
P−セレクチンリガンド蛋白
【課題】内皮細胞への白血球の付着を阻害し、炎症反応を阻害しうる、P−セレクチンリガンド糖蛋白、該蛋白をコードするDNA配列の提供。
【解決手段】(a)特定のアミノ酸配列を有するP−セレクチンリガンド糖蛋白、および(b)抗体に由来する第2のアミノ酸配列を含む、融合蛋白、および該蛋白をコードするDNA配列。
【解決手段】(a)特定のアミノ酸配列を有するP−セレクチンリガンド糖蛋白、および(b)抗体に由来する第2のアミノ酸配列を含む、融合蛋白、および該蛋白をコードするDNA配列。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、内皮細胞への白血球の付着を阻害することにより作用する抗炎症物質の分野に関する。より詳細には、本発明は、セレクチンとして知られる哺乳動物付着蛋白の新規リガンドに指向される。
【0002】
本願は、現在放棄されている1992年10月23日出願の米国出願第07/965662号の一部継続出願であった1993年8月26日出願の同時係属出願第08/112608号の一部継続出願であった1994年4月28日出願の同時係属出願第08/235398号の一部継続出願であった1994年9月30日出願の同時係属出願第08/316305号の一部継続出願であった1995年4月25日出願の同時係属出願第08/428734号の一部継続出願である。
【背景技術】
【0003】
炎症の間、白血球は血管内皮に付着し、内皮下組織に侵入するが、これはセレクチンまたはLECCAMクラスの蛋白の標的細胞上のリガンドへの特異的結合により媒介される相互作用である。かかるセレクチンにより媒介される細胞付着は、血栓性疾患および寄生虫性疾患においても起こり、腫瘍細胞の転移性拡張に関与している可能性もある。
【0004】
セレクチン蛋白はN末端レクチン様ドメイン、表皮増殖因子様ドメイン、および補体結合蛋白に相同性を有する領域により特徴づけられる。これまで3種のヒトセレクチン蛋白が同定されており、それらはE−セレクチン(以前はELAM−1と呼ばれた)、L−セレクチン(以前はLAM−1と呼ばれた)およびP−セレクチン(以前はPADGEMまたはGMP−140と呼ばれた)である。
【0005】
E−セレクチンは、サイトカインによる活性化から数時間後に内皮細胞上に誘導され、好中球と内皮との間のカルシウム依存性相互作用を媒介する。L−セレクチンはリンパ球誘導受容体であり、P−セレクチンは、血小板が活性化された場合、血小板細胞表面に急激に出現し、好中球または単球の血小板へのカルシウム依存性付着を媒介する。P−セレクチンは内皮細胞のWeibl-Palade体中にも見出される。P−セレクチンはこれらの小胞から放出されて、ヒスタミンまたはスロンビンにより刺激された内皮への好中球の初期の結合を媒介する。
【0006】
セレクチンは、標的細胞表面上に存在するリガンドとの特異的相互作用により付着を媒介すると考えられている。一般的には、セレクチンのリガンドは、少なくとも一部には炭水化物部分を含む。例えば、E−セレクチンは、末端構造
【化1】

を有する炭水化物に結合し、また、末端構造
【化2】
を有する炭水化物にも結合する。式中、Rは炭水化物鎖の残りの部分である。これらの炭水化物は血液型抗原として知られており、通常には、それぞれシアリル・ルイスxおよびシアリル・ルイスaと呼ばれている。内皮細胞表面上にシアリル・ルイスx抗原のみが存在することは、E−セレクチン発現細胞への結合を促進するには十分であるかもしれない。E−セレクチンは、末端構造
【化3】
を有する炭水化物にも結合する。
【0007】
E−セレクチンと同様、各セレクチンは種々の親和性で広範な炭水化物に結合するように思われる。セレクチンにより媒介される付着の強度(結合親和性)は、細胞表面上の炭水化物密度およびセレクチン密度にも依存する。
【0008】
P−セレクチンは、非シアル化形態のルイスx血液型抗原を含む炭水化物に結合し、シアリル・ルイスxに対しては親和性が高い。また、P−セレクチンは、異種性3−硫酸化ガラクトシルセラミドでありミエロイドおよび腫瘍細胞から脂質抽出により単離されたスルファチド類を認識する可能性がある。しかしながら、P−セレクチン担持細胞のP−セレクチンリガンド担持細胞への結合は、リガンド担持細胞をプロテアーゼ処理した場合には見られず、P−セレクチンリガンドが糖蛋白であることが示される。
【0009】
P−セレクチンに関する2種の推定上の糖蛋白リガンドが最近同定され、その1つは部分精製されている(Moore et al., J. Cell Biol. 118, 445-456 (1992))(非特許文献1)。しかしながら、これらの糖蛋白のアミノ酸組成もアミノ酸配列も開示されていない。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Moore et al., J. Cell Biol. 118, 445-456 (1992)
【発明の概要】
【発明が解決しようとする課題】
【0011】
したがって、本発明の解決すべき課題は、P−セレクチンのリガンドとして作用する蛋白およびDNAを同定および単離することである。加えて、本発明の解決すべき課題は、P−セレクチンリガンド糖蛋白、該蛋白をコードするDNA配列、該DNAで形質転換された宿主細胞、およびP−セレクチンリガンド蛋白の製造方法を提供することである。
【課題を解決するための手段】
【0012】
1の具体例において、本発明は、P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2に示すアミノ酸1からアミノ酸402までのアミノ酸配列を含む。可溶性P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物も提供され、該蛋白は配列番号:2のアミノ酸1からアミノ酸310までを含む。さらに本発明は、成熟P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列を含む。もう1つの具体例において、本発明は、可溶性成熟P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列を含む。もう1つの具体例において、本発明は、P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:4に示すアミノ酸配列を含む。さらに本発明は、本発明単離DNAのいずれか1つを含む発現ベクターを含む組成物を提供し、該DNAは発現制御配列に作動可能に連結されている。上記DNAのいずれか1つを含む発現ベクターで形質転換された宿主細胞も提供される。さらに、P−セレクチンリガンド蛋白の製造方法も提供され、該方法は:
(a)本発明DNAのいずれか1つを含む発現ベクターで形質転換された宿主細胞を適当な倍地中で培養し、ついで、
(b)P−セレクチンリガンド蛋白を培地から精製する
ことを含む。
【0013】
もう1つの具体例において、本発明は、配列番号:2のアミノ酸21からアミノ酸402までのアミノ酸配列を含む蛋白を含む組成物を提供し、該蛋白は実質的に他の哺乳動物蛋白を含まない。さらに本発明は、配列番号:2のアミノ酸21からアミノ酸310までのアミノ酸配列を含む可溶性P−セレクチンリガンド蛋白を含み、該蛋白は実質的に他の哺乳動物蛋白を含まない。もう1つの具体例において、本発明は、配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列を含むP−セレクチンリガンド蛋白を含み、該蛋白は実質的に他の哺乳動物蛋白を含まない。また本発明は、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列を含む成熟P−セレクチンリガンド蛋白を含む組成物を提供し、該蛋白は実質的に他の哺乳動物蛋白を含まない。さらに、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列を含む可溶性成熟P−セレクチンリガンド蛋白を含む組成物が提供され、該蛋白は実質的に他の哺乳動物蛋白を含まない。もう1つの具体例において、本発明は、配列番号:4に示すアミノ酸配列を含む蛋白を含む組成物を提供する。
【0014】
さらにもう1つの具体例において、本発明は、P−セレクチンリガンド蛋白に対して特異的な抗体を含む組成物を提供する。
【0015】
もう1つの具体例において、本発明は、下記工程:
(a)配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列、および配列番号:4に示すアミノ酸配列からなる群より選択されるアミノ酸配列を含むP−セレクチンリガンド蛋白にP−セレクチンを結合させ(該結合は第1の結合混合物を生じる);
(b)第1の結合混合物中のP−セレクチン蛋白とP−セレクチンリガンド蛋白との間の結合量を測定し;
(c)P−セレクチン蛋白およびP−セレクチンリガンド蛋白に化合物を結合させて第2の結合混合物を得て;
(d)第2の結合混合物中の結合量を測定し;ついで
(e)第1の結合混合物中の結合量を第2の結合混合物中の結合量と比較する
ことを含み、第2の結合混合物の結合量の減少か起こった場合、化合物がP−セレクチンにより媒介される細胞間付着を阻害しうるものである、P−セレクチンにより媒介される細胞間付着の阻害剤の同定方法を提供する。
【0016】
もう1つの具体例において、本発明は、下記工程:
(a)配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列、および配列番号:4に示すアミノ酸配列からなる群より選択されるアミノ酸配列を含むE−セレクチンリガンド蛋白にE−セレクチンを結合させ(該結合は第1の結合混合物を生じる);
(b)第1の結合混合物中のE−セレクチン蛋白とE−セレクチンリガンド蛋白との間の結合量を測定し;
(c)E−セレクチン蛋白およびE−セレクチンリガンド蛋白に化合物を結合させて第2の結合混合物を得て;
(d)第2の結合混合物中の結合量を測定し;ついで
(e)第1の結合混合物中の結合量を第2の結合混合物中の結合量と比較する
を含み、第2の結合混合物の結合量の減少が起こった場合、化合物がE−セレクチンにより媒介される細胞間付着を阻害しうるものである、E−セレクチンにより媒介される細胞間付着の阻害剤の同定方法を提供する。これらの方法を用いて、E−セレクチンをL−セレクチンに置き換えるとL−セレクチン阻害剤を探すこともできる。
【0017】
また本発明は、(a)P−セレクチンリガンド蛋白をコードするDNAおよびシアリル・ルイスX(sLex)またはシアリル・ルイスA(sLea)を合成できるフコシルトランスフェラーゼ((α1,3/α1,4)フコシルトランスフェラーゼまたは(α1,3)フコシルトランスフェラーゼのごときフコシルトランスフェラーゼ)をコードするDNAで宿主細胞を同時形質転換し;(b)適当な培地中で宿主細胞を培養し;ついで(c)P−セレクチンリガンド蛋白を培地から精製することを含む、P−セレクチンリガンド蛋白の製造方法を包含する。他の特定の具体例において、対になった塩基性アミノ酸変換酵素(paired basic amino acid converting enzyme)をコードするDNAおよび/またはGlcNAcトランスフェラーゼ(好ましくは「コア2トランスフェラーゼ」)をコードするDNAで宿主細胞を同時形質転換する。好ましい具体例において、P−セレクチンリガンド蛋白は全長または可溶性形態である。
【0018】
他の具体例において、本発明は、P−セレクチンリガンド蛋白活性を有するP−蛋白リガンド蛋白を包含する。好ましい具体例において、リガンド蛋白は、配列番号:2のアミノ酸42からアミノ酸60までのアミノ酸配列を含む蛋白、配列番号:2のアミノ酸42からアミノ酸60までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸88までのアミノ酸配列を含む蛋白、配列番号:2のアミノ酸42からアミノ酸88までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸118までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸189までのアミノ酸配列を必須として含む蛋白である。他の好ましい具体例において、配列番号:2の位置65、111および292におけるアスパラギン残基の少なくとも1つが欠失または置換される。リガンド蛋白の特定の好ましい具体例は、配列番号:2の位置46、48および51において少なくとも1つのチロシン残基を含む。これらのP−セレクチンリガンド蛋白をコードするDNA、かかるDNAで形質転換された宿主細胞、かかる宿主細胞を培養することによる蛋白の製造方法、蛋白を含む医薬組成物、蛋白を用いるセレクチン結合阻害剤の同定方法、蛋白に対する抗体、ならびに蛋白を用いる、セレクチンにより媒介される結合の阻害方法も本発明により包含される。
【0019】
さらなる他の具体例において、本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のアミノ酸配列、および(b)P−セレクチンリガンド以外の蛋白の配列に由来する第2のアミノ酸配列を含む融合蛋白をコードする単離DNAを提供する。好ましくは、発現制御配列はそれらのヌクレオチド配列に作動可能に連結される。かかるDNAで形質転換された宿主細胞も提供される。また本発明は、(a)融合蛋白の発現に適した条件下で宿主細胞を培養し;ついで(b)融合蛋白を培地から精製することを含む、融合蛋白の製造方法を提供する。かかる方法により製造される融合蛋白も提供される。
【0020】
特定の好ましい具体例において、かかる融合蛋白の第1のアミノ酸配列は、配列番号:2のアミノ酸42からアミノ酸402まで、配列番号:2のアミノ酸42からアミノ酸301まで、配列番号:2のアミノ酸42からアミノ酸88まで、配列番号:2のアミノ酸42からアミノ酸118まで、または配列番号:2のアミノ酸42からアミノ酸189までを含む。
【0021】
他の好ましい具体例において、DNAは、配列番号:35のヌクレオチド123からヌクレオチド939までのヌクレオチド配列、配列番号:35のヌクレオチド配列、配列番号:37のヌクレオチド123からヌクレオチド807までのヌクレオチド配列、配列番号:37のヌクレオチド配列、配列番号:39のヌクレオチド123からヌクレオチド1311までのヌクレオチド配列、配列番号:39のヌクレオチド配列、配列番号:41のヌクレオチド123からヌクレオチド792までのヌクレオチド配列、配列番号:41のヌクレオチド配列を含む。
【0022】
また本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のアミノ酸配列、および(b)P−セレクチンリガンド以外の蛋白の配列に由来する第2のアミノ酸配列を含む融合蛋白を提供する。好ましくは、第1のアミノ酸配列は配列番号:2のアミノ酸42からアミノ酸402まで、配列番号:2のアミノ酸42からアミノ酸310まで、配列番号:2のアミノ酸42からアミノ酸88まで、配列番号:2のアミノ酸42からアミノ酸118まで、配列番号:2のアミノ酸42からアミノ酸189までを含む。
【0023】
ある種の特に好ましい具体例において、融合蛋白は、配列番号:36アミノ酸42からアミノ酸313までのアミノ酸配列、配列番号:36のアミノ酸配列、配列番号:38のアミノ酸42からアミノ酸269までのアミノ酸配列、配列番号:38のアミノ酸配列、配列番号:40のアミノ酸42からアミノ酸437までのアミノ酸配列、配列番号:40のアミノ酸配列、配列番号:42のアミノ酸42からアミノ酸264までのアミノ酸配列、配列番号:42のアミノ酸配列を含む。
【0024】
他の好ましい具体例において、第2のアミノ酸配列は第1のアミノ酸配列のC末端またはN末端に連結され、結合配列により結合されても、されなくてもよい。
【0025】
さらなる他の具体例において、第2のアミノ酸配列は、抗体、サイトカイン、増殖因子、分化因子、ホルモン、酵素、受容体またはそれらのフラグメントならびにリガンドからなる群より選択される蛋白に由来するものである。好ましくは、第2のアミノ酸配列は、抗体配列由来、抗体のFc部分由来であり、あるいはまた抗体由来の配列の変異体である。
【0026】
さらなる具体例において、本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のペプチド、およびO−セレクチンリガンド以外の蛋白の配列に由来する第2のペプチドを含む組成物であって、第1のペプチドおよび第2のペプチドがペプチド結合以外の部分により化学結合されている組成物を提供する。本発明P−セレクチンリガンド蛋白をかかる組成物に使用することができる。
【図面の簡単な説明】
【0027】
【図1】コア2ありおよびなしで発現されたP−セレクチンリガンド蛋白の結合を比較したグラフである。
【図2】コア2ありおよびなしで発現されたP−セレクチンリガンド蛋白の免疫沈降のオートラジオグラフである。
【図3】P−セレクチンリガンド蛋白(コア2ありおよびなしで発現)のP−セレクチン/IgGキメラ(LEC−γ1)および抗P−セレクチンリガンド蛋白モノクローナル抗体(MAb275)への結合のフローサイトメトリー分析の結果を示す。
【図4】P−およびE−セレクチン/IgGキメラに結合する、P−セレクチンリガンド蛋白を含む蛋白のオートラジオグラフである。
【図5】配列番号:2の全長P−セレクチンリガンド蛋白の構造的特徴を図式的に示したものである。
【図6】P−セレクチンリガンド蛋白のセレクチン類への結合におけるN−結合糖鎖付加部位の役割を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図7】P−セレクチンリガンド蛋白のセレクチン類への結合におけるN−結合糖鎖付加部位の役割を調べるための実験結果を示す。
【図8】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図9】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図10】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図11】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図12】P−セレクチンリガンド蛋白のセレクチン類への結合における種々の欠失の影響を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図13】実施例4(c)の定量的プレート結合アッセイを図式的に示す。
【図14】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図15】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図16】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図17】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図18】P−セレクチンリガンド蛋白のアニオン性領域のセレクチン類への結合におけるチロシン残基の変化の影響を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図19】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図20】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図21】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図22】P−セレクチンリガンド蛋白のP−およびE−セレクチンへの結合についての提案されたモデルを示す。
【図23】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチンへの結合を比較する実験の結果を示す。
【図24】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチンへの結合を比較する実験の結果を示す。
【図25】すでに3/4フコシルトランスフェラーゼおよびコア2トランスフェラーゼを発現しているCHO細胞の発現産物の分析であり、CHO細胞はpsPSL.T7、ΔTM、I316またはpsPSL.QCでトランスフェクションされており、メトトレキセートを用いて増幅された。ならし培地を直接分析するか、あるいはまずLEC−γ1で沈降させ、ついで非還元的条件下および還元的条件下でSDS−PAGEにより分析した。
【図26】P−およびE−セレクチンによりアフィニティー捕捉されたミエロイド細胞膜蛋白のSDS−PAGE分離である。3H−グルコサミンで代謝的に標識したU937細胞から膜溶解物を調製し、固定化P−およびE−セレクチンならびに対照ヒトIgG1とともにアフィニティー沈降させた。ゲル電気泳動前に溶離蛋白をDTT存在下(「還元」)または不存在下(「非還元」)で処理した。レーン:レーン1はヒトIgG1によるアフィニティー捕捉;レーン2はP−セレクチンによるアフィニティー捕捉;レーン3はE−セレクチンによるアフィニティー捕捉。
【図27】連続的アフィニティー捕捉実験である。3H−標識U937溶解物種をP−またはE−セレクチンによりアフィニティー捕捉し、溶離し、ついで、抗PSGL−1抗血清Rb3443との免疫沈降に供した。レーン:1および2は、免疫前のウサギの血清(レーン1)およびRb3443(レーン2)を用いる新鮮ミエロイド細胞膜溶解物の対照免疫沈降である。レーン3〜5は、すでにP−セレクチン(レーン3)、E−セレクチン(レーン4)、およびヒトIgG1(レーン5)によりアフィニティー捕捉され、ついで溶離されたミエロイド細胞膜溶解物のRb3443との免疫沈降である。
【図28】ミエロイド膜抽出物のCD43およびPSGL−1含量の比較である。標識U937細胞抽出物を抗PSGL−1ウサギポリクローナル抗体Rb3443または抗CD43マウスMabとともに免疫沈降させ、ついでSDS−PAGE/オートラジオグラフィーに供した。レーン:1は、対照である免疫前のウサギ血清との免疫沈降である。2はRb3443との免疫沈降である。3は対照イソタイプに合致するマウス抗体との免疫沈降である。4は抗CD43抗体との免疫沈降である。
【図29】COSトランスフェクション実験である。PSGL−2またはCD43をコードするプラスミド、ならびにFuc−TIIIまたはFucTVIIをコードするプラスミドを用いてトランスフェクションしたCOS M6細胞を35S−メチオニンで代謝的に標識し、材料および方法において説明するように、膜をアフィニティー捕捉実験用に調製した。トランスフェクションに使用したcDNAをレーンの上に示す。(A)E−セレクチン、(B)P−セレクチン、および(C)抗PSGL−1抗血清Rb3443および抗CD43 Mabを用いて沈降を行った。
【図30】P−およびE−セレクチン結合の阻害に関する種々のP−セレクチンリガンド蛋白のスクリーニングをまとめたものである(実施例13参照)。
【発明を実施するための形態】
【0028】
上記の図のいくつかには、それらが示す構造中に配列番号:2の番号付けとは異なる番号づけを便利のために使用する。それらの図において、可溶性成熟P−セレクチンリガンドの1番目のアミノ酸を出発点として用いて残基の番号付けを行う。それゆえ、それらの図に使用する残基の番号は配列番号:2のものよりも41少ない。例えば、それらの図中の残基19は配列番号:2の残基60に対応する。
【0029】
本発明者らは、ヒト内皮細胞および血小板上のP−セレクチンのリガンドとして作用する蛋白をコードする新規DNAを初めて同定し、単離した。DNA配列を配列番号:1に示す。P−セレクチンリガンド蛋白の完全なアミノ酸配列(すなわち、成熟ペプチドにリーダー配列が付加したもの)は、配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列により特徴付けられる。疎水性分析および既知開裂パターンとの比較により、20ないし22個のアミノ酸、すなわち配列番号:2のアミノ酸1から20までまたはアミノ酸1から22までのシグナル配列が予想された。P−セレクチンリガンド蛋白は、PACE(対になった塩基性アミノ酸変換酵素)開裂部位(−Arg−Asp−Arg−Arg−)を配列番号:2のアミノ酸38〜41に有する。本発明成熟P−セレクチンリガンド蛋白は配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列により特徴付けられる。可溶性形態のP−セレクチンリガンド蛋白は配列番号:2のアミノ酸21からアミノ酸310を含むことにより特徴付けられる。もう1つの可溶性形態の成熟P−セレクチンリガンド蛋白は配列番号:2のアミノ酸42からアミノ酸310に示すアミノ酸配列により特徴付けられる。可溶性形態のP−セレクチンリガンド蛋白は、さらに室温において水溶液中に可溶性であることにより特徴付けられる。もちろん、これらの蛋白をコードする、配列番号:1に示す対応DNA配列も本発明に包含される。
【0030】
本発明P−セレクチンリガンドは糖蛋白であり、以下の末端炭水化物の1つまたはそれ以上を含む可能性がある:
【化4】
【0031】
式中、Rは炭水化物鎖の残りの部分であり、直接P−セレクチンリガンド蛋白に共有結合しているか、あるいはP−セレクチンリガンド蛋白に共有結合した脂質部分に共有結合している。本発明P−セレクチンリガンド糖蛋白はさらに硫酸化されていてもよく、あるいは翻訳後修飾されていてもよい。COSおよびCHO細胞中で発現される場合、全長のP−セレクチンリガンド蛋白(配列番号:2のアミノ酸1から402まで、または配列番号:2のアミノ酸42から402まで)は、非還元的ADA−ポリアクリルアミドゲル電気泳動によれば220kDaの見かけの分子量を有するホモダイマー蛋白である。
【0032】
全長のP−セレクチンリガンド蛋白の構造を図5に図式的に示す。配列番号:2のP−セレクチンリガンド蛋白の3つの領域は以下のとおりである:細胞外ドメイン(配列番号:2のアミノ酸21から310まで)、膜貫通ドメイン(配列番号:2のアミノ酸311から322まで)、および細胞内、細胞質ドメイン(配列番号:2のアミノ酸333から402まで)。細胞外ドメインは、N−結合糖鎖付加が可能な3つのコンセンサストリペプチド部位(Asn−X−Ser/Thr)を有し、それらはAsn残基65、111、および292から開始する。細胞外ドメインはさらに、チロシン硫酸化可能な3つの部位を残基46、48、および51に有する。残基55〜267からなる領域は高いパーセンテージのプロリンおよびスレオニンを含み、アミノ酸コンセンサス配列Ala−Thr/Met−Glu−Ala−Gln−Thr−Thr−X−Pro/Leu−Ala/Thrからなる10量体の15個のリピートを含む。ここにXはPro、Ala、Gln、Glu、またはArgのいずれであってもよい。これらの領域のごとき領域は高度にO−糖鎖付加された蛋白の特徴を有する。
【0033】
P−セレクチンリガンド蛋白をコードする遺伝子およびフコシルトランスフェラーゼ(以下、FTという)、好ましくは(α1,3/α1,4)フコシルランスフェラーゼ(「3/4FT」という)をコードする遺伝子を用いて同時トランスフェクションされたCOSまたはCHO細胞は、表面上にP−セレクチンを発現するCHO細胞に結合しうるが、表面上にP−セレクチンを発現しないCHO細胞には結合しない。精製形態またはCHO細胞表面上に発現されたセレクチンに結合するためには、P−セレクチンリガンド蛋白をコードする遺伝子をFTをコードする遺伝子とともに同時トランスフェクションしなければならない。なぜなら、一方のみの遺伝子でのトランスフェクションではP−セレクチン結合活性が無いかまたは著しく低下しているからである。本発明P−セレクチンリガンド蛋白のP−セレクチンへの結合は、EDTAにより、あるいはP−セレクチン特異的中和モノクローナル抗体により阻害されうる。本発明P−セレクチンリガンド蛋白のP−セレクチンへの結合は、P−セレクチン特異的非中和モノクローナル抗体により、あるいはイソタイプ対照によっては阻害されない。これらの結果は、本発明P−セレクチンリガンド蛋白の結合特異性を特徴付ける。
【0034】
本発明の目的のために、実施例4のCHO−P−セレクチン結合アッセイにおいて細胞表面に存在するP−セレクチンに、あるいは別の表面に固定されたP−セレクチン、例えば実施例4のペトリ皿に固定化されたキメラP−セレクチン−IgGγ1蛋白にカルシウム依存的に結合する場合、「P−セレクチンリガンド蛋白活性」を有すると定義される蛋白であると定義され、「P−セレクチンリガンド蛋白」または「P−セレクチンリガンド糖蛋白」または単に「P−セレクチンリガンド」と呼ばれる。
【0035】
実施例5(C)に詳述されsPSL.T7と命名されたキメラな可溶性形態のP−セレクチンリガンド蛋白を用いて本発明P−セレクチンリガンド蛋白の糖鎖付加状態を研究した。3/4FTとともに同時トランスフェクションされたCOS細胞から産生されるsPSL.T7蛋白を、実施例6(C)に詳述するように翻訳後糖鎖付加により徹底的に修飾した。かくして、少なくともそのいくつかがシアル化されているN−およびO−結合オリゴ糖鎖が本発明P−セレクチンリガンド蛋白上に存在すると考えられる。
【0036】
本発明P−セレクチンリガンド蛋白はE−セレクチンおよびL−セレクチンにも結合する。sPSL.T7またはP−セレクチンリガンド−Ig融合物をコードするDNAおよび3/4FTをコードするDNAで同時トランスフェクションされたCOS細胞により得られたならし培地をプラスチックマイクロタイタープレートのウェルにコーティングすると、E−セレクチンを発現するCHO細胞がプレートに結合する。しかしながら、E−セレクチンを発現しないCHO細胞はかかるプレートに結合しない。sPSL.T7をコードするDNAおよび3/4FTをコードするDNAで同時トランスフェクションされたCOS細胞によるならし培地でコーティングされたマイクロタイタープレートへのE−セレクチン発現CHO細胞の結合は、EDTAの存在またはE−セレクチン特異的中和抗体の存在下で消滅する。sPSL.T7 DNAのみでトランスフェクションされたCOS細胞によるならし培地は、マイクロタイタープレートのウェルにコーティングされた場合、E−セレクチン発現CHO細胞の結合を引き起こさない。これらの理由により、本発明P−セレクチンリガンド蛋白は、P−セレクチンにより媒介される細胞間付着のほかにも、E−セレクチンにより媒介される細胞間付着の阻害剤として有用であると考えられる。
【0037】
固定化されたP−セレクチンのFcキメラを用いるアフィニティー捕捉により決定されるように、COS細胞により産生された可溶性P−セレクチンリガンド蛋白に対して生成された抗体は、P−セレクチンに特異的に結合する主要HL−60糖蛋白と免疫反応する。U937細胞は類似の免疫反応性糖蛋白リガンドを有する。よって、3H−グルコサミン標識U937細胞の界面活性剤抽出物とともに前以てインキュベーションされた固定化P−セレクチンのEDTAでの溶離により、単一の糖蛋白種が観察される。この主要種は、非還元的条件下のSDS−PAGEによれば220kDの見かけの分子量を示し、還元的条件下では100kDの見かけの分子量を示す。H−60細胞から単離された相当種の場合、U937リガンドは、COS組み換えP−セレクチンリガンド蛋白に対して生成されたポリクローナル抗体と免疫反応する。さらに、固定化されたE−セレクチンのFcキメラを用いる、U937細胞および細胞膜標品からのE−セレクチンリガンドのアフィニティー捕捉により、主要U937 P−セレクチンリガンドと同一の分子量および電気泳動上の挙動を示す単一の主要種が得られる。よって、E−およびP−セレクチンはU937細胞の同じ主要糖蛋白リガンドを認識し、その糖蛋白リガンドは抗P−セレクチンリガンド蛋白抗体と免疫反応し、全長の組み換えP−セレクチンリガンド蛋白と同じ見かけの分子量および電気泳動上の挙動を有する。
【0038】
P−セレクチンと相互作用可能な、あるいはP−セレクチンにより媒介される細胞間付着を阻害しうるP−セレクチンリガンド蛋白のフラグメントも、本発明に包含される。かかるフラグメントは、セリンおよびスレオニン残基の頻度が少ないP−セレクチンリガンド蛋白の領域である配列番号:2のアミノ酸21から54まで;プロリン、セリンおよびスレオニンの頻度が多く、さらにアスパラギン結合糖蛋白のコンセンサス配列(Asn−XSer/Thr)2個を有する配列番号:2のアミノ酸55から127まで;プロリン、セリン、およびスレオニンいずれの頻度も高く、10個のアミノ酸のコンセンサス配列:Ala−(Thr/Met)−Glu−Ala−Gln−Thr−Thr−(Pro/Arg/Gln/Ala/Glu)−(Leu/Pro)−(Ala/Thr)の15個のリピートを含む配列番号:2のアミノ酸128から267までのもう1つの大きなフラグメント(この大きなフラグメント中の小さいフラグメントも、P−セレクチンとの相互作用能を保持し、あるいはP−セレクチンにより媒介される細胞間付着の阻害剤として作用しうる。);アスパラギン結合糖鎖付加のコンセンサスを含み、配列番号:2のアミノ酸268から308までのアミノ酸を含む領域;配列番号:2のアミノ酸309から333までにより示される蛋白の疎水性領域;ならびに配列番号:2のアミノ酸334から402までのP−セレクチンリガンド蛋白の両親媒性領域を含む。さらなるフラグメントは、配列番号:2のアミノ酸43からアミノ酸56まで、配列番号:2のアミノ酸42からアミノ酸60までを含んで射てもよく、アミノ酸46、アミノ酸48、および/またはアミノ酸51において1個またはそれ以上の硫酸化またはリン酸化(Domcheck et al., Biochemistry 31:9865-9870 (1992))チロシンを有していてもよい。P−セレクチンリガンド蛋白のフラグメントは直鎖状であってもよく、あるいは既知方法、例えばH. U. Saragovi, et al., Bio/Technology 10, 773-778 (1992)およびR. S. McDowell, et al., J. Amer.Chem. Soc. 114, 9245-9253 (1992)に記載の方法(それらを参照により本明細書に記載されているものとみなす)を用いて環化されていてもよい。本発明の目的のため、本明細書で「P−セレクチンリガンド蛋白」という場合はすべて、P−セレクチンに結合しうるフラグメントを含むものとする。
【0039】
かかるフラグメントを免疫グロブリンのごときキャリア分子に融合させてP−セレクチンリガンド結合部位の価数を増加させてもよい。例えば、配列番号:2のアミノ酸42からアミノ酸295までまたはアミノ酸42からアミノ酸88のフラグメントのごとき可溶性形態のP−セレクチンリガンド蛋白を、「リンカー」配列を介して免疫グロブリンのFc部分(得られるキメラに所望品質(例えば、長い半減期または免疫原性の低下)を付与するための無処理配列または変異配列)に融合させてもよい。2価形態のP−セレクチンリガンド蛋白に関しては、実施例5(D)および配列番号:6に示すように、かかる融合はIgG分子のFc部分に対するものであってよい。他の免疫グロブリンイソタイプを用いてかかる融合物を得てもよい。例えば、P−セレクチンリガンド蛋白−IgM融合物は10価形態の本発明P−セレクチンリガンド蛋白を生じるであろう。
【0040】
他の蛋白由来のアミノ酸配列への本発明P−セレクチンリガンド蛋白の融合物を構築してもよい。かかる目的に好ましいP−セレクチンリガンド蛋白は、配列番号:2のアミノ酸42からアミノ酸295までのフラグメントまたはアミノ酸42からアミノ酸88までのフラグメントを包含する。望ましい融合蛋白は、例えばサイトカイン、増殖因子および分化因子(骨形態形成蛋白、例えばBMP類))、ホルモン、酵素、受容体成分またはフラグメントならびに他のリガンドのようなP−セレクチンリガンドとは異なる生物学的活性を有する蛋白由来のアミノ酸配列を含んでいてもよい。また、P−セレクチンリガンド蛋白を他の蛋白または薬剤に化学結合させることもできる。かかる使用において、P−セレクチンリガンド蛋白は、セレクチン分子との相互作用能によって、治療効果を増強することにより融合または結合された薬剤の薬剤動力学および/または生体分配を変化させる。例えば、サイトカイン配列へのP−セレクチンリガンド蛋白の融合は、サイトカイン活性を炎症部位に指向することができる。かかる場合、融合蛋白のP−セレクチンリガンド蛋白部分は炎症部位において発現されたセレクチンに結合するであろう。この結合は、融合蛋白のサイトカイン部分を局在化させ、その同族の受容体または近くの細胞表面への結合を可能にするであろう。同様に、他のリガンドをかかる融合蛋白において使用して、対応受容体を発現する細胞をP−セレクチン発現部位に引き寄せることができる。かかる融合物の好ましい例を実施例15において説明する。
【0041】
P−セレクチンリガンド蛋白を含む融合蛋白において、P−セレクチンリガンド以外の蛋白由来のアミノ酸配列をP−セレクチンリガンド由来の配列のN末端またはC末端に連結することができる。連結は直接的(すなわち、いずれの蛋白にも由来しない介在結合配列を含まない)であってもよく、あるいは連結配列を介するものであってもよい。
【0042】
かかる融合蛋白を用いる哺乳動物の治療方法も本発明により企図される。かかる場合、融合蛋白を用いて、P−セレクチンリガンド蛋白が融合している蛋白により効果が発揮される症状を治療する。例えば、IL−11へのP−セレクチンリガンド蛋白の融合物を用いて、セレクチンをその表面に発現する骨髄内皮細胞にIL−11活性を局在化させることができる。局在化されると、融合蛋白のIL−11部分は巨核細胞の前駆細胞を刺激するであろう。同様に、BMPへのPセレクチンリガンド蛋白の融合物を用いて、障害部位における骨または軟骨の形成を刺激することもできる。障害組織はP−セレクチンを発現し、そのことにより融合蛋白が結合する。局在化されると、融合蛋白のBMP部分は障害部位において骨または軟骨の産生を刺激するであろう。
【0043】
下記実施例に詳述するように、本発明P−セレクチンリガンド蛋白を、最初は発現クローニング法(Clarkら、米国特許第4675285号)を用いて得た。ヒトプロミエロサイト細胞系HL−60(S. J. Collins, et al., Nature 270, 347-349 (1977), ATCC番号CCL 240)からcDNAライブラリーを構築した。このライブラリーを3/4FTをコードするDNAとともにCOS細胞中に同時トランスフェクションし、P−セレクチンの細胞外部分およびヒトIgGγ1モノクローナル抗体のFc部分からなるキメラ分子への結合に関してトランスフェクション体をスクリーニングした。キメラP−セレクチンに結合する同時トランスフェクションを、P−セレクチンリガンド蛋白をコードする蛋白に関して豊富化した。このスクリーニング工程を数回繰り返して、P−セレクチンリガンド蛋白をコードするcDNAに関してさらにプラスミド集団豊富化した。第2のクローニング段階において、豊富化したプラスミド集団を、3/4FT遺伝子とともに再度COS細胞中に同時トランスフェクションし、表面上にP−セレクチンを発現する蛍光標識CHO細胞系への結合に関してスクリーニングした。この方法により単一のcDNAクローンを得て、pMT21:PL85と命名した。1992年10月16日にpMT21:PL85プラスミドをAmerican Type Culture Collectionに寄託し、受託番号ATCC 69096を付与された。
【0044】
本発明新規DNAを配列番号:1に示す。本発明DNAは種々の形態のP−セレクチンリガンド蛋白をコードしうる。例えば、1の具体例において、本発明DNAは配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列を有する全P−セレクチンリガンド蛋白をコードする。もう1つの具体例において、本発明DNAは配列番号:2のアミノ酸21からアミノ酸402までのアミノ酸配列により特徴づけられる、シグナル配列を欠いたP−セレクチンリガンド蛋白の1形態をコードする。さらなる具体例において、本発明DNAは、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列により特徴づけられる成熟P−セレクチンリガンド蛋白をコードする。本発明DNAのもう1つの具体例は、配列番号:2のアミノ酸1からアミノ酸310までのアミノ酸配列により特徴づけられる可溶性形態のP−セレクチンリガンド蛋白をコードする。また、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列により特徴づけられる可溶性形態の成熟P−セレクチンリガンド蛋白をコードするDNAも本発明の具体例である。さらに、配列番号:2のアミノ酸21からアミノ酸310までのアミノ酸配列により特徴づけられる、シグナル配列を欠いた可溶性形態のP−セレクチンリガンド蛋白をコードするDNAも本発明の具体例である。本発明DNAは他のヒトDNAを含まず、それゆえ、単離DNAとして特徴づけられる。上に詳述したように、P−セレクチンと相互作用するP−セレクチンリガンドフラグメントをコードするDNAも本発明に包含される。
【0045】
P−セレクチンリガンド蛋白cDNAプローブを用いるノーザン分析により、種々のヒト細胞系(HL−60、THP−1、U937)およびヒト単球ならびに多形核白血球においてP−セレクチンリガンド蛋白mRNA転写物の発現が観察された。それらの細胞系すべてにおいて、2.5kbの主要転写物が観察された。HL60およびU937細胞系ならびに多形核白血球において約4kbのマイナーな種が観察された。対照的に、ヒト肝芽腫細胞系HepG2においてP−セレクチンリガンドmRNA発現が検出された。
【0046】
サザンブロット分析によりわかるように、本発明P−セレクチンリガンド蛋白は単一コピー遺伝子によりコードされており、多遺伝子ファミリーの一部ではない。本発明P−セレクチンリガンド蛋白のゲノム形態は、5'非翻訳領域中のヌクレオチド54に局在する約9kbの大きなイントロンを含む。多形核白血球および単球において、本発明P−セレクチンリガンド蛋白は配列番号:3に示すDNA配列によりコードされている。この具体例において、P−セレクチンリガンド蛋白は16個のリピート領域を含む。同様に本発明単離DNAは配列番号:3に示すDNA配列においても具体化され、1993年10月22日にAmerican Type Culture Collectionに寄託され、受託番号ATCC 75577を付与されたプラスミドpPL85R16に含まれている。
【0047】
また本発明は、配列番号:1に示す単離DNAまたは配列番号:3に示す単離DNAの対立遺伝子変種、すなわちP−セレクチンリガンド活性を有する蛋白をコードする配列番号:1または配列番号:3の単離DNAの天然の別形態を包含する。厳密な条件下(例えば、65℃で4xSSC、または42℃で50%ホルムアミドおよび4xSSC)または弛緩した条件下(50℃で4xSSC、または42℃で30〜40%ホルムアミド)において配列番号:1に示すDNAまたは配列番号:3に示すDNAにハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する単離DNAも本発明に包含される。P−セレクチンリガンド蛋白をコードし、遺伝学的コードの縮重により配列番号:1に示す単離DNAまたは配列番号:3に示すDNAとは異なっており、やはりP−セレクチンリガンド蛋白活性を有する単離DNAも本発明に包含される。点突然変異による、あるいはP−セレクチンリガンド活性、半減期または生成レベルを増強する修飾を導入されたことによる、配列番号:1に示すDNAまたは配列番号:3に示すDNAの変種も本発明に包含される。本発明の目的のため、本明細書の「配列番号:1のDNA」は、配列番号:1に示す特定のDNA配列を含むDNAのほかに、配列番号:2の成熟P−セレクチンリガンド蛋白をコードするDNA;P−セレクチンに結合しうる配列番号:2のP−セレクチンリガンド蛋白のフラグメントをコードするDNA;可溶性形態の配列番号:2のP−セレクチンリガンド蛋白をコードするDNA;配列番号:1のDNA配列にハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する蛋白をコードしており、遺伝学的コードの縮重により配列番号:1のDNAとは異なっているDNA;ならびに上記配列番号:1のDNA配列の変種も包含する。同様に、「配列番号:3のDNA」は、配列番号:3に示す特定のDNA配列を含むDNAのほかに、配列番号:4の成熟P−セレクチンリガンド蛋白をコードするDNA;P−セレクチンに結合しうる配列番号:4のP−セレクチンリガンド蛋白のフラグメントをコードするDNA;可溶性形態の配列番号:4のP−セレクチンリガンド蛋白をコードするDNA;配列番号:3のDNA配列にハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する蛋白をコードしており、遺伝学的コードの縮重により配列番号:3のDNAとは異なっているDNA;ならびに上記配列番号:3のDNA配列の変種も包含する。
【0048】
P−セレクチンリガンド蛋白の膜貫通および細胞質ドメインをコードする領域が欠失され、さらに/あるいは細胞外ドメインのカルボキシ末端のアミノ酸に対するコドンの3'側にストップコドンが導入された修飾DNAの発現により、可溶性形態のP−セレクチンリガンド蛋白をコードするDNAを調製してもよい。例えば、疎水性分析により、配列番号:2に示すP−セレクチンリガンド蛋白が配列番号:2のアミノ酸311から332までを含む膜貫通ドメインならびに配列番号:2のアミノ酸333から402までを含む細胞質ドメインを有することが予想される。当該分野で知られた特異的突然変異法を包含する標準的な分子生物学的手法により、あるいは適当なオリゴヌクレオチドプライマーを用いるポリメラーゼ連鎖反応により上記修飾DNAを得てもよい。種々の可溶性P−セレクチンリガンド蛋白をコードするいくつかのDNAを得る方法を実施例5に示す。
【0049】
全長配列の一部が欠失または変化している修飾DNAの発現により、他のフラグメントおよび別形態のP−セレクチンリガンド蛋白をコードするDNAを調製してもよい。P−セレクチンリガンド活性を保持させつつ、P−セレクチンリガンド蛋白配列の実質的な欠失を行うことができる。例えば、配列番号:2のアミノ酸42からアミノ酸189までの配列、配列番号:2のアミノ酸42からアミノ酸118までの配列、または配列番号:2のアミノ酸42からアミノ酸89までの配列を含むP−セレクチンリガンド蛋白はそれぞれP−セレクチン蛋白結合活性およびE−セレクチン結合能を有する。1個またはそれ以上のN−結合糖鎖付加部位(例えば、配列番号:2のアミノ酸65、111および292のごとき部位)が他のアミノ酸に変化させられ、あるいは欠失されたP−セレクチンリガンド蛋白もやはりP−セレクチン蛋白結合活性およびE−セレクチン結合能を保持している。配列番号:2のアミノ酸42からアミノ酸60までを含むP−セレクチンリガンド蛋白(配列番号:2のアミノ酸45からアミノ酸58までの非常にアニオン性の領域を含む)もまたP−セレクチンリガンド蛋白活性を保持している。しかしながら、かかる配列に限定されたP−セレクチンリガンド蛋白はE−セレクチンに結合しない。好ましくは、P−セレクチンリガンド蛋白は、配列番号:2のアミノ酸46、48および51に見出されるチロシン残基のうち少なくとも1個(より好ましくは少なくとも2個、最も好ましくは3個すべて)を保持しているものであり、それらの残基の硫酸化はP−セレクチンリガンド蛋白活性に貢献しうる。これらの活性フラグメントおよび他の活性フラグメントまたは別形態のP−セレクチンリガンド蛋白をコードするDNAの構築を、当業者に知られた方法に従って行ってもよい。
【0050】
本発明単離DNAを、Kaufman et al., Nucleic Acids Res. 19, 4485-4490 (1991)に開示されたpMT2またはpED発現ベクターのごとき発現制御配列に作動可能に連結して、P−セレクチンリガンドを組み換え的に製造してもよい。多くの適当な発現制御配列が当該分野において知られている。組み換え蛋白発現の一般的方法も知られており、R. Kaufman, Methods in Enzymology 185, 537-566 (1990)に説明がある。本明細書の「作動可能に連結」は、酵素的または化学的な連結により本発明単離DNAおよび発現制御配列間に共有結合が形成され、その結果、連結されたDNA/発現制御配列を用いて形質転換(トランスフェクション)された宿主細胞によりP−セレクチンリガンド蛋白が発現されることを意味する。
【0051】
対になったアミノ酸配列(例えば、−Lys−Arg−および−Arg−Arg−)のカルボキシル側で前駆体ペプチドを開裂して成熟蛋白を生じる、いくつかのエンド蛋白分解酵素が知られている。一般的には、かかる酵素は、対になった塩基性アミノ酸変換酵素またはPACEとして知られ、成熟蛋白の組み換え製造方法におけるそれらの使用はWO92/09698および米国特許出願第07/885972号に十分に開示されており、参照によりそれらを本明細書に記載されているものとみなす。PACEファミリーの酵素は、組み換え宿主細胞における前駆体ポリペプチドの蛋白分解的プロセッシングの効率を高めることが知られている。上記のごとく、本発明P−セレクチンリガンド蛋白はかかる
PACE開裂部位を含んでいる。
【0052】
本明細書記載の可溶性P−セレクチンリガンド蛋白をコードするDNA配列およびWO92/09698および米国特許出願第07/885972号(参照によりそれらを本明細書に記載されているものとみなす)に記載のPACEをコードするDNA配列または配列番号:5のDNA配列を含む宿主細胞により、本発明の可溶性成熟P−セレクチンリガンド蛋白を製造してもよい。かかる宿主細胞は、可溶性P−セレクチンリガンド蛋白DNAおよびPACE DNAをそれぞれ含む別個の発現ベクターでの同時形質転換または逐次形質転換の結果としてのDNAを含んでいてもよい。3/4FTをコードする第3のDNAを、P−セレクチンリガンド蛋白およびPACEをコードするDNAとともに同時形質転換してもよい。別法として、宿主細胞は、可溶性P−セレクチンリガンド蛋白DNAおよびPACE DNAの両方を含む単一の発現ベクターでの形質転換の結果としてのDNAを含んでいてもよい。かかる発現ベクターの構築は分子生物学の当業者のレベル内である。同時形質転換および形質転換の方法も知られている。
【0053】
PACEをコードする多くのDNA配列が知られている。例えば、フリン(furin)として知られる1の形態のPACEをコードするDNAがA. M. W. van den Ouweland et al., Nucl. Acids Res. 18, 664 (1990)に開示されており、参照により本明細書に記載されているものとみなす。PACESOLとして知られる可溶性形態のPACEをコードするcDNAを配列番号:5に示す。他の形態のPACEをコードするDNAも存在し、PACEがアミノ酸38〜41においてP−セレクチンリガンド蛋白を開裂しうるかぎり、かかるPACEをコードするDNAを用いて本発明の可溶性成熟P−セレクチンリガンド蛋白を得てもよい。好ましくは、可溶性形態のPACEをコードするDNAを用いて本発明の可溶性成熟P−セレクチンリガンド蛋白を得る。
【0054】
可溶性形態のP−セレクチンリガンド蛋白およびPACEをそれぞれコードするDNAを、別個にあるいは一緒にして、上記pMT2またはpED発現ベクター中に含まれるような発現制御配列に作動可能に連結して、PACEにより開裂された可溶性P−セレクチンリガンドを組み換え的に製造してもよい。さらなる適当な発現制御配列が当該分野において知られている。下記実施例3(C)および3(D)は本発明の可溶性成熟P−セレクチンリガンド蛋白の製造方法を説明する。
【0055】
多くのタイプの細胞がP−セレクチンリガンド蛋白発現のための適当な宿主細胞として作動しうる。適当な宿主細胞は、P−セレクチンリガンド蛋白に特徴的な炭水化物側鎖を結合しうるものである。かかる能力は、天然に存在するかどうか、化学的突然変異により誘導されたかどうか、あるいは糖鎖付加酵素をコードするDNA配列を含む適当な発現プラスミドで宿主細胞がトランスフェクションされたかどうかに関係なく、宿主細胞中の適当な糖鎖付加酵素の存在により生じるものである。例えば、宿主細胞は、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト腎臓293細胞、ヒト上皮A431細胞、Colo 205細胞、3T3細胞、CV−1細胞、他の形質転換された霊長類細胞系、正常2倍体細胞、1次組織、1次移植物のインビトロ培養から得られる細胞株、HeLa細胞、マウスL細胞、BHK、HL−60、U937、またはHak細胞を包含する。
【0056】
本発明単離DNAおよび適当な糖鎖付加酵素をコードする1またはそれ以上のDNAを、1またはそれ以上の昆虫発現ベクター中の適当な制御配列に作動可能に連結し、昆虫発現系を用いることにより蛋白を得てもよい。バキュロウイルス/昆虫細胞発現系のための材料および方法は、例えばInvitrogen, San Diego, California, USAからのキット形態(MaxBacRキット)で市販されており、かかる方法は当該分野においてよく知られており、SummersおよびSmith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987)(参照により本明細書に記載されているものとみなす)に記載のようなものがある。上記の適当な単離DNAを用いて、昆虫細胞において可溶性形態のP−セレクチンリガンド蛋白を得てもよい。1形態のPACEをコードするDNAを昆虫細胞中でさらに同時発現させて、PACEにより開裂された形態のP−セレクチンリガンド蛋白を得てもよい。
【0057】
別法として、酵母のごとき下等真核細胞または細菌のごとき原核細胞においてP−セレクチンリガンド蛋白を製造することも可能である。潜在的に適当な酵母株は、Saccharomyces cerevisiae、Schizosaccharomyces pombe、Kluyveromyces株、Candida、または異種蛋白の発現能を有する酵母株を包含する。潜在的に適当な細菌株は、Escherichia coli、Bacillus subtilis、Salmonella typhimurium、または異種蛋白の発現能を有する細菌株を包含する。P−セレクチンリガンド蛋白が酵母または細菌において生成される場合、蛋白の適当な部分に適当な炭水化物を共有結合させる必要がある。既知化学的または酵素的方法を用いてかかる共有結合を行ってもよい。
【0058】
本発明P−セレクチンリガンド蛋白をトランスジェニック動物の産生物として、例えばP−セレクチンリガンド蛋白をコードするDNA配列を含む体細胞または生殖細胞により特徴づけられるトランスジェニックなウシ、ヤギ、ブタ、またはヒツジの乳の成分として発現させてもよい。
【0059】
GlcNacトランスフェラーゼ、好ましくはコア2トランスフェラーゼとしても知られるUDP−GlcNac:Gal β1−3GalNac−R(GlcNac to GalNAc)β1−6GlcNAcトランスフェラーゼ(EC 2.4.1.102)での宿主細胞の形質転換により、P−セレクチンリガンド蛋白のP−セレクチン結合活性を増大させてもよい。
【0060】
P−セレクチンリガンド蛋白上に存在するO−結合グリカンはP−セレクチンへの結合に重要であることが示されている(D. Sako et al., Cell 75, 1175-1186 (1993))。ミエロイド細胞のO−結合グリカン上のシアリルLexが複合分枝構造上に存在することが報告されている(Maemura, K. and Fukuda, M., J. Biol. Chem. 267, 24379-24386 (1992))。かかるオリゴサッカライド構造の生成に関与する酵素が「コア2」である。コア2酵素活性はCOS細胞中に非常に低レベルで見出され、CHO細胞中には痕跡レベルで見出される。P−セレクチンリガンド蛋白、(α1,3/α1,4)フコシルトランスフェラーゼおよびコア2をコードするDNAで同時形質転換された宿主細胞は、P−セレクチンに対する結合が20〜30倍増強されたP−セレクチンリガンド蛋白を生じうる。
【0061】
ある種の好ましい具体例において、可溶性P−セレクチンリガンド蛋白、3/4FT、コア2およびPACEをコードするDNAで同時形質転換した宿主細胞によりP−セレクチンリガンド蛋白を製造する。
【0062】
P−セレクチン結合糖蛋白の発現に適した培養条件下で形質転換宿主細胞を培養することにより本発明P−セレクチンリガンド蛋白を製造してもよい。次いで、得られた発現糖蛋白を、ゲル濾過およびイオン交換クロマトグラフィーのごとき既知精製プロセスを用いて培地または細胞抽出物から精製してもよい。可溶性形態の本発明P−セレクチンリガンド蛋白を、レンズマメレクチン−セファロース(Sepharose)(登録商標)によるクロマトグラフィー、ついで0.5M α−メチルーマンノースでの溶離により生成することができる。ついで、溶離した可溶性P−セレクチンリガンド蛋白を0〜70%硫酸アンモニウム工程によりさらに精製し、濃縮してもよい。ついで、蛋白を回収し、再懸濁し、TSK G4000SWXLによるサイズ排除クロマトグラフィーによりさらに精製する。別法として、発現細胞から全膜フラクションを調製し、Triton X-100のごとき非イオン性界面活性剤で膜を抽出することにより本発明の全長のP−セレクチンリガンド蛋白を精製することができる。ついで、界面活性剤抽出物を、固定化P−セレクチンを含むアフィニティーカラムに通し、0.1%界面活性剤を含有するバッファー中の10mM EDTAを用いてP−セレクチンリガンド蛋白をカラムから溶離することができる。ついで、アフィニティーカラムから溶離した物質を透析してEDTAを除去し、レンズマメ-Sepharose(登録商標)アフィニティーカラムで精製し、再度.05M α−メチル−マンノシドで溶離する。
【0063】
別法として、市販蛋白濃縮フィルター、例えばAmiconまたはMillipore Pellicon限外濾過ユニットを用いて本発明P−セレクチンリガンド蛋白を濃縮してもよい。濃縮工程後、濃縮物をゲル濾過媒体のごとき精製マトリックスに負荷する。別法として、アニオン交換樹脂、例えば懸垂ジエチルアミノエチル(DEAE)基を有するマトリックスまたは基材を用いることができる。マトリックスはアクリルアミド、アガロース、デキストラン、セルロースまたは蛋白精製に通常使用される他のタイプのものがある。別法として、カチオン交換工程を用いることができる。適当なカチオン交換体は、スルホプロピルまたはカルボキシメチル基を含む種々の不溶性マトリックスを包含する。スルホプロピル基が好ましい(例えば、S-Sepharose(登録商標)カラム)。コンカナバリンA−アガロース、ヘパリン−トヨパール(toyopearl)(登録商標)またはCibacrom blue 3GA Sepharose(登録商標)のごときアフィニティー樹脂による、あるいはフェニルエーテル、ブルエーテル、またはプロピルエーテルのごとき樹脂を用いる疎水性相互作用クロマトグラフィーによる、あるいは免疫アフィニティークロマトグラフィーによる1またはそれ以上のカラム工程により、培養上清からP−セレクチンリガンド蛋白を精製してもよい。
【0064】
最後に、疎水性RP−HPLC媒体、例えば懸垂メチルまたは他の脂肪族基を有するシリカゲルを用いる1またはそれ以上の逆相高品質液体クロマトグラフィー(RP−PLC)工程を用いてさらにP−セレクチンリガンド蛋白を精製することができる。いくつかまたはすべての上記精製工程を種々組み合わせて用いて実質的に均一な単離組み換え蛋白を得ることもできる。かくして精製されたP−セレクチンリガンド蛋白は他の哺乳動物蛋白を実質的に含まず、本発明により「単離蛋白」と定義される。
【0065】
単離P−セレクチンリガンド蛋白は、P−、E−またはL−セレクチンにより媒介される細胞間付着により特徴づけられる症状の治療において有用でありうる。かかる症状は、心筋梗塞、細菌もしくはウイルス感染、転移性の症状、関節炎のごとき炎症性疾患、痛風、葡萄膜炎、急性呼吸困難症候群、喘息、気腫、晩発型高血圧反応、全身性紅斑狼瘡、火傷もしくはしもやけのごとき熱的傷害、自己免疫性甲状腺炎、実験的なアレルギー性脳脊髄炎、多発性硬化症、外傷後の多発性器官傷害症候群、糖尿病、Reynaud症候群、好中球性皮膚病(Sweet症候群)、炎症性腸症候群、Grave病、糸球体腎炎、歯肉炎、歯周炎、溶血性の尿毒性症候群、潰瘍性大腸炎、Crohn病、壊死性全腸炎、顆粒球輸血関連症候群、サイトカインにより誘導される毒性等を包含する(これらに限らない)。単離P−セレクチンリガンド蛋白は器官移植にも有用で、移植用に器官を調製すること、および器官移植拒絶反応を抑制することの両方において有用である。したがって、器官切除前にP−セレクチンリガンド蛋白を生きた器官ドナーまたは生きていない器官ドナーに投与してもよい。さらに、P−セレクチンリガンド蛋白をエクスビボで器官保存溶液中のドナーの器官に投与してもよく、レシピエントに融着前、および/またはレシピエントに融着後に投与してもよい。単離P−セレクチンリガンド蛋白を用いて血液透析およびロイコフォレシスの患者を治療してもよい。さらに、単離P−セレクチンリガンド蛋白を抗転移剤として使用してもよい。単離P−セレクチンリガンド蛋白をそれ自体、P−、E−またはL−セレクチンにより媒介される細胞間付着の阻害剤として用いてもよく、あるいはP−、E−またはL−セレクチンにより媒介される細胞間付着の阻害剤を設計するために使用してもよい。本発明は、単離P−セレクチンリガンド蛋白を含有する医薬組成物ならびに単離P−セレクチンリガンド蛋白を用いる治療方法または単離P−セレクチンリガンド蛋白の使用方法を包含する。
【0066】
細胞から精製された、または組み換え的に製造された単離P−セレクチンリガンド蛋白を医薬上許容される担体と混合して医薬組成物中に用いてもよい。かかる組成物は、P−セレクチンリガンド蛋白および担体、希釈剤、充填剤、塩類、バッファー、安定化剤、可溶化剤、および当該分野でよく知られた他の物質を含有していてもよい。用語「医薬上許容される」は、有効成分の生物学的活性の有効性を妨害しない無毒の物質を意味する。担体の特性は投与経路に依存する。本発明医薬組成物は、M−CSF、GM−CSF、TNF、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−13、IL−14、IL−15、IFN、TNF0、TNF1、TNF2、G−CSF、Meg−CSF、スロンボポイエチン、幹細胞因子、およびエリスロポイエチンのごときサイトカイン、リンホカイン、または他の造血因子を含有していてもよい。医薬組成物はさらに、プラスミノーゲンアクチベーターおよび因子VIIIのごとき血栓溶解因子または抗血栓因子を含有していてもよい。医薬組成物はさらに他の抗炎症剤を含有していてもよい。かかるさらなる因子および/または作用剤を医薬組成物に含有させて、単離P−セレクチンリガンド蛋白との相乗効果を発揮させ、あるいは単離P−セレクチンリガンド蛋白により引き起こされる副作用を最小化してもよい。逆に、特定のサイトカイン、リンホカイン、他の造血因子、血栓溶解因子または抗血栓因子、または抗炎症剤の処方に単離P−セレクチンリガンド蛋白を含有させて、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子または抗血栓因子、または抗炎症剤の副作用を最小化してもよい。
【0067】
本発明医薬組成物はリポソーム形態であってもよい。リポソーム形態においては、単離P−セレクチンリガンド蛋白は、水溶液中のミセル、不溶性単層、液状結晶、またはラメラ層として凝集形態となって存在する脂質のごとき両親媒性物質と混合されて、他の医薬上許容される担体とともに存在している。リポソーム処方に適する脂質は、モノグリセリド、ジグリセリド、スルファチド、リゾレシチン、リン脂質、サポニン、胆汁酸等を包含するが、これらに限らない。かかるリポソーム処方の調製は当業者のレベル内であり、例えば、米国特許第4235871号、第4501728号、第4837028号および第4737323号に開示されており、参照によりこれらを本明細書に記載されているものとみなす。
【0068】
本明細書の用語「治療上有効量」は、医薬組成物または方法の各有効成分の合計量が有意な患者の利益、すなわちP−セレクチンまたはE−セレクチンにより媒介される細胞付着の治療、治癒、予防または改善、あるいはかかる症状の治療、治癒、予防または改善の速度の増加を示すに十分であることを意味する。単独投与される個々の有効成分について用いる場合、該用語は当該成分のみに関するものである。組み合わせた成分について用いる場合、該用語は、混合して、逐次または同時に投与するかどうかにかかわらず、治療効果を生じる有効成分の合計量に関するものである。
【0069】
本発明治療方法または使用方法の実施に際して、治療上有効量の単離P−セレクチンリガンド蛋白を、P−セレクチンにより媒介される疾病状態を有する哺乳動物に投与する。本発明方法に従って単離P−セレクチンリガンド蛋白を単独で、あるいは受容体アンタゴニスト、リガンドアンタゴニスト、サイトカイン、リンホカインまたは他の造血因子を用いる治療のごとき他の治療とともに投与することができる。1またはそれ以上のサイトカイン、リンホカインまたは他の造血因子とともに共投与する場合、単離P−セレクチンリガンド蛋白をサイトカイン、リンホカインまたは他の造血因子、血栓溶解因子または抗血栓因子と同時に、あるいは逐次投与することができる。逐次投与の場合、担当医は他の抗炎症剤、サイトカイン、リンホカインまたは他の造血因子、血栓溶解因子または抗血栓因子と単離P−セレクチンリガンド蛋白との適当な順序を決定するであろう。
【0070】
医薬組成物中に使用される、あるいは本発明方法の実施に用いられる単離P−セレクチンリガンド蛋白の投与を、摂食、吸入、または皮膚、皮下、または静脈注射のごとき種々の慣用的経路で行うことができる。
【0071】
治療上有効量の単離P−セレクチンリガンド蛋白を経口投与する場合、単離P−セレクチンリガンド蛋白は錠剤、カプセル、粉末、溶液またはエリキシルの形態であろう。錠剤形態で投与する場合、本発明医薬組成物はさらにゼラチンのごとき固体担体またはアジュバントを含有してもよい。錠剤、カプセル、および粉末は約5ないし95%、好ましくは約25%ないし90%の単離P−セレクチンリガンド蛋白を含有する。液体形態で投与する場合、水、鉱油、ピーナッツ油、ミネラルオイル、大豆油もしくはゴマ油のごとき動物もしくは植物の油脂、または合成油脂のごとき液体担体を添加してもよい。液体形態の医薬組成物はさらに、生理学的セイライン溶液、ブドウ糖もしくは他の糖溶液、またはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールのごときグリコール類を含有してもよい。液体形態で投与する場合、医薬組成物は約0.5ないし90重量%、好ましくは約1ないし50重量%の単離P−セレクチンリガンド蛋白を含む。
【0072】
治療上有効量の単離P−セレクチンリガンド蛋白を静脈、皮膚または皮下注射により投与する場合、単離P−セレクチンリガンド蛋白はパイロジェン不含の非経口的に許容される水溶液の形態であろう。かかる非経口的に許容される蛋白溶液の調製は、pH、等張性、安定性等に関して当該分野の技術の範囲内である。静脈、皮膚、または皮下注射に好ましい医薬組成物は、本発明化合物のほかに、注射用塩化ナトリウム、注射用リンゲル溶液、注射用ブドウ糖および塩化ナトリウム、注射用乳酸加リンゲル溶液のごとき等張性の担体、または当該分野で知られた他の担体を含有すべきである。本発明医薬組成物は安定化剤、保存料、バッファー、抗酸化剤、または当業者に知られた他の添加物を含有していてもよい。
【0073】
本発明医薬組成物中の単離P−セレクチンリガンド蛋白の量は、治療すべき症状の性質および重さ、ならびに患者の治療前の性質に依存するであろう。最終的には、担当医が個々の患者の治療に用いる単離P−セレクチンリガンド蛋白の量を決定するであろう。最初に、担当医は単離P−セレクチンリガンド蛋白を低用量で投与して患者の応答を観察する。患者に関して最適治療効果が得られるまで、より高用量の単離P−セレクチンリガンド蛋白を投与し、最適効果が得られた時点で用量をもはや増加させない。本発明方法の実施に用いる種々の医薬組成物は、体重1kgあたり約0.1μgないし約100mgの単離P−セレクチンリガンド蛋白を含有すべきである。
【0074】
本発明医薬組成物を用いた静脈注射による治療期間は、治療すべき疾病の重さおよび個々の患者それぞれの潜在的応答に依存するであろう。単離P−セレクチンリガンド蛋白の1回の適用期間は連続静脈投与の場合12ないし24時間の範囲であろう。最終的に、担当医が、本発明医薬組成物を用いた静脈注射による適当な治療期間を決定するであろう。
【0075】
単離P−セレクチンリガンド蛋白を用いて動物を免役して、P−セレクチンリガンド蛋白と特異的に反応し、P−セレクチンにより媒介される細胞付着を阻害するポリクローナルおよびモノクローナル抗体を得てもよい。P−セレクチンリガンド蛋白全体または可溶性成熟P−セレクチンリガンド蛋白のごときそのフラグメントを免疫原として用いてかかる抗体を得てもよい。P−セレクチンリガンド蛋白の小型のフラグメント、例えば下記フラグメントを用いて動物を免疫してもよい。小型フラグメントとしては、例えば、配列番号:2のアミノ酸42からアミノ酸56まで、配列番号:2のアミノ酸127からアミノ酸138までがある。さらなるペプチド免疫原は配列番号:2のアミノ酸238からアミノ酸248までを含み、ペプチドのアミノ末端にアラニン残基が付加されたものである。もう1つのペプチド免疫原は、配列番号:2のアミノ酸43からアミノ酸56までを含み、位置46、48または51のいずれかまたはすべてにおいて硫酸化チロシンを有するものである。ペプチド免疫源はカルボキシ末端にシステイン残基をさらに含んでいてもよく、キーホールリムペット・ヘモシアニン(KLH)のごときハプテンに結合する。かかるペプチドを合成する方法は当該分野において知られており、例えば、R.P.Merrifield,J.Amer.Chem.Soc.85,2149-2154 (1963);J.L.Krstenansky,et.al.,FEBS Lett.211,10(1987)に記載のごときものがある。
【0076】
P−セレクチンリガンド糖蛋白またはP−セレクチンリガンド糖蛋白の特徴である複合型炭水化物部分に結合するモノクローナル抗体は、炎症性疾患およびある種の形態の癌の免疫検出用診断薬でありうる。小胞肺癌腫のごときいくつかの癌細胞は検出可能なレベルのP−セレクチンリガンド蛋白を発現しうる。癌細胞によるこのP−セレクチンリガンド蛋白の異常な発現はこれらの細胞の転移において役割を果たしているかもしれない。
【0077】
P−セレクチンリガンド糖蛋白またはP−セレクチンリガンド糖蛋白の特徴である複合型炭水化物部分に結合する中和抗体は、P−セレクチンリガンド蛋白の異常発現が関与している炎症性疾患およびある種の形態の癌の治療薬として有用でありうる。これらの中和モノクローナル抗体は、P−セレクチンリガンド蛋白のセレクチンにより媒介される細胞間付着機能をブロックしうる。P−セレクチンリガンド蛋白の結合をブロックすることにより、不適切な炎症部位への白血球の付着が消滅または著しく減少させられる。癌細胞または白血病性細胞の場合、P−セレクチンリガンド蛋白に対する中和モノクローナル抗体は、P−セレクチンリガンド蛋白により媒介されうる癌細胞の転移による蔓延の検出および防止において有用でありうる。さらに、これらの細胞に結合するモノクローナル抗体は、抗体依存性の細胞により媒介される細胞毒性(ADCC)に関する癌細胞を標的とする可能性があり、よって、癌細胞の除去を助ける。P−セレクチンリガンド蛋白と反応するヒト抗体を、ヒト免疫グロブリンをコードする遺伝を生殖細胞系中に含むトランスジェニック動物において産生させてもよい。下記実施例7は、P−セレクチンリガンド蛋白フラグメントに特異的なウサギポリクローナル抗体の製造を説明する。
【0078】
本発明P−セレクチンリガンド蛋白を用いて、P−セレクチンリガンド蛋白と結合しうる、よってP−セレクチンまたはE−セレクチンにより媒介される細胞間付着の阻害剤として作用しうる作用剤をスクリーニングしてもよい。固定化された、あるいは固定化されていない所望結合蛋白を用いる結合アッセイは当該分野においてよく知られており、本発明P−セレクチンリガンド蛋白を用いる目的に使用可能である。適当なスクリーニングアッセイは、下記実施例3および9のような細胞を用いるものであってもよい。別法として、精製蛋白を用いるスクリーニングアッセイを用いてかかる作用剤を同定してもよい。例えば、精製形態のP−セレクチンリガンド蛋白を担体に固定化し、潜在的阻害剤の存在下および不存在下において精製P−セレクチンへの結合を測定してもよい。別法として、適当な結合アッセイは担体に固定化された精製P−セレクチンならびに可溶性形態の本発明P−セレクチンリガンド蛋白を用いるものであってもよい。
【0079】
いずれのP−セレクチンリガンド蛋白を上記スクリーニングアッセイにおいて使用してもよい。例えば、配列番号:2のアミノ酸1からアミノ酸402までの全長P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよく、あるいは配列番号:2のアミノ酸42からアミノ酸402までの成熟P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよく、あるいは配列番号:2のアミノ酸42からアミノ酸310までの可溶性成熟P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよい。別法として、配列番号:4のアミノ酸1からアミノ酸412までのP−セレクチンリガンド蛋白、または配列番号:4のアミノ酸42からアミノ酸412までの成熟形態のP−セレクチンリガンド蛋白、または配列番号:4のアミノ酸42からアミノ酸320までの可溶性成熟形態のP−セレクチンリガンド蛋白を用いて、本発明に従って、細胞間付着の阻害剤をスクリーニングしてもよい。
【0080】
かかるスクリーニングアッセイにおいて、P−セレクチンまたはE−セレクチンとP−セレクチンリガンド蛋白とを結合させることにより第1の結合混合物を得て、第1の結合混合物中の結合量(B0)を測定する。P−、E−、またはL−セレクチン、P−セレクチンリガンド蛋白、ならびにスクリーニングすべき化合物または作用剤を結合させることにより第2の結合混合物を得て、第2の結合混合物中の結合量(B)を測定する。第1および第2の結合混合物中の結合量を、例えばB/B0を計算することにより比較する。第1の結合混合物中の結合と比較して第2の結合混合物中の結合の減少が観察される場合、化合物または作用剤はP−、E−、またはL−セレクチンにより媒介される細胞間付着を阻害しうると考えられる。結合混合物の処方および最適化は当業者のレベル内であり、かかる結合混合物は結合を促進または最適化するバッファーおよび塩類を含んでいてもよく、さらなる対照アッセイを本発明スクリーニングアッセイにおいて行ってもよい。
【0081】
P−、E−、またはL−セレクチンに対するP−セレクチンリガンド蛋白の結合活性を少なくとも約10%、好ましくは約50%以上低下させることがわかった化合物を同定し、ついで、L−セクレチンへの結合のアッセイおよびインビボアッセイを包含する他のセクレチン結合アッセイにおいて2次スクリーニングしてもよい。これらの手段により、セレクチンにより媒介される細胞間付着に対する阻害活性を有し、抗炎症剤として適当な化合物を同定してもよい。
【実施例1】
【0082】
P−セレクチンリガンド蛋白遺伝子のクローニング
A.HL60 cDNAライブラリーの構築
P−セレクチンリガンドの発現クローニングのためにHL60 cDNAライブラリーを構築した。Fast Track mRNA Isolation Kit (Invitrogen; San Diego, CA)を用いてヒトプロミエロサイト細胞系HL60(S. J. Collins, et al.,上記文献)からポリA+RNAを単離した。ポリA+RNAフラクションから2本鎖cDNAを合成し、平滑末端をEcoRIアダプター(5'-AATTCCGTCGACTCTAGAG-3', 配列番号:7; 5'-CTCTAGAGTCGACGG-3', 配列番号:8)に連結した。EcoRIエンドヌクレアーゼとともにインキュベーションされ、ついで、ウシ腸アルカリ性ホスファターゼとともにインキュベーションされ、その後精製された発現ベクターpMT21(R. Kaufman et al., J. Mol. Cell. Biol. 9, 946-958 (1989))中にcDNAを連結した。連結生成物から2μ取ってイー・コリ(E. coli) DH5α細胞中にエレクトロポレーションし、10mM MgCl2、10mM
MgSO4、および2%グリセロールを補足した1mlのSOB培地(J. Sambrook et al., Molecular Cloning:A Laboratory Manual, New York, Cold Spring Harbor Press, p1.90 (1989))中、37℃で1時間増殖させた。ライブラリーを小さい部分に分けるために、細菌懸濁液1mlずつをアンピシリン存在下の寒天プレートに撒き、1mlあたりのコロニー数を計算した。各コロニーが1のcDNAクローンを有すると仮定すると、600000クローンが得られ、1プールあたり約16000クローンの部分に分かれた。38のプールそれぞれを、アンピシリン存在下のLブロス中で一晩増殖させ、CsClグラジエントによりプラスミドを精製した。
【0083】
B.P−セレクチンリガンド蛋白遺伝子のスクリーニング
第1段階において、実施例4(A)のLEC−γ1結合アッセイを用いて、HL60 cDNAライブラリーをパンニングし、そのことにより目的プラスミドを豊富化させた。HL60 cDNAライブラリープール各6μgを2μgの3/4FT遺伝子(実施例2)とともにCHO細胞中に同時トランスフェクションした。トランスフェクションから約45時間後、1mM EGTA中、37℃で15分間細胞をインキュベーションし、ついで、セルリフターでかき取ることによりCOS細胞をプレートから取った。1mMカルシウムを含有するHanks緩衝化セイライン(HBSS)で細胞を2回洗浄した。細胞を4mlのHBSSに再懸濁した。懸濁された、トランスフェクションされたCOS細胞を、実施例4(A)において説明するLEC−γ1結合アッセイを用いてスクリーニングした。
【0084】
付着性COS細胞をHirts抽出物(B. Hirts, J. Mol. Biol., 26, 365-369 (1967))から回収し、ついで、増幅のためにイー・コリ DH5α細胞中にエレクトロポレーションした。豊富化されたプラスミド集団をCsClグラジエントで精製し、3/4FT遺伝子(実施例2)をCOS細胞中に再トランスフェクションした。トランスフェクション、スクリーニング、ついで、プラスミド増幅工程を全部で3回繰り返し、ついで、LEC−γ1でコーティングされたプレートに結合するプールを視覚的に検出した。その後、陽性プラスミドプールを部分に分けた。これには陽性抽出物からのHirts抽出物のイー・コリDH5α細胞中へのエレクトロポレーション、ついで、上記のごとく1mlあたりのコロニー数の計算を用いた。一定数のコロニーをアンピシリン存在下の寒天プレートに撒くことにより種々のプールサイズが得られた。ニトロセルロースリフトを行い、フィルターを新たな寒天プレート上に置くことにより2系のプレートを調製した。2系のプレートは、陽性であると同定されたプール由来の各コロニーおよびコロニー群を選択するためのリファレンスプレートとして役立った。
【0085】
クローニングの第2段階において、最初のスクリーニング段階に使用したのと同じ手順により、COS細胞をライブラリーから分けたプールおよび3/4FT遺伝子を用いて同時トランスフェクションした。トランスフェクションから48時間後、実施例4(B)の蛍光CHO:P−セレクチンアッセイを用いてトランスフェクションされた細胞をスクリーニングした。上記のごとく、陽性プールをさらに分けて、最後に各コロニーをスクリーニングし、陽性クローンを同定した。この方法を用いて、1個の陽性クローンpMT21:PL85を得て、P−セレクチンリガンド蛋白をコードしていることがわかった。pMT21:PL85中に含まれるP−セレクチンリガンドのDNA配列を配列番号:1に示し、pMT21:PL85によりコードされるP−セレクチンリガンド蛋白の結合特性を下記実施例4(C)において説明する。
【実施例2】
【0086】
1,3/1,4フコシルトランスフェラーゼ遺伝子のクローニング
ヒト全ゲノムDNA(Clontech Laboratories)からPCR手段を用いて1,3/1,4フコシルトランスフェラーゼ遺伝子(3/4FT)をクローン化した。センスオリゴヌクレオチドプライマーは遺伝子のXbaI部位および5'末端を含み(5'-TAGCATACGCTCTAGAGCATGGATCCCCTGGGTGCAGCCAAGC-3', 配列番号:9)、アンチセンスオリゴヌクレオチドプライマーは遺伝子のEcoRI部位および3'末端を含んでいた(5'-CCGGAATTCTCAGGTGAACCAAGCCGC-3', 配列番号:10)。PCR生成物をXbaI、ついでEcoRIで逐次消化し、標準的なゲル精製法により精製した。ついで、XbaI、ついでEcoRIで逐次消化され、標準的なゲル精製法により精製されたベクターpMT3Sv2ADA(R. Kaufman, Methods in Enzymology,上記)中にこの遺伝子を連結した。コンピテントHB101細胞(Biorad)をこの連結生成物でトランスフェクションし、ついで、アンピシリン存在下の寒天プレートに撒いた。アンピシリン耐性形質転換体のニトロセルロースフィルターリフトを、遺伝子の中央部の506〜530の領域に対して相捕的な放射性標識オリゴヌクレオチド(5'-AAGTATCTGTCCAGGGCTTCCAGGT-3', 配列番号:11)でプローブした(J. Sambrook et al., 上記)。
【0087】
20個の陽性クローンからプラスミドDNA微量標品を調製した。ついで、精製DNAをEcoRIおよびXbaIで消化して正しいサイズのインサートを有する正しいクローンを同定した。ついで、このクローン(pEA.3/4FT)を大スケールまで増殖させ、CsCl密度グラジエントバンド法によりDNAを単離した。下記の細胞−細胞結合アッセイにおいて遺伝子の機能を評価した。DEAEデキストラン処理、DMSOショック処理、ついでクロロキンインキュベーションを用いてCOS−1サル細胞(クローンM6; M. Horwitz et al., Mol. Appl. Genet., 2:147-149 (1983))を3/4FTでトランスフェクションした(L. Sompeyrac and K. Dana, Proc. Natl. Acad. Sci. 78:7575-7578 (1981); M. Lopata et al., Nucleic Acids Res., 12:5707-5717 (1984); H. Luthman and G. Magnuson, Nucleic Acids Res., 11:1295-1308 (1983))。トランスフェクションされたCOS細胞を懸濁し、E−セレクチン発現CHO細胞系への結合を定量した(G. Larsen et al., J. Biol. Chem., 267:11104-11110 (1992))。このアッセイにより、3/4FTでトランスフェクションされたCOS細胞がシアル化Lewisxエピトープを細胞表面に発現しうることが確認された。
【実施例3】
【0088】
P−セレクチンリガンド蛋白の発現
A.LEC11細胞におけるP−セレクチンリガンドの発現
以下のようにして、機能的P−セレクチンリガンドをSLex陽性チャイニーズハムスター卵巣(CHO)細胞系LEC11(Campbell, C. and Stanley, P. Cell 35:303-309 (1983))において発現させた。約8μgのP−セレクチンリガンド遺伝子含有プラスミド8pMT21:PL85、実施例1)をLEC11細胞中にトランスフェクションした。トランスフェクションから68時間後、細胞を2.5mMの酪酸ナトリウムで4時間処理した。6−CFD標識CHO:P−セレクチン細胞結合アッセイ(実施例4、セクションBにおいて説明)を用いて調べたところ、細胞はP−セレクチン付着を誘導することがわかった。対照的に、LEC11細胞単独および対照プラスミドでトランスフェクションされたLEC11細胞はいずれもP−セレクチン付着を誘導しなかった。
【0089】
B.COS細胞における可溶性P−セレクチンリガンドの発現
8μgのpED.sPSL.T7(実施例5C参照)および実施例2の4μgのpEA.3/4FTプラスミド、8μgのpED.sPSL.T7のみ、あるいは8μgのプラスミドベクター(pMT21)および4μgのpEA.3/4FT遺伝子を用いてCOS細胞をトランスフェクションした。トランスフェクションから45時間後、細胞をPBSで2回すすぎ、2mM L−グルタミン、100U/mlペニシリンおよび100μg/mlストレプトマイシンを補足した無血清DMEM(フェノールレッド不含)(JRH Biosciences)中、37℃で一晩インキュベーションした。フッ化フェニルメチルスルホニル、アプロチニンおよびNaN3を添加して最終濃度をそれぞれ1mM、2μg/mlおよび0.02%とし、ならし培地を遠心分離してすべての残渣を除去した。
【0090】
免疫沈降実験のために、COS細胞をpED.sPSL.T7およびpEA.3/4FTで同時トランスフェクションすることにより標識可溶性P−セレクチンリガンド蛋白を得た。トランスフェクションから45時間後、COS細胞を250μCi/mlの35Sメチオニン(NEN)で5時間標識し、培地を集めた。抗T7抗体との免疫沈降によりsPSL.T7蛋白の発現を確認した。
【0091】
C.COS細胞におけるPACEにより開裂されたP−セレクチンリガンドの発現
COS細胞を実施例5(C)のpED.sPSL.T7プラスミド、実施例2のpEA.3/4FT cDNA、および配列番号:5のPACE cDNAを含むプラスミドで同時トランスフェクションした。pED.sPSL.T7プラスミドおよびpEA.3/4FTプラスミドのみを用いて対照同時トランスフェクションを並行して行った。45時間後、これらのトランスフェクションCOS細胞からのならし培地をプラスチックディッシュ上にコーティングし、CHO:P−セレクチン細胞への結合(実施例4)を調べた。PACEとともに同時発現されたP−セレクチンリガンドを含む培地でコーティングされたディッシュについては結合したCHO:P−セレクチン細胞の約20倍の増加が確認された(PACEとともに同時発現されなかったP−セレクチンリガンドを含む培地と比較)。
【0092】
PACEとともに同時トランスフェクションされた精製pPSL.T7蛋白のN末端のアミノ酸配列決定により、すべてのリガンドがPACEコンセンサス部位(配列番号:1のアミノ酸38〜41)において開裂されたこが示された。同時トランスフェクションされたCOS細胞の35S−メチオニンでの放射性標識、ついでオートラジオグラフィーを行い、かなりの量のP−セレクチンリガンドが両方のトランスフェクションにおいて分泌されたことが示された。
【0093】
D.CHO細胞におけるP−セレクチンリガンド蛋白の発現
以下のようにして、全長形態(アミノ酸1〜402)のP−セレクチンリガンド蛋白をCHO(DUKX)細胞系(Urlaub & Chasin, Proc. natl. Acad. Sci. USA 77, 4216-4220 (1980))において発現させた。約25μgのpMT21:PL85プラスミドおよび約8μgのpED.3.4FT(pEA3/4FTをEcoRIおよびXbaIで制限的に消化し、得られたフラグメントをpEDプラスミドに挿入することにより得た)を、リン酸カルシウム法を用いてCHO(DUKX)細胞中に同時トランスフェクションした。トランスフェクション体をメトトレキセート耐性に関して選択した。2週間後、抗SLex抗体(CSLEX−1,米国特許第4752569号)およびヤギ赤血球(sRBC)の抱合体(クロミッククロリド法(Goding, J. W., J. Immunol. Methods 10:61-66 (1976))により調製)を用いることにより、下記のごとく個々のコロニーをSLex発現についてスクリーニングした。洗液が透明になるまで、sRBCを0.15M NaClで洗浄し、ついで、sRBC50%懸濁液を0.15M NaCl中に調製した。50μgのCSLEX−1を含有する0.2mlのsRBC懸濁液に1mlの0.01%クロミッククロリド溶液を滴下し、その間ボルテックスで撹拌した。37℃で30分インキュベーション後、10mlのリン酸塩緩衝化セイライン(PBS)溶液を反応物に添加した。抱合体を
PBSで1回洗浄し、ついで、10mlのPBSに懸濁した。トランスフェクションを入れたプレートをPBSで洗浄し、ついで、3mlのPBSおよび1mlのsRBC/CSLEX−1抱合体を各プレートに添加した。陽性コロニーはトランスルミネーターで赤色を呈し、これらを、10%ウシ胎児血清含有アルファ培地中に拾った。2週間後、コロニーをステップワイズ増幅(2、10、25、100、150nMの濃度のメトトレキセートを用いる)に供した。得られた安定な細胞系をCD−PSGL−1(R3.4)と命名した。実施例7(A)のポリクローナル抗P−セレクチンリガンド蛋白抗体を用いる免疫沈降研究によりP−セレクチンリガンド蛋白の発現を確認した。CD−PSGL−1(R3.4)細胞系により産生されるP−セレクチンリガンド蛋白の機能を、実施例4(A)のごとくLEC−γ1への結合についてトランスフェクション体をアッセイすることにより試験した。
【0094】
アデノシンデアミナーゼ選択(Kaufman, et al., PNAS (USA) 83:3136-3140 (1986))の下で、配列番号:5に示すPACEをコードするcDNAをすでに発現している安定なCHO−PACE系においてsPSL.T7蛋白を発現させた。リン酸カルシウム法を用いてpsPSL.T7(25μg)およびpED.3/4FT(8μg)をCHO−PACE細胞中に同時トランスフェクションした。メトトレキセート耐性に関してトランスフェクション体を選択し、sRBC/CSLEX−1抱合体に結合する個々のコロニーを拾った。2週間培養後、コロニーを上記ステップワイズ増幅に供した。得られた安定な細胞系をCP/PSL−T7(R4.1)と命名した。T7特異的モノクローナル抗体または実施例4(A)のLEC−γ1キメラのいずれかを用いる標準的な免疫沈降法によりpPSL.T7蛋白の発現を確認した。同様のやり方で、pMT21:PL85およびpED.3/4FTをCHO−PACEc系に同時トランスフェクションすることにより成熟全長形態(アミノ酸42〜402)のP−セレクチンリガンド蛋白発現する安定な細胞系を得た。
【0095】
実施例5(B)のsPSL.Q蛋白および実施例5(D)のsPSL.Fc蛋白を発現する安定な細胞系を以下のようにして構築した。プラスミドpED.sPSL.Q(25μg)またはpED.sPSL.Fc(25μg)を、上記の約25μgのpED.3/4FTプラスミドおよび配列番号:5のPACE cDNAとネオマイシン耐性遺伝子を含む約20μgのプラスミドとともに、リン酸カルシウム法を用いてCHO(DUKX)細胞中に同時トランスフェクションした。トランスフェクション体をメトトレキセートおよびG418抗生物質耐性に関して選択した。約2週間後、sRBC/CSLEX−1抱合体結合を用いて個々のコロニーをSLEx発現についてスクリーニングした。陽性コロニーを、1mg/mlの濃度のG418培地中に拾った。2〜3週間培養後、細胞をステップワイズメトトレキセート増幅させて選択した。得られた安定な細胞系をCD−sPSL.Q(R8.2)およびCD−sPSL.Fc(R8.1)と命名した。sPSL.QおよびsPSL.Fc蛋白の発現を、実施例7(A)の抗P−セレクチンリガンド蛋白ポリクローナル抗体を用いる標準的な免疫沈降法により確認した。
【実施例4】
【0096】
P−セレクチンにより媒介される細胞間付着のアッセイ
A.LEC−γ1結合アッセイ
キメラ形態のヒトIgGγ1(LEC−γ1)のFc部分に抱合したP−セレクチンをコードするDNAを、既知方法(Aruffo et al. Cell 67, 35-44 (1991))を用いて構築し、高レベルのキメラLEC−γ1蛋白の発現に関してdhfr−CHO細胞中に安定にトランスフェクションし、下記結合アッセイに使用するためにキメラLEC−γ1蛋白を精製した。
【0097】
ペトリ皿を、まず、ポリクローナル抗ヒトIgGγ1 Fc抗体でコーティングし、ついで、LEC−γ1蛋白でコーティングした。この方法は、キメラ分子のP−セレクチン部分がプレート表面に存在するようにLEC−γ1構築物を配向させる。カルシウムの存在下および不存在下において、配向LEC−γ1へのHL60の付着を定量した。HL60付着はカルシウム依存的であることが示され、キメラ分子がHL60細胞上のそのリガンドへのP−セレクチンの結合機能を保持していることが示された。さらに配向LEC−γ1へのHL60細胞の結合は、P−セレクチンに対する中和モノクローナル抗体によりブロックされることも示され、P−セレクチン結合の特異性が示された。
【0098】
B.蛍光CHO−P−セレクチン結合アッセイ
当該アッセイには、P−セレクチンリガンド遺伝子および3/4FT遺伝子で同時トランスフェクションされたCOS細胞表面に結合し、クラスターを形成しうる蛍光標識CHO:P−セレクチン細胞系(Laesen et al., J. Biol. Chem., 267, 11104-11110 (1992))を用いた。CHO:P−セレクチン細胞を1%ウシ胎児血清含有DME中1.5x106個/mlとなるよう懸濁し、6−カルボキシフルオレセイン二酢酸(6−CFD)を最終濃度100μg/mlとなるよう添加することにより標識した。37℃で15分インキュベーション後、細胞を培地で洗浄し、1x105個/mlとなるよう再懸濁した。洗浄されたCOSトランスフェクション体の入ったアッセイすべき各プレートに標識細胞5mlを添加し、室温で10分間インキュベーションした。培地で4回洗浄することにより非付着細胞除去した。ついで、付着CHO:P−セレクチン細胞のロゼットを求めて蛍光顕微鏡でプレートをスキャンした。
【0099】
C.放射性標識CHO:P−セレクチン細胞を用いる定量的付着アッセイ
スクリーニングの最初の段階に用いたのと同じ手順により、実施例1のpMT21:PL85プラスミドおよび実施例2のpEA.3/4FTプラスミドでCOS細胞を同時トランスフェクションした。対照として、COS細胞をpMT21:PL85のみ、あるいはpEA.3/4FTのみ、あるいはインサートを含まない同様のプラスミド(「模擬」)でトランスフェクションした。トランスフェクションから24時間後、トランスフェクション細胞をトリプシン処理し、Costar6ウェル組織培養プレート中に分配した。既知方法を用いてCHO:P−セレクチン細胞を3H−チミジンで16時間標識し、ついで、1% BSAを含有するα培地(対照);1% BSA、5mM EDTAおよび5mM EGTAを含有するα培地;1% BSAおよび10μg/mlの中和抗P−セレクチンモノクローナル抗体を含有するα培地;ならびに1% BSAおよび非中和抗P−セレクチンモノクローナル抗体を含有するα培地中、1x106/mlとして個4℃で30分プレインキュベーションした。ついで、プレインキュベーションした細胞をトランスフェクションしたCOS細胞を含むウェルに添加した。インキュベーションから10分後、培地を4回注入することにより未結合細胞を除去した。トリプシン処理により結合CHO:P−セレクチン細胞を遊離させ、シンチレーションカウンティングにより定量した。
【0100】
P−セレクチンリガンドおよび3/4FTで同時トランスフェクションされたCOS細胞は、COS模擬細胞と比較して5.4倍のCHO:P−セレクチン細胞結合を誘導した。EGTAおよびEDTA存在下でのアッセイは、模擬トランスフェクションCOS細胞の結合レベルが低下した。同様に、中和抗P−セレクチン抗体とのインキュベーションによっても特異的結合が減少し、非中和抗体は効果がなかった。対照的に、P−セレクチンのみでトランスフェクションされたCHO細胞へのCHO:P−セレクチンの結合は、EDTAおよびEGTAの存在下または不存在下の両方において、模擬トランスフェクションCOS細胞への結合と統計学的に相異はなかった。3/4FTのみでトランスフェクションされたCOS細胞へのCHO:P−セレクチン細胞の結合は、模擬トランスフェクションCOSに対する結合よりも約2倍促進されたが、EDTAおよびEGTAの存在または不存在によっては影響されなかった。
【実施例5】
【0101】
可溶性P−セレクチンリガンドの構築
実施例1においてHL60細部からのcDNAライブラリーの製造に用いた
EcoRIアダプターはXbaI制限部位(TCTAGA)を配列番号:1の開始5'側に有し、それはpMT21:PL85プラスミド中にも存在する。可溶性形態のPSLを得るために、pMT21:PL85プラスミドをXbaIおよびHincII(配列番号:1のヌクレオチド944の後ろを開裂)で制限消化した。かくして約950bpのフラグメントが得られ、それはバリン295のコドンを含め、バリン295までのコードされるリガンドの細胞外セグメント全体を含んでいた。これを単離し、下記セクションAからDまでに示すようにして、可溶性形態のP−セレクチンリガンド蛋白をコードするDNAを得るために使用した。
【0102】
A.psPSL.QCの構築
フラグメントを精製し、Asn296からCys310までのコドンが再生成され、Cys310の直後に新たなストップコドンを導入された2本鎖合成オリゴヌクレオチドDNAとともに、哺乳動物発現ベクターpEDのXbaIおよびEcoRI部位間に連結した。当該オリゴヌクレオチドの配列は以下のごとし:
5'-AACTACCCAGTGGGAGCACCAGACCACATCTCTGTGAAGCAGTGCTAG (配列番号:12)
5'-AATTCTAGCACTGCTTCACAGAGATGTGGTCTGGTGCTCCCACTGGGTAGTT (配列番号:13)
得られたプラスミドをpED.sPSL.QCと命名し、そのプラスミドから発現される蛋白をsPSL.QCと命名した。
【0103】
B.psPSL.Qの構築
フラグメントを精製し、Asn296からGln309までのコドンが再生成され、Gln309の直後に新たなストップコドンを導入された2本鎖合成オリゴヌクレオチドDNAとともに、pEDプラスミド(Kaufman et al., 1991)のXbaIおよびEcoRI部位間に連結した。オリゴヌクレオチドの配列は以下のごとし:
5'-AACTACCCAGTGGGAGCACCAGACCACATCTCTGTGAAGCAGTAG (配列番号:14)
5'-AATTCTACTGCTTCACAGAGATGTGGTCTGGTGCTCCCACTGGGTAGTT (配列番号:15)
得られたプラスミドをpED.sPSL.Qと命名し、そのプラスミドから発現される蛋白をsPSL.Qと命名した。
【0104】
C.psPSL.T7の構築
ファージT7大カプシド蛋白由来のエピトープを含む14個のアミノ酸をコードするオリゴヌクレオチドを合成し、さらにベクター配列由来の32個のアミノ酸を用いてエピトープ「タグ」をC末端に融合させた。2つのオリゴヌクレオチドは下記配列:
5'-CTAGACCCGGGATGGCATCCATGACAGGAGGACAACAAATGGTAGGCCGTAG (配列番号:16) および
5'-AATTCTACGGCCTACCCATTTGTTGTCCTCCTGTCATGGATGCCATCCCGGGT (配列番号:17)
を有し、それらを2本鎖化し、哺乳動物発現プラスミドpEDの大XbaI−EcoRIフラグメントと連結した。得られたプラスミドpED.T7をXbaIおよびSmaIで制限消化し、上記950bpのXbaI−HincIIフラグメントに連結し、プラスミドpED.sPSL.T7を得た。
【0105】
pED.sPSL.T7の発現により得られる蛋白をsPSL.T7と命名した。
【0106】
D.可溶性P−セレクチンリガンド−IgGFcキメラの構築
ヒト免疫グロブリンIgG1のFc部分に融合した可溶性の細胞外形態のP−セレクチンリガンド蛋白をコードするプラスミドDNAを以下のようにして構築した。哺乳動物発現ベクターpED.Fcは、ヒトIgG1のFc領域をコードする配列を含み、該配列はユニークなXbaI制限部位を介してヒンジ領域へのコーディング配列のアミノ末端の融合を可能にする新規リンカー配列を伴っている。3つのフラグメントの連結を行った。pED.FcをXbaIで制限消化し、直鎖状にしてゲル精製した。上記pMT21:PL85由来の950bpのフラグメントは第2のフラグメントを含んでいた。第3のフラグメントは、以下の配列を有するアニーリングされた合成オリゴヌクレオチドDNAからなっていた。
5' - CTGCGGCCGCAGT (配列番号:18)
5' - CTAGACTGCGGCCGCAG (配列番号:19)
【0107】
連結生成物をプラスミドDNAとして増幅し、正しい配置を有する個々のクローンをDNA配列決定により同定した。プラスミドをpED.PSL.Fcと命名した。得られた可溶性P−セレクチンリガンド/Fc融合蛋白のDNAコーディング領域を配列番号:6に示す。
【実施例6】
【0108】
発現されたP−セレクチンリガンドの特徴付け
A.COS細胞上に発現された全長P−セレクチンリガンド蛋白の結合特性
実施例2のpEA.3/4FTプラスミドおよび実施例1のpMT21:PL85プラスミドでのCOS細胞の同時トランスフェクションにより、CHO:P−セレクチン細胞に特異的に結合するCOS細胞を得る。この結合は、pEA.3/4FTおよびpMT21:PL85で同時トランスフェクションした場合にのみ観察される。いずれかのプラスミドのみの使用はCHO:P−セレクチン細胞に結合しないCHO細胞を生じる。P−セレクチンを発現しない親のCHO(DUKX)細胞系およびpEA.3/4FTおよびpMT21:PL85で同時トランスフェクションされたCOS細胞の間には結合は観察されない。同時トランスフェクション細胞およびCHO:P−セレクチン細胞の間の結合は、EDTAおよびEGTAのごとき2価イオンのキレーターに対して感受性があり、P−セレクチンにより媒介される細胞付着のCa++依存性と矛盾しない。中和抗P−セレクチンモノクローナル抗体は、CHO:P−セレクチン細胞とpEA.3/4FTおよびpMT21:PL85で同時トランスフェクションされたCHO細胞との間の結合をブロックしたが、非中和抗P−セレクチンモノクローナル抗体は結合に影響しなかった。抗体の結果は、COS細胞表面に発現されるP−セレクチンリガンド蛋白への結合にはP−セレクチンの機能的ドメインが必要であることを示す。
【0109】
B.COS細胞において発現された全長P−セレクチンリガンドの電気泳動特性
同時トランスフェクションCOS細胞の界面活性剤抽出物を以下のようにして調製した。同時トランスフェクションから45時間後、約1.5x107個の細胞を5mlの溶解バッファー(10mMピペラジン−N,N'−ビス[2−エタンスルホン酸](PIPES)pH7.5、100mM KCl、3mM MgCl2、1mMベンズアミジン、0.5μg/mlロイペプチン、0.75μg/mlペプスタチン、1mMエチレンジアミン、および1μg/mlアプロチニン)に懸濁した。低速遠心分離(500xg、10分)により細胞残渣を除去し、超遠心(100000xg、60分)により膜フラクションを集めた。高速遠心分離により得た膜ペレットを抽出バッファー(10mM 3−[N−モルホリノ]プロパンスルホン酸(MOPS)pH7.5、0.1M NaCl、0.02% NaN3、1% Thesit(登録商標)(Sigma),1mMベンズアミジン、0.5μg/mlロイペプチン、0.75μg/mlペプスタチン、1mMエチレンジアミン、および1μg/mlアプロチニン)に再懸濁した。ついで、以下のようにして試料をSDSポリアクリルアミドゲル電気泳動に供し、ニトロセルロースブロットに移した。界面活性剤抽出物の一部を1% SDSローディングバッファーに懸濁し、100℃で5分間加熱し、ついで、8〜16%ポリアクリルアミドゲル(還元的)または6%ゲル(非還元的)にロードし、Laemmliバッファー系中で泳動した。Immobilon-P(登録商標)トランスファー膜を用いてブロットを調製した。ブロットを10mM MOPS pH7.5、0.1M NaCl、0.02% NaN3、1mM MgCl2、1mM CaCl2、および10%脱脂乳に、4℃で一晩浸した。ブロットを上記バッファー(乳不含)で1回すすぎ、ブロティングバッファー(10mM MOPS pH7.5、0.1M NaCl、1% ウシ血清アルブミン、0.05% Thesit、1mM MgCl2、1mM CaCl2)中、室温で30分インキュベーションした。
【0110】
ついで、下記のごとくブロットをP−セレクチンリガンドを求めてプローブした。50ngのP−セレクチン/Fcキメラを、3μCiの125I−プロテインAとともにブロッティングバッファー中室温で30分インキュベーションした。この時点で、さらなる賦形剤(例えば、EDTA、EGTA、モノクローナル抗体)をプレインキュベーション混合物に添加して、P−セレクチンリガンドへのキメラの結合の対する影響を評価することができる。ついで、プレインキュベーションされた混合物をブロットとともに室温で60分インキュベーションし、ついで、ブロットを同じブロッティングバッファー(ウシ血清アルブミン不含)で4回洗浄し、風乾し、ついで、−70℃でオートラジオグラフを行った。
【0111】
非還元条件下において、同時トランスフェクションCOS細胞から調製した膜抽出物に関してこの方法により2つのバンドが観察された。主要バンドは約220kDの分子量のところまで移動したが、マイナーバンドは110kDのところまで移動した。還元条件下においては、約110kDの分子量のただ1つのバンドが観察され、非還元条件下ではP−セレクチンリガンドはホモダイマーとして存在することが示された。還元されたモノマーの大体の分子量は、cDNAクローンの推定アミノ酸配列から予想されるもの(45kD)よりも大きく、発現された蛋白がさらなる翻訳後修飾を受けたことが示される(実施例6(C)参照)。非特異的IgG1プローブがブロット上にバンドを生じないことにより、P−セレクチン/Fcキメラの特異性を確認した。さらに、P−セレクチン/Fcキメラのブロットへの結合は、EDTA、EGTA、および中和抗P−セレクチンモノクローナル抗体によって消滅した。pEA.3/4FTおよびpMT:PL85プラスミドで同時トランスフェクションされたCOS細胞の膜抽出物からのみ、ブロット上の特異的バンドが観察された。対照トランスフェクション体(pEA.3/4FTまたはpMT21:PL85)からの膜抽出物はブロット上にバンドを生じなかった。
【0112】
C.P−セレクチンリガンド蛋白の糖鎖付加
組み換えP−セレクチンリガンド上に共有結合した炭水化物の存在およびP−セレクチンへの結合におけるその役割を以下のようにして調べた。実施例5(C)のpED.sPSL.T7および実施例2のpEA.3/4FTプラスミドでCOS細胞を同時トランスフェクションした。48時間後、細胞に35S−メチオニンのパルスを与えた。200μlの35Sメチオニン標識sPSL.T7ならし培地を5μgのLEC−γ1とともに、2mM CaCl2および1mg/mlウシ血清アルブミン(BSA)の存在下でインキュベーションした。4℃で2時間回転撹拌した後、プロテインA−Sepharoseビーズ(Pharmacia)を4℃において1時間かけて添加し、遠心分離によりペレット化させ、2mM CaCl2および1mg/mlBSAを含有するTris緩衝化セイライン(20mM Tris−HCl、150mM NaCl pH7.5、以下TBSという)で2回洗浄した。ついで、ペレットを再懸濁し、ノイラミニダーゼ(Streptococcus pneumoniae)、O−グリカナーゼ、およびN−グリカナーゼ(すべてGenzymeから得た)で以下のようにして処理した。すべてのグリコシダーゼ消化を37℃で一晩行った。ノイラミニダーゼ消化には、ペレットを50μlの2−(モルホリノ)−エタンスルホン酸(MES)バッファー,pH6.5(Calbiochem)および0.1% SDS中に再懸濁し、95℃で5分間加熱し、ついで、ペレット化させた。上清を、1.4% n−オクチルβ−D−グルコピラノシド(OGP)、10mM酢酸カルシウム、20mMカコジル酸ナトリウムおよび2.5mM PMSFを含むようにし、最終pH7.0とした。8μlのノイラミニダーゼを添加して最終濃度1ユニット/mlとした。ノイラミニダーゼ/O−グリカナーゼ消化には、試料をノイラミニダーゼとともに上記のごとく調製し、さらにO−グリカナーゼを添加して最終濃度0.1ユニット/mlとした。N−グリカナーゼ消化には、ペレットを54μlのMESバッファーおよび1% SDSに再懸濁し、95℃で5分間加熱し、ついでペレット化させた。上清を、0.2Mリン酸ナトリウム、3.5% OGP、および2.5mM PMSFを含むようにし、最終pHを8.5とした。N−グリカナーゼを添加して最終濃度12ユニット/mlとし、上記のごとくインキュベーションした。
【0113】
sPSL.T7に対するグリコシダーゼ処理の影響を2とおりのやり方で評価した。これを行うために、各消化蛋白試料を2つの等しいフラクションに分けた。一方のフラクションを、実施例7(A)のP−セレクチンポリクローナル抗体を用いて沈降させて電気泳動度に対する消化の影響を示した。他方のフラクションを、実施例4(A)のLEC−γ1キメラを用いて沈降させて消化後に残存するP−セレクチンリガンド結合活性を評価した。免疫沈降した試料を、還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動およびオートラジオグラフィーにより分析した。
【0114】
グリコシダーゼ処理なしの場合、オートラジオグラフィーは各沈降物に対応するバンド(分子量110kD)を示した。P−セレクチンリガンド蛋白をノイラミニダーゼで処理した場合、抗P−セレクチンリガンドポリクローナル抗体沈降物は泳動度がわずかに減少し、シアル酸除去の除去と矛盾しなかった。LEC−γ1により沈降したP−セレクチンリガンド蛋白の量はノイラミニダーゼ処理後に有意に減少し、P−セレクチン/P−セレクチンリガンド相互作用におけるシアル酸残基の役割と矛盾しなかった。P−セレクチンリガンド蛋白をノイラミニダーゼおよびO−グリカナーゼの両方で処理した場合、抗P−セレクチンリガンドポリクローナル抗体との沈降後に電気泳動度の実質的な増加が観察され、多くのO−結合オリゴサッカライド鎖が除去されたことが示された。しかしながら、P−セレクチンリガンド蛋白からのO−結合オリゴサッカライドの除去が不完全であるかもしれない。なぜなら、電気泳動度が分子量38kDの蛋白(配列番号:1のアミノ酸配列から推定)に対応しなかったからである。ノイラミニダーゼ/O−グリカナーゼは、LEC−γ1に結合したP−セレクチンリガンド蛋白をわずかしか消化せず、さらに、P−セレクチン/P−セレクチンリガンド相互作用におけるオリゴサッカライドの役割が示された。N−グリカナーゼで精製P−セレクチンリガンドを処理すると電気泳動度がわずかに増加し、N−結合糖鎖付加に関するいくつかのコンセンサス部位が占領されていることが示された。LEC−γ1により沈降したP−セレクチンリガンド蛋白の量はわずかに減少し、N−結合糖鎖付加もP−セレクチン/P−セレクチンリガンド相互作用に関与しているが、シアル化およびO−結合糖鎖付加のような劇的なものではないことが示された。
【実施例7】
【0115】
P−セレクチンリガンドに特異的なポリクローナル抗体
A.ポリクローナルウサギ抗P−セレクチンリガンド蛋白/マルトース結合蛋白融合蛋白
イー・コリにおいて得られた融合蛋白でウサギを免疫することにより抗P−セレクチンリガンドポリクローナル抗体を得た。融合蛋白は、P−セレクチンリガンドのアミノ末端側1/3(配列番号:1のアミノ酸1から110まで)がイン・フレームでマルトース結合蛋白に融合している(Maina, C. V. et al., Gene 74, 365-373 (1988); Riggs, P., in Current Protocols in Molecular Biology, F. M. Ausbel et al., Eds., Greene Associates/Wiley Interscience (New York, 1990) chapter 16.6)。ここに用いた条件下において、融合蛋白抗体はP−セレクチンリガンド蛋白を認識する。
【0116】
B.ポリクローナルウサギ抗sPSL.T7蛋白
下記スキームに従って、可溶性形態の本発明蛋白(sPSL.T7;実施例5(C)参照)を見かけ上均一にまで精製した。COS細胞を3つのプラスミド(それぞれsPSL.T7(実施例5(C))、3/4FT(実施例2)および可溶性形態のPACE(配列番号:5に示す)をコードしている)でトランスフェクションした。72時間後、ならし培地を集め、組み換えsPSL.T7を以下のようにして精製した。
【0117】
ならし培地を50mM MOPS、150mM NaCl、0.5mM CaCl2および0.5mM MnCl2,pH7.2で2倍に希釈し、同じバッファーで平衡化しておいたレンズマメ−Sepharose 4Bカラムに負荷した。負荷後、280nmの吸光度が安定なベースラインまで低下するまでカラムを同じバッファーで洗浄した。ついで、カラムを、0.5M α−メチル−マンノシドおよび0.3M NaClに調節した同じバッファーで溶離した。カラムの5〜15倍体積のこの溶離バッファー中に組み換えsPSL.T7を集めた。ついで、レンズマメレクチン溶離物を、4℃においてカラム溶離物1リットルあたり472gの硫酸アンモニウムを添加することにより0〜70%硫酸アンモニウム沈殿させた。30分撹拌後、沈殿を最小量のTBS(20mM Tris−HCl、150mM NaCl,pH7.5)に再懸濁し、TBSで平衡化したTSK G4000SWXLゲル濾過カラムに負荷した。カラム流速0.5ml/分とし、ガードカラムを使用した。再懸濁硫酸アンモニウムペレットを250μl未満の部分試料としてカラムに注入し、SDS−PAGEおよびウェスタン分析によりフラクションを分析した。sPSL.T7含有フラクションをプールし、ついで、ウサギの免疫に使用した。
【0118】
抗原プライミングおよびその後のブーストにより3カ月にわたって標準的方法でsPSL.T7に対する抗体を得た。詳細には、50μgのsPSL.T7(0.1% SDS中に混合し、100℃で10分加熱することにより変性)を完全フロイントのアジュバントと混合し、5カ所に皮下注射することにより最初の免疫を行った。25μgのsPSL.T7(0.1% SDS中に混合し、100℃で10分加熱することにより変性)[3回目およびそれ以後のブーストには12.5μg]を不完全フロイントのアジュバントと混合し、2カ所に皮下注射することにより(あるいは後に筋肉内注射)2回目以降のブースト(すべて皮下注射)を行った(2週間ごとに注射)。2週間ごとに試験採血を行って抗体力価をモニターした。抗体力価が適当なレベルに達したら、大量採血を行い、全血清フラクションを調製した。このポリクローナル抗体調合物を用いて、実施例4記載の方法と同様の方法で、CHO:P−セレクチン細胞へのHL60細胞の特異的結合を阻害した。
【0119】
このアッセイには、マイクロタイタープレートの底に撒かれたCHO細胞に結合した蛍光標識HL60細胞(BCECFAM;2',7'−ビス−(2−カルボキシメチル)−5−(および−6)−カルボキシフルイレセイン,アセトキシメチルエステルで標識)を使用した。標識HL60細胞を、ポリクローナル抗体含有血清または免疫前の血清のいずれかとともに4℃で30分プレインキュベーションした。次いで、細胞を洗浄し、CHO:P−セレクチン細胞とともに10分間インキュベーションした。ついで、プレートを洗浄し、蛍光マイクロタイタープレートリーダーを用いて蛍光を読んだ。このアッセイを用いると、1:15希釈の抗−sPSL.T7ポリクローナル血清はCHO:P−セレクチンへのHL60細胞の結合を本質的に完全に阻害した。1:150の抗血清希釈ではCHO:P−セレクチンへのHL60細胞の結合がまだ示された。免疫前の血清はCHO:P−セレクチンへのHL60細胞の結合に影響しなかった。
【実施例8】
【0120】
コア2との同時形質転換
A.コア2GlcNAcトランスフェラーゼをコードするcDNAの単離
コア2GlcNAcトランスフェラーゼをコードしているcDNAを、標準的な生物学的方法により単離した。公表されているヒトコア2配列(Bierhuizen, M. F. A., Fukuda, M., Proc. natl. Acad. Sci. 89, 9326-9330 (1992))に基づいて5'および3'末端における2つのオリゴヌクレオチドを設計した。HL60 cDNAライブラリーのプール(Sako, D., Cell 75, 1179-1186 (1993))を鋳型として用いて、標準的PCRプロトコールによりコア2コーディング配列を増幅した。PCR増幅されたフラグメントを精製し、pEDベクター中にサブクローンした。cDNAを単離するために、PCR反応において陽性のシグナルを発したプールをイー・コリ中に形質転換し、プレートに撒いた。形質転換体をニトロセルロースフィルター上に移し、標準的プロトコールに従って32P放射性標識PCRフラグメントとハイブリダイゼーションさせた。溶液クローンを拾い、再度プレートに撒くことにより精製した。cDNAおよびPCRクローンの配列をジデオキシ配列決定により確認した。
【0121】
B.コア2酵素を発現する安定なPSGL−1チャイニーズハムスター卵巣細胞系の取得
実施例3に従って作成した、全長のP−セレクチンリガンド蛋白および3/4FTフコシルトランスフェラーゼを発現する細胞系を、リン酸カルシウム法により、コア2 cDNAおよびネオマイシン耐性遺伝子(pMT4Neo)で同時トランスフェクションした。約2週間後、安定なG418耐性トランスフェクション体を単一単離体としてあるいはプールとして拾った。これらのトランスフェクション体を、1mg/mlのG418含有完全DMEM培地中で増殖させ、コア2酵素活性につき分析した(Higgins, E. A., et al., J. Biol. Chem. 266, 6280-6290 (1991))。コア2活性に関して陽性のコロニーまたはプールを、種々の方法によりP−セレクチンへのP−セレクチンリガンド結合に関して分析した。同様の方法で、P−セレクチンリガンド蛋白を発現する、あるいは3/4フコシルトランスフェラーゼおよびPACE酵素の両方とともに可溶性P−セレクチンリガンド蛋白を発現する細胞系(実施例3参照)を用いて、上記のごとく安定なコア2の同時トランスフェクション体を単離した。
【0122】
C.P−セレクチン結合活性に対するコア2の影響
P−セレクチン結合活性に対するコア2の影響を、3つの異なる方法により評価した。
【0123】
1.固定化可溶性P−セレクチンまたはP−セレクチン/IgGキメラへのmPSGL−1トランスフェクション体の結合
4℃において、50mM Tris pH9.5中の1μg/mlの抗ヒトFc抗体で48ウェルプレートをコーティングした。HBSSバッファーで2回洗浄後、P−セレクチン/IgGキメラ(濃度0.1〜1μg/ml、実施例5)を4℃で一晩HBSSバッファー中にプレーティングした。4℃で3ないし4時間プレートをBSAでブロックした。可溶性P−セレクチンリガンド蛋白の場合、同じバッファー中の蛋白を直接プレートにコーティングした。3H標識CHO細胞を2mM EGTAでリフトし、PBSで3回洗浄し、最終濃度106個/mlとして再懸濁した。この懸濁液の300μlの部分試料を各ウェルに添加した(300000個/ウェル)。室温で12時間インキュベーション後、ウェルを無血清DMEMで4回洗浄して未結合細胞を除去した。結合細胞を5mM EGTAでリフトし、シンチレーションカウンターでカウントした。無処理のP−セレクチンリガンド蛋白結合に関する陽性対照として使用するU937細胞をガンマグロブリン(5mg/ml)で前処理して内在性Fc受容体をブロックし、ついで、P−セレクチンIgGキメラに結合させた。比較結合データを図1に示す。
【0124】
2.P−セレクチン/IgGキメラを用いる免疫沈降
形質転換体から調製された組み換え全長または可溶性P−セレクチンリガンド蛋白(さらにコア2を伴うもの、あるいは伴わないもの)を35S−メチオニンで標識し、ついで、実施例7および5ならびにSako, D., Cell 175, 1179-1186 (1993)にすでに記載のごとく、抗P−セレクチンリガンド蛋白ポリクローナル抗体またはP−セレクチン/IgGキメラのいずれかと免疫沈降させた。データを図2に示す。
【0125】
3.フローサイトメトリー
安定なネズミP−セレクチンリガンド蛋白トランスフェクション体(コア2を伴うもの、あるいは伴わないもの)を、P−セレクチン/IgGキメラ(LecY1)(実施例5)または抗P−セレクチンリガンド蛋白モノクローナル抗体(Mab275、配列番号:2のアミノ酸42からアミノ酸56までの配列を有するペプチドに対して生成)のいずれかを用いる標準的なFACS法により分析した。両方の試薬をFITC標識プロテインAに前以て抱合させた。2mM CaCl2存在下においてこの抱合体とともに4℃で30分インキュベーション後に、細胞を分析した。データを図3に示す。
【実施例9】
【0126】
P−セレクチンリガンド蛋白のE−セレクチン結合
Bevilacqua et al., Science, 243:1160 (1989)に報告されている配列のアミノ酸−21から536までを含むE−セレクチンをコードしているDNAを用いて、P−セレクチン/IgGキメラに関して実施例5で説明したようにしてE−セレクチン/IgGキメラを作成した。
【0127】
U937細胞(約6.5x107個)を組織培養プレートから回収、新鮮完全RPMI培地および50μCi/mlの3H−グルコサミン塩酸塩(糖蛋白の蛋白結合炭水化物部分を標識)[Varki, FASEB 5:226-235 (1991)]を含む50mlの培地に二等分した。48時間インキュベーション後、両方の培養物から遠心分離により細胞を回収し、PBSで3回洗浄した。ペレット化した細胞をそれぞれ2.5mlの溶解バッファー(1% Triton X-100を含有)に懸濁し、プローブソニケーションを2分間行うことにより細胞を破壊した。界面活性剤溶解物を氷上に3時間置き、ついで、さらに2分間再ソニケーションした。溶解物を16000rpmで5分間遠心分離し、上清を回収し、界面活性剤不含の溶解バッファーでそれぞれ12mlとした。。2つの希釈細胞溶解物のうち1つに、P−セレクチン/IgGキメラ(実施例5参照)に前以てカップリングさせた100μlのプロテインAセファロースを添加し、もう1つには、E−セレクチン/IgGキメラに前以てカップリングさせた100μlのプロテインAセファロースを添加した。往復運動により混合しながら結合反応を4℃で一晩進行させた。場合によっては、U937細胞由来の精製膜が標識蛋白の界面活性剤抽出のための出発物質として役立った。これらの場合、界面活性剤抽出およびアフィニティー沈殿工程は上記のものと本質的に同じであった。
【0128】
インキュベーション後、2つの並行反応混合物をそれぞれ2000rpmで遠心分離し、上清を捨てた。樹脂ペレットをバッファー(10mM MOPS、100mM NaCl、1mM CaCl2、1mM MgCl2、0.02% NaN3,pH7.5、Triton X-100含有[1回目および2回目の洗浄には0.25%、3回目の洗浄には0.1%、4回目の洗浄には0.01%])で4回洗浄した。0.01% Triton X-100含有バッファー1mlを用いて溶離前の最後の洗浄を行い、これらの洗液を液体シンチレーションカウンテイング(LSC)による放射活性カウント定量のためにとっておいた。ついで、樹脂を、0.01% Triton X-100および10mM EDTAを含有するバッファー1mlを用い、4℃で一晩往復運動により混合しながら溶離を行った。上清を遠心分離により回収し、ついで、LSCにより定量した。
【0129】
EDTAにより樹脂から遊離した物質のオートラジオグラフィーを、10%架橋SDS−PAGEゲルで電気泳動し(必要ならばCentricon-10ユニットにより試料を約10000cpmに濃縮)、ついで、製造者の説明に従ってゲルをEN3HANCE(DuPont)で処理し、その後市販ゲルドライヤーでゲルを2時間乾燥させることにより行った。−80℃で最低3日間乾燥ゲルをX線フィルムにさらした。
【0130】
前以てU937細胞の界面活性剤抽出物にさらした固定化E−またはP−セレクチンのEDTAでの溶離、ついで、EDTAでの徹底的な洗浄により、遊離状態の3H−グルコサミン標識蛋白が得られた。EDTA溶離液からの放射性標識回収量は、EDTA処理前の最終洗液において観察されるカウントよりも少なくとも10倍多かった。この観察結果は、P−およびE−セレクチンキメラは両方ともU937全細胞溶解物由来のリガンドをEDTA依存的な様式でアフィニティー捕捉し、ついで、樹脂をEDTAで処理することにより、捕捉されたリガンドが遊離されたことを示唆する。
【0131】
EDTAにより2種のキメラから遊離された蛋白についての評価を、非還元的条件下でのSDS−PAGEおよびオートラジオグラフィーにより行った(市販14C−標識分子量標準物質を用いた)。図4のオートラジオグラフにより示されるように、P−セレクチンキメラ(レーン2および10)で処理した全細胞溶解物、およびE−セレクチンキメラ(レーン4および8)で処理した全細胞溶解物からの遊離カウントは、非還元的条件においては200kDの主要種(レーン2および4)に、還元的条件下では100kDの種(レーン8および10)に対応していた。図4に示す異なる実験においては精製膜抽出物が全細胞のかわりに出発物質として使用され、E−セレクチンキメラ(レーン3(非還元的)、およびレーン9(還元的))およびP−セレクチンキメラ(示さず)は同様の結果を示した。他の実験は、P−セレクチンに結合する主要U937糖蛋白がRb3026(組み換えsPSGL1.T7に対して生成したポリクローナル抗体)と免疫反応することを示す。それゆえ、P−およびE−セレクチンは、各場合において同じ特性で、単一の主要糖蛋白種を特異的に認識する。
【実施例10】
【0132】
欠失または変化した形態の可溶性P−セレクチンリガンド蛋白の製造および分析
A.DNA構築物の製造
以下のようにして、末端切断形態のP−セレクチンリガンド蛋白−IgGキメラを得た。プラスミドpED.PSL.FcをPstIおよびNotIで制限消化し、Fc部分を含む6kbのフラグメントおよびベクターpEDFc6kbをゲル精製した。プラスミド構築物pED.149.Fc、pED.47.FcおよびpED.19.Fcを、標準的なPCR法により、下記のオリゴヌクレオチドプライマーのペアーを用いて作成した。
すべての構築物用の「上流」プライマー:
5'-CCAGGTCCAACTGCAGGTCGACTCTAGAGGGCACTTCTTCTGGGCCCACG-3' (配列番号:20)
148Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCCCTCCAGAACCCATGGCTGCTGGTTGCAGTGG-3' (配列番号:21)
47Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCGCAGCAGGCTCCACAGTGGTAG-3' (配列番号:22)
19Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCGGAGGCTCCGTTTCTGGCAG-3' (配列番号:23)
【0133】
PCR反応用鋳型DNAはpED.PSL.Fcであった。PCR条件は、Perkin-Elmer Thermocyclerを用いて94℃で1分;42℃で1分;72℃で3分を25サイクルである。最後のサイクル完了後、反応物を25℃においてKlenow酵素で30分処理した。ついで、フェノールクロロホルム抽出を行い、酢酸ナトリウムを添加して0.3Mとし、ついで、PCR生成物であるDNAを2.5倍体積のエタノールで沈殿させた。DNAペレットを70%エタノールですすぎ、残存エタノールを蒸発させた。再懸濁したDNAをPstIおよびNotIで消化し、ゲル精製し、上記pEDFc6kbフラグメントに連結した。制限分析により正しい構築物を同定し、DNA配列決定により確認した。
【0134】
pED.148Fcを鋳型として用い、下記突然変異オリゴヌクレオチドを用いて部位特異的突然変異法(Maniatis et al., 1989, MolecularCloning:A Laboratory Manual, Cold Spring Harbor Laboratories)を行うことによりプラスミドpED.ΔY148.Fc、pED.H24.Q70.148.Fcを作成した。
ΔY148用:5'-CGGAGACAGGCCACCGAATTCCTGCCAGAAACG-3' (配列番号:24)
H24用:5'-CCTCCAGAAATGCTGAGGCACAGCACTGACACCACTCCTC-3' (配列番号:25)
Q70用:5'-GAGCTGGCCAACATGGGGCAACTGTCCACGGATTCAGCAG-3' (配列番号:26)
【0135】
コロニーハイブリダイゼーション(Maniatis et al.上記)により陽性クローンを同定した。
【0136】
pED.ΔY148.FcをEcoRIで制限消化し、下記の2本鎖オリゴヌクレオチドに連結することによりpED.FFFE.148.Fcを構築した。
5'-AATTCGAGTTCCTAGATTTTG-3' (配列番号:27) および
5'-AATTCAAAATCTAGGAACTCG-3' (配列番号:28).
【0137】
pED.ΔY148.FcをEcoRIおよびNotIで制限消化し、ついで、下記の2本鎖オリゴヌクレオチドに連結することによりシリーズ
pED.FYYD.19.Fc、pED.FFYD.19.FcおよびpED.FFFD.19.Fcの構築物を作成した。
【0138】
pED.FYYD.19.Fc用:
5'-AATTCGAGTACCTAGATTATGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:29) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCATAATCTAGGTACTCG-3' (配列番号:30);
pED.FFYD.19.Fc用:
5'-AATTCGAGTTCCTAGATTATGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:31) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCATAATCTAGGAACTCG-3' (配列番号:32);
pED.FFFD.19.Fc用:
5'-AATTCGAGTTCCTAGATTTCGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:33) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCGAAATCTAGGAACTCG-3' (配列番号:34).
【0139】
B.欠失または変化した形態の可溶性P−セレクチンリガンド蛋白の分析のためのプレート結合アッセイ
種々の変異形態の可溶性PSGL−1/Fcキメラをコードする個々のDNAを、実施例3(C)に記載のごとく、pEA.3/4FTおよびPACE cDNAとともにCOS細胞中に同時トランスフェクションした。約107個のCOS細胞からトランスフェクション40〜64時間後に集めた無血清培地50mlを、2mM CaCl2を捕捉したTBSで平衡化した0.25mlのプロテインAセファロース(Pharmacia)カラムにより精製した。20mlのTBS/CaCl2で洗浄後、0.5mlの0.1M酢酸、0.15M NaCl、2mM CaCl2で結合物質を溶離させた。溶離した物質を1/20倍体積の3M Tris pH9.0で中和した。280nmの吸光度を測定し、PAGE/SDS/Laemmliゲルのクーマシーブルー染色により溶離物質を定量した。
【0140】
硫酸化されていない形態の可溶性PSGL−1を得るために、上記のごとく(トランスフェクション後に細胞を50mMコール酸(Sigma)存在下で培養すること以外は)適当なFcキメラでのCOS細胞のトランスフェクションを行った。
【0141】
実施例4(C)に記載のごとく(下記の変更はあるが)、CHO:P−セレクチン、CHO:E−セレクチンおよびCHO:L−セレクチン発現細胞の定量的な付着を行った:COS細胞および抗体を省略した。そのかわり、種々の量のプロテインAにより精製された可溶性PSGL−1/Fcキメラを用いて48ウェルマイクロタイタープレート(Costar)を4℃で16時間コーティングした。未結合物質を除去し、コーティングしたウェルを、1mg/mlのBSAおよび2mMのCaCl2を含有するHankの緩衝化セイライン(HBS)で4℃において1時間処理した。トリチウム標識したセレクチン発現CHO細胞を添加し、実施例4(C)に記載のごとく結合を定量した。
【0142】
C.N−結合糖鎖付加部位の変化の効果
3種のP−セレクチンリガンド−IgGキメラを発現する構築物を構築して、セレクチン結合に対するN−結合糖鎖付加部位の影響を調べた。これらの構築物は下記配列を有する。
【0143】
148.Fc 配列番号:2のアミノ酸42〜189。
【0144】
Q70.148.Fc 配列番号:2のアミノ酸42〜189、配列番号:2の位置111のアスパラギン残基がグルタミン残基に置き換わっている。
【0145】
H24.Q70.148.Fc 配列番号:2のアミノ酸42〜189、配列番号:2の位置65のアスパラギン残基がヒスチジン残基に、配列番号:2の位置111のアスパラギン残基がグルタミン残基に置き換わっている。
【0146】
これらの構築物を図式的に図6に示す。
【0147】
プロテインAおよびP−セレクチン−IgGキメラへのこれらの構築物の結合を比較した。これらの実験結果を図7に示す。オートラジオグラフのレーン4、5および6を比較すると、可溶性P−セレクチンリガンド蛋白中の最初の2つのN−糖鎖付加部位の1つまたは両方の除去によってはP−セレクチンへの結合に対する有為な影響がないことが示される。
【0148】
D.チロシンの影響
可溶性蛋白のアニオン性領域の変化によるP−セレクチンリガンド蛋白のセレクチンへの結合におけるチロシンの役割を調べるために構築物を作成した。下記構築物を作成した。
【0149】
ΔY148.Fc 配列番号:2のアミノ酸42〜189、アミノ酸46〜52が欠失。
【0150】
FFFE.148.Fc 配列番号:2のアミノ酸42〜189、位置46、48および51のチロシン残基がフェニルアラニン残基に、位置52のアスパラギン酸残基がグルタミン酸残基に置き換わっている。
【0151】
これらの構築物を図8に図式的に示す。
【0152】
標識硫酸存在下で対応する構築物を発現させることにより、P−セレクチンリガンド蛋白の硫酸化の程度および部位を調べた。148.FcおよびΔY.148.Fcの硫酸化の程度を、硫酸化されていないP−セレクチン−IgGキメラの硫酸かの程度と比較した。結果を図9に示す。これらのデータは、硫酸取り込みの大部分がP−セレクチンリガンド蛋白のアニオン性領域中へのものであることを示す。
【0153】
アニオン性領域の硫酸化がチロシン残基において起こるかどうかを調べるためにさらなる構築物作成した。下記構築物を作成した。
【0154】
FYYD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46のチロシン残基がフェニルアラニン残基に置き換わっている。
【0155】
FFYD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46および48のチロシン残基がフェニルアラニン残基に置き換わっている。
【0156】
FFFD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46、48および51のチロシン残基がフェニルアラニン残基に置き換わっている。
【0157】
これらの構築物を次式的に図9に示す。
【0158】
これらの構築物の硫酸化の程度を19.Fc(「YYYD.19.Fc」)と比較した。結果を図10に示す。FTTD.19.Fcは有意な硫酸化を示したが、FFFD.19.Fcは実質的にほとんど硫酸化されなかった。よって、P−セレクチンリガンド蛋白のアニオン性領域のチロシン残基が硫酸化の主要部位である。
【0159】
P−セレクチンリガンド蛋白から硫酸を除去すると、P−セレクチンへの結合が実質的に減少した。コール酸処理された148.FcのP−セレクチンへの結合を試験した。図11に示すように、コール酸処理による硫酸化の阻害により、P−セレクチンへのP−セレクチンリガンド蛋白の結合が減少した。
【0160】
E.C末端欠失の影響
いくつかのさらなるC末端欠失構築物を下記のように作成した。
254.Fc 配列番号:2のアミノ酸42〜295。
47.Fc 配列番号:2のアミノ酸42〜88。
19.Fc 配列番号:2のアミノ酸42〜60。
これらの構築物を図式的に図12に示す。
【0161】
254.Fc、148.Fc、47.Fcおよび19.FcのP−セレクチン、E−セレクチンおよびL−セレクチンへの結合を試験した。図23および24は、これらの欠失キメラのセレクチンへの結合および対照を比較したものである。結果を図12にもまとめた。
【0162】
F.P−セレクチンおよびE−セレクチン発現細胞への結合
実施例4(C)の定量的プレート結合アッセイを用いて、P−セレクチンおよびE−セレクチンを発現する細胞への上記の種々の構築物の結合を比較した(図13において図式的に説明する)。
【0163】
図14は、P−セレクチン発現CHO細胞への148.Fc、ΔY.148.Fc、FFFE.148.FcおよびヒトIgG1の結合を比較する。ΔY.148.Fcのアニオン性領域中のすべてのチロシン残基を欠失させると結合が消滅した。FFFE.148.Fcのチロシン残基をフェニルアラニン残基に置き換えると、148.Fcと比較して実質的に結合が減少した。よって、全長のアニオン性領域の存在はP−セレクチン結合に必須であり、P−セレクチン結合はこの領域の硫酸化により促進されることが示された。図14は、148.Fc、ΔY.148.FcおよびFFFE.148.Fcは、セレクチンを発現しないCHO細胞には結合しないことを示す対照実験を示す。
【0164】
図15は、148.Fc、ΔY.148.Fc、FFFE.148.FcおよびヒトIgG1のE−セレクチン発現細胞への結合を比較する。E−セレクチン結合は、もとの配列の欠失または変化によっては影響されなかった。よって、アニオン性領域はE−セレクチン結合には必要ないことが示された。
【0165】
図16は、図14および15の結果をまとめたものである。
【0166】
図17は、P−およびE−セレクチン発現CHO細胞への47.Fcの結合を比較する。47.Fcは、配列番号:2の位置111および292におけるN−結合糖鎖付加部位の欠失にもかかわらず、両セレクチンへの実質的な結合を示した。
【0167】
図19は、P−セレクチン発現CHO細胞へのFYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fc、148.FcおよびヒトIgG1の結合を比較する。FFFD.19.Fcのアニオン性領域中のすべてのチロシン残基の置換により結合が消滅した。FYYD.19.Fcの位置46のチロシン残基をフェニルアラニン残基に置き換えると、148.Fcと比較して実質的に結合が減少した。H42.Q70.148.FcのN−結合糖鎖付加部位の変更は結合に影響しなかった。よって、P−セレクチン結合はアニオン性領域における硫酸化により促進され、N−結合糖鎖付加はP−セレクチン結合には必要ないことが示された。図19は、FYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fcおよび148.Fcは、ヒトIgG1単独以上にはセレクチンを発現しないCHO細胞に結合しないことを示す。
【0168】
図20は、E−P−セレクチン発現CHO細胞へのFYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fc、148.FcおよびヒトIgG1の結合を比較する。FYYD.19.FcおよびFFFD.19.Fcの程度までリガンド蛋白を末端切断すると、実質的にE−セレクチン結合が減少した。H24.Q70.148.FcにおけるN−糖鎖付加部位の変化は、実質的にはE−セレクチン結合に影響しなかった。よって、配列番号:2のアミノ酸42から60までを含むP−セレクチンリガンド蛋白は選択的にP−セレクチンに結合でき、E−セレクチンには実質的にほとんど結合しないことが示された。
【0169】
図21は、図19および20の結果をまとめたものである。
【0170】
G.P−およびE−セレクチン結合に関する結論
これらのデータから、P−セレクチンリガンド蛋白によるP−セレクチン結合およびE−セレクチン結合の間の関係に関するいくつかの結論が導かれる。N−結合炭水化物はP−またはE−セレクチンへのP−セレクチンリガンド蛋白の結合には必要ない。配列番号:2のアミノ酸42〜60程度の小さいP−セレクチンリガンド蛋白はP−セレクチンへの結合能を有し、E−セレクチンには実質的にほとんど結合しない。
【0171】
図22は、P−およびE−セレクチンへのP−セレクチンリガンド蛋白の結合に関して提案された図式的モデルを示す。O−結合sLex炭水化物は、P−セレクチン結合およびE−セレクチン結合の両方に必要であることが示された。ここに示すデータは、硫酸化チロシン残基はP−セレクチン結合には関与するが、E−セレクチン結合には関与しないことを示す。出願人のデータは、N−結合糖鎖付加結合部位は不要であることも示す。
【実施例11】
【0172】
PSGL−1形態の凝集現象およびダイマー形成に関する試験
部位特異的突然変異法および/またはストップコドンを導入されたプライマーを用いるPCR増幅により一群のPSGL−1変異体を構築した。すべての変異体実験のための鋳型はpPL85.R16(ATCC 75577、出願人により寄託)であった。
【0173】
第1群の変異体(C310SおよびC327S)は、野生型と比較すると1種のアミノ酸変化のみを有する全長のPSGL−1.R16(それぞれ位置310または327においてCysがSerになっている)。pEA.3/4FTおよびへ変異体C3010SまたはC327Sで同時トランスフェクションされたCOS細胞を35S−メチオニンで標識した。細胞溶解物を調製し、変異体蛋白を実施例7(A)のP−セレクチンリガンドポリクローナル抗体で免疫沈降させ、非還元的条件下および還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動により分析した。
【0174】
変異体C327Sならびに野生型PSGL−1.R16は非還元的条件下ではホモダイマーとして移動し、還元的条件下ではモノマーとして移動した。対照的に、変異体C310Sは非還元的条件下および還元的条件下いずれにおいてもモノマーとして移動し、位置310のシステインがPSGL−1のダイマー形成に必要であることが示された。
【0175】
両方の変異体を、P−セレクチンへの結合能についても分析した。同時トランスフェクションCOS細胞の界面活性剤抽出物を、実施例4(A)のLEC−γ1キメラで沈殿させた。沈殿物を非還元的条件下および還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動およびオートラジオグラフィーにより分析した。PSGL−1.R16およびC327Sは両方ともLEC−γ1により効果的に沈殿したが、LEC−γ1へのC310Sの結合はいちじるしく減少し、ダイマー形態のPSGL−1はモノマー形態よりも強固にP−セレクチンに結合することが示された。
【0176】
第2のセットの変異体は可溶性形態のPSGL−1.R16をコードしており、それらを表1に示す。部位特異的突然変異法により変異体ΔTMが得られ、これは膜貫通ドメインの欠失を有し、その後にRLSRKAが続いている。部位特異的突然変異法または所望位置にストップコドンを導入されたPCRプライマーを用いるPCR増幅により変異体L311、L312、A313、I314、L315、A318およびT322を得た。変異体の名称は各末端切断形態のPSGL−1.R16のC末端アミノ酸を示す。
【0177】
下記判断基準に従って変異体を分析した。
1.トランスフェクションCOS細胞からの発現および分泌
2.モノマー形成対ダイマー形成
3.凝集形態の欠如
4.P−セレクチン結合(LEC−γ1キメラ)
【0178】
変異体ΔTMおよびI316は4つの判断基準すべてを満たした。実施例5(A)のsPSL.QC、L311、L312、A313、I314およびL315のごとき短い可溶性形態のPSGL−1はダイマーを形成せず、実施例5(C)のsPSL.T7、A318およびT322のごとき長い可溶性形態のPSGL−1はあまり望ましくない高分子凝集体を形成した。
【0179】
すでに3/4フコシルトランスフェラーゼおよびコア2トランスフェラーゼを発現しているCHO細胞を、psPSL.T7、ΔTM、I316またはpsPSL.QCでトランスフェクションし、メトトレキセートを用いて増幅した。安定なクローンを単離し、35S−メチオニンで標識した。ならし培地を直接分析するか、あるいはまずLEC−γ1で沈殿させ、ついで、非還元的条件下および還元的条件下でSDS−PAGEにより分析した(図25)。結果は、ΔTMおよびI316がダイマー形成およびP−セレクチン結合において最も効果的であることを示した。
【0180】
表I
【表1】
【実施例12】
【0181】
P−およびE−セレクチンへのPSGL−1結合の特異性
材料. ヒトE−セレクチンの細胞外ドメインおよびヒトIgG1のFc部分を含むキメラ蛋白を、先に説明したP−セレクチンキメラ、LEC−γ1と同様にして構築した。バキュロウイルスに感染したトリコプルシア・ニ(Trichoplusia ni)ハイ・ファイブ(high five)細胞(Invitrogen)において可溶性E−セレクチンキメラを発現させ、プロテインAセファロースクロマトグラフィーにより均一に精製した。それぞれα(1,3/1,4)−フコシルトランスフェラーゼ(Fuc−TIII)、PSGL−1、CD43(ロイコシアリン)、および可溶性の対になったアミノ酸変換酵素(PACE)をCOSで発現させるためのプラスミドベクターpEQA.3/4FT、pPL85、pFCD43、およびpEA.sPACEは本明細書ならびに文献においても記載されている(Sako et al. (1993) Cell 75, 1179-1186; Rehemtulla, A. & Kaufman, R. J. (1992) Curr. Opin. Biotechnol. 3, 560-565; Wasley et al. (1993) J. Biol. Chem. 268, 8458-8465)。公表された配列(Natsuka et al. (1994) Journal of Biological Chemistry 269, 16789-16794; Sasaki et al. (1994) J. Biol. Chem. 269, 14730-14737)由来のオリゴヌクレオチドプローブを用いて、Fuc−TVII cDNA(プラスミドpMT.FT7)をHL60 cDNA発現ライブラリーからクローン化した。成熟(PACE開裂)PSGL−1のN末端の最初の15個のアミノ酸を含むペプチドに対してポリクローナル中和ウサギ抗体Rb3443を生成させた。モノクローナル抗CD43抗体(Becton DickinsonまたはBiodesign Internationalから得た)およびイソタイプ対照抗体を、アフィニティー精製されたヤギの抗体(マウスIgGに対するもの)(Cappel, Organon Teknik Corporation)が共有結合しているセファロースTM−4Bからなる固体支持体にカップリングさせた。セレクチンキメラおよびネズミ抗体の、プロテインAセファロース4ファストフロー(Pharmacia)および抗マウスIgG樹脂へのそれぞれのアフィニティーカップリングを、樹脂1mlあたり2mgの蛋白の割合で行った。抗血清Rb3443をプロテインAセファロースに、樹脂1mlに1mgの割合でカップリングさせた。反応後の上清についてのミクロ−BCAアッセイ(Pierce)により示されるカップリング効率は少なくとも95%であった。アプロチニンおよびペプスタチンはBoehringer Mannheimから得て、ベンズアミジン、ロイペプチン、およびフッ化フェニルメチルスルホニル(PMSF)はSigmaから得た。
【0182】
ミエロイド細胞の標識および膜抽出. 浮遊状態で〜1.3x106個/mlまで増殖したU937またはHL60細胞を、10%ウシ胎児血清および2.5mCiの3H−グルコサミンHCl(Dupont/NEN)を補足したRPMI 1640培地50ml中で48時間標識した。この方法により、細胞1個あたり1cpmよりも高い活性が通常的に得られた。標識細胞をPBSで洗浄し、細胞溶解バッファー(10mM MOPS、150mM NaCl、4mM CaCl2および4mM MgCl2,pH7.5、プロテアーゼ阻害剤アプロチニン20mg/ml、ベンズアミジン10mM、ロイペプチン20mg/ml、ペプスタチン8mg/mlおよびPMSF10mMを含有)に懸濁し、氷上で数サイクルのプローブソニケーションに供した。低速遠心分離により核および細胞残渣を除去し、100000g、1時間の遠心分離により細胞膜を上清から回収し、1M NaClを含有する細胞溶解バッファーに懸濁することにより洗浄し、高速遠心分離し、最後に3mlの膜可溶化バッファー(1% Triton-100を含有する細胞溶解バッファー)に再懸濁した。数サイクルのソニケーションおよび氷上でのインキュベーションを行って膜フラクションを可溶化させた。最後に、低速遠心分離工程を行って不溶性膜残渣を除去した。
【0183】
トランスフェクションCOS細胞の標識および膜抽出. 8μgのプラスミドpPL85またはpFCD43および4μgのpES.sPACE、ならびに4μgのpEA.3/4FTまたはpMT.FT7を用い、DEAE−デキストランおよびクロロキン(25)を用いてCOS M6細胞をトランスフェクションした。回収してから40〜45時間後、トランスフェクション細胞を血清およびメチオニンを含まない培地中で30分飢餓状態とし、ついで、無血清DME中[35S]−メチオニンを5時間与えた。標識細胞を洗浄し、EGTAとともにインキュベーションしてディッシュ表面への付着を緩め、ディッシュからかき取り、ペレット化させ、ついで、冷10mM PIPESバッファー,pH7.5(100mM KCl、3mM NaCl、3.5mM MgCl2、およびプロテアーゼ阻害剤(上記参照)を含有)中に再懸濁した。ついで、ソニケーションにより膜抽出を行い、低速遠心分離、高速遠心分離、ついで、標識ミエロイド細胞について上で説明したようにして膜溶解バッファー中で可溶化を行った。
【0184】
アフィニティー沈殿. 膜抽出物を細胞溶解バッファーまたはTBSCバッファー(20mM Tris−HCl、150mM NaCl、2mM CaCl2,pH7.5、5mg/mlのウシ血清アルブミン(純度約99%、Sigma)を補足)で1:4または1:5に希釈した。かくして、Triton X-100を0.2〜0.25%にまで希釈した抽出物を、往復運動により混合しながらヒトIgG1−プロテインAセファロースとともに4℃で一晩インキュベーションした。ついで、前以て清澄化した上清を、E−またはP−セレクチンキメラ、対照ヒトIgG1、Rb3443またはウサギの免疫前の血清、または抗CD43抗体、またはヤギ抗マウスIgGセファロースに前以てカップリングされたイソタイプ対照と前以てカップリングされたプロテインAセファロースと、4℃で6〜12時間反応させた。洗液上清の放射活性がバックグラウンドレベルに低下するまで0.1〜0.5% Triton X-100を含有するバッファー中で5回またはそれ以上樹脂を洗浄した。P−またはE−セレクチン樹脂に特異的に結合した蛋白の溶離を、室温において10mM EDTAまたは5mM EDTA/5mM EGTAを用いて、あるいはSDS−PAGE試料バッファー(Laemmli, U. E. (1970) Nature 227, 680-685)中で煮沸することにより行った。一方、抗体樹脂に結合した蛋白の溶離を後者の方法により徹底的に行った。還元的条件下での分離のために、ジチオスレイトールを試料バッファーに添加し、最終濃度100mMとした。かくして調製した試料を7.5%ゲルを用いるSDS−PAGEにより分離し、En3Hance(Dupont)で処理し、乾燥させ、ついで、オートラジオグラフィーフィルムにさらした。
【0185】
引き続き行うアフィニティー捕捉実験のために、上記のごとく膜抽出物を前以て清澄化させ、P−もしくはE−セレクチンまたはヒトIgG1とともにアフィニティー沈殿させ、洗浄した。ついで、4℃において10mM MOPS、150mM NaCl,pH7.5中5mM EDTAで試料を樹脂から溶離させた(1時間転がしながら撹拌、溶離は2回行った)。最初および2回目の溶離液を一緒にし、ついで、すでに説明したプロトコールに従って固定化Rb3443とともに免疫沈降させた。
【0186】
結果:
対照ヒトIgG1と並行して、可溶性E−およびP−セレクチンキメラを用いて、下記の「方法」において説明するようにして3H−グルコサミン標識U937細胞の界面活性剤可溶化膜抽出物をプローブした。固定化セレクチンからの溶離物についてのSDS−PAGE/オートラジオグラフィーによる試験(図26)により、P−セレクチン溶離物およびE−セレクチン溶離物の両方における主要蛋白種の存在が明らかとなり、その電気泳動特性は下記のごとし。非還元的条件下では分子量200kDaであり、還元により分子量120kDaの種に変換される(図26、それぞれレーン2および3)。場合によっては、P−セレクチン溶離物およびE−セレクチン溶離物の両方においてさらなるバンドが観察され、おそらく、この主要バンドに対応するものであり、自然に還元された物質の存在(非還元試料中の120kDa種)および不完全な還元(還元試料中の200kDa)を反映していた。さらに、DTTでの還元により影響されないE−セレクチン溶離物中の分子量150kDaの痕跡バンドが場合によって観察された。固定化ヒトIgG1を用いた対照実験(図26、レーン1)、あるいはセレクチン樹脂の溶離をEDTAまたはSDSの不存在下で行った実験(データ示さず)においてはバンドは観察されなかった。HL60細胞を用いた場合にも本質的に同一の結果が得られた(データ示さず)。それゆえ、これらの認識の性質は、これらの蛋白と各セレクチンとのレクチンドメインを介する特異的な金属依存性相互作用であると解釈される(Lasky, L. A. (1992) Science 258, 964-969; Drickamer, K. (1988) J. Biol. Chem. 263, 9557)。
【0187】
E−セレクチンとともに沈殿する主要バンドの金属依存性認識および電気泳動上の挙動は、すでに同定されているP−セレクチンカウンターリセプター、P−セレクチン糖蛋白リガンドまたはPSGL−1の特性と矛盾しない(Moore et al. (1994) J. Biol. Chem. 269, 23318-23327; Moore et al. (1992) J. Cell Biol. 118, 445-456; Sako et al.)。
【0188】
この種が実際にPSGL−1であるかどうかを評価するために、E−セレクチン沈殿およびP−セレクチン沈殿両方のEDTA溶離物を、PSGL−1特異的ポリクローナル抗血清Rb3443と反応させた。図27に示すように、いずれのセレクチンとのアフィニティー捕捉により単離された主要バンドも、この抗血清を用いると免疫沈降した(それぞれレーン3および4)。対照IgG1 EDTA溶離物の免疫沈降後に検出された種はなかった(図27、レーン5)。新鮮3H−標識U937膜抽出物を用いる直接免疫沈降によりRb3443の特異性が確認された。Rb3443との沈降によりPSGL−1の電気泳動特性(非還元的条件下で分子量200kDa、還元的条件下で分子量120kDa;図27、レーン2)を有する単一バンドが回収された。一方、免疫前の抗血清との沈降は物質を捕捉しなかった(図27、レーン1)。これらの結果は、E−セレクチンおよびP−セレクチンの両方により特異的に捕捉される、ミエロイド細胞由来の主要蛋白種がPSGL−1であることを示す。
【0189】
PSGL−1に対するE−セレクチンの特異性をさらに評価するために、CD43(ロイコシアリンともいう)(ミエロイド細胞SLex残基の主要部分を有することが知られている豊富な細胞表面シアロ糖蛋白(Maemura, K. & Fukuda, M. (1992) J. Biol. Chem. 267, 24379-24386))の存在について、U937膜溶解物を直接プローブした。かくして、「方法」に記載のごとく、3H−グルコサミン標識U937細胞の膜抽出物を抗CD43抗体でプローブし、並行してPSGL−1特異的抗血清Rb3443および対照抗体でプローブした。同量の膜溶解物から、CD43抗体はPSGL−1抗血清よりも30倍過剰な放射カウントにおいて沈降させた。SDS−PAGE/オートラジオグラフィーによる免疫沈降の評価(図28)により、各抗体に対して1本の特異的バンドが明らかとなった。Rb3443は、PSGL−1の電気泳動特性を有する単一種を捕捉した(図28、レーン2)。対照的に、CD43抗体は、還元に感受性のない分子量120kDaの種を沈降させ(図28、レーン4)、CD43中にシステインが存在しないことと矛盾しない。対照抗体との免疫沈降(図28、レーン1および3)は予想通り陰性であることがわかった。U937細胞中にはPSGL−1よりもかなり多量のCD43が存在すると思われ、HL60細胞中のこれらの蛋白の定量結果と矛盾しない(Ushiyama et al. (1993) J. Biol. Chem. 268, 15229-15237)。よって、E−セレクチンがミエロイド細胞由来のCD43を沈殿させることができないのは、それらの細胞系におけるその不存在によるものではないように思われる。E−セレクチンが痕跡量のCD43を捕捉するという可能性(すなわち、図26の非還元的条件下レーン3における強度の小さい分子量120kDaのバンド(モノマーPSGL−1とも矛盾しない))も排除できないが、PSGL−1はミエロイド膜抽出物から沈殿する主要蛋白であると思われる。
【0190】
PSGL−1 cDNAとα(1,3/1,4)フコシルトランスフェラーゼ(Fuc−TIII)との同時トランスフェクションによりCOS細胞において発現された組み換えPSGL−1は、最もよくP−セレクチンに結合に関した(Sako et al)。興味深いことに、組み換えPSGL−1に対するE−セレクチンの認識を示すための初期の努力は失敗に終わった。P−セレクチン捕捉がうまく行く条件下において、E−セレクチンは同時トランスフェクションされたCOS細胞膜溶解物由来のカウンターリセプターを捕捉できなかった。この結果に対する1の解釈は、Fuc−TIIIは組み換えPSGL−1をP−セレクチンにより認識されるように修飾できたが、E−セレクチン認識に必要なミエロイドPSGL−1においてはそのような適当な修飾をすることができなかったというものである。SLex炭水化物構造を生成することのできるミエロイドのフコシルトランスフェラーゼFuc−TVIIについての最近のクローニング(Natsuka et al (1994) Journal of Biological Chemistry 269, 16789-12764; Sasaki et al (1994) J. Biol. Chem. 269, 14730-14737)は、この解釈を価値あるものとする。
【0191】
PSGL−1またはCD43をコードするcDNAおよびFuc−TIIIまたはFuc−TVIIのいずれかをコードするcDNAでCOS細胞を同時形質転換した。トランスフェクションCOS細胞から膜溶解物を調製し、固定化E−セレクチンキメラまたはP−セレクチンキメラあるいはPSGL−1またはCD43に対する抗体とともにこれらを沈降させた。沈降生成物をSDS−PAGE/オートラジオグラフィーにより評価し、ついで、EDTA/EGTAによるそれらの遊離(セレクチンにより媒介される結合に関して)またはSDS中での煮沸(免疫沈降物に関して)により評価した。結果を図29に示す。
【0192】
図29Aに示すように、COSにより発現されたPSGL−1のE−セレクチン捕捉はトランスフェクションに使用したフコシルトランスフェラーゼの性質に依存した。3つの別個の実験において、Fuc−TVIIはE−セレクチンによるPSGL−1の沈殿を促進したが、Fuc−TIIIは促進しなかった。Fuc−TIIIがPSGL−1に対してE−セレクチンとの反応性を付与しないことは、PSGL−1の発現不足によるものとすることはできない。なぜなら、特異的抗血清Rb3443は、有意なかなりの量のPSGL−1をFuc−TIIIおよびFuc−TVIIトランスフェクション対から免疫沈降させたからである(図29C)。さらにそのうえ、P−セレクチンは、いずれのフコシルトランスフェラーゼとも等量のPSGL−1を沈殿させることができ(図29B)、これらのトランスフェクションにおいてFuc−TIIIおよびFuc−TVIIは発現され、活性を有していたことが示される。
【0193】
COS組み換え発現系において、高アフィニティーE−セレクチン認識もまた、適当なポリペプチドの存在に依存していた。ポリペプチドの長さ、見かけの分子量、および高頻度かつ特異的なタイプの翻訳後修飾はCD43およびPSGL−1において類似しているが(Maemura, K. &Fukuda, M. (1992) J. Biol. Chem. 267, 24379-24386)、いずれのフコシルトランスフェラーゼも、同時トランスフェクションCOS細胞中の組み換えロイコシアリンに対して高アフィニティーE−セレクチン(またはP−セレクチン)認識を付与することはできなかった(図29A)。抗CD43抗体との免疫沈降は、かなりの量のロイコシアリンがFuc−TIII同時トランスフェクション体およびFuc−TVII同時トランスフェクション体の両方において発現されたことを示す(図29C)。E−セレクチンがCD43を捕捉できないことは、同時トランスフェクション細胞中のフコシルトランスフェラーゼ活性の不足によるものではなかった。PSGL−1またはCD43およびFuc−TIIIまたはFuc−TVIIのいずれかでトランスフェクションされたCOS細胞についてのFACS分析はすべて、SLex特異的抗体CSLEX−1との高レベルの反応性を示す。それゆえ、これらの結果から、高アフィニティーE−セレクチン認識には、特異的フコシルトランスフェラーゼにより適当に修飾された特異的ポリペプチドの存在が必要であることが示唆される。
【実施例13】
【0194】
PSGL−1由来のペプチドによるP−セレクチン/PSGL−1結合の阻害
PSGL−1の配列(配列番号:2)由来の多くのペプチドを、P−セレクチン/PSGL−1結合を阻害する能力に関して試験した。試験したペプチドを図30に記載する。
【0195】
以下のプロトコールに従って阻害を試験した。96ウェルプレートのウェルを、4℃において、50μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2(pH7.5)中のPSGL−1でコーティングした。ウェルから液体を除去した後、ウェル1個あたり150μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2、0.05%ゼラチン(pH7.5)を添加して未占領部位をブロックした。1/2時間ないし2時間後、ブロックバッファーをウェルから除去し、Lec−γ1(P−セレクチン−ヒトIgG Fcキメラ)(2μg/ml)、ビオチン化ヤギ抗ヒト抗体、およびストレプトアビジン−抱合アルカリ性ホスファターゼ(30分ないし1時間室温で拡散させた)ならびに潜在的阻害剤からなる混合物100μlをウェル1個あたりにつき添加した。各プレートを振盪し、プレートからブロックを除去するために強くたたいた。暗所で1時間、回転させながらインキュベーションを継続した。150μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2、0.05%ツイン−20で2回洗浄し、ついで、150μlの1M ジエタノールアミン、0.5mM MgCl2で洗浄して未結合混合物をプレートから洗い流した。10mM DEA/0.5mM MgCl2中のアルカリ性ホスファターゼ用発色基質PNPPを添加し、ついで、プレートを405nmにおいて読んだ。
【0196】
これらのアッセイの結果を図30に示す。配列番号:2のアミノ酸48〜51(チロシン残基がリン酸化されている)およびアミノ酸42〜56を含むペプチドは特に望ましい結果を示した。
【実施例14】
【0197】
可溶性形態のPSGL−1の精製
可溶性形態のP−セレクチンリガンド蛋白の実質的な精製を下記プロトコールに従って行った。
【0198】
可溶性P−セレクチンリガンド蛋白I316(配列番号:2のアミノ酸42からアミノ酸316まで)を、本明細書記載のごとくCHO細胞において発現させた。CHO細胞のならし培地を、分子量10000または30000のいずれかのカットオフのPellicon限外濾過膜ユニット(Millipore)で濃縮して、もとの体積の約1/10とした。ついで、バッファーを25mM Tris、1mM CaCl2,pH7.4と交換した。
【0199】
バッファー交換した濃縮物をToyopearl QAE 550C(TosoHaas)カラムに負荷した。別法として、バッファー交換工程を省略し、濃縮物1部を3部の25mM Tris、1mM CaCl2,pH7.4で希釈し、ついで、カラムに負荷することもできる。4℃においてカラムをその5〜10倍体積の25mM Tris、1mM CaCl2,pH7.4で洗浄した。
【0200】
25mM Tris、1mM CaCl2,pH7.4バッファー中の直線的グラジエント(0M NaClから1.0M NaClまで)を用い、カラムの約5倍の体積のところでP−セレクチンリガンド蛋白が溶離した。2つのピークがカラムから溶離した。第2のピークがP−セレクチンリガンド蛋白を含んでおり、バルクとして集めた。
【0201】
QAEカラムからのピークを分子量30000カットオフの十字流限外濾過膜(Millipore)を用いて濃縮し、ついで、バッファーを25mM Tris、150mM NaCl、1mM CaCl2,pH7.4と交換した。
【0202】
バッファー交換された濃縮物を、4℃においてJacalinアガロースカラムに一晩かけて負荷した。カラムを透析濾過バッファーで洗浄し、P−セレクチンリガンド蛋白は、20℃におけるメチルα−D−ガラクトピラノシド(0〜100mMまたは0〜50mM)のグラジエントで溶離した。JacalinカラムからのフラクションをSDS−PAGEにより分析し、最も純度の高いフラクションをプールした。
【実施例15】
【0203】
P−セレクチンリガンド蛋白融合物
異なるアミノ酸配列を有する、4種のP−セレクチンリガンド蛋白融合物を構築した。それらは47.Fc、47.AGP、47.BMPおよび47.IL11であった。
【0204】
47.Fc:シグナルペプチド、PACE開裂部位、およびもとのFc配列のHis224において変異したヒトIgG1のFc領域に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:35に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:36に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:36のアミノ酸42から開始する。Fc部分における変異は、もとのFc配列のLeu234およびGly237がAlaに変化したものである。
【0205】
47.AGP:シグナルペプチド、PACE開裂部位、および成熟ヒトAGPの最初のロイシン残基に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:37に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:38に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:38のアミノ酸42から開始する。
【0206】
47.BMP:シグナルペプチド、PACE開裂部位、および成熟ヒトBMP−2の配列(その最初の8個のアミノ酸が欠失されている)に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:39に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:40に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:40のアミノ酸42から開始する。
【0207】
47.IL11:シグナルペプチド、PACE開裂部位、および成熟ヒトIL−11の配列に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:41に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:42に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:42のアミノ酸42から開始する。
【0208】
本明細書において引用した特許および文献を、本発明に十分に記載されているものとみなす。
【技術分野】
【0001】
本発明は、内皮細胞への白血球の付着を阻害することにより作用する抗炎症物質の分野に関する。より詳細には、本発明は、セレクチンとして知られる哺乳動物付着蛋白の新規リガンドに指向される。
【0002】
本願は、現在放棄されている1992年10月23日出願の米国出願第07/965662号の一部継続出願であった1993年8月26日出願の同時係属出願第08/112608号の一部継続出願であった1994年4月28日出願の同時係属出願第08/235398号の一部継続出願であった1994年9月30日出願の同時係属出願第08/316305号の一部継続出願であった1995年4月25日出願の同時係属出願第08/428734号の一部継続出願である。
【背景技術】
【0003】
炎症の間、白血球は血管内皮に付着し、内皮下組織に侵入するが、これはセレクチンまたはLECCAMクラスの蛋白の標的細胞上のリガンドへの特異的結合により媒介される相互作用である。かかるセレクチンにより媒介される細胞付着は、血栓性疾患および寄生虫性疾患においても起こり、腫瘍細胞の転移性拡張に関与している可能性もある。
【0004】
セレクチン蛋白はN末端レクチン様ドメイン、表皮増殖因子様ドメイン、および補体結合蛋白に相同性を有する領域により特徴づけられる。これまで3種のヒトセレクチン蛋白が同定されており、それらはE−セレクチン(以前はELAM−1と呼ばれた)、L−セレクチン(以前はLAM−1と呼ばれた)およびP−セレクチン(以前はPADGEMまたはGMP−140と呼ばれた)である。
【0005】
E−セレクチンは、サイトカインによる活性化から数時間後に内皮細胞上に誘導され、好中球と内皮との間のカルシウム依存性相互作用を媒介する。L−セレクチンはリンパ球誘導受容体であり、P−セレクチンは、血小板が活性化された場合、血小板細胞表面に急激に出現し、好中球または単球の血小板へのカルシウム依存性付着を媒介する。P−セレクチンは内皮細胞のWeibl-Palade体中にも見出される。P−セレクチンはこれらの小胞から放出されて、ヒスタミンまたはスロンビンにより刺激された内皮への好中球の初期の結合を媒介する。
【0006】
セレクチンは、標的細胞表面上に存在するリガンドとの特異的相互作用により付着を媒介すると考えられている。一般的には、セレクチンのリガンドは、少なくとも一部には炭水化物部分を含む。例えば、E−セレクチンは、末端構造
【化1】
を有する炭水化物に結合し、また、末端構造
【化2】
を有する炭水化物にも結合する。式中、Rは炭水化物鎖の残りの部分である。これらの炭水化物は血液型抗原として知られており、通常には、それぞれシアリル・ルイスxおよびシアリル・ルイスaと呼ばれている。内皮細胞表面上にシアリル・ルイスx抗原のみが存在することは、E−セレクチン発現細胞への結合を促進するには十分であるかもしれない。E−セレクチンは、末端構造
【化3】
を有する炭水化物にも結合する。
【0007】
E−セレクチンと同様、各セレクチンは種々の親和性で広範な炭水化物に結合するように思われる。セレクチンにより媒介される付着の強度(結合親和性)は、細胞表面上の炭水化物密度およびセレクチン密度にも依存する。
【0008】
P−セレクチンは、非シアル化形態のルイスx血液型抗原を含む炭水化物に結合し、シアリル・ルイスxに対しては親和性が高い。また、P−セレクチンは、異種性3−硫酸化ガラクトシルセラミドでありミエロイドおよび腫瘍細胞から脂質抽出により単離されたスルファチド類を認識する可能性がある。しかしながら、P−セレクチン担持細胞のP−セレクチンリガンド担持細胞への結合は、リガンド担持細胞をプロテアーゼ処理した場合には見られず、P−セレクチンリガンドが糖蛋白であることが示される。
【0009】
P−セレクチンに関する2種の推定上の糖蛋白リガンドが最近同定され、その1つは部分精製されている(Moore et al., J. Cell Biol. 118, 445-456 (1992))(非特許文献1)。しかしながら、これらの糖蛋白のアミノ酸組成もアミノ酸配列も開示されていない。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Moore et al., J. Cell Biol. 118, 445-456 (1992)
【発明の概要】
【発明が解決しようとする課題】
【0011】
したがって、本発明の解決すべき課題は、P−セレクチンのリガンドとして作用する蛋白およびDNAを同定および単離することである。加えて、本発明の解決すべき課題は、P−セレクチンリガンド糖蛋白、該蛋白をコードするDNA配列、該DNAで形質転換された宿主細胞、およびP−セレクチンリガンド蛋白の製造方法を提供することである。
【課題を解決するための手段】
【0012】
1の具体例において、本発明は、P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2に示すアミノ酸1からアミノ酸402までのアミノ酸配列を含む。可溶性P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物も提供され、該蛋白は配列番号:2のアミノ酸1からアミノ酸310までを含む。さらに本発明は、成熟P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列を含む。もう1つの具体例において、本発明は、可溶性成熟P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列を含む。もう1つの具体例において、本発明は、P−セレクチンリガンド蛋白をコードする単離DNAを含む組成物を提供し、該蛋白は配列番号:4に示すアミノ酸配列を含む。さらに本発明は、本発明単離DNAのいずれか1つを含む発現ベクターを含む組成物を提供し、該DNAは発現制御配列に作動可能に連結されている。上記DNAのいずれか1つを含む発現ベクターで形質転換された宿主細胞も提供される。さらに、P−セレクチンリガンド蛋白の製造方法も提供され、該方法は:
(a)本発明DNAのいずれか1つを含む発現ベクターで形質転換された宿主細胞を適当な倍地中で培養し、ついで、
(b)P−セレクチンリガンド蛋白を培地から精製する
ことを含む。
【0013】
もう1つの具体例において、本発明は、配列番号:2のアミノ酸21からアミノ酸402までのアミノ酸配列を含む蛋白を含む組成物を提供し、該蛋白は実質的に他の哺乳動物蛋白を含まない。さらに本発明は、配列番号:2のアミノ酸21からアミノ酸310までのアミノ酸配列を含む可溶性P−セレクチンリガンド蛋白を含み、該蛋白は実質的に他の哺乳動物蛋白を含まない。もう1つの具体例において、本発明は、配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列を含むP−セレクチンリガンド蛋白を含み、該蛋白は実質的に他の哺乳動物蛋白を含まない。また本発明は、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列を含む成熟P−セレクチンリガンド蛋白を含む組成物を提供し、該蛋白は実質的に他の哺乳動物蛋白を含まない。さらに、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列を含む可溶性成熟P−セレクチンリガンド蛋白を含む組成物が提供され、該蛋白は実質的に他の哺乳動物蛋白を含まない。もう1つの具体例において、本発明は、配列番号:4に示すアミノ酸配列を含む蛋白を含む組成物を提供する。
【0014】
さらにもう1つの具体例において、本発明は、P−セレクチンリガンド蛋白に対して特異的な抗体を含む組成物を提供する。
【0015】
もう1つの具体例において、本発明は、下記工程:
(a)配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列、および配列番号:4に示すアミノ酸配列からなる群より選択されるアミノ酸配列を含むP−セレクチンリガンド蛋白にP−セレクチンを結合させ(該結合は第1の結合混合物を生じる);
(b)第1の結合混合物中のP−セレクチン蛋白とP−セレクチンリガンド蛋白との間の結合量を測定し;
(c)P−セレクチン蛋白およびP−セレクチンリガンド蛋白に化合物を結合させて第2の結合混合物を得て;
(d)第2の結合混合物中の結合量を測定し;ついで
(e)第1の結合混合物中の結合量を第2の結合混合物中の結合量と比較する
ことを含み、第2の結合混合物の結合量の減少か起こった場合、化合物がP−セレクチンにより媒介される細胞間付着を阻害しうるものである、P−セレクチンにより媒介される細胞間付着の阻害剤の同定方法を提供する。
【0016】
もう1つの具体例において、本発明は、下記工程:
(a)配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列、および配列番号:4に示すアミノ酸配列からなる群より選択されるアミノ酸配列を含むE−セレクチンリガンド蛋白にE−セレクチンを結合させ(該結合は第1の結合混合物を生じる);
(b)第1の結合混合物中のE−セレクチン蛋白とE−セレクチンリガンド蛋白との間の結合量を測定し;
(c)E−セレクチン蛋白およびE−セレクチンリガンド蛋白に化合物を結合させて第2の結合混合物を得て;
(d)第2の結合混合物中の結合量を測定し;ついで
(e)第1の結合混合物中の結合量を第2の結合混合物中の結合量と比較する
を含み、第2の結合混合物の結合量の減少が起こった場合、化合物がE−セレクチンにより媒介される細胞間付着を阻害しうるものである、E−セレクチンにより媒介される細胞間付着の阻害剤の同定方法を提供する。これらの方法を用いて、E−セレクチンをL−セレクチンに置き換えるとL−セレクチン阻害剤を探すこともできる。
【0017】
また本発明は、(a)P−セレクチンリガンド蛋白をコードするDNAおよびシアリル・ルイスX(sLex)またはシアリル・ルイスA(sLea)を合成できるフコシルトランスフェラーゼ((α1,3/α1,4)フコシルトランスフェラーゼまたは(α1,3)フコシルトランスフェラーゼのごときフコシルトランスフェラーゼ)をコードするDNAで宿主細胞を同時形質転換し;(b)適当な培地中で宿主細胞を培養し;ついで(c)P−セレクチンリガンド蛋白を培地から精製することを含む、P−セレクチンリガンド蛋白の製造方法を包含する。他の特定の具体例において、対になった塩基性アミノ酸変換酵素(paired basic amino acid converting enzyme)をコードするDNAおよび/またはGlcNAcトランスフェラーゼ(好ましくは「コア2トランスフェラーゼ」)をコードするDNAで宿主細胞を同時形質転換する。好ましい具体例において、P−セレクチンリガンド蛋白は全長または可溶性形態である。
【0018】
他の具体例において、本発明は、P−セレクチンリガンド蛋白活性を有するP−蛋白リガンド蛋白を包含する。好ましい具体例において、リガンド蛋白は、配列番号:2のアミノ酸42からアミノ酸60までのアミノ酸配列を含む蛋白、配列番号:2のアミノ酸42からアミノ酸60までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸88までのアミノ酸配列を含む蛋白、配列番号:2のアミノ酸42からアミノ酸88までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸118までのアミノ酸配列を必須として含む蛋白、配列番号:2のアミノ酸42からアミノ酸189までのアミノ酸配列を必須として含む蛋白である。他の好ましい具体例において、配列番号:2の位置65、111および292におけるアスパラギン残基の少なくとも1つが欠失または置換される。リガンド蛋白の特定の好ましい具体例は、配列番号:2の位置46、48および51において少なくとも1つのチロシン残基を含む。これらのP−セレクチンリガンド蛋白をコードするDNA、かかるDNAで形質転換された宿主細胞、かかる宿主細胞を培養することによる蛋白の製造方法、蛋白を含む医薬組成物、蛋白を用いるセレクチン結合阻害剤の同定方法、蛋白に対する抗体、ならびに蛋白を用いる、セレクチンにより媒介される結合の阻害方法も本発明により包含される。
【0019】
さらなる他の具体例において、本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のアミノ酸配列、および(b)P−セレクチンリガンド以外の蛋白の配列に由来する第2のアミノ酸配列を含む融合蛋白をコードする単離DNAを提供する。好ましくは、発現制御配列はそれらのヌクレオチド配列に作動可能に連結される。かかるDNAで形質転換された宿主細胞も提供される。また本発明は、(a)融合蛋白の発現に適した条件下で宿主細胞を培養し;ついで(b)融合蛋白を培地から精製することを含む、融合蛋白の製造方法を提供する。かかる方法により製造される融合蛋白も提供される。
【0020】
特定の好ましい具体例において、かかる融合蛋白の第1のアミノ酸配列は、配列番号:2のアミノ酸42からアミノ酸402まで、配列番号:2のアミノ酸42からアミノ酸301まで、配列番号:2のアミノ酸42からアミノ酸88まで、配列番号:2のアミノ酸42からアミノ酸118まで、または配列番号:2のアミノ酸42からアミノ酸189までを含む。
【0021】
他の好ましい具体例において、DNAは、配列番号:35のヌクレオチド123からヌクレオチド939までのヌクレオチド配列、配列番号:35のヌクレオチド配列、配列番号:37のヌクレオチド123からヌクレオチド807までのヌクレオチド配列、配列番号:37のヌクレオチド配列、配列番号:39のヌクレオチド123からヌクレオチド1311までのヌクレオチド配列、配列番号:39のヌクレオチド配列、配列番号:41のヌクレオチド123からヌクレオチド792までのヌクレオチド配列、配列番号:41のヌクレオチド配列を含む。
【0022】
また本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のアミノ酸配列、および(b)P−セレクチンリガンド以外の蛋白の配列に由来する第2のアミノ酸配列を含む融合蛋白を提供する。好ましくは、第1のアミノ酸配列は配列番号:2のアミノ酸42からアミノ酸402まで、配列番号:2のアミノ酸42からアミノ酸310まで、配列番号:2のアミノ酸42からアミノ酸88まで、配列番号:2のアミノ酸42からアミノ酸118まで、配列番号:2のアミノ酸42からアミノ酸189までを含む。
【0023】
ある種の特に好ましい具体例において、融合蛋白は、配列番号:36アミノ酸42からアミノ酸313までのアミノ酸配列、配列番号:36のアミノ酸配列、配列番号:38のアミノ酸42からアミノ酸269までのアミノ酸配列、配列番号:38のアミノ酸配列、配列番号:40のアミノ酸42からアミノ酸437までのアミノ酸配列、配列番号:40のアミノ酸配列、配列番号:42のアミノ酸42からアミノ酸264までのアミノ酸配列、配列番号:42のアミノ酸配列を含む。
【0024】
他の好ましい具体例において、第2のアミノ酸配列は第1のアミノ酸配列のC末端またはN末端に連結され、結合配列により結合されても、されなくてもよい。
【0025】
さらなる他の具体例において、第2のアミノ酸配列は、抗体、サイトカイン、増殖因子、分化因子、ホルモン、酵素、受容体またはそれらのフラグメントならびにリガンドからなる群より選択される蛋白に由来するものである。好ましくは、第2のアミノ酸配列は、抗体配列由来、抗体のFc部分由来であり、あるいはまた抗体由来の配列の変異体である。
【0026】
さらなる具体例において、本発明は、(a)配列番号:2のアミノ酸42からアミノ酸60までを含む第1のペプチド、およびO−セレクチンリガンド以外の蛋白の配列に由来する第2のペプチドを含む組成物であって、第1のペプチドおよび第2のペプチドがペプチド結合以外の部分により化学結合されている組成物を提供する。本発明P−セレクチンリガンド蛋白をかかる組成物に使用することができる。
【図面の簡単な説明】
【0027】
【図1】コア2ありおよびなしで発現されたP−セレクチンリガンド蛋白の結合を比較したグラフである。
【図2】コア2ありおよびなしで発現されたP−セレクチンリガンド蛋白の免疫沈降のオートラジオグラフである。
【図3】P−セレクチンリガンド蛋白(コア2ありおよびなしで発現)のP−セレクチン/IgGキメラ(LEC−γ1)および抗P−セレクチンリガンド蛋白モノクローナル抗体(MAb275)への結合のフローサイトメトリー分析の結果を示す。
【図4】P−およびE−セレクチン/IgGキメラに結合する、P−セレクチンリガンド蛋白を含む蛋白のオートラジオグラフである。
【図5】配列番号:2の全長P−セレクチンリガンド蛋白の構造的特徴を図式的に示したものである。
【図6】P−セレクチンリガンド蛋白のセレクチン類への結合におけるN−結合糖鎖付加部位の役割を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図7】P−セレクチンリガンド蛋白のセレクチン類への結合におけるN−結合糖鎖付加部位の役割を調べるための実験結果を示す。
【図8】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図9】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図10】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図11】P−セレクチンリガンド蛋白のセレクチン類への結合における硫酸化チロシン残基の役割を調べる実験の結果を示す。
【図12】P−セレクチンリガンド蛋白のセレクチン類への結合における種々の欠失の影響を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図13】実施例4(c)の定量的プレート結合アッセイを図式的に示す。
【図14】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図15】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図16】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図17】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図18】P−セレクチンリガンド蛋白のアニオン性領域のセレクチン類への結合におけるチロシン残基の変化の影響を調べる目的で構築されたいくつかのP−セレクチンリガンド蛋白フラグメントを図式的に示す。
【図19】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図20】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図21】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチン類への結合を比較する実験の結果を示す。
【図22】P−セレクチンリガンド蛋白のP−およびE−セレクチンへの結合についての提案されたモデルを示す。
【図23】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチンへの結合を比較する実験の結果を示す。
【図24】種々の欠失および変化したP−セレクチンリガンド蛋白のセレクチンへの結合を比較する実験の結果を示す。
【図25】すでに3/4フコシルトランスフェラーゼおよびコア2トランスフェラーゼを発現しているCHO細胞の発現産物の分析であり、CHO細胞はpsPSL.T7、ΔTM、I316またはpsPSL.QCでトランスフェクションされており、メトトレキセートを用いて増幅された。ならし培地を直接分析するか、あるいはまずLEC−γ1で沈降させ、ついで非還元的条件下および還元的条件下でSDS−PAGEにより分析した。
【図26】P−およびE−セレクチンによりアフィニティー捕捉されたミエロイド細胞膜蛋白のSDS−PAGE分離である。3H−グルコサミンで代謝的に標識したU937細胞から膜溶解物を調製し、固定化P−およびE−セレクチンならびに対照ヒトIgG1とともにアフィニティー沈降させた。ゲル電気泳動前に溶離蛋白をDTT存在下(「還元」)または不存在下(「非還元」)で処理した。レーン:レーン1はヒトIgG1によるアフィニティー捕捉;レーン2はP−セレクチンによるアフィニティー捕捉;レーン3はE−セレクチンによるアフィニティー捕捉。
【図27】連続的アフィニティー捕捉実験である。3H−標識U937溶解物種をP−またはE−セレクチンによりアフィニティー捕捉し、溶離し、ついで、抗PSGL−1抗血清Rb3443との免疫沈降に供した。レーン:1および2は、免疫前のウサギの血清(レーン1)およびRb3443(レーン2)を用いる新鮮ミエロイド細胞膜溶解物の対照免疫沈降である。レーン3〜5は、すでにP−セレクチン(レーン3)、E−セレクチン(レーン4)、およびヒトIgG1(レーン5)によりアフィニティー捕捉され、ついで溶離されたミエロイド細胞膜溶解物のRb3443との免疫沈降である。
【図28】ミエロイド膜抽出物のCD43およびPSGL−1含量の比較である。標識U937細胞抽出物を抗PSGL−1ウサギポリクローナル抗体Rb3443または抗CD43マウスMabとともに免疫沈降させ、ついでSDS−PAGE/オートラジオグラフィーに供した。レーン:1は、対照である免疫前のウサギ血清との免疫沈降である。2はRb3443との免疫沈降である。3は対照イソタイプに合致するマウス抗体との免疫沈降である。4は抗CD43抗体との免疫沈降である。
【図29】COSトランスフェクション実験である。PSGL−2またはCD43をコードするプラスミド、ならびにFuc−TIIIまたはFucTVIIをコードするプラスミドを用いてトランスフェクションしたCOS M6細胞を35S−メチオニンで代謝的に標識し、材料および方法において説明するように、膜をアフィニティー捕捉実験用に調製した。トランスフェクションに使用したcDNAをレーンの上に示す。(A)E−セレクチン、(B)P−セレクチン、および(C)抗PSGL−1抗血清Rb3443および抗CD43 Mabを用いて沈降を行った。
【図30】P−およびE−セレクチン結合の阻害に関する種々のP−セレクチンリガンド蛋白のスクリーニングをまとめたものである(実施例13参照)。
【発明を実施するための形態】
【0028】
上記の図のいくつかには、それらが示す構造中に配列番号:2の番号付けとは異なる番号づけを便利のために使用する。それらの図において、可溶性成熟P−セレクチンリガンドの1番目のアミノ酸を出発点として用いて残基の番号付けを行う。それゆえ、それらの図に使用する残基の番号は配列番号:2のものよりも41少ない。例えば、それらの図中の残基19は配列番号:2の残基60に対応する。
【0029】
本発明者らは、ヒト内皮細胞および血小板上のP−セレクチンのリガンドとして作用する蛋白をコードする新規DNAを初めて同定し、単離した。DNA配列を配列番号:1に示す。P−セレクチンリガンド蛋白の完全なアミノ酸配列(すなわち、成熟ペプチドにリーダー配列が付加したもの)は、配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列により特徴付けられる。疎水性分析および既知開裂パターンとの比較により、20ないし22個のアミノ酸、すなわち配列番号:2のアミノ酸1から20までまたはアミノ酸1から22までのシグナル配列が予想された。P−セレクチンリガンド蛋白は、PACE(対になった塩基性アミノ酸変換酵素)開裂部位(−Arg−Asp−Arg−Arg−)を配列番号:2のアミノ酸38〜41に有する。本発明成熟P−セレクチンリガンド蛋白は配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列により特徴付けられる。可溶性形態のP−セレクチンリガンド蛋白は配列番号:2のアミノ酸21からアミノ酸310を含むことにより特徴付けられる。もう1つの可溶性形態の成熟P−セレクチンリガンド蛋白は配列番号:2のアミノ酸42からアミノ酸310に示すアミノ酸配列により特徴付けられる。可溶性形態のP−セレクチンリガンド蛋白は、さらに室温において水溶液中に可溶性であることにより特徴付けられる。もちろん、これらの蛋白をコードする、配列番号:1に示す対応DNA配列も本発明に包含される。
【0030】
本発明P−セレクチンリガンドは糖蛋白であり、以下の末端炭水化物の1つまたはそれ以上を含む可能性がある:
【化4】
【0031】
式中、Rは炭水化物鎖の残りの部分であり、直接P−セレクチンリガンド蛋白に共有結合しているか、あるいはP−セレクチンリガンド蛋白に共有結合した脂質部分に共有結合している。本発明P−セレクチンリガンド糖蛋白はさらに硫酸化されていてもよく、あるいは翻訳後修飾されていてもよい。COSおよびCHO細胞中で発現される場合、全長のP−セレクチンリガンド蛋白(配列番号:2のアミノ酸1から402まで、または配列番号:2のアミノ酸42から402まで)は、非還元的ADA−ポリアクリルアミドゲル電気泳動によれば220kDaの見かけの分子量を有するホモダイマー蛋白である。
【0032】
全長のP−セレクチンリガンド蛋白の構造を図5に図式的に示す。配列番号:2のP−セレクチンリガンド蛋白の3つの領域は以下のとおりである:細胞外ドメイン(配列番号:2のアミノ酸21から310まで)、膜貫通ドメイン(配列番号:2のアミノ酸311から322まで)、および細胞内、細胞質ドメイン(配列番号:2のアミノ酸333から402まで)。細胞外ドメインは、N−結合糖鎖付加が可能な3つのコンセンサストリペプチド部位(Asn−X−Ser/Thr)を有し、それらはAsn残基65、111、および292から開始する。細胞外ドメインはさらに、チロシン硫酸化可能な3つの部位を残基46、48、および51に有する。残基55〜267からなる領域は高いパーセンテージのプロリンおよびスレオニンを含み、アミノ酸コンセンサス配列Ala−Thr/Met−Glu−Ala−Gln−Thr−Thr−X−Pro/Leu−Ala/Thrからなる10量体の15個のリピートを含む。ここにXはPro、Ala、Gln、Glu、またはArgのいずれであってもよい。これらの領域のごとき領域は高度にO−糖鎖付加された蛋白の特徴を有する。
【0033】
P−セレクチンリガンド蛋白をコードする遺伝子およびフコシルトランスフェラーゼ(以下、FTという)、好ましくは(α1,3/α1,4)フコシルランスフェラーゼ(「3/4FT」という)をコードする遺伝子を用いて同時トランスフェクションされたCOSまたはCHO細胞は、表面上にP−セレクチンを発現するCHO細胞に結合しうるが、表面上にP−セレクチンを発現しないCHO細胞には結合しない。精製形態またはCHO細胞表面上に発現されたセレクチンに結合するためには、P−セレクチンリガンド蛋白をコードする遺伝子をFTをコードする遺伝子とともに同時トランスフェクションしなければならない。なぜなら、一方のみの遺伝子でのトランスフェクションではP−セレクチン結合活性が無いかまたは著しく低下しているからである。本発明P−セレクチンリガンド蛋白のP−セレクチンへの結合は、EDTAにより、あるいはP−セレクチン特異的中和モノクローナル抗体により阻害されうる。本発明P−セレクチンリガンド蛋白のP−セレクチンへの結合は、P−セレクチン特異的非中和モノクローナル抗体により、あるいはイソタイプ対照によっては阻害されない。これらの結果は、本発明P−セレクチンリガンド蛋白の結合特異性を特徴付ける。
【0034】
本発明の目的のために、実施例4のCHO−P−セレクチン結合アッセイにおいて細胞表面に存在するP−セレクチンに、あるいは別の表面に固定されたP−セレクチン、例えば実施例4のペトリ皿に固定化されたキメラP−セレクチン−IgGγ1蛋白にカルシウム依存的に結合する場合、「P−セレクチンリガンド蛋白活性」を有すると定義される蛋白であると定義され、「P−セレクチンリガンド蛋白」または「P−セレクチンリガンド糖蛋白」または単に「P−セレクチンリガンド」と呼ばれる。
【0035】
実施例5(C)に詳述されsPSL.T7と命名されたキメラな可溶性形態のP−セレクチンリガンド蛋白を用いて本発明P−セレクチンリガンド蛋白の糖鎖付加状態を研究した。3/4FTとともに同時トランスフェクションされたCOS細胞から産生されるsPSL.T7蛋白を、実施例6(C)に詳述するように翻訳後糖鎖付加により徹底的に修飾した。かくして、少なくともそのいくつかがシアル化されているN−およびO−結合オリゴ糖鎖が本発明P−セレクチンリガンド蛋白上に存在すると考えられる。
【0036】
本発明P−セレクチンリガンド蛋白はE−セレクチンおよびL−セレクチンにも結合する。sPSL.T7またはP−セレクチンリガンド−Ig融合物をコードするDNAおよび3/4FTをコードするDNAで同時トランスフェクションされたCOS細胞により得られたならし培地をプラスチックマイクロタイタープレートのウェルにコーティングすると、E−セレクチンを発現するCHO細胞がプレートに結合する。しかしながら、E−セレクチンを発現しないCHO細胞はかかるプレートに結合しない。sPSL.T7をコードするDNAおよび3/4FTをコードするDNAで同時トランスフェクションされたCOS細胞によるならし培地でコーティングされたマイクロタイタープレートへのE−セレクチン発現CHO細胞の結合は、EDTAの存在またはE−セレクチン特異的中和抗体の存在下で消滅する。sPSL.T7 DNAのみでトランスフェクションされたCOS細胞によるならし培地は、マイクロタイタープレートのウェルにコーティングされた場合、E−セレクチン発現CHO細胞の結合を引き起こさない。これらの理由により、本発明P−セレクチンリガンド蛋白は、P−セレクチンにより媒介される細胞間付着のほかにも、E−セレクチンにより媒介される細胞間付着の阻害剤として有用であると考えられる。
【0037】
固定化されたP−セレクチンのFcキメラを用いるアフィニティー捕捉により決定されるように、COS細胞により産生された可溶性P−セレクチンリガンド蛋白に対して生成された抗体は、P−セレクチンに特異的に結合する主要HL−60糖蛋白と免疫反応する。U937細胞は類似の免疫反応性糖蛋白リガンドを有する。よって、3H−グルコサミン標識U937細胞の界面活性剤抽出物とともに前以てインキュベーションされた固定化P−セレクチンのEDTAでの溶離により、単一の糖蛋白種が観察される。この主要種は、非還元的条件下のSDS−PAGEによれば220kDの見かけの分子量を示し、還元的条件下では100kDの見かけの分子量を示す。H−60細胞から単離された相当種の場合、U937リガンドは、COS組み換えP−セレクチンリガンド蛋白に対して生成されたポリクローナル抗体と免疫反応する。さらに、固定化されたE−セレクチンのFcキメラを用いる、U937細胞および細胞膜標品からのE−セレクチンリガンドのアフィニティー捕捉により、主要U937 P−セレクチンリガンドと同一の分子量および電気泳動上の挙動を示す単一の主要種が得られる。よって、E−およびP−セレクチンはU937細胞の同じ主要糖蛋白リガンドを認識し、その糖蛋白リガンドは抗P−セレクチンリガンド蛋白抗体と免疫反応し、全長の組み換えP−セレクチンリガンド蛋白と同じ見かけの分子量および電気泳動上の挙動を有する。
【0038】
P−セレクチンと相互作用可能な、あるいはP−セレクチンにより媒介される細胞間付着を阻害しうるP−セレクチンリガンド蛋白のフラグメントも、本発明に包含される。かかるフラグメントは、セリンおよびスレオニン残基の頻度が少ないP−セレクチンリガンド蛋白の領域である配列番号:2のアミノ酸21から54まで;プロリン、セリンおよびスレオニンの頻度が多く、さらにアスパラギン結合糖蛋白のコンセンサス配列(Asn−XSer/Thr)2個を有する配列番号:2のアミノ酸55から127まで;プロリン、セリン、およびスレオニンいずれの頻度も高く、10個のアミノ酸のコンセンサス配列:Ala−(Thr/Met)−Glu−Ala−Gln−Thr−Thr−(Pro/Arg/Gln/Ala/Glu)−(Leu/Pro)−(Ala/Thr)の15個のリピートを含む配列番号:2のアミノ酸128から267までのもう1つの大きなフラグメント(この大きなフラグメント中の小さいフラグメントも、P−セレクチンとの相互作用能を保持し、あるいはP−セレクチンにより媒介される細胞間付着の阻害剤として作用しうる。);アスパラギン結合糖鎖付加のコンセンサスを含み、配列番号:2のアミノ酸268から308までのアミノ酸を含む領域;配列番号:2のアミノ酸309から333までにより示される蛋白の疎水性領域;ならびに配列番号:2のアミノ酸334から402までのP−セレクチンリガンド蛋白の両親媒性領域を含む。さらなるフラグメントは、配列番号:2のアミノ酸43からアミノ酸56まで、配列番号:2のアミノ酸42からアミノ酸60までを含んで射てもよく、アミノ酸46、アミノ酸48、および/またはアミノ酸51において1個またはそれ以上の硫酸化またはリン酸化(Domcheck et al., Biochemistry 31:9865-9870 (1992))チロシンを有していてもよい。P−セレクチンリガンド蛋白のフラグメントは直鎖状であってもよく、あるいは既知方法、例えばH. U. Saragovi, et al., Bio/Technology 10, 773-778 (1992)およびR. S. McDowell, et al., J. Amer.Chem. Soc. 114, 9245-9253 (1992)に記載の方法(それらを参照により本明細書に記載されているものとみなす)を用いて環化されていてもよい。本発明の目的のため、本明細書で「P−セレクチンリガンド蛋白」という場合はすべて、P−セレクチンに結合しうるフラグメントを含むものとする。
【0039】
かかるフラグメントを免疫グロブリンのごときキャリア分子に融合させてP−セレクチンリガンド結合部位の価数を増加させてもよい。例えば、配列番号:2のアミノ酸42からアミノ酸295までまたはアミノ酸42からアミノ酸88のフラグメントのごとき可溶性形態のP−セレクチンリガンド蛋白を、「リンカー」配列を介して免疫グロブリンのFc部分(得られるキメラに所望品質(例えば、長い半減期または免疫原性の低下)を付与するための無処理配列または変異配列)に融合させてもよい。2価形態のP−セレクチンリガンド蛋白に関しては、実施例5(D)および配列番号:6に示すように、かかる融合はIgG分子のFc部分に対するものであってよい。他の免疫グロブリンイソタイプを用いてかかる融合物を得てもよい。例えば、P−セレクチンリガンド蛋白−IgM融合物は10価形態の本発明P−セレクチンリガンド蛋白を生じるであろう。
【0040】
他の蛋白由来のアミノ酸配列への本発明P−セレクチンリガンド蛋白の融合物を構築してもよい。かかる目的に好ましいP−セレクチンリガンド蛋白は、配列番号:2のアミノ酸42からアミノ酸295までのフラグメントまたはアミノ酸42からアミノ酸88までのフラグメントを包含する。望ましい融合蛋白は、例えばサイトカイン、増殖因子および分化因子(骨形態形成蛋白、例えばBMP類))、ホルモン、酵素、受容体成分またはフラグメントならびに他のリガンドのようなP−セレクチンリガンドとは異なる生物学的活性を有する蛋白由来のアミノ酸配列を含んでいてもよい。また、P−セレクチンリガンド蛋白を他の蛋白または薬剤に化学結合させることもできる。かかる使用において、P−セレクチンリガンド蛋白は、セレクチン分子との相互作用能によって、治療効果を増強することにより融合または結合された薬剤の薬剤動力学および/または生体分配を変化させる。例えば、サイトカイン配列へのP−セレクチンリガンド蛋白の融合は、サイトカイン活性を炎症部位に指向することができる。かかる場合、融合蛋白のP−セレクチンリガンド蛋白部分は炎症部位において発現されたセレクチンに結合するであろう。この結合は、融合蛋白のサイトカイン部分を局在化させ、その同族の受容体または近くの細胞表面への結合を可能にするであろう。同様に、他のリガンドをかかる融合蛋白において使用して、対応受容体を発現する細胞をP−セレクチン発現部位に引き寄せることができる。かかる融合物の好ましい例を実施例15において説明する。
【0041】
P−セレクチンリガンド蛋白を含む融合蛋白において、P−セレクチンリガンド以外の蛋白由来のアミノ酸配列をP−セレクチンリガンド由来の配列のN末端またはC末端に連結することができる。連結は直接的(すなわち、いずれの蛋白にも由来しない介在結合配列を含まない)であってもよく、あるいは連結配列を介するものであってもよい。
【0042】
かかる融合蛋白を用いる哺乳動物の治療方法も本発明により企図される。かかる場合、融合蛋白を用いて、P−セレクチンリガンド蛋白が融合している蛋白により効果が発揮される症状を治療する。例えば、IL−11へのP−セレクチンリガンド蛋白の融合物を用いて、セレクチンをその表面に発現する骨髄内皮細胞にIL−11活性を局在化させることができる。局在化されると、融合蛋白のIL−11部分は巨核細胞の前駆細胞を刺激するであろう。同様に、BMPへのPセレクチンリガンド蛋白の融合物を用いて、障害部位における骨または軟骨の形成を刺激することもできる。障害組織はP−セレクチンを発現し、そのことにより融合蛋白が結合する。局在化されると、融合蛋白のBMP部分は障害部位において骨または軟骨の産生を刺激するであろう。
【0043】
下記実施例に詳述するように、本発明P−セレクチンリガンド蛋白を、最初は発現クローニング法(Clarkら、米国特許第4675285号)を用いて得た。ヒトプロミエロサイト細胞系HL−60(S. J. Collins, et al., Nature 270, 347-349 (1977), ATCC番号CCL 240)からcDNAライブラリーを構築した。このライブラリーを3/4FTをコードするDNAとともにCOS細胞中に同時トランスフェクションし、P−セレクチンの細胞外部分およびヒトIgGγ1モノクローナル抗体のFc部分からなるキメラ分子への結合に関してトランスフェクション体をスクリーニングした。キメラP−セレクチンに結合する同時トランスフェクションを、P−セレクチンリガンド蛋白をコードする蛋白に関して豊富化した。このスクリーニング工程を数回繰り返して、P−セレクチンリガンド蛋白をコードするcDNAに関してさらにプラスミド集団豊富化した。第2のクローニング段階において、豊富化したプラスミド集団を、3/4FT遺伝子とともに再度COS細胞中に同時トランスフェクションし、表面上にP−セレクチンを発現する蛍光標識CHO細胞系への結合に関してスクリーニングした。この方法により単一のcDNAクローンを得て、pMT21:PL85と命名した。1992年10月16日にpMT21:PL85プラスミドをAmerican Type Culture Collectionに寄託し、受託番号ATCC 69096を付与された。
【0044】
本発明新規DNAを配列番号:1に示す。本発明DNAは種々の形態のP−セレクチンリガンド蛋白をコードしうる。例えば、1の具体例において、本発明DNAは配列番号:2のアミノ酸1からアミノ酸402までのアミノ酸配列を有する全P−セレクチンリガンド蛋白をコードする。もう1つの具体例において、本発明DNAは配列番号:2のアミノ酸21からアミノ酸402までのアミノ酸配列により特徴づけられる、シグナル配列を欠いたP−セレクチンリガンド蛋白の1形態をコードする。さらなる具体例において、本発明DNAは、配列番号:2のアミノ酸42からアミノ酸402までのアミノ酸配列により特徴づけられる成熟P−セレクチンリガンド蛋白をコードする。本発明DNAのもう1つの具体例は、配列番号:2のアミノ酸1からアミノ酸310までのアミノ酸配列により特徴づけられる可溶性形態のP−セレクチンリガンド蛋白をコードする。また、配列番号:2のアミノ酸42からアミノ酸310までのアミノ酸配列により特徴づけられる可溶性形態の成熟P−セレクチンリガンド蛋白をコードするDNAも本発明の具体例である。さらに、配列番号:2のアミノ酸21からアミノ酸310までのアミノ酸配列により特徴づけられる、シグナル配列を欠いた可溶性形態のP−セレクチンリガンド蛋白をコードするDNAも本発明の具体例である。本発明DNAは他のヒトDNAを含まず、それゆえ、単離DNAとして特徴づけられる。上に詳述したように、P−セレクチンと相互作用するP−セレクチンリガンドフラグメントをコードするDNAも本発明に包含される。
【0045】
P−セレクチンリガンド蛋白cDNAプローブを用いるノーザン分析により、種々のヒト細胞系(HL−60、THP−1、U937)およびヒト単球ならびに多形核白血球においてP−セレクチンリガンド蛋白mRNA転写物の発現が観察された。それらの細胞系すべてにおいて、2.5kbの主要転写物が観察された。HL60およびU937細胞系ならびに多形核白血球において約4kbのマイナーな種が観察された。対照的に、ヒト肝芽腫細胞系HepG2においてP−セレクチンリガンドmRNA発現が検出された。
【0046】
サザンブロット分析によりわかるように、本発明P−セレクチンリガンド蛋白は単一コピー遺伝子によりコードされており、多遺伝子ファミリーの一部ではない。本発明P−セレクチンリガンド蛋白のゲノム形態は、5'非翻訳領域中のヌクレオチド54に局在する約9kbの大きなイントロンを含む。多形核白血球および単球において、本発明P−セレクチンリガンド蛋白は配列番号:3に示すDNA配列によりコードされている。この具体例において、P−セレクチンリガンド蛋白は16個のリピート領域を含む。同様に本発明単離DNAは配列番号:3に示すDNA配列においても具体化され、1993年10月22日にAmerican Type Culture Collectionに寄託され、受託番号ATCC 75577を付与されたプラスミドpPL85R16に含まれている。
【0047】
また本発明は、配列番号:1に示す単離DNAまたは配列番号:3に示す単離DNAの対立遺伝子変種、すなわちP−セレクチンリガンド活性を有する蛋白をコードする配列番号:1または配列番号:3の単離DNAの天然の別形態を包含する。厳密な条件下(例えば、65℃で4xSSC、または42℃で50%ホルムアミドおよび4xSSC)または弛緩した条件下(50℃で4xSSC、または42℃で30〜40%ホルムアミド)において配列番号:1に示すDNAまたは配列番号:3に示すDNAにハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する単離DNAも本発明に包含される。P−セレクチンリガンド蛋白をコードし、遺伝学的コードの縮重により配列番号:1に示す単離DNAまたは配列番号:3に示すDNAとは異なっており、やはりP−セレクチンリガンド蛋白活性を有する単離DNAも本発明に包含される。点突然変異による、あるいはP−セレクチンリガンド活性、半減期または生成レベルを増強する修飾を導入されたことによる、配列番号:1に示すDNAまたは配列番号:3に示すDNAの変種も本発明に包含される。本発明の目的のため、本明細書の「配列番号:1のDNA」は、配列番号:1に示す特定のDNA配列を含むDNAのほかに、配列番号:2の成熟P−セレクチンリガンド蛋白をコードするDNA;P−セレクチンに結合しうる配列番号:2のP−セレクチンリガンド蛋白のフラグメントをコードするDNA;可溶性形態の配列番号:2のP−セレクチンリガンド蛋白をコードするDNA;配列番号:1のDNA配列にハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する蛋白をコードしており、遺伝学的コードの縮重により配列番号:1のDNAとは異なっているDNA;ならびに上記配列番号:1のDNA配列の変種も包含する。同様に、「配列番号:3のDNA」は、配列番号:3に示す特定のDNA配列を含むDNAのほかに、配列番号:4の成熟P−セレクチンリガンド蛋白をコードするDNA;P−セレクチンに結合しうる配列番号:4のP−セレクチンリガンド蛋白のフラグメントをコードするDNA;可溶性形態の配列番号:4のP−セレクチンリガンド蛋白をコードするDNA;配列番号:3のDNA配列にハイブリダイゼーションし、P−セレクチンリガンド蛋白活性を有する蛋白をコードしており、遺伝学的コードの縮重により配列番号:3のDNAとは異なっているDNA;ならびに上記配列番号:3のDNA配列の変種も包含する。
【0048】
P−セレクチンリガンド蛋白の膜貫通および細胞質ドメインをコードする領域が欠失され、さらに/あるいは細胞外ドメインのカルボキシ末端のアミノ酸に対するコドンの3'側にストップコドンが導入された修飾DNAの発現により、可溶性形態のP−セレクチンリガンド蛋白をコードするDNAを調製してもよい。例えば、疎水性分析により、配列番号:2に示すP−セレクチンリガンド蛋白が配列番号:2のアミノ酸311から332までを含む膜貫通ドメインならびに配列番号:2のアミノ酸333から402までを含む細胞質ドメインを有することが予想される。当該分野で知られた特異的突然変異法を包含する標準的な分子生物学的手法により、あるいは適当なオリゴヌクレオチドプライマーを用いるポリメラーゼ連鎖反応により上記修飾DNAを得てもよい。種々の可溶性P−セレクチンリガンド蛋白をコードするいくつかのDNAを得る方法を実施例5に示す。
【0049】
全長配列の一部が欠失または変化している修飾DNAの発現により、他のフラグメントおよび別形態のP−セレクチンリガンド蛋白をコードするDNAを調製してもよい。P−セレクチンリガンド活性を保持させつつ、P−セレクチンリガンド蛋白配列の実質的な欠失を行うことができる。例えば、配列番号:2のアミノ酸42からアミノ酸189までの配列、配列番号:2のアミノ酸42からアミノ酸118までの配列、または配列番号:2のアミノ酸42からアミノ酸89までの配列を含むP−セレクチンリガンド蛋白はそれぞれP−セレクチン蛋白結合活性およびE−セレクチン結合能を有する。1個またはそれ以上のN−結合糖鎖付加部位(例えば、配列番号:2のアミノ酸65、111および292のごとき部位)が他のアミノ酸に変化させられ、あるいは欠失されたP−セレクチンリガンド蛋白もやはりP−セレクチン蛋白結合活性およびE−セレクチン結合能を保持している。配列番号:2のアミノ酸42からアミノ酸60までを含むP−セレクチンリガンド蛋白(配列番号:2のアミノ酸45からアミノ酸58までの非常にアニオン性の領域を含む)もまたP−セレクチンリガンド蛋白活性を保持している。しかしながら、かかる配列に限定されたP−セレクチンリガンド蛋白はE−セレクチンに結合しない。好ましくは、P−セレクチンリガンド蛋白は、配列番号:2のアミノ酸46、48および51に見出されるチロシン残基のうち少なくとも1個(より好ましくは少なくとも2個、最も好ましくは3個すべて)を保持しているものであり、それらの残基の硫酸化はP−セレクチンリガンド蛋白活性に貢献しうる。これらの活性フラグメントおよび他の活性フラグメントまたは別形態のP−セレクチンリガンド蛋白をコードするDNAの構築を、当業者に知られた方法に従って行ってもよい。
【0050】
本発明単離DNAを、Kaufman et al., Nucleic Acids Res. 19, 4485-4490 (1991)に開示されたpMT2またはpED発現ベクターのごとき発現制御配列に作動可能に連結して、P−セレクチンリガンドを組み換え的に製造してもよい。多くの適当な発現制御配列が当該分野において知られている。組み換え蛋白発現の一般的方法も知られており、R. Kaufman, Methods in Enzymology 185, 537-566 (1990)に説明がある。本明細書の「作動可能に連結」は、酵素的または化学的な連結により本発明単離DNAおよび発現制御配列間に共有結合が形成され、その結果、連結されたDNA/発現制御配列を用いて形質転換(トランスフェクション)された宿主細胞によりP−セレクチンリガンド蛋白が発現されることを意味する。
【0051】
対になったアミノ酸配列(例えば、−Lys−Arg−および−Arg−Arg−)のカルボキシル側で前駆体ペプチドを開裂して成熟蛋白を生じる、いくつかのエンド蛋白分解酵素が知られている。一般的には、かかる酵素は、対になった塩基性アミノ酸変換酵素またはPACEとして知られ、成熟蛋白の組み換え製造方法におけるそれらの使用はWO92/09698および米国特許出願第07/885972号に十分に開示されており、参照によりそれらを本明細書に記載されているものとみなす。PACEファミリーの酵素は、組み換え宿主細胞における前駆体ポリペプチドの蛋白分解的プロセッシングの効率を高めることが知られている。上記のごとく、本発明P−セレクチンリガンド蛋白はかかる
PACE開裂部位を含んでいる。
【0052】
本明細書記載の可溶性P−セレクチンリガンド蛋白をコードするDNA配列およびWO92/09698および米国特許出願第07/885972号(参照によりそれらを本明細書に記載されているものとみなす)に記載のPACEをコードするDNA配列または配列番号:5のDNA配列を含む宿主細胞により、本発明の可溶性成熟P−セレクチンリガンド蛋白を製造してもよい。かかる宿主細胞は、可溶性P−セレクチンリガンド蛋白DNAおよびPACE DNAをそれぞれ含む別個の発現ベクターでの同時形質転換または逐次形質転換の結果としてのDNAを含んでいてもよい。3/4FTをコードする第3のDNAを、P−セレクチンリガンド蛋白およびPACEをコードするDNAとともに同時形質転換してもよい。別法として、宿主細胞は、可溶性P−セレクチンリガンド蛋白DNAおよびPACE DNAの両方を含む単一の発現ベクターでの形質転換の結果としてのDNAを含んでいてもよい。かかる発現ベクターの構築は分子生物学の当業者のレベル内である。同時形質転換および形質転換の方法も知られている。
【0053】
PACEをコードする多くのDNA配列が知られている。例えば、フリン(furin)として知られる1の形態のPACEをコードするDNAがA. M. W. van den Ouweland et al., Nucl. Acids Res. 18, 664 (1990)に開示されており、参照により本明細書に記載されているものとみなす。PACESOLとして知られる可溶性形態のPACEをコードするcDNAを配列番号:5に示す。他の形態のPACEをコードするDNAも存在し、PACEがアミノ酸38〜41においてP−セレクチンリガンド蛋白を開裂しうるかぎり、かかるPACEをコードするDNAを用いて本発明の可溶性成熟P−セレクチンリガンド蛋白を得てもよい。好ましくは、可溶性形態のPACEをコードするDNAを用いて本発明の可溶性成熟P−セレクチンリガンド蛋白を得る。
【0054】
可溶性形態のP−セレクチンリガンド蛋白およびPACEをそれぞれコードするDNAを、別個にあるいは一緒にして、上記pMT2またはpED発現ベクター中に含まれるような発現制御配列に作動可能に連結して、PACEにより開裂された可溶性P−セレクチンリガンドを組み換え的に製造してもよい。さらなる適当な発現制御配列が当該分野において知られている。下記実施例3(C)および3(D)は本発明の可溶性成熟P−セレクチンリガンド蛋白の製造方法を説明する。
【0055】
多くのタイプの細胞がP−セレクチンリガンド蛋白発現のための適当な宿主細胞として作動しうる。適当な宿主細胞は、P−セレクチンリガンド蛋白に特徴的な炭水化物側鎖を結合しうるものである。かかる能力は、天然に存在するかどうか、化学的突然変異により誘導されたかどうか、あるいは糖鎖付加酵素をコードするDNA配列を含む適当な発現プラスミドで宿主細胞がトランスフェクションされたかどうかに関係なく、宿主細胞中の適当な糖鎖付加酵素の存在により生じるものである。例えば、宿主細胞は、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト腎臓293細胞、ヒト上皮A431細胞、Colo 205細胞、3T3細胞、CV−1細胞、他の形質転換された霊長類細胞系、正常2倍体細胞、1次組織、1次移植物のインビトロ培養から得られる細胞株、HeLa細胞、マウスL細胞、BHK、HL−60、U937、またはHak細胞を包含する。
【0056】
本発明単離DNAおよび適当な糖鎖付加酵素をコードする1またはそれ以上のDNAを、1またはそれ以上の昆虫発現ベクター中の適当な制御配列に作動可能に連結し、昆虫発現系を用いることにより蛋白を得てもよい。バキュロウイルス/昆虫細胞発現系のための材料および方法は、例えばInvitrogen, San Diego, California, USAからのキット形態(MaxBacRキット)で市販されており、かかる方法は当該分野においてよく知られており、SummersおよびSmith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987)(参照により本明細書に記載されているものとみなす)に記載のようなものがある。上記の適当な単離DNAを用いて、昆虫細胞において可溶性形態のP−セレクチンリガンド蛋白を得てもよい。1形態のPACEをコードするDNAを昆虫細胞中でさらに同時発現させて、PACEにより開裂された形態のP−セレクチンリガンド蛋白を得てもよい。
【0057】
別法として、酵母のごとき下等真核細胞または細菌のごとき原核細胞においてP−セレクチンリガンド蛋白を製造することも可能である。潜在的に適当な酵母株は、Saccharomyces cerevisiae、Schizosaccharomyces pombe、Kluyveromyces株、Candida、または異種蛋白の発現能を有する酵母株を包含する。潜在的に適当な細菌株は、Escherichia coli、Bacillus subtilis、Salmonella typhimurium、または異種蛋白の発現能を有する細菌株を包含する。P−セレクチンリガンド蛋白が酵母または細菌において生成される場合、蛋白の適当な部分に適当な炭水化物を共有結合させる必要がある。既知化学的または酵素的方法を用いてかかる共有結合を行ってもよい。
【0058】
本発明P−セレクチンリガンド蛋白をトランスジェニック動物の産生物として、例えばP−セレクチンリガンド蛋白をコードするDNA配列を含む体細胞または生殖細胞により特徴づけられるトランスジェニックなウシ、ヤギ、ブタ、またはヒツジの乳の成分として発現させてもよい。
【0059】
GlcNacトランスフェラーゼ、好ましくはコア2トランスフェラーゼとしても知られるUDP−GlcNac:Gal β1−3GalNac−R(GlcNac to GalNAc)β1−6GlcNAcトランスフェラーゼ(EC 2.4.1.102)での宿主細胞の形質転換により、P−セレクチンリガンド蛋白のP−セレクチン結合活性を増大させてもよい。
【0060】
P−セレクチンリガンド蛋白上に存在するO−結合グリカンはP−セレクチンへの結合に重要であることが示されている(D. Sako et al., Cell 75, 1175-1186 (1993))。ミエロイド細胞のO−結合グリカン上のシアリルLexが複合分枝構造上に存在することが報告されている(Maemura, K. and Fukuda, M., J. Biol. Chem. 267, 24379-24386 (1992))。かかるオリゴサッカライド構造の生成に関与する酵素が「コア2」である。コア2酵素活性はCOS細胞中に非常に低レベルで見出され、CHO細胞中には痕跡レベルで見出される。P−セレクチンリガンド蛋白、(α1,3/α1,4)フコシルトランスフェラーゼおよびコア2をコードするDNAで同時形質転換された宿主細胞は、P−セレクチンに対する結合が20〜30倍増強されたP−セレクチンリガンド蛋白を生じうる。
【0061】
ある種の好ましい具体例において、可溶性P−セレクチンリガンド蛋白、3/4FT、コア2およびPACEをコードするDNAで同時形質転換した宿主細胞によりP−セレクチンリガンド蛋白を製造する。
【0062】
P−セレクチン結合糖蛋白の発現に適した培養条件下で形質転換宿主細胞を培養することにより本発明P−セレクチンリガンド蛋白を製造してもよい。次いで、得られた発現糖蛋白を、ゲル濾過およびイオン交換クロマトグラフィーのごとき既知精製プロセスを用いて培地または細胞抽出物から精製してもよい。可溶性形態の本発明P−セレクチンリガンド蛋白を、レンズマメレクチン−セファロース(Sepharose)(登録商標)によるクロマトグラフィー、ついで0.5M α−メチルーマンノースでの溶離により生成することができる。ついで、溶離した可溶性P−セレクチンリガンド蛋白を0〜70%硫酸アンモニウム工程によりさらに精製し、濃縮してもよい。ついで、蛋白を回収し、再懸濁し、TSK G4000SWXLによるサイズ排除クロマトグラフィーによりさらに精製する。別法として、発現細胞から全膜フラクションを調製し、Triton X-100のごとき非イオン性界面活性剤で膜を抽出することにより本発明の全長のP−セレクチンリガンド蛋白を精製することができる。ついで、界面活性剤抽出物を、固定化P−セレクチンを含むアフィニティーカラムに通し、0.1%界面活性剤を含有するバッファー中の10mM EDTAを用いてP−セレクチンリガンド蛋白をカラムから溶離することができる。ついで、アフィニティーカラムから溶離した物質を透析してEDTAを除去し、レンズマメ-Sepharose(登録商標)アフィニティーカラムで精製し、再度.05M α−メチル−マンノシドで溶離する。
【0063】
別法として、市販蛋白濃縮フィルター、例えばAmiconまたはMillipore Pellicon限外濾過ユニットを用いて本発明P−セレクチンリガンド蛋白を濃縮してもよい。濃縮工程後、濃縮物をゲル濾過媒体のごとき精製マトリックスに負荷する。別法として、アニオン交換樹脂、例えば懸垂ジエチルアミノエチル(DEAE)基を有するマトリックスまたは基材を用いることができる。マトリックスはアクリルアミド、アガロース、デキストラン、セルロースまたは蛋白精製に通常使用される他のタイプのものがある。別法として、カチオン交換工程を用いることができる。適当なカチオン交換体は、スルホプロピルまたはカルボキシメチル基を含む種々の不溶性マトリックスを包含する。スルホプロピル基が好ましい(例えば、S-Sepharose(登録商標)カラム)。コンカナバリンA−アガロース、ヘパリン−トヨパール(toyopearl)(登録商標)またはCibacrom blue 3GA Sepharose(登録商標)のごときアフィニティー樹脂による、あるいはフェニルエーテル、ブルエーテル、またはプロピルエーテルのごとき樹脂を用いる疎水性相互作用クロマトグラフィーによる、あるいは免疫アフィニティークロマトグラフィーによる1またはそれ以上のカラム工程により、培養上清からP−セレクチンリガンド蛋白を精製してもよい。
【0064】
最後に、疎水性RP−HPLC媒体、例えば懸垂メチルまたは他の脂肪族基を有するシリカゲルを用いる1またはそれ以上の逆相高品質液体クロマトグラフィー(RP−PLC)工程を用いてさらにP−セレクチンリガンド蛋白を精製することができる。いくつかまたはすべての上記精製工程を種々組み合わせて用いて実質的に均一な単離組み換え蛋白を得ることもできる。かくして精製されたP−セレクチンリガンド蛋白は他の哺乳動物蛋白を実質的に含まず、本発明により「単離蛋白」と定義される。
【0065】
単離P−セレクチンリガンド蛋白は、P−、E−またはL−セレクチンにより媒介される細胞間付着により特徴づけられる症状の治療において有用でありうる。かかる症状は、心筋梗塞、細菌もしくはウイルス感染、転移性の症状、関節炎のごとき炎症性疾患、痛風、葡萄膜炎、急性呼吸困難症候群、喘息、気腫、晩発型高血圧反応、全身性紅斑狼瘡、火傷もしくはしもやけのごとき熱的傷害、自己免疫性甲状腺炎、実験的なアレルギー性脳脊髄炎、多発性硬化症、外傷後の多発性器官傷害症候群、糖尿病、Reynaud症候群、好中球性皮膚病(Sweet症候群)、炎症性腸症候群、Grave病、糸球体腎炎、歯肉炎、歯周炎、溶血性の尿毒性症候群、潰瘍性大腸炎、Crohn病、壊死性全腸炎、顆粒球輸血関連症候群、サイトカインにより誘導される毒性等を包含する(これらに限らない)。単離P−セレクチンリガンド蛋白は器官移植にも有用で、移植用に器官を調製すること、および器官移植拒絶反応を抑制することの両方において有用である。したがって、器官切除前にP−セレクチンリガンド蛋白を生きた器官ドナーまたは生きていない器官ドナーに投与してもよい。さらに、P−セレクチンリガンド蛋白をエクスビボで器官保存溶液中のドナーの器官に投与してもよく、レシピエントに融着前、および/またはレシピエントに融着後に投与してもよい。単離P−セレクチンリガンド蛋白を用いて血液透析およびロイコフォレシスの患者を治療してもよい。さらに、単離P−セレクチンリガンド蛋白を抗転移剤として使用してもよい。単離P−セレクチンリガンド蛋白をそれ自体、P−、E−またはL−セレクチンにより媒介される細胞間付着の阻害剤として用いてもよく、あるいはP−、E−またはL−セレクチンにより媒介される細胞間付着の阻害剤を設計するために使用してもよい。本発明は、単離P−セレクチンリガンド蛋白を含有する医薬組成物ならびに単離P−セレクチンリガンド蛋白を用いる治療方法または単離P−セレクチンリガンド蛋白の使用方法を包含する。
【0066】
細胞から精製された、または組み換え的に製造された単離P−セレクチンリガンド蛋白を医薬上許容される担体と混合して医薬組成物中に用いてもよい。かかる組成物は、P−セレクチンリガンド蛋白および担体、希釈剤、充填剤、塩類、バッファー、安定化剤、可溶化剤、および当該分野でよく知られた他の物質を含有していてもよい。用語「医薬上許容される」は、有効成分の生物学的活性の有効性を妨害しない無毒の物質を意味する。担体の特性は投与経路に依存する。本発明医薬組成物は、M−CSF、GM−CSF、TNF、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−13、IL−14、IL−15、IFN、TNF0、TNF1、TNF2、G−CSF、Meg−CSF、スロンボポイエチン、幹細胞因子、およびエリスロポイエチンのごときサイトカイン、リンホカイン、または他の造血因子を含有していてもよい。医薬組成物はさらに、プラスミノーゲンアクチベーターおよび因子VIIIのごとき血栓溶解因子または抗血栓因子を含有していてもよい。医薬組成物はさらに他の抗炎症剤を含有していてもよい。かかるさらなる因子および/または作用剤を医薬組成物に含有させて、単離P−セレクチンリガンド蛋白との相乗効果を発揮させ、あるいは単離P−セレクチンリガンド蛋白により引き起こされる副作用を最小化してもよい。逆に、特定のサイトカイン、リンホカイン、他の造血因子、血栓溶解因子または抗血栓因子、または抗炎症剤の処方に単離P−セレクチンリガンド蛋白を含有させて、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子または抗血栓因子、または抗炎症剤の副作用を最小化してもよい。
【0067】
本発明医薬組成物はリポソーム形態であってもよい。リポソーム形態においては、単離P−セレクチンリガンド蛋白は、水溶液中のミセル、不溶性単層、液状結晶、またはラメラ層として凝集形態となって存在する脂質のごとき両親媒性物質と混合されて、他の医薬上許容される担体とともに存在している。リポソーム処方に適する脂質は、モノグリセリド、ジグリセリド、スルファチド、リゾレシチン、リン脂質、サポニン、胆汁酸等を包含するが、これらに限らない。かかるリポソーム処方の調製は当業者のレベル内であり、例えば、米国特許第4235871号、第4501728号、第4837028号および第4737323号に開示されており、参照によりこれらを本明細書に記載されているものとみなす。
【0068】
本明細書の用語「治療上有効量」は、医薬組成物または方法の各有効成分の合計量が有意な患者の利益、すなわちP−セレクチンまたはE−セレクチンにより媒介される細胞付着の治療、治癒、予防または改善、あるいはかかる症状の治療、治癒、予防または改善の速度の増加を示すに十分であることを意味する。単独投与される個々の有効成分について用いる場合、該用語は当該成分のみに関するものである。組み合わせた成分について用いる場合、該用語は、混合して、逐次または同時に投与するかどうかにかかわらず、治療効果を生じる有効成分の合計量に関するものである。
【0069】
本発明治療方法または使用方法の実施に際して、治療上有効量の単離P−セレクチンリガンド蛋白を、P−セレクチンにより媒介される疾病状態を有する哺乳動物に投与する。本発明方法に従って単離P−セレクチンリガンド蛋白を単独で、あるいは受容体アンタゴニスト、リガンドアンタゴニスト、サイトカイン、リンホカインまたは他の造血因子を用いる治療のごとき他の治療とともに投与することができる。1またはそれ以上のサイトカイン、リンホカインまたは他の造血因子とともに共投与する場合、単離P−セレクチンリガンド蛋白をサイトカイン、リンホカインまたは他の造血因子、血栓溶解因子または抗血栓因子と同時に、あるいは逐次投与することができる。逐次投与の場合、担当医は他の抗炎症剤、サイトカイン、リンホカインまたは他の造血因子、血栓溶解因子または抗血栓因子と単離P−セレクチンリガンド蛋白との適当な順序を決定するであろう。
【0070】
医薬組成物中に使用される、あるいは本発明方法の実施に用いられる単離P−セレクチンリガンド蛋白の投与を、摂食、吸入、または皮膚、皮下、または静脈注射のごとき種々の慣用的経路で行うことができる。
【0071】
治療上有効量の単離P−セレクチンリガンド蛋白を経口投与する場合、単離P−セレクチンリガンド蛋白は錠剤、カプセル、粉末、溶液またはエリキシルの形態であろう。錠剤形態で投与する場合、本発明医薬組成物はさらにゼラチンのごとき固体担体またはアジュバントを含有してもよい。錠剤、カプセル、および粉末は約5ないし95%、好ましくは約25%ないし90%の単離P−セレクチンリガンド蛋白を含有する。液体形態で投与する場合、水、鉱油、ピーナッツ油、ミネラルオイル、大豆油もしくはゴマ油のごとき動物もしくは植物の油脂、または合成油脂のごとき液体担体を添加してもよい。液体形態の医薬組成物はさらに、生理学的セイライン溶液、ブドウ糖もしくは他の糖溶液、またはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールのごときグリコール類を含有してもよい。液体形態で投与する場合、医薬組成物は約0.5ないし90重量%、好ましくは約1ないし50重量%の単離P−セレクチンリガンド蛋白を含む。
【0072】
治療上有効量の単離P−セレクチンリガンド蛋白を静脈、皮膚または皮下注射により投与する場合、単離P−セレクチンリガンド蛋白はパイロジェン不含の非経口的に許容される水溶液の形態であろう。かかる非経口的に許容される蛋白溶液の調製は、pH、等張性、安定性等に関して当該分野の技術の範囲内である。静脈、皮膚、または皮下注射に好ましい医薬組成物は、本発明化合物のほかに、注射用塩化ナトリウム、注射用リンゲル溶液、注射用ブドウ糖および塩化ナトリウム、注射用乳酸加リンゲル溶液のごとき等張性の担体、または当該分野で知られた他の担体を含有すべきである。本発明医薬組成物は安定化剤、保存料、バッファー、抗酸化剤、または当業者に知られた他の添加物を含有していてもよい。
【0073】
本発明医薬組成物中の単離P−セレクチンリガンド蛋白の量は、治療すべき症状の性質および重さ、ならびに患者の治療前の性質に依存するであろう。最終的には、担当医が個々の患者の治療に用いる単離P−セレクチンリガンド蛋白の量を決定するであろう。最初に、担当医は単離P−セレクチンリガンド蛋白を低用量で投与して患者の応答を観察する。患者に関して最適治療効果が得られるまで、より高用量の単離P−セレクチンリガンド蛋白を投与し、最適効果が得られた時点で用量をもはや増加させない。本発明方法の実施に用いる種々の医薬組成物は、体重1kgあたり約0.1μgないし約100mgの単離P−セレクチンリガンド蛋白を含有すべきである。
【0074】
本発明医薬組成物を用いた静脈注射による治療期間は、治療すべき疾病の重さおよび個々の患者それぞれの潜在的応答に依存するであろう。単離P−セレクチンリガンド蛋白の1回の適用期間は連続静脈投与の場合12ないし24時間の範囲であろう。最終的に、担当医が、本発明医薬組成物を用いた静脈注射による適当な治療期間を決定するであろう。
【0075】
単離P−セレクチンリガンド蛋白を用いて動物を免役して、P−セレクチンリガンド蛋白と特異的に反応し、P−セレクチンにより媒介される細胞付着を阻害するポリクローナルおよびモノクローナル抗体を得てもよい。P−セレクチンリガンド蛋白全体または可溶性成熟P−セレクチンリガンド蛋白のごときそのフラグメントを免疫原として用いてかかる抗体を得てもよい。P−セレクチンリガンド蛋白の小型のフラグメント、例えば下記フラグメントを用いて動物を免疫してもよい。小型フラグメントとしては、例えば、配列番号:2のアミノ酸42からアミノ酸56まで、配列番号:2のアミノ酸127からアミノ酸138までがある。さらなるペプチド免疫原は配列番号:2のアミノ酸238からアミノ酸248までを含み、ペプチドのアミノ末端にアラニン残基が付加されたものである。もう1つのペプチド免疫原は、配列番号:2のアミノ酸43からアミノ酸56までを含み、位置46、48または51のいずれかまたはすべてにおいて硫酸化チロシンを有するものである。ペプチド免疫源はカルボキシ末端にシステイン残基をさらに含んでいてもよく、キーホールリムペット・ヘモシアニン(KLH)のごときハプテンに結合する。かかるペプチドを合成する方法は当該分野において知られており、例えば、R.P.Merrifield,J.Amer.Chem.Soc.85,2149-2154 (1963);J.L.Krstenansky,et.al.,FEBS Lett.211,10(1987)に記載のごときものがある。
【0076】
P−セレクチンリガンド糖蛋白またはP−セレクチンリガンド糖蛋白の特徴である複合型炭水化物部分に結合するモノクローナル抗体は、炎症性疾患およびある種の形態の癌の免疫検出用診断薬でありうる。小胞肺癌腫のごときいくつかの癌細胞は検出可能なレベルのP−セレクチンリガンド蛋白を発現しうる。癌細胞によるこのP−セレクチンリガンド蛋白の異常な発現はこれらの細胞の転移において役割を果たしているかもしれない。
【0077】
P−セレクチンリガンド糖蛋白またはP−セレクチンリガンド糖蛋白の特徴である複合型炭水化物部分に結合する中和抗体は、P−セレクチンリガンド蛋白の異常発現が関与している炎症性疾患およびある種の形態の癌の治療薬として有用でありうる。これらの中和モノクローナル抗体は、P−セレクチンリガンド蛋白のセレクチンにより媒介される細胞間付着機能をブロックしうる。P−セレクチンリガンド蛋白の結合をブロックすることにより、不適切な炎症部位への白血球の付着が消滅または著しく減少させられる。癌細胞または白血病性細胞の場合、P−セレクチンリガンド蛋白に対する中和モノクローナル抗体は、P−セレクチンリガンド蛋白により媒介されうる癌細胞の転移による蔓延の検出および防止において有用でありうる。さらに、これらの細胞に結合するモノクローナル抗体は、抗体依存性の細胞により媒介される細胞毒性(ADCC)に関する癌細胞を標的とする可能性があり、よって、癌細胞の除去を助ける。P−セレクチンリガンド蛋白と反応するヒト抗体を、ヒト免疫グロブリンをコードする遺伝を生殖細胞系中に含むトランスジェニック動物において産生させてもよい。下記実施例7は、P−セレクチンリガンド蛋白フラグメントに特異的なウサギポリクローナル抗体の製造を説明する。
【0078】
本発明P−セレクチンリガンド蛋白を用いて、P−セレクチンリガンド蛋白と結合しうる、よってP−セレクチンまたはE−セレクチンにより媒介される細胞間付着の阻害剤として作用しうる作用剤をスクリーニングしてもよい。固定化された、あるいは固定化されていない所望結合蛋白を用いる結合アッセイは当該分野においてよく知られており、本発明P−セレクチンリガンド蛋白を用いる目的に使用可能である。適当なスクリーニングアッセイは、下記実施例3および9のような細胞を用いるものであってもよい。別法として、精製蛋白を用いるスクリーニングアッセイを用いてかかる作用剤を同定してもよい。例えば、精製形態のP−セレクチンリガンド蛋白を担体に固定化し、潜在的阻害剤の存在下および不存在下において精製P−セレクチンへの結合を測定してもよい。別法として、適当な結合アッセイは担体に固定化された精製P−セレクチンならびに可溶性形態の本発明P−セレクチンリガンド蛋白を用いるものであってもよい。
【0079】
いずれのP−セレクチンリガンド蛋白を上記スクリーニングアッセイにおいて使用してもよい。例えば、配列番号:2のアミノ酸1からアミノ酸402までの全長P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよく、あるいは配列番号:2のアミノ酸42からアミノ酸402までの成熟P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよく、あるいは配列番号:2のアミノ酸42からアミノ酸310までの可溶性成熟P−セレクチンリガンド蛋白を用いて阻害剤をスクリーニングしてもよい。別法として、配列番号:4のアミノ酸1からアミノ酸412までのP−セレクチンリガンド蛋白、または配列番号:4のアミノ酸42からアミノ酸412までの成熟形態のP−セレクチンリガンド蛋白、または配列番号:4のアミノ酸42からアミノ酸320までの可溶性成熟形態のP−セレクチンリガンド蛋白を用いて、本発明に従って、細胞間付着の阻害剤をスクリーニングしてもよい。
【0080】
かかるスクリーニングアッセイにおいて、P−セレクチンまたはE−セレクチンとP−セレクチンリガンド蛋白とを結合させることにより第1の結合混合物を得て、第1の結合混合物中の結合量(B0)を測定する。P−、E−、またはL−セレクチン、P−セレクチンリガンド蛋白、ならびにスクリーニングすべき化合物または作用剤を結合させることにより第2の結合混合物を得て、第2の結合混合物中の結合量(B)を測定する。第1および第2の結合混合物中の結合量を、例えばB/B0を計算することにより比較する。第1の結合混合物中の結合と比較して第2の結合混合物中の結合の減少が観察される場合、化合物または作用剤はP−、E−、またはL−セレクチンにより媒介される細胞間付着を阻害しうると考えられる。結合混合物の処方および最適化は当業者のレベル内であり、かかる結合混合物は結合を促進または最適化するバッファーおよび塩類を含んでいてもよく、さらなる対照アッセイを本発明スクリーニングアッセイにおいて行ってもよい。
【0081】
P−、E−、またはL−セレクチンに対するP−セレクチンリガンド蛋白の結合活性を少なくとも約10%、好ましくは約50%以上低下させることがわかった化合物を同定し、ついで、L−セクレチンへの結合のアッセイおよびインビボアッセイを包含する他のセクレチン結合アッセイにおいて2次スクリーニングしてもよい。これらの手段により、セレクチンにより媒介される細胞間付着に対する阻害活性を有し、抗炎症剤として適当な化合物を同定してもよい。
【実施例1】
【0082】
P−セレクチンリガンド蛋白遺伝子のクローニング
A.HL60 cDNAライブラリーの構築
P−セレクチンリガンドの発現クローニングのためにHL60 cDNAライブラリーを構築した。Fast Track mRNA Isolation Kit (Invitrogen; San Diego, CA)を用いてヒトプロミエロサイト細胞系HL60(S. J. Collins, et al.,上記文献)からポリA+RNAを単離した。ポリA+RNAフラクションから2本鎖cDNAを合成し、平滑末端をEcoRIアダプター(5'-AATTCCGTCGACTCTAGAG-3', 配列番号:7; 5'-CTCTAGAGTCGACGG-3', 配列番号:8)に連結した。EcoRIエンドヌクレアーゼとともにインキュベーションされ、ついで、ウシ腸アルカリ性ホスファターゼとともにインキュベーションされ、その後精製された発現ベクターpMT21(R. Kaufman et al., J. Mol. Cell. Biol. 9, 946-958 (1989))中にcDNAを連結した。連結生成物から2μ取ってイー・コリ(E. coli) DH5α細胞中にエレクトロポレーションし、10mM MgCl2、10mM
MgSO4、および2%グリセロールを補足した1mlのSOB培地(J. Sambrook et al., Molecular Cloning:A Laboratory Manual, New York, Cold Spring Harbor Press, p1.90 (1989))中、37℃で1時間増殖させた。ライブラリーを小さい部分に分けるために、細菌懸濁液1mlずつをアンピシリン存在下の寒天プレートに撒き、1mlあたりのコロニー数を計算した。各コロニーが1のcDNAクローンを有すると仮定すると、600000クローンが得られ、1プールあたり約16000クローンの部分に分かれた。38のプールそれぞれを、アンピシリン存在下のLブロス中で一晩増殖させ、CsClグラジエントによりプラスミドを精製した。
【0083】
B.P−セレクチンリガンド蛋白遺伝子のスクリーニング
第1段階において、実施例4(A)のLEC−γ1結合アッセイを用いて、HL60 cDNAライブラリーをパンニングし、そのことにより目的プラスミドを豊富化させた。HL60 cDNAライブラリープール各6μgを2μgの3/4FT遺伝子(実施例2)とともにCHO細胞中に同時トランスフェクションした。トランスフェクションから約45時間後、1mM EGTA中、37℃で15分間細胞をインキュベーションし、ついで、セルリフターでかき取ることによりCOS細胞をプレートから取った。1mMカルシウムを含有するHanks緩衝化セイライン(HBSS)で細胞を2回洗浄した。細胞を4mlのHBSSに再懸濁した。懸濁された、トランスフェクションされたCOS細胞を、実施例4(A)において説明するLEC−γ1結合アッセイを用いてスクリーニングした。
【0084】
付着性COS細胞をHirts抽出物(B. Hirts, J. Mol. Biol., 26, 365-369 (1967))から回収し、ついで、増幅のためにイー・コリ DH5α細胞中にエレクトロポレーションした。豊富化されたプラスミド集団をCsClグラジエントで精製し、3/4FT遺伝子(実施例2)をCOS細胞中に再トランスフェクションした。トランスフェクション、スクリーニング、ついで、プラスミド増幅工程を全部で3回繰り返し、ついで、LEC−γ1でコーティングされたプレートに結合するプールを視覚的に検出した。その後、陽性プラスミドプールを部分に分けた。これには陽性抽出物からのHirts抽出物のイー・コリDH5α細胞中へのエレクトロポレーション、ついで、上記のごとく1mlあたりのコロニー数の計算を用いた。一定数のコロニーをアンピシリン存在下の寒天プレートに撒くことにより種々のプールサイズが得られた。ニトロセルロースリフトを行い、フィルターを新たな寒天プレート上に置くことにより2系のプレートを調製した。2系のプレートは、陽性であると同定されたプール由来の各コロニーおよびコロニー群を選択するためのリファレンスプレートとして役立った。
【0085】
クローニングの第2段階において、最初のスクリーニング段階に使用したのと同じ手順により、COS細胞をライブラリーから分けたプールおよび3/4FT遺伝子を用いて同時トランスフェクションした。トランスフェクションから48時間後、実施例4(B)の蛍光CHO:P−セレクチンアッセイを用いてトランスフェクションされた細胞をスクリーニングした。上記のごとく、陽性プールをさらに分けて、最後に各コロニーをスクリーニングし、陽性クローンを同定した。この方法を用いて、1個の陽性クローンpMT21:PL85を得て、P−セレクチンリガンド蛋白をコードしていることがわかった。pMT21:PL85中に含まれるP−セレクチンリガンドのDNA配列を配列番号:1に示し、pMT21:PL85によりコードされるP−セレクチンリガンド蛋白の結合特性を下記実施例4(C)において説明する。
【実施例2】
【0086】
1,3/1,4フコシルトランスフェラーゼ遺伝子のクローニング
ヒト全ゲノムDNA(Clontech Laboratories)からPCR手段を用いて1,3/1,4フコシルトランスフェラーゼ遺伝子(3/4FT)をクローン化した。センスオリゴヌクレオチドプライマーは遺伝子のXbaI部位および5'末端を含み(5'-TAGCATACGCTCTAGAGCATGGATCCCCTGGGTGCAGCCAAGC-3', 配列番号:9)、アンチセンスオリゴヌクレオチドプライマーは遺伝子のEcoRI部位および3'末端を含んでいた(5'-CCGGAATTCTCAGGTGAACCAAGCCGC-3', 配列番号:10)。PCR生成物をXbaI、ついでEcoRIで逐次消化し、標準的なゲル精製法により精製した。ついで、XbaI、ついでEcoRIで逐次消化され、標準的なゲル精製法により精製されたベクターpMT3Sv2ADA(R. Kaufman, Methods in Enzymology,上記)中にこの遺伝子を連結した。コンピテントHB101細胞(Biorad)をこの連結生成物でトランスフェクションし、ついで、アンピシリン存在下の寒天プレートに撒いた。アンピシリン耐性形質転換体のニトロセルロースフィルターリフトを、遺伝子の中央部の506〜530の領域に対して相捕的な放射性標識オリゴヌクレオチド(5'-AAGTATCTGTCCAGGGCTTCCAGGT-3', 配列番号:11)でプローブした(J. Sambrook et al., 上記)。
【0087】
20個の陽性クローンからプラスミドDNA微量標品を調製した。ついで、精製DNAをEcoRIおよびXbaIで消化して正しいサイズのインサートを有する正しいクローンを同定した。ついで、このクローン(pEA.3/4FT)を大スケールまで増殖させ、CsCl密度グラジエントバンド法によりDNAを単離した。下記の細胞−細胞結合アッセイにおいて遺伝子の機能を評価した。DEAEデキストラン処理、DMSOショック処理、ついでクロロキンインキュベーションを用いてCOS−1サル細胞(クローンM6; M. Horwitz et al., Mol. Appl. Genet., 2:147-149 (1983))を3/4FTでトランスフェクションした(L. Sompeyrac and K. Dana, Proc. Natl. Acad. Sci. 78:7575-7578 (1981); M. Lopata et al., Nucleic Acids Res., 12:5707-5717 (1984); H. Luthman and G. Magnuson, Nucleic Acids Res., 11:1295-1308 (1983))。トランスフェクションされたCOS細胞を懸濁し、E−セレクチン発現CHO細胞系への結合を定量した(G. Larsen et al., J. Biol. Chem., 267:11104-11110 (1992))。このアッセイにより、3/4FTでトランスフェクションされたCOS細胞がシアル化Lewisxエピトープを細胞表面に発現しうることが確認された。
【実施例3】
【0088】
P−セレクチンリガンド蛋白の発現
A.LEC11細胞におけるP−セレクチンリガンドの発現
以下のようにして、機能的P−セレクチンリガンドをSLex陽性チャイニーズハムスター卵巣(CHO)細胞系LEC11(Campbell, C. and Stanley, P. Cell 35:303-309 (1983))において発現させた。約8μgのP−セレクチンリガンド遺伝子含有プラスミド8pMT21:PL85、実施例1)をLEC11細胞中にトランスフェクションした。トランスフェクションから68時間後、細胞を2.5mMの酪酸ナトリウムで4時間処理した。6−CFD標識CHO:P−セレクチン細胞結合アッセイ(実施例4、セクションBにおいて説明)を用いて調べたところ、細胞はP−セレクチン付着を誘導することがわかった。対照的に、LEC11細胞単独および対照プラスミドでトランスフェクションされたLEC11細胞はいずれもP−セレクチン付着を誘導しなかった。
【0089】
B.COS細胞における可溶性P−セレクチンリガンドの発現
8μgのpED.sPSL.T7(実施例5C参照)および実施例2の4μgのpEA.3/4FTプラスミド、8μgのpED.sPSL.T7のみ、あるいは8μgのプラスミドベクター(pMT21)および4μgのpEA.3/4FT遺伝子を用いてCOS細胞をトランスフェクションした。トランスフェクションから45時間後、細胞をPBSで2回すすぎ、2mM L−グルタミン、100U/mlペニシリンおよび100μg/mlストレプトマイシンを補足した無血清DMEM(フェノールレッド不含)(JRH Biosciences)中、37℃で一晩インキュベーションした。フッ化フェニルメチルスルホニル、アプロチニンおよびNaN3を添加して最終濃度をそれぞれ1mM、2μg/mlおよび0.02%とし、ならし培地を遠心分離してすべての残渣を除去した。
【0090】
免疫沈降実験のために、COS細胞をpED.sPSL.T7およびpEA.3/4FTで同時トランスフェクションすることにより標識可溶性P−セレクチンリガンド蛋白を得た。トランスフェクションから45時間後、COS細胞を250μCi/mlの35Sメチオニン(NEN)で5時間標識し、培地を集めた。抗T7抗体との免疫沈降によりsPSL.T7蛋白の発現を確認した。
【0091】
C.COS細胞におけるPACEにより開裂されたP−セレクチンリガンドの発現
COS細胞を実施例5(C)のpED.sPSL.T7プラスミド、実施例2のpEA.3/4FT cDNA、および配列番号:5のPACE cDNAを含むプラスミドで同時トランスフェクションした。pED.sPSL.T7プラスミドおよびpEA.3/4FTプラスミドのみを用いて対照同時トランスフェクションを並行して行った。45時間後、これらのトランスフェクションCOS細胞からのならし培地をプラスチックディッシュ上にコーティングし、CHO:P−セレクチン細胞への結合(実施例4)を調べた。PACEとともに同時発現されたP−セレクチンリガンドを含む培地でコーティングされたディッシュについては結合したCHO:P−セレクチン細胞の約20倍の増加が確認された(PACEとともに同時発現されなかったP−セレクチンリガンドを含む培地と比較)。
【0092】
PACEとともに同時トランスフェクションされた精製pPSL.T7蛋白のN末端のアミノ酸配列決定により、すべてのリガンドがPACEコンセンサス部位(配列番号:1のアミノ酸38〜41)において開裂されたこが示された。同時トランスフェクションされたCOS細胞の35S−メチオニンでの放射性標識、ついでオートラジオグラフィーを行い、かなりの量のP−セレクチンリガンドが両方のトランスフェクションにおいて分泌されたことが示された。
【0093】
D.CHO細胞におけるP−セレクチンリガンド蛋白の発現
以下のようにして、全長形態(アミノ酸1〜402)のP−セレクチンリガンド蛋白をCHO(DUKX)細胞系(Urlaub & Chasin, Proc. natl. Acad. Sci. USA 77, 4216-4220 (1980))において発現させた。約25μgのpMT21:PL85プラスミドおよび約8μgのpED.3.4FT(pEA3/4FTをEcoRIおよびXbaIで制限的に消化し、得られたフラグメントをpEDプラスミドに挿入することにより得た)を、リン酸カルシウム法を用いてCHO(DUKX)細胞中に同時トランスフェクションした。トランスフェクション体をメトトレキセート耐性に関して選択した。2週間後、抗SLex抗体(CSLEX−1,米国特許第4752569号)およびヤギ赤血球(sRBC)の抱合体(クロミッククロリド法(Goding, J. W., J. Immunol. Methods 10:61-66 (1976))により調製)を用いることにより、下記のごとく個々のコロニーをSLex発現についてスクリーニングした。洗液が透明になるまで、sRBCを0.15M NaClで洗浄し、ついで、sRBC50%懸濁液を0.15M NaCl中に調製した。50μgのCSLEX−1を含有する0.2mlのsRBC懸濁液に1mlの0.01%クロミッククロリド溶液を滴下し、その間ボルテックスで撹拌した。37℃で30分インキュベーション後、10mlのリン酸塩緩衝化セイライン(PBS)溶液を反応物に添加した。抱合体を
PBSで1回洗浄し、ついで、10mlのPBSに懸濁した。トランスフェクションを入れたプレートをPBSで洗浄し、ついで、3mlのPBSおよび1mlのsRBC/CSLEX−1抱合体を各プレートに添加した。陽性コロニーはトランスルミネーターで赤色を呈し、これらを、10%ウシ胎児血清含有アルファ培地中に拾った。2週間後、コロニーをステップワイズ増幅(2、10、25、100、150nMの濃度のメトトレキセートを用いる)に供した。得られた安定な細胞系をCD−PSGL−1(R3.4)と命名した。実施例7(A)のポリクローナル抗P−セレクチンリガンド蛋白抗体を用いる免疫沈降研究によりP−セレクチンリガンド蛋白の発現を確認した。CD−PSGL−1(R3.4)細胞系により産生されるP−セレクチンリガンド蛋白の機能を、実施例4(A)のごとくLEC−γ1への結合についてトランスフェクション体をアッセイすることにより試験した。
【0094】
アデノシンデアミナーゼ選択(Kaufman, et al., PNAS (USA) 83:3136-3140 (1986))の下で、配列番号:5に示すPACEをコードするcDNAをすでに発現している安定なCHO−PACE系においてsPSL.T7蛋白を発現させた。リン酸カルシウム法を用いてpsPSL.T7(25μg)およびpED.3/4FT(8μg)をCHO−PACE細胞中に同時トランスフェクションした。メトトレキセート耐性に関してトランスフェクション体を選択し、sRBC/CSLEX−1抱合体に結合する個々のコロニーを拾った。2週間培養後、コロニーを上記ステップワイズ増幅に供した。得られた安定な細胞系をCP/PSL−T7(R4.1)と命名した。T7特異的モノクローナル抗体または実施例4(A)のLEC−γ1キメラのいずれかを用いる標準的な免疫沈降法によりpPSL.T7蛋白の発現を確認した。同様のやり方で、pMT21:PL85およびpED.3/4FTをCHO−PACEc系に同時トランスフェクションすることにより成熟全長形態(アミノ酸42〜402)のP−セレクチンリガンド蛋白発現する安定な細胞系を得た。
【0095】
実施例5(B)のsPSL.Q蛋白および実施例5(D)のsPSL.Fc蛋白を発現する安定な細胞系を以下のようにして構築した。プラスミドpED.sPSL.Q(25μg)またはpED.sPSL.Fc(25μg)を、上記の約25μgのpED.3/4FTプラスミドおよび配列番号:5のPACE cDNAとネオマイシン耐性遺伝子を含む約20μgのプラスミドとともに、リン酸カルシウム法を用いてCHO(DUKX)細胞中に同時トランスフェクションした。トランスフェクション体をメトトレキセートおよびG418抗生物質耐性に関して選択した。約2週間後、sRBC/CSLEX−1抱合体結合を用いて個々のコロニーをSLEx発現についてスクリーニングした。陽性コロニーを、1mg/mlの濃度のG418培地中に拾った。2〜3週間培養後、細胞をステップワイズメトトレキセート増幅させて選択した。得られた安定な細胞系をCD−sPSL.Q(R8.2)およびCD−sPSL.Fc(R8.1)と命名した。sPSL.QおよびsPSL.Fc蛋白の発現を、実施例7(A)の抗P−セレクチンリガンド蛋白ポリクローナル抗体を用いる標準的な免疫沈降法により確認した。
【実施例4】
【0096】
P−セレクチンにより媒介される細胞間付着のアッセイ
A.LEC−γ1結合アッセイ
キメラ形態のヒトIgGγ1(LEC−γ1)のFc部分に抱合したP−セレクチンをコードするDNAを、既知方法(Aruffo et al. Cell 67, 35-44 (1991))を用いて構築し、高レベルのキメラLEC−γ1蛋白の発現に関してdhfr−CHO細胞中に安定にトランスフェクションし、下記結合アッセイに使用するためにキメラLEC−γ1蛋白を精製した。
【0097】
ペトリ皿を、まず、ポリクローナル抗ヒトIgGγ1 Fc抗体でコーティングし、ついで、LEC−γ1蛋白でコーティングした。この方法は、キメラ分子のP−セレクチン部分がプレート表面に存在するようにLEC−γ1構築物を配向させる。カルシウムの存在下および不存在下において、配向LEC−γ1へのHL60の付着を定量した。HL60付着はカルシウム依存的であることが示され、キメラ分子がHL60細胞上のそのリガンドへのP−セレクチンの結合機能を保持していることが示された。さらに配向LEC−γ1へのHL60細胞の結合は、P−セレクチンに対する中和モノクローナル抗体によりブロックされることも示され、P−セレクチン結合の特異性が示された。
【0098】
B.蛍光CHO−P−セレクチン結合アッセイ
当該アッセイには、P−セレクチンリガンド遺伝子および3/4FT遺伝子で同時トランスフェクションされたCOS細胞表面に結合し、クラスターを形成しうる蛍光標識CHO:P−セレクチン細胞系(Laesen et al., J. Biol. Chem., 267, 11104-11110 (1992))を用いた。CHO:P−セレクチン細胞を1%ウシ胎児血清含有DME中1.5x106個/mlとなるよう懸濁し、6−カルボキシフルオレセイン二酢酸(6−CFD)を最終濃度100μg/mlとなるよう添加することにより標識した。37℃で15分インキュベーション後、細胞を培地で洗浄し、1x105個/mlとなるよう再懸濁した。洗浄されたCOSトランスフェクション体の入ったアッセイすべき各プレートに標識細胞5mlを添加し、室温で10分間インキュベーションした。培地で4回洗浄することにより非付着細胞除去した。ついで、付着CHO:P−セレクチン細胞のロゼットを求めて蛍光顕微鏡でプレートをスキャンした。
【0099】
C.放射性標識CHO:P−セレクチン細胞を用いる定量的付着アッセイ
スクリーニングの最初の段階に用いたのと同じ手順により、実施例1のpMT21:PL85プラスミドおよび実施例2のpEA.3/4FTプラスミドでCOS細胞を同時トランスフェクションした。対照として、COS細胞をpMT21:PL85のみ、あるいはpEA.3/4FTのみ、あるいはインサートを含まない同様のプラスミド(「模擬」)でトランスフェクションした。トランスフェクションから24時間後、トランスフェクション細胞をトリプシン処理し、Costar6ウェル組織培養プレート中に分配した。既知方法を用いてCHO:P−セレクチン細胞を3H−チミジンで16時間標識し、ついで、1% BSAを含有するα培地(対照);1% BSA、5mM EDTAおよび5mM EGTAを含有するα培地;1% BSAおよび10μg/mlの中和抗P−セレクチンモノクローナル抗体を含有するα培地;ならびに1% BSAおよび非中和抗P−セレクチンモノクローナル抗体を含有するα培地中、1x106/mlとして個4℃で30分プレインキュベーションした。ついで、プレインキュベーションした細胞をトランスフェクションしたCOS細胞を含むウェルに添加した。インキュベーションから10分後、培地を4回注入することにより未結合細胞を除去した。トリプシン処理により結合CHO:P−セレクチン細胞を遊離させ、シンチレーションカウンティングにより定量した。
【0100】
P−セレクチンリガンドおよび3/4FTで同時トランスフェクションされたCOS細胞は、COS模擬細胞と比較して5.4倍のCHO:P−セレクチン細胞結合を誘導した。EGTAおよびEDTA存在下でのアッセイは、模擬トランスフェクションCOS細胞の結合レベルが低下した。同様に、中和抗P−セレクチン抗体とのインキュベーションによっても特異的結合が減少し、非中和抗体は効果がなかった。対照的に、P−セレクチンのみでトランスフェクションされたCHO細胞へのCHO:P−セレクチンの結合は、EDTAおよびEGTAの存在下または不存在下の両方において、模擬トランスフェクションCOS細胞への結合と統計学的に相異はなかった。3/4FTのみでトランスフェクションされたCOS細胞へのCHO:P−セレクチン細胞の結合は、模擬トランスフェクションCOSに対する結合よりも約2倍促進されたが、EDTAおよびEGTAの存在または不存在によっては影響されなかった。
【実施例5】
【0101】
可溶性P−セレクチンリガンドの構築
実施例1においてHL60細部からのcDNAライブラリーの製造に用いた
EcoRIアダプターはXbaI制限部位(TCTAGA)を配列番号:1の開始5'側に有し、それはpMT21:PL85プラスミド中にも存在する。可溶性形態のPSLを得るために、pMT21:PL85プラスミドをXbaIおよびHincII(配列番号:1のヌクレオチド944の後ろを開裂)で制限消化した。かくして約950bpのフラグメントが得られ、それはバリン295のコドンを含め、バリン295までのコードされるリガンドの細胞外セグメント全体を含んでいた。これを単離し、下記セクションAからDまでに示すようにして、可溶性形態のP−セレクチンリガンド蛋白をコードするDNAを得るために使用した。
【0102】
A.psPSL.QCの構築
フラグメントを精製し、Asn296からCys310までのコドンが再生成され、Cys310の直後に新たなストップコドンを導入された2本鎖合成オリゴヌクレオチドDNAとともに、哺乳動物発現ベクターpEDのXbaIおよびEcoRI部位間に連結した。当該オリゴヌクレオチドの配列は以下のごとし:
5'-AACTACCCAGTGGGAGCACCAGACCACATCTCTGTGAAGCAGTGCTAG (配列番号:12)
5'-AATTCTAGCACTGCTTCACAGAGATGTGGTCTGGTGCTCCCACTGGGTAGTT (配列番号:13)
得られたプラスミドをpED.sPSL.QCと命名し、そのプラスミドから発現される蛋白をsPSL.QCと命名した。
【0103】
B.psPSL.Qの構築
フラグメントを精製し、Asn296からGln309までのコドンが再生成され、Gln309の直後に新たなストップコドンを導入された2本鎖合成オリゴヌクレオチドDNAとともに、pEDプラスミド(Kaufman et al., 1991)のXbaIおよびEcoRI部位間に連結した。オリゴヌクレオチドの配列は以下のごとし:
5'-AACTACCCAGTGGGAGCACCAGACCACATCTCTGTGAAGCAGTAG (配列番号:14)
5'-AATTCTACTGCTTCACAGAGATGTGGTCTGGTGCTCCCACTGGGTAGTT (配列番号:15)
得られたプラスミドをpED.sPSL.Qと命名し、そのプラスミドから発現される蛋白をsPSL.Qと命名した。
【0104】
C.psPSL.T7の構築
ファージT7大カプシド蛋白由来のエピトープを含む14個のアミノ酸をコードするオリゴヌクレオチドを合成し、さらにベクター配列由来の32個のアミノ酸を用いてエピトープ「タグ」をC末端に融合させた。2つのオリゴヌクレオチドは下記配列:
5'-CTAGACCCGGGATGGCATCCATGACAGGAGGACAACAAATGGTAGGCCGTAG (配列番号:16) および
5'-AATTCTACGGCCTACCCATTTGTTGTCCTCCTGTCATGGATGCCATCCCGGGT (配列番号:17)
を有し、それらを2本鎖化し、哺乳動物発現プラスミドpEDの大XbaI−EcoRIフラグメントと連結した。得られたプラスミドpED.T7をXbaIおよびSmaIで制限消化し、上記950bpのXbaI−HincIIフラグメントに連結し、プラスミドpED.sPSL.T7を得た。
【0105】
pED.sPSL.T7の発現により得られる蛋白をsPSL.T7と命名した。
【0106】
D.可溶性P−セレクチンリガンド−IgGFcキメラの構築
ヒト免疫グロブリンIgG1のFc部分に融合した可溶性の細胞外形態のP−セレクチンリガンド蛋白をコードするプラスミドDNAを以下のようにして構築した。哺乳動物発現ベクターpED.Fcは、ヒトIgG1のFc領域をコードする配列を含み、該配列はユニークなXbaI制限部位を介してヒンジ領域へのコーディング配列のアミノ末端の融合を可能にする新規リンカー配列を伴っている。3つのフラグメントの連結を行った。pED.FcをXbaIで制限消化し、直鎖状にしてゲル精製した。上記pMT21:PL85由来の950bpのフラグメントは第2のフラグメントを含んでいた。第3のフラグメントは、以下の配列を有するアニーリングされた合成オリゴヌクレオチドDNAからなっていた。
5' - CTGCGGCCGCAGT (配列番号:18)
5' - CTAGACTGCGGCCGCAG (配列番号:19)
【0107】
連結生成物をプラスミドDNAとして増幅し、正しい配置を有する個々のクローンをDNA配列決定により同定した。プラスミドをpED.PSL.Fcと命名した。得られた可溶性P−セレクチンリガンド/Fc融合蛋白のDNAコーディング領域を配列番号:6に示す。
【実施例6】
【0108】
発現されたP−セレクチンリガンドの特徴付け
A.COS細胞上に発現された全長P−セレクチンリガンド蛋白の結合特性
実施例2のpEA.3/4FTプラスミドおよび実施例1のpMT21:PL85プラスミドでのCOS細胞の同時トランスフェクションにより、CHO:P−セレクチン細胞に特異的に結合するCOS細胞を得る。この結合は、pEA.3/4FTおよびpMT21:PL85で同時トランスフェクションした場合にのみ観察される。いずれかのプラスミドのみの使用はCHO:P−セレクチン細胞に結合しないCHO細胞を生じる。P−セレクチンを発現しない親のCHO(DUKX)細胞系およびpEA.3/4FTおよびpMT21:PL85で同時トランスフェクションされたCOS細胞の間には結合は観察されない。同時トランスフェクション細胞およびCHO:P−セレクチン細胞の間の結合は、EDTAおよびEGTAのごとき2価イオンのキレーターに対して感受性があり、P−セレクチンにより媒介される細胞付着のCa++依存性と矛盾しない。中和抗P−セレクチンモノクローナル抗体は、CHO:P−セレクチン細胞とpEA.3/4FTおよびpMT21:PL85で同時トランスフェクションされたCHO細胞との間の結合をブロックしたが、非中和抗P−セレクチンモノクローナル抗体は結合に影響しなかった。抗体の結果は、COS細胞表面に発現されるP−セレクチンリガンド蛋白への結合にはP−セレクチンの機能的ドメインが必要であることを示す。
【0109】
B.COS細胞において発現された全長P−セレクチンリガンドの電気泳動特性
同時トランスフェクションCOS細胞の界面活性剤抽出物を以下のようにして調製した。同時トランスフェクションから45時間後、約1.5x107個の細胞を5mlの溶解バッファー(10mMピペラジン−N,N'−ビス[2−エタンスルホン酸](PIPES)pH7.5、100mM KCl、3mM MgCl2、1mMベンズアミジン、0.5μg/mlロイペプチン、0.75μg/mlペプスタチン、1mMエチレンジアミン、および1μg/mlアプロチニン)に懸濁した。低速遠心分離(500xg、10分)により細胞残渣を除去し、超遠心(100000xg、60分)により膜フラクションを集めた。高速遠心分離により得た膜ペレットを抽出バッファー(10mM 3−[N−モルホリノ]プロパンスルホン酸(MOPS)pH7.5、0.1M NaCl、0.02% NaN3、1% Thesit(登録商標)(Sigma),1mMベンズアミジン、0.5μg/mlロイペプチン、0.75μg/mlペプスタチン、1mMエチレンジアミン、および1μg/mlアプロチニン)に再懸濁した。ついで、以下のようにして試料をSDSポリアクリルアミドゲル電気泳動に供し、ニトロセルロースブロットに移した。界面活性剤抽出物の一部を1% SDSローディングバッファーに懸濁し、100℃で5分間加熱し、ついで、8〜16%ポリアクリルアミドゲル(還元的)または6%ゲル(非還元的)にロードし、Laemmliバッファー系中で泳動した。Immobilon-P(登録商標)トランスファー膜を用いてブロットを調製した。ブロットを10mM MOPS pH7.5、0.1M NaCl、0.02% NaN3、1mM MgCl2、1mM CaCl2、および10%脱脂乳に、4℃で一晩浸した。ブロットを上記バッファー(乳不含)で1回すすぎ、ブロティングバッファー(10mM MOPS pH7.5、0.1M NaCl、1% ウシ血清アルブミン、0.05% Thesit、1mM MgCl2、1mM CaCl2)中、室温で30分インキュベーションした。
【0110】
ついで、下記のごとくブロットをP−セレクチンリガンドを求めてプローブした。50ngのP−セレクチン/Fcキメラを、3μCiの125I−プロテインAとともにブロッティングバッファー中室温で30分インキュベーションした。この時点で、さらなる賦形剤(例えば、EDTA、EGTA、モノクローナル抗体)をプレインキュベーション混合物に添加して、P−セレクチンリガンドへのキメラの結合の対する影響を評価することができる。ついで、プレインキュベーションされた混合物をブロットとともに室温で60分インキュベーションし、ついで、ブロットを同じブロッティングバッファー(ウシ血清アルブミン不含)で4回洗浄し、風乾し、ついで、−70℃でオートラジオグラフを行った。
【0111】
非還元条件下において、同時トランスフェクションCOS細胞から調製した膜抽出物に関してこの方法により2つのバンドが観察された。主要バンドは約220kDの分子量のところまで移動したが、マイナーバンドは110kDのところまで移動した。還元条件下においては、約110kDの分子量のただ1つのバンドが観察され、非還元条件下ではP−セレクチンリガンドはホモダイマーとして存在することが示された。還元されたモノマーの大体の分子量は、cDNAクローンの推定アミノ酸配列から予想されるもの(45kD)よりも大きく、発現された蛋白がさらなる翻訳後修飾を受けたことが示される(実施例6(C)参照)。非特異的IgG1プローブがブロット上にバンドを生じないことにより、P−セレクチン/Fcキメラの特異性を確認した。さらに、P−セレクチン/Fcキメラのブロットへの結合は、EDTA、EGTA、および中和抗P−セレクチンモノクローナル抗体によって消滅した。pEA.3/4FTおよびpMT:PL85プラスミドで同時トランスフェクションされたCOS細胞の膜抽出物からのみ、ブロット上の特異的バンドが観察された。対照トランスフェクション体(pEA.3/4FTまたはpMT21:PL85)からの膜抽出物はブロット上にバンドを生じなかった。
【0112】
C.P−セレクチンリガンド蛋白の糖鎖付加
組み換えP−セレクチンリガンド上に共有結合した炭水化物の存在およびP−セレクチンへの結合におけるその役割を以下のようにして調べた。実施例5(C)のpED.sPSL.T7および実施例2のpEA.3/4FTプラスミドでCOS細胞を同時トランスフェクションした。48時間後、細胞に35S−メチオニンのパルスを与えた。200μlの35Sメチオニン標識sPSL.T7ならし培地を5μgのLEC−γ1とともに、2mM CaCl2および1mg/mlウシ血清アルブミン(BSA)の存在下でインキュベーションした。4℃で2時間回転撹拌した後、プロテインA−Sepharoseビーズ(Pharmacia)を4℃において1時間かけて添加し、遠心分離によりペレット化させ、2mM CaCl2および1mg/mlBSAを含有するTris緩衝化セイライン(20mM Tris−HCl、150mM NaCl pH7.5、以下TBSという)で2回洗浄した。ついで、ペレットを再懸濁し、ノイラミニダーゼ(Streptococcus pneumoniae)、O−グリカナーゼ、およびN−グリカナーゼ(すべてGenzymeから得た)で以下のようにして処理した。すべてのグリコシダーゼ消化を37℃で一晩行った。ノイラミニダーゼ消化には、ペレットを50μlの2−(モルホリノ)−エタンスルホン酸(MES)バッファー,pH6.5(Calbiochem)および0.1% SDS中に再懸濁し、95℃で5分間加熱し、ついで、ペレット化させた。上清を、1.4% n−オクチルβ−D−グルコピラノシド(OGP)、10mM酢酸カルシウム、20mMカコジル酸ナトリウムおよび2.5mM PMSFを含むようにし、最終pH7.0とした。8μlのノイラミニダーゼを添加して最終濃度1ユニット/mlとした。ノイラミニダーゼ/O−グリカナーゼ消化には、試料をノイラミニダーゼとともに上記のごとく調製し、さらにO−グリカナーゼを添加して最終濃度0.1ユニット/mlとした。N−グリカナーゼ消化には、ペレットを54μlのMESバッファーおよび1% SDSに再懸濁し、95℃で5分間加熱し、ついでペレット化させた。上清を、0.2Mリン酸ナトリウム、3.5% OGP、および2.5mM PMSFを含むようにし、最終pHを8.5とした。N−グリカナーゼを添加して最終濃度12ユニット/mlとし、上記のごとくインキュベーションした。
【0113】
sPSL.T7に対するグリコシダーゼ処理の影響を2とおりのやり方で評価した。これを行うために、各消化蛋白試料を2つの等しいフラクションに分けた。一方のフラクションを、実施例7(A)のP−セレクチンポリクローナル抗体を用いて沈降させて電気泳動度に対する消化の影響を示した。他方のフラクションを、実施例4(A)のLEC−γ1キメラを用いて沈降させて消化後に残存するP−セレクチンリガンド結合活性を評価した。免疫沈降した試料を、還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動およびオートラジオグラフィーにより分析した。
【0114】
グリコシダーゼ処理なしの場合、オートラジオグラフィーは各沈降物に対応するバンド(分子量110kD)を示した。P−セレクチンリガンド蛋白をノイラミニダーゼで処理した場合、抗P−セレクチンリガンドポリクローナル抗体沈降物は泳動度がわずかに減少し、シアル酸除去の除去と矛盾しなかった。LEC−γ1により沈降したP−セレクチンリガンド蛋白の量はノイラミニダーゼ処理後に有意に減少し、P−セレクチン/P−セレクチンリガンド相互作用におけるシアル酸残基の役割と矛盾しなかった。P−セレクチンリガンド蛋白をノイラミニダーゼおよびO−グリカナーゼの両方で処理した場合、抗P−セレクチンリガンドポリクローナル抗体との沈降後に電気泳動度の実質的な増加が観察され、多くのO−結合オリゴサッカライド鎖が除去されたことが示された。しかしながら、P−セレクチンリガンド蛋白からのO−結合オリゴサッカライドの除去が不完全であるかもしれない。なぜなら、電気泳動度が分子量38kDの蛋白(配列番号:1のアミノ酸配列から推定)に対応しなかったからである。ノイラミニダーゼ/O−グリカナーゼは、LEC−γ1に結合したP−セレクチンリガンド蛋白をわずかしか消化せず、さらに、P−セレクチン/P−セレクチンリガンド相互作用におけるオリゴサッカライドの役割が示された。N−グリカナーゼで精製P−セレクチンリガンドを処理すると電気泳動度がわずかに増加し、N−結合糖鎖付加に関するいくつかのコンセンサス部位が占領されていることが示された。LEC−γ1により沈降したP−セレクチンリガンド蛋白の量はわずかに減少し、N−結合糖鎖付加もP−セレクチン/P−セレクチンリガンド相互作用に関与しているが、シアル化およびO−結合糖鎖付加のような劇的なものではないことが示された。
【実施例7】
【0115】
P−セレクチンリガンドに特異的なポリクローナル抗体
A.ポリクローナルウサギ抗P−セレクチンリガンド蛋白/マルトース結合蛋白融合蛋白
イー・コリにおいて得られた融合蛋白でウサギを免疫することにより抗P−セレクチンリガンドポリクローナル抗体を得た。融合蛋白は、P−セレクチンリガンドのアミノ末端側1/3(配列番号:1のアミノ酸1から110まで)がイン・フレームでマルトース結合蛋白に融合している(Maina, C. V. et al., Gene 74, 365-373 (1988); Riggs, P., in Current Protocols in Molecular Biology, F. M. Ausbel et al., Eds., Greene Associates/Wiley Interscience (New York, 1990) chapter 16.6)。ここに用いた条件下において、融合蛋白抗体はP−セレクチンリガンド蛋白を認識する。
【0116】
B.ポリクローナルウサギ抗sPSL.T7蛋白
下記スキームに従って、可溶性形態の本発明蛋白(sPSL.T7;実施例5(C)参照)を見かけ上均一にまで精製した。COS細胞を3つのプラスミド(それぞれsPSL.T7(実施例5(C))、3/4FT(実施例2)および可溶性形態のPACE(配列番号:5に示す)をコードしている)でトランスフェクションした。72時間後、ならし培地を集め、組み換えsPSL.T7を以下のようにして精製した。
【0117】
ならし培地を50mM MOPS、150mM NaCl、0.5mM CaCl2および0.5mM MnCl2,pH7.2で2倍に希釈し、同じバッファーで平衡化しておいたレンズマメ−Sepharose 4Bカラムに負荷した。負荷後、280nmの吸光度が安定なベースラインまで低下するまでカラムを同じバッファーで洗浄した。ついで、カラムを、0.5M α−メチル−マンノシドおよび0.3M NaClに調節した同じバッファーで溶離した。カラムの5〜15倍体積のこの溶離バッファー中に組み換えsPSL.T7を集めた。ついで、レンズマメレクチン溶離物を、4℃においてカラム溶離物1リットルあたり472gの硫酸アンモニウムを添加することにより0〜70%硫酸アンモニウム沈殿させた。30分撹拌後、沈殿を最小量のTBS(20mM Tris−HCl、150mM NaCl,pH7.5)に再懸濁し、TBSで平衡化したTSK G4000SWXLゲル濾過カラムに負荷した。カラム流速0.5ml/分とし、ガードカラムを使用した。再懸濁硫酸アンモニウムペレットを250μl未満の部分試料としてカラムに注入し、SDS−PAGEおよびウェスタン分析によりフラクションを分析した。sPSL.T7含有フラクションをプールし、ついで、ウサギの免疫に使用した。
【0118】
抗原プライミングおよびその後のブーストにより3カ月にわたって標準的方法でsPSL.T7に対する抗体を得た。詳細には、50μgのsPSL.T7(0.1% SDS中に混合し、100℃で10分加熱することにより変性)を完全フロイントのアジュバントと混合し、5カ所に皮下注射することにより最初の免疫を行った。25μgのsPSL.T7(0.1% SDS中に混合し、100℃で10分加熱することにより変性)[3回目およびそれ以後のブーストには12.5μg]を不完全フロイントのアジュバントと混合し、2カ所に皮下注射することにより(あるいは後に筋肉内注射)2回目以降のブースト(すべて皮下注射)を行った(2週間ごとに注射)。2週間ごとに試験採血を行って抗体力価をモニターした。抗体力価が適当なレベルに達したら、大量採血を行い、全血清フラクションを調製した。このポリクローナル抗体調合物を用いて、実施例4記載の方法と同様の方法で、CHO:P−セレクチン細胞へのHL60細胞の特異的結合を阻害した。
【0119】
このアッセイには、マイクロタイタープレートの底に撒かれたCHO細胞に結合した蛍光標識HL60細胞(BCECFAM;2',7'−ビス−(2−カルボキシメチル)−5−(および−6)−カルボキシフルイレセイン,アセトキシメチルエステルで標識)を使用した。標識HL60細胞を、ポリクローナル抗体含有血清または免疫前の血清のいずれかとともに4℃で30分プレインキュベーションした。次いで、細胞を洗浄し、CHO:P−セレクチン細胞とともに10分間インキュベーションした。ついで、プレートを洗浄し、蛍光マイクロタイタープレートリーダーを用いて蛍光を読んだ。このアッセイを用いると、1:15希釈の抗−sPSL.T7ポリクローナル血清はCHO:P−セレクチンへのHL60細胞の結合を本質的に完全に阻害した。1:150の抗血清希釈ではCHO:P−セレクチンへのHL60細胞の結合がまだ示された。免疫前の血清はCHO:P−セレクチンへのHL60細胞の結合に影響しなかった。
【実施例8】
【0120】
コア2との同時形質転換
A.コア2GlcNAcトランスフェラーゼをコードするcDNAの単離
コア2GlcNAcトランスフェラーゼをコードしているcDNAを、標準的な生物学的方法により単離した。公表されているヒトコア2配列(Bierhuizen, M. F. A., Fukuda, M., Proc. natl. Acad. Sci. 89, 9326-9330 (1992))に基づいて5'および3'末端における2つのオリゴヌクレオチドを設計した。HL60 cDNAライブラリーのプール(Sako, D., Cell 75, 1179-1186 (1993))を鋳型として用いて、標準的PCRプロトコールによりコア2コーディング配列を増幅した。PCR増幅されたフラグメントを精製し、pEDベクター中にサブクローンした。cDNAを単離するために、PCR反応において陽性のシグナルを発したプールをイー・コリ中に形質転換し、プレートに撒いた。形質転換体をニトロセルロースフィルター上に移し、標準的プロトコールに従って32P放射性標識PCRフラグメントとハイブリダイゼーションさせた。溶液クローンを拾い、再度プレートに撒くことにより精製した。cDNAおよびPCRクローンの配列をジデオキシ配列決定により確認した。
【0121】
B.コア2酵素を発現する安定なPSGL−1チャイニーズハムスター卵巣細胞系の取得
実施例3に従って作成した、全長のP−セレクチンリガンド蛋白および3/4FTフコシルトランスフェラーゼを発現する細胞系を、リン酸カルシウム法により、コア2 cDNAおよびネオマイシン耐性遺伝子(pMT4Neo)で同時トランスフェクションした。約2週間後、安定なG418耐性トランスフェクション体を単一単離体としてあるいはプールとして拾った。これらのトランスフェクション体を、1mg/mlのG418含有完全DMEM培地中で増殖させ、コア2酵素活性につき分析した(Higgins, E. A., et al., J. Biol. Chem. 266, 6280-6290 (1991))。コア2活性に関して陽性のコロニーまたはプールを、種々の方法によりP−セレクチンへのP−セレクチンリガンド結合に関して分析した。同様の方法で、P−セレクチンリガンド蛋白を発現する、あるいは3/4フコシルトランスフェラーゼおよびPACE酵素の両方とともに可溶性P−セレクチンリガンド蛋白を発現する細胞系(実施例3参照)を用いて、上記のごとく安定なコア2の同時トランスフェクション体を単離した。
【0122】
C.P−セレクチン結合活性に対するコア2の影響
P−セレクチン結合活性に対するコア2の影響を、3つの異なる方法により評価した。
【0123】
1.固定化可溶性P−セレクチンまたはP−セレクチン/IgGキメラへのmPSGL−1トランスフェクション体の結合
4℃において、50mM Tris pH9.5中の1μg/mlの抗ヒトFc抗体で48ウェルプレートをコーティングした。HBSSバッファーで2回洗浄後、P−セレクチン/IgGキメラ(濃度0.1〜1μg/ml、実施例5)を4℃で一晩HBSSバッファー中にプレーティングした。4℃で3ないし4時間プレートをBSAでブロックした。可溶性P−セレクチンリガンド蛋白の場合、同じバッファー中の蛋白を直接プレートにコーティングした。3H標識CHO細胞を2mM EGTAでリフトし、PBSで3回洗浄し、最終濃度106個/mlとして再懸濁した。この懸濁液の300μlの部分試料を各ウェルに添加した(300000個/ウェル)。室温で12時間インキュベーション後、ウェルを無血清DMEMで4回洗浄して未結合細胞を除去した。結合細胞を5mM EGTAでリフトし、シンチレーションカウンターでカウントした。無処理のP−セレクチンリガンド蛋白結合に関する陽性対照として使用するU937細胞をガンマグロブリン(5mg/ml)で前処理して内在性Fc受容体をブロックし、ついで、P−セレクチンIgGキメラに結合させた。比較結合データを図1に示す。
【0124】
2.P−セレクチン/IgGキメラを用いる免疫沈降
形質転換体から調製された組み換え全長または可溶性P−セレクチンリガンド蛋白(さらにコア2を伴うもの、あるいは伴わないもの)を35S−メチオニンで標識し、ついで、実施例7および5ならびにSako, D., Cell 175, 1179-1186 (1993)にすでに記載のごとく、抗P−セレクチンリガンド蛋白ポリクローナル抗体またはP−セレクチン/IgGキメラのいずれかと免疫沈降させた。データを図2に示す。
【0125】
3.フローサイトメトリー
安定なネズミP−セレクチンリガンド蛋白トランスフェクション体(コア2を伴うもの、あるいは伴わないもの)を、P−セレクチン/IgGキメラ(LecY1)(実施例5)または抗P−セレクチンリガンド蛋白モノクローナル抗体(Mab275、配列番号:2のアミノ酸42からアミノ酸56までの配列を有するペプチドに対して生成)のいずれかを用いる標準的なFACS法により分析した。両方の試薬をFITC標識プロテインAに前以て抱合させた。2mM CaCl2存在下においてこの抱合体とともに4℃で30分インキュベーション後に、細胞を分析した。データを図3に示す。
【実施例9】
【0126】
P−セレクチンリガンド蛋白のE−セレクチン結合
Bevilacqua et al., Science, 243:1160 (1989)に報告されている配列のアミノ酸−21から536までを含むE−セレクチンをコードしているDNAを用いて、P−セレクチン/IgGキメラに関して実施例5で説明したようにしてE−セレクチン/IgGキメラを作成した。
【0127】
U937細胞(約6.5x107個)を組織培養プレートから回収、新鮮完全RPMI培地および50μCi/mlの3H−グルコサミン塩酸塩(糖蛋白の蛋白結合炭水化物部分を標識)[Varki, FASEB 5:226-235 (1991)]を含む50mlの培地に二等分した。48時間インキュベーション後、両方の培養物から遠心分離により細胞を回収し、PBSで3回洗浄した。ペレット化した細胞をそれぞれ2.5mlの溶解バッファー(1% Triton X-100を含有)に懸濁し、プローブソニケーションを2分間行うことにより細胞を破壊した。界面活性剤溶解物を氷上に3時間置き、ついで、さらに2分間再ソニケーションした。溶解物を16000rpmで5分間遠心分離し、上清を回収し、界面活性剤不含の溶解バッファーでそれぞれ12mlとした。。2つの希釈細胞溶解物のうち1つに、P−セレクチン/IgGキメラ(実施例5参照)に前以てカップリングさせた100μlのプロテインAセファロースを添加し、もう1つには、E−セレクチン/IgGキメラに前以てカップリングさせた100μlのプロテインAセファロースを添加した。往復運動により混合しながら結合反応を4℃で一晩進行させた。場合によっては、U937細胞由来の精製膜が標識蛋白の界面活性剤抽出のための出発物質として役立った。これらの場合、界面活性剤抽出およびアフィニティー沈殿工程は上記のものと本質的に同じであった。
【0128】
インキュベーション後、2つの並行反応混合物をそれぞれ2000rpmで遠心分離し、上清を捨てた。樹脂ペレットをバッファー(10mM MOPS、100mM NaCl、1mM CaCl2、1mM MgCl2、0.02% NaN3,pH7.5、Triton X-100含有[1回目および2回目の洗浄には0.25%、3回目の洗浄には0.1%、4回目の洗浄には0.01%])で4回洗浄した。0.01% Triton X-100含有バッファー1mlを用いて溶離前の最後の洗浄を行い、これらの洗液を液体シンチレーションカウンテイング(LSC)による放射活性カウント定量のためにとっておいた。ついで、樹脂を、0.01% Triton X-100および10mM EDTAを含有するバッファー1mlを用い、4℃で一晩往復運動により混合しながら溶離を行った。上清を遠心分離により回収し、ついで、LSCにより定量した。
【0129】
EDTAにより樹脂から遊離した物質のオートラジオグラフィーを、10%架橋SDS−PAGEゲルで電気泳動し(必要ならばCentricon-10ユニットにより試料を約10000cpmに濃縮)、ついで、製造者の説明に従ってゲルをEN3HANCE(DuPont)で処理し、その後市販ゲルドライヤーでゲルを2時間乾燥させることにより行った。−80℃で最低3日間乾燥ゲルをX線フィルムにさらした。
【0130】
前以てU937細胞の界面活性剤抽出物にさらした固定化E−またはP−セレクチンのEDTAでの溶離、ついで、EDTAでの徹底的な洗浄により、遊離状態の3H−グルコサミン標識蛋白が得られた。EDTA溶離液からの放射性標識回収量は、EDTA処理前の最終洗液において観察されるカウントよりも少なくとも10倍多かった。この観察結果は、P−およびE−セレクチンキメラは両方ともU937全細胞溶解物由来のリガンドをEDTA依存的な様式でアフィニティー捕捉し、ついで、樹脂をEDTAで処理することにより、捕捉されたリガンドが遊離されたことを示唆する。
【0131】
EDTAにより2種のキメラから遊離された蛋白についての評価を、非還元的条件下でのSDS−PAGEおよびオートラジオグラフィーにより行った(市販14C−標識分子量標準物質を用いた)。図4のオートラジオグラフにより示されるように、P−セレクチンキメラ(レーン2および10)で処理した全細胞溶解物、およびE−セレクチンキメラ(レーン4および8)で処理した全細胞溶解物からの遊離カウントは、非還元的条件においては200kDの主要種(レーン2および4)に、還元的条件下では100kDの種(レーン8および10)に対応していた。図4に示す異なる実験においては精製膜抽出物が全細胞のかわりに出発物質として使用され、E−セレクチンキメラ(レーン3(非還元的)、およびレーン9(還元的))およびP−セレクチンキメラ(示さず)は同様の結果を示した。他の実験は、P−セレクチンに結合する主要U937糖蛋白がRb3026(組み換えsPSGL1.T7に対して生成したポリクローナル抗体)と免疫反応することを示す。それゆえ、P−およびE−セレクチンは、各場合において同じ特性で、単一の主要糖蛋白種を特異的に認識する。
【実施例10】
【0132】
欠失または変化した形態の可溶性P−セレクチンリガンド蛋白の製造および分析
A.DNA構築物の製造
以下のようにして、末端切断形態のP−セレクチンリガンド蛋白−IgGキメラを得た。プラスミドpED.PSL.FcをPstIおよびNotIで制限消化し、Fc部分を含む6kbのフラグメントおよびベクターpEDFc6kbをゲル精製した。プラスミド構築物pED.149.Fc、pED.47.FcおよびpED.19.Fcを、標準的なPCR法により、下記のオリゴヌクレオチドプライマーのペアーを用いて作成した。
すべての構築物用の「上流」プライマー:
5'-CCAGGTCCAACTGCAGGTCGACTCTAGAGGGCACTTCTTCTGGGCCCACG-3' (配列番号:20)
148Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCCCTCCAGAACCCATGGCTGCTGGTTGCAGTGG-3' (配列番号:21)
47Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCGCAGCAGGCTCCACAGTGGTAG-3' (配列番号:22)
19Fc用の「下流」プライマー:
5'-TATTATCTGTGCGGCCGCGGAGGCTCCGTTTCTGGCAG-3' (配列番号:23)
【0133】
PCR反応用鋳型DNAはpED.PSL.Fcであった。PCR条件は、Perkin-Elmer Thermocyclerを用いて94℃で1分;42℃で1分;72℃で3分を25サイクルである。最後のサイクル完了後、反応物を25℃においてKlenow酵素で30分処理した。ついで、フェノールクロロホルム抽出を行い、酢酸ナトリウムを添加して0.3Mとし、ついで、PCR生成物であるDNAを2.5倍体積のエタノールで沈殿させた。DNAペレットを70%エタノールですすぎ、残存エタノールを蒸発させた。再懸濁したDNAをPstIおよびNotIで消化し、ゲル精製し、上記pEDFc6kbフラグメントに連結した。制限分析により正しい構築物を同定し、DNA配列決定により確認した。
【0134】
pED.148Fcを鋳型として用い、下記突然変異オリゴヌクレオチドを用いて部位特異的突然変異法(Maniatis et al., 1989, MolecularCloning:A Laboratory Manual, Cold Spring Harbor Laboratories)を行うことによりプラスミドpED.ΔY148.Fc、pED.H24.Q70.148.Fcを作成した。
ΔY148用:5'-CGGAGACAGGCCACCGAATTCCTGCCAGAAACG-3' (配列番号:24)
H24用:5'-CCTCCAGAAATGCTGAGGCACAGCACTGACACCACTCCTC-3' (配列番号:25)
Q70用:5'-GAGCTGGCCAACATGGGGCAACTGTCCACGGATTCAGCAG-3' (配列番号:26)
【0135】
コロニーハイブリダイゼーション(Maniatis et al.上記)により陽性クローンを同定した。
【0136】
pED.ΔY148.FcをEcoRIで制限消化し、下記の2本鎖オリゴヌクレオチドに連結することによりpED.FFFE.148.Fcを構築した。
5'-AATTCGAGTTCCTAGATTTTG-3' (配列番号:27) および
5'-AATTCAAAATCTAGGAACTCG-3' (配列番号:28).
【0137】
pED.ΔY148.FcをEcoRIおよびNotIで制限消化し、ついで、下記の2本鎖オリゴヌクレオチドに連結することによりシリーズ
pED.FYYD.19.Fc、pED.FFYD.19.FcおよびpED.FFFD.19.Fcの構築物を作成した。
【0138】
pED.FYYD.19.Fc用:
5'-AATTCGAGTACCTAGATTATGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:29) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCATAATCTAGGTACTCG-3' (配列番号:30);
pED.FFYD.19.Fc用:
5'-AATTCGAGTTCCTAGATTATGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:31) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCATAATCTAGGAACTCG-3' (配列番号:32);
pED.FFFD.19.Fc用:
5'-AATTCGAGTTCCTAGATTTCGATTTCCTGCCAGAAACTGAGCCTCCGC-3' (配列番号:33) および
5'-GGCCGCGGAGGCTCAGTTTCTGGCAGGAAATCGAAATCTAGGAACTCG-3' (配列番号:34).
【0139】
B.欠失または変化した形態の可溶性P−セレクチンリガンド蛋白の分析のためのプレート結合アッセイ
種々の変異形態の可溶性PSGL−1/Fcキメラをコードする個々のDNAを、実施例3(C)に記載のごとく、pEA.3/4FTおよびPACE cDNAとともにCOS細胞中に同時トランスフェクションした。約107個のCOS細胞からトランスフェクション40〜64時間後に集めた無血清培地50mlを、2mM CaCl2を捕捉したTBSで平衡化した0.25mlのプロテインAセファロース(Pharmacia)カラムにより精製した。20mlのTBS/CaCl2で洗浄後、0.5mlの0.1M酢酸、0.15M NaCl、2mM CaCl2で結合物質を溶離させた。溶離した物質を1/20倍体積の3M Tris pH9.0で中和した。280nmの吸光度を測定し、PAGE/SDS/Laemmliゲルのクーマシーブルー染色により溶離物質を定量した。
【0140】
硫酸化されていない形態の可溶性PSGL−1を得るために、上記のごとく(トランスフェクション後に細胞を50mMコール酸(Sigma)存在下で培養すること以外は)適当なFcキメラでのCOS細胞のトランスフェクションを行った。
【0141】
実施例4(C)に記載のごとく(下記の変更はあるが)、CHO:P−セレクチン、CHO:E−セレクチンおよびCHO:L−セレクチン発現細胞の定量的な付着を行った:COS細胞および抗体を省略した。そのかわり、種々の量のプロテインAにより精製された可溶性PSGL−1/Fcキメラを用いて48ウェルマイクロタイタープレート(Costar)を4℃で16時間コーティングした。未結合物質を除去し、コーティングしたウェルを、1mg/mlのBSAおよび2mMのCaCl2を含有するHankの緩衝化セイライン(HBS)で4℃において1時間処理した。トリチウム標識したセレクチン発現CHO細胞を添加し、実施例4(C)に記載のごとく結合を定量した。
【0142】
C.N−結合糖鎖付加部位の変化の効果
3種のP−セレクチンリガンド−IgGキメラを発現する構築物を構築して、セレクチン結合に対するN−結合糖鎖付加部位の影響を調べた。これらの構築物は下記配列を有する。
【0143】
148.Fc 配列番号:2のアミノ酸42〜189。
【0144】
Q70.148.Fc 配列番号:2のアミノ酸42〜189、配列番号:2の位置111のアスパラギン残基がグルタミン残基に置き換わっている。
【0145】
H24.Q70.148.Fc 配列番号:2のアミノ酸42〜189、配列番号:2の位置65のアスパラギン残基がヒスチジン残基に、配列番号:2の位置111のアスパラギン残基がグルタミン残基に置き換わっている。
【0146】
これらの構築物を図式的に図6に示す。
【0147】
プロテインAおよびP−セレクチン−IgGキメラへのこれらの構築物の結合を比較した。これらの実験結果を図7に示す。オートラジオグラフのレーン4、5および6を比較すると、可溶性P−セレクチンリガンド蛋白中の最初の2つのN−糖鎖付加部位の1つまたは両方の除去によってはP−セレクチンへの結合に対する有為な影響がないことが示される。
【0148】
D.チロシンの影響
可溶性蛋白のアニオン性領域の変化によるP−セレクチンリガンド蛋白のセレクチンへの結合におけるチロシンの役割を調べるために構築物を作成した。下記構築物を作成した。
【0149】
ΔY148.Fc 配列番号:2のアミノ酸42〜189、アミノ酸46〜52が欠失。
【0150】
FFFE.148.Fc 配列番号:2のアミノ酸42〜189、位置46、48および51のチロシン残基がフェニルアラニン残基に、位置52のアスパラギン酸残基がグルタミン酸残基に置き換わっている。
【0151】
これらの構築物を図8に図式的に示す。
【0152】
標識硫酸存在下で対応する構築物を発現させることにより、P−セレクチンリガンド蛋白の硫酸化の程度および部位を調べた。148.FcおよびΔY.148.Fcの硫酸化の程度を、硫酸化されていないP−セレクチン−IgGキメラの硫酸かの程度と比較した。結果を図9に示す。これらのデータは、硫酸取り込みの大部分がP−セレクチンリガンド蛋白のアニオン性領域中へのものであることを示す。
【0153】
アニオン性領域の硫酸化がチロシン残基において起こるかどうかを調べるためにさらなる構築物作成した。下記構築物を作成した。
【0154】
FYYD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46のチロシン残基がフェニルアラニン残基に置き換わっている。
【0155】
FFYD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46および48のチロシン残基がフェニルアラニン残基に置き換わっている。
【0156】
FFFD.19.Fc 配列番号:2のアミノ酸42〜60、配列番号:2の位置46、48および51のチロシン残基がフェニルアラニン残基に置き換わっている。
【0157】
これらの構築物を次式的に図9に示す。
【0158】
これらの構築物の硫酸化の程度を19.Fc(「YYYD.19.Fc」)と比較した。結果を図10に示す。FTTD.19.Fcは有意な硫酸化を示したが、FFFD.19.Fcは実質的にほとんど硫酸化されなかった。よって、P−セレクチンリガンド蛋白のアニオン性領域のチロシン残基が硫酸化の主要部位である。
【0159】
P−セレクチンリガンド蛋白から硫酸を除去すると、P−セレクチンへの結合が実質的に減少した。コール酸処理された148.FcのP−セレクチンへの結合を試験した。図11に示すように、コール酸処理による硫酸化の阻害により、P−セレクチンへのP−セレクチンリガンド蛋白の結合が減少した。
【0160】
E.C末端欠失の影響
いくつかのさらなるC末端欠失構築物を下記のように作成した。
254.Fc 配列番号:2のアミノ酸42〜295。
47.Fc 配列番号:2のアミノ酸42〜88。
19.Fc 配列番号:2のアミノ酸42〜60。
これらの構築物を図式的に図12に示す。
【0161】
254.Fc、148.Fc、47.Fcおよび19.FcのP−セレクチン、E−セレクチンおよびL−セレクチンへの結合を試験した。図23および24は、これらの欠失キメラのセレクチンへの結合および対照を比較したものである。結果を図12にもまとめた。
【0162】
F.P−セレクチンおよびE−セレクチン発現細胞への結合
実施例4(C)の定量的プレート結合アッセイを用いて、P−セレクチンおよびE−セレクチンを発現する細胞への上記の種々の構築物の結合を比較した(図13において図式的に説明する)。
【0163】
図14は、P−セレクチン発現CHO細胞への148.Fc、ΔY.148.Fc、FFFE.148.FcおよびヒトIgG1の結合を比較する。ΔY.148.Fcのアニオン性領域中のすべてのチロシン残基を欠失させると結合が消滅した。FFFE.148.Fcのチロシン残基をフェニルアラニン残基に置き換えると、148.Fcと比較して実質的に結合が減少した。よって、全長のアニオン性領域の存在はP−セレクチン結合に必須であり、P−セレクチン結合はこの領域の硫酸化により促進されることが示された。図14は、148.Fc、ΔY.148.FcおよびFFFE.148.Fcは、セレクチンを発現しないCHO細胞には結合しないことを示す対照実験を示す。
【0164】
図15は、148.Fc、ΔY.148.Fc、FFFE.148.FcおよびヒトIgG1のE−セレクチン発現細胞への結合を比較する。E−セレクチン結合は、もとの配列の欠失または変化によっては影響されなかった。よって、アニオン性領域はE−セレクチン結合には必要ないことが示された。
【0165】
図16は、図14および15の結果をまとめたものである。
【0166】
図17は、P−およびE−セレクチン発現CHO細胞への47.Fcの結合を比較する。47.Fcは、配列番号:2の位置111および292におけるN−結合糖鎖付加部位の欠失にもかかわらず、両セレクチンへの実質的な結合を示した。
【0167】
図19は、P−セレクチン発現CHO細胞へのFYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fc、148.FcおよびヒトIgG1の結合を比較する。FFFD.19.Fcのアニオン性領域中のすべてのチロシン残基の置換により結合が消滅した。FYYD.19.Fcの位置46のチロシン残基をフェニルアラニン残基に置き換えると、148.Fcと比較して実質的に結合が減少した。H42.Q70.148.FcのN−結合糖鎖付加部位の変更は結合に影響しなかった。よって、P−セレクチン結合はアニオン性領域における硫酸化により促進され、N−結合糖鎖付加はP−セレクチン結合には必要ないことが示された。図19は、FYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fcおよび148.Fcは、ヒトIgG1単独以上にはセレクチンを発現しないCHO細胞に結合しないことを示す。
【0168】
図20は、E−P−セレクチン発現CHO細胞へのFYYD.19.Fc、FFFD.19.Fc、H24.Q70.148.Fc、148.FcおよびヒトIgG1の結合を比較する。FYYD.19.FcおよびFFFD.19.Fcの程度までリガンド蛋白を末端切断すると、実質的にE−セレクチン結合が減少した。H24.Q70.148.FcにおけるN−糖鎖付加部位の変化は、実質的にはE−セレクチン結合に影響しなかった。よって、配列番号:2のアミノ酸42から60までを含むP−セレクチンリガンド蛋白は選択的にP−セレクチンに結合でき、E−セレクチンには実質的にほとんど結合しないことが示された。
【0169】
図21は、図19および20の結果をまとめたものである。
【0170】
G.P−およびE−セレクチン結合に関する結論
これらのデータから、P−セレクチンリガンド蛋白によるP−セレクチン結合およびE−セレクチン結合の間の関係に関するいくつかの結論が導かれる。N−結合炭水化物はP−またはE−セレクチンへのP−セレクチンリガンド蛋白の結合には必要ない。配列番号:2のアミノ酸42〜60程度の小さいP−セレクチンリガンド蛋白はP−セレクチンへの結合能を有し、E−セレクチンには実質的にほとんど結合しない。
【0171】
図22は、P−およびE−セレクチンへのP−セレクチンリガンド蛋白の結合に関して提案された図式的モデルを示す。O−結合sLex炭水化物は、P−セレクチン結合およびE−セレクチン結合の両方に必要であることが示された。ここに示すデータは、硫酸化チロシン残基はP−セレクチン結合には関与するが、E−セレクチン結合には関与しないことを示す。出願人のデータは、N−結合糖鎖付加結合部位は不要であることも示す。
【実施例11】
【0172】
PSGL−1形態の凝集現象およびダイマー形成に関する試験
部位特異的突然変異法および/またはストップコドンを導入されたプライマーを用いるPCR増幅により一群のPSGL−1変異体を構築した。すべての変異体実験のための鋳型はpPL85.R16(ATCC 75577、出願人により寄託)であった。
【0173】
第1群の変異体(C310SおよびC327S)は、野生型と比較すると1種のアミノ酸変化のみを有する全長のPSGL−1.R16(それぞれ位置310または327においてCysがSerになっている)。pEA.3/4FTおよびへ変異体C3010SまたはC327Sで同時トランスフェクションされたCOS細胞を35S−メチオニンで標識した。細胞溶解物を調製し、変異体蛋白を実施例7(A)のP−セレクチンリガンドポリクローナル抗体で免疫沈降させ、非還元的条件下および還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動により分析した。
【0174】
変異体C327Sならびに野生型PSGL−1.R16は非還元的条件下ではホモダイマーとして移動し、還元的条件下ではモノマーとして移動した。対照的に、変異体C310Sは非還元的条件下および還元的条件下いずれにおいてもモノマーとして移動し、位置310のシステインがPSGL−1のダイマー形成に必要であることが示された。
【0175】
両方の変異体を、P−セレクチンへの結合能についても分析した。同時トランスフェクションCOS細胞の界面活性剤抽出物を、実施例4(A)のLEC−γ1キメラで沈殿させた。沈殿物を非還元的条件下および還元的条件下でのSDS−ポリアクリルアミドゲル電気泳動およびオートラジオグラフィーにより分析した。PSGL−1.R16およびC327Sは両方ともLEC−γ1により効果的に沈殿したが、LEC−γ1へのC310Sの結合はいちじるしく減少し、ダイマー形態のPSGL−1はモノマー形態よりも強固にP−セレクチンに結合することが示された。
【0176】
第2のセットの変異体は可溶性形態のPSGL−1.R16をコードしており、それらを表1に示す。部位特異的突然変異法により変異体ΔTMが得られ、これは膜貫通ドメインの欠失を有し、その後にRLSRKAが続いている。部位特異的突然変異法または所望位置にストップコドンを導入されたPCRプライマーを用いるPCR増幅により変異体L311、L312、A313、I314、L315、A318およびT322を得た。変異体の名称は各末端切断形態のPSGL−1.R16のC末端アミノ酸を示す。
【0177】
下記判断基準に従って変異体を分析した。
1.トランスフェクションCOS細胞からの発現および分泌
2.モノマー形成対ダイマー形成
3.凝集形態の欠如
4.P−セレクチン結合(LEC−γ1キメラ)
【0178】
変異体ΔTMおよびI316は4つの判断基準すべてを満たした。実施例5(A)のsPSL.QC、L311、L312、A313、I314およびL315のごとき短い可溶性形態のPSGL−1はダイマーを形成せず、実施例5(C)のsPSL.T7、A318およびT322のごとき長い可溶性形態のPSGL−1はあまり望ましくない高分子凝集体を形成した。
【0179】
すでに3/4フコシルトランスフェラーゼおよびコア2トランスフェラーゼを発現しているCHO細胞を、psPSL.T7、ΔTM、I316またはpsPSL.QCでトランスフェクションし、メトトレキセートを用いて増幅した。安定なクローンを単離し、35S−メチオニンで標識した。ならし培地を直接分析するか、あるいはまずLEC−γ1で沈殿させ、ついで、非還元的条件下および還元的条件下でSDS−PAGEにより分析した(図25)。結果は、ΔTMおよびI316がダイマー形成およびP−セレクチン結合において最も効果的であることを示した。
【0180】
表I
【表1】
【実施例12】
【0181】
P−およびE−セレクチンへのPSGL−1結合の特異性
材料. ヒトE−セレクチンの細胞外ドメインおよびヒトIgG1のFc部分を含むキメラ蛋白を、先に説明したP−セレクチンキメラ、LEC−γ1と同様にして構築した。バキュロウイルスに感染したトリコプルシア・ニ(Trichoplusia ni)ハイ・ファイブ(high five)細胞(Invitrogen)において可溶性E−セレクチンキメラを発現させ、プロテインAセファロースクロマトグラフィーにより均一に精製した。それぞれα(1,3/1,4)−フコシルトランスフェラーゼ(Fuc−TIII)、PSGL−1、CD43(ロイコシアリン)、および可溶性の対になったアミノ酸変換酵素(PACE)をCOSで発現させるためのプラスミドベクターpEQA.3/4FT、pPL85、pFCD43、およびpEA.sPACEは本明細書ならびに文献においても記載されている(Sako et al. (1993) Cell 75, 1179-1186; Rehemtulla, A. & Kaufman, R. J. (1992) Curr. Opin. Biotechnol. 3, 560-565; Wasley et al. (1993) J. Biol. Chem. 268, 8458-8465)。公表された配列(Natsuka et al. (1994) Journal of Biological Chemistry 269, 16789-16794; Sasaki et al. (1994) J. Biol. Chem. 269, 14730-14737)由来のオリゴヌクレオチドプローブを用いて、Fuc−TVII cDNA(プラスミドpMT.FT7)をHL60 cDNA発現ライブラリーからクローン化した。成熟(PACE開裂)PSGL−1のN末端の最初の15個のアミノ酸を含むペプチドに対してポリクローナル中和ウサギ抗体Rb3443を生成させた。モノクローナル抗CD43抗体(Becton DickinsonまたはBiodesign Internationalから得た)およびイソタイプ対照抗体を、アフィニティー精製されたヤギの抗体(マウスIgGに対するもの)(Cappel, Organon Teknik Corporation)が共有結合しているセファロースTM−4Bからなる固体支持体にカップリングさせた。セレクチンキメラおよびネズミ抗体の、プロテインAセファロース4ファストフロー(Pharmacia)および抗マウスIgG樹脂へのそれぞれのアフィニティーカップリングを、樹脂1mlあたり2mgの蛋白の割合で行った。抗血清Rb3443をプロテインAセファロースに、樹脂1mlに1mgの割合でカップリングさせた。反応後の上清についてのミクロ−BCAアッセイ(Pierce)により示されるカップリング効率は少なくとも95%であった。アプロチニンおよびペプスタチンはBoehringer Mannheimから得て、ベンズアミジン、ロイペプチン、およびフッ化フェニルメチルスルホニル(PMSF)はSigmaから得た。
【0182】
ミエロイド細胞の標識および膜抽出. 浮遊状態で〜1.3x106個/mlまで増殖したU937またはHL60細胞を、10%ウシ胎児血清および2.5mCiの3H−グルコサミンHCl(Dupont/NEN)を補足したRPMI 1640培地50ml中で48時間標識した。この方法により、細胞1個あたり1cpmよりも高い活性が通常的に得られた。標識細胞をPBSで洗浄し、細胞溶解バッファー(10mM MOPS、150mM NaCl、4mM CaCl2および4mM MgCl2,pH7.5、プロテアーゼ阻害剤アプロチニン20mg/ml、ベンズアミジン10mM、ロイペプチン20mg/ml、ペプスタチン8mg/mlおよびPMSF10mMを含有)に懸濁し、氷上で数サイクルのプローブソニケーションに供した。低速遠心分離により核および細胞残渣を除去し、100000g、1時間の遠心分離により細胞膜を上清から回収し、1M NaClを含有する細胞溶解バッファーに懸濁することにより洗浄し、高速遠心分離し、最後に3mlの膜可溶化バッファー(1% Triton-100を含有する細胞溶解バッファー)に再懸濁した。数サイクルのソニケーションおよび氷上でのインキュベーションを行って膜フラクションを可溶化させた。最後に、低速遠心分離工程を行って不溶性膜残渣を除去した。
【0183】
トランスフェクションCOS細胞の標識および膜抽出. 8μgのプラスミドpPL85またはpFCD43および4μgのpES.sPACE、ならびに4μgのpEA.3/4FTまたはpMT.FT7を用い、DEAE−デキストランおよびクロロキン(25)を用いてCOS M6細胞をトランスフェクションした。回収してから40〜45時間後、トランスフェクション細胞を血清およびメチオニンを含まない培地中で30分飢餓状態とし、ついで、無血清DME中[35S]−メチオニンを5時間与えた。標識細胞を洗浄し、EGTAとともにインキュベーションしてディッシュ表面への付着を緩め、ディッシュからかき取り、ペレット化させ、ついで、冷10mM PIPESバッファー,pH7.5(100mM KCl、3mM NaCl、3.5mM MgCl2、およびプロテアーゼ阻害剤(上記参照)を含有)中に再懸濁した。ついで、ソニケーションにより膜抽出を行い、低速遠心分離、高速遠心分離、ついで、標識ミエロイド細胞について上で説明したようにして膜溶解バッファー中で可溶化を行った。
【0184】
アフィニティー沈殿. 膜抽出物を細胞溶解バッファーまたはTBSCバッファー(20mM Tris−HCl、150mM NaCl、2mM CaCl2,pH7.5、5mg/mlのウシ血清アルブミン(純度約99%、Sigma)を補足)で1:4または1:5に希釈した。かくして、Triton X-100を0.2〜0.25%にまで希釈した抽出物を、往復運動により混合しながらヒトIgG1−プロテインAセファロースとともに4℃で一晩インキュベーションした。ついで、前以て清澄化した上清を、E−またはP−セレクチンキメラ、対照ヒトIgG1、Rb3443またはウサギの免疫前の血清、または抗CD43抗体、またはヤギ抗マウスIgGセファロースに前以てカップリングされたイソタイプ対照と前以てカップリングされたプロテインAセファロースと、4℃で6〜12時間反応させた。洗液上清の放射活性がバックグラウンドレベルに低下するまで0.1〜0.5% Triton X-100を含有するバッファー中で5回またはそれ以上樹脂を洗浄した。P−またはE−セレクチン樹脂に特異的に結合した蛋白の溶離を、室温において10mM EDTAまたは5mM EDTA/5mM EGTAを用いて、あるいはSDS−PAGE試料バッファー(Laemmli, U. E. (1970) Nature 227, 680-685)中で煮沸することにより行った。一方、抗体樹脂に結合した蛋白の溶離を後者の方法により徹底的に行った。還元的条件下での分離のために、ジチオスレイトールを試料バッファーに添加し、最終濃度100mMとした。かくして調製した試料を7.5%ゲルを用いるSDS−PAGEにより分離し、En3Hance(Dupont)で処理し、乾燥させ、ついで、オートラジオグラフィーフィルムにさらした。
【0185】
引き続き行うアフィニティー捕捉実験のために、上記のごとく膜抽出物を前以て清澄化させ、P−もしくはE−セレクチンまたはヒトIgG1とともにアフィニティー沈殿させ、洗浄した。ついで、4℃において10mM MOPS、150mM NaCl,pH7.5中5mM EDTAで試料を樹脂から溶離させた(1時間転がしながら撹拌、溶離は2回行った)。最初および2回目の溶離液を一緒にし、ついで、すでに説明したプロトコールに従って固定化Rb3443とともに免疫沈降させた。
【0186】
結果:
対照ヒトIgG1と並行して、可溶性E−およびP−セレクチンキメラを用いて、下記の「方法」において説明するようにして3H−グルコサミン標識U937細胞の界面活性剤可溶化膜抽出物をプローブした。固定化セレクチンからの溶離物についてのSDS−PAGE/オートラジオグラフィーによる試験(図26)により、P−セレクチン溶離物およびE−セレクチン溶離物の両方における主要蛋白種の存在が明らかとなり、その電気泳動特性は下記のごとし。非還元的条件下では分子量200kDaであり、還元により分子量120kDaの種に変換される(図26、それぞれレーン2および3)。場合によっては、P−セレクチン溶離物およびE−セレクチン溶離物の両方においてさらなるバンドが観察され、おそらく、この主要バンドに対応するものであり、自然に還元された物質の存在(非還元試料中の120kDa種)および不完全な還元(還元試料中の200kDa)を反映していた。さらに、DTTでの還元により影響されないE−セレクチン溶離物中の分子量150kDaの痕跡バンドが場合によって観察された。固定化ヒトIgG1を用いた対照実験(図26、レーン1)、あるいはセレクチン樹脂の溶離をEDTAまたはSDSの不存在下で行った実験(データ示さず)においてはバンドは観察されなかった。HL60細胞を用いた場合にも本質的に同一の結果が得られた(データ示さず)。それゆえ、これらの認識の性質は、これらの蛋白と各セレクチンとのレクチンドメインを介する特異的な金属依存性相互作用であると解釈される(Lasky, L. A. (1992) Science 258, 964-969; Drickamer, K. (1988) J. Biol. Chem. 263, 9557)。
【0187】
E−セレクチンとともに沈殿する主要バンドの金属依存性認識および電気泳動上の挙動は、すでに同定されているP−セレクチンカウンターリセプター、P−セレクチン糖蛋白リガンドまたはPSGL−1の特性と矛盾しない(Moore et al. (1994) J. Biol. Chem. 269, 23318-23327; Moore et al. (1992) J. Cell Biol. 118, 445-456; Sako et al.)。
【0188】
この種が実際にPSGL−1であるかどうかを評価するために、E−セレクチン沈殿およびP−セレクチン沈殿両方のEDTA溶離物を、PSGL−1特異的ポリクローナル抗血清Rb3443と反応させた。図27に示すように、いずれのセレクチンとのアフィニティー捕捉により単離された主要バンドも、この抗血清を用いると免疫沈降した(それぞれレーン3および4)。対照IgG1 EDTA溶離物の免疫沈降後に検出された種はなかった(図27、レーン5)。新鮮3H−標識U937膜抽出物を用いる直接免疫沈降によりRb3443の特異性が確認された。Rb3443との沈降によりPSGL−1の電気泳動特性(非還元的条件下で分子量200kDa、還元的条件下で分子量120kDa;図27、レーン2)を有する単一バンドが回収された。一方、免疫前の抗血清との沈降は物質を捕捉しなかった(図27、レーン1)。これらの結果は、E−セレクチンおよびP−セレクチンの両方により特異的に捕捉される、ミエロイド細胞由来の主要蛋白種がPSGL−1であることを示す。
【0189】
PSGL−1に対するE−セレクチンの特異性をさらに評価するために、CD43(ロイコシアリンともいう)(ミエロイド細胞SLex残基の主要部分を有することが知られている豊富な細胞表面シアロ糖蛋白(Maemura, K. & Fukuda, M. (1992) J. Biol. Chem. 267, 24379-24386))の存在について、U937膜溶解物を直接プローブした。かくして、「方法」に記載のごとく、3H−グルコサミン標識U937細胞の膜抽出物を抗CD43抗体でプローブし、並行してPSGL−1特異的抗血清Rb3443および対照抗体でプローブした。同量の膜溶解物から、CD43抗体はPSGL−1抗血清よりも30倍過剰な放射カウントにおいて沈降させた。SDS−PAGE/オートラジオグラフィーによる免疫沈降の評価(図28)により、各抗体に対して1本の特異的バンドが明らかとなった。Rb3443は、PSGL−1の電気泳動特性を有する単一種を捕捉した(図28、レーン2)。対照的に、CD43抗体は、還元に感受性のない分子量120kDaの種を沈降させ(図28、レーン4)、CD43中にシステインが存在しないことと矛盾しない。対照抗体との免疫沈降(図28、レーン1および3)は予想通り陰性であることがわかった。U937細胞中にはPSGL−1よりもかなり多量のCD43が存在すると思われ、HL60細胞中のこれらの蛋白の定量結果と矛盾しない(Ushiyama et al. (1993) J. Biol. Chem. 268, 15229-15237)。よって、E−セレクチンがミエロイド細胞由来のCD43を沈殿させることができないのは、それらの細胞系におけるその不存在によるものではないように思われる。E−セレクチンが痕跡量のCD43を捕捉するという可能性(すなわち、図26の非還元的条件下レーン3における強度の小さい分子量120kDaのバンド(モノマーPSGL−1とも矛盾しない))も排除できないが、PSGL−1はミエロイド膜抽出物から沈殿する主要蛋白であると思われる。
【0190】
PSGL−1 cDNAとα(1,3/1,4)フコシルトランスフェラーゼ(Fuc−TIII)との同時トランスフェクションによりCOS細胞において発現された組み換えPSGL−1は、最もよくP−セレクチンに結合に関した(Sako et al)。興味深いことに、組み換えPSGL−1に対するE−セレクチンの認識を示すための初期の努力は失敗に終わった。P−セレクチン捕捉がうまく行く条件下において、E−セレクチンは同時トランスフェクションされたCOS細胞膜溶解物由来のカウンターリセプターを捕捉できなかった。この結果に対する1の解釈は、Fuc−TIIIは組み換えPSGL−1をP−セレクチンにより認識されるように修飾できたが、E−セレクチン認識に必要なミエロイドPSGL−1においてはそのような適当な修飾をすることができなかったというものである。SLex炭水化物構造を生成することのできるミエロイドのフコシルトランスフェラーゼFuc−TVIIについての最近のクローニング(Natsuka et al (1994) Journal of Biological Chemistry 269, 16789-12764; Sasaki et al (1994) J. Biol. Chem. 269, 14730-14737)は、この解釈を価値あるものとする。
【0191】
PSGL−1またはCD43をコードするcDNAおよびFuc−TIIIまたはFuc−TVIIのいずれかをコードするcDNAでCOS細胞を同時形質転換した。トランスフェクションCOS細胞から膜溶解物を調製し、固定化E−セレクチンキメラまたはP−セレクチンキメラあるいはPSGL−1またはCD43に対する抗体とともにこれらを沈降させた。沈降生成物をSDS−PAGE/オートラジオグラフィーにより評価し、ついで、EDTA/EGTAによるそれらの遊離(セレクチンにより媒介される結合に関して)またはSDS中での煮沸(免疫沈降物に関して)により評価した。結果を図29に示す。
【0192】
図29Aに示すように、COSにより発現されたPSGL−1のE−セレクチン捕捉はトランスフェクションに使用したフコシルトランスフェラーゼの性質に依存した。3つの別個の実験において、Fuc−TVIIはE−セレクチンによるPSGL−1の沈殿を促進したが、Fuc−TIIIは促進しなかった。Fuc−TIIIがPSGL−1に対してE−セレクチンとの反応性を付与しないことは、PSGL−1の発現不足によるものとすることはできない。なぜなら、特異的抗血清Rb3443は、有意なかなりの量のPSGL−1をFuc−TIIIおよびFuc−TVIIトランスフェクション対から免疫沈降させたからである(図29C)。さらにそのうえ、P−セレクチンは、いずれのフコシルトランスフェラーゼとも等量のPSGL−1を沈殿させることができ(図29B)、これらのトランスフェクションにおいてFuc−TIIIおよびFuc−TVIIは発現され、活性を有していたことが示される。
【0193】
COS組み換え発現系において、高アフィニティーE−セレクチン認識もまた、適当なポリペプチドの存在に依存していた。ポリペプチドの長さ、見かけの分子量、および高頻度かつ特異的なタイプの翻訳後修飾はCD43およびPSGL−1において類似しているが(Maemura, K. &Fukuda, M. (1992) J. Biol. Chem. 267, 24379-24386)、いずれのフコシルトランスフェラーゼも、同時トランスフェクションCOS細胞中の組み換えロイコシアリンに対して高アフィニティーE−セレクチン(またはP−セレクチン)認識を付与することはできなかった(図29A)。抗CD43抗体との免疫沈降は、かなりの量のロイコシアリンがFuc−TIII同時トランスフェクション体およびFuc−TVII同時トランスフェクション体の両方において発現されたことを示す(図29C)。E−セレクチンがCD43を捕捉できないことは、同時トランスフェクション細胞中のフコシルトランスフェラーゼ活性の不足によるものではなかった。PSGL−1またはCD43およびFuc−TIIIまたはFuc−TVIIのいずれかでトランスフェクションされたCOS細胞についてのFACS分析はすべて、SLex特異的抗体CSLEX−1との高レベルの反応性を示す。それゆえ、これらの結果から、高アフィニティーE−セレクチン認識には、特異的フコシルトランスフェラーゼにより適当に修飾された特異的ポリペプチドの存在が必要であることが示唆される。
【実施例13】
【0194】
PSGL−1由来のペプチドによるP−セレクチン/PSGL−1結合の阻害
PSGL−1の配列(配列番号:2)由来の多くのペプチドを、P−セレクチン/PSGL−1結合を阻害する能力に関して試験した。試験したペプチドを図30に記載する。
【0195】
以下のプロトコールに従って阻害を試験した。96ウェルプレートのウェルを、4℃において、50μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2(pH7.5)中のPSGL−1でコーティングした。ウェルから液体を除去した後、ウェル1個あたり150μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2、0.05%ゼラチン(pH7.5)を添加して未占領部位をブロックした。1/2時間ないし2時間後、ブロックバッファーをウェルから除去し、Lec−γ1(P−セレクチン−ヒトIgG Fcキメラ)(2μg/ml)、ビオチン化ヤギ抗ヒト抗体、およびストレプトアビジン−抱合アルカリ性ホスファターゼ(30分ないし1時間室温で拡散させた)ならびに潜在的阻害剤からなる混合物100μlをウェル1個あたりにつき添加した。各プレートを振盪し、プレートからブロックを除去するために強くたたいた。暗所で1時間、回転させながらインキュベーションを継続した。150μlの10mM MOPS、150mM NaCl、1mM CaCl2、1mM MgCl2、0.05%ツイン−20で2回洗浄し、ついで、150μlの1M ジエタノールアミン、0.5mM MgCl2で洗浄して未結合混合物をプレートから洗い流した。10mM DEA/0.5mM MgCl2中のアルカリ性ホスファターゼ用発色基質PNPPを添加し、ついで、プレートを405nmにおいて読んだ。
【0196】
これらのアッセイの結果を図30に示す。配列番号:2のアミノ酸48〜51(チロシン残基がリン酸化されている)およびアミノ酸42〜56を含むペプチドは特に望ましい結果を示した。
【実施例14】
【0197】
可溶性形態のPSGL−1の精製
可溶性形態のP−セレクチンリガンド蛋白の実質的な精製を下記プロトコールに従って行った。
【0198】
可溶性P−セレクチンリガンド蛋白I316(配列番号:2のアミノ酸42からアミノ酸316まで)を、本明細書記載のごとくCHO細胞において発現させた。CHO細胞のならし培地を、分子量10000または30000のいずれかのカットオフのPellicon限外濾過膜ユニット(Millipore)で濃縮して、もとの体積の約1/10とした。ついで、バッファーを25mM Tris、1mM CaCl2,pH7.4と交換した。
【0199】
バッファー交換した濃縮物をToyopearl QAE 550C(TosoHaas)カラムに負荷した。別法として、バッファー交換工程を省略し、濃縮物1部を3部の25mM Tris、1mM CaCl2,pH7.4で希釈し、ついで、カラムに負荷することもできる。4℃においてカラムをその5〜10倍体積の25mM Tris、1mM CaCl2,pH7.4で洗浄した。
【0200】
25mM Tris、1mM CaCl2,pH7.4バッファー中の直線的グラジエント(0M NaClから1.0M NaClまで)を用い、カラムの約5倍の体積のところでP−セレクチンリガンド蛋白が溶離した。2つのピークがカラムから溶離した。第2のピークがP−セレクチンリガンド蛋白を含んでおり、バルクとして集めた。
【0201】
QAEカラムからのピークを分子量30000カットオフの十字流限外濾過膜(Millipore)を用いて濃縮し、ついで、バッファーを25mM Tris、150mM NaCl、1mM CaCl2,pH7.4と交換した。
【0202】
バッファー交換された濃縮物を、4℃においてJacalinアガロースカラムに一晩かけて負荷した。カラムを透析濾過バッファーで洗浄し、P−セレクチンリガンド蛋白は、20℃におけるメチルα−D−ガラクトピラノシド(0〜100mMまたは0〜50mM)のグラジエントで溶離した。JacalinカラムからのフラクションをSDS−PAGEにより分析し、最も純度の高いフラクションをプールした。
【実施例15】
【0203】
P−セレクチンリガンド蛋白融合物
異なるアミノ酸配列を有する、4種のP−セレクチンリガンド蛋白融合物を構築した。それらは47.Fc、47.AGP、47.BMPおよび47.IL11であった。
【0204】
47.Fc:シグナルペプチド、PACE開裂部位、およびもとのFc配列のHis224において変異したヒトIgG1のFc領域に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:35に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:36に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:36のアミノ酸42から開始する。Fc部分における変異は、もとのFc配列のLeu234およびGly237がAlaに変化したものである。
【0205】
47.AGP:シグナルペプチド、PACE開裂部位、および成熟ヒトAGPの最初のロイシン残基に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:37に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:38に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:38のアミノ酸42から開始する。
【0206】
47.BMP:シグナルペプチド、PACE開裂部位、および成熟ヒトBMP−2の配列(その最初の8個のアミノ酸が欠失されている)に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:39に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:40に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:40のアミノ酸42から開始する。
【0207】
47.IL11:シグナルペプチド、PACE開裂部位、および成熟ヒトIL−11の配列に融合した成熟P−セレクチンリガンド配列の最初の47個のアミノ酸をコードするcDNAを構築した。cDNA構築物の配列を配列番号:41に示す。融合ポイントはヌクレオチド261における新規なNotI部位である。cDNA構築物によりコードされるアミノ酸配列を配列番号:42に示す。コードされる融合蛋白の成熟アミノ酸配列は配列番号:42のアミノ酸42から開始する。
【0208】
本明細書において引用した特許および文献を、本発明に十分に記載されているものとみなす。
【特許請求の範囲】
【請求項1】
(a)配列番号:2のアミノ酸42からアミノ酸88までを含む第1のアミノ酸配列、ここで、該第1のアミノ酸配列はP−セレクチンに結合するものであり;および
(b)抗体のFc配列を含む第2のアミノ酸配列、ここで、Fc配列のアミノ酸Leu234およびGly237はアラニンに変化している;
を含む融合蛋白をコードする単離DNA。
【請求項1】
(a)配列番号:2のアミノ酸42からアミノ酸88までを含む第1のアミノ酸配列、ここで、該第1のアミノ酸配列はP−セレクチンに結合するものであり;および
(b)抗体のFc配列を含む第2のアミノ酸配列、ここで、Fc配列のアミノ酸Leu234およびGly237はアラニンに変化している;
を含む融合蛋白をコードする単離DNA。
【図1】

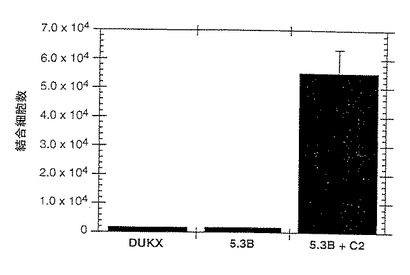
【図2】

【図3】
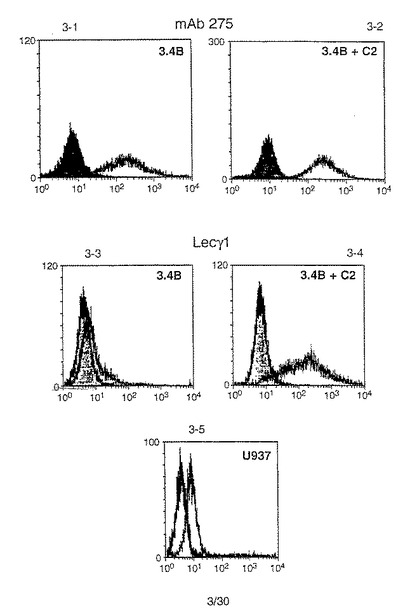
【図4】
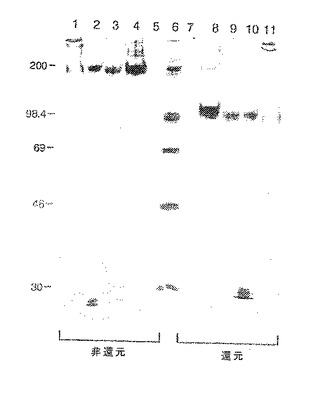
【図5】
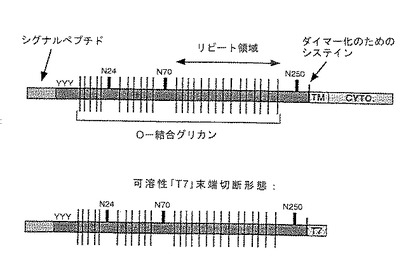
【図6】
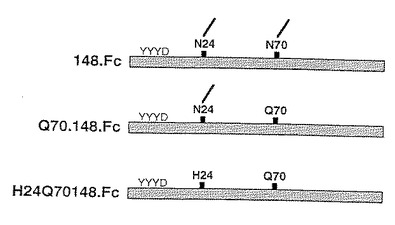
【図7】

【図8】
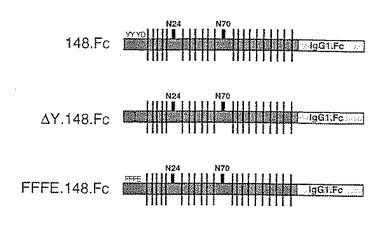
【図9】

【図10】
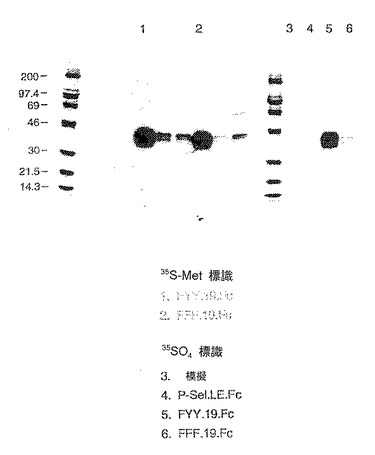
【図11】
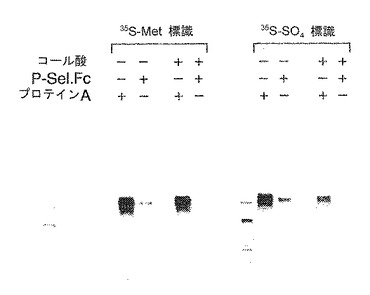
【図12】
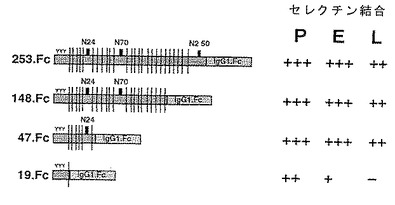
【図13】
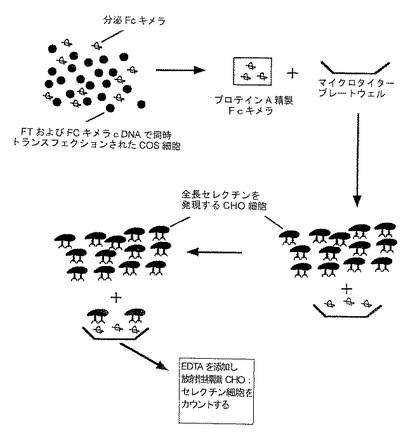
【図14】
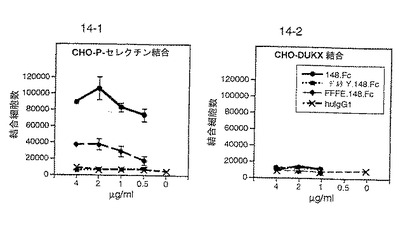
【図15】
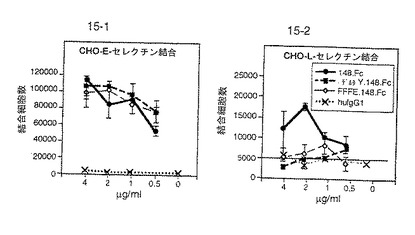
【図16】
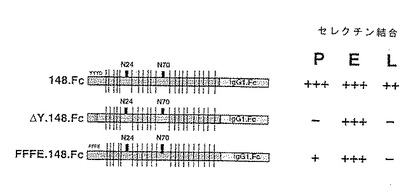
【図17】
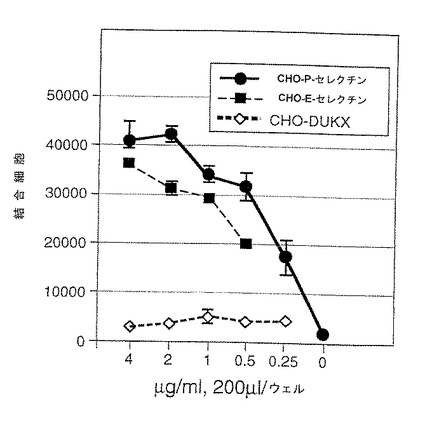
【図18】
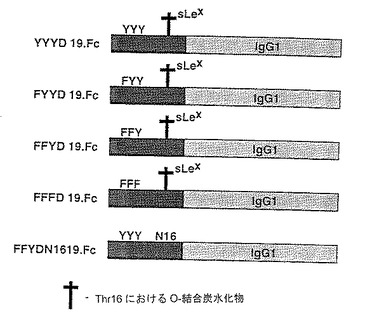
【図19】
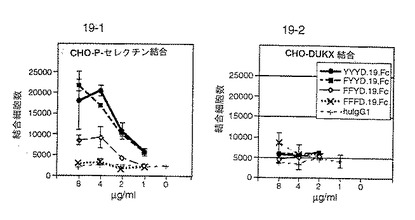
【図20】
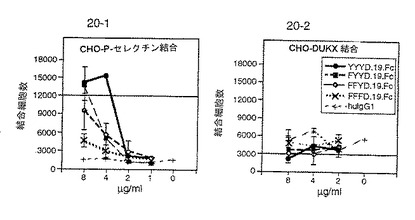
【図21】
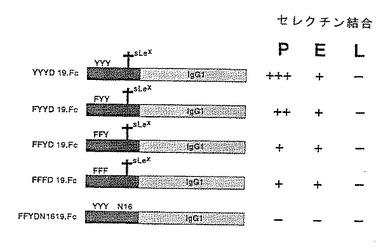
【図22】
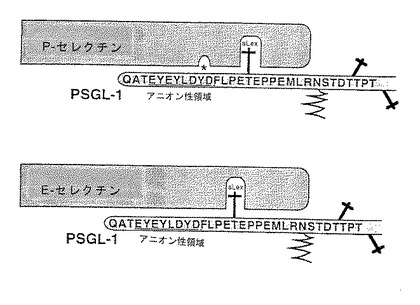
【図23】
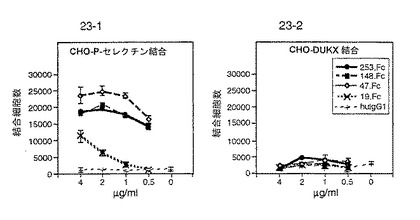
【図24】
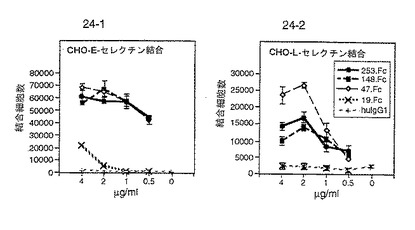
【図25】
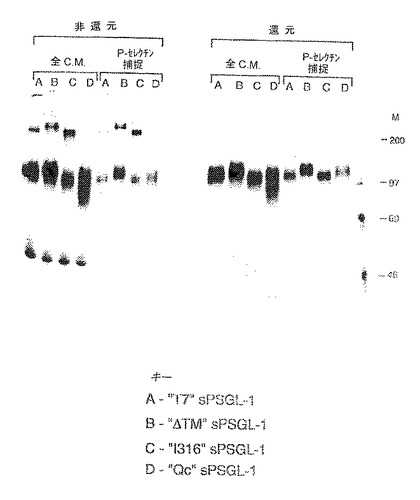
【図26】
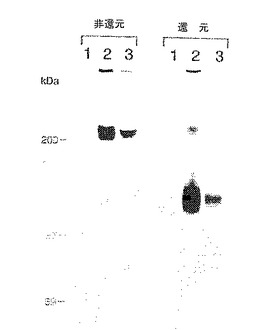
【図27】
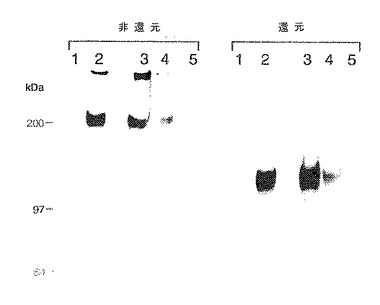
【図28】
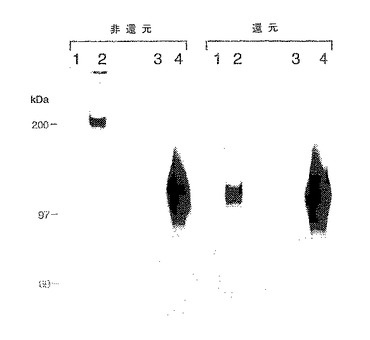
【図29】
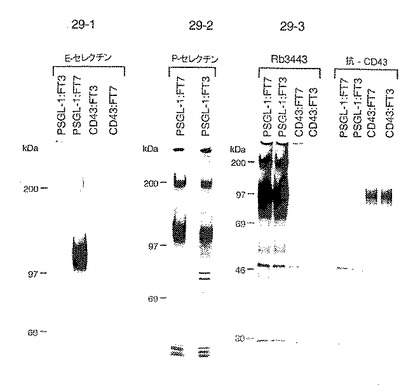
【図30】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【公開番号】特開2012−110338(P2012−110338A)
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願番号】特願2012−14311(P2012−14311)
【出願日】平成24年1月26日(2012.1.26)
【分割の表示】特願2008−266326(P2008−266326)の分割
【原出願日】平成9年8月29日(1997.8.29)
【出願人】(501418214)ジェネティクス インスティテュート,エルエルシー (35)
【Fターム(参考)】
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願日】平成24年1月26日(2012.1.26)
【分割の表示】特願2008−266326(P2008−266326)の分割
【原出願日】平成9年8月29日(1997.8.29)
【出願人】(501418214)ジェネティクス インスティテュート,エルエルシー (35)
【Fターム(参考)】
[ Back to top ]