
NADH/NAD比の制御による2−デオキシ−シロ−イノソースの製造方法
【課題】2−デオキシ−シロ−イノソース(DOI)生産性を飛躍的に向上させうる細胞内代謝産物制御技術を提供すること。
【解決手段】2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
【解決手段】2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、2−デオキシ−シロ−イノソース合成酵素を用いて2−デオキシ−シロ−イノソースを製造する方法においてNADH/NAD比を制御することを特徴とする2−デオキシ−シロ−イノソースの製造方法に関する。
【背景技術】
【0002】
現在、カテコールなどの芳香族化合物の多くは、石油を原料として生産されているが、石油資源の枯渇問題や二酸化炭素排出量の削減などの観点から、バイオマスを利用した環境調和型の新規製造プロセスの開発が求められている。
【0003】
一方、2−デオキシ−シロ−イノソース(以下、DOI)が、産業上有用とされる芳香族化合物(カテコール等)へと変換できることが見出され(非特許文献1)、さらには、このDOIを、バイオマスの構成要素の一つであるグルコースから合成できることが報告された(非特許文献1)。本DOI製造プロセスで重要となるのがグルコース−6−リン酸からDOIへと変換する2−デオキシ−シロ−イノソース合成酵素(以下、DOI合成酵素)であり、DOIは上記芳香族化合物への変換のみならず各種有用化合物の中間体ともなり得ることから、DOI合成酵素は非常に大きな注目を浴びている。
【0004】
DOI合成酵素はブチロシン生産菌であるBacillus circulans に属する微生物から1997年に単離精製(非特許文献2)され、続いて、その遺伝子配列も公開されている(特許文献1)。この他にもStreptoalloteichus hindustanus JCM3268株(非特許文献3)由来のDOI合成酵素などがこれまでに発見されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2000−236881号公報
【非特許文献】
【0006】
【非特許文献1】Tetrahedron Letters 41,1935−1938,2000
【非特許文献2】The Journal of Antibiotics 50(5), 424−428, 1997
【非特許文献3】The Journal of Antibiotics 59(6), 358−361, 2006
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、前記DOI合成酵素の安定性と比活性などの各種特性は、満足のいくものではなく、また、数千トン規模の工業化製法に向けたDOI発酵生産に関する研究開発、特に細胞内代謝物の制御に関するものはほとんどなされてこなかった。
本発明は、従来法と比較して、DOI発酵生産性を飛躍的に向上させうる細胞内代謝産物制御技術を提供することを解決すべき課題とする。
【課題を解決するための手段】
【0008】
これらの課題を解決するため鋭意検討を重ねた結果、本発明者らは、DOI合成酵素の活性が反応系中のNADH/NAD比に予想し得ない領域で影響を受けることを見出し、従来法と比較して、DOI発酵生産性を飛躍的に向上させうるDOI生産技術を見出し、本発明を完成するに至った。
【0009】
すなわち、本発明は以下の[1]〜[7]に示すDOI製造方法に関するものである。
[1] 2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
[2] グルコースを構成成分とする多糖類あるいはグルコースを含有する原料を用いることを特徴とする、[1]に記載の方法。
[3] 2−デオキシ−シロ−イノソース合成酵素が、安定温度範囲が少なくとも46℃まで安定である耐熱性酵素である、[1]又は[2]に記載の方法。
[4] 2−デオキシ−シロ−イノソース合成酵素が、pH6.0〜 8.0で安定な酵素である、[1]から[3]の何れか記載の方法。
[5] 2−デオキシ−シロ−イノソース合成酵素が下記の特性を有する酵素である、[1]又は[2]に記載の方法。
(1)作用:本酵素は、グルコース−6−リン酸を2−デオキシ−シロ−イノソースへと変換する機能を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
[6] 2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12に記載のアミノ酸配列を有する2−デオキシ−シロ−イノソース合成酵素である、[1]から[2]の何れか記載の方法。
[7] 2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12で示されるアミノ酸配列において1個または複数個のアミノ酸の欠失、付加、及び/又は置換を含み、配列番号2、4、6、8、10又は12で示されるアミノ酸配列に対して80%以上の相同性を有して且つ、高温安定性及び/又は広範囲pH安定性を持つ2−デオキシ−シロ−イノソース合成酵素である、[1]から[5]の何れか記載の方法。
【発明の効果】
【0010】
本発明によると、菌体あたりのDOI発酵生産性の向上に寄与することから、短時間で大量のDOIを合成することができる。また、触媒となる菌体量の低減も可能となり、原料利用効率を向上させることもできる。
【図面の簡単な説明】
【0011】
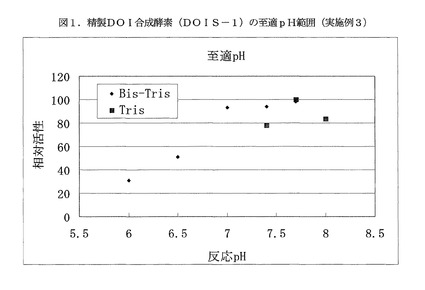
【図1】図1は、精製DOI合成酵素(DOIS−1)の至適pH範囲を示す(実施例3)。
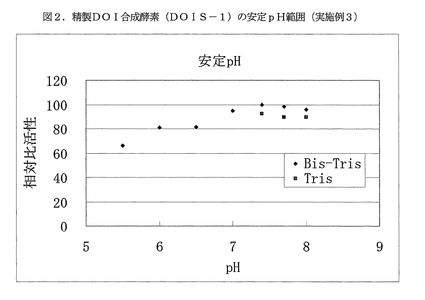
【図2】図2は、精製DOI合成酵素(DOIS−1)の安定pH範囲を示す(実施例3)。
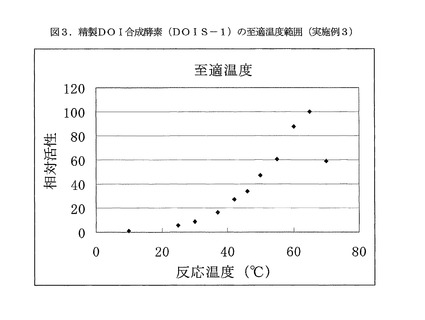
【図3】図3は、精製DOI合成酵素(DOIS−1)の至適温度範囲を示す(実施例3)。
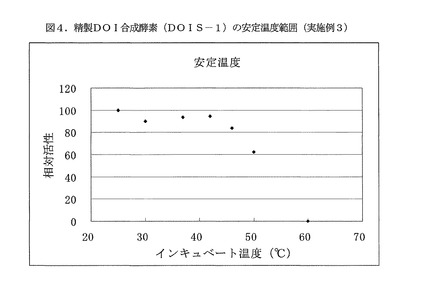
【図4】図4は、精製DOI合成酵素(DOIS−1)の安定温度範囲を示す(実施例3)。
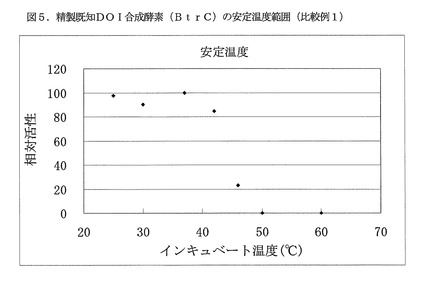
【図5】図5は、精製既知DOI合成酵素(BtrC)の安定温度範囲を示す(比較例1)。
【発明を実施するための形態】
【0012】
以下、本発明について具体的に説明する。
[DOI合成酵素]
本発明を実施するための形態(以下、本実施の形態という)のDOI合成酵素は、下記の特性を有し、
(1)作用:本酵素は、グルコース−6−リン酸をDOIへと変換する機能を有する。
さらには、下記に示す一部またはすべての特性を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
【0013】
配列番号2、4、6、8、10及び12に従来のDOI合成酵素と比較して、耐熱性やpH安定性などの面で優れた新規DOI合成酵素のアミノ酸配列を例示するが、このアミノ酸配列からなるタンパク質がDOI合成酵素活性を有する限り、当該アミノ酸配列において少なくとも1個のアミノ酸に欠失、置換、付加等の変異が生じてもよい。例えば、配列番号2、4、6、8、10又は12に示されるアミノ酸配列に対して80%以上、好ましくは85%以上、さらに好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは99%以上の相同性の範囲内で、配列番号2、4、6、8、10又は12で表されるアミノ酸配列の1個または複数個(例えば1から50個、好ましくは1から30個、さらに好ましくは1から20個、さらに好ましくは1から10個、さらに好ましくは1から5個、さらに好ましくは1から3個))のアミノ酸の欠失、付加、及び/又は置換してもよい。
DOI合成酵素中のアミノ酸に欠失、置換、付加等の変異を生じさせる方法としては、PCR法、エラープローンPCR法、DNAシャッフリング法やキメラ酵素を作製する手法等の公知の方法が利用できる。
上記に、新規DOI合成酵素を示したが、もちろん本発明は従来のDOI合成酵素を用いても良い。
DOI合成酵素活性を測定する方法としては、例えば、1000mMグルコース−6−リン酸溶液20μL、100mMNAD+溶液50μL、100mM塩化コバルト六水和物溶液50μLを含むpH7.0の150mMBis−Tris緩衝液900μLに適当な濃度のDOI合成酵素液100μLを混合し、5〜60分間程度反応させた後、酵素を失活させ、DOIを定量する方法などが挙げられる。
酵素の至適温度範囲あるいは至適pH範囲は、反応温度あるいは反応pHを変化させて酵素活性を測定すればよい。さらにDOI合成酵素を種々のpH条件下又は温度条件下に一定時間さらした後に酵素活性を測定することにより、安定pH範囲及び安定温度範囲を調べることができる。例えば、各pH条件におけるDOI合成酵素の評価には、Bis−Tris緩衝液(pH5.5〜8.0)およびTris緩衝液(pH7.4〜8.0)を用いることができる。
至適pH範囲とは、最大活性を100とするときの活性値が70以上の範囲であり、安定pH範囲とは最大活性を100とするときの活性値が70以上の範囲である。また、至適温度範囲とは、最大活性を100とするときの活性値が50以上の範囲であり、安定温度範囲は最大活性を100とするときの活性値が50以上の範囲である。
本実施の形態におけるDOI合成酵素の高温安定性とは、安定温度範囲が46℃以下、より好ましくは、50℃以下、さらにより好ましくは60℃以下、さらに特に好ましくは95℃以下まで安定なことである。高温安定性を有する2−デオキシ−シロ−イノソース合成酵素としては、例えば、50℃で1時間インキュベートした後の残存酵素活性(最大活性を100とする相対活性)が50以上である酵素を選抜することができる。
本実施の形態における広範囲pH安定性を持つDOI合成酵素は、安定pH範囲が少なくともpH6.0〜7.0、好ましくはpH6.0〜7.4、より好ましくはpH6.0〜7.7、より好ましくはpH6.0〜8.0、特により好ましくはpH4.0〜9.0である特性を有する。
【0014】
[DOI合成酵素遺伝子]
本実施の形態のDOI合成酵素遺伝子は、メタゲノムや微生物から単離・抽出した天然のものでも良く、また、その塩基配列に従ってPCR法、人工合成法等の公知の方法によって合成したものでも良い。また、新規DOI酵素キメラ遺伝子の作成には、既知DOI合成酵素遺伝子を組み合わせてもよく、例示される遺伝子としてはPaenibacillus sp.NBRC13157株、Streptoalloteichus hindustanus JCM3268、Streptomyces fradiae NBRC12773株などに由来するDOI合成酵素遺伝子が挙げられる。
【0015】
配列番号1、3、5、7、9及び11に新規DOI合成酵素遺伝子の塩基配列を例示するが、もちろん本発明は従来のDOI合成酵素遺伝子を用いても良い。
【0016】
[組み換えベクター及び形質転換体]
本実施の形態の組換えベクターは、プラスミド等の公知のベクターに本発明の遺伝子を連結(挿入)して得ることができる。前記ベクターは宿主中で複製可能なものであれば特に限定されず、例えばプラスミドDNA、ファージDNA等が挙げられる。
【0017】
前記プラスミドDNAとしては、大腸菌由来のプラスミド(例えば pBR322, pBR325, pUC18, pUC119, pTrcHis, pBlueBacHis 等)、枯草菌由来のプラスミド(例えば pUB110, pTP5 等)、酵母由来のプラスミド(例えば YEp13, YEp24, YCp50, pYE52 等)などが、ファージ DNAとしてはλファージ等が挙げられる。
【0018】
前記ベクターへの本発明の遺伝子の挿入は、まず、精製されたDNAを適当な制限酵素で切断し、ベクターDNAの適当な制限酵素部位またはマルチクローニングサイトに挿入してベクターに連結する方法が採用される。
【0019】
宿主内で外来遺伝子を発現させるためには、構造遺伝子の前に、適当なプロモーターを配置させる必要がある。前記プロモーターは特に限定されず、宿主内で機能することが知られている任意のものを用いることができる。なおプロモーターについては、後述する形質転換体において、宿主ごとに詳述する。また、必要であればエンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、リボソーム結合配列(SD配列)、ターミネーター配列等を配置させてもよい。
【0020】
本実施の形態の形質転換体は、本発明の組換えベクターを目的遺伝子が発現しうるように宿主中に導入することによって得ることができる。ここで宿主としては、本発明のDNAを発現できるものであれば特に限定されず、例えば、エッシェリヒア・コリ(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロテイ(Rhizobium meliloti)等のリゾビウム属に属する細菌、またサッカロミセス・セルビシエ(Saccharomyces cerevisiae)等の酵母、その他COS細胞、CHO細胞等の動物細胞、あるいはSf19、Sf21等の昆虫細胞を挙げることができる。
【0021】
また、本実施の形態の形質転換体として利用する宿主は、グルコース−6−リン酸をグルコース、フルクトース、ガラクトース、キシロースなどの単糖類から合成する機能を有するものが好ましい。また、必要に応じて、二つ以上の単糖が連結した多糖類を分解する機能を有することも好ましい。
【0022】
大腸菌等の細菌を宿主とする場合は、本発明の組換えベクターが各細菌中で自律複製可能であるとともにプロモーター、リボゾーム結合配列、本発明遺伝子、転写終結配列により構成されていることが望ましい。また、プロモーターを制御する遺伝子が含まれていても良い。大腸菌としてはエッシェリヒア・コリ(E. coli)K12、DH1、DH10B(Invitrogen社)、BL21-CodonPlus(DE3)-RIL(ストラタジーン社)、TOP10F等が挙げられ、枯草菌としてはバチルス・ズブチリス(B. subtilis)MI114、207-21 等が挙げられる。
【0023】
プロモーターとしては大腸菌等の宿主で機能するものであれば特に限定されず、例えばgapAプロモーター、gadAプロモーター、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の大腸菌由来のプロモーターや、T7プロモーター等のファージ由来のプロモーターを用いることができる。
【0024】
細菌への組換えベクターの導入方法は、細菌にDNAを導入できる方法であれば特に限定されないが、例えばカルシウムイオンを用いる方法(Cohen, SN et al. Proc. Natl. Acad. Sci. USA, 69 : 2110 (1972))、エレクトロポレーション法等が挙げられる。
【0025】
酵母を宿主とする場合は、例えばサッカロミセス・セルビシエ(S. cerevisiae)、ピヒア・パストリス(Pichia pastoris)等が用いられる。この場合、プロモーターとしては酵母で発現しうるもの、例えばgal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα 1プロモーター、PHO5プロモーター、AOXプロモーター等を挙げることができる。
【0026】
酵母への組換えベクターの導入方法としては、例えばエレクトロポレーション法(Becker, D.M. et al. : Methods. Enzymol., 194 : 180 (1990))、スフェロプラスト法(Hinnen, A. et al. : Proc Natl. Acad. Sci. USA, 75 : 1929 (1978))、酢酸リチウム法(Itoh, H. : J. Bacteriol., 153 : 163 (1983))等を挙げることができる。
【0027】
[DOI生産法]
本実施の形態のDOI生産法では、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする。なお、本実施の形態で言う反応系とは、細胞内でもよいし、インビトロでもよい。DOI合成酵素を発現可能な微生物を利用する場合、本実施の形態のDOI生産法では、反応系(細胞内)のNADH/NAD比を制御することを除いては、DOI合成酵素産生微生物を常法に従って培養して得られる培養物からDOIを採取することでえられる。
【0028】
本実施の形態における反応系(細胞内)のNADH/NAD比の制御は、菌の培養開始と同時に行っても良いが、より好ましくは、菌の生育およびDOI合成酵素の発現などの効率性を考慮し、DOIの生成濃度が2g/Lに到達してから開始する。また、DOIの最終到達濃度が50g/Lを超えるようなDOI製造プロセスにおいては、DOIの生成濃度が30g/Lに到達してから反応系(細胞内)のNADH/NAD比の制御を開始してもよい。
【0029】
反応系(細胞内)のNADH/NAD比は、好ましくは1.4以下、より好ましくは1.0以下、さらにより好ましくは0.5以下、さらにより好ましくは0.1以下、特により好ましくは0.02以下である。
【0030】
反応系(細胞内)のNADH/NAD比は、制御開始から少なくとも1.4以下に1分間保てばよく、より好ましくは1時間、より好ましくは10時間、さらにより好ましくは24時間、特により好ましくは48時間保てばよい。
【0031】
本実施の形態における反応系(細胞内)のNADH/NAD比を1.4以下に保つ効果的な手法としては、常法に従えばよく、例えば、非特許文献(Metabolic Engineering Volume 7, Issue 2, March 2005, Pages 104-115)に知られているピルビン酸キナーゼ関連遺伝子欠損が利用でき、本手法を用いれば反応系(細胞内)のNADH/NAD比が0.010にできることが知られている。本発明における反応系(細胞内)のNADH/NAD比の測定は、非特許文献(Metabolic Engineering Volume 7, Issue 2, March 2005, Pages 104-115)に記載の手法などで行うことができる。
【0032】
また、本実施の形態における反応系(細胞内)のNADH/NAD比を1.4以下に保つその他の効果的な手法としては、非特許文献(Appl Microbiol Biotechnol. 2004 Apr;64(3):367-75. Epub 2003 Dec 12.)に記載のグルコースを炭素源とした嫌気(微好気)条件下での培養技術を利用することができる。本技術を用いれば、好気条件で、反応系(細胞内)のNADH/NAD比が1.44にも関わらず、これを嫌気培養することにより0.13まで低下させることができる。
【0033】
培養に用いる炭素源としては、形質転換体が資化できるものであればグルコースを構成成分とする多糖類を直接用いてもよく、より好ましくは、単糖類もしくはでんぷんや米ぬかや廃糖蜜などの多糖類を含む原料に由来する単糖類が挙げられ、具体的にはD−グルコースなどが挙げられる。また、プロモーターの発現が誘導型である場合には、適時誘導物質を添加すればよい。炭素源以外の栄養源としては、窒素源、無機塩類、その他有機栄養源を含む培地を使用できる。培養温度はその菌の生育可能な温度であれば良い。培養時間には特に制限はないが、1日から7日程度でよい。その後、得られた培養菌体又は培養上清液からDOIを回収すればよい。
【0034】
炭素源としては、D−グルコース、ガラクトース、マルトース、サッカロース、トレハロース等の糖類や油脂類や脂肪酸類さらにはn−パラフィン等を用いることができる。油脂としては、例えば、ナタネ油、ヤシ油、パーム油、パーム核油などがあげられる。脂肪酸としてはヘキサン酸、オクタン酸、デカン酸、ラウリン酸、オレイン酸、パルミチン酸、リノール酸、リノレン酸、ミリスチン酸などの飽和・不飽和脂肪酸、あるいはこれら脂肪酸のエステルや塩など脂肪酸誘導体などがあげられる。
【0035】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、リン酸アンモニウムなどのアンモニウム塩の他、ペプトン、肉エキス、酵母エキスなどが挙げられる。無機塩類としては、例えば、リン酸水素ナトリウム、リン酸二水素カリウム、塩化マグネシウム、硫酸マグネシウム、塩化ナトリウム、塩化コバルトなどが挙げられる。
【0036】
その他の有機栄養源としては、アミノ酸類、例えば、アデニン、ヒスチジン、ロイシン、ウラシル、トリプトファンなどが挙げられる。
【0037】
生成するDOIの確認は、非特許文献(Journal of Biotechnology 129, 502-509 (2007))に記載の方法などを用いることができる。
【0038】
本実施の形態において、DOIの培養液からの回収は,例えば、次のような方法が使用できる。培養終了後、培養液から遠心分離器や濾過装置などで菌体を除き、培養上清液を得る。この培養上清液に対してさらに濾過処理を行い、菌体等の固形物を除き、その濾液にイオン交換樹脂を添加し、蒸留水で溶出を行う。ICP発光分析、pHなどを測定しながら不純物を含まないフラクションを分取して、その水溶液の溶媒を取り除くことでDOIを回収することができる。得られたDOIの分析は、例えば、高速液体クロマトグラフィーや核磁気共鳴法などにより行う。
【0039】
本実施の形態のインビトロ下でのDOI製造温度は、反応速度や酵素の安定性の点から、好ましくは10〜95℃、より好ましくは20〜70℃であり、さらに好ましくは30〜60℃である。反応pHは広範囲で調整可能であり、酵素の安定性の点から、好ましくはpH2.0〜10.0、より好ましくはpH4.0〜8.0、さらに好ましくは6.0〜8.0である。
【0040】
しかしながら、本発明は以上の各種条件に限定されるものではなく、適宜選択することができる。
【0041】
以下、実施例により具体的に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【実施例】
【0042】
[実施例1]
(1)染色体DNAの調製
常法に従ってPaenibacillus sp.NBRC13157株の染色体DNAを調製した。
Paenibacillus sp.NBRC13157株をNR寒天プレート(1% Bacto Tryptone、0.2% Yeast Extract、1% エルリッヒカツオエキス、1.5% Bacto Agar、pH7.0)で、30℃、1日間培養してコロニーを形成させた。その1白金耳を、NR培地(1% Bacto Tryptone、0.2% Yeast Extract、1% エルリッヒカツオエキス、pH7.0)30mLを150mL三角フラスコに分注したものに接種して、30℃、180rpmで1日間培養した。この培養液を、4℃で、1,2000g×1分間遠心分離して上清を除去し、菌体を回収した。
得られた菌体をLysisバッファー(50mM Tris−HCl(pH 8.0)、20mM EDTA、50mM グルコース)に懸濁し、よく洗浄した。遠心分離して菌体を回収した後、Lysisバッファーに再懸濁し、これにリゾチームを加え37℃で45分間インキュベートした。次いでSDSとRNaseを添加し、37℃で45分間インキュベートした。その後Proteinase Kを添加し、50℃で60分間穏やかに振盪した。ここで得られた溶液をフェノール−クロロホルム、クロロホルムで処理後、エタノール沈殿し、析出した核酸をガラスピペットに巻きつけて回収した。この核酸を70%エタノールで洗浄後、乾燥し、TEに再溶解した。この操作により、約100μgの染色体DNAを調製した。
【0043】
(2)DOI合成酵素遺伝子の単離
上記(1)において調製した染色体DNAからDOI合成酵素遺伝子を増幅するためのPCRプライマーとして、センスプライマーは配列番号15の配列を有するオリゴDNAを、アンチセンスプライマーは配列番号16の配列を有するオリゴDNAをそれぞれ合成した。
ここで得られたPCRプライマーを用い、上記(1)において調製した染色体DNAを鋳型としてPCR法によるDOI合成酵素遺伝子の増幅を行い、1107塩基対からなるPCR産物を取得した。
【0044】
ここで得られたPCR産物の遺伝子配列をDNAシークエンサーで解析することで確認し、配列番号13の塩基配列を有する既知DOI合成酵素btrC遺伝子を得た。また、配列番号14には、既知DOI合成酵素BtrCのアミノ酸配列を示した。
【0045】
(3)DOI合成酵素遺伝子の発現プラスミドベクターの構築及び形質転換
上記(2)で取得したPCR産物の平滑末端化、リン酸化を行い、pUC19に大腸菌由来のgapAプロモーター、SD配列、ターミネーターを連結したプラスミドにライゲーションした。このプラスミドベクターには大腸菌中で外来遺伝子として連結された遺伝子を効率的に転写できるgapAプロモーターが導入されており、グルコースを含む培地で組換え微生物を培養した場合においてもDOI合成酵素遺伝子を効率的に発現・製造させることができる。
ここで得られたプラスミドベクターを、塩化カルシウム法で調製した大腸菌JM109株のコンピテントセルにヒートショック法で形質転換し、組換え微生物を作製した。
【0046】
(4)耐熱性DOI合成酵素遺伝子の取得
上記(3)で取得したプラスミドベクターに対し、常法による変異導入を行うことで、配列番号1(DOIS−1)、配列番号3(DOIS−2)、配列番号5(DOIS−3)、配列番号7(DOIS−4)に記載の耐熱性DOI合成酵素遺伝子を得ることができる。また、これらの遺伝子配列に対応するそれぞれのアミノ酸配列を配列番号2、4、6及び8に示した。ここで取得したDOI合成酵素遺伝子(DOIS−1)についても(3)と同様に形質転換体の取得を行った。
【0047】
(5)広範囲pH安定型DOI合成酵素遺伝子の取得
上記(3)で取得したプラスミドベクターに対し、常法による変異導入を行うことで、配列番号9(DOIS−5)、配列番号11(DOIS−6)、に記載の広範囲pH安定型DOI合成酵素遺伝子を得ることができる。また、これらの遺伝子配列に対応するそれぞれのアミノ酸配列を配列番号10及び12に示した。
【0048】
[実施例2]
[精製酵素の取得]
実施例1で作製したDOI合成酵素(DOIS−1)の形質転換体を100mg/Lのアンピシリンを含むLBプレートで、37℃、一日間培養して、コロニーを形成させた。
次いで100mg/Lのアンピシリンを含むLB培地30mLを150mL容の三角フラスコに入れ、上記プレートからコロニーを白金耳で植菌し、37℃で、3〜8時間、OD(600nm)が0.5程度になるまで180rpmで回転振盪培養を行い、これを本培養の前培養液とした。
【0049】
500mL容の三角フラスコ36本に、2g/Lのグルコースと100mg/Lのアンピシリンを含むLB培地を100mLずつ入れ、それぞれの三角フラスコに0.5mLの前培養液を添加し、37℃で、16時間、180rpmで回転振盪培養を行った。
次いでこの培養液を、4℃で、10,000g×30分間遠心分離して上清を除去し、0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)で数回洗浄しながら菌体を回収した。回収した菌体は、−80℃で凍結保存した。
【0050】
この凍結保存菌体を0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)で懸濁し、リゾチーム(Sigma社製 卵白由来)を180mgとデオキシリボヌクレアーゼI(WaKo社製 ウシ由来 組換え体溶液)を60μL添加して、37℃で、5時間、120rpmで振盪し、菌体破砕を行った。
菌体破砕後の溶液を、4℃で、10,000g×30分間遠心分離して菌体残渣を除去し、上清を回収した。
【0051】
この上清に硫酸アンモニウムを加えて30%飽和とし、4℃でしばらく攪拌した後に、生じた沈殿を、4℃で、10,000g×30分間遠心分離して除去し、上清を回収した。
この上清にさらに硫酸アンモニウムを加えて40.0%飽和とし、4℃でしばらく攪拌した後に、生じた沈殿を、4℃で、10,000g×30分間遠心分離して上清を除去し、沈殿を回収した。次いでこの沈殿を0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)に溶解させた。
【0052】
この溶解液を4℃で、平均分画分子量10,000の限外濾過膜を用いて濃縮し、0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)を加え、再度、濃縮するという脱塩操作を2〜3回繰り返した。
上記で得られた酵素溶液を、50mM Tris−HCl緩衝液(pH7.7)で平衡化した「DEAE Sepharose FF」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、0〜0.4Mの塩化ナトリウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
【0053】
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、10重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)で懸濁し、10重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)で平衡化した「HiTrap Phenyl FF (high sub)」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、10〜0重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、50mM Tris−HCl緩衝液(pH7.7)で平衡化した「MonoQ 5/50 GL」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、0〜0.2Mの塩化ナトリウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
【0054】
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、0.2mg/L塩化コバルト六水和物と0.1MNaClを含む50mM Tris−HCl緩衝液(pH7.7)で平衡化した「HiLoad 16/60 Superdex 200」(商標)(GEヘルスケア バイオサイエンス株式会社)に充填した後、同様の緩衝液で溶出を行った。
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、精製DOI合成酵素(DOIS−1)を得た。
【0055】
上記、一連の精製酵素取得実験と同様にして、精製DOI合成酵素(DOIS−2)、精製DOI合成酵素(DOIS−3)、精製DOI合成酵素(DOI−4)、精製DOI合成酵素(DOI−5)および精製DOI合成酵素(DOI−6)、精製既知DOI合成酵素(BtrC)を取得した。
【0056】
[実施例3]
[新規DOI合成酵素の評価]
実施例2で得られた精製DOI合成酵素を用いて、その作用に関する実験を行った。
【0057】
(1)至適pH範囲
各pHにおける活性測定では、反応液に精製DOI合成酵素(DOIS−1)を適量添加し、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mM、各種緩衝液が100mMとなるようにして反応液を調整した。緩衝液としては、Bis−Tris緩衝液(pH6.0〜7.7)およびTris緩衝液(pH7.4〜8.0) を使用した。反応温度は30℃とし、生成するDOIを定量することで活性を測定した。最大活性を100とする相対活性を求め、この結果を図1に示す。本発明酵素における至適pH範囲は、pH7.0〜 7.7であった。
【0058】
(2)安定pH範囲
pH5.5〜8.0の範囲の100mMの緩衝液を用いて、精製DOI合成酵素(DOIS−1)を各pHで30℃、60分間インキュベートし、その残存酵素活性を測定した。インキュベートに用いた緩衝液としては、Bis−Tris緩衝液(pH5.5〜8.0)およびTris緩衝液(pH7.4〜8.0)を使用した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図2に示した。本酵素の安定pH範囲は、pH6.0〜 8.0であり、非常に幅広いpH範囲で安定であった。精製DOI合成酵素(DOIS−2)および精製DOI合成酵素(DOIS−3)も同様の結果を示した。
【0059】
(3)至適温度範囲
反応温度10〜70℃の各反応温度条件において、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、精製DOI合成酵素(DOIS−1)の酵素活性測定を実施した。最大活性を100とする相対活性を求め、この結果を図3に示す。至適温度範囲は55〜70℃であり、反応温度65℃における比活性は1.8μmol(DOI)/min/mg(酵素)と非常に高い値を示した。
【0060】
(4)安定温度範囲
精製DOI合成酵素(DOIS−1)を添加して且つ、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなるように調整した100mMBis−Tris緩衝液(pH7.0)を、温度範囲25〜60℃の各温度で1時間インキュベートした後、それぞれの残存酵素活性を測定した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図4に示した。精製DOI合成酵素(DOIS−1)の安定温度範囲は50℃以下であり、精製DOI合成酵素(DOIS−2)および精製DOI合成酵素(DOIS−3)も同様の結果を示した。これらの高温安定性は、既知酵素にはない新規特性であった。
【0061】
また、安定化剤として作用することが知られているNAD+およびコバルトイオン非存在化でインキュベートした安定温度範囲試験を実施したところ、精製DOI合成酵素(DOIS−1)のインキュベート温度46℃における相対活性は38、インキュベート温度42℃における相対活性は89、インキュベート温度37℃における相対活性は100であった。
【0062】
(5)分子量
分離ゲル濃度が10%の「レディーゲルJ」(商標)(日本バイオ・ラッド ラボラトリーズ株式会社)を用いるSDS−ポリアクリルアミドゲル電気泳動により、分子量を求めたところ、精製DOI合成酵素(DOIS−1)の分子量は約40,000であり、アミノ酸配列から推定される分子量40,656とほぼ一致した。
【0063】
[比較例1]
[既知DOI合成酵素の評価]
配列番号14に記載のアミノ酸配列を有する精製既知DOI合成酵素(BtrC)に関して、実施例3−(4)安定温度範囲と同様にして以下実験を行った。
精製既知DOI合成酵素(BtrC)を添加して且つ、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなるように調整した100mMBis−Tris緩衝液(pH7.0)を、温度範囲25〜60℃の各温度で1時間インキュベートした後、それぞれの残存酵素活性を測定した。
【0064】
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図5に示した。精製既知DOI合成酵素(BtrC)の安定温度範囲は42℃以下であり、46℃での相対活性は23、50℃での相対活性は0と著しく低い熱安定性を示した。
【0065】
また、安定化剤として作用することが知られているNAD+およびコバルトイオン非存在化でインキュベートした安定温度範囲試験を実施したところ、精製既知DOI合成酵素(BtrC)のインキュベート温度42℃および46℃における相対活性は0、インキュベート温度37℃における相対活性は14であった。本実験からも既存酵素が著しく低い熱安定性を有することが示された。
【0066】
配列番号14に記載のアミノ酸配列を有する精製既知DOI合成酵素(BtrC)に関して、実施例3−(2)安定pH範囲と同様にして以下実験を行った。
精製既知DOI合成酵素(BtrC)を各pHで30℃、60分間インキュベートし、その残存酵素活性を測定した。インキュベートに用いた緩衝液としては、Bis−Tris緩衝液(pH5.5〜7.0)およびTris緩衝液(pH7.4〜8.0)を使用した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求めたところ、pH6における相対活性は27と非常に低い値であった。
【0067】
[実施例4]
[NADH/NAD比の酵素活性依存性]
NAD+およびNADHを表1と表2に記載の濃度となるように調整し且つ、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、精製DOI合成酵素(DOIS−1)の酵素活性測定を実施した。NADH/NAD比が0となる際の活性を100とするときの活性値をそれぞれ求め、この結果を表1および表2に示した。
表1および表2に示すように、NADH/NAD比にDOI合成酵素活性は非常に大きな影響を受けることが明らかとなった。
【0068】
【表1】

【0069】
【表2】
【産業上の利用可能性】
【0070】
本発明を用いることにより、利用価値の高いDOIを効率的な製造プロセスを用いて生産することができる。
【技術分野】
【0001】
本発明は、2−デオキシ−シロ−イノソース合成酵素を用いて2−デオキシ−シロ−イノソースを製造する方法においてNADH/NAD比を制御することを特徴とする2−デオキシ−シロ−イノソースの製造方法に関する。
【背景技術】
【0002】
現在、カテコールなどの芳香族化合物の多くは、石油を原料として生産されているが、石油資源の枯渇問題や二酸化炭素排出量の削減などの観点から、バイオマスを利用した環境調和型の新規製造プロセスの開発が求められている。
【0003】
一方、2−デオキシ−シロ−イノソース(以下、DOI)が、産業上有用とされる芳香族化合物(カテコール等)へと変換できることが見出され(非特許文献1)、さらには、このDOIを、バイオマスの構成要素の一つであるグルコースから合成できることが報告された(非特許文献1)。本DOI製造プロセスで重要となるのがグルコース−6−リン酸からDOIへと変換する2−デオキシ−シロ−イノソース合成酵素(以下、DOI合成酵素)であり、DOIは上記芳香族化合物への変換のみならず各種有用化合物の中間体ともなり得ることから、DOI合成酵素は非常に大きな注目を浴びている。
【0004】
DOI合成酵素はブチロシン生産菌であるBacillus circulans に属する微生物から1997年に単離精製(非特許文献2)され、続いて、その遺伝子配列も公開されている(特許文献1)。この他にもStreptoalloteichus hindustanus JCM3268株(非特許文献3)由来のDOI合成酵素などがこれまでに発見されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2000−236881号公報
【非特許文献】
【0006】
【非特許文献1】Tetrahedron Letters 41,1935−1938,2000
【非特許文献2】The Journal of Antibiotics 50(5), 424−428, 1997
【非特許文献3】The Journal of Antibiotics 59(6), 358−361, 2006
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、前記DOI合成酵素の安定性と比活性などの各種特性は、満足のいくものではなく、また、数千トン規模の工業化製法に向けたDOI発酵生産に関する研究開発、特に細胞内代謝物の制御に関するものはほとんどなされてこなかった。
本発明は、従来法と比較して、DOI発酵生産性を飛躍的に向上させうる細胞内代謝産物制御技術を提供することを解決すべき課題とする。
【課題を解決するための手段】
【0008】
これらの課題を解決するため鋭意検討を重ねた結果、本発明者らは、DOI合成酵素の活性が反応系中のNADH/NAD比に予想し得ない領域で影響を受けることを見出し、従来法と比較して、DOI発酵生産性を飛躍的に向上させうるDOI生産技術を見出し、本発明を完成するに至った。
【0009】
すなわち、本発明は以下の[1]〜[7]に示すDOI製造方法に関するものである。
[1] 2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
[2] グルコースを構成成分とする多糖類あるいはグルコースを含有する原料を用いることを特徴とする、[1]に記載の方法。
[3] 2−デオキシ−シロ−イノソース合成酵素が、安定温度範囲が少なくとも46℃まで安定である耐熱性酵素である、[1]又は[2]に記載の方法。
[4] 2−デオキシ−シロ−イノソース合成酵素が、pH6.0〜 8.0で安定な酵素である、[1]から[3]の何れか記載の方法。
[5] 2−デオキシ−シロ−イノソース合成酵素が下記の特性を有する酵素である、[1]又は[2]に記載の方法。
(1)作用:本酵素は、グルコース−6−リン酸を2−デオキシ−シロ−イノソースへと変換する機能を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
[6] 2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12に記載のアミノ酸配列を有する2−デオキシ−シロ−イノソース合成酵素である、[1]から[2]の何れか記載の方法。
[7] 2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12で示されるアミノ酸配列において1個または複数個のアミノ酸の欠失、付加、及び/又は置換を含み、配列番号2、4、6、8、10又は12で示されるアミノ酸配列に対して80%以上の相同性を有して且つ、高温安定性及び/又は広範囲pH安定性を持つ2−デオキシ−シロ−イノソース合成酵素である、[1]から[5]の何れか記載の方法。
【発明の効果】
【0010】
本発明によると、菌体あたりのDOI発酵生産性の向上に寄与することから、短時間で大量のDOIを合成することができる。また、触媒となる菌体量の低減も可能となり、原料利用効率を向上させることもできる。
【図面の簡単な説明】
【0011】
【図1】図1は、精製DOI合成酵素(DOIS−1)の至適pH範囲を示す(実施例3)。
【図2】図2は、精製DOI合成酵素(DOIS−1)の安定pH範囲を示す(実施例3)。
【図3】図3は、精製DOI合成酵素(DOIS−1)の至適温度範囲を示す(実施例3)。
【図4】図4は、精製DOI合成酵素(DOIS−1)の安定温度範囲を示す(実施例3)。
【図5】図5は、精製既知DOI合成酵素(BtrC)の安定温度範囲を示す(比較例1)。
【発明を実施するための形態】
【0012】
以下、本発明について具体的に説明する。
[DOI合成酵素]
本発明を実施するための形態(以下、本実施の形態という)のDOI合成酵素は、下記の特性を有し、
(1)作用:本酵素は、グルコース−6−リン酸をDOIへと変換する機能を有する。
さらには、下記に示す一部またはすべての特性を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
【0013】
配列番号2、4、6、8、10及び12に従来のDOI合成酵素と比較して、耐熱性やpH安定性などの面で優れた新規DOI合成酵素のアミノ酸配列を例示するが、このアミノ酸配列からなるタンパク質がDOI合成酵素活性を有する限り、当該アミノ酸配列において少なくとも1個のアミノ酸に欠失、置換、付加等の変異が生じてもよい。例えば、配列番号2、4、6、8、10又は12に示されるアミノ酸配列に対して80%以上、好ましくは85%以上、さらに好ましくは90%以上、さらに好ましくは95%以上、さらに好ましくは99%以上の相同性の範囲内で、配列番号2、4、6、8、10又は12で表されるアミノ酸配列の1個または複数個(例えば1から50個、好ましくは1から30個、さらに好ましくは1から20個、さらに好ましくは1から10個、さらに好ましくは1から5個、さらに好ましくは1から3個))のアミノ酸の欠失、付加、及び/又は置換してもよい。
DOI合成酵素中のアミノ酸に欠失、置換、付加等の変異を生じさせる方法としては、PCR法、エラープローンPCR法、DNAシャッフリング法やキメラ酵素を作製する手法等の公知の方法が利用できる。
上記に、新規DOI合成酵素を示したが、もちろん本発明は従来のDOI合成酵素を用いても良い。
DOI合成酵素活性を測定する方法としては、例えば、1000mMグルコース−6−リン酸溶液20μL、100mMNAD+溶液50μL、100mM塩化コバルト六水和物溶液50μLを含むpH7.0の150mMBis−Tris緩衝液900μLに適当な濃度のDOI合成酵素液100μLを混合し、5〜60分間程度反応させた後、酵素を失活させ、DOIを定量する方法などが挙げられる。
酵素の至適温度範囲あるいは至適pH範囲は、反応温度あるいは反応pHを変化させて酵素活性を測定すればよい。さらにDOI合成酵素を種々のpH条件下又は温度条件下に一定時間さらした後に酵素活性を測定することにより、安定pH範囲及び安定温度範囲を調べることができる。例えば、各pH条件におけるDOI合成酵素の評価には、Bis−Tris緩衝液(pH5.5〜8.0)およびTris緩衝液(pH7.4〜8.0)を用いることができる。
至適pH範囲とは、最大活性を100とするときの活性値が70以上の範囲であり、安定pH範囲とは最大活性を100とするときの活性値が70以上の範囲である。また、至適温度範囲とは、最大活性を100とするときの活性値が50以上の範囲であり、安定温度範囲は最大活性を100とするときの活性値が50以上の範囲である。
本実施の形態におけるDOI合成酵素の高温安定性とは、安定温度範囲が46℃以下、より好ましくは、50℃以下、さらにより好ましくは60℃以下、さらに特に好ましくは95℃以下まで安定なことである。高温安定性を有する2−デオキシ−シロ−イノソース合成酵素としては、例えば、50℃で1時間インキュベートした後の残存酵素活性(最大活性を100とする相対活性)が50以上である酵素を選抜することができる。
本実施の形態における広範囲pH安定性を持つDOI合成酵素は、安定pH範囲が少なくともpH6.0〜7.0、好ましくはpH6.0〜7.4、より好ましくはpH6.0〜7.7、より好ましくはpH6.0〜8.0、特により好ましくはpH4.0〜9.0である特性を有する。
【0014】
[DOI合成酵素遺伝子]
本実施の形態のDOI合成酵素遺伝子は、メタゲノムや微生物から単離・抽出した天然のものでも良く、また、その塩基配列に従ってPCR法、人工合成法等の公知の方法によって合成したものでも良い。また、新規DOI酵素キメラ遺伝子の作成には、既知DOI合成酵素遺伝子を組み合わせてもよく、例示される遺伝子としてはPaenibacillus sp.NBRC13157株、Streptoalloteichus hindustanus JCM3268、Streptomyces fradiae NBRC12773株などに由来するDOI合成酵素遺伝子が挙げられる。
【0015】
配列番号1、3、5、7、9及び11に新規DOI合成酵素遺伝子の塩基配列を例示するが、もちろん本発明は従来のDOI合成酵素遺伝子を用いても良い。
【0016】
[組み換えベクター及び形質転換体]
本実施の形態の組換えベクターは、プラスミド等の公知のベクターに本発明の遺伝子を連結(挿入)して得ることができる。前記ベクターは宿主中で複製可能なものであれば特に限定されず、例えばプラスミドDNA、ファージDNA等が挙げられる。
【0017】
前記プラスミドDNAとしては、大腸菌由来のプラスミド(例えば pBR322, pBR325, pUC18, pUC119, pTrcHis, pBlueBacHis 等)、枯草菌由来のプラスミド(例えば pUB110, pTP5 等)、酵母由来のプラスミド(例えば YEp13, YEp24, YCp50, pYE52 等)などが、ファージ DNAとしてはλファージ等が挙げられる。
【0018】
前記ベクターへの本発明の遺伝子の挿入は、まず、精製されたDNAを適当な制限酵素で切断し、ベクターDNAの適当な制限酵素部位またはマルチクローニングサイトに挿入してベクターに連結する方法が採用される。
【0019】
宿主内で外来遺伝子を発現させるためには、構造遺伝子の前に、適当なプロモーターを配置させる必要がある。前記プロモーターは特に限定されず、宿主内で機能することが知られている任意のものを用いることができる。なおプロモーターについては、後述する形質転換体において、宿主ごとに詳述する。また、必要であればエンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、リボソーム結合配列(SD配列)、ターミネーター配列等を配置させてもよい。
【0020】
本実施の形態の形質転換体は、本発明の組換えベクターを目的遺伝子が発現しうるように宿主中に導入することによって得ることができる。ここで宿主としては、本発明のDNAを発現できるものであれば特に限定されず、例えば、エッシェリヒア・コリ(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロテイ(Rhizobium meliloti)等のリゾビウム属に属する細菌、またサッカロミセス・セルビシエ(Saccharomyces cerevisiae)等の酵母、その他COS細胞、CHO細胞等の動物細胞、あるいはSf19、Sf21等の昆虫細胞を挙げることができる。
【0021】
また、本実施の形態の形質転換体として利用する宿主は、グルコース−6−リン酸をグルコース、フルクトース、ガラクトース、キシロースなどの単糖類から合成する機能を有するものが好ましい。また、必要に応じて、二つ以上の単糖が連結した多糖類を分解する機能を有することも好ましい。
【0022】
大腸菌等の細菌を宿主とする場合は、本発明の組換えベクターが各細菌中で自律複製可能であるとともにプロモーター、リボゾーム結合配列、本発明遺伝子、転写終結配列により構成されていることが望ましい。また、プロモーターを制御する遺伝子が含まれていても良い。大腸菌としてはエッシェリヒア・コリ(E. coli)K12、DH1、DH10B(Invitrogen社)、BL21-CodonPlus(DE3)-RIL(ストラタジーン社)、TOP10F等が挙げられ、枯草菌としてはバチルス・ズブチリス(B. subtilis)MI114、207-21 等が挙げられる。
【0023】
プロモーターとしては大腸菌等の宿主で機能するものであれば特に限定されず、例えばgapAプロモーター、gadAプロモーター、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の大腸菌由来のプロモーターや、T7プロモーター等のファージ由来のプロモーターを用いることができる。
【0024】
細菌への組換えベクターの導入方法は、細菌にDNAを導入できる方法であれば特に限定されないが、例えばカルシウムイオンを用いる方法(Cohen, SN et al. Proc. Natl. Acad. Sci. USA, 69 : 2110 (1972))、エレクトロポレーション法等が挙げられる。
【0025】
酵母を宿主とする場合は、例えばサッカロミセス・セルビシエ(S. cerevisiae)、ピヒア・パストリス(Pichia pastoris)等が用いられる。この場合、プロモーターとしては酵母で発現しうるもの、例えばgal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα 1プロモーター、PHO5プロモーター、AOXプロモーター等を挙げることができる。
【0026】
酵母への組換えベクターの導入方法としては、例えばエレクトロポレーション法(Becker, D.M. et al. : Methods. Enzymol., 194 : 180 (1990))、スフェロプラスト法(Hinnen, A. et al. : Proc Natl. Acad. Sci. USA, 75 : 1929 (1978))、酢酸リチウム法(Itoh, H. : J. Bacteriol., 153 : 163 (1983))等を挙げることができる。
【0027】
[DOI生産法]
本実施の形態のDOI生産法では、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする。なお、本実施の形態で言う反応系とは、細胞内でもよいし、インビトロでもよい。DOI合成酵素を発現可能な微生物を利用する場合、本実施の形態のDOI生産法では、反応系(細胞内)のNADH/NAD比を制御することを除いては、DOI合成酵素産生微生物を常法に従って培養して得られる培養物からDOIを採取することでえられる。
【0028】
本実施の形態における反応系(細胞内)のNADH/NAD比の制御は、菌の培養開始と同時に行っても良いが、より好ましくは、菌の生育およびDOI合成酵素の発現などの効率性を考慮し、DOIの生成濃度が2g/Lに到達してから開始する。また、DOIの最終到達濃度が50g/Lを超えるようなDOI製造プロセスにおいては、DOIの生成濃度が30g/Lに到達してから反応系(細胞内)のNADH/NAD比の制御を開始してもよい。
【0029】
反応系(細胞内)のNADH/NAD比は、好ましくは1.4以下、より好ましくは1.0以下、さらにより好ましくは0.5以下、さらにより好ましくは0.1以下、特により好ましくは0.02以下である。
【0030】
反応系(細胞内)のNADH/NAD比は、制御開始から少なくとも1.4以下に1分間保てばよく、より好ましくは1時間、より好ましくは10時間、さらにより好ましくは24時間、特により好ましくは48時間保てばよい。
【0031】
本実施の形態における反応系(細胞内)のNADH/NAD比を1.4以下に保つ効果的な手法としては、常法に従えばよく、例えば、非特許文献(Metabolic Engineering Volume 7, Issue 2, March 2005, Pages 104-115)に知られているピルビン酸キナーゼ関連遺伝子欠損が利用でき、本手法を用いれば反応系(細胞内)のNADH/NAD比が0.010にできることが知られている。本発明における反応系(細胞内)のNADH/NAD比の測定は、非特許文献(Metabolic Engineering Volume 7, Issue 2, March 2005, Pages 104-115)に記載の手法などで行うことができる。
【0032】
また、本実施の形態における反応系(細胞内)のNADH/NAD比を1.4以下に保つその他の効果的な手法としては、非特許文献(Appl Microbiol Biotechnol. 2004 Apr;64(3):367-75. Epub 2003 Dec 12.)に記載のグルコースを炭素源とした嫌気(微好気)条件下での培養技術を利用することができる。本技術を用いれば、好気条件で、反応系(細胞内)のNADH/NAD比が1.44にも関わらず、これを嫌気培養することにより0.13まで低下させることができる。
【0033】
培養に用いる炭素源としては、形質転換体が資化できるものであればグルコースを構成成分とする多糖類を直接用いてもよく、より好ましくは、単糖類もしくはでんぷんや米ぬかや廃糖蜜などの多糖類を含む原料に由来する単糖類が挙げられ、具体的にはD−グルコースなどが挙げられる。また、プロモーターの発現が誘導型である場合には、適時誘導物質を添加すればよい。炭素源以外の栄養源としては、窒素源、無機塩類、その他有機栄養源を含む培地を使用できる。培養温度はその菌の生育可能な温度であれば良い。培養時間には特に制限はないが、1日から7日程度でよい。その後、得られた培養菌体又は培養上清液からDOIを回収すればよい。
【0034】
炭素源としては、D−グルコース、ガラクトース、マルトース、サッカロース、トレハロース等の糖類や油脂類や脂肪酸類さらにはn−パラフィン等を用いることができる。油脂としては、例えば、ナタネ油、ヤシ油、パーム油、パーム核油などがあげられる。脂肪酸としてはヘキサン酸、オクタン酸、デカン酸、ラウリン酸、オレイン酸、パルミチン酸、リノール酸、リノレン酸、ミリスチン酸などの飽和・不飽和脂肪酸、あるいはこれら脂肪酸のエステルや塩など脂肪酸誘導体などがあげられる。
【0035】
窒素源としては、例えば、アンモニア、塩化アンモニウム、硫酸アンモニウム、リン酸アンモニウムなどのアンモニウム塩の他、ペプトン、肉エキス、酵母エキスなどが挙げられる。無機塩類としては、例えば、リン酸水素ナトリウム、リン酸二水素カリウム、塩化マグネシウム、硫酸マグネシウム、塩化ナトリウム、塩化コバルトなどが挙げられる。
【0036】
その他の有機栄養源としては、アミノ酸類、例えば、アデニン、ヒスチジン、ロイシン、ウラシル、トリプトファンなどが挙げられる。
【0037】
生成するDOIの確認は、非特許文献(Journal of Biotechnology 129, 502-509 (2007))に記載の方法などを用いることができる。
【0038】
本実施の形態において、DOIの培養液からの回収は,例えば、次のような方法が使用できる。培養終了後、培養液から遠心分離器や濾過装置などで菌体を除き、培養上清液を得る。この培養上清液に対してさらに濾過処理を行い、菌体等の固形物を除き、その濾液にイオン交換樹脂を添加し、蒸留水で溶出を行う。ICP発光分析、pHなどを測定しながら不純物を含まないフラクションを分取して、その水溶液の溶媒を取り除くことでDOIを回収することができる。得られたDOIの分析は、例えば、高速液体クロマトグラフィーや核磁気共鳴法などにより行う。
【0039】
本実施の形態のインビトロ下でのDOI製造温度は、反応速度や酵素の安定性の点から、好ましくは10〜95℃、より好ましくは20〜70℃であり、さらに好ましくは30〜60℃である。反応pHは広範囲で調整可能であり、酵素の安定性の点から、好ましくはpH2.0〜10.0、より好ましくはpH4.0〜8.0、さらに好ましくは6.0〜8.0である。
【0040】
しかしながら、本発明は以上の各種条件に限定されるものではなく、適宜選択することができる。
【0041】
以下、実施例により具体的に説明するが、本発明はこれらの実施例により何ら限定されるものではない。
【実施例】
【0042】
[実施例1]
(1)染色体DNAの調製
常法に従ってPaenibacillus sp.NBRC13157株の染色体DNAを調製した。
Paenibacillus sp.NBRC13157株をNR寒天プレート(1% Bacto Tryptone、0.2% Yeast Extract、1% エルリッヒカツオエキス、1.5% Bacto Agar、pH7.0)で、30℃、1日間培養してコロニーを形成させた。その1白金耳を、NR培地(1% Bacto Tryptone、0.2% Yeast Extract、1% エルリッヒカツオエキス、pH7.0)30mLを150mL三角フラスコに分注したものに接種して、30℃、180rpmで1日間培養した。この培養液を、4℃で、1,2000g×1分間遠心分離して上清を除去し、菌体を回収した。
得られた菌体をLysisバッファー(50mM Tris−HCl(pH 8.0)、20mM EDTA、50mM グルコース)に懸濁し、よく洗浄した。遠心分離して菌体を回収した後、Lysisバッファーに再懸濁し、これにリゾチームを加え37℃で45分間インキュベートした。次いでSDSとRNaseを添加し、37℃で45分間インキュベートした。その後Proteinase Kを添加し、50℃で60分間穏やかに振盪した。ここで得られた溶液をフェノール−クロロホルム、クロロホルムで処理後、エタノール沈殿し、析出した核酸をガラスピペットに巻きつけて回収した。この核酸を70%エタノールで洗浄後、乾燥し、TEに再溶解した。この操作により、約100μgの染色体DNAを調製した。
【0043】
(2)DOI合成酵素遺伝子の単離
上記(1)において調製した染色体DNAからDOI合成酵素遺伝子を増幅するためのPCRプライマーとして、センスプライマーは配列番号15の配列を有するオリゴDNAを、アンチセンスプライマーは配列番号16の配列を有するオリゴDNAをそれぞれ合成した。
ここで得られたPCRプライマーを用い、上記(1)において調製した染色体DNAを鋳型としてPCR法によるDOI合成酵素遺伝子の増幅を行い、1107塩基対からなるPCR産物を取得した。
【0044】
ここで得られたPCR産物の遺伝子配列をDNAシークエンサーで解析することで確認し、配列番号13の塩基配列を有する既知DOI合成酵素btrC遺伝子を得た。また、配列番号14には、既知DOI合成酵素BtrCのアミノ酸配列を示した。
【0045】
(3)DOI合成酵素遺伝子の発現プラスミドベクターの構築及び形質転換
上記(2)で取得したPCR産物の平滑末端化、リン酸化を行い、pUC19に大腸菌由来のgapAプロモーター、SD配列、ターミネーターを連結したプラスミドにライゲーションした。このプラスミドベクターには大腸菌中で外来遺伝子として連結された遺伝子を効率的に転写できるgapAプロモーターが導入されており、グルコースを含む培地で組換え微生物を培養した場合においてもDOI合成酵素遺伝子を効率的に発現・製造させることができる。
ここで得られたプラスミドベクターを、塩化カルシウム法で調製した大腸菌JM109株のコンピテントセルにヒートショック法で形質転換し、組換え微生物を作製した。
【0046】
(4)耐熱性DOI合成酵素遺伝子の取得
上記(3)で取得したプラスミドベクターに対し、常法による変異導入を行うことで、配列番号1(DOIS−1)、配列番号3(DOIS−2)、配列番号5(DOIS−3)、配列番号7(DOIS−4)に記載の耐熱性DOI合成酵素遺伝子を得ることができる。また、これらの遺伝子配列に対応するそれぞれのアミノ酸配列を配列番号2、4、6及び8に示した。ここで取得したDOI合成酵素遺伝子(DOIS−1)についても(3)と同様に形質転換体の取得を行った。
【0047】
(5)広範囲pH安定型DOI合成酵素遺伝子の取得
上記(3)で取得したプラスミドベクターに対し、常法による変異導入を行うことで、配列番号9(DOIS−5)、配列番号11(DOIS−6)、に記載の広範囲pH安定型DOI合成酵素遺伝子を得ることができる。また、これらの遺伝子配列に対応するそれぞれのアミノ酸配列を配列番号10及び12に示した。
【0048】
[実施例2]
[精製酵素の取得]
実施例1で作製したDOI合成酵素(DOIS−1)の形質転換体を100mg/Lのアンピシリンを含むLBプレートで、37℃、一日間培養して、コロニーを形成させた。
次いで100mg/Lのアンピシリンを含むLB培地30mLを150mL容の三角フラスコに入れ、上記プレートからコロニーを白金耳で植菌し、37℃で、3〜8時間、OD(600nm)が0.5程度になるまで180rpmで回転振盪培養を行い、これを本培養の前培養液とした。
【0049】
500mL容の三角フラスコ36本に、2g/Lのグルコースと100mg/Lのアンピシリンを含むLB培地を100mLずつ入れ、それぞれの三角フラスコに0.5mLの前培養液を添加し、37℃で、16時間、180rpmで回転振盪培養を行った。
次いでこの培養液を、4℃で、10,000g×30分間遠心分離して上清を除去し、0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)で数回洗浄しながら菌体を回収した。回収した菌体は、−80℃で凍結保存した。
【0050】
この凍結保存菌体を0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)で懸濁し、リゾチーム(Sigma社製 卵白由来)を180mgとデオキシリボヌクレアーゼI(WaKo社製 ウシ由来 組換え体溶液)を60μL添加して、37℃で、5時間、120rpmで振盪し、菌体破砕を行った。
菌体破砕後の溶液を、4℃で、10,000g×30分間遠心分離して菌体残渣を除去し、上清を回収した。
【0051】
この上清に硫酸アンモニウムを加えて30%飽和とし、4℃でしばらく攪拌した後に、生じた沈殿を、4℃で、10,000g×30分間遠心分離して除去し、上清を回収した。
この上清にさらに硫酸アンモニウムを加えて40.0%飽和とし、4℃でしばらく攪拌した後に、生じた沈殿を、4℃で、10,000g×30分間遠心分離して上清を除去し、沈殿を回収した。次いでこの沈殿を0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)に溶解させた。
【0052】
この溶解液を4℃で、平均分画分子量10,000の限外濾過膜を用いて濃縮し、0.2mg/L塩化コバルト六水和物を含む50mM Tris−HCl緩衝液(pH7.7)を加え、再度、濃縮するという脱塩操作を2〜3回繰り返した。
上記で得られた酵素溶液を、50mM Tris−HCl緩衝液(pH7.7)で平衡化した「DEAE Sepharose FF」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、0〜0.4Mの塩化ナトリウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
【0053】
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、10重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)で懸濁し、10重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)で平衡化した「HiTrap Phenyl FF (high sub)」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、10〜0重量%の硫酸アンモニウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、50mM Tris−HCl緩衝液(pH7.7)で平衡化した「MonoQ 5/50 GL」(商標)(GEヘルスケア バイオサイエンス株式会社)に吸着させた後、0〜0.2Mの塩化ナトリウムを含有する50mM Tris−HCl緩衝液(pH7.7)の濃度勾配法によって酵素を溶出させた。
【0054】
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、0.2mg/L塩化コバルト六水和物と0.1MNaClを含む50mM Tris−HCl緩衝液(pH7.7)で平衡化した「HiLoad 16/60 Superdex 200」(商標)(GEヘルスケア バイオサイエンス株式会社)に充填した後、同様の緩衝液で溶出を行った。
上記で溶出した活性画分を集め、平均分画分子量10,000の限外濾過膜を用いて濃縮し、精製DOI合成酵素(DOIS−1)を得た。
【0055】
上記、一連の精製酵素取得実験と同様にして、精製DOI合成酵素(DOIS−2)、精製DOI合成酵素(DOIS−3)、精製DOI合成酵素(DOI−4)、精製DOI合成酵素(DOI−5)および精製DOI合成酵素(DOI−6)、精製既知DOI合成酵素(BtrC)を取得した。
【0056】
[実施例3]
[新規DOI合成酵素の評価]
実施例2で得られた精製DOI合成酵素を用いて、その作用に関する実験を行った。
【0057】
(1)至適pH範囲
各pHにおける活性測定では、反応液に精製DOI合成酵素(DOIS−1)を適量添加し、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mM、各種緩衝液が100mMとなるようにして反応液を調整した。緩衝液としては、Bis−Tris緩衝液(pH6.0〜7.7)およびTris緩衝液(pH7.4〜8.0) を使用した。反応温度は30℃とし、生成するDOIを定量することで活性を測定した。最大活性を100とする相対活性を求め、この結果を図1に示す。本発明酵素における至適pH範囲は、pH7.0〜 7.7であった。
【0058】
(2)安定pH範囲
pH5.5〜8.0の範囲の100mMの緩衝液を用いて、精製DOI合成酵素(DOIS−1)を各pHで30℃、60分間インキュベートし、その残存酵素活性を測定した。インキュベートに用いた緩衝液としては、Bis−Tris緩衝液(pH5.5〜8.0)およびTris緩衝液(pH7.4〜8.0)を使用した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図2に示した。本酵素の安定pH範囲は、pH6.0〜 8.0であり、非常に幅広いpH範囲で安定であった。精製DOI合成酵素(DOIS−2)および精製DOI合成酵素(DOIS−3)も同様の結果を示した。
【0059】
(3)至適温度範囲
反応温度10〜70℃の各反応温度条件において、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、精製DOI合成酵素(DOIS−1)の酵素活性測定を実施した。最大活性を100とする相対活性を求め、この結果を図3に示す。至適温度範囲は55〜70℃であり、反応温度65℃における比活性は1.8μmol(DOI)/min/mg(酵素)と非常に高い値を示した。
【0060】
(4)安定温度範囲
精製DOI合成酵素(DOIS−1)を添加して且つ、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなるように調整した100mMBis−Tris緩衝液(pH7.0)を、温度範囲25〜60℃の各温度で1時間インキュベートした後、それぞれの残存酵素活性を測定した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図4に示した。精製DOI合成酵素(DOIS−1)の安定温度範囲は50℃以下であり、精製DOI合成酵素(DOIS−2)および精製DOI合成酵素(DOIS−3)も同様の結果を示した。これらの高温安定性は、既知酵素にはない新規特性であった。
【0061】
また、安定化剤として作用することが知られているNAD+およびコバルトイオン非存在化でインキュベートした安定温度範囲試験を実施したところ、精製DOI合成酵素(DOIS−1)のインキュベート温度46℃における相対活性は38、インキュベート温度42℃における相対活性は89、インキュベート温度37℃における相対活性は100であった。
【0062】
(5)分子量
分離ゲル濃度が10%の「レディーゲルJ」(商標)(日本バイオ・ラッド ラボラトリーズ株式会社)を用いるSDS−ポリアクリルアミドゲル電気泳動により、分子量を求めたところ、精製DOI合成酵素(DOIS−1)の分子量は約40,000であり、アミノ酸配列から推定される分子量40,656とほぼ一致した。
【0063】
[比較例1]
[既知DOI合成酵素の評価]
配列番号14に記載のアミノ酸配列を有する精製既知DOI合成酵素(BtrC)に関して、実施例3−(4)安定温度範囲と同様にして以下実験を行った。
精製既知DOI合成酵素(BtrC)を添加して且つ、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなるように調整した100mMBis−Tris緩衝液(pH7.0)を、温度範囲25〜60℃の各温度で1時間インキュベートした後、それぞれの残存酵素活性を測定した。
【0064】
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求め、この結果を図5に示した。精製既知DOI合成酵素(BtrC)の安定温度範囲は42℃以下であり、46℃での相対活性は23、50℃での相対活性は0と著しく低い熱安定性を示した。
【0065】
また、安定化剤として作用することが知られているNAD+およびコバルトイオン非存在化でインキュベートした安定温度範囲試験を実施したところ、精製既知DOI合成酵素(BtrC)のインキュベート温度42℃および46℃における相対活性は0、インキュベート温度37℃における相対活性は14であった。本実験からも既存酵素が著しく低い熱安定性を有することが示された。
【0066】
配列番号14に記載のアミノ酸配列を有する精製既知DOI合成酵素(BtrC)に関して、実施例3−(2)安定pH範囲と同様にして以下実験を行った。
精製既知DOI合成酵素(BtrC)を各pHで30℃、60分間インキュベートし、その残存酵素活性を測定した。インキュベートに用いた緩衝液としては、Bis−Tris緩衝液(pH5.5〜7.0)およびTris緩衝液(pH7.4〜8.0)を使用した。
残存酵素活性測定は、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、NAD+(オリエンタル酵母製)が5mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、反応温度30℃で実施した。最大活性を100とする相対活性を求めたところ、pH6における相対活性は27と非常に低い値であった。
【0067】
[実施例4]
[NADH/NAD比の酵素活性依存性]
NAD+およびNADHを表1と表2に記載の濃度となるように調整し且つ、グルコース−6−リン酸二ナトリウム塩(オリエンタル酵母製)が20mM、塩化コバルト六水和物が5mMとなる100mMBis−Tris緩衝液(pH7.0)中で、精製DOI合成酵素(DOIS−1)の酵素活性測定を実施した。NADH/NAD比が0となる際の活性を100とするときの活性値をそれぞれ求め、この結果を表1および表2に示した。
表1および表2に示すように、NADH/NAD比にDOI合成酵素活性は非常に大きな影響を受けることが明らかとなった。
【0068】
【表1】
【0069】
【表2】
【産業上の利用可能性】
【0070】
本発明を用いることにより、利用価値の高いDOIを効率的な製造プロセスを用いて生産することができる。
【特許請求の範囲】
【請求項1】
2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
【請求項2】
グルコースを構成成分とする多糖類あるいはグルコースを含有する原料を用いることを特徴とする、請求項1に記載の方法。
【請求項3】
2−デオキシ−シロ−イノソース合成酵素が、安定温度範囲が少なくとも46℃まで安定である耐熱性酵素である、請求項1又は2に記載の方法。
【請求項4】
2−デオキシ−シロ−イノソース合成酵素が、pH6.0〜 8.0で安定な酵素である、請求項1から3の何れか1項記載の方法。
【請求項5】
2−デオキシ−シロ−イノソース合成酵素が下記の特性を有する酵素である、請求項1または2に記載の方法。
(1)作用:本酵素は、グルコース−6−リン酸を2−デオキシ−シロ−イノソースへと変換する機能を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
【請求項6】
2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12に記載のアミノ酸配列を有する2−デオキシ−シロ−イノソース合成酵素である、請求項1から2の何れか1項記載の方法。
【請求項7】
2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12で示されるアミノ酸配列において1個または複数個のアミノ酸の欠失、付加、及び/又は置換を含み、配列番号2、4、6、8、10又は12で示されるアミノ酸配列に対して80%以上の相同性を有して且つ、高温安定性及び/又は広範囲pH安定性を持つ2−デオキシ−シロ−イノソース合成酵素である、請求項1から5の何れか1項に記載の方法。
【請求項1】
2−デオキシ−シロ−イノソース合成酵素又は2−デオキシ−シロ−イノソース合成酵素を発現可能な微生物を利用してグルコース−6−リン酸から2−デオキシ−シロ−イノソースを製造する方法において、反応系におけるNADH/NAD比を1.4以下に制御することを特徴とする、2−デオキシ−シロ−イノソースの製造方法。
【請求項2】
グルコースを構成成分とする多糖類あるいはグルコースを含有する原料を用いることを特徴とする、請求項1に記載の方法。
【請求項3】
2−デオキシ−シロ−イノソース合成酵素が、安定温度範囲が少なくとも46℃まで安定である耐熱性酵素である、請求項1又は2に記載の方法。
【請求項4】
2−デオキシ−シロ−イノソース合成酵素が、pH6.0〜 8.0で安定な酵素である、請求項1から3の何れか1項記載の方法。
【請求項5】
2−デオキシ−シロ−イノソース合成酵素が下記の特性を有する酵素である、請求項1または2に記載の方法。
(1)作用:本酵素は、グルコース−6−リン酸を2−デオキシ−シロ−イノソースへと変換する機能を有する。
(2)安定温度範囲:少なくとも46℃まで安定である。
(3)補酵素としてNAD+を利用。
(4)至適pH範囲:7.0〜 7.7
(5)安定pH範囲:少なくとも6.0〜 8.0の範囲で安定である。
(6)至適温度範囲:55 〜 70 ℃
(7)分子量:39000〜42000
(8)補因子:Co2+イオンの添加により活性向上。
(9)比活性:1.0μmol/min/mg以上(反応温度65℃)。
【請求項6】
2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12に記載のアミノ酸配列を有する2−デオキシ−シロ−イノソース合成酵素である、請求項1から2の何れか1項記載の方法。
【請求項7】
2−デオキシ−シロ−イノソース合成酵素が、配列番号2、4、6、8、10又は12で示されるアミノ酸配列において1個または複数個のアミノ酸の欠失、付加、及び/又は置換を含み、配列番号2、4、6、8、10又は12で示されるアミノ酸配列に対して80%以上の相同性を有して且つ、高温安定性及び/又は広範囲pH安定性を持つ2−デオキシ−シロ−イノソース合成酵素である、請求項1から5の何れか1項に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2010−227023(P2010−227023A)
【公開日】平成22年10月14日(2010.10.14)
【国際特許分類】
【出願番号】特願2009−78714(P2009−78714)
【出願日】平成21年3月27日(2009.3.27)
【出願人】(303046314)旭化成ケミカルズ株式会社 (2,513)
【Fターム(参考)】
【公開日】平成22年10月14日(2010.10.14)
【国際特許分類】
【出願日】平成21年3月27日(2009.3.27)
【出願人】(303046314)旭化成ケミカルズ株式会社 (2,513)
【Fターム(参考)】
[ Back to top ]