
O−アセチル−ホモセリン生産菌株およびこれを用いてO−アセチチル−ホモセリンを生産する方法
【課題】L−メチオニン前駆体としてのO−アセチルホモセリンを高収率で生産する菌株、およびこれを用いてO−アセチルホモセリンを生産する方法を提供する。
【解決手段】前記菌株は、アセチル−CoAシンターゼ遺伝子(acs)、および/またはCoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)が導入および増進された、エシェリキア属菌株である。
【解決手段】前記菌株は、アセチル−CoAシンターゼ遺伝子(acs)、および/またはCoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)が導入および増進された、エシェリキア属菌株である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、L−メチオニン前駆体であるO−アセチル−ホモセリンを高収率で生産することが可能な菌株に関する。また、本発明は、前記菌株を用いてO−アセチル−ホモセリンを高収率で生産する方法に関する。
【背景技術】
【0002】
メチオニン(methionine)は、動物飼料、食品および医薬品に適用するために、化学的または生物学的に合成できる。
化学合成において、メチオニンは、主に、5−(β−メチルメルカプトエチル)ヒダントインを加水分解させる反応によって生産される。ところが、このような化学合成によって生産されたメチオニンは、L型とD型の混合形態で生産され、それぞれを分離する難しい追加的工程があるという欠点がある。そこで、本発明者は、かかる問題点を解決するために、生物学的方法を用いてL−メチオニンを選択的に生産することが可能な技術を開発して特許を出願したことがある(国際特許公開公報第2008/013432号)。この方法は、簡便に「2段階工法」と命名し、発酵によるL−メチオニン前駆体生産工程、および酵素による前記L−メチオニン前駆体のL−メチオニンへの転換工程を含む。前記L−メチオニン前駆体は、好ましくはO−アセチルホモセリン(O-acetylhomoserine)およびO−スクシニルホモセリン(O-succinylhomoserine)を含む。このような2段階工法を開発することにより、既存の問題となった、硫化物特有の基質毒性問題、メチオニンとSAMeによるメチオニン合成におけるフィードバック調節問題、およびシスタチオニンガンマシンターゼ(cystathionine gamma synthase)、O−スクシニルホモセリンスルフヒドリラーゼ(O-succinylhomoserine sulfhydrylase)およびO−アセチルホモセリンスルフヒドリラーゼ(O-acetylhomoserine sulfhydrylase)特有の中間産物分解活性問題を全て解決することができた。また、L−メチオニンのみを選択的に生産することができるため、DL−メチオニンを同時に生産する既存の化学合成工程に比べて優れた工程であり、さらに同一の反応によって副産物として有機酸、例えば、コハク酸および酢酸を同時に生産することが可能な非常に優れた工程である。
【0003】
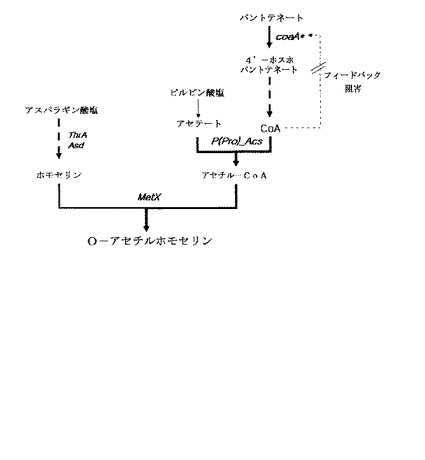
O−アセチルホモセリンは、メチオニン生産の前駆体として使用される物質であって、メチオニン生合成経路上にある中間体である(国際特許公開公報第2008/013432号)。O−アセチルホモセリンは、下記反応式のようにホモセリンO−アセチルトランスフェラーゼ(O-acetyl transferase)によってL−ホモセリンおよびアセチル−CoAを基質として合成される。
L−ホモセリン+アセチル−CoA → O−アセチル−ホモセリン
本発明者の米国公開特許第2009−253186号では、thrAの強化によってL−ホモセリンの生合成を増加させ、metXの強化によってO−アセチル−ホモセリンの生合成を増加させる菌株、および前記菌株を用いて高収率でO−アセチルホモセリンを生産する方法を開示した。これに関連し、本発明者は、O−アセチルホモセリンの生合成に用いられる二つの基質の一つであるアセチル−CoAを増加させることにより、よりさらに高い収率でO−アセチルホモセリンを生産することができることを確認した。
【0004】
一般に、大腸菌において、アセチル−CoAはピルビン酸塩から生合成される。しかし、過度な糖が存在する場合、アセチル−CoAは培地中に蓄積された酢酸によって生成できる。酢酸からアセチル−CoAが合成される場合、2つの互いに異なる生合成経路によって合成できる。酢酸からアセチル−CoAを生合成する一つの経路は、アセチル−CoAシンターゼ(ACS)によってアセチルアデニレート(acetyladenylate、AcAMP)を中間体としてアセチル−CoAを生合成する経路であり、もう一つの経路は、アセテートキナーゼ(acetate kinase、ACK)およびホスホトランスアセチラーゼ(phosphotransacetylase、PTA)によってアセチルホスフェート(acetylphosphate)を介してアセチル−CoAを生合成する経路である(JOURNAL OF BACTERIOLOGY, May 1995, p. 2878-2886)。特に、アセチル−CoAシンターゼによる生合成経路の場合、アセチル−CoAシンターゼ酵素自体がアセテートに対する高い親和度を持っているため、細胞内または外部に存在する低濃度の酢酸をアセチル−CoAに生合成する特徴を持つ。これに対し、ACK−PTA経路によるアセチル−CoA生合成の場合、酵素のアセテートに対する低い親和度を有する特性上、混合酸発酵(mixed acid fermentation)によって生成された高濃度のアセテートによってのみその活性を持つ特徴を示す(J. Gen. Microbiol. 102:327-336.)。
【0005】
前記アセチル−CoAを生合成する2つの経路のうち、前記アセチル−CoAシンターゼの場合、プロモーターのアップストリームに位置したCRP−結合部位が存在し、このような理由により、転写段階で分解代謝物阻害(Catabolite repression)によって指数的成長期まで発現が阻害され、以後、静止期では発現が増加する様相を示す(Mol Microbiol. 2004 Jan;51(1):241-54.)。
したがって、発酵中盤に培地中に蓄積されるアセテートは、ACK−PTA経路を用いてアセチル−CoAを合成することができる。しかし、ACK−PTA経路の場合、両方向性を有するため、アセテートの濃度が低くないときにアセチル−CoAがアセテートに転換できる。すなわち、ACK−TPA経路の間にアセチル−CoAが消耗され、O−アセチルホモセリンの合成に否定的影響を与える。
【0006】
アセチル−CoAの生合成において、アセテート以外に別の基質として使用されるCoA(Coenzyme A)は、代表的な細胞内のアシル基伝達体である。CoAは、ビタミンパントテネートから一連の生合成過程によって生合成される。パントテネートからCoAを生合成する酵素は次のとおりである。その第1段階として、パントテネートキナーゼ(Pantothenate Kinase、coaA)によってパントテネート(ビタミンB5)を4’−ホスホパントテネート(4’-phosphopantothenate)に合成し、第2段階として、P−PanCysシンターゼ/P−PanCysデカルボキシラーゼ(P-PanCys synthase/P-PanCys decarboxylase、coaBC)によって4’−ホスホパントテネートにシステインを結合して4’−ホスホパントデノイル−L−システイン(4’-phosphopantothenoyl-L-cysteine)を合成し、これを脱炭酸化して4’−ホスホパンテテイン(4’-phosphopantetheine)を合成し、第3段階として、P−PanSHアデニリルトランスフェラーゼ(P-PanSH adenylyltransferase、coaD)によって3−デホスホ−CoAを合成し、最後の段階として、デホスホ−CoAキナーゼ(dephospho-CoA(deP-CoA) kinase、coaE)によってATPを用いてリン酸化することにより、CoAを生合成する。
【0007】
一般に、CoAは、細胞内で多様な代謝反応はもちろん、多くの合成反応の基質として使用される。このような理由により、細胞内のCoA濃度は一定量以上存在しないように調節される。CoA poolの濃度を調節する段階は、CoA生合成経路上で第1の生合成段階である、パントテネートから4’−ホスホパントテネートを合成する段階に関与し、速度調節酵素としてのパントテネートキナーゼがCoA濃度によるフィードバック阻害(feedback inhibition)を受け、その活性阻害を受けて細胞内のCoA濃度を調節する(J Biol Chem. 1994 Oct 28; 269(43):27051-8)。ところが、常に細胞内で一定の水準に維持されるCoAの濃度はアセチル−CoAによるO−アセチルホモセリンを高収率で生産するのに問題として作用することができる。文献によれば、細胞内に存在するCoA poolの濃度によってフィードバック阻害を受けるパントテネートキナーゼの106番目のアミノ酸をR(arginine)からA(alanine)へ置換する場合(JOURNAL OF BACTERIOLOGY, 185, June 2003, p. 3410-3415)、CoAによるフィードバック阻害が解除できるものと確認されたことがある。野生型タンパク質の場合、40mMのCoA存在の際に残存活性が約20%である。これに対し、R106A変異タンパク質の場合、同一濃度のCoA存在の際にも全く活性の阻害が起こらなかった。また、変異タンパク質を発現した菌株において、CoAの細胞内濃度が野生型に比べて増加する現象を示すことを確認した。
【0008】
本発明者は、O−アセチルホモセリンの生産を増加させるために鋭意努力した結果、図1に示すように、(a)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子を導入および増進させる場合、および(b)アセチル−CoAシンターゼ遺伝子を導入および増進させる場合、O−アセチルホモセリンの生産能力が向上することを見出し、本発明を完成した。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】国際特許公開公報第2008/013432号
【特許文献2】国際特許公開公報第2008/013432号
【特許文献3】米国公開特許第2009−253186号
【非特許文献1】JOURNAL OF BACTERIOLOGY, May 1995, p. 2878-2886
【非特許文献2】J. Gen. Microbiol. 102:327-336.
【非特許文献3】Mol Microbiol. 2004 Jan;51(1):241-54.
【非特許文献4】J Biol Chem. 1994 Oct 28; 269(43):27051-8
【非特許文献5】JOURNAL OF BACTERIOLOGY, 185, June 2003, p. 3410-3415
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明の目的は、より高い収率でO−アセチルホモセリンを生産するために、O−アセチルホモセリンの生産を増加させることが可能なacs遺伝子の過発現およびCoAによってフィードバック阻害を受けるcoaA遺伝子のフィードバック阻害解除によってアセチル−CoAの生合成経路を強化し、これにより高収率のO−アセチルホモセリンを生産する微生物菌株を提供することにある。
また、本発明の他の目的は、 前記菌株を用いてO−アセチルホモセリンを高収率で生産する方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明のある観点によれば、(a)アセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)、または(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株を提供する。
本発明の他の観点によれば、前記菌株を培養培地で発酵する段階を含む、当該培地内でO−アセチルホモセリンを生産する方法を提供する。
本発明の別の観点によれば、(a)前記O−アセチルホモセリンを生産することが可能なエシェリキア属菌株を培養して発酵によってO−アセチルホモセリンを生産する段階と、(b)生産されたO−アセチルホモセリンを分離する段階と、(c)前記分離されたO−アセチルホモセリンをメチルメルカプタンと共に、シスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼ、およびO−スクシニルホモセリンスルフヒドリラーゼよりなる群から選ばれた酵素を用いて酵素反応を起し、L−メチオニンおよびアセテートを生産する方法を提供する。
【発明の効果】
【0012】
したがって、本発明によれば、O−アセチルホモセリンを高収率で生産することが可能な菌株を使用する場合、高収率でメチオニンを化学的合成方法より環境親和的に生産することができる。また、生産されたO−アセチル−L−ホモセリンは、O−アセチル−ホモセリンスルフヒドリラーゼによってメチオニンと酢酸との合成の前駆体として用いて高収率でL−メチオニンを生物転換することができる。転換された前記L−メチオニンは動物飼料または動物飼料添加剤だけでなく、ヒトの食品または食品添加剤の生産に有用である。
本発明の前記および他の目的、特徴および利点は、添付図面を参照する次の説明からさらに明確に理解されるであろう。
【図面の簡単な説明】
【0013】
【図1】図1はO−アセチル−ホモセリンを高収率で生産することが可能なO−アセチル−ホモセリン生合成経路を示す図である。
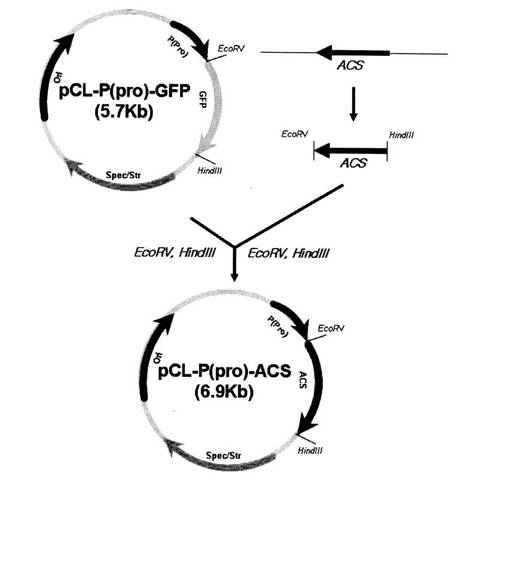
【図2】図2はアセチル−CoAシンターゼ遺伝子(acs)発現用ベクターpCL−P(pro)−acsの遺伝子の地図及び構成を示す図である。
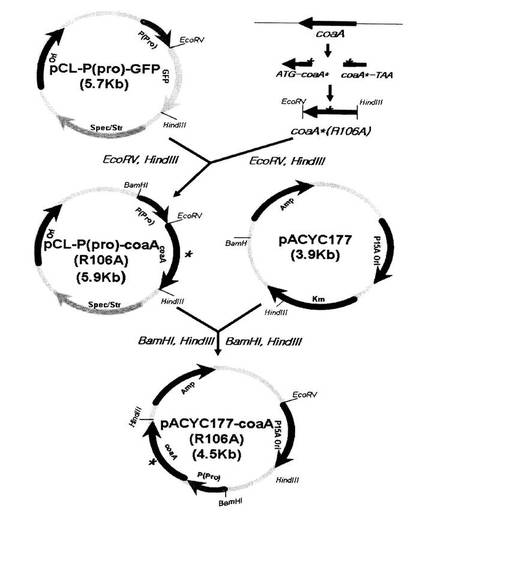
【図3】図3はフィードバック阻害が解除されたパントテネートキナーゼをコードするcoaA(R106A)遺伝子発現用ベクターpCL−P(pro)−coaA(R106A)とpACYC−coaA(R106A)の遺伝子の地図及び構成を示す図である。
【発明を実施するための形態】
【0014】
一つの様態として、本発明は、(a)アセチル−CoAシンターゼ遺伝子(acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)、または(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株を提供する。
本発明で使用される用語「L−メチオニン前駆体」とは、メチオニン生合成経路の一部分である代謝物質、またはこの代謝物質から派生した物質をいう。具体的に、本発明におけるL−メチオニン前駆体とはO−アセチルホモセリンを意味する。
【0015】
本発明で使用される用語「O−アセチルホモセリン生産菌株」とは、O−アセチルホモセリンを生物体内で生産することが可能な原核または真核微生物菌株であって、本発明に係る操作によってO−アセチルホモセリンを菌株内に蓄積することが可能な菌株をいう。例えば、本発明で有用な前記菌株は、エシェリキア属(Escherichia sp.)、エルウィニア属(Erwinia sp.)、セラシア属(Serratia sp.)、プロビデンシア属(Providencia sp.)、コリネバクテリウム属(Corynebacteria sp.)、シュードモナス属(Pseudomonas sp.)、レプトスピラ属(Leptospira sp.)、サルモネラ属(Salmonellar sp.)、ブレビバクテリア属(Brevibacteria sp.)、ヒポモナス属(Hypomononas sp.)、クロモバクテリウム属(Chromobacterium sp.)、ノカルジア属(Norcardia sp.)、カビ類(fungi)または酵母類(yeast)に属する微生物菌株が含まれ得る。好ましくはエシェリキア属、コリネバクテリウム属、レプトスピラ属の微生物菌株と酵母である。さらに好ましくはエシェリキア属の微生物菌株であり、さらに好ましくはリシン、トレオニン、イソロイシンまたはメチオニンを生産する菌株由来であり、最も好ましくは本発明によって提供された米国特許第12/062835号(米国公開特許第2009−253186号)に記載されているthrAおよびmetX遺伝子を導入したトレオニン生産菌株由来のO−アセチル−L−ホモセリン生合成経路が強化された大腸菌菌株(寄託番号KCCM10921P)である。
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、アセチル−CoAの生合成に関与するアセチルCoAシンターゼ遺伝子を前記菌株に導入及び増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
【0016】
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、CoA生合成経路上のCoA蓄積によるフィードバック阻害を解除したパントテネートキナーゼ遺伝子を前記菌株に導入および増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、アセチル−CoAの生合成に関与するアセチルCoAシンターゼ遺伝子を前記菌株に導入および増進させ、同時にCoA生合成経路上のCoA蓄積によるフィードバック阻害を解除したパントテネートキナーゼ遺伝子を導入および増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
本発明で使用される用語「導入および増進」は相応する遺伝子によってコードされる酵素の細胞内活性の増加を意味し、これは一般的に遺伝子の過発現によって行われ得る。目的遺伝子の過発現は、前記遺伝子のプロモーター部位および/または5’−UTR地域の塩基配列を変形させることにより、タンパク質発現を増進させることができ、目的遺伝子を染色体上に追加導入することにより、発現を強化させることができ、目的遺伝子をベクター上に自己プロモーターまたは強化された別個のプロモーターと共に導入して菌株に形質転換させることにより、タンパク質の発現量を強化させることができる。また、目的遺伝子のORF(open reading frame)地域に突然変異を導入することにより、目的遺伝子を過発現させることができる。過発現の測定によれば、相応するタンパク質の活性または濃度が、野生型タンパク質または初期の微生物菌株における活性または濃度を基準として、一般に最小10%、25%、50%、75%、100%、150%、200%、300%、400%または500%、最大1000%または2000%まで増加する。
【0017】
好ましくは、前記遺伝子を導入および増進させるためには強力なプロモーターへの置換、プロモーターの変異誘発、または遺伝子コピー数の増加による方法を使用することができ、さらに好ましくは、強力なプロモーターへの置換によって前記遺伝子を導入および増進させることができる。前記強力なプロモーターは、当業界において発現を常に増加させることができると知られているプロモーターであればその制限はないが、pTac、pTrc、pPro、pR、pLなどのプロモーターが使用でき、最も好ましくは配列番号9のpProプロモーターであり、前記遺伝子は配列番号9のpProプロモーターを全体または一部含むことができる。
酵素のアセチル−CoAシンターゼおよびパントテネートキナーゼは多様な微生物種に由来できる。このような活性を持つタンパク質をコードする遺伝子は、本発明においてそれぞれacsおよびcoaAと通称できる。
より具体的に、本発明では、アセチル−CoAシンターゼ活性が強化されるように変形してO−アセチル−ホモセリン生産能が向上した菌株、および前記菌株を用いてO−アセチルホモセリンを生産する方法を提供する。
本発明の具体的な実施態様として、O−アセチルホモセリン生産菌株を製造する方法は次のとおりである。
O−アセチル−L−ホモセリンの生産を増加させることを可能とするために、菌株をアセチル−CoAシンターゼタンパク質をコードする遺伝子のプロモーター部分を、配列番号9の構成的プロモーターP(pro)で置換して代謝物質阻害のない常時発現形態のプラスミド形態に製作して目的遺伝子の過発現を誘導した。前記アセチル−CoAシンターゼをコードする遺伝子を通称「acs」と表記する。これは文献(Mol Syst Biol. 2006;2:2006.0007. Epub 2006 Feb 21.)によって公開された大腸菌のゲノム配列から得ることができる(gi:89110790)。また、前記遺伝子配列は、米国生物工学情報センター(NCBI)および日本DNAデータバンク(DDBJ)などのデータベースからも得ることができる。
前記アセチル−CoAシンターゼは、下記反応を触媒する活性を持っている。よって、このような活性を有する遺伝子の発現を強化する場合、細胞内へのアセチル−CoAの蓄積を誘導することができる。
アセテート+CoA ⇔ アセチル−CoA
【0018】
次に、構成的発現プラスミドを含む菌株においてアセチル−CoAの合成をさらに増加させるための操作を行うことができる。CoA生合成経路の第1の段階であり且つ速度調節酵素であるパントテネートキナーゼを、CoAによるフィードバック阻害を解除するために突然変異させる。このために、パントテネートキナーゼの遺伝子配列上に突然変異を導入する。フィードバック阻害の解除されたパントテネートキナーゼをコードする遺伝子coaA(R106A)の発現を強化させる。この遺伝子は文献(Mol Syst Biol. 2006;2:2006.0007. Epub 2006 Feb 21.)によって公開された大腸菌のゲノム配列から得ることができる(gi:89110060)。また、前記遺伝子配列は米国生物工学情報センター(NCBI)および日本DNAデータバンク(DDBJ)などのデータベースからも得ることができる。
前記フィードバック阻害の解除されたパントテネートキナーゼは、下記反応式に示すように、パントテネートを4’−ホスホパントテネートに合成する活性を持っており、細胞内に蓄積されたCoAの濃度による活性阻害を受けない特性を持っている。よって、このようなフィードバック阻害を受けない活性を持つ遺伝子の発現を強化する場合、細胞内にCoA poolの増加を誘導することができる。
パントテネート+ATP ⇔ 4’−ホスホパントネート+ADP
前記方法によって製作されたO−アセチルホモセリン生産菌株は、細胞内に多量のCoAとアセチル−CoAを蓄積し、metXによりコードされる酵素によってホモセリンとアセチル−CoAを基質としてO−アセチル−L−ホモセリンを高収率で生産することが可能な菌株を製作することができる。
前述した方法によって製作したO−アセチルホモセリン生産菌株CJM−XにpCL−P(pro)−Acs、pCL−P(pro)−coaA(R106A)、およびpCL−P(pro)−acsとpACYC−coaA(R106A)を形質転換した菌株をそれぞれ「大腸菌CA05−0565」、「大腸菌CA05−0564」および「大腸菌CA05−0566」と命名し、KCCM(Korean Culture Center of Microorganism、韓国ソウル市西大門区弘済1洞ユリムビル361−221)に2009年8月11日付でそれぞれ寄託番号KCCM11023P、KCCM11022PおよびKCCM11024Pで寄託した。
【0019】
好適な一実施態様として、本発明は、アセチル−CoAの生合成に関与する遺伝子acsのプロモーターを構成的プロモーターProに置換して過発現させたO−アセチルホモセリン生産菌株を提供する。より具体的に、pProプロモーターは配列番号9の全体または一部が使用できる。
好適な一実施態様として、本発明は、CoA生合成経路上の主要段階を担当するパントテネートキナーゼのCoA蓄積によるフィードバック阻害が解除されたO−アセチルホモセリン生産能が向上した菌株を提供する。また、高収率でO−アセチルホモセリンを生産する方法を提供する。好ましくは、前記パントテネートキナーゼのアミノ酸配列において106番目のアミノ酸が他のアミノ酸に置換されることにより、CoA蓄積によるフィードバック阻害が解除され、O−アセチルホモセリン生産能を向上させることができる。さらに好ましくは、前記パントテネートキナーゼのアミノ酸配列において106番目のアルギニンがアラニンに置換されることにより、CoA蓄積によるフィードバック阻害が解除され、O−アセチルホモセリン生産能を向上させることができる。最も好ましくは、前記CoA蓄積によるフィードバック阻害の解除されたパントテネートキナーゼのアミノ酸配列が配列番号8のパントテネートキナーゼを用いることにより、O−アセチルホモセリンの生産能を向上させることができる。
【0020】
また、好適な一実施態様として、前記2つの方法を併行して使用し、アセチル−CoAシンターゼ活性及びパントテネートキナーゼ活性を強化してO−アセチルホモセリン生産能を向上させる方法を用いて製作されたO−アセチルホモセリン生産大腸菌を提供する。
好ましくは、O−アセチルホモセリン生産菌株の製造に有用な前記大腸菌はリシン、トレオニン、イソロイシンまたはメチオニン生産菌株のいずれか一つを用いることができる。さらに好ましくは、前記大腸菌は(a)ホモセリンアセチルトランスフェラーゼの活性、(b)アスパルトキナーゼまたはホモセリンデヒドロゲナーゼの活性、または(c)前記(a)および(b)の両方ともが導入された菌株を用いて製造することができる。最も好ましくは大腸菌CJM−X/pthrA(M)−CL(寄託番号KCCM10921P)由来の菌株を用いて製造することができる。
本発明の具体的な実施例では、acs遺伝子をクローニングして、配列番号9の構成的プロモーターpProの発現調節を受けるacs発現用ベクターpCL−P(pro)−acsを製作し(図2)、CoA蓄積によるフィードバック阻害が解除されるようにアミノ酸配列の106番目のアルギニンがアラニンに置換された配列番号8の変異coaA遺伝子(R106A)をクローニングして、 配列番号9の構成的プロモーターpProの発現調節を受ける変異coaA遺伝子発現用ベクターpCL−P(pro)−coaA(R106A)およびpACYC−coaA(R106A)を製作した(図3)。本発明者の米国特許第12/062835号(即ち、米国公開特許第2009−253186号)に開示されたthrAおよびmetXの強化菌株CJM−X/pthrA(M)−CL(寄託番号KCCM10921P)からpthrA(M)−CLプラスミドを除去して製作されたCJM−X菌株に、pCL−P(pro)−acs、pCL−P(pro)−coaA(R106A)、並びにpCL−P(pro)−acsおよびpACYC−coaA(R106A)を形質転換することにより、それぞれ(a)アセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)、および(c)前記(a)および(b)の両方ともが導入および増進される特徴を有する、O−アセチルホモセリン生産能が向上した菌株を製造した(実施例1〜3)。これら菌株のO−アセチルホモセリン生産能をフラスコ培養によって比較した結果、対照群であるCJM−Xに比べて、前記(a)のアセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)を有するpCL−P(pro)−acsが形質転換された菌株はO−アセチルホモセリン生産量が2.8g/L、収率が4.7%向上したことを確認した。また、前記(b)のCoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子(coaA)を有するpCL−P(pro)−coaA(R106A)の形質転換された菌株は、対照群に比べてO−アセチルホモセリン生産量が2.1g/L、収率が3.5%向上することを確認した。また、pCL−P(pro)−acsおよびpACYC−coaA(R106A)が形質転換された前記(a)および(b)の両方ともが導入および増進される特徴を有する菌株は、対照群に比べて生産量が6.8g/L、収率が11.4%向上することを確認した(実施例2、表2)。また、本発明の方法で製造された菌株のO−アセチルホモセリン生産能が向上することを確認した。
【0021】
別の様態として、本発明は、前記方法によって製造されたO−アセチル−ホモセリン生産能が向上した大腸菌を培養培地で発酵し、培地にO−アセチル−ホモセリンを蓄積させることにより、O−アセチル−ホモセリンを生産する方法に関する。
別の様態として、本発明は、(a)前記方法によって製造されたO−アセチルホモセリン生産能が向上したエシェリキア属由来の菌株を培養し、発酵によってO−アセチルホモセリンを生産する段階、(b)生産されたO−アセチルホモセリンを分離する段階、および(c)前記分離されたO−アセチルホモセリンおよびメチルメルカプタンを基質として、シスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼおよびO−スクシニルホモセリンスルフヒドリラーゼの活性を有する酵素よりなる群から選ばれた転換酵素を用いて酵素反応を起し、L−メチオニンおよびアセテートを生産する方法に関する。
前記方法は、本発明者が2007年に出願した特許(国際特許公開公報第2008/013432号に基づき、メチオニン転換酵素としてのシスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼ、O−スクシニルホモセリンスルフヒドリラーゼの活性を用いてL−メチオニンを生産する方法であって、本発明で提供する菌株を用いる場合、さらに高い収率でL−メチオニンを生産することができる。
【0022】
前記で製造されたO−アセチル−L−ホモセリン生産菌株の培養過程は、当業界に知られている適切な培地と培養条件に応じて行われ得る。このような培養過程は、当業者であれば、選択される菌株に応じて容易に調整して使用することができる。前記培養方法の例には回分式、連続式および流加式培養が含まれるが、これに限定されない。このような各種培養方法は、例えば文献『“Biochemical Engineering” by James M. Lee, Prentice-Hall International Editions, pp 138-176』に開示されている。
培養に使用される培地は、特定の菌株の要求条件を満足させなければならない。多様な微生物の培地は、例えば文献『“Manual of Methods for General Bacteriology” by the American Society for Bacteriology, Washington D. C., USA, 1981』に開示されている。一般的に前記培地は多様な炭素源、窒素源、および微量元素成分を含む。炭素源は、ブドウ糖、乳糖、蔗糖、果糖、麦芽糖、澱粉およびセルロースなどの炭水化物;大豆油、ヒマワリ油、ヒマシ油、ヤシ油などの脂肪;パルミチン酸、ステアリン酸およびリノール酸などの脂肪酸;グリセロールおよびエタノールなどのアルコールと酢酸などの有機酸を含む。これらの炭素源は単独でまたは組み合わせて使用できる。窒素源としては、ペプトン、酵母抽出液、肉汁、麦芽抽出液、コーンスティープリカー((corn steep liquor)CSL)およびベスン粉(bean flour)などの有機窒素源、および尿素、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウムおよび硝酸アンモニウムなどの無機窒素源を含む。これらの窒素源は、単独で或いは組み合わせて使用できる。前記培地にはリン酸源としてさらにリン酸二水素カリウム(potassium dihydrogen phosphate)、リン酸水素カリウム(dipotassium hydrogen phosphate)および相応するナトリウム含有塩(sodium-containing salts)を含むことができる。また、培地は硫酸マグネシウムまたは硫酸鉄などの金属を含むことができる。また、アミノ酸、ビタミン、および適した前駆体などが添加できる。これらの培地または前駆体は、培養物に回分式または連続式で添加できる。
【0023】
また、培養中に水酸化アンモニウム、水酸化カリウム、アンモニア、リン酸および硫酸などの化合物を適切な方式で添加することにより、培養物のpHを調整することができる。また、培養中に脂肪酸ポリグリコールエステルなどの消泡剤を使用することにより、培養中の気泡生成を抑制することができる。また、培養液の好気性条件を保つために、培養液内に酸素または酸素含有ガス(例えば、空気)が注入できる。培養物の温度は一般に20〜45℃、好ましくは25〜40℃である。培養はL−メチオニン前駆体の生産が所望の水準に到達するまで続けることが可能であり、好ましい培養時間は10〜160時間である。
【0024】
以下、実施例によって本発明をより詳しく説明する。ところが、これらの実施例は本発明を例示的に説明するためのもので、本発明の範囲を限定するものではない。
【0025】
実施例1:O−アセチルホモセリン生産菌株の製造
<1−1>acs遺伝子のクローニング
acs遺伝子のクローニングは、大腸菌W3110の染色体DNA(ATCC27325)を鋳型としたPCRによって、アセチル−CoAシンターゼであるacs遺伝子を確保した。米国国立保健院の遺伝子銀行(NIH GenBank)に基づいてacs遺伝子の塩基配列情報(NCBI登録番号gi:89110790)を確保した。その配列は配列番号7に示した。前記配列に基づいてacs遺伝子のATG部分とTAAを含有するORF部分を増幅し、制限酵素EcoRVとHindIII認識部位を含んでいるプライマー(配列番号1および2)を合成した。大腸菌W3110の染色体DNAを鋳型とし、前記配列番号1および2のプライマーを用いてPCRを行った。重合酵素はPfuUltraTM高信頼DNAポリメラーゼを使用した。PCRは、変性96℃、30秒;アニーリング50℃、30秒;および重合反応72℃、2分を1サイクルとして30サイクル繰返し行った。その結果、asc遺伝子と制限酵素EcoRVとHindIIIを含有した約2.0kbの増幅された遺伝子を獲得した。
前記PCRによって獲得したacs遺伝子の末端に含まれた制限酵素EcoRV、HindIIIを処理し、制限酵素EcoRV、HindIIIが処理されたpPro−GFPベクターに接合によってクローニングし、最終的に構成的プロモーターProによって発現調節されるacs遺伝子のクローニングされたpCL−P(pro)−acs組み換えベクターを製作した。図2はacs発現用ベクターpCL−P(pro)−acsを示す図である。
【0026】
<1−2>フィードバック抵抗性coaA遺伝子のクローニング
フィードバック阻害が解除されたcoaA遺伝子のクローニングは、大腸菌W3110の染色体DNA(ATCC27325)を鋳型としたPCRによって、パントテネートキナーゼであるcoaA遺伝子を確保した。米国国立保健院の遺伝子銀行(NIH GenBank)に基づいてcoaA遺伝子の塩基配列情報(NCBI登録番号gi:89110060)を確保し、これに基づいてcoaA遺伝子のATG部分からフィードバック阻害の解除が可能な316番目〜318番目の遺伝子配列をCGTからGCCに変更し、制限酵素EcoRVが含まれたプライマー(配列番号3および4)を合成した。また、フィードバック阻害の解除が可能な316番目〜318番目の遺伝子配列をCGTからGCCに変更し、TAA部分までを含み、制限酵素HindIIIを含むように増幅可能なプライマー(配列番号5および6)を合成した。
【0027】
大腸菌W3110の染色体DNAを鋳型とし、前記配列番号3および4と配列番号5および6のプライマーを用いてPCRを行った。重合酵素は、PfuUltraTM高信頼DNAポリメラーゼを使用した。PCRは、変性96℃、30秒;アニーリング50℃、30秒;および重合反応72℃、2分を1サイクルとして30サイクル繰返し行った。その結果、coaA遺伝子のATG部分から316番目〜318番目の遺伝子配列がCGTからGCCに変更された部分を含むように突然変異を導入した約344bpの増幅された遺伝子断片と、突然変異を導入して316番目〜318番目の遺伝子配列がCGTからGCCに変更された部分を含み、TAA部分を含む約642bpの増幅された遺伝子を獲得した。
前記で獲得した、増幅された2つの遺伝子を鋳型としてPCRを行った。PCRは、変性96℃、60秒;アニーリング50℃、60秒;および重合反応72℃、2分を1サイクルとして10サイクル繰返し行った後、配列番号3および6のプライマーを添加し、しかる後に、20サイクル繰返し行った。その結果、coaA遺伝子の316番目〜318番目の遺伝子配列がCGTからGCCに変更された963bpの増幅されたcoaA(R106A)遺伝子を獲得した。
【0028】
前記PCRによって獲得したcoaA遺伝子の末端に含まれた制限酵素EcoRVおよびHindIIIを処理し、制限酵素EcoRVおよびHindIIIが処理されたpPro−GFPベクターに接合によってクローニングし、最終的に構成的プロモーターPro(配列番号9)によって発現調節を受け、106番目のアミノ酸のコドンがCGTからGCCに突然変異されたcoaA遺伝子がクローニングされたpCL−P(pro)−coaA(R106A)組み換えベクターを製作した。前記変異coaAのアミノ酸配列は配列番号8に示した。
前記で製作されたpCL−P(pro)−coaA(R106A)組み換えベクターをpACYC177ベクターにクローニングするために、pCL−P(pro)−coaA(R106A)組み換えベクターに制限酵素BamHIおよびHindIIIを処理することにより、1.45kbのProプロモーターを含むcoaA(R106A)切片を得た。こうして得られたProプロモーターを含むcoaA(R106A)切片を、BamHIおよびHindIIIを処理したpACYC177ベクターと接合することにより、pACYC−coaA(R106A)ベクターを製作した。図3はcoaA(R106A)発現用ベクターpCL−P(pro)−coaA(R106A)およびpACYC−coaA(R106A)を示す図である。
【0029】
<1−3>O−アセチルホモセリン生産菌株の製作
AcsとcoaAの発現が強化されたO−アセチルホモセリン生産菌株の製作のために、米国公開特許第2009−253186号に開示されたCJM−X/pthrA(M)−CL(寄託番号KCCM10921P)菌株からpthrA(M)−CLプラスミドを除去し、pthrA(M)−CLの除去された菌株CJM−Xを製作した。こうして製作されたCJM−X菌株に、実施例<1−1>および<1−2>で製造されたpCL−P(pro)−acsおよびpCL−P(pro)−coaA(R106A)プラスミドを取り込んでそれぞれ形質転換(KCCM11023PおよびKCCM11022P)し、LB−Sp(酵母抽出10g/L、NaCl5g/L、トリプトン10g/L、ストレプトマイシン25μg/L)培地で培養した後、ストレプトマイシン耐性を有するコロニー10株ずつを選抜した。さらに、前記で製作したpCL−P(pro)−acsを含むCJM−X菌株に、実施例<1−2>で製作したpACYC−CoaA(R106A)を取り込んで形質転換(KCCM11024P)し、LB−Sp−Ap(酵母抽出10g/L、NaCl5g/L、トリプトン10g/L、ストレプトマイシン25μg/L、アンピシリン50μg/L)培地で培養した後、ストレプトマイシンとアンピシリンに対して耐性を示すコロニー10株を選抜した。選抜されたコロニーのO−アセチルホモセリン生産性を互いに比較した。
【0030】
実施例2:O−アセチルホモセリン生産のための発酵
実施例1で製作された菌株のメチオニン前駆体としてのO−アセチルホモセリンの生産能力を確保するために、三角フラスコ培養を行った。
表1に示したO−アセチル−ホモセリン力価培地を用いて三角フラスコでO−アセチルホモセリンの生産性を比較した。
【0031】
【表1】

【0032】
32℃の培養器でLB固体培地内に一晩培養した単一コロニーを25mLのO−アセチルホモセリン力価培地に1白金耳ずつ接種して、250rpmで攪拌しながら32℃で42〜64時間培養した。培養物からO−アセチルホモセリンをHPLCによって定量的に分析した。分析結果を表2に示した。
下記表2に示した結果のように、acs遺伝子のプロモーターを構成的発現プロモーターP(pro)に置換したacs遺伝子を過発現する場合、対照群菌株としてのCJM−X菌株に比べてO−アセチルホモセリンの生産量が2.8g/L、収率は4.7%向上することを確認することができた。また、フィードバック阻害の解除されたcoaA(R106A)遺伝子を過発現させる場合、対照群菌株であるCJM−X菌株に比べてO−アセチルホモセリンの生産量が2.1g/L、収率は3.5%向上することを確認することができた。acsとcoaA(R106A)を同時に過発現した場合、対照群菌株としてのCJM−X菌株に比べてO−アセチルホモセリンの生産量が6.8g/L、収率は11.4%が向上することを確認した。
このようなフラスコ培養の結果からみて、acs遺伝子の発現強化、フィードバック阻害の解除されたcoaA遺伝子の発現強化、またはacsおよびフィードバック阻害の解除されたcoaA遺伝子の発現強化がO−アセチルホモセリンの生産性および収率の向上に効果的であることを確認することができた。
【0033】
【表2】
【産業上の利用可能性】
【0034】
上述したように、本発明は、培地内で発酵するときに培地で高収率でO−アセチルホモセリンを生産するエシェリキア属菌株を提供する。また、前記O−アセチルホモセリンとメチルカプタンを基質として用いて、メチオニン転換酵素の存在下に転換反応を行うことにより、高収率でL−メチオニンと酢酸を同時に生産することができる。
以上、本発明の好適な実施例について説明の目的で開示したが、当業者であれば、添付した請求の範囲に開示された本発明の精神と範囲から逸脱することなく、様々な変形、追加または置換を加え得ることを理解するであろう。
【技術分野】
【0001】
本発明は、L−メチオニン前駆体であるO−アセチル−ホモセリンを高収率で生産することが可能な菌株に関する。また、本発明は、前記菌株を用いてO−アセチル−ホモセリンを高収率で生産する方法に関する。
【背景技術】
【0002】
メチオニン(methionine)は、動物飼料、食品および医薬品に適用するために、化学的または生物学的に合成できる。
化学合成において、メチオニンは、主に、5−(β−メチルメルカプトエチル)ヒダントインを加水分解させる反応によって生産される。ところが、このような化学合成によって生産されたメチオニンは、L型とD型の混合形態で生産され、それぞれを分離する難しい追加的工程があるという欠点がある。そこで、本発明者は、かかる問題点を解決するために、生物学的方法を用いてL−メチオニンを選択的に生産することが可能な技術を開発して特許を出願したことがある(国際特許公開公報第2008/013432号)。この方法は、簡便に「2段階工法」と命名し、発酵によるL−メチオニン前駆体生産工程、および酵素による前記L−メチオニン前駆体のL−メチオニンへの転換工程を含む。前記L−メチオニン前駆体は、好ましくはO−アセチルホモセリン(O-acetylhomoserine)およびO−スクシニルホモセリン(O-succinylhomoserine)を含む。このような2段階工法を開発することにより、既存の問題となった、硫化物特有の基質毒性問題、メチオニンとSAMeによるメチオニン合成におけるフィードバック調節問題、およびシスタチオニンガンマシンターゼ(cystathionine gamma synthase)、O−スクシニルホモセリンスルフヒドリラーゼ(O-succinylhomoserine sulfhydrylase)およびO−アセチルホモセリンスルフヒドリラーゼ(O-acetylhomoserine sulfhydrylase)特有の中間産物分解活性問題を全て解決することができた。また、L−メチオニンのみを選択的に生産することができるため、DL−メチオニンを同時に生産する既存の化学合成工程に比べて優れた工程であり、さらに同一の反応によって副産物として有機酸、例えば、コハク酸および酢酸を同時に生産することが可能な非常に優れた工程である。
【0003】
O−アセチルホモセリンは、メチオニン生産の前駆体として使用される物質であって、メチオニン生合成経路上にある中間体である(国際特許公開公報第2008/013432号)。O−アセチルホモセリンは、下記反応式のようにホモセリンO−アセチルトランスフェラーゼ(O-acetyl transferase)によってL−ホモセリンおよびアセチル−CoAを基質として合成される。
L−ホモセリン+アセチル−CoA → O−アセチル−ホモセリン
本発明者の米国公開特許第2009−253186号では、thrAの強化によってL−ホモセリンの生合成を増加させ、metXの強化によってO−アセチル−ホモセリンの生合成を増加させる菌株、および前記菌株を用いて高収率でO−アセチルホモセリンを生産する方法を開示した。これに関連し、本発明者は、O−アセチルホモセリンの生合成に用いられる二つの基質の一つであるアセチル−CoAを増加させることにより、よりさらに高い収率でO−アセチルホモセリンを生産することができることを確認した。
【0004】
一般に、大腸菌において、アセチル−CoAはピルビン酸塩から生合成される。しかし、過度な糖が存在する場合、アセチル−CoAは培地中に蓄積された酢酸によって生成できる。酢酸からアセチル−CoAが合成される場合、2つの互いに異なる生合成経路によって合成できる。酢酸からアセチル−CoAを生合成する一つの経路は、アセチル−CoAシンターゼ(ACS)によってアセチルアデニレート(acetyladenylate、AcAMP)を中間体としてアセチル−CoAを生合成する経路であり、もう一つの経路は、アセテートキナーゼ(acetate kinase、ACK)およびホスホトランスアセチラーゼ(phosphotransacetylase、PTA)によってアセチルホスフェート(acetylphosphate)を介してアセチル−CoAを生合成する経路である(JOURNAL OF BACTERIOLOGY, May 1995, p. 2878-2886)。特に、アセチル−CoAシンターゼによる生合成経路の場合、アセチル−CoAシンターゼ酵素自体がアセテートに対する高い親和度を持っているため、細胞内または外部に存在する低濃度の酢酸をアセチル−CoAに生合成する特徴を持つ。これに対し、ACK−PTA経路によるアセチル−CoA生合成の場合、酵素のアセテートに対する低い親和度を有する特性上、混合酸発酵(mixed acid fermentation)によって生成された高濃度のアセテートによってのみその活性を持つ特徴を示す(J. Gen. Microbiol. 102:327-336.)。
【0005】
前記アセチル−CoAを生合成する2つの経路のうち、前記アセチル−CoAシンターゼの場合、プロモーターのアップストリームに位置したCRP−結合部位が存在し、このような理由により、転写段階で分解代謝物阻害(Catabolite repression)によって指数的成長期まで発現が阻害され、以後、静止期では発現が増加する様相を示す(Mol Microbiol. 2004 Jan;51(1):241-54.)。
したがって、発酵中盤に培地中に蓄積されるアセテートは、ACK−PTA経路を用いてアセチル−CoAを合成することができる。しかし、ACK−PTA経路の場合、両方向性を有するため、アセテートの濃度が低くないときにアセチル−CoAがアセテートに転換できる。すなわち、ACK−TPA経路の間にアセチル−CoAが消耗され、O−アセチルホモセリンの合成に否定的影響を与える。
【0006】
アセチル−CoAの生合成において、アセテート以外に別の基質として使用されるCoA(Coenzyme A)は、代表的な細胞内のアシル基伝達体である。CoAは、ビタミンパントテネートから一連の生合成過程によって生合成される。パントテネートからCoAを生合成する酵素は次のとおりである。その第1段階として、パントテネートキナーゼ(Pantothenate Kinase、coaA)によってパントテネート(ビタミンB5)を4’−ホスホパントテネート(4’-phosphopantothenate)に合成し、第2段階として、P−PanCysシンターゼ/P−PanCysデカルボキシラーゼ(P-PanCys synthase/P-PanCys decarboxylase、coaBC)によって4’−ホスホパントテネートにシステインを結合して4’−ホスホパントデノイル−L−システイン(4’-phosphopantothenoyl-L-cysteine)を合成し、これを脱炭酸化して4’−ホスホパンテテイン(4’-phosphopantetheine)を合成し、第3段階として、P−PanSHアデニリルトランスフェラーゼ(P-PanSH adenylyltransferase、coaD)によって3−デホスホ−CoAを合成し、最後の段階として、デホスホ−CoAキナーゼ(dephospho-CoA(deP-CoA) kinase、coaE)によってATPを用いてリン酸化することにより、CoAを生合成する。
【0007】
一般に、CoAは、細胞内で多様な代謝反応はもちろん、多くの合成反応の基質として使用される。このような理由により、細胞内のCoA濃度は一定量以上存在しないように調節される。CoA poolの濃度を調節する段階は、CoA生合成経路上で第1の生合成段階である、パントテネートから4’−ホスホパントテネートを合成する段階に関与し、速度調節酵素としてのパントテネートキナーゼがCoA濃度によるフィードバック阻害(feedback inhibition)を受け、その活性阻害を受けて細胞内のCoA濃度を調節する(J Biol Chem. 1994 Oct 28; 269(43):27051-8)。ところが、常に細胞内で一定の水準に維持されるCoAの濃度はアセチル−CoAによるO−アセチルホモセリンを高収率で生産するのに問題として作用することができる。文献によれば、細胞内に存在するCoA poolの濃度によってフィードバック阻害を受けるパントテネートキナーゼの106番目のアミノ酸をR(arginine)からA(alanine)へ置換する場合(JOURNAL OF BACTERIOLOGY, 185, June 2003, p. 3410-3415)、CoAによるフィードバック阻害が解除できるものと確認されたことがある。野生型タンパク質の場合、40mMのCoA存在の際に残存活性が約20%である。これに対し、R106A変異タンパク質の場合、同一濃度のCoA存在の際にも全く活性の阻害が起こらなかった。また、変異タンパク質を発現した菌株において、CoAの細胞内濃度が野生型に比べて増加する現象を示すことを確認した。
【0008】
本発明者は、O−アセチルホモセリンの生産を増加させるために鋭意努力した結果、図1に示すように、(a)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子を導入および増進させる場合、および(b)アセチル−CoAシンターゼ遺伝子を導入および増進させる場合、O−アセチルホモセリンの生産能力が向上することを見出し、本発明を完成した。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】国際特許公開公報第2008/013432号
【特許文献2】国際特許公開公報第2008/013432号
【特許文献3】米国公開特許第2009−253186号
【非特許文献1】JOURNAL OF BACTERIOLOGY, May 1995, p. 2878-2886
【非特許文献2】J. Gen. Microbiol. 102:327-336.
【非特許文献3】Mol Microbiol. 2004 Jan;51(1):241-54.
【非特許文献4】J Biol Chem. 1994 Oct 28; 269(43):27051-8
【非特許文献5】JOURNAL OF BACTERIOLOGY, 185, June 2003, p. 3410-3415
【発明の概要】
【発明が解決しようとする課題】
【0010】
本発明の目的は、より高い収率でO−アセチルホモセリンを生産するために、O−アセチルホモセリンの生産を増加させることが可能なacs遺伝子の過発現およびCoAによってフィードバック阻害を受けるcoaA遺伝子のフィードバック阻害解除によってアセチル−CoAの生合成経路を強化し、これにより高収率のO−アセチルホモセリンを生産する微生物菌株を提供することにある。
また、本発明の他の目的は、 前記菌株を用いてO−アセチルホモセリンを高収率で生産する方法を提供することにある。
【課題を解決するための手段】
【0011】
本発明のある観点によれば、(a)アセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)、または(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株を提供する。
本発明の他の観点によれば、前記菌株を培養培地で発酵する段階を含む、当該培地内でO−アセチルホモセリンを生産する方法を提供する。
本発明の別の観点によれば、(a)前記O−アセチルホモセリンを生産することが可能なエシェリキア属菌株を培養して発酵によってO−アセチルホモセリンを生産する段階と、(b)生産されたO−アセチルホモセリンを分離する段階と、(c)前記分離されたO−アセチルホモセリンをメチルメルカプタンと共に、シスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼ、およびO−スクシニルホモセリンスルフヒドリラーゼよりなる群から選ばれた酵素を用いて酵素反応を起し、L−メチオニンおよびアセテートを生産する方法を提供する。
【発明の効果】
【0012】
したがって、本発明によれば、O−アセチルホモセリンを高収率で生産することが可能な菌株を使用する場合、高収率でメチオニンを化学的合成方法より環境親和的に生産することができる。また、生産されたO−アセチル−L−ホモセリンは、O−アセチル−ホモセリンスルフヒドリラーゼによってメチオニンと酢酸との合成の前駆体として用いて高収率でL−メチオニンを生物転換することができる。転換された前記L−メチオニンは動物飼料または動物飼料添加剤だけでなく、ヒトの食品または食品添加剤の生産に有用である。
本発明の前記および他の目的、特徴および利点は、添付図面を参照する次の説明からさらに明確に理解されるであろう。
【図面の簡単な説明】
【0013】
【図1】図1はO−アセチル−ホモセリンを高収率で生産することが可能なO−アセチル−ホモセリン生合成経路を示す図である。
【図2】図2はアセチル−CoAシンターゼ遺伝子(acs)発現用ベクターpCL−P(pro)−acsの遺伝子の地図及び構成を示す図である。
【図3】図3はフィードバック阻害が解除されたパントテネートキナーゼをコードするcoaA(R106A)遺伝子発現用ベクターpCL−P(pro)−coaA(R106A)とpACYC−coaA(R106A)の遺伝子の地図及び構成を示す図である。
【発明を実施するための形態】
【0014】
一つの様態として、本発明は、(a)アセチル−CoAシンターゼ遺伝子(acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)、または(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株を提供する。
本発明で使用される用語「L−メチオニン前駆体」とは、メチオニン生合成経路の一部分である代謝物質、またはこの代謝物質から派生した物質をいう。具体的に、本発明におけるL−メチオニン前駆体とはO−アセチルホモセリンを意味する。
【0015】
本発明で使用される用語「O−アセチルホモセリン生産菌株」とは、O−アセチルホモセリンを生物体内で生産することが可能な原核または真核微生物菌株であって、本発明に係る操作によってO−アセチルホモセリンを菌株内に蓄積することが可能な菌株をいう。例えば、本発明で有用な前記菌株は、エシェリキア属(Escherichia sp.)、エルウィニア属(Erwinia sp.)、セラシア属(Serratia sp.)、プロビデンシア属(Providencia sp.)、コリネバクテリウム属(Corynebacteria sp.)、シュードモナス属(Pseudomonas sp.)、レプトスピラ属(Leptospira sp.)、サルモネラ属(Salmonellar sp.)、ブレビバクテリア属(Brevibacteria sp.)、ヒポモナス属(Hypomononas sp.)、クロモバクテリウム属(Chromobacterium sp.)、ノカルジア属(Norcardia sp.)、カビ類(fungi)または酵母類(yeast)に属する微生物菌株が含まれ得る。好ましくはエシェリキア属、コリネバクテリウム属、レプトスピラ属の微生物菌株と酵母である。さらに好ましくはエシェリキア属の微生物菌株であり、さらに好ましくはリシン、トレオニン、イソロイシンまたはメチオニンを生産する菌株由来であり、最も好ましくは本発明によって提供された米国特許第12/062835号(米国公開特許第2009−253186号)に記載されているthrAおよびmetX遺伝子を導入したトレオニン生産菌株由来のO−アセチル−L−ホモセリン生合成経路が強化された大腸菌菌株(寄託番号KCCM10921P)である。
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、アセチル−CoAの生合成に関与するアセチルCoAシンターゼ遺伝子を前記菌株に導入及び増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
【0016】
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、CoA生合成経路上のCoA蓄積によるフィードバック阻害を解除したパントテネートキナーゼ遺伝子を前記菌株に導入および増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
好適な一実施態様として、本発明は、アセチル−CoA生合成経路を強化してO−アセチルホモセリン生産能が向上するように、アセチル−CoAの生合成に関与するアセチルCoAシンターゼ遺伝子を前記菌株に導入および増進させ、同時にCoA生合成経路上のCoA蓄積によるフィードバック阻害を解除したパントテネートキナーゼ遺伝子を導入および増進させた菌株をO−アセチルホモセリン生産菌株として提供する。
本発明で使用される用語「導入および増進」は相応する遺伝子によってコードされる酵素の細胞内活性の増加を意味し、これは一般的に遺伝子の過発現によって行われ得る。目的遺伝子の過発現は、前記遺伝子のプロモーター部位および/または5’−UTR地域の塩基配列を変形させることにより、タンパク質発現を増進させることができ、目的遺伝子を染色体上に追加導入することにより、発現を強化させることができ、目的遺伝子をベクター上に自己プロモーターまたは強化された別個のプロモーターと共に導入して菌株に形質転換させることにより、タンパク質の発現量を強化させることができる。また、目的遺伝子のORF(open reading frame)地域に突然変異を導入することにより、目的遺伝子を過発現させることができる。過発現の測定によれば、相応するタンパク質の活性または濃度が、野生型タンパク質または初期の微生物菌株における活性または濃度を基準として、一般に最小10%、25%、50%、75%、100%、150%、200%、300%、400%または500%、最大1000%または2000%まで増加する。
【0017】
好ましくは、前記遺伝子を導入および増進させるためには強力なプロモーターへの置換、プロモーターの変異誘発、または遺伝子コピー数の増加による方法を使用することができ、さらに好ましくは、強力なプロモーターへの置換によって前記遺伝子を導入および増進させることができる。前記強力なプロモーターは、当業界において発現を常に増加させることができると知られているプロモーターであればその制限はないが、pTac、pTrc、pPro、pR、pLなどのプロモーターが使用でき、最も好ましくは配列番号9のpProプロモーターであり、前記遺伝子は配列番号9のpProプロモーターを全体または一部含むことができる。
酵素のアセチル−CoAシンターゼおよびパントテネートキナーゼは多様な微生物種に由来できる。このような活性を持つタンパク質をコードする遺伝子は、本発明においてそれぞれacsおよびcoaAと通称できる。
より具体的に、本発明では、アセチル−CoAシンターゼ活性が強化されるように変形してO−アセチル−ホモセリン生産能が向上した菌株、および前記菌株を用いてO−アセチルホモセリンを生産する方法を提供する。
本発明の具体的な実施態様として、O−アセチルホモセリン生産菌株を製造する方法は次のとおりである。
O−アセチル−L−ホモセリンの生産を増加させることを可能とするために、菌株をアセチル−CoAシンターゼタンパク質をコードする遺伝子のプロモーター部分を、配列番号9の構成的プロモーターP(pro)で置換して代謝物質阻害のない常時発現形態のプラスミド形態に製作して目的遺伝子の過発現を誘導した。前記アセチル−CoAシンターゼをコードする遺伝子を通称「acs」と表記する。これは文献(Mol Syst Biol. 2006;2:2006.0007. Epub 2006 Feb 21.)によって公開された大腸菌のゲノム配列から得ることができる(gi:89110790)。また、前記遺伝子配列は、米国生物工学情報センター(NCBI)および日本DNAデータバンク(DDBJ)などのデータベースからも得ることができる。
前記アセチル−CoAシンターゼは、下記反応を触媒する活性を持っている。よって、このような活性を有する遺伝子の発現を強化する場合、細胞内へのアセチル−CoAの蓄積を誘導することができる。
アセテート+CoA ⇔ アセチル−CoA
【0018】
次に、構成的発現プラスミドを含む菌株においてアセチル−CoAの合成をさらに増加させるための操作を行うことができる。CoA生合成経路の第1の段階であり且つ速度調節酵素であるパントテネートキナーゼを、CoAによるフィードバック阻害を解除するために突然変異させる。このために、パントテネートキナーゼの遺伝子配列上に突然変異を導入する。フィードバック阻害の解除されたパントテネートキナーゼをコードする遺伝子coaA(R106A)の発現を強化させる。この遺伝子は文献(Mol Syst Biol. 2006;2:2006.0007. Epub 2006 Feb 21.)によって公開された大腸菌のゲノム配列から得ることができる(gi:89110060)。また、前記遺伝子配列は米国生物工学情報センター(NCBI)および日本DNAデータバンク(DDBJ)などのデータベースからも得ることができる。
前記フィードバック阻害の解除されたパントテネートキナーゼは、下記反応式に示すように、パントテネートを4’−ホスホパントテネートに合成する活性を持っており、細胞内に蓄積されたCoAの濃度による活性阻害を受けない特性を持っている。よって、このようなフィードバック阻害を受けない活性を持つ遺伝子の発現を強化する場合、細胞内にCoA poolの増加を誘導することができる。
パントテネート+ATP ⇔ 4’−ホスホパントネート+ADP
前記方法によって製作されたO−アセチルホモセリン生産菌株は、細胞内に多量のCoAとアセチル−CoAを蓄積し、metXによりコードされる酵素によってホモセリンとアセチル−CoAを基質としてO−アセチル−L−ホモセリンを高収率で生産することが可能な菌株を製作することができる。
前述した方法によって製作したO−アセチルホモセリン生産菌株CJM−XにpCL−P(pro)−Acs、pCL−P(pro)−coaA(R106A)、およびpCL−P(pro)−acsとpACYC−coaA(R106A)を形質転換した菌株をそれぞれ「大腸菌CA05−0565」、「大腸菌CA05−0564」および「大腸菌CA05−0566」と命名し、KCCM(Korean Culture Center of Microorganism、韓国ソウル市西大門区弘済1洞ユリムビル361−221)に2009年8月11日付でそれぞれ寄託番号KCCM11023P、KCCM11022PおよびKCCM11024Pで寄託した。
【0019】
好適な一実施態様として、本発明は、アセチル−CoAの生合成に関与する遺伝子acsのプロモーターを構成的プロモーターProに置換して過発現させたO−アセチルホモセリン生産菌株を提供する。より具体的に、pProプロモーターは配列番号9の全体または一部が使用できる。
好適な一実施態様として、本発明は、CoA生合成経路上の主要段階を担当するパントテネートキナーゼのCoA蓄積によるフィードバック阻害が解除されたO−アセチルホモセリン生産能が向上した菌株を提供する。また、高収率でO−アセチルホモセリンを生産する方法を提供する。好ましくは、前記パントテネートキナーゼのアミノ酸配列において106番目のアミノ酸が他のアミノ酸に置換されることにより、CoA蓄積によるフィードバック阻害が解除され、O−アセチルホモセリン生産能を向上させることができる。さらに好ましくは、前記パントテネートキナーゼのアミノ酸配列において106番目のアルギニンがアラニンに置換されることにより、CoA蓄積によるフィードバック阻害が解除され、O−アセチルホモセリン生産能を向上させることができる。最も好ましくは、前記CoA蓄積によるフィードバック阻害の解除されたパントテネートキナーゼのアミノ酸配列が配列番号8のパントテネートキナーゼを用いることにより、O−アセチルホモセリンの生産能を向上させることができる。
【0020】
また、好適な一実施態様として、前記2つの方法を併行して使用し、アセチル−CoAシンターゼ活性及びパントテネートキナーゼ活性を強化してO−アセチルホモセリン生産能を向上させる方法を用いて製作されたO−アセチルホモセリン生産大腸菌を提供する。
好ましくは、O−アセチルホモセリン生産菌株の製造に有用な前記大腸菌はリシン、トレオニン、イソロイシンまたはメチオニン生産菌株のいずれか一つを用いることができる。さらに好ましくは、前記大腸菌は(a)ホモセリンアセチルトランスフェラーゼの活性、(b)アスパルトキナーゼまたはホモセリンデヒドロゲナーゼの活性、または(c)前記(a)および(b)の両方ともが導入された菌株を用いて製造することができる。最も好ましくは大腸菌CJM−X/pthrA(M)−CL(寄託番号KCCM10921P)由来の菌株を用いて製造することができる。
本発明の具体的な実施例では、acs遺伝子をクローニングして、配列番号9の構成的プロモーターpProの発現調節を受けるacs発現用ベクターpCL−P(pro)−acsを製作し(図2)、CoA蓄積によるフィードバック阻害が解除されるようにアミノ酸配列の106番目のアルギニンがアラニンに置換された配列番号8の変異coaA遺伝子(R106A)をクローニングして、 配列番号9の構成的プロモーターpProの発現調節を受ける変異coaA遺伝子発現用ベクターpCL−P(pro)−coaA(R106A)およびpACYC−coaA(R106A)を製作した(図3)。本発明者の米国特許第12/062835号(即ち、米国公開特許第2009−253186号)に開示されたthrAおよびmetXの強化菌株CJM−X/pthrA(M)−CL(寄託番号KCCM10921P)からpthrA(M)−CLプラスミドを除去して製作されたCJM−X菌株に、pCL−P(pro)−acs、pCL−P(pro)−coaA(R106A)、並びにpCL−P(pro)−acsおよびpACYC−coaA(R106A)を形質転換することにより、それぞれ(a)アセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)、(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)、および(c)前記(a)および(b)の両方ともが導入および増進される特徴を有する、O−アセチルホモセリン生産能が向上した菌株を製造した(実施例1〜3)。これら菌株のO−アセチルホモセリン生産能をフラスコ培養によって比較した結果、対照群であるCJM−Xに比べて、前記(a)のアセチル−CoAシンターゼ遺伝子(Acetyl-CoA synthase、acs)を有するpCL−P(pro)−acsが形質転換された菌株はO−アセチルホモセリン生産量が2.8g/L、収率が4.7%向上したことを確認した。また、前記(b)のCoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼ遺伝子(coaA)を有するpCL−P(pro)−coaA(R106A)の形質転換された菌株は、対照群に比べてO−アセチルホモセリン生産量が2.1g/L、収率が3.5%向上することを確認した。また、pCL−P(pro)−acsおよびpACYC−coaA(R106A)が形質転換された前記(a)および(b)の両方ともが導入および増進される特徴を有する菌株は、対照群に比べて生産量が6.8g/L、収率が11.4%向上することを確認した(実施例2、表2)。また、本発明の方法で製造された菌株のO−アセチルホモセリン生産能が向上することを確認した。
【0021】
別の様態として、本発明は、前記方法によって製造されたO−アセチル−ホモセリン生産能が向上した大腸菌を培養培地で発酵し、培地にO−アセチル−ホモセリンを蓄積させることにより、O−アセチル−ホモセリンを生産する方法に関する。
別の様態として、本発明は、(a)前記方法によって製造されたO−アセチルホモセリン生産能が向上したエシェリキア属由来の菌株を培養し、発酵によってO−アセチルホモセリンを生産する段階、(b)生産されたO−アセチルホモセリンを分離する段階、および(c)前記分離されたO−アセチルホモセリンおよびメチルメルカプタンを基質として、シスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼおよびO−スクシニルホモセリンスルフヒドリラーゼの活性を有する酵素よりなる群から選ばれた転換酵素を用いて酵素反応を起し、L−メチオニンおよびアセテートを生産する方法に関する。
前記方法は、本発明者が2007年に出願した特許(国際特許公開公報第2008/013432号に基づき、メチオニン転換酵素としてのシスタチオニンガンマシンターゼ、O−アセチルホモセリンスルフヒドリラーゼ、O−スクシニルホモセリンスルフヒドリラーゼの活性を用いてL−メチオニンを生産する方法であって、本発明で提供する菌株を用いる場合、さらに高い収率でL−メチオニンを生産することができる。
【0022】
前記で製造されたO−アセチル−L−ホモセリン生産菌株の培養過程は、当業界に知られている適切な培地と培養条件に応じて行われ得る。このような培養過程は、当業者であれば、選択される菌株に応じて容易に調整して使用することができる。前記培養方法の例には回分式、連続式および流加式培養が含まれるが、これに限定されない。このような各種培養方法は、例えば文献『“Biochemical Engineering” by James M. Lee, Prentice-Hall International Editions, pp 138-176』に開示されている。
培養に使用される培地は、特定の菌株の要求条件を満足させなければならない。多様な微生物の培地は、例えば文献『“Manual of Methods for General Bacteriology” by the American Society for Bacteriology, Washington D. C., USA, 1981』に開示されている。一般的に前記培地は多様な炭素源、窒素源、および微量元素成分を含む。炭素源は、ブドウ糖、乳糖、蔗糖、果糖、麦芽糖、澱粉およびセルロースなどの炭水化物;大豆油、ヒマワリ油、ヒマシ油、ヤシ油などの脂肪;パルミチン酸、ステアリン酸およびリノール酸などの脂肪酸;グリセロールおよびエタノールなどのアルコールと酢酸などの有機酸を含む。これらの炭素源は単独でまたは組み合わせて使用できる。窒素源としては、ペプトン、酵母抽出液、肉汁、麦芽抽出液、コーンスティープリカー((corn steep liquor)CSL)およびベスン粉(bean flour)などの有機窒素源、および尿素、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、炭酸アンモニウムおよび硝酸アンモニウムなどの無機窒素源を含む。これらの窒素源は、単独で或いは組み合わせて使用できる。前記培地にはリン酸源としてさらにリン酸二水素カリウム(potassium dihydrogen phosphate)、リン酸水素カリウム(dipotassium hydrogen phosphate)および相応するナトリウム含有塩(sodium-containing salts)を含むことができる。また、培地は硫酸マグネシウムまたは硫酸鉄などの金属を含むことができる。また、アミノ酸、ビタミン、および適した前駆体などが添加できる。これらの培地または前駆体は、培養物に回分式または連続式で添加できる。
【0023】
また、培養中に水酸化アンモニウム、水酸化カリウム、アンモニア、リン酸および硫酸などの化合物を適切な方式で添加することにより、培養物のpHを調整することができる。また、培養中に脂肪酸ポリグリコールエステルなどの消泡剤を使用することにより、培養中の気泡生成を抑制することができる。また、培養液の好気性条件を保つために、培養液内に酸素または酸素含有ガス(例えば、空気)が注入できる。培養物の温度は一般に20〜45℃、好ましくは25〜40℃である。培養はL−メチオニン前駆体の生産が所望の水準に到達するまで続けることが可能であり、好ましい培養時間は10〜160時間である。
【0024】
以下、実施例によって本発明をより詳しく説明する。ところが、これらの実施例は本発明を例示的に説明するためのもので、本発明の範囲を限定するものではない。
【0025】
実施例1:O−アセチルホモセリン生産菌株の製造
<1−1>acs遺伝子のクローニング
acs遺伝子のクローニングは、大腸菌W3110の染色体DNA(ATCC27325)を鋳型としたPCRによって、アセチル−CoAシンターゼであるacs遺伝子を確保した。米国国立保健院の遺伝子銀行(NIH GenBank)に基づいてacs遺伝子の塩基配列情報(NCBI登録番号gi:89110790)を確保した。その配列は配列番号7に示した。前記配列に基づいてacs遺伝子のATG部分とTAAを含有するORF部分を増幅し、制限酵素EcoRVとHindIII認識部位を含んでいるプライマー(配列番号1および2)を合成した。大腸菌W3110の染色体DNAを鋳型とし、前記配列番号1および2のプライマーを用いてPCRを行った。重合酵素はPfuUltraTM高信頼DNAポリメラーゼを使用した。PCRは、変性96℃、30秒;アニーリング50℃、30秒;および重合反応72℃、2分を1サイクルとして30サイクル繰返し行った。その結果、asc遺伝子と制限酵素EcoRVとHindIIIを含有した約2.0kbの増幅された遺伝子を獲得した。
前記PCRによって獲得したacs遺伝子の末端に含まれた制限酵素EcoRV、HindIIIを処理し、制限酵素EcoRV、HindIIIが処理されたpPro−GFPベクターに接合によってクローニングし、最終的に構成的プロモーターProによって発現調節されるacs遺伝子のクローニングされたpCL−P(pro)−acs組み換えベクターを製作した。図2はacs発現用ベクターpCL−P(pro)−acsを示す図である。
【0026】
<1−2>フィードバック抵抗性coaA遺伝子のクローニング
フィードバック阻害が解除されたcoaA遺伝子のクローニングは、大腸菌W3110の染色体DNA(ATCC27325)を鋳型としたPCRによって、パントテネートキナーゼであるcoaA遺伝子を確保した。米国国立保健院の遺伝子銀行(NIH GenBank)に基づいてcoaA遺伝子の塩基配列情報(NCBI登録番号gi:89110060)を確保し、これに基づいてcoaA遺伝子のATG部分からフィードバック阻害の解除が可能な316番目〜318番目の遺伝子配列をCGTからGCCに変更し、制限酵素EcoRVが含まれたプライマー(配列番号3および4)を合成した。また、フィードバック阻害の解除が可能な316番目〜318番目の遺伝子配列をCGTからGCCに変更し、TAA部分までを含み、制限酵素HindIIIを含むように増幅可能なプライマー(配列番号5および6)を合成した。
【0027】
大腸菌W3110の染色体DNAを鋳型とし、前記配列番号3および4と配列番号5および6のプライマーを用いてPCRを行った。重合酵素は、PfuUltraTM高信頼DNAポリメラーゼを使用した。PCRは、変性96℃、30秒;アニーリング50℃、30秒;および重合反応72℃、2分を1サイクルとして30サイクル繰返し行った。その結果、coaA遺伝子のATG部分から316番目〜318番目の遺伝子配列がCGTからGCCに変更された部分を含むように突然変異を導入した約344bpの増幅された遺伝子断片と、突然変異を導入して316番目〜318番目の遺伝子配列がCGTからGCCに変更された部分を含み、TAA部分を含む約642bpの増幅された遺伝子を獲得した。
前記で獲得した、増幅された2つの遺伝子を鋳型としてPCRを行った。PCRは、変性96℃、60秒;アニーリング50℃、60秒;および重合反応72℃、2分を1サイクルとして10サイクル繰返し行った後、配列番号3および6のプライマーを添加し、しかる後に、20サイクル繰返し行った。その結果、coaA遺伝子の316番目〜318番目の遺伝子配列がCGTからGCCに変更された963bpの増幅されたcoaA(R106A)遺伝子を獲得した。
【0028】
前記PCRによって獲得したcoaA遺伝子の末端に含まれた制限酵素EcoRVおよびHindIIIを処理し、制限酵素EcoRVおよびHindIIIが処理されたpPro−GFPベクターに接合によってクローニングし、最終的に構成的プロモーターPro(配列番号9)によって発現調節を受け、106番目のアミノ酸のコドンがCGTからGCCに突然変異されたcoaA遺伝子がクローニングされたpCL−P(pro)−coaA(R106A)組み換えベクターを製作した。前記変異coaAのアミノ酸配列は配列番号8に示した。
前記で製作されたpCL−P(pro)−coaA(R106A)組み換えベクターをpACYC177ベクターにクローニングするために、pCL−P(pro)−coaA(R106A)組み換えベクターに制限酵素BamHIおよびHindIIIを処理することにより、1.45kbのProプロモーターを含むcoaA(R106A)切片を得た。こうして得られたProプロモーターを含むcoaA(R106A)切片を、BamHIおよびHindIIIを処理したpACYC177ベクターと接合することにより、pACYC−coaA(R106A)ベクターを製作した。図3はcoaA(R106A)発現用ベクターpCL−P(pro)−coaA(R106A)およびpACYC−coaA(R106A)を示す図である。
【0029】
<1−3>O−アセチルホモセリン生産菌株の製作
AcsとcoaAの発現が強化されたO−アセチルホモセリン生産菌株の製作のために、米国公開特許第2009−253186号に開示されたCJM−X/pthrA(M)−CL(寄託番号KCCM10921P)菌株からpthrA(M)−CLプラスミドを除去し、pthrA(M)−CLの除去された菌株CJM−Xを製作した。こうして製作されたCJM−X菌株に、実施例<1−1>および<1−2>で製造されたpCL−P(pro)−acsおよびpCL−P(pro)−coaA(R106A)プラスミドを取り込んでそれぞれ形質転換(KCCM11023PおよびKCCM11022P)し、LB−Sp(酵母抽出10g/L、NaCl5g/L、トリプトン10g/L、ストレプトマイシン25μg/L)培地で培養した後、ストレプトマイシン耐性を有するコロニー10株ずつを選抜した。さらに、前記で製作したpCL−P(pro)−acsを含むCJM−X菌株に、実施例<1−2>で製作したpACYC−CoaA(R106A)を取り込んで形質転換(KCCM11024P)し、LB−Sp−Ap(酵母抽出10g/L、NaCl5g/L、トリプトン10g/L、ストレプトマイシン25μg/L、アンピシリン50μg/L)培地で培養した後、ストレプトマイシンとアンピシリンに対して耐性を示すコロニー10株を選抜した。選抜されたコロニーのO−アセチルホモセリン生産性を互いに比較した。
【0030】
実施例2:O−アセチルホモセリン生産のための発酵
実施例1で製作された菌株のメチオニン前駆体としてのO−アセチルホモセリンの生産能力を確保するために、三角フラスコ培養を行った。
表1に示したO−アセチル−ホモセリン力価培地を用いて三角フラスコでO−アセチルホモセリンの生産性を比較した。
【0031】
【表1】
【0032】
32℃の培養器でLB固体培地内に一晩培養した単一コロニーを25mLのO−アセチルホモセリン力価培地に1白金耳ずつ接種して、250rpmで攪拌しながら32℃で42〜64時間培養した。培養物からO−アセチルホモセリンをHPLCによって定量的に分析した。分析結果を表2に示した。
下記表2に示した結果のように、acs遺伝子のプロモーターを構成的発現プロモーターP(pro)に置換したacs遺伝子を過発現する場合、対照群菌株としてのCJM−X菌株に比べてO−アセチルホモセリンの生産量が2.8g/L、収率は4.7%向上することを確認することができた。また、フィードバック阻害の解除されたcoaA(R106A)遺伝子を過発現させる場合、対照群菌株であるCJM−X菌株に比べてO−アセチルホモセリンの生産量が2.1g/L、収率は3.5%向上することを確認することができた。acsとcoaA(R106A)を同時に過発現した場合、対照群菌株としてのCJM−X菌株に比べてO−アセチルホモセリンの生産量が6.8g/L、収率は11.4%が向上することを確認した。
このようなフラスコ培養の結果からみて、acs遺伝子の発現強化、フィードバック阻害の解除されたcoaA遺伝子の発現強化、またはacsおよびフィードバック阻害の解除されたcoaA遺伝子の発現強化がO−アセチルホモセリンの生産性および収率の向上に効果的であることを確認することができた。
【0033】
【表2】
【産業上の利用可能性】
【0034】
上述したように、本発明は、培地内で発酵するときに培地で高収率でO−アセチルホモセリンを生産するエシェリキア属菌株を提供する。また、前記O−アセチルホモセリンとメチルカプタンを基質として用いて、メチオニン転換酵素の存在下に転換反応を行うことにより、高収率でL−メチオニンと酢酸を同時に生産することができる。
以上、本発明の好適な実施例について説明の目的で開示したが、当業者であれば、添付した請求の範囲に開示された本発明の精神と範囲から逸脱することなく、様々な変形、追加または置換を加え得ることを理解するであろう。
【特許請求の範囲】
【請求項1】
(a)アセチル−CoAシンターゼ遺伝子(acs)、
(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)、または
(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株。
【請求項2】
前記遺伝子の導入および増進はプロモーターの置換、プロモーターの変異、または遺伝子コピー数の増加によることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項3】
前記プロモーターの置換はpTrc、pPro、pRおよびpLプロモーターよりなる群から選ばれたプロモーターの置換によることを特徴とする、請求項2に記載のエシェリキア属菌株。
【請求項4】
前記プロモーターは配列番号9のpProプロモーター全体または一部を含むように置換されたことを特徴とする、請求項3に記載のエシェリキア属菌株。
【請求項5】
前記パントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)がコードする、CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼは、野生型酵素のアミノ酸配列(NCBI−GeneID 948479)の106番目のアミノ酸が別のアミノ酸に置換されたことを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項6】
前記106番目のアミノ酸が、野生型酵素のアミノ酸であるアルギニンの代わりにアラニンで置換されたことを特徴とする、請求項5に記載のエシェリキア属菌株。
【請求項7】
前記アミノ酸配列は配列番号8であることを特徴とする、請求項6に記載のエシェリキア属菌株。
【請求項8】
前記菌株はリシン、トレオニン、イソロイシンまたはメチオニンを生産することが可能な菌株に由来することを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項9】
前記菌株は、(a)ホモセリンアセチルトランスフェラーゼの活性、(b)アスパルトキナーゼまたはホモセリンデヒドロゲナーゼの活性、または(c)前記(a)および(b)の両方ともが導入および増進された菌株に由来することを特徴とする、請求項8に記載のエシェリキア属菌株。
【請求項10】
前記菌株は寄託番号KCCM10921Pの菌株に由来することを特徴とする、請求項9に記載のエシェリキア属菌株。
【請求項11】
前記(a)の菌株は大腸菌(Escherichia sp.)CA05−0565(寄託番号KCCM11023P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項12】
前記(b)の菌株は大腸菌CA05−0564(寄託番号KCCM11022P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項13】
前記(c)の菌株は大腸菌CA05−0566(寄託番号KCCM11024P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項14】
前記菌株は大腸菌(Escherichia coli.)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項1】
(a)アセチル−CoAシンターゼ遺伝子(acs)、
(b)CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼをコードするパントテネートキナーゼ遺伝子(coaA)、または
(c)前記(a)のアセチル−CoAシンターゼ遺伝子(acs)および前記(b)のパントテネートキナーゼ遺伝子(coaA)の両方ともが導入および増進された、O−アセチルホモセリンを生産することが可能なエシェリキア属(Escherichia sp.)菌株。
【請求項2】
前記遺伝子の導入および増進はプロモーターの置換、プロモーターの変異、または遺伝子コピー数の増加によることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項3】
前記プロモーターの置換はpTrc、pPro、pRおよびpLプロモーターよりなる群から選ばれたプロモーターの置換によることを特徴とする、請求項2に記載のエシェリキア属菌株。
【請求項4】
前記プロモーターは配列番号9のpProプロモーター全体または一部を含むように置換されたことを特徴とする、請求項3に記載のエシェリキア属菌株。
【請求項5】
前記パントテネートキナーゼ遺伝子(Panthothenate kinase、coaA)がコードする、CoA蓄積によるフィードバック阻害が解除されたパントテネートキナーゼは、野生型酵素のアミノ酸配列(NCBI−GeneID 948479)の106番目のアミノ酸が別のアミノ酸に置換されたことを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項6】
前記106番目のアミノ酸が、野生型酵素のアミノ酸であるアルギニンの代わりにアラニンで置換されたことを特徴とする、請求項5に記載のエシェリキア属菌株。
【請求項7】
前記アミノ酸配列は配列番号8であることを特徴とする、請求項6に記載のエシェリキア属菌株。
【請求項8】
前記菌株はリシン、トレオニン、イソロイシンまたはメチオニンを生産することが可能な菌株に由来することを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項9】
前記菌株は、(a)ホモセリンアセチルトランスフェラーゼの活性、(b)アスパルトキナーゼまたはホモセリンデヒドロゲナーゼの活性、または(c)前記(a)および(b)の両方ともが導入および増進された菌株に由来することを特徴とする、請求項8に記載のエシェリキア属菌株。
【請求項10】
前記菌株は寄託番号KCCM10921Pの菌株に由来することを特徴とする、請求項9に記載のエシェリキア属菌株。
【請求項11】
前記(a)の菌株は大腸菌(Escherichia sp.)CA05−0565(寄託番号KCCM11023P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項12】
前記(b)の菌株は大腸菌CA05−0564(寄託番号KCCM11022P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項13】
前記(c)の菌株は大腸菌CA05−0566(寄託番号KCCM11024P)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【請求項14】
前記菌株は大腸菌(Escherichia coli.)であることを特徴とする、請求項1に記載のエシェリキア属菌株。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2011−45359(P2011−45359A)
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−54800(P2010−54800)
【出願日】平成22年3月11日(2010.3.11)
【出願人】(508081075)シージェイチェイルジェダンコーポレーション (6)
【Fターム(参考)】
【公開日】平成23年3月10日(2011.3.10)
【国際特許分類】
【出願番号】特願2010−54800(P2010−54800)
【出願日】平成22年3月11日(2010.3.11)
【出願人】(508081075)シージェイチェイルジェダンコーポレーション (6)
【Fターム(参考)】
[ Back to top ]