5g/L及びAmpicillin(SIGMA社製) 50mg/L)で一晩培養(37℃)した後、100mlの同組成培地に1%植菌し、好気的に振盪培養した。OD600の値が0.5〜0.9に達したところで対数増殖期を過ぎたと判断し、最終濃度1mMとなるようにIPTG(イソプロピル-1-チオ-β-D-ガラクトシド)(TaKaRa社製)を添加することでギ酸脱水素酵素遺伝子の発現誘導を開始した。その後、培養温度を20℃として、更に16時間培養した。
【0049】
次に、得られた培養液を遠心分離(13krpm,4℃,10min)により集菌後、1ml/0.1g(菌体湿重量)の0.1Mリン酸カリウムバッファーで懸濁し、1g/0.1g(菌体湿重量)の0.1mm径のガラスビーズを加えマルチビーズショッカー(安井器械社製)を用いて菌体を破砕し、粗酵素液を得た。粗酵素液のギ酸脱水素酵素活性の測定は、162mM ギ酸ナトリウム、1.62mM NADを含む0.1Mリン酸カリウムバッファー(pH7.5)中で、37℃でのNADHの生成に伴う340nmの吸光度の増加を測定することによりおこなった。また、タンパク質の定量はBovine serum albumin(BSA)(BioRad社製)を標準タンパク質としてBradford法によりおこなった。その結果を図2に示した。
【0050】
図2に示すように、アカカビ由来のギ酸脱水素酵素遺伝子を含む形質転換体由来の粗酵素液ではNADHの生成に伴う340nmの吸光度の増加が確認された。一方、Emericella nidulans由来のギ酸脱水素酵素遺伝子を含む形質転換体由来の粗酵素液では340nmの吸光度増加は確認されなかった。同様にギ酸脱水素酵素遺伝子を含まないベクターのみによって形質転換された大腸菌粗酵素液も340nmの吸光度の増加が確認されなかった。以上の結果から、本実施例に示した発現誘導方法によれば、アカカビ由来のギ酸脱水素酵素を、大腸菌発現系において生産可能であることが確認された。
【0051】
〔実施例3〕
本実施例では、実施例2で得られたアカカビ由来のギ酸脱水素酵素を精製し、精製されたアカカビ由来のギ酸脱水素酵素の比活性を従来公知のギ酸脱水素酵素との間で比較した。
【0052】
先ず実施例2で得られたアカカビ由来のギ酸脱水素酵素を含む粗破砕液をサンプルとして、HiTrap Q FFカラム 5ml(アマシャムバイオサイエンス社製)を使用し、図3に示す手順でカラム精製を実施した。溶出画分のうちギ酸脱水素酵素を含む10フラクション(全体の1/3)を2次精製のサンプルとした。
【0053】
そして、上記で得られたギ酸脱水素酵素を含む画分は、Amicon Ultra-15(30kDaカット)を用いて脱塩後、10mM KPB(pH7.2)で適量(3bed=15ml程度)に希釈した。二次精製要カラムとしてEcono-Pac CHT-IIカラム 5mlを使用し、図4の手順でカラム精製を実施した。二次精製後に精製度80%以上の画分を得た。
【0054】
得られた画分は、アカカビ由来のギ酸脱水素酵素(GzFDH)のギ酸分解反応における活性比較を行うため濃度調整した。対照として性状が公知であるCandida boidinii由来FDH(CbFDH)(シグマ社製)を用いた。CbFDHはリン酸カリウム緩衝液に溶解後、本実施例で上述した方法と同様の方法で精製したものを標品とした。精製したGzFDHとCbFDHのそれぞれについて、BSAを標準タンパク質としたBradford法で定量後、濃度が一定となるよう調製した。Bradford法染色液としてBio-Safe CBB G-250ステイン(BioRad社製)を使用した。
【0055】
精製して濃度調整済みのGzFDH、CbFDHの2種についてギ酸分解反応のエンドポイント法による活性測定を行い、反応速度を比較した。FDH触媒によるギ酸分解反応は次式で示される。
HCOO- + NAD+ → CO2 + NADH
【0056】
ここに電子伝達物質であるMethoxy PMS(mPMS)と、酸化還元発色指示薬であるWST1(共にDOJINDO社製)を加えることで次式のように反応が進むため、波長438nmの吸光度で黄色ホルマザンを測定することで、ギ酸分解量の定量が可能となる。なお黄色ホルマザンの吸光係数はNADHの約6倍であり、NADHの直接測定よりも高感度での定量が可能となる。
NADH + mPMS → NAD+ + mPMS(還元型)
mPMS(還元型) + WST1 → mPMS + 黄色ホルマザン(37000 / M・cm、438 nm)
【0057】
反応液の組成は、162mMのHCOONa、1.62mMのNAD、3.33μg/mlのmPMS、0.33mg/mlのWST1、0.02mg/mlのGzFDH又はCbFDH及び100mMのリン酸カリウム緩衝液(pH7.5)とし、全量を100μlとした。反応液を混和後、25℃で30分間の反応を行った後、プレートリーダー(TECAN社製)によってA430を測定した。反応後の黄色ホルマザン生成量(反応速度)の比較を図5に示す。また従来公知のギ酸脱水素酵素の性状を網羅した論文データ(Biochemistry vol.69, No.11 2004 991252-1267)との比較を図6に示す。これらの結果および公知データとの比較から、今回取得したアカカビ由来のギ酸脱水素酵素は、従来公知のギ酸脱水素酵素と比較すると世界最高レベルの比活性を有していることが示された。
【0058】
〔比較例1〕
比較例1では、実施例1で作製した発現ベクターを含む大腸菌形質転換体を用いて、対数増殖期に発現誘導を行う以外は実施例2と同様にしてアカカビ由来のギ酸脱水素酵素の生産を試みる。比較例1では、上記発現有の結果としてタンパク質の発現は確認されるが、ギ酸脱水素酵素活性を検出することができない。
【0059】
〔比較例2〕
比較例2では、実施例1で作製した発現ベクターを含む大腸菌形質転換体を用いて、対数増殖期を過ぎた時点で発現誘導し、その後、37℃で培養を継続する以外は実施例2と同様にしてアカカビ由来のギ酸脱水素酵素の生産を試みる。比較例2では、上記発現有の結果としてタンパク質の発現は確認されるが、ギ酸脱水素酵素活性を検出することができない。
【図面の簡単な説明】
【0060】
【図1】GzFDH/pET23b(+)及びEnFDH/pET23b(+)の構築の工程を示す模式図である。
【図2】実施例2で調製した粗酵素液を用いて酵素反応を行いNADHの生成に伴う340nmの吸光度を検出した結果を示す特性図である。
【図3】実施例3で行ったカラム精製の手順を示すフローチャートである。
【図4】実施例3で行った二次精製の手順を示すフローチャートである。
【図5】精製したGzFDH及びCbFDHについて反応速度を比較した結果を示す特性図である。
【図6】精製したGzFDH及びCbFDHについて、従来公知のギ酸脱水素酵素の性状を網羅した論文データ(Biochemistry vol.69, No.11 2004 991252-1267)と比較した結果を示す特性図である。
【特許請求の範囲】
【請求項1】
ギ酸及びNAD+を含む反応系にアカカビ由来のギ酸脱水素酵素を作用させる、
NADHの製造方法。
【請求項2】
上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項1記載のNADHの製造方法。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
【請求項3】
上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることを特徴とする請求項1記載のNADHの製造方法。
【請求項4】
アカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導し、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養し、
上記ギ酸脱水素酵素を取得することを特徴とする、ギ酸脱水素酵素の製造方法。
【請求項5】
上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
【請求項6】
上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【請求項7】
上記対数増殖を過ぎた時は定常期であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【請求項8】
上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度は15〜37℃であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【請求項1】
ギ酸及びNAD+を含む反応系にアカカビ由来のギ酸脱水素酵素を作用させる、
NADHの製造方法。
【請求項2】
上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項1記載のNADHの製造方法。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
【請求項3】
上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることを特徴とする請求項1記載のNADHの製造方法。
【請求項4】
アカカビ由来のギ酸脱水素酵素遺伝子を誘導型プロモーターの制御下に配置したベクターを導入した宿主を培養し、対数増殖期を過ぎた時に上記ギ酸脱水素酵素遺伝子の発現を誘導し、上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度で培養し、
上記ギ酸脱水素酵素を取得することを特徴とする、ギ酸脱水素酵素の製造方法。
【請求項5】
上記アカカビ由来のギ酸脱水素酵素は、以下の(a)〜(c)のいずれかのタンパク質であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
(a)配列番号2に示すアミノ酸配列を含むタンパク質
(b)配列番号2に示すアミノ酸配列において1又は複数個のアミノ酸が欠失、置換、付加又は挿入されたアミノ酸配列を含み、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
(c)配列番号1に示す塩基配列の相補的な塩基配列からなるポリヌクレオチドに対してストリンジェントな条件下においてハイブリダイズするポリヌクレオチドによってコードされ、ギ酸及びNAD+を基質として二酸化炭素及びNADHを生産物とする反応における触媒活性を有するタンパク質
【請求項6】
上記アカカビ由来のギ酸脱水素酵素は、配列番号2に示すアミノ酸配列に対して85%以上の相同性を有するアミノ酸配列を含むタンパク質であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【請求項7】
上記対数増殖を過ぎた時は定常期であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【請求項8】
上記宿主の生育至適温度よりも低く且つ上記宿主が生存可能な温度は15〜37℃であることを特徴とする請求項4記載のギ酸脱水素酵素の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2009−247296(P2009−247296A)
【公開日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願番号】特願2008−100448(P2008−100448)
【出願日】平成20年4月8日(2008.4.8)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
【公開日】平成21年10月29日(2009.10.29)
【国際特許分類】
【出願日】平成20年4月8日(2008.4.8)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【Fターム(参考)】
[ Back to top ]