
O6−アルキルグアニン−DNAアルキルトランスフェラーゼの突然変異体
本発明は、野生型ヒトAGTと比較したときに、(a)DNA相互作用の低下;(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;(d)酸化条件下での安定性の向上;(e)基質との反応後の細胞内での安定性の向上;(f)基質との反応前後の細胞外部での安定性の向上;(g)試験管内溶解度の向上;(h)O6−アルキルグアニン基質に対する反応性の向上;(i)DNAベース基質に対する反応性の低下;および(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;から選択された2つ以上の利点を示す、AGT突然変異体に関する。上記の向上した特性を備えたこのようなAGT突然変異体は、野生型ヒトAGTの1〜25個のアミノ酸が他のアミノ酸によって置換され、場合により連続鎖からの1〜5個のアミノ酸が1、2、または3つの位置で欠損または付加され、および/またはN末端の1〜4個のアミノ酸またはC末端の1〜40個のアミノ酸が欠損している突然変異体である。本発明は更に、本発明のAGT突然変異体を有する融合タンパク質に組み込まれている対象となるタンパク質を検出および/または操作する方法に関する。本発明の別の目的は、このようなAGT突然変異体および対象となるタンパク質を含むAGT融合タンパク質である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、野生型ヒトO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(hAGT)の突然変異体、および標識を基質からこれらのO6−アルキルグアニン−DNAアルキルトランスフェラーゼ突然変異体および対象となるタンパク質からなる融合タンパク質へ転移させる方法に関するものである。
【背景技術】
【0002】
N−メチル−N−ニトロソ尿素などの求電子物質の変異原性および発癌性作用は、主に、DNAにおけるグアニンのO6−アルキル化による。哺乳類および細菌は、DNAアルキル化からそれら自体を保護するために、これらの損傷を修復するタンパク質であるO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を所有する。AGTは、アルキル基をアルキル化グアニンおよびグアニン誘導体の位置O−6から、自身のシステインの1つのメルカプト基に転移させて、非可逆的にアルキル化されたAGTを生じさせる。基礎を成すメカニズムは、SN2タイプの求核反応であり、これはメチル基だけでなく、ベンジル基も容易に転用される理由を説明する。腫瘍細胞におけるヒトAGT(hAGT、配列番号:1)の過剰発現は、アルキル化薬、例えばプロカルバジン、ダカルバジン、テモゾロミド、およびビス−2−クロロエチル−N−ニトロソ尿素に対する耐性の主な理由であるため、AGTの阻害剤は、化学療法での増感剤として使用するために提案されてきた(Peggら, Prog Nucleic Acid Res Mol Biol 51: 167-223,1995)。米国特許第5,691,307号は、ベンジル基内に各種の置換基を有するO6−ベンジルグアニンおよび腫瘍細胞でのAGTレベルを枯渇させ、それによりアルキル化抗腫瘍薬に対する反応性を上昇させるためのその使用について記載している。同様に国際公開公報第97/20843号は、O6−ベンジル−およびO6−ヘテロアリールメチル−ピリミジン誘導体に相当するAGT枯渇剤を更に開示している。
【0003】
ドイツ特許第199 03 895号は、AGTのビオチン化を引き起こす、ビオチン化O6−アルキルグアニン誘導体とヒトAGTとの間の反応に依存するAGTのレベルを測定するアッセイを開示している。これは次に、ストレプトアビジンコーティングプレート上のAGTの分離および例えばELISAアッセイでのその検出を可能にする。腫瘍細胞中のAGTのレベルを監視するための、そしてAGT阻害剤のスクリーニングで使用するためのアッセイが提案されている。
【0004】
Damoiseauxら、Chem Biochem 4: 285-287,2001は、研究および化学療法での支援のために癌細胞におけるこの酵素のレベルの検出を再び容易なものとするために、ヒトAGTを標識する化学プローブとして使用するための、オリゴデオキシリボヌクレオチドに組み込まれた修飾O6−アルキル化グアニン誘導体を開示している。
【0005】
国際公開公報第02/083937号は、対象となるタンパク質を検出および/または操作する方法を開示し、該方法ではタンパク質を、AGTに融合し、標識を担持するAGT基質とAGT融合タンパク質を接触し、標識を使用してAGT融合タンパク質を検出し、場合により更に操作する。使用される複数のAGT融合タンパク質、AGT基質および広範に渡る標識の一般的構造原理、ならびに該方法で有用な標識を検出する方法が記載されている。AGTの他の形に触れられているが、ヒトAGTのみが例示されている。
【0006】
PCT/EP03/10859(国際公開公報第2004/031404号)は、対象となるタンパク質を検出および/または操作するための言及された方法で使用される特定のAGT融合タンパク質と、この方法によって得られる標識融合タンパク質と、特定のAGT融合タンパク質を使用する方法について記載している。
【0007】
PCT/EP03/10889(国際公開公報第2004/031405号)は、対象となるタンパク質を検出および/または操作するための言及した方法で特に適切な標識を担持する追加のAGT基質と、このような特に標識された基質の応用について開示している。この特許出願は、この追加のAGT基質の製造方法についても記載している。
【0008】
ヒトAGT突然変異型Gly160Trp(Xu-Welliverら、Biochemical Pharmacology 58: 1279-1285,1999)は、ベンジルグアニン誘導体に対して、野生型ヒトAGTよりも多少反応性である。Juilleratら, Chem Biol 10: 313-317,2003は、AGT融合タンパク質の合成基質による生体内(細胞内)標識に有効な更に反応性のパートナーを求めて、ヒトAGTの多数の突然変異体を作成した。位置140、157、159、および160における突然変異体が報告された。突然変異型Asn157GlySer159Gluは、野生型AGTと比較しておおよそ20倍の、ベンジルグアニン誘導体に対する活性の上昇を示している。
【0009】
hAGTにおける以下の追加の突然変異体は、hAGTのDNA結合を妨害するが、ベンジルグアニン誘導体に対する活性に著しく干渉しないことが示されている:Lys125Ala、Ala127ThrおよびArg128A1a。Limら、EMBO J 15:4050−4060,1996,およびDanielsら,EMBO J 19:1719−1730,2000を参照。
【0010】
ヒトAGT(pdb−ID 1EH6、Danielsら、EMBO J 19:1719,2000)の結晶構造は、Asn207の後で切断された機能性タンパク質を使用して解明された。
【0011】
発明の概要
本発明は、野生型ヒトAGTと比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の利点を示す、AGT突然変異体に関する。
【0012】
本発明のAGT突然変異体は、野生型ヒトAGTの1〜25個のアミノ酸が他のアミノ酸によって置換され、場合により連続鎖からの1〜5個のアミノ酸が1、2、または3つの位置で欠損または付加され、および/またはN末端の1〜4個のアミノ酸またはC末端の1〜40個のアミノ酸が欠損している、上記の向上した特性を備えた突然変異体である。
【0013】
本発明は更に、本発明のAGT突然変異体を有する融合タンパク質に組み込まれている対象となるタンパク質を検出および/または操作する方法に関する。本発明の別の目的は、このようなAGT突然変異体および対象となるタンパク質を含むAGT融合タンパク質である。
【0014】
発明の詳細な説明
対象となるタンパク質を、AGT融合タンパク質に組み込み、AGT融合タンパク質を、標識を担持する特定のAGT基質と接触させ、AGT融合タンパク質を、標識を認識および/処理するために設計された系において、標識を使用して検出し、場合により更に操作する、対象となるタンパク質を検出および/または操作するための前に記載の方法において、AGTの性能を、野生型ヒトAGTを突然変異型AGTと置換することによって更に向上できる。突然変異型AGTを包含するこのような向上方法は、本発明の目的である。別の目的は上記方法に特に適したAGT突然変異体であり、AGT融合タンパク質は、このようなAGT突然変異体および少なくとも1種の対象となるタンパク質を含む1種以上の他のタンパク質を含む。対象となるタンパク質は、いかなるタンパク質であってもよい。
【0015】
本発明のAGT突然変異体は、例えば、野生型ヒトAGTまたは既知のAGT突然変異体と比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の特性を示す突然変異体を含む。
【0016】
(a)DNA相互作用の低下
「DNA相互作用の低下」のある本発明の突然変異型AGTは、野生型ヒトAGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、20%未満のDNA結合、好ましくは2%未満のDNA結合を示し、最も好ましくは検出可能なDNA結合を示さない。DNAとの相互作用は例えば、低塩およびDNase非存在の条件下でE.coli抽出物から同時精製されたDNAの量を評価することによって定量される。これは野生型(ヒト)および突然変異型AGTの融合タンパク質(例えばGSTへの融合)の並行精製の間で分光法によって比較される(260および280nmでの吸収率の比)。あるいはAGTのDNAとの相互作用は、DNAの存在下での、基質としてのBG−Cy3(Cy3に結合されたO6−(4−アミノエチル−ベンジル)−グアニン)とのAGT反応性の阻害として測定される。この手法は、BG−Cy3のAGTとの反応時におけるCy3蛍光物質の蛍光の著しい増加を利用する。AGTのCy3−標識ベンジルグアニン誘導体との反応動力学には、サケ精子DNAの各種の濃度での数時間に渡る蛍光強度測定が続く。野生型hAGTは、DNAにテトラマーとして結合するのに対して、未結合hAGTタンパク質はモノマーのままであることが示された(Rasimasら、J Biol Chem 278(10): 7973-80,2003)。DNAに結合できないAGT突然変異体もテトラマー化しない。
【0017】
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化
「もはや核に限定されない局在化」を伴う本発明の突然変異型AGTは、真核細胞、例えば哺乳類細胞での発現時に細胞での突然変異型AGTの実質的に均一な局在化を示す。AGT突然変異体の細胞内局在化は、AGT欠損HeLa細胞またはCHO細胞を構成的発現のための構築物をヒトサイトメガロウィルス前初期プロモータの下で用いて一過性に形質移入することによって調査する。細胞を細胞膜透過性ジアセチルフルオレセイン修飾O6−ベンジルグアニンによって染色して(Juilleratら、Chem Biol 10: 313-317,2003の物質4)、共焦点レーザ顕微鏡法によって解析する。細胞質および核の蛍光強度を突然変異型AGTと野生型AGTで比較する。野生型ヒトAGTは、核での優先的局在化と、細胞質でのもっぱらわずかな局在化を示す。
【0018】
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
「溶解性タンパク質(すなわち細胞溶解後に溶解性画分に見出され、封入体には見出されないタンパク質)としての発現収率の向上」を伴う本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、溶解タンパク質として3倍を超える発現収率、好ましくは5倍を超える発現収率、最も好ましくは10倍を超える発現収率を示す。この発現収率の向上は、同時に、発現に使用される「宿主での安定性」の尺度となる。発現収率は、E.coliまたは遺伝子組換えタンパク質用の他のいずれかの標準的生産細胞、例えば酵母、あるいは好ましくは昆虫細胞、CHO細胞もしくはHeLa細胞において測定する。発現収率を定量するために、融合パートナーが容易な精製および定量を可能にするように、AGT融合タンパク質を選択できる。例えばE.coliでの発現収率は、E.coliの並行させた発現培養からの溶解性および非溶解性GST野生型AGT融合タンパク質ならびにGST突然変異型AGT融合タンパク質の収率を測定および比較することによって決定する。細胞溶解後の溶解性画分および非溶解性画分(すなわち封入体)のサンプルをSDS−PAGEにかけて、相当するAGT融合タンパク質のバンド染色強度を比較する。溶解性タンパク質は、アフィニティクロマトグラフィーによる細胞抽出物からの融合タンパク質(例えばグルタチオンセファロースによるGST−AGT融合タンパク質の場合)の精製後、精製した画分にタンパク質濃度のアッセイを行うことによって定量する(Bradford,Anal Biochem 72: 248-54,1976)。溶解性画分中の同様のタンパク質の発現収率(点突然変異での相違)は、精製状態と同様に、未精製状態でのタンパク質安定性の尺度として使用できる(Ohageら、J Mol Biol 291: 1119-1128,1999、Wirzら、Protein Sci 8: 2245-50,1999)。したがって、これは折畳み安定性および凝集傾向の尺度として見なされる。
【0019】
(c’)実用的な見地から、E.coliにおける溶解性タンパク質としての発現収率の向上が特に重要である。
【0020】
(d)酸化条件下での安定性の向上
「酸化条件下での安定性の向上」を伴う本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、2倍を超える活性タンパク質の収率、好ましくは5倍を超える活性タンパク質の収率、最も好ましくは10倍を超える活性タンパク質の収率を示し、すなわちAGTタンパク質は、緩衝水溶液(例えば100mM NaCl、10mM HEPES、pH7.4、ジチオスレイトールまたはベータ−メルカプトエタノール無添加)中、酸化条件下で1時間以上のインキュベーション時間の後に、AGT基質に対してその活性を維持する。AGT基質に対する活性は、ジチオスレイトールまたはベータ−メルカプトエタノールなどの還元剤を添加せずに精製後に測定する。あるいは突然変異型AGTおよび野生型ヒトの活性を、適切なシグナル配列へのそれらの融合による酸化性酸化還元電位を有する細胞コンパートメント(例えばE.coliのペリプラズム)内へのエクスポート後に比較する。還元および酸化条件下でのAGT基質に対する活性は、ジチオスレイトールまたはベータ−メルカプトエタノールなどの還元剤の存在下または非存在下で反応を実施することによって比較する。
【0021】
(e)基質との反応後の細胞内での安定性の向上
「細胞内での向上した安定性」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、細胞(例えば哺乳類細胞)内での細胞透過性基質との反応後に、2倍を超える安定性、好ましくは3倍を超える安定性、最も好ましくは6倍を超える安定性を示す。安定性は、突然変異型AGT融合タンパク質について、AGT基質との反応後に、AGT融合タンパク質の強度および局在化を、共焦点レーザ走査顕微鏡法を用いて解析することによって決定する。
【0022】
(f)基質との反応前後の細胞外部での安定性の向上
「細胞外部での向上した安定性」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、2倍を超える安定性、好ましくは4倍を超える安定性、最も好ましくは6倍を超える安定性を示す。反応前の安定性は、突然変異型AGTまたは突然変異型AGT融合タンパク質について、精製サンプルを緩衝水溶液中で4℃にて2週間まで、−20℃にて6ヶ月までインキュベートすることによって決定する。複数の時点にてアリコートを取り、Juilleratら、Chem Biol 10: 313-317,2003によって記載されているように反応性AGTの濃度を概算する。標識基質との反応後の、そして未反応基質からの続いての分離後の野生型AGTおよびAGT突然変異体の安定性は、4℃にて2週間および−20℃にて3ヶ月に渡って溶解性画分中の標識の濃度を定量することによって決定される。
【0023】
(f’)実際的な観点から、標識基質との反応後の安定性は特に重要である。
【0024】
(g)試験管内溶解度の向上
「試験管内溶解度の向上」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、2倍を超える溶解度、好ましくは5倍を超える溶解度、最も好ましくは10倍を超える溶解度を示す。野生型AGTおよびAGT突然変異体の試験管内溶解度は、適切な緩衝液(例えば100mM NaCl、20mM Tris、pH8.0、20%グリセロール、1mM DTT)にて確立された1つ以上の濃度での4℃または最高37℃での精製サンプルの一晩に渡るインキュベーション後に残存するタンパク質の量を決定することによって測定する。
【0025】
(h)O6−アルキルグアニン基質に対する反応性の向上
「向上した反応性」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、3倍を超える反応性、好ましくは5倍を超える反応性、最も好ましくは10倍を超える反応性を示す。O6−ベンジルグアニン基質に対する活性は、Juilleratら、Chem Biol 10: 313-317,2003によって記載されているように測定する。
【0026】
(i)DNAベース基質に対する反応性の低下
「DNAベース基質に対する反応性の低下」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、DNAベース基質に対する10%未満の反応性、好ましくはDNAベース基質に対する1%未満の反応性を示し、最も好ましくはDNAベース基質に対する検出可能な反応性を示さない。野生型AGTまたは突然変異型AGTがアルキル化DNA基質と反応する能力は、O6−ベンジルグアニンを含有する合成オリゴヌクレオチド(配列番号:2、位置14にて修飾)によるその不活性化の反応として測定する。次に反応をビオチン化O6−アルキルグアニンによるインキュベーションにより失活させる。サンプルにウェスタンブロッティングおよびストレプトアビジン誘導体により検出させて、これらの基質の速度定数を得る。あるいは野生型または突然変異型AGTがアルキル化DNA基質と反応する能力をAGT蛍光基質と競合するその標識効率によって測定する。BG−Cy3(Cy3に結合したO6−ベンジルグアニン)によるAGTの反応速度論の後には、O6−ベンジルグアニンを含有する競合する合成オリゴヌクレオチド(配列番号:2、位置14にて修飾)の各種濃度での蛍光強度測定が続く。
【0027】
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下
「N9−置換O6−アルキルグアニン基質に対する反応性の低下」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、N9−置換O6−アルキルグアニン基質に対して10%未満の反応性、好ましくは2%未満の反応性を示し、最も好ましくは検出可能な反応性を示さない。このようなN9−置換O6−アルキルグアニン基質は、野生型AGTの天然基質である。野生型AGTまたは突然変異型AGTがN9−修飾O6−アルキルグアニン基質と反応する能力は、競合実験でのビオチン化N9−非置換O6−アルキルグアニン基質の存在下でのこのような基質との、例えば低分子量N9−シクロペンチル−O6−ベンジルグアニンとのその反応の速度として測定する。次にサンプルにウェスタンブロッティングおよびストレプトアビジン誘導体による検出を受けさせて、これらの基質の速度定数を得る。あるいはこのような基質に対する反応性を、AGTとBG−Cy3との反応の阻害として測定する。AGTのBG−Cy3との反応の後に、プレインキュベーション後の、またはN9−修飾O6−アルキルグアニン基質との直接競合におけるAGTへの結合時のCy3蛍光強度の上昇が続く。
【0028】
本発明の好ましいAGT突然変異体は、共通する複数の好ましい特性を有する突然変異体、例えば
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f)基質との反応前後の細胞外部での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
を示す突然変異体である。
【0029】
更に好ましいAGT突然変異体は、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での5倍を超える安定性、
(f)基質との反応前の、細胞外での4倍を超える安定性、特に基質との反応後の(f’)、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
を示す突然変異体である。
【0030】
最も好ましいAGT突然変異体は
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
を示す突然変異体である。
【0031】
当技術水準のAGT突然変異体は、多くの所望の特性の一部のみを有する。例えばLimら、EMBO J 15: 4050-4060,1996によって記載されている突然変異体は、哺乳類細胞内での発現時に、DNA結合の減少および細胞質内への局在化を示す。Juilleratら、Chem Biol 10: 313-317,2003で記載されている突然変異体は、O6−ベンジルグアニン誘導体に対する反応性の上昇を示す。
【0032】
本発明のAGT突然変異体は、野生型ヒトAGTと比較して上述の向上した特性を備えた突然変異体であり、そこでは野生型ヒトAGTの1〜25個、好ましくは6〜22個、特に特に7、8、9、10、11、12、13、14、15、16、17、18、または19個のアミノ酸が、他のアミノ酸によって置換され、場合により連続鎖中の1〜5個のアミノ酸が、1、2、または3つの位置にて欠損もしくは付加され、および/またはN末端にて1〜4個のアミノ酸が、またはC末端にて1〜40個の、好ましくは20〜35個の、特に25〜30個のアミノ酸が欠損している。
【0033】
好ましくは、以下の位置のアミノ酸が置換される:
(A)E.coliにおける発現収率を上昇させ、タンパク質に酸化をより受けにくくする、AlaまたはValによって、好ましくはAlaによって置換されたCys62。
(B)他のアミノ酸置換、好ましくはCys150−Ser151−Ser152の置換、特にCys150の置換と組合された、E.coliにおける発現を上昇させて、野生型ヒトAGTに匹敵する基質反応性を維持する、Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Ser、好ましくはSer−Hisによって置換されたGln115−Gln116。
(C)他のアミノ酸置換、好ましくはGln115−Gln116/Cys150−Ser151−Ser152の置換またはGly131−Gly132/Met134−Arg135の置換、特にCys62/Gln115−Gln116/Gly131−Gly132/Met134−Arg135/Cys150−Ser151−Ser152の置換および182の後での切断と組合された、E.coliにおける発現収率を上昇させ、DNA結合を減少させて、哺乳類細胞での核局在化を無効にする(Limら、EMBO J 15: 4050-4060,1996)、Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128。
(D)E.coliのペリプラズムおよび細胞質における発現収率を上昇させ、DNA結合を減少させて、オリゴヌクレオチド(O6−アルキル−N9−デオキシリボシルグアニンを含有する)、O6−アルキル−N9−デオキシリボシルグアニンおよびN9−シクロペンチル−O6−ベンジルグアニンまたは他のN9置換O6−ベンジルグアニンとの反応性を無効にするのに対して、N9位置で置換されていないO6−アルキルグアニン基質に対する反応性を上昇させる、Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Val、好ましくはLys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134。
(E)野生型hAGTと比較してE.coliのペリプラズムにおいてより効率的な発現を可能にして、O6−アルキルグアニン基質に対する反応性を維持し、タンパク質に酸化をより受けにくくして、DNA結合を減少させる、Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−、好ましくはAsn−Ile−Asnによって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、またはSerもしくはThrによって置換されたCys150。
(F)N9位置で置換されていないO6−アルキルグアニン基質に対する反応速度を上昇させる(Juilleratら、Chem Biol 10: 313-317,2003)のに対して、E.coliのペリプラズムおよび細胞質における発現収率を上昇させ、DNA結合を減少させて、オリゴヌクレオチド(O6−アルキル−N9−デオキシリボシルグアニンを含有する)、O6−アルキル−N9−デオキシリボシルグアニンおよびN9−シクロペンチル−O6−ベンジルグアニンとの反応性を無効にする、他のアミノ酸置換、好ましくはGln115−Gln116/Cys150−Ser151−Ser152の置換またはGly131−Gly132/Met134−Arg135の置換、特にCys62/Gln115−Gln116/Lys125/Ala127−Arg128/Gly131−Gly132/Met134−Arg135/Cys150−Ser151−Ser152の置換および182の後での切断と組合された、Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trp、最も好ましくはGly−Glu−Alaによって置換されたAsn157/Ser159−Gly160、あるいはGly/Glu(特に好ましい)によって置換されたAsn157/Ser159、GlyまたはArgによって置換されたAsn157。
(G)発現収率をやや上昇させる、Gly182の後の切断(アミノ酸183−207の欠損)。
【0034】
本発明の突然変異体は、特に、修飾(A)、(B)、(D)、(E)、(F)および(G)のうち2つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体、および修飾(A)〜(G)のうちの3つ以上、例えば3または4つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体である。他の好ましい突然変異体は、修飾(A)、(B)、(C)、(E)、(F)、および(G)のうち3つ以上、例えば3または4つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体である。
【0035】
特に好ましいのは、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞を通じての細胞質全体への分布、DNA結合の減少、およびO6−ベンジルグアニン基質に対する反応性の上昇を示す、突然変異型Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu、Gly182の後の切断、
E.coliにおける発現収率の上昇、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇、およびO6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性の実質的な低下を示す、突然変異型Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu、
E.coliにおける発現収率の実質的な上昇を示すが、O6−ベンジルグアニジン基質に対する活性を維持する、突然変異型Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞における細胞質全体への分布、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇、およびO6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する少なくとも100分の1に減少した反応性を示す、突然変異型Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断、および、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞における細胞質全体への分布、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇を示すが、O6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性を維持する、突然変異型Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断
である。
【0036】
検討される更なる突然変異体は、修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断、および場合により1〜15個の、好ましくは1〜10個の、特に3〜7個の追加のアミノ酸修飾、例えば
Glu115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、AGT突然変異体である。
【0037】
同様に検討される突然変異体は、修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断、および場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾、例えば
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Asn150Asp、Ile151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Asn150Val、Ile151Ser、Asn152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Asn150Gly、Ile151Gly、Asn152Asp、Ala154Asp;
Asn150Val、Ala154Asp;
Asn150Glu、Ile151Gly、Asn152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ile151Gly;
Asn150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、AGT突然変異体である。
【0038】
同様に好ましいのは、修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異体;および修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異体である。
【0039】
特に好ましいのは、実施例の、特に実施例2〜7および表1の化合物である。
【0040】
突然変異体は、当業者に公知の技法によって得られる。AGT変異体は、好ましくは、新しいO6−アルキルグアニン−DNAアルキルトランスフェラーゼを産生および選択するために、当業者に既知のタンパク質工学技法を使用して、および/または分子進化を使用して生成できる。このような技法は、例えば、部位特異的変異誘発、飽和変異誘発、変異を配列の任意の箇所に導入するためのエラープローン(error prone)PCR、および飽和変異誘発の後に使用されるDNAシャッフリングである。ファージ提示法を活用すると、突然変異体は、本発明のO6−ベンジルグアニンおよびAGT基質に対する活性の著しい向上ならびに酸化条件下での安定性の向上と共に見出される。AGTは、繊維状ファージ上にファージカプシドタンパク質pIIIを有する融合タンパク質として機能的に提示することが可能であり、AGTの異常なメカニズムを使用して、向上した特性、反応速度の上昇を備えたAGTを提示するファージを選択できる、例えばJuilleratら, Chem Biol 10: 313-317,2003を参照。
【0041】
本発明において、対象となるタンパク質またはペプチドは、上記のAGT突然変異体に融合される。対象となるタンパク質またはペプチドは、いかなる長さでもよく、二次、三次、または四次構造を有していても、有していなくてもよく、好ましくは少なくとも12個のアミノ酸および最大2000個のアミノ酸からなる。このような対象となるタンパク質またはペプチドの例を以下に与えるが、例えば酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタタンパク質、核局在化シグナルタンパク質、タンパク質補助因子、小型モノマー性GTPase、ATP結合カセットタンパク質、細胞内構造タンパク質、タンパク質を特定の細胞コンパートメントに標的化させることに関与する配列を備えたタンパク質、ラベルまたは親和性タグとして一般に使用されるタンパク質および上述のタンパク質のドメインまたはサブドメインである。対象となるタンパク質またはペプチドは好ましくはAGT突然変異体に、酵素によって開裂できるリンカーによって、例えばDNA段階にて適切な制限酵素、例えばBglIIによって開裂できるAGATCTによって、および/またはタンパク質段階にて適切な酵素、例えばタバコエッチウィルスNla(TEV)プロテアーゼによって開裂可能なリンカーによって融合される。融合タンパク質は、原核宿主、好ましくはE.coliにて、または真核宿主、例えば酵母、昆虫、または哺乳類細胞にて発現できる。
【0042】
AGT突然変異体は、以下に記載する適切な基質上に存在する標識を融合タンパク質のAGT部分のシステイン残基の1つに標識を転移する特性を有する。
【0043】
対象となるタンパク質およびAGT突然変異体を含む融合タンパク質を、標識を有する特定の基質に以下に記載するように接触させる。AGT突然変異体が基質と反応して、基質の標識を転移するような反応条件を選択する。通常の条件は、室温、例えば約25℃で約pH7の緩衝溶液である。しかしながら、AGT突然変異体は、多様な他の条件の下でも反応し、本明細書で挙げるこれらの条件は、本発明の範囲を制限しないことが理解される。
【0044】
基質の標識部分は、融合タンパク質が対象とする用途に応じて当業者が選択できる。AGT突然変異体を含む融合タンパク質を基質に接触させた後に、標識は融合タンパク質に共有結合する。次に標識AGT突然変異型融合タンパク質は、転移された標識によって更に操作および/または検出される。標識は、複数の同じまたは異なる標識からなる場合がある。基質が1つを超える標識を含有する場合、対応する標識AGT突然変異型融合タンパク質も、標識融合タンパク質を更に操作および/または検出するための更なる選択肢を与える、1つを超える標識を含むことになる。
【0045】
「操作」の下では、いずれの物理または化学処理も理解される。例えば操作は、細胞からの単離、標準精製技法、例えばクロマトグラフィーを用いた精製、化学試薬との、または特に結合パートナーが固相に固定されている場合の結合対の結合パートナーとの反応などを意味し得る。このような操作は、標識Lに依存することがあり、標識融合タンパク質の「検出」に加えて行うことができる。標識融合タンパク質に操作および検出の両方を行う場合、検出は、操作の前または後でもよく、あるいは本明細書で定義されたような操作の間に実施できる。
【0046】
特定のAGT基質は、特許出願PCT/EP03/10889(国際公開公報第2004/031405号)に開示されているようなAGT基質、例えば式(I)
【0047】
【化1】

【0048】
〔式中、
R1−R2は、AGTによって基質として認識される基であり;
Xは、酸素または硫黄であり;
R3は、芳香族またはヘテロ芳香族基、あるいは場合によりCH2に結合された二重結合を有する、置換不飽和アルキル、シクロアルキル、またはヘテロシクリル基であり;
R4は、リンカーであり;そして
Lは、標識、複数の同じまたは異なる標識、R4をR1に結合して環式基質を生成する結合、または更なる基−R3−CH2−X−R1−R2である。〕
の化合物であり、
【0049】
基R1−R2において、残基R1は、好ましくは、AGTによって基質として認識される、窒素原子1〜5個を含有するヘテロ芳香族基、好ましくは式(II)
【0050】
【化2】
【0051】
〔式中、
R2は、水素、炭素原子1〜10個のアルキル、または糖部分であり;
R5は、水素、ハロゲン、例えばクロロまたはブロモ、トリフルオロメチル、またはヒドロキシであり;そして
R6は、水素、ヒドロキシ、または非置換もしくは置換のアミノである。〕
のプリンラジカルであり、
【0052】
R5またはR6がヒドロキシである場合、プリンラジカルは主にその互変異性形で存在し、互変異性形ではR5またはR6を保持する炭素原子に隣接する窒素が水素原子を有し、この窒素原子とR5またはR6を保持する炭素原子との間の二重結合が単結合であり、そしてR5またはR6がそれぞれ二重結合された酸素である。
【0053】
R6が非置換であるか、または置換のアミノであり、そしてプリンラジカルの結合に接続された残基Xが酸素である場合、式(II)の残基は、グアニン誘導体である。
【0054】
本発明の方法で有用な他の特定の基質は、R1が、式(II)のプリンラジカルであり、R2が、シクロアルキル、例えばシクロペンチル、あるいはアルキル、例えばプロピルまたはイソブチル、あるいは置換アルキル、例えばシアノメチルである、式(I)の化合物である。
【0055】
本発明は、生体内(細胞内)および試験管内の両方での標識AGT突然変異型融合タンパク質のための方法を提供する。AGT突然変異型融合タンパク質の生体内標識化という用語は、細胞のコンパートメントすべてはもちろんのこと、細胞外スペースを指示するAGT突然変異型融合タンパク質のコンパートメントすべてでの標識化を含む。AGT突然変異型融合タンパク質の標識化が、生体内で実施され、そしてAGT突然変異体に融合したタンパク質が膜タンパク質、より詳細には原形質膜タンパク質である場合、融合タンパク質のAGT部分は膜のどちら側にも結合することが可能であり、例えば原形質膜の細胞質側または細胞外側に結合することが可能である。
【0056】
標識化を試験管内で実施する場合、融合タンパク質の標識化は、AGT突然変異型融合タンパク質の細胞抽出物中で、あるいはその精製または濃縮形を用いてのどちらかで実施できる。
【0057】
標識化を生体内(細胞内)で、または細胞抽出物中で実施する場合、宿主の内因性AGTの標識化を好都合に考慮できる。宿主の内因性AGTがO6−アルキルグアニン誘導体または関連化合物を基質として受入れない場合、融合タンパク質の標識化は、特異的である。哺乳類細胞において、例えばヒト、マウス、またはラット細胞において、内因性AGTの標識化は可能である。これらの実験において、内因性AGTはもちろんのこと、AGT突然変異型融合タンパク質の同時標識化が問題を引き起こす、このような実験においては、既知のAGT欠損細胞株を使用できる。
【0058】
特定の基質に対して非反応性であり、次に内因性AGTによって認識される突然変異型AGT融合タンパク質を使用する場合、内因性AGTの活性を遮断するために、特定の突然変異型AGT融合タンパク質と反応するように設計された基質を用いて細胞をインキュベートする前またはその間に、このような非反応性基質を使用できる。例えばN9−置換O6−アルキルグアニン誘導体と、例えばN9−シクロペンチル−O6−ベンジルグアニンと反応しない突然変異型AGTを使用できる。内因性AGTを含有する哺乳類細胞において、この野生型AGTは、次に、標識化の前に、または突然変異型AGT融合タンパク質をこの突然変異型AGTによって認識された別の基質によって標識化する間に、N9−シクロペンチル−O6−ベンジルグアニンによって遮断できる。
【0059】
有効濃度の内因性野生型AGTが特定の生体サンプル中に存在しない場合、実験用に選択された突然変異型AGTが反応性でない基質を使用して、内因性AGTを事前に不活性化する必要はないであろう。このような条件下では、野生型AGTの特定の基質と反応しない突然変異型AGT融合タンパク質を利用することにより、AGTの2つの異なる突然変異体(あるいはAGTの1つの突然変異体および野生型AGT)を2つの異なる基質によって選択的に標識化できる。このことは、野生型hAGTタンパク質と比較して低分子量の基質に対して選択性を示す、ここでは「AGT−A」と呼ぶ突然変異型AGT、および特定の基質と野生型hAGTの10分の1、好ましくは100分の1の速さで反応する、「AGT−B」と呼ぶ別の突然変異型AGTを使用することによって実施される。突然変異型「AGT−A」(または野生型AGT)および突然変異型「AGT−B」の両方を生体サンプルに存在させて、その生体サンプル−例えば細胞抽出物または無傷の細胞−を最初に突然変異型「AGT−A」(または野生型AGT)によって選択的に認識される基質と一定時間反応させて、基質「A」による突然変異型「AGT−A」タンパク質(または野生型AGT)の完全な、またはほぼ完全なターンオーバーを引き起こす。しかし、突然変異型「AGT−B」タンパク質は未反応またはほぼ未反応のまま残し、次いで突然変異型「AGT−B」と反応性である(そして突然変異型「AGT−A」または野生型AGTも反応性であり得る)基質「B」を用いてインキュベーションし、突然変異型「AGT−B」タンパク質と基質「B」との優先的反応も引き起こす。なぜならば、突然変異型「AGT−A」タンパク質(または野生型AGT)が基質「A」によって既に不活性化されているためである。同様に、基質「B」の選択した濃度の存在下でさえ、選択した濃度での突然変異型「AGT−A」(または野生型AGT)の基質「A」に対する反応性が突然変異型「AGT−A」(または野生型AGT)の基質「A」との優先的反応を引き起こすという条件で、突然変異型「AGT−B」および突然変異型「AGT−A」(または野生型AGT)を含有する混合物を、基質「A」および基質「B」と同時にインキュベートし、突然変異型「AGT−A」(または野生型AGT)の基質「A」との優先的反応および突然変異型「AGT−B」の基質「B」との優先的反応を引き起すことができる。2つの異なる基質「A」および「B」は例えば、蛍光共鳴エネルギー移動対(FRET)の2つの化合物、または近接アッセイのための1つの蛍光物質および1つのクエンチャーを保持してもよい。
【0060】
本発明は、対象となるタンパク質を検出および/または操作する方法にも関し、該方法では対象となるタンパク質を融合タンパク質内にAGT突然変異体と共に組み込み、標識を有する特定のAGT基質とAGT融合タンパク質を接触させ、標識を認識および/または処理するために設計された系において標識を使用してAGT融合タンパク質を検出し、場合により操作する。このことは前の段落に記載するように、AGT突然変異体と1つの基質との優先的な(または特に低い)反応性を利用している。特に本発明は、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質、ならびに更なるAGT融合タンパク質を含有するAGT融合タンパク質混合物を、AGT突然変異体または更なるAGTのどちらかに対して選択的である特定の基質と接触させて、混合物を更なる基質と処理して、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質を、標識を認識および/または処理するために設計された系において標識を使用して検出し、場合により更に操作する方法に関する。更なる基質は、混合物と特定の基質の反応終了後に、または特定の基質と共にAGT融合タンパク質混合物に添加できる。
【0061】
より詳細には、本発明は、特定の基質の標識が更なる基質の標識と相互作用し、例えば標識が蛍光共鳴エネルギー移動対(FRET)の化合物あるいは近接アッセイのための1つの蛍光物質および1つのクエンチャーである、これらの方法に関する。
【実施例】
【0062】
使用した省略形:
AGTM=修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異型
BG−Bt=ビオチンに結合されたO6−(4−アミノメチル−ベンジル)−グアニン
BG−Cy3=Cy3に結合されたO6−(4−アミノメチル−ベンジル)−グアニン
DTT=1,4−ジチオスレイトール
GST=グルタチオン−S−トランスフェラーゼ(Schistosoma japonicum 由来)
HEPES=2−[4−(2−ヒドロキシエチル)−1−ピペラジニル]エタンスルホン酸
IPTG=イソプロピルβ−D−1−チオガラクトピラノシド
PEG=ポリエチレングリコール
PMSF=フェニルメタンスルホニルフルオリド
SDS−PAGE=ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動
【0063】
〔実施例1〕
突然変異Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子の2つの特に重複する領域である、突然変異Asn157Gly、Ser159Gluを含有するAGT(Juilleratら、Chem Biol 10: 313-317,2003)を配列番号:3、配列番号:6および配列番号:4、配列番号:5のプライマーによって別個の反応にて増幅した。その部分相補性に関して、これらの2つの部分を更なるPCR反応にて構築し、配列番号:3および配列番号:4のプライマーによって増幅して、ここで突然変異Lys125Ala、Ala127Thr、Arg128Alaを更に含有する完全な遺伝子を産生させた。続いて遺伝子を発現ベクターpGEX−2T(Pharmacia)BamHI部位とEcoRI部位との間でクローニングした。これにより遺伝子がベクターによって提供されたGSTタンパク質のC末端融合物として挿入遺伝子の発現が可能となる。
【0064】
このベクターからのタンパク質発現をE.coli菌株BL21で実施した。指数関数的に増殖する培養は、0.5mM IPTGによって誘起して、発現を24℃にて3.5時間実施した。
【0065】
精製:収穫した細胞を1mM PMSFおよび2μg/mlアプロチニンを添加した、50mMホスフェート、0.5M NaCl、1mM DTTを含有する緩衝液で再懸濁させて、リゾチームおよび超音波処理によって破壊した。細胞破片を40000xgでの遠心分離によって分離した。抽出物を事前に平衡にしたグルタチオンセファロース(Amersham)に加え、次に20ベッドボリューム(50mMホスフェート、0.5M NaCl、1mM DTT)で洗浄した。変異したGST−AGT融合タンパク質を50mM Tris・HCl、pH7.9中の10mM還元グルタチオンで溶出させた。精製したタンパク質を50mM HEPES、pH7.2、1mM DTT、30%グリセロールで透析して、次に−80℃にて保存した。
【0066】
タンパク質収率の決定:GST−AGT融合タンパク質の純度および相対量をSDS−PAGEにサンプルをかけることによって比較した。純粋画分のUVスペクトルは、Perkin Elmer Lambda 10を使用して記録した。大量のDNAを含有しないタンパク質サンプルでは、タンパク質含有率の尺度として280nmでの吸光度を同じ条件下で生成した野生型hAGTまたはPGEG−hAGTの吸光度と比較した。
【0067】
試験管内でタンパク質のDNA結合特性を比較するために、収穫した細胞を再懸濁させ、同じ方法の後に、しかし50mMホスフェート、0.1M NaCl、1mM DTTを含有する緩衝液中でGST融合タンパク質を精製した。透析サンプルにおいて、次に同時精製されたDNAの量をUV分光法によって見積もった。スペクトルは、Perkin Elmer Lambda 10を使用して記録した。DNAの含有率は、280および260nmにおける吸光度の比に相当する。この値を、野生型hAGTまたはPGEG−hAGTを精製して得られた値とそれぞれ比較した。
【0068】
活性アッセイ:精製した変異GST−AGTは、ビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら, Chem Biol 10: 313-317,2003の物質3a)を用いて試験管内でインキュベートした。総反応物量80μlにて、0.2μM GST−AGTを50mM HEPES pH7.2および1mM DTT中の1μM基質を用いて室温にてインキュベートした。複数の時点にてアリコートをSDS−Laemmli緩衝液中の1mM O6−ベンジルグアニン(Sigma)で失活させて、ウェスタンブロッティング分析を受けさせた(ニュートラアビジン−ペルオキシダーゼコンジュゲート(PIERCE),Renaissance reagent plus(NEN))。対応するバンドの強度は、Kodak Image Station 440によって定量した。
【0069】
生体内局在化:変異AGT(Lys125Ala、Ala127Thr、Argl28Ala、Asn157Gly、Ser159Glu)遺伝子を配列番号:7、配列番号:8のプライマーで増幅して、ベクターpEGFP−Nuc(Clontech)のNheI部位とBamHI部位との間でクローニングした。この構築物を内因性AGTが欠損したCHO細胞に形質移入した(B.Kainaら、Carcinogenesis 12,1857-1867,1991)。24時間のAGT一過性発現の後に、細胞をJuilleratら、Chem Biol 10: 313-317,2003の物質4の0.5μM で5分間インキュベートして、PBSで30分間洗浄した。細胞を、水(1.2開口数)対物レンズを備えたZeiss LSM 510顕微鏡(Carl Zeiss AG)での488nmアルゴン/クリプトンレーザ線を使用して、レーザ走査共焦顕微鏡法によって撮像した。
【0070】
E.coliにおいて、突然変異型AGT Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159GluのGST融合は、PGEG−hAGTの少なくとも3倍の溶解性タンパク質を生じる。それは少なくとも10分の1に減少したDNA結合を示し、O6−ベンジルグアニン基質に対する活性を維持している。CHO細胞では、蛍光標識変異AGTは、細胞質全体に分布しており、優先的な核局在化は検出できない。
【0071】
〔実施例2〕
突然変異Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu
実施例1の突然変異型AGT遺伝子を、配列番号:3、配列番号:10および配列番号:4、配列番号:9のオリゴヌクレオチドを用いて実施例1に記載するように増幅およびクローニングすると、PCRによるその取込み時に遺伝子内に更なる突然変異Cys62Alaを引き起こさせた。タンパク質発現、精製および収率の決定は、実施例1に記載するように実施した。突然変異型遺伝子は、ベクターpAK100内の繊維状ファージのg3タンパク質へ融合させて遺伝子をサブクローニングするために、SfiI制限部位を含有する配列番号:11、配列番号:12のプライマーによってPCR増幅した(Krebberら、J Immunol Methods 201: 35-55,1997)。非サプレッサ菌株E.coli BL21内で遺伝子を発現させるときに、アンバー停止コドンは、AGT遺伝子後の翻訳を終止させる。したがって、このベクターからの突然変異型AGTタンパク質のペリプラズム発現を実施例1で記載するように実施した。収穫した細胞を1mM PMSFおよび2μg/mlアプロチニンを添加した、50mMホスフェート、1M NaCl、1mM DTTを含有する緩衝液で再懸濁させて、リゾチームおよび超音波処理によって破壊した。細胞破片を40000xgでの遠心分離によって分離した。実施例1で記載するように、上清をSDS−PAGEによるタンパク質収率の定量および活性アッセイに直接供した。
【0072】
突然変異型Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Gluは、O6−ベンジルグアニン基質に対する活性を保持している。この突然変異型AGTからの溶解性GST融合タンパク質の収率は、実施例1の突然変異型Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Gluの収率の少なくとも2倍である。E.coliのペリプラズムでの発現による活性AGT突然変異体の収率は、PGEG−hAGTの収率の少なくとも3倍である。
【0073】
〔実施例3〕
182での切断、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子(Asn157Gly、Ser159Glu、Juilleratら、Chem Biol 10:313−317,2003を参照)を配列番号:3、配列番号:13のプライマーによって増幅して、コドン182の後に停止コドンおよびEcoRIを導入して、次に発現ベクターpGEX2T(Pharmacia)のBamHI部位とEcoRI部位との間でクローニングした。タンパク質発現、精製および発現収率の見積りは、実施例1に記載するように実施した。
【0074】
切断PGEG−hAGT突然変異体は、O6−ベンジルグアニン基質に対する活性を保持している。溶解性突然変異型GST−AGT融合タンパク質の収率は、突然変異型PGEG−hAGTの収率の少なくとも2倍である。
【0075】
〔実施例4〕
突然変異Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子(Asn157Gly、Ser159Glu、Juilleratら、Chem Biol 10:313−317,2003を参照)を、実施例1に記載するように配列番号:11、配列番号:15および配列番号:12、配列番号:14のプライマーでPCR増幅して、構築した。プライマーは、ヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)をhAGT遺伝子のコドン131、132、134、135に相当する位置に含有する。遺伝子を繊維状ファージのg3タンパク質へ融合させて、SfiI制限部位を介してベクターpAK100内にクローニングした。生じた遺伝子ライブラリをファージ提示に使用した。
【0076】
このライブラリのファージの産生をE.coli JM101細胞にて実施した。指数関数的培養物にヘルパーファージを重感染させて、24℃にて一晩培養した。この培養物の上清を1μMジゴキシゲニン化O6−ベンジルグアニン(Juilleratら, Chem Biol 10: 313-317,2003の物質2)によって6分間インキュベートした。次の選択ラウンドでは、反応時間がそれぞれ90秒および45秒まで短縮し、基質の濃度は、10nMまで低下して選択圧を上昇させた。この反応からのファージを4% PEG/3% NaClを用いた沈殿によって精製した。ジゴキシゲニンによって今や共有結合的に標識された突然変異型AGTを有するファージを、抗ジゴキシゲニン抗体(Roche Diagnostics)でコーティングした磁気ビーズを用いたインキュベーションによって単離して、細菌の再感染に使用した。
【0077】
選択したAGT突然変異体を実施例1に記載するように、pGEX2T内にサブクローニングして、発現させて、精製およびキャラクタリゼーションした。N9−置換およびN9−非置換O6−ベンジルグアニン基質間の識別は、N9−置換およびN9−非置換O6−ベンジルグアニンを用いた精製された変異GST−AGTのインビトロでの競合実験によって評価した。0.2μM GST−AGTのアリコートを、50mM HEPES、pH7.3、および1mM DTT中の様々な濃度のN9−置換基質(0、0.5〜100μMまで)および0.5μMビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら, Chem Biol 10: 313-317, 2003の物質3a)を用いて室温にてインキュベートした。45分後、SDS−Laemmli緩衝液の添加および熱変性によって反応を停止させた。サンプルにSDS−PAGEおよびウェスタンブロッティング分析に供した(ニュートラアビジン−ペルオキシダーゼコンジュゲート(PIERCE),Renaissance reagent plus(NEN))。対応するバンドの強度は、Kodak Image Station 440によって定量した。
【0078】
この突然変異型AGTからの溶解性GST融合タンパク質の収率は、PGEG−hAGTの収率の少なくとも2倍である。突然変異型AGTは、O6−ベンジルグアニン基質に対して少なくとも2倍更に上昇した活性を示す。O6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性は、PGEG−hAGTと比較して、少なくとも10分の1に、特定のクローンでは少なくとも100分の1に低下する。オリゴヌクレオチド、例えば位置14のグアニンがO6−ベンジル基を有する配列番号:2のオリゴヌクレオチドとの反応性は、PGEG−hAGTと比較して、少なくとも1000分の1に低下する。
【0079】
〔実施例5〕
突然変異Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu
実施例4に記載したように、3つのPCR断片から配列番号:11、配列番号:17、配列番号:16、配列番号:19および配列番号:12、配列番号:18のプライマーの組合せを使用して、3回の別個のPCR反応で、AGT突然変異体のライブラリを構築した。遺伝子を部分的に重複する断片から構築して、断片のうち2つは、コドン115−116および150−152にそれぞれ対応する位置に、ランダムヌクレオチド混合物NNKを含有している。ファージ提示選択は、Juilleratら, Chem Biol 10: 313-317,2003に記載されているように実施した。選択したタンパク質を実施例1に記載するように、pGEX内にサブクローニングして、発現させ、精製およびキャラクタリゼーションした。ペリプラズム発現は実施例2に記載するように実施した。
【0080】
この突然変異型AGTからの溶解性GST融合タンパク質の収率は、PGEG−hAGTからの収率の少なくとも2倍である。突然変異型AGTは、O6−ベンジルグアニン基質に対する活性を保持している。E.coliのペリプラズムでの発現による活性AGT突然変異体の収率は、PGEG−hAGTの収率の少なくとも2倍である。
【0081】
〔実施例6〕
突然変異Cys62Ala、GIn115Ser、Gln116His、Lys125Ala、Ala127Thr、Ara128Ala、Gly131Lys、Gly132Thr、Met134Leu、Ara135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、182での切断、「AGTM」
実施例2、3、4、5による突然変異を組合せた。実施例1からの突然変異型AGT遺伝子の4つの重複断片(Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu)を、配列番号:3、配列番号:23;配列番号:21、配列番号:22;配列番号:20、配列番号:25および配列番号:24、配列番号:13のプライマー組合せを使用して増幅し、遺伝子へと構築して、実施例1に記載するようにpGEX2T内へサブクローニングした。実施例1に記載するように、突然変異型AGT(「AGTM」)を発現させ、精製およびキャラクタリゼーションした。
【0082】
適切な緩衝液(例えば100mM NaCl、20mM Tris、pH8.0、20%グリセロール)中で確立された1つ以上の濃度において4℃または最高37℃での精製サンプルの一晩に渡るインキュベーション後に溶解性画分中に残存するタンパク質の量を決定することによって、AGTMまたはAGTM融合タンパク質の試験管内での溶解度を測定して、野生型AGTまたは既知のAGT突然変異体、例えば「PGEG−hAGT」の溶解度と比較した(Juilleratら、Chem Biol 10: 313-317,2003)。
【0083】
突然変異型AGTMは、実施例1〜5に記載する突然変異体と比較して少なくとも2倍の発現速度の更なる上昇を示し、野生型AGTと比較したときに少なくとも4倍の試験管内での溶解度の上昇を有する。それはO6−ベンジルグアニンに対する反応性を保持しているが、N9−置換O6−アルキルグアニン誘導体との少なくとも10分の1までの反応性の著しい低下、DNA基質との少なくとも100分の1までの反応性の低下を示す。
【0084】
酸化条件下での安定性:配列番号:11および配列番号:28のプライマーを使用して突然変異型遺伝子をPCR増幅して、実施例2で記載するように、該遺伝子をpAK100内にサブクローニングした。実施例2に記載するようにタンパク質発現および細胞溶解を行って、透明な細胞溶解物を得た。PGEA−hAGT(Juilleratら,Chem Biol 10: 313−317,2003)および突然変異体の反応性を比較するために、1μM BG−Btを用いてサンプルを50分間インキュベートして、SDS−Laemmli緩衝液で失活させ、ニュートラアビジン−ペルオキシダーゼコンジュゲート(Pierce)またはモノクローナルマウス抗FLAG IgG抗体(M2、Sigma)および二次抗マウスIgG::HRPコンジュゲート、ならびにRenaissance reagent plus(NEN)を用いてウェスタンブロッティングによって分析した。ブロットをKodak Image station 440で視覚化した。抗FLAG IgG抗体を使用して、溶解性AGTタンパク質の総量を見積もったが、ニュートラアビジンは、その反応性部分のみを定量した。AGT突然変異体の予想分子量は、それぞれ21および24kDaである。
【0085】
ペリプラズム発現は2〜3倍のAGTMをもたらすが、突然変異型PGEA−hAGTでは、少なくとも10倍のこの突然変異体の活性タンパク質をもたらす(タンパク質収率へ正規化)。
【0086】
BG−Cy3アッセイで測定したDNAベース基質に対する反応性:前に記載したO6−(4−アミノエチル−ベンジル)グアニン(Kepplerら、Nature Biotechnol.21: 86-89,2003)をCy3の市販のN−ヒドロキシスクシンイミドエステル(Cy3−NHS、Amersham)と反応させることによって、BG−Cy3を調製した。カップリング反応は周囲温度にて、N,N−ジメチルホルムアミド中でトリエチルアミン1当量の存在下で実施した。BG−Cy3コンジュゲートは、C18カラムでの逆相HPLCによって、0.1%トリフルオロ酢酸〜アセトニトリルの線形勾配を使用して精製した。溶媒蒸発後、BG−Cy3をジメチルスルホキシドに溶解させ、−20℃にて保存した。DNAベース基質との反応性を測定するために、反応緩衝液(50mM HEPES、pH7.2、1mM DTT、200mg/ml BSA)中に各種の濃度(0〜5μM)の配列番号:2のオリゴヌクレオチド(位置14で修飾)、0.2μM GST−AGT融合タンパク質、および0.5μM BG−Cy3を含有する反応混合物をマイクロタイタープレートでインキュベートした。Cy3誘導体の蛍光(励起519nm、発光572nm)をSpectramax Geminiプレートリーダー(Molecular Devices)で測定した。
【0087】
突然変異型AGTMは、5μMまでのBG修飾オリゴヌクレオチドの存在下でBG−Cy3とのその反応の低下を示していない(図1A)。PGEG−hAGTの活性は、0.5μM(またはそれ以上)BG修飾オリゴヌクレオチドの存在下でバックグラウンドレベル以下(3%未満)である(図1B)。
【0088】
DNA結合:DNA結合は、AGT突然変異体とBG誘導体との反応のDNAによる阻害として測定した。オリゴヌクレオチドの代わりにサケ精子DNA(25μg/ml)を含有する反応混合物中のAGT突然変異体の反応速度をBG−Cy3アッセイによって測定した。反応速度は、得られたデータを単指数上昇に最大まで適合させることによって決定した。
【0089】
AGTMのBG−Cy3に対する反応速度は、DNAの存在下で不変であるのに対して、PGEG−hAGTのBG−Cy3に対する速度は、DNAの存在下で5分の1に低下している。
【0090】
反応後の安定性:12xHis−タグならびにSbfIおよびAscIクローニング部位を含めるために合成オリゴヌクレオチド(配列番号:29および配列番号:30)NcoI部位とHindIII部位との間でクローニングすることによって、ベクターpBAD/His A(Invitrogen)を修飾した。AGTMの遺伝子は、配列番号:31および配列番号:32のプライマーによって増幅して、SbfI/AscI部位間にてこのベクター内へクローニングした。野生型AGTおよび突然変異型AGT(Lys125Ala、Alal27Thr、Arg128Ala、Asn157Gly、Ser159Glu)を配列番号:31および配列番号:33のプライマーによって増幅して、このベクター内へクローニングした。E.coli菌株Top10においてタンパク質発現を実施した。指数関数的に増殖する培養物を0.2%アラビノースによって誘起し、発現を28℃にて16時間に渡って実施した。0.5M NaCl、80mMイミダゾール、1% Triton、250μM PMSF、1mM β−メルカプトエタノール、5mg/mlリゾチーム、20μg/ml DNaseを含有する緩衝液を10mg/gまで添加することによって細胞を溶解させ、45分間撹拌した。10,000gにて30分間の遠心分離後の上清を平衡したNi−NTA(Qiagen)カラムに装填して0.5M NaCl、100mMイミダゾール、1mM β−メルカプトエタノールの20カラム体積で、続いて0.25M NaCl、100mMイミダゾール、20%グリセロールの3カラム体積で洗浄した。タンパク質を0.25M NaCl、400mMイミダゾール、20%グリセロールで溶出させた。1mM DTTを添加した後、タンパク質を−80℃で保存した。
【0091】
AGTタンパク質(10μMにて)をフルオレセイン修飾O6−ベンジルグアニン(Juilleratら、Chem Biol 10: 313-317,2003の物質5、BG−FL、20μM)と定量的に反応させて、サイズ排除カラム(Econopac 10DG、Bio−Rad)で精製し、7日間インキュベートした。各種の時点にて、アリコートを25,000gにて遠心分離して、1% SDSを上清に添加した。上清の蛍光をVictor2蛍光プレートリーダー(PerkinElmer、485/535nmにて励起/発光)で測定した。
【0092】
7日間に渡って溶解性蛍光タンパク質の約60%を失うhAGTまたは突然変異型(Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu)とは対照的に、標識突然変異型AGTMは、7日間を超えて溶液中にて安定なままである。
【0093】
N9−修飾O6−ベンジルグアニン誘導体に対するAGTの反応性:実験は、BG−Cy3アッセイに基づく。AGTのBG−Cy3との反応時に、Cy3の蛍光放出強度の2〜3倍の上昇が認められる。化合物N9−イソブチル−O6−ベンジルグアニン、N9−プロピル−O6−ベンジルグアニン、またはN9−ニトリル−O6−ベンジルグアニンに対する、AGTM(実施例6)対PGEG−hAGT(Juilleratら2003、突然変異N157G、S159Eを含有する)の反応性を、GST−AGT融合タンパク質10μMのサンプルを反応緩衝液(50mM HEPES、pH7.2、100mM NaCl、0.1% Tween−20、1mM DTT)中のこれらのサンプル100μMによって室温にて15分間に渡って事前にインキュベートすることによって調査した。次にBG−Cy3(20μM最終濃度)を添加して、反応物を室温にて更に1時間インキュベートした。サンプルを反応緩衝液によって1:20で希釈して、マイクロタイタープレートに移動した。Cy3蛍光物質の蛍光強度(励起:519nm、発光:572nm)をSpectra Max Geminiマイクロプレートリーダー(Molecular Devices)によって測定した。
【0094】
両方のAGT突然変異体をBG(100μM、15分の反応時間)によって効率的に予備遮断した(図3、No.3)。AGTMの5%は、BGでの予備遮断後に活性のままである(灰色棒)。N9−修飾BG誘導体に対するAGTMとPGEG−hAGTとの反応性の著しい差が見られる。AGTMは、1%〜10%の範囲でN9−修飾BG誘導体と反応する(図3、No.4、5、6、灰色棒)。AGTMとは対照的に、PGEG−hAGTは、3つすべてのN9−修飾BG誘導体と効率的に反応する。BG−Cy3による次のインキュベーション時にはバックグラウンドを超える蛍光の増加は認められず、N9−置換化合物との完全な反応を示す(図3、No.4、5、6、黒色棒)。
【0095】
〔実施例7〕
1つの生体サンプル中に存在する2つの異なるAGTの変異体と2つの異なる基質との反応
実施例6(AGTM)の突然変異型AGTを配列番号:26および配列番号:27のプライマーによってPCR増幅して、NdeIおよびBamHIを介してpET15b内へサブクローニングした。このベクターからの遺伝子発現を実施例1に記載するようにE.coli菌株BL21(DE3)にて実施し、ベクターによってコードされるN末端融合されたHis−タグを有するタンパク質を生じさせた。細胞を収穫して、実施例1に記載するように、0.5M NaCl、10mMイミダゾール、50mMホスフェート、pH8.0を含有する抽出緩衝液中に抽出した。タンパク質を含有する抽出物を事前に平衡にされたNi−NTA−セファロース(Qiagen)に加え、次にこれを、20mMイミダゾールを含有する緩衝液20ベッドボリュームで洗浄した。250mMイミダゾールを含有する緩衝液を用いてHis−タグ化タンパク質を溶出させた。実施例1に記載するように精製したタンパク質を透析し、保存して、後でキャラクタリゼーションした。
【0096】
実験では、2つのAGTタンパク質には異なるサイズの2つのタグを使用した:His−タグ化 AGTM(約20kDa)およびGST−hAGT(実施例1に記載するようにクローニングおよび調製、しかし野生型hAGT遺伝子から開始、約48kDa)を、50mMHEPES、1mMDTT、200mg/mLBSAを含有する反応緩衝液(pH7.3)でそれぞれ0.2μMおよび1.2μM最終濃度まで希釈した。サンプルを基質N9−シクロペンチル−O6−ブロモテニル−グアニン(CPTG、0、5および10μM最終濃度)およびビオチン化O6−ベンジルグアニン(BG−Bt、5μM最終濃度)の混合物で30分間インキュベートした。反応物をSDS−Laemmli緩衝液の添加によって失活させ、ウェスタンブロッティング分析に供した(実施例1)。2つのAGT変異体は異なる質量を有するため、2つのタンパク質はアクリルアミドゲルで分離して、独立して分析できる。GST−hAGTに対するCPTGの高い特異性が認められる。同じ濃度では(どちらの基質も5μM最終濃度)、GST−hAGTの95%がCPTGと反応するのに対して、His−タグ化 AGTMはわずか5%である。
【0097】
基質CPTGは、シクロペンテニルメチルカーボネートによる6−クロログアニンの位置N−9のシクロペンテニル化と、6−クロロ−N9−シクロペンチル−グアニンへの還元と、3級アミンの存在下での4−ブロモ−2−ヒドロキシメチルチオフェンとの反応によって入手できる。
【0098】
【化3】
【0099】
更なる実験において、実施例6に記載する突然変異型AGTMを配列番号:34および配列番号:36のプライマーによって増幅し、ベクターpEGFP−NucのNheI部位とBglII部位との間へクローニングした。酵母β−ガラクトシダーゼ遺伝子を配列番号:37および配列番号:38のプライマーによって増幅して、BglII/BamHIを介してこのベクターへサブクローニングして、AGTM−β−GaI融合遺伝子を有するベクターを生成させた。hAGT遺伝子を実施例1に記載する原理に基づいて、配列番号:39、配列番号:40、配列番号:34、および配列番号:35のプライマーによって増幅し、NheI/BglIIを介してpEGFP−Nuc内へクローニングして、突然変異Gly160TrpをコードするAGT−NLS3融合遺伝子を生成させた。AGT突然変異体両方の同時の一過性発現後に、細胞を5μM CPTGで10分間、次にJuilleratら、Chem Biol 10: 313-317,2003の物質45μMで20分間インキュベートした。実施例1に記載するように細胞を洗浄および撮像した。
【0100】
CHO細胞において、細胞質に局在化したAGTM−β−ガラクトシダーゼ融合タンパク質を蛍光基質物質4との反応によって、選択的に蛍光標識化した。核内の他のAGT融合タンパク質の著しい標識化は認められない。AGT(Gly160Trp)が細胞内でCPTGと効率的に反応して、それゆえその後、蛍光基質によって標識できないことが結論付けられる。実施例6(AGTM)の突然変異体は、細胞内でCPTGによる事前のインキュベーション後に蛍光基質に対して反応性のままであり、したがって第1のステップでCPTGと反応しない。
【0101】
〔実施例8〕
化合物Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、182での切断における更なる突然変異
A)エラープローンPCR:ベクターpAK100からの実施例6のAGTMの遺伝子を、配列番号:42のプライマーおよび配列番号:41のビオチン化プライマーによってTaq−ポリメラーゼを使用して、120μM(6−(2−デオキシ−β−D−エリスロ−ペント−フラノシル)−3,4−ジヒドロ−6H,8H−ピリミド[4,5−c][1,2]オキサジン−2−オン)−5’−トリホスフェートおよび480μM 2’−デオキシ−8−オキソ−グアノシン−トリホスフェート(どちらもTrilink Biotechnologiesより)でスパイクしたエラープローンPCR反応によって増幅した。生成物をストレプトアビジンコーティングビーズ(Dynabeads M−280、Dynal)上に捕捉して、次に配列番号:43および配列番号:44のプライマーを用いて標準PCRによって増幅した。
【0102】
B)飽和突然変異誘発、領域150−154:AGTM(実施例6の突然変異体)の遺伝子を配列番号:11、配列番号:45、配列番号:46および配列番号:28のプライマーを用いてPCRによって実施例1に記載するように増幅して、SfiIを介してpAK100内へサブクローニングした。プライマーは、hAGT遺伝子のコドン150−154に相当する位置にヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)を含有する。
【0103】
C)飽和突然変異誘発、領域31−35:AGTM(実施例6の突然変異体)をコードする遺伝子を配列番号:11、配列番号:48、配列番号:47および配列番号:28のプライマーを用いてPCRによって実施例1に記載するように増幅して、SfiIを介してpAK100内へサブクローニングした。プライマー47は、hAGT遺伝子のコドン31−35に相当する位置にヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)を含有し プライマー48は、アンチセンスストランドのための混合物MNN(M=CまたはA)を含有する。
【0104】
増幅生成物を繊維状ファージのg3遺伝子に融合させて、SfiI制限部位を介してベクターpAK100へクローニングした。実施例4に記載したように、生じた遺伝子ライブラリをファージ提示に使用した。ライブラリの突然変異体をBG−Bt 5〜50nMによって10分間に渡って選択した。選択された突然変異体を配列番号:3および配列番号:13のプライマーを使用してpGEX内へサブクローニングした。突然変異型AGTの反応速度を実施例6、DNAベース基質に対する反応性(BG−Cy3アッセイ)に記載するように決定した。
【0105】
突然変異の組合せ:BG誘導体との反応速度を向上するために見出された選択からの突然変異を、鋳型として1つの突然変異型遺伝子を使用して、そして突然変異の点にてミスマッチであるプライマー配列での更なる突然変異を含めて、配列番号:3および配列番号:13のプライマーにより実施例1に記載するようにPCRによって組合せた。突然変異を重複し60℃以下でハイブリダイズするのに十分な長さを有する2つの各変異原性プライマーを選択して、相補性ストランドの一方へそれぞれアニーリングさせた。アニーリング温度は、各G/C対では4℃および各A/T対では2℃の寄与を想定して計算した。生じた遺伝子を実施例1に記載するように、pGEX内へサブクローニングした。実施例1、4および6に記載するように、突然変異型AGTタンパク質を調製およびキャラクタリゼーションした。
【0106】
これらの選択からの組合せ突然変異を有するAGT突然変異体は、実施例1〜6の突然変異体について記載した好都合な特性を示す。突然変異体は、実施例1〜5に記載する突然変異体に匹敵する発現速度を有し、野生型AGTと比較したときに試験管内で少なくとも4倍の溶解度の上昇を有する。それらは少なくとも10分の1へのN9−置換O6−アルキルグアニン誘導体との反応性の低下と、少なくとも100分の1へのDNAまたはDNA基質との相互作用および反応性それぞれの低下を示す。E.coliのペリプラズムにおける発現からの活性AGT突然変異体の収率は、PGEG−hAGTの収率よりも少なくとも5倍の高さである。CHO細胞において、蛍光標識化変異AGTは細胞質全体に分布しており、優先的な核局在化は検出できない。その上、これらのAGT突然変異体は野生型hAGTと比較したときに、O6−ベンジルグアニン基質に対する反応性のかなりの上昇を示す(表1を参照)。
【0107】
【表1】
【0108】
BG−Cy3活性アッセイによって反応速度を決定して、野生型hAGTについてこのアッセイで得られた値(kobs=116M−1cm−1)で割った。活性アッセイでは、精製したGST融合タンパク質を、「p.」のマークが付いた(実施例1に記載するように調製した)突然変異体について使用した;ペリプラズム抽出物を、「x.」のマークが付いた(実施例2に記載するように調製した)突然変異体について使用した。
【図面の簡単な説明】
【0109】
【図1】表示濃度でのBG修飾オリゴヌクレオチド(配列番号:2)の存在下でのAGTM(実施例6、Aの突然変異体)またはPGEG−hAGT(B)のBG−Cy3との反応。Spectra Max Geminiプレートリーダー(Molecular Devices)で検出した任意単位[a.u.]の蛍光FL(励起519nm、572nmにて放出)は、時間(t)(分[min])にプロットされている。
【図2】実施例7に記載するような、野生型hAGT(GST−hAGT融合タンパク質)およびAGTMと、2つの基質N9−シクロペンチル−O6−ブロモテニル−グアニン(CPTG)およびビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら2003の物質3a)との反応のウェスタンブロット。BG−Btは、すべてのサンプル中に5μMで存在する。BG−Btと反応したAGTを検出する、ニュートラアビジン−ペルオキシダーゼコンジュゲートおよび発光ペルオキシダーゼ基質を用いた検出。同じ濃度のCPTGおよびBG−Btを用いた同様のインキュベーションの下でさえ、GST−hAGTのわずか5%が基質BG−Btと反応する。
【図3】N9−修飾BG誘導体とのAGT突然変異体の反応性。10μM AGTM(灰色バー)またはPGEG−hAGT(黒色バー)を100μM N9−修飾BG誘導体(以下のリストを参照)で予備遮断して、次に20μM BG−Cy3で標識した。1時間の反応時間の後、595nmにて任意単位[a.u.]で示した蛍光強度(FI)を測定する。黒色実線は、タンパク質に結合していない蛍光物質のバックグラウンド蛍光に相当する条件1の測定値に設定されている。この線より上の値は、N9−修飾BG誘導体との低速または不完全な反応を示す。1 タンパク質を含まないBG−Cy3の蛍光、バックグラウンド蛍光。2 BG−Cy3との反応;予備遮断なし、正の対照。3 BG−Cy3との反応、100μM BGによって予備遮断。4 BG−Cy3との反応、100μM N9−イソブチル−O6−ベンジルグアニンによって予備遮断。5 BG−Cy3との反応、100μM N9−プロピル−O6−ベンジルグアニンによって予備遮断。6 BG−Cy3との反応、100μM N9−シアノメチル−O6−ベンジルグアニンによって予備遮断。
【技術分野】
【0001】
本発明は、野生型ヒトO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(hAGT)の突然変異体、および標識を基質からこれらのO6−アルキルグアニン−DNAアルキルトランスフェラーゼ突然変異体および対象となるタンパク質からなる融合タンパク質へ転移させる方法に関するものである。
【背景技術】
【0002】
N−メチル−N−ニトロソ尿素などの求電子物質の変異原性および発癌性作用は、主に、DNAにおけるグアニンのO6−アルキル化による。哺乳類および細菌は、DNAアルキル化からそれら自体を保護するために、これらの損傷を修復するタンパク質であるO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を所有する。AGTは、アルキル基をアルキル化グアニンおよびグアニン誘導体の位置O−6から、自身のシステインの1つのメルカプト基に転移させて、非可逆的にアルキル化されたAGTを生じさせる。基礎を成すメカニズムは、SN2タイプの求核反応であり、これはメチル基だけでなく、ベンジル基も容易に転用される理由を説明する。腫瘍細胞におけるヒトAGT(hAGT、配列番号:1)の過剰発現は、アルキル化薬、例えばプロカルバジン、ダカルバジン、テモゾロミド、およびビス−2−クロロエチル−N−ニトロソ尿素に対する耐性の主な理由であるため、AGTの阻害剤は、化学療法での増感剤として使用するために提案されてきた(Peggら, Prog Nucleic Acid Res Mol Biol 51: 167-223,1995)。米国特許第5,691,307号は、ベンジル基内に各種の置換基を有するO6−ベンジルグアニンおよび腫瘍細胞でのAGTレベルを枯渇させ、それによりアルキル化抗腫瘍薬に対する反応性を上昇させるためのその使用について記載している。同様に国際公開公報第97/20843号は、O6−ベンジル−およびO6−ヘテロアリールメチル−ピリミジン誘導体に相当するAGT枯渇剤を更に開示している。
【0003】
ドイツ特許第199 03 895号は、AGTのビオチン化を引き起こす、ビオチン化O6−アルキルグアニン誘導体とヒトAGTとの間の反応に依存するAGTのレベルを測定するアッセイを開示している。これは次に、ストレプトアビジンコーティングプレート上のAGTの分離および例えばELISAアッセイでのその検出を可能にする。腫瘍細胞中のAGTのレベルを監視するための、そしてAGT阻害剤のスクリーニングで使用するためのアッセイが提案されている。
【0004】
Damoiseauxら、Chem Biochem 4: 285-287,2001は、研究および化学療法での支援のために癌細胞におけるこの酵素のレベルの検出を再び容易なものとするために、ヒトAGTを標識する化学プローブとして使用するための、オリゴデオキシリボヌクレオチドに組み込まれた修飾O6−アルキル化グアニン誘導体を開示している。
【0005】
国際公開公報第02/083937号は、対象となるタンパク質を検出および/または操作する方法を開示し、該方法ではタンパク質を、AGTに融合し、標識を担持するAGT基質とAGT融合タンパク質を接触し、標識を使用してAGT融合タンパク質を検出し、場合により更に操作する。使用される複数のAGT融合タンパク質、AGT基質および広範に渡る標識の一般的構造原理、ならびに該方法で有用な標識を検出する方法が記載されている。AGTの他の形に触れられているが、ヒトAGTのみが例示されている。
【0006】
PCT/EP03/10859(国際公開公報第2004/031404号)は、対象となるタンパク質を検出および/または操作するための言及された方法で使用される特定のAGT融合タンパク質と、この方法によって得られる標識融合タンパク質と、特定のAGT融合タンパク質を使用する方法について記載している。
【0007】
PCT/EP03/10889(国際公開公報第2004/031405号)は、対象となるタンパク質を検出および/または操作するための言及した方法で特に適切な標識を担持する追加のAGT基質と、このような特に標識された基質の応用について開示している。この特許出願は、この追加のAGT基質の製造方法についても記載している。
【0008】
ヒトAGT突然変異型Gly160Trp(Xu-Welliverら、Biochemical Pharmacology 58: 1279-1285,1999)は、ベンジルグアニン誘導体に対して、野生型ヒトAGTよりも多少反応性である。Juilleratら, Chem Biol 10: 313-317,2003は、AGT融合タンパク質の合成基質による生体内(細胞内)標識に有効な更に反応性のパートナーを求めて、ヒトAGTの多数の突然変異体を作成した。位置140、157、159、および160における突然変異体が報告された。突然変異型Asn157GlySer159Gluは、野生型AGTと比較しておおよそ20倍の、ベンジルグアニン誘導体に対する活性の上昇を示している。
【0009】
hAGTにおける以下の追加の突然変異体は、hAGTのDNA結合を妨害するが、ベンジルグアニン誘導体に対する活性に著しく干渉しないことが示されている:Lys125Ala、Ala127ThrおよびArg128A1a。Limら、EMBO J 15:4050−4060,1996,およびDanielsら,EMBO J 19:1719−1730,2000を参照。
【0010】
ヒトAGT(pdb−ID 1EH6、Danielsら、EMBO J 19:1719,2000)の結晶構造は、Asn207の後で切断された機能性タンパク質を使用して解明された。
【0011】
発明の概要
本発明は、野生型ヒトAGTと比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の利点を示す、AGT突然変異体に関する。
【0012】
本発明のAGT突然変異体は、野生型ヒトAGTの1〜25個のアミノ酸が他のアミノ酸によって置換され、場合により連続鎖からの1〜5個のアミノ酸が1、2、または3つの位置で欠損または付加され、および/またはN末端の1〜4個のアミノ酸またはC末端の1〜40個のアミノ酸が欠損している、上記の向上した特性を備えた突然変異体である。
【0013】
本発明は更に、本発明のAGT突然変異体を有する融合タンパク質に組み込まれている対象となるタンパク質を検出および/または操作する方法に関する。本発明の別の目的は、このようなAGT突然変異体および対象となるタンパク質を含むAGT融合タンパク質である。
【0014】
発明の詳細な説明
対象となるタンパク質を、AGT融合タンパク質に組み込み、AGT融合タンパク質を、標識を担持する特定のAGT基質と接触させ、AGT融合タンパク質を、標識を認識および/処理するために設計された系において、標識を使用して検出し、場合により更に操作する、対象となるタンパク質を検出および/または操作するための前に記載の方法において、AGTの性能を、野生型ヒトAGTを突然変異型AGTと置換することによって更に向上できる。突然変異型AGTを包含するこのような向上方法は、本発明の目的である。別の目的は上記方法に特に適したAGT突然変異体であり、AGT融合タンパク質は、このようなAGT突然変異体および少なくとも1種の対象となるタンパク質を含む1種以上の他のタンパク質を含む。対象となるタンパク質は、いかなるタンパク質であってもよい。
【0015】
本発明のAGT突然変異体は、例えば、野生型ヒトAGTまたは既知のAGT突然変異体と比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の特性を示す突然変異体を含む。
【0016】
(a)DNA相互作用の低下
「DNA相互作用の低下」のある本発明の突然変異型AGTは、野生型ヒトAGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、20%未満のDNA結合、好ましくは2%未満のDNA結合を示し、最も好ましくは検出可能なDNA結合を示さない。DNAとの相互作用は例えば、低塩およびDNase非存在の条件下でE.coli抽出物から同時精製されたDNAの量を評価することによって定量される。これは野生型(ヒト)および突然変異型AGTの融合タンパク質(例えばGSTへの融合)の並行精製の間で分光法によって比較される(260および280nmでの吸収率の比)。あるいはAGTのDNAとの相互作用は、DNAの存在下での、基質としてのBG−Cy3(Cy3に結合されたO6−(4−アミノエチル−ベンジル)−グアニン)とのAGT反応性の阻害として測定される。この手法は、BG−Cy3のAGTとの反応時におけるCy3蛍光物質の蛍光の著しい増加を利用する。AGTのCy3−標識ベンジルグアニン誘導体との反応動力学には、サケ精子DNAの各種の濃度での数時間に渡る蛍光強度測定が続く。野生型hAGTは、DNAにテトラマーとして結合するのに対して、未結合hAGTタンパク質はモノマーのままであることが示された(Rasimasら、J Biol Chem 278(10): 7973-80,2003)。DNAに結合できないAGT突然変異体もテトラマー化しない。
【0017】
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化
「もはや核に限定されない局在化」を伴う本発明の突然変異型AGTは、真核細胞、例えば哺乳類細胞での発現時に細胞での突然変異型AGTの実質的に均一な局在化を示す。AGT突然変異体の細胞内局在化は、AGT欠損HeLa細胞またはCHO細胞を構成的発現のための構築物をヒトサイトメガロウィルス前初期プロモータの下で用いて一過性に形質移入することによって調査する。細胞を細胞膜透過性ジアセチルフルオレセイン修飾O6−ベンジルグアニンによって染色して(Juilleratら、Chem Biol 10: 313-317,2003の物質4)、共焦点レーザ顕微鏡法によって解析する。細胞質および核の蛍光強度を突然変異型AGTと野生型AGTで比較する。野生型ヒトAGTは、核での優先的局在化と、細胞質でのもっぱらわずかな局在化を示す。
【0018】
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
「溶解性タンパク質(すなわち細胞溶解後に溶解性画分に見出され、封入体には見出されないタンパク質)としての発現収率の向上」を伴う本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、溶解タンパク質として3倍を超える発現収率、好ましくは5倍を超える発現収率、最も好ましくは10倍を超える発現収率を示す。この発現収率の向上は、同時に、発現に使用される「宿主での安定性」の尺度となる。発現収率は、E.coliまたは遺伝子組換えタンパク質用の他のいずれかの標準的生産細胞、例えば酵母、あるいは好ましくは昆虫細胞、CHO細胞もしくはHeLa細胞において測定する。発現収率を定量するために、融合パートナーが容易な精製および定量を可能にするように、AGT融合タンパク質を選択できる。例えばE.coliでの発現収率は、E.coliの並行させた発現培養からの溶解性および非溶解性GST野生型AGT融合タンパク質ならびにGST突然変異型AGT融合タンパク質の収率を測定および比較することによって決定する。細胞溶解後の溶解性画分および非溶解性画分(すなわち封入体)のサンプルをSDS−PAGEにかけて、相当するAGT融合タンパク質のバンド染色強度を比較する。溶解性タンパク質は、アフィニティクロマトグラフィーによる細胞抽出物からの融合タンパク質(例えばグルタチオンセファロースによるGST−AGT融合タンパク質の場合)の精製後、精製した画分にタンパク質濃度のアッセイを行うことによって定量する(Bradford,Anal Biochem 72: 248-54,1976)。溶解性画分中の同様のタンパク質の発現収率(点突然変異での相違)は、精製状態と同様に、未精製状態でのタンパク質安定性の尺度として使用できる(Ohageら、J Mol Biol 291: 1119-1128,1999、Wirzら、Protein Sci 8: 2245-50,1999)。したがって、これは折畳み安定性および凝集傾向の尺度として見なされる。
【0019】
(c’)実用的な見地から、E.coliにおける溶解性タンパク質としての発現収率の向上が特に重要である。
【0020】
(d)酸化条件下での安定性の向上
「酸化条件下での安定性の向上」を伴う本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、2倍を超える活性タンパク質の収率、好ましくは5倍を超える活性タンパク質の収率、最も好ましくは10倍を超える活性タンパク質の収率を示し、すなわちAGTタンパク質は、緩衝水溶液(例えば100mM NaCl、10mM HEPES、pH7.4、ジチオスレイトールまたはベータ−メルカプトエタノール無添加)中、酸化条件下で1時間以上のインキュベーション時間の後に、AGT基質に対してその活性を維持する。AGT基質に対する活性は、ジチオスレイトールまたはベータ−メルカプトエタノールなどの還元剤を添加せずに精製後に測定する。あるいは突然変異型AGTおよび野生型ヒトの活性を、適切なシグナル配列へのそれらの融合による酸化性酸化還元電位を有する細胞コンパートメント(例えばE.coliのペリプラズム)内へのエクスポート後に比較する。還元および酸化条件下でのAGT基質に対する活性は、ジチオスレイトールまたはベータ−メルカプトエタノールなどの還元剤の存在下または非存在下で反応を実施することによって比較する。
【0021】
(e)基質との反応後の細胞内での安定性の向上
「細胞内での向上した安定性」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、細胞(例えば哺乳類細胞)内での細胞透過性基質との反応後に、2倍を超える安定性、好ましくは3倍を超える安定性、最も好ましくは6倍を超える安定性を示す。安定性は、突然変異型AGT融合タンパク質について、AGT基質との反応後に、AGT融合タンパク質の強度および局在化を、共焦点レーザ走査顕微鏡法を用いて解析することによって決定する。
【0022】
(f)基質との反応前後の細胞外部での安定性の向上
「細胞外部での向上した安定性」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、2倍を超える安定性、好ましくは4倍を超える安定性、最も好ましくは6倍を超える安定性を示す。反応前の安定性は、突然変異型AGTまたは突然変異型AGT融合タンパク質について、精製サンプルを緩衝水溶液中で4℃にて2週間まで、−20℃にて6ヶ月までインキュベートすることによって決定する。複数の時点にてアリコートを取り、Juilleratら、Chem Biol 10: 313-317,2003によって記載されているように反応性AGTの濃度を概算する。標識基質との反応後の、そして未反応基質からの続いての分離後の野生型AGTおよびAGT突然変異体の安定性は、4℃にて2週間および−20℃にて3ヶ月に渡って溶解性画分中の標識の濃度を定量することによって決定される。
【0023】
(f’)実際的な観点から、標識基質との反応後の安定性は特に重要である。
【0024】
(g)試験管内溶解度の向上
「試験管内溶解度の向上」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、2倍を超える溶解度、好ましくは5倍を超える溶解度、最も好ましくは10倍を超える溶解度を示す。野生型AGTおよびAGT突然変異体の試験管内溶解度は、適切な緩衝液(例えば100mM NaCl、20mM Tris、pH8.0、20%グリセロール、1mM DTT)にて確立された1つ以上の濃度での4℃または最高37℃での精製サンプルの一晩に渡るインキュベーション後に残存するタンパク質の量を決定することによって測定する。
【0025】
(h)O6−アルキルグアニン基質に対する反応性の向上
「向上した反応性」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、3倍を超える反応性、好ましくは5倍を超える反応性、最も好ましくは10倍を超える反応性を示す。O6−ベンジルグアニン基質に対する活性は、Juilleratら、Chem Biol 10: 313-317,2003によって記載されているように測定する。
【0026】
(i)DNAベース基質に対する反応性の低下
「DNAベース基質に対する反応性の低下」を備えた本発明の突然変異型AGTは、野生型ヒトAGTと比較したときに、DNAベース基質に対する10%未満の反応性、好ましくはDNAベース基質に対する1%未満の反応性を示し、最も好ましくはDNAベース基質に対する検出可能な反応性を示さない。野生型AGTまたは突然変異型AGTがアルキル化DNA基質と反応する能力は、O6−ベンジルグアニンを含有する合成オリゴヌクレオチド(配列番号:2、位置14にて修飾)によるその不活性化の反応として測定する。次に反応をビオチン化O6−アルキルグアニンによるインキュベーションにより失活させる。サンプルにウェスタンブロッティングおよびストレプトアビジン誘導体により検出させて、これらの基質の速度定数を得る。あるいは野生型または突然変異型AGTがアルキル化DNA基質と反応する能力をAGT蛍光基質と競合するその標識効率によって測定する。BG−Cy3(Cy3に結合したO6−ベンジルグアニン)によるAGTの反応速度論の後には、O6−ベンジルグアニンを含有する競合する合成オリゴヌクレオチド(配列番号:2、位置14にて修飾)の各種濃度での蛍光強度測定が続く。
【0027】
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下
「N9−置換O6−アルキルグアニン基質に対する反応性の低下」を備えた本発明の突然変異型AGTは、野生型AGTまたは「PGEG−hAGT」(Juilleratら、Chem Biol 10: 313-317,2003)などの既知のAGT突然変異体と比較したときに、N9−置換O6−アルキルグアニン基質に対して10%未満の反応性、好ましくは2%未満の反応性を示し、最も好ましくは検出可能な反応性を示さない。このようなN9−置換O6−アルキルグアニン基質は、野生型AGTの天然基質である。野生型AGTまたは突然変異型AGTがN9−修飾O6−アルキルグアニン基質と反応する能力は、競合実験でのビオチン化N9−非置換O6−アルキルグアニン基質の存在下でのこのような基質との、例えば低分子量N9−シクロペンチル−O6−ベンジルグアニンとのその反応の速度として測定する。次にサンプルにウェスタンブロッティングおよびストレプトアビジン誘導体による検出を受けさせて、これらの基質の速度定数を得る。あるいはこのような基質に対する反応性を、AGTとBG−Cy3との反応の阻害として測定する。AGTのBG−Cy3との反応の後に、プレインキュベーション後の、またはN9−修飾O6−アルキルグアニン基質との直接競合におけるAGTへの結合時のCy3蛍光強度の上昇が続く。
【0028】
本発明の好ましいAGT突然変異体は、共通する複数の好ましい特性を有する突然変異体、例えば
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f)基質との反応前後の細胞外部での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
を示す突然変異体である。
【0029】
更に好ましいAGT突然変異体は、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での5倍を超える安定性、
(f)基質との反応前の、細胞外での4倍を超える安定性、特に基質との反応後の(f’)、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
を示す突然変異体である。
【0030】
最も好ましいAGT突然変異体は
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、特にE.coliにおける(c’)、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前の、細胞外での6倍を超える安定性、特に基質との反応後の(f’)、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
を示す突然変異体である。
【0031】
当技術水準のAGT突然変異体は、多くの所望の特性の一部のみを有する。例えばLimら、EMBO J 15: 4050-4060,1996によって記載されている突然変異体は、哺乳類細胞内での発現時に、DNA結合の減少および細胞質内への局在化を示す。Juilleratら、Chem Biol 10: 313-317,2003で記載されている突然変異体は、O6−ベンジルグアニン誘導体に対する反応性の上昇を示す。
【0032】
本発明のAGT突然変異体は、野生型ヒトAGTと比較して上述の向上した特性を備えた突然変異体であり、そこでは野生型ヒトAGTの1〜25個、好ましくは6〜22個、特に特に7、8、9、10、11、12、13、14、15、16、17、18、または19個のアミノ酸が、他のアミノ酸によって置換され、場合により連続鎖中の1〜5個のアミノ酸が、1、2、または3つの位置にて欠損もしくは付加され、および/またはN末端にて1〜4個のアミノ酸が、またはC末端にて1〜40個の、好ましくは20〜35個の、特に25〜30個のアミノ酸が欠損している。
【0033】
好ましくは、以下の位置のアミノ酸が置換される:
(A)E.coliにおける発現収率を上昇させ、タンパク質に酸化をより受けにくくする、AlaまたはValによって、好ましくはAlaによって置換されたCys62。
(B)他のアミノ酸置換、好ましくはCys150−Ser151−Ser152の置換、特にCys150の置換と組合された、E.coliにおける発現を上昇させて、野生型ヒトAGTに匹敵する基質反応性を維持する、Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Ser、好ましくはSer−Hisによって置換されたGln115−Gln116。
(C)他のアミノ酸置換、好ましくはGln115−Gln116/Cys150−Ser151−Ser152の置換またはGly131−Gly132/Met134−Arg135の置換、特にCys62/Gln115−Gln116/Gly131−Gly132/Met134−Arg135/Cys150−Ser151−Ser152の置換および182の後での切断と組合された、E.coliにおける発現収率を上昇させ、DNA結合を減少させて、哺乳類細胞での核局在化を無効にする(Limら、EMBO J 15: 4050-4060,1996)、Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128。
(D)E.coliのペリプラズムおよび細胞質における発現収率を上昇させ、DNA結合を減少させて、オリゴヌクレオチド(O6−アルキル−N9−デオキシリボシルグアニンを含有する)、O6−アルキル−N9−デオキシリボシルグアニンおよびN9−シクロペンチル−O6−ベンジルグアニンまたは他のN9置換O6−ベンジルグアニンとの反応性を無効にするのに対して、N9位置で置換されていないO6−アルキルグアニン基質に対する反応性を上昇させる、Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Val、好ましくはLys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134。
(E)野生型hAGTと比較してE.coliのペリプラズムにおいてより効率的な発現を可能にして、O6−アルキルグアニン基質に対する反応性を維持し、タンパク質に酸化をより受けにくくして、DNA結合を減少させる、Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−、好ましくはAsn−Ile−Asnによって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、またはSerもしくはThrによって置換されたCys150。
(F)N9位置で置換されていないO6−アルキルグアニン基質に対する反応速度を上昇させる(Juilleratら、Chem Biol 10: 313-317,2003)のに対して、E.coliのペリプラズムおよび細胞質における発現収率を上昇させ、DNA結合を減少させて、オリゴヌクレオチド(O6−アルキル−N9−デオキシリボシルグアニンを含有する)、O6−アルキル−N9−デオキシリボシルグアニンおよびN9−シクロペンチル−O6−ベンジルグアニンとの反応性を無効にする、他のアミノ酸置換、好ましくはGln115−Gln116/Cys150−Ser151−Ser152の置換またはGly131−Gly132/Met134−Arg135の置換、特にCys62/Gln115−Gln116/Lys125/Ala127−Arg128/Gly131−Gly132/Met134−Arg135/Cys150−Ser151−Ser152の置換および182の後での切断と組合された、Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trp、最も好ましくはGly−Glu−Alaによって置換されたAsn157/Ser159−Gly160、あるいはGly/Glu(特に好ましい)によって置換されたAsn157/Ser159、GlyまたはArgによって置換されたAsn157。
(G)発現収率をやや上昇させる、Gly182の後の切断(アミノ酸183−207の欠損)。
【0034】
本発明の突然変異体は、特に、修飾(A)、(B)、(D)、(E)、(F)および(G)のうち2つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体、および修飾(A)〜(G)のうちの3つ以上、例えば3または4つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体である。他の好ましい突然変異体は、修飾(A)、(B)、(C)、(E)、(F)、および(G)のうち3つ以上、例えば3または4つが、そして場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾が存在する突然変異体である。
【0035】
特に好ましいのは、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞を通じての細胞質全体への分布、DNA結合の減少、およびO6−ベンジルグアニン基質に対する反応性の上昇を示す、突然変異型Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu、Gly182の後の切断、
E.coliにおける発現収率の上昇、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇、およびO6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性の実質的な低下を示す、突然変異型Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu、
E.coliにおける発現収率の実質的な上昇を示すが、O6−ベンジルグアニジン基質に対する活性を維持する、突然変異型Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞における細胞質全体への分布、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇、およびO6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する少なくとも100分の1に減少した反応性を示す、突然変異型Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断、および、
E.coliにおける発現収率の上昇、酸化に対する感受性の低下、CHO細胞における細胞質全体への分布、少なくとも1000分の1に減少したDNA結合、O6−ベンジルグアニン基質に対する反応性の上昇を示すが、O6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性を維持する、突然変異型Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断
である。
【0036】
検討される更なる突然変異体は、修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断、および場合により1〜15個の、好ましくは1〜10個の、特に3〜7個の追加のアミノ酸修飾、例えば
Glu115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、AGT突然変異体である。
【0037】
同様に検討される突然変異体は、修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断、および場合により1〜10個の、特に3〜7個の追加のアミノ酸修飾、例えば
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Asn150Asp、Ile151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Asn150Val、Ile151Ser、Asn152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Asn150Gly、Ile151Gly、Asn152Asp、Ala154Asp;
Asn150Val、Ala154Asp;
Asn150Glu、Ile151Gly、Asn152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ile151Gly;
Asn150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、AGT突然変異体である。
【0038】
同様に好ましいのは、修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異体;および修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異体である。
【0039】
特に好ましいのは、実施例の、特に実施例2〜7および表1の化合物である。
【0040】
突然変異体は、当業者に公知の技法によって得られる。AGT変異体は、好ましくは、新しいO6−アルキルグアニン−DNAアルキルトランスフェラーゼを産生および選択するために、当業者に既知のタンパク質工学技法を使用して、および/または分子進化を使用して生成できる。このような技法は、例えば、部位特異的変異誘発、飽和変異誘発、変異を配列の任意の箇所に導入するためのエラープローン(error prone)PCR、および飽和変異誘発の後に使用されるDNAシャッフリングである。ファージ提示法を活用すると、突然変異体は、本発明のO6−ベンジルグアニンおよびAGT基質に対する活性の著しい向上ならびに酸化条件下での安定性の向上と共に見出される。AGTは、繊維状ファージ上にファージカプシドタンパク質pIIIを有する融合タンパク質として機能的に提示することが可能であり、AGTの異常なメカニズムを使用して、向上した特性、反応速度の上昇を備えたAGTを提示するファージを選択できる、例えばJuilleratら, Chem Biol 10: 313-317,2003を参照。
【0041】
本発明において、対象となるタンパク質またはペプチドは、上記のAGT突然変異体に融合される。対象となるタンパク質またはペプチドは、いかなる長さでもよく、二次、三次、または四次構造を有していても、有していなくてもよく、好ましくは少なくとも12個のアミノ酸および最大2000個のアミノ酸からなる。このような対象となるタンパク質またはペプチドの例を以下に与えるが、例えば酵素、DNA結合タンパク質、転写調節タンパク質、膜タンパク質、核レセプタタンパク質、核局在化シグナルタンパク質、タンパク質補助因子、小型モノマー性GTPase、ATP結合カセットタンパク質、細胞内構造タンパク質、タンパク質を特定の細胞コンパートメントに標的化させることに関与する配列を備えたタンパク質、ラベルまたは親和性タグとして一般に使用されるタンパク質および上述のタンパク質のドメインまたはサブドメインである。対象となるタンパク質またはペプチドは好ましくはAGT突然変異体に、酵素によって開裂できるリンカーによって、例えばDNA段階にて適切な制限酵素、例えばBglIIによって開裂できるAGATCTによって、および/またはタンパク質段階にて適切な酵素、例えばタバコエッチウィルスNla(TEV)プロテアーゼによって開裂可能なリンカーによって融合される。融合タンパク質は、原核宿主、好ましくはE.coliにて、または真核宿主、例えば酵母、昆虫、または哺乳類細胞にて発現できる。
【0042】
AGT突然変異体は、以下に記載する適切な基質上に存在する標識を融合タンパク質のAGT部分のシステイン残基の1つに標識を転移する特性を有する。
【0043】
対象となるタンパク質およびAGT突然変異体を含む融合タンパク質を、標識を有する特定の基質に以下に記載するように接触させる。AGT突然変異体が基質と反応して、基質の標識を転移するような反応条件を選択する。通常の条件は、室温、例えば約25℃で約pH7の緩衝溶液である。しかしながら、AGT突然変異体は、多様な他の条件の下でも反応し、本明細書で挙げるこれらの条件は、本発明の範囲を制限しないことが理解される。
【0044】
基質の標識部分は、融合タンパク質が対象とする用途に応じて当業者が選択できる。AGT突然変異体を含む融合タンパク質を基質に接触させた後に、標識は融合タンパク質に共有結合する。次に標識AGT突然変異型融合タンパク質は、転移された標識によって更に操作および/または検出される。標識は、複数の同じまたは異なる標識からなる場合がある。基質が1つを超える標識を含有する場合、対応する標識AGT突然変異型融合タンパク質も、標識融合タンパク質を更に操作および/または検出するための更なる選択肢を与える、1つを超える標識を含むことになる。
【0045】
「操作」の下では、いずれの物理または化学処理も理解される。例えば操作は、細胞からの単離、標準精製技法、例えばクロマトグラフィーを用いた精製、化学試薬との、または特に結合パートナーが固相に固定されている場合の結合対の結合パートナーとの反応などを意味し得る。このような操作は、標識Lに依存することがあり、標識融合タンパク質の「検出」に加えて行うことができる。標識融合タンパク質に操作および検出の両方を行う場合、検出は、操作の前または後でもよく、あるいは本明細書で定義されたような操作の間に実施できる。
【0046】
特定のAGT基質は、特許出願PCT/EP03/10889(国際公開公報第2004/031405号)に開示されているようなAGT基質、例えば式(I)
【0047】
【化1】
【0048】
〔式中、
R1−R2は、AGTによって基質として認識される基であり;
Xは、酸素または硫黄であり;
R3は、芳香族またはヘテロ芳香族基、あるいは場合によりCH2に結合された二重結合を有する、置換不飽和アルキル、シクロアルキル、またはヘテロシクリル基であり;
R4は、リンカーであり;そして
Lは、標識、複数の同じまたは異なる標識、R4をR1に結合して環式基質を生成する結合、または更なる基−R3−CH2−X−R1−R2である。〕
の化合物であり、
【0049】
基R1−R2において、残基R1は、好ましくは、AGTによって基質として認識される、窒素原子1〜5個を含有するヘテロ芳香族基、好ましくは式(II)
【0050】
【化2】
【0051】
〔式中、
R2は、水素、炭素原子1〜10個のアルキル、または糖部分であり;
R5は、水素、ハロゲン、例えばクロロまたはブロモ、トリフルオロメチル、またはヒドロキシであり;そして
R6は、水素、ヒドロキシ、または非置換もしくは置換のアミノである。〕
のプリンラジカルであり、
【0052】
R5またはR6がヒドロキシである場合、プリンラジカルは主にその互変異性形で存在し、互変異性形ではR5またはR6を保持する炭素原子に隣接する窒素が水素原子を有し、この窒素原子とR5またはR6を保持する炭素原子との間の二重結合が単結合であり、そしてR5またはR6がそれぞれ二重結合された酸素である。
【0053】
R6が非置換であるか、または置換のアミノであり、そしてプリンラジカルの結合に接続された残基Xが酸素である場合、式(II)の残基は、グアニン誘導体である。
【0054】
本発明の方法で有用な他の特定の基質は、R1が、式(II)のプリンラジカルであり、R2が、シクロアルキル、例えばシクロペンチル、あるいはアルキル、例えばプロピルまたはイソブチル、あるいは置換アルキル、例えばシアノメチルである、式(I)の化合物である。
【0055】
本発明は、生体内(細胞内)および試験管内の両方での標識AGT突然変異型融合タンパク質のための方法を提供する。AGT突然変異型融合タンパク質の生体内標識化という用語は、細胞のコンパートメントすべてはもちろんのこと、細胞外スペースを指示するAGT突然変異型融合タンパク質のコンパートメントすべてでの標識化を含む。AGT突然変異型融合タンパク質の標識化が、生体内で実施され、そしてAGT突然変異体に融合したタンパク質が膜タンパク質、より詳細には原形質膜タンパク質である場合、融合タンパク質のAGT部分は膜のどちら側にも結合することが可能であり、例えば原形質膜の細胞質側または細胞外側に結合することが可能である。
【0056】
標識化を試験管内で実施する場合、融合タンパク質の標識化は、AGT突然変異型融合タンパク質の細胞抽出物中で、あるいはその精製または濃縮形を用いてのどちらかで実施できる。
【0057】
標識化を生体内(細胞内)で、または細胞抽出物中で実施する場合、宿主の内因性AGTの標識化を好都合に考慮できる。宿主の内因性AGTがO6−アルキルグアニン誘導体または関連化合物を基質として受入れない場合、融合タンパク質の標識化は、特異的である。哺乳類細胞において、例えばヒト、マウス、またはラット細胞において、内因性AGTの標識化は可能である。これらの実験において、内因性AGTはもちろんのこと、AGT突然変異型融合タンパク質の同時標識化が問題を引き起こす、このような実験においては、既知のAGT欠損細胞株を使用できる。
【0058】
特定の基質に対して非反応性であり、次に内因性AGTによって認識される突然変異型AGT融合タンパク質を使用する場合、内因性AGTの活性を遮断するために、特定の突然変異型AGT融合タンパク質と反応するように設計された基質を用いて細胞をインキュベートする前またはその間に、このような非反応性基質を使用できる。例えばN9−置換O6−アルキルグアニン誘導体と、例えばN9−シクロペンチル−O6−ベンジルグアニンと反応しない突然変異型AGTを使用できる。内因性AGTを含有する哺乳類細胞において、この野生型AGTは、次に、標識化の前に、または突然変異型AGT融合タンパク質をこの突然変異型AGTによって認識された別の基質によって標識化する間に、N9−シクロペンチル−O6−ベンジルグアニンによって遮断できる。
【0059】
有効濃度の内因性野生型AGTが特定の生体サンプル中に存在しない場合、実験用に選択された突然変異型AGTが反応性でない基質を使用して、内因性AGTを事前に不活性化する必要はないであろう。このような条件下では、野生型AGTの特定の基質と反応しない突然変異型AGT融合タンパク質を利用することにより、AGTの2つの異なる突然変異体(あるいはAGTの1つの突然変異体および野生型AGT)を2つの異なる基質によって選択的に標識化できる。このことは、野生型hAGTタンパク質と比較して低分子量の基質に対して選択性を示す、ここでは「AGT−A」と呼ぶ突然変異型AGT、および特定の基質と野生型hAGTの10分の1、好ましくは100分の1の速さで反応する、「AGT−B」と呼ぶ別の突然変異型AGTを使用することによって実施される。突然変異型「AGT−A」(または野生型AGT)および突然変異型「AGT−B」の両方を生体サンプルに存在させて、その生体サンプル−例えば細胞抽出物または無傷の細胞−を最初に突然変異型「AGT−A」(または野生型AGT)によって選択的に認識される基質と一定時間反応させて、基質「A」による突然変異型「AGT−A」タンパク質(または野生型AGT)の完全な、またはほぼ完全なターンオーバーを引き起こす。しかし、突然変異型「AGT−B」タンパク質は未反応またはほぼ未反応のまま残し、次いで突然変異型「AGT−B」と反応性である(そして突然変異型「AGT−A」または野生型AGTも反応性であり得る)基質「B」を用いてインキュベーションし、突然変異型「AGT−B」タンパク質と基質「B」との優先的反応も引き起こす。なぜならば、突然変異型「AGT−A」タンパク質(または野生型AGT)が基質「A」によって既に不活性化されているためである。同様に、基質「B」の選択した濃度の存在下でさえ、選択した濃度での突然変異型「AGT−A」(または野生型AGT)の基質「A」に対する反応性が突然変異型「AGT−A」(または野生型AGT)の基質「A」との優先的反応を引き起こすという条件で、突然変異型「AGT−B」および突然変異型「AGT−A」(または野生型AGT)を含有する混合物を、基質「A」および基質「B」と同時にインキュベートし、突然変異型「AGT−A」(または野生型AGT)の基質「A」との優先的反応および突然変異型「AGT−B」の基質「B」との優先的反応を引き起すことができる。2つの異なる基質「A」および「B」は例えば、蛍光共鳴エネルギー移動対(FRET)の2つの化合物、または近接アッセイのための1つの蛍光物質および1つのクエンチャーを保持してもよい。
【0060】
本発明は、対象となるタンパク質を検出および/または操作する方法にも関し、該方法では対象となるタンパク質を融合タンパク質内にAGT突然変異体と共に組み込み、標識を有する特定のAGT基質とAGT融合タンパク質を接触させ、標識を認識および/または処理するために設計された系において標識を使用してAGT融合タンパク質を検出し、場合により操作する。このことは前の段落に記載するように、AGT突然変異体と1つの基質との優先的な(または特に低い)反応性を利用している。特に本発明は、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質、ならびに更なるAGT融合タンパク質を含有するAGT融合タンパク質混合物を、AGT突然変異体または更なるAGTのどちらかに対して選択的である特定の基質と接触させて、混合物を更なる基質と処理して、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質を、標識を認識および/または処理するために設計された系において標識を使用して検出し、場合により更に操作する方法に関する。更なる基質は、混合物と特定の基質の反応終了後に、または特定の基質と共にAGT融合タンパク質混合物に添加できる。
【0061】
より詳細には、本発明は、特定の基質の標識が更なる基質の標識と相互作用し、例えば標識が蛍光共鳴エネルギー移動対(FRET)の化合物あるいは近接アッセイのための1つの蛍光物質および1つのクエンチャーである、これらの方法に関する。
【実施例】
【0062】
使用した省略形:
AGTM=修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断を有するAGT突然変異型
BG−Bt=ビオチンに結合されたO6−(4−アミノメチル−ベンジル)−グアニン
BG−Cy3=Cy3に結合されたO6−(4−アミノメチル−ベンジル)−グアニン
DTT=1,4−ジチオスレイトール
GST=グルタチオン−S−トランスフェラーゼ(Schistosoma japonicum 由来)
HEPES=2−[4−(2−ヒドロキシエチル)−1−ピペラジニル]エタンスルホン酸
IPTG=イソプロピルβ−D−1−チオガラクトピラノシド
PEG=ポリエチレングリコール
PMSF=フェニルメタンスルホニルフルオリド
SDS−PAGE=ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動
【0063】
〔実施例1〕
突然変異Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子の2つの特に重複する領域である、突然変異Asn157Gly、Ser159Gluを含有するAGT(Juilleratら、Chem Biol 10: 313-317,2003)を配列番号:3、配列番号:6および配列番号:4、配列番号:5のプライマーによって別個の反応にて増幅した。その部分相補性に関して、これらの2つの部分を更なるPCR反応にて構築し、配列番号:3および配列番号:4のプライマーによって増幅して、ここで突然変異Lys125Ala、Ala127Thr、Arg128Alaを更に含有する完全な遺伝子を産生させた。続いて遺伝子を発現ベクターpGEX−2T(Pharmacia)BamHI部位とEcoRI部位との間でクローニングした。これにより遺伝子がベクターによって提供されたGSTタンパク質のC末端融合物として挿入遺伝子の発現が可能となる。
【0064】
このベクターからのタンパク質発現をE.coli菌株BL21で実施した。指数関数的に増殖する培養は、0.5mM IPTGによって誘起して、発現を24℃にて3.5時間実施した。
【0065】
精製:収穫した細胞を1mM PMSFおよび2μg/mlアプロチニンを添加した、50mMホスフェート、0.5M NaCl、1mM DTTを含有する緩衝液で再懸濁させて、リゾチームおよび超音波処理によって破壊した。細胞破片を40000xgでの遠心分離によって分離した。抽出物を事前に平衡にしたグルタチオンセファロース(Amersham)に加え、次に20ベッドボリューム(50mMホスフェート、0.5M NaCl、1mM DTT)で洗浄した。変異したGST−AGT融合タンパク質を50mM Tris・HCl、pH7.9中の10mM還元グルタチオンで溶出させた。精製したタンパク質を50mM HEPES、pH7.2、1mM DTT、30%グリセロールで透析して、次に−80℃にて保存した。
【0066】
タンパク質収率の決定:GST−AGT融合タンパク質の純度および相対量をSDS−PAGEにサンプルをかけることによって比較した。純粋画分のUVスペクトルは、Perkin Elmer Lambda 10を使用して記録した。大量のDNAを含有しないタンパク質サンプルでは、タンパク質含有率の尺度として280nmでの吸光度を同じ条件下で生成した野生型hAGTまたはPGEG−hAGTの吸光度と比較した。
【0067】
試験管内でタンパク質のDNA結合特性を比較するために、収穫した細胞を再懸濁させ、同じ方法の後に、しかし50mMホスフェート、0.1M NaCl、1mM DTTを含有する緩衝液中でGST融合タンパク質を精製した。透析サンプルにおいて、次に同時精製されたDNAの量をUV分光法によって見積もった。スペクトルは、Perkin Elmer Lambda 10を使用して記録した。DNAの含有率は、280および260nmにおける吸光度の比に相当する。この値を、野生型hAGTまたはPGEG−hAGTを精製して得られた値とそれぞれ比較した。
【0068】
活性アッセイ:精製した変異GST−AGTは、ビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら, Chem Biol 10: 313-317,2003の物質3a)を用いて試験管内でインキュベートした。総反応物量80μlにて、0.2μM GST−AGTを50mM HEPES pH7.2および1mM DTT中の1μM基質を用いて室温にてインキュベートした。複数の時点にてアリコートをSDS−Laemmli緩衝液中の1mM O6−ベンジルグアニン(Sigma)で失活させて、ウェスタンブロッティング分析を受けさせた(ニュートラアビジン−ペルオキシダーゼコンジュゲート(PIERCE),Renaissance reagent plus(NEN))。対応するバンドの強度は、Kodak Image Station 440によって定量した。
【0069】
生体内局在化:変異AGT(Lys125Ala、Ala127Thr、Argl28Ala、Asn157Gly、Ser159Glu)遺伝子を配列番号:7、配列番号:8のプライマーで増幅して、ベクターpEGFP−Nuc(Clontech)のNheI部位とBamHI部位との間でクローニングした。この構築物を内因性AGTが欠損したCHO細胞に形質移入した(B.Kainaら、Carcinogenesis 12,1857-1867,1991)。24時間のAGT一過性発現の後に、細胞をJuilleratら、Chem Biol 10: 313-317,2003の物質4の0.5μM で5分間インキュベートして、PBSで30分間洗浄した。細胞を、水(1.2開口数)対物レンズを備えたZeiss LSM 510顕微鏡(Carl Zeiss AG)での488nmアルゴン/クリプトンレーザ線を使用して、レーザ走査共焦顕微鏡法によって撮像した。
【0070】
E.coliにおいて、突然変異型AGT Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159GluのGST融合は、PGEG−hAGTの少なくとも3倍の溶解性タンパク質を生じる。それは少なくとも10分の1に減少したDNA結合を示し、O6−ベンジルグアニン基質に対する活性を維持している。CHO細胞では、蛍光標識変異AGTは、細胞質全体に分布しており、優先的な核局在化は検出できない。
【0071】
〔実施例2〕
突然変異Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu
実施例1の突然変異型AGT遺伝子を、配列番号:3、配列番号:10および配列番号:4、配列番号:9のオリゴヌクレオチドを用いて実施例1に記載するように増幅およびクローニングすると、PCRによるその取込み時に遺伝子内に更なる突然変異Cys62Alaを引き起こさせた。タンパク質発現、精製および収率の決定は、実施例1に記載するように実施した。突然変異型遺伝子は、ベクターpAK100内の繊維状ファージのg3タンパク質へ融合させて遺伝子をサブクローニングするために、SfiI制限部位を含有する配列番号:11、配列番号:12のプライマーによってPCR増幅した(Krebberら、J Immunol Methods 201: 35-55,1997)。非サプレッサ菌株E.coli BL21内で遺伝子を発現させるときに、アンバー停止コドンは、AGT遺伝子後の翻訳を終止させる。したがって、このベクターからの突然変異型AGTタンパク質のペリプラズム発現を実施例1で記載するように実施した。収穫した細胞を1mM PMSFおよび2μg/mlアプロチニンを添加した、50mMホスフェート、1M NaCl、1mM DTTを含有する緩衝液で再懸濁させて、リゾチームおよび超音波処理によって破壊した。細胞破片を40000xgでの遠心分離によって分離した。実施例1で記載するように、上清をSDS−PAGEによるタンパク質収率の定量および活性アッセイに直接供した。
【0072】
突然変異型Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Gluは、O6−ベンジルグアニン基質に対する活性を保持している。この突然変異型AGTからの溶解性GST融合タンパク質の収率は、実施例1の突然変異型Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Gluの収率の少なくとも2倍である。E.coliのペリプラズムでの発現による活性AGT突然変異体の収率は、PGEG−hAGTの収率の少なくとも3倍である。
【0073】
〔実施例3〕
182での切断、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子(Asn157Gly、Ser159Glu、Juilleratら、Chem Biol 10:313−317,2003を参照)を配列番号:3、配列番号:13のプライマーによって増幅して、コドン182の後に停止コドンおよびEcoRIを導入して、次に発現ベクターpGEX2T(Pharmacia)のBamHI部位とEcoRI部位との間でクローニングした。タンパク質発現、精製および発現収率の見積りは、実施例1に記載するように実施した。
【0074】
切断PGEG−hAGT突然変異体は、O6−ベンジルグアニン基質に対する活性を保持している。溶解性突然変異型GST−AGT融合タンパク質の収率は、突然変異型PGEG−hAGTの収率の少なくとも2倍である。
【0075】
〔実施例4〕
突然変異Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu
PGEG−hAGT遺伝子(Asn157Gly、Ser159Glu、Juilleratら、Chem Biol 10:313−317,2003を参照)を、実施例1に記載するように配列番号:11、配列番号:15および配列番号:12、配列番号:14のプライマーでPCR増幅して、構築した。プライマーは、ヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)をhAGT遺伝子のコドン131、132、134、135に相当する位置に含有する。遺伝子を繊維状ファージのg3タンパク質へ融合させて、SfiI制限部位を介してベクターpAK100内にクローニングした。生じた遺伝子ライブラリをファージ提示に使用した。
【0076】
このライブラリのファージの産生をE.coli JM101細胞にて実施した。指数関数的培養物にヘルパーファージを重感染させて、24℃にて一晩培養した。この培養物の上清を1μMジゴキシゲニン化O6−ベンジルグアニン(Juilleratら, Chem Biol 10: 313-317,2003の物質2)によって6分間インキュベートした。次の選択ラウンドでは、反応時間がそれぞれ90秒および45秒まで短縮し、基質の濃度は、10nMまで低下して選択圧を上昇させた。この反応からのファージを4% PEG/3% NaClを用いた沈殿によって精製した。ジゴキシゲニンによって今や共有結合的に標識された突然変異型AGTを有するファージを、抗ジゴキシゲニン抗体(Roche Diagnostics)でコーティングした磁気ビーズを用いたインキュベーションによって単離して、細菌の再感染に使用した。
【0077】
選択したAGT突然変異体を実施例1に記載するように、pGEX2T内にサブクローニングして、発現させて、精製およびキャラクタリゼーションした。N9−置換およびN9−非置換O6−ベンジルグアニン基質間の識別は、N9−置換およびN9−非置換O6−ベンジルグアニンを用いた精製された変異GST−AGTのインビトロでの競合実験によって評価した。0.2μM GST−AGTのアリコートを、50mM HEPES、pH7.3、および1mM DTT中の様々な濃度のN9−置換基質(0、0.5〜100μMまで)および0.5μMビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら, Chem Biol 10: 313-317, 2003の物質3a)を用いて室温にてインキュベートした。45分後、SDS−Laemmli緩衝液の添加および熱変性によって反応を停止させた。サンプルにSDS−PAGEおよびウェスタンブロッティング分析に供した(ニュートラアビジン−ペルオキシダーゼコンジュゲート(PIERCE),Renaissance reagent plus(NEN))。対応するバンドの強度は、Kodak Image Station 440によって定量した。
【0078】
この突然変異型AGTからの溶解性GST融合タンパク質の収率は、PGEG−hAGTの収率の少なくとも2倍である。突然変異型AGTは、O6−ベンジルグアニン基質に対して少なくとも2倍更に上昇した活性を示す。O6−アルキル−N9−デオキシリボシルグアニンまたはN9−シクロペンチル−O6−ベンジルグアニンに対する反応性は、PGEG−hAGTと比較して、少なくとも10分の1に、特定のクローンでは少なくとも100分の1に低下する。オリゴヌクレオチド、例えば位置14のグアニンがO6−ベンジル基を有する配列番号:2のオリゴヌクレオチドとの反応性は、PGEG−hAGTと比較して、少なくとも1000分の1に低下する。
【0079】
〔実施例5〕
突然変異Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu
実施例4に記載したように、3つのPCR断片から配列番号:11、配列番号:17、配列番号:16、配列番号:19および配列番号:12、配列番号:18のプライマーの組合せを使用して、3回の別個のPCR反応で、AGT突然変異体のライブラリを構築した。遺伝子を部分的に重複する断片から構築して、断片のうち2つは、コドン115−116および150−152にそれぞれ対応する位置に、ランダムヌクレオチド混合物NNKを含有している。ファージ提示選択は、Juilleratら, Chem Biol 10: 313-317,2003に記載されているように実施した。選択したタンパク質を実施例1に記載するように、pGEX内にサブクローニングして、発現させ、精製およびキャラクタリゼーションした。ペリプラズム発現は実施例2に記載するように実施した。
【0080】
この突然変異型AGTからの溶解性GST融合タンパク質の収率は、PGEG−hAGTからの収率の少なくとも2倍である。突然変異型AGTは、O6−ベンジルグアニン基質に対する活性を保持している。E.coliのペリプラズムでの発現による活性AGT突然変異体の収率は、PGEG−hAGTの収率の少なくとも2倍である。
【0081】
〔実施例6〕
突然変異Cys62Ala、GIn115Ser、Gln116His、Lys125Ala、Ala127Thr、Ara128Ala、Gly131Lys、Gly132Thr、Met134Leu、Ara135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、182での切断、「AGTM」
実施例2、3、4、5による突然変異を組合せた。実施例1からの突然変異型AGT遺伝子の4つの重複断片(Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu)を、配列番号:3、配列番号:23;配列番号:21、配列番号:22;配列番号:20、配列番号:25および配列番号:24、配列番号:13のプライマー組合せを使用して増幅し、遺伝子へと構築して、実施例1に記載するようにpGEX2T内へサブクローニングした。実施例1に記載するように、突然変異型AGT(「AGTM」)を発現させ、精製およびキャラクタリゼーションした。
【0082】
適切な緩衝液(例えば100mM NaCl、20mM Tris、pH8.0、20%グリセロール)中で確立された1つ以上の濃度において4℃または最高37℃での精製サンプルの一晩に渡るインキュベーション後に溶解性画分中に残存するタンパク質の量を決定することによって、AGTMまたはAGTM融合タンパク質の試験管内での溶解度を測定して、野生型AGTまたは既知のAGT突然変異体、例えば「PGEG−hAGT」の溶解度と比較した(Juilleratら、Chem Biol 10: 313-317,2003)。
【0083】
突然変異型AGTMは、実施例1〜5に記載する突然変異体と比較して少なくとも2倍の発現速度の更なる上昇を示し、野生型AGTと比較したときに少なくとも4倍の試験管内での溶解度の上昇を有する。それはO6−ベンジルグアニンに対する反応性を保持しているが、N9−置換O6−アルキルグアニン誘導体との少なくとも10分の1までの反応性の著しい低下、DNA基質との少なくとも100分の1までの反応性の低下を示す。
【0084】
酸化条件下での安定性:配列番号:11および配列番号:28のプライマーを使用して突然変異型遺伝子をPCR増幅して、実施例2で記載するように、該遺伝子をpAK100内にサブクローニングした。実施例2に記載するようにタンパク質発現および細胞溶解を行って、透明な細胞溶解物を得た。PGEA−hAGT(Juilleratら,Chem Biol 10: 313−317,2003)および突然変異体の反応性を比較するために、1μM BG−Btを用いてサンプルを50分間インキュベートして、SDS−Laemmli緩衝液で失活させ、ニュートラアビジン−ペルオキシダーゼコンジュゲート(Pierce)またはモノクローナルマウス抗FLAG IgG抗体(M2、Sigma)および二次抗マウスIgG::HRPコンジュゲート、ならびにRenaissance reagent plus(NEN)を用いてウェスタンブロッティングによって分析した。ブロットをKodak Image station 440で視覚化した。抗FLAG IgG抗体を使用して、溶解性AGTタンパク質の総量を見積もったが、ニュートラアビジンは、その反応性部分のみを定量した。AGT突然変異体の予想分子量は、それぞれ21および24kDaである。
【0085】
ペリプラズム発現は2〜3倍のAGTMをもたらすが、突然変異型PGEA−hAGTでは、少なくとも10倍のこの突然変異体の活性タンパク質をもたらす(タンパク質収率へ正規化)。
【0086】
BG−Cy3アッセイで測定したDNAベース基質に対する反応性:前に記載したO6−(4−アミノエチル−ベンジル)グアニン(Kepplerら、Nature Biotechnol.21: 86-89,2003)をCy3の市販のN−ヒドロキシスクシンイミドエステル(Cy3−NHS、Amersham)と反応させることによって、BG−Cy3を調製した。カップリング反応は周囲温度にて、N,N−ジメチルホルムアミド中でトリエチルアミン1当量の存在下で実施した。BG−Cy3コンジュゲートは、C18カラムでの逆相HPLCによって、0.1%トリフルオロ酢酸〜アセトニトリルの線形勾配を使用して精製した。溶媒蒸発後、BG−Cy3をジメチルスルホキシドに溶解させ、−20℃にて保存した。DNAベース基質との反応性を測定するために、反応緩衝液(50mM HEPES、pH7.2、1mM DTT、200mg/ml BSA)中に各種の濃度(0〜5μM)の配列番号:2のオリゴヌクレオチド(位置14で修飾)、0.2μM GST−AGT融合タンパク質、および0.5μM BG−Cy3を含有する反応混合物をマイクロタイタープレートでインキュベートした。Cy3誘導体の蛍光(励起519nm、発光572nm)をSpectramax Geminiプレートリーダー(Molecular Devices)で測定した。
【0087】
突然変異型AGTMは、5μMまでのBG修飾オリゴヌクレオチドの存在下でBG−Cy3とのその反応の低下を示していない(図1A)。PGEG−hAGTの活性は、0.5μM(またはそれ以上)BG修飾オリゴヌクレオチドの存在下でバックグラウンドレベル以下(3%未満)である(図1B)。
【0088】
DNA結合:DNA結合は、AGT突然変異体とBG誘導体との反応のDNAによる阻害として測定した。オリゴヌクレオチドの代わりにサケ精子DNA(25μg/ml)を含有する反応混合物中のAGT突然変異体の反応速度をBG−Cy3アッセイによって測定した。反応速度は、得られたデータを単指数上昇に最大まで適合させることによって決定した。
【0089】
AGTMのBG−Cy3に対する反応速度は、DNAの存在下で不変であるのに対して、PGEG−hAGTのBG−Cy3に対する速度は、DNAの存在下で5分の1に低下している。
【0090】
反応後の安定性:12xHis−タグならびにSbfIおよびAscIクローニング部位を含めるために合成オリゴヌクレオチド(配列番号:29および配列番号:30)NcoI部位とHindIII部位との間でクローニングすることによって、ベクターpBAD/His A(Invitrogen)を修飾した。AGTMの遺伝子は、配列番号:31および配列番号:32のプライマーによって増幅して、SbfI/AscI部位間にてこのベクター内へクローニングした。野生型AGTおよび突然変異型AGT(Lys125Ala、Alal27Thr、Arg128Ala、Asn157Gly、Ser159Glu)を配列番号:31および配列番号:33のプライマーによって増幅して、このベクター内へクローニングした。E.coli菌株Top10においてタンパク質発現を実施した。指数関数的に増殖する培養物を0.2%アラビノースによって誘起し、発現を28℃にて16時間に渡って実施した。0.5M NaCl、80mMイミダゾール、1% Triton、250μM PMSF、1mM β−メルカプトエタノール、5mg/mlリゾチーム、20μg/ml DNaseを含有する緩衝液を10mg/gまで添加することによって細胞を溶解させ、45分間撹拌した。10,000gにて30分間の遠心分離後の上清を平衡したNi−NTA(Qiagen)カラムに装填して0.5M NaCl、100mMイミダゾール、1mM β−メルカプトエタノールの20カラム体積で、続いて0.25M NaCl、100mMイミダゾール、20%グリセロールの3カラム体積で洗浄した。タンパク質を0.25M NaCl、400mMイミダゾール、20%グリセロールで溶出させた。1mM DTTを添加した後、タンパク質を−80℃で保存した。
【0091】
AGTタンパク質(10μMにて)をフルオレセイン修飾O6−ベンジルグアニン(Juilleratら、Chem Biol 10: 313-317,2003の物質5、BG−FL、20μM)と定量的に反応させて、サイズ排除カラム(Econopac 10DG、Bio−Rad)で精製し、7日間インキュベートした。各種の時点にて、アリコートを25,000gにて遠心分離して、1% SDSを上清に添加した。上清の蛍光をVictor2蛍光プレートリーダー(PerkinElmer、485/535nmにて励起/発光)で測定した。
【0092】
7日間に渡って溶解性蛍光タンパク質の約60%を失うhAGTまたは突然変異型(Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu)とは対照的に、標識突然変異型AGTMは、7日間を超えて溶液中にて安定なままである。
【0093】
N9−修飾O6−ベンジルグアニン誘導体に対するAGTの反応性:実験は、BG−Cy3アッセイに基づく。AGTのBG−Cy3との反応時に、Cy3の蛍光放出強度の2〜3倍の上昇が認められる。化合物N9−イソブチル−O6−ベンジルグアニン、N9−プロピル−O6−ベンジルグアニン、またはN9−ニトリル−O6−ベンジルグアニンに対する、AGTM(実施例6)対PGEG−hAGT(Juilleratら2003、突然変異N157G、S159Eを含有する)の反応性を、GST−AGT融合タンパク質10μMのサンプルを反応緩衝液(50mM HEPES、pH7.2、100mM NaCl、0.1% Tween−20、1mM DTT)中のこれらのサンプル100μMによって室温にて15分間に渡って事前にインキュベートすることによって調査した。次にBG−Cy3(20μM最終濃度)を添加して、反応物を室温にて更に1時間インキュベートした。サンプルを反応緩衝液によって1:20で希釈して、マイクロタイタープレートに移動した。Cy3蛍光物質の蛍光強度(励起:519nm、発光:572nm)をSpectra Max Geminiマイクロプレートリーダー(Molecular Devices)によって測定した。
【0094】
両方のAGT突然変異体をBG(100μM、15分の反応時間)によって効率的に予備遮断した(図3、No.3)。AGTMの5%は、BGでの予備遮断後に活性のままである(灰色棒)。N9−修飾BG誘導体に対するAGTMとPGEG−hAGTとの反応性の著しい差が見られる。AGTMは、1%〜10%の範囲でN9−修飾BG誘導体と反応する(図3、No.4、5、6、灰色棒)。AGTMとは対照的に、PGEG−hAGTは、3つすべてのN9−修飾BG誘導体と効率的に反応する。BG−Cy3による次のインキュベーション時にはバックグラウンドを超える蛍光の増加は認められず、N9−置換化合物との完全な反応を示す(図3、No.4、5、6、黒色棒)。
【0095】
〔実施例7〕
1つの生体サンプル中に存在する2つの異なるAGTの変異体と2つの異なる基質との反応
実施例6(AGTM)の突然変異型AGTを配列番号:26および配列番号:27のプライマーによってPCR増幅して、NdeIおよびBamHIを介してpET15b内へサブクローニングした。このベクターからの遺伝子発現を実施例1に記載するようにE.coli菌株BL21(DE3)にて実施し、ベクターによってコードされるN末端融合されたHis−タグを有するタンパク質を生じさせた。細胞を収穫して、実施例1に記載するように、0.5M NaCl、10mMイミダゾール、50mMホスフェート、pH8.0を含有する抽出緩衝液中に抽出した。タンパク質を含有する抽出物を事前に平衡にされたNi−NTA−セファロース(Qiagen)に加え、次にこれを、20mMイミダゾールを含有する緩衝液20ベッドボリュームで洗浄した。250mMイミダゾールを含有する緩衝液を用いてHis−タグ化タンパク質を溶出させた。実施例1に記載するように精製したタンパク質を透析し、保存して、後でキャラクタリゼーションした。
【0096】
実験では、2つのAGTタンパク質には異なるサイズの2つのタグを使用した:His−タグ化 AGTM(約20kDa)およびGST−hAGT(実施例1に記載するようにクローニングおよび調製、しかし野生型hAGT遺伝子から開始、約48kDa)を、50mMHEPES、1mMDTT、200mg/mLBSAを含有する反応緩衝液(pH7.3)でそれぞれ0.2μMおよび1.2μM最終濃度まで希釈した。サンプルを基質N9−シクロペンチル−O6−ブロモテニル−グアニン(CPTG、0、5および10μM最終濃度)およびビオチン化O6−ベンジルグアニン(BG−Bt、5μM最終濃度)の混合物で30分間インキュベートした。反応物をSDS−Laemmli緩衝液の添加によって失活させ、ウェスタンブロッティング分析に供した(実施例1)。2つのAGT変異体は異なる質量を有するため、2つのタンパク質はアクリルアミドゲルで分離して、独立して分析できる。GST−hAGTに対するCPTGの高い特異性が認められる。同じ濃度では(どちらの基質も5μM最終濃度)、GST−hAGTの95%がCPTGと反応するのに対して、His−タグ化 AGTMはわずか5%である。
【0097】
基質CPTGは、シクロペンテニルメチルカーボネートによる6−クロログアニンの位置N−9のシクロペンテニル化と、6−クロロ−N9−シクロペンチル−グアニンへの還元と、3級アミンの存在下での4−ブロモ−2−ヒドロキシメチルチオフェンとの反応によって入手できる。
【0098】
【化3】
【0099】
更なる実験において、実施例6に記載する突然変異型AGTMを配列番号:34および配列番号:36のプライマーによって増幅し、ベクターpEGFP−NucのNheI部位とBglII部位との間へクローニングした。酵母β−ガラクトシダーゼ遺伝子を配列番号:37および配列番号:38のプライマーによって増幅して、BglII/BamHIを介してこのベクターへサブクローニングして、AGTM−β−GaI融合遺伝子を有するベクターを生成させた。hAGT遺伝子を実施例1に記載する原理に基づいて、配列番号:39、配列番号:40、配列番号:34、および配列番号:35のプライマーによって増幅し、NheI/BglIIを介してpEGFP−Nuc内へクローニングして、突然変異Gly160TrpをコードするAGT−NLS3融合遺伝子を生成させた。AGT突然変異体両方の同時の一過性発現後に、細胞を5μM CPTGで10分間、次にJuilleratら、Chem Biol 10: 313-317,2003の物質45μMで20分間インキュベートした。実施例1に記載するように細胞を洗浄および撮像した。
【0100】
CHO細胞において、細胞質に局在化したAGTM−β−ガラクトシダーゼ融合タンパク質を蛍光基質物質4との反応によって、選択的に蛍光標識化した。核内の他のAGT融合タンパク質の著しい標識化は認められない。AGT(Gly160Trp)が細胞内でCPTGと効率的に反応して、それゆえその後、蛍光基質によって標識できないことが結論付けられる。実施例6(AGTM)の突然変異体は、細胞内でCPTGによる事前のインキュベーション後に蛍光基質に対して反応性のままであり、したがって第1のステップでCPTGと反応しない。
【0101】
〔実施例8〕
化合物Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、182での切断における更なる突然変異
A)エラープローンPCR:ベクターpAK100からの実施例6のAGTMの遺伝子を、配列番号:42のプライマーおよび配列番号:41のビオチン化プライマーによってTaq−ポリメラーゼを使用して、120μM(6−(2−デオキシ−β−D−エリスロ−ペント−フラノシル)−3,4−ジヒドロ−6H,8H−ピリミド[4,5−c][1,2]オキサジン−2−オン)−5’−トリホスフェートおよび480μM 2’−デオキシ−8−オキソ−グアノシン−トリホスフェート(どちらもTrilink Biotechnologiesより)でスパイクしたエラープローンPCR反応によって増幅した。生成物をストレプトアビジンコーティングビーズ(Dynabeads M−280、Dynal)上に捕捉して、次に配列番号:43および配列番号:44のプライマーを用いて標準PCRによって増幅した。
【0102】
B)飽和突然変異誘発、領域150−154:AGTM(実施例6の突然変異体)の遺伝子を配列番号:11、配列番号:45、配列番号:46および配列番号:28のプライマーを用いてPCRによって実施例1に記載するように増幅して、SfiIを介してpAK100内へサブクローニングした。プライマーは、hAGT遺伝子のコドン150−154に相当する位置にヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)を含有する。
【0103】
C)飽和突然変異誘発、領域31−35:AGTM(実施例6の突然変異体)をコードする遺伝子を配列番号:11、配列番号:48、配列番号:47および配列番号:28のプライマーを用いてPCRによって実施例1に記載するように増幅して、SfiIを介してpAK100内へサブクローニングした。プライマー47は、hAGT遺伝子のコドン31−35に相当する位置にヌクレオチド混合物NNK(N=A、C、GまたはT;K=GまたはT)を含有し プライマー48は、アンチセンスストランドのための混合物MNN(M=CまたはA)を含有する。
【0104】
増幅生成物を繊維状ファージのg3遺伝子に融合させて、SfiI制限部位を介してベクターpAK100へクローニングした。実施例4に記載したように、生じた遺伝子ライブラリをファージ提示に使用した。ライブラリの突然変異体をBG−Bt 5〜50nMによって10分間に渡って選択した。選択された突然変異体を配列番号:3および配列番号:13のプライマーを使用してpGEX内へサブクローニングした。突然変異型AGTの反応速度を実施例6、DNAベース基質に対する反応性(BG−Cy3アッセイ)に記載するように決定した。
【0105】
突然変異の組合せ:BG誘導体との反応速度を向上するために見出された選択からの突然変異を、鋳型として1つの突然変異型遺伝子を使用して、そして突然変異の点にてミスマッチであるプライマー配列での更なる突然変異を含めて、配列番号:3および配列番号:13のプライマーにより実施例1に記載するようにPCRによって組合せた。突然変異を重複し60℃以下でハイブリダイズするのに十分な長さを有する2つの各変異原性プライマーを選択して、相補性ストランドの一方へそれぞれアニーリングさせた。アニーリング温度は、各G/C対では4℃および各A/T対では2℃の寄与を想定して計算した。生じた遺伝子を実施例1に記載するように、pGEX内へサブクローニングした。実施例1、4および6に記載するように、突然変異型AGTタンパク質を調製およびキャラクタリゼーションした。
【0106】
これらの選択からの組合せ突然変異を有するAGT突然変異体は、実施例1〜6の突然変異体について記載した好都合な特性を示す。突然変異体は、実施例1〜5に記載する突然変異体に匹敵する発現速度を有し、野生型AGTと比較したときに試験管内で少なくとも4倍の溶解度の上昇を有する。それらは少なくとも10分の1へのN9−置換O6−アルキルグアニン誘導体との反応性の低下と、少なくとも100分の1へのDNAまたはDNA基質との相互作用および反応性それぞれの低下を示す。E.coliのペリプラズムにおける発現からの活性AGT突然変異体の収率は、PGEG−hAGTの収率よりも少なくとも5倍の高さである。CHO細胞において、蛍光標識化変異AGTは細胞質全体に分布しており、優先的な核局在化は検出できない。その上、これらのAGT突然変異体は野生型hAGTと比較したときに、O6−ベンジルグアニン基質に対する反応性のかなりの上昇を示す(表1を参照)。
【0107】
【表1】
【0108】
BG−Cy3活性アッセイによって反応速度を決定して、野生型hAGTについてこのアッセイで得られた値(kobs=116M−1cm−1)で割った。活性アッセイでは、精製したGST融合タンパク質を、「p.」のマークが付いた(実施例1に記載するように調製した)突然変異体について使用した;ペリプラズム抽出物を、「x.」のマークが付いた(実施例2に記載するように調製した)突然変異体について使用した。
【図面の簡単な説明】
【0109】
【図1】表示濃度でのBG修飾オリゴヌクレオチド(配列番号:2)の存在下でのAGTM(実施例6、Aの突然変異体)またはPGEG−hAGT(B)のBG−Cy3との反応。Spectra Max Geminiプレートリーダー(Molecular Devices)で検出した任意単位[a.u.]の蛍光FL(励起519nm、572nmにて放出)は、時間(t)(分[min])にプロットされている。
【図2】実施例7に記載するような、野生型hAGT(GST−hAGT融合タンパク質)およびAGTMと、2つの基質N9−シクロペンチル−O6−ブロモテニル−グアニン(CPTG)およびビオチン化O6−ベンジルグアニン(BG−Bt、Juilleratら2003の物質3a)との反応のウェスタンブロット。BG−Btは、すべてのサンプル中に5μMで存在する。BG−Btと反応したAGTを検出する、ニュートラアビジン−ペルオキシダーゼコンジュゲートおよび発光ペルオキシダーゼ基質を用いた検出。同じ濃度のCPTGおよびBG−Btを用いた同様のインキュベーションの下でさえ、GST−hAGTのわずか5%が基質BG−Btと反応する。
【図3】N9−修飾BG誘導体とのAGT突然変異体の反応性。10μM AGTM(灰色バー)またはPGEG−hAGT(黒色バー)を100μM N9−修飾BG誘導体(以下のリストを参照)で予備遮断して、次に20μM BG−Cy3で標識した。1時間の反応時間の後、595nmにて任意単位[a.u.]で示した蛍光強度(FI)を測定する。黒色実線は、タンパク質に結合していない蛍光物質のバックグラウンド蛍光に相当する条件1の測定値に設定されている。この線より上の値は、N9−修飾BG誘導体との低速または不完全な反応を示す。1 タンパク質を含まないBG−Cy3の蛍光、バックグラウンド蛍光。2 BG−Cy3との反応;予備遮断なし、正の対照。3 BG−Cy3との反応、100μM BGによって予備遮断。4 BG−Cy3との反応、100μM N9−イソブチル−O6−ベンジルグアニンによって予備遮断。5 BG−Cy3との反応、100μM N9−プロピル−O6−ベンジルグアニンによって予備遮断。6 BG−Cy3との反応、100μM N9−シアノメチル−O6−ベンジルグアニンによって予備遮断。
【特許請求の範囲】
【請求項1】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)突然変異体であって、
野生型ヒトAGTと比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の利点を示す、AGT突然変異体。
【請求項2】
好都合な特性が
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f)基質との反応前後の細胞外部での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
である、請求項1記載のAGT突然変異体。
【請求項3】
好都合な特性が
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f’)基質との反応後の、細胞外での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
である、請求項1記載のAGT突然変異体。
【請求項4】
好都合な特性が
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(f)基質との反応前および後の、細胞外での4倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
である、請求項1または2記載のAGT突然変異体。
【請求項5】
好都合な特性が
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(f’)基質との反応後の、細胞外での4倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
である、請求項1または3記載のAGT突然変異体。
【請求項6】
好都合な特性が
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
である、請求項1または2記載のAGT突然変異体。
【請求項7】
好都合な特性が
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
である、請求項1または3記載のAGT突然変異体。
【請求項8】
野生型ヒトAGTの1〜25個のアミノ酸が、他のアミノ酸によって置換され、場合により連続鎖からの1〜5個のアミノ酸が1、2、または3つの位置で欠損または付加され、および/またはN末端の1〜4個のアミノ酸またはC末端の1〜40個のアミノ酸が欠損している、請求項1〜7のいずれか一項記載のAGT突然変異体。
【請求項9】
2つ以上の修飾が
(A)AlaまたはValによって置換されたCys62;
(B)Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Serによって置換されたGln115−Gln116;
(D)Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Valによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−によって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、あるいはSerまたはThrによって置換されたCys150;
(F)Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trpによって置換されたAsn157/Ser159−Gly160、あるいはGly/Gluによって置換されたAsn157/Ser159、あるいはGlyまたはArgによって置換されたAsn157;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項10】
3つ以上の修飾が
(A)AlaまたはValによって置換されたCys62;
(B)Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Serによって置換されたGln115−Gln116;
(c)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(D)Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Valによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−によって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、あるいはSerまたはThrによって置換されたCys150;
(F)Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trpによって置換されたAsn157/Ser159−Gly160、あるいはGly/Gluによって置換されたAsn157/Ser159、あるいはGlyまたはArgによって置換されたAsn157;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項11】
2つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(D)Lys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、またはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項12】
3つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(C)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(D)Lys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、またはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項13】
3つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(C)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項14】
修飾
Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu、Gly182の後の切断;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu;
Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu;ならびに
Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断;
を有する突然変異体より選択される、請求項8記載のAGT突然変異体。
【請求項15】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項16】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により1〜10個の追加のアミノ酸修飾を有する、請求項8記載のAGT突然変異体。
【請求項17】
修飾Cys62Ala、Glnl15Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により3〜7個の追加のアミノ酸修飾を有する、請求項16記載のAGT突然変異体。
【請求項18】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、請求項16記載のAGT突然変異体。
【請求項19】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により1〜10個の追加のアミノ酸修飾を有する、請求項8記載のAGT突然変異体。
【請求項20】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により3〜7個の追加のアミノ酸修飾を有する、請求項19記載のAGT突然変異体。
【請求項21】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157G1y、Ser159G1u、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項22】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項23】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項24】
修飾Lys32Ile、Leu33Phe,Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Gly、Ser151Gly、Ser152Asp、Ala154Asp、Asn157Gly、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項25】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Val、Ser152Arg、Gly153Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項26】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Ala154Thr、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項27】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Ala154Thr、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項28】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Val、Ser152Arg、Gly153Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
を有する、請求項8記載のAGT突然変異体。
【請求項29】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Gly、Ser151Gly、Ser152Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項30】
修飾Cys62Ala、Lysl25Ala,Ala127Thr、Arg128Ala、Cys150Glu、Ser151Gly、Ser152Glu、Ala154Arg、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Metl34Leu、Argl35Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項31】
対象となるタンパク質を検出および/または操作する方法であって、対象となるタンパク質を、請求項1〜30のいずれか一項記載のAGT突然変異体を有する融合タンパク質に組み込み、AGT融合タンパク質を、標識を担持する特定のAGT基質と接触させ、AGT融合タンパク質を、標識を認識および/処理するために設計された系において標識を使用して検出し、場合により更に操作する方法。
【請求項32】
対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質、ならびに更なるAGT融合タンパク質を含有するAGT融合タンパク質混合物を、AGT突然変異体または更なるAGTのいずれかに対して、選択性を有する特定の基質と接触させ、混合物を更なる基質によって処理し、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質を、標識を認識および/処理するために設計された系において標識を使用して検出し、場合により更に操作する、請求項31記載の方法。
【請求項33】
混合物と特定の基質との反応終了後に、更なる基質をAGT融合タンパク質混合物に添加する、請求項32記載の方法。
【請求項34】
更なる基質を、AGT融合タンパク質混合物に特定の基質と共に添加する、請求項32記載の方法。
【請求項35】
標識を認識および/処理するために設計された系において、特定の基質の標識を、更なる基質の標識と相互作用させる、請求項34記載の方法。
【請求項36】
特定の基質の標識およびさらなる基質の標識が、蛍光共鳴エネルギー移動対(FRET)の化合物または近接アッセイのための1つの蛍光物質および1つのクエンチャーである、請求項35記載の方法。
【請求項37】
請求項1〜30のいずれか一項記載のAGT突然変異体および対象となるタンパク質を含む、AGT融合タンパク質。
【請求項1】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)突然変異体であって、
野生型ヒトAGTと比較したときに、
(a)DNA相互作用の低下;
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化;
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上;
(d)酸化条件下での安定性の向上;
(e)基質との反応後の細胞内での安定性の向上;
(f)基質との反応前後の細胞外部での安定性の向上;
(g)試験管内溶解度の向上;
(h)O6−アルキルグアニン基質に対する反応性の向上;
(i)DNAベース基質に対する反応性の低下;ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
より選択される2つ以上の利点を示す、AGT突然変異体。
【請求項2】
好都合な特性が
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f)基質との反応前後の細胞外部での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種宿主における溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
である、請求項1記載のAGT突然変異体。
【請求項3】
好都合な特性が
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(d)酸化条件下での安定性の向上、
(f’)基質との反応後の、細胞外での安定性の向上、
(g)試験管内溶解度の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、ならびに
(i)DNAベース基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける溶解性タンパク質としての発現収率の向上および安定性の向上、
(e)基質との反応後の細胞内での安定性の向上、
(h)O6−アルキルグアニン基質に対する反応性の向上、
(i)DNAベース基質に対する反応性の低下、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する反応性の低下;
である、請求項1記載のAGT突然変異体。
【請求項4】
好都合な特性が
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(f)基質との反応前および後の、細胞外での4倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
である、請求項1または2記載のAGT突然変異体。
【請求項5】
好都合な特性が
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する5倍を超える反応性;
あるいは
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(d)酸化条件下での5倍を超える安定性、
(f’)基質との反応後の、細胞外での4倍を超える安定性、
(g)5倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する反応性の向上;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、ならびに
(i)DNAベース基質に対する1%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
あるいは
(a)2%未満のDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての5倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での3倍を超える安定性、
(h)O6−アルキルグアニン基質に対する5倍を超える反応性、
(i)DNAベース基質に対する1%未満の反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する2%未満の反応性;
である、請求項1または3記載のAGT突然変異体。
【請求項6】
好都合な特性が
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c)各種の宿主における、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f)基質との反応前および後の、細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
である、請求項1または2記載のAGT突然変異体。
【請求項7】
好都合な特性が
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、ならびに
(h)O6−アルキルグアニン基質に対する10倍を超える反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)検出不能なDNA結合、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(e)基質との反応後の細胞内での6倍を超える安定性、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、ならびに
(i)DNAベース基質に対する検出不能な反応性;
あるいは
(a)DNA相互作用の低下、
(b)もはや核に限定されない、真核細胞での発現タンパク質の局在化、
(c’)E.coliにおける、溶解性タンパク質としての10倍を超える発現収率および安定性の向上、
(d)酸化条件下での10倍を超える安定性、
(e)基質との反応後の細胞内での6倍を超える安定性、
(f’)基質との反応後の細胞外での6倍を超える安定性、
(g)10倍を超える試験管内溶解度、
(h)O6−アルキルグアニン基質に対する10倍を超える反応性、
(i)DNAベース基質に対する検出不能な反応性、ならびに
(j)N9−置換O6−アルキルグアニン基質に対する検出不能な反応性;
である、請求項1または3記載のAGT突然変異体。
【請求項8】
野生型ヒトAGTの1〜25個のアミノ酸が、他のアミノ酸によって置換され、場合により連続鎖からの1〜5個のアミノ酸が1、2、または3つの位置で欠損または付加され、および/またはN末端の1〜4個のアミノ酸またはC末端の1〜40個のアミノ酸が欠損している、請求項1〜7のいずれか一項記載のAGT突然変異体。
【請求項9】
2つ以上の修飾が
(A)AlaまたはValによって置換されたCys62;
(B)Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Serによって置換されたGln115−Gln116;
(D)Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Valによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−によって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、あるいはSerまたはThrによって置換されたCys150;
(F)Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trpによって置換されたAsn157/Ser159−Gly160、あるいはGly/Gluによって置換されたAsn157/Ser159、あるいはGlyまたはArgによって置換されたAsn157;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項10】
3つ以上の修飾が
(A)AlaまたはValによって置換されたCys62;
(B)Ala−Asn、Asn−Asn、Ser−His、Ser−Ser、Pro−Pro、Pro−Ser、Pro−Thr、またはThr−Serによって置換されたGln115−Gln116;
(c)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(D)Val−His/Leu−Arg、Lys−Thr/Leu−Ser、Gln−Val/Leu−Ser、またはMet−Thr/Met−Valによって置換されたGly131−Gly132/Met134−Arg135、あるいはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asn、Pro−Leu−Pro、Pro−Arg−Thr、Ser−Phe−Pro−、またはSer−His−Thr−によって置換されたCys150−Ser151−Ser152、あるいはPhe−AsnまたはArg−Asnによって置換されたCys150−Ser151、あるいはHis/Thr、Leu/Asn、Leu/Asn、Leu/ProまたはPro/Leuによって置換されたCys150/Ser152、あるいはSerまたはThrによって置換されたCys150;
(F)Phe/Arg/Gluによって置換されたPro140/Asn157/Ser159、あるいはMet/Trp/Valによって置換されたPro140/Asn157/Gly160、あるいはGly/Glu−Ala、Gly/Asn−Trp、Pro/Gln−CysまたはGly−Gln−Trpによって置換されたAsn157/Ser159−Gly160、あるいはGly/Gluによって置換されたAsn157/Ser159、あるいはGlyまたはArgによって置換されたAsn157;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項11】
2つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(D)Lys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、またはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項12】
3つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(C)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(D)Lys−Thr/Leu−Serによって置換されたGly131−Gly132/Met134−Arg135、またはVal−His/Leuによって置換されたGly131−Gly132/Met134;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項13】
3つ以上の修飾が
(A)Alaによって置換されたCys62;
(B)Ser−Hisによって置換されたGln115−Gln116;
(C)Alaによって置換されたLys125およびThr−Alaによって置換されたAla127−Arg128;
(E)Asn−Ile−Asnによって置換されたCys150−Ser151−Ser152、またはSerもしくはThrによって置換されたCys150;
(F)あるいはGly/Gluによって置換されたAsn157/Ser159;ならびに
(G)Gly182の後の切断;
そして、場合により、1〜10個の追加のアミノ酸修飾
より選択される、請求項8記載のAGT突然変異体。
【請求項14】
修飾
Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Asn157Gly、Ser159Glu、Gly182の後の切断;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Asn157Gly、Ser159Glu;
Gln115Ser、Gln116His、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu;ならびに
Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断;
を有する突然変異体より選択される、請求項8記載のAGT突然変異体。
【請求項15】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項16】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により1〜10個の追加のアミノ酸修飾を有する、請求項8記載のAGT突然変異体。
【請求項17】
修飾Cys62Ala、Glnl15Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により3〜7個の追加のアミノ酸修飾を有する、請求項16記載のAGT突然変異体。
【請求項18】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、請求項16記載のAGT突然変異体。
【請求項19】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により1〜10個の追加のアミノ酸修飾を有する、請求項8記載のAGT突然変異体。
【請求項20】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により3〜7個の追加のアミノ酸修飾を有する、請求項19記載のAGT突然変異体。
【請求項21】
修飾Cys62Ala、Gln115Ser、Gln116His、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157G1y、Ser159G1u、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Ser150Asn、Ser151Ile、Ser152Asn;
Lys8Thr、Lys32Ile、Leu33Phe、Thr127Ala、Ser150Asp、Ser151Gly、Ala154Thr;
Lys32Ile、Leu33Phe、Ser150Val、Ser152Arg、Gly153Asp、Ala154Asp;
Lys32Ile、Leu33Phe、Ser150Gly、Ser151Gly、Ser152Asp、Ala154Asp;
Ser150Val、Ala154Asp;
Ser150Glu、Ser151Gly、Ser152Glu、Ala154Arg;
Lys8Thr、Thr127Ala、Ala154Thr;
Lys32Ile、Leu33Phe;
Ala154Thr;
Leu33Phe;
Ser151Gly;
Ser150Asp;
Thr127Ala;ならびに
Lys32Ile、Leu33Phe、およびLeu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項22】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項23】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser、Cys150Ser、Asn157Gly、Ser159Glu、Gly182の後の切断を有する、請求項8記載のAGT突然変異体。
【請求項24】
修飾Lys32Ile、Leu33Phe,Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Gly、Ser151Gly、Ser152Asp、Ala154Asp、Asn157Gly、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項25】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Val、Ser152Arg、Gly153Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断および場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項26】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Asn、Ser151Ile、Ser152Asn、Ala154Thr、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項27】
修飾Lys32Ile、Leu33Phe、Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Ser、Ala154Thr、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項28】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Val、Ser152Arg、Gly153Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;ならびに
Leu34の欠損;
を有する、請求項8記載のAGT突然変異体。
【請求項29】
修飾Cys62Ala、Lys125Ala、Ala127Thr、Arg128Ala、Cys150Gly、Ser151Gly、Ser152Asp、Ala154Asp、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Met134Leu、Arg135Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項30】
修飾Cys62Ala、Lysl25Ala,Ala127Thr、Arg128Ala、Cys150Glu、Ser151Gly、Ser152Glu、Ala154Arg、Asn157Gly、Ser159Glu、Gly182の後の切断、ならびに場合により
Gln115Ser、Gln116His;
Gly131Lys、Gly132Thr、Metl34Leu、Argl35Ser;および
Leu34の欠損;
より選択される更なる突然変異を有する、請求項8記載のAGT突然変異体。
【請求項31】
対象となるタンパク質を検出および/または操作する方法であって、対象となるタンパク質を、請求項1〜30のいずれか一項記載のAGT突然変異体を有する融合タンパク質に組み込み、AGT融合タンパク質を、標識を担持する特定のAGT基質と接触させ、AGT融合タンパク質を、標識を認識および/処理するために設計された系において標識を使用して検出し、場合により更に操作する方法。
【請求項32】
対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質、ならびに更なるAGT融合タンパク質を含有するAGT融合タンパク質混合物を、AGT突然変異体または更なるAGTのいずれかに対して、選択性を有する特定の基質と接触させ、混合物を更なる基質によって処理し、対象となるタンパク質およびAGT突然変異体のAGT融合タンパク質を、標識を認識および/処理するために設計された系において標識を使用して検出し、場合により更に操作する、請求項31記載の方法。
【請求項33】
混合物と特定の基質との反応終了後に、更なる基質をAGT融合タンパク質混合物に添加する、請求項32記載の方法。
【請求項34】
更なる基質を、AGT融合タンパク質混合物に特定の基質と共に添加する、請求項32記載の方法。
【請求項35】
標識を認識および/処理するために設計された系において、特定の基質の標識を、更なる基質の標識と相互作用させる、請求項34記載の方法。
【請求項36】
特定の基質の標識およびさらなる基質の標識が、蛍光共鳴エネルギー移動対(FRET)の化合物または近接アッセイのための1つの蛍光物質および1つのクエンチャーである、請求項35記載の方法。
【請求項37】
請求項1〜30のいずれか一項記載のAGT突然変異体および対象となるタンパク質を含む、AGT融合タンパク質。
【図1】

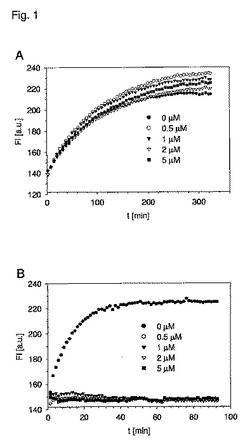
【図2】
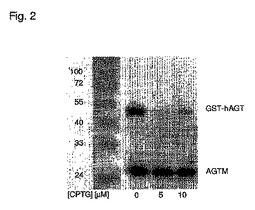
【図3】
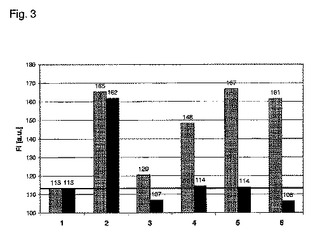
【図2】
【図3】
【公表番号】特表2007−525988(P2007−525988A)
【公表日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願番号】特願2007−501284(P2007−501284)
【出願日】平成17年3月1日(2005.3.1)
【国際出願番号】PCT/EP2005/050899
【国際公開番号】WO2005/085431
【国際公開日】平成17年9月15日(2005.9.15)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
【公表日】平成19年9月13日(2007.9.13)
【国際特許分類】
【出願日】平成17年3月1日(2005.3.1)
【国際出願番号】PCT/EP2005/050899
【国際公開番号】WO2005/085431
【国際公開日】平成17年9月15日(2005.9.15)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
[ Back to top ]