
Nogo−66受容体(NGR)に対する中和モノクローナル抗体及びその使用
本発明は、Nogo−66受容体に結合する単離蛋白質、特にモノクローナル抗体に関する。具体的には、これらの抗体はNogo−66受容体の天然リガンドの結合を阻害し、Nogo−66受容体を中和することができる。本発明のこれらの抗体又はその部分は例えばNgR又はNogo−66活性が有害である障害に罹患したヒトにおいて、NgRを検出し、NgR活性を阻害するために有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本願はNogo−66受容体結合性蛋白質、特にNogo−66受容体と結合し、Nogo−66受容体の機能を中和することが可能なモノクローナル抗体について記載する。従って、これらの抗体は限定されないが、哺乳動物脳損傷、脊髄損傷、脳卒中、神経変性疾患及び統合失調症等の数種の状態の治療に有用であると思われる。
【背景技術】
【0002】
哺乳動物中枢神経系(CNS)内の損傷後の軸索再生は殆どの場合に不可能であり、転帰はCNS内の神経線維の固有の再成長能と、損傷部位の微細環境に局在しており、損傷した線維区域の再成長、従って再生を積極的に妨害するCNS内の阻害因子とのバランスに依存する。
【0003】
希突起膠細胞により産生されるCNSミエリンがインビトロ及びインビボで成長円錐崩壊を引き起こし、その結果、軸索伸長の直接阻害を生じることにより損傷の初期段階における軸索成長に最も関係の深い非許容因子であることは定説である(研究報告については、Leeら,2003参照)。CNSミエリンの主要阻害因子として希突起膠細胞ミエリン糖蛋白(OMgp)、ミエリン関連糖蛋白(MAG)及びNogo−Aがつい最近同定された(Domeniconiら,2002;研究報告:Woolf & Bloechinger,2002;McGee & Strittmatter,2003;Leeら,2003)。最後に挙げた蛋白質は主要阻害機能を発揮するドメインNogo−66を含む(GrandPreら,2000)。興味深いことに、全3種の阻害蛋白質はCNSで高い発現レベルを示し、同一のニューロングリコシルホスファチジルイノシトール(GPI)部分により固定された受容体であるNogo−66受容体ないしNgRと相互作用する(Fournierら,2001)。Nogo−66受容体NgRは473アミノ酸グリコシルホスファチジルイノシトール結合蛋白質である。この蛋白質はN末端シグナル配列とそれに続く8個のロイシンリッチリピートドメイン、(一緒になって所謂細胞外ドメインを形成する)ロイシンリッチリピートC末端ドメイン及びGPI固定ドメインから構成される。GPIアンカーを介して、NgRは外部ニューロン原形質膜と結合している。
【0004】
NgR自体は3個のCNS集積GPI固定蛋白質(NgR、NgR2及びNgR3と言う。)のファミリーに属しており、ファミリーの配列一致度は約40%であるが、全体の構造編成は非常によく似ている(Bartonら,2003;Laurenら,2003;Pignotら,2003)。NgRは複数のミエリン関連阻害分子と相互作用することが知られている唯一のメンバーであるが、MAGもNgR2と相互作用することが最近報告されている(Venkateshら,2005)。NgRホモログの機能は現時点では不明である。NgR自体は齧歯類又はニワトリで初期発生中には発現されないが、成体動物では高い発現レベルを示し、NgRは脊髄を含むCNS領域の全部ではないとしても大半で発現される(Huntら,2002a,b)。ニワトリ(Fournierら,2001)、ラット(Huntら,2002a)及びマウス(Wangら,2002b)ではmRNAと蛋白質レベルの両者で脊髄発現が報告されている。成体CNS組織内において、NgR蛋白質はその軸索突起を含む全成熟ニューロンで発現される。リガンドがNgRと結合すると、細胞内シグナル伝達カスケードが開始し、その結果、軸索伸長阻害と成長円錐崩壊が生じる。NgRは膜貫通ドメインを含まないので、シグナル伝達にはNgR/リガンド相互作用シグナルを細胞に導入する共受容体が必要である。NgRシグナル伝達の初期段階は共受容体p75又はTROYとのその相互作用である(Wongら,2002;Shaoら,2005;Parkら,2005)。Lingo−1と呼ばれる第2の共受容体も同定されている。NgRとP75又はTROYとLingo−1の三元複合体のみが機能的シグナル伝達複合体を構成する(Miら,2004;Parkら,2005)。このシグナル伝達の結果はアクチン細胞骨格の再構成である。ニューロンにおいて、このアクチン細胞骨格変化は軸索伸長の阻害と成長円錐崩壊の誘導をもたらす。
【0005】
インビトロにおいて、NgR(−/−)マウスに由来する後根神経節細胞はNogo66結合能が低下し、成長円錐崩壊アッセイにおいてNogo66、Fc−MAG、OMgp又はミエリンの阻害作用に対して低応答性である(Kimら,2004)。NgR(−/−)マウスは部分的又は完全な脊髄損傷後に赤核脊髄路と縫線核脊髄路を含む脳幹路の再生増加を示した。脊髄の完全な実験的離断後でも、NgR(−/−)マウスはオープンフィールド試験で機能回復の増加を示した。脊髄の半側切断及び完全離断後に、NgR(−/−)マウスの回復はホモ接合(+/+)及びヘテロ接合同腹子よりも有意に良好であった(Kimら,2004)。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Leeら,2003
【非特許文献2】Domeniconiら,2002
【非特許文献3】Woolf & Bloechinger,2002
【非特許文献4】McGee & Strittmatter,2003
【非特許文献5】GrandPreら,2000
【非特許文献6】Fournierら,2001
【非特許文献7】Bartonら,2003
【非特許文献8】Laurenら,2003
【非特許文献9】Pignotら,2003
【非特許文献10】Venkateshら,2005
【非特許文献11】Huntら,2002a,b
【非特許文献12】Wangら,2002b
【非特許文献13】Wongら,2002
【非特許文献14】Shaoら,2005
【非特許文献15】Parkら,2005
【非特許文献16】Miら,2004
【非特許文献17】Kimら,2004
【発明の概要】
【0007】
本願はNogo−66結合について選択的に競合し、NgR活性が有害であり得る障害を改善すると予想される抗NgR中和モノクローナル抗体の作製について記載する。本発明の中和モノクローナル抗体は例えば、特に急性脊髄損傷、脳損傷又は神経変性疾患(例えばハンチントン舞踏病、パーキンソン病、アルツハイマー病又は多発性硬化症)後の損傷したCNSにおいてニューロン再生を促進すると予想される。
【図面の簡単な説明】
【0008】
【図1】図1a及びbは抗体mAb50及びmAb51とヒト及びラットNgRとの結合を示す。
【図2】AP−Nogo66とNgR−Fcとの結合のmAb50及びmAb51による競合。
【図3】HEK293f細胞で発現させたヒト及びラットNgRとNogo66との結合のmAB50及びmAB51による競合。
【図4】mAB50及びmAB51とNTera2細胞との結合。
【図5】NTera2細胞凝集物からの神経突起伸長の定量。
【図6】hNgRの欠失変異体。
【図7】MAG−FcのNgR−Fcへの結合の競合。
【図8】許容条件下及び阻害条件下におけるラット後根神経節ニューロン及びNogo66により誘導される神経突起伸長阻害のmAb50による中和。
【図9】ラットDRG細胞におけるNogo66により誘導される神経突起伸長阻害のmAB50及びmAb51による中和。
【0009】
配列表
配列番号1a:ヒトNgR蛋白質
配列番号1b:ヒトNgRヌクレオチド
配列番号2a:ラットNgR蛋白質
配列番号2b:ラットNgRヌクレオチド
配列番号3:抗体クローン50VH
配列番号4:抗体クローン50VL
配列番号5:抗体クローン51VH
配列番号6:抗体クローン51VL
配列番号7a:AP−Nogo−66蛋白質
配列番号7b:AP−Nogo−66ヌクレオチド。
【0010】
詳細な説明
本願はNogo受容体(NgR)と相互作用する単離結合性蛋白質、特にヒト及びラットNgRと結合してこれを中和する中和モノクローナル抗体に関する。これらの抗体はNgRとの結合についてNogo−66と競合することができる。本願の他の側面としては、このような結合性蛋白質の作製方法、前記蛋白質を使用した医薬組成物、及び前記蛋白質の使用方法が挙げられる。
【0011】
抗体
本願の主要な態様はNogo−66受容体(NgR)の少なくとも1個のエピトープと特異的に結合する単離された蛋白質又はポリペプチドを含む。「単離された蛋白質」又は「単離されたポリペプチド」なる用語はその起源又は誘導源によりその天然状態でこれに付随する天然結合成分と結合していない蛋白質又はポリペプチド、同一種に由来する他の蛋白質を実質的に含まない蛋白質又はポリペプチド、別の種に由来する細胞により発現される蛋白質又はポリペプチド、あるいは自然界に存在しない蛋白質又はポリペプチドである。従って、化学的に合成されるポリペプチド又はその天然起源である細胞とは異なる細胞系で合成されるポリペプチドはその天然結合成分から「単離」されることになる。当分野で周知の蛋白質精製技術を使用して、単離により蛋白質から天然結合成分を実質的に除去することもできる。本明細書で使用する「ポリペプチド」なる用語はアミノ酸の任意ポリマー鎖を意味する。「ペプチド」及び「蛋白質」なる用語はポリペプチドなる用語と交換可能に使用し、同様にアミノ酸のポリマー鎖を意味する。「ポリペプチド」なる用語は天然又は人工蛋白質、蛋白質フラグメント及び蛋白質配列のポリペプチドアナログを包含する。ポリペプチドはモノマーでもポリマーでもよい。
【0012】
Nogo−66受容体(NgR)の少なくとも1個のエピトープと特異的に結合する単離蛋白質又はポリペプチドはリガンドと前記NgRの結合を阻害することができる。Nogo−66受容体NgRは473アミノ酸グリコシルホスファチジルイノシトール結合蛋白質である。この蛋白質はN末端シグナル配列とそれに続く8個のロイシンリッチリピートドメイン、(一緒になって所謂細胞外ドメインを形成する)ロイシンリッチリピートC末端ドメイン及びGPI固定ドメインから構成される。GPIアンカーを介して、NgRは外部ニューロン原形質膜と結合している。本発明の好ましい蛋白質はヒトNgRの少なくとも1個のエピトープと結合するモノクローナル中和抗体又はその抗原結合フラグメントである。Nogo66は軸索伸長阻害を誘導すると共に成長円錐崩壊を促進するCNSミエリンの数種の主要阻害因子の1つである。
【0013】
本明細書で使用する「抗体」なる用語はジスルフィド結合により相互に結合された2本の重(H)鎖と2本の軽(L)鎖の4本のポリペプチド鎖から構成される免疫グロブリン分子を意味する。抗体はある分子と特異的に反応することにより分子を抗体と結合することができる場合に、この分子と「結合することが可能」であると言う。「エピトープ」なる用語は抗体と結合することが可能であり、更にこの抗体により認識することができる任意分子の部分を意味する。エピトープないし「抗原決定基」は通常、アミノ酸や糖側鎖等の化学的に活性な表面分子群から構成され、特定の三次元構造特性と特定の電荷特性をもつ。
【0014】
本明細書で使用する抗体の「抗原結合フラグメント」(又は単に「抗原フラグメント」)なる用語は受容体と特異的に結合し、これを夫々活性化又は調節する能力を保持する抗体の1個以上の部分を意味する。抗体の抗原結合機能は全長抗体のフラグメントにより実現可能であることが分かっている。抗体の「抗原結合部分」なる用語に含まれる結合フラグメントの例としては、(i)VL、VH、CL及びCH1ドメインから構成される1価フラグメントであるFabフラグメント;(ii)ヒンジ領域でジスルフィド橋により結合された2個のFabフラグメントからなる2価フラグメントであるF(ab’)2フラグメント;(iii)VHドメインとCH1ドメインから構成されるFdフラグメント;(iv)抗体の単一アームのVLドメインとVHドメインから構成されるFvフラグメント;(v)VHドメインから構成されるdAbフラグメント(Wardら,(1989)Nature 341:544−546);並びに(vi)単離CDRが挙げられる。更に、Fvフラグメントの2個のドメインVL及びVHは別々の遺伝子によりコードされるが、VL領域とVH領域が対合して1本鎖Fv(scFv)抗体と呼ばれる1価分子を形成する単一蛋白質としての作製を可能にする合成リンカーにより組換え法を使用して結合することができる。(例えばBirdら,(1988)Science 242:423−426;Hustonら,(1988)Proceedings of the National Academy of Science USA 85:5879−5883参照。)このようなscFv抗体も抗体の抗原結合部分なる用語に含むものとする。ダイアボディ等の他の形態の1本鎖抗体もこの用語に含む。ダイアボディはVH及びVLドメインが1本のポリペプチド鎖で発現される2価の二重特異性抗体であるが、非常に短いリンカーを使用するため、同一鎖の2領域間で対合することができず、これらの領域は別の鎖の相補性領域と対合し、同一受容体上又は2個の受容体分子に2個の抗原結合部位を形成する。(例えばHolligerら,(1993)Proceedings of the National Academy of Science USA 90:6444−6448;Poljakら,(1994)Structure 2:1121−1123参照)。
【0015】
本明細書で使用する「モノクローナル抗体」とは異なる抗体の混合物を含む「ポリクローナル」抗体調製物と異なり、共通重鎖及び共通軽鎖アミノ酸配列を有する抗体分子の調製物を意味するものとする。モノクローナル抗体はファージ、細菌、酵母又はリボソームディスプレイ等の数種の新規技術や、ハイブリドーマ由来抗体(例えばケーラーとミルスタインの標準ハイブリドーマ法((1975)Nature 256:495−497)等のハイブリドーマ技術により作製したハイブリドーマにより分泌される抗体)等の旧来の方法により作製することができる。本発明の抗体は哺乳動物細胞株で作製したNgR蛋白質を使用して標準免疫/ハイブリドーマ技術により作製した。
【0016】
本明細書で使用する「中和モノクローナル抗体」とは特定抗原と結合すると、前記抗原に対するリガンドの結合を競合及び阻害することが可能な抗体分子調製物を意味するものとする。本願の特定ケースにおいて、本発明の中和抗体はNgRとの結合についてNogo66と競合し、Nogo66とNgRの結合に起因するNogo66生物活性又は機能を妨害することができる。
【0017】
好ましくは、本願のモノクローナル中和抗体はヒト抗体である。「ヒト抗体」なる用語はヒト生殖細胞系列免疫グロブリン配列に対応又は由来する可変領域と定常領域を有する抗体を意味する(例えばKabatら,Sequences of Proteins of Immunological Interest,第5版,U.S.Department of Health and Human Services,NIH Publication No.91−3242,1991参照)。本願のヒト抗体はヒト生殖細胞系列免疫グロブリン配列によりコードされないアミノ酸残基(例えばランダムもしくは部位特異的変異誘発によりインビトロ又は体細胞変異によりインビボで導入される変異)を例えばCDR、特にCDR3に含んでいてよい。本明細書で使用する「CDR」なる用語は抗体可変配列内の相補性決定領域を意味する。重鎖及び軽鎖可変領域の各々に3個のCDRがあり、可変領域の各々についてCDR1、CDR2及びCDR3と呼ぶ。各種態様において、抗体は組換え抗体又はモノクローナル抗体である。本願の最も好ましい中和抗体を本明細書ではmAb50及びmAb51(夫々ATCC No.PTA−8383及びPTA−8384)と呼ぶ。mAb50及びmAb51抗体と機能的抗体フラグメント、mAb50及びmAb51関連抗体と機能的抗体フラグメント、並びにmAb50及びmAb51に等価の特性(例えばNgRに対する高い結合親和性と低い解離反応速度と高い中和能)を有する他の抗体と機能的抗体フラグメントを本発明に含むものとする。
【0018】
免疫原性NgRポリペプチドに対する本願の抗NgR抗体又はそのフラグメントの結合親和性と解離速度は当分野で公知のいずれかの方法により測定することができる。例えば、結合親和性は競合ELISA、cRIA、BIAcore又はKinExA技術により測定することができる。解離速度もBIAcore又はKinExA技術により測定することができる。結合親和性と解離速度は例えばBIAcoreを使用して表面プラズモン共鳴法により測定される。
【0019】
本願の好ましい抗体の1例は配列番号3の配列を含む重鎖可変領域(VH領域)と配列番号4の配列を含む軽鎖可変領域(L領域)を含む配列(mAB50抗体)に対して少なくとも90%のアミノ酸配列一致度を有する。別の好ましい態様は配列番号5の配列を含む重鎖可変領域(VH領域)と配列番号6の配列を含む軽鎖可変領域(L領域)を含む配列(mAB51抗体)に対して少なくとも90%のアミノ酸配列一致度を有する。
【0020】
好ましくは、mAb50抗体とmAb51抗体は1×10−9M未満のEC50でヒトNgRと結合し、より好ましくは、前記抗体は1×10−10M未満のEC50でNgRと結合し、最も好ましくは、前記抗体は4×10−11M未満のEC50でNgRと結合する。
【0021】
抗NgR抗体mAb50及びmAb51はプロNgR、成熟NgR及び短縮型NgR等の各種形態のヒトNgRと結合するものとする。抗体mAb50及びmAb51はNgR2やNgR3等の他のNgRホモログ、又は他のLRR含有蛋白質と特異的に結合しない。他方、抗体mAb50及びmAb51は他の種、特に齧歯類に由来するNgR、より具体的にはラットNgRに対して交差反応性を示す。MAb52〜62は更にマウスNgRに対しても交差反応性である。例えば、抗体はラットに由来するNgRと結合する(ラットNgRに対する両者抗体のIC50は約3×10−11Mである)。
【0022】
本願の(NgR)と相互作用する単離結合性蛋白質は抗体又はその抗原結合部分が1個以上の糖鎖残基を含むグリコシル化結合性蛋白質でもよい。初期インビボ蛋白質産生後に翻訳後修飾と呼ばれるプロセシングを受ける場合がある。特に、糖鎖(グリコシル)残基が酵素により付加される場合があり、このプロセスをグリコシル化と言う。その結果として得られた蛋白質は共有結合オリゴ糖側鎖をもち、グリコシル化蛋白質又は糖蛋白質と呼ばれる。蛋白質グリコシル化は該当蛋白質のアミノ酸配列と、蛋白質が発現される宿主細胞に依存する。生物によって産生されるグリコシル化酵素(例えばグリコシルトランスフェラーゼやグリコシダーゼ)は異なり、利用可能な基質(ヌクレオチド糖)も異なる。このような因子により、蛋白質グリコシル化パターンとグリコシル残基の組成は特定蛋白質が発現される宿主系により異なる。本発明で有用なグリコシル残基としては限定されないが、グルコース、ガラクトース、マンノース、フコース、n−アセチルグルコサミン及びシアル酸が挙げられる。グリコシル化結合性蛋白質はグリコシル化パターンがヒトであるようなグリコシル残基を含むことが好ましい。
【0023】
本願の抗体はIgG1、IgG2、IgG3、IgG4、IgA、IgE、IgM又はIgD定常領域等の重鎖定常領域を含む。更に、抗体はκ軽鎖定常領域又はλ軽鎖定常領域のいずれかの軽鎖定常領域を含むことができる。抗体はκ軽鎖定常領域を含むことが好ましい。あるいは、抗体部分は例えばFabフラグメント又は1本鎖Fvフラグメントとすることができる。抗体エフェクター機能を改変するためにFc部分のアミノ酸残基を置換することは当分野で公知である(Winterら,米国特許第5,648,260号;5,624,821号明細書)。抗体のFc部分は数種の重要なエフェクター機能(例えばサイトカイン誘導、ADCC、貪食作用、補体依存性細胞傷害作用(CDC)並びに抗体及び抗原−抗体複合体の半減期/クリアランス率)を媒介する。これらのエフェクター機能は治療目的に応じて治療用抗体に望ましい場合もあるが、不要又は有害な場合もある。所定のヒトIgGアイソタイプ、特にIgG1及びIgG3は夫々FcγRs及び補体C1qとの結合を介してADCCとCDCを媒介する。胎児性Fc受容体(FcRn)は抗体の循環半減期を決定する必須成分である。更に別の態様では、抗体のエフェクター機能を改変するように、抗体の定常領域(例えば抗体のFc領域)の少なくとも1個のアミノ酸を置換する。
【0024】
組換え蛋白質の作製
免疫及びELISAアッセイと、(「実施例」のセクションに記載する)神経突起伸長アッセイのために、可溶性ヒト及びラットNgRを作製した。アルカリホスファターゼタグと融合したリガンドNogo66(AP−Nogo66)も作製し、神経突起伸長アッセイの阻害因子及び(同様に「実施例」のセクションに記載する)ELISA試験とFACS試験におけるリガンドとして利用した。
【0025】
ヒト及びラットNgR
ヒトNgR蛋白質はアクセション番号AAG53612に基づくものとした。(アミノ酸27〜450の)蛋白質DNAをpSecベクター(Ambion)にクローニングし、CHO−K1細胞で安定に発現させることにより蛋白質を作製した。発現させた受容体は配列番号1(ヒトNgR蛋白質)及び配列番号1b(ヒトNgRヌクレオチド)に従い、C末端にMycと6×Hisタグを結合した全長蛋白質の424アミノ酸(アミノ酸27〜450)から構成された。セルファクトリー当たり5000mlのUltraCHO無血清培地(Cambrex Bio Science)を加えた40チャンバーセルファクトリー8個でヒトsecNgR 27−450 aa_D6_CHO−K1細胞をコンフルエントまで培養した(約5日間)。その後、上清40リットルを遠心し、Hemoflow Fカラム(Fresenius Medical Care)で500mlまで濃縮した。濃縮液を−80℃で凍結させた。蛋白質精製のために、濃縮蛋白質上清に20mM NaH2PO4;140mM NaCl(pH7.4)500mlを加えて1000mlとし、再び300mlまで濃縮した。濃縮工程をもう一度繰返し、最後に20mM NaH2PO4;140mM NaCl(pH7.4)300mlを濃縮液に加えた。20mM NaH2PO4;300mM NaCl;pH8.0で平衡化しておいたNi−NTA−Superflow Fa.(Qiagen catalog #30430)50mlを濃縮液600mlに加え、6℃で1時間撹拌し、6℃で沈降させ、上清を捨て、Ni−NTAビーズをカラムに充填した。カラムを室温にて20mM NaH2PO4;300mM NaCl(pH8.0)10CVで洗浄した後、20mM NaH2PO4;300mM NaCl;10mMイミダゾール(pH8.0)5〜10CVで洗浄した。カラムを20mM NaH2PO4;300mM NaCl;100mMイミダゾール;pH8.0で溶出させ、UV−280nm活性ピークを採取した。次に溶出液を6℃にて一晩25mM Tris/HCl(pH7.0)5Lで透析し、透析液を室温でQ−Sepharoseカラム(カラムサイズ1.6cm×3cm;容量6ml;Amersham Biosciences catalog #17−0510−01)にロードした。緩衝液Aは50mM Tris/HCl;pH7.0とした。緩衝液Bは50mM Tris/HCl;1M NaCl;pH7.0とし、流速2ml/分とした。勾配は0%B保持5CV;0−50%B12CV;50−100%B2CV;100%B保持5CVとした。フラクションサイズは2.5mlとした。その後、フラクションをSDS−PAGで分析し、SDS−page上のサイズと純度(高グリコシル化NgR−HisでNgR−His最高純度;低グリコシル化NgR−Hisで最高NgR−His純度)によりフラクションプールした。プールしたフラクションを12〜14kDa透析チューブで6℃にて20mM NaH2PO4;140mM NaCl;pH7.4で1回透析し、フラクションを0.2μm滅菌フィルターで濾過し、その後の使用に備えて6℃で保存した。長期間保存用として、受容体フラクションを分取し、−80℃で保存した。
【0026】
ラットNgR DNA(アクセション番号AAM46772)をpcDNA3.1(Invitrogen)にクローニングし、HEK293F細胞で一過性発現システムにより発現及び生産させた。蛋白質は配列番号2a(ラットNgR蛋白質)及び配列番号2b(ラットNgRヌクレオチド)に従い、6×Hisタグと結合したアミノ酸27〜450を含んでいた。ラット蛋白質の生産はHEK293F細胞で48〜72時間標準一過性発現により実施した。細胞上清を回収し、蛋白質の精製はヒト蛋白質について上述したと同様の工程に従った。所定の実験では、R&D Systemsから入手した蛋白質を使用した。これらの蛋白質は、ヒト組換えNgR/Fcキメラ,カタログ番号1208−NG及び組換えマウスNogo受容体/Fcキメラ,カタログ番号1440−NGであった。
【0027】
NgRの細胞表面発現
細胞表面で発現されるNgRには、アミノ酸1〜473の完全オープンリーディングフレームを含む全長受容体配列(夫々ラットAAM46772及びヒトAAG53612)をpcDNA4にクローニングした。プラスミドを標準手順に従ってCHO−K1又はHEK293細胞に形質移入した。要約すると、細胞をペトリ皿でMEM培地に播種し、Fugene 6(Roche)を製造業者の指示に従って形質移入した。150μg/mlゼオシンを使用して2〜3週間選択を実施し、蛋白質発現をFACSにより確認した(下記実施例4参照)。
【0028】
HEK293F細胞で一過性発現させるために、製造業者(Invitrogen;Free Style System)の指示に従って細胞を懸濁培養し、48〜72時間後に回収し、FACS試験に使用した(下記実施例のセクション参照)。
【0029】
AP−Nogo66の生産
AP−Nogo66(配列番号7a及び配列番号7b)を標準条件下で生産した。要約すると、Nogo66をpAPTag5ベクターにクローニングし、HEK293細胞に構築物を形質移入し、RPMI Glutamax+10%FCS,150μg/mlゼオシンで選択した。蛋白質生産のために、セルファクトリー当たり1200mlのRPMI(Invitrogen)+10%FCSを加えた10チャンバーセルファクトリー6個でHEK293細胞をコンフルエントまで培養した(約3日間)。その後、上清を捨て、Pro293a−CDM(Cambrex Bio Science)1200mlを各セルファクトリーに充填した。細胞を更に3日間培養した。その後、上清7200mlを遠心し、Hemoflow Fカラム(Fresenius Medical Care)で350mlまで濃縮した。1mM PefablocSC(ROCHE)の添加後、濃縮液を分取し、−80℃で凍結させた。
【0030】
抗体及び抗体産生細胞株の作製
本願の抗体は適切な宿主(例えばヒト、マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ、ウマ等の脊椎動物、爬虫類、魚類、両生類、並びに鳥類、爬虫類及び魚類の卵)に免疫により作製することができる。このような抗体はポリクローナル抗体でもモノクローナル抗体でもよい。本願の抗体を作製するためには、本発明の免疫原性NgRポリペプチド又はそのフラグメントにより宿主を免疫する。「免疫」なる用語は本明細書では抗原を免疫レパートリーに提示するプロセスを意味し、レパートリーは遺伝子改変されていない天然生物に存在するか又は人工ヒト免疫レパートリーを示すように改変されたものを含むトランスジェニック生物に存在するかを問わない。同様に、「免疫原製剤」とはアジュバント又は抗原の免疫原性を強化する他の添加剤を加えた抗原製剤である。
【0031】
動物の免疫は当分野で公知のいずれかの方法により実施することができる。例えばHarlow and Lane,Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Press,1990参照。マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ及びウマ等の非ヒト動物の免疫方法は当分野で周知である。例えばHarlow and Lane及び米国特許第5,994,619号明細書参照。好ましい1態様では、NgR抗原をアジュバントと共に投与し、免疫応答を刺激する。このようなアジュバントとしては完全もしくは不完全フロイントアジュバント、RIBI(ムラミルジペプチド)又はISCOM(免疫刺激複合体)が挙げられる。このようなアジュバントはポリペプチドを局所沈着物中に分離することにより迅速な分散から保護することもできるし、マクロファージや免疫系の他の成分に対して化学走化性の因子を分泌するように宿主を刺激する物質を添加してもよい。ポリペプチドを投与している場合には、免疫スケジュールは数週間にわたってポリペプチドを2回以上投与することが好ましい。
【0032】
無傷又は破壊した細胞の細胞膜に伴われたNgRにより動物宿主を免疫することが考えられ、本願の抗体は本発明の免疫原性NgRポリペプチドと結合することにより同定される。
【0033】
動物宿主をNgR抗原により免疫後、動物から抗体及び/又は抗体産生細胞を得ることができる。動物から採血する又は動物を屠殺することにより動物から抗NgR抗体含有血清が得られる。血清は動物から採取したまま使用してもよいし、血清から免疫グロブリンフラクションを採取してもよいし、血清から抗NgR抗体を精製してもよい。こうして得られた血清又は免疫グロブリンはポリクローナルであるため、一連の異種特性をもつ。
【0034】
抗体産生細胞株
本願は更に免疫動物から作製可能な抗体産生不死化ハイブリドーマについて記載する。好ましくは、免疫動物はヒト免疫グロブリン遺伝子を発現する非ヒト動物であり、脾臓B細胞を非ヒト動物と同一種に由来する骨髄腫と融合させる。
【0035】
免疫後、動物を屠殺し、脾臓B細胞を当分野で周知のように不死化骨髄腫細胞と融合させる。例えばHarlow and Lane,前出参照。骨髄腫細胞は免疫グロブリンポリペプチドを分泌しないことが好ましい(非分泌型細胞株)。融合及び抗体選択後、NgRもしくはその一部又はNgRを発現する細胞を使用してハイブリドーマをスクリーニングする。好ましくは、初期スクリーニングは酵素免疫測定法(ELISA)又は放射免疫測定法(RIA)、好ましくはELISAを使用して実施される(実施例のセクションにELISAスクリーニングの1例を記載する)。
【0036】
抗NgR抗体を産生するハイブリドーマを選択し、クローニングし、以下に詳述するように、活発なハイブリドーマ増殖、高い抗体産生能及び望ましい抗体特性等の望ましい特性について更にスクリーニングする。ハイブリドーマを培養し、同系動物、免疫系を欠損する動物(例えばヌードマウス)でインビボ増殖させる、あるいは細胞培養でインビトロ増殖させる。ハイブリドーマの選択、クローニング及び増殖方法は当業者に周知である。好ましい1態様において、ハイブリドーマは上記のようにマウスハイブリドーマである。別の好ましい態様において、ハイブリドーマは非ヒト非マウス種(例えばラット、ヒツジ、ブタ、ヤギ、ウシ又はウマ)で作製される。別の態様において、ハイブリドーマは抗NgR抗体を発現するヒト細胞にヒト非分泌型骨髄腫が融合されたヒトハイブリドーマである。
【0037】
本願は、米国特許第5,627,052号明細書、国際公開第92/02551号パンフレット及びBabcock,J.S.ら(1996)Proc.Natl.Acad.Sci.USA 93:7843−7848に記載されているような選択リンパ球抗体法(SLAM)と当分野で呼ばれる手順を使用して単一単離リンパ球から作製される組換え抗体についても記載する。この方法では、リンカー(例えばビオチン)を使用して抗原NgR又はそのフラグメントをヒツジ赤血球と結合させ、これを使用してNgRに対する特異性をもつ抗体を分泌する単一細胞を同定する抗原特異的溶血プラークアッセイを使用することにより、該当抗体を分泌する単一細胞(例えば任意免疫動物に由来するリンパ球)をスクリーニングする。該当抗体分泌細胞の同定後、重鎖及び軽鎖可変領域cDNAを逆転写酵素−PCRにより細胞から得た後、COS又はCHO細胞等の哺乳動物宿主細胞で適切な免疫グロブリン定常領域(例えばヒト定常領域)のコンテキストにおいてこれらの可変領域を発現させることができる。インビボ選択したリンパ球に由来する増幅免疫グロブリン配列を形質移入した宿主細胞をその後、例えば形質移入した細胞のパニングにより更にインビトロ分析及び選択し、NgRに対する抗体を発現する細胞を単離することができる。
【0038】
インビトロ作製した抗体
本願に記載する抗体を作製するためには、抗体ライブラリーをスクリーニングして所望の結合特異性をもつ抗体を同定するインビトロ法を使用することもできる。組換え抗体ライブラリーのこのようなスクリーニング方法は当分野で周知である。
【0039】
組換え抗体ライブラリーはNgR又はNgRの一部により免疫された対象に由来することができる。あるいは、組換え抗体ライブラリーはナイーブ対象、即ちNgRにより免疫されていない対象に由来することができる(例えばヒトNgRにより免疫されていないヒト対象に由来するヒト抗体ライブラリー)。本願の抗体はヒトNgRを含むペプチド(例えばhNgRの一部に対応するペプチド)で組換え抗体ライブラリーをスクリーニングし、NgRを認識する抗体を選択することにより選択される。このようなスクリーニングと選択を実施するための方法は前段落に挙げた文献に記載されているように、当分野で周知である。hNgRに対する特定結合親和性をもつ本願の抗体(例えば特定koff速度定数でヒトNgRから解離するもの)を選択するためには、当分野で公知の表面プラズモン共鳴法を使用して所望のkoff速度定数をもつ抗体を選択することができる。hNgRに対して特定中和活性をもつ本願の抗体(例えば特定IC50をもつもの)を選択するためには、hNgR活性の阻害を評価する方法として当分野で公知の標準方法を使用することができる。
【0040】
抗体の特性
本願のmAB50及びmAB51又はその抗原結合部分はヒトNgRと結合し、表面プラズモン共鳴法により測定した場合に約0.1s−1以下、好ましくは1×10−2s−1以下、より好ましくは1×10−3s−1以下、更に好ましくは1×10−4s−1以下、最も好ましくは1×10−5s−1以下のkoff速度定数でヒトNgRから解離する。本明細書で使用する「表面プラズモン共鳴」なる用語は例えばBIAcoreシステム(Pharmacia Biosensor AB,Uppsala,Sweden及びPiscataway,NJ)を使用してバイオセンサーマトリックス内の蛋白濃度の変化の検出によりリアルタイム生体特異的相互作用の分析を可能にする光学現象を意味する。更に詳細な記載については、Jonsson,U.ら(1993)Ann.Biol.Clin.51:19−26;Jonsson,U.ら(1991)Biotechniques 11:620−627;Johnsson,B.ら(1995)J.Mol Recognit.8:125−131;及びJohnnson,B.ら(1991)Anal.Biochem.198:268−277参照。
【0041】
【表1】

【0042】
本明細書で使用する「Kon」なる用語は当分野で公知の通り、抗体と抗原が抗体/抗原複合体を形成するための会合速度定数を意味するものとする。
【0043】
本明細書で使用する「Koff」なる用語は当分野で公知の通り、抗体/抗原複合体から抗体が解離する解離速度定数を意味する。本明細書で使用する「Kd」なる用語は当分野で公知の通り、特定抗体−抗原相互作用の解離定数を意味するものとする。
【0044】
あるいは、本願のmAb50及びmAb51又はその抗原結合部分は約1×10−6M以下、好ましくは1×10−7M以下、好ましくは1×10−8M以下、より好ましくは1×10−9M以下、より好ましくは1×10−10M以下、最も好ましくは1×10−11M以下のIC50でヒトNgRを阻害することができる。本明細書で使用するIC50なる用語はリガンドNogo66とNgRの結合について競合する抗体の濃度を意味するものとする。
【0045】
融合抗体及びイムノアドヘジン
本願は別のポリペプチドに連結した本願の抗Nogo受容体−1抗体の全部又は一部を含む作製可能な融合抗体又はイムノアドヘジンについても記載する。ある態様において、抗Nogo受容体−1抗体の可変領域のみをポリペプチドに連結する。他の態様において、本願の抗Nogo受容体−1抗体のVHドメインを第1のポリペプチドに連結し、VHドメインとVLドメインを相互作用させて抗体結合部位を形成するように第1のポリペプチドと会合する第2のポリペプチドに抗体のVLドメインを連結する。他の態様において、VHドメインとVLドメインを相互作用させるリンカーによりVHドメインをVLドメインから分離する(下記1本鎖抗体の項参照)。その後、VH−リンカー−VL抗体を該当ポリペプチドに連結する。融合抗体はNogo受容体−1リガンドを発現する細胞又は組織にポリペプチドを誘導するのに有用である。該当ポリペプチドは毒素等の治療剤でもよいし、可視化可能な酵素(例えば西洋ワサビペルオキシダーゼ)等の診断剤でもよい。更に、2個(以上)の1本鎖抗体を相互に連結した融合抗体を作製することもできる。これは、1本のポリペプチド鎖上に2価又は多価抗体を作製したい場合や、二重特異性抗体を作製したい場合に有用である。
【0046】
1態様は本願の抗体又は抗体部分を誘導体化した又は別の機能的分子(例えば別のペプチド又は蛋白質)と連結した標識結合性蛋白質を提供する。例えば、核酸、別の抗体(例えば二重特異性抗体又はダイアボディ)、検出可能な物質、細胞傷害性物質、薬剤、及び/又は抗体もしくは抗体部分と別の分子(例えばストレプトアビジンコア領域やポリヒスチジンタグ)との結合を媒介することができる蛋白質もしくはペプチド等の1個以上の他の分子部分と本願の抗体又は抗体部分を(化学的カップリング、遺伝子融合、非共有的結合又は他の方法により)機能的に連結することにより、本願の標識結合性蛋白質を誘導することができる。
【0047】
本願の抗体又は抗体部分を誘導体化することができる有用な検出可能な物質としては蛍光化合物が挙げられる。検出可能な蛍光物質の例としてはフルオレセイン、フルオレセインイソチオシアネート、ローダミン、5−ジメチルアミン−1−ナフタレンスルホニルクロリド、フィコエリスリン等が挙げられる。アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、グルコースオキシダーゼ等の検出可能な酵素で抗体を誘導体化することもできる。検出可能な酵素で抗体を誘導体化する場合には、検出可能な反応生成物を生成するために酵素により使用される他の試薬を加えることにより抗体を検出する。例えば、検出可能な物質として西洋ワサビペルオキシダーゼを使用する場合には、過酸化水素とジアミノベンジジンを加えると、検出可能な着色反応生成物が得られる。抗体を核酸、ビオチンで誘導体化し、アビジン又はストレプトアビジン結合の間接測定により検出することもできる。
【0048】
本願の別の態様は結晶化結合性蛋白質を提供する。本明細書で使用する「結晶化」なる用語は結晶形態で存在する抗体又はその抗原結合部分を意味する。結晶は物質の固体状態の1形態であり、非晶質固体状態や液体結晶状態等の他の形態から区別される。結晶は原子、イオン、分子(例えば抗体等の蛋白質)、又は分子集合(例えば抗原/抗体複合体)の規則的な反復三次元配列から構成される。これらの三次元配列は当分野で周知の特定の数学的関係に従って配置されている。結晶中で反復している基本単位ないし構成単位を非対称単位と言う。所定の明確な結晶対称性に一致する配置で非対称単位が反復することにより、結晶の「単位格子」を構成する。単位格子が全3方向に規則的平行移動により反復することにより、結晶を構成する。Giege,R.and Ducruix,A.Barrett,Crystallization of Nucleic Acids and Proteins,a Practical Approach,第2版,pp.20 1−16,Oxford University Press,New York,New York,(1999)参照。
【0049】
好ましくは、本願は本明細書に開示するような全長抗NgR抗体及びそのフラグメントの結晶と、このような結晶を含有する製剤及び組成物について記載する。1態様において、結晶化結合性蛋白質は結合性蛋白質の可溶性部分よりも長いインビボ半減期をもつ。別の態様において、結合性蛋白質は結晶化後に生物活性を保持する。
【0050】
本発明の結晶化結合性蛋白質は当分野で公知の方法に従って作製することができる。
【0051】
1本鎖抗体
本願は本発明の免疫原性NgRと結合する1本鎖抗体(scFv)を含む。scFvを作製するためには、VH配列とVL配列を連続した1本鎖蛋白質として発現させ、VL領域とVH領域を可撓性リンカーにより結合できるように、可撓性リンカーをコードするDNA、例えばアミノ酸配列(G1Y4−Ser)をコードするDNAにVHとVをコードするDNAを機能的に連結する(例えばBirdら,(1988)Science 242:423−42 6;Hustonら,(1988)Proc.Natl.Acad.Sci.USA 85:5879−5883;McCaffertyら,30 Nature(1 99 0)34 8:552−554参照)。1本鎖抗体はただ1個のVHとVLを使用する場合には1価とすることができ、2個のVHとVLを使用する場合には2価とすることができ、3個以上のVHとVLを使用する場合には多価とすることができる。
【0052】
キメラ抗体
本願は更に一方の特異性が本願の免疫原性Nogo受容体−1ポリペプチドに対するものである二重特異性抗体又はその抗原結合フラグメントを含む。例えば、第1の結合ドメインを介して本発明の免疫原性NgRポリペプチドと特異的に結合し、第2の結合ドメインを介して第2の分子と結合するキメラ抗体を作製することができる。「キメラ抗体」なる用語はある種に由来する重鎖及び軽鎖可変領域配列と、別の種に由来する定常領域配列を含む抗体(例えばヒト定常領域に連結したマウス重鎖及び軽鎖可変領域をもつ抗体)を意味する。キメラ抗体は組換え分子生物学技術により作製することもできるし、相互に物理的に結合させてもよい。更に、2個以上のVHとVLを含み、本発明の免疫原性ポリペプチドと、ミエリンによる成長円錐崩壊と神経突起伸長及び発芽の阻害を低減する働きがあるとされる別の分子とに特異的に結合する1本鎖抗体を作製することもできる。このような二重特異性抗体は、例えばFangerら,Immunol Methods 4:72−81(1994)及びWright and Harris,20(前出)等の周知技術を使用して作製することができる。所定態様では、キメラ抗体は本発明の抗体に由来する可変領域の1個以上を使用して作製される。別の態様では、キメラ抗体は前記抗体に由来する1個以上のCDR領域を使用して作製される。「ヒト化抗体」なる用語は非ヒト種(例えばマウス)に由来する重鎖及び軽鎖可変領域配列を含むが、VH及び/又はVL配列の少なくとも一部をより「ヒト様」即ちヒト生殖細胞系列可変配列に類似するように改変した抗体を意味する。ヒト化抗体の1例はヒトCDR配列を非ヒトVH及びVL配列に導入して対応する非ヒトCDR配列を置換したCDRグラフト抗体である。
【0053】
ヒト化抗体
ヒト化抗体は所望抗原と結合する非ヒト種に由来する抗体分子であり、非ヒト種に由来する1個以上の相補性決定領域(CDR)とヒト免疫グロブリン分子に由来するフレームワーク領域をもつ。ヒトIg配列は当分野で公知である。当分野で公知の通り、このようなインポート配列を使用して免疫原性を低下させたり、結合性、親和性、会合速度、解離速度、アビディティ、特異性、半減期又は他の適切な特性のいずれかを低下させ、増加させ又は改変することができる。
【0054】
抗原結合を改変する、好ましくは改善するためには、ヒトフレームワーク領域のフレームワーク残基をCDRドナー抗体に由来する対応する残基で置換すればよい。これらのフレームワーク置換は当分野で周知の方法により同定され、例えば抗原結合に重要なフレームワーク残基を同定するにはCDR残基とフレームワーク残基の相互作用のモデル化を使用し、特定位置の異常なフレームワーク残基を同定するには配列比較を使用する。(例えばQueenら,米国特許第5,585,089号明細書;Riechmannら,Nature 332:323(1988)参照)。三次元免疫グロブリンモデルは広く入手可能であり、当業者に周知である。選択された候補免疫グロブリン配列の推定三次元配座構造を図解表示するコンピュータープログラムも入手可能である。これらの表示を検討すると、候補免疫グロブリン配列の機能に残基が果たすと予想される役割を分析することができ、即ち候補免疫グロブリンがその抗原と結合する能力に影響を与える残基を分析することができる。こうして、ターゲット抗原に対する親和性の増加等の所望抗体特性が得られるようにコンセンサス配列とインポート配列からFR残基を選択し、組み合わせることができる。一般に、CDR残基は抗原結合の改変に直接的且つ最も実質的に関与している。当分野で公知の各種技術を使用して抗体をヒト化することができ、限定されないが、Carterら,Proc.Natl.Acad.Sci.U.S.A.89:4285(1992);Prestaら,J.Immunol.151:2623(1993),Padlan,Molecular Immunology 28(4/5):489−498(1991);Studnickaら,Protein Engineering 7(6):805−814(1994);Roguskaら,PNAS 91:969−973(1994)に記載されている技術が挙げられる。
【0055】
誘導体化及び標識抗体
本願の抗体又は抗原結合フラグメントは誘導体化又は別の機能的分子(例えば別のペプチド又は蛋白質)と連結することができる。一般に、抗体又は抗原結合フラグメントは本発明の抗原性ペプチドとの結合が誘導体化又は標識により悪影響を受けないように誘導体化される。例えば、別の抗体(例えば二重特異性抗体又はダイアボディ)、検出可能な物質、細胞傷害性物質、薬剤及び/又は抗体もしくは抗体部分と別の分子(例えばストレプトアビジンコア領域又はポリヒスチジンタグ)との結合を媒介することができる蛋白質もしくはペプチド等の1個以上の他の分子部分と本願の抗体又は抗体部分を(化学的カップリング、遺伝子融合、非共有的結合又は他の方法により)機能的に連結することができる。更に、抗体又はその抗原結合部分は、抗体又は抗体部分と1種以上の他の蛋白質又はペプチドとの共有又は非共有的結合により形成される大きな免疫接着分子の一部でもよい。このような免疫接着分子の例としてはストレプトアビジンコア領域を使用して四量体scFv分子を作製する場合(Kipriyanovら(1995)Human Antibodies and Hybridomas 6:93−101)や、システイン残基、マーカーペプチド及びC末端ポリヒスチジンタグを使用して2価ビオチン化scFv分子を作製する場合(Kipriyanovら(1994)Molecular Immunology 31:1047−1058)が挙げられる。Fab及びF(ab’)2フラグメント等の抗体部分は夫々全長抗体のパパイン又はペプシン消化等の慣用技術を使用して全長抗体から作製することができる。更に、抗体、抗体部分及び免疫接着分子は標準組換えDNA技術を使用して得ることができる。
【0056】
誘導体化抗体は(同一型又は例えば二重特異性抗体を作製するためには異なる型の)2個以上の抗体を架橋することにより作製することができる。適切な架橋剤としては適切なスペーサーにより分離された2個の別個の反応基をもつヘテロ2官能性のもの(例えばm−マレイミドベンゾイル−N−ヒドロキシスクシンイミドエステル)又はホモ2官能性のもの(例えばスベリン酸ジスクシンイミジル)が挙げられる。このようなリンカーはPierce Chemical Company,Rockford,Illから入手可能である。
【0057】
誘導体化抗体は標識抗体でもよい。例えば、本発明の抗体又は抗体部分を誘導体化することができる検出剤はフルオレセイン、フルオレセインイソチオシアネート、ローダミン、5−ジメチルアミン−1−ナフタレンスルホニルクロリド、フィコエリスリン、ランタニド蛍光体等の蛍光化合物である。西洋ワサビペルオキシダーゼ、ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ、グルコースオキシダーゼ等の検出に有用な酵素で抗体を標識してもよい。検出可能な酵素で標識する態様では、検出可能な反応生成物を生成するために酵素により使用される他の試薬を加えることにより抗体を検出する。例えば、西洋ワサビペルオキシダーゼを過酸化水素及びジアミノベンジジンと併用する。抗体をビオチンで標識し、アビジン又はストレプトアビジン結合の間接測定により検出することもできる。二次レポーターにより認識される所定ポリペプチドエピトープ(例えばロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)で抗体を標識してもよい。抗Nogo受容体−1抗体又はその抗原フラグメントを放射性標識アミノ酸で標識してもよい。放射性ラベルは診断目的と治療目的の両者に使用することができる。放射性標識抗Nogo受容体−1抗体は例えば対象におけるNogo受容体−1濃度を測定するために診断用として使用することができる。更に、放射性標識抗Nogo受容体−1抗体は脊髄損傷を治療するために治療用として使用することもできる。
【0058】
ポリペプチド用ラベルの例としては限定されないが、放射性同位体ないし放射性核種である15N、35S、90Y、99Tc、111In、125I、131I、177Lu、166Ho、153Smが挙げられる。ポリエチレングリコール(PEG)、メチルもしくはエチル基又は糖鎖基等の化学基で抗Nogo受容体−1抗体又はその抗原フラグメントを誘導体化してもよい。これらの基は抗体の生物学的特性を改善するため、例えば血清半減期を延長するため又は組織結合を増加するために有用であると思われる。更に、ポリペプチド用ラベルとしては核酸も挙げられ、例えばPCRによる検出用又は遺伝子発現強化用のDNAや、NgRを保持する細胞又は組織における遺伝子発現を抑制するためのsiRNAも挙げられる。
【0059】
抗Nogo受容体−1抗体のクラス及びサブクラスは当分野で公知の方法のいずれかにより決定することができる。一般に、抗体のクラス及びサブクラスは特定クラス及びサブクラスの抗体に特異的な抗体を使用して決定することができる。このような抗体は市販されている。クラス及びサブクラスはELISA、ウェスタンブロット及び他の技術により決定することができる。あるいは、クラス及びサブクラスは抗体の重鎖及び/又は軽鎖の定常領域の全部又は一部を配列決定し、そのアミノ酸配列を種々のクラス及びサブクラスの免疫グロブリンの既知アミノ酸配列と比較し、抗体のクラス及びサブクラスを決定することにより決定することができる。
【0060】
抗NgR抗体によるNgR活性の阻害
本願の抗Nogo受容体−1抗体又はその抗原結合フラグメントはリガンドとNgRの結合を阻害する。このような阻害のIC50は当分野で公知のいずれかの方法、例えばELISA、RIA又は機能的拮抗により測定することができる。IC50は0.01〜100nMとすることができる。好ましくは、IC50は1〜10nMである。より好ましくは、本発明の抗Nogo受容体−1抗体又はその抗原結合フラグメントのIC50は0.1nM〜1nMである。最も好ましくは、IC50は0.1nM未満である。
【0061】
二重可変領域抗体
本明細書で使用する二重可変領(DVD)結合性抗体とは2個以上の抗原結合部位を含む結合性蛋白質であり、4価又は多価結合性抗体である。「多価結合性抗体」なる用語は本明細書では2個以上の抗原結合部位を含む結合性蛋白質の意味で使用する。多価結合性抗体は3個以上の抗原結合部位をもつように構築することが好ましく、一般に天然抗体以外のものである。「多重特異性結合性抗体」なる用語は2個以上の関連又は無関連ターゲットと結合することが可能な結合性蛋白質を意味する。このようなDVDは単一特異性、即ち1種類の抗原としか結合できないものでもよいし、多重特異性、即ち2種類以上の抗原と結合できるものでもよい。2個の重鎖DVDポリペプチドと2個の軽鎖DVDポリペプチドを含むDVD結合性抗体をDVD Igと言う。DVD Igの各半分は重鎖DVDポリペプチドと軽鎖DVDポリペプチドと2個の抗原結合部位を含む。各結合部位は重鎖可変領域と軽鎖可変領域を含み、抗原結合部位1個当たり合計6個のCDRが抗原結合に関与している。DVD結合性蛋白質とDVD結合性蛋白質の作製方法は米国特許出願第11/507,050号に開示されている。本発明はNgRと結合することが可能な結合性蛋白質を含むDVD結合性蛋白質を含むものとする。好ましくは、DVD結合性蛋白質はNgRと第2のターゲットとに結合することが可能である。第2のターゲットは反発性ガイダンス分子(RGM)、Nogo−A、MAG、OMgp、CSPGから構成される群から選択される。CSPGとしては、アグリカン、ブレビカン、バーシカン、ニューロカン、ホスファカン又はTe38から選択することができる。従って、これらの例はミエリン由来阻害剤と、NgRの公知ニューロン共受容体を含む。
【0062】
二重特異性抗体
本願は「二重特異性抗体」技術についても記載する。二重特異性抗体はアゴニスト、アンタゴニスト又は両者の各種組み合わせとして機能することができる。本明細書で使用する「アゴニスト」なる用語は該当分子と接触した場合に、アゴニストの不在下で観察される活性又は機能の強さに比較して分子の所定活性又は機能の強さを増加するモジュレーターを意味する。本明細書で使用する「アンタゴニスト」又は「阻害剤」なる用語は該当分子と接触した場合に、アンタゴニストの不在下で観察される活性又は機能の強さに比較して分子の所定活性又は機能の強さを低下させるモジュレーターを意味する。特定該当アンタゴニストとしては、Nogo−66の生物活性を阻害又は調節するものが挙げられる。Nogo−66のアンタゴニスト及び阻害剤としては限定されないが、Nogo−66受容体(NgR)と相互作用する任意分子、好ましくはモノクローナル抗体が挙げられる。
【0063】
なお、NgRとの相互作用の結果、受容体又は他のリガンド/細胞膜成分の結合と中和を生じる場合があり、複数の疾患に対する付加的又は相乗的作用に有用であると思われる。
【0064】
本願はNgRとp75、NgRとTROY、NgRとLINGO−1等のNgR抗体とNgR共受容体の組み合わせについても記載する。本願はNgRとそのリガンド及びNgRとミエリン由来阻害因子に交差反応する抗体も含む。これらの抗体としては、NgRと反発性ガイダンス分子(RGM)、NGRとNogo−A、NGRとMAG、NgRとOMpg、NgRとCSPGに交差反応する抗体が挙げられる。CSPGとしては、アグリカン、ブレビカン、バーシカン、ニューロカン、ホスファカン又はTe38から選択することができる。従って、これらの例はミエリン由来阻害剤と、NgRの公知ニューロン共受容体を含む。
【0065】
本願はNgRと成長因子受容体の二重特異性抗体についても記載し、このような成長因子としては限定されないが、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、上皮成長因子(EGF)、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、ニューロトロフィン、血小板由来成長因子(PDGF)、エリスロポエチン(EPO)、トロンボポエチン(TPO)、ミオスタチン(GDF−8)、増殖分化因子−9(GDF9)、塩基性線維芽細胞増殖因子(bFGF又はFGF2)、グリア細胞由来神経栄養因子(GDNF)、毛様体神経栄養因子(CNTF)が挙げられる。
【0066】
抗体の使用
本願の中和抗体又はその部分はヒトNgRと結合することができるので、酵素免疫測定法(ELISA)、放射免疫測定法(RIA)又は組織免疫組織化学等の慣用イムノアッセイを使用して(例えば血清や血漿等の生体サンプル中で)ヒトNgRを検出するために使用することができる。本願は、生体サンプル中のヒトNgRの検出方法として、生体サンプルを本発明の抗体又は抗体部分と接触させる段階と、及び、ヒトNgRと結合した抗体(又は抗体部分)又は未結合抗体(又は抗体部分)を検出することにより、生体サンプル中のヒトNgRを検出する段階とを含む方法を提供する。抗体は結合抗体又は未結合抗体の検出を容易にするために検出可能な物質で直接又は間接的に標識する。適切な検出可能な物質としては各種酵素、補欠分子族、蛍光材料、発光材料及び放射性材料が挙げられる。適切な酵素の例としては西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−ガラクトシダーゼ又はアセチルコリンエステラーゼが挙げられ;適切な補欠分子族複合体の例としてはストレプトアビジン/ビオチンとアビジン/ビオチンが挙げられ;適切な蛍光材料の例としてはウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド又はフィコエリスリンが挙げられ;発光材料の1例としてはルミノールが挙げられ;適切な放射性材料の例としては3H、14C、35S、90Y、99Tc、111In、125I、131I、177Lu、166Ho、153Smが挙げられる。
【0067】
本願の抗体及び抗体部分はヒトNgR活性をインビトロ及びインビボの両者で中和できることが好ましい。従って、本発明のこのような抗体及び抗体部分はNogo−66とNgRの結合又はそれに起因する活性を阻害するために使用することができる。
【0068】
別の態様において、本願は対象、有利にはNgRに起因する活性が有害である疾患又は障害に罹患した対象におけるNogo−66活性又はNgR活性を低下させる方法を提供する。本願はこのような疾患又は障害に罹患した対象におけるNgR活性を低下させる方法として、対象のNgR活性を低下させるように本願の抗体又は抗体部分を対象に投与する段階を含む方法を提供する。好ましくは、NgRはヒトNgRであり、対象はヒト対象である。あるいは、対象は本発明の抗体が結合することが可能なNgRを発現する哺乳動物でもよい。更に、対象はNgRを導入した哺乳動物でもよい。本願の抗体は治療目的でヒト対象に投与することができる。更に、本願の抗体は獣医学的目的のため又はヒト疾患の動物モデルとして抗体が結合することが可能なNgRを発現する非ヒト哺乳動物に投与することもできる。後者に関して、このような動物モデルは本発明の抗体の治療効果の評価(例えば投与量及び投与時間の試験)に有用であると思われる。
【0069】
本明細書で使用する「NgR活性が有害である障害」なる用語はこの障害に罹患した対象におけるNgR又はそれに起因する活性の存在が障害の病理生理の原因である又は障害の増悪に寄与する因子であることが分かっている又はその疑いのある疾病及び他の障害を包含するものとする。従って、NgR活性が有害である障害はNgR活性の低下が障害の症状及び/又は進行を緩和すると予想される障害である。本発明の抗体で治療することができる障害の非限定的な例としては本発明の抗体の医薬組成物に関する下記セクションに記載する障害が挙げられる。
【0070】
NgRは神経変性又は神経変性プロセスの阻害に関連する神経疾患を伴うため、麻痺を生じる各種疾患に関連する病理において重要な役割を果たすことが認められている。このような疾患としては、筋委縮性側索硬化症、腕神経叢損傷、外傷性脳損傷を含む脳損傷、脳性麻痺、フリードライヒ運動失調症、ギラン・バレー症候群、白質ジストロフィー、多発性硬化症、ポリオ後遺症、二分脊椎、脊髄損傷、脊髄性筋萎縮症、脊髄腫瘍、脳卒中、横断性脊髄炎が挙げられる。更に、NgRは認知症、老人性認知症、軽度認知障害、アルツハイマー関連認知症、ハンチントン舞踏病、遅発性ジスキネジア、運動亢進症、躁病、梅毒性パーキンソン病、スチール・リチャードソン症候群、ダウン症候群、重症筋無力症にも役割を果たすことが認められている。
【0071】
NgRとそのリガンドは既知炎症因子を伴う炎症又は自己免疫状態の発生と進行にも関与していると思われる(Teng & Tang,2005;Fontoura & Steinmann,2006)。これらの疾患として、限定されないが、関節リウマチ、変形性関節症、若年性慢性関節炎、敗血症性関節炎、ライム関節炎、乾癬性関節炎、反応性関節炎、脊椎関節症、全身性エリテマトーデス、クローン病、潰瘍性大腸炎、炎症性腸疾患、インスリン依存性糖尿病、甲状腺炎、喘息、アレルギー疾患、乾癬、皮膚炎、強皮症、移植片対宿主病、臓器移植拒絶反応、臓器移植に伴う急性又は慢性免疫疾患、サルコイドーシス、アテローム性動脈硬化症、播種性血管内凝固、川崎病、グレーブス病、、ネフローゼ症候群、慢性疲労症候群、ウェゲナー肉芽腫症、シェーンライン・ヘノッホ紫斑病、顕微鏡的腎血管炎、慢性活動性肝炎、ブドウ膜炎、敗血性ショック、毒素性ショック症候群、敗血症症候群、悪液質、感染性疾患、寄生虫症、後天性免疫不全症候群、急性横断性脊髄炎、ハンチントン舞踏病、パーキンソン病、アルツハイマー病、脳卒中、原発性胆汁性肝硬変、溶血性貧血、悪性腫瘍、心不全、心筋梗塞、アジソン病、散発性I型多腺性内分泌不全症及びII型多腺性内分泌不全症、シュミット症候群、成人(急性)呼吸窮迫症候群、脱毛症、先天性脱毛症、血清反応陰性関節症、関節症、ライター病、乾癬性関節症、潰瘍性大腸炎性関節症、腸病性滑膜炎、クラミジア、エルシニア及びサルモネラ関連関節症、脊椎関節症、アテローム性疾患/動脈硬化症、アトピー性アレルギー、自己免疫性水疱症、尋常性天疱瘡、紅斑性天疱瘡、類天疱瘡、線状IgA疾患、自己免疫性溶血性貧血、クームス陽性溶血性貧血、後天性悪性貧血、若年性悪性貧血、筋痛性脳炎/ロイヤルフリー病、慢性粘膜皮膚カンジダ症、巨細胞性動脈炎、原発性硬化性肝炎、特発性自己免疫性肝炎、後天性免疫不全症候群、後天性免疫不全症関連疾患、B型肝炎、C型肝炎、分類不能型免疫不全症(分類不能型低ガンマグロブリン血症)、拡張型心筋症、女性不妊症、卵巣不全、早発卵巣不全、線維性肺疾患、特発性線維化性肺胞炎、炎症後間質性肺疾患、間質性肺炎、結合組織病関連間質性肺疾患、混合性結合組織病関連肺疾患、全身性硬化症関連間質性肺疾患、関節リウマチ関連間質性肺疾患、全身性エリテマトーデス関連肺疾患、皮膚筋炎/多発性筋炎関連肺疾患、ショーグレン病関連肺疾患、強直性脊椎炎関連肺疾患、血管炎性びまん性肺疾患、ヘモジデリン沈着症関連肺疾患、薬物誘発性間質性肺疾患、線維症、放射線線維症、閉塞性細気管支炎、慢性好酸球性肺炎、リンパ球浸潤性肺疾患、感染後間質性肺疾患、通風性関節炎、自己免疫性肝炎、1型自己免疫性肝炎(旧称自己免疫性又はルポイド肝炎)、2型自己免疫性肝炎(抗LKM抗体肝炎)、自己免疫性低血糖症、黒色表皮症を伴うB型インスリン抵抗性、副甲状腺機能低下症、臓器移植に伴う急性免疫疾患、臓器移植に伴う慢性免疫疾患、変形性関節症、原発性硬化性胆管炎、1型乾癬、2型乾癬、特発性白血球減少症、自己免疫性好中球減少症、腎疾患NOS、糸球体腎炎、腎臓の顕微鏡的血管炎、ライム病、円板状エリテマトーデス、特発性男性不妊症ないしNOS、精子自己免疫、多発性硬化症(全サブタイプ)、交感性眼炎、結合組織病続発性肺高血圧症、グッドパスチャー症候群、結節性多発性動脈炎の肺症状、急性リウマチ熱、リウマチ性脊椎炎、スティル病、全身性硬化症、ショーグレン症候群、高安動脈炎、自己免疫性血小板減少症、特発性血小板減少症、自己免疫性甲状腺疾患、甲状腺機能亢進症、甲状腺腫性自己免疫性甲状腺機能低下症(橋本病)、萎縮性自己免疫性甲状腺機能低下症、原発性粘液水腫、水晶体起因性ブドウ膜炎、原発性血管炎、白斑急性肝疾患、慢性肝疾患、アルコール性肝硬変、アルコール性肝傷害、胆汁鬱滞症、特異体質性肝疾患、薬物誘発性肝炎、非アルコール性脂肪肝炎、アレルギー及び喘息、B群連鎖球菌(GBS)感染症、精神障害(例えば鬱病及び統合失調症)、Th2型及びTh1型疾患、急性及び慢性疼痛(各種疼痛)、及び癌(例えば肺癌、乳癌、胃癌、膀胱癌、結腸癌、膵臓癌、卵巣癌、前立腺癌及び直腸癌)及び血液悪性疾患(白血病及びリンパ腫)が挙げられる。本願のヒト抗体又は抗体部分は自己免疫疾患、特に炎症に関連するもの(例えばリウマチ性脊椎炎、アレルギー、自己免疫性糖尿病、自己免疫性ブドウ膜炎)に罹患したヒトを治療するために使用することができる。
【0072】
NgRが他の蛋白質と相互作用することも知られており、このような蛋白質として、限定されないが、細胞接着、細胞移動、細胞トラッキング、軸索経路検出に関連する蛋白質及び細胞外マトリックス蛋白質が挙げられる。本願に含まれる可能な治療薬組み合わせとしては、NgRに対する抗体とセマフォリン(特にSema−1a,1b;Sema−2a;Sema3A,B,C,D,E,F;Sema4A,D;Sema5A;Sema6D;Sema7A;Sema VA)、プレキシン(プレキシン−A1−4,プレキシン−B1−3,プレキシン−C1,プレキシン−D1,Tim−2)、ニューロピリン(ニューロピリン−1及びニューロピリン−2)、カドヘリン(E−カドヘリン及びN−カドヘリン)、ネトリン(ネトリン−1)、エフリン(EphA3,4,6,7,8;B2,B3)Eph受容体、Ephリガンド、Ig CAM、テナスシン−C、CSPG、テナスシン、Sema3A、フィブロネクチン、ラミニン−1、コラーゲン(例えばコラーゲン−IV)、Robo、Abl、N−カドヘリン、L1、NCAMが挙げられる。
【0073】
NgRは細胞外アミロイド前駆体蛋白質フラグメント(Aβ)と相互作用することも記載されている。従って、本願はNgRとAβ種(Aβ1−40、Aβ1−42、Aβオリゴマー、Aβマルチマー、Aβグロブロマー)に対する抗体間の組み合わせを含む。この型の併用療法はアルツハイマー病の治療に有用であると思われる。この組み合わせは神経突起伸長/神経保護に加え、AD患者におけるプラーク沈着と認知能力の改善という二重の効果があると思われる。軸索及び樹状突起の萎縮はアルツハイマー病の非常に初期の特徴であり、併用治療が非常に有効であると思われる。
【0074】
更に、従来記載されているように、上記パートナーの任意1種間の二重特異性抗体も使用することができる。上記のようなこのような抗体製剤は、アルツハイマー病、パーキンソン病、脊髄損傷、外傷性脳損傷、多発性硬化症、末梢神経損傷、統合失調症、鬱病、不安症、加えて上記の可塑性及び神経突起成長及び神経毒性に関連する疾患のいずれものの治療に有用であると思われる。
【0075】
本願の抗体は細胞内標的蛋白質に到達するように膜貫通導入を可能にするペプチドと併用してもよい。このようなペプチド配列として、限定されないが、tat、アンテナペディア、ポリアルギニン、所定の抗菌ペプチドが挙げられる。このようなペプチドは細胞原形質膜のみならず、血液脳関門、腸粘膜、髄膜等の上皮及び内皮膜も含めた膜を通して導入を可能にする。
【0076】
このようなペプチドは細胞シグナル伝達阻害剤を細胞に導入することもでき、このような阻害剤としてはROCK、低分子量GTPアーゼ、アクチン及びミエリン安定剤等のNgRシグナル伝達分子に対する抗体又は小分子が挙げられる。
【0077】
本願の抗体又は抗体部分は前段落に記載したようなNgR活性が関与する障害の治療に有用な1種以上の補助的な小分子治療剤と併用投与することもできる。当然のことながら、本願の抗体又はその抗原結合部分は単独で使用することもできるし、補助剤(例えば,治療剤)と併用することもでき、前記補助剤はその所期目的に合わせて当業者により選択される。例えば、補助剤は本発明の抗体により治療される疾患又は病態を治療するために有用であると当業者に認められている治療剤とすることができる。補助剤は治療用組成物に有益な属性を付与する物質(例えば組成物の粘性を変化させる物質)でもよい。好ましい組み合わせとしては限定されないが、抗精神病薬が挙げられ、限定されないが、例えばリスペリドン、オランザピン、クエチアピン、フェノチアジン(クロルプロマジン、フルフェナジン、レボメプロマジン、ペリシアニン、ペルフェナジン、プロクロルペラジン、プロマジン、チオリダジン、トリフルオペラジン)、ブチロフェノン(ベンペリドール、ハロペリドール)、ゾテピン、ロキサピン、アリピプラゾール、セルトラリン、ジプラシドン、Rhoキナーゼ活性の小分子阻害剤(ROCK)(例えばファスジル、ジメチルファスジル又は他の任意ROCK阻害剤)、GABA A受容体又は代謝型グルタミン酸受容体(mGluRs)に対する低分子量受容体リガンド、非ステロイド性抗炎症薬(NSAIDS)、抗炎症性コルチコステロイド(例えばメチルプレドニゾロン)が挙げられる。
【0078】
本発明の医薬組成物
本願の抗体及び抗体部分は対象への投与に適した医薬組成物に配合することができる。本願の医薬組成物は「治療有効量」又は「予防有効量」の本発明の抗体又は抗体部分を含有することができる。「治療有効量」とは必要な用量と期間で所望治療結果を達成するために有効な量を意味する。抗体又は抗体部分の治療有効量は、当業者が決定することができ、個体の疾患状態、年齢、性別及び体重並びに抗体又は抗体部分が個体に所望応答を誘発する能力等の因子により異なる。治療有効量は、抗体又は抗体部分の有益な治療作用が毒性又は有害作用を上回る量でもある。「予防有効量」とは、必要な用量と期間で所望予防結果を達成するために有効な量を意味する。一般に、予防用量は、初期疾患ステージ又はそれ以前の対象に使用するので、予防有効量は治療有効量よりも少なくなる。
【0079】
一般に、医薬組成物は本発明の抗体又は抗体部分と医薬的に許容可能なキャリヤーを含有する。本明細書で使用する「医薬的に許容可能なキャリヤー」は生理的に適合可能な全溶媒、分散媒、コーティング、抗細菌及び抗真菌剤、等張及び吸収遅延剤等を包含する。医薬的に許容可能なキャリヤーの例としては水、食塩水、リン酸緩衝食塩水、デキストロース、グリセロール、エタノール等の1種以上とその組み合わせが挙げられる。多くの場合には、例えば糖類、多価アルコール(例えばマンニトール、ソルビトール)又は塩化ナトリウム等の等張化剤を組成物に加えることが好ましい。医薬的に許容可能なキャリヤーとしては更に抗体又は抗体部分の保存期間又は効力を増す微量の補助物質(例えば湿潤剤、乳化剤、防腐剤又は緩衝液)も挙げられる。
【0080】
本願の組成物は各種形態を取ることができる。これらの形態としては、例えば液体、半固体及び固体剤形が挙げられ、例えば溶液(例えば注射溶液及び輸液溶液)、分散液、懸濁液、錠剤、ピル、散剤、リポソーム及び座剤が挙げられる。好ましい剤形は所期投与方法及び治療用途により異なる。典型的な好ましい組成物は他の抗体をヒトに受動免疫するために使用されているものと同様の組成物等の注射溶液又は輸液溶液の形態である。好ましい投与方法は非経口投与(例えば静脈内、皮下、腹膜内、筋肉内)である。好ましい1態様では、静脈内輸液又は注射により抗体を投与する。別の好ましい態様では、筋肉内又は皮下注射により抗体を投与する。更に別の好ましい態様としては抗体の髄腔内投与が挙げられる。
【0081】
治療用組成物は一般に製造及び保存条件下で無菌安定でなければならない。組成物は溶液、マイクロエマルション、分散液、リポソーム又は高薬剤濃度に適した他の規則的構造として製剤化することができる。滅菌注射溶液は必要量の活性化合物(即ち抗体又は抗体部分)を必要に応じて上記成分の1種又はその組み合わせと共に適切な溶媒に加えた後に濾過滅菌することにより製造することができる。一般に、分散液は塩基性分散媒と上記から選択される必要な他の成分を含有する滅菌ビークルに活性化合物を加えることにより製造される。滅菌注射溶液の調製用滅菌凍結乾燥粉末の場合には、好ましい製造方法は予め滅菌濾過しておいたその溶液から活性成分と所望の付加成分のいずれかの粉末を生成する真空乾燥と噴霧乾燥である。例えばレシチン等のコーティングの使用、分散液の場合には必要な粒度の維持及び界面活性剤の使用により、溶液の適正な流動性を維持することができる。吸収を遅らせる物質(例えばモノステアリン酸塩やゼラチン)を組成物に加えることにより、注射用組成物の長期吸収が可能になる。
【0082】
本発明の抗体及び抗体部分は当分野で公知の各種方法により投与することができるが、多くの治療用途に好ましい投与経路/方法は皮下注射、静脈内注射又は輸液である。当業者に自明の通り、投与経路及び/又は方法は所望結果により異なる。ある態様において、化合物の迅速な放出を防止するキャリヤーを活性化合物に配合することができ、例えばインプラント、経皮パッチ及びマイクロカプセル送達システム等の制御放出製剤が挙げられる。エチレン酢酸ビニル、ポリ酸無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル及びポリ乳酸等の生分解性生体適合性ポリマーを使用することができる。このような製剤の多数の製造方法が特許登録されており、あるいは一般に当業者に公知である。例えばRobinson編,Sustained and Controlled Release Drug Delivery Systems,Marcel Dekker,Inc.,New York,1978参照。
【0083】
ある態様において、本願の抗体又は抗体部分を例えば不活性希釈剤又は同化性可食性キャリヤーと共に経口投与することができる。化合物(及び所望により他の成分)を硬又は軟シェルゼラチンカプセルに封入してもよいし、錠剤に圧縮してもよいし、対象の食事に直接添加してもよい。経口治療投与には、化合物に賦形剤を添加し、内服錠、口腔錠、トローチ剤、カプセル剤、エリキシル剤、懸濁液剤、シロップ剤、ウェハース剤等の形態で使用することができる。本発明の化合物を非経口投与以外の方法で投与するためには、その不活性化を防止するための材料を化合物に被覆したり、化合物と併用投与することが必要な場合がある。
【実施例】
【0084】
以下、実施例により本願を更に説明するが、これらの実施例は本願の例証のみを目的とし、その範囲を限定するものではない。
【0085】
(実施例1)
抗体の作製
A/Jマウス(The Jackson Laboratories,Bar Harbor Me)が組換えヒト及びラットNgR蛋白質(夫々配列番号1a及び配列番号2a)により免疫された。組換えヒト又はラットNgR蛋白質50ugを初回注射では完全フロイントアジュバントに加え、2回目以降はImmuneasy(登録商標)(Qiagen)に加えてマウスに4回皮下免疫した。融合の4日前に、マウスに抗原10ugを静脈内注射した。融合については、ケーラーとミルスタインの標準技術(Kohler G and Milstein C;Nature Vol.256,495−497頁(1975))を使用して免疫動物からの脾細胞をSP2/0−Ag14骨髄腫細胞と5:1の比で融合させた。融合から7〜10日後に、顕微鏡でハイブリドーマコロニーが観察されたので、上清をELISAアッセイにより試験した。PBS中1ug/mlの組換えヒト又はラットNgR蛋白質をELISAプレートに4℃で一晩被覆し、室温で1時間ブロックした。希釈した上清を温置し、結合をヤギ抗マウスIgFc−HRPコンジュゲートで検出した。ELISAで陽性の抗体産生ハイブリドーマ細胞をスケールアップし、限界希釈法によりサブクローニングした。Zymed EIAアイソタイプキットを使用して抗体のアイソタイプを決定した。
【0086】
各種ELISAフォーマットが確立されており、ヒト、ラット又はマウスNgRと結合する抗体を同定するための最初のスクリーニングとして日常的に使用されている。ELISAアッセイで反応した抗体を次に、組換えヒトNgR又は組換えラットNgRを安定に発現するHEK又はCHO細胞と結合させ、形質移入していない細胞又は対照細胞と結合しないものについて試験した。ELISAフォーマット用として、可溶性受容体を作製した(上記参照)。FACS試験には、全長NgR蛋白質を組換え細胞株で発現させた。ELISA結合アッセイとFACSアッセイにおけるMab50(配列番号3及び配列番号4)とMab51(配列番号5及び配列番号6)の結果を表2に示す。
【0087】
【表2】
【0088】
同一実験プロトコールを使用して他の抗体Mab1、Mab52、Mab53、Mab54、Mab55、Mab56、Mab57、Mab58、Mab59、Mab60、Mab61及びMab62を得た。
【0089】
(実施例2)
ELISAによる抗体特異性と結合親和性の測定
実施例1で作製したモノクローナル抗体の特異性を測定するために、NgR−Fc(ヒトNgRのアミノ酸Met1〜Ser447とヒトFcフラグメントを含む融合蛋白質(R&D Systems))とラットNgR(配列番号2a)を使用してELISAを実施した。2種のリガンドを96ウェルマイクロタイタープレート(Nunc Maxisorb;0.2μg/ウェル)に固定化した。ブロッキング試薬として、Tris−HCL,pH7.2中2%ウシ血清アルブミン(BSA)を室温で2時間使用した。モノクローナル抗体は10,000ng/mlの濃度から出発して使用した。西洋ワサビペルオキシダーゼ(Sigma)で標識した二次抗マウス抗体で結合抗体を検出し、3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して発色させた。mAb50とmAb51はヒトNgRと特異的に結合した。ヒトNgR−Fc 0.2μg/ウェルを使用した場合には、どちらのモノクローナル抗体でも20ng/ml未満の濃度で50%結合が認められた(図1A)。同様の実験でmAb50とmAb51はラットNgRのアミノ酸から構成されるポリペプチドにも特異的に結合した。96ウェルマイクロタイタープレートのウェル当たり0.2μgのラットNgRを使用した場合には、どちらのモノクローナル抗体でも20ng/ml未満の濃度で50%結合が認められた(図1B)。本発明のその他のモノクローナル抗体(mAB)もヒト及びラットNgRと結合した。各抗体で得られたシグナルの差から結合親和性に相違があると思われた(表3参照)。
【0090】
【表3】
【0091】
(実施例3)
ドットブロットとウェスタンブロットを使用した可溶性ヒト及びラットNgRとの抗体結合の特性決定
ドットブロットのために、異なる濃度の蛋白質2μlを乾燥ニトロセルロース膜上のTTBS緩衝液にスポットした。ウェスタンブロットのために、20%メタノールを添加したNovex転写用緩衝液に濾紙とニトロセルロースを10分間浸漬した。ブロッティングはNovexチャンバーで定電流(100mA)下に2時間室温で実施した。
【0092】
スポット当たりの蛋白質の添加量は、
a)100μg/ml〜〜200ng/スポット
b)50μg/ml〜〜100ng/スポット
c)10μg/ml〜〜20ng/スポット
d)5μg/ml〜〜10ng/スポット
e)1μg/ml〜〜2ng/スポット
f)500ng/ml〜〜1ng/スポット
とした。プローブをスポット後、膜を10分間室温で乾燥した後、免疫検出プロトコールを開始した。
【0093】
試験した全モノクローナル抗体はヒト及びラットNgRと結合した。異なる抗体で得られたシグナル間の差から結合親和性に相違があると思われた。(MAB61はニトロセルロース膜のプローブに使用した条件下でブロット上に高いバックグラウンドを示した。)NgR「対照」に対する抗体を除去するとシグナルを示さなかったので、結合は抗体依存性であった。
【0094】
モノクローナル抗体はウェスタンブロットにて変性NgRと様々に反応した。MAB1のみがヒト及びラットNgRに顕著なシグナルを示した。抗体の結合能の対照として、蛋白質をSDSゲルから転写させた後に非変性ヒトNgRと非変性ラットNgRを含むドットをニトロセルロース膜にスポットした。これらのドットを陽性対照として利用した処、全モノクローナル抗体は同一ニトロセルロース膜上の非変性蛋白質と結合することが確認された。結果を表4にまとめる。
【0095】
【表4】
【0096】
(実施例4)
AP−Nogo66と可溶性ヒトNogo受容体(hNgR−Fc)の結合の競合
モノクローナル抗体を更に特性決定するために、実施例2の競合アッセイと同様の結合アッセイを使用し、実施例1で作製したmAbの、AP−Nogo66のヒトNgR−Fcへの結合を阻害する能力を試験した。ヒトNgR−Fc(0.2μg/ウェル)を50mM炭酸Na緩衝液(pH9)中で96ウェルマイクロタイタープレート(Nunc Maxisorb)に4℃で一晩固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。ウェルに一定濃度のAP−Nogo66(終濃度Tris−HCL,pH7.2+0.1%BSA中0.15nM)を加え、Ab濃度を徐々に増加させた。プレートを90分間室温で温置した。各温置段階中に、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05%Tween20)で洗浄した。AP−Nogo66の結合をAttoPhos基質(Roche)で検出し、蛍光単位をPolarstar(BMG)計器で測定した。図2はmAb50とmAb51について0分と30分で行った測定の相対蛍光単位(RFU)を示す。上記結合はmAb50とmAb51により完全に阻害され、AP−Nogo66とヒトNgR−Fcの結合を50%阻害するためにはどちらの抗体も10倍モル過剰量が必要であった。mAb1を除く他の抗体mab52、mAb53、mAb54、mAb55、mAb56、mAb57、mAb58、mAb59、mAb60、mAb61及びmAb62もmAb50及び51で認められたと同様の濃度範囲でAP−Nogo66と可溶性Nogo受容体の結合を阻害した。
【0097】
(実施例5)
HEK293f細胞におけるAP−Nogo66とヒト及びラットNgRの結合のmAB50及びmAb51による競合
実施例1に記載したように作製したmAbの結合特性と競合特性を更に特性決定するために、ヒト又はラットNgRを一過的に発現するHEK293f細胞の懸濁液を使用した。形質移入から48時間後に細胞を96ウェルマイクロタイタープレート(Nunc Maxisorb)に撒き、リン酸緩衝食塩水(1%BSAを添加したPBS)で洗浄した。1μg/200μlのAP−Nogo66(終濃度40nM)と種々の濃度のモノクローナル抗体(20μg、4、0.8及び0.16μg/200μl)を加え、4℃で1時間温置した。同一アイソタイプのモノクローナル抗体を陰性対照として使用した。ZenonマウスIgG2a標識キット(Invitrogen)を使用してAlexa Fluorで標識した抗アルカリホスファターゼ抗体(Sigma)でAP−Nogo66の結合を検出した。二次抗体を4℃で1時間温置した。温置後に、細胞をPBSで洗浄し、FACS分析した。20μg/200μl濃度(20倍モル過剰)では、ポリクローナルNgR抗体とNgR−Fc(いずれもR&D Systems)はAP−Nogo66結合を60%〜70%阻害し、mAb50及び51はAP−Nogo66結合を約90%阻害した。mAb50及びmAB51をラット及びヒトNgR中に20μgの濃度で使用した場合の抗体アイソタイプ対照を図3aに示す。抗体アイソタイプ対照は黒線(左端のシグナル)で示し、抗体不在下のAP−Nogo66結合は赤線(右端シグナル)で示す。ヒト及びラットNgRはいずれも同等にAP−Nogo66と結合した。AP−Nogo66蛍光はヒト及びラットNgRを発現するHEK293f細胞で両者抗体の存在下では低強度側に移動する(斜線部;緑色)。異なる抗体濃度を使用すると、コルモゴロフ・スミルノフ(K−S)アッセイにより測定した場合にヒト及びラットNgRの両者でAP−Nogo66に対して2倍モル過剰のモノクローナル抗体50及び51でIC50が認められた。
【0098】
図3bは種々の濃度、即ち20.0μg,4.0,0.8及び0.16μg/200μlのmAb50及びmAb51による結果を示す。本発明の他のmAbはmAb50及び51で認められたと同様の濃度範囲でAP−Nogo66とHEK293f細胞で発現させたNogo受容体の結合を阻害した。AP−Nogo66とHEK293f細胞で発現させたヒトNgRの結合の50%競合に必要な抗体量を示す表5参照。
【0099】
【表5】
【0100】
(実施例6)
NTera2細胞中でAP−Nogo66により誘導された神経突起伸長阻害のmAB50及びmAb51による中和
ヒトNTera−2細胞及び齧歯類(マウス皮質ニューロン、ラット皮質ニューロン及びラット小脳顆粒細胞ニューロン)細胞型と、リガンドとしてAP−Nogo66及びミエリンを使用してインビボ状況に近似する機能システムで神経突起伸長に及ぼす本発明の抗体の効果を試験した。
【0101】
NTera2細胞(ヒト奇形癌細胞株)はレチノイン酸を使用して神経突起様細胞に分化させることができ、この細胞はポリクローナル抗体とNgRの適度の結合(FACS結合)により示されるようにNgR mRNA(PCR)と細胞表面蛋白質を発現する。夫々アイソタイプ対照抗体(非斜線領域)とmAb50及びmAb51(斜線領域)(図4)を使用してFACS分析により細胞表面上のNTera−2細胞による天然ヒトNgRの発現を検出した。両者抗体とNtera−2細胞の表面で発現されるNgR細胞の結合を斜線領域により示す。
【0102】
これらの分化細胞における神経突起伸長又は成長円錐崩壊の分析はヒトニューロンに可能な限り近似するインビトロシステムである。NTera2細胞(German National Resource Center for Biologicals,DMSZ,Braunschweigから入手)を融解し、DMEM培地(Gibco #31966−021+10%ウシ胎仔血清(FCS)+5%ウマ血清(PS))を加えた175cm2培養フラスコ(Greiner bio−one #660175)に撒いた。数日間培養後、細胞を再培養した。このために、細胞をPBS(Gibco #14190−094)で1回洗浄し、トリプシン/EDTA(Gibco #25300−054)で洗浄し、トリプシン/EDTAの存在下で5分間温置した。分化のために、細胞を再懸濁し、175cm2フラスコでDMEM(Gibco 31966−021,Lot.Nr.3092594)+10%FCS+5%PS+1%ペニシリン/ストレプトマイシン(5000/5000単位/mL)+レチノイン酸(SIGMA #R2625)終濃度10μMに細胞2.5×106個を撒いた。
【0103】
分化のために、終濃度10μMのレチノイン酸(SIGMA #R2625)を3週間にわたって週2回NTera2細胞に加えた。21日間分化後に、細胞を再培養した。このために、細胞をPBSで1回洗浄し、トリプシン/EDTAで洗浄し、トリプシン/EDTAの存在下で5分間温置した。細胞を再懸濁し、1:6に分割し、175cm2フラスコ6本でDMEM(Gibco 31966−021,Lot.Nr.3092594)+10%FCS+5%PS+1%PenStrep)に撒き、2〜3日間培養した。2〜3日後に細胞をPBSで洗浄し、物理的に剥がし、5分間1000rpmで遠心し、Neurobasal培地(Gibco #21103−049)+2mM L−グルタミン(Gibco #25030−024)+ペニシリン/ストレプトマイシン+B27−サプリメントに再懸濁し、三角フラスコ(Corning #431143)で予備凝集させた。こうして、細胞106個/mLを2×15mL凝集培地(Neurobsalmedium(Gibco #21103−049)+2mM L−グルタミン(Gibco #25030−024)+ペニシリン/ストレプトマイシン+B27−サプリメント)に加え、37℃,5%CO2下に一晩温和に撹拌し、阻害基質と対照基質を事前被覆した96ウェルプレート(Biocoat Poly−D−Lysin Cellware 96−Well Black/Clear Plate Becton Dickinson #35 4640(35 6640))に撒いた。阻害基質では、96ウェルプレートの半数に滅菌PBS中AP−Nogo66(AP−Nogo66濃度は15μg/mLとした)+ラミニン(Sigma,L−2020,Lot 014K4060,(ストック溶液.1mg/mL);ウェル当たり最終ラミニン量20μg)100μLを事前被覆した。許容基質では、プレートの残りの半数にラミニン100μl(20μg)を被覆した。2時間温置後に、プレートをPBSで2回洗浄し、培地40μlを補充した各ウェルに予め凝集させた懸濁液50μlを撒いた。プレートを37℃で2時間温置し、最後に、予め希釈したmAb50又はmAb51溶液10μlを加え、最終抗体濃度1〜100μg/mLとした。細胞を37℃,5%CO2下に一晩温置し、翌日、2%パラホルムアルデヒド(SIGMA #P−6148)で固定し、その後の分析に備えて4℃で保存した。ソフトウェアAxioVision LE Rel.4.1を使用し、標準評価パラメーター(凝集面積及び凝集面積&神経突起成長面積)を使用して神経突起伸長の分析を実施した。
【0104】
上記方法によりNTera2凝集物からの神経突起伸長を定量した。結果を図5に示す通り、mAb50では2μg/ml、mAb51では1μg/mlで神経突起伸長の有意な改善を観測することができる。Nogo66処理に対する有意差:*=p値<0.05;***=p値<0.00.1。
【0105】
抗体Mab52、53、54、55、56、57、58、59、60、61及び62でも神経突起伸長阻害の改善が得られた。Mab1とMab4はAP−Nogo66による神経突起伸長の阻害を示さなかった。
【0106】
(実施例7)
hNgRの欠失変異体:抗体の発現、精製及び結合。
【0107】
GPIリンカー形成(450最終アミノ酸)と膜結合を阻止するようにC末端アミノ酸を欠失させると共に、分泌蛋白質量を潜在的に増加させるように分泌シグナルを使用することにより、hNgRを可溶性蛋白質として発現させた。使用した発現システムは293F細胞における発現プラスミドの一過的形質移入に基づくものとした。Ni−NTA(ニッケル−ニトリロ三酢酸)ビーズを使用することによりHisタグ付き分泌蛋白質を捕獲した。hisタグを付けた無関係の蛋白質(RGM A−His)を陰性対照として使用してPAGEとウェスタンブロットにより溶出後の蛋白質の純度とサイズを分析した。これらの調製物におけるNgR−蛋白質をより良好に調整するために、全欠失変異体を検出すると予想されるhNgRに対するポリクローナル抗体(AF1208)を使用してドットブロットによりNgRの量を測定した。異なるNgR欠失変異体に調整量の蛋白質を使用するドットブロットを実施した。上記抗体による免疫検出にドットブロットを使用した。
【0108】
NgR欠失変異体を発現する293F細胞の細胞培養上清を非精製NgR源として使用して抗体とNgR欠失変異体の結合を更に特性決定した。
(i)フルニエらによるNgR欠失変異体の作製(Truncated Soluble Nogo Receptor Binds Nogo−66 and Blocks Inhibition of Axon Growth by Myelin;Alyson E.Fournier,Graham C.Gould,Betty P.Liu,Stephen M.Strittmatter;The Journal of Neuroscience,October 15,2002,22(20):8876−8883)。ヒトNgRの7種類の欠失変異体が文献に記載されている(Fournierら)。これらの7種類の変異体構築物がpSecTag2A IgK/hNgR 27−450/Myc/Hisから作製された。この構築物はpSecTag2AベクターにIgKリーダーペプチドとC末端Myc及びHisタグに融合したヒトNgRのアミノ酸27−450のコーディング領域を含む。以下の構築物を作製した:
【0109】
【化1】
【0110】
pSecTag2AigK/27−310/Myc/Hisを除く全構築物はQuikChange II XL Site Directed Mutagenesis Kit(Stratagene,#200521)を使用して作製され、形質転換は大腸菌XL10 Gold細胞中にて実施された。pSecTag2AigK/NgR/27−310/Myc/Hisについて、コーディング配列の相当領域を増幅し、pSecTag2Aにクローニングした。NgRコーディング領域の異なる部分を欠失させるために以下の変異誘発及び増幅プライマーを使用した。
【0111】
【化2】
【0112】
図6の模式図参照。
【0113】
(ii)NgR欠失変異体の発現
ヒトNgRのC末端アミノ酸を欠失させると、膜固定性及び細胞上清中の可溶性受容体蛋白質の存在が失われる。N末端において、hNgRのシグナルペプチド(アミノ酸1−27)をベクター中にコードされているシグナルペプチドで置換した。従って、hNgRのアミノ酸27から開始し、hNgRのアミノ酸450を終点とするN末端及びC末端欠失変異体(hNgR27−450)をこれらの全変異体の基礎として使用し、細胞上清中に可溶性蛋白質の存在を可能にした。更に、これらの変異体にはこれらの蛋白質の精製を可能にするための短いアミノ酸タグを加えた。hNgR変異体のDNAを293F細胞で一過的に発現させた。72時間後に遠心により細胞上清を回収した。下記方法により蛋白質精製を実施した。ヒトNgRの7種類の異なる変異体をコードする以下のDNAによる形質移入を実施した。
【0114】
【化3】
【0115】
hNgR変異体のDNAを293F細胞で一過的に発現させた。
【0116】
(iii)Ni−キレート親和性(Ni−NTA)を使用したNgR蛋白質の精製
Ni−NTAスーパーフロービーズ(Qiagen Qiagen #1018611)を使用した。ビーズ上清を13500rpmで遠心し、上清を捨て、ビーズを新鮮なPBSに再懸濁することにより、ビーズをPBS(リン酸緩衝食塩水,Invitrogen)で3回洗浄した。ビーズ懸濁液200μlを細胞培養上清30mlに使用した。ビーズを細胞培養上清と共に4℃でローテータで60rpmにて温置し、温置後に遠心(10分間,3000rpm)し、ビーズをペレット化した。上清を捨て、ビーズをPBSで3回洗浄した。溶出用緩衝液(PBS,160mM NaCl,150mMイミダゾール)250μlを使用して結合性蛋白質をビーズから溶出させた。室温のローテータで30分間温置後、13.500rpmで3分間遠心することによりビーズをペレット化した。上清を除去した。溶出した蛋白質をその後の分析に備えて−20℃で凍結させた。hNgRの欠失変異体によるドットブロット実験からのデータを表6にまとめる。
【0117】
【表6】
【0118】
(実施例8A)
MAG−FcとNgR−Fcの結合の競合
モノクローナル抗体を更に特性決定するために、実施例2の競合アッセイと同様の結合アッセイを使用して実施例1で作製した2種類のmAbがMAG−FcとヒトNgR−Fcの結合を阻害する能力を試験した。NgR−Fc(0.2μg/ウェル,R&D Systems)を4℃の50mM炭酸Na緩衝液(pH9)中、96ウェルマイクロタイタープレート(Nunc Maxisorb)に固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。MAG−Fc(R&D Systems recombinant Rat MAG/Fc Chimera,Catalog #538−MG)をZenonヒトIgG標識キット(Molecular Probes)により西洋ワサビペルオキシダーゼで標識した。ウェルに一定濃度の標識MAG−Fc(Tris−HCL,pH7.2+0.1%BSA中終濃度50ng/ml)と、指定濃度のmAb50及びmAb51を加えた。未標識MAG−FcをNgR−Fc対照として使用してこれを60分間室温で温置した。各温置段階中、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05% Tween 20)で洗浄した。3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して標識MAG−Fcを発色させた。図7に示すように、mAb50(黒三角)とmAb51(白三角)はMAG−FcとヒトNgRの結合を阻害しなかった。MAG−FcとNgR−Fcは標識MAG−Fc(黒円)とヒトNgR−Fcの結合に競合した(ヒトNgRのアミノ酸Met1〜Ser447とヒトFcフラグメントを含む融合蛋白質(R&D Systems);黒四角)。従って、mAb50とmAb51はこれらの条件下でNgRとの結合に関してMAGと競合しなかった(図7)。他の抗体はいずれもこれらの条件下でNgRとの結合に関してMAGと競合しなかった。
【0119】
(実施例8B)
OMgpとNgR−Fcの結合の競合
モノクローナル抗体を更に特性決定するために、実施例8aの競合アッセイと同様の結合アッセイを使用して実施例1で作製した2種類のmAbの、oMgpのヒトNgR−Fcへの結合を阻害する能力を試験した。NgR−Fc(0.2μg/ウェル,R&D Systems)を4℃の50mM炭酸Na緩衝液(pH9)中、96ウェルマイクロタイタープレート(Nunc Maxisorb)に固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。ウェルに一定濃度のヒトOMgp(R&D Systems/1673−OM;Tris−HCL,pH7.2+0.1%BSA中終濃度500ng/ml)と、指定濃度のmAb50及びmAb51を加えた。この混合物を60分間室温で温置した。西洋ワサビペルオキシダーゼ(Roche)で標識した抗His抗体を使用してOMgpを検出した。各温置段階中、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05% Tween 20)で洗浄した。3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して標識抗His抗体を発色させた。
【0120】
表7に示すように、mAb50とmAb51はOMgpとヒトNgRの結合を部分的に阻害した(30〜40%範囲)。mAb52とmAb59以外の他の抗体も、100倍モル過剰を上回る80μg/mlまでの濃度でもOMgpとヒトNgRの結合を部分的に阻害した(表7)。
【0121】
【表7】
【0122】
(実施例9)
ラットDRG細胞におけるAP−Nogo66により誘導した神経突起伸長阻害のmAB50及びmAb51による中和
生後3〜6日の子ラットからの後根神経節(DRG)を使用して神経突起伸長に及ぼすNgR抗体の作用を調べた。
【0123】
DRGを作製するために、子ラット6〜8匹を外科用鋏で断頭した。腹側臓器と椎骨を除去することにより腹側から脊柱を摘出した。次に、精密鋏により脊柱を縦方向に切開した。DRGを付けたまま脊髄を摘出し、PBSを加えた10cmペトリ皿に移した。2本の精密鉗子を使用することによりDRGから脊髄と結合神経線維を分離し、PBS 1mlを加えた35mmペトリ皿に移した。全DRGを採取後、コラゲナーゼのPBS溶液(4mg/mlコラゲナーゼタイプI,Worthington #CLS−1)0,5mlを加え、DRGを20〜30分間37℃で温置した。トリプシン溶液(PBS中0,5%トリプシン(SERVA #37290))0,5mlを加え、DRGを更に15〜25分間37℃で温置した。DRGを15mlチューブに移し、培地(DMEM Nut Mix F12(Gibco #31330−038)+5%FCS(Gibco,熱不活化)+5%ウマ血清(Sigma,熱不活化)+1%ペニシリン/ストレプトマイシン(Gibco #15140−122))10mlを加えた。DRGがチューブの底に沈降後、上清を除去した。パスツールピペットに3〜5回通した後、口径の小さいパスツールピペットに更に2〜3回通すことによりDRGを培地2ml中で解離させた。細胞塊の沈降後、解離した細胞を含む上清を新しいチューブに移した。5分間1000rpmで遠心することにより細胞を採取し、培地2mlに再懸濁し、カウントし、神経成長因子(NGF;終濃度62,5μg/ml,Roche #1014331)を加えた培地で所望細胞密度まで希釈した。ポリリジンを被覆し、更にコーティング溶液100μl(滅菌水50mlで希釈したラミニン(1mg/ml Sigma #L−2020)バイアル1本)を被覆しておいた96ウェルプレート(例えばBeckton Dickinson #356640)のウェル当たり80μlの容量で細胞4000〜7000個を撒いた後、30分間〜3時間37℃で温置した。細胞培養後、濃度を増加しながら容量10μlの抗体を加えた。CO2インキュベーターで37℃にて2時間温置後、10μlのAP−Nogo66を加えた。細胞を18〜30時間増殖させ、4%パラホルムアルデヒドのPBS(リン酸緩衝食塩水;Gibco #14190−094)溶液100μlを加えた後に4℃で少なくとも12時間温置することにより固定させた。96ウェルプレートの代わりに、ポリリジンを被覆したカバースリップ(例えばBecton Dickinson #354085)と24ウェルプレート(例えばFalcon #353047)を併用し、コーティング溶液500μlを加えることにより被覆した。抗βIIIチューブリン抗体(例えばAbeam #abl4545)とCy3標識二次抗体(例えばJackson ImmunoResearch #715−165−151)を併用して間接免疫蛍光により神経突起伸長を可視化した。ビスベンズイミド(H33258)を二次抗体に加えることにより神経を染色した。BDTM Pathway Bioimager(Beckton Dickinson)で10倍の倍率で顕微鏡写真を撮影し、AttoNO(Beckton Dickinson)ソフトウェアを使用して神経突起長を測定した。神経突起伸長を画像1枚当たりのDRG数に正規化した。図8は1例としてAP−Nogo66により誘導した神経突起伸長阻害のmAb50による中和を示す。図9は夫々抗体mAb50及びmAb51を使用した実験の結果を示す。AP−Nogo66を加えると、AP−Nogo66を加えない対照条件(1本目の棒グラフ)に比較して神経突起伸長の長さは著しく減少する(2本目の棒グラフ)。どちらの抗体もAP−Nogo66により誘導した神経突起伸長の阻害を用量依存的に中和した。DRG数に正規化した神経突起長は抗体を加えずにAP−Nogo66を加えた場合(2本目の棒グラフ)に比較してどちらの抗体でも50μg/mlで統計的に有意差を示す(最後の棒グラフ)。
【0124】
抗体Mab52、53、54、55、56、57、58、59、60、61及び62でも神経突起伸長阻害の改善が得られた。Mab1とMab4はAP−Nogo66による神経突起伸長の阻害を改善しなかった。
【0125】
これらの結果から、このような抗体は成長阻害環境で神経突起成長を刺激する潜在能力があると結論することができる。
【技術分野】
【0001】
本願はNogo−66受容体結合性蛋白質、特にNogo−66受容体と結合し、Nogo−66受容体の機能を中和することが可能なモノクローナル抗体について記載する。従って、これらの抗体は限定されないが、哺乳動物脳損傷、脊髄損傷、脳卒中、神経変性疾患及び統合失調症等の数種の状態の治療に有用であると思われる。
【背景技術】
【0002】
哺乳動物中枢神経系(CNS)内の損傷後の軸索再生は殆どの場合に不可能であり、転帰はCNS内の神経線維の固有の再成長能と、損傷部位の微細環境に局在しており、損傷した線維区域の再成長、従って再生を積極的に妨害するCNS内の阻害因子とのバランスに依存する。
【0003】
希突起膠細胞により産生されるCNSミエリンがインビトロ及びインビボで成長円錐崩壊を引き起こし、その結果、軸索伸長の直接阻害を生じることにより損傷の初期段階における軸索成長に最も関係の深い非許容因子であることは定説である(研究報告については、Leeら,2003参照)。CNSミエリンの主要阻害因子として希突起膠細胞ミエリン糖蛋白(OMgp)、ミエリン関連糖蛋白(MAG)及びNogo−Aがつい最近同定された(Domeniconiら,2002;研究報告:Woolf & Bloechinger,2002;McGee & Strittmatter,2003;Leeら,2003)。最後に挙げた蛋白質は主要阻害機能を発揮するドメインNogo−66を含む(GrandPreら,2000)。興味深いことに、全3種の阻害蛋白質はCNSで高い発現レベルを示し、同一のニューロングリコシルホスファチジルイノシトール(GPI)部分により固定された受容体であるNogo−66受容体ないしNgRと相互作用する(Fournierら,2001)。Nogo−66受容体NgRは473アミノ酸グリコシルホスファチジルイノシトール結合蛋白質である。この蛋白質はN末端シグナル配列とそれに続く8個のロイシンリッチリピートドメイン、(一緒になって所謂細胞外ドメインを形成する)ロイシンリッチリピートC末端ドメイン及びGPI固定ドメインから構成される。GPIアンカーを介して、NgRは外部ニューロン原形質膜と結合している。
【0004】
NgR自体は3個のCNS集積GPI固定蛋白質(NgR、NgR2及びNgR3と言う。)のファミリーに属しており、ファミリーの配列一致度は約40%であるが、全体の構造編成は非常によく似ている(Bartonら,2003;Laurenら,2003;Pignotら,2003)。NgRは複数のミエリン関連阻害分子と相互作用することが知られている唯一のメンバーであるが、MAGもNgR2と相互作用することが最近報告されている(Venkateshら,2005)。NgRホモログの機能は現時点では不明である。NgR自体は齧歯類又はニワトリで初期発生中には発現されないが、成体動物では高い発現レベルを示し、NgRは脊髄を含むCNS領域の全部ではないとしても大半で発現される(Huntら,2002a,b)。ニワトリ(Fournierら,2001)、ラット(Huntら,2002a)及びマウス(Wangら,2002b)ではmRNAと蛋白質レベルの両者で脊髄発現が報告されている。成体CNS組織内において、NgR蛋白質はその軸索突起を含む全成熟ニューロンで発現される。リガンドがNgRと結合すると、細胞内シグナル伝達カスケードが開始し、その結果、軸索伸長阻害と成長円錐崩壊が生じる。NgRは膜貫通ドメインを含まないので、シグナル伝達にはNgR/リガンド相互作用シグナルを細胞に導入する共受容体が必要である。NgRシグナル伝達の初期段階は共受容体p75又はTROYとのその相互作用である(Wongら,2002;Shaoら,2005;Parkら,2005)。Lingo−1と呼ばれる第2の共受容体も同定されている。NgRとP75又はTROYとLingo−1の三元複合体のみが機能的シグナル伝達複合体を構成する(Miら,2004;Parkら,2005)。このシグナル伝達の結果はアクチン細胞骨格の再構成である。ニューロンにおいて、このアクチン細胞骨格変化は軸索伸長の阻害と成長円錐崩壊の誘導をもたらす。
【0005】
インビトロにおいて、NgR(−/−)マウスに由来する後根神経節細胞はNogo66結合能が低下し、成長円錐崩壊アッセイにおいてNogo66、Fc−MAG、OMgp又はミエリンの阻害作用に対して低応答性である(Kimら,2004)。NgR(−/−)マウスは部分的又は完全な脊髄損傷後に赤核脊髄路と縫線核脊髄路を含む脳幹路の再生増加を示した。脊髄の完全な実験的離断後でも、NgR(−/−)マウスはオープンフィールド試験で機能回復の増加を示した。脊髄の半側切断及び完全離断後に、NgR(−/−)マウスの回復はホモ接合(+/+)及びヘテロ接合同腹子よりも有意に良好であった(Kimら,2004)。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Leeら,2003
【非特許文献2】Domeniconiら,2002
【非特許文献3】Woolf & Bloechinger,2002
【非特許文献4】McGee & Strittmatter,2003
【非特許文献5】GrandPreら,2000
【非特許文献6】Fournierら,2001
【非特許文献7】Bartonら,2003
【非特許文献8】Laurenら,2003
【非特許文献9】Pignotら,2003
【非特許文献10】Venkateshら,2005
【非特許文献11】Huntら,2002a,b
【非特許文献12】Wangら,2002b
【非特許文献13】Wongら,2002
【非特許文献14】Shaoら,2005
【非特許文献15】Parkら,2005
【非特許文献16】Miら,2004
【非特許文献17】Kimら,2004
【発明の概要】
【0007】
本願はNogo−66結合について選択的に競合し、NgR活性が有害であり得る障害を改善すると予想される抗NgR中和モノクローナル抗体の作製について記載する。本発明の中和モノクローナル抗体は例えば、特に急性脊髄損傷、脳損傷又は神経変性疾患(例えばハンチントン舞踏病、パーキンソン病、アルツハイマー病又は多発性硬化症)後の損傷したCNSにおいてニューロン再生を促進すると予想される。
【図面の簡単な説明】
【0008】
【図1】図1a及びbは抗体mAb50及びmAb51とヒト及びラットNgRとの結合を示す。
【図2】AP−Nogo66とNgR−Fcとの結合のmAb50及びmAb51による競合。
【図3】HEK293f細胞で発現させたヒト及びラットNgRとNogo66との結合のmAB50及びmAB51による競合。
【図4】mAB50及びmAB51とNTera2細胞との結合。
【図5】NTera2細胞凝集物からの神経突起伸長の定量。
【図6】hNgRの欠失変異体。
【図7】MAG−FcのNgR−Fcへの結合の競合。
【図8】許容条件下及び阻害条件下におけるラット後根神経節ニューロン及びNogo66により誘導される神経突起伸長阻害のmAb50による中和。
【図9】ラットDRG細胞におけるNogo66により誘導される神経突起伸長阻害のmAB50及びmAb51による中和。
【0009】
配列表
配列番号1a:ヒトNgR蛋白質
配列番号1b:ヒトNgRヌクレオチド
配列番号2a:ラットNgR蛋白質
配列番号2b:ラットNgRヌクレオチド
配列番号3:抗体クローン50VH
配列番号4:抗体クローン50VL
配列番号5:抗体クローン51VH
配列番号6:抗体クローン51VL
配列番号7a:AP−Nogo−66蛋白質
配列番号7b:AP−Nogo−66ヌクレオチド。
【0010】
詳細な説明
本願はNogo受容体(NgR)と相互作用する単離結合性蛋白質、特にヒト及びラットNgRと結合してこれを中和する中和モノクローナル抗体に関する。これらの抗体はNgRとの結合についてNogo−66と競合することができる。本願の他の側面としては、このような結合性蛋白質の作製方法、前記蛋白質を使用した医薬組成物、及び前記蛋白質の使用方法が挙げられる。
【0011】
抗体
本願の主要な態様はNogo−66受容体(NgR)の少なくとも1個のエピトープと特異的に結合する単離された蛋白質又はポリペプチドを含む。「単離された蛋白質」又は「単離されたポリペプチド」なる用語はその起源又は誘導源によりその天然状態でこれに付随する天然結合成分と結合していない蛋白質又はポリペプチド、同一種に由来する他の蛋白質を実質的に含まない蛋白質又はポリペプチド、別の種に由来する細胞により発現される蛋白質又はポリペプチド、あるいは自然界に存在しない蛋白質又はポリペプチドである。従って、化学的に合成されるポリペプチド又はその天然起源である細胞とは異なる細胞系で合成されるポリペプチドはその天然結合成分から「単離」されることになる。当分野で周知の蛋白質精製技術を使用して、単離により蛋白質から天然結合成分を実質的に除去することもできる。本明細書で使用する「ポリペプチド」なる用語はアミノ酸の任意ポリマー鎖を意味する。「ペプチド」及び「蛋白質」なる用語はポリペプチドなる用語と交換可能に使用し、同様にアミノ酸のポリマー鎖を意味する。「ポリペプチド」なる用語は天然又は人工蛋白質、蛋白質フラグメント及び蛋白質配列のポリペプチドアナログを包含する。ポリペプチドはモノマーでもポリマーでもよい。
【0012】
Nogo−66受容体(NgR)の少なくとも1個のエピトープと特異的に結合する単離蛋白質又はポリペプチドはリガンドと前記NgRの結合を阻害することができる。Nogo−66受容体NgRは473アミノ酸グリコシルホスファチジルイノシトール結合蛋白質である。この蛋白質はN末端シグナル配列とそれに続く8個のロイシンリッチリピートドメイン、(一緒になって所謂細胞外ドメインを形成する)ロイシンリッチリピートC末端ドメイン及びGPI固定ドメインから構成される。GPIアンカーを介して、NgRは外部ニューロン原形質膜と結合している。本発明の好ましい蛋白質はヒトNgRの少なくとも1個のエピトープと結合するモノクローナル中和抗体又はその抗原結合フラグメントである。Nogo66は軸索伸長阻害を誘導すると共に成長円錐崩壊を促進するCNSミエリンの数種の主要阻害因子の1つである。
【0013】
本明細書で使用する「抗体」なる用語はジスルフィド結合により相互に結合された2本の重(H)鎖と2本の軽(L)鎖の4本のポリペプチド鎖から構成される免疫グロブリン分子を意味する。抗体はある分子と特異的に反応することにより分子を抗体と結合することができる場合に、この分子と「結合することが可能」であると言う。「エピトープ」なる用語は抗体と結合することが可能であり、更にこの抗体により認識することができる任意分子の部分を意味する。エピトープないし「抗原決定基」は通常、アミノ酸や糖側鎖等の化学的に活性な表面分子群から構成され、特定の三次元構造特性と特定の電荷特性をもつ。
【0014】
本明細書で使用する抗体の「抗原結合フラグメント」(又は単に「抗原フラグメント」)なる用語は受容体と特異的に結合し、これを夫々活性化又は調節する能力を保持する抗体の1個以上の部分を意味する。抗体の抗原結合機能は全長抗体のフラグメントにより実現可能であることが分かっている。抗体の「抗原結合部分」なる用語に含まれる結合フラグメントの例としては、(i)VL、VH、CL及びCH1ドメインから構成される1価フラグメントであるFabフラグメント;(ii)ヒンジ領域でジスルフィド橋により結合された2個のFabフラグメントからなる2価フラグメントであるF(ab’)2フラグメント;(iii)VHドメインとCH1ドメインから構成されるFdフラグメント;(iv)抗体の単一アームのVLドメインとVHドメインから構成されるFvフラグメント;(v)VHドメインから構成されるdAbフラグメント(Wardら,(1989)Nature 341:544−546);並びに(vi)単離CDRが挙げられる。更に、Fvフラグメントの2個のドメインVL及びVHは別々の遺伝子によりコードされるが、VL領域とVH領域が対合して1本鎖Fv(scFv)抗体と呼ばれる1価分子を形成する単一蛋白質としての作製を可能にする合成リンカーにより組換え法を使用して結合することができる。(例えばBirdら,(1988)Science 242:423−426;Hustonら,(1988)Proceedings of the National Academy of Science USA 85:5879−5883参照。)このようなscFv抗体も抗体の抗原結合部分なる用語に含むものとする。ダイアボディ等の他の形態の1本鎖抗体もこの用語に含む。ダイアボディはVH及びVLドメインが1本のポリペプチド鎖で発現される2価の二重特異性抗体であるが、非常に短いリンカーを使用するため、同一鎖の2領域間で対合することができず、これらの領域は別の鎖の相補性領域と対合し、同一受容体上又は2個の受容体分子に2個の抗原結合部位を形成する。(例えばHolligerら,(1993)Proceedings of the National Academy of Science USA 90:6444−6448;Poljakら,(1994)Structure 2:1121−1123参照)。
【0015】
本明細書で使用する「モノクローナル抗体」とは異なる抗体の混合物を含む「ポリクローナル」抗体調製物と異なり、共通重鎖及び共通軽鎖アミノ酸配列を有する抗体分子の調製物を意味するものとする。モノクローナル抗体はファージ、細菌、酵母又はリボソームディスプレイ等の数種の新規技術や、ハイブリドーマ由来抗体(例えばケーラーとミルスタインの標準ハイブリドーマ法((1975)Nature 256:495−497)等のハイブリドーマ技術により作製したハイブリドーマにより分泌される抗体)等の旧来の方法により作製することができる。本発明の抗体は哺乳動物細胞株で作製したNgR蛋白質を使用して標準免疫/ハイブリドーマ技術により作製した。
【0016】
本明細書で使用する「中和モノクローナル抗体」とは特定抗原と結合すると、前記抗原に対するリガンドの結合を競合及び阻害することが可能な抗体分子調製物を意味するものとする。本願の特定ケースにおいて、本発明の中和抗体はNgRとの結合についてNogo66と競合し、Nogo66とNgRの結合に起因するNogo66生物活性又は機能を妨害することができる。
【0017】
好ましくは、本願のモノクローナル中和抗体はヒト抗体である。「ヒト抗体」なる用語はヒト生殖細胞系列免疫グロブリン配列に対応又は由来する可変領域と定常領域を有する抗体を意味する(例えばKabatら,Sequences of Proteins of Immunological Interest,第5版,U.S.Department of Health and Human Services,NIH Publication No.91−3242,1991参照)。本願のヒト抗体はヒト生殖細胞系列免疫グロブリン配列によりコードされないアミノ酸残基(例えばランダムもしくは部位特異的変異誘発によりインビトロ又は体細胞変異によりインビボで導入される変異)を例えばCDR、特にCDR3に含んでいてよい。本明細書で使用する「CDR」なる用語は抗体可変配列内の相補性決定領域を意味する。重鎖及び軽鎖可変領域の各々に3個のCDRがあり、可変領域の各々についてCDR1、CDR2及びCDR3と呼ぶ。各種態様において、抗体は組換え抗体又はモノクローナル抗体である。本願の最も好ましい中和抗体を本明細書ではmAb50及びmAb51(夫々ATCC No.PTA−8383及びPTA−8384)と呼ぶ。mAb50及びmAb51抗体と機能的抗体フラグメント、mAb50及びmAb51関連抗体と機能的抗体フラグメント、並びにmAb50及びmAb51に等価の特性(例えばNgRに対する高い結合親和性と低い解離反応速度と高い中和能)を有する他の抗体と機能的抗体フラグメントを本発明に含むものとする。
【0018】
免疫原性NgRポリペプチドに対する本願の抗NgR抗体又はそのフラグメントの結合親和性と解離速度は当分野で公知のいずれかの方法により測定することができる。例えば、結合親和性は競合ELISA、cRIA、BIAcore又はKinExA技術により測定することができる。解離速度もBIAcore又はKinExA技術により測定することができる。結合親和性と解離速度は例えばBIAcoreを使用して表面プラズモン共鳴法により測定される。
【0019】
本願の好ましい抗体の1例は配列番号3の配列を含む重鎖可変領域(VH領域)と配列番号4の配列を含む軽鎖可変領域(L領域)を含む配列(mAB50抗体)に対して少なくとも90%のアミノ酸配列一致度を有する。別の好ましい態様は配列番号5の配列を含む重鎖可変領域(VH領域)と配列番号6の配列を含む軽鎖可変領域(L領域)を含む配列(mAB51抗体)に対して少なくとも90%のアミノ酸配列一致度を有する。
【0020】
好ましくは、mAb50抗体とmAb51抗体は1×10−9M未満のEC50でヒトNgRと結合し、より好ましくは、前記抗体は1×10−10M未満のEC50でNgRと結合し、最も好ましくは、前記抗体は4×10−11M未満のEC50でNgRと結合する。
【0021】
抗NgR抗体mAb50及びmAb51はプロNgR、成熟NgR及び短縮型NgR等の各種形態のヒトNgRと結合するものとする。抗体mAb50及びmAb51はNgR2やNgR3等の他のNgRホモログ、又は他のLRR含有蛋白質と特異的に結合しない。他方、抗体mAb50及びmAb51は他の種、特に齧歯類に由来するNgR、より具体的にはラットNgRに対して交差反応性を示す。MAb52〜62は更にマウスNgRに対しても交差反応性である。例えば、抗体はラットに由来するNgRと結合する(ラットNgRに対する両者抗体のIC50は約3×10−11Mである)。
【0022】
本願の(NgR)と相互作用する単離結合性蛋白質は抗体又はその抗原結合部分が1個以上の糖鎖残基を含むグリコシル化結合性蛋白質でもよい。初期インビボ蛋白質産生後に翻訳後修飾と呼ばれるプロセシングを受ける場合がある。特に、糖鎖(グリコシル)残基が酵素により付加される場合があり、このプロセスをグリコシル化と言う。その結果として得られた蛋白質は共有結合オリゴ糖側鎖をもち、グリコシル化蛋白質又は糖蛋白質と呼ばれる。蛋白質グリコシル化は該当蛋白質のアミノ酸配列と、蛋白質が発現される宿主細胞に依存する。生物によって産生されるグリコシル化酵素(例えばグリコシルトランスフェラーゼやグリコシダーゼ)は異なり、利用可能な基質(ヌクレオチド糖)も異なる。このような因子により、蛋白質グリコシル化パターンとグリコシル残基の組成は特定蛋白質が発現される宿主系により異なる。本発明で有用なグリコシル残基としては限定されないが、グルコース、ガラクトース、マンノース、フコース、n−アセチルグルコサミン及びシアル酸が挙げられる。グリコシル化結合性蛋白質はグリコシル化パターンがヒトであるようなグリコシル残基を含むことが好ましい。
【0023】
本願の抗体はIgG1、IgG2、IgG3、IgG4、IgA、IgE、IgM又はIgD定常領域等の重鎖定常領域を含む。更に、抗体はκ軽鎖定常領域又はλ軽鎖定常領域のいずれかの軽鎖定常領域を含むことができる。抗体はκ軽鎖定常領域を含むことが好ましい。あるいは、抗体部分は例えばFabフラグメント又は1本鎖Fvフラグメントとすることができる。抗体エフェクター機能を改変するためにFc部分のアミノ酸残基を置換することは当分野で公知である(Winterら,米国特許第5,648,260号;5,624,821号明細書)。抗体のFc部分は数種の重要なエフェクター機能(例えばサイトカイン誘導、ADCC、貪食作用、補体依存性細胞傷害作用(CDC)並びに抗体及び抗原−抗体複合体の半減期/クリアランス率)を媒介する。これらのエフェクター機能は治療目的に応じて治療用抗体に望ましい場合もあるが、不要又は有害な場合もある。所定のヒトIgGアイソタイプ、特にIgG1及びIgG3は夫々FcγRs及び補体C1qとの結合を介してADCCとCDCを媒介する。胎児性Fc受容体(FcRn)は抗体の循環半減期を決定する必須成分である。更に別の態様では、抗体のエフェクター機能を改変するように、抗体の定常領域(例えば抗体のFc領域)の少なくとも1個のアミノ酸を置換する。
【0024】
組換え蛋白質の作製
免疫及びELISAアッセイと、(「実施例」のセクションに記載する)神経突起伸長アッセイのために、可溶性ヒト及びラットNgRを作製した。アルカリホスファターゼタグと融合したリガンドNogo66(AP−Nogo66)も作製し、神経突起伸長アッセイの阻害因子及び(同様に「実施例」のセクションに記載する)ELISA試験とFACS試験におけるリガンドとして利用した。
【0025】
ヒト及びラットNgR
ヒトNgR蛋白質はアクセション番号AAG53612に基づくものとした。(アミノ酸27〜450の)蛋白質DNAをpSecベクター(Ambion)にクローニングし、CHO−K1細胞で安定に発現させることにより蛋白質を作製した。発現させた受容体は配列番号1(ヒトNgR蛋白質)及び配列番号1b(ヒトNgRヌクレオチド)に従い、C末端にMycと6×Hisタグを結合した全長蛋白質の424アミノ酸(アミノ酸27〜450)から構成された。セルファクトリー当たり5000mlのUltraCHO無血清培地(Cambrex Bio Science)を加えた40チャンバーセルファクトリー8個でヒトsecNgR 27−450 aa_D6_CHO−K1細胞をコンフルエントまで培養した(約5日間)。その後、上清40リットルを遠心し、Hemoflow Fカラム(Fresenius Medical Care)で500mlまで濃縮した。濃縮液を−80℃で凍結させた。蛋白質精製のために、濃縮蛋白質上清に20mM NaH2PO4;140mM NaCl(pH7.4)500mlを加えて1000mlとし、再び300mlまで濃縮した。濃縮工程をもう一度繰返し、最後に20mM NaH2PO4;140mM NaCl(pH7.4)300mlを濃縮液に加えた。20mM NaH2PO4;300mM NaCl;pH8.0で平衡化しておいたNi−NTA−Superflow Fa.(Qiagen catalog #30430)50mlを濃縮液600mlに加え、6℃で1時間撹拌し、6℃で沈降させ、上清を捨て、Ni−NTAビーズをカラムに充填した。カラムを室温にて20mM NaH2PO4;300mM NaCl(pH8.0)10CVで洗浄した後、20mM NaH2PO4;300mM NaCl;10mMイミダゾール(pH8.0)5〜10CVで洗浄した。カラムを20mM NaH2PO4;300mM NaCl;100mMイミダゾール;pH8.0で溶出させ、UV−280nm活性ピークを採取した。次に溶出液を6℃にて一晩25mM Tris/HCl(pH7.0)5Lで透析し、透析液を室温でQ−Sepharoseカラム(カラムサイズ1.6cm×3cm;容量6ml;Amersham Biosciences catalog #17−0510−01)にロードした。緩衝液Aは50mM Tris/HCl;pH7.0とした。緩衝液Bは50mM Tris/HCl;1M NaCl;pH7.0とし、流速2ml/分とした。勾配は0%B保持5CV;0−50%B12CV;50−100%B2CV;100%B保持5CVとした。フラクションサイズは2.5mlとした。その後、フラクションをSDS−PAGで分析し、SDS−page上のサイズと純度(高グリコシル化NgR−HisでNgR−His最高純度;低グリコシル化NgR−Hisで最高NgR−His純度)によりフラクションプールした。プールしたフラクションを12〜14kDa透析チューブで6℃にて20mM NaH2PO4;140mM NaCl;pH7.4で1回透析し、フラクションを0.2μm滅菌フィルターで濾過し、その後の使用に備えて6℃で保存した。長期間保存用として、受容体フラクションを分取し、−80℃で保存した。
【0026】
ラットNgR DNA(アクセション番号AAM46772)をpcDNA3.1(Invitrogen)にクローニングし、HEK293F細胞で一過性発現システムにより発現及び生産させた。蛋白質は配列番号2a(ラットNgR蛋白質)及び配列番号2b(ラットNgRヌクレオチド)に従い、6×Hisタグと結合したアミノ酸27〜450を含んでいた。ラット蛋白質の生産はHEK293F細胞で48〜72時間標準一過性発現により実施した。細胞上清を回収し、蛋白質の精製はヒト蛋白質について上述したと同様の工程に従った。所定の実験では、R&D Systemsから入手した蛋白質を使用した。これらの蛋白質は、ヒト組換えNgR/Fcキメラ,カタログ番号1208−NG及び組換えマウスNogo受容体/Fcキメラ,カタログ番号1440−NGであった。
【0027】
NgRの細胞表面発現
細胞表面で発現されるNgRには、アミノ酸1〜473の完全オープンリーディングフレームを含む全長受容体配列(夫々ラットAAM46772及びヒトAAG53612)をpcDNA4にクローニングした。プラスミドを標準手順に従ってCHO−K1又はHEK293細胞に形質移入した。要約すると、細胞をペトリ皿でMEM培地に播種し、Fugene 6(Roche)を製造業者の指示に従って形質移入した。150μg/mlゼオシンを使用して2〜3週間選択を実施し、蛋白質発現をFACSにより確認した(下記実施例4参照)。
【0028】
HEK293F細胞で一過性発現させるために、製造業者(Invitrogen;Free Style System)の指示に従って細胞を懸濁培養し、48〜72時間後に回収し、FACS試験に使用した(下記実施例のセクション参照)。
【0029】
AP−Nogo66の生産
AP−Nogo66(配列番号7a及び配列番号7b)を標準条件下で生産した。要約すると、Nogo66をpAPTag5ベクターにクローニングし、HEK293細胞に構築物を形質移入し、RPMI Glutamax+10%FCS,150μg/mlゼオシンで選択した。蛋白質生産のために、セルファクトリー当たり1200mlのRPMI(Invitrogen)+10%FCSを加えた10チャンバーセルファクトリー6個でHEK293細胞をコンフルエントまで培養した(約3日間)。その後、上清を捨て、Pro293a−CDM(Cambrex Bio Science)1200mlを各セルファクトリーに充填した。細胞を更に3日間培養した。その後、上清7200mlを遠心し、Hemoflow Fカラム(Fresenius Medical Care)で350mlまで濃縮した。1mM PefablocSC(ROCHE)の添加後、濃縮液を分取し、−80℃で凍結させた。
【0030】
抗体及び抗体産生細胞株の作製
本願の抗体は適切な宿主(例えばヒト、マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ、ウマ等の脊椎動物、爬虫類、魚類、両生類、並びに鳥類、爬虫類及び魚類の卵)に免疫により作製することができる。このような抗体はポリクローナル抗体でもモノクローナル抗体でもよい。本願の抗体を作製するためには、本発明の免疫原性NgRポリペプチド又はそのフラグメントにより宿主を免疫する。「免疫」なる用語は本明細書では抗原を免疫レパートリーに提示するプロセスを意味し、レパートリーは遺伝子改変されていない天然生物に存在するか又は人工ヒト免疫レパートリーを示すように改変されたものを含むトランスジェニック生物に存在するかを問わない。同様に、「免疫原製剤」とはアジュバント又は抗原の免疫原性を強化する他の添加剤を加えた抗原製剤である。
【0031】
動物の免疫は当分野で公知のいずれかの方法により実施することができる。例えばHarlow and Lane,Antibodies:A Laboratory Manual,New York:Cold Spring Harbor Press,1990参照。マウス、ラット、ヒツジ、ヤギ、ブタ、ウシ及びウマ等の非ヒト動物の免疫方法は当分野で周知である。例えばHarlow and Lane及び米国特許第5,994,619号明細書参照。好ましい1態様では、NgR抗原をアジュバントと共に投与し、免疫応答を刺激する。このようなアジュバントとしては完全もしくは不完全フロイントアジュバント、RIBI(ムラミルジペプチド)又はISCOM(免疫刺激複合体)が挙げられる。このようなアジュバントはポリペプチドを局所沈着物中に分離することにより迅速な分散から保護することもできるし、マクロファージや免疫系の他の成分に対して化学走化性の因子を分泌するように宿主を刺激する物質を添加してもよい。ポリペプチドを投与している場合には、免疫スケジュールは数週間にわたってポリペプチドを2回以上投与することが好ましい。
【0032】
無傷又は破壊した細胞の細胞膜に伴われたNgRにより動物宿主を免疫することが考えられ、本願の抗体は本発明の免疫原性NgRポリペプチドと結合することにより同定される。
【0033】
動物宿主をNgR抗原により免疫後、動物から抗体及び/又は抗体産生細胞を得ることができる。動物から採血する又は動物を屠殺することにより動物から抗NgR抗体含有血清が得られる。血清は動物から採取したまま使用してもよいし、血清から免疫グロブリンフラクションを採取してもよいし、血清から抗NgR抗体を精製してもよい。こうして得られた血清又は免疫グロブリンはポリクローナルであるため、一連の異種特性をもつ。
【0034】
抗体産生細胞株
本願は更に免疫動物から作製可能な抗体産生不死化ハイブリドーマについて記載する。好ましくは、免疫動物はヒト免疫グロブリン遺伝子を発現する非ヒト動物であり、脾臓B細胞を非ヒト動物と同一種に由来する骨髄腫と融合させる。
【0035】
免疫後、動物を屠殺し、脾臓B細胞を当分野で周知のように不死化骨髄腫細胞と融合させる。例えばHarlow and Lane,前出参照。骨髄腫細胞は免疫グロブリンポリペプチドを分泌しないことが好ましい(非分泌型細胞株)。融合及び抗体選択後、NgRもしくはその一部又はNgRを発現する細胞を使用してハイブリドーマをスクリーニングする。好ましくは、初期スクリーニングは酵素免疫測定法(ELISA)又は放射免疫測定法(RIA)、好ましくはELISAを使用して実施される(実施例のセクションにELISAスクリーニングの1例を記載する)。
【0036】
抗NgR抗体を産生するハイブリドーマを選択し、クローニングし、以下に詳述するように、活発なハイブリドーマ増殖、高い抗体産生能及び望ましい抗体特性等の望ましい特性について更にスクリーニングする。ハイブリドーマを培養し、同系動物、免疫系を欠損する動物(例えばヌードマウス)でインビボ増殖させる、あるいは細胞培養でインビトロ増殖させる。ハイブリドーマの選択、クローニング及び増殖方法は当業者に周知である。好ましい1態様において、ハイブリドーマは上記のようにマウスハイブリドーマである。別の好ましい態様において、ハイブリドーマは非ヒト非マウス種(例えばラット、ヒツジ、ブタ、ヤギ、ウシ又はウマ)で作製される。別の態様において、ハイブリドーマは抗NgR抗体を発現するヒト細胞にヒト非分泌型骨髄腫が融合されたヒトハイブリドーマである。
【0037】
本願は、米国特許第5,627,052号明細書、国際公開第92/02551号パンフレット及びBabcock,J.S.ら(1996)Proc.Natl.Acad.Sci.USA 93:7843−7848に記載されているような選択リンパ球抗体法(SLAM)と当分野で呼ばれる手順を使用して単一単離リンパ球から作製される組換え抗体についても記載する。この方法では、リンカー(例えばビオチン)を使用して抗原NgR又はそのフラグメントをヒツジ赤血球と結合させ、これを使用してNgRに対する特異性をもつ抗体を分泌する単一細胞を同定する抗原特異的溶血プラークアッセイを使用することにより、該当抗体を分泌する単一細胞(例えば任意免疫動物に由来するリンパ球)をスクリーニングする。該当抗体分泌細胞の同定後、重鎖及び軽鎖可変領域cDNAを逆転写酵素−PCRにより細胞から得た後、COS又はCHO細胞等の哺乳動物宿主細胞で適切な免疫グロブリン定常領域(例えばヒト定常領域)のコンテキストにおいてこれらの可変領域を発現させることができる。インビボ選択したリンパ球に由来する増幅免疫グロブリン配列を形質移入した宿主細胞をその後、例えば形質移入した細胞のパニングにより更にインビトロ分析及び選択し、NgRに対する抗体を発現する細胞を単離することができる。
【0038】
インビトロ作製した抗体
本願に記載する抗体を作製するためには、抗体ライブラリーをスクリーニングして所望の結合特異性をもつ抗体を同定するインビトロ法を使用することもできる。組換え抗体ライブラリーのこのようなスクリーニング方法は当分野で周知である。
【0039】
組換え抗体ライブラリーはNgR又はNgRの一部により免疫された対象に由来することができる。あるいは、組換え抗体ライブラリーはナイーブ対象、即ちNgRにより免疫されていない対象に由来することができる(例えばヒトNgRにより免疫されていないヒト対象に由来するヒト抗体ライブラリー)。本願の抗体はヒトNgRを含むペプチド(例えばhNgRの一部に対応するペプチド)で組換え抗体ライブラリーをスクリーニングし、NgRを認識する抗体を選択することにより選択される。このようなスクリーニングと選択を実施するための方法は前段落に挙げた文献に記載されているように、当分野で周知である。hNgRに対する特定結合親和性をもつ本願の抗体(例えば特定koff速度定数でヒトNgRから解離するもの)を選択するためには、当分野で公知の表面プラズモン共鳴法を使用して所望のkoff速度定数をもつ抗体を選択することができる。hNgRに対して特定中和活性をもつ本願の抗体(例えば特定IC50をもつもの)を選択するためには、hNgR活性の阻害を評価する方法として当分野で公知の標準方法を使用することができる。
【0040】
抗体の特性
本願のmAB50及びmAB51又はその抗原結合部分はヒトNgRと結合し、表面プラズモン共鳴法により測定した場合に約0.1s−1以下、好ましくは1×10−2s−1以下、より好ましくは1×10−3s−1以下、更に好ましくは1×10−4s−1以下、最も好ましくは1×10−5s−1以下のkoff速度定数でヒトNgRから解離する。本明細書で使用する「表面プラズモン共鳴」なる用語は例えばBIAcoreシステム(Pharmacia Biosensor AB,Uppsala,Sweden及びPiscataway,NJ)を使用してバイオセンサーマトリックス内の蛋白濃度の変化の検出によりリアルタイム生体特異的相互作用の分析を可能にする光学現象を意味する。更に詳細な記載については、Jonsson,U.ら(1993)Ann.Biol.Clin.51:19−26;Jonsson,U.ら(1991)Biotechniques 11:620−627;Johnsson,B.ら(1995)J.Mol Recognit.8:125−131;及びJohnnson,B.ら(1991)Anal.Biochem.198:268−277参照。
【0041】
【表1】
【0042】
本明細書で使用する「Kon」なる用語は当分野で公知の通り、抗体と抗原が抗体/抗原複合体を形成するための会合速度定数を意味するものとする。
【0043】
本明細書で使用する「Koff」なる用語は当分野で公知の通り、抗体/抗原複合体から抗体が解離する解離速度定数を意味する。本明細書で使用する「Kd」なる用語は当分野で公知の通り、特定抗体−抗原相互作用の解離定数を意味するものとする。
【0044】
あるいは、本願のmAb50及びmAb51又はその抗原結合部分は約1×10−6M以下、好ましくは1×10−7M以下、好ましくは1×10−8M以下、より好ましくは1×10−9M以下、より好ましくは1×10−10M以下、最も好ましくは1×10−11M以下のIC50でヒトNgRを阻害することができる。本明細書で使用するIC50なる用語はリガンドNogo66とNgRの結合について競合する抗体の濃度を意味するものとする。
【0045】
融合抗体及びイムノアドヘジン
本願は別のポリペプチドに連結した本願の抗Nogo受容体−1抗体の全部又は一部を含む作製可能な融合抗体又はイムノアドヘジンについても記載する。ある態様において、抗Nogo受容体−1抗体の可変領域のみをポリペプチドに連結する。他の態様において、本願の抗Nogo受容体−1抗体のVHドメインを第1のポリペプチドに連結し、VHドメインとVLドメインを相互作用させて抗体結合部位を形成するように第1のポリペプチドと会合する第2のポリペプチドに抗体のVLドメインを連結する。他の態様において、VHドメインとVLドメインを相互作用させるリンカーによりVHドメインをVLドメインから分離する(下記1本鎖抗体の項参照)。その後、VH−リンカー−VL抗体を該当ポリペプチドに連結する。融合抗体はNogo受容体−1リガンドを発現する細胞又は組織にポリペプチドを誘導するのに有用である。該当ポリペプチドは毒素等の治療剤でもよいし、可視化可能な酵素(例えば西洋ワサビペルオキシダーゼ)等の診断剤でもよい。更に、2個(以上)の1本鎖抗体を相互に連結した融合抗体を作製することもできる。これは、1本のポリペプチド鎖上に2価又は多価抗体を作製したい場合や、二重特異性抗体を作製したい場合に有用である。
【0046】
1態様は本願の抗体又は抗体部分を誘導体化した又は別の機能的分子(例えば別のペプチド又は蛋白質)と連結した標識結合性蛋白質を提供する。例えば、核酸、別の抗体(例えば二重特異性抗体又はダイアボディ)、検出可能な物質、細胞傷害性物質、薬剤、及び/又は抗体もしくは抗体部分と別の分子(例えばストレプトアビジンコア領域やポリヒスチジンタグ)との結合を媒介することができる蛋白質もしくはペプチド等の1個以上の他の分子部分と本願の抗体又は抗体部分を(化学的カップリング、遺伝子融合、非共有的結合又は他の方法により)機能的に連結することにより、本願の標識結合性蛋白質を誘導することができる。
【0047】
本願の抗体又は抗体部分を誘導体化することができる有用な検出可能な物質としては蛍光化合物が挙げられる。検出可能な蛍光物質の例としてはフルオレセイン、フルオレセインイソチオシアネート、ローダミン、5−ジメチルアミン−1−ナフタレンスルホニルクロリド、フィコエリスリン等が挙げられる。アルカリホスファターゼ、西洋ワサビペルオキシダーゼ、グルコースオキシダーゼ等の検出可能な酵素で抗体を誘導体化することもできる。検出可能な酵素で抗体を誘導体化する場合には、検出可能な反応生成物を生成するために酵素により使用される他の試薬を加えることにより抗体を検出する。例えば、検出可能な物質として西洋ワサビペルオキシダーゼを使用する場合には、過酸化水素とジアミノベンジジンを加えると、検出可能な着色反応生成物が得られる。抗体を核酸、ビオチンで誘導体化し、アビジン又はストレプトアビジン結合の間接測定により検出することもできる。
【0048】
本願の別の態様は結晶化結合性蛋白質を提供する。本明細書で使用する「結晶化」なる用語は結晶形態で存在する抗体又はその抗原結合部分を意味する。結晶は物質の固体状態の1形態であり、非晶質固体状態や液体結晶状態等の他の形態から区別される。結晶は原子、イオン、分子(例えば抗体等の蛋白質)、又は分子集合(例えば抗原/抗体複合体)の規則的な反復三次元配列から構成される。これらの三次元配列は当分野で周知の特定の数学的関係に従って配置されている。結晶中で反復している基本単位ないし構成単位を非対称単位と言う。所定の明確な結晶対称性に一致する配置で非対称単位が反復することにより、結晶の「単位格子」を構成する。単位格子が全3方向に規則的平行移動により反復することにより、結晶を構成する。Giege,R.and Ducruix,A.Barrett,Crystallization of Nucleic Acids and Proteins,a Practical Approach,第2版,pp.20 1−16,Oxford University Press,New York,New York,(1999)参照。
【0049】
好ましくは、本願は本明細書に開示するような全長抗NgR抗体及びそのフラグメントの結晶と、このような結晶を含有する製剤及び組成物について記載する。1態様において、結晶化結合性蛋白質は結合性蛋白質の可溶性部分よりも長いインビボ半減期をもつ。別の態様において、結合性蛋白質は結晶化後に生物活性を保持する。
【0050】
本発明の結晶化結合性蛋白質は当分野で公知の方法に従って作製することができる。
【0051】
1本鎖抗体
本願は本発明の免疫原性NgRと結合する1本鎖抗体(scFv)を含む。scFvを作製するためには、VH配列とVL配列を連続した1本鎖蛋白質として発現させ、VL領域とVH領域を可撓性リンカーにより結合できるように、可撓性リンカーをコードするDNA、例えばアミノ酸配列(G1Y4−Ser)をコードするDNAにVHとVをコードするDNAを機能的に連結する(例えばBirdら,(1988)Science 242:423−42 6;Hustonら,(1988)Proc.Natl.Acad.Sci.USA 85:5879−5883;McCaffertyら,30 Nature(1 99 0)34 8:552−554参照)。1本鎖抗体はただ1個のVHとVLを使用する場合には1価とすることができ、2個のVHとVLを使用する場合には2価とすることができ、3個以上のVHとVLを使用する場合には多価とすることができる。
【0052】
キメラ抗体
本願は更に一方の特異性が本願の免疫原性Nogo受容体−1ポリペプチドに対するものである二重特異性抗体又はその抗原結合フラグメントを含む。例えば、第1の結合ドメインを介して本発明の免疫原性NgRポリペプチドと特異的に結合し、第2の結合ドメインを介して第2の分子と結合するキメラ抗体を作製することができる。「キメラ抗体」なる用語はある種に由来する重鎖及び軽鎖可変領域配列と、別の種に由来する定常領域配列を含む抗体(例えばヒト定常領域に連結したマウス重鎖及び軽鎖可変領域をもつ抗体)を意味する。キメラ抗体は組換え分子生物学技術により作製することもできるし、相互に物理的に結合させてもよい。更に、2個以上のVHとVLを含み、本発明の免疫原性ポリペプチドと、ミエリンによる成長円錐崩壊と神経突起伸長及び発芽の阻害を低減する働きがあるとされる別の分子とに特異的に結合する1本鎖抗体を作製することもできる。このような二重特異性抗体は、例えばFangerら,Immunol Methods 4:72−81(1994)及びWright and Harris,20(前出)等の周知技術を使用して作製することができる。所定態様では、キメラ抗体は本発明の抗体に由来する可変領域の1個以上を使用して作製される。別の態様では、キメラ抗体は前記抗体に由来する1個以上のCDR領域を使用して作製される。「ヒト化抗体」なる用語は非ヒト種(例えばマウス)に由来する重鎖及び軽鎖可変領域配列を含むが、VH及び/又はVL配列の少なくとも一部をより「ヒト様」即ちヒト生殖細胞系列可変配列に類似するように改変した抗体を意味する。ヒト化抗体の1例はヒトCDR配列を非ヒトVH及びVL配列に導入して対応する非ヒトCDR配列を置換したCDRグラフト抗体である。
【0053】
ヒト化抗体
ヒト化抗体は所望抗原と結合する非ヒト種に由来する抗体分子であり、非ヒト種に由来する1個以上の相補性決定領域(CDR)とヒト免疫グロブリン分子に由来するフレームワーク領域をもつ。ヒトIg配列は当分野で公知である。当分野で公知の通り、このようなインポート配列を使用して免疫原性を低下させたり、結合性、親和性、会合速度、解離速度、アビディティ、特異性、半減期又は他の適切な特性のいずれかを低下させ、増加させ又は改変することができる。
【0054】
抗原結合を改変する、好ましくは改善するためには、ヒトフレームワーク領域のフレームワーク残基をCDRドナー抗体に由来する対応する残基で置換すればよい。これらのフレームワーク置換は当分野で周知の方法により同定され、例えば抗原結合に重要なフレームワーク残基を同定するにはCDR残基とフレームワーク残基の相互作用のモデル化を使用し、特定位置の異常なフレームワーク残基を同定するには配列比較を使用する。(例えばQueenら,米国特許第5,585,089号明細書;Riechmannら,Nature 332:323(1988)参照)。三次元免疫グロブリンモデルは広く入手可能であり、当業者に周知である。選択された候補免疫グロブリン配列の推定三次元配座構造を図解表示するコンピュータープログラムも入手可能である。これらの表示を検討すると、候補免疫グロブリン配列の機能に残基が果たすと予想される役割を分析することができ、即ち候補免疫グロブリンがその抗原と結合する能力に影響を与える残基を分析することができる。こうして、ターゲット抗原に対する親和性の増加等の所望抗体特性が得られるようにコンセンサス配列とインポート配列からFR残基を選択し、組み合わせることができる。一般に、CDR残基は抗原結合の改変に直接的且つ最も実質的に関与している。当分野で公知の各種技術を使用して抗体をヒト化することができ、限定されないが、Carterら,Proc.Natl.Acad.Sci.U.S.A.89:4285(1992);Prestaら,J.Immunol.151:2623(1993),Padlan,Molecular Immunology 28(4/5):489−498(1991);Studnickaら,Protein Engineering 7(6):805−814(1994);Roguskaら,PNAS 91:969−973(1994)に記載されている技術が挙げられる。
【0055】
誘導体化及び標識抗体
本願の抗体又は抗原結合フラグメントは誘導体化又は別の機能的分子(例えば別のペプチド又は蛋白質)と連結することができる。一般に、抗体又は抗原結合フラグメントは本発明の抗原性ペプチドとの結合が誘導体化又は標識により悪影響を受けないように誘導体化される。例えば、別の抗体(例えば二重特異性抗体又はダイアボディ)、検出可能な物質、細胞傷害性物質、薬剤及び/又は抗体もしくは抗体部分と別の分子(例えばストレプトアビジンコア領域又はポリヒスチジンタグ)との結合を媒介することができる蛋白質もしくはペプチド等の1個以上の他の分子部分と本願の抗体又は抗体部分を(化学的カップリング、遺伝子融合、非共有的結合又は他の方法により)機能的に連結することができる。更に、抗体又はその抗原結合部分は、抗体又は抗体部分と1種以上の他の蛋白質又はペプチドとの共有又は非共有的結合により形成される大きな免疫接着分子の一部でもよい。このような免疫接着分子の例としてはストレプトアビジンコア領域を使用して四量体scFv分子を作製する場合(Kipriyanovら(1995)Human Antibodies and Hybridomas 6:93−101)や、システイン残基、マーカーペプチド及びC末端ポリヒスチジンタグを使用して2価ビオチン化scFv分子を作製する場合(Kipriyanovら(1994)Molecular Immunology 31:1047−1058)が挙げられる。Fab及びF(ab’)2フラグメント等の抗体部分は夫々全長抗体のパパイン又はペプシン消化等の慣用技術を使用して全長抗体から作製することができる。更に、抗体、抗体部分及び免疫接着分子は標準組換えDNA技術を使用して得ることができる。
【0056】
誘導体化抗体は(同一型又は例えば二重特異性抗体を作製するためには異なる型の)2個以上の抗体を架橋することにより作製することができる。適切な架橋剤としては適切なスペーサーにより分離された2個の別個の反応基をもつヘテロ2官能性のもの(例えばm−マレイミドベンゾイル−N−ヒドロキシスクシンイミドエステル)又はホモ2官能性のもの(例えばスベリン酸ジスクシンイミジル)が挙げられる。このようなリンカーはPierce Chemical Company,Rockford,Illから入手可能である。
【0057】
誘導体化抗体は標識抗体でもよい。例えば、本発明の抗体又は抗体部分を誘導体化することができる検出剤はフルオレセイン、フルオレセインイソチオシアネート、ローダミン、5−ジメチルアミン−1−ナフタレンスルホニルクロリド、フィコエリスリン、ランタニド蛍光体等の蛍光化合物である。西洋ワサビペルオキシダーゼ、ガラクトシダーゼ、ルシフェラーゼ、アルカリホスファターゼ、グルコースオキシダーゼ等の検出に有用な酵素で抗体を標識してもよい。検出可能な酵素で標識する態様では、検出可能な反応生成物を生成するために酵素により使用される他の試薬を加えることにより抗体を検出する。例えば、西洋ワサビペルオキシダーゼを過酸化水素及びジアミノベンジジンと併用する。抗体をビオチンで標識し、アビジン又はストレプトアビジン結合の間接測定により検出することもできる。二次レポーターにより認識される所定ポリペプチドエピトープ(例えばロイシンジッパー対配列、二次抗体の結合部位、金属結合ドメイン、エピトープタグ)で抗体を標識してもよい。抗Nogo受容体−1抗体又はその抗原フラグメントを放射性標識アミノ酸で標識してもよい。放射性ラベルは診断目的と治療目的の両者に使用することができる。放射性標識抗Nogo受容体−1抗体は例えば対象におけるNogo受容体−1濃度を測定するために診断用として使用することができる。更に、放射性標識抗Nogo受容体−1抗体は脊髄損傷を治療するために治療用として使用することもできる。
【0058】
ポリペプチド用ラベルの例としては限定されないが、放射性同位体ないし放射性核種である15N、35S、90Y、99Tc、111In、125I、131I、177Lu、166Ho、153Smが挙げられる。ポリエチレングリコール(PEG)、メチルもしくはエチル基又は糖鎖基等の化学基で抗Nogo受容体−1抗体又はその抗原フラグメントを誘導体化してもよい。これらの基は抗体の生物学的特性を改善するため、例えば血清半減期を延長するため又は組織結合を増加するために有用であると思われる。更に、ポリペプチド用ラベルとしては核酸も挙げられ、例えばPCRによる検出用又は遺伝子発現強化用のDNAや、NgRを保持する細胞又は組織における遺伝子発現を抑制するためのsiRNAも挙げられる。
【0059】
抗Nogo受容体−1抗体のクラス及びサブクラスは当分野で公知の方法のいずれかにより決定することができる。一般に、抗体のクラス及びサブクラスは特定クラス及びサブクラスの抗体に特異的な抗体を使用して決定することができる。このような抗体は市販されている。クラス及びサブクラスはELISA、ウェスタンブロット及び他の技術により決定することができる。あるいは、クラス及びサブクラスは抗体の重鎖及び/又は軽鎖の定常領域の全部又は一部を配列決定し、そのアミノ酸配列を種々のクラス及びサブクラスの免疫グロブリンの既知アミノ酸配列と比較し、抗体のクラス及びサブクラスを決定することにより決定することができる。
【0060】
抗NgR抗体によるNgR活性の阻害
本願の抗Nogo受容体−1抗体又はその抗原結合フラグメントはリガンドとNgRの結合を阻害する。このような阻害のIC50は当分野で公知のいずれかの方法、例えばELISA、RIA又は機能的拮抗により測定することができる。IC50は0.01〜100nMとすることができる。好ましくは、IC50は1〜10nMである。より好ましくは、本発明の抗Nogo受容体−1抗体又はその抗原結合フラグメントのIC50は0.1nM〜1nMである。最も好ましくは、IC50は0.1nM未満である。
【0061】
二重可変領域抗体
本明細書で使用する二重可変領(DVD)結合性抗体とは2個以上の抗原結合部位を含む結合性蛋白質であり、4価又は多価結合性抗体である。「多価結合性抗体」なる用語は本明細書では2個以上の抗原結合部位を含む結合性蛋白質の意味で使用する。多価結合性抗体は3個以上の抗原結合部位をもつように構築することが好ましく、一般に天然抗体以外のものである。「多重特異性結合性抗体」なる用語は2個以上の関連又は無関連ターゲットと結合することが可能な結合性蛋白質を意味する。このようなDVDは単一特異性、即ち1種類の抗原としか結合できないものでもよいし、多重特異性、即ち2種類以上の抗原と結合できるものでもよい。2個の重鎖DVDポリペプチドと2個の軽鎖DVDポリペプチドを含むDVD結合性抗体をDVD Igと言う。DVD Igの各半分は重鎖DVDポリペプチドと軽鎖DVDポリペプチドと2個の抗原結合部位を含む。各結合部位は重鎖可変領域と軽鎖可変領域を含み、抗原結合部位1個当たり合計6個のCDRが抗原結合に関与している。DVD結合性蛋白質とDVD結合性蛋白質の作製方法は米国特許出願第11/507,050号に開示されている。本発明はNgRと結合することが可能な結合性蛋白質を含むDVD結合性蛋白質を含むものとする。好ましくは、DVD結合性蛋白質はNgRと第2のターゲットとに結合することが可能である。第2のターゲットは反発性ガイダンス分子(RGM)、Nogo−A、MAG、OMgp、CSPGから構成される群から選択される。CSPGとしては、アグリカン、ブレビカン、バーシカン、ニューロカン、ホスファカン又はTe38から選択することができる。従って、これらの例はミエリン由来阻害剤と、NgRの公知ニューロン共受容体を含む。
【0062】
二重特異性抗体
本願は「二重特異性抗体」技術についても記載する。二重特異性抗体はアゴニスト、アンタゴニスト又は両者の各種組み合わせとして機能することができる。本明細書で使用する「アゴニスト」なる用語は該当分子と接触した場合に、アゴニストの不在下で観察される活性又は機能の強さに比較して分子の所定活性又は機能の強さを増加するモジュレーターを意味する。本明細書で使用する「アンタゴニスト」又は「阻害剤」なる用語は該当分子と接触した場合に、アンタゴニストの不在下で観察される活性又は機能の強さに比較して分子の所定活性又は機能の強さを低下させるモジュレーターを意味する。特定該当アンタゴニストとしては、Nogo−66の生物活性を阻害又は調節するものが挙げられる。Nogo−66のアンタゴニスト及び阻害剤としては限定されないが、Nogo−66受容体(NgR)と相互作用する任意分子、好ましくはモノクローナル抗体が挙げられる。
【0063】
なお、NgRとの相互作用の結果、受容体又は他のリガンド/細胞膜成分の結合と中和を生じる場合があり、複数の疾患に対する付加的又は相乗的作用に有用であると思われる。
【0064】
本願はNgRとp75、NgRとTROY、NgRとLINGO−1等のNgR抗体とNgR共受容体の組み合わせについても記載する。本願はNgRとそのリガンド及びNgRとミエリン由来阻害因子に交差反応する抗体も含む。これらの抗体としては、NgRと反発性ガイダンス分子(RGM)、NGRとNogo−A、NGRとMAG、NgRとOMpg、NgRとCSPGに交差反応する抗体が挙げられる。CSPGとしては、アグリカン、ブレビカン、バーシカン、ニューロカン、ホスファカン又はTe38から選択することができる。従って、これらの例はミエリン由来阻害剤と、NgRの公知ニューロン共受容体を含む。
【0065】
本願はNgRと成長因子受容体の二重特異性抗体についても記載し、このような成長因子としては限定されないが、神経成長因子(NGF)、脳由来神経栄養因子(BDNF)、上皮成長因子(EGF)、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、ニューロトロフィン、血小板由来成長因子(PDGF)、エリスロポエチン(EPO)、トロンボポエチン(TPO)、ミオスタチン(GDF−8)、増殖分化因子−9(GDF9)、塩基性線維芽細胞増殖因子(bFGF又はFGF2)、グリア細胞由来神経栄養因子(GDNF)、毛様体神経栄養因子(CNTF)が挙げられる。
【0066】
抗体の使用
本願の中和抗体又はその部分はヒトNgRと結合することができるので、酵素免疫測定法(ELISA)、放射免疫測定法(RIA)又は組織免疫組織化学等の慣用イムノアッセイを使用して(例えば血清や血漿等の生体サンプル中で)ヒトNgRを検出するために使用することができる。本願は、生体サンプル中のヒトNgRの検出方法として、生体サンプルを本発明の抗体又は抗体部分と接触させる段階と、及び、ヒトNgRと結合した抗体(又は抗体部分)又は未結合抗体(又は抗体部分)を検出することにより、生体サンプル中のヒトNgRを検出する段階とを含む方法を提供する。抗体は結合抗体又は未結合抗体の検出を容易にするために検出可能な物質で直接又は間接的に標識する。適切な検出可能な物質としては各種酵素、補欠分子族、蛍光材料、発光材料及び放射性材料が挙げられる。適切な酵素の例としては西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β−ガラクトシダーゼ又はアセチルコリンエステラーゼが挙げられ;適切な補欠分子族複合体の例としてはストレプトアビジン/ビオチンとアビジン/ビオチンが挙げられ;適切な蛍光材料の例としてはウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド又はフィコエリスリンが挙げられ;発光材料の1例としてはルミノールが挙げられ;適切な放射性材料の例としては3H、14C、35S、90Y、99Tc、111In、125I、131I、177Lu、166Ho、153Smが挙げられる。
【0067】
本願の抗体及び抗体部分はヒトNgR活性をインビトロ及びインビボの両者で中和できることが好ましい。従って、本発明のこのような抗体及び抗体部分はNogo−66とNgRの結合又はそれに起因する活性を阻害するために使用することができる。
【0068】
別の態様において、本願は対象、有利にはNgRに起因する活性が有害である疾患又は障害に罹患した対象におけるNogo−66活性又はNgR活性を低下させる方法を提供する。本願はこのような疾患又は障害に罹患した対象におけるNgR活性を低下させる方法として、対象のNgR活性を低下させるように本願の抗体又は抗体部分を対象に投与する段階を含む方法を提供する。好ましくは、NgRはヒトNgRであり、対象はヒト対象である。あるいは、対象は本発明の抗体が結合することが可能なNgRを発現する哺乳動物でもよい。更に、対象はNgRを導入した哺乳動物でもよい。本願の抗体は治療目的でヒト対象に投与することができる。更に、本願の抗体は獣医学的目的のため又はヒト疾患の動物モデルとして抗体が結合することが可能なNgRを発現する非ヒト哺乳動物に投与することもできる。後者に関して、このような動物モデルは本発明の抗体の治療効果の評価(例えば投与量及び投与時間の試験)に有用であると思われる。
【0069】
本明細書で使用する「NgR活性が有害である障害」なる用語はこの障害に罹患した対象におけるNgR又はそれに起因する活性の存在が障害の病理生理の原因である又は障害の増悪に寄与する因子であることが分かっている又はその疑いのある疾病及び他の障害を包含するものとする。従って、NgR活性が有害である障害はNgR活性の低下が障害の症状及び/又は進行を緩和すると予想される障害である。本発明の抗体で治療することができる障害の非限定的な例としては本発明の抗体の医薬組成物に関する下記セクションに記載する障害が挙げられる。
【0070】
NgRは神経変性又は神経変性プロセスの阻害に関連する神経疾患を伴うため、麻痺を生じる各種疾患に関連する病理において重要な役割を果たすことが認められている。このような疾患としては、筋委縮性側索硬化症、腕神経叢損傷、外傷性脳損傷を含む脳損傷、脳性麻痺、フリードライヒ運動失調症、ギラン・バレー症候群、白質ジストロフィー、多発性硬化症、ポリオ後遺症、二分脊椎、脊髄損傷、脊髄性筋萎縮症、脊髄腫瘍、脳卒中、横断性脊髄炎が挙げられる。更に、NgRは認知症、老人性認知症、軽度認知障害、アルツハイマー関連認知症、ハンチントン舞踏病、遅発性ジスキネジア、運動亢進症、躁病、梅毒性パーキンソン病、スチール・リチャードソン症候群、ダウン症候群、重症筋無力症にも役割を果たすことが認められている。
【0071】
NgRとそのリガンドは既知炎症因子を伴う炎症又は自己免疫状態の発生と進行にも関与していると思われる(Teng & Tang,2005;Fontoura & Steinmann,2006)。これらの疾患として、限定されないが、関節リウマチ、変形性関節症、若年性慢性関節炎、敗血症性関節炎、ライム関節炎、乾癬性関節炎、反応性関節炎、脊椎関節症、全身性エリテマトーデス、クローン病、潰瘍性大腸炎、炎症性腸疾患、インスリン依存性糖尿病、甲状腺炎、喘息、アレルギー疾患、乾癬、皮膚炎、強皮症、移植片対宿主病、臓器移植拒絶反応、臓器移植に伴う急性又は慢性免疫疾患、サルコイドーシス、アテローム性動脈硬化症、播種性血管内凝固、川崎病、グレーブス病、、ネフローゼ症候群、慢性疲労症候群、ウェゲナー肉芽腫症、シェーンライン・ヘノッホ紫斑病、顕微鏡的腎血管炎、慢性活動性肝炎、ブドウ膜炎、敗血性ショック、毒素性ショック症候群、敗血症症候群、悪液質、感染性疾患、寄生虫症、後天性免疫不全症候群、急性横断性脊髄炎、ハンチントン舞踏病、パーキンソン病、アルツハイマー病、脳卒中、原発性胆汁性肝硬変、溶血性貧血、悪性腫瘍、心不全、心筋梗塞、アジソン病、散発性I型多腺性内分泌不全症及びII型多腺性内分泌不全症、シュミット症候群、成人(急性)呼吸窮迫症候群、脱毛症、先天性脱毛症、血清反応陰性関節症、関節症、ライター病、乾癬性関節症、潰瘍性大腸炎性関節症、腸病性滑膜炎、クラミジア、エルシニア及びサルモネラ関連関節症、脊椎関節症、アテローム性疾患/動脈硬化症、アトピー性アレルギー、自己免疫性水疱症、尋常性天疱瘡、紅斑性天疱瘡、類天疱瘡、線状IgA疾患、自己免疫性溶血性貧血、クームス陽性溶血性貧血、後天性悪性貧血、若年性悪性貧血、筋痛性脳炎/ロイヤルフリー病、慢性粘膜皮膚カンジダ症、巨細胞性動脈炎、原発性硬化性肝炎、特発性自己免疫性肝炎、後天性免疫不全症候群、後天性免疫不全症関連疾患、B型肝炎、C型肝炎、分類不能型免疫不全症(分類不能型低ガンマグロブリン血症)、拡張型心筋症、女性不妊症、卵巣不全、早発卵巣不全、線維性肺疾患、特発性線維化性肺胞炎、炎症後間質性肺疾患、間質性肺炎、結合組織病関連間質性肺疾患、混合性結合組織病関連肺疾患、全身性硬化症関連間質性肺疾患、関節リウマチ関連間質性肺疾患、全身性エリテマトーデス関連肺疾患、皮膚筋炎/多発性筋炎関連肺疾患、ショーグレン病関連肺疾患、強直性脊椎炎関連肺疾患、血管炎性びまん性肺疾患、ヘモジデリン沈着症関連肺疾患、薬物誘発性間質性肺疾患、線維症、放射線線維症、閉塞性細気管支炎、慢性好酸球性肺炎、リンパ球浸潤性肺疾患、感染後間質性肺疾患、通風性関節炎、自己免疫性肝炎、1型自己免疫性肝炎(旧称自己免疫性又はルポイド肝炎)、2型自己免疫性肝炎(抗LKM抗体肝炎)、自己免疫性低血糖症、黒色表皮症を伴うB型インスリン抵抗性、副甲状腺機能低下症、臓器移植に伴う急性免疫疾患、臓器移植に伴う慢性免疫疾患、変形性関節症、原発性硬化性胆管炎、1型乾癬、2型乾癬、特発性白血球減少症、自己免疫性好中球減少症、腎疾患NOS、糸球体腎炎、腎臓の顕微鏡的血管炎、ライム病、円板状エリテマトーデス、特発性男性不妊症ないしNOS、精子自己免疫、多発性硬化症(全サブタイプ)、交感性眼炎、結合組織病続発性肺高血圧症、グッドパスチャー症候群、結節性多発性動脈炎の肺症状、急性リウマチ熱、リウマチ性脊椎炎、スティル病、全身性硬化症、ショーグレン症候群、高安動脈炎、自己免疫性血小板減少症、特発性血小板減少症、自己免疫性甲状腺疾患、甲状腺機能亢進症、甲状腺腫性自己免疫性甲状腺機能低下症(橋本病)、萎縮性自己免疫性甲状腺機能低下症、原発性粘液水腫、水晶体起因性ブドウ膜炎、原発性血管炎、白斑急性肝疾患、慢性肝疾患、アルコール性肝硬変、アルコール性肝傷害、胆汁鬱滞症、特異体質性肝疾患、薬物誘発性肝炎、非アルコール性脂肪肝炎、アレルギー及び喘息、B群連鎖球菌(GBS)感染症、精神障害(例えば鬱病及び統合失調症)、Th2型及びTh1型疾患、急性及び慢性疼痛(各種疼痛)、及び癌(例えば肺癌、乳癌、胃癌、膀胱癌、結腸癌、膵臓癌、卵巣癌、前立腺癌及び直腸癌)及び血液悪性疾患(白血病及びリンパ腫)が挙げられる。本願のヒト抗体又は抗体部分は自己免疫疾患、特に炎症に関連するもの(例えばリウマチ性脊椎炎、アレルギー、自己免疫性糖尿病、自己免疫性ブドウ膜炎)に罹患したヒトを治療するために使用することができる。
【0072】
NgRが他の蛋白質と相互作用することも知られており、このような蛋白質として、限定されないが、細胞接着、細胞移動、細胞トラッキング、軸索経路検出に関連する蛋白質及び細胞外マトリックス蛋白質が挙げられる。本願に含まれる可能な治療薬組み合わせとしては、NgRに対する抗体とセマフォリン(特にSema−1a,1b;Sema−2a;Sema3A,B,C,D,E,F;Sema4A,D;Sema5A;Sema6D;Sema7A;Sema VA)、プレキシン(プレキシン−A1−4,プレキシン−B1−3,プレキシン−C1,プレキシン−D1,Tim−2)、ニューロピリン(ニューロピリン−1及びニューロピリン−2)、カドヘリン(E−カドヘリン及びN−カドヘリン)、ネトリン(ネトリン−1)、エフリン(EphA3,4,6,7,8;B2,B3)Eph受容体、Ephリガンド、Ig CAM、テナスシン−C、CSPG、テナスシン、Sema3A、フィブロネクチン、ラミニン−1、コラーゲン(例えばコラーゲン−IV)、Robo、Abl、N−カドヘリン、L1、NCAMが挙げられる。
【0073】
NgRは細胞外アミロイド前駆体蛋白質フラグメント(Aβ)と相互作用することも記載されている。従って、本願はNgRとAβ種(Aβ1−40、Aβ1−42、Aβオリゴマー、Aβマルチマー、Aβグロブロマー)に対する抗体間の組み合わせを含む。この型の併用療法はアルツハイマー病の治療に有用であると思われる。この組み合わせは神経突起伸長/神経保護に加え、AD患者におけるプラーク沈着と認知能力の改善という二重の効果があると思われる。軸索及び樹状突起の萎縮はアルツハイマー病の非常に初期の特徴であり、併用治療が非常に有効であると思われる。
【0074】
更に、従来記載されているように、上記パートナーの任意1種間の二重特異性抗体も使用することができる。上記のようなこのような抗体製剤は、アルツハイマー病、パーキンソン病、脊髄損傷、外傷性脳損傷、多発性硬化症、末梢神経損傷、統合失調症、鬱病、不安症、加えて上記の可塑性及び神経突起成長及び神経毒性に関連する疾患のいずれものの治療に有用であると思われる。
【0075】
本願の抗体は細胞内標的蛋白質に到達するように膜貫通導入を可能にするペプチドと併用してもよい。このようなペプチド配列として、限定されないが、tat、アンテナペディア、ポリアルギニン、所定の抗菌ペプチドが挙げられる。このようなペプチドは細胞原形質膜のみならず、血液脳関門、腸粘膜、髄膜等の上皮及び内皮膜も含めた膜を通して導入を可能にする。
【0076】
このようなペプチドは細胞シグナル伝達阻害剤を細胞に導入することもでき、このような阻害剤としてはROCK、低分子量GTPアーゼ、アクチン及びミエリン安定剤等のNgRシグナル伝達分子に対する抗体又は小分子が挙げられる。
【0077】
本願の抗体又は抗体部分は前段落に記載したようなNgR活性が関与する障害の治療に有用な1種以上の補助的な小分子治療剤と併用投与することもできる。当然のことながら、本願の抗体又はその抗原結合部分は単独で使用することもできるし、補助剤(例えば,治療剤)と併用することもでき、前記補助剤はその所期目的に合わせて当業者により選択される。例えば、補助剤は本発明の抗体により治療される疾患又は病態を治療するために有用であると当業者に認められている治療剤とすることができる。補助剤は治療用組成物に有益な属性を付与する物質(例えば組成物の粘性を変化させる物質)でもよい。好ましい組み合わせとしては限定されないが、抗精神病薬が挙げられ、限定されないが、例えばリスペリドン、オランザピン、クエチアピン、フェノチアジン(クロルプロマジン、フルフェナジン、レボメプロマジン、ペリシアニン、ペルフェナジン、プロクロルペラジン、プロマジン、チオリダジン、トリフルオペラジン)、ブチロフェノン(ベンペリドール、ハロペリドール)、ゾテピン、ロキサピン、アリピプラゾール、セルトラリン、ジプラシドン、Rhoキナーゼ活性の小分子阻害剤(ROCK)(例えばファスジル、ジメチルファスジル又は他の任意ROCK阻害剤)、GABA A受容体又は代謝型グルタミン酸受容体(mGluRs)に対する低分子量受容体リガンド、非ステロイド性抗炎症薬(NSAIDS)、抗炎症性コルチコステロイド(例えばメチルプレドニゾロン)が挙げられる。
【0078】
本発明の医薬組成物
本願の抗体及び抗体部分は対象への投与に適した医薬組成物に配合することができる。本願の医薬組成物は「治療有効量」又は「予防有効量」の本発明の抗体又は抗体部分を含有することができる。「治療有効量」とは必要な用量と期間で所望治療結果を達成するために有効な量を意味する。抗体又は抗体部分の治療有効量は、当業者が決定することができ、個体の疾患状態、年齢、性別及び体重並びに抗体又は抗体部分が個体に所望応答を誘発する能力等の因子により異なる。治療有効量は、抗体又は抗体部分の有益な治療作用が毒性又は有害作用を上回る量でもある。「予防有効量」とは、必要な用量と期間で所望予防結果を達成するために有効な量を意味する。一般に、予防用量は、初期疾患ステージ又はそれ以前の対象に使用するので、予防有効量は治療有効量よりも少なくなる。
【0079】
一般に、医薬組成物は本発明の抗体又は抗体部分と医薬的に許容可能なキャリヤーを含有する。本明細書で使用する「医薬的に許容可能なキャリヤー」は生理的に適合可能な全溶媒、分散媒、コーティング、抗細菌及び抗真菌剤、等張及び吸収遅延剤等を包含する。医薬的に許容可能なキャリヤーの例としては水、食塩水、リン酸緩衝食塩水、デキストロース、グリセロール、エタノール等の1種以上とその組み合わせが挙げられる。多くの場合には、例えば糖類、多価アルコール(例えばマンニトール、ソルビトール)又は塩化ナトリウム等の等張化剤を組成物に加えることが好ましい。医薬的に許容可能なキャリヤーとしては更に抗体又は抗体部分の保存期間又は効力を増す微量の補助物質(例えば湿潤剤、乳化剤、防腐剤又は緩衝液)も挙げられる。
【0080】
本願の組成物は各種形態を取ることができる。これらの形態としては、例えば液体、半固体及び固体剤形が挙げられ、例えば溶液(例えば注射溶液及び輸液溶液)、分散液、懸濁液、錠剤、ピル、散剤、リポソーム及び座剤が挙げられる。好ましい剤形は所期投与方法及び治療用途により異なる。典型的な好ましい組成物は他の抗体をヒトに受動免疫するために使用されているものと同様の組成物等の注射溶液又は輸液溶液の形態である。好ましい投与方法は非経口投与(例えば静脈内、皮下、腹膜内、筋肉内)である。好ましい1態様では、静脈内輸液又は注射により抗体を投与する。別の好ましい態様では、筋肉内又は皮下注射により抗体を投与する。更に別の好ましい態様としては抗体の髄腔内投与が挙げられる。
【0081】
治療用組成物は一般に製造及び保存条件下で無菌安定でなければならない。組成物は溶液、マイクロエマルション、分散液、リポソーム又は高薬剤濃度に適した他の規則的構造として製剤化することができる。滅菌注射溶液は必要量の活性化合物(即ち抗体又は抗体部分)を必要に応じて上記成分の1種又はその組み合わせと共に適切な溶媒に加えた後に濾過滅菌することにより製造することができる。一般に、分散液は塩基性分散媒と上記から選択される必要な他の成分を含有する滅菌ビークルに活性化合物を加えることにより製造される。滅菌注射溶液の調製用滅菌凍結乾燥粉末の場合には、好ましい製造方法は予め滅菌濾過しておいたその溶液から活性成分と所望の付加成分のいずれかの粉末を生成する真空乾燥と噴霧乾燥である。例えばレシチン等のコーティングの使用、分散液の場合には必要な粒度の維持及び界面活性剤の使用により、溶液の適正な流動性を維持することができる。吸収を遅らせる物質(例えばモノステアリン酸塩やゼラチン)を組成物に加えることにより、注射用組成物の長期吸収が可能になる。
【0082】
本発明の抗体及び抗体部分は当分野で公知の各種方法により投与することができるが、多くの治療用途に好ましい投与経路/方法は皮下注射、静脈内注射又は輸液である。当業者に自明の通り、投与経路及び/又は方法は所望結果により異なる。ある態様において、化合物の迅速な放出を防止するキャリヤーを活性化合物に配合することができ、例えばインプラント、経皮パッチ及びマイクロカプセル送達システム等の制御放出製剤が挙げられる。エチレン酢酸ビニル、ポリ酸無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル及びポリ乳酸等の生分解性生体適合性ポリマーを使用することができる。このような製剤の多数の製造方法が特許登録されており、あるいは一般に当業者に公知である。例えばRobinson編,Sustained and Controlled Release Drug Delivery Systems,Marcel Dekker,Inc.,New York,1978参照。
【0083】
ある態様において、本願の抗体又は抗体部分を例えば不活性希釈剤又は同化性可食性キャリヤーと共に経口投与することができる。化合物(及び所望により他の成分)を硬又は軟シェルゼラチンカプセルに封入してもよいし、錠剤に圧縮してもよいし、対象の食事に直接添加してもよい。経口治療投与には、化合物に賦形剤を添加し、内服錠、口腔錠、トローチ剤、カプセル剤、エリキシル剤、懸濁液剤、シロップ剤、ウェハース剤等の形態で使用することができる。本発明の化合物を非経口投与以外の方法で投与するためには、その不活性化を防止するための材料を化合物に被覆したり、化合物と併用投与することが必要な場合がある。
【実施例】
【0084】
以下、実施例により本願を更に説明するが、これらの実施例は本願の例証のみを目的とし、その範囲を限定するものではない。
【0085】
(実施例1)
抗体の作製
A/Jマウス(The Jackson Laboratories,Bar Harbor Me)が組換えヒト及びラットNgR蛋白質(夫々配列番号1a及び配列番号2a)により免疫された。組換えヒト又はラットNgR蛋白質50ugを初回注射では完全フロイントアジュバントに加え、2回目以降はImmuneasy(登録商標)(Qiagen)に加えてマウスに4回皮下免疫した。融合の4日前に、マウスに抗原10ugを静脈内注射した。融合については、ケーラーとミルスタインの標準技術(Kohler G and Milstein C;Nature Vol.256,495−497頁(1975))を使用して免疫動物からの脾細胞をSP2/0−Ag14骨髄腫細胞と5:1の比で融合させた。融合から7〜10日後に、顕微鏡でハイブリドーマコロニーが観察されたので、上清をELISAアッセイにより試験した。PBS中1ug/mlの組換えヒト又はラットNgR蛋白質をELISAプレートに4℃で一晩被覆し、室温で1時間ブロックした。希釈した上清を温置し、結合をヤギ抗マウスIgFc−HRPコンジュゲートで検出した。ELISAで陽性の抗体産生ハイブリドーマ細胞をスケールアップし、限界希釈法によりサブクローニングした。Zymed EIAアイソタイプキットを使用して抗体のアイソタイプを決定した。
【0086】
各種ELISAフォーマットが確立されており、ヒト、ラット又はマウスNgRと結合する抗体を同定するための最初のスクリーニングとして日常的に使用されている。ELISAアッセイで反応した抗体を次に、組換えヒトNgR又は組換えラットNgRを安定に発現するHEK又はCHO細胞と結合させ、形質移入していない細胞又は対照細胞と結合しないものについて試験した。ELISAフォーマット用として、可溶性受容体を作製した(上記参照)。FACS試験には、全長NgR蛋白質を組換え細胞株で発現させた。ELISA結合アッセイとFACSアッセイにおけるMab50(配列番号3及び配列番号4)とMab51(配列番号5及び配列番号6)の結果を表2に示す。
【0087】
【表2】
【0088】
同一実験プロトコールを使用して他の抗体Mab1、Mab52、Mab53、Mab54、Mab55、Mab56、Mab57、Mab58、Mab59、Mab60、Mab61及びMab62を得た。
【0089】
(実施例2)
ELISAによる抗体特異性と結合親和性の測定
実施例1で作製したモノクローナル抗体の特異性を測定するために、NgR−Fc(ヒトNgRのアミノ酸Met1〜Ser447とヒトFcフラグメントを含む融合蛋白質(R&D Systems))とラットNgR(配列番号2a)を使用してELISAを実施した。2種のリガンドを96ウェルマイクロタイタープレート(Nunc Maxisorb;0.2μg/ウェル)に固定化した。ブロッキング試薬として、Tris−HCL,pH7.2中2%ウシ血清アルブミン(BSA)を室温で2時間使用した。モノクローナル抗体は10,000ng/mlの濃度から出発して使用した。西洋ワサビペルオキシダーゼ(Sigma)で標識した二次抗マウス抗体で結合抗体を検出し、3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して発色させた。mAb50とmAb51はヒトNgRと特異的に結合した。ヒトNgR−Fc 0.2μg/ウェルを使用した場合には、どちらのモノクローナル抗体でも20ng/ml未満の濃度で50%結合が認められた(図1A)。同様の実験でmAb50とmAb51はラットNgRのアミノ酸から構成されるポリペプチドにも特異的に結合した。96ウェルマイクロタイタープレートのウェル当たり0.2μgのラットNgRを使用した場合には、どちらのモノクローナル抗体でも20ng/ml未満の濃度で50%結合が認められた(図1B)。本発明のその他のモノクローナル抗体(mAB)もヒト及びラットNgRと結合した。各抗体で得られたシグナルの差から結合親和性に相違があると思われた(表3参照)。
【0090】
【表3】
【0091】
(実施例3)
ドットブロットとウェスタンブロットを使用した可溶性ヒト及びラットNgRとの抗体結合の特性決定
ドットブロットのために、異なる濃度の蛋白質2μlを乾燥ニトロセルロース膜上のTTBS緩衝液にスポットした。ウェスタンブロットのために、20%メタノールを添加したNovex転写用緩衝液に濾紙とニトロセルロースを10分間浸漬した。ブロッティングはNovexチャンバーで定電流(100mA)下に2時間室温で実施した。
【0092】
スポット当たりの蛋白質の添加量は、
a)100μg/ml〜〜200ng/スポット
b)50μg/ml〜〜100ng/スポット
c)10μg/ml〜〜20ng/スポット
d)5μg/ml〜〜10ng/スポット
e)1μg/ml〜〜2ng/スポット
f)500ng/ml〜〜1ng/スポット
とした。プローブをスポット後、膜を10分間室温で乾燥した後、免疫検出プロトコールを開始した。
【0093】
試験した全モノクローナル抗体はヒト及びラットNgRと結合した。異なる抗体で得られたシグナル間の差から結合親和性に相違があると思われた。(MAB61はニトロセルロース膜のプローブに使用した条件下でブロット上に高いバックグラウンドを示した。)NgR「対照」に対する抗体を除去するとシグナルを示さなかったので、結合は抗体依存性であった。
【0094】
モノクローナル抗体はウェスタンブロットにて変性NgRと様々に反応した。MAB1のみがヒト及びラットNgRに顕著なシグナルを示した。抗体の結合能の対照として、蛋白質をSDSゲルから転写させた後に非変性ヒトNgRと非変性ラットNgRを含むドットをニトロセルロース膜にスポットした。これらのドットを陽性対照として利用した処、全モノクローナル抗体は同一ニトロセルロース膜上の非変性蛋白質と結合することが確認された。結果を表4にまとめる。
【0095】
【表4】
【0096】
(実施例4)
AP−Nogo66と可溶性ヒトNogo受容体(hNgR−Fc)の結合の競合
モノクローナル抗体を更に特性決定するために、実施例2の競合アッセイと同様の結合アッセイを使用し、実施例1で作製したmAbの、AP−Nogo66のヒトNgR−Fcへの結合を阻害する能力を試験した。ヒトNgR−Fc(0.2μg/ウェル)を50mM炭酸Na緩衝液(pH9)中で96ウェルマイクロタイタープレート(Nunc Maxisorb)に4℃で一晩固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。ウェルに一定濃度のAP−Nogo66(終濃度Tris−HCL,pH7.2+0.1%BSA中0.15nM)を加え、Ab濃度を徐々に増加させた。プレートを90分間室温で温置した。各温置段階中に、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05%Tween20)で洗浄した。AP−Nogo66の結合をAttoPhos基質(Roche)で検出し、蛍光単位をPolarstar(BMG)計器で測定した。図2はmAb50とmAb51について0分と30分で行った測定の相対蛍光単位(RFU)を示す。上記結合はmAb50とmAb51により完全に阻害され、AP−Nogo66とヒトNgR−Fcの結合を50%阻害するためにはどちらの抗体も10倍モル過剰量が必要であった。mAb1を除く他の抗体mab52、mAb53、mAb54、mAb55、mAb56、mAb57、mAb58、mAb59、mAb60、mAb61及びmAb62もmAb50及び51で認められたと同様の濃度範囲でAP−Nogo66と可溶性Nogo受容体の結合を阻害した。
【0097】
(実施例5)
HEK293f細胞におけるAP−Nogo66とヒト及びラットNgRの結合のmAB50及びmAb51による競合
実施例1に記載したように作製したmAbの結合特性と競合特性を更に特性決定するために、ヒト又はラットNgRを一過的に発現するHEK293f細胞の懸濁液を使用した。形質移入から48時間後に細胞を96ウェルマイクロタイタープレート(Nunc Maxisorb)に撒き、リン酸緩衝食塩水(1%BSAを添加したPBS)で洗浄した。1μg/200μlのAP−Nogo66(終濃度40nM)と種々の濃度のモノクローナル抗体(20μg、4、0.8及び0.16μg/200μl)を加え、4℃で1時間温置した。同一アイソタイプのモノクローナル抗体を陰性対照として使用した。ZenonマウスIgG2a標識キット(Invitrogen)を使用してAlexa Fluorで標識した抗アルカリホスファターゼ抗体(Sigma)でAP−Nogo66の結合を検出した。二次抗体を4℃で1時間温置した。温置後に、細胞をPBSで洗浄し、FACS分析した。20μg/200μl濃度(20倍モル過剰)では、ポリクローナルNgR抗体とNgR−Fc(いずれもR&D Systems)はAP−Nogo66結合を60%〜70%阻害し、mAb50及び51はAP−Nogo66結合を約90%阻害した。mAb50及びmAB51をラット及びヒトNgR中に20μgの濃度で使用した場合の抗体アイソタイプ対照を図3aに示す。抗体アイソタイプ対照は黒線(左端のシグナル)で示し、抗体不在下のAP−Nogo66結合は赤線(右端シグナル)で示す。ヒト及びラットNgRはいずれも同等にAP−Nogo66と結合した。AP−Nogo66蛍光はヒト及びラットNgRを発現するHEK293f細胞で両者抗体の存在下では低強度側に移動する(斜線部;緑色)。異なる抗体濃度を使用すると、コルモゴロフ・スミルノフ(K−S)アッセイにより測定した場合にヒト及びラットNgRの両者でAP−Nogo66に対して2倍モル過剰のモノクローナル抗体50及び51でIC50が認められた。
【0098】
図3bは種々の濃度、即ち20.0μg,4.0,0.8及び0.16μg/200μlのmAb50及びmAb51による結果を示す。本発明の他のmAbはmAb50及び51で認められたと同様の濃度範囲でAP−Nogo66とHEK293f細胞で発現させたNogo受容体の結合を阻害した。AP−Nogo66とHEK293f細胞で発現させたヒトNgRの結合の50%競合に必要な抗体量を示す表5参照。
【0099】
【表5】
【0100】
(実施例6)
NTera2細胞中でAP−Nogo66により誘導された神経突起伸長阻害のmAB50及びmAb51による中和
ヒトNTera−2細胞及び齧歯類(マウス皮質ニューロン、ラット皮質ニューロン及びラット小脳顆粒細胞ニューロン)細胞型と、リガンドとしてAP−Nogo66及びミエリンを使用してインビボ状況に近似する機能システムで神経突起伸長に及ぼす本発明の抗体の効果を試験した。
【0101】
NTera2細胞(ヒト奇形癌細胞株)はレチノイン酸を使用して神経突起様細胞に分化させることができ、この細胞はポリクローナル抗体とNgRの適度の結合(FACS結合)により示されるようにNgR mRNA(PCR)と細胞表面蛋白質を発現する。夫々アイソタイプ対照抗体(非斜線領域)とmAb50及びmAb51(斜線領域)(図4)を使用してFACS分析により細胞表面上のNTera−2細胞による天然ヒトNgRの発現を検出した。両者抗体とNtera−2細胞の表面で発現されるNgR細胞の結合を斜線領域により示す。
【0102】
これらの分化細胞における神経突起伸長又は成長円錐崩壊の分析はヒトニューロンに可能な限り近似するインビトロシステムである。NTera2細胞(German National Resource Center for Biologicals,DMSZ,Braunschweigから入手)を融解し、DMEM培地(Gibco #31966−021+10%ウシ胎仔血清(FCS)+5%ウマ血清(PS))を加えた175cm2培養フラスコ(Greiner bio−one #660175)に撒いた。数日間培養後、細胞を再培養した。このために、細胞をPBS(Gibco #14190−094)で1回洗浄し、トリプシン/EDTA(Gibco #25300−054)で洗浄し、トリプシン/EDTAの存在下で5分間温置した。分化のために、細胞を再懸濁し、175cm2フラスコでDMEM(Gibco 31966−021,Lot.Nr.3092594)+10%FCS+5%PS+1%ペニシリン/ストレプトマイシン(5000/5000単位/mL)+レチノイン酸(SIGMA #R2625)終濃度10μMに細胞2.5×106個を撒いた。
【0103】
分化のために、終濃度10μMのレチノイン酸(SIGMA #R2625)を3週間にわたって週2回NTera2細胞に加えた。21日間分化後に、細胞を再培養した。このために、細胞をPBSで1回洗浄し、トリプシン/EDTAで洗浄し、トリプシン/EDTAの存在下で5分間温置した。細胞を再懸濁し、1:6に分割し、175cm2フラスコ6本でDMEM(Gibco 31966−021,Lot.Nr.3092594)+10%FCS+5%PS+1%PenStrep)に撒き、2〜3日間培養した。2〜3日後に細胞をPBSで洗浄し、物理的に剥がし、5分間1000rpmで遠心し、Neurobasal培地(Gibco #21103−049)+2mM L−グルタミン(Gibco #25030−024)+ペニシリン/ストレプトマイシン+B27−サプリメントに再懸濁し、三角フラスコ(Corning #431143)で予備凝集させた。こうして、細胞106個/mLを2×15mL凝集培地(Neurobsalmedium(Gibco #21103−049)+2mM L−グルタミン(Gibco #25030−024)+ペニシリン/ストレプトマイシン+B27−サプリメント)に加え、37℃,5%CO2下に一晩温和に撹拌し、阻害基質と対照基質を事前被覆した96ウェルプレート(Biocoat Poly−D−Lysin Cellware 96−Well Black/Clear Plate Becton Dickinson #35 4640(35 6640))に撒いた。阻害基質では、96ウェルプレートの半数に滅菌PBS中AP−Nogo66(AP−Nogo66濃度は15μg/mLとした)+ラミニン(Sigma,L−2020,Lot 014K4060,(ストック溶液.1mg/mL);ウェル当たり最終ラミニン量20μg)100μLを事前被覆した。許容基質では、プレートの残りの半数にラミニン100μl(20μg)を被覆した。2時間温置後に、プレートをPBSで2回洗浄し、培地40μlを補充した各ウェルに予め凝集させた懸濁液50μlを撒いた。プレートを37℃で2時間温置し、最後に、予め希釈したmAb50又はmAb51溶液10μlを加え、最終抗体濃度1〜100μg/mLとした。細胞を37℃,5%CO2下に一晩温置し、翌日、2%パラホルムアルデヒド(SIGMA #P−6148)で固定し、その後の分析に備えて4℃で保存した。ソフトウェアAxioVision LE Rel.4.1を使用し、標準評価パラメーター(凝集面積及び凝集面積&神経突起成長面積)を使用して神経突起伸長の分析を実施した。
【0104】
上記方法によりNTera2凝集物からの神経突起伸長を定量した。結果を図5に示す通り、mAb50では2μg/ml、mAb51では1μg/mlで神経突起伸長の有意な改善を観測することができる。Nogo66処理に対する有意差:*=p値<0.05;***=p値<0.00.1。
【0105】
抗体Mab52、53、54、55、56、57、58、59、60、61及び62でも神経突起伸長阻害の改善が得られた。Mab1とMab4はAP−Nogo66による神経突起伸長の阻害を示さなかった。
【0106】
(実施例7)
hNgRの欠失変異体:抗体の発現、精製及び結合。
【0107】
GPIリンカー形成(450最終アミノ酸)と膜結合を阻止するようにC末端アミノ酸を欠失させると共に、分泌蛋白質量を潜在的に増加させるように分泌シグナルを使用することにより、hNgRを可溶性蛋白質として発現させた。使用した発現システムは293F細胞における発現プラスミドの一過的形質移入に基づくものとした。Ni−NTA(ニッケル−ニトリロ三酢酸)ビーズを使用することによりHisタグ付き分泌蛋白質を捕獲した。hisタグを付けた無関係の蛋白質(RGM A−His)を陰性対照として使用してPAGEとウェスタンブロットにより溶出後の蛋白質の純度とサイズを分析した。これらの調製物におけるNgR−蛋白質をより良好に調整するために、全欠失変異体を検出すると予想されるhNgRに対するポリクローナル抗体(AF1208)を使用してドットブロットによりNgRの量を測定した。異なるNgR欠失変異体に調整量の蛋白質を使用するドットブロットを実施した。上記抗体による免疫検出にドットブロットを使用した。
【0108】
NgR欠失変異体を発現する293F細胞の細胞培養上清を非精製NgR源として使用して抗体とNgR欠失変異体の結合を更に特性決定した。
(i)フルニエらによるNgR欠失変異体の作製(Truncated Soluble Nogo Receptor Binds Nogo−66 and Blocks Inhibition of Axon Growth by Myelin;Alyson E.Fournier,Graham C.Gould,Betty P.Liu,Stephen M.Strittmatter;The Journal of Neuroscience,October 15,2002,22(20):8876−8883)。ヒトNgRの7種類の欠失変異体が文献に記載されている(Fournierら)。これらの7種類の変異体構築物がpSecTag2A IgK/hNgR 27−450/Myc/Hisから作製された。この構築物はpSecTag2AベクターにIgKリーダーペプチドとC末端Myc及びHisタグに融合したヒトNgRのアミノ酸27−450のコーディング領域を含む。以下の構築物を作製した:
【0109】
【化1】
【0110】
pSecTag2AigK/27−310/Myc/Hisを除く全構築物はQuikChange II XL Site Directed Mutagenesis Kit(Stratagene,#200521)を使用して作製され、形質転換は大腸菌XL10 Gold細胞中にて実施された。pSecTag2AigK/NgR/27−310/Myc/Hisについて、コーディング配列の相当領域を増幅し、pSecTag2Aにクローニングした。NgRコーディング領域の異なる部分を欠失させるために以下の変異誘発及び増幅プライマーを使用した。
【0111】
【化2】
【0112】
図6の模式図参照。
【0113】
(ii)NgR欠失変異体の発現
ヒトNgRのC末端アミノ酸を欠失させると、膜固定性及び細胞上清中の可溶性受容体蛋白質の存在が失われる。N末端において、hNgRのシグナルペプチド(アミノ酸1−27)をベクター中にコードされているシグナルペプチドで置換した。従って、hNgRのアミノ酸27から開始し、hNgRのアミノ酸450を終点とするN末端及びC末端欠失変異体(hNgR27−450)をこれらの全変異体の基礎として使用し、細胞上清中に可溶性蛋白質の存在を可能にした。更に、これらの変異体にはこれらの蛋白質の精製を可能にするための短いアミノ酸タグを加えた。hNgR変異体のDNAを293F細胞で一過的に発現させた。72時間後に遠心により細胞上清を回収した。下記方法により蛋白質精製を実施した。ヒトNgRの7種類の異なる変異体をコードする以下のDNAによる形質移入を実施した。
【0114】
【化3】
【0115】
hNgR変異体のDNAを293F細胞で一過的に発現させた。
【0116】
(iii)Ni−キレート親和性(Ni−NTA)を使用したNgR蛋白質の精製
Ni−NTAスーパーフロービーズ(Qiagen Qiagen #1018611)を使用した。ビーズ上清を13500rpmで遠心し、上清を捨て、ビーズを新鮮なPBSに再懸濁することにより、ビーズをPBS(リン酸緩衝食塩水,Invitrogen)で3回洗浄した。ビーズ懸濁液200μlを細胞培養上清30mlに使用した。ビーズを細胞培養上清と共に4℃でローテータで60rpmにて温置し、温置後に遠心(10分間,3000rpm)し、ビーズをペレット化した。上清を捨て、ビーズをPBSで3回洗浄した。溶出用緩衝液(PBS,160mM NaCl,150mMイミダゾール)250μlを使用して結合性蛋白質をビーズから溶出させた。室温のローテータで30分間温置後、13.500rpmで3分間遠心することによりビーズをペレット化した。上清を除去した。溶出した蛋白質をその後の分析に備えて−20℃で凍結させた。hNgRの欠失変異体によるドットブロット実験からのデータを表6にまとめる。
【0117】
【表6】
【0118】
(実施例8A)
MAG−FcとNgR−Fcの結合の競合
モノクローナル抗体を更に特性決定するために、実施例2の競合アッセイと同様の結合アッセイを使用して実施例1で作製した2種類のmAbがMAG−FcとヒトNgR−Fcの結合を阻害する能力を試験した。NgR−Fc(0.2μg/ウェル,R&D Systems)を4℃の50mM炭酸Na緩衝液(pH9)中、96ウェルマイクロタイタープレート(Nunc Maxisorb)に固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。MAG−Fc(R&D Systems recombinant Rat MAG/Fc Chimera,Catalog #538−MG)をZenonヒトIgG標識キット(Molecular Probes)により西洋ワサビペルオキシダーゼで標識した。ウェルに一定濃度の標識MAG−Fc(Tris−HCL,pH7.2+0.1%BSA中終濃度50ng/ml)と、指定濃度のmAb50及びmAb51を加えた。未標識MAG−FcをNgR−Fc対照として使用してこれを60分間室温で温置した。各温置段階中、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05% Tween 20)で洗浄した。3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して標識MAG−Fcを発色させた。図7に示すように、mAb50(黒三角)とmAb51(白三角)はMAG−FcとヒトNgRの結合を阻害しなかった。MAG−FcとNgR−Fcは標識MAG−Fc(黒円)とヒトNgR−Fcの結合に競合した(ヒトNgRのアミノ酸Met1〜Ser447とヒトFcフラグメントを含む融合蛋白質(R&D Systems);黒四角)。従って、mAb50とmAb51はこれらの条件下でNgRとの結合に関してMAGと競合しなかった(図7)。他の抗体はいずれもこれらの条件下でNgRとの結合に関してMAGと競合しなかった。
【0119】
(実施例8B)
OMgpとNgR−Fcの結合の競合
モノクローナル抗体を更に特性決定するために、実施例8aの競合アッセイと同様の結合アッセイを使用して実施例1で作製した2種類のmAbの、oMgpのヒトNgR−Fcへの結合を阻害する能力を試験した。NgR−Fc(0.2μg/ウェル,R&D Systems)を4℃の50mM炭酸Na緩衝液(pH9)中、96ウェルマイクロタイタープレート(Nunc Maxisorb)に固定化した後、Tris−HCL,pH7.2中2%BSAで室温にて2時間ブロッキング段階を実施した。ウェルに一定濃度のヒトOMgp(R&D Systems/1673−OM;Tris−HCL,pH7.2+0.1%BSA中終濃度500ng/ml)と、指定濃度のmAb50及びmAb51を加えた。この混合物を60分間室温で温置した。西洋ワサビペルオキシダーゼ(Roche)で標識した抗His抗体を使用してOMgpを検出した。各温置段階中、プレートを洗浄用緩衝液(10mM Tris−HCl,pH7.2+0.05% Tween 20)で洗浄した。3,3’,5,5’−テトラメチルベンジジン基質(TMB,Pierce)を標準条件下で使用して標識抗His抗体を発色させた。
【0120】
表7に示すように、mAb50とmAb51はOMgpとヒトNgRの結合を部分的に阻害した(30〜40%範囲)。mAb52とmAb59以外の他の抗体も、100倍モル過剰を上回る80μg/mlまでの濃度でもOMgpとヒトNgRの結合を部分的に阻害した(表7)。
【0121】
【表7】
【0122】
(実施例9)
ラットDRG細胞におけるAP−Nogo66により誘導した神経突起伸長阻害のmAB50及びmAb51による中和
生後3〜6日の子ラットからの後根神経節(DRG)を使用して神経突起伸長に及ぼすNgR抗体の作用を調べた。
【0123】
DRGを作製するために、子ラット6〜8匹を外科用鋏で断頭した。腹側臓器と椎骨を除去することにより腹側から脊柱を摘出した。次に、精密鋏により脊柱を縦方向に切開した。DRGを付けたまま脊髄を摘出し、PBSを加えた10cmペトリ皿に移した。2本の精密鉗子を使用することによりDRGから脊髄と結合神経線維を分離し、PBS 1mlを加えた35mmペトリ皿に移した。全DRGを採取後、コラゲナーゼのPBS溶液(4mg/mlコラゲナーゼタイプI,Worthington #CLS−1)0,5mlを加え、DRGを20〜30分間37℃で温置した。トリプシン溶液(PBS中0,5%トリプシン(SERVA #37290))0,5mlを加え、DRGを更に15〜25分間37℃で温置した。DRGを15mlチューブに移し、培地(DMEM Nut Mix F12(Gibco #31330−038)+5%FCS(Gibco,熱不活化)+5%ウマ血清(Sigma,熱不活化)+1%ペニシリン/ストレプトマイシン(Gibco #15140−122))10mlを加えた。DRGがチューブの底に沈降後、上清を除去した。パスツールピペットに3〜5回通した後、口径の小さいパスツールピペットに更に2〜3回通すことによりDRGを培地2ml中で解離させた。細胞塊の沈降後、解離した細胞を含む上清を新しいチューブに移した。5分間1000rpmで遠心することにより細胞を採取し、培地2mlに再懸濁し、カウントし、神経成長因子(NGF;終濃度62,5μg/ml,Roche #1014331)を加えた培地で所望細胞密度まで希釈した。ポリリジンを被覆し、更にコーティング溶液100μl(滅菌水50mlで希釈したラミニン(1mg/ml Sigma #L−2020)バイアル1本)を被覆しておいた96ウェルプレート(例えばBeckton Dickinson #356640)のウェル当たり80μlの容量で細胞4000〜7000個を撒いた後、30分間〜3時間37℃で温置した。細胞培養後、濃度を増加しながら容量10μlの抗体を加えた。CO2インキュベーターで37℃にて2時間温置後、10μlのAP−Nogo66を加えた。細胞を18〜30時間増殖させ、4%パラホルムアルデヒドのPBS(リン酸緩衝食塩水;Gibco #14190−094)溶液100μlを加えた後に4℃で少なくとも12時間温置することにより固定させた。96ウェルプレートの代わりに、ポリリジンを被覆したカバースリップ(例えばBecton Dickinson #354085)と24ウェルプレート(例えばFalcon #353047)を併用し、コーティング溶液500μlを加えることにより被覆した。抗βIIIチューブリン抗体(例えばAbeam #abl4545)とCy3標識二次抗体(例えばJackson ImmunoResearch #715−165−151)を併用して間接免疫蛍光により神経突起伸長を可視化した。ビスベンズイミド(H33258)を二次抗体に加えることにより神経を染色した。BDTM Pathway Bioimager(Beckton Dickinson)で10倍の倍率で顕微鏡写真を撮影し、AttoNO(Beckton Dickinson)ソフトウェアを使用して神経突起長を測定した。神経突起伸長を画像1枚当たりのDRG数に正規化した。図8は1例としてAP−Nogo66により誘導した神経突起伸長阻害のmAb50による中和を示す。図9は夫々抗体mAb50及びmAb51を使用した実験の結果を示す。AP−Nogo66を加えると、AP−Nogo66を加えない対照条件(1本目の棒グラフ)に比較して神経突起伸長の長さは著しく減少する(2本目の棒グラフ)。どちらの抗体もAP−Nogo66により誘導した神経突起伸長の阻害を用量依存的に中和した。DRG数に正規化した神経突起長は抗体を加えずにAP−Nogo66を加えた場合(2本目の棒グラフ)に比較してどちらの抗体でも50μg/mlで統計的に有意差を示す(最後の棒グラフ)。
【0124】
抗体Mab52、53、54、55、56、57、58、59、60、61及び62でも神経突起伸長阻害の改善が得られた。Mab1とMab4はAP−Nogo66による神経突起伸長の阻害を改善しなかった。
【0125】
これらの結果から、このような抗体は成長阻害環境で神経突起成長を刺激する潜在能力があると結論することができる。
【特許請求の範囲】
【請求項1】
Nogo−66受容体の少なくとも1個のエピトープと特異的に相互作用する単離された結合性蛋白質。
【請求項2】
単離された蛋白質がモノクローナル中和抗体又はその抗原結合フラグメントである、請求項1に記載の単離された結合性蛋白質。
【請求項3】
前記抗原結合フラグメントがVH及びVL領域を含む、請求項2に記載の結合性蛋白質。
【請求項4】
前記中和抗体が、Nogo−66の、その受容体への結合能を低下させる、請求項2に記載の中和抗体。
【請求項5】
前記中和抗体がNogo−66を中和することができる、請求項2に記載の中和抗体。
【請求項6】
Nogo66受容体がヒト、カニクイザル、ラット、ニワトリ、カエル及び魚類から選択される、請求項2に記載の抗体。
【請求項7】
Nogo66受容体ポリペプチドが配列番号1a及び配列番号2aから選択されるアミノ酸配列に対して90%の相同度を有する、請求項2に記載の抗体。
【請求項8】
Nogo66受容体が配列番号1b及び配列番号2bの核酸配列に対して90%の相同度を有する核酸によりコードされている、請求項2に記載の抗体。
【請求項9】
配列番号3の配列を含む重鎖可変領域(VH領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項10】
配列番号4の配列を含む軽鎖可変領域(VL領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項11】
配列番号5の配列を含む重鎖可変領域(VH領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項12】
配列番号6の配列を含む軽鎖可変領域(VL領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項13】
10−7Mから10−13Mの解離定数(Kd)を有する、請求項3に記載の抗体。
【請求項14】
103M−1s−1から107M−1s−1の会合速度(Kon)を有する、請求項3に記載の抗体。
【請求項15】
10−3s−1から少なくとも約10−6s−1の解離速度(Koff)を有する、請求項3に記載の抗体。
【請求項16】
10−6s−1から少なくとも約10−12s−1のIC50値を有する、請求項3に記載の抗体。
【請求項17】
1×10−9Mから1×10−11MのEC50にてヒトNgRに結合する、請求項3に記載の抗体。
【請求項18】
ヒトNgRに結合し、抗体がグリコシル化されている、請求項3に記載の抗体。
【請求項19】
前記抗体又は抗原結合フラグメントがマウス抗体、ヒト化抗体、完全ヒト、キメラ抗体、ヒト化抗体の抗原結合フラグメント又はキメラ抗体の抗原結合フラグメントである、請求項3に記載の抗体又は抗原結合フラグメント。
【請求項20】
前記抗体又は抗原結合フラグメントがFabフラグメント、Fab’フラグメント、F(ab’)2フラグメント及びFvフラグメントから構成される群から選択される抗原結合フラグメントである、請求項3に記載の抗体又は抗原結合フラグメント。
【請求項21】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体であり、前記モノクローナル抗体がハイブリドーマ細胞株ATCC NO.PTA−8383により分泌されるモノクローナル抗体である前記モノクローナル抗体。
【請求項22】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体であり、前記モノクローナル抗体がハイブリドーマ細胞株ATCC NO.PTA−8384により分泌されるモノクローナル抗体である前記モノクローナル抗体。
【請求項23】
結合がNogo−66受容体の不活性化をもたらす、請求項21に記載のモノクローナル抗体。
【請求項24】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体を産生するハイブリドーマ細胞株。
【請求項25】
ハイブリドーマがマウス及びヒトハイブリドーマから構成される群から選択される、請求項24に記載のハイブリドーマ細胞株。
【請求項26】
ハイブリドーマがラット、ヒツジ、ブタ、ウシ、ヤギ及びウマハイブリドーマから構成される群から選択される、請求項24に記載のハイブリドーマ細胞株。
【請求項27】
結合がNogo−66受容体の不活性化をもたらす、請求項22に記載のハイブリドーマ細胞株。
【請求項28】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体を産生するハイブリドーマ細胞株。
【請求項29】
ハイブリドーマがマウス及びヒトハイブリドーマから構成される群から選択される、請求項27に記載のハイブリドーマ細胞株。
【請求項30】
ハイブリドーマがラット、ヒツジ、ブタ、ウシ、ヤギ及びウマハイブリドーマから構成される群から選択される、請求項27に記載のハイブリドーマ細胞株。
【請求項31】
a)nM範囲以下の親和性で哺乳動物NgRに結合する特性;
b)神経突起伸長アッセイにおいてnM以下の効力にてインビトロのNogo−66の結合を機能的に阻害する特性;
c)神経突起伸長アッセイにおいてnM以下の効力にてインビトロのNogo−66の結合を機能的に阻害する特性;
d)眼神経挫滅モデルにて出芽をインビボ誘導する特性;又は
e)インビボ実験的脊髄損傷を緩和する特性
から構成される群から選択される少なくとも1種の特性を有するモノクローナル中和抗体又はその抗原結合フラグメント。
【請求項32】
請求項3に記載のモノクローナル中和抗体又は抗原結合フラグメントをコードする単離された核酸。
【請求項33】
pcDNA、pTT、pTT3、pEFBOS、pBV、pJV及びpBJから構成される群から選択される、請求項28に記載の単離された核酸を含むベクター。
【請求項34】
原生動物細胞、動物細胞、植物細胞及び真菌細胞から構成される群から選択される、請求項33に記載のベクターで形質転換された宿主細胞。
【請求項35】
動物細胞がHEK293、CHO及びCOSを含む群から選択される哺乳動物細胞である、請求項30に記載の宿主細胞。
【請求項36】
真核細胞である、請求項33に記載のベクターで形質転換された宿主細胞。
【請求項37】
ヒト及び/又はラットNgRに結合する結合性蛋白質を生産するために十分な条件下に宿主細胞を培地中に培養する段階を含む、ヒト及び/又はラットNgRに結合する請求項1に記載の結合性蛋白質の生産方法。
【請求項38】
請求項3から5のいずれか一項に記載のモノクローナル抗体又は抗原結合部分と及び医薬的に許容可能なキャリヤーとを含有する医薬組成物。
【請求項39】
請求項3から5のいずれか一項に記載の単離蛋白質を対象に投与する段階を含む、Nogo−66受容体に結合するNogo−66を低減させることが必要な対象においてNogo−66受容体に結合するNogo−66を低減させる方法。
【請求項40】
請求項3から5のいずれか一項に記載の単離蛋白質を使用する段階を含む、Nogo−66により誘導される神経突起伸長阻害を回復させる方法。
【請求項41】
請求項3から5のいずれか一項に記載の抗体を単独投与又は他の治療剤と併用投与する段階を含む、NgR活性に関連する障害の対象の治療方法。請求項3から5のいずれか一項に記載の抗体を単独投与する段階又は対象におけるヒトNgR活性を低下させるような他の治療剤と併用投与する段階を含む、NgR活性が有害である障害に罹患した対象においてヒトNgR活性を低下させる方法。
【請求項42】
障害が筋委縮性側索硬化症、腕神経叢損傷、外傷性脳損傷を含む脳損傷、脳性麻痺、フリードライヒ運動失調症、ギラン・バレー症候群、白質ジストロフィー、多発性硬化症、ポリオ後遺症、二分脊椎、脊髄損傷、脊髄性筋萎縮症、脊髄腫瘍、脳卒中及び横断性脊髄炎を含む群から選択される神経疾患を含む、請求項41に記載の方法。
【請求項43】
障害が認知症、老人性認知症、軽度認知障害、アルツハイマー関連認知症、ハンチントン舞踏病、遅発性ジスキネジア、運動亢進症、躁病、梅毒性パーキンソン病、スチール・リチャードソン症候群、ダウン症候群、重症筋無力症、神経外傷、血管アミロイドーシス、アミロイドーシスを伴う脳出血I、脳炎症、フリードライヒ運動失調症、急性錯乱、筋委縮性側索硬化症、緑内障及びアルツハイマー病を含む群から選択される神経疾患を含む、請求項41に記載の方法。
【請求項1】
Nogo−66受容体の少なくとも1個のエピトープと特異的に相互作用する単離された結合性蛋白質。
【請求項2】
単離された蛋白質がモノクローナル中和抗体又はその抗原結合フラグメントである、請求項1に記載の単離された結合性蛋白質。
【請求項3】
前記抗原結合フラグメントがVH及びVL領域を含む、請求項2に記載の結合性蛋白質。
【請求項4】
前記中和抗体が、Nogo−66の、その受容体への結合能を低下させる、請求項2に記載の中和抗体。
【請求項5】
前記中和抗体がNogo−66を中和することができる、請求項2に記載の中和抗体。
【請求項6】
Nogo66受容体がヒト、カニクイザル、ラット、ニワトリ、カエル及び魚類から選択される、請求項2に記載の抗体。
【請求項7】
Nogo66受容体ポリペプチドが配列番号1a及び配列番号2aから選択されるアミノ酸配列に対して90%の相同度を有する、請求項2に記載の抗体。
【請求項8】
Nogo66受容体が配列番号1b及び配列番号2bの核酸配列に対して90%の相同度を有する核酸によりコードされている、請求項2に記載の抗体。
【請求項9】
配列番号3の配列を含む重鎖可変領域(VH領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項10】
配列番号4の配列を含む軽鎖可変領域(VL領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項11】
配列番号5の配列を含む重鎖可変領域(VH領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項12】
配列番号6の配列を含む軽鎖可変領域(VL領域)を含む配列に対して少なくとも90%のアミノ酸配列一致度を有する、請求項3から5のいずれか一項に記載の抗体。
【請求項13】
10−7Mから10−13Mの解離定数(Kd)を有する、請求項3に記載の抗体。
【請求項14】
103M−1s−1から107M−1s−1の会合速度(Kon)を有する、請求項3に記載の抗体。
【請求項15】
10−3s−1から少なくとも約10−6s−1の解離速度(Koff)を有する、請求項3に記載の抗体。
【請求項16】
10−6s−1から少なくとも約10−12s−1のIC50値を有する、請求項3に記載の抗体。
【請求項17】
1×10−9Mから1×10−11MのEC50にてヒトNgRに結合する、請求項3に記載の抗体。
【請求項18】
ヒトNgRに結合し、抗体がグリコシル化されている、請求項3に記載の抗体。
【請求項19】
前記抗体又は抗原結合フラグメントがマウス抗体、ヒト化抗体、完全ヒト、キメラ抗体、ヒト化抗体の抗原結合フラグメント又はキメラ抗体の抗原結合フラグメントである、請求項3に記載の抗体又は抗原結合フラグメント。
【請求項20】
前記抗体又は抗原結合フラグメントがFabフラグメント、Fab’フラグメント、F(ab’)2フラグメント及びFvフラグメントから構成される群から選択される抗原結合フラグメントである、請求項3に記載の抗体又は抗原結合フラグメント。
【請求項21】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体であり、前記モノクローナル抗体がハイブリドーマ細胞株ATCC NO.PTA−8383により分泌されるモノクローナル抗体である前記モノクローナル抗体。
【請求項22】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体であり、前記モノクローナル抗体がハイブリドーマ細胞株ATCC NO.PTA−8384により分泌されるモノクローナル抗体である前記モノクローナル抗体。
【請求項23】
結合がNogo−66受容体の不活性化をもたらす、請求項21に記載のモノクローナル抗体。
【請求項24】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体を産生するハイブリドーマ細胞株。
【請求項25】
ハイブリドーマがマウス及びヒトハイブリドーマから構成される群から選択される、請求項24に記載のハイブリドーマ細胞株。
【請求項26】
ハイブリドーマがラット、ヒツジ、ブタ、ウシ、ヤギ及びウマハイブリドーマから構成される群から選択される、請求項24に記載のハイブリドーマ細胞株。
【請求項27】
結合がNogo−66受容体の不活性化をもたらす、請求項22に記載のハイブリドーマ細胞株。
【請求項28】
Nogo−66受容体の少なくとも1個のエピトープに特異的に結合するモノクローナル抗体を産生するハイブリドーマ細胞株。
【請求項29】
ハイブリドーマがマウス及びヒトハイブリドーマから構成される群から選択される、請求項27に記載のハイブリドーマ細胞株。
【請求項30】
ハイブリドーマがラット、ヒツジ、ブタ、ウシ、ヤギ及びウマハイブリドーマから構成される群から選択される、請求項27に記載のハイブリドーマ細胞株。
【請求項31】
a)nM範囲以下の親和性で哺乳動物NgRに結合する特性;
b)神経突起伸長アッセイにおいてnM以下の効力にてインビトロのNogo−66の結合を機能的に阻害する特性;
c)神経突起伸長アッセイにおいてnM以下の効力にてインビトロのNogo−66の結合を機能的に阻害する特性;
d)眼神経挫滅モデルにて出芽をインビボ誘導する特性;又は
e)インビボ実験的脊髄損傷を緩和する特性
から構成される群から選択される少なくとも1種の特性を有するモノクローナル中和抗体又はその抗原結合フラグメント。
【請求項32】
請求項3に記載のモノクローナル中和抗体又は抗原結合フラグメントをコードする単離された核酸。
【請求項33】
pcDNA、pTT、pTT3、pEFBOS、pBV、pJV及びpBJから構成される群から選択される、請求項28に記載の単離された核酸を含むベクター。
【請求項34】
原生動物細胞、動物細胞、植物細胞及び真菌細胞から構成される群から選択される、請求項33に記載のベクターで形質転換された宿主細胞。
【請求項35】
動物細胞がHEK293、CHO及びCOSを含む群から選択される哺乳動物細胞である、請求項30に記載の宿主細胞。
【請求項36】
真核細胞である、請求項33に記載のベクターで形質転換された宿主細胞。
【請求項37】
ヒト及び/又はラットNgRに結合する結合性蛋白質を生産するために十分な条件下に宿主細胞を培地中に培養する段階を含む、ヒト及び/又はラットNgRに結合する請求項1に記載の結合性蛋白質の生産方法。
【請求項38】
請求項3から5のいずれか一項に記載のモノクローナル抗体又は抗原結合部分と及び医薬的に許容可能なキャリヤーとを含有する医薬組成物。
【請求項39】
請求項3から5のいずれか一項に記載の単離蛋白質を対象に投与する段階を含む、Nogo−66受容体に結合するNogo−66を低減させることが必要な対象においてNogo−66受容体に結合するNogo−66を低減させる方法。
【請求項40】
請求項3から5のいずれか一項に記載の単離蛋白質を使用する段階を含む、Nogo−66により誘導される神経突起伸長阻害を回復させる方法。
【請求項41】
請求項3から5のいずれか一項に記載の抗体を単独投与又は他の治療剤と併用投与する段階を含む、NgR活性に関連する障害の対象の治療方法。請求項3から5のいずれか一項に記載の抗体を単独投与する段階又は対象におけるヒトNgR活性を低下させるような他の治療剤と併用投与する段階を含む、NgR活性が有害である障害に罹患した対象においてヒトNgR活性を低下させる方法。
【請求項42】
障害が筋委縮性側索硬化症、腕神経叢損傷、外傷性脳損傷を含む脳損傷、脳性麻痺、フリードライヒ運動失調症、ギラン・バレー症候群、白質ジストロフィー、多発性硬化症、ポリオ後遺症、二分脊椎、脊髄損傷、脊髄性筋萎縮症、脊髄腫瘍、脳卒中及び横断性脊髄炎を含む群から選択される神経疾患を含む、請求項41に記載の方法。
【請求項43】
障害が認知症、老人性認知症、軽度認知障害、アルツハイマー関連認知症、ハンチントン舞踏病、遅発性ジスキネジア、運動亢進症、躁病、梅毒性パーキンソン病、スチール・リチャードソン症候群、ダウン症候群、重症筋無力症、神経外傷、血管アミロイドーシス、アミロイドーシスを伴う脳出血I、脳炎症、フリードライヒ運動失調症、急性錯乱、筋委縮性側索硬化症、緑内障及びアルツハイマー病を含む群から選択される神経疾患を含む、請求項41に記載の方法。
【図1】

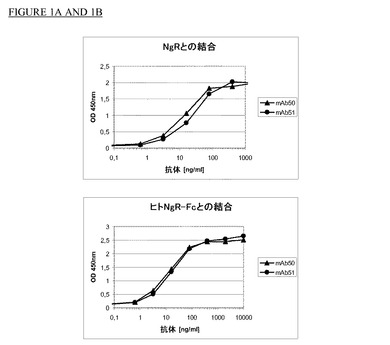
【図2】
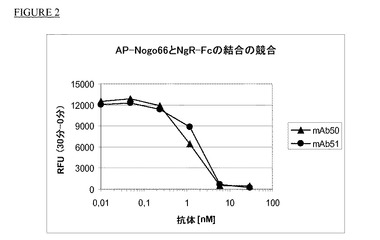
【図3】
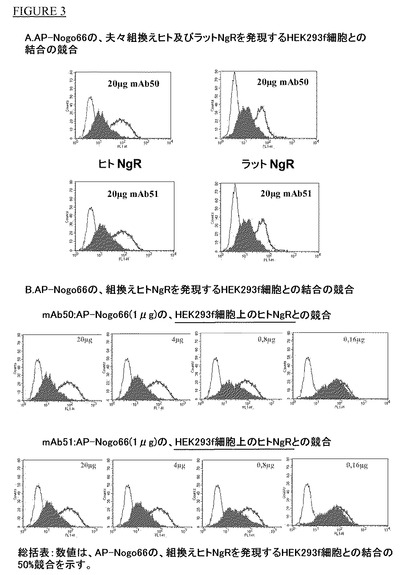
【図4】
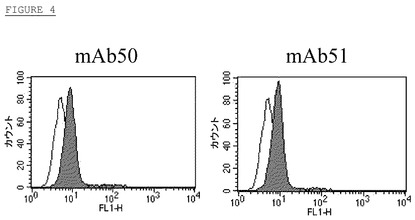
【図5】
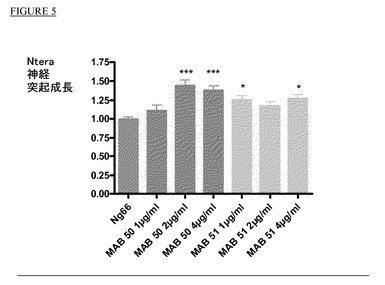
【図6】
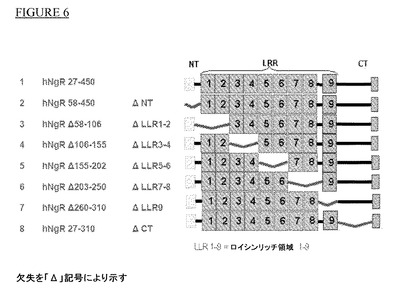
【図7】
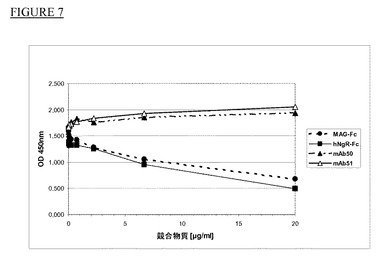
【図8】
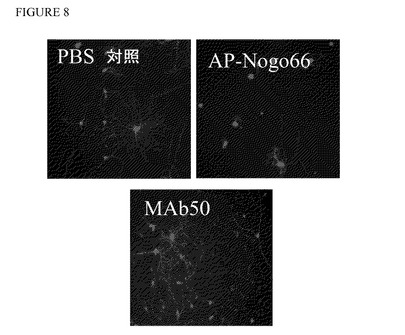
【図9】
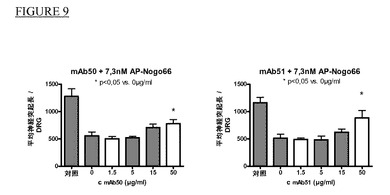
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2010−509939(P2010−509939A)
【公表日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願番号】特願2009−538504(P2009−538504)
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/US2007/085349
【国際公開番号】WO2008/064292
【国際公開日】平成20年5月29日(2008.5.29)
【出願人】(391008788)アボット・ラボラトリーズ (650)
【氏名又は名称原語表記】ABBOTT LABORATORIES
【出願人】(502104228)アボット ゲーエムベーハー ウント カンパニー カーゲー (89)
【Fターム(参考)】
【公表日】平成22年4月2日(2010.4.2)
【国際特許分類】
【出願日】平成19年11月21日(2007.11.21)
【国際出願番号】PCT/US2007/085349
【国際公開番号】WO2008/064292
【国際公開日】平成20年5月29日(2008.5.29)
【出願人】(391008788)アボット・ラボラトリーズ (650)
【氏名又は名称原語表記】ABBOTT LABORATORIES
【出願人】(502104228)アボット ゲーエムベーハー ウント カンパニー カーゲー (89)
【Fターム(参考)】
[ Back to top ]