
PPARδリガンドのスクリーニング方法
【課題】PPARδリガンド(アゴニスト又はアンタゴニスト)としての有効性をハイスループットに評価できる手段を提供すること
【解決手段】PPARδのリガンド結合領域を含む第1領域と、転写因子のDNA結合領域又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子;PPARγコアクチベーター1−αの受容体相互作用領域を含む第3領域と、前記第2領域がDNA結合領域を含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDNA結合領域を含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子;及び前記DNA結合領域が結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト;を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
【解決手段】PPARδのリガンド結合領域を含む第1領域と、転写因子のDNA結合領域又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子;PPARγコアクチベーター1−αの受容体相互作用領域を含む第3領域と、前記第2領域がDNA結合領域を含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDNA結合領域を含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子;及び前記DNA結合領域が結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト;を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ペルオキシソームプロリフェレータ活性化受容体δ(PPARδ)のリガンドのスクリーニング方法に関する。
【背景技術】
【0002】
肥満は生活習慣病を始めとする様々な疾患の原因であり、北米においては重大な社会問題となっている。治療法としては食餌療法や運動療法などがあるが、重度の肥満に対しては即効性に乏しく、十分な効果が得られないのが現状である。PPARδは、脂肪酸およびその誘導体などをリガンドとする核内受容体で、糖代謝、脂質代謝、エネルギー産生に関与することが知られている。PPARδは脂肪の燃焼を促進させる作用があり、恒常的に活性化したPPARδを脂肪細胞特異的に過剰発現させたマウスでは、高脂肪食等の肥満誘因負荷を与えてもほとんど肥満にならない(Wang YX. et al., Cell. 113. 159-170, 2003)。また驚いたことにこのマウスでは、十分な運動を行った動物のような筋繊維の変化が認められることから(Wang YX. et al., Cell. 113. 159-170, 2003)、PPARδは単なる抗肥満薬としてだけでなく、運動効果も期待できるような抗肥満薬の分子標的として有望視されており、そのアゴニストは即効性の高い抗肥満薬としての作用が期待されている。PPARδは開拓されたばかりの標的分子であるため、今後、そのリガンドの探索や開発競争が激化し、それに伴い、PPARδリガンドをハイスループットにスクリーニングできる手法に対するニーズが高まると予想される。尚、PPARδと同様にPPARのサブタイプの一つであるPPARγについては、リガンド依存的にPPARγと相互作用するタンパク質のスクリーニング法(特許文献1)や、アゴニスト及びアンタゴニストのスクリーニング法(特許文献2)が知られている。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】国際公開第2003/062427号パンフレット
【特許文献2】特開平11−56369号公報
【非特許文献】
【0004】
【非特許文献1】J. Nishikawa, K. Saito, J. Goto, F. Dakeyama, M. Matsuo, T. Nishihara: Toxicol. Appl. Pharmacol., 154, 76(1999)
【非特許文献2】T. Kanayama, S. Mamiya, T. Nishihara, J. Nishikawa: J Biocliem., 133. 791(2003)
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の課題は、PPARδリガンド(アゴニスト又はアンタゴニスト)としての被験物質の有効性を、ハイスループットに評価できる手段を提供することにある。
【課題を解決するための手段】
【0006】
これまでに、PPARδについてはアゴニストに良好な反応を示す酵母two-hybrid(ツーハイブリッド)スクリーニング系は構築されていない。PPARのように内因性のアゴニストが未知であるいわゆるオーファン受容体の多くは、ステロイドホルモン受容体等とは異なり、下等生物にも存在し、また宿主自体に内因性のリガンドを有している可能性が示唆されている。PPARδについても例外ではないと考えられ、実はセオリー通り酵母two-hybrid系を構築しても、アゴニストに依存した反応は見られない。酵母内には、もともとPPARのアゴニストになるような内因性の脂溶性生理活性物質が豊富に存在していることが指摘されており、これが一因となっていると考えられる。尚、核内ホルモン受容体とそのコアクチベーターとの相互作用を利用した酵母two-hybrid系によって、内分泌撹乱物質をスクリーニングする方法が報告されている(非特許文献1)。
【0007】
このような背景の下、本発明者らはエストロゲン受容体やレチノイン酸受容体等で良好な反応性を示した主要なコアクチベーター7種の部分配列を用いて、PPARδリガンドスクリーニング系の構築を試みた。その結果、PPARγコアクチベーター1−α(PGC1α)が最も良好な反応性を示した。しかも、全長を用いた場合よりも、適度な長さの部分PGC1α(受容体相互作用領域(RID)を含む)を用いることで良好な反応性を示すことも見出した。更には、PGC1αをコアクチベーターとしたスクリーニング系によれば、アゴニスト活性とアンタゴニスト活性の両者を評価可能であることが確認された。
【0008】
以上の通り、本発明者の検討によってPGC1αの有用性が判明し、それを利用したPPARδリガンド評価系(スクリーニング系)が完成した。ところで、従来のPPARδリガンド評価系として、マイクロイムノプレートと精製タンパク質を用いた無細胞検出系が先行して存在する(非特許文献2)。基本的に無細胞検出系においても応用するイベントは酵母two-hybrid系と同様である。無細胞検出系は、酵母two-hybrid系よりもハイスループットに検討することが可能であるという利点を有する上、酵母を用いた場合には、被験物質の酵母に対する毒性を考慮した上で検討を行う必要があるが、無細胞系においては毒性を気にせずに濃度を設定できる利点も有する。しかしその一方で、無細胞系は天然物や食品成分等の疎水性粗抽出分画のような混合物では擬陽性が非常に多く、スクリーニングできない。これは無細胞系がリガンド依存的、非依存的にかかわらず、タンパク質間相互作用が起こりさえすれば陽性と判定されるためで、このようなサンプルにはタンパク質変性作用を有していたり、タンパク質に非特異的に接着することでタンパク質同士を結合させるような化合物が含まれていたりするためであると考えられる。したがってサンプルの種類を問わない酵母two-hybrid系は、無細胞系よりもスクリーニングできる検体が幅広く、アドバンテージを有している。即ち、無細胞系に比して汎用性が高い点も利点といえる。
【0009】
一方、動物細胞内で核内受容体のアゴニスト活性を評価する実験系として、酵母の転写因子であるGAL4のDNA結合領域(DNA binding domain:DBD)と核内受容体のリガンド結合領域(ligand binding domain:LBD)の融合タンパク質と転写開始点上流にガラクトース代謝遺伝子の上流域活性化配列(upstream activation sequence:UAS)が存在しているルシフェラーゼを利用した系が存在する。この実験系は、核内受容体リガンドの核内受容体LBDへの結合から標的遺伝子の転写活性化までの、核内受容体によるリガンド依存的な遺伝子転写制御のイベントを一通り網羅しているため、酵母による核内受容体アゴニスト活性評価法よりも正確な評価が可能であるといえる。本発明者らが完成させた上記評価系と当該実験系を比較した結果、両者の間に相関を認め上記評価系の有効性が裏付けられた。また、上記評価系による結果(データ)を評価する上で重要且つ有用な情報が得られた。
以下に列挙する本願発明は、以上の成果ないし知見に基づく。
[1]PPARδのLBDを含む第1領域と、転写因子のDBD又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子、
PGC1αのRIDを含む第3領域と、前記第2領域がDBDを含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDBDを含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子、及び
前記DBDが結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト、
を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
[2]前記第3領域が、PGC1αの一部分である、[1]に記載のスクリーニング方法。
[3]前記第3領域が、PGC1αのN末端側から1番目のアミノ酸〜188番目のアミノ酸からなる、[2]に記載のスクリーニング方法。
[4]前記第3領域が、配列番号5のアミノ酸配列からなる、[3]に記載のスクリーニング方法。
[5]前記細胞が酵母細胞である、[1]〜[4]のいずれか一項に記載のスクリーニング方法。
[6]前記転写因子が酵母のGAL4である、[5]に記載のスクリーニング方法。
[7]前記第2領域が前記転写因子のDBDを含み、前記第4領域が前記転写因子の転写活性化領域を含む、[1]〜[6]のいずれか一項に記載のスクリーニング方法。
【図面の簡単な説明】
【0010】
【図1】PPARδアゴニスト活性評価系に使用するコアクチベーターの検討。
【図2】PGC1α全長を発現する酵母を用いた酵母による検討。
【図3】PGC1αおよびPPARδ LBDを発現する酵母を用いたPPARδアゴニスト活性評価
【図4】PGC1αおよびPPARδ LBDを発現する酵母を用いたPPARδアンタゴニスト活性評価。
【図5】レポーターアッセイによるPPARδアゴニスト活性評価。
【図6】レポーターアッセイによるPPARδアンタゴニスト活性評価。
【発明を実施するための形態】
【0011】
本発明はPPARδのリガンド、即ちアゴニスト(作動薬)またはアンタゴニスト(拮抗薬)をスクリーニングする方法を提供する。本発明のスクリーニング方法ではPPARδと転写共因子とのリガンド依存的な相互作用が生ずるように操作した細胞を用い、当該相互作用に伴うレポーター遺伝子の発現を指標として、被験物質の有効性を評価する。PPARδと転写共因子とのリガンド依存的な相互作用が生ずるように、本発明のスクリーニング方法に用いる細胞は、3要素、即ち第1遺伝子、第2遺伝子及びレポーターコンストラクトを含む。以下、各要素について詳細に説明する。尚、ヒトPPARδのアミノ酸配列を配列番号1に示す。
【0012】
<第1遺伝子>
本発明では、PPARδのLBDを含む第1領域と、転写因子のDBD又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする遺伝子のことを第1遺伝子と呼ぶ。PPARδのLBDは、PPARδのアミノ酸配列のN末端側から254番目〜441番目までのアミノ酸を含む領域と考えられている。少なくとも当該領域を含む限り、第1領域の長さは特に限定されない。第1領域の具体例を配列番号2(PPARδの140a.a.〜441a.a.)に示す。
【0013】
第2領域は転写因子のDBD又は転写活性化領域を含む。好ましい転写因子として酵母GAL4を用いることができる。即ち、好ましい一態様では、GAL4のDBDとGAL4の転写活性化領域を用いる。一方、LexAのDBDとVP16の転写活性化領域の組合せや、LexAのDBDと大腸菌由来のB42転写活性化領域の組合せを用いることにしてもよい。
【0014】
一態様では、第2領域が転写因子のDBDを含む。DBDは、後述の応答配列に対する結合能を示す一方で、転写活性化能を示さない。別の一態様では、第2領域が転写因子の転写活性化領域を含む。転写活性化領域とは、転写活性化能を示す一方、DNA結合能を有しない領域である。転写因子のDBDは、例えばGAL4の場合、N末端側(およそ第1番目から147番目までのアミノ酸を含む領域)に存在することが知られている。同様に転写活性化領域は、例えばGAL4の場合、C末端側(およそ第768番目から881番目までのアミノ酸を含む領域)に存在することが知られている。少なくとも当該領域を含む限り、第2領域の長さは特に限定されない。DBDを採用した場合の第2領域の具体例を配列番号3(GAL4のDBD)に示す。
【0015】
上記の如き構成の第1領域と第2領域が連結されてなる融合タンパク質が第1遺伝子にコードされる。第1領域の機能と第2領域の機能に支障のない限りにおいて、第1領域と第2領域の間に別の配列が介在していてもよい。好ましい態様では、第1領域と第2領域が直接連結されている。第1領域と第2領域の間ではなく、第1領域又は第2領域の外側に他の配列が付加されていてもよい。また、第1領域と第2領域の順序は特に問わず、いずれが上流側に配置されていてもよい。尚、第1遺伝子の配列の例を配列番号4に示す。
【0016】
<第2遺伝子>
本発明では、PGC1αのRIDを含む第3領域と、前記第2領域がDBDを含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDBDを含む第4領域とが連結されてなる融合タンパク質をコードする遺伝子のことを第2遺伝子と呼ぶ。
【0017】
PGC1αでは、そのRIDはN末端側から88番目〜92番目までのアミノ酸、144番目〜148番目、または210番目〜214番目までのアミノ酸を含む領域と考えられている。少なくともいずれかの当該領域を1つ以上含む限り、第3領域の長さは特に限定されない。本発明者らの検討の結果、PGC1α全長を用いた場合には、良好な相互作用が認められなかった。この知見に基づき、好ましくは、PGC1αの一部を第3領域に使用する。PGC1αの全長は798アミノ酸残基であるが、例えば、1番目〜188番目のアミノ酸の部分PGC1α(但し、RIDを含む)を用いるとよい。好ましい第3領域の具体例を配列番号5(1a.a.〜188a.a.)に示す。
【0018】
第4領域は転写因子のDBD又は転写活性化領域を含むが、第2領域との間でDBDと転写活性化領域からなるペアーが形成されるように、第2領域にDBDを使用した場合には、第4領域には転写因子の転写活性化領域を使用し、これとは逆に、第2領域に転写活性化領域を使用した場合には、第4領域には転写因子のDBDを使用する。このように構成することによって、第2領域が供給する転写因子の要素と、第4領域が提供する転写因子の要素が一体となることで、応答配列への結合及びそれに続くレポーター遺伝子の転写活性化が引き起こされる。第2領域と第4領域による当該作用・効果が生ずる限りにおいて、第4領域が含む転写因子の要素の由来が、第2領域が含む転写因子の要素の由来と同一である必要はない。尚、転写活性化領域を採用した場合の第4領域の具体例を配列番号6(GAL4の転写活性化領域)に示す。
【0019】
上記の如き構成の第3領域と第4領域が連結されてなる融合タンパク質が第2遺伝子にコードされる。第3領域の機能と第4領域の機能に支障のない限りにおいて、第3領域と第4領域の間に別の配列が介在していてもよい。好ましい態様では、第3領域と第4領域が直接連結されている。第3領域と第4領域の間ではなく、第3領域又は第4領域の外側に他の配列が付加されていてもよい。また、第3領域と第4領域の順序は特に問わず、いずれが上流側に配置されていてもよい。尚、第2遺伝子の配列の例を配列番号7に示す。
【0020】
<レポーターコンストラクト>
本発明では、上記DBDが結合し得る応答配列及び当該応答配列に連結されたレポーター遺伝子を含むものをレポーターコンストラクトと呼ぶ。使用する転写因子に対応した応答配列が採用される。例えば、転写因子としてGAL4を採用した場合、応答配列としてUASを用いることができる。
【0021】
レポーター遺伝子には、例えば、出芽酵母の栄養要求性遺伝子(ADE2、HIS3、URA3)、大腸菌由来のβ−ガラクトシダーゼ遺伝子(lacZ)、バクテリアトランスポゾン由来のクロラムフェニコールアセチルトランスフェラーゼ遺伝子(CAT)、ホタル由来のルシフェラーゼ遺伝子(Luc)を用いることができる。レポーター遺伝子は、応答配列の制御下に配置される。即ち、応答配列に対して作動的にレポーター遺伝子が連結される。
【0022】
第1遺伝子、第2遺伝子及びレポーターコンストラクトは、遺伝子組換え技術を用いて調製できる。第1遺伝子に含まれる、PPARδのLBDをコードするDNAや転写因子の一部をコードするDNA、或いは第2遺伝子に含まれる、PGC1αのRIDをコードするDNAや転写因子の一部をコードするDNAなどは、既知のアミノ酸配列や塩基配列の情報などをもとに設計・合成したプライマーやプローブを用いて、例えばcDNAライブラリーから単離することができる。単離したDNAを制限酵素やPCR法などを用いて連結することにより、第1遺伝子及び第2遺伝子を構築することができる。
【0023】
以上で説明した3要素を細胞に導入することにより、本発明に使用するスクリーニング用細胞を得ることができる。例えば第1遺伝子、第2遺伝子、レポーターコンストラクトをそれぞれ適当なベクター(例えばプラスミドベクター)に組み込み、当該ベクターで宿主細胞を形質転換すればよい。二つ以上の要素を一つのベクターで導入することにしてもよい。尚、各種レポーター遺伝子に関して、それが応答配列とともに組み込まれた細胞を利用可能な状況にある。そのような細胞を利用すれば、第1遺伝子及び第2遺伝子のみを導入することによって、本発明に必要なスクリーニング用細胞が得られる。
【0024】
好ましい態様では、酵母two-hybrid系として本発明のスクリーニング方法を構成する。酵母two-hybrid系とは、酵母を宿主細胞として用い、細胞内で生じるタンパク質相互作用の存在を検出するものである。基本的には真核細胞の転写因子の多くがDBDと転写活性化領域に分割が可能であり、これらの領域に調べようとする2種類のタンパク質をそれぞれに連結して酵母内で発現させたときに転写因子の活性化が起これば、これらのタンパク質は会合していることが分かる。酵母two-hybrid系を核内受容体のリガンドスクリーニング系に応用する場合は、一般に、核内受容体と転写因子(例えば酵母転写因子GAL4)のDBDとの融合タンパク質及びコアクチベーターと転写因子の転写活性化領域の融合タンパク質を酵母内で発現させる。また、これらとともに転写因子の応答配列をレポーター遺伝子の上流に挿入したレポータープラスミドも酵母内に導入する。例えば、被験物質がアゴニスト活性を有している場合には、核内受容体(本発明の場合はPPARδ)のLBDのコンフォメーションが変化し、コアクチベーターをリクルートするため、両融合タンパク質は結合し転写因子の働きを持つタンパク質複合体が形成される。このタンパク質複合体は転写因子として機能し、レポーター遺伝子の発現を誘導する。したがって、被験物質のリガンド活性をレポーター遺伝子の発現量で評価することができる。被験物質のリガンド活性の強弱は、活性化された核内受存体のコアクチベーターのリクルート能に反映されることから、このレポーター遺伝子の発現の度合いを指標にリガンド活性を定量的に検出することが可能となる。典型的なアゴニストを試験系に加えておいて、同時に被験物質を共存させることによりアンタゴニスト活性も検出できる。酵母two-hybrid系には典型的にはサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)が用いられる。尚、酵母two-hybrid系の原理や構成等については、J. Nishikawa, K. Saito, J. Goto, F. Dakeyama, M. Matsuo, T. Nishihara: Toxicol. Appl. Pharmacol., 154, 76(1999)が参考になる。
【0025】
DBDと転写活性化領域に分割された転写因子の活性化を評価できるのであれば、酵母細胞以外の真核細胞(例えば哺乳動物細胞や昆虫細胞)を用いることも可能である。
【0026】
本発明のスクリーニング方法では、上記のように3要素を含むスクリーニング用細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析する。具体的には、例えば次のステップ(1)〜(3)を行えばよい。
(1)スクリーニング用細胞に被験物質を接触させるステップ
(2)レポーター遺伝子の発現を検出するステップ
(3)検出結果に基づき、被験物質による、リガンド依存的な相互作用の変化の有無及び/又は程度を調べるステップ
【0027】
ステップ(1)では、用意したスクリーニング用細胞に対して、被験物質を接触させる。例えば、スクリーニング用細胞の培養液中に被験物質を添加する。或いは、スクリーニング用細胞の培養液を、被験物質を含有するものに交換する。培養方法は常法に従えばよく、スクリーニング用細胞の種類に応じて決定できる。被験物質との接触時間、即ち被験物質を添加などすることによって、培養液中に被験物質が共存するようになってから、ステップ(2)に至るまでの時間は予備実験を通して設定すればよいが、例えば30分〜6時間である。
【0028】
試験物質としては様々な分子サイズの有機化合物又は無機化合物を用いることができる。有機化合物の例として、核酸、ペプチド、タンパク質、脂質(単純脂質、複合脂質(ホスホグリセリド、スフィンゴ脂質、グリコシルグリセリド、セレブロシド等)、プロスタグランジン、イソプレノイド、テルペン、ステロイド、ポリフェノール、カテキン、ビタミン(B1、B2、B3、B5、B6、B7、B9、B12、C、A、D、E等)を例示できる。試験物質は天然物由来であっても、或いは合成によるものであってもよい。後者の場合には例えばコンビナトリアル合成の手法を利用して効率的なスクリーニング系を構築することができる。尚、細胞抽出液、培養上清などを試験物質として用いてもよい。また、既存の薬剤を試験物質としてもよい。2種類以上の被験物質を同時に添加することにより、被験物質間の相互作用、相乗作用などを調べることにしてもよい。
【0029】
ステップ(1)に続くステップ(2)では、レポーター遺伝子の発現を検出する。検出は常法に従えばよい。本発明で用いることが可能な汎用的なレポーター遺伝子に関しては、基本的な検出法が確立されており、成書や過去の論文等を参考にして、適切な方法や条件を容易に選択ないし設定することが可能である。
【0030】
ステップ(3)では、ステップ(2)の検出結果に基づき、リガンド依存的な相互作用に及ぼす被験物質の影響・効果を調べる。具体的には、スクリーニング用細胞に被験物質を接触させたことにより、リガンド依存的な相互作用の変化が生じたか、又はリガンド依存的な相互作用の変化はどの程度か、或いはこれらの両方を調べ、被験物質のアゴニスト又はアンタゴニストとしての有効性を判定する。例えば、被験物質の接触(共存)によってレポーター遺伝子の発現の増強を認めた場合(発現を検出できるようになった場合も含む)、即ち、リガンド依存的な相互作用を正に調節した場合、被験物質がアゴニスト活性を有すると判定できる。通常は、スクリーニング用細胞に被験物質を接触させない場合(陰性対照)と比較し、被験物質のアゴニスト活性を判定する。また、このような陰性対照の利用と併せて或いはその代わりに、アゴニスト活性を示すことが判明している物質(既知アゴニスト)をスクリーニング用細胞に接触させた場合(陽性対照)と比較し、被験物質のアゴニスト活性の程度を評価してもよい。以上のように陰性対照及び/又は陽性対照との比較に基づき被験物質の有効性を判定・評価すれば、より客観的且つ信頼性の高い判定結果が得られる。
【0031】
一方、被験物質のアンタゴニスト活性を評価する場合には、原則、アゴニストの共存下でステップ(1)を行い(即ち、被験物質とアゴニストをスクリーニング用細胞に接触させる)、その場合のレポーター遺伝子の発現を、アゴニストのみをスクリーニング用細胞に接触させた場合のレポーター遺伝子の発現と比較する。後者に比較して前者にレポーター遺伝子の発現の減弱を認めた場合、即ち、アゴニストに起因するリガンド依存的な相互作用を被験物質が負に調節した場合、被験物質がアンタゴニスト活性を有すると判定できる。従って、アンタゴニスト候補として当該被験物質が選抜される。
【0032】
発現増強又は減弱の程度に基づき、被験物質のアゴニスト又はアンタゴニストとしての有効性を定量的に判断してもよい。尚、PPARδのアゴニストとしてGW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)が知られており、同様にPPARδのアンタゴニストとしてGSK0660が知られている。
【0033】
PPARδのアゴニストは、即効性の高い抗肥満薬の有効成分として期待される。他方、PPARδのアンタゴニストは、ヒトの肺ガン、肝ガン、乳ガン由来細胞株の増殖を抑制し、アポトーシスを誘導すること(Zaveri NT. et al., Cancer Biol Ther. 8. 1252-1261, 2009)から、抗ガン剤の有効成分として期待される。
【0034】
本発明のスクリーニング方法によって選択された物質が十分な薬効を有する場合には、当該物質をそのまま医薬の有効成分として使用することができる。一方で十分な薬効を有しない場合には化学的修飾などの改変を施してその薬効を高めた上で、医薬の有効成分として使用することができる。勿論、十分な薬効を有する場合であっても、更なる薬効の増大を目的として同様の改変を施してもよい。
【実施例】
【0035】
以下の説明では次の略号を使用する。
AD : activation domain(転写活性化領域)
RID : Receptor Interaction Domain(受容体相互作用領域)
DBD : DNA binding domain(DAN結合領域)
LBD : ligand binding domain(リガンド結合領域)
UAS : upstream activating sequence(ガラクトース代謝遺伝子の上流域活性化配列)
【0036】
1.実験材料
(1)酵母two-hybrid法で使用する試薬および組成
・YPDA培地
20 g/L ペプトン
10 g/L 酵母エキス(Yeast extract)
20 g/L グルコース
30 mg/L L-アデニンヘミ硫酸塩
・SD-L/-T培地
6.7 g/L Difco yeast nitrogen base without amino acid
100 mL/L 10x Dropout溶液(-Leu, -Trp)
20 g/L グルコース
・Dropout溶液(-Leu, -Trp)
300 mg/L L-イソロイシン
1500 mg/L L-バリン
200 mg/L L-アデニンヘミ硫酸塩
200 mg/L L-アルギニンHCl
200 mg/L L-ヒスチジンHCl・H2O
300 mg/L L-リジンHCl
200 mg/L L-メチオニン
500 mg/L L-フェニルアラニン
2000 mg/L L-スレオニン
300 mg/L L-チロシン
200 mg/L L-ウラシル
・SD-L/-Tプレート
SD-L/-T培地に寒天を2%(w/v)となるように加えたもの
・10x TE
100 mM Tris-HCl(pH 7.5)
10 mM EDTA
・10x LiAc溶液
1 M 酢酸リチウム(pH 7.5)
・50%(w/v) PEG 4000溶液
・LiAc/TE溶液
10x TE 1 mL
10x LiAc 1 mL
超純水 8 mL
・LiAc/TE/PEG溶液
10x TE 1 mL
10x LiAc 1 mL
50%(w/v) PEG 4000溶液 8 mL
・Z-バッファー
60 mM Na2HPO4
40 mM NaH2PO4
10 mM KCl
1 mM MgSO4
【0037】
(2)実験に使用したPPARδリガンド
【化1】

アゴニスト:GW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)
アンタゴニスト:GSK0660
【0038】
(3)酵母two-hybrid法で使用するプラスミドの構築
(a)酵母内でGAL4-ADとTIF2の融合タンパク質を発現するプラスミド(pGAD424-TIF2)
ヒトTIF2のRID(483-1090 a.a.(配列番号8))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaq(Applied Biosystems)を用いてPCRを行うことで増幅し、pBluescript KS-(stratagene)に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424(clontech)に挿入し、プラスミドを構築した。
【0039】
(b)酵母内でGAL4-ADとPGC1αRID(1-188 a.a.(配列番号5))の融合タンパク質を発現するプラスミド(pGAD424-PGC1α)
ヒトPGC1αのRID(1-188 a.a.(配列番号5))をコードする配列をHepG2細胞由来cDNAを鋳型とし、LA taq(TAKARA BIO)を用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、Sma I、Sal Iを用いて配列を切り出し、pGAD424に挿入し、プラスミドを構築した。
【0040】
(c)酵母内でGAL4-ADとPGC1α(798 a.a.(配列番号9))の融合タンパク質を発現するプラスミド(pGAD424-PGC1αfull)
ヒトPGC1αをコードする配列をHepG2細胞由来cDNAを鋳型とし、KOD-Plus(TOYOBO)を用いてPCRを行うことで増幅した後、Sma I、Sal Iで処理した。これをSma I、Sal Iで処理したpGAD424に挿入し、プラスミドを構築した。またDNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した。
【0041】
(d)酵母内でGAL4-ADとPGC1βの融合タンパク質を発現するプラスミド(pGAD424-PGC1β)
ヒトPGC1βのRID(1-264 a.a.(配列番号10))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、BamH I、Sal Iを用いて配列を切り出し、pGAD424に挿入し、プラスミドを構築した。
【0042】
(e)酵母内でGAL4-ADとSRC1の融合タンパク質を発現する
プラスミド(pGAD424-SRC1)
ヒトSRC1のRID(570-783 a.a.(配列番号11))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0043】
(f)酵母内でGAL4-ADとp300/CBPの融合タンパク質を発現するプラスミド(pGAD424-CBP)
ヒトp300/CBPのRID(61-522 a.a.(配列番号12))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0044】
(g)酵母内でGAL4-ADとACTRの融合タンパク質を発現するプラスミド(pGAD424-ACTR)
ヒトACTRのRID(625-820 a.a.(配列番号13))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0045】
(h)酵母内でGAL4-ADとRIP140の融合タンパク質を発現するプラスミド(pGAD424-RIP140)
ヒトRIP140のRID(752-1158 a.a.(配列番号14))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0046】
(i)酵母内でGAL4-DBDとPPARδ LBD(140-441 a.a.)の融合タンパク質を発現するプラスミド(pGBT9-PPARδ LBD)
ヒトPPARδ LBD(140-441 a.a.(配列番号2))をコードする配列をHepG2細胞由来のcDNAを鋳型とし、KOD-Plusを用いてPCRを行うことで増幅した後、Sma I、Sal Iで処理した。これをSma I、Sal Iで処理したpGBT9(clontech)に挿入し、プラスミドを構築した。またDNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した。
【0047】
(4)レポーターアッセイで使用するプラスミドの構築
(A)培養細胞内でホタルルシフェラーゼを発現するレポータープラスミド(p4xUAS-tk-Luc)
4コピーのGAL4DBD結合配列であるUAS 配列(5’-CGACGG AGTACTGTCCTCCGAGCT-3’(配列番号15))をSma Iで処理したptk-luc(clontech)に挿入し、プラスミドを構築した。
【0048】
(B)培養細胞内でGAL4-DBDとPPARδ LBD(140-441 a.a.(配列番号2))の融合タンパク質を発現するプラスミド(pM-PPARδ LBD)
pGBT9-PPARδ LBDをEcoR I、Sal I処理することでヒトPPARδ LBD(140-441 a.a.(配列番号2))をコードする配列を切り出し、これをEcoR I、Sal I処理したpM(clontech)に組み込みプラスミドを構築した。
【0049】
2.実験方法
(1)酵母へのトランスフォーメーション
酵母(Y190株)をYPDA培地15 mL(1トランスフォーメーションあたり)で30℃、16〜24時間培養後、遠心分離(室温、3,000 rpm、5分)により集菌し、1xTE、LiAc/TE溶液で洗浄後、150μLのLiAc/TE溶液で懸濁した。懸濁した酵母を1.5 mLチューブに移し、pGAD424プラスミド(TIF2、PGC1α、PGC1αfull、PGC1β、SRC1)およびpGBT9-PPARδ LBDプラスミドをそれぞれ1μg相当量、キャリアーDNA溶液(ウシ胸腺DNA、SIGMA D-8661)を7μL添加し、LiAc/TE/PEG溶液を850μL加え、よく混和した。混和後、30℃で30分、42℃で15分インキュベートした後、遠心分離(室温、3,000 rpm、3分)を行い、LiAc/TE/PEG溶液を除去した。沈殿を500μLの超純水で懸濁し、再度遠心分離(室温、3,000 rpm、3分)を行い、上清を除去した後、200μLのLiAc/TEで再懸濁し、SD-L/-Tプレートに塗布した。プレートは適当なコロニーが大きさのコロニーが得られるまで(2〜4晩)30℃でインキュベートし、コロニーが生えたプレートはβ-ガラクトシダーゼ活性測定時まで4℃で保存した。
【0050】
(2)β-ガラクトシダーゼ活性測定
pGAD424プラスミドとpGBT9プラスミドを導入した酵母を15 mLのSD-L/-T培地に植菌した後、30℃で一晩培養し、前培養液とした。前培養液を150μLとり、96穴プレートに移し、プレートリーダーで600 nmの吸光度(OD600)を測定した上で、前培養液をOD600=0.05となるようにSD-L/-T培地で希釈し、希釈液250μLを1.5 mLチューブに移した。さらに試料溶液を2.5μL加え、良く混和した後、30℃で4時間培養した。培養後、培養液を150μLとり、96穴プレートに移し、プレートリーダーでOD600を測定した。残りの培養液100μLを遠心(室温、15,000 rpm、5分)し、上清を除き、1 mg/mLのZymolyase-20T(生化学バイオビジネス株式会社)を含むZバッファーを200μL加え、良く混和した後、37℃で30分静置し、酵母を溶解した。酵母の溶解を確認した後、4 mg/mLのo-nitrophenyl-β-galactoside(Sigma)を含むZバッファーを40μL加えて良く混和し、30℃で発色反応を開始させ、30分後に1M Na2CO3溶液を100μL加えて反応を停止させた。遠心(室温、15,000 rpm、5分)後、上清150μLを96穴プレートに移し415 nmの吸光度(OD415)を測定し、次式を用いてβ-ガラクトシダーゼ活性を算出した。
【数1】
【0051】
(3)細胞培養
ヒト胎盤絨毛細胞株であるJEG-3細胞(American Type Culture Collection, ATCC)の培養には10%ウシ胎児血清、2 mM L-グルタミン、1 mMピルビン酸ナトリウム(Nacalai Tesque)及び0.1 mM MEM非必須アミノ酸溶液(GIBCO)を加えたMEM培地(Nacalai Tesque)を用いた。培養は37℃、5% CO2存在下で行った。遺伝子導入時には無血清培地で、遺伝子導入以降の培養は活性炭処理したウシ胎児血清を1%加えた培地を用いた。
【0052】
(4)遺伝子導入および化合物処理
JEG-3細胞を3.0x104細胞/ウェルとなるように24穴平底プレート(corning)に播種し、24時間後に遺伝子導入を行った。遺伝子導入には、Lipofectamine Reagent(Invitrogen)およびPlus Reagent(Invitrogen)を用いて、その標準プロトコールに従って操作を行った。プラスミド量は、1ウェルあたりp4xUAS-tk-Luc 50 ng、pM-PPARδ LBD 20 ng、pGL 4.75(Promega)10 pgとした。遺伝子導入3時間後に、1%活性炭処理ウシ胎児血清含有培地に交換し、培地交換の21時間後に化合物の処理を行った。
【0053】
(5)ルシフェラーゼアッセイ
化合物処理24時間後に細胞をPBSで2回洗浄した後、Passive Lysis Buffer(Promega)で溶解した。20分静置後、細胞溶解液20μLを96穴白色平底プレート(NUNC)に加え、Dual-Luciferase Reporter Assay System(Promega)を用いてルミノメーター(Berthold Japan)によりホタルルシフェラーゼ活性を測定した。内部標準として、pGL 4.75のウミシイタケルシフェラーゼ活性を測定し、これを用いて遺伝子導入効率を補正した。
【0054】
3.実験結果
(1)酵母によるPPARδアゴニスト活性評価系に使用するコアクチベーターの選別
酵母two-hybrid法は、酵母を宿主細胞に用いて、任意の2つのタンパク質の結合をレポーター遺伝子の発現を指標に検出するシステムである。核内受容体にリガンドが結合していない状態では、コリプレッサー複合体(標的遺伝子の転写を抑制する働きをする)が核内受容体に結合し、標的遺伝子の転写を抑制しているが、リガンドが結合すると核内受容体の立体構造が変化し、それに伴って、コリプレッサーが解離し、コアクチベーター複合体(標的遺伝子の転写を促進する働きをする)が結合して、標的遺伝子の転写を促進する。酵母two-hybrid法を用いた核内受容体アゴニスト活性評価系はこのイベントを応用している。すなわち、酵母内で酵母の転写制御因子であるGAL4と核内受容体LBD融合タンパク質にアゴニストが結合すると、核内受容体LBDが前述の立体構造変化が起こることでコアクチベーターを迎え入れる状態となり、そこにGAL4の活性化領域とコアクチベーター融合タンパク質のコアクチベーター部分が結合することで、レポーター遺伝子が発現する。またアゴニスト作用の強弱は、活性化された核内受容体のコアクチベーターのリクルート能に反映されることから、このレポーター遺伝子の発現の度合いを指標にアゴニスト作用を定量的に検出することが可能となる。
【0055】
この系を用いてPPARδアゴニスト活性を評価するにあたっては、リガンド添加によりレポーター遺伝子発現が大きく上昇させられるようなコアクチベーターを使用する必要がある。そこで、PPARδアゴニスト活性に適したコアクチベーターの選別を行った。検討にはTIF2、PGC1α、PGC1β、SRC-1の4つのコアクチベーターを用いた。コアクチベーターを発現するプラスミド、PPARδ LBDを発現するプラスミドを酵母に導入した後これを培養し、PPARδアゴニストであるGW0742と一定時間反応させた。その後酵母を回収し、細胞壁を破壊した後に、レポーター遺伝子であるβ-ガラクトシダーゼ(β-gal)の活性を測定した。
【0056】
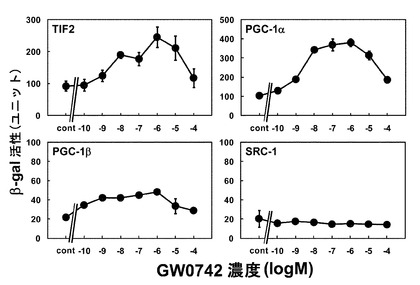
検討の結果、TIF2、PGC1αを用いた場合にはアゴニスト添加により、β-gal活性の上昇が認められ、上昇の程度は、PGC1αの方が大きかった(図1)。一方でPGC1βを用いた場合には、アゴニスト添加によりβ-gal活性が若干上昇したが、全体的にその活性は低く、SRC-1を用いた場合には、アゴニストを添加してもβ-gal活性は上昇しなかった(図1)。この結果から、酵母によるPPARδアゴニスト活性評価に使用するコアクチベーターはPGC1αが適していることが明らかとなった。
【0057】
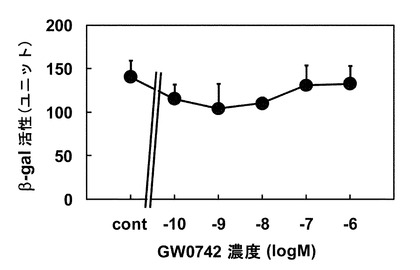
この検討でPGC1αは核内受容体LBDと相互作用する領域のみを発現させたが、PGC1α全長を発現させることで、アゴニスト添加によるβ-gal活性上昇の程度が大きくなるか検討を行った。検討の結果、PGC1α全長を用いた場合には、アゴニスト添加してもβ-gal活性は全く上昇せず(図2)、PPARδアゴニスト活性を評価する場合にはPGC1αの核内受容体LBDと相互作用する領域のみを発現させた方がよいことが明らかとなった。
【0058】
(2)酵母を用いたPPARδアゴニスト、アンタゴニストの評価
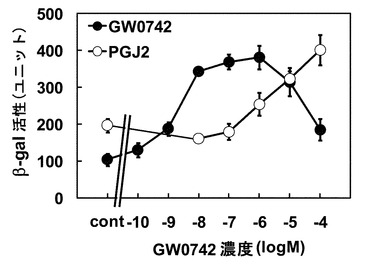
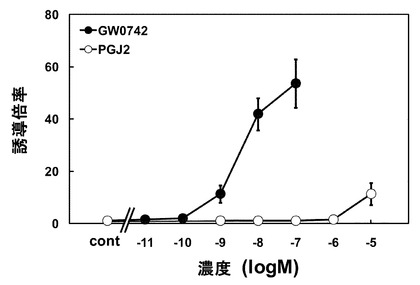
検討の結果から、酵母によるPPARδアゴニスト活性評価に使用するコアクチベーターにはPGC1αが適していることが明らかとなったため、PGC1α発現プラスミド及びPPARδ LBD発現するプラスミドを導入した酵母を用いて検討を行った。まず、アゴニスト活性の評価を行った。PPARδアゴニストとして機能することがしられているGW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)について検討を行ったところ、GW0742は、1 nM以上で、PGJ2は1μM以上で濃度依存的なβ-gal活性の上昇が確認された(図3)。
【0059】
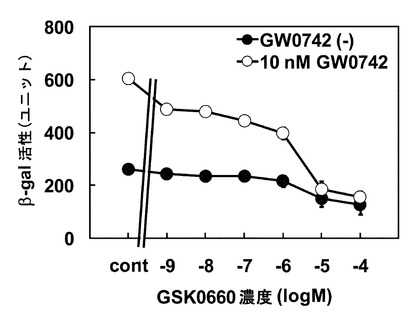
続いて、PPARδアンタゴニストであるGSK0660が、酵母の系においてもアンタゴニストとして機能して、GW0742によるβ-gal活性上昇を抑制するか検討を行った。10 nM GW0742および1 nM~100 μM GSK0660を酵母に添加し検討を行ったところ、GSK0660濃度依存的にβ-gal活性上昇作用は抑制され、10μM以上の濃度において顕著に抑制された(図4)。PPARδアゴニストによりβ-gal活性が上昇し、PPARδアンタゴニストによりその活性上昇が抑制されたことから、この酵母によりPPARδアゴニスト活性およびアンタゴニスト活性を評価できることが確認された。
【0060】
(3)レポーターアッセイによるPPARδアゴニスト、アンタゴニスト活性評価
動物細胞内で核内受容体のアゴニスト活性を評価する実験系として、酵母の転写因子であるGAL4 DBDと核内受容体のLBDの融合タンパク質と転写開始点上流にUASが存在しているルシフェラーゼを利用した系が存在する。この実験系は、GAL4 DBDと核内受容体LBD融合タンパク質のLBD部分に核内受容体リガンドとなる物質が結合すると、DBD部分がUASに結合し、レポーター遺伝子であるルシフェラーゼの転写が開始されるという仕組みになっている。この実験系は、核内受容体リガンドの核内受容体LBDへの結合から標的遺伝子の転写活性化までの、核内受容体によるリガンド依存的な遺伝子転写制御のイベントを一通り網羅しているため、酵母による核内受容体アゴニスト活性評価法よりも正確な評価が可能である。そこで、この実験系を用いて酵母を用いた評価系での結果を検証した。レポーターアッセイによりGW0742、PGJ2のPPARδアゴニスト活性を評価した。その結果、GW0742は1nM以上でレポーター遺伝子であるルシフェラーゼの顕著な活性上昇が認められ、PGJ2は10μMでルシフェラーゼ活性の上昇が認められた(図5)。
【0061】
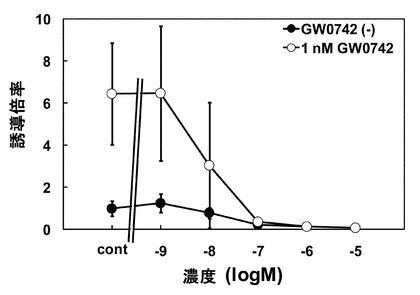
酵母には細胞壁があるため、培養細胞と酵母とでは物質の透過性が異なる。このことにより、レポーターアッセイで反応が認められる濃度よりも1オーダー高くなる傾向が認められている。GW0742はこの傾向と合致していたが、PGJ2は、活性上昇が認められる濃度が酵母の方が低くなっていた。このことから、酵母の系では、PGJ2のPPARδアゴニスト活性を実際よりも強く評価する可能性が示唆された。続いて、酵母の系と同様にPPARδアンタゴニストであるGSK0660の評価を行った。1nM GW0742および1nM〜10μM GSK0660を培養細胞に処理し検討を行ったところ、GSK0660 10nMでルシフェラーゼ活性の減少が認められ、100nMでGW0742によるルシフェラーゼ活性上昇作用をほぼ完全に抑制した(図6)。レポーターアッセイ、酵母two-hybrid系共にGSK0660によりGW0742のアゴニスト作用が抑制されたが、完全に抑制する濃度がレポーターアッセイでは100nMであったのに対し、酵母two-hybrid系では10μMであった。このことから、酵母による評価ではアンタゴニスト活性を過小評価する可能性が示唆された。
【産業上の利用可能性】
【0062】
本発明のスクリーニング方法は、PPARδアゴニスト又はアンタゴニストを探索、同定するツールとして有用である。本発明のスクリーニング方法はハイスループット化に有利である。従って、例えば、抗肥満薬として有望視されているPPARδアゴニストを効率的に同定する手段として利用されることが多いに期待できる。
【0063】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【技術分野】
【0001】
本発明は、ペルオキシソームプロリフェレータ活性化受容体δ(PPARδ)のリガンドのスクリーニング方法に関する。
【背景技術】
【0002】
肥満は生活習慣病を始めとする様々な疾患の原因であり、北米においては重大な社会問題となっている。治療法としては食餌療法や運動療法などがあるが、重度の肥満に対しては即効性に乏しく、十分な効果が得られないのが現状である。PPARδは、脂肪酸およびその誘導体などをリガンドとする核内受容体で、糖代謝、脂質代謝、エネルギー産生に関与することが知られている。PPARδは脂肪の燃焼を促進させる作用があり、恒常的に活性化したPPARδを脂肪細胞特異的に過剰発現させたマウスでは、高脂肪食等の肥満誘因負荷を与えてもほとんど肥満にならない(Wang YX. et al., Cell. 113. 159-170, 2003)。また驚いたことにこのマウスでは、十分な運動を行った動物のような筋繊維の変化が認められることから(Wang YX. et al., Cell. 113. 159-170, 2003)、PPARδは単なる抗肥満薬としてだけでなく、運動効果も期待できるような抗肥満薬の分子標的として有望視されており、そのアゴニストは即効性の高い抗肥満薬としての作用が期待されている。PPARδは開拓されたばかりの標的分子であるため、今後、そのリガンドの探索や開発競争が激化し、それに伴い、PPARδリガンドをハイスループットにスクリーニングできる手法に対するニーズが高まると予想される。尚、PPARδと同様にPPARのサブタイプの一つであるPPARγについては、リガンド依存的にPPARγと相互作用するタンパク質のスクリーニング法(特許文献1)や、アゴニスト及びアンタゴニストのスクリーニング法(特許文献2)が知られている。
【先行技術文献】
【特許文献】
【0003】
【特許文献1】国際公開第2003/062427号パンフレット
【特許文献2】特開平11−56369号公報
【非特許文献】
【0004】
【非特許文献1】J. Nishikawa, K. Saito, J. Goto, F. Dakeyama, M. Matsuo, T. Nishihara: Toxicol. Appl. Pharmacol., 154, 76(1999)
【非特許文献2】T. Kanayama, S. Mamiya, T. Nishihara, J. Nishikawa: J Biocliem., 133. 791(2003)
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明の課題は、PPARδリガンド(アゴニスト又はアンタゴニスト)としての被験物質の有効性を、ハイスループットに評価できる手段を提供することにある。
【課題を解決するための手段】
【0006】
これまでに、PPARδについてはアゴニストに良好な反応を示す酵母two-hybrid(ツーハイブリッド)スクリーニング系は構築されていない。PPARのように内因性のアゴニストが未知であるいわゆるオーファン受容体の多くは、ステロイドホルモン受容体等とは異なり、下等生物にも存在し、また宿主自体に内因性のリガンドを有している可能性が示唆されている。PPARδについても例外ではないと考えられ、実はセオリー通り酵母two-hybrid系を構築しても、アゴニストに依存した反応は見られない。酵母内には、もともとPPARのアゴニストになるような内因性の脂溶性生理活性物質が豊富に存在していることが指摘されており、これが一因となっていると考えられる。尚、核内ホルモン受容体とそのコアクチベーターとの相互作用を利用した酵母two-hybrid系によって、内分泌撹乱物質をスクリーニングする方法が報告されている(非特許文献1)。
【0007】
このような背景の下、本発明者らはエストロゲン受容体やレチノイン酸受容体等で良好な反応性を示した主要なコアクチベーター7種の部分配列を用いて、PPARδリガンドスクリーニング系の構築を試みた。その結果、PPARγコアクチベーター1−α(PGC1α)が最も良好な反応性を示した。しかも、全長を用いた場合よりも、適度な長さの部分PGC1α(受容体相互作用領域(RID)を含む)を用いることで良好な反応性を示すことも見出した。更には、PGC1αをコアクチベーターとしたスクリーニング系によれば、アゴニスト活性とアンタゴニスト活性の両者を評価可能であることが確認された。
【0008】
以上の通り、本発明者の検討によってPGC1αの有用性が判明し、それを利用したPPARδリガンド評価系(スクリーニング系)が完成した。ところで、従来のPPARδリガンド評価系として、マイクロイムノプレートと精製タンパク質を用いた無細胞検出系が先行して存在する(非特許文献2)。基本的に無細胞検出系においても応用するイベントは酵母two-hybrid系と同様である。無細胞検出系は、酵母two-hybrid系よりもハイスループットに検討することが可能であるという利点を有する上、酵母を用いた場合には、被験物質の酵母に対する毒性を考慮した上で検討を行う必要があるが、無細胞系においては毒性を気にせずに濃度を設定できる利点も有する。しかしその一方で、無細胞系は天然物や食品成分等の疎水性粗抽出分画のような混合物では擬陽性が非常に多く、スクリーニングできない。これは無細胞系がリガンド依存的、非依存的にかかわらず、タンパク質間相互作用が起こりさえすれば陽性と判定されるためで、このようなサンプルにはタンパク質変性作用を有していたり、タンパク質に非特異的に接着することでタンパク質同士を結合させるような化合物が含まれていたりするためであると考えられる。したがってサンプルの種類を問わない酵母two-hybrid系は、無細胞系よりもスクリーニングできる検体が幅広く、アドバンテージを有している。即ち、無細胞系に比して汎用性が高い点も利点といえる。
【0009】
一方、動物細胞内で核内受容体のアゴニスト活性を評価する実験系として、酵母の転写因子であるGAL4のDNA結合領域(DNA binding domain:DBD)と核内受容体のリガンド結合領域(ligand binding domain:LBD)の融合タンパク質と転写開始点上流にガラクトース代謝遺伝子の上流域活性化配列(upstream activation sequence:UAS)が存在しているルシフェラーゼを利用した系が存在する。この実験系は、核内受容体リガンドの核内受容体LBDへの結合から標的遺伝子の転写活性化までの、核内受容体によるリガンド依存的な遺伝子転写制御のイベントを一通り網羅しているため、酵母による核内受容体アゴニスト活性評価法よりも正確な評価が可能であるといえる。本発明者らが完成させた上記評価系と当該実験系を比較した結果、両者の間に相関を認め上記評価系の有効性が裏付けられた。また、上記評価系による結果(データ)を評価する上で重要且つ有用な情報が得られた。
以下に列挙する本願発明は、以上の成果ないし知見に基づく。
[1]PPARδのLBDを含む第1領域と、転写因子のDBD又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子、
PGC1αのRIDを含む第3領域と、前記第2領域がDBDを含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDBDを含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子、及び
前記DBDが結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト、
を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
[2]前記第3領域が、PGC1αの一部分である、[1]に記載のスクリーニング方法。
[3]前記第3領域が、PGC1αのN末端側から1番目のアミノ酸〜188番目のアミノ酸からなる、[2]に記載のスクリーニング方法。
[4]前記第3領域が、配列番号5のアミノ酸配列からなる、[3]に記載のスクリーニング方法。
[5]前記細胞が酵母細胞である、[1]〜[4]のいずれか一項に記載のスクリーニング方法。
[6]前記転写因子が酵母のGAL4である、[5]に記載のスクリーニング方法。
[7]前記第2領域が前記転写因子のDBDを含み、前記第4領域が前記転写因子の転写活性化領域を含む、[1]〜[6]のいずれか一項に記載のスクリーニング方法。
【図面の簡単な説明】
【0010】
【図1】PPARδアゴニスト活性評価系に使用するコアクチベーターの検討。
【図2】PGC1α全長を発現する酵母を用いた酵母による検討。
【図3】PGC1αおよびPPARδ LBDを発現する酵母を用いたPPARδアゴニスト活性評価
【図4】PGC1αおよびPPARδ LBDを発現する酵母を用いたPPARδアンタゴニスト活性評価。
【図5】レポーターアッセイによるPPARδアゴニスト活性評価。
【図6】レポーターアッセイによるPPARδアンタゴニスト活性評価。
【発明を実施するための形態】
【0011】
本発明はPPARδのリガンド、即ちアゴニスト(作動薬)またはアンタゴニスト(拮抗薬)をスクリーニングする方法を提供する。本発明のスクリーニング方法ではPPARδと転写共因子とのリガンド依存的な相互作用が生ずるように操作した細胞を用い、当該相互作用に伴うレポーター遺伝子の発現を指標として、被験物質の有効性を評価する。PPARδと転写共因子とのリガンド依存的な相互作用が生ずるように、本発明のスクリーニング方法に用いる細胞は、3要素、即ち第1遺伝子、第2遺伝子及びレポーターコンストラクトを含む。以下、各要素について詳細に説明する。尚、ヒトPPARδのアミノ酸配列を配列番号1に示す。
【0012】
<第1遺伝子>
本発明では、PPARδのLBDを含む第1領域と、転写因子のDBD又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする遺伝子のことを第1遺伝子と呼ぶ。PPARδのLBDは、PPARδのアミノ酸配列のN末端側から254番目〜441番目までのアミノ酸を含む領域と考えられている。少なくとも当該領域を含む限り、第1領域の長さは特に限定されない。第1領域の具体例を配列番号2(PPARδの140a.a.〜441a.a.)に示す。
【0013】
第2領域は転写因子のDBD又は転写活性化領域を含む。好ましい転写因子として酵母GAL4を用いることができる。即ち、好ましい一態様では、GAL4のDBDとGAL4の転写活性化領域を用いる。一方、LexAのDBDとVP16の転写活性化領域の組合せや、LexAのDBDと大腸菌由来のB42転写活性化領域の組合せを用いることにしてもよい。
【0014】
一態様では、第2領域が転写因子のDBDを含む。DBDは、後述の応答配列に対する結合能を示す一方で、転写活性化能を示さない。別の一態様では、第2領域が転写因子の転写活性化領域を含む。転写活性化領域とは、転写活性化能を示す一方、DNA結合能を有しない領域である。転写因子のDBDは、例えばGAL4の場合、N末端側(およそ第1番目から147番目までのアミノ酸を含む領域)に存在することが知られている。同様に転写活性化領域は、例えばGAL4の場合、C末端側(およそ第768番目から881番目までのアミノ酸を含む領域)に存在することが知られている。少なくとも当該領域を含む限り、第2領域の長さは特に限定されない。DBDを採用した場合の第2領域の具体例を配列番号3(GAL4のDBD)に示す。
【0015】
上記の如き構成の第1領域と第2領域が連結されてなる融合タンパク質が第1遺伝子にコードされる。第1領域の機能と第2領域の機能に支障のない限りにおいて、第1領域と第2領域の間に別の配列が介在していてもよい。好ましい態様では、第1領域と第2領域が直接連結されている。第1領域と第2領域の間ではなく、第1領域又は第2領域の外側に他の配列が付加されていてもよい。また、第1領域と第2領域の順序は特に問わず、いずれが上流側に配置されていてもよい。尚、第1遺伝子の配列の例を配列番号4に示す。
【0016】
<第2遺伝子>
本発明では、PGC1αのRIDを含む第3領域と、前記第2領域がDBDを含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDBDを含む第4領域とが連結されてなる融合タンパク質をコードする遺伝子のことを第2遺伝子と呼ぶ。
【0017】
PGC1αでは、そのRIDはN末端側から88番目〜92番目までのアミノ酸、144番目〜148番目、または210番目〜214番目までのアミノ酸を含む領域と考えられている。少なくともいずれかの当該領域を1つ以上含む限り、第3領域の長さは特に限定されない。本発明者らの検討の結果、PGC1α全長を用いた場合には、良好な相互作用が認められなかった。この知見に基づき、好ましくは、PGC1αの一部を第3領域に使用する。PGC1αの全長は798アミノ酸残基であるが、例えば、1番目〜188番目のアミノ酸の部分PGC1α(但し、RIDを含む)を用いるとよい。好ましい第3領域の具体例を配列番号5(1a.a.〜188a.a.)に示す。
【0018】
第4領域は転写因子のDBD又は転写活性化領域を含むが、第2領域との間でDBDと転写活性化領域からなるペアーが形成されるように、第2領域にDBDを使用した場合には、第4領域には転写因子の転写活性化領域を使用し、これとは逆に、第2領域に転写活性化領域を使用した場合には、第4領域には転写因子のDBDを使用する。このように構成することによって、第2領域が供給する転写因子の要素と、第4領域が提供する転写因子の要素が一体となることで、応答配列への結合及びそれに続くレポーター遺伝子の転写活性化が引き起こされる。第2領域と第4領域による当該作用・効果が生ずる限りにおいて、第4領域が含む転写因子の要素の由来が、第2領域が含む転写因子の要素の由来と同一である必要はない。尚、転写活性化領域を採用した場合の第4領域の具体例を配列番号6(GAL4の転写活性化領域)に示す。
【0019】
上記の如き構成の第3領域と第4領域が連結されてなる融合タンパク質が第2遺伝子にコードされる。第3領域の機能と第4領域の機能に支障のない限りにおいて、第3領域と第4領域の間に別の配列が介在していてもよい。好ましい態様では、第3領域と第4領域が直接連結されている。第3領域と第4領域の間ではなく、第3領域又は第4領域の外側に他の配列が付加されていてもよい。また、第3領域と第4領域の順序は特に問わず、いずれが上流側に配置されていてもよい。尚、第2遺伝子の配列の例を配列番号7に示す。
【0020】
<レポーターコンストラクト>
本発明では、上記DBDが結合し得る応答配列及び当該応答配列に連結されたレポーター遺伝子を含むものをレポーターコンストラクトと呼ぶ。使用する転写因子に対応した応答配列が採用される。例えば、転写因子としてGAL4を採用した場合、応答配列としてUASを用いることができる。
【0021】
レポーター遺伝子には、例えば、出芽酵母の栄養要求性遺伝子(ADE2、HIS3、URA3)、大腸菌由来のβ−ガラクトシダーゼ遺伝子(lacZ)、バクテリアトランスポゾン由来のクロラムフェニコールアセチルトランスフェラーゼ遺伝子(CAT)、ホタル由来のルシフェラーゼ遺伝子(Luc)を用いることができる。レポーター遺伝子は、応答配列の制御下に配置される。即ち、応答配列に対して作動的にレポーター遺伝子が連結される。
【0022】
第1遺伝子、第2遺伝子及びレポーターコンストラクトは、遺伝子組換え技術を用いて調製できる。第1遺伝子に含まれる、PPARδのLBDをコードするDNAや転写因子の一部をコードするDNA、或いは第2遺伝子に含まれる、PGC1αのRIDをコードするDNAや転写因子の一部をコードするDNAなどは、既知のアミノ酸配列や塩基配列の情報などをもとに設計・合成したプライマーやプローブを用いて、例えばcDNAライブラリーから単離することができる。単離したDNAを制限酵素やPCR法などを用いて連結することにより、第1遺伝子及び第2遺伝子を構築することができる。
【0023】
以上で説明した3要素を細胞に導入することにより、本発明に使用するスクリーニング用細胞を得ることができる。例えば第1遺伝子、第2遺伝子、レポーターコンストラクトをそれぞれ適当なベクター(例えばプラスミドベクター)に組み込み、当該ベクターで宿主細胞を形質転換すればよい。二つ以上の要素を一つのベクターで導入することにしてもよい。尚、各種レポーター遺伝子に関して、それが応答配列とともに組み込まれた細胞を利用可能な状況にある。そのような細胞を利用すれば、第1遺伝子及び第2遺伝子のみを導入することによって、本発明に必要なスクリーニング用細胞が得られる。
【0024】
好ましい態様では、酵母two-hybrid系として本発明のスクリーニング方法を構成する。酵母two-hybrid系とは、酵母を宿主細胞として用い、細胞内で生じるタンパク質相互作用の存在を検出するものである。基本的には真核細胞の転写因子の多くがDBDと転写活性化領域に分割が可能であり、これらの領域に調べようとする2種類のタンパク質をそれぞれに連結して酵母内で発現させたときに転写因子の活性化が起これば、これらのタンパク質は会合していることが分かる。酵母two-hybrid系を核内受容体のリガンドスクリーニング系に応用する場合は、一般に、核内受容体と転写因子(例えば酵母転写因子GAL4)のDBDとの融合タンパク質及びコアクチベーターと転写因子の転写活性化領域の融合タンパク質を酵母内で発現させる。また、これらとともに転写因子の応答配列をレポーター遺伝子の上流に挿入したレポータープラスミドも酵母内に導入する。例えば、被験物質がアゴニスト活性を有している場合には、核内受容体(本発明の場合はPPARδ)のLBDのコンフォメーションが変化し、コアクチベーターをリクルートするため、両融合タンパク質は結合し転写因子の働きを持つタンパク質複合体が形成される。このタンパク質複合体は転写因子として機能し、レポーター遺伝子の発現を誘導する。したがって、被験物質のリガンド活性をレポーター遺伝子の発現量で評価することができる。被験物質のリガンド活性の強弱は、活性化された核内受存体のコアクチベーターのリクルート能に反映されることから、このレポーター遺伝子の発現の度合いを指標にリガンド活性を定量的に検出することが可能となる。典型的なアゴニストを試験系に加えておいて、同時に被験物質を共存させることによりアンタゴニスト活性も検出できる。酵母two-hybrid系には典型的にはサッカロマイセス・セレビシエ(Saccharomyces cerevisiae)が用いられる。尚、酵母two-hybrid系の原理や構成等については、J. Nishikawa, K. Saito, J. Goto, F. Dakeyama, M. Matsuo, T. Nishihara: Toxicol. Appl. Pharmacol., 154, 76(1999)が参考になる。
【0025】
DBDと転写活性化領域に分割された転写因子の活性化を評価できるのであれば、酵母細胞以外の真核細胞(例えば哺乳動物細胞や昆虫細胞)を用いることも可能である。
【0026】
本発明のスクリーニング方法では、上記のように3要素を含むスクリーニング用細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析する。具体的には、例えば次のステップ(1)〜(3)を行えばよい。
(1)スクリーニング用細胞に被験物質を接触させるステップ
(2)レポーター遺伝子の発現を検出するステップ
(3)検出結果に基づき、被験物質による、リガンド依存的な相互作用の変化の有無及び/又は程度を調べるステップ
【0027】
ステップ(1)では、用意したスクリーニング用細胞に対して、被験物質を接触させる。例えば、スクリーニング用細胞の培養液中に被験物質を添加する。或いは、スクリーニング用細胞の培養液を、被験物質を含有するものに交換する。培養方法は常法に従えばよく、スクリーニング用細胞の種類に応じて決定できる。被験物質との接触時間、即ち被験物質を添加などすることによって、培養液中に被験物質が共存するようになってから、ステップ(2)に至るまでの時間は予備実験を通して設定すればよいが、例えば30分〜6時間である。
【0028】
試験物質としては様々な分子サイズの有機化合物又は無機化合物を用いることができる。有機化合物の例として、核酸、ペプチド、タンパク質、脂質(単純脂質、複合脂質(ホスホグリセリド、スフィンゴ脂質、グリコシルグリセリド、セレブロシド等)、プロスタグランジン、イソプレノイド、テルペン、ステロイド、ポリフェノール、カテキン、ビタミン(B1、B2、B3、B5、B6、B7、B9、B12、C、A、D、E等)を例示できる。試験物質は天然物由来であっても、或いは合成によるものであってもよい。後者の場合には例えばコンビナトリアル合成の手法を利用して効率的なスクリーニング系を構築することができる。尚、細胞抽出液、培養上清などを試験物質として用いてもよい。また、既存の薬剤を試験物質としてもよい。2種類以上の被験物質を同時に添加することにより、被験物質間の相互作用、相乗作用などを調べることにしてもよい。
【0029】
ステップ(1)に続くステップ(2)では、レポーター遺伝子の発現を検出する。検出は常法に従えばよい。本発明で用いることが可能な汎用的なレポーター遺伝子に関しては、基本的な検出法が確立されており、成書や過去の論文等を参考にして、適切な方法や条件を容易に選択ないし設定することが可能である。
【0030】
ステップ(3)では、ステップ(2)の検出結果に基づき、リガンド依存的な相互作用に及ぼす被験物質の影響・効果を調べる。具体的には、スクリーニング用細胞に被験物質を接触させたことにより、リガンド依存的な相互作用の変化が生じたか、又はリガンド依存的な相互作用の変化はどの程度か、或いはこれらの両方を調べ、被験物質のアゴニスト又はアンタゴニストとしての有効性を判定する。例えば、被験物質の接触(共存)によってレポーター遺伝子の発現の増強を認めた場合(発現を検出できるようになった場合も含む)、即ち、リガンド依存的な相互作用を正に調節した場合、被験物質がアゴニスト活性を有すると判定できる。通常は、スクリーニング用細胞に被験物質を接触させない場合(陰性対照)と比較し、被験物質のアゴニスト活性を判定する。また、このような陰性対照の利用と併せて或いはその代わりに、アゴニスト活性を示すことが判明している物質(既知アゴニスト)をスクリーニング用細胞に接触させた場合(陽性対照)と比較し、被験物質のアゴニスト活性の程度を評価してもよい。以上のように陰性対照及び/又は陽性対照との比較に基づき被験物質の有効性を判定・評価すれば、より客観的且つ信頼性の高い判定結果が得られる。
【0031】
一方、被験物質のアンタゴニスト活性を評価する場合には、原則、アゴニストの共存下でステップ(1)を行い(即ち、被験物質とアゴニストをスクリーニング用細胞に接触させる)、その場合のレポーター遺伝子の発現を、アゴニストのみをスクリーニング用細胞に接触させた場合のレポーター遺伝子の発現と比較する。後者に比較して前者にレポーター遺伝子の発現の減弱を認めた場合、即ち、アゴニストに起因するリガンド依存的な相互作用を被験物質が負に調節した場合、被験物質がアンタゴニスト活性を有すると判定できる。従って、アンタゴニスト候補として当該被験物質が選抜される。
【0032】
発現増強又は減弱の程度に基づき、被験物質のアゴニスト又はアンタゴニストとしての有効性を定量的に判断してもよい。尚、PPARδのアゴニストとしてGW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)が知られており、同様にPPARδのアンタゴニストとしてGSK0660が知られている。
【0033】
PPARδのアゴニストは、即効性の高い抗肥満薬の有効成分として期待される。他方、PPARδのアンタゴニストは、ヒトの肺ガン、肝ガン、乳ガン由来細胞株の増殖を抑制し、アポトーシスを誘導すること(Zaveri NT. et al., Cancer Biol Ther. 8. 1252-1261, 2009)から、抗ガン剤の有効成分として期待される。
【0034】
本発明のスクリーニング方法によって選択された物質が十分な薬効を有する場合には、当該物質をそのまま医薬の有効成分として使用することができる。一方で十分な薬効を有しない場合には化学的修飾などの改変を施してその薬効を高めた上で、医薬の有効成分として使用することができる。勿論、十分な薬効を有する場合であっても、更なる薬効の増大を目的として同様の改変を施してもよい。
【実施例】
【0035】
以下の説明では次の略号を使用する。
AD : activation domain(転写活性化領域)
RID : Receptor Interaction Domain(受容体相互作用領域)
DBD : DNA binding domain(DAN結合領域)
LBD : ligand binding domain(リガンド結合領域)
UAS : upstream activating sequence(ガラクトース代謝遺伝子の上流域活性化配列)
【0036】
1.実験材料
(1)酵母two-hybrid法で使用する試薬および組成
・YPDA培地
20 g/L ペプトン
10 g/L 酵母エキス(Yeast extract)
20 g/L グルコース
30 mg/L L-アデニンヘミ硫酸塩
・SD-L/-T培地
6.7 g/L Difco yeast nitrogen base without amino acid
100 mL/L 10x Dropout溶液(-Leu, -Trp)
20 g/L グルコース
・Dropout溶液(-Leu, -Trp)
300 mg/L L-イソロイシン
1500 mg/L L-バリン
200 mg/L L-アデニンヘミ硫酸塩
200 mg/L L-アルギニンHCl
200 mg/L L-ヒスチジンHCl・H2O
300 mg/L L-リジンHCl
200 mg/L L-メチオニン
500 mg/L L-フェニルアラニン
2000 mg/L L-スレオニン
300 mg/L L-チロシン
200 mg/L L-ウラシル
・SD-L/-Tプレート
SD-L/-T培地に寒天を2%(w/v)となるように加えたもの
・10x TE
100 mM Tris-HCl(pH 7.5)
10 mM EDTA
・10x LiAc溶液
1 M 酢酸リチウム(pH 7.5)
・50%(w/v) PEG 4000溶液
・LiAc/TE溶液
10x TE 1 mL
10x LiAc 1 mL
超純水 8 mL
・LiAc/TE/PEG溶液
10x TE 1 mL
10x LiAc 1 mL
50%(w/v) PEG 4000溶液 8 mL
・Z-バッファー
60 mM Na2HPO4
40 mM NaH2PO4
10 mM KCl
1 mM MgSO4
【0037】
(2)実験に使用したPPARδリガンド
【化1】
アゴニスト:GW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)
アンタゴニスト:GSK0660
【0038】
(3)酵母two-hybrid法で使用するプラスミドの構築
(a)酵母内でGAL4-ADとTIF2の融合タンパク質を発現するプラスミド(pGAD424-TIF2)
ヒトTIF2のRID(483-1090 a.a.(配列番号8))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaq(Applied Biosystems)を用いてPCRを行うことで増幅し、pBluescript KS-(stratagene)に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424(clontech)に挿入し、プラスミドを構築した。
【0039】
(b)酵母内でGAL4-ADとPGC1αRID(1-188 a.a.(配列番号5))の融合タンパク質を発現するプラスミド(pGAD424-PGC1α)
ヒトPGC1αのRID(1-188 a.a.(配列番号5))をコードする配列をHepG2細胞由来cDNAを鋳型とし、LA taq(TAKARA BIO)を用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、Sma I、Sal Iを用いて配列を切り出し、pGAD424に挿入し、プラスミドを構築した。
【0040】
(c)酵母内でGAL4-ADとPGC1α(798 a.a.(配列番号9))の融合タンパク質を発現するプラスミド(pGAD424-PGC1αfull)
ヒトPGC1αをコードする配列をHepG2細胞由来cDNAを鋳型とし、KOD-Plus(TOYOBO)を用いてPCRを行うことで増幅した後、Sma I、Sal Iで処理した。これをSma I、Sal Iで処理したpGAD424に挿入し、プラスミドを構築した。またDNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した。
【0041】
(d)酵母内でGAL4-ADとPGC1βの融合タンパク質を発現するプラスミド(pGAD424-PGC1β)
ヒトPGC1βのRID(1-264 a.a.(配列番号10))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、BamH I、Sal Iを用いて配列を切り出し、pGAD424に挿入し、プラスミドを構築した。
【0042】
(e)酵母内でGAL4-ADとSRC1の融合タンパク質を発現する
プラスミド(pGAD424-SRC1)
ヒトSRC1のRID(570-783 a.a.(配列番号11))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0043】
(f)酵母内でGAL4-ADとp300/CBPの融合タンパク質を発現するプラスミド(pGAD424-CBP)
ヒトp300/CBPのRID(61-522 a.a.(配列番号12))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0044】
(g)酵母内でGAL4-ADとACTRの融合タンパク質を発現するプラスミド(pGAD424-ACTR)
ヒトACTRのRID(625-820 a.a.(配列番号13))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0045】
(h)酵母内でGAL4-ADとRIP140の融合タンパク質を発現するプラスミド(pGAD424-RIP140)
ヒトRIP140のRID(752-1158 a.a.(配列番号14))をコードする配列をヒト肝臓由来cDNAを鋳型とし、DNA Polymerase AmpliTaqを用いてPCRを行うことで増幅し、pBluescript KS-に挿入した。DNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した後、pGAD424に挿入し、プラスミドを構築した。
【0046】
(i)酵母内でGAL4-DBDとPPARδ LBD(140-441 a.a.)の融合タンパク質を発現するプラスミド(pGBT9-PPARδ LBD)
ヒトPPARδ LBD(140-441 a.a.(配列番号2))をコードする配列をHepG2細胞由来のcDNAを鋳型とし、KOD-Plusを用いてPCRを行うことで増幅した後、Sma I、Sal Iで処理した。これをSma I、Sal Iで処理したpGBT9(clontech)に挿入し、プラスミドを構築した。またDNAシークエンス解析により、挿入した配列がGenBankに登録されているものと一致することを確認した。
【0047】
(4)レポーターアッセイで使用するプラスミドの構築
(A)培養細胞内でホタルルシフェラーゼを発現するレポータープラスミド(p4xUAS-tk-Luc)
4コピーのGAL4DBD結合配列であるUAS 配列(5’-CGACGG AGTACTGTCCTCCGAGCT-3’(配列番号15))をSma Iで処理したptk-luc(clontech)に挿入し、プラスミドを構築した。
【0048】
(B)培養細胞内でGAL4-DBDとPPARδ LBD(140-441 a.a.(配列番号2))の融合タンパク質を発現するプラスミド(pM-PPARδ LBD)
pGBT9-PPARδ LBDをEcoR I、Sal I処理することでヒトPPARδ LBD(140-441 a.a.(配列番号2))をコードする配列を切り出し、これをEcoR I、Sal I処理したpM(clontech)に組み込みプラスミドを構築した。
【0049】
2.実験方法
(1)酵母へのトランスフォーメーション
酵母(Y190株)をYPDA培地15 mL(1トランスフォーメーションあたり)で30℃、16〜24時間培養後、遠心分離(室温、3,000 rpm、5分)により集菌し、1xTE、LiAc/TE溶液で洗浄後、150μLのLiAc/TE溶液で懸濁した。懸濁した酵母を1.5 mLチューブに移し、pGAD424プラスミド(TIF2、PGC1α、PGC1αfull、PGC1β、SRC1)およびpGBT9-PPARδ LBDプラスミドをそれぞれ1μg相当量、キャリアーDNA溶液(ウシ胸腺DNA、SIGMA D-8661)を7μL添加し、LiAc/TE/PEG溶液を850μL加え、よく混和した。混和後、30℃で30分、42℃で15分インキュベートした後、遠心分離(室温、3,000 rpm、3分)を行い、LiAc/TE/PEG溶液を除去した。沈殿を500μLの超純水で懸濁し、再度遠心分離(室温、3,000 rpm、3分)を行い、上清を除去した後、200μLのLiAc/TEで再懸濁し、SD-L/-Tプレートに塗布した。プレートは適当なコロニーが大きさのコロニーが得られるまで(2〜4晩)30℃でインキュベートし、コロニーが生えたプレートはβ-ガラクトシダーゼ活性測定時まで4℃で保存した。
【0050】
(2)β-ガラクトシダーゼ活性測定
pGAD424プラスミドとpGBT9プラスミドを導入した酵母を15 mLのSD-L/-T培地に植菌した後、30℃で一晩培養し、前培養液とした。前培養液を150μLとり、96穴プレートに移し、プレートリーダーで600 nmの吸光度(OD600)を測定した上で、前培養液をOD600=0.05となるようにSD-L/-T培地で希釈し、希釈液250μLを1.5 mLチューブに移した。さらに試料溶液を2.5μL加え、良く混和した後、30℃で4時間培養した。培養後、培養液を150μLとり、96穴プレートに移し、プレートリーダーでOD600を測定した。残りの培養液100μLを遠心(室温、15,000 rpm、5分)し、上清を除き、1 mg/mLのZymolyase-20T(生化学バイオビジネス株式会社)を含むZバッファーを200μL加え、良く混和した後、37℃で30分静置し、酵母を溶解した。酵母の溶解を確認した後、4 mg/mLのo-nitrophenyl-β-galactoside(Sigma)を含むZバッファーを40μL加えて良く混和し、30℃で発色反応を開始させ、30分後に1M Na2CO3溶液を100μL加えて反応を停止させた。遠心(室温、15,000 rpm、5分)後、上清150μLを96穴プレートに移し415 nmの吸光度(OD415)を測定し、次式を用いてβ-ガラクトシダーゼ活性を算出した。
【数1】
【0051】
(3)細胞培養
ヒト胎盤絨毛細胞株であるJEG-3細胞(American Type Culture Collection, ATCC)の培養には10%ウシ胎児血清、2 mM L-グルタミン、1 mMピルビン酸ナトリウム(Nacalai Tesque)及び0.1 mM MEM非必須アミノ酸溶液(GIBCO)を加えたMEM培地(Nacalai Tesque)を用いた。培養は37℃、5% CO2存在下で行った。遺伝子導入時には無血清培地で、遺伝子導入以降の培養は活性炭処理したウシ胎児血清を1%加えた培地を用いた。
【0052】
(4)遺伝子導入および化合物処理
JEG-3細胞を3.0x104細胞/ウェルとなるように24穴平底プレート(corning)に播種し、24時間後に遺伝子導入を行った。遺伝子導入には、Lipofectamine Reagent(Invitrogen)およびPlus Reagent(Invitrogen)を用いて、その標準プロトコールに従って操作を行った。プラスミド量は、1ウェルあたりp4xUAS-tk-Luc 50 ng、pM-PPARδ LBD 20 ng、pGL 4.75(Promega)10 pgとした。遺伝子導入3時間後に、1%活性炭処理ウシ胎児血清含有培地に交換し、培地交換の21時間後に化合物の処理を行った。
【0053】
(5)ルシフェラーゼアッセイ
化合物処理24時間後に細胞をPBSで2回洗浄した後、Passive Lysis Buffer(Promega)で溶解した。20分静置後、細胞溶解液20μLを96穴白色平底プレート(NUNC)に加え、Dual-Luciferase Reporter Assay System(Promega)を用いてルミノメーター(Berthold Japan)によりホタルルシフェラーゼ活性を測定した。内部標準として、pGL 4.75のウミシイタケルシフェラーゼ活性を測定し、これを用いて遺伝子導入効率を補正した。
【0054】
3.実験結果
(1)酵母によるPPARδアゴニスト活性評価系に使用するコアクチベーターの選別
酵母two-hybrid法は、酵母を宿主細胞に用いて、任意の2つのタンパク質の結合をレポーター遺伝子の発現を指標に検出するシステムである。核内受容体にリガンドが結合していない状態では、コリプレッサー複合体(標的遺伝子の転写を抑制する働きをする)が核内受容体に結合し、標的遺伝子の転写を抑制しているが、リガンドが結合すると核内受容体の立体構造が変化し、それに伴って、コリプレッサーが解離し、コアクチベーター複合体(標的遺伝子の転写を促進する働きをする)が結合して、標的遺伝子の転写を促進する。酵母two-hybrid法を用いた核内受容体アゴニスト活性評価系はこのイベントを応用している。すなわち、酵母内で酵母の転写制御因子であるGAL4と核内受容体LBD融合タンパク質にアゴニストが結合すると、核内受容体LBDが前述の立体構造変化が起こることでコアクチベーターを迎え入れる状態となり、そこにGAL4の活性化領域とコアクチベーター融合タンパク質のコアクチベーター部分が結合することで、レポーター遺伝子が発現する。またアゴニスト作用の強弱は、活性化された核内受容体のコアクチベーターのリクルート能に反映されることから、このレポーター遺伝子の発現の度合いを指標にアゴニスト作用を定量的に検出することが可能となる。
【0055】
この系を用いてPPARδアゴニスト活性を評価するにあたっては、リガンド添加によりレポーター遺伝子発現が大きく上昇させられるようなコアクチベーターを使用する必要がある。そこで、PPARδアゴニスト活性に適したコアクチベーターの選別を行った。検討にはTIF2、PGC1α、PGC1β、SRC-1の4つのコアクチベーターを用いた。コアクチベーターを発現するプラスミド、PPARδ LBDを発現するプラスミドを酵母に導入した後これを培養し、PPARδアゴニストであるGW0742と一定時間反応させた。その後酵母を回収し、細胞壁を破壊した後に、レポーター遺伝子であるβ-ガラクトシダーゼ(β-gal)の活性を測定した。
【0056】
検討の結果、TIF2、PGC1αを用いた場合にはアゴニスト添加により、β-gal活性の上昇が認められ、上昇の程度は、PGC1αの方が大きかった(図1)。一方でPGC1βを用いた場合には、アゴニスト添加によりβ-gal活性が若干上昇したが、全体的にその活性は低く、SRC-1を用いた場合には、アゴニストを添加してもβ-gal活性は上昇しなかった(図1)。この結果から、酵母によるPPARδアゴニスト活性評価に使用するコアクチベーターはPGC1αが適していることが明らかとなった。
【0057】
この検討でPGC1αは核内受容体LBDと相互作用する領域のみを発現させたが、PGC1α全長を発現させることで、アゴニスト添加によるβ-gal活性上昇の程度が大きくなるか検討を行った。検討の結果、PGC1α全長を用いた場合には、アゴニスト添加してもβ-gal活性は全く上昇せず(図2)、PPARδアゴニスト活性を評価する場合にはPGC1αの核内受容体LBDと相互作用する領域のみを発現させた方がよいことが明らかとなった。
【0058】
(2)酵母を用いたPPARδアゴニスト、アンタゴニストの評価
検討の結果から、酵母によるPPARδアゴニスト活性評価に使用するコアクチベーターにはPGC1αが適していることが明らかとなったため、PGC1α発現プラスミド及びPPARδ LBD発現するプラスミドを導入した酵母を用いて検討を行った。まず、アゴニスト活性の評価を行った。PPARδアゴニストとして機能することがしられているGW0742、15-デオキシ-Δ12,14-プロスタグランジンJ2(PGJ2)について検討を行ったところ、GW0742は、1 nM以上で、PGJ2は1μM以上で濃度依存的なβ-gal活性の上昇が確認された(図3)。
【0059】
続いて、PPARδアンタゴニストであるGSK0660が、酵母の系においてもアンタゴニストとして機能して、GW0742によるβ-gal活性上昇を抑制するか検討を行った。10 nM GW0742および1 nM~100 μM GSK0660を酵母に添加し検討を行ったところ、GSK0660濃度依存的にβ-gal活性上昇作用は抑制され、10μM以上の濃度において顕著に抑制された(図4)。PPARδアゴニストによりβ-gal活性が上昇し、PPARδアンタゴニストによりその活性上昇が抑制されたことから、この酵母によりPPARδアゴニスト活性およびアンタゴニスト活性を評価できることが確認された。
【0060】
(3)レポーターアッセイによるPPARδアゴニスト、アンタゴニスト活性評価
動物細胞内で核内受容体のアゴニスト活性を評価する実験系として、酵母の転写因子であるGAL4 DBDと核内受容体のLBDの融合タンパク質と転写開始点上流にUASが存在しているルシフェラーゼを利用した系が存在する。この実験系は、GAL4 DBDと核内受容体LBD融合タンパク質のLBD部分に核内受容体リガンドとなる物質が結合すると、DBD部分がUASに結合し、レポーター遺伝子であるルシフェラーゼの転写が開始されるという仕組みになっている。この実験系は、核内受容体リガンドの核内受容体LBDへの結合から標的遺伝子の転写活性化までの、核内受容体によるリガンド依存的な遺伝子転写制御のイベントを一通り網羅しているため、酵母による核内受容体アゴニスト活性評価法よりも正確な評価が可能である。そこで、この実験系を用いて酵母を用いた評価系での結果を検証した。レポーターアッセイによりGW0742、PGJ2のPPARδアゴニスト活性を評価した。その結果、GW0742は1nM以上でレポーター遺伝子であるルシフェラーゼの顕著な活性上昇が認められ、PGJ2は10μMでルシフェラーゼ活性の上昇が認められた(図5)。
【0061】
酵母には細胞壁があるため、培養細胞と酵母とでは物質の透過性が異なる。このことにより、レポーターアッセイで反応が認められる濃度よりも1オーダー高くなる傾向が認められている。GW0742はこの傾向と合致していたが、PGJ2は、活性上昇が認められる濃度が酵母の方が低くなっていた。このことから、酵母の系では、PGJ2のPPARδアゴニスト活性を実際よりも強く評価する可能性が示唆された。続いて、酵母の系と同様にPPARδアンタゴニストであるGSK0660の評価を行った。1nM GW0742および1nM〜10μM GSK0660を培養細胞に処理し検討を行ったところ、GSK0660 10nMでルシフェラーゼ活性の減少が認められ、100nMでGW0742によるルシフェラーゼ活性上昇作用をほぼ完全に抑制した(図6)。レポーターアッセイ、酵母two-hybrid系共にGSK0660によりGW0742のアゴニスト作用が抑制されたが、完全に抑制する濃度がレポーターアッセイでは100nMであったのに対し、酵母two-hybrid系では10μMであった。このことから、酵母による評価ではアンタゴニスト活性を過小評価する可能性が示唆された。
【産業上の利用可能性】
【0062】
本発明のスクリーニング方法は、PPARδアゴニスト又はアンタゴニストを探索、同定するツールとして有用である。本発明のスクリーニング方法はハイスループット化に有利である。従って、例えば、抗肥満薬として有望視されているPPARδアゴニストを効率的に同定する手段として利用されることが多いに期待できる。
【0063】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【特許請求の範囲】
【請求項1】
PPARδのリガンド結合領域を含む第1領域と、転写因子のDNA結合領域又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子、
PPARγコアクチベーター1−αの受容体相互作用領域を含む第3領域と、前記第2領域がDNA結合領域を含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDNA結合領域を含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子、及び
前記DNA結合領域が結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト、
を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
【請求項2】
前記第3領域が、PPARγコアクチベーター1−αの一部分である、請求項1に記載のスクリーニング方法。
【請求項3】
前記第3領域が、PPARγコアクチベーター1−αのN末端側から1番目のアミノ酸〜188番目のアミノ酸からなる、請求項2に記載のスクリーニング方法。
【請求項4】
前記第3領域が、配列番号5のアミノ酸配列からなる、請求項3に記載のスクリーニング方法。
【請求項5】
前記細胞が酵母細胞である、請求項1〜4のいずれか一項に記載のスクリーニング方法。
【請求項6】
前記転写因子が酵母のGAL4である、請求項5に記載のスクリーニング方法。
【請求項7】
前記第2領域が前記転写因子のDNA結合領域を含み、前記第4領域が前記転写因子の転写活性化領域を含む、請求項1〜6のいずれか一項に記載のスクリーニング方法。
【請求項1】
PPARδのリガンド結合領域を含む第1領域と、転写因子のDNA結合領域又は転写活性化領域を含む第2領域とが連結されてなる融合タンパク質をコードする第1遺伝子、
PPARγコアクチベーター1−αの受容体相互作用領域を含む第3領域と、前記第2領域がDNA結合領域を含む場合には転写因子の転写活性化領域を含み、前記第2領域が転写活性化領域を含む場合には転写因子のDNA結合領域を含む第4領域とが連結されてなる融合タンパク質をコードする第2遺伝子、及び
前記DNA結合領域が結合し得る応答配列及び該応答配列に連結されたレポーター遺伝子を含むレポーターコンストラクト、
を含む細胞に被験物質を接触させ、前記レポーター遺伝子の発現を指標として、前記被験物質による、リガンド依存的な相互作用の変化を分析することを特徴とする、PPARδのアゴニスト又はアンタゴニストのスクリーニング方法。
【請求項2】
前記第3領域が、PPARγコアクチベーター1−αの一部分である、請求項1に記載のスクリーニング方法。
【請求項3】
前記第3領域が、PPARγコアクチベーター1−αのN末端側から1番目のアミノ酸〜188番目のアミノ酸からなる、請求項2に記載のスクリーニング方法。
【請求項4】
前記第3領域が、配列番号5のアミノ酸配列からなる、請求項3に記載のスクリーニング方法。
【請求項5】
前記細胞が酵母細胞である、請求項1〜4のいずれか一項に記載のスクリーニング方法。
【請求項6】
前記転写因子が酵母のGAL4である、請求項5に記載のスクリーニング方法。
【請求項7】
前記第2領域が前記転写因子のDNA結合領域を含み、前記第4領域が前記転写因子の転写活性化領域を含む、請求項1〜6のいずれか一項に記載のスクリーニング方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2012−205546(P2012−205546A)
【公開日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願番号】特願2011−73838(P2011−73838)
【出願日】平成23年3月30日(2011.3.30)
【出願人】(591060289)岐阜市 (15)
【Fターム(参考)】
【公開日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願日】平成23年3月30日(2011.3.30)
【出願人】(591060289)岐阜市 (15)
【Fターム(参考)】
[ Back to top ]