
NOGO受容体アンタゴニスト
【課題】Nogo受容体-1のそのリガンドに対する結合を阻害し、ミエリン-媒介成長円錐崩壊および神経突起伸長阻害を減弱する分子を提供すること。
【解決手段】免疫原性Nogo受容体-1ポリペプチド、Nogo受容体-1抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸が開示される。かかるNogo受容体抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸を含む組成物およびその製造及び使用方法も開示される。
【解決手段】免疫原性Nogo受容体-1ポリペプチド、Nogo受容体-1抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸が開示される。かかるNogo受容体抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸を含む組成物およびその製造及び使用方法も開示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、神経生物学および分子生物学に関する。より具体的には、本発明は、免疫原性Nogo受容体-1ポリペプチド、Nogo受容体-1抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質およびそれをコードする核酸に関する。本発明はさらにかかるNogo受容体抗体、その抗原結合性フラグメント、免疫原性Nogo受容体-1ポリペプチド、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸を含む組成物およびその製造及び使用方法にも関する。
【背景技術】
【0002】
神経細胞の軸索および樹状突起は、神経細胞からの長い細胞の伸長物である。伸長する軸索の遠位先端または神経突起は、成長円錐として知られる特殊な領域を含む。成長円錐は局所環境を感知し、神経細胞の標的細胞に向かった軸索成長を案内する。成長円錐はいくつかの環境的な合図、例えば、表面接着性、成長因子、神経伝達物質および電場に応答する。円錐における成長の誘導には様々なクラスの接着分子、細胞間シグナル、および成長円錐を刺激および阻害する因子が関与する。成長する神経突起の成長円錐は、様々な速度で進行するが、典型的には1日あたり1〜2mmの速度である。
【0003】
成長円錐は、手のような形であり、胚における表面に差次的に接着する広く平らな膨張部を備える(微小突起または糸状仮足)。糸状仮足は絶えず活動性であり、糸状仮足のなかには成長円錐内へ退縮するものもあり、基層に沿って伸長を続けるものもある。異なる糸状仮足間の伸長は膜状仮足を形成する。
【0004】
成長円錐は、膜状仮足および糸状仮足によってその前方およびいずれかの側面の領域を探索する。伸長物が成長に望ましくない表面に接触するとそれは後退する。伸長物が成長に好ましい表面に接触すると、それは伸長し続けその方向に成長円錐をガイドする。成長円錐は基層の表面の特徴において小さい変動でガイドされうる。成長円錐が適当な標的細胞に到達すると、シナプス結合が生じる。
【0005】
神経細胞の機能は、神経細胞とその直近の環境におけるその他の細胞の間の接触に大幅に影響される(U. Rutishauser, T. M. Jessell, Physiol. Rev. 1988, 68, p. 819)。かかる細胞には、特別(specialized)グリア細胞、中枢神経系 (CNS)における乏突起膠細胞、末梢神経系(PNS)におけるシュワン細胞が含まれ、これは神経細胞の軸索をミエリン(多層膜の絶縁体構造)鞘で覆う(G. Lemke, in An Introduction to Molecular Neurobiology, Z. Hall, Ed. [Sinauer, Sunderland, Mass., 1992], p. 281)。
【0006】
CNS神経細胞は傷害後の再生能を有するが、それらはミエリンに存在する阻害性タンパク質およびおそらくその局所環境に通常みられるその他のタイプの分子の存在により再生を阻害される(Brittis and Flanagan, Neuron, 2001, 30, pp. 11-14; Jones et al., J. Neurosci. 2002, 22, pp. 2792-2803; Grimpe et al., J. Neurosci. 2002, 22, pp. 3144-3160)。
【0007】
乏突起膠細胞上にみられるいくつかのミエリン阻害性タンパク質が特徴づけられており、例えば以下が挙げられる:NogoA (Chen et al., Nature, 2000, 403, 434-439; Grandpre et al., Nature 2000, 403, 439-444)、ミエリン関連糖タンパク質(MAG, McKerracher et al, Neuron 1994, 13, 805-811; Mukhopadhyay et al, Neuron 1994, 13, 757-767)および乏突起膠細胞糖タンパク質(OM-gp, Mikol and Stefansson, J. Cell. Biol. 1988, 106, 1273-1279)。これらタンパク質はそれぞれ別々に神経細胞Nogo受容体-1のリガンドであることが示された(Wang et al., Nature 2002, 417, 941-944; Liu et al., Science, 2002, 297, 1190-93; Grandpre et al., Nature 2000, 403, 439-444; Chen et al., Nature, 2000, 403, 434-439; Domeniconi et al., Nuron, 2002, 35, 283-90)。
【0008】
Nogo受容体-1は8つのロイシンリッチリピートを含むGPI-アンカー(anchored)膜タンパク質である(Fournier et al., Nature 2001, 409, 341-346)。阻害性タンパク質(例えば、NogoA, MAGおよびOM-gp)との相互作用時に、Nogo受容体-1複合体は、成長円錐の崩壊および神経突起伸長の阻害を導くシグナルを伝達する。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】U. Rutishauser, T. M. Jessell, Physiol. Rev. 1988, 68, p. 819
【非特許文献2】G. Lemke, in An Introduction to Molecular Neurobiology, Z. Hall, Ed. [Sinauer, Sunderland, Mass., 1992], p. 281
【非特許文献3】Brittis and Flanagan, Neuron, 2001, 30, pp. 11-14
【非特許文献4】Jones et al., J. Neurosci. 2002, 22, pp. 2792-2803
【非特許文献5】Grimpe et al., J. Neurosci. 2002, 22, pp. 3144-3160
【非特許文献6】Chen et al., Nature, 2000, 403, 434-439
【非特許文献7】Grandpre et al., Nature 2000, 403, 439-444
【非特許文献8】McKerracher et al, Neuron 1994, 13, 805-811
【非特許文献9】Mukhopadhyay et al, Neuron 1994, 13, 757-767
【非特許文献10】Mikol and Stefansson, J. Cell. Biol. 1988, 106, 1273-1279
【非特許文献11】Wang et al., Nature 2002, 417, 941-944
【非特許文献12】Liu et al., Science, 2002, 297, 1190-93
【非特許文献13】Domeniconi et al., Nuron, 2002, 35, 283-90
【非特許文献14】Fournier et al., Nature 2001, 409, 341-346
【発明の概要】
【発明が解決しようとする課題】
【0010】
Nogo受容体-1のそのリガンドに対する結合を阻害し、ミエリン-媒介成長円錐崩壊および神経突起伸長阻害を減弱する分子が緊急に要求されている。
【課題を解決するための手段】
【0011】
(発明の概要)
本発明は、可溶性Nogo受容体-1ポリペプチドおよびそれを含む融合タンパク質、ならびにNogo受容体-1の特定の免疫原性領域に対する抗体およびその抗原性フラグメントに関する。本発明はまた、本発明の抗体に結合する免疫原性Nogo受容体-1ポリペプチドに関する。本発明はさらに、本発明のポリペプチドをコードする核酸、かかる核酸を含むベクターおよび宿主細胞、ならびに該ペプチドの製造方法に関する。本発明の抗体、可溶性受容体および受容体融合タンパク質はNogo受容体-1を拮抗またはブロックし、Nogo受容体-1のそのリガンドへの結合の阻害、神経細胞における成長円錐崩壊の阻害、および神経細胞における神経突起伸長または出芽の阻害の抑制に有用である。
【0012】
ある態様において、本発明は、配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択される免疫原性ポリペプチドを提供する。
【0013】
ある態様において、本発明は、該免疫原性ポリペプチドをコードする核酸、該核酸を含むベクターおよび該核酸またはベクターを含む宿主細胞を提供する。ある態様において、核酸は、発現制御配列に作動可能に連結している。
【0014】
ある態様において、本発明は以下の工程を含む免疫原性ポリペプチドの産生方法を提供する:
(a)免疫原性ペプチドをコードする核酸またはそれをコードするベクターを含む宿主細胞を培養する工程;および、
(b)宿主細胞または培地からポリペプチドを回収する工程。
【0015】
ある態様において、本発明は以下の工程を含む、Nogo受容体-1に特異的に結合する抗体の産生方法を提供する:
(a) 配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチドまたは該ポリペプチドを発現する宿主細胞で宿主を免疫する工程;および、
(b)抗体を回収する工程。
ある態様において、抗体またはその抗原結合性フラグメントはこの方法によって産生される。ある態様において、抗体またはその抗原結合性フラグメントは、配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチドに特異的に結合する。ある態様において、抗体または抗原結合性フラグメントは、(a) 神経細胞の成長円錐崩壊を阻害する;(b)神経突起伸長および神経細胞における出芽の阻害を減少させる;そして(c) Nogo受容体-1のリガンドへの結合を阻害する。ある態様において、抗体または抗原結合性フラグメントは死の危険にある神経細胞の生存を促進する。ある態様において、死の危険にある神経細胞は動物、例えば、哺乳類におけるものである。ある態様において、神経突起伸長および出芽は軸索成長である。ある態様において、神経細胞は、中枢神経系(CNS)神経細胞である。
【0016】
ある態様において、抗体または抗原結合性フラグメントはモノクローナル抗体である。ある態様において、抗体または抗原結合性フラグメントはマウス抗体である。ある態様において、抗体はヒト化抗体、キメラ抗体あるいは一本鎖抗体である。
【0017】
ある態様において、本発明は以下からなる群から選択されるハイブリドーマ細胞株を提供する:HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585)。ある態様において、抗体またはその抗原結合性フラグメントは、ハイブリドーマ細胞株によって産生される。
【0018】
ある態様において、抗体またはその抗原結合性フラグメントは、以下からなる群から選択されるアミノ酸配列を含む軽鎖を有する(a) 配列番号15のアミノ酸配列;(b) 配列番号16のアミノ酸配列;および(c) 配列番号22、23および24のCDR1、CDR2およびCDR3アミノ酸配列を含むアミノ酸配列。ある態様において、抗体またはその抗原結合性フラグメントは以下からなる群から選択されるアミノ酸配列を含む重鎖を有する (a)配列番号17のアミノ酸配列; (b) 配列番号18のアミノ酸配列;および、(c) 配列番号19、20および21のCDR1、CDR2およびCDR3アミノ酸配列を含むアミノ酸配列。ある態様において、本発明は、該抗体またはその抗原結合性フラグメントをコードする核酸を提供する。ある態様において、核酸は発現制御配列に作動可能に連結している。ある態様において、本発明は該核酸を含むベクターを提供する。ある態様において、本発明は該核酸または該核酸を含むベクターを含む宿主細胞を提供する。
【0019】
ある態様において、抗体またはその抗原結合性フラグメントは、ハイブリドーマ細胞株によって産生される抗体の Nogo受容体-1または配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択される免疫原性ポリペプチドに対する結合を競合的に阻害する。
【0020】
ある態様において、本発明は以下の工程を含むリガンドに対するNogo受容体-1の結合の阻害方法を提供する:Nogo受容体-1を本発明の抗体または抗原結合性フラグメントと接触させる工程。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。
【0021】
ある態様において、本発明は以下の工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する:神経細胞を本発明の抗体またはその抗原結合性フラグメントと接触させる工程。ある態様において、本発明は、以下の工程を含む神経細胞における神経突起伸長または出芽の阻害の減少方法を提供する:神経細胞を本発明の抗体またはその抗原結合性フラグメントと接触させる工程。ある態様において、神経細胞はCNS 神経細胞である。これら方法いくつかにおいて、神経突起伸長または出芽は軸索成長である。
【0022】
ある態様において、本発明は以下の工程を含む死の危険にある神経細胞の生存を促進する方法を提供する:神経細胞を有効量の(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメントまたは(b)可溶性Nogo受容体-1ポリペプチドと接触させる工程。ある態様において、可溶性Nogo受容体-1ポリペプチドは、 融合タンパク質、例えば、Fc-融合タンパク質である。ある態様において、融合タンパク質は、sNogoR344-Fcタンパク質である。ある態様において、神経細胞はインビトロである。ある態様において、神経細胞は例えば、多発性硬化症、ALS、ハンチントン病、アルツハイマー病、パーキンソン病、糖尿病性神経障害、脳卒中、外傷性脳障害および脊髄損傷等の徴候または症状を示す哺乳類におけるものである。
【0023】
ある態様において、本発明は、以下の工程を含み、神経細胞が哺乳類におけるものである、死の危険にある神経細胞の生存を促進する方法を提供する:
(a)(i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(ii) 可溶性Nogo受容体-1ポリペプチドを発現する培養宿主細胞を提供する工程;および、
(b)宿主細胞を哺乳類に神経細胞においてまたはその近くの部位にて導入する工程。
【0024】
ある態様において、本発明は以下の工程を含み、神経細胞が死の危険にある、哺乳類における神経細胞の生存を促進する遺伝子治療方法を提供する:以下の(a)または(b)をコードするヌクレオチド配列を含むウイルスベクターを神経細胞においてまたはその近くの部位にて投与する工程:(a)抗-Nogo受容体-1抗体または抗原結合性フラグメント ; または(b)可溶性Nogo受容体-1ポリペプチド、ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは、神経細胞の生存を促進するのに十分な量にて哺乳類においてヌクレオチド配列から発現する。
【0025】
ある態様において、本発明は、Nogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる可溶性Nogo受容体-1ポリペプチドを提供する。ある態様において、該可溶性Nogo受容体-1ポリペプチドはシグナル配列に連結している。ある態様において、LRRは異種LRRを含む。ある態様において、本発明は以下からなる群から選択される可溶性Nogo受容体-1ポリペプチドを提供する: 配列番号6のアミノ酸残基26-344; 配列番号7のアミノ酸残基26-310; 配列番号8のアミノ酸残基26-344; 配列番号9のアミノ酸残基 26-310; 配列番号8のアミノ酸残基 27-344;および配列番号9のアミノ酸残基 27-310。ある態様において、本発明は、該可溶性Nogo受容体-1ポリペプチドをコードする核酸を提供する。ある態様において、核酸は発現制御配列に作動可能に連結している。ある態様において、本発明は該核酸を含むベクターを提供する。ある態様において、本発明は、該核酸または該核酸を含むベクターを含む宿主細胞を提供する。
【0026】
ある態様において、本発明は以下の工程を含む本発明の可溶性Nogo受容体-1ポリペプチドの産生方法を提供する:
(a) 可溶性Nogo受容体-1ポリペプチドをコードする核酸または該核酸を含むベクターを含む宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程。
【0027】
ある態様において、本発明は、可溶性Nogo受容体-1および異種ポリペプチドを含むNogo受容体-1融合タンパク質を提供する。ある態様において、可溶性Nogo受容体-1ポリペプチドはNogo受容体-1のN-末端ドメイン (NT)、8 ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる。ある態様において、可溶性Nogo受容体-1ポリペプチドはシグナル配列に連結している。ある態様において、Nogo受容体-1融合タンパク質は異種LRRを含む。ある態様において、Nogo受容体-1融合タンパク質は以下からなる群から選択されるポリペプチドを含む: 配列番号6のアミノ酸残基26-344; 配列番号7のアミノ酸残基26-310; 配列番号8のアミノ酸残基26-344; 配列番号9のアミノ酸残基26-310; 配列番号8のアミノ酸残基 27-344; 配列番号9のアミノ酸残基27-310。ある態様において、異種ポリペプチドは、免疫グロブリン定常領域を含む。ある態様において、免疫グロブリン定常領域は、免疫グロブリン重鎖定常領域である。ある態様において、免疫グロブリン重鎖定常領域はIgG 重鎖定常領域である。ある態様において、異種ポリペプチドはFc領域である。ある態様において、Nogo受容体-1融合タンパク質はダイマーである。
【0028】
ある態様において、本発明は、Nogo受容体-1融合タンパク質をコードする核酸を提供する。ある態様において、Nogo受容体-1融合タンパク質をコードする核酸は、発現制御配列に作動可能に連結している。
【0029】
ある態様において、本発明はNogo受容体-1融合タンパク質をコードする核酸を含むベクターを提供する。
【0030】
ある態様において、本発明は、Nogo受容体-1融合タンパク質をコードする核酸をまたはNogo受容体-1融合タンパク質をコードする核酸を含むベクターを含む宿主細胞を提供する。
【0031】
ある態様において、本発明は以下の工程を含むNogo受容体-1融合タンパク質の産生方法を提供する:
(a) Nogo受容体-1融合タンパク質をコードする核酸または該核酸を含むベクターを含む宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からNogo受容体-1融合タンパク質を回収する工程。
【0032】
ある態様において、本発明は以下の工程を含む、リガンドに対するNogo受容体-1の結合を阻害する方法を提供する:リガンドを本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質に接触させる工程。
【0033】
ある態様において、本発明は以下の工程を含むNogo受容体-1リガンドの活性の調節方法を提供する: Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。
【0034】
ある態様において、本発明は以下の工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する:Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。ある態様において、本発明は以下の工程を含む、神経細胞における神経突起伸長または出芽の阻害の減少方法を提供する:Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。ある態様において、神経細胞は CNS 神経細胞である。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。ある態様において、神経突起伸長または出芽は軸索成長である。
【0035】
ある態様において、本発明は、医薬上許容される担体および以下から選択される成分を含む組成物を提供する(a) 本発明による抗体または抗原結合性フラグメント; (b) 本発明による可溶性Nogo受容体-1ポリペプチド;および(c) 本発明によるNogo受容体-1融合タンパク質。ある態様において、組成物はさらに1または複数のさらなる治療薬を含む。
【図面の簡単な説明】
【0036】
【図1】図1は、Nogo受容体-1の構造の模式図である。ヒト sNogoR310は残基 26-310を含み、sNogoR344は残基 26-344を含む。ラットsNogoR310は残基 27-310を含み、 sNogoR344は残基 27-344を含む。
【図2】図2は、ベクターNti(商標)ソフトウェアを用いたNogo受容体-1 タンパク質の抗原性プロットを示す。ラットP-617は配列番号10であり、ラットP-618 は配列番号11である。
【図3】図3Aは、抗-Nogo受容体-1抗体, 7E11の結合活性を示すグラフである。グラフはNogo66のNogo受容体-1への結合に対する7E11濃度の効果を表す。図3Bは抗-Nogo受容体-1抗体, 1H2の結合活性を示す。グラフは、1H2 濃度のNogo66の sNogoR344-Fcに対する結合への効果を表す(本明細書そして米国特許出願 60/402,866 においてFc-sNogoR344またはIg-sNogoR344とも称される)。Fc-MAGはsNogoR344-Fc への結合についてNogo66と競合しなかった。
【図4】図4は、抗-Nogo-R-1抗体 1H2, 3G5および2F7についての ELISAの結果を示す。固定化抗原の存在下の抗体のOD450に対する効果を測定した。固定化抗原は、sNogoR310-Fc (本明細書そして米国特許出願 60/402,866において Fc-sNogoR310またはIg-sNogoR310とも称される)、sNogoR344-Fc、p-617、p-618、p-4、p-5および卵白アルブミンおよびBSA。
【図5】図5は、様々な量のミエリンの存在下でのモノクローナル抗体、7E11、のラットDRG神経突起伸長に対する効果を示すグラフである。
【図6】図6Aは以下のコンペティターの存在下でのsNogoR310の125I-Nogo66および125I-Nogo40への結合に対する効果を示すグラフである: Nogo66、Nogo40および抗-Nogo受容体-1モノクローナル抗体上清。図6Bは、125I-Nogo66のsNogoR310への結合活性を示す。
【図7】図7は、sNogoR310-Fc の、sNogoR310に対する125I-Nogo40の結合への効果を示すグラフである。
【図8】図8は、sNogoR310-Fcの125I-Nogo40に対する結合活性を示すグラフである。
【図9A】図9Aはミエリンの存在下または非存在下でのsNogoR310の神経突起伸長/細胞に対する効果を示すグラフである。
【図9B】図9Bはミエリンの存在下または非存在下での神経突起伸長に対するsNogoR310の効果を示すグラフである。
【図10A】図10Aは様々な量のミエリンの存在下または非存在下でのP4ラットDRG神経突起伸長に対するsNogoR310-Fcの効果を示すグラフである。
【図10B】図10Bは、PBS、PBS + sNogoR310-Fc、20ng ミエリンおよびミエリン + sNogoR310-Fcでの処理後の神経突起数/細胞を示す。
【図11】図11は、様々な量のミエリンの存在下でのモノクローナル抗体 5B10 の DRG神経突起伸長/細胞に対する効果を示すグラフである。
【図12】図12は、ラット脊髄離断モデルにおける損傷の誘導後30日までのBBBスコアに対するsNogoR310-Fcの効果を示すグラフである。
【図13】図13Aおよび13BはWTまたは系統 08または系統 01からのトランスジェニックマウスにおける歩行運動BBBスコアを背部片側切断後の時間に対する関数として報告する。図13Cは、最大許容傾斜平面角をWTおよびトランスジェニックマウスについての損傷後時間の関数として示すグラフである。図13Dは、傾斜格子登上間の後肢エラーを損傷後時間の関数として示す。すべてのグラフにおいて、各群において7-9匹のマウスからの平均±s.e.m.を報告する。トランスジェニック群からの値はWTマウスからの値と有意差がある(**、P < 0.01; スチューデントt検定)。
【図14】図14Aは、媒体またはsNogoR310-Fc 処理動物における背部片側切断後の時間の関数として歩行運動BBBスコアを示す。図14Bは格子歩行の際の後肢エラーを損傷後時間の関数として示す。図14Cは、無傷または損傷 + sNogoR310-Fcラットよりコントロールマウスにおいてストライド長が短く、ストライド幅が広いことを明らかにする足跡分析である。すべてのグラフにおいて、各群において7-9匹のラットからの平均±s.e.m.を報告する。sNogoR310-Fc群の値は、コントロールと有意差がある(図14A-B)。コントロール値は図14Cにおける非-SCIまたはSCI + sNogoR310-Fcラットと有意差がある(*、p< 0.05; **、p < 0.01;スチューデントt検定)。
【発明を実施するための形態】
【0037】
(発明の詳細な説明)
定義および一般的教示
特に断りのない限り、本明細書において用いるすべての技術及び科学用語は本発明の属する分野の当業者に通常理解される意味と同義である。争いのある場合、定義を含む本出願が優先する。また文脈により要求されない限り、単数形の語は、複数も含み、複数形の語は、単数も含む。本明細書において言及する、すべての刊行物、特許およびその他の文献はあらゆる目的のためその全体を引用により本出願に含める。
【0038】
本明細書に記載するものと類似または均等の方法及び材料が本発明の実施または試験に利用できるが、好適な方法および材料を以下に記載する。材料、方法および実施例は単に例示のものであり、限定の意図ではない。本発明のその他の特徴および利点は、詳細な説明および特許請求の範囲から明らかであろう。
【0039】
本明細書および特許請求の範囲を通して「含む」なる語またはその同義語は、言及した要素または要素群を含む意味であるがその他の要素または要素群を排除するものではない。
【0040】
本発明をさらに明確にするため以下の用語および定義を本明細書において提供する。
【0041】
本明細書において用いる、「抗体」とはインタクトな免疫グロブリンまたはその抗原結合性フラグメントを意味する。本発明の抗体はいずれのアイソタイプまたはクラスでもよく(例えば、M、D、G、EおよびA)、いずれのサブクラスでもよく(例えば、G1-4, A1-2) そしてカッパ(κ)またはラムダ(λ)軽鎖のいずれをもつものでのよい。
【0042】
本明細書において用いる「Fc」とは、パパイン消化によって得られる抗体の重鎖定常領域の部分を意味する。
【0043】
本明細書において用いる、「NogoR融合タンパク質」とは、異種ポリペプチドに融合した可溶性Nogo受容体-1部分を含むタンパク質を意味する。
【0044】
本明細書において用いる、「ヒト化抗体」とは、非-ヒト配列の少なくとも一部がヒト配列で置換された抗体を意味する。ヒト化抗体の作成法の例は米国特許第6054297、5886152および5877293号にみることができる。
【0045】
本明細書において用いる「キメラ抗体」とは、第一の抗体からの1または複数の領域および少なくとも1つのその他の抗体からの1または複数の領域を含む抗体を意味する。第一の抗体およびさらなる抗体は同種由来のものでも異種由来のものでもよい。
【0046】
本明細書および米国特許出願第60/402,866号において用いる、「Nogo受容体」、「NogoR」、「NogoR-1」、「NgR」、および「NgR-1」はそれぞれNogo受容体-1を意味する。
【0047】
免疫原性Nogo受容体-1ポリペプチド
一つの態様において、本発明は免疫原性のNogo受容体-1ポリペプチドに関する。本発明のある態様において、免疫原性ポリペプチドは実質的に以下からなる群から選択されるアミノ酸配列からなる:LDLSDNAQLRVVDPTT (ラット)(配列番号1); LDLSDNAQLRSVDPAT (ヒト)(配列番号2); AVASGPFRPFQTNQLTDEELLGLPKCCQPDAADKA (ラット)(配列番号3); AVATGPYHPIWTGRATDEEPLGLPKCCQPDAADKA (ヒト)(配列番号4);およびCRLGQAGSGA (マウス)(配列番号5)。
【0048】
ある態様において、本発明は配列番号1-5のポリペプチドをコードする核酸に関する。本発明のある態様において、核酸分子は発現制御配列(例えば、pCDNA(I))に連結している。
【0049】
本発明はまた、本発明の免疫原性ポリペプチドをコードする核酸を含むベクターに関する。本発明のある態様において、ベクターはクローニングベクターである。本発明のある態様において、ベクターは発現ベクターである。本発明のある態様において、ベクターは少なくとも1つの選抜可能マーカーを含む。
【0050】
本発明はまた、上記の核酸またはベクターを含む宿主細胞に関する。
【0051】
本発明はまた、宿主細胞を培養する工程を含む、本発明の免疫原性ポリペプチドの産生方法に関する。ある態様において、宿主細胞は原核生物である。ある態様において、宿主細胞は真核生物である。ある態様において、宿主細胞は酵母である。
【0052】
抗体
本発明はさらに、本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗体またはその抗原結合性フラグメントに関する。ある態様において、抗体または抗原結合性フラグメントは実質的に配列番号1-5からなる群から選択されるアミノ酸配列からなるポリペプチドに結合する。本発明の抗体または抗原結合性フラグメントはインビボでもインビトロでも産生できる。抗体または抗原結合性フラグメントの産生を以下に説明する。
【0053】
本発明の抗体またはその抗原結合性フラグメントはNogo受容体-1のリガンド(例えば、NogoA, NogoB, NogoC, MAG, OM-gp)に対する結合を阻害し、ミエリンに媒介される神経突起伸長および出芽、特に軸索成長の阻害を減少させ、ミエリンに媒介される成長円錐崩壊を減弱させる。
【0054】
ある態様において、抗-Nogo受容体-1抗体またはその抗原結合性フラグメントはマウス由来である。ある態様において、Nogo受容体-1はラット由来である。別の態様において、Nogo受容体-1はヒト由来である。ある態様において、抗-Nogo受容体-1抗体またはその抗原結合性フラグメントは組換え、遺伝子操作、ヒト化および/またはキメラである。
【0055】
ある態様において、抗体は以下からなる群から選択される: モノクローナル7E11(ATCC受託番号、PTA-4587); モノクローナル 1H2 (ATCC受託番号、PTA-4584); モノクローナル 2F7 (ATCC受託番号、PTA-4585); モノクローナル 3G5 (ATCC受託番号、PTA-4586);およびモノクローナル 5B10 (ATCC受託番号、PTA-4588)。ある態様において、抗体はポリクローナル抗体46である。
【0056】
例示的な抗原結合性フラグメントは、Fab、Fab'、F(ab')2、Fv、Fd、dAbおよび相補性決定領域 (CDR)フラグメントを含むフラグメント、一本鎖抗体 (scFv)、キメラ抗体、二価抗体(diabodies)およびポリペプチドに特異的な抗原結合性を付与するのに十分な免疫グロブリンの少なくとも一部を含むポリペプチド(例えば、イムノアドヘシン)である。
【0057】
本明細書において用いる、FdはVHおよびCH1ドメインからなるフラグメントを意味する;Fvは抗体の1つの腕のVLおよびVHドメインからなるフラグメントを意味する;そしてdAbはVHドメインからなるフラグメントを意味する(Ward et al., Nature 341:544 546, 1989)。本明細書において用いる、一本鎖抗体(scFv)はVL領域およびVH領域が対となってそれらを一つのタンパク質鎖にさせる合成リンカーを介して一価分子を形成する抗体を意味する(Bird et al., Science 242:423 426, 1988およびHuston et al., Proc. Natl. Acad. Sci. USA 85:5879 5883, 1988)。本明細書において用いる、二価抗体は二重特異性抗体であって、VHおよびVL ドメインが一本のポリペプチド鎖上に発現するが、同じ鎖上の該2つのドメイン間の対合を可能とするのには短すぎるリンカーを用いてドメインを別の鎖の相補的ドメインと対合させ2つの抗原結合性部位が形成されるものを意味する(例えば、Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90:6444 6448, 1993,およびPoljak, R. J., et al., Structure 2:1121 1123, 1994を参照されたい)。本明細書において用いる、目的の抗原に特異的に結合するイムノアドヘシンとは、1または複数のCDRが共有結合または非共有結合によって導入された分子を意味する。
【0058】
ある態様において、本発明は本発明のNogo受容体-1抗体のサブユニットポリペプチドを提供し、ここでサブユニットポリペプチドは以下からなる群から選択される:(a)重鎖またはその可変領域;および(b)軽鎖またはその可変領域。
【0059】
ある態様において、本発明は本発明のNogo受容体-1抗体のサブユニットポリペプチドの重鎖またはその可変領域あるいは軽鎖およびその可変領域をコードする核酸を提供する。
【0060】
ある態様において、本発明は本発明のNogo受容体-1抗体の超可変領域 (CDR)またはCDRをコードする核酸を提供する。
【0061】
免疫
本発明の抗体は好適な宿主 (例えば、脊椎動物、例えばヒト、マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ、ウマ、爬虫類、魚類、両生類および鳥類、爬虫類および魚類の卵中)の免疫により生成させうる。かかる抗体はポリクローナルであってもモノクローナルであってもよい。
【0062】
ある態様においては、宿主を本発明の免疫原性Nogo受容体-1ポリペプチドで免疫する。別の態様において、宿主をインタクトまたは崩壊した細胞の細胞膜に結合したNogo受容体-1で免疫し、本発明の抗体を本発明の免疫原性Nogo受容体-1ポリペプチドへの結合によって同定する。
【0063】
ある態様において、Nogo受容体-1抗原を免疫応答を刺激するアジュバントとともに投与する。アジュバントはしばしば抗原への免疫応答を誘発するために抗原に加えて投与する必要がある。かかるアジュバントは通常は、免疫部位において単核食細胞を集めて非特異的炎症を促進する不溶性または非崩壊性物質である。アジュバントの例としては、これらに限定されないが以下が挙げられる:フロイントアジュバント、RIBI(ムラミルジペプチド)、ISCOM (免疫刺激複合体)またはそれらのフラグメント。
【0064】
抗体の製造方法の概説としては、例えば、Harlow and Lane (1988), Antibodies, A Laboratory Manual, Yelton, D.E. et al. (1981); Ann. Rev. of Biochem., 50, pp. 657-80., およびAusubel et al. (1989); Current Protocols in Molecular Biology (New York: John Wiley & Sons)を参照されたい。本発明の免疫原性Nogo受容体-1ポリペプチドによる免疫反応性の測定は当該技術分野で周知の様々な方法によって行うことが出来、例えば、イムノブロットアッセイおよびELISAが挙げられる。
【0065】
抗体および抗体産生細胞株の産生
本発明のモノクローナル抗体は例えば前掲のHarlow and Lane (1988)に記載の標準的手順によって作ることが出来る。
【0066】
簡単に説明すると、適当な時期に動物を屠殺し、リンパ節および/または脾臓のB-細胞を当該技術分野で周知のいずれかの技術によって不死化する。かかる技術としては、これらに限定されないが、例えば、EBVなどによる形質転換またはミエローマ細胞等の不死化細胞株との融合が挙げられる。その後、細胞をクローン分離し、各クローンの上清を本発明の免疫原性Nogo受容体-1ポリペプチドに特異的な抗体の産生について試験する。ハイブリドーマの選抜、クローニングおよび拡張方法は当該技術分野で周知である。同様に、免疫グロブリン遺伝子のヌクレオチドおよびアミノ酸配列決定方法は当該技術分野で知られている。
【0067】
本発明の抗体のその他の好適な産生技術には、本発明のNogo受容体-1または免疫原性ポリペプチドへリンパ球をインビトロで曝すこと、あるいは、ファージまたは類似のベクターにおける抗体ライブラリーの選抜が含まれる。Huse et al., Science, 246, pp. 1275-81 (1989)を参照されたい。本発明に有用な抗体は改変してもしなくても利用できる。
【0068】
抗原(この場合、Nogo受容体-1または本発明の免疫原性ポリペプチド)および抗体は共有結合または非共有結合によって検出可能なシグナルを提供する物質を結合させることにより標識できる。様々な標識および結合(conjugation)技術が当該技術分野で知られており、本発明の実施に用いることが出来る。好適な標識にはこれらに限定されないが以下が含まれる:放射性ヌクレオチド、酵素、基質、補因子、阻害剤、蛍光試薬、化学発光試薬、磁性粒子等。かかる標識の使用を教示する特許としては以下が挙げられる:米国特許3817837号; 3850752号; 3939350号; 3996345号; 4277437号; 4275149号および4366241号。また、組換え免疫グロブリンも産生できる(米国特許4816567号を参照されたい)。
【0069】
本発明のある態様において、抗体は複数の結合特異性を有し、例えば、二機能性抗体は当業者に知られている多くの技術のいずれかによって調製される。かかる技術としては例えば、雑種ハイブリドーマの産生、ジスルフィド交換、化学的架橋、2つのモノクローナル抗体間のペプチドリンカーの付加、特定の細胞株への2セットの免疫グロブリン重鎖および軽鎖の導入などが挙げられる (より詳細には以下を参照されたい)。
【0070】
本発明の抗体は、例えば不死化ヒト細胞によって、SCID-hu マウスによって、あるいはその他の「ヒト」抗体を産生することが出来る非ヒト動物によって産生されたヒトモノクローナル抗体であってもよい。
【0071】
ファージディスプレーライブラリー
本発明の抗-Nogo受容体-1抗体は組換えコンビナトリアル抗体ライブラリーのスクリーニングによって単離することが出来る。例示的なコンビナトリアルライブラリーは本発明の免疫原性Nogo受容体-1ポリペプチドへの結合についてのものであり、例えばscFvファージディスプレーライブラリーが挙げられ、これは本発明の免疫原性Nogo受容体-1ポリペプチドで免疫した動物由来のmRNAから調製したVLおよびVHcDNAを用いて調製される。かかるライブラリーの調製方法およびスクリーニング方法は当該技術分野で知られている。ファージディスプレーライブラリーを作成するための市販の方法および材料がある (例えば、Pharmacia組換えファージ抗体システム、カタログ番号27 9400 01; Stratagene SurfZAP(商標)ファージディスプレーキット、カタログ番号240612;およびMorphoSysのその他のもの)。また、抗体ディスプレーライブラリーの作成およびスクリーニングに使用できるその他の方法及び試薬もある(例えば、Ladner et al. 米国特許第5223409号; Kang et al. PCT公開WO 92/18619号; Dower et al. PCT 公開 WO 91/17271; Winter et al. PCT公開 WO 92/20791; Markland et al. PCT公開WO 92/15679; Breitling et al. PCT 公開 WO 93/01288; McCafferty et al. PCT 公開 WO 92/01047; Garrard et al. PCT公開WO 92/09690; Fuchs et al. (1991) Bio/Technology 9:1370 1372; Hay et al. (1992) Hum.Antibod. Hybdiromas 3:81 85; Huse et al. (1989) Science 246:1275 1281; McCafferty et al., Nature (1990) 348:552 554; Griffiths et al. (1993) EMBO J. 12:725 734; Hawkins et al. (1992) J. Mol. Biol. 226:889 896; Clackson et al. (1991) Nature 352:624 628; Gram et al. (1992) Proc. Natl. Acad. Sci. USA 89:3576 3580; Garrad et al. (1991) Bio/Technology 9:1373 1377; Hoogenboom et al. (1991) Nucl. Acids Res. 19:4133 4137;およびBarbas et al. (1991) Proc. Natl. Acad. Sci. USA 88:7978 7982を参照)。
【0072】
組換え免疫グロブリンディスプレーライブラリーからの本発明の抗-Nogo受容体-1抗体のスクリーニングおよび単離後、選択した抗体をコードする核酸をディスプレーパッケージから回収し(例えば、ファージゲノムから)、標準的組換えDNA技術によってその他の発現ベクターにサブクローニングすればよい。所望の場合、核酸をさらに操作して以下に記載のように本発明のその他の抗体形態を作成してもよい。コンビナトリアルライブラリーのスクリーニングによって単離された抗体を発現するために、抗体重鎖および軽鎖またはその可変領域をコードするDNAを組換え発現ベクターにクローニングし、上記のように哺乳類宿主細胞に導入する。
【0073】
クラススイッチ
本発明の抗-Nogo受容体-1抗体はいずれのアイソタイプでもよい。あらゆる所望のアイソタイプの抗体がクラススイッチによって産生できる。クラススイッチのために、CLまたはCHをコードするヌクレオチド配列を含まないVLまたはVHをコードする核酸を当該技術分野で周知の方法を用いて単離する。VLまたはVHをコードする核酸を所望のクラスの免疫グロブリン分子からのCLまたはCHをコードするヌクレオチド配列と作動可能に連結する。これは上記のようにCLまたはCH鎖を含むベクターまたは核酸を用いて達成できる。例えば、元はIgMであった本発明の抗-Nogo受容体-1抗体をIgGへとクラススイッチ出来る。さらに、クラススイッチを用いて1つのIgG サブクラスをその他に、例えばIgG1からIgG2に変換できる。
【0074】
突然変異抗体
別の態様において本発明の抗体または抗原結合性フラグメントを重鎖および/または軽鎖の可変ドメインにおいて突然変異させて抗体の結合特性を変化させてもよい。例えば、突然変異は1または複数のCDR領域において導入してNogo受容体-1についての抗体のKdを増減させてもよいし、Koffを増減させてもよいし、抗体の結合特異性を変化させてもよい。部位特異的突然変異誘発技術は当該技術分野で周知である。例えば、Sambrook et al. and Ausubel et al., 前掲を参照されたい。好適な態様において本発明の抗-Nogo受容体-1抗体の可変領域において生殖系列と比較して変化していることが知られているアミノ酸残基において突然変異を導入する。ある態様において、本発明の抗-Nogo受容体-1抗体の可変領域において生殖系列と比較して変化していることが知られている1または複数のアミノ酸残基に突然変異を導入する。別の態様において、抗体重鎖または軽鎖可変領域をコードする核酸を1または複数のフレームワーク領域において突然変異させる。突然変異はフレームワーク領域または定常ドメインに導入して半減期を上昇させてもよい。フレームワーク領域または定常ドメインにおける突然変異はまた、抗体の免疫原性を変化させ、別の分子への共有または非共有結合のための部位を提供するか、補体固定としての性質を変化させることもある。突然変異は1つの突然変異抗体におけるフレームワーク領域、定常ドメインおよび可変領域のそれぞれにおいて導入してもよい。あるいは、突然変異は1つの突然変異抗体においてフレームワーク領域、可変領域または定常ドメインのいずれか1つのみにおいて導入してもよい。
【0075】
融合抗体およびイムノアドヘシン
別の態様において、融合抗体またはイムノアドヘシンを作成してもよく、これは別のポリペプチドに連結した本発明の抗-Nogo受容体-1抗体の全部または一部を含む。ある態様において、抗-Nogo受容体-1抗体の可変領域のみをポリペプチドに連結する。別の態様において、本発明の抗-Nogo受容体-1抗体のVHドメインを第一のポリペプチドに連結し、抗体のVLドメインを第二のポリペプチドに連結させる。第二のポリペプチドは第一のポリペプチドとVHおよびVLドメインが互いに相互作用して抗体結合部位を形成するように結合する。別の態様において、VHドメインとVLドメインとは、VHおよびVLドメインが互いに相互作用するのを可能にするリンカーによって分離する(以下の一本鎖抗体を参照されたい)。VH -リンカー- VL 抗体を次いで目的のポリペプチドと連結する。融合抗体はNogo受容体-1リガンドを発現する細胞または組織に対してポリペプチドを標的化するのに有用である。目的のポリペプチドは例えば毒素などの治療薬であってもよく、例えば容易に可視化できる酵素などの診断薬であってもよく、例えばセイヨウワサビペルオキシダーゼが挙げられる。さらに、2(または3以上)の一本鎖抗体が互いに連結する融合抗体を作成してもよい。これは1つのポリペプチド鎖上に二価または多価抗体を作る場合や、二重特異性抗体を作る場合に有用である。
【0076】
一本鎖抗体
本発明は、本発明の免疫原性Nogo受容体-1ポリペプチドに結合する一本鎖抗体(scFv)を含む。ScFvを産生するためには、VHおよびVLをコードするDNAを屈曲性リンカー、例えば、アミノ酸配列 (Gly4 Ser)3(配列番号10)をコードするDNAと作動可能に連結し、VHおよびVL配列が連続した一本鎖タンパク質として発現するようにし、ここで、VLおよびVH 領域は屈曲性リンカーによって連結している(例えば、Bird et al. (1988) Science 242:423 426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879 5883; McCafferty et al., Nature (1990) 348:552 554を参照されたい)。一本鎖抗体は1つのVHおよびVLのみを用いる場合、一価であってもよいし、2つのVHおよびVLを用いる場合、二価であってもよいし、3以上のVHおよびVLを用いる場合、多価であってもよい。
【0077】
キメラ抗体
本発明はさらにその1つの特異性が本発明の免疫原性Nogo受容体-1ポリペプチドに対するものである二重特異性抗体またはその抗原結合性フラグメントを含む。1つの態様において、本発明の免疫原性Nogo受容体-1ポリペプチドに1つの結合ドメインを介して、そして第二の分子に第二の結合ドメインを介して特異的に結合するキメラ抗体を作成してもよい。キメラ抗体は組換え分子生物学技術によって産生してもよいし、互いに物理的に結合させてもよい。さらに、2以上のVHおよびVLを含み、本発明の免疫原性ポリペプチドと、ミエリン媒介成長円錐崩壊と神経突起伸長および出芽の阻害の減弱に関連するその他の分子に特異的に結合する一本鎖抗体を作成してもよい。かかる二重特異性抗体は周知技術によって作成でき、例えば、 Fanger et al. Immunol Methods 4: 72-81 (1994)およびWright and Harris,前掲を参照されたい。また、(iii)に関しては例えば、Traunecker et al. Int. J. Cancer (Suppl.) 7: 51-52 (1992)を参照されたい。
【0078】
ある態様において、キメラ抗体は本発明の抗体由来の1または複数の可変領域を用いて調製する。別の態様において、キメラ抗体は該抗体からの1または複数のCDR領域を用いて調製する。
【0079】
誘導体化および標識化抗体
本発明の抗体または抗原結合性フラグメントは誘導体化または別の分子に連結してもよい(例えば、別のペプチドまたはタンパク質)。一般に、抗体または抗原結合性フラグメントは本発明の免疫原性ポリペプチドへの結合が誘導体化または標識によって阻害されないように誘導体化する。例えば、本発明の抗体または抗体部分を機能的に(化学的カップリング、遺伝子融合、非共有的結合その他によって)1または複数のその他の分子的実体に結合させるとよい。分子的実体としては例えば、別の抗体(例えば、二重特異性抗体または二価抗体)、検出試薬、細胞毒性試薬、医薬および/または抗体または抗原結合性フラグメントのその他の分子への結合を媒介しうるタンパク質またはペプチドが挙げられる(例えば、ストレプトアビジンコア領域またはポリヒスチジンタグ)。
【0080】
ある態様において、誘導体化抗体は2以上の抗体(同じタイプでも異なるタイプでもよく、例えば二重特異性抗体を作るもの)の架橋によって産生する。好適なクロスリンカーにはヘテロ二機能性のもの、即ち2種類の反応性の基が適当なスペーサーによって分離されているもの(例えば、m- マレイミドベンゾイル-N-ヒドロキシスクシンイミドエステル)またはホモ二機能性のもの(例えば ジスクシンイミジルスベレート)がある。かかるリンカーはPierce Chemical Company, Rockford, Illから入手できる。
【0081】
ある態様において、誘導体化抗体は標識化抗体である。例えば、本発明の抗体または抗体部分が誘導体化される検出試薬は蛍光化合物であり、例えば、フルオレセイン、フルオレセインイソチアシアナート、ローダミン、5-ジメチルアミン-1-ナフタレンスルホニルクロリド、フィコエリトリン、ランタニドリン等が挙げられる。抗体は検出に有用な酵素で標識してもよく、かかる酵素としては、例えば、セイヨウワサビペルオキシダーゼ、β-ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ、グルコースオキシダーゼ等が挙げられる。検出可能な酵素で標識する態様において、抗体は、酵素が検出可能な反応産物を産生するのに用いるさらなる試薬の添加によって検出する。例えば、セイヨウワサビペルオキシダーゼでは過酸化水素およびジアミノベンジジンを用いる。抗体はビオチンで標識してもよく、この場合アビジンまたはストレプトアビジン結合の間接的測定によって検出する。抗体は二次レポーター(例えば、ロイシンジッパーペア配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)によって認識されるあらかじめ決定したポリペプチドエピトープで標識してもよい。
【0082】
抗-Nogo受容体-1抗体またはその抗原-フラグメントを放射標識アミノ酸で標識してもよい。放射標識は診断および治療用途の両方に用いられる。放射標識抗-Nogo受容体-1抗体は診断に用いることが出来、例えば、対象におけるNogo受容体-1レベルの判定に用いられる。さらに、放射標識抗-Nogo受容体-1抗体は脊髄損傷の治療に治療的に用いられる。ポリペプチドのための標識の例は、これらに限定されないが以下の放射性同位体または放射性ヌクレオチドが挙げられる- 3H、14C、15N、35S、90Y、99Tc、111In、125I、131I。
【0083】
抗-Nogo受容体-1抗体またはその抗原-フラグメントは以下のような化学基で誘導体化してもよい:ポリエチレングリコール(PEG)、メチルまたはエチル基または炭水化物基。これらの基は抗体の生物学的特性の向上、例えば、血清半減期の上昇または組織結合性の上昇、に有用である。
【0084】
抗-Nogo受容体-1抗体の特徴付け
抗-Nogo受容体-1抗体のクラスおよびサブクラス
抗-Nogo受容体-1抗体のクラスおよびサブクラスは当該技術分野で知られているいずれの方法で判定してもよい。一般に、抗体のクラスおよびサブクラスは、抗体の特定のクラスおよびサブクラスに特異的な抗体を用いて判定できる。かかる抗体は市販されている。クラスおよびサブクラスはELISA、ウェスタンブロットおよびその他の技術によって判定できる。あるいは、クラスおよびサブクラスは、抗体の重鎖および/または軽鎖の定常ドメインの全部または一部の配列決定、そのアミノ酸配列と免疫グロブリンの様々なクラスおよびサブクラスの公知のアミノ酸配列との比較、および抗体のクラスおよびサブクラスの決定によって判定できる。
【0085】
抗-Nogo受容体-1抗体のNogo受容体-1への結合アフィニティー
本発明の抗-Nogo受容体-1抗体の本発明の免疫原性Nogo受容体-1ポリペプチドに対する結合アフィニティーおよび解離速度は当該技術分野で知られているいずれの方法でも測定できる。例えば、結合アフィニティーは、競合的ELISA、RIA、BIAcoreまたはKinExA技術によって測定できる。解離速度もまた、BIAcoreまたは KinExA 技術によって測定できる。結合アフィニティーおよび解離速度は例えばBIAcoreを用いた表面プラズモン共鳴によって測定される。
【0086】
7E11および1H2のKdを測定したところ、それぞれ1 x 10-7 Mおよび2 x 10-8 Mであった。
【0087】
抗-Nogo受容体-1抗体によるNogo受容体-1活性の阻害
ある態様において、本発明の抗-Nogo受容体-1抗体または抗原結合性フラグメントは、Nogo受容体-1のリガンドに対する結合を阻害する。かかる阻害のIC50 は当該技術分野で知られているいずれの方法でも測定でき、例えば、ELISA、RIAまたは機能アンタゴニズムが挙げられる。ある態様において、IC50は0.1〜500 nMである。ある態様において、IC50は10〜400 nMである。さらに別の態様において、 抗体またはその部分のIC50は60 nM〜400 nMである。7E11および1H2のIC50を結合アッセイにおいて測定したところそれぞれ400 nM および60 nMであった。以下の表3を参照されたい。
【0088】
要約すると、当業者であれば、本発明の教示に接した場合、本発明の抗体の生物学的特性を変化させるのに用いられる多くの方法を利用することが出来る。例えば、所与の抗体分子の安定性、半減期、免疫原性、毒性、アフィニティーまたは収率を増減する方法等が挙げられる。あるいはその他の方法によって抗体の生物学的特性を特定の用途により好適なように変化させることができる。
【0089】
本発明の抗体を含む組成物および用途を以下に記載する。
可溶性Nogo受容体-1ポリペプチド
タンパク質
全長Nogo受容体-1は、シグナル配列、N-末端領域(NT)、8つのロイシンリッチリピート(LRR)、LRRCT領域(8つのロイシンリッチリピートのC-末端の1つのロイシンリッチリピートドメイン)、C-末端領域 (CT)および GPI アンカー (図1参照)からなる。
【0090】
本発明のいくつかの態様は可溶性Nogo受容体-1ポリペプチドを提供する。本発明の可溶性Nogo受容体-1ポリペプチドは、NT ドメイン; 8 LRRおよびLRRCT ドメインを含み、シグナル配列および機能性GPIアンカーを欠く(即ち、 GPI アンカーがないか、細胞膜と有効に結合する能力を欠くGPI アンカーがある)。
【0091】
ある態様において、可溶性Nogo受容体-1ポリペプチドは異種LRRを含む。ある態様において、可溶性Nogo受容体-1ポリペプチドは2、3、4、5、6、7または8の異種LRRを含む。異種LRRとは、Nogo受容体-1以外のタンパク質から得たLRRを意味する。異種LRRの起源となりうる例示的なタンパク質は、toll-様受容体(TLR1.2); T-細胞活性化ロイシンリピートリッチタンパク質; デセオリン; OM-gp; インスリン様成長因子結合タンパク質酸不安定性サブユニット slitおよびrobo; そしてtoll-様受容体4である。
【0092】
ある態様において、本発明は319 アミノ酸の可溶性Nogo受容体-1ポリペプチド(可溶性Nogo受容体-1 344、sNogoR1-344またはsNogoR344)( 配列番号6および8の残基 26-344 または配列番号8の残基 27-344)を提供する。ある態様において、本発明は285 アミノ酸の可溶性Nogo受容体-1ポリペプチド (可溶性Nogo受容体-1 310、sNogoR1-310またはsNogoR310)(配列番号7および9の残基 26-310または配列番号9の残基27-310)を提供する。図1を参照されたい。
【0093】
【表1】

[全長および1-310 マウス配列をこの表に追加すべきか?]
【0094】
本発明のある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは、リガンドのNogo受容体-1への結合の阻害に用いられ、Nogo受容体-1リガンドのアンタゴニストとして作用する。本発明のある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは神経細胞の例えば軸索成長における神経突起伸長および出芽の阻害の減少および神経細胞におけるミエリン媒介成長円錐崩壊の阻害に用いられる。ある態様において、神経細胞はCNS 神経細胞である。
【0095】
sNogoR310およびsNogoR344は驚くべきことにNogoA、NogoB、NogoC、MAGおよびOM-gpのNogo受容体-1に対する結合をブロックする。
【0096】
ある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは、さらに異種ポリペプチドを含む融合タンパク質の構成成分である。ある態様において、異種ポリペプチドは免疫グロブリン定常ドメインである。ある態様において、免疫グロブリン定常ドメインは重鎖定常ドメインである。ある態様において、異種ポリペプチドはFc フラグメントである。ある態様において、Fcは本発明の可溶性Nogo受容体-1ポリペプチドのC-末端に連結する。ある態様において、融合Nogo受容体-1タンパク質はダイマーである。
【0097】
本発明の核酸分子
本発明は本発明のポリペプチドをコードする核酸を提供し、本発明のポリペプチドには配列番号1-9のいずれかのポリペプチドが含まれる。ある態様において、核酸は以下からなる群から選択されるポリペプチドをコードする:配列番号6および8に示すNogo受容体-1のアミノ酸残基 26-344または配列番号8に示すNogo受容体-1のアミノ酸残基 27-344。ある態様において、核酸分子は以下からなる群から選択されるポリペプチドをコードする:配列番号7および9に示すNogo受容体-1のアミノ酸残基 26-310または配列番号9に示すNogo受容体-1のアミノ酸残基 27-310。本明細書において用いる、「核酸」とは、ゲノムDNA、cDNA、mRNAおよびアンチセンス分子ならびに以下のような核酸を意味する:別の骨格に基づく核酸または天然または合成の別の塩基を含む核酸。ある態様において、核酸はさらに転写プロモーターおよび所望によりシグナル配列を含み、そのいずれも本発明のポリペプチドをコードするヌクレオチド配列に作動可能に連結している。
【0098】
ある態様において、本発明は本発明のNogo受容体-1融合タンパク質をコードする核酸を提供する。ある態様において、核酸は本発明のNogo受容体-1融合タンパク質をコードし、本発明のNogo受容体-1融合タンパク質には、以下からなる群から選択されるポリペプチドを含む融合タンパク質が含まれる:配列番号6および8に示すNogo受容体-1のアミノ酸残基 26-344または配列番号8のアミノ酸残基 27-344および配列番号7および9に示すNogo受容体-1のアミノ酸残基 26-310または配列番号9のアミノ酸残基27-310。ある態様において、Nogo受容体-1融合タンパク質をコードする核酸はさらに、転写プロモーターおよび所望によりシグナル配列を含む。ある態様において、ヌクレオチド配列はさらに免疫グロブリン定常領域をコードする。ある態様において、免疫グロブリン定常領域は重鎖定常領域である。ある態様において、ヌクレオチド配列はさらにヒンジ領域に連結した免疫グロブリン重鎖定常領域をコードする。ある態様において、核酸はさらにFcをコードする。ある態様において、Nogo受容体-1融合タンパク質はFc フラグメントを含む。
【0099】
本発明のコード核酸はさらに診断およびプローブ用の検出可能な標識を含むように改変してもよい。様々なかかる標識は当該技術分野で知られており、本明細書に記載するコード分子に容易に適用できる。好適な標識としては、これらに限定されないが、ビオチン、放射標識ヌクレオチド等が挙げられる。当業者であれば標識化コード核酸分子を得るために当該技術分野で知られたいずれの標識も利用することが出来る。
【0100】
組成物
ある態様において、本発明は以下からなる群から選択される免疫原性ポリペプチドを含む組成物を提供する:配列番号1、配列番号2、配列番号3、配列番号4および配列番号5。
【0101】
ある態様において、本発明は本発明の抗-Nogo受容体-1抗体またはその抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質を含む組成物を提供する。
【0102】
ある態様において、本発明の組成物は賦形剤および補助剤を含む好適な医薬上許容される担体を含んでいてもよく、かかる担体は活性化合物を作用部位に送達するために医薬上使用されうる調製物への加工を促進する。非経口投与に好適な製剤には、水可溶性形態、例えば、水-可溶性塩における活性化合物の水溶液が挙げられる。適当に油性の注射用懸濁液などの活性化合物の懸濁液を投与してもよい。好適な親油性溶媒または媒体としては、脂肪油、例えば、ごま油または合成脂肪酸エステル、例えば、エチルオレアートまたはトリグリセリドが挙げられる。水性注射用懸濁液は懸濁液の粘度を上昇させる物質を含んでいてもよく、かかる物質としては、例えば、カルボキシメチルセルロースナトリウム、ソルビトールおよびデキストランが挙げられる。所望により、懸濁液はまた安定剤を含んでいてもよい。リポソームも本発明の分子を被覆して細胞へと送達するのに利用できる。例示的な「医薬上許容される担体」はあらゆる溶媒、分散媒、被覆、抗細菌剤および抗真菌剤、生理的に許容される等張および吸収遅延剤等、水、生理的食塩水、リン酸緩衝食塩水、デキストロース、グリセロール、エタノール等およびそれらの組み合わせである。ある態様において、組成物は等張剤を含み、例えば、糖、ポリアルコール、例えば、マンニトール、ソルビトールまたは塩化ナトリウムが挙げられる。ある態様において、組成物は医薬上許容される物質、例えば、少量の補助物質、例えば湿潤剤または乳化剤、保存料または緩衝剤を含み、これらは本発明の抗体、抗原結合性フラグメント、可溶性Nogo受容体または融合タンパク質の有効期間または有効性を改善する。
【0103】
本発明の組成物は様々な形態であってよく、例えば、液体、半流動性および固体剤形が含まれ、例えば液体溶液(例えば、注射可能および注入可能溶液)、分散液または懸濁液が挙げられる。好ましい形態は意図する投与方法および治療用途に依存する。1つの態様において、組成物は注射可能または注入可能溶液形態であり、例えば、ヒトのその他の抗体による受動免疫に用いられるような組成物等が挙げられる。
【0104】
組成物は溶液、マイクロエマルジョン、分散液、リポソームまたはその他の高薬剤濃度に好適な構造に製剤するとよい。無菌注射可能溶液は、要求される量の抗-Nogo受容体-1抗体を所望により上記の1または複数の成分とともに適当な溶媒に導入し、次いでフィルター滅菌することによって調製される。一般に、分散液は活性化合物を基本分散媒を含む無菌媒体に所望により上記のその他の成分とともに導入することによって調製される。無菌注射可能溶液の調製物のための無菌粉末の場合、好ましい調製方法は減圧乾燥および凍結乾燥であり、これにより先に無菌-ろ過した溶液から活性成分の粉末とさらなる所望の成分が得られる。溶液の適当な流動性は例えば以下のように維持されうる。レシチン等の被覆剤の使用、分散液の場合要求される粒径の維持、そして界面活性剤の使用。注射可能組成物の吸収の延長は吸収を遅延させる薬剤、例えば、モノステアリン酸塩およびゼラチンを組成物に含めることによってもたらされうる。
【0105】
ある態様において、活性化合物は化合物が迅速に放出するのを保護する担体とともに調製すればよく、例えば、徐放性製剤とすればよい。かかる徐放性製剤としては、インプラント、経皮パッチおよびマイクロカプセル化送達系が挙げられる。生分解性、生体適合性ポリマーを用いることが出来、例えば、酢酸エチレンビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステルおよびポリ乳酸が挙げられる。かかる製剤の多くの調製方法が特許されているかあるいは当業者に周知である。例えば、Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978を参照されたい。
【0106】
補足性活性化合物を組成物に含めてもよい。ある態様において、本発明のNogo受容体-1抗体またはその抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質は1または複数のさらなる治療薬と共製剤および/または共投与される。
【0107】
本発明の医薬組成物は、「治療上有効量」または「予防的に有効量」の本発明の抗体、抗原結合性フラグメント、ポリペプチドまたは融合タンパク質を含んでいるとよい。「治療上有効量」とは、投与および必要な期間において所望の治療結果を達成するのに有効な量を意味する。Nogo受容体-1抗体またはその抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはNogo受容体融合タンパク質の治療上有効量は、様々な因子に応じて変動し得、かかる因子としては例えば、個人の疾患状態、年齢、性別および体重が挙げられる。治療上有効量は抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはNogo受容体融合タンパク質のあらゆる毒性や有害作用よりも治療的に有用な効果が上回るものでもある。「予防的に有効量」とは投与および必要な期間において所望の予防結果を達成するのに有効な量を意味する。典型的には予防用量は対象に疾患前または疾患のより早期の段階にて用いられるために、予防的に有効量は治療上有効量より少ない。
【0108】
用法・用量は最適な望ましい応答(例えば治療または予防応答)が得られるよう調整すればよい。例えば、1のボーラスを投与してもよいし、いくつかに分割された用量を長期に投与してもよいし、治療状況の切迫に応じて比例的に増減してもよい。投与の便宜のために用量単位形態における非経口組成物を製剤するのが特に好ましく、本明細書において用いる用量単位形態とは、治療すべき哺乳類対象にとっての単位用量として好適な物理的に分離された単位であり、各単位は、必要な医薬担体とともに所望の治療効果が得られるよう計算されたあらかじめ決定した量の活性化合物を含む。本発明の用量単位形態の特定は以下によって示され直接依存する:(a)抗体、抗原結合性フラグメントおよび可溶性受容体-1ポリペプチドまたはNogo受容体融合タンパク質の固有の特性および達成すべき特定の治療または予防効果そして(b)かかる抗体、抗原結合性フラグメントおよび可溶性受容体-1ポリペプチドまたはNogo受容体融合タンパク質の配合における個人の感受性の処置についての当該技術分野に固有の制限。ある態様において、治療上有効用量はNogo受容体-1抗体またはその抗原結合性フラグメントについて1日あたり 0.1 - 4 mg/Kgの範囲である。ある態様において、 Nogo受容体-1抗体またはその抗原結合性フラグメントの治療上有効用量範囲は1日あたり0.2 - 4 mg/Kgである。ある態様において、Nogo受容体-1抗体またはその抗原結合性フラグメントの治療上有効用量範囲は1日あたり0.2 mg/Kgである。
【0109】
抗体、抗原結合性フラグメント、可溶性受容体および融合タンパク質の使用
ある態様において、本発明は必要とする哺乳類に抗-Nogo受容体-1抗体、かかる抗体の抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはかかるポリペプチドを含む融合タンパク質を投与することによるNogo受容体-1活性の阻害方法を提供する。
【0110】
ある態様において、本発明はNogo受容体-1と本発明の抗体または抗原結合性フラグメントを接触させる工程を含むNogo受容体-1のリガンドへの結合の阻害方法を提供する。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。
【0111】
ある態様において、本発明は、神経細胞と本発明の抗体またはその抗原結合性フラグメントとを接触させる工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する。ある態様において、本発明は、神経細胞と本発明の抗体または抗原結合性フラグメントとを接触させる工程を含む神経細胞における神経突起伸長または出芽の阻害を減少させる方法を提供する。ある態様において、神経細胞はCNS 神経細胞である。かかる方法のいくつかにおいて神経突起伸長または出芽は軸索成長である。
【0112】
ある態様において、本発明は、以下の工程を含む、神経細胞が死の危険にある哺乳類における、神経細胞の生存を促進させる方法を提供する:(a) (i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(ii)可溶性Nogo受容体-1ポリペプチド、を発現する培養宿主細胞を提供する工程; および、(b) 神経細胞においてまたはその近くの部位にて哺乳類に宿主細胞を導入する工程。 Almudena Ramon-Cueto, M Isabel Cordero, Fernando F Santos-Benito and Jesus Avila (2000) Functional recovery of paralegic rats and motor axon regneration in their spinal cords by olfactory ensheathing cells. Neuron 25, 425-435。
【0113】
ある態様において、本発明は以下の工程を含む神経細胞が哺乳類のものである死の危険にある神経細胞の生存を促進するための遺伝子治療方法を提供する:神経細胞においてまたはその近くの部位にて(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(b)可溶性Nogo受容体-1ポリペプチド、をコードするヌクレオチド配列を含むウイルスベクターを投与する工程、ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは哺乳類において神経細胞の生存を促進するのに十分な量にてヌクレオチド配列から発現する。かかる態様にとって有用なウイルスベクターおよび方法は、例えば、Noel et al., Human Gene Therapy, 13, 1483-93 (2002)に記載されている。
【0114】
ある態様において、本発明は、リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含むリガンドへのNogo受容体-1の結合の阻害方法を提供する。
【0115】
ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、Nogo受容体-1リガンドの活性を調節する方法を提供する。
【0116】
ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における成長円錐崩壊の阻害方法を提供する。ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法を提供する。ある態様において、神経細胞はCNS 神経細胞である。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。ある態様において、神経突起伸長または出芽は軸索成長である。
【0117】
本明細書に記載するあらゆるタイプの抗体または受容体を治療的に用いることが出来る。ある態様において、抗-Nogo受容体-1抗体はヒト抗体である。ある態様において、哺乳類はヒト患者である。ある態様において、抗体またはその抗原結合性フラグメントは獣医用またはヒト疾患の動物モデルとして、抗体と交差反応するNogo受容体-1を発現する非-ヒト哺乳類(例えば、霊長類、カニクイザルまたはアカゲザル)に投与される。かかる動物モデルは本発明の抗体の治療効果の評価に有用である。
【0118】
ある態様において、抗-Nogo受容体-1抗体または抗原結合性フラグメントあるいは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質の投与を用いて損傷部位への軸索成長を促進することにより脊髄損傷を治療する。
【0119】
本発明の抗-Nogo受容体-1抗体または抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質は単独で、その他の試薬と組み合わせて、または逐次に組み合わせて提供するとよい。その他の試薬は特定の病理プロセスを調節するものである。例えば、抗炎症薬をさらなる神経細胞損傷および軸索再生の阻害のブロックの手段として脳卒中の後に共投与するとよい。本明細書において用いる、Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1およびNogo受容体融合タンパク質は、同時、逐次または別々に投与される場合、1または複数のさらなる治療薬と組み合わせて投与されるという。
【0120】
本発明の抗-Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチド、Nogo受容体-1融合タンパク質は、非経口、皮下、静脈内、筋肉内、腹腔内、経皮、吸入または頬側経路にて投与するとよい。例えば、薬剤は損傷部位へ微量注入により局所投与すればよい。典型的な部位としては、これに限定されないが、損傷に起因する脊髄の損傷領域が挙げられる。投与用量はレシピエントの年齢、健康状態、体重、あるとすれば併用療法の種類、治療頻度および所望の効果の性質に依存する。
【0121】
本発明の化合物はインビボにて、通常、哺乳類、例えば、ヒト、ヒツジ、ウマ、ウシ、ブタ、イヌ、ネコ、ラットおよびマウスにおいて、またはインビトロにて利用できる。
【0122】
本発明のベクター
ある態様において、本発明は、コード配列を含む組換えDNA分子(rDNA)を提供する。本明細書において用いる、rDNA分子とは、分子操作を受けたDNA分子である。rDNA分子の作成方法は当該技術分野で周知であり、例えば、Sambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory Pressを参照されたい。あるrDNA分子においては、コードDNA配列は発現制御配列およびベクター配列に作動可能に連結している。
【0123】
ある態様において、本発明は、本発明のポリペプチドをコードする核酸を含むベクターを提供する。本発明の核酸が作動可能に連結するベクターおよび発現制御配列の選択は直接、当該技術分野で周知のように所望の機能特性に依存する(例えば、タンパク質発現および形質転換すべき宿主細胞)。本発明のベクターは少なくとも複製または宿主染色体への挿入が可能なものであり、好ましくは、rDNA分子に含まれる構造遺伝子の発現も可能にするものである。
【0124】
作動可能に連結したタンパク質コード配列の発現を制御するのに用いられる発現制御要素は当該技術分野で知られており、これらに限定されないが、例えば、誘導性プロモーター、構成的プロモーター、選抜シグナルおよびその他の制御要素が挙げられる。好ましくは、誘導性プロモーターは迅速に制御できるものであり、例えば、宿主細胞の培地の栄養成分に応答性のものなどである。
【0125】
1つの態様において、コード核酸分子を含むベクターは原核生物レプリコンを含み、レプリコンとは、それによって形質転換された細菌宿主細胞等の原核生物宿主細胞において染色体外で組換えDNA分子の自己複製と維持を可能にする能力を有するDNA配列である。かかるレプリコンは当該技術分野で周知である。さらに、原核生物レプリコンを含むベクターは、その発現が検出可能または選抜可能マーカー、例えば薬剤耐性マーカーを付与する遺伝子を含んでいてもよい。細菌の薬剤耐性遺伝子に典型的なものは、アンピシリンまたはテトラサイクリンへの耐性を付与するものである。
【0126】
原核生物レプリコンを含むベクターは、細菌宿主細胞、例えば、大腸菌におけるコード遺伝子配列の発現(転写および翻訳)の誘導を可能にする原核生物またはバクテリオファージプロモーターを含んでいてもよい。プロモーターとは、RNA ポリメラーゼに結合し転写を起こすDNA配列によって形成される発現制御要素である。細菌宿主に適合するプロモーター配列は典型的には、本発明のDNAセグメントの挿入のために便宜な制限部位を含むプラスミドベクターにおいて提供される。かかるベクタープラスミドの例は、pUC8、pUC9、pBR322およびpBR329 (Bio-Rad(登録商標) Laboratories)、pPLおよびpKK223 (Pharmacia)である。好適な原核生物宿主を用いて本発明のタンパク質をコードする組換えDNA分子を発現させることが出来る。
【0127】
真核生物細胞、好ましくは脊椎動物細胞に適合する発現ベクターもコード配列を含むrDNA分子の形成に利用できる。真核生物細胞発現ベクターは当該技術分野で周知であり、いくつか市販されている。典型的にはかかるベクターは所望のDNAセグメントの挿入のための便宜な制限部位を含むように提供される。かかるベクターの例は、pSVLおよびpKSV-10 (Pharmacia)、pBPV-1、pML2d (International Biotechnologies)、pTDT1 (ATCC31255)およびその他の真核生物発現ベクターである。
【0128】
本発明のrDNA分子の構築に用いられる真核生物細胞発現ベクターはさらに真核生物細胞において有効な、好ましくは薬剤耐性選抜マーカーといった選抜可能マーカーを含んでいてもよい。好ましい薬剤耐性マーカーは、その発現の結果、ネオマイシン耐性をもたらす遺伝子、即ちネオマイシンホスホトランスフェラーゼ (neo)遺伝子である(Southern et al., (1982) J. Mol. Anal. Genet. 1, 327-341)。あるいは選抜可能マーカーは別のプラスミドに存在してもよく、2つのベクターは宿主細胞に共形質移入によって導入され、トランスフェクタントは、選抜可能マーカーのための適当な薬剤中で培養することによって選抜する。
【0129】
本発明の抗体または抗体部分の発現のために、部分または全長の軽鎖および重鎖をコードするDNAを発現ベクターに挿入し、それら遺伝子を転写および翻訳制御配列に作動可能に連結する。発現ベクターとしては、プラスミド、レトロウイルス、コスミド、YAC、EBV-由来エピソームなどが挙げられる。抗体遺伝子をベクターに連結してベクター中の転写および翻訳制御配列が抗体遺伝子の転写および翻訳の制御の所望の機能を果たすようにする。発現ベクターおよび発現制御配列は用いる発現宿主細胞に適合するものを選択する。抗体軽鎖遺伝子と抗体重鎖遺伝子を別々のベクターに挿入してもよい。ある態様において、両方の遺伝子を同じ発現ベクターに挿入する。抗体遺伝子は標準的方法(例えば抗体遺伝子フラグメントおよびベクターの相補的制限部位のライゲーションまたは制限部位が無い場合は平滑末端ライゲーション)によって発現ベクターに挿入する。
【0130】
便宜なベクターは機能的に完全なヒトCHまたはCL 免疫グロブリン配列をコードするものであり、適当な制限部位を含み、操作によってあらゆるVHまたはVL 配列が上記のように容易に挿入され発現するものである。かかるベクターにおいて、スプライシングが通常、挿入されたJ 領域におけるスプライス供与部位とヒト C 領域の前にあるスプライス受容部位の間で起こり、ヒト CHエキソンにおけるスプライス領域においても起こる。ポリアデニル化および転写終結は、コード領域の下流のネイティブな染色体部位にて起こる。組換え発現ベクターはまた、宿主細胞からの抗体鎖の分泌を容易にするシグナルペプチドをコードしていてもよい。抗体鎖遺伝子をベクターにクローニングし、シグナルペプチドが抗体鎖遺伝子のアミノ末端にインフレームにて連結するようにするとよい。シグナルペプチドは免疫グロブリンシグナルペプチドであっても異種シグナルペプチド(即ち、非免疫グロブリンタンパク質からのシグナルペプチド)であってもよい。
【0131】
本発明の免疫原性ポリペプチド、Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質に加えて、本発明の組換え発現ベクターは、宿主細胞におけるその発現を制御する調節配列を担持する。当業者であれば調節配列の選択を含む発現ベクターの設計は形質転換する宿主細胞の選択、所望のタンパク質発現レベルなどの因子に依存することを理解するであろう。好ましい哺乳類宿主細胞発現のための調節配列としては哺乳類細胞における高レベルのタンパク質発現をもららすウイルス性要素、例えば、レトロウイルスLTR、サイトメガロウイルス(CMV) (例えば、CMV プロモーター/エンハンサー)、サルウイルス 40 (SV40) (例えば、SV40 プロモーター/エンハンサー)、アデノウイルス、(例えば、アデノウイルス主要後期プロモーター (AdMLP))、ポリオーマ由来のプロモーターおよび/またはエンハンサーおよび強力な哺乳類プロモーター、例えば、ネイティブな免疫グロブリンおよびアクチンプロモーターが挙げられる。ウイルス性制御要素についての更に詳細な説明およびその配列については例えば以下を参照されたい:Stinskiの米国特許第5168062号、Bell et alの米国特許第4510245号およびSchaffner et alの米国特許第4968615号。
【0132】
異種遺伝子および調節配列に加えて、本発明の組換え発現ベクターはさらなる配列を担持していてもよく、例えば、宿主細胞におけるベクターの複製を制御する配列(例えば、複製起点)および選抜可能マーカー遺伝子が挙げられる。選抜可能マーカー遺伝子は、ベクターが導入された宿主細胞の選抜を容易にする(例えば、米国特許第4399216号、4634665号および5179017号、これらはすべてAxel et alによる)。例えば典型的には、選抜可能マーカー遺伝子はベクターが導入された宿主細胞に薬剤耐性を付与する。薬剤としては例えば、G418、ハイグロマイシンまたはメトトレキサートが挙げられる。好ましい選抜可能マーカー遺伝子としてはジヒドロ葉酸レダクターゼ (DHFR)遺伝子(メトトレキサート選抜/増幅を用いるdhfr- 宿主細胞における使用)およびneo遺伝子(G418 選抜用)が挙げられる。
【0133】
本発明のタンパク質の組換え産生のための宿主細胞および方法
本発明の抗-Nogo受容体-1抗体、免疫原性ペプチド、可溶性Nogo受容体-1ポリペプチド、可溶性Nogo受容体-1融合タンパク質をコードする核酸分子およびかかる核酸分子を含むベクターを用いて好適な宿主細胞を形質転換するとよい。形質転換は宿主細胞にポリヌクレオチドを導入するいずれの公知方法によってもよい。異種ポリヌクレオチドの哺乳類細胞への導入方法は当該技術分野で周知あり、デキストラン-媒介形質移入、リン酸カルシウム沈降、ポリブレン-媒介形質移入、プロトプラスト融合、エレクトロポレーション、リポソームへのポリヌクレオチドのカプセル封入および直接のDNAの核へのマイクロインジェクションが挙げられる。さらに核酸分子は哺乳類細胞にウイルスベクターによって導入してもよい。
【0134】
適当な細胞宿主の本発明のrDNA分子による形質転換は、周知方法により達成され、かかる方法は典型的には使用するベクターのタイプおよび宿主系に依存する。原核生物宿主細胞の形質転換については、エレクトロポレーションおよび塩処理方法を用いるとよい(例えば、Sambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory Press; Cohen et al., (1972) Proc. Natl. Acad. Sci. USA 69, 2110-2114を参照されたい)。脊椎動物細胞のrDNAを含むベクターによる形質転換については、エレクトロポレーション、カチオン脂質または塩処理方法を用いるとよい(例えば、Graham et al., (1973) Virology 52, 456-467; Wigler et al., (1979) Proc. Natl. Acad. Sci. USA 76, 1373-1376を参照されたい)。
【0135】
形質転換が成功した細胞、即ち本発明のrDNA分子を含む細胞は選抜可能マーカーについての選抜などの周知技術によって同定できる。例えば、本発明のrDNAの導入の結果得られる細胞をクローニングしてシングルコロニーを得ることが出来る。かかるコロニーからの細胞を回収し、溶解し、そのDNA内容物をrDNAの存在について、例えばSouthern, (1975) J. Mol. Biol. 98, 503-517に記載の方法で調べることが出来、あるいは細胞から産生されたタンパク質を免疫学的方法でアッセイすることが出来る。
【0136】
発現用宿主として利用できる哺乳類細胞株は当該技術分野で周知であり、アメリカンタイプカルチャーコレクション(ATCC)から得られる多くの不死化細胞株が含まれる。これらにはとりわけ、 チャイニーズハムスター卵巣(CHO)細胞、NSO、SP2 細胞、HeLa 細胞、仔ハムスター腎臓 (BHK) 細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)、A549 細胞、およびその他の多くの細胞株が含まれる。特に好ましい細胞株はどの細胞株の発現レベルが高いかを測定することによって選択される。その他の使用可能な細胞株は昆虫細胞株、例えば、Sf9 細胞である。本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質をコードする組換え発現ベクターは哺乳類宿主細胞に導入され、宿主細胞における抗体、ポリペプチドおよび融合ポリペプチドの発現を可能にするのに十分な期間宿主細胞を培養することにより産生され、あるいはより好ましくは本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質を宿主細胞が生育する培地で選抜する。本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質は、標準的タンパク質精製方法を用いて培地から回収できる。
【0137】
さらに本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質(またはそれら由来の部分)の産生細胞株からの発現は多くの公知技術を用いて促進できる。例えば、グルタミンシンゼターゼ遺伝子発現系 (GS系)は特定の条件下での発現促進の常套の手段である。GS系については欧州特許第0216846号、0256055号および0323997号および欧州特許出願第89303964.4号に記載されている。
【0138】
宿主細胞
本発明はさらに、本発明のNogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび/または可溶性Nogo受容体-1融合タンパク質をコードする核酸分子で形質転換された宿主細胞を提供する。宿主細胞は原核生物でも真核生物でもよい。本発明のタンパク質の発現に有用な真核生物細胞は、細胞株が細胞培養方法に適合し、発現ベクターの増殖および遺伝子産物の発現に適合する限り特に限定されない。好ましい真核生物宿主細胞としては、これらに限定されないが、酵母、昆虫および哺乳類細胞、好ましくは脊椎動物細胞、例えば、マウス、ラット、サルまたはヒト細胞株が挙げられる。有用な真核生物宿主細胞の例としては、ATCCからCCL61として入手できるチャイニーズハムスター卵巣 (CHO) 細胞、ATCCからCRL1658として入手できるNIH Swiss マウス 胚細胞NIH-3T3、仔ハムスター腎臓細胞(BHK)、その他の真核生物組織細胞細胞株が挙げられる。
【0139】
rDNA分子を用いる組換えタンパク質産生
本発明はさらに、本明細書に記載した核酸分子を用いる本発明のNogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび/または可溶性Nogo受容体-1融合タンパク質の産生方法を提供する。一般に組換え形態のタンパク質の産生は典型的には以下の工程を含む:
【0140】
第一に、本発明のタンパク質をコードする核酸分子を得る。コード配列がイントロンで分断されていない場合、それはあらゆる宿主における発現に直ちに好適である。
【0141】
核酸分子を次いで所望により、上記の好適な制御配列と作動可能に連結させ、タンパク質オープンリーディングフレームを含む発現単位を形成する。発現単位を用いて好適な宿主を形質転換し、形質転換した宿主を組換えタンパク質の産生を可能とする条件下で培養する。所望により、組換えタンパク質を培地または細胞から単離する;タンパク質の回収および精製はいくらかの不純物が許容される場合には特に必要ではない。
【0142】
上記の各工程は様々な方法で行うことが出来る。例えば、所望のコード配列をゲノム断片から得て直接適当な宿主において用いることが出来る。様々な宿主において作動可能な発現ベクターの構築は上記の適当なレプリコンおよび制御配列を用いて達成される。制御配列、発現ベクターおよび形質転換方法は、遺伝子発現に用いられる宿主細胞のタイプに依存し、先に詳細に説明した。好適な制限部位を、通常は入手可能ではないとしても、コード配列の末端に付加して、かかるベクターへの挿入用の切り出し可能な遺伝子を提供することが出来る。当業者であれば当該技術分野で知られている宿主/発現系を本発明の核酸分子とともに組換えタンパク質の産生に容易に用いることが出来る。
【0143】
本発明の理解をさらに容易にするために。以下に実施例を示す。かかる実施例は単に例示の目的であり、本発明の範囲を限定する意図のものではない。
【実施例1】
【0144】
マウスモノクローナル抗-Nogo受容体-1抗体の産生
本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗-Nogo受容体-1抗体を以下の方法および手順を用いて作製した。
【0145】
免疫
2種類の免疫アプローチを用いた:
1. 免疫原としてNogo受容体-1 (NogoR-1)を含むCOS-7細胞または細胞膜
ラットNogo受容体-1遺伝子(GenBank番号 AF 462390)を、CMV プロモーターおよび薬剤選抜用のジェネティシン耐性遺伝子を含む哺乳類発現ベクター pEAG1256 (Biogen)にサブクローニングした。組換えプラスミドをSuperfect (Qiagen)を用いてCOS-7細胞にトランスフェクトした。トランスフェクタントをジェネティシン (Gibco, 2 mg/ml)を用いて選抜し、クローニングし、FACSによってNogo受容体-1タンパク質の表面発現を確認した。COS-7膜を記載された手順[Wang et al., J. Neurochem.,75:1155-1161 (2000)]にしたがってかかる細胞から調製し、2回洗浄し、1 mg/ml [タンパク質濃度]にて10% グリセロール中、-70℃で保存した。
【0146】
8週齢の雌性RBF マウス (Jackson Labs, Bar Harbor, ME)を50μg ラットNogo受容体-1-COS-7 膜またはNogo受容体-1を表面に発現する全COS-7細胞のいずれかおよび50μl RIBI MPL+TDM+CWS アジュバント (Sigma Chemical Co., St. Louis, MO)を含む乳濁液で腹腔内に2週間毎に免疫した(Lipman et al., 1992)。免疫したマウスからの血清を1回目の免疫前、2回目および3回目の免疫の7日後、および3回目の免疫の38日後に回収し、抗-Nogo受容体-1抗体力価を以下のようにELISAによって測定した。
【0147】
2. 免疫原としての特定のNogo受容体-1 ペプチド
ラットNogo受容体-1遺伝子配列をベクター NTiソフトウェア (図2)を用いた抗原性分析にかけた。分析によって同定された抗原ペプチドをキーホール・リンペット・ヘモシアニン (KLH)と標準的グルタールデヒド法を用いて結合させた。
【0148】
8週齡の雌性RBF マウス(Jackson Labs, Bar Harbor, ME)を50 μg KLH-結合ペプチドおよび50 μl 完全フロイントアジュバント(Sigma Chemical Co., St. Louis, MO) を含む乳濁液で2週間毎に腹腔内に免疫した。免疫したマウスからの血清を1回目の免疫前、2回目および3回目の免疫の1週間後に回収し、抗-Nogo受容体-1抗体力価を測定した。追加抗原投与量を3回目の免疫後に与えた。この追加抗原投与量免疫の3日後、融合実験を開始した。
【0149】
ハイブリドーマ産生およびスクリーニング
抗原性Nogo受容体-1ペプチドで免疫したマウスからの血清をELISAにてスクリーニングし、Nogo受容体-1を発現するCOS-7細胞で免疫したマウスからの血清をフローサイトメトリーによってスクリーニングした。Nogo受容体-1-COS-7細胞に特異的に結合する抗体についてフローサイトメトリーで同定したところ陽性であったマウスを屠殺した。脾細胞をマウスから単離しFL653ミエローマ(Ig-/HGPRT- Balb/c マウスミエローマのAPRT- 誘導体、10% FBS、4500 mg/L グルコース、4 mM L-グルタミンおよび20 mg/ml 8-アザグアニンを含有するDMEM中で維持)と以前に記載されているようにして(Kennett et al., 1993. Monoclonal Antibodies: A New Dimension in Biological Analysis. Plenum Press, New York)融合させた。融合細胞を24-または48-ウェルプレート(Corning Glass Works, Corning, NY)に播き、アデニン、アミノプテリンおよびチミジン含有培地を与えた。AAT耐性培養物を以下に説明するようにしてNogo受容体-1-COS-7細胞またはNogo受容体-1抗原ペプチドへの結合についてELISAまたはフローサイトメトリーでスクリーニングした。陽性ウェル中の細胞をさらに限界希釈によってサブクローニングした。
【0150】
抗体のNogo受容体-1抗原ペプチドへの結合をスクリーニングするために、免疫原として用いたペプチドをBSAと結合させた。50 μlの0.1 M 炭酸水素ナトリウム緩衝液, pH 9.0 中の0.5μgの結合ペプチドを96-ウェルMaxiSorp プレート(Nunc)の各ウェルに添加した。プレートを37℃で1時間または4℃で16時間インキュベートし、非特異的結合部位を0.1% BSA、0.1%卵白アルブミン、0.1%ブロット(blotto)および0.001% アジドを含む25 mM HEPES, pH 7.4を用いてブロックした。ハイブリドーマ上清を添加し、25℃で1時間インキュベートした。PBSでの3回の洗浄後、 50μlの1:10,000希釈のセイヨウワサビペルオキシダーゼ-結合ヤギ抗-マウス二次抗体(Jackson ImmunoResearch Inc.)を各ウェルに添加し、さらに1時間インキュベートした。3回の洗浄後、色をTMB (Pierce)で現像し、2 M硫酸で停止した。着色強度を分光光度計で450 nmにてモニターした。
【0151】
抗体を全長Nogo受容体-1への結合について以下のようにスクリーニングした。COS-7細胞を製造業者の指示に従って0.1 uM CellTracker Green CMFDA (Molecular Probes, Eugene, OR)で標識した。等量のCellTracker標識コントロール細胞を洗浄したNogo受容体-1-COS-7細胞と混合し、抗-Nogo受容体-1被験血清とのインキュベーションに供した。15μlの細胞混合物を96-ウェルV底ポリスチレンプレート(Costar 3877, Corning, NY)の各ウェルに分配し、100μlのハイブリドーマ上清またはコントロール抗-Nogo受容体-1抗体を添加した。4℃で30分のインキュベーション後、細胞を洗浄し、PBS中の50 μlのR-フィコエリトリン-結合アフィニティー精製 F(ab')2 フラグメントヤギ抗-マウス IgG Fcガンマ特異的二次抗体(1:200, Jackson ImmunoResearch Laboratory, West Grove, PA)とともにインキュベートした。インキュベーションの最後に、細胞をPBSで2回洗浄し、1% FBSを含む200μlのPBSに懸濁し、FACS分析にかけた。あるいは、Nogo受容体-1-COS-7細胞をハイブリドーマ上清と混合し、R-フィコエリトリン-結合ヤギ抗-マウス二次抗体で処理し、直接標準的FACS分析にかけた。
【0152】
本発明者らは様々な免疫原を用いて25の抗-Nogo受容体-1抗体を作成した。本発明者らは免疫原としてラットNogo受容体-1 残基 110-125に対応するペプチド配列を用いて2つの抗体、7E11および5B10を作成した。本発明者らは免疫原として全長ラットNogo受容体-1でトランスフェクトしたCOS-7細胞から調製した膜を用いて3つの抗体、1H2、3G5および2F7を作成した。本発明者らは免疫原としてsNogoR310-Fcを用いて13の抗体を作成し(1D9.3、1E4.7、1B4.3、2C4.3、1F10.3、2H1.4、1H3.3、1G4.1、1E4.1、2G7.1、2C4.1、2F11.1および1H4.1)、免疫原としてラットNogo受容体-1 残基 423-434に対応するペプチド配列を用いて7つの抗体を作成した(2E8.1、2G11.2および1B5.1)。
【0153】
モノクローナル抗体7E11および5B10の配列分析
本発明者らはトータルRNAを Qiagen RNeasy ミニキットを用いて抽出し、単離したRNA からcDNAを作成した。本発明者らは、軽鎖配列をPCRによってプライマー、5'-TGAGGAGACGGTGACCGTGGTCCCTTGGCCCCAG-3'(配列番号12)および5'-AGGTSMARCTGCAGSAGTCWGG-3'(配列番号25)を用いて増幅した。本発明者らは、重鎖配列をPCRによってプライマー、5'-GGGGATATCCACCATGAAGTTGCCTGTTAGGCTGTTG-3'(配列番号13)および5'-GGGGATATCCACCATGAGGKCCCCWGCTCAGYTYCTKGGA-3'(配列番号14)を用いて増幅した。かかるプライマーは以下の縮重ヌクレオチドを含む: SはGまたはC;MはAまたはC、RはGまたはA;WはAまたはT;KはGまたはT;そしてYはTまたはCを表す。本発明者らはPCRフラグメントを配列決定ベクターにクローニングし、配列決定ベクターに特異的なプライマーを用いてジデオキシ鎖終結によってCDRのDNA配列を決定した。本発明者らは理論的にDNA配列を翻訳した結果、モノクローナル抗体 7E11および5B10の重鎖および軽鎖のCDR領域の部分アミノ酸配列を表2に示す。mAbの重鎖および軽鎖からの3つのCDRを表2において下線にて示す。7E11および5B10の軽鎖は94%のアミノ酸配列同一性を、重鎖は91%のアミノ酸配列同一性を有していた。mAb、7E11、5B10および1H2はIgG1 アイソタイプであり、mAb、3G5および2F7はIgG2a アイソタイプである。これら5つのmAbはそれぞれカッパアイソタイプの軽鎖を有する。本発明者らはこのアプローチによってその他のモノクローナル抗体の配列を分析した。
【0154】
【表2】
【0155】
モノクローナル抗-Nogo受容体-1抗体による可溶性Nogo受容体-1へのリガンド結合の阻害
本発明者らは上記のように産生した抗-Nogo受容体-1 モノクローナル抗体をそれらがNogo受容体-1へのリガンド結合を阻害するかを調べるべく試験した。
【0156】
下記のように産生した0.5μgの、ラットNogo受容体-1のアミノ酸残基 26-344およびラットIgG1分子のヒンジおよびFc領域を含む可溶性Nogo受容体-1融合タンパク質(sNogoR344-Fc)を、250μgのタンパク質-A- またはコムギ胚芽凝集素-結合SPAビーズ(Amersham Pharmacia Biotech)に2時間25℃で固定化した。50μlのHEPES-緩衝インキュベーション培地(10 mM HEPES, pH 7.4、0.1% ウシ血清アルブミン、0.1% 卵白アルブミン、2 mM MgCl2、2 mM CaCl2およびプロテアーゼ阻害剤)中のFc-sNogoR-1[これはどのsNOGO?]と結合したSPAビーズ、抗-Nogo受容体-1 mAbおよび1μl 125I-Nogo66 (Amersham, 2000 Ci/mmol, 1nM)を各サンプルウェルに添加した。16時間後、放射能を四連のサンプルにおいてTopCount (Packard)を用いて測定した。IC50値をカーブフィッティッング分析(図3)から算出した(PRISM, GraphPad Software, NJ)。いくつかの実験において、本発明者らは、AP-リガンド結合体(例えば、AP-Nogo66)も使用し、結合をアルカリホスファターゼ活性をモニターすることにより検出した。本発明者らはまた、mAbの、リガンドMAG-FcおよびAP-OM-gpのNogo受容体-1への結合をブロックする能力もアッセイした。
【0157】
モノクローナル抗体7E11、5B10、1H2、3G5および2F7はすべてNogo66、MAGおよびOM-gpのsNogoR344-Fcへの結合を阻害した。7E11および1H2について計算したNogo66についての IC50はそれぞれ400 nMおよび60 nMであった。3つのリガンドのNogo受容体-1への結合のmAb-媒介阻害をモニターするELISAからのデータを表3に要約する。
【0158】
【表3】
【0159】
パーセント置換は30 nM 抗体にて示され、特定のmAbについてのEC50 は記載されているカーブフィッティッング分析から決定した。「−」は、検出可能な活性が無いこと、そして「ND」は測定していないことを示す。
【実施例2】
【0160】
Fab-ファージ抗-Nogo受容体-1抗体の産生
本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗-Nogo受容体-1 Fab-ファージ抗体も、以下のようにFab-ファージライブラリーをスクリーニングすることによって得た。
【0161】
MorphoSys Fab-ファージライブラリー HuCAL(登録商標)GOLDを、組換えラット可溶性sNogoR310-Fcタンパク質およびラットNogo受容体-1を発現するCOS-7細胞に対してスクリーニングした。Nogo受容体-1に特異的に結合したFab-ファージを精製し、特徴づけた。14D5の重鎖はVH2遺伝子由来であり、軽鎖はVK1遺伝子由来である。これらFab-ファージの1つ、14D5の重鎖および軽鎖のCDRのアミノ酸配列を表4に示す。
【0162】
【表4】
【0163】
14D5は一価および二価の両方の形態においてラットNogo受容体-1に結合する。さらに、14D5はマウスおよびヒトNogo受容体-1およびヒトNogo受容体-2に結合するがマウスNogo受容体-3には結合しない。
【実施例3】
【0164】
抗-Nogo受容体-1モノクローナル抗体によるNogo受容体-1の免疫沈降
免疫沈降を行うために、100μlの溶解した細胞または50μlのPiPLC処理細胞を、400または450μlの抽出バッファー [10 mM Tris-HCl, pH 7.2、0.5% Tween-20、0.2 mM PMSF]またはRIPAバッファーと30μlのタンパク質AまたはGおよび1-2μgの抗体の存在下でそれぞれ混合した。混合物をシェーカーで4℃で16 時間インキュベートした。
【0165】
サンプルを穏やかに遠心してタンパク質AまたはG結合ビーズをペレットとした。ビーズを3回1 ml洗浄バッファー (10 mM Tris-HCl, pH 7.2、0.1% Tween-20)で洗浄した。最終洗浄は10%の元の洗浄バッファーで行った。
【0166】
ビーズを100μlの10% ベータ・メルカプトエタノールを含む2X SDSに再懸濁した。サンプルを室温でインキュベートした後、4-20% Tris-グリシンゲルのSDS-PAGEで泳動した。SDS-PAGEゲル分析によって判定したところ、モノクローナル抗体、3G5および2F7は、Nogo受容体-1を免疫沈降する。
【実施例4】
【0167】
抗体特異性のELISAによる判定
実施例1および2で産生したモノクローナルおよびFab-ファージ抗体の特異性を調べるために、本発明者らはNogo受容体-1ポリペプチドのパネルを用いてELISAを行った。パネルは、sNogoR310-Fc (ラットNogo受容体-1のアミノ酸26-310およびラットFcフラグメントを含む融合タンパク質)、sNogoR344-Fc(前掲)、ポリペプチド p-617 (配列番号1)、ポリペプチド p-618 (ラットNogo受容体-1のLRR7領域からの19-アミノ酸ポリペプチド;図2;配列番号11)およびポリペプチド p-4およびp-5 (それぞれNogo受容体-1のLRR5およびLRRCT領域からのポリペプチド)からなるものであった。卵白アルブミンおよびBSAをコントロールとして用いた。図4に示すように、mAb、1H2、3G5および2F7はすべてsNogoR344-Fcに特異的に結合した。類似の実験において、これら抗体はまた、ラットNogo受容体-1のアミノ酸310-344 (配列番号3)からなるポリペプチドに特異的に結合し、mAb、7E11および5B10はポリペプチド p-617 (配列番号1)に特異的に結合した。
【0168】
sNogoR310-Fc免疫からの10の抗体(1D9.3、1E4.7、1B4.3、2C4.3、1F10.3、2H1.4、1H3.3、1G4.1、1E4.1および2G7.1)は結合について互いに置換し、これはそれらがsNogoR310-Fc上の類似のまたはオーバーラップするエピトープを認識することを示す。sNogoR310-Fc 免疫からの他の3つの抗体(2C4.1、2F11.1および1H4.1)はアミノ酸残基 26-310に位置する異なるエピトープを認識する。
【0169】
本発明者らはまた、Fab-ファージ 14D5を用いてELISA結合アッセイを行った。AP-Nogo66、AP-OM-gpおよびMAG-Fc リガンドを固定化したsNogoR344-Fcに結合させたところ、1μMの14D5が完全にNogoおよびMAG結合を阻害した。10 μMの14D5がOM-gpのsNogoR344-Fcに対する結合の完全な阻害に要求された。
【実施例5】
【0170】
神経突起伸長アッセイ
先に産生したモノクローナルおよびFab-ファージ抗体の神経細胞に対するCNSミエリンの阻害効果を低減させる能力を試験するため、Lab-Tek 培養スライド(4ウェル)を0.l mg/mlのポリ-D-リジン(Sigma)でコーティングした。CNSミエリンまたはPBSを3 μlの滴としてスポットした。蛍光ミクロスフェア(Polysciences)をミエリン/PBSに添加して後の滴の同定を可能にした(Grandpre et al, Nature 403, 2000)。Lab-Tekスライドをすすぎ、次いで10 μg/mlラミニン(Gibco)でコーティングした。P3-4 SDラット子から後根神経節(DRG)を1 mg/ml コラゲナーゼタイプ1 (Worthington)で分離し、先端熱加工パスツールピペットで粉砕し、あらかじめプレートして神経細胞を豊富にし、最終的にはあらかじめコーティングした Lab-Tek 培養スライドに23,000細胞/ウェルにてプレーティングした。培地は5% 熱不活性化ドナーウマ血清、5% 熱不活性化胎児ウシ血清および50 ng/ml mNGF を含有するF12であり、37℃、5% CO2にて6時間インキュベートした。15μg/mlのmAb 7E11をプレーティングの直後に添加した。
【0171】
スライドを20分間、20% スクロースを含む4% パラホルムアルデヒドで固定し、1:500に希釈した神経細胞マーカー抗ベータ-III-チューブリン(Covance TUJ1)で染色した。二次抗体として抗-マウス Alexa Fluor 594 (Molecular Probes)を1:300に希釈し、スライドをGel/Mount(Biomed)でカバースリップした。5x デジタル画像を OpenLabソフトウェアで得てMetaMorphソフトウェアを用いて神経突起伸長を定量分析した。
【0172】
MAb 7E11は神経突起伸長のミエリン-媒介阻害からDRG神経細胞を保護した(図5)。類似の結果がmAb、1H2および3G5によっても観察された。
【0173】
ラットP7 DRG 神経細胞をCNSミエリン基質で培養する神経突起伸長保護アッセイにおいて、二価 14D5も効率的に神経突起伸長を促した。
【実施例6】
【0174】
Nogo受容体-1をトランスフェクトした細胞での7E11による免疫組織化学
実施例1に記載のように産生した抗-Nogo受容体-1 mAbの結合特性をさらに特徴づけるために、本発明者らはラットまたはヒトNogo受容体-1を発現する固定および生COS-7または293細胞への結合を比較した。
【0175】
固定細胞:
Nogo受容体-1でトランスフェクトした細胞および非-トランスフェクト細胞を8-ウェル Lab-Tek培養スライドに播き、4% パラホルムアルデヒドで15分間固定し、PBS中の10% 正常ヤギ血清、0.1% Triton X-100で1時間ブロッキングした。Mab 7E11をブロッキング溶液中15μg/mlおよび1.5μg/mlで添加し、室温で2時間インキュベートした; Alexa-結合二次抗体 抗-マウス(Molecular Probes)をブロッキング溶液中1:300希釈にて1時間インキュベートした; すべての核を標識するためにDAPI を5μg/mlにて二次抗体に添加した。
【0176】
生細胞:
トランスフェクト細胞および非-トランスフェクト細胞を8ウェル Lab-Tek培養スライドに播き、FACSバッファー(4% ドナーウマ血清含有)で30 分間4℃にてブロッキングし、FACS バッファー中15 μg/mlおよび1.5 μg/mlにて7E11と1時間4℃でインキュベートし、すすぎ、二次抗体抗-マウス-Alexa(FACSバッファー中1:300)と30分間4℃でインキュベートした。
【0177】
免疫組織化学染色実験は、すべてのmAbがラットNogo受容体-1を発現する細胞に結合したことを示した。mAb、7E11、2G7.1および2C4.1はヒトNogo受容体-1を発現する固定細胞と生細胞との両方に結合した。
【実施例7】
【0178】
脊髄挫傷のマウスモデル
神経細胞に対するインビボでの実施例1で産生した抗-Nogo受容体-1 mAbの効果を試験するために、本発明者らはマウス脊髄挫傷モデルを用いた。
【0179】
雌性マウス(18-22 g)を鎮痛薬および抗生剤で予防処置した。マウスを麻酔し、立体顕微鏡下で脊柱固定して定位固定装置に入れた。脊髄への外傷を重量ドロップ法(M. Li et al., Functional role and therapeutic implications of neuronal caspase-1 and -3 in a mouse model of traumatic spinal cord injury. Neuroscience Vol. 99, pp. 333-342, 2000)の改変法によって導入した。
【0180】
簡単に説明すると、T9およびT10椎弓切除術を行い脊柱をT9-T10横断方法を両側的に支持する1対のマウス横断クランプを用いて安定化した。直径1.4 mm、重さ2 gのステンレス鋼衝撃棒を、硬膜の2.5 cm上に位置させ、T10レベルにて脊髄に落下させた。手術中、マウスを37℃の毛布上に維持し、手術後に脱水を避けるために1 mlの昇温無菌生理的食塩水をそれぞれのマウスに皮下投与した。膀胱を手作業で反射的膀胱制御が回復するまで1日1回絞った。
【0181】
すべての動物に手術の8-12時間後おきに術後鎮痛薬を与え、その後7日間1日2回抗生物質処理を行った。動物には試験期間餌と水を自由に与えた。抗-Nogo受容体-1抗体を以下のラット脊髄離断モデルにて記載するように28日間くも膜下腔内注射によって損傷部位に送達した。
【実施例8】
【0182】
可溶性Nogo受容体-1融合タンパク質の特徴付け
可溶性Nogo受容体-1ポリペプチド(sNogoR-1)および融合タンパク質(Fc-sNogoR-1)を特徴づけるために、本発明者らは以下の実験を行った。
【0183】
3μgの可溶性Nogo受容体(sNogoR310-FcおよびsNogoR344-Fc)を250 μgのWGA-SPA ビーズに固定化させ、最終容量100 μLの結合バッファー(20 mM HEPES, pH 7.4, 2 mM Ca, 2 mM Mg, 0.1% BSA, 0.1% 卵白アルブミンおよびプロテアーゼ阻害剤)中の0.5 μL の放射性リガンド(終濃度 0.5 nM)を与えた。リガンドには、10 μMのNogo66、10 μMの125I-Nogo40 (NogoAのアミノ酸1-40)が含まれ、各リガンドセットは10 μLの抗-Nogo受容体-1抗体上清を含む。Nogo40上の3つのチロシンを別々にヨウ素化し、それぞれNogo40-A,-Bおよび -Cと命名した。三連の平均値を正規化%結合放射能として示す(図6、7および8)。エラーバーはSEMを示す。阻害剤の非存在下での結合放射能を100%とし、10 μM Nogo40存在下での最小結合放射能を0%としてデータを正規化した。
【実施例9】
【0184】
可溶性Nogo受容体-1融合タンパク質へのリガンド結合の阻害
実施例8の結合アッセイに類似の結合アッセイを用いて実施例1で産生した2つのmAbが125I-Nogo66のsNogoR344-Fcへの結合を阻害する能力を調べた。Mab、2F7および3G5は、125I-Nogo66のsNogoR344-Fcへの結合を阻害した。
【実施例10】
【0185】
神経突起伸長アッセイ
Lab-Tek培養スライド(4ウェル)を0.1 mg/ml ポリ-D-リジン (Sigma)でコーティングした。CNSミエリンを単独でまたはsNogoR310、sNogoR310-Fc 融合タンパク質、mAb 5B10またはコントロールPBSと混合して別々に3μlの滴としてスポットした。蛍光ミクロスフェア(Polysciences)をミエリン/PBSに添加して後の滴の同定を可能とした(Grandpre et al, Nature 403, 2000)。Lab-Tekスライドをすすぎ、10μg/ml ラミニン (Gibco)でコーティングした。
【0186】
P3-4 SDラット子からの後根神経節(DRG)を1 mg/ml コラゲナーゼタイプ1(Worthington)で単離し、先端熱加工パスツールピペットで粉砕し、あらかじめプレーティングして神経細胞を豊富にし、最終的に23,000 細胞/ウェルであらかじめコーティングしたLabtek 培養スライドにプレーティングした。培地は5% 熱不活性化ドナーウマ血清、5% 熱不活性化胎児ウシ血清および50 ng/ml mNGFを含有するF12であり、37℃、5% CO2にて6時間インキュベートした。
【0187】
スライドを20% スクロースを含有する4% パラホルムアルデヒドで20分間固定し、1:500希釈神経細胞マーカー抗ベータ-III-チューブリン(Covance TUJ1)にて染色した。二次抗体として、抗-マウス Alexa Fluor 594 (Molecular Probes)を1:300に希釈し、スライドをGel/Mount(Biomeda)でカバースリップした。5x デジタル画像をOpenLabソフトウェアで得て、MetaMorphソフトウェアを用いて神経突起伸長を定量分析した。
【0188】
sNogoR310、sNogoR310-FcおよびmAb 5B10はすべて神経突起伸長のミエリン-媒介阻害からDRG神経細胞を保護した(図9-11)。sNogoR310を用いてニワトリ神経細胞を用いた類似のアッセイを行ったところ保護することが判明した。
【0189】
本発明者らはまた、ラミニンの存在下および非存在下での細胞成長による実験を行うことにより、可溶性Nogo受容体の神経保護効果を調べた。ラミニン非含有培地での神経細胞成長は悪く、神経細胞ストレス状態のモデルである。
【0190】
DRGを出生後6-7日のラット子(P6-7)から解体し、単一細胞に分離し、上記のようにポリ-D-リジンであらかじめコーティングした96-ウェルプレートに播いた。いくつかのウェルにおいては、2 μg/mlラミニンを2-3 時間添加し、すすいだ後、細胞を播いた。18-20 時間のインキュベーションの後、プレートを4% パラホルムアルデヒドで固定し、1:500希釈のウサギ抗-ベータ-III-チューブリン抗体(Covance)および1:100希釈の抗-HuC/D (Molecular Probes)で染色し、蛍光二次抗体 (Molecular Probes)を1:200希釈にて添加した。ArrayScan II (Cellomics)を用いて5x デジタル画像を得、神経突起伸長をウェルあたりの平均神経突起伸長/神経細胞として神経突起伸長アプリケーションを用いて定量した。3 ウェル/条件からの9つの5x 画像を分析した。
【0191】
いくつかの実験において、PC12細胞(Neuroscreen)のサブクローンを用いた(Cellomics)。Neuroscreen細胞をあらかじめ7日間200 ng/ml NGFによって分化させ、剥離し、ポリ-D-リジンであらかじめコーティングしておいた96-ウェルプレートに播いた。いくつかのウェルにおいては、5 μg/mlのラミニンを2-3 時間添加し、すすぎ、その後細胞を播いた。2日間のインキュベーション後、プレートを4% パラホルムアルデヒドで固定し、1:500希釈ウサギ 抗-ベータ-III-チューブリン抗体(Covance)およびHoechst (核染色)で染色した。ArrayScan IIを用いてDRG 細胞における神経突起伸長を定量した。
【0192】
sNogoR344-FcまたはラットIgGを溶液中でP6-7 DRG 神経細胞および分化Neuroscreen細胞にプレーティング時に添加した。
【0193】
sNogoR344-Fcの神経保護効果をP6 DRG 神経細胞をラミニンの非存在下で培養した際に、1 μMおよび10 μMにて観察した。神経突起伸長の定量は、sNogoR344-Fc添加による用量依存的上昇を示した。ラミニン基質上で培養しているDRG神経細胞への同じ濃度のsNogoR344-Fc の添加では、異常な効果はみられず、sNogoR344-Fcはストレス細胞に対してのみ活性であることが示された。ラミニンの非存在下での同じ濃度のsNogoR344-Fcの神経保護効果もまた、Neuroscreen細胞でみられた。
【実施例11】
【0194】
Fc-sNogoR-1融合タンパク質の産生および精製
ラットNogo受容体-1のアミノ酸1-310をコードするcDNAコンストラクトを哺乳類発現ベクターに含まれるラット IgG1 Fcと融合し、このベクターをチャイニーズハムスター卵巣(CHO) (DG44) 細胞にエレクトロポレーションした。細胞を10% 透析胎児ウシ血清、2 mM グルタミンおよび抗細菌-抗真菌薬を補充したアルファ-MEMで維持した。形質移入の2日後、培養上清を回収し還元条件下でウェスタンブロット分析を行った。約60 kDaのタンパク質バンドをポリクローナルウサギ抗-Nogo受容体-1抗体を用いて検出した。細胞を拡張し、R-PE 結合ヤギ抗-ラット IgG 抗体を用いてソートした。2回目のソートの後、細胞を96-ウェルプレートにて1細胞/ウェルの密度で播いた。個々のウェルからの分泌された可溶性Nogo受容体-1タンパク質レベルをサンドイッチELISAを用いて試験及び比較した。ELISA プレートはヤギ抗-ラット IgG Fcκ特異的抗体でコーティングした。培養上清を入れた。結合可溶性Nogo受容体-1タンパク質をHRP結合ロバ抗-ラット IgG Fab、Fc-特異的抗体で検出した。クローン4C12が最大の分泌レベルであった。4C12を拡張し、撹拌フラスコ中CHO-M7培地で培養した。分泌レベルは37℃で約10 mg/Lであった。
【0195】
sNogoR310-Fc 融合タンパク質を発現するCHO細胞をラージスケールで培養した。1.7 L の濃縮培養上清を10 Lのバイオリアクターランから得た。pHを1/10量の1.0 M Tris-HCl, pH 8.9の添加により上昇させた。固体塩化ナトリウムおよびグリシンをそれぞれ3.0 M および1.5 M添加した。10 mM Tris-HCl、3 M 塩化ナトリウム、1.5 M グリシン, pH 8.9で平衡化した60 mLのタンパク質 A-Sepharoseカラムを調製した。蠕動ポンプを用いて濃縮培養上清をカラムに1.5 mL/分にてアプライした。カラムを300 mLの10 mM Tris-HCl、3 M 塩化ナトリウム、1.5 M グリシン, pH 8.9、次いで120 mL 5 mM Tris-HCl、3 M 塩化ナトリウム, pH 8.9で洗浄した。タンパク質を25 mM リン酸ナトリウム、100 mM 塩化ナトリウム, pH 2.8で溶出した。10 mL フラクションを1.0 mL の1.0 M HEPES, pH 8.5を含む試験管に回収した。タンパク質フラクションをプールし3 x 2 Lの5 mM リン酸ナトリウム、300 mM NaCl, pH 7.4で透析した。
【実施例12】
【0196】
脊髄横切アッセイ
インビボでの機能回復を促進する能力を調べるため、sNogoR-1融合タンパク質をラット脊髄横切アッセイにおいて試験した。
【0197】
Alzet 浸透ポンプを使用日に新たに調製した被験溶液(PBS中sNogoR310-Fc)にロードした。ローディング濃度を計算したところ5および50 μMであった。ポンプは動物への移植の40時間以上前に37℃で開始した。雌性ロングエバンスラットに手術前鎮痛薬および精神安定薬を与え、イソフルラン(O2中3%)を用いて麻酔した。
【0198】
ラットを定位固定枠におき、運動野を路(tract)追跡薬 BDA (10,000 MW)の注入のために両側的に曝した。次いでラットをT5-T6にて脊髄の背部を片側切断し、次いで被験化合物の送達のためにくも膜下腔内カテーテルおよびポンプシステムを移植した(1群あたりn=11)。
【0199】
ラットを回復させ、手術後28日間生存させた。BBBシステムを用いた挙動スコアリングを損傷の導入後28日間記録した後、生存相の試験を終結させた。灌流および固定の後、脊髄を除き、凍結保護し、切片として染色し、軸索を計数した。
【0200】
Basso-Beattie-Bresnahan (BBB) 歩行運動強度スケール(Basso et al., 1996, Neurotrauma 13, 343-359)という、傾斜平面試験および傾斜格子歩行試験 (Li and Strittmatter, 2003, J Neurosci. 2003, 23, 4219-27)を損傷後ラットおよびマウスにてモニターした。傾斜平面試験のために、本発明者らは、50 cm x 60 cm ボードが5秒間マウスが滑り落ちずに傾けることが出来る最大角を測定した。傾斜格子歩行のために、マウスを訓練して45度のスロープの針金格子 (35 cm長、2.54 cm格子)を登らせた。格子平面の下に後肢が落ちる例の数を下から上までの各運動にて測定した。ラット挙動試験のために、BBB歩行運動スケール、格子歩行および足跡分析を行った。格子歩行のために、ラットを針金格子 (70 cm長、2.54 cm格子)上を歩行するように訓練し、格子平面下に後肢が落ちる例数を計数した。足跡分析のために、ラット後肢の歩行パターンを90 cm の路を横切る連続移動運動にわたってインクで記録し、各側のストライド長およびストライド幅を計算した(Metz et al., 2000, Brain Res., 883, 165-177)。これら挙動試験はすべて少なくとも2個体で行った。手術、挙動試験および組織学的分析において、研究者には、ミニポンプ内の化合物が何かを教えなかった。
【0201】
sNogoR310-Fcは機能回復を促進した(図12)。
【実施例13】
【0202】
ラット脊髄挫傷アッセイ
インビボでの神経細胞に対する可溶性Nogo受容体-1ポリペプチドおよび融合タンパク質の効果をラット脊髄挫傷アッセイで試験した。
【0203】
雌性ズキンロングエバンスラット(170-190 g)を鎮痛薬と抗生剤で予防処置した。手術の10分前に、動物を2.5 mg/kgのミダゾラム(腹腔内)で鎮静化し、O2中の2-3% イソフルランで麻酔した。ラットを次いで剪毛し、アルコールとベタジンで拭き、眼球潤滑剤を点眼した。次いで、切開を正中線下で行い、T7−T12の脊椎を曝した。
【0204】
背部椎弓切除術ををT9 1/2およびT10にて行って索を曝した。ラットを衝撃装置に載せた。T7およびT8セグメントをまずクランプし、次いでT11およびT12 セグメントを尾クランプに結合させた。軟材料をラットの胸部の下に敷いた。衝撃棒を0位に合わせ、電気的アースクリップを創傷縁につけた。衝撃棒を25.0 mmに上げ、適宜曝された脊髄の真上になるよう調節した。次ぎに衝撃棒を曝された脊髄に落とし、衝撃棒をすぐに上げた。
【0205】
ラットをおろし、Gelfoam(登録商標)を傷に当てた。傷の上の筋肉を縫合し、切開を外科的に閉じた。動物を麻酔から覚めるまでインキュベーターに入れた。ラットに抗生物質、鎮痛薬および要求に応じて生理的食塩水を与えた。膀胱を機能が回復するまで朝晩絞った。
【0206】
可溶性Nogo受容体-1融合タンパク質(例えば、sNogoR310-Fc)を上記ラット脊髄離断モデルに記載のようにくも膜下腔内投与した。BBBスコアリングを手術の1日後、次いでその後4-6週間、2週間毎に行った。
【実施例14】
【0207】
トランスジェニックマウスにおけるsNogoR310の発現
本発明者らは、インビボで発現させた場合の効果を試験するために、可溶性Nogo受容体-1タンパク質を発現するトランスジェニックマウスを作製した。
【0208】
本発明者らはマウス sNogoR310 cDNA (Nogo受容体-1のアミノ酸1-310に対応)をC-3123 ベクターのNotI 部位にクローニングした。このベクターにおいて、sNogoR310 発現はグリア細胞繊維性酸性タンパク質 (gfap) 遺伝子調節要素の制御下にあり、損傷部位の反応性星状細胞からの分泌を促進し、高レベルの発現が可能である。本発明者らは結果として得られたベクターを順次AatIIおよびSfiIで消化し、3.4 kb フラグメント上にgfap::sNogoR310 コンストラクトを単離した。本発明者らはこのフラグメントを胚にマイクロインジェクションし、トランスジェニックマウスを作製した。本発明者らは、導入遺伝子が組み込まれていることをPCRによって確認し、5つの創始系統を同定した。本発明者らは発現レベルが最も高い2つの創始系統のヘテロ接合性雄と雌性 C57BL/6J マウスとを交配した。本発明者らはNogo受容体-1に対する抗体を用いたウェスタンブロット分析によってGFAP-陽性細胞がヘテロ接合性トランスジェニックマウスにおいてsNogoR310を発現および分泌していることを確認した。
【0209】
本発明者らはプロテアーゼ阻害剤(Roche)を追加したTris-緩衝食塩水中で皮質および脊髄をホモゲナイズし、ホモジネートを40,000 rpmで20分間、4℃で遠心分離した。本発明者らは、上清を4% パラホルムアルデヒドで20分間処理して抗体特異性を強化し、イムノブロッティングの前に透析した。本発明者らは、粒状フラクションをRIPA バッファー (PBS 中1% Triton X-100、0.5% デオキシコール酸ナトリウム、0.1% SDS)中で超音波処理してホモゲナイズし、その結果得られたホモジネートを遠心分離し、上記のようにこの上清(界面活性剤-可溶性粒状フラクション)を処理した。本発明者らは、1:2000希釈のNogo受容体-1に対するウサギ抗血清を用いたイムノブロットによって20 μgの脳または脊髄タンパク質を分析した。本発明者らは、免疫反応性をAP-結合抗-ウサギ IgGおよびNBT/BCIP AP 基質とのインキュベーションによって可視化した。
【0210】
本発明者らは、2つのトランスジェニック系統、Tg08およびTg01からの皮質および脊髄の界面活性剤-フリー可溶性抽出物において分泌された37 kDa sNogoR310を検出したが、同腹仔野生型(WT)マウスにおいてはあるにしてもわずかしか、可溶性Nogo受容体-1 タンパク質は37または81 kDaにて存在しなかった。粒状フラクションを調べたところ、WTとトランスジェニックマウスの両方において内在性Nogo受容体-1が同レベル存在することが確認された。
【実施例15】
【0211】
損傷後のトランスジェニックマウスにおけるsNogoR310の発現
本発明者らは背部上-片側切断損傷を行うことにより、トランスジェニックマウスにおけるsNogoR310 発現に対するCNS損傷の効果を調べた。本発明者らは、ヘテロ接合性雄とC57/BL6 雌とを実施例14に記載のように交配することにより、sNogoR310 トランスジェニックおよび非トランスジェニックコントロール動物を得た。
【0212】
本発明者らは、成体雌性ヘテロ接合性トランスジェニックまたは同腹仔WTマウス (10-16週齡)を深く麻酔し、完全椎弓切除術を行い、脊髄のT6およびT7 レベルの背部を完全に曝した。本発明者らは、背部上-片側切断をT6にて30-ゲージ針および1対の小型はさみを用いて行い、完全に背部および背外側皮質脊髄路(CST)を切断した。本発明者らは、脊髄の背部に数回印をした針を通し、損傷の深さが1.0 mmであることを確かめた。本発明者らは椎弓切除術上の筋肉層を縫合し、手術用とじ金で背中の皮膚を閉じた。皮質脊髄路を追跡するために、本発明者らは、脊髄損傷の14日後に頭蓋骨内の右側脳皮質に刻み孔を開けた。本発明者らはトレーサーBDA (PBS 中10% 、MW 10,000) (Molecular Probes, Eugene, OR)を皮質表面から0.7 mm の深さにて4つの注射部位に適用した。損傷の4週間後、マウスを心臓を介してPBS、次いで4% パラホルムアルデヒドで灌流した。sNogoR310 発現実験に用いたマウスはトレーサー注射を受けなかった。
【0213】
ウェスタンブロット分析に用いるマウスについてT3〜L3レベルの脊髄を灌流せずに損傷の14日後に回収した。Nogo受容体-1 免疫組織化学染色に用いるマウスは片側切断の10日後に4% パラホルムアルデヒドで灌流し、損傷脊髄を切片とするために取り出した。トランスジェニックおよびWTマウスの損傷した脳におけるsNogoR310 発現を調べるために、定位固定装置(David Kopf, Tujunga, CA)に備えた11番手術用メス刃によって皮質穿刺損傷を行った。十字縫合の0.5 mm 後側、正中線から1.5 mm 側方、3.5 mm 深さの4 mm 傍矢状切断部を作った。
【0214】
本発明者らは、トランスジェニックマウスにおいて損傷の10日後に脊髄の可溶性抽出物におけるsNogoR310レベルの上昇を検出したが、WTマウスでは検出されず、これは損傷のまわりのGFAPの損傷後の上方制御と一致する。これがNogo-Aの代償的上方制御に起因するのではないことを確認するために、本発明者らはその発現を試験し、WTおよびトランスジェニックマウスからの無傷または損傷の皮質および脊髄のいずれにおいても同様であることを見いだした。
【0215】
本発明者らはNogo受容体-1およびGFAPに対する抗体による、損傷領域を含む損傷脳および脊髄の免疫染色により、損傷CNSにおけるsNogoR310の細胞性発現を調べた。反応性アストロサイトグリアの概ねの形態はWTとトランスジェニックマウスで異なることはなかったが、細胞内および細胞外空間の両方におけるNogo受容体-1について染色された密度はWTマウスと比較してgfap::sNogoR310 トランスジェニックマウスの方が顕著に高く、これは、トランスジェニックマウスにおける損傷のまわりのsNogoR310 発現の上昇を示す。トランスジェニックマウスにおいてのみ、Nogo受容体-1 タンパク質はアストロサイトマーカーGFAPと共に局在していた。トランスジェニックサンプルにおいて、散在性非細胞性染色が強くなっていたが、これは細胞外空間におけるsNogoR310と一致する。神経細胞の細胞体 Nogo受容体-1染色はWTとトランスジェニックマウスの両方で検出された。
【実施例16】
【0216】
トランスジェニックマウスにおいて分泌sNogoR310はCST出芽を誘導する
本発明者らはトランスジェニックマウスの損傷周囲のsNogoR310の発現上昇が損傷軸索の再生をもたらすかを試験した。
【0217】
本発明者らは、下行皮質脊髄路(CST)の統合性を、順行性トレーサー、ビオチンデキストランアミン(BDA) を右運動野に、Li and Strittmatter, 2003, J. Neurosci., 23, 4219-27に記載のように注入することによって調べた。同腹仔WTマウスにおいて、明らかな背部CST(dCST)が損傷の頭側で緊密に束になっており、数本の背外側CST繊維が同側方に観察された。少数のBDA-標識した短い側枝出芽が灰白質、特に前索における灰白質に突出したが、出芽は主にトレーサー注射の反対側の索にて確認された。しかし、損傷sNogoR310 トランスジェニックマウスからの背部片側切断の頭側の切片は、まったく異なるBDA 標識パターンを示した。高密度のBDA-標識CST線維が系統Tg08または系統Tg01からのすべてのトランスジェニックマウスにおける突出したdCSTの外側にみられた。異所性の線維が灰白質領域にわたって伸長しており、いくらかの線維は、外側および背外側白質に到達していた。いくつかの線維(横断切片あたり4-12の出芽)が脊髄の逆側にみられた(トレーサー注入部位の同側)。側枝出芽のマイクロデンシトメトリー測定は、sNogoR310 トランスジェニックマウスにおけるおよそ10倍の出芽密度の上昇を示した。損傷の1〜4 mm 頭側からの傍矢状長軸方向切片を調べたところ、sNogoR310 トランスジェニックマウスにおいてdCST線維が腹側灰白質領域に分岐する出芽を多数伸長しており、これは同腹仔WT動物ではみられなかった。一般にトランスジェニックマウスにおける損傷の頭側の出芽のパターンおよび程度は、Nogo受容体-1アンタゴニストペプチド NEP1-40で全身的に処置されたマウスにおいて観察されるものと同様であった(Li and Strittmatter, 2003)。
【0218】
これらの結果は分泌sNogoR310がトランスジェニックマウスにおけるCST出芽を誘導することを示す。
【実施例17】
【0219】
sNogoR310 トランスジェニックマウスにおいて再生するCST軸索は損傷部位を迂回して遠位脊髄に向かう
本発明者らはトランスジェニックマウスから損傷部位(8 mm全長)に対して4 mm 頭側、4 mm 尾側の脊髄を単離し、それをグルタルアルデヒド-重合アルブミンマトリックスに包埋し、ビブラトーム上で傍矢状に切断した(30μm厚)。本発明者らは損傷部位から5-7 mm 頭側、5-7 mm 尾側の脊髄から横断切片(50 μm)を集めた。ラットにおけるsNogoR310-Fc 注射実験のために、損傷部位の10 mm 頭側から10 mm 尾側に伸長する脊髄を振動マイクロトーム上で傍矢状に(50 μm)切断した。横断切片を損傷部位の11-16 mm 頭側、11-16 mm 尾側の脊髄から集めた。本発明者らは切片をアビジン-ビオチン-ペルオキシダーゼ複合体とインキュベートし、ニッケル促進化ジアミノベンジジンHRP反応によりBDAトレーサーを可視化した(Grandpre, 2002, Nature, 417, 547-551)。本発明者らは、間接的免疫蛍光による、セロトニン免疫組織化学(抗-5-HT 抗体)のためにいくつかの切片を加工した。損傷領域を可視化するために、本発明者らはGFAPに対する抗体を用いていくらかの切片を二重染色した(Sigma, St. Louis, MO)。 本発明者らは切片を標本にし、脱水し、封入剤で被覆した。
【0220】
本発明者らは損傷後にトランスジェニックマウスにおいて発現したsNogoR310によって誘導された線維が、機能回復させるために損傷領域を横切って尾脊髄へと向かうかを調べた(実施例16参照)。
【0221】
明箱に入れた損傷部位を横切る連続的傍矢状切片は、再生CST繊維の損傷から数ミリの全体的な分布パターンを示した。WTマウスからの切片には損傷部位の上に伸長するCST線維はみられなかった。sNogoR310 トランスジェニックマウスからの類似の切片は、横切領域を横切り、遠位灰白質および白質領域に高度に分岐したパターンにて突出する多くのCST線維を示した。片側切断に近接する頭側に、高密度のBDA-標識CST出芽が損傷領域へと突出するdCSTから生じたが、ほとんどのCST出芽は瘢痕形成および組織空洞化が顕著である横切領域を横切ることはできなかった。少ないがかなり有意な部分の再生軸索が腹側および腹外側灰白質および白質の残っている組織架橋を介して損傷部位を迂回していた。さらに、いくらかのCST線維は損傷背部および背外側脊髄を介して横切領域自体を横切って遠位領域へ向かっているようであった。損傷の近傍においては、再生する線維の経路は典型的には蛇行状であり、頭側のCSTにおける正常に直線上の線維とはかなり異なっていた。側枝および分岐した線維は遠位脊髄の灰白質領域にもっとも頻繁にみられた。再構築は、各トランスジェニックマウスにおける損傷の1-4 mm 尾側のあらゆるレベルにおいて頭側-尾側軸を経路とする5-15 BDA-標識化再生線維を示した。背部片側切断の5-7 mm 尾側の横断切片については、BDA-標識CST軸索が各トランスジェニックマウスにおいて灰白質と白質領域の両方においてみられた。トランスジェニックマウスの線維数は、矢状切片の近位レベルと類似の数のBDA-標識CST線維を示した。
【0222】
CST線維に加えて、その他の下行路、例えば、縫線核脊髄線維もマウスの歩行運動機能に寄与している。このマウス背部上-片側切断モデルにおいて、横切は大部分のセロトニン作動性線維を損傷し、これら線維密度は、腹側角において約80%減少していた。尾脊髄の腹側角におけるセロトニン線維の全長の分析は、WT群と比較してトランスジェニックマウスにおける方がかなり多数のかかる線維を示し、これはトランスジェニックマウスにおけるsNogoR310の成長促進効果が、軸索下行性経路に限定されないことを示す。
【実施例18】
【0223】
sNogoR310のトランスジェニック発現は歩行運動回復を向上させる
CST軸索追跡およびセロトニン作動性線維分析は、トランスジェニックマウスにおける星状細胞から放出されるsNogoR310が、脊髄における損傷下行性軸索の広範な解剖学的再生を刺激することを示した。本発明者らは実施例12に記載のいくつかの挙動試験を行って、かかる再生線維が機能回復に有益であるかを調べた。
【0224】
BBB試験によりアッセイしたところ、WTマウスは生存の4週間の間に部分的に歩行運動機能を回復した。損傷の4週間後には、ほとんどのWTマウスは体重を支えられる一貫した足底歩進によって特徴づけられるレベルに回復したが、時折しか前肢-後肢協調を示さず、表面に最初に接触させると片側の肢が回転した。一方、系統Tg08およびTg 01の両方からのsNogoR310 トランスジェニックマウスのBBBスコアは観察の7-28日間にわたってコントロール群より有意に高かった(図13Aおよび13B)。損傷の28日後には、ほとんどのトランスジェニックマウスが一貫した前肢-後肢協調を示し、片側の肢の位置は体と平行であった。
【0225】
本発明者らは2つのさらなる挙動試験を行ってsNogoR310 トランスジェニックマウスの性能を特徴づけた。まず、本発明者らはマウスが握力を5秒間は失うことなくボードを傾斜させられる最大角を測定した。背部片側切断損傷の前に、トランスジェニックおよびWTマウスの両方は55度に傾けたボードにて姿勢を維持することが出来た。損傷の7-28日後、維持可能な角度はすべてのマウスにおいて低下したが、トランスジェニックマウスによって維持可能な角度はコントロール群より有意に高かった(図13C)。もう1つの挙動試験において、マウスを垂直線に対して45度傾斜させた格子を登らせて格子平面下に落ちる後肢運動を計数した(Metz et al., 2000)。損傷前の訓練においてはいずれのマウスもこの試験でエラーを起こさなかった。損傷後の2-6週間においてはWTマウスにおいては多数の肢欠陥エラーがみられほとんど改善されなかった。一方、sNogoR310 トランスジェニックマウスはこの期間にわたって格子登上が徐々に改善し、損傷後1-3週間後に改善の大部分が起こった(図13D)。したがって、星状細胞からsNogoR310を分泌するトランスジェニックマウスは、CST再生、縫線核脊髄出芽および胸髄片側切断後の運動機能の改善を示した。
【実施例19】
【0226】
sNogoR310-Fcタンパク質のくも膜下腔内投与はCST出芽を誘導する
脊髄損傷後の可溶性Nogo受容体-1の成長促進性の効果の第二の試験として、本発明者らは精製したタンパク質をくも膜下腔内投与した。
【0227】
本発明者らはラットNogo受容体-1のリガンド結合ドメイン(27-310)とラット IgG1 Fc ドメインを融合させ安定性および精製を向上させた。本発明者らは安定にトランスフェクトしたCHO細胞からタンパク質を精製した。このタンパク質はNogo-66、MAGおよびミエリン作用をインビトロでブロックするということが以前のマウス sNogoR310-Myc His についての研究で示されている(Fournier et al., 2002, J Neurosci., 22, 8876-8883; Liu et al., 2002, Science, 297, 1190-1193)。 本発明者らは、sNogoR310-Fc タンパク質を浸透ミニポンプを用いて胸部背部中央片側切断損傷を有するラットのくも膜下腔内に送達した。損傷後4週間の生存期間において、1.2 mgのsNogoR310-Fc タンパク質を各ラットに局所投与した。媒体処置(1.2 mg ラット IgG)を受けたラットにおいて、片側切断の頭側の切片は緊密に束になった顕著な背部CSTを示し、損傷部位上の異所性BDA-標識CST線維は非常にわずかであった。sNogoR310-Fc タンパク質を与えられた損傷ラットからの損傷の頭側の切片は、全く異なる標識パターンを示した。BDA-標識CSTから出芽した多数の異所性の線維が横断および傍矢状切片から観察された。いくつかの場合には、突出は正中線近くのdCST領域を横切って索の外周に伸びており、背外側CSTと混ざっていた。出芽軸索は白質よりも灰白質に多く伸びていた。dCST近くの異所性出芽線維の測定(横断切片において100 μm以上、矢状切片において200 μm以上)によりsNogoR310-Fc-処理ラットにおける方がより多いことが明らかとなった。
【実施例20】
【0228】
sNogoR310-Fc処理ラットにおいてCST軸索は遠位脊髄に向かって再生する
本発明者らは、雌性SDラット(190-250 g)を深く麻酔し、脊髄レベルT6-7において椎弓切除術を行って脊髄を露出させた。本発明者らは脊髄の背部半分を30-ゲージ針と1対の小型はさみで切断してCSGT路の背部を分断し、損傷の深さ(1.8 mm)を11番メスの鋭利部分を索の背部半分に通すことにより確認した(Grandpre et al., 2002, Nature, 417, 547-551)。PBS 中の1.2 mg ラット IgGまたはPBS中の1.2 mg sNogoR310-Fc 融合タンパク質で満たした浸透ミニポンプ (Alzet 2ML4, 2 ml 容量、2.5 μl/時、28日間送達)を動物の背の筋肉下に縫合した。ミニポンプの出口に連結したカテーテルを硬膜の小孔を介して脊髄のT7-8 レベルにてくも膜下腔内空間に挿入した。
【0229】
Nogo受容体-1アンタゴニストタンパク質の注入は広範なラット片側切断の頭側の出芽を誘導したが、より重要なことは遠位脊髄に突出した出芽CST線維が歩行運動回復に寄与するか否かである。媒体-処理ラットからの損傷部位を横切る長軸方向切片では、BDA-標識腹側CST線維が検出不可能であるか損傷レベルを下回る非常に少量であった(GrandPre et al., 2002; Weidner et al., 2001, Proc. Natl. Acad. Sci. USA, 98, 3513-3518)。sNogoR310-Fc 処理ラットからの類似の切片においては、多くのBDA-標識線維が横切部位を迂回して主に腹側および腹外側脊髄の架橋組織を介して尾脊髄に突出していることが示された。アストロサイトマーカーGFAPの免疫染色により、横切の程度が中心管領域より深く達していることが示された。突出した背部CSTにおける頭側の線維が直線上であるのと異なり、再生CST線維は通常、遠位脊髄、特に灰白質領域において高度に分岐した軌道にしたがった。かかる線維は脊髄の多くの領域で観察されたが、脊髄を通じて脊髄の中央部および背部半分においてより容易に観察された。矢状切片からのCST線維数は各sNogoR310-Fc-処理ラットにおいて、損傷の尾側1-2 mmにおいておよそ20のBDA-標識軸索、そして損傷の遠位7-8 mmにおいて15の追跡軸索を示した。
【0230】
一般にこれら線維の分岐パターンは局所NEP 1-40 ペプチド処理動物のものと類似しているが、各出芽においてより多くの側枝分岐がsNogoR310-Fc タンパク質で処理した切片から観察される。遠位脊髄からの出芽の測定により、sNogoR310-Fc-処理ラットにおける各出芽の側枝全長はNEP 1-40-処理動物の2倍多いことが示された。両Nogo受容体-1 アンタゴニスト-処理群における脊髄の尾側1-10 mmにおける出芽数(長さ200 μm以上)はコントロール群と比較しておよそ20-40倍多かった。sNogoR310-Fc処理ラットからの方が局所NEP 1-40 処理からよりも多くの出芽が観察されたが(〜50対25 出芽/ラット)、この差は統計的に有意ではなかった(p=0.1713, t-検定)。
【0231】
再生CST軸索がsNogoR310-Fc 処理を受けたラットの片側切断の11-15 mm 尾側の脊髄の横断切片において観察された。これら線維は脊髄の灰白質と白質の両方で観察された。灰白質において検出された線維はしばしば白質領域においてより多くの側枝分岐を示した。一方、媒体処理群からの横断切片では、時折のみBDA-標識が腹側白質領域にみられ、これは無傷腹側CST軸索と一致する。このレベルの遠位脊髄において、Nogo受容体-1 アンタゴニスト-処理群[sNogoR310-FcおよびNEP 1-40]からのBDA-標識CST線維の平均数は、媒体-処理ラットのおよそ20倍であった。即ち、両方Nogo受容体アンタゴニストである、sNogoR310-Fc タンパク質およびNEP 1-40ペプチドにより、遠位脊髄において多くのCST軸索が再生したが、前者によって誘導される出芽の方がより高度な分岐パターンを示した。
【実施例21】
【0232】
局所sNogoR310-Fcは損傷ラット脊髄における赤核脊髄(Rubropinal)およびセロトニン作動性軸索の出芽を誘導する
片側切断の14日後、CST線維を追跡するために下肢の感覚野に横わたる頭蓋骨の各側面に刻み孔を開けた。順行性神経細胞トレーサーBDA(PBS中10%、皮質あたり3.5 μl)を各側面の硬膜から1.5 mm の深さの7つの注入部位に与えた(Grandpre, 2002)。ラットにおける赤核脊髄路追跡のために、トレーサーBDA (1 μl; MW 10,000; PBS 中10%)を左側の赤核に注射した(十字縫合から5.8 mm 後側、頭蓋骨表面から0.7 mm 外側、7.0 mm 腹側)。BDA注射の2週間後、これら動物をPBS、次いで4% パラホルムアルデヒドで灌流し、組織を組織化学のために収集した。
【0233】
損傷赤核脊髄路(RST)線維の修復は脊髄損傷後の機能向上に寄与する(Liu et al., 1999, J. Neurosci., 19, 4370-4387)。CNS 神経細胞における広範なNogo受容体-1の分布(Wang et al., 2002, J. Neurosci., 22, 5505-5515)により、アンタゴニストによるNogo受容体-1の阻害の結果、損傷後のRST 軸索の再生を導くことが可能となる。sNogoR310-Fcの損傷RSTへの効果を調べるために、この経路の統合性を左赤核にBDAを注射することにより追跡した。脊髄レベルにおいて、RST線維は脊髄の背外側白質領域に通常位置し、この研究の背部片側切断によって横断される。コントロールラットからの損傷の11-15 mm 頭側の横断切片において、少量の短いBDA-標識線維が突出RSTと背部角灰白質の間にみられた。sNogoR310-Fcで処理された同レベルの切片は主要RSTと背部角灰白質の間の多くの連結する線維を示した。媒体-処理ラットのSCIの11-15 mm 遠位の横断切片ではBDA-標識RST線維はみられなかった。一方、同レベルのsNogoR310-Fc処理を受けた切片は多くのBDA-標識RST 線維をトレーサー注射の反対側の灰白質と白質の両方で示した。分岐パターンを示すいくつかの出芽がBDA注射の同側の灰白質にみられた。
【0234】
縫線核脊髄線維もsNogoR310-Fc処理脊髄損傷ラットにおいて調べた。免疫染色により、損傷の11-15 mm頭側のセロトニン作動性線維密度は媒体処理群とsNogoR310-Fc処理群で同様であることが示された。損傷の11-15 mm 下の切片においては、sNogoR310-Fc処理ラットにおけるセロトニン線維はコントロール群の2倍多かった。これらの結果はsNogoR310-Fc タンパク質によるNogo受容体-1阻害への応答性はCST線維に限定されず、その他の下行路、例えば、赤核脊髄およびセロトニン作動性軸索もまたNogo受容体-1アンタゴニズムに応答性であることを示した。
【実施例22】
【0235】
sNogoR310-Fcによる局所処理はラットにおける機能回復を改善する
sNogoR310-Fc タンパク質のくも膜下腔内は、脊髄損傷後にいくつかの下行路における軸索再生を刺激する。本発明者らは、該タンパク質が損傷脊髄における機能回復も向上させるかどうかを調べた。
【0236】
片側切断の2週間後、媒体-処理ラットにおける歩行運動BBBスコアは安定レベル、12に達した(図14A)。損傷の4週間後、ほとんどのコントロール(7匹のうち6匹)は、頻度が一定の体重を支持する足底歩行および頻度が一定の前肢-後肢協調を示したが、表面と接触した際に片方の肢位置が回転した。一方、sNogoR310-Fc タンパク質処理を受けたラットにおいては、歩行運動スコアは損傷後2-4週間向上し続けた。損傷の4週間後、9匹全てのsNogoR310-Fc 処理動物は一定した前肢-後肢協調を示し、試験表面と接触した際、平行な肢位置であった。
【0237】
格子歩行を用いて脊髄損傷後の下行性の細かい動作の制御の欠陥を評価した(Metz et al., 2000)。格子歩行を行うためには、前肢-後肢協調および腹外側、皮質脊髄および赤核脊髄線維に媒介される随意運動制御が必要である。損傷前の訓練において、すべてのラットは格子バーのうえにその後肢を正確においた。損傷の2-4週間後、コントロールラットはセッションあたり8-9のエラーをおこし、その期間にわたって改善されなかった。一方、sNogoR310-Fcで処理したラットは格子歩行において徐々に向上し、エラーは有意に少なかった(平均4-7/セッション)。この向上のほとんどは損傷の2-3週間後に起こった。コントロール群における後肢足跡の分析により、片側切断の4週間後において、無傷ラットまたはsNogoR310-Fc処理を受けた損傷動物と比較してストライド長が有意に減少し、ストライド幅は増加していた(図14C)。それゆえ、これらの複数の挙動試験によりアンタゴニストタンパク質の局所注射によるNogo受容体-1機能のブロックが損傷後の歩行運動回復を促進することが示された。
【0238】
生物寄託
ハイブリドーマ、HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585)をAmerican Type Culture Collection (ATCC), 10801 University Boulevard, Manassas, VA 20110-2209, USA, on August 9, 2002に寄託した。
【0239】
当業者であれば理解するように、様々な変更および改変が本発明の範囲を逸脱することなく本発明の好ましい態様に対して行うことが出来る。かかる改変はすべて本発明の範囲に含まれる。
【0240】
本発明の具体的態様を示す:
(1)配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチド、
(2)項目(1)のポリペプチドをコードする核酸、
(3)発現制御配列に作動可能に連結した項目(2)の核酸、
(4)項目(2)または(3)の核酸を含むベクター、
(5)項目(2)または(3)の核酸または項目(4)のベクターを含む宿主細胞、
(6)以下の工程を含む項目(1)のポリペプチドの産生方法:
(a) 項目(5)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程、
(7)以下の工程を含む抗体の産生方法:
(a) 項目(1)のポリペプチドまたは項目(5)の宿主細胞で宿主を免疫する工程;および、(b)抗体を回収する工程、
(8)項目(7)の方法によって産生される抗体または該抗体の抗原結合性フラグメント、
(9)項目(1)のポリペプチドに特異的に結合する抗体またはその抗原結合性フラグメント、
(10)抗体が、
(a) 神経細胞の成長円錐崩壊を阻害し;
(b) 神経細胞における神経突起伸長および出芽の阻害を減少させ;そして、
(c) リガンドに対するNogo受容体-1の結合を阻害する、
項目(8)または(9)の抗体または抗原結合性フラグメント、
(11)神経突起伸長および出芽が軸索成長である、項目(10)の抗体または抗原結合性フラグメント、
(12)神経細胞が中枢神経系神経細胞である、項目(10)の抗体または抗原結合性フラグメント、
(13)抗体がモノクローナル抗体である項目(8)または(9)の抗体または抗原結合性フラグメント、
(14)抗体がマウス抗体である項目(8)または(9)の抗体または抗原結合性フラグメント、
(15)抗体がヒト化抗体、キメラ抗体および一本鎖抗体からなる群から選択される項目(8)または(9)の抗体、
(16)HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585) からなる群から選択されるハイブリドーマ細胞株、
(17)項目(16)のハイブリドーマ細胞株によって産生される抗体またはその抗原結合性フラグメント、
(18)項目(17)の抗体の、項目(1)のポリペプチドまたはNogo受容体-1への結合を競合的に阻害する抗体またはその抗原結合性フラグメント、
(19)Nogo受容体-1と項目(10)または(17)の抗体または抗原結合性フラグメントとを接触させる工程を含む、リガンドに対するNogo受容体-1結合を阻害する方法、
(20)リガンドがNogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される、項目(19)の方法、
(21)神経細胞と項目(10)または(17)の抗体またはその抗原結合性フラグメントとを接触させる工程を含む、神経細胞における成長円錐崩壊を阻害する方法、
(22)神経細胞と項目(10)または(17)の抗体またはその抗原結合性フラグメントとを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法、
(23)神経突起伸長または出芽が軸索成長である項目(22)の方法、
(24)神経細胞が中枢神経神経細胞である項目(21)または(22)の方法、
(25)Nogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる可溶性Nogo受容体-1ポリペプチド、
(26)シグナル配列に連結した項目(25)の可溶性Nogo受容体-1ポリペプチド、
(27)LRRが異種LRRを含む項目(25)または(26)の可溶性Nogo受容体-1ポリペプチド、
(28)配列番号6のアミノ酸残基 26-344、配列番号7のアミノ酸残基 26-310、配列番号8のアミノ酸残基 26-344、配列番号9のアミノ酸残基 26-310、配列番号8のアミノ酸残基 27-344および配列番号9のアミノ酸残基 27-310からなる群から選択される可溶性Nogo受容体-1ポリペプチド、
(29)項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドをコードする核酸、
(30)発現制御配列に作動可能に連結した項目(28)の核酸、
(31)項目(29)または(30)の核酸を含むベクター、
(32)項目(29)または(30)の核酸または項目(31)のベクターを含む宿主細胞、
(33)以下の工程を含む項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドの産生方法:
(a) 項目(32)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程、
(34)可溶性Nogo受容体-1ポリペプチドおよび異種ポリペプチドを含むNogo受容体-1融合タンパク質、
(35)可溶性Nogo受容体-1ポリペプチドがNogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる、項目(34)のNogo受容体-1融合タンパク質、
(36)シグナル配列に連結した項目(35)のNogo受容体-1融合タンパク質、
(37)LRRが異種LRRを含む項目(34)または(35)のNogo受容体-1融合タンパク質、
(38)可溶性Nogo受容体-1ポリペプチドが項目(25)−(28)のいずれかのポリペプチドを含む、項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(39)異種ポリペプチドが免疫グロブリン定常領域を含む項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(40)免疫グロブリン定常領域が免疫グロブリン重鎖定常領域である項目(39)のNogo受容体-1融合タンパク質、
(41)免疫グロブリン重鎖定常領域がIgG 重鎖定常領域である項目(40)のNogo受容体-1融合タンパク質、
(42)異種ポリペプチドがFcである項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(43)Nogo受容体-1融合タンパク質がダイマーである項目(34)−(37のいずれかのNogo受容体-1融合タンパク質、
(44)項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質をコードする核酸、
(45)発現制御配列に作動可能に連結した項目(44)の核酸、
(46)項目(44)または(45)の核酸を含むベクター、
(47)項目(44)または(45)の核酸または項目(46)のベクターを含む宿主細胞、
(48)以下の工程を含む項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質の産生方法:
(a) 項目(47)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からNogo受容体-1ポリペプチドを回収する工程、
(49)リガンドと項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、リガンドに対するNogo受容体-1の結合を阻害する方法、
(50)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、Nogo受容体-1リガンドの活性を調節する方法、
(51)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における成長円錐崩壊の阻害方法、
(52)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法、
(53)リガンドがNogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される項目(49)−(52)のいずれかの方法、
(54)神経突起伸長または出芽が軸索成長である項目(52)の方法、
(55)神経細胞がCNS神経細胞である項目(51)または(52)の方法、
(56)医薬上許容される担体および以下から選択される成分を含む組成物:
(a) 項目(8)、(9)および(16)のいずれかの抗体または抗原結合性フラグメント;
(b) 項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチド;および、
(c) 項目(34)−(37)のいずれかの融合タンパク質、
(57)さらに1または複数のさらなる治療薬を含む項目(56)の組成物、
(58)神経細胞と有効量の以下の(a)または(b)とを接触させる工程を含む死の危険にある神経細胞の生存を促進する方法:
(a)抗-Nogo受容体-1抗体または抗原結合性フラグメント;または、
(b)可溶性Nogo受容体-1ポリペプチド、
(59)神経細胞がインビトロである項目(58)の方法、
(60)可溶性Nogo受容体-1ポリペプチドが融合タンパク質である項目(58)の方法、
(61)融合タンパク質がFc 融合タンパク質である項目(60)の方法、
(62)Fc 融合タンパク質がIg-sNogoR344である項目(61)の方法、
(63)神経細胞が哺乳類におけるものである項目(58)の方法、
(64)哺乳類が多発性硬化症、ALS、ハンチントン病、アルツハイマー病、パーキンソン病、糖尿病性神経障害、脳卒中、外傷性脳障害または脊髄損傷の徴候または症状を示す項目(63)の方法、
(65)神経細胞が死の危険にある、以下の工程を含む哺乳類における神経細胞の生存を促進する方法:
(a)以下の(i)または(ii)を発現する培養宿主細胞を提供する工程、
(i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または、
(ii)可溶性Nogo受容体-1ポリペプチド;および、
(b) 神経細胞においてまたはその近くの部位にて哺乳類に該宿主細胞を導入する工程、
(66)神経細胞が哺乳類におけるものである、以下の工程を含む死の危険にある神経細胞の生存を促進する遺伝子治療方法:
以下の(a)または(b)をコードするヌクレオチド配列を含むウイルスベクターを神経細胞においてまたはその近くの部位にて投与する工程、
(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または、
(b)可溶性Nogo受容体-1ポリペプチド、
ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは哺乳類において神経細胞の生存を促進するのに十分な量にてヌクレオチド配列から発現する、
(67)Nogo受容体-1に特異的に結合する抗体またはその抗原結合性フラグメントであって、該抗体またはフラグメントが以下からなる群から選択されるアミノ酸配列を含む軽鎖を含む抗体またはその抗原結合性フラグメント:
(a) 配列番号15のアミノ酸配列;
(b) 配列番号16のアミノ酸配列;および、
(c) 配列番号22、23および24のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(68)さらに以下からなる群から選択されるアミノ酸配列を含む重鎖を含む項目(67)の抗体または抗原結合性フラグメント:
(a) 配列番号17のアミノ酸配列;
(b) 配列番号18のアミノ酸配列;および、
(c) 配列番号19、20および21のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(69)Nogo受容体-1に特異的に結合する抗体またはその抗原結合性フラグメントであって該抗体またはフラグメントが以下からなる群から選択されるアミノ酸配列を含む重鎖を含む抗体またはその抗原結合性フラグメント:
(a) 配列番号17のアミノ酸配列;
(b) 配列番号18のアミノ酸配列;および、
(c) 配列番号19、20および21のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(70)さらに以下からなる群から選択されるアミノ酸配列を含む軽鎖を含む項目(69)の抗体または抗原結合性フラグメント:
(a) 配列番号15のアミノ酸配列;
(b) 配列番号16のアミノ酸配列;および、
(c) 配列番号22、23および24のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列。
【技術分野】
【0001】
本発明は、神経生物学および分子生物学に関する。より具体的には、本発明は、免疫原性Nogo受容体-1ポリペプチド、Nogo受容体-1抗体、その抗原結合性フラグメント、可溶性Nogo受容体およびその融合タンパク質およびそれをコードする核酸に関する。本発明はさらにかかるNogo受容体抗体、その抗原結合性フラグメント、免疫原性Nogo受容体-1ポリペプチド、可溶性Nogo受容体およびその融合タンパク質ならびにそれをコードする核酸を含む組成物およびその製造及び使用方法にも関する。
【背景技術】
【0002】
神経細胞の軸索および樹状突起は、神経細胞からの長い細胞の伸長物である。伸長する軸索の遠位先端または神経突起は、成長円錐として知られる特殊な領域を含む。成長円錐は局所環境を感知し、神経細胞の標的細胞に向かった軸索成長を案内する。成長円錐はいくつかの環境的な合図、例えば、表面接着性、成長因子、神経伝達物質および電場に応答する。円錐における成長の誘導には様々なクラスの接着分子、細胞間シグナル、および成長円錐を刺激および阻害する因子が関与する。成長する神経突起の成長円錐は、様々な速度で進行するが、典型的には1日あたり1〜2mmの速度である。
【0003】
成長円錐は、手のような形であり、胚における表面に差次的に接着する広く平らな膨張部を備える(微小突起または糸状仮足)。糸状仮足は絶えず活動性であり、糸状仮足のなかには成長円錐内へ退縮するものもあり、基層に沿って伸長を続けるものもある。異なる糸状仮足間の伸長は膜状仮足を形成する。
【0004】
成長円錐は、膜状仮足および糸状仮足によってその前方およびいずれかの側面の領域を探索する。伸長物が成長に望ましくない表面に接触するとそれは後退する。伸長物が成長に好ましい表面に接触すると、それは伸長し続けその方向に成長円錐をガイドする。成長円錐は基層の表面の特徴において小さい変動でガイドされうる。成長円錐が適当な標的細胞に到達すると、シナプス結合が生じる。
【0005】
神経細胞の機能は、神経細胞とその直近の環境におけるその他の細胞の間の接触に大幅に影響される(U. Rutishauser, T. M. Jessell, Physiol. Rev. 1988, 68, p. 819)。かかる細胞には、特別(specialized)グリア細胞、中枢神経系 (CNS)における乏突起膠細胞、末梢神経系(PNS)におけるシュワン細胞が含まれ、これは神経細胞の軸索をミエリン(多層膜の絶縁体構造)鞘で覆う(G. Lemke, in An Introduction to Molecular Neurobiology, Z. Hall, Ed. [Sinauer, Sunderland, Mass., 1992], p. 281)。
【0006】
CNS神経細胞は傷害後の再生能を有するが、それらはミエリンに存在する阻害性タンパク質およびおそらくその局所環境に通常みられるその他のタイプの分子の存在により再生を阻害される(Brittis and Flanagan, Neuron, 2001, 30, pp. 11-14; Jones et al., J. Neurosci. 2002, 22, pp. 2792-2803; Grimpe et al., J. Neurosci. 2002, 22, pp. 3144-3160)。
【0007】
乏突起膠細胞上にみられるいくつかのミエリン阻害性タンパク質が特徴づけられており、例えば以下が挙げられる:NogoA (Chen et al., Nature, 2000, 403, 434-439; Grandpre et al., Nature 2000, 403, 439-444)、ミエリン関連糖タンパク質(MAG, McKerracher et al, Neuron 1994, 13, 805-811; Mukhopadhyay et al, Neuron 1994, 13, 757-767)および乏突起膠細胞糖タンパク質(OM-gp, Mikol and Stefansson, J. Cell. Biol. 1988, 106, 1273-1279)。これらタンパク質はそれぞれ別々に神経細胞Nogo受容体-1のリガンドであることが示された(Wang et al., Nature 2002, 417, 941-944; Liu et al., Science, 2002, 297, 1190-93; Grandpre et al., Nature 2000, 403, 439-444; Chen et al., Nature, 2000, 403, 434-439; Domeniconi et al., Nuron, 2002, 35, 283-90)。
【0008】
Nogo受容体-1は8つのロイシンリッチリピートを含むGPI-アンカー(anchored)膜タンパク質である(Fournier et al., Nature 2001, 409, 341-346)。阻害性タンパク質(例えば、NogoA, MAGおよびOM-gp)との相互作用時に、Nogo受容体-1複合体は、成長円錐の崩壊および神経突起伸長の阻害を導くシグナルを伝達する。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】U. Rutishauser, T. M. Jessell, Physiol. Rev. 1988, 68, p. 819
【非特許文献2】G. Lemke, in An Introduction to Molecular Neurobiology, Z. Hall, Ed. [Sinauer, Sunderland, Mass., 1992], p. 281
【非特許文献3】Brittis and Flanagan, Neuron, 2001, 30, pp. 11-14
【非特許文献4】Jones et al., J. Neurosci. 2002, 22, pp. 2792-2803
【非特許文献5】Grimpe et al., J. Neurosci. 2002, 22, pp. 3144-3160
【非特許文献6】Chen et al., Nature, 2000, 403, 434-439
【非特許文献7】Grandpre et al., Nature 2000, 403, 439-444
【非特許文献8】McKerracher et al, Neuron 1994, 13, 805-811
【非特許文献9】Mukhopadhyay et al, Neuron 1994, 13, 757-767
【非特許文献10】Mikol and Stefansson, J. Cell. Biol. 1988, 106, 1273-1279
【非特許文献11】Wang et al., Nature 2002, 417, 941-944
【非特許文献12】Liu et al., Science, 2002, 297, 1190-93
【非特許文献13】Domeniconi et al., Nuron, 2002, 35, 283-90
【非特許文献14】Fournier et al., Nature 2001, 409, 341-346
【発明の概要】
【発明が解決しようとする課題】
【0010】
Nogo受容体-1のそのリガンドに対する結合を阻害し、ミエリン-媒介成長円錐崩壊および神経突起伸長阻害を減弱する分子が緊急に要求されている。
【課題を解決するための手段】
【0011】
(発明の概要)
本発明は、可溶性Nogo受容体-1ポリペプチドおよびそれを含む融合タンパク質、ならびにNogo受容体-1の特定の免疫原性領域に対する抗体およびその抗原性フラグメントに関する。本発明はまた、本発明の抗体に結合する免疫原性Nogo受容体-1ポリペプチドに関する。本発明はさらに、本発明のポリペプチドをコードする核酸、かかる核酸を含むベクターおよび宿主細胞、ならびに該ペプチドの製造方法に関する。本発明の抗体、可溶性受容体および受容体融合タンパク質はNogo受容体-1を拮抗またはブロックし、Nogo受容体-1のそのリガンドへの結合の阻害、神経細胞における成長円錐崩壊の阻害、および神経細胞における神経突起伸長または出芽の阻害の抑制に有用である。
【0012】
ある態様において、本発明は、配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択される免疫原性ポリペプチドを提供する。
【0013】
ある態様において、本発明は、該免疫原性ポリペプチドをコードする核酸、該核酸を含むベクターおよび該核酸またはベクターを含む宿主細胞を提供する。ある態様において、核酸は、発現制御配列に作動可能に連結している。
【0014】
ある態様において、本発明は以下の工程を含む免疫原性ポリペプチドの産生方法を提供する:
(a)免疫原性ペプチドをコードする核酸またはそれをコードするベクターを含む宿主細胞を培養する工程;および、
(b)宿主細胞または培地からポリペプチドを回収する工程。
【0015】
ある態様において、本発明は以下の工程を含む、Nogo受容体-1に特異的に結合する抗体の産生方法を提供する:
(a) 配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチドまたは該ポリペプチドを発現する宿主細胞で宿主を免疫する工程;および、
(b)抗体を回収する工程。
ある態様において、抗体またはその抗原結合性フラグメントはこの方法によって産生される。ある態様において、抗体またはその抗原結合性フラグメントは、配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチドに特異的に結合する。ある態様において、抗体または抗原結合性フラグメントは、(a) 神経細胞の成長円錐崩壊を阻害する;(b)神経突起伸長および神経細胞における出芽の阻害を減少させる;そして(c) Nogo受容体-1のリガンドへの結合を阻害する。ある態様において、抗体または抗原結合性フラグメントは死の危険にある神経細胞の生存を促進する。ある態様において、死の危険にある神経細胞は動物、例えば、哺乳類におけるものである。ある態様において、神経突起伸長および出芽は軸索成長である。ある態様において、神経細胞は、中枢神経系(CNS)神経細胞である。
【0016】
ある態様において、抗体または抗原結合性フラグメントはモノクローナル抗体である。ある態様において、抗体または抗原結合性フラグメントはマウス抗体である。ある態様において、抗体はヒト化抗体、キメラ抗体あるいは一本鎖抗体である。
【0017】
ある態様において、本発明は以下からなる群から選択されるハイブリドーマ細胞株を提供する:HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585)。ある態様において、抗体またはその抗原結合性フラグメントは、ハイブリドーマ細胞株によって産生される。
【0018】
ある態様において、抗体またはその抗原結合性フラグメントは、以下からなる群から選択されるアミノ酸配列を含む軽鎖を有する(a) 配列番号15のアミノ酸配列;(b) 配列番号16のアミノ酸配列;および(c) 配列番号22、23および24のCDR1、CDR2およびCDR3アミノ酸配列を含むアミノ酸配列。ある態様において、抗体またはその抗原結合性フラグメントは以下からなる群から選択されるアミノ酸配列を含む重鎖を有する (a)配列番号17のアミノ酸配列; (b) 配列番号18のアミノ酸配列;および、(c) 配列番号19、20および21のCDR1、CDR2およびCDR3アミノ酸配列を含むアミノ酸配列。ある態様において、本発明は、該抗体またはその抗原結合性フラグメントをコードする核酸を提供する。ある態様において、核酸は発現制御配列に作動可能に連結している。ある態様において、本発明は該核酸を含むベクターを提供する。ある態様において、本発明は該核酸または該核酸を含むベクターを含む宿主細胞を提供する。
【0019】
ある態様において、抗体またはその抗原結合性フラグメントは、ハイブリドーマ細胞株によって産生される抗体の Nogo受容体-1または配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択される免疫原性ポリペプチドに対する結合を競合的に阻害する。
【0020】
ある態様において、本発明は以下の工程を含むリガンドに対するNogo受容体-1の結合の阻害方法を提供する:Nogo受容体-1を本発明の抗体または抗原結合性フラグメントと接触させる工程。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。
【0021】
ある態様において、本発明は以下の工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する:神経細胞を本発明の抗体またはその抗原結合性フラグメントと接触させる工程。ある態様において、本発明は、以下の工程を含む神経細胞における神経突起伸長または出芽の阻害の減少方法を提供する:神経細胞を本発明の抗体またはその抗原結合性フラグメントと接触させる工程。ある態様において、神経細胞はCNS 神経細胞である。これら方法いくつかにおいて、神経突起伸長または出芽は軸索成長である。
【0022】
ある態様において、本発明は以下の工程を含む死の危険にある神経細胞の生存を促進する方法を提供する:神経細胞を有効量の(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメントまたは(b)可溶性Nogo受容体-1ポリペプチドと接触させる工程。ある態様において、可溶性Nogo受容体-1ポリペプチドは、 融合タンパク質、例えば、Fc-融合タンパク質である。ある態様において、融合タンパク質は、sNogoR344-Fcタンパク質である。ある態様において、神経細胞はインビトロである。ある態様において、神経細胞は例えば、多発性硬化症、ALS、ハンチントン病、アルツハイマー病、パーキンソン病、糖尿病性神経障害、脳卒中、外傷性脳障害および脊髄損傷等の徴候または症状を示す哺乳類におけるものである。
【0023】
ある態様において、本発明は、以下の工程を含み、神経細胞が哺乳類におけるものである、死の危険にある神経細胞の生存を促進する方法を提供する:
(a)(i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(ii) 可溶性Nogo受容体-1ポリペプチドを発現する培養宿主細胞を提供する工程;および、
(b)宿主細胞を哺乳類に神経細胞においてまたはその近くの部位にて導入する工程。
【0024】
ある態様において、本発明は以下の工程を含み、神経細胞が死の危険にある、哺乳類における神経細胞の生存を促進する遺伝子治療方法を提供する:以下の(a)または(b)をコードするヌクレオチド配列を含むウイルスベクターを神経細胞においてまたはその近くの部位にて投与する工程:(a)抗-Nogo受容体-1抗体または抗原結合性フラグメント ; または(b)可溶性Nogo受容体-1ポリペプチド、ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは、神経細胞の生存を促進するのに十分な量にて哺乳類においてヌクレオチド配列から発現する。
【0025】
ある態様において、本発明は、Nogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる可溶性Nogo受容体-1ポリペプチドを提供する。ある態様において、該可溶性Nogo受容体-1ポリペプチドはシグナル配列に連結している。ある態様において、LRRは異種LRRを含む。ある態様において、本発明は以下からなる群から選択される可溶性Nogo受容体-1ポリペプチドを提供する: 配列番号6のアミノ酸残基26-344; 配列番号7のアミノ酸残基26-310; 配列番号8のアミノ酸残基26-344; 配列番号9のアミノ酸残基 26-310; 配列番号8のアミノ酸残基 27-344;および配列番号9のアミノ酸残基 27-310。ある態様において、本発明は、該可溶性Nogo受容体-1ポリペプチドをコードする核酸を提供する。ある態様において、核酸は発現制御配列に作動可能に連結している。ある態様において、本発明は該核酸を含むベクターを提供する。ある態様において、本発明は、該核酸または該核酸を含むベクターを含む宿主細胞を提供する。
【0026】
ある態様において、本発明は以下の工程を含む本発明の可溶性Nogo受容体-1ポリペプチドの産生方法を提供する:
(a) 可溶性Nogo受容体-1ポリペプチドをコードする核酸または該核酸を含むベクターを含む宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程。
【0027】
ある態様において、本発明は、可溶性Nogo受容体-1および異種ポリペプチドを含むNogo受容体-1融合タンパク質を提供する。ある態様において、可溶性Nogo受容体-1ポリペプチドはNogo受容体-1のN-末端ドメイン (NT)、8 ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる。ある態様において、可溶性Nogo受容体-1ポリペプチドはシグナル配列に連結している。ある態様において、Nogo受容体-1融合タンパク質は異種LRRを含む。ある態様において、Nogo受容体-1融合タンパク質は以下からなる群から選択されるポリペプチドを含む: 配列番号6のアミノ酸残基26-344; 配列番号7のアミノ酸残基26-310; 配列番号8のアミノ酸残基26-344; 配列番号9のアミノ酸残基26-310; 配列番号8のアミノ酸残基 27-344; 配列番号9のアミノ酸残基27-310。ある態様において、異種ポリペプチドは、免疫グロブリン定常領域を含む。ある態様において、免疫グロブリン定常領域は、免疫グロブリン重鎖定常領域である。ある態様において、免疫グロブリン重鎖定常領域はIgG 重鎖定常領域である。ある態様において、異種ポリペプチドはFc領域である。ある態様において、Nogo受容体-1融合タンパク質はダイマーである。
【0028】
ある態様において、本発明は、Nogo受容体-1融合タンパク質をコードする核酸を提供する。ある態様において、Nogo受容体-1融合タンパク質をコードする核酸は、発現制御配列に作動可能に連結している。
【0029】
ある態様において、本発明はNogo受容体-1融合タンパク質をコードする核酸を含むベクターを提供する。
【0030】
ある態様において、本発明は、Nogo受容体-1融合タンパク質をコードする核酸をまたはNogo受容体-1融合タンパク質をコードする核酸を含むベクターを含む宿主細胞を提供する。
【0031】
ある態様において、本発明は以下の工程を含むNogo受容体-1融合タンパク質の産生方法を提供する:
(a) Nogo受容体-1融合タンパク質をコードする核酸または該核酸を含むベクターを含む宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からNogo受容体-1融合タンパク質を回収する工程。
【0032】
ある態様において、本発明は以下の工程を含む、リガンドに対するNogo受容体-1の結合を阻害する方法を提供する:リガンドを本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質に接触させる工程。
【0033】
ある態様において、本発明は以下の工程を含むNogo受容体-1リガンドの活性の調節方法を提供する: Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。
【0034】
ある態様において、本発明は以下の工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する:Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。ある態様において、本発明は以下の工程を含む、神経細胞における神経突起伸長または出芽の阻害の減少方法を提供する:Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程。ある態様において、神経細胞は CNS 神経細胞である。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。ある態様において、神経突起伸長または出芽は軸索成長である。
【0035】
ある態様において、本発明は、医薬上許容される担体および以下から選択される成分を含む組成物を提供する(a) 本発明による抗体または抗原結合性フラグメント; (b) 本発明による可溶性Nogo受容体-1ポリペプチド;および(c) 本発明によるNogo受容体-1融合タンパク質。ある態様において、組成物はさらに1または複数のさらなる治療薬を含む。
【図面の簡単な説明】
【0036】
【図1】図1は、Nogo受容体-1の構造の模式図である。ヒト sNogoR310は残基 26-310を含み、sNogoR344は残基 26-344を含む。ラットsNogoR310は残基 27-310を含み、 sNogoR344は残基 27-344を含む。
【図2】図2は、ベクターNti(商標)ソフトウェアを用いたNogo受容体-1 タンパク質の抗原性プロットを示す。ラットP-617は配列番号10であり、ラットP-618 は配列番号11である。
【図3】図3Aは、抗-Nogo受容体-1抗体, 7E11の結合活性を示すグラフである。グラフはNogo66のNogo受容体-1への結合に対する7E11濃度の効果を表す。図3Bは抗-Nogo受容体-1抗体, 1H2の結合活性を示す。グラフは、1H2 濃度のNogo66の sNogoR344-Fcに対する結合への効果を表す(本明細書そして米国特許出願 60/402,866 においてFc-sNogoR344またはIg-sNogoR344とも称される)。Fc-MAGはsNogoR344-Fc への結合についてNogo66と競合しなかった。
【図4】図4は、抗-Nogo-R-1抗体 1H2, 3G5および2F7についての ELISAの結果を示す。固定化抗原の存在下の抗体のOD450に対する効果を測定した。固定化抗原は、sNogoR310-Fc (本明細書そして米国特許出願 60/402,866において Fc-sNogoR310またはIg-sNogoR310とも称される)、sNogoR344-Fc、p-617、p-618、p-4、p-5および卵白アルブミンおよびBSA。
【図5】図5は、様々な量のミエリンの存在下でのモノクローナル抗体、7E11、のラットDRG神経突起伸長に対する効果を示すグラフである。
【図6】図6Aは以下のコンペティターの存在下でのsNogoR310の125I-Nogo66および125I-Nogo40への結合に対する効果を示すグラフである: Nogo66、Nogo40および抗-Nogo受容体-1モノクローナル抗体上清。図6Bは、125I-Nogo66のsNogoR310への結合活性を示す。
【図7】図7は、sNogoR310-Fc の、sNogoR310に対する125I-Nogo40の結合への効果を示すグラフである。
【図8】図8は、sNogoR310-Fcの125I-Nogo40に対する結合活性を示すグラフである。
【図9A】図9Aはミエリンの存在下または非存在下でのsNogoR310の神経突起伸長/細胞に対する効果を示すグラフである。
【図9B】図9Bはミエリンの存在下または非存在下での神経突起伸長に対するsNogoR310の効果を示すグラフである。
【図10A】図10Aは様々な量のミエリンの存在下または非存在下でのP4ラットDRG神経突起伸長に対するsNogoR310-Fcの効果を示すグラフである。
【図10B】図10Bは、PBS、PBS + sNogoR310-Fc、20ng ミエリンおよびミエリン + sNogoR310-Fcでの処理後の神経突起数/細胞を示す。
【図11】図11は、様々な量のミエリンの存在下でのモノクローナル抗体 5B10 の DRG神経突起伸長/細胞に対する効果を示すグラフである。
【図12】図12は、ラット脊髄離断モデルにおける損傷の誘導後30日までのBBBスコアに対するsNogoR310-Fcの効果を示すグラフである。
【図13】図13Aおよび13BはWTまたは系統 08または系統 01からのトランスジェニックマウスにおける歩行運動BBBスコアを背部片側切断後の時間に対する関数として報告する。図13Cは、最大許容傾斜平面角をWTおよびトランスジェニックマウスについての損傷後時間の関数として示すグラフである。図13Dは、傾斜格子登上間の後肢エラーを損傷後時間の関数として示す。すべてのグラフにおいて、各群において7-9匹のマウスからの平均±s.e.m.を報告する。トランスジェニック群からの値はWTマウスからの値と有意差がある(**、P < 0.01; スチューデントt検定)。
【図14】図14Aは、媒体またはsNogoR310-Fc 処理動物における背部片側切断後の時間の関数として歩行運動BBBスコアを示す。図14Bは格子歩行の際の後肢エラーを損傷後時間の関数として示す。図14Cは、無傷または損傷 + sNogoR310-Fcラットよりコントロールマウスにおいてストライド長が短く、ストライド幅が広いことを明らかにする足跡分析である。すべてのグラフにおいて、各群において7-9匹のラットからの平均±s.e.m.を報告する。sNogoR310-Fc群の値は、コントロールと有意差がある(図14A-B)。コントロール値は図14Cにおける非-SCIまたはSCI + sNogoR310-Fcラットと有意差がある(*、p< 0.05; **、p < 0.01;スチューデントt検定)。
【発明を実施するための形態】
【0037】
(発明の詳細な説明)
定義および一般的教示
特に断りのない限り、本明細書において用いるすべての技術及び科学用語は本発明の属する分野の当業者に通常理解される意味と同義である。争いのある場合、定義を含む本出願が優先する。また文脈により要求されない限り、単数形の語は、複数も含み、複数形の語は、単数も含む。本明細書において言及する、すべての刊行物、特許およびその他の文献はあらゆる目的のためその全体を引用により本出願に含める。
【0038】
本明細書に記載するものと類似または均等の方法及び材料が本発明の実施または試験に利用できるが、好適な方法および材料を以下に記載する。材料、方法および実施例は単に例示のものであり、限定の意図ではない。本発明のその他の特徴および利点は、詳細な説明および特許請求の範囲から明らかであろう。
【0039】
本明細書および特許請求の範囲を通して「含む」なる語またはその同義語は、言及した要素または要素群を含む意味であるがその他の要素または要素群を排除するものではない。
【0040】
本発明をさらに明確にするため以下の用語および定義を本明細書において提供する。
【0041】
本明細書において用いる、「抗体」とはインタクトな免疫グロブリンまたはその抗原結合性フラグメントを意味する。本発明の抗体はいずれのアイソタイプまたはクラスでもよく(例えば、M、D、G、EおよびA)、いずれのサブクラスでもよく(例えば、G1-4, A1-2) そしてカッパ(κ)またはラムダ(λ)軽鎖のいずれをもつものでのよい。
【0042】
本明細書において用いる「Fc」とは、パパイン消化によって得られる抗体の重鎖定常領域の部分を意味する。
【0043】
本明細書において用いる、「NogoR融合タンパク質」とは、異種ポリペプチドに融合した可溶性Nogo受容体-1部分を含むタンパク質を意味する。
【0044】
本明細書において用いる、「ヒト化抗体」とは、非-ヒト配列の少なくとも一部がヒト配列で置換された抗体を意味する。ヒト化抗体の作成法の例は米国特許第6054297、5886152および5877293号にみることができる。
【0045】
本明細書において用いる「キメラ抗体」とは、第一の抗体からの1または複数の領域および少なくとも1つのその他の抗体からの1または複数の領域を含む抗体を意味する。第一の抗体およびさらなる抗体は同種由来のものでも異種由来のものでもよい。
【0046】
本明細書および米国特許出願第60/402,866号において用いる、「Nogo受容体」、「NogoR」、「NogoR-1」、「NgR」、および「NgR-1」はそれぞれNogo受容体-1を意味する。
【0047】
免疫原性Nogo受容体-1ポリペプチド
一つの態様において、本発明は免疫原性のNogo受容体-1ポリペプチドに関する。本発明のある態様において、免疫原性ポリペプチドは実質的に以下からなる群から選択されるアミノ酸配列からなる:LDLSDNAQLRVVDPTT (ラット)(配列番号1); LDLSDNAQLRSVDPAT (ヒト)(配列番号2); AVASGPFRPFQTNQLTDEELLGLPKCCQPDAADKA (ラット)(配列番号3); AVATGPYHPIWTGRATDEEPLGLPKCCQPDAADKA (ヒト)(配列番号4);およびCRLGQAGSGA (マウス)(配列番号5)。
【0048】
ある態様において、本発明は配列番号1-5のポリペプチドをコードする核酸に関する。本発明のある態様において、核酸分子は発現制御配列(例えば、pCDNA(I))に連結している。
【0049】
本発明はまた、本発明の免疫原性ポリペプチドをコードする核酸を含むベクターに関する。本発明のある態様において、ベクターはクローニングベクターである。本発明のある態様において、ベクターは発現ベクターである。本発明のある態様において、ベクターは少なくとも1つの選抜可能マーカーを含む。
【0050】
本発明はまた、上記の核酸またはベクターを含む宿主細胞に関する。
【0051】
本発明はまた、宿主細胞を培養する工程を含む、本発明の免疫原性ポリペプチドの産生方法に関する。ある態様において、宿主細胞は原核生物である。ある態様において、宿主細胞は真核生物である。ある態様において、宿主細胞は酵母である。
【0052】
抗体
本発明はさらに、本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗体またはその抗原結合性フラグメントに関する。ある態様において、抗体または抗原結合性フラグメントは実質的に配列番号1-5からなる群から選択されるアミノ酸配列からなるポリペプチドに結合する。本発明の抗体または抗原結合性フラグメントはインビボでもインビトロでも産生できる。抗体または抗原結合性フラグメントの産生を以下に説明する。
【0053】
本発明の抗体またはその抗原結合性フラグメントはNogo受容体-1のリガンド(例えば、NogoA, NogoB, NogoC, MAG, OM-gp)に対する結合を阻害し、ミエリンに媒介される神経突起伸長および出芽、特に軸索成長の阻害を減少させ、ミエリンに媒介される成長円錐崩壊を減弱させる。
【0054】
ある態様において、抗-Nogo受容体-1抗体またはその抗原結合性フラグメントはマウス由来である。ある態様において、Nogo受容体-1はラット由来である。別の態様において、Nogo受容体-1はヒト由来である。ある態様において、抗-Nogo受容体-1抗体またはその抗原結合性フラグメントは組換え、遺伝子操作、ヒト化および/またはキメラである。
【0055】
ある態様において、抗体は以下からなる群から選択される: モノクローナル7E11(ATCC受託番号、PTA-4587); モノクローナル 1H2 (ATCC受託番号、PTA-4584); モノクローナル 2F7 (ATCC受託番号、PTA-4585); モノクローナル 3G5 (ATCC受託番号、PTA-4586);およびモノクローナル 5B10 (ATCC受託番号、PTA-4588)。ある態様において、抗体はポリクローナル抗体46である。
【0056】
例示的な抗原結合性フラグメントは、Fab、Fab'、F(ab')2、Fv、Fd、dAbおよび相補性決定領域 (CDR)フラグメントを含むフラグメント、一本鎖抗体 (scFv)、キメラ抗体、二価抗体(diabodies)およびポリペプチドに特異的な抗原結合性を付与するのに十分な免疫グロブリンの少なくとも一部を含むポリペプチド(例えば、イムノアドヘシン)である。
【0057】
本明細書において用いる、FdはVHおよびCH1ドメインからなるフラグメントを意味する;Fvは抗体の1つの腕のVLおよびVHドメインからなるフラグメントを意味する;そしてdAbはVHドメインからなるフラグメントを意味する(Ward et al., Nature 341:544 546, 1989)。本明細書において用いる、一本鎖抗体(scFv)はVL領域およびVH領域が対となってそれらを一つのタンパク質鎖にさせる合成リンカーを介して一価分子を形成する抗体を意味する(Bird et al., Science 242:423 426, 1988およびHuston et al., Proc. Natl. Acad. Sci. USA 85:5879 5883, 1988)。本明細書において用いる、二価抗体は二重特異性抗体であって、VHおよびVL ドメインが一本のポリペプチド鎖上に発現するが、同じ鎖上の該2つのドメイン間の対合を可能とするのには短すぎるリンカーを用いてドメインを別の鎖の相補的ドメインと対合させ2つの抗原結合性部位が形成されるものを意味する(例えば、Holliger, P., et al., Proc. Natl. Acad. Sci. USA 90:6444 6448, 1993,およびPoljak, R. J., et al., Structure 2:1121 1123, 1994を参照されたい)。本明細書において用いる、目的の抗原に特異的に結合するイムノアドヘシンとは、1または複数のCDRが共有結合または非共有結合によって導入された分子を意味する。
【0058】
ある態様において、本発明は本発明のNogo受容体-1抗体のサブユニットポリペプチドを提供し、ここでサブユニットポリペプチドは以下からなる群から選択される:(a)重鎖またはその可変領域;および(b)軽鎖またはその可変領域。
【0059】
ある態様において、本発明は本発明のNogo受容体-1抗体のサブユニットポリペプチドの重鎖またはその可変領域あるいは軽鎖およびその可変領域をコードする核酸を提供する。
【0060】
ある態様において、本発明は本発明のNogo受容体-1抗体の超可変領域 (CDR)またはCDRをコードする核酸を提供する。
【0061】
免疫
本発明の抗体は好適な宿主 (例えば、脊椎動物、例えばヒト、マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ、ウマ、爬虫類、魚類、両生類および鳥類、爬虫類および魚類の卵中)の免疫により生成させうる。かかる抗体はポリクローナルであってもモノクローナルであってもよい。
【0062】
ある態様においては、宿主を本発明の免疫原性Nogo受容体-1ポリペプチドで免疫する。別の態様において、宿主をインタクトまたは崩壊した細胞の細胞膜に結合したNogo受容体-1で免疫し、本発明の抗体を本発明の免疫原性Nogo受容体-1ポリペプチドへの結合によって同定する。
【0063】
ある態様において、Nogo受容体-1抗原を免疫応答を刺激するアジュバントとともに投与する。アジュバントはしばしば抗原への免疫応答を誘発するために抗原に加えて投与する必要がある。かかるアジュバントは通常は、免疫部位において単核食細胞を集めて非特異的炎症を促進する不溶性または非崩壊性物質である。アジュバントの例としては、これらに限定されないが以下が挙げられる:フロイントアジュバント、RIBI(ムラミルジペプチド)、ISCOM (免疫刺激複合体)またはそれらのフラグメント。
【0064】
抗体の製造方法の概説としては、例えば、Harlow and Lane (1988), Antibodies, A Laboratory Manual, Yelton, D.E. et al. (1981); Ann. Rev. of Biochem., 50, pp. 657-80., およびAusubel et al. (1989); Current Protocols in Molecular Biology (New York: John Wiley & Sons)を参照されたい。本発明の免疫原性Nogo受容体-1ポリペプチドによる免疫反応性の測定は当該技術分野で周知の様々な方法によって行うことが出来、例えば、イムノブロットアッセイおよびELISAが挙げられる。
【0065】
抗体および抗体産生細胞株の産生
本発明のモノクローナル抗体は例えば前掲のHarlow and Lane (1988)に記載の標準的手順によって作ることが出来る。
【0066】
簡単に説明すると、適当な時期に動物を屠殺し、リンパ節および/または脾臓のB-細胞を当該技術分野で周知のいずれかの技術によって不死化する。かかる技術としては、これらに限定されないが、例えば、EBVなどによる形質転換またはミエローマ細胞等の不死化細胞株との融合が挙げられる。その後、細胞をクローン分離し、各クローンの上清を本発明の免疫原性Nogo受容体-1ポリペプチドに特異的な抗体の産生について試験する。ハイブリドーマの選抜、クローニングおよび拡張方法は当該技術分野で周知である。同様に、免疫グロブリン遺伝子のヌクレオチドおよびアミノ酸配列決定方法は当該技術分野で知られている。
【0067】
本発明の抗体のその他の好適な産生技術には、本発明のNogo受容体-1または免疫原性ポリペプチドへリンパ球をインビトロで曝すこと、あるいは、ファージまたは類似のベクターにおける抗体ライブラリーの選抜が含まれる。Huse et al., Science, 246, pp. 1275-81 (1989)を参照されたい。本発明に有用な抗体は改変してもしなくても利用できる。
【0068】
抗原(この場合、Nogo受容体-1または本発明の免疫原性ポリペプチド)および抗体は共有結合または非共有結合によって検出可能なシグナルを提供する物質を結合させることにより標識できる。様々な標識および結合(conjugation)技術が当該技術分野で知られており、本発明の実施に用いることが出来る。好適な標識にはこれらに限定されないが以下が含まれる:放射性ヌクレオチド、酵素、基質、補因子、阻害剤、蛍光試薬、化学発光試薬、磁性粒子等。かかる標識の使用を教示する特許としては以下が挙げられる:米国特許3817837号; 3850752号; 3939350号; 3996345号; 4277437号; 4275149号および4366241号。また、組換え免疫グロブリンも産生できる(米国特許4816567号を参照されたい)。
【0069】
本発明のある態様において、抗体は複数の結合特異性を有し、例えば、二機能性抗体は当業者に知られている多くの技術のいずれかによって調製される。かかる技術としては例えば、雑種ハイブリドーマの産生、ジスルフィド交換、化学的架橋、2つのモノクローナル抗体間のペプチドリンカーの付加、特定の細胞株への2セットの免疫グロブリン重鎖および軽鎖の導入などが挙げられる (より詳細には以下を参照されたい)。
【0070】
本発明の抗体は、例えば不死化ヒト細胞によって、SCID-hu マウスによって、あるいはその他の「ヒト」抗体を産生することが出来る非ヒト動物によって産生されたヒトモノクローナル抗体であってもよい。
【0071】
ファージディスプレーライブラリー
本発明の抗-Nogo受容体-1抗体は組換えコンビナトリアル抗体ライブラリーのスクリーニングによって単離することが出来る。例示的なコンビナトリアルライブラリーは本発明の免疫原性Nogo受容体-1ポリペプチドへの結合についてのものであり、例えばscFvファージディスプレーライブラリーが挙げられ、これは本発明の免疫原性Nogo受容体-1ポリペプチドで免疫した動物由来のmRNAから調製したVLおよびVHcDNAを用いて調製される。かかるライブラリーの調製方法およびスクリーニング方法は当該技術分野で知られている。ファージディスプレーライブラリーを作成するための市販の方法および材料がある (例えば、Pharmacia組換えファージ抗体システム、カタログ番号27 9400 01; Stratagene SurfZAP(商標)ファージディスプレーキット、カタログ番号240612;およびMorphoSysのその他のもの)。また、抗体ディスプレーライブラリーの作成およびスクリーニングに使用できるその他の方法及び試薬もある(例えば、Ladner et al. 米国特許第5223409号; Kang et al. PCT公開WO 92/18619号; Dower et al. PCT 公開 WO 91/17271; Winter et al. PCT公開 WO 92/20791; Markland et al. PCT公開WO 92/15679; Breitling et al. PCT 公開 WO 93/01288; McCafferty et al. PCT 公開 WO 92/01047; Garrard et al. PCT公開WO 92/09690; Fuchs et al. (1991) Bio/Technology 9:1370 1372; Hay et al. (1992) Hum.Antibod. Hybdiromas 3:81 85; Huse et al. (1989) Science 246:1275 1281; McCafferty et al., Nature (1990) 348:552 554; Griffiths et al. (1993) EMBO J. 12:725 734; Hawkins et al. (1992) J. Mol. Biol. 226:889 896; Clackson et al. (1991) Nature 352:624 628; Gram et al. (1992) Proc. Natl. Acad. Sci. USA 89:3576 3580; Garrad et al. (1991) Bio/Technology 9:1373 1377; Hoogenboom et al. (1991) Nucl. Acids Res. 19:4133 4137;およびBarbas et al. (1991) Proc. Natl. Acad. Sci. USA 88:7978 7982を参照)。
【0072】
組換え免疫グロブリンディスプレーライブラリーからの本発明の抗-Nogo受容体-1抗体のスクリーニングおよび単離後、選択した抗体をコードする核酸をディスプレーパッケージから回収し(例えば、ファージゲノムから)、標準的組換えDNA技術によってその他の発現ベクターにサブクローニングすればよい。所望の場合、核酸をさらに操作して以下に記載のように本発明のその他の抗体形態を作成してもよい。コンビナトリアルライブラリーのスクリーニングによって単離された抗体を発現するために、抗体重鎖および軽鎖またはその可変領域をコードするDNAを組換え発現ベクターにクローニングし、上記のように哺乳類宿主細胞に導入する。
【0073】
クラススイッチ
本発明の抗-Nogo受容体-1抗体はいずれのアイソタイプでもよい。あらゆる所望のアイソタイプの抗体がクラススイッチによって産生できる。クラススイッチのために、CLまたはCHをコードするヌクレオチド配列を含まないVLまたはVHをコードする核酸を当該技術分野で周知の方法を用いて単離する。VLまたはVHをコードする核酸を所望のクラスの免疫グロブリン分子からのCLまたはCHをコードするヌクレオチド配列と作動可能に連結する。これは上記のようにCLまたはCH鎖を含むベクターまたは核酸を用いて達成できる。例えば、元はIgMであった本発明の抗-Nogo受容体-1抗体をIgGへとクラススイッチ出来る。さらに、クラススイッチを用いて1つのIgG サブクラスをその他に、例えばIgG1からIgG2に変換できる。
【0074】
突然変異抗体
別の態様において本発明の抗体または抗原結合性フラグメントを重鎖および/または軽鎖の可変ドメインにおいて突然変異させて抗体の結合特性を変化させてもよい。例えば、突然変異は1または複数のCDR領域において導入してNogo受容体-1についての抗体のKdを増減させてもよいし、Koffを増減させてもよいし、抗体の結合特異性を変化させてもよい。部位特異的突然変異誘発技術は当該技術分野で周知である。例えば、Sambrook et al. and Ausubel et al., 前掲を参照されたい。好適な態様において本発明の抗-Nogo受容体-1抗体の可変領域において生殖系列と比較して変化していることが知られているアミノ酸残基において突然変異を導入する。ある態様において、本発明の抗-Nogo受容体-1抗体の可変領域において生殖系列と比較して変化していることが知られている1または複数のアミノ酸残基に突然変異を導入する。別の態様において、抗体重鎖または軽鎖可変領域をコードする核酸を1または複数のフレームワーク領域において突然変異させる。突然変異はフレームワーク領域または定常ドメインに導入して半減期を上昇させてもよい。フレームワーク領域または定常ドメインにおける突然変異はまた、抗体の免疫原性を変化させ、別の分子への共有または非共有結合のための部位を提供するか、補体固定としての性質を変化させることもある。突然変異は1つの突然変異抗体におけるフレームワーク領域、定常ドメインおよび可変領域のそれぞれにおいて導入してもよい。あるいは、突然変異は1つの突然変異抗体においてフレームワーク領域、可変領域または定常ドメインのいずれか1つのみにおいて導入してもよい。
【0075】
融合抗体およびイムノアドヘシン
別の態様において、融合抗体またはイムノアドヘシンを作成してもよく、これは別のポリペプチドに連結した本発明の抗-Nogo受容体-1抗体の全部または一部を含む。ある態様において、抗-Nogo受容体-1抗体の可変領域のみをポリペプチドに連結する。別の態様において、本発明の抗-Nogo受容体-1抗体のVHドメインを第一のポリペプチドに連結し、抗体のVLドメインを第二のポリペプチドに連結させる。第二のポリペプチドは第一のポリペプチドとVHおよびVLドメインが互いに相互作用して抗体結合部位を形成するように結合する。別の態様において、VHドメインとVLドメインとは、VHおよびVLドメインが互いに相互作用するのを可能にするリンカーによって分離する(以下の一本鎖抗体を参照されたい)。VH -リンカー- VL 抗体を次いで目的のポリペプチドと連結する。融合抗体はNogo受容体-1リガンドを発現する細胞または組織に対してポリペプチドを標的化するのに有用である。目的のポリペプチドは例えば毒素などの治療薬であってもよく、例えば容易に可視化できる酵素などの診断薬であってもよく、例えばセイヨウワサビペルオキシダーゼが挙げられる。さらに、2(または3以上)の一本鎖抗体が互いに連結する融合抗体を作成してもよい。これは1つのポリペプチド鎖上に二価または多価抗体を作る場合や、二重特異性抗体を作る場合に有用である。
【0076】
一本鎖抗体
本発明は、本発明の免疫原性Nogo受容体-1ポリペプチドに結合する一本鎖抗体(scFv)を含む。ScFvを産生するためには、VHおよびVLをコードするDNAを屈曲性リンカー、例えば、アミノ酸配列 (Gly4 Ser)3(配列番号10)をコードするDNAと作動可能に連結し、VHおよびVL配列が連続した一本鎖タンパク質として発現するようにし、ここで、VLおよびVH 領域は屈曲性リンカーによって連結している(例えば、Bird et al. (1988) Science 242:423 426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879 5883; McCafferty et al., Nature (1990) 348:552 554を参照されたい)。一本鎖抗体は1つのVHおよびVLのみを用いる場合、一価であってもよいし、2つのVHおよびVLを用いる場合、二価であってもよいし、3以上のVHおよびVLを用いる場合、多価であってもよい。
【0077】
キメラ抗体
本発明はさらにその1つの特異性が本発明の免疫原性Nogo受容体-1ポリペプチドに対するものである二重特異性抗体またはその抗原結合性フラグメントを含む。1つの態様において、本発明の免疫原性Nogo受容体-1ポリペプチドに1つの結合ドメインを介して、そして第二の分子に第二の結合ドメインを介して特異的に結合するキメラ抗体を作成してもよい。キメラ抗体は組換え分子生物学技術によって産生してもよいし、互いに物理的に結合させてもよい。さらに、2以上のVHおよびVLを含み、本発明の免疫原性ポリペプチドと、ミエリン媒介成長円錐崩壊と神経突起伸長および出芽の阻害の減弱に関連するその他の分子に特異的に結合する一本鎖抗体を作成してもよい。かかる二重特異性抗体は周知技術によって作成でき、例えば、 Fanger et al. Immunol Methods 4: 72-81 (1994)およびWright and Harris,前掲を参照されたい。また、(iii)に関しては例えば、Traunecker et al. Int. J. Cancer (Suppl.) 7: 51-52 (1992)を参照されたい。
【0078】
ある態様において、キメラ抗体は本発明の抗体由来の1または複数の可変領域を用いて調製する。別の態様において、キメラ抗体は該抗体からの1または複数のCDR領域を用いて調製する。
【0079】
誘導体化および標識化抗体
本発明の抗体または抗原結合性フラグメントは誘導体化または別の分子に連結してもよい(例えば、別のペプチドまたはタンパク質)。一般に、抗体または抗原結合性フラグメントは本発明の免疫原性ポリペプチドへの結合が誘導体化または標識によって阻害されないように誘導体化する。例えば、本発明の抗体または抗体部分を機能的に(化学的カップリング、遺伝子融合、非共有的結合その他によって)1または複数のその他の分子的実体に結合させるとよい。分子的実体としては例えば、別の抗体(例えば、二重特異性抗体または二価抗体)、検出試薬、細胞毒性試薬、医薬および/または抗体または抗原結合性フラグメントのその他の分子への結合を媒介しうるタンパク質またはペプチドが挙げられる(例えば、ストレプトアビジンコア領域またはポリヒスチジンタグ)。
【0080】
ある態様において、誘導体化抗体は2以上の抗体(同じタイプでも異なるタイプでもよく、例えば二重特異性抗体を作るもの)の架橋によって産生する。好適なクロスリンカーにはヘテロ二機能性のもの、即ち2種類の反応性の基が適当なスペーサーによって分離されているもの(例えば、m- マレイミドベンゾイル-N-ヒドロキシスクシンイミドエステル)またはホモ二機能性のもの(例えば ジスクシンイミジルスベレート)がある。かかるリンカーはPierce Chemical Company, Rockford, Illから入手できる。
【0081】
ある態様において、誘導体化抗体は標識化抗体である。例えば、本発明の抗体または抗体部分が誘導体化される検出試薬は蛍光化合物であり、例えば、フルオレセイン、フルオレセインイソチアシアナート、ローダミン、5-ジメチルアミン-1-ナフタレンスルホニルクロリド、フィコエリトリン、ランタニドリン等が挙げられる。抗体は検出に有用な酵素で標識してもよく、かかる酵素としては、例えば、セイヨウワサビペルオキシダーゼ、β-ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ、グルコースオキシダーゼ等が挙げられる。検出可能な酵素で標識する態様において、抗体は、酵素が検出可能な反応産物を産生するのに用いるさらなる試薬の添加によって検出する。例えば、セイヨウワサビペルオキシダーゼでは過酸化水素およびジアミノベンジジンを用いる。抗体はビオチンで標識してもよく、この場合アビジンまたはストレプトアビジン結合の間接的測定によって検出する。抗体は二次レポーター(例えば、ロイシンジッパーペア配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)によって認識されるあらかじめ決定したポリペプチドエピトープで標識してもよい。
【0082】
抗-Nogo受容体-1抗体またはその抗原-フラグメントを放射標識アミノ酸で標識してもよい。放射標識は診断および治療用途の両方に用いられる。放射標識抗-Nogo受容体-1抗体は診断に用いることが出来、例えば、対象におけるNogo受容体-1レベルの判定に用いられる。さらに、放射標識抗-Nogo受容体-1抗体は脊髄損傷の治療に治療的に用いられる。ポリペプチドのための標識の例は、これらに限定されないが以下の放射性同位体または放射性ヌクレオチドが挙げられる- 3H、14C、15N、35S、90Y、99Tc、111In、125I、131I。
【0083】
抗-Nogo受容体-1抗体またはその抗原-フラグメントは以下のような化学基で誘導体化してもよい:ポリエチレングリコール(PEG)、メチルまたはエチル基または炭水化物基。これらの基は抗体の生物学的特性の向上、例えば、血清半減期の上昇または組織結合性の上昇、に有用である。
【0084】
抗-Nogo受容体-1抗体の特徴付け
抗-Nogo受容体-1抗体のクラスおよびサブクラス
抗-Nogo受容体-1抗体のクラスおよびサブクラスは当該技術分野で知られているいずれの方法で判定してもよい。一般に、抗体のクラスおよびサブクラスは、抗体の特定のクラスおよびサブクラスに特異的な抗体を用いて判定できる。かかる抗体は市販されている。クラスおよびサブクラスはELISA、ウェスタンブロットおよびその他の技術によって判定できる。あるいは、クラスおよびサブクラスは、抗体の重鎖および/または軽鎖の定常ドメインの全部または一部の配列決定、そのアミノ酸配列と免疫グロブリンの様々なクラスおよびサブクラスの公知のアミノ酸配列との比較、および抗体のクラスおよびサブクラスの決定によって判定できる。
【0085】
抗-Nogo受容体-1抗体のNogo受容体-1への結合アフィニティー
本発明の抗-Nogo受容体-1抗体の本発明の免疫原性Nogo受容体-1ポリペプチドに対する結合アフィニティーおよび解離速度は当該技術分野で知られているいずれの方法でも測定できる。例えば、結合アフィニティーは、競合的ELISA、RIA、BIAcoreまたはKinExA技術によって測定できる。解離速度もまた、BIAcoreまたは KinExA 技術によって測定できる。結合アフィニティーおよび解離速度は例えばBIAcoreを用いた表面プラズモン共鳴によって測定される。
【0086】
7E11および1H2のKdを測定したところ、それぞれ1 x 10-7 Mおよび2 x 10-8 Mであった。
【0087】
抗-Nogo受容体-1抗体によるNogo受容体-1活性の阻害
ある態様において、本発明の抗-Nogo受容体-1抗体または抗原結合性フラグメントは、Nogo受容体-1のリガンドに対する結合を阻害する。かかる阻害のIC50 は当該技術分野で知られているいずれの方法でも測定でき、例えば、ELISA、RIAまたは機能アンタゴニズムが挙げられる。ある態様において、IC50は0.1〜500 nMである。ある態様において、IC50は10〜400 nMである。さらに別の態様において、 抗体またはその部分のIC50は60 nM〜400 nMである。7E11および1H2のIC50を結合アッセイにおいて測定したところそれぞれ400 nM および60 nMであった。以下の表3を参照されたい。
【0088】
要約すると、当業者であれば、本発明の教示に接した場合、本発明の抗体の生物学的特性を変化させるのに用いられる多くの方法を利用することが出来る。例えば、所与の抗体分子の安定性、半減期、免疫原性、毒性、アフィニティーまたは収率を増減する方法等が挙げられる。あるいはその他の方法によって抗体の生物学的特性を特定の用途により好適なように変化させることができる。
【0089】
本発明の抗体を含む組成物および用途を以下に記載する。
可溶性Nogo受容体-1ポリペプチド
タンパク質
全長Nogo受容体-1は、シグナル配列、N-末端領域(NT)、8つのロイシンリッチリピート(LRR)、LRRCT領域(8つのロイシンリッチリピートのC-末端の1つのロイシンリッチリピートドメイン)、C-末端領域 (CT)および GPI アンカー (図1参照)からなる。
【0090】
本発明のいくつかの態様は可溶性Nogo受容体-1ポリペプチドを提供する。本発明の可溶性Nogo受容体-1ポリペプチドは、NT ドメイン; 8 LRRおよびLRRCT ドメインを含み、シグナル配列および機能性GPIアンカーを欠く(即ち、 GPI アンカーがないか、細胞膜と有効に結合する能力を欠くGPI アンカーがある)。
【0091】
ある態様において、可溶性Nogo受容体-1ポリペプチドは異種LRRを含む。ある態様において、可溶性Nogo受容体-1ポリペプチドは2、3、4、5、6、7または8の異種LRRを含む。異種LRRとは、Nogo受容体-1以外のタンパク質から得たLRRを意味する。異種LRRの起源となりうる例示的なタンパク質は、toll-様受容体(TLR1.2); T-細胞活性化ロイシンリピートリッチタンパク質; デセオリン; OM-gp; インスリン様成長因子結合タンパク質酸不安定性サブユニット slitおよびrobo; そしてtoll-様受容体4である。
【0092】
ある態様において、本発明は319 アミノ酸の可溶性Nogo受容体-1ポリペプチド(可溶性Nogo受容体-1 344、sNogoR1-344またはsNogoR344)( 配列番号6および8の残基 26-344 または配列番号8の残基 27-344)を提供する。ある態様において、本発明は285 アミノ酸の可溶性Nogo受容体-1ポリペプチド (可溶性Nogo受容体-1 310、sNogoR1-310またはsNogoR310)(配列番号7および9の残基 26-310または配列番号9の残基27-310)を提供する。図1を参照されたい。
【0093】
【表1】
[全長および1-310 マウス配列をこの表に追加すべきか?]
【0094】
本発明のある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは、リガンドのNogo受容体-1への結合の阻害に用いられ、Nogo受容体-1リガンドのアンタゴニストとして作用する。本発明のある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは神経細胞の例えば軸索成長における神経突起伸長および出芽の阻害の減少および神経細胞におけるミエリン媒介成長円錐崩壊の阻害に用いられる。ある態様において、神経細胞はCNS 神経細胞である。
【0095】
sNogoR310およびsNogoR344は驚くべきことにNogoA、NogoB、NogoC、MAGおよびOM-gpのNogo受容体-1に対する結合をブロックする。
【0096】
ある態様において、本発明の可溶性Nogo受容体-1ポリペプチドは、さらに異種ポリペプチドを含む融合タンパク質の構成成分である。ある態様において、異種ポリペプチドは免疫グロブリン定常ドメインである。ある態様において、免疫グロブリン定常ドメインは重鎖定常ドメインである。ある態様において、異種ポリペプチドはFc フラグメントである。ある態様において、Fcは本発明の可溶性Nogo受容体-1ポリペプチドのC-末端に連結する。ある態様において、融合Nogo受容体-1タンパク質はダイマーである。
【0097】
本発明の核酸分子
本発明は本発明のポリペプチドをコードする核酸を提供し、本発明のポリペプチドには配列番号1-9のいずれかのポリペプチドが含まれる。ある態様において、核酸は以下からなる群から選択されるポリペプチドをコードする:配列番号6および8に示すNogo受容体-1のアミノ酸残基 26-344または配列番号8に示すNogo受容体-1のアミノ酸残基 27-344。ある態様において、核酸分子は以下からなる群から選択されるポリペプチドをコードする:配列番号7および9に示すNogo受容体-1のアミノ酸残基 26-310または配列番号9に示すNogo受容体-1のアミノ酸残基 27-310。本明細書において用いる、「核酸」とは、ゲノムDNA、cDNA、mRNAおよびアンチセンス分子ならびに以下のような核酸を意味する:別の骨格に基づく核酸または天然または合成の別の塩基を含む核酸。ある態様において、核酸はさらに転写プロモーターおよび所望によりシグナル配列を含み、そのいずれも本発明のポリペプチドをコードするヌクレオチド配列に作動可能に連結している。
【0098】
ある態様において、本発明は本発明のNogo受容体-1融合タンパク質をコードする核酸を提供する。ある態様において、核酸は本発明のNogo受容体-1融合タンパク質をコードし、本発明のNogo受容体-1融合タンパク質には、以下からなる群から選択されるポリペプチドを含む融合タンパク質が含まれる:配列番号6および8に示すNogo受容体-1のアミノ酸残基 26-344または配列番号8のアミノ酸残基 27-344および配列番号7および9に示すNogo受容体-1のアミノ酸残基 26-310または配列番号9のアミノ酸残基27-310。ある態様において、Nogo受容体-1融合タンパク質をコードする核酸はさらに、転写プロモーターおよび所望によりシグナル配列を含む。ある態様において、ヌクレオチド配列はさらに免疫グロブリン定常領域をコードする。ある態様において、免疫グロブリン定常領域は重鎖定常領域である。ある態様において、ヌクレオチド配列はさらにヒンジ領域に連結した免疫グロブリン重鎖定常領域をコードする。ある態様において、核酸はさらにFcをコードする。ある態様において、Nogo受容体-1融合タンパク質はFc フラグメントを含む。
【0099】
本発明のコード核酸はさらに診断およびプローブ用の検出可能な標識を含むように改変してもよい。様々なかかる標識は当該技術分野で知られており、本明細書に記載するコード分子に容易に適用できる。好適な標識としては、これらに限定されないが、ビオチン、放射標識ヌクレオチド等が挙げられる。当業者であれば標識化コード核酸分子を得るために当該技術分野で知られたいずれの標識も利用することが出来る。
【0100】
組成物
ある態様において、本発明は以下からなる群から選択される免疫原性ポリペプチドを含む組成物を提供する:配列番号1、配列番号2、配列番号3、配列番号4および配列番号5。
【0101】
ある態様において、本発明は本発明の抗-Nogo受容体-1抗体またはその抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質を含む組成物を提供する。
【0102】
ある態様において、本発明の組成物は賦形剤および補助剤を含む好適な医薬上許容される担体を含んでいてもよく、かかる担体は活性化合物を作用部位に送達するために医薬上使用されうる調製物への加工を促進する。非経口投与に好適な製剤には、水可溶性形態、例えば、水-可溶性塩における活性化合物の水溶液が挙げられる。適当に油性の注射用懸濁液などの活性化合物の懸濁液を投与してもよい。好適な親油性溶媒または媒体としては、脂肪油、例えば、ごま油または合成脂肪酸エステル、例えば、エチルオレアートまたはトリグリセリドが挙げられる。水性注射用懸濁液は懸濁液の粘度を上昇させる物質を含んでいてもよく、かかる物質としては、例えば、カルボキシメチルセルロースナトリウム、ソルビトールおよびデキストランが挙げられる。所望により、懸濁液はまた安定剤を含んでいてもよい。リポソームも本発明の分子を被覆して細胞へと送達するのに利用できる。例示的な「医薬上許容される担体」はあらゆる溶媒、分散媒、被覆、抗細菌剤および抗真菌剤、生理的に許容される等張および吸収遅延剤等、水、生理的食塩水、リン酸緩衝食塩水、デキストロース、グリセロール、エタノール等およびそれらの組み合わせである。ある態様において、組成物は等張剤を含み、例えば、糖、ポリアルコール、例えば、マンニトール、ソルビトールまたは塩化ナトリウムが挙げられる。ある態様において、組成物は医薬上許容される物質、例えば、少量の補助物質、例えば湿潤剤または乳化剤、保存料または緩衝剤を含み、これらは本発明の抗体、抗原結合性フラグメント、可溶性Nogo受容体または融合タンパク質の有効期間または有効性を改善する。
【0103】
本発明の組成物は様々な形態であってよく、例えば、液体、半流動性および固体剤形が含まれ、例えば液体溶液(例えば、注射可能および注入可能溶液)、分散液または懸濁液が挙げられる。好ましい形態は意図する投与方法および治療用途に依存する。1つの態様において、組成物は注射可能または注入可能溶液形態であり、例えば、ヒトのその他の抗体による受動免疫に用いられるような組成物等が挙げられる。
【0104】
組成物は溶液、マイクロエマルジョン、分散液、リポソームまたはその他の高薬剤濃度に好適な構造に製剤するとよい。無菌注射可能溶液は、要求される量の抗-Nogo受容体-1抗体を所望により上記の1または複数の成分とともに適当な溶媒に導入し、次いでフィルター滅菌することによって調製される。一般に、分散液は活性化合物を基本分散媒を含む無菌媒体に所望により上記のその他の成分とともに導入することによって調製される。無菌注射可能溶液の調製物のための無菌粉末の場合、好ましい調製方法は減圧乾燥および凍結乾燥であり、これにより先に無菌-ろ過した溶液から活性成分の粉末とさらなる所望の成分が得られる。溶液の適当な流動性は例えば以下のように維持されうる。レシチン等の被覆剤の使用、分散液の場合要求される粒径の維持、そして界面活性剤の使用。注射可能組成物の吸収の延長は吸収を遅延させる薬剤、例えば、モノステアリン酸塩およびゼラチンを組成物に含めることによってもたらされうる。
【0105】
ある態様において、活性化合物は化合物が迅速に放出するのを保護する担体とともに調製すればよく、例えば、徐放性製剤とすればよい。かかる徐放性製剤としては、インプラント、経皮パッチおよびマイクロカプセル化送達系が挙げられる。生分解性、生体適合性ポリマーを用いることが出来、例えば、酢酸エチレンビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステルおよびポリ乳酸が挙げられる。かかる製剤の多くの調製方法が特許されているかあるいは当業者に周知である。例えば、Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978を参照されたい。
【0106】
補足性活性化合物を組成物に含めてもよい。ある態様において、本発明のNogo受容体-1抗体またはその抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質は1または複数のさらなる治療薬と共製剤および/または共投与される。
【0107】
本発明の医薬組成物は、「治療上有効量」または「予防的に有効量」の本発明の抗体、抗原結合性フラグメント、ポリペプチドまたは融合タンパク質を含んでいるとよい。「治療上有効量」とは、投与および必要な期間において所望の治療結果を達成するのに有効な量を意味する。Nogo受容体-1抗体またはその抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはNogo受容体融合タンパク質の治療上有効量は、様々な因子に応じて変動し得、かかる因子としては例えば、個人の疾患状態、年齢、性別および体重が挙げられる。治療上有効量は抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはNogo受容体融合タンパク質のあらゆる毒性や有害作用よりも治療的に有用な効果が上回るものでもある。「予防的に有効量」とは投与および必要な期間において所望の予防結果を達成するのに有効な量を意味する。典型的には予防用量は対象に疾患前または疾患のより早期の段階にて用いられるために、予防的に有効量は治療上有効量より少ない。
【0108】
用法・用量は最適な望ましい応答(例えば治療または予防応答)が得られるよう調整すればよい。例えば、1のボーラスを投与してもよいし、いくつかに分割された用量を長期に投与してもよいし、治療状況の切迫に応じて比例的に増減してもよい。投与の便宜のために用量単位形態における非経口組成物を製剤するのが特に好ましく、本明細書において用いる用量単位形態とは、治療すべき哺乳類対象にとっての単位用量として好適な物理的に分離された単位であり、各単位は、必要な医薬担体とともに所望の治療効果が得られるよう計算されたあらかじめ決定した量の活性化合物を含む。本発明の用量単位形態の特定は以下によって示され直接依存する:(a)抗体、抗原結合性フラグメントおよび可溶性受容体-1ポリペプチドまたはNogo受容体融合タンパク質の固有の特性および達成すべき特定の治療または予防効果そして(b)かかる抗体、抗原結合性フラグメントおよび可溶性受容体-1ポリペプチドまたはNogo受容体融合タンパク質の配合における個人の感受性の処置についての当該技術分野に固有の制限。ある態様において、治療上有効用量はNogo受容体-1抗体またはその抗原結合性フラグメントについて1日あたり 0.1 - 4 mg/Kgの範囲である。ある態様において、 Nogo受容体-1抗体またはその抗原結合性フラグメントの治療上有効用量範囲は1日あたり0.2 - 4 mg/Kgである。ある態様において、Nogo受容体-1抗体またはその抗原結合性フラグメントの治療上有効用量範囲は1日あたり0.2 mg/Kgである。
【0109】
抗体、抗原結合性フラグメント、可溶性受容体および融合タンパク質の使用
ある態様において、本発明は必要とする哺乳類に抗-Nogo受容体-1抗体、かかる抗体の抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドまたはかかるポリペプチドを含む融合タンパク質を投与することによるNogo受容体-1活性の阻害方法を提供する。
【0110】
ある態様において、本発明はNogo受容体-1と本発明の抗体または抗原結合性フラグメントを接触させる工程を含むNogo受容体-1のリガンドへの結合の阻害方法を提供する。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。
【0111】
ある態様において、本発明は、神経細胞と本発明の抗体またはその抗原結合性フラグメントとを接触させる工程を含む神経細胞における成長円錐崩壊の阻害方法を提供する。ある態様において、本発明は、神経細胞と本発明の抗体または抗原結合性フラグメントとを接触させる工程を含む神経細胞における神経突起伸長または出芽の阻害を減少させる方法を提供する。ある態様において、神経細胞はCNS 神経細胞である。かかる方法のいくつかにおいて神経突起伸長または出芽は軸索成長である。
【0112】
ある態様において、本発明は、以下の工程を含む、神経細胞が死の危険にある哺乳類における、神経細胞の生存を促進させる方法を提供する:(a) (i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(ii)可溶性Nogo受容体-1ポリペプチド、を発現する培養宿主細胞を提供する工程; および、(b) 神経細胞においてまたはその近くの部位にて哺乳類に宿主細胞を導入する工程。 Almudena Ramon-Cueto, M Isabel Cordero, Fernando F Santos-Benito and Jesus Avila (2000) Functional recovery of paralegic rats and motor axon regneration in their spinal cords by olfactory ensheathing cells. Neuron 25, 425-435。
【0113】
ある態様において、本発明は以下の工程を含む神経細胞が哺乳類のものである死の危険にある神経細胞の生存を促進するための遺伝子治療方法を提供する:神経細胞においてまたはその近くの部位にて(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または(b)可溶性Nogo受容体-1ポリペプチド、をコードするヌクレオチド配列を含むウイルスベクターを投与する工程、ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは哺乳類において神経細胞の生存を促進するのに十分な量にてヌクレオチド配列から発現する。かかる態様にとって有用なウイルスベクターおよび方法は、例えば、Noel et al., Human Gene Therapy, 13, 1483-93 (2002)に記載されている。
【0114】
ある態様において、本発明は、リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含むリガンドへのNogo受容体-1の結合の阻害方法を提供する。
【0115】
ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、Nogo受容体-1リガンドの活性を調節する方法を提供する。
【0116】
ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における成長円錐崩壊の阻害方法を提供する。ある態様において、本発明は、Nogo受容体-1リガンドと本発明の可溶性Nogo受容体-1ポリペプチドまたはNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法を提供する。ある態様において、神経細胞はCNS 神経細胞である。ある態様において、リガンドは、NogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される。ある態様において、神経突起伸長または出芽は軸索成長である。
【0117】
本明細書に記載するあらゆるタイプの抗体または受容体を治療的に用いることが出来る。ある態様において、抗-Nogo受容体-1抗体はヒト抗体である。ある態様において、哺乳類はヒト患者である。ある態様において、抗体またはその抗原結合性フラグメントは獣医用またはヒト疾患の動物モデルとして、抗体と交差反応するNogo受容体-1を発現する非-ヒト哺乳類(例えば、霊長類、カニクイザルまたはアカゲザル)に投与される。かかる動物モデルは本発明の抗体の治療効果の評価に有用である。
【0118】
ある態様において、抗-Nogo受容体-1抗体または抗原結合性フラグメントあるいは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質の投与を用いて損傷部位への軸索成長を促進することにより脊髄損傷を治療する。
【0119】
本発明の抗-Nogo受容体-1抗体または抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドまたは融合タンパク質は単独で、その他の試薬と組み合わせて、または逐次に組み合わせて提供するとよい。その他の試薬は特定の病理プロセスを調節するものである。例えば、抗炎症薬をさらなる神経細胞損傷および軸索再生の阻害のブロックの手段として脳卒中の後に共投与するとよい。本明細書において用いる、Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1およびNogo受容体融合タンパク質は、同時、逐次または別々に投与される場合、1または複数のさらなる治療薬と組み合わせて投与されるという。
【0120】
本発明の抗-Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチド、Nogo受容体-1融合タンパク質は、非経口、皮下、静脈内、筋肉内、腹腔内、経皮、吸入または頬側経路にて投与するとよい。例えば、薬剤は損傷部位へ微量注入により局所投与すればよい。典型的な部位としては、これに限定されないが、損傷に起因する脊髄の損傷領域が挙げられる。投与用量はレシピエントの年齢、健康状態、体重、あるとすれば併用療法の種類、治療頻度および所望の効果の性質に依存する。
【0121】
本発明の化合物はインビボにて、通常、哺乳類、例えば、ヒト、ヒツジ、ウマ、ウシ、ブタ、イヌ、ネコ、ラットおよびマウスにおいて、またはインビトロにて利用できる。
【0122】
本発明のベクター
ある態様において、本発明は、コード配列を含む組換えDNA分子(rDNA)を提供する。本明細書において用いる、rDNA分子とは、分子操作を受けたDNA分子である。rDNA分子の作成方法は当該技術分野で周知であり、例えば、Sambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory Pressを参照されたい。あるrDNA分子においては、コードDNA配列は発現制御配列およびベクター配列に作動可能に連結している。
【0123】
ある態様において、本発明は、本発明のポリペプチドをコードする核酸を含むベクターを提供する。本発明の核酸が作動可能に連結するベクターおよび発現制御配列の選択は直接、当該技術分野で周知のように所望の機能特性に依存する(例えば、タンパク質発現および形質転換すべき宿主細胞)。本発明のベクターは少なくとも複製または宿主染色体への挿入が可能なものであり、好ましくは、rDNA分子に含まれる構造遺伝子の発現も可能にするものである。
【0124】
作動可能に連結したタンパク質コード配列の発現を制御するのに用いられる発現制御要素は当該技術分野で知られており、これらに限定されないが、例えば、誘導性プロモーター、構成的プロモーター、選抜シグナルおよびその他の制御要素が挙げられる。好ましくは、誘導性プロモーターは迅速に制御できるものであり、例えば、宿主細胞の培地の栄養成分に応答性のものなどである。
【0125】
1つの態様において、コード核酸分子を含むベクターは原核生物レプリコンを含み、レプリコンとは、それによって形質転換された細菌宿主細胞等の原核生物宿主細胞において染色体外で組換えDNA分子の自己複製と維持を可能にする能力を有するDNA配列である。かかるレプリコンは当該技術分野で周知である。さらに、原核生物レプリコンを含むベクターは、その発現が検出可能または選抜可能マーカー、例えば薬剤耐性マーカーを付与する遺伝子を含んでいてもよい。細菌の薬剤耐性遺伝子に典型的なものは、アンピシリンまたはテトラサイクリンへの耐性を付与するものである。
【0126】
原核生物レプリコンを含むベクターは、細菌宿主細胞、例えば、大腸菌におけるコード遺伝子配列の発現(転写および翻訳)の誘導を可能にする原核生物またはバクテリオファージプロモーターを含んでいてもよい。プロモーターとは、RNA ポリメラーゼに結合し転写を起こすDNA配列によって形成される発現制御要素である。細菌宿主に適合するプロモーター配列は典型的には、本発明のDNAセグメントの挿入のために便宜な制限部位を含むプラスミドベクターにおいて提供される。かかるベクタープラスミドの例は、pUC8、pUC9、pBR322およびpBR329 (Bio-Rad(登録商標) Laboratories)、pPLおよびpKK223 (Pharmacia)である。好適な原核生物宿主を用いて本発明のタンパク質をコードする組換えDNA分子を発現させることが出来る。
【0127】
真核生物細胞、好ましくは脊椎動物細胞に適合する発現ベクターもコード配列を含むrDNA分子の形成に利用できる。真核生物細胞発現ベクターは当該技術分野で周知であり、いくつか市販されている。典型的にはかかるベクターは所望のDNAセグメントの挿入のための便宜な制限部位を含むように提供される。かかるベクターの例は、pSVLおよびpKSV-10 (Pharmacia)、pBPV-1、pML2d (International Biotechnologies)、pTDT1 (ATCC31255)およびその他の真核生物発現ベクターである。
【0128】
本発明のrDNA分子の構築に用いられる真核生物細胞発現ベクターはさらに真核生物細胞において有効な、好ましくは薬剤耐性選抜マーカーといった選抜可能マーカーを含んでいてもよい。好ましい薬剤耐性マーカーは、その発現の結果、ネオマイシン耐性をもたらす遺伝子、即ちネオマイシンホスホトランスフェラーゼ (neo)遺伝子である(Southern et al., (1982) J. Mol. Anal. Genet. 1, 327-341)。あるいは選抜可能マーカーは別のプラスミドに存在してもよく、2つのベクターは宿主細胞に共形質移入によって導入され、トランスフェクタントは、選抜可能マーカーのための適当な薬剤中で培養することによって選抜する。
【0129】
本発明の抗体または抗体部分の発現のために、部分または全長の軽鎖および重鎖をコードするDNAを発現ベクターに挿入し、それら遺伝子を転写および翻訳制御配列に作動可能に連結する。発現ベクターとしては、プラスミド、レトロウイルス、コスミド、YAC、EBV-由来エピソームなどが挙げられる。抗体遺伝子をベクターに連結してベクター中の転写および翻訳制御配列が抗体遺伝子の転写および翻訳の制御の所望の機能を果たすようにする。発現ベクターおよび発現制御配列は用いる発現宿主細胞に適合するものを選択する。抗体軽鎖遺伝子と抗体重鎖遺伝子を別々のベクターに挿入してもよい。ある態様において、両方の遺伝子を同じ発現ベクターに挿入する。抗体遺伝子は標準的方法(例えば抗体遺伝子フラグメントおよびベクターの相補的制限部位のライゲーションまたは制限部位が無い場合は平滑末端ライゲーション)によって発現ベクターに挿入する。
【0130】
便宜なベクターは機能的に完全なヒトCHまたはCL 免疫グロブリン配列をコードするものであり、適当な制限部位を含み、操作によってあらゆるVHまたはVL 配列が上記のように容易に挿入され発現するものである。かかるベクターにおいて、スプライシングが通常、挿入されたJ 領域におけるスプライス供与部位とヒト C 領域の前にあるスプライス受容部位の間で起こり、ヒト CHエキソンにおけるスプライス領域においても起こる。ポリアデニル化および転写終結は、コード領域の下流のネイティブな染色体部位にて起こる。組換え発現ベクターはまた、宿主細胞からの抗体鎖の分泌を容易にするシグナルペプチドをコードしていてもよい。抗体鎖遺伝子をベクターにクローニングし、シグナルペプチドが抗体鎖遺伝子のアミノ末端にインフレームにて連結するようにするとよい。シグナルペプチドは免疫グロブリンシグナルペプチドであっても異種シグナルペプチド(即ち、非免疫グロブリンタンパク質からのシグナルペプチド)であってもよい。
【0131】
本発明の免疫原性ポリペプチド、Nogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質に加えて、本発明の組換え発現ベクターは、宿主細胞におけるその発現を制御する調節配列を担持する。当業者であれば調節配列の選択を含む発現ベクターの設計は形質転換する宿主細胞の選択、所望のタンパク質発現レベルなどの因子に依存することを理解するであろう。好ましい哺乳類宿主細胞発現のための調節配列としては哺乳類細胞における高レベルのタンパク質発現をもららすウイルス性要素、例えば、レトロウイルスLTR、サイトメガロウイルス(CMV) (例えば、CMV プロモーター/エンハンサー)、サルウイルス 40 (SV40) (例えば、SV40 プロモーター/エンハンサー)、アデノウイルス、(例えば、アデノウイルス主要後期プロモーター (AdMLP))、ポリオーマ由来のプロモーターおよび/またはエンハンサーおよび強力な哺乳類プロモーター、例えば、ネイティブな免疫グロブリンおよびアクチンプロモーターが挙げられる。ウイルス性制御要素についての更に詳細な説明およびその配列については例えば以下を参照されたい:Stinskiの米国特許第5168062号、Bell et alの米国特許第4510245号およびSchaffner et alの米国特許第4968615号。
【0132】
異種遺伝子および調節配列に加えて、本発明の組換え発現ベクターはさらなる配列を担持していてもよく、例えば、宿主細胞におけるベクターの複製を制御する配列(例えば、複製起点)および選抜可能マーカー遺伝子が挙げられる。選抜可能マーカー遺伝子は、ベクターが導入された宿主細胞の選抜を容易にする(例えば、米国特許第4399216号、4634665号および5179017号、これらはすべてAxel et alによる)。例えば典型的には、選抜可能マーカー遺伝子はベクターが導入された宿主細胞に薬剤耐性を付与する。薬剤としては例えば、G418、ハイグロマイシンまたはメトトレキサートが挙げられる。好ましい選抜可能マーカー遺伝子としてはジヒドロ葉酸レダクターゼ (DHFR)遺伝子(メトトレキサート選抜/増幅を用いるdhfr- 宿主細胞における使用)およびneo遺伝子(G418 選抜用)が挙げられる。
【0133】
本発明のタンパク質の組換え産生のための宿主細胞および方法
本発明の抗-Nogo受容体-1抗体、免疫原性ペプチド、可溶性Nogo受容体-1ポリペプチド、可溶性Nogo受容体-1融合タンパク質をコードする核酸分子およびかかる核酸分子を含むベクターを用いて好適な宿主細胞を形質転換するとよい。形質転換は宿主細胞にポリヌクレオチドを導入するいずれの公知方法によってもよい。異種ポリヌクレオチドの哺乳類細胞への導入方法は当該技術分野で周知あり、デキストラン-媒介形質移入、リン酸カルシウム沈降、ポリブレン-媒介形質移入、プロトプラスト融合、エレクトロポレーション、リポソームへのポリヌクレオチドのカプセル封入および直接のDNAの核へのマイクロインジェクションが挙げられる。さらに核酸分子は哺乳類細胞にウイルスベクターによって導入してもよい。
【0134】
適当な細胞宿主の本発明のrDNA分子による形質転換は、周知方法により達成され、かかる方法は典型的には使用するベクターのタイプおよび宿主系に依存する。原核生物宿主細胞の形質転換については、エレクトロポレーションおよび塩処理方法を用いるとよい(例えば、Sambrook et al., (1989) Molecular Cloning - A Laboratory Manual, Cold Spring Harbor Laboratory Press; Cohen et al., (1972) Proc. Natl. Acad. Sci. USA 69, 2110-2114を参照されたい)。脊椎動物細胞のrDNAを含むベクターによる形質転換については、エレクトロポレーション、カチオン脂質または塩処理方法を用いるとよい(例えば、Graham et al., (1973) Virology 52, 456-467; Wigler et al., (1979) Proc. Natl. Acad. Sci. USA 76, 1373-1376を参照されたい)。
【0135】
形質転換が成功した細胞、即ち本発明のrDNA分子を含む細胞は選抜可能マーカーについての選抜などの周知技術によって同定できる。例えば、本発明のrDNAの導入の結果得られる細胞をクローニングしてシングルコロニーを得ることが出来る。かかるコロニーからの細胞を回収し、溶解し、そのDNA内容物をrDNAの存在について、例えばSouthern, (1975) J. Mol. Biol. 98, 503-517に記載の方法で調べることが出来、あるいは細胞から産生されたタンパク質を免疫学的方法でアッセイすることが出来る。
【0136】
発現用宿主として利用できる哺乳類細胞株は当該技術分野で周知であり、アメリカンタイプカルチャーコレクション(ATCC)から得られる多くの不死化細胞株が含まれる。これらにはとりわけ、 チャイニーズハムスター卵巣(CHO)細胞、NSO、SP2 細胞、HeLa 細胞、仔ハムスター腎臓 (BHK) 細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)、A549 細胞、およびその他の多くの細胞株が含まれる。特に好ましい細胞株はどの細胞株の発現レベルが高いかを測定することによって選択される。その他の使用可能な細胞株は昆虫細胞株、例えば、Sf9 細胞である。本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質をコードする組換え発現ベクターは哺乳類宿主細胞に導入され、宿主細胞における抗体、ポリペプチドおよび融合ポリペプチドの発現を可能にするのに十分な期間宿主細胞を培養することにより産生され、あるいはより好ましくは本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質を宿主細胞が生育する培地で選抜する。本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質は、標準的タンパク質精製方法を用いて培地から回収できる。
【0137】
さらに本発明の免疫原性ポリペプチド、Nogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび可溶性Nogo受容体-1融合タンパク質(またはそれら由来の部分)の産生細胞株からの発現は多くの公知技術を用いて促進できる。例えば、グルタミンシンゼターゼ遺伝子発現系 (GS系)は特定の条件下での発現促進の常套の手段である。GS系については欧州特許第0216846号、0256055号および0323997号および欧州特許出願第89303964.4号に記載されている。
【0138】
宿主細胞
本発明はさらに、本発明のNogo受容体-1抗体、抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび/または可溶性Nogo受容体-1融合タンパク質をコードする核酸分子で形質転換された宿主細胞を提供する。宿主細胞は原核生物でも真核生物でもよい。本発明のタンパク質の発現に有用な真核生物細胞は、細胞株が細胞培養方法に適合し、発現ベクターの増殖および遺伝子産物の発現に適合する限り特に限定されない。好ましい真核生物宿主細胞としては、これらに限定されないが、酵母、昆虫および哺乳類細胞、好ましくは脊椎動物細胞、例えば、マウス、ラット、サルまたはヒト細胞株が挙げられる。有用な真核生物宿主細胞の例としては、ATCCからCCL61として入手できるチャイニーズハムスター卵巣 (CHO) 細胞、ATCCからCRL1658として入手できるNIH Swiss マウス 胚細胞NIH-3T3、仔ハムスター腎臓細胞(BHK)、その他の真核生物組織細胞細胞株が挙げられる。
【0139】
rDNA分子を用いる組換えタンパク質産生
本発明はさらに、本明細書に記載した核酸分子を用いる本発明のNogo受容体-1抗体または抗原結合性フラグメント、可溶性Nogo受容体-1ポリペプチドおよび/または可溶性Nogo受容体-1融合タンパク質の産生方法を提供する。一般に組換え形態のタンパク質の産生は典型的には以下の工程を含む:
【0140】
第一に、本発明のタンパク質をコードする核酸分子を得る。コード配列がイントロンで分断されていない場合、それはあらゆる宿主における発現に直ちに好適である。
【0141】
核酸分子を次いで所望により、上記の好適な制御配列と作動可能に連結させ、タンパク質オープンリーディングフレームを含む発現単位を形成する。発現単位を用いて好適な宿主を形質転換し、形質転換した宿主を組換えタンパク質の産生を可能とする条件下で培養する。所望により、組換えタンパク質を培地または細胞から単離する;タンパク質の回収および精製はいくらかの不純物が許容される場合には特に必要ではない。
【0142】
上記の各工程は様々な方法で行うことが出来る。例えば、所望のコード配列をゲノム断片から得て直接適当な宿主において用いることが出来る。様々な宿主において作動可能な発現ベクターの構築は上記の適当なレプリコンおよび制御配列を用いて達成される。制御配列、発現ベクターおよび形質転換方法は、遺伝子発現に用いられる宿主細胞のタイプに依存し、先に詳細に説明した。好適な制限部位を、通常は入手可能ではないとしても、コード配列の末端に付加して、かかるベクターへの挿入用の切り出し可能な遺伝子を提供することが出来る。当業者であれば当該技術分野で知られている宿主/発現系を本発明の核酸分子とともに組換えタンパク質の産生に容易に用いることが出来る。
【0143】
本発明の理解をさらに容易にするために。以下に実施例を示す。かかる実施例は単に例示の目的であり、本発明の範囲を限定する意図のものではない。
【実施例1】
【0144】
マウスモノクローナル抗-Nogo受容体-1抗体の産生
本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗-Nogo受容体-1抗体を以下の方法および手順を用いて作製した。
【0145】
免疫
2種類の免疫アプローチを用いた:
1. 免疫原としてNogo受容体-1 (NogoR-1)を含むCOS-7細胞または細胞膜
ラットNogo受容体-1遺伝子(GenBank番号 AF 462390)を、CMV プロモーターおよび薬剤選抜用のジェネティシン耐性遺伝子を含む哺乳類発現ベクター pEAG1256 (Biogen)にサブクローニングした。組換えプラスミドをSuperfect (Qiagen)を用いてCOS-7細胞にトランスフェクトした。トランスフェクタントをジェネティシン (Gibco, 2 mg/ml)を用いて選抜し、クローニングし、FACSによってNogo受容体-1タンパク質の表面発現を確認した。COS-7膜を記載された手順[Wang et al., J. Neurochem.,75:1155-1161 (2000)]にしたがってかかる細胞から調製し、2回洗浄し、1 mg/ml [タンパク質濃度]にて10% グリセロール中、-70℃で保存した。
【0146】
8週齢の雌性RBF マウス (Jackson Labs, Bar Harbor, ME)を50μg ラットNogo受容体-1-COS-7 膜またはNogo受容体-1を表面に発現する全COS-7細胞のいずれかおよび50μl RIBI MPL+TDM+CWS アジュバント (Sigma Chemical Co., St. Louis, MO)を含む乳濁液で腹腔内に2週間毎に免疫した(Lipman et al., 1992)。免疫したマウスからの血清を1回目の免疫前、2回目および3回目の免疫の7日後、および3回目の免疫の38日後に回収し、抗-Nogo受容体-1抗体力価を以下のようにELISAによって測定した。
【0147】
2. 免疫原としての特定のNogo受容体-1 ペプチド
ラットNogo受容体-1遺伝子配列をベクター NTiソフトウェア (図2)を用いた抗原性分析にかけた。分析によって同定された抗原ペプチドをキーホール・リンペット・ヘモシアニン (KLH)と標準的グルタールデヒド法を用いて結合させた。
【0148】
8週齡の雌性RBF マウス(Jackson Labs, Bar Harbor, ME)を50 μg KLH-結合ペプチドおよび50 μl 完全フロイントアジュバント(Sigma Chemical Co., St. Louis, MO) を含む乳濁液で2週間毎に腹腔内に免疫した。免疫したマウスからの血清を1回目の免疫前、2回目および3回目の免疫の1週間後に回収し、抗-Nogo受容体-1抗体力価を測定した。追加抗原投与量を3回目の免疫後に与えた。この追加抗原投与量免疫の3日後、融合実験を開始した。
【0149】
ハイブリドーマ産生およびスクリーニング
抗原性Nogo受容体-1ペプチドで免疫したマウスからの血清をELISAにてスクリーニングし、Nogo受容体-1を発現するCOS-7細胞で免疫したマウスからの血清をフローサイトメトリーによってスクリーニングした。Nogo受容体-1-COS-7細胞に特異的に結合する抗体についてフローサイトメトリーで同定したところ陽性であったマウスを屠殺した。脾細胞をマウスから単離しFL653ミエローマ(Ig-/HGPRT- Balb/c マウスミエローマのAPRT- 誘導体、10% FBS、4500 mg/L グルコース、4 mM L-グルタミンおよび20 mg/ml 8-アザグアニンを含有するDMEM中で維持)と以前に記載されているようにして(Kennett et al., 1993. Monoclonal Antibodies: A New Dimension in Biological Analysis. Plenum Press, New York)融合させた。融合細胞を24-または48-ウェルプレート(Corning Glass Works, Corning, NY)に播き、アデニン、アミノプテリンおよびチミジン含有培地を与えた。AAT耐性培養物を以下に説明するようにしてNogo受容体-1-COS-7細胞またはNogo受容体-1抗原ペプチドへの結合についてELISAまたはフローサイトメトリーでスクリーニングした。陽性ウェル中の細胞をさらに限界希釈によってサブクローニングした。
【0150】
抗体のNogo受容体-1抗原ペプチドへの結合をスクリーニングするために、免疫原として用いたペプチドをBSAと結合させた。50 μlの0.1 M 炭酸水素ナトリウム緩衝液, pH 9.0 中の0.5μgの結合ペプチドを96-ウェルMaxiSorp プレート(Nunc)の各ウェルに添加した。プレートを37℃で1時間または4℃で16時間インキュベートし、非特異的結合部位を0.1% BSA、0.1%卵白アルブミン、0.1%ブロット(blotto)および0.001% アジドを含む25 mM HEPES, pH 7.4を用いてブロックした。ハイブリドーマ上清を添加し、25℃で1時間インキュベートした。PBSでの3回の洗浄後、 50μlの1:10,000希釈のセイヨウワサビペルオキシダーゼ-結合ヤギ抗-マウス二次抗体(Jackson ImmunoResearch Inc.)を各ウェルに添加し、さらに1時間インキュベートした。3回の洗浄後、色をTMB (Pierce)で現像し、2 M硫酸で停止した。着色強度を分光光度計で450 nmにてモニターした。
【0151】
抗体を全長Nogo受容体-1への結合について以下のようにスクリーニングした。COS-7細胞を製造業者の指示に従って0.1 uM CellTracker Green CMFDA (Molecular Probes, Eugene, OR)で標識した。等量のCellTracker標識コントロール細胞を洗浄したNogo受容体-1-COS-7細胞と混合し、抗-Nogo受容体-1被験血清とのインキュベーションに供した。15μlの細胞混合物を96-ウェルV底ポリスチレンプレート(Costar 3877, Corning, NY)の各ウェルに分配し、100μlのハイブリドーマ上清またはコントロール抗-Nogo受容体-1抗体を添加した。4℃で30分のインキュベーション後、細胞を洗浄し、PBS中の50 μlのR-フィコエリトリン-結合アフィニティー精製 F(ab')2 フラグメントヤギ抗-マウス IgG Fcガンマ特異的二次抗体(1:200, Jackson ImmunoResearch Laboratory, West Grove, PA)とともにインキュベートした。インキュベーションの最後に、細胞をPBSで2回洗浄し、1% FBSを含む200μlのPBSに懸濁し、FACS分析にかけた。あるいは、Nogo受容体-1-COS-7細胞をハイブリドーマ上清と混合し、R-フィコエリトリン-結合ヤギ抗-マウス二次抗体で処理し、直接標準的FACS分析にかけた。
【0152】
本発明者らは様々な免疫原を用いて25の抗-Nogo受容体-1抗体を作成した。本発明者らは免疫原としてラットNogo受容体-1 残基 110-125に対応するペプチド配列を用いて2つの抗体、7E11および5B10を作成した。本発明者らは免疫原として全長ラットNogo受容体-1でトランスフェクトしたCOS-7細胞から調製した膜を用いて3つの抗体、1H2、3G5および2F7を作成した。本発明者らは免疫原としてsNogoR310-Fcを用いて13の抗体を作成し(1D9.3、1E4.7、1B4.3、2C4.3、1F10.3、2H1.4、1H3.3、1G4.1、1E4.1、2G7.1、2C4.1、2F11.1および1H4.1)、免疫原としてラットNogo受容体-1 残基 423-434に対応するペプチド配列を用いて7つの抗体を作成した(2E8.1、2G11.2および1B5.1)。
【0153】
モノクローナル抗体7E11および5B10の配列分析
本発明者らはトータルRNAを Qiagen RNeasy ミニキットを用いて抽出し、単離したRNA からcDNAを作成した。本発明者らは、軽鎖配列をPCRによってプライマー、5'-TGAGGAGACGGTGACCGTGGTCCCTTGGCCCCAG-3'(配列番号12)および5'-AGGTSMARCTGCAGSAGTCWGG-3'(配列番号25)を用いて増幅した。本発明者らは、重鎖配列をPCRによってプライマー、5'-GGGGATATCCACCATGAAGTTGCCTGTTAGGCTGTTG-3'(配列番号13)および5'-GGGGATATCCACCATGAGGKCCCCWGCTCAGYTYCTKGGA-3'(配列番号14)を用いて増幅した。かかるプライマーは以下の縮重ヌクレオチドを含む: SはGまたはC;MはAまたはC、RはGまたはA;WはAまたはT;KはGまたはT;そしてYはTまたはCを表す。本発明者らはPCRフラグメントを配列決定ベクターにクローニングし、配列決定ベクターに特異的なプライマーを用いてジデオキシ鎖終結によってCDRのDNA配列を決定した。本発明者らは理論的にDNA配列を翻訳した結果、モノクローナル抗体 7E11および5B10の重鎖および軽鎖のCDR領域の部分アミノ酸配列を表2に示す。mAbの重鎖および軽鎖からの3つのCDRを表2において下線にて示す。7E11および5B10の軽鎖は94%のアミノ酸配列同一性を、重鎖は91%のアミノ酸配列同一性を有していた。mAb、7E11、5B10および1H2はIgG1 アイソタイプであり、mAb、3G5および2F7はIgG2a アイソタイプである。これら5つのmAbはそれぞれカッパアイソタイプの軽鎖を有する。本発明者らはこのアプローチによってその他のモノクローナル抗体の配列を分析した。
【0154】
【表2】
【0155】
モノクローナル抗-Nogo受容体-1抗体による可溶性Nogo受容体-1へのリガンド結合の阻害
本発明者らは上記のように産生した抗-Nogo受容体-1 モノクローナル抗体をそれらがNogo受容体-1へのリガンド結合を阻害するかを調べるべく試験した。
【0156】
下記のように産生した0.5μgの、ラットNogo受容体-1のアミノ酸残基 26-344およびラットIgG1分子のヒンジおよびFc領域を含む可溶性Nogo受容体-1融合タンパク質(sNogoR344-Fc)を、250μgのタンパク質-A- またはコムギ胚芽凝集素-結合SPAビーズ(Amersham Pharmacia Biotech)に2時間25℃で固定化した。50μlのHEPES-緩衝インキュベーション培地(10 mM HEPES, pH 7.4、0.1% ウシ血清アルブミン、0.1% 卵白アルブミン、2 mM MgCl2、2 mM CaCl2およびプロテアーゼ阻害剤)中のFc-sNogoR-1[これはどのsNOGO?]と結合したSPAビーズ、抗-Nogo受容体-1 mAbおよび1μl 125I-Nogo66 (Amersham, 2000 Ci/mmol, 1nM)を各サンプルウェルに添加した。16時間後、放射能を四連のサンプルにおいてTopCount (Packard)を用いて測定した。IC50値をカーブフィッティッング分析(図3)から算出した(PRISM, GraphPad Software, NJ)。いくつかの実験において、本発明者らは、AP-リガンド結合体(例えば、AP-Nogo66)も使用し、結合をアルカリホスファターゼ活性をモニターすることにより検出した。本発明者らはまた、mAbの、リガンドMAG-FcおよびAP-OM-gpのNogo受容体-1への結合をブロックする能力もアッセイした。
【0157】
モノクローナル抗体7E11、5B10、1H2、3G5および2F7はすべてNogo66、MAGおよびOM-gpのsNogoR344-Fcへの結合を阻害した。7E11および1H2について計算したNogo66についての IC50はそれぞれ400 nMおよび60 nMであった。3つのリガンドのNogo受容体-1への結合のmAb-媒介阻害をモニターするELISAからのデータを表3に要約する。
【0158】
【表3】
【0159】
パーセント置換は30 nM 抗体にて示され、特定のmAbについてのEC50 は記載されているカーブフィッティッング分析から決定した。「−」は、検出可能な活性が無いこと、そして「ND」は測定していないことを示す。
【実施例2】
【0160】
Fab-ファージ抗-Nogo受容体-1抗体の産生
本発明の免疫原性Nogo受容体-1ポリペプチドに特異的に結合する抗-Nogo受容体-1 Fab-ファージ抗体も、以下のようにFab-ファージライブラリーをスクリーニングすることによって得た。
【0161】
MorphoSys Fab-ファージライブラリー HuCAL(登録商標)GOLDを、組換えラット可溶性sNogoR310-Fcタンパク質およびラットNogo受容体-1を発現するCOS-7細胞に対してスクリーニングした。Nogo受容体-1に特異的に結合したFab-ファージを精製し、特徴づけた。14D5の重鎖はVH2遺伝子由来であり、軽鎖はVK1遺伝子由来である。これらFab-ファージの1つ、14D5の重鎖および軽鎖のCDRのアミノ酸配列を表4に示す。
【0162】
【表4】
【0163】
14D5は一価および二価の両方の形態においてラットNogo受容体-1に結合する。さらに、14D5はマウスおよびヒトNogo受容体-1およびヒトNogo受容体-2に結合するがマウスNogo受容体-3には結合しない。
【実施例3】
【0164】
抗-Nogo受容体-1モノクローナル抗体によるNogo受容体-1の免疫沈降
免疫沈降を行うために、100μlの溶解した細胞または50μlのPiPLC処理細胞を、400または450μlの抽出バッファー [10 mM Tris-HCl, pH 7.2、0.5% Tween-20、0.2 mM PMSF]またはRIPAバッファーと30μlのタンパク質AまたはGおよび1-2μgの抗体の存在下でそれぞれ混合した。混合物をシェーカーで4℃で16 時間インキュベートした。
【0165】
サンプルを穏やかに遠心してタンパク質AまたはG結合ビーズをペレットとした。ビーズを3回1 ml洗浄バッファー (10 mM Tris-HCl, pH 7.2、0.1% Tween-20)で洗浄した。最終洗浄は10%の元の洗浄バッファーで行った。
【0166】
ビーズを100μlの10% ベータ・メルカプトエタノールを含む2X SDSに再懸濁した。サンプルを室温でインキュベートした後、4-20% Tris-グリシンゲルのSDS-PAGEで泳動した。SDS-PAGEゲル分析によって判定したところ、モノクローナル抗体、3G5および2F7は、Nogo受容体-1を免疫沈降する。
【実施例4】
【0167】
抗体特異性のELISAによる判定
実施例1および2で産生したモノクローナルおよびFab-ファージ抗体の特異性を調べるために、本発明者らはNogo受容体-1ポリペプチドのパネルを用いてELISAを行った。パネルは、sNogoR310-Fc (ラットNogo受容体-1のアミノ酸26-310およびラットFcフラグメントを含む融合タンパク質)、sNogoR344-Fc(前掲)、ポリペプチド p-617 (配列番号1)、ポリペプチド p-618 (ラットNogo受容体-1のLRR7領域からの19-アミノ酸ポリペプチド;図2;配列番号11)およびポリペプチド p-4およびp-5 (それぞれNogo受容体-1のLRR5およびLRRCT領域からのポリペプチド)からなるものであった。卵白アルブミンおよびBSAをコントロールとして用いた。図4に示すように、mAb、1H2、3G5および2F7はすべてsNogoR344-Fcに特異的に結合した。類似の実験において、これら抗体はまた、ラットNogo受容体-1のアミノ酸310-344 (配列番号3)からなるポリペプチドに特異的に結合し、mAb、7E11および5B10はポリペプチド p-617 (配列番号1)に特異的に結合した。
【0168】
sNogoR310-Fc免疫からの10の抗体(1D9.3、1E4.7、1B4.3、2C4.3、1F10.3、2H1.4、1H3.3、1G4.1、1E4.1および2G7.1)は結合について互いに置換し、これはそれらがsNogoR310-Fc上の類似のまたはオーバーラップするエピトープを認識することを示す。sNogoR310-Fc 免疫からの他の3つの抗体(2C4.1、2F11.1および1H4.1)はアミノ酸残基 26-310に位置する異なるエピトープを認識する。
【0169】
本発明者らはまた、Fab-ファージ 14D5を用いてELISA結合アッセイを行った。AP-Nogo66、AP-OM-gpおよびMAG-Fc リガンドを固定化したsNogoR344-Fcに結合させたところ、1μMの14D5が完全にNogoおよびMAG結合を阻害した。10 μMの14D5がOM-gpのsNogoR344-Fcに対する結合の完全な阻害に要求された。
【実施例5】
【0170】
神経突起伸長アッセイ
先に産生したモノクローナルおよびFab-ファージ抗体の神経細胞に対するCNSミエリンの阻害効果を低減させる能力を試験するため、Lab-Tek 培養スライド(4ウェル)を0.l mg/mlのポリ-D-リジン(Sigma)でコーティングした。CNSミエリンまたはPBSを3 μlの滴としてスポットした。蛍光ミクロスフェア(Polysciences)をミエリン/PBSに添加して後の滴の同定を可能にした(Grandpre et al, Nature 403, 2000)。Lab-Tekスライドをすすぎ、次いで10 μg/mlラミニン(Gibco)でコーティングした。P3-4 SDラット子から後根神経節(DRG)を1 mg/ml コラゲナーゼタイプ1 (Worthington)で分離し、先端熱加工パスツールピペットで粉砕し、あらかじめプレートして神経細胞を豊富にし、最終的にはあらかじめコーティングした Lab-Tek 培養スライドに23,000細胞/ウェルにてプレーティングした。培地は5% 熱不活性化ドナーウマ血清、5% 熱不活性化胎児ウシ血清および50 ng/ml mNGF を含有するF12であり、37℃、5% CO2にて6時間インキュベートした。15μg/mlのmAb 7E11をプレーティングの直後に添加した。
【0171】
スライドを20分間、20% スクロースを含む4% パラホルムアルデヒドで固定し、1:500に希釈した神経細胞マーカー抗ベータ-III-チューブリン(Covance TUJ1)で染色した。二次抗体として抗-マウス Alexa Fluor 594 (Molecular Probes)を1:300に希釈し、スライドをGel/Mount(Biomed)でカバースリップした。5x デジタル画像を OpenLabソフトウェアで得てMetaMorphソフトウェアを用いて神経突起伸長を定量分析した。
【0172】
MAb 7E11は神経突起伸長のミエリン-媒介阻害からDRG神経細胞を保護した(図5)。類似の結果がmAb、1H2および3G5によっても観察された。
【0173】
ラットP7 DRG 神経細胞をCNSミエリン基質で培養する神経突起伸長保護アッセイにおいて、二価 14D5も効率的に神経突起伸長を促した。
【実施例6】
【0174】
Nogo受容体-1をトランスフェクトした細胞での7E11による免疫組織化学
実施例1に記載のように産生した抗-Nogo受容体-1 mAbの結合特性をさらに特徴づけるために、本発明者らはラットまたはヒトNogo受容体-1を発現する固定および生COS-7または293細胞への結合を比較した。
【0175】
固定細胞:
Nogo受容体-1でトランスフェクトした細胞および非-トランスフェクト細胞を8-ウェル Lab-Tek培養スライドに播き、4% パラホルムアルデヒドで15分間固定し、PBS中の10% 正常ヤギ血清、0.1% Triton X-100で1時間ブロッキングした。Mab 7E11をブロッキング溶液中15μg/mlおよび1.5μg/mlで添加し、室温で2時間インキュベートした; Alexa-結合二次抗体 抗-マウス(Molecular Probes)をブロッキング溶液中1:300希釈にて1時間インキュベートした; すべての核を標識するためにDAPI を5μg/mlにて二次抗体に添加した。
【0176】
生細胞:
トランスフェクト細胞および非-トランスフェクト細胞を8ウェル Lab-Tek培養スライドに播き、FACSバッファー(4% ドナーウマ血清含有)で30 分間4℃にてブロッキングし、FACS バッファー中15 μg/mlおよび1.5 μg/mlにて7E11と1時間4℃でインキュベートし、すすぎ、二次抗体抗-マウス-Alexa(FACSバッファー中1:300)と30分間4℃でインキュベートした。
【0177】
免疫組織化学染色実験は、すべてのmAbがラットNogo受容体-1を発現する細胞に結合したことを示した。mAb、7E11、2G7.1および2C4.1はヒトNogo受容体-1を発現する固定細胞と生細胞との両方に結合した。
【実施例7】
【0178】
脊髄挫傷のマウスモデル
神経細胞に対するインビボでの実施例1で産生した抗-Nogo受容体-1 mAbの効果を試験するために、本発明者らはマウス脊髄挫傷モデルを用いた。
【0179】
雌性マウス(18-22 g)を鎮痛薬および抗生剤で予防処置した。マウスを麻酔し、立体顕微鏡下で脊柱固定して定位固定装置に入れた。脊髄への外傷を重量ドロップ法(M. Li et al., Functional role and therapeutic implications of neuronal caspase-1 and -3 in a mouse model of traumatic spinal cord injury. Neuroscience Vol. 99, pp. 333-342, 2000)の改変法によって導入した。
【0180】
簡単に説明すると、T9およびT10椎弓切除術を行い脊柱をT9-T10横断方法を両側的に支持する1対のマウス横断クランプを用いて安定化した。直径1.4 mm、重さ2 gのステンレス鋼衝撃棒を、硬膜の2.5 cm上に位置させ、T10レベルにて脊髄に落下させた。手術中、マウスを37℃の毛布上に維持し、手術後に脱水を避けるために1 mlの昇温無菌生理的食塩水をそれぞれのマウスに皮下投与した。膀胱を手作業で反射的膀胱制御が回復するまで1日1回絞った。
【0181】
すべての動物に手術の8-12時間後おきに術後鎮痛薬を与え、その後7日間1日2回抗生物質処理を行った。動物には試験期間餌と水を自由に与えた。抗-Nogo受容体-1抗体を以下のラット脊髄離断モデルにて記載するように28日間くも膜下腔内注射によって損傷部位に送達した。
【実施例8】
【0182】
可溶性Nogo受容体-1融合タンパク質の特徴付け
可溶性Nogo受容体-1ポリペプチド(sNogoR-1)および融合タンパク質(Fc-sNogoR-1)を特徴づけるために、本発明者らは以下の実験を行った。
【0183】
3μgの可溶性Nogo受容体(sNogoR310-FcおよびsNogoR344-Fc)を250 μgのWGA-SPA ビーズに固定化させ、最終容量100 μLの結合バッファー(20 mM HEPES, pH 7.4, 2 mM Ca, 2 mM Mg, 0.1% BSA, 0.1% 卵白アルブミンおよびプロテアーゼ阻害剤)中の0.5 μL の放射性リガンド(終濃度 0.5 nM)を与えた。リガンドには、10 μMのNogo66、10 μMの125I-Nogo40 (NogoAのアミノ酸1-40)が含まれ、各リガンドセットは10 μLの抗-Nogo受容体-1抗体上清を含む。Nogo40上の3つのチロシンを別々にヨウ素化し、それぞれNogo40-A,-Bおよび -Cと命名した。三連の平均値を正規化%結合放射能として示す(図6、7および8)。エラーバーはSEMを示す。阻害剤の非存在下での結合放射能を100%とし、10 μM Nogo40存在下での最小結合放射能を0%としてデータを正規化した。
【実施例9】
【0184】
可溶性Nogo受容体-1融合タンパク質へのリガンド結合の阻害
実施例8の結合アッセイに類似の結合アッセイを用いて実施例1で産生した2つのmAbが125I-Nogo66のsNogoR344-Fcへの結合を阻害する能力を調べた。Mab、2F7および3G5は、125I-Nogo66のsNogoR344-Fcへの結合を阻害した。
【実施例10】
【0185】
神経突起伸長アッセイ
Lab-Tek培養スライド(4ウェル)を0.1 mg/ml ポリ-D-リジン (Sigma)でコーティングした。CNSミエリンを単独でまたはsNogoR310、sNogoR310-Fc 融合タンパク質、mAb 5B10またはコントロールPBSと混合して別々に3μlの滴としてスポットした。蛍光ミクロスフェア(Polysciences)をミエリン/PBSに添加して後の滴の同定を可能とした(Grandpre et al, Nature 403, 2000)。Lab-Tekスライドをすすぎ、10μg/ml ラミニン (Gibco)でコーティングした。
【0186】
P3-4 SDラット子からの後根神経節(DRG)を1 mg/ml コラゲナーゼタイプ1(Worthington)で単離し、先端熱加工パスツールピペットで粉砕し、あらかじめプレーティングして神経細胞を豊富にし、最終的に23,000 細胞/ウェルであらかじめコーティングしたLabtek 培養スライドにプレーティングした。培地は5% 熱不活性化ドナーウマ血清、5% 熱不活性化胎児ウシ血清および50 ng/ml mNGFを含有するF12であり、37℃、5% CO2にて6時間インキュベートした。
【0187】
スライドを20% スクロースを含有する4% パラホルムアルデヒドで20分間固定し、1:500希釈神経細胞マーカー抗ベータ-III-チューブリン(Covance TUJ1)にて染色した。二次抗体として、抗-マウス Alexa Fluor 594 (Molecular Probes)を1:300に希釈し、スライドをGel/Mount(Biomeda)でカバースリップした。5x デジタル画像をOpenLabソフトウェアで得て、MetaMorphソフトウェアを用いて神経突起伸長を定量分析した。
【0188】
sNogoR310、sNogoR310-FcおよびmAb 5B10はすべて神経突起伸長のミエリン-媒介阻害からDRG神経細胞を保護した(図9-11)。sNogoR310を用いてニワトリ神経細胞を用いた類似のアッセイを行ったところ保護することが判明した。
【0189】
本発明者らはまた、ラミニンの存在下および非存在下での細胞成長による実験を行うことにより、可溶性Nogo受容体の神経保護効果を調べた。ラミニン非含有培地での神経細胞成長は悪く、神経細胞ストレス状態のモデルである。
【0190】
DRGを出生後6-7日のラット子(P6-7)から解体し、単一細胞に分離し、上記のようにポリ-D-リジンであらかじめコーティングした96-ウェルプレートに播いた。いくつかのウェルにおいては、2 μg/mlラミニンを2-3 時間添加し、すすいだ後、細胞を播いた。18-20 時間のインキュベーションの後、プレートを4% パラホルムアルデヒドで固定し、1:500希釈のウサギ抗-ベータ-III-チューブリン抗体(Covance)および1:100希釈の抗-HuC/D (Molecular Probes)で染色し、蛍光二次抗体 (Molecular Probes)を1:200希釈にて添加した。ArrayScan II (Cellomics)を用いて5x デジタル画像を得、神経突起伸長をウェルあたりの平均神経突起伸長/神経細胞として神経突起伸長アプリケーションを用いて定量した。3 ウェル/条件からの9つの5x 画像を分析した。
【0191】
いくつかの実験において、PC12細胞(Neuroscreen)のサブクローンを用いた(Cellomics)。Neuroscreen細胞をあらかじめ7日間200 ng/ml NGFによって分化させ、剥離し、ポリ-D-リジンであらかじめコーティングしておいた96-ウェルプレートに播いた。いくつかのウェルにおいては、5 μg/mlのラミニンを2-3 時間添加し、すすぎ、その後細胞を播いた。2日間のインキュベーション後、プレートを4% パラホルムアルデヒドで固定し、1:500希釈ウサギ 抗-ベータ-III-チューブリン抗体(Covance)およびHoechst (核染色)で染色した。ArrayScan IIを用いてDRG 細胞における神経突起伸長を定量した。
【0192】
sNogoR344-FcまたはラットIgGを溶液中でP6-7 DRG 神経細胞および分化Neuroscreen細胞にプレーティング時に添加した。
【0193】
sNogoR344-Fcの神経保護効果をP6 DRG 神経細胞をラミニンの非存在下で培養した際に、1 μMおよび10 μMにて観察した。神経突起伸長の定量は、sNogoR344-Fc添加による用量依存的上昇を示した。ラミニン基質上で培養しているDRG神経細胞への同じ濃度のsNogoR344-Fc の添加では、異常な効果はみられず、sNogoR344-Fcはストレス細胞に対してのみ活性であることが示された。ラミニンの非存在下での同じ濃度のsNogoR344-Fcの神経保護効果もまた、Neuroscreen細胞でみられた。
【実施例11】
【0194】
Fc-sNogoR-1融合タンパク質の産生および精製
ラットNogo受容体-1のアミノ酸1-310をコードするcDNAコンストラクトを哺乳類発現ベクターに含まれるラット IgG1 Fcと融合し、このベクターをチャイニーズハムスター卵巣(CHO) (DG44) 細胞にエレクトロポレーションした。細胞を10% 透析胎児ウシ血清、2 mM グルタミンおよび抗細菌-抗真菌薬を補充したアルファ-MEMで維持した。形質移入の2日後、培養上清を回収し還元条件下でウェスタンブロット分析を行った。約60 kDaのタンパク質バンドをポリクローナルウサギ抗-Nogo受容体-1抗体を用いて検出した。細胞を拡張し、R-PE 結合ヤギ抗-ラット IgG 抗体を用いてソートした。2回目のソートの後、細胞を96-ウェルプレートにて1細胞/ウェルの密度で播いた。個々のウェルからの分泌された可溶性Nogo受容体-1タンパク質レベルをサンドイッチELISAを用いて試験及び比較した。ELISA プレートはヤギ抗-ラット IgG Fcκ特異的抗体でコーティングした。培養上清を入れた。結合可溶性Nogo受容体-1タンパク質をHRP結合ロバ抗-ラット IgG Fab、Fc-特異的抗体で検出した。クローン4C12が最大の分泌レベルであった。4C12を拡張し、撹拌フラスコ中CHO-M7培地で培養した。分泌レベルは37℃で約10 mg/Lであった。
【0195】
sNogoR310-Fc 融合タンパク質を発現するCHO細胞をラージスケールで培養した。1.7 L の濃縮培養上清を10 Lのバイオリアクターランから得た。pHを1/10量の1.0 M Tris-HCl, pH 8.9の添加により上昇させた。固体塩化ナトリウムおよびグリシンをそれぞれ3.0 M および1.5 M添加した。10 mM Tris-HCl、3 M 塩化ナトリウム、1.5 M グリシン, pH 8.9で平衡化した60 mLのタンパク質 A-Sepharoseカラムを調製した。蠕動ポンプを用いて濃縮培養上清をカラムに1.5 mL/分にてアプライした。カラムを300 mLの10 mM Tris-HCl、3 M 塩化ナトリウム、1.5 M グリシン, pH 8.9、次いで120 mL 5 mM Tris-HCl、3 M 塩化ナトリウム, pH 8.9で洗浄した。タンパク質を25 mM リン酸ナトリウム、100 mM 塩化ナトリウム, pH 2.8で溶出した。10 mL フラクションを1.0 mL の1.0 M HEPES, pH 8.5を含む試験管に回収した。タンパク質フラクションをプールし3 x 2 Lの5 mM リン酸ナトリウム、300 mM NaCl, pH 7.4で透析した。
【実施例12】
【0196】
脊髄横切アッセイ
インビボでの機能回復を促進する能力を調べるため、sNogoR-1融合タンパク質をラット脊髄横切アッセイにおいて試験した。
【0197】
Alzet 浸透ポンプを使用日に新たに調製した被験溶液(PBS中sNogoR310-Fc)にロードした。ローディング濃度を計算したところ5および50 μMであった。ポンプは動物への移植の40時間以上前に37℃で開始した。雌性ロングエバンスラットに手術前鎮痛薬および精神安定薬を与え、イソフルラン(O2中3%)を用いて麻酔した。
【0198】
ラットを定位固定枠におき、運動野を路(tract)追跡薬 BDA (10,000 MW)の注入のために両側的に曝した。次いでラットをT5-T6にて脊髄の背部を片側切断し、次いで被験化合物の送達のためにくも膜下腔内カテーテルおよびポンプシステムを移植した(1群あたりn=11)。
【0199】
ラットを回復させ、手術後28日間生存させた。BBBシステムを用いた挙動スコアリングを損傷の導入後28日間記録した後、生存相の試験を終結させた。灌流および固定の後、脊髄を除き、凍結保護し、切片として染色し、軸索を計数した。
【0200】
Basso-Beattie-Bresnahan (BBB) 歩行運動強度スケール(Basso et al., 1996, Neurotrauma 13, 343-359)という、傾斜平面試験および傾斜格子歩行試験 (Li and Strittmatter, 2003, J Neurosci. 2003, 23, 4219-27)を損傷後ラットおよびマウスにてモニターした。傾斜平面試験のために、本発明者らは、50 cm x 60 cm ボードが5秒間マウスが滑り落ちずに傾けることが出来る最大角を測定した。傾斜格子歩行のために、マウスを訓練して45度のスロープの針金格子 (35 cm長、2.54 cm格子)を登らせた。格子平面の下に後肢が落ちる例の数を下から上までの各運動にて測定した。ラット挙動試験のために、BBB歩行運動スケール、格子歩行および足跡分析を行った。格子歩行のために、ラットを針金格子 (70 cm長、2.54 cm格子)上を歩行するように訓練し、格子平面下に後肢が落ちる例数を計数した。足跡分析のために、ラット後肢の歩行パターンを90 cm の路を横切る連続移動運動にわたってインクで記録し、各側のストライド長およびストライド幅を計算した(Metz et al., 2000, Brain Res., 883, 165-177)。これら挙動試験はすべて少なくとも2個体で行った。手術、挙動試験および組織学的分析において、研究者には、ミニポンプ内の化合物が何かを教えなかった。
【0201】
sNogoR310-Fcは機能回復を促進した(図12)。
【実施例13】
【0202】
ラット脊髄挫傷アッセイ
インビボでの神経細胞に対する可溶性Nogo受容体-1ポリペプチドおよび融合タンパク質の効果をラット脊髄挫傷アッセイで試験した。
【0203】
雌性ズキンロングエバンスラット(170-190 g)を鎮痛薬と抗生剤で予防処置した。手術の10分前に、動物を2.5 mg/kgのミダゾラム(腹腔内)で鎮静化し、O2中の2-3% イソフルランで麻酔した。ラットを次いで剪毛し、アルコールとベタジンで拭き、眼球潤滑剤を点眼した。次いで、切開を正中線下で行い、T7−T12の脊椎を曝した。
【0204】
背部椎弓切除術ををT9 1/2およびT10にて行って索を曝した。ラットを衝撃装置に載せた。T7およびT8セグメントをまずクランプし、次いでT11およびT12 セグメントを尾クランプに結合させた。軟材料をラットの胸部の下に敷いた。衝撃棒を0位に合わせ、電気的アースクリップを創傷縁につけた。衝撃棒を25.0 mmに上げ、適宜曝された脊髄の真上になるよう調節した。次ぎに衝撃棒を曝された脊髄に落とし、衝撃棒をすぐに上げた。
【0205】
ラットをおろし、Gelfoam(登録商標)を傷に当てた。傷の上の筋肉を縫合し、切開を外科的に閉じた。動物を麻酔から覚めるまでインキュベーターに入れた。ラットに抗生物質、鎮痛薬および要求に応じて生理的食塩水を与えた。膀胱を機能が回復するまで朝晩絞った。
【0206】
可溶性Nogo受容体-1融合タンパク質(例えば、sNogoR310-Fc)を上記ラット脊髄離断モデルに記載のようにくも膜下腔内投与した。BBBスコアリングを手術の1日後、次いでその後4-6週間、2週間毎に行った。
【実施例14】
【0207】
トランスジェニックマウスにおけるsNogoR310の発現
本発明者らは、インビボで発現させた場合の効果を試験するために、可溶性Nogo受容体-1タンパク質を発現するトランスジェニックマウスを作製した。
【0208】
本発明者らはマウス sNogoR310 cDNA (Nogo受容体-1のアミノ酸1-310に対応)をC-3123 ベクターのNotI 部位にクローニングした。このベクターにおいて、sNogoR310 発現はグリア細胞繊維性酸性タンパク質 (gfap) 遺伝子調節要素の制御下にあり、損傷部位の反応性星状細胞からの分泌を促進し、高レベルの発現が可能である。本発明者らは結果として得られたベクターを順次AatIIおよびSfiIで消化し、3.4 kb フラグメント上にgfap::sNogoR310 コンストラクトを単離した。本発明者らはこのフラグメントを胚にマイクロインジェクションし、トランスジェニックマウスを作製した。本発明者らは、導入遺伝子が組み込まれていることをPCRによって確認し、5つの創始系統を同定した。本発明者らは発現レベルが最も高い2つの創始系統のヘテロ接合性雄と雌性 C57BL/6J マウスとを交配した。本発明者らはNogo受容体-1に対する抗体を用いたウェスタンブロット分析によってGFAP-陽性細胞がヘテロ接合性トランスジェニックマウスにおいてsNogoR310を発現および分泌していることを確認した。
【0209】
本発明者らはプロテアーゼ阻害剤(Roche)を追加したTris-緩衝食塩水中で皮質および脊髄をホモゲナイズし、ホモジネートを40,000 rpmで20分間、4℃で遠心分離した。本発明者らは、上清を4% パラホルムアルデヒドで20分間処理して抗体特異性を強化し、イムノブロッティングの前に透析した。本発明者らは、粒状フラクションをRIPA バッファー (PBS 中1% Triton X-100、0.5% デオキシコール酸ナトリウム、0.1% SDS)中で超音波処理してホモゲナイズし、その結果得られたホモジネートを遠心分離し、上記のようにこの上清(界面活性剤-可溶性粒状フラクション)を処理した。本発明者らは、1:2000希釈のNogo受容体-1に対するウサギ抗血清を用いたイムノブロットによって20 μgの脳または脊髄タンパク質を分析した。本発明者らは、免疫反応性をAP-結合抗-ウサギ IgGおよびNBT/BCIP AP 基質とのインキュベーションによって可視化した。
【0210】
本発明者らは、2つのトランスジェニック系統、Tg08およびTg01からの皮質および脊髄の界面活性剤-フリー可溶性抽出物において分泌された37 kDa sNogoR310を検出したが、同腹仔野生型(WT)マウスにおいてはあるにしてもわずかしか、可溶性Nogo受容体-1 タンパク質は37または81 kDaにて存在しなかった。粒状フラクションを調べたところ、WTとトランスジェニックマウスの両方において内在性Nogo受容体-1が同レベル存在することが確認された。
【実施例15】
【0211】
損傷後のトランスジェニックマウスにおけるsNogoR310の発現
本発明者らは背部上-片側切断損傷を行うことにより、トランスジェニックマウスにおけるsNogoR310 発現に対するCNS損傷の効果を調べた。本発明者らは、ヘテロ接合性雄とC57/BL6 雌とを実施例14に記載のように交配することにより、sNogoR310 トランスジェニックおよび非トランスジェニックコントロール動物を得た。
【0212】
本発明者らは、成体雌性ヘテロ接合性トランスジェニックまたは同腹仔WTマウス (10-16週齡)を深く麻酔し、完全椎弓切除術を行い、脊髄のT6およびT7 レベルの背部を完全に曝した。本発明者らは、背部上-片側切断をT6にて30-ゲージ針および1対の小型はさみを用いて行い、完全に背部および背外側皮質脊髄路(CST)を切断した。本発明者らは、脊髄の背部に数回印をした針を通し、損傷の深さが1.0 mmであることを確かめた。本発明者らは椎弓切除術上の筋肉層を縫合し、手術用とじ金で背中の皮膚を閉じた。皮質脊髄路を追跡するために、本発明者らは、脊髄損傷の14日後に頭蓋骨内の右側脳皮質に刻み孔を開けた。本発明者らはトレーサーBDA (PBS 中10% 、MW 10,000) (Molecular Probes, Eugene, OR)を皮質表面から0.7 mm の深さにて4つの注射部位に適用した。損傷の4週間後、マウスを心臓を介してPBS、次いで4% パラホルムアルデヒドで灌流した。sNogoR310 発現実験に用いたマウスはトレーサー注射を受けなかった。
【0213】
ウェスタンブロット分析に用いるマウスについてT3〜L3レベルの脊髄を灌流せずに損傷の14日後に回収した。Nogo受容体-1 免疫組織化学染色に用いるマウスは片側切断の10日後に4% パラホルムアルデヒドで灌流し、損傷脊髄を切片とするために取り出した。トランスジェニックおよびWTマウスの損傷した脳におけるsNogoR310 発現を調べるために、定位固定装置(David Kopf, Tujunga, CA)に備えた11番手術用メス刃によって皮質穿刺損傷を行った。十字縫合の0.5 mm 後側、正中線から1.5 mm 側方、3.5 mm 深さの4 mm 傍矢状切断部を作った。
【0214】
本発明者らは、トランスジェニックマウスにおいて損傷の10日後に脊髄の可溶性抽出物におけるsNogoR310レベルの上昇を検出したが、WTマウスでは検出されず、これは損傷のまわりのGFAPの損傷後の上方制御と一致する。これがNogo-Aの代償的上方制御に起因するのではないことを確認するために、本発明者らはその発現を試験し、WTおよびトランスジェニックマウスからの無傷または損傷の皮質および脊髄のいずれにおいても同様であることを見いだした。
【0215】
本発明者らはNogo受容体-1およびGFAPに対する抗体による、損傷領域を含む損傷脳および脊髄の免疫染色により、損傷CNSにおけるsNogoR310の細胞性発現を調べた。反応性アストロサイトグリアの概ねの形態はWTとトランスジェニックマウスで異なることはなかったが、細胞内および細胞外空間の両方におけるNogo受容体-1について染色された密度はWTマウスと比較してgfap::sNogoR310 トランスジェニックマウスの方が顕著に高く、これは、トランスジェニックマウスにおける損傷のまわりのsNogoR310 発現の上昇を示す。トランスジェニックマウスにおいてのみ、Nogo受容体-1 タンパク質はアストロサイトマーカーGFAPと共に局在していた。トランスジェニックサンプルにおいて、散在性非細胞性染色が強くなっていたが、これは細胞外空間におけるsNogoR310と一致する。神経細胞の細胞体 Nogo受容体-1染色はWTとトランスジェニックマウスの両方で検出された。
【実施例16】
【0216】
トランスジェニックマウスにおいて分泌sNogoR310はCST出芽を誘導する
本発明者らはトランスジェニックマウスの損傷周囲のsNogoR310の発現上昇が損傷軸索の再生をもたらすかを試験した。
【0217】
本発明者らは、下行皮質脊髄路(CST)の統合性を、順行性トレーサー、ビオチンデキストランアミン(BDA) を右運動野に、Li and Strittmatter, 2003, J. Neurosci., 23, 4219-27に記載のように注入することによって調べた。同腹仔WTマウスにおいて、明らかな背部CST(dCST)が損傷の頭側で緊密に束になっており、数本の背外側CST繊維が同側方に観察された。少数のBDA-標識した短い側枝出芽が灰白質、特に前索における灰白質に突出したが、出芽は主にトレーサー注射の反対側の索にて確認された。しかし、損傷sNogoR310 トランスジェニックマウスからの背部片側切断の頭側の切片は、まったく異なるBDA 標識パターンを示した。高密度のBDA-標識CST線維が系統Tg08または系統Tg01からのすべてのトランスジェニックマウスにおける突出したdCSTの外側にみられた。異所性の線維が灰白質領域にわたって伸長しており、いくらかの線維は、外側および背外側白質に到達していた。いくつかの線維(横断切片あたり4-12の出芽)が脊髄の逆側にみられた(トレーサー注入部位の同側)。側枝出芽のマイクロデンシトメトリー測定は、sNogoR310 トランスジェニックマウスにおけるおよそ10倍の出芽密度の上昇を示した。損傷の1〜4 mm 頭側からの傍矢状長軸方向切片を調べたところ、sNogoR310 トランスジェニックマウスにおいてdCST線維が腹側灰白質領域に分岐する出芽を多数伸長しており、これは同腹仔WT動物ではみられなかった。一般にトランスジェニックマウスにおける損傷の頭側の出芽のパターンおよび程度は、Nogo受容体-1アンタゴニストペプチド NEP1-40で全身的に処置されたマウスにおいて観察されるものと同様であった(Li and Strittmatter, 2003)。
【0218】
これらの結果は分泌sNogoR310がトランスジェニックマウスにおけるCST出芽を誘導することを示す。
【実施例17】
【0219】
sNogoR310 トランスジェニックマウスにおいて再生するCST軸索は損傷部位を迂回して遠位脊髄に向かう
本発明者らはトランスジェニックマウスから損傷部位(8 mm全長)に対して4 mm 頭側、4 mm 尾側の脊髄を単離し、それをグルタルアルデヒド-重合アルブミンマトリックスに包埋し、ビブラトーム上で傍矢状に切断した(30μm厚)。本発明者らは損傷部位から5-7 mm 頭側、5-7 mm 尾側の脊髄から横断切片(50 μm)を集めた。ラットにおけるsNogoR310-Fc 注射実験のために、損傷部位の10 mm 頭側から10 mm 尾側に伸長する脊髄を振動マイクロトーム上で傍矢状に(50 μm)切断した。横断切片を損傷部位の11-16 mm 頭側、11-16 mm 尾側の脊髄から集めた。本発明者らは切片をアビジン-ビオチン-ペルオキシダーゼ複合体とインキュベートし、ニッケル促進化ジアミノベンジジンHRP反応によりBDAトレーサーを可視化した(Grandpre, 2002, Nature, 417, 547-551)。本発明者らは、間接的免疫蛍光による、セロトニン免疫組織化学(抗-5-HT 抗体)のためにいくつかの切片を加工した。損傷領域を可視化するために、本発明者らはGFAPに対する抗体を用いていくらかの切片を二重染色した(Sigma, St. Louis, MO)。 本発明者らは切片を標本にし、脱水し、封入剤で被覆した。
【0220】
本発明者らは損傷後にトランスジェニックマウスにおいて発現したsNogoR310によって誘導された線維が、機能回復させるために損傷領域を横切って尾脊髄へと向かうかを調べた(実施例16参照)。
【0221】
明箱に入れた損傷部位を横切る連続的傍矢状切片は、再生CST繊維の損傷から数ミリの全体的な分布パターンを示した。WTマウスからの切片には損傷部位の上に伸長するCST線維はみられなかった。sNogoR310 トランスジェニックマウスからの類似の切片は、横切領域を横切り、遠位灰白質および白質領域に高度に分岐したパターンにて突出する多くのCST線維を示した。片側切断に近接する頭側に、高密度のBDA-標識CST出芽が損傷領域へと突出するdCSTから生じたが、ほとんどのCST出芽は瘢痕形成および組織空洞化が顕著である横切領域を横切ることはできなかった。少ないがかなり有意な部分の再生軸索が腹側および腹外側灰白質および白質の残っている組織架橋を介して損傷部位を迂回していた。さらに、いくらかのCST線維は損傷背部および背外側脊髄を介して横切領域自体を横切って遠位領域へ向かっているようであった。損傷の近傍においては、再生する線維の経路は典型的には蛇行状であり、頭側のCSTにおける正常に直線上の線維とはかなり異なっていた。側枝および分岐した線維は遠位脊髄の灰白質領域にもっとも頻繁にみられた。再構築は、各トランスジェニックマウスにおける損傷の1-4 mm 尾側のあらゆるレベルにおいて頭側-尾側軸を経路とする5-15 BDA-標識化再生線維を示した。背部片側切断の5-7 mm 尾側の横断切片については、BDA-標識CST軸索が各トランスジェニックマウスにおいて灰白質と白質領域の両方においてみられた。トランスジェニックマウスの線維数は、矢状切片の近位レベルと類似の数のBDA-標識CST線維を示した。
【0222】
CST線維に加えて、その他の下行路、例えば、縫線核脊髄線維もマウスの歩行運動機能に寄与している。このマウス背部上-片側切断モデルにおいて、横切は大部分のセロトニン作動性線維を損傷し、これら線維密度は、腹側角において約80%減少していた。尾脊髄の腹側角におけるセロトニン線維の全長の分析は、WT群と比較してトランスジェニックマウスにおける方がかなり多数のかかる線維を示し、これはトランスジェニックマウスにおけるsNogoR310の成長促進効果が、軸索下行性経路に限定されないことを示す。
【実施例18】
【0223】
sNogoR310のトランスジェニック発現は歩行運動回復を向上させる
CST軸索追跡およびセロトニン作動性線維分析は、トランスジェニックマウスにおける星状細胞から放出されるsNogoR310が、脊髄における損傷下行性軸索の広範な解剖学的再生を刺激することを示した。本発明者らは実施例12に記載のいくつかの挙動試験を行って、かかる再生線維が機能回復に有益であるかを調べた。
【0224】
BBB試験によりアッセイしたところ、WTマウスは生存の4週間の間に部分的に歩行運動機能を回復した。損傷の4週間後には、ほとんどのWTマウスは体重を支えられる一貫した足底歩進によって特徴づけられるレベルに回復したが、時折しか前肢-後肢協調を示さず、表面に最初に接触させると片側の肢が回転した。一方、系統Tg08およびTg 01の両方からのsNogoR310 トランスジェニックマウスのBBBスコアは観察の7-28日間にわたってコントロール群より有意に高かった(図13Aおよび13B)。損傷の28日後には、ほとんどのトランスジェニックマウスが一貫した前肢-後肢協調を示し、片側の肢の位置は体と平行であった。
【0225】
本発明者らは2つのさらなる挙動試験を行ってsNogoR310 トランスジェニックマウスの性能を特徴づけた。まず、本発明者らはマウスが握力を5秒間は失うことなくボードを傾斜させられる最大角を測定した。背部片側切断損傷の前に、トランスジェニックおよびWTマウスの両方は55度に傾けたボードにて姿勢を維持することが出来た。損傷の7-28日後、維持可能な角度はすべてのマウスにおいて低下したが、トランスジェニックマウスによって維持可能な角度はコントロール群より有意に高かった(図13C)。もう1つの挙動試験において、マウスを垂直線に対して45度傾斜させた格子を登らせて格子平面下に落ちる後肢運動を計数した(Metz et al., 2000)。損傷前の訓練においてはいずれのマウスもこの試験でエラーを起こさなかった。損傷後の2-6週間においてはWTマウスにおいては多数の肢欠陥エラーがみられほとんど改善されなかった。一方、sNogoR310 トランスジェニックマウスはこの期間にわたって格子登上が徐々に改善し、損傷後1-3週間後に改善の大部分が起こった(図13D)。したがって、星状細胞からsNogoR310を分泌するトランスジェニックマウスは、CST再生、縫線核脊髄出芽および胸髄片側切断後の運動機能の改善を示した。
【実施例19】
【0226】
sNogoR310-Fcタンパク質のくも膜下腔内投与はCST出芽を誘導する
脊髄損傷後の可溶性Nogo受容体-1の成長促進性の効果の第二の試験として、本発明者らは精製したタンパク質をくも膜下腔内投与した。
【0227】
本発明者らはラットNogo受容体-1のリガンド結合ドメイン(27-310)とラット IgG1 Fc ドメインを融合させ安定性および精製を向上させた。本発明者らは安定にトランスフェクトしたCHO細胞からタンパク質を精製した。このタンパク質はNogo-66、MAGおよびミエリン作用をインビトロでブロックするということが以前のマウス sNogoR310-Myc His についての研究で示されている(Fournier et al., 2002, J Neurosci., 22, 8876-8883; Liu et al., 2002, Science, 297, 1190-1193)。 本発明者らは、sNogoR310-Fc タンパク質を浸透ミニポンプを用いて胸部背部中央片側切断損傷を有するラットのくも膜下腔内に送達した。損傷後4週間の生存期間において、1.2 mgのsNogoR310-Fc タンパク質を各ラットに局所投与した。媒体処置(1.2 mg ラット IgG)を受けたラットにおいて、片側切断の頭側の切片は緊密に束になった顕著な背部CSTを示し、損傷部位上の異所性BDA-標識CST線維は非常にわずかであった。sNogoR310-Fc タンパク質を与えられた損傷ラットからの損傷の頭側の切片は、全く異なる標識パターンを示した。BDA-標識CSTから出芽した多数の異所性の線維が横断および傍矢状切片から観察された。いくつかの場合には、突出は正中線近くのdCST領域を横切って索の外周に伸びており、背外側CSTと混ざっていた。出芽軸索は白質よりも灰白質に多く伸びていた。dCST近くの異所性出芽線維の測定(横断切片において100 μm以上、矢状切片において200 μm以上)によりsNogoR310-Fc-処理ラットにおける方がより多いことが明らかとなった。
【実施例20】
【0228】
sNogoR310-Fc処理ラットにおいてCST軸索は遠位脊髄に向かって再生する
本発明者らは、雌性SDラット(190-250 g)を深く麻酔し、脊髄レベルT6-7において椎弓切除術を行って脊髄を露出させた。本発明者らは脊髄の背部半分を30-ゲージ針と1対の小型はさみで切断してCSGT路の背部を分断し、損傷の深さ(1.8 mm)を11番メスの鋭利部分を索の背部半分に通すことにより確認した(Grandpre et al., 2002, Nature, 417, 547-551)。PBS 中の1.2 mg ラット IgGまたはPBS中の1.2 mg sNogoR310-Fc 融合タンパク質で満たした浸透ミニポンプ (Alzet 2ML4, 2 ml 容量、2.5 μl/時、28日間送達)を動物の背の筋肉下に縫合した。ミニポンプの出口に連結したカテーテルを硬膜の小孔を介して脊髄のT7-8 レベルにてくも膜下腔内空間に挿入した。
【0229】
Nogo受容体-1アンタゴニストタンパク質の注入は広範なラット片側切断の頭側の出芽を誘導したが、より重要なことは遠位脊髄に突出した出芽CST線維が歩行運動回復に寄与するか否かである。媒体-処理ラットからの損傷部位を横切る長軸方向切片では、BDA-標識腹側CST線維が検出不可能であるか損傷レベルを下回る非常に少量であった(GrandPre et al., 2002; Weidner et al., 2001, Proc. Natl. Acad. Sci. USA, 98, 3513-3518)。sNogoR310-Fc 処理ラットからの類似の切片においては、多くのBDA-標識線維が横切部位を迂回して主に腹側および腹外側脊髄の架橋組織を介して尾脊髄に突出していることが示された。アストロサイトマーカーGFAPの免疫染色により、横切の程度が中心管領域より深く達していることが示された。突出した背部CSTにおける頭側の線維が直線上であるのと異なり、再生CST線維は通常、遠位脊髄、特に灰白質領域において高度に分岐した軌道にしたがった。かかる線維は脊髄の多くの領域で観察されたが、脊髄を通じて脊髄の中央部および背部半分においてより容易に観察された。矢状切片からのCST線維数は各sNogoR310-Fc-処理ラットにおいて、損傷の尾側1-2 mmにおいておよそ20のBDA-標識軸索、そして損傷の遠位7-8 mmにおいて15の追跡軸索を示した。
【0230】
一般にこれら線維の分岐パターンは局所NEP 1-40 ペプチド処理動物のものと類似しているが、各出芽においてより多くの側枝分岐がsNogoR310-Fc タンパク質で処理した切片から観察される。遠位脊髄からの出芽の測定により、sNogoR310-Fc-処理ラットにおける各出芽の側枝全長はNEP 1-40-処理動物の2倍多いことが示された。両Nogo受容体-1 アンタゴニスト-処理群における脊髄の尾側1-10 mmにおける出芽数(長さ200 μm以上)はコントロール群と比較しておよそ20-40倍多かった。sNogoR310-Fc処理ラットからの方が局所NEP 1-40 処理からよりも多くの出芽が観察されたが(〜50対25 出芽/ラット)、この差は統計的に有意ではなかった(p=0.1713, t-検定)。
【0231】
再生CST軸索がsNogoR310-Fc 処理を受けたラットの片側切断の11-15 mm 尾側の脊髄の横断切片において観察された。これら線維は脊髄の灰白質と白質の両方で観察された。灰白質において検出された線維はしばしば白質領域においてより多くの側枝分岐を示した。一方、媒体処理群からの横断切片では、時折のみBDA-標識が腹側白質領域にみられ、これは無傷腹側CST軸索と一致する。このレベルの遠位脊髄において、Nogo受容体-1 アンタゴニスト-処理群[sNogoR310-FcおよびNEP 1-40]からのBDA-標識CST線維の平均数は、媒体-処理ラットのおよそ20倍であった。即ち、両方Nogo受容体アンタゴニストである、sNogoR310-Fc タンパク質およびNEP 1-40ペプチドにより、遠位脊髄において多くのCST軸索が再生したが、前者によって誘導される出芽の方がより高度な分岐パターンを示した。
【実施例21】
【0232】
局所sNogoR310-Fcは損傷ラット脊髄における赤核脊髄(Rubropinal)およびセロトニン作動性軸索の出芽を誘導する
片側切断の14日後、CST線維を追跡するために下肢の感覚野に横わたる頭蓋骨の各側面に刻み孔を開けた。順行性神経細胞トレーサーBDA(PBS中10%、皮質あたり3.5 μl)を各側面の硬膜から1.5 mm の深さの7つの注入部位に与えた(Grandpre, 2002)。ラットにおける赤核脊髄路追跡のために、トレーサーBDA (1 μl; MW 10,000; PBS 中10%)を左側の赤核に注射した(十字縫合から5.8 mm 後側、頭蓋骨表面から0.7 mm 外側、7.0 mm 腹側)。BDA注射の2週間後、これら動物をPBS、次いで4% パラホルムアルデヒドで灌流し、組織を組織化学のために収集した。
【0233】
損傷赤核脊髄路(RST)線維の修復は脊髄損傷後の機能向上に寄与する(Liu et al., 1999, J. Neurosci., 19, 4370-4387)。CNS 神経細胞における広範なNogo受容体-1の分布(Wang et al., 2002, J. Neurosci., 22, 5505-5515)により、アンタゴニストによるNogo受容体-1の阻害の結果、損傷後のRST 軸索の再生を導くことが可能となる。sNogoR310-Fcの損傷RSTへの効果を調べるために、この経路の統合性を左赤核にBDAを注射することにより追跡した。脊髄レベルにおいて、RST線維は脊髄の背外側白質領域に通常位置し、この研究の背部片側切断によって横断される。コントロールラットからの損傷の11-15 mm 頭側の横断切片において、少量の短いBDA-標識線維が突出RSTと背部角灰白質の間にみられた。sNogoR310-Fcで処理された同レベルの切片は主要RSTと背部角灰白質の間の多くの連結する線維を示した。媒体-処理ラットのSCIの11-15 mm 遠位の横断切片ではBDA-標識RST線維はみられなかった。一方、同レベルのsNogoR310-Fc処理を受けた切片は多くのBDA-標識RST 線維をトレーサー注射の反対側の灰白質と白質の両方で示した。分岐パターンを示すいくつかの出芽がBDA注射の同側の灰白質にみられた。
【0234】
縫線核脊髄線維もsNogoR310-Fc処理脊髄損傷ラットにおいて調べた。免疫染色により、損傷の11-15 mm頭側のセロトニン作動性線維密度は媒体処理群とsNogoR310-Fc処理群で同様であることが示された。損傷の11-15 mm 下の切片においては、sNogoR310-Fc処理ラットにおけるセロトニン線維はコントロール群の2倍多かった。これらの結果はsNogoR310-Fc タンパク質によるNogo受容体-1阻害への応答性はCST線維に限定されず、その他の下行路、例えば、赤核脊髄およびセロトニン作動性軸索もまたNogo受容体-1アンタゴニズムに応答性であることを示した。
【実施例22】
【0235】
sNogoR310-Fcによる局所処理はラットにおける機能回復を改善する
sNogoR310-Fc タンパク質のくも膜下腔内は、脊髄損傷後にいくつかの下行路における軸索再生を刺激する。本発明者らは、該タンパク質が損傷脊髄における機能回復も向上させるかどうかを調べた。
【0236】
片側切断の2週間後、媒体-処理ラットにおける歩行運動BBBスコアは安定レベル、12に達した(図14A)。損傷の4週間後、ほとんどのコントロール(7匹のうち6匹)は、頻度が一定の体重を支持する足底歩行および頻度が一定の前肢-後肢協調を示したが、表面と接触した際に片方の肢位置が回転した。一方、sNogoR310-Fc タンパク質処理を受けたラットにおいては、歩行運動スコアは損傷後2-4週間向上し続けた。損傷の4週間後、9匹全てのsNogoR310-Fc 処理動物は一定した前肢-後肢協調を示し、試験表面と接触した際、平行な肢位置であった。
【0237】
格子歩行を用いて脊髄損傷後の下行性の細かい動作の制御の欠陥を評価した(Metz et al., 2000)。格子歩行を行うためには、前肢-後肢協調および腹外側、皮質脊髄および赤核脊髄線維に媒介される随意運動制御が必要である。損傷前の訓練において、すべてのラットは格子バーのうえにその後肢を正確においた。損傷の2-4週間後、コントロールラットはセッションあたり8-9のエラーをおこし、その期間にわたって改善されなかった。一方、sNogoR310-Fcで処理したラットは格子歩行において徐々に向上し、エラーは有意に少なかった(平均4-7/セッション)。この向上のほとんどは損傷の2-3週間後に起こった。コントロール群における後肢足跡の分析により、片側切断の4週間後において、無傷ラットまたはsNogoR310-Fc処理を受けた損傷動物と比較してストライド長が有意に減少し、ストライド幅は増加していた(図14C)。それゆえ、これらの複数の挙動試験によりアンタゴニストタンパク質の局所注射によるNogo受容体-1機能のブロックが損傷後の歩行運動回復を促進することが示された。
【0238】
生物寄託
ハイブリドーマ、HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585)をAmerican Type Culture Collection (ATCC), 10801 University Boulevard, Manassas, VA 20110-2209, USA, on August 9, 2002に寄託した。
【0239】
当業者であれば理解するように、様々な変更および改変が本発明の範囲を逸脱することなく本発明の好ましい態様に対して行うことが出来る。かかる改変はすべて本発明の範囲に含まれる。
【0240】
本発明の具体的態様を示す:
(1)配列番号1、配列番号2、配列番号3、配列番号4および配列番号5からなる群から選択されるポリペプチド、
(2)項目(1)のポリペプチドをコードする核酸、
(3)発現制御配列に作動可能に連結した項目(2)の核酸、
(4)項目(2)または(3)の核酸を含むベクター、
(5)項目(2)または(3)の核酸または項目(4)のベクターを含む宿主細胞、
(6)以下の工程を含む項目(1)のポリペプチドの産生方法:
(a) 項目(5)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程、
(7)以下の工程を含む抗体の産生方法:
(a) 項目(1)のポリペプチドまたは項目(5)の宿主細胞で宿主を免疫する工程;および、(b)抗体を回収する工程、
(8)項目(7)の方法によって産生される抗体または該抗体の抗原結合性フラグメント、
(9)項目(1)のポリペプチドに特異的に結合する抗体またはその抗原結合性フラグメント、
(10)抗体が、
(a) 神経細胞の成長円錐崩壊を阻害し;
(b) 神経細胞における神経突起伸長および出芽の阻害を減少させ;そして、
(c) リガンドに対するNogo受容体-1の結合を阻害する、
項目(8)または(9)の抗体または抗原結合性フラグメント、
(11)神経突起伸長および出芽が軸索成長である、項目(10)の抗体または抗原結合性フラグメント、
(12)神経細胞が中枢神経系神経細胞である、項目(10)の抗体または抗原結合性フラグメント、
(13)抗体がモノクローナル抗体である項目(8)または(9)の抗体または抗原結合性フラグメント、
(14)抗体がマウス抗体である項目(8)または(9)の抗体または抗原結合性フラグメント、
(15)抗体がヒト化抗体、キメラ抗体および一本鎖抗体からなる群から選択される項目(8)または(9)の抗体、
(16)HB 7E11 (ATCC受託番号、PTA-4587)、HB 1H2 (ATCC受託番号、PTA-4584)、HB 3G5 (ATCC受託番号、PTA-4586)、HB 5B10 (ATCC受託番号、PTA-4588)およびHB 2F7 (ATCC受託番号、PTA-4585) からなる群から選択されるハイブリドーマ細胞株、
(17)項目(16)のハイブリドーマ細胞株によって産生される抗体またはその抗原結合性フラグメント、
(18)項目(17)の抗体の、項目(1)のポリペプチドまたはNogo受容体-1への結合を競合的に阻害する抗体またはその抗原結合性フラグメント、
(19)Nogo受容体-1と項目(10)または(17)の抗体または抗原結合性フラグメントとを接触させる工程を含む、リガンドに対するNogo受容体-1結合を阻害する方法、
(20)リガンドがNogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される、項目(19)の方法、
(21)神経細胞と項目(10)または(17)の抗体またはその抗原結合性フラグメントとを接触させる工程を含む、神経細胞における成長円錐崩壊を阻害する方法、
(22)神経細胞と項目(10)または(17)の抗体またはその抗原結合性フラグメントとを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法、
(23)神経突起伸長または出芽が軸索成長である項目(22)の方法、
(24)神経細胞が中枢神経神経細胞である項目(21)または(22)の方法、
(25)Nogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる可溶性Nogo受容体-1ポリペプチド、
(26)シグナル配列に連結した項目(25)の可溶性Nogo受容体-1ポリペプチド、
(27)LRRが異種LRRを含む項目(25)または(26)の可溶性Nogo受容体-1ポリペプチド、
(28)配列番号6のアミノ酸残基 26-344、配列番号7のアミノ酸残基 26-310、配列番号8のアミノ酸残基 26-344、配列番号9のアミノ酸残基 26-310、配列番号8のアミノ酸残基 27-344および配列番号9のアミノ酸残基 27-310からなる群から選択される可溶性Nogo受容体-1ポリペプチド、
(29)項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドをコードする核酸、
(30)発現制御配列に作動可能に連結した項目(28)の核酸、
(31)項目(29)または(30)の核酸を含むベクター、
(32)項目(29)または(30)の核酸または項目(31)のベクターを含む宿主細胞、
(33)以下の工程を含む項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドの産生方法:
(a) 項目(32)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からポリペプチドを回収する工程、
(34)可溶性Nogo受容体-1ポリペプチドおよび異種ポリペプチドを含むNogo受容体-1融合タンパク質、
(35)可溶性Nogo受容体-1ポリペプチドがNogo受容体-1のN-末端ドメイン(NT)、8ロイシンリッチリピートドメイン(LRR)およびLRR C-末端ドメイン(LRRCT)から実質的になる、項目(34)のNogo受容体-1融合タンパク質、
(36)シグナル配列に連結した項目(35)のNogo受容体-1融合タンパク質、
(37)LRRが異種LRRを含む項目(34)または(35)のNogo受容体-1融合タンパク質、
(38)可溶性Nogo受容体-1ポリペプチドが項目(25)−(28)のいずれかのポリペプチドを含む、項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(39)異種ポリペプチドが免疫グロブリン定常領域を含む項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(40)免疫グロブリン定常領域が免疫グロブリン重鎖定常領域である項目(39)のNogo受容体-1融合タンパク質、
(41)免疫グロブリン重鎖定常領域がIgG 重鎖定常領域である項目(40)のNogo受容体-1融合タンパク質、
(42)異種ポリペプチドがFcである項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質、
(43)Nogo受容体-1融合タンパク質がダイマーである項目(34)−(37のいずれかのNogo受容体-1融合タンパク質、
(44)項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質をコードする核酸、
(45)発現制御配列に作動可能に連結した項目(44)の核酸、
(46)項目(44)または(45)の核酸を含むベクター、
(47)項目(44)または(45)の核酸または項目(46)のベクターを含む宿主細胞、
(48)以下の工程を含む項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質の産生方法:
(a) 項目(47)の宿主細胞を培養する工程;および、
(b) 宿主細胞または培地からNogo受容体-1ポリペプチドを回収する工程、
(49)リガンドと項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、リガンドに対するNogo受容体-1の結合を阻害する方法、
(50)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、Nogo受容体-1リガンドの活性を調節する方法、
(51)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における成長円錐崩壊の阻害方法、
(52)Nogo受容体-1リガンドと項目(25)−(28)のいずれかのポリペプチドまたは項目(34)−(37)のいずれかのNogo受容体-1融合タンパク質とを接触させる工程を含む、神経細胞における神経突起伸長または出芽の阻害を減少させる方法、
(53)リガンドがNogoA、NogoB、NogoC、MAGおよびOM-gpからなる群から選択される項目(49)−(52)のいずれかの方法、
(54)神経突起伸長または出芽が軸索成長である項目(52)の方法、
(55)神経細胞がCNS神経細胞である項目(51)または(52)の方法、
(56)医薬上許容される担体および以下から選択される成分を含む組成物:
(a) 項目(8)、(9)および(16)のいずれかの抗体または抗原結合性フラグメント;
(b) 項目(25)−(28)のいずれかの可溶性Nogo受容体-1ポリペプチド;および、
(c) 項目(34)−(37)のいずれかの融合タンパク質、
(57)さらに1または複数のさらなる治療薬を含む項目(56)の組成物、
(58)神経細胞と有効量の以下の(a)または(b)とを接触させる工程を含む死の危険にある神経細胞の生存を促進する方法:
(a)抗-Nogo受容体-1抗体または抗原結合性フラグメント;または、
(b)可溶性Nogo受容体-1ポリペプチド、
(59)神経細胞がインビトロである項目(58)の方法、
(60)可溶性Nogo受容体-1ポリペプチドが融合タンパク質である項目(58)の方法、
(61)融合タンパク質がFc 融合タンパク質である項目(60)の方法、
(62)Fc 融合タンパク質がIg-sNogoR344である項目(61)の方法、
(63)神経細胞が哺乳類におけるものである項目(58)の方法、
(64)哺乳類が多発性硬化症、ALS、ハンチントン病、アルツハイマー病、パーキンソン病、糖尿病性神経障害、脳卒中、外傷性脳障害または脊髄損傷の徴候または症状を示す項目(63)の方法、
(65)神経細胞が死の危険にある、以下の工程を含む哺乳類における神経細胞の生存を促進する方法:
(a)以下の(i)または(ii)を発現する培養宿主細胞を提供する工程、
(i)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または、
(ii)可溶性Nogo受容体-1ポリペプチド;および、
(b) 神経細胞においてまたはその近くの部位にて哺乳類に該宿主細胞を導入する工程、
(66)神経細胞が哺乳類におけるものである、以下の工程を含む死の危険にある神経細胞の生存を促進する遺伝子治療方法:
以下の(a)または(b)をコードするヌクレオチド配列を含むウイルスベクターを神経細胞においてまたはその近くの部位にて投与する工程、
(a)抗-Nogo受容体-1抗体またはその抗原結合性フラグメント;または、
(b)可溶性Nogo受容体-1ポリペプチド、
ここで、抗-Nogo受容体-1抗体、抗原結合性フラグメントまたは可溶性Nogo受容体-1ポリペプチドは哺乳類において神経細胞の生存を促進するのに十分な量にてヌクレオチド配列から発現する、
(67)Nogo受容体-1に特異的に結合する抗体またはその抗原結合性フラグメントであって、該抗体またはフラグメントが以下からなる群から選択されるアミノ酸配列を含む軽鎖を含む抗体またはその抗原結合性フラグメント:
(a) 配列番号15のアミノ酸配列;
(b) 配列番号16のアミノ酸配列;および、
(c) 配列番号22、23および24のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(68)さらに以下からなる群から選択されるアミノ酸配列を含む重鎖を含む項目(67)の抗体または抗原結合性フラグメント:
(a) 配列番号17のアミノ酸配列;
(b) 配列番号18のアミノ酸配列;および、
(c) 配列番号19、20および21のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(69)Nogo受容体-1に特異的に結合する抗体またはその抗原結合性フラグメントであって該抗体またはフラグメントが以下からなる群から選択されるアミノ酸配列を含む重鎖を含む抗体またはその抗原結合性フラグメント:
(a) 配列番号17のアミノ酸配列;
(b) 配列番号18のアミノ酸配列;および、
(c) 配列番号19、20および21のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列、
(70)さらに以下からなる群から選択されるアミノ酸配列を含む軽鎖を含む項目(69)の抗体または抗原結合性フラグメント:
(a) 配列番号15のアミノ酸配列;
(b) 配列番号16のアミノ酸配列;および、
(c) 配列番号22、23および24のCDR1、CDR2およびCDR3 アミノ酸配列を含むアミノ酸配列。
【特許請求の範囲】
【請求項1】
以下からなる群から選択される単離可溶性Nogo受容体-1 ポリペプチド: 配列番号6のアミノ酸 26-344; 配列番号7のアミノ酸 26-310; 配列番号8のアミノ酸 26-344; 配列番号9のアミノ酸 26-310; 配列番号8のアミノ酸 27-344 ;および配列番号9のアミノ酸 27-310;ここで、該可溶性Nogo受容体-1 ポリペプチドは神経突起伸長阻害を阻害する。
【請求項1】
以下からなる群から選択される単離可溶性Nogo受容体-1 ポリペプチド: 配列番号6のアミノ酸 26-344; 配列番号7のアミノ酸 26-310; 配列番号8のアミノ酸 26-344; 配列番号9のアミノ酸 26-310; 配列番号8のアミノ酸 27-344 ;および配列番号9のアミノ酸 27-310;ここで、該可溶性Nogo受容体-1 ポリペプチドは神経突起伸長阻害を阻害する。
【図1】

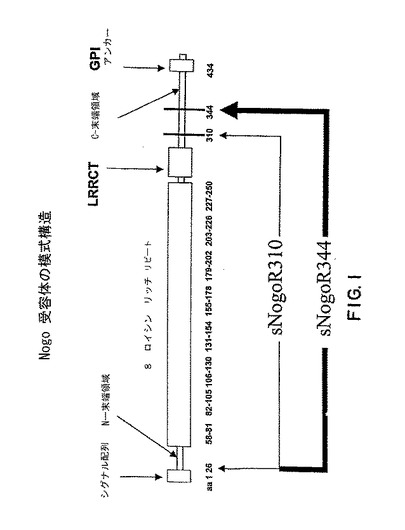
【図2】
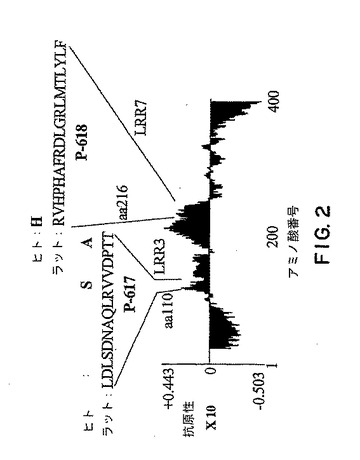
【図3】
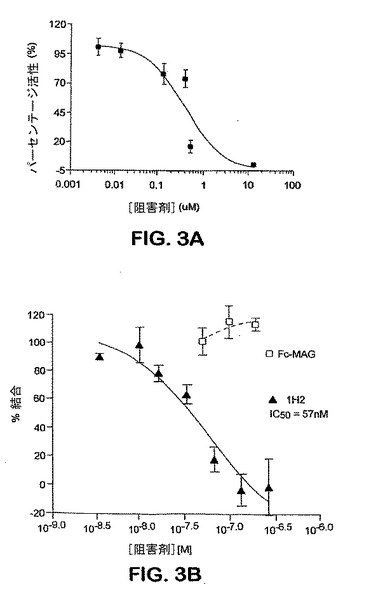
【図4】
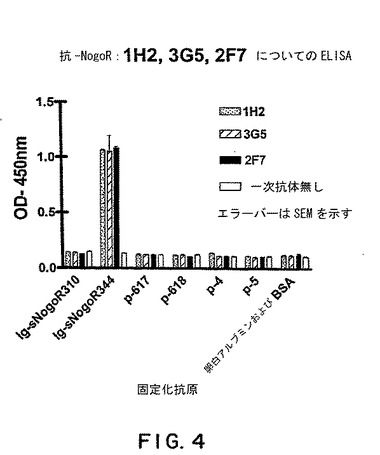
【図5】
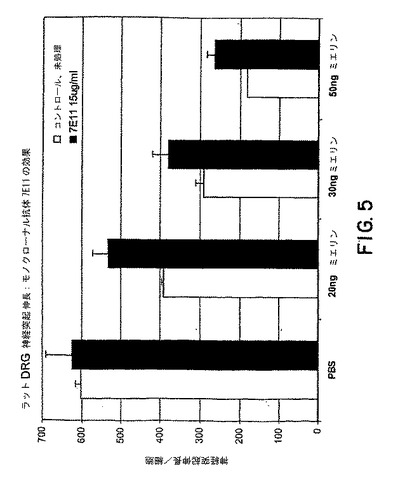
【図6】
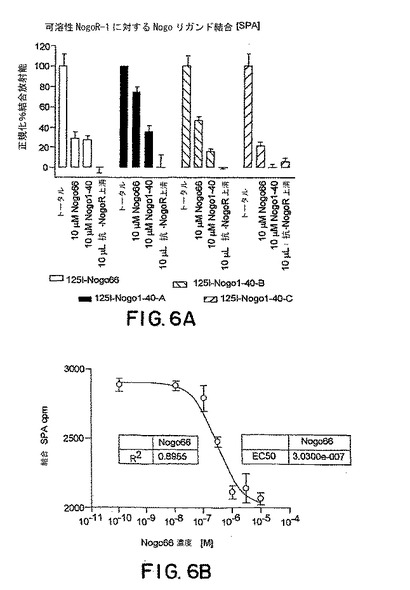
【図7】
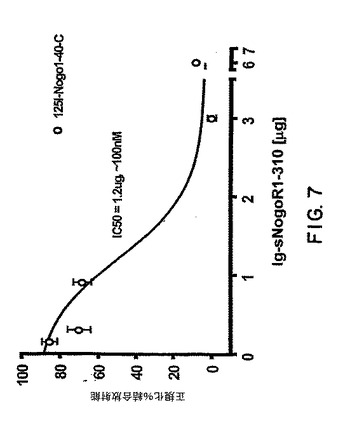
【図8】
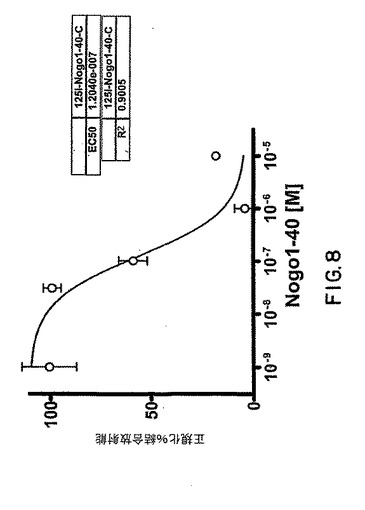
【図9A】
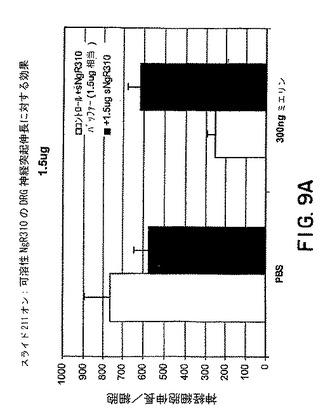
【図9B】
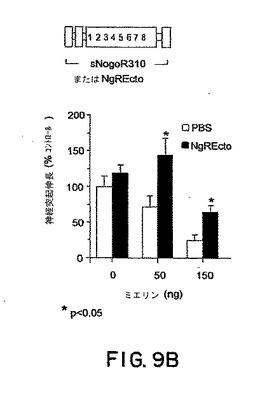
【図10A】
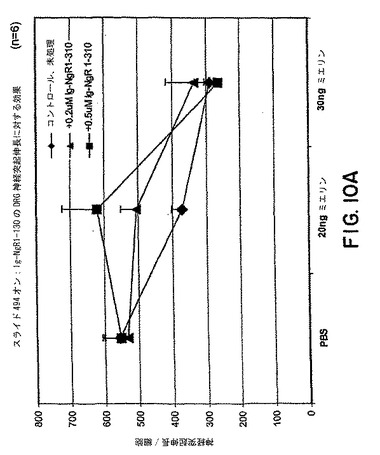
【図10B】
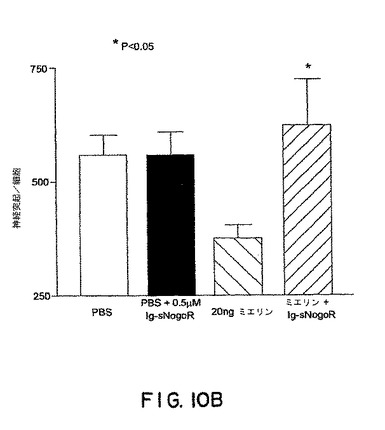
【図11】
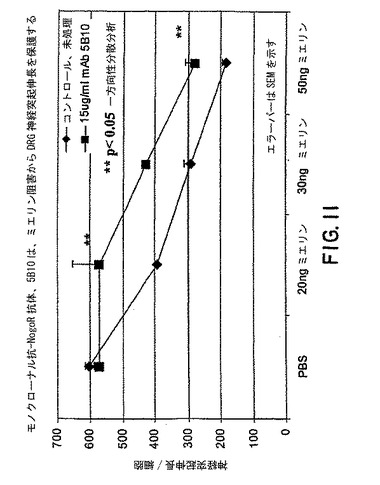
【図12】
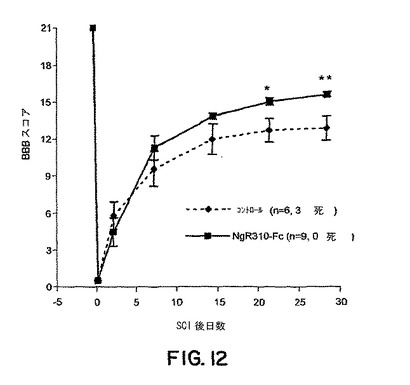
【図13】
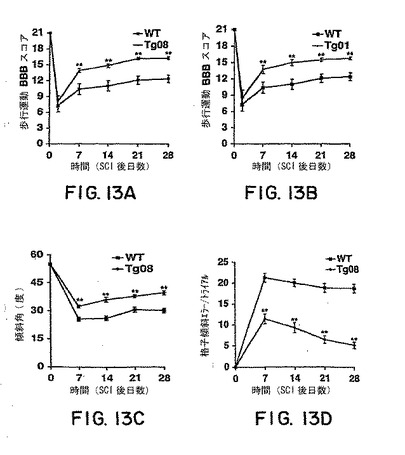
【図14】
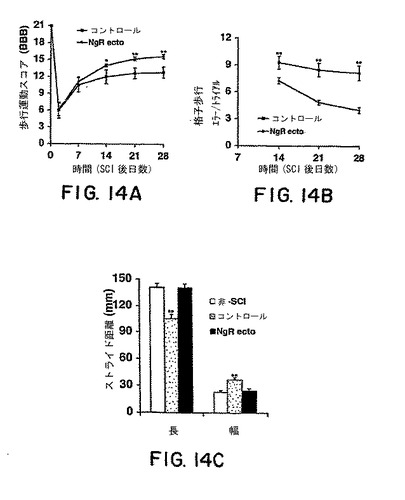
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2011−173899(P2011−173899A)
【公開日】平成23年9月8日(2011.9.8)
【国際特許分類】
【出願番号】特願2011−85613(P2011−85613)
【出願日】平成23年4月7日(2011.4.7)
【分割の表示】特願2004−527960(P2004−527960)の分割
【原出願日】平成15年8月7日(2003.8.7)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【出願人】(592221528)バイオジェン・アイデック・エムエイ・インコーポレイテッド (224)
【Fターム(参考)】
【公開日】平成23年9月8日(2011.9.8)
【国際特許分類】
【出願日】平成23年4月7日(2011.4.7)
【分割の表示】特願2004−527960(P2004−527960)の分割
【原出願日】平成15年8月7日(2003.8.7)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【出願人】(592221528)バイオジェン・アイデック・エムエイ・インコーポレイテッド (224)
【Fターム(参考)】
[ Back to top ]