
PDX−1発現神経内分泌腫瘍中のPDX−1オンコジーンを標的とする新規な治療的RNA干渉技術
標的細胞中におけるPDX−1オンコジーンの発現をノックダウンするための、二機能性shRNAに基づく組成物及び方法を本明細書中に記載する。本発明はまた、PDX−1オンコジーンを過剰発現する組織を標的とするshRNA含有発現ベクターを送達する方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般に、遺伝子を標的とするがん療法の分野に、より詳しくは、膵臓神経内分泌腫瘍の治療のための新規な治療法及び薬物送達系の開発に関する。
【背景技術】
【0002】
本発明の範囲を限定することなく、本発明の背景を、膵臓神経内分泌腫瘍の治療としての、PDX−1オンコジーンの発現を標的とするshRNAに基づく治療法の設計及び送達に関連して記載する。
【0003】
膵島新生物(islet neoplasia)は、膵臓神経内分泌腫瘍(NET)、インスリノーマ及びカルチノイドなどの、膵島細胞から生じる障害の総称である。膵島細胞腫瘍の5年生存率は35〜50%である(Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72、Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17)。膵島新生物は、比較的若年の患者(診断時年齢50才)に発症する(Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511、Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14)。インスリノーマ患者は特に、効果的な治療がない、コントロール不能な低血糖症に非常に苦しむ。低血糖症は、高インスリン血症によって起こり、神経低糖症の症状、例えば、頭痛、めまい感、嗜眠、複視、発作及び交感神経活性化と共に現れる(Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511)。外科手術が依然として最も効果的な治療法であり、効果的な術後補助療法はない。原発腫瘍及び転移の同時摘除が可能であるとはいえ、再発率は依然として高い。さらに、生存率は全身療法で改善されていない(House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9、Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67)。したがって、NETを治療するための新規で効果的な治療方法が必要である。
【0004】
膵臓十二指腸ホメオボックス−1(PDX−1,pancreatic duodenal homeobox-1)は、膵臓器官形成の胚転写因子であり、大部分のNET及び全ての膵島細胞インスリノーマにおいて過剰発現される。転写因子PDX−1遺伝子は、良性及び悪性インスリノーマにおけるインスリン分泌の制御に直接関与している(Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6、Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75、Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25)。これは、種々のがんでも認められ、増殖、分化及び移動と関係していると考えられ、NETの発生及び膵臓胚形成において主要な役割を担っている(Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6、Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75、Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25、Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6、Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075、Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225)。成人膵臓において、PDX−1はβ細胞の90%によって発現され、インスリン、グルコキナーゼ及び2型グルコーストランスポーター(GLUT2)の発現を制御することが示されている(Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31、Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502、Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93、Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20、Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51)。これらの遺伝子は、急性高血糖症及び慢性高血糖症のいずれにおいても、グルコースホメオスタシスの維持に重要な役割を担う。急性高血糖症においては、PDX−1はβ−細胞の細胞質から核に移動し、インスリンの産生及び分泌の増加の第1ステップとして、インスリン遺伝子を活性化する(Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206、Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8)。
【0005】
PDX−1は多くのがんにおいて過剰発現されることが、本発明者らなどによって示された(Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8、Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30、Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40、Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8)。特に、本発明者らは、PDX−1が80を超える膵臓がん及び膵島細胞新生物において過剰発現されることを発見した(Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego)。また、インスリノーマの100%においてPDX−1過剰発現が認められる。本発明者らは最近になって、PDX−1が膵臓がん及びインスリノーマ細胞株の増殖及び浸潤を制御することを実証した(Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20)。当業者ならば、がんにおけるPDX−1の過剰発現を調節する組成物及び方法が提供されれば、このような組成物及び方法は有益であり、当技術分野への多大な貢献といえることがわかるであろう。同様に、薬物送達技術に精通した者ならば、非標的器官を迂回し、PDX−1を過剰発現している標的器官への治療薬の送達を目標とする送達系が提供されれば、このような送達系は医療の実践において臨床的に有用であるといえることがわかるであろう。PDX−1を標的とすることによる膵臓腺癌の治療戦略は、Brunicardiに対して発行された米国特許第6,716,824号明細書(2004年)に示されている。この特許は、インスリン分泌細胞、例えば、β−細胞、PDX−1陽性ヒト膵管癌及び特定の転写因子を含有する他の細胞を選択的に標的とする、RIP−tk(ラットインスリンプロモーター-チミジンキナーゼ)コンストラクトのための組換え核酸に関する。Brunicardiの発明は、膵臓がん、例えば、β−細胞インスリノーマの治療に有用である。
【0006】
前臨床試験は、がん関連標的のサイレンシングにRNA干渉技術(RNAi)を使用できることを裏付けている(Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9、Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7、Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54、Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8、Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7、Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45、Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205、Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4、Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16、Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33、Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7)。インビボの研究もまた、腫瘍細胞の成長(Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7、Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7、Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300、Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84、Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71)、転移(Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9、Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62、Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6)、血管新生(Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22、Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70)及び化学療法抵抗性(Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9、Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84、Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63)に重要な成分のRNAi標的化によって有益な結果を示した。RNAi技術の適用には、2つの型の分子、化学的に合成された二本鎖低分子干渉RNA(siRNA)又はベクターに基づく低分子ヘアピン型RNA(shRNA)が最初に使用された。siRNA及びshRNAは同様なノックダウン機能を達成できるが、siRNAとshRNAとは本質的に異なる分子である。天然起源のマイクロRNA(miRNA)と同じ生物発生経路を共有するshRNAは、プロセシングされ、細胞質に輸送され、細胞質においてRNA干渉サイレンシング複合体(RISC)にロードされる(Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5)。内因性miRNAの一次転写産物は、核内でゲノムDNAからヘアピン構造を有する長いRNA鎖として合成され、それが、一連の内因性リボヌクレアーゼ(RNase)III酵素によってプロセシングされて、21〜23塩基対の成熟二重鎖miRNAとなる。少なくとも2つのリボヌクレアーゼIII酵素複合体が成熟プロセスに関与する。最初に、Drosha/DGCR8複合体が、プレmiRNAヘアピン構造を形成し、それが、核外輸送後にRLCに組み込まれる。RLCにおいて、第2の酵素、Dicerが、ループを切り取り、成熟miRNAを生じる(Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23)。これは、RNAiのためにRISCにロードされるエフェクター分子である(Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8)。
【0007】
アルゴノート(Ago,argonaute)ファミリーのタンパク質は一般に、細胞質RISCと関連する。これらのタンパク質は、siRNA又はmiRNAのロードに関与し、転写型(ヘテロクロマチンを標的とする)遺伝子サイレンシング及び転写後遺伝子サイレンシングにも関わる。Agoと複合体形成したRISCにロードされたsiRNA又はmiRNAのガイド鎖は、配列相補性を有する標的mRNAを探し出す。エンドヌクレアーゼ活性のあるAgo−2はmRNAを切断して、mRNA分解を開始する(Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60、Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6)。或いは、標的mRNAの3’UTRへの部分的な相補的結合が、プロセシングボディ(P−ボディ)中への隔離によって翻訳抑制(translational repression)を行う(Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25)。標的mRNAの不安定化をもたらす脱アデニル化が、P−ボディにおいて観察されている(Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24、Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46)。現在、多数の動物モデルにおいて、RNAi媒介遺伝子ノックダウンが強力であり、特異的であり、忍容性が高いという確たる証拠がある。これらの知見により、RNAi療法を臨床に移すための科学的論拠が得られる。現在のところ、少なくとも10種のRNAiに基づく薬が前期の相の臨床試験中であり((2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.)、そのうち4種はがんに関連している(Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15、Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73、Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7、Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84)。別のsiRNAもまた、動物モデルにおいて効力を示し(Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15、Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73、Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84)、ヒト以外の霊長類において安全性(Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21)を示した。
【0008】
米国特許出願公開第2009/0163431号明細書(Kazhdanら、2009年)は、PDX−1を阻害するための方法及び組成物を開示している。本発明の方法及び組成物に含まれている抗PDX−1剤は、抗体、アプタマー、アンチセンスオリゴヌクレオチド、リボザイム及び/又は阻害化合物を含む。本発明は、PDX−1をコードするRNA分子の特定の領域と非常に又は完全に相補的な有効量の抗PDX−1剤と腫瘍細胞を接触させるステップを含む、腫瘍細胞におけるPDX−1発現の阻害方法を提供する。このような作用剤は、PDX−1をコードするRNA分子と特異的にハイブリダイズし、腫瘍細胞におけるPDX−1遺伝子の発現を阻害する。抗PDX−1 siRNA及び/又はshRNAを含む組成物が記載されている。本発明による抗PDX−1 siRNA又はshRNAをコードする組換え型の発現コンストラクトが、記載されている。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許第6,716,824号明細書
【特許文献2】米国特許出願公開第2009/0163431号明細書
【非特許文献】
【0010】
【非特許文献1】Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72
【非特許文献2】Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17
【非特許文献3】Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511
【非特許文献4】Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14
【非特許文献5】House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9
【非特許文献6】Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67
【非特許文献7】Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6
【非特許文献8】Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75
【非特許文献9】Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2
【非特許文献10】Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25
【非特許文献11】Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6
【非特許文献12】Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075
【非特許文献13】Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352
【非特許文献14】Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225
【非特許文献15】Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31
【非特許文献16】Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502
【非特許文献17】Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93
【非特許文献18】Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20
【非特許文献19】Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51
【非特許文献20】Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206
【非特許文献21】Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8
【非特許文献22】Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8
【非特許文献23】Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30
【非特許文献24】Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40
【非特許文献25】Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8
【非特許文献26】Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego
【非特許文献27】Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20
【非特許文献28】Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9
【非特許文献29】Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7
【非特許文献30】Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54
【非特許文献31】Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8
【非特許文献32】Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7
【非特許文献33】Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45
【非特許文献34】Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205
【非特許文献35】Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4
【非特許文献36】Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16
【非特許文献37】Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33
【非特許文献38】Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7
【非特許文献39】Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7
【非特許文献40】Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300
【非特許文献41】Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84
【非特許文献42】Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71
【非特許文献43】Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9
【非特許文献44】Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62
【非特許文献45】Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6
【非特許文献46】Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22
【非特許文献47】Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70
【非特許文献48】Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9
【非特許文献49】Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84
【非特許文献50】Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63
【非特許文献51】Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5
【非特許文献52】Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23
【非特許文献53】Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8
【非特許文献54】Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60
【非特許文献55】Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6
【非特許文献56】Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25
【非特許文献57】Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24
【非特許文献58】Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46
【非特許文献59】(2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.
【非特許文献60】Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15
【非特許文献61】Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73
【非特許文献62】Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7
【非特許文献63】Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84
【非特許文献64】Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84
【非特許文献65】Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、標的の翻訳抑制及び転写後mRNA分解を同時に誘発するためのPDX bi−shRNAを含む。PDX bi−shRNAコンストラクトは、同じmRNA標的特異性を有するsiRNAと比較して、高い効力及び長期間にわたる活性を達成する。本発明者らは、PDX−1ノックダウンがアポトーシスを誘発し、増殖を停止し、NETがん細胞の侵襲性を低下させ、癌腫異種移植片を有するSCIDマウスの生存率を改善することを示した。本発明者らはまた、IV(静脈内)BIVリポソームPDX−1 shRNAの複数回処理が、SCIDマウスにおいてヒトPC異種移植片をほぼ完全に消失させたことを示し、マウスインスリノーマモデルにおいて応答及び延命効果を確認している。
【0012】
2層中空小胞(BIV,Bilamellar invaginated vesicle)は、一時的にマスクされた標的リポソームを使用して、細胞膜との融合による細胞への治療薬の送達を行う送達系である。これらのマスクされたステルスリポソーム(stealthed liposome)は、エンドサイトーシス経路を回避することによって、より高いトランスフェクション効率を達成する。Copernicusの凝縮核酸送達技術は、別の非ウイルス性の合成的モジュラープラットフォームであり、単一分子又はDNA若しくはRNAが、ポリカチオンによって凝縮されて、非常に高いトランスフェクション効率を有するナノ粒子が生じる。本発明者らは、マスクされたステルスBIVリポソーム中への凝縮shRNA含有DNAナノ粒子のパッケージングによる新規ながん治療法を開発した。
【課題を解決するための手段】
【0013】
本発明は、プロモーターと、プロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターであって、前記核酸インサートが、PDX−1転写因子をコードする転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする発現ベクターを含む。転写物へのshRNAの結合は、RNA干渉によってPDX−1発現を阻害する。shRNAは二機能性であって、siRNA(切断依存性)モチーフとmiRNA(切断非依存性)モチーフの両方が同時に組み込まれていてもよい。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。一態様において、1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態(modification)からなる群から選択される配列を含む。
【0014】
本発明はまた、shRNAをPDX−1mRNA標的に送達する系を提供する。この送達系は、プロモーターとプロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターにカップリングされているリポソームなどの治療薬担体を含み、該インサートは、PDX−1転写因子をコードする転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする。1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。転写物へのshRNAの結合は、RNA干渉によってPDX−1発現を阻害する。shRNAは二機能性であって、siRNA(切断依存性)モチーフとmiRNA(切断非依存性)モチーフの両方を組み込むことができる。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。治療薬担体は、凝縮されたDNAナノ粒子とすることができる。一実施形態において、発現ベクターのDNAは、ポリカチオンと組み合わせることによって凝縮できる。より詳しくは、DNAは、N末端システイン残基及び30リジン残基を含むペプチドで修飾された10kDポリエチレングリコール(PEG)であるCK30PEG10kと組み合わせることによって、凝縮できる。治療薬担体はリポソームであってもよく、その中で、DNAは、30merリジン縮合ペプチドと組み合わされることによって凝縮され得る。一実施形態において、リポソームは、2層中空小胞(BIV)とすることができる。リポソームは、「スマート」レセプター標的化部分で装飾することによって、標的組織を特異的に標的とすることができる。一例において、標的化部分は、低分子二価βターン模倣体とすることができる。リポソームは、非標的器官を迂回させるために、可逆的にマスクすることができる。一例において、可逆的マスキングは、リポソームをn−ドデシル−β−D−マルトピラノシドなどの低分子量脂質(分子量約500以下)でコーティングすることによって実施できる。本発明の別の実施形態において、凝縮DNAナノ粒子とリポソーム送達系とを組み合わせることができる。したがって、shRNA発現ベクターを含有する凝縮DNAナノ粒子を、リポソームに封入することができる。これらの凝縮DNA−ナノ粒子含有リポソームは、BIVとすることができる。リポソームは、標的化部分で装飾することができ、可逆的にマスクすることもできる。
【0015】
本発明の別の実施形態は、PDX−1を発現する標的組織にshRNAを送達する方法であって、(i)プロモーターと、PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAをコードする、プロモーターに作動可能に連結されている核酸のインサートとを含む発現ベクターを調製するステップと、(ii)前記発現ベクターを治療薬担体と組み合わせるステップと、(iii)治療有効量の発現ベクターと治療薬担体との複合体を、それを必要とする患者に投与するステップとを含む方法である。治療薬担体は、凝縮DNAナノ粒子とすることができる。一実施形態において、発現ベクターのDNAは、ポリカチオンと組み合わせることによって凝縮することができる。より詳しくは、DNAは、CK30PEG10kと組み合わせることによって凝縮することができる。治療薬担体は、リポソームであってもよい。一実施形態において、リポソームはBIVとすることができる。リポソームは、「スマート」レセプター標的化部分で装飾することによって、標的組織を特異的に標的とすることができる。一例において、標的化部分は、低分子二価βターン模倣体とすることができる。リポソームは、非標的器官を迂回させるために、可逆的にマスクすることができる。一例において、可逆的マスキングは、リポソームをn−ドデシル−D−マルトピラノシドなどの低分子量脂質(分子量約500以下)でコーティングすることによって実施できる。本発明の別の実施形態において、凝縮DNAナノ粒子とリポソーム送達系とを組み合わせることができる。したがって、shRNA発現ベクターを含有する凝縮DNAナノ粒子を、リポソームに封入することができる。これらの凝縮DNAナノ粒子含有リポソームは、BIVとすることができる。リポソームは、標的化部分で装飾することができ、可逆的にマスクすることもできる。一態様において、1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。
【0016】
本発明はまた、標的細胞中のPDX−1発現をサイレンシングする方法であって、(i)標的細胞を選択するステップと、(ii)PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAを発現するベクターを標的細胞にトランスフェクトするステップとを含み、がん細胞への前記ベクターのトランスフェクションがRNA干渉によってPDX−1発現を阻害する方法を提供する。shRNAは二機能性であって、切断依存性モチーフと切断非依存性モチーフの両方が同時に組み込まれていてもよい。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。
【0017】
本発明はまた、がん患者における腫瘍細胞成長の抑制方法であって、(i)腫瘍の治療を必要とする患者を特定するステップと、(ii)PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAを発現するベクターを前記腫瘍細胞にトランスフェクトするステップとを含み、前記ベクターがトランスフェクトされたがん細胞が、アポトーシス、増殖停止、又は侵襲性の低下をもたらす方法を含む。shRNAは、二機能性であって、切断依存性モチーフと切断非依存性モチーフの両方が同時に組み込まれていてもよく、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。より詳しくは、標的のがんは、膵臓神経内分泌腫瘍(NET)とすることができる。
【0018】
本発明の特徴及び利点のより完全な理解のために、添付図面と共に本発明の詳細な説明に言及する。
【図面の簡単な説明】
【0019】
【図1A−1B】bi−shRNASTMN1ベクターのインビボ成長阻害活性を示すグラフである。図1Aは、CCL−247腫瘍異種移植片に対して、bi−shRNASTMN1リポプレックス(pGBI2)若しくはスクランブル対照リポプレックス(pGBI5)又は懸濁液(D5W)の単回腫瘍内注射を行った場合に観察される阻害を示す。値は平均±SEMを表す。図1Bは、SCIDマウスにおける原発骨肉腫異種移植片に対するbi−shRNASTMN1の成長阻害活性を示す。bi−shRNASTMN1リポプレックス又は対照の連日腫瘍内注射を6回行った。
【図2】bi−sh−STMN1とsiRNASTMN1とを比較する、STMN1mRNA標的切断と細胞成長阻害との相関を示すグラフである。bi−shRNASTMN1(赤色)及びsiRNASTMN1(黄色)の複合用量反応曲線は、STMN1mRNA切断と相関した。x軸は、左から右へと増加するプラスミド/siRNA濃度の用量である。y軸は、24時間処理後のパーセント細胞生存率である。各データ点は、三重反復試料の平均を表す。bi−shRNASTMN1の濃度範囲は、1.44×10−12M〜5.63×10−15Mであった。siRNASTMN1の濃度範囲は、5×10−7M〜1×10−10Mであった。電気泳動図挿入部分は、トランスフェクトされたCCL−247細胞から検出される5’RACE産物を示し、これは切断生成物を示している。
【図3A−3B】過剰発現されたヒト(図3A)及びマウス(図3B)PDX1mRNA(配列番号1及び2)に対する二機能性shRNAの効果を示すグラフである。結腸がんCCL−247細胞に、PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又はPDX−1 cDNA及びbi−shRNA発現ベクターを同時トランスフェクトした。pUMVC3は、インサートを含まない発現ベクター対照(配列番号9)である。トランスフェクションの24,48,72及び96時間後に、qRT−PCRのために全RNAを単離した。PDX−1mRNAレベルは、GAPDH mRNAに基準化した。各試料の比較PDX−1mRNAレベルは、培地のみの試料(トランスフェクションなし)に規準化した。
【図4】ヒトPDX−1(配列番号1)に対する二機能性shRNAのサイレンシング効果を示す図である。結腸がんCCL−247細胞に、種特異的PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又は種特異的PDX−1 cDNAとbisRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない対照発現ベクターである。トランスフェクションの48、72及び96時間後に、細胞を収集し、PDX−1及びβ−アクチンに特異なモノクローナル抗体によるウエスタン免疫ブロットを行った。
【図5A−5B】RACE−PCRによるヒト(図5A)PDX1及びマウス(図5B)PDX1(配列番号1及び2)の標的特異的切断の同定を示す図である。結腸がんCCL−247細胞に、PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又はPDX−1 cDNAとbi−shRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない発現ベクター対照である。トランスフェクションの24及び96時間後に、5’RACEアッセイのために全RNAを単離して、標的部位切断産物を検出した。RT−PCRで増幅された切断産物が、アガロースゲル電気泳動図上に示された。
【図6A−6C】CATレポーター遺伝子産生に対するマスキング剤の効果を示すグラフである。図6Aは肺に対する効果を示し、図6Bは心臓に対する効果を示し、図6Cは肝臓に対する効果を示す。BALB/cマウスへの静脈内注射後に、マスキングの増加につれて、肺(図6A)及び心臓(図6B)における非特異的局在化の減少が見られる。マスキングは、目的とする標的部位へのCATレポーター遺伝子の局在化の増加と関連する(肝臓;図6C)。
【図7A−7C】凝縮DNAナノ粒子の構造及び機能を示す図とグラフである。図7Aは、楕円体又は棒状として凝縮されたDNAナノ粒子の電子顕微鏡写真を示す。バーは、100nmである。図7Bは、対数期の又は成長が停止した神経芽細胞腫細胞に、裸のDNA又は凝縮DNAのリポソーム混合物を加えたことを示している。凝縮DNAは、遺伝子発現を、有糸分裂後細胞で1000倍超、対数期細胞で6〜8倍改善する。図7Cは、IV投与によって又は腫瘍内(IT)注射として投与された、結腸がん隣接外植片(colon cancer flank explant)を有するSCIDマウスにおける、凝縮DNA遺伝子導入(gene transfer)を示す。IT注射は、高レベルのルシフェラーゼ活性を生じたが、IV投与は生じなかった。
【図8】トランスフェクションの48時間後におけるPDX−1タンパク質発現の減少に対する配列特異性を示す図である。結腸がんCCL−247細胞に、種特異的PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又は種特異的PDX−1 cDNAとbisRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない対照発現ベクターである。トランスフェクションの48時間後に、細胞を収集して、PDX−1及びβ−アクチンに特異的なモノクローナル抗体によるウエスタン免疫ブロットを行った。
【図9A−9B】インビボにおけるbi−shRNA−PDX−1リポプレックスによる処理後のSCIDマウス異種移植片モデル:β−TC−6細胞インスリノーマモデル(図9A)及びPANC−1膵臓がんモデル(図9B)の血糖値を経時的に示すグラフである。SCIDマウスに腫瘍細胞を移植後に2週間、bi−shRNA−PDX−1リポプレックス50μgによる処理を3サイクル行った。処理及び非処理マウスの空腹時血糖値を、処理後90日まで一定時間間隔で監視した。
【図10A−10B】PANC−1膵臓がんモデル(図10A)及びβ−TC−6細胞インスリノーマモデル(図10B)のカプラン−マイヤー(Kaplan-Meier)生存率分析を示すグラフである。腫瘍細胞を移植したSCIDマウスは、処理を行わないか、又は対照ベクターリポプレックス(pUMVC3)若しくはbi−shRNA−PDX−1リポプレックスで処理する(30μg/サイクルで3サイクル)かのいずれかとした。処理後90日まで、マウスを生存について監視した。
【図11】ヒトインスリノーマ腫瘍及びマウスTC−6インスリノーマ細胞におけるPDX−1の過剰発現率を示す図である。
【図12】mPDX−1 RNAサイレンシング後のβ−TC−6細胞周期タンパク質発現率を示すグラフである。
【図13】マウスインスリノーマSCIDモデルにおける、L−mPDX−1 shRNA処理されたマウスの生存率を示すグラフである。
【図14】腫瘍体積の比較データを示すグラフである。
【図15】本発明の新規bi−shRNAPDX−1を製造するためのプラスミドコンストラクトを示す図である。
【発明を実施するための形態】
【0020】
本発明の種々の実施形態の製造及び使用を以下に詳細に記載するが、本発明は、種々の具体的状況で具体化し得る多くの応用可能な発明概念を提供するものと理解すべきである。本明細書中に記載した具体的な実施形態は、本発明の具体的な製造及び使用方法の単なる実例であって、本発明の範囲の限界を定めるものではない。
【0021】
本発明の理解を容易にするために、多くの用語を以下に定義する。本明細書中で定義する用語は、本発明に関連する分野の当業者によって一般に理解される意味を有する。「a」、「an」及び「the」などの用語は、単一の構成要素を指すだけでなく、その一般的種類を含み、その具体例を説明するために使用できるものとする。本明細書中の専門用語を使用して、本発明の具体的な実施形態を説明するが、それらの使用は、特許請求の範囲に概説される場合を除いて、本発明の範囲の限界を定めるものではない。
【0022】
本明細書中で使用する「核酸」又は「核酸分子」という用語は、ポリヌクレオチド、例えば、デオキシリボ核酸(DNA)又はリボ核酸(RNA)、オリゴヌクレオチド、ポリメラーゼ連鎖反応(PCR)によって生成されるフラグメント、並びにライゲーション、切断、エンドヌクレアーゼ作用及びエキソヌクレアーゼ作用のいずれかによって生成されるフラグメントを指す。核酸分子は、天然起源のヌクレオチドであるモノマー(例えば、DNA及びRNA)、若しくは天然起源のヌクレオチドのアナログ(例えば、天然起源のヌクレオチドのα−エナンチオマー型)、又は両方の組合せから構成され得る。修飾ヌクレオチドは、糖部分及び/又はピリミジン若しくはプリン塩基部分が変更されているものとすることができる。糖修飾形態は、例えば、ハロゲン、アルキル基、アミン及びアジド基による、1若しくは2以上のヒドロキシル基の置換を含み、又は糖はエーテル若しくはエステルとして機能化されていてもよい。さらに、糖部分全体が、立体配置的及び電子的に類似した構造、例えば、アザ糖及び炭素環式糖アナログと置き換えられることもできる。塩基部分の修飾形態の例としては、アルキル化プリン及びピリミジン、アシル化プリン若しくはピリミジン又は他の周知の複素環代替物が挙げられる。核酸モノマーは、ホスホジエステル結合又はこのような結合のアナログによって連結され得る。ホスホジエステル結合のアナログとしては、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニリデート、ホスホルアミデートなどが挙げられる。「核酸分子」という用語はまた、ポリアミド主鎖に天然起源又は修飾核酸塩基が付着している、いわゆる「ペプチド核酸」を含む。核酸は一本鎖でも、二重鎖でもよい。
【0023】
本明細書及び特許請求の範囲中で使用する「発現ベクター」という用語は、宿主細胞で発現される遺伝子をコードする核酸分子を含む。典型的には、発現ベクターは、転写プロモーター、遺伝子及び転写ターミネーターを含む。遺伝子発現は通常、プロモーターにコントロールされ、このような遺伝子は、プロモーターに「作動可能に連結されている」と言われる。同様に、制御エレメントがコアプロモーターの活性を調節する場合には、制御エレメントとコアプロモーターとは、作動可能に連結されている。「プロモーター」という用語は、宿主酵母細胞中で構造遺伝子と関連する場合に、適切な成長条件下で1)転写、2)翻訳又は3)mRNA安定性の1又は2以上を、プロモーター配列の非存在下での転写、翻訳又はmRNA安定性(mRNAのより長い半減期)と比較して増加させる、その構造遺伝子に関する任意のDNA配列を指す。
【0024】
本明細書中で使用する「オンコジーン」という用語は、悪性新生物細胞の形成及び生存を可能にする遺伝子を指す(Bradshaw, T.K.: Mutagenesis 1, 91-97(1986)。
【0025】
本明細書中で使用する「レセプター」という用語は、「リガンド」と称される生物活性分子と結合する細胞関連タンパク質(cell-associated protein)を意味する。この相互作用が、細胞に対するリガンドの効果を媒介する。レセプターは、膜結合レセプター、細胞質レセプター又は核内レセプター;モノマーレセプター(例えば、甲状腺刺激ホルモンレセプター、βアドレナリン作動性レセプター)又は多量体レセプター(例えば、PDGFレセプター、成長ホルモンレセプター、IL−3レセプター、GM−CSFレセプター、G−CSFレセプター、エリスロポエチンレセプター及びIL−6レセプター)とすることができる。膜結合型レセプターは、細胞外リガンド結合ドメイン及び細胞内エフェクタードメインを含むマルチドメイン構造を特徴とし、シグナル変換に典型的に関与する。特定の膜結合レセプターにおいては、細胞外リガンド結合ドメインと細胞内エフェクタードメインとは、完全な機能性レセプターを含む別個のポリペプチド中に位置する。
【0026】
「ハイブリダイズする」という用語は、核酸の鎖が塩基対形成によって相補鎖と結合する任意のプロセスを指す。
【0027】
「トランスフェクション」という用語は、真核細胞中への外来DNAの導入を指す。トランスフェクションは、当技術分野で知られている種々の手段、例えば、リン酸カルシウム−DNA共沈、DEAE−デキストラン媒介トランスフェクション、ポリブレン媒介トランスフェクション、エレクトロポレーション、マイクロインジェクション、リポソーム融合、リポフェクション、プロトプラスト融合、レトロウイルス感染及び微粒子銃によって行うことができる。
【0028】
本明細書中で使用する「リポソーム」という用語は、内部水性空間を取り囲む脂質二重層からなる閉鎖構造を指す。本明細書中で使用する「ポリカチオン」という用語は、同一分子内に第4級アンモニウムラジカルなどの複数のカチオン部分を有する物質を意味し、遊離塩基及びそれらの薬学的に許容される塩を含む。
【0029】
本発明は、標的の翻訳抑制と転写後mRNA分解とを同時に誘発するために開発されたPDX bi−shRNAを含む。プロトタイプのコンストラクトは、同じmRNA標的特異性を有するsiRNAと比較して高い効力及び長期間にわたる活性を達成する。本発明者らは、PDX−1ノックダウンがNETがん細胞のアポトーシスを誘発し、増殖を停止し、侵襲性を低下させ、癌腫異種移植片を有するSCIDマウスの生存率を改善することを示している。本発明者はまた、IV(静脈内)BIVリポソームPDX−1 shRNAの複数の処理が、SCIDマウスにおいてヒトPC異種移植片をほぼ完全に消失させたことを示し、マウスインスリノーマモデルにおいて応答及び延命効果を確認している。
【0030】
bi−shRNA:本発明者らは、二機能性shRNAとして知られている第3の独特のRNAiプラットフォームを開発した。概念的には、shRNAがロードされたRISCによってRNAiを達成して、切断依存性又は切断非依存性mRNAノックダウンを促進できる。両立体配置のshRNAの同時発現(それゆえに、二機能性shRNAと命名されている)は、siRNAと比較して、サイレンシングのより急速な開始時に、より効果的な標的遺伝子ノックダウンを達成する(mRNA及びタンパク質の代謝回転率にもかかわらず)と共に、耐久性が大きいことを本発明者らは示した。二機能性shRNA発現単位の基本設計は、2つのステムループshRNA構造を含む。これらのステムループの1つは、切断依存性RISCロードのための完全にマッチしたパッセンジャー鎖及びガイド鎖から構成され、もう1つのステムループは、切断非依存性RISCロードのためのミスマッチパッセンジャー鎖(9〜12位)を有する。この二機能性設計は、第1に、二機能性設計は、異なるRISC型へのガイド鎖のロードを促進し、したがって、mRNAターゲティングを促進する;第2に、同一標的mRNAに対して切断依存性RISC及び切断非依存性RISCが存在することにより、分解と翻訳阻害/隔離プロセスの両方によってサイレンシングが促進されるという2つの理由で、はるかに効率的である。強力な遺伝子ノックダウンエフェクターは、単一Pol IIプロモーターのコントロール下で多重化shRNAによって空間的及び時間的コントロールを達成する。本発明者らが設計したプラットフォームは、天然のプロセスを模倣している。他の研究者による多数の研究及び文献は、本発明者らのアプローチを裏付けている(Okamura, K., et al., Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev, 2004. 18(14): pp. 1655-66、Miyoshi, K., et al., Slicer function of Drosophila Argonautes and its involvement in RISC formation. Genes Dev, 2005. 19(23): pp. 2837-48、Tomari, Y., T. Du, and P.D. Zamore, Sorting of Drosophila small silencing RNAs. Cell, 2007. 130(2): pp. 299-308、Forstemann, K., et al., Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell, 2007. 130(2): pp. 287-97、Landthaler, M., et al., Molecular characterization of human Argonaute-containing ribonucleoprotein complexes and their bound target mRNAs. RNA, 2008. 14(12): pp. 2580-96)。
【0031】
本発明者らは、miR30骨格を用いて、オンコプロテインであるスタスミン(stathmin)(STMN1、オンコプロテイン18、プロソリン(prosolin)、p19、op18)を再構築する微小管に対する新規な二機能性(bi−shRNA)を調製した。bi−shRNASTMN1は、同じ標的部位に対して、siRNAと比較して、より効果的なサイレンシング活性を示した。STMN1は、有糸分裂紡錘体形成に大きく関与する(Rana, S., et al., Stathmin 1: a novel therapeutic target for anticancer activity. Expert Rev Anticancer Ther, 2008. 8(9): pp. 1461-70、Iancu, C., et al., Effects of stathmin inhibition on the mitotic spindle. J Cell Sci, 2001. 114(Pt 5): pp. 909-16)。本発明のbi−shRNASTMN1コンストラクトは、同じ標的に対するsiRNAと比較した場合に、安全で効果的な標的ノックダウン及び腫瘍細胞死滅における有意な用量有利性を示している。さらに、インビボにおいてbi−shRNASTMN1リポプレックスによって、有意な腫瘍成長コントロール及び延命効果が示された(図1A及び1B)。本発明者らは、マッチパッセンジャー鎖とミスマッチパッセンジャー鎖とを区別できるRT−PCR方法を使用して、成熟分子及びエフェクター分子(完全マッチ鎖を有するdsRNA及び特定のミスマッチを有するdsRNA)の両方の細胞内転写及びプロセシングを検証した(Chen, C., et al., Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res, 2005. 33(20): pp. e179)。ほとんどのがん細胞には、高いDrosha及びDicer発現が見られる。がん細胞の内因性Dicerレベルに関しては議論がある(Merritt, W.M., et al., Dicer, Drosha, and outcomes in patients with ovarian cancer. N Engl J Med, 2008. 359(25): pp. 2641-50)。しかし、大部分の研究は、Dicer発現が低レベルであっても、sh又はbi−shRNAiノックダウンはがん細胞において非常に効果的であることを示している(Senzer, N., D. Rao, and J. Nemunaitis, Letter to the Editor: Does Dicer expression affect shRNA processing? Gene Regulation and Systems Biology (in press))。本発明者らは、対照siRNASTMN1処理細胞中では完全マッチパッセンジャー鎖のみがより合わされるのとは対照的に、bi−shRNASTMN1に対する成熟miRNA/siRNA成分に対応する、予測されるマッチ及びミスマッチshRNAの発現を確認した(Rao, D.R., Maples, P.B., Senzer, N., Kumar, P., Wang, Z., Pappen, B.O., Yu, Y., Haddock, C., Tong, A., Nemunaitis, J., Bifunctional shRNA: A novel approach of RNA interference. (submitted), 2009)。bi−shRNASTMN1の機構をさらに裏付けるために、5’RACE法による本発明の研究の成果から、bi−shRNASTMN1のsiRNA(マッチ)成分の標的切断部位に相当すると予想される配列を有するSTMN1切断産物の存在が確認された。STMN1発現腫瘍細胞の効果的なノックダウン(93%)が観察された。これは、bi−shRNASTMN1の切断依存性及び非依存性媒介ノックダウンの結果を反映している。さらに、bi−shRNASTMN1(GBI−2)と比較して、別個の成分である切断依存性(GBI−1)及び切断非依存性(GBI−3)ベクターによるノックダウン後に観察されるSTMN1mRNA動態は、予測される機構と一致していた。
【0032】
bi−shRNASTMN1ベクターのエフェクター機構及び機能的効力を実証後、本発明者らは、インビトロでの抗がん活性をさらに探求し、いくつかのがん細胞株において効果的な細胞死滅の実証に成功した。同じ配列を標的とするにもかかわらず、bi−shRNASTMN1はsiRNASTMN1よりも5log低いモル濃度でIC50を達成した(図2)。
【0033】
インスリノーマは、最も一般的な型の膵島細胞腫瘍である。悪性インスリノーマは、重度の形態の高インスリン血症であり、コントロールできない低血糖症につながる。前述のように、膵臓十二指腸ホメオボックス−1(PDX−1)は、ホメオドメイン含有転写因子ファミリーに属し、膵臓器官形成において主要な役割を担う。PDX−1は、インスリン、グルコキナーゼ及び2型グルコーストランスポーターの転写を制御することによって、β細胞機能を維持する。PDX−1は、インスリノーマにおいて過剰発現され、高インスリン血症をもたらす。PDX−1は、膵臓腫瘍において一般に過剰発現されることも認められている。転移性膵臓がんは、診断からの生存期間が4〜6カ月である。
【0034】
マウスインスリノーマモデルにおけるshRNAノックダウンPDX−1:インスリノーマにおけるPDX−1の発現を実証するために、Dr.Brunicardiの研究室は、PDX−1に対する強力な抗体を開発した。本発明者らは、この抗体を用いて、PDX−1がマウスインスリノーマにおいてヒトインスリノーマと同様なレベルまで顕著に過剰発現されることを実証した(図11)。本発明者らは、マウスモデルにおけるノックダウン効果を試験するために、マウス(mu)shRNAを調製した。しかし、インスリノーマのサイズが増加するので、コントロールできない低血糖症によりこれらのマウスの寿命は2〜3カ月であるため、研究は制限される。最初の研究においては、インビトロにおいてβTC−6細胞をmu−shPDX−1 shRNAで処理し、MTSアッセイを行って、細胞生存率を評価した。空ベクターと比較して、mu−shPDX−1処理細胞は、複数の時点にわたって細胞増殖のかなりの減少を示した。本発明者らは、DNA合成がノックダウンによって複数の時点において有意に減少することをさらに実証し、PDX−1ノックダウンに応答するいくつかの細胞周期関連タンパク質のウエスタンブロットによってノックダウンを実証した(図12)。
【0035】
PDX−1発現のサイレンシングは、腫瘍成長を阻害するための魅力的な方法といえる。本発明者らは、PDX−1遺伝子発現のサイレンシングのための二機能性shRNAを設計した。ヒトPDX−1(配列番号1)又はマウスPDX−1(配列番号1)のいずれかに対するMiR30をベースとする二機能性shRNAカセットを、pUMVC3ベクター中にクローニングした。図15は、新規bi−shRNAPDX−1を製造するためのプラスミドコンストラクトを示す。マウス又はヒトPDX−1を標的とする二機能性shRNAを、マウス又はヒトPDX−1のいずれかを発現する発現ベクターと共に、ヒトの結腸がん細胞株に同時にエレクトロポレートした。RACE−PCRを用いて、ヒト及びマウスPDX1mRNAの、可能性のある切断産物を調べた。RT−QPCR及び免疫ブロット法を使用して、ヒトPDX−1又はマウスPDX−1のいずれかの発現のノックダウンを調べた。
【0036】
ヒト膵臓十二指腸ホメオボックス1(PDX1)mRNA(配列番号1):
【0037】
【0038】
前記配列において、コード領域はボールドイタリック体で示され、第1及び第2標的部位はそれぞれ、下線及び太字で示されている。
【0039】
マウス膵臓十二指腸ホメオボックス1(PDX1)mRNA(配列番号2):
【0040】
【0041】
前記配列において、コード領域は太字のイタリック体で示され、標的部位は下線で示されている。
【0042】
図3A及び3Bはそれぞれ、強制的に過剰発現されたヒト及びマウスPDX1に対する二機能性shRNAの効果を示す。トランスフェクションの24時間後にヒトPDX−1に対する二機能性shRNAのサイレンシング効果が観察され、少なくとも96時間持続した(図4)。最大のサイレンシング効果、ヒトPDX−1の80%ノックダウンは、トランスフェクションの72時間後に達成された。さらに、マウスPDX−1を標的とする二機能性shRNA(ヒトの配列との相同性が68%である)は、ヒトPDX−1の発現に影響を及ぼさなかった(図8)。同様に、マウスPDX−1の発現は、トランスフェクションの24時間後にサイレンシングを受け、このサイレンシング効果は少なくとも96時間持続した(図4)。最大のサイレンシング効果、マウスPDX−1発現の95%が、トランスフェクションの48時間後に観察された。さらに、ヒトPDX−1を標的とする二機能性shRNA(マウスの配列との配列相同性が79%)もまた、マウスPDX−1の発現を変化させなかった(図8)。
【0043】
RACE−PCRによって同定されたヒト及びマウスPDX−1mRNAの切断産物はそれぞれ、図5A及び5Bに示されている。図5A及び5Bは、ヒトPDX1mRNA及びマウスPDX1mRNAがいずれも、予測された標的領域の中央で正確に切断されることを示した。加えて、ヒトPDX−1を標的とする二機能性shRNAは、マウスPDX−1mRNAの切断を引き起こさず、逆の場合も同じであった。
【0044】
前述の研究は、ヒト又はマウスPDX1のいずれかを標的とする二機能性shRNAの効力及び種特異性を実証した。bi−shRNAhPDX1は、ヒトPDX1の発現をノックダウンしたが、マウスPDX1の発現に対するサイレンシング効果を示さなかった。同様に、bi−shRNAmPDX1はマウスPDX1の発現をノックダウンしたが、ヒトPDX1の発現に対するサイレンシング効果を示さなかった。
【0045】
リポソーム送達系:本発明者らが以前に検証したリポソーム送達系は、1,2−ジオレイル−3−トリメチル-アンモニオプロパン(DOTAP)及びコレステロールを含んでいた(Ruponen M, H.P., Ronkko S, Pelkonen J, Tammi M, Urtti A, Extracellular and intracellular barriers in non-viral gene delivery J Control Release, 2003. 93(2): pp. 213-217)。この配合物は、DNAと組み合わされて、2層中空小胞(リポソームBIV)内に核酸を封入する複合体を形成する。本発明者らのうちの一人は、RNA、DNA及びRNAiプラスミドの送達を改善するために、BIV送達系のいくつかの特徴を最適化した。リポソームBIVは融合性であるので、核酸分解(Simberg D, W.A., Barenholz Y, Reversible mode of binding of serum proteins to DOTAP/cholesterol Lipoplexes: a possible explanation for intravenous lipofection efficiency. Hum Gene Ther, 2005. 16(9): pp. 1087-1096)及びTLR媒介オフターゲット効果をもたらし得るエンドサイトーシス媒介DNA細胞移入を回避する。このリポソーム送達系は、本発明者らなどが臨床試験での使用に成功した(Jay, C., et al., Preclinical Assessment of wt GNE Gene Plasmid for Management of Hereditary Inclusion Body Myopathy 2 (HIBM2). Gene Regulation & Systems Biology, 2008. 2: pp. 243-52、Frank O, R.C., Heberlein C, Neuhoff Nv, Schrock E, Schambach A, et al., Tumor cells escape suicide gene therapy by genetic and epigenetic instability. Blood 2004. 104(12): pp. 3543-3539、Ito I, J.L., Tanaka F, Saito Y, Gopalan B, Branch CD, Xu K, Atkinson EN, Bekele BN, Stephens LC, Minna JD, Roth JA, Ramesh R Liposomal vector mediated delivery of the 3p FUS1 gene demonstrates potent antitumor activity against human lung cancer in vivo. Cancer Gene Ther, 2004. 11(11): pp. 733-9、Templeton, N.S., Reversible masking of liposomal complexes for targeted delivery, USPTO, Editor. 2006, Baylor College of Medicine: USA、Phadke, A.P., et al., Safety and in vivo expression of a GNE-transgene: A novel treatment approach for Hereditary Inclusion Body Myopathy-2. Gene Regulation & Systems Biology, 2009. 3: pp. 89-101)。過去10年にわたる累積的な研究は、血清アルブミンなどの負電荷を持つ血清タンパク質への非特異的結合(オプソニン化)を最小化する(Tong, A.W., et al., Systemic therapeutic gene delivery for cancer: crafting paris' arrow. Curr Gene Ther, 2009. 9(1): pp. 45-60)ためには、最適化送達媒体が、効率的な血管内輸送のためにゼータ電位10mV以下のステルス化(一般にPEG化によって達成される)ナノ粒子である必要がある(Pirollo, K.F. and E.H. Chang, Targeted delivery of small interfering RNA: approaching effective cancer therapies. Cancer Res, 2008. 68(5): pp. 1247-50、Khalil, I.A., et al., Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev, 2006. 58(1): pp. 32-45、Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9)ことを示している。標的化部分、例えば、抗体及びそれらの単鎖誘導体(scFv)、糖又はペプチドの組込みは、導入遺伝子の標的細胞への局在化をさらに増加できる。
【0046】
本発明者らは、インビボでのPEGを用いない複合体の標的化送達を創出し、それによって、過度に長期の循環半減期を回避した(Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9、Simoes, S., et al., Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands or pH-sensitive peptides. Gene Ther, 1999. 6(11): pp. 1798-807、Simoes, S., et al., Cationic liposomes for gene delivery. Expert Opin Drug Deliv, 2005. 2(2): pp. 237-54、Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81)。PEG化は、膜透過性を改善するDNA又はsiRNAオリゴヌクレオチド送達に関連するが、本発明者らは、この方法はBIVリポソーム構造に立体障害をもたらし、結果としてDNA封入が非効率的になり、遺伝子発現が減少することを示した。さらにまた、PEG化された複合体は、主にエンドサイトーシス経路を通って細胞に入り、リソソーム中で核酸の大部分を分解する。PEGは循環において極めて長い半減期を示すが、これは、細胞毒性薬ドキソルビシンを封入するPEG化リポソーム製剤ドキシルによって例証されるように、患者に対して問題を引き起こした(Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81、Sapra, P., B., et al., Ligand-targeted liposomes for cancer treatmentLiposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. Curr Drug DelivJ Clin Oncol, 20051995. 213(47): pp. 369-811777-85、Uziely, B., et al., Liposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. J Clin Oncol, 1995. 13(7): pp. 1777-85)。特異的細胞表面レセプター(例えば、HER2/neu)への送達のためにドキシルにリガンドを付加する試みは、腫瘍特異的送達を増加しなかった(Park, J.W., et al., Tumor targeting using anti-her2 immunoliposomes. J Control Release, 2001. 74(1-3): pp. 95-113)。
【0047】
この推論に基づき、本発明のBIVは、DOTAP及び合成コレステロールを用い、専有の手動押出プロセスを使用して調製した(Bartlett, D.W., N. S., et al., Impact of tumor-specific targetingImproved DNA: liposome complexes for increased systemic delivery and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticlesgene expression. Nat Biotechnol Bioeng, 20081997. 9915(47): pp. 975-85647-52)。さらにまた、可逆的マスキング技術を使用して送達を最適化した。可逆的マスキングは、荷電してないためにBIV複合体の表面と緩く結合する低分子量脂質(分子量約500以下;例えば、n−ドデシル−β−D−マルトピラノシド)を使用し、結果として正電荷を持つBIV複合体を一時的に遮蔽して、非標的化器官を迂回する。これらの小脂質は、血流において剪断力によって除去される。それらが標的細胞に到達するまでに、電荷が再曝露されて(最適には約45mV)、移入が促進される。図6Aは、BALB/cマウスにおけるCAT−封入プラスミドを封入しているBIVを用いた、静脈内投与による肺のトランスフェクションの最適化を、図6Bは、静脈内投与による心臓のトランスフェクションの最適化を示す。逆マスキング複合体が全身的に循環する場合には、ゼータ電位アナライザーで測定される電荷が4.8mVであるにもかかわらず、トランスフェクションは最小であった。マスキングがBIVから複合体を可逆的に解離するので、これらの複合体は本来の荷電状態(45.5mV)に戻り、結果として肝臓の肝細胞において膜融合による細胞移入及び高レベルの導入遺伝子(CAT)発現を促進した(図6C)。「ジェネリック」BIV複合体を使用して、導入遺伝子の発現を100倍超増加させ、ほぼ一致したCATレベルを肺で生じさせた。可逆的マスキングを使用して、他の器官ではCAT産生は観察されなかった。
【0048】
本発明のBIV送達系が他に類を見ないほど効率的な理由は、1つには、複合体が治療薬を細胞膜との融合によって細胞中に送達し、エンドサイトーシス経路を回避するためである。リポソーム移入の2つの主要な移入機構は、エンドサイトーシス又は細胞膜との直接融合による。マウス肺胞マクロファージにおけるポリエチレン−アミン(PEI)を用いた比較研究によって示されるように、本発明者らは、インビトロ及びインビボの両方において送達されるBIV複合体に封入された核酸が、直接融合によって細胞に移入すること、並びにBIVが主にエンドソーム取込みを回避することを発見した。トランスフェクション後2〜3時間以内におけるローダミン標識オリゴヌクレオチドの95%以上の局在化によって示される(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40、Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40、Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54)ように、PEIは、エンドソーム中に急速且つ旺盛に取り込まれることが知られている。
【0049】
装飾BIVを用いたがん標的化送達:最近になって、Bartlett及びDavisは、腫瘍を標的とするナノ粒子(NP)によって全身に送達されるsiRNAが、皮下腫瘍の成長阻害において、非装飾NPと比較して有意に効果的であることを示した(Bartlett, D.W. and M.E. Davis, Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol Bioeng, 2008. 99(4): pp. 975-85)。標的化送達は、薬物動態又は生体内分布に大きく影響を与えず、これは依然として大部分がEPR(血管透過性−滞留性亢進)効果の結果であった(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40)が、細胞取込みの増加によって導入遺伝子発現を改善するようであった(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40、Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40、Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54)。
【0050】
実のところ、治療用BIVの開発において重要な「欠けている部分(missing piece)」は、BIV−複合体の表面に位置してそれらを標的細胞に誘導することができるような非免疫原性リガンドの同定であった。これは、リポソーム表面で多量体化される小ペプチドによって行える可能性もあるが、小ペプチドは、反復注射後に免疫応答を発生する可能性がある。抗体、抗体フラグメント、タンパク質、部分タンパク質などを含む比較的大きい他のリガンドは、リポソーム表面における標的化送達に関しては、小ペプチドを使用する場合よりもはるかに扱いにくい。よって、本発明の複合体は、腫瘍脈管構造の内皮孔及び固形腫瘍の間質圧勾配を含む厳重なバリアを透過する(Ramesh, R., et al., Successful treatment of primary and disseminated human lung cancers by systemic delivery of tumor suppressor genes using an improved liposome vector. Mol Ther, 2001. 3(3): pp. 337-50)だけでなく、腫瘍細胞を直接的に標的とする限りにおいて、独特である。したがって、本発明の治療方法は、EPR効果にのみ依存する送達に限定されず、腫瘍を直接的に標的とする(Hashizume, H., et al., Openings between defective endothelial cells explain tumor vessel leakiness. Am J Pathol, 2000. 156(4): pp. 1363-80、Netti, P.A., et al., Effect of transvascular fluid exchange on pressure-flow relationship in tumors: a proposed mechanism for tumor blood flow heterogeneity. Microvasc Res, 1996. 52(1): pp. 27-46、Yuan, F., et al., Vascular permeability and microcirculation of gliomas and mammary carcinomas transplanted in rat and mouse cranial windows. Cancer Res, 1994. 54(17): pp. 4564-8)。
【0051】
本発明には、タンパク質を選択的に結合するように設計された小分子を使用できる。重要なことに、調製される小分子は「二価」であるので、細胞表面レセプターを結合するのに特に適し、タンパク質−リガンド相互作用においてホットスポットに見られる二次構造モチーフに類似している。Burgessのグループは、細胞表面レセプターに対して親和性を有する二価βターン模倣体の設計に成功した(Feng, Y., et al., Solid-phase SN2 macrocyclization reactions to form beta-turn mimics. Org Lett, 1999. 1(1): pp. 121-4、Bruno, M.A., et al., Long-lasting rescue of age-associated deficits in cognition and the CNS cholinergic phenotype by a partial agonist peptidomimetic ligand of TrkA. J Neurosci, 2004. 24(37): pp. 8009-18、Zaccaro, M.C., et al., Selective small molecule peptidomimetic ligands of TrkC and TrkA receptors afford discrete or complete neurotrophic activities. Chem Biol, 2005. 12(9): pp. 1015-28)。本発明者らは、糖尾部を有する二価分子を生成するようにこの戦略を適合させ、これらの適合小分子から機能化BIV複合体を調製した。ライブラリーをスクリーニングする効率的なハイスループット技術を開発し、実施した。
【0052】
凝縮DNAナノ粒子:有糸分裂後細胞における安全で効率的なDNA送達:Copernicusの核酸送達技術は、非ウイルス性の合成的モジュラープラットフォームであり、DNA又はsiRNAの単一分子がポリカチオンによって凝縮されて、最小可能体積を有するナノ粒子を生成する(Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86)。インビボ送達のために最適化されたポリカチオンは、N末端システイン残基及び30リジン残基を含むペプチドで修飾された10kDaポリエチレングリコール(PEG)(CK30PEG10k)である。これらの複合体の形状は、DNA凝縮時のリジン対イオンにある程度左右される(Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51)。棒状ナノ粒子の最小断面直径は、ペイロードプラスミドの寸法に関係なく8〜11nmであるが、楕円体の最小直径は、典型的な発現プラスミドの場合、20〜22nmである(図7A)(Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51)。重要なことに、これらのDNAナノ粒子は、培養中の非分裂細胞に大量にトランスフェクトすることができる。凝縮DNAのリポソーム混合物は、リポソームの裸のDNA混合物と比較して、遺伝子発現レベルを1,000倍超増加させる(図7B)。インビボ投与後、凝縮DNAは、肺(Ziady, A.G., et al., Transfection of airway epithelium by stable PEGylated poly-L-lysine DNA nanoparticles in vivo. Mol Ther, 2003. 8(6): pp. 936-47)、脳(Yurek, D.M., et al., Long-term transgene expression in the central nervous system using DNA nanoparticles. Mol Ther, 2009. 17(4): pp. 641-50、Yurek, D.M., et al., Compacted DNA nanoparticle gene transfer of GDNF to the rat striatum enhances the survival of grafted fetal dopamine neurons. Cell Transplant, 2009)及び眼(Farjo, R., et al., Efficient non-viral ocular gene transfer with compacted DNA nanoparticles. PLoS One, 2006. 1: pp. e38、Cai, X., et al., A partial structural and functional rescue of a retinitis pigmentosa model with compacted DNA nanoparticles. PLoS One, 2009. 4(4): pp. e5290)において有糸分裂後細胞に大量にトランスフェクトする。これらの各系において、有糸分裂後細胞にトランスフェクトする凝縮DNAの注目すべき能力は、これらのナノ粒子のサイズが小さく、ナノ粒子が25nmの核膜細孔を通過できることによると考えられる(Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86)。
【0053】
これらのDNAナノ粒子に関する1つの取込み機構は、細胞表面ヌクレオリン(26nm KD)との結合に基づく。DNAナノ粒子は、その後の細胞質輸送によって非分解経路を経て核に入り、核内でほどけて、生物活性DNAを放出する(Chen, X., et al., Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Mol Ther, 2008. 16(2): pp. 333-42)。遺伝子導入後1年にもわたる長期インビボ発現が実証された。これらのナノ粒子は、無害の毒性プロフィールを有し、トール様レセプターを刺激しないので毒性サイトカイン応答を回避し、凝縮DNAが何百ものCpGアイランドを有し、リポソームと混合される場合であっても、毒性作用は観察されていない(Ziady, A.G., et al., Minimal toxicity of stabilized compacted DNA nanoparticles in the murine lung. Mol Ther, 2003. 8(6): pp. 948-56、Ding, X.-Q., et al., Ocular delivery of compacted DNA-nanoparticles does not elicit toxicity in the mouse retina. Plos One, 2009. In press)。DNAナノ粒子が、嚢胞性線維症試験においてヒトに投与されたところ、結果は期待の持てるものであり、ナノ粒子に起因する有害事象なく、ほとんどの患者はCFTRタンパク質の生物活性を示した(Konstan, M.W., et al., Compacted DNA nanoparticles administered to the nasal mucosa of cystic fibrosis subjects are safe and demonstrate partial to complete cystic fibrosis transmembrane regulator reconstitution. Hum Gene Ther, 2004. 15(12): pp. 1255-69)。
【0054】
本発明者らとCopernicus(Cleaveland、OH)との初期の共同研究において、結腸がん腫瘍外植片を有するSCIDマウスに、尾静脈又は腫瘍内(IT)注射によって凝縮DNAを投与した(図7C)。IV注射は遺伝子導入をもたらさなかったが、IT投与後には著しく高いレベルの遺伝子導入が認められた。これは、これらのDNAナノ粒子が、局所送達後における腫瘍へのトランスフェクトに高い活性を有することを示している。
【0055】
新規送達系:原発腫瘍及び転移部位への全身的遺伝子導入が望ましいので、凝縮DNAの腫瘍への送達は、30merリジン縮合ペプチドと組み合わせることによって凝縮されたDNAからなるこれらのナノ粒子を、有標BIVリポソーム中にパッケージングすることによって促進できる。凝縮DNAとマスクされたステルスリポソームとのこの新規なカップリングは、新規ながん遺伝子治療法といえる。
【0056】
前述のように、膵臓がん細胞株PANC−1におけるRNA干渉によるPDX−1発現のサイレンシングは、インビトロでは細胞増殖を阻害し、インビボでは腫瘍成長を抑制すると共に腫瘍細胞アポトーシスを増加させた。この戦略に基づき、本発明者らは、インスリノーマ、膵臓がん及び神経内分泌腫瘍の治療への臨床応用のための二機能性shRNA(bi−shRNA−PDX−1)を開発した。さらに、本発明者らは、インビトロでの効力並びにインスリノーマ/SCIDモデル及びPANC−1/SCID膵臓腫瘍モデルを用いたインビボでの効力を実証する。
【0057】
インビトロでのノックダウン効力は、PDX−1及びbi−shRNA発現ベクターの同時トランスフェクションによる、PDX−1陰性ヒトがん細胞株(HCT116)におけるPDX−1発現のウエスタン免疫ブロット及びqRT−PCR分析によって評価した(図3A、3B、4及び8)。動物モデルは、インビボインスリノーマモデルに関しては1×106個のβ−TC−6細胞の、又はPCモデルに関しては5×105個のPANC−1細胞の腹腔内(IP)注射によって調製した。接種2週間後に、マウスを、各モデルについて無作為に2群に分け;尾静脈注射によって、1つの群には35μgの対照ベクター(shRNAインサートを含まないプラスミド)を含有するリポプレックス(DOTAP:コレステロールリポソームプラスミド複合体)を、第2の群にはマウス特異的bi−msh−PDX−1(インスリノーマモデルの場合)又はヒト特異的bi−hsh−PDX−1(PCモデルの場合)を35μg含有するリポプレックスを与えた。2つの反復送達を、2週間毎に合計3回の注射で行った。血中インスリン値及び血糖値を、定期的な時間間隔で監視した。3回目の注射の2、10及び18週間後に、病理評価及び腫瘍評価のために、各群6匹のマウスを屠殺した。
【0058】
HCT116ヒトがん細胞に、プラスミドを1:1の比率でエレクトロポレートした。PDX−1mRNAを、qRT−PCRを用いてΔΔCt法によって定量的に比較した(図3A及び3B)。mRNAレベルを空ベクター対照と比較して、表1に要約する%発現率を得た。PDX−1mRNAは、トランスフェクションの24、48及び72時間後に分析した。
【0059】
【表1】
【0060】
ヒト及びマウス特異的bi−shRNA−PDX−1は、インビトロにおいてPDX−1mRNA発現の効果的で種特異的なノックダウンを示す。
【0061】
絶食マウスの血糖値を、最初の2サイクルの各処理の7日後、45日目及び90日目に測定した。マウスインスリノーマモデルにおいて、mbi−shRNA−PDX−1による処理は、β−TC−6細胞によって誘発された低血糖症からマウスを効果的にレスキューした(図9A)が、hbi−shRNA−PDX−1は、膵臓がんモデルの場合に血糖値に対して効果がなかった(図9B)。β−TC−6インスリノーマモデルにおいて、未処理マウスは全て、90日目の測定の前に死亡した。PANC−1膵臓がんモデルの結果を、図9Bに示す。
【0062】
カプラン−マイヤー生存率分析は、PANC−1モデル(図10A)及びβ−TC−6インスリノーマモデル(図10B)に対するbi−shRNA−PDX−1処理の有益性をさらに示す。インスリノーマモデルについては、処理群のマウス(mbi−shRNA−PDX−1リポプレックス各30μgで3サイクル処理)10匹及び対照群のマウス(bi−shRNAインサートを含まないpUMVC3ベクターを含有するリポプレックス各30μgで3サイクル処理)10匹を、生存について監視した。生存は95日間監視した。PCモデルについては、PANC−1接種マウス5匹は処理せず、マウス10匹は、bi−shRNAインサートを含まないpUMVC3ベクターを含有するリポプレックス各30μgで3サイクル処理し、マウス15匹は、hbi−shRNA−PDX−1リポプレックス各30μgで3サイクル処理した。腫瘍体積の比較のために、各群5匹のマウスを、60及び90日目に屠殺した。処理の60日後に、非処理マウスは100%が平均1330.75mm3のサイズの腫瘍を発現したが、処理マウスは50%のみが、平均50.5mm3のサイズの目視可能な腫瘍を有していた。処理の90日後に、処理マウスの37.5%は目視可能な腫瘍を有さず、残りのマウスは平均サイズ36.7mm3の腫瘍を有していた。腫瘍体積の比較研究の結果を、表2及び図14に示す。
【0063】
生存率は、SCIDマウスでも、BIVリポソーム送達されたshRNAPDX−1(L−mu−shRNAPDX−1)のIV注入後に向上し、L−mu−shRNAPDX−1注射の回数と相関するようであった(図13)。リポプレックス(即ち、インビボ研究のために使用される脂質−プラスミド複合体)を、ベクターそのものと区別するために接頭語「L−」で表す。インスリン値の減少及び血糖値の増加は、インスリノーマ成長阻害というL−mu−shRNAPDX−1処理の結果に対応する。L−mu−shPDX−1による処理後の動物の腫瘍応答は、フローサイトメトリーによるインスリノーマ細胞のPDX−1ノックダウンの検出並びに細胞周期タンパク質(サイクリンE、cdk4、cdk2)のノックダウン及びサイクリン依存性キナーゼ阻害因子(p27)の上方制御と相関した。アポトーシスも示された。本発明者らはさらに、PDX−1発現性ヒト膵臓がんマウス異種移植片モデルにおいて、shRNAPDX−1リポプレックスのIV注入の用量依存的な延命効果をさらに実証した。
【0064】
【表2】
【0065】
新規bi−shRNAPDX−1:二機能性shRNA戦略を適用するために、本発明者らは、マウス(mu)−bi−shRNAPDX−1を調製し、検証した。mu−bi−shRNAPDX−1は、miR−30a主鎖を有する2つのステムループ構造からなる。第1のステムループ構造は、完全相補的なガイド鎖及びパッセンジャー鎖を有し、第2のステムループ構造は、パッセンジャー鎖の10及び11位に2つの塩基対ミスマッチを有する。mu−bi−shRNAPDX−1は、マウスPDX−1mRNA(NM_008814)のヌクレオチド#723〜#741を標的とする。
【0066】
mu−bi−shRNAPDX−1発現単位を、オリゴヌクレオチドの組合せと共に構築した。5’及び3’末端それぞれにSalI及びNotI部位を有する構築発現単位を、pUMVC3のSalI及びNotI部位に挿入した。インサートに隣接するプライマーで両方向から配列決定することによって、インサート配列を確認した。マイクロRNAの検出のために開発されたステムループRT−PCR法を用いて、mu−bi−shRNAPDX−1をトランスフェクトした細胞中の成熟shRNAを検出した。本発明者らはさらに、5’RACEアッセイによってmu−bi−shRNAPDX−1の活性を検証して、標的部位切断を検出した。
【0067】
本発明のbi−shRNAプラスミドのインサート配列を以下に示す。
【0068】
pGBI-20:ヒトbi−shRNA−PDX1(配列番号3):
【0069】
TCGACTGCTGTTGAAGTGAGCGCCAGTTCCTATTCAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCAGTTCCTATCTAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0070】
配列番号3の下線部分(25〜43の核酸残渣を含む)は、主要標的部位を表す。
【0071】
pGBI-21:ヒトbi−shRNA−PDX−1(配列番号4):
【0072】
TCGACTGCTGTTGAAGTGAGCGCCAGTTCCTATTCAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0073】
pGBI-22:ヒトbi−shRNA−PDX−1(配列番号5):
【0074】
TCGACTGCTGTTGAAGTGAGCGCCCAGTTATTTACAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCCAGTTATTTCTAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0075】
pGBI-23:ヒトbi−shRNA−PDX−1(配列番号6):
【0076】
TCGACTGCTGTTGAAGTGAGCGCCCAGTTATTTACAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0077】
pGBI-24:マウスbi−shRNA−PDX−1(配列番号7):
【0078】
TCGACTGCTGTTGAAGTGAGCGCCGGAAGATAAGAAACGTAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCGGAAGATAAACAACATAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0079】
配列番号4の下線部分(25〜43の核酸残渣を含む)は、主要標的部位を表す。
【0080】
pGBI-25:マウスbi−shRNA−PDX−1(配列番号8):
【0081】
TCGACTGCTGTTGAAGTGAGCGCCGGAAGATAAGAAACGTAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0082】
本発明の新規bi−shRNA治療薬の調製は、転写後の遺伝子ノックダウンによって効果的な治療結果を達成するのに必要な有効全身用量を減少させることができる最先端の方法といえる。PDX−1過剰発現腫瘍の全身標的化へと迅速に移行できる効果的で臨床的に応用可能な送達法が整っている。
【0083】
本明細書中に記載した実施形態はいずれも、本発明の任意の方法、キット、試薬又は組成物に関して実施可能であり、逆の場合も同様であると考えられる。さらにまた、本発明の組成物は、本発明の方法の実施に使用できる。
【0084】
本明細書中に記載した特定の実施形態は一例として示すのであって、本発明を限定するものとして示すのではないことがわかるであろう。この発明の主要な特徴は、本発明の範囲から逸脱せずに、種々の実施形態で実施可能である。当業者ならば、常法にすぎない実験を使用すれば、本明細書中に記載した具体的な方法の多数の均等物を認識、又は確認できるであろう。このような均等物を、本発明の範囲内であるとみなし、特許請求の範囲によって保護する。
【0085】
本明細書中に記載した全ての公表文献及び特許出願は、本発明が関連する技術分野の当業者の技術レベルを示す。全ての公表文献及び特許出願は、個々の公表文献又は特許出願が参照によって明確且つ個別に組み込まれていることが示された場合と同程度まで、参照によって本明細書中に組み入れる。
【0086】
単語「a」又は「an」の使用は、特許請求の範囲及び/又は明細書中で用語「含む(comprising)」と共に使用する場合、「1の」を意味することができるが、「1又は2以上の」、「少なくとも1の」及び「1又は1超の」の意味とも矛盾しない。特許請求の範囲における用語「又は(or)」の使用は、選択肢のみを指すことが明確に示されているか又は選択肢が相互排他的である場合を除いて、「及び/又は(and/or)」を意味して使われるものであるが、本開示は選択肢のみ及び「及び/又は」を指す定義も支持する。本出願の全体を通じて、「約」という用語は、値が、その値の測定に用いられた装置、方法の誤差の固有変動、又は研究対象間に存在する変動を含むことを示すのに用いる。
【0087】
本明細書及び特許請求の範囲中で使用する「含む(comprising)」(並びにcomprisingの任意の形、例えば、「comprise」及び「comprises」)、「有する(having)」(並びにhavingの任意の形、例えば、「have」及び「has」)、「含む(including)」(並びにincludingの任意の形、例えば、「includes」及び「include」)又は「含有する(containing)」(並びにcontainingの任意の形、例えば、「contains」及び「contain」)という用語は、包括的又は非限定的であり、列挙していない追加の要素又は方法のステップを排除しない。
【0088】
本明細書中で使用する「又はそれらの組合せ」という用語は、この用語の前に記載された用語の全ての順列及び組合せを指す。例えば、「A、B、C又はそれらの組合せ」は、A、B、C、AB、AC、BC又はABCの少なくとも1つを含むことを意図し、特定の状況で順序が重要である場合には、BA、CA、CB、CBA、BCA、ACB、BAC又はCABも含むことを意図する。この例に続いて、1又は2以上のアイテム又は用語の反復、例えば、BB、AAA、MB、BBC、AAABCCCC、CBBAAA、CABABBなどを含む組合せも明白に含まれる。当業者は、状況から明らかでない限り、典型的には、任意の組合せ中のアイテム又は用語の数に制限がないことがわかるであろう。
【0089】
本明細書中で使用する近似の単語、例えば、限定するものではないが、「約」、「実質的な」又は「実質的に」が指す状態は、それによって修飾される場合に、必ずしも絶対的又は完全でないと理解されるが、その状態が存在しているとするに値する、当業者にとっては十分に近いとみなされるであろう状態を指す。記載が変動し得る程度は、可能な変化の大きさと、修飾された特徴が修飾されていない特徴に必須の特性及び能力を依然として有すると当業者が認めることができる変化の大きさによるであろう。しかし、一般に、前記説明を受けて、「約」などの近似の単語で修飾された本明細書中の数値は、記載された値から少なくとも±1、2、3、4、5、6、7、10、12又は15%変動してもよい。
【0090】
本明細書中に開示され且つ特許請求の範囲に記載された組成物及び/又は方法は全て、必要以上の実験を行わずに本開示を踏まえて製造及び実施可能である。本発明の組成物及び方法を好ましい実施形態に関して記載したが、本発明の概念、精神及び範囲から逸脱せずに、本明細書中に記載した組成物及び/又は方法に対して、並びに方法のステップ又は一連のステップにおいて、変形形態が適用されてもよいことは、当業者には明白であろう。当業者に明白なこのような類似した置換形態及び変更形態は全て、添付した特許請求の範囲によって定義される本発明の精神、範囲及び概念の範囲内とみなされる。
【0091】
(参考文献)
U.S.Patent Application No. 2009/0163431: Compositions and Methods for Modulation of PDX-1
U.S.Patent No. 6,716,824: Treatment of Pancreatic Adenocarcinoma by Cytotoxic Gene Therapy
1. Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72
2. Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17
3. Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511
4. Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14
5. House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9
6. Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67
7. Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6
8. Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75
9. Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2
10. Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25
11. Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6
12. Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075
13. Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352
14. Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225
15. Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31
16. Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502
17. Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93
18. Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20
19. Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51
20. Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206
21. Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8
22. Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8
23. Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30
24. Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40
25. Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8
26. Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego
27. Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20
28. Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9
29. Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7
30. Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54
31. Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8
32. Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7
33. Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45
34. Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205
35. Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4
36. Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16
37. Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33
38. Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7
39. Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7
40. Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300
41. Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84
42. Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71
43. Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9
44. Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62
45. Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6
46. Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22
47. Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70
48. Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9
49. Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84
50. Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63
51. Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5
52. Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23
53. Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8
54. Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60
55. Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6
56. Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25
57. Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24
58. Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46
59. (2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.
60. Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15
61. Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73
62. Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7
63. Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84
64. Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84
65. Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21
66. Okamura, K., et al., Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev, 2004. 18(14): pp. 1655-66
67. Miyoshi, K., et al., Slicer function of Drosophila Argonautes and its involvement in RISC formation. Genes Dev, 2005. 19(23): pp. 2837-48
68. Tomari, Y., T. Du, and P.D. Zamore, Sorting of Drosophila small silencing RNAs. Cell, 2007. 130(2): pp. 299-308
69. Forstemann, K., et al., Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell, 2007. 130(2): pp. 287-97
70. Landthaler, M., et al., Molecular characterization of human Argonaute-containing ribonucleoprotein complexes and their bound target mRNAs. RNA, 2008. 14(12): pp. 2580-96
71. Rana, S., et al., Stathmin 1: a novel therapeutic target for anticancer activity. Expert Rev Anticancer Ther, 2008. 8(9): pp. 1461-70
72. Iancu, C., et al., Effects of stathmin inhibition on the mitotic spindle. J Cell Sci, 2001. 114(Pt 5): pp. 909-16
73. Chen, C., et al., Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res, 2005. 33(20): pp. e179
74. Merritt, W.M., et al., Dicer, Drosha, and outcomes in patients with ovarian cancer. N EnglJ Med, 2008. 359(25): pp. 2641-50
75. Senzer, N., D. Rao, and J. Nemunaitis, Letter to the Editor: Does Dicer expression affect shRNA processing? Gene Regulation and Systems Biology (in press)
76. Rao, D.R., Maples, P.B., Senzer, N., Kumar, P., Wang, Z., Pappen, B.O., Yu, Y., Haddock, C., Tong, A., Nemunaitis, J., Bifunctional shRNA: A novel approach of RNA interference. (submitted), 2009
77. Ruponen M, H.P., Ronkko S, Pelkonen J, Tammi M, Urtti A, Extracellular and intracellular barriers in non-viral gene delivery J Control Release, 2003. 93(2): pp. 213-217
78. Simberg D, W.A., Barenholz Y, Reversible mode of binding of serum proteins to DOTAP/cholesterol Lipoplexes: a possible explanation for intravenous lipofection efficiency. Hum Gene Ther, 2005. 16(9): pp. 1087-1096
79. Jay, C., et al., Preclinical Assessment of wt GNE Gene Plasmid for Management of Hereditary Inclusion Body Myopathy 2 (HIBM2). Gene Regulation & Systems Biology, 2008. 2: pp. 243-52
80. Frank O, R.C., Heberlein C, Neuhoff Nv, Schrock E, Schambach A, et al., Tumor cells escape suicide gene therapy by genetic and epigenetic instability. Blood 2004. 104(12): pp. 3543-3539
81. Ito I, J.L., Tanaka F, Saito Y, Gopalan B, Branch CD, Xu K, Atkinson EN, Bekele BN, Stephens LC, Minna JD, Roth JA, Ramesh R Liposomal vector mediated delivery of the 3p FUS1 gene demonstrates potent antitumor activity against human lung cancer in vivo. Cancer Gene Ther, 2004. 11(11): pp. 733-9
82. Templeton, N.S., Reversible masking of liposomal complexes for targeted delivery, USPTO, Editor. 2006, Baylor Collegeof Medicine: USA
83. Phadke, A.P., et al., Safety and in vivo expression of a GNE-transgene: A novel treatment approach for Hereditary Inclusion Body Myopathy-2. Gene Regulation & Systems Biology, 2009. 3: pp. 89-101
84. Pirollo, K.F. and E.H. Chang, Targeted delivery of small interfering RNA: approaching effective cancer therapies. Cancer Res, 2008. 68(5): pp. 1247-50
85. Khalil, I.A., et al., Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev, 2006. 58(1): pp. 32-45
86. Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9
87. Tong, A.W., et al., Systemic therapeutic gene delivery for cancer: crafting paris' arrow. Curr Gene Ther, 2009. 9(1): pp. 45-60
88. Simoes, S., et al., Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands or pH-sensitive peptides. Gene Ther, 1999. 6(11): pp. 1798-807
89. Simoes, S., et al., Cationic liposomes for gene delivery. Expert Opin Drug Deliv, 2005. 2(2): pp. 237-54
90. Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81
91. Sapra, P., B., et al., Ligand-targeted liposomes for cancer treatmentLiposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. Curr Drug DelivJ Clin Oncol, 20051995. 213(47): pp. 369-811777-85
92. Uziely, B., et al., Liposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. J Clin Oncol, 1995. 13(7): pp. 1777-85
93. Park, J.W., et al., Tumor targeting using anti-her2 immunoliposomes. J Control Release, 2001. 74(1-3): pp. 95-113
94. Bartlett, D.W., N. S., et al., Impact of tumor-specific targetingImproved DNA: liposome complexes for increased systemic delivery and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticlesgene expression. Nat Biotechnol Bioeng, 20081997. 9915(47): pp. 975-85647-52
95. Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40
96. Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40
97. Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54
98. Bartlett, D.W. and M.E. Davis, Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol Bioeng, 2008. 99(4): pp. 975-85
99. Ramesh, R., et al., Successful treatment of primary and disseminated human lung cancers by systemic delivery of tumor suppressor genes using an improved liposome vector. Mol Ther, 2001. 3(3): pp. 337-50
100. Hashizume, H., et al., Openings between defective endothelial cells explain tumor vessel leakiness. Am J Pathol, 2000. 156(4): pp. 1363-80
101. Netti, P.A., et al., Effect of transvascular fluid exchange on pressure-flow relationship in tumors: a proposed mechanism for tumor blood flow heterogeneity. Microvasc Res, 1996. 52(1): pp. 27-46
102. Yuan, F., et al., Vascular permeability and microcirculation of gliomas and mammary carcinomas transplanted in rat and mouse cranial windows. Cancer Res, 1994. 54(17): pp. 4564-8
103. Feng, Y., et al., Solid-phase SN2 macrocyclization reactions to form beta-turn mimics. Org Lett, 1999. 1(1): pp. 121-4
104. Bruno, M.A., et al., Long-lasting rescue of age-associated deficits in cognition and the CNS cholinergic phenotype by a partial agonist peptidomimetic ligand of TrkA. J Neurosci, 2004. 24(37): pp. 8009-18
105. Zaccaro, M.C., et al., Selective small molecule peptidomimetic ligands of TrkC and TrkA receptors afford discrete or complete neurotrophic activities. Chem Biol, 2005. 12(9): pp. 1015-28
106. Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86
107. Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51
108. Ziady, A.G., et al., Transfection of airway epithelium by stable PEGylated poly-L-lysine DNA nanoparticles in vivo. Mol Ther, 2003. 8(6): pp. 936-47
109. Yurek, D.M., et al., Long-term transgene expression in the central nervous system using DNA nanoparticles. Mol Ther, 2009. 17(4): pp. 641-50
110. Yurek, D.M., et al., Compacted DNA nanoparticle gene transfer of GDNF to the rat striatum enhances the survival of grafted fetal dopamine neurons. Cell Transplant, 2009
111. Farjo, R., et al., Efficient non-viral ocular gene transfer with compacted DNA nanoparticles. PLoS One, 2006. 1: pp. e38
112. Cai, X., et al., A partial structural and functional rescue of a retinitis pigmentosa model with compacted DNA nanoparticles. PLoS One, 2009. 4(4): pp. e5290
113. Chen, X., et al., Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Mol Ther, 2008. 16(2): pp. 333-42
114. Ziady, A.G., et al., Minimal toxicity of stabilized compacted DNA nanoparticles in the murine lung. Mol Ther, 2003. 8(6): pp. 948-56
115. Ding, X.-Q., et al., Ocular delivery of compacted DNA-nanoparticles does not elicit toxicity in the mouse retina. Plos One, 2009. In press
116. Konstan, M.W., et al., Compacted DNA nanoparticles administered to the nasal mucosa of cystic fibrosis subjects are safe and demonstrate partial to complete cystic fibrosis transmembrane regulator reconstitution. Hum Gene Ther, 2004. 15(12): pp. 1255-69
【図1A】
【図1B】
【技術分野】
【0001】
本発明は、一般に、遺伝子を標的とするがん療法の分野に、より詳しくは、膵臓神経内分泌腫瘍の治療のための新規な治療法及び薬物送達系の開発に関する。
【背景技術】
【0002】
本発明の範囲を限定することなく、本発明の背景を、膵臓神経内分泌腫瘍の治療としての、PDX−1オンコジーンの発現を標的とするshRNAに基づく治療法の設計及び送達に関連して記載する。
【0003】
膵島新生物(islet neoplasia)は、膵臓神経内分泌腫瘍(NET)、インスリノーマ及びカルチノイドなどの、膵島細胞から生じる障害の総称である。膵島細胞腫瘍の5年生存率は35〜50%である(Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72、Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17)。膵島新生物は、比較的若年の患者(診断時年齢50才)に発症する(Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511、Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14)。インスリノーマ患者は特に、効果的な治療がない、コントロール不能な低血糖症に非常に苦しむ。低血糖症は、高インスリン血症によって起こり、神経低糖症の症状、例えば、頭痛、めまい感、嗜眠、複視、発作及び交感神経活性化と共に現れる(Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511)。外科手術が依然として最も効果的な治療法であり、効果的な術後補助療法はない。原発腫瘍及び転移の同時摘除が可能であるとはいえ、再発率は依然として高い。さらに、生存率は全身療法で改善されていない(House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9、Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67)。したがって、NETを治療するための新規で効果的な治療方法が必要である。
【0004】
膵臓十二指腸ホメオボックス−1(PDX−1,pancreatic duodenal homeobox-1)は、膵臓器官形成の胚転写因子であり、大部分のNET及び全ての膵島細胞インスリノーマにおいて過剰発現される。転写因子PDX−1遺伝子は、良性及び悪性インスリノーマにおけるインスリン分泌の制御に直接関与している(Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6、Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75、Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25)。これは、種々のがんでも認められ、増殖、分化及び移動と関係していると考えられ、NETの発生及び膵臓胚形成において主要な役割を担っている(Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6、Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75、Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25、Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6、Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075、Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352、Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225)。成人膵臓において、PDX−1はβ細胞の90%によって発現され、インスリン、グルコキナーゼ及び2型グルコーストランスポーター(GLUT2)の発現を制御することが示されている(Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31、Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502、Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93、Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20、Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51)。これらの遺伝子は、急性高血糖症及び慢性高血糖症のいずれにおいても、グルコースホメオスタシスの維持に重要な役割を担う。急性高血糖症においては、PDX−1はβ−細胞の細胞質から核に移動し、インスリンの産生及び分泌の増加の第1ステップとして、インスリン遺伝子を活性化する(Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206、Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8)。
【0005】
PDX−1は多くのがんにおいて過剰発現されることが、本発明者らなどによって示された(Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8、Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30、Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40、Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8)。特に、本発明者らは、PDX−1が80を超える膵臓がん及び膵島細胞新生物において過剰発現されることを発見した(Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego)。また、インスリノーマの100%においてPDX−1過剰発現が認められる。本発明者らは最近になって、PDX−1が膵臓がん及びインスリノーマ細胞株の増殖及び浸潤を制御することを実証した(Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20)。当業者ならば、がんにおけるPDX−1の過剰発現を調節する組成物及び方法が提供されれば、このような組成物及び方法は有益であり、当技術分野への多大な貢献といえることがわかるであろう。同様に、薬物送達技術に精通した者ならば、非標的器官を迂回し、PDX−1を過剰発現している標的器官への治療薬の送達を目標とする送達系が提供されれば、このような送達系は医療の実践において臨床的に有用であるといえることがわかるであろう。PDX−1を標的とすることによる膵臓腺癌の治療戦略は、Brunicardiに対して発行された米国特許第6,716,824号明細書(2004年)に示されている。この特許は、インスリン分泌細胞、例えば、β−細胞、PDX−1陽性ヒト膵管癌及び特定の転写因子を含有する他の細胞を選択的に標的とする、RIP−tk(ラットインスリンプロモーター-チミジンキナーゼ)コンストラクトのための組換え核酸に関する。Brunicardiの発明は、膵臓がん、例えば、β−細胞インスリノーマの治療に有用である。
【0006】
前臨床試験は、がん関連標的のサイレンシングにRNA干渉技術(RNAi)を使用できることを裏付けている(Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9、Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7、Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54、Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8、Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7、Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45、Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205、Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4、Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16、Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33、Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7)。インビボの研究もまた、腫瘍細胞の成長(Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7、Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7、Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300、Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84、Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71)、転移(Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9、Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62、Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6)、血管新生(Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22、Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70)及び化学療法抵抗性(Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9、Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84、Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63)に重要な成分のRNAi標的化によって有益な結果を示した。RNAi技術の適用には、2つの型の分子、化学的に合成された二本鎖低分子干渉RNA(siRNA)又はベクターに基づく低分子ヘアピン型RNA(shRNA)が最初に使用された。siRNA及びshRNAは同様なノックダウン機能を達成できるが、siRNAとshRNAとは本質的に異なる分子である。天然起源のマイクロRNA(miRNA)と同じ生物発生経路を共有するshRNAは、プロセシングされ、細胞質に輸送され、細胞質においてRNA干渉サイレンシング複合体(RISC)にロードされる(Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5)。内因性miRNAの一次転写産物は、核内でゲノムDNAからヘアピン構造を有する長いRNA鎖として合成され、それが、一連の内因性リボヌクレアーゼ(RNase)III酵素によってプロセシングされて、21〜23塩基対の成熟二重鎖miRNAとなる。少なくとも2つのリボヌクレアーゼIII酵素複合体が成熟プロセスに関与する。最初に、Drosha/DGCR8複合体が、プレmiRNAヘアピン構造を形成し、それが、核外輸送後にRLCに組み込まれる。RLCにおいて、第2の酵素、Dicerが、ループを切り取り、成熟miRNAを生じる(Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23)。これは、RNAiのためにRISCにロードされるエフェクター分子である(Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8)。
【0007】
アルゴノート(Ago,argonaute)ファミリーのタンパク質は一般に、細胞質RISCと関連する。これらのタンパク質は、siRNA又はmiRNAのロードに関与し、転写型(ヘテロクロマチンを標的とする)遺伝子サイレンシング及び転写後遺伝子サイレンシングにも関わる。Agoと複合体形成したRISCにロードされたsiRNA又はmiRNAのガイド鎖は、配列相補性を有する標的mRNAを探し出す。エンドヌクレアーゼ活性のあるAgo−2はmRNAを切断して、mRNA分解を開始する(Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60、Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6)。或いは、標的mRNAの3’UTRへの部分的な相補的結合が、プロセシングボディ(P−ボディ)中への隔離によって翻訳抑制(translational repression)を行う(Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25)。標的mRNAの不安定化をもたらす脱アデニル化が、P−ボディにおいて観察されている(Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24、Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46)。現在、多数の動物モデルにおいて、RNAi媒介遺伝子ノックダウンが強力であり、特異的であり、忍容性が高いという確たる証拠がある。これらの知見により、RNAi療法を臨床に移すための科学的論拠が得られる。現在のところ、少なくとも10種のRNAiに基づく薬が前期の相の臨床試験中であり((2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.)、そのうち4種はがんに関連している(Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15、Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73、Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7、Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84)。別のsiRNAもまた、動物モデルにおいて効力を示し(Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15、Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73、Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84)、ヒト以外の霊長類において安全性(Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21)を示した。
【0008】
米国特許出願公開第2009/0163431号明細書(Kazhdanら、2009年)は、PDX−1を阻害するための方法及び組成物を開示している。本発明の方法及び組成物に含まれている抗PDX−1剤は、抗体、アプタマー、アンチセンスオリゴヌクレオチド、リボザイム及び/又は阻害化合物を含む。本発明は、PDX−1をコードするRNA分子の特定の領域と非常に又は完全に相補的な有効量の抗PDX−1剤と腫瘍細胞を接触させるステップを含む、腫瘍細胞におけるPDX−1発現の阻害方法を提供する。このような作用剤は、PDX−1をコードするRNA分子と特異的にハイブリダイズし、腫瘍細胞におけるPDX−1遺伝子の発現を阻害する。抗PDX−1 siRNA及び/又はshRNAを含む組成物が記載されている。本発明による抗PDX−1 siRNA又はshRNAをコードする組換え型の発現コンストラクトが、記載されている。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】米国特許第6,716,824号明細書
【特許文献2】米国特許出願公開第2009/0163431号明細書
【非特許文献】
【0010】
【非特許文献1】Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72
【非特許文献2】Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17
【非特許文献3】Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511
【非特許文献4】Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14
【非特許文献5】House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9
【非特許文献6】Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67
【非特許文献7】Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6
【非特許文献8】Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75
【非特許文献9】Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2
【非特許文献10】Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25
【非特許文献11】Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6
【非特許文献12】Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075
【非特許文献13】Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352
【非特許文献14】Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225
【非特許文献15】Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31
【非特許文献16】Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502
【非特許文献17】Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93
【非特許文献18】Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20
【非特許文献19】Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51
【非特許文献20】Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206
【非特許文献21】Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8
【非特許文献22】Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8
【非特許文献23】Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30
【非特許文献24】Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40
【非特許文献25】Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8
【非特許文献26】Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego
【非特許文献27】Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20
【非特許文献28】Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9
【非特許文献29】Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7
【非特許文献30】Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54
【非特許文献31】Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8
【非特許文献32】Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7
【非特許文献33】Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45
【非特許文献34】Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205
【非特許文献35】Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4
【非特許文献36】Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16
【非特許文献37】Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33
【非特許文献38】Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7
【非特許文献39】Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7
【非特許文献40】Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300
【非特許文献41】Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84
【非特許文献42】Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71
【非特許文献43】Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9
【非特許文献44】Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62
【非特許文献45】Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6
【非特許文献46】Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22
【非特許文献47】Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70
【非特許文献48】Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9
【非特許文献49】Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84
【非特許文献50】Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63
【非特許文献51】Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5
【非特許文献52】Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23
【非特許文献53】Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8
【非特許文献54】Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60
【非特許文献55】Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6
【非特許文献56】Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25
【非特許文献57】Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24
【非特許文献58】Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46
【非特許文献59】(2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.
【非特許文献60】Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15
【非特許文献61】Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73
【非特許文献62】Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7
【非特許文献63】Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84
【非特許文献64】Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84
【非特許文献65】Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、標的の翻訳抑制及び転写後mRNA分解を同時に誘発するためのPDX bi−shRNAを含む。PDX bi−shRNAコンストラクトは、同じmRNA標的特異性を有するsiRNAと比較して、高い効力及び長期間にわたる活性を達成する。本発明者らは、PDX−1ノックダウンがアポトーシスを誘発し、増殖を停止し、NETがん細胞の侵襲性を低下させ、癌腫異種移植片を有するSCIDマウスの生存率を改善することを示した。本発明者らはまた、IV(静脈内)BIVリポソームPDX−1 shRNAの複数回処理が、SCIDマウスにおいてヒトPC異種移植片をほぼ完全に消失させたことを示し、マウスインスリノーマモデルにおいて応答及び延命効果を確認している。
【0012】
2層中空小胞(BIV,Bilamellar invaginated vesicle)は、一時的にマスクされた標的リポソームを使用して、細胞膜との融合による細胞への治療薬の送達を行う送達系である。これらのマスクされたステルスリポソーム(stealthed liposome)は、エンドサイトーシス経路を回避することによって、より高いトランスフェクション効率を達成する。Copernicusの凝縮核酸送達技術は、別の非ウイルス性の合成的モジュラープラットフォームであり、単一分子又はDNA若しくはRNAが、ポリカチオンによって凝縮されて、非常に高いトランスフェクション効率を有するナノ粒子が生じる。本発明者らは、マスクされたステルスBIVリポソーム中への凝縮shRNA含有DNAナノ粒子のパッケージングによる新規ながん治療法を開発した。
【課題を解決するための手段】
【0013】
本発明は、プロモーターと、プロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターであって、前記核酸インサートが、PDX−1転写因子をコードする転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする発現ベクターを含む。転写物へのshRNAの結合は、RNA干渉によってPDX−1発現を阻害する。shRNAは二機能性であって、siRNA(切断依存性)モチーフとmiRNA(切断非依存性)モチーフの両方が同時に組み込まれていてもよい。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。一態様において、1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態(modification)からなる群から選択される配列を含む。
【0014】
本発明はまた、shRNAをPDX−1mRNA標的に送達する系を提供する。この送達系は、プロモーターとプロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターにカップリングされているリポソームなどの治療薬担体を含み、該インサートは、PDX−1転写因子をコードする転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする。1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。転写物へのshRNAの結合は、RNA干渉によってPDX−1発現を阻害する。shRNAは二機能性であって、siRNA(切断依存性)モチーフとmiRNA(切断非依存性)モチーフの両方を組み込むことができる。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。治療薬担体は、凝縮されたDNAナノ粒子とすることができる。一実施形態において、発現ベクターのDNAは、ポリカチオンと組み合わせることによって凝縮できる。より詳しくは、DNAは、N末端システイン残基及び30リジン残基を含むペプチドで修飾された10kDポリエチレングリコール(PEG)であるCK30PEG10kと組み合わせることによって、凝縮できる。治療薬担体はリポソームであってもよく、その中で、DNAは、30merリジン縮合ペプチドと組み合わされることによって凝縮され得る。一実施形態において、リポソームは、2層中空小胞(BIV)とすることができる。リポソームは、「スマート」レセプター標的化部分で装飾することによって、標的組織を特異的に標的とすることができる。一例において、標的化部分は、低分子二価βターン模倣体とすることができる。リポソームは、非標的器官を迂回させるために、可逆的にマスクすることができる。一例において、可逆的マスキングは、リポソームをn−ドデシル−β−D−マルトピラノシドなどの低分子量脂質(分子量約500以下)でコーティングすることによって実施できる。本発明の別の実施形態において、凝縮DNAナノ粒子とリポソーム送達系とを組み合わせることができる。したがって、shRNA発現ベクターを含有する凝縮DNAナノ粒子を、リポソームに封入することができる。これらの凝縮DNA−ナノ粒子含有リポソームは、BIVとすることができる。リポソームは、標的化部分で装飾することができ、可逆的にマスクすることもできる。
【0015】
本発明の別の実施形態は、PDX−1を発現する標的組織にshRNAを送達する方法であって、(i)プロモーターと、PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAをコードする、プロモーターに作動可能に連結されている核酸のインサートとを含む発現ベクターを調製するステップと、(ii)前記発現ベクターを治療薬担体と組み合わせるステップと、(iii)治療有効量の発現ベクターと治療薬担体との複合体を、それを必要とする患者に投与するステップとを含む方法である。治療薬担体は、凝縮DNAナノ粒子とすることができる。一実施形態において、発現ベクターのDNAは、ポリカチオンと組み合わせることによって凝縮することができる。より詳しくは、DNAは、CK30PEG10kと組み合わせることによって凝縮することができる。治療薬担体は、リポソームであってもよい。一実施形態において、リポソームはBIVとすることができる。リポソームは、「スマート」レセプター標的化部分で装飾することによって、標的組織を特異的に標的とすることができる。一例において、標的化部分は、低分子二価βターン模倣体とすることができる。リポソームは、非標的器官を迂回させるために、可逆的にマスクすることができる。一例において、可逆的マスキングは、リポソームをn−ドデシル−D−マルトピラノシドなどの低分子量脂質(分子量約500以下)でコーティングすることによって実施できる。本発明の別の実施形態において、凝縮DNAナノ粒子とリポソーム送達系とを組み合わせることができる。したがって、shRNA発現ベクターを含有する凝縮DNAナノ粒子を、リポソームに封入することができる。これらの凝縮DNAナノ粒子含有リポソームは、BIVとすることができる。リポソームは、標的化部分で装飾することができ、可逆的にマスクすることもできる。一態様において、1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。
【0016】
本発明はまた、標的細胞中のPDX−1発現をサイレンシングする方法であって、(i)標的細胞を選択するステップと、(ii)PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAを発現するベクターを標的細胞にトランスフェクトするステップとを含み、がん細胞への前記ベクターのトランスフェクションがRNA干渉によってPDX−1発現を阻害する方法を提供する。shRNAは二機能性であって、切断依存性モチーフと切断非依存性モチーフの両方が同時に組み込まれていてもよい。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。1又は2以上のshRNAは、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。
【0017】
本発明はまた、がん患者における腫瘍細胞成長の抑制方法であって、(i)腫瘍の治療を必要とする患者を特定するステップと、(ii)PDX−1をコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上のshRNAを発現するベクターを前記腫瘍細胞にトランスフェクトするステップとを含み、前記ベクターがトランスフェクトされたがん細胞が、アポトーシス、増殖停止、又は侵襲性の低下をもたらす方法を含む。shRNAは、二機能性であって、切断依存性モチーフと切断非依存性モチーフの両方が同時に組み込まれていてもよく、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む。本発明の一実施形態において、shRNAは、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある。shRNAが標的とするmRNA配列は、PDX−1mRNA転写物の3’非翻訳(UTR)領域に限定されず、本発明の一実施形態において、shRNAはコード配列を標的とすることができる。より詳しくは、標的のがんは、膵臓神経内分泌腫瘍(NET)とすることができる。
【0018】
本発明の特徴及び利点のより完全な理解のために、添付図面と共に本発明の詳細な説明に言及する。
【図面の簡単な説明】
【0019】
【図1A−1B】bi−shRNASTMN1ベクターのインビボ成長阻害活性を示すグラフである。図1Aは、CCL−247腫瘍異種移植片に対して、bi−shRNASTMN1リポプレックス(pGBI2)若しくはスクランブル対照リポプレックス(pGBI5)又は懸濁液(D5W)の単回腫瘍内注射を行った場合に観察される阻害を示す。値は平均±SEMを表す。図1Bは、SCIDマウスにおける原発骨肉腫異種移植片に対するbi−shRNASTMN1の成長阻害活性を示す。bi−shRNASTMN1リポプレックス又は対照の連日腫瘍内注射を6回行った。
【図2】bi−sh−STMN1とsiRNASTMN1とを比較する、STMN1mRNA標的切断と細胞成長阻害との相関を示すグラフである。bi−shRNASTMN1(赤色)及びsiRNASTMN1(黄色)の複合用量反応曲線は、STMN1mRNA切断と相関した。x軸は、左から右へと増加するプラスミド/siRNA濃度の用量である。y軸は、24時間処理後のパーセント細胞生存率である。各データ点は、三重反復試料の平均を表す。bi−shRNASTMN1の濃度範囲は、1.44×10−12M〜5.63×10−15Mであった。siRNASTMN1の濃度範囲は、5×10−7M〜1×10−10Mであった。電気泳動図挿入部分は、トランスフェクトされたCCL−247細胞から検出される5’RACE産物を示し、これは切断生成物を示している。
【図3A−3B】過剰発現されたヒト(図3A)及びマウス(図3B)PDX1mRNA(配列番号1及び2)に対する二機能性shRNAの効果を示すグラフである。結腸がんCCL−247細胞に、PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又はPDX−1 cDNA及びbi−shRNA発現ベクターを同時トランスフェクトした。pUMVC3は、インサートを含まない発現ベクター対照(配列番号9)である。トランスフェクションの24,48,72及び96時間後に、qRT−PCRのために全RNAを単離した。PDX−1mRNAレベルは、GAPDH mRNAに基準化した。各試料の比較PDX−1mRNAレベルは、培地のみの試料(トランスフェクションなし)に規準化した。
【図4】ヒトPDX−1(配列番号1)に対する二機能性shRNAのサイレンシング効果を示す図である。結腸がんCCL−247細胞に、種特異的PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又は種特異的PDX−1 cDNAとbisRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない対照発現ベクターである。トランスフェクションの48、72及び96時間後に、細胞を収集し、PDX−1及びβ−アクチンに特異なモノクローナル抗体によるウエスタン免疫ブロットを行った。
【図5A−5B】RACE−PCRによるヒト(図5A)PDX1及びマウス(図5B)PDX1(配列番号1及び2)の標的特異的切断の同定を示す図である。結腸がんCCL−247細胞に、PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又はPDX−1 cDNAとbi−shRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない発現ベクター対照である。トランスフェクションの24及び96時間後に、5’RACEアッセイのために全RNAを単離して、標的部位切断産物を検出した。RT−PCRで増幅された切断産物が、アガロースゲル電気泳動図上に示された。
【図6A−6C】CATレポーター遺伝子産生に対するマスキング剤の効果を示すグラフである。図6Aは肺に対する効果を示し、図6Bは心臓に対する効果を示し、図6Cは肝臓に対する効果を示す。BALB/cマウスへの静脈内注射後に、マスキングの増加につれて、肺(図6A)及び心臓(図6B)における非特異的局在化の減少が見られる。マスキングは、目的とする標的部位へのCATレポーター遺伝子の局在化の増加と関連する(肝臓;図6C)。
【図7A−7C】凝縮DNAナノ粒子の構造及び機能を示す図とグラフである。図7Aは、楕円体又は棒状として凝縮されたDNAナノ粒子の電子顕微鏡写真を示す。バーは、100nmである。図7Bは、対数期の又は成長が停止した神経芽細胞腫細胞に、裸のDNA又は凝縮DNAのリポソーム混合物を加えたことを示している。凝縮DNAは、遺伝子発現を、有糸分裂後細胞で1000倍超、対数期細胞で6〜8倍改善する。図7Cは、IV投与によって又は腫瘍内(IT)注射として投与された、結腸がん隣接外植片(colon cancer flank explant)を有するSCIDマウスにおける、凝縮DNA遺伝子導入(gene transfer)を示す。IT注射は、高レベルのルシフェラーゼ活性を生じたが、IV投与は生じなかった。
【図8】トランスフェクションの48時間後におけるPDX−1タンパク質発現の減少に対する配列特異性を示す図である。結腸がんCCL−247細胞に、種特異的PDX−1 cDNA発現ベクターのみをトランスフェクトするか、又は種特異的PDX−1 cDNAとbisRNA発現ベクターとを同時トランスフェクトした。pUMVC3は、インサートを含まない対照発現ベクターである。トランスフェクションの48時間後に、細胞を収集して、PDX−1及びβ−アクチンに特異的なモノクローナル抗体によるウエスタン免疫ブロットを行った。
【図9A−9B】インビボにおけるbi−shRNA−PDX−1リポプレックスによる処理後のSCIDマウス異種移植片モデル:β−TC−6細胞インスリノーマモデル(図9A)及びPANC−1膵臓がんモデル(図9B)の血糖値を経時的に示すグラフである。SCIDマウスに腫瘍細胞を移植後に2週間、bi−shRNA−PDX−1リポプレックス50μgによる処理を3サイクル行った。処理及び非処理マウスの空腹時血糖値を、処理後90日まで一定時間間隔で監視した。
【図10A−10B】PANC−1膵臓がんモデル(図10A)及びβ−TC−6細胞インスリノーマモデル(図10B)のカプラン−マイヤー(Kaplan-Meier)生存率分析を示すグラフである。腫瘍細胞を移植したSCIDマウスは、処理を行わないか、又は対照ベクターリポプレックス(pUMVC3)若しくはbi−shRNA−PDX−1リポプレックスで処理する(30μg/サイクルで3サイクル)かのいずれかとした。処理後90日まで、マウスを生存について監視した。
【図11】ヒトインスリノーマ腫瘍及びマウスTC−6インスリノーマ細胞におけるPDX−1の過剰発現率を示す図である。
【図12】mPDX−1 RNAサイレンシング後のβ−TC−6細胞周期タンパク質発現率を示すグラフである。
【図13】マウスインスリノーマSCIDモデルにおける、L−mPDX−1 shRNA処理されたマウスの生存率を示すグラフである。
【図14】腫瘍体積の比較データを示すグラフである。
【図15】本発明の新規bi−shRNAPDX−1を製造するためのプラスミドコンストラクトを示す図である。
【発明を実施するための形態】
【0020】
本発明の種々の実施形態の製造及び使用を以下に詳細に記載するが、本発明は、種々の具体的状況で具体化し得る多くの応用可能な発明概念を提供するものと理解すべきである。本明細書中に記載した具体的な実施形態は、本発明の具体的な製造及び使用方法の単なる実例であって、本発明の範囲の限界を定めるものではない。
【0021】
本発明の理解を容易にするために、多くの用語を以下に定義する。本明細書中で定義する用語は、本発明に関連する分野の当業者によって一般に理解される意味を有する。「a」、「an」及び「the」などの用語は、単一の構成要素を指すだけでなく、その一般的種類を含み、その具体例を説明するために使用できるものとする。本明細書中の専門用語を使用して、本発明の具体的な実施形態を説明するが、それらの使用は、特許請求の範囲に概説される場合を除いて、本発明の範囲の限界を定めるものではない。
【0022】
本明細書中で使用する「核酸」又は「核酸分子」という用語は、ポリヌクレオチド、例えば、デオキシリボ核酸(DNA)又はリボ核酸(RNA)、オリゴヌクレオチド、ポリメラーゼ連鎖反応(PCR)によって生成されるフラグメント、並びにライゲーション、切断、エンドヌクレアーゼ作用及びエキソヌクレアーゼ作用のいずれかによって生成されるフラグメントを指す。核酸分子は、天然起源のヌクレオチドであるモノマー(例えば、DNA及びRNA)、若しくは天然起源のヌクレオチドのアナログ(例えば、天然起源のヌクレオチドのα−エナンチオマー型)、又は両方の組合せから構成され得る。修飾ヌクレオチドは、糖部分及び/又はピリミジン若しくはプリン塩基部分が変更されているものとすることができる。糖修飾形態は、例えば、ハロゲン、アルキル基、アミン及びアジド基による、1若しくは2以上のヒドロキシル基の置換を含み、又は糖はエーテル若しくはエステルとして機能化されていてもよい。さらに、糖部分全体が、立体配置的及び電子的に類似した構造、例えば、アザ糖及び炭素環式糖アナログと置き換えられることもできる。塩基部分の修飾形態の例としては、アルキル化プリン及びピリミジン、アシル化プリン若しくはピリミジン又は他の周知の複素環代替物が挙げられる。核酸モノマーは、ホスホジエステル結合又はこのような結合のアナログによって連結され得る。ホスホジエステル結合のアナログとしては、ホスホロチオエート、ホスホロジチオエート、ホスホロセレノエート、ホスホロジセレノエート、ホスホロアニロチオエート、ホスホルアニリデート、ホスホルアミデートなどが挙げられる。「核酸分子」という用語はまた、ポリアミド主鎖に天然起源又は修飾核酸塩基が付着している、いわゆる「ペプチド核酸」を含む。核酸は一本鎖でも、二重鎖でもよい。
【0023】
本明細書及び特許請求の範囲中で使用する「発現ベクター」という用語は、宿主細胞で発現される遺伝子をコードする核酸分子を含む。典型的には、発現ベクターは、転写プロモーター、遺伝子及び転写ターミネーターを含む。遺伝子発現は通常、プロモーターにコントロールされ、このような遺伝子は、プロモーターに「作動可能に連結されている」と言われる。同様に、制御エレメントがコアプロモーターの活性を調節する場合には、制御エレメントとコアプロモーターとは、作動可能に連結されている。「プロモーター」という用語は、宿主酵母細胞中で構造遺伝子と関連する場合に、適切な成長条件下で1)転写、2)翻訳又は3)mRNA安定性の1又は2以上を、プロモーター配列の非存在下での転写、翻訳又はmRNA安定性(mRNAのより長い半減期)と比較して増加させる、その構造遺伝子に関する任意のDNA配列を指す。
【0024】
本明細書中で使用する「オンコジーン」という用語は、悪性新生物細胞の形成及び生存を可能にする遺伝子を指す(Bradshaw, T.K.: Mutagenesis 1, 91-97(1986)。
【0025】
本明細書中で使用する「レセプター」という用語は、「リガンド」と称される生物活性分子と結合する細胞関連タンパク質(cell-associated protein)を意味する。この相互作用が、細胞に対するリガンドの効果を媒介する。レセプターは、膜結合レセプター、細胞質レセプター又は核内レセプター;モノマーレセプター(例えば、甲状腺刺激ホルモンレセプター、βアドレナリン作動性レセプター)又は多量体レセプター(例えば、PDGFレセプター、成長ホルモンレセプター、IL−3レセプター、GM−CSFレセプター、G−CSFレセプター、エリスロポエチンレセプター及びIL−6レセプター)とすることができる。膜結合型レセプターは、細胞外リガンド結合ドメイン及び細胞内エフェクタードメインを含むマルチドメイン構造を特徴とし、シグナル変換に典型的に関与する。特定の膜結合レセプターにおいては、細胞外リガンド結合ドメインと細胞内エフェクタードメインとは、完全な機能性レセプターを含む別個のポリペプチド中に位置する。
【0026】
「ハイブリダイズする」という用語は、核酸の鎖が塩基対形成によって相補鎖と結合する任意のプロセスを指す。
【0027】
「トランスフェクション」という用語は、真核細胞中への外来DNAの導入を指す。トランスフェクションは、当技術分野で知られている種々の手段、例えば、リン酸カルシウム−DNA共沈、DEAE−デキストラン媒介トランスフェクション、ポリブレン媒介トランスフェクション、エレクトロポレーション、マイクロインジェクション、リポソーム融合、リポフェクション、プロトプラスト融合、レトロウイルス感染及び微粒子銃によって行うことができる。
【0028】
本明細書中で使用する「リポソーム」という用語は、内部水性空間を取り囲む脂質二重層からなる閉鎖構造を指す。本明細書中で使用する「ポリカチオン」という用語は、同一分子内に第4級アンモニウムラジカルなどの複数のカチオン部分を有する物質を意味し、遊離塩基及びそれらの薬学的に許容される塩を含む。
【0029】
本発明は、標的の翻訳抑制と転写後mRNA分解とを同時に誘発するために開発されたPDX bi−shRNAを含む。プロトタイプのコンストラクトは、同じmRNA標的特異性を有するsiRNAと比較して高い効力及び長期間にわたる活性を達成する。本発明者らは、PDX−1ノックダウンがNETがん細胞のアポトーシスを誘発し、増殖を停止し、侵襲性を低下させ、癌腫異種移植片を有するSCIDマウスの生存率を改善することを示している。本発明者はまた、IV(静脈内)BIVリポソームPDX−1 shRNAの複数の処理が、SCIDマウスにおいてヒトPC異種移植片をほぼ完全に消失させたことを示し、マウスインスリノーマモデルにおいて応答及び延命効果を確認している。
【0030】
bi−shRNA:本発明者らは、二機能性shRNAとして知られている第3の独特のRNAiプラットフォームを開発した。概念的には、shRNAがロードされたRISCによってRNAiを達成して、切断依存性又は切断非依存性mRNAノックダウンを促進できる。両立体配置のshRNAの同時発現(それゆえに、二機能性shRNAと命名されている)は、siRNAと比較して、サイレンシングのより急速な開始時に、より効果的な標的遺伝子ノックダウンを達成する(mRNA及びタンパク質の代謝回転率にもかかわらず)と共に、耐久性が大きいことを本発明者らは示した。二機能性shRNA発現単位の基本設計は、2つのステムループshRNA構造を含む。これらのステムループの1つは、切断依存性RISCロードのための完全にマッチしたパッセンジャー鎖及びガイド鎖から構成され、もう1つのステムループは、切断非依存性RISCロードのためのミスマッチパッセンジャー鎖(9〜12位)を有する。この二機能性設計は、第1に、二機能性設計は、異なるRISC型へのガイド鎖のロードを促進し、したがって、mRNAターゲティングを促進する;第2に、同一標的mRNAに対して切断依存性RISC及び切断非依存性RISCが存在することにより、分解と翻訳阻害/隔離プロセスの両方によってサイレンシングが促進されるという2つの理由で、はるかに効率的である。強力な遺伝子ノックダウンエフェクターは、単一Pol IIプロモーターのコントロール下で多重化shRNAによって空間的及び時間的コントロールを達成する。本発明者らが設計したプラットフォームは、天然のプロセスを模倣している。他の研究者による多数の研究及び文献は、本発明者らのアプローチを裏付けている(Okamura, K., et al., Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev, 2004. 18(14): pp. 1655-66、Miyoshi, K., et al., Slicer function of Drosophila Argonautes and its involvement in RISC formation. Genes Dev, 2005. 19(23): pp. 2837-48、Tomari, Y., T. Du, and P.D. Zamore, Sorting of Drosophila small silencing RNAs. Cell, 2007. 130(2): pp. 299-308、Forstemann, K., et al., Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell, 2007. 130(2): pp. 287-97、Landthaler, M., et al., Molecular characterization of human Argonaute-containing ribonucleoprotein complexes and their bound target mRNAs. RNA, 2008. 14(12): pp. 2580-96)。
【0031】
本発明者らは、miR30骨格を用いて、オンコプロテインであるスタスミン(stathmin)(STMN1、オンコプロテイン18、プロソリン(prosolin)、p19、op18)を再構築する微小管に対する新規な二機能性(bi−shRNA)を調製した。bi−shRNASTMN1は、同じ標的部位に対して、siRNAと比較して、より効果的なサイレンシング活性を示した。STMN1は、有糸分裂紡錘体形成に大きく関与する(Rana, S., et al., Stathmin 1: a novel therapeutic target for anticancer activity. Expert Rev Anticancer Ther, 2008. 8(9): pp. 1461-70、Iancu, C., et al., Effects of stathmin inhibition on the mitotic spindle. J Cell Sci, 2001. 114(Pt 5): pp. 909-16)。本発明のbi−shRNASTMN1コンストラクトは、同じ標的に対するsiRNAと比較した場合に、安全で効果的な標的ノックダウン及び腫瘍細胞死滅における有意な用量有利性を示している。さらに、インビボにおいてbi−shRNASTMN1リポプレックスによって、有意な腫瘍成長コントロール及び延命効果が示された(図1A及び1B)。本発明者らは、マッチパッセンジャー鎖とミスマッチパッセンジャー鎖とを区別できるRT−PCR方法を使用して、成熟分子及びエフェクター分子(完全マッチ鎖を有するdsRNA及び特定のミスマッチを有するdsRNA)の両方の細胞内転写及びプロセシングを検証した(Chen, C., et al., Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res, 2005. 33(20): pp. e179)。ほとんどのがん細胞には、高いDrosha及びDicer発現が見られる。がん細胞の内因性Dicerレベルに関しては議論がある(Merritt, W.M., et al., Dicer, Drosha, and outcomes in patients with ovarian cancer. N Engl J Med, 2008. 359(25): pp. 2641-50)。しかし、大部分の研究は、Dicer発現が低レベルであっても、sh又はbi−shRNAiノックダウンはがん細胞において非常に効果的であることを示している(Senzer, N., D. Rao, and J. Nemunaitis, Letter to the Editor: Does Dicer expression affect shRNA processing? Gene Regulation and Systems Biology (in press))。本発明者らは、対照siRNASTMN1処理細胞中では完全マッチパッセンジャー鎖のみがより合わされるのとは対照的に、bi−shRNASTMN1に対する成熟miRNA/siRNA成分に対応する、予測されるマッチ及びミスマッチshRNAの発現を確認した(Rao, D.R., Maples, P.B., Senzer, N., Kumar, P., Wang, Z., Pappen, B.O., Yu, Y., Haddock, C., Tong, A., Nemunaitis, J., Bifunctional shRNA: A novel approach of RNA interference. (submitted), 2009)。bi−shRNASTMN1の機構をさらに裏付けるために、5’RACE法による本発明の研究の成果から、bi−shRNASTMN1のsiRNA(マッチ)成分の標的切断部位に相当すると予想される配列を有するSTMN1切断産物の存在が確認された。STMN1発現腫瘍細胞の効果的なノックダウン(93%)が観察された。これは、bi−shRNASTMN1の切断依存性及び非依存性媒介ノックダウンの結果を反映している。さらに、bi−shRNASTMN1(GBI−2)と比較して、別個の成分である切断依存性(GBI−1)及び切断非依存性(GBI−3)ベクターによるノックダウン後に観察されるSTMN1mRNA動態は、予測される機構と一致していた。
【0032】
bi−shRNASTMN1ベクターのエフェクター機構及び機能的効力を実証後、本発明者らは、インビトロでの抗がん活性をさらに探求し、いくつかのがん細胞株において効果的な細胞死滅の実証に成功した。同じ配列を標的とするにもかかわらず、bi−shRNASTMN1はsiRNASTMN1よりも5log低いモル濃度でIC50を達成した(図2)。
【0033】
インスリノーマは、最も一般的な型の膵島細胞腫瘍である。悪性インスリノーマは、重度の形態の高インスリン血症であり、コントロールできない低血糖症につながる。前述のように、膵臓十二指腸ホメオボックス−1(PDX−1)は、ホメオドメイン含有転写因子ファミリーに属し、膵臓器官形成において主要な役割を担う。PDX−1は、インスリン、グルコキナーゼ及び2型グルコーストランスポーターの転写を制御することによって、β細胞機能を維持する。PDX−1は、インスリノーマにおいて過剰発現され、高インスリン血症をもたらす。PDX−1は、膵臓腫瘍において一般に過剰発現されることも認められている。転移性膵臓がんは、診断からの生存期間が4〜6カ月である。
【0034】
マウスインスリノーマモデルにおけるshRNAノックダウンPDX−1:インスリノーマにおけるPDX−1の発現を実証するために、Dr.Brunicardiの研究室は、PDX−1に対する強力な抗体を開発した。本発明者らは、この抗体を用いて、PDX−1がマウスインスリノーマにおいてヒトインスリノーマと同様なレベルまで顕著に過剰発現されることを実証した(図11)。本発明者らは、マウスモデルにおけるノックダウン効果を試験するために、マウス(mu)shRNAを調製した。しかし、インスリノーマのサイズが増加するので、コントロールできない低血糖症によりこれらのマウスの寿命は2〜3カ月であるため、研究は制限される。最初の研究においては、インビトロにおいてβTC−6細胞をmu−shPDX−1 shRNAで処理し、MTSアッセイを行って、細胞生存率を評価した。空ベクターと比較して、mu−shPDX−1処理細胞は、複数の時点にわたって細胞増殖のかなりの減少を示した。本発明者らは、DNA合成がノックダウンによって複数の時点において有意に減少することをさらに実証し、PDX−1ノックダウンに応答するいくつかの細胞周期関連タンパク質のウエスタンブロットによってノックダウンを実証した(図12)。
【0035】
PDX−1発現のサイレンシングは、腫瘍成長を阻害するための魅力的な方法といえる。本発明者らは、PDX−1遺伝子発現のサイレンシングのための二機能性shRNAを設計した。ヒトPDX−1(配列番号1)又はマウスPDX−1(配列番号1)のいずれかに対するMiR30をベースとする二機能性shRNAカセットを、pUMVC3ベクター中にクローニングした。図15は、新規bi−shRNAPDX−1を製造するためのプラスミドコンストラクトを示す。マウス又はヒトPDX−1を標的とする二機能性shRNAを、マウス又はヒトPDX−1のいずれかを発現する発現ベクターと共に、ヒトの結腸がん細胞株に同時にエレクトロポレートした。RACE−PCRを用いて、ヒト及びマウスPDX1mRNAの、可能性のある切断産物を調べた。RT−QPCR及び免疫ブロット法を使用して、ヒトPDX−1又はマウスPDX−1のいずれかの発現のノックダウンを調べた。
【0036】
ヒト膵臓十二指腸ホメオボックス1(PDX1)mRNA(配列番号1):
【0037】
【0038】
前記配列において、コード領域はボールドイタリック体で示され、第1及び第2標的部位はそれぞれ、下線及び太字で示されている。
【0039】
マウス膵臓十二指腸ホメオボックス1(PDX1)mRNA(配列番号2):
【0040】
【0041】
前記配列において、コード領域は太字のイタリック体で示され、標的部位は下線で示されている。
【0042】
図3A及び3Bはそれぞれ、強制的に過剰発現されたヒト及びマウスPDX1に対する二機能性shRNAの効果を示す。トランスフェクションの24時間後にヒトPDX−1に対する二機能性shRNAのサイレンシング効果が観察され、少なくとも96時間持続した(図4)。最大のサイレンシング効果、ヒトPDX−1の80%ノックダウンは、トランスフェクションの72時間後に達成された。さらに、マウスPDX−1を標的とする二機能性shRNA(ヒトの配列との相同性が68%である)は、ヒトPDX−1の発現に影響を及ぼさなかった(図8)。同様に、マウスPDX−1の発現は、トランスフェクションの24時間後にサイレンシングを受け、このサイレンシング効果は少なくとも96時間持続した(図4)。最大のサイレンシング効果、マウスPDX−1発現の95%が、トランスフェクションの48時間後に観察された。さらに、ヒトPDX−1を標的とする二機能性shRNA(マウスの配列との配列相同性が79%)もまた、マウスPDX−1の発現を変化させなかった(図8)。
【0043】
RACE−PCRによって同定されたヒト及びマウスPDX−1mRNAの切断産物はそれぞれ、図5A及び5Bに示されている。図5A及び5Bは、ヒトPDX1mRNA及びマウスPDX1mRNAがいずれも、予測された標的領域の中央で正確に切断されることを示した。加えて、ヒトPDX−1を標的とする二機能性shRNAは、マウスPDX−1mRNAの切断を引き起こさず、逆の場合も同じであった。
【0044】
前述の研究は、ヒト又はマウスPDX1のいずれかを標的とする二機能性shRNAの効力及び種特異性を実証した。bi−shRNAhPDX1は、ヒトPDX1の発現をノックダウンしたが、マウスPDX1の発現に対するサイレンシング効果を示さなかった。同様に、bi−shRNAmPDX1はマウスPDX1の発現をノックダウンしたが、ヒトPDX1の発現に対するサイレンシング効果を示さなかった。
【0045】
リポソーム送達系:本発明者らが以前に検証したリポソーム送達系は、1,2−ジオレイル−3−トリメチル-アンモニオプロパン(DOTAP)及びコレステロールを含んでいた(Ruponen M, H.P., Ronkko S, Pelkonen J, Tammi M, Urtti A, Extracellular and intracellular barriers in non-viral gene delivery J Control Release, 2003. 93(2): pp. 213-217)。この配合物は、DNAと組み合わされて、2層中空小胞(リポソームBIV)内に核酸を封入する複合体を形成する。本発明者らのうちの一人は、RNA、DNA及びRNAiプラスミドの送達を改善するために、BIV送達系のいくつかの特徴を最適化した。リポソームBIVは融合性であるので、核酸分解(Simberg D, W.A., Barenholz Y, Reversible mode of binding of serum proteins to DOTAP/cholesterol Lipoplexes: a possible explanation for intravenous lipofection efficiency. Hum Gene Ther, 2005. 16(9): pp. 1087-1096)及びTLR媒介オフターゲット効果をもたらし得るエンドサイトーシス媒介DNA細胞移入を回避する。このリポソーム送達系は、本発明者らなどが臨床試験での使用に成功した(Jay, C., et al., Preclinical Assessment of wt GNE Gene Plasmid for Management of Hereditary Inclusion Body Myopathy 2 (HIBM2). Gene Regulation & Systems Biology, 2008. 2: pp. 243-52、Frank O, R.C., Heberlein C, Neuhoff Nv, Schrock E, Schambach A, et al., Tumor cells escape suicide gene therapy by genetic and epigenetic instability. Blood 2004. 104(12): pp. 3543-3539、Ito I, J.L., Tanaka F, Saito Y, Gopalan B, Branch CD, Xu K, Atkinson EN, Bekele BN, Stephens LC, Minna JD, Roth JA, Ramesh R Liposomal vector mediated delivery of the 3p FUS1 gene demonstrates potent antitumor activity against human lung cancer in vivo. Cancer Gene Ther, 2004. 11(11): pp. 733-9、Templeton, N.S., Reversible masking of liposomal complexes for targeted delivery, USPTO, Editor. 2006, Baylor College of Medicine: USA、Phadke, A.P., et al., Safety and in vivo expression of a GNE-transgene: A novel treatment approach for Hereditary Inclusion Body Myopathy-2. Gene Regulation & Systems Biology, 2009. 3: pp. 89-101)。過去10年にわたる累積的な研究は、血清アルブミンなどの負電荷を持つ血清タンパク質への非特異的結合(オプソニン化)を最小化する(Tong, A.W., et al., Systemic therapeutic gene delivery for cancer: crafting paris' arrow. Curr Gene Ther, 2009. 9(1): pp. 45-60)ためには、最適化送達媒体が、効率的な血管内輸送のためにゼータ電位10mV以下のステルス化(一般にPEG化によって達成される)ナノ粒子である必要がある(Pirollo, K.F. and E.H. Chang, Targeted delivery of small interfering RNA: approaching effective cancer therapies. Cancer Res, 2008. 68(5): pp. 1247-50、Khalil, I.A., et al., Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev, 2006. 58(1): pp. 32-45、Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9)ことを示している。標的化部分、例えば、抗体及びそれらの単鎖誘導体(scFv)、糖又はペプチドの組込みは、導入遺伝子の標的細胞への局在化をさらに増加できる。
【0046】
本発明者らは、インビボでのPEGを用いない複合体の標的化送達を創出し、それによって、過度に長期の循環半減期を回避した(Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9、Simoes, S., et al., Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands or pH-sensitive peptides. Gene Ther, 1999. 6(11): pp. 1798-807、Simoes, S., et al., Cationic liposomes for gene delivery. Expert Opin Drug Deliv, 2005. 2(2): pp. 237-54、Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81)。PEG化は、膜透過性を改善するDNA又はsiRNAオリゴヌクレオチド送達に関連するが、本発明者らは、この方法はBIVリポソーム構造に立体障害をもたらし、結果としてDNA封入が非効率的になり、遺伝子発現が減少することを示した。さらにまた、PEG化された複合体は、主にエンドサイトーシス経路を通って細胞に入り、リソソーム中で核酸の大部分を分解する。PEGは循環において極めて長い半減期を示すが、これは、細胞毒性薬ドキソルビシンを封入するPEG化リポソーム製剤ドキシルによって例証されるように、患者に対して問題を引き起こした(Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81、Sapra, P., B., et al., Ligand-targeted liposomes for cancer treatmentLiposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. Curr Drug DelivJ Clin Oncol, 20051995. 213(47): pp. 369-811777-85、Uziely, B., et al., Liposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. J Clin Oncol, 1995. 13(7): pp. 1777-85)。特異的細胞表面レセプター(例えば、HER2/neu)への送達のためにドキシルにリガンドを付加する試みは、腫瘍特異的送達を増加しなかった(Park, J.W., et al., Tumor targeting using anti-her2 immunoliposomes. J Control Release, 2001. 74(1-3): pp. 95-113)。
【0047】
この推論に基づき、本発明のBIVは、DOTAP及び合成コレステロールを用い、専有の手動押出プロセスを使用して調製した(Bartlett, D.W., N. S., et al., Impact of tumor-specific targetingImproved DNA: liposome complexes for increased systemic delivery and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticlesgene expression. Nat Biotechnol Bioeng, 20081997. 9915(47): pp. 975-85647-52)。さらにまた、可逆的マスキング技術を使用して送達を最適化した。可逆的マスキングは、荷電してないためにBIV複合体の表面と緩く結合する低分子量脂質(分子量約500以下;例えば、n−ドデシル−β−D−マルトピラノシド)を使用し、結果として正電荷を持つBIV複合体を一時的に遮蔽して、非標的化器官を迂回する。これらの小脂質は、血流において剪断力によって除去される。それらが標的細胞に到達するまでに、電荷が再曝露されて(最適には約45mV)、移入が促進される。図6Aは、BALB/cマウスにおけるCAT−封入プラスミドを封入しているBIVを用いた、静脈内投与による肺のトランスフェクションの最適化を、図6Bは、静脈内投与による心臓のトランスフェクションの最適化を示す。逆マスキング複合体が全身的に循環する場合には、ゼータ電位アナライザーで測定される電荷が4.8mVであるにもかかわらず、トランスフェクションは最小であった。マスキングがBIVから複合体を可逆的に解離するので、これらの複合体は本来の荷電状態(45.5mV)に戻り、結果として肝臓の肝細胞において膜融合による細胞移入及び高レベルの導入遺伝子(CAT)発現を促進した(図6C)。「ジェネリック」BIV複合体を使用して、導入遺伝子の発現を100倍超増加させ、ほぼ一致したCATレベルを肺で生じさせた。可逆的マスキングを使用して、他の器官ではCAT産生は観察されなかった。
【0048】
本発明のBIV送達系が他に類を見ないほど効率的な理由は、1つには、複合体が治療薬を細胞膜との融合によって細胞中に送達し、エンドサイトーシス経路を回避するためである。リポソーム移入の2つの主要な移入機構は、エンドサイトーシス又は細胞膜との直接融合による。マウス肺胞マクロファージにおけるポリエチレン−アミン(PEI)を用いた比較研究によって示されるように、本発明者らは、インビトロ及びインビボの両方において送達されるBIV複合体に封入された核酸が、直接融合によって細胞に移入すること、並びにBIVが主にエンドソーム取込みを回避することを発見した。トランスフェクション後2〜3時間以内におけるローダミン標識オリゴヌクレオチドの95%以上の局在化によって示される(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40、Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40、Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54)ように、PEIは、エンドソーム中に急速且つ旺盛に取り込まれることが知られている。
【0049】
装飾BIVを用いたがん標的化送達:最近になって、Bartlett及びDavisは、腫瘍を標的とするナノ粒子(NP)によって全身に送達されるsiRNAが、皮下腫瘍の成長阻害において、非装飾NPと比較して有意に効果的であることを示した(Bartlett, D.W. and M.E. Davis, Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol Bioeng, 2008. 99(4): pp. 975-85)。標的化送達は、薬物動態又は生体内分布に大きく影響を与えず、これは依然として大部分がEPR(血管透過性−滞留性亢進)効果の結果であった(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40)が、細胞取込みの増加によって導入遺伝子発現を改善するようであった(Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40、Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40、Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54)。
【0050】
実のところ、治療用BIVの開発において重要な「欠けている部分(missing piece)」は、BIV−複合体の表面に位置してそれらを標的細胞に誘導することができるような非免疫原性リガンドの同定であった。これは、リポソーム表面で多量体化される小ペプチドによって行える可能性もあるが、小ペプチドは、反復注射後に免疫応答を発生する可能性がある。抗体、抗体フラグメント、タンパク質、部分タンパク質などを含む比較的大きい他のリガンドは、リポソーム表面における標的化送達に関しては、小ペプチドを使用する場合よりもはるかに扱いにくい。よって、本発明の複合体は、腫瘍脈管構造の内皮孔及び固形腫瘍の間質圧勾配を含む厳重なバリアを透過する(Ramesh, R., et al., Successful treatment of primary and disseminated human lung cancers by systemic delivery of tumor suppressor genes using an improved liposome vector. Mol Ther, 2001. 3(3): pp. 337-50)だけでなく、腫瘍細胞を直接的に標的とする限りにおいて、独特である。したがって、本発明の治療方法は、EPR効果にのみ依存する送達に限定されず、腫瘍を直接的に標的とする(Hashizume, H., et al., Openings between defective endothelial cells explain tumor vessel leakiness. Am J Pathol, 2000. 156(4): pp. 1363-80、Netti, P.A., et al., Effect of transvascular fluid exchange on pressure-flow relationship in tumors: a proposed mechanism for tumor blood flow heterogeneity. Microvasc Res, 1996. 52(1): pp. 27-46、Yuan, F., et al., Vascular permeability and microcirculation of gliomas and mammary carcinomas transplanted in rat and mouse cranial windows. Cancer Res, 1994. 54(17): pp. 4564-8)。
【0051】
本発明には、タンパク質を選択的に結合するように設計された小分子を使用できる。重要なことに、調製される小分子は「二価」であるので、細胞表面レセプターを結合するのに特に適し、タンパク質−リガンド相互作用においてホットスポットに見られる二次構造モチーフに類似している。Burgessのグループは、細胞表面レセプターに対して親和性を有する二価βターン模倣体の設計に成功した(Feng, Y., et al., Solid-phase SN2 macrocyclization reactions to form beta-turn mimics. Org Lett, 1999. 1(1): pp. 121-4、Bruno, M.A., et al., Long-lasting rescue of age-associated deficits in cognition and the CNS cholinergic phenotype by a partial agonist peptidomimetic ligand of TrkA. J Neurosci, 2004. 24(37): pp. 8009-18、Zaccaro, M.C., et al., Selective small molecule peptidomimetic ligands of TrkC and TrkA receptors afford discrete or complete neurotrophic activities. Chem Biol, 2005. 12(9): pp. 1015-28)。本発明者らは、糖尾部を有する二価分子を生成するようにこの戦略を適合させ、これらの適合小分子から機能化BIV複合体を調製した。ライブラリーをスクリーニングする効率的なハイスループット技術を開発し、実施した。
【0052】
凝縮DNAナノ粒子:有糸分裂後細胞における安全で効率的なDNA送達:Copernicusの核酸送達技術は、非ウイルス性の合成的モジュラープラットフォームであり、DNA又はsiRNAの単一分子がポリカチオンによって凝縮されて、最小可能体積を有するナノ粒子を生成する(Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86)。インビボ送達のために最適化されたポリカチオンは、N末端システイン残基及び30リジン残基を含むペプチドで修飾された10kDaポリエチレングリコール(PEG)(CK30PEG10k)である。これらの複合体の形状は、DNA凝縮時のリジン対イオンにある程度左右される(Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51)。棒状ナノ粒子の最小断面直径は、ペイロードプラスミドの寸法に関係なく8〜11nmであるが、楕円体の最小直径は、典型的な発現プラスミドの場合、20〜22nmである(図7A)(Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51)。重要なことに、これらのDNAナノ粒子は、培養中の非分裂細胞に大量にトランスフェクトすることができる。凝縮DNAのリポソーム混合物は、リポソームの裸のDNA混合物と比較して、遺伝子発現レベルを1,000倍超増加させる(図7B)。インビボ投与後、凝縮DNAは、肺(Ziady, A.G., et al., Transfection of airway epithelium by stable PEGylated poly-L-lysine DNA nanoparticles in vivo. Mol Ther, 2003. 8(6): pp. 936-47)、脳(Yurek, D.M., et al., Long-term transgene expression in the central nervous system using DNA nanoparticles. Mol Ther, 2009. 17(4): pp. 641-50、Yurek, D.M., et al., Compacted DNA nanoparticle gene transfer of GDNF to the rat striatum enhances the survival of grafted fetal dopamine neurons. Cell Transplant, 2009)及び眼(Farjo, R., et al., Efficient non-viral ocular gene transfer with compacted DNA nanoparticles. PLoS One, 2006. 1: pp. e38、Cai, X., et al., A partial structural and functional rescue of a retinitis pigmentosa model with compacted DNA nanoparticles. PLoS One, 2009. 4(4): pp. e5290)において有糸分裂後細胞に大量にトランスフェクトする。これらの各系において、有糸分裂後細胞にトランスフェクトする凝縮DNAの注目すべき能力は、これらのナノ粒子のサイズが小さく、ナノ粒子が25nmの核膜細孔を通過できることによると考えられる(Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86)。
【0053】
これらのDNAナノ粒子に関する1つの取込み機構は、細胞表面ヌクレオリン(26nm KD)との結合に基づく。DNAナノ粒子は、その後の細胞質輸送によって非分解経路を経て核に入り、核内でほどけて、生物活性DNAを放出する(Chen, X., et al., Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Mol Ther, 2008. 16(2): pp. 333-42)。遺伝子導入後1年にもわたる長期インビボ発現が実証された。これらのナノ粒子は、無害の毒性プロフィールを有し、トール様レセプターを刺激しないので毒性サイトカイン応答を回避し、凝縮DNAが何百ものCpGアイランドを有し、リポソームと混合される場合であっても、毒性作用は観察されていない(Ziady, A.G., et al., Minimal toxicity of stabilized compacted DNA nanoparticles in the murine lung. Mol Ther, 2003. 8(6): pp. 948-56、Ding, X.-Q., et al., Ocular delivery of compacted DNA-nanoparticles does not elicit toxicity in the mouse retina. Plos One, 2009. In press)。DNAナノ粒子が、嚢胞性線維症試験においてヒトに投与されたところ、結果は期待の持てるものであり、ナノ粒子に起因する有害事象なく、ほとんどの患者はCFTRタンパク質の生物活性を示した(Konstan, M.W., et al., Compacted DNA nanoparticles administered to the nasal mucosa of cystic fibrosis subjects are safe and demonstrate partial to complete cystic fibrosis transmembrane regulator reconstitution. Hum Gene Ther, 2004. 15(12): pp. 1255-69)。
【0054】
本発明者らとCopernicus(Cleaveland、OH)との初期の共同研究において、結腸がん腫瘍外植片を有するSCIDマウスに、尾静脈又は腫瘍内(IT)注射によって凝縮DNAを投与した(図7C)。IV注射は遺伝子導入をもたらさなかったが、IT投与後には著しく高いレベルの遺伝子導入が認められた。これは、これらのDNAナノ粒子が、局所送達後における腫瘍へのトランスフェクトに高い活性を有することを示している。
【0055】
新規送達系:原発腫瘍及び転移部位への全身的遺伝子導入が望ましいので、凝縮DNAの腫瘍への送達は、30merリジン縮合ペプチドと組み合わせることによって凝縮されたDNAからなるこれらのナノ粒子を、有標BIVリポソーム中にパッケージングすることによって促進できる。凝縮DNAとマスクされたステルスリポソームとのこの新規なカップリングは、新規ながん遺伝子治療法といえる。
【0056】
前述のように、膵臓がん細胞株PANC−1におけるRNA干渉によるPDX−1発現のサイレンシングは、インビトロでは細胞増殖を阻害し、インビボでは腫瘍成長を抑制すると共に腫瘍細胞アポトーシスを増加させた。この戦略に基づき、本発明者らは、インスリノーマ、膵臓がん及び神経内分泌腫瘍の治療への臨床応用のための二機能性shRNA(bi−shRNA−PDX−1)を開発した。さらに、本発明者らは、インビトロでの効力並びにインスリノーマ/SCIDモデル及びPANC−1/SCID膵臓腫瘍モデルを用いたインビボでの効力を実証する。
【0057】
インビトロでのノックダウン効力は、PDX−1及びbi−shRNA発現ベクターの同時トランスフェクションによる、PDX−1陰性ヒトがん細胞株(HCT116)におけるPDX−1発現のウエスタン免疫ブロット及びqRT−PCR分析によって評価した(図3A、3B、4及び8)。動物モデルは、インビボインスリノーマモデルに関しては1×106個のβ−TC−6細胞の、又はPCモデルに関しては5×105個のPANC−1細胞の腹腔内(IP)注射によって調製した。接種2週間後に、マウスを、各モデルについて無作為に2群に分け;尾静脈注射によって、1つの群には35μgの対照ベクター(shRNAインサートを含まないプラスミド)を含有するリポプレックス(DOTAP:コレステロールリポソームプラスミド複合体)を、第2の群にはマウス特異的bi−msh−PDX−1(インスリノーマモデルの場合)又はヒト特異的bi−hsh−PDX−1(PCモデルの場合)を35μg含有するリポプレックスを与えた。2つの反復送達を、2週間毎に合計3回の注射で行った。血中インスリン値及び血糖値を、定期的な時間間隔で監視した。3回目の注射の2、10及び18週間後に、病理評価及び腫瘍評価のために、各群6匹のマウスを屠殺した。
【0058】
HCT116ヒトがん細胞に、プラスミドを1:1の比率でエレクトロポレートした。PDX−1mRNAを、qRT−PCRを用いてΔΔCt法によって定量的に比較した(図3A及び3B)。mRNAレベルを空ベクター対照と比較して、表1に要約する%発現率を得た。PDX−1mRNAは、トランスフェクションの24、48及び72時間後に分析した。
【0059】
【表1】
【0060】
ヒト及びマウス特異的bi−shRNA−PDX−1は、インビトロにおいてPDX−1mRNA発現の効果的で種特異的なノックダウンを示す。
【0061】
絶食マウスの血糖値を、最初の2サイクルの各処理の7日後、45日目及び90日目に測定した。マウスインスリノーマモデルにおいて、mbi−shRNA−PDX−1による処理は、β−TC−6細胞によって誘発された低血糖症からマウスを効果的にレスキューした(図9A)が、hbi−shRNA−PDX−1は、膵臓がんモデルの場合に血糖値に対して効果がなかった(図9B)。β−TC−6インスリノーマモデルにおいて、未処理マウスは全て、90日目の測定の前に死亡した。PANC−1膵臓がんモデルの結果を、図9Bに示す。
【0062】
カプラン−マイヤー生存率分析は、PANC−1モデル(図10A)及びβ−TC−6インスリノーマモデル(図10B)に対するbi−shRNA−PDX−1処理の有益性をさらに示す。インスリノーマモデルについては、処理群のマウス(mbi−shRNA−PDX−1リポプレックス各30μgで3サイクル処理)10匹及び対照群のマウス(bi−shRNAインサートを含まないpUMVC3ベクターを含有するリポプレックス各30μgで3サイクル処理)10匹を、生存について監視した。生存は95日間監視した。PCモデルについては、PANC−1接種マウス5匹は処理せず、マウス10匹は、bi−shRNAインサートを含まないpUMVC3ベクターを含有するリポプレックス各30μgで3サイクル処理し、マウス15匹は、hbi−shRNA−PDX−1リポプレックス各30μgで3サイクル処理した。腫瘍体積の比較のために、各群5匹のマウスを、60及び90日目に屠殺した。処理の60日後に、非処理マウスは100%が平均1330.75mm3のサイズの腫瘍を発現したが、処理マウスは50%のみが、平均50.5mm3のサイズの目視可能な腫瘍を有していた。処理の90日後に、処理マウスの37.5%は目視可能な腫瘍を有さず、残りのマウスは平均サイズ36.7mm3の腫瘍を有していた。腫瘍体積の比較研究の結果を、表2及び図14に示す。
【0063】
生存率は、SCIDマウスでも、BIVリポソーム送達されたshRNAPDX−1(L−mu−shRNAPDX−1)のIV注入後に向上し、L−mu−shRNAPDX−1注射の回数と相関するようであった(図13)。リポプレックス(即ち、インビボ研究のために使用される脂質−プラスミド複合体)を、ベクターそのものと区別するために接頭語「L−」で表す。インスリン値の減少及び血糖値の増加は、インスリノーマ成長阻害というL−mu−shRNAPDX−1処理の結果に対応する。L−mu−shPDX−1による処理後の動物の腫瘍応答は、フローサイトメトリーによるインスリノーマ細胞のPDX−1ノックダウンの検出並びに細胞周期タンパク質(サイクリンE、cdk4、cdk2)のノックダウン及びサイクリン依存性キナーゼ阻害因子(p27)の上方制御と相関した。アポトーシスも示された。本発明者らはさらに、PDX−1発現性ヒト膵臓がんマウス異種移植片モデルにおいて、shRNAPDX−1リポプレックスのIV注入の用量依存的な延命効果をさらに実証した。
【0064】
【表2】
【0065】
新規bi−shRNAPDX−1:二機能性shRNA戦略を適用するために、本発明者らは、マウス(mu)−bi−shRNAPDX−1を調製し、検証した。mu−bi−shRNAPDX−1は、miR−30a主鎖を有する2つのステムループ構造からなる。第1のステムループ構造は、完全相補的なガイド鎖及びパッセンジャー鎖を有し、第2のステムループ構造は、パッセンジャー鎖の10及び11位に2つの塩基対ミスマッチを有する。mu−bi−shRNAPDX−1は、マウスPDX−1mRNA(NM_008814)のヌクレオチド#723〜#741を標的とする。
【0066】
mu−bi−shRNAPDX−1発現単位を、オリゴヌクレオチドの組合せと共に構築した。5’及び3’末端それぞれにSalI及びNotI部位を有する構築発現単位を、pUMVC3のSalI及びNotI部位に挿入した。インサートに隣接するプライマーで両方向から配列決定することによって、インサート配列を確認した。マイクロRNAの検出のために開発されたステムループRT−PCR法を用いて、mu−bi−shRNAPDX−1をトランスフェクトした細胞中の成熟shRNAを検出した。本発明者らはさらに、5’RACEアッセイによってmu−bi−shRNAPDX−1の活性を検証して、標的部位切断を検出した。
【0067】
本発明のbi−shRNAプラスミドのインサート配列を以下に示す。
【0068】
pGBI-20:ヒトbi−shRNA−PDX1(配列番号3):
【0069】
TCGACTGCTGTTGAAGTGAGCGCCAGTTCCTATTCAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCAGTTCCTATCTAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0070】
配列番号3の下線部分(25〜43の核酸残渣を含む)は、主要標的部位を表す。
【0071】
pGBI-21:ヒトbi−shRNA−PDX−1(配列番号4):
【0072】
TCGACTGCTGTTGAAGTGAGCGCCAGTTCCTATTCAACAAGTATAGTGAAGCCACAGATGTATACTTGTTGAATAGGAACTGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0073】
pGBI-22:ヒトbi−shRNA−PDX−1(配列番号5):
【0074】
TCGACTGCTGTTGAAGTGAGCGCCCAGTTATTTACAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCCAGTTATTTCTAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0075】
pGBI-23:ヒトbi−shRNA−PDX−1(配列番号6):
【0076】
TCGACTGCTGTTGAAGTGAGCGCCCAGTTATTTACAAACAGGTTAGTGAAGCCACAGATGTAACCTGTTTGTAAATAACTGGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0077】
pGBI-24:マウスbi−shRNA−PDX−1(配列番号7):
【0078】
TCGACTGCTGTTGAAGTGAGCGCCGGAAGATAAGAAACGTAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCAGCTCACTACATTACTCAGCTGTTGAAGTGAGCGCCGGAAGATAAACAACATAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0079】
配列番号4の下線部分(25〜43の核酸残渣を含む)は、主要標的部位を表す。
【0080】
pGBI-25:マウスbi−shRNA−PDX−1(配列番号8):
【0081】
TCGACTGCTGTTGAAGTGAGCGCCGGAAGATAAGAAACGTAGTTAGTGAAGCCACAGATGTAACTACGTTTCTTATCTTCCGTTGCCTACTGCCTCGGAAGCTTAATAAAGGATCTTTTATTTTCATTGGC
【0082】
本発明の新規bi−shRNA治療薬の調製は、転写後の遺伝子ノックダウンによって効果的な治療結果を達成するのに必要な有効全身用量を減少させることができる最先端の方法といえる。PDX−1過剰発現腫瘍の全身標的化へと迅速に移行できる効果的で臨床的に応用可能な送達法が整っている。
【0083】
本明細書中に記載した実施形態はいずれも、本発明の任意の方法、キット、試薬又は組成物に関して実施可能であり、逆の場合も同様であると考えられる。さらにまた、本発明の組成物は、本発明の方法の実施に使用できる。
【0084】
本明細書中に記載した特定の実施形態は一例として示すのであって、本発明を限定するものとして示すのではないことがわかるであろう。この発明の主要な特徴は、本発明の範囲から逸脱せずに、種々の実施形態で実施可能である。当業者ならば、常法にすぎない実験を使用すれば、本明細書中に記載した具体的な方法の多数の均等物を認識、又は確認できるであろう。このような均等物を、本発明の範囲内であるとみなし、特許請求の範囲によって保護する。
【0085】
本明細書中に記載した全ての公表文献及び特許出願は、本発明が関連する技術分野の当業者の技術レベルを示す。全ての公表文献及び特許出願は、個々の公表文献又は特許出願が参照によって明確且つ個別に組み込まれていることが示された場合と同程度まで、参照によって本明細書中に組み入れる。
【0086】
単語「a」又は「an」の使用は、特許請求の範囲及び/又は明細書中で用語「含む(comprising)」と共に使用する場合、「1の」を意味することができるが、「1又は2以上の」、「少なくとも1の」及び「1又は1超の」の意味とも矛盾しない。特許請求の範囲における用語「又は(or)」の使用は、選択肢のみを指すことが明確に示されているか又は選択肢が相互排他的である場合を除いて、「及び/又は(and/or)」を意味して使われるものであるが、本開示は選択肢のみ及び「及び/又は」を指す定義も支持する。本出願の全体を通じて、「約」という用語は、値が、その値の測定に用いられた装置、方法の誤差の固有変動、又は研究対象間に存在する変動を含むことを示すのに用いる。
【0087】
本明細書及び特許請求の範囲中で使用する「含む(comprising)」(並びにcomprisingの任意の形、例えば、「comprise」及び「comprises」)、「有する(having)」(並びにhavingの任意の形、例えば、「have」及び「has」)、「含む(including)」(並びにincludingの任意の形、例えば、「includes」及び「include」)又は「含有する(containing)」(並びにcontainingの任意の形、例えば、「contains」及び「contain」)という用語は、包括的又は非限定的であり、列挙していない追加の要素又は方法のステップを排除しない。
【0088】
本明細書中で使用する「又はそれらの組合せ」という用語は、この用語の前に記載された用語の全ての順列及び組合せを指す。例えば、「A、B、C又はそれらの組合せ」は、A、B、C、AB、AC、BC又はABCの少なくとも1つを含むことを意図し、特定の状況で順序が重要である場合には、BA、CA、CB、CBA、BCA、ACB、BAC又はCABも含むことを意図する。この例に続いて、1又は2以上のアイテム又は用語の反復、例えば、BB、AAA、MB、BBC、AAABCCCC、CBBAAA、CABABBなどを含む組合せも明白に含まれる。当業者は、状況から明らかでない限り、典型的には、任意の組合せ中のアイテム又は用語の数に制限がないことがわかるであろう。
【0089】
本明細書中で使用する近似の単語、例えば、限定するものではないが、「約」、「実質的な」又は「実質的に」が指す状態は、それによって修飾される場合に、必ずしも絶対的又は完全でないと理解されるが、その状態が存在しているとするに値する、当業者にとっては十分に近いとみなされるであろう状態を指す。記載が変動し得る程度は、可能な変化の大きさと、修飾された特徴が修飾されていない特徴に必須の特性及び能力を依然として有すると当業者が認めることができる変化の大きさによるであろう。しかし、一般に、前記説明を受けて、「約」などの近似の単語で修飾された本明細書中の数値は、記載された値から少なくとも±1、2、3、4、5、6、7、10、12又は15%変動してもよい。
【0090】
本明細書中に開示され且つ特許請求の範囲に記載された組成物及び/又は方法は全て、必要以上の実験を行わずに本開示を踏まえて製造及び実施可能である。本発明の組成物及び方法を好ましい実施形態に関して記載したが、本発明の概念、精神及び範囲から逸脱せずに、本明細書中に記載した組成物及び/又は方法に対して、並びに方法のステップ又は一連のステップにおいて、変形形態が適用されてもよいことは、当業者には明白であろう。当業者に明白なこのような類似した置換形態及び変更形態は全て、添付した特許請求の範囲によって定義される本発明の精神、範囲及び概念の範囲内とみなされる。
【0091】
(参考文献)
U.S.Patent Application No. 2009/0163431: Compositions and Methods for Modulation of PDX-1
U.S.Patent No. 6,716,824: Treatment of Pancreatic Adenocarcinoma by Cytotoxic Gene Therapy
1. Hirshberg, B., et al., Malignant insulinoma: spectrum of unusual clinical features. Cancer, 2005. 104(2): pp. 264-72
2. Pavelic, K., et al., Molecular genetics of malignant insulinoma. Anticancer Res, 1996. 16(4A): pp. 1707-17
3. Kaltsas, G.A., G.M. Besser, and A.B. Grossman, The diagnosis and medical management of advanced neuroendocrine tumors. Endocr Rev, 2004. 25(3): pp. 458-511
4. Pelengaris, S. and M. Khan, Oncogenic co-operation in beta-cell tumorigenesis. Endocr Relat Cancer, 2001. 8(4): pp. 307-14
5. House, M.G. and R.D. Schulick, Endocrine tumors of the pancreas. Curr Opin Oncol, 2006. 18(1): pp. 23-9
6. Vezzosi, D., et al., Octreotide in insulinoma patients: efficacy on hypoglycemia, relationships with Octreoscan scintigraphy and immunostaining with anti-sst2A and anti-sst5 antibodies. Eur J Endocrinol, 2005. 152(5): pp. 757-67
7. Brennan, M.F., Pancreatic cancer. J Gastroenterol Hepatol, 2000. 15 Suppl: pp. G13-6
8. Urgell, E., et al., Prospective evaluation of the contribution of K-ras mutational analysis and CA 19.9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer. Eur J Cancer, 2000. 36(16): pp. 2069-75
9. Willett, C.G., W.J. Daly, and A.L. Warshaw, CA 19-9 is an index of response to neoadjunctive chemoradiation therapy in pancreatic cancer. Am J Surg, 1996. 172(4): pp. 350-2
10. Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front Biosci, 1998. 3: pp. E214-25
11. Brennan, M., Pancreatic cancer. J. Gastroenterol. Hepatol, 2000. 15: pp. G13-6
12. Urgell E, P.P., Boadas J, Capella G, Queralto MJM, Boluda R, et al, Prospective evaluation of the contribution of K-ras mutational analysis and CA 19-9 measurement to cytological diagnosis in patients with clinical suspicion of pancreatic cancer Eur. J. Cancer, 2000. 36(16): pp. 2069-2075
13. Willet C, D.W., Warshaw A., CA19-9 is an index of response to neoadjuvant chemoradiation in pancreatic cancer. Am. J. Surg, 1996. 172: pp. 350-352
14. Blaszkowsky, L., Treatment of advanced and metastatic pancreatic cancer. Front. Biosci 1998. 3: pp. 214-225
15. Watada, H., et al., PDX-1 induces insulin and glucokinase gene expressions in alphaTC1 clone 6 cells in the presence of betacellulin. Diabetes, 1996. 45(12): pp. 1826-31
16. Bretherton-Watt, D., N. Gore, and D.S. Boam, Insulin upstream factor 1 and a novel ubiquitous factor bind to the human islet amyloid polypeptide/amylin gene promoter. Biochem J, 1996. 313 ( Pt 2): pp. 495-502
17. Carty, M.D., et al., Identification of cis- and trans-active factors regulating human islet amyloid polypeptide gene expression in pancreatic beta-cells. J Biol Chem, 1997. 272(18): pp. 11986-93
18. Serup, P., et al., Induction of insulin and islet amyloid polypeptide production in pancreatic islet glucagonoma cells by insulin promoter factor 1. Proc Natl Acad Sci U S A, 1996. 93(17): pp. 9015-20
19. Watada, H., et al., Involvement of the homeodomain-containing transcription factor PDX-1 in islet amyloid polypeptide gene transcription. Biochem Biophys Res Commun, 1996. 229(3): pp. 746-51
20. Ballian, N., et al., Proliferation, hyperplasia, neogenesis, and neoplasia in the islets of Langerhans. Pancreas, 2007. 35(3): pp. 199-206
21. Hagman, D.K., et al., Palmitate inhibits insulin gene expression by altering PDX-1 nuclear localization and reducing MafA expression in isolated rat islets of Langerhans. J Biol Chem, 2005. 280(37): pp. 32413-8
22. Leys CM, N.S., Rudzinski E, Kaminishi M, Montgomery E, Washington MK, Goldenring JR, Expression of PDX-1 in human gastric metaplasia and gastric adenocarcinoma. Hum Pathol., 2006. 37(9): pp. 1162-8
23. Sakai H, E.Y., Li XL, Akiyama Y, Miyake S, Takizawa T, Konishi N, Tatematsu M, Koike M, Yuasa Y, PDX-1 homeobox protein expression in pseudopyloric glands and gastric carcinomas. Gut, 2004. 53(3): pp. 323-30
24. Miyatsuka T, K.H., Shiraiwa T, Matsuoka TA, Yamamoto K, et al, Persistent expression of PDX-1 in the pancreas causes acinar-to-ductal metaplasia through Stat3 activation. Genes Dev, 2006. 20(11): pp. 1435-40
25. Wang, X.P., et al., Tissue MicroArray analyses of pancreatic duodenal homeobox-1 in human cancers. World J Surg, 2005. 29(3): pp. 334-8
26. Ayala, G., Prognostic Value of Akt-1 in Prostate Cancer: A Computerized Quantitative Approach with Quantum Dot Technology, in Abstract and presentation. 2007, United States Academy of Pathology: San Diego
27. Liu, S., et al., PDX-1 acts as a potential molecular target for treatment of human pancreatic cancer. Pancreas, 2008. 37(2): pp. 210-20
28. Scherr, M., et al., Specific inhibition of bcr-abl gene expression by small interfering RNA. Blood, 2003. 101(4): pp. 1566-9
29. Brummelkamp, T.R., R. Bernards, and R. Agami, Stable suppression of tumorigenicity by virus-mediated RNA interference. Cancer Cell, 2002. 2(3): pp. 243-7
30. Martinez, L.A., et al., Synthetic small inhibiting RNAs: efficient tools to inactivate oncogenic mutations and restore p53 pathways. Proc Natl Acad Sci U S A, 2002. 99(23): pp. 14849-54
31. Yoshinouchi, M., et al., In vitro and in vivo growth suppression of human papillomavirus 16-positive cervical cancer cells by E6 siRNA. Mol Ther, 2003. 8(5): pp. 762-8
32. Choudhury, A., et al., Small interfering RNA (siRNA) inhibits the expression of the Her2/neu gene, upregulates HLA class I and induces apoptosis of Her2/neu positive tumor cell lines. Int J Cancer, 2004. 108(1): pp. 71-7
33. Yang, G., et al., Inhibition of breast and ovarian tumor growth through multiple signaling pathways by using retrovirus-mediated small interfering RNA against Her-2/neu gene expression. J Biol Chem, 2004. 279(6): pp. 4339-45
34. Farrow, B., et al., Inhibition of pancreatic cancer cell growth and induction of apoptosis with novel therapies directed against protein kinase A. Surgery, 2003. 134(2): pp. 197-205
35. Yague, E., C.F. Higgins, and S. Raguz, Complete reversal of multidrug resistance by stable expression of small interfering RNAs targeting MDR1. Gene Ther, 2004. 11(14): pp. 1170-4
36. Kosciolek, B.A., et al., Inhibition of telomerase activity in human cancer cells by RNA interference. Mol Cancer Ther, 2003. 2(3): pp. 209-16
37. Cioca, D.P., Y. Aoki, and K. Kiyosawa, RNA interference is a functional pathway with therapeutic potential in human myeloid leukemia cell lines. Cancer Gene Ther, 2003. 10(2): pp. 125-33
38. Kawasaki, H. and K. Taira, Short hairpin type of dsRNAs that are controlled by tRNA(Val) promoter significantly induce RNAi-mediated gene silencing in the cytoplasm of human cells. Nucleic Acids Res, 2003. 31(2): pp. 700-7
39. Li, K., et al., Use of RNA interference to target cyclin E-overexpressing hepatocellular carcinoma. Cancer Res, 2003. 63(13): pp. 3593-7
40. Verma, U.N., et al., Small interfering RNAs directed against beta-catenin inhibit the in vitro and in vivo growth of colon cancer cells. Clin Cancer Res, 2003. 9(4): pp. 1291-300
41. Aharinejad, S., et al., Colony-stimulating factor-1 blockade by antisense oligonucleotides and small interfering RNAs suppresses growth of human mammary tumor xenografts in mice. Cancer Res, 2004. 64(15): pp. 5378-84
42. Uchida, H., et al., Adenovirus-mediated transfer of siRNA against survivin induced apoptosis and attenuated tumor cell growth in vitro and in vivo. Mol Ther, 2004. 10(1): pp. 162-71
43. Salisbury, A.J. and V.M. Macaulay, Development of molecular agents for IGF receptor targeting. Horm Metab Res, 2003. 35(11-12): pp. 843-9
44. Duxbury, M.S., et al., Focal adhesion kinase gene silencing promotes anoikis and suppresses metastasis of human pancreatic adenocarcinoma cells. Surgery, 2004. 135(5): pp. 555-62
45. Duxbury, M.S., et al., Systemic siRNA-mediated gene silencing: a new approach to targeted therapy of cancer. Ann Surg, 2004. 240(4): pp. 667-74; discussion 675-6
46. Filleur, S., et al., SiRNA-mediated inhibition of vascular endothelial growth factor severely limits tumor resistance to antiangiogenic thrombospondin-1 and slows tumor vascularization and growth. Cancer Res, 2003. 63(14): pp. 3919-22
47. Takei, Y., et al., A small interfering RNA targeting vascular endothelial growth factor as cancer therapeutics. Cancer Res, 2004. 64(10): pp. 3365-70
48. Lakka, S.S., et al., Inhibition of cathepsin B and MMP-9 gene expression in glioblastoma cell line via RNA interference reduces tumor cell invasion, tumor growth and angiogenesis. Oncogene, 2004. 23(27): pp. 4681-9
49. Singh, A., et al., RNAi-mediated silencing of nuclear factor erythroid-2-related factor 2 gene expression in non-small cell lung cancer inhibits tumor growth and increases efficacy of chemotherapy. Cancer Res, 2008. 68(19): pp. 7975-84
50. Nakahira, S., et al., Involvement of ribonucleotide reductase M1 subunit overexpression in gemcitabine resistance of human pancreatic cancer. Int J Cancer, 2007. 120(6): pp. 1355-63
51. Cullen, B.R., RNAi the natural way. Nat Genet, 2005. 37(11): pp. 1163-5
52. Zeng, Y. and B.R. Cullen, Sequence requirements for micro RNA processing and function in human cells. RNA, 2003. 9(1): pp. 112-23
53. Silva, J.M., et al., Second-generation shRNA libraries covering the mouse and human genomes. Nat Genet, 2005. 37(11): pp. 1281-8
54. Hutvagner, G. and P.D. Zamore, A microRNA in a multiple-turnover RNAi enzyme complex. Science, 2002. 297(5589): pp. 2056-60
55. Yekta, S., I.H. Shih, and D.P. Bartel, MicroRNA-directed cleavage of HOXB8 mRNA. Science, 2004. 304(5670): pp. 594-6
56. Pillai, R.S., C.G. Artus, and W. Filipowicz, Tethering of human Ago proteins to mRNA mimics the miRNA-mediated repression of protein synthesis. Rna, 2004. 10(10): pp. 1518-25
57. Valencia-Sanchez, M.A., et al., Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev, 2006. 20(5): pp. 515-24
58. Parker, R. and U. Sheth, P bodies and the control of mRNA translation and degradation. Mol Cell, 2007. 25(5): pp. 635-46
59. (2008 ) Breakdown of RNAi-Based Drugs in the Clinic. RNAi News Volume.
60. Heidel, J.D., et al., Potent siRNA inhibitors of ribonucleotide reductase subunit RRM2 reduce cell proliferation in vitro and in vivo. Clin Cancer Res, 2007. 13(7): pp. 2207-15
61. Judge, A.D., et al., Confirming the RNAi-mediated mechanism of action of siRNA-based cancer therapeutics in mice. J Clin Invest, 2009. 119(3): pp. 661-73
62. Zukiel, R., et al., Suppression of human brain tumor with interference RNA specific for tenascin-C. Cancer Biol Ther, 2006. 5(8): pp. 1002-7
63. Wyszko, E., et al., A multivariate analysis of patients with brain tumors treated with ATN-RNA. Acta Pol Pharm, 2008. 65(6): pp. 677-84
64. Calvo, A., et al., Identification of VEGF-regulated genes associated with increased lung metastatic potential: functional involvement of tenascin-C in tumor growth and lung metastasis. Oncogene, 2008. 27(40): pp. 5373-84
65. Heidel, J.D., et al., Administration in non-human primates of escalating intravenous doses of targeted nanoparticles containing ribonucleotide reductase subunit M2 siRNA. Proc Natl Acad Sci U S A, 2007. 104(14): pp. 5715-21
66. Okamura, K., et al., Distinct roles for Argonaute proteins in small RNA-directed RNA cleavage pathways. Genes Dev, 2004. 18(14): pp. 1655-66
67. Miyoshi, K., et al., Slicer function of Drosophila Argonautes and its involvement in RISC formation. Genes Dev, 2005. 19(23): pp. 2837-48
68. Tomari, Y., T. Du, and P.D. Zamore, Sorting of Drosophila small silencing RNAs. Cell, 2007. 130(2): pp. 299-308
69. Forstemann, K., et al., Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell, 2007. 130(2): pp. 287-97
70. Landthaler, M., et al., Molecular characterization of human Argonaute-containing ribonucleoprotein complexes and their bound target mRNAs. RNA, 2008. 14(12): pp. 2580-96
71. Rana, S., et al., Stathmin 1: a novel therapeutic target for anticancer activity. Expert Rev Anticancer Ther, 2008. 8(9): pp. 1461-70
72. Iancu, C., et al., Effects of stathmin inhibition on the mitotic spindle. J Cell Sci, 2001. 114(Pt 5): pp. 909-16
73. Chen, C., et al., Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Res, 2005. 33(20): pp. e179
74. Merritt, W.M., et al., Dicer, Drosha, and outcomes in patients with ovarian cancer. N EnglJ Med, 2008. 359(25): pp. 2641-50
75. Senzer, N., D. Rao, and J. Nemunaitis, Letter to the Editor: Does Dicer expression affect shRNA processing? Gene Regulation and Systems Biology (in press)
76. Rao, D.R., Maples, P.B., Senzer, N., Kumar, P., Wang, Z., Pappen, B.O., Yu, Y., Haddock, C., Tong, A., Nemunaitis, J., Bifunctional shRNA: A novel approach of RNA interference. (submitted), 2009
77. Ruponen M, H.P., Ronkko S, Pelkonen J, Tammi M, Urtti A, Extracellular and intracellular barriers in non-viral gene delivery J Control Release, 2003. 93(2): pp. 213-217
78. Simberg D, W.A., Barenholz Y, Reversible mode of binding of serum proteins to DOTAP/cholesterol Lipoplexes: a possible explanation for intravenous lipofection efficiency. Hum Gene Ther, 2005. 16(9): pp. 1087-1096
79. Jay, C., et al., Preclinical Assessment of wt GNE Gene Plasmid for Management of Hereditary Inclusion Body Myopathy 2 (HIBM2). Gene Regulation & Systems Biology, 2008. 2: pp. 243-52
80. Frank O, R.C., Heberlein C, Neuhoff Nv, Schrock E, Schambach A, et al., Tumor cells escape suicide gene therapy by genetic and epigenetic instability. Blood 2004. 104(12): pp. 3543-3539
81. Ito I, J.L., Tanaka F, Saito Y, Gopalan B, Branch CD, Xu K, Atkinson EN, Bekele BN, Stephens LC, Minna JD, Roth JA, Ramesh R Liposomal vector mediated delivery of the 3p FUS1 gene demonstrates potent antitumor activity against human lung cancer in vivo. Cancer Gene Ther, 2004. 11(11): pp. 733-9
82. Templeton, N.S., Reversible masking of liposomal complexes for targeted delivery, USPTO, Editor. 2006, Baylor Collegeof Medicine: USA
83. Phadke, A.P., et al., Safety and in vivo expression of a GNE-transgene: A novel treatment approach for Hereditary Inclusion Body Myopathy-2. Gene Regulation & Systems Biology, 2009. 3: pp. 89-101
84. Pirollo, K.F. and E.H. Chang, Targeted delivery of small interfering RNA: approaching effective cancer therapies. Cancer Res, 2008. 68(5): pp. 1247-50
85. Khalil, I.A., et al., Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol Rev, 2006. 58(1): pp. 32-45
86. Li, S.D., et al., Tumor-targeted delivery of siRNA by self-assembled nanoparticles. Mol Ther, 2008. 16(1): pp. 163-9
87. Tong, A.W., et al., Systemic therapeutic gene delivery for cancer: crafting paris' arrow. Curr Gene Ther, 2009. 9(1): pp. 45-60
88. Simoes, S., et al., Mechanisms of gene transfer mediated by lipoplexes associated with targeting ligands or pH-sensitive peptides. Gene Ther, 1999. 6(11): pp. 1798-807
89. Simoes, S., et al., Cationic liposomes for gene delivery. Expert Opin Drug Deliv, 2005. 2(2): pp. 237-54
90. Sapra, P., P. Tyagi, and T.M. Allen, Ligand-targeted liposomes for cancer treatment. Curr Drug Deliv, 2005. 2(4): pp. 369-81
91. Sapra, P., B., et al., Ligand-targeted liposomes for cancer treatmentLiposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. Curr Drug DelivJ Clin Oncol, 20051995. 213(47): pp. 369-811777-85
92. Uziely, B., et al., Liposomal doxorubicin: antitumor activity and unique toxicities during two complementary phase I studies. J Clin Oncol, 1995. 13(7): pp. 1777-85
93. Park, J.W., et al., Tumor targeting using anti-her2 immunoliposomes. J Control Release, 2001. 74(1-3): pp. 95-113
94. Bartlett, D.W., N. S., et al., Impact of tumor-specific targetingImproved DNA: liposome complexes for increased systemic delivery and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticlesgene expression. Nat Biotechnol Bioeng, 20081997. 9915(47): pp. 975-85647-52
95. Kirpotin, D.B., et al., Antibody targeting of long-circulating lipidic nanoparticles does not increase tumor localization but does increase internalization in animal models. Cancer Res, 2006. 66(13): pp. 6732-40
96. Maeda, N., et al., Enhancement of anticancer activity in antineovascular therapy is based on the intratumoral distribution of the active targeting carrier for anticancer drugs. Biol Pharm Bull, 2006. 29(9): pp. 1936-40
97. Bartlett, D.W., et al., Impact of tumor-specific targeting on the biodistribution and efficacy of siRNA nanoparticles measured by multimodality in vivo imaging. Proc Natl Acad Sci U S A, 2007. 104(39): pp. 15549-54
98. Bartlett, D.W. and M.E. Davis, Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol Bioeng, 2008. 99(4): pp. 975-85
99. Ramesh, R., et al., Successful treatment of primary and disseminated human lung cancers by systemic delivery of tumor suppressor genes using an improved liposome vector. Mol Ther, 2001. 3(3): pp. 337-50
100. Hashizume, H., et al., Openings between defective endothelial cells explain tumor vessel leakiness. Am J Pathol, 2000. 156(4): pp. 1363-80
101. Netti, P.A., et al., Effect of transvascular fluid exchange on pressure-flow relationship in tumors: a proposed mechanism for tumor blood flow heterogeneity. Microvasc Res, 1996. 52(1): pp. 27-46
102. Yuan, F., et al., Vascular permeability and microcirculation of gliomas and mammary carcinomas transplanted in rat and mouse cranial windows. Cancer Res, 1994. 54(17): pp. 4564-8
103. Feng, Y., et al., Solid-phase SN2 macrocyclization reactions to form beta-turn mimics. Org Lett, 1999. 1(1): pp. 121-4
104. Bruno, M.A., et al., Long-lasting rescue of age-associated deficits in cognition and the CNS cholinergic phenotype by a partial agonist peptidomimetic ligand of TrkA. J Neurosci, 2004. 24(37): pp. 8009-18
105. Zaccaro, M.C., et al., Selective small molecule peptidomimetic ligands of TrkC and TrkA receptors afford discrete or complete neurotrophic activities. Chem Biol, 2005. 12(9): pp. 1015-28
106. Liu, G., et al., Nanoparticles of compacted DNA transfect postmitotic cells. J Biol Chem, 2003. 278(35): pp. 32578-86
107. Fink, T.L., et al., Plasmid size up to 20 kbp does not limit effective in vivo lung gene transfer using compacted DNA nanoparticles. Gene Ther, 2006. 13(13): pp. 1048-51
108. Ziady, A.G., et al., Transfection of airway epithelium by stable PEGylated poly-L-lysine DNA nanoparticles in vivo. Mol Ther, 2003. 8(6): pp. 936-47
109. Yurek, D.M., et al., Long-term transgene expression in the central nervous system using DNA nanoparticles. Mol Ther, 2009. 17(4): pp. 641-50
110. Yurek, D.M., et al., Compacted DNA nanoparticle gene transfer of GDNF to the rat striatum enhances the survival of grafted fetal dopamine neurons. Cell Transplant, 2009
111. Farjo, R., et al., Efficient non-viral ocular gene transfer with compacted DNA nanoparticles. PLoS One, 2006. 1: pp. e38
112. Cai, X., et al., A partial structural and functional rescue of a retinitis pigmentosa model with compacted DNA nanoparticles. PLoS One, 2009. 4(4): pp. e5290
113. Chen, X., et al., Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Mol Ther, 2008. 16(2): pp. 333-42
114. Ziady, A.G., et al., Minimal toxicity of stabilized compacted DNA nanoparticles in the murine lung. Mol Ther, 2003. 8(6): pp. 948-56
115. Ding, X.-Q., et al., Ocular delivery of compacted DNA-nanoparticles does not elicit toxicity in the mouse retina. Plos One, 2009. In press
116. Konstan, M.W., et al., Compacted DNA nanoparticles administered to the nasal mucosa of cystic fibrosis subjects are safe and demonstrate partial to complete cystic fibrosis transmembrane regulator reconstitution. Hum Gene Ther, 2004. 15(12): pp. 1255-69
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
プロモーターと、
前記プロモーターに作動可能に連結されている核酸インサートと
を含む発現ベクターであって、前記核酸インサートが、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができ、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する、1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする、発現ベクター。
【請求項2】
shRNAに1又は2以上のsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項1に記載の発現ベクター。
【請求項3】
shRNAが、PDX−1オンコジーン発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項1に記載の発現ベクター。
【請求項4】
shRNAが二機能性shRNAとしてさらに定義される、請求項1に記載の発現ベクター。
【請求項5】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項1に記載の発現ベクター。
【請求項6】
標的とされる領域が、PDX−1オンコジーン転写物の3’UTR領域配列である、請求項1に記載の発現ベクター。
【請求項7】
治療薬担体と、
プロモーター、及びPDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズし、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする、前記プロモーターに作動可能に連結されている核酸インサートを含む発現ベクターと
を含む治療薬送達系。
【請求項8】
治療薬担体が凝縮DNAナノ粒子である、請求項7に記載の送達系。
【請求項9】
DNAナノ粒子が、1又は2以上のポリカチオンで凝縮されている、請求項8に記載の送達系。
【請求項10】
1又は2以上のポリカチオンが、10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)である、請求項8に記載の送達系。
【請求項11】
凝縮DNAナノ粒子がさらにリポソーム中に封入されている、請求項8に記載の送達系。
【請求項12】
リポソームが2層中空小胞(BIV)である、請求項11に記載の送達系。
【請求項13】
リポソームが、可逆的にマスクされたリポソームである、請求項11に記載の送達系。
【請求項14】
リポソームが、1又は2以上の「スマート」レセプター標的化部分で装飾されている、請求項11に記載の送達系。
【請求項15】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項14に記載の送達系。
【請求項16】
治療薬担体がリポソームである、請求項7に記載の送達系。
【請求項17】
リポソームが、1又は2以上の「スマート」レセプター標的化部分で装飾されている2層中空小胞(BIV)であり、前記リポソームが可逆的にマスクされているリポソームである、請求項16に記載の送達系。
【請求項18】
「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項17に記載の送達系。
【請求項19】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項7に記載の送達系。
【請求項20】
PDX−1オンコジーンを発現する標的組織に1又は2以上のshRNAを送達する方法であって、
プロモーターと、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズし、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する前記1又は2以上のshRNAをコードする、前記プロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターを調製するステップと、
前記発現ベクターを治療薬担体と組み合わせるステップであって、前記治療薬担体が1又は2以上の「スマート」レセプター標的化部分で装飾されているリポソームであるステップと、
治療有効量の前記発現ベクターと治療薬担体との複合体を、それを必要とする患者に投与するステップと
を含む方法。
【請求項21】
治療薬担体が凝縮DNAナノ粒子である、請求項20に記載の方法。
【請求項22】
DNAナノ粒子が1又は2以上のポリカチオンで凝縮されており、前記1又は2以上のポリカチオンが10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)又は30merリジン縮合ペプチドを含む、請求項21に記載の方法。
【請求項23】
凝縮DNAナノ粒子がリポソーム中にさらに封入されており、前記リポソームが、2層中空小胞(BIV)であり、1又は2以上の「スマート」レセプター標的化部分で装飾されている、請求項21に記載の方法。
【請求項24】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項23に記載の方法。
【請求項25】
リポソームが、可逆的にマスクされているリポソームである、請求項23に記載の方法。
【請求項26】
リポソームが2層中空小胞(BIV)である、請求項20に記載の方法。
【請求項27】
リポソームが、可逆的にマスクされているリポソームである、請求項20に記載の方法。
【請求項28】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項20に記載の方法。
【請求項29】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項20に記載の方法。
【請求項30】
1又は2以上の標的細胞中のPDX−1オンコジーンの発現をサイレンシングする方法であって、
前記1又は2以上の標的細胞を選択するステップと、
前記PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)を発現するベクターを前記標的細胞にトランスフェクトするステップと
を含み、前記1又は2以上の標的細胞への前記ベクターのトランスフェクションがRNA干渉によってPDX−1オンコジーンの発現を阻害する方法。
【請求項31】
shRNAにsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項30に記載の方法。
【請求項32】
shRNAが、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項30に記載の方法。
【請求項33】
shRNAが二機能性shRNAとしてさらに定義される、請求項30に記載の方法。
【請求項34】
標的とされる領域が、PDX−1オンコジーン転写物の3’UTR領域配列である、請求項30に記載の方法。
【請求項35】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項30に記載の方法。
【請求項36】
ヒト対象における、腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方の方法であって、
腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方を必要とする前記ヒト対象を特定するステップと、
治療薬担体複合体中の発現ベクターを、腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方に十分な量で、前記ヒト対象に投与するステップと
を含み、前記発現ベクターが、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上の二機能性低分子ヘアピン型RNA(shRNA)を発現し、前記ハイブリダイゼーションが腫瘍細胞のアポトーシス、増殖停止、又は侵襲性の低下をもたらす方法。
【請求項37】
膵島新生物障害が、膵臓神経内分泌腫瘍(NET)、インスリノーマ及びカルチノイドを含む、請求項36に記載の方法。
【請求項38】
二機能性shRNAにsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項36に記載の方法。
【請求項39】
二機能性shRNAが、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項36に記載の方法。
【請求項40】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項36に記載の方法。
【請求項41】
標的とされる領域が、PDX−1転写物の3’UTR領域配列である、請求項36に記載の方法。
【請求項42】
膵島新生物障害が、膵臓神経内分泌腫瘍(NET)である、請求項36に記載の方法。
【請求項43】
治療薬担体が、凝縮DNAナノ粒子、又は1若しくは2以上の「スマート」レセプター標的化部分で装飾された、可逆的にマスクされたリポソームである、請求項36に記載の方法。
【請求項44】
DNAナノ粒子が、1又は2以上のポリカチオンで凝縮されており、前記1又は2以上のポリカチオンが、10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)又は30merリジン縮合ペプチドである、請求項43に記載の方法。
【請求項45】
可逆的にマスクされたリポソームが2層中空小胞(BIV)である、請求項43に記載の方法。
【請求項46】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項43に記載の方法。
【請求項47】
凝縮DNAナノ粒子がリポソーム中にさらに封入されている、請求項43に記載の方法。
【請求項1】
プロモーターと、
前記プロモーターに作動可能に連結されている核酸インサートと
を含む発現ベクターであって、前記核酸インサートが、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができ、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する、1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする、発現ベクター。
【請求項2】
shRNAに1又は2以上のsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項1に記載の発現ベクター。
【請求項3】
shRNAが、PDX−1オンコジーン発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項1に記載の発現ベクター。
【請求項4】
shRNAが二機能性shRNAとしてさらに定義される、請求項1に記載の発現ベクター。
【請求項5】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項1に記載の発現ベクター。
【請求項6】
標的とされる領域が、PDX−1オンコジーン転写物の3’UTR領域配列である、請求項1に記載の発現ベクター。
【請求項7】
治療薬担体と、
プロモーター、及びPDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズし、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する1又は2以上の低分子ヘアピン型RNA(shRNA)をコードする、前記プロモーターに作動可能に連結されている核酸インサートを含む発現ベクターと
を含む治療薬送達系。
【請求項8】
治療薬担体が凝縮DNAナノ粒子である、請求項7に記載の送達系。
【請求項9】
DNAナノ粒子が、1又は2以上のポリカチオンで凝縮されている、請求項8に記載の送達系。
【請求項10】
1又は2以上のポリカチオンが、10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)である、請求項8に記載の送達系。
【請求項11】
凝縮DNAナノ粒子がさらにリポソーム中に封入されている、請求項8に記載の送達系。
【請求項12】
リポソームが2層中空小胞(BIV)である、請求項11に記載の送達系。
【請求項13】
リポソームが、可逆的にマスクされたリポソームである、請求項11に記載の送達系。
【請求項14】
リポソームが、1又は2以上の「スマート」レセプター標的化部分で装飾されている、請求項11に記載の送達系。
【請求項15】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項14に記載の送達系。
【請求項16】
治療薬担体がリポソームである、請求項7に記載の送達系。
【請求項17】
リポソームが、1又は2以上の「スマート」レセプター標的化部分で装飾されている2層中空小胞(BIV)であり、前記リポソームが可逆的にマスクされているリポソームである、請求項16に記載の送達系。
【請求項18】
「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項17に記載の送達系。
【請求項19】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項7に記載の送達系。
【請求項20】
PDX−1オンコジーンを発現する標的組織に1又は2以上のshRNAを送達する方法であって、
プロモーターと、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズし、かつ、RNA干渉によってPDX−1オンコジーン発現を阻害する前記1又は2以上のshRNAをコードする、前記プロモーターに作動可能に連結されている核酸インサートとを含む発現ベクターを調製するステップと、
前記発現ベクターを治療薬担体と組み合わせるステップであって、前記治療薬担体が1又は2以上の「スマート」レセプター標的化部分で装飾されているリポソームであるステップと、
治療有効量の前記発現ベクターと治療薬担体との複合体を、それを必要とする患者に投与するステップと
を含む方法。
【請求項21】
治療薬担体が凝縮DNAナノ粒子である、請求項20に記載の方法。
【請求項22】
DNAナノ粒子が1又は2以上のポリカチオンで凝縮されており、前記1又は2以上のポリカチオンが10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)又は30merリジン縮合ペプチドを含む、請求項21に記載の方法。
【請求項23】
凝縮DNAナノ粒子がリポソーム中にさらに封入されており、前記リポソームが、2層中空小胞(BIV)であり、1又は2以上の「スマート」レセプター標的化部分で装飾されている、請求項21に記載の方法。
【請求項24】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項23に記載の方法。
【請求項25】
リポソームが、可逆的にマスクされているリポソームである、請求項23に記載の方法。
【請求項26】
リポソームが2層中空小胞(BIV)である、請求項20に記載の方法。
【請求項27】
リポソームが、可逆的にマスクされているリポソームである、請求項20に記載の方法。
【請求項28】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項20に記載の方法。
【請求項29】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項20に記載の方法。
【請求項30】
1又は2以上の標的細胞中のPDX−1オンコジーンの発現をサイレンシングする方法であって、
前記1又は2以上の標的細胞を選択するステップと、
前記PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上の低分子ヘアピン型RNA(shRNA)を発現するベクターを前記標的細胞にトランスフェクトするステップと
を含み、前記1又は2以上の標的細胞への前記ベクターのトランスフェクションがRNA干渉によってPDX−1オンコジーンの発現を阻害する方法。
【請求項31】
shRNAにsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項30に記載の方法。
【請求項32】
shRNAが、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項30に記載の方法。
【請求項33】
shRNAが二機能性shRNAとしてさらに定義される、請求項30に記載の方法。
【請求項34】
標的とされる領域が、PDX−1オンコジーン転写物の3’UTR領域配列である、請求項30に記載の方法。
【請求項35】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項30に記載の方法。
【請求項36】
ヒト対象における、腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方の方法であって、
腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方を必要とする前記ヒト対象を特定するステップと、
治療薬担体複合体中の発現ベクターを、腫瘍細胞成長の抑制、膵島新生物障害の治療又は両方に十分な量で、前記ヒト対象に投与するステップと
を含み、前記発現ベクターが、PDX−1オンコジーンをコードするmRNA転写物の領域とハイブリダイズすることができる1又は2以上の二機能性低分子ヘアピン型RNA(shRNA)を発現し、前記ハイブリダイゼーションが腫瘍細胞のアポトーシス、増殖停止、又は侵襲性の低下をもたらす方法。
【請求項37】
膵島新生物障害が、膵臓神経内分泌腫瘍(NET)、インスリノーマ及びカルチノイドを含む、請求項36に記載の方法。
【請求項38】
二機能性shRNAにsiRNA(切断依存性)モチーフ及びmiRNA(切断非依存性)モチーフが組み込まれている、請求項36に記載の方法。
【請求項39】
二機能性shRNAが、PDX−1発現の切断依存性阻害因子でもあり、切断非依存性阻害因子でもある、請求項36に記載の方法。
【請求項40】
1又は2以上のshRNAが、配列番号3、配列番号4、配列番号5、配列番号6、配列番号7、配列番号8及びそれらの組合せ又は修飾形態からなる群から選択される配列を含む、請求項36に記載の方法。
【請求項41】
標的とされる領域が、PDX−1転写物の3’UTR領域配列である、請求項36に記載の方法。
【請求項42】
膵島新生物障害が、膵臓神経内分泌腫瘍(NET)である、請求項36に記載の方法。
【請求項43】
治療薬担体が、凝縮DNAナノ粒子、又は1若しくは2以上の「スマート」レセプター標的化部分で装飾された、可逆的にマスクされたリポソームである、請求項36に記載の方法。
【請求項44】
DNAナノ粒子が、1又は2以上のポリカチオンで凝縮されており、前記1又は2以上のポリカチオンが、10kDAポリエチレングリコール(PEG)置換システイン−リジン3merペプチド(CK30PEG10k)又は30merリジン縮合ペプチドである、請求項43に記載の方法。
【請求項45】
可逆的にマスクされたリポソームが2層中空小胞(BIV)である、請求項43に記載の方法。
【請求項46】
1又は2以上の「スマート」レセプター標的化部分が低分子二価βターン模倣体である、請求項43に記載の方法。
【請求項47】
凝縮DNAナノ粒子がリポソーム中にさらに封入されている、請求項43に記載の方法。
【図2】

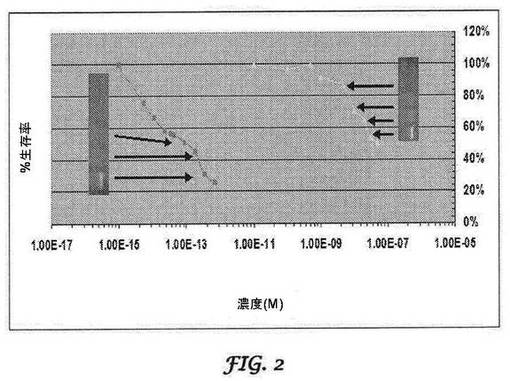
【図3A】
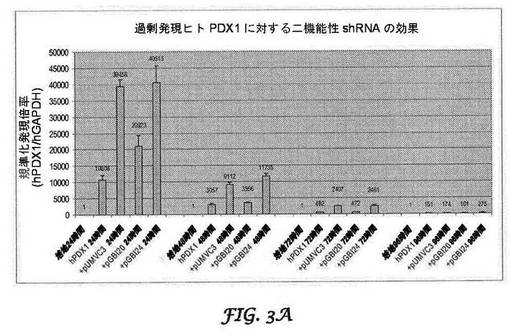
【図3B】
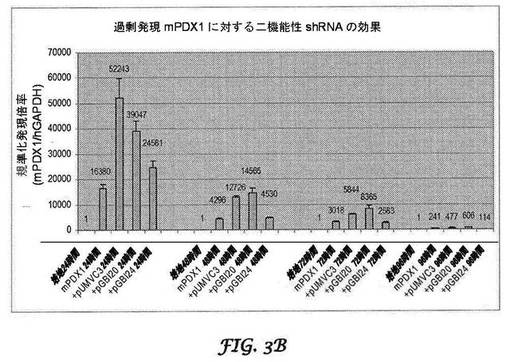
【図4】
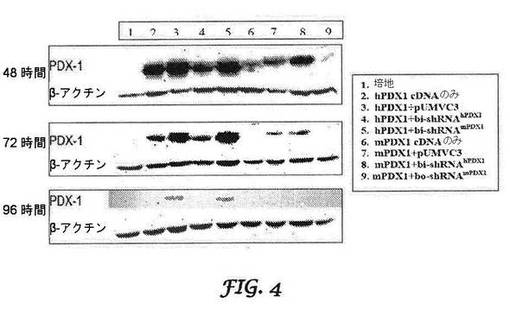
【図5A】
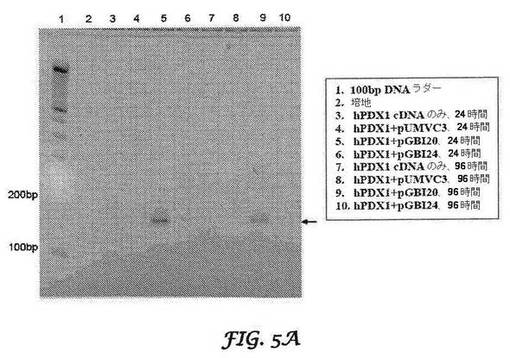
【図5B】

【図6A】
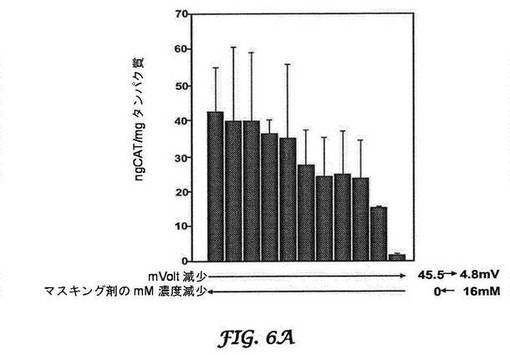
【図6B】
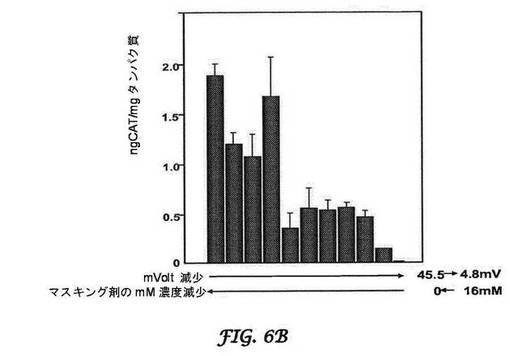
【図6C】
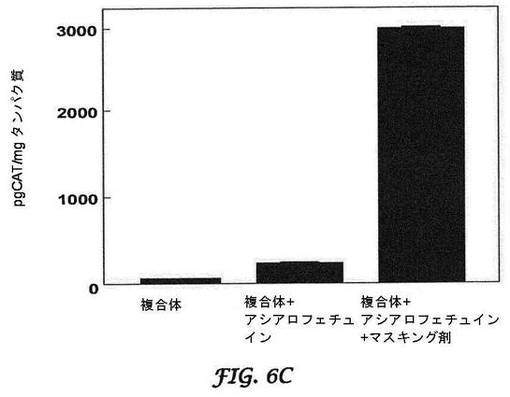
【図7A】

【図7B】
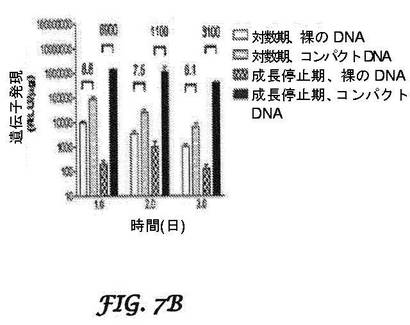
【図7C】
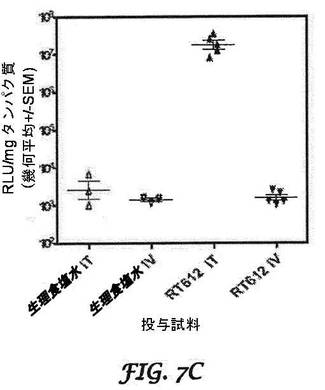
【図8】
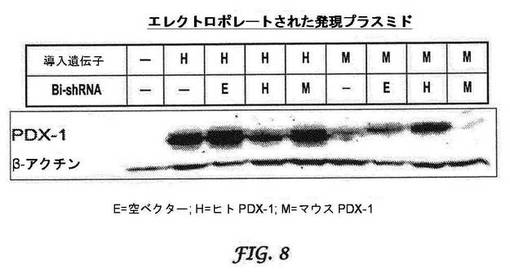
【図9A】
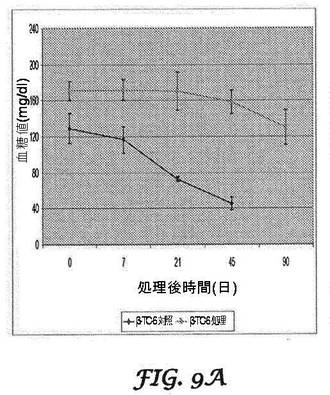
【図9B】
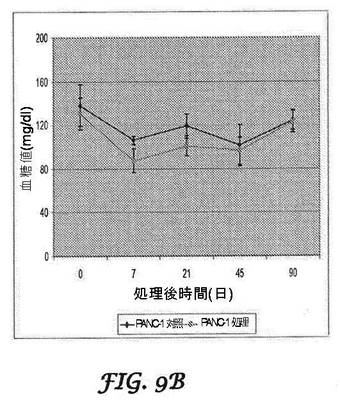
【図10A】
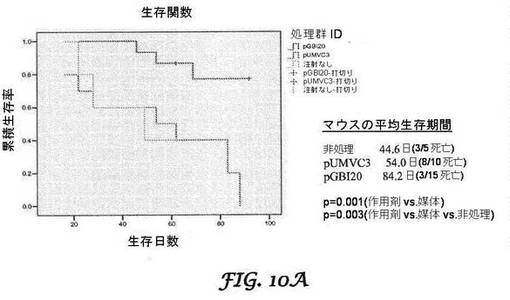
【図10B】
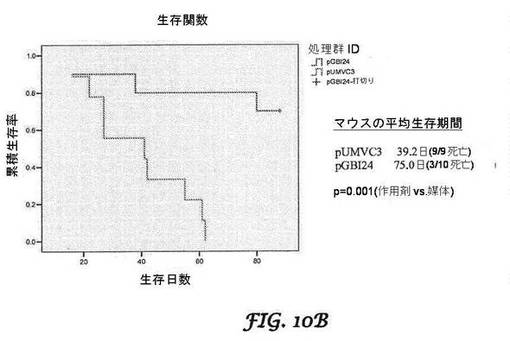
【図11】
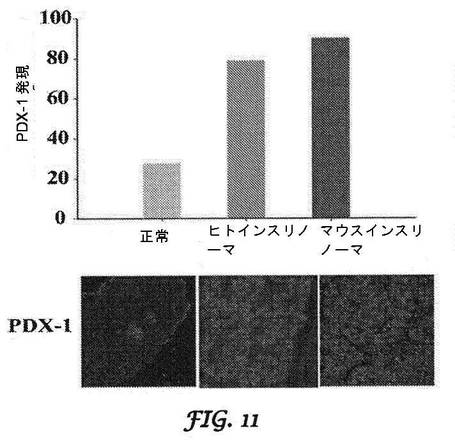
【図12】
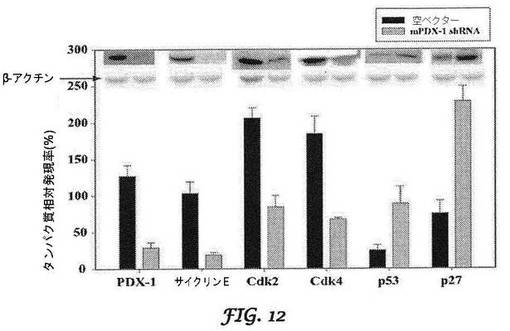
【図13】
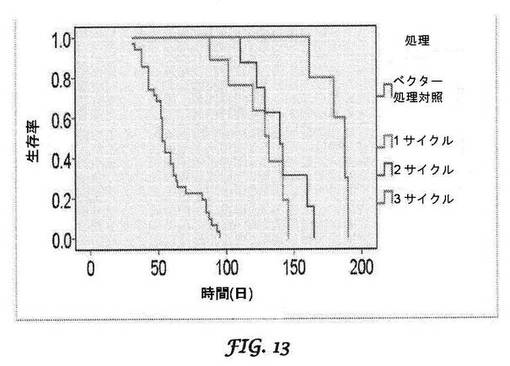
【図14】
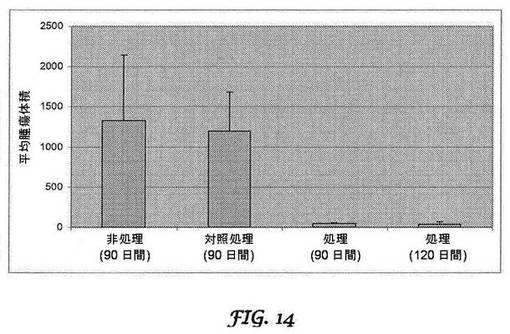
【図15】
【図3A】
【図3B】
【図4】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図7A】
【図7B】
【図7C】
【図8】
【図9A】
【図9B】
【図10A】
【図10B】
【図11】
【図12】
【図13】
【図14】
【図15】
【公表番号】特表2013−509183(P2013−509183A)
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願番号】特願2012−537017(P2012−537017)
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/US2010/054350
【国際公開番号】WO2011/053660
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(510285779)グラダリス インク. (4)
【氏名又は名称原語表記】GRADALIS,INC.
【Fターム(参考)】
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/US2010/054350
【国際公開番号】WO2011/053660
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(510285779)グラダリス インク. (4)
【氏名又は名称原語表記】GRADALIS,INC.
【Fターム(参考)】
[ Back to top ]