
P450遺伝子クラスターに関するノックアウトマウス
本発明は、薬物代謝に関与する遺伝子のクラスターがノックアウトされた薬物代謝マウスモデルの作出に関する。治療用途や他の目的のための新しい薬物や化学薬品の開発は非常に複雑である。特に重要なのは、このような化学薬品が、適切な薬物動態を有していようと、代謝の結果として如何なる安全性の問題が生じようと、体内でどのように処理されるか把握することである。薬物の代謝や処理、排出に関与するタンパク質の多くは、遺伝子の数や機能、調節において非常に顕著な種差を示す多重遺伝子族のメンバーである。このような理由から、代謝経路や毒性を確かめるために実験動物で行う実験は、大幅に妥協したものになることがあり、その結果、ヒトの状況を忠実に示すことがなくなる。このような複雑さの一例は、哺乳動物シトクロムP450系(即ち、特定の代謝機能を実施するタンパク質の多重遺伝子族のサイズが種間で大きく異なる)において反映される。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、薬物代謝に関与する遺伝子のクラスターがノックアウトされた薬物代謝マウスモデルの作出に関する。
【背景技術】
【0002】
治療用途や他の目的のための新しい薬物や化学薬品の開発は非常に複雑である。特に重要なのは、このような化学薬品が、適切な薬物動態を有していようと代謝の結果として何らかの安全性の問題を生じさせるものであろうと、体内でどのように処理されるか把握することである。薬物の代謝や処理(disposition)、排出に関与するタンパク質の多くは、遺伝子の数や機能、調節において非常に顕著な種差を示す多重遺伝子族のメンバーである。このような理由から、代謝経路や毒性を確かめるために実験動物で行う実験は、大幅に妥協したものになることがあり、その結果、ヒトの状況を忠実に示すことがなくなる。
【0003】
このような複雑さの一例は、哺乳動物シトクロムP450系で見られる。即ち、この系では特定の代謝機能を担うタンパク質の多重遺伝子族のサイズが種によって大きく異なる。ヒトP450遺伝子をマウスゲノムに導入することによってヒト化マウスモデルを作出する提案が文献報告されている。しかし、導入と同時にマウス遺伝子を無効化しないと、該遺伝子の活性からの干渉が大きくなり、最終的には、その系からの如何なる読み出しも無意味になる。より高度なアプローチにおいては、ヒト化の際に等価なマウス遺伝子を無効化することが提案されており(WO2006/064197参照)、これによってマウスの機能をヒト配列で置換することができる。
【0004】
しかし、Nelsonら、2004年(Pharmacogenomics 14(1): 1)によると、マウスには約72種類の機能性遺伝子が存在するが、ヒトにおいては約27種類しか存在しない。従って、多くの遺伝子に対し、直接的にオルソロガスな遺伝子は少なく(例えば、CYP2E1やCYP1A1、CYP1A2)、ヒトにおいてはごく僅かな遺伝子しか存在しないが、マウスにおいて多数の等価な遺伝子が存在する。その例としては、CYP2A/2B遺伝子ファミリーやCYP2Dが挙げられる。WO2006/064197では、個々のヒト遺伝子をクラスター内の複数の等価なマウス遺伝子と交換することが提案されており、これによって、マウスモデルでヒト薬物代謝の特性を詳細に解析することが可能になることが期待されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
しかし、マウスP450遺伝子のクラスターの機能をヒト等価物で置換せずに該クラスターを欠失させる試みは未だになされていない。このようなP450酵素によって行われる酵素反応は、非常に重要な生命プロセスに関与する。実際、多数のCYP遺伝子における変異は先天性代謝異常に関与し、また、数々の重要な臨床的に価値のある疾患に寄与する。
【0006】
しかし、上述の重要な代謝機能の検討を可能にするマウスを作出できるよう、全シトクロムP450遺伝子クラスターを個々に組み合わせて欠失させることができれば非常に有利であろう。複数の遺伝子クラスターが欠失したマウスによって、化学/薬物動態や体内分布、毒性を定める上でのシトクロムP450系の一般的な役割を確認することができるであろう。このようなモデルはそれ自身でも、また、特定の遺伝子ファミリー又はクラスターに関してヒト化したモデルと組み合わせても非常に有用で多くの用途があると考えられる。
【課題を解決するための手段】
【0007】
本発明によると、薬物代謝酵素をコードする多重遺伝子クラスターの全てのメンバーが機能しないようにされており(rendered inoperable)、欠失クラスターの中の遺伝子と等価な機能を有するヒト置換遺伝子を含まないマウスが提供される。
【0008】
本明細書においては、マウスの全組織内の特定のP450クラスターからシトクロムP450遺伝子全てを欠失させることによって、見かけ上正常で生存可能なマウス系が得られることが示される。この知見から、上述のシトクロムP450酵素に起因するハウスキーピング的な役割が、予測される表現型の結果と著しく違うことにおいて、結局は生存や受精に必須ではあり得ないことが暗示され、マウスにおける個々の複数のP450遺伝子クラスターノックアウトの開発への道が開かれる。
【0009】
これはかなり驚くべきことである。現在まで文献においては、P450薬物代謝酵素をコードする遺伝子クラスターが欠失したマウスについては報告されていない。この一理由としては、シトクロムP450系が、外来化学薬品の代謝や処理において主な役割を果たすことに加え、胚発生や正常なホメオスタシスに必須の多数のホルモンの代謝や処理に関与することが分かっていることが考えられる。例としては、レチノイン酸やステロイドホルモンが挙げられる。全シトクロムP450遺伝子クラスターの欠失によって胚や新生児の致死がもたらされることが予想されよう。結局はこのようなことが生じないのは、かなり驚くべきことである。
【0010】
こうして作出したモデルにおいては、上述の薬物代謝機能が完全に欠失している。従って、このようなモデルは幅広い潜在的用途を有する。第1の例においては、例えば、このようなモデルを用いて、薬物や化学薬品の有効性や毒性、薬物動態、分布におけるP450代謝の役割を確かめることができる。他の可能性は、第2相薬物代謝酵素のクラスター(例えば、UGT1A及び/又はGST遺伝子)の完全な除去である。
【0011】
本発明者らは、このような研究を行う際には、関連機能を有するタンパク質をコードする遺伝子を試験動物においてできるだけ多く欠失させることが最も重要であると考えている。この一理由は、各種薬物代謝遺伝子同士の間(特に、P450遺伝子間)で高度な冗長性が見られるためである。一例としてはCYP2D6P450クラスターを挙げることができるが、該クラスターにおいては、ヒトにおける唯一の機能性遺伝子に対応して、基質特異性が重複する9種類の機能性遺伝子が存在する。この理由から、マウスクラスター内の遺伝子の1個のみを欠失させ、残りを機能性モジュールとして保持するのは殆ど意味がない。
【0012】
「多重遺伝子クラスター」とは、薬物代謝酵素をコードする二を超える遺伝子のクラスターを意味する。本明細書において「クラスター」とは、二を超える遺伝子産物を産生する能力を有し、同一遺伝子の複数のスプライシングを含むゲノムの如何なる領域をも指す。
【0013】
例えば、マウスには7種類のP450遺伝子クラスター、即ち、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターが存在すると考えられる。
【0014】
現在の知見によると、Cyp2abfgstクラスターは次の機能性遺伝子、即ち、2s1、2b10、2b13、2b9、2a4、2b23、2b19、2g1、2a5、2a22、2a12、2f2及び2t4を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子13種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8、9、10、11又は12種のみ)が欠失の対象とされ得る状況であってもよい。
【0015】
現在の知見によると、Cyp2cクラスターは次の機能性遺伝子、即ち、2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。
【0016】
これらの遺伝子15種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8、9、10、11、12、13又は14種のみ)が欠失の対象とされ得る状況であってもよい。例えば、実施例2に記載のノックアウトから2c44遺伝子を特異的に排除したが、これは、該遺伝子が主要マウスCyp2cクラスターの4.1Mb下流に位置し、Cyp遺伝子と何ら関係の無い遺伝子数個分だけ主要クラスターから離れていることに基づく。結果として、これらの無関係な遺伝子の除去は望ましくなく、得られる表現型を複雑にするであろう。
【0017】
現在の知見によると、Cyp2dクラスターは次の機能性遺伝子、即ち、2d22、2d11、2d10、2d9、2d12、2d34、2d13、2d40及び2d26を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子9種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7又は8種のみ)が欠失の対象とされ得る状況であってもよい。コーカソイドの約6%においては等価なヒト遺伝子クラスター発現からのCYP2D6の発現が無いため、このマウスクラスター内の遺伝子の欠失は実際に真のヒト多型を反映する。このタンパク質は神経遮断薬(例えば、抗うつ薬や統合失調症治療用薬物)の代謝において主要な役割を果たすため、かなり重要である。製薬会社はCYP2D6によって代謝される薬物を撤退(back)させたがらないため、薬物を開発している間にその薬物のCYP2D6ライアビリティーの有無をできるだけ早く知る必要がある。従って、CYP2D6レベルの存在下及び非存在下の両方で薬物(特に抗うつ薬)の代謝について研究し得ることは大きな利益となる。
【0018】
現在の知見によると、Cyp2jクラスターは次の機能性遺伝子、即ち、2j13、2j12、2j7、2j11、2j8、2j6、2j9及び2j5を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子8種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば3、4、5、6又は7種のみ)が欠失の対象とされ得る状況であってもよい。
【0019】
現在の知見によると、Cyp3aクラスターは次の機能性遺伝子、即ち、3a13、3a57、3a16、3a41、3a11、3a25及び3a59を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子7種全てが欠失していてもよいが、これらの遺伝子の一部のみ(例えば、3、4、5又は6種のみ)が欠失の対象とされ得る状況であってもよい。例えば、本発明の好ましい実施形態は、3a57、3a16、3a41、3a11及び3a25遺伝子が欠失しているが、3a13及び3a59遺伝子の一方又は両方が無傷のまま残っているマウスである。これらの遺伝子の両方は物理的に該クラスターの外側に位置し、Cypファミリーに属さない遺伝子によってCyp3a遺伝子の残りから離れている。Cypと無関係の遺伝子の欠失は本発明のモデルの価値を分かりにくくするため、これは理想的ではなく、よってあまり好ましくない。また、これらの遺伝子も含むクラスターのノックアウトは技術的に難しく、該クラスターの残りの欠失も含む細胞においてES細胞レベルでこれらの特定遺伝子を更にノックアウトすることによってのみ成し得る。
【0020】
現在の知見によると、Cyp4abxクラスターは次の機能性遺伝子、即ち、4x1、4a29、4a12a、4a12b、4a30b、4a14、4a10、4a31、4a32及び4b1を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子10種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8又は9種のみ)が欠失の対象とされ得る状況であってもよい。
【0021】
現在の知見によると、Cyp4fクラスターは次の機能性遺伝子、即ち、4f39、4f17、4f16、4f37、4f40、4f15、4f14及び4f13を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子8種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6又は7種のみ)が欠失の対象とされ得る状況であってもよい。
【0022】
UGT1Aクラスターは次の機能性遺伝子、即ち、1a1、1a2、1a5、1a6、1a6a、1a7c、1a8、1a9及び1a10を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子9種全てが欠失しているが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7又は8種のみ)が欠失の対象とされ得る状況であってもよい。UGT1a1遺伝子の欠失によって疾患が生じるため、この遺伝子を欠失させないのが好ましい。
【0023】
本発明の方法においては、クラスター欠失の組合せも考えられる。従って、本発明の或る実施形態においては、複数のP450クラスターを機能しないようにすることができる。例えば、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターのいずれか2種を同一動物内で欠失させることができる。他の実施形態においては、完全ヌルP450バックグラウンドのマウスを作出するために、該遺伝子クラスターの3種、4種、5種、6種又は全7種を欠失させることができる。ノックアウトするクラスターの好ましい組合せとしては、Cyp3a、2c及び2dの内の2種又は全3種を含むもの、例えば、Cyp3aとCyp2c、Cyp2cとCyp2d、及びCyp3aとCyp2dが挙げられる。Cyp3a及びCyp2cクラスターの欠失の組合せが好ましく、これによってP450代謝の大部分がノックアウトされる。
【0024】
これらのモデルは、同一遺伝子ファミリーのメンバー間(実際には、遺伝子ファミリー間)の機能的冗長性のため、個々のシトクロムP450アイソザイムの欠失に比べて大きな利益をもたらす。
【0025】
本発明の動物の全組織においてP450の機能性が欠失しているのが好ましい。肝臓が薬物代謝に関して真に重要な唯一の組織であるという間違った考えが広く受け入れられている。真実はこれとは程遠く、実際、P450の機能は、肝臓以外の様々な組織(特に、消化管や血液脳関門等が挙げられる)で発現される。従って、全組織での機能の完全抑止は、P450クラスターノックアウトの作用を適切に評価するために必要である。現在利用可能な技術を用いる場合、このようなことは、胚性幹細胞段階でP450クラスター内の全遺伝子の発現をノックアウトする技術によってのみ実際に成し遂げられる。P450酵素に割り当てられている発生学的役割のため、その大規模なノックアウトは不可能であろうことが常に推測される。
【0026】
従って、本発明のモデルは、特定のP450系が不活性化された他のモデルよりも利益をもたらす。他のモデルの一例としては、肝臓でしか生じないシトクロムP450レダクターゼの条件的不活性化が挙げられる。これに対し、本発明のモデルにおいては、好ましいことに該酵素の欠失が全組織で生じ得る。
【0027】
ある状況下では、全クラスター又はクラスターの組合せのノックアウトが致死の原因となる可能性もあるが、本発明によると、このシナリオにおいては、一組織のみにおいて遺伝子、遺伝子の組合せ又はクラスターをノックアウトするのが好ましい場合もある。このような状況下でノックアウトを限定すべき最も適切な組織は、薬物代謝における役割を考慮すれば肝臓となるであろう。
【0028】
また、多くの理由から、本発明の実験的アプローチもシトクロムP450阻害剤の使用に比べて大きな利益をもたらす。第一に、特異性を欠く阻害剤はそれ自体が薬理学的作用や毒物学的作用を有する。第二に、このような分子は慢性的な投薬状況下では用いることができない。更に、阻害剤は、試験対象の化合物の治療特性や毒物学的特性を調節することが多い。また、阻害剤は、特定の遺伝子ファミリー内の全シトクロムP450を等価な効力レベルで同時に阻害する能力を有しない。最後に、阻害剤を用いた実験は遥かに面倒且つ複雑で解釈が難しい。
【0029】
本発明に係るマウス系の作出によって、ヒトにおける薬物反応や化学反応を決める因子に対する我々の理解が顕著に増すであろう。このようなモデルは、多くの異なるスクリーニングシナリオ(例えば、効力スクリーニングやPK/PDモデリング、薬物安全性試験等)に適用することができる。本発明のノックアウトモデルは、ヒトにおいて見られないげっ歯類特異性代謝産物が生成し、従来の動物試験から得た効力、PK/PD又は安全性の結果を分かりにくくしているような場合には特に注目され得る。
【0030】
本発明のマウスモデルの有用性に関する根拠は、一般にP450代謝速度が遅いため、マウスが多くの薬物をヒトよりも遥かに速く(ほぼ10倍速く)代謝することである。従って、等価な薬物の半減期がヒトにおいて数時間である場合には、マウスでは半減期は恐らく30分間となるであろう。当然のことながら、マウスでは正常な処理経路がマスクされるという作用が得られるが、これは、該経路が有効になる機会が無いためである。従って、一以上のマウスP450クラスター(特に、支配的なCyp3a及び2cクラスター)を欠失させることによって、このような薬物処理経路を論点から排除する効果が得られる。このようなノックアウトマウスをヒトP450クラスターに関してヒト化することによって、ヒト薬物代謝経路を内在性マウス酵素からの干渉無しに評価することができるであろう。
【0031】
本発明の動物モデルはそれ自体が薬物モデルとして興味深い。例えば、該動物モデルによって、欠失していなければ薬物を代謝する機能を有していたであろう酵素の非存在下で薬物代謝を評価することができる。従って、本発明の動物モデルによって、P450系内の冗長性の程度を評価することができ、第1の選択酵素が存在しない場合には他のP450酵素が引き継ぐことができる。また、これによって、第2の選択酵素が恐らく異なる反応経路を用いて作用し、通常は産生しないであろう毒性の二次代謝物を生じさせるかどうかを評価することもできる。
【0032】
本発明のマウスの特徴は、P450クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないことである。これは、該クラスターによってコードされるP450機能が真に機能しないようにされており、他の生体由来の相同又はオルソロガスなクラスターで置換されていないことを意味する。このような動物はそれ自身が有用であり、下流においても作出することができる(例えば、ヒトP450遺伝子クラスターをマウスに導入した動物)。しかし、このような動物については本発明の範囲から意図的に除外する。
【0033】
「等価な機能」とは、機能しないようにするP450機能に対して機能的に取って代わることのできる如何なる遺伝子又は遺伝子クラスターをも意味する。従って、その例としては、他の生体における認知されたオルソロガスなカウンターパートが挙げられる。ヒト及びマウスのオルソログの例は当業者には公知であり、その多くはNelsonら、2004年(Pharmacogenomics 14(1): 1)に記載されている。
【0034】
本発明に係る好ましい薬物モデルは、Cyp3aクラスターが全組織で機能しないようにされており、Cyp3aクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp3aマウスクラスターに対するヒト等価遺伝子は、CYP3A4、3A5、3A7及び3A43である。遺伝子3a57、3a16、3a41、3a11、3a25及び3a59の全てが欠失した、本発明のこの様相に係るマウスの作出及び部分的特徴付けは実施例に記載されているが、このマウスは本発明の一様相を構成する。既存の定説によると、このようなホモ接合的欠失は胚の致死の原因となり得るため、上述のように、この遺伝子型の生存可能なマウスが作出されたということは大いに驚くべきことであると共に非常に意義のある結果である。
【0035】
本発明のこの様相に係るCyp3aクラスターKOの最初の特徴付けから、以下の表現型の特徴が明らかになった。
【0036】
A.一実施形態においては、本発明のこの様相に係るマウスの血漿臨床化学パラメータ(トリグリセリド、アラニントランスフェラーゼ、アルカリホスファターゼ、アスパラギン酸アミノトランスフェラーゼ、アルブミン、総ビリルビン、クレアチンキナーゼ、高密度リポタンパク質及び低密度リポタンパク質)は、未処理野生型マウス(例えば、C57BL/6J)の公知の正常範囲内である。
【0037】
B.第二の実施形態においては、本発明のこの様相に係るマウスの、IP投与後のミダゾラムの薬物動態プロファイルは、野生型マウス(例えば、C57BL/6J)で見られるのと同様の濃度/時間曲線下総面積である(即ち、野生型マウスの場合の80%〜120%)。
【0038】
C.他の実施形態においては、本発明のこの様相に係るマウスに対しリファンピシンで前処理(例えば、60mg/kg、3日間投与)した上でミダゾラムをIP投与した後の算出濃度/時間曲線下総面積が、未処理Cyp3aクラスターKOマウスの場合と比べて、好ましくは170%〜370%、より好ましくは220%〜320%、より好ましくは240%〜300%、より好ましくは250%〜290%、より好ましくは260%〜280%、より好ましくは265%〜275%減少している。
【0039】
D.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによる7−ベンジルオキシキノリンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベルまでの低下が見られる。
【0040】
E.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによるミダゾラムの1’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベルまでの低下が見られる。
【0041】
F.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるミダゾラムの1’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベル、より好ましくは<10%のレベルまでの低下が見られる。
【0042】
G.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによるミダゾラムの4’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<80%のレベル、好ましくは<70%のレベル、より好ましくは<65%のレベルまでの低下が見られる。
【0043】
H.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるミダゾラムの4’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、好ましくは<40%のレベル、より好ましくは<35%のレベル、好ましくは<30%のレベル、より好ましくは<25%のレベルまでの低下が見られる。
【0044】
I.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる処理(例えば、60mg/kg、3日間投与)後のミダゾラムの水酸化が未処理Cyp3aクラスターKOマウスと比べて僅かに増加、即ち、好ましくは101%〜150%、より好ましくは110%〜140%、より好ましくは120〜130%増加する。また、水酸化の増加は、野生型マウス(例えば、C57BL/6J)においてリファンピシン処理時の水酸化で見られる増加に比べて有意に少なく、水酸化の増加は野生型マウスで見られる増加の<60%が好ましく、野生型マウスで見られる増加の<50%が好ましく、野生型マウスで見られる増加の<40%がより好ましく、野生型マウスで見られる増加の<35%がより好ましい。
【0045】
J.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)に続くミダゾラムのIV投与後の1’−ヒドロキシミダゾラムの薬物動態プロファイルにおいて、AUCが野生型マウス(例えば、C57BL/6J)で記録されたものに比べて低く、即ち、AUCは野生型マウスで記録されたものの<60%が好ましく、<50%がより好ましく、<40%がより好ましく、<35%がより好ましい。
【0046】
K.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)に続くミダゾラムの経口投与後の1’−ヒドロキシミダゾラムの薬物動態プロファイルにおいて、AUCが野生型マウス(例えば、C57BL/6J)で記録されたものに比べて低く、即ち、AUCは野生型マウスで記録されたものの<90%が好ましく、<85%がより好ましく、<80%がより好ましい。
【0047】
L.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによる7−ベンジルオキシキノリンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベル、より好ましくは<10%のレベルまでの低下が見られる。
【0048】
M.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるトリアゾラムの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<25%のレベル、より好ましくは<10%のレベル、より好ましくは<5%のレベル、より好ましくは<2.5%のレベルまでの低下が見られる。
【0049】
N.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の腸ミクロソームによるトリアゾラムの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<25%のレベル、より好ましくは<10%のレベル、より好ましくは<5%のレベル、より好ましくは<2.5%のレベルまでの低下が見られる。
【0050】
O.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるニフェジピンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくはC57BL/6Jの<30%のレベル、より好ましくは<25%のレベルまでの低下が見られる。
【0051】
P.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるテストステロンの6β水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型で記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくは<30%のレベルまでの低下が見られる。
【0052】
Q.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるテストステロンの16β水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な増加、即ち、野生型マウスで記録されたものに対して>150%のレベル、より好ましくは>200%のレベル、より好ましくは>250%のレベル、より好ましくは>300%のレベルまでの増加が見られる。
【0053】
R.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるジベンジルフルオレシンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<25%のレベル、より好ましくは<20%のレベル、より好ましくは<15%のレベル、より好ましくは<12.5%のレベルまでの低下が見られる。
【0054】
S.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の腸ミクロソームによる4−トリフルオロメチルクマリンの野生型マウス(例えば、C57BL/6J)と比べての有意な増加、即ち、野生型マウスで記録されたものに対して>150%のレベル、より好ましくは>175%のレベル、より好ましくは>200%のレベルまでの増加が見られる。
【0055】
T.他の実施形態においては、本発明のこの様相に係るマウスでは、野生型マウス(例えば、C57BL/6J)と比べてベンゾキシレゾルフィンO−脱ベンジル化活性の低下、即ち、野生型マウスで記録されたものに対して好ましくは<90%のレベル、より好ましくは<80%のレベル、より好ましくは<70%のレベル、より好ましくは<60%のレベル、より好ましくは<50%のレベルまでのO−脱ベンジル化活性の低下が見られる。
【0056】
U.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンで前処理(例えば、60mg/kg、3日間投与)を行った際、野生型マウス(例えば、C57BL/6J)と比べてベンゾキシレゾルフィンO−脱ベンジル化活性の低下、即ち、野生型マウスで記録されたものに対して好ましくは<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくは<30%のレベルまでのO−脱ベンジル化活性の低下が見られる。
【0057】
V.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態A〜Uの代謝表現型の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20又は全21種を示す。
【0058】
W.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される検出可能レベルのCyp3aは肝ミクロソームにおいては発現されない。
【0059】
X.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームでのCyp1aの発現レベルがC57BL/6Jマウスと比べて高い。
【0060】
Y.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームにおけるCyp2cタンパク質の発現が野生型マウス(例えば、C57BL/6J)と比べて増加する。
【0061】
Z.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームにおけるCyp2dタンパク質のレベルが野生型マウス(例えば、C57BL/6J)と比べて正常であり、肝ミクロソームにおけるCyp2dタンパク質のレベルが野生型マウス(例えば、C57BL/6J)と比べて高い。
【0062】
AA.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソーム及び腸ミクロソームの両方におけるCyp2eタンパク質のレベルに関し、野生型マウス(例えば、C57BL/6J)と比べて有意差が無い。
【0063】
AB.他の実施形態においては、本発明のこの様相に係るマウスでは、シトクロムCレダクターゼ活性で確認されるP450オキシドレダクターゼ(POR)タンパク質の発現レベルに関し、野生型マウス(例えば、C57BL/6J)と比べて有意差が無い。
【0064】
AC.他の実施形態においては、本発明のこの様相に係るマウスでは、総P450レベルは野生型マウス(例えば、C57BL/6J)で記録されたものに比べて有意に高いが、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)の際には有意に変化しない。
【0065】
AD.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態W〜ACのタンパク質発現表現型の内の1、2、3、4、5、6又は全7種を示す。
【0066】
AE.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp3a11、Cyp3a16及びCyp3a25の肝転写産物(liver transcript)レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>50倍、より好ましくは>100倍、より好ましくは>100倍抑制されている。
【0067】
AF.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp4a13の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、より好ましくは>5倍、より好ましくは>10倍、より好ましくは>12.5倍、より好ましくは>15倍抑制されている。
【0068】
AG.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c38の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.5倍アップレギュレートされている。
【0069】
AH.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2a4の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0070】
AI.他の実施形態においては、本発明のこの様相に係るマウスでは、GSTP1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.4倍アップレギュレートされている。
【0071】
AJ.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍抑制されている。
【0072】
AK.他の実施形態においては、本発明のこの様相に係るマウスでは、UGT2B17の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0073】
AL.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCB10の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.2倍アップレギュレートされている。
【0074】
AM.他の実施形態においては、本発明のこの様相に係るマウスでは、CAMK2Bの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.6倍アップレギュレートされている。
【0075】
AN.他の実施形態においては、本発明のこの様相に係るマウスでは、CITED2の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.2倍アップレギュレートされている。
【0076】
AO.他の実施形態においては、本発明のこの様相に係るマウスでは、DNAJC12の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4.1倍抑制されている。
【0077】
AP.他の実施形態においては、本発明のこの様相に係るマウスでは、FMO5の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍アップレギュレートされている。
【0078】
AQ.他の実施形態においては、本発明のこの様相に係るマウスでは、HSD2B3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍アップレギュレートされている。
【0079】
AR.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.1倍抑制されている。
【0080】
AS.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNBの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>3倍、より好ましくは>4倍、より好ましくは>5倍、より好ましくは>5.1倍抑制されている。
【0081】
AT.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP2K6の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、好ましくは>2.5倍、好ましくは>3倍、好ましくは>3.3倍アップレギュレートされている。
【0082】
AU.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍抑制されている。
【0083】
AV.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍抑制されている。
【0084】
AW.他の実施形態においては、本発明のこの様相に係るマウスでは、STIP1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍アップレギュレートされている。
【0085】
AX.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2a4の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.4倍アップレギュレートされている。
【0086】
AY.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b13の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2.5倍、より好ましくは>4倍、より好ましくは>4.5倍、より好ましくは>5倍、より好ましくは>5.5倍アップレギュレートされている。
【0087】
AZ.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b10の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>4倍、好ましくは>5倍、好ましくは>6倍、好ましくは>7倍、好ましくは>7.2倍アップレギュレートされている。
【0088】
BA.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b9の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.2倍アップレギュレートされている。
【0089】
BB.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c54の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0090】
BC.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c54の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0091】
BD.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp4a2bの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.3倍アップレギュレートされている。
【0092】
BE.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0093】
BF.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.4倍抑制されている。
【0094】
BG.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCD1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0095】
BH.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCD3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.5倍アップレギュレートされている。
【0096】
BI.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCG8の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0097】
BJ.他の実施形態においては、本発明のこの様相に係るマウスでは、ARK1C14の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0098】
BK.他の実施形態においては、本発明のこの様相に係るマウスでは、CAMK2Bの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>3.7倍アップレギュレートされている。
【0099】
BL.他の実施形態においては、本発明のこの様相に係るマウスでは、CITED2の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0100】
BM.他の実施形態においては、本発明のこの様相に係るマウスでは、FMO5の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.7倍アップレギュレートされている。
【0101】
BN.他の実施形態においては、本発明のこの様相に係るマウスでは、HSD3B3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後にC57BL/6Jと比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.9倍アップレギュレートされている。
【0102】
BO.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNBの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍抑制されている。
【0103】
BP.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP2K6の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.6倍アップレギュレートされている。
【0104】
BQ.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、好ましくは>2.5倍、好ましくは>2.7倍抑制されている。
【0105】
BR.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態AE〜BQのトランスクリプトーム表現型の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38又は全39種を示す。
【0106】
BS.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態A〜BRの表現型の特徴の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70又は全71種を示す。
【0107】
本発明に係る他の好ましいモデルは、Cyp2cクラスターが全組織で機能しないようにされており、Cyp2cクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp2cマウスクラスターに対するヒト等価遺伝子は2C8、2C9、2C18及び2C19である。遺伝子2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44の全てが全組織で機能しないようにされている本発明のこの様相に係るマウスを作出するための戦略は実施例に記載される。ホモ接合体Cyp2cノックアウトマウスが作出された。
【0108】
本発明に係る他の好ましいモデルは、Cyp2dクラスターが全組織で機能しないようにされており、Cyp2dクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp2dマウスクラスターに対するヒト等価遺伝子は2D6、2D7及び2D8である。遺伝子2d22、2d11、2d10、2d9、2d12、2d34、2d13、2d40及び2d26の全てが全組織で機能しないようにされている本発明のこの様相に係るマウスを作出するための戦略は実施例に記載されている。ホモ接合体Cyp2dノックアウトマウスが作出された。
【0109】
一実施形態においては、本発明のこの様相に係るマウスでは、ブフラロールIP投与後の算出した濃度/時間曲線下総面積が、野生型マウス(例えば、C57BL/6J)の場合と比べて、好ましくは>5倍、より好ましくは>10倍、より好ましくは>15倍、より好ましくは>20倍増加する。
【0110】
本発明に係る更に好ましいモデルは、Cyp3aクラスター及びCyp2cクラスターの両方が全組織で機能しないようにされているマウスである。遺伝子3a57、3a16、3a41、3a11、3a25、3a59、2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44の全てが欠失した、本発明のこの様相に係るマウスの作出及び部分的特徴付けは実施例に記載されているが、このマウスは本発明の一様相を構成する。
【0111】
本発明においては、如何なるP450クラスターをも機能しないようにすることができる。「機能しないようにする」とは、遺伝子又は遺伝子機能を欠失させることを意味する。これは、遺伝子のコード配列をマウスゲノムから完全に欠失させることによって行うことができる。或いは、コード配列を他の配列の挿入、欠失又は置換によって変異させて欠失させることができる。例えば、一以上の変異(フレームシフト変異等)は、得られるRNA転写産物が非機能性タンパク質又は切断型タンパク質をコードするように生じさせることができる。或いは、染色体配列に挿入を行ってアミノ酸コードを分断することができる。
【0112】
同様に、欠失対象のP450遺伝子配列をある配列(例えば、成功した欠失体をスクリーニングするための基準として用いることができる選択配列やマーカー配列)と交換することができる。このような一戦略はWallaceら(Cell 128, 197-209 2007)によって考案されており、遺伝子交換のコンテクスト内ではあるが、本発明の方法に適用可能である。この方法では、ベクターを用いた置換配列が内在性ゲノムマウス配列に取って代わるのに2種の分子間相同組換えイベントが必要となるような、マウス染色体とBAC又はYACベクターとの間での配列の交換が想定されている。
【0113】
好ましい一システムにおいては、相同組換えのメカニズムを用いて遺伝子クラスターを該クラスターが存在しない他の配列と交換する。このような方法は、次の段階:
a)置換対象のマウス標的遺伝子クラスターが各々の端で組換え部位と隣接するように相同組換えによってマウス染色体に一対の部位特異的組換え部位を組込むことと、
b)部位特異的組換え部位間で組換えを行い、マウス標的遺伝子クラスターが染色体から除去され、残存部位特異的組換え部位で置換されるようにすることと、を含むのが好ましい。
【0114】
相同組換えを行うための方法は当該技術分野では公知であり、外部から供給されたDNA分子と標的染色体との間で相同領域を用いてRT部位を導入する。この方法においては、置換配列内の5’及び3’相同アームによって置換配列と標的との間の組換えが促進され、遺伝子クラスターが欠失されるようになる。この戦略を用いた方法は、本出願人による2007年9月14日出願の同時係属特許出願No.GB0718029.2「発明の名称:二段階のクラスター欠失及びヒト化」に報告されている。この戦略は上述のWallaceの技術に比べて遥かに効率的であり、分子内組換えイベントが約106分の1の頻度で生じる。
【0115】
しかし、多くのクラスターは数百キロベースのゲノムの相当のストレッチに及ぶ。一例としてはマウスCYP3a遺伝子クラスターが挙げられ、Cyp3a13が含まれていなくても約800kbに及ぶ。このようなシナリオにおいては、2種の別個の置換核酸をその間にクラスター全体が広がるようにマウスゲノム内に導入し、その後、この2種の置換核酸間での組換えによって該核酸間にある遺伝子クラスターを除去する戦略を用いるのが遥かに効率的である。
【0116】
この構成においては、置換配列の各々は、5’相同アームと3’相同アームとの間に選択マーカー及び少なくとも1種のリコンビナーゼ標的(RT)部位(例えば、loxPやlox5171、FRT、F3)があるようにデザインする。このように、両方の置換核酸をうまく組込んで、欠失対象クラスターに隣接するように選択することが可能である。RT部位を認識する適切な部位特異的リコンビナーゼ(SSR)にマウス細胞を曝露することによってクラスターを除去するのは技術的に容易である。「SSR」とは、特定のDNA遺伝子座におけるDNA再構成を仲介する組換え系の如何なるタンパク質成分をも意味し、例えば、インテグラーゼ又はレゾルバーゼ/インベルターゼクラスのSSR(Abremski, K.E. and Hoess, R.H. (1992) Protein Engineering 5, 87-91; Khan, et al, (1991) Nucleic acids Res. 19, 851-860; Nunes-Duby et al, (1998) Nucleic Acids Res 26 391-406; Thorpe and Smith, (1998) P.N.A.S USA 95 5505-10)及びイントロンコード化エンドヌクレアーゼによって仲介される部位特異的組換え(Perrin et al, (1993) EMBO J. 12, 2939-2947)が挙げられる。SSR酵素活性に曝露することによって、RT部位の配置で確認されるDNA再構成が生じ、その結果、直線DNA分子内で介在配列が除去又は切除される。
【0117】
当該技術分野で周知であり、以下に更に説明する方法に従って、上述の組換え段階はマウス胚性幹細胞内で行うのが好ましい。胚性幹細胞は全能性細胞の培養細胞株であり、該細胞を初期胚に導入すると発育して生体の全組織に定着するようになる。
【0118】
本発明の方法の第1の段階は、一対のRT部位をマウス染色体内に組込むことである。置換対象のマウス標的遺伝子クラスター配列は、各々の端においてRT部位と隣接する必要がある。2個のRT部位間での組換えによって介在遺伝子クラスターが切除される。RT部位を染色体内に組込むための方法は当業者には公知であろうが、相同組換えプロセスを用いて行うのが好ましい。
【0119】
RT対は、LoxP、FRT、attP/attB及びrox(Sauer and McDermott. Nucleic Acids Res. 32(20): 6086-95, 2004)から成る群から選択されるのが好ましい。このリストはほんの一例として挙げたに過ぎず、これらに限定されるものではない。この様相においては、前記部位特異的組換え部位対間の組換えは、対応する部位特異的リコンビナーゼ(即ち、Cre、FLP、PhiC31及びDre)によって行われる。前記部位特異的組換えはインビボ又はインビトロで実施し得ることは理解されよう。
【0120】
実際には、各RT部位は、選択可能マーカーと連結する(好ましくは、連続する)ように構築する必要がある。選択可能マーカーを組込むことによってマウス染色体内のRT部位を検出することができ、それによって、RT部位対の挿入が成功したかどうかをモニターすることができる。各選択可能マーカーは、マウス標的配列とRT部位との間の空間にあるように配置するのが好ましい。これは、組換えが成功した際には、選択可能マーカーが染色体から除去され、本発明の方法においてはこれ以上参加しないことを意味する。このように、ヒト化後に染色体内に残るヒト配列の調節を選択可能マーカーが乱すことがないことは確実であり得る。選択可能マーカーの各々は互いに異なっているのが好ましい。これによって、各組換え部位の挿入を互いに別々に独立してモニターすることができる。選択可能マーカーは、成長ES細胞が曝露され得る化学化合物(例えば、抗生物質)に対するある種の耐性をコードする遺伝子であるのが好ましい。その例としては、細胞成長培地に添加される抗生物質に対する耐性を付与する選択可能マーカー(例えば、G418、ハイグロマイシン又はピューロマイシンに対する耐性を付与するネオマイシン耐性マーカー)の使用が挙げられる。更なる例としては、本発明の上述の様相における核酸配列(又はその一部)に相補的であり、それに対してハイブリダイズする核酸配列を用いた検出が挙げられる。例えば、サザンブロット解析やノーザンブロット解析、PCRが挙げられる。
【0121】
好ましい戦略においては、変化したマウス胚性幹細胞の作出(好ましくは、別々の段階での作出)を伴い、これによって、各RT部位がその選択可能マーカーと共に別々に組込まれる。その後、変化した胚性幹細胞を胚盤胞に挿入することができる。従来、胚盤胞は雌性マウスから交配の約3日後に単離している。最大20個の変化した胚性幹細胞(好ましくは、約16個)をそのような胚盤胞に同時に挿入し得ることは理解されよう。変化した胚性幹細胞を胚盤胞に挿入することにより、好ましくは、妊娠マウスの特性を反映するように誘導された偽妊娠マウスに移植することによって、胚性幹細胞は発育初期胚に組込まれるようになる。この方法によると、変化した胚性幹細胞を含む胚盤胞は偽妊娠マウスの子宮壁内に移植され、妊娠期間が終了するまで該マウス内で発育し続ける。変化した胚性幹細胞は増殖し、分裂して発育トランスジェニックマウスの全組織(生殖系列を含む)に定着するようになる。
【0122】
本発明の方法の一様相においては、作出したトランスジェニックマウスは、各体細胞組織及び生殖系列内に変化した細胞と未変化の細胞を含むキメラとなり得る。
【0123】
大きいDNA断片(200kb〜数メガベース)の欠失に適したCre/lox介在欠失を仲介するための方法は以下の論文に記載されている(Li ZW, Stark G, Gotz J, Rulicke T, Gschwind M, Huber G, Muller U, Weissmann C. Generation of mice with a 200-kb amyloid precursor protein gene deletion by Cre recombinase-mediated site-specific recombination in embryonic stem cells Proc Natl Acad Sci U S A. 1996 Jun 11;93(12):6158-62. Erratum in: Proc Natl Acad Sci U S A 1996 Oct 15;93(21):12052; in Su H, Wang X, Bradley A. Nested chromosomal deletions induced with retroviral vectors in mice. Nat Genet. 2000 Jan;24(l):92-5); Call LM, Moore CS, Stetten G, Gearhart JD. A cre-lox recombination system for the targeted integration of circular yeast artificial chromosomes into embryonic stem cells. Hum Mol Genet. 2000 JuI 22;9(12):1745-51)。
【0124】
RT部位間で組換えを行うためには、ゲノムをSSR酵素形態のSSR活性に曝露する必要がある。上述のように、部位特異的組換えイベントはインビトロ又はインビボで行うことができる。部位特異的組換えは、トランスジェニックマウス内でSSRの活性を誘導することによって行うことができる。実際、生体系で遺伝子型を変化させるために部位特異的組換えをうまく利用するには、組換えイベントを調節するための戦略が必要である。これは、得られる発現パターンが上述の組織特異的な要素が活性となる時間及び場所に制限されるように、リコンビナーゼmRNA又はタンパク質の発現を制御することによって行うことができる(Baubonis and Sauer (1993) Nucl Acids Res. 21, 2025-2029; Sauer B, (1994) Curr Opin Biotechnol 5:521-7; Rajewsky et al., (1996) J Clin Invest 98, 600-3; Metzger and Feil, (1999) Curr. Opinions Biotechnology 10, 470-476)。
【0125】
研究者らは、SSR酵素を発現させるために、直接トランスフェクションや組換えウイルスによる感染、SSRタンパク質をコードするDNAやmRNA又は該タンパク質自身の注入を用いてきた(Konsolaki et al, (1992) New Biol. 4: 551-557)。このようなSSR酵素の発現ではなく活性を調節することによって、より正確に制御することができる。一戦略においては、SSR酵素をステロイド受容体のリガンド結合ドメイン(LBD)に融合させた融合タンパク質を用いてSSR−LBDタンパク質を得る(EP-B-0 707 599;Logie and Stewart (1995) P.N.A.S. USA 92: 5940-5944; Brocard et al, (1997) P.N.A.S. USA 94: 14559-14563; Akagi et al, (1997) Nucleic Acids Res 25, 1766-73参照)。この戦略は、SSR活性を活性化するステロイド受容体のリガンドをリガンドが受容体部分に結合している場合のみ用いることに依存している。該受容体のLBDは、同族リガンドの非存在下ではSSRの活性を抑制する。同族リガンドの送達によってSSRの抑制を軽減し、RT部位間での組換えを行うことができる。
【0126】
このように、誘導については、前記SSRの転写を誘導するか、SSRの翻訳を誘導するか、又はSSRから阻害剤を除去することによって行うことができる。或いは、SSRを前記トランスジェニックマウスに人工的に導入することができる。本発明の方法の一要素は、トランスジェニックマウス内で部位特異的組換えを行うことによってマウス標的遺伝子の除去と同時にヒト化マウスの作出が可能であることである。
【0127】
トランスジェニックマウスを欠失株マウスと交雑させることによって部位特異的組換えを実施し得るのが好ましい。本明細書において「欠失株」とは、生殖系列で部位特異的リコンビナーゼを発現するマウスに関するが、これを、トランスジェニックマウスと交雑させてマウス標的遺伝子配列を除去することができる。このように、インビボ組換えによって着目遺伝子に対してヘテロ接合性の子孫が産生する。こうして、トランスジェニックマウスを欠失株と交雑させることによって、後代が産生すると共に、ヒト置換遺伝子配列と部位特異的リコンビナーゼを含有するように変化したマウス染色体を含む細胞が産生し、その結果、マウス標的遺伝子が除去され、細胞の機能的ヒト化が生じる。従って、このようなトランスジェニックマウスは、特定の遺伝子又は遺伝子クラスターに関するヒト化に対してヘテロ接合性となる。
【0128】
ある実施形態においては、部位特異的リコンビナーゼがリコンビナーゼ株マウスのある組織でのみ発現するのが望ましいことがある。ある遺伝子又は遺伝子クラスターの欠失が致死的であるか又は亜致死性の表現型作用をもたらし得ることは、当該技術分野では知られている。また、このような遺伝子をそのヒト等価物で置換しても致死を防止できない場合がある。このような状況下では、部位特異的リコンビナーゼをある組織(例えば、肝臓)でのみ発現させることによって、致死に関する如何なるこのような問題をも克服し得る。これは、特定の遺伝子がある組織において必須である場合、このように部位特異的リコンビナーゼを発現させることによってマウス遺伝子が該組織において存続し得るため、特に有利となる。
【0129】
本発明のこの様相においては、SSRはアルブミン−Creであり得る。アルブミン−Creは、RT部位LoxPに作用するSSR Creの特定のバリアントである。アルブミン−Creは肝臓においてのみ発現するため、マウス標的配列を肝臓以外の全ての組織で存続させることができ、致死に関する潜在的な問題を克服する一方、機能的にヒト化された肝臓が得られる。
【0130】
最終的には、上述の方法によって作出した2種のヘテロ接合体マウスを交雑させて、着目遺伝子のヒト対立遺伝子に対してホモ接合性のトランスジェニックマウスを作出することができる。2種のヘテロ接合体トランスジェニックマウスの交雑によって、欠失に対してホモ接合性の後代の一部が産生するであろう。
【0131】
本発明の更なる実施形態においては、以降に開示の方法によって、トランスジェニック非ヒト動物を上述の特徴を全て有するように新しく作出する。
【0132】
また、部位特異的組換えイベントを体細胞において行うことも可能であり、その後、該細胞をWO00/51424の方法に記載のクローン化マウス又はそのバリアントのコロニーを形成するために核移植ドナーとして用いることができる。
【0133】
本発明の他の実施形態においては、本発明のマウスを交雑によって作出する。例えば、特定のクラスターの遺伝子の一部が欠失した部分欠失体を他の部分欠失体と交雑させて、特定のクラスター内の遺伝子機能が全て欠失した動物を作出することができるであろう。
【0134】
本発明の更なる実施形態においては、以降に開示の方法によって、トランスジェニックマウスを上述の特徴を全て有するように新しく作出する。
【0135】
トランスジェニックな組合せ
最終的には、本発明の動物モデルをヌルバックグラウンドとして用い、げっ歯類酵素の機能に取って代わり得るヒト遺伝子を同一の染色体領域に直接組込むか、又は他の部位に組込むことによって該遺伝子の導入を行うことができる。ヒト遺伝子を同一の染色体領域に組込むのが好ましいが、これは、染色体の完全性が保持されるため、発現や組織分布の生理学的パターンが同様に保持される可能性が高いからである。従って、上述の細胞やマウスの調製に関する本発明の実施形態においては、該細胞や動物を更なる遺伝子組換えに付すことができるが、該遺伝子組換えは更なる遺伝子又はタンパク質コード核酸配列の導入である。該遺伝子組換えは、細胞又は細胞株の一過性又は安定トランスフェクション、細胞又は細胞株におけるエピソーム発現系、又は、前核マイクロインジェクションや非胚性幹(ES)細胞における組換えイベント、非ヒト胚性幹(ES)細胞におけるランダムな遺伝子組換え、又は核移植クローニング手順においてドナー核として用いる核を有する細胞のトランスフェクションによるトランスジェニック非ヒト動物の調製であり得る。
【0136】
特に、一以上のP450遺伝子クラスターが機能しないようにされている本発明に係るマウスを一以上のヒト遺伝子に関してヒト化し得ることが予想される。欠失したクラスターはCyp3aクラスター、Cyp2cクラスター及び/又はCyp2dクラスターであるのが好ましい。後述のように、これらのクラスターとヒト化薬物輸送体、ヒト化転写因子又はヒト化第II相薬物代謝酵素との如何なる単一又は複数の組合せも好ましい。
【0137】
例えば、このようなマウスを転写因子に関してヒト化することができる。好適な転写因子の例としては、プレグナンX受容体(PXR)や構成的アンドロスタン受容体(CAR)、PPAR(α、δ及びγ)、NRF2、Ah受容体、HNF1、HNF4が挙げられる。これらの転写因子の1種、2種又はそれ以上に関してマウスをヒト化することができる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたのも好ましい。ヌルバックグラウンドに関してP450クラスター欠失と二重ヒト化hCAR及び/又はhPXRマウスの組合せは、本発明に係る好ましいモデルである。
【0138】
また、本発明に係るマウスを第2相薬物代謝酵素に関してヒト化することもできる。このような酵素の例としては、グルクロニルトランスフェラーゼ(例えば、UGT1A遺伝子又は遺伝子クラスター)やグルタチオントランスフェラーゼ(例えば、GST(グルタチオンS−トランスフェラーゼ)(GST−m1及び/又はt1クラスターを含む))、スルホニルトランスフェラーゼ、アセチルトランスフェラーゼが挙げられる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたものも好ましい。
【0139】
また、本発明に係るマウスを薬物輸送タンパク質に関してヒト化することもできるが、その例としては、多剤耐性タンパク質(例えば、mdr1やmdr3)や多剤耐性関連タンパク質(MRP)(例えば、MRP1及び/又はMRP2及び/又はMRP6)、有機アニオン輸送ポリペプチド(OATP)由来のものが挙げられる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたのも好ましい。
【0140】
細胞
本発明の他の様相は、本発明の上述のいずれか一様相における特性を有するように改変された細胞に関する。本発明に係る好ましい細胞型は肝細胞及び神経細胞である。該細胞はマウス細胞であることができる。
【0141】
本発明のこの様相における細胞は、本発明の知見を十分体得した当業者には明らかなように、本発明に係るトランスジェニックマウスから標準的な技法を用いて作出することができる。好適な方法は多くの標準的な実験マニュアル、例えば、Davisら、Basic Methods in Molecular Biology (1986); Sambrook Molecular Cloning; A Laboratory Manual, Third Edition (2000); Ausubel et al., 1991 [supra]; Spector, Goldman & Leinwald, 1998)に記載されている。
【0142】
このような細胞を作出する好ましい一方法は、上述のようにヒト化マウスをSV40不死化マウス(例えば、不死マウス(タコニック))と交雑させることである。次に、当該技術分野で周知の技法によってこのような動物から細胞を単離することができる。肝臓でのみ活性であるため肝細胞しか産生することができないアルブミンプロモーターを用いる先行技術のトランスジェニック系とは対照的に、本発明において作出されるトランスジェニック動物由来の細胞は、薬物動態解析にかなり重要な細胞(例えば、肝細胞や神経細胞)等の様々な細胞型から選択することができる。
【0143】
上述の特性を有する、本発明に係るトランスジェニック動物から単離された幹細胞も本発明の有用な様相である。このような細胞は多能性であってもよく、部分的に分化していてもよい。幹細胞は成体幹細胞であってもよく、胚性幹細胞であってもよい。より一般には、用いる幹細胞は後胚発達段階(例えば、胎生期や新生期、幼若期、成体期)で得られることができる。このように単離された幹細胞を用いて特定の細胞型(例えば、肝細胞や神経細胞)を作出することができる。このような細胞も本発明の一様相を構成する。
本発明の方法によって作出した細胞又は動物をモデル系として用いて、他の生体(特にヒト)における薬物や他の生体異物化合物の代謝を確認することができる。
【0144】
アッセイ
本発明の動物、組織及び細胞を用いて、薬物化合物がどのように代謝されるか確認することができる。単一又は複数のシトクロムP450遺伝子クラスターが欠失したマウス系、或いは、シトクロムP450遺伝子クラスター交換を薬物の代謝や処理に関連する他の遺伝子と組合わせたマウス系の作出によって、ヒトにおける薬物反応や化学反応を決める因子、更にはこのような遺伝子と化学毒性の関連性についての我々の理解が顕著に増すであろう。このようなモデルは、効力スクリーニングやPK/PDモデリング、薬物安全性試験に適用することができる。
【0145】
本発明のクラスターノックアウトによってヒト化及び更なるノックアウト実験を行うことができる。作出する動物の性質に応じて、多数の異なるアッセイプロセスに着目することができる。特に、特定の薬物代謝遺伝子クラスターの如何なる薬物化合物の代謝への関与も確認することができる。また、特定のクラスターが欠失した際に第2の選択経路による代謝への代償作用があるとすれば、それを評価することによって如何なる薬物代謝系における冗長性の程度も確認することができる。また、如何なる毒性二次代謝物の生成も別々の代謝酵素に帰属させることができる。また、体組織内での転写因子や薬物代謝酵素、薬物輸送タンパク質の処理や分布に薬物化合物が影響を及ぼすかどうか調べることができる。また、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現の持続期間に薬物化合物が影響を及ぼすかどうか調べることができる。有用なアッセイプロセスの他の例は当業者には明らかであろう。
【0146】
動物における表現型の変化(例えば、生理学的作用)を測定することができる。このような生理学的作用としては、例えば、病態(胆管壊死等)や中毒性副作用を挙げることができる。
【0147】
薬物化合物の代謝速度を調べることができる。この代謝速度は、該化合物の投与によってもたらされる毒性や活性を測定するか、該化合物の半減期を測定するか、又は薬物代謝酵素のレベルを測定することによって求めることができる。例えば、該化合物の代謝速度は、酸化生成物の生成速度や酸化中間体から次に産生される生成物の生成速度として測定することができる。或いは、この代謝速度は、最初の化合物(initial compound)の半減期や消失速度として、又は最初の化合物や最初の化合物から産生される代謝産物の毒性や活性の変化として表わすことができる。半減期は、様々な時点で採取したサンプルに存在する薬物化合物の量を求めることによって測定することができる。薬物化合物の量は、標準的な方法(例えば、高速液体クロマトグラフィーや質量分析、化合物特異的抗体を用いたウェスタンブロット解析、他の適切な方法)によって測定することができる。
【0148】
また、薬物化合物が特定の状況下で毒性代謝物や発癌性代謝産物に代謝されるかどうかを、例えば、該化合物の組織やタンパク質、DNAへの共有結合を測定するか、又はグルタチオンの欠乏を測定することによって調べることもできる。
【0149】
上述の種の測定は、薬物化合物投与後、1を越える時点、3時点、5時点又は10以上の時点で行うのが好ましい。
【0150】
従って、本発明の更なる様相は、転写因子や薬物代謝酵素、薬物輸送タンパク質の活性や発現レベルに対する薬物化合物の作用を確認するために提供されるスクリーニング方法に関する。このような方法は、本発明の上述のいずれか一様相におけるトランスジェニック動物、又は該動物由来の組織又は細胞に薬物化合物を投与することを含む。
【0151】
スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質をコードする遺伝子の誘導を測定することを含むことができる。また、スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現のレベル、又はこのような発現の持続期間を測定することを含むことができる。また、スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現の分布を測定することを含むことができる。
【0152】
薬物化合物の存在下及び非存在下でアッセイを行って、分布や代謝、毒性の差を確認することができる。様々なトランスジェニック動物や細胞、組織に対する薬物化合物の作用を評価することによって、特定の遺伝子の存在下及び非存在下における薬物化合物の作用を確認することができる。例えば、ヌルバックグラウンドを有する動物と着目遺伝子(例えば、PXRやCAR、MDR1、第1相代謝酵素、第2相代謝酵素)に関してヒト化した動物との間で薬物化合物の作用を評価することも可能である。
【0153】
従って、更なる様相において、本発明は、本明細書に記載の生体異物の代謝又は毒性を調べるための方法であって、本明細書に記載の非ヒト動物、組織又は細胞の2種以上、3種以上、4種以上、5種以上、6種以上、7種以上、8種以上、9種以上又は10種以上に対して薬物化合物を投与することを含む方法を提供する。このような方法は、異なる非ヒト動物、組織又は細胞に関して得られた実験結果を比較する段階を更に含むのが好ましい。
【0154】
二種以上の薬物化合物を投与することができる。例えば、ある薬物化合物がCAR遺伝子の誘導をもたらす場合、該化合物はCAR転写因子を活性化すると判断される。また、対照としてのヒトCAR受容体を発現する動物にCAR受容体インバースアゴニスト(例えば、クロトリマゾール)を投与することもできる。
【0155】
本発明の更なる様相におけるアッセイによって、第1の薬物化合物の代謝が第2の薬物化合物で調節されるかどうか確認するためのスクリーニング方法を提供することができる。この方法は、第2の化合物の存在下及び非存在下で第1の化合物を本発明の上述のいずれか一様相におけるトランスジェニック動物、又は該動物由来の組織又は細胞に投与することと、表現型効果をモニタリングすることとを含む。或いは、上述のように、スクリーニング段階は、遺伝子の誘導、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現のレベルや持続期間、分布を測定することを含むことができる。第2の化合物がこれらの試験因子のいずれか一因子の変化をもたらす場合、第2の化合物は第1の化合物の代謝を調節することが確認される。例えば、第1の化合物の投与によってもたらされる毒性や活性を測定するか、又は第1の薬物化合物の半減期を測定することによって生理学的作用をアッセイすることができる。
【0156】
このように、アッセイを用いて、特定のタンパク質の発現を活性化又は誘導する能力が低いか又は検出できないために、副作用が少ないか又はインビボでの半減期が長いことが期待される薬物化合物の類似体を容易に同定することができる。
【0157】
以下、本発明の様々な様相や実施形態を実施例によって更に詳細に説明する。本発明の範囲を逸脱することなく詳細な変更を実施し得ることは理解されよう。
【図面の簡単な説明】
【0158】
【図1a】マウスCyp3aクラスター欠失のための戦略。(A)マウスCyp3aクラスター内の機能性遺伝子の染色体の構成及び方向の概略図(出典:Nelsonら、2004年)。偽遺伝子は示されていない。(B)Cyp3a57及びCyp3a59のエクソン/イントロン構造。エクソンは黒い棒で示し、ATGは両遺伝子の翻訳開始点を示す。相同組換えのターゲティングアームの位置をそれぞれ、薄灰色線(Cyp3a57)及び濃灰色線(Cyp3a59)でハイライトする。(C)相同組換えによりCyp3a57(左)及びCyp3a59(右)のターゲティングに用いたベクター。LoxP部位、lox5171部位、frt部位及びf3部位をそれぞれ、白い三角、縞模様の三角、点模様の三角、及び黒い三角で示す。(D)Cyp3a57及びCyp3a59遺伝子座における同一対立遺伝子上での相同組換え後の二重標的ES細胞内のCyp3aクラスターのゲノム構成。(E)loxP部位におけるCre介在組換え後のマウスCyp3aクラスターの欠失。Cyp3a57、Cyp3a16、Cyp3a41、Cyp3a11及びCyp3a25由来の全てのエクソンとイントロンは完全に欠失し、エクソン1〜4及びCyp3a59のプロモーターも欠失する。従って、Cre介在欠失後に残存する唯一の機能性Cyp3a遺伝子はCyp3a13であり、これは、>800kbのゲノムDNAやCypと無関係の多数の機能性遺伝子によってクラスターの残りから離れている。マウスCyp3aクラスターの欠失が成功したことを実証するのに用いたプライマーを矢印で示す。 分かり易くするため、配列は一定の縮尺率では示していない。TK=チミジンキナーゼ発現カセット、Hygro=ハイグロマイシン発現カセット、ZsGreen=ZsGreen発現カセット、P=ネオマイシンの発現を促進するプロモーター、5’ΔNeo=ATG欠損ネオマイシン。
【図1b−1】Cyp3aクラスター欠失に対するマウスホモ接合性のPCR確認。
【図1b−2】Cyp3aクラスター欠失に対するマウスホモ接合性のPCR確認。
【図1c】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な肝臓切片のヘマトキシリン及びエオシン染色。20倍の対物レンズを用いてCyp3aクラスターKO(Con)の画像を得た。他の画像の全てについては10倍の対物レンズを用いた。
【図1d】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な小腸切片のヘマトキシリン及びエオシン染色。10倍の対物レンズを用いて画像を得た。
【図1e】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な腎臓切片のヘマトキシリン及びエオシン染色。10倍の対物レンズを用いて画像を得た。
【図1f】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な肝臓切片のオイルレッドO染色。40倍の対物レンズを用いて画像を得た。
【図1g】対照とリファンピシン(60mg/kg、3日間)で処理したC57BL/6J及びCyp3aクラスターKOマウスの血漿の臨床化学解析:トリグリセリド、低密度リポタンパク(LDL)、高密度リポタンパク質(HDL)、クレアチンキナーゼ(CRE)、総ビリルビン(BIL−T)、アスパラギン酸アミノトランスフェラーゼ(AST)、アラニントランスフェラーゼ(ALT)、アルカリホスファターゼ(ALP)、アルブミン。データは平均±S.D.で示す(n=4)。
【図1h】図1h:コーン油(3日間投与)(A)又はリファンピシン(60mg/kg、3日間投与)(B)で処理後4日目の野生型及びCyp3aクラスターKOマウスにおけるミダゾラム(1mg/kg)薬物動態(誤差棒はSDを示す)。N.B.:リファンピシン処理C57BL/6Jマウスの10分時点でのSDは45ng/mLであった。グラフAとBのスケールを同じに保つため、対応する誤差棒は一部のみを示した。
【図1i】WinNonlinソフトウェアの非コンパートメント解析を用いたミダゾラムPKから算出した濃度/時間曲線下総面積(AUC)。
【図1j】C57BL/6J及びCyp3aクラスターKOマウスにおけるCyp3aアイソフォームの発現及び活性。A)ウェスタンブロッティング;肝ミクロソーム由来のタンパク質1μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.1pmolのCyp3a11組換えタンパク質(hisタグ)を用いた。一次抗体としてはウサギ抗CYP3A2(Colin Henderson, Ninewells Hospital, Dundee)を1:2000の濃度で用いた。B)7−BQの酸化、C)ミダゾラムの1’−水酸化及びD)ミダゾラムの4−水酸化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(**:P<0.01、***:P<0.001)。
【図1k】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp1aタンパク質の発現及び活性。(A)−ウェスタンブロッティング;肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.1pmolのCyp1a1組換えタンパク質を用いた。一次抗体としてはヤギ抗Cyp1a1/1a2(BDゲンテスト、cat#458131)を1:2000の濃度で用いた。(B)−エトシキレゾルフィン脱エチル化、(C)−メトキシレゾルフィン脱メチル化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05、**:P<0.01)。
【図1l】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの(A)肝ミクロソーム及び(B)腸ミクロソームにおけるCyp2bタンパク質の発現。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.01pmolのCyp2b20組換えタンパク質(hisタグ)を用いた。一次抗体としてはウサギ抗Cyp2b(Colin Henderson, Ninewells Hospital, Dundee)を1:2000の濃度で用いた。(C)−ペントキシレゾルフィン脱ペンチル化。(D)−ベンゾキシレゾルフィン脱ベンジル化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05)。
【図1m】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの肝ミクロソームにおけるCyp2cタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手したCyp2c29(0.5pmol)組換えタンパク質を用いた。一次抗体としてはウサギ抗Cyp2c(Colin Henderson, Ninewells Hospital, Dundee)を1:50000の濃度で用いた。
【図1n】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの(A)肝ミクロソーム及び(B)腸ミクロソーム。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した組換えCyp2d22(0.2pmol)を用いた。一次抗体としてはヒツジ抗Cyp2d(Colin Henderson, Ninewells Hospital, Dundee)を1:20000の濃度で用いた。
【図1o】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの(A)肝ミクロソーム及び(B)腸ミクロソームにおけるCyp2eの発現。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.2pmolのCyp2e1組換えタンパク質を用いた。一次抗体としてはヒツジ抗Cyp2e(SAPU BRC)を1:4000の濃度で用いた。
【図1p】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp4aタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した1μgのAPFO誘導ラットミクロソームを用いた。一次抗体としてはウサギ抗Cyp4a(Colin Henderson, Ninewells Hospital, Dundee)を1:20000の濃度で用いた。
【図1q】(A)媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウス(KO)の肝ミクロソームにおけるP450オキシドレダクターゼタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはBDゲンテスト(cat#452172)から入手した3μgの雄性ヒト肝ミクロソーム(プール)を用いた。一次抗体としてはウサギ抗P450レダクターゼ(CXRバイオサイエンシーズ)を1:5000の濃度で用いた。(B)−C57BL/6J及びCyp3aクラスターKOマウスのマウス肝ミクロソームにおけるシトクロムCレダクターゼ活性。データは平均±SDで示す(C57BL/6J(n=3)、Cyp3aクラスターKO(n=4))。
【図1r】媒体とリファンピシン(60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおける総P450量。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05、**:P<0.01、***:P<0.001)。
【図1s】(A)−リファンピシン(60mg/kg、3日間投与)による処理後4日目にミダゾラム(1mg/kg)をIV投与した後の野生型、Cyp3aクラスターKO及びCyp3a11KOマウスにおける1’−ヒドロキシミダゾラム薬物動態(誤差棒はSDを示す)。(B)1’−ヒドロキシミダゾラムPKから算出した曲線下面積(AUC)。
【図1t】リファンピシン(60mg/kg、3日間投与)による処理後4日目にミダゾラム(2mg/kg)を経口投与した後の野生型、Cyp3aクラスターKO及びCyp3a11KOマウスにおける1’−ヒドロキシミダゾラム薬物動態(誤差棒はSDを示す)。(B)1’−ヒドロキシミダゾラムPKから算出した曲線下面積(AUC)。
【図1u】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるトリアゾラムのα水酸化。データは平均+SDで示す(C57BL/6J及びCyp3a11KOの肝ミクロソーム(n=4)、Cyp3aクラスターKOの肝ミクロソーム及びC57BL/6Jの腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001)。Cyp3aクラスターKO及びCyp3a11KOの腸ミクロソームの活性は定量下限未満であった。
【図1v】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるニフェジピンの酸化。データは平均+SDで示す(C57BL/16J及びCyp3a11KOのミクロソーム(n=4)、Cyp3aクラスターKO(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001)。
【図1w】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるテストステロンの6β水酸化。データは平均+SDで示す(肝ミクロソーム(n=4)、腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001、**:p<0.01)。
【図1x】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソームによるテストステロンの16β水酸化。データは平均+SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(**:p<0.01)。
【図1y】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるDBFの酸化。データは平均+SDで示す(肝ミクロソーム(n=4)、腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001、*:p<0.05)。
【図1z】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるBFCの酸化。データは平均+/−SDで示す(肝ミクロソーム(n=4)、C57BL/6Jの腸ミクロソーム(n=4)、Cyp3a11KOの腸ミクロソーム(n=3)、Cyp3aクラスターKOの腸ミクロソーム(n=2))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(*:p<0.05)。
【図2a】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2b】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2c】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2d−1】Cyp2cクラスター欠失に対するマウスホモ接合性のPCR確認。
【図2d−2】Cyp2cクラスター欠失に対するマウスホモ接合性のPCR確認。
【図2e】Cyp3a/Cyp2s複合クラスターKO株の構築のPCR確認。
【図2f】Cyp3a/Cyp2s複合クラスターKO株の構築のPCR確認。
【図3a】Cyp2dクラスター交換のための方法を示す。A)マウスCyp2d26の3’ターゲティング。
【図3b】Cyp2dクラスター交換のための方法を示す。B)マウスCyp2dクラスターの5’ターゲティング。
【図3c】Cyp2dクラスター交換のための方法を示す。C)概要:Cyp2dノックアウト。
【図3d−1】Cyp2dクラスター欠失に対するマウスホモ接合性のPCR確認。
【図3d−2】Cyp2dクラスター欠失に対するマウスホモ接合性のPCR確認。
【図3e】野生型(例えば、C75BL/6J)及びCyp2dクラスターKOマウスにおけるブフラロール(10mg/kg、単回経口投与)の薬物動態。データは、投与後240分まで全血中で検出されたブフラロール濃度の平均±SD(n=4)で示す。
【図4a】CYP1Aノックアウトのための方法を示す。A)マウスCyp1aの5’ターゲティング。
【図4b】CYP1Aノックアウトのための方法を示す。B)マウスCyp1aの3’ターゲティング。
【図4c】CYP1Aノックアウトのための方法を示す。C)概要:Cyp1aノックアウト。
【発明を実施するための形態】
【実施例】
【0159】
実施例1:Cyp3aクラスター交換
相同組換えによってマウスCyp3aクラスターの5’末端を標的にして、図1aに示すように、frt、lox5171及び野生型loxP部位がCyp3a57遺伝子のエクソン1の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットとチミジンキナーゼ(TK)発現カセットを共導入した。
【0160】
マウスCyp3aクラスターの3’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、野生型loxP及びf3部位がCyp3a59遺伝子のエクソン4の下流に挿入されるようにした。図1bに示すように、5’欠損ネオマイシン耐性カセットとTK発現カセットを共導入した。翻訳開始ATGは、loxP部位によって5’欠損ネオマイシン耐性カセットからインフレームで分離させて、loxP部位によってコードされる他のアミノ酸がネオマイシンタンパク質のN末端に付着するようにする。
【0161】
Cre介在組換えによってマウスCyp3aクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。これについては、Creを用いた二重標的ES細胞のトランスフェクションとそれに続くガンシクロビルを用いた選択によってインビトロで試験した。図1eに示すように、同一対立遺伝子上でターゲティングを行った細胞においてのみ、TK遺伝子はクラスターの両端で欠失し、ヌクレオシド類似体ガンシクロビルから毒性三リン酸類似体へのTK介在転化が防止される。従って、同一対立遺伝子上でターゲティングを行ったES細胞のみがガンシクロビル選択後も生存する。
【0162】
生存ES細胞クローンをPCRで更に解析し、マウスCyp3aクラスターが欠失し、PCR陽性クローンが胚盤胞に注入されてキメラマウスが作出されたことを確認した。その後の交雑によって、Cyp3aクラスターがホモ欠損したマウスを最終的に作出した。
【0163】
その後、ES細胞に残存する野生型loxP及びlox5171部位を用い、Cre介在挿入によって置換遺伝子(例えば、ヒトCYP3Aクラスター)をマウスクラスターの前の位置において挿入することができる。残存プロモーター、ATG及びインフレーム野生型loxP部位を用いて、5’欠損選択カセット(例えば、5’欠損ネオマイシン)を再度挿入し、高効率で正しく挿入されたクローンを選択することができる。更なるfrt部位及びf3部位が正しい方向に置換カセットと共に共導入されている場合には、両端におけるfrt部位及びf3部位を用い、Flp介在欠失によって、置換遺伝子と共に導入された選択カセットを欠失させることができる。
【0164】
Cyp3a57の800bp上流に位置し、Cypと無関係の多数の遺伝子によってマウスCyp3aクラスターから離れているCyp3a13遺伝子は、上述のマウスCyp3aクラスターの欠失に影響されない。
【0165】
図1bは、IDが219678、219679、219680、219681及び219682のマウスのゲノムからのマウスCyp3aクラスター欠如のPCR確認を示す。
【0166】
C57BL/6Jとの比較によるCyp3aクラスターKOマウスの解析
研究は4群(4匹/群)で行った。マウスには、リファンピシン(60mg/kg)を3日間、腹腔内(IP)投与した。研究4日目に、ミダゾラム(1mg/kg)をIP注射により投与し、10μLの血液サンプルを10分、20分、40分、60分、120分、240分及び360分で採取してPK解析を行った(表1)。
【0167】
【表1】
【0168】
病理学
肝腫
媒体処理Cyp3aクラスターKOマウスにおいては、生物学的に無関係と思われる媒体処理C57BL/6Jマウスと比べて、肝臓/身体重量比が僅か(〜6%)ではあるが統計的に有意に増加した。リファンピシン処理の場合には、いずれのマウス株においても肝腫は生じなかった(表2)。
【0169】
【表2】
【0170】
病理組織学
ヘマトキシリン及びエオシン染色
全群のマウス全ての腎臓サンプル、肝臓サンプル及び小腸サンプルに対して病理組織学的検討を行った。処理C57BL/6Jマウスにおいては軽度の小葉中心性肝細胞肥大が認められた。Cyp3aクラスターKOマウスの肝臓には微細な小葉中心性肥大の徴候もあった。このような変化はリファンピシンによる処理に関連すると考えられた。肝臓で記録された他の微視的な知見は全て、処理マウスと対照マウスとの間で有意差が無かった。いずれの差もランダムな事象であり、自然発生的であり、マウスで通常見られる正常なバックグラウンド病理の範囲内であると見なされた(図1c)。腎臓及び小腸においてはリファンピシン処理に関連する微視的知見は記録されなかった(図1d〜1e)。この研究の条件下では、2種の異なるマウス株に対してリファンピシン(60mg/kg体重)を3日間反復投与することによって、両方の株で肝細胞肥大の病理学的証拠が得られた。トランスジェニックマウス(Cyp3aクラスターKO)ではあまり大きな影響を受けないように思われた。
【0171】
オイルレッドO染色
オイルレッドO染色により脂質沈着について肝臓サンプルを調べた。媒体処理マウス肝臓のいずれにおいても脂肪蓄積の徴候は無かった(図1f)。リファンピシン処理C57BL/6Jマウスの肝臓においては、核周辺に中程度(グレード1)の滴模様のオイルレッドO染色が観察された。これは、リファンピシン処理Cyp3aクラスターKOマウスの肝臓切片では検出できなかった。
【0172】
臨床化学
全てのマウスから血漿サンプルを採取し、トリグリセリド、アラニントランスフェラーゼ、アルカリホスファターゼ、アスパラギン酸アミノトランスフェラーゼ、アルブミン、ビリルビン(総ビリルビン及び直接ビリルビン)、クレアチンキナーゼ、高密度リポタンパク質、低密度リポタンパク質及びγ−グルタミルトランスフェラーゼについて解析した。γ−グルタミルトランスフェラーゼ活性及び直接ビリルビンレベルは定量限界未満であった。他の血漿臨床化学パラメータの値は全て、未処理C57BL/6Jマウスの公知の正常範囲内であった(図1g)。
【0173】
分子/生化学的表現型
ミダゾラムの薬物動態
野生型及びCyp3aクラスターKOマウスにおけるミダゾラムの薬物動態を図1hに示す。
【0174】
両方のマウス株はIP投与後に基質の速やかな吸収を示した。これは、3日間のリファンピシンの先行投与によっては影響を受けなかった。ミダゾラムのAUCは、野生型及びCyp3aクラスターKO対照マウスにおいて同様であった(図1i)。リファンピシン処理によって両方のマウス株で基質の排除が促進されたが、C57BL/6Jマウスにおいてより高い作用が示された。アイソフォーム特異的基質が腹腔内投与された場合、肝臓における代謝シトクロムP450の発現レベルは該基質のAUCに反比例することが先に示されている。従って、ミダゾラムのAUCは、Cyp3aクラスターKOマウスではその主要代謝酵素のクラスターが欠失しているため、より高いことが予測された。しかし、この場合にはそうではなく、恐らく、次の文献(van Waterschoot et al., 2008)に記載ものと同様の何らかの代償性変化が関与していることが示唆される。
【0175】
Cyp3aタンパク質の発現及び活性
Cyp3aクラスターKOマウスの肝ミクロソームにおけるCyp3aの発現は、処理マウス及び対照マウスの両方においてウェスタンブロッティングによる検出限界未満のレベルまで低下した(図1j)。これは、対応するC57BL/6Jマウス実験群と比べてCyp3a特異的基質7−BQの酸化が有意に低下(対照マウスにおいては18%、リファンピシン処理マウスにおいては6.7%まで低下)したことと関連付けられた。Cyp3aアイソフォーム及びCyp2cアイソフォームの両方によって触媒されるミダゾラムの1’−水酸化は、対照マウス及び処理マウスでそれぞれ53%、18%まで低下した。ミダゾラムの4−水酸化の低下(65%まで)は小さく、対照C57BL/6Jと比べてCyp3aクラスターKO対照群では統計的に有意ではなかった。これは、4−水酸化がマウスCyp3aによって支配的に触媒されることが示されている(Perl off et al., 2000)ことを考慮すると予期せぬ結果であった。ミダゾラムの4−水酸化は、Cyp3aクラスターKOリファンピシン処理群においては野生型マウスと比べて23%まで低下した。
【0176】
Cyp3aクラスターKOマウスのリファンピシンによる前処理によって、7−BQの酸化は僅かではあるが(168%)、統計的に有意に(P<0.05)増加した。Cyp3aクラスターKOマウスにおけるミダゾラムの水酸化に対するリファンピシンの作用は小さく(1’−水酸化では123%増加、4−水酸化では112%)、統計的に有意ではなかった。
【0177】
腸ミクロソームのウェスタンブロッティングでは如何なるCyp3aバンドも得られなかった(データは示さず)。
【0178】
腸ミクロソームの比活性の測定によって得たデータの不整合性が高かったが、これは、上皮細胞を擦り取って腸ミクロソームを調製する際にむらがあることを反映しているように思われる(Bonkovsky et al., 1985; Mohri and Uesawa, 2001)(データは示さず)。
【0179】
本研究に用いた抗体は、試験したサブファミリー由来の主要な組換えシトクロムP450との相互作用後にウェスタンブロット上で明確なバンドを示した。しかし、1種のP450サブファミリー内の異なるアイソフォームに対する抗体の特異性については検討しなかった。
【0180】
Cyp1aタンパク質の発現及び活性
ウェスタンブロッティングで確認したCyp1aの発現は、Cyp3aクラスターKOマウスの肝ミクロソームにおいては野生型に比べて僅かに高かった(図1k)。リファンピシン処理によって、両方のマウス株でCyp1aが僅かにダウンレギュレートされた。エトキシレゾルフィン脱エチル化活性(EROD)は、Cyp3aクラスターKOマウスの肝ミクロソーム及び野生型マウスの肝ミクロソームの両方において同様であった。ERODはCyp1a1アイソフォーム及びCyp1a2アイソフォームの両方によって触媒されるが、Cyp1a1の方が主に寄与する。リファンピシン処理によって、両方の株でEROD代謝が僅かに誘導された。野生型マウスとCyp3aクラスターKOマウスとの間でメトキシレゾルフィン脱メチル化(MROD)活性の差は無かった。MRODは主にCyp1a2によって触媒される。
【0181】
Cyp2bタンパク質の発現及び活性
マウス肝ミクロソームでのCyp2bの発現は、C57BL/6Jマウス及びCyp3aクラスターKOマウスの両方において低かった(図1l)。リファンピシン処理によって、両方の株でCyp2bが僅かに誘導された。これはPROD活性の増加が少ないことと関連付けられた。腸ミクロソームでは顕著に高いCyp2bの発現が示されたが、リファンピシン処理に対して感受性が低かった。媒体処理Cyp3aクラスターKOマウスの肝ミクロソームでは、ベンゾキシレゾルフィンO−脱アルキル化(BROD)活性が野生型に比べて低いことが示された。BRODは先にマウスCyp2bの基質として用いられた(Donato et al., 2003)。リファンピシン処理によって、試験した両方の株においてBRODが誘導された。これらのデータから、Cyp3a(更に、恐らくはCyp2cアイソフォーム)由来の酵素がBRODに関与し得ることが示唆される。
【0182】
Cyp2cタンパク質の発現
C57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp2cタンパク質の発現はウェスタンブロッティングによって評価した(図1m)。タンパク質バンド強度は、野生型と比べて、対照Cyp3aクラスターKOマウス由来のサンプルにおいて僅かに増加した。両方のマウス株では、リファンピシン処理後に顕著なCyp2c誘導が示された。
【0183】
Cyp2dタンパク質の発現
Cyp2dタンパク質のバンドは、肝臓サンプル及び腸サンプルの両方において検出された(図1n)。未処理野生型マウスの肝臓サンプルとCyp3aクラスターKOマウスの肝臓サンプルとの間でCyp2dタンパク質バンド強度に顕著な差は無かった。リファンピシン処理によって、C57BL/6JマウスではCyp2dの発現が僅かに低下したが、Cyp3aクラスターKOマウスでは僅かに増加した。野生型に比べて、Cyp3aクラスターKOでは腸Cyp2dレベルが僅かに増加した。腸ミクロソームでのCyp2dレベルに対するリファンピシン処理の顕著な作用はいずれのマウス株でも見られなかった。
【0184】
Cyp2eタンパク質の発現
Cyp2eタンパク質バンド強度は、野生型と比べてCyp3aクラスターKOマウスの肝ミクロソームでは僅かに高かった(図1o)。肝ミクロソームにおけるCyp2eレベルは、いずれのマウス株においてもリファンピシン処理によっては顕著に変化しなかった。同様に、Cyp2e発現のレベルは、野生型と比べてCyp3aクラスターKOマウスの腸においては高かった。リファンピシン処理によって、Cyp3aクラスターKOマウスでは腸Cyp2eが僅かに誘導されたが、野生型マウスでは腸Cyp2eが僅かにダウンレギュレートされた。
【0185】
Cyp4aタンパク質の発現
Cyp4aタンパク質は両方のマウス株の肝ミクロソームで検出された(図1p)。野生型マウス由来サンプルにおけるCyp4a発現レベルはリファンピシン処理後に顕著に増加した。
【0186】
マウスP450オキシドレダクターゼ(POR)タンパク質の発現
シトクロムP450オキシドレダクターゼレベルは、C57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおいては同様であった(図1q)。両方のマウス株は、PORのタンパク質発現レベルの顕著な増加によってリファンピシン処理に反応した。リファンピシン処理マウスの肝ミクロソームでは、野生型マウスと比べてシトクロムCの還元における活性が僅かに高いことが示された。この増加は統計的に有意ではなかった。
【0187】
総P450量
Cyp3aクラスターKOマウスでは、C57BL/6Jマウスと比べて、媒体処理マウスの肝ミクロソームの総P450量が僅かではあるが統計的に有意に増加することが示された(図1r)。リファンピシン処理によって、野生型マウスでは総P450量が顕著に増加した。しかし、Cyp3aクラスターKOマウスの肝ミクロソームのP450量は変化しなかった。
【0188】
結論
Cyp3aクラスターKOマウスの場合、血漿、肝臓、腎臓又は小腸においては、如何なる遺伝子型誘導の病理学的変化も示されなかった。このモデルで示されたCyp3a発現のレベルは、ウェスタンブロッティングによる検出限界未満であった。Cyp3a特異的基質7−BQに対する活性が顕著に低下(野生型マウスと比べて20%未満のレベルまで)した。ミダゾラムクリアランスに関するインビトロデータから、Cyp3aサブファミリー由来のP450はこの基質の排除に関与する唯一の代謝酵素ではないことが示唆された。未処理Cyp3aクラスターKOマウスの肝ミクロソームのミダゾラム水酸化活性は、野生型株のものに対して〜60%であった。このことは、その酸化における他のP450(例えば、Cyp2c)の関与によって説明できよう(Perloff et al., 2000; van Waterschoot et al., 2008)。リファンピシン処理によって野生型ではCyp3aが誘導されたが、Cyp3aクラスターKOでは誘導されなかった。これによって、処理Cyp3aクラスターKOマウスの肝ミクロソームにおけるミダゾラム水酸化活性にはより顕著な差(野生型に比べて20%)がもたらされた。ミダゾラムPKデータによって、ミダゾラム排除への複数の代謝酵素の関与についての更なる証拠が得られた。未処理Cyp3aクラスターKOマウスとC57BL/6Jマウスとの間ではミダゾラムPKの差は無かったが、これは先に公表されたデータと関連付けられ、Cyp3aクラスターKO株におけるP450(例えば、Cyp2cサブファミリー由来)の多少のアップレギュレーションに起因し得る(van Waterschoot et al., 2008)。リファンピシンの投与によって両方のマウス株でミダゾラムの排除が促進された。これは代謝酵素の誘導を示し、野生型マウス及びCyp3aクラスターKOマウスにおいてそれぞれ、最大470%及び270%であった(概算はAUCの変化に基づいて為された)。誘導C57BL/6Jマウスの肝ミクロソームでもミダゾラム水酸化活性の高い増加(最大380%)を示したが、Cyp3aクラスターKO株の肝ミクロソームで示されたミダゾラム水酸化の誘導はほんの僅かであった(最大125%)。インビトロ及びインビボでのミダゾラムクリアランスに対するリファンピシンの作用の差は、Cyp3aクラスターKOマウスのインビボでのミダゾラム排除の代替経路の誘導を暗示する。
【0189】
更なる実験を行い、1’−ヒドロキシミダゾラムのPKプロファイル及び他のCyp3a基質の酵素修飾について比較した。
【0190】
研究は9群で行った(表3)。マウスにはリファンピシン(60mg/kg)を3日間腹腔内(ip)投与した。研究4日目に、ミダゾラムを1〜6群のマウスにはIV注射によって投与(1mg/kg)し、7〜9群のマウスには経口投与(2mg/kg)した。血液サンプルをIV投与後2分、5分、10分、20分、30分、45分、60分、90分、120分及び150分(連続出血)で採取し、また、経口投与後10分、20分、40分、60分、90分、120分、240分及び360分で採取し、PK解析を行った(表3)。
【0191】
【表3】
【0192】
1’−ヒドロキシミダゾラムの薬物動態
先のリファンピシン処理によって野生型マウスではCyp3aが誘導されたが、Cyp3aクラスターKOマウスでは誘導されなかった。これによって、2種類のマウス株間ではインビトロでのミダゾラム水酸化活性に顕著な差がもたらされた。しかし、インビボでのミダゾラム排除における差はあまり顕著ではなく、これは、Cyp3aクラスターKOマウスにおけるインビボでのミダゾラム排除の代替経路の誘導を示唆している。本研究では、P450発現レベル概算のためのより高感度で具体的な方法として、リファンピシン誘導マウスへのミダゾラムのIV投与及び経口投与後に1’−ヒドロキシミダゾラムの薬物代謝(一次代謝産物)をモニターした。血液をB−グルクロニダーゼで前処理した後に1’−ヒドロキシミダゾラムの総濃度を測定した。親化合物をIV注入した後の1’−ヒドロキシミダゾラムの薬物動態を図1sに示す。
【0193】
曲線の終末段階はC57BL/6Jマウス及びCyp3a11KOマウスの両方において同様であるが、これは、この代謝産物のクリアランスにおける株差が非常に小さいことを示唆する。Cyp3aクラスターKOマウスPKの終末段階は、1’−ヒドロキシミダゾラムの排除が他の株と比べて僅かに遅いことを示す。1’−ヒドロキシミダゾラムのAUCは、Cyp3aクラスターKOマウスでは野生型株と比べて顕著に(〜3倍)低下したが、これは、Cyp3aサブファミリー由来のシトクロムP450がこの代謝産物生成の主要な経路であることを示す。Cyp3a11KOマウスにおけるAUCの低下はあまり顕著ではなかった(〜2倍)が、これは、1’−ヒドロキシミダゾラムの生成にCyp3aサブファミリーの他のメンバーが多少関与していることを示唆する。ミダゾラム経口投与後の1’−ヒドロキシミダゾラムの薬物動態を図1tに示す。
【0194】
曲線の終末段階は全ての株で同様であった。しかし、初期の吸収段階では差が見られた。野生型マウスと比べて、Cyp3aクラスターKOマウスやCyp3a11KOマウスではバイオアベイラビリティが低かった。これは、ミダゾラム経口投与後の1’−ヒドロキシミダゾラムの生成に腸シトクロムP450が大きく寄与していることを示す。このことは、IV注入後に野生型マウスとKOマウスとの間で1’−ヒドロキシミダゾラムのAUCが非常に大きく異なっていることを示したIVデータによって支持された。本研究においては、曲線の吸収段階を定めるデータ点が非常に少なかったため、C57BL/6JにおけるCyp3a酵素からの影響は過小評価されているように思われる。
【0195】
Cyp3a酵素活性
肝ミクロソーム及び腸ミクロソームによるCyp3a特異的基質の酸化は、トリアゾラム、ニフェジピン、テストステロン、ジベンジルフルオレセイン(DBF)及び7−ベンジルオキシ−4−トリフルオロメチルクマリン(BFC)を用いて試験した。トリアゾラム、ニフェジピン及びテストステロンは、10種のCYP3A4基質の様々なCYP3A4リガンドとの相互作用の統計的解析に基づき、CYP3A4内の異なる基質グループに帰属された(Kenworthy et al., 1999)。DBFとBFCは蛍光代謝物を産生し、CYP3A4阻害剤のハイスループットスクリーニングにおいて広く応用されている。他の蛍光基質である7−ベンジルオキシキノリン(BQ)は、Cyp3aに対して良好な特異性を示している。
【0196】
関連するグラフの全てにおいて、小腸由来サンプルの活性は右のY軸スケールを参照し、肝ミクロソームの活性は左のY軸スケールを参照されたい。
【0197】
トリアゾラムα−水酸化
肝ミクロソーム及び腸ミクロソームによるトリアゾラム酸化の結果を図1uに示す。Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方においてトリアゾラムの水酸化は顕著に低下した。このことは、公表データ(Perloff et a1.,200O)とよく関連付けられ、トリアゾラムはCyp3aサブファミリー由来のマウスシトクロムP450に対して高い特異性を有する基質であることが示された。また、このデータから、Cyp3aクラスターKO及びCyp3a11KOの活性は非常に低く、統計的に違いが無かったため、水酸化にはCyp3a11が大きく寄与し、他のCyp3a酵素からの影響は無視できることも分かった。
【0198】
ニフェジピン酸化
Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソームによるニフェジピン酸化は、野生型マウスと比べて有意に(P<0.001)低下した(図1v)。この低下はトリアゾラムの場合と比べてあまり顕著ではなかったが、これは、異なるサブファミリー由来の酵素の潜在的な関与を示す。Cyp3aクラスターKO及びCyp3a11KOの平均値は異なったが、統計的な有意差は無かった。腸ミクロソームによるニフェジピン酸化においては統計的な有意差は無かった。
【0199】
テストステロン水酸化
テストステロンは、数種の異なるシトクロムP450によって水酸化される。6p−水酸化は通常、Cyp3aサブファミリー由来の酵素に起因するが、16B−水酸化はCyp2b10及びCyp2c29によって触媒される(Honkakoski et al., 1992)。Cyp3aクラスターノックアウトマウスにおけるCyp2cサブファミリー由来酵素のアップレギュレーションは先に報告されている(van Waterschoot et al., 2008)。従って、両方の水酸化については肝ミクロソーム及び腸ミクロソームを用いて検討した(図1w〜1x)。
【0200】
Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方においてテストステロンの6β−水酸化は有意に低下した。トランスジェニックマウス株の肝ミクロソームによる6β−テストステロンの生成においては差が無かった。Cyp3aクラスターKOマウスの腸ミクロソームの活性はCyp3a11KOマウスの場合と比べて低かった(P<0.05)が、これは、マウス小腸によるテストステロン6β−水酸化に他のCyp3a酵素が関与していることを示し得る。テストステロンの16β−水酸化は、野生型マウスと比べて両方のトランスジェニック株の肝ミクロソームで増加したが、これは何らかの代償性変化を示唆し、この変化は、Cyp2cサブファミリー由来の酵素の過剰発現であると思われる(van Waterschoot et al., 2008)。Cyp3aクラスターKOの平均値はCyp3a11KOの場合と比べて高かったが、統計的な有意差は無かった。
【0201】
DBFの酸化
DBFの酸化の結果を図1yに示す。肝ミクロソームによるDBF酸化は、Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方に由来するサンプルにおいては顕著な低下を示した。2種のノックアウト株間では統計的な有意差は無かったが、これは、Cyp3a11が主要な代謝酵素であることを示唆する。腸ミクロソームによるDBF酸化においては、野生型株とノックアウト株との間で統計的な有意差は無かった。
【0202】
BFCの酸化
BFCの酸化の結果を図1zに示す。
肝ミクロソームサンプル間でBFC活性に差は無かった。腸ミクロソームにおいては、僅かではあるが統計的に有意な増加が見られたが、これは、BFCを酸化する、トランスジェニックマウスにおける他のサブファミリー由来のシトクロムP450のアップレギュレーションを示唆する。
【0203】
結論
Cyp3aクラスターKOモデルは先に基質特異的挙動を示した。このモデルと野生型マウスとの明らかな差は7−BQの酸化に見られた。しかし、ミダゾラムの水酸化では差が見られず、これは、この反応における他のP450(例えば、Cyp2c)の関与を示唆する(Perloff et al., 2000; van Waterschoot et al., 2008)。リファンピシン処理によって野生型ではCyp3aが誘導されたが、Cyp3aクラスターKOでは誘導されなかった。この結果、処理Cyp3aクラスターKOマウスの肝ミクロソームにおけるミダゾラム水酸化活性は顕著に低下した。ミダゾラムAUCで示されたように、インビボでの差はあまり顕著ではなかったが、これは、Cyp3aクラスターKOマウスにおけるミダゾラム排除の代替経路の誘導を示唆する。このことは、ミダゾラム代謝産物の薬物動態によって、Cyp3a発現レベルをより具体的且つ高感度に概算し得ることを暗示した。
【0204】
親化合物をIV投与した後の1’−ヒドロキシミダゾラムのAUCはCyp3aクラスターKOマウスにおいて顕著に(〜3倍)低下したが、これは、この代謝産物の生成にCyp3aサブファミリーが大きく寄与することを示す。この方法は、先に用いられた方法の改良であり、ミダゾラムAUCの測定(1.6倍)に基づいた。しかし、インビトロで見られた差(6倍)には完全に対応しなかった。この違いは、親化合物及び代謝産物自身の両方の分布量やクリアランスからの代謝産物AUCの複合依存性によって説明し得る(Rowland and Tozer, 1995)。腸代謝や吸収が代謝産物の処理に関与する場合には、親薬物の経口投与後に他の複雑な因子が存在する。マウス株間の基質酸化における差は、肝ミクロソームと腸ミクロソームとの間でテストステロンやトリアゾラムに関してはよく対応した。しかし、ニフェジピン、DBF及びBFCの酸化に関してはパターンが異なった。これは、後者の反応に様々な酵素が関与し、これらの酵素の相対濃度が肝臓や小腸で異なることを示し得る。このことは、シトクロムP450の発現レベルの概算において、経口投与後の代謝物AUCがIV注入後の代謝物AUCと比べて有益ではないことを示唆した。本研究に用いたCyp3a基質は、その酸化におけるCyp3aサブファミリーの影響の点で次のようにランク付けすることができる(トリアゾラム>DBF:テストステロン>ニフェジピン>>BFC)。
【0205】
CYP3A4基質であるBFCの酸化に関するデータから、Cyp3aサブファミリー由来のマウスシトクロムP450に対するこの化合物の特異性が低いことが分かる。このことは、先に報告されたBFC酸化におけるヒトシトクロムP450とラットシトクロムP450との間の差(Stresser et al., 2002)と一致する。
【0206】
Cyp3aクラスターノックアウトの作用に関しては、全ゲノムマイクロアレイ及び肝臓から調製したRNAを用いて転写レベルでも評価した。本研究においては、Agilent Whole Mouse Genome Oligo Microarrayスライド(G4122−60510)及びAgilent 44KガスケットスライドG2534−60005を用いた。
【0207】
マウスには、表1に示すように、リファンピシン/コーン油(60mg/kg/3日間)又は媒体のみを腹腔内注射によって投与した。遺伝子発現の変化倍数(Fold changes)を表4に示す。
【0208】
【表4−1】
【0209】
【表4−2】
【0210】
【表4−3】
【0211】
実施例2:Cyp2cクラスター交換
相同組換えによってマウスCyp2cクラスターの5’末端を標的にして、野生型loxP及びf3部位がCyp2c55遺伝子のエクソン1の下流に挿入されるようにした。図2aに示すように、5’欠損ネオマイシン耐性カセットとTK発現カセットを共導入した。翻訳開始ATGは、loxP部位によって5’欠損ネオマイシン耐性カセットからインフレームで分離させて、loxP部位によってコードされる他のアミノ酸がネオマイシンタンパク質のN末端に付着するようにする。
【0212】
マウスCyp2cクラスターの3’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、図2bに示すように、frt、lox5171及び野生型loxP部位がCyp2c70遺伝子のエクソン5の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットとチミジンキナーゼ(TK)発現カセットを共導入した。
【0213】
Cre介在組換えによってマウスCyp2cクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。これについては、Creを用いた二重標的ES細胞のトランスフェクションとそれに続くガンシクロビルを用いた選択によってインビトロで試験した。図2cに示すように、同一対立遺伝子上でターゲティングを行った細胞においてのみ、TK遺伝子はクラスターの両端で欠失し、ヌクレオシド類似体ガンシクロビルから毒性三リン酸類似体へのTK介在転化が防止される。従って、同一対立遺伝子上でターゲティングを行ったES細胞のみがガンシクロビル選択後も生存する。生存ES細胞クローンをPCRで更に解析し、マウスCyp2cクラスターが欠失し、PCR陽性クローンが胚盤胞に注入されてキメラマウスが作出されたことを確認した。その後の交雑によって、Cyp2cクラスターがホモ欠損したマウスを作出した。
【0214】
その後、ES細胞に残存する野生型loxP及びlox5171部位を用い、Cre介在挿入によって置換遺伝子(例えば、ヒトCYP2Cクラスター)をマウスクラスターの前の位置において挿入することができる。残存プロモーター、ATG及びインフレーム野生型loxP部位を用いて、5’欠損選択カセット(例えば、5’欠損ネオマイシン)を再度挿入し、高効率で正しく挿入されたクローンを選択することができる。更なるfrt部位及びf3部位が正しい方向に置換カセットと共に共導入されている場合には、両端におけるfrt部位及びf3部位を用い、Flp介在欠失によって、置換遺伝子と共に導入された選択カセットを欠失させることができる。このホモ接合性クラスターKO作出のPCRによる実証を図2dに示す。
【0215】
実施例3:ホモ接合体Cyp3aクラスターノックアウト/Cyp2cクラスターノックアウトマウス
Cyp3aKOマウスとCyp2cKOマウスを交雑させて生存可能なマウス(ID234254)を繁殖させた。このマウスは、マウスCyp3aクラスター(7個の遺伝子が欠失)及びCyp2cクラスター(14個の遺伝子が欠失)の両方がホモ欠損している。
【0216】
図2eは、マウスID234254のゲノムからマウスCyp3aクラスターが欠如していることのPCR確認を示す。図2fは、同一マウスのゲノムからマウスCyp2cクラスターが欠如していることのPCR確認を示す。
【0217】
ホモ接合体Cyp3aクラスターKO/Cyp2cクラスターKOマウスは、実施例1で詳述するように、その病理組織学、生化学、タンパク質発現及びmRNA転写レベルに関して特徴付けされよう。また、公知で仮定のCyp2cP450アイソフォーム基質を用いた該株の更なる特徴付けも実施されよう。
【0218】
実施例4:Cyp2dクラスター欠失
相同組換えによってマウスCyp2dクラスターの3’末端を標的にして、図3aに示すように、frt、lox5171及び野生型loxP部位がCyp2d26遺伝子のエクソン1の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットを共導入した。続くマウスクラスターの欠失によって無傷のCyp2d26プロモーターとCyp2d26遺伝子のエクソン1が残るが、これらは、後述のようにクラスターの5’末端において挿入されるヒトCYP2D6の発現を妨げる可能性があるため、Cyp2d26プロモーターからの発現を終結させる3’ターゲティングベクターにスプライスアクセプターポリアデニル化モチーフ(sApA)を含有させた。
【0219】
マウスCyp2dクラスターの5’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、図3bに示すように、frt部位、野生型loxP部位及びネオマイシン耐性カセット3’によってloxP部位に隣接させたヒトCYP2D6発現カセットがCyp2d22遺伝子のエクソン3の下流に挿入されるようにした。
【0220】
loxP部位におけるCre介在組換えによってヒト発現カセットを維持しながらマウスCyp2dクラスターを欠失させることができるが、Cyp2d遺伝子座の完全なノックアウト(図3cに示す)をもたらすマウスクラスターとヒト発現カセットの同時欠失は、frt部位におけるFlp介在組換えの結果である。
【0221】
Cre介在又はFlp介在組換えによってマウスCyp2dクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。従って、Creを用いたインビトロ欠失によって二重標的ESクローンを更に解析した後、クラスターの欠失を可能とするプライマー対を用いてPCR解析を行った。インビトロCre欠失はトランスフェクト細胞の一部でしか生じず、また、欠失が成功した個々の細胞を選択することができないため、PCR解析は欠失クローン及び未欠失クローンのプールに対して行った。PCR陽性未欠失親クローンを胚盤胞に注入してキメラマウスを作出した。Cre又はFlp欠失株、或いは各リコンビナーゼを組織特異的に発現する株と交雑させた後に交雑受精させることによって、全身又は特定の組織において欠失し、ヒト発現カセットを含む(又は含まない)ホモ接合Cyp2dクラスター欠失マウスが作出されよう。
【0222】
このホモ接合性クラスターKO作出のPCRによる実証を図3dに示す。
【0223】
Cyp2dKOマウスにおけるブフラロールPK
Cyp2d特異的基質であるブフラロールの薬物動態をCyp2dKOマウス(Cyp2dアイソフォーム全てが無効化されたマウス)において検討した。野生型であるC57BL/6Jマウスを対照として用いた。ブフラロールのAUCは、野生型マウスと比べるとCyp2dKOマウスでは顕著に増加したことが示された(図3e)。このことは、インビトロデータ(Bogaards et al., 2000)(即ち、ブフラロール水酸化の速度は、ヒト肝ミクロソームと比べてマウス肝ミクロソームでは〜10倍高いことが示された)とよく関連付けられた。
【0224】
参考文献
Anderson, S. P., Dunn, C., Laughter, A., Yoon, L., Swanson, C., Stulnig, T. M., Steffensen, K. R., Chandraratna, R. A., Gustafsson, J. A., and Corton, J. C. (2004). Overlapping transcriptional programs regulated by the nuclear receptors peroxisome proliferator-activated receptor alpha, retinoid X receptor, and liver X receptor in mouse liver. Mol Pharmacol 66, 1440-52.
Bogaards JJ, Bertrand M, Jackson P, Oudshoorn MJ, Weaver RJ, van Bladeren PJ and Walther B (2000) Determining the best animal model for human cytochrome P450 activities: a comparison of mouse, rat, rabbit, dog, micropig, monkey and
Bonkovsky HL, Hauri HP, Marti U, Gasser R and Meyer UA (1985) Cytochrome P450 of small intestinal epithelial cells. Immunochemical characterization of the increase in cytochrome P450 caused by phenobarbital. Gastroenterology 88:458-467.
Donato MT, Klocke R, Castell JV, Stenzel K, Paul D and Gomez-Lechon MJ (2003) Constitutive and inducible expression of CYP enzymes in immortal hepatocytes derived from SV40 transgenic mice. Xenobiotica 33:459-473
Honkakoski, P, Auriola S and Lang MA (1992) Distinct induction profiles of three phenobarbital-responsive mouse liver cytochrome P450 isozymes. Biochem Pharmacol
43:2121-2128.
Kenworthy KE, Bloomer JC, Clark SE and Houston JB (1999) CYP3A4 drug interactions: correlation of 10 in vitro probe substrates. Br J Clin Pharmacol 48:716-727.
Ide, T., Shimano, H., Yoshikawa, T., Yahagi, N., Amemiya-Kudo, M., Matsuzaka, T., Nakakuki, M., Yatoh, S., Iizuka, Y., Tomita, S., Ohashi, K., Takahashi, A., Sone, H., Gotoda, T., Osuga, J., Ishibashi, S., and Yamada, N. (2003). Cross-talk between peroxisome proliferator-activated receptor (PPAR) alpha and liver X receptor (LXR) in nutritional regulation of fatty acid metabolism. II. LXRs suppress lipid degradation gene promoters through inhibition of PPAR signaling. Mol Endocrinol 17, 1255-67.
Inc, A. t. (2007). Agilent Feature Extraction Software v9.5 User Guide. Agilent Publication number g4460-90005, 1-250.
Mohri K and Uesawa Y (2001) Enzymatic activities in the microsomes prepared from rat small intestinal epithelial cells by differential procedures. Pharm Res 18:1232-1236.
Perloff MD, von Moltke LL, Court MH, Kotegawa T, Shader RI and Greenblatt DJ (2000) Midazolam and triazolam biotransformation in mouse and human liver microsomes: relative contribution of CYP3A and CYP2C isoforms. J Pharmacol Exp Ther 292:618-628.
Rowland M and Tozer N (1995) Clinical Pharmacokinetics: Concepts and Applications. Lippincot Williams & Wilkins, Philiadelphia, US.
Saini, S. P., Sonoda, J., Xu, L., Toma, D., Uppal, H., Mu, Y., Ren, S., Moore, D. D., Evans, R. M., and Xie, W. (2004). A novel constitutive androstane receptor-mediated and CYP3A-independent pathway of bile acid detoxification. Mol Pharmacol 65, 292-300.
Stresser DM, Turner SD, Blanchard AP, Miller VP and Crespi CL (2002). Cytochrome P450 fluorimetric substrates: identification of isoform-selective probes for rat CYP2D2 and human CYP3A4. Drug Metab Dispos 30:845-852.
van Waterschoot, R. A., van Herwaarden, A. E., Lagas, J. S., Sparidans, R. W., Wagenaar, E., van der Kruijssen, C. M., Goldstein, J. A., Zeldin, D. C., Beijnen, J. H., and Schinkel, A. H. (2007). Midazolam metabolism in Cytochrome P450 3A knockout mice can be attributed to upregulated CYP2C enzymes. Mol Pharmacol. 73, 1029-36.
Weng, L., Dai, H., Zhan, Y., He, Y., Stepaniants, S. B., and Bassett, D. E. (2006). Rosetta error model for gene expression analysis. Bioinformatics 22, 1111-21.
Xu, C., Li, C. Y., and Kong, A. N. (2005). Induction of phase I, II and III drug
metabolism/transport by xenobiotics. Arch Pharm Res 28, 249-68.
Zhou, J., Zhang, J., and Xie, W. (2005). Xenobiotic nuclear receptor-mediated regulation of UDP-glucuronosyl-transferases. Curr Drug Metab. 6, 289-98.
【技術分野】
【0001】
本発明は、薬物代謝に関与する遺伝子のクラスターがノックアウトされた薬物代謝マウスモデルの作出に関する。
【背景技術】
【0002】
治療用途や他の目的のための新しい薬物や化学薬品の開発は非常に複雑である。特に重要なのは、このような化学薬品が、適切な薬物動態を有していようと代謝の結果として何らかの安全性の問題を生じさせるものであろうと、体内でどのように処理されるか把握することである。薬物の代謝や処理(disposition)、排出に関与するタンパク質の多くは、遺伝子の数や機能、調節において非常に顕著な種差を示す多重遺伝子族のメンバーである。このような理由から、代謝経路や毒性を確かめるために実験動物で行う実験は、大幅に妥協したものになることがあり、その結果、ヒトの状況を忠実に示すことがなくなる。
【0003】
このような複雑さの一例は、哺乳動物シトクロムP450系で見られる。即ち、この系では特定の代謝機能を担うタンパク質の多重遺伝子族のサイズが種によって大きく異なる。ヒトP450遺伝子をマウスゲノムに導入することによってヒト化マウスモデルを作出する提案が文献報告されている。しかし、導入と同時にマウス遺伝子を無効化しないと、該遺伝子の活性からの干渉が大きくなり、最終的には、その系からの如何なる読み出しも無意味になる。より高度なアプローチにおいては、ヒト化の際に等価なマウス遺伝子を無効化することが提案されており(WO2006/064197参照)、これによってマウスの機能をヒト配列で置換することができる。
【0004】
しかし、Nelsonら、2004年(Pharmacogenomics 14(1): 1)によると、マウスには約72種類の機能性遺伝子が存在するが、ヒトにおいては約27種類しか存在しない。従って、多くの遺伝子に対し、直接的にオルソロガスな遺伝子は少なく(例えば、CYP2E1やCYP1A1、CYP1A2)、ヒトにおいてはごく僅かな遺伝子しか存在しないが、マウスにおいて多数の等価な遺伝子が存在する。その例としては、CYP2A/2B遺伝子ファミリーやCYP2Dが挙げられる。WO2006/064197では、個々のヒト遺伝子をクラスター内の複数の等価なマウス遺伝子と交換することが提案されており、これによって、マウスモデルでヒト薬物代謝の特性を詳細に解析することが可能になることが期待されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
しかし、マウスP450遺伝子のクラスターの機能をヒト等価物で置換せずに該クラスターを欠失させる試みは未だになされていない。このようなP450酵素によって行われる酵素反応は、非常に重要な生命プロセスに関与する。実際、多数のCYP遺伝子における変異は先天性代謝異常に関与し、また、数々の重要な臨床的に価値のある疾患に寄与する。
【0006】
しかし、上述の重要な代謝機能の検討を可能にするマウスを作出できるよう、全シトクロムP450遺伝子クラスターを個々に組み合わせて欠失させることができれば非常に有利であろう。複数の遺伝子クラスターが欠失したマウスによって、化学/薬物動態や体内分布、毒性を定める上でのシトクロムP450系の一般的な役割を確認することができるであろう。このようなモデルはそれ自身でも、また、特定の遺伝子ファミリー又はクラスターに関してヒト化したモデルと組み合わせても非常に有用で多くの用途があると考えられる。
【課題を解決するための手段】
【0007】
本発明によると、薬物代謝酵素をコードする多重遺伝子クラスターの全てのメンバーが機能しないようにされており(rendered inoperable)、欠失クラスターの中の遺伝子と等価な機能を有するヒト置換遺伝子を含まないマウスが提供される。
【0008】
本明細書においては、マウスの全組織内の特定のP450クラスターからシトクロムP450遺伝子全てを欠失させることによって、見かけ上正常で生存可能なマウス系が得られることが示される。この知見から、上述のシトクロムP450酵素に起因するハウスキーピング的な役割が、予測される表現型の結果と著しく違うことにおいて、結局は生存や受精に必須ではあり得ないことが暗示され、マウスにおける個々の複数のP450遺伝子クラスターノックアウトの開発への道が開かれる。
【0009】
これはかなり驚くべきことである。現在まで文献においては、P450薬物代謝酵素をコードする遺伝子クラスターが欠失したマウスについては報告されていない。この一理由としては、シトクロムP450系が、外来化学薬品の代謝や処理において主な役割を果たすことに加え、胚発生や正常なホメオスタシスに必須の多数のホルモンの代謝や処理に関与することが分かっていることが考えられる。例としては、レチノイン酸やステロイドホルモンが挙げられる。全シトクロムP450遺伝子クラスターの欠失によって胚や新生児の致死がもたらされることが予想されよう。結局はこのようなことが生じないのは、かなり驚くべきことである。
【0010】
こうして作出したモデルにおいては、上述の薬物代謝機能が完全に欠失している。従って、このようなモデルは幅広い潜在的用途を有する。第1の例においては、例えば、このようなモデルを用いて、薬物や化学薬品の有効性や毒性、薬物動態、分布におけるP450代謝の役割を確かめることができる。他の可能性は、第2相薬物代謝酵素のクラスター(例えば、UGT1A及び/又はGST遺伝子)の完全な除去である。
【0011】
本発明者らは、このような研究を行う際には、関連機能を有するタンパク質をコードする遺伝子を試験動物においてできるだけ多く欠失させることが最も重要であると考えている。この一理由は、各種薬物代謝遺伝子同士の間(特に、P450遺伝子間)で高度な冗長性が見られるためである。一例としてはCYP2D6P450クラスターを挙げることができるが、該クラスターにおいては、ヒトにおける唯一の機能性遺伝子に対応して、基質特異性が重複する9種類の機能性遺伝子が存在する。この理由から、マウスクラスター内の遺伝子の1個のみを欠失させ、残りを機能性モジュールとして保持するのは殆ど意味がない。
【0012】
「多重遺伝子クラスター」とは、薬物代謝酵素をコードする二を超える遺伝子のクラスターを意味する。本明細書において「クラスター」とは、二を超える遺伝子産物を産生する能力を有し、同一遺伝子の複数のスプライシングを含むゲノムの如何なる領域をも指す。
【0013】
例えば、マウスには7種類のP450遺伝子クラスター、即ち、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターが存在すると考えられる。
【0014】
現在の知見によると、Cyp2abfgstクラスターは次の機能性遺伝子、即ち、2s1、2b10、2b13、2b9、2a4、2b23、2b19、2g1、2a5、2a22、2a12、2f2及び2t4を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子13種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8、9、10、11又は12種のみ)が欠失の対象とされ得る状況であってもよい。
【0015】
現在の知見によると、Cyp2cクラスターは次の機能性遺伝子、即ち、2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。
【0016】
これらの遺伝子15種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8、9、10、11、12、13又は14種のみ)が欠失の対象とされ得る状況であってもよい。例えば、実施例2に記載のノックアウトから2c44遺伝子を特異的に排除したが、これは、該遺伝子が主要マウスCyp2cクラスターの4.1Mb下流に位置し、Cyp遺伝子と何ら関係の無い遺伝子数個分だけ主要クラスターから離れていることに基づく。結果として、これらの無関係な遺伝子の除去は望ましくなく、得られる表現型を複雑にするであろう。
【0017】
現在の知見によると、Cyp2dクラスターは次の機能性遺伝子、即ち、2d22、2d11、2d10、2d9、2d12、2d34、2d13、2d40及び2d26を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子9種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7又は8種のみ)が欠失の対象とされ得る状況であってもよい。コーカソイドの約6%においては等価なヒト遺伝子クラスター発現からのCYP2D6の発現が無いため、このマウスクラスター内の遺伝子の欠失は実際に真のヒト多型を反映する。このタンパク質は神経遮断薬(例えば、抗うつ薬や統合失調症治療用薬物)の代謝において主要な役割を果たすため、かなり重要である。製薬会社はCYP2D6によって代謝される薬物を撤退(back)させたがらないため、薬物を開発している間にその薬物のCYP2D6ライアビリティーの有無をできるだけ早く知る必要がある。従って、CYP2D6レベルの存在下及び非存在下の両方で薬物(特に抗うつ薬)の代謝について研究し得ることは大きな利益となる。
【0018】
現在の知見によると、Cyp2jクラスターは次の機能性遺伝子、即ち、2j13、2j12、2j7、2j11、2j8、2j6、2j9及び2j5を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子8種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば3、4、5、6又は7種のみ)が欠失の対象とされ得る状況であってもよい。
【0019】
現在の知見によると、Cyp3aクラスターは次の機能性遺伝子、即ち、3a13、3a57、3a16、3a41、3a11、3a25及び3a59を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子7種全てが欠失していてもよいが、これらの遺伝子の一部のみ(例えば、3、4、5又は6種のみ)が欠失の対象とされ得る状況であってもよい。例えば、本発明の好ましい実施形態は、3a57、3a16、3a41、3a11及び3a25遺伝子が欠失しているが、3a13及び3a59遺伝子の一方又は両方が無傷のまま残っているマウスである。これらの遺伝子の両方は物理的に該クラスターの外側に位置し、Cypファミリーに属さない遺伝子によってCyp3a遺伝子の残りから離れている。Cypと無関係の遺伝子の欠失は本発明のモデルの価値を分かりにくくするため、これは理想的ではなく、よってあまり好ましくない。また、これらの遺伝子も含むクラスターのノックアウトは技術的に難しく、該クラスターの残りの欠失も含む細胞においてES細胞レベルでこれらの特定遺伝子を更にノックアウトすることによってのみ成し得る。
【0020】
現在の知見によると、Cyp4abxクラスターは次の機能性遺伝子、即ち、4x1、4a29、4a12a、4a12b、4a30b、4a14、4a10、4a31、4a32及び4b1を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子10種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7、8又は9種のみ)が欠失の対象とされ得る状況であってもよい。
【0021】
現在の知見によると、Cyp4fクラスターは次の機能性遺伝子、即ち、4f39、4f17、4f16、4f37、4f40、4f15、4f14及び4f13を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子8種全てが欠失しているのが好ましいが、これらの遺伝子の一部のみ(例えば、3、4、5、6又は7種のみ)が欠失の対象とされ得る状況であってもよい。
【0022】
UGT1Aクラスターは次の機能性遺伝子、即ち、1a1、1a2、1a5、1a6、1a6a、1a7c、1a8、1a9及び1a10を含む。従って、本発明はその一様相において、これらの遺伝子が全組織で機能しないようにされており、該クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスを提供する。これらの遺伝子9種全てが欠失しているが、これらの遺伝子の一部のみ(例えば、3、4、5、6、7又は8種のみ)が欠失の対象とされ得る状況であってもよい。UGT1a1遺伝子の欠失によって疾患が生じるため、この遺伝子を欠失させないのが好ましい。
【0023】
本発明の方法においては、クラスター欠失の組合せも考えられる。従って、本発明の或る実施形態においては、複数のP450クラスターを機能しないようにすることができる。例えば、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターのいずれか2種を同一動物内で欠失させることができる。他の実施形態においては、完全ヌルP450バックグラウンドのマウスを作出するために、該遺伝子クラスターの3種、4種、5種、6種又は全7種を欠失させることができる。ノックアウトするクラスターの好ましい組合せとしては、Cyp3a、2c及び2dの内の2種又は全3種を含むもの、例えば、Cyp3aとCyp2c、Cyp2cとCyp2d、及びCyp3aとCyp2dが挙げられる。Cyp3a及びCyp2cクラスターの欠失の組合せが好ましく、これによってP450代謝の大部分がノックアウトされる。
【0024】
これらのモデルは、同一遺伝子ファミリーのメンバー間(実際には、遺伝子ファミリー間)の機能的冗長性のため、個々のシトクロムP450アイソザイムの欠失に比べて大きな利益をもたらす。
【0025】
本発明の動物の全組織においてP450の機能性が欠失しているのが好ましい。肝臓が薬物代謝に関して真に重要な唯一の組織であるという間違った考えが広く受け入れられている。真実はこれとは程遠く、実際、P450の機能は、肝臓以外の様々な組織(特に、消化管や血液脳関門等が挙げられる)で発現される。従って、全組織での機能の完全抑止は、P450クラスターノックアウトの作用を適切に評価するために必要である。現在利用可能な技術を用いる場合、このようなことは、胚性幹細胞段階でP450クラスター内の全遺伝子の発現をノックアウトする技術によってのみ実際に成し遂げられる。P450酵素に割り当てられている発生学的役割のため、その大規模なノックアウトは不可能であろうことが常に推測される。
【0026】
従って、本発明のモデルは、特定のP450系が不活性化された他のモデルよりも利益をもたらす。他のモデルの一例としては、肝臓でしか生じないシトクロムP450レダクターゼの条件的不活性化が挙げられる。これに対し、本発明のモデルにおいては、好ましいことに該酵素の欠失が全組織で生じ得る。
【0027】
ある状況下では、全クラスター又はクラスターの組合せのノックアウトが致死の原因となる可能性もあるが、本発明によると、このシナリオにおいては、一組織のみにおいて遺伝子、遺伝子の組合せ又はクラスターをノックアウトするのが好ましい場合もある。このような状況下でノックアウトを限定すべき最も適切な組織は、薬物代謝における役割を考慮すれば肝臓となるであろう。
【0028】
また、多くの理由から、本発明の実験的アプローチもシトクロムP450阻害剤の使用に比べて大きな利益をもたらす。第一に、特異性を欠く阻害剤はそれ自体が薬理学的作用や毒物学的作用を有する。第二に、このような分子は慢性的な投薬状況下では用いることができない。更に、阻害剤は、試験対象の化合物の治療特性や毒物学的特性を調節することが多い。また、阻害剤は、特定の遺伝子ファミリー内の全シトクロムP450を等価な効力レベルで同時に阻害する能力を有しない。最後に、阻害剤を用いた実験は遥かに面倒且つ複雑で解釈が難しい。
【0029】
本発明に係るマウス系の作出によって、ヒトにおける薬物反応や化学反応を決める因子に対する我々の理解が顕著に増すであろう。このようなモデルは、多くの異なるスクリーニングシナリオ(例えば、効力スクリーニングやPK/PDモデリング、薬物安全性試験等)に適用することができる。本発明のノックアウトモデルは、ヒトにおいて見られないげっ歯類特異性代謝産物が生成し、従来の動物試験から得た効力、PK/PD又は安全性の結果を分かりにくくしているような場合には特に注目され得る。
【0030】
本発明のマウスモデルの有用性に関する根拠は、一般にP450代謝速度が遅いため、マウスが多くの薬物をヒトよりも遥かに速く(ほぼ10倍速く)代謝することである。従って、等価な薬物の半減期がヒトにおいて数時間である場合には、マウスでは半減期は恐らく30分間となるであろう。当然のことながら、マウスでは正常な処理経路がマスクされるという作用が得られるが、これは、該経路が有効になる機会が無いためである。従って、一以上のマウスP450クラスター(特に、支配的なCyp3a及び2cクラスター)を欠失させることによって、このような薬物処理経路を論点から排除する効果が得られる。このようなノックアウトマウスをヒトP450クラスターに関してヒト化することによって、ヒト薬物代謝経路を内在性マウス酵素からの干渉無しに評価することができるであろう。
【0031】
本発明の動物モデルはそれ自体が薬物モデルとして興味深い。例えば、該動物モデルによって、欠失していなければ薬物を代謝する機能を有していたであろう酵素の非存在下で薬物代謝を評価することができる。従って、本発明の動物モデルによって、P450系内の冗長性の程度を評価することができ、第1の選択酵素が存在しない場合には他のP450酵素が引き継ぐことができる。また、これによって、第2の選択酵素が恐らく異なる反応経路を用いて作用し、通常は産生しないであろう毒性の二次代謝物を生じさせるかどうかを評価することもできる。
【0032】
本発明のマウスの特徴は、P450クラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないことである。これは、該クラスターによってコードされるP450機能が真に機能しないようにされており、他の生体由来の相同又はオルソロガスなクラスターで置換されていないことを意味する。このような動物はそれ自身が有用であり、下流においても作出することができる(例えば、ヒトP450遺伝子クラスターをマウスに導入した動物)。しかし、このような動物については本発明の範囲から意図的に除外する。
【0033】
「等価な機能」とは、機能しないようにするP450機能に対して機能的に取って代わることのできる如何なる遺伝子又は遺伝子クラスターをも意味する。従って、その例としては、他の生体における認知されたオルソロガスなカウンターパートが挙げられる。ヒト及びマウスのオルソログの例は当業者には公知であり、その多くはNelsonら、2004年(Pharmacogenomics 14(1): 1)に記載されている。
【0034】
本発明に係る好ましい薬物モデルは、Cyp3aクラスターが全組織で機能しないようにされており、Cyp3aクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp3aマウスクラスターに対するヒト等価遺伝子は、CYP3A4、3A5、3A7及び3A43である。遺伝子3a57、3a16、3a41、3a11、3a25及び3a59の全てが欠失した、本発明のこの様相に係るマウスの作出及び部分的特徴付けは実施例に記載されているが、このマウスは本発明の一様相を構成する。既存の定説によると、このようなホモ接合的欠失は胚の致死の原因となり得るため、上述のように、この遺伝子型の生存可能なマウスが作出されたということは大いに驚くべきことであると共に非常に意義のある結果である。
【0035】
本発明のこの様相に係るCyp3aクラスターKOの最初の特徴付けから、以下の表現型の特徴が明らかになった。
【0036】
A.一実施形態においては、本発明のこの様相に係るマウスの血漿臨床化学パラメータ(トリグリセリド、アラニントランスフェラーゼ、アルカリホスファターゼ、アスパラギン酸アミノトランスフェラーゼ、アルブミン、総ビリルビン、クレアチンキナーゼ、高密度リポタンパク質及び低密度リポタンパク質)は、未処理野生型マウス(例えば、C57BL/6J)の公知の正常範囲内である。
【0037】
B.第二の実施形態においては、本発明のこの様相に係るマウスの、IP投与後のミダゾラムの薬物動態プロファイルは、野生型マウス(例えば、C57BL/6J)で見られるのと同様の濃度/時間曲線下総面積である(即ち、野生型マウスの場合の80%〜120%)。
【0038】
C.他の実施形態においては、本発明のこの様相に係るマウスに対しリファンピシンで前処理(例えば、60mg/kg、3日間投与)した上でミダゾラムをIP投与した後の算出濃度/時間曲線下総面積が、未処理Cyp3aクラスターKOマウスの場合と比べて、好ましくは170%〜370%、より好ましくは220%〜320%、より好ましくは240%〜300%、より好ましくは250%〜290%、より好ましくは260%〜280%、より好ましくは265%〜275%減少している。
【0039】
D.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによる7−ベンジルオキシキノリンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベルまでの低下が見られる。
【0040】
E.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによるミダゾラムの1’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベルまでの低下が見られる。
【0041】
F.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるミダゾラムの1’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベル、より好ましくは<10%のレベルまでの低下が見られる。
【0042】
G.他の実施形態においては、本発明のこの様相に係るマウスでは、肝ミクロソームによるミダゾラムの4’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<80%のレベル、好ましくは<70%のレベル、より好ましくは<65%のレベルまでの低下が見られる。
【0043】
H.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるミダゾラムの4’−水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、好ましくは<40%のレベル、より好ましくは<35%のレベル、好ましくは<30%のレベル、より好ましくは<25%のレベルまでの低下が見られる。
【0044】
I.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる処理(例えば、60mg/kg、3日間投与)後のミダゾラムの水酸化が未処理Cyp3aクラスターKOマウスと比べて僅かに増加、即ち、好ましくは101%〜150%、より好ましくは110%〜140%、より好ましくは120〜130%増加する。また、水酸化の増加は、野生型マウス(例えば、C57BL/6J)においてリファンピシン処理時の水酸化で見られる増加に比べて有意に少なく、水酸化の増加は野生型マウスで見られる増加の<60%が好ましく、野生型マウスで見られる増加の<50%が好ましく、野生型マウスで見られる増加の<40%がより好ましく、野生型マウスで見られる増加の<35%がより好ましい。
【0045】
J.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)に続くミダゾラムのIV投与後の1’−ヒドロキシミダゾラムの薬物動態プロファイルにおいて、AUCが野生型マウス(例えば、C57BL/6J)で記録されたものに比べて低く、即ち、AUCは野生型マウスで記録されたものの<60%が好ましく、<50%がより好ましく、<40%がより好ましく、<35%がより好ましい。
【0046】
K.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)に続くミダゾラムの経口投与後の1’−ヒドロキシミダゾラムの薬物動態プロファイルにおいて、AUCが野生型マウス(例えば、C57BL/6J)で記録されたものに比べて低く、即ち、AUCは野生型マウスで記録されたものの<90%が好ましく、<85%がより好ましく、<80%がより好ましい。
【0047】
L.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによる7−ベンジルオキシキノリンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<30%のレベル、より好ましくは<20%のレベル、より好ましくは<10%のレベルまでの低下が見られる。
【0048】
M.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるトリアゾラムの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<25%のレベル、より好ましくは<10%のレベル、より好ましくは<5%のレベル、より好ましくは<2.5%のレベルまでの低下が見られる。
【0049】
N.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の腸ミクロソームによるトリアゾラムの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<25%のレベル、より好ましくは<10%のレベル、より好ましくは<5%のレベル、より好ましくは<2.5%のレベルまでの低下が見られる。
【0050】
O.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるニフェジピンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくはC57BL/6Jの<30%のレベル、より好ましくは<25%のレベルまでの低下が見られる。
【0051】
P.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるテストステロンの6β水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型で記録されたものに対して<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくは<30%のレベルまでの低下が見られる。
【0052】
Q.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるテストステロンの16β水酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な増加、即ち、野生型マウスで記録されたものに対して>150%のレベル、より好ましくは>200%のレベル、より好ましくは>250%のレベル、より好ましくは>300%のレベルまでの増加が見られる。
【0053】
R.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の肝ミクロソームによるジベンジルフルオレシンの酸化に関して野生型マウス(例えば、C57BL/6J)と比べて有意な低下、即ち、野生型マウスで記録されたものに対して<25%のレベル、より好ましくは<20%のレベル、より好ましくは<15%のレベル、より好ましくは<12.5%のレベルまでの低下が見られる。
【0054】
S.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)後の腸ミクロソームによる4−トリフルオロメチルクマリンの野生型マウス(例えば、C57BL/6J)と比べての有意な増加、即ち、野生型マウスで記録されたものに対して>150%のレベル、より好ましくは>175%のレベル、より好ましくは>200%のレベルまでの増加が見られる。
【0055】
T.他の実施形態においては、本発明のこの様相に係るマウスでは、野生型マウス(例えば、C57BL/6J)と比べてベンゾキシレゾルフィンO−脱ベンジル化活性の低下、即ち、野生型マウスで記録されたものに対して好ましくは<90%のレベル、より好ましくは<80%のレベル、より好ましくは<70%のレベル、より好ましくは<60%のレベル、より好ましくは<50%のレベルまでのO−脱ベンジル化活性の低下が見られる。
【0056】
U.他の実施形態においては、本発明のこの様相に係るマウスでは、リファンピシンで前処理(例えば、60mg/kg、3日間投与)を行った際、野生型マウス(例えば、C57BL/6J)と比べてベンゾキシレゾルフィンO−脱ベンジル化活性の低下、即ち、野生型マウスで記録されたものに対して好ましくは<50%のレベル、より好ましくは<40%のレベル、より好ましくは<35%のレベル、より好ましくは<30%のレベルまでのO−脱ベンジル化活性の低下が見られる。
【0057】
V.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態A〜Uの代謝表現型の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20又は全21種を示す。
【0058】
W.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される検出可能レベルのCyp3aは肝ミクロソームにおいては発現されない。
【0059】
X.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームでのCyp1aの発現レベルがC57BL/6Jマウスと比べて高い。
【0060】
Y.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームにおけるCyp2cタンパク質の発現が野生型マウス(例えば、C57BL/6J)と比べて増加する。
【0061】
Z.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソームにおけるCyp2dタンパク質のレベルが野生型マウス(例えば、C57BL/6J)と比べて正常であり、肝ミクロソームにおけるCyp2dタンパク質のレベルが野生型マウス(例えば、C57BL/6J)と比べて高い。
【0062】
AA.他の実施形態においては、本発明のこの様相に係るマウスでは、ウェスタンブロッティングで確認される肝ミクロソーム及び腸ミクロソームの両方におけるCyp2eタンパク質のレベルに関し、野生型マウス(例えば、C57BL/6J)と比べて有意差が無い。
【0063】
AB.他の実施形態においては、本発明のこの様相に係るマウスでは、シトクロムCレダクターゼ活性で確認されるP450オキシドレダクターゼ(POR)タンパク質の発現レベルに関し、野生型マウス(例えば、C57BL/6J)と比べて有意差が無い。
【0064】
AC.他の実施形態においては、本発明のこの様相に係るマウスでは、総P450レベルは野生型マウス(例えば、C57BL/6J)で記録されたものに比べて有意に高いが、リファンピシンによる前処理(例えば、60mg/kg、3日間投与)の際には有意に変化しない。
【0065】
AD.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態W〜ACのタンパク質発現表現型の内の1、2、3、4、5、6又は全7種を示す。
【0066】
AE.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp3a11、Cyp3a16及びCyp3a25の肝転写産物(liver transcript)レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>50倍、より好ましくは>100倍、より好ましくは>100倍抑制されている。
【0067】
AF.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp4a13の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、より好ましくは>5倍、より好ましくは>10倍、より好ましくは>12.5倍、より好ましくは>15倍抑制されている。
【0068】
AG.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c38の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.5倍アップレギュレートされている。
【0069】
AH.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2a4の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0070】
AI.他の実施形態においては、本発明のこの様相に係るマウスでは、GSTP1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.4倍アップレギュレートされている。
【0071】
AJ.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍抑制されている。
【0072】
AK.他の実施形態においては、本発明のこの様相に係るマウスでは、UGT2B17の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0073】
AL.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCB10の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.2倍アップレギュレートされている。
【0074】
AM.他の実施形態においては、本発明のこの様相に係るマウスでは、CAMK2Bの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.6倍アップレギュレートされている。
【0075】
AN.他の実施形態においては、本発明のこの様相に係るマウスでは、CITED2の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.2倍アップレギュレートされている。
【0076】
AO.他の実施形態においては、本発明のこの様相に係るマウスでは、DNAJC12の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4.1倍抑制されている。
【0077】
AP.他の実施形態においては、本発明のこの様相に係るマウスでは、FMO5の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍アップレギュレートされている。
【0078】
AQ.他の実施形態においては、本発明のこの様相に係るマウスでは、HSD2B3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍アップレギュレートされている。
【0079】
AR.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.1倍抑制されている。
【0080】
AS.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNBの肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>3倍、より好ましくは>4倍、より好ましくは>5倍、より好ましくは>5.1倍抑制されている。
【0081】
AT.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP2K6の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、好ましくは>2.5倍、好ましくは>3倍、好ましくは>3.3倍アップレギュレートされている。
【0082】
AU.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍抑制されている。
【0083】
AV.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K3の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍抑制されている。
【0084】
AW.他の実施形態においては、本発明のこの様相に係るマウスでは、STIP1の肝転写産物レベルが野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.3倍アップレギュレートされている。
【0085】
AX.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2a4の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.4倍アップレギュレートされている。
【0086】
AY.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b13の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2.5倍、より好ましくは>4倍、より好ましくは>4.5倍、より好ましくは>5倍、より好ましくは>5.5倍アップレギュレートされている。
【0087】
AZ.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b10の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>4倍、好ましくは>5倍、好ましくは>6倍、好ましくは>7倍、好ましくは>7.2倍アップレギュレートされている。
【0088】
BA.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2b9の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.2倍アップレギュレートされている。
【0089】
BB.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c54の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0090】
BC.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp2c54の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0091】
BD.他の実施形態においては、本発明のこの様相に係るマウスでは、Cyp4a2bの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.3倍アップレギュレートされている。
【0092】
BE.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0093】
BF.他の実施形態においては、本発明のこの様相に係るマウスでは、MGST3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>1.5倍、より好ましくは>2倍、より好ましくは>2.4倍抑制されている。
【0094】
BG.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCD1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0095】
BH.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCD3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.5倍アップレギュレートされている。
【0096】
BI.他の実施形態においては、本発明のこの様相に係るマウスでは、ABCG8の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0097】
BJ.他の実施形態においては、本発明のこの様相に係るマウスでは、ARK1C14の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.7倍アップレギュレートされている。
【0098】
BK.他の実施形態においては、本発明のこの様相に係るマウスでは、CAMK2Bの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>3.7倍アップレギュレートされている。
【0099】
BL.他の実施形態においては、本発明のこの様相に係るマウスでは、CITED2の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>2.8倍アップレギュレートされている。
【0100】
BM.他の実施形態においては、本発明のこの様相に係るマウスでは、FMO5の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.7倍アップレギュレートされている。
【0101】
BN.他の実施形態においては、本発明のこの様相に係るマウスでは、HSD3B3の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後にC57BL/6Jと比べて、好ましくは>2倍、より好ましくは>3倍、より好ましくは>4倍、より好ましくは>4.9倍アップレギュレートされている。
【0102】
BO.他の実施形態においては、本発明のこの様相に係るマウスでは、JUNBの肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍抑制されている。
【0103】
BP.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP2K6の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、より好ましくは>2.5倍、より好ましくは>3倍、より好ましくは>3.6倍アップレギュレートされている。
【0104】
BQ.他の実施形態においては、本発明のこの様相に係るマウスでは、MAP3K1の肝転写産物レベルがリファンピシンによる株の前処理(例えば、60mg/kg、3日間投与)後に野生型マウス(例えば、C57BL/6J)と比べて、好ましくは>2倍、好ましくは>2.5倍、好ましくは>2.7倍抑制されている。
【0105】
BR.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態AE〜BQのトランスクリプトーム表現型の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38又は全39種を示す。
【0106】
BS.他の実施形態においては、本発明のこの様相に係るマウスは、実施形態A〜BRの表現型の特徴の内の1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70又は全71種を示す。
【0107】
本発明に係る他の好ましいモデルは、Cyp2cクラスターが全組織で機能しないようにされており、Cyp2cクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp2cマウスクラスターに対するヒト等価遺伝子は2C8、2C9、2C18及び2C19である。遺伝子2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44の全てが全組織で機能しないようにされている本発明のこの様相に係るマウスを作出するための戦略は実施例に記載される。ホモ接合体Cyp2cノックアウトマウスが作出された。
【0108】
本発明に係る他の好ましいモデルは、Cyp2dクラスターが全組織で機能しないようにされており、Cyp2dクラスター内の遺伝子の機能に対して等価な機能を有する置換遺伝子を含まないマウスである。Cyp2dマウスクラスターに対するヒト等価遺伝子は2D6、2D7及び2D8である。遺伝子2d22、2d11、2d10、2d9、2d12、2d34、2d13、2d40及び2d26の全てが全組織で機能しないようにされている本発明のこの様相に係るマウスを作出するための戦略は実施例に記載されている。ホモ接合体Cyp2dノックアウトマウスが作出された。
【0109】
一実施形態においては、本発明のこの様相に係るマウスでは、ブフラロールIP投与後の算出した濃度/時間曲線下総面積が、野生型マウス(例えば、C57BL/6J)の場合と比べて、好ましくは>5倍、より好ましくは>10倍、より好ましくは>15倍、より好ましくは>20倍増加する。
【0110】
本発明に係る更に好ましいモデルは、Cyp3aクラスター及びCyp2cクラスターの両方が全組織で機能しないようにされているマウスである。遺伝子3a57、3a16、3a41、3a11、3a25、3a59、2c55、2c65、2c66、2c29、2c38、2c39、2c67、2c68、2c40、2c69、2c37、2c54、2c50、2c70及び2c44の全てが欠失した、本発明のこの様相に係るマウスの作出及び部分的特徴付けは実施例に記載されているが、このマウスは本発明の一様相を構成する。
【0111】
本発明においては、如何なるP450クラスターをも機能しないようにすることができる。「機能しないようにする」とは、遺伝子又は遺伝子機能を欠失させることを意味する。これは、遺伝子のコード配列をマウスゲノムから完全に欠失させることによって行うことができる。或いは、コード配列を他の配列の挿入、欠失又は置換によって変異させて欠失させることができる。例えば、一以上の変異(フレームシフト変異等)は、得られるRNA転写産物が非機能性タンパク質又は切断型タンパク質をコードするように生じさせることができる。或いは、染色体配列に挿入を行ってアミノ酸コードを分断することができる。
【0112】
同様に、欠失対象のP450遺伝子配列をある配列(例えば、成功した欠失体をスクリーニングするための基準として用いることができる選択配列やマーカー配列)と交換することができる。このような一戦略はWallaceら(Cell 128, 197-209 2007)によって考案されており、遺伝子交換のコンテクスト内ではあるが、本発明の方法に適用可能である。この方法では、ベクターを用いた置換配列が内在性ゲノムマウス配列に取って代わるのに2種の分子間相同組換えイベントが必要となるような、マウス染色体とBAC又はYACベクターとの間での配列の交換が想定されている。
【0113】
好ましい一システムにおいては、相同組換えのメカニズムを用いて遺伝子クラスターを該クラスターが存在しない他の配列と交換する。このような方法は、次の段階:
a)置換対象のマウス標的遺伝子クラスターが各々の端で組換え部位と隣接するように相同組換えによってマウス染色体に一対の部位特異的組換え部位を組込むことと、
b)部位特異的組換え部位間で組換えを行い、マウス標的遺伝子クラスターが染色体から除去され、残存部位特異的組換え部位で置換されるようにすることと、を含むのが好ましい。
【0114】
相同組換えを行うための方法は当該技術分野では公知であり、外部から供給されたDNA分子と標的染色体との間で相同領域を用いてRT部位を導入する。この方法においては、置換配列内の5’及び3’相同アームによって置換配列と標的との間の組換えが促進され、遺伝子クラスターが欠失されるようになる。この戦略を用いた方法は、本出願人による2007年9月14日出願の同時係属特許出願No.GB0718029.2「発明の名称:二段階のクラスター欠失及びヒト化」に報告されている。この戦略は上述のWallaceの技術に比べて遥かに効率的であり、分子内組換えイベントが約106分の1の頻度で生じる。
【0115】
しかし、多くのクラスターは数百キロベースのゲノムの相当のストレッチに及ぶ。一例としてはマウスCYP3a遺伝子クラスターが挙げられ、Cyp3a13が含まれていなくても約800kbに及ぶ。このようなシナリオにおいては、2種の別個の置換核酸をその間にクラスター全体が広がるようにマウスゲノム内に導入し、その後、この2種の置換核酸間での組換えによって該核酸間にある遺伝子クラスターを除去する戦略を用いるのが遥かに効率的である。
【0116】
この構成においては、置換配列の各々は、5’相同アームと3’相同アームとの間に選択マーカー及び少なくとも1種のリコンビナーゼ標的(RT)部位(例えば、loxPやlox5171、FRT、F3)があるようにデザインする。このように、両方の置換核酸をうまく組込んで、欠失対象クラスターに隣接するように選択することが可能である。RT部位を認識する適切な部位特異的リコンビナーゼ(SSR)にマウス細胞を曝露することによってクラスターを除去するのは技術的に容易である。「SSR」とは、特定のDNA遺伝子座におけるDNA再構成を仲介する組換え系の如何なるタンパク質成分をも意味し、例えば、インテグラーゼ又はレゾルバーゼ/インベルターゼクラスのSSR(Abremski, K.E. and Hoess, R.H. (1992) Protein Engineering 5, 87-91; Khan, et al, (1991) Nucleic acids Res. 19, 851-860; Nunes-Duby et al, (1998) Nucleic Acids Res 26 391-406; Thorpe and Smith, (1998) P.N.A.S USA 95 5505-10)及びイントロンコード化エンドヌクレアーゼによって仲介される部位特異的組換え(Perrin et al, (1993) EMBO J. 12, 2939-2947)が挙げられる。SSR酵素活性に曝露することによって、RT部位の配置で確認されるDNA再構成が生じ、その結果、直線DNA分子内で介在配列が除去又は切除される。
【0117】
当該技術分野で周知であり、以下に更に説明する方法に従って、上述の組換え段階はマウス胚性幹細胞内で行うのが好ましい。胚性幹細胞は全能性細胞の培養細胞株であり、該細胞を初期胚に導入すると発育して生体の全組織に定着するようになる。
【0118】
本発明の方法の第1の段階は、一対のRT部位をマウス染色体内に組込むことである。置換対象のマウス標的遺伝子クラスター配列は、各々の端においてRT部位と隣接する必要がある。2個のRT部位間での組換えによって介在遺伝子クラスターが切除される。RT部位を染色体内に組込むための方法は当業者には公知であろうが、相同組換えプロセスを用いて行うのが好ましい。
【0119】
RT対は、LoxP、FRT、attP/attB及びrox(Sauer and McDermott. Nucleic Acids Res. 32(20): 6086-95, 2004)から成る群から選択されるのが好ましい。このリストはほんの一例として挙げたに過ぎず、これらに限定されるものではない。この様相においては、前記部位特異的組換え部位対間の組換えは、対応する部位特異的リコンビナーゼ(即ち、Cre、FLP、PhiC31及びDre)によって行われる。前記部位特異的組換えはインビボ又はインビトロで実施し得ることは理解されよう。
【0120】
実際には、各RT部位は、選択可能マーカーと連結する(好ましくは、連続する)ように構築する必要がある。選択可能マーカーを組込むことによってマウス染色体内のRT部位を検出することができ、それによって、RT部位対の挿入が成功したかどうかをモニターすることができる。各選択可能マーカーは、マウス標的配列とRT部位との間の空間にあるように配置するのが好ましい。これは、組換えが成功した際には、選択可能マーカーが染色体から除去され、本発明の方法においてはこれ以上参加しないことを意味する。このように、ヒト化後に染色体内に残るヒト配列の調節を選択可能マーカーが乱すことがないことは確実であり得る。選択可能マーカーの各々は互いに異なっているのが好ましい。これによって、各組換え部位の挿入を互いに別々に独立してモニターすることができる。選択可能マーカーは、成長ES細胞が曝露され得る化学化合物(例えば、抗生物質)に対するある種の耐性をコードする遺伝子であるのが好ましい。その例としては、細胞成長培地に添加される抗生物質に対する耐性を付与する選択可能マーカー(例えば、G418、ハイグロマイシン又はピューロマイシンに対する耐性を付与するネオマイシン耐性マーカー)の使用が挙げられる。更なる例としては、本発明の上述の様相における核酸配列(又はその一部)に相補的であり、それに対してハイブリダイズする核酸配列を用いた検出が挙げられる。例えば、サザンブロット解析やノーザンブロット解析、PCRが挙げられる。
【0121】
好ましい戦略においては、変化したマウス胚性幹細胞の作出(好ましくは、別々の段階での作出)を伴い、これによって、各RT部位がその選択可能マーカーと共に別々に組込まれる。その後、変化した胚性幹細胞を胚盤胞に挿入することができる。従来、胚盤胞は雌性マウスから交配の約3日後に単離している。最大20個の変化した胚性幹細胞(好ましくは、約16個)をそのような胚盤胞に同時に挿入し得ることは理解されよう。変化した胚性幹細胞を胚盤胞に挿入することにより、好ましくは、妊娠マウスの特性を反映するように誘導された偽妊娠マウスに移植することによって、胚性幹細胞は発育初期胚に組込まれるようになる。この方法によると、変化した胚性幹細胞を含む胚盤胞は偽妊娠マウスの子宮壁内に移植され、妊娠期間が終了するまで該マウス内で発育し続ける。変化した胚性幹細胞は増殖し、分裂して発育トランスジェニックマウスの全組織(生殖系列を含む)に定着するようになる。
【0122】
本発明の方法の一様相においては、作出したトランスジェニックマウスは、各体細胞組織及び生殖系列内に変化した細胞と未変化の細胞を含むキメラとなり得る。
【0123】
大きいDNA断片(200kb〜数メガベース)の欠失に適したCre/lox介在欠失を仲介するための方法は以下の論文に記載されている(Li ZW, Stark G, Gotz J, Rulicke T, Gschwind M, Huber G, Muller U, Weissmann C. Generation of mice with a 200-kb amyloid precursor protein gene deletion by Cre recombinase-mediated site-specific recombination in embryonic stem cells Proc Natl Acad Sci U S A. 1996 Jun 11;93(12):6158-62. Erratum in: Proc Natl Acad Sci U S A 1996 Oct 15;93(21):12052; in Su H, Wang X, Bradley A. Nested chromosomal deletions induced with retroviral vectors in mice. Nat Genet. 2000 Jan;24(l):92-5); Call LM, Moore CS, Stetten G, Gearhart JD. A cre-lox recombination system for the targeted integration of circular yeast artificial chromosomes into embryonic stem cells. Hum Mol Genet. 2000 JuI 22;9(12):1745-51)。
【0124】
RT部位間で組換えを行うためには、ゲノムをSSR酵素形態のSSR活性に曝露する必要がある。上述のように、部位特異的組換えイベントはインビトロ又はインビボで行うことができる。部位特異的組換えは、トランスジェニックマウス内でSSRの活性を誘導することによって行うことができる。実際、生体系で遺伝子型を変化させるために部位特異的組換えをうまく利用するには、組換えイベントを調節するための戦略が必要である。これは、得られる発現パターンが上述の組織特異的な要素が活性となる時間及び場所に制限されるように、リコンビナーゼmRNA又はタンパク質の発現を制御することによって行うことができる(Baubonis and Sauer (1993) Nucl Acids Res. 21, 2025-2029; Sauer B, (1994) Curr Opin Biotechnol 5:521-7; Rajewsky et al., (1996) J Clin Invest 98, 600-3; Metzger and Feil, (1999) Curr. Opinions Biotechnology 10, 470-476)。
【0125】
研究者らは、SSR酵素を発現させるために、直接トランスフェクションや組換えウイルスによる感染、SSRタンパク質をコードするDNAやmRNA又は該タンパク質自身の注入を用いてきた(Konsolaki et al, (1992) New Biol. 4: 551-557)。このようなSSR酵素の発現ではなく活性を調節することによって、より正確に制御することができる。一戦略においては、SSR酵素をステロイド受容体のリガンド結合ドメイン(LBD)に融合させた融合タンパク質を用いてSSR−LBDタンパク質を得る(EP-B-0 707 599;Logie and Stewart (1995) P.N.A.S. USA 92: 5940-5944; Brocard et al, (1997) P.N.A.S. USA 94: 14559-14563; Akagi et al, (1997) Nucleic Acids Res 25, 1766-73参照)。この戦略は、SSR活性を活性化するステロイド受容体のリガンドをリガンドが受容体部分に結合している場合のみ用いることに依存している。該受容体のLBDは、同族リガンドの非存在下ではSSRの活性を抑制する。同族リガンドの送達によってSSRの抑制を軽減し、RT部位間での組換えを行うことができる。
【0126】
このように、誘導については、前記SSRの転写を誘導するか、SSRの翻訳を誘導するか、又はSSRから阻害剤を除去することによって行うことができる。或いは、SSRを前記トランスジェニックマウスに人工的に導入することができる。本発明の方法の一要素は、トランスジェニックマウス内で部位特異的組換えを行うことによってマウス標的遺伝子の除去と同時にヒト化マウスの作出が可能であることである。
【0127】
トランスジェニックマウスを欠失株マウスと交雑させることによって部位特異的組換えを実施し得るのが好ましい。本明細書において「欠失株」とは、生殖系列で部位特異的リコンビナーゼを発現するマウスに関するが、これを、トランスジェニックマウスと交雑させてマウス標的遺伝子配列を除去することができる。このように、インビボ組換えによって着目遺伝子に対してヘテロ接合性の子孫が産生する。こうして、トランスジェニックマウスを欠失株と交雑させることによって、後代が産生すると共に、ヒト置換遺伝子配列と部位特異的リコンビナーゼを含有するように変化したマウス染色体を含む細胞が産生し、その結果、マウス標的遺伝子が除去され、細胞の機能的ヒト化が生じる。従って、このようなトランスジェニックマウスは、特定の遺伝子又は遺伝子クラスターに関するヒト化に対してヘテロ接合性となる。
【0128】
ある実施形態においては、部位特異的リコンビナーゼがリコンビナーゼ株マウスのある組織でのみ発現するのが望ましいことがある。ある遺伝子又は遺伝子クラスターの欠失が致死的であるか又は亜致死性の表現型作用をもたらし得ることは、当該技術分野では知られている。また、このような遺伝子をそのヒト等価物で置換しても致死を防止できない場合がある。このような状況下では、部位特異的リコンビナーゼをある組織(例えば、肝臓)でのみ発現させることによって、致死に関する如何なるこのような問題をも克服し得る。これは、特定の遺伝子がある組織において必須である場合、このように部位特異的リコンビナーゼを発現させることによってマウス遺伝子が該組織において存続し得るため、特に有利となる。
【0129】
本発明のこの様相においては、SSRはアルブミン−Creであり得る。アルブミン−Creは、RT部位LoxPに作用するSSR Creの特定のバリアントである。アルブミン−Creは肝臓においてのみ発現するため、マウス標的配列を肝臓以外の全ての組織で存続させることができ、致死に関する潜在的な問題を克服する一方、機能的にヒト化された肝臓が得られる。
【0130】
最終的には、上述の方法によって作出した2種のヘテロ接合体マウスを交雑させて、着目遺伝子のヒト対立遺伝子に対してホモ接合性のトランスジェニックマウスを作出することができる。2種のヘテロ接合体トランスジェニックマウスの交雑によって、欠失に対してホモ接合性の後代の一部が産生するであろう。
【0131】
本発明の更なる実施形態においては、以降に開示の方法によって、トランスジェニック非ヒト動物を上述の特徴を全て有するように新しく作出する。
【0132】
また、部位特異的組換えイベントを体細胞において行うことも可能であり、その後、該細胞をWO00/51424の方法に記載のクローン化マウス又はそのバリアントのコロニーを形成するために核移植ドナーとして用いることができる。
【0133】
本発明の他の実施形態においては、本発明のマウスを交雑によって作出する。例えば、特定のクラスターの遺伝子の一部が欠失した部分欠失体を他の部分欠失体と交雑させて、特定のクラスター内の遺伝子機能が全て欠失した動物を作出することができるであろう。
【0134】
本発明の更なる実施形態においては、以降に開示の方法によって、トランスジェニックマウスを上述の特徴を全て有するように新しく作出する。
【0135】
トランスジェニックな組合せ
最終的には、本発明の動物モデルをヌルバックグラウンドとして用い、げっ歯類酵素の機能に取って代わり得るヒト遺伝子を同一の染色体領域に直接組込むか、又は他の部位に組込むことによって該遺伝子の導入を行うことができる。ヒト遺伝子を同一の染色体領域に組込むのが好ましいが、これは、染色体の完全性が保持されるため、発現や組織分布の生理学的パターンが同様に保持される可能性が高いからである。従って、上述の細胞やマウスの調製に関する本発明の実施形態においては、該細胞や動物を更なる遺伝子組換えに付すことができるが、該遺伝子組換えは更なる遺伝子又はタンパク質コード核酸配列の導入である。該遺伝子組換えは、細胞又は細胞株の一過性又は安定トランスフェクション、細胞又は細胞株におけるエピソーム発現系、又は、前核マイクロインジェクションや非胚性幹(ES)細胞における組換えイベント、非ヒト胚性幹(ES)細胞におけるランダムな遺伝子組換え、又は核移植クローニング手順においてドナー核として用いる核を有する細胞のトランスフェクションによるトランスジェニック非ヒト動物の調製であり得る。
【0136】
特に、一以上のP450遺伝子クラスターが機能しないようにされている本発明に係るマウスを一以上のヒト遺伝子に関してヒト化し得ることが予想される。欠失したクラスターはCyp3aクラスター、Cyp2cクラスター及び/又はCyp2dクラスターであるのが好ましい。後述のように、これらのクラスターとヒト化薬物輸送体、ヒト化転写因子又はヒト化第II相薬物代謝酵素との如何なる単一又は複数の組合せも好ましい。
【0137】
例えば、このようなマウスを転写因子に関してヒト化することができる。好適な転写因子の例としては、プレグナンX受容体(PXR)や構成的アンドロスタン受容体(CAR)、PPAR(α、δ及びγ)、NRF2、Ah受容体、HNF1、HNF4が挙げられる。これらの転写因子の1種、2種又はそれ以上に関してマウスをヒト化することができる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたのも好ましい。ヌルバックグラウンドに関してP450クラスター欠失と二重ヒト化hCAR及び/又はhPXRマウスの組合せは、本発明に係る好ましいモデルである。
【0138】
また、本発明に係るマウスを第2相薬物代謝酵素に関してヒト化することもできる。このような酵素の例としては、グルクロニルトランスフェラーゼ(例えば、UGT1A遺伝子又は遺伝子クラスター)やグルタチオントランスフェラーゼ(例えば、GST(グルタチオンS−トランスフェラーゼ)(GST−m1及び/又はt1クラスターを含む))、スルホニルトランスフェラーゼ、アセチルトランスフェラーゼが挙げられる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたものも好ましい。
【0139】
また、本発明に係るマウスを薬物輸送タンパク質に関してヒト化することもできるが、その例としては、多剤耐性タンパク質(例えば、mdr1やmdr3)や多剤耐性関連タンパク質(MRP)(例えば、MRP1及び/又はMRP2及び/又はMRP6)、有機アニオン輸送ポリペプチド(OATP)由来のものが挙げられる。また、WO2006/064197に記載のように、内在性等価マウス遺伝子が無効化されたのも好ましい。
【0140】
細胞
本発明の他の様相は、本発明の上述のいずれか一様相における特性を有するように改変された細胞に関する。本発明に係る好ましい細胞型は肝細胞及び神経細胞である。該細胞はマウス細胞であることができる。
【0141】
本発明のこの様相における細胞は、本発明の知見を十分体得した当業者には明らかなように、本発明に係るトランスジェニックマウスから標準的な技法を用いて作出することができる。好適な方法は多くの標準的な実験マニュアル、例えば、Davisら、Basic Methods in Molecular Biology (1986); Sambrook Molecular Cloning; A Laboratory Manual, Third Edition (2000); Ausubel et al., 1991 [supra]; Spector, Goldman & Leinwald, 1998)に記載されている。
【0142】
このような細胞を作出する好ましい一方法は、上述のようにヒト化マウスをSV40不死化マウス(例えば、不死マウス(タコニック))と交雑させることである。次に、当該技術分野で周知の技法によってこのような動物から細胞を単離することができる。肝臓でのみ活性であるため肝細胞しか産生することができないアルブミンプロモーターを用いる先行技術のトランスジェニック系とは対照的に、本発明において作出されるトランスジェニック動物由来の細胞は、薬物動態解析にかなり重要な細胞(例えば、肝細胞や神経細胞)等の様々な細胞型から選択することができる。
【0143】
上述の特性を有する、本発明に係るトランスジェニック動物から単離された幹細胞も本発明の有用な様相である。このような細胞は多能性であってもよく、部分的に分化していてもよい。幹細胞は成体幹細胞であってもよく、胚性幹細胞であってもよい。より一般には、用いる幹細胞は後胚発達段階(例えば、胎生期や新生期、幼若期、成体期)で得られることができる。このように単離された幹細胞を用いて特定の細胞型(例えば、肝細胞や神経細胞)を作出することができる。このような細胞も本発明の一様相を構成する。
本発明の方法によって作出した細胞又は動物をモデル系として用いて、他の生体(特にヒト)における薬物や他の生体異物化合物の代謝を確認することができる。
【0144】
アッセイ
本発明の動物、組織及び細胞を用いて、薬物化合物がどのように代謝されるか確認することができる。単一又は複数のシトクロムP450遺伝子クラスターが欠失したマウス系、或いは、シトクロムP450遺伝子クラスター交換を薬物の代謝や処理に関連する他の遺伝子と組合わせたマウス系の作出によって、ヒトにおける薬物反応や化学反応を決める因子、更にはこのような遺伝子と化学毒性の関連性についての我々の理解が顕著に増すであろう。このようなモデルは、効力スクリーニングやPK/PDモデリング、薬物安全性試験に適用することができる。
【0145】
本発明のクラスターノックアウトによってヒト化及び更なるノックアウト実験を行うことができる。作出する動物の性質に応じて、多数の異なるアッセイプロセスに着目することができる。特に、特定の薬物代謝遺伝子クラスターの如何なる薬物化合物の代謝への関与も確認することができる。また、特定のクラスターが欠失した際に第2の選択経路による代謝への代償作用があるとすれば、それを評価することによって如何なる薬物代謝系における冗長性の程度も確認することができる。また、如何なる毒性二次代謝物の生成も別々の代謝酵素に帰属させることができる。また、体組織内での転写因子や薬物代謝酵素、薬物輸送タンパク質の処理や分布に薬物化合物が影響を及ぼすかどうか調べることができる。また、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現の持続期間に薬物化合物が影響を及ぼすかどうか調べることができる。有用なアッセイプロセスの他の例は当業者には明らかであろう。
【0146】
動物における表現型の変化(例えば、生理学的作用)を測定することができる。このような生理学的作用としては、例えば、病態(胆管壊死等)や中毒性副作用を挙げることができる。
【0147】
薬物化合物の代謝速度を調べることができる。この代謝速度は、該化合物の投与によってもたらされる毒性や活性を測定するか、該化合物の半減期を測定するか、又は薬物代謝酵素のレベルを測定することによって求めることができる。例えば、該化合物の代謝速度は、酸化生成物の生成速度や酸化中間体から次に産生される生成物の生成速度として測定することができる。或いは、この代謝速度は、最初の化合物(initial compound)の半減期や消失速度として、又は最初の化合物や最初の化合物から産生される代謝産物の毒性や活性の変化として表わすことができる。半減期は、様々な時点で採取したサンプルに存在する薬物化合物の量を求めることによって測定することができる。薬物化合物の量は、標準的な方法(例えば、高速液体クロマトグラフィーや質量分析、化合物特異的抗体を用いたウェスタンブロット解析、他の適切な方法)によって測定することができる。
【0148】
また、薬物化合物が特定の状況下で毒性代謝物や発癌性代謝産物に代謝されるかどうかを、例えば、該化合物の組織やタンパク質、DNAへの共有結合を測定するか、又はグルタチオンの欠乏を測定することによって調べることもできる。
【0149】
上述の種の測定は、薬物化合物投与後、1を越える時点、3時点、5時点又は10以上の時点で行うのが好ましい。
【0150】
従って、本発明の更なる様相は、転写因子や薬物代謝酵素、薬物輸送タンパク質の活性や発現レベルに対する薬物化合物の作用を確認するために提供されるスクリーニング方法に関する。このような方法は、本発明の上述のいずれか一様相におけるトランスジェニック動物、又は該動物由来の組織又は細胞に薬物化合物を投与することを含む。
【0151】
スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質をコードする遺伝子の誘導を測定することを含むことができる。また、スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現のレベル、又はこのような発現の持続期間を測定することを含むことができる。また、スクリーニング段階は、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現の分布を測定することを含むことができる。
【0152】
薬物化合物の存在下及び非存在下でアッセイを行って、分布や代謝、毒性の差を確認することができる。様々なトランスジェニック動物や細胞、組織に対する薬物化合物の作用を評価することによって、特定の遺伝子の存在下及び非存在下における薬物化合物の作用を確認することができる。例えば、ヌルバックグラウンドを有する動物と着目遺伝子(例えば、PXRやCAR、MDR1、第1相代謝酵素、第2相代謝酵素)に関してヒト化した動物との間で薬物化合物の作用を評価することも可能である。
【0153】
従って、更なる様相において、本発明は、本明細書に記載の生体異物の代謝又は毒性を調べるための方法であって、本明細書に記載の非ヒト動物、組織又は細胞の2種以上、3種以上、4種以上、5種以上、6種以上、7種以上、8種以上、9種以上又は10種以上に対して薬物化合物を投与することを含む方法を提供する。このような方法は、異なる非ヒト動物、組織又は細胞に関して得られた実験結果を比較する段階を更に含むのが好ましい。
【0154】
二種以上の薬物化合物を投与することができる。例えば、ある薬物化合物がCAR遺伝子の誘導をもたらす場合、該化合物はCAR転写因子を活性化すると判断される。また、対照としてのヒトCAR受容体を発現する動物にCAR受容体インバースアゴニスト(例えば、クロトリマゾール)を投与することもできる。
【0155】
本発明の更なる様相におけるアッセイによって、第1の薬物化合物の代謝が第2の薬物化合物で調節されるかどうか確認するためのスクリーニング方法を提供することができる。この方法は、第2の化合物の存在下及び非存在下で第1の化合物を本発明の上述のいずれか一様相におけるトランスジェニック動物、又は該動物由来の組織又は細胞に投与することと、表現型効果をモニタリングすることとを含む。或いは、上述のように、スクリーニング段階は、遺伝子の誘導、転写因子や薬物代謝酵素、薬物輸送タンパク質の発現のレベルや持続期間、分布を測定することを含むことができる。第2の化合物がこれらの試験因子のいずれか一因子の変化をもたらす場合、第2の化合物は第1の化合物の代謝を調節することが確認される。例えば、第1の化合物の投与によってもたらされる毒性や活性を測定するか、又は第1の薬物化合物の半減期を測定することによって生理学的作用をアッセイすることができる。
【0156】
このように、アッセイを用いて、特定のタンパク質の発現を活性化又は誘導する能力が低いか又は検出できないために、副作用が少ないか又はインビボでの半減期が長いことが期待される薬物化合物の類似体を容易に同定することができる。
【0157】
以下、本発明の様々な様相や実施形態を実施例によって更に詳細に説明する。本発明の範囲を逸脱することなく詳細な変更を実施し得ることは理解されよう。
【図面の簡単な説明】
【0158】
【図1a】マウスCyp3aクラスター欠失のための戦略。(A)マウスCyp3aクラスター内の機能性遺伝子の染色体の構成及び方向の概略図(出典:Nelsonら、2004年)。偽遺伝子は示されていない。(B)Cyp3a57及びCyp3a59のエクソン/イントロン構造。エクソンは黒い棒で示し、ATGは両遺伝子の翻訳開始点を示す。相同組換えのターゲティングアームの位置をそれぞれ、薄灰色線(Cyp3a57)及び濃灰色線(Cyp3a59)でハイライトする。(C)相同組換えによりCyp3a57(左)及びCyp3a59(右)のターゲティングに用いたベクター。LoxP部位、lox5171部位、frt部位及びf3部位をそれぞれ、白い三角、縞模様の三角、点模様の三角、及び黒い三角で示す。(D)Cyp3a57及びCyp3a59遺伝子座における同一対立遺伝子上での相同組換え後の二重標的ES細胞内のCyp3aクラスターのゲノム構成。(E)loxP部位におけるCre介在組換え後のマウスCyp3aクラスターの欠失。Cyp3a57、Cyp3a16、Cyp3a41、Cyp3a11及びCyp3a25由来の全てのエクソンとイントロンは完全に欠失し、エクソン1〜4及びCyp3a59のプロモーターも欠失する。従って、Cre介在欠失後に残存する唯一の機能性Cyp3a遺伝子はCyp3a13であり、これは、>800kbのゲノムDNAやCypと無関係の多数の機能性遺伝子によってクラスターの残りから離れている。マウスCyp3aクラスターの欠失が成功したことを実証するのに用いたプライマーを矢印で示す。 分かり易くするため、配列は一定の縮尺率では示していない。TK=チミジンキナーゼ発現カセット、Hygro=ハイグロマイシン発現カセット、ZsGreen=ZsGreen発現カセット、P=ネオマイシンの発現を促進するプロモーター、5’ΔNeo=ATG欠損ネオマイシン。
【図1b−1】Cyp3aクラスター欠失に対するマウスホモ接合性のPCR確認。
【図1b−2】Cyp3aクラスター欠失に対するマウスホモ接合性のPCR確認。
【図1c】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な肝臓切片のヘマトキシリン及びエオシン染色。20倍の対物レンズを用いてCyp3aクラスターKO(Con)の画像を得た。他の画像の全てについては10倍の対物レンズを用いた。
【図1d】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な小腸切片のヘマトキシリン及びエオシン染色。10倍の対物レンズを用いて画像を得た。
【図1e】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な腎臓切片のヘマトキシリン及びエオシン染色。10倍の対物レンズを用いて画像を得た。
【図1f】対照(Con)とリファンピシン(RIF、60mg/kg、3日間)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウスから得た代表的な肝臓切片のオイルレッドO染色。40倍の対物レンズを用いて画像を得た。
【図1g】対照とリファンピシン(60mg/kg、3日間)で処理したC57BL/6J及びCyp3aクラスターKOマウスの血漿の臨床化学解析:トリグリセリド、低密度リポタンパク(LDL)、高密度リポタンパク質(HDL)、クレアチンキナーゼ(CRE)、総ビリルビン(BIL−T)、アスパラギン酸アミノトランスフェラーゼ(AST)、アラニントランスフェラーゼ(ALT)、アルカリホスファターゼ(ALP)、アルブミン。データは平均±S.D.で示す(n=4)。
【図1h】図1h:コーン油(3日間投与)(A)又はリファンピシン(60mg/kg、3日間投与)(B)で処理後4日目の野生型及びCyp3aクラスターKOマウスにおけるミダゾラム(1mg/kg)薬物動態(誤差棒はSDを示す)。N.B.:リファンピシン処理C57BL/6Jマウスの10分時点でのSDは45ng/mLであった。グラフAとBのスケールを同じに保つため、対応する誤差棒は一部のみを示した。
【図1i】WinNonlinソフトウェアの非コンパートメント解析を用いたミダゾラムPKから算出した濃度/時間曲線下総面積(AUC)。
【図1j】C57BL/6J及びCyp3aクラスターKOマウスにおけるCyp3aアイソフォームの発現及び活性。A)ウェスタンブロッティング;肝ミクロソーム由来のタンパク質1μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.1pmolのCyp3a11組換えタンパク質(hisタグ)を用いた。一次抗体としてはウサギ抗CYP3A2(Colin Henderson, Ninewells Hospital, Dundee)を1:2000の濃度で用いた。B)7−BQの酸化、C)ミダゾラムの1’−水酸化及びD)ミダゾラムの4−水酸化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(**:P<0.01、***:P<0.001)。
【図1k】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp1aタンパク質の発現及び活性。(A)−ウェスタンブロッティング;肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.1pmolのCyp1a1組換えタンパク質を用いた。一次抗体としてはヤギ抗Cyp1a1/1a2(BDゲンテスト、cat#458131)を1:2000の濃度で用いた。(B)−エトシキレゾルフィン脱エチル化、(C)−メトキシレゾルフィン脱メチル化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05、**:P<0.01)。
【図1l】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの(A)肝ミクロソーム及び(B)腸ミクロソームにおけるCyp2bタンパク質の発現。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.01pmolのCyp2b20組換えタンパク質(hisタグ)を用いた。一次抗体としてはウサギ抗Cyp2b(Colin Henderson, Ninewells Hospital, Dundee)を1:2000の濃度で用いた。(C)−ペントキシレゾルフィン脱ペンチル化。(D)−ベンゾキシレゾルフィン脱ベンジル化。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05)。
【図1m】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの肝ミクロソームにおけるCyp2cタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手したCyp2c29(0.5pmol)組換えタンパク質を用いた。一次抗体としてはウサギ抗Cyp2c(Colin Henderson, Ninewells Hospital, Dundee)を1:50000の濃度で用いた。
【図1n】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの(A)肝ミクロソーム及び(B)腸ミクロソーム。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した組換えCyp2d22(0.2pmol)を用いた。一次抗体としてはヒツジ抗Cyp2d(Colin Henderson, Ninewells Hospital, Dundee)を1:20000の濃度で用いた。
【図1o】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKO(KO)マウスの(A)肝ミクロソーム及び(B)腸ミクロソームにおけるCyp2eの発現。肝ミクロソーム及び腸ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した0.2pmolのCyp2e1組換えタンパク質を用いた。一次抗体としてはヒツジ抗Cyp2e(SAPU BRC)を1:4000の濃度で用いた。
【図1p】媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp4aタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはCXRバイオサイエンシーズから入手した1μgのAPFO誘導ラットミクロソームを用いた。一次抗体としてはウサギ抗Cyp4a(Colin Henderson, Ninewells Hospital, Dundee)を1:20000の濃度で用いた。
【図1q】(A)媒体(対照)とリファンピシン(RIF;60mg/kg、3日間投与)で処理したC57BL/6J(WT)及びCyp3aクラスターKOマウス(KO)の肝ミクロソームにおけるP450オキシドレダクターゼタンパク質の発現。肝ミクロソーム由来のタンパク質2μgを7.5%SDSpageゲルに添加した。正の対照にはBDゲンテスト(cat#452172)から入手した3μgの雄性ヒト肝ミクロソーム(プール)を用いた。一次抗体としてはウサギ抗P450レダクターゼ(CXRバイオサイエンシーズ)を1:5000の濃度で用いた。(B)−C57BL/6J及びCyp3aクラスターKOマウスのマウス肝ミクロソームにおけるシトクロムCレダクターゼ活性。データは平均±SDで示す(C57BL/6J(n=3)、Cyp3aクラスターKO(n=4))。
【図1r】媒体とリファンピシン(60mg/kg、3日間投与)で処理したC57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおける総P450量。データは平均±SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:対照との有意差有り(*:P<0.05、**:P<0.01、***:P<0.001)。
【図1s】(A)−リファンピシン(60mg/kg、3日間投与)による処理後4日目にミダゾラム(1mg/kg)をIV投与した後の野生型、Cyp3aクラスターKO及びCyp3a11KOマウスにおける1’−ヒドロキシミダゾラム薬物動態(誤差棒はSDを示す)。(B)1’−ヒドロキシミダゾラムPKから算出した曲線下面積(AUC)。
【図1t】リファンピシン(60mg/kg、3日間投与)による処理後4日目にミダゾラム(2mg/kg)を経口投与した後の野生型、Cyp3aクラスターKO及びCyp3a11KOマウスにおける1’−ヒドロキシミダゾラム薬物動態(誤差棒はSDを示す)。(B)1’−ヒドロキシミダゾラムPKから算出した曲線下面積(AUC)。
【図1u】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるトリアゾラムのα水酸化。データは平均+SDで示す(C57BL/6J及びCyp3a11KOの肝ミクロソーム(n=4)、Cyp3aクラスターKOの肝ミクロソーム及びC57BL/6Jの腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001)。Cyp3aクラスターKO及びCyp3a11KOの腸ミクロソームの活性は定量下限未満であった。
【図1v】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるニフェジピンの酸化。データは平均+SDで示す(C57BL/16J及びCyp3a11KOのミクロソーム(n=4)、Cyp3aクラスターKO(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001)。
【図1w】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるテストステロンの6β水酸化。データは平均+SDで示す(肝ミクロソーム(n=4)、腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001、**:p<0.01)。
【図1x】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソームによるテストステロンの16β水酸化。データは平均+SDで示す(n=4)。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(**:p<0.01)。
【図1y】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるDBFの酸化。データは平均+SDで示す(肝ミクロソーム(n=4)、腸ミクロソーム(n=3))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(***:P<0.001、*:p<0.05)。
【図1z】C57BL/6J、Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソーム及び腸ミクロソームによるBFCの酸化。データは平均+/−SDで示す(肝ミクロソーム(n=4)、C57BL/6Jの腸ミクロソーム(n=4)、Cyp3a11KOの腸ミクロソーム(n=3)、Cyp3aクラスターKOの腸ミクロソーム(n=2))。活性は対応のないt検定(両側P値)で比較した。*:有意差有り(*:p<0.05)。
【図2a】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2b】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2c】図2a〜2cはCyp2cクラスター交換のための方法を示す。A)マウスCyp2cクラスターの5’ターゲティング、B)マウスCyp2cクラスターの3’ターゲティング、C)概要:Cyp2cクラスターノックアウト、D)マウスID234254のゲノムからのマウスCyp3a クラスター欠如のPCR確認、E)マウスID234254のゲノムからのマウス3a クラスター欠如のPCR確認。
【図2d−1】Cyp2cクラスター欠失に対するマウスホモ接合性のPCR確認。
【図2d−2】Cyp2cクラスター欠失に対するマウスホモ接合性のPCR確認。
【図2e】Cyp3a/Cyp2s複合クラスターKO株の構築のPCR確認。
【図2f】Cyp3a/Cyp2s複合クラスターKO株の構築のPCR確認。
【図3a】Cyp2dクラスター交換のための方法を示す。A)マウスCyp2d26の3’ターゲティング。
【図3b】Cyp2dクラスター交換のための方法を示す。B)マウスCyp2dクラスターの5’ターゲティング。
【図3c】Cyp2dクラスター交換のための方法を示す。C)概要:Cyp2dノックアウト。
【図3d−1】Cyp2dクラスター欠失に対するマウスホモ接合性のPCR確認。
【図3d−2】Cyp2dクラスター欠失に対するマウスホモ接合性のPCR確認。
【図3e】野生型(例えば、C75BL/6J)及びCyp2dクラスターKOマウスにおけるブフラロール(10mg/kg、単回経口投与)の薬物動態。データは、投与後240分まで全血中で検出されたブフラロール濃度の平均±SD(n=4)で示す。
【図4a】CYP1Aノックアウトのための方法を示す。A)マウスCyp1aの5’ターゲティング。
【図4b】CYP1Aノックアウトのための方法を示す。B)マウスCyp1aの3’ターゲティング。
【図4c】CYP1Aノックアウトのための方法を示す。C)概要:Cyp1aノックアウト。
【発明を実施するための形態】
【実施例】
【0159】
実施例1:Cyp3aクラスター交換
相同組換えによってマウスCyp3aクラスターの5’末端を標的にして、図1aに示すように、frt、lox5171及び野生型loxP部位がCyp3a57遺伝子のエクソン1の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットとチミジンキナーゼ(TK)発現カセットを共導入した。
【0160】
マウスCyp3aクラスターの3’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、野生型loxP及びf3部位がCyp3a59遺伝子のエクソン4の下流に挿入されるようにした。図1bに示すように、5’欠損ネオマイシン耐性カセットとTK発現カセットを共導入した。翻訳開始ATGは、loxP部位によって5’欠損ネオマイシン耐性カセットからインフレームで分離させて、loxP部位によってコードされる他のアミノ酸がネオマイシンタンパク質のN末端に付着するようにする。
【0161】
Cre介在組換えによってマウスCyp3aクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。これについては、Creを用いた二重標的ES細胞のトランスフェクションとそれに続くガンシクロビルを用いた選択によってインビトロで試験した。図1eに示すように、同一対立遺伝子上でターゲティングを行った細胞においてのみ、TK遺伝子はクラスターの両端で欠失し、ヌクレオシド類似体ガンシクロビルから毒性三リン酸類似体へのTK介在転化が防止される。従って、同一対立遺伝子上でターゲティングを行ったES細胞のみがガンシクロビル選択後も生存する。
【0162】
生存ES細胞クローンをPCRで更に解析し、マウスCyp3aクラスターが欠失し、PCR陽性クローンが胚盤胞に注入されてキメラマウスが作出されたことを確認した。その後の交雑によって、Cyp3aクラスターがホモ欠損したマウスを最終的に作出した。
【0163】
その後、ES細胞に残存する野生型loxP及びlox5171部位を用い、Cre介在挿入によって置換遺伝子(例えば、ヒトCYP3Aクラスター)をマウスクラスターの前の位置において挿入することができる。残存プロモーター、ATG及びインフレーム野生型loxP部位を用いて、5’欠損選択カセット(例えば、5’欠損ネオマイシン)を再度挿入し、高効率で正しく挿入されたクローンを選択することができる。更なるfrt部位及びf3部位が正しい方向に置換カセットと共に共導入されている場合には、両端におけるfrt部位及びf3部位を用い、Flp介在欠失によって、置換遺伝子と共に導入された選択カセットを欠失させることができる。
【0164】
Cyp3a57の800bp上流に位置し、Cypと無関係の多数の遺伝子によってマウスCyp3aクラスターから離れているCyp3a13遺伝子は、上述のマウスCyp3aクラスターの欠失に影響されない。
【0165】
図1bは、IDが219678、219679、219680、219681及び219682のマウスのゲノムからのマウスCyp3aクラスター欠如のPCR確認を示す。
【0166】
C57BL/6Jとの比較によるCyp3aクラスターKOマウスの解析
研究は4群(4匹/群)で行った。マウスには、リファンピシン(60mg/kg)を3日間、腹腔内(IP)投与した。研究4日目に、ミダゾラム(1mg/kg)をIP注射により投与し、10μLの血液サンプルを10分、20分、40分、60分、120分、240分及び360分で採取してPK解析を行った(表1)。
【0167】
【表1】
【0168】
病理学
肝腫
媒体処理Cyp3aクラスターKOマウスにおいては、生物学的に無関係と思われる媒体処理C57BL/6Jマウスと比べて、肝臓/身体重量比が僅か(〜6%)ではあるが統計的に有意に増加した。リファンピシン処理の場合には、いずれのマウス株においても肝腫は生じなかった(表2)。
【0169】
【表2】
【0170】
病理組織学
ヘマトキシリン及びエオシン染色
全群のマウス全ての腎臓サンプル、肝臓サンプル及び小腸サンプルに対して病理組織学的検討を行った。処理C57BL/6Jマウスにおいては軽度の小葉中心性肝細胞肥大が認められた。Cyp3aクラスターKOマウスの肝臓には微細な小葉中心性肥大の徴候もあった。このような変化はリファンピシンによる処理に関連すると考えられた。肝臓で記録された他の微視的な知見は全て、処理マウスと対照マウスとの間で有意差が無かった。いずれの差もランダムな事象であり、自然発生的であり、マウスで通常見られる正常なバックグラウンド病理の範囲内であると見なされた(図1c)。腎臓及び小腸においてはリファンピシン処理に関連する微視的知見は記録されなかった(図1d〜1e)。この研究の条件下では、2種の異なるマウス株に対してリファンピシン(60mg/kg体重)を3日間反復投与することによって、両方の株で肝細胞肥大の病理学的証拠が得られた。トランスジェニックマウス(Cyp3aクラスターKO)ではあまり大きな影響を受けないように思われた。
【0171】
オイルレッドO染色
オイルレッドO染色により脂質沈着について肝臓サンプルを調べた。媒体処理マウス肝臓のいずれにおいても脂肪蓄積の徴候は無かった(図1f)。リファンピシン処理C57BL/6Jマウスの肝臓においては、核周辺に中程度(グレード1)の滴模様のオイルレッドO染色が観察された。これは、リファンピシン処理Cyp3aクラスターKOマウスの肝臓切片では検出できなかった。
【0172】
臨床化学
全てのマウスから血漿サンプルを採取し、トリグリセリド、アラニントランスフェラーゼ、アルカリホスファターゼ、アスパラギン酸アミノトランスフェラーゼ、アルブミン、ビリルビン(総ビリルビン及び直接ビリルビン)、クレアチンキナーゼ、高密度リポタンパク質、低密度リポタンパク質及びγ−グルタミルトランスフェラーゼについて解析した。γ−グルタミルトランスフェラーゼ活性及び直接ビリルビンレベルは定量限界未満であった。他の血漿臨床化学パラメータの値は全て、未処理C57BL/6Jマウスの公知の正常範囲内であった(図1g)。
【0173】
分子/生化学的表現型
ミダゾラムの薬物動態
野生型及びCyp3aクラスターKOマウスにおけるミダゾラムの薬物動態を図1hに示す。
【0174】
両方のマウス株はIP投与後に基質の速やかな吸収を示した。これは、3日間のリファンピシンの先行投与によっては影響を受けなかった。ミダゾラムのAUCは、野生型及びCyp3aクラスターKO対照マウスにおいて同様であった(図1i)。リファンピシン処理によって両方のマウス株で基質の排除が促進されたが、C57BL/6Jマウスにおいてより高い作用が示された。アイソフォーム特異的基質が腹腔内投与された場合、肝臓における代謝シトクロムP450の発現レベルは該基質のAUCに反比例することが先に示されている。従って、ミダゾラムのAUCは、Cyp3aクラスターKOマウスではその主要代謝酵素のクラスターが欠失しているため、より高いことが予測された。しかし、この場合にはそうではなく、恐らく、次の文献(van Waterschoot et al., 2008)に記載ものと同様の何らかの代償性変化が関与していることが示唆される。
【0175】
Cyp3aタンパク質の発現及び活性
Cyp3aクラスターKOマウスの肝ミクロソームにおけるCyp3aの発現は、処理マウス及び対照マウスの両方においてウェスタンブロッティングによる検出限界未満のレベルまで低下した(図1j)。これは、対応するC57BL/6Jマウス実験群と比べてCyp3a特異的基質7−BQの酸化が有意に低下(対照マウスにおいては18%、リファンピシン処理マウスにおいては6.7%まで低下)したことと関連付けられた。Cyp3aアイソフォーム及びCyp2cアイソフォームの両方によって触媒されるミダゾラムの1’−水酸化は、対照マウス及び処理マウスでそれぞれ53%、18%まで低下した。ミダゾラムの4−水酸化の低下(65%まで)は小さく、対照C57BL/6Jと比べてCyp3aクラスターKO対照群では統計的に有意ではなかった。これは、4−水酸化がマウスCyp3aによって支配的に触媒されることが示されている(Perl off et al., 2000)ことを考慮すると予期せぬ結果であった。ミダゾラムの4−水酸化は、Cyp3aクラスターKOリファンピシン処理群においては野生型マウスと比べて23%まで低下した。
【0176】
Cyp3aクラスターKOマウスのリファンピシンによる前処理によって、7−BQの酸化は僅かではあるが(168%)、統計的に有意に(P<0.05)増加した。Cyp3aクラスターKOマウスにおけるミダゾラムの水酸化に対するリファンピシンの作用は小さく(1’−水酸化では123%増加、4−水酸化では112%)、統計的に有意ではなかった。
【0177】
腸ミクロソームのウェスタンブロッティングでは如何なるCyp3aバンドも得られなかった(データは示さず)。
【0178】
腸ミクロソームの比活性の測定によって得たデータの不整合性が高かったが、これは、上皮細胞を擦り取って腸ミクロソームを調製する際にむらがあることを反映しているように思われる(Bonkovsky et al., 1985; Mohri and Uesawa, 2001)(データは示さず)。
【0179】
本研究に用いた抗体は、試験したサブファミリー由来の主要な組換えシトクロムP450との相互作用後にウェスタンブロット上で明確なバンドを示した。しかし、1種のP450サブファミリー内の異なるアイソフォームに対する抗体の特異性については検討しなかった。
【0180】
Cyp1aタンパク質の発現及び活性
ウェスタンブロッティングで確認したCyp1aの発現は、Cyp3aクラスターKOマウスの肝ミクロソームにおいては野生型に比べて僅かに高かった(図1k)。リファンピシン処理によって、両方のマウス株でCyp1aが僅かにダウンレギュレートされた。エトキシレゾルフィン脱エチル化活性(EROD)は、Cyp3aクラスターKOマウスの肝ミクロソーム及び野生型マウスの肝ミクロソームの両方において同様であった。ERODはCyp1a1アイソフォーム及びCyp1a2アイソフォームの両方によって触媒されるが、Cyp1a1の方が主に寄与する。リファンピシン処理によって、両方の株でEROD代謝が僅かに誘導された。野生型マウスとCyp3aクラスターKOマウスとの間でメトキシレゾルフィン脱メチル化(MROD)活性の差は無かった。MRODは主にCyp1a2によって触媒される。
【0181】
Cyp2bタンパク質の発現及び活性
マウス肝ミクロソームでのCyp2bの発現は、C57BL/6Jマウス及びCyp3aクラスターKOマウスの両方において低かった(図1l)。リファンピシン処理によって、両方の株でCyp2bが僅かに誘導された。これはPROD活性の増加が少ないことと関連付けられた。腸ミクロソームでは顕著に高いCyp2bの発現が示されたが、リファンピシン処理に対して感受性が低かった。媒体処理Cyp3aクラスターKOマウスの肝ミクロソームでは、ベンゾキシレゾルフィンO−脱アルキル化(BROD)活性が野生型に比べて低いことが示された。BRODは先にマウスCyp2bの基質として用いられた(Donato et al., 2003)。リファンピシン処理によって、試験した両方の株においてBRODが誘導された。これらのデータから、Cyp3a(更に、恐らくはCyp2cアイソフォーム)由来の酵素がBRODに関与し得ることが示唆される。
【0182】
Cyp2cタンパク質の発現
C57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおけるCyp2cタンパク質の発現はウェスタンブロッティングによって評価した(図1m)。タンパク質バンド強度は、野生型と比べて、対照Cyp3aクラスターKOマウス由来のサンプルにおいて僅かに増加した。両方のマウス株では、リファンピシン処理後に顕著なCyp2c誘導が示された。
【0183】
Cyp2dタンパク質の発現
Cyp2dタンパク質のバンドは、肝臓サンプル及び腸サンプルの両方において検出された(図1n)。未処理野生型マウスの肝臓サンプルとCyp3aクラスターKOマウスの肝臓サンプルとの間でCyp2dタンパク質バンド強度に顕著な差は無かった。リファンピシン処理によって、C57BL/6JマウスではCyp2dの発現が僅かに低下したが、Cyp3aクラスターKOマウスでは僅かに増加した。野生型に比べて、Cyp3aクラスターKOでは腸Cyp2dレベルが僅かに増加した。腸ミクロソームでのCyp2dレベルに対するリファンピシン処理の顕著な作用はいずれのマウス株でも見られなかった。
【0184】
Cyp2eタンパク質の発現
Cyp2eタンパク質バンド強度は、野生型と比べてCyp3aクラスターKOマウスの肝ミクロソームでは僅かに高かった(図1o)。肝ミクロソームにおけるCyp2eレベルは、いずれのマウス株においてもリファンピシン処理によっては顕著に変化しなかった。同様に、Cyp2e発現のレベルは、野生型と比べてCyp3aクラスターKOマウスの腸においては高かった。リファンピシン処理によって、Cyp3aクラスターKOマウスでは腸Cyp2eが僅かに誘導されたが、野生型マウスでは腸Cyp2eが僅かにダウンレギュレートされた。
【0185】
Cyp4aタンパク質の発現
Cyp4aタンパク質は両方のマウス株の肝ミクロソームで検出された(図1p)。野生型マウス由来サンプルにおけるCyp4a発現レベルはリファンピシン処理後に顕著に増加した。
【0186】
マウスP450オキシドレダクターゼ(POR)タンパク質の発現
シトクロムP450オキシドレダクターゼレベルは、C57BL/6J及びCyp3aクラスターKOマウスの肝ミクロソームにおいては同様であった(図1q)。両方のマウス株は、PORのタンパク質発現レベルの顕著な増加によってリファンピシン処理に反応した。リファンピシン処理マウスの肝ミクロソームでは、野生型マウスと比べてシトクロムCの還元における活性が僅かに高いことが示された。この増加は統計的に有意ではなかった。
【0187】
総P450量
Cyp3aクラスターKOマウスでは、C57BL/6Jマウスと比べて、媒体処理マウスの肝ミクロソームの総P450量が僅かではあるが統計的に有意に増加することが示された(図1r)。リファンピシン処理によって、野生型マウスでは総P450量が顕著に増加した。しかし、Cyp3aクラスターKOマウスの肝ミクロソームのP450量は変化しなかった。
【0188】
結論
Cyp3aクラスターKOマウスの場合、血漿、肝臓、腎臓又は小腸においては、如何なる遺伝子型誘導の病理学的変化も示されなかった。このモデルで示されたCyp3a発現のレベルは、ウェスタンブロッティングによる検出限界未満であった。Cyp3a特異的基質7−BQに対する活性が顕著に低下(野生型マウスと比べて20%未満のレベルまで)した。ミダゾラムクリアランスに関するインビトロデータから、Cyp3aサブファミリー由来のP450はこの基質の排除に関与する唯一の代謝酵素ではないことが示唆された。未処理Cyp3aクラスターKOマウスの肝ミクロソームのミダゾラム水酸化活性は、野生型株のものに対して〜60%であった。このことは、その酸化における他のP450(例えば、Cyp2c)の関与によって説明できよう(Perloff et al., 2000; van Waterschoot et al., 2008)。リファンピシン処理によって野生型ではCyp3aが誘導されたが、Cyp3aクラスターKOでは誘導されなかった。これによって、処理Cyp3aクラスターKOマウスの肝ミクロソームにおけるミダゾラム水酸化活性にはより顕著な差(野生型に比べて20%)がもたらされた。ミダゾラムPKデータによって、ミダゾラム排除への複数の代謝酵素の関与についての更なる証拠が得られた。未処理Cyp3aクラスターKOマウスとC57BL/6Jマウスとの間ではミダゾラムPKの差は無かったが、これは先に公表されたデータと関連付けられ、Cyp3aクラスターKO株におけるP450(例えば、Cyp2cサブファミリー由来)の多少のアップレギュレーションに起因し得る(van Waterschoot et al., 2008)。リファンピシンの投与によって両方のマウス株でミダゾラムの排除が促進された。これは代謝酵素の誘導を示し、野生型マウス及びCyp3aクラスターKOマウスにおいてそれぞれ、最大470%及び270%であった(概算はAUCの変化に基づいて為された)。誘導C57BL/6Jマウスの肝ミクロソームでもミダゾラム水酸化活性の高い増加(最大380%)を示したが、Cyp3aクラスターKO株の肝ミクロソームで示されたミダゾラム水酸化の誘導はほんの僅かであった(最大125%)。インビトロ及びインビボでのミダゾラムクリアランスに対するリファンピシンの作用の差は、Cyp3aクラスターKOマウスのインビボでのミダゾラム排除の代替経路の誘導を暗示する。
【0189】
更なる実験を行い、1’−ヒドロキシミダゾラムのPKプロファイル及び他のCyp3a基質の酵素修飾について比較した。
【0190】
研究は9群で行った(表3)。マウスにはリファンピシン(60mg/kg)を3日間腹腔内(ip)投与した。研究4日目に、ミダゾラムを1〜6群のマウスにはIV注射によって投与(1mg/kg)し、7〜9群のマウスには経口投与(2mg/kg)した。血液サンプルをIV投与後2分、5分、10分、20分、30分、45分、60分、90分、120分及び150分(連続出血)で採取し、また、経口投与後10分、20分、40分、60分、90分、120分、240分及び360分で採取し、PK解析を行った(表3)。
【0191】
【表3】
【0192】
1’−ヒドロキシミダゾラムの薬物動態
先のリファンピシン処理によって野生型マウスではCyp3aが誘導されたが、Cyp3aクラスターKOマウスでは誘導されなかった。これによって、2種類のマウス株間ではインビトロでのミダゾラム水酸化活性に顕著な差がもたらされた。しかし、インビボでのミダゾラム排除における差はあまり顕著ではなく、これは、Cyp3aクラスターKOマウスにおけるインビボでのミダゾラム排除の代替経路の誘導を示唆している。本研究では、P450発現レベル概算のためのより高感度で具体的な方法として、リファンピシン誘導マウスへのミダゾラムのIV投与及び経口投与後に1’−ヒドロキシミダゾラムの薬物代謝(一次代謝産物)をモニターした。血液をB−グルクロニダーゼで前処理した後に1’−ヒドロキシミダゾラムの総濃度を測定した。親化合物をIV注入した後の1’−ヒドロキシミダゾラムの薬物動態を図1sに示す。
【0193】
曲線の終末段階はC57BL/6Jマウス及びCyp3a11KOマウスの両方において同様であるが、これは、この代謝産物のクリアランスにおける株差が非常に小さいことを示唆する。Cyp3aクラスターKOマウスPKの終末段階は、1’−ヒドロキシミダゾラムの排除が他の株と比べて僅かに遅いことを示す。1’−ヒドロキシミダゾラムのAUCは、Cyp3aクラスターKOマウスでは野生型株と比べて顕著に(〜3倍)低下したが、これは、Cyp3aサブファミリー由来のシトクロムP450がこの代謝産物生成の主要な経路であることを示す。Cyp3a11KOマウスにおけるAUCの低下はあまり顕著ではなかった(〜2倍)が、これは、1’−ヒドロキシミダゾラムの生成にCyp3aサブファミリーの他のメンバーが多少関与していることを示唆する。ミダゾラム経口投与後の1’−ヒドロキシミダゾラムの薬物動態を図1tに示す。
【0194】
曲線の終末段階は全ての株で同様であった。しかし、初期の吸収段階では差が見られた。野生型マウスと比べて、Cyp3aクラスターKOマウスやCyp3a11KOマウスではバイオアベイラビリティが低かった。これは、ミダゾラム経口投与後の1’−ヒドロキシミダゾラムの生成に腸シトクロムP450が大きく寄与していることを示す。このことは、IV注入後に野生型マウスとKOマウスとの間で1’−ヒドロキシミダゾラムのAUCが非常に大きく異なっていることを示したIVデータによって支持された。本研究においては、曲線の吸収段階を定めるデータ点が非常に少なかったため、C57BL/6JにおけるCyp3a酵素からの影響は過小評価されているように思われる。
【0195】
Cyp3a酵素活性
肝ミクロソーム及び腸ミクロソームによるCyp3a特異的基質の酸化は、トリアゾラム、ニフェジピン、テストステロン、ジベンジルフルオレセイン(DBF)及び7−ベンジルオキシ−4−トリフルオロメチルクマリン(BFC)を用いて試験した。トリアゾラム、ニフェジピン及びテストステロンは、10種のCYP3A4基質の様々なCYP3A4リガンドとの相互作用の統計的解析に基づき、CYP3A4内の異なる基質グループに帰属された(Kenworthy et al., 1999)。DBFとBFCは蛍光代謝物を産生し、CYP3A4阻害剤のハイスループットスクリーニングにおいて広く応用されている。他の蛍光基質である7−ベンジルオキシキノリン(BQ)は、Cyp3aに対して良好な特異性を示している。
【0196】
関連するグラフの全てにおいて、小腸由来サンプルの活性は右のY軸スケールを参照し、肝ミクロソームの活性は左のY軸スケールを参照されたい。
【0197】
トリアゾラムα−水酸化
肝ミクロソーム及び腸ミクロソームによるトリアゾラム酸化の結果を図1uに示す。Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方においてトリアゾラムの水酸化は顕著に低下した。このことは、公表データ(Perloff et a1.,200O)とよく関連付けられ、トリアゾラムはCyp3aサブファミリー由来のマウスシトクロムP450に対して高い特異性を有する基質であることが示された。また、このデータから、Cyp3aクラスターKO及びCyp3a11KOの活性は非常に低く、統計的に違いが無かったため、水酸化にはCyp3a11が大きく寄与し、他のCyp3a酵素からの影響は無視できることも分かった。
【0198】
ニフェジピン酸化
Cyp3aクラスターKO及びCyp3a11KOマウスの肝ミクロソームによるニフェジピン酸化は、野生型マウスと比べて有意に(P<0.001)低下した(図1v)。この低下はトリアゾラムの場合と比べてあまり顕著ではなかったが、これは、異なるサブファミリー由来の酵素の潜在的な関与を示す。Cyp3aクラスターKO及びCyp3a11KOの平均値は異なったが、統計的な有意差は無かった。腸ミクロソームによるニフェジピン酸化においては統計的な有意差は無かった。
【0199】
テストステロン水酸化
テストステロンは、数種の異なるシトクロムP450によって水酸化される。6p−水酸化は通常、Cyp3aサブファミリー由来の酵素に起因するが、16B−水酸化はCyp2b10及びCyp2c29によって触媒される(Honkakoski et al., 1992)。Cyp3aクラスターノックアウトマウスにおけるCyp2cサブファミリー由来酵素のアップレギュレーションは先に報告されている(van Waterschoot et al., 2008)。従って、両方の水酸化については肝ミクロソーム及び腸ミクロソームを用いて検討した(図1w〜1x)。
【0200】
Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方においてテストステロンの6β−水酸化は有意に低下した。トランスジェニックマウス株の肝ミクロソームによる6β−テストステロンの生成においては差が無かった。Cyp3aクラスターKOマウスの腸ミクロソームの活性はCyp3a11KOマウスの場合と比べて低かった(P<0.05)が、これは、マウス小腸によるテストステロン6β−水酸化に他のCyp3a酵素が関与していることを示し得る。テストステロンの16β−水酸化は、野生型マウスと比べて両方のトランスジェニック株の肝ミクロソームで増加したが、これは何らかの代償性変化を示唆し、この変化は、Cyp2cサブファミリー由来の酵素の過剰発現であると思われる(van Waterschoot et al., 2008)。Cyp3aクラスターKOの平均値はCyp3a11KOの場合と比べて高かったが、統計的な有意差は無かった。
【0201】
DBFの酸化
DBFの酸化の結果を図1yに示す。肝ミクロソームによるDBF酸化は、Cyp3aクラスターKOマウス及びCyp3a11KOマウスの両方に由来するサンプルにおいては顕著な低下を示した。2種のノックアウト株間では統計的な有意差は無かったが、これは、Cyp3a11が主要な代謝酵素であることを示唆する。腸ミクロソームによるDBF酸化においては、野生型株とノックアウト株との間で統計的な有意差は無かった。
【0202】
BFCの酸化
BFCの酸化の結果を図1zに示す。
肝ミクロソームサンプル間でBFC活性に差は無かった。腸ミクロソームにおいては、僅かではあるが統計的に有意な増加が見られたが、これは、BFCを酸化する、トランスジェニックマウスにおける他のサブファミリー由来のシトクロムP450のアップレギュレーションを示唆する。
【0203】
結論
Cyp3aクラスターKOモデルは先に基質特異的挙動を示した。このモデルと野生型マウスとの明らかな差は7−BQの酸化に見られた。しかし、ミダゾラムの水酸化では差が見られず、これは、この反応における他のP450(例えば、Cyp2c)の関与を示唆する(Perloff et al., 2000; van Waterschoot et al., 2008)。リファンピシン処理によって野生型ではCyp3aが誘導されたが、Cyp3aクラスターKOでは誘導されなかった。この結果、処理Cyp3aクラスターKOマウスの肝ミクロソームにおけるミダゾラム水酸化活性は顕著に低下した。ミダゾラムAUCで示されたように、インビボでの差はあまり顕著ではなかったが、これは、Cyp3aクラスターKOマウスにおけるミダゾラム排除の代替経路の誘導を示唆する。このことは、ミダゾラム代謝産物の薬物動態によって、Cyp3a発現レベルをより具体的且つ高感度に概算し得ることを暗示した。
【0204】
親化合物をIV投与した後の1’−ヒドロキシミダゾラムのAUCはCyp3aクラスターKOマウスにおいて顕著に(〜3倍)低下したが、これは、この代謝産物の生成にCyp3aサブファミリーが大きく寄与することを示す。この方法は、先に用いられた方法の改良であり、ミダゾラムAUCの測定(1.6倍)に基づいた。しかし、インビトロで見られた差(6倍)には完全に対応しなかった。この違いは、親化合物及び代謝産物自身の両方の分布量やクリアランスからの代謝産物AUCの複合依存性によって説明し得る(Rowland and Tozer, 1995)。腸代謝や吸収が代謝産物の処理に関与する場合には、親薬物の経口投与後に他の複雑な因子が存在する。マウス株間の基質酸化における差は、肝ミクロソームと腸ミクロソームとの間でテストステロンやトリアゾラムに関してはよく対応した。しかし、ニフェジピン、DBF及びBFCの酸化に関してはパターンが異なった。これは、後者の反応に様々な酵素が関与し、これらの酵素の相対濃度が肝臓や小腸で異なることを示し得る。このことは、シトクロムP450の発現レベルの概算において、経口投与後の代謝物AUCがIV注入後の代謝物AUCと比べて有益ではないことを示唆した。本研究に用いたCyp3a基質は、その酸化におけるCyp3aサブファミリーの影響の点で次のようにランク付けすることができる(トリアゾラム>DBF:テストステロン>ニフェジピン>>BFC)。
【0205】
CYP3A4基質であるBFCの酸化に関するデータから、Cyp3aサブファミリー由来のマウスシトクロムP450に対するこの化合物の特異性が低いことが分かる。このことは、先に報告されたBFC酸化におけるヒトシトクロムP450とラットシトクロムP450との間の差(Stresser et al., 2002)と一致する。
【0206】
Cyp3aクラスターノックアウトの作用に関しては、全ゲノムマイクロアレイ及び肝臓から調製したRNAを用いて転写レベルでも評価した。本研究においては、Agilent Whole Mouse Genome Oligo Microarrayスライド(G4122−60510)及びAgilent 44KガスケットスライドG2534−60005を用いた。
【0207】
マウスには、表1に示すように、リファンピシン/コーン油(60mg/kg/3日間)又は媒体のみを腹腔内注射によって投与した。遺伝子発現の変化倍数(Fold changes)を表4に示す。
【0208】
【表4−1】
【0209】
【表4−2】
【0210】
【表4−3】
【0211】
実施例2:Cyp2cクラスター交換
相同組換えによってマウスCyp2cクラスターの5’末端を標的にして、野生型loxP及びf3部位がCyp2c55遺伝子のエクソン1の下流に挿入されるようにした。図2aに示すように、5’欠損ネオマイシン耐性カセットとTK発現カセットを共導入した。翻訳開始ATGは、loxP部位によって5’欠損ネオマイシン耐性カセットからインフレームで分離させて、loxP部位によってコードされる他のアミノ酸がネオマイシンタンパク質のN末端に付着するようにする。
【0212】
マウスCyp2cクラスターの3’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、図2bに示すように、frt、lox5171及び野生型loxP部位がCyp2c70遺伝子のエクソン5の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットとチミジンキナーゼ(TK)発現カセットを共導入した。
【0213】
Cre介在組換えによってマウスCyp2cクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。これについては、Creを用いた二重標的ES細胞のトランスフェクションとそれに続くガンシクロビルを用いた選択によってインビトロで試験した。図2cに示すように、同一対立遺伝子上でターゲティングを行った細胞においてのみ、TK遺伝子はクラスターの両端で欠失し、ヌクレオシド類似体ガンシクロビルから毒性三リン酸類似体へのTK介在転化が防止される。従って、同一対立遺伝子上でターゲティングを行ったES細胞のみがガンシクロビル選択後も生存する。生存ES細胞クローンをPCRで更に解析し、マウスCyp2cクラスターが欠失し、PCR陽性クローンが胚盤胞に注入されてキメラマウスが作出されたことを確認した。その後の交雑によって、Cyp2cクラスターがホモ欠損したマウスを作出した。
【0214】
その後、ES細胞に残存する野生型loxP及びlox5171部位を用い、Cre介在挿入によって置換遺伝子(例えば、ヒトCYP2Cクラスター)をマウスクラスターの前の位置において挿入することができる。残存プロモーター、ATG及びインフレーム野生型loxP部位を用いて、5’欠損選択カセット(例えば、5’欠損ネオマイシン)を再度挿入し、高効率で正しく挿入されたクローンを選択することができる。更なるfrt部位及びf3部位が正しい方向に置換カセットと共に共導入されている場合には、両端におけるfrt部位及びf3部位を用い、Flp介在欠失によって、置換遺伝子と共に導入された選択カセットを欠失させることができる。このホモ接合性クラスターKO作出のPCRによる実証を図2dに示す。
【0215】
実施例3:ホモ接合体Cyp3aクラスターノックアウト/Cyp2cクラスターノックアウトマウス
Cyp3aKOマウスとCyp2cKOマウスを交雑させて生存可能なマウス(ID234254)を繁殖させた。このマウスは、マウスCyp3aクラスター(7個の遺伝子が欠失)及びCyp2cクラスター(14個の遺伝子が欠失)の両方がホモ欠損している。
【0216】
図2eは、マウスID234254のゲノムからマウスCyp3aクラスターが欠如していることのPCR確認を示す。図2fは、同一マウスのゲノムからマウスCyp2cクラスターが欠如していることのPCR確認を示す。
【0217】
ホモ接合体Cyp3aクラスターKO/Cyp2cクラスターKOマウスは、実施例1で詳述するように、その病理組織学、生化学、タンパク質発現及びmRNA転写レベルに関して特徴付けされよう。また、公知で仮定のCyp2cP450アイソフォーム基質を用いた該株の更なる特徴付けも実施されよう。
【0218】
実施例4:Cyp2dクラスター欠失
相同組換えによってマウスCyp2dクラスターの3’末端を標的にして、図3aに示すように、frt、lox5171及び野生型loxP部位がCyp2d26遺伝子のエクソン1の下流に挿入されるようにした。更に、図示のように、ハイグロマイシン耐性カセットを共導入した。続くマウスクラスターの欠失によって無傷のCyp2d26プロモーターとCyp2d26遺伝子のエクソン1が残るが、これらは、後述のようにクラスターの5’末端において挿入されるヒトCYP2D6の発現を妨げる可能性があるため、Cyp2d26プロモーターからの発現を終結させる3’ターゲティングベクターにスプライスアクセプターポリアデニル化モチーフ(sApA)を含有させた。
【0219】
マウスCyp2dクラスターの5’末端における次の相同組換え段階によって、正しくターゲティングされたES細胞を更に改変して、図3bに示すように、frt部位、野生型loxP部位及びネオマイシン耐性カセット3’によってloxP部位に隣接させたヒトCYP2D6発現カセットがCyp2d22遺伝子のエクソン3の下流に挿入されるようにした。
【0220】
loxP部位におけるCre介在組換えによってヒト発現カセットを維持しながらマウスCyp2dクラスターを欠失させることができるが、Cyp2d遺伝子座の完全なノックアウト(図3cに示す)をもたらすマウスクラスターとヒト発現カセットの同時欠失は、frt部位におけるFlp介在組換えの結果である。
【0221】
Cre介在又はFlp介在組換えによってマウスCyp2dクラスターの欠失を可能とするためには、クラスター両端におけるターゲティングを同一対立遺伝子上で行う必要がある。従って、Creを用いたインビトロ欠失によって二重標的ESクローンを更に解析した後、クラスターの欠失を可能とするプライマー対を用いてPCR解析を行った。インビトロCre欠失はトランスフェクト細胞の一部でしか生じず、また、欠失が成功した個々の細胞を選択することができないため、PCR解析は欠失クローン及び未欠失クローンのプールに対して行った。PCR陽性未欠失親クローンを胚盤胞に注入してキメラマウスを作出した。Cre又はFlp欠失株、或いは各リコンビナーゼを組織特異的に発現する株と交雑させた後に交雑受精させることによって、全身又は特定の組織において欠失し、ヒト発現カセットを含む(又は含まない)ホモ接合Cyp2dクラスター欠失マウスが作出されよう。
【0222】
このホモ接合性クラスターKO作出のPCRによる実証を図3dに示す。
【0223】
Cyp2dKOマウスにおけるブフラロールPK
Cyp2d特異的基質であるブフラロールの薬物動態をCyp2dKOマウス(Cyp2dアイソフォーム全てが無効化されたマウス)において検討した。野生型であるC57BL/6Jマウスを対照として用いた。ブフラロールのAUCは、野生型マウスと比べるとCyp2dKOマウスでは顕著に増加したことが示された(図3e)。このことは、インビトロデータ(Bogaards et al., 2000)(即ち、ブフラロール水酸化の速度は、ヒト肝ミクロソームと比べてマウス肝ミクロソームでは〜10倍高いことが示された)とよく関連付けられた。
【0224】
参考文献
Anderson, S. P., Dunn, C., Laughter, A., Yoon, L., Swanson, C., Stulnig, T. M., Steffensen, K. R., Chandraratna, R. A., Gustafsson, J. A., and Corton, J. C. (2004). Overlapping transcriptional programs regulated by the nuclear receptors peroxisome proliferator-activated receptor alpha, retinoid X receptor, and liver X receptor in mouse liver. Mol Pharmacol 66, 1440-52.
Bogaards JJ, Bertrand M, Jackson P, Oudshoorn MJ, Weaver RJ, van Bladeren PJ and Walther B (2000) Determining the best animal model for human cytochrome P450 activities: a comparison of mouse, rat, rabbit, dog, micropig, monkey and
Bonkovsky HL, Hauri HP, Marti U, Gasser R and Meyer UA (1985) Cytochrome P450 of small intestinal epithelial cells. Immunochemical characterization of the increase in cytochrome P450 caused by phenobarbital. Gastroenterology 88:458-467.
Donato MT, Klocke R, Castell JV, Stenzel K, Paul D and Gomez-Lechon MJ (2003) Constitutive and inducible expression of CYP enzymes in immortal hepatocytes derived from SV40 transgenic mice. Xenobiotica 33:459-473
Honkakoski, P, Auriola S and Lang MA (1992) Distinct induction profiles of three phenobarbital-responsive mouse liver cytochrome P450 isozymes. Biochem Pharmacol
43:2121-2128.
Kenworthy KE, Bloomer JC, Clark SE and Houston JB (1999) CYP3A4 drug interactions: correlation of 10 in vitro probe substrates. Br J Clin Pharmacol 48:716-727.
Ide, T., Shimano, H., Yoshikawa, T., Yahagi, N., Amemiya-Kudo, M., Matsuzaka, T., Nakakuki, M., Yatoh, S., Iizuka, Y., Tomita, S., Ohashi, K., Takahashi, A., Sone, H., Gotoda, T., Osuga, J., Ishibashi, S., and Yamada, N. (2003). Cross-talk between peroxisome proliferator-activated receptor (PPAR) alpha and liver X receptor (LXR) in nutritional regulation of fatty acid metabolism. II. LXRs suppress lipid degradation gene promoters through inhibition of PPAR signaling. Mol Endocrinol 17, 1255-67.
Inc, A. t. (2007). Agilent Feature Extraction Software v9.5 User Guide. Agilent Publication number g4460-90005, 1-250.
Mohri K and Uesawa Y (2001) Enzymatic activities in the microsomes prepared from rat small intestinal epithelial cells by differential procedures. Pharm Res 18:1232-1236.
Perloff MD, von Moltke LL, Court MH, Kotegawa T, Shader RI and Greenblatt DJ (2000) Midazolam and triazolam biotransformation in mouse and human liver microsomes: relative contribution of CYP3A and CYP2C isoforms. J Pharmacol Exp Ther 292:618-628.
Rowland M and Tozer N (1995) Clinical Pharmacokinetics: Concepts and Applications. Lippincot Williams & Wilkins, Philiadelphia, US.
Saini, S. P., Sonoda, J., Xu, L., Toma, D., Uppal, H., Mu, Y., Ren, S., Moore, D. D., Evans, R. M., and Xie, W. (2004). A novel constitutive androstane receptor-mediated and CYP3A-independent pathway of bile acid detoxification. Mol Pharmacol 65, 292-300.
Stresser DM, Turner SD, Blanchard AP, Miller VP and Crespi CL (2002). Cytochrome P450 fluorimetric substrates: identification of isoform-selective probes for rat CYP2D2 and human CYP3A4. Drug Metab Dispos 30:845-852.
van Waterschoot, R. A., van Herwaarden, A. E., Lagas, J. S., Sparidans, R. W., Wagenaar, E., van der Kruijssen, C. M., Goldstein, J. A., Zeldin, D. C., Beijnen, J. H., and Schinkel, A. H. (2007). Midazolam metabolism in Cytochrome P450 3A knockout mice can be attributed to upregulated CYP2C enzymes. Mol Pharmacol. 73, 1029-36.
Weng, L., Dai, H., Zhan, Y., He, Y., Stepaniants, S. B., and Bassett, D. E. (2006). Rosetta error model for gene expression analysis. Bioinformatics 22, 1111-21.
Xu, C., Li, C. Y., and Kong, A. N. (2005). Induction of phase I, II and III drug
metabolism/transport by xenobiotics. Arch Pharm Res 28, 249-68.
Zhou, J., Zhang, J., and Xie, W. (2005). Xenobiotic nuclear receptor-mediated regulation of UDP-glucuronosyl-transferases. Curr Drug Metab. 6, 289-98.
【特許請求の範囲】
【請求項1】
薬物代謝酵素をコードする遺伝子クラスターの全てのメンバーが機能しないようにされており、欠失したクラスター内の遺伝子と等価な機能を有するヒト置換遺伝子を含まないマウス。
【請求項2】
該遺伝子クラスターは全ての組織において機能しないようにされている、請求項1に記載のマウス。
【請求項3】
薬物代謝酵素をコードする遺伝子クラスターは、P450遺伝子クラスター等の第I相薬物代謝酵素遺伝子クラスターである、請求項1又は請求項2に記載のマウス。
【請求項4】
薬物代謝酵素をコードする遺伝子クラスターは、UGT遺伝子クラスター等の第II相薬物代謝酵素遺伝子クラスターである、請求項1又は請求項2に記載のマウス。
【請求項5】
第I相薬物代謝酵素遺伝子クラスターは、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターから成る群から選択される、請求項3に記載のマウス。
【請求項6】
第I相薬物代謝酵素遺伝子クラスターは、Cyp3aクラスター、Cyp2cクラスター、Cyp2dクラスター、又はCyp3aクラスターとCyp2cクラスターの多重ノックアウトである、請求項5に記載のマウス。
【請求項7】
第I相、第II相及び/又は第III相薬物代謝酵素及び/又は薬物輸送体をコードする多重遺伝子クラスターが機能しないようにされている、先の請求項のいずれか1項に記載のマウス。
【請求項8】
該遺伝子クラスターは、コード配列の欠失によって機能しないようにされている、先の請求項のいずれか1項に記載のマウス。
【請求項9】
コード配列の欠失は、相同組換えのメカニズムによって為されている、請求項8に記載のマウス。
【請求項10】
マウス染色体への更なるヒト遺伝子の導入を伴う更なる遺伝子組換えが行われている、先の請求項のいずれか1項に記載のマウス。
【請求項11】
前記更なるヒト遺伝子は、プレグナンX受容体(PXR)、構成的アンドロスタン受容体(CAR)、PPAR(α、δ及びγ)、NRF2、Ah受容体、HNF1及びHNF4の一以上から選択される、請求項10に記載のマウス。
【請求項12】
前記更なるヒト遺伝子は、グルクロニルトランスフェラーゼ(例えば、UGT1A遺伝子)、グルタチオントランスフェラーゼ(例えば、GST(グルタチオンS−トランスフェラーゼ))、スルホトランスフェラーゼ及びアセチルトランスフェラーゼから選択される、請求項10又は請求項11に記載のマウス。
【請求項13】
前記更なるヒト遺伝子は、多剤耐性タンパク質(例えば、mdr1及びmdr3)及び多剤耐性関連タンパク質(MRP)(例えば、MRP1及び/又はMRP2及び/又はMRP6)、又は有機アニオン輸送ポリペプチド(OATP)から選択される、請求項10〜12のいずれか1項に記載のマウス。
【請求項14】
先の請求項のいずれか1項に記載のマウスから単離された細胞。
【請求項15】
請求項1〜13のいずれか1項に記載のマウス又は請求項14に記載の細胞の集団を用いるアッセイ方法。
【請求項16】
請求項1〜13のいずれか1項に記載のマウスを作出する方法であって、
a)置換対象のマウス標的遺伝子クラスターが各々の端で組換え部位と隣接するように相同組換えによってマウス染色体に一対の部位特異的組換え部位を組込むことと、
b)部位特異的組換え部位間で組換えを行い、マウス標的遺伝子クラスターが染色体から除去され、残存部位特異的組換え部位で置換されるようにすることとを含む方法。
【請求項1】
薬物代謝酵素をコードする遺伝子クラスターの全てのメンバーが機能しないようにされており、欠失したクラスター内の遺伝子と等価な機能を有するヒト置換遺伝子を含まないマウス。
【請求項2】
該遺伝子クラスターは全ての組織において機能しないようにされている、請求項1に記載のマウス。
【請求項3】
薬物代謝酵素をコードする遺伝子クラスターは、P450遺伝子クラスター等の第I相薬物代謝酵素遺伝子クラスターである、請求項1又は請求項2に記載のマウス。
【請求項4】
薬物代謝酵素をコードする遺伝子クラスターは、UGT遺伝子クラスター等の第II相薬物代謝酵素遺伝子クラスターである、請求項1又は請求項2に記載のマウス。
【請求項5】
第I相薬物代謝酵素遺伝子クラスターは、Cyp2abfgstクラスター、Cyp2cクラスター、Cyp2dクラスター、Cyp2jクラスター、Cyp3aクラスター、Cyp4abxクラスター及びCyp4fクラスターから成る群から選択される、請求項3に記載のマウス。
【請求項6】
第I相薬物代謝酵素遺伝子クラスターは、Cyp3aクラスター、Cyp2cクラスター、Cyp2dクラスター、又はCyp3aクラスターとCyp2cクラスターの多重ノックアウトである、請求項5に記載のマウス。
【請求項7】
第I相、第II相及び/又は第III相薬物代謝酵素及び/又は薬物輸送体をコードする多重遺伝子クラスターが機能しないようにされている、先の請求項のいずれか1項に記載のマウス。
【請求項8】
該遺伝子クラスターは、コード配列の欠失によって機能しないようにされている、先の請求項のいずれか1項に記載のマウス。
【請求項9】
コード配列の欠失は、相同組換えのメカニズムによって為されている、請求項8に記載のマウス。
【請求項10】
マウス染色体への更なるヒト遺伝子の導入を伴う更なる遺伝子組換えが行われている、先の請求項のいずれか1項に記載のマウス。
【請求項11】
前記更なるヒト遺伝子は、プレグナンX受容体(PXR)、構成的アンドロスタン受容体(CAR)、PPAR(α、δ及びγ)、NRF2、Ah受容体、HNF1及びHNF4の一以上から選択される、請求項10に記載のマウス。
【請求項12】
前記更なるヒト遺伝子は、グルクロニルトランスフェラーゼ(例えば、UGT1A遺伝子)、グルタチオントランスフェラーゼ(例えば、GST(グルタチオンS−トランスフェラーゼ))、スルホトランスフェラーゼ及びアセチルトランスフェラーゼから選択される、請求項10又は請求項11に記載のマウス。
【請求項13】
前記更なるヒト遺伝子は、多剤耐性タンパク質(例えば、mdr1及びmdr3)及び多剤耐性関連タンパク質(MRP)(例えば、MRP1及び/又はMRP2及び/又はMRP6)、又は有機アニオン輸送ポリペプチド(OATP)から選択される、請求項10〜12のいずれか1項に記載のマウス。
【請求項14】
先の請求項のいずれか1項に記載のマウスから単離された細胞。
【請求項15】
請求項1〜13のいずれか1項に記載のマウス又は請求項14に記載の細胞の集団を用いるアッセイ方法。
【請求項16】
請求項1〜13のいずれか1項に記載のマウスを作出する方法であって、
a)置換対象のマウス標的遺伝子クラスターが各々の端で組換え部位と隣接するように相同組換えによってマウス染色体に一対の部位特異的組換え部位を組込むことと、
b)部位特異的組換え部位間で組換えを行い、マウス標的遺伝子クラスターが染色体から除去され、残存部位特異的組換え部位で置換されるようにすることとを含む方法。
【図1a】

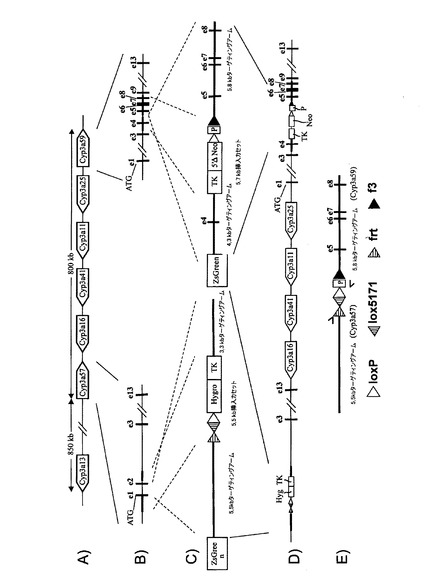
【図1b−1】
【図1b−2】
【図1c】

【図1d】
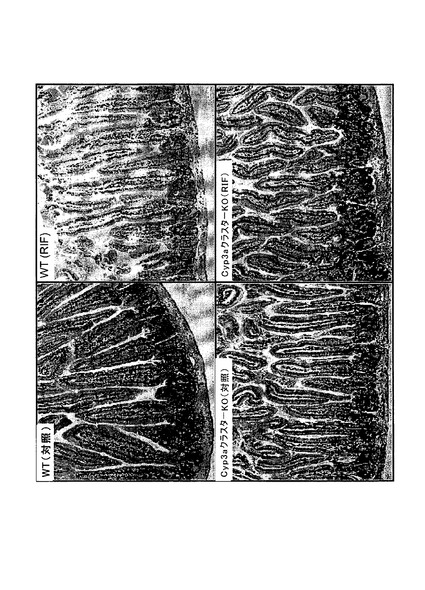
【図1e】
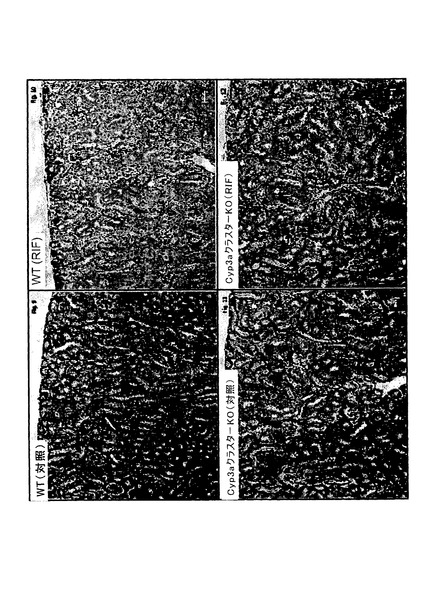
【図1f】
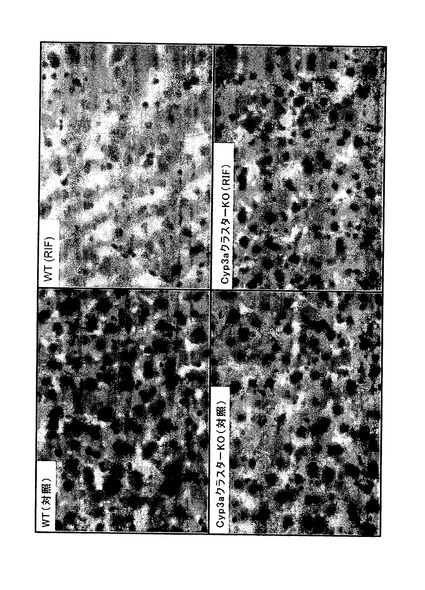
【図1g】
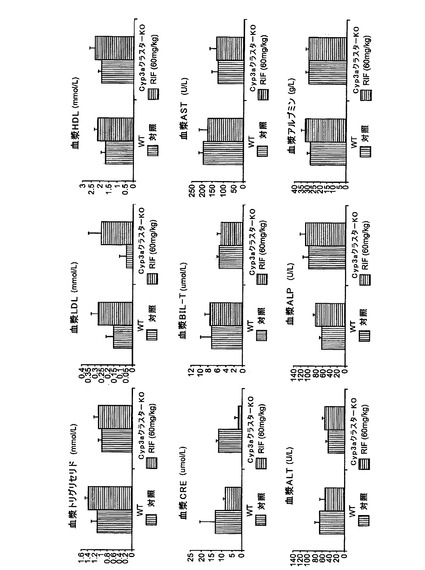
【図1h】
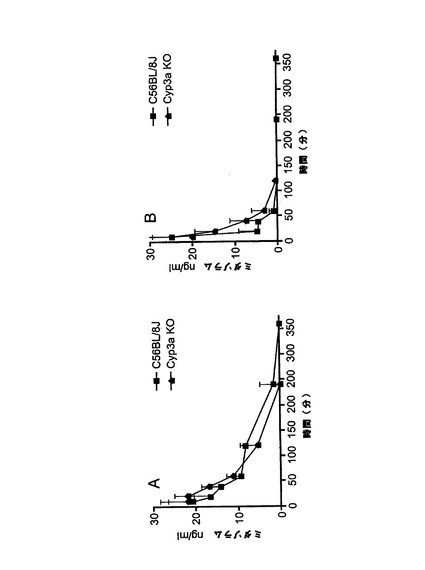
【図1i】
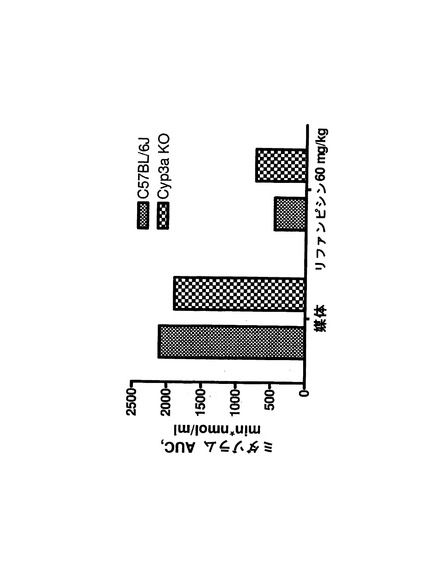
【図1j】

【図1k】
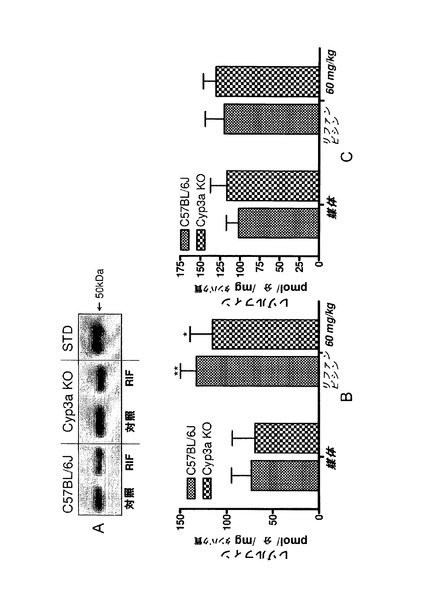
【図1l】
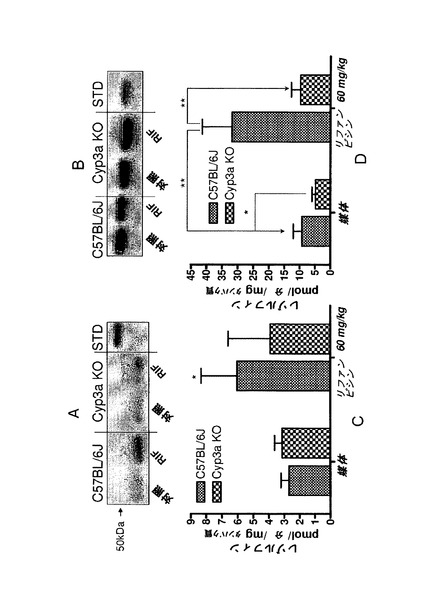
【図1m】
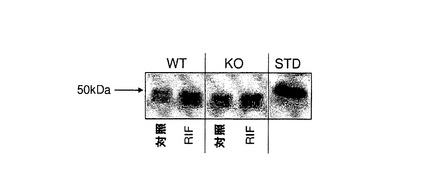
【図1n】
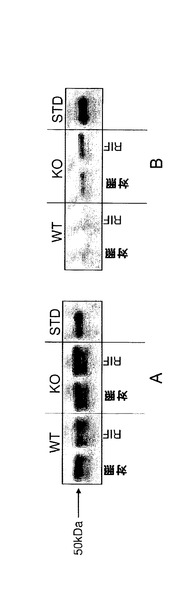
【図1o】
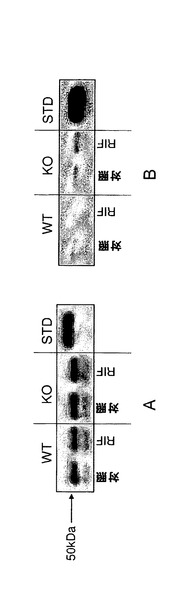
【図1p】
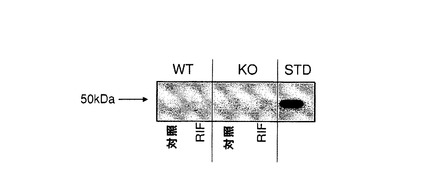
【図1q】
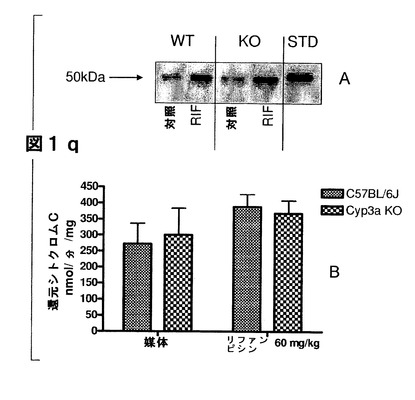
【図1r】
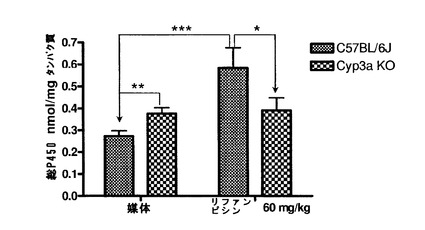
【図1s】
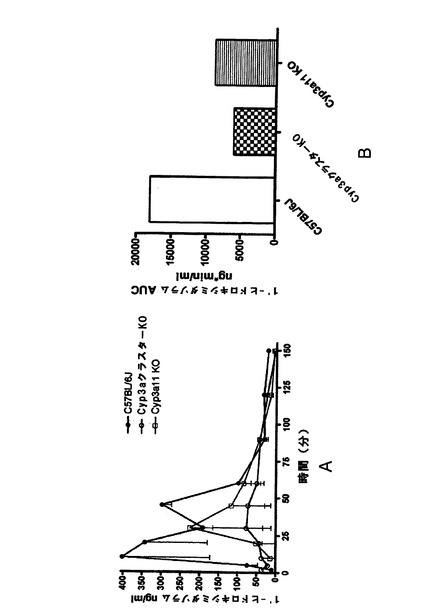
【図1t】
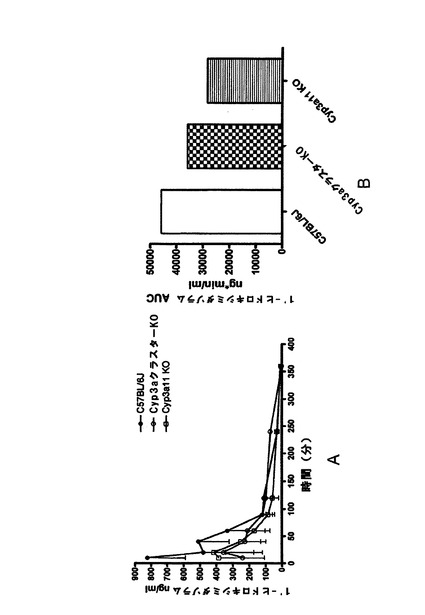
【図1u】
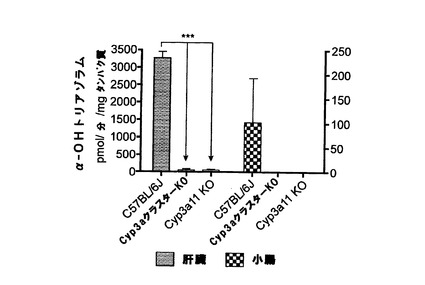
【図1v】
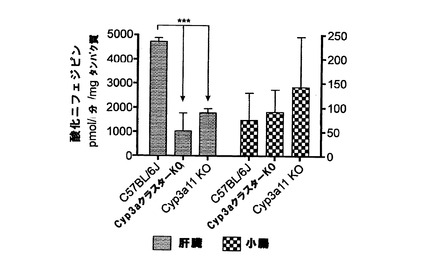
【図1w】
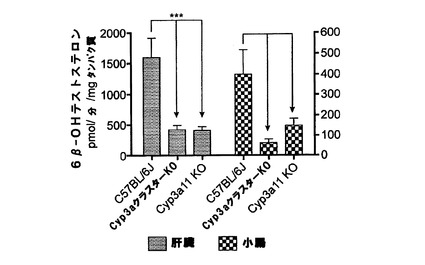
【図1x】
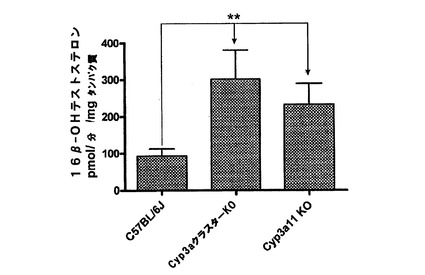
【図1y】
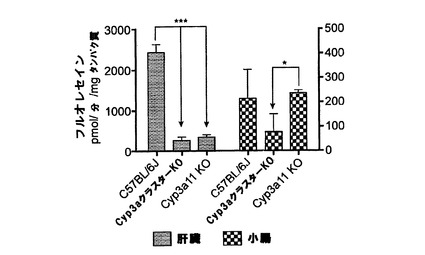
【図1z】
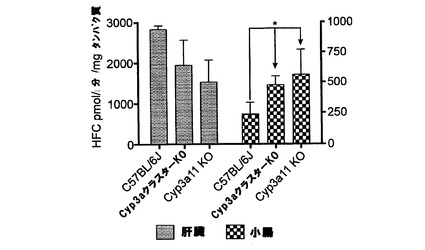
【図2a】
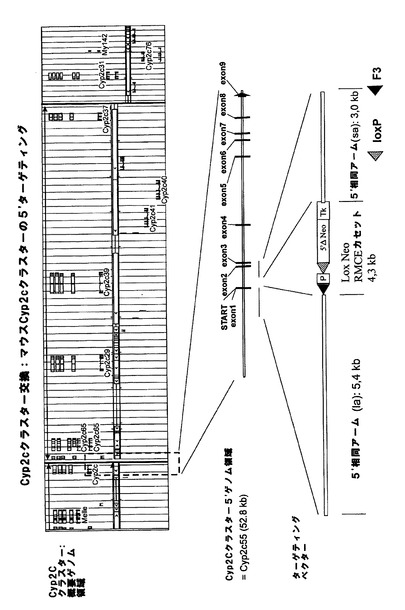
【図2b】
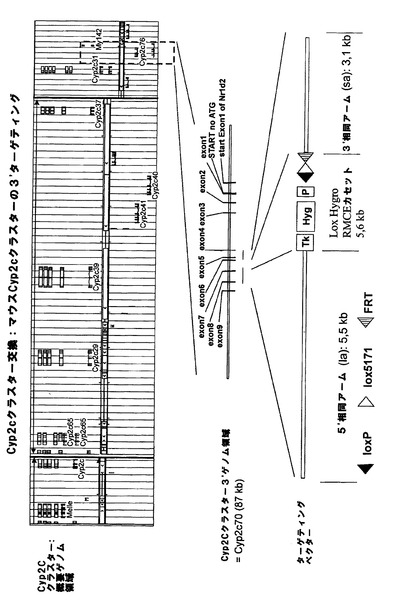
【図2c】
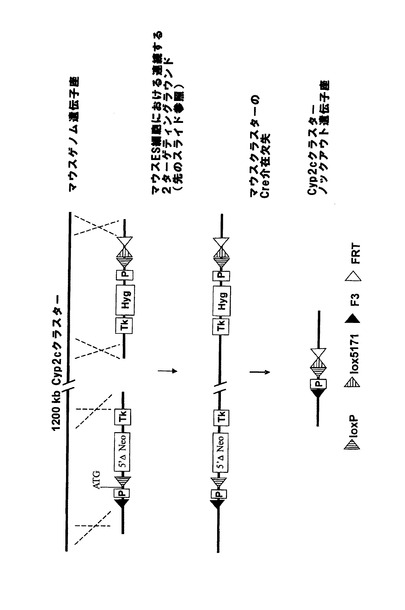
【図2d−1】

【図2d−2】

【図2e】
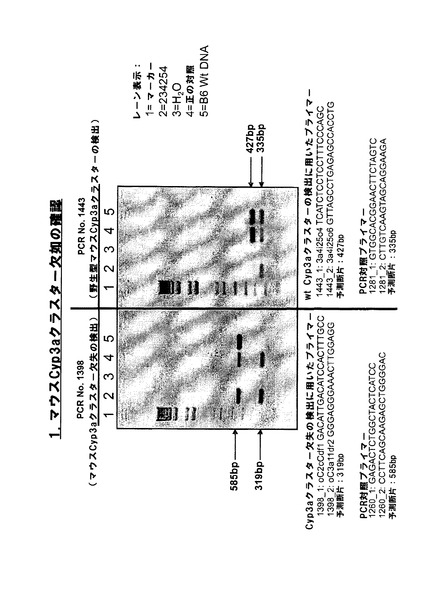
【図2f】
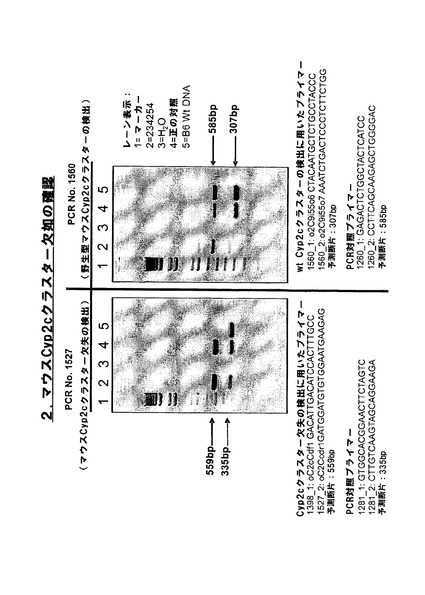
【図3a】
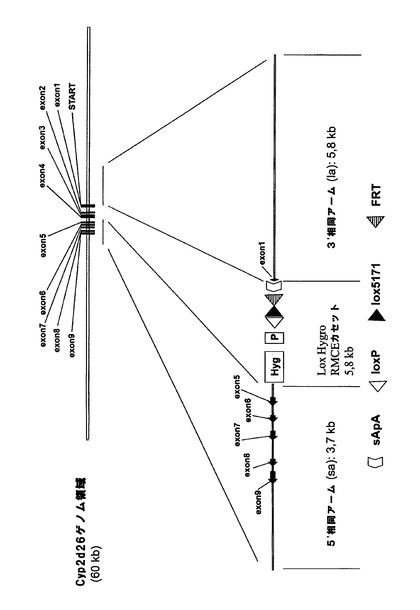
【図3b】
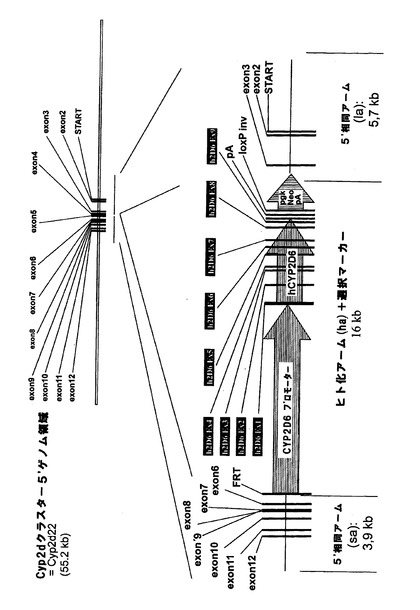
【図3c】
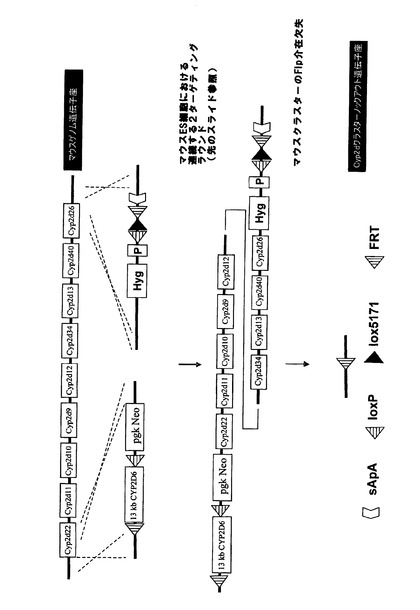
【図3d−1】

【図3d−2】
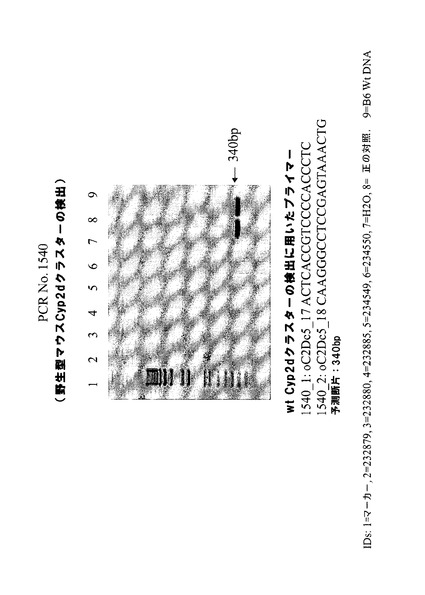
【図3e】
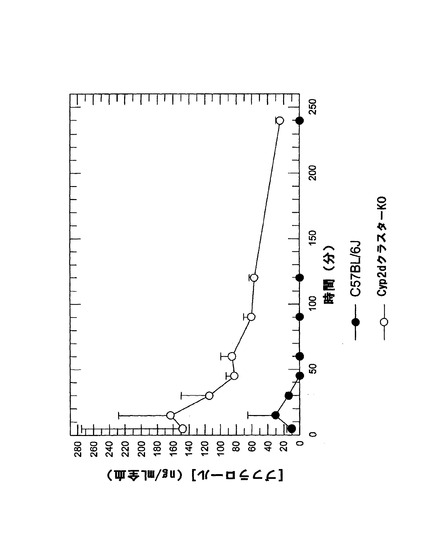
【図4a】
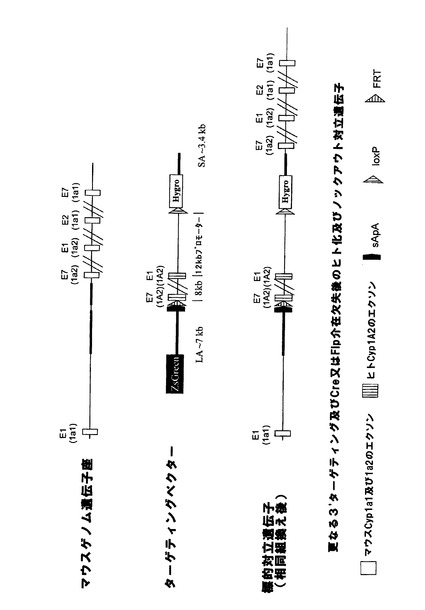
【図4b】
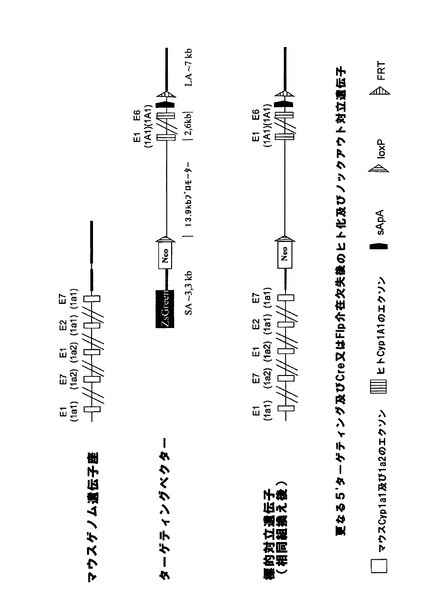
【図4c】
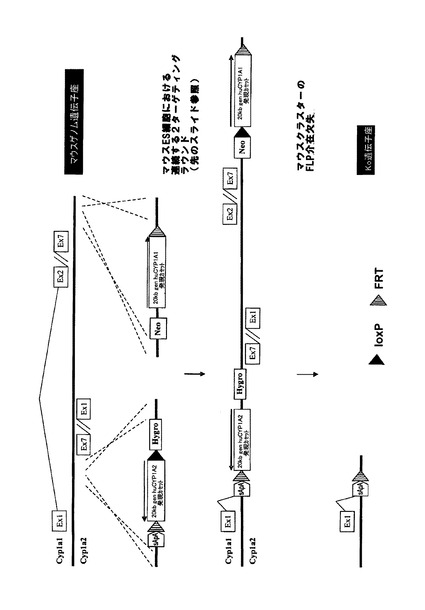
【図1b−1】
【図1b−2】
【図1c】
【図1d】
【図1e】
【図1f】
【図1g】
【図1h】
【図1i】
【図1j】
【図1k】
【図1l】
【図1m】
【図1n】
【図1o】
【図1p】
【図1q】
【図1r】
【図1s】
【図1t】
【図1u】
【図1v】
【図1w】
【図1x】
【図1y】
【図1z】
【図2a】
【図2b】
【図2c】
【図2d−1】
【図2d−2】
【図2e】
【図2f】
【図3a】
【図3b】
【図3c】
【図3d−1】
【図3d−2】
【図3e】
【図4a】
【図4b】
【図4c】
【公表番号】特表2011−500048(P2011−500048A)
【公表日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願番号】特願2010−529451(P2010−529451)
【出願日】平成20年10月17日(2008.10.17)
【国際出願番号】PCT/GB2008/003531
【国際公開番号】WO2009/050484
【国際公開日】平成21年4月23日(2009.4.23)
【出願人】(507194084)アイティーアイ・スコットランド・リミテッド (30)
【Fターム(参考)】
【公表日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願日】平成20年10月17日(2008.10.17)
【国際出願番号】PCT/GB2008/003531
【国際公開番号】WO2009/050484
【国際公開日】平成21年4月23日(2009.4.23)
【出願人】(507194084)アイティーアイ・スコットランド・リミテッド (30)
【Fターム(参考)】
[ Back to top ]