
PAN−DR結合ポリペプチドおよびその使用
本発明は、複数の対立遺伝子によってコードされるHLAクラスII分子を結合することのできる新規の人工的オリゴペプチドを提供する。このオリゴペプチドは、配列AX1FVAAX2TLX3AX3AX4A(配列番号1)を含み、ここでX1は、W、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択され;X3はHおよびKからなる群から選択され;X4はA、DおよびEからなる群から選択されており、ただしここでオリゴペプチド配列がAKFV AAWTLKAAAでないことが条件となっている。本発明は同様に、オリゴペプチドを含むより大きなペプチド、オリゴペプチドおよびより大きなペプチドをコードするポリヌクレオチド、ならびにオリゴペプチド、ペプチドまたはポリヌクレオチドを含む組成物にも関する。同様に開示されているのは、免疫応答を誘発するための方法である。
【発明の詳細な説明】
【背景技術】
【0001】
Tヘルパー細胞により認識されるエピトープを含む免疫原性ペプチドが、免疫応答を誘発する上で有用であることが発見されてきた。特定の決定基に対する抗体応答を増強するためのヘルパーペプチドの使用は、例えばHervas−Stubbsら、Vaccine12:867−871(1994)中に記載されている。
【0002】
MHC対立遺伝子のペプチド結合ポケットをライニングする対立遺伝子多型残基は各々の対立遺伝子に固有のペプチドセットを結合させる能力を付与する傾向にあるが、所与のペプチドが2つ以上のMHC対立遺伝子に結合することが実証されたケースも存在する。例えば、複数の研究者が、多数のDRタイプに関連していくつかのエピトープの変性結合および/または認識を報告し、或る種のエピトープが「万能」エピトープであるかもしれないという概念を導いている(Buschら、Int.Immunol.2:443−451(1990);Panina−Bordignonら、Eur.J.Immunol.19:2237−2242(1989);Sinigagliaら、Nature336:778−780(1988);O’Sullivanら、J.Immunol.147:2663−2669(1991);Roacheら、J.Immunol.144:1849−1856(1991);Hillら、J.Immunol.147:189−197(1991))。Pan−DR結合ペプチドは、例えば米国特許第6,413,935号(US6,413,935);国際公開第95/07707号(WO95/07707);国際公開第2005/120563号(WO/2005/120563);およびAlexanderら、Immunity 1:751−761(1994)中で記述されてきた)。これらのペプチドは、抗原に対するさまざまな免疫応答の生成を助けることが示されてきた。
【発明の概要】
【0003】
本発明は、タンパク質分解酵素に対する安定性が改善された(短ペプチド、オリゴペプチドおよびポリペプチドの両方の形での)免疫原性ペプチドの提供という観点から見たPADRE配列(配列番号3)内の選択された変異の効果の研究に基づいている。
【0004】
したがって本発明は、100nM未満、例えば少なくとも50nMのIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むまたはこれで構成されている単離ポリペプチドであって、オリゴペプチド配列がAX1FVAAX2TLX3AX4A(配列番号1)を含み、
− X1がW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;
− X3がHおよびKからなる群から選択され;
− X4がA、DおよびEからなる群から選択され;
ただしここでオリゴペプチド配列がAKFVAAWTLKAAA(配列番号3)でないことが条件となっている、
単離ポリペプチドに関する。
【0005】
本発明はさらに、本発明のポリペプチドをコードするポリヌクレオチドおよびこのようなポリヌクレオチドを含むベクターにも関する。同様に本発明が包含しているのは、本発明のポリヌクレオチドを含む細胞である。本発明はさらに、本発明のポリペプチドまたはオリゴペプチドまたはポリヌクレオチドまたはベクターまたは細胞を含む、医薬組成物を含めた組成物に関する。最後に本発明は、それを必要としている対象に対して、本発明のポリペプチドまたはオリゴペプチドまたはポリヌクレオチドまたはベクターまたは細胞または組成物を投与することによって、免疫応答を刺激する方法に関する。
【図面の簡単な説明】
【0006】
【図1−1】さまざまなHLA−DR分子に対する本発明のペプチドのHLA−DR結合を決定する結合実験からのデータを示す。詳細については実施例1を参照のこと。
【図1−2】図1−1に同じ。
【図1−3】図1−1に同じ。
【図1−4】図1−1に同じ。
【図1−5】図1−1に同じ。
【0007】
定義
別段の定義がない限り、本明細書中で使用されている全ての技術的および科学的用語は、本発明が属する技術分野の当業者が一般的に理解するものと同じ意味を有する。本発明は、組換え遺伝学の分野における常用手技に依存するものである。本発明中で使用される一般的な方法を開示する基本的教書としては、Sambrookら、Molecular Cloning、A Laboratory Manual(第3版2001);Kriegler、Gene Transfer and Expression:ALaboratory Manual(1990);およびCurrent Protocols in Molecular Biology (Ausubelら、編、1994))がある。
【0008】
本明細書中で使用される「オリゴペプチド」または「ペプチド」という用語は、少なくとも4つのアミノ酸残基またはアミノ酸ミメティックの連鎖を意味する。オリゴペプチドまたはペプチドは、その中性(未荷電)形態または塩形態のいずれであれ、また、グリコシル化、側鎖酸化またはリン酸化を含めた(ただしこれらに限定されない)修飾が無い状態かまたはこれらの修飾を1つ以上含む状態のいずれであれ、さまざまな長さであり得る。任意の修飾は本明細書中で記述されるポリペプチドの生物活性を破壊するものではないが、本発明は、(例えば生体内で修飾が除去されるまで活性を限定するために)修飾が生物活性を削減または削除する選択肢を含むものである。
【0009】
「ポリペプチド」および「タンパク質」という用語は、本明細書において互換的に使用されて、アミノ酸残基のポリマーを意味する。これらの用語は、1つ以上のアミノ酸残基が対応する天然に発生するアミノ酸の人工的化学ミメティックであるアミノ酸ポリマーならびに天然に発生するアミノ酸ポリマーおよび天然に発生しないアミノ酸ポリマーに適用される。本明細書中で使用されるこれらの用語は、アミノ酸残基が共有ペプチド結合によって連結されている、全長タンパク質(すなわち抗原)を含むあらゆる長さのアミノ酸鎖を包含する。これらの用語はさらに、例えばグリコシル化、アセチル化、リン酸化、アミド化、公知の保護/封鎖基による誘導体化、タンパク質分解による分割または非天然発生アミノ酸による修飾などの翻訳後修飾を受けたポリペプチドを含んでいる。
【0010】
本発明のポリペプチドとして同様に含まれるのは、前述のポリペプチドのフラグメント、誘導体、類似体またはバリアント、およびそれらの任意の組合せである。ポリペプチドおよびそのフラグメント、誘導体、類似体またはバリアントは、抗原性および免疫原性ポリペプチドであり得る。
【0011】
「アミノ酸残基」、「アミノ酸」および「残基」は、ペプチド、オリゴペプチドまたはタンパク質中のアミノ酸残基に言及している場合、互換的に使用され、本明細書中で使用されている通り、アミド結合またはアミド結合ミメティックを通して少なくとも1つの他のアミノ酸またはアミノ酸ミメティックに共有結合により接合されたアミノ酸またはアミノ酸ミメティックを意味する。
【0012】
本明細書中で使用される「アミノ酸」という用語は、無条件の場合、「L−アミノ酸」またはL−アミノ酸ミメティックを意味する。
【0013】
本明細書中で使用される通り、ペプチド化合物を記述するために使用される命名法は、各アミノ酸残基の左側(N末端)にアミノ基が提示され右側(C末端)にアルボキシル基が提示されている従来の実践法にしたがっている。エピトープ内でアミノ酸残基の位置が言及されている場合、それらは、アミノからカルボキシル方向に付番されており、1位は、エピトープまたはそれが一部を成していてよいペプチドまたはタンパク質のアミノ末端に基も近い位置である。アミノ酸構造式においては、各残基は一般に、標準的な三文字または一文字の呼称で表わされる。アミノ酸残基のL形態は、大文字一文字かまたは三文字記号の最初の大文字によって表わされ、D−形態を有するこれらのアミノ酸のD形態は、小文字一文字かまたは小文字三文字記号により表わされる。グリシンは、非対称炭素原子を全く有しておらず、単に「Gly」または「G」と称される。
【0014】
本明細書中で使用されているアミノ酸の記号を以下に示す。
【0015】
【表1】

【0016】
本明細書中で使用される「panDR結合ペプチド」または「pan−DR結合エピトープ」という用語は、2つ以上のMHCクラスIIDR分子を結合させる(例えば100nM未満例えば少なくとも50nMのIC50を有する2つ以上のMHC分子の各々を結合させる)分子ファミリーの一成員を意味する。一部の実施形態においては、本発明のpan−DR結合オリゴペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11または全12個の最も一般的なDR対立遺伝子(DR1、2w2b、2w2a、3、4w4、4w14、5、7、52a、52b、52cおよび53)を結合させることのできるペプチドである。本発明のpan−DR結合オリゴペプチドは、第2の決定基に対する免疫応答を促進することに加えて、標的免疫原自体としても役立つことができる。こうして、例えばpan−DR結合ペプチド自体が炭水化物エピトープに連結される場合、免疫応答は、pan−DR結合ペプチドと炭水化物エピトープの両方に対するものであってよい。
【0017】
本明細書中で使用される「PADRE」という用語は、アミノ酸配列AKFVAAWTLKAAA(配列番号3)を有するpanDR結合ペプチドを意味する。
【0018】
本明細書中で使用される「PADRE類似体」という表現は、PADREと比べて多くとも1、2、3または4個のアミノ酸変更を含むアミノ酸配列を含み、うち少なくとも1つがK2、W7、K10またはA12内で行なわれているアミノ酸配列を含むpanDRペプチドを意味する。
【0019】
本明細書中で使用される「IC50」という用語は、基準ペプチドの結合の50%の阻害が観察される結合検定中のペプチドの濃度を意味する。検定が実施される条件(すなわち限定的MHCタンパク質および標識ペプチド濃度)に応じて、これらの値はKD値に接近してよい。結合を決定するための検定は、例えばその各々の開示が参照により本明細書に援用されている国際公開第94/20127号(WO94/20127)および国際公開第94/03205号(WO94/03205)の中で詳述されている。検定条件が変動した場合そして使用される特定の試薬(例えばMHC調製物など)に応じて、IC50値は、多くの場合劇的に変化し得る。例えば、MHC分子の過度の濃度は、所与のリガンドの見かけのIC50測定値を増大させる。
【0020】
あるいは、結合は、例えばPADREなどの基準ペプチドとの関係において表現される。特定の検定の感度が増減するにつれて試験対象のペプチドのIC50値は幾分か変化するかもしれないが、基準ペプチドとの関係における結合は有意に変化しない。例えば、基準ペプチドのIC50が10倍増加するような条件下で実施される検定においては、被検ペプチドのIC50値もおよそ10倍シフトする。したがって、あいまいさを避けるため、ペプチドが良好な結合性を有するか、あるいは中度の、弱いまたは不良な結合性を有するかについての査定は一般に、標準ペプチドのIC50との関係における対象ペプチドのIC50に基づく。結合は同様に、生細胞(例えば、Ceppelliniら、Nature339:392、1989;Christnickら、Nature352:67、1991;Buschら、Int.Immunol.2:443、19990;Hillら、J.Immunol.147:189、1991;del Guercioら、J.Immunol.154:685、1995))、洗浄剤溶解物を用いた無細胞系((例えば、Cerundoloら、J.Immunol.21:2069、1991)、固定化された精製済みMHC(例えば、Hillら、J.Immunol.152、2890、1994;Marshallら、J.Immunol.152:4946、1994);ELISA系(例えばReayら、EMBO J.11:2829、1992)、表面プラズモン共鳴(例えば、Khilkoら、J.Biol.Chem.268:15425、1993)、高流量可溶性相検定(Hammerら、J.Exp.Med.180:2353、1994)およびクラスIMHC安定化または会合の測定(例えば、Ljunggrenら、Nature346:476、1990;Schumacherら、Cell62:563、1990;Townsendら、Cell62:285、1990;Parkerら、J.Immunol.149:1896、1992)を使用するものを含めた他の検定系を用いて判定されてよい。
【0021】
「発現」という用語は、コーディング配列によってコードされる産物の生物学的産生を意味する。大部分の場合において、コーディング配列を含めたDNA配列は転写されてメッセンジャーRNA(mRNA)を形成する。メッセンジャーRNAは次に翻訳されて、関連する生物活性を有するポリペプチド産物を形成する。同様に、発現プロセスには、転写のRNA産物へのさらなるプロセッシングステップ、例えばイントロンを除去するためのスプライシング、および/またはポリペプチド産物の翻訳後プロセッシングが関与してよい。
【0022】
本明細書中で使用される「構築物」という用語は、一般に、天然に発生しない組成物を一般に表わす。構築物は合成技術、例えば組換え型DNAの調製および発現または核酸またはアミノ酸用の化学合成技術によって生成可能である。構築物は同様に、結果がその形態では天然に見出されないような形で1つの材料を別の材料と添加または親和させることによっても生成可能である。「マルチ−エピトープ構築物」という用語は、「ミニ遺伝子」または「マルチ−エピトープ核酸ワクチン」という用語と互換的に使用可能であり、例えば一部の実施形態ではMHCクラスIおよびT細胞受容体および/またはMHCクラスIIおよびT細胞受容体などの、免疫系内で機能する分子に結合し得る任意の長さのペプチドエピトープをコードする多数のエピトープ核酸を含む。マルチエピトープ構築物中のエピトープ核酸は、例えばクラスIIMHCエピトープまたはクラスIMHCエピトープとクラスIIMHCエピトープの組合せをコードできる。
【0023】
特定のアミノ酸配列に関して、「エピトープ」は、特定の免疫グロブリンによる認識に関与する一組のアミノ酸残基、またはT細胞に関連しては、T細胞受容体タンパク質および/または主要組織適合複合体(MHC)受容体による認識に必要な残基である。試験管内または生体内における免疫系の設定において、エピトープは、免疫グロブリン、T細胞受容体またはMHC分子により認識される部位を合わさって形成している、例えば一次、二次および三次ペプチド構造、および電荷などの分子の集合的特徴である。したがって、「エピトープ」という用語には、適切なMHC分子に結合しその後細胞傷害性T細胞応答またはヘルパーT細胞応答を誘発できる、あるいは代替的に、抗体を結合させその後、免疫原性ペプチドを誘導する抗原に対する抗体応答を誘発することのできる、本発明の免疫原性ペプチドが含まれるが、これらに限定されない。
【0024】
「フランキング残基」とは、エピトープの隣りに位置づけされている残基である。フランキング残基は、エピトープのN末端またはC末端に隣接する位置に導入または挿入され得る。
【0025】
「免疫原」および「抗原」という用語は互換的に使用され、細胞または体液性免疫応答の対象となるべきあらゆる化合物を意味する。さらに、本発明の抗原または免疫原性ペプチドは、線状であるすなわちポリペプチド中の隣接するアミノ酸で構成されていてよく、あるいは抗体−エピトープの場合には三次元または立体配座のもの、すなわち、機能的エピトープがポリペプチドの二次または三次構造に起因して一緒になって1つのエピトープを形成する非隣接アミノ酸で構成されているものであってよい。
【0026】
本明細書中で使用される「抗原決定基」という用語は、免疫応答を惹起する、促す、または生成するように誘発され得るあらゆる構造、例えば炭水化物エピトープ、脂質、タンパク質、ペプチドまたはそれらの組合せである。
【0027】
「抗体」という用語は、1つ以上のタンパク質および/または抗原を含む(ただしこれらに限定されない)組成物の中に含まれる特定されたタンパク質または抗原と相互作用しかつこれに結合することのできる1つまたは複数の免疫グロブリン遺伝子によって実質的にコードされるポリペプチドを意味する。
【0028】
「CTLエピトープ」という用語は、特定のMHCクラスI分子により認識され結合され、かつ特定のMHCクラスI分子と複合体化された場合にTリンパ球により認識されるペプチドを意味する。一部の実施形態においては、CTLエピトープは、長さがアミノ酸約8〜約13個、約9〜約11個または約9〜約10個であり得る。
【0029】
「HTLエピトープ」という用語は、特定のMHCクラスII分子により認識され結合され、かつ特定のMHCクラスII分子と複合体化された場合にTリンパ球により認識されるペプチドを意味する。一部の実施形態においては、HTLエピトープは、長さがアミノ酸約6〜約30個、約8〜約30個、約10〜約30個、約12〜約30個、約6〜約25個、約8〜約25個、約10〜約25個、約12〜約25個、約6〜約18個、約8〜約18個、約10〜約18個または約12〜約18個であり得る。
【0030】
Tリンパ球がMHC分子とCTLまたはHTLエピトープ間の複合体を「認識する」という記載は、本明細書、Tリンパ球上のT細胞受容体が複合体に結合し、その効果でTリンパ球が活性化されることを意味する。
【0031】
2つ以上のペプチド配列に関連して「同一の」または「同一性」百分率という用語は、配列比較アルゴリズムを用いてまたは手作業によるアラインメントおよび目視によって測定されるように比較ウインドウ全体にわたる最大限の一致について比較され整列させられた場合に、同じであるかまたは同じであるアミノ酸残基を規定の百分率で有する2つ以上の配列または下位配列を意味する。
【0032】
配列同一性および配列類似性百分率を決定するのに適したアルゴリズムの一例は、それぞれAltschulら、(1977)Nuc.Acids Res.25:3389−3402、およびAltschulら、(1990)J.Mol.Biol.215:403−410に記述されているBLASTおよびBLAST2.0アルゴリズムである。BLAST解析を実施するためのソフトウェアは、国立生物工学情報センター(http://www.ncbi.nlm.nih.gov/)を通して公開されている。このアルゴリズムには、データベース配列中の同じ長さのワードと整列させた場合に或る正値の閾値スコアTと整合するかまたはそれを満たす問い合わせ配列内の長さWの短かいワードを識別することによって、高スコアの配列対(HSP)をまず最初に識別するステップが関与する。Tは、近傍ワードスコア閾値と呼ばれる(Altschulら、上掲書)。これらの初期近傍ワードヒットは、それらを含むより長いHSPを発見するための検索を開始するためのシードとして作用する。ワードヒットは、累積アラインメントスコアを増大できるかぎり、各々の配列に沿って両方向に拡張される。累積スコアは、ヌクレオチド配列について、パラメータM(一対の整合する残基のための報酬スコア;つねに0超)およびN(不整合残基のためのペナルティスコア;つねに0未満)を用いて計算される。アミノ酸配列については、累積スコアを計算するのにスコアリングマトリクスが使用される。各方向でのワードヒットの拡張は、次の場合に中断される:累積アラインメントスコアが、その最大達成値からXという数量だけ下落した場合;累積スコアが、1つ以上の負のスコアの残基アラインメントの累積に起因してゼロ以下となった場合;またはいずれかの配列の終りに達した場合。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は、デフォルトとして、11のワート長(W)、10という期待値、M=5、N=−4および両ストランドの比較を使用する。アミノ酸配列については、BLASTPプログラムはデフォルトとして、3というワード長、10という期待値(E)を使用し、BLOSUM62スコアリングマトリクス(HenikoffおよびHenikoff(1989)Proc.Natl.Acad.Sci.USA89:10915参照)は、50というアラインメント(B)、10という期待値(E)、M=5、N=−4そして両ストランドの比較を使用する。
【0033】
「単離(された)」または「生物学的に純粋な」という語句は、天然の状態で発見された場合に通常それに随伴する構成要素を実質的にまたは本質的に含んでいない材料を意味する。
【0034】
本明細書中で使用されている「リンカー」という用語は、2つの機能群(例えばpan−DR結合ペプチドと所望の免疫原)の間の共有結合およびスペーシングを提供するために用いられる任意の化合物のことである。一部の実施形態において、リンカーは、生理学的条件下で実質的に非電荷で線状または分岐側鎖を有していてよい脂肪族炭素鎖、アミノ酸またはアミノ酸ミメティックなどの中性分子を含む。一部の場合において、リンカーはそれ自体免疫原性であり得る。本発明において有用であるさまざまなリンカーについて以下でより詳細に記述する。さらに「連結する(link)」および「コンジュゲートする(conjugate)」という動詞は、本明細書において互換的に使用され、2つ以上の種の共有結合付着を意味する。
【0035】
「連結する」または「接合する」という用語は、組換え融合、共有結合、ジスルフィド結合、イオン結合、水素結合および静電結合を含めた(ただしこれらに限定されない)、ペプチドを機能的に結合するための当該技術分野において公知のあらゆる方法を意味する。
【0036】
「直接連結された」または「直接接合された」という用語は、何も介在することなく接合されていることを意味する。例えば、直接接合されている2つのペプチドの場合、1つのペプチドは、上述の通り別のペプチドに対して、2つのペプチドの間にいかなる配列、分子、スペーサー、リンカーなども介在することなく接合または結合されると考えられる。直接接合されたペプチドは、共通のアミノ酸を共有していてもいなくてもよい。すなわち2つのペプチドの配列は、重複しなくてもしてもよい。後者は例えば、本発明のPADRE類似体が挿入を用いて別の(ポリ)ペプチド内に導入される場合に該当し、PADRE類似体のアミノ酸配列の存在を保証するためには、挿入点にあるアミノ酸残基と共にPADRE類似体のための配列全体を提供するアミノ酸残基を挿入することしか必要ではない。
【0037】
「間接的に連結された」という用語は、何かが介在している状態で接合されていることを意味する。例えば、2つのペプチドが間接的に接合されている場合、1つのペプチドは上述の通り別のペプチドに対し、2つのペプチド間に配列、分子、スペーサー、リンカーなどが介在している状態で接合または結合されると考えられる。
【0038】
「主要組織適合複合体」または「MHC」は、生理学的免疫応答に関与する細胞相互作用の制御において役割を果たす遺伝子クラスタである。ヒトにおいては、MHC複合体はHLA複合体としても公知である。MHCおよびHLA複合体の詳細な説明については、例えばPaul、Fundamental Immunology、第3版、Raven Press、New York、1993を参照のこと。
【0039】
「医薬的に許容される」という語句は、一般に無毒性、不活性および/または生理学的に適合性ある組成物を意味する。
【発明を実施するための形態】
【0040】
発明者らは、pan−DR結合性をもつ新しい部類のペプチド、すなわちペプチドが数多くの異なるDR対立遺伝子によりコードされるMHCクラスII分子に結合して、ペプチドが広範囲の個体において免疫応答を刺激できるようにすることを発見した。さらに発明者らは、この新しいペプチドの多くが、著しく改善されたプロテアーゼ耐性を有し、こうしてより長い生体内半減期を可能にすることを発見した。意外にも、これらのペプチドの一部は、公知のpan−DRペプチドに比べて免疫応答を高める大幅に増大した能力を示す。
【0041】
本発明のポリペプチド(またはそのフラグメント)の生物活性は、適切なMHC分子を結合し、標的免疫原または免疫原ミメティックに対する免疫応答を誘発するのを任意に補助するTヘルパー応答を誘発する能力である。抗体応答を刺激するために有用なペプチドの場合、本発明のペプチドはTヘルパー応答を誘発し、このことが今度は標的免疫原に対する体液性応答の誘発を補助する。
【0042】
II. pan−DR結合ペプチド
本発明は、pan−DR結合ペプチドである新しい部類のペプチドを提供し、その使用を提供する。本発明のペプチドは、数多くの異なるDR対立遺伝子の2つ以上を結合でき、こうしてさまざまな抗原/免疫原に対する免疫応答を増大させるのに用いられる。本発明のペプチドはさらに、例えば米国特許第5,736,142号(US5,736,142)で開示されているような公知のpan−DR結合ペプチドと比べて増強した免疫応答を惹起するのに用いられる。
【0043】
本明細書中で実証される通り、本発明者らは、多数のHLA対立遺伝子によりコードされるMHC分子に結合する一連のペプチド配列を同定した。一部にはこのデータを考慮して、本発明者らは、以下の配列モチーフが、活性および免疫原性の改善されたpan−DR結合ペプチドを表わすものと判定した。一部の実施形態において、本発明のポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLX3AX4A(配列番号1)を含み、式中X1はW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択され;X3はHおよびKからなる群から選択され;X4はA、DおよびEからなる群から選択される。一部の実施形態において、ポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLHAAA(配列番号2)を含み、式中X1はY、H、I、E、N、QおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択される。本発明によると、オリゴペプチド配列はAKFVAAWTLKAAA(配列番号3)を含まない。
【0044】
一部の実施形態において、本発明のポリペプチドは、配列番号1、配列番号2、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号50、配列番号51、配列番号52、配列番号53、配列番号54、配列番号55、配列番号56、配列番号57、配列番号58、配列番号59、配列番号60、配列番号61、配列番号62、配列番号63、配列番号64、配列番号65、配列番号66、配列番号67、配列番号68、配列番号69、配列番号70、配列番号71または配列番号72から選択されたオリゴペプチドの13アミノ酸C末端フラグメントを構成する1つ以上のアミノ酸配列の1つ以上のコピーを含む。
【0045】
本発明のpan−DR結合ペプチドは、多数の異なるMHC対立遺伝子を結合することができ、「ヘルパーペプチド」とも呼ばれ得る。一部の実施形態において、pan−DR結合ペプチドは、2、3、4、5、6、7、8、9、10、11、12以上の異なるMHCクラスII対立遺伝子によりコードされるMHC分子を結合することができる。一部の実施形態において、誘発される応答は、増強された免疫応答である。一部の実施形態において、免疫応答は、本発明のpan−DR結合オリゴペプチドに向けられている。代替的に、または付加的に、MHC分子に対するpan−DR結合オリゴペプチドの結合は、例えば1つ以上のCTLペプチドを含むポリペプチドなどの第2の抗原に対する免疫応答および/またはタンパク質または非タンパク質抗原に対する体液性(すなわち抗体)応答をさらに増強する。
【0046】
本明細書中で記述されているpan−DRポリペプチド配列は単独で、またはより大きな融合タンパク質またはコンシュゲートの一部として使用可能である。こうして、本発明のポリペプチドは、本明細書中で記されているpan−DRペプチド配列に加えていずれかのまたは両方の末端に追加のアミノ酸を含み得る。例えば、タグまたはマーカーとして他のアミノ酸配列を含み入れるかまたは生体内または試験管内の安定性を促進するかまたそうでなければ当該技術分野において一般に理解される通りの追加のメリットを含み入れることが望ましい可能性がある。さらに、本発明のポリペプチドは、ポリペプチド内に1つ以上のpan−DRオリゴペプチド配列の1、2、3個以上のコピーを含むことができかつ/または本発明のポリペプチド内に多数の異なるpan−DRオリゴペプチド配列を含むことができる。その上、ポリペプチドは、例えばこのような配列またはこのような配列を含むポリペプチドに対する免疫応答を増強することが必要である場合に、少なくとも1つの追加のHTLオリゴペプチドおよび/またはCTLオリゴペプチドおよび/または抗体誘発性ポリペプチドを含むことができる。このような実施形態において、本明細書中に記述されているpan−DR配列は、キーホールリンペットヘモシアニン、破傷風トキソイドおよびジフテリアトキソイドなどの従来の担体タンパク質に対する代替物として、または先行技術の万能T−ヘルパーエピトープの代替物として機能する。
【0047】
本発明のポリペプチドは任意の長さのものであり得る。一部の実施形態において、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列および任意には他のアミノ酸配列を含むポリペプチドは、例えばアミノ酸1000、900、800、700、600、500、400、300、200、100、80、60、50、40、30、20または15個以下の長さを有するが、多くの場合、1つまたは複数のpan−DR結合オリゴペプチドを含むポリペプチドは、(ポリペプチドが未変性ポリペプチド配列の大部分を含むように)本発明のpan−DR結合オリゴペプチドの1つまたは数個を導入することによって修飾されている未変性ポリペプチドとほぼ同じ長さを有する。一方、本発明のpan−DR結合オリゴペプチドを含む本発明の一部のポリペプチドは、「マルチエピトープ構築物」、すなわち1つ以上の抗原から誘導された複数のエピトープを含み、構築物の抗原性または免疫原性を最適化するためにエピトープが非天然発生手順で組織されていないポリペプチドである。したがって、本発明のポリヌクレオチドの開示の下で以下に論述するマルチエピトープ発現産物も同様に、本発明のペプチドの実施形態である。
【0048】
このようなマルチエピトープ構築物において、複数のエピトープは典型的に、複数のCTLエピトープ、複数のB細胞エピトープ、複数のTヘルパーリンパ球エピトープ、複数のCTLおよびB細胞エピトープ、複数のCTLおよびTヘルパーリンパ球エピトープ、複数のB細胞およびTヘルパーリンパ球エピトープ、ならびに複数のB細胞、CTLおよびTヘルパーリンパ球エピトープから選択される。これらのエピトープは、単一の抗原タンパク質から誘導されてよいが、少なくとも2つの異なるポリペプチド抗原から誘導されてもよく、同様にこれらは同じまたは異なる種(例えば細菌、ウイルスおよび寄生虫の種)から誘導されてよい。
【0049】
一部の実施形態では、pan−DR結合オリゴペプチド配列は、末端NH2アシル化、例えばアルカノイル(C1−C20)またはチオグリコリルアセチル化、末端カルボキシアミド化、例えばアンモニア、メチルアミンなどによって修飾されていることにより、原初の配列と異なっている可能性がある。一部の実施形態において、これらの修飾は、支持体またはその他の分子に対する連結のための部位を提供してよい。
【0050】
以上で同定されたペプチドの生物活性は、さまざまなシステム内で検定されてよい。典型的には、抗原特異的T細胞活性を阻害する能力が試験される。1つの例示的プロトコルにおいては、余剰のペプチドが、公知のMHC発現(例えばDR1)を有する抗原提示細胞および公知の抗原特異性(例えば破傷風毒素830−843)およびMHC制限(ここでもDR1)を有するT細胞クローン、そして免疫原性ペプチド自体(例えば破傷風毒素830−843)と共にインキュベートされる。検定培養は、T細胞増殖のために充分な時間、例えば4日間インキュベートされ、その後、最後の18時間のインキューベーションの間[3H]−チミジンでパルス処理することなどの標準的な手順を用いて増殖が測定される。ペプチドが与えられなかった対照と比べた阻害百分率が、次に計算される。
【0051】
試験管内検定における抗原提示を阻害するペプチドの能力は、生体内での免疫応答を阻害するペプチドの能力と相関されてきた。生体内活性は、例えば免疫調節ペプチドそしてペプチドにより認識される特定のMHC分子に制限されるものとして公知である免疫原を投与することなどによって、動物モデルにおいて判定されてよい。その後、動物からTリンパ球が取り出され、一用量範囲の免疫原と共に培養される。従来の手段、例えば[3H]−チミジンでのパルス処理および適切な対照に対する比較により、刺激の阻害が測定される。当然のことながら、或る種の実験上の詳細は、当業者には明白である。同様に、参照により本明細書に援用されているAdoriniら、Nature334:623−625(1988)も参照のこと。
【0052】
定義されたMHC分子、特にMHCクラスII分子を伴う多数の細胞が公知であり、例えばthe American Type Culture Collection(例えば、「Catalogue of Cell Lines and Hybridomas」、第6版(1988)を参照)Rockville、Maryland、U.S.A.から容易に入手可能である。
【0053】
本発明のオリゴペプチドの一部の実施形態は、NおよびC末端残基に対する修飾を含む。当業者であれば充分理解できるように、NおよびC末端を修飾して、ペプチドの物理的または化学的特性を改変し、例えば結合、安定性、生物学的利用能、連結の容易さなどに影響を与えてもよい。
【0054】
任意には、例えばNまたはC末端におけるさまざまなアミノ酸ミメティックまたはD−アミノ酸でのペプチドの修飾が使用可能であり、例えば生体内でのペプチドの安定性を増大させる上で有用である。このようなペプチドは、「inverso」型または「retroinverso」型として、すなわち一配列のLアミノ酸をDアミノ酸で置換することによってかまたはアミノ酸の配列を逆転させDアミノ酸でL−アミノ酸を置換することによって合成されてよい。Dペプチドは実質的にペプチダーゼに対する耐性がより高く、したがってそのL−ペプチド対応物と比べて血清および組織中でより安定していることから、生理学的条件下でのDペプチドの安定性は、対応するLペプチドと比べた親和性の差異を補償する以上のものであるかもしれない。さらに、置換を伴うまたは伴わないLアミノ酸含有ペプチドをDアミノ酸でキャッピングして、免疫原性ペプチドのエクソペプチダーゼ破壊を阻害することができる。
【0055】
ポリペプチドの生体内安定性は、数多くの方法で検定できる。例えば、ペプチダーゼおよびさまざまな生体媒質例えばヒトの血漿および血清を使用して、安定性が試験されてきた。例えば、全て参照により本明細書に援用されているVerhoefら、Eur.J.Drug Metab.Pharmacokin.11:291−302(1986);Walterら、Proc.Soc.Exp.Biol.Med.148:98−103(1975);Witterら、Neuroendocrinology30:377−381(1980);Verhoefら、J.Endocrinology110:557−562(1986);Handaら、Eur.J.Pharmacol.70:531−540(1981);Bizzozeroら、Eur.J.Biochem.122:251−258(1982);Chang、Eur.J.Biochem.151:217−224(1985)を参照のこと。
【0056】
生物活性にとって不可欠である一部のアミノ酸残基、例えば重要な接触部位にあるアミノ酸残基は一般に生物活性に対する不利な効果無く改変され得ないことは容易に認識されることから、本発明のペプチドまたは類似体は、一部の残基の順序または組成を改変させることによって修飾可能である。重要でないアミノ酸は、L−α−アミノ酸またはそのD−異性体などのタンパク質内で自然に発生するアミノ酸に限定される必要はなく、非タンパク質アミノ酸例えばβ−γ−δアミノ酸、ならびにL−α−アミノ酸の数多くの誘導体も含んでいてよい。本発明のオリゴペプチドはLアミノ酸かDアミノ酸のいずれかを含み得るが、通常、コア結合領域内部にDアミノ酸を含むことはできない。
【0057】
本発明のペプチドは、多様な方法で調製可能である。一部の実施形態では、組換えDNA技術が利用され、この場合本発明の免疫原性ポリペプチドをコードするヌクレオチド配列が発現ベクター内に挿入され、適切な宿主細胞へと形質転換またはトランスフェクトされ、発現に適した条件下で培養される。これらの手順は、Sambrookら、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、Cold Spring Harbor、New York(1989)中に一般的に記載されている通り、当該技術分野において一般に公知である。あるいは、そのサイズに応じて、従来の技術にしたがって、溶液中かまたは固体支持体上でポリペプチドを合成することができる。さまざまな自動合成装置が市販されており、公知のプロトコルにしたがって使用可能である。例えばStewartおよびYoung、Solid Phase Peptide Synthesis、第2版、Pierce Chemical Co.(1984)、上掲書。
を参照のこと。
【0058】
III. ポリヌクレオチド
本発明は、本明細書中で記述されている通りの本発明のポリペプチドをコードするポリヌクレオチドを提供する。例えば、ポリペプチドをコードするポリヌクレオチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLX3AX4A(配列番号1)を含み、式中X1はW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択され;X3はHおよびKからなる群から選択され;X4はA、DおよびEから選択される。一部の実施形態において、ポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLHAAA(配列番号2)を含み、式中X1はY、H、I、E、N、QおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択される。本発明によると、オリゴペプチド配列はAKFVAAWTLKAAA(配列番号3)を含まない。一部の実施形態において、本発明のポリヌクレオチドは、配列番号1、配列番号2、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号50、配列番号51、配列番号52、配列番号53、配列番号54、配列番号55、配列番号56、配列番号57、配列番号58、配列番号59、配列番号60、配列番号61、配列番号62、配列番号63、配列番号64、配列番号65、配列番号66、配列番号67、配列番号68、配列番号69、配列番号70、配列番号71または配列番号72から選択されたオリゴペプチドの13アミノ酸C末端フラグメントを各々構成する1つ以上のアミノ酸配列をコードし、ポリヌクレオチドは、他のアミノ酸配列と融合して配列または切形を含むポリペプチドをコードしてよい。したがって、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列を含めた2つ以上のCTLおよび/またはHTLオリゴペプチド配列を含む、本明細書中で開示されている通りのポリペプチドをコードする本発明のポリヌクレオチドが提供される。
【0059】
本発明のポリヌクレオチド構築物は、上述のペプチドまたはポリペプチドのいずれでもコードすることができる。例えば多数のクラスIおよび/またはクラスIIMHCエピトープおよび/またはB細胞エピトープ、例えば2つ以上のクラスIIMHCエピトープ(すなわち本明細書中に記述されている少なくとも1つのpan−DR結合オリゴペプチド、例えば配列番号1、2および4〜72)またはクラスIMHCエピトープとクラスIIMHCエピトープおよびB細胞エピトープの組合せなど。クラスIMHCコーディングエピトープ核酸は、「CTLエピトープ核酸」とも呼ばれ、クラスIIMHCコーディングエピトープ核酸は「HTLエピトープ核酸」とも呼ばれる。「B細胞エピトープ」は、抗体またはB細胞受容体を結合させる線状または立体構造エピトープである。一部のマルチエピトープ構築物は、クラスIMHCエピトープおよび/またはB細胞エピトープをコードするその配列の一部分と、クラスIIMHCエピトープをコードする別の部分とを有することができる。一部の実施形態において、CTLエピトープ核酸は、長さがアミノ酸約8〜約15個、例えばアミノ酸約8〜約11個、例えばアミノ酸約9個のエピトープペプチドをコードする。HTLエピトープ核酸は、本明細書中に記述されている通り少なくとも1つのpan−DRオリゴペプチドをコードするが、2つ以上のHTLエピトープが含まれる場合、他のHTLエピトープも同様に使用可能である。一部の実施形態において、ポリヌクレオチド構築物は、例えば5個以上、10個以上、15個以上、20個以上または25個以上のエピトープ核酸を含む。マルチエピトープポリヌクレオチド構築物中のCTLエピトープ核酸は全て、1つの生体に由来するものであり得(例えば全てのエピトープ核酸のヌクレオチド配列はHIV株内に存在するかもしれない)、またはマルチエピトープ構築物は2つ以上の異なる生体内に存在するエピトープ核酸を含み得る(例えば一部のエピトープがHIV由来で一部がHCV由来)。以下で記述する通り、マルチエピトープ構築物内の1つ以上のエピトープ核酸は、スペーサー核酸によりフランキングされていてよい。
【0060】
この先で理解されるように、本発明のPADRE類似体に連結されているかまたはその他の形でこの類似体を含むポリペプチドを含むB細胞エピトープをコードする核酸構築物が、本発明の一実施形態である。このような核酸構築物は典型的に、発現産物中に存在するPADRE類似体がポリペプチド由来のB細胞エピトープの立体構造とマイナスに干渉しない場合に融合構築物をコードする。B細胞エピトープが線状である場合、B細胞エピトープは単純にPADRE類似体に融合されてよく、したがってコーディング核酸の提供は比較的簡単である。しかしながら、B細胞エピトープが立体構造エピトープである場合、PADRE類似体コーディング核酸の導入点の典型的な例は、ポリペプチドのフレキシブルループ内かまたは、フレキシブル末端内(例えばタンパク質ドメイン全体かさらにはポリペプチド全体を使用することによる立体構造エピトープの3D構造の保存が求められる場合)であるが、例えば問題のポリペプチドの細胞内に閉じ込められた部分をコードする領域は生体内での免疫応答と関連性をもたないことから、これらの領域内にPADRE類似体コーディング核酸を導入することも同様に可能である。このような核酸構築物の発現産物は、導入されたPADRE類似体配列が発現産物に対する免疫応答の惹起において増大したTリンパ球ヘルプを提供することになるため、抗体誘発用免疫原として有用である。こうして、発現産物は免疫原として直接使用されてよく(例えば動物における抗体産生のために使用される免疫原性組成物またはワクチン内の抗体誘発用)、あるいは、コーディング核酸での核酸免疫付与に付された動物において生体内で発現された後、その効果を及ぼしてもよい。
【0061】
「スペーサー」とは、接合エピトープの発生を防止するためおよび/またはプロセッシングの効率を増大させるためマルチエピトープ構築物内の2つのエピトープの間に挿入される配列を意味する。マルチエピトープ構築物は、1つ以上のスペーサー核酸を有する場合がある。スペーサー核酸は構築物内で各エピトープ核酸にフランキングしてよく、あるいはスペーサー核酸対エピトープ核酸の比率は約2対10、約5対10、約7対10、約8対10、または約9対10であってよく、ここで約8対10という比率が一部の構築物にとって有利な結果を生み出すものと判定されている。
【0062】
スペーサー核酸は1つ以上のアミノ酸をコードしてよい。一部の実施形態において、マルチエピトープ構築物内のクラスIMHCエピトープにフランキングするスペーサー核酸は、長さがアミノ酸1〜約8個、アミノ酸2〜8個、アミノ酸3〜8個、アミノ酸4〜8個、アミノ酸5〜8個、アミノ酸6〜8個、またはアミノ酸7〜8個である。マルチエピトープ構築物内でクラスIIMHCエピトープにフランキングするスペーサー核酸は、一部の実施形態において長さがアミノ酸5、6、7個以上、そして一部の実施形態においてアミノ酸5個または6個超である。
【0063】
1構築物内のスペーサーの数、スペーサポリヌクレオチド内でコードされるアミノ酸の数およびスペーサーのアミノ酸組成を、エピトープのプロセッシングを最適化しかつ/または接合エピトープを最小限に抑えるように選択することができる。一部の実施形態では、スペーサーは、エピトーププロセッシングおよび接合モチーフを同時に最適化することにより選択される。本明細書では、エピトーププロセッシングを最適化するための適切なアミノ酸について記述されている。同様に、構築物内の接合エピトープ数を最小限にするための適切なアミノ酸スペーシングが、本明細書中に、クラスIおよびクラスIIHLAについて記述されている。例えば、クラスIIMHCエピトープにフランキングするスペーサーは、一部の実施形態において、一般にG、Pおよび/またはN残基が一次アンカー残基であるものとして公知でないことからこれらを含むことができる(例えばPCT米国特許第00/19774号(PCT/US00/19774)を参照のこと)。一部の実施形態において、クラスIMHCエピトープにフランキングするためのスペーサーは、交互のGおよびP残基例えば(GP)n、(PG)n、(GP)nG、(PG)nPなどを含み、ここでnは1〜10、2〜約2の間の整数であり、一部の実施形態において、このようなスペーサーの具体例はGPGPGまたはPGPGPである。クラスIMHCエピトープの一部の実施形態において、スペーサーは、任意にはK、NまたはGが先行する1、2、3個以上の連続するアラニン(A)残基を含む。
【0064】
一部のマルチエピトープ構築物において、各スペーサー核酸が同じアミノ酸配列をコードすることで充分である。同じアミノ酸配列をコードする2つのスペーサー核酸を有するマルチエピトープ構築物において、これらのスペーサーをコードするスペーサー核酸は同じまたは異なるヌクレオチド配列を有していてよく、ここで、細胞内にマルチエピトープ構築物が挿入された場合、異なるヌクレオチド配列が、意図されない組換え事象の発生確率を減少させるかもしれない。
【0065】
他のマルチエピトープ構築物においては、スペーサー核酸の1つ以上が異なるアミノ酸配列をコードしてよい。スペーサー核酸の多くがマルチエピトープ構築物内の同じアミノ酸配列をコードしてよいが、1、2、3、4、5個以上のスペーサー核酸が異なるアミノ酸配列をコードする場合があり、マルチエピトープ構築物内のスペーサー核酸の全てが異なるアミノ酸配列をコードすることが可能である。スペーサー核酸は、本明細書中で記述されている通り、スペーサー配列がエピトーププロセッシングを最大限にするかおよび/または接合エピトープを最小限にするかを判定することによって、自らフランキングするエピトープ核酸に関して最適化されてよい。
【0066】
マルチエピトープ構築物は、1つの構築物内のスペーサーが別の構築物に比べてエピトーププロセッシングを最適化するかまたは接合エピトープを最小限に抑えるかによって、互いに区別されてよく、一部の実施形態においては、1つの構築物がもう1つの構築物に比べてエピトーププロセッシングおよび接合エピトープについて同時に最適化されている場合、構築物を区別してよい。1つの構築物がエピトーププロセッシングおよび接合モチーフについて最適化されているか否かを決定するためのコンピュータ援用方法および試験管内および生体内実験室方法が、本明細書において記述されている。
【0067】
一部の実施形態において、本発明のポリヌクレオチド構築物は、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列そして任意にはポリペプチドに関連して本明細書中で記述された配列のいずれかまたは全てをコードする核酸を含む発現ベクターとして提供されている。このような発現ベクターの構成は、例えばその開示が参照により本明細書に援用されているPCT米国特許第99/10646号(PCT/US99/10646)の中に記載されている。発現ベクターは、抗原が発現され適切なMHC分子に標的化されるように生体の適切な細胞内で核酸をコードする転写単位を発現することのできる少なくとも1つのプロモータ要素を含んでいる。例えば、ヒトへの投与のためには、ヒト細胞内で機能するプロモータ要素が発現ベクター内に取込まれる。
【0068】
本発明において使用される一般的方法を開示する基本的教書には、Sambrookら、Molecular Cloning、A Laboratory Manual(第2版、1989);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990);および Current Protocols in Molecular Biology(Ausubelら、編、1994);Oligonucleotide Synthesis.A Practical Approach(Gait編、1984);Kuijpers、Nucleic Acids Research 18(17):5197(1994);Dueholm、J.Org.Chem.59:5767−5773(1994);Methods in Molecular Biology、第20巻(Agrawal、編);およびTijssen、Laboratory Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Acid Probes、例えば、第I部、第2章、「Overview of principles of hybridization and the strategy of nucleic acid probe assays」(1993))が含まれる。
【0069】
関連するオリゴペプチド配列(例えばエピトープ)をコードする核酸は、標準的な技術にしたがって構築物の中に組立てることができる。一部の実施形態において、pan−DR結合オリゴペプチドそして任意にはマルチエピトープポリペプチドをコードする核酸配列は、オリゴヌクレオチドプライマを用いる増幅技術を用いて単離されるかまたは化学的に合成される。該当する場合には、組換えクローニング技術も使用可能である。所望のエピトープを増幅する(構築物を組立てるのにPCRを使用する場合)またはコードする(構築物を組立てるのに合成オリゴヌクレオチドを使用する場合)オリゴヌクレオチド配列が選択される。
【0070】
典型的にはプライマを使用する増幅技術を用いて、DNAまたはRNAからの選択されたエピトープをコードする配列を増幅し単離する。(米国特許第4,683,195号(U.S.4,683,195)および4,683,202号(4,683,202);PCR Protocols:A Guide to Methods and Applications(Innisら、編、1990))を参照のこと)。ポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR)などの方法を使用して、mRNAから、cDNAから、ゲノムライブラリからまたはcDNAライブラリから直接エピトープ核酸配列を増幅することができる。プライマ内に制限エンドヌクレアーゼ部位を取込むことが可能である。PCR反応により増幅されたマルチエピトープ構築物をアガロースゲルから精製し、適切なベクター内にクローニングすることが可能である。
【0071】
本発明のポリヌクレオチドを構成するために、合成オリゴヌクレオチドも使用することができる。一部の実施形態において、この方法は、遺伝子のセンスおよびナンセンスストランドを表わす一連の重複するオリゴヌクレオチドを用いて実施される。これらのDNAフラグメントは次にアニールされ、ライゲートされ、クローニングされる。市販されていないオリゴヌクレオチドは、Van Devanterら、Nucleic Acids Res.、72:6159−6168(1984)中に記載されている通りの自動合成装置を用いてBeaucageおよびCaruthers、Tetrahedron Letts.22:1859−1862(1981)が最初に記述した固相ホスホラミジットトリエステル法にしたがって化学的に合成可能である。オリゴヌクレオチドの精製は、PearsonおよびReanier、J.Chrom.255:137−149(1983)内で記述されているようなアニオン交換HPLCまたは未変成アクリルアミドゲル電気泳動法による。
【0072】
一部の実施形態において、本発明のポリヌクレオチド構築物のエピトープは、転写を導くための強力なプロモータならびにエンハンサやポリアデニル化部位などの他の調節配列を含む発現ベクターの形にサブクローニングされる。適切なプロモータは当該技術分野において周知であり、例えばSambrookら、およびAusubelら、の中に記載されている。哺乳動物細胞のための真核生物発現系は当該技術分野において周知であり、市販されている。このようなプロモータ要素には例えば、サイトメガウイルス(CMV)、ラウス肉腫ウイルスLTRおよびSV40が含まれる。
【0073】
発現ベクターは典型的には、宿主細胞内でのポリヌクレオチド構築物の発現に必要とされる追加の要素を全て含む転写単位または発現カセットを含む。例えば、発現カセットは写しの効率的ポリアデニル化のために必要なシグナルおよびマルチエピトープ構築物に作動的に連結されたプロモータを含み得る。カセットの追加の要素は、機能的スプライスドナーおよびアクセプタ部位を伴うイントロンおよびエンハンサを含んでいてよい。
【0074】
プロモータ配列に加えて、発現カセットは、効率的終結を提供するために構造遺伝子の下流側に転写終結領域も含むことができる。終結領域は、プロモータ配列と同じ遺伝子から得られてもよいし、あるいは異なる遺伝子から得られてもよい。
【0075】
本発明のペプチドを発現するために使用される発現ベクター内に選択可能なマーカーを取込むことができる。これらの遺伝子は、選択培地中で成長した形質転換済み宿主細胞の存続または成長に必要な、タンパク質などの遺伝子産物をコードすることができる。選択遺伝子を含む遺伝子で形質転換されていない宿主細胞は培地中で存続しない。典型的な選択遺伝子は、抗生物質またはその他の毒素、例えばアンピシリン、ネオマイシン、カナマイシン、クロラムフェニコールまたはテトラサイクリンに対する耐性を付与するタンパク質をコードする。代替的には、選択可能なマーカーは、栄養要求性欠乏症を補足するかまたは複合培地からは入手できない重要な栄養素を供給するタンパク質をコードしてよい。一部の実施形態において、ベクターは、例えばE.coli(大腸菌)内、または宿主細胞内への導入に先立ちベクターがその中で複製されるその他の細胞内で機能的である1つの選択可能なマーカーを有するものである。当業者にとっては、数多くの選択可能なマーカーが公知である。
【0076】
細胞内に遺伝情報を輸送するために、さまざまな発現ベクターを使用することができる。真核細胞内での発現のために使用される従来のベクターのうち任意のものを使用してよい。真核ウイルス由来の調節要素を含む発現ベクターが、典型的に真核細胞発現ベクター、例えばSV40ベクター、CMVベクター、乳頭腫ウイルスベクターおよびエプスタイン・バー・ウイルス由来のベクター内で使用される。
【0077】
本発明のポリヌクレオチド構築物は、プラスミドベクターならびにウイルスまたは細菌ベクターを含めたさまざまなベクターから発現可能である。ウイルス発現ベクターの例としては、弱毒化ウイルス宿主例えば牛痘または鶏痘が含まれる。このアブローチの例として、本発明の(ポリ)ペプチドをコードするヌクレオチド配列を発現するためのベクターとしてワクチニアウイルスが使用される。腫瘍を担持する宿主内への導入時点で、組換えワクチニアウイルスは免疫原性ペプチドを発現し、こうして宿主CTLおよび/またはHTL応答を惹起する。ワクチニアベクターおよび免疫付与プロトコルにおいて有用な方法は、例えば米国特許第4,722,848号(US4,722,848)の中で記述されている。
【0078】
本発明のポリヌクレオチドは、E.coli、その他の細菌宿主、酵母および昆虫細胞を含めたさまざまな宿主細胞中で発現されて、本明細書中で記述されているタンパク質発現産物を提供することができる。宿主細胞は、例えば酵母細胞、細菌細胞または糸状菌細胞などの微生物である。適切な宿主細胞の例としては、他の数多くのもののうち例えばシュードモナス属、エシュリキア属(例えばE.coli)、バチラス属が含まれる。適切な酵母細胞は、例えばサッカロミセス属(例えばS.cerevisiae)およびカンジタ属を含めた複数の属のうちのいずれかのものであり得る。適切な真核細胞は、ピチア属、アスペルギルス属そして組換え産生に適したその他の真菌から選択される。適切な昆虫細胞は、例えばS2、Sf9およびHi−5細胞を含めた複数の亜種の細胞であり得る。
【0079】
当業者にとっては、例えばアデノおよびアデノ関連ウイルスベクター、レトロウイルスベクター、非ウイルスベクター例えばBCG(Bacillus Calmette Guerin)、チフス菌ベクター、無毒化炭疽菌毒素ベクターなどの、治療的投与または免疫付与に有用な多様な他のベクターが明白である。したがって、本発明は同様に、必要としている個体に対してこのようなベクターを投与することによる、このようなベクターの予防的または治療的使用にも関する。
【0080】
マルチエピトープ構築物の免疫原性および抗原性は、本明細書中で記述されている通りに評価される。
【0081】
以上で記したように、本発明は同様に、本明細書中で記述されている通りの本発明のポリヌクレオチドのいずれかによりコードされる単離ポリペプチドにも関し、したがって、これらのポリペプチドも同様に本発明の「ペプチド」またはポリペプチドである。
【0082】
IV. さらなる抗原
以上で説明したように、本発明のpan−DRオリゴペプチドは、さまざまな抗原に対する細胞または体液性免疫応答を増強するために使用することができる。このような抗原に由来する抗原決定基または抗原自体は、本発明のpan−DR結合ペプチドと混和した形、またはそれに連結させた形で投与可能である。例えば、免疫応答を惹起するまたは増強するために、抗原決定基を本発明のヘルパーペプチドと連結または混合するか、またはそれと共に連続して投与することができる。本質的に、本発明のpan−DR結合オリゴペプチド配列と組合せた形でいかなる抗原でも使用することができる(例えば多糖類、タンパク質、糖タンパク質、脂質、糖脂質、リポ多糖類など)。タンパク質抗原は、細菌、真菌、ウイルス、原生動物、蠕虫および他の寄生虫などの感染性因子から誘導されてよいが、或る種の疾病状態において過剰発現されているかまたはその他の形で不適当である抗原あるいは癌抗原などの疾病関連抗原であってもよい。国際公開第00/20027号(WO00/20027)に列挙されている癌抗原は全て、関連性あるタンパク質抗原であり、さらなる抗原は炎症に付随する抗原である(ここではTNFがその好例である)。本発明は、全てが本発明の一部を成している数多くのTNF構築物を開示しており、一般にこれらの構築物は全て、それらをコードする核酸フラグメント、核酸フラグメントを含むベクターおよび核酸フラグメントまたはベクターを含む宿主細胞(またはベクターで形質転換されている宿主細胞)と同様、本発明の一部を成すものである。
【0083】
一部の実施形態において、本発明のオリゴペプチドは、単独で投与される。一部の実施形態において、本発明のオリゴペプチドは、第2の抗原決定基と併用して投与される。一部の実施形態において、本発明のオリゴペプチドは、抗原決定基と混和される。一部の実施形態において、本発明のペプチドは、抗原決定基に連結される。例えば、タンパク質抗原を、(ペプチド結合を介して)融合タンパク質としてpan−DR結合オリゴペプチドに対して間接的に、または直接的に連結させることができる。非タンパク質抗原を含めた抗原を、他の共有結合コンジュゲーション方法を介して、pan−DR結合オリゴペプチドまたはオリゴペプチド配列を含むポリペプチドに対し連結させることができる。
【0084】
一部の実施形態において、本発明のペプチドと共に投与される抗原決定基はタンパク質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は多糖類である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は糖タンパク質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は脂質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は糖脂質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基はリポ多糖類である。
【0085】
炭水化物エピトープは炭水化物構造を含むが、コンジュゲート例えば糖タンパク質、糖ペプチド、糖脂質など、DNA、RNA、または多糖類オリゴ糖または単糖として存在することができ、これらに対する免疫応答が所望される。炭水化物エピトープは、広範囲の免疫応答を誘発してよい。当業者であれば、本明細書中で例示されているさまざまな炭水化物構造を、抗原性に不利な影響を及ぼすことなく標準的な方法にしたがってさまざまな形で修飾することができる、ということを認識するものである。例えば、サッカリドの単糖単位は、さまざまな形で置換されるかまたさらには小有機分子と置換されてよく、これは単糖類のためのミメティックとして役立つ。
【0086】
適切な抗原の例としては、炭水化物ベースのワクチンの中で使用可能である細菌表面多糖類から誘導されたものが含まれる。細菌は典型的には、糖タンパク質、糖脂質、リポ多糖類の0特異的側鎖、莢膜多糖類などの一部としてその細胞表面上で炭水化物を発現する。例示的細菌菌株としては、肺炎連鎖球菌(例えば国際公開第2005/120563号(WO/2005/120563)およびその中の炭水化物エピトープを参照のこと)、ナイセリア・メニンギチジス(Neisseria meningitidis)、ヘモフィラス・インフルエンザ(Haemophilus influenza)、クレブシエラ菌株(Klebsiella spp.)、シュードモナス菌株(Pseudomonas spp.)、サルモネラ菌株(Salmonella spp.)、シゲラ菌株(Shigella spp.)およびグループBストレプトコッキィ(streptococci)が含まれる。
【0087】
数多くの適切な細菌炭水化物エピトープが、先行技術(例えばSandersら、Pediatr.Res.37:812−819(1995);Bartoloniら、Vaccine13:463−470(1995);Pirofski、ら、Infect.Immun.63:2906−2911(1995)および国際公開第WO93/21948号(WO93/21948)))中で記述されており、例えば米国特許第6,413,935号(US6,413,935)に記載の通りである。
【0088】
一般に、以下の国際特許出願のいずれか1つにおいて開示されているHLA結合エピトープは、本発明によると有用であるかもしれず、したがってそれらの内容は参照により本明細書に援用される;国際公開第93/03764号(WO93/03764)、国際公開第95/22317号(WO95/22317)、国際公開第94/03205号(WO94/03205)、国際公開第95/19783号(WO95/19783)、国際公開第97/34617号(WO97/34617)、国際公開第02/20053号(WO02/20053)、国際公開第94/20127号(WO94/20127)、国際公開第97/34621号(WO97/34621)、国際公開第02/20616号(WO02/20616)、国際公開第95/07707号(WO95/07707)、国際公開第95/04817号(WO95/04817)、国際公開第98/33888号(WO98/33888)、国際公開第94/26774号(WO94/26774)、国際公開第96/03140号(WO96/03140)、国際公開第02/20035号(WO02/20035)、国際公開第96/40213号(WO96/40213)、国際公開第97/26784号(WO97/26784)、国際公開第97/33602号(WO97/33602)、国際公開第98/32456号(WO98/32456)、国際公開第99/61916号(WO99/61916)、国際公開第99/45954号(WO99/45954)、国際公開第99/65522号(WO99/65522)、国際公開第01/62776号(WO01/62776)、国際公開第01/00225号(WO01/00225)、国際公開第2004/031211号(WO2004/031211)、国際公開第99/58658号(WO99/58658)、国際公開第2005/012502号(WO2005/012502)、国際公開第00/44775号(WO00/44775)、国際公開第02/019986号(WO02/019986)、国際公開第2004/031210号(WO2004/031210)、国際公開第01/21189号(WO01/21189)、国際公開第01/24810号(WO01/24810)、国際公開第2005/033265号(WO2005/033265)、国際公開第01/42270号(WO01/42270)、国際公開第01/41788号(WO01/41788)、国際公開第01/42267号(WO01/42267)、国際公開第01/45728号(WO01/45728)、国際公開第01/41787号(WO01/41787)、国際公開第02/061435号(WO02/061435)、国際公開第02/061435号(WO02/061435)、国際公開第01/41741号(WO01/41741)、国際公開第2004/052917号(WO2004/052917)、国際公開第01/36452号(WO01/36452)、国際公開第03/087126号(WO03/087126)、国際公開第01/47541号(WO01/47541)、国際公開第02/083714号(WO02/083714)、国際公開第01/41799号(WO01/41799)、国際公開第2005/089164号(WO2005/089164)、国際公開第2003/040165号(WO)、国際公開第2005/120563号(WO2005/120563)、国際公開第2004/094454号(WO2004/094454)、国際公開第2004/053086号(WO2004/053086)、国際公開第2004/089973号(WO2004/089973)、国際公開第2008/054540号(WO2008/054540)、国際公開第2008/039267号(WO2008/039267)、国際公開第99/19478号(WO99/19478)、国際公開第92/21033号(WO92/21033)、国際公開第94/11738号(WO94/11738)、国際公開第93/22338号(WO93/22338)、国際公開第95/25530号(WO95/25530)、国際公開第95/25739号(WO95/25739)、国際公開第2005/118626号(WO2005/118626)、国際公開第93/03753号(WO93/03753)、国際公開第94/19011号(WO94/19011)、国際公開第95/03777号(WO95/03777)、国際公開第95/19783号(WO95/19783)、国際公開第95/04542号(WO95/04542)、国際公開第01/42270号(WO01/42270)、国際公開第01/41788号(WO01/41788)、国際公開第01/42267号(WO01/42267)、国際公開第01/45728号(WO01/45728)、国際公開第01/41787号(WO01/41787)、国際公開第01/41741号(WO01/41741)、国際公開第2004/052917号(WO2004/052917)、国際公開第2004/094454号(WO2004/094454)および国際公開第2004/089973号(WO2004/089973)。
【0089】
V. コンジュゲートの調製
本発明のpan−DR結合ペプチドは、さまざまな形で少なくとも1つのさらなる抗原決定基に連結され得る。末端を介したまたはリジンのε−アミノ基を介したイオン相互作用が可能である。残基の側基と抗原決定基の間の水素結合も同様に可能である。その他の実施形態においては、pan−DR結合ペプチドと抗原決定基の間の立体構造相互作用が、安定した付着を発生させるかもしれない。
【0090】
以上で指摘した通り、抗原決定基は、本発明のコンジュゲートを調製するためにpan−DR結合ペプチドに対し共有結合されてよい。一部の実施形態においては、抗原決定基/pan−DR結合ペプチドが、スペーサー分子またはリンカーにより連結される。一部の実施形態において、抗原決定基はリンカー無しでpan−DR結合ペプチドに付着されてよい。
【0091】
スペーサーまたはリンカーは典型的には、生理学的条件下で実質的に非荷電性であり線状または分岐側鎖を有していてよい中性分子例えば脂肪族炭素鎖、アミノ酸またはアミノ酸ミメティックで構成されている。当業者にとっては、さまざまな生体分子を連結するための数多くの組成物および方法が公知である。例えば、炭水化物エピトープに対してpan−DR結合ペプチドを共有結合させるための数多くの方法が可能である。pan−DR結合ペプチドを炭水化物抗原に連結するのに適した方法は、例えば国際公開第93/21948号(WO93/21948)中で開示されている。
【0092】
数多くのリンカーが周知であり、市販されているかまたは科学的文献中に記載されている。本発明において使用される連結分子は、分子の2つの部分が、それらに曝露された分子と独立してかつ自由に相互作用できるようにするのに充分な長さを有する。炭水化物エピトープの場合、連結分子の長さは典型的に1〜50原子である。一部の実施形態において、連結分子はアリールアセチレン、2〜14個のモノマー単位を含むエチレングリコールオリゴマー、ジアミン、二塩基酸、アミノ酸またはそれらの組合せである。他の適切なリンカーとしては、アミノ酸側鎖を通して異なる炭水化物部分が連結されるセラミドおよびアミノ酸残基などの脂質分子が含まれる。
【0093】
使用される特定の連結分子は、その化学的/物理的特性に基づいて選択されてよい。連結分子は、各端部に適切な官能基を有し、一基は炭水化物部分上の反応性部位に対する付着に適切であり、もう一方の基はアミノ酸/ペプチド部分に対する付着に適切である。例えば、炭水化物部分に対する付着に適切な基は、カルボン酸、エステル、イソシアネート、ハロゲン化アルキル、ハロゲン化アシルおよびイソチオシアネートである。アミノ酸部分に対する付着のためには、類似の基が有用であると考えられる。官能基の適切な選択は、アミノ酸またはペプチドの反応性部分の性質によって左右されるものである。
【0094】
1群の実施形態においては、アルキルまたはアルキレン基が連結基として有用であり、1〜20個の炭素原子を有し、一部の実施形態においては3〜6個の炭素原子を含む。例えば、ポリエチレングリコールおよび関係する構造を含むリンカーを使用することができる。「ポリエチレングリコール」という用語は、例えばヘキサエチレングリコール(HO−(CH2CH2O)5−CH2CH2OH)などのエチレングリコールの反復単位を有する分子に言及するために用いられる。連結基に言及するために「ポリエチレングリコール」という用語が使用される場合、当業者は、その他のポリエーテルまたはポリオールも同様に使用できる(すなわちポリプロピレングリコールまたはエチレンとプロピレングリコールの混合物)ということを理解すると考えられる。
【0095】
別の実施形態群において、アルキルまたはアルキレン連結基は過フッ素化され、生物学的分解を受ける可能性が低くなっている。米国特許第5,055,562号(US.5,055,562)を参照のこと。一部の実施形態において、連結基は、アミノカプロン酸、4−ヒドロキシ酪酸、4−メルカプト酪酸、3−アミノ−1−プロパノール、エタノールアミン、ペルフルオロエタノールアミンおよびペルフルオロヒドロキシ酪酸を含む。一部の実施形態において、2つの部分はポリエチレングリコール部分を介して連結されている。
【0096】
一部の実施形態において、pan−DR結合ペプチドとその他のペプチド(例えばpan−DR結合ペプチドとCTLまたはB細胞エピトープ(ただしこれらに限定されない))の間のリンカーは、Ala、Glyまたは非極性アミノ酸または中性極性アミノ酸のその他のスペーサーから選択され得る。本明細書中の一部の実施形態において、中性スペーサーはAlaである。任意に存在するスペーサーは同じ残基で構成されている必要はなく、したがってヘテロまたはホモオリゴマーであってよい、ということが理解される。一部の実施形態において、例示的スペーサーは、Alaのホモオリゴマーである。存在する場合、スペーサーは通常少なくとも1つまたは2つの残基、さらに通常は3〜6個の残基である。一部の実施形態において、pan−DR結合ペプチドは、CTLまたは抗体誘発ペプチドにコンジュゲートされる。一部の実施形態において、pan−DR結合ペプチドはアミノ末端に位置づけされる。ペプチドは、Ala−Ala−Alaなどの中性リンカーによって接合され得、一部の実施形態においては、典型的にはSer−Ser連結などを介してペプチドコンジュゲートのアミノ末端に付着される、Lys残基((PAM)2Lys)のアルファおよびイプシロンアミノ基に付着されているパルミチン酸などの脂質残基を含み得る。
【0097】
CTLまたは抗体誘発ペプチドは、直接、あるいはCTLペプチドのアミノまたはカルボキシ末端のいずれかにあるスペーサーを介して、pan−DR結合ペプチドに連結されてよい。CTLまたは抗体誘発ペプチドまたはpan−DR結合ペプチドのいずれかのアミノ末端はアシル化可能である。一部の実施形態において、CTLペプチド/pan−DR結合ペプチドコンジュゲートは、以下で記述する通り、Gly、Gly−Gly、Ser、Ser−Serなどの1つ以上の連結残基を介して、一部のアルカノイル(C1−C20)脂質に連結され得る。一部の実施形態において、脂質部分はコレステロール、脂肪酸などを含む。
【0098】
一部の実施形態において、本発明の医薬組成物は、CTLのプライミングを補助する少なくとも1つの成分を含み得る。脂質が、ウイルス抗原に対する生体内でのCTLプライミングを補助できる作用物質として同定されてきた。例えば、ステロイド例えばコレステロール、脂肪酸例えばパルミチン酸残基(ただしこれらに限定されない)を、システイン残基のスルフヒドリル基、Lys残基のアルファおよびイプシロンアミノ基に付着させ、次に例えば1つ以上の連結残基例えばGly、Gly−Gly−、Ser、Ser−Serなどを介して、免疫原性ペプチド例えばpan−DR結合ペプチドに対して連結することができる。一部の実施形態において、脂肪酸の代りに、最終的アミノ酸(例えばシステイン残基)に対し、エーテル連結を通して長鎖アルキル基を連結することができる。
【0099】
脂質化されたペプチドは、リポソーム中に取り込まれたミセル型で直接か、または例えば不完全フロインドアジュバントなどのアジュバント中で乳化された状態のいずれかで、注射され得る。一部の実施形態において、特に有効な免疫原は、免疫原性ペプチドのアミノ末端に対して例えばSer−Serなどの連結を介して付着されているLysのアルファおよびイプシロンアミノ基に付着されたパルミチン酸を含む。
【0100】
CTL応答の脂質プライミングの別の例として、適切なペプチドに対し共有結合により付着されている場合にウイルス特異的CTLをプライミングするために、E.coliリポタンパク質例えばトリパルミトイル−S−グリセリルシステンリセリル−セリン(P3CSS)を使用することができる。Deresら、Nature342:561−564(1989)を参照のこと。本発明のペプチドは例えばP3CSSにカップリングさせることができ、リポペプチドは個体に投与されて標的抗原に対するCTL応答を特異的にプライミングすることができる。一部の実施形態において、中和抗体の誘発も同様に、適切なエピトープを表示するペプチドにコンジュゲートされたP3CSSを用いてプライミングされ得、2つの組成物を組合せて、感染に対する体液性応答と細胞媒介応答の両方をより効果的に惹起することができる。
【0101】
炭水化物エピトープにコンジュゲートされたpan−DR結合ペプチドの場合、脂質部分は、ペプチドの反対側の末端に連結されてよい(例えば炭水化物はC末端に連結され、脂質はN末端に連結される)。一部の実施形態において、脂質および炭水化物部分は両方共、ペプチドの同じ端部に連結されてよい。一部の実施形態において、2つの部分は、N末端上の同じリンカーに連結されてよい。
【0102】
VI. 免疫応答を刺激する方法
本発明のポリペプチド免疫原は、当業者にとっては公知のさまざまな方法を用いて、E.coliなどの発現宿主(または以上で論述した他の宿主細胞)から発現され、濃縮され、精製されてよく、代替的には、免疫原は液相または固相ペプチド合成により調製されてよい。精製されたポリペプチドまたはペプチド免疫原は、例えば免疫応答を刺激するかまたは増強し、任意には保護および/または治療的免疫応答を生成する目的で、個体の体内に投与するための医薬的に許容される賦形剤と共に調合可能である。「保護および/または治療的免疫応答」という語句は、何らかの形で疾病の症候、副作用または進行を防止するかまたは少なくとも部分的に阻み感染性因子を一掃する、例えば感染性因子から誘導された疾病関連抗原に対するCTLおよび/またはHTLおよび/または抗体応答を意味する。一部の実施形態において、本発明のポリペプチドまたはポリヌクレオチドは、免疫学的に有効な量の本発明のペプチドのうちの1つ以上と適切な薬学的担体とを含むワクチンの形に調合される。したがって、本発明のペプチドは、個別にかまたは単一組成物または多数の組成物のいずれかに組合わされた状態で投与可能である。
【0103】
本発明はさらに、免疫応答を刺激するため、本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物を投与する方法に関する。発現ベクターは、例えばDonnellyら、(Ann.Rev.Immunol.、15:617−648(1997));Felgnerら、(米国特許第5,580,859号(US.5,580,859)、1996年12月3日発行);Felgner(米国特許第5,703,055号(US.5,703,055)、1997年12月30日発行);およびCarsonら、(米国特許第5,679,647号(US.5,679,647)、1997年10月21日発行)中に記載の当該技術分野において周知の方法によって投与される。一実施形態においては、マルチエピトープ構築物は、ネイキッド核酸として投与される。
【0104】
本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物は、例えば経口、膣内、直腸または非経口経路例えば静脈内、筋内、皮下、眼窩内、関節包内、腹腔内、嚢内経路を含めたさまざまな経路によって、または例えばそれぞれ皮膚パッチまたは経皮イオントフォレシスを用いた経皮的な受動吸収または吸収促進によって、対象の体内で免疫応答を刺激するために投与することができる。さらにこの組成物を注射、挿管法によってかまたは局所的に投与することもでき、この局所的投与は、軟こうまたは粉末の直接塗布などによる受動的なもの、あるいは鼻腔用スプレーまたは吸入薬などを用いた能動的なものであり得る。発現ベクターは同様に局所的スプレーとして投与することができ、この場合、組成物の一成分は適切な高圧ガスである。医薬組成物は同様に、所望される場合、例えばFelgnerら、米国特許第5,703,055号(US.5,703,055);Gregoriadis、Liposome Technology、第I巻〜第III巻(第2版、1993)中に記載されているように、リポソーム、ミクロスフェアまたはその他のポリマーマトリクス内に取り込むこともできる。例えば、リン脂質または他の脂質で構成されたリポソームは、比較的簡単に製造し投与できる無毒性で生理学的に許容される代謝性担体である。
【0105】
本発明の発現ベクターまたはそれから誘導されたポリペプチドは、例えばFelgnerら、米国特許第5,580,859号(US.5,580,859)および5,703,055号(US.5,703,055)中に記載されている通りに、動物の体組織の間質腔に送出され得る。本発明の発現ベクターの筋肉への投与は、皮内および皮下注射および経皮投与を含め、特に有効な投与方法である。イオントフォレシスによるものなどの経皮投与も同様に、筋肉に対して本発明の発現ベクターを送出する有効な方法である。本発明の発現ベクターの表皮投与も同様に利用可能である。表皮投与には、刺激物に対する免疫応答を刺激するために表皮の最も外側の層を機械的または化学的に刺激することが関与する(Carsonら、米国特許第5,679,647号(US.5,679,647))。
【0106】
免疫応答を刺激するために本発明の発現ベクターまたはそれから誘導されたポリペプチドを投与する他の効果的方法には、例えばCarsonら、米国特許第5,679,647号(US.5,679,647)中に記載されている通りの粘膜投与が含まれる。粘膜投与について、最も有効な投与方法には、発現ベクターおよび医薬組成物を含む適切なエアロゾルの鼻腔内投与が含まれる。生殖器、膣および眼部位の粘膜組織に対する発現ベクターの送出のためには、座薬および局所的調製物も同様に有効である。追加的には、発現ベクターを粒子に複合体化させ、ワクチンガンにより投与することもできる。
【0107】
結論として、本発明のペプチドは、予防目的(すなわち後の疾病の危険性を減少させる目的)および治療目的の両方の目的で免疫応答を誘発するために有用であるが、本発明のペプチドは、実験的にかつ抗体および/またはBリンパ球を単離できる動物の体内で抗体を誘発する目的で、免疫応答を誘発するためにも有用であり、このことはそれ自体、後にモノクローナル抗体および抗体誘導体の産生を可能にする。
【0108】
VII. 医薬組成物
本発明のポリペプチド、本発明のポリヌクレオチドおよびその医薬組成物およびワクチン組成物は、予防および/または治療目的で哺乳動物、特にヒトに対し投与可能である。本発明のポリペプチドは、病原体または癌関連または癌特異的生体分子(例えばタンパク質、炭水化物など)を含めた(ただしこれらに限定されない)抗原に対する免疫応答を惹起および/または増強するために使用可能である。本発明を用いて治療可能な疾病の例としては、さまざまな細菌感染、ウイルス感染、真菌感染、寄生虫感染および癌が含まれる。タンパク性抗原に関して以上で論述した通り、国際公開第00/20027号(WO00/20027)からの癌抗原に関連する癌も、関節リウマチおよび炎症性腸疾患などのさまざまな炎症性疾患と同様の疾病例である。
【0109】
一部の治療的利用分野において、本発明は、すでに癌、炎症性疾患を患っているかまたは問題のウイルスまたは微生物に感染している個体に対して投与される。疾病の潜伏期または急性期にある個体は、適宜本発明を用いて別個に、または他の治療と併用して治療されてよい。
【0110】
一部の治療的利用分野において、本発明の組成物は、微生物または腫瘍抗原に対する有効なCTL応答または体液性応答を惹起し、かつ症候および/または合併症を治ゆするかあるいは少なくとも部分的に阻むのに充分な量で患者に投与される。これを達成するために適切な量が「治療上有効な量」として定義される。この用途にとって有効な量は、一部にはペプチド組成物、投与方法、治療対象の疾病の病期および重症度、患者の体重および全身的健康状態、および処方医師の判断に左右される。
【0111】
本発明の組成物の治療上有効な量は一般に、初期免疫付与すなわち治療的または予防的治療のためには、体重70kgの患者に対しポリペプチド約1.0μg〜約10,000μg、例えば約100〜約8000μg、例えば約200〜約6000μgである。一部の実施形態において、これらの用量には、特異的免疫応答を測定することにより患者の応答および身体条件に応じて数週間乃至数カ月にわたり、追加免疫処方計画に準じて約1.0μg〜約3000μgのペプチドの追加免疫投薬量が後続する。
【0112】
本発明の組成物は一般に重篤な疾病状態すなわち生命にかかわるかまたは潜在的に生命にかかわる状況において利用可能であるということを念頭に置いておくべきである。このような場合、異物の最少化およびコンジュゲートの相対的無毒性を考慮して、これらの組成物を実質的に余剰に投与することが可能であり、治療医師がそれを望ましいと感じるかもしれない。
【0113】
一部の実施形態において、本発明は、細菌感染、ウイルス感染、真菌感染、寄生虫感染および癌を含めた(ただしこれらに限定されない)特定の疾病を予防および/または改善するために、予防的に使用可能である。有効量は、以上で記述した通りである。追加的に、ワクチン技術の当業者であれば、追加免疫することおよび投薬量および投薬計画を調整することなどによって適宜、予防的処置をいかに調整または修正すべきかも認識していると考えられる。
【0114】
本発明の調合ワクチンは、医薬的に許容されるアジュバントと組合わせることができる。調合ワクチンは、水溶液、懸濁液またはエマルジョンであり得る。本発明のワクチン中の各免疫原の免疫学的に有効な量は、過度の実験なく当該技術分野において公知の方法により決定可能である。
【0115】
アジュバントは、医薬的に許容されるあらゆるアジュバントであり得る。一部の実施形態において、アジュバントはミョウバンベースの化合物である。一部の実施形態において、アジュバントは水酸化アルミニウムである。他の実施形態においては、アジュバントはリン酸アルミニウムである。一部の実施形態において、アジュバントはEmunade(登録商標)である。Emunade(登録商標)は、油、水および水酸化アルミニウムの組合せで構成されたアジュバントである。一部の実施形態において、アジュバントはQuil Aである。一部の実施形態において、アジュバントはQuil Aにコレステロールを加えたものである。一部の実施形態において、アジュバントは、MF59およびProvaxなど(ただしこれらに限定されない)のエマルジョンである。
【0116】
一部の実施形態において、ISCOMがアジュバントとして用いられる。ISCOMは、Immune Stimulating Complex(免疫刺激複合体)の頭字語であり、その技法は例えばMoreinら、(Nature308:457−460(1984))の中に記載されている。ISCOMは、以下の通りに形成される新油性免疫刺激複合体である。ポリペプチドは、非イオン洗浄剤(例えばMega−9、TritonX−I00、オクチルグリコシド、Digitonin、NonidetP−40、C12E8、Lubrol、Tween−80)などを用いて、標準的方法により可溶化される。ISCOM形成を補助するために、脂質混合物が添加される。脂質混合物は、ホスファチジルコリンおよび合成コレステロールを含み得る。一部の実施形態において、混合物はまず最初に、室温で撹拌しながら非イオン洗浄剤で処理され、次に脂質混合物(例えば等量のホスファチジルコリンとコレステロール)が添加され、撹拌が続行される。ポリペプチド組成物にQuil A(精製サポニン配糖体)が添加され、撹拌が続行される。その後、非イオン洗浄剤が除去される(例えば酢酸アンモニウムでの膜分離法による)。ISCOMのマトリクスがQuil Aにより形成される。電子顕微鏡で見たISCOM粒子の形態は、約35nmのサイズの典型的なカゴ様構造を示す。ISCOM形成期は、接線流膜分離法を用いて精製可能である。ISCOMは、臨界濃度でミセルを自然発生的に形成するQuil Aの能力に基づいて、かつ精製された抗原を封入する疎水性/親水性連結によって、マルチマー形状で精製抗原を提示する。ISCOMの形成は、典型的なかご様構造が形成されたことを確認するための電子顕微鏡によって確認される。約0.01〜0.1%の最終的濃度を得るためにQuil Aを添加することができる。一部の実施形態において、最終的濃度は約0.05%である。
【0117】
本発明は同様に、医薬的に許容される担体と本発明の発現ベクターまたは本発明のポリペプチドとを含む医薬組成物にも関する。医薬的に許容される担体は、当該技術分野において周知であり、生理的緩衝食塩水、アルコール/水溶液またはその他の溶媒またはビヒクル例えばグリコール、グリセロール、油例えばオリーブ油または注射用有機エステル、脂質またはリポソームを含めた、水溶液または非水性溶液、懸濁液およびエマルジョンを含む。
【0118】
医薬的に許容される担体は、例えば発現ベクターを安定化するかまたは発現ベクターの吸収を増大させるために作用する生理学的に許容される化合物を含むことができる。このような生理学的に許容可能な化合物には、例えばグルコース、スクロースまたはデキストランなどの炭水化物、アスコルビン酸またはグルタチオンなどの酸化防止剤、キレート剤、低分子量ポリペプチド、抗菌薬、不活性ガスまたはその他の安定化剤または賦形剤が含まれる。発現ベクターは、さらに、ペプチド、ポリペプチドおよび炭水化物などのその他の成分と複合体化され得る。発現ベクターは同様に、例えばワクチンガンを用いて個体に投与できる粒子またはビーズへと複合体化され得る。当業者であれば、生理学的に許容される化合物を含めた医薬的に許容される担体の選択が、例えば発現ベクターの投与経路により左右されるということを認識していると考えられる。
【0119】
本発明はさらに、本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物を投与して免疫応答を刺激する方法にも関する。発現ベクターは、例えばDonnellyら、{Ann.Rev.Immunol.、15:617−648(1997));Felgnerら、(米国特許第5,580,859号(US.5,580,859)、1996年12月3日発行);Felgner(米国特許第5,703,055号(US.5,703,055)、1997年12月30日発行);およびCarsonら、(米国特許第5,679,647号(US.5,679,647)、1997年10月21日発行)中で記述されている通りの当該技術分野において周知の方法により投与される。一実施形態において、本発明のポリヌクレオチドは、ネイキッド核酸として投与される。
【0120】
本発明の組成物は、抗体または抗体誘導体を調製することを目的として、例えば実験動物または動物の体内で免疫応答を誘発するためにも有用である。このような組成物においては、組成物が医薬的に許容されるものでなくてはならないという要件は、重要性が低く、例えばヒトにおいて使用するのに適しているとみなされていないアジュバントを使用することが可能である。
【実施例】
【0121】
実施例の前文
以下の実施例は、請求対象の発明を限定するためではなく、例証するために提供されるものである。
【0122】
PADRE(配列番号3)内部のタンパク質分解分割が、PADRE含有タンパク質内、特に2つのリジン残基K2およびK10のまわりで、ただしW7のまわりにも観察された。PADRE内部の分割は、組換えにより産生されたタンパク質内で不均質、生産収量の低下および安定性の改変を導くことから望ましくない。上流側および下流側のプロセスの間に添加剤を使用することによって分割をうまく回避できるにせよ、それでも、結果として得られるPADRE含有ワクチンはなお低い免疫原性および薬物動態特性例えば半減期の短縮および1つまたは複数のTヘルパーエピトープの不適切な提示をなおも示すかもしれない。したがって、PADRE配列からプロテアーゼ感受性アミノ酸/配列を除去する必要がある。
【0123】
プロテアーゼ活性に対する感受性が低くなったPADRE類似体を提示することで、特にE.coliおよびショウジョウバエ発現系などの発現系においてプロテアーゼに対する組換え発現産物の感受性が削減されて、タンパク質産生の上流側および/または下流側プロセスの間のpan−DR結合ペプチドアミノ酸配列内部のタンパク質分解分割が回避されることになる。さらに、PADRE類似体が、単離形態でまたは免疫原性ポリペプチドの一部としてヒトに投与される場合、感受性の削減によって、薬物動態特性の改善(すなわちより長い血清半減期)そして場合によっては免疫原性の改善というさらなる利点がもたらされる。
【0124】
これに基づいて、配列番号3の数多くの類似体を調製し以下の一連の同定ステップに付して、タンパク質分解を比較的受けにくい改良型pan−DR結合ペプチドを同定した:
1)合成PADRE類似体のライブラリの調製ステップ;
2)優れたHLA結合特性について、1)に由来する類似体を検定し選択するステップ;
3)生体内原位置プロテアーゼ耐性について、2)で選択した類似体を検定し選択するステップ;
4)優れた免疫原特性について、3)で選択した類似体を検定し選択するステップ。
【0125】
実施例1
PADRE類似体のHLA−DR結合についての試験
標準的な固相ペプチド合成により、合計69個の異なるPADRE類似体(配列番号:4〜72)を合成した。これらのPADRE類似体は全て、N−からC−末端に向かって、2つのアラニン残基とそれに続く残基3〜15により構成され、これは(2つのアラニン残基が先行するPADREである配列番号4以外)配列番号3の類似体である。2つの追加のN末端アラニンを含み入れる理由は、PADRE類似体の免疫原特性が13C末端アミノ酸内に存在する一方で、単離ペプチドがプロテアーゼ耐性についての試験管内検定においてエンドプロテアーゼ耐性を示すことができるようになることを保証するためであった。
【0126】
ペプチドを、以下の手順にしたがってHLA−DRに対する結合について試験した:
【0127】
ペプチド/HLA−DR結合検定
クラスIIMHC結合検定のための標準的な作業手順は、Sidneyら、(1998)、Current Protocols in Immunology、18.3.1−18.3.19、1998中に記載されている。簡潔に言うと、精製ヒトクラスII分子[5〜500nM]を、さまざまな未標識ペプチド結合阻害物質および1〜10nM125I−放射性標識プローブペプチドと共に48時間、プロテアーゼ阻害物質カクテルの存在下で5%のDMSを含むPBS中でインキュベートした。インキューベーション混合物中の最終的洗浄剤濃度は、0.05%NonidetP−40であった。pH4.5で実施したDR3およびpH5.0で実施したDRw53を除いて、検定をpH7.0で実施した。pHは、Setteら、(1992)、J Immunol、148:844−51で記述されている通りに調整した。
【0128】
Sidneyらの中に記載されている通りにHPLC方法を使用してMHC分子に対するペプチド結合を測定する代りに、抗MHCクラスII抗体がコーティングされた抗体平板ベースの捕獲検定を利用した。この検定は、大量平板内放射分析検定用に特定的に設計された96ウェルの白色ポリスチレンマイクロタイター平板を使用するように開発されたものである。MHCに結合した125I標識ペプチドの測定は、高速大量処理性能を得るためにきわめて感度の高い検定を可能にするTOPCOUNT(Perkin−Elmer Instruments)ベンチトップマイクロプレートシンチレーションおよびルミネセンスカウンタを用いて実施される。
【0129】
ペプチド結合阻害物質を、30μg/ml〜300pg/mlの範囲内の濃度で試験した。市販の曲線適合アルゴリズムを用いて、データをコンピュータ内でプロットし、50%の阻害を生成する用量(IC50)を決定した。適切な化学量論的条件下で、精製DR分子に対する未標識被験ペプチドのIC50は、相互作用の親和性(KD)の適正な近似である。2〜4個の完全に独立した実験においてペプチドを試験した。
【0130】
結果
IC50値を介して表わされる結合親和性を、各ペプチドについて図1に列挙する。これらのIC50値に基づいて、類似体(38PADRE類似体)の画分が充分に優れたHLA結合能力(図1にボールド体と下線で示す)を示すという結論が下された。これらの優れた結合性を有するもののうち、当初10個を選択してプロテアーゼ耐性についてさらに試験した:配列番号;4、10、19、23、29、38、46、52、67および69。
【0131】
配列番号3および4が示す類似の結合特性により証明されるように、配列番号4〜69内に2つのN末端アラニンを含み入れることでHLA結合特性に影響が及ぶことはなかったという点を指摘しておくことも重要である。
【0132】
実施例2
選択されたPADRE類似体のプロテアーゼ耐性試験
選択された優れた結合性を有するPADRE類似体のプロテアーゼ耐性を試験するために、クローニング技術により、ヒトTNFα内のループ(ループEE)内にその配列を挿入した。修飾されたTNFα分子を発現するE.coli由来の無細胞タンパク質粗製抽出物を調製し、PADRE類似体配列内部のタンパク質分解分割を、SDS−PAGEとそれに続く免疫ブロット法によって分析した。
【0133】
全長、膜結合TNFαは、アミノ酸233個の長さをもつタンパク質である。アミノ末端フラグメント1〜76は、TNFαのアミノ酸157個の長さの可溶形態(77〜233)中には存在しない。
【0134】
ヒトTNFαワクチン候補の設計のために用いられる鋳型は、E.coli中での発現用のN末端Metが先行する、可溶形態のTNFα(フラグメント77〜233、配列番号73)である。さらに、TNFαのその2つの受容体に対する結合を取り除くために点変異を行ない(Y875)、こうしてタンパク質の細胞傷害性を根絶した。したがってワクチン候補は、長さがアミノ酸158個の配列番号74に基づくものである。
【0135】
TNF37.87(配列番号75)は、A185とE186の間にPADREが挿入されているTNFバリアント(…PEGA−EAK…)である。PADREはAで始まることから、Aは反復されず、その結果13個ではなく12個のアミノ酸が挿入された:
...PEGA−AKFVAAWTLKAAA−EAK... → PEG−A−KFVAAWTLKAAAEAK...
【0136】
したがって、当初プロテアーゼ耐性について試験したTNFバリアントは、以下の通りである:
配列番号75−TNF37.87−「従来の」PADREを含む基準タンパク質:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAKFVAAWTLKAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
− ならびに、下線位置においてそれぞれ配列番号4、10、19、23、29、38、46、52、67および69の12のC末端アミノ酸残基を各々同じように有するTNFバリアントを含む9個のPADRE類似体すなわち:
配列番号76−TNF37.87−007配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAWFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号77−TNF37.87−016配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAFFVAANTLKADAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号78−TNF37.87−020配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAYFVAAFTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号79−TNF37.87−026配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAHFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号80−TNF37.87−035配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAHFVAAFTLKAEAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号81−TNF37.87−043配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAEFVAAWTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号82−TNF37.87−049配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGANFVAAYTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号83−TNF37.87−064配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAQFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号84−TNF37.87−066配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAIFVAAWTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
【0137】
全ての配列中、2重下線は初期メチオニルおよびY87S変異をそれぞれ示し、1本下線は、挿入されたpanDR結合配列を示し;ボールド体は、PADRE(配列番号3)に比べたpan−DR結合ペプチド中のアミノ酸変化を表わす。
【0138】
組換えTNFαバリアントのクローニング、発現、精製および特徴づけ
鋳型として、TNF(Y87S)をコードする合成cDNAを使用した。PADREまたはPADRE類似体をコードするオリゴヌクレオチドを、PCRによりA185とE186の間で鋳型内に挿入した(上記参照)。結果として得たcDNAをpET28b+(Novagen)内にクローニングし、E.coli菌株HMS174へと形質転換させた。組換え発現のため、結果として得たE.coli菌株を発酵槽内において37℃にて合成培地中で培養した。組換え発現の誘発は、1mMのIPTGを添加することで開始し、温度を25℃まで低下させた。IPTG誘発から4、8または24時間後に培養を収穫した。Nielsenら、(2004)、J Biol.Chem.279:33593−33600中で記述されているようにTNFα特異的モノクローナル抗体を用いて親和性クロマトグラフィにより90%超の均質性までTNFαバリアントを精製した。精製したTNFαバリアントをMALDI−TOF質量分析法によって特徴づけしてそれらの同一性と無欠性を確認した。
【0139】
生体内原位置プロテアーゼ検定:ウエスタンブロット法を用いたTNF−αバリアントのプロテアーゼ耐性の分析:
試験すべきさまざまなTNF−αバリアントを発現するE.coli細胞培養のアリコート(1ml)を遠心分離(5000g/5分/5℃)し、培地を廃棄した。pH6.0で20mMのBis−Tris1ml中に細胞ペレットを再懸濁させた。超音波処理(各超音波処理ラウンド間で20秒間休止して、11μmの振幅で4×15秒)により、細胞破壊を達成した。結果として得た懸濁液を遠心分離し(20000g/30分/8℃)、上清(粗製抽出物)を新しい管に移した。次に、ブラッドフォードタンパク質分析(Biorad)を用いて、タンパク質含有量について粗製抽出物を分析した。4〜12%のNuPAGEゲル(Novex)上に等量のタンパク質を投入した。SDS−PAGEの後、セミドライブロット法によりニトロセルロース膜上にタンパク質を移した。免疫反応については、Tris50mM、NaCl150mM、EDTA5mM、Igepal0.1%、ゼラチン0.5%中で、ウサギTNFα抗血清(1時間;希釈度1:10,000)およびホースラディッシュペルオキシダーゼでコンシュゲートされた二次抗体(Dako P448)と共にインキューベーションを行なった。検出にはECLTM試薬(GE Healthcare)を用いた。
【0140】
結果
被験TNFバリアントのうち、3つ(配列番号78、79および84)はプロテアーゼ検定においてプロテアーゼ耐性があることが示され(データ示さず)、これらのTNFバリアントが、PADRE類似体配列の内部でのタンパク質分解分割に対する感受性をもたないということを証明した。
【0141】
配列番号23、29および69の各々の少なくとも13C末端アミノ酸残基により画定されるPADRE類似体がタンパク質分解に対する耐性の所望の増加を示し、かつ配列番号23、29および69の13C末端アミノ酸残基を含み入れることで修飾されたタンパク質は、組換えにより生成された場合により優れた収量を示し(これは、発現産物のフラグメント化が全くまたは限定的にしか発生しないことに起因する)、均質な発現産物を提供し、場合によってはプロテアーゼ耐性免疫原の生物学的半減期の延長を原因として免疫原性の増大を提供するものであるという結論が下された。
【0142】
実施例3
選択されたPADRE類似体の免疫原性試験
実施例2内で同定した3つのプロテアーゼ耐性PADRE類似体をその免疫原特性について試験した。1本の実験ライン上で、ヒトのドナー由来のPBMC(末梢血単核細胞)内および2つのマウス株(DR4およびbxd)内で遊離ペプチドとしてペプチドを試験した。第2の実験ライン内では、実施例2で同定したプロテアーゼ耐性TNFバリアントに関連して、PADRE類似体を試験した。
【0143】
ヒトPBMCにおけるペプチドの試験
ヒトPBMC(末梢血単核細胞)を、配列番号23、29および69の13アミノ酸残基と同一の配列をもつペプチドで刺激した(以下では、これら3つの13量体をそれぞれPADRE.Y2.F7.H10、PADRE.H2.N7.H10およびPADRE.12.H10と呼ぶ)。1日目および4日目に、IL−2を添加した。7日目に、he細胞の一部分を取り出してINF−γELI−POT検定に付した。残りの細胞を再刺激した。8日目および11日目に、IL−2を添加した。14日目にCD4細胞を精製し、INF−γELISPOT検定において試験した。
【0144】
INF−γELISPOT検定は、本質的にTangri Sら、(2005)、J Immunol 174、3187−96中で記述されている通りに実施した。
【0145】
マウスにおけるペプチドの試験
HLA−DR4遺伝子導入マウスとbxdマウス株の各々の3匹の個体からなるグループを、CFA中の20μg(6μlの20mg/mlペプチドおよび294μlのPBS1×+300μlのCFA)または2μg(6μlの2mg/mlペプチド+294μlのPBS1×+300μlのCFA)のペプチドを用いて尾の付け根で免疫付与した。10〜14日後に、脾臓を収穫し、CD4細胞を精製し、INF−γELISPOT検定(McKinneyら、(2004)、J Immunol 173、1941−50に記載の通り)で試験した。被験ペプチドは、ヒトPBMCで試験されたものと同一であった。
【0146】
マウスにおけるTNFαバリアントの試験
3匹のHLA−DR4遺伝子導入マウスからなるグループを、マウス一匹あたり100μlの完全フロインドアジュバント(CFA)中10μgのTNF由来の抗原を用いて、尾の付け根で免疫付与した。最初の免疫付与から14日目に、100μlのIFA(不完全フロインドアジュバント)中の同じ用量の抗原を用いて、動物に追加免疫を付与した。TNFα特異的抗体力価を決定するため、各々の免疫付与から14日後に、血清を収集した。その後動物を屠殺して、INF−γELISPOT検定に付した。
【0147】
マウスの抗血清中のTNFα特異的抗体力価の決定
直接ELISAを用いて、TNFα特異的抗体力価を決定した。96ウェル平板(Maxisorb、Nunc)を組換えTNFαバリアントTNFY87S(pH9.6で炭酸緩衝液中5μg/ml、100μl/ウェル、4℃で一晩)を用いてコーティングした。その後平板を3回洗浄し、37℃で2時間200μl/ウェルのブロッキング緩衝液(1%のウシ血清アルブミンおよび0.05%のTween20を含むリン酸緩衝生理食塩水)を用いてインキュベートした。平板を3回洗浄し、ヒトTNFαバリアントで免疫付与したマウス由来のプールされた血清を、合計100μl/ウェルのブロッキング緩衝液中での1/10希釈ステップ(出発希釈度1/1000)を用いて、6ステップで滴定した。全ての試料をデュプリケートで滴定した。血清と対照を2時間37℃でインキュベートした。平板を3回洗浄し、100μlのビオチニル化ヤギ抗マウスIgG(ブロッキング緩衝液中で1/10000希釈)を各ウェルに移し、37℃で1時間インキュベートした。最後に、平板を3回洗浄し、100μl/ウェルのアビジン−ペルオキシダーゼ複合体(Vectastain Elite Vector PK−6100)を用いて室温で45分間インキュベートした。平板を再び洗浄し、TMBSを用いて現像した。10〜20分後、100μlの4NのH2SO4で反応を停止させ、ELISA読取り装置を用いてA450値を決定した。0.5のA450値を生成する抗血清希釈度として抗体力価を定義づけた。
【0148】
結果
ヒトPBMC中のペプチドの試験から得られた結果は、PADRE.Y2.F7.H10およびPADRE.I2.H10が、配列番号3に比べ7日目と14日目の両日にINF−γELISPOT検定においてより優れた免疫応答を提供し、一方PADRE.H2.N7.H10は、配列番号3と同じ規模の免疫応答を提供したことを明らかにした。
【0149】
マウスにおけるPADRE類似体ペプチドの試験では、3個の被験類似体の全てが、DR4マウスにおいて20μg用量でPADREよりも良い性能を示した一方、ELISPOT検定により測定された通り2μg用量でDR4遺伝子導入マウスにおいて、PADRE.I2.H10ペプチドのみが免疫応答の改善を提供した。bxdマウスモデルでは、3つの被験PADRE類似体の全てが、両方の用量でELISPOT検定内でPADREに比べて有意な免疫応答を提供したが、PADREに比べた免疫応答の改善は得られなかった。
【0150】
PADRE類似体を含むTNFαバリアントでの免疫付与は、以下のような結果を提供した:すなわち
INF−γELISPOTにより測定されたHLADR4遺伝子導入マウスにおけるHTL誘発は、全てのTNFα−バリアントが、TNFα内に導入されたPADRE類似体に対するHTLを誘発したこと、そしてこの点においてPADRE.Y2.F7.H10およびPADRE.I2.H10がPADREよりも優れていたこと(ここでPADRE.H2.N7.H10はPADREとほぼ同じHTL誘発を提供した)を明らかにした。さらに全てのバリアントは、野生型TNFαにより誘発されたものよりも著しく高い抗体力価を提供し、PADRE.Y2.F7.H10およびPADRE.I2.H10を含むバリアントは、PADREに比べ優れており、ここでもまた、PADRE.H2.N7.H10はPADREに匹敵する免疫応答を提供した。実際、抗体力価とHTL誘発を相関させた場合、明らかに線形相関が存在していた。
【0151】
実施例4
8個のさらなるPADRE類似体の選択および試験
panDR結合性でプロテアーゼ耐性を有する免疫原性PADRE類似体PADRE.Y2.F7.H10、PADRE.H2.N7.H10およびPADRE.I2.H10の同定に基づいて、実施例1において優れた結合性を有するものとして同定されたさらなるPADRE類似体を試験することが決定された。被験ペプチドは、配列番号15(PADRE.F2.N7.H10)、21(PADRE.Y2.H10)、24(PADRE.Y2.N7.H10)、26(PADRE.H2.H10)、47(PADRE.E2.Y7.H10)、54(PADRE.N2.N7.H10)、66(PADRE.Q2.F7.H10)、および72(PADRE.I2.N7.H10)からの13C末端アミノ酸残基を有するペプチドであった。
【0152】
これらの配列のうち3つすなわちPADRE.E2.Y7.H10、PADRE.N2.N7.H10およびPADRE.Q2.N7.H10は、実施例2に記された手順に従ってプロテアーゼ耐性を有することが示された。
【0153】
結論
上述の実施例は、プロテアーゼ耐性および/または免疫原性の観点から見て改善された特性を有する数多くのPADRE類似体の存在を実証した。配列番号3内で入念に修飾を行なうことで、組換え発現された場合および生体内免疫原として使用された場合の両方において、PADREよりも安定した免疫原が提供される、ということが実証されている。
【0154】
本明細書中で記述されている実施例および実施形態は単に例証を目的としており、それに照らしたさまざまな修正または変更が当業者には示唆されるものであり、それらは本出願の精神および視野そして添付のクレームの範囲内含み入れられるべきものである。本明細書中で引用される全ての刊行物、特許および特許出願は、あらゆる目的でその全体が参照により本明細書に援用される。
【0155】
【表2】
【表3】
【背景技術】
【0001】
Tヘルパー細胞により認識されるエピトープを含む免疫原性ペプチドが、免疫応答を誘発する上で有用であることが発見されてきた。特定の決定基に対する抗体応答を増強するためのヘルパーペプチドの使用は、例えばHervas−Stubbsら、Vaccine12:867−871(1994)中に記載されている。
【0002】
MHC対立遺伝子のペプチド結合ポケットをライニングする対立遺伝子多型残基は各々の対立遺伝子に固有のペプチドセットを結合させる能力を付与する傾向にあるが、所与のペプチドが2つ以上のMHC対立遺伝子に結合することが実証されたケースも存在する。例えば、複数の研究者が、多数のDRタイプに関連していくつかのエピトープの変性結合および/または認識を報告し、或る種のエピトープが「万能」エピトープであるかもしれないという概念を導いている(Buschら、Int.Immunol.2:443−451(1990);Panina−Bordignonら、Eur.J.Immunol.19:2237−2242(1989);Sinigagliaら、Nature336:778−780(1988);O’Sullivanら、J.Immunol.147:2663−2669(1991);Roacheら、J.Immunol.144:1849−1856(1991);Hillら、J.Immunol.147:189−197(1991))。Pan−DR結合ペプチドは、例えば米国特許第6,413,935号(US6,413,935);国際公開第95/07707号(WO95/07707);国際公開第2005/120563号(WO/2005/120563);およびAlexanderら、Immunity 1:751−761(1994)中で記述されてきた)。これらのペプチドは、抗原に対するさまざまな免疫応答の生成を助けることが示されてきた。
【発明の概要】
【0003】
本発明は、タンパク質分解酵素に対する安定性が改善された(短ペプチド、オリゴペプチドおよびポリペプチドの両方の形での)免疫原性ペプチドの提供という観点から見たPADRE配列(配列番号3)内の選択された変異の効果の研究に基づいている。
【0004】
したがって本発明は、100nM未満、例えば少なくとも50nMのIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むまたはこれで構成されている単離ポリペプチドであって、オリゴペプチド配列がAX1FVAAX2TLX3AX4A(配列番号1)を含み、
− X1がW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;
− X3がHおよびKからなる群から選択され;
− X4がA、DおよびEからなる群から選択され;
ただしここでオリゴペプチド配列がAKFVAAWTLKAAA(配列番号3)でないことが条件となっている、
単離ポリペプチドに関する。
【0005】
本発明はさらに、本発明のポリペプチドをコードするポリヌクレオチドおよびこのようなポリヌクレオチドを含むベクターにも関する。同様に本発明が包含しているのは、本発明のポリヌクレオチドを含む細胞である。本発明はさらに、本発明のポリペプチドまたはオリゴペプチドまたはポリヌクレオチドまたはベクターまたは細胞を含む、医薬組成物を含めた組成物に関する。最後に本発明は、それを必要としている対象に対して、本発明のポリペプチドまたはオリゴペプチドまたはポリヌクレオチドまたはベクターまたは細胞または組成物を投与することによって、免疫応答を刺激する方法に関する。
【図面の簡単な説明】
【0006】
【図1−1】さまざまなHLA−DR分子に対する本発明のペプチドのHLA−DR結合を決定する結合実験からのデータを示す。詳細については実施例1を参照のこと。
【図1−2】図1−1に同じ。
【図1−3】図1−1に同じ。
【図1−4】図1−1に同じ。
【図1−5】図1−1に同じ。
【0007】
定義
別段の定義がない限り、本明細書中で使用されている全ての技術的および科学的用語は、本発明が属する技術分野の当業者が一般的に理解するものと同じ意味を有する。本発明は、組換え遺伝学の分野における常用手技に依存するものである。本発明中で使用される一般的な方法を開示する基本的教書としては、Sambrookら、Molecular Cloning、A Laboratory Manual(第3版2001);Kriegler、Gene Transfer and Expression:ALaboratory Manual(1990);およびCurrent Protocols in Molecular Biology (Ausubelら、編、1994))がある。
【0008】
本明細書中で使用される「オリゴペプチド」または「ペプチド」という用語は、少なくとも4つのアミノ酸残基またはアミノ酸ミメティックの連鎖を意味する。オリゴペプチドまたはペプチドは、その中性(未荷電)形態または塩形態のいずれであれ、また、グリコシル化、側鎖酸化またはリン酸化を含めた(ただしこれらに限定されない)修飾が無い状態かまたはこれらの修飾を1つ以上含む状態のいずれであれ、さまざまな長さであり得る。任意の修飾は本明細書中で記述されるポリペプチドの生物活性を破壊するものではないが、本発明は、(例えば生体内で修飾が除去されるまで活性を限定するために)修飾が生物活性を削減または削除する選択肢を含むものである。
【0009】
「ポリペプチド」および「タンパク質」という用語は、本明細書において互換的に使用されて、アミノ酸残基のポリマーを意味する。これらの用語は、1つ以上のアミノ酸残基が対応する天然に発生するアミノ酸の人工的化学ミメティックであるアミノ酸ポリマーならびに天然に発生するアミノ酸ポリマーおよび天然に発生しないアミノ酸ポリマーに適用される。本明細書中で使用されるこれらの用語は、アミノ酸残基が共有ペプチド結合によって連結されている、全長タンパク質(すなわち抗原)を含むあらゆる長さのアミノ酸鎖を包含する。これらの用語はさらに、例えばグリコシル化、アセチル化、リン酸化、アミド化、公知の保護/封鎖基による誘導体化、タンパク質分解による分割または非天然発生アミノ酸による修飾などの翻訳後修飾を受けたポリペプチドを含んでいる。
【0010】
本発明のポリペプチドとして同様に含まれるのは、前述のポリペプチドのフラグメント、誘導体、類似体またはバリアント、およびそれらの任意の組合せである。ポリペプチドおよびそのフラグメント、誘導体、類似体またはバリアントは、抗原性および免疫原性ポリペプチドであり得る。
【0011】
「アミノ酸残基」、「アミノ酸」および「残基」は、ペプチド、オリゴペプチドまたはタンパク質中のアミノ酸残基に言及している場合、互換的に使用され、本明細書中で使用されている通り、アミド結合またはアミド結合ミメティックを通して少なくとも1つの他のアミノ酸またはアミノ酸ミメティックに共有結合により接合されたアミノ酸またはアミノ酸ミメティックを意味する。
【0012】
本明細書中で使用される「アミノ酸」という用語は、無条件の場合、「L−アミノ酸」またはL−アミノ酸ミメティックを意味する。
【0013】
本明細書中で使用される通り、ペプチド化合物を記述するために使用される命名法は、各アミノ酸残基の左側(N末端)にアミノ基が提示され右側(C末端)にアルボキシル基が提示されている従来の実践法にしたがっている。エピトープ内でアミノ酸残基の位置が言及されている場合、それらは、アミノからカルボキシル方向に付番されており、1位は、エピトープまたはそれが一部を成していてよいペプチドまたはタンパク質のアミノ末端に基も近い位置である。アミノ酸構造式においては、各残基は一般に、標準的な三文字または一文字の呼称で表わされる。アミノ酸残基のL形態は、大文字一文字かまたは三文字記号の最初の大文字によって表わされ、D−形態を有するこれらのアミノ酸のD形態は、小文字一文字かまたは小文字三文字記号により表わされる。グリシンは、非対称炭素原子を全く有しておらず、単に「Gly」または「G」と称される。
【0014】
本明細書中で使用されているアミノ酸の記号を以下に示す。
【0015】
【表1】
【0016】
本明細書中で使用される「panDR結合ペプチド」または「pan−DR結合エピトープ」という用語は、2つ以上のMHCクラスIIDR分子を結合させる(例えば100nM未満例えば少なくとも50nMのIC50を有する2つ以上のMHC分子の各々を結合させる)分子ファミリーの一成員を意味する。一部の実施形態においては、本発明のpan−DR結合オリゴペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11または全12個の最も一般的なDR対立遺伝子(DR1、2w2b、2w2a、3、4w4、4w14、5、7、52a、52b、52cおよび53)を結合させることのできるペプチドである。本発明のpan−DR結合オリゴペプチドは、第2の決定基に対する免疫応答を促進することに加えて、標的免疫原自体としても役立つことができる。こうして、例えばpan−DR結合ペプチド自体が炭水化物エピトープに連結される場合、免疫応答は、pan−DR結合ペプチドと炭水化物エピトープの両方に対するものであってよい。
【0017】
本明細書中で使用される「PADRE」という用語は、アミノ酸配列AKFVAAWTLKAAA(配列番号3)を有するpanDR結合ペプチドを意味する。
【0018】
本明細書中で使用される「PADRE類似体」という表現は、PADREと比べて多くとも1、2、3または4個のアミノ酸変更を含むアミノ酸配列を含み、うち少なくとも1つがK2、W7、K10またはA12内で行なわれているアミノ酸配列を含むpanDRペプチドを意味する。
【0019】
本明細書中で使用される「IC50」という用語は、基準ペプチドの結合の50%の阻害が観察される結合検定中のペプチドの濃度を意味する。検定が実施される条件(すなわち限定的MHCタンパク質および標識ペプチド濃度)に応じて、これらの値はKD値に接近してよい。結合を決定するための検定は、例えばその各々の開示が参照により本明細書に援用されている国際公開第94/20127号(WO94/20127)および国際公開第94/03205号(WO94/03205)の中で詳述されている。検定条件が変動した場合そして使用される特定の試薬(例えばMHC調製物など)に応じて、IC50値は、多くの場合劇的に変化し得る。例えば、MHC分子の過度の濃度は、所与のリガンドの見かけのIC50測定値を増大させる。
【0020】
あるいは、結合は、例えばPADREなどの基準ペプチドとの関係において表現される。特定の検定の感度が増減するにつれて試験対象のペプチドのIC50値は幾分か変化するかもしれないが、基準ペプチドとの関係における結合は有意に変化しない。例えば、基準ペプチドのIC50が10倍増加するような条件下で実施される検定においては、被検ペプチドのIC50値もおよそ10倍シフトする。したがって、あいまいさを避けるため、ペプチドが良好な結合性を有するか、あるいは中度の、弱いまたは不良な結合性を有するかについての査定は一般に、標準ペプチドのIC50との関係における対象ペプチドのIC50に基づく。結合は同様に、生細胞(例えば、Ceppelliniら、Nature339:392、1989;Christnickら、Nature352:67、1991;Buschら、Int.Immunol.2:443、19990;Hillら、J.Immunol.147:189、1991;del Guercioら、J.Immunol.154:685、1995))、洗浄剤溶解物を用いた無細胞系((例えば、Cerundoloら、J.Immunol.21:2069、1991)、固定化された精製済みMHC(例えば、Hillら、J.Immunol.152、2890、1994;Marshallら、J.Immunol.152:4946、1994);ELISA系(例えばReayら、EMBO J.11:2829、1992)、表面プラズモン共鳴(例えば、Khilkoら、J.Biol.Chem.268:15425、1993)、高流量可溶性相検定(Hammerら、J.Exp.Med.180:2353、1994)およびクラスIMHC安定化または会合の測定(例えば、Ljunggrenら、Nature346:476、1990;Schumacherら、Cell62:563、1990;Townsendら、Cell62:285、1990;Parkerら、J.Immunol.149:1896、1992)を使用するものを含めた他の検定系を用いて判定されてよい。
【0021】
「発現」という用語は、コーディング配列によってコードされる産物の生物学的産生を意味する。大部分の場合において、コーディング配列を含めたDNA配列は転写されてメッセンジャーRNA(mRNA)を形成する。メッセンジャーRNAは次に翻訳されて、関連する生物活性を有するポリペプチド産物を形成する。同様に、発現プロセスには、転写のRNA産物へのさらなるプロセッシングステップ、例えばイントロンを除去するためのスプライシング、および/またはポリペプチド産物の翻訳後プロセッシングが関与してよい。
【0022】
本明細書中で使用される「構築物」という用語は、一般に、天然に発生しない組成物を一般に表わす。構築物は合成技術、例えば組換え型DNAの調製および発現または核酸またはアミノ酸用の化学合成技術によって生成可能である。構築物は同様に、結果がその形態では天然に見出されないような形で1つの材料を別の材料と添加または親和させることによっても生成可能である。「マルチ−エピトープ構築物」という用語は、「ミニ遺伝子」または「マルチ−エピトープ核酸ワクチン」という用語と互換的に使用可能であり、例えば一部の実施形態ではMHCクラスIおよびT細胞受容体および/またはMHCクラスIIおよびT細胞受容体などの、免疫系内で機能する分子に結合し得る任意の長さのペプチドエピトープをコードする多数のエピトープ核酸を含む。マルチエピトープ構築物中のエピトープ核酸は、例えばクラスIIMHCエピトープまたはクラスIMHCエピトープとクラスIIMHCエピトープの組合せをコードできる。
【0023】
特定のアミノ酸配列に関して、「エピトープ」は、特定の免疫グロブリンによる認識に関与する一組のアミノ酸残基、またはT細胞に関連しては、T細胞受容体タンパク質および/または主要組織適合複合体(MHC)受容体による認識に必要な残基である。試験管内または生体内における免疫系の設定において、エピトープは、免疫グロブリン、T細胞受容体またはMHC分子により認識される部位を合わさって形成している、例えば一次、二次および三次ペプチド構造、および電荷などの分子の集合的特徴である。したがって、「エピトープ」という用語には、適切なMHC分子に結合しその後細胞傷害性T細胞応答またはヘルパーT細胞応答を誘発できる、あるいは代替的に、抗体を結合させその後、免疫原性ペプチドを誘導する抗原に対する抗体応答を誘発することのできる、本発明の免疫原性ペプチドが含まれるが、これらに限定されない。
【0024】
「フランキング残基」とは、エピトープの隣りに位置づけされている残基である。フランキング残基は、エピトープのN末端またはC末端に隣接する位置に導入または挿入され得る。
【0025】
「免疫原」および「抗原」という用語は互換的に使用され、細胞または体液性免疫応答の対象となるべきあらゆる化合物を意味する。さらに、本発明の抗原または免疫原性ペプチドは、線状であるすなわちポリペプチド中の隣接するアミノ酸で構成されていてよく、あるいは抗体−エピトープの場合には三次元または立体配座のもの、すなわち、機能的エピトープがポリペプチドの二次または三次構造に起因して一緒になって1つのエピトープを形成する非隣接アミノ酸で構成されているものであってよい。
【0026】
本明細書中で使用される「抗原決定基」という用語は、免疫応答を惹起する、促す、または生成するように誘発され得るあらゆる構造、例えば炭水化物エピトープ、脂質、タンパク質、ペプチドまたはそれらの組合せである。
【0027】
「抗体」という用語は、1つ以上のタンパク質および/または抗原を含む(ただしこれらに限定されない)組成物の中に含まれる特定されたタンパク質または抗原と相互作用しかつこれに結合することのできる1つまたは複数の免疫グロブリン遺伝子によって実質的にコードされるポリペプチドを意味する。
【0028】
「CTLエピトープ」という用語は、特定のMHCクラスI分子により認識され結合され、かつ特定のMHCクラスI分子と複合体化された場合にTリンパ球により認識されるペプチドを意味する。一部の実施形態においては、CTLエピトープは、長さがアミノ酸約8〜約13個、約9〜約11個または約9〜約10個であり得る。
【0029】
「HTLエピトープ」という用語は、特定のMHCクラスII分子により認識され結合され、かつ特定のMHCクラスII分子と複合体化された場合にTリンパ球により認識されるペプチドを意味する。一部の実施形態においては、HTLエピトープは、長さがアミノ酸約6〜約30個、約8〜約30個、約10〜約30個、約12〜約30個、約6〜約25個、約8〜約25個、約10〜約25個、約12〜約25個、約6〜約18個、約8〜約18個、約10〜約18個または約12〜約18個であり得る。
【0030】
Tリンパ球がMHC分子とCTLまたはHTLエピトープ間の複合体を「認識する」という記載は、本明細書、Tリンパ球上のT細胞受容体が複合体に結合し、その効果でTリンパ球が活性化されることを意味する。
【0031】
2つ以上のペプチド配列に関連して「同一の」または「同一性」百分率という用語は、配列比較アルゴリズムを用いてまたは手作業によるアラインメントおよび目視によって測定されるように比較ウインドウ全体にわたる最大限の一致について比較され整列させられた場合に、同じであるかまたは同じであるアミノ酸残基を規定の百分率で有する2つ以上の配列または下位配列を意味する。
【0032】
配列同一性および配列類似性百分率を決定するのに適したアルゴリズムの一例は、それぞれAltschulら、(1977)Nuc.Acids Res.25:3389−3402、およびAltschulら、(1990)J.Mol.Biol.215:403−410に記述されているBLASTおよびBLAST2.0アルゴリズムである。BLAST解析を実施するためのソフトウェアは、国立生物工学情報センター(http://www.ncbi.nlm.nih.gov/)を通して公開されている。このアルゴリズムには、データベース配列中の同じ長さのワードと整列させた場合に或る正値の閾値スコアTと整合するかまたはそれを満たす問い合わせ配列内の長さWの短かいワードを識別することによって、高スコアの配列対(HSP)をまず最初に識別するステップが関与する。Tは、近傍ワードスコア閾値と呼ばれる(Altschulら、上掲書)。これらの初期近傍ワードヒットは、それらを含むより長いHSPを発見するための検索を開始するためのシードとして作用する。ワードヒットは、累積アラインメントスコアを増大できるかぎり、各々の配列に沿って両方向に拡張される。累積スコアは、ヌクレオチド配列について、パラメータM(一対の整合する残基のための報酬スコア;つねに0超)およびN(不整合残基のためのペナルティスコア;つねに0未満)を用いて計算される。アミノ酸配列については、累積スコアを計算するのにスコアリングマトリクスが使用される。各方向でのワードヒットの拡張は、次の場合に中断される:累積アラインメントスコアが、その最大達成値からXという数量だけ下落した場合;累積スコアが、1つ以上の負のスコアの残基アラインメントの累積に起因してゼロ以下となった場合;またはいずれかの配列の終りに達した場合。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTNプログラム(ヌクレオチド配列用)は、デフォルトとして、11のワート長(W)、10という期待値、M=5、N=−4および両ストランドの比較を使用する。アミノ酸配列については、BLASTPプログラムはデフォルトとして、3というワード長、10という期待値(E)を使用し、BLOSUM62スコアリングマトリクス(HenikoffおよびHenikoff(1989)Proc.Natl.Acad.Sci.USA89:10915参照)は、50というアラインメント(B)、10という期待値(E)、M=5、N=−4そして両ストランドの比較を使用する。
【0033】
「単離(された)」または「生物学的に純粋な」という語句は、天然の状態で発見された場合に通常それに随伴する構成要素を実質的にまたは本質的に含んでいない材料を意味する。
【0034】
本明細書中で使用されている「リンカー」という用語は、2つの機能群(例えばpan−DR結合ペプチドと所望の免疫原)の間の共有結合およびスペーシングを提供するために用いられる任意の化合物のことである。一部の実施形態において、リンカーは、生理学的条件下で実質的に非電荷で線状または分岐側鎖を有していてよい脂肪族炭素鎖、アミノ酸またはアミノ酸ミメティックなどの中性分子を含む。一部の場合において、リンカーはそれ自体免疫原性であり得る。本発明において有用であるさまざまなリンカーについて以下でより詳細に記述する。さらに「連結する(link)」および「コンジュゲートする(conjugate)」という動詞は、本明細書において互換的に使用され、2つ以上の種の共有結合付着を意味する。
【0035】
「連結する」または「接合する」という用語は、組換え融合、共有結合、ジスルフィド結合、イオン結合、水素結合および静電結合を含めた(ただしこれらに限定されない)、ペプチドを機能的に結合するための当該技術分野において公知のあらゆる方法を意味する。
【0036】
「直接連結された」または「直接接合された」という用語は、何も介在することなく接合されていることを意味する。例えば、直接接合されている2つのペプチドの場合、1つのペプチドは、上述の通り別のペプチドに対して、2つのペプチドの間にいかなる配列、分子、スペーサー、リンカーなども介在することなく接合または結合されると考えられる。直接接合されたペプチドは、共通のアミノ酸を共有していてもいなくてもよい。すなわち2つのペプチドの配列は、重複しなくてもしてもよい。後者は例えば、本発明のPADRE類似体が挿入を用いて別の(ポリ)ペプチド内に導入される場合に該当し、PADRE類似体のアミノ酸配列の存在を保証するためには、挿入点にあるアミノ酸残基と共にPADRE類似体のための配列全体を提供するアミノ酸残基を挿入することしか必要ではない。
【0037】
「間接的に連結された」という用語は、何かが介在している状態で接合されていることを意味する。例えば、2つのペプチドが間接的に接合されている場合、1つのペプチドは上述の通り別のペプチドに対し、2つのペプチド間に配列、分子、スペーサー、リンカーなどが介在している状態で接合または結合されると考えられる。
【0038】
「主要組織適合複合体」または「MHC」は、生理学的免疫応答に関与する細胞相互作用の制御において役割を果たす遺伝子クラスタである。ヒトにおいては、MHC複合体はHLA複合体としても公知である。MHCおよびHLA複合体の詳細な説明については、例えばPaul、Fundamental Immunology、第3版、Raven Press、New York、1993を参照のこと。
【0039】
「医薬的に許容される」という語句は、一般に無毒性、不活性および/または生理学的に適合性ある組成物を意味する。
【発明を実施するための形態】
【0040】
発明者らは、pan−DR結合性をもつ新しい部類のペプチド、すなわちペプチドが数多くの異なるDR対立遺伝子によりコードされるMHCクラスII分子に結合して、ペプチドが広範囲の個体において免疫応答を刺激できるようにすることを発見した。さらに発明者らは、この新しいペプチドの多くが、著しく改善されたプロテアーゼ耐性を有し、こうしてより長い生体内半減期を可能にすることを発見した。意外にも、これらのペプチドの一部は、公知のpan−DRペプチドに比べて免疫応答を高める大幅に増大した能力を示す。
【0041】
本発明のポリペプチド(またはそのフラグメント)の生物活性は、適切なMHC分子を結合し、標的免疫原または免疫原ミメティックに対する免疫応答を誘発するのを任意に補助するTヘルパー応答を誘発する能力である。抗体応答を刺激するために有用なペプチドの場合、本発明のペプチドはTヘルパー応答を誘発し、このことが今度は標的免疫原に対する体液性応答の誘発を補助する。
【0042】
II. pan−DR結合ペプチド
本発明は、pan−DR結合ペプチドである新しい部類のペプチドを提供し、その使用を提供する。本発明のペプチドは、数多くの異なるDR対立遺伝子の2つ以上を結合でき、こうしてさまざまな抗原/免疫原に対する免疫応答を増大させるのに用いられる。本発明のペプチドはさらに、例えば米国特許第5,736,142号(US5,736,142)で開示されているような公知のpan−DR結合ペプチドと比べて増強した免疫応答を惹起するのに用いられる。
【0043】
本明細書中で実証される通り、本発明者らは、多数のHLA対立遺伝子によりコードされるMHC分子に結合する一連のペプチド配列を同定した。一部にはこのデータを考慮して、本発明者らは、以下の配列モチーフが、活性および免疫原性の改善されたpan−DR結合ペプチドを表わすものと判定した。一部の実施形態において、本発明のポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLX3AX4A(配列番号1)を含み、式中X1はW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択され;X3はHおよびKからなる群から選択され;X4はA、DおよびEからなる群から選択される。一部の実施形態において、ポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLHAAA(配列番号2)を含み、式中X1はY、H、I、E、N、QおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択される。本発明によると、オリゴペプチド配列はAKFVAAWTLKAAA(配列番号3)を含まない。
【0044】
一部の実施形態において、本発明のポリペプチドは、配列番号1、配列番号2、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号50、配列番号51、配列番号52、配列番号53、配列番号54、配列番号55、配列番号56、配列番号57、配列番号58、配列番号59、配列番号60、配列番号61、配列番号62、配列番号63、配列番号64、配列番号65、配列番号66、配列番号67、配列番号68、配列番号69、配列番号70、配列番号71または配列番号72から選択されたオリゴペプチドの13アミノ酸C末端フラグメントを構成する1つ以上のアミノ酸配列の1つ以上のコピーを含む。
【0045】
本発明のpan−DR結合ペプチドは、多数の異なるMHC対立遺伝子を結合することができ、「ヘルパーペプチド」とも呼ばれ得る。一部の実施形態において、pan−DR結合ペプチドは、2、3、4、5、6、7、8、9、10、11、12以上の異なるMHCクラスII対立遺伝子によりコードされるMHC分子を結合することができる。一部の実施形態において、誘発される応答は、増強された免疫応答である。一部の実施形態において、免疫応答は、本発明のpan−DR結合オリゴペプチドに向けられている。代替的に、または付加的に、MHC分子に対するpan−DR結合オリゴペプチドの結合は、例えば1つ以上のCTLペプチドを含むポリペプチドなどの第2の抗原に対する免疫応答および/またはタンパク質または非タンパク質抗原に対する体液性(すなわち抗体)応答をさらに増強する。
【0046】
本明細書中で記述されているpan−DRポリペプチド配列は単独で、またはより大きな融合タンパク質またはコンシュゲートの一部として使用可能である。こうして、本発明のポリペプチドは、本明細書中で記されているpan−DRペプチド配列に加えていずれかのまたは両方の末端に追加のアミノ酸を含み得る。例えば、タグまたはマーカーとして他のアミノ酸配列を含み入れるかまたは生体内または試験管内の安定性を促進するかまたそうでなければ当該技術分野において一般に理解される通りの追加のメリットを含み入れることが望ましい可能性がある。さらに、本発明のポリペプチドは、ポリペプチド内に1つ以上のpan−DRオリゴペプチド配列の1、2、3個以上のコピーを含むことができかつ/または本発明のポリペプチド内に多数の異なるpan−DRオリゴペプチド配列を含むことができる。その上、ポリペプチドは、例えばこのような配列またはこのような配列を含むポリペプチドに対する免疫応答を増強することが必要である場合に、少なくとも1つの追加のHTLオリゴペプチドおよび/またはCTLオリゴペプチドおよび/または抗体誘発性ポリペプチドを含むことができる。このような実施形態において、本明細書中に記述されているpan−DR配列は、キーホールリンペットヘモシアニン、破傷風トキソイドおよびジフテリアトキソイドなどの従来の担体タンパク質に対する代替物として、または先行技術の万能T−ヘルパーエピトープの代替物として機能する。
【0047】
本発明のポリペプチドは任意の長さのものであり得る。一部の実施形態において、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列および任意には他のアミノ酸配列を含むポリペプチドは、例えばアミノ酸1000、900、800、700、600、500、400、300、200、100、80、60、50、40、30、20または15個以下の長さを有するが、多くの場合、1つまたは複数のpan−DR結合オリゴペプチドを含むポリペプチドは、(ポリペプチドが未変性ポリペプチド配列の大部分を含むように)本発明のpan−DR結合オリゴペプチドの1つまたは数個を導入することによって修飾されている未変性ポリペプチドとほぼ同じ長さを有する。一方、本発明のpan−DR結合オリゴペプチドを含む本発明の一部のポリペプチドは、「マルチエピトープ構築物」、すなわち1つ以上の抗原から誘導された複数のエピトープを含み、構築物の抗原性または免疫原性を最適化するためにエピトープが非天然発生手順で組織されていないポリペプチドである。したがって、本発明のポリヌクレオチドの開示の下で以下に論述するマルチエピトープ発現産物も同様に、本発明のペプチドの実施形態である。
【0048】
このようなマルチエピトープ構築物において、複数のエピトープは典型的に、複数のCTLエピトープ、複数のB細胞エピトープ、複数のTヘルパーリンパ球エピトープ、複数のCTLおよびB細胞エピトープ、複数のCTLおよびTヘルパーリンパ球エピトープ、複数のB細胞およびTヘルパーリンパ球エピトープ、ならびに複数のB細胞、CTLおよびTヘルパーリンパ球エピトープから選択される。これらのエピトープは、単一の抗原タンパク質から誘導されてよいが、少なくとも2つの異なるポリペプチド抗原から誘導されてもよく、同様にこれらは同じまたは異なる種(例えば細菌、ウイルスおよび寄生虫の種)から誘導されてよい。
【0049】
一部の実施形態では、pan−DR結合オリゴペプチド配列は、末端NH2アシル化、例えばアルカノイル(C1−C20)またはチオグリコリルアセチル化、末端カルボキシアミド化、例えばアンモニア、メチルアミンなどによって修飾されていることにより、原初の配列と異なっている可能性がある。一部の実施形態において、これらの修飾は、支持体またはその他の分子に対する連結のための部位を提供してよい。
【0050】
以上で同定されたペプチドの生物活性は、さまざまなシステム内で検定されてよい。典型的には、抗原特異的T細胞活性を阻害する能力が試験される。1つの例示的プロトコルにおいては、余剰のペプチドが、公知のMHC発現(例えばDR1)を有する抗原提示細胞および公知の抗原特異性(例えば破傷風毒素830−843)およびMHC制限(ここでもDR1)を有するT細胞クローン、そして免疫原性ペプチド自体(例えば破傷風毒素830−843)と共にインキュベートされる。検定培養は、T細胞増殖のために充分な時間、例えば4日間インキュベートされ、その後、最後の18時間のインキューベーションの間[3H]−チミジンでパルス処理することなどの標準的な手順を用いて増殖が測定される。ペプチドが与えられなかった対照と比べた阻害百分率が、次に計算される。
【0051】
試験管内検定における抗原提示を阻害するペプチドの能力は、生体内での免疫応答を阻害するペプチドの能力と相関されてきた。生体内活性は、例えば免疫調節ペプチドそしてペプチドにより認識される特定のMHC分子に制限されるものとして公知である免疫原を投与することなどによって、動物モデルにおいて判定されてよい。その後、動物からTリンパ球が取り出され、一用量範囲の免疫原と共に培養される。従来の手段、例えば[3H]−チミジンでのパルス処理および適切な対照に対する比較により、刺激の阻害が測定される。当然のことながら、或る種の実験上の詳細は、当業者には明白である。同様に、参照により本明細書に援用されているAdoriniら、Nature334:623−625(1988)も参照のこと。
【0052】
定義されたMHC分子、特にMHCクラスII分子を伴う多数の細胞が公知であり、例えばthe American Type Culture Collection(例えば、「Catalogue of Cell Lines and Hybridomas」、第6版(1988)を参照)Rockville、Maryland、U.S.A.から容易に入手可能である。
【0053】
本発明のオリゴペプチドの一部の実施形態は、NおよびC末端残基に対する修飾を含む。当業者であれば充分理解できるように、NおよびC末端を修飾して、ペプチドの物理的または化学的特性を改変し、例えば結合、安定性、生物学的利用能、連結の容易さなどに影響を与えてもよい。
【0054】
任意には、例えばNまたはC末端におけるさまざまなアミノ酸ミメティックまたはD−アミノ酸でのペプチドの修飾が使用可能であり、例えば生体内でのペプチドの安定性を増大させる上で有用である。このようなペプチドは、「inverso」型または「retroinverso」型として、すなわち一配列のLアミノ酸をDアミノ酸で置換することによってかまたはアミノ酸の配列を逆転させDアミノ酸でL−アミノ酸を置換することによって合成されてよい。Dペプチドは実質的にペプチダーゼに対する耐性がより高く、したがってそのL−ペプチド対応物と比べて血清および組織中でより安定していることから、生理学的条件下でのDペプチドの安定性は、対応するLペプチドと比べた親和性の差異を補償する以上のものであるかもしれない。さらに、置換を伴うまたは伴わないLアミノ酸含有ペプチドをDアミノ酸でキャッピングして、免疫原性ペプチドのエクソペプチダーゼ破壊を阻害することができる。
【0055】
ポリペプチドの生体内安定性は、数多くの方法で検定できる。例えば、ペプチダーゼおよびさまざまな生体媒質例えばヒトの血漿および血清を使用して、安定性が試験されてきた。例えば、全て参照により本明細書に援用されているVerhoefら、Eur.J.Drug Metab.Pharmacokin.11:291−302(1986);Walterら、Proc.Soc.Exp.Biol.Med.148:98−103(1975);Witterら、Neuroendocrinology30:377−381(1980);Verhoefら、J.Endocrinology110:557−562(1986);Handaら、Eur.J.Pharmacol.70:531−540(1981);Bizzozeroら、Eur.J.Biochem.122:251−258(1982);Chang、Eur.J.Biochem.151:217−224(1985)を参照のこと。
【0056】
生物活性にとって不可欠である一部のアミノ酸残基、例えば重要な接触部位にあるアミノ酸残基は一般に生物活性に対する不利な効果無く改変され得ないことは容易に認識されることから、本発明のペプチドまたは類似体は、一部の残基の順序または組成を改変させることによって修飾可能である。重要でないアミノ酸は、L−α−アミノ酸またはそのD−異性体などのタンパク質内で自然に発生するアミノ酸に限定される必要はなく、非タンパク質アミノ酸例えばβ−γ−δアミノ酸、ならびにL−α−アミノ酸の数多くの誘導体も含んでいてよい。本発明のオリゴペプチドはLアミノ酸かDアミノ酸のいずれかを含み得るが、通常、コア結合領域内部にDアミノ酸を含むことはできない。
【0057】
本発明のペプチドは、多様な方法で調製可能である。一部の実施形態では、組換えDNA技術が利用され、この場合本発明の免疫原性ポリペプチドをコードするヌクレオチド配列が発現ベクター内に挿入され、適切な宿主細胞へと形質転換またはトランスフェクトされ、発現に適した条件下で培養される。これらの手順は、Sambrookら、Molecular Cloning、A Laboratory Manual、Cold Spring Harbor Press、Cold Spring Harbor、New York(1989)中に一般的に記載されている通り、当該技術分野において一般に公知である。あるいは、そのサイズに応じて、従来の技術にしたがって、溶液中かまたは固体支持体上でポリペプチドを合成することができる。さまざまな自動合成装置が市販されており、公知のプロトコルにしたがって使用可能である。例えばStewartおよびYoung、Solid Phase Peptide Synthesis、第2版、Pierce Chemical Co.(1984)、上掲書。
を参照のこと。
【0058】
III. ポリヌクレオチド
本発明は、本明細書中で記述されている通りの本発明のポリペプチドをコードするポリヌクレオチドを提供する。例えば、ポリペプチドをコードするポリヌクレオチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLX3AX4A(配列番号1)を含み、式中X1はW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択され;X3はHおよびKからなる群から選択され;X4はA、DおよびEから選択される。一部の実施形態において、ポリペプチドは、少なくとも2、3、4、5、6、7、8、9、10、11、12以上の異なるDR対立遺伝子によりコードされるMHC分子を結合できるオリゴペプチド配列を含み、ここでオリゴペプチド配列はAX1FVAAX2TLHAAA(配列番号2)を含み、式中X1はY、H、I、E、N、QおよびKからなる群から選択され;X2はF、N、YおよびWからなる群から選択される。本発明によると、オリゴペプチド配列はAKFVAAWTLKAAA(配列番号3)を含まない。一部の実施形態において、本発明のポリヌクレオチドは、配列番号1、配列番号2、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8、配列番号9、配列番号10、配列番号11、配列番号12、配列番号13、配列番号14、配列番号15、配列番号16、配列番号17、配列番号18、配列番号19、配列番号20、配列番号21、配列番号22、配列番号23、配列番号24、配列番号25、配列番号26、配列番号27、配列番号28、配列番号29、配列番号30、配列番号31、配列番号32、配列番号33、配列番号34、配列番号35、配列番号36、配列番号37、配列番号38、配列番号39、配列番号40、配列番号41、配列番号42、配列番号43、配列番号44、配列番号45、配列番号46、配列番号47、配列番号48、配列番号49、配列番号50、配列番号51、配列番号52、配列番号53、配列番号54、配列番号55、配列番号56、配列番号57、配列番号58、配列番号59、配列番号60、配列番号61、配列番号62、配列番号63、配列番号64、配列番号65、配列番号66、配列番号67、配列番号68、配列番号69、配列番号70、配列番号71または配列番号72から選択されたオリゴペプチドの13アミノ酸C末端フラグメントを各々構成する1つ以上のアミノ酸配列をコードし、ポリヌクレオチドは、他のアミノ酸配列と融合して配列または切形を含むポリペプチドをコードしてよい。したがって、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列を含めた2つ以上のCTLおよび/またはHTLオリゴペプチド配列を含む、本明細書中で開示されている通りのポリペプチドをコードする本発明のポリヌクレオチドが提供される。
【0059】
本発明のポリヌクレオチド構築物は、上述のペプチドまたはポリペプチドのいずれでもコードすることができる。例えば多数のクラスIおよび/またはクラスIIMHCエピトープおよび/またはB細胞エピトープ、例えば2つ以上のクラスIIMHCエピトープ(すなわち本明細書中に記述されている少なくとも1つのpan−DR結合オリゴペプチド、例えば配列番号1、2および4〜72)またはクラスIMHCエピトープとクラスIIMHCエピトープおよびB細胞エピトープの組合せなど。クラスIMHCコーディングエピトープ核酸は、「CTLエピトープ核酸」とも呼ばれ、クラスIIMHCコーディングエピトープ核酸は「HTLエピトープ核酸」とも呼ばれる。「B細胞エピトープ」は、抗体またはB細胞受容体を結合させる線状または立体構造エピトープである。一部のマルチエピトープ構築物は、クラスIMHCエピトープおよび/またはB細胞エピトープをコードするその配列の一部分と、クラスIIMHCエピトープをコードする別の部分とを有することができる。一部の実施形態において、CTLエピトープ核酸は、長さがアミノ酸約8〜約15個、例えばアミノ酸約8〜約11個、例えばアミノ酸約9個のエピトープペプチドをコードする。HTLエピトープ核酸は、本明細書中に記述されている通り少なくとも1つのpan−DRオリゴペプチドをコードするが、2つ以上のHTLエピトープが含まれる場合、他のHTLエピトープも同様に使用可能である。一部の実施形態において、ポリヌクレオチド構築物は、例えば5個以上、10個以上、15個以上、20個以上または25個以上のエピトープ核酸を含む。マルチエピトープポリヌクレオチド構築物中のCTLエピトープ核酸は全て、1つの生体に由来するものであり得(例えば全てのエピトープ核酸のヌクレオチド配列はHIV株内に存在するかもしれない)、またはマルチエピトープ構築物は2つ以上の異なる生体内に存在するエピトープ核酸を含み得る(例えば一部のエピトープがHIV由来で一部がHCV由来)。以下で記述する通り、マルチエピトープ構築物内の1つ以上のエピトープ核酸は、スペーサー核酸によりフランキングされていてよい。
【0060】
この先で理解されるように、本発明のPADRE類似体に連結されているかまたはその他の形でこの類似体を含むポリペプチドを含むB細胞エピトープをコードする核酸構築物が、本発明の一実施形態である。このような核酸構築物は典型的に、発現産物中に存在するPADRE類似体がポリペプチド由来のB細胞エピトープの立体構造とマイナスに干渉しない場合に融合構築物をコードする。B細胞エピトープが線状である場合、B細胞エピトープは単純にPADRE類似体に融合されてよく、したがってコーディング核酸の提供は比較的簡単である。しかしながら、B細胞エピトープが立体構造エピトープである場合、PADRE類似体コーディング核酸の導入点の典型的な例は、ポリペプチドのフレキシブルループ内かまたは、フレキシブル末端内(例えばタンパク質ドメイン全体かさらにはポリペプチド全体を使用することによる立体構造エピトープの3D構造の保存が求められる場合)であるが、例えば問題のポリペプチドの細胞内に閉じ込められた部分をコードする領域は生体内での免疫応答と関連性をもたないことから、これらの領域内にPADRE類似体コーディング核酸を導入することも同様に可能である。このような核酸構築物の発現産物は、導入されたPADRE類似体配列が発現産物に対する免疫応答の惹起において増大したTリンパ球ヘルプを提供することになるため、抗体誘発用免疫原として有用である。こうして、発現産物は免疫原として直接使用されてよく(例えば動物における抗体産生のために使用される免疫原性組成物またはワクチン内の抗体誘発用)、あるいは、コーディング核酸での核酸免疫付与に付された動物において生体内で発現された後、その効果を及ぼしてもよい。
【0061】
「スペーサー」とは、接合エピトープの発生を防止するためおよび/またはプロセッシングの効率を増大させるためマルチエピトープ構築物内の2つのエピトープの間に挿入される配列を意味する。マルチエピトープ構築物は、1つ以上のスペーサー核酸を有する場合がある。スペーサー核酸は構築物内で各エピトープ核酸にフランキングしてよく、あるいはスペーサー核酸対エピトープ核酸の比率は約2対10、約5対10、約7対10、約8対10、または約9対10であってよく、ここで約8対10という比率が一部の構築物にとって有利な結果を生み出すものと判定されている。
【0062】
スペーサー核酸は1つ以上のアミノ酸をコードしてよい。一部の実施形態において、マルチエピトープ構築物内のクラスIMHCエピトープにフランキングするスペーサー核酸は、長さがアミノ酸1〜約8個、アミノ酸2〜8個、アミノ酸3〜8個、アミノ酸4〜8個、アミノ酸5〜8個、アミノ酸6〜8個、またはアミノ酸7〜8個である。マルチエピトープ構築物内でクラスIIMHCエピトープにフランキングするスペーサー核酸は、一部の実施形態において長さがアミノ酸5、6、7個以上、そして一部の実施形態においてアミノ酸5個または6個超である。
【0063】
1構築物内のスペーサーの数、スペーサポリヌクレオチド内でコードされるアミノ酸の数およびスペーサーのアミノ酸組成を、エピトープのプロセッシングを最適化しかつ/または接合エピトープを最小限に抑えるように選択することができる。一部の実施形態では、スペーサーは、エピトーププロセッシングおよび接合モチーフを同時に最適化することにより選択される。本明細書では、エピトーププロセッシングを最適化するための適切なアミノ酸について記述されている。同様に、構築物内の接合エピトープ数を最小限にするための適切なアミノ酸スペーシングが、本明細書中に、クラスIおよびクラスIIHLAについて記述されている。例えば、クラスIIMHCエピトープにフランキングするスペーサーは、一部の実施形態において、一般にG、Pおよび/またはN残基が一次アンカー残基であるものとして公知でないことからこれらを含むことができる(例えばPCT米国特許第00/19774号(PCT/US00/19774)を参照のこと)。一部の実施形態において、クラスIMHCエピトープにフランキングするためのスペーサーは、交互のGおよびP残基例えば(GP)n、(PG)n、(GP)nG、(PG)nPなどを含み、ここでnは1〜10、2〜約2の間の整数であり、一部の実施形態において、このようなスペーサーの具体例はGPGPGまたはPGPGPである。クラスIMHCエピトープの一部の実施形態において、スペーサーは、任意にはK、NまたはGが先行する1、2、3個以上の連続するアラニン(A)残基を含む。
【0064】
一部のマルチエピトープ構築物において、各スペーサー核酸が同じアミノ酸配列をコードすることで充分である。同じアミノ酸配列をコードする2つのスペーサー核酸を有するマルチエピトープ構築物において、これらのスペーサーをコードするスペーサー核酸は同じまたは異なるヌクレオチド配列を有していてよく、ここで、細胞内にマルチエピトープ構築物が挿入された場合、異なるヌクレオチド配列が、意図されない組換え事象の発生確率を減少させるかもしれない。
【0065】
他のマルチエピトープ構築物においては、スペーサー核酸の1つ以上が異なるアミノ酸配列をコードしてよい。スペーサー核酸の多くがマルチエピトープ構築物内の同じアミノ酸配列をコードしてよいが、1、2、3、4、5個以上のスペーサー核酸が異なるアミノ酸配列をコードする場合があり、マルチエピトープ構築物内のスペーサー核酸の全てが異なるアミノ酸配列をコードすることが可能である。スペーサー核酸は、本明細書中で記述されている通り、スペーサー配列がエピトーププロセッシングを最大限にするかおよび/または接合エピトープを最小限にするかを判定することによって、自らフランキングするエピトープ核酸に関して最適化されてよい。
【0066】
マルチエピトープ構築物は、1つの構築物内のスペーサーが別の構築物に比べてエピトーププロセッシングを最適化するかまたは接合エピトープを最小限に抑えるかによって、互いに区別されてよく、一部の実施形態においては、1つの構築物がもう1つの構築物に比べてエピトーププロセッシングおよび接合エピトープについて同時に最適化されている場合、構築物を区別してよい。1つの構築物がエピトーププロセッシングおよび接合モチーフについて最適化されているか否かを決定するためのコンピュータ援用方法および試験管内および生体内実験室方法が、本明細書において記述されている。
【0067】
一部の実施形態において、本発明のポリヌクレオチド構築物は、本発明の少なくとも1つのpan−DR結合オリゴペプチド配列そして任意にはポリペプチドに関連して本明細書中で記述された配列のいずれかまたは全てをコードする核酸を含む発現ベクターとして提供されている。このような発現ベクターの構成は、例えばその開示が参照により本明細書に援用されているPCT米国特許第99/10646号(PCT/US99/10646)の中に記載されている。発現ベクターは、抗原が発現され適切なMHC分子に標的化されるように生体の適切な細胞内で核酸をコードする転写単位を発現することのできる少なくとも1つのプロモータ要素を含んでいる。例えば、ヒトへの投与のためには、ヒト細胞内で機能するプロモータ要素が発現ベクター内に取込まれる。
【0068】
本発明において使用される一般的方法を開示する基本的教書には、Sambrookら、Molecular Cloning、A Laboratory Manual(第2版、1989);Kriegler、Gene Transfer and Expression:A Laboratory Manual(1990);および Current Protocols in Molecular Biology(Ausubelら、編、1994);Oligonucleotide Synthesis.A Practical Approach(Gait編、1984);Kuijpers、Nucleic Acids Research 18(17):5197(1994);Dueholm、J.Org.Chem.59:5767−5773(1994);Methods in Molecular Biology、第20巻(Agrawal、編);およびTijssen、Laboratory Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Acid Probes、例えば、第I部、第2章、「Overview of principles of hybridization and the strategy of nucleic acid probe assays」(1993))が含まれる。
【0069】
関連するオリゴペプチド配列(例えばエピトープ)をコードする核酸は、標準的な技術にしたがって構築物の中に組立てることができる。一部の実施形態において、pan−DR結合オリゴペプチドそして任意にはマルチエピトープポリペプチドをコードする核酸配列は、オリゴヌクレオチドプライマを用いる増幅技術を用いて単離されるかまたは化学的に合成される。該当する場合には、組換えクローニング技術も使用可能である。所望のエピトープを増幅する(構築物を組立てるのにPCRを使用する場合)またはコードする(構築物を組立てるのに合成オリゴヌクレオチドを使用する場合)オリゴヌクレオチド配列が選択される。
【0070】
典型的にはプライマを使用する増幅技術を用いて、DNAまたはRNAからの選択されたエピトープをコードする配列を増幅し単離する。(米国特許第4,683,195号(U.S.4,683,195)および4,683,202号(4,683,202);PCR Protocols:A Guide to Methods and Applications(Innisら、編、1990))を参照のこと)。ポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR)などの方法を使用して、mRNAから、cDNAから、ゲノムライブラリからまたはcDNAライブラリから直接エピトープ核酸配列を増幅することができる。プライマ内に制限エンドヌクレアーゼ部位を取込むことが可能である。PCR反応により増幅されたマルチエピトープ構築物をアガロースゲルから精製し、適切なベクター内にクローニングすることが可能である。
【0071】
本発明のポリヌクレオチドを構成するために、合成オリゴヌクレオチドも使用することができる。一部の実施形態において、この方法は、遺伝子のセンスおよびナンセンスストランドを表わす一連の重複するオリゴヌクレオチドを用いて実施される。これらのDNAフラグメントは次にアニールされ、ライゲートされ、クローニングされる。市販されていないオリゴヌクレオチドは、Van Devanterら、Nucleic Acids Res.、72:6159−6168(1984)中に記載されている通りの自動合成装置を用いてBeaucageおよびCaruthers、Tetrahedron Letts.22:1859−1862(1981)が最初に記述した固相ホスホラミジットトリエステル法にしたがって化学的に合成可能である。オリゴヌクレオチドの精製は、PearsonおよびReanier、J.Chrom.255:137−149(1983)内で記述されているようなアニオン交換HPLCまたは未変成アクリルアミドゲル電気泳動法による。
【0072】
一部の実施形態において、本発明のポリヌクレオチド構築物のエピトープは、転写を導くための強力なプロモータならびにエンハンサやポリアデニル化部位などの他の調節配列を含む発現ベクターの形にサブクローニングされる。適切なプロモータは当該技術分野において周知であり、例えばSambrookら、およびAusubelら、の中に記載されている。哺乳動物細胞のための真核生物発現系は当該技術分野において周知であり、市販されている。このようなプロモータ要素には例えば、サイトメガウイルス(CMV)、ラウス肉腫ウイルスLTRおよびSV40が含まれる。
【0073】
発現ベクターは典型的には、宿主細胞内でのポリヌクレオチド構築物の発現に必要とされる追加の要素を全て含む転写単位または発現カセットを含む。例えば、発現カセットは写しの効率的ポリアデニル化のために必要なシグナルおよびマルチエピトープ構築物に作動的に連結されたプロモータを含み得る。カセットの追加の要素は、機能的スプライスドナーおよびアクセプタ部位を伴うイントロンおよびエンハンサを含んでいてよい。
【0074】
プロモータ配列に加えて、発現カセットは、効率的終結を提供するために構造遺伝子の下流側に転写終結領域も含むことができる。終結領域は、プロモータ配列と同じ遺伝子から得られてもよいし、あるいは異なる遺伝子から得られてもよい。
【0075】
本発明のペプチドを発現するために使用される発現ベクター内に選択可能なマーカーを取込むことができる。これらの遺伝子は、選択培地中で成長した形質転換済み宿主細胞の存続または成長に必要な、タンパク質などの遺伝子産物をコードすることができる。選択遺伝子を含む遺伝子で形質転換されていない宿主細胞は培地中で存続しない。典型的な選択遺伝子は、抗生物質またはその他の毒素、例えばアンピシリン、ネオマイシン、カナマイシン、クロラムフェニコールまたはテトラサイクリンに対する耐性を付与するタンパク質をコードする。代替的には、選択可能なマーカーは、栄養要求性欠乏症を補足するかまたは複合培地からは入手できない重要な栄養素を供給するタンパク質をコードしてよい。一部の実施形態において、ベクターは、例えばE.coli(大腸菌)内、または宿主細胞内への導入に先立ちベクターがその中で複製されるその他の細胞内で機能的である1つの選択可能なマーカーを有するものである。当業者にとっては、数多くの選択可能なマーカーが公知である。
【0076】
細胞内に遺伝情報を輸送するために、さまざまな発現ベクターを使用することができる。真核細胞内での発現のために使用される従来のベクターのうち任意のものを使用してよい。真核ウイルス由来の調節要素を含む発現ベクターが、典型的に真核細胞発現ベクター、例えばSV40ベクター、CMVベクター、乳頭腫ウイルスベクターおよびエプスタイン・バー・ウイルス由来のベクター内で使用される。
【0077】
本発明のポリヌクレオチド構築物は、プラスミドベクターならびにウイルスまたは細菌ベクターを含めたさまざまなベクターから発現可能である。ウイルス発現ベクターの例としては、弱毒化ウイルス宿主例えば牛痘または鶏痘が含まれる。このアブローチの例として、本発明の(ポリ)ペプチドをコードするヌクレオチド配列を発現するためのベクターとしてワクチニアウイルスが使用される。腫瘍を担持する宿主内への導入時点で、組換えワクチニアウイルスは免疫原性ペプチドを発現し、こうして宿主CTLおよび/またはHTL応答を惹起する。ワクチニアベクターおよび免疫付与プロトコルにおいて有用な方法は、例えば米国特許第4,722,848号(US4,722,848)の中で記述されている。
【0078】
本発明のポリヌクレオチドは、E.coli、その他の細菌宿主、酵母および昆虫細胞を含めたさまざまな宿主細胞中で発現されて、本明細書中で記述されているタンパク質発現産物を提供することができる。宿主細胞は、例えば酵母細胞、細菌細胞または糸状菌細胞などの微生物である。適切な宿主細胞の例としては、他の数多くのもののうち例えばシュードモナス属、エシュリキア属(例えばE.coli)、バチラス属が含まれる。適切な酵母細胞は、例えばサッカロミセス属(例えばS.cerevisiae)およびカンジタ属を含めた複数の属のうちのいずれかのものであり得る。適切な真核細胞は、ピチア属、アスペルギルス属そして組換え産生に適したその他の真菌から選択される。適切な昆虫細胞は、例えばS2、Sf9およびHi−5細胞を含めた複数の亜種の細胞であり得る。
【0079】
当業者にとっては、例えばアデノおよびアデノ関連ウイルスベクター、レトロウイルスベクター、非ウイルスベクター例えばBCG(Bacillus Calmette Guerin)、チフス菌ベクター、無毒化炭疽菌毒素ベクターなどの、治療的投与または免疫付与に有用な多様な他のベクターが明白である。したがって、本発明は同様に、必要としている個体に対してこのようなベクターを投与することによる、このようなベクターの予防的または治療的使用にも関する。
【0080】
マルチエピトープ構築物の免疫原性および抗原性は、本明細書中で記述されている通りに評価される。
【0081】
以上で記したように、本発明は同様に、本明細書中で記述されている通りの本発明のポリヌクレオチドのいずれかによりコードされる単離ポリペプチドにも関し、したがって、これらのポリペプチドも同様に本発明の「ペプチド」またはポリペプチドである。
【0082】
IV. さらなる抗原
以上で説明したように、本発明のpan−DRオリゴペプチドは、さまざまな抗原に対する細胞または体液性免疫応答を増強するために使用することができる。このような抗原に由来する抗原決定基または抗原自体は、本発明のpan−DR結合ペプチドと混和した形、またはそれに連結させた形で投与可能である。例えば、免疫応答を惹起するまたは増強するために、抗原決定基を本発明のヘルパーペプチドと連結または混合するか、またはそれと共に連続して投与することができる。本質的に、本発明のpan−DR結合オリゴペプチド配列と組合せた形でいかなる抗原でも使用することができる(例えば多糖類、タンパク質、糖タンパク質、脂質、糖脂質、リポ多糖類など)。タンパク質抗原は、細菌、真菌、ウイルス、原生動物、蠕虫および他の寄生虫などの感染性因子から誘導されてよいが、或る種の疾病状態において過剰発現されているかまたはその他の形で不適当である抗原あるいは癌抗原などの疾病関連抗原であってもよい。国際公開第00/20027号(WO00/20027)に列挙されている癌抗原は全て、関連性あるタンパク質抗原であり、さらなる抗原は炎症に付随する抗原である(ここではTNFがその好例である)。本発明は、全てが本発明の一部を成している数多くのTNF構築物を開示しており、一般にこれらの構築物は全て、それらをコードする核酸フラグメント、核酸フラグメントを含むベクターおよび核酸フラグメントまたはベクターを含む宿主細胞(またはベクターで形質転換されている宿主細胞)と同様、本発明の一部を成すものである。
【0083】
一部の実施形態において、本発明のオリゴペプチドは、単独で投与される。一部の実施形態において、本発明のオリゴペプチドは、第2の抗原決定基と併用して投与される。一部の実施形態において、本発明のオリゴペプチドは、抗原決定基と混和される。一部の実施形態において、本発明のペプチドは、抗原決定基に連結される。例えば、タンパク質抗原を、(ペプチド結合を介して)融合タンパク質としてpan−DR結合オリゴペプチドに対して間接的に、または直接的に連結させることができる。非タンパク質抗原を含めた抗原を、他の共有結合コンジュゲーション方法を介して、pan−DR結合オリゴペプチドまたはオリゴペプチド配列を含むポリペプチドに対し連結させることができる。
【0084】
一部の実施形態において、本発明のペプチドと共に投与される抗原決定基はタンパク質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は多糖類である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は糖タンパク質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は脂質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基は糖脂質である。一部の実施形態において、本発明のペプチドと共に投与される抗原決定基はリポ多糖類である。
【0085】
炭水化物エピトープは炭水化物構造を含むが、コンジュゲート例えば糖タンパク質、糖ペプチド、糖脂質など、DNA、RNA、または多糖類オリゴ糖または単糖として存在することができ、これらに対する免疫応答が所望される。炭水化物エピトープは、広範囲の免疫応答を誘発してよい。当業者であれば、本明細書中で例示されているさまざまな炭水化物構造を、抗原性に不利な影響を及ぼすことなく標準的な方法にしたがってさまざまな形で修飾することができる、ということを認識するものである。例えば、サッカリドの単糖単位は、さまざまな形で置換されるかまたさらには小有機分子と置換されてよく、これは単糖類のためのミメティックとして役立つ。
【0086】
適切な抗原の例としては、炭水化物ベースのワクチンの中で使用可能である細菌表面多糖類から誘導されたものが含まれる。細菌は典型的には、糖タンパク質、糖脂質、リポ多糖類の0特異的側鎖、莢膜多糖類などの一部としてその細胞表面上で炭水化物を発現する。例示的細菌菌株としては、肺炎連鎖球菌(例えば国際公開第2005/120563号(WO/2005/120563)およびその中の炭水化物エピトープを参照のこと)、ナイセリア・メニンギチジス(Neisseria meningitidis)、ヘモフィラス・インフルエンザ(Haemophilus influenza)、クレブシエラ菌株(Klebsiella spp.)、シュードモナス菌株(Pseudomonas spp.)、サルモネラ菌株(Salmonella spp.)、シゲラ菌株(Shigella spp.)およびグループBストレプトコッキィ(streptococci)が含まれる。
【0087】
数多くの適切な細菌炭水化物エピトープが、先行技術(例えばSandersら、Pediatr.Res.37:812−819(1995);Bartoloniら、Vaccine13:463−470(1995);Pirofski、ら、Infect.Immun.63:2906−2911(1995)および国際公開第WO93/21948号(WO93/21948)))中で記述されており、例えば米国特許第6,413,935号(US6,413,935)に記載の通りである。
【0088】
一般に、以下の国際特許出願のいずれか1つにおいて開示されているHLA結合エピトープは、本発明によると有用であるかもしれず、したがってそれらの内容は参照により本明細書に援用される;国際公開第93/03764号(WO93/03764)、国際公開第95/22317号(WO95/22317)、国際公開第94/03205号(WO94/03205)、国際公開第95/19783号(WO95/19783)、国際公開第97/34617号(WO97/34617)、国際公開第02/20053号(WO02/20053)、国際公開第94/20127号(WO94/20127)、国際公開第97/34621号(WO97/34621)、国際公開第02/20616号(WO02/20616)、国際公開第95/07707号(WO95/07707)、国際公開第95/04817号(WO95/04817)、国際公開第98/33888号(WO98/33888)、国際公開第94/26774号(WO94/26774)、国際公開第96/03140号(WO96/03140)、国際公開第02/20035号(WO02/20035)、国際公開第96/40213号(WO96/40213)、国際公開第97/26784号(WO97/26784)、国際公開第97/33602号(WO97/33602)、国際公開第98/32456号(WO98/32456)、国際公開第99/61916号(WO99/61916)、国際公開第99/45954号(WO99/45954)、国際公開第99/65522号(WO99/65522)、国際公開第01/62776号(WO01/62776)、国際公開第01/00225号(WO01/00225)、国際公開第2004/031211号(WO2004/031211)、国際公開第99/58658号(WO99/58658)、国際公開第2005/012502号(WO2005/012502)、国際公開第00/44775号(WO00/44775)、国際公開第02/019986号(WO02/019986)、国際公開第2004/031210号(WO2004/031210)、国際公開第01/21189号(WO01/21189)、国際公開第01/24810号(WO01/24810)、国際公開第2005/033265号(WO2005/033265)、国際公開第01/42270号(WO01/42270)、国際公開第01/41788号(WO01/41788)、国際公開第01/42267号(WO01/42267)、国際公開第01/45728号(WO01/45728)、国際公開第01/41787号(WO01/41787)、国際公開第02/061435号(WO02/061435)、国際公開第02/061435号(WO02/061435)、国際公開第01/41741号(WO01/41741)、国際公開第2004/052917号(WO2004/052917)、国際公開第01/36452号(WO01/36452)、国際公開第03/087126号(WO03/087126)、国際公開第01/47541号(WO01/47541)、国際公開第02/083714号(WO02/083714)、国際公開第01/41799号(WO01/41799)、国際公開第2005/089164号(WO2005/089164)、国際公開第2003/040165号(WO)、国際公開第2005/120563号(WO2005/120563)、国際公開第2004/094454号(WO2004/094454)、国際公開第2004/053086号(WO2004/053086)、国際公開第2004/089973号(WO2004/089973)、国際公開第2008/054540号(WO2008/054540)、国際公開第2008/039267号(WO2008/039267)、国際公開第99/19478号(WO99/19478)、国際公開第92/21033号(WO92/21033)、国際公開第94/11738号(WO94/11738)、国際公開第93/22338号(WO93/22338)、国際公開第95/25530号(WO95/25530)、国際公開第95/25739号(WO95/25739)、国際公開第2005/118626号(WO2005/118626)、国際公開第93/03753号(WO93/03753)、国際公開第94/19011号(WO94/19011)、国際公開第95/03777号(WO95/03777)、国際公開第95/19783号(WO95/19783)、国際公開第95/04542号(WO95/04542)、国際公開第01/42270号(WO01/42270)、国際公開第01/41788号(WO01/41788)、国際公開第01/42267号(WO01/42267)、国際公開第01/45728号(WO01/45728)、国際公開第01/41787号(WO01/41787)、国際公開第01/41741号(WO01/41741)、国際公開第2004/052917号(WO2004/052917)、国際公開第2004/094454号(WO2004/094454)および国際公開第2004/089973号(WO2004/089973)。
【0089】
V. コンジュゲートの調製
本発明のpan−DR結合ペプチドは、さまざまな形で少なくとも1つのさらなる抗原決定基に連結され得る。末端を介したまたはリジンのε−アミノ基を介したイオン相互作用が可能である。残基の側基と抗原決定基の間の水素結合も同様に可能である。その他の実施形態においては、pan−DR結合ペプチドと抗原決定基の間の立体構造相互作用が、安定した付着を発生させるかもしれない。
【0090】
以上で指摘した通り、抗原決定基は、本発明のコンジュゲートを調製するためにpan−DR結合ペプチドに対し共有結合されてよい。一部の実施形態においては、抗原決定基/pan−DR結合ペプチドが、スペーサー分子またはリンカーにより連結される。一部の実施形態において、抗原決定基はリンカー無しでpan−DR結合ペプチドに付着されてよい。
【0091】
スペーサーまたはリンカーは典型的には、生理学的条件下で実質的に非荷電性であり線状または分岐側鎖を有していてよい中性分子例えば脂肪族炭素鎖、アミノ酸またはアミノ酸ミメティックで構成されている。当業者にとっては、さまざまな生体分子を連結するための数多くの組成物および方法が公知である。例えば、炭水化物エピトープに対してpan−DR結合ペプチドを共有結合させるための数多くの方法が可能である。pan−DR結合ペプチドを炭水化物抗原に連結するのに適した方法は、例えば国際公開第93/21948号(WO93/21948)中で開示されている。
【0092】
数多くのリンカーが周知であり、市販されているかまたは科学的文献中に記載されている。本発明において使用される連結分子は、分子の2つの部分が、それらに曝露された分子と独立してかつ自由に相互作用できるようにするのに充分な長さを有する。炭水化物エピトープの場合、連結分子の長さは典型的に1〜50原子である。一部の実施形態において、連結分子はアリールアセチレン、2〜14個のモノマー単位を含むエチレングリコールオリゴマー、ジアミン、二塩基酸、アミノ酸またはそれらの組合せである。他の適切なリンカーとしては、アミノ酸側鎖を通して異なる炭水化物部分が連結されるセラミドおよびアミノ酸残基などの脂質分子が含まれる。
【0093】
使用される特定の連結分子は、その化学的/物理的特性に基づいて選択されてよい。連結分子は、各端部に適切な官能基を有し、一基は炭水化物部分上の反応性部位に対する付着に適切であり、もう一方の基はアミノ酸/ペプチド部分に対する付着に適切である。例えば、炭水化物部分に対する付着に適切な基は、カルボン酸、エステル、イソシアネート、ハロゲン化アルキル、ハロゲン化アシルおよびイソチオシアネートである。アミノ酸部分に対する付着のためには、類似の基が有用であると考えられる。官能基の適切な選択は、アミノ酸またはペプチドの反応性部分の性質によって左右されるものである。
【0094】
1群の実施形態においては、アルキルまたはアルキレン基が連結基として有用であり、1〜20個の炭素原子を有し、一部の実施形態においては3〜6個の炭素原子を含む。例えば、ポリエチレングリコールおよび関係する構造を含むリンカーを使用することができる。「ポリエチレングリコール」という用語は、例えばヘキサエチレングリコール(HO−(CH2CH2O)5−CH2CH2OH)などのエチレングリコールの反復単位を有する分子に言及するために用いられる。連結基に言及するために「ポリエチレングリコール」という用語が使用される場合、当業者は、その他のポリエーテルまたはポリオールも同様に使用できる(すなわちポリプロピレングリコールまたはエチレンとプロピレングリコールの混合物)ということを理解すると考えられる。
【0095】
別の実施形態群において、アルキルまたはアルキレン連結基は過フッ素化され、生物学的分解を受ける可能性が低くなっている。米国特許第5,055,562号(US.5,055,562)を参照のこと。一部の実施形態において、連結基は、アミノカプロン酸、4−ヒドロキシ酪酸、4−メルカプト酪酸、3−アミノ−1−プロパノール、エタノールアミン、ペルフルオロエタノールアミンおよびペルフルオロヒドロキシ酪酸を含む。一部の実施形態において、2つの部分はポリエチレングリコール部分を介して連結されている。
【0096】
一部の実施形態において、pan−DR結合ペプチドとその他のペプチド(例えばpan−DR結合ペプチドとCTLまたはB細胞エピトープ(ただしこれらに限定されない))の間のリンカーは、Ala、Glyまたは非極性アミノ酸または中性極性アミノ酸のその他のスペーサーから選択され得る。本明細書中の一部の実施形態において、中性スペーサーはAlaである。任意に存在するスペーサーは同じ残基で構成されている必要はなく、したがってヘテロまたはホモオリゴマーであってよい、ということが理解される。一部の実施形態において、例示的スペーサーは、Alaのホモオリゴマーである。存在する場合、スペーサーは通常少なくとも1つまたは2つの残基、さらに通常は3〜6個の残基である。一部の実施形態において、pan−DR結合ペプチドは、CTLまたは抗体誘発ペプチドにコンジュゲートされる。一部の実施形態において、pan−DR結合ペプチドはアミノ末端に位置づけされる。ペプチドは、Ala−Ala−Alaなどの中性リンカーによって接合され得、一部の実施形態においては、典型的にはSer−Ser連結などを介してペプチドコンジュゲートのアミノ末端に付着される、Lys残基((PAM)2Lys)のアルファおよびイプシロンアミノ基に付着されているパルミチン酸などの脂質残基を含み得る。
【0097】
CTLまたは抗体誘発ペプチドは、直接、あるいはCTLペプチドのアミノまたはカルボキシ末端のいずれかにあるスペーサーを介して、pan−DR結合ペプチドに連結されてよい。CTLまたは抗体誘発ペプチドまたはpan−DR結合ペプチドのいずれかのアミノ末端はアシル化可能である。一部の実施形態において、CTLペプチド/pan−DR結合ペプチドコンジュゲートは、以下で記述する通り、Gly、Gly−Gly、Ser、Ser−Serなどの1つ以上の連結残基を介して、一部のアルカノイル(C1−C20)脂質に連結され得る。一部の実施形態において、脂質部分はコレステロール、脂肪酸などを含む。
【0098】
一部の実施形態において、本発明の医薬組成物は、CTLのプライミングを補助する少なくとも1つの成分を含み得る。脂質が、ウイルス抗原に対する生体内でのCTLプライミングを補助できる作用物質として同定されてきた。例えば、ステロイド例えばコレステロール、脂肪酸例えばパルミチン酸残基(ただしこれらに限定されない)を、システイン残基のスルフヒドリル基、Lys残基のアルファおよびイプシロンアミノ基に付着させ、次に例えば1つ以上の連結残基例えばGly、Gly−Gly−、Ser、Ser−Serなどを介して、免疫原性ペプチド例えばpan−DR結合ペプチドに対して連結することができる。一部の実施形態において、脂肪酸の代りに、最終的アミノ酸(例えばシステイン残基)に対し、エーテル連結を通して長鎖アルキル基を連結することができる。
【0099】
脂質化されたペプチドは、リポソーム中に取り込まれたミセル型で直接か、または例えば不完全フロインドアジュバントなどのアジュバント中で乳化された状態のいずれかで、注射され得る。一部の実施形態において、特に有効な免疫原は、免疫原性ペプチドのアミノ末端に対して例えばSer−Serなどの連結を介して付着されているLysのアルファおよびイプシロンアミノ基に付着されたパルミチン酸を含む。
【0100】
CTL応答の脂質プライミングの別の例として、適切なペプチドに対し共有結合により付着されている場合にウイルス特異的CTLをプライミングするために、E.coliリポタンパク質例えばトリパルミトイル−S−グリセリルシステンリセリル−セリン(P3CSS)を使用することができる。Deresら、Nature342:561−564(1989)を参照のこと。本発明のペプチドは例えばP3CSSにカップリングさせることができ、リポペプチドは個体に投与されて標的抗原に対するCTL応答を特異的にプライミングすることができる。一部の実施形態において、中和抗体の誘発も同様に、適切なエピトープを表示するペプチドにコンジュゲートされたP3CSSを用いてプライミングされ得、2つの組成物を組合せて、感染に対する体液性応答と細胞媒介応答の両方をより効果的に惹起することができる。
【0101】
炭水化物エピトープにコンジュゲートされたpan−DR結合ペプチドの場合、脂質部分は、ペプチドの反対側の末端に連結されてよい(例えば炭水化物はC末端に連結され、脂質はN末端に連結される)。一部の実施形態において、脂質および炭水化物部分は両方共、ペプチドの同じ端部に連結されてよい。一部の実施形態において、2つの部分は、N末端上の同じリンカーに連結されてよい。
【0102】
VI. 免疫応答を刺激する方法
本発明のポリペプチド免疫原は、当業者にとっては公知のさまざまな方法を用いて、E.coliなどの発現宿主(または以上で論述した他の宿主細胞)から発現され、濃縮され、精製されてよく、代替的には、免疫原は液相または固相ペプチド合成により調製されてよい。精製されたポリペプチドまたはペプチド免疫原は、例えば免疫応答を刺激するかまたは増強し、任意には保護および/または治療的免疫応答を生成する目的で、個体の体内に投与するための医薬的に許容される賦形剤と共に調合可能である。「保護および/または治療的免疫応答」という語句は、何らかの形で疾病の症候、副作用または進行を防止するかまたは少なくとも部分的に阻み感染性因子を一掃する、例えば感染性因子から誘導された疾病関連抗原に対するCTLおよび/またはHTLおよび/または抗体応答を意味する。一部の実施形態において、本発明のポリペプチドまたはポリヌクレオチドは、免疫学的に有効な量の本発明のペプチドのうちの1つ以上と適切な薬学的担体とを含むワクチンの形に調合される。したがって、本発明のペプチドは、個別にかまたは単一組成物または多数の組成物のいずれかに組合わされた状態で投与可能である。
【0103】
本発明はさらに、免疫応答を刺激するため、本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物を投与する方法に関する。発現ベクターは、例えばDonnellyら、(Ann.Rev.Immunol.、15:617−648(1997));Felgnerら、(米国特許第5,580,859号(US.5,580,859)、1996年12月3日発行);Felgner(米国特許第5,703,055号(US.5,703,055)、1997年12月30日発行);およびCarsonら、(米国特許第5,679,647号(US.5,679,647)、1997年10月21日発行)中に記載の当該技術分野において周知の方法によって投与される。一実施形態においては、マルチエピトープ構築物は、ネイキッド核酸として投与される。
【0104】
本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物は、例えば経口、膣内、直腸または非経口経路例えば静脈内、筋内、皮下、眼窩内、関節包内、腹腔内、嚢内経路を含めたさまざまな経路によって、または例えばそれぞれ皮膚パッチまたは経皮イオントフォレシスを用いた経皮的な受動吸収または吸収促進によって、対象の体内で免疫応答を刺激するために投与することができる。さらにこの組成物を注射、挿管法によってかまたは局所的に投与することもでき、この局所的投与は、軟こうまたは粉末の直接塗布などによる受動的なもの、あるいは鼻腔用スプレーまたは吸入薬などを用いた能動的なものであり得る。発現ベクターは同様に局所的スプレーとして投与することができ、この場合、組成物の一成分は適切な高圧ガスである。医薬組成物は同様に、所望される場合、例えばFelgnerら、米国特許第5,703,055号(US.5,703,055);Gregoriadis、Liposome Technology、第I巻〜第III巻(第2版、1993)中に記載されているように、リポソーム、ミクロスフェアまたはその他のポリマーマトリクス内に取り込むこともできる。例えば、リン脂質または他の脂質で構成されたリポソームは、比較的簡単に製造し投与できる無毒性で生理学的に許容される代謝性担体である。
【0105】
本発明の発現ベクターまたはそれから誘導されたポリペプチドは、例えばFelgnerら、米国特許第5,580,859号(US.5,580,859)および5,703,055号(US.5,703,055)中に記載されている通りに、動物の体組織の間質腔に送出され得る。本発明の発現ベクターの筋肉への投与は、皮内および皮下注射および経皮投与を含め、特に有効な投与方法である。イオントフォレシスによるものなどの経皮投与も同様に、筋肉に対して本発明の発現ベクターを送出する有効な方法である。本発明の発現ベクターの表皮投与も同様に利用可能である。表皮投与には、刺激物に対する免疫応答を刺激するために表皮の最も外側の層を機械的または化学的に刺激することが関与する(Carsonら、米国特許第5,679,647号(US.5,679,647))。
【0106】
免疫応答を刺激するために本発明の発現ベクターまたはそれから誘導されたポリペプチドを投与する他の効果的方法には、例えばCarsonら、米国特許第5,679,647号(US.5,679,647)中に記載されている通りの粘膜投与が含まれる。粘膜投与について、最も有効な投与方法には、発現ベクターおよび医薬組成物を含む適切なエアロゾルの鼻腔内投与が含まれる。生殖器、膣および眼部位の粘膜組織に対する発現ベクターの送出のためには、座薬および局所的調製物も同様に有効である。追加的には、発現ベクターを粒子に複合体化させ、ワクチンガンにより投与することもできる。
【0107】
結論として、本発明のペプチドは、予防目的(すなわち後の疾病の危険性を減少させる目的)および治療目的の両方の目的で免疫応答を誘発するために有用であるが、本発明のペプチドは、実験的にかつ抗体および/またはBリンパ球を単離できる動物の体内で抗体を誘発する目的で、免疫応答を誘発するためにも有用であり、このことはそれ自体、後にモノクローナル抗体および抗体誘導体の産生を可能にする。
【0108】
VII. 医薬組成物
本発明のポリペプチド、本発明のポリヌクレオチドおよびその医薬組成物およびワクチン組成物は、予防および/または治療目的で哺乳動物、特にヒトに対し投与可能である。本発明のポリペプチドは、病原体または癌関連または癌特異的生体分子(例えばタンパク質、炭水化物など)を含めた(ただしこれらに限定されない)抗原に対する免疫応答を惹起および/または増強するために使用可能である。本発明を用いて治療可能な疾病の例としては、さまざまな細菌感染、ウイルス感染、真菌感染、寄生虫感染および癌が含まれる。タンパク性抗原に関して以上で論述した通り、国際公開第00/20027号(WO00/20027)からの癌抗原に関連する癌も、関節リウマチおよび炎症性腸疾患などのさまざまな炎症性疾患と同様の疾病例である。
【0109】
一部の治療的利用分野において、本発明は、すでに癌、炎症性疾患を患っているかまたは問題のウイルスまたは微生物に感染している個体に対して投与される。疾病の潜伏期または急性期にある個体は、適宜本発明を用いて別個に、または他の治療と併用して治療されてよい。
【0110】
一部の治療的利用分野において、本発明の組成物は、微生物または腫瘍抗原に対する有効なCTL応答または体液性応答を惹起し、かつ症候および/または合併症を治ゆするかあるいは少なくとも部分的に阻むのに充分な量で患者に投与される。これを達成するために適切な量が「治療上有効な量」として定義される。この用途にとって有効な量は、一部にはペプチド組成物、投与方法、治療対象の疾病の病期および重症度、患者の体重および全身的健康状態、および処方医師の判断に左右される。
【0111】
本発明の組成物の治療上有効な量は一般に、初期免疫付与すなわち治療的または予防的治療のためには、体重70kgの患者に対しポリペプチド約1.0μg〜約10,000μg、例えば約100〜約8000μg、例えば約200〜約6000μgである。一部の実施形態において、これらの用量には、特異的免疫応答を測定することにより患者の応答および身体条件に応じて数週間乃至数カ月にわたり、追加免疫処方計画に準じて約1.0μg〜約3000μgのペプチドの追加免疫投薬量が後続する。
【0112】
本発明の組成物は一般に重篤な疾病状態すなわち生命にかかわるかまたは潜在的に生命にかかわる状況において利用可能であるということを念頭に置いておくべきである。このような場合、異物の最少化およびコンジュゲートの相対的無毒性を考慮して、これらの組成物を実質的に余剰に投与することが可能であり、治療医師がそれを望ましいと感じるかもしれない。
【0113】
一部の実施形態において、本発明は、細菌感染、ウイルス感染、真菌感染、寄生虫感染および癌を含めた(ただしこれらに限定されない)特定の疾病を予防および/または改善するために、予防的に使用可能である。有効量は、以上で記述した通りである。追加的に、ワクチン技術の当業者であれば、追加免疫することおよび投薬量および投薬計画を調整することなどによって適宜、予防的処置をいかに調整または修正すべきかも認識していると考えられる。
【0114】
本発明の調合ワクチンは、医薬的に許容されるアジュバントと組合わせることができる。調合ワクチンは、水溶液、懸濁液またはエマルジョンであり得る。本発明のワクチン中の各免疫原の免疫学的に有効な量は、過度の実験なく当該技術分野において公知の方法により決定可能である。
【0115】
アジュバントは、医薬的に許容されるあらゆるアジュバントであり得る。一部の実施形態において、アジュバントはミョウバンベースの化合物である。一部の実施形態において、アジュバントは水酸化アルミニウムである。他の実施形態においては、アジュバントはリン酸アルミニウムである。一部の実施形態において、アジュバントはEmunade(登録商標)である。Emunade(登録商標)は、油、水および水酸化アルミニウムの組合せで構成されたアジュバントである。一部の実施形態において、アジュバントはQuil Aである。一部の実施形態において、アジュバントはQuil Aにコレステロールを加えたものである。一部の実施形態において、アジュバントは、MF59およびProvaxなど(ただしこれらに限定されない)のエマルジョンである。
【0116】
一部の実施形態において、ISCOMがアジュバントとして用いられる。ISCOMは、Immune Stimulating Complex(免疫刺激複合体)の頭字語であり、その技法は例えばMoreinら、(Nature308:457−460(1984))の中に記載されている。ISCOMは、以下の通りに形成される新油性免疫刺激複合体である。ポリペプチドは、非イオン洗浄剤(例えばMega−9、TritonX−I00、オクチルグリコシド、Digitonin、NonidetP−40、C12E8、Lubrol、Tween−80)などを用いて、標準的方法により可溶化される。ISCOM形成を補助するために、脂質混合物が添加される。脂質混合物は、ホスファチジルコリンおよび合成コレステロールを含み得る。一部の実施形態において、混合物はまず最初に、室温で撹拌しながら非イオン洗浄剤で処理され、次に脂質混合物(例えば等量のホスファチジルコリンとコレステロール)が添加され、撹拌が続行される。ポリペプチド組成物にQuil A(精製サポニン配糖体)が添加され、撹拌が続行される。その後、非イオン洗浄剤が除去される(例えば酢酸アンモニウムでの膜分離法による)。ISCOMのマトリクスがQuil Aにより形成される。電子顕微鏡で見たISCOM粒子の形態は、約35nmのサイズの典型的なカゴ様構造を示す。ISCOM形成期は、接線流膜分離法を用いて精製可能である。ISCOMは、臨界濃度でミセルを自然発生的に形成するQuil Aの能力に基づいて、かつ精製された抗原を封入する疎水性/親水性連結によって、マルチマー形状で精製抗原を提示する。ISCOMの形成は、典型的なかご様構造が形成されたことを確認するための電子顕微鏡によって確認される。約0.01〜0.1%の最終的濃度を得るためにQuil Aを添加することができる。一部の実施形態において、最終的濃度は約0.05%である。
【0117】
本発明は同様に、医薬的に許容される担体と本発明の発現ベクターまたは本発明のポリペプチドとを含む医薬組成物にも関する。医薬的に許容される担体は、当該技術分野において周知であり、生理的緩衝食塩水、アルコール/水溶液またはその他の溶媒またはビヒクル例えばグリコール、グリセロール、油例えばオリーブ油または注射用有機エステル、脂質またはリポソームを含めた、水溶液または非水性溶液、懸濁液およびエマルジョンを含む。
【0118】
医薬的に許容される担体は、例えば発現ベクターを安定化するかまたは発現ベクターの吸収を増大させるために作用する生理学的に許容される化合物を含むことができる。このような生理学的に許容可能な化合物には、例えばグルコース、スクロースまたはデキストランなどの炭水化物、アスコルビン酸またはグルタチオンなどの酸化防止剤、キレート剤、低分子量ポリペプチド、抗菌薬、不活性ガスまたはその他の安定化剤または賦形剤が含まれる。発現ベクターは、さらに、ペプチド、ポリペプチドおよび炭水化物などのその他の成分と複合体化され得る。発現ベクターは同様に、例えばワクチンガンを用いて個体に投与できる粒子またはビーズへと複合体化され得る。当業者であれば、生理学的に許容される化合物を含めた医薬的に許容される担体の選択が、例えば発現ベクターの投与経路により左右されるということを認識していると考えられる。
【0119】
本発明はさらに、本発明の発現ベクターまたはそれから誘導されたポリペプチドを含む医薬組成物を投与して免疫応答を刺激する方法にも関する。発現ベクターは、例えばDonnellyら、{Ann.Rev.Immunol.、15:617−648(1997));Felgnerら、(米国特許第5,580,859号(US.5,580,859)、1996年12月3日発行);Felgner(米国特許第5,703,055号(US.5,703,055)、1997年12月30日発行);およびCarsonら、(米国特許第5,679,647号(US.5,679,647)、1997年10月21日発行)中で記述されている通りの当該技術分野において周知の方法により投与される。一実施形態において、本発明のポリヌクレオチドは、ネイキッド核酸として投与される。
【0120】
本発明の組成物は、抗体または抗体誘導体を調製することを目的として、例えば実験動物または動物の体内で免疫応答を誘発するためにも有用である。このような組成物においては、組成物が医薬的に許容されるものでなくてはならないという要件は、重要性が低く、例えばヒトにおいて使用するのに適しているとみなされていないアジュバントを使用することが可能である。
【実施例】
【0121】
実施例の前文
以下の実施例は、請求対象の発明を限定するためではなく、例証するために提供されるものである。
【0122】
PADRE(配列番号3)内部のタンパク質分解分割が、PADRE含有タンパク質内、特に2つのリジン残基K2およびK10のまわりで、ただしW7のまわりにも観察された。PADRE内部の分割は、組換えにより産生されたタンパク質内で不均質、生産収量の低下および安定性の改変を導くことから望ましくない。上流側および下流側のプロセスの間に添加剤を使用することによって分割をうまく回避できるにせよ、それでも、結果として得られるPADRE含有ワクチンはなお低い免疫原性および薬物動態特性例えば半減期の短縮および1つまたは複数のTヘルパーエピトープの不適切な提示をなおも示すかもしれない。したがって、PADRE配列からプロテアーゼ感受性アミノ酸/配列を除去する必要がある。
【0123】
プロテアーゼ活性に対する感受性が低くなったPADRE類似体を提示することで、特にE.coliおよびショウジョウバエ発現系などの発現系においてプロテアーゼに対する組換え発現産物の感受性が削減されて、タンパク質産生の上流側および/または下流側プロセスの間のpan−DR結合ペプチドアミノ酸配列内部のタンパク質分解分割が回避されることになる。さらに、PADRE類似体が、単離形態でまたは免疫原性ポリペプチドの一部としてヒトに投与される場合、感受性の削減によって、薬物動態特性の改善(すなわちより長い血清半減期)そして場合によっては免疫原性の改善というさらなる利点がもたらされる。
【0124】
これに基づいて、配列番号3の数多くの類似体を調製し以下の一連の同定ステップに付して、タンパク質分解を比較的受けにくい改良型pan−DR結合ペプチドを同定した:
1)合成PADRE類似体のライブラリの調製ステップ;
2)優れたHLA結合特性について、1)に由来する類似体を検定し選択するステップ;
3)生体内原位置プロテアーゼ耐性について、2)で選択した類似体を検定し選択するステップ;
4)優れた免疫原特性について、3)で選択した類似体を検定し選択するステップ。
【0125】
実施例1
PADRE類似体のHLA−DR結合についての試験
標準的な固相ペプチド合成により、合計69個の異なるPADRE類似体(配列番号:4〜72)を合成した。これらのPADRE類似体は全て、N−からC−末端に向かって、2つのアラニン残基とそれに続く残基3〜15により構成され、これは(2つのアラニン残基が先行するPADREである配列番号4以外)配列番号3の類似体である。2つの追加のN末端アラニンを含み入れる理由は、PADRE類似体の免疫原特性が13C末端アミノ酸内に存在する一方で、単離ペプチドがプロテアーゼ耐性についての試験管内検定においてエンドプロテアーゼ耐性を示すことができるようになることを保証するためであった。
【0126】
ペプチドを、以下の手順にしたがってHLA−DRに対する結合について試験した:
【0127】
ペプチド/HLA−DR結合検定
クラスIIMHC結合検定のための標準的な作業手順は、Sidneyら、(1998)、Current Protocols in Immunology、18.3.1−18.3.19、1998中に記載されている。簡潔に言うと、精製ヒトクラスII分子[5〜500nM]を、さまざまな未標識ペプチド結合阻害物質および1〜10nM125I−放射性標識プローブペプチドと共に48時間、プロテアーゼ阻害物質カクテルの存在下で5%のDMSを含むPBS中でインキュベートした。インキューベーション混合物中の最終的洗浄剤濃度は、0.05%NonidetP−40であった。pH4.5で実施したDR3およびpH5.0で実施したDRw53を除いて、検定をpH7.0で実施した。pHは、Setteら、(1992)、J Immunol、148:844−51で記述されている通りに調整した。
【0128】
Sidneyらの中に記載されている通りにHPLC方法を使用してMHC分子に対するペプチド結合を測定する代りに、抗MHCクラスII抗体がコーティングされた抗体平板ベースの捕獲検定を利用した。この検定は、大量平板内放射分析検定用に特定的に設計された96ウェルの白色ポリスチレンマイクロタイター平板を使用するように開発されたものである。MHCに結合した125I標識ペプチドの測定は、高速大量処理性能を得るためにきわめて感度の高い検定を可能にするTOPCOUNT(Perkin−Elmer Instruments)ベンチトップマイクロプレートシンチレーションおよびルミネセンスカウンタを用いて実施される。
【0129】
ペプチド結合阻害物質を、30μg/ml〜300pg/mlの範囲内の濃度で試験した。市販の曲線適合アルゴリズムを用いて、データをコンピュータ内でプロットし、50%の阻害を生成する用量(IC50)を決定した。適切な化学量論的条件下で、精製DR分子に対する未標識被験ペプチドのIC50は、相互作用の親和性(KD)の適正な近似である。2〜4個の完全に独立した実験においてペプチドを試験した。
【0130】
結果
IC50値を介して表わされる結合親和性を、各ペプチドについて図1に列挙する。これらのIC50値に基づいて、類似体(38PADRE類似体)の画分が充分に優れたHLA結合能力(図1にボールド体と下線で示す)を示すという結論が下された。これらの優れた結合性を有するもののうち、当初10個を選択してプロテアーゼ耐性についてさらに試験した:配列番号;4、10、19、23、29、38、46、52、67および69。
【0131】
配列番号3および4が示す類似の結合特性により証明されるように、配列番号4〜69内に2つのN末端アラニンを含み入れることでHLA結合特性に影響が及ぶことはなかったという点を指摘しておくことも重要である。
【0132】
実施例2
選択されたPADRE類似体のプロテアーゼ耐性試験
選択された優れた結合性を有するPADRE類似体のプロテアーゼ耐性を試験するために、クローニング技術により、ヒトTNFα内のループ(ループEE)内にその配列を挿入した。修飾されたTNFα分子を発現するE.coli由来の無細胞タンパク質粗製抽出物を調製し、PADRE類似体配列内部のタンパク質分解分割を、SDS−PAGEとそれに続く免疫ブロット法によって分析した。
【0133】
全長、膜結合TNFαは、アミノ酸233個の長さをもつタンパク質である。アミノ末端フラグメント1〜76は、TNFαのアミノ酸157個の長さの可溶形態(77〜233)中には存在しない。
【0134】
ヒトTNFαワクチン候補の設計のために用いられる鋳型は、E.coli中での発現用のN末端Metが先行する、可溶形態のTNFα(フラグメント77〜233、配列番号73)である。さらに、TNFαのその2つの受容体に対する結合を取り除くために点変異を行ない(Y875)、こうしてタンパク質の細胞傷害性を根絶した。したがってワクチン候補は、長さがアミノ酸158個の配列番号74に基づくものである。
【0135】
TNF37.87(配列番号75)は、A185とE186の間にPADREが挿入されているTNFバリアント(…PEGA−EAK…)である。PADREはAで始まることから、Aは反復されず、その結果13個ではなく12個のアミノ酸が挿入された:
...PEGA−AKFVAAWTLKAAA−EAK... → PEG−A−KFVAAWTLKAAAEAK...
【0136】
したがって、当初プロテアーゼ耐性について試験したTNFバリアントは、以下の通りである:
配列番号75−TNF37.87−「従来の」PADREを含む基準タンパク質:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAKFVAAWTLKAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
− ならびに、下線位置においてそれぞれ配列番号4、10、19、23、29、38、46、52、67および69の12のC末端アミノ酸残基を各々同じように有するTNFバリアントを含む9個のPADRE類似体すなわち:
配列番号76−TNF37.87−007配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAWFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号77−TNF37.87−016配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAFFVAANTLKADAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号78−TNF37.87−020配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAYFVAAFTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号79−TNF37.87−026配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAHFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号80−TNF37.87−035配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAHFVAAFTLKAEAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号81−TNF37.87−043配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAEFVAAWTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号82−TNF37.87−049配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGANFVAAYTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号83−TNF37.87−064配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAQFVAANTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
配列番号84−TNF37.87−066配列:
MVRSSSRTPSDKPVAHVVANPQAEGQLQWLNRRANALLANGVELRDNQLVVPSEGLYLIYSQVLFKGQGCPSTHVLLTHTISRIAVSSQTKVNLLSAIKSPCQRETPEGAIFVAAWTLHAAAEAKPWYEPIYLGGVFQLEKGDRLSAEINRPDYLDFAESGQVYFGIIAL
【0137】
全ての配列中、2重下線は初期メチオニルおよびY87S変異をそれぞれ示し、1本下線は、挿入されたpanDR結合配列を示し;ボールド体は、PADRE(配列番号3)に比べたpan−DR結合ペプチド中のアミノ酸変化を表わす。
【0138】
組換えTNFαバリアントのクローニング、発現、精製および特徴づけ
鋳型として、TNF(Y87S)をコードする合成cDNAを使用した。PADREまたはPADRE類似体をコードするオリゴヌクレオチドを、PCRによりA185とE186の間で鋳型内に挿入した(上記参照)。結果として得たcDNAをpET28b+(Novagen)内にクローニングし、E.coli菌株HMS174へと形質転換させた。組換え発現のため、結果として得たE.coli菌株を発酵槽内において37℃にて合成培地中で培養した。組換え発現の誘発は、1mMのIPTGを添加することで開始し、温度を25℃まで低下させた。IPTG誘発から4、8または24時間後に培養を収穫した。Nielsenら、(2004)、J Biol.Chem.279:33593−33600中で記述されているようにTNFα特異的モノクローナル抗体を用いて親和性クロマトグラフィにより90%超の均質性までTNFαバリアントを精製した。精製したTNFαバリアントをMALDI−TOF質量分析法によって特徴づけしてそれらの同一性と無欠性を確認した。
【0139】
生体内原位置プロテアーゼ検定:ウエスタンブロット法を用いたTNF−αバリアントのプロテアーゼ耐性の分析:
試験すべきさまざまなTNF−αバリアントを発現するE.coli細胞培養のアリコート(1ml)を遠心分離(5000g/5分/5℃)し、培地を廃棄した。pH6.0で20mMのBis−Tris1ml中に細胞ペレットを再懸濁させた。超音波処理(各超音波処理ラウンド間で20秒間休止して、11μmの振幅で4×15秒)により、細胞破壊を達成した。結果として得た懸濁液を遠心分離し(20000g/30分/8℃)、上清(粗製抽出物)を新しい管に移した。次に、ブラッドフォードタンパク質分析(Biorad)を用いて、タンパク質含有量について粗製抽出物を分析した。4〜12%のNuPAGEゲル(Novex)上に等量のタンパク質を投入した。SDS−PAGEの後、セミドライブロット法によりニトロセルロース膜上にタンパク質を移した。免疫反応については、Tris50mM、NaCl150mM、EDTA5mM、Igepal0.1%、ゼラチン0.5%中で、ウサギTNFα抗血清(1時間;希釈度1:10,000)およびホースラディッシュペルオキシダーゼでコンシュゲートされた二次抗体(Dako P448)と共にインキューベーションを行なった。検出にはECLTM試薬(GE Healthcare)を用いた。
【0140】
結果
被験TNFバリアントのうち、3つ(配列番号78、79および84)はプロテアーゼ検定においてプロテアーゼ耐性があることが示され(データ示さず)、これらのTNFバリアントが、PADRE類似体配列の内部でのタンパク質分解分割に対する感受性をもたないということを証明した。
【0141】
配列番号23、29および69の各々の少なくとも13C末端アミノ酸残基により画定されるPADRE類似体がタンパク質分解に対する耐性の所望の増加を示し、かつ配列番号23、29および69の13C末端アミノ酸残基を含み入れることで修飾されたタンパク質は、組換えにより生成された場合により優れた収量を示し(これは、発現産物のフラグメント化が全くまたは限定的にしか発生しないことに起因する)、均質な発現産物を提供し、場合によってはプロテアーゼ耐性免疫原の生物学的半減期の延長を原因として免疫原性の増大を提供するものであるという結論が下された。
【0142】
実施例3
選択されたPADRE類似体の免疫原性試験
実施例2内で同定した3つのプロテアーゼ耐性PADRE類似体をその免疫原特性について試験した。1本の実験ライン上で、ヒトのドナー由来のPBMC(末梢血単核細胞)内および2つのマウス株(DR4およびbxd)内で遊離ペプチドとしてペプチドを試験した。第2の実験ライン内では、実施例2で同定したプロテアーゼ耐性TNFバリアントに関連して、PADRE類似体を試験した。
【0143】
ヒトPBMCにおけるペプチドの試験
ヒトPBMC(末梢血単核細胞)を、配列番号23、29および69の13アミノ酸残基と同一の配列をもつペプチドで刺激した(以下では、これら3つの13量体をそれぞれPADRE.Y2.F7.H10、PADRE.H2.N7.H10およびPADRE.12.H10と呼ぶ)。1日目および4日目に、IL−2を添加した。7日目に、he細胞の一部分を取り出してINF−γELI−POT検定に付した。残りの細胞を再刺激した。8日目および11日目に、IL−2を添加した。14日目にCD4細胞を精製し、INF−γELISPOT検定において試験した。
【0144】
INF−γELISPOT検定は、本質的にTangri Sら、(2005)、J Immunol 174、3187−96中で記述されている通りに実施した。
【0145】
マウスにおけるペプチドの試験
HLA−DR4遺伝子導入マウスとbxdマウス株の各々の3匹の個体からなるグループを、CFA中の20μg(6μlの20mg/mlペプチドおよび294μlのPBS1×+300μlのCFA)または2μg(6μlの2mg/mlペプチド+294μlのPBS1×+300μlのCFA)のペプチドを用いて尾の付け根で免疫付与した。10〜14日後に、脾臓を収穫し、CD4細胞を精製し、INF−γELISPOT検定(McKinneyら、(2004)、J Immunol 173、1941−50に記載の通り)で試験した。被験ペプチドは、ヒトPBMCで試験されたものと同一であった。
【0146】
マウスにおけるTNFαバリアントの試験
3匹のHLA−DR4遺伝子導入マウスからなるグループを、マウス一匹あたり100μlの完全フロインドアジュバント(CFA)中10μgのTNF由来の抗原を用いて、尾の付け根で免疫付与した。最初の免疫付与から14日目に、100μlのIFA(不完全フロインドアジュバント)中の同じ用量の抗原を用いて、動物に追加免疫を付与した。TNFα特異的抗体力価を決定するため、各々の免疫付与から14日後に、血清を収集した。その後動物を屠殺して、INF−γELISPOT検定に付した。
【0147】
マウスの抗血清中のTNFα特異的抗体力価の決定
直接ELISAを用いて、TNFα特異的抗体力価を決定した。96ウェル平板(Maxisorb、Nunc)を組換えTNFαバリアントTNFY87S(pH9.6で炭酸緩衝液中5μg/ml、100μl/ウェル、4℃で一晩)を用いてコーティングした。その後平板を3回洗浄し、37℃で2時間200μl/ウェルのブロッキング緩衝液(1%のウシ血清アルブミンおよび0.05%のTween20を含むリン酸緩衝生理食塩水)を用いてインキュベートした。平板を3回洗浄し、ヒトTNFαバリアントで免疫付与したマウス由来のプールされた血清を、合計100μl/ウェルのブロッキング緩衝液中での1/10希釈ステップ(出発希釈度1/1000)を用いて、6ステップで滴定した。全ての試料をデュプリケートで滴定した。血清と対照を2時間37℃でインキュベートした。平板を3回洗浄し、100μlのビオチニル化ヤギ抗マウスIgG(ブロッキング緩衝液中で1/10000希釈)を各ウェルに移し、37℃で1時間インキュベートした。最後に、平板を3回洗浄し、100μl/ウェルのアビジン−ペルオキシダーゼ複合体(Vectastain Elite Vector PK−6100)を用いて室温で45分間インキュベートした。平板を再び洗浄し、TMBSを用いて現像した。10〜20分後、100μlの4NのH2SO4で反応を停止させ、ELISA読取り装置を用いてA450値を決定した。0.5のA450値を生成する抗血清希釈度として抗体力価を定義づけた。
【0148】
結果
ヒトPBMC中のペプチドの試験から得られた結果は、PADRE.Y2.F7.H10およびPADRE.I2.H10が、配列番号3に比べ7日目と14日目の両日にINF−γELISPOT検定においてより優れた免疫応答を提供し、一方PADRE.H2.N7.H10は、配列番号3と同じ規模の免疫応答を提供したことを明らかにした。
【0149】
マウスにおけるPADRE類似体ペプチドの試験では、3個の被験類似体の全てが、DR4マウスにおいて20μg用量でPADREよりも良い性能を示した一方、ELISPOT検定により測定された通り2μg用量でDR4遺伝子導入マウスにおいて、PADRE.I2.H10ペプチドのみが免疫応答の改善を提供した。bxdマウスモデルでは、3つの被験PADRE類似体の全てが、両方の用量でELISPOT検定内でPADREに比べて有意な免疫応答を提供したが、PADREに比べた免疫応答の改善は得られなかった。
【0150】
PADRE類似体を含むTNFαバリアントでの免疫付与は、以下のような結果を提供した:すなわち
INF−γELISPOTにより測定されたHLADR4遺伝子導入マウスにおけるHTL誘発は、全てのTNFα−バリアントが、TNFα内に導入されたPADRE類似体に対するHTLを誘発したこと、そしてこの点においてPADRE.Y2.F7.H10およびPADRE.I2.H10がPADREよりも優れていたこと(ここでPADRE.H2.N7.H10はPADREとほぼ同じHTL誘発を提供した)を明らかにした。さらに全てのバリアントは、野生型TNFαにより誘発されたものよりも著しく高い抗体力価を提供し、PADRE.Y2.F7.H10およびPADRE.I2.H10を含むバリアントは、PADREに比べ優れており、ここでもまた、PADRE.H2.N7.H10はPADREに匹敵する免疫応答を提供した。実際、抗体力価とHTL誘発を相関させた場合、明らかに線形相関が存在していた。
【0151】
実施例4
8個のさらなるPADRE類似体の選択および試験
panDR結合性でプロテアーゼ耐性を有する免疫原性PADRE類似体PADRE.Y2.F7.H10、PADRE.H2.N7.H10およびPADRE.I2.H10の同定に基づいて、実施例1において優れた結合性を有するものとして同定されたさらなるPADRE類似体を試験することが決定された。被験ペプチドは、配列番号15(PADRE.F2.N7.H10)、21(PADRE.Y2.H10)、24(PADRE.Y2.N7.H10)、26(PADRE.H2.H10)、47(PADRE.E2.Y7.H10)、54(PADRE.N2.N7.H10)、66(PADRE.Q2.F7.H10)、および72(PADRE.I2.N7.H10)からの13C末端アミノ酸残基を有するペプチドであった。
【0152】
これらの配列のうち3つすなわちPADRE.E2.Y7.H10、PADRE.N2.N7.H10およびPADRE.Q2.N7.H10は、実施例2に記された手順に従ってプロテアーゼ耐性を有することが示された。
【0153】
結論
上述の実施例は、プロテアーゼ耐性および/または免疫原性の観点から見て改善された特性を有する数多くのPADRE類似体の存在を実証した。配列番号3内で入念に修飾を行なうことで、組換え発現された場合および生体内免疫原として使用された場合の両方において、PADREよりも安定した免疫原が提供される、ということが実証されている。
【0154】
本明細書中で記述されている実施例および実施形態は単に例証を目的としており、それに照らしたさまざまな修正または変更が当業者には示唆されるものであり、それらは本出願の精神および視野そして添付のクレームの範囲内含み入れられるべきものである。本明細書中で引用される全ての刊行物、特許および特許出願は、あらゆる目的でその全体が参照により本明細書に援用される。
【0155】
【表2】
【表3】
【特許請求の範囲】
【請求項1】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含む単離ポリペプチドであって、オリゴペプチド配列がAX1FVAAX2TLX3AX4A(配列番号1)を含み、
− X1がW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;
− X3がHおよびKからなる群から選択され;
− X4がA、DおよびEからなる群から選択され;
ただしここでオリゴペプチド配列がAKFVAAWTLKAAAでないことが条件となっている、
単離ポリペプチド。
【請求項2】
− X1がY、H、I、E、N、QおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;かつ
− X3がHであり、
− X4がAである、
請求項1に記載の単離ポリペプチド。
【請求項3】
前記オリゴペプチドが配列番号4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71および72からなる群から選択されたオリゴペプチド中の13個のアミノ酸残基C末端フラグメントと同一のアミノ酸配列を有する、請求項1に記載の単離ポリペプチド。
【請求項4】
前記ポリペプチドがオリゴペプチドに加えて未変性ポリペプチド配列の大部分を含んでいる、請求項1〜3のいずれか一項に記載の単離ポリペプチド。
【請求項5】
前記未変性ポリペプチド配列がヒトTNFα由来のものである、請求項4に記載の単離ポリペプチド。
【請求項6】
前記ポリペプチドが、配列番号76::77、78、79、80、81、82、83および84のいずれか1つから選択されたアミノ酸配列を有する、請求項5に記載の単離ポリペプチド。
【請求項7】
前記オリゴペプチドに加えて、少なくとも1つの抗原由来の複数のエピトープを含む、請求項1〜3のいずれか一項に記載の単離ポリペプチド。
【請求項8】
前記複数のエピトープが、複数のCTLエピトープ、複数のB−細胞エピトープ、複数のTヘルパーリンパ球エピトープ、複数のCTLおよびB細胞エピトープ、複数のCTLおよびTヘルパーリンパ球エピトープ、複数のB細胞およびTヘルパーリンパ球エピトープおよび複数のB細胞、CTLおよびTヘルパーリンパ球エピトープから選択されている、請求項7に記載の単離ポリペプチド。
【請求項9】
前記エピトープが1つの単一抗原タンパク質に由来するものである、請求項7または8に記載の単離ポリペプチド。
【請求項10】
前記エピトープが少なくとも2つの異なる抗原タンパク質に由来するものである、請求項7または8に記載の単離ポリペプチド。
【請求項11】
前記少なくとも2つの異なる抗原タンパク質が同じ種に由来するものである、請求項10に記載の単離ポリペプチド。
【請求項12】
前記ポリペプチドが100個以下のアミノ酸を有する、請求項1〜3および6〜11のいずれか一項に記載の単離ポリペプチド。
【請求項13】
前記ポリペプチドが、MHC分子を結合できるオリゴペプチド配列と重複しない第2のオリゴペプチド配列を含む、請求項1または4または6〜11に記載の単離ポリペプチド。
【請求項14】
前記第2のオリゴペプチド配列がCTL配列またはB細胞エピトープを含む、請求項13に記載の単離ポリペプチド。
【請求項15】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードする単離ポリヌクレオチドであって、オリゴペプチド配列が請求項1〜4のいずれか一項に規定された通りである、単離ポリヌクレオチド。
【請求項16】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドに対して作動的に連結されたプロモータを含む核酸を含む単離ベクターであって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである単離ベクター。
【請求項17】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドに対して作動的に連結されたプロモータを含む核酸を含む細胞であって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである細胞。
【請求項18】
単離細胞である、請求項17に記載の細胞。
【請求項19】
ポリペプチドと抗原を含む組成物であって、ポリペプチドが100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含み、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである組成物。
【請求項20】
前記抗原が第2のポリペプチドである、請求項19に記載の組成物。
【請求項21】
前記抗原が非タンパク性抗原である、請求項19に記載の組成物。
【請求項22】
前記抗原が炭水化物である、請求項19に記載の組成物。
【請求項23】
生理学的に許容される賦形剤と100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドとを含む医薬組成物であって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである医薬組成物。
【請求項24】
抗原をさらに含む、請求項23に記載の医薬組成物。
【請求項25】
免疫応答を刺激する方法であって、人間などの動物に対して有効量のポリペプチドを投与するステップを含む方法であって、ポリペプチドが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含み、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである方法。
【請求項26】
免疫応答を刺激する方法であって、人間などの動物に対して有効量のポリヌクレオチドを投与するステップを含む方法であって、ポリヌクレオチドが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードし、配列が請求項1〜14のいずれか一項に規定された通りである方法。
【請求項27】
免疫応答を刺激する方法であって、人間などの動物に対して有効量の非病原性ウイルスまたは微生物を投与するステップを含む方法であって、微生物またはウイルスが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドで形質転換され、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである方法。
【請求項1】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含む単離ポリペプチドであって、オリゴペプチド配列がAX1FVAAX2TLX3AX4A(配列番号1)を含み、
− X1がW、F、Y、H、D、E、N、Q、IおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;
− X3がHおよびKからなる群から選択され;
− X4がA、DおよびEからなる群から選択され;
ただしここでオリゴペプチド配列がAKFVAAWTLKAAAでないことが条件となっている、
単離ポリペプチド。
【請求項2】
− X1がY、H、I、E、N、QおよびKからなる群から選択され;
− X2がF、N、YおよびWからなる群から選択され;かつ
− X3がHであり、
− X4がAである、
請求項1に記載の単離ポリペプチド。
【請求項3】
前記オリゴペプチドが配列番号4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71および72からなる群から選択されたオリゴペプチド中の13個のアミノ酸残基C末端フラグメントと同一のアミノ酸配列を有する、請求項1に記載の単離ポリペプチド。
【請求項4】
前記ポリペプチドがオリゴペプチドに加えて未変性ポリペプチド配列の大部分を含んでいる、請求項1〜3のいずれか一項に記載の単離ポリペプチド。
【請求項5】
前記未変性ポリペプチド配列がヒトTNFα由来のものである、請求項4に記載の単離ポリペプチド。
【請求項6】
前記ポリペプチドが、配列番号76::77、78、79、80、81、82、83および84のいずれか1つから選択されたアミノ酸配列を有する、請求項5に記載の単離ポリペプチド。
【請求項7】
前記オリゴペプチドに加えて、少なくとも1つの抗原由来の複数のエピトープを含む、請求項1〜3のいずれか一項に記載の単離ポリペプチド。
【請求項8】
前記複数のエピトープが、複数のCTLエピトープ、複数のB−細胞エピトープ、複数のTヘルパーリンパ球エピトープ、複数のCTLおよびB細胞エピトープ、複数のCTLおよびTヘルパーリンパ球エピトープ、複数のB細胞およびTヘルパーリンパ球エピトープおよび複数のB細胞、CTLおよびTヘルパーリンパ球エピトープから選択されている、請求項7に記載の単離ポリペプチド。
【請求項9】
前記エピトープが1つの単一抗原タンパク質に由来するものである、請求項7または8に記載の単離ポリペプチド。
【請求項10】
前記エピトープが少なくとも2つの異なる抗原タンパク質に由来するものである、請求項7または8に記載の単離ポリペプチド。
【請求項11】
前記少なくとも2つの異なる抗原タンパク質が同じ種に由来するものである、請求項10に記載の単離ポリペプチド。
【請求項12】
前記ポリペプチドが100個以下のアミノ酸を有する、請求項1〜3および6〜11のいずれか一項に記載の単離ポリペプチド。
【請求項13】
前記ポリペプチドが、MHC分子を結合できるオリゴペプチド配列と重複しない第2のオリゴペプチド配列を含む、請求項1または4または6〜11に記載の単離ポリペプチド。
【請求項14】
前記第2のオリゴペプチド配列がCTL配列またはB細胞エピトープを含む、請求項13に記載の単離ポリペプチド。
【請求項15】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードする単離ポリヌクレオチドであって、オリゴペプチド配列が請求項1〜4のいずれか一項に規定された通りである、単離ポリヌクレオチド。
【請求項16】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドに対して作動的に連結されたプロモータを含む核酸を含む単離ベクターであって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである単離ベクター。
【請求項17】
100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドに対して作動的に連結されたプロモータを含む核酸を含む細胞であって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである細胞。
【請求項18】
単離細胞である、請求項17に記載の細胞。
【請求項19】
ポリペプチドと抗原を含む組成物であって、ポリペプチドが100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含み、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである組成物。
【請求項20】
前記抗原が第2のポリペプチドである、請求項19に記載の組成物。
【請求項21】
前記抗原が非タンパク性抗原である、請求項19に記載の組成物。
【請求項22】
前記抗原が炭水化物である、請求項19に記載の組成物。
【請求項23】
生理学的に許容される賦形剤と100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドとを含む医薬組成物であって、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである医薬組成物。
【請求項24】
抗原をさらに含む、請求項23に記載の医薬組成物。
【請求項25】
免疫応答を刺激する方法であって、人間などの動物に対して有効量のポリペプチドを投与するステップを含む方法であって、ポリペプチドが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含み、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである方法。
【請求項26】
免疫応答を刺激する方法であって、人間などの動物に対して有効量のポリヌクレオチドを投与するステップを含む方法であって、ポリヌクレオチドが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードし、配列が請求項1〜14のいずれか一項に規定された通りである方法。
【請求項27】
免疫応答を刺激する方法であって、人間などの動物に対して有効量の非病原性ウイルスまたは微生物を投与するステップを含む方法であって、微生物またはウイルスが、100nM未満のIC50値を有する少なくとも3つの異なるHLA−DR対立遺伝子によりコードされるMHCクラスII分子を結合できるオリゴペプチド配列を含むポリペプチドをコードするポリヌクレオチドで形質転換され、オリゴペプチド配列が請求項1〜14のいずれか一項に規定された通りである方法。
【図1−1】

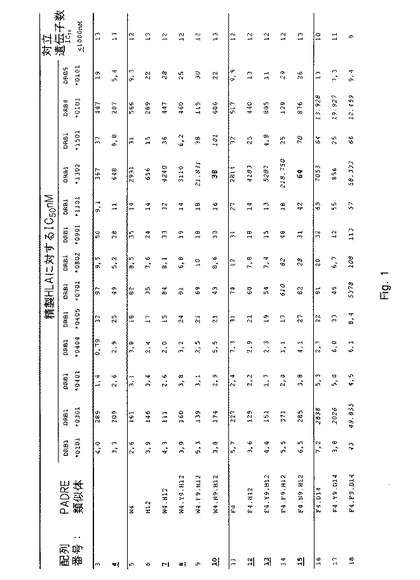
【図1−2】
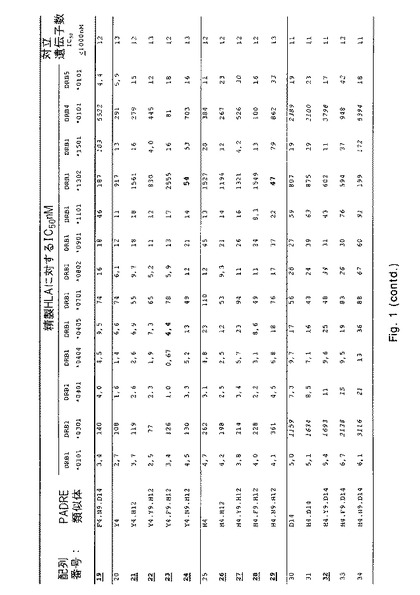
【図1−3】
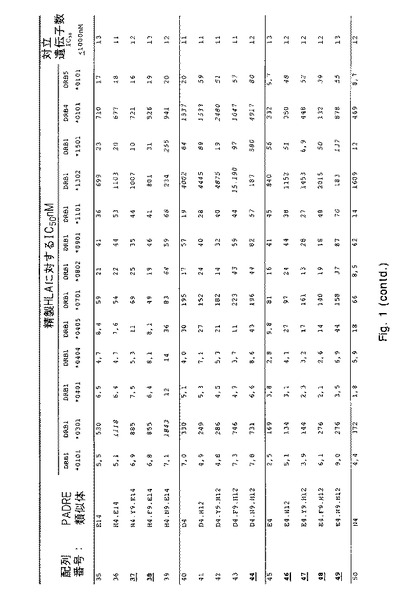
【図1−4】
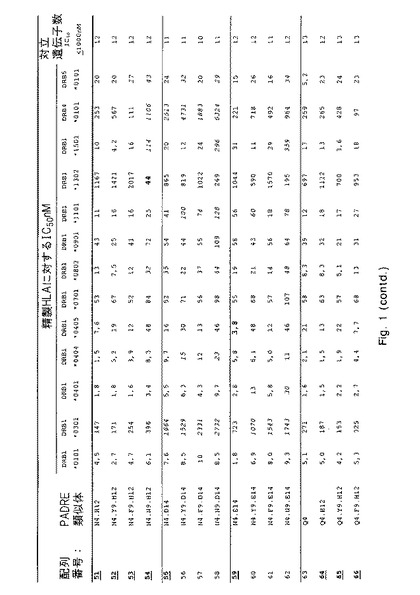
【図1−5】
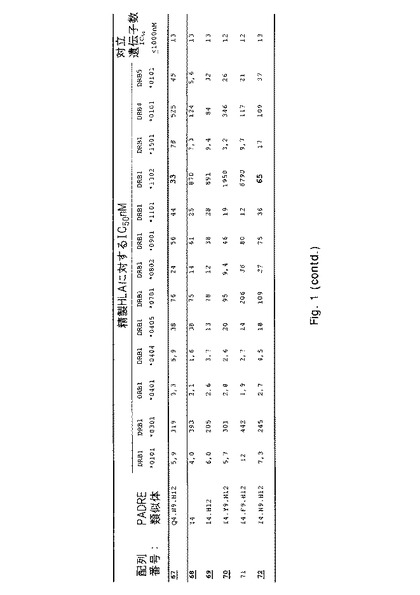
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【公表番号】特表2012−516140(P2012−516140A)
【公表日】平成24年7月19日(2012.7.19)
【国際特許分類】
【出願番号】特願2011−546814(P2011−546814)
【出願日】平成22年1月26日(2010.1.26)
【国際出願番号】PCT/EP2010/050839
【国際公開番号】WO2010/086294
【国際公開日】平成22年8月5日(2010.8.5)
【出願人】(500430660)エピミューン,インコーポレイティド (2)
【Fターム(参考)】
【公表日】平成24年7月19日(2012.7.19)
【国際特許分類】
【出願日】平成22年1月26日(2010.1.26)
【国際出願番号】PCT/EP2010/050839
【国際公開番号】WO2010/086294
【国際公開日】平成22年8月5日(2010.8.5)
【出願人】(500430660)エピミューン,インコーポレイティド (2)
【Fターム(参考)】
[ Back to top ]