
ORF7欠損水痘ウイルス、該ウイルスを含むワクチン、及びその使用
ORF7欠損水痘ウイルス、そのウイルスを含むワクチン、及びそれらの使用、並びにそのウイルスを製造する方法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルス学及び生物医薬の分野に属する。とりわけ、本発明は、ORF7欠損水痘ウイルス、このウイルスを含有するワクチン、及びその使用を提供する。本発明は、水痘帯状疱疹ウイルスを予防するための候補ワクチンを提供する。
【背景技術】
【0002】
水痘帯状疱疹ウイルス(「水痘ウイルス」又は「VZV」と称する)は、厳密に種特異的なウイルスであり、ヒトにおいて水痘又は帯状疱疹を引き起こす病原性因子である。水痘ウイルスのP−Oka株は、典型的な水痘を有する、「岡(Oka)」という姓の3歳の日本人男児の水疱液から、1971年に高橋(Takahashi)により初めて単離され、ヒト胎児肺細胞においてこのウイルスを継代培養することにより得られた。P−Okaは、水痘及び帯状疱疹を引き起こす能力を有すると考えられている(Takahashi M、Otsuka T、Okuno Yら、「院内小児における水痘の拡大を防止するために用いられる生ワクチン(Live vaccine used to prevent the spread of varicella in children in hospital)」、Lancet 1974、2:1288〜90)。
【0003】
米国特許第3,985,615号では、モルモット胎仔組織(GPEC)及びヒト胎児肺細胞においてこのウイルスを連続的に継代培養することにより、弱毒化P−Oka株が得られること、水痘を予防するためにV−Oka生ワクチンを調製することが開示された。米国特許第4,000,256号では、ヒト胎児線維芽細胞WI−38株においてVZVを10〜80回にわたり継代培養して、弱毒化VZV生ワクチンを製造することが記載された。これまでのところ、細胞内でP−Oka株を継代培養することにより得られる「弱毒化生水痘ウイルスOka株」(臨時速報、53号−41202Kimiaki、Lancet2:1288〜1290、1974)だけが、水痘又は帯状疱疹を予防するためのワクチンの調製についてWHOにより承認されており、この株から調製されたワクチンは、世界中で広く用いられている[水痘ワクチン(生)採用の要件(Requirement for Varicella Vaccine(Live)Adopted)、1984:WHO Technical Report Series、725号、102〜104ページ、1985]。水痘ワクチンを接種された集団は、自然感染した集団より帯状疱疹の発生率が低く、大部分の小児は免疫不全であることが報告されているが、実際のところ、ワクチン接種を受けた集団においてもなお、帯状疱疹は出現し、また、ワクチン接種を受けた集団における帯状疱疹の水疱液からもV−Oka株が単離された(Schmid D.Sら、「水痘帯状疱疹ウイルスの動態に対する水痘ワクチンの影響(Impact of varicella vaccine on varicella−zoster virus dynamics)」、Clinical Microbiology Reviews、2010年1月、202〜217ページ)。
【0004】
中国特許第CN1163604C号は、Oka株による弱毒化水痘ワクチンの同定に関するが、これにより、V−Oka株が混合株であることが分子レベルで明らかにされ、これは多数の異なる配列で構成されていた。V−Okaワクチンを接種された患者が、V−Oka株を含有する帯状疱疹を発症したと報告されているので、弱毒化水痘ウイルス株には、野生型の株も存在しうる。これは、弱毒化ウイルス株にもなお、何らかの潜在的有害性が存在することを意味する。しかし、これまでのところ、VZVに対する他のワクチンは市場に出ていない。このウイルスは、種特異性が厳密であり、天然の宿主はヒトに限られる。また、VZVの遺伝子機能を研究するのに用いるために利用可能な動物モデルは存在しない。さらに、このウイルスは、宿主細胞に対する結合親和性が強い。遊離ウイルスは、容易に不活化される。したがって、形質転換に用いることができるVZVウイルスのDNAを得ることは極めて困難である。結果として、このウイルスの遺伝子機能についての研究過程の進展は、長期にわたって非常に遅々としたものである。
【0005】
研究に細菌人工染色体(BAC)技術が適用され、組織培養法が進歩し、異種生物間における臓器移植の障壁が突破された(複合型免疫不全マウスモデルの開発)ことにより、種特異的VZVの遺伝子機能についての研究領域が拡大されている。VZVの遺伝子機能についての研究は、in vitroにおいて実施することもでき、動物への移植後に行うこともできる。2004年に、Kazuhiro N.らは、「大腸菌における感染性BACとしての水痘帯状疱疹ウイルスゲノムのクローニング(Cloning of the varicella−zoster virus genome as an infectious bacterial artificial chromosome in Escherichia coli)」において、BACベクター中にクローニングされたVZV Oka株の全ゲノムが、大腸菌(Escherichia coli)及びヒト胎児肺細胞において増殖しうることを明らかにした。このBACは、ヒト胎児肺細胞へと共形質転換される酵素であるCreにより、正確に切り出すことができる。結果として得られる組換えVZVウイルスは、親ウイルスであるP−Oka株と同じ構造及び特徴を有する。BACを適用することにより、VZVの遺伝子機能についての研究が大幅に促進されている。各機能性遺伝子について研究し、これにより、VZVの遺伝子機能についての研究過程を加速化させることが可能である。
【0006】
ウイルスゲノムの8607〜9368位に位置するVZVのORF7は、29kDaのタンパク質をコードし、テグメント内に位置すると考えられている。しかし、その機能は、依然として不明である。
【0007】
本発明者らは、驚くべきことであるが、広範な研究の後に、VZVに感染した患者における皮膚及び感覚神経節の機能をORF7が決定することを見出した。ORF7は、ORF7において以下のうちのいずれかが生じる場合、機能を喪失する:完全又は部分的な欠失、停止コドンの挿入、1塩基又は2塩基の置換、とりわけORF7の欠失又は反転逆フレームシフト突然変異。すなわち、VZVはもはや、ヒトの皮膚及び感覚神経節に感染しなくなる。このため、水痘を発生させる潜在的な危険性、又は帯状疱疹を引き起こす再活性化の危険性は生じない(本明細書において、本発明者らは、これを、ORF7が欠損することにより引き起こされるORF7の機能喪失と規定するが、この機能とは、ヒトの皮膚及び感覚神経節に接触することを意味する)。単一クローン株を得るために、大腸菌(E.coli)内で、抗生物質又はガラクトースにより、ORF7欠失VZV株(VZV−7D BAC又はVZV−7DRM BAC)をさらにスクリーニングすることができる。このため、混合型V−Oka株の潜在的な危険性は存在しない。ORF7欠失VZV株は、ORF7が欠失したウイルス又はORF7の突然変異体ウイルスによる、より安全でより有効なワクチンを開発するための扉を開く。
【発明の概要】
【課題を解決するための手段】
【0008】
一態様では、本発明は、ORF7が完全に若しくは部分的に欠失している、停止コドンが挿入されている、又は1塩基若しくは2塩基が置換若しくは付加されている水痘ウイルス(例えば、P−Oka株)に関し、記載されているVZVは、ヒトの皮膚及び感覚神経節に感染することができない。
【0009】
ORF7の配列(8607〜9386bp)を、以下に示す。
【化1】

【0010】
本発明の一実施形態では、ORF7の全体又は部分が、抗生物質耐性遺伝子及び/又は非機能性核酸配列により置換されている。前記非機能性核酸配列とは、それがORF7の機能を回復しないことを意味する。すなわち、上記のVZVウイルスはやはり、ヒトの皮膚及び感覚神経節に感染することが不可能である。該非機能性核酸配列は、融合タンパク質をコードしていなくてもよく、コードされるタンパク質がORF7機能を回復することができない条件で融合タンパク質をコードしていてもよい。本発明の一実施形態では、上記の非機能性核酸配列が、反転逆フレームシフト突然変異を有するORF7である。
【0011】
「ORF7」断片という用語は、ORF7核酸配列の全体又は部分を指す。
【0012】
本発明の一実施形態では、該記載されている反転逆フレームシフト突然変異を有するORF7断片の配列が、配列番号13に示されている。
【0013】
本発明の一実施形態では、抗生物質耐性遺伝子が、ペニシリン耐性遺伝子、ストレプトマイシン耐性遺伝子、及びカナマイシン耐性遺伝子から選択される。
【0014】
本発明の一実施形態では、本発明者らは、相同組換えの原理を適用することにより、スタンフォード大学医学部のAnn Arvin博士の研究室により臨床で単離されたP−Oka株に基づく組換えクローン、並びにVZV−BAC(P−Oka)のフレームワークを作製した。全長ORF7を、カナマイシン耐性遺伝子により置換してVZV−7D BACを作製し、又は反転逆フレームシフト突然変異を有するORF7断片により置換してVZV−7DRM BACを作製した。前記Kanrの配列は、以下に示される。
【化2】
【0015】
特に、本発明は、2009年7月28日に中国微生物株寄託管理委員会普通微生物センター(China General Microbiological Culture Collection Center)に寄託されている水痘帯状疱疹ウイルス株を提供し、その寄託番号は、CGMCC第3207号である。
【0016】
このウイルス株が、ヒトの皮膚及び感覚神経節に感染しないことは、in vitro及びin vivoでの研究により確認されている。そのため、水痘を引き起こす潜在的な危険性、又は帯状疱疹を引き起こす再活性化の危険性は見られない可能性が極めて高い。このウイルスは、MRC−5細胞、MeWo細胞、及びARPE細胞において、P−Oka株と同じ増殖特性で増殖する。その一方で、胸腺組織培養物における生育及び増殖特性も変化しない。このウイルス株は、P−Oka株のすべての糖タンパク質を保持し、そのため、P−Oka株と同じ免疫原性を示すはずである。このウイルス株は、カナマイシン又はガラクトースによりスクリーニングされる、該ウイルスの単一クローンである。したがって、V−Oka株のクローンと混在する潜在的な危険性は存在しない。ORF7全体の欠失(VZV−7D BAC)、又はORF7の反転逆フレームシフト突然変異(VZV−7DRM BAC)により、本発明のウイルス株に逆突然変異が導入される可能性はない。VZV−7D BAC又はVZV−7DRM BACから調製される弱毒性生ワクチン株は、より安全で、より有効である。
【0017】
別の態様では、本発明は、上記の水痘ウイルスを調製する方法であって、
1)野生型VZVのゲノムを、細菌人工染色体(BAC)ベクター中にクローニングして、ORF7の完全又は部分的な欠失、1塩基又は複数塩基の置換又は挿入を有する組換え水痘ウイルスVZV−BACを作製するステップであって、前記組換え水痘ウイルスが、ヒトの皮膚及び感覚神経節に感染することができないステップと、
2)1)で得られたVZVからDNAを単離し、単離されたDNA及び酵素Creを含有する組換えプラスミドにより、MRC−5細胞、Mewo細胞、又はARPE細胞を共形質転換するステップと、
3)酵素Creにより、MRC−5細胞、Mewo細胞、又はARPE細胞からBACを切り出すステップと
を含む方法に関する。
【0018】
本発明の一実施形態では、野生型VZVが、P−Oka株であり、ステップ1)におけるORF7が、Kanr、又は反転逆フレームシフト突然変異を有するORF7断片により置換されている。本発明の特定の一実施形態では、反転逆フレームシフト突然変異を有するORF7断片が、配列番号13として示される。
【0019】
別の態様ではまた、本発明は、上記の任意の水痘ウイルスを含む組成物にも関する。
【0020】
さらに別の態様ではまた、本発明は、水痘及び/又は帯状疱疹を予防するためのワクチンであって、上記の任意の水痘ウイルス、及びワクチンにおいて許容される賦形剤又は許容される担体を含有するワクチンにも関する。
【0021】
本発明で用いられる「水痘又は帯状疱疹を予防するためのワクチン」という用語は、水痘ワクチン、帯状疱疹ワクチン、並びに水痘及び帯状疱疹を予防するためのワクチンを指す。
【0022】
別の態様ではまた、本発明は、水痘ワクチン及び/又は帯状疱疹ワクチンの調製における、上記の水痘ウイルスのうちのいずれか1つの使用にも関する。
【0023】
別の態様ではまた、本発明は、発現ベクターの調製における、上記の水痘ウイルスのうちのいずれか1つの使用にも関する。
【0024】
本発明はまた、BACと、前記BAC中に挿入されている上記の水痘ウイルスのうちのいずれか1つのDNAとで構成される発現ベクターにも関する。発現ベクターは、上記の水痘ウイルスのうちのいずれか1つのDNAを含む。ベクターの機能に影響を及ぼすことなく、少なくとも10kbの外来遺伝子を上記の発現ベクターに挿入することができる。これはまた外来遺伝子も発現することができる。加えて、該発現ベクターは、ORF7の欠失の結果として安全である。該発現ベクターはまた、in vitro及びin vivoにおけるVZV−7DRM BACの機能をモニタリングする目的で、ルシフェラーゼ遺伝子による標識化(Zhangら、Journal of Virology、2007年9月、9024〜9033ページ)にも用いることができる。
【0025】
別の態様ではまた、本発明は、上記の発現ベクターと、挿入された外来遺伝子とを含む組換えベクターにも関する。
【0026】
別の態様ではまた、本発明は、上記の発現ベクター又は組換えベクターを含有する組換え細胞にも関する。
【0027】
本発明の一実施形態では、組換え細胞として用いられる宿主細胞は、MRC−5細胞、Mewo細胞、ARPE細胞、2BS細胞、WI−38細胞、及びKMB17細胞から選択される。
【0028】
別の態様ではまた、本発明は、水痘及び/又は帯状疱疹を予防する方法であって、それを必要とする患者に有効量の上記のワクチンを接種するステップを含む方法にも関する。
【0029】
本明細書では、「DPI」という用語は、「感染後日数」を表し、感染後の日数を指す。
【図面の簡単な説明】
【0030】
【図1】P−Oka BAC(VZV BAC)ベクターの構築を示す図である。
【図2】ORF7欠失VZV BAC株(VZV−7D BAC)の調製を示す図である。
【図3】ORF7欠失VZVの電気泳動を示す図である。
【図4A】後根神経節におけるVZV−7Dの培養を示す図である。MeWo細胞において培養した、野生型ウイルス(WT)、逆突然変異体ウイルス(7R)、欠損ウイルス(7D)についての増殖曲線を示す図である。
【図4B】後根神経節におけるVZV−7Dの培養を示す図である。後根神経節(DRG)において培養した、野生型ウイルス(WT)、逆突然変異体ウイルス(7R)、欠損ウイルス(7D)についての増殖曲線を示す図である。
【図5】ORF−7逆突然変異体VZV(VZV−7R)の調製について示す図である。
【図6】ヒトMeWo細胞において培養した、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
【図7】ヒト胸腺組織培養物(TOC)において培養した、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
【図8】ヒト皮膚組織培養物(SOC)中の、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
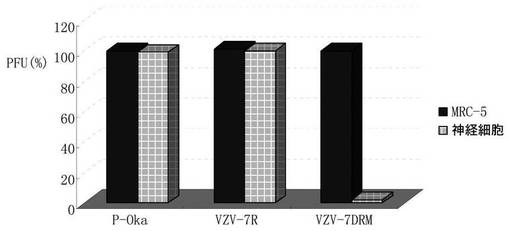
【図9A】WT、VZV−7DRM、及びVZV−7Rに由来するDNAをトランスフェクトしたMeWo細胞及び神経芽腫(SH−SY5Y)細胞を示す図である。神経芽腫細胞において、VZV−7DRMは増殖できない。
【図9B】ヒト胚の脊髄から摘出した後根神経節(DRG)を示す図である。
【図9C】DRGを、重症複合型免疫欠損マウスモデル(SCID−hu)の腎被膜へと移植し、次いで、WT又はVZV−7DRMを感染させた。IVIS計測器によりウイルスの増殖を測定する。ヒト組織を移植した重症複合型免疫欠損マウス(SCID−hu)において、VZV−7DRMは増殖できない。
【図9D】感染後、GFP強度の増大により、DRG培養物において、WT VZVが増殖できることが示された。
【発明を実施するための形態】
【0031】
以下の実施例に言及することにより、本発明の実施形態を詳細に説明する。これらの実施例が、本発明の実施を例示することを意図するものであり、本発明の範囲を限定することを意図するものではないことを、当業者は理解するであろう。具体的に例示されない技法又は条件については、当技術分野において説明されている(例えば、J.Sambrookら、「Molecular Cloning:A Laboratory Manual」、第3版、Science Press、Peitang Huang訳)技法及び条件に従って、又は製造元の指示書に従って、該手順を実施する。別段に指定しない限り、試薬及び装置は市販されている。
【実施例】
【0032】
(例1)
p−Oka BAC(VZV BAC)の構築
(1)BACベクター:BACベクターであるpUSF−6は、原核細胞の複製起点(ori)、該複製遺伝子及び分配遺伝子(repE、parA、及びparB)、Camr遺伝子、緑色蛍光タンパク質(gfp)遺伝子、2つの500bpのVZV断片a及びb(グレー)、並びに2つのloxP部位(白)を含有する。BamHIによるpUSF−6の消化の結果として直鎖断片を得、これを、相同組換えにより、VZVを含有するコスミドであるpvSpe23に挿入した。
【0033】
(2)pOkaゲノムについての概略図は、VZVが、固有の長いセグメント(UL)と、固有の短いセグメント(US)とを伴う、125kbのヌクレオチド配列を含有することを示す。全ゲノムを、重複領域を含む4つのVZVセグメントへと消化し、コスミド中にクローニングし、その結果として、4つの組換えクローン、すなわち、pvFsp73、pvSpe14、pvPme19、及びpvSpe23を得た。相同組換えにより、pUSF−6を、VZVコスミドであるpvSpe23のORF60とORF61との間に挿入した。
【0034】
(3)BACベクター(pUSF−6)を含有するpvSpe23を、他の3つのVZVコスミドと共にMeWo細胞へと共トランスフェクトしたところ、その結果として得られる組換えウイルス(VZV BAC)は、MeWo細胞内で複製されることが可能であり、緑色蛍光プラークを生成した。
【0035】
図1にもまた、該手順を示す。
【0036】
VZV BACの増殖曲線を、野生型pOkaの増殖曲線と比較した。各dpiにおけるプラーク形成単位(PFU)を記録した。結果は、VZV BAC(緑)が、検出可能な増殖の欠陥を示さず、これが、in vitroにおける親ウイルスの増殖と符合することを示唆する。
【0037】
(例2)
ORF7欠損VZV BAC(VZV−7D BAC)の作製及び検証
既に説明されているluc VZV BAC(Zhangら、Journal of Virology、2007年9月、9024〜9033ページ)を参照することにより、ORF7欠失突然変異体を構築した。手順を図2及び下記で説明した。
【0038】
(1)カナマイシン耐性遺伝子に相同な20bpのヌクレオチドと、欠失の標的とされるORF7の開始コドン又は停止コドンに近接する隣接配列に相同な40bpのヌクレオチドとを含有するように2つのプライマーをデザインした。プライマーF1及びR1の配列を以下に示す。
【化3】
【0039】
プラスミドpGEM−oriV/kan1(Netterwald、Yangら、2005)により、KanR遺伝子を増幅した。そのPCR産物をDpnIで消化し、ゲル抽出キット(Qiagen)により回収した。
【0040】
(2)BioRad Gene Pulser II装置を用いて、1.6kV及び250uFでエレクトロポレーションにより、約200ngずつの各PCR産物を、VZV Luc BACを保有するDY380株へと形質転換した。
【0041】
(3)高効率の相同組換え系として機能するDY380株により、40bpという短い相同配列の組換えが可能となる。42℃で活性化され、32℃で抑制される組換え酵素により、カナマイシン耐性遺伝子(kanr)とORF7との相同組換えを実現する。結果として、ORF7が、Kanrにより置換される。32℃で30μg/mlのカナマイシンを含有する寒天プレート上で、単一組換え体を選択した。VZV突然変異体(ORF7欠失による)のDNAを、大腸菌から単離した。
【0042】
(4)HindIII消化によりウイルスゲノムの完全性を検証し、PCR解析により組換えDNAを確認した。KanrによりORF7が置換された、ORF7欠損BAC(VZV−7D BAC)が得られたことが確認される。
【0043】
(例3)
反転逆フレームシフト突然変異を有するVZV BAC(VZV−7DRM BAC)の作製及び検証
戦略:野生型ウイルスのORF7を、galk遺伝子で置換した。galk遺伝子を有する組換えVZVは、ガラクトースを唯一の炭素供給源とするマッコンキー寒天培地上において、明赤色のプラークを生成させるはずである。この方法によるならば、単一クローンが選択されるであろう。次いで、galk遺伝子を、反転逆フレームシフト突然変異配列で置換し、完全最小培地プレート上において単一クローンを選択して、VZV−7DRM BACを得る。
【0044】
詳細なプロトコールを以下に示す。
【0045】
(1)pGalK(1〜82bp:プロモーター;83〜1231bp:GalKのオープンリーディングフレーム)により、galk遺伝子を増幅するための増幅プライマーF2及びR2をデザインする。これらのフォワードプライマー及びリバースプライマーは各々が、5'側に60bpの、ORF7に隣接する相同配列を有する。GalK配列を以下(配列番号5)に示す。
【化4】
【0046】
プライマーF2及びR2の配列を以下に示す。
【化5】
【0047】
(2)これらのプライマーを用いて、GalKカセットを増幅する。この産物を、DpnIにより終夜にわたり消化し、精製する。
【0048】
(3)VZV BACを、大腸菌のSW102株へと形質転換する。
【0049】
(4)単一コロニーを5mlのLB−CM培地(クロラムフェニコールを含むLB培地)へと接種し、32℃で終夜にわたり増殖させる。
【0050】
(5)終夜にわたる培養物を、250mlのLB−CMol培地へと接種し、ODが0.4〜0.6となるまで、振とうしながら32℃で増殖させる。
【0051】
(6)エレクトロコンピテント細胞(VZV BACで形質転換したSW102細胞)を調製する。
【0052】
(7)ステップ(6)で調製されたエレクトロコンピテント細胞(VZV BACで形質転換したSW102細胞)を、ステップ(2)による1〜2μl(約200ng)の精製されたPCR産物により、例2において説明したエレクトロポレーションパラメータで形質転換する。
【0053】
(8)32℃で1mlのLB培地中で、1時間にわたり、形質転換体を回収する。
【0054】
(9)回収時間の後、1×M9塩中で、以下の通り、3回にわたり細菌を洗浄する。培養物を13,200RPMで15秒間にわたり遠心分離にかけ、ピペットで上清を除去する。ペレットを1×M9塩中に再懸濁させ、上記の通り、再度遠心分離にかける。洗浄ステップを再度繰り返す。3回にわたり洗浄した後、上清を除去し、ペレットを1mlの1倍濃度M9塩中に再懸濁させる。次いで、ペレットの懸濁液、10倍の希釈系列、及び100倍の希釈系列を、ガラクトース、ロイシン、ビオチン、及びクロラムフェニコールを含むM63最小培地プレートへと播種する。
【0055】
(10)32℃で3日間にわたりプレートをインキュベートする。
【0056】
(11)数個のコロニーを、ガラクトース+クロラムフェニコールを含むマッコンキー寒天培地プレート上へと画線培養して、単一コロニーを得る。3日間にわたるインキュベーション後に出現するコロニーはGal+コロニーのはずであるが、Gal−混入を除去するためには、明赤色の単一コロニーを得てから、次のステップへと進むことが重要である。
【0057】
(12)単一の明赤色の(Gal+)コロニーを採取して、5mlのLB−CM培地へと接種し、32℃で終夜にわたり培養する。該終夜にわたる培養物からBAC DNAを調製し、PCRによりgalk遺伝子を検証する。用いられるプライマーは、
【化6】
である。
【0058】
(13)プライマーF4(配列番号10)及びR4(配列番号11)を用いて、ORF7停止コドンの3'側に近接する457bpの配列(457bpのORF配列を、配列番号12に示す)を増幅する。プライマーF4及びR4は、1塩基のフレームシフトを導入するようにデザインされているので、フレーシフト突然変異を有するPCR産物が生成される。
【0059】
F4及びR4の配列を、以下に示す。
【化7】
【0060】
457bpのオープンリーディングフレームは、
【化8】
である。
【0061】
上記のステップ4〜10を繰り返して、エレクトロコンピテントSW102細胞を得る。
【0062】
(14)ステップ(13)の熱ショック処理した細菌50μlを、ステップ(2)のgalk遺伝子に隣接する領域と相同的なPCR産物200ngにより形質転換する。振とうしながら、32℃の水浴中で4.5時間にわたりインキュベートすることにより、50mlのバッフル付き三角フラスコ内のLB培地10ml中に回収する。この長い回収時間は、対象の配列だけを含有する組換えVZV BACを得る(したがって、GalKカセットを含有するVZV BACを除去する)のに役立つ。プライマーF4は、ORF7の5'側に隣接する60bpの相同配列と、ORF7の3'側に近接する457bpの逆配列とで構成され、プライマーR4は、ORF7の3’側に隣接する60bpの相同配列と、ORF7の3’側に近接する457bpのフォワード配列とで構成されるので、増幅されたPCR産物と、ORF7遺伝子座内のgalk遺伝子との相同組換えにより、3'側457bpの反転逆リンクがもたらされ、その結果として、機能を発現しない反転逆フレームシフト突然変異(配列番号13)が得られる。反転逆フレームシフト突然変異配列を、以下に示す。
【化9】
【0063】
(15)ステップ9の通り、1mlの培養物をスピニングし、1×M9塩中で3回にわたり洗浄し、1mlの1倍濃度M9塩中に再懸濁させる。ペレットの懸濁液及び10倍の希釈系列(各々100μlずつ)を、グリセロール、ロイシン、ビオチン、2−デオキシ−ガラクトース、及びクロラムフェニコールを含むM63最小培地プレートへと播種する。
【0064】
(16)32℃で3〜4日間にわたりインキュベートする。
【0065】
(17)コロニーを採取し、それらをLB−CMプレート及びGalK−M63 CM最小培地プレート上に画線培養する。LB−CMプレート上では増殖するが、最小培地プレート上では増殖しない単一コロニーを選択する。ウイルスDNAを抽出し、HindIII消化により完全性を検証し、PCRにより組換えDNAを確認する。ORF7を含むVZV BACが逆フレームシフト型突然変異(VZV−7DRM BAC)を反転させることを確認する。
【0066】
上記の手順で用いた試薬は、表1の通りに調製する。
【表1】
【0067】
(例4)
VZV−7Dウイルス及びVZV−7DRMウイルスの作製
(1)例2で得たVZV−7D BACからDNAを抽出する。
【0068】
(2)精製されたDNAを、エレクトロポレーションを介して、MRC−5細胞(Marchini、Liuら、2001)へと個別にトランスフェクトする(BACにおいて保有されるGFPにより、ウイルスの増殖をモニタリングすることができる)か、又は精製されたDNAを、Cre発現ベクターと共に、MRC−5細胞へと共トランスフェクトした(Marchini、Liuら、2001)。Creにより、BACに隣接するloxP部位で、BACを正確に切り出し、VZV 7Dウイルスを得た。
【0069】
同様にまた、ORF7の反転逆フレームシフト突然変異を有するVZV−7DRMウイルスも得た。VZV 7DRMは、反転逆配列であるORF7の一部だけを含有するので、発現することができず、ORF7機能を示さない。VZV−7DRM及びVZV−7Dはいずれも、示す機能が同じである。
【0070】
(例5)
VZV−7Dの検証
単層MRC−5細胞に、例4で調製され、2%ウシ血清を補充したMEM培地中で培養されたVZV−7Dを感染させた。細胞のほぼ80%が細胞変性効果を呈示したところで、培地を廃棄し、細胞を採取し、DNAを抽出する。P−Okaは対照としての役割を果たす。PCRによりORF−10遺伝子及びORF−7遺伝子を増幅した。VZV−7D及びP−OkaのDNAをHindIIIにより消化した。PCR産物と、HindIII消化生成物とを、0.5%アガロースゲル中の電気泳動にかけた。結果を図3に示した。VZV−7DにおけるORF−10のPCR産物はP−Okaと同じであるが、VZV−7DにおけるORF−7バンドは不在である。これにより、VZV−7DにおけるORF−7が欠失されていることが確認された。VZV−7D及びP−Okaとで、HindIIIにより消化されたウイルスDNAのプロファイルの差違は見られなかった。
【0071】
(例6)
ヒト後根神経節内におけるVZV−7Dの培養
ヒト胎児脊椎(妊娠20週齢)から、頸椎及び胸椎の後根神経節(DRG)を摘出した。1%のペニシリン/ストレプトマイシンを含む1×氷冷PBS中でこのDRGを簡単に洗浄し、次いで、氷冷組織培養培地(15%の熱不活化ウシ胎仔血清と、1%のペニシリン/ストレプトマイシンとを含むDEM)へと移した。0.5mlの培地で覆われた6ウェルプレート内の、コラーゲンでコーティングされたカバースリップ上に各DRGを個別に入れ、培地を1回交換して、5日間にわたり培養した。6ウェルプレート内で50%のMeWo細胞を調製し、滴定及び増殖曲線(図4A)の作成を行った。6日目に、P−Oka、VZV−7D、及びVZV−7Rを感染させたばかりのMewo細胞を採取し、培地中で1mlの最終容量へと再懸濁させた。各DRGは、6ウェルプレート内のコラーゲンでコーティングされたカバースリップ上で個別に増殖させる。10μlの懸濁液を用いて、ウイルス力価を決定し、2μlの懸濁液を用いて、感染MeWo細胞の増殖曲線を作成した。細胞懸濁液の残りを、20回にわたり27ゲージ針に通し、4000rpmで10分間にわたり遠心分離することにより、細胞破砕物をペレット化させた。上清を等量に分割し、これらを用いてDRGに感染させ(約120μl/試料1例)、加えて、2滴(20μl)の細胞懸濁液を用いて、別のDRGセットに感染させた。37℃/5%CO2中で3時間にわたるインキュベーション後、1×PBSでDRGを洗浄し、新鮮な培地を添加した。24時間間隔でルシフェラーゼ活性を測定した。試料を、150μg/mlのD−ルシフェリンと共に、37℃で10分間にわたりインキュベートした。IVIS測定器により光子をカウントし、ソフトウェアにより解析した。結果は、図4Bに示した。
【0072】
結果は、VZV−7Dが、MeWo細胞内では良好に増殖するが、DRG内ではほとんど増殖しないことを示す。VZV−7DがDRGに感染しないという事実は、安全な弱毒化生ワクチンを開発するための基盤をもたらす。
【0073】
(例7)
VZV−7Dに対する逆突然変異体(VZV−7R)の構築及び検証
ORF7欠失突然変異体は、HindIII消化によるだけでなく、相同組換えにより得られる完全VZVウイルスによっても同定することができる。すなわち、完全VZVは、相同組換えの機構によりレスキューされる、図5を参照されたい)。Invitrogen製のHigh Fidelity Platinum Taq DNAポリメラーゼキットを用いるPCRにより、ORF7断片を増幅し、プラスミドpGEM−lox−zeoのNotI部位とBglII部位との間にクローニングして、pGEM−zeo−ORF7プラスミドを形成した。配列決定解析により、クローニングされた断片を検証したところ、PCR増幅により、不適正なコドンがORF7へと導入されることはなかったことが示される。次いで、2つのプライマーを用いて、pGEM−zeo−ORF7から、PCRによりzeoR−ORF7カセットを増幅した。
【化10】
【0074】
結果として得られるPCR産物は、各末端に40bpの相同VZVセグメントを含有し、前記セグメントは、kanR遺伝子に近接し、欠失ORF7遺伝子内に位置する。結果として得られるキメラPCR産物を、上記のエレクトロポレーションを介して、ORF7D BACクローンを保有するDY380株へと形質転換する。50μg/mlのZeocin(Invitrogen)及び12.5μg/mlのクロラムフェニコールを含有する寒天プレート上、32℃で組換え体を選択した。それらの抗生物質感受性により、適正なVZVクローンを確認した。適正なクローンは、クロラムフェニコール、ハイグロマイシン、Zeocinに耐性であり、カナマイシン及びアンピシリンに感受性のはずである。制限酵素消化及びPCR解析により、これらのクローンをさらに検証した。
【0075】
検証されたクローン(ORF7が欠失している、又はレスキューluc VZV BACを有する、大腸菌DY380株によるクローン)を、12.5μg/mlのクロラムフェニコールを含む500mlのLB培地中、32℃で20時間にわたりインキュベートした。Nucleobond Maxiprep BAC DNA単離キット(BD Biosciences、Palo Alto、CA)を用いて、VZV BAC DNAを単離し、これらを、製造元の指示書に従い、FuGene6トランスフェクション試薬(Roche、Indianapolis、IN)を用いてMRC−5細胞にトランスフェクトするのに用いた。6ウェルプレートの各ウェル(35mm)について、DNA対トランスフェクション試薬の比は、1.5μg:6μlであった。トランスフェクションの3日後には、VZVプラークが正常に見て取れた。VZVゲノムから、BACベクター(2つのloxP部位により挟まれる)を除去するために、Cre発現ベクターを、VZV BAC DNAと共に、MRC−5細胞へと共トランスフェクトした(Marchini、Liuら、2001)。Cre酵素により、BACは正確に欠失され、完全VZVを生成させた。
【0076】
ORF−7レスキューウイルスは、P−Oka株と同じ生育及び増殖特性を示す。これらの結果は、逆突然変異により、ORF−7の機能を回復させうることを示した。
【0077】
(例8)
Mewo細胞におけるVZV−7DRMの培養
6ウェルプレート内の単層MeWo細胞にモック感染させ、又はlucVZV−7DRM(Lucを有するVZV−7DRM)、luc7VZV−7R(Lucを有するVZ−7R)、及びlucVZV(Lucを有するP−Oka株)を感染させた。2%ウシ血清を含むMEM培地中、37℃でMewo細胞を培養した。感染の24時間後に、D−ルシフェリンを150μg/mlの最終濃度まで、培養ウェルに添加した。37℃で10分間にわたるインキュベーション後、IVIS Imaging System(50 Series;Xenogen Corporation、Alameda、CA)を用いて、異なるウェル内の発光を同時に3連で記録した。測定後、ルシフェリン含有培地を、通常の細胞培地で置換した。24時間ごとに7日間にわたり、この測定を繰り返した。平均データ対時間をプロットすること(図6)により、増殖曲線を作成した。
【0078】
ヒトMeWo細胞にモック感染させ、又は野生型VZV(lucVZV)、又はVZV−7DRM、又はVZV−7Dレスキュー株(luc7R)を感染させ、7日間にわたり培養した。感染後毎日、IVISシステムにより3連で光子カウントを記録した。平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に誤差バーを表示した。この実験の結果は、VZV−7DRM及びVZV−7Rが、野生型ウイルスと同様に十分にMeWo細胞内で増殖することを示した。一方、BACを挿入した後、VZV−7DRMが正常に増殖しうることが示された。BACにより保有される遺伝子もまた、発現することが可能である。すなわち、VZV−7DRMは、異種遺伝子の発現ベクターとして用いることができる。
【0079】
(例9)
ヒト胸腺組織におけるVZV−7DRMの培養
ヒト胸腺−肝臓組織を、CB−17SCID/ベージュマウスへの異種移植片として同時移植した。ヒト胸腺−肝臓組織の組合せ移植片は、文献(Jennifer F.M.、Journal of Virology、1995年9月、5236〜5242ページ)で説明されている方法に従い構築する。移植の3カ月後、手術によりヒト胸腺−肝臓異種移植片を露出させ、これらに3連で、モック感染させ、又は10〜20μlのMeWo細胞により培養した、2×103〜4×103PFUのlucVZV−7DRM(ルシフェラーゼを有するVZV−7DRM)及びlucVZV(ルシフェラーゼを有するP−Oka)を感染させた。感染後、毎日蛍光強度を測定した。腹腔内(i.p.)注射により、ルシフェラーゼ基質であるD−ルシフェリン250μlを添加した。10分後、例8で説明した方法に従い、IVIS Imaging Systemを用いて、3連で、異種移植片中の蛍光を記録した。結果を図7に示した。
【0080】
7日間にわたり、培養されたヒト胸腺組織に、モック感染させ、又は野生型VZV、若しくはVZV−7DRMを感染させた。感染後毎日(DPI)、IVISシステムを用いて光子カウントを測定し、平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に、3回にわたるデータ測定による誤差バーを表示する。この実験により、VZV−7DRM突然変異体が、胸腺組織(T細胞)内で、野生型VZVと同様に増殖しうることが示された。
【0081】
同様の実験によりまた、VZV−7Dも、胸腺組織(T細胞)内で、野生型VZVと同様に増殖しうることが裏付けられた。
【0082】
(例10)
皮膚におけるVZV−7DRMの培養
文献(Jennifer F.M.、Journal of Virology、1995年9月、5236〜5242ページ)で説明されている方法に従い、ヒト胎児皮膚を、完全移植片としてSCIDマウスへと皮下移植した。移植の4週間後、マウスの皮膚に3連で、モック感染させ、又はlucVZV−7DRM(Lucを有するVZV−7DRM)、lucVZV−7R(Lucを有するVZ−7R)及びlucVZV(Lucを有するP−Oka株)を感染させた。感染後、毎日蛍光強度を測定した。i.p.注射により、ルシフェラーゼ基質であるD−ルシフェリン250μlを添加した。10分後、例8で説明した方法に従い、IVIS Imaging Systemを用いて、3連で、異種移植片中の蛍光を記録した。結果を図8に示した。
【0083】
7日間にわたり、培養されたヒト皮膚組織に、モック感染させるか、野生型VZV(P−Oka株)、若しくはORF7欠損突然変異体(VZV−7DRM)、又はORF7逆突然変異体(VZV−7R)を感染させた。感染後毎日(DPI)、IVISシステムを用いて光子カウントを測定し、平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に、3回にわたるデータ測定による誤差バーを表示する。この実験により、VZV−7DRM突然変異体が、皮膚における増殖で大きな欠陥を示し、この機能の欠陥が、VZV−7DRMにおける逆突然変異により回復されうることが示された。
【0084】
同様の実験によりまた、VZV−7D突然変異体も、皮膚における増殖で大きな欠陥を示し、この機能の欠陥が、VZV−7Dにおける逆突然変異により回復されうることが見出される。
【0085】
(例11)
神経細胞にVZVが感染するのにORF7が必須であることが実験により確認された
1.神経細胞腫細胞への感染
MeWo細胞並びに神経細胞腫細胞系であるSH−SY5Yに、野生型(WT)、ORF7欠失突然変異体(VZV−7DRM)BAC、及びORF7逆突然変異体(VZV−7R)BACに由来するDNAをトランスフェクトした。結果は、WT、VZV−7Rが、MeWo細胞及び神経細胞腫細胞において良好に増殖しうるが、VZV−7DRMが、神経細胞腫細胞においては増殖しえないことを示した(図9A)。これにより、神経細胞にVZVが感染するのに、ORF7が本質的である可能性が極めて高いことが示唆される。
【0086】
2.DRG移植片への感染
この例における試験1の結果をさらに検証するため、ヒト胎児DRGをSCIDマウスへと移植し、ウイルス感染の動物モデルとして用いた。結果は、野生型ウイルスが、短い複製周期の後に潜在化することを示す(蛍光現象による)一方、VZV−7DがDRG移植片中では複製されえないことを示した。これらの結果は、ORF7が、VZVの向神経性因子である可能性が極めて高いことを示した(図9B〜D)。
【0087】
上記の2つの試験により、ニューロンにVZVが感染するのに、ORF7が本質的であることが裏付けられた。
【0088】
(例12)
VZV−7DRMによるワクチンの調製並びにVZV−7DRMについての免疫原性アッセイ
単層ARPE細胞又はMRC−5細胞に、0.1の感染多重度(MOI)でVZV−7DRMを感染させた。40分間にわたりウイルスを細胞に付着させた後、フラスコに培地を添加し、これを35℃でインキュベートした。細胞のうちの約80%が細胞症を提示したら(一般に、3日後)、0.1%EDTAで消化することにより、細胞を採取した。採取された細胞を遠心分離し、保護剤を添加し、破砕及び清明化の後にワクチンを得た。
【0089】
1回のワクチン接種当たり5匹ずつのモルモット(体重約250g)に、投与1回当たり5000pfu/0.5mlで、復元されたワクチンを接種した。初回免疫化の4週間後に、追加免疫化を施した。P−Oka及びV−Okaを対照として用いる。
【0090】
追加免疫化の2週間後、心臓穿刺により血液を回収した。gp−ELISAにより、血清抗体力価を検出した。免疫阻害アッセイにより、ウイルスに対する抗体の中和能を決定した。ワクチン接種の前後に、10倍の血清希釈液を、1:1の比でウイルスと混合した。37℃で60分間にわたるインキュベーションの後、混合物を用いて、MRC−5細胞に感染させた。5%CO2中37℃における7日間のインキュベーション後、培地を除去し、クーマシーブルーで細胞を染色した。プラーク形成単位をカウントすることにより、ウイルス力価を計算した。100×([ワクチン接種前に血清により中和されるウイルス力価]−[ワクチン接種後に血清により中和されるウイルス力価])/[ワクチン接種前に血清により中和されるウイルス力価]として、抗体によるウイルス中和度を計算する。結果を表2に示す。
【表2】
【0091】
結果により、VZV−7DRMが、良好な免疫原性を保有し、それに対する抗体による中和度は、V−Okaの場合と同等であることが示された。VZV−7DRMによるワクチン接種後の血清は、3種のウイルスすべてに対して良好な中和効果を有し、ワクチンとして開発される可能性を示す。
【0092】
同様の実験によりまた、VZV−7Dも良好な中和効果を有し、ワクチンとして開発される可能性を有することが示された。
【0093】
(例13)
VZV−7DRMをベクターとして用いる生物学的活性物質の調製
ORF7に対する隣接相同性を含有するEV71−VP1プライマーによるPCRを介して、pT−VP1プラスミドから、EV71−VP1カセットを増幅した。
【0094】
EV71−VP1カセットは、以下に示される。
【化11】
【0095】
PCRプライマー配列:
F6(ORF7の5'側隣接領域に対して相同な配列;開始コドン;EV71−VP1に相同な配列):
【化12】
【0096】
R6(ORF7の3’側隣接領域に対して相同な配列;EV71−VP1に相同な配列):
【化13】
【0097】
VZV−7DRM BACを含有する大腸菌のエレクトロコンピテントSW102株を、精製されたPCR産物により形質転換する。例2に従い、組換えSW102株をスクリーニング及び同定した。適切な培養の後、VZV−7DRM−VP1 BACを単離し、次いで、エレクトロポレーションを介して、Cre発現ベクターと共に、ARPE細胞へと共トランスフェクトした。BACの両端におけるloxP部位から、BACの全体を切り出し、その後、組換えVZV−7DRM−VP1ウイルスを得た。単層ARPE細胞に組換えウイルスを感染させ、35℃で3〜4日間にわたり培養した。そこで、細胞は細胞症を提示した。組換えウイルスの培養物上清、並びに細胞溶解(凍結融解又は超音波処理を繰り返した後の)に由来する遠心分離上清におけるEV71特異的サンドイッチELISAにより、VP1の活性を検出した。EV71及びVZV−7DRMを、対照として用いる。結果(表3)は、VZV−7DRM−VP1ウイルスを感染させたARPE細胞においてVP1活性が検出されたことを示す。これは、VP1が、VZV−7DRMウイルス内に組み換えられていることを示唆する。この例により、外来遺伝子を発現させるベクターとして、VZV−7DRMを用いうることが示された。
【表3】
【0098】
同様の実験により、VZV−7Dは外来遺伝子を発現させる発現ベクターとしても用いることができることが裏付けられた。
【0099】
本発明の実施形態を詳細に説明してきたが、本発明の範囲を逸脱しない限りにおいて、本開示の教示に従い、これらの詳細に対して各種の改変及び置換を行いうることを、当業者は理解するであろう。付属の特許請求の範囲、及びそれらの同等物により、本発明の全範囲が規定される。
【図1A】
【図1B】
【図1C】
【図1D】
【技術分野】
【0001】
本発明は、ウイルス学及び生物医薬の分野に属する。とりわけ、本発明は、ORF7欠損水痘ウイルス、このウイルスを含有するワクチン、及びその使用を提供する。本発明は、水痘帯状疱疹ウイルスを予防するための候補ワクチンを提供する。
【背景技術】
【0002】
水痘帯状疱疹ウイルス(「水痘ウイルス」又は「VZV」と称する)は、厳密に種特異的なウイルスであり、ヒトにおいて水痘又は帯状疱疹を引き起こす病原性因子である。水痘ウイルスのP−Oka株は、典型的な水痘を有する、「岡(Oka)」という姓の3歳の日本人男児の水疱液から、1971年に高橋(Takahashi)により初めて単離され、ヒト胎児肺細胞においてこのウイルスを継代培養することにより得られた。P−Okaは、水痘及び帯状疱疹を引き起こす能力を有すると考えられている(Takahashi M、Otsuka T、Okuno Yら、「院内小児における水痘の拡大を防止するために用いられる生ワクチン(Live vaccine used to prevent the spread of varicella in children in hospital)」、Lancet 1974、2:1288〜90)。
【0003】
米国特許第3,985,615号では、モルモット胎仔組織(GPEC)及びヒト胎児肺細胞においてこのウイルスを連続的に継代培養することにより、弱毒化P−Oka株が得られること、水痘を予防するためにV−Oka生ワクチンを調製することが開示された。米国特許第4,000,256号では、ヒト胎児線維芽細胞WI−38株においてVZVを10〜80回にわたり継代培養して、弱毒化VZV生ワクチンを製造することが記載された。これまでのところ、細胞内でP−Oka株を継代培養することにより得られる「弱毒化生水痘ウイルスOka株」(臨時速報、53号−41202Kimiaki、Lancet2:1288〜1290、1974)だけが、水痘又は帯状疱疹を予防するためのワクチンの調製についてWHOにより承認されており、この株から調製されたワクチンは、世界中で広く用いられている[水痘ワクチン(生)採用の要件(Requirement for Varicella Vaccine(Live)Adopted)、1984:WHO Technical Report Series、725号、102〜104ページ、1985]。水痘ワクチンを接種された集団は、自然感染した集団より帯状疱疹の発生率が低く、大部分の小児は免疫不全であることが報告されているが、実際のところ、ワクチン接種を受けた集団においてもなお、帯状疱疹は出現し、また、ワクチン接種を受けた集団における帯状疱疹の水疱液からもV−Oka株が単離された(Schmid D.Sら、「水痘帯状疱疹ウイルスの動態に対する水痘ワクチンの影響(Impact of varicella vaccine on varicella−zoster virus dynamics)」、Clinical Microbiology Reviews、2010年1月、202〜217ページ)。
【0004】
中国特許第CN1163604C号は、Oka株による弱毒化水痘ワクチンの同定に関するが、これにより、V−Oka株が混合株であることが分子レベルで明らかにされ、これは多数の異なる配列で構成されていた。V−Okaワクチンを接種された患者が、V−Oka株を含有する帯状疱疹を発症したと報告されているので、弱毒化水痘ウイルス株には、野生型の株も存在しうる。これは、弱毒化ウイルス株にもなお、何らかの潜在的有害性が存在することを意味する。しかし、これまでのところ、VZVに対する他のワクチンは市場に出ていない。このウイルスは、種特異性が厳密であり、天然の宿主はヒトに限られる。また、VZVの遺伝子機能を研究するのに用いるために利用可能な動物モデルは存在しない。さらに、このウイルスは、宿主細胞に対する結合親和性が強い。遊離ウイルスは、容易に不活化される。したがって、形質転換に用いることができるVZVウイルスのDNAを得ることは極めて困難である。結果として、このウイルスの遺伝子機能についての研究過程の進展は、長期にわたって非常に遅々としたものである。
【0005】
研究に細菌人工染色体(BAC)技術が適用され、組織培養法が進歩し、異種生物間における臓器移植の障壁が突破された(複合型免疫不全マウスモデルの開発)ことにより、種特異的VZVの遺伝子機能についての研究領域が拡大されている。VZVの遺伝子機能についての研究は、in vitroにおいて実施することもでき、動物への移植後に行うこともできる。2004年に、Kazuhiro N.らは、「大腸菌における感染性BACとしての水痘帯状疱疹ウイルスゲノムのクローニング(Cloning of the varicella−zoster virus genome as an infectious bacterial artificial chromosome in Escherichia coli)」において、BACベクター中にクローニングされたVZV Oka株の全ゲノムが、大腸菌(Escherichia coli)及びヒト胎児肺細胞において増殖しうることを明らかにした。このBACは、ヒト胎児肺細胞へと共形質転換される酵素であるCreにより、正確に切り出すことができる。結果として得られる組換えVZVウイルスは、親ウイルスであるP−Oka株と同じ構造及び特徴を有する。BACを適用することにより、VZVの遺伝子機能についての研究が大幅に促進されている。各機能性遺伝子について研究し、これにより、VZVの遺伝子機能についての研究過程を加速化させることが可能である。
【0006】
ウイルスゲノムの8607〜9368位に位置するVZVのORF7は、29kDaのタンパク質をコードし、テグメント内に位置すると考えられている。しかし、その機能は、依然として不明である。
【0007】
本発明者らは、驚くべきことであるが、広範な研究の後に、VZVに感染した患者における皮膚及び感覚神経節の機能をORF7が決定することを見出した。ORF7は、ORF7において以下のうちのいずれかが生じる場合、機能を喪失する:完全又は部分的な欠失、停止コドンの挿入、1塩基又は2塩基の置換、とりわけORF7の欠失又は反転逆フレームシフト突然変異。すなわち、VZVはもはや、ヒトの皮膚及び感覚神経節に感染しなくなる。このため、水痘を発生させる潜在的な危険性、又は帯状疱疹を引き起こす再活性化の危険性は生じない(本明細書において、本発明者らは、これを、ORF7が欠損することにより引き起こされるORF7の機能喪失と規定するが、この機能とは、ヒトの皮膚及び感覚神経節に接触することを意味する)。単一クローン株を得るために、大腸菌(E.coli)内で、抗生物質又はガラクトースにより、ORF7欠失VZV株(VZV−7D BAC又はVZV−7DRM BAC)をさらにスクリーニングすることができる。このため、混合型V−Oka株の潜在的な危険性は存在しない。ORF7欠失VZV株は、ORF7が欠失したウイルス又はORF7の突然変異体ウイルスによる、より安全でより有効なワクチンを開発するための扉を開く。
【発明の概要】
【課題を解決するための手段】
【0008】
一態様では、本発明は、ORF7が完全に若しくは部分的に欠失している、停止コドンが挿入されている、又は1塩基若しくは2塩基が置換若しくは付加されている水痘ウイルス(例えば、P−Oka株)に関し、記載されているVZVは、ヒトの皮膚及び感覚神経節に感染することができない。
【0009】
ORF7の配列(8607〜9386bp)を、以下に示す。
【化1】
【0010】
本発明の一実施形態では、ORF7の全体又は部分が、抗生物質耐性遺伝子及び/又は非機能性核酸配列により置換されている。前記非機能性核酸配列とは、それがORF7の機能を回復しないことを意味する。すなわち、上記のVZVウイルスはやはり、ヒトの皮膚及び感覚神経節に感染することが不可能である。該非機能性核酸配列は、融合タンパク質をコードしていなくてもよく、コードされるタンパク質がORF7機能を回復することができない条件で融合タンパク質をコードしていてもよい。本発明の一実施形態では、上記の非機能性核酸配列が、反転逆フレームシフト突然変異を有するORF7である。
【0011】
「ORF7」断片という用語は、ORF7核酸配列の全体又は部分を指す。
【0012】
本発明の一実施形態では、該記載されている反転逆フレームシフト突然変異を有するORF7断片の配列が、配列番号13に示されている。
【0013】
本発明の一実施形態では、抗生物質耐性遺伝子が、ペニシリン耐性遺伝子、ストレプトマイシン耐性遺伝子、及びカナマイシン耐性遺伝子から選択される。
【0014】
本発明の一実施形態では、本発明者らは、相同組換えの原理を適用することにより、スタンフォード大学医学部のAnn Arvin博士の研究室により臨床で単離されたP−Oka株に基づく組換えクローン、並びにVZV−BAC(P−Oka)のフレームワークを作製した。全長ORF7を、カナマイシン耐性遺伝子により置換してVZV−7D BACを作製し、又は反転逆フレームシフト突然変異を有するORF7断片により置換してVZV−7DRM BACを作製した。前記Kanrの配列は、以下に示される。
【化2】
【0015】
特に、本発明は、2009年7月28日に中国微生物株寄託管理委員会普通微生物センター(China General Microbiological Culture Collection Center)に寄託されている水痘帯状疱疹ウイルス株を提供し、その寄託番号は、CGMCC第3207号である。
【0016】
このウイルス株が、ヒトの皮膚及び感覚神経節に感染しないことは、in vitro及びin vivoでの研究により確認されている。そのため、水痘を引き起こす潜在的な危険性、又は帯状疱疹を引き起こす再活性化の危険性は見られない可能性が極めて高い。このウイルスは、MRC−5細胞、MeWo細胞、及びARPE細胞において、P−Oka株と同じ増殖特性で増殖する。その一方で、胸腺組織培養物における生育及び増殖特性も変化しない。このウイルス株は、P−Oka株のすべての糖タンパク質を保持し、そのため、P−Oka株と同じ免疫原性を示すはずである。このウイルス株は、カナマイシン又はガラクトースによりスクリーニングされる、該ウイルスの単一クローンである。したがって、V−Oka株のクローンと混在する潜在的な危険性は存在しない。ORF7全体の欠失(VZV−7D BAC)、又はORF7の反転逆フレームシフト突然変異(VZV−7DRM BAC)により、本発明のウイルス株に逆突然変異が導入される可能性はない。VZV−7D BAC又はVZV−7DRM BACから調製される弱毒性生ワクチン株は、より安全で、より有効である。
【0017】
別の態様では、本発明は、上記の水痘ウイルスを調製する方法であって、
1)野生型VZVのゲノムを、細菌人工染色体(BAC)ベクター中にクローニングして、ORF7の完全又は部分的な欠失、1塩基又は複数塩基の置換又は挿入を有する組換え水痘ウイルスVZV−BACを作製するステップであって、前記組換え水痘ウイルスが、ヒトの皮膚及び感覚神経節に感染することができないステップと、
2)1)で得られたVZVからDNAを単離し、単離されたDNA及び酵素Creを含有する組換えプラスミドにより、MRC−5細胞、Mewo細胞、又はARPE細胞を共形質転換するステップと、
3)酵素Creにより、MRC−5細胞、Mewo細胞、又はARPE細胞からBACを切り出すステップと
を含む方法に関する。
【0018】
本発明の一実施形態では、野生型VZVが、P−Oka株であり、ステップ1)におけるORF7が、Kanr、又は反転逆フレームシフト突然変異を有するORF7断片により置換されている。本発明の特定の一実施形態では、反転逆フレームシフト突然変異を有するORF7断片が、配列番号13として示される。
【0019】
別の態様ではまた、本発明は、上記の任意の水痘ウイルスを含む組成物にも関する。
【0020】
さらに別の態様ではまた、本発明は、水痘及び/又は帯状疱疹を予防するためのワクチンであって、上記の任意の水痘ウイルス、及びワクチンにおいて許容される賦形剤又は許容される担体を含有するワクチンにも関する。
【0021】
本発明で用いられる「水痘又は帯状疱疹を予防するためのワクチン」という用語は、水痘ワクチン、帯状疱疹ワクチン、並びに水痘及び帯状疱疹を予防するためのワクチンを指す。
【0022】
別の態様ではまた、本発明は、水痘ワクチン及び/又は帯状疱疹ワクチンの調製における、上記の水痘ウイルスのうちのいずれか1つの使用にも関する。
【0023】
別の態様ではまた、本発明は、発現ベクターの調製における、上記の水痘ウイルスのうちのいずれか1つの使用にも関する。
【0024】
本発明はまた、BACと、前記BAC中に挿入されている上記の水痘ウイルスのうちのいずれか1つのDNAとで構成される発現ベクターにも関する。発現ベクターは、上記の水痘ウイルスのうちのいずれか1つのDNAを含む。ベクターの機能に影響を及ぼすことなく、少なくとも10kbの外来遺伝子を上記の発現ベクターに挿入することができる。これはまた外来遺伝子も発現することができる。加えて、該発現ベクターは、ORF7の欠失の結果として安全である。該発現ベクターはまた、in vitro及びin vivoにおけるVZV−7DRM BACの機能をモニタリングする目的で、ルシフェラーゼ遺伝子による標識化(Zhangら、Journal of Virology、2007年9月、9024〜9033ページ)にも用いることができる。
【0025】
別の態様ではまた、本発明は、上記の発現ベクターと、挿入された外来遺伝子とを含む組換えベクターにも関する。
【0026】
別の態様ではまた、本発明は、上記の発現ベクター又は組換えベクターを含有する組換え細胞にも関する。
【0027】
本発明の一実施形態では、組換え細胞として用いられる宿主細胞は、MRC−5細胞、Mewo細胞、ARPE細胞、2BS細胞、WI−38細胞、及びKMB17細胞から選択される。
【0028】
別の態様ではまた、本発明は、水痘及び/又は帯状疱疹を予防する方法であって、それを必要とする患者に有効量の上記のワクチンを接種するステップを含む方法にも関する。
【0029】
本明細書では、「DPI」という用語は、「感染後日数」を表し、感染後の日数を指す。
【図面の簡単な説明】
【0030】
【図1】P−Oka BAC(VZV BAC)ベクターの構築を示す図である。
【図2】ORF7欠失VZV BAC株(VZV−7D BAC)の調製を示す図である。
【図3】ORF7欠失VZVの電気泳動を示す図である。
【図4A】後根神経節におけるVZV−7Dの培養を示す図である。MeWo細胞において培養した、野生型ウイルス(WT)、逆突然変異体ウイルス(7R)、欠損ウイルス(7D)についての増殖曲線を示す図である。
【図4B】後根神経節におけるVZV−7Dの培養を示す図である。後根神経節(DRG)において培養した、野生型ウイルス(WT)、逆突然変異体ウイルス(7R)、欠損ウイルス(7D)についての増殖曲線を示す図である。
【図5】ORF−7逆突然変異体VZV(VZV−7R)の調製について示す図である。
【図6】ヒトMeWo細胞において培養した、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
【図7】ヒト胸腺組織培養物(TOC)において培養した、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
【図8】ヒト皮膚組織培養物(SOC)中の、蛍光タグ(luc)を有するVZV−7DRMについての増殖曲線を示す図である。
【図9A】WT、VZV−7DRM、及びVZV−7Rに由来するDNAをトランスフェクトしたMeWo細胞及び神経芽腫(SH−SY5Y)細胞を示す図である。神経芽腫細胞において、VZV−7DRMは増殖できない。
【図9B】ヒト胚の脊髄から摘出した後根神経節(DRG)を示す図である。
【図9C】DRGを、重症複合型免疫欠損マウスモデル(SCID−hu)の腎被膜へと移植し、次いで、WT又はVZV−7DRMを感染させた。IVIS計測器によりウイルスの増殖を測定する。ヒト組織を移植した重症複合型免疫欠損マウス(SCID−hu)において、VZV−7DRMは増殖できない。
【図9D】感染後、GFP強度の増大により、DRG培養物において、WT VZVが増殖できることが示された。
【発明を実施するための形態】
【0031】
以下の実施例に言及することにより、本発明の実施形態を詳細に説明する。これらの実施例が、本発明の実施を例示することを意図するものであり、本発明の範囲を限定することを意図するものではないことを、当業者は理解するであろう。具体的に例示されない技法又は条件については、当技術分野において説明されている(例えば、J.Sambrookら、「Molecular Cloning:A Laboratory Manual」、第3版、Science Press、Peitang Huang訳)技法及び条件に従って、又は製造元の指示書に従って、該手順を実施する。別段に指定しない限り、試薬及び装置は市販されている。
【実施例】
【0032】
(例1)
p−Oka BAC(VZV BAC)の構築
(1)BACベクター:BACベクターであるpUSF−6は、原核細胞の複製起点(ori)、該複製遺伝子及び分配遺伝子(repE、parA、及びparB)、Camr遺伝子、緑色蛍光タンパク質(gfp)遺伝子、2つの500bpのVZV断片a及びb(グレー)、並びに2つのloxP部位(白)を含有する。BamHIによるpUSF−6の消化の結果として直鎖断片を得、これを、相同組換えにより、VZVを含有するコスミドであるpvSpe23に挿入した。
【0033】
(2)pOkaゲノムについての概略図は、VZVが、固有の長いセグメント(UL)と、固有の短いセグメント(US)とを伴う、125kbのヌクレオチド配列を含有することを示す。全ゲノムを、重複領域を含む4つのVZVセグメントへと消化し、コスミド中にクローニングし、その結果として、4つの組換えクローン、すなわち、pvFsp73、pvSpe14、pvPme19、及びpvSpe23を得た。相同組換えにより、pUSF−6を、VZVコスミドであるpvSpe23のORF60とORF61との間に挿入した。
【0034】
(3)BACベクター(pUSF−6)を含有するpvSpe23を、他の3つのVZVコスミドと共にMeWo細胞へと共トランスフェクトしたところ、その結果として得られる組換えウイルス(VZV BAC)は、MeWo細胞内で複製されることが可能であり、緑色蛍光プラークを生成した。
【0035】
図1にもまた、該手順を示す。
【0036】
VZV BACの増殖曲線を、野生型pOkaの増殖曲線と比較した。各dpiにおけるプラーク形成単位(PFU)を記録した。結果は、VZV BAC(緑)が、検出可能な増殖の欠陥を示さず、これが、in vitroにおける親ウイルスの増殖と符合することを示唆する。
【0037】
(例2)
ORF7欠損VZV BAC(VZV−7D BAC)の作製及び検証
既に説明されているluc VZV BAC(Zhangら、Journal of Virology、2007年9月、9024〜9033ページ)を参照することにより、ORF7欠失突然変異体を構築した。手順を図2及び下記で説明した。
【0038】
(1)カナマイシン耐性遺伝子に相同な20bpのヌクレオチドと、欠失の標的とされるORF7の開始コドン又は停止コドンに近接する隣接配列に相同な40bpのヌクレオチドとを含有するように2つのプライマーをデザインした。プライマーF1及びR1の配列を以下に示す。
【化3】
【0039】
プラスミドpGEM−oriV/kan1(Netterwald、Yangら、2005)により、KanR遺伝子を増幅した。そのPCR産物をDpnIで消化し、ゲル抽出キット(Qiagen)により回収した。
【0040】
(2)BioRad Gene Pulser II装置を用いて、1.6kV及び250uFでエレクトロポレーションにより、約200ngずつの各PCR産物を、VZV Luc BACを保有するDY380株へと形質転換した。
【0041】
(3)高効率の相同組換え系として機能するDY380株により、40bpという短い相同配列の組換えが可能となる。42℃で活性化され、32℃で抑制される組換え酵素により、カナマイシン耐性遺伝子(kanr)とORF7との相同組換えを実現する。結果として、ORF7が、Kanrにより置換される。32℃で30μg/mlのカナマイシンを含有する寒天プレート上で、単一組換え体を選択した。VZV突然変異体(ORF7欠失による)のDNAを、大腸菌から単離した。
【0042】
(4)HindIII消化によりウイルスゲノムの完全性を検証し、PCR解析により組換えDNAを確認した。KanrによりORF7が置換された、ORF7欠損BAC(VZV−7D BAC)が得られたことが確認される。
【0043】
(例3)
反転逆フレームシフト突然変異を有するVZV BAC(VZV−7DRM BAC)の作製及び検証
戦略:野生型ウイルスのORF7を、galk遺伝子で置換した。galk遺伝子を有する組換えVZVは、ガラクトースを唯一の炭素供給源とするマッコンキー寒天培地上において、明赤色のプラークを生成させるはずである。この方法によるならば、単一クローンが選択されるであろう。次いで、galk遺伝子を、反転逆フレームシフト突然変異配列で置換し、完全最小培地プレート上において単一クローンを選択して、VZV−7DRM BACを得る。
【0044】
詳細なプロトコールを以下に示す。
【0045】
(1)pGalK(1〜82bp:プロモーター;83〜1231bp:GalKのオープンリーディングフレーム)により、galk遺伝子を増幅するための増幅プライマーF2及びR2をデザインする。これらのフォワードプライマー及びリバースプライマーは各々が、5'側に60bpの、ORF7に隣接する相同配列を有する。GalK配列を以下(配列番号5)に示す。
【化4】
【0046】
プライマーF2及びR2の配列を以下に示す。
【化5】
【0047】
(2)これらのプライマーを用いて、GalKカセットを増幅する。この産物を、DpnIにより終夜にわたり消化し、精製する。
【0048】
(3)VZV BACを、大腸菌のSW102株へと形質転換する。
【0049】
(4)単一コロニーを5mlのLB−CM培地(クロラムフェニコールを含むLB培地)へと接種し、32℃で終夜にわたり増殖させる。
【0050】
(5)終夜にわたる培養物を、250mlのLB−CMol培地へと接種し、ODが0.4〜0.6となるまで、振とうしながら32℃で増殖させる。
【0051】
(6)エレクトロコンピテント細胞(VZV BACで形質転換したSW102細胞)を調製する。
【0052】
(7)ステップ(6)で調製されたエレクトロコンピテント細胞(VZV BACで形質転換したSW102細胞)を、ステップ(2)による1〜2μl(約200ng)の精製されたPCR産物により、例2において説明したエレクトロポレーションパラメータで形質転換する。
【0053】
(8)32℃で1mlのLB培地中で、1時間にわたり、形質転換体を回収する。
【0054】
(9)回収時間の後、1×M9塩中で、以下の通り、3回にわたり細菌を洗浄する。培養物を13,200RPMで15秒間にわたり遠心分離にかけ、ピペットで上清を除去する。ペレットを1×M9塩中に再懸濁させ、上記の通り、再度遠心分離にかける。洗浄ステップを再度繰り返す。3回にわたり洗浄した後、上清を除去し、ペレットを1mlの1倍濃度M9塩中に再懸濁させる。次いで、ペレットの懸濁液、10倍の希釈系列、及び100倍の希釈系列を、ガラクトース、ロイシン、ビオチン、及びクロラムフェニコールを含むM63最小培地プレートへと播種する。
【0055】
(10)32℃で3日間にわたりプレートをインキュベートする。
【0056】
(11)数個のコロニーを、ガラクトース+クロラムフェニコールを含むマッコンキー寒天培地プレート上へと画線培養して、単一コロニーを得る。3日間にわたるインキュベーション後に出現するコロニーはGal+コロニーのはずであるが、Gal−混入を除去するためには、明赤色の単一コロニーを得てから、次のステップへと進むことが重要である。
【0057】
(12)単一の明赤色の(Gal+)コロニーを採取して、5mlのLB−CM培地へと接種し、32℃で終夜にわたり培養する。該終夜にわたる培養物からBAC DNAを調製し、PCRによりgalk遺伝子を検証する。用いられるプライマーは、
【化6】
である。
【0058】
(13)プライマーF4(配列番号10)及びR4(配列番号11)を用いて、ORF7停止コドンの3'側に近接する457bpの配列(457bpのORF配列を、配列番号12に示す)を増幅する。プライマーF4及びR4は、1塩基のフレームシフトを導入するようにデザインされているので、フレーシフト突然変異を有するPCR産物が生成される。
【0059】
F4及びR4の配列を、以下に示す。
【化7】
【0060】
457bpのオープンリーディングフレームは、
【化8】
である。
【0061】
上記のステップ4〜10を繰り返して、エレクトロコンピテントSW102細胞を得る。
【0062】
(14)ステップ(13)の熱ショック処理した細菌50μlを、ステップ(2)のgalk遺伝子に隣接する領域と相同的なPCR産物200ngにより形質転換する。振とうしながら、32℃の水浴中で4.5時間にわたりインキュベートすることにより、50mlのバッフル付き三角フラスコ内のLB培地10ml中に回収する。この長い回収時間は、対象の配列だけを含有する組換えVZV BACを得る(したがって、GalKカセットを含有するVZV BACを除去する)のに役立つ。プライマーF4は、ORF7の5'側に隣接する60bpの相同配列と、ORF7の3'側に近接する457bpの逆配列とで構成され、プライマーR4は、ORF7の3’側に隣接する60bpの相同配列と、ORF7の3’側に近接する457bpのフォワード配列とで構成されるので、増幅されたPCR産物と、ORF7遺伝子座内のgalk遺伝子との相同組換えにより、3'側457bpの反転逆リンクがもたらされ、その結果として、機能を発現しない反転逆フレームシフト突然変異(配列番号13)が得られる。反転逆フレームシフト突然変異配列を、以下に示す。
【化9】
【0063】
(15)ステップ9の通り、1mlの培養物をスピニングし、1×M9塩中で3回にわたり洗浄し、1mlの1倍濃度M9塩中に再懸濁させる。ペレットの懸濁液及び10倍の希釈系列(各々100μlずつ)を、グリセロール、ロイシン、ビオチン、2−デオキシ−ガラクトース、及びクロラムフェニコールを含むM63最小培地プレートへと播種する。
【0064】
(16)32℃で3〜4日間にわたりインキュベートする。
【0065】
(17)コロニーを採取し、それらをLB−CMプレート及びGalK−M63 CM最小培地プレート上に画線培養する。LB−CMプレート上では増殖するが、最小培地プレート上では増殖しない単一コロニーを選択する。ウイルスDNAを抽出し、HindIII消化により完全性を検証し、PCRにより組換えDNAを確認する。ORF7を含むVZV BACが逆フレームシフト型突然変異(VZV−7DRM BAC)を反転させることを確認する。
【0066】
上記の手順で用いた試薬は、表1の通りに調製する。
【表1】
【0067】
(例4)
VZV−7Dウイルス及びVZV−7DRMウイルスの作製
(1)例2で得たVZV−7D BACからDNAを抽出する。
【0068】
(2)精製されたDNAを、エレクトロポレーションを介して、MRC−5細胞(Marchini、Liuら、2001)へと個別にトランスフェクトする(BACにおいて保有されるGFPにより、ウイルスの増殖をモニタリングすることができる)か、又は精製されたDNAを、Cre発現ベクターと共に、MRC−5細胞へと共トランスフェクトした(Marchini、Liuら、2001)。Creにより、BACに隣接するloxP部位で、BACを正確に切り出し、VZV 7Dウイルスを得た。
【0069】
同様にまた、ORF7の反転逆フレームシフト突然変異を有するVZV−7DRMウイルスも得た。VZV 7DRMは、反転逆配列であるORF7の一部だけを含有するので、発現することができず、ORF7機能を示さない。VZV−7DRM及びVZV−7Dはいずれも、示す機能が同じである。
【0070】
(例5)
VZV−7Dの検証
単層MRC−5細胞に、例4で調製され、2%ウシ血清を補充したMEM培地中で培養されたVZV−7Dを感染させた。細胞のほぼ80%が細胞変性効果を呈示したところで、培地を廃棄し、細胞を採取し、DNAを抽出する。P−Okaは対照としての役割を果たす。PCRによりORF−10遺伝子及びORF−7遺伝子を増幅した。VZV−7D及びP−OkaのDNAをHindIIIにより消化した。PCR産物と、HindIII消化生成物とを、0.5%アガロースゲル中の電気泳動にかけた。結果を図3に示した。VZV−7DにおけるORF−10のPCR産物はP−Okaと同じであるが、VZV−7DにおけるORF−7バンドは不在である。これにより、VZV−7DにおけるORF−7が欠失されていることが確認された。VZV−7D及びP−Okaとで、HindIIIにより消化されたウイルスDNAのプロファイルの差違は見られなかった。
【0071】
(例6)
ヒト後根神経節内におけるVZV−7Dの培養
ヒト胎児脊椎(妊娠20週齢)から、頸椎及び胸椎の後根神経節(DRG)を摘出した。1%のペニシリン/ストレプトマイシンを含む1×氷冷PBS中でこのDRGを簡単に洗浄し、次いで、氷冷組織培養培地(15%の熱不活化ウシ胎仔血清と、1%のペニシリン/ストレプトマイシンとを含むDEM)へと移した。0.5mlの培地で覆われた6ウェルプレート内の、コラーゲンでコーティングされたカバースリップ上に各DRGを個別に入れ、培地を1回交換して、5日間にわたり培養した。6ウェルプレート内で50%のMeWo細胞を調製し、滴定及び増殖曲線(図4A)の作成を行った。6日目に、P−Oka、VZV−7D、及びVZV−7Rを感染させたばかりのMewo細胞を採取し、培地中で1mlの最終容量へと再懸濁させた。各DRGは、6ウェルプレート内のコラーゲンでコーティングされたカバースリップ上で個別に増殖させる。10μlの懸濁液を用いて、ウイルス力価を決定し、2μlの懸濁液を用いて、感染MeWo細胞の増殖曲線を作成した。細胞懸濁液の残りを、20回にわたり27ゲージ針に通し、4000rpmで10分間にわたり遠心分離することにより、細胞破砕物をペレット化させた。上清を等量に分割し、これらを用いてDRGに感染させ(約120μl/試料1例)、加えて、2滴(20μl)の細胞懸濁液を用いて、別のDRGセットに感染させた。37℃/5%CO2中で3時間にわたるインキュベーション後、1×PBSでDRGを洗浄し、新鮮な培地を添加した。24時間間隔でルシフェラーゼ活性を測定した。試料を、150μg/mlのD−ルシフェリンと共に、37℃で10分間にわたりインキュベートした。IVIS測定器により光子をカウントし、ソフトウェアにより解析した。結果は、図4Bに示した。
【0072】
結果は、VZV−7Dが、MeWo細胞内では良好に増殖するが、DRG内ではほとんど増殖しないことを示す。VZV−7DがDRGに感染しないという事実は、安全な弱毒化生ワクチンを開発するための基盤をもたらす。
【0073】
(例7)
VZV−7Dに対する逆突然変異体(VZV−7R)の構築及び検証
ORF7欠失突然変異体は、HindIII消化によるだけでなく、相同組換えにより得られる完全VZVウイルスによっても同定することができる。すなわち、完全VZVは、相同組換えの機構によりレスキューされる、図5を参照されたい)。Invitrogen製のHigh Fidelity Platinum Taq DNAポリメラーゼキットを用いるPCRにより、ORF7断片を増幅し、プラスミドpGEM−lox−zeoのNotI部位とBglII部位との間にクローニングして、pGEM−zeo−ORF7プラスミドを形成した。配列決定解析により、クローニングされた断片を検証したところ、PCR増幅により、不適正なコドンがORF7へと導入されることはなかったことが示される。次いで、2つのプライマーを用いて、pGEM−zeo−ORF7から、PCRによりzeoR−ORF7カセットを増幅した。
【化10】
【0074】
結果として得られるPCR産物は、各末端に40bpの相同VZVセグメントを含有し、前記セグメントは、kanR遺伝子に近接し、欠失ORF7遺伝子内に位置する。結果として得られるキメラPCR産物を、上記のエレクトロポレーションを介して、ORF7D BACクローンを保有するDY380株へと形質転換する。50μg/mlのZeocin(Invitrogen)及び12.5μg/mlのクロラムフェニコールを含有する寒天プレート上、32℃で組換え体を選択した。それらの抗生物質感受性により、適正なVZVクローンを確認した。適正なクローンは、クロラムフェニコール、ハイグロマイシン、Zeocinに耐性であり、カナマイシン及びアンピシリンに感受性のはずである。制限酵素消化及びPCR解析により、これらのクローンをさらに検証した。
【0075】
検証されたクローン(ORF7が欠失している、又はレスキューluc VZV BACを有する、大腸菌DY380株によるクローン)を、12.5μg/mlのクロラムフェニコールを含む500mlのLB培地中、32℃で20時間にわたりインキュベートした。Nucleobond Maxiprep BAC DNA単離キット(BD Biosciences、Palo Alto、CA)を用いて、VZV BAC DNAを単離し、これらを、製造元の指示書に従い、FuGene6トランスフェクション試薬(Roche、Indianapolis、IN)を用いてMRC−5細胞にトランスフェクトするのに用いた。6ウェルプレートの各ウェル(35mm)について、DNA対トランスフェクション試薬の比は、1.5μg:6μlであった。トランスフェクションの3日後には、VZVプラークが正常に見て取れた。VZVゲノムから、BACベクター(2つのloxP部位により挟まれる)を除去するために、Cre発現ベクターを、VZV BAC DNAと共に、MRC−5細胞へと共トランスフェクトした(Marchini、Liuら、2001)。Cre酵素により、BACは正確に欠失され、完全VZVを生成させた。
【0076】
ORF−7レスキューウイルスは、P−Oka株と同じ生育及び増殖特性を示す。これらの結果は、逆突然変異により、ORF−7の機能を回復させうることを示した。
【0077】
(例8)
Mewo細胞におけるVZV−7DRMの培養
6ウェルプレート内の単層MeWo細胞にモック感染させ、又はlucVZV−7DRM(Lucを有するVZV−7DRM)、luc7VZV−7R(Lucを有するVZ−7R)、及びlucVZV(Lucを有するP−Oka株)を感染させた。2%ウシ血清を含むMEM培地中、37℃でMewo細胞を培養した。感染の24時間後に、D−ルシフェリンを150μg/mlの最終濃度まで、培養ウェルに添加した。37℃で10分間にわたるインキュベーション後、IVIS Imaging System(50 Series;Xenogen Corporation、Alameda、CA)を用いて、異なるウェル内の発光を同時に3連で記録した。測定後、ルシフェリン含有培地を、通常の細胞培地で置換した。24時間ごとに7日間にわたり、この測定を繰り返した。平均データ対時間をプロットすること(図6)により、増殖曲線を作成した。
【0078】
ヒトMeWo細胞にモック感染させ、又は野生型VZV(lucVZV)、又はVZV−7DRM、又はVZV−7Dレスキュー株(luc7R)を感染させ、7日間にわたり培養した。感染後毎日、IVISシステムにより3連で光子カウントを記録した。平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に誤差バーを表示した。この実験の結果は、VZV−7DRM及びVZV−7Rが、野生型ウイルスと同様に十分にMeWo細胞内で増殖することを示した。一方、BACを挿入した後、VZV−7DRMが正常に増殖しうることが示された。BACにより保有される遺伝子もまた、発現することが可能である。すなわち、VZV−7DRMは、異種遺伝子の発現ベクターとして用いることができる。
【0079】
(例9)
ヒト胸腺組織におけるVZV−7DRMの培養
ヒト胸腺−肝臓組織を、CB−17SCID/ベージュマウスへの異種移植片として同時移植した。ヒト胸腺−肝臓組織の組合せ移植片は、文献(Jennifer F.M.、Journal of Virology、1995年9月、5236〜5242ページ)で説明されている方法に従い構築する。移植の3カ月後、手術によりヒト胸腺−肝臓異種移植片を露出させ、これらに3連で、モック感染させ、又は10〜20μlのMeWo細胞により培養した、2×103〜4×103PFUのlucVZV−7DRM(ルシフェラーゼを有するVZV−7DRM)及びlucVZV(ルシフェラーゼを有するP−Oka)を感染させた。感染後、毎日蛍光強度を測定した。腹腔内(i.p.)注射により、ルシフェラーゼ基質であるD−ルシフェリン250μlを添加した。10分後、例8で説明した方法に従い、IVIS Imaging Systemを用いて、3連で、異種移植片中の蛍光を記録した。結果を図7に示した。
【0080】
7日間にわたり、培養されたヒト胸腺組織に、モック感染させ、又は野生型VZV、若しくはVZV−7DRMを感染させた。感染後毎日(DPI)、IVISシステムを用いて光子カウントを測定し、平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に、3回にわたるデータ測定による誤差バーを表示する。この実験により、VZV−7DRM突然変異体が、胸腺組織(T細胞)内で、野生型VZVと同様に増殖しうることが示された。
【0081】
同様の実験によりまた、VZV−7Dも、胸腺組織(T細胞)内で、野生型VZVと同様に増殖しうることが裏付けられた。
【0082】
(例10)
皮膚におけるVZV−7DRMの培養
文献(Jennifer F.M.、Journal of Virology、1995年9月、5236〜5242ページ)で説明されている方法に従い、ヒト胎児皮膚を、完全移植片としてSCIDマウスへと皮下移植した。移植の4週間後、マウスの皮膚に3連で、モック感染させ、又はlucVZV−7DRM(Lucを有するVZV−7DRM)、lucVZV−7R(Lucを有するVZ−7R)及びlucVZV(Lucを有するP−Oka株)を感染させた。感染後、毎日蛍光強度を測定した。i.p.注射により、ルシフェラーゼ基質であるD−ルシフェリン250μlを添加した。10分後、例8で説明した方法に従い、IVIS Imaging Systemを用いて、3連で、異種移植片中の蛍光を記録した。結果を図8に示した。
【0083】
7日間にわたり、培養されたヒト皮膚組織に、モック感染させるか、野生型VZV(P−Oka株)、若しくはORF7欠損突然変異体(VZV−7DRM)、又はORF7逆突然変異体(VZV−7R)を感染させた。感染後毎日(DPI)、IVISシステムを用いて光子カウントを測定し、平均光子カウント対時間をプロットすることにより、増殖曲線を作成した。図中に、3回にわたるデータ測定による誤差バーを表示する。この実験により、VZV−7DRM突然変異体が、皮膚における増殖で大きな欠陥を示し、この機能の欠陥が、VZV−7DRMにおける逆突然変異により回復されうることが示された。
【0084】
同様の実験によりまた、VZV−7D突然変異体も、皮膚における増殖で大きな欠陥を示し、この機能の欠陥が、VZV−7Dにおける逆突然変異により回復されうることが見出される。
【0085】
(例11)
神経細胞にVZVが感染するのにORF7が必須であることが実験により確認された
1.神経細胞腫細胞への感染
MeWo細胞並びに神経細胞腫細胞系であるSH−SY5Yに、野生型(WT)、ORF7欠失突然変異体(VZV−7DRM)BAC、及びORF7逆突然変異体(VZV−7R)BACに由来するDNAをトランスフェクトした。結果は、WT、VZV−7Rが、MeWo細胞及び神経細胞腫細胞において良好に増殖しうるが、VZV−7DRMが、神経細胞腫細胞においては増殖しえないことを示した(図9A)。これにより、神経細胞にVZVが感染するのに、ORF7が本質的である可能性が極めて高いことが示唆される。
【0086】
2.DRG移植片への感染
この例における試験1の結果をさらに検証するため、ヒト胎児DRGをSCIDマウスへと移植し、ウイルス感染の動物モデルとして用いた。結果は、野生型ウイルスが、短い複製周期の後に潜在化することを示す(蛍光現象による)一方、VZV−7DがDRG移植片中では複製されえないことを示した。これらの結果は、ORF7が、VZVの向神経性因子である可能性が極めて高いことを示した(図9B〜D)。
【0087】
上記の2つの試験により、ニューロンにVZVが感染するのに、ORF7が本質的であることが裏付けられた。
【0088】
(例12)
VZV−7DRMによるワクチンの調製並びにVZV−7DRMについての免疫原性アッセイ
単層ARPE細胞又はMRC−5細胞に、0.1の感染多重度(MOI)でVZV−7DRMを感染させた。40分間にわたりウイルスを細胞に付着させた後、フラスコに培地を添加し、これを35℃でインキュベートした。細胞のうちの約80%が細胞症を提示したら(一般に、3日後)、0.1%EDTAで消化することにより、細胞を採取した。採取された細胞を遠心分離し、保護剤を添加し、破砕及び清明化の後にワクチンを得た。
【0089】
1回のワクチン接種当たり5匹ずつのモルモット(体重約250g)に、投与1回当たり5000pfu/0.5mlで、復元されたワクチンを接種した。初回免疫化の4週間後に、追加免疫化を施した。P−Oka及びV−Okaを対照として用いる。
【0090】
追加免疫化の2週間後、心臓穿刺により血液を回収した。gp−ELISAにより、血清抗体力価を検出した。免疫阻害アッセイにより、ウイルスに対する抗体の中和能を決定した。ワクチン接種の前後に、10倍の血清希釈液を、1:1の比でウイルスと混合した。37℃で60分間にわたるインキュベーションの後、混合物を用いて、MRC−5細胞に感染させた。5%CO2中37℃における7日間のインキュベーション後、培地を除去し、クーマシーブルーで細胞を染色した。プラーク形成単位をカウントすることにより、ウイルス力価を計算した。100×([ワクチン接種前に血清により中和されるウイルス力価]−[ワクチン接種後に血清により中和されるウイルス力価])/[ワクチン接種前に血清により中和されるウイルス力価]として、抗体によるウイルス中和度を計算する。結果を表2に示す。
【表2】
【0091】
結果により、VZV−7DRMが、良好な免疫原性を保有し、それに対する抗体による中和度は、V−Okaの場合と同等であることが示された。VZV−7DRMによるワクチン接種後の血清は、3種のウイルスすべてに対して良好な中和効果を有し、ワクチンとして開発される可能性を示す。
【0092】
同様の実験によりまた、VZV−7Dも良好な中和効果を有し、ワクチンとして開発される可能性を有することが示された。
【0093】
(例13)
VZV−7DRMをベクターとして用いる生物学的活性物質の調製
ORF7に対する隣接相同性を含有するEV71−VP1プライマーによるPCRを介して、pT−VP1プラスミドから、EV71−VP1カセットを増幅した。
【0094】
EV71−VP1カセットは、以下に示される。
【化11】
【0095】
PCRプライマー配列:
F6(ORF7の5'側隣接領域に対して相同な配列;開始コドン;EV71−VP1に相同な配列):
【化12】
【0096】
R6(ORF7の3’側隣接領域に対して相同な配列;EV71−VP1に相同な配列):
【化13】
【0097】
VZV−7DRM BACを含有する大腸菌のエレクトロコンピテントSW102株を、精製されたPCR産物により形質転換する。例2に従い、組換えSW102株をスクリーニング及び同定した。適切な培養の後、VZV−7DRM−VP1 BACを単離し、次いで、エレクトロポレーションを介して、Cre発現ベクターと共に、ARPE細胞へと共トランスフェクトした。BACの両端におけるloxP部位から、BACの全体を切り出し、その後、組換えVZV−7DRM−VP1ウイルスを得た。単層ARPE細胞に組換えウイルスを感染させ、35℃で3〜4日間にわたり培養した。そこで、細胞は細胞症を提示した。組換えウイルスの培養物上清、並びに細胞溶解(凍結融解又は超音波処理を繰り返した後の)に由来する遠心分離上清におけるEV71特異的サンドイッチELISAにより、VP1の活性を検出した。EV71及びVZV−7DRMを、対照として用いる。結果(表3)は、VZV−7DRM−VP1ウイルスを感染させたARPE細胞においてVP1活性が検出されたことを示す。これは、VP1が、VZV−7DRMウイルス内に組み換えられていることを示唆する。この例により、外来遺伝子を発現させるベクターとして、VZV−7DRMを用いうることが示された。
【表3】
【0098】
同様の実験により、VZV−7Dは外来遺伝子を発現させる発現ベクターとしても用いることができることが裏付けられた。
【0099】
本発明の実施形態を詳細に説明してきたが、本発明の範囲を逸脱しない限りにおいて、本開示の教示に従い、これらの詳細に対して各種の改変及び置換を行いうることを、当業者は理解するであろう。付属の特許請求の範囲、及びそれらの同等物により、本発明の全範囲が規定される。
【図1A】
【図1B】
【図1C】
【図1D】
【特許請求の範囲】
【請求項1】
そのORF7が完全に若しくは部分的に欠失している、又は1塩基若しくは2塩基が置換若しくは付加されていることを特徴とし、ヒトの皮膚及び感覚神経節に感染することができない水痘ウイルス。
【請求項2】
前記ORF7が、抗生物質耐性遺伝子及び/又は非機能性核酸配列により完全に又は部分的に置換されている、請求項1に記載の水痘ウイルス。
【請求項3】
前記抗生物質耐性遺伝子が、ペニシリン耐性遺伝子、ストレプトマイシン耐性遺伝子、及びカナマイシン耐性遺伝子から選択される、請求項2に記載の水痘ウイルス。
【請求項4】
前記非機能性核酸配列が、反転逆フレームシフト突然変異を有するORF7断片を指す、請求項2に記載の水痘ウイルス。
【請求項5】
前記反転逆フレームシフト突然変異を有するORF7断片の配列が、配列番号13に示されている、請求項4に記載の水痘ウイルス。
【請求項6】
2009年7月28日に、寄託番号CGMCC第3207号で中国微生物株寄託管理委員会普通微生物センター(China General Microbiological Culture Collection Center)に寄託されている水痘ウイルス。
【請求項7】
請求項1に記載の水痘ウイルスを調製する方法であって、
1)野生型水痘ウイルスのゲノムを、細菌人工染色体(BAC)ベクター中にクローニングして、ORF7の完全又は部分的な欠失、1塩基又は複数塩基の置換又は挿入を有する組換え水痘ウイルスVZV−BACを作製するステップであって、前記組換え水痘ウイルスが、ヒトの皮膚及び感覚神経節に感染することができない上記ステップと、
2)1)で得られたVZVからDNAを単離し、単離されたDNA及び酵素Creを含有する組換えプラスミドにより、Mewo細胞、MRC−5細胞、ARPE細胞又は2BS細胞又はWI−38細胞又はKMB17細胞を共形質転換するステップと、
3)酵素Creにより、Mewo細胞、MRC−5細胞、ARPE細胞又は2BS細胞又はWI−38細胞又はKMB17細胞からBACを切り出すステップと
を含む、上記方法。
【請求項8】
前記野生型水痘ウイルスが、P−Oka株であり、ステップ1)における前記ORF7が、カナマイシン耐性遺伝子、又はその配列が配列番号13に示されている反転逆フレームシフト突然変異を有するORF7断片により置換されている、請求項7に記載の方法。
【請求項9】
請求項1〜6のいずれか一項に記載の水痘ウイルスを含む組合せ。
【請求項10】
水痘及び/又は帯状疱疹を予防するためのワクチンであって、請求項1〜6のいずれか一項に記載の水痘ウイルス、ワクチンにおいて許容される賦形剤又は担体を含む、上記ワクチン。
【請求項11】
水痘及び/又は帯状疱疹を予防するためのワクチンを調製するための、請求項1〜6のいずれか一項に記載の水痘ウイルスの使用。
【請求項12】
発現ベクターを調製するための、請求項1〜6のいずれか一項に記載の水痘ウイルスの使用。
【請求項13】
BACと、前記BAC中に挿入されている請求項1〜6のいずれか一項に記載の水痘ウイルスのDNAとで構成される発現ベクター。
【請求項14】
請求項1〜6のいずれか一項に記載の水痘ウイルスDNAのDNAである発現ベクター。
【請求項15】
請求項13又は14に記載の発現ベクターと、挿入外来遺伝子とを含む組換えベクター。
【請求項16】
請求項13若しくは14に記載の発現ベクター、又は請求項15に記載の組換えベクターを含む組換え細胞。
【請求項17】
前記宿主細胞が、以下の細胞:MRC−5細胞、Mewo細胞、ARPE細胞、2BS細胞、WI−38細胞、及びKMB17細胞から選択される、請求項16に記載の組換え細胞。
【請求項18】
水痘及び/又は帯状疱疹を予防する方法であって、それを必要とする患者に有効量の請求項10に記載のワクチンを接種するステップを含む、上記方法。
【請求項1】
そのORF7が完全に若しくは部分的に欠失している、又は1塩基若しくは2塩基が置換若しくは付加されていることを特徴とし、ヒトの皮膚及び感覚神経節に感染することができない水痘ウイルス。
【請求項2】
前記ORF7が、抗生物質耐性遺伝子及び/又は非機能性核酸配列により完全に又は部分的に置換されている、請求項1に記載の水痘ウイルス。
【請求項3】
前記抗生物質耐性遺伝子が、ペニシリン耐性遺伝子、ストレプトマイシン耐性遺伝子、及びカナマイシン耐性遺伝子から選択される、請求項2に記載の水痘ウイルス。
【請求項4】
前記非機能性核酸配列が、反転逆フレームシフト突然変異を有するORF7断片を指す、請求項2に記載の水痘ウイルス。
【請求項5】
前記反転逆フレームシフト突然変異を有するORF7断片の配列が、配列番号13に示されている、請求項4に記載の水痘ウイルス。
【請求項6】
2009年7月28日に、寄託番号CGMCC第3207号で中国微生物株寄託管理委員会普通微生物センター(China General Microbiological Culture Collection Center)に寄託されている水痘ウイルス。
【請求項7】
請求項1に記載の水痘ウイルスを調製する方法であって、
1)野生型水痘ウイルスのゲノムを、細菌人工染色体(BAC)ベクター中にクローニングして、ORF7の完全又は部分的な欠失、1塩基又は複数塩基の置換又は挿入を有する組換え水痘ウイルスVZV−BACを作製するステップであって、前記組換え水痘ウイルスが、ヒトの皮膚及び感覚神経節に感染することができない上記ステップと、
2)1)で得られたVZVからDNAを単離し、単離されたDNA及び酵素Creを含有する組換えプラスミドにより、Mewo細胞、MRC−5細胞、ARPE細胞又は2BS細胞又はWI−38細胞又はKMB17細胞を共形質転換するステップと、
3)酵素Creにより、Mewo細胞、MRC−5細胞、ARPE細胞又は2BS細胞又はWI−38細胞又はKMB17細胞からBACを切り出すステップと
を含む、上記方法。
【請求項8】
前記野生型水痘ウイルスが、P−Oka株であり、ステップ1)における前記ORF7が、カナマイシン耐性遺伝子、又はその配列が配列番号13に示されている反転逆フレームシフト突然変異を有するORF7断片により置換されている、請求項7に記載の方法。
【請求項9】
請求項1〜6のいずれか一項に記載の水痘ウイルスを含む組合せ。
【請求項10】
水痘及び/又は帯状疱疹を予防するためのワクチンであって、請求項1〜6のいずれか一項に記載の水痘ウイルス、ワクチンにおいて許容される賦形剤又は担体を含む、上記ワクチン。
【請求項11】
水痘及び/又は帯状疱疹を予防するためのワクチンを調製するための、請求項1〜6のいずれか一項に記載の水痘ウイルスの使用。
【請求項12】
発現ベクターを調製するための、請求項1〜6のいずれか一項に記載の水痘ウイルスの使用。
【請求項13】
BACと、前記BAC中に挿入されている請求項1〜6のいずれか一項に記載の水痘ウイルスのDNAとで構成される発現ベクター。
【請求項14】
請求項1〜6のいずれか一項に記載の水痘ウイルスDNAのDNAである発現ベクター。
【請求項15】
請求項13又は14に記載の発現ベクターと、挿入外来遺伝子とを含む組換えベクター。
【請求項16】
請求項13若しくは14に記載の発現ベクター、又は請求項15に記載の組換えベクターを含む組換え細胞。
【請求項17】
前記宿主細胞が、以下の細胞:MRC−5細胞、Mewo細胞、ARPE細胞、2BS細胞、WI−38細胞、及びKMB17細胞から選択される、請求項16に記載の組換え細胞。
【請求項18】
水痘及び/又は帯状疱疹を予防する方法であって、それを必要とする患者に有効量の請求項10に記載のワクチンを接種するステップを含む、上記方法。
【図5】

【図9B】

【図9D】

【図2】
【図3】
【図4A】
【図4B】
【図6】
【図7】
【図8】
【図9A】
【図9C】
【図9B】
【図9D】
【図2】
【図3】
【図4A】
【図4B】
【図6】
【図7】
【図8】
【図9A】
【図9C】
【公表番号】特表2013−500024(P2013−500024A)
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願番号】特願2012−521930(P2012−521930)
【出願日】平成22年7月27日(2010.7.27)
【国際出願番号】PCT/CN2010/001139
【国際公開番号】WO2011/011971
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(512022686)ニュージャージー メディカル スクール (1)
【出願人】(508198258)ベイジン ワンタイ バイオロジカル ファーマシー エンタープライズ カンパニー, リミテッド (3)
【出願人】(510183280)シャアメン ユニバーシティ (2)
【Fターム(参考)】
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願日】平成22年7月27日(2010.7.27)
【国際出願番号】PCT/CN2010/001139
【国際公開番号】WO2011/011971
【国際公開日】平成23年2月3日(2011.2.3)
【出願人】(512022686)ニュージャージー メディカル スクール (1)
【出願人】(508198258)ベイジン ワンタイ バイオロジカル ファーマシー エンタープライズ カンパニー, リミテッド (3)
【出願人】(510183280)シャアメン ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]