
O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)の使用方法
【課題】O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を用いた、標識を基質からAGTを含む融合タンパク質に移転する方法の提供。
【解決手段】新たな物理学的または化学的特性を融合タンパク質に導入する分子を融合タンパク質に結合させることによって、インビトロとインビボとの双方での融合タンパク質の検出および/または操作を可能にする。そのような分子の例は、とりわけ、分光学的プローブもしくはリポーター分子、親和性タグ、反応性ラジカルを発生できる分子、架橋結合剤、タンパク質−タンパク質相互作用を仲介するリガンド、または融合タンパク質の固定化に適した分子である。
【解決手段】新たな物理学的または化学的特性を融合タンパク質に導入する分子を融合タンパク質に結合させることによって、インビトロとインビボとの双方での融合タンパク質の検出および/または操作を可能にする。そのような分子の例は、とりわけ、分光学的プローブもしくはリポーター分子、親和性タグ、反応性ラジカルを発生できる分子、架橋結合剤、タンパク質−タンパク質相互作用を仲介するリガンド、または融合タンパク質の固定化に適した分子である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、標識を基質から、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質へと移転する方法、特に、標識付けられた融合タンパク質を検出かつ/または操作することをさらに含む方法に関するものである。
【背景技術】
【0002】
複雑な生物系の理解における進歩は、裏面にある生体分子、特にタンパク質の相互作用を特性評価することに依存する。増加する数の生物のDNA配列決定は、それらの読み枠(ORF)を同定したが、対応するタンパク質の生細胞内での挙動を調べ、インビボおよびインビトロにおける多数のタンパク質の相互作用を特性評価できる可能性は、限られている。この目的の実現を目指すほとんどの方針は、結合したタンパク質の環境内での変化に応じ、物理学的、生理学的または化学的応答を引き出す融合タンパク質を構成することに基づく。その例は、酵母ツーハイブリッドシステム、分割ユビキチン、および緑色蛍光タンパク質(GFP)融合タンパク質を含む。しかし、これらの手法は、すべて、様々な限定または短所を有する。
【0003】
ドイツ国特許願第199 03 895A(Kai Johnsson)は、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)の検出のためのELISAアッセイを記載している。N−メチル−N−ニトロソ尿素のような求電子剤の突然変異誘発および発癌効果は、主として、DNA中のグアニンのO6−アルキル化による。自身をDNAアルキル化から防護するために、哺乳動物および細菌は、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)というタンパク質を保有していて、それがこれらの損傷を修復する[Pegg et al., 1995]。AGTは、アルキル基を、SN2反応で、それ自身のシステインの一つに移転し、その結果、不可逆的にアルキル化された酵素を生じる。腫瘍細胞内でのAGTの過剰発現は、薬物耐性、特にプロカルバジン、ダカルバジン、テモゾロミドおよびビス−2−クロロエチル−N−ニトロソ尿素のようなアルキル化薬物に対するそれを獲得するのを可能にするため、AGTの阻害剤が、増感剤として化学療法に用いるために提唱されている[Pegg et al., 1995]。DE 199 03 895Aは、ビオチニル化されたO6−アルキルグアニン誘導体とAGTとの間の、AGTのビオチニル化へと導く反応に依拠する、AGTレベルを測定するアッセイを開示している。次いで、これは、ストレプトアビジン被覆プレート上でのAGTの分離と、例えばELISAアッセイでの、その検出とを可能にする。このアッセイは、腫瘍組織中のAGTレベルを追跡すること、AGT阻害剤を化学療法における増感剤として用いて処置を調整すること、およびAGT阻害剤についてスクリーニングするのに用いることが示唆されている。
【0004】
Damoiseaux、KepplerおよびJohnsson[ChemBiochem., 4: 285-287, 2001]は、やはり癌細胞内のこの酵素のレベルを検出するのを容易にして、研究および化学療法の際の助けにするために、AGT標識付けのための化学プローブのように用いるための、オリゴデオキシリボヌクレオチドに組み込んだ改質O6−アルキル化グアニン誘導体を開示している。2種類の変種AGT基質、およびビオチンで標識付けするAGT(DE 199 03 895 Aに記載されたのと同じ)についてのアッセイが開示されている。加えて、AGTの指向性進化にこれらのO6−アルキル化誘導体を用いることが示唆されている。
【0005】
発明の要約
広義には、本発明は、ある系内の対象となるタンパク質またはペプチドを標識付けし、場合により、次いで操作および/もしくは検出する方法に、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)をさらに用いることに関するものであって、該方法では、該タンパク質またはペプチドとAGTとの融合を、標識付けられた基質に接触させる結果、AGTが該標識を基質からAGT融合に移転し、そのため、移転された標識により、標識付けられたAGT−タンパク質融合が操作かつ/または検出されるのを可能にする。これは、AGTが融合タンパク質として存在しない、AGTレベル測定用アッセイの従来の用い方とは対照的である。
【0006】
上記により、第一の態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法を提供する。融合タンパク質への標識の移転後は、該方法は、標識付けられた融合タンパク質を検出および/または操作する、その後の工程をさらに包含してもよい。
【0007】
一実施態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を標識付けする方法であって、融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法を提供する。
【0008】
いくつかの実施態様では、該方法は、例えば、標識付けられた融合タンパク質を検出および/または操作する、その後の一つもしくはそれ以上の工程を含む。
【0009】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を検出する方法であって、該融合タンパク質と、標識を有する基質とを接触させる結果、AGTが、ラベルを、それが該融合タンパク質に共有結合させるように移転すること、および該標識を用いて、タンパク質構成体を検出することを含む方法を提供する。
【0010】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を操作する方法であって、該融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転すること、および融合タンパク質を、標識によって融合タンパク質に導入された物理学的および/または化学的特性を用いて操作することを含む方法を提供する。
【0011】
本発明のこの態様のいくつかの実施態様では、該方法は、標識を用いてタンパク質構成体を検出することを含んでよい。
【0012】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を固体支持体に固定化する方法であって、該融合タンパク質を、固体支持体に結合されるか、または結合され得る、標識を有する基質に接触させ、AGTが標識を、それが該融合タンパク質に共有結合させるように移転し、そのため、該融合タンパク質が固体支持体に結合されるか、またはその後に結合することができることを含む方法を提供する。本発明の、標識を初めに固体支持体に結合させない実施態様では、該方法は、標識付けられた融合タンパク質を固体支持体に接触させる結果、それが固体支持体上で固定化される工程をさらに含んでよい。本発明のこの態様のこの好ましい実施態様では、標識が移転されたときか、またはその後の反応の際のいずれかに、標識は、共有結合で固体支持体に結合してよいか、あるいは特異的結合対の一方の成員であってよくて、その他方の成員が、共有結合によってか、またはその他の手段によって(例えば、ビオチンおよびアビジンまたはストレプトアビジンの結合対を用いて)該固体支持体に結合されるか、もしくは結合され得る。
【0013】
さらに一つの態様では、本発明は、AGT融合タンパク質をインビボとインビトロとの双方で標識付けする方法を提供する。AGT融合タンパク質のインビボ標識付けという用語は、細胞のすべての区画内での標識付けはもとより、細胞外空間を指向するAGT融合タンパク質の標識付けも含む。AGT融合タンパク質の標識付けをインビボで実施し、AGTに融合されたタンパク質が原形質膜タンパク質であるならば、融合タンパク質のAGT部分は、原形質膜の細胞質側または細胞外側のいずれにも結合することができる。標識付けをインビトロで実施するならば、融合タンパク質の標識付けは、細胞抽出物中でか、またはAGT融合タンパク質の精製された形態もしくは富化された形態のいずれで実施することもできる。
【0014】
さらに一つの態様では、本発明は、候補化合物または候補化合物のライブラリーと、標的物質または標的物質のライブラリーとの相互作用を決定する方法を提供する。該化合物および物質の例は、リガンドおよびタンパク質、薬物および該薬物の標的、または小分子およびタンパク質を含む。この方法では、AGTに融合した対象となるタンパク質は、転写因子のDNA結合ドメイン、または転写因子の活性化ドメインを含み、標的物質または標的物質のライブラリーを、該DNA結合ドメインの他方、または該転写因子の活性化ドメインに結合させ、かつ標識が、標的物質と作用し合うと想定される候補化合物または候補化合物のライブラリーである。
【0015】
好ましい実施態様では、該方法は、候補化合物または候補化合物のライブラリーを、AGTタンパク質融合に移転すること、および候補化合物で標識付けしたAGT融合タンパク質と、標的物質とを接触させる結果、AGT融合タンパク質に結合した候補化合物と、標的物質との相互作用が、転写因子を活性化することをさらに含む。そうして、活性化された転写因子は、リポーターの発現を駆動し、それは、該方法を細胞内で実施するならば、リポーターの発現が細胞に選択的な利点を与えた場合に、検出することができる。いくつかの実施態様では、該方法は、候補化合物または標的物質を検出、単離、同定または特性評価することのような、追加的な一つもしくはそれ以上の工程を包含してもよい。
【0016】
本願では、O6−アルキルグアニン−DNAアルキルトランスフェラーゼまたは「AGT」は、基質に存在する標識を、融合タンパク質のAGT形成部分のシステイン残基の一つに移転するという特性を有する。好ましい実施態様では、AGTは、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ、例えば、Peggら(1995)およびその中の引用文献に記載されている、ヒトO6−アルキルグアニン−DNAアルキルトランスフェラーゼである。しかし、他のアルキルグアニン−DNAアルキルトランスフェラーゼ、例えばRoyら(1995)に記載されているこの酵素のマウスまたはラット形態も公知であって、それらが上に定義された特性を有することを条件に、本発明に用いることができる。本発明では、O6−アルキルグアニン−DNAアルキルトランスフェラーゼは、一つまたはそれ以上のアミノ酸の置換、欠失もしくは付加によって異なり得るが、基質に存在する標識を、融合を形成しているAGTまたはタンパク質もしくはペプチドに移転するという特性は依然として保持している、野生型AGTの変種も含む。AGTのその他の変種は、当業者には周知の手法を用いて化学的に改質してもよい。AGTの変種は、当業者に公知のタンパク質工学の手法を用い、かつ/または新たなO6−アルキルグアニン−DNAアルキルトランスフェラーゼを発生かつ選択する分子的進化を用いて、生成してよい。
【0017】
本発明では、融合タンパク質のAGTを有するタンパク質部分の基準は、いかなる長さのタンパク質、ポリペプチドおよびペプチドを含み、かつ二次、三次または四次構造を保有するものと保有しないものとの双方を含む。本発明の適用の例を下記に示す。
【0018】
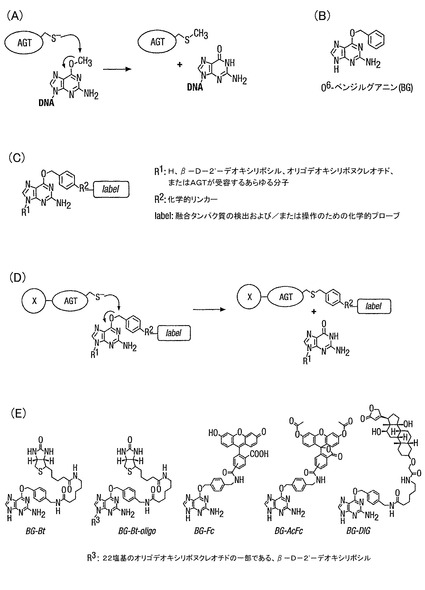
本発明では、標識付け基質は、好ましくは、標識付けベンジルグアニン基質であり、より好ましくは、O6−ベンジルグアニン誘導体である。そのような誘導体の例は、下記の一般式:
【0019】
【化3】

【0020】
[式中、
R1は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であり、好ましくは2〜99ヌクレオチドの長さを有するβ−D−2′−デオキシリボシルであり;
【0021】
R2は、リンカー基、例えば、置換または非置換アルキル鎖、ポリエチレングリコールのような柔軟なリンカーであり;そして
【0022】
labelは、本明細書に記載されたとおりの、融合タンパク質を検出かつ/または操作する原因となる分子である]
を有する、ベンジル環の4位が誘導体化された、O6−ベンジルグアニン誘導体である。
【0023】
本発明に従って用いるのに適した改質O6−ベンジルグアニン誘導体の例を、図1に示す。さらに、本発明者らは、AGTが、基質の同一性におけるかなりの程度の自由度を許容して、下記の一般式:
【0024】
【化4】
【0025】
[式中、
R1は、AGTによって受容されて、AGTが標識をタンパク質の融合へと移転するのを許す基、例えば、置換もしくは非置換アルキル鎖、環の大きさが3〜10炭素の置換もしくは非置換シクロアルキル基、環の大きさが3〜10炭素の置換もしくは非置換複素環、または環の大きさが3〜10炭素の置換もしくは非置換芳香族複素環であり;
【0026】
R2は、リンカー基、例えば、置換もしくは非置換アルキル鎖、またはポリエチレングリコールのような、様々な長さの柔軟なリンカーであり;そして
【0027】
R3は、プロトン、β−D−2′−デオキシリボシル、または好ましくは2〜99ヌクレオチドの長さを有する、オリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;
【0028】
labelは、本明細書に記載されたとおりの、融合タンパク質を検出かつ/または操作する原因となる分子である]
を有する、広範囲の基質が用いられるのを可能とすることができることを見出した。
【0029】
基質の標識部分は、融合タンパク質が意図される適用に応じて、当業者が選ぶことができる。標識の例は、下記のものを含む:
【0030】
(1)発蛍光団、発色団、磁気プローブまたは造量剤のような、分光学的プローブ;
【0031】
(2)放射性標識付け分子;
【0032】
(3)パートナーに特異的に結合できる特異的結合対の一方の部分である分子。そのような特異的結合対は、当技術に周知であり、例えば、アビジンまたはストレプトアビジンに結合することができるビオチンを含む;
【0033】
(4)その他の生体分子と作用し合うと思われる分子;
【0034】
(5)その他の生体分子と作用し合うと思われる分子のライブラリー;
【0035】
(6)当業者に公知のとおり[Nadeau et al., 2002]、その他の生体分子と架橋結合できる分子;
【0036】
(7)束縛された金属キレートのような[Hori et al., 2002]、H2O2およびアスコルビン酸塩に接触するとヒドロキシルラジカルを発生できる分子(最終原稿を確認されたい);
【0037】
(8)マラカイトグリーンのような[Jay et al., 1999]、光を照射すると反応性ラジカルを発生できる分子;
【0038】
(9)スライドグラス、微量滴定プレート、または一般に当業者に公知のいかなる重合体であってもよい、固体支持体に共有結合させる分子;
【0039】
(10)その相補鎖との塩基対合をなし得る核酸またはその誘導体;
【0040】
(11)脂質、または他の膜挿入特性を有する疎水性分子;
【0041】
(12)望ましい酵素的、化学的または物理的特性を有する生体分子;
【0042】
(13)上記に列挙された特性のいずれかの組合せを保有する分子。
【0043】
例示のために、ここで、本発明の実施態様を、添付の図面を参照して、より詳しく説明する。
【0044】
詳細な説明
本発明の下記の適用は、例示のために提供され、限定のためではない。本明細書に開示された方法は、一般的には、ある範囲の応用に適用することができ、融合タンパク質を、(1)標識付けられた融合タンパク質の環境における変化を感知かつ誘導できる標識、ならびに/または(2)標識が融合タンパク質に特異的に導入した物理および/もしくは化学的特性によって融合タンパク質を操作する助けとなる標識で、融合タンパク質を特異的に、かつ共有結合で標識化することができる。本明細書に開示された方法は、インビボとインビトロとの双方で、AGT融合タンパク質を標識付けするのに用いることができる。
【0045】
本発明は、所望のタンパク質への標識の特異的な結合は、問題のそのタンパク質と、ヒトのDNA修復酵素であるO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(hAGT)のようなO6−アルキルグアニン−DNAアルキルトランスフェラーゼとの間の融合タンパク質を構成し、後者の機序を利用することによって、実施することができるとの理解に基づく。この酵素は、アルキル基を、その基質、すなわちO6−アルキルグアニン−DNAからそのシステイン残基の一つに不可逆的に移転する(図1)。hAGTと急速に反応する基質類似体は、O6−ベンジルグアニンであって、二次速度定数は、約103秒-1M-1である(図1)。本発明者らは、O6−ベンジルグアニンのベンジル環のC4での置換が、O6−ベンジルグアニン誘導体に対するhAGTの反応性に有意には影響しないことを示した。これは、標識付けられたAGT融合タンパク質をインビボまたはインビトロで、共有結合によって特異的に結合するのに、標識がベンジル環のC4に結合したO6−ベンジルグアニン誘導体を用いることを可能にする(図1D)。標識付けは、融合タンパク質の性質には無関係である。
【0046】
標識付けをインビボまたは細胞抽出物中で実施するならば、宿主の内在AGTの標識付けを考慮に入れるのが好都合である。宿主の内在AGTがO6−ベンジルグアニン誘導体または関連化合物を基質として受容しないならば、融合タンパク質の標識付けは特異的である。哺乳動物細胞(ヒト、マウス、ラット)では、内在AGTの標識付けは可能である。内在AGTとAGT融合問題との同時標識付けが問題を提起する、これらの実験では、以前に記載したAGT欠乏細胞系を用いることができる[Kaina et al., 1991]。一般的には、本発明は、融合タンパク質との標識の共有結合による特異的な結合を用いて、AGT融合タンパク質の挙動を監視するか、もしくはそれに影響を与えるか、または導入された標識によってAGT融合タンパク質を操作する手法のすべての適用に利用することができる。この技術を用いるための適用の例を、以下に述べる。
【0047】
(1)分光学的プローブまたはリポーター基としての標識
AGTに移転できる、検出可能な標識、例えば発蛍光団、発色団、磁気プローブ、放射性標識付け分子またはその他のいかなる分光学的プローブをも基質が担持する、標識付けAGT基質、例えばO6−ベンジルグアニン誘導体の使用は、本発明を、検出できる標識を細胞内、細胞表面(インビボ)またはインビトロのいずれかでAGT融合タンパク質に特異的に共有結合によって結合するのに用いることを可能とする。これは、AGT融合タンパク質のインビボまたはインビトロでの検出および特性評価を可能とする。インビボという用語は、細胞のすべての区画内での標識付けはもとより、細胞外空間を指向するAGT融合タンパク質の標識付けも含む。この方法は、対象となるタンパク質にやはり遺伝学的に融合され、生細胞内でのその探索を可能とする、緑色蛍光タンパク質(GFP)の適用と対比することができる。GFPおよびその突然変異体の短所は、それが、主として、自然に産する発蛍光団の使用に限られることである。
【0048】
(2)AGT融合タンパク質を検出かつ単離するためのタグとしての標識
ビオチンのような親和性タグで標識付けしたAGT基質、例えばO6−ベンジルグアニン誘導体の使用は、親和性タグをAGT−タンパク質融合に移転し、そのため親和性タグの結合パートナーによって、融合タンパク質を結合するのに、本発明を用いることを可能にする。例としては、ビオチンのような親和性タグで標識付けしたAGT基質を、AGT融合タンパク質を発現する(細菌性または真核性の)細胞にか、またはそのような細胞の細胞抽出物、もしくは精製されたAGT融合タンパク質に付加することは、親和性タグによる融合タンパク質の共有結合的修飾へと導くことになる。そうして、これは、親和性タグとその結合パートナー、例えばビオチンの場合、固定化されたアビジンまたはストレプトアビジンとの間の相互作用を用いる、融合タンパク質の単離を可能にする。標識を、切断できる結合、例えばジスルフィド架橋を含有するリンカーを介してAGT融合タンパク質に結合するか、またはリンカーが光切断できるならば、AGT融合タンパク質は、親和性タグから、その単離後に遊離させることができる。
【0049】
(3)反応性ラジカルの源泉としての標識
AGT基質、例えばO6−ベンジルグアニン誘導体は、外部刺激に触れると反応性ラジカル、例えばヒドロキシルラジカルを発生できる、AGT融合タンパク質に、標識を導入するのに用いることができる。そうして、発生したラジカルは、AGT融合タンパク質はもとより、AGT融合タンパク質のごく近傍にあるようなタンパク質も不活性化することができて、これらのタンパク質の役割の研究を可能にする。そのような標識の例は、H2O2やアスコルビン酸塩に接触するとヒドロキシルラジカルを生ずる、束縛された金属−キレート錯体、およびレーザー照射されるとヒドロキシルラジカルを生ずるマラカイトグリーンのような、発色団である。ヒドロキシルラジカルを発生させるのに発色団やレーザーを用いることは、レーザー分子不活性化法(CALI)として当技術で公知である[Jay et al., 1998]。CALIは、細胞内の一定のタンパク質を、時間的に制御され、空間的に分解された方式で、特異的に不活性化するのに用いられる方法であり、発色団およびタンパク質の空間的近傍を基礎にしている。レーザー照射されると、発色団は、ヒドロキシルラジカルを発生し、それが、発色団内、およびそのわずか約0.1nm以内のすべてのタンパク質を不活性化する。従来、発色団は、対象となるタンパク質の空間的近傍に、対象となるタンパク質に特異的な発色団標識付け抗体を微量注入することによって近付けている。本発明では、マラカイトグリーンのような発色団によるAGT融合タンパク質の標識付け、およびその後のレーザー照射は、AGT融合タンパク質はもとより、AGT融合タンパク質と作用し合うようなタンパク質も、時間的に制御され、空間的に分解された方式で不活性化するのを可能にすると思われる。この方法は、インビボまたはインビトロの双方で適用することができる。
【0050】
同様にして、AGT融合タンパク質は、束縛された金属−キレートで標識付けすることができ、AGT融合タンパク質、およびAGT融合タンパク質と作用し合うようなタンパク質は、H2O2およびアスコルビン酸塩に接触させると、特異的な方式で不活性化することができる。この方法は、AGT融合タンパク質、またはAGT融合タンパク質のごく近傍にあるそれの機能を調べることばかりでなく、AGT融合タンパク質のごく近傍にあるようなタンパク質を同定することにも用いることができる。ここで、AGT融合タンパク質のごく近傍にあるタンパク質は、該タンパク質のフラグメントの特異的な抗体による検出、高分離能2D電気泳動ゲル上のこれらのタンパク質の消滅、または切断されたタンパク質フラグメントの、分離、および質量分析法もしくはN末端分解によるタンパク質配列決定のような配列決定手法による同定のいずれかによって同定されるようなものとして同定することができる。
【0051】
(4)タンパク質−タンパク質相互作用を仲介するリガンドとしての標識
標識付けられたAGT基質、例えばO6−ベンジルグアニン誘導体は、リガンドをAGT−タンパク質融合に移転するのに用いることができる。これは、リガンドのパートナー、例えばタンパク質をAGT−タンパク質融合に結合するのを可能にする。例えば、標識が、結合パートナーに結合できるリガンドである場合、そのような基質とAGT融合タンパク質とを接触させることは、融合タンパク質とのリガンドの特異的結合へと導くことになる。リガンドが、もう一つのタンパク質Yに結合し、タンパク質Yと標識付けられたAGT融合タンパク質との二量体化が、生物学的機能、または測定できるシグナルへと導くならば、この生物学的機能または測定可能シグナルは、標識を担持するAGT基質の付加に依存する。具体的な例は、小分子で遺伝子発現を調節するための、Hoら、1996が記載した、いわゆる三雑種系におけるAGT基質およびAGT融合タンパク質の使用であると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。リガンド、例えばFK506に結合するタンパク質Y、例えばFKBPを、転写因子の活性化ドメインに結合させる。リガンド、この例ではFK506を担持するAGT基質を細胞に供給することが、機能的な転写因子の形成、および遺伝子発現へと導くことになる。
【0052】
(5)その標的が未知である薬物または生物学的活性分子としての標識
標識を担持し、その標識が、未だ同定されていないタンパク質Yに結合する薬物または生物学的活性小分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。ここでは、目標は、生物学的活性分子の標的であるタンパク質Yを同定することになると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。未知の標的タンパク質Yを発現する生物のcDNAライブラリーを、転写因子の活性化ドメインに融合させる。標識を担持し、その標識が薬物または生物学的活性小分子である、O6−ベンジルグアニン誘導体または関連AGT基質の付加が、この分子が、cDNAライブラリー内に存在し、活性化ドメインに融合した、その標的タンパク質Yに結合する場合にのみ、機能的転写因子の形成、および遺伝子発現へと導くことになる。遺伝子発現を選択的な利点に結び付けるならば、該薬物または生体活性分子の標的遺伝子Yを有するプラスミドを担持する、対応する宿主を同定することができる。
【0053】
(6)タンパク質Yに結合する分子を同定するための小分子のライブラリーとしての標識
標識を担持し、その標識が、化学的分子のライブラリーである、O6−ベンジルグアニンまたは関連AGT基質の使用:ここでは、目標は、インビボ条件下でタンパク質Yに結合する、潜在的な薬物標的であり得る小分子を同定することになると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。標的タンパク質Yを、転写因子の活性化ドメインに融合させる。標識として結合させた小分子のライブラリーを、O6−ベンジルグアニン誘導体に付加することが、標識(すなわち小分子)が、活性化ドメインに融合したその標的タンパク質Yに結合する場合にのみ、機能的転写因子の形成、および遺伝子発現へと導くことになる。遺伝子発現を選択的な利点に結び付けるならば、ライブラリーの、宿主の成長へと導くような分子を同定することができる。
【0054】
(7)AGT融合タンパク質を固定化し、かつ/またはAGT融合タンパク質のタンパク質アレーを創出するためのBG誘導体の使用
標識を担持し、その標識が、担体の表面に共有結合で結合されているか、または該標識が、それ自体該表面に結合されたもう一つの分子によって非共有結合で結合され得る分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。後者の取組み方についての例は、標識がビオチンであり、表面に結合された分子がストレプトアビジンまたはアビジンである場合である。担体についてのあり得る例は、スライドガラス、微量滴定プレート、または機能化された任意の重合体のいずれかでもあると思われる。標識によるAGT基質の固定化は、標識の融合タンパク質への移転によって、担体へのAGT融合タンパク質のその後の固定化を可能にする。空間的に分解された方式での(異なる)AGT融合タンパク質の担体でのスポット形成は、タンパク質アレーの生成を可能にする。
【0055】
(8)AGT融合タンパク質と作用し合うタンパク質を検出するための架橋結合剤としての標識
標識を担持し、その標識が、他のタンパク質と架橋結合することができる分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。そのような架橋結合剤の例は、マレイミド、活性エステルまたはアジ化物およびその他の、当業者に公知であり、Nadeauら、2002に記載されたもののような、官能基を含有する分子である。そのようなAGT基質を、他のタンパク質と作用し合うAGT融合タンパク質と(インビボまたはインビトロで)接触させることは、AGT融合タンパク質と、標識を介して作用し合うそのタンパク質との共有結合での架橋結合へと導くことができる。これは、AGT融合タンパク質と作用し合うタンパク質の同定を可能にする。
【0056】
実施例
下記の実施例は、本発明を実施する方法の完全な開示および記載を当業者に提供するために説明するもので、本発明の対象範囲を限定することを意図するものではない。
【0057】
実施例A:大腸菌におけるAGT融合タンパク質の共有結合による標識付け
下記の例、すなわちBG−Btを用いた、大腸菌におけるPep−hAGTの標識付けは、大腸菌におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。hAGTのN末端に融合したペプチド(Pep−hAGTを生じる)の配列は、(一字コードでは)MHHHHHHSSAであって、hAGTの最初のアミノ酸である、メチオニンがこれに続く。N末端融合ペプチドを有するhAGT、すなわちPep−hAGTをコーディングしている、pET−15b(Novagen)に基づく発現ベクターを含有するXL−1Blueという大腸菌細胞の液体培養物を、0.6の光学的密度OD600nmまで増殖させた。IPTGを1mMの最終濃度までを加えることによって、Pep−hAGTの発現を誘導した。同時に、BG−Btを10μMの最終濃度まで加え、細菌を37℃で2時間温置した。細胞を遠心分離によって採集し、ペレットを2回洗浄して、過剰なBG−Btを除去した。再懸濁させた細胞のアリコートを、ウエスタンブロット分析によって分析した。ビオチニル化タンパク質を、ストレプトアビジン−ペルオキシダーゼ複合体(NEN)および化学発光性ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用いて検出した(図2)。
【0058】
実施例B:酵母におけるAGT融合タンパク質の共有結合による標識付け
下記の実施例は、酵母におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、BG−Btを用いて、hAGT−DHFR−HAという融合タンパク質を酵母中でビオチニル化した。この融合タンパク質は、標準的な分子生物学の手順を用いて、DNAレベルで構成される。略述すると、hAGTの停止コドンを、アミノ酸RSGIに対するコドンで置き換え、次いで、マウスからのDHFRの第1アミノ酸、すなわちメチオニンに対するコドンで後続させた[Nunberg et al., 1980]。hAGTとDHFRとの間のリンカーに対するコドンは、BglII部位もコード化していて、そのDNA配列は、AGATCTである。DHFRとHAタグとの間の融合を構成するために、DHFRの停止コドンを、HA−タグの第1アミノ酸に対するコドンで置き換えた[Kolodziej, 1991]。HA−タグに停止コドンを後続させた。hAGT−DHFR−HAというタンパク質がpcup1プロモーターの制御下にある、発現ベクターp314AK1を含有する、L40という酵母細胞の培養を、0.6のOD600まで増殖させた。hAGT−DHFR−HAの発現は、CuSO4を100μMの濃度まで加えることによって誘導し、BG−Btを、10μMの濃度まで同時に加えた。2.5時間および5時間後にアリコートを採取し、遠心分離によって、細胞を採集した。ペレットを2回洗浄して、残留BG−Btを除去した。凍結/融解周期の循環によって酵母細胞を溶解した後、細胞抽出物を、ELISAを用いて、ビオチニル化hAGT−DHFR−HA融合タンパク質の存在について分析した。略述すると、ビオチニル化hAGT−DHFR−HAを、ストレプトアビジン被覆した微量滴定ウェル内に固定化し、抗HA抗体(Babco)を一次抗体として、かつ抗マウスHRP複合体(Sigma)を二次抗体として用いることによって検出した(図1)[Kolodziej, 1991]。ELISAは、ペルオキシダーゼ基質ABTSを用いて展開し、シグナル(吸光度)をOD405nmで測定した。インビボビオチニル化hAGT−DHFR−HA融合タンパク質についてのシグナルは、バックグラウンドを少なくとも5倍上回った。バックグラウンドシグナルは、厳密に上記のとおり処理したが、BG−Btの添加を省いた細胞から得られた、細胞溶解物のOD405nmとして定義した。
【0059】
実施例C:酵母におけるhAGT融合タンパク質の共有結合による標識付け
下記の実施例は、酵母におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、BG−DIGを用いて、hAGT−DHFR−HA融合タンパク質を酵母中でジゴキシゲニンで標識付けした。hAGT−DHFR−HA融合の構成は、実施例Bに記載されている。hAGT−DHFR融合タンパク質の遺伝子がpcup1プロモーターの制御下にある、発現ベクターp314AK1を含有する、L40という酵母細胞の培養を、1.2のOD600nmまで増殖させた。hAGT−DHFR融合タンパク質の発現は、CuSO4を100μMの濃度まで加えることによって誘導し、BG−DIGを、20μMの濃度まで同時に加えた。2時間後、振盪フラスコ培養体1mlからの細胞を、遠心分離によって採集した。ペレットを培地で3回洗浄して、残留BIG−DIGを除去した。凍結/融解周期の循環によって酵母細胞を溶解した後、細胞抽出物を、ジゴキシゲニン化hAGT−DHFR融合タンパク質の存在について、ウエスタンブロット分析によって分析した。ジゴキシゲニン化タンパク質は、抗ジゴキシゲニン−ペルオキシダーゼ複合体(Roche)、および化学発光性ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用いて検出した(図2)。
【0060】
実施例D:ヒト細胞系におけるhAGT融合タンパク質の共有結合による標識付け
下記の実施例は、哺乳動物細胞におけるAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、ヒトの細胞(HEK293)内の内在性hAGTを、BG−AcFcを用いてフルオレセインで標識付けした。HEK293細胞を、5μMのBG−AcFcとともにPBS中に5分間温置した。アセチル化されたフルオレセイン誘導体であるBG−AcFcは、細胞浸透性であり、非蛍光性であるが、細胞内で急速に水解されて、蛍光性のBG−Fcを生じることが期待できる。次いで、細胞を、PBSを変えることによって洗浄して、いかなる過剰な基質BG−AcFcも除去し、PBS中に20分間温置した。次いで、共焦点蛍光顕微鏡で画像を得た(励起492nm;発光510nm)。対照実験として、HEK293細胞を上記のとおり処理したが、BG−AcFcを加える前に、O6−ベンジルグアニン(1μM)とともに終夜温置した。これは、内在性hAGTを不活性化し、そのため核内での蛍光の蓄積を防止するはずであった。期待されたとおり、細胞をO6−ベンジルグアニンとともに前温置したとき、核内での蛍光蓄積は、全く観察されなかった。hAGTがBG−Fcを基質として受容することを、独立に確認するため、組換えPep−hAGT(10μM、実施例Aに記載のとおり)を、100μMのBG−Fcとともに25℃で、50mMトリス−Cl、10mMDTT、1mMEDTA、200μg/mlのBSA、10%グリセリン、pH7.4中で10分間温置した後、PBS(リン酸緩衝化生理食塩水:137mMNaCl、2.7mMKCl、10mMNa2HPO4、1.8mMKH2PO4、pH7.4)900μlを加えた。過剰な基質BG−Fcの分離は、NAP(登録商標)−10カラム(Pharmacia)で、供給者の指示によるゲル濾過によって達成した。次いで、Pep−hAGTを、標準的な蛍光分光光度計で特性評価した。サンプルは、それぞれ、222、238および490nmで励起し、波長523nmで最大発光を示し、このタンパク質が、フルオレセインで標識付けられたことを確認した。PBS中20nMのBG−Fcの溶液を、参照として測定した。基質の発光波長は、519nmであった(励起は、それぞれ、237、323および490nm)。
【0061】
実施例E:細胞抽出物中でのAGT融合タンパク質の共有結合による標識付け
AGTとの融合タンパク質は、細胞抽出物中では直接標識付けし、かつ操作(ここでは固定化)することができることを立証するため、下記のhAGTとのN−およびC−末端融合タンパク質を、標準的な分子クローニングの手順によって構成し、酵母の発現ベクター内にクローニングした:
【0062】
(i)V5−NLS−B42−hAGT(ここで、V5はV5エピトープを、NLSはSV40の大T抗原核局在配列を、またB42は、人工転写活性化因子であるB42[Ma et al., 1987]を意味する)。B42のトランス活性化ドメインの最後のコドンに、21アミノ酸の配列ASKKGTELGSTTSNGRQCAGILを後続させた。最後の3コドンは、B42にhAGTをC末端クローニングするためのEcoRI部位を含んでいる。NotI部位は、hAGTに対するC末端制限部位であって、その配列は、停止コドンを含む;
【0063】
(ii)hAGT−HA−Ura3(ここで、Ura3は、酵母の酵素オロチン酸デカルボキシラーゼを意味し、HAは、Haエピトープを意味する)。ここでは、hAGTに対する停止コドンを、RSリンカーで置き換え、HA−タグの最初のアミノ酸を後続させた。HA−タグは、Ura3遺伝子が直接後続する;
【0064】
(iii)hAGT−DHFR−HA(ここで、DHFRは、マウスのジヒドロ葉酸レダクターゼを意味し、HAは、Haエピトープを意味する)。構成については、実施例Bを参照されたい;および
【0065】
(iv)SSN6−hAGT(ここで、SSN6は、酵母のDNA転写リプレッサー[Schultz et al., 1987]を意味する)。ここでは、hAGTの停止コドンを、アミノ酸RSGSGに対するコドンで置き換えて、そうして、酵母のSSN6の最初のアミノ酸、すなわちメチオニンに対するコドンが後続する。
【0066】
遺伝子の発現は、すべて、pcup1プロモーターによって制御した。融合タンパク質の一つをコーディングしている発現ベクターを含有するL40酵母細胞を、0.6のOD600まで増殖させ、融合タンパク質の発現は、CuSO4を100μMの濃度まで加えることによって誘導した。アリコート(2ml)を5時間後に採取し、遠心分離によって細胞を採集した。凍結/融解周期の循環によって細胞を溶解した後、酵母抽出物を、BG−Bt−oligo(10pmol)とともに室温で20分間温置して、融合タンパク質のビオチニル化へと導いた。次いで、懸濁液を、ストレプトアビジン被覆した微量滴定プレート(Roche molecular biochemicals)内に移し、1時間温置した。ウェルをPBSで徹底的に洗浄した後、固定化された融合タンパク質を、抗HA抗体(Babco)または抗hAGT抗体(SSN6−hAGT融合タンパク質の場合)のいずれかを一次抗体として、かつ抗マウスペルオキシダーゼ複合体(Sigma、#A4416)を二次抗体として用い、次いで、標準的な生化学手順を用いて、ペルオキシダーゼの基質ABTSとともに温置することによって、検出した。すべての場合に、OD405nmとして測定されたシグナルは、バックグラウンドを少なくとも5倍上回った。バックグラウンドは、各融合タンパク質について、BG−Bt−oligoの細胞抽出物への添加を省くことによって測定した。
【0067】
参考文献
本明細書に引用された参考文献は、すべて、特に参照として組み込まれる。参考文献は、アルファベット順である。
【0068】
【表1】
【図面の簡単な説明】
【0069】
【図1】(A)O6−アルキルグアニン−DNAアルキルトランスフェラーゼの機序を示す図である。(B)O6−ベンジルグアニンの構造を示す図である。(C)実施例に用いたO6−ベンジルグアニン誘導体の一般的構造を示す図である。(D)AGT融合タンパク質を標識付けする一般的スキームであって、Xは、AGTに融合させたタンパク質である。(E)実施例に用いたAGT基質の構造を示す図である。オリゴヌクレオチド(22量体)の配列は、5′-GTGGTGGGCGCTGXAGGCGTGG-3′であって、X=BG−Btである。
【図2】インビボでAGT融合タンパク質を標識付けした後のウエスタンブロット分析を示す図である。(A)培地中にBG−Btを含む場合と含まない場合の、Pep−hAGTを発現する大腸菌の全細胞抽出物のウエスタンブロット分析。ストレプトアビジン−ペルオキシダーゼ複合体を用いて、ビオチニル化タンパク質を検出。20kDの帯域は、BG−Btの不在下での大腸菌内でビオチニル化タンパク質に相当する。(B)培地中にBG−DIGを含む場合と含まない場合の、hAGT−DHFR−HA融合タンパク質を発現する酵母の全細胞抽出物のウエスタンブロット分析。抗ジゴキシゲニン−ペルオキシダーゼ複合体を用いて、ジゴキシゲニン標識付けタンパク質を検出。
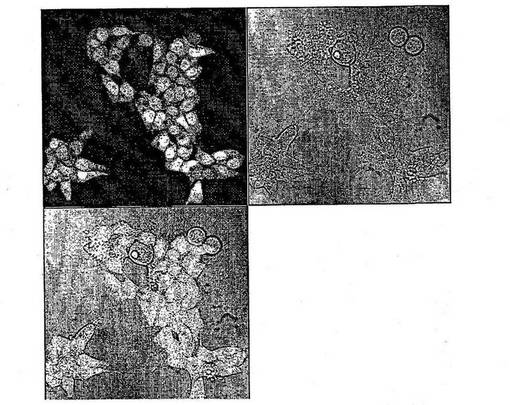
【図3】BG−AcFcとともに温置し、蛍光および光学顕微鏡測定によって分析したHEK293細胞を示す図である。蛍光が細胞核に蓄積されているのを認めることができる。これは、hAGTが核に局在するのが知られていることから[Ali et al., 1998]、期待された結果である。第三の図面は、蛍光および光学顕微鏡測定から得られた画像を重ね合わせたものである。
【技術分野】
【0001】
本発明は、標識を基質から、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質へと移転する方法、特に、標識付けられた融合タンパク質を検出かつ/または操作することをさらに含む方法に関するものである。
【背景技術】
【0002】
複雑な生物系の理解における進歩は、裏面にある生体分子、特にタンパク質の相互作用を特性評価することに依存する。増加する数の生物のDNA配列決定は、それらの読み枠(ORF)を同定したが、対応するタンパク質の生細胞内での挙動を調べ、インビボおよびインビトロにおける多数のタンパク質の相互作用を特性評価できる可能性は、限られている。この目的の実現を目指すほとんどの方針は、結合したタンパク質の環境内での変化に応じ、物理学的、生理学的または化学的応答を引き出す融合タンパク質を構成することに基づく。その例は、酵母ツーハイブリッドシステム、分割ユビキチン、および緑色蛍光タンパク質(GFP)融合タンパク質を含む。しかし、これらの手法は、すべて、様々な限定または短所を有する。
【0003】
ドイツ国特許願第199 03 895A(Kai Johnsson)は、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)の検出のためのELISAアッセイを記載している。N−メチル−N−ニトロソ尿素のような求電子剤の突然変異誘発および発癌効果は、主として、DNA中のグアニンのO6−アルキル化による。自身をDNAアルキル化から防護するために、哺乳動物および細菌は、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)というタンパク質を保有していて、それがこれらの損傷を修復する[Pegg et al., 1995]。AGTは、アルキル基を、SN2反応で、それ自身のシステインの一つに移転し、その結果、不可逆的にアルキル化された酵素を生じる。腫瘍細胞内でのAGTの過剰発現は、薬物耐性、特にプロカルバジン、ダカルバジン、テモゾロミドおよびビス−2−クロロエチル−N−ニトロソ尿素のようなアルキル化薬物に対するそれを獲得するのを可能にするため、AGTの阻害剤が、増感剤として化学療法に用いるために提唱されている[Pegg et al., 1995]。DE 199 03 895Aは、ビオチニル化されたO6−アルキルグアニン誘導体とAGTとの間の、AGTのビオチニル化へと導く反応に依拠する、AGTレベルを測定するアッセイを開示している。次いで、これは、ストレプトアビジン被覆プレート上でのAGTの分離と、例えばELISAアッセイでの、その検出とを可能にする。このアッセイは、腫瘍組織中のAGTレベルを追跡すること、AGT阻害剤を化学療法における増感剤として用いて処置を調整すること、およびAGT阻害剤についてスクリーニングするのに用いることが示唆されている。
【0004】
Damoiseaux、KepplerおよびJohnsson[ChemBiochem., 4: 285-287, 2001]は、やはり癌細胞内のこの酵素のレベルを検出するのを容易にして、研究および化学療法の際の助けにするために、AGT標識付けのための化学プローブのように用いるための、オリゴデオキシリボヌクレオチドに組み込んだ改質O6−アルキル化グアニン誘導体を開示している。2種類の変種AGT基質、およびビオチンで標識付けするAGT(DE 199 03 895 Aに記載されたのと同じ)についてのアッセイが開示されている。加えて、AGTの指向性進化にこれらのO6−アルキル化誘導体を用いることが示唆されている。
【0005】
発明の要約
広義には、本発明は、ある系内の対象となるタンパク質またはペプチドを標識付けし、場合により、次いで操作および/もしくは検出する方法に、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)をさらに用いることに関するものであって、該方法では、該タンパク質またはペプチドとAGTとの融合を、標識付けられた基質に接触させる結果、AGTが該標識を基質からAGT融合に移転し、そのため、移転された標識により、標識付けられたAGT−タンパク質融合が操作かつ/または検出されるのを可能にする。これは、AGTが融合タンパク質として存在しない、AGTレベル測定用アッセイの従来の用い方とは対照的である。
【0006】
上記により、第一の態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法を提供する。融合タンパク質への標識の移転後は、該方法は、標識付けられた融合タンパク質を検出および/または操作する、その後の工程をさらに包含してもよい。
【0007】
一実施態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を標識付けする方法であって、融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法を提供する。
【0008】
いくつかの実施態様では、該方法は、例えば、標識付けられた融合タンパク質を検出および/または操作する、その後の一つもしくはそれ以上の工程を含む。
【0009】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を検出する方法であって、該融合タンパク質と、標識を有する基質とを接触させる結果、AGTが、ラベルを、それが該融合タンパク質に共有結合させるように移転すること、および該標識を用いて、タンパク質構成体を検出することを含む方法を提供する。
【0010】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を操作する方法であって、該融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転すること、および融合タンパク質を、標識によって融合タンパク質に導入された物理学的および/または化学的特性を用いて操作することを含む方法を提供する。
【0011】
本発明のこの態様のいくつかの実施態様では、該方法は、標識を用いてタンパク質構成体を検出することを含んでよい。
【0012】
さらに一つの態様では、本発明は、対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質を固体支持体に固定化する方法であって、該融合タンパク質を、固体支持体に結合されるか、または結合され得る、標識を有する基質に接触させ、AGTが標識を、それが該融合タンパク質に共有結合させるように移転し、そのため、該融合タンパク質が固体支持体に結合されるか、またはその後に結合することができることを含む方法を提供する。本発明の、標識を初めに固体支持体に結合させない実施態様では、該方法は、標識付けられた融合タンパク質を固体支持体に接触させる結果、それが固体支持体上で固定化される工程をさらに含んでよい。本発明のこの態様のこの好ましい実施態様では、標識が移転されたときか、またはその後の反応の際のいずれかに、標識は、共有結合で固体支持体に結合してよいか、あるいは特異的結合対の一方の成員であってよくて、その他方の成員が、共有結合によってか、またはその他の手段によって(例えば、ビオチンおよびアビジンまたはストレプトアビジンの結合対を用いて)該固体支持体に結合されるか、もしくは結合され得る。
【0013】
さらに一つの態様では、本発明は、AGT融合タンパク質をインビボとインビトロとの双方で標識付けする方法を提供する。AGT融合タンパク質のインビボ標識付けという用語は、細胞のすべての区画内での標識付けはもとより、細胞外空間を指向するAGT融合タンパク質の標識付けも含む。AGT融合タンパク質の標識付けをインビボで実施し、AGTに融合されたタンパク質が原形質膜タンパク質であるならば、融合タンパク質のAGT部分は、原形質膜の細胞質側または細胞外側のいずれにも結合することができる。標識付けをインビトロで実施するならば、融合タンパク質の標識付けは、細胞抽出物中でか、またはAGT融合タンパク質の精製された形態もしくは富化された形態のいずれで実施することもできる。
【0014】
さらに一つの態様では、本発明は、候補化合物または候補化合物のライブラリーと、標的物質または標的物質のライブラリーとの相互作用を決定する方法を提供する。該化合物および物質の例は、リガンドおよびタンパク質、薬物および該薬物の標的、または小分子およびタンパク質を含む。この方法では、AGTに融合した対象となるタンパク質は、転写因子のDNA結合ドメイン、または転写因子の活性化ドメインを含み、標的物質または標的物質のライブラリーを、該DNA結合ドメインの他方、または該転写因子の活性化ドメインに結合させ、かつ標識が、標的物質と作用し合うと想定される候補化合物または候補化合物のライブラリーである。
【0015】
好ましい実施態様では、該方法は、候補化合物または候補化合物のライブラリーを、AGTタンパク質融合に移転すること、および候補化合物で標識付けしたAGT融合タンパク質と、標的物質とを接触させる結果、AGT融合タンパク質に結合した候補化合物と、標的物質との相互作用が、転写因子を活性化することをさらに含む。そうして、活性化された転写因子は、リポーターの発現を駆動し、それは、該方法を細胞内で実施するならば、リポーターの発現が細胞に選択的な利点を与えた場合に、検出することができる。いくつかの実施態様では、該方法は、候補化合物または標的物質を検出、単離、同定または特性評価することのような、追加的な一つもしくはそれ以上の工程を包含してもよい。
【0016】
本願では、O6−アルキルグアニン−DNAアルキルトランスフェラーゼまたは「AGT」は、基質に存在する標識を、融合タンパク質のAGT形成部分のシステイン残基の一つに移転するという特性を有する。好ましい実施態様では、AGTは、O6−アルキルグアニン−DNAアルキルトランスフェラーゼ、例えば、Peggら(1995)およびその中の引用文献に記載されている、ヒトO6−アルキルグアニン−DNAアルキルトランスフェラーゼである。しかし、他のアルキルグアニン−DNAアルキルトランスフェラーゼ、例えばRoyら(1995)に記載されているこの酵素のマウスまたはラット形態も公知であって、それらが上に定義された特性を有することを条件に、本発明に用いることができる。本発明では、O6−アルキルグアニン−DNAアルキルトランスフェラーゼは、一つまたはそれ以上のアミノ酸の置換、欠失もしくは付加によって異なり得るが、基質に存在する標識を、融合を形成しているAGTまたはタンパク質もしくはペプチドに移転するという特性は依然として保持している、野生型AGTの変種も含む。AGTのその他の変種は、当業者には周知の手法を用いて化学的に改質してもよい。AGTの変種は、当業者に公知のタンパク質工学の手法を用い、かつ/または新たなO6−アルキルグアニン−DNAアルキルトランスフェラーゼを発生かつ選択する分子的進化を用いて、生成してよい。
【0017】
本発明では、融合タンパク質のAGTを有するタンパク質部分の基準は、いかなる長さのタンパク質、ポリペプチドおよびペプチドを含み、かつ二次、三次または四次構造を保有するものと保有しないものとの双方を含む。本発明の適用の例を下記に示す。
【0018】
本発明では、標識付け基質は、好ましくは、標識付けベンジルグアニン基質であり、より好ましくは、O6−ベンジルグアニン誘導体である。そのような誘導体の例は、下記の一般式:
【0019】
【化3】
【0020】
[式中、
R1は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であり、好ましくは2〜99ヌクレオチドの長さを有するβ−D−2′−デオキシリボシルであり;
【0021】
R2は、リンカー基、例えば、置換または非置換アルキル鎖、ポリエチレングリコールのような柔軟なリンカーであり;そして
【0022】
labelは、本明細書に記載されたとおりの、融合タンパク質を検出かつ/または操作する原因となる分子である]
を有する、ベンジル環の4位が誘導体化された、O6−ベンジルグアニン誘導体である。
【0023】
本発明に従って用いるのに適した改質O6−ベンジルグアニン誘導体の例を、図1に示す。さらに、本発明者らは、AGTが、基質の同一性におけるかなりの程度の自由度を許容して、下記の一般式:
【0024】
【化4】
【0025】
[式中、
R1は、AGTによって受容されて、AGTが標識をタンパク質の融合へと移転するのを許す基、例えば、置換もしくは非置換アルキル鎖、環の大きさが3〜10炭素の置換もしくは非置換シクロアルキル基、環の大きさが3〜10炭素の置換もしくは非置換複素環、または環の大きさが3〜10炭素の置換もしくは非置換芳香族複素環であり;
【0026】
R2は、リンカー基、例えば、置換もしくは非置換アルキル鎖、またはポリエチレングリコールのような、様々な長さの柔軟なリンカーであり;そして
【0027】
R3は、プロトン、β−D−2′−デオキシリボシル、または好ましくは2〜99ヌクレオチドの長さを有する、オリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;
【0028】
labelは、本明細書に記載されたとおりの、融合タンパク質を検出かつ/または操作する原因となる分子である]
を有する、広範囲の基質が用いられるのを可能とすることができることを見出した。
【0029】
基質の標識部分は、融合タンパク質が意図される適用に応じて、当業者が選ぶことができる。標識の例は、下記のものを含む:
【0030】
(1)発蛍光団、発色団、磁気プローブまたは造量剤のような、分光学的プローブ;
【0031】
(2)放射性標識付け分子;
【0032】
(3)パートナーに特異的に結合できる特異的結合対の一方の部分である分子。そのような特異的結合対は、当技術に周知であり、例えば、アビジンまたはストレプトアビジンに結合することができるビオチンを含む;
【0033】
(4)その他の生体分子と作用し合うと思われる分子;
【0034】
(5)その他の生体分子と作用し合うと思われる分子のライブラリー;
【0035】
(6)当業者に公知のとおり[Nadeau et al., 2002]、その他の生体分子と架橋結合できる分子;
【0036】
(7)束縛された金属キレートのような[Hori et al., 2002]、H2O2およびアスコルビン酸塩に接触するとヒドロキシルラジカルを発生できる分子(最終原稿を確認されたい);
【0037】
(8)マラカイトグリーンのような[Jay et al., 1999]、光を照射すると反応性ラジカルを発生できる分子;
【0038】
(9)スライドグラス、微量滴定プレート、または一般に当業者に公知のいかなる重合体であってもよい、固体支持体に共有結合させる分子;
【0039】
(10)その相補鎖との塩基対合をなし得る核酸またはその誘導体;
【0040】
(11)脂質、または他の膜挿入特性を有する疎水性分子;
【0041】
(12)望ましい酵素的、化学的または物理的特性を有する生体分子;
【0042】
(13)上記に列挙された特性のいずれかの組合せを保有する分子。
【0043】
例示のために、ここで、本発明の実施態様を、添付の図面を参照して、より詳しく説明する。
【0044】
詳細な説明
本発明の下記の適用は、例示のために提供され、限定のためではない。本明細書に開示された方法は、一般的には、ある範囲の応用に適用することができ、融合タンパク質を、(1)標識付けられた融合タンパク質の環境における変化を感知かつ誘導できる標識、ならびに/または(2)標識が融合タンパク質に特異的に導入した物理および/もしくは化学的特性によって融合タンパク質を操作する助けとなる標識で、融合タンパク質を特異的に、かつ共有結合で標識化することができる。本明細書に開示された方法は、インビボとインビトロとの双方で、AGT融合タンパク質を標識付けするのに用いることができる。
【0045】
本発明は、所望のタンパク質への標識の特異的な結合は、問題のそのタンパク質と、ヒトのDNA修復酵素であるO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(hAGT)のようなO6−アルキルグアニン−DNAアルキルトランスフェラーゼとの間の融合タンパク質を構成し、後者の機序を利用することによって、実施することができるとの理解に基づく。この酵素は、アルキル基を、その基質、すなわちO6−アルキルグアニン−DNAからそのシステイン残基の一つに不可逆的に移転する(図1)。hAGTと急速に反応する基質類似体は、O6−ベンジルグアニンであって、二次速度定数は、約103秒-1M-1である(図1)。本発明者らは、O6−ベンジルグアニンのベンジル環のC4での置換が、O6−ベンジルグアニン誘導体に対するhAGTの反応性に有意には影響しないことを示した。これは、標識付けられたAGT融合タンパク質をインビボまたはインビトロで、共有結合によって特異的に結合するのに、標識がベンジル環のC4に結合したO6−ベンジルグアニン誘導体を用いることを可能にする(図1D)。標識付けは、融合タンパク質の性質には無関係である。
【0046】
標識付けをインビボまたは細胞抽出物中で実施するならば、宿主の内在AGTの標識付けを考慮に入れるのが好都合である。宿主の内在AGTがO6−ベンジルグアニン誘導体または関連化合物を基質として受容しないならば、融合タンパク質の標識付けは特異的である。哺乳動物細胞(ヒト、マウス、ラット)では、内在AGTの標識付けは可能である。内在AGTとAGT融合問題との同時標識付けが問題を提起する、これらの実験では、以前に記載したAGT欠乏細胞系を用いることができる[Kaina et al., 1991]。一般的には、本発明は、融合タンパク質との標識の共有結合による特異的な結合を用いて、AGT融合タンパク質の挙動を監視するか、もしくはそれに影響を与えるか、または導入された標識によってAGT融合タンパク質を操作する手法のすべての適用に利用することができる。この技術を用いるための適用の例を、以下に述べる。
【0047】
(1)分光学的プローブまたはリポーター基としての標識
AGTに移転できる、検出可能な標識、例えば発蛍光団、発色団、磁気プローブ、放射性標識付け分子またはその他のいかなる分光学的プローブをも基質が担持する、標識付けAGT基質、例えばO6−ベンジルグアニン誘導体の使用は、本発明を、検出できる標識を細胞内、細胞表面(インビボ)またはインビトロのいずれかでAGT融合タンパク質に特異的に共有結合によって結合するのに用いることを可能とする。これは、AGT融合タンパク質のインビボまたはインビトロでの検出および特性評価を可能とする。インビボという用語は、細胞のすべての区画内での標識付けはもとより、細胞外空間を指向するAGT融合タンパク質の標識付けも含む。この方法は、対象となるタンパク質にやはり遺伝学的に融合され、生細胞内でのその探索を可能とする、緑色蛍光タンパク質(GFP)の適用と対比することができる。GFPおよびその突然変異体の短所は、それが、主として、自然に産する発蛍光団の使用に限られることである。
【0048】
(2)AGT融合タンパク質を検出かつ単離するためのタグとしての標識
ビオチンのような親和性タグで標識付けしたAGT基質、例えばO6−ベンジルグアニン誘導体の使用は、親和性タグをAGT−タンパク質融合に移転し、そのため親和性タグの結合パートナーによって、融合タンパク質を結合するのに、本発明を用いることを可能にする。例としては、ビオチンのような親和性タグで標識付けしたAGT基質を、AGT融合タンパク質を発現する(細菌性または真核性の)細胞にか、またはそのような細胞の細胞抽出物、もしくは精製されたAGT融合タンパク質に付加することは、親和性タグによる融合タンパク質の共有結合的修飾へと導くことになる。そうして、これは、親和性タグとその結合パートナー、例えばビオチンの場合、固定化されたアビジンまたはストレプトアビジンとの間の相互作用を用いる、融合タンパク質の単離を可能にする。標識を、切断できる結合、例えばジスルフィド架橋を含有するリンカーを介してAGT融合タンパク質に結合するか、またはリンカーが光切断できるならば、AGT融合タンパク質は、親和性タグから、その単離後に遊離させることができる。
【0049】
(3)反応性ラジカルの源泉としての標識
AGT基質、例えばO6−ベンジルグアニン誘導体は、外部刺激に触れると反応性ラジカル、例えばヒドロキシルラジカルを発生できる、AGT融合タンパク質に、標識を導入するのに用いることができる。そうして、発生したラジカルは、AGT融合タンパク質はもとより、AGT融合タンパク質のごく近傍にあるようなタンパク質も不活性化することができて、これらのタンパク質の役割の研究を可能にする。そのような標識の例は、H2O2やアスコルビン酸塩に接触するとヒドロキシルラジカルを生ずる、束縛された金属−キレート錯体、およびレーザー照射されるとヒドロキシルラジカルを生ずるマラカイトグリーンのような、発色団である。ヒドロキシルラジカルを発生させるのに発色団やレーザーを用いることは、レーザー分子不活性化法(CALI)として当技術で公知である[Jay et al., 1998]。CALIは、細胞内の一定のタンパク質を、時間的に制御され、空間的に分解された方式で、特異的に不活性化するのに用いられる方法であり、発色団およびタンパク質の空間的近傍を基礎にしている。レーザー照射されると、発色団は、ヒドロキシルラジカルを発生し、それが、発色団内、およびそのわずか約0.1nm以内のすべてのタンパク質を不活性化する。従来、発色団は、対象となるタンパク質の空間的近傍に、対象となるタンパク質に特異的な発色団標識付け抗体を微量注入することによって近付けている。本発明では、マラカイトグリーンのような発色団によるAGT融合タンパク質の標識付け、およびその後のレーザー照射は、AGT融合タンパク質はもとより、AGT融合タンパク質と作用し合うようなタンパク質も、時間的に制御され、空間的に分解された方式で不活性化するのを可能にすると思われる。この方法は、インビボまたはインビトロの双方で適用することができる。
【0050】
同様にして、AGT融合タンパク質は、束縛された金属−キレートで標識付けすることができ、AGT融合タンパク質、およびAGT融合タンパク質と作用し合うようなタンパク質は、H2O2およびアスコルビン酸塩に接触させると、特異的な方式で不活性化することができる。この方法は、AGT融合タンパク質、またはAGT融合タンパク質のごく近傍にあるそれの機能を調べることばかりでなく、AGT融合タンパク質のごく近傍にあるようなタンパク質を同定することにも用いることができる。ここで、AGT融合タンパク質のごく近傍にあるタンパク質は、該タンパク質のフラグメントの特異的な抗体による検出、高分離能2D電気泳動ゲル上のこれらのタンパク質の消滅、または切断されたタンパク質フラグメントの、分離、および質量分析法もしくはN末端分解によるタンパク質配列決定のような配列決定手法による同定のいずれかによって同定されるようなものとして同定することができる。
【0051】
(4)タンパク質−タンパク質相互作用を仲介するリガンドとしての標識
標識付けられたAGT基質、例えばO6−ベンジルグアニン誘導体は、リガンドをAGT−タンパク質融合に移転するのに用いることができる。これは、リガンドのパートナー、例えばタンパク質をAGT−タンパク質融合に結合するのを可能にする。例えば、標識が、結合パートナーに結合できるリガンドである場合、そのような基質とAGT融合タンパク質とを接触させることは、融合タンパク質とのリガンドの特異的結合へと導くことになる。リガンドが、もう一つのタンパク質Yに結合し、タンパク質Yと標識付けられたAGT融合タンパク質との二量体化が、生物学的機能、または測定できるシグナルへと導くならば、この生物学的機能または測定可能シグナルは、標識を担持するAGT基質の付加に依存する。具体的な例は、小分子で遺伝子発現を調節するための、Hoら、1996が記載した、いわゆる三雑種系におけるAGT基質およびAGT融合タンパク質の使用であると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。リガンド、例えばFK506に結合するタンパク質Y、例えばFKBPを、転写因子の活性化ドメインに結合させる。リガンド、この例ではFK506を担持するAGT基質を細胞に供給することが、機能的な転写因子の形成、および遺伝子発現へと導くことになる。
【0052】
(5)その標的が未知である薬物または生物学的活性分子としての標識
標識を担持し、その標識が、未だ同定されていないタンパク質Yに結合する薬物または生物学的活性小分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。ここでは、目標は、生物学的活性分子の標的であるタンパク質Yを同定することになると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。未知の標的タンパク質Yを発現する生物のcDNAライブラリーを、転写因子の活性化ドメインに融合させる。標識を担持し、その標識が薬物または生物学的活性小分子である、O6−ベンジルグアニン誘導体または関連AGT基質の付加が、この分子が、cDNAライブラリー内に存在し、活性化ドメインに融合した、その標的タンパク質Yに結合する場合にのみ、機能的転写因子の形成、および遺伝子発現へと導くことになる。遺伝子発現を選択的な利点に結び付けるならば、該薬物または生体活性分子の標的遺伝子Yを有するプラスミドを担持する、対応する宿主を同定することができる。
【0053】
(6)タンパク質Yに結合する分子を同定するための小分子のライブラリーとしての標識
標識を担持し、その標識が、化学的分子のライブラリーである、O6−ベンジルグアニンまたは関連AGT基質の使用:ここでは、目標は、インビボ条件下でタンパク質Yに結合する、潜在的な薬物標的であり得る小分子を同定することになると思われる。この場合は、AGTを、転写因子のDNA結合ドメインに融合させる。標的タンパク質Yを、転写因子の活性化ドメインに融合させる。標識として結合させた小分子のライブラリーを、O6−ベンジルグアニン誘導体に付加することが、標識(すなわち小分子)が、活性化ドメインに融合したその標的タンパク質Yに結合する場合にのみ、機能的転写因子の形成、および遺伝子発現へと導くことになる。遺伝子発現を選択的な利点に結び付けるならば、ライブラリーの、宿主の成長へと導くような分子を同定することができる。
【0054】
(7)AGT融合タンパク質を固定化し、かつ/またはAGT融合タンパク質のタンパク質アレーを創出するためのBG誘導体の使用
標識を担持し、その標識が、担体の表面に共有結合で結合されているか、または該標識が、それ自体該表面に結合されたもう一つの分子によって非共有結合で結合され得る分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。後者の取組み方についての例は、標識がビオチンであり、表面に結合された分子がストレプトアビジンまたはアビジンである場合である。担体についてのあり得る例は、スライドガラス、微量滴定プレート、または機能化された任意の重合体のいずれかでもあると思われる。標識によるAGT基質の固定化は、標識の融合タンパク質への移転によって、担体へのAGT融合タンパク質のその後の固定化を可能にする。空間的に分解された方式での(異なる)AGT融合タンパク質の担体でのスポット形成は、タンパク質アレーの生成を可能にする。
【0055】
(8)AGT融合タンパク質と作用し合うタンパク質を検出するための架橋結合剤としての標識
標識を担持し、その標識が、他のタンパク質と架橋結合することができる分子である、O6−ベンジルグアニン誘導体または関連AGT基質の使用。そのような架橋結合剤の例は、マレイミド、活性エステルまたはアジ化物およびその他の、当業者に公知であり、Nadeauら、2002に記載されたもののような、官能基を含有する分子である。そのようなAGT基質を、他のタンパク質と作用し合うAGT融合タンパク質と(インビボまたはインビトロで)接触させることは、AGT融合タンパク質と、標識を介して作用し合うそのタンパク質との共有結合での架橋結合へと導くことができる。これは、AGT融合タンパク質と作用し合うタンパク質の同定を可能にする。
【0056】
実施例
下記の実施例は、本発明を実施する方法の完全な開示および記載を当業者に提供するために説明するもので、本発明の対象範囲を限定することを意図するものではない。
【0057】
実施例A:大腸菌におけるAGT融合タンパク質の共有結合による標識付け
下記の例、すなわちBG−Btを用いた、大腸菌におけるPep−hAGTの標識付けは、大腸菌におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。hAGTのN末端に融合したペプチド(Pep−hAGTを生じる)の配列は、(一字コードでは)MHHHHHHSSAであって、hAGTの最初のアミノ酸である、メチオニンがこれに続く。N末端融合ペプチドを有するhAGT、すなわちPep−hAGTをコーディングしている、pET−15b(Novagen)に基づく発現ベクターを含有するXL−1Blueという大腸菌細胞の液体培養物を、0.6の光学的密度OD600nmまで増殖させた。IPTGを1mMの最終濃度までを加えることによって、Pep−hAGTの発現を誘導した。同時に、BG−Btを10μMの最終濃度まで加え、細菌を37℃で2時間温置した。細胞を遠心分離によって採集し、ペレットを2回洗浄して、過剰なBG−Btを除去した。再懸濁させた細胞のアリコートを、ウエスタンブロット分析によって分析した。ビオチニル化タンパク質を、ストレプトアビジン−ペルオキシダーゼ複合体(NEN)および化学発光性ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用いて検出した(図2)。
【0058】
実施例B:酵母におけるAGT融合タンパク質の共有結合による標識付け
下記の実施例は、酵母におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、BG−Btを用いて、hAGT−DHFR−HAという融合タンパク質を酵母中でビオチニル化した。この融合タンパク質は、標準的な分子生物学の手順を用いて、DNAレベルで構成される。略述すると、hAGTの停止コドンを、アミノ酸RSGIに対するコドンで置き換え、次いで、マウスからのDHFRの第1アミノ酸、すなわちメチオニンに対するコドンで後続させた[Nunberg et al., 1980]。hAGTとDHFRとの間のリンカーに対するコドンは、BglII部位もコード化していて、そのDNA配列は、AGATCTである。DHFRとHAタグとの間の融合を構成するために、DHFRの停止コドンを、HA−タグの第1アミノ酸に対するコドンで置き換えた[Kolodziej, 1991]。HA−タグに停止コドンを後続させた。hAGT−DHFR−HAというタンパク質がpcup1プロモーターの制御下にある、発現ベクターp314AK1を含有する、L40という酵母細胞の培養を、0.6のOD600まで増殖させた。hAGT−DHFR−HAの発現は、CuSO4を100μMの濃度まで加えることによって誘導し、BG−Btを、10μMの濃度まで同時に加えた。2.5時間および5時間後にアリコートを採取し、遠心分離によって、細胞を採集した。ペレットを2回洗浄して、残留BG−Btを除去した。凍結/融解周期の循環によって酵母細胞を溶解した後、細胞抽出物を、ELISAを用いて、ビオチニル化hAGT−DHFR−HA融合タンパク質の存在について分析した。略述すると、ビオチニル化hAGT−DHFR−HAを、ストレプトアビジン被覆した微量滴定ウェル内に固定化し、抗HA抗体(Babco)を一次抗体として、かつ抗マウスHRP複合体(Sigma)を二次抗体として用いることによって検出した(図1)[Kolodziej, 1991]。ELISAは、ペルオキシダーゼ基質ABTSを用いて展開し、シグナル(吸光度)をOD405nmで測定した。インビボビオチニル化hAGT−DHFR−HA融合タンパク質についてのシグナルは、バックグラウンドを少なくとも5倍上回った。バックグラウンドシグナルは、厳密に上記のとおり処理したが、BG−Btの添加を省いた細胞から得られた、細胞溶解物のOD405nmとして定義した。
【0059】
実施例C:酵母におけるhAGT融合タンパク質の共有結合による標識付け
下記の実施例は、酵母におけるhAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、BG−DIGを用いて、hAGT−DHFR−HA融合タンパク質を酵母中でジゴキシゲニンで標識付けした。hAGT−DHFR−HA融合の構成は、実施例Bに記載されている。hAGT−DHFR融合タンパク質の遺伝子がpcup1プロモーターの制御下にある、発現ベクターp314AK1を含有する、L40という酵母細胞の培養を、1.2のOD600nmまで増殖させた。hAGT−DHFR融合タンパク質の発現は、CuSO4を100μMの濃度まで加えることによって誘導し、BG−DIGを、20μMの濃度まで同時に加えた。2時間後、振盪フラスコ培養体1mlからの細胞を、遠心分離によって採集した。ペレットを培地で3回洗浄して、残留BIG−DIGを除去した。凍結/融解周期の循環によって酵母細胞を溶解した後、細胞抽出物を、ジゴキシゲニン化hAGT−DHFR融合タンパク質の存在について、ウエスタンブロット分析によって分析した。ジゴキシゲニン化タンパク質は、抗ジゴキシゲニン−ペルオキシダーゼ複合体(Roche)、および化学発光性ペルオキシダーゼ基質(Renaissance reagent plus, NEN)を用いて検出した(図2)。
【0060】
実施例D:ヒト細胞系におけるhAGT融合タンパク質の共有結合による標識付け
下記の実施例は、哺乳動物細胞におけるAGT融合タンパク質の共有結合による標識付けの実施可能性を立証した。ここでは、ヒトの細胞(HEK293)内の内在性hAGTを、BG−AcFcを用いてフルオレセインで標識付けした。HEK293細胞を、5μMのBG−AcFcとともにPBS中に5分間温置した。アセチル化されたフルオレセイン誘導体であるBG−AcFcは、細胞浸透性であり、非蛍光性であるが、細胞内で急速に水解されて、蛍光性のBG−Fcを生じることが期待できる。次いで、細胞を、PBSを変えることによって洗浄して、いかなる過剰な基質BG−AcFcも除去し、PBS中に20分間温置した。次いで、共焦点蛍光顕微鏡で画像を得た(励起492nm;発光510nm)。対照実験として、HEK293細胞を上記のとおり処理したが、BG−AcFcを加える前に、O6−ベンジルグアニン(1μM)とともに終夜温置した。これは、内在性hAGTを不活性化し、そのため核内での蛍光の蓄積を防止するはずであった。期待されたとおり、細胞をO6−ベンジルグアニンとともに前温置したとき、核内での蛍光蓄積は、全く観察されなかった。hAGTがBG−Fcを基質として受容することを、独立に確認するため、組換えPep−hAGT(10μM、実施例Aに記載のとおり)を、100μMのBG−Fcとともに25℃で、50mMトリス−Cl、10mMDTT、1mMEDTA、200μg/mlのBSA、10%グリセリン、pH7.4中で10分間温置した後、PBS(リン酸緩衝化生理食塩水:137mMNaCl、2.7mMKCl、10mMNa2HPO4、1.8mMKH2PO4、pH7.4)900μlを加えた。過剰な基質BG−Fcの分離は、NAP(登録商標)−10カラム(Pharmacia)で、供給者の指示によるゲル濾過によって達成した。次いで、Pep−hAGTを、標準的な蛍光分光光度計で特性評価した。サンプルは、それぞれ、222、238および490nmで励起し、波長523nmで最大発光を示し、このタンパク質が、フルオレセインで標識付けられたことを確認した。PBS中20nMのBG−Fcの溶液を、参照として測定した。基質の発光波長は、519nmであった(励起は、それぞれ、237、323および490nm)。
【0061】
実施例E:細胞抽出物中でのAGT融合タンパク質の共有結合による標識付け
AGTとの融合タンパク質は、細胞抽出物中では直接標識付けし、かつ操作(ここでは固定化)することができることを立証するため、下記のhAGTとのN−およびC−末端融合タンパク質を、標準的な分子クローニングの手順によって構成し、酵母の発現ベクター内にクローニングした:
【0062】
(i)V5−NLS−B42−hAGT(ここで、V5はV5エピトープを、NLSはSV40の大T抗原核局在配列を、またB42は、人工転写活性化因子であるB42[Ma et al., 1987]を意味する)。B42のトランス活性化ドメインの最後のコドンに、21アミノ酸の配列ASKKGTELGSTTSNGRQCAGILを後続させた。最後の3コドンは、B42にhAGTをC末端クローニングするためのEcoRI部位を含んでいる。NotI部位は、hAGTに対するC末端制限部位であって、その配列は、停止コドンを含む;
【0063】
(ii)hAGT−HA−Ura3(ここで、Ura3は、酵母の酵素オロチン酸デカルボキシラーゼを意味し、HAは、Haエピトープを意味する)。ここでは、hAGTに対する停止コドンを、RSリンカーで置き換え、HA−タグの最初のアミノ酸を後続させた。HA−タグは、Ura3遺伝子が直接後続する;
【0064】
(iii)hAGT−DHFR−HA(ここで、DHFRは、マウスのジヒドロ葉酸レダクターゼを意味し、HAは、Haエピトープを意味する)。構成については、実施例Bを参照されたい;および
【0065】
(iv)SSN6−hAGT(ここで、SSN6は、酵母のDNA転写リプレッサー[Schultz et al., 1987]を意味する)。ここでは、hAGTの停止コドンを、アミノ酸RSGSGに対するコドンで置き換えて、そうして、酵母のSSN6の最初のアミノ酸、すなわちメチオニンに対するコドンが後続する。
【0066】
遺伝子の発現は、すべて、pcup1プロモーターによって制御した。融合タンパク質の一つをコーディングしている発現ベクターを含有するL40酵母細胞を、0.6のOD600まで増殖させ、融合タンパク質の発現は、CuSO4を100μMの濃度まで加えることによって誘導した。アリコート(2ml)を5時間後に採取し、遠心分離によって細胞を採集した。凍結/融解周期の循環によって細胞を溶解した後、酵母抽出物を、BG−Bt−oligo(10pmol)とともに室温で20分間温置して、融合タンパク質のビオチニル化へと導いた。次いで、懸濁液を、ストレプトアビジン被覆した微量滴定プレート(Roche molecular biochemicals)内に移し、1時間温置した。ウェルをPBSで徹底的に洗浄した後、固定化された融合タンパク質を、抗HA抗体(Babco)または抗hAGT抗体(SSN6−hAGT融合タンパク質の場合)のいずれかを一次抗体として、かつ抗マウスペルオキシダーゼ複合体(Sigma、#A4416)を二次抗体として用い、次いで、標準的な生化学手順を用いて、ペルオキシダーゼの基質ABTSとともに温置することによって、検出した。すべての場合に、OD405nmとして測定されたシグナルは、バックグラウンドを少なくとも5倍上回った。バックグラウンドは、各融合タンパク質について、BG−Bt−oligoの細胞抽出物への添加を省くことによって測定した。
【0067】
参考文献
本明細書に引用された参考文献は、すべて、特に参照として組み込まれる。参考文献は、アルファベット順である。
【0068】
【表1】
【図面の簡単な説明】
【0069】
【図1】(A)O6−アルキルグアニン−DNAアルキルトランスフェラーゼの機序を示す図である。(B)O6−ベンジルグアニンの構造を示す図である。(C)実施例に用いたO6−ベンジルグアニン誘導体の一般的構造を示す図である。(D)AGT融合タンパク質を標識付けする一般的スキームであって、Xは、AGTに融合させたタンパク質である。(E)実施例に用いたAGT基質の構造を示す図である。オリゴヌクレオチド(22量体)の配列は、5′-GTGGTGGGCGCTGXAGGCGTGG-3′であって、X=BG−Btである。
【図2】インビボでAGT融合タンパク質を標識付けした後のウエスタンブロット分析を示す図である。(A)培地中にBG−Btを含む場合と含まない場合の、Pep−hAGTを発現する大腸菌の全細胞抽出物のウエスタンブロット分析。ストレプトアビジン−ペルオキシダーゼ複合体を用いて、ビオチニル化タンパク質を検出。20kDの帯域は、BG−Btの不在下での大腸菌内でビオチニル化タンパク質に相当する。(B)培地中にBG−DIGを含む場合と含まない場合の、hAGT−DHFR−HA融合タンパク質を発現する酵母の全細胞抽出物のウエスタンブロット分析。抗ジゴキシゲニン−ペルオキシダーゼ複合体を用いて、ジゴキシゲニン標識付けタンパク質を検出。
【図3】BG−AcFcとともに温置し、蛍光および光学顕微鏡測定によって分析したHEK293細胞を示す図である。蛍光が細胞核に蓄積されているのを認めることができる。これは、hAGTが核に局在するのが知られていることから[Ali et al., 1998]、期待された結果である。第三の図面は、蛍光および光学顕微鏡測定から得られた画像を重ね合わせたものである。
【特許請求の範囲】
【請求項1】
対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法。
【請求項2】
さらに、融合タンパク質を標識を用いて検出することを含む、請求項1に記載の方法。
【請求項3】
標識付けられた融合タンパク質をインビトロ系内で検出することを含む、請求項2に記載の方法。
【請求項4】
インビトロ系内で、標識付けを細胞抽出物中でか、または融合タンパク質の精製もしくは富化された形態を用いて実施する、請求項3に記載の方法。
【請求項5】
標識付けられた融合タンパク質をインビボ系内で検出することを含む、請求項2に記載の方法。
【請求項6】
インビボ系が細胞である、請求項5に記載の方法。
【請求項7】
さらに、対照配列に連結された融合タンパク質をコーディングしている核酸を含む発現ベクターで細胞を形質転換し、その発現を導くという最初の工程を含む、請求項6に記載の方法。
【請求項8】
さらに、標識付けられた融合タンパク質を、標識によって融合タンパク質に導入された特性を用いて操作することを含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
対象となるタンパク質を、AGTのN−末端またはC−末端に融合させる、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、ヒトのO6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、マウスまたはラットのO6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、一つ以上のアミノ酸の置換、欠失もしくは付加によってヒト、ラットまたはマウスO6−アルキルグアニン−DNAアルキルトランスフェラーゼと異なるアミノ酸配列を有するが、基質に存在する標識をAGT−タンパク質融合に移転するという特性は保持している突然変異O6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
標識を融合タンパク質のシステイン残基に共有結合で結合させる、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
標識付けられた基質を核酸分子に組み込む、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
核酸分子が2〜99ヌクレオチドの長さのオリゴヌクレオチドである、請求項14に記載の方法。
【請求項16】
標識付けられた基質がベンジルグアニン基質である、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
ベンジルグアニン基質が、ベンジル環のC4位が標識で置換されている、請求項16に記載の方法。
【請求項18】
標識付けベンジルグアニン基質が、一般式:
【化1】
[式中、
R1は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;
R2は、リンカー基であり;そして
labelは、融合タンパク質を検出かつ/または操作するのに用いられる基を表す]
で示される、請求項16または17に記載の方法。
【請求項19】
リンカー基R2が、置換もしくは非置換アルキル鎖またはポリエチレングリコールである、請求項18に記載の方法。
【請求項20】
標識付けられたベンジルグアニン基質が、一般式:
【化2】
[式中、
R1は、AGTによって受容されて、AGTが標識をタンパク質融合へと移転するのを許す基であり;
R2は、リンカー基であり;
R3は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;そして
labelは、融合タンパク質を検出かつ/または操作するのに用いられる基を表す]
で示される、請求項18または19に記載の方法。
【請求項21】
R1が、置換もしくは非置換アルキル鎖、環の大きさが3〜10炭素の置換もしくは非置換シクロアルキル基、環の大きさが3〜10炭素の置換もしくは非置換複素環、または環の大きさが3〜10炭素の置換もしくは非置換芳香族複素環である、請求項20に記載の方法。
【請求項22】
リンカー基R2が、置換もしくは非置換アルキル鎖またはポリエチレングリコールである、請求項20または21に記載の方法。
【請求項23】
オリゴデオキシリボヌクレオチドが2〜99ヌクレオチドの長さを有する、請求項18〜22のいずれか1項に記載の方法。
【請求項24】
標識が、分光学的プローブ、親和性タグ、特異的な結合対の第二の成員に特異的に結合できる、特異的結合対の第一の成員、反応性ラジカルを発生できる分子、固相であるか、または固相に結合されるか、もしくは固相に結合され得る分子、候補化合物もしくは候補化合物のライブラリー、他の生体分子に架橋結合できる分子、その相補鎖と塩基対合をなし得る核酸もしくはその誘導体、脂質、あるいは膜挿入特性を有する疎水性分子のうち一つ以上である、請求項1〜23のいずれか1項に記載の方法。
【請求項25】
分光学的プローブが、発蛍光団、染料もしくは造量剤、放射性標識付け分子、または磁気プローブである、請求項24に記載の方法。
【請求項26】
分光学的プローブを用いて、融合タンパク質をインビトロもしくはインビボで検出するか、または特性評価する、請求項24または25に記載の方法。
【請求項27】
親和性タグがビオチン、アビジンまたはストレプトアビジンである、請求項24に記載の方法。
【請求項28】
親和性タグを、切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を該親和性タグから遊離させることができる、請求項24または27に記載の方法。
【請求項29】
親和性タグを、光切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を親和性タグから遊離させることができる、請求項24または27に記載の方法。
【請求項30】
融合タンパク質を、固体支持体に結合されるか、または結合され得る標識付け基質に接触させることによって、融合タンパク質を固定化することをさらに含んで、AGTが標識を、それが該融合タンパク質に共有結合で結合され、そのため、標識付けられた融合タンパク質が固体支持体に結合されるか、またはその後に結合され得るように移転する、請求項1〜29のいずれか1項に記載の方法。
【請求項31】
さらに、標識を初めに固体支持体に結合させず、該方法が、標識付けられた融合タンパク質を固体支持体に接触させる結果、それが固体支持体上で固定化される工程を含む、請求項30に記載の方法。
【請求項32】
標識が特異的結合対の一方の成員であり、その他方の成員を、固体支持体に結合させるか、または結合させ得る、請求項30または31に記載の方法。
【請求項33】
特異的結合対がビオチンおよびアビジンまたはストレプトアビジンである、請求項32に記載の方法。
【請求項34】
標識を、該標識の移転によってか、またはその後の反応中に、固体支持体に共有結合で結合させる、請求項30に記載の方法。
【請求項35】
標識を、切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を、固体支持体への固定化後に標識から遊離させることができる、請求項30〜34のいずれか1項に記載の方法。
【請求項36】
標識を、光切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を、固体支持体への固定化後に標識から遊離させることができる、請求項30〜34のいずれか1項に記載の方法。
【請求項37】
対象となる複数のタンパク質を固体支持体に固定化することを含む、請求項30〜36のいずれか1項に記載の方法。
【請求項38】
対象となるタンパク質を、整列させて固体支持体に固定化する、請求項37に記載の方法。
【請求項39】
アレーが、複数の空間分解位置を含む、請求項38に記載の方法。
【請求項40】
固体支持体が、ガラスまたは重合体を含む、請求項30〜39のいずれか1項に記載の方法。
【請求項41】
反応性ラジカルを発生できる分子が、反応性ヒドロキシルラジカルを発生する、請求項24に記載の方法。
【請求項42】
反応性ヒドロキシルラジカルを発生できる分子が、H2O2およびアスコルビン酸塩に接触するとラジカルを発生する、束縛された金属キレートである、請求項41に記載の方法。
【請求項43】
反応性ヒドロキシルラジカルを発生できる分子が、光を照射するとラジカルを発生する、請求項41に記載の方法。
【請求項44】
反応性ヒドロキシルラジカルを発生できる分子がマラカイトグリーンである、請求項43に記載の方法。
【請求項45】
ラジカルが、融合タンパク質、または融合タンパク質の近傍に位置するもう一つのタンパク質を不活性化する、請求項24または41〜44のいずれか1項に記載の方法。
【請求項46】
ラジカルが、タンパク質のフラグメント、または融合タンパク質の近傍に位置するもう一つのタンパク質を発生する、請求項41〜45のいずれか1項に記載の方法。
【請求項47】
フラグメントの発生を、融合タンパク質のごく近傍のタンパク質の同定に用いる、請求項46記載の方法。
【請求項48】
タンパク質またはもう一つのタンパク質の不活性化が、レーザー分子不活性化法(CALI)を用いる、請求項45に記載の方法。
【請求項49】
標識が、特異的結合対の第一の成員である、請求項24に記載の方法。
【請求項50】
さらに、融合タンパク質を、特異的結合対の第二の成員を含む分子に接触させることを含む、請求項49に記載の方法。
【請求項51】
特異的結合対の第一および第二の成員の相互作用または結合が、検出できる結果を生じる、請求項49または50に記載の方法。
【請求項52】
特異的結合対の第一および第二の成員が、第一および第二のタンパク質、抗体および抗原、酵素および基質、またはリガンドおよび受容体である、請求項49〜51のいずれか1項に記載の方法。
【請求項53】
AGTに融合した対象となるタンパク質が、転写因子のDNA結合ドメイン、または転写因子の活性化ドメインを含み、標的物質または標的物質のライブラリーを、該DNA結合ドメインまたは該転写因子の活性化ドメインの他方に結合させ、かつ標識が、標的物質と作用し合うと想定される候補化合物または候補化合物のライブラリーである、請求項1〜24のいずれか1項に記載の方法。
【請求項54】
候補化合物または候補化合物のライブラリー、および標的物質または標的物質のライブラリーが、リガンドおよびタンパク質、薬物および該薬物の標的、小分子およびタンパク質、ならびに酵素およびその基質または阻害剤から選択される、請求項53に記載の方法。
【請求項55】
候補化合物または候補化合物のライブラリーを、AGTタンパク質融合に移転することを含む、請求項53または54に記載の方法。
【請求項56】
候補化合物で標識付けしたAGT融合タンパク質と、標的物質とを接触させる結果、AGT融合タンパク質に結合した候補化合物と、標的物質との相互作用が、転写因子を再構成かつ活性化することをさらに含む、請求項55に記載の方法。
【請求項57】
活性化された転写因子が、リポーターの発現を発動する、請求項56に記載の方法。
【請求項58】
リポーターが、測定できるシグナルへとその活性が導くタンパク質、酵素または核酸である、請求項57に記載の方法。
【請求項59】
該方法が、細胞内で実施され、リポーターの発現が、選択的な利点を細胞に与える、請求項53〜58のいずれか1項に記載の方法。
【請求項60】
AGT融合タンパク質、および/またはAGTタンパク質融合と作用し合う標的物質を単離することをさらに含む、請求項53〜59のいずれか1項に記載の方法。
【請求項61】
標的物質と作用し合う候補化合物を検出かつ/または同定することをさらに含む、請求項53〜60のいずれか1項に記載の方法。
【請求項62】
標識が、タンパク質を架橋結合することができる、請求項24に記載の方法。
【請求項63】
架橋結合する標識が、マレイミド基、エステル基またはアジド基を含む、請求項62記載の方法。
【請求項64】
融合タンパク質を、候補タンパク質の結合パートナーに接触させ、架橋結合する標識を反応させる結果、作用し合う融合タンパク質と候補タンパク質とが互いに共有結合させる、請求項62または63に記載の方法。
【請求項65】
架橋結合した融合タンパク質および/または候補タンパク質を検出かつ/もしくは同定することをさらに含む、請求項62〜64のいずれか1項に記載の方法。
【請求項1】
対象となるタンパク質およびO6−アルキルグアニン−DNAアルキルトランスフェラーゼ(AGT)を含む融合タンパク質と、標識を有する基質とを接触させる結果、AGTが標識を、それが該融合タンパク質に共有結合させるように移転することを含む方法。
【請求項2】
さらに、融合タンパク質を標識を用いて検出することを含む、請求項1に記載の方法。
【請求項3】
標識付けられた融合タンパク質をインビトロ系内で検出することを含む、請求項2に記載の方法。
【請求項4】
インビトロ系内で、標識付けを細胞抽出物中でか、または融合タンパク質の精製もしくは富化された形態を用いて実施する、請求項3に記載の方法。
【請求項5】
標識付けられた融合タンパク質をインビボ系内で検出することを含む、請求項2に記載の方法。
【請求項6】
インビボ系が細胞である、請求項5に記載の方法。
【請求項7】
さらに、対照配列に連結された融合タンパク質をコーディングしている核酸を含む発現ベクターで細胞を形質転換し、その発現を導くという最初の工程を含む、請求項6に記載の方法。
【請求項8】
さらに、標識付けられた融合タンパク質を、標識によって融合タンパク質に導入された特性を用いて操作することを含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
対象となるタンパク質を、AGTのN−末端またはC−末端に融合させる、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、ヒトのO6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、マウスまたはラットのO6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
O6−アルキルグアニン−DNAアルキルトランスフェラーゼが、一つ以上のアミノ酸の置換、欠失もしくは付加によってヒト、ラットまたはマウスO6−アルキルグアニン−DNAアルキルトランスフェラーゼと異なるアミノ酸配列を有するが、基質に存在する標識をAGT−タンパク質融合に移転するという特性は保持している突然変異O6−アルキルグアニン−DNAアルキルトランスフェラーゼである、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
標識を融合タンパク質のシステイン残基に共有結合で結合させる、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
標識付けられた基質を核酸分子に組み込む、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
核酸分子が2〜99ヌクレオチドの長さのオリゴヌクレオチドである、請求項14に記載の方法。
【請求項16】
標識付けられた基質がベンジルグアニン基質である、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
ベンジルグアニン基質が、ベンジル環のC4位が標識で置換されている、請求項16に記載の方法。
【請求項18】
標識付けベンジルグアニン基質が、一般式:
【化1】
[式中、
R1は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;
R2は、リンカー基であり;そして
labelは、融合タンパク質を検出かつ/または操作するのに用いられる基を表す]
で示される、請求項16または17に記載の方法。
【請求項19】
リンカー基R2が、置換もしくは非置換アルキル鎖またはポリエチレングリコールである、請求項18に記載の方法。
【請求項20】
標識付けられたベンジルグアニン基質が、一般式:
【化2】
[式中、
R1は、AGTによって受容されて、AGTが標識をタンパク質融合へと移転するのを許す基であり;
R2は、リンカー基であり;
R3は、プロトン、β−D−2′−デオキシリボシル、またはオリゴデオキシリボヌクレオチドの一部であるβ−D−2′−デオキシリボシルであり;そして
labelは、融合タンパク質を検出かつ/または操作するのに用いられる基を表す]
で示される、請求項18または19に記載の方法。
【請求項21】
R1が、置換もしくは非置換アルキル鎖、環の大きさが3〜10炭素の置換もしくは非置換シクロアルキル基、環の大きさが3〜10炭素の置換もしくは非置換複素環、または環の大きさが3〜10炭素の置換もしくは非置換芳香族複素環である、請求項20に記載の方法。
【請求項22】
リンカー基R2が、置換もしくは非置換アルキル鎖またはポリエチレングリコールである、請求項20または21に記載の方法。
【請求項23】
オリゴデオキシリボヌクレオチドが2〜99ヌクレオチドの長さを有する、請求項18〜22のいずれか1項に記載の方法。
【請求項24】
標識が、分光学的プローブ、親和性タグ、特異的な結合対の第二の成員に特異的に結合できる、特異的結合対の第一の成員、反応性ラジカルを発生できる分子、固相であるか、または固相に結合されるか、もしくは固相に結合され得る分子、候補化合物もしくは候補化合物のライブラリー、他の生体分子に架橋結合できる分子、その相補鎖と塩基対合をなし得る核酸もしくはその誘導体、脂質、あるいは膜挿入特性を有する疎水性分子のうち一つ以上である、請求項1〜23のいずれか1項に記載の方法。
【請求項25】
分光学的プローブが、発蛍光団、染料もしくは造量剤、放射性標識付け分子、または磁気プローブである、請求項24に記載の方法。
【請求項26】
分光学的プローブを用いて、融合タンパク質をインビトロもしくはインビボで検出するか、または特性評価する、請求項24または25に記載の方法。
【請求項27】
親和性タグがビオチン、アビジンまたはストレプトアビジンである、請求項24に記載の方法。
【請求項28】
親和性タグを、切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を該親和性タグから遊離させることができる、請求項24または27に記載の方法。
【請求項29】
親和性タグを、光切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を親和性タグから遊離させることができる、請求項24または27に記載の方法。
【請求項30】
融合タンパク質を、固体支持体に結合されるか、または結合され得る標識付け基質に接触させることによって、融合タンパク質を固定化することをさらに含んで、AGTが標識を、それが該融合タンパク質に共有結合で結合され、そのため、標識付けられた融合タンパク質が固体支持体に結合されるか、またはその後に結合され得るように移転する、請求項1〜29のいずれか1項に記載の方法。
【請求項31】
さらに、標識を初めに固体支持体に結合させず、該方法が、標識付けられた融合タンパク質を固体支持体に接触させる結果、それが固体支持体上で固定化される工程を含む、請求項30に記載の方法。
【請求項32】
標識が特異的結合対の一方の成員であり、その他方の成員を、固体支持体に結合させるか、または結合させ得る、請求項30または31に記載の方法。
【請求項33】
特異的結合対がビオチンおよびアビジンまたはストレプトアビジンである、請求項32に記載の方法。
【請求項34】
標識を、該標識の移転によってか、またはその後の反応中に、固体支持体に共有結合で結合させる、請求項30に記載の方法。
【請求項35】
標識を、切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を、固体支持体への固定化後に標識から遊離させることができる、請求項30〜34のいずれか1項に記載の方法。
【請求項36】
標識を、光切断できるリンカーを介して融合タンパク質に結合する結果、該融合タンパク質を、固体支持体への固定化後に標識から遊離させることができる、請求項30〜34のいずれか1項に記載の方法。
【請求項37】
対象となる複数のタンパク質を固体支持体に固定化することを含む、請求項30〜36のいずれか1項に記載の方法。
【請求項38】
対象となるタンパク質を、整列させて固体支持体に固定化する、請求項37に記載の方法。
【請求項39】
アレーが、複数の空間分解位置を含む、請求項38に記載の方法。
【請求項40】
固体支持体が、ガラスまたは重合体を含む、請求項30〜39のいずれか1項に記載の方法。
【請求項41】
反応性ラジカルを発生できる分子が、反応性ヒドロキシルラジカルを発生する、請求項24に記載の方法。
【請求項42】
反応性ヒドロキシルラジカルを発生できる分子が、H2O2およびアスコルビン酸塩に接触するとラジカルを発生する、束縛された金属キレートである、請求項41に記載の方法。
【請求項43】
反応性ヒドロキシルラジカルを発生できる分子が、光を照射するとラジカルを発生する、請求項41に記載の方法。
【請求項44】
反応性ヒドロキシルラジカルを発生できる分子がマラカイトグリーンである、請求項43に記載の方法。
【請求項45】
ラジカルが、融合タンパク質、または融合タンパク質の近傍に位置するもう一つのタンパク質を不活性化する、請求項24または41〜44のいずれか1項に記載の方法。
【請求項46】
ラジカルが、タンパク質のフラグメント、または融合タンパク質の近傍に位置するもう一つのタンパク質を発生する、請求項41〜45のいずれか1項に記載の方法。
【請求項47】
フラグメントの発生を、融合タンパク質のごく近傍のタンパク質の同定に用いる、請求項46記載の方法。
【請求項48】
タンパク質またはもう一つのタンパク質の不活性化が、レーザー分子不活性化法(CALI)を用いる、請求項45に記載の方法。
【請求項49】
標識が、特異的結合対の第一の成員である、請求項24に記載の方法。
【請求項50】
さらに、融合タンパク質を、特異的結合対の第二の成員を含む分子に接触させることを含む、請求項49に記載の方法。
【請求項51】
特異的結合対の第一および第二の成員の相互作用または結合が、検出できる結果を生じる、請求項49または50に記載の方法。
【請求項52】
特異的結合対の第一および第二の成員が、第一および第二のタンパク質、抗体および抗原、酵素および基質、またはリガンドおよび受容体である、請求項49〜51のいずれか1項に記載の方法。
【請求項53】
AGTに融合した対象となるタンパク質が、転写因子のDNA結合ドメイン、または転写因子の活性化ドメインを含み、標的物質または標的物質のライブラリーを、該DNA結合ドメインまたは該転写因子の活性化ドメインの他方に結合させ、かつ標識が、標的物質と作用し合うと想定される候補化合物または候補化合物のライブラリーである、請求項1〜24のいずれか1項に記載の方法。
【請求項54】
候補化合物または候補化合物のライブラリー、および標的物質または標的物質のライブラリーが、リガンドおよびタンパク質、薬物および該薬物の標的、小分子およびタンパク質、ならびに酵素およびその基質または阻害剤から選択される、請求項53に記載の方法。
【請求項55】
候補化合物または候補化合物のライブラリーを、AGTタンパク質融合に移転することを含む、請求項53または54に記載の方法。
【請求項56】
候補化合物で標識付けしたAGT融合タンパク質と、標的物質とを接触させる結果、AGT融合タンパク質に結合した候補化合物と、標的物質との相互作用が、転写因子を再構成かつ活性化することをさらに含む、請求項55に記載の方法。
【請求項57】
活性化された転写因子が、リポーターの発現を発動する、請求項56に記載の方法。
【請求項58】
リポーターが、測定できるシグナルへとその活性が導くタンパク質、酵素または核酸である、請求項57に記載の方法。
【請求項59】
該方法が、細胞内で実施され、リポーターの発現が、選択的な利点を細胞に与える、請求項53〜58のいずれか1項に記載の方法。
【請求項60】
AGT融合タンパク質、および/またはAGTタンパク質融合と作用し合う標的物質を単離することをさらに含む、請求項53〜59のいずれか1項に記載の方法。
【請求項61】
標的物質と作用し合う候補化合物を検出かつ/または同定することをさらに含む、請求項53〜60のいずれか1項に記載の方法。
【請求項62】
標識が、タンパク質を架橋結合することができる、請求項24に記載の方法。
【請求項63】
架橋結合する標識が、マレイミド基、エステル基またはアジド基を含む、請求項62記載の方法。
【請求項64】
融合タンパク質を、候補タンパク質の結合パートナーに接触させ、架橋結合する標識を反応させる結果、作用し合う融合タンパク質と候補タンパク質とが互いに共有結合させる、請求項62または63に記載の方法。
【請求項65】
架橋結合した融合タンパク質および/または候補タンパク質を検出かつ/もしくは同定することをさらに含む、請求項62〜64のいずれか1項に記載の方法。
【図1】

【図2】

【図3】
【図2】
【図3】
【公開番号】特開2008−154587(P2008−154587A)
【公開日】平成20年7月10日(2008.7.10)
【国際特許分類】
【外国語出願】
【出願番号】特願2007−328757(P2007−328757)
【出願日】平成19年12月20日(2007.12.20)
【分割の表示】特願2002−582274(P2002−582274)の分割
【原出願日】平成14年4月5日(2002.4.5)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
【公開日】平成20年7月10日(2008.7.10)
【国際特許分類】
【出願番号】特願2007−328757(P2007−328757)
【出願日】平成19年12月20日(2007.12.20)
【分割の表示】特願2002−582274(P2002−582274)の分割
【原出願日】平成14年4月5日(2002.4.5)
【出願人】(503183293)ウペエフエル・エコル・ポリテクニック・フェデラル・ドゥ・ローザンヌ (11)
【氏名又は名称原語表記】EPFL ECOLE POLYTECHNIQUEFEDERALE DE LAUSANNE
【Fターム(参考)】
[ Back to top ]