
PD−1アンタゴニストおよび感染性疾患を処置するための方法
(1)迅速T細胞媒介応答の惹起不全、(2)T細胞の消耗、T細胞の無応答もしくはその双方の誘導、または(3)単球、マクロファージ、樹状細胞および/または例えば細胞内病原体を死滅させるのに必要とされるような他のAPCの活性化不全から起こる感染または疾患を処置するための方法および組成物。該方法および組成物は、PD−1と結合してそれを遮断し、阻害的シグナル伝達を回避または低減することによるか、またはPD−L1などのPD−1のリガンド(Hgands)と結合し、それにより、そのリガンドがPD−1と結合して(このにまたは部分的に)阻害シグナルを送達しないようにすることにより、望ましくないT細胞阻害の問題を解決する。この免疫応答は種々の親和性(すなわち、必要とされるものよりも高いまたは低い)で結合するアンタゴニストを提供することにより、また、投与される薬剤の用量を変えることにより、また、投与計画中、間欠的投与を行うことにより、また、その組合せにより(再び投与される前にそれが結合されている分子から薬剤の解離をもたらす(初回刺激と追加免疫を用いる抗原惹起で見られるものに類似))調節され得る。いくつかの場合では、免疫系を刺激した後に、その刺激を取り去ることが特に望ましいことがある。

【発明の詳細な説明】
【発明の背景】
【0001】
関連出願の相互参照
本願は2008年8月25日に出願された米国仮出願第61/091,502号公報、2008年8月25日に出願された米国仮出願第61/091,694号公報、2008年8月25日に出願された米国仮出願第61/091,709号公報、2008年8月25日に出願された米国仮出願第61/091,705号公報、2009年1月5日に出願された米国仮出願第61/142,548号公報、および2009年4月1日に出願された米国仮出願第61/165,652号公報の優先権および利益を主張するものであり、これらは許容される場合には引用することにより本発明の開示の一部とされる。
【0002】
発明の分野
本発明は一般に、癌または感染などの疾患、特に、T細胞の消耗、T細胞の無応答もしくはその双方または細胞内病原体、すなわち例えばリーシュマニアがAPC(例えば、単球、樹状細胞、マクロファージ)または上皮細胞上でPD−1リガンドをアップレギュレートすることにより免疫応答を逃れる疾患を含む疾患を処置するための免疫調節組成物および方法に関する。
【0003】
背景技術
微生物感染に対する宿主の耐性には、二つの主要な重複する防御系、先天免疫、および適応免疫が組み込まれている。ウイルス、細菌、および寄生虫を包含する細胞内病原体は、感染後の初期に即座に、感染宿主において非特異的体液性応答と細胞エフェクター応答を刺激する活性化シグナルを中継する。これらの先天的防御応答の助けで、微生物の増殖速度が数日間遅れ、その間に適応系の免疫が誘導され、長期間病原体と対抗するよう促される(適応的/長期免疫)。これらの免疫応答はT細胞により媒介される。多くの細胞内病原体に関して、感染防御免疫は、前もって感染と闘う手助けをするように他の免疫細胞を刺激するサイトカインなどの化合物を産生するCD4+ヘルパーT細胞の生成、病原体に感染した宿主細胞を排除するCD8+細胞傷害性Tリンパ球(CTL)により主として媒介される細胞媒介応答、およびTヘルパー細胞により媒介される抗体応答の双方を必要とする。しかしながら、それらの生物が初期の免疫活性化を迂回し、エフェクター免疫応答および長期メモリー応答を損なえば、感染が確立され、持続する。これが急性および慢性感染を招く。
【0004】
研究によれば、初期の免疫破壊は多くの場合、クラスI MHC分子による抗原プロセシングおよび/または提示に関与する細胞内経路を標的とすることが証明されている。これの結果、初期の免疫活性化が不十分となり、また、その生物に対する一次応答がほとんど、または全く無くなる。これによりこれらの生物は確立され、細胞内病原体では、免疫系から「隠れた」ままとなる。さらに最近の研究では、多くの場合でこれらの病原が低いが測定可能な特異的免疫応答を刺激することが示されている。しかしながら、T細胞が病原体と闘うことにより「消耗」し、時間が経つにつれ段階的に有効性が低くなるという顕著な変化を受ける場合には、慢性感染が起こる。これはT細胞の消耗として知られる現象である。
【0005】
B7タンパク質は、免疫細胞(例えば、T細胞)に免疫応答を刺激または阻害する第二のシグナルを与える働きをする。PD−L1(B7−H1)およびPD−L2(PD−DC)は、共通の受容体PD−1と結合するB7ファミリーの分子の阻害メンバーである。PD−L1は多様な組織および細胞種で広く発現されるが、PD−L2の発現は主として活性化された樹状細胞(DC)およびマクロファージに限られる。CD28ファミリー受容体のメンバーであるPD−1は、活性化されたT細胞、B細胞、ナチュラルキラー(NK)細胞、単球、DCおよびマクロファージで誘導的に発現される。T細胞の消耗は、T細胞機能に負の調節を行う、PD−1受容体を介した阻害的T細胞シグナル伝達により生じることが示されている。
【0006】
そのリガンドによるPD−1結合の主要な結果は、T細胞受容体(TCR)下流でのシグナル伝達の阻害である。よって、PD−1を介したシグナル伝達は通常、T細胞に対して、T細胞増殖の低下またはT細胞活性におけるその他の低下をもたらす抑制または阻害シグナルを与える。PD−1のシグナル伝達は、主要組織適合性複合体(MHC)により提示されるペプチド抗原と近接したリガンドとの結合を必要とすると考えられ、これはTCRと結合されている(Freeman Proc. Natl. Acad. Sci. U. S. A 105:10275-10276 (2008))。PD−L1はT細胞において阻害的シグナル伝達を引き起こす主要なPD−1リガンドである。
【0007】
多くの細胞内病原体に対する一次免疫応答およびエフェクター免疫応答が不十分な結果として、とりわけヒト免疫不全ウイルス(HIV)、C型肝炎ウイルス(HCV)、単純ヘルペスウイルス(HSV)、結核菌(M. tuberculosis)、C.トラコマチス(C. trachomatis)、マラリアなどのこれら多くの生物に対する効果的なワクチンは存在しない。慢性感染が定着し、宿主免疫系がこれらの慢性または潜在感染を排除できない場合に重大な問題となる。抗原/ワクチンに対する不十分な一次応答およびエフェクター応答はまた、急性免疫が必要とされる場合(それ以外の点で有効なワクチンが作製可能な場合であっても)、例えば、インフルエンザなどの地域流行/汎流行的大発生または感染性病原体(例えば、炭疽)によるバイオテロ攻撃の、ならびに免疫系が未発達または弱まっている小児および高齢者集団において問題となる。
【0008】
ワクチンの免疫原性および保護を改善するための一つのアプローチがアジュバントの使用である。アジュバントは免疫応答を改善するためにワクチンに加えられる成分である。開発され、試験されているほとんどのアジュバントは主として先天的免疫応答(抗原特異的でない)、抗体応答および極めて少ない場合であるが、弱いT細胞応答を惹起する。利用可能なアジュバントに、一次応答を増大させ、細胞内病原体に対する防御免疫を惹起するのに必要とされるものである、強力なエフェクター応答または迅速なT細胞増殖応答を誘導するものはない。
【0009】
よって、本発明の一つの目的は、一次免疫応答およびエフェクター免疫応答の双方を増強するワクチンアジュバントを提供することである。
【0010】
もう一つの目的は、より迅速な防御誘導ならびに慢性感染に対する強力なエフェクター応答をもたらす組成物を提供することである。
【0011】
別の目的は、T細胞の消耗、T細胞の無応答またはその双方を誘導する感染を処置するための組成物および方法を提供することである。
【0012】
本発明のさらに別の目的は、単球、樹状細胞、マクロファージを含む抗原提示細胞の細胞内感染を処置するための組成物および方法を提供することである。
【発明の概要】
【0013】
(1)迅速T細胞媒介応答の惹起不全、(2)T細胞の消耗、T細胞の無応答もしくはその双方の誘導、または(3)単球、マクロファージ、樹状細胞、および/または例えば細胞内病原体を死滅させるのに必要とされるような他のAPCの活性化不全から起こる感染または疾患を処置するための方法および組成物。これらは急性(例えば、毒素誘発性)、慢性、遅発性、または潜在感染により引き起こされ得る。本発明の方法および組成物は、PD−1と結合してそれを遮断し、阻害的シグナル伝達を回避または低減することによるか、またはPD−L1などのPD−1のリガンドと結合してそれを遮断し、それにより、そのリガンドがPD−1と結合して(完全にまたは部分的に)阻害シグナルを送達しないようにすることにより、望ましくないT細胞阻害の問題を解決する。これらの分子は一般にPD−1アンタゴニストと呼ばれ、PD−1またはPD−L1などのリガンドと直接結合する両化合物を含む。いずれの場合でも、T細胞の増殖または活性化などのT細胞応答が増強される。さらに、PD−1アンタゴニストは、細胞内病原体によりアップレギュレートされている、抗原提示細胞(単球、マクロファージ、樹状細胞、上皮細胞などのAPC)で発現されるPD−1リガンドと結合してそれを遮断することもできる。
【0014】
免疫応答を増強する、または増大させ得るには、1)T細胞活性を阻害する分子に干渉すること、例えば、分子がPD−1であり、a)受容体(PD−1)を遮断するか、もしくはb)リガンド(B7−H1もしくはB7−DC)を遮断する場合、または2)T細胞活性を活性化させる分子を増大させる、例えば、分子がCD28であり、アゴニストが加えられる場合の、二つの機構がある。免疫応答は、投与される薬剤の用量を変えることにより、投与計画中、間欠的投与を行うことにより、またその組合せにより(再び投与される前に分子から薬剤の解離をもたらす(初回刺激と追加免疫を用いる抗原惹起で見られるものに類似))、種々の親和性(すなわち、必要とされるものよりも高いまたは低い)で結合するアンタゴニストを提供することにより調節され得る。いくつかの場合では、免疫系を刺激した後にその刺激を取り去ることが特に望ましいことがある。その結合相手に対するアンタゴニストの親和性を用いて、解離に必要な時間を決定することができる(親和性の高い薬剤は親和性の低い薬剤よりも解離に時間がかかる)。また、PD−1またはリガンドのいずれかと結合する、また同じ分子に異なる親和性で結合するアンタゴニストの組合せを用いて、免疫刺激の程度を調節することもできる。
【0015】
これらの組成物は、(i)PD−1を介した阻害的シグナル伝達を誘導することなくPD−1と結合してそれを遮断し、PD−L1およびPD−L2などのリガンドの結合を妨げ、それにより、PD−1により媒介される阻害シグナルの活性化を妨げるか、または(ii)PD−1のリガンドと結合し、PD−1受容体との結合を回避し、それにより、PD−1のより媒介される阻害シグナルの活性化を妨げるPD−1アンタゴニストを含む。
【0016】
好ましい組成物は、十分なT細胞応答の欠如、T細胞の消耗、T細胞の無応答、ならびに単球、マクロファージ、樹状細胞および他のAPCの活性化、または被験体におけるこれらの影響の総てを低減するまたは克服するのに有効な量の非抗体PD−1アンタゴニスト、例えば、PD−L2融合タンパク質(PD−L2−Ig)を含む。PD−1アンタゴニストはまた、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合するPD−L1タンパク質、断片、変異体または融合物も含む。これらのPD−L1断片は非機能的PD−L1断片とも呼ばれる。PD−L2ポリペプチド、融合タンパク質、および非機能的PD−L1断片は、PD−1の内因性リガンドがPD−1と相互作用しないようにすることにより、T細胞においてPD−1を介して起こる阻害的シグナル伝達を阻害または低減することができる。さらなる好ましいPD−1アンタゴニストとしては、PD−1のリガンドと結合し、T細胞上の内因性PD−1受容体との結合を妨げるPD−1またはその可溶性断片が含まれる。これらのPD−1断片は可溶性PD−1断片とも呼ばれる。その他のPD−1アンタゴニストとしては、PD−L1と結合し、PD−L1とPD−1の結合を妨げるB7.1またはその可溶性断片が含まれる。
【0017】
さらなる実施形態としては、阻害的シグナル伝達を生じることなくPD−1受容体と、またはPD−L1およびPD−L2などのPD−1受容体のリガンドのいずれかと結合してそれを遮断する抗体が含まれる。PD−L2ポリペプチド、融合タンパク質および非機能的PD−L1断片はまた、T細胞またはAPC上の別の受容体と結合することによってT細胞を活性化することもできる。
【0018】
アンタゴニストの作用は、T細胞の消耗、T細胞の無応答、またはその双方を克服する手助けをするとともに、感染または癌により誘導される単球、マクロファージ、樹状細胞、および他のAPCを活性化する。PD−L2ポリペプチドまたは融合タンパク質で処置可能な代表的感染としては、限定されるものではないが、ウイルス、細菌、寄生虫、原虫、または真菌により引き起こされる感染が挙げられる。処置可能な例示的ウイルス感染としては、限定されるものではないが、肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、ヒトTリンパ球向性ウイルス(HTLV)、ヘルペスウイルス、インフルエンザ、エプスタイン・バーウイルス、フィロウイルス、またはヒト乳頭腫ウイルスにより引き起こされる感染が挙げられる。処置可能な他の感染としては、プラスモジウム、マイコプラズマ、結核菌(M. tuberculosis)、炭疽菌(Bacillus anthracis)、ブドウ状球菌(Staphylococcus)、およびC.トラコマチス(C. trachomatis)により引き起こされるものが挙げられる。
【0019】
PD−1アンタゴニストは、ウイルス抗原、細菌抗原、原虫抗原、および腫瘍特異的抗原などの1以上の抗原を含有するワクチンと組み合わせて、または交互に投与することができる。PD−1アンタゴニストは、被験体において一次免疫応答およびエフェクター細胞応答を増強するためにワクチンとともに有効なアジュバントとして使用することができる。処置される好ましい被験体は免疫系が弱まっているか、または免疫不全であり、65歳を超えるか、または2歳未満である。
【図面の簡単な説明】
【0020】
【図1】図1A−Bは、PD−1結合ELISAにおけるB7−DC−IgのPD−1への結合を示すグラフである。
【図2】図2は、B7−DC−IgがPD−1発現CHO細胞と結合することを示すグラフである。
【図3】図3は、B7−DC−IgがPD−1との結合に関してB7−H1と競合することを示すグラフである。
【図4】図4は、B7−DC−Igの組合せ処置が腫瘍モデルにおいて抗原特異的メモリーCTLの生成をもたらしたことを示す。
【図5】図5は、B7−DC−IgがHSV−2ウイルス粒子の放出を低減し、HSV−2ワクチンの存在下でマウスの生存率を高めたことを示す。
【発明の具体的説明】
【0021】
I.定義
本明細書において「単離された」とは、その化合物が天然に存在する場合と異なる環境にある、例えば、ペプチドをそれが天然には見られない濃度まで濃縮することなどによってその天然環境から分離された目的化合物(例えば、ポリヌクレオチドまたはポリペプチドのいずれか)を表すことを意味する。「単離された」とは、目的化合物が有意に富化された、かつ/または目的化合物が部分的または有意に精製されたサンプル内にある化合物を含むことを意味する。「有意に」とは、統計学的に有意に大きいことを意味する。
【0022】
本明細書において「ポリペプチド」とは、修飾(例えば、リン酸化またはグリコシル化)とは無関係に任意の長さのアミノ酸鎖を意味する。
【0023】
本明細書において「変異体」ポリペプチドは、対応する野生型ポリペプチドのアミノ酸配列に比べて少なくとも一つのアミノ酸配列の変更を含む。
【0024】
本明細書において「アミノ酸配列の変更」とは、例えば、1以上のアミノ酸の置換、欠失または挿入であり得る。
【0025】
本明細書において「ベクター」とは、別のDNAセグメントが、その挿入されたセグメントの複製を生じるように挿入することができるプラスミド、ファージ、またはコスミドなどのレプリコンである。本明細書に記載のベクターは発現ベクターであり得る。
【0026】
本明細書において「発現ベクター」とは、1以上の発現制御配列を含むベクターである。
【0027】
本明細書において「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。
【0028】
本明細書において「機能的に連結される」とは、発現制御配列が目的のコード配列の発現を有効に制御するように遺伝構築物に組み込まれることを意味する。
【0029】
本明細書においてポリペプチドの「断片」とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。一般に、断片はアミノ酸5個以上の長さである。
【0030】
本明細書において「価数」とは、分子当たりに利用可能な結合部位の数を意味する。
【0031】
本明細書において「保存的」アミノ酸置換とは、置換されたアミノ酸が類似の構造的または化学的特性を有する置換である。
【0032】
本明細書において「非保存的」アミノ酸置換とは、置換されたアミノ酸の電荷、疎水性または嵩が有意に変化するものである。
【0033】
本明細書において「単離された核酸」とは、哺乳類ゲノムにおいてその核酸の一端または両端に通常隣接している核酸をはじめ、哺乳類ゲノムに存在する他の核酸分子から分離された核酸を意味する。
【0034】
本明細書において、核酸に関して「単離された」は任意の非天然核酸配列を含む(このような非天然配列は天然には見られず、天然ゲノムには直接連続する配列を持たないため)。
【0035】
本明細書において「宿主細胞」とは、組換え発現ベクターが導入できる原核細胞および真核細胞を意味する。
【0036】
本明細書において「形質転換させる」および「トランスフェクトする」とは、当該技術分野で知られているいくつかの技術によって核酸(例えばベクター)を細胞に導入することを包含する。
【0037】
本明細書において「抗体」とは、完全分子ならびに抗原−結合部位を含むその断片の双方を含むことを意味する。これらには完全抗体のFc断片を欠いたFabおよびF(ab’)2断片が含まれる。
【0038】
「免疫細胞」とは、免疫応答に役割を果たす造血系起源の細胞を意味する。免疫細胞は、リンパ球(例えば、B細胞およびT細胞)、ナチュラルキラー細胞および骨髄細胞(例えば、単球、マクロファージ、好酸球、肥満細胞、好塩基球、および顆粒球)を含む。
【0039】
「T細胞」とは、CD4+T細胞またはCD8+T細胞を意味する。T細胞とは、TH1細胞、TH2細胞、およびTh17細胞を含む。
【0040】
「T細胞細胞傷害性」とは、CD8+T細胞の活性化により媒介されるいずれの免疫応答も含む。例示的免疫応答としては、サイトカインの産生、CD8+T細胞の増殖、グランザイムまたはパーフォリンの産生、および感染性病原体の排除が含まれる。
【0041】
「免疫細胞」とは、T細胞、B細胞、およびリンパ球を意味する。
【0042】
「阻害的シグナル伝達」とは、T細胞の増殖を低減することによるものであれ、他のいずれかの阻害機構によるものであれ、T細胞応答を抑制する、またはそうでなければ低減する作用を有するPD−L1または他のリガンドによるPD−1受容体を介したシグナル伝達を意味する。
【0043】
II.PD−1アンタゴニスト
PD−1とPD−L1の間の相互作用に干渉するのに好ましいPD−1アンタゴニスト化合物は、PD−L2(B7−DCとしても知られる)、PD−L2の細胞外ドメイン、PD−L2の融合タンパク質、およびPD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合してそれを遮断し、するPD−L1とPD−1の結合を妨げるその変異体である。さらなるPD−1アンタゴニストとしては、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合するPD−L1の断片、PD−1のリガンドと結合し、T細胞上の内因性PD−1受容体と結合するのを妨げるPD−1または可溶性その断片、ならびにPD−L1と結合し、PD−L1とPD−1の結合を妨げることができるB7.1または可溶性その断片が含まれる。ある特定の実施形態では、PD−1アンタゴニストは被験体におけるT細胞傷害性を高める。この多機能性PD−1アンタゴニストは被験体において強力な免疫応答を誘導し、T細胞の消耗およびT細胞の無応答を克服する助けをする。
【0044】
PD−1アンタゴニストはPD−1のリガンドと結合し、それと相互作用するか、またはリガンドとPD−1受容体の結合を阻害するか、またはPD−1受容体を介したシグナル伝達に結びつくことなくPD−1受容体と直接結合する。好ましい実施形態では、PD−1アンタゴニストはPD−1と直接結合し、PD−1阻害的シグナル伝達を遮断する。他の実施形態では、PD−1アンタゴニストはPD−1のリガンドと結合し、リガンドがPD−1を介した阻害的シグナル伝達を誘発するのを低減または阻害する。さらに別の実施形態では、PD−1アンタゴニストは、PD−1受容体以外の受容体と結合することによりT細胞を活性化することができる。
【0045】
PD−1アンタゴニストは小分子アンタゴニストであり得る。「小分子」とは、100ダルトンを超え、約2,500ダルトン未満、好ましくは100〜2000ダルトンの間、より好ましくは約100〜約1250ダルトンの間、より好ましくは約100〜約1000ダルトンの間、より好ましくは約100〜約750ダルトンの間、より好ましくは約200〜約500ダルトンの間の分子量を有する小有機化合物を意味する。これらの小分子は多くの場合、環状炭素または複素環式構造および/または1以上の官能基で置換された芳香族または多環芳香族構造を含む。これらの小分子アンタゴニストは、PD−L1およびPD−L2などのPD−1のリガンドと結合し、そのリガンドがPD−1と相互作用しないようにすることにより、またはPD−1受容体を介したシグナル伝達を誘発することなくPD−1受容体と直接結合することにより、PD−1受容体シグナル伝達を低減するか、またはそれと干渉する。
【0046】
例示的PD−1アンタゴニストとしては、限定されるものではないが、PD−L2、PD−L1、PD−1またはB7−1ポリペプチド、およびその変異体、断片または融合タンパク質が挙げられる。さらなる実施形態としては、これらのタンパク質のいずれかと結合する抗体が挙げられる。
【0047】
A.PD−L2に基づくPD−1アンタゴニスト
1.PD−1と結合する、PD−L2に基づくPD−1アンタゴニスト
PD−1アンタゴニストは免疫細胞上のPD−1と結合し、阻害的PD−1のシグナル伝達を遮断する。PD−1のシグナル伝達は、PD−1リガンド(PD−L2またはPD−L1、一般に、PD−L1)が免疫シナプス内でTCR:MHC複合体と近接してPD−1と結合する必要があると思われる。よって、PD−1を介した阻害的シグナル伝達を遮断し、場合により、T細胞膜上でのPD−1とTCRの同時結合を妨げるタンパク質、抗体または小分子は有用なPD−1アンタゴニストである。
【0048】
代表的なポリペプチドアンタゴニストとしては、限定されるものではないが、PD−L2ポリペプチド、その断片、その融合タンパク質、およびその変異体が挙げられる。PD−1と結合し、PD−1を介した阻害的シグナル伝達を遮断するPD−L2ポリペプチドは好ましい実施形態の一つである。他の実施形態としては、PD−1の天然リガンドが結合しないように、かつ、シグナル伝達を誘発しないようにするPD−1アンタゴニストが含まれる。ある特定の実施形態では、開示されたPD−L2ポリペプチドは、免疫シナプスに関してペプチド−MHC複合体によるTCRの同時結合が存在しないので、PD−1受容体を介したシグナル伝達が低減されているか、またはそれを誘発する能力がないと考えられる。PD−1受容体を介したシグナル伝達は、T細胞の活性化およびT細胞の増殖を減衰する負のシグナルを伝達するので、PD−1のシグナル伝達経路を阻害すれば、そうでなければ減衰される細胞を活性化させることができる。
【0049】
2.例示的PD−L2ポリペプチドPD−1アンタゴニスト
ネズミPD−L2ポリペプチドは、
【化1】
(配列番号1)または
【化2】
(配列番号2)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0050】
ヒトPD−L2ポリペプチドは、
【化3】
(配列番号3)または
【化4】
(配列番号4)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0051】
非ヒト霊長類(カニクイザル)PD−L2ポリペプチドは、
【化5】
(配列番号5)または
【化6】
(配列番号6)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号1、3、および5はそれぞれシグナルペプチドを含む。
【0052】
B.PD−L1に基づくPD−1アンタゴニスト
1.PD−1受容体と結合するPD−L1に基づくPD−1アンタゴニスト
PD−1受容体と結合する他のPD−1アンタゴニストとしては、限定されるものではないが、PD−L1ポリペプチド、その断片、その融合タンパク質およびその変異体が挙げられる。これらのPD−1ポリペプチドアンタゴニストはPD−1受容体と結合してそれを遮断し、PD−1受容体を介した阻害的シグナル伝達が低減されているか、またはそれを誘発する能力がない。一つの実施形態では、PD−L1ポリペプチドは、免疫シナプスに関してペプチド−MHC複合体によるTCRの同時結合が存在しないので、PD−1受容体を介したシグナル伝達が低減されているか、またはそれを誘発する能力がないと考えられる。PD−1受容体を介したシグナル伝達は、T細胞の活性化およびT細胞の増殖を減衰する負のシグナルを伝達するので、PD−L1ポリペプチドを用いてPD−1のシグナル伝達を阻害すれば、そうでなければ減衰される細胞を活性化することができる。
【0053】
2.例示的PD−L1ポリペプチドPD−1アンタゴニスト
ネズミPD−L1ポリペプチドは、
【化7】
(配列番号7)または
【化8】
(配列番号8)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0054】
ヒトPD−L1ポリペプチドは、
【化9】
(配列番号9)
【化10】
(配列番号10)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0055】
配列番号7および9はそれぞれシグナルペプチドを含む。
【0056】
C.B7.1およびPD−1に基づくPD−1アンタゴニスト
1.PD−L1およびPD−L2と結合するB7.1およびPD−1に基づくPD−1アンタゴニスト
他の有用なポリペプチドとしては、PD−L1またはPD−L2などのPD−1リガンドと結合し、内因性PD−1受容体との結合を妨げ、それにより阻害的シグナル伝達を妨げることができるPD−1受容体タンパク質、またはその可溶性断片が含まれる。このような断片はまた、場合により天然リガンドとの結合を増大させるA99L突然変異などの突然変異を含む、PD−1タンパク質の可溶性ECD部分も含む。PD−L1はまた、タンパク質B7.1と結合することも示されている(Butte, et al., Immunity, 27(1): 111-122 (2007))。よって、PD−L1リガンドと結合し、内因性PD−1受容体との結合を妨げ、それにより阻害的シグナル伝達を妨げることができるB7.1またはその可溶性断片もまた有用である。
【0057】
2.例示的B7.1ポリペプチドPD−1アンタゴニスト
ネズミB7.1ポリペプチドは、
【化11】
(配列番号11)または
【化12】
(配列番号12)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0058】
ヒトB7.1ポリペプチドは、
【化13】
(配列番号13)または
【化14】
(配列番号14)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号11および13はそれぞれシグナルペプチドを含む。
【0059】
3.例示的PD−1ポリペプチドPD−1アンタゴニスト
ヒトPD−1ポリペプチドは、
【化15】
(配列番号15)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0060】
非ヒト霊長類(カニクイザル)PD−1ポリペプチドは、
【化16】
(配列番号16)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号15および16はそれぞれシグナルペプチドを含む。
【0061】
D.PD−1アンタゴニストポリペプチドの断片
PD−1アンタゴニストポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。本明細書において、PD−1アンタゴニストポリペプチドの断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0062】
有用な断片は、それらの天然リガンドと結合する能力を保持するものである。全長PD−1アンタゴニストポリペプチドの断片であるPD−1アンタゴニストポリペプチドは一般に、全長PD−1アンタゴニストポリペプチドに比べてその天然リガンドと結合する能力の少なくとも20パーセント、30パーセント、40パーセント、50パーセント、60パーセント、70パーセント、80パーセント、90パーセント、95パーセント、98パーセント、99パーセント、100パーセントを有する、またはさらには100パーセントを超える。
【0063】
例えば、PD−L2およびPD−L1の有用な断片は、PD−1と結合する能力を保持するものである。PD−L2およびPD−L1断片は一般に、全長PD−L2およびPD−L1に比べてPD−1と結合する能力の少なくとも20パーセント、30パーセント、40パーセント、50パーセント、60パーセント、70パーセント、80パーセント、90パーセント、95パーセント、98パーセント、99パーセント、100パーセントを有する、またはさらには100パーセントを超えるものである。
【0064】
PD−1アンタゴニストポリペプチドの断片は可溶性断片を含む。可溶性PD−1アンタゴニストポリペプチド断片は、産生細胞から放出、分泌またはそうでなければ抽出され得るPD−1アンタゴニストポリペプチドの断片である。PD−1アンタゴニストポリペプチドの可溶性断片は、該ポリペプチドの細胞外ドメインの一部または全部を含み、細胞内ドメインおよび/または膜貫通ドメインの一部または全部を欠く。一実施形態では、PD−1アンタゴニストポリペプチド断片はPD−1アンタゴニストポリペプチドの全細胞外ドメインを含む。細胞外ドメインは膜貫通ドメイン由来の1、2、3、4または5個のアミノ酸を含み得ると考えられる。あるいは、細胞外ドメインはC末端、N末端または双方から除去された1、2、3、4、または5個のアミノ酸を有し得る。
【0065】
一般に、PD−1アンタゴニストポリペプチドまたはその断片は、シグナル配列をコードする配列を含む核酸から発現される。このシグナル配列が一般に未熟なポリペプチドから切断されて、シグナル配列を欠く成熟ポリペプチドが生じる。PD−1アンタゴニストポリペプチドのこのシグナル配列は、そのポリペプチドの発現レベル、分泌、溶解度または他の特性に影響を与えるために標準的な分子生物学的技術を用いて別のポリペプチドのシグナル配列で置換することができる。PD−1アンタゴニストポリペプチドシグナル配列を置換するために用いられるシグナル配列は当該技術分野で知られているいずれのものであってもよい。
【0066】
1.PD−L2細胞外ドメイン
a.ヒトPD−L2細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはヒトPD−L2またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化17】
(配列番号17)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0067】
別の実施形態では、PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化18】
(配列番号18)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0068】
このシグナル配列は成熟タンパク質では除去されると考えられる。さらに、産生中に宿主からのタンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができると考えられる。配列番号19は、シグナル配列を含まない配列番号18のヒトアミノ酸配列を示す。
【化19】
(配列番号19)
【0069】
別の実施形態では、PD−1アンタゴニストポリペプチドはヒトPD−L2のIgVドメインを含む。第一の融合相手は、
【化20】
(配列番号20)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0070】
PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化21】
(配列番号21)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0071】
b.非ヒト霊長類PD−L2細胞外ドメイン
一つの実施形態では、PD−1アンタゴニストポリペプチドは非ヒト霊長類(カニクイザル)PD−L2の細胞外ドメインまたはその断片を含む。PD−1アンタゴニストポリペプチドは、
【化22】
(配列番号22)
と少なくとも80%、85%、90%、95%、99%または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0072】
別の実施形態では、PD−1アンタゴニストポリペプチドは、非ヒト霊長類アミノ酸配列:
【化23】
(配列番号23)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0073】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からの融合タンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができる。配列番号24は、シグナル配列を含まない配列番号23の非ヒト霊長類アミノ酸配列を示す。
【化24】
(配列番号24)
【0074】
別の実施形態では、PD−1アンタゴニストポリペプチドは非ヒト霊長類PD−L2のIgVドメインを含む。第一の融合相手は、
【化25】
(配列番号25)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0075】
PD−1アンタゴニストポリペプチドは、非ヒト霊長類アミノ酸配列:
【化26】
(配列番号26)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0076】
d.ネズミPD−L2細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはネズミPD−L2またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化27】
(配列番号27)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0077】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化28】
(配列番号28)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0078】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができる。配列番号29は、シグナル配列を含まない配列番号28のネズミアミノ酸配列を示す。
【化29】
(配列番号29)
【0079】
別の実施形態においては、PD−1アンタゴニストポリペプチドはネズミPD−L2のIgVドメインを含む。第一の融合相手は、
【化30】
(配列番号30)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0080】
PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化31】
(配列番号31)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0081】
d.PD−L2細胞外ドメイン断片
PD−L2細胞外ドメインは、シグナルペプチドまたはPD−L2の推定膜貫通ドメイン由来の1以上のアミノ酸を含み得る。分泌の際に切断されるシグナルペプチドのアミノ酸の数は発現系および宿主によって異なり得る。さらに、PD−1と結合する能力を保持するC末端またはN末端から1以上のアミノ酸を欠くPD−L2細胞外ドメインの断片を使用することもできる。
【0082】
第一の融合相手として使用することができるネズミPD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号53の
【化32】
が挙げられる。
【0083】
ネズミPD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号1の
【化33】
が挙げられる。シグナルペプチドは、配列番号1内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0084】
第一の融合相手として使用することができるヒトPD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号56の
【化34】
が挙げられる。
【0085】
ヒトPD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号3の
【化35】
が挙げられる。シグナルペプチドは、配列番号3内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0086】
第一の融合相手として使用することができる非ヒト霊長類PD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号5の
【化36】
が挙げられる。
【0087】
非ヒト霊長類PD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号5の
【化37】
が挙げられる。シグナルペプチドは、配列番号5内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0088】
PD−L2タンパク質はまた、配列番号3(ヒト全長)のアミノ酸20−121または配列番号23(細胞外ドメインまたはECD)のアミノ酸1−102のPD−1結合断片も含む。その特定の実施形態では、PD−L2ポリペプチドまたはPD−1結合断片はまた、配列番号3の残基110−114にアミノ酸WDYKYを、または配列番号23の残基91−95にWDYKYを組み込んでいる。限定されない例として、このようなPD−1結合断片は、配列番号3のアミノ酸20−121の配列の少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも60、少なくとも70、少なくとも75、少なくとも80、少なくとも85、少なくとも90、少なくとも95または少なくとも100個の連続するアミノ酸を含んでなり、このようなPD−1結合断片のそれぞれの好ましい実施形態は、部分断片として配列番号3の残基110−114に見られるアミノ酸WDYKY、または配列番号23の残基91−95に見られるWDYKYを含んでなる。
【0089】
2.PD−L1細胞外ドメイン
一つの実施形態では、変異体PD−L1ポリペプチドは細胞外ドメインの全部または一部を含む。PD−L1の代表的な細胞外ドメインのアミノ酸配列は、
【化38】
(配列番号32)
と80%、85%、90%、95%、または99%の配列同一性を有し得る。
【0090】
PD−L1の膜貫通ドメインは配列番号9の239番のアミノ酸で始まる。PD−L1の好適な断片は、シグナルペプチド配列、例えば、配列番号9またはその変異体の1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、膜貫通ドメインの1、2、3、4、5、6、7、8、9、もしくは10個のアミノ酸、またはその組合せを含み得る。
【0091】
ネズミPD−L1の細胞外ドメインは以下のアミノ酸配列:
【化39】
(配列番号33)
を有する。
【0092】
ネズミPD−L1の膜貫通ドメインは配列番号7の240番のアミノ酸で始まる。ある特定の実施形態では、PD−L1ポリペプチドは、シグナルペプチドの1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、膜貫通ドメインの1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、またはその組合せを有するネズミPD−L1の細胞外ドメインを含む。
【0093】
3.B7.1細胞外ドメイン
a.ネズミB7.1細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミB7.1またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化40】
(配列番号34)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0094】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化41】
(配列番号35)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0095】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために、他の生物由来のシグナルペプチドを使用することができる。配列番号36は、シグナル配列を含まない配列番号35のネズミアミノ酸配列を示す。
【化42】
(配列番号36)
【0096】
別の実施形態においては、PD−1アンタゴニストポリペプチドはネズミB7.1のIgVドメインを含む。第一の融合相手は、
【化43】
(配列番号37)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0097】
PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化44】
(配列番号38)(B7.1Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0098】
b.ヒトB7.1細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはヒトB7.1またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化45】
(配列番号39)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0099】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化46】
(配列番号40)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0100】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために、他の生物由来のシグナルペプチドを使用することができる。配列番号41は、シグナル配列を含まない配列番号40のヒトアミノ酸配列を示す。
【化47】
(配列番号41)
【0101】
別の実施形態においては、PD−1アンタゴニストポリペプチドはヒトB7.1のIgVドメインを含む。第一の融合相手は、
【化48】
(配列番号42)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0102】
PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化49】
(配列番号43)(B7.1Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0103】
3.B7.1細胞外ドメイン断片
共刺激ポリペプチドドメインとして使用することができるネズミB7.1の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号11の
【化50】
が挙げられる。
【0104】
ネズミB7.1のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号11の
【化51】
が挙げられる。シグナルペプチドは、配列番号11内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0105】
共刺激ポリペプチドドメインとして使用することができるヒトB7.1の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号13の
【化52】
が挙げられる。
【0106】
ヒトB7.1のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号13の
【化53】
が挙げられる。シグナルペプチドは、配列番号13内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0107】
E.変異体
1.変異体PD−L2およびPD−L1 PD−1アンタゴニスト
さらなるPD−1アンタゴニストとしては、生理学的条件下でPD−1と結合する能力を保持し、PD−1に対する結合が増強されるか、またはPD−1受容体を介したシグナル伝達を促進する能力が低減されるように突然変異誘発されたPD−L2およびPD−L1、ポリペプチドおよびその断片が含まれる。一つの実施形態は、PD−1を活性化し、T細胞へ阻害シグナルを伝達するポリペプチドの能力を非突然変異PD−L2またはPD−L1に比べて阻害または低減する1以上のアミノ酸置換、欠失、または挿入を含む単離されたPD−L2およびPD−L1ポリペプチドを提供する。該PD−L2およびPD−L1ポリペプチドはいずれの起源種のものであってもよい。一つの実施形態においては、PD−L2またはPD−L1ポリペプチドは哺乳類種に由来する。好ましい実施形態では、PD−L2またはPD−L1ポリペプチドはヒトまたは非ヒト霊長類起源のものである。
【0108】
別の実施形態においては、変異型PD−L2またはPD−L1ポリペプチドは、PD−1に対して野生型または非変異型PD−L2またはPD−L1と同じ結合活性を有するが、非突然変異PD−L2またはPD−L1ポリペプチドに比べてPD−1受容体を介したシグナル伝達を刺激する能力を持たないか、または10%未満である。他の実施形態では、該変異型PD−L2またはPD−L1ポリペプチドは野生型PD−L2またはPD−L1よりもPD−1に対して10%、20%、30%、40%、50%、60%、70%、80%、90%、100%またはそれを超える結合活性を有し、非突然変異PD−L2またはPD−L1ポリペプチドに比べてPD−1受容体を介したシグナル伝達を刺激する能力が50%、40%、30%、20%、または10%未満である。
【0109】
変異型PD−L2またはPD−L1ポリペプチドは、アミノ酸置換、欠失、または挿入の任意の組合せを有し得る。一つの実施形態では、単離されたPD−L2またはPD−L1変異型ポリペプチドは、それらのアミノ酸配列が野生型PD−L2またはPD−L1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するように、ある整数のアミノ酸変化を有する。好ましい実施形態では、B7−H1変異型ポリペプチドは、野生型ネズミ、非ヒト霊長類またはヒトPD−L2またはPD−L1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するアミノ酸配列を有する。
【0110】
配列同一性%は、コンピュータープログラムまたは直接的配列比較を用いて計算することができる。2配列間の同一性を決定するための好ましいコンピュータープログラム法としては、限定されるものではないが、GCGプログラムパッケージ、FASTA、BLASTP、およびTBLASTNが挙げられる(例えば、D. W. Mount, 2001, Bioinformatics: Sequence and Genome Analysis, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 参照)。BLASTPおよびTBLASTNプログラムはNCBIおよび他のソースから公的に入手可能である。周知のSmith Watermanアルゴリズムも同一性を決定するために使用可能である。
【0111】
アミノ酸配列比較のためのパラメーター例としては以下のものが挙げられる:1)Needleman and Wunsch (J. Mol. Biol., 48:443-453 (1970))からのアルゴリズム、2)Hentikoff and Hentikoff (Proc. Natl. Acad. Sci. U.S.A., 89:10915-10919 (1992))からのBLOSSUM62比較マトリックス、3)ギャップペナルティー=12、および4)ギャップレングスペナルティー=4。これらのパラメーターを用いる有用なプログラムは、「ギャップ」プログラム(Genetics Computer Group, Madison, Wis.)として公的に入手可能である。上述のパラメーターはポリペプチド比較のためのデフォルトパラメーターである(エンドギャップに対するペナルティーは無い)。
【0112】
あるいは、ポリペプチド配列同一性は、次式:同一性%=(同一の残基の数)/(アミノ酸残基のアライメント長)*100を用いて計算することができる。この計算では、配列長は内部ギャップを含むが、末端ギャップは含まない。
【0113】
PD−L2またはPD−L1ポリペプチドのアミノ酸置換は「保存的」または「非保存的」であり得る。本明細書において、「保存的」アミノ酸置換は、置換されたアミノ酸が類似の構造的または化学的特性を有する置換であり、「非保存的」アミノ酸置換は、置換されたアミノ酸の電荷、疎水性、または嵩が有意に変更されるものである。非保存的置換は、(a)例えば、シートまたはらせんコンフォメーションのような置換領域におけるペプチド主鎖の構造、(b)標的部位における分子の電荷または疎水性、または(c)側鎖の嵩の維持に対するそれらの効果がより有意に異なる。
【0114】
保存的アミノ酸置換の例としては、置換が以下の5群中のものが挙げられる:1)小さな脂肪族、非極性またはやや極性の残基(Ala、Ser、Thr、Pro、Gly)、2)極性、負電荷を有する残基およびそれらのアミド(Asp、Asn、Glu、Gln)、極性、正電荷を有する残基(His、Arg、Lys)、大きな脂肪族、非極性残基(Met、Leu、Ile、Val、Cys)、および大きな芳香族残基(Phe、Tyr、Trp)。非保存的アミノ酸置換の例としては、1)親水性残基(例えば、セリルまたはトレオニル)が疎水性残基(例えば、ロイシル、イソロイシル、フェニルアラニル、バリルまたはアラニル)から(またはにより)置換される場合、2)システインまたはプロリンが他の残基から(またはにより)置換される場合、3)電気的に陽性の側鎖を有する残基(例えば、リシル、アルギニルまたはヒスチジル)が電気的に陰性の残基(例えば、グルタミルまたはアスパルチル)から(またはにより)置換される場合、または4)嵩高の側鎖を有する残基(例えば、フェニルアラニン)が側鎖を持たない残基(例えば、グリシン)から(またはにより)置換される場合が挙げられる。
【0115】
しかしながら、挙げられたアミノ酸位置における置換は、任意のアミノ酸またはアミノ酸類似体を用いて行うことができると理解される。例えば、挙げられた位置における置換は、任意の天然アミノ酸(例えば、アラニン、アスパラギン酸、アスパラギン、アルギニン、システイン、グリシン、グルタミン酸、グルタミン、ヒスチジン、ロイシン、バリン、イソロイシン、リシン、メチオニン、プロリン、トレオニン、セリン、フェニルアラニン、トリプトファン、またはチロシン)を用いて行うことができる。
【0116】
本明細書に記載される置換はマウス、非ヒト霊長類およびヒトPD−L2またはPD−L1に関するものであるが、当業者ならば他種(例えば、ラット、ハムスター、モルモット、アレチネズミ、ウサギ、イヌ、ネコ、ウマ、ブタ、ヒツジ、またはウシ)由来の相当するポリペプチドにおいても等価の変更を容易に行うことができることを注記する。しかしながら、結合は種特異的な成分であることから、PD−1アンタゴニストをヒト投与する場合にはヒトを使用するのが好ましい。
【0117】
一つの実施形態においては、開示されている単離された変異型PD−L2またはPD−L1ポリペプチドはPD−1のアンタゴニストであり、PD−1を介したシグナル伝達を誘発することなくPD−1と結合してそれを遮断する。PD−1シグナル伝達によるT細胞の減衰を妨げることにより、より多くのT細胞の活性化に利用可能である。T細胞阻害を妨げると、PD−1アンタゴニストと接触していないT細胞に比べて、T細胞応答が高まるか、T細胞の増殖が増大し、T細胞によるサイトカインの産生および/または分泌が高まるか、T細胞の分化およびエフェクター機能が刺激されるか、またはT細胞の生存が促進される。この相互作用から生じるT細胞応答は一般に、PD−1アンタゴニストポリペプチドの非存在下での応答よりも大きい。PD−1アンタゴニストポリペプチドの非存在下でのT細胞の応答は応答性がないか、またはPD−1アンタゴニストポリペプチドの存在下よりも有意に低いものであり得る。このT細胞の応答は、エフェクター(例えば、CTLまたは抗体産生B細胞)応答、1以上のエフェクター(例えば、CTLまたは抗体産生B細胞)応答を助けるヘルパー応答、または抑制的応答であり得る。
【0118】
2分子間の結合親和性を測定する方法は技術分野でよく知られている。PD−1に対する変異型PD−L2またはPD−L1ポリペプチドの結合親和性を測定する方法としては、限定されるものではないが、蛍光活性化細胞選別(FACS)、表面プラズモン共鳴、蛍光異方性、アフィニティークロマトグラフィー、および親和性選択−質量分析が挙げられる。
【0119】
本明細書に開示される変異型ポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。好ましい断片は、PD−1と結合するのに有効な細胞外ドメインの全部または一部を含む。本明細書において、断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0120】
2.変異型B7.1およびPD−1アンタゴニスト
さらなるPD−1アンタゴニストとしては、生理学的条件下でPD−L2および/またはPD−L1と結合する能力を保持するか、またはPD−L2および/またはPD−L1に対する結合が増強されるように改変されているB7.1およびPD−1ポリペプチドおよびその断片が含まれる。このような変異体PD−1タンパク質としては、天然リガンドに対する結合を増強する、A99L突然変異などの突然変異を含むPD−1タンパク質の可溶性ECD部分が挙げられる(Molnar et al., Crystal structure of the complex between programmed death-1 (PD-1) and its ligand PD-L2, PNAS, Vol. 105, pp. 10483-10488 (29 July 2008))。B7.1およびPD−1ポリペプチドはいずれの起源種のものであってもよい。一つの実施形態では、B7.1またはPD−1ポリペプチドは哺乳類種に由来する。好ましい実施形態では、B7.1またはPD−1ポリペプチドはヒトまたは非ヒト霊長類起源のものである。
【0121】
変異型B7.1またはPD−1ポリペプチドは、アミノ酸置換、欠失、または挿入の任意の組合せを有し得る。一つの実施形態では、単離されたB7.1またはPD−1変異型ポリペプチドは、それらのアミノ酸配列が野生型B7.1またはPD−1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するように、ある整数のアミノ酸変化を有する。好ましい実施形態では、B7.1またはPD−1変異型ポリペプチドは、野生型ネズミ、非ヒト霊長類、またはヒトB7.1またはPD−1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するアミノ酸配列を有する。
【0122】
B7.1またはPD−1ポリペプチドにおけるアミノ酸置換は「保存的」または「非保存的」であり得る。保存的および非保存的置換は上記に定義されている。
【0123】
一つの実施形態においては、開示されている単離された変異型B7.1またはPD−1ポリペプチドはPD−1のアンタゴニストであり、PD−L2および/またはPD−L1と結合し、それによりそれらの内因性PD−1との結合を遮断する。PD−1シグナル伝達によるT細胞の減衰を妨げることにより、より多くのT細胞を活性化に利用可能とする。T細胞阻害を妨げると、PD−1アンタゴニストと接触していないT細胞に比べて、T細胞応答が高まるか、T細胞の増殖が増大するか、T細胞によるサイトカインの産生および/または分泌が高まるか、T細胞の分化およびエフェクター機能が刺激されるか、またはT細胞の生存が促進される。この相互作用から生じるT細胞応答は一般に、PD−1アンタゴニストポリペプチドの非存在下での応答よりも大きい。PD−1アンタゴニストポリペプチドの非存在下でのT細胞の応答は、応答性がないか、またはPD−1アンタゴニストポリペプチドの存在下よりも有意に低いものであり得る。このT細胞の応答は、エフェクター(例えば、CTLまたは抗体産生B細胞)応答、1以上のエフェクター(例えば、CTLまたは抗体産生B細胞)応答を助けるヘルパー応答、または抑制的応答であり得る。
【0124】
変異体ポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。好ましい断片は、PD−L2および/またはPD−L1と結合するのに有効な細胞外ドメインの全部または一部を含む。本明細書において、断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0125】
F.融合タンパク質
いくつかの実施形態では、PD−1アンタゴニストは、第一のポリペプチドドメインと第二のドメインとを含む融合タンパク質である。この融合タンパク質はT細胞受容体に結合することができるか、あるいは好ましくはこの融合タンパク質は、例えばPD−1と競合的に結合することによって、T細胞と結合して、T細胞への阻害的シグナル伝達を遮断することができる。開示されている組成物は、天然阻害リガンドとPD−1との結合に干渉することにより、PD−1を介したシグナル伝達を有効に遮断する。好適な共刺激ポリペプチドとしては、阻害性T細胞シグナル伝達受容体に対する結合親和性が増強または低下されたPD−1などの変異型ポリペプチドおよび/またはその断片が含まれる。
【0126】
融合タンパク質はまた場合により、第一のポリペプチドドメインを抗原結合ドメインから分離するペプチドまたはポリペプチドリンカードメインを含んでもよい。
【0127】
本明細書に開示される融合タンパク質は式I:
N−R1−R2−R3−C
(式中、「N」は融合タンパク質のN末端を表し、「C」は融合タンパク質のC末端を表し、「R1」はPD−L2、PD−L1、B7.1、もしくはPD−1ポリペプチドまたは抗原結合標的化ドメインであり、「R2」はペプチド/ポリペプチドリンカードメインであり、「R3」は標的化ドメインまたは抗原結合標的化ドメインであり、ここで、「R1」が抗原結合標的化ドメインである場合には、「R3」はポリペプチドドメインであり、「R1」がPD−L2、PD−L1、B7.1またはPD−1ポリペプチドドメインである場合には、「R3」は抗原結合標的化ドメインである)
で表されるものである。好ましい実施形態では、「R1」はPD−L2、PD−L1、B7.1、またはPD−1ポリペプチドドメインであり、「R3」は抗原結合標的化ドメインである。
【0128】
場合により、融合タンパク質はさらに、2以上の融合タンパク質を二量体化または多量体化する働きをするドメインを含む。該融合タンパク質を二量体化または多量体化する働きをするドメインは別個のドメインであってもよいし、あるいはまたその融合タンパク質の他のドメイン(PD−L2、PD−L1、B7.1もしくはPD−1ポリペプチドドメイン、抗原結合標的化ドメインまたはペプチド/ポリペプチドリンカードメイン)の一つに含まれていてもよい。
【0129】
融合タンパク質は二量体化または多量体化されていてもよい。二量体化または多量体化は二量体化または多量体化ドメインを介して2以上の融合タンパク質間で生じ得る。あるいは、融合タンパク質の二量体化または多量体化は化学的架橋によっても生じする。形成される二量体または多量体は、ホモ二量体/ホモ多量体またはヘテロ二量体/ヘテロ多量体であり得る。
【0130】
融合タンパク質のモジュール性およびそれらの種々の組合せでの二量体化または多量体化能は、腫瘍細胞の微小環境または免疫調節組織に対する免疫応答を増強する働きをする標的化分子に豊富な選択肢を与える。
【0131】
1.抗原結合標的化ドメイン
融合タンパク質はまた抗原結合標的化ドメインを含む。いくつかの実施形態では、標的化ドメインは、感染性疾患を引き起こす病原体に応答したT細胞の活性化の調節に関与する免疫組織に特異的な抗原、リガンドまたは受容体と結合する。
【0132】
標的化ドメイン
標的化のための抗原、リガンド、および受容体
一つの実施形態においては、融合タンパク質は、感染性疾患を引き起こす病原体に応答したT細胞の活性化の調節に関与する免疫組織により発現される抗原と特異的に結合するドメインを含む。
【0133】
標的化ドメインの分子種
リガンドおよび受容体
一つの実施形態においては、疾病標的化ドメインは、罹患細胞で特異的に発現されるか、または正常組織に比べて罹患細胞で過剰発現される細胞表面抗原もしくは受容体と結合するリガンドである。罹患細胞はまた、増殖および発達に影響を及ぼす微小環境中に多数のリガンドを分泌する。限定されるものではないが、増殖因子、サイトカインおよびケモカイン(上記に示されるケモカインを含む)をはじめとする、罹患細胞により分泌されるリガンドと結合する受容体は、開示されている融合タンパク質に用いるのに好適である。罹患細胞により分泌されるリガンドは、分泌されたリガンドと結合する受容体の可溶性断片を用いて標的化され得る。可溶性受容体断片は産生細胞から放出、分泌またはそうでなければ抽出され得る断片ポリペプチドであり、全細胞外ドメインまたはその断片を含む。
【0134】
単鎖ポリペプチド抗体
別の実施形態では、疾病関連標的化ドメインは、罹患細胞で特異的に発現されるか、または正常組織に比べて罹患細胞で過剰発現される細胞表面抗原または受容体と結合する単鎖抗体である。単鎖ドメイン抗体は共阻害性受容体アンタゴニストドメインに関して上記されている。
【0135】
Fcドメイン
別の実施形態では、疾病または疾病関連標的化ドメインは、罹患細胞で発現されるFc受容体と結合する免疫グロブリン重鎖のFcドメインである。Fc領域は、第一の定常領域免疫グロブリンドメインを除く抗体の定常領域を含むポリペプチドを含む。よって、FcはIgA、IgDおよびIgGの最後の二つの定常領域免疫グロブリンドメインとIgEおよびIgMの最後の3つの定常領域免疫グロブリンドメインを意味する。好ましい実施形態では、Fcドメインはヒトまたはネズミ免疫グロブリンに由来する。より好ましい実施形態では、Fcドメインは、CH2およびCH3領域を含むヒトIgG1またはネズミIgG2aに由来する。
【0136】
一つの実施形態においては、ヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3域は、
【化54】
(配列番号44)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0137】
配列番号44によりコードされるヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3領域は以下のアミノ酸配列:
【化55】
(配列番号45)
を有する。
【0138】
別の実施形態においては、ネズミ免疫グロブリンCγ2a鎖のヒンジ、CH2およびCH3領域は、
【化56】
(配列番号46)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0139】
配列番号46によりコードされるネズミ免疫グロブリンCγ2a鎖のヒンジ、CH2およびCH3領域は以下のアミノ酸配列:
【化57】
(配列番号47)
を有する。
【0140】
一つの実施形態においては、Fcドメインは、腫瘍もしくは腫瘍関連新生血管系で特異的に発現されるか、または正常組織に比べて腫瘍もしくは腫瘍関連新生ポリペプチド血管系で過剰発現される特異的Fc受容体との結合を増強する1以上のアミノ酸挿入、欠失または置換を含み得る。好適なアミノ酸置換は上記のように保存的および非保存的置換を含む。
【0141】
非ホジキンリンパ腫またはワルデンストロームマクログロブリン血症に対してリツキシマブ(キメラマウス/CD20に対するヒトIgG1モノクローナル抗体)で処置された患者における治療結果は、ヒトIgG1のFcドメインに対して明瞭な内因的親和性を有するFcγ受容体の対立遺伝子変異体の個々の発現と相関していた。特に、Fc受容体CD16A(FcγRIIIA)を活性化する低親和性の高親和性対立遺伝子を有する患者はより高い奏功率を示し、非ホジキンリンパ腫の場合には、無進行生存を改善した。別の実施形態では、Fcドメインは、低親和性阻害Fc受容体CD32B(FcγRIIB)との結合を低減し、かつ低親和性活性化Fc受容体CD16A(FcγRIIIA)との野生型レベルの結合を保持するか、またはそれとの結合を増強する以上のアミノ酸挿入、欠失、または置換を含み得る。
【0142】
好ましい実施形態では、Fcドメインは、CD16Aとの結合を増強するアミノ酸挿入、欠失、または置換を含む。CD16Aとの結合を増強し、CD32Bとの結合を低減するヒトIgG1のFcドメインの多数の置換が当該技術分野に知られており、Stavenhagen, et al., Cancer Res., 57(18):8882-90 (2007)に記載されている。CD32Bとの結合が低減され、かつ/またはCD16Aとの結合が増強されたヒトIgG1 Fcドメインの変異体の例は、F243L、R929P、Y300L、V305IまたはP296L置換を含む。これらのアミノ酸置換はヒトIgG1 Fcドメインにいずれの組合せで存在してもよい。一実施形態では、ヒトIgG1 Fcドメイン変異体はF243L、R929P、およびY300L置換を含む。別の実施形態においては、ヒトIgG1 Fcドメイン変異体はF243L、R929P、Y300L、V305I、およびP296L置換を含む。
【0143】
グリコホスファチジルイノシトールアンカードメイン
別の実施形態においては、疾病または疾病関連新生血管系標的化ドメインは、グリコシルホスファチジルイノシトール(GPI)アンカーの翻訳後付加のためのシグナルを与えるポリペプチドである。GPIアンカーは、多くの真核生物のタンパク質のC末端に翻訳後に付加される糖脂質構造である。この修飾は細胞膜の外片に付着タンパク質をしっかり固定する。GPIアンカーは、T細胞に対する提示のために細胞表面にT細胞受容体結合ドメインを付着させるのに使用することができる。この実施形態では、GPIアンカードメインはT細胞受容体結合ドメインに対してC末端にある。
【0144】
一つの実施形態においては、GPIアンカードメインは、ポリペプチドが真核生物系で発現される場合にGPIアンカーの翻訳後付加のためにシグナルを伝達するポリペプチドである。アンカーの付加はGPIアンカーシグナル配列により決定され、アンカー付加の部位における小アミノ酸のセット(ω部位)とそれに続く親水性スペーサーからなり、疎水性ストレッチで終わる(Low, FASEB J., 3:1600-1608 (1989))。このシグナル配列の切断は、保存された中央成分(Low, FASEB J., 3:1600-1608 (1989))を有するが、可変の末梢部分(Homans et al., Nature, 333:269-272 (1988))を有するアンカーを付加する前にERで起こる。GPIアンカータンパク質のC末端は、ホスホエタノールアミン橋を介して保存性の高いコアグリカン、マンノース(α1−2)マンノース(α1−6)マンノース(α1−4)グルコサミン(α1−6)ミオイノシトールに連結される。リン脂質テールはGPIアンカーを細胞膜の付着させる。このグリカンコアは、ホスホエタノールアミン基、マンノース、ガラクトース、シアル酸または他の糖などの側鎖で様々に修飾することができる。第一のマンノース残基に付着されている最も一般的な側鎖は別のマンノースである。グリカンコアの第三のマンノースに付着されているN−アセチルガラクトサミン含有多糖などの複合側鎖が哺乳類アンカー構造に見られる。コアグルコサミンの修飾はまれである。タンパク質および起源種によって、ホスホイノシトール環の脂質アンカーはジアシルグリセロール、アルキルアシルグリセロールまたはセラミドである。これらの脂質種は炭素14〜28個の範囲で長さが異なり、飽和であっても不飽和であってもよい。多くのGPIアンカーはまた、イノシトール環の2−ヒドロキシルにパルミチン酸などの付加的脂肪酸を含む。この余分な脂肪酸により、GPIアンカーはPI−PLCによる切断に耐性となる。
【0145】
GPIアンカーは、GPI翻訳後修飾を行うことができる真核生物系でGPIアンカードメインを含む融合タンパク質を発現させることよって達成することができる。GPIアンカードメインは腫瘍または腫瘍血管系標的化ドメインとして使用することもできるし、あるいはすでに別個の腫瘍または腫瘍血管系標的化ドメインを含む融合タンパク質に追加することもできる。
【0146】
別の実施形態においては、GPIアンカー部分は、インビトロ酵素的または化学的工程を介して、単離されたT細胞受容体結合ドメインに直接付加される。この実施形態では、GPIアンカーは、GPIアンカードメインを必要とすることなくポリペプチドに付加することができる。GPIアンカー部分は、T細胞受容体結合ドメインおよび腫瘍または腫瘍血管系標的化ドメインを有する、本明細書に記載の融合タンパク質に付加することができる。あるいは、GPIアンカーは、腫瘍または腫瘍血管系標的化ドメインをコードする融合相手を必要とすることなく、T細胞受容体結合ドメインポリペプチドに直接付加することができる。
【0147】
2.ペプチドまたはポリペプチドリンカードメイン
融合タンパク質は場合により、共刺激ポリペプチドドメインを抗原結合標的化ドメインから分離するペプチドまたはポリペプチドリンカードメインを含む。
【0148】
抗体のヒンジ領域
一つの実施形態においては、リンカードメインは免疫グロブリンのヒンジ領域を含む。好ましい実施形態では、ヒンジ領域はヒト免疫グロブリンに由来する。ヒンジが由来し得る好適なヒト免疫グロブリンとしては、IgG、IgD、およびIgAが挙げられる。好ましい実施形態では、ヒンジ領域はヒトIgGに由来する。
【0149】
別の実施形態においては、リンカードメインは、上記のような免疫グロブリンのヒンジ領域を含み、さらに1以上の付加的な免疫グロブリンドメインを含む。一つの実施形態では、付加的ドメインは免疫グロブリンのFcドメインを含む。本明細書においてFc領域は、第一の定常領域免疫グロブリンドメインを除く抗体の定常領域を含むポリペプチドを含む。よって、FcIgA、IgD、およびIgGの最後の二つ二つの定常領域免疫グロブリンドメインとIgEおよびIgMの最後の3つの定常領域免疫グロブリンドメインを意味する。好ましい実施形態では、Fcドメインはヒト免疫グロブリンに由来する。より好ましい実施形態では、FcドメインはCH2およびCH3領域を含むヒトIgGに由来する。
【0150】
別の実施形態においては、リンカードメインは免疫グロブリンのヒンジ領域と、免疫グロブリン重鎖のCH1ドメインまたは免疫グロブリン軽鎖のCLドメインのいずれかを含む。好ましい実施形態では、CH1またはCLドメインはヒト免疫グロブリンに由来する。CLドメインはκ軽鎖またはλ軽鎖のいずれかに由来するものであり得る。より好ましい実施形態では、CH1またはCLドメインはヒトIgGに由来する。
【0151】
免疫グロブリンヒンジ領域および他のドメインのアミノ酸配列は当該技術分野でよく知られている。
【0152】
他のペプチド/ポリペプチドリンカードメイン
他の好適なペプチド/ポリペプチドリンカードメインは、天然または非天然ペプチドまたはポリペプチドを含む。ペプチドリンカー配列は少なくともアミノ酸2個の長さである。好ましくは、ペプチドまたはポリペプチドドメインは柔軟なペプチドまたはポリペプチドである。「柔軟なリンカー」とは、柔軟なリンカーの非存在下で二つの連結されたポリペプチドが持つものよりも、それにより連結された二つのポリペプチドの回転自由度を高めるペプチド結合により連結された2以上のアミノ酸残基を含むペプチドまたはポリペプチドを意味する。このような回転自由度は、柔軟なリンカーによって各接近標的抗原に連結された2以上の抗原結合部位をより有効とする。柔軟なペプチド/ポリペプチドの例としては、限定されるものではないが、アミノ酸配列Gly−Ser、Gly−Ser−Gly−Ser(配列番号74)、Ala−Ser、Gly−Gly−Gly−Ser(配列番号75)、(Gly4−Ser)3(配列番号76)、および(Gly4−Ser)4(配列番号77)が挙げられる。さらなる柔軟なペプチド/ポリペプチド配列も当該技術分野でよく知られている。
【0153】
3.二量体化および多量体化ドメイン
融合タンパク質は場合により、2以上の融合タンパク質を二量体化または多量体化する働きをする二量体化または多量体化ドメインを含む。融合タンパク質を二量体化または多量体化する働きをするドメインは分離ドメインであってもよいし、あるいはまた融合タンパク質の他のドメイン(T細胞共刺激/共阻害受容体結合ドメイン、腫瘍/腫瘍新生血管系抗原結合ドメインまたはペプチド/ポリペプチドリンカードメイン)の一つの中に含まれてもよい。
【0154】
二量体化ドメイン
「二量体化ドメイン」とは、少なくとも二つのアミノ酸残基または少なくとも二つのペプチドもしくはポリペプチド(同じまたは異なるアミノ酸配列を持ち得る)の会合によって形成される。これらのペプチドまたはポリペプチドは共有結合的会合および/または非共有結合的会合によって互いに相互作用し得る。好ましい二量体化ドメインは、融合タンパク質相手上のシステインと分子間ジスルフィド結合を形成することができる少なくとも一つのシステインを含む。二量体化ドメインは、融合タンパク質相手の間でジスルフィド結合が形成できるように1以上のシステイン残基を含み得る。一つの実施形態では、二量体化ドメインは一つ、二つ、または3つ〜約10のシステイン残基を含む。好ましい実施形態では、二量体化ドメインは免疫グロブリンのヒンジ領域である。この特定の実施形態では、二量体化ドメインは融合タンパク質のリンカーペプチド/ポリペプチド内に含まれる。
【0155】
さらなる二量体化ドメインの例も当該技術分野で知られており、限定されるものではないが、コイルドコイル、アシッドパッチ(acid patches)、ジンクフィンガー、カルシウムハンド、CH1−CL対、米国特許第5,821,333号公報に記載されている操作された「こぶ(knob)」および/または「突起(protruberance)」を有する「境界」、ロイシンジッパー(例えば、junおよび/またはfos由来)(米国特許第5,932,448号公報)、SH2(srcホモロジー2)、SH3(srcホモロジー3)(Vidal, et al., Biochemistry, 43, 7336-44 ((2004))、ホスホチロシン結合(PTB)(Zhou, et al., Nature, 378:584-592 (1995))、WW(Sudol, Prog. Biochys. Mol. Bio., 65:113-132 (1996))、PDZ(Kim, et al., Nature, 378: 85-88 (1995)、 Komau, et al., Science, 269:1737-1740 (1995))14−3−3、WD40(Hu, et al., J Biol Chem., 273, 33489-33494 (1998))EH、Lim、イソロイシンジッパー、受容体二量体対(例えば、インターロイキン−8受容体(IL−8R)、ならびにインテグリンヘテロ二量体、例えば、LFA−1およびGPIIIb/IIIaまたはその二量体化領域、二量体リガンドポリペプチド(例えば、神経成長因子(NGF)、ニューロトロフィン−3(NT−3)、インターロイキン−8(IL−8)、血管内皮増殖因子(VEGF)、VEGF−C、VEGF−D、PDGFメンバー、および脳由来神経栄養因子(BDNF)(Arakawa, et al., J. Biol. Chem., 269(45): 27833-27839 (1994)およびRadziejewski, et al., Biochem., 32(48): 1350 (1993))が挙げられ、親和性が変更されたこれらのドメインの変異体であってもよい。これらのポリペプチド対は、酵母二つのハイブリッドスクリーンを含む当該技術分野で知られている方法によって同定することができる。酵母ツーハイブリッドスクリーンは米国特許第5,283,173号公報および同第6,562,576号公報に記載されており、双方とも引用することにより本明細書の開示の一部とされる。相互作用ドメイン対の間の親和性はKatahira, et al., J. Biol. Chem., 277, 9242-9246 (2002) の記載を含む、当該技術分野で知られている方法を用いて決定することができる。あるいは、ペプチド配列のライブラリーを、例えばWO01/00814号公報に記載されている方法を用いてヘテロ二量体形成に関してスクリーニングすることもできる。タンパク質−タンパク質相互作用に関する有用な方法はまた米国特許第6,790,624号公報にも記載されている。
【0156】
多量体化ドメイン
「多量体化ドメイン」とは、三つ以上のペプチドまたはポリペプチドを共有結合的会合および/または非共有結合的会合によって互いに相互作用させるドメインである。好適な多量体化ドメインとしては、限定されるものではないが、コイルドコイルドメインが挙げられる。コイルドコイルは、通常、7つのアミノ酸(ヘプタッドリピート)または11のアミノ酸(ウンデカドリピート)の配列に、3および4残基の間隔を置いた主として疎水性の残基の連続パターンを有し、組み立てられて(折りたたまれて)多量性のらせん束を形成するペプチド配列を意味する。3および4残基おきのいくつかの不規則な分布を含む配列を有するコイルドコイルも意図される。疎水性残基は特に、疎水性アミノ酸Val、Ile、Leu、Met、Tyr、Phe、およびTrpである。主として疎水性とは、残基の少なくとも50%が記載の疎水性アミノ酸から選択されるべきであることを意味する。
【0157】
コイルドコイルドメインはラミニンに由来してもよい。細胞外間隙で、ヘテロ三量体コイルドコイルタンパク質ラミニンは基底膜の形成に重要な役割を果たす。明らかに、多機能オリゴマー構造はラミニン機能に必要である。コイルドコイルドメインはまた、3つ(TSP−1およびTSP−2)または5つ(TSP−3、TSP−4およびTSP−5)の鎖が接続されたトロンボスポンジンまたは平行5鎖コイルドコイル(Malashkevich ,et al., Science, 274: 761-765 (1996))へと折りたたまれるCOMP(COMPcc)(Guo, et at., EMBO J., 1998, 17: 5265-5272)に由来してもよい。
【0158】
他のタンパク質に由来するさらなるコイルドコイルドメインおよびポリペプチドの多量体化を媒介する他のドメインも当該技術分野で知られており、開示されている融合タンパク質で用いるのに好適である。
【0159】
4.融合タンパク質例
PD−L2
代表的なネズミPD−L2融合タンパク質は、
【化58】
(配列番号52)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0160】
配列番号79によりコードされているネズミPD−L2融合タンパク質は以下のアミノ酸配列:
【化59】
(配列番号53)
を有する。
【0161】
シグナル配列を含まない配列番号53のネズミPD−L2融合タンパク質のアミノ酸配列は、
【化60】
(配列番号54)
である。
【0162】
代表的なヒトPD−L2融合タンパク質は、
【化61】
(配列番号55)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0163】
配列番号82によりコードされているヒトPD−L2融合タンパク質は以下のアミノ酸配列:
【化62】
(配列番号56)
を有する。
【0164】
シグナル配列を含まない配列番号83のヒトPD−L2融合タンパク質のアミノ酸配列は
【化63】
(配列番号57)
である。
【0165】
代表的な非ヒト霊長類(カニクイザル)PD−L2融合タンパク質タンパク質は以下のアミノ酸配列:
【化64】
(配列番号86)
を有する。
【0166】
シグナル配列を含まない配列番号86の非ヒト霊長類(カニクイザル)PD−L2 融合タンパク質のアミノ酸配列は
【化65】
(配列番号87)
である。
【0167】
G.PD−1受容体アンタゴニストをコードする単離された核酸分子
PD−1アンタゴニストポリペプチド、その変異体およびその融合タンパク質をコードする単離された核酸配列が開示される。本明細書において「単離された核酸」とは、哺乳類ゲノムにおいてその核酸の一端または両端に通常隣接している核酸をはじめ、哺乳類ゲノムに存在する他の核酸分子から分離された核酸を意味する。
【0168】
単離された核酸は、例えば、天然ゲノムにおいてそのDNA分子にすぐ隣接して通常見られる核酸配列の一つが除去されているか、または存在しない限り、DNA分子であってもよい。よって、単離された核酸としては、限定されるものではないが、他の配列とは独立した分離分子として存在するDNA分子(例えば、化学的に合成された核酸またはcDNAまたはPCRまたは制限エンドヌクレアーゼ処理によって生成されるゲノムDNA断片)ならびにベクター、自律的に複製するプラスミド、ウイルス(例えば、レトロウイルス、レンチウイルス、アデノウイルスまたはヘルペスウイルス)に、または原核生物または真核生物のゲノムDNAに組み込まれた組換えDNAが挙げられる。さらに、単離された核酸としては、ハイブリッドまたは融合核酸の一部である組換えDNA分子などの操作核酸が挙げられる。例えば、cDNAライブラリーもしくはゲノムライブラリーの中の数百から数百万の他の核酸に存在する核酸、またはゲノムDNA制限消化を含むゲルスライスは単離された核酸とはみなさない。
【0169】
核酸はセンス配向であってもアンチセンス配向であってもよく、あるいはPD−L2、PD−L1、PD−1またはB7.1ポリペプチドまたはその変異体をコードする参照配列と相補的であってもよい。参照配列としては、例えば、当該技術分野で知られており、上記に述べられているヒトPD−L2、ヒトPD−L1、またはネズミPD−L2およびネズミPD−L1のヌクレオチド配列が含まれる。
【0170】
核酸はDNA、RNA、または核酸類似体であり得る。核酸類似体は塩基部分、糖部分またはリン酸主鎖において修飾することができる。このような修飾は例えば、核酸の安定性、ハイブリダイゼーションまたは溶解度を改善することができる。塩基部分における修飾としては、デオキシチミジンをデオキシウリジン、デオキシシチジンを5−メチル−2’−デオキシシチジンまたは5−ブロモ−2’−デオキシシチジンにいうものが挙げられる。糖部分の修飾としては、リボース糖の2’ヒドロキシルの修飾による2’−O−メチルまたは2’−O−アリル糖の形成が挙げられる。デオキシリボースリン酸主鎖は、各塩基部分が6員のモルホリノ環に連結されているモルホリノ核酸、またはデオキシリン酸主鎖がシュードペプチド主鎖で置換され、4つの塩基が保持されているペプチド核酸を生成するように修飾することができる。例えば、Summerton and Weller (1997) Antisence Nucleic Acid Drug Dev. 7:187-195、およびHyrup et al. (1996) Bioorgan. Med. Chem. 4:5-23参照。さらに、デオキシリン酸主鎖は、例えば、ホスホロチオエートまたはホスホロジチオエート主鎖、ホスホロアミダイトまたはアルキルホスホトリエステル主鎖で置換することができる。
【0171】
H.PD−1受容体アンタゴニストを発現するベクターおよび宿主細胞
上記の核酸は、細胞で発現させるためにベクターに挿入することができる。本明細書において「ベクター」とは、別のDNAセグメントが、挿入されたセグメントの複製が起こるように挿入され得る、プラスミド、ファージまたはコスミドなどのレプリコンである。ベクターは発現ベクターであり得る。「発現ベクター」とは、1以上の発現制御配列を含むベクターであり、「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。
【0172】
ベクター中の核酸は1以上の発現制御配列に機能的に連結させることができる。本明細書において「機能的に連結される」とは、発現制御配列が目的のコード配列の発現を有効に制御するように遺伝子構築物に組み込まれることを意味する。発現制御配列の例としては、プロモーター、エンハンサー、および転写終結領域が挙げられる。プロモーターは、一般に、転写が開始する点の上流100ヌクレオチド以内(一般に、RNAポリメラーゼIIの開始部位付近)のDNA分子の領域からなる発現制御配列である。コード配列をプロモーターの制御下に置くためには、そのポリペプチドの翻訳リーディングフレームの翻訳開始部位をプロモーターの下流1〜約15ヌクレオチドの間に置く必要がある。エンハンサーは、時間、場所、およびレベルに関して発現特異性を与える。プロモーターとは異なり、エンハンサーは転写部位から様々な距離に位置する場合にも機能し得る。エンハンサーはまた、転写開始部位から下流に位置してもよい。コード配列は、RNAポリメラーゼがコード配列をmRNAへ転写することができ、それがコード配列によりコードされているタンパク質へ翻訳され得る場合に、「機能的に連結され」かつ細胞内の発現制御配列の「制御下」にある。
【0173】
好適な発現ベクターとしては、限定されるものではないが、例えば、バクテリオファージ、バキュロウイルス、タバコモザイクウイルス、ヘルペスウイルス、サイトメガロウイルス、レトロウイルス、ワクシニアウイルス、アデノウイルスおよびアデノ随伴ウイルスに由来するプラスミドおよびウイルスベクターが挙げられる。多くのベクターおよび発現系がNovagen (Madison, WI)、Clontech (Palo Alto, CA)、Stratagene (La Jolla, CA)およびInvitrogen Life Technologies (Carlsbad, CA)などの会社から市販されている。
【0174】
発現ベクターはタグ配列を含み得る。タグ配列は一般に、コードされているポリペプチドとの融合物として発現される。このようなタグは、カルボキシル末端またはアミノ末端を含むポリペプチド内のいずれの位置にでも挿入することができる。有用なタグの例としては、限定されるものではないが、緑色蛍光タンパク質(GFP)、グルタチオンS−トランスフェラーゼ(GST)、ポリヒスチジン、c−myc、血球凝集素、Flag(商標)タグ(Kodak, New Haven, CT)、マルトースE結合タンパク質およびタンパク質Aが挙げられる。一実施形態では、変異体PD−L2融合タンパク質は、Ig重鎖定常領域の1以上のドメインをコードする、好ましくは、ヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3領域に相当するアミノ酸配列を有する核酸を含むベクター内に存在する。
【0175】
発現される核酸を含むベクターを宿主細胞に導入することができる。「宿主細胞」とは、組換え発現ベクターが導入可能な原核生物のおよび真核細胞を含むものとする。本明細書において「形質転換される」および「トランスフェクトされる」は、多くの技術の一つによって核酸分子(例えばベクター)を細胞に導入することを包含する。特定の技術に限定されるものではないが、これらの技術の多くは当該技術分野で十分に確立されている。
【0176】
原核細胞は、例えば、エレクトロポレーションまたは塩化カルシウム媒介形質転換により核酸で形質転換させることができる。核酸は、例えば、リン酸カルシウム共沈殿、DEAE−デキストラン媒介トランスフェクション、リポフェクション、エレクトロポレーションまたはマイクロインジェクションを含む技術によって哺乳類細胞へトランスフェクトすることができる。宿主細胞(例えば、原核細胞または真核細胞、例えばCHO細胞)を用いて、例えば、本明細書に記載のPD−1アンタゴニストポリペプチドを生産することができる。
【0177】
I.抗体PD−1アンタゴニスト
PD−1アンタゴニストまたはPD−1のエピトープと反応性のあるモノクローナルおよびポリクローナル抗体が開示されている。モノクローナル抗体(mAbs)およびそれらの製造および使用方法は、Kohler and Milstein, Nature 256:495-497 (1975)、米国特許第4,376,110号公報、Hartlow, E. et al., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1988)、 Monoclonal Antibodies and Hybridomas: A New Dimension in Biological Analyses, Plenum Press, New York, N.Y. (1980)、 H. Zola et al., Monoclonal Hybridoma Antibodies: Techniques and Applications, CRC Press, 1982に記載されている。
【0178】
PD−1と結合し、PD−1を介したシグナル伝達を遮断し、かつ、現行使用されているものよりも親和性が低く、投与後3か月、2か月、1か月、3週間、2週間、1週間、または数日未満の期間で抗体を解離させる抗体が免疫応答の増強、増進、または刺激に好ましい。
【0179】
本発明の別の実施形態は、B7−HなどのPD−1のリガンドと結合する架橋したPD−1受容体と結合する抗体を含んでなる二重特異性抗体を含む。好ましい実施形態では、このPD−1結合部分は、PD−1受容体を介したシグナル伝達を低減または阻害する。
【0180】
イムノアッセイ法は、Coligan, J. E. et al., eds., Current Protocols in Immunology, Wiley-Interscience, New York 1991 (または最新版)、 Butt, W. R. (ed.) Practical Immunoassay: The State of the Art, Dekker, N.Y., 1984、 Bizollon, Ch. A., ed., Monoclonal Antibodies and New Trends in Immunoassays, Elsevier, N.Y., 1984、 Butler, J. E., ELISA (Chapter 29), In: van Oss, C. J. et al., (eds), Immunochemistry, Marcel Dekker, Inc., New York, 1994, pp. 759-803、 Butler, J. E. (ed.), Immunochemistry of Solid-Phase Immunoassay, CRC Press, Boca Raton, 1991、 Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986、 Work, T. S. et al., Laboratory Techniques and Biochemistry in Molecular Biology, North Holland Publishing Company, NY, (1978) (Chapter by Chard, T., "An Introduction to Radioimmune Assay and Related Techniques")に記載されている。
【0181】
抗イディオタイプ抗体は、例えば、Idiotypy in Biology and Medicine, Academic Press, New York, 1984、 Immunological Reviews Volume 79, 1984、 Immunological Reviews Volume 90, 1986、 Curr. Top. Microbiol., Immunol. Volume 119, 1985、 Bona, C. et al., CRC Crit. Rev. Immunol., pp. 33-81 (1981)、 Jerme, N K, Ann. Immunol. 125C:373-389 (1974)、 Jerne, N K, In: Idiotypes--Antigens on the Inside, Westen-Schnurr, I., ed., Editiones Roche, Basel, 1982, Urbain, J. et al., Ann. Immunol. 133D:179-(1982)、 Rajewsky, K. et al., Ann. Rev. Immunol. 1:569-607 (1983)に記載されている。
【0182】
これらの抗体は異種、同種異系、同系、またはその改変形態(ヒト化またはキメラ抗体など)であり得る。特定の抗体、例えば、抗PD−L2抗体のイディオタイプに特異的な抗イディオタイプ抗体も含まれる。「抗体」との用語は、完全分子ならびに抗原結合部位を含み、かつ、およびPD−1アンタゴニストエピトープと結合することができるその断片の双方を含むことを意味する。これらには、完全抗体のFc断片を欠き、循環からより迅速に排除され、完全抗体よりも低い非特異的組織結合性を持ち得るFabおよびF(ab’)2断片が含まれる(Wahl et al., J. Nuc. Med. 24:316-325 (1983))。また、Fv断片も含まれる(Hochman, J. et al. (1973) Biochemistry 12:1130-1135、 Sharon, J. et al.(1976) Biochemistry 15:1591-1594)。これらの種々の断片は、プロテアーゼ切断または化学的切断などの常法を用いて作製される(例えば、Rousseaux et al., Meth. Enzymol., 121:663-69 (1986)参照)。
【0183】
ポリクローナル抗体は、ウサギ、ヤギ、齧歯類などの免疫動物から血清として得られ、それ以上処理することなくそのまま用いてもよく、あるいは硫酸アンモニウム沈殿、イオン交換クロマトグラフィーおよびアフィニティークロマトグラフィーなどの慣例の富化または精製方法を施してもよい。
【0184】
免疫原は、完全なPD−1アンタゴニスト、PD−1または断片またはその誘導体を含む。好ましい免疫原は、PD−1アンタゴニストまたはPD−1の細胞外ドメイン(ECD)の全部または一部を含み、ここで、これらの残基はグリコシル化などの翻訳後調節を含む。細胞外ドメインを含む免疫原は、例えば、慣例の組換え法を用いたクローン化遺伝子の発現または起源細胞からの単離などの当該技術分野で知られている種々の方法で生産される。
【0185】
モノクローナル抗体は、Kohler and Milstein, Nature, 256:495-97 (1975)により紹介された手順およびその改変法(上記の参照を参照)などの慣例のハイブリドーマ技術を用いて生産され得る。動物、好ましくはマウスを上記のような免疫原に感作させることにより初回刺激し、刺激動物に所望の抗体応答を惹起させる。初回刺激動物のリンパ節、脾臓、または末梢血からのBリンパ球を、一般にポリエチレングリコール(PEG)などの誘導促進剤の存在下で骨髄腫細胞と融合させる。多くのネズミ骨髄腫細胞系統はいずれもがこのような使用に利用できる:P3−NS1/1−Ag4−1、P3−x63−k0Ag8.653、Sp2/0−Ag14またはHL1−653骨髄腫系統(ATCC, Rockville, Md.から入手可能)。その後の工程は、融合されてない親骨髄腫細胞と、ドナーリンパ球細胞とがやがて死に至り、ハイブリドーマ細胞だけが生き残るような選択培地で増殖させることを含む。これらをクローニングし、増殖させ、それらの上清を、例えば、PD−L2またはPD−L1融合タンパク質を用いたイムノアッセイ技術によって所望の特異性の抗体が存在するか否かをスクリーニングする。例えば、制限希釈によって陽性クローンをサブクローニングし、モノクローナル抗体を単離する。
【0186】
これらの方法に従って生産されたハイブリドーマは、当該技術分野で知られている技術(一般に、Fink et al., Prog. Clin. Pathol., 9:121-33 (1984)参照)を用いて、インビトロまたはインビボ(腹水中)で増殖させることができる。一般に、個々の細胞系統を培養で増殖させ、高濃度の単一モノクローナル抗体を含む培養培地を、デカンテーション、濾過または遠心分離によって採取することができる。
【0187】
抗体は、正常な多量体構造の代わりに単鎖抗体またはscFvとして生産してもよい。単鎖抗体は、着目するIgに由来する超可変領域を含み、完全なIgのサイズの一部でありつつ、天然Igの抗原結合部位を再形成する(Skerra, A. et al. Science, 240: 1038-1041 (1988)、 Pluckthun, A. et al. Methods Enzymol. 178: 497-515 (1989)、 Winter, G. et al. Nature, 349: 293-299 (1991))。好ましい実施形態では、抗体は慣例の分子生物学的技術を用いて生産される。
【0188】
III.製造方法
PD−1アンタゴニストの生産方法
ポリペプチドおよびその変異体
単離されたPD−1アンタゴニストまたはその変異体は、例えば、化学的合成または宿主細胞での組換え生産によって得ることができる。PD−1アンタゴニストポリペプチドを組換え生産するためには、そのポリペプチドをコードするヌクレオチド配列を含む核酸を用いて、細菌または真核生物宿主細胞(例えば、昆虫、酵母、または哺乳類細胞)に形質転換、形質導入またはトランスフェクションを行うことができる。一般に、核酸構築物は、PD−1アンタゴニストポリペプチドをコードするヌクレオチド配列に機能的に連結された調節配列を含む。調節配列(本明細書では発現制御配列とも呼ばれる)は一般に遺伝子産物をコードしないが、それらが機能的に連結されている核酸配列の発現に影響を与える。
【0189】
ポリペプチドを発現および生産するのに有用な原核生物系および真核生物系は当該技術分野でよく知られており、例えば、BL−21などの大腸菌(Escherichia coli)株、およびCHO細胞などの培養哺乳類細胞が挙げられる。
【0190】
真核生物宿主細胞では、PD−1アンタゴニストポリペプチドを発現させるために多くのウイルスに基づく発現系が利用可能である。ウイルスに基づく発現系は当該技術分野でよく知られており、限定されるものではないが、バキュロウイルス、SV40、レトロウイルス、またはワクシニアに基づくウイルスベクターが挙げられる。
【0191】
PD−1アンタゴニストポリペプチドを安定発現する哺乳類細胞系統は、適当な制御エレメントと選択マーカーを備えた発現ベクターを用いて作出することができる。例えば、真核生物発現ベクターpCR3.1(Invitrogen Life Technologies) およびp91023(B)(Wong et al. (1985) Science 228:810-815参照)は、例えば、チャイニーズハムスター卵巣(CHO)細胞、COS−1細胞、ヒト胎児腎臓293細胞、NIH3T3細胞、BHK21細胞、MDCK細胞、およびヒト血管内皮細胞(HUVEC)での変異体共刺激ポリペプチドの発現に好適である。エレクトロポレーション、リポフェクション、リン酸カルシウムまたは塩化カルシウム共沈殿、DEAEデキストランまたは他の好適なトランスフェクション方法による発現ベクターの導入の後、安定な細胞系統を選択することができる(例えば、G418、カナマイシンまたはハイグロマイシンに対する抗生物質耐性による)。トランスフェクト細胞は、目的のポリペプチドが発現されるように培養することができ、このポリペプチドを例えば、細胞培養上清または溶解細胞から回収することができる。あるいは、PD−1アンタゴニストポリペプチドは、(a)増幅された配列をpcDNA3(Invitrogen Life Technologies)などの哺乳類発現ベクターに連結し、(b)インビトロにおいてコムギ胚芽抽出液またはウサギ網状赤血球溶解液を用いて転写および翻訳することによって生産することもできる。
【0192】
PD−1アンタゴニストポリペプチドは、例えば、DEAEイオン交換、ゲル濾過、およびヒドロキシルアパタイトクロマトグラフィーなどのクロマトグラフィー法を用いて単離することができる。例えば、細胞培養上清または細胞質抽出液中のPD−1アンタゴニストポリペプチドは、Gタンパク質カラムを用いて単離することができる。いくつかの実施形態では、変異体PD−1アンタゴニストポリペプチドは、ポリペプチドを親和性マトリックスに捕捉させるアミノ酸配列を含むように「操作」することができる。ポリペプチドの精製を助けるために、例えば、c−myc、血球凝集素、ポリヒスチジン、またはFlag(商標)(Kodak)などのタグを使用することができる。このようなタグは、カルボキシル末端またはアミノ末端を含むポリペプチド内のいずれの位置にも挿入することができる。有用であり得る他の融合物としては、アルカリ性ホスファターゼなど、ポリペプチドの検出を助ける酵素を含むことができる。共刺激ポリペプチドを精製するのに、イムノアフィニティークロマトグラフィーを使用することもできる。
【0193】
変異体ポリペプチドを作出するためにランダム突然変異を導入する方法は当該技術分野で知られている。PD−1、PD−L1、またはPD−L2と相互作用するペプチドをスクリーニングするためにランダムペプチドディスプレーライブラリーを使用することができる。このようなランダムペプチドディスプレーライブラリーを作出およびスクリーニングするための技術は当該技術分野で知られており(Ladnerら、米国特許第5,223,409号公報、Ladnerら、米国特許第4,946,778号公報、Ladnerら、米国特許第5,403,484号公報、およびLadnerら、米国特許第5,571,698号)、ランダムペプチドディスプレーライブラリーおよびこのようなライブラリーをスクリーニングするためのキットが市販されている。
【0194】
B.PD−1アンタゴニストポリペプチドをコードする単離された核酸分子の生産方法
PD−1アンタゴニストポリペプチドをコードする単離された核酸分子は、限定されるものではないが、一般的分子クローニングおよび化学的核酸合成技術を含む標準的技術によって生産することができる。例えば、変異体共刺激ポリペプチドをコードする単離された核酸を得るために、ポリメラーゼ連鎖反応(PCR)技術を使用することができる。PCRは、標的核酸が酵素的に増幅される技術である。一般に、着目する領域の末端から、またはそれを超える配列情報を用いれば、増幅される鋳型の反対側の鎖と配列が同一であるオリゴヌクレオチドプライマーを設計することができる。PCRを用いて、全ゲノムDNAまたは全細胞RNA由来の配列を含め、DNAならびにRNAから特定の配列を増幅させることができる。プライマーは一般に14〜40ヌクレオチドの長さであるが、10ヌクレオチド〜数百ヌクレオチドの範囲の長さであってもよい。一般的なPCR技術は、例えば、PCR Primaer: A Laboratory Manual, Dieffenbach and Dveksler編, Cold Spring Harbor Laboratory Press, 1995に記載されている。鋳型の供給源としてRNAを用いる場合には、逆転写酵素を用いて相補的DNA(cDNA)鎖を合成することができる。単離された核酸を得るために、リガーゼ連鎖反応、鎖置換増幅、自己持続配列複製または核酸配列に基づく増幅を使用することもできる。例えば、Lewis (1992) Genetic Engineering News 12:1、 Guatelli et al. (1990) Proc. Natl. Acad. Sci. USA 87:1874-1878、およびWeiss (1991) Science 254:1292-1293参照。
【0195】
単離された核酸は、単一の核酸分子としてか、またはオリゴヌクレオチド系列としてのいずれかで化学的に合成することができる(例えば、3’から5’方向への自動DNA合成のためのホスホルアミダイト技術を使用)。例えば、所望の配列を含み、各対が短い相補的セグメント(例えば約15ヌクレオチド)を含み、その結果、オリゴヌクレオチド対がアニーリングされる際に二重らせんが形成される、長いオリゴヌクレオチド(例えば>100ヌクレオチド)の1以上の対を合成することができる。DNAポリメラーゼオリゴヌクレオチドを延長するのにも使用することができ、その結果、オリゴヌクレオチド対につき一つの一本鎖、二本鎖核酸分子が得られ、次にこれをベクターに連結することができる。また、単離された核酸は突然変異誘発により得ることもできる。PD−1アンタゴニストポリペプチドをコードする核酸は、PCRを介したオリゴヌクレオチド指定突然変異誘発および/または部位特異的突然変異誘発を含む標準的技術を用いて変異させることができる。Short Protocols in Molecular Biology. Chapter 8, Green Publishing Associates and John Wiley & Sons, edited by Ausubel et al, 1992参照。修飾可能なアミノ酸位置の例としては、本明細書に記載されているものが含まれる。
【0196】
IV.処方物
A.PD−1アンタゴニスト処方物
PD−1アンタゴニストを含有する医薬組成物が提供される。ペプチドまたはポリペプチドを含有する医薬組成物は非経口(筋肉内、腹腔内、静脈内(IV)または皮下注射)、経皮(受動的またはイオン泳動もしくはエレクトロポレーションを使用)または経粘膜(鼻腔、膣、直腸または舌下)投与経路による投与用であり得る。また、組成物は生浸食性インサートを用いて投与してもよいし、適当なリンパ系組織(例えば、脾臓、リンパ節もしくは粘膜に関連するリンパ系組織)に直接送達、または器官もしくは腫瘍に直接送達してもよい。これらの組成物は各投与経路に適当な剤形で処方することができる。ペプチドまたはポリペプチドでないPD−1受容体のアンタゴニストを含有する組成物はさらに腸内投与用に処方することもできる。
【0197】
本明細書において「有効量」または「治療上有効な量」とは、処置される障害の1以上の症状を処置、阻害または緩和するのに、またはそうでなければされる所望の薬理学的および/または生理学的作用を提供するのに十分な用量を意味する。厳密な用量は、被験体に依存する変数(例えば、年齢、免疫系の健全性など)、疾患および行われる処置などの種々の要因によって異なる。PD−1アンタゴニストの治療上有効な量は、免疫応答を活性化、増強、増大または持続させ、かつ/またはT細胞の消耗および/またはT細胞の無応答を克服または緩和し、かつ/または単球、マクロファージ、樹状細胞および他の抗原提示細胞(APC)を活性化する。
【0198】
好ましい実施形態では、PD−1アンタゴニストは、腫瘍モデリングおよびバイオアベイラビリティからの外挿に基づき、0.1〜20mg/kgの範囲で投与される。最も好ましい範囲は5〜20mgPD−1アンタゴニスト/kgである。一般に、静脈注射または注入では、別の経路によって投与する場合よりも用量は低くなり得る。
【0199】
1.非経口投与用処方物
好ましい実施形態においては、ペプチドおよびポリペプチドを含有するものを含む開示されている組成物は、非経口注射により水溶液として投与される。処方物はまた懸濁液またはエマルションの形態であってもよい。一般に、有効量のペプチドまたはポリペプチドを含み、場合により薬学上許容される希釈剤、保存剤、可溶化剤、乳化剤、アジュバントおよび/または担体を含む医薬組成物が提供される。このような組成物は無菌水、緩衝生理食塩水(例えば、Tris−HCl、酢酸、リン酸)、pHおよびイオン強度、ならびに場合により、洗剤および可溶化剤(例えば、TWEEN(商標)20、TWEEN80、ポリソルベート80)、抗酸化剤(例えば、アスコルビン酸、メタ重亜硫酸ナトリウム)、および保存剤(例えば、Thimersol、ベンジルアルコール)および増量剤(例えば、ラクトース、マンニトール)などの添加剤を含む。非水性溶媒またはビヒクルの例としては、プロピレングリコール、ポリエチレングリコール、オリーブ油およびコーン油などの植物油、ゼラチン、およびオレイン酸エチルなどの注射可能な有機エステルがある。処方物は凍結乾燥させ、使用直前に再溶解/再懸濁させてもよい。処方物は例えば、細菌保持フィルターによる濾過、除菌剤の組成物への配合、組成物の照射または組成物の加熱によって除菌してもよい。
【0200】
2.制御送達ポリマーマトリックス
1以上のPD−1アンタゴニストまたはPD−1アンタゴニストをコードする核酸を含有する組成物は徐放性処方物として投与することができる。徐放性ポリマーデバイスはポリマーデバイス(ロッド、シリンダー、フィルム、ディスク)の移植または注射(微粒子)後に長時間全身への放出を行うことができる。マトリックスは、ペプチドが固体ポリマーマトリックス内に分散されているマイクロスフェア、またはコアがポリマー剤皮とは異なる材料のものであり、そのコア(本質的に液体であっても固体であってもよい)内にペプチドが分散もしくは懸濁されているマイクロカプセルなどの微粒子の形態であってもよい。本明細書で特に定義されない限り、微粒子、マイクロスフェアおよびマイクロカプセルは互換的に用いられる。あるいは、ポリマーは数ナノメートルから4センチメートルの範囲の薄スラブもしくはフィルムとしての鋳物、摩砕もしくは他の標準的な技術によって製造された粉末、またはさらにはヒドロゲルなどのゲルであってもよい。マトリックスはまた、免疫応答を調節するため、免疫不全患者(カテーテルが挿入されている高齢者または未熟児など)における感染予防のため、またはただれ、床ずれ潰瘍などの治癒を促進するために用いられるマトリックスの場合と同様に治癒における補助とするために医療機器の内外に組み込むことができる。
【0201】
非生分解性マトリックスでも生分解性マトリックスでもPD−1アンタゴニストまたはそれらをコードする核酸の送達に使用することができるが、生分解性マトリックスが好ましい。これらは天然ポリマーであっても合成ポリマーであってもよいが、分解および放出特性の特徴付けがより良いため、合成ポリマーが好ましい。ポリマーは、放出が望まれる期間に基づいて選択される。直線的放出が最も有用である場合もあるが、パルス放出または「バルク放出」がより有効な結果をもたらす場合もある。ポリマーはヒドロゲル(一般に最大約90重量%の水を吸収する)の形態であってもよく、場合により多価イオンまたはポリマーで架橋されていてもよい。
【0202】
マトリックスは、溶媒蒸発、噴霧乾燥、溶媒抽出、および当業者に知られている他の方法によって形成させることができる。生浸食性マイクロスフェアは、例えば、Mathiowitz and Langer, J. Controlled Release, 5:13-22 (1987)、 Mathiowitz, et al., Reactive Polymers, 6:275-283 (1987)、およびMathiowitz, et al., J. Appl. Polymer Sci., 35:755-774 (1988)に記載されているような、薬物送達用のマイクロスフェアを作製するために開発されたいずれかの方法を用いて製造することができる。
【0203】
徐放性経口処方物が望ましい場合がある。PD−1阻害的シグナル伝達のアンタゴニストは、拡散または溶脱機構のいずれかによる放出を可能とする不活性マトリックス、例えば、フィルムまたはガム中に配合することができる。また、緩徐崩壊マトリックスを処方物に配合してもよい。別の徐放性形態としては、水を入り込ませて浸透圧作用により単一の小開口部から薬剤を押し出す半透膜に薬剤が封入されるものがある。経口処方物では、放出の場所は胃、小腸(十二指腸、空腸もしくは回腸)または大腸であり得る。好ましくは、この放出は有効剤(もしくは誘導体)の保護によるか、または腸などの胃環境を超えた有効剤の放出によるかのいずれかで、胃環境の有害な影響を回避する完全な胃耐性を確保するためには、腸溶コーティング(すなわち、少なくともpH5.0までは不透性)が不可欠である。これらのコーティングは混合フィルム、またはBanner Pharmacapsから市販されているものなどのカプセル剤として使用可能である。
【0204】
これらのデバイスは移植または注射領域を処置するために局所放出用に処方することができ、一般に、全身処置用の用量よりもはるかに低い用量を送達する。これらのデバイスはまた、全身送達用に処方することもできる。これらは埋植または皮下注射することができる。
【0205】
3.腸内投与用処方物
PD−1のアンタゴニストはまた、経口送達用にも処方することができる。経口固体剤形は当業者に知られている。固体剤形としては、錠剤、カプセル剤、丸剤、トローチ剤またはロゼンジ剤、カシェ剤、ペレット、粉末もしくは顆粒、またはポリ乳酸、ポリグリコール酸などのポリマー化合物の特定の調製物、もしくはリポソームへの材料の組み込みが挙げられる。このような組成物は本発明のタンパク質および誘導体の物理的状態、安定性、インビボ放出速度、およびインビボクリアランス速度に影響を及ぼし得る。例えば、Remington's Pharmaceutical Sciences, 21st Ed. (2005, Lippincott, Williams & Wilins, Baltimore, Md. 21201) pages 889-964参照。組成物は液体形態で調製してもよいし、あるいは乾燥粉末(例えば、凍結乾燥)形態であってもよい。リポソームまたはポリマーカプセル封入を組成物の処方に用いてよい。Marshall, K. In: Modern Pharmaceutics, G. S. Banker and C. T. Rhodes編 Chapter 10, 1979も参照。一般に、処方物は有効剤と、胃環境でPD−1アンタゴニストを保護し、腸で生活性材料を放出する不活性成分を含む。
【0206】
薬学上許容されるエマルション、溶液、懸濁液、およびシロップを含む経口投与用の液体剤形は、不活性希釈剤、湿潤剤、乳化剤および沈殿防止剤などのアジュバント、ならびに甘味剤、香味剤、および芳香剤を含む他の成分を含み得る。
【0207】
B.PD−1アンタゴニストを含むワクチン
ワクチンは、感染細胞を排除するために強いT細胞応答を必要とする。PD−1アンタゴニストは、一次免疫応答およびエフェクター細胞の活性と、数とを促進、増大、または増強するためにワクチンの成分として投与することができる。ワクチンは抗原、PD−1アンタゴニスト(またはその供給源)、および場合により他のアジュバントおよび標的化分子を含む。PD−1アンタゴニストの供給源は、開示されているPD−L2ポリペプチド、PD−L2融合タンパク質、その変異体、PD−L1断片、PD−1断片、PD−L2ポリペプチドをコードする核酸、PD−L2融合タンパク質、その変異体、PD−L1断片またはPD−1断片、または上記のPD−1のポリペプチドリガンドを発現するベクターを含む宿主細胞のいずれかを含む。
【0208】
1.抗原
抗原は、ペプチド、タンパク質、多糖類、糖類、脂質、核酸、またはその組合せであり得る。抗原は、ウイルス、細菌、寄生虫、原虫、真菌、ヒストプラズマ、組織、または形質転換細胞に由来するものであり得、全細胞またはその免疫原成分、例えば、細胞壁成分またはその分子成分であり得る。
【0209】
好適な抗原は当該技術分野で知られており、商業的官業および科学ソースから入手可能である。一つの実施形態においては、抗原は不活性化または弱毒された完全な生物である。これらの生物はウイルス、寄生虫および細菌などの感染性生物であり得る。これらの生物は腫瘍細胞または淋病もしくはマラリアなどのウイルスまたは細胞内病原体に感染した細胞であり得る。抗原は腫瘍または腫瘍またはウイルスもしくは細菌源に由来する精製または部分精製ポリペプチドであり得る。抗原は、異種発現系においそのポリペプチド抗原をコードするDNAを発現させることにより産生された組換えポリペプチドであってもよい。抗原は、抗原タンパク質の全部または一部をコードするDNAであってもよい。DNAはプラスミドDNAなどのベクターDNAの形態であってもよい。
【0210】
抗原は単一の抗原として提供されてもよいし、あるいは組合せで提供されてもよい。抗原はまた、ポリペプチドまたは核酸の複合体混合物として提供されてもよい。
【0211】
i.ウイルス抗原
ウイルス抗原は、限定されるものではないが、以下のウイルス科:アレナウイルス科、アルテリウイルス科、アストロウイルス科、バキュロウイルス科、バドナウイルス科、バルナウイルス科、ビルナウイルス科、ブロモウイルス科、ブンヤウイルス科、カリシウイルス科、カピロウイルス科、カルラウイルス科、カリモウイルス科、サーコウイルス科、コモウイルス科、コロナウイルス科(例えば、重症急性呼吸器症候群(SARS)ウイルスなどのコロナウイルス)、コルチコウイルス科、シストウイルス科、デルタウイルス科、ディアンソウイルス科(Dianthoviridae)、エナノウイルス科、フィロウイルス科(例えば、マールブルグウイルスおよびエボラウイルス(例えば、ザイール株、レストン株、コートジボワール株またはスーダン株))、フラビウイルス科(例えば、C型肝炎ウイルス、デング熱ウイルス1、デング熱ウイルス2、デング熱ウイルス3およびデング熱ウイルス4)、ヘパドナウイルス科、ヘルペスウイルス科(例えば、ヒトヘルペスウイルス1、3、4、5および6、ならびにサイトメガロウイルス)、ヒポウイルス科、イリドウイルス科、レビウイルス科、ミクロウイルス科、オルトミクソウイルス科(例えば、A型およびB型およびC型インフルエンザウイルス)、パポバウイルス科、パラミクソウイルス科(例えば、麻疹ウイルス、流行性耳下腺炎ウイルスおよびヒト呼吸器多核体ウイルス(respiratory synticial virus))、パルボウイルス科、ピコルナウイルス科(例えば、ポリオウイルス、ライノウイルス、ヘパトウイルスおよびアフトウイルス)、ポックスウイルス科(例えば、ワクシニアウイルスおよびスモールポックスウイルス)、レオウイルス科(例えば、ロタウイルス)、レトロウイルス科(例えば、レンチウイルス、例えば、ヒト免疫不全ウイルス(HIV)1およびHIV 2)、ラブドウイルス科(例えば、狂犬病ウイルス、麻疹ウイルス、呼吸器多核体ウイルスなど)、トガウイルス科(例えば、風疹ウイルス、デング熱ウイルスなど)、トリウイルス科のいずれかに由来するウイルスを含む任意のウイルスから単離することができる。好適なウイルス抗原はまた、デングタンパク質M、デングタンパク質E、デングD1NS1、デングD1NS2、およびデングD1NS3の全部または一部を含む。
【0212】
ウイルス抗原は、乳頭腫ウイルス、ヘルペスウイルス、すなわち、単純ヘルペス1および2、肝炎ウイルス、例えば、A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、D型肝炎ウイルス(HDV)、E型肝炎ウイルス(HEV)およびG型肝炎ウイルス(HGV)、ダニ媒介性脳炎ウイルス、パラインフルエンザ、水痘帯状疱疹ウイルス、サイトメガロウイルス(cytomeglavirus)、エプスタイン・バーウイルス、ロタウイルス、ライノウイルス、アデノウイルス、コクサッキーウイルス、ウマ脳炎ウイルス、日本脳炎ウイルス、黄熱ウイルス、リフトバレー熱ウイルス、およびリンパ球性脈絡髄膜炎ウイルスなどの特定の株または株の組合せに由来し得る。
【0213】
ii.細菌抗原
細菌抗原は、限定されるものではないが、アクチノミセス属、アナベナ属、バチルス属、バクテロイデス属、デロビブリオ属、ボルデテラ属、ボレリア属、カンピロバクター属、カウロバクター属、クラミジア属、クロロビウム属、クロマチウム属、クロストリジウム属、コリネバクテリウム属、サイトファガ属、ディノコッカス属、エシェリキア属、フランシセラ属、ハロバクテリア属、ヘリコバクター属、ヘモフィルス属、B型ヘモフィルス・インフルエンザ(HIB)、ハイフォミクロビウム属、レジオネラ属、レプトスピラ属、リステリア属、A型、B型およびC型髄膜炎球菌、メタノバクテリウム属、球菌属、ミオバクテリウム属(Myobacterium)、マイコプラズマ属、ミクソコッカス属、ナイセリア属、ニトロバクター属、オシラトリア属、プロクロロン属、プロテウス属、シュードモナス属、ロドスピリルム属、リケッチア属、サルモネラ菌属、赤痢菌属、スピリルム属、スピロヘータ属、ブドウ状球菌属、連鎖球菌属、ストレプトミセス属、スルフォロブス属、サーモプラズマ属、チオバチルス属およびトレポネーマ属、ビブリオ属、およびエルシニア属を含む任意の細菌に由来し得る。
【0214】
iii.寄生虫抗原
寄生虫の抗原は、限定されるものではないが、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツム(Histoplasma capsulatum)、カンジダ・アルビカンス(Candida albicans)、カンジダ・トロピカリス(Candida tropicalis)、ノカルジア・アステロイデス(Nocardia asteroides)、リケッチア・リケッチイ(Rickettsia ricketsii)、発疹熱リケッチア(Rickettsia typhi)、肺炎マイコプラズマ(Mycoplasma pneumoniae)、クラミジア・シッタシ(Chlamydial psittaci)、クラミジア・トラコマチス(Chlamydial trachomatis)、熱帯熱マラリア原虫(Plasmodium falciparum)、ブルーストリパノソーマ(Trypanosoma brucei)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ゴンディ(Toxoplasma gondii)、膣トリコモナス(Trichomonas vaginalis)、およびマンソン住血吸虫(Schistosoma mansoni)などの寄生虫から得ることができる。これらには胞子虫抗原、マラリア原虫抗原、例えば、スポロゾイト周囲タンパク質、スポロゾイト表面タンパク質、肝臓段階抗原、頂端膜関連タンパク質またはメロゾイト表面タンパク質の全部または一部が含まれる。
【0215】
iv.腫瘍抗原
抗原は、限定されるものではないが、α−アクチニン−4、Bcr−Abl融合タンパク質、Casp−8、β−カテニン、cdc27、cdk4、cdkn2a、coa−1、dek−can融合タンパク質、EF2、ETV6−AML1融合タンパク質、LDLR−フコシルトランスフェラーゼAS融合タンパク質、HLA−A2、HLA−A11、hsp70−2、KIAAO205、Mart2、Mum−1、2および3、ネオ−PAP、ミオシンクラスI、OS−9、pml−RARα融合タンパク質、PTPRK、K−ras、N−ras、トリオースリン酸イソメラーゼ、Bage−1、Gage3、4、5、6、7、GnTV、Herv−K−mel、Lage−1、Mage−A1、2、3、4、6、10、12、Mage−C2、NA−88、NY−Eso−1/Lage−2、SP17、SSX−2、およびTRP2−Int2、MelanA(MART−I)、gp100(Pmel17)、チロシナーゼ、TRP−1、TRP−2、MAGE−1、MAGE−3、BAGE、GAGE−1、GAGE−2、p15(58)、CEA、RAGE、NY−ESO(LAGE)、SCP−1、Hom/Mel−40、PRAME、p53、H−Ras、HER−2/neu、BCR−ABL、E2A−PRL、H4−RET、IGH−IGK、MYL−RAR、エプスタイン・バーウイルス抗原、EBNA、ヒト乳頭腫ウイルス(HPV)抗原E6およびE7、TSP−180、MAGE−4、MAGE−5、MAGE−6、p185erbB2、p180erbB−3、c−met、nm−23H1、PSA、TAG−72−4、CA19−9、CA72−4、CAM17.1、NuMa、K−ras、β−カテニン、CDK4、Mum−1、p16、TAGE、PSMA、PSCA、CT7、テロメラーゼ、43−9F、5T4、791Tgp72、α−フェトタンパク質、13HCG、BCA225、BTAA、CA125、CA15−3(CA 27.29\BCAA)、CA195、CA242、CA−50、CAM43、CD68\KP1、CO−029、FGF−5、G250、Ga733(EpCAM)、HTgp−175、M344、MA−50、MG7−Ag、MOV18、NB\70K、NY−CO−1、RCAS1、SDCCAG16、TA−90(Mac−2結合タンパク質\シクロフィリンC関連タンパク質)、TAAL6、TAG72、TLP、およびTPSなどの腫瘍関連高原または腫瘍特異的抗原を含む腫瘍抗原であり得る。また、BCGなどの腫瘍抗原は、アジュバントに対する免疫刺激剤として使用することもできる。
【0216】
2.アジュバント
場合により、ワクチンはアジュバントを含み得る。アジュバントは、限定されるものではないが、以下の1以上であり得る:オイルエマルション(例えば、フロイントのアジュバント)、サポニン処方物、ウィロソーム(virosomes)およびウイルス様粒子、細菌および細菌誘導体、免疫刺激性オリゴヌクレオチド、ADP−リボシル化毒素および解毒誘導体、ミョウバン、BCG、無機物含有組成物(例えば、アルミニウム塩およびカルシウム塩などの無機塩)、水酸化物、リン酸塩、硫酸塩など)、生体接着剤および/または粘膜接着剤、微粒子、リポソーム、ポリオキシエチレンエーテルおよびポリオキシエチレンエステル処方物、ポリホスファゼン、ムラミルペプチド、イミダゾキノロン化合物、および界面活性物質(例えば、リゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、オイルエマルション、キーホールリンペットヘモシアニン、およびジニトロフェノール)。
【0217】
アジュバントはまたサイトカイン、インターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12など)、インターフェロン(例えば、インターフェロンγ)、マクロファージコロニー刺激因子、および腫瘍壊死因子などの免疫調節剤を含み得る。変異体PD−L2ポリペプチドの他、B7ファミリーの他のポリペプチドを含む他の共刺激分子を投与してもよい。このようなタンパク質性アジュバントは全長ポリペプチドもしくはその活性断片として、またはプラスミドDNAなどのDNAの形態で提供されてよい。
【0218】
IV.使用方法
PD−1アンタゴニストポリペプチドおよびその断片、ならびにこれらのポリペプチドおよび融合タンパク質をコードする核酸、またはPD−1アンタゴニストポリペプチドを発現する細胞は、抗原に対する一次免疫応答を増強するとともにT細胞の抗原特異的増殖の増強、T細胞によるサイトカイン生産の増強および分化刺激などのエフェクター機能を増強するために使用することができる。PD−1アンタゴニスト組成物は、それを必要とする被験体に、T細胞の消耗および/またはT細胞の無応答を克服するのに有効な量で投与することができる。T細胞の消耗またはT細胞の無応答の克服は、既知の技術を用いてT細胞機能を測定することにより決定することができる。好ましいPD−1アンタゴニストポリペプチドは、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合し、かつT細胞を共刺激する能力を保持するように操作される。
【0219】
PD−1アンタゴニストのインビトロ適用は、例えば、免疫機構の基礎的科学研究において、またはT細胞機能の研究もしくは例えば受動免疫療法に用いる活性化T細胞の生産のために有用であり得る。さらに、PD−1アンタゴニストは、T細胞が得られた被験体において着目する抗原に対する免疫性を試験すべく設計されたインビトロアッセイ(例えば、T細胞増殖アッセイ)に加えることができる。このようなアッセイへのPD−1アンタゴニストの付加は、より強力な、従ってより検出の容易なインビトロ応答をもたらすと考えられる。
【0220】
A.免疫増強のためのPD−1アンタゴニストの投与
PD−1アンタゴニストは一般に、インビボおよびエクスビボにおいて免疫応答刺激治療薬として有用である。好ましい実施形態においては、これらの組成物は、T細胞の消耗またはT細胞の無応答が既に起こり、その感染が長期にわたって宿主に残存する感染を処置するのに有用である。処置される例示的感染は、肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、ヒトTリンパ球向性ウイルス(HTLV)、ヘルペスウイルス、エプスタイン・バーウイルス、またはヒト乳頭腫ウイルスにより引き起こされる慢性感染である。他の感染もPD−1アンタゴニストを用いて処置可能であると考えられる。開示されている組成物はワクチンの一部としても有用である。好ましい実施形態では、治療または予防される疾患の種類は、細菌、ウイルス、原虫、蠕虫、または細胞内に侵入する、すなわち細胞傷害性Tリンパ球により攻撃される、他の微生物病原体により引き起こされる慢性感染性疾患である。
【0221】
ヒトおよび動物モデルにおける慢性感染は、宿主免疫応答の、機能的なCD8+およびCD4+T細胞集団の生成および維持不全に関連し、その結果として、感染性を中和するための抗体応答も不十分となる。この機能の欠損はT細胞の消耗と呼ばれる。T細胞の無応答は、リンパ球が抗原と遭遇した後に内因的に機能的に活性化されるが、長期間、低応答状態で生存を維持するという耐性機構である。慢性感染を処置するための一つの方法は、消耗したT細胞を活性化すること、または被験体におけるT細胞の消耗を逆転させるとともにT細胞の無応答を克服することである。T細胞の消耗の逆転は、PD−1とそのリガンドPD−L1(B7−H1)およびPD−L2(PD−L2)の間の相互作用に干渉することによって達成することができる。急性の、多くの場合致死的な病原体の作用は、毒素、またはその毒素により引き起こされる損傷の前に十分な免疫応答を惹起することができない他の因子によって媒介され得る。これは、PD−1とそのリガンドの間の相互作用に干渉し、より有効な迅速免疫応答を可能とすることにより克服され得る。
【0222】
ウイルス感染は主としてT細胞により排除されるので、感染力のあるウイルス病原体のより迅速な、または十分な排除がヒトまたは動物被験体に有益となる状況では、T細胞活性の増強が治療上有用である。よって、PD−1アンタゴニストは、限定されるものではないが、免疫不全(例えば、HIV)、乳頭腫(例えば、HPV)、ヘルペス(例えば、HSV)、脳炎、インフルエンザ(例えば、ヒトA型インフルエンザウイルス)、および風邪(例えば、ヒトライノウイルス)ウイルス感染を含む、局所または全身ウイルス感染の処置のために投与することができる。例えば、PD−1アンタゴニスト組成物を含む医薬処方物は、ヘルペス病巣または帯状疱疹または性器疣贅などのウイルス性皮膚疾患を処置するために局所投与することができる。PD−1アンタゴニスト組成物の医薬処方物はまた、限定されるものではないが、AIDS、インフルエンザ、風邪または脳炎を含む全身性ウイルス性疾患を処置するためにも投与することができる。
【0223】
処置可能な代表的感染としては、限定されるものではないが、アクチノミセス属、アナベナ属、バチルス属、バクテロイデス属、デロビブリオ属、ボルデテラ属、ボレリア属、カンピロバクター属、カウロバクター属、クラミジア属、クロロビウム属、クロマチウム属、クロストリジウム属、コリネバクテリウム属、サイトファガ属、ディノコッカス属、エシェリキア属、フランシセラ属、ハロバクテリア属、ヘリコバクター属、ヘモフィルス属、B型ヘモフィルス・インフルエンザ(HIB)、ヒトプラズマ属、ハイフォミクロビウム属、レジオネラ属、リーシュマニア属、レプトスピラ属、リステリア属、A型、B型およびC型髄膜炎球菌、メタノバクテリウム属、球菌属、ミオバクテリウム属(Myobacterium)、マイコプラズマ属、ミクソコッカス属、ナイセリア属、ニトロバクター属、オシラトリア属、プロクロロン属、プロテウス属、シュードモナス属、ロドスピリルム属、リケッチア属、サルモネラ菌属、赤痢菌属、スピリルム属、スピロヘータ属、ブドウ状球菌属、連鎖球菌属、ストレプトミセス属、スルフォロブス属、サーモプラズマ属、チオバチルス属およびトレポネーマ属、ビブリオ属、エルシニア属、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツム(Histoplasma capsulatum)、カンジダ・アルビカンス(Candida albicans)、カンジダ・トロピカリス(Candida tropicalis)、ノカルジア・アステロイデス(Nocardia asteroides)、リケッチア・リケッチイ(Rickettsia ricketsii)、発疹熱リケッチア(Rickettsia typhi)、肺炎マイコプラズマ(Mycoplasma pneumoniae)、クラミジア・シッタシ(Chlamydial psittaci)、クラミジア・トラコマチス(Chlamydial trachomatis)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(Plasmodium vivax)、ブルーストリパノソーマ(Trypanosoma brucei)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ゴンディ(Toxoplas
ma gondii)、膣トリコモナス(Trichomonas vaginalis)およびマンソン住血吸虫(Schistosoma mansoni)を含む微生物により引き起こされる感染が挙げられる。
【0224】
B.ワクチンにおけるPD−1アンタゴニストの使用
PD−1アンタゴニストまたはそれをコードする核酸は単独で投与してもよいし、あるいは他の好適な処置と組み合わせて投与してもよい。一つの実施形態においては、PD−1アンタゴニストは上記のようなワクチン組成物とともに、またはワクチン阻害剤の一成分として投与することができる。ワクチン組成物の好適な成分は上記されている。開示されているPD−1アンタゴニストはワクチン投与の前、ワクチン投与と同時、またはワクチン投与の後に投与することができる。一つの実施形態においては、PD−1アンタゴニスト組成物はワクチン投与と同時に投与される。
【0225】
PD−1アンタゴニスト組成物は、被験体において次に感染性病原体に曝された際の耐性を付与する予防ワクチンとともに、またはウイルスに感染した被験体におけるウイルス抗原などの既存の抗原に対する被験体の免疫応答を誘発または増強するために使用できる治療ワクチンとともに投与してもよい。
【0226】
予防的、治療的または脱感作免疫応答の所望の結果は、当該技術分野でよく知られている原理に従い、疾患によって異なり得る。例えば、感染性病原体に対する免疫応答は感染性病原体の定着および複製を完全に防ぐことができ、「無菌免疫(sterile immunity)」および疾病症状の非存在に影響を及ぼす。しかしながら、感染性病原体に対するワクチンは、それが症状の数、重篤度または持続時間を軽減するか、症状を有する集団中の個体数を少なくするか、または感染性病原体の伝染を軽減すれば、有効であると見なせる。同様に、癌、アレルゲン、または感染性病原体に対する免疫応答は疾患を完全に治療する場合、症状を緩和する場合、または疾患に対する全体的な治療的介入の一つの態様である場合がある。
【0227】
PD−1アンタゴニストは、少なくとも一つの成分抗原または抗原組成物に対し、PD−1アンタゴニストを用いずに対応する組成物で得られるエフェクター細胞応答に比べて向上したエフェクター細胞応答(CD4T細胞免疫応答など)を誘導する。「向上したエフェクター細胞応答」との用語は、ワクチン組成物の投与後にヒト患者で得られる、PD−1アンタゴニストを用いずに同じ組成物の投与後に得られるものよりも高いエフェクター細胞応答(CD4応答など)を意味する。例えば、ヒト患者においてPD−1アンタゴニスト、好ましくはPD−L2−Igと抗原調製物を含有する免疫原組成物を投与した際に、アジュバントを含まないその抗原調製物を含有する免疫原組成物の投与後に誘導される応答に比べて高いCD4T細胞応答が得られる。このような処方物は有利には、MHCクラスII分子により提示される抗原エピトープを検出することができる抗抗原エフェクター細胞応答を誘導するために用いられる。
【0228】
この向上したエフェクター細胞応答は、免疫刺激されていない患者、すなわち、その抗原に対して血清陰性である患者で得ることができる。この血清陰性は、患者がその抗原に直面したことがない(いわゆる「ナイーブ」患者)か、あるいはまた、かつて遭遇したその抗原に応答できなかった結果であり得る。この向上したエフェクター細胞応答は、高齢者、一般に65歳以上、または高リスクな医学的状態を有する65歳より若い成人(「高リスク」成人)、または2歳未満の小児など、免疫不全被験体で得られるのが好ましい。
【0229】
向上したエフェクター細胞応答は、以下のサイトカインのいずれかを産生する細胞:(1)少なくとも二つの異なるサイトカイン(CD40L、IL−2、IFN−γ、TNF−α)を産生する細胞、(2)少なくともCD40Lと別のサイトカイン(IL−2、TNF−α、IFN−γ)を産生する細胞、(3)少なくともIL−2と別のサイトカイン(CD40L、TNF−α、IFN−γ)を産生する細胞、(4)少なくともIFN−γと別のサイトカイン(IL−2、TNF−α、CD40L)を産生する細胞、および(5)少なくともTNF−αと別のサイトカイン(IL−2、CD40L、IFN−γ)の数を測定することにより評価することができる。
【0230】
向上したエフェクター細胞応答は、上記のサイトカインのいずれかを産生する細胞がワクチン組成物の投与後に、PD−1アンタゴニストを含まない組成物を投与した場合に比べて多い量である際に存在する。一般に、上記の5つの条件のうち少なくとも一つ、好ましくは二つが満たされる。特定の実施形態では、4つ総てのサイトカインを産生する細胞が、ワクチン接種群で非ワクチン接種群に比べて多い数で存在する。
【0231】
免疫原組成物は、皮内、粘膜、例えば、鼻内、経口、筋肉内、または皮下などのいずれの好適な送達経路によって投与してもよい。当該技術分野では他の送達経路もよく知られている。免疫原組成物にとっては筋肉内送達経路が好ましい。皮内送達はもう一つの好適な経路である。皮内送達には、例えば短針デバイスなどの任意の好適なデバイスが使用可能である。皮内ワクチンはまた、針の皮膚への有効侵入長を制限するデバイスにより投与してもよい。また、液体ジェット式注射器によって、または角質層を穿孔し、真皮に達するジェットを生じる針によって、液体ワクチンを真皮へ送達するジェット注射デバイスを使用することもできる。ジェット注射デバイスは当該技術分野で既知である。皮膚の外層から真皮へと粉末形態のワクチンを加速化するために圧縮ガスを用いる弾道粉末/粒子送達デバイスも使用可能である。さらに、皮内投与の従来のMantoux法で、通常のシリンジを使用することもできる。
【0232】
もう一つの好適な投与経路は皮下経路である。皮下送達には、例えば、従来の針などの好適なデバイスを使用することができる。好ましくは、無針ジェット式注射器が用いられる。無針注射器は当該技術分野で既知である。より好ましくは、デバイスは液体ワクチン処方物が予め充填されている。
【0233】
あるいは、ワクチンは鼻腔内投与される。一般に、ワクチンは鼻咽頭領域に、好ましくは肺へ吸入されずに、局所投与される。ワクチン処方物を、肺に入ることなく、または実質的に入ることなく鼻咽頭領域へ送達する鼻腔内送達デバイスを用いるのが好ましい。ワクチンの鼻腔内投与に好ましいデバイスはスプレーデバイスである。鼻腔スプレーデバイスは市販されている。ネブライザーは容易に肺へ吸入される可能性のある極めて微細なスプレーを生じるので、鼻腔粘膜に十分に達しない。従って、ネブライザーは好ましくない。鼻腔用の好ましいスプレーデバイスは、デバイスの性能が、使用者がかける圧力に依存しないデバイスである。これらのデバイスは圧力閾値デバイスとして知られる。閾値圧力がかけられた場合にのみノズルから液体が放出される。これらのデバイスは規則的な液滴サイズを有するスプレーを達成することを容易とする。本発明とともに用いるのに好適な圧力閾値デバイスは当該技術分野で公知であり、市販されている。
【0234】
好ましい鼻腔デバイスは、1〜200μm、好ましくは10〜120μmの範囲の液滴(液体として水を用いて測定)を生じる。10μmを下回ると吸入のリスクがあるので、10μm未満の液滴が約5%を超えないことが望ましい。120μmを超える液滴は、120μmを超える液滴ならびにそれより小さい液滴は拡散しないので、120μmを超える液体が約5%を超えないことが望ましい。
【0235】
二用量送達(bi-dose delivery)は、ワクチンとともに用いるための鼻腔内送達系のもう一つの特徴である。二用量デバイスは、単回ワクチン用量の二つの分割量を含み、一つの分割用量は各鼻孔に投与するためのものである。一般に、二つの分割用量は単一のチャンバー内に存在し、このデバイスの構成は単一の分割用量を一度に効率的に送達することを可能とする。あるいは、ワクチンの投与に一用量デバイスを用いてもよい。
【0236】
免疫原組成物は数日、数週間または数ヶ月の間にわたって2回以上で与えることができる。一つの実施形態では、異なる投与経路が用いられ、最初に筋肉内投与を行ってよく、場合によりPD−1アンタゴニストを含んでもよい追加免疫組成物が、例えば皮内、皮下、または鼻腔内などの異なる経路により投与してよい。
【0237】
免疫原組成物により付与された向上したエフェクター細胞応答は1回の投与の後に得られるのが理想的であり得る。この単回用量アプローチは、バイオテロリストの攻撃および流行を含む、急速に進行する大発生に極めて関連が強い。ある特定の状況では、特に、高齢者集団、または特定の抗原に対して初めてワクチン接種される年少の小児(9歳未満)では、2回用量の同じ組成物を投与することが有益であり得る。同じ組成物の2回目の用量(なお、最初のワクチン接種用の組成物とみなされる)は、進行中の一次免疫応答中に投与することができ、初回投与から十分な時間間隔が置かれる。一般に、2回目の用量の組成物は、未応答または応答が不十分な個体において免疫系の刺激を助けるために、初回用量の数週間、または約1か月、例えば、2週間、3週間、4週間、5週間、または6週間後に投与される。
【0238】
特定の実施形態においては、免疫原組成物の投与は、代わりにまたはその上に、アジュバントを含む免疫原組成物を投与した患者において、アジュバントを含まない組成物で免疫された個体において誘導されたBメモリー細胞応答に比べて向上したBメモリー細胞応答を誘導する。向上したBメモリー細胞応答は、インビトロ分化の刺激により測定される、抗原遭遇時に抗体分泌血漿細胞へと分化し得る末梢血Bリンパ球の頻度の増加を意味するものとする(実施例の節、例えば、Elispot B細胞記憶の方法を参照)。
【0239】
さらに別の実施形態では、免疫原組成物は一次免疫応答ならびにCD8応答を増強する。初回ワクチン接種用の免疫原組成物の1回用量を投与すると、良好な血清保護がもたらされ、アジュバントを含まない処方物で得られるものに比べて、特定の抗原に対して向上したCD4T細胞またはCD8T細胞免疫応答が誘導される。この結果、全体的な罹患率および死亡率が引き下げられ、また、肺炎および他のインフルエンザ様疾患のための緊急入院が防げる。この方法により、−アジュバントを含まない処方物で誘導される応答に比べて、より長い時間持続する、例えば、初回ワクチン接種後なお1年存在するCD4T細胞応答を誘導することができる。
【0240】
好ましくは、無刺激被験体で得られる向上したCD4T細胞免疫応答などのCD4T細胞免疫応答は、交差反応性CD4Tヘルパー応答の誘導を含む。特に、交差反応性CD4T細胞の量が増す。「交差反応性」CD4応答とは、例えばインフルエンザ株の間で共有されているエピトープを標的化するCD4T細胞を意味する。
【0241】
この用量のPD−1アンタゴニストはヒトにおいて抗原に対する免疫応答を増強する。特に、好適なPD−1アンタゴニスト量は、アジュバントを含まない組成物に比べて、または別のPD−1アンタゴニスト量のアジュバントを含む組成物に比べて、組成物の免疫能を向上させるものである。通常、免疫原組成物の用量は約0.5ml〜約1mlの範囲である。一般的なワクチン用量は0.5ml、0.6ml、0.7ml、0.8ml、0.9ml、または1mlである。好ましい実施形態では、ワクチン組成物1ml当たりに終濃度50μgのPD−1アンタゴニスト、好ましくはPD−L2−Ig、ワクチン用量0.5ml当たり25μgが含まれる。他の好ましい実施形態では、ワクチン組成物1ml当たり終濃度35.7μgまたは71.4μgのPD−1アンタゴニストが含まれる。具体的には、0.5mlのワクチン用量は、1用量当たり25μgまたは50μgのPD−1アンタゴニストを含む。さらに別の実施形態では、この用量は100μg以上である。免疫原組成物は通常、一元放射免疫拡散法(SRD)(J. M. Wood et al.: J. Biol. Stand. 5 (1977) 237-247、 J. M. Wood et al., J. Biol. Stand. 9 (1981) 317-330)により測定した場合、15μgの抗原成分を含む。
【0242】
被験体に免疫原組成物を再接種することができる。一般に、再接種は初回接種の少なくとも6か月後、好ましくは8〜14か月後、より好ましくは10〜12か月後前後で行われる。
【0243】
再接種用の免疫原組成物(追加免疫組成物)は、不活性されているか、または生きた状態で弱毒されている、いずれの種類の抗原調製物も含み得る。それは、初回接種用に用いた免疫原組成物と同じ種類の抗原調製物、例えば、スプリットインフルエンザウイルスまたはそのスプリットインフルエンザウイルス抗原調製物、完全ビリオン、精製サブユニットワクチンまたはウィロソーム(virosome)を含み得る。あるいは、追加免疫組成物は、初回接種用に用いたものと別の種類の抗原、すなわち、スプリットインフルエンザウイルスまたはそのスプリットインフルエンザウイルス抗原調製物、完全なビリオン、精製サブユニットワクチン、またはウィロソームを含み得る。
【0244】
ウイルスに対するワクチンに関して、使用する場合、追加免疫組成物は一般に次のウイルスシーズン、例えば、最初の免疫原組成物のおよそ1年後に投与する。追加免疫組成物はまた、その後毎年(3回目、4回目、5回目の接種など)与えてもよい。追加免疫組成物は初回接種に用いた組成物と同じであってもよい。
【0245】
好ましくは、再接種は以下のいずれか、好ましくは二つまたは総てを誘導する:PD−1アンタゴニストを含まない抗原調製物での初回接種の後に誘導された同等の応答に比べて(i)抗原調製物に対する向上したエフェクター細胞応答、または(ii)向上したB細胞メモリー応答、または(iii)向上した体液性応答。好ましくは、PD−1アンタゴニストを含有する免疫原性抗原調製物を再接種した後に誘導される免疫応答は、アジュバントを含まない組成物を再接種した後に誘導される対応する応答よりも高い。
【0246】
免疫原組成物は一価または多価、すなわち、二価、三価、または四価であり得る。好ましくは、その免疫原組成物は三価または四価である。多価とは、一般に異なる種または株からの抗原の供給源の数を意味する。ウイルスに関しては、少なくとも一つの株が汎流行的大発生に関連するか、または汎流行的大発生に関連する可能性がある。
【0247】
C.標的化抗原提示細胞
別の実施形態は、抗原提示細胞(APC)を1以上の開示されているPD−1アンタゴニストと、APCにおけるPD−1シグナル伝達を阻害、低減または遮断するのに有効な量で接触させることを提供する。APCにおけるPD−1シグナル伝達の遮断は、細胞内病原体または細胞内病原体に感染した細胞の排除を増強するAPCを活性化する。
【0248】
D.組合せ療法
PD−1アンタゴニスト組成物はそれを必要とする被験体に単独で、または1以上の付加的治療薬と組み合わせて投与することができる。付加的治療薬は処置される症状、障害、または疾患に基づいて選択される。例えば、PD−1アンタゴニストは免疫応答を増強または促進する働きをする1以上の付加的薬剤と同時に投与することができる。
【0249】
E.結合特性の調節
PD−1アンタゴニストの結合特性は投与される用量および用量計画に関連する。MDX−1106などの既存の抗体PD−1アンタゴニストは、1回投与の後少なくとも3ヶ月間、T細胞上のPD−1分子の60〜80%を持続的に占有することを示す(Brahmer, et al. J. Clin. Oncology, 27:(155) 3018 (2009))。好ましい実施形態では、開示されているPD−1アンタゴニストは、免疫細胞上のPD−1分子の、より短期またはより低パーセンテージの占有を示すPD−1との結合特性を有する。例えば、開示されているPD−1アンタゴニストは一般に、1回用量の投与後1週間、2週間、3週間またはさらには1か月で、免疫細胞上のPD−1分子の占有率5、10、15、20、25、30、35、40、45、50%未満を示す。他の実施形態では、開示されているPD−1アンタゴニストはPD−1に対してMDX−1106よりも低い結合親和性を有する。MDX−1106などの抗体に関して、PD−1−Ig融合タンパク質はその受容体に対して比較的低い親和性を有し、従って、比較的速い解離速度を有するはずである。
【0250】
他の実施形態では、PD−1アンタゴニストは、次の投与の前に減衰される周期的に増強される免疫応答を惹起するために数日、数週間、または数ヶ月間にわたって間欠的に投与され、これにより、免疫応答を誘導する、免疫応答を刺激する、または免疫応答を増強する働きをすることができる。
【実施例】
【0251】
本発明は、以下の非限定的実施例を参照すればさらに理解される。
【0252】
実施例1:B7−DCのPD−1への結合
ヒトB7−DC−IgのPD−1結合活性を、ELISAにより評価した。96ウェルELISAプレートを、BupH Carbonate/Bicarbonate pH9.4バッファー(Pierce)で希釈した0.75μg/mLの組換えヒトPD−1/Fc(R&D Systems)100μLで2時間コーティングした後、BSA溶液(Jackson ImmunoResearch)を用いて90〜120分間ブロックした。連続誘導体希釈したヒトB7−DC−IgならびにヒトIgG1イソ型対照を90分間結合させた。結合したB7−DC−Igは、100μLの0.5μg/mLビオチン結合抗ヒトB7−DCクローンMIH18(eBioscience)、次いで1:1000希釈HRP−ストレプトアビジン(BD Bioscience)およびTMB基質(BioFX)を用いて検出した。プレートリーダー(Molecular Devices)を用いて450nmの吸光度を読み取り、データを、SoftMaxで4パラメーターロジスティック適合を用いて解析した。
【0253】
ネズミB7−DC−IgのPD−1結合活性を、ELISAにより評価した。96ウェルのELISAプレートを、BupH Carbonate/Bicarbonate pH9.4バッファー(Pierce)で希釈した0.75μg/mLの組換えマウスPD−1/Fc(R&D Systems) 100μLを用いて2時間コーティングし、次いでBSA溶液(Candor-Bioscience)を用いて90分間ブロックした。連続希釈したネズミB7−DC−Ig(野生型、ならびにPD−1への低減された結合のために選択されたD111SおよびK113S変異株)、ならびにネズミIgG2aイソ型対照を90分間結合させた。結合したB7−DC−Igは、100μLの0.25μg/mLビオチン結合抗マウスB7−DCクローン112(eBioscience)、次いで1:2000希釈HRP−ストレプトアビジン(BD Bioscience)およびTMB基質(BioFX)を用いて検出した。プレートリーダー(Molecular Devices)を用いて450nmの吸光度を読み取り、データを、SoftMaxで4−パラメーターロジスティック適合を用いて解析した。
【0254】
図1Aおよび1Bは、PD−1結合ELISAにおける、OD450対B7−DC−Ig(μg/ml)の量の線グラフを示す。図1Aは、4つの異なるロットのヒトB7−DC−Igの結合を示す。図4Bは、野生型ネズミB7−DC−Ig(円)、DS変異株(D111Sを置換したB7−DC−Ig、三角)、およびKS変異株(K113Sを置換したB7−DC−Ig、四角)、およびネズミIgG2aイソ型対照(ひし形)の結合を示す。
【0255】
実施例2:B7−DCのPD−1発現CHO細胞への結合
B7−DC−Igを、最初にアロフィコシアニン(APC)と結合させ、次いでPD−1を構成的に発現するCHO細胞系統またはPD−1を発現しない親CHO細胞とともに種々な濃度でインキュベートした。結合は、フローサイトメトリーにより分析した。図2は、プローブ濃度(x軸)の関数としての、B7−DC−Ig−APCの蛍光強度の中央値(MFI)(y軸)を示す。B7−DC−Ig−APCはCHO.PD−1細胞(黒円)に結合するが、トランスフェクトされていないCHO細胞には結合しない(灰色三角)。
【0256】
実施例3:PD−1への結合に関するB7−DC−IgのB7−H1との競合
B7−H1−Igを、最初にアロフィコシアニン(APC)と結合させた。B7−H1−Ig−APCをプローブおよび細胞混合物に加える前に、種々の濃度の非標識B7−DC−Igを、最初に、PD−1を構成的に発現するCHO細胞系統とインキュベートした。図3は、加えた非標識B7−DC−Igコンペティターの濃度(x軸)の関数としての、B7−H1−Ig−APCの蛍光強度の中央値(MFI)(y軸)を示す。非標識B7−DC−Ig濃度が増加するにつれて、CHO細胞に結合するB7−H1−Ig−APC量は減少し、このことは、PD−1への結合に関してB7−DC−IgがB7−H1と競合することを実証している。
【0257】
実施例4:シクロホスファミドとB7−DC−Igの組合せが腫瘍特異的メモリー細胞傷害性Tリンパ球を生成する
9〜11週齢のBalb/Cマウスの皮下に1.0×105のCT26結腸直腸腫瘍細胞を移植した。腫瘍移植後10日目に、マウスに100mg/kgのシクロホスファミドを投与した。B7−DC−Ig処置は1日後である11日目に開始した。マウスを、100μgのB7−DC−Igを用いて、1週間に2用量、4週間にわたり合計8用量で処置した。CTX+B7−DC−Ig処置投与計画を受けたマウスの75%において44日までに定着腫瘍が根絶され、一方、対照CTX単独群のマウスは全例が、腫瘍増殖の結果死亡したか、または腫瘍がIACUCにより承認されたサイズを超えたため安楽死させた。
【0258】
上記実験でCT26結腸直腸定着腫瘍を根絶したマウスに、44日目および70日目に1×105個のCT26細胞を再負荷した。再負荷により腫瘍が増殖しなかったことは、シクロホスファミドおよびB7−DC−Igの組合せ処置が、長期にわたる抗腫瘍免疫を発展させたことを示唆している。ビヒクル対照群のマウスは全例が腫瘍を発現した。これにより、定着腫瘍に対する処置の有効性、ならびにB7−DCIgの併用処置が腫瘍抗原に対する記憶応答をもたらしたことが示された。
【0259】
上記実験でCT26結腸直腸定着腫瘍を根絶したマウスに、44日目に2.5×105個のCT26細胞を再負荷した。7日後、マウスの脾臓を単離した。マウス脾細胞を、Golgi遮断薬(BD BioScience)の存在下、5または50μg/mLのオバアルブミン(OVA)またはAH1ペプチドとともに6時間パルス処理し、CD8+/IFNγ+T細胞を評価することによりメモリーTエフェクター細胞を分析した。図4の結果は、CT26腫瘍根絶マウスにおいて多量のCT26特異的Tエフェクター細胞が存在することを示している。
【0260】
実施例5:B7−DC−IgはHSVウイルス粒子の放出を低減し、マウスの生存率を高めた
8〜10週齢のBalb/Cマウスをまず、ビヒクル(白四角)または300μgのB7−DC−Ig(黒四角)とともに4×104用量の弱毒HSV−2生ワクチンで免疫した(図5Aおよび5B)。1か月後、総てのマウスを5×105PFUのHSV−2 G−6株を腟内投与した。図5Aは、ウイルス投与後9時間、1日、2日、3日、4日および5日目における膣領域のスワブのウイルス粒子力価を示す。図5Bは、ウイルス投与後12日目におけるマウスの生存率を示す。これにより、ワクチンとともにB7−DC−Iが存在すると、ウイルス量を低減し、動物の生存率を高めることができる。
【図1A】
【図1B】
【発明の背景】
【0001】
関連出願の相互参照
本願は2008年8月25日に出願された米国仮出願第61/091,502号公報、2008年8月25日に出願された米国仮出願第61/091,694号公報、2008年8月25日に出願された米国仮出願第61/091,709号公報、2008年8月25日に出願された米国仮出願第61/091,705号公報、2009年1月5日に出願された米国仮出願第61/142,548号公報、および2009年4月1日に出願された米国仮出願第61/165,652号公報の優先権および利益を主張するものであり、これらは許容される場合には引用することにより本発明の開示の一部とされる。
【0002】
発明の分野
本発明は一般に、癌または感染などの疾患、特に、T細胞の消耗、T細胞の無応答もしくはその双方または細胞内病原体、すなわち例えばリーシュマニアがAPC(例えば、単球、樹状細胞、マクロファージ)または上皮細胞上でPD−1リガンドをアップレギュレートすることにより免疫応答を逃れる疾患を含む疾患を処置するための免疫調節組成物および方法に関する。
【0003】
背景技術
微生物感染に対する宿主の耐性には、二つの主要な重複する防御系、先天免疫、および適応免疫が組み込まれている。ウイルス、細菌、および寄生虫を包含する細胞内病原体は、感染後の初期に即座に、感染宿主において非特異的体液性応答と細胞エフェクター応答を刺激する活性化シグナルを中継する。これらの先天的防御応答の助けで、微生物の増殖速度が数日間遅れ、その間に適応系の免疫が誘導され、長期間病原体と対抗するよう促される(適応的/長期免疫)。これらの免疫応答はT細胞により媒介される。多くの細胞内病原体に関して、感染防御免疫は、前もって感染と闘う手助けをするように他の免疫細胞を刺激するサイトカインなどの化合物を産生するCD4+ヘルパーT細胞の生成、病原体に感染した宿主細胞を排除するCD8+細胞傷害性Tリンパ球(CTL)により主として媒介される細胞媒介応答、およびTヘルパー細胞により媒介される抗体応答の双方を必要とする。しかしながら、それらの生物が初期の免疫活性化を迂回し、エフェクター免疫応答および長期メモリー応答を損なえば、感染が確立され、持続する。これが急性および慢性感染を招く。
【0004】
研究によれば、初期の免疫破壊は多くの場合、クラスI MHC分子による抗原プロセシングおよび/または提示に関与する細胞内経路を標的とすることが証明されている。これの結果、初期の免疫活性化が不十分となり、また、その生物に対する一次応答がほとんど、または全く無くなる。これによりこれらの生物は確立され、細胞内病原体では、免疫系から「隠れた」ままとなる。さらに最近の研究では、多くの場合でこれらの病原が低いが測定可能な特異的免疫応答を刺激することが示されている。しかしながら、T細胞が病原体と闘うことにより「消耗」し、時間が経つにつれ段階的に有効性が低くなるという顕著な変化を受ける場合には、慢性感染が起こる。これはT細胞の消耗として知られる現象である。
【0005】
B7タンパク質は、免疫細胞(例えば、T細胞)に免疫応答を刺激または阻害する第二のシグナルを与える働きをする。PD−L1(B7−H1)およびPD−L2(PD−DC)は、共通の受容体PD−1と結合するB7ファミリーの分子の阻害メンバーである。PD−L1は多様な組織および細胞種で広く発現されるが、PD−L2の発現は主として活性化された樹状細胞(DC)およびマクロファージに限られる。CD28ファミリー受容体のメンバーであるPD−1は、活性化されたT細胞、B細胞、ナチュラルキラー(NK)細胞、単球、DCおよびマクロファージで誘導的に発現される。T細胞の消耗は、T細胞機能に負の調節を行う、PD−1受容体を介した阻害的T細胞シグナル伝達により生じることが示されている。
【0006】
そのリガンドによるPD−1結合の主要な結果は、T細胞受容体(TCR)下流でのシグナル伝達の阻害である。よって、PD−1を介したシグナル伝達は通常、T細胞に対して、T細胞増殖の低下またはT細胞活性におけるその他の低下をもたらす抑制または阻害シグナルを与える。PD−1のシグナル伝達は、主要組織適合性複合体(MHC)により提示されるペプチド抗原と近接したリガンドとの結合を必要とすると考えられ、これはTCRと結合されている(Freeman Proc. Natl. Acad. Sci. U. S. A 105:10275-10276 (2008))。PD−L1はT細胞において阻害的シグナル伝達を引き起こす主要なPD−1リガンドである。
【0007】
多くの細胞内病原体に対する一次免疫応答およびエフェクター免疫応答が不十分な結果として、とりわけヒト免疫不全ウイルス(HIV)、C型肝炎ウイルス(HCV)、単純ヘルペスウイルス(HSV)、結核菌(M. tuberculosis)、C.トラコマチス(C. trachomatis)、マラリアなどのこれら多くの生物に対する効果的なワクチンは存在しない。慢性感染が定着し、宿主免疫系がこれらの慢性または潜在感染を排除できない場合に重大な問題となる。抗原/ワクチンに対する不十分な一次応答およびエフェクター応答はまた、急性免疫が必要とされる場合(それ以外の点で有効なワクチンが作製可能な場合であっても)、例えば、インフルエンザなどの地域流行/汎流行的大発生または感染性病原体(例えば、炭疽)によるバイオテロ攻撃の、ならびに免疫系が未発達または弱まっている小児および高齢者集団において問題となる。
【0008】
ワクチンの免疫原性および保護を改善するための一つのアプローチがアジュバントの使用である。アジュバントは免疫応答を改善するためにワクチンに加えられる成分である。開発され、試験されているほとんどのアジュバントは主として先天的免疫応答(抗原特異的でない)、抗体応答および極めて少ない場合であるが、弱いT細胞応答を惹起する。利用可能なアジュバントに、一次応答を増大させ、細胞内病原体に対する防御免疫を惹起するのに必要とされるものである、強力なエフェクター応答または迅速なT細胞増殖応答を誘導するものはない。
【0009】
よって、本発明の一つの目的は、一次免疫応答およびエフェクター免疫応答の双方を増強するワクチンアジュバントを提供することである。
【0010】
もう一つの目的は、より迅速な防御誘導ならびに慢性感染に対する強力なエフェクター応答をもたらす組成物を提供することである。
【0011】
別の目的は、T細胞の消耗、T細胞の無応答またはその双方を誘導する感染を処置するための組成物および方法を提供することである。
【0012】
本発明のさらに別の目的は、単球、樹状細胞、マクロファージを含む抗原提示細胞の細胞内感染を処置するための組成物および方法を提供することである。
【発明の概要】
【0013】
(1)迅速T細胞媒介応答の惹起不全、(2)T細胞の消耗、T細胞の無応答もしくはその双方の誘導、または(3)単球、マクロファージ、樹状細胞、および/または例えば細胞内病原体を死滅させるのに必要とされるような他のAPCの活性化不全から起こる感染または疾患を処置するための方法および組成物。これらは急性(例えば、毒素誘発性)、慢性、遅発性、または潜在感染により引き起こされ得る。本発明の方法および組成物は、PD−1と結合してそれを遮断し、阻害的シグナル伝達を回避または低減することによるか、またはPD−L1などのPD−1のリガンドと結合してそれを遮断し、それにより、そのリガンドがPD−1と結合して(完全にまたは部分的に)阻害シグナルを送達しないようにすることにより、望ましくないT細胞阻害の問題を解決する。これらの分子は一般にPD−1アンタゴニストと呼ばれ、PD−1またはPD−L1などのリガンドと直接結合する両化合物を含む。いずれの場合でも、T細胞の増殖または活性化などのT細胞応答が増強される。さらに、PD−1アンタゴニストは、細胞内病原体によりアップレギュレートされている、抗原提示細胞(単球、マクロファージ、樹状細胞、上皮細胞などのAPC)で発現されるPD−1リガンドと結合してそれを遮断することもできる。
【0014】
免疫応答を増強する、または増大させ得るには、1)T細胞活性を阻害する分子に干渉すること、例えば、分子がPD−1であり、a)受容体(PD−1)を遮断するか、もしくはb)リガンド(B7−H1もしくはB7−DC)を遮断する場合、または2)T細胞活性を活性化させる分子を増大させる、例えば、分子がCD28であり、アゴニストが加えられる場合の、二つの機構がある。免疫応答は、投与される薬剤の用量を変えることにより、投与計画中、間欠的投与を行うことにより、またその組合せにより(再び投与される前に分子から薬剤の解離をもたらす(初回刺激と追加免疫を用いる抗原惹起で見られるものに類似))、種々の親和性(すなわち、必要とされるものよりも高いまたは低い)で結合するアンタゴニストを提供することにより調節され得る。いくつかの場合では、免疫系を刺激した後にその刺激を取り去ることが特に望ましいことがある。その結合相手に対するアンタゴニストの親和性を用いて、解離に必要な時間を決定することができる(親和性の高い薬剤は親和性の低い薬剤よりも解離に時間がかかる)。また、PD−1またはリガンドのいずれかと結合する、また同じ分子に異なる親和性で結合するアンタゴニストの組合せを用いて、免疫刺激の程度を調節することもできる。
【0015】
これらの組成物は、(i)PD−1を介した阻害的シグナル伝達を誘導することなくPD−1と結合してそれを遮断し、PD−L1およびPD−L2などのリガンドの結合を妨げ、それにより、PD−1により媒介される阻害シグナルの活性化を妨げるか、または(ii)PD−1のリガンドと結合し、PD−1受容体との結合を回避し、それにより、PD−1のより媒介される阻害シグナルの活性化を妨げるPD−1アンタゴニストを含む。
【0016】
好ましい組成物は、十分なT細胞応答の欠如、T細胞の消耗、T細胞の無応答、ならびに単球、マクロファージ、樹状細胞および他のAPCの活性化、または被験体におけるこれらの影響の総てを低減するまたは克服するのに有効な量の非抗体PD−1アンタゴニスト、例えば、PD−L2融合タンパク質(PD−L2−Ig)を含む。PD−1アンタゴニストはまた、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合するPD−L1タンパク質、断片、変異体または融合物も含む。これらのPD−L1断片は非機能的PD−L1断片とも呼ばれる。PD−L2ポリペプチド、融合タンパク質、および非機能的PD−L1断片は、PD−1の内因性リガンドがPD−1と相互作用しないようにすることにより、T細胞においてPD−1を介して起こる阻害的シグナル伝達を阻害または低減することができる。さらなる好ましいPD−1アンタゴニストとしては、PD−1のリガンドと結合し、T細胞上の内因性PD−1受容体との結合を妨げるPD−1またはその可溶性断片が含まれる。これらのPD−1断片は可溶性PD−1断片とも呼ばれる。その他のPD−1アンタゴニストとしては、PD−L1と結合し、PD−L1とPD−1の結合を妨げるB7.1またはその可溶性断片が含まれる。
【0017】
さらなる実施形態としては、阻害的シグナル伝達を生じることなくPD−1受容体と、またはPD−L1およびPD−L2などのPD−1受容体のリガンドのいずれかと結合してそれを遮断する抗体が含まれる。PD−L2ポリペプチド、融合タンパク質および非機能的PD−L1断片はまた、T細胞またはAPC上の別の受容体と結合することによってT細胞を活性化することもできる。
【0018】
アンタゴニストの作用は、T細胞の消耗、T細胞の無応答、またはその双方を克服する手助けをするとともに、感染または癌により誘導される単球、マクロファージ、樹状細胞、および他のAPCを活性化する。PD−L2ポリペプチドまたは融合タンパク質で処置可能な代表的感染としては、限定されるものではないが、ウイルス、細菌、寄生虫、原虫、または真菌により引き起こされる感染が挙げられる。処置可能な例示的ウイルス感染としては、限定されるものではないが、肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、ヒトTリンパ球向性ウイルス(HTLV)、ヘルペスウイルス、インフルエンザ、エプスタイン・バーウイルス、フィロウイルス、またはヒト乳頭腫ウイルスにより引き起こされる感染が挙げられる。処置可能な他の感染としては、プラスモジウム、マイコプラズマ、結核菌(M. tuberculosis)、炭疽菌(Bacillus anthracis)、ブドウ状球菌(Staphylococcus)、およびC.トラコマチス(C. trachomatis)により引き起こされるものが挙げられる。
【0019】
PD−1アンタゴニストは、ウイルス抗原、細菌抗原、原虫抗原、および腫瘍特異的抗原などの1以上の抗原を含有するワクチンと組み合わせて、または交互に投与することができる。PD−1アンタゴニストは、被験体において一次免疫応答およびエフェクター細胞応答を増強するためにワクチンとともに有効なアジュバントとして使用することができる。処置される好ましい被験体は免疫系が弱まっているか、または免疫不全であり、65歳を超えるか、または2歳未満である。
【図面の簡単な説明】
【0020】
【図1】図1A−Bは、PD−1結合ELISAにおけるB7−DC−IgのPD−1への結合を示すグラフである。
【図2】図2は、B7−DC−IgがPD−1発現CHO細胞と結合することを示すグラフである。
【図3】図3は、B7−DC−IgがPD−1との結合に関してB7−H1と競合することを示すグラフである。
【図4】図4は、B7−DC−Igの組合せ処置が腫瘍モデルにおいて抗原特異的メモリーCTLの生成をもたらしたことを示す。
【図5】図5は、B7−DC−IgがHSV−2ウイルス粒子の放出を低減し、HSV−2ワクチンの存在下でマウスの生存率を高めたことを示す。
【発明の具体的説明】
【0021】
I.定義
本明細書において「単離された」とは、その化合物が天然に存在する場合と異なる環境にある、例えば、ペプチドをそれが天然には見られない濃度まで濃縮することなどによってその天然環境から分離された目的化合物(例えば、ポリヌクレオチドまたはポリペプチドのいずれか)を表すことを意味する。「単離された」とは、目的化合物が有意に富化された、かつ/または目的化合物が部分的または有意に精製されたサンプル内にある化合物を含むことを意味する。「有意に」とは、統計学的に有意に大きいことを意味する。
【0022】
本明細書において「ポリペプチド」とは、修飾(例えば、リン酸化またはグリコシル化)とは無関係に任意の長さのアミノ酸鎖を意味する。
【0023】
本明細書において「変異体」ポリペプチドは、対応する野生型ポリペプチドのアミノ酸配列に比べて少なくとも一つのアミノ酸配列の変更を含む。
【0024】
本明細書において「アミノ酸配列の変更」とは、例えば、1以上のアミノ酸の置換、欠失または挿入であり得る。
【0025】
本明細書において「ベクター」とは、別のDNAセグメントが、その挿入されたセグメントの複製を生じるように挿入することができるプラスミド、ファージ、またはコスミドなどのレプリコンである。本明細書に記載のベクターは発現ベクターであり得る。
【0026】
本明細書において「発現ベクター」とは、1以上の発現制御配列を含むベクターである。
【0027】
本明細書において「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。
【0028】
本明細書において「機能的に連結される」とは、発現制御配列が目的のコード配列の発現を有効に制御するように遺伝構築物に組み込まれることを意味する。
【0029】
本明細書においてポリペプチドの「断片」とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。一般に、断片はアミノ酸5個以上の長さである。
【0030】
本明細書において「価数」とは、分子当たりに利用可能な結合部位の数を意味する。
【0031】
本明細書において「保存的」アミノ酸置換とは、置換されたアミノ酸が類似の構造的または化学的特性を有する置換である。
【0032】
本明細書において「非保存的」アミノ酸置換とは、置換されたアミノ酸の電荷、疎水性または嵩が有意に変化するものである。
【0033】
本明細書において「単離された核酸」とは、哺乳類ゲノムにおいてその核酸の一端または両端に通常隣接している核酸をはじめ、哺乳類ゲノムに存在する他の核酸分子から分離された核酸を意味する。
【0034】
本明細書において、核酸に関して「単離された」は任意の非天然核酸配列を含む(このような非天然配列は天然には見られず、天然ゲノムには直接連続する配列を持たないため)。
【0035】
本明細書において「宿主細胞」とは、組換え発現ベクターが導入できる原核細胞および真核細胞を意味する。
【0036】
本明細書において「形質転換させる」および「トランスフェクトする」とは、当該技術分野で知られているいくつかの技術によって核酸(例えばベクター)を細胞に導入することを包含する。
【0037】
本明細書において「抗体」とは、完全分子ならびに抗原−結合部位を含むその断片の双方を含むことを意味する。これらには完全抗体のFc断片を欠いたFabおよびF(ab’)2断片が含まれる。
【0038】
「免疫細胞」とは、免疫応答に役割を果たす造血系起源の細胞を意味する。免疫細胞は、リンパ球(例えば、B細胞およびT細胞)、ナチュラルキラー細胞および骨髄細胞(例えば、単球、マクロファージ、好酸球、肥満細胞、好塩基球、および顆粒球)を含む。
【0039】
「T細胞」とは、CD4+T細胞またはCD8+T細胞を意味する。T細胞とは、TH1細胞、TH2細胞、およびTh17細胞を含む。
【0040】
「T細胞細胞傷害性」とは、CD8+T細胞の活性化により媒介されるいずれの免疫応答も含む。例示的免疫応答としては、サイトカインの産生、CD8+T細胞の増殖、グランザイムまたはパーフォリンの産生、および感染性病原体の排除が含まれる。
【0041】
「免疫細胞」とは、T細胞、B細胞、およびリンパ球を意味する。
【0042】
「阻害的シグナル伝達」とは、T細胞の増殖を低減することによるものであれ、他のいずれかの阻害機構によるものであれ、T細胞応答を抑制する、またはそうでなければ低減する作用を有するPD−L1または他のリガンドによるPD−1受容体を介したシグナル伝達を意味する。
【0043】
II.PD−1アンタゴニスト
PD−1とPD−L1の間の相互作用に干渉するのに好ましいPD−1アンタゴニスト化合物は、PD−L2(B7−DCとしても知られる)、PD−L2の細胞外ドメイン、PD−L2の融合タンパク質、およびPD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合してそれを遮断し、するPD−L1とPD−1の結合を妨げるその変異体である。さらなるPD−1アンタゴニストとしては、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合するPD−L1の断片、PD−1のリガンドと結合し、T細胞上の内因性PD−1受容体と結合するのを妨げるPD−1または可溶性その断片、ならびにPD−L1と結合し、PD−L1とPD−1の結合を妨げることができるB7.1または可溶性その断片が含まれる。ある特定の実施形態では、PD−1アンタゴニストは被験体におけるT細胞傷害性を高める。この多機能性PD−1アンタゴニストは被験体において強力な免疫応答を誘導し、T細胞の消耗およびT細胞の無応答を克服する助けをする。
【0044】
PD−1アンタゴニストはPD−1のリガンドと結合し、それと相互作用するか、またはリガンドとPD−1受容体の結合を阻害するか、またはPD−1受容体を介したシグナル伝達に結びつくことなくPD−1受容体と直接結合する。好ましい実施形態では、PD−1アンタゴニストはPD−1と直接結合し、PD−1阻害的シグナル伝達を遮断する。他の実施形態では、PD−1アンタゴニストはPD−1のリガンドと結合し、リガンドがPD−1を介した阻害的シグナル伝達を誘発するのを低減または阻害する。さらに別の実施形態では、PD−1アンタゴニストは、PD−1受容体以外の受容体と結合することによりT細胞を活性化することができる。
【0045】
PD−1アンタゴニストは小分子アンタゴニストであり得る。「小分子」とは、100ダルトンを超え、約2,500ダルトン未満、好ましくは100〜2000ダルトンの間、より好ましくは約100〜約1250ダルトンの間、より好ましくは約100〜約1000ダルトンの間、より好ましくは約100〜約750ダルトンの間、より好ましくは約200〜約500ダルトンの間の分子量を有する小有機化合物を意味する。これらの小分子は多くの場合、環状炭素または複素環式構造および/または1以上の官能基で置換された芳香族または多環芳香族構造を含む。これらの小分子アンタゴニストは、PD−L1およびPD−L2などのPD−1のリガンドと結合し、そのリガンドがPD−1と相互作用しないようにすることにより、またはPD−1受容体を介したシグナル伝達を誘発することなくPD−1受容体と直接結合することにより、PD−1受容体シグナル伝達を低減するか、またはそれと干渉する。
【0046】
例示的PD−1アンタゴニストとしては、限定されるものではないが、PD−L2、PD−L1、PD−1またはB7−1ポリペプチド、およびその変異体、断片または融合タンパク質が挙げられる。さらなる実施形態としては、これらのタンパク質のいずれかと結合する抗体が挙げられる。
【0047】
A.PD−L2に基づくPD−1アンタゴニスト
1.PD−1と結合する、PD−L2に基づくPD−1アンタゴニスト
PD−1アンタゴニストは免疫細胞上のPD−1と結合し、阻害的PD−1のシグナル伝達を遮断する。PD−1のシグナル伝達は、PD−1リガンド(PD−L2またはPD−L1、一般に、PD−L1)が免疫シナプス内でTCR:MHC複合体と近接してPD−1と結合する必要があると思われる。よって、PD−1を介した阻害的シグナル伝達を遮断し、場合により、T細胞膜上でのPD−1とTCRの同時結合を妨げるタンパク質、抗体または小分子は有用なPD−1アンタゴニストである。
【0048】
代表的なポリペプチドアンタゴニストとしては、限定されるものではないが、PD−L2ポリペプチド、その断片、その融合タンパク質、およびその変異体が挙げられる。PD−1と結合し、PD−1を介した阻害的シグナル伝達を遮断するPD−L2ポリペプチドは好ましい実施形態の一つである。他の実施形態としては、PD−1の天然リガンドが結合しないように、かつ、シグナル伝達を誘発しないようにするPD−1アンタゴニストが含まれる。ある特定の実施形態では、開示されたPD−L2ポリペプチドは、免疫シナプスに関してペプチド−MHC複合体によるTCRの同時結合が存在しないので、PD−1受容体を介したシグナル伝達が低減されているか、またはそれを誘発する能力がないと考えられる。PD−1受容体を介したシグナル伝達は、T細胞の活性化およびT細胞の増殖を減衰する負のシグナルを伝達するので、PD−1のシグナル伝達経路を阻害すれば、そうでなければ減衰される細胞を活性化させることができる。
【0049】
2.例示的PD−L2ポリペプチドPD−1アンタゴニスト
ネズミPD−L2ポリペプチドは、
【化1】
(配列番号1)または
【化2】
(配列番号2)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0050】
ヒトPD−L2ポリペプチドは、
【化3】
(配列番号3)または
【化4】
(配列番号4)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0051】
非ヒト霊長類(カニクイザル)PD−L2ポリペプチドは、
【化5】
(配列番号5)または
【化6】
(配列番号6)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号1、3、および5はそれぞれシグナルペプチドを含む。
【0052】
B.PD−L1に基づくPD−1アンタゴニスト
1.PD−1受容体と結合するPD−L1に基づくPD−1アンタゴニスト
PD−1受容体と結合する他のPD−1アンタゴニストとしては、限定されるものではないが、PD−L1ポリペプチド、その断片、その融合タンパク質およびその変異体が挙げられる。これらのPD−1ポリペプチドアンタゴニストはPD−1受容体と結合してそれを遮断し、PD−1受容体を介した阻害的シグナル伝達が低減されているか、またはそれを誘発する能力がない。一つの実施形態では、PD−L1ポリペプチドは、免疫シナプスに関してペプチド−MHC複合体によるTCRの同時結合が存在しないので、PD−1受容体を介したシグナル伝達が低減されているか、またはそれを誘発する能力がないと考えられる。PD−1受容体を介したシグナル伝達は、T細胞の活性化およびT細胞の増殖を減衰する負のシグナルを伝達するので、PD−L1ポリペプチドを用いてPD−1のシグナル伝達を阻害すれば、そうでなければ減衰される細胞を活性化することができる。
【0053】
2.例示的PD−L1ポリペプチドPD−1アンタゴニスト
ネズミPD−L1ポリペプチドは、
【化7】
(配列番号7)または
【化8】
(配列番号8)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0054】
ヒトPD−L1ポリペプチドは、
【化9】
(配列番号9)
【化10】
(配列番号10)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0055】
配列番号7および9はそれぞれシグナルペプチドを含む。
【0056】
C.B7.1およびPD−1に基づくPD−1アンタゴニスト
1.PD−L1およびPD−L2と結合するB7.1およびPD−1に基づくPD−1アンタゴニスト
他の有用なポリペプチドとしては、PD−L1またはPD−L2などのPD−1リガンドと結合し、内因性PD−1受容体との結合を妨げ、それにより阻害的シグナル伝達を妨げることができるPD−1受容体タンパク質、またはその可溶性断片が含まれる。このような断片はまた、場合により天然リガンドとの結合を増大させるA99L突然変異などの突然変異を含む、PD−1タンパク質の可溶性ECD部分も含む。PD−L1はまた、タンパク質B7.1と結合することも示されている(Butte, et al., Immunity, 27(1): 111-122 (2007))。よって、PD−L1リガンドと結合し、内因性PD−1受容体との結合を妨げ、それにより阻害的シグナル伝達を妨げることができるB7.1またはその可溶性断片もまた有用である。
【0057】
2.例示的B7.1ポリペプチドPD−1アンタゴニスト
ネズミB7.1ポリペプチドは、
【化11】
(配列番号11)または
【化12】
(配列番号12)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0058】
ヒトB7.1ポリペプチドは、
【化13】
(配列番号13)または
【化14】
(配列番号14)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号11および13はそれぞれシグナルペプチドを含む。
【0059】
3.例示的PD−1ポリペプチドPD−1アンタゴニスト
ヒトPD−1ポリペプチドは、
【化15】
(配列番号15)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0060】
非ヒト霊長類(カニクイザル)PD−1ポリペプチドは、
【化16】
(配列番号16)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
配列番号15および16はそれぞれシグナルペプチドを含む。
【0061】
D.PD−1アンタゴニストポリペプチドの断片
PD−1アンタゴニストポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。本明細書において、PD−1アンタゴニストポリペプチドの断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0062】
有用な断片は、それらの天然リガンドと結合する能力を保持するものである。全長PD−1アンタゴニストポリペプチドの断片であるPD−1アンタゴニストポリペプチドは一般に、全長PD−1アンタゴニストポリペプチドに比べてその天然リガンドと結合する能力の少なくとも20パーセント、30パーセント、40パーセント、50パーセント、60パーセント、70パーセント、80パーセント、90パーセント、95パーセント、98パーセント、99パーセント、100パーセントを有する、またはさらには100パーセントを超える。
【0063】
例えば、PD−L2およびPD−L1の有用な断片は、PD−1と結合する能力を保持するものである。PD−L2およびPD−L1断片は一般に、全長PD−L2およびPD−L1に比べてPD−1と結合する能力の少なくとも20パーセント、30パーセント、40パーセント、50パーセント、60パーセント、70パーセント、80パーセント、90パーセント、95パーセント、98パーセント、99パーセント、100パーセントを有する、またはさらには100パーセントを超えるものである。
【0064】
PD−1アンタゴニストポリペプチドの断片は可溶性断片を含む。可溶性PD−1アンタゴニストポリペプチド断片は、産生細胞から放出、分泌またはそうでなければ抽出され得るPD−1アンタゴニストポリペプチドの断片である。PD−1アンタゴニストポリペプチドの可溶性断片は、該ポリペプチドの細胞外ドメインの一部または全部を含み、細胞内ドメインおよび/または膜貫通ドメインの一部または全部を欠く。一実施形態では、PD−1アンタゴニストポリペプチド断片はPD−1アンタゴニストポリペプチドの全細胞外ドメインを含む。細胞外ドメインは膜貫通ドメイン由来の1、2、3、4または5個のアミノ酸を含み得ると考えられる。あるいは、細胞外ドメインはC末端、N末端または双方から除去された1、2、3、4、または5個のアミノ酸を有し得る。
【0065】
一般に、PD−1アンタゴニストポリペプチドまたはその断片は、シグナル配列をコードする配列を含む核酸から発現される。このシグナル配列が一般に未熟なポリペプチドから切断されて、シグナル配列を欠く成熟ポリペプチドが生じる。PD−1アンタゴニストポリペプチドのこのシグナル配列は、そのポリペプチドの発現レベル、分泌、溶解度または他の特性に影響を与えるために標準的な分子生物学的技術を用いて別のポリペプチドのシグナル配列で置換することができる。PD−1アンタゴニストポリペプチドシグナル配列を置換するために用いられるシグナル配列は当該技術分野で知られているいずれのものであってもよい。
【0066】
1.PD−L2細胞外ドメイン
a.ヒトPD−L2細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはヒトPD−L2またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化17】
(配列番号17)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0067】
別の実施形態では、PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化18】
(配列番号18)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0068】
このシグナル配列は成熟タンパク質では除去されると考えられる。さらに、産生中に宿主からのタンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができると考えられる。配列番号19は、シグナル配列を含まない配列番号18のヒトアミノ酸配列を示す。
【化19】
(配列番号19)
【0069】
別の実施形態では、PD−1アンタゴニストポリペプチドはヒトPD−L2のIgVドメインを含む。第一の融合相手は、
【化20】
(配列番号20)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0070】
PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化21】
(配列番号21)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0071】
b.非ヒト霊長類PD−L2細胞外ドメイン
一つの実施形態では、PD−1アンタゴニストポリペプチドは非ヒト霊長類(カニクイザル)PD−L2の細胞外ドメインまたはその断片を含む。PD−1アンタゴニストポリペプチドは、
【化22】
(配列番号22)
と少なくとも80%、85%、90%、95%、99%または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0072】
別の実施形態では、PD−1アンタゴニストポリペプチドは、非ヒト霊長類アミノ酸配列:
【化23】
(配列番号23)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0073】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からの融合タンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができる。配列番号24は、シグナル配列を含まない配列番号23の非ヒト霊長類アミノ酸配列を示す。
【化24】
(配列番号24)
【0074】
別の実施形態では、PD−1アンタゴニストポリペプチドは非ヒト霊長類PD−L2のIgVドメインを含む。第一の融合相手は、
【化25】
(配列番号25)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0075】
PD−1アンタゴニストポリペプチドは、非ヒト霊長類アミノ酸配列:
【化26】
(配列番号26)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0076】
d.ネズミPD−L2細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはネズミPD−L2またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化27】
(配列番号27)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0077】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化28】
(配列番号28)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0078】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために他の生物由来のシグナルペプチドを使用することができる。配列番号29は、シグナル配列を含まない配列番号28のネズミアミノ酸配列を示す。
【化29】
(配列番号29)
【0079】
別の実施形態においては、PD−1アンタゴニストポリペプチドはネズミPD−L2のIgVドメインを含む。第一の融合相手は、
【化30】
(配列番号30)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0080】
PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化31】
(配列番号31)(PD−L2Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0081】
d.PD−L2細胞外ドメイン断片
PD−L2細胞外ドメインは、シグナルペプチドまたはPD−L2の推定膜貫通ドメイン由来の1以上のアミノ酸を含み得る。分泌の際に切断されるシグナルペプチドのアミノ酸の数は発現系および宿主によって異なり得る。さらに、PD−1と結合する能力を保持するC末端またはN末端から1以上のアミノ酸を欠くPD−L2細胞外ドメインの断片を使用することもできる。
【0082】
第一の融合相手として使用することができるネズミPD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号53の
【化32】
が挙げられる。
【0083】
ネズミPD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号1の
【化33】
が挙げられる。シグナルペプチドは、配列番号1内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0084】
第一の融合相手として使用することができるヒトPD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号56の
【化34】
が挙げられる。
【0085】
ヒトPD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号3の
【化35】
が挙げられる。シグナルペプチドは、配列番号3内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0086】
第一の融合相手として使用することができる非ヒト霊長類PD−L2の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号5の
【化36】
が挙げられる。
【0087】
非ヒト霊長類PD−L2のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号5の
【化37】
が挙げられる。シグナルペプチドは、配列番号5内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0088】
PD−L2タンパク質はまた、配列番号3(ヒト全長)のアミノ酸20−121または配列番号23(細胞外ドメインまたはECD)のアミノ酸1−102のPD−1結合断片も含む。その特定の実施形態では、PD−L2ポリペプチドまたはPD−1結合断片はまた、配列番号3の残基110−114にアミノ酸WDYKYを、または配列番号23の残基91−95にWDYKYを組み込んでいる。限定されない例として、このようなPD−1結合断片は、配列番号3のアミノ酸20−121の配列の少なくとも10、少なくとも20、少なくとも30、少なくとも40、少なくとも50、少なくとも60、少なくとも70、少なくとも75、少なくとも80、少なくとも85、少なくとも90、少なくとも95または少なくとも100個の連続するアミノ酸を含んでなり、このようなPD−1結合断片のそれぞれの好ましい実施形態は、部分断片として配列番号3の残基110−114に見られるアミノ酸WDYKY、または配列番号23の残基91−95に見られるWDYKYを含んでなる。
【0089】
2.PD−L1細胞外ドメイン
一つの実施形態では、変異体PD−L1ポリペプチドは細胞外ドメインの全部または一部を含む。PD−L1の代表的な細胞外ドメインのアミノ酸配列は、
【化38】
(配列番号32)
と80%、85%、90%、95%、または99%の配列同一性を有し得る。
【0090】
PD−L1の膜貫通ドメインは配列番号9の239番のアミノ酸で始まる。PD−L1の好適な断片は、シグナルペプチド配列、例えば、配列番号9またはその変異体の1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、膜貫通ドメインの1、2、3、4、5、6、7、8、9、もしくは10個のアミノ酸、またはその組合せを含み得る。
【0091】
ネズミPD−L1の細胞外ドメインは以下のアミノ酸配列:
【化39】
(配列番号33)
を有する。
【0092】
ネズミPD−L1の膜貫通ドメインは配列番号7の240番のアミノ酸で始まる。ある特定の実施形態では、PD−L1ポリペプチドは、シグナルペプチドの1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、膜貫通ドメインの1、2、3、4、5、6、7、8、9、もしくは10個の連続するアミノ酸、またはその組合せを有するネズミPD−L1の細胞外ドメインを含む。
【0093】
3.B7.1細胞外ドメイン
a.ネズミB7.1細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミB7.1またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化40】
(配列番号34)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0094】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化41】
(配列番号35)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0095】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために、他の生物由来のシグナルペプチドを使用することができる。配列番号36は、シグナル配列を含まない配列番号35のネズミアミノ酸配列を示す。
【化42】
(配列番号36)
【0096】
別の実施形態においては、PD−1アンタゴニストポリペプチドはネズミB7.1のIgVドメインを含む。第一の融合相手は、
【化43】
(配列番号37)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0097】
PD−1アンタゴニストポリペプチドは、ネズミアミノ酸配列:
【化44】
(配列番号38)(B7.1Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0098】
b.ヒトB7.1細胞外ドメイン
一つの実施形態においては、PD−1アンタゴニストポリペプチドはヒトB7.1またはその断片の細胞外ドメインを含む。PD−1アンタゴニストポリペプチドは、
【化45】
(配列番号39)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0099】
別の実施形態においては、PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化46】
(配列番号40)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0100】
シグナル配列は成熟タンパク質では除去される。さらに、産生中に宿主からのタンパク質の分泌を増強するために、他の生物由来のシグナルペプチドを使用することができる。配列番号41は、シグナル配列を含まない配列番号40のヒトアミノ酸配列を示す。
【化47】
(配列番号41)
【0101】
別の実施形態においては、PD−1アンタゴニストポリペプチドはヒトB7.1のIgVドメインを含む。第一の融合相手は、
【化48】
(配列番号42)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有するヌクレオチド配列によりコードされ得る。
【0102】
PD−1アンタゴニストポリペプチドは、ヒトアミノ酸配列:
【化49】
(配列番号43)(B7.1Vとも呼ばれる)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有し得る。
【0103】
3.B7.1細胞外ドメイン断片
共刺激ポリペプチドドメインとして使用することができるネズミB7.1の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号11の
【化50】
が挙げられる。
【0104】
ネズミB7.1のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号11の
【化51】
が挙げられる。シグナルペプチドは、配列番号11内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0105】
共刺激ポリペプチドドメインとして使用することができるヒトB7.1の好適な断片の例としては、限定されるものではないが、以下のもの:
配列番号13の
【化52】
が挙げられる。
【0106】
ヒトB7.1のさらなる好適な断片としては、限定されるものではないが、場合によりシグナルペプチドの1〜5個のアミノ酸がN末端に付加された、以下のもの:
配列番号13の
【化53】
が挙げられる。シグナルペプチドは、配列番号13内に含まれるシグナルペプチドを含め、本明細書に開示されているいずれのものであってもよく、または当該技術分野で知られているいずれのシグナルペプチドであってもよい。
【0107】
E.変異体
1.変異体PD−L2およびPD−L1 PD−1アンタゴニスト
さらなるPD−1アンタゴニストとしては、生理学的条件下でPD−1と結合する能力を保持し、PD−1に対する結合が増強されるか、またはPD−1受容体を介したシグナル伝達を促進する能力が低減されるように突然変異誘発されたPD−L2およびPD−L1、ポリペプチドおよびその断片が含まれる。一つの実施形態は、PD−1を活性化し、T細胞へ阻害シグナルを伝達するポリペプチドの能力を非突然変異PD−L2またはPD−L1に比べて阻害または低減する1以上のアミノ酸置換、欠失、または挿入を含む単離されたPD−L2およびPD−L1ポリペプチドを提供する。該PD−L2およびPD−L1ポリペプチドはいずれの起源種のものであってもよい。一つの実施形態においては、PD−L2またはPD−L1ポリペプチドは哺乳類種に由来する。好ましい実施形態では、PD−L2またはPD−L1ポリペプチドはヒトまたは非ヒト霊長類起源のものである。
【0108】
別の実施形態においては、変異型PD−L2またはPD−L1ポリペプチドは、PD−1に対して野生型または非変異型PD−L2またはPD−L1と同じ結合活性を有するが、非突然変異PD−L2またはPD−L1ポリペプチドに比べてPD−1受容体を介したシグナル伝達を刺激する能力を持たないか、または10%未満である。他の実施形態では、該変異型PD−L2またはPD−L1ポリペプチドは野生型PD−L2またはPD−L1よりもPD−1に対して10%、20%、30%、40%、50%、60%、70%、80%、90%、100%またはそれを超える結合活性を有し、非突然変異PD−L2またはPD−L1ポリペプチドに比べてPD−1受容体を介したシグナル伝達を刺激する能力が50%、40%、30%、20%、または10%未満である。
【0109】
変異型PD−L2またはPD−L1ポリペプチドは、アミノ酸置換、欠失、または挿入の任意の組合せを有し得る。一つの実施形態では、単離されたPD−L2またはPD−L1変異型ポリペプチドは、それらのアミノ酸配列が野生型PD−L2またはPD−L1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するように、ある整数のアミノ酸変化を有する。好ましい実施形態では、B7−H1変異型ポリペプチドは、野生型ネズミ、非ヒト霊長類またはヒトPD−L2またはPD−L1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するアミノ酸配列を有する。
【0110】
配列同一性%は、コンピュータープログラムまたは直接的配列比較を用いて計算することができる。2配列間の同一性を決定するための好ましいコンピュータープログラム法としては、限定されるものではないが、GCGプログラムパッケージ、FASTA、BLASTP、およびTBLASTNが挙げられる(例えば、D. W. Mount, 2001, Bioinformatics: Sequence and Genome Analysis, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. 参照)。BLASTPおよびTBLASTNプログラムはNCBIおよび他のソースから公的に入手可能である。周知のSmith Watermanアルゴリズムも同一性を決定するために使用可能である。
【0111】
アミノ酸配列比較のためのパラメーター例としては以下のものが挙げられる:1)Needleman and Wunsch (J. Mol. Biol., 48:443-453 (1970))からのアルゴリズム、2)Hentikoff and Hentikoff (Proc. Natl. Acad. Sci. U.S.A., 89:10915-10919 (1992))からのBLOSSUM62比較マトリックス、3)ギャップペナルティー=12、および4)ギャップレングスペナルティー=4。これらのパラメーターを用いる有用なプログラムは、「ギャップ」プログラム(Genetics Computer Group, Madison, Wis.)として公的に入手可能である。上述のパラメーターはポリペプチド比較のためのデフォルトパラメーターである(エンドギャップに対するペナルティーは無い)。
【0112】
あるいは、ポリペプチド配列同一性は、次式:同一性%=(同一の残基の数)/(アミノ酸残基のアライメント長)*100を用いて計算することができる。この計算では、配列長は内部ギャップを含むが、末端ギャップは含まない。
【0113】
PD−L2またはPD−L1ポリペプチドのアミノ酸置換は「保存的」または「非保存的」であり得る。本明細書において、「保存的」アミノ酸置換は、置換されたアミノ酸が類似の構造的または化学的特性を有する置換であり、「非保存的」アミノ酸置換は、置換されたアミノ酸の電荷、疎水性、または嵩が有意に変更されるものである。非保存的置換は、(a)例えば、シートまたはらせんコンフォメーションのような置換領域におけるペプチド主鎖の構造、(b)標的部位における分子の電荷または疎水性、または(c)側鎖の嵩の維持に対するそれらの効果がより有意に異なる。
【0114】
保存的アミノ酸置換の例としては、置換が以下の5群中のものが挙げられる:1)小さな脂肪族、非極性またはやや極性の残基(Ala、Ser、Thr、Pro、Gly)、2)極性、負電荷を有する残基およびそれらのアミド(Asp、Asn、Glu、Gln)、極性、正電荷を有する残基(His、Arg、Lys)、大きな脂肪族、非極性残基(Met、Leu、Ile、Val、Cys)、および大きな芳香族残基(Phe、Tyr、Trp)。非保存的アミノ酸置換の例としては、1)親水性残基(例えば、セリルまたはトレオニル)が疎水性残基(例えば、ロイシル、イソロイシル、フェニルアラニル、バリルまたはアラニル)から(またはにより)置換される場合、2)システインまたはプロリンが他の残基から(またはにより)置換される場合、3)電気的に陽性の側鎖を有する残基(例えば、リシル、アルギニルまたはヒスチジル)が電気的に陰性の残基(例えば、グルタミルまたはアスパルチル)から(またはにより)置換される場合、または4)嵩高の側鎖を有する残基(例えば、フェニルアラニン)が側鎖を持たない残基(例えば、グリシン)から(またはにより)置換される場合が挙げられる。
【0115】
しかしながら、挙げられたアミノ酸位置における置換は、任意のアミノ酸またはアミノ酸類似体を用いて行うことができると理解される。例えば、挙げられた位置における置換は、任意の天然アミノ酸(例えば、アラニン、アスパラギン酸、アスパラギン、アルギニン、システイン、グリシン、グルタミン酸、グルタミン、ヒスチジン、ロイシン、バリン、イソロイシン、リシン、メチオニン、プロリン、トレオニン、セリン、フェニルアラニン、トリプトファン、またはチロシン)を用いて行うことができる。
【0116】
本明細書に記載される置換はマウス、非ヒト霊長類およびヒトPD−L2またはPD−L1に関するものであるが、当業者ならば他種(例えば、ラット、ハムスター、モルモット、アレチネズミ、ウサギ、イヌ、ネコ、ウマ、ブタ、ヒツジ、またはウシ)由来の相当するポリペプチドにおいても等価の変更を容易に行うことができることを注記する。しかしながら、結合は種特異的な成分であることから、PD−1アンタゴニストをヒト投与する場合にはヒトを使用するのが好ましい。
【0117】
一つの実施形態においては、開示されている単離された変異型PD−L2またはPD−L1ポリペプチドはPD−1のアンタゴニストであり、PD−1を介したシグナル伝達を誘発することなくPD−1と結合してそれを遮断する。PD−1シグナル伝達によるT細胞の減衰を妨げることにより、より多くのT細胞の活性化に利用可能である。T細胞阻害を妨げると、PD−1アンタゴニストと接触していないT細胞に比べて、T細胞応答が高まるか、T細胞の増殖が増大し、T細胞によるサイトカインの産生および/または分泌が高まるか、T細胞の分化およびエフェクター機能が刺激されるか、またはT細胞の生存が促進される。この相互作用から生じるT細胞応答は一般に、PD−1アンタゴニストポリペプチドの非存在下での応答よりも大きい。PD−1アンタゴニストポリペプチドの非存在下でのT細胞の応答は応答性がないか、またはPD−1アンタゴニストポリペプチドの存在下よりも有意に低いものであり得る。このT細胞の応答は、エフェクター(例えば、CTLまたは抗体産生B細胞)応答、1以上のエフェクター(例えば、CTLまたは抗体産生B細胞)応答を助けるヘルパー応答、または抑制的応答であり得る。
【0118】
2分子間の結合親和性を測定する方法は技術分野でよく知られている。PD−1に対する変異型PD−L2またはPD−L1ポリペプチドの結合親和性を測定する方法としては、限定されるものではないが、蛍光活性化細胞選別(FACS)、表面プラズモン共鳴、蛍光異方性、アフィニティークロマトグラフィー、および親和性選択−質量分析が挙げられる。
【0119】
本明細書に開示される変異型ポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。好ましい断片は、PD−1と結合するのに有効な細胞外ドメインの全部または一部を含む。本明細書において、断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0120】
2.変異型B7.1およびPD−1アンタゴニスト
さらなるPD−1アンタゴニストとしては、生理学的条件下でPD−L2および/またはPD−L1と結合する能力を保持するか、またはPD−L2および/またはPD−L1に対する結合が増強されるように改変されているB7.1およびPD−1ポリペプチドおよびその断片が含まれる。このような変異体PD−1タンパク質としては、天然リガンドに対する結合を増強する、A99L突然変異などの突然変異を含むPD−1タンパク質の可溶性ECD部分が挙げられる(Molnar et al., Crystal structure of the complex between programmed death-1 (PD-1) and its ligand PD-L2, PNAS, Vol. 105, pp. 10483-10488 (29 July 2008))。B7.1およびPD−1ポリペプチドはいずれの起源種のものであってもよい。一つの実施形態では、B7.1またはPD−1ポリペプチドは哺乳類種に由来する。好ましい実施形態では、B7.1またはPD−1ポリペプチドはヒトまたは非ヒト霊長類起源のものである。
【0121】
変異型B7.1またはPD−1ポリペプチドは、アミノ酸置換、欠失、または挿入の任意の組合せを有し得る。一つの実施形態では、単離されたB7.1またはPD−1変異型ポリペプチドは、それらのアミノ酸配列が野生型B7.1またはPD−1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するように、ある整数のアミノ酸変化を有する。好ましい実施形態では、B7.1またはPD−1変異型ポリペプチドは、野生型ネズミ、非ヒト霊長類、またはヒトB7.1またはPD−1ポリペプチドのアミノ酸配列と少なくとも60、70、80、85、90、95、97、98、99、99.5、または100%の同一性を有するアミノ酸配列を有する。
【0122】
B7.1またはPD−1ポリペプチドにおけるアミノ酸置換は「保存的」または「非保存的」であり得る。保存的および非保存的置換は上記に定義されている。
【0123】
一つの実施形態においては、開示されている単離された変異型B7.1またはPD−1ポリペプチドはPD−1のアンタゴニストであり、PD−L2および/またはPD−L1と結合し、それによりそれらの内因性PD−1との結合を遮断する。PD−1シグナル伝達によるT細胞の減衰を妨げることにより、より多くのT細胞を活性化に利用可能とする。T細胞阻害を妨げると、PD−1アンタゴニストと接触していないT細胞に比べて、T細胞応答が高まるか、T細胞の増殖が増大するか、T細胞によるサイトカインの産生および/または分泌が高まるか、T細胞の分化およびエフェクター機能が刺激されるか、またはT細胞の生存が促進される。この相互作用から生じるT細胞応答は一般に、PD−1アンタゴニストポリペプチドの非存在下での応答よりも大きい。PD−1アンタゴニストポリペプチドの非存在下でのT細胞の応答は、応答性がないか、またはPD−1アンタゴニストポリペプチドの存在下よりも有意に低いものであり得る。このT細胞の応答は、エフェクター(例えば、CTLまたは抗体産生B細胞)応答、1以上のエフェクター(例えば、CTLまたは抗体産生B細胞)応答を助けるヘルパー応答、または抑制的応答であり得る。
【0124】
変異体ポリペプチドは全長ポリペプチドであってもよいし、あるいは全長ポリペプチドの断片であってもよい。好ましい断片は、PD−L2および/またはPD−L1と結合するのに有効な細胞外ドメインの全部または一部を含む。本明細書において、断片とは、全長タンパク質のより短いポリペプチドであるポリペプチドの任意のサブセットを意味する。
【0125】
F.融合タンパク質
いくつかの実施形態では、PD−1アンタゴニストは、第一のポリペプチドドメインと第二のドメインとを含む融合タンパク質である。この融合タンパク質はT細胞受容体に結合することができるか、あるいは好ましくはこの融合タンパク質は、例えばPD−1と競合的に結合することによって、T細胞と結合して、T細胞への阻害的シグナル伝達を遮断することができる。開示されている組成物は、天然阻害リガンドとPD−1との結合に干渉することにより、PD−1を介したシグナル伝達を有効に遮断する。好適な共刺激ポリペプチドとしては、阻害性T細胞シグナル伝達受容体に対する結合親和性が増強または低下されたPD−1などの変異型ポリペプチドおよび/またはその断片が含まれる。
【0126】
融合タンパク質はまた場合により、第一のポリペプチドドメインを抗原結合ドメインから分離するペプチドまたはポリペプチドリンカードメインを含んでもよい。
【0127】
本明細書に開示される融合タンパク質は式I:
N−R1−R2−R3−C
(式中、「N」は融合タンパク質のN末端を表し、「C」は融合タンパク質のC末端を表し、「R1」はPD−L2、PD−L1、B7.1、もしくはPD−1ポリペプチドまたは抗原結合標的化ドメインであり、「R2」はペプチド/ポリペプチドリンカードメインであり、「R3」は標的化ドメインまたは抗原結合標的化ドメインであり、ここで、「R1」が抗原結合標的化ドメインである場合には、「R3」はポリペプチドドメインであり、「R1」がPD−L2、PD−L1、B7.1またはPD−1ポリペプチドドメインである場合には、「R3」は抗原結合標的化ドメインである)
で表されるものである。好ましい実施形態では、「R1」はPD−L2、PD−L1、B7.1、またはPD−1ポリペプチドドメインであり、「R3」は抗原結合標的化ドメインである。
【0128】
場合により、融合タンパク質はさらに、2以上の融合タンパク質を二量体化または多量体化する働きをするドメインを含む。該融合タンパク質を二量体化または多量体化する働きをするドメインは別個のドメインであってもよいし、あるいはまたその融合タンパク質の他のドメイン(PD−L2、PD−L1、B7.1もしくはPD−1ポリペプチドドメイン、抗原結合標的化ドメインまたはペプチド/ポリペプチドリンカードメイン)の一つに含まれていてもよい。
【0129】
融合タンパク質は二量体化または多量体化されていてもよい。二量体化または多量体化は二量体化または多量体化ドメインを介して2以上の融合タンパク質間で生じ得る。あるいは、融合タンパク質の二量体化または多量体化は化学的架橋によっても生じする。形成される二量体または多量体は、ホモ二量体/ホモ多量体またはヘテロ二量体/ヘテロ多量体であり得る。
【0130】
融合タンパク質のモジュール性およびそれらの種々の組合せでの二量体化または多量体化能は、腫瘍細胞の微小環境または免疫調節組織に対する免疫応答を増強する働きをする標的化分子に豊富な選択肢を与える。
【0131】
1.抗原結合標的化ドメイン
融合タンパク質はまた抗原結合標的化ドメインを含む。いくつかの実施形態では、標的化ドメインは、感染性疾患を引き起こす病原体に応答したT細胞の活性化の調節に関与する免疫組織に特異的な抗原、リガンドまたは受容体と結合する。
【0132】
標的化ドメイン
標的化のための抗原、リガンド、および受容体
一つの実施形態においては、融合タンパク質は、感染性疾患を引き起こす病原体に応答したT細胞の活性化の調節に関与する免疫組織により発現される抗原と特異的に結合するドメインを含む。
【0133】
標的化ドメインの分子種
リガンドおよび受容体
一つの実施形態においては、疾病標的化ドメインは、罹患細胞で特異的に発現されるか、または正常組織に比べて罹患細胞で過剰発現される細胞表面抗原もしくは受容体と結合するリガンドである。罹患細胞はまた、増殖および発達に影響を及ぼす微小環境中に多数のリガンドを分泌する。限定されるものではないが、増殖因子、サイトカインおよびケモカイン(上記に示されるケモカインを含む)をはじめとする、罹患細胞により分泌されるリガンドと結合する受容体は、開示されている融合タンパク質に用いるのに好適である。罹患細胞により分泌されるリガンドは、分泌されたリガンドと結合する受容体の可溶性断片を用いて標的化され得る。可溶性受容体断片は産生細胞から放出、分泌またはそうでなければ抽出され得る断片ポリペプチドであり、全細胞外ドメインまたはその断片を含む。
【0134】
単鎖ポリペプチド抗体
別の実施形態では、疾病関連標的化ドメインは、罹患細胞で特異的に発現されるか、または正常組織に比べて罹患細胞で過剰発現される細胞表面抗原または受容体と結合する単鎖抗体である。単鎖ドメイン抗体は共阻害性受容体アンタゴニストドメインに関して上記されている。
【0135】
Fcドメイン
別の実施形態では、疾病または疾病関連標的化ドメインは、罹患細胞で発現されるFc受容体と結合する免疫グロブリン重鎖のFcドメインである。Fc領域は、第一の定常領域免疫グロブリンドメインを除く抗体の定常領域を含むポリペプチドを含む。よって、FcはIgA、IgDおよびIgGの最後の二つの定常領域免疫グロブリンドメインとIgEおよびIgMの最後の3つの定常領域免疫グロブリンドメインを意味する。好ましい実施形態では、Fcドメインはヒトまたはネズミ免疫グロブリンに由来する。より好ましい実施形態では、Fcドメインは、CH2およびCH3領域を含むヒトIgG1またはネズミIgG2aに由来する。
【0136】
一つの実施形態においては、ヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3域は、
【化54】
(配列番号44)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0137】
配列番号44によりコードされるヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3領域は以下のアミノ酸配列:
【化55】
(配列番号45)
を有する。
【0138】
別の実施形態においては、ネズミ免疫グロブリンCγ2a鎖のヒンジ、CH2およびCH3領域は、
【化56】
(配列番号46)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0139】
配列番号46によりコードされるネズミ免疫グロブリンCγ2a鎖のヒンジ、CH2およびCH3領域は以下のアミノ酸配列:
【化57】
(配列番号47)
を有する。
【0140】
一つの実施形態においては、Fcドメインは、腫瘍もしくは腫瘍関連新生血管系で特異的に発現されるか、または正常組織に比べて腫瘍もしくは腫瘍関連新生ポリペプチド血管系で過剰発現される特異的Fc受容体との結合を増強する1以上のアミノ酸挿入、欠失または置換を含み得る。好適なアミノ酸置換は上記のように保存的および非保存的置換を含む。
【0141】
非ホジキンリンパ腫またはワルデンストロームマクログロブリン血症に対してリツキシマブ(キメラマウス/CD20に対するヒトIgG1モノクローナル抗体)で処置された患者における治療結果は、ヒトIgG1のFcドメインに対して明瞭な内因的親和性を有するFcγ受容体の対立遺伝子変異体の個々の発現と相関していた。特に、Fc受容体CD16A(FcγRIIIA)を活性化する低親和性の高親和性対立遺伝子を有する患者はより高い奏功率を示し、非ホジキンリンパ腫の場合には、無進行生存を改善した。別の実施形態では、Fcドメインは、低親和性阻害Fc受容体CD32B(FcγRIIB)との結合を低減し、かつ低親和性活性化Fc受容体CD16A(FcγRIIIA)との野生型レベルの結合を保持するか、またはそれとの結合を増強する以上のアミノ酸挿入、欠失、または置換を含み得る。
【0142】
好ましい実施形態では、Fcドメインは、CD16Aとの結合を増強するアミノ酸挿入、欠失、または置換を含む。CD16Aとの結合を増強し、CD32Bとの結合を低減するヒトIgG1のFcドメインの多数の置換が当該技術分野に知られており、Stavenhagen, et al., Cancer Res., 57(18):8882-90 (2007)に記載されている。CD32Bとの結合が低減され、かつ/またはCD16Aとの結合が増強されたヒトIgG1 Fcドメインの変異体の例は、F243L、R929P、Y300L、V305IまたはP296L置換を含む。これらのアミノ酸置換はヒトIgG1 Fcドメインにいずれの組合せで存在してもよい。一実施形態では、ヒトIgG1 Fcドメイン変異体はF243L、R929P、およびY300L置換を含む。別の実施形態においては、ヒトIgG1 Fcドメイン変異体はF243L、R929P、Y300L、V305I、およびP296L置換を含む。
【0143】
グリコホスファチジルイノシトールアンカードメイン
別の実施形態においては、疾病または疾病関連新生血管系標的化ドメインは、グリコシルホスファチジルイノシトール(GPI)アンカーの翻訳後付加のためのシグナルを与えるポリペプチドである。GPIアンカーは、多くの真核生物のタンパク質のC末端に翻訳後に付加される糖脂質構造である。この修飾は細胞膜の外片に付着タンパク質をしっかり固定する。GPIアンカーは、T細胞に対する提示のために細胞表面にT細胞受容体結合ドメインを付着させるのに使用することができる。この実施形態では、GPIアンカードメインはT細胞受容体結合ドメインに対してC末端にある。
【0144】
一つの実施形態においては、GPIアンカードメインは、ポリペプチドが真核生物系で発現される場合にGPIアンカーの翻訳後付加のためにシグナルを伝達するポリペプチドである。アンカーの付加はGPIアンカーシグナル配列により決定され、アンカー付加の部位における小アミノ酸のセット(ω部位)とそれに続く親水性スペーサーからなり、疎水性ストレッチで終わる(Low, FASEB J., 3:1600-1608 (1989))。このシグナル配列の切断は、保存された中央成分(Low, FASEB J., 3:1600-1608 (1989))を有するが、可変の末梢部分(Homans et al., Nature, 333:269-272 (1988))を有するアンカーを付加する前にERで起こる。GPIアンカータンパク質のC末端は、ホスホエタノールアミン橋を介して保存性の高いコアグリカン、マンノース(α1−2)マンノース(α1−6)マンノース(α1−4)グルコサミン(α1−6)ミオイノシトールに連結される。リン脂質テールはGPIアンカーを細胞膜の付着させる。このグリカンコアは、ホスホエタノールアミン基、マンノース、ガラクトース、シアル酸または他の糖などの側鎖で様々に修飾することができる。第一のマンノース残基に付着されている最も一般的な側鎖は別のマンノースである。グリカンコアの第三のマンノースに付着されているN−アセチルガラクトサミン含有多糖などの複合側鎖が哺乳類アンカー構造に見られる。コアグルコサミンの修飾はまれである。タンパク質および起源種によって、ホスホイノシトール環の脂質アンカーはジアシルグリセロール、アルキルアシルグリセロールまたはセラミドである。これらの脂質種は炭素14〜28個の範囲で長さが異なり、飽和であっても不飽和であってもよい。多くのGPIアンカーはまた、イノシトール環の2−ヒドロキシルにパルミチン酸などの付加的脂肪酸を含む。この余分な脂肪酸により、GPIアンカーはPI−PLCによる切断に耐性となる。
【0145】
GPIアンカーは、GPI翻訳後修飾を行うことができる真核生物系でGPIアンカードメインを含む融合タンパク質を発現させることよって達成することができる。GPIアンカードメインは腫瘍または腫瘍血管系標的化ドメインとして使用することもできるし、あるいはすでに別個の腫瘍または腫瘍血管系標的化ドメインを含む融合タンパク質に追加することもできる。
【0146】
別の実施形態においては、GPIアンカー部分は、インビトロ酵素的または化学的工程を介して、単離されたT細胞受容体結合ドメインに直接付加される。この実施形態では、GPIアンカーは、GPIアンカードメインを必要とすることなくポリペプチドに付加することができる。GPIアンカー部分は、T細胞受容体結合ドメインおよび腫瘍または腫瘍血管系標的化ドメインを有する、本明細書に記載の融合タンパク質に付加することができる。あるいは、GPIアンカーは、腫瘍または腫瘍血管系標的化ドメインをコードする融合相手を必要とすることなく、T細胞受容体結合ドメインポリペプチドに直接付加することができる。
【0147】
2.ペプチドまたはポリペプチドリンカードメイン
融合タンパク質は場合により、共刺激ポリペプチドドメインを抗原結合標的化ドメインから分離するペプチドまたはポリペプチドリンカードメインを含む。
【0148】
抗体のヒンジ領域
一つの実施形態においては、リンカードメインは免疫グロブリンのヒンジ領域を含む。好ましい実施形態では、ヒンジ領域はヒト免疫グロブリンに由来する。ヒンジが由来し得る好適なヒト免疫グロブリンとしては、IgG、IgD、およびIgAが挙げられる。好ましい実施形態では、ヒンジ領域はヒトIgGに由来する。
【0149】
別の実施形態においては、リンカードメインは、上記のような免疫グロブリンのヒンジ領域を含み、さらに1以上の付加的な免疫グロブリンドメインを含む。一つの実施形態では、付加的ドメインは免疫グロブリンのFcドメインを含む。本明細書においてFc領域は、第一の定常領域免疫グロブリンドメインを除く抗体の定常領域を含むポリペプチドを含む。よって、FcIgA、IgD、およびIgGの最後の二つ二つの定常領域免疫グロブリンドメインとIgEおよびIgMの最後の3つの定常領域免疫グロブリンドメインを意味する。好ましい実施形態では、Fcドメインはヒト免疫グロブリンに由来する。より好ましい実施形態では、FcドメインはCH2およびCH3領域を含むヒトIgGに由来する。
【0150】
別の実施形態においては、リンカードメインは免疫グロブリンのヒンジ領域と、免疫グロブリン重鎖のCH1ドメインまたは免疫グロブリン軽鎖のCLドメインのいずれかを含む。好ましい実施形態では、CH1またはCLドメインはヒト免疫グロブリンに由来する。CLドメインはκ軽鎖またはλ軽鎖のいずれかに由来するものであり得る。より好ましい実施形態では、CH1またはCLドメインはヒトIgGに由来する。
【0151】
免疫グロブリンヒンジ領域および他のドメインのアミノ酸配列は当該技術分野でよく知られている。
【0152】
他のペプチド/ポリペプチドリンカードメイン
他の好適なペプチド/ポリペプチドリンカードメインは、天然または非天然ペプチドまたはポリペプチドを含む。ペプチドリンカー配列は少なくともアミノ酸2個の長さである。好ましくは、ペプチドまたはポリペプチドドメインは柔軟なペプチドまたはポリペプチドである。「柔軟なリンカー」とは、柔軟なリンカーの非存在下で二つの連結されたポリペプチドが持つものよりも、それにより連結された二つのポリペプチドの回転自由度を高めるペプチド結合により連結された2以上のアミノ酸残基を含むペプチドまたはポリペプチドを意味する。このような回転自由度は、柔軟なリンカーによって各接近標的抗原に連結された2以上の抗原結合部位をより有効とする。柔軟なペプチド/ポリペプチドの例としては、限定されるものではないが、アミノ酸配列Gly−Ser、Gly−Ser−Gly−Ser(配列番号74)、Ala−Ser、Gly−Gly−Gly−Ser(配列番号75)、(Gly4−Ser)3(配列番号76)、および(Gly4−Ser)4(配列番号77)が挙げられる。さらなる柔軟なペプチド/ポリペプチド配列も当該技術分野でよく知られている。
【0153】
3.二量体化および多量体化ドメイン
融合タンパク質は場合により、2以上の融合タンパク質を二量体化または多量体化する働きをする二量体化または多量体化ドメインを含む。融合タンパク質を二量体化または多量体化する働きをするドメインは分離ドメインであってもよいし、あるいはまた融合タンパク質の他のドメイン(T細胞共刺激/共阻害受容体結合ドメイン、腫瘍/腫瘍新生血管系抗原結合ドメインまたはペプチド/ポリペプチドリンカードメイン)の一つの中に含まれてもよい。
【0154】
二量体化ドメイン
「二量体化ドメイン」とは、少なくとも二つのアミノ酸残基または少なくとも二つのペプチドもしくはポリペプチド(同じまたは異なるアミノ酸配列を持ち得る)の会合によって形成される。これらのペプチドまたはポリペプチドは共有結合的会合および/または非共有結合的会合によって互いに相互作用し得る。好ましい二量体化ドメインは、融合タンパク質相手上のシステインと分子間ジスルフィド結合を形成することができる少なくとも一つのシステインを含む。二量体化ドメインは、融合タンパク質相手の間でジスルフィド結合が形成できるように1以上のシステイン残基を含み得る。一つの実施形態では、二量体化ドメインは一つ、二つ、または3つ〜約10のシステイン残基を含む。好ましい実施形態では、二量体化ドメインは免疫グロブリンのヒンジ領域である。この特定の実施形態では、二量体化ドメインは融合タンパク質のリンカーペプチド/ポリペプチド内に含まれる。
【0155】
さらなる二量体化ドメインの例も当該技術分野で知られており、限定されるものではないが、コイルドコイル、アシッドパッチ(acid patches)、ジンクフィンガー、カルシウムハンド、CH1−CL対、米国特許第5,821,333号公報に記載されている操作された「こぶ(knob)」および/または「突起(protruberance)」を有する「境界」、ロイシンジッパー(例えば、junおよび/またはfos由来)(米国特許第5,932,448号公報)、SH2(srcホモロジー2)、SH3(srcホモロジー3)(Vidal, et al., Biochemistry, 43, 7336-44 ((2004))、ホスホチロシン結合(PTB)(Zhou, et al., Nature, 378:584-592 (1995))、WW(Sudol, Prog. Biochys. Mol. Bio., 65:113-132 (1996))、PDZ(Kim, et al., Nature, 378: 85-88 (1995)、 Komau, et al., Science, 269:1737-1740 (1995))14−3−3、WD40(Hu, et al., J Biol Chem., 273, 33489-33494 (1998))EH、Lim、イソロイシンジッパー、受容体二量体対(例えば、インターロイキン−8受容体(IL−8R)、ならびにインテグリンヘテロ二量体、例えば、LFA−1およびGPIIIb/IIIaまたはその二量体化領域、二量体リガンドポリペプチド(例えば、神経成長因子(NGF)、ニューロトロフィン−3(NT−3)、インターロイキン−8(IL−8)、血管内皮増殖因子(VEGF)、VEGF−C、VEGF−D、PDGFメンバー、および脳由来神経栄養因子(BDNF)(Arakawa, et al., J. Biol. Chem., 269(45): 27833-27839 (1994)およびRadziejewski, et al., Biochem., 32(48): 1350 (1993))が挙げられ、親和性が変更されたこれらのドメインの変異体であってもよい。これらのポリペプチド対は、酵母二つのハイブリッドスクリーンを含む当該技術分野で知られている方法によって同定することができる。酵母ツーハイブリッドスクリーンは米国特許第5,283,173号公報および同第6,562,576号公報に記載されており、双方とも引用することにより本明細書の開示の一部とされる。相互作用ドメイン対の間の親和性はKatahira, et al., J. Biol. Chem., 277, 9242-9246 (2002) の記載を含む、当該技術分野で知られている方法を用いて決定することができる。あるいは、ペプチド配列のライブラリーを、例えばWO01/00814号公報に記載されている方法を用いてヘテロ二量体形成に関してスクリーニングすることもできる。タンパク質−タンパク質相互作用に関する有用な方法はまた米国特許第6,790,624号公報にも記載されている。
【0156】
多量体化ドメイン
「多量体化ドメイン」とは、三つ以上のペプチドまたはポリペプチドを共有結合的会合および/または非共有結合的会合によって互いに相互作用させるドメインである。好適な多量体化ドメインとしては、限定されるものではないが、コイルドコイルドメインが挙げられる。コイルドコイルは、通常、7つのアミノ酸(ヘプタッドリピート)または11のアミノ酸(ウンデカドリピート)の配列に、3および4残基の間隔を置いた主として疎水性の残基の連続パターンを有し、組み立てられて(折りたたまれて)多量性のらせん束を形成するペプチド配列を意味する。3および4残基おきのいくつかの不規則な分布を含む配列を有するコイルドコイルも意図される。疎水性残基は特に、疎水性アミノ酸Val、Ile、Leu、Met、Tyr、Phe、およびTrpである。主として疎水性とは、残基の少なくとも50%が記載の疎水性アミノ酸から選択されるべきであることを意味する。
【0157】
コイルドコイルドメインはラミニンに由来してもよい。細胞外間隙で、ヘテロ三量体コイルドコイルタンパク質ラミニンは基底膜の形成に重要な役割を果たす。明らかに、多機能オリゴマー構造はラミニン機能に必要である。コイルドコイルドメインはまた、3つ(TSP−1およびTSP−2)または5つ(TSP−3、TSP−4およびTSP−5)の鎖が接続されたトロンボスポンジンまたは平行5鎖コイルドコイル(Malashkevich ,et al., Science, 274: 761-765 (1996))へと折りたたまれるCOMP(COMPcc)(Guo, et at., EMBO J., 1998, 17: 5265-5272)に由来してもよい。
【0158】
他のタンパク質に由来するさらなるコイルドコイルドメインおよびポリペプチドの多量体化を媒介する他のドメインも当該技術分野で知られており、開示されている融合タンパク質で用いるのに好適である。
【0159】
4.融合タンパク質例
PD−L2
代表的なネズミPD−L2融合タンパク質は、
【化58】
(配列番号52)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0160】
配列番号79によりコードされているネズミPD−L2融合タンパク質は以下のアミノ酸配列:
【化59】
(配列番号53)
を有する。
【0161】
シグナル配列を含まない配列番号53のネズミPD−L2融合タンパク質のアミノ酸配列は、
【化60】
(配列番号54)
である。
【0162】
代表的なヒトPD−L2融合タンパク質は、
【化61】
(配列番号55)
と少なくとも80%、85%、90%、95%、99%、または100%の配列同一性を有する核酸によりコードされる。
【0163】
配列番号82によりコードされているヒトPD−L2融合タンパク質は以下のアミノ酸配列:
【化62】
(配列番号56)
を有する。
【0164】
シグナル配列を含まない配列番号83のヒトPD−L2融合タンパク質のアミノ酸配列は
【化63】
(配列番号57)
である。
【0165】
代表的な非ヒト霊長類(カニクイザル)PD−L2融合タンパク質タンパク質は以下のアミノ酸配列:
【化64】
(配列番号86)
を有する。
【0166】
シグナル配列を含まない配列番号86の非ヒト霊長類(カニクイザル)PD−L2 融合タンパク質のアミノ酸配列は
【化65】
(配列番号87)
である。
【0167】
G.PD−1受容体アンタゴニストをコードする単離された核酸分子
PD−1アンタゴニストポリペプチド、その変異体およびその融合タンパク質をコードする単離された核酸配列が開示される。本明細書において「単離された核酸」とは、哺乳類ゲノムにおいてその核酸の一端または両端に通常隣接している核酸をはじめ、哺乳類ゲノムに存在する他の核酸分子から分離された核酸を意味する。
【0168】
単離された核酸は、例えば、天然ゲノムにおいてそのDNA分子にすぐ隣接して通常見られる核酸配列の一つが除去されているか、または存在しない限り、DNA分子であってもよい。よって、単離された核酸としては、限定されるものではないが、他の配列とは独立した分離分子として存在するDNA分子(例えば、化学的に合成された核酸またはcDNAまたはPCRまたは制限エンドヌクレアーゼ処理によって生成されるゲノムDNA断片)ならびにベクター、自律的に複製するプラスミド、ウイルス(例えば、レトロウイルス、レンチウイルス、アデノウイルスまたはヘルペスウイルス)に、または原核生物または真核生物のゲノムDNAに組み込まれた組換えDNAが挙げられる。さらに、単離された核酸としては、ハイブリッドまたは融合核酸の一部である組換えDNA分子などの操作核酸が挙げられる。例えば、cDNAライブラリーもしくはゲノムライブラリーの中の数百から数百万の他の核酸に存在する核酸、またはゲノムDNA制限消化を含むゲルスライスは単離された核酸とはみなさない。
【0169】
核酸はセンス配向であってもアンチセンス配向であってもよく、あるいはPD−L2、PD−L1、PD−1またはB7.1ポリペプチドまたはその変異体をコードする参照配列と相補的であってもよい。参照配列としては、例えば、当該技術分野で知られており、上記に述べられているヒトPD−L2、ヒトPD−L1、またはネズミPD−L2およびネズミPD−L1のヌクレオチド配列が含まれる。
【0170】
核酸はDNA、RNA、または核酸類似体であり得る。核酸類似体は塩基部分、糖部分またはリン酸主鎖において修飾することができる。このような修飾は例えば、核酸の安定性、ハイブリダイゼーションまたは溶解度を改善することができる。塩基部分における修飾としては、デオキシチミジンをデオキシウリジン、デオキシシチジンを5−メチル−2’−デオキシシチジンまたは5−ブロモ−2’−デオキシシチジンにいうものが挙げられる。糖部分の修飾としては、リボース糖の2’ヒドロキシルの修飾による2’−O−メチルまたは2’−O−アリル糖の形成が挙げられる。デオキシリボースリン酸主鎖は、各塩基部分が6員のモルホリノ環に連結されているモルホリノ核酸、またはデオキシリン酸主鎖がシュードペプチド主鎖で置換され、4つの塩基が保持されているペプチド核酸を生成するように修飾することができる。例えば、Summerton and Weller (1997) Antisence Nucleic Acid Drug Dev. 7:187-195、およびHyrup et al. (1996) Bioorgan. Med. Chem. 4:5-23参照。さらに、デオキシリン酸主鎖は、例えば、ホスホロチオエートまたはホスホロジチオエート主鎖、ホスホロアミダイトまたはアルキルホスホトリエステル主鎖で置換することができる。
【0171】
H.PD−1受容体アンタゴニストを発現するベクターおよび宿主細胞
上記の核酸は、細胞で発現させるためにベクターに挿入することができる。本明細書において「ベクター」とは、別のDNAセグメントが、挿入されたセグメントの複製が起こるように挿入され得る、プラスミド、ファージまたはコスミドなどのレプリコンである。ベクターは発現ベクターであり得る。「発現ベクター」とは、1以上の発現制御配列を含むベクターであり、「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。
【0172】
ベクター中の核酸は1以上の発現制御配列に機能的に連結させることができる。本明細書において「機能的に連結される」とは、発現制御配列が目的のコード配列の発現を有効に制御するように遺伝子構築物に組み込まれることを意味する。発現制御配列の例としては、プロモーター、エンハンサー、および転写終結領域が挙げられる。プロモーターは、一般に、転写が開始する点の上流100ヌクレオチド以内(一般に、RNAポリメラーゼIIの開始部位付近)のDNA分子の領域からなる発現制御配列である。コード配列をプロモーターの制御下に置くためには、そのポリペプチドの翻訳リーディングフレームの翻訳開始部位をプロモーターの下流1〜約15ヌクレオチドの間に置く必要がある。エンハンサーは、時間、場所、およびレベルに関して発現特異性を与える。プロモーターとは異なり、エンハンサーは転写部位から様々な距離に位置する場合にも機能し得る。エンハンサーはまた、転写開始部位から下流に位置してもよい。コード配列は、RNAポリメラーゼがコード配列をmRNAへ転写することができ、それがコード配列によりコードされているタンパク質へ翻訳され得る場合に、「機能的に連結され」かつ細胞内の発現制御配列の「制御下」にある。
【0173】
好適な発現ベクターとしては、限定されるものではないが、例えば、バクテリオファージ、バキュロウイルス、タバコモザイクウイルス、ヘルペスウイルス、サイトメガロウイルス、レトロウイルス、ワクシニアウイルス、アデノウイルスおよびアデノ随伴ウイルスに由来するプラスミドおよびウイルスベクターが挙げられる。多くのベクターおよび発現系がNovagen (Madison, WI)、Clontech (Palo Alto, CA)、Stratagene (La Jolla, CA)およびInvitrogen Life Technologies (Carlsbad, CA)などの会社から市販されている。
【0174】
発現ベクターはタグ配列を含み得る。タグ配列は一般に、コードされているポリペプチドとの融合物として発現される。このようなタグは、カルボキシル末端またはアミノ末端を含むポリペプチド内のいずれの位置にでも挿入することができる。有用なタグの例としては、限定されるものではないが、緑色蛍光タンパク質(GFP)、グルタチオンS−トランスフェラーゼ(GST)、ポリヒスチジン、c−myc、血球凝集素、Flag(商標)タグ(Kodak, New Haven, CT)、マルトースE結合タンパク質およびタンパク質Aが挙げられる。一実施形態では、変異体PD−L2融合タンパク質は、Ig重鎖定常領域の1以上のドメインをコードする、好ましくは、ヒト免疫グロブリンCγ1鎖のヒンジ、CH2およびCH3領域に相当するアミノ酸配列を有する核酸を含むベクター内に存在する。
【0175】
発現される核酸を含むベクターを宿主細胞に導入することができる。「宿主細胞」とは、組換え発現ベクターが導入可能な原核生物のおよび真核細胞を含むものとする。本明細書において「形質転換される」および「トランスフェクトされる」は、多くの技術の一つによって核酸分子(例えばベクター)を細胞に導入することを包含する。特定の技術に限定されるものではないが、これらの技術の多くは当該技術分野で十分に確立されている。
【0176】
原核細胞は、例えば、エレクトロポレーションまたは塩化カルシウム媒介形質転換により核酸で形質転換させることができる。核酸は、例えば、リン酸カルシウム共沈殿、DEAE−デキストラン媒介トランスフェクション、リポフェクション、エレクトロポレーションまたはマイクロインジェクションを含む技術によって哺乳類細胞へトランスフェクトすることができる。宿主細胞(例えば、原核細胞または真核細胞、例えばCHO細胞)を用いて、例えば、本明細書に記載のPD−1アンタゴニストポリペプチドを生産することができる。
【0177】
I.抗体PD−1アンタゴニスト
PD−1アンタゴニストまたはPD−1のエピトープと反応性のあるモノクローナルおよびポリクローナル抗体が開示されている。モノクローナル抗体(mAbs)およびそれらの製造および使用方法は、Kohler and Milstein, Nature 256:495-497 (1975)、米国特許第4,376,110号公報、Hartlow, E. et al., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1988)、 Monoclonal Antibodies and Hybridomas: A New Dimension in Biological Analyses, Plenum Press, New York, N.Y. (1980)、 H. Zola et al., Monoclonal Hybridoma Antibodies: Techniques and Applications, CRC Press, 1982に記載されている。
【0178】
PD−1と結合し、PD−1を介したシグナル伝達を遮断し、かつ、現行使用されているものよりも親和性が低く、投与後3か月、2か月、1か月、3週間、2週間、1週間、または数日未満の期間で抗体を解離させる抗体が免疫応答の増強、増進、または刺激に好ましい。
【0179】
本発明の別の実施形態は、B7−HなどのPD−1のリガンドと結合する架橋したPD−1受容体と結合する抗体を含んでなる二重特異性抗体を含む。好ましい実施形態では、このPD−1結合部分は、PD−1受容体を介したシグナル伝達を低減または阻害する。
【0180】
イムノアッセイ法は、Coligan, J. E. et al., eds., Current Protocols in Immunology, Wiley-Interscience, New York 1991 (または最新版)、 Butt, W. R. (ed.) Practical Immunoassay: The State of the Art, Dekker, N.Y., 1984、 Bizollon, Ch. A., ed., Monoclonal Antibodies and New Trends in Immunoassays, Elsevier, N.Y., 1984、 Butler, J. E., ELISA (Chapter 29), In: van Oss, C. J. et al., (eds), Immunochemistry, Marcel Dekker, Inc., New York, 1994, pp. 759-803、 Butler, J. E. (ed.), Immunochemistry of Solid-Phase Immunoassay, CRC Press, Boca Raton, 1991、 Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986、 Work, T. S. et al., Laboratory Techniques and Biochemistry in Molecular Biology, North Holland Publishing Company, NY, (1978) (Chapter by Chard, T., "An Introduction to Radioimmune Assay and Related Techniques")に記載されている。
【0181】
抗イディオタイプ抗体は、例えば、Idiotypy in Biology and Medicine, Academic Press, New York, 1984、 Immunological Reviews Volume 79, 1984、 Immunological Reviews Volume 90, 1986、 Curr. Top. Microbiol., Immunol. Volume 119, 1985、 Bona, C. et al., CRC Crit. Rev. Immunol., pp. 33-81 (1981)、 Jerme, N K, Ann. Immunol. 125C:373-389 (1974)、 Jerne, N K, In: Idiotypes--Antigens on the Inside, Westen-Schnurr, I., ed., Editiones Roche, Basel, 1982, Urbain, J. et al., Ann. Immunol. 133D:179-(1982)、 Rajewsky, K. et al., Ann. Rev. Immunol. 1:569-607 (1983)に記載されている。
【0182】
これらの抗体は異種、同種異系、同系、またはその改変形態(ヒト化またはキメラ抗体など)であり得る。特定の抗体、例えば、抗PD−L2抗体のイディオタイプに特異的な抗イディオタイプ抗体も含まれる。「抗体」との用語は、完全分子ならびに抗原結合部位を含み、かつ、およびPD−1アンタゴニストエピトープと結合することができるその断片の双方を含むことを意味する。これらには、完全抗体のFc断片を欠き、循環からより迅速に排除され、完全抗体よりも低い非特異的組織結合性を持ち得るFabおよびF(ab’)2断片が含まれる(Wahl et al., J. Nuc. Med. 24:316-325 (1983))。また、Fv断片も含まれる(Hochman, J. et al. (1973) Biochemistry 12:1130-1135、 Sharon, J. et al.(1976) Biochemistry 15:1591-1594)。これらの種々の断片は、プロテアーゼ切断または化学的切断などの常法を用いて作製される(例えば、Rousseaux et al., Meth. Enzymol., 121:663-69 (1986)参照)。
【0183】
ポリクローナル抗体は、ウサギ、ヤギ、齧歯類などの免疫動物から血清として得られ、それ以上処理することなくそのまま用いてもよく、あるいは硫酸アンモニウム沈殿、イオン交換クロマトグラフィーおよびアフィニティークロマトグラフィーなどの慣例の富化または精製方法を施してもよい。
【0184】
免疫原は、完全なPD−1アンタゴニスト、PD−1または断片またはその誘導体を含む。好ましい免疫原は、PD−1アンタゴニストまたはPD−1の細胞外ドメイン(ECD)の全部または一部を含み、ここで、これらの残基はグリコシル化などの翻訳後調節を含む。細胞外ドメインを含む免疫原は、例えば、慣例の組換え法を用いたクローン化遺伝子の発現または起源細胞からの単離などの当該技術分野で知られている種々の方法で生産される。
【0185】
モノクローナル抗体は、Kohler and Milstein, Nature, 256:495-97 (1975)により紹介された手順およびその改変法(上記の参照を参照)などの慣例のハイブリドーマ技術を用いて生産され得る。動物、好ましくはマウスを上記のような免疫原に感作させることにより初回刺激し、刺激動物に所望の抗体応答を惹起させる。初回刺激動物のリンパ節、脾臓、または末梢血からのBリンパ球を、一般にポリエチレングリコール(PEG)などの誘導促進剤の存在下で骨髄腫細胞と融合させる。多くのネズミ骨髄腫細胞系統はいずれもがこのような使用に利用できる:P3−NS1/1−Ag4−1、P3−x63−k0Ag8.653、Sp2/0−Ag14またはHL1−653骨髄腫系統(ATCC, Rockville, Md.から入手可能)。その後の工程は、融合されてない親骨髄腫細胞と、ドナーリンパ球細胞とがやがて死に至り、ハイブリドーマ細胞だけが生き残るような選択培地で増殖させることを含む。これらをクローニングし、増殖させ、それらの上清を、例えば、PD−L2またはPD−L1融合タンパク質を用いたイムノアッセイ技術によって所望の特異性の抗体が存在するか否かをスクリーニングする。例えば、制限希釈によって陽性クローンをサブクローニングし、モノクローナル抗体を単離する。
【0186】
これらの方法に従って生産されたハイブリドーマは、当該技術分野で知られている技術(一般に、Fink et al., Prog. Clin. Pathol., 9:121-33 (1984)参照)を用いて、インビトロまたはインビボ(腹水中)で増殖させることができる。一般に、個々の細胞系統を培養で増殖させ、高濃度の単一モノクローナル抗体を含む培養培地を、デカンテーション、濾過または遠心分離によって採取することができる。
【0187】
抗体は、正常な多量体構造の代わりに単鎖抗体またはscFvとして生産してもよい。単鎖抗体は、着目するIgに由来する超可変領域を含み、完全なIgのサイズの一部でありつつ、天然Igの抗原結合部位を再形成する(Skerra, A. et al. Science, 240: 1038-1041 (1988)、 Pluckthun, A. et al. Methods Enzymol. 178: 497-515 (1989)、 Winter, G. et al. Nature, 349: 293-299 (1991))。好ましい実施形態では、抗体は慣例の分子生物学的技術を用いて生産される。
【0188】
III.製造方法
PD−1アンタゴニストの生産方法
ポリペプチドおよびその変異体
単離されたPD−1アンタゴニストまたはその変異体は、例えば、化学的合成または宿主細胞での組換え生産によって得ることができる。PD−1アンタゴニストポリペプチドを組換え生産するためには、そのポリペプチドをコードするヌクレオチド配列を含む核酸を用いて、細菌または真核生物宿主細胞(例えば、昆虫、酵母、または哺乳類細胞)に形質転換、形質導入またはトランスフェクションを行うことができる。一般に、核酸構築物は、PD−1アンタゴニストポリペプチドをコードするヌクレオチド配列に機能的に連結された調節配列を含む。調節配列(本明細書では発現制御配列とも呼ばれる)は一般に遺伝子産物をコードしないが、それらが機能的に連結されている核酸配列の発現に影響を与える。
【0189】
ポリペプチドを発現および生産するのに有用な原核生物系および真核生物系は当該技術分野でよく知られており、例えば、BL−21などの大腸菌(Escherichia coli)株、およびCHO細胞などの培養哺乳類細胞が挙げられる。
【0190】
真核生物宿主細胞では、PD−1アンタゴニストポリペプチドを発現させるために多くのウイルスに基づく発現系が利用可能である。ウイルスに基づく発現系は当該技術分野でよく知られており、限定されるものではないが、バキュロウイルス、SV40、レトロウイルス、またはワクシニアに基づくウイルスベクターが挙げられる。
【0191】
PD−1アンタゴニストポリペプチドを安定発現する哺乳類細胞系統は、適当な制御エレメントと選択マーカーを備えた発現ベクターを用いて作出することができる。例えば、真核生物発現ベクターpCR3.1(Invitrogen Life Technologies) およびp91023(B)(Wong et al. (1985) Science 228:810-815参照)は、例えば、チャイニーズハムスター卵巣(CHO)細胞、COS−1細胞、ヒト胎児腎臓293細胞、NIH3T3細胞、BHK21細胞、MDCK細胞、およびヒト血管内皮細胞(HUVEC)での変異体共刺激ポリペプチドの発現に好適である。エレクトロポレーション、リポフェクション、リン酸カルシウムまたは塩化カルシウム共沈殿、DEAEデキストランまたは他の好適なトランスフェクション方法による発現ベクターの導入の後、安定な細胞系統を選択することができる(例えば、G418、カナマイシンまたはハイグロマイシンに対する抗生物質耐性による)。トランスフェクト細胞は、目的のポリペプチドが発現されるように培養することができ、このポリペプチドを例えば、細胞培養上清または溶解細胞から回収することができる。あるいは、PD−1アンタゴニストポリペプチドは、(a)増幅された配列をpcDNA3(Invitrogen Life Technologies)などの哺乳類発現ベクターに連結し、(b)インビトロにおいてコムギ胚芽抽出液またはウサギ網状赤血球溶解液を用いて転写および翻訳することによって生産することもできる。
【0192】
PD−1アンタゴニストポリペプチドは、例えば、DEAEイオン交換、ゲル濾過、およびヒドロキシルアパタイトクロマトグラフィーなどのクロマトグラフィー法を用いて単離することができる。例えば、細胞培養上清または細胞質抽出液中のPD−1アンタゴニストポリペプチドは、Gタンパク質カラムを用いて単離することができる。いくつかの実施形態では、変異体PD−1アンタゴニストポリペプチドは、ポリペプチドを親和性マトリックスに捕捉させるアミノ酸配列を含むように「操作」することができる。ポリペプチドの精製を助けるために、例えば、c−myc、血球凝集素、ポリヒスチジン、またはFlag(商標)(Kodak)などのタグを使用することができる。このようなタグは、カルボキシル末端またはアミノ末端を含むポリペプチド内のいずれの位置にも挿入することができる。有用であり得る他の融合物としては、アルカリ性ホスファターゼなど、ポリペプチドの検出を助ける酵素を含むことができる。共刺激ポリペプチドを精製するのに、イムノアフィニティークロマトグラフィーを使用することもできる。
【0193】
変異体ポリペプチドを作出するためにランダム突然変異を導入する方法は当該技術分野で知られている。PD−1、PD−L1、またはPD−L2と相互作用するペプチドをスクリーニングするためにランダムペプチドディスプレーライブラリーを使用することができる。このようなランダムペプチドディスプレーライブラリーを作出およびスクリーニングするための技術は当該技術分野で知られており(Ladnerら、米国特許第5,223,409号公報、Ladnerら、米国特許第4,946,778号公報、Ladnerら、米国特許第5,403,484号公報、およびLadnerら、米国特許第5,571,698号)、ランダムペプチドディスプレーライブラリーおよびこのようなライブラリーをスクリーニングするためのキットが市販されている。
【0194】
B.PD−1アンタゴニストポリペプチドをコードする単離された核酸分子の生産方法
PD−1アンタゴニストポリペプチドをコードする単離された核酸分子は、限定されるものではないが、一般的分子クローニングおよび化学的核酸合成技術を含む標準的技術によって生産することができる。例えば、変異体共刺激ポリペプチドをコードする単離された核酸を得るために、ポリメラーゼ連鎖反応(PCR)技術を使用することができる。PCRは、標的核酸が酵素的に増幅される技術である。一般に、着目する領域の末端から、またはそれを超える配列情報を用いれば、増幅される鋳型の反対側の鎖と配列が同一であるオリゴヌクレオチドプライマーを設計することができる。PCRを用いて、全ゲノムDNAまたは全細胞RNA由来の配列を含め、DNAならびにRNAから特定の配列を増幅させることができる。プライマーは一般に14〜40ヌクレオチドの長さであるが、10ヌクレオチド〜数百ヌクレオチドの範囲の長さであってもよい。一般的なPCR技術は、例えば、PCR Primaer: A Laboratory Manual, Dieffenbach and Dveksler編, Cold Spring Harbor Laboratory Press, 1995に記載されている。鋳型の供給源としてRNAを用いる場合には、逆転写酵素を用いて相補的DNA(cDNA)鎖を合成することができる。単離された核酸を得るために、リガーゼ連鎖反応、鎖置換増幅、自己持続配列複製または核酸配列に基づく増幅を使用することもできる。例えば、Lewis (1992) Genetic Engineering News 12:1、 Guatelli et al. (1990) Proc. Natl. Acad. Sci. USA 87:1874-1878、およびWeiss (1991) Science 254:1292-1293参照。
【0195】
単離された核酸は、単一の核酸分子としてか、またはオリゴヌクレオチド系列としてのいずれかで化学的に合成することができる(例えば、3’から5’方向への自動DNA合成のためのホスホルアミダイト技術を使用)。例えば、所望の配列を含み、各対が短い相補的セグメント(例えば約15ヌクレオチド)を含み、その結果、オリゴヌクレオチド対がアニーリングされる際に二重らせんが形成される、長いオリゴヌクレオチド(例えば>100ヌクレオチド)の1以上の対を合成することができる。DNAポリメラーゼオリゴヌクレオチドを延長するのにも使用することができ、その結果、オリゴヌクレオチド対につき一つの一本鎖、二本鎖核酸分子が得られ、次にこれをベクターに連結することができる。また、単離された核酸は突然変異誘発により得ることもできる。PD−1アンタゴニストポリペプチドをコードする核酸は、PCRを介したオリゴヌクレオチド指定突然変異誘発および/または部位特異的突然変異誘発を含む標準的技術を用いて変異させることができる。Short Protocols in Molecular Biology. Chapter 8, Green Publishing Associates and John Wiley & Sons, edited by Ausubel et al, 1992参照。修飾可能なアミノ酸位置の例としては、本明細書に記載されているものが含まれる。
【0196】
IV.処方物
A.PD−1アンタゴニスト処方物
PD−1アンタゴニストを含有する医薬組成物が提供される。ペプチドまたはポリペプチドを含有する医薬組成物は非経口(筋肉内、腹腔内、静脈内(IV)または皮下注射)、経皮(受動的またはイオン泳動もしくはエレクトロポレーションを使用)または経粘膜(鼻腔、膣、直腸または舌下)投与経路による投与用であり得る。また、組成物は生浸食性インサートを用いて投与してもよいし、適当なリンパ系組織(例えば、脾臓、リンパ節もしくは粘膜に関連するリンパ系組織)に直接送達、または器官もしくは腫瘍に直接送達してもよい。これらの組成物は各投与経路に適当な剤形で処方することができる。ペプチドまたはポリペプチドでないPD−1受容体のアンタゴニストを含有する組成物はさらに腸内投与用に処方することもできる。
【0197】
本明細書において「有効量」または「治療上有効な量」とは、処置される障害の1以上の症状を処置、阻害または緩和するのに、またはそうでなければされる所望の薬理学的および/または生理学的作用を提供するのに十分な用量を意味する。厳密な用量は、被験体に依存する変数(例えば、年齢、免疫系の健全性など)、疾患および行われる処置などの種々の要因によって異なる。PD−1アンタゴニストの治療上有効な量は、免疫応答を活性化、増強、増大または持続させ、かつ/またはT細胞の消耗および/またはT細胞の無応答を克服または緩和し、かつ/または単球、マクロファージ、樹状細胞および他の抗原提示細胞(APC)を活性化する。
【0198】
好ましい実施形態では、PD−1アンタゴニストは、腫瘍モデリングおよびバイオアベイラビリティからの外挿に基づき、0.1〜20mg/kgの範囲で投与される。最も好ましい範囲は5〜20mgPD−1アンタゴニスト/kgである。一般に、静脈注射または注入では、別の経路によって投与する場合よりも用量は低くなり得る。
【0199】
1.非経口投与用処方物
好ましい実施形態においては、ペプチドおよびポリペプチドを含有するものを含む開示されている組成物は、非経口注射により水溶液として投与される。処方物はまた懸濁液またはエマルションの形態であってもよい。一般に、有効量のペプチドまたはポリペプチドを含み、場合により薬学上許容される希釈剤、保存剤、可溶化剤、乳化剤、アジュバントおよび/または担体を含む医薬組成物が提供される。このような組成物は無菌水、緩衝生理食塩水(例えば、Tris−HCl、酢酸、リン酸)、pHおよびイオン強度、ならびに場合により、洗剤および可溶化剤(例えば、TWEEN(商標)20、TWEEN80、ポリソルベート80)、抗酸化剤(例えば、アスコルビン酸、メタ重亜硫酸ナトリウム)、および保存剤(例えば、Thimersol、ベンジルアルコール)および増量剤(例えば、ラクトース、マンニトール)などの添加剤を含む。非水性溶媒またはビヒクルの例としては、プロピレングリコール、ポリエチレングリコール、オリーブ油およびコーン油などの植物油、ゼラチン、およびオレイン酸エチルなどの注射可能な有機エステルがある。処方物は凍結乾燥させ、使用直前に再溶解/再懸濁させてもよい。処方物は例えば、細菌保持フィルターによる濾過、除菌剤の組成物への配合、組成物の照射または組成物の加熱によって除菌してもよい。
【0200】
2.制御送達ポリマーマトリックス
1以上のPD−1アンタゴニストまたはPD−1アンタゴニストをコードする核酸を含有する組成物は徐放性処方物として投与することができる。徐放性ポリマーデバイスはポリマーデバイス(ロッド、シリンダー、フィルム、ディスク)の移植または注射(微粒子)後に長時間全身への放出を行うことができる。マトリックスは、ペプチドが固体ポリマーマトリックス内に分散されているマイクロスフェア、またはコアがポリマー剤皮とは異なる材料のものであり、そのコア(本質的に液体であっても固体であってもよい)内にペプチドが分散もしくは懸濁されているマイクロカプセルなどの微粒子の形態であってもよい。本明細書で特に定義されない限り、微粒子、マイクロスフェアおよびマイクロカプセルは互換的に用いられる。あるいは、ポリマーは数ナノメートルから4センチメートルの範囲の薄スラブもしくはフィルムとしての鋳物、摩砕もしくは他の標準的な技術によって製造された粉末、またはさらにはヒドロゲルなどのゲルであってもよい。マトリックスはまた、免疫応答を調節するため、免疫不全患者(カテーテルが挿入されている高齢者または未熟児など)における感染予防のため、またはただれ、床ずれ潰瘍などの治癒を促進するために用いられるマトリックスの場合と同様に治癒における補助とするために医療機器の内外に組み込むことができる。
【0201】
非生分解性マトリックスでも生分解性マトリックスでもPD−1アンタゴニストまたはそれらをコードする核酸の送達に使用することができるが、生分解性マトリックスが好ましい。これらは天然ポリマーであっても合成ポリマーであってもよいが、分解および放出特性の特徴付けがより良いため、合成ポリマーが好ましい。ポリマーは、放出が望まれる期間に基づいて選択される。直線的放出が最も有用である場合もあるが、パルス放出または「バルク放出」がより有効な結果をもたらす場合もある。ポリマーはヒドロゲル(一般に最大約90重量%の水を吸収する)の形態であってもよく、場合により多価イオンまたはポリマーで架橋されていてもよい。
【0202】
マトリックスは、溶媒蒸発、噴霧乾燥、溶媒抽出、および当業者に知られている他の方法によって形成させることができる。生浸食性マイクロスフェアは、例えば、Mathiowitz and Langer, J. Controlled Release, 5:13-22 (1987)、 Mathiowitz, et al., Reactive Polymers, 6:275-283 (1987)、およびMathiowitz, et al., J. Appl. Polymer Sci., 35:755-774 (1988)に記載されているような、薬物送達用のマイクロスフェアを作製するために開発されたいずれかの方法を用いて製造することができる。
【0203】
徐放性経口処方物が望ましい場合がある。PD−1阻害的シグナル伝達のアンタゴニストは、拡散または溶脱機構のいずれかによる放出を可能とする不活性マトリックス、例えば、フィルムまたはガム中に配合することができる。また、緩徐崩壊マトリックスを処方物に配合してもよい。別の徐放性形態としては、水を入り込ませて浸透圧作用により単一の小開口部から薬剤を押し出す半透膜に薬剤が封入されるものがある。経口処方物では、放出の場所は胃、小腸(十二指腸、空腸もしくは回腸)または大腸であり得る。好ましくは、この放出は有効剤(もしくは誘導体)の保護によるか、または腸などの胃環境を超えた有効剤の放出によるかのいずれかで、胃環境の有害な影響を回避する完全な胃耐性を確保するためには、腸溶コーティング(すなわち、少なくともpH5.0までは不透性)が不可欠である。これらのコーティングは混合フィルム、またはBanner Pharmacapsから市販されているものなどのカプセル剤として使用可能である。
【0204】
これらのデバイスは移植または注射領域を処置するために局所放出用に処方することができ、一般に、全身処置用の用量よりもはるかに低い用量を送達する。これらのデバイスはまた、全身送達用に処方することもできる。これらは埋植または皮下注射することができる。
【0205】
3.腸内投与用処方物
PD−1のアンタゴニストはまた、経口送達用にも処方することができる。経口固体剤形は当業者に知られている。固体剤形としては、錠剤、カプセル剤、丸剤、トローチ剤またはロゼンジ剤、カシェ剤、ペレット、粉末もしくは顆粒、またはポリ乳酸、ポリグリコール酸などのポリマー化合物の特定の調製物、もしくはリポソームへの材料の組み込みが挙げられる。このような組成物は本発明のタンパク質および誘導体の物理的状態、安定性、インビボ放出速度、およびインビボクリアランス速度に影響を及ぼし得る。例えば、Remington's Pharmaceutical Sciences, 21st Ed. (2005, Lippincott, Williams & Wilins, Baltimore, Md. 21201) pages 889-964参照。組成物は液体形態で調製してもよいし、あるいは乾燥粉末(例えば、凍結乾燥)形態であってもよい。リポソームまたはポリマーカプセル封入を組成物の処方に用いてよい。Marshall, K. In: Modern Pharmaceutics, G. S. Banker and C. T. Rhodes編 Chapter 10, 1979も参照。一般に、処方物は有効剤と、胃環境でPD−1アンタゴニストを保護し、腸で生活性材料を放出する不活性成分を含む。
【0206】
薬学上許容されるエマルション、溶液、懸濁液、およびシロップを含む経口投与用の液体剤形は、不活性希釈剤、湿潤剤、乳化剤および沈殿防止剤などのアジュバント、ならびに甘味剤、香味剤、および芳香剤を含む他の成分を含み得る。
【0207】
B.PD−1アンタゴニストを含むワクチン
ワクチンは、感染細胞を排除するために強いT細胞応答を必要とする。PD−1アンタゴニストは、一次免疫応答およびエフェクター細胞の活性と、数とを促進、増大、または増強するためにワクチンの成分として投与することができる。ワクチンは抗原、PD−1アンタゴニスト(またはその供給源)、および場合により他のアジュバントおよび標的化分子を含む。PD−1アンタゴニストの供給源は、開示されているPD−L2ポリペプチド、PD−L2融合タンパク質、その変異体、PD−L1断片、PD−1断片、PD−L2ポリペプチドをコードする核酸、PD−L2融合タンパク質、その変異体、PD−L1断片またはPD−1断片、または上記のPD−1のポリペプチドリガンドを発現するベクターを含む宿主細胞のいずれかを含む。
【0208】
1.抗原
抗原は、ペプチド、タンパク質、多糖類、糖類、脂質、核酸、またはその組合せであり得る。抗原は、ウイルス、細菌、寄生虫、原虫、真菌、ヒストプラズマ、組織、または形質転換細胞に由来するものであり得、全細胞またはその免疫原成分、例えば、細胞壁成分またはその分子成分であり得る。
【0209】
好適な抗原は当該技術分野で知られており、商業的官業および科学ソースから入手可能である。一つの実施形態においては、抗原は不活性化または弱毒された完全な生物である。これらの生物はウイルス、寄生虫および細菌などの感染性生物であり得る。これらの生物は腫瘍細胞または淋病もしくはマラリアなどのウイルスまたは細胞内病原体に感染した細胞であり得る。抗原は腫瘍または腫瘍またはウイルスもしくは細菌源に由来する精製または部分精製ポリペプチドであり得る。抗原は、異種発現系においそのポリペプチド抗原をコードするDNAを発現させることにより産生された組換えポリペプチドであってもよい。抗原は、抗原タンパク質の全部または一部をコードするDNAであってもよい。DNAはプラスミドDNAなどのベクターDNAの形態であってもよい。
【0210】
抗原は単一の抗原として提供されてもよいし、あるいは組合せで提供されてもよい。抗原はまた、ポリペプチドまたは核酸の複合体混合物として提供されてもよい。
【0211】
i.ウイルス抗原
ウイルス抗原は、限定されるものではないが、以下のウイルス科:アレナウイルス科、アルテリウイルス科、アストロウイルス科、バキュロウイルス科、バドナウイルス科、バルナウイルス科、ビルナウイルス科、ブロモウイルス科、ブンヤウイルス科、カリシウイルス科、カピロウイルス科、カルラウイルス科、カリモウイルス科、サーコウイルス科、コモウイルス科、コロナウイルス科(例えば、重症急性呼吸器症候群(SARS)ウイルスなどのコロナウイルス)、コルチコウイルス科、シストウイルス科、デルタウイルス科、ディアンソウイルス科(Dianthoviridae)、エナノウイルス科、フィロウイルス科(例えば、マールブルグウイルスおよびエボラウイルス(例えば、ザイール株、レストン株、コートジボワール株またはスーダン株))、フラビウイルス科(例えば、C型肝炎ウイルス、デング熱ウイルス1、デング熱ウイルス2、デング熱ウイルス3およびデング熱ウイルス4)、ヘパドナウイルス科、ヘルペスウイルス科(例えば、ヒトヘルペスウイルス1、3、4、5および6、ならびにサイトメガロウイルス)、ヒポウイルス科、イリドウイルス科、レビウイルス科、ミクロウイルス科、オルトミクソウイルス科(例えば、A型およびB型およびC型インフルエンザウイルス)、パポバウイルス科、パラミクソウイルス科(例えば、麻疹ウイルス、流行性耳下腺炎ウイルスおよびヒト呼吸器多核体ウイルス(respiratory synticial virus))、パルボウイルス科、ピコルナウイルス科(例えば、ポリオウイルス、ライノウイルス、ヘパトウイルスおよびアフトウイルス)、ポックスウイルス科(例えば、ワクシニアウイルスおよびスモールポックスウイルス)、レオウイルス科(例えば、ロタウイルス)、レトロウイルス科(例えば、レンチウイルス、例えば、ヒト免疫不全ウイルス(HIV)1およびHIV 2)、ラブドウイルス科(例えば、狂犬病ウイルス、麻疹ウイルス、呼吸器多核体ウイルスなど)、トガウイルス科(例えば、風疹ウイルス、デング熱ウイルスなど)、トリウイルス科のいずれかに由来するウイルスを含む任意のウイルスから単離することができる。好適なウイルス抗原はまた、デングタンパク質M、デングタンパク質E、デングD1NS1、デングD1NS2、およびデングD1NS3の全部または一部を含む。
【0212】
ウイルス抗原は、乳頭腫ウイルス、ヘルペスウイルス、すなわち、単純ヘルペス1および2、肝炎ウイルス、例えば、A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、C型肝炎ウイルス(HCV)、D型肝炎ウイルス(HDV)、E型肝炎ウイルス(HEV)およびG型肝炎ウイルス(HGV)、ダニ媒介性脳炎ウイルス、パラインフルエンザ、水痘帯状疱疹ウイルス、サイトメガロウイルス(cytomeglavirus)、エプスタイン・バーウイルス、ロタウイルス、ライノウイルス、アデノウイルス、コクサッキーウイルス、ウマ脳炎ウイルス、日本脳炎ウイルス、黄熱ウイルス、リフトバレー熱ウイルス、およびリンパ球性脈絡髄膜炎ウイルスなどの特定の株または株の組合せに由来し得る。
【0213】
ii.細菌抗原
細菌抗原は、限定されるものではないが、アクチノミセス属、アナベナ属、バチルス属、バクテロイデス属、デロビブリオ属、ボルデテラ属、ボレリア属、カンピロバクター属、カウロバクター属、クラミジア属、クロロビウム属、クロマチウム属、クロストリジウム属、コリネバクテリウム属、サイトファガ属、ディノコッカス属、エシェリキア属、フランシセラ属、ハロバクテリア属、ヘリコバクター属、ヘモフィルス属、B型ヘモフィルス・インフルエンザ(HIB)、ハイフォミクロビウム属、レジオネラ属、レプトスピラ属、リステリア属、A型、B型およびC型髄膜炎球菌、メタノバクテリウム属、球菌属、ミオバクテリウム属(Myobacterium)、マイコプラズマ属、ミクソコッカス属、ナイセリア属、ニトロバクター属、オシラトリア属、プロクロロン属、プロテウス属、シュードモナス属、ロドスピリルム属、リケッチア属、サルモネラ菌属、赤痢菌属、スピリルム属、スピロヘータ属、ブドウ状球菌属、連鎖球菌属、ストレプトミセス属、スルフォロブス属、サーモプラズマ属、チオバチルス属およびトレポネーマ属、ビブリオ属、およびエルシニア属を含む任意の細菌に由来し得る。
【0214】
iii.寄生虫抗原
寄生虫の抗原は、限定されるものではないが、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツム(Histoplasma capsulatum)、カンジダ・アルビカンス(Candida albicans)、カンジダ・トロピカリス(Candida tropicalis)、ノカルジア・アステロイデス(Nocardia asteroides)、リケッチア・リケッチイ(Rickettsia ricketsii)、発疹熱リケッチア(Rickettsia typhi)、肺炎マイコプラズマ(Mycoplasma pneumoniae)、クラミジア・シッタシ(Chlamydial psittaci)、クラミジア・トラコマチス(Chlamydial trachomatis)、熱帯熱マラリア原虫(Plasmodium falciparum)、ブルーストリパノソーマ(Trypanosoma brucei)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ゴンディ(Toxoplasma gondii)、膣トリコモナス(Trichomonas vaginalis)、およびマンソン住血吸虫(Schistosoma mansoni)などの寄生虫から得ることができる。これらには胞子虫抗原、マラリア原虫抗原、例えば、スポロゾイト周囲タンパク質、スポロゾイト表面タンパク質、肝臓段階抗原、頂端膜関連タンパク質またはメロゾイト表面タンパク質の全部または一部が含まれる。
【0215】
iv.腫瘍抗原
抗原は、限定されるものではないが、α−アクチニン−4、Bcr−Abl融合タンパク質、Casp−8、β−カテニン、cdc27、cdk4、cdkn2a、coa−1、dek−can融合タンパク質、EF2、ETV6−AML1融合タンパク質、LDLR−フコシルトランスフェラーゼAS融合タンパク質、HLA−A2、HLA−A11、hsp70−2、KIAAO205、Mart2、Mum−1、2および3、ネオ−PAP、ミオシンクラスI、OS−9、pml−RARα融合タンパク質、PTPRK、K−ras、N−ras、トリオースリン酸イソメラーゼ、Bage−1、Gage3、4、5、6、7、GnTV、Herv−K−mel、Lage−1、Mage−A1、2、3、4、6、10、12、Mage−C2、NA−88、NY−Eso−1/Lage−2、SP17、SSX−2、およびTRP2−Int2、MelanA(MART−I)、gp100(Pmel17)、チロシナーゼ、TRP−1、TRP−2、MAGE−1、MAGE−3、BAGE、GAGE−1、GAGE−2、p15(58)、CEA、RAGE、NY−ESO(LAGE)、SCP−1、Hom/Mel−40、PRAME、p53、H−Ras、HER−2/neu、BCR−ABL、E2A−PRL、H4−RET、IGH−IGK、MYL−RAR、エプスタイン・バーウイルス抗原、EBNA、ヒト乳頭腫ウイルス(HPV)抗原E6およびE7、TSP−180、MAGE−4、MAGE−5、MAGE−6、p185erbB2、p180erbB−3、c−met、nm−23H1、PSA、TAG−72−4、CA19−9、CA72−4、CAM17.1、NuMa、K−ras、β−カテニン、CDK4、Mum−1、p16、TAGE、PSMA、PSCA、CT7、テロメラーゼ、43−9F、5T4、791Tgp72、α−フェトタンパク質、13HCG、BCA225、BTAA、CA125、CA15−3(CA 27.29\BCAA)、CA195、CA242、CA−50、CAM43、CD68\KP1、CO−029、FGF−5、G250、Ga733(EpCAM)、HTgp−175、M344、MA−50、MG7−Ag、MOV18、NB\70K、NY−CO−1、RCAS1、SDCCAG16、TA−90(Mac−2結合タンパク質\シクロフィリンC関連タンパク質)、TAAL6、TAG72、TLP、およびTPSなどの腫瘍関連高原または腫瘍特異的抗原を含む腫瘍抗原であり得る。また、BCGなどの腫瘍抗原は、アジュバントに対する免疫刺激剤として使用することもできる。
【0216】
2.アジュバント
場合により、ワクチンはアジュバントを含み得る。アジュバントは、限定されるものではないが、以下の1以上であり得る:オイルエマルション(例えば、フロイントのアジュバント)、サポニン処方物、ウィロソーム(virosomes)およびウイルス様粒子、細菌および細菌誘導体、免疫刺激性オリゴヌクレオチド、ADP−リボシル化毒素および解毒誘導体、ミョウバン、BCG、無機物含有組成物(例えば、アルミニウム塩およびカルシウム塩などの無機塩)、水酸化物、リン酸塩、硫酸塩など)、生体接着剤および/または粘膜接着剤、微粒子、リポソーム、ポリオキシエチレンエーテルおよびポリオキシエチレンエステル処方物、ポリホスファゼン、ムラミルペプチド、イミダゾキノロン化合物、および界面活性物質(例えば、リゾレシチン、プルロニックポリオール、ポリアニオン、ペプチド、オイルエマルション、キーホールリンペットヘモシアニン、およびジニトロフェノール)。
【0217】
アジュバントはまたサイトカイン、インターロイキン(例えば、IL−1、IL−2、IL−4、IL−5、IL−6、IL−7、IL−12など)、インターフェロン(例えば、インターフェロンγ)、マクロファージコロニー刺激因子、および腫瘍壊死因子などの免疫調節剤を含み得る。変異体PD−L2ポリペプチドの他、B7ファミリーの他のポリペプチドを含む他の共刺激分子を投与してもよい。このようなタンパク質性アジュバントは全長ポリペプチドもしくはその活性断片として、またはプラスミドDNAなどのDNAの形態で提供されてよい。
【0218】
IV.使用方法
PD−1アンタゴニストポリペプチドおよびその断片、ならびにこれらのポリペプチドおよび融合タンパク質をコードする核酸、またはPD−1アンタゴニストポリペプチドを発現する細胞は、抗原に対する一次免疫応答を増強するとともにT細胞の抗原特異的増殖の増強、T細胞によるサイトカイン生産の増強および分化刺激などのエフェクター機能を増強するために使用することができる。PD−1アンタゴニスト組成物は、それを必要とする被験体に、T細胞の消耗および/またはT細胞の無応答を克服するのに有効な量で投与することができる。T細胞の消耗またはT細胞の無応答の克服は、既知の技術を用いてT細胞機能を測定することにより決定することができる。好ましいPD−1アンタゴニストポリペプチドは、PD−1を介した阻害的シグナル伝達を誘発することなくPD−1と結合し、かつT細胞を共刺激する能力を保持するように操作される。
【0219】
PD−1アンタゴニストのインビトロ適用は、例えば、免疫機構の基礎的科学研究において、またはT細胞機能の研究もしくは例えば受動免疫療法に用いる活性化T細胞の生産のために有用であり得る。さらに、PD−1アンタゴニストは、T細胞が得られた被験体において着目する抗原に対する免疫性を試験すべく設計されたインビトロアッセイ(例えば、T細胞増殖アッセイ)に加えることができる。このようなアッセイへのPD−1アンタゴニストの付加は、より強力な、従ってより検出の容易なインビトロ応答をもたらすと考えられる。
【0220】
A.免疫増強のためのPD−1アンタゴニストの投与
PD−1アンタゴニストは一般に、インビボおよびエクスビボにおいて免疫応答刺激治療薬として有用である。好ましい実施形態においては、これらの組成物は、T細胞の消耗またはT細胞の無応答が既に起こり、その感染が長期にわたって宿主に残存する感染を処置するのに有用である。処置される例示的感染は、肝炎ウイルス、ヒト免疫不全ウイルス(HIV)、ヒトTリンパ球向性ウイルス(HTLV)、ヘルペスウイルス、エプスタイン・バーウイルス、またはヒト乳頭腫ウイルスにより引き起こされる慢性感染である。他の感染もPD−1アンタゴニストを用いて処置可能であると考えられる。開示されている組成物はワクチンの一部としても有用である。好ましい実施形態では、治療または予防される疾患の種類は、細菌、ウイルス、原虫、蠕虫、または細胞内に侵入する、すなわち細胞傷害性Tリンパ球により攻撃される、他の微生物病原体により引き起こされる慢性感染性疾患である。
【0221】
ヒトおよび動物モデルにおける慢性感染は、宿主免疫応答の、機能的なCD8+およびCD4+T細胞集団の生成および維持不全に関連し、その結果として、感染性を中和するための抗体応答も不十分となる。この機能の欠損はT細胞の消耗と呼ばれる。T細胞の無応答は、リンパ球が抗原と遭遇した後に内因的に機能的に活性化されるが、長期間、低応答状態で生存を維持するという耐性機構である。慢性感染を処置するための一つの方法は、消耗したT細胞を活性化すること、または被験体におけるT細胞の消耗を逆転させるとともにT細胞の無応答を克服することである。T細胞の消耗の逆転は、PD−1とそのリガンドPD−L1(B7−H1)およびPD−L2(PD−L2)の間の相互作用に干渉することによって達成することができる。急性の、多くの場合致死的な病原体の作用は、毒素、またはその毒素により引き起こされる損傷の前に十分な免疫応答を惹起することができない他の因子によって媒介され得る。これは、PD−1とそのリガンドの間の相互作用に干渉し、より有効な迅速免疫応答を可能とすることにより克服され得る。
【0222】
ウイルス感染は主としてT細胞により排除されるので、感染力のあるウイルス病原体のより迅速な、または十分な排除がヒトまたは動物被験体に有益となる状況では、T細胞活性の増強が治療上有用である。よって、PD−1アンタゴニストは、限定されるものではないが、免疫不全(例えば、HIV)、乳頭腫(例えば、HPV)、ヘルペス(例えば、HSV)、脳炎、インフルエンザ(例えば、ヒトA型インフルエンザウイルス)、および風邪(例えば、ヒトライノウイルス)ウイルス感染を含む、局所または全身ウイルス感染の処置のために投与することができる。例えば、PD−1アンタゴニスト組成物を含む医薬処方物は、ヘルペス病巣または帯状疱疹または性器疣贅などのウイルス性皮膚疾患を処置するために局所投与することができる。PD−1アンタゴニスト組成物の医薬処方物はまた、限定されるものではないが、AIDS、インフルエンザ、風邪または脳炎を含む全身性ウイルス性疾患を処置するためにも投与することができる。
【0223】
処置可能な代表的感染としては、限定されるものではないが、アクチノミセス属、アナベナ属、バチルス属、バクテロイデス属、デロビブリオ属、ボルデテラ属、ボレリア属、カンピロバクター属、カウロバクター属、クラミジア属、クロロビウム属、クロマチウム属、クロストリジウム属、コリネバクテリウム属、サイトファガ属、ディノコッカス属、エシェリキア属、フランシセラ属、ハロバクテリア属、ヘリコバクター属、ヘモフィルス属、B型ヘモフィルス・インフルエンザ(HIB)、ヒトプラズマ属、ハイフォミクロビウム属、レジオネラ属、リーシュマニア属、レプトスピラ属、リステリア属、A型、B型およびC型髄膜炎球菌、メタノバクテリウム属、球菌属、ミオバクテリウム属(Myobacterium)、マイコプラズマ属、ミクソコッカス属、ナイセリア属、ニトロバクター属、オシラトリア属、プロクロロン属、プロテウス属、シュードモナス属、ロドスピリルム属、リケッチア属、サルモネラ菌属、赤痢菌属、スピリルム属、スピロヘータ属、ブドウ状球菌属、連鎖球菌属、ストレプトミセス属、スルフォロブス属、サーモプラズマ属、チオバチルス属およびトレポネーマ属、ビブリオ属、エルシニア属、クリプトコッカス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツム(Histoplasma capsulatum)、カンジダ・アルビカンス(Candida albicans)、カンジダ・トロピカリス(Candida tropicalis)、ノカルジア・アステロイデス(Nocardia asteroides)、リケッチア・リケッチイ(Rickettsia ricketsii)、発疹熱リケッチア(Rickettsia typhi)、肺炎マイコプラズマ(Mycoplasma pneumoniae)、クラミジア・シッタシ(Chlamydial psittaci)、クラミジア・トラコマチス(Chlamydial trachomatis)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(Plasmodium vivax)、ブルーストリパノソーマ(Trypanosoma brucei)、赤痢アメーバ(Entamoeba histolytica)、トキソプラズマ・ゴンディ(Toxoplas
ma gondii)、膣トリコモナス(Trichomonas vaginalis)およびマンソン住血吸虫(Schistosoma mansoni)を含む微生物により引き起こされる感染が挙げられる。
【0224】
B.ワクチンにおけるPD−1アンタゴニストの使用
PD−1アンタゴニストまたはそれをコードする核酸は単独で投与してもよいし、あるいは他の好適な処置と組み合わせて投与してもよい。一つの実施形態においては、PD−1アンタゴニストは上記のようなワクチン組成物とともに、またはワクチン阻害剤の一成分として投与することができる。ワクチン組成物の好適な成分は上記されている。開示されているPD−1アンタゴニストはワクチン投与の前、ワクチン投与と同時、またはワクチン投与の後に投与することができる。一つの実施形態においては、PD−1アンタゴニスト組成物はワクチン投与と同時に投与される。
【0225】
PD−1アンタゴニスト組成物は、被験体において次に感染性病原体に曝された際の耐性を付与する予防ワクチンとともに、またはウイルスに感染した被験体におけるウイルス抗原などの既存の抗原に対する被験体の免疫応答を誘発または増強するために使用できる治療ワクチンとともに投与してもよい。
【0226】
予防的、治療的または脱感作免疫応答の所望の結果は、当該技術分野でよく知られている原理に従い、疾患によって異なり得る。例えば、感染性病原体に対する免疫応答は感染性病原体の定着および複製を完全に防ぐことができ、「無菌免疫(sterile immunity)」および疾病症状の非存在に影響を及ぼす。しかしながら、感染性病原体に対するワクチンは、それが症状の数、重篤度または持続時間を軽減するか、症状を有する集団中の個体数を少なくするか、または感染性病原体の伝染を軽減すれば、有効であると見なせる。同様に、癌、アレルゲン、または感染性病原体に対する免疫応答は疾患を完全に治療する場合、症状を緩和する場合、または疾患に対する全体的な治療的介入の一つの態様である場合がある。
【0227】
PD−1アンタゴニストは、少なくとも一つの成分抗原または抗原組成物に対し、PD−1アンタゴニストを用いずに対応する組成物で得られるエフェクター細胞応答に比べて向上したエフェクター細胞応答(CD4T細胞免疫応答など)を誘導する。「向上したエフェクター細胞応答」との用語は、ワクチン組成物の投与後にヒト患者で得られる、PD−1アンタゴニストを用いずに同じ組成物の投与後に得られるものよりも高いエフェクター細胞応答(CD4応答など)を意味する。例えば、ヒト患者においてPD−1アンタゴニスト、好ましくはPD−L2−Igと抗原調製物を含有する免疫原組成物を投与した際に、アジュバントを含まないその抗原調製物を含有する免疫原組成物の投与後に誘導される応答に比べて高いCD4T細胞応答が得られる。このような処方物は有利には、MHCクラスII分子により提示される抗原エピトープを検出することができる抗抗原エフェクター細胞応答を誘導するために用いられる。
【0228】
この向上したエフェクター細胞応答は、免疫刺激されていない患者、すなわち、その抗原に対して血清陰性である患者で得ることができる。この血清陰性は、患者がその抗原に直面したことがない(いわゆる「ナイーブ」患者)か、あるいはまた、かつて遭遇したその抗原に応答できなかった結果であり得る。この向上したエフェクター細胞応答は、高齢者、一般に65歳以上、または高リスクな医学的状態を有する65歳より若い成人(「高リスク」成人)、または2歳未満の小児など、免疫不全被験体で得られるのが好ましい。
【0229】
向上したエフェクター細胞応答は、以下のサイトカインのいずれかを産生する細胞:(1)少なくとも二つの異なるサイトカイン(CD40L、IL−2、IFN−γ、TNF−α)を産生する細胞、(2)少なくともCD40Lと別のサイトカイン(IL−2、TNF−α、IFN−γ)を産生する細胞、(3)少なくともIL−2と別のサイトカイン(CD40L、TNF−α、IFN−γ)を産生する細胞、(4)少なくともIFN−γと別のサイトカイン(IL−2、TNF−α、CD40L)を産生する細胞、および(5)少なくともTNF−αと別のサイトカイン(IL−2、CD40L、IFN−γ)の数を測定することにより評価することができる。
【0230】
向上したエフェクター細胞応答は、上記のサイトカインのいずれかを産生する細胞がワクチン組成物の投与後に、PD−1アンタゴニストを含まない組成物を投与した場合に比べて多い量である際に存在する。一般に、上記の5つの条件のうち少なくとも一つ、好ましくは二つが満たされる。特定の実施形態では、4つ総てのサイトカインを産生する細胞が、ワクチン接種群で非ワクチン接種群に比べて多い数で存在する。
【0231】
免疫原組成物は、皮内、粘膜、例えば、鼻内、経口、筋肉内、または皮下などのいずれの好適な送達経路によって投与してもよい。当該技術分野では他の送達経路もよく知られている。免疫原組成物にとっては筋肉内送達経路が好ましい。皮内送達はもう一つの好適な経路である。皮内送達には、例えば短針デバイスなどの任意の好適なデバイスが使用可能である。皮内ワクチンはまた、針の皮膚への有効侵入長を制限するデバイスにより投与してもよい。また、液体ジェット式注射器によって、または角質層を穿孔し、真皮に達するジェットを生じる針によって、液体ワクチンを真皮へ送達するジェット注射デバイスを使用することもできる。ジェット注射デバイスは当該技術分野で既知である。皮膚の外層から真皮へと粉末形態のワクチンを加速化するために圧縮ガスを用いる弾道粉末/粒子送達デバイスも使用可能である。さらに、皮内投与の従来のMantoux法で、通常のシリンジを使用することもできる。
【0232】
もう一つの好適な投与経路は皮下経路である。皮下送達には、例えば、従来の針などの好適なデバイスを使用することができる。好ましくは、無針ジェット式注射器が用いられる。無針注射器は当該技術分野で既知である。より好ましくは、デバイスは液体ワクチン処方物が予め充填されている。
【0233】
あるいは、ワクチンは鼻腔内投与される。一般に、ワクチンは鼻咽頭領域に、好ましくは肺へ吸入されずに、局所投与される。ワクチン処方物を、肺に入ることなく、または実質的に入ることなく鼻咽頭領域へ送達する鼻腔内送達デバイスを用いるのが好ましい。ワクチンの鼻腔内投与に好ましいデバイスはスプレーデバイスである。鼻腔スプレーデバイスは市販されている。ネブライザーは容易に肺へ吸入される可能性のある極めて微細なスプレーを生じるので、鼻腔粘膜に十分に達しない。従って、ネブライザーは好ましくない。鼻腔用の好ましいスプレーデバイスは、デバイスの性能が、使用者がかける圧力に依存しないデバイスである。これらのデバイスは圧力閾値デバイスとして知られる。閾値圧力がかけられた場合にのみノズルから液体が放出される。これらのデバイスは規則的な液滴サイズを有するスプレーを達成することを容易とする。本発明とともに用いるのに好適な圧力閾値デバイスは当該技術分野で公知であり、市販されている。
【0234】
好ましい鼻腔デバイスは、1〜200μm、好ましくは10〜120μmの範囲の液滴(液体として水を用いて測定)を生じる。10μmを下回ると吸入のリスクがあるので、10μm未満の液滴が約5%を超えないことが望ましい。120μmを超える液滴は、120μmを超える液滴ならびにそれより小さい液滴は拡散しないので、120μmを超える液体が約5%を超えないことが望ましい。
【0235】
二用量送達(bi-dose delivery)は、ワクチンとともに用いるための鼻腔内送達系のもう一つの特徴である。二用量デバイスは、単回ワクチン用量の二つの分割量を含み、一つの分割用量は各鼻孔に投与するためのものである。一般に、二つの分割用量は単一のチャンバー内に存在し、このデバイスの構成は単一の分割用量を一度に効率的に送達することを可能とする。あるいは、ワクチンの投与に一用量デバイスを用いてもよい。
【0236】
免疫原組成物は数日、数週間または数ヶ月の間にわたって2回以上で与えることができる。一つの実施形態では、異なる投与経路が用いられ、最初に筋肉内投与を行ってよく、場合によりPD−1アンタゴニストを含んでもよい追加免疫組成物が、例えば皮内、皮下、または鼻腔内などの異なる経路により投与してよい。
【0237】
免疫原組成物により付与された向上したエフェクター細胞応答は1回の投与の後に得られるのが理想的であり得る。この単回用量アプローチは、バイオテロリストの攻撃および流行を含む、急速に進行する大発生に極めて関連が強い。ある特定の状況では、特に、高齢者集団、または特定の抗原に対して初めてワクチン接種される年少の小児(9歳未満)では、2回用量の同じ組成物を投与することが有益であり得る。同じ組成物の2回目の用量(なお、最初のワクチン接種用の組成物とみなされる)は、進行中の一次免疫応答中に投与することができ、初回投与から十分な時間間隔が置かれる。一般に、2回目の用量の組成物は、未応答または応答が不十分な個体において免疫系の刺激を助けるために、初回用量の数週間、または約1か月、例えば、2週間、3週間、4週間、5週間、または6週間後に投与される。
【0238】
特定の実施形態においては、免疫原組成物の投与は、代わりにまたはその上に、アジュバントを含む免疫原組成物を投与した患者において、アジュバントを含まない組成物で免疫された個体において誘導されたBメモリー細胞応答に比べて向上したBメモリー細胞応答を誘導する。向上したBメモリー細胞応答は、インビトロ分化の刺激により測定される、抗原遭遇時に抗体分泌血漿細胞へと分化し得る末梢血Bリンパ球の頻度の増加を意味するものとする(実施例の節、例えば、Elispot B細胞記憶の方法を参照)。
【0239】
さらに別の実施形態では、免疫原組成物は一次免疫応答ならびにCD8応答を増強する。初回ワクチン接種用の免疫原組成物の1回用量を投与すると、良好な血清保護がもたらされ、アジュバントを含まない処方物で得られるものに比べて、特定の抗原に対して向上したCD4T細胞またはCD8T細胞免疫応答が誘導される。この結果、全体的な罹患率および死亡率が引き下げられ、また、肺炎および他のインフルエンザ様疾患のための緊急入院が防げる。この方法により、−アジュバントを含まない処方物で誘導される応答に比べて、より長い時間持続する、例えば、初回ワクチン接種後なお1年存在するCD4T細胞応答を誘導することができる。
【0240】
好ましくは、無刺激被験体で得られる向上したCD4T細胞免疫応答などのCD4T細胞免疫応答は、交差反応性CD4Tヘルパー応答の誘導を含む。特に、交差反応性CD4T細胞の量が増す。「交差反応性」CD4応答とは、例えばインフルエンザ株の間で共有されているエピトープを標的化するCD4T細胞を意味する。
【0241】
この用量のPD−1アンタゴニストはヒトにおいて抗原に対する免疫応答を増強する。特に、好適なPD−1アンタゴニスト量は、アジュバントを含まない組成物に比べて、または別のPD−1アンタゴニスト量のアジュバントを含む組成物に比べて、組成物の免疫能を向上させるものである。通常、免疫原組成物の用量は約0.5ml〜約1mlの範囲である。一般的なワクチン用量は0.5ml、0.6ml、0.7ml、0.8ml、0.9ml、または1mlである。好ましい実施形態では、ワクチン組成物1ml当たりに終濃度50μgのPD−1アンタゴニスト、好ましくはPD−L2−Ig、ワクチン用量0.5ml当たり25μgが含まれる。他の好ましい実施形態では、ワクチン組成物1ml当たり終濃度35.7μgまたは71.4μgのPD−1アンタゴニストが含まれる。具体的には、0.5mlのワクチン用量は、1用量当たり25μgまたは50μgのPD−1アンタゴニストを含む。さらに別の実施形態では、この用量は100μg以上である。免疫原組成物は通常、一元放射免疫拡散法(SRD)(J. M. Wood et al.: J. Biol. Stand. 5 (1977) 237-247、 J. M. Wood et al., J. Biol. Stand. 9 (1981) 317-330)により測定した場合、15μgの抗原成分を含む。
【0242】
被験体に免疫原組成物を再接種することができる。一般に、再接種は初回接種の少なくとも6か月後、好ましくは8〜14か月後、より好ましくは10〜12か月後前後で行われる。
【0243】
再接種用の免疫原組成物(追加免疫組成物)は、不活性されているか、または生きた状態で弱毒されている、いずれの種類の抗原調製物も含み得る。それは、初回接種用に用いた免疫原組成物と同じ種類の抗原調製物、例えば、スプリットインフルエンザウイルスまたはそのスプリットインフルエンザウイルス抗原調製物、完全ビリオン、精製サブユニットワクチンまたはウィロソーム(virosome)を含み得る。あるいは、追加免疫組成物は、初回接種用に用いたものと別の種類の抗原、すなわち、スプリットインフルエンザウイルスまたはそのスプリットインフルエンザウイルス抗原調製物、完全なビリオン、精製サブユニットワクチン、またはウィロソームを含み得る。
【0244】
ウイルスに対するワクチンに関して、使用する場合、追加免疫組成物は一般に次のウイルスシーズン、例えば、最初の免疫原組成物のおよそ1年後に投与する。追加免疫組成物はまた、その後毎年(3回目、4回目、5回目の接種など)与えてもよい。追加免疫組成物は初回接種に用いた組成物と同じであってもよい。
【0245】
好ましくは、再接種は以下のいずれか、好ましくは二つまたは総てを誘導する:PD−1アンタゴニストを含まない抗原調製物での初回接種の後に誘導された同等の応答に比べて(i)抗原調製物に対する向上したエフェクター細胞応答、または(ii)向上したB細胞メモリー応答、または(iii)向上した体液性応答。好ましくは、PD−1アンタゴニストを含有する免疫原性抗原調製物を再接種した後に誘導される免疫応答は、アジュバントを含まない組成物を再接種した後に誘導される対応する応答よりも高い。
【0246】
免疫原組成物は一価または多価、すなわち、二価、三価、または四価であり得る。好ましくは、その免疫原組成物は三価または四価である。多価とは、一般に異なる種または株からの抗原の供給源の数を意味する。ウイルスに関しては、少なくとも一つの株が汎流行的大発生に関連するか、または汎流行的大発生に関連する可能性がある。
【0247】
C.標的化抗原提示細胞
別の実施形態は、抗原提示細胞(APC)を1以上の開示されているPD−1アンタゴニストと、APCにおけるPD−1シグナル伝達を阻害、低減または遮断するのに有効な量で接触させることを提供する。APCにおけるPD−1シグナル伝達の遮断は、細胞内病原体または細胞内病原体に感染した細胞の排除を増強するAPCを活性化する。
【0248】
D.組合せ療法
PD−1アンタゴニスト組成物はそれを必要とする被験体に単独で、または1以上の付加的治療薬と組み合わせて投与することができる。付加的治療薬は処置される症状、障害、または疾患に基づいて選択される。例えば、PD−1アンタゴニストは免疫応答を増強または促進する働きをする1以上の付加的薬剤と同時に投与することができる。
【0249】
E.結合特性の調節
PD−1アンタゴニストの結合特性は投与される用量および用量計画に関連する。MDX−1106などの既存の抗体PD−1アンタゴニストは、1回投与の後少なくとも3ヶ月間、T細胞上のPD−1分子の60〜80%を持続的に占有することを示す(Brahmer, et al. J. Clin. Oncology, 27:(155) 3018 (2009))。好ましい実施形態では、開示されているPD−1アンタゴニストは、免疫細胞上のPD−1分子の、より短期またはより低パーセンテージの占有を示すPD−1との結合特性を有する。例えば、開示されているPD−1アンタゴニストは一般に、1回用量の投与後1週間、2週間、3週間またはさらには1か月で、免疫細胞上のPD−1分子の占有率5、10、15、20、25、30、35、40、45、50%未満を示す。他の実施形態では、開示されているPD−1アンタゴニストはPD−1に対してMDX−1106よりも低い結合親和性を有する。MDX−1106などの抗体に関して、PD−1−Ig融合タンパク質はその受容体に対して比較的低い親和性を有し、従って、比較的速い解離速度を有するはずである。
【0250】
他の実施形態では、PD−1アンタゴニストは、次の投与の前に減衰される周期的に増強される免疫応答を惹起するために数日、数週間、または数ヶ月間にわたって間欠的に投与され、これにより、免疫応答を誘導する、免疫応答を刺激する、または免疫応答を増強する働きをすることができる。
【実施例】
【0251】
本発明は、以下の非限定的実施例を参照すればさらに理解される。
【0252】
実施例1:B7−DCのPD−1への結合
ヒトB7−DC−IgのPD−1結合活性を、ELISAにより評価した。96ウェルELISAプレートを、BupH Carbonate/Bicarbonate pH9.4バッファー(Pierce)で希釈した0.75μg/mLの組換えヒトPD−1/Fc(R&D Systems)100μLで2時間コーティングした後、BSA溶液(Jackson ImmunoResearch)を用いて90〜120分間ブロックした。連続誘導体希釈したヒトB7−DC−IgならびにヒトIgG1イソ型対照を90分間結合させた。結合したB7−DC−Igは、100μLの0.5μg/mLビオチン結合抗ヒトB7−DCクローンMIH18(eBioscience)、次いで1:1000希釈HRP−ストレプトアビジン(BD Bioscience)およびTMB基質(BioFX)を用いて検出した。プレートリーダー(Molecular Devices)を用いて450nmの吸光度を読み取り、データを、SoftMaxで4パラメーターロジスティック適合を用いて解析した。
【0253】
ネズミB7−DC−IgのPD−1結合活性を、ELISAにより評価した。96ウェルのELISAプレートを、BupH Carbonate/Bicarbonate pH9.4バッファー(Pierce)で希釈した0.75μg/mLの組換えマウスPD−1/Fc(R&D Systems) 100μLを用いて2時間コーティングし、次いでBSA溶液(Candor-Bioscience)を用いて90分間ブロックした。連続希釈したネズミB7−DC−Ig(野生型、ならびにPD−1への低減された結合のために選択されたD111SおよびK113S変異株)、ならびにネズミIgG2aイソ型対照を90分間結合させた。結合したB7−DC−Igは、100μLの0.25μg/mLビオチン結合抗マウスB7−DCクローン112(eBioscience)、次いで1:2000希釈HRP−ストレプトアビジン(BD Bioscience)およびTMB基質(BioFX)を用いて検出した。プレートリーダー(Molecular Devices)を用いて450nmの吸光度を読み取り、データを、SoftMaxで4−パラメーターロジスティック適合を用いて解析した。
【0254】
図1Aおよび1Bは、PD−1結合ELISAにおける、OD450対B7−DC−Ig(μg/ml)の量の線グラフを示す。図1Aは、4つの異なるロットのヒトB7−DC−Igの結合を示す。図4Bは、野生型ネズミB7−DC−Ig(円)、DS変異株(D111Sを置換したB7−DC−Ig、三角)、およびKS変異株(K113Sを置換したB7−DC−Ig、四角)、およびネズミIgG2aイソ型対照(ひし形)の結合を示す。
【0255】
実施例2:B7−DCのPD−1発現CHO細胞への結合
B7−DC−Igを、最初にアロフィコシアニン(APC)と結合させ、次いでPD−1を構成的に発現するCHO細胞系統またはPD−1を発現しない親CHO細胞とともに種々な濃度でインキュベートした。結合は、フローサイトメトリーにより分析した。図2は、プローブ濃度(x軸)の関数としての、B7−DC−Ig−APCの蛍光強度の中央値(MFI)(y軸)を示す。B7−DC−Ig−APCはCHO.PD−1細胞(黒円)に結合するが、トランスフェクトされていないCHO細胞には結合しない(灰色三角)。
【0256】
実施例3:PD−1への結合に関するB7−DC−IgのB7−H1との競合
B7−H1−Igを、最初にアロフィコシアニン(APC)と結合させた。B7−H1−Ig−APCをプローブおよび細胞混合物に加える前に、種々の濃度の非標識B7−DC−Igを、最初に、PD−1を構成的に発現するCHO細胞系統とインキュベートした。図3は、加えた非標識B7−DC−Igコンペティターの濃度(x軸)の関数としての、B7−H1−Ig−APCの蛍光強度の中央値(MFI)(y軸)を示す。非標識B7−DC−Ig濃度が増加するにつれて、CHO細胞に結合するB7−H1−Ig−APC量は減少し、このことは、PD−1への結合に関してB7−DC−IgがB7−H1と競合することを実証している。
【0257】
実施例4:シクロホスファミドとB7−DC−Igの組合せが腫瘍特異的メモリー細胞傷害性Tリンパ球を生成する
9〜11週齢のBalb/Cマウスの皮下に1.0×105のCT26結腸直腸腫瘍細胞を移植した。腫瘍移植後10日目に、マウスに100mg/kgのシクロホスファミドを投与した。B7−DC−Ig処置は1日後である11日目に開始した。マウスを、100μgのB7−DC−Igを用いて、1週間に2用量、4週間にわたり合計8用量で処置した。CTX+B7−DC−Ig処置投与計画を受けたマウスの75%において44日までに定着腫瘍が根絶され、一方、対照CTX単独群のマウスは全例が、腫瘍増殖の結果死亡したか、または腫瘍がIACUCにより承認されたサイズを超えたため安楽死させた。
【0258】
上記実験でCT26結腸直腸定着腫瘍を根絶したマウスに、44日目および70日目に1×105個のCT26細胞を再負荷した。再負荷により腫瘍が増殖しなかったことは、シクロホスファミドおよびB7−DC−Igの組合せ処置が、長期にわたる抗腫瘍免疫を発展させたことを示唆している。ビヒクル対照群のマウスは全例が腫瘍を発現した。これにより、定着腫瘍に対する処置の有効性、ならびにB7−DCIgの併用処置が腫瘍抗原に対する記憶応答をもたらしたことが示された。
【0259】
上記実験でCT26結腸直腸定着腫瘍を根絶したマウスに、44日目に2.5×105個のCT26細胞を再負荷した。7日後、マウスの脾臓を単離した。マウス脾細胞を、Golgi遮断薬(BD BioScience)の存在下、5または50μg/mLのオバアルブミン(OVA)またはAH1ペプチドとともに6時間パルス処理し、CD8+/IFNγ+T細胞を評価することによりメモリーTエフェクター細胞を分析した。図4の結果は、CT26腫瘍根絶マウスにおいて多量のCT26特異的Tエフェクター細胞が存在することを示している。
【0260】
実施例5:B7−DC−IgはHSVウイルス粒子の放出を低減し、マウスの生存率を高めた
8〜10週齢のBalb/Cマウスをまず、ビヒクル(白四角)または300μgのB7−DC−Ig(黒四角)とともに4×104用量の弱毒HSV−2生ワクチンで免疫した(図5Aおよび5B)。1か月後、総てのマウスを5×105PFUのHSV−2 G−6株を腟内投与した。図5Aは、ウイルス投与後9時間、1日、2日、3日、4日および5日目における膣領域のスワブのウイルス粒子力価を示す。図5Bは、ウイルス投与後12日目におけるマウスの生存率を示す。これにより、ワクチンとともにB7−DC−Iが存在すると、ウイルス量を低減し、動物の生存率を高めることができる。
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
免疫応答を調節する方法であって、疾病病原体または感染細胞に対する免疫応答誘導する、増大させる、または増強するのに有効な量のPD−1アンタゴニストを投与することを含んでなり、その分子の用量、その分子の投与時期、および/またはその分子の親和性によりリガンドのPD−1受容体への間欠的接近を可能にする、方法。
【請求項2】
PD−1アンタゴニストが内因性PD−L1と、PD−1との結合を阻害または低減する、請求項1に記載の方法。
【請求項3】
PD−1アンタゴニストが内因性PD−L2と、PD−1との結合を阻害または低減する、請求項1に記載の方法。
【請求項4】
PD−1アンタゴニストがPD−1と結合する、請求項1に記載の方法。
【請求項5】
PD−1アンタゴニストがPD−1、PD−L1、PD−L2、B7.1、およびその断片からなる群から選択される、請求項1に記載の方法。
【請求項6】
前記分子がPD−1またはそのリガンドとインビボ投与後、3か月以内の期間結合する、請求項1に記載の方法。
【請求項7】
1を超えるPD−1アンタゴニストが投与される、請求項1に記載の方法。
【請求項8】
感染が慢性ウイルス感染、細菌感染、真菌感染、マイコプラズマ感染、寄生虫感染であり、感染の急性期に毒素により媒介される疾患を誘発するか、または感染がT細胞応答の低下を特徴とする、請求項1に記載の方法。
【請求項9】
ウイルス感染が、肝炎ウイルス、ヒト免疫不全ウイルス、ヒトTリンパ球向性ウイルス、ヘルペスウイルス、エプスタイン・バーウイルス、フィロウイルス、ヒト乳頭腫ウイルス、エプスタイン・バーウイルス、インフルエンザウイルス、呼吸器多核体ウイルス(respiratory synticial virus)、脳炎ウイルス、デング熱ウイルス、および乳頭腫ウイルスによる感染である、請求項8に記載の方法。
【請求項10】
寄生虫感染がマラリアまたはリーシュマニアである、請求項4に記載の方法。
【請求項11】
細菌感染が、結核菌(Mycobacterium tuberculosis)、炭疽菌(Bacillus anthracis)、ブドウ状球菌(Staphylococcus)、リステリア菌(Listeria)、およびクラミジア・トラコマチス(Clamydia trachomatis)からなる群から選択される細菌により引き起こされる、請求項8に記載の方法。
【請求項12】
疾病に対する免疫応答を増強するためにPD−1アンタゴニストと組み合わせて疾病抗原を投与することさらに含んでなる、請求項1に記載の方法。
【請求項13】
PD−1アンタゴニストがPD−1リガンドの融合タンパク質である、請求項1に記載の方法。
【請求項14】
融合タンパク質が、PD−1と結合し得るPD−L2の細胞外ドメインまたはその断片を含んでなる、請求項13に記載の方法。
【請求項15】
融合タンパク質が配列番号83のアミノ酸配列を有する、請求項14に記載の方法。
【請求項16】
PD−1アンタゴニストとともに、免疫調節剤、Tregの機能を枯渇させる、または阻害する薬剤、および共刺激分子からなる群から選択される付加的な有効薬剤を投与することをさらに含んでなる、請求項1に記載の方法。
【請求項17】
付加的な有効薬剤がCD4+CD25+Tregの機能を枯渇させる、または阻害する薬剤である、請求項17に記載の方法。
【請求項18】
CD4+CD25+Tregの機能を枯渇させる、または阻害する薬剤がシクロホスファミドである、請求項17に記載の方法。
【請求項19】
抗原提示細胞機能を増強させるための請求項1の方法であり、APCをPD−1アンタゴニストと、APCにおけるPD−1のシグナル伝達を阻害、低減、もしくは遮断する、または罹患細胞または感染細胞の除去を増強するのに有効な量で接触させることを含んでなる、方法。
【請求項20】
PD−1アンタゴニストを1以上の疾病抗原と組み合わせて含んでなる、組成物。
【請求項21】
PD−1アンタゴニストをワクチンと組み合わせて含んでなる、組成物。
【請求項1】
免疫応答を調節する方法であって、疾病病原体または感染細胞に対する免疫応答誘導する、増大させる、または増強するのに有効な量のPD−1アンタゴニストを投与することを含んでなり、その分子の用量、その分子の投与時期、および/またはその分子の親和性によりリガンドのPD−1受容体への間欠的接近を可能にする、方法。
【請求項2】
PD−1アンタゴニストが内因性PD−L1と、PD−1との結合を阻害または低減する、請求項1に記載の方法。
【請求項3】
PD−1アンタゴニストが内因性PD−L2と、PD−1との結合を阻害または低減する、請求項1に記載の方法。
【請求項4】
PD−1アンタゴニストがPD−1と結合する、請求項1に記載の方法。
【請求項5】
PD−1アンタゴニストがPD−1、PD−L1、PD−L2、B7.1、およびその断片からなる群から選択される、請求項1に記載の方法。
【請求項6】
前記分子がPD−1またはそのリガンドとインビボ投与後、3か月以内の期間結合する、請求項1に記載の方法。
【請求項7】
1を超えるPD−1アンタゴニストが投与される、請求項1に記載の方法。
【請求項8】
感染が慢性ウイルス感染、細菌感染、真菌感染、マイコプラズマ感染、寄生虫感染であり、感染の急性期に毒素により媒介される疾患を誘発するか、または感染がT細胞応答の低下を特徴とする、請求項1に記載の方法。
【請求項9】
ウイルス感染が、肝炎ウイルス、ヒト免疫不全ウイルス、ヒトTリンパ球向性ウイルス、ヘルペスウイルス、エプスタイン・バーウイルス、フィロウイルス、ヒト乳頭腫ウイルス、エプスタイン・バーウイルス、インフルエンザウイルス、呼吸器多核体ウイルス(respiratory synticial virus)、脳炎ウイルス、デング熱ウイルス、および乳頭腫ウイルスによる感染である、請求項8に記載の方法。
【請求項10】
寄生虫感染がマラリアまたはリーシュマニアである、請求項4に記載の方法。
【請求項11】
細菌感染が、結核菌(Mycobacterium tuberculosis)、炭疽菌(Bacillus anthracis)、ブドウ状球菌(Staphylococcus)、リステリア菌(Listeria)、およびクラミジア・トラコマチス(Clamydia trachomatis)からなる群から選択される細菌により引き起こされる、請求項8に記載の方法。
【請求項12】
疾病に対する免疫応答を増強するためにPD−1アンタゴニストと組み合わせて疾病抗原を投与することさらに含んでなる、請求項1に記載の方法。
【請求項13】
PD−1アンタゴニストがPD−1リガンドの融合タンパク質である、請求項1に記載の方法。
【請求項14】
融合タンパク質が、PD−1と結合し得るPD−L2の細胞外ドメインまたはその断片を含んでなる、請求項13に記載の方法。
【請求項15】
融合タンパク質が配列番号83のアミノ酸配列を有する、請求項14に記載の方法。
【請求項16】
PD−1アンタゴニストとともに、免疫調節剤、Tregの機能を枯渇させる、または阻害する薬剤、および共刺激分子からなる群から選択される付加的な有効薬剤を投与することをさらに含んでなる、請求項1に記載の方法。
【請求項17】
付加的な有効薬剤がCD4+CD25+Tregの機能を枯渇させる、または阻害する薬剤である、請求項17に記載の方法。
【請求項18】
CD4+CD25+Tregの機能を枯渇させる、または阻害する薬剤がシクロホスファミドである、請求項17に記載の方法。
【請求項19】
抗原提示細胞機能を増強させるための請求項1の方法であり、APCをPD−1アンタゴニストと、APCにおけるPD−1のシグナル伝達を阻害、低減、もしくは遮断する、または罹患細胞または感染細胞の除去を増強するのに有効な量で接触させることを含んでなる、方法。
【請求項20】
PD−1アンタゴニストを1以上の疾病抗原と組み合わせて含んでなる、組成物。
【請求項21】
PD−1アンタゴニストをワクチンと組み合わせて含んでなる、組成物。
【図2】

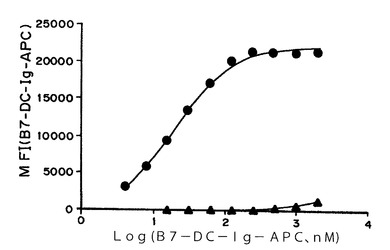
【図3】
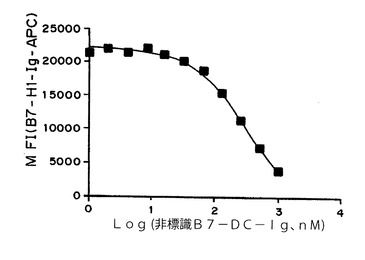
【図4】
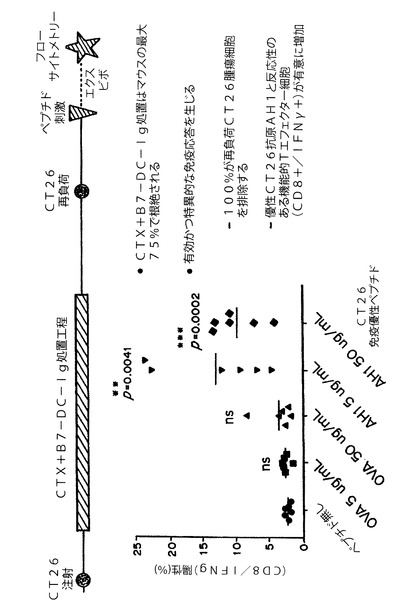
【図5】
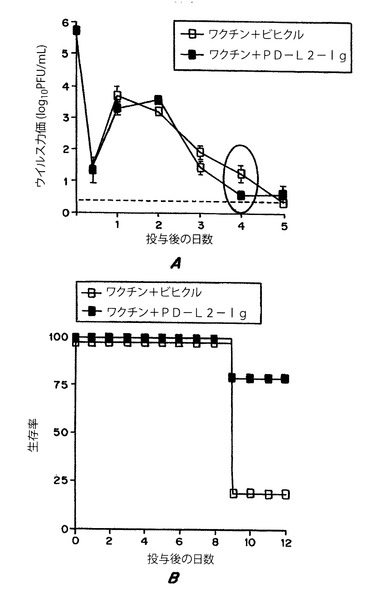
【図3】
【図4】
【図5】
【公表番号】特表2012−500855(P2012−500855A)
【公表日】平成24年1月12日(2012.1.12)
【国際特許分類】
【出願番号】特願2011−525158(P2011−525158)
【出願日】平成21年8月25日(2009.8.25)
【国際出願番号】PCT/US2009/054970
【国際公開番号】WO2010/098788
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(511049439)アンプリミューン、インコーポレーテッド (5)
【氏名又は名称原語表記】AMPLIMMUNE, INC.
【Fターム(参考)】
【公表日】平成24年1月12日(2012.1.12)
【国際特許分類】
【出願日】平成21年8月25日(2009.8.25)
【国際出願番号】PCT/US2009/054970
【国際公開番号】WO2010/098788
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(511049439)アンプリミューン、インコーポレーテッド (5)
【氏名又は名称原語表記】AMPLIMMUNE, INC.
【Fターム(参考)】
[ Back to top ]