
RNAバクテリオファージのウイルス様粒子でのペプチドディスプレイ及びアフィニティー選択のためのプラスミド及び方法
本発明は、ウイルス様粒子(VLP)、特にMS2 VLPを含むVLPでペプチドディスプレイ価数を制御するためのシステム及び方法に関する。この方法では、大量の野生型及び少量の一本鎖二量体コートタンパク質を、単一のRNAから生成できる。価数は、単一のRNAから大量の野生型及び少量の一本鎖二量体コーティングタンパク質の産生を可能にするシステムを提供することによって免疫原(ワクチン)産生において制御され、これによってVLP、特にMS2 VLPで広い範囲にわたって、1粒子当たり平均で1未満から90程度まで、提示(ディスプレイ)価数レベルの容易な調節が可能になる。これによって、低い価数を示すVLPを含めて、免疫原及びワクチンの産生が容易になる。ウイルス様粒子の発現に有用な核酸構築物が開示されて、これは異種ペプチドの挿入によって改変されるMS2のコートポリペプチドから構成され、ここでこの異種ペプチドは、ウイルス様粒子上で提示され、MS2 niRNAをカプシドで包む。異種ペプチドであって、ウイルス様粒子上で提示され、PP7 mRNAをカプシドで包む異種ペプチドの挿入によって改変されたPP7のコートポリペプチドから構成されるウイルス様粒子の発現において有用な核酸構築物もまた開示される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願及び支援助成金
本出願は、発明の名称「VLPでのペプチドディスプレイ価数の制御(Control of Peptide Display Valency on VLP)」である2009年12月31日出願の米国特許仮出願第61/335,122号、発明の名称「バクテリオファージMS2 VLPでのランダム配列ペプチドライブラリーの容易な構築のためのプラスミドベクター、並びに関連の構築物、ライブラリー及び方法(Plasmid Vectors for Facile Construction of Random Sequence Peptide Libraries on Bacteriophage MS2 VLPs and Related Constructs, Libraries,and Methods」である2009年12月31日出願の米国特許仮出願第61/335,120号、及び発明の名称「バクテリオファージPP7のウイルス様粒子でのペプチドディスプレイ(Peptide Display on Virus−Like Particles of Bacteriophage PP7)」である2009年12月31日出願の米国特許仮出願第61/335,121号から優先権の利益を主張し、それぞれの出願の内容は、その全体が本明細書に援用される。
【0002】
本特許出願は、国立衛生研究所によって助成金番号GM042901及びR01 AI065240により支援された。米国政府は、本発明において一定の権利を有する。
【0003】
本発明の分野
本発明は、RNAバクテリオファージ、特にMS2及びPP7のウイルス様粒子(virus-like particule, VLP)でのペプチドのディスプレイ(提示)のためのシステム及び方法に関する。所望の結合機能を有するペプチドがアフィニティー選択によって単離される、高い複雑性のランダム配列及び抗原フラグメントライブラリーの構築を容易にする方法及びプラスミドベクターが記載されている。ペプチドディスプレイの密度は、アフィニティー選択のストリンジェンシーにおける重要な決定因子であるので、VLPでペプチドディスプレイ価数を制御するための方法及びプラスミドもまた記載される。本発明の方法によって、VLP、特にMS2 VLPでの広い範囲(すなわち、1粒子当たり平均で1未満から90まで)にわたるディスプレイ価数レベルの容易な調節が可能になり、これによって、低価数を示すVLPを含めて、免疫原及びワクチンの特定及び産生が容易になる。このシステムは、主にワクチン開発を視野に入れて開発されてきたが、薬物及び造影剤の細胞又は組織型特異的な標的送達に有用性を有するペプチド−VLPの特定を含め、様々な他の適用において、並びに新規な物質の鋳型合成において、有用性を有する。
【0004】
本出願は、VLPディスプレイシステムの実現を容易にする方法及びプラスミドベクターを記載する。本発明は、ウイルス様粒子上で提示され、VLPの合成及びVLPで提示されるペプチドの合成を指向する特定のRNA(MS2又はPP7のいずれか)をカプシドで包む異種ペプチドの挿入によってそれぞれが改変される、MS2又はPP7のコートポリペプチドから構成されるウイルス様粒子の発現において有用な核酸構築物を提供する。関連のウイルス様粒子、方法及び免疫原性組成物もまた提供される。
【背景技術】
【0005】
発明の背景
ワクチンとしてのVLP。近年の組み換えDNA技術の進歩によって、動物及びヒトの両方において有効な防御免疫を可能にするための十分な量のタンパク質が得られるように、免疫原性タンパク質が特定され、クローニングされ、適切な宿主中で発現されたワクチンの導入がもたらされた。ほとんどの有効なワクチンの多くは、ビリオン表面が中和抗体を誘発する強力な能力に基づく。これらとしては、防御抗体の応答を効果的に誘導する、ポリオ、インフルエンザ及び狂犬病などの認可された不活化ウイルスワクチン又は弱毒化ウイルスワクチンが挙げられる。さらに最近では、ヒトパピローマウイルス(HPV)及びB型肝炎ウイルス(HBV)の構造タンパク質の自己アセンブルに基づくサブユニットワクチンが、食品医薬品局によって承認された。このサブユニットは適切な宿主で発現され、次いで本物のウイルスを構造的に模倣するが、ウイルスのゲノムを欠いているので非感染性である粒子へと自己アセンブルされる。これらのいわゆるウイルス様粒子(VLP)は、個々の粒子それぞれで、ウイルス様粒子を含む構造タンパク質が多数のコピーにおいて存在するので、一般に極めて免疫原性である。この高密度の抗原提示は、それらの粒子が強固な抗体応答を誘発するのを特に有効にする。HBVワクチン及びHPVワクチンは、それぞれのウイルス自体の構造タンパク質からアセンブルされたVLPに基づくが、VLPはまた、異種エピトープの高密度提示のための足場として利用されてもよい。VLPは一般に高度反復性を示し、従って極めて免疫原性の構造であるので、それらは多数の異なるウイルス種に由来し得る。本発明の方法は、ファージディスプレイと類似の方法によって、免疫原性提示及びエピトープ開発の両方のために、RNAバクテリオファージ(特にMS2及びPP7)のVLPを利用することを目的に行われる[1,2]。
【0006】
RNAバクテリオファージ。一本鎖RNAバクテリオファージは、天然に広く分布して見られるウイルスのグループである。いくつかは、ゲノム配列、分子生物学、並びにカプシド構造及び集合に関して詳細に特徴付けられた。MS2はおそらく、最も研究されたグループのメンバーであって、本発明者の研究室で行われたほとんどの研究の焦点であったが、近年の研究はまた、PP7と呼ばれる関連のファージも開拓している。MS2は、4つのタンパク質(マチュラーゼ、コート、溶解(lysis)及びレプリカーゼ)のみをコードする3569ヌクレオチドの一本鎖RNAゲノムを有する。このウイルス粒子は、180個のコートポリペプチド、1分子のマチュラーゼ、及び1コピーのRNAゲノムから構成される。コートタンパク質自体が完全に正二十面体のシェルの形成を担うので、MS2 VLPは、プラスミドから単独の遺伝子の生成物として生成され得る。従って、ペプチドディスプレイのために用いられる他のファージと比較して、RNA VLPは著しく簡易である。ペプチドディスプレイ及びアフィニティー選択のためのMS2及びPP7のVLPの操作は、本願の発明者によって近年示されており[1,2]、また本明細書の後段にも記載される。
【0007】
従来のファージディスプレイによるエピトープ特定。ファージディスプレイは、ランダムなアミノ酸配列の大きいライブラリーの提示を可能にするいくつかの技術のうちの1つであり、それらから特定の専門的な機能(例えば、特定の抗体に結合する能力)を有するペプチドを選択する目的を備える。最も一般的に用いられるファージディスプレイ法は、繊維状ファージ(例えば、M13)に基づく。この基本的な概念は、ウイルス構造タンパク質のうちの1つに対して、ファージのDNAゲノム中に遺伝子的に融合されたランダムな配列のライブラリーを含んでいる組み換えバクテリオファージゲノムを作成することである。このような組み換え体が細菌にトランスフェクトされる時、それぞれがウイルス粒子を生成し、そのウイルス粒子は、その表面上に特定のペプチドを提示し、そのペプチドをコードする同じ組み換えゲノムをパッケージングする。これによって、この方法に必須の遺伝子型及び表現型の連鎖が確立される。任意関数(例えば、レセプターの結合、免疫原性)は、アフィニティー選択の使用と、続く大腸菌での増殖による選択物の増幅とによって、ペプチドディスプレイファージの複雑なライブラリーから選択され得る。ペプチドディスプレイファージの膨大なライブラリーでは、特定のレセプター(例えば、モノクローナル抗体)に結合できるごく少数が、アフィニティー精製されて、次いで大腸菌中での増殖によって増幅され得る。比較的単純な集団を得るには、通常数回の反復回数の選択及び増幅で十分であり、それから所望の活性を有するペプチドを提示する個々のファージをクローニングし、次いで特徴付けすることができる。選択する分子が抗体である場合、このようにして特定されたペプチドは、抗体によって認識されるエピトープであり、そして適切な条件下では、免疫された患者又は動物において、その天然の抗原中でそのエピトープに特異的な抗体応答を誘発し得る。
【0008】
しかし、繊維状ファージディスプレイには不利な点がある。最も重要なことに、繊維状ファージの分子生物学の特異な様式によって、本当に強力な免疫原性について必要な高密度でペプチドを提示することが困難又は不可能となる場合が多い。このことは、免疫原(すなわち、ワクチン)として有用であるべきペプチドエピトープはアフィニティー選択によって特定され得るが、これらのペプチドエピトープは、通常は化学的に合成されて、次いでさらに免疫原性の担体に結合されなければならないということを意味する。不幸にも、ペプチドは、そのアフィニティー選択及び最適化の間にそのペプチドが存在する構造的状況からこのように分離された場合、活性を失う場合が多い。ここで記載したRNAファージVLPディスプレイシステムは、他方では、アフィニティー選択及び免疫原性エピトープの提示を単一のプラットフォーム上で行う能力を生み出す。これは、高密度ペプチドエピトープディスプレイとアフィニティー選択能力とを組み合わせた結果であって、エピトープのアフィニティー選択の間に存在する構造的制約が、免疫過程の間維持され得るということを意味し、エピトープが所望の抗体応答を誘発するのに必要な構造を保持する確率が増大する。
【0009】
RNAファージVLPディスプレイ法の概説。本発明者は、MS2及びPP7を含めて、RNAバクテリオファージのVLPに基づくペプチドディスプレイ及びアフィニティー選択のための技術を以前に記載したが[1,2]、ここでその技術を簡略に説明する。VLPディスプレイ法の開発には、2つの前提条件が満足されることを必要とする:第一に、RNAファージコートタンパク質の形態、及びその中の適切に折り畳んでVLPにアセンブルする能力を破壊することなく外来ペプチドの挿入を許容する部位を特定する必要があった。コートタンパク質の表面上のAB−ループを、ペプチド挿入のための部位として選択した。ここに挿入されたペプチドは、VLPの表面上で顕著に提示される。不幸にも、コートタンパク質の野生型は、AB−ループ中のペプチド挿入に極めて耐性がなく、その圧倒的多数(通常は98%を超える)が折り畳みの失敗に至る。コートタンパク質は、通常は二量体として折り畳まれるが、そのうちの90が正二十面体VLPへとアセンブルされる。本発明者は、新規な形態のコートタンパク質を操作して、それを安定化し、さらなるAB−ループ挿入の耐性を与えた。そうするために、本発明者は、二量体中で2つの同一のポリペプチド鎖のN末端及びC末端の近接性を利用した(図1)。コートタンパク質コード配列を複製すること、次いで2つのコピーを単一のリーディングフレームに融合することによって、本発明者は、いわゆる一本鎖の二量体を生成した。この形態のタンパク質は、劇的にさらに熱力学的に安定であり、その折り畳みは一本鎖二量体の下流のコピーのAB−ループに挿入されたペプチドにかなり耐性である[1,2]。得られたVLPは、1つの二量体当たり1つのペプチド、又は1つのVLP当たり90のペプチドを提示する。ペプチドディスプレイ/アフィニティー選択能力についての第二の前提条件は、アフィニティー選択された配列を増幅するための手段を提供することが必須であることから、遺伝子型に対する表現型の連鎖である。この要件は、本発明者が、RNAファージVLPがその合成を指示するメッセンジャーRNAをカプシドで包むということを示した際に満たされた[1,2]。このことは、アフィニティー選択されたペプチド−VLPの配列が、逆転写及びポリメラーゼ連鎖反応によって増幅され得ることを意味する。選択の標的がモノクローナル抗体である場合、得られたアフィニティー選択されたVLPは、動物又は患者において、その活性が選択抗体の活性を模倣する抗体を引き出すためのワクチン候補物である。
【0010】
ペプチドディスプレイ価数及びその制御に関する考慮。本出願は、RNAファージVLPでの複雑なランダム配列及び抗原フラグメントのライブラリーの構築、並びに特定のモノクローナル抗体(又は他の任意のレセプター)に結合するペプチドのこのようなライブラリーからのアフィニティー選択を容易にするプラスミドベクターを記載する。元々記載されるとおり、RNAファージVLPディスプレイ技術は、それぞれのVLPに90個のペプチドを提示することに注意されたい。これは、ペプチドが一本鎖二量体の1つのAB−ループに挿入され、90個の二量体がVLPを作成するからである[1,2]。これらの粒子の多価数は、MS2 VLPのほとんどの適用について望ましい。例えば、この粒子の高い免疫原性は、提示されるペプチドの高密度に関連し、従ってワクチン中で価値のある特性である。しかし、複数の同時の弱い相互作用という理由のみにより、アフィニティー選択過程の間、多価数によって、選択標的について内因性の高い結合親和性を有するペプチドを提示する粒子を、密接に結合する粒子から識別することが困難になる。この「結合活性対親和性」のジレンマは、繊維状ファージディスプレイを用いての高い親和性のペプチドリガンドの選択において十分立証された問題である[3−5]。本発明は、広い範囲にわたって、すなわち1粒子当たり1未満から90まで、平均のペプチドディスプレイ価数を調節する手段を導入することによって、VLPディスプレイシステムにおいてこの問題に取り組む。これによって、アフィニティー選択過程の間にペプチドディスプレイの密度を変更することが可能になる。選択は数回行われ、初回は通常は多価数の提示を用いて行われ、これによって標的に関してある程度の最小の親和性を有する全てのペプチドを含む比較的複雑な集団を得る。その後の回では、ペプチドディスプレイの価数は減少されることができ、それによって選択のストリンジェンシーが増大し、結果として抗体標的に関して高い親和性を有するペプチドの単離及び好ましいエピトープの良好な分子模倣物が得られる。
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明の目的は、低価数の異種ペプチドを提示するVLPを提供することである。
【0012】
本発明のさらなる目的は、低価数又は高価数の異種ペプチドを提示するVLPなどのVLPを産生するための核酸構築物を提供することである。
【0013】
本発明のさらに別の目的は、低価数又は高価数の異種ペプチドの提示を制御し得るVLPなどのVLPを産生するための核酸構築物を提供することである。
【0014】
本発明のさらに別の目的は、高い親和性の免疫原を特定し、これらの免疫原を治療及び他の製剤又は適用において用いるために、低価数又は高価数の異種ペプチドの提示を制御し得るVLPを産生するための核酸構築物を提供することである。
【0015】
本発明のさらなる目的は、選択された抗体に対して高い親和性を示す免疫原性ペプチドを特定するための方法を提供することである。
【0016】
本発明のさらに別の目的は、高い親和性のペプチドをVLPに組み込むことである。
【0017】
本発明のさらなる目的は、本発明によるVLPを用いて免疫原性の方法及び組成物を提供することである。
【0018】
本発明のこれら及び/又は他の目的のいずれか1つ以上は、以下の本発明の説明から容易に得られる。
【課題を解決するための手段】
【0019】
発明の要旨
本発明は、選択的に免疫原性のペプチドを組み込むVLPを特定及び提供することのさらに効果的な手段を提供するために、高い複雑性のランダム配列及び抗原フラグメントペプチドライブラリーの構築を容易にし、かつMS2 VLPでのペプチドディスプレイの価数(すなわち、密度)を制御することを可能にする、組成物、構造(プラスミドを含む)、システム及び方法に関する。本発明は、その内容全体がここに援用され、上記及び[1,2,6,7]において簡略に記載された米国特許出願公開第2009/0054246号及び出願番号PCT/US2007/018614(WO08/024427として公開)において別に開示された、方法、組成物、粒子、単位及びその他の開示の拡張を示す。
【0020】
本発明は、所定のモノクローナル抗体に最も高い親和性を有するペプチドの選択が、天然の抗原の最良の分子模倣物を提供すること、そしてこれらのペプチドが、関係する抗体応答を提供又は誘導する可能性が最も高いことをもたらす。これらのペプチドは、患者において免疫原性を誘導し、防御応答を提供するために特に適しているものとして提案される。これらのペプチドから調製されるワクチン及び/又はこれらのペプチドを組み込むワクチンは、特にアジュバント無しで投与される本発明によるワクチンを含み、低減された副作用を有し、より効果的である。
【0021】
RNAファージMS2(pDSP1及びpDSP62)のVLP及びRNAファージPP7(pET2P7K32及びpDSP7)由来のVLPにおいて、ランダム配列又は抗原フラグメントペプチドライブラリーの構築を容易にするプラスミドベクターが記載される。これらのベクターは、1011〜1012個を超える個々のメンバーを有するライブラリーの作製を可能にする。しかし、これらのベクターは、高密度で(すなわち、1つのVLP当たり90個)の外来ペプチドを均一に提示するVLPを生じる。上記で説明したとおり、このことは、ペプチドに対して高レベルの免疫原性を与えるが、多価数が選択のストリンジェンシーを低下して、集団の中で最も緊密に結合する種を優先的に選択することを困難にすることでアフィニティー選択の間に問題を生じることが多いので、ワクチン適用のための明白な利点である。
【0022】
本発明は、ペプチドディスプレイ価数制御の問題に対する簡易な解決である、単一のRNAからの少なくとも四(4)アミノ酸長の異種ペプチドを含む大量の野生型及び少量の一本鎖(好ましくは、二量体)のコートタンパク質の産生を可能にするシステムを示す。このアプローチは、pDSP1(am)、pDSP62(am)、pET2P7K32(am)及びpDSP7(am)のようなプラスミドを構築することを含む。これらは、例えば、一本鎖二量体中でコーティングタンパク質の下流の第一のアミノ酸を通常コードするコドン(アラニン)の代わりに、終止コドン(好ましくは、アンバー終止コドン)を含むように改変された、上記のプラスミドの単純な改変体である。これらのプラスミドは、一本鎖二量体の二等分の接合部に終止コドンを有するので(例えば、pDSP1を参照)、それらのプラスミドは正常には、単位長さの野生型コートタンパク質しか産生せず、このタンパク質は当然にVLPにアセンブルする。しかし、本発明のアプローチでは、このプラスミド又は第二のプラスミド(例えば、pNMsupA)は、コート配列を翻訳するわずかな割合のリボソームに終止コドンを読み飛ばしさせ、異種ペプチドを含む一本鎖二量体を生じさせる量でサプレッサーtRNAがもたらされるようにして、プロモーター(例えば、異なる不適合性群からのクロラムフェニコール耐性プラスミド上のlacプロモーター)の制御下で発現されるアラニン挿入サプレッサーtRNA遺伝子などのtRNA遺伝子[8,9]を挿入して改変される。得られたタンパク質は、ゲストの異種ペプチドが、好ましくは、例えば第二のAB−ループ中若しくはカルボキシル末端又は下流サブユニット内の他の位置に挿入されており、同じmRNAから発現された野生型タンパク質とコアセンブル(共集合)して、低い価数を呈するモザイクのVLPを形成する。
【0023】
本発明によれば、VLPは、1つのVLP当たり1未満から九十(90)の異種ペプチド、好ましくは1つのVLP当たり約1〜約十(10)の異種ペプチド、さらに好ましくは1つのVLP当たり約1〜約5の異種ペプチド、さらに好ましくは1つのVLP当たり約1〜約3の異種ペプチド、最も好ましくは1つのVLP当たり約2〜約4の異種ペプチドを提示するように制御された方式で生成され得る。本発明によるペプチド密度の低下は、アフィニティー選択のストリンジェンシーが増大したVLPを生じ、後に高い提示密度の形式に戻った場合に強力に免疫原性となる高い親和性のペプチドの容易な特定が可能になる。アガロースゲル電気泳動及びノーザンブロットによって、本発明によって生じた粒子が関連のRNAをカプシドで包むことを確認する。本発明の方法はさらに、価数(1つのVLP当たりに生じるペプチドの数)が、サプレッサーtRNAの発現レベルを制御することによって、例えばサプレッサーtRNA合成のレベルを調節することによって、広い範囲にわたって調節され得ることになり、これは、例えば誘導因子濃度の関数としてその活性が調節され得るプロモーター(例えば、proB又は他の適切なプロモーター)からtRNAを発現することによって、適宜達成され得る。価数のレベルはまた、異なるサプレッサーtRNA又はその変異体の利用を通じて制御されることができ、より大きい又はより小さい内因性の抑制の効率を有する。
【0024】
当業者は、価数の制御が過剰な野生型コートタンパク質と一緒に組み換えタンパク質を同時発現することによってより簡単に達成されることができ、これによって外来のペプチドの含量が低下されたモザイクカプシドが生じると考えるかもしれない。不幸にも、このアプローチは、価数を適切なレベルまで(例えば、平均で1つのVLP当たり1未満から2〜3のペプチドまで)低下することが、野生型タンパク質が組み換えペプチドよりもかなり過剰に発現されることを必要とするので実用的でない。このような同時発現の方法は、過剰な関連のない(すなわち、非外来ペプチドをコードする)RNAを生じ、これがかなり多量であることによって、ペプチドコードRNAよりも優先してVLP内にパッケージングされる。特異的なカプシド形成は、RNA中のいかなる簡易なパッケージングシグナルの存在からも独立して出現するので(1)、選択的なカプシド形成のために少数派のペプチドコード種を選び出すことは簡単な方法ではないと考えられる。この同時発現アプローチの実行可能な改変型が存在することは想像できるが、これらの改変型は、あまりにも複雑になりがちである。本発明者のシステムは、単一のmRNAから両方の型のタンパク質を生成することによって、この問題を解決する。
【0025】
例として、以下の実施形態は、本発明をさらに実証するために用いられ得る。以下に記載される実施形態のそれぞれでは、MS2コートポリペプチド又はPP7コートポリペプチドが個々の核酸構築物において用いられることができるが、MS2又はPP7コートポリペプチドへの限定は、このような限定が記載の文脈の中で適当でない限り、考慮されるべきではないことに注意されたい。
【0026】
本発明の一実施形態は、次のものを含む核酸構築物(例えば、pDSP1を参照)を提供する:
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーター(例えば、本発明の詳細な説明において後述されるような)であって、このコートポリペプチド二量体のコード配列は、コートポリペプチド二量体のABループを規定する配列のその部分に対して、5’側に配置された第一の制限部位(例えば、SalI又はKpnI)を規定するように改変されている;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位(例えば、BamHI);
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)。
(d)抗生物質耐性遺伝子(例えば、カナマイシン)、及び
(e)原核生物細胞での複製のための複製起点(例えば、プラスミドColE1由来の複製起点)。
【0027】
別の実施形態では、本発明は、次のものを含む核酸構築物(たとえば、pDSP62を参照)を提供する:
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、この一本鎖二量体配列の二分の一が、改変された二量体を生成するために複数のサイレントヌクレオチド置換体で改変されて(すなわち、「コドンジャグリングされて(codon juggled)」)、突然変異のオリゴヌクレオチドプライマーが、この改変された二量体(好ましくは、2つの配列がプライマーのハイブリダイゼーションによって識別されることを可能にするAB−ループのいずれかの側における20ヌクレオチド単位内の十分な数の変異)の改変された半分又は未改変の半分に対して、特異的にアニールされ得る、プロモーターと;
(b)ABループを特定するコート配列のその部分に対して5’側に配置された第一の制限部位(例えば、SalI)と、
(c)コートポリペプチド二量体のコード配列に対して3’側の第二の制限部位(例えば、BamHI)と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と;
(e)第一の抗生物質に対して耐性の遺伝子と;
(f)原核生物細胞での複製のための複製起点と;
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点(例えば、M13又はfd)と;
(h)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージ(例えば、M13、fd)。
【0028】
別の実施形態では、本発明の核酸構築物に関して、バクテリオファージMS2(又はPP7)一本鎖コートポリペプチド二量体のコード配列はさらに、異種ペプチドをコードする核酸配列を含み、この構築物は必要に応じて、第二の制限部位に対して3’側に配置された転写ターミネーターを含む。そのような核酸構築物は、異種ペプチドの挿入によって改変されたMS2(又はPP7)のコートポリペプチドから構成されるウイルス様粒子の発現に有用であり、ここでは異種ペプチドがウイルス様粒子に提示されて、MS2(又はPP7)のmRNAをカプシドで包む。
【0029】
別の実施形態では(例えば、pET2P7K32)、核酸構築物は次のものを含む:
(a)バクテリオファージPP7(又はMS2)一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、このコートポリペプチド二量体のコード配列は、コートポリペプチド二量体のコード配列の下流部分に位置され、かつコートポリペプチド二量体のABループを規定する配列に対して5’側に、又はその中に配置された、第一の制限部位を規定するように改変されている、プロモーターと;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と;
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と、
(d)抗生物質耐性遺伝子と;
(e)原核生物細胞での複製のための複製起点。
【0030】
別の実施形態(例えば、pDSP7)では、核酸構築物は次のものを含む:
(a)バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、この一本鎖二量体配列の半分が、複数のサイレントヌクレオチド置換体で改変されて(すなわち、「コドンジャグリングされて(codon juggled)」)、変異原性のオリゴヌクレオチドプライマーが、一方又はもう一方の半分に対して特異的にアニールされ得る(好ましくは、2つの配列がプライマーのハイブリダイゼーションによって識別されることを可能にするAB−ループのいずれかの側における20ヌクレオチド単位内の十分な数の変異である)、プロモーターと;
(b)コートポリペプチド二量体のコード配列の下流部分に位置し、かつコートポリペプチド二量体のABループを規定する配列に対して5’側に、又はその中に配置された第一の制限部位と;
(c)コートポリペプチド二量体のコード配列に対して3’側に配置された制限部位と;
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と、
(e)第一の抗生物質に対して耐性の遺伝子と;
(f)原核生物細胞での複製のための複製起点と;
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点(例えば、M13、fd)と;
(h)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージ(例えば、M13、fd)。
【0031】
本発明の別の実施形態では、核酸構築物は次のものを含む:
(a)バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、このコートポリペプチド二量体のコード配列は、(1)コートポリペプチド二量体のコード配列の下流部分に位置され、コートポリペプチド二量体のABループを規定する配列に対して5’側に又はその中に配置された第一の制限部位を規定し、及び(2)ヌクレオチド配列(NNS)xを含み、ここでNは任意のヌクレオチドであり、Sはグアノシンヌクレオチド(G)又はシチジンヌクレオチド(C)であり、xは1〜500の整数である、ように改変されている、プロモーターと;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と;
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと;
(d)抗生物質耐性遺伝子と;
(e)原核生物細胞での複製のための複製起点。
【0032】
本明細書に記載される本発明の特定の実施形態では、本明細書に記載されるような、上記の特定の構成要素は、選択的ではあるが、構築物内に含まれ得ることが好ましい。
【0033】
他の実施形態(例えば、DSP1(am)、pDSP62(am)、pET2P7K32(am)及びpDSP7(am)と呼ばれる上記のプラスミドの誘導体)では、提示価数の制御は、以下の追加の特徴を含むことによってもたらされる:
(a)一本鎖二量体の上流半分と下流半分との間の接合部のナンセンスコドン(例えば、本明細書で他に記載されるようなアンバーコドン);
(b)上記の(a)に記載されるナンセンスコドンで翻訳終結を部分的に抑制することができるサプレッサーtRNA(例えば、アラニン−挿入アンバー−サプレッサー)を生成するプラスミド(例えば、pNMsupA)。このプラスミドは、第二の不適合性群由来の複製起点を有し、第二の抗生物質に対する耐性を付与し、これによって上記のコートタンパク質産生プラスミドの1つもまた含む細菌中で、そのプラスミドの安定な維持が可能になる。
【0034】
ワクチン候補物のライブラリー構築及びアフィニティー選択の概略
ここではMS2 VLPシステムを例として用いるが、同様の方法が、本出願でやはり示されるPP7システムにもあてはまることが理解されるべきである。
【0035】
1.ライブラリー構築。ランダムな配列ライブラリーは、2つの方法のいずれかによって生成され得る。これらは、後にさらに詳細に記載され、図9b及び図15に図示される。(A)PCR産物は、pDSP1へ、ランダムな配列でalIとBamHIとの間にクローニングされ、このランダムな配列は、AB−ループ中にそれを導入するような方法で結合される。この方法は、比較的低い複雑性のライブラリー(典型的には107〜108個のメンバー)の便利な構築物について適切であるが、より高レベルまで大規模化するには不便である。(B)より複雑性が高いライブラリー(例えば、109〜1011個以上)については、合成オリゴヌクレオチドプライマーは、pDSP62の一本鎖バージョンにアニールされ、DNAポリメラーゼで伸長されて、二本鎖の環状分子を生じ、次にこれがDNAリガーゼの作用によって共有結合的に閉じられる(図12を参照)。いずれかの方法によって産生される組み換えDNA分子は、大腸菌の適切な発現株(例えば、BL21(DE3))に導入され、そこでそれらの大腸菌がVLPを合成する。pDSP1又はpDSP62から産生される粒子は、1粒子当たり90という密度(高密度)で外来ペプチドを提示する。
【0036】
2.アフィニティー選択は、例えば、ある手順(例えば、バイオパニング、又は抗体に高い親和性を示すペプチドを迅速に特定及び分離するための他のアプローチ)にVLPライブラリーを供することによって行われ、ここでは、モノクローナル抗体はマルチウェルのプラスチックプレートの1つ以上のウェルの表面に吸着される。VLPライブラリー溶液を、免疫された抗体とともにインキュベートし、次いで未結合のVLPを洗い流して廃棄し、いずれかの結合したVLPを、通常は溶液のpHを低下することによって溶出する。続いて、溶出したVLPは熱変性され、それが含むRNAは、RNAに対してその3’末端(BamHI部位の十分下流)付近でアニールするオリゴヌクレオチドプライマーを用いて逆転写によってDNAにコピーされる。次いで、得られたcDNAを、第一の制限部位(例えば、SalI)の上流及び第二の制限部位(例えば、BamHI)の下流に特異的にアニールするプライマーを用いてPCRによって増幅する。
【0037】
3.再クローニング(recloning)。上記で得られたPCR産物を、第一及び第二の制限酵素(例えば、SalI及びBamHI)で分解し、次いで適切な発現ベクター中に再クローニングする。2回目の選択回を低ペプチド価数で行うべき場合、PCR産物は、pDSP1(am)又はpDSP62(am)中にクローニングされ、選択された集団全体を、pNMsupAを含む大腸菌の発現株中に導入する(図7を参照)。このように生成されたVLPは、一旦アフィニティー選択されれば、最初のライブラリーに存在したペプチドの集団よりもかなり簡素な集団しか提示しないので、平均して、その外来ペプチドの2〜3のコピーしか提示しない(すなわち、低価数である)。
【0038】
4.アフィニティー選択及び再クローニングのさらなる反復回は、上記のとおり行う。2〜4回又は好ましくは3回若しくは4回(高い価数で1又は2回、続いて低い価数で1又は2回)が典型的には、選択のモノクローナル抗体に緊密に結合するペプチドを提示するVLPの簡素な集団を生じるのには十分である。選択が完了したと考えられる場合、配列をpDSP1又はpDSP62中に再度クローニングし、ここでペプチドは、最大の免疫原性について高密度(1カプシド当たり最大90ペプチドまで)で再度提示される。個々のクローンが得られて、DNA配列分析による特徴付けに供される。選択抗体についてのインビトロにおけるそれらの親和性を評価し、それらが所望の抗体応答を引き出す能力を決定する。十分特徴付けられたモノクローナル抗体標的を使用する実証実験では、本発明者は、その配列が以前に特定されたエピトープの配列を模倣するペプチドを回収した。得られたVLPで免疫された動物は、もとの抗原のエピトープを認識する抗体を生じる。
【0039】
本発明のプラスミド及び他の側面の詳細は、以下の発明の詳細な説明にさらに記載される。
【0040】
本発明による上記の実施形態及び/又は他の実施形態のいずれか1つ以上が、以下の本発明の説明からさらに容易に得られる。
【図面の簡単な説明】
【0041】
【図1】MS2コートタンパク質二量体の構造を図示する。左側上の写真は、2つのサブユニット鎖のN末端及びC末端の近接性を強調する。この特徴によって、外来ペプチド挿入の高い耐性を有する、一本鎖二量体の構築が容易になった。また、図示(左下)されるのは、二量体及びインタクトなVLPの両方における、AB−ループの高い接近性である。垂直方向には、VLP自体の構造が示される。VLPが構成されるコートタンパク質は、準同一によって3つのわずかに異なる配置をとり、赤、青及び緑で示される。AB−ループは黄色で示される。VLP表面上でのそれらの反復性の性質及びそれらの露出に注目されたい。
【図2】VLPのランダム配列ペプチドライブラリーの提示の基礎を示す。ランダム配列ペプチド挿入物のライブラリーは、組み換えDNA方法によって作成され、得られたプラスミド集団は、形質転換によって大腸菌に導入され、VLPのライブラリーが生成される。集団中の個々のVLPのそれぞれは、その表面上に異なるペプチドを提示し、そしてその中に(mRNAの形態で)、その合成のための遺伝情報を含む。
【図3】アフィニティー選択の過程を図示する。ランダム配列ペプチドライブラリーを提示するVLPの集団を、表面で免疫されたモノクローナル抗体とともにインキュベートする。VLPのほとんどは、通常は抗体に結合できず、洗い流されて廃棄される。次いで、ペプチドが抗体との結合を示すいずれかのVLPを特異的に溶出し、それらが含むRNAを逆転写によってDNAにコピーし、PCRによって増幅し、そしてアフィニティー選択されたVLPの産生のために発現プラスミド(例えば、pDSP62又はpDSP62(am))中に再クローニングする。選択は、典型的には反復して(すなわち、2回を超えて)、好ましくは3〜5回行われる。
【図4】ペプチドディスプレイ価数の重要性を示す。多価数のディスプレイによって、本質的に緊密な結合因子を複数のレセプターと同時に相互作用する弱い結合因子と識別することが困難になり得る。通常は、初回のアフィニティー選択は高い価数で行うが、その後の回では、価数は低レベルまで低下され、それによって選択のストリンジェンシーを増大し、選択抗体について最高の親和性を有するペプチドの単離を確実にする。
【図5】AB−ループ中に挿入するために都合のよいクローニング部位を有するプラスミドpDSP1を示す。pDSP1は、例えば1つだけのSalI及びKpnI制限部位を含むように改変された、一本鎖二量体のコード配列を発現する。この二量体は、AB−ループへの外来配列の簡易なクローニングを容易にする。これらの部位を1つだけにするには、ベクター中及び上流のコート配列中で、多数のSalI及びKpnI部位を破壊することが必要であった。
【図6】プラスミドpDSP62を示す。一本鎖のDNAの産生を容易にするために、M13複製起点をプラスミド中に導入し、M13起点の配向に応じてpDSP61及びpDSP62として特定した。このプラスミドはまた、いわゆる「コドンジャグリングされた(codon juggled)」コート配列を、一本鎖二量体の上流半分に含む。コドンジャグリングされた半分は、下流の半分と同じアミノ酸配列をコードするが、可能な最大数のサイレント置換を含むことによってその配列とは異なり、2つの半分をオリゴヌクレオチドのアニールのために識別可能とする。
【図7】単一のRNAから大量の野生型及び少量のAB−ループ組み換えタンパク質を産生するための例示的なシステムを示す。一本鎖二量体においてコートタンパク質の下流のコピーの第一のアミノ酸を通常はコードするアラニンコドンの代わりにアンバー終止コドンを含む、pDSP1の改変体(pDSP1(am))を構築した。さらに、アラニン挿入サプレッサーtRNA遺伝子を合成して、クローニングした。この遺伝子は、コート配列を翻訳する少ない割合のリボソームに終止コドンを読み飛ばしさせて一本鎖二量体を生じさせる量で産生される。得られたタンパク質(とそのゲストペプチド)は、同じmRNAから発現される野生型タンパク質とコアセンブル(共集合)して、本明細書の他で記載されるようなモザイクカプシド及び低減された価数を生じる。
【図8】下に列挙するプラスミドで産生される精製VLPのSDSゲル電気泳動を示す。実験のポイントは、精製VLPにおける、上の図7について及び本出願の実施例の項において記載するような、一本鎖「読み飛ばし」(リードスルー、readthrough)生成物の含量を示すことである。精製されたVLPは、以下のプラスミドから生成された(左から右へ): pDSP1は、コートタンパク質一本鎖二量体のみを含むVLPを生成する。 pDSP1−Flagは、その第二のAB−ループ中に挿入されたFlagエピトープを有する一本鎖二量体のみを含むVLPを生成する。 pDSP1(am)は、一本鎖二量体中のコート配列の接合部にナンセンス変異を有し、これによってpDSP1(am)は、大量の野生型コートタンパク質を生じ、そしてサプレッサー−tRNA(pNMsupAから提供される)の存在下では、少量の一本鎖二量体を生じる。ナンセンス抑制は、この場合低い効率を有するので、ごくわずかなリボソームが、終止コドンを読み飛ばして一本鎖二量体を生成する。この2つの形態のコートタンパク質は、コアセンブルしてモザイク又はハイブリッドのVLPになり、これは大部分の野生型コートタンパク質及び少量の一本鎖二量体(野生型コートタンパク質のレベルの約3%と推定される)からなる。 pDSP1(am)−Flagは、pDSP1(am)のようであるが、一本鎖二量体の第二のAB−ループにフラグエピトープが挿入されている。それは、大部分の野生型コートタンパク質、及びそのAB−ループにフラグペプチドが挿入されている少量の一本鎖二量体を生成する。 図の右の2つの矢印は、pDSP1(am)及びpDSP1(am)−Flagの終止コドンの抑制によって生成される2つのタンパク質の位置を示す。
【図9】図9a及び図9bは、異種ペプチドをコードする核酸配列をそのプラスミド中に挿入するためのpDSP1プラスミド(9a)及び技術(9b)を示す。
【図10】pDSP62プラスミドを表し、一本鎖二量体の下流コピーのAB−ループ中へ配列を特異的に挿入するために、どのように合成オリゴヌクレオチドプライマーが設計され得るかを模式的に示す。オリゴヌクレオチドは、AB−ループに隣接するコート配列に対して完全にアニールするように設計され、それらの間に外来配列を挿入する。
【図11】図11aは、pDSP1プラスミドの核酸配列(配列番号1)を含む。図11bは、pDSP1(am)の核酸配列(配列番号2)を示す。図11cは、pDSP62プラスミドの核酸配列(配列番号3)を示す。図11dは、pDSP62(am)の核酸配列(配列番号4)を示す。pDSP1(am)及びpDSP62(am)のプラスミド配列は、アンバーコドンを導入するヌクレオチド置換によってのみ、pDSP1及びpDSP62と異なる。図11eは、M13K07のクロラムフェニコール耐性誘導体であるM13CM1の核酸配列(配列番号5)である。
【図12】ランダムなペプチド配列ライブラリーを生成するためにpDSP62及びM13CM1を利用する方法を示す。M13CM1は、pDSP62がそのM13起点から複製することを可能にするヘルパーウイルスである。従って、pDSP62を含む大腸菌細胞のM13CM1による感染によって、一本鎖の環状pDSP62の生成が可能になる。dut−、ung−細菌株中で増殖した場合、DNAは、DNA中に通常存在するdTTPのいくつかに代えてdUTPで置換される。ペプチドコード配列の部位特異的な挿入は、DNAポリメラーゼでの合成オリゴヌクレオチドの伸長によって達成され、その環は、DNAリガーゼの作用によって閉じられる。この得られた共有結合的に閉じられた環状DNAを、大腸菌のdut+、ung+株中にエレクトロポレーションによって導入し、そこでdUTP置換鎖が変性され、挿入の高頻度(通常は85〜90%)の組み込みを生じる[10]。
【図13】図13a及び図13bは、pP7Kプラスミド(13a)及びp2P7K32プラスミド(13b)を示す。これらのプラスミドは、翻訳抑制及びVLPアセンブリ試験によってPP7コートタンパク質AB−ループの挿入耐性を試験するためだけに、PP7 VLPプラットフォームの開発の間に利用された(実施例の項を参照)。それらは、後述されたアフィニティー選択のためのVLPライブラリーの構築に用いられる、pET2P7K32又はpDSP7と混同されるべきではない。
【図14】PP7コートタンパク質AB−ループ付近のヌクレオチド及びアミノ酸の配列を示し、また特異的なペプチド挿入物の挿入のために用いたプライマー配列を示す。多数の特異的及びランダムなペプチド配列を挿入して、AB−ループ挿入に対するPP7一本鎖二量体の一般的な耐性を試験し、特定の挿入されたペプチドの免疫原性を決定した。
【図15】PP7一本鎖コートタンパク質のAB−ループの挿入耐性を実証するために、p2P7K32中のランダムな配列ペプチドライブラリーの構築のために用いたスキームを図示する。この方法は、pDSP1のMS2一本鎖二量体中への挿入について図5bで示される方法と同様である。
【図16】本明細書に記載される種々のライブラリーそれぞれからの24個のクローンの細胞溶解液すべてのアガロースゲル電気泳動を示す。得られたクローンのほぼ100%が翻訳抑制をすることができ、このことは、それぞれのタンパク質が適切に折り畳まれた確率を示している。タンパク質が正確に折り畳まれたことをさらに確認するために、それぞれのVLPを形成する能力を、クローンのランダムな選択をこの図に示されるように電気泳動及びウエスタンブロットに供することによって評価した。それぞれのセットの上半分は、エチジウムブロマイド染色したゲルであり、下の半分は、複製のゲルのウエスタンブロットである。それぞれのセットにおける最も左側のレーンは、p2P7K32対照である。それぞれのクローンは、ランダムに生成された異なるペプチド配列を含み、ほぼ全てがVLPを生じるという事実によって、PP7コートタンパク質一本鎖二量体の高レベルの挿入耐性が実証される。
【図17】図17aはpETP7Kプラスミドを示し、図17bはpETP7K32プラスミドを示し、図17cはpDSP7プラスミドを示す。これらのプラスミドは、PP7コートタンパク質の高レベルの過剰発現のために生成された。pDSP7プラスミドは、PP7コートタンパク質の一本鎖二量体を含み、ここでは配列の二分の一が、AB−ループコード配列の近傍に十分な数のヌクレオチド置換を含んで、変異原性のオリゴヌクレオチドをアニールするために二つの半分を識別可能にする。ここで示される実施例では、上流の配列全体が、可能な最大数のサイレント変異を含むように改変される。pET2P7K32及びpDSP7は、MS2コートタンパク質プロデューサー、pDSP1及びpDSP62のPP7アナログとみなされるべきである。
【図18】pETP7K又はpET2P7K32を含む細菌中で生成されたVLPから抽出されたRNAのホルムアルデヒド/アガロースゲル上での電気泳動によって、それらが生成するVLPが、それらの合成を指向するmRNAをカプシドで包むことが示される。これは、逆転写及びPCRによってアフィニティー選択された配列を回収する手段を提供する。別のレーンは、同じプラスミドのインビトロでの転写によって生成されたRNAを含む。左の写真は、エチジウム染色ゲルを示す。右側は、PP7コートタンパク質センス鎖に特異的なオリゴヌクレオチドでプローブされた複製ゲルのブロットである。このプローブは、MS2又はQβコートタンパク質配列(示さず)のインビトロ転写に由来する同様の量のRNAとは反応できない。
【図19】PP7コートタンパク質中にクローニングされたいくつかの特異的なペプチド配列の列挙を示す。
【図20】抗−L2 mAb(RG−1)が、PP7 L2−VLPには結合するが、PP7 V3−VLPには結合しないことを示す。mAb RG−1の希釈物を、500ng/ウェルのL2−VLP又はV3−VLPと反応させた。結合は、西洋ワサビペルオキシダーゼ標識したヤギ抗−マウスIgG二次を用い、続いてABTSでの発色によって検出した。反応性は、405nmでの吸光度(OD405)の測定によって決定した。
【図21】V3ペプチド提示のPP7 VLPが免疫の際に抗−V3 IgG応答を誘導することを図示する。PP7 V3−VLPで、又は対照としてL2−VLPで免疫されたマウスでの抗−V3 IgG抗体応答を示す。マウスを、10μgのVLPと不完全フロイントアジュバントとを用いて3回免疫し、次いで血清を最終の追加免疫の2週後に収集した。別々のマウス7匹から得た希釈血清(6匹はV3−VLPで免疫し、1匹はL2−VLPで免疫した)を、HIVLAI由来のV3ループの一部を提示するペプチドとの反応性についてELISAによって試験した。結合は、西洋ワサビペルオキシダーゼ標識したヤギ抗−マウスIgG二次を用い、続いてABTSでの発色によって検出した。反応性は、405nmでの吸光度(OD405)の測定によって決定した。
【図22】抗Flag M2抗体でのMS2 VLPディスプレイを用いてのアフィニティー選択の結果を示す。抗Flag M2抗体のエピトープ配列は、これ以前に他者により十分に明らかにされている。4回のそれぞれにおけるいくつかのペプチドの配列は、選択の進行を明らかにするために示される。1回及び2回で見い出されるペプチドは、天然のエピトープの認識可能なエレメントを含むが、3回及び4回では類似性は明白であり、配列は野性型エピトープの配列に密接にマッチしている。実際に、4回目の配列は、十分に確立された繊維状ファージディスプレイの方法によって以前に得られた配列よりも密接に天然のエピトープにマッチする(「NEB Transcript,Summer,1006(New England Biolabsのウェブサイト、www.neb.comで入手可能)を参照)。
【図23】図23a〜23dは、pET2P7K32(配列番号6)、pET2P7K32(am)(配列番号7)、pDSP7(配列番号8)、及びpDSP7(am)(配列番号9)のプラスミドのヌクレオチド配列を示す。これらは、図11a〜11dに示される対応するMS2VLPプロデューサーのPP7コートタンパク質−産生アナログである。
【発明を実施するための形態】
【0042】
発明の詳細な説明
本発明によれば、当該分野の技術の範囲内の従来の分子生物学、微生物学及び組み換えDNA技術が使用され得る。このような技術は、文献中で詳細に説明される。例えば、Sambrookら、2001年,「Molecular Cloning:A Laboratory Manual」;Ausubel編集,1994年,「Current Protocols in Molecular Biology」第I〜III巻;Celis編集,1994年,「Cell Biology:A Laboratory Handbook」第I〜III巻;Coligan編集,1994年,「Current Protocols in Immunology」第I〜III巻;Gait編集、1984年,「Oligonucleotide Synthesis」;Hames & Higgins編集、1985年,「Nucleic Acid Hybridization」;Hames & Higgins編集、1984年,「Transcription And Translation」;Freshney編集、1986年,「Animal Cell Culture」;IRL Press,1986年,「Immobilized Cells And Enzymes」;Perbal,1984年,「A Practical Guide To Molecular Cloning」を参照のこと。
【0043】
ある範囲の値が示される場合、その範囲の上限と下限との間の、その状況が他を明確に示すのでない場合における下限の単位の十分の一までのそれぞれのその間の値、及びいずれかの他の言及された範囲における言及された値又はその間の値は、本発明の範囲内に包含されることが理解される。これらのより小さい範囲の上限及び下限は、独立してそのより小さい範囲内に含まれることができ、そしてその言及された範囲でいずれかの特に除外される限界が付された本発明の範囲内にもまた包含される。その言及される範囲が限界の一方又は両方を含む場合、それらの含まれる限界のいずれか又は両方を除外した範囲もまた本発明に含まれる。
【0044】
別段規定しない限り、本明細書において用いられるすべての技術及び科学的用語は、本発明が属する技術分野の当業者によって通常理解されるのと同じ意味を有する。本明細書に記載される方法及び材料と同様又は等価の任意の方法及び材料もまた、本発明の実行又は試験において用いられ得るが、好ましい方法及び材料がここに記載される。
【0045】
本明細書及び添付の特許請求の範囲で用いる場合、単数形(a、an及びthe)は、その状況が明らかに他を示すのでない限り、複数形の言及を包含する。
【0046】
さらに、次の用語は、下記の定義を有するものとする。
【0047】
本明細書において、「ポリヌクレオチド」という用語は、多量体形の任意の長さのヌクレオチド、リボヌクレオチド又はデオキシヌクレオチドのいずれかを指し、そして二本鎖及び一本鎖のDNA及びRNAの両方を包含する。ポリヌクレオチドは、例えばコード領域及び調節性配列(例えば、プロモーター又は転写ターミネーター)などの非コード領域といった、異なる機能を有するヌクレオチド配列を含んでもよい。ポリヌクレオチドは、天然の供給源から直接得られてもよいし、又は組み換え技術、酵素技術若しくは化学的技術の補助によって調製されてもよい。ポリヌクレオチドは、直線状であっても又は位相幾何学における環状であってもよい。ポリヌクレオチドは、例えば、発現ベクター若しくはクローニングベクターなどのプラスミド若しくはバクテリオファージベクターの一部、又はフラグメントであってもよい。
【0048】
制限エンドヌクレアーゼは、明確に規定された配列でDNAを切断する酵素である。それらは、例えば、同じ制限エンドヌクレアーゼでの消化によって生成される他のDNAフラグメントに、DNAリガーゼの作用を通じて容易に連結される特定のDNAフラグメントを生成するために、組み換えDNA技術で用いられる。本出願では、その認識配列がそれぞれGTCGAC、GGTACC、及びGGATCCであるSalI、KpnI、及びBamHIを含め、いくつかの特異的な制限エンドヌクレアーゼに対して言及する。
【0049】
本明細書において、「ポリペプチド」という用語は広義には、ペプチド結合によって一緒に連結される2つ以上のアミノ酸のポリマーを指す。「ポリペプチド」という用語はまた、ジスルフィド結合によって結合された2つ以上のポリペプチドを含む分子、又は多量体(例えば、二量体、四量体)として共有結合若しくは非共有結合的に結合されるポリペプチドの複合体を包含する。したがって、ペプチド、オリゴペプチド及びタンパク質という用語は、すべてがポリペプチドの定義内に包含され、そしてこれらの用語は交換可能に用いられる。これらの用語は、アミノ酸のポリマーの特定の長さを暗示するものでも、ポリペプチドが組み換え技術、化学的若しくは酵素的な合成を用いて生成されるか、又は天然に存在するかを暗示したり識別したりすることを意図するものでもないことが理解されるべきである。
【0050】
「一本鎖二量体」という用語は、通常は二量体のタンパク質であって、その2つのサブユニットが遺伝子的に(化学的に、共有結合を通して)単一のポリペプチド鎖に融合されたタンパク質を指す。具体的には、本発明では、MS2及びPP7コートタンパク質の両方の一本鎖二量体が構築された。これらのタンパク質のそれぞれは、天然には同一のポリペプチド鎖の二量体である。MS2及びPP7コートタンパク質二量体の両方において、1つのサブユニットのN末端は、コンパニオンサブユニットのC末端に対して物理的に近接して存在する(図1を参照)。一本鎖コートタンパク質二量体は、コートタンパク質のDNAコード配列を複製し、次いでそれらを尾−頭方式で互いに融合することによる組み換えDNA方法を用いて生成された。この結果は、単一のポリペプチド鎖であって、ここではコートタンパク質アミノ酸は2回存在し、上流コピーのC末端が、下流コピーのN末端に共有結合されている。通常の(野性型)の2つのサブユニットは、2つの鎖の間の非共有結合的な相互作用を通じてのみ会合される。一本鎖二量体では、これらの非共有結合的な相互作用が維持されるが、2つのサブユニットは、互いにさらに共有結合的に連結された。これは、タンパク質の折り畳み構造をかなり安定化し、そのタンパク質に対して上記のようなペプチド挿入の高い耐性を付与する。
【0051】
本出願は、コートタンパク質の「AB−ループ」に言及することが多い。RNAファージコートタンパク質は、保存された三級構造を保有する。MS2及びPP7コートタンパク質は、例えば、それぞれが図1で示されるMS2コートタンパク質の構造によって例示される構造を有する。それぞれのポリペプチド鎖は、AからGの文字で示される多数のβ鎖に折り畳まれる。β鎖のA及びBは、VLPの表面に露出されるヘアピンの頂部に2つの鎖を接続する3アミノ酸鎖のループを有するヘアピンを形成する。本出願に示されるように、ABループに挿入されるペプチドは、VLPの表面上に露出され、強力な免疫原性である。
【0052】
本明細書に記載されるアミノ酸残基は、「L」異性体型であることが好ましい。しかし、「D」異性体型の残基は、そのポリペプチドによって所望の機能が保持される限り、いずれのLアミノ酸残基とも代えることができる。NH2は、ポリペプチドのアミノ末端に存在する遊離のアミノ基を指す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離のカルボキシル基を指す。
【0053】
「コード配列」の用語は、本明細書では、そのタンパク質生成物のアミノ酸配列を直接特定する核酸配列の部分として規定される。コード配列の境界は、一般に原核生物においては、リボソーム結合(又はシャイン・ダルガノ)部位及び翻訳開始コドン(通常はAUG)によって決定される。真核生物においては、オープンリーディングフレームの開始部位に位置するAUG開始コドン(mRNAの5’末端に通常は近い)、及びオープンリーディングフレームの末端、通常はmRNAの3’末端付近に位置してそれを特定する翻訳終結配列(ナンセンスコドン:UAG、UGA、又はUAAのうちの1つ)によって決定される。限定するものではないが、コード配列としては、DNA、cDNA、及び組み換え核酸配列などを挙げることができる。
【0054】
上記で簡略に注記したとおり、「終止コドン」又は「終結コドン」とは、翻訳の終結をシグナル伝達するメッセンジャーRNA内のヌクレオチドトリプレットである。タンパク質は、アミノ酸の固有の配列であって、メッセンジャーRNAにおけるほとんどのコドンは、成長しているタンパク質鎖へのアミノ酸の付加に相当する−終止コドンは、このプロセスの終結をシグナル伝達し、アミノ酸鎖を遊離する。標準的な遺伝子コードでは、3つの終止コドンがある:UAG(RNA中)/TAG(DNA中)(「アンバー」終止コドンとしても知られる)、UAA/TAA(「オーカー」終止コドンとしても知られる)、及びUGA/TGA(「オパール」又は「アンバー」終止コドンとしても知られる)。この主要な群に対するいくつかの改変が知られている。本発明における終止コドンの使用は、通常はタンパク質合成を終止又は終結させる。しかし、tRNA中には、終止コドンを認識することを可能にさせ、リボソームに終止コドンを読み飛ばさせ、終止コドンの下流のコードされるペプチドの合成を可能にする変異がある[11−13]。例えば、アンバー終止コドンを認識するtRNA中の変異は、翻訳がコドンを「読み飛ばし」て全長タンパク質を生じることを可能にし、それによって、正常な形態のタンパク質を回収し、終止コドンを「抑制する」。ほとんどの場合、終止コドンの抑制は、部分的にしか効果的でない−わずかな割合のリボソームしか終止コドンを読み飛ばすことができないことが多い。しかし、ある場合には、抑制はかなりより効果的であり得る。2〜3のサプレッサーtRNAは、簡単に高度な内因性の抑制効率を保有する。他の場合には、弱いサプレッサーは、単にそれをより高レベルで発現することによってさらに効率的にされ得る。本発明の特定の実施形態では、終止コドンは、その終止コドンの下流の転写単位内にコードされるペプチドの合成を制御するために転写単位内に組み込まれる。終止コドンを認識し、終止コドンの下流のペプチドの合成を可能にするtRNAの制御された合成を提供することによって、そのほとんどが異種ペプチドを含まないコートタンパク質の集団内に、異種ペプチドを含むコートタンパク質が生成され得る。野生型(異種ペプチドが存在しない)コートタンパク質を含む異種ペプチドのこの混合物からアセンブルされるこの得られたVLPでは、異種提示の価数がかなり低くなる。
【0055】
組み換え細胞の「異種」領域とは、より大きい分子と関連して見られることが全く無い、より大きい核酸分子内の核酸の特定可能なセグメントである。「異種」ペプチドとは、より大きいポリペプチドと関連して見られることが全く無い、ポリペプチドの特定可能なセグメントであるペプチドである。
【0056】
VLPの価数とは、粒子に提示される異種ペプチドのコピー数を指す。「低い価数」の異種ペプチド、好ましくは少なくとも4のペプチド単位の免疫原性ペプチドを示すウイルス粒子は、このウイルス粒子を含むコートポリペプチド二量体中で、1未満から最大で約10以上の異種ペプチドを提示する粒子である。低い価数を示すウイルス粒子は、異種ペプチド(好ましくは、野生型コートポリペプチド)を含まない複数のコートポリペプチド二量体、及び異種ペプチドを、好ましくはコートポリペプチドの下流サブユニットのA−Bループ内又は一本鎖二量体コートポリペプチドのカルボキシ末端に含む少数のコートポリペプチド二量体から形成され、従ってモザイクVLPを形成する。
【0057】
文脈内で用いられる「複製起点」とは、通常はDNA複製開始領域を特定することによってDNA合成に関与するDNA配列を指す。必要な因子(DNAポリメラーゼなど)の存在下で、複製起点は、それに会合するDNAを複製させる。例えば、ColE1複製起点(pDSP1などのようなプラスミド中で用いられる)は、細菌の染色体とは独立して複製する能力を、多くの通常用いられるプラスミドクローニングベクターに与える。別の例はp15A複製起点であり、それは本願の他の箇所で記載されるプラスミドpNMsupAで用いらる(図7を参照)。ファージM13由来の追加の複製起点がプラスミド上に存在すること(例えば、pDSP62と同様)によって、大腸菌細胞がいわゆるヘルパーファージ(例えば、本出願中で記載されるM13CM1)で感染される場合にその起点を用いて複製する追加の能力が付与され、これによって必須のタンパク質因子がもたらされる。M13は、細胞内で二本鎖の環状DNAとして複製するが、一本鎖環状型も生じ、これはファージ粒子内にそれをパッケージングする。これらの粒子によって、図12に図示される方法を用いたライブラリー構築のために有用である、pDSP62及びpDSP7(本出願中の他の箇所に記載される)のようなプラスミドのための一本鎖環状DNAの都合のよい供給源が得られる。
【0058】
「プロモーター配列」とは、細胞中でRNAポリメラーゼを結合し、下流(3’方向)のコード配列の転写を開始することができるDNA調節領域である。本発明を規定するためには、このプロモーター配列は、バックグラウンドを超える検出可能なレベルで転写を開始するために必要な最小数の塩基又はエレメントを含む。プロモーター内では、RNAポリメラーゼの結合を担うDNA配列、及び転写開始に必須であるあらゆる関連の因子が見られる。細菌中では、プロモーターは通常、−35及び−10のコンセンサス配列と、多かれ少なかれ特定の転写開始部位とからなる。真核生物プロモーターは、常にではないが、「TATA」ボックス及び「CAT」ボックスを含む場合が多い。細菌の発現ベクター(通常はプラスミド又はファージ)は、一般的には大腸菌のLactose、Arabinose、Tryptophan及びProBオペロン由来のプロモーター、並びにバクテリオファージ供給源由来の他のプロモーターなどの、天然の供給源由来のプロモーターを利用する。例としては、バクテリオファージλ、T7、T3及びSP6由来のプロモーターが挙げられる。
【0059】
細菌では、転写は、一般的に細菌のrho因子の作用をその活性のために必要とするか否かに応じてrho依存性及びrho非依存性(又は内因性)ターミネーターに分類される、特定の転写終結配列で通常は終わる。これらのターミネーターは、RNAポリメラーゼにその転写活性を停止させる部位を特定し、そのためそれらは主としてRNAの3’末端を規定するが、リボヌクレアーゼの続いての作用がRNAをさらに整える場合もある。
【0060】
「抗生物質耐性遺伝子」とは、所定の抗生物質に対する耐性を細菌に付与するタンパク質をコードする遺伝子を指す。例えば、カナマイシン耐性遺伝子は、薬物を改変して不活性化するホスホトランスフェラーゼの合成を指向する。カナマイシン耐性遺伝子がプラスミド(例えば、pDSP1)上に存在すれば、形質転換された細菌内のプラスミドの存在について選択する機序が提供される。同様に、クロラムフェニコール耐性遺伝子は、アセチル化を通じて抗生物質を不活性化するアセチルトランスフェラーゼ酵素を生成することによって、細菌が薬物の存在下で増殖することを可能にする。本出願では、クロラムフェニコール耐性を用いて、pNMsupA及びM13CM1の細菌内での維持を確実にする。
【0061】
「逆転写及びPCR」は、アフィニティー選択されたVLPの核酸配列を増幅する手段として本出願で示される。「逆転写」とは、RNA分子のDNAコピー(又はcDNA)が、逆転写酵素の作用によって生成されるプロセスを指す。本出願では、逆転写を用いて、アフィニティー選択されたVLPにおいてカプシドで包まれたRNA配列のDNAコピーを生成する。逆転写酵素は、プライマーがRNAにアニールされることを必要とする(以下を参照)。
【0062】
「PCR」という用語は、インビトロで特定のDNA配列の増幅のために用いられる技術である、ポリメラーゼ連鎖反応を指す。「PCRプライマー」という用語は、標的DNAにアニールすることを可能にし、それによりDNAポリメラーゼ(例えば、Taq DNAポリメラーゼ)がDNA合成を開始することができるDNA配列(通常は合成オリゴヌクレオチド)を指す。PCRプライマーの対をポリメラーゼ連鎖反応で用いて、DNAの2つの鎖のそれぞれでDNA合成を開始し、これによって、例えば図9b及び図15に図示されるように2つのプライマーの間でDNAセグメントを増幅する。
【0063】
逆転写及びPCRに用いられるプライマーの例をここに示す。
E2:5’ TCA GCG GTG GCA GCA GCC AA 3’−本出願に記載されるVLPによってカプシドで包まれるRNAの3’末端付近にアニールする;逆転写をプライムするために用いられる。
E3.2:5’ CGG GCT TTG TTA GCA GCC GG 3’−E2プライマー部位のすぐ上流の部位において、上述の逆転写によって生成されるcDNAの3’末端付近にアニールする;PCR反応中で3’−プライマーとして機能して、アフィニティー選択された配列を増幅する。
E2及びE3.2プライマーは、pDSP1、pDSP1(am)、pDSP62、pDSP62(am)、pET2P7K32、pET2P7K32(am)、pDSP7、及びpDSP7(am)と共通の配列を含み、従ってこれらの供給源のそれぞれに由来するVLPによってカプシドで包まれるRNAの逆転写及びPCRに有用である。PCRは、下に記載されるプラスミド特異5’プライマーのうちの1つとのE3.2の対合に依存する。簡略のために、pDSP1及びpDSP1(am)についてのプライマーのみを示すが、他のVLPで見られる配列に特異的な類似のプライマーが必要に応じて利用されることも理解されるべきである。
J2:5’ACT CCG GCC TCT ACG GCA AC 3’−pDSP1の一本鎖二量体配列の間の接合部で配列に特異的にアニールするプライマー。E3.2とのPCR反応では、一本鎖二量体の下流の半分を含むDNAセグメントが増幅される(AB−ループ中に挿入されたペプチドの配列を含む)。SalI及びBamHIによるPCR産物の消化によって、関連のプラスミドのSalIとBamHIとの間に挿入され得るフラグメントを生成する(例えば、図9aを参照)。
J2(アンバー):5’ AC TCC GGC ATC TAC TAG AAC TTT AC 3’−このプライマーは、上記J2と全く同じように機能するが、pDSP1(am)から生成される配列に特異的である。
【0064】
「発現制御配列」とは、別のDNA配列の転写及び翻訳を制御及び調節するDNA配列である。コード配列は、RNAポリメラーゼがコード配列をmRNA(これは、次にコード配列によってコードされるタンパク質に翻訳される)に転写する場合、細胞中で転写及び翻訳制御配列の「制御下」にある。転写制御配列とは、宿主細胞中でコード配列の発現をもたらす、例えばプロモーター、エンハンサー、ポリアデニル化シグナル、ターミネーターなどのDNA調節性配列である。翻訳制御配列は、メッセンジャーRNAの翻訳の効率を、通常はリボソーム結合及び翻訳開始の効率を制御することによって決定する。例えば、本出願の他の箇所で考察されるように、RNAファージのコートタンパク質は、周知のファージレプリカーゼの翻訳リプレッサーである。コートタンパク質は、感染した細胞中において十分に高濃度で蓄積することで、ファージレプリカーゼ遺伝子の翻訳開始領域(シャイン・ダルガーノ及びイニシエーターAUG)を含むRNAヘアピンに結合する。これは、複製からウイルスアセンブリまでの移行が起きるウイルスのライフサイクルのある時点で、リボソーム結合を妨げて、レプリカーゼの合成を遮断する。
【0065】
細胞は、外因性又は異種のDNAによって、このようなDNAが細胞の内側に導入された場合に、「形質転換される」。形質転換するDNAは、細胞のゲノムを構成している染色体DNAに組み込まれ(共有結合され)てもよいし、又は組み込まれなくてもよい。例えば原核生物、酵母及び哺乳動物細胞では、形質転換するDNAは、複製起点のプラスミド上に存在するおかげで通常は細菌染色体とは独立に複製する、プラスミドのようなエピソームエレメント上で維持されてもよい。真核生物細胞に関して、安定に形質転換された細胞は、形質転換するDNAが染色体中に組み込まれることで、染色体複製を通じて娘細胞によってDNAが受け継がれる細胞である。この安定性は、形質転換するDNAを含んでいる娘細胞の集団から構成される細胞株又はクローンを樹立する真核生物細胞の能力によって実証される。
【0066】
本明細書に開示される配列と同じアミノ酸配列を有するポリペプチドをコードするが、本明細書に開示される核酸に縮重する、本発明のポリペプチド(単数又は複数)をコードする核酸配列もまた本発明の範囲内であることが理解されるべきである。「縮重する(degenerate to)」とは、異なる3文字コドンを用いて個別のアミノ酸を特定することを意味する。
【0067】
本明細書において、「エピトープ」とは、ポリペプチドの抗原決定基を指す。エピトープは、エピトープに特有の空間的高次構造中に3個のアミノ酸を含み得る。一般的には、エピトープは、少なくとも5個のこのようなアミノ酸からなり、さらに一般的には、少なくとも8〜10個のこのようなアミノ酸からなる。アミノ酸の空間的高次構造を決定する方法は、当該分野で公知であり、例えばX線結晶学及び二次元核磁気共鳴が挙げられる。
【0068】
本明細書において、「ミモトープ(mimotope)」とは、本物の抗原性エピトープを模倣するペプチドである。場合によっては、そのアミノ酸配列は、もとの抗原のエピトープとある程度の類似を示し得るが、場合によっては、配列の類似性はほとんど又は全く存在しない。そのような場合、ミモトープは、異なるアミノ酸配列を用いるエピトープの3D構造を模倣する。糖鎖抗原のエピトープのような、非ペプチドのエピトープさえも模倣するミモトープが確認され得る。
【0069】
本明細書において、「コートタンパク質(単数又は複数)」という用語は、バクテリオファージ又はRNAファージのカプシドアセンブリ内に組み込まれ得る、このバクテリオファージ又はRNA−ファージのタンパク質(単数又は複数)を指す。
【0070】
本明細書において、「コートポリペプチド」とは、コートタンパク質の機能を保有し、さらに全長のコートタンパク質及び/又はその一本鎖改変体を包含する、コートタンパク質のポリペプチドフラグメントとして本明細書において規定される。
【0071】
本明細書において、「免疫応答」という用語は、B−リンパ球、T−リンパ球及び/又は抗原提示細胞の活性化若しくは増殖をもたらす体液性免疫応答及び/又は細胞性免疫応答を指す。しかし、ある場合には、免疫応答は、低い強度であって、本発明による少なくとも1つの物質を用いる場合にのみ検出可能になることがある。「免疫原性」とは、生きている生物体の免疫系を刺激するために用いられる作用物であり、それによりその免疫系の1つ以上の機能が増大されて、免疫原に向けられる。「免疫原性ポリペプチド」とは、細胞性及び/又は体液性の免疫応答を引き出すポリペプチドであり、アジュバントの有無にかかわらず、単独か又は担体に対して連結されている。好ましくは、抗原提示細胞は活性化され得る。
【0072】
本明細書において、「自己抗原」という用語は、宿主のDNAによってコードされるタンパク質、及び宿主のDNAによってコードされるタンパク質又はRNAによって生成される生成物を指す。さらに、2つの若しくはいくつかの自己分子の組み合わせから生じるか又は自己分子の画分に相当するタンパク質、及び高い相同性又は上記のように2つの自己分子を有する(>95%、好ましくは>97%、さらに好ましくは>99%)タンパク質もまた、自己とみなしてもよい。自己抗原の例としては、限定するものではないが、ErbB−2、アミロイド−β、免疫グロブリンE(IgE)、ガストリン、グレリン、血管内皮成長因子(VEGF)、インターロイキン(IL)−17、IL−23、IL−13、CCR5、CXCR4、神経成長因子(NGF)、アンジオテンシンII、TRANCE/RANKL及びMUC−1が挙げられる。
【0073】
本明細書において、「ワクチン」という用語は、本発明の組成物を含みかつ動物に投与され得る形態である処方物を指す。
【0074】
本明細書において、「バクテリオファージのウイルス様粒子」という用語は、バクテリオファージの構造を模倣し、非複製的及び非感染性であり、通常は感染性ウイルスとしてそのバクテリオファージの増殖に必要な1つ以上のウイルス遺伝子を欠いているウイルス様粒子(VLP)を指す。RNAバクテリオファージのVLPは、通常はまた、宿主に対するウイルスの結合又は進入を担う1又は複数のタンパク質をコードする1又は複数の遺伝子を欠いている。
【0075】
この定義はまた、前述の1又は複数の遺伝子が依然として存在しているが不活性であり、従ってやはりバクテリオファージの非複製的及び非感染性のウイルス様粒子ももたらす、バクテリオファージのウイルス様粒子を包含する。
【0076】
「RNAバクテリオファージコートタンパク質のVLP」とは、RNAバクテリオファージコートタンパク質の1つ以上のサブユニットの自己アセンブリから形成されるカプシド構造として規定され、通常はそのコートタンパク質自体のmRNAを含み、さらに任意で宿主RNAを含む。本出願の目的のために、RNAファージVLPは通常、180コピーの野生型コートタンパク質二量体から、若しくは90コピーの一本鎖二量体コートタンパク質から、又は可変数の野生型二量体及び1粒子当たり全部で90個の一本鎖二量体コートタンパク質を含むモザイクVLPとして、アセンブルされる。
【0077】
核酸分子は、発現制御配列が核酸配列の転写及び翻訳を制御及び調節する場合、その発現制御配列に対して「作動可能に連結される」か、又は「作動可能に会合される」。「作動可能に連結される」という用語は、発現されるべき核酸配列の前に適切な開始シグナル(例えば、ATG)を有すること、並びに発現制御配列の制御下で核酸配列の発現及びこの核酸配列によってコードされる所望の生成物の産生を可能にするために正確なリーディングフレームを維持することを包含する。組み換えDNA分子に挿入することが望ましい遺伝子が適切な開始シグナルを含まない場合、このような開始シグナルを遺伝子の前に挿入してもよい。
【0078】
「ストリンジェントなハイブリダイゼーション条件」という用語は、当業者に知られており、Current Protocols in Molecular Biology,John Wiley & Sons.N.Y.(1989年),6.3.1−6.3.6.に見られる。好ましくは、ストリンジェントなハイブリダイゼーション条件の非限定的な例は、約45℃で6×の塩化ナトリウム/クエン酸ナトリウム(SSC)中でのハイブリダイゼーション、続いて0.2.×SSC、0.1%SDSにおいて50℃で、好ましくは55℃で、さらに好ましくは60℃又は65℃での1回以上の洗浄である。
【0079】
ウイルス様粒子の生成
本発明は、ウイルス様ファージ粒子、並びにこれらの粒子をインビボ及びインビトロで生成するための方法に関する。通常、これらの粒子はインビボで生成されるが、これらの粒子の使用はインビトロ状況で適用されてもよく、このアプローチは、本発明によって予想される。本発明によって、実験の複雑性を増大して、相互作用選択に必要な時間を短縮することが可能になる。この方法は典型的には、インビトロでビリオンを産生する工程、及びこのビリオンを回収する工程を包含する。本明細書において、ビリオンを「インビトロで」生成するとは、細胞の外側で、例えば無細胞系で、ビリオンを生成する工程を指し、一方で「インビボで」ビリオンを生成するとは、細胞、例えば大腸菌又は緑膿菌細胞の内側でビリオンを生成する工程を指す。
【0080】
バクテリオファージ
本明細書で想定される系(システム)は、一本鎖RNAバクテリオファージの特性に基づく(The BacteriophagesでのRNA Bacteriophages, Calendar,RL編集.Oxford University Press.2005年を参照)。この群の公知のウイルスは、大腸菌、緑膿菌及びアシネトバクターのなど多様な細菌を攻撃する。それぞれ、極めて類似のゲノム構成、複製ストラテジー及びビリオン構造を保有している。特に、バクテリオファージは、一本鎖の(+)−センスRNAゲノムを含み、マチュラーゼ、コート及びレプリカーゼの遺伝子を含み、かつ小型(300オングストローム未満)の正二重面体のカプシドを有する。これらとしては、限定するものではないが、MS2、Qβ、R17、SP、PP7、GA、M11、MX1、f4、Cb5、Cb12r、Cb23r、7s及びf2 RNAバクテリオファージが挙げられる。
【0081】
PP7は、緑膿菌の一本鎖RNAバクテリオファージであって、MS2及びQβのような大腸菌ファージとは遠縁である。PP7バクテリオファージは、通常は緑膿菌に感染するが、PP7コートタンパク質が大腸菌中で図9及び図13に記載されるプラスミドのようなプラスミドから発現される場合、ウイルス様粒子が容易に生成される。PP7コートタンパク質は、特異的なRNA結合活性を備え、PP7レプリカーゼの翻訳開始領域に融合された配列の翻訳を抑制し得る、特異的なRNA−結合タンパク質であるということが確認された。これは、PP7の翻訳オペレーターを抑制するが、MS2又はQβファージのオペレーターは抑制しないためである。脱凝集したウイルス様粒子からのコートタンパク質の精製及びそのRNA結合活性を再構成するための条件は、確立されている。インビトロにおけるPP7オペレーターRNAの解離定数は、約1nMであることが確認された。コートタンパク質がレプリカーゼ−β−ガラクトシダーゼ融合タンパク質の翻訳を抑制する遺伝子系を用いて、PP7 RNAの結合に重要なアミノ酸残基が特定された。Peabodyら、Translational repression and specific RNA binding by the coat protein of the Pseudomonas phage PP72001,J.Biol.Chem.,Jun 22;276(25):22507−13.Epub 2001年4月16日。
【0082】
いくつかの一本鎖RNAバクテリオファージのコートタンパク質は、公知の翻訳リプレッサーであり、レプリカーゼリボソーム結合部位を含むRNAヘアピンを結合することによってウイルスレプリカーゼ合成を遮断する。RNAファージのX線構造決定によって、コートタンパク質アミノ酸配列の比較から明らかになる相同性が、三次構造において反映されることが示される。リプレッサー及びウイルス粒子の基本的構築ブロックの両方であるコートタンパク質二量体は、一緒になってRNAが結合される大きいβシート表面を形成する2つの結び付けられた単量体からなる。コートタンパク質のそれぞれは、異なるRNAを結合するために共通の構造的なフレームワークを用い、それによって特定のRNA−タンパク質認識の根拠を調査する機会を与える。そのコートタンパク質がMS2のアミノ酸配列に対してわずか13%のアミノ酸配列同一性しか示さない緑膿菌のRNAバクテリオファージである、PP7のコートタンパク質のRNA結合特性を本明細書に記載する。以下の知見もまた示される:(1)PP7のコートタンパク質は、翻訳リプレッサーである;(2)PP7レプリカーゼ翻訳開始部位を含むRNAヘアピンは、インビボ及びインビトロの両方でPP7コートタンパク質によって特異的に結合され、このことはこの構造が翻訳オペレーターであることを示す;並びに(3)RNA結合部位がコートタンパク質β−シートに存在する。この部位のマップは上に示した。
【0083】
例示のために、この群での特によく特徴付けられたメンバー、MS2のゲノムを利用し、これはわずか4個のタンパク質(そのうち2つがビリオンの構造的な成分である)をコードする一本鎖の(+)−センスRNAの3569ヌクレオチド長である。ウイルス粒子は、180コピーのコートタンパク質から形成される正二重面体のカプシド及び1分子のマチュラーゼタンパク質と一緒になった1分子のRNAゲノムから構成される。コートタンパク質はまた、特異的なRNA結合タンパク質である。アセンブリは、コートタンパク質が、レプリカーゼシストロンの5’末端に近いRNAヘアピンである、その特異的な認識標識と会合する場合に開始され得る(その全体が本明細書に援用される、2009年2月26日公開の米国特許出願公開第20090054246の図1B及び配列番号1を参照)。次いでウイルス粒子は、細胞がウイルス溶解タンパク質の影響下で破裂する時に培地中に遊離される。感染性ウイルスの形成は、少なくとも3つの構成要素、すなわちコートタンパク質、マチュラーゼ及びウイルスゲノムRNAを必要とするが、実験では、正二重面体のカプシドのシェルのアセンブリに必要な情報は、コートタンパク質自体の中に完全に含まれることが示される。例えば、精製されたコートタンパク質は、RNAの存在によって刺激されるプロセスにおいてインビトロでカプシドを形成し得る(Beckettら、,1988年,J.Mol Biol 204:939〜47)。さらに、細胞中でプラスミドから発現されるコートタンパク質は、インビボでウイルス様粒子にアセンブルする(Peabody,D.S.,1990年,J Biol Chem 265:5684〜5689)。
【0084】
コートポリペプチド
コード領域によってコードされるコートポリペプチドは、典型的には、少なくとも120、好ましくは少なくとも125アミノ酸長であり、約135アミノ酸長以下、好ましくは130アミノ酸長以下である。本質的にいずれかの一本鎖のRNAバクテリオファージ由来のコートポリペプチドが用いられ得ることが予想される。コートポリペプチドの例としては、限定するものではないが、MS2コートポリペプチド(例えば、米国特許出願公開第20090054246号の配列番号2を参照)、R17コートポリペプチド(例えば、ジーンバンク・アクセッション番号(Genbank Accession No.)P03612を参照)、PRR1コートポリペプチド(例えば、ジーンバンク・アクセッション番号ABH03627を参照)、frファージコートポリペプチド(例えば、ジーンバンク・アクセッション番号NP_039624を参照)、GAコートポリペプチド(例えば、ジーンバンク・アクセッション番号P07234を参照)、Qβコートポリペプチド(例えば、ジーンバンク・アクセッション番号P03615を参照)、SPコートポリペプチド(例えば、ジーンバンク・アクセッション番号P09673を参照)、f4コートポリペプチド(例えば、ジーンバンク・アクセッション番号M37979.1を参照)及びPP7コートポリペプチド(例えば、ジーンバンク・アクセッション番号P0363 0を参照)が挙げられる。
【0085】
PP7コートポリペプチドの例としては、限定するものではないが、RNAヘアピンを有する複合体でのPP7コートタンパク質二量体の種々の鎖が挙げられる(例えば、ジーンバンク・アクセッション番号2QUXR;2QUXO;2QUX_L;2QUX_I;2QUX_F;及び2QUX_C)。また、本明細書の実施例1、及びPeabodyら、RNA recognition site of PP7 coat protein,Nucleic Acids Research.2002年,第30巻,第19号、4138〜4144.[14,15]も参照のこと。
【0086】
本発明において有用なコートポリペプチドとしてはまた、上記で開示されたコートポリペプチドの1つ以上と類似性を有するポリペプチドが挙げられる。この類似性は、構造的類似性と見なされる。構造的類似性は、2つのアミノ酸配列(すなわち、候補アミノ酸配列及びアミノ酸配列)の残基を整列させて、それらの配列の長さに沿った同一のアミノ酸の数を最大限に利用することによって決定され得る;いずれか又は両方の配列におけるギャップは、同一のアミノ酸の数を最大限に利用するために整列させることにおいて許されるが、それぞれの配列におけるアミノ酸は、それにかかわらずその適切な順序のままでなければならない。候補アミノ酸配列とは、例えば米国特許出願公開第2009/0054246号の配列番号2に示されるアミノ酸配列と比較されるアミノ酸配列である。候補アミノ酸配列は、一本鎖RNAウイルスから単離されることも、組み換え技術を用いて産生されることも、又は化学的若しくは酵素的に合成されることもできる。好ましくは、2つのアミノ酸配列は、Tatusovaら(FEMS Microbial Lett 1999年,174:247〜250)に記載されたような、またhttp://www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html.で入手可能である、GCGパッケージ(バージョン10.2,Madison WI)でのBESTFITアルゴリズム、又はBLAST2サーチアルゴリズムのBlastpプログラムを用いて比較される。好ましくは、全てのBLAST 2サーチパラメーターのデフォールト値を用い、これはマトリックス=BLOSUM62;オープンギャップペナルティー=11、エクステンションギャップペナルティー=1、ギャップ xドロップオフ=50、期待値=10、ワードサイズ=3、及び任意でフィルターオンを含む。BLASTサーチアルゴリズムを用いる2つのアミノ酸配列の比較において、構造的類似性は、「同一性」と呼ばれる。
【0087】
また好ましくは、コートポリペプチドとしては、上記で開示されるアミノ酸配列の1つ以上に対して、少なくとも80%のアミノ酸同一性、少なくとも85%のアミノ酸同一性、少なくとも90%のアミノ酸同一性、又は少なくとも95%のアミノ酸同一性を有するアミノ酸配列を有するポリペプチドが挙げられる。好ましくは、コートポリペプチドは活性である。コートポリペプチドが活性であるか否かは、カプシドを形成して一本鎖RNA分子をパッケージングするポリペプチドの能力を評価することによって決定され得る。このような評価は、インビボ又はインビトロの系を用いて行われることができ、このような方法は当該分野及び通常作業において公知である。あるいは、ポリペプチドは、言及されたコートポリペプチド及び/又は機能的活性のように類似の3次元構造を有する場合に、構造的に類似であるとみなされ得る。
【0088】
コートポリペプチド又はポリペプチド中に挿入された異種ペプチド配列は、ランダムなペプチド配列であり得る。特定の実施形態では、ランダム配列は配列Xaanを有し、ここでnは、少なくとも4、少なくとも6、又は少なくとも8及び20以下、18以下、又は16以下であり、そしてそれぞれのXaaは、独立したランダムなアミノ酸である。あるいは、ペプチドフラグメントは、規定された配列を有し、公知の機能(例えば、抗原性、免疫原性)を保有し得る。異種配列は、コートポリペプチドのアミノ末端、コートポリペプチドのカルボキシ末端に存在してもよいし、又はコートポリペプチド内の他の箇所に存在してもよい。好ましくは、異種配列は、挿入配列がカプシドの外面に発現されるようにして、コートポリペプチド中のある位置に存在する。特定の実施形態において、及び本明細書の後述の実施例に記載されるように、ペプチド配列は、上述のコートポリペプチドのABループ領域に挿入されてもよい。このような位置の例としては、例えば、コートポリペプチドのアミノ酸11〜17、又はアミノ酸13〜17の直後のコートポリペプチド中への挿入配列の挿入が挙げられる。最も特殊な実施形態では、異種ペプチドが、MS−2のアミノ酸11〜17、又は特に13〜17に対応する部位に挿入される。
【0089】
本発明による特定の実施形態では、異種ペプチドが、以下に対応する部位に挿入される:
(a)MS−2、R17及びfrコートポリペプチドのアミノ酸11〜17、又は特に13〜17;
(b)GAコートポリペプチドのアミノ酸10〜16
(c)Qβ及びSPコートポリペプチドのアミノ酸10〜17;
(d)PP7コートポリペプチドのアミノ酸8〜11、及び
(e)PRR1コートポリペプチドのアミノ酸9〜17。
【0090】
あるいは、異種ペプチドは、コートポリペプチドのN−末端に挿入されても、又はC−末端に挿入されてもよい。
【0091】
異種ペプチドは、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択できる。
【0092】
この異種ペプチドとしては、限定するものではないが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択されるペプチドが挙げられる。
【0093】
構造的に類似のコートポリペプチドにおいて対応する位置を決定するために、この構造的に類似のコートポリペプチドのアミノ酸配列を、上記で特定されたような名前を挙げられたコートポリペプチドの配列と整列させる。例えば、MS−2コートポリペプチドに対して構造的に類似のコートポリペプチドの対応する位置を、配列番号2(本明細書に援用される米国特許出願公開第2009/0054246号の)と整列させる。この整列から、配列番号1(やはり米国特許出願公開第2009/0054246号の)の所定の位置に対応する他のコートポリペプチドにおける位置を決定し得る。
【0094】
特定の実施形態では、コートポリペプチドは、上流及び下流のサブユニットを含む一本鎖二量体である。それぞれのサブユニットは、機能的なコートポリペプチド配列を含む。異種ペプチドは、本明細書において上述された部位で上流及び/又は下流のサブユニットに挿入されることができ、例えば、好ましくは下流サブユニットのA−Bループ領域に挿入されることができる。特定の実施形態では、このコートポリペプチドは、本明細書に援用される米国特許出願公開第2009/0054246号の配列番号12に示される配列を有し得るMS2コートポリペプチドの一本鎖二量体である。
【0095】
特定の実施形態では、コートポリペプチドは、上流及び下流のサブユニットを含む一本鎖二量体である。それぞれのサブユニットは、機能的なコートポリペプチド配列を含む。異種ペプチドは、本明細書において上述された部位で上流及び/又は下流のサブユニットに挿入されることができ、例えば、下流サブユニットのA−B領域に挿入されることができる。特定の実施形態では、このコートポリペプチドは、PP7コートポリペプチドの一本鎖二量体である。
【0096】
転写単位の調製
本発明の転写単位は、発現調節性領域(例えばプロモーター)、コートポリペプチド
をコードする配列、及び転写ターミネーターを含む。RNAポリヌクレオチドは、任意でコート認識部位(「パッケージングシグナル」、「翻訳オペレーター配列」、「コート認識部位」とも呼ばれる)を含んでもよい。あるいは、転写単位は、翻訳オペレーター配列を含まなくてもよい。プロモーター、コード領域、転写ターミネーター、及び存在する場合はコート認識部位は、一般的に作動可能に連結される。「作動可能に連結される」又は「〜と作動可能に会合される」とは、そのように記載された構成要素が、それらの意図する方法で機能することを可能にする関係にある並列状態を指す。調節性配列は、コード領域の発現が、調節性配列と両立する条件下で達成されるような方法で連結される場合に、そのコード領域と「作動可能に連結される」か、又は「作動可能に会合される」。コート認識部位は、存在する場合、それが意図する方法で機能するならば、RNAポリヌクレオチド内の任意の位置でよい。
【0097】
本発明は、いずれかの特定のプロモーターの使用によって限定されず、広範な種々のプロモーターが公知である。本発明で用いられるプロモーターは、構成的プロモーターであっても、誘導プロモーターであってもよい。好ましいプロモーターは、コートポリペプチドをコードするコード領域によってコードされた高レベルのRNAを駆動することができる。このようなプロモーターの例は、当該分野で公知であり、例えばlacプロモーターT7、T3、及びSP6プロモーターなどが挙げられる。
【0098】
本明細書に記載されるコートポリペプチドをコードするコード領域のヌクレオチド配列は、容易に決定される。本明細書に記載されるコートポリペプチドの1つをコードするヌクレオチド配列の分類の例は、配列番号3(本明細書に援用される米国特許出願公開第2009/0054246の)のヌクレオチド4080〜4470である。これらの分類のヌクレオチド配列は、大型だが有限であり、この分類のそれぞれのメンバーのヌクレオチド配列は、標準的な遺伝子コードを参照して当業者によって容易に決定され得る。
【0099】
さらに、RNAバクテリオファージ一本鎖コートポリペプチドのコード配列は、異種ペプチドの挿入のための部位、及びその異種ペプチドのコード配列自体を含む。特定の実施形態では、その異種ペプチドの挿入のための部位は、制限酵素部位である。
【0100】
特定の実施形態では、コード領域は、コートポリペプチドの一本鎖二量体をコードする。最も特殊な実施形態では、コード領域は、改変された一本鎖コートポリペプチド二量体をコードし、ここでこの改変は、挿入部位に少なくとも4個のアミノ酸のコード配列の挿入を含む。このような転写単位の特定の実施形態の概略図は、米国特許出願公開第2009/0054246号の図3に示される。転写単位は、lacプロモーターのような細菌のプロモーターを含んでもよく、又はT7プロモーターのようなバクテリオファージプロモーター、及び任意でT7転写ターミネーターを含んでもよい。
【0101】
プロモーター及び融合ポリペプチドをコードするコード領域を含むことに加えて、RNAポリヌクレオチドは、通常は転写ターミネーター及び任意でコート認識部位を含む。コート認識部位は、RNAとして存在する場合、ヘアピンを形成するヌクレオチド配列である。これはまた、当該分野では、翻訳オペレーター、パッケージングシグナル、及びRNA結合部位とも呼ばれる。限定する意図はないが、この構造は、翻訳リプレッサー(例えば、コートポリペプチド)によって認識される結合部位として機能し、RNAパッケージングを開始すると考えられる。コート認識部位のヌクレオチド配列は当該分野で公知であり、これには例えば配列番号1(米国特許出願公開第2009/0054246号の図1Bを参照)のヌクレオチドが挙げられる。他のコート認識配列は、一本鎖RNAバクテリオファージR17、GA、Qβ、SP、及びPP7において特徴付けられており、そして当業者にとって容易に利用可能である。本質的に任意の転写ターミネーターが、それがプロモーターとともに機能するならば、RNAポリヌクレオチド中で用いられ得る。転写ターミネーターは、当業者に公知であり、容易に利用可能であり、そしてごく普通に用いられる。
【0102】
合成
後にさらに詳細に記載されるように、本発明のVLPは、特に転写単位が細菌のプロモーターを含むならば、転写単位を細菌に導入することによってインビボで合成され得る。あるいは、VLPは、カップリングされた無細胞の転写/翻訳システムにおいてインビトロで生成されることができる。
【0103】
異種の物質をカプシドで包むVLPのアセンブリ
合成の間、VLPは通常、VLPが翻訳によって生成されるメッセンジャー−RNAと会合する。これは、本出願に記載されるシステムのアフィニティー選択能力にとって重要である。しかし、いくつかの他の適用(例えば、薬物又は造影剤の標的化送達)では、VLPへの他の物質の導入が所望され得る。これらのVLPは、異種物質の存在下でインビトロのVLPアセンブリ反応を行うことによってアセンブルされ得る。具体的には、精製されたコートタンパク質サブユニットは、変性剤(通常は酢酸)で脱凝集されたVLPから得られる。タンパク質サブユニットは、異種の物質と混合される。特定の実施形態では、この物質は、VLPの内部に対してある程度の親和性を有し、好ましくは負に荷電されている。
【0104】
別の方法は、翻訳オペレーターの合成RNAバージョンに対して異種の物質を結合する工程を含む。インビトロのアセンブリ反応の間、RNAは、その認識部位に緊密に結合し、得られたVLP中に効率的に組み込まれ、それと一緒に外来物質を保持する。
【0105】
別の実施形態では、この物質は、VLP表面に自然に存在する細孔を通じてVLP中に受動的に拡散される。特定の実施形態では、この物質はこれらの細孔を通過するのに十分小さく(MSでは、それは直径約10オングストロームである)、かつVLPの内部に高い親和性を有する。
【0106】
VLP集団
上記のとおり、本発明は、VLP集団又はライブラリーに関する。本明細書における「集団」及び「ライブラリー」という用語は、交換可能に用いられ、従って同義であるとみなされる。1つの特定の実施形態では、ライブラリーはランダムなライブラリーであってもよい;別の実施形態では、このライブラリーは、抗原性ポリペプチド由来のフラグメントのライブラリーである、抗原フラグメントライブラリーである。
【0107】
ランダムライブラリー(集団)
ペプチドをコードするオリゴヌクレオチドが調製され得る。1つの特定の実施形態では、特定のアミノ酸をコードするトリプレットは、組成NNS(ここでNはA、G、C又はTであり、SはG又はTである)か、あるいはNNY(ここでNはA、G、C、又はTであり、YはC又はTである)を有する。複数のトリプレットがコートタンパク質遺伝子に挿入されて、タンパク質生成物への対応するペプチドの挿入がもたらされる。終止コドンの存在を最小化するために、ペプチドライブラリーは、天然のアミノ酸組成をより正確に反映するように設計された特注の三ヌクレオチドのホスホラミダイト混合物(Glen Research,Inc.から入手可能)から合成され、終止コドンを完全に欠いているオリゴヌクレオチドを用いて構築されることができる。
【0108】
コートタンパク質へのこのようなランダム配列の挿入によって、VLPの集団(又はライブラリー)の合成がもたらされ、それぞれの粒子は、その表面上で異なるペプチドを提示する。このような集団は、極度に大きくてもよく、数十億の個々のメンバーからなってもよい。実質的に任意のレセプター(例えば、モノクローナル抗体)に特異的なリガンドがこのようなライブラリーに存在することが一般に観察されるが、それらは通常、集団全体のわずかな画分に相当する。アフィニティー選択とその後の増幅(本発明の場合は、カプシドで包まれたRNAの逆転写及びPCRによる)によって、このような希少な種の回収、分析及び開発が可能になる。
【実施例】
【0109】
本発明は、本発明の例示として示される以下の非限定的な例を参照することによってさらによく理解され得る。以下の実施例は、本発明の好ましい実施形態をさらに詳細に例示するために示しているが、決して本発明の範囲を限定するものとして解釈されるべきではない。
【0110】
低い価数の異種ペプチドを示すVLPの生成
本出願は、MS2 VLPのペプチドディスプレイのための技術の開発における近年の進歩を詳述しており、これはランダム配列ペプチドライブラリーの容易な生成のため及びペプチドディスプレイの価数の制御のための新規なプラスミドベクターの説明を含む。これは、米国特許出願公開20090054246号に記載される方法の進歩を表す。
【0111】
本発明は、繊維状ファージディスプレイの能力に類似するアフィニティー選択能力を有するMS2ウイルス様粒子(VLP)プラットフォームの開発に関する。本発明は、部分的には、RNAバクテリオファージのコートタンパク質の、外来のペプチドを提示する能力及びそれらの合成のためにテンプレートとして働く同じmRNAをカプシドで包む能力の両方に基づく。このプロセスは、VLPを形成する細菌細胞中のプラスミドからのコートタンパク質のランダムなペプチド融合物の合成を必要とする。VLPは、細胞から抽出されて、特定の抗体に対する結合のためのアフィニティー選択に供される。最終的に、RNAは、選択されたVLPから抽出されて、カプシドで包まれた配列を回収及び増幅するために逆転写及びPCRに供され、この配列が次にクローニングされて細菌中に再導入され、ここでそれらの配列は、別の回の合成、アセンブリ及び選択のためのテンプレートとして機能する。このプロセスは、必要な回数だけサイクルが繰り返され、最後に、選択された配列を選択されたVLPの高レベルの細菌発現のためにクローニングする。
【0112】
いくつかの他の用途の中でも、ワクチン開発を容易にするために開発されている本発明の方法は、単一のプラットフォームでは一緒に利用できなかった重要な特徴を有する。第一に、本発明の方法は、ウイルス様粒子(VLP)の表面上の高密度反復性アレイとして外来ペプチドを提示することによって高い免疫原性を確実なものにする。これは、外来抗原に対する活発な免疫応答を生じるだけでなく、免疫寛容を克服し、さらに自己抗原に対する抗体を誘導することもできる。第二に、ファージディスプレイと類似のプロセスでは、本発明のプラットフォームは、複雑なランダム配列ライブラリーからアフィニティー選択された配列の回収及び増幅を可能にする。従って、本発明は、単一のプラットフォームにおいて、価数制限及びアフィニティー選択によって関連のエピトープを特定する能力と、その次に免疫系に対してそれらのエピトープをワクチンとして示す能力とを合わせ持つ。エピトープは、抗体標的に対するアフィニティー選択によって特定され、最適化され、次いでプラットフォームを変化することなく、免疫系に対して直接ワクチンとして提示され得る。このペプチドは、それらのもともとの選択の間に存在する構造的連結を変更することなく高密度で提示され、これによって天然のエピトープの忠実な分子模倣物が単離され、それらの最適構造が免疫プロセスの間に維持され、関係する抗体応答が誘導される確率が増大する。
【0113】
MS2 VLPでの高い複雑性のランダム配列ペプチドライブラリー構築物のためのプラスミドベクター
MS2 VLPのランダム配列ペプチドライブラリーを利用する最初の実験は、T7転写単位を有するアンピシリン耐性プラスミドであるpET3dの単純な誘導体で実施された。これは、高い複雑性のライブラリーの作成に便利な最適なシステムよりはいくらか劣ると判明したが、そのため本発明者は、重要であると思われる多数の最適特徴を組み合わせた一連のベクターを作成した。
これらとしては以下が挙げられる:
【0114】
A.AB−ループでの挿入のために従来のクローニング部位を有する一本鎖二量体。
pDSP1(図1)は、AB−ループ−コード配列又はその近位において、例えば1つだけのSalI及びKpnI制限部位を含むように改変された、一本鎖二量体のコード配列を発現する。典型的には、BamHI部位は、コート配列の下流に含まれる。この形態の二量体は、外来配列のAB−ループ中への簡易なクローニングを容易にする。これらの部位を1つだけにするため、ベクター中、上流コート配列中及びプラスミド配列中で、多数のSalI及びKpnI部位を破壊する必要があった。
【0115】
B.カナマイシン耐性。pDSP1は、カナマイシンに対する耐性を付与する。これによって、形質転換されてない細胞の最初の高いバックグラウンドに対して、液体培養物中の形質転換された細菌の選択を可能にすることにより、ライブラリー構築がかなり容易になる。最初の生成プラスミドベクターによって付与されたアンピシリン耐性は、この目的に対して不適当であった。これは、形質転換された大腸菌による抗生物質の急速な分解によって、培養物中で驚くほど短時間後に選択の損失が生じるからである。これは形質転換されていない細胞の過剰増殖を可能にした。当然ながらこの細胞は、効率的な形質転換方法が利用される場合でさえ、通常はこの集団の大部分に相当する。
【0116】
C.M13の複製起点。今日に至るまで、本発明者は、PCRプライマーを利用して一方の末端にランダムな配列が結合されたフラグメントを生成する手順によって、ランダム配列ライブラリーを構築している。次いで、このフラグメントを適切な制限エンドヌクレアーゼによって消化して、上述される1つだけの部位(典型的にはSalIとBamHIとの間、図1を参照)においてpDSP1中にクローニングする。このような方法を用いて、本発明者は、最大109までの個々の組み換え体からなるライブラリーを作成した。しかし、このサイズのライブラリーは、これらの方法を用いて構築するには不便である;かなり大きいライブラリーの構築は、典型的なライゲーション反応で見られるよりもさらに高収率の組み換えDNAを効果的に生じる方法によって容易にされる。具体的には、本発明者は、部位特異的な突然変異誘発(2)のための方法のバリエーションを利用するが、この方法は、1011個の複雑性の範囲で繊維状ファージライブラリーを生成するために他者によって既に用いられている(3)。この方法は、dUTP置換一本鎖環状テンプレート上のミスマッチのプライマーの伸長に依拠する(dUによるdTの置換は、CJ236又はBW313のようなdut−、ung−宿主におけるテンプレートDNAの成長によって達成される)。ランダム配列ペプチドライブラリーを作成するために、プライマーは、AB−ループのいずれかの側でのコート配列に対する相補的な配列が、それぞれの側に配置されたランダムなDNA配列を含む。このプライマーは、DNAポリメラーゼで伸長されて、共有結合的に閉鎖された二本鎖環が、DNAリガーゼの作用によって生成される。ung+株の形質転換(例えば、エレクトロポレーションによる)は、変異鎖の選択的増殖のための強力な選択をもたらし、そして挿入された配列を保有する組み換え体を高収率で生じる。これらの挿入の変異原性反応は、エレクトロポレーションによっておよそ1011個以上の個々の組み換え体を容易に生成するのに十分な比較的大量のDNA(例えば、20ug)で行われることができる。
【0117】
一本鎖DNAの生成を容易にするために、M13の複製起点がプラスミド中に導入され、そしてM13起点の方向に応じて、pDSP61及びpDSP62と呼ばれる(図2)。また、M13CM1と呼ばれるヘルパーファージを構築した。これは、クロラムフェニコールでカナマイシン耐性を置き換えるM13K07の誘導体であり、これによってカナマイシン耐性を付与するM13cm1ヘルパー及びpDSP6の同時の存在が選択可能になる。プラスミドを含むdut-、ung-株(例えば、CJ236)のM13ヘルパーファージによる重複感染によって、dUTP−置換一本鎖DNAの容易な生成がもたらされる。
【0118】
D.合成の「コドンジャグリングされた」コート遺伝子。ライブラリー構築のためにプライマー伸長突然変異を用いる要望によって、新しい問題が導入され、いわゆる「コドンジャグリングされた」コート配列をpDSP6へ導入することが必要になった。本発明のペプチドディスプレイ法は、外来のペプチドを一本鎖二量体の2つのAB−ループのうちの一方にのみ特異的に導入する能力に依拠する。上記のスキームでは、変異原性プライマーは、通常は一本鎖二量体の両方の半分での配列にアニールして、二重挿入が生じる。しかし、AB−ループの両方における同時挿入は、タンパク質折り畳みの失敗を高頻度に生じる。この理由のために、コートタンパク質のコドンジャグリングされたバージョンの合成が完成された。このコドンジャグリングされたバージョンは、可能な最大数のサイレントヌクレオチド置換を一本鎖二量体の上流側半分に導入し、これによって野生型コートタンパク質のアミノ酸配列を有するポリペプチドを生じる。しかし、多くの変異の存在によって、ジャグリングされた配列は、変異原性のオリゴヌクレオチドに対して効率的にアニールできなくなる。この方法では、挿入は、一本鎖二量体の下流側半分を標的にする。
【0119】
それぞれpDSP1(am)及びpDSP62(am)と呼ばれるpDSP1及びpDSP62の改変体は、pDSP1及びPDSP62の特徴の全てを保持するが、部位指向性の突然変異によって改変されて、アラニンコドン(一本鎖二量体中のコートタンパク質の下流のコピーの最初のアミノ酸を特定する)が、UAG(アンバーコドンとして公知の特異的なナンセンスコドン又は終止コドン)に変換される。この終止コドンの抑制によって、単一のmRNAから、野性型及び一本鎖二量体のコートタンパク質の両方の合成が可能になる。これによって、後でさらに詳細に記載されるような平均のペプチドディスプレイ価数を制御する能力が得られる。
【0120】
ナンセンスコドンの読み飛ばしは、通常は抑制されている特定の終止コドンに特異的なサプレッサーtRNAの存在を必要とする。3つの終止コドン全てに活性なサプレッサーtRNAが記載されたが、分子生物学者には周知である。本明細書に記載される研究は、アンバーコドンを利用し、従ってこの場合、アラニンを挿入するために特別に設計されたアンバー−サプレッサーtRNAの使用を必要とする。しかし、他の終止コドン(UAA及びUGA)のそれぞれについて特異的なサプレッサーtRNA、及び種々のアミノ酸を挿入することが同様に利用され得ることが理解されるべきである。
【0121】
細菌細胞中でサプレッサーtRNAを生成するために、pNMsupAと呼ばれるプラスミドを生成した(図3を参照)。これは、pACYC18の誘導体であり、p15Aプラスミド不適合性群由来の複製起点を含む。これはまた、クロラムフェニコールに対する耐性も提供する。このことは、このプラスミドが、ColE1起点を有しかつカナマイシンに対する耐性を付与するpDSP1(am)又はpDSP62(am)をすでに含んでいる細菌細胞では、安定に維持され得るということを意味する。プラスミドpNMsupAはまた、lacプロモーター、及び合成のサプレッサーtRNA遺伝子が挿入されたpUC18のポリリンカー領域を含む。tRNA遺伝子の転写は、lacプロモーターの制御下であり、このことは、サプレッサーtRNAの合成が、lacオペロンの誘導因子(例えば、IPTG)によって制御されるということを意味する。合成tRNA遺伝子の配列は、Kleinaら(4)によって以前に記載された配列を基にしており、下に示される。これには、pNMsupA中でそのクローニングを容易にするEcoRI及びPstI部位が隣接する。
【0122】
【化1】

【0123】
サプレッサーtRNAのレベルを変更することは、ナンセンス抑制のレベルを変更するためにも知られている。pDSP1(am)又はpDSP62(am)からの一本鎖二量体の合成のレベルをこのように変更することによって、1個のVLP当たりに提示されるペプチドの平均数を制御することが可能になり、これによって広い範囲にわたって提示価数の制御が可能になると考えられた。pNMsupAはlacプロモーターを使用するが、サプレッサーtRNA合成の制御のために種々のプロモーターが用いられ得ること、及びいくつか(例えば、プロピオン酸塩オペロン(5)のプロモーター)は、実際に、それらのそれぞれの誘導因子の種々の濃度に対して、適切に管理された、段階的な応答を良好に提供できるということは明らかであろう。
【0124】
III.提示価数の制御
MS2 VLPで提示されたペプチドの数を制御する手段があることが望ましい。多価数のMS2 VLPディスプレイ(1粒子当たり90コピーのエピトープ)によって、抗体について高い内因性の親和性を有するペプチドを提示するVLPを、低い内因性の親和性を有するが、複数の弱い相互作用をする能力のおかげで依然として緊密に結合するものから、区別することが困難になるはずである(すなわち、結合活性対親和性)。これは、繊維状ファージディスプレイについての周知の問題であり、ここでは高い親和性の相互作用の選択は、通常は低い提示価数の使用を必要とする。最高の親和性のペプチドを選択する場合に、適切な分子模倣物の発見はかなり増加される。以下は、選択のストリンジェンシーを改善するために提示価数を減少するための方法の説明である。
【0125】
価数の制御。所定のモノクローナル抗体に対する最高の親和性を有するペプチドの選択が天然の抗原の最良の分子模倣物を提供すること、及びこれらは関係する抗体応答を誘導する可能性が最も高いことが仮定される。理想的には、このアプローチは、多価提示を用いて初回の選択を行い、これにより標的に対していくつかの最少の親和性を有する全てのペプチドを含む比較的複雑な集団を得る。次いで、親和性選択のストリンジェンシーを増大するために、その後の回では提示価数を少なくすることが望ましい。
【0126】
このアプローチでは、単一のRNAから大量の野性型及び少量のAB−ループ組み換えタンパク質の生成を可能にするシステム(系)が提供される。一本鎖二量体中のコートタンパク質の下流のコピーの最初のアミノ酸を普通はコードするアラニンコドンの代わりにアンバー終止コドンを含む、pDSP1の改変体(pDSP1(am))を構築した(図3を参照)。従って、pDSP1(am)は、普通は野性型コートタンパク質のみを産生し、これは当然ながら普通はVLPにアセンブルされる。さらに、本発明者は、アラニン−挿入サプレッサーtRNA遺伝子を合成してクローニングした。異なる不適合性群由来のクロラムフェニコール耐性プラスミド上でのlacプロモーターの制御下で発現されて、サプレッサーtRNAは、コート配列を翻訳するわずかな割合のリボソームにアンバー(終止)コドンを読み飛ばしさせて、一本鎖二量体を産生する量で産生される。得られたタンパク質は(そのゲストペプチドとともに)、同じmRNAから発現される野性型タンパク質とアセンブルして、モザイクカプシドを形成する。精製されたVLPはこのベクターから産生され、それらは平均して、1個のVLP当たり約3個のペプチドを提示すると予測された。SDSゲル電気泳動(図4を参照)によって、精製されたVLP中の「読み飛ばし」生成物の内容を示す。これらを試験して、予想どおりアフィニティー選択のストリンジェンシーの増大を確認した。アガロースゲル電気泳動及びノーザンブロットによって、粒子が関係するRNAをカプシドで包むことが確認される。必要であれば、価数はさらに、サプレッサーtRNAの発現レベルを減少することによって減少されることができる。実際に、サプレッサーtRNA合成のレベルを調節することによって、広い範囲にわたって読み飛ばし生成物の発現(及び提示価数)を制御することが可能である。これは、活性が誘導因子の濃度に応じて正確に調節され得るプロモーター(例えば、proB)からtRNAを発現することによって達成される。
【0127】
抗原フラグメントライブラリー
別の方法では、クローニングされた抗原遺伝子又は表現型ゲノムの存在を利用してランダムな抗原フラグメントライブラリーを作成する。このようなライブラリーの作製のためには、いくつかの方法が存在する。1つは、例えば適切な平均サイズ(例えば、30bp)のフラグメントを生成するためにDNaseIで処理することによる、抗原遺伝子のランダムなフラグメントを要件とする。これらは、コートポリペプチドをコードする遺伝子中の適切な部位に対して平滑末端で連結(ライゲート)される(例えば、一本鎖コートタンパク質二量体のAB−ループ中において)。次いで得られたライブラリーを、アフィニティー選択に供して、抗体によって認識されるペプチドを提示するVLPを回収する。特定の実施形態では、制限部位を、コートポリペプチドのAB−ループ又はN−末端に挿入してもよい。
【0128】
合成
RNAファージVLPは、溶解されてVLPが抽出される生きている大腸菌細胞中のプラスミドから通常は生成される。本発明者によって今まで行われ、本出願の別の箇所で記載された実験では、MS2及びPP7のVLPのランダム配列ペプチドライブラリーは、この方法で正確に生成された。しかし、特定の別の実施形態では、本発明の集団は、当該分野で公知の手順を用いて、結合されたインビトロの転写/翻訳システムにおいて合成されてもよい(例えば、米国特許第7,008,651号;Krarnerら、1999年,Cell−free Coupled Transcription−translation Systems From E.coli.In Protein Expression.A Practical Approach,Higgins及びHames(編集),Oxford University Pressを参照)。特定の実施形態では、バクテリオファージT7の(又は関連の)RNAポリメラーゼを用いて、このようにして生成された大量のRNAを効率的に翻訳するために最適化されたシステム(系)において、T7プロモーターの制御下で、クローニングされた遺伝子の高レベル転写を指向する(例えば、Kimら、1996年,Eur J Biochem 239:881〜886;Jewettら、2004,Biotech及びBioeng 86:19〜26を参照)。
【0129】
VLPがインビボで生成される場合、大腸菌の細胞自体が区画化を提供して、これが所定のコートタンパク質−ペプチド組み換え体の複数のコピーがそのmRNAと特異的にアセンブルすることを確実にする。同様の形態の区画化が提供されないと、特に本発明の集団において、テンプレートの混合物(すなわち、ライブラリー)からインビトロで合成の間に、異なるペプチドに対するそれらの融合によって識別される異なる個々のコートポリペプチドは、恐らく互いのmRNAをパッケージングし、それによって有効なファージディスプレイに必要な遺伝子型/表現型の連鎖を破壊する可能性がある。さらに、それぞれのVLPは、多数のサブユニットからアセンブルされるので、ハイブリッドVLPの形成が生じ得る。従って、1つの好ましい実施形態では、本発明の集団又はライブラリーを調製する場合、1回以上のサイクルの転写/翻訳反応を、水/オイルのエマルジョン中で行う(Tawfikら,1998年,Nat Biotechnol 16:652〜6)。この現在では十分に確立された方法では、個々のテンプレートを、水/オイルのエマルジョンの水の部分に隔離する。適切な条件下で、多数の水性の微小液滴が形成されることができ、それぞれが平均して単一のDNAテンプレート分子及び転写/翻訳の機構を含む。それらはオイルに囲まれるので、これらの部分は互いに通じない。このような液滴中で合成されたコートポリペプチドは、それらをコードする同じmRNAと特異的に会合するはずであり、1つのペプチドのみを提示するVLPへとアセンブルするはずである。合成後に、このエマルジョンは破壊され、VLPは回収されて選択に供されることができる。1つの特定の実施形態では、転写/翻訳反応の全てが水/オイルのエマルジョン中で行われる。1つの特定の実施形態では、1つの液滴当たり1つのテンプレートのみを含む液滴(1つのペプチドのみを提示するVLP)だけが単離される。別の実施形態では、混合VLPを含む液滴が、1回以上のサイクルの転写/翻訳反応で単離され(1つの液滴当たり複数のテンプレート)、その後にペプチドを1つだけ提示するVLP(1つの液滴当たり1個のテンプレート)が単離される。
【0130】
VLP及びVLP集団の使用
本発明のVLP及びVLP集団は、多くの用途の可能性がある。以下にさらに詳細に記載されるように、VLPは、免疫原性組成物、特にワクチンとして、薬物送達デバイスとして、生体医学造影剤として、又は自己アセンブリナノデバイスとして用いられ得る。本発明のVLP集団は、適切なワクチン候補を選択するために用いられ得る。
【0131】
ワクチン候補の選択
本発明のVLP集団又はライブラリーは、ワクチン候補物を選択するために用いられ得る。このライブラリーは、ランダムライブラリーであっても、又は抗原性ライブラリーであってもよい。ランダムあるいは抗原由来ペプチドの配列ライブラリーは、VLPの表面及び特異的な標的エピトープ、又はおそらくミモトープ上に提示され、次いで抗体を用いるアフィニティー選択によって単離される。VLPは、それら自体のmRNAをカプシドで包むので、それら(及びそれらのゲストペプチド)をコードする配列は、逆転写及びPCRによって回収され得る。個々のアフィニティー選択されたVLPは、続いてクローニング、過剰発現及び精製される。
【0132】
ファージディスプレイにおけるアフィニティー選択のための技術は十分開発されており、本発明のVLPディスプレイシステムに直接適用可能である。つまり、抗体(又は抗血清)は、ランダム配列又は抗原フラグメント提示ライブラリー中のVLPのペプチドとの複合体を形成することができる。典型的には、抗体はビオチンで標識され、それによりこの複合体は、ストレプトアビジンでコーティングされた表面、磁気ビーズ又は他の適切な固定媒体に対する結合によって捕獲されることができる。洗浄後、結合されたVLPが溶出され、RNAは、アフィニティー選択された集団から抽出されて、逆転写及びPCRに供されてコートをコードする配列を回収し、この配列が次に再クローニングされて、最良結合の改変体が得られるまで、発現及びアフィニティー選択のさらなる回に供される。VLPからRNAを回収するための多数のスキームが、容易に想定される。1つの魅力的な可能性は、ストレプトアビジンコーティングされたPCRチューブ中でビオチン−mAb−VLP複合体を単に捕獲し、次にVLPを熱的に変性し、それらのRNA内容物を直接RT−PCRに供することである。多くの明らかな代替手段が存在し、種々の固定媒体の結合能力などの考慮に応じて調節が必要な場合もある。一旦、選択された配列がRT−PCRによって回収されれば、それらをクローニングして大腸菌中に再導入することは簡単なことであり、それぞれの段階で注意して必須のライブラリー多様性(これは当然ながら各回の選択で減少する)を保存する。選択が完了したら、それぞれのクローンは、VLPワクチン候補を産生するために過剰発現されることができる。
【0133】
MS2 VLPプラットフォーム上でのアフィニティー選択の効率を確立するため、エピトープが十分特徴付けされたモノクローナル抗体標的を用いて上記の方法によって選択が行われた。M2 抗−Flagモノクローナル抗体、及びライブラリーは、pDSP1中で構築され、アミノ酸13及び16についてのコドンの間に挿入された(すなわち、13/16挿入モード)10個のNNSトリプレットを含む。この特定のライブラリーは、約108個の独立したクローンを含み、かつ外来のペプチドを高い価数で提示した。それ以降、さらに複雑なライブラリーをpDSP62(上記参照)を用いて構築した。しかし、pDSP1ライブラリーが、Flagエピトープに遭遇する合理的な可能性を得るために十分複雑であると考えられた。第一回の選択を、抗体分子に対して10倍過剰であると推測されたVLPを用いて、プラスチックウェルに対する吸着によって固定された250ngの抗体に対して行った。過剰の洗浄後に、結合したVLPを溶出し、次に逆転写及びPCRに供した。PCR生成物をSalI及びBamHIで消化し、2回目で使用されるVLPの産生のためにpDSP62中にクローニングした。ここにおいて、及び全ての引き続く回において、選択物のクローニングによって、少なくとも5×106個の独立したクローンを得た。2回目の選択は、1回目と同じ条件下で行われた。2回目及び3回目の生成物を、pDSP62(am)にクローニングして、VLPを上記のアンバーサプレッサー(pNMsupA)の存在下で生成した。このことは、3回目及び4回目では、ペプチドが低価数で提示されたことを意味する。4回目では、抗体の量は50ngまで減少され、それによりVLPは、抗体に比較して約50倍過剰で存在する。4回目の後、VLPの過剰産生及び精製を期待して、高価数の提示のために生成物をpDSP62中にクローニングした。それぞれの回からの2〜3の選択物の配列を図22に示す。初期の回に得られた配列は、公知のFlagエピトープDYKDDDDKLに対して限られた類似性を示すが、特にYKジペプチドなどの、特定の重要なエレメントが既に明白である。3回目まで、全ての配列は、少なくとも1つの下流DとともにDYKエレメントを示す。4回目までに、1個の配列のみを、配列分析に供された7個のクローンから得た。野生型フラグエピトープの配列に対するその類似性は明白である。Flagエピトープは、従来の繊維状ファージディスプレイ方法を用いて以前にマッピングされており、その結果は、NEB Transcript,1996年 夏で報告された(The NEB Transcriptは、ファージディスプレイライブラリー及びアフィニティー選択キットの商業的供給業者である、New England Biolabsの刊行物である(彼らのウェブサイト、www.Neb.comで入手可能))。図22に報告される配列は、現実にNEBの実験で得られた配列よりも実際のエピトープに対してよく適合しており、このことは、MS2 VLPディスプレイシステムが、アフィニティー選択によるエピトープ特定のための繊維状ファージディスプレイと少なくとも同じ程度に効率的であることを示す。
【0134】
免疫原性組成物
上述したとおり、本発明のスクリーニング手順によって特定されるVLPを用いて、免疫原性組成物、特にワクチンを処方してもよい。ワクチンは、動物に対して投与され得る形態でなければならない。典型的には、ワクチンは、本発明の組成物が懸濁又は溶解されている、従来の生理食塩水又は緩衝化水溶液媒体を含む。この形態では、本発明の組成物は、ある状態又は障害を予防、緩和又は処置するために都合よく用いられ得る。宿主への導入の際、ワクチンは、これらに限定されないが、抗体及び/若しくはサイトカインの産生、細胞傷害性T細胞、抗原提示細胞、ヘルパーT細胞、樹状細胞及び/若しくは他の細胞の活性化などの免疫応答、並びに/又は他の細胞性応答を誘発することができる。
【0135】
任意で、本発明のワクチンはさらに、本発明の化合物に対してわずかな割合でも又は大きな割合でも存在することができるアジュバントを含む。「アジュバント」という用語は、本明細書において、免疫応答の非特異的な刺激因子又は宿主中でデポの生成を可能にする物質であって、本発明のワクチンと組み合された場合、さらにより増強された免疫応答を提供するものを指す。種々のアジュバントが用いられることができる。例としては、完全及び不完全フロイントアジュバント、水酸化アルミニウム及び改変ムラミルジペプチドが挙げられる。
【0136】
任意で、本発明のワクチンは、本発明の化合物に対してわずかな割合でも又は大きな割合でも存在することができるアジュバントをさらに含む。
【0137】
標的化薬物送達
特定の細胞レセプターに結合するペプチドを特定し、それによって標的細胞に結合かつ進入する能力を持った(例えば、エンドサイトーシスによる)粒子を生成するために、アフィニティー選択を用いることができる。MS2 VLPは、20nmの大きさの内径を有する中空の球体である。特定の実施形態では、VLPは、薬物、例えば送達されるべきタンパク質毒素及び任意で細胞型特異的なレセプターに結合するリガンドを含む。このような粒子の内部組成は、例えば、リシンのようなタンパク質毒素をそれに特異的に充填することによって、又は合成の翻訳オペレーター模倣物に対してカップリングすることによって調整され得る。このような粒子の外面に対して細胞特異的なレセプターを結合する能力を付与することによって、選択される細胞型を毒素(又は他の薬物)の送達の標的とすることができる。関連する側面では、コートポリペプチド二量体を含むVLPは、細菌毒素、アジュバント又は免疫刺激性核酸などの異種物質を実際にカプシドで包み得る。
【0138】
生体医学造影剤
薬物が特定の細胞型を標的とし得る同じ方法では、核磁気共鳴画像化のための造影剤が、特定の細胞又は組織に対して送達されることができると考えられ、可能性としては、MRIの診断力を大いに増大する。実際に、MS2粒子は、MRIのコントラストを大きく増大するために、既にガンドリニウムで標識されている(Andersonら,2006年,Nano Letters 6(6),1160〜1164)。従って、特定の実施形態では、このような粒子は、それらの表面上で適切なレセプター特異的ペプチドを提示することによって、特定の部位を標的とすることができるであろう。関連の側面では、コートポリペプチド二量体を含んでいるVLPは、実際に造影剤をカプシドで包み得る。
【0139】
自己アセンブリするナノデバイス
本発明のVLPは、金属結合タンパク質を提示する繊維状ファージ粒子のいずれかの末端に親和性を有するペプチドを含み得る。繊維状ファージ粒子のいずれかの末端に親和性を有するVLPは、繊維状ファージナノワイヤの末端に対してこれらの球体(及びそれらが含むものなら何でも)を連結する可能性を生じる。あるいは、VLPは、金属結合ペプチド(例えば、金及び亜鉛)を提示でき、それにより通常ではない電気的及び光学的な特性を有するアレイが得られる場合がある。あるいは、これらのアレイに自己アセンブリする能力が改善されたVLPは、特定の表面に対する親和性を有するペプチド、又はVLP自体の自己会合の特性を変更するペプチドを提示することによって産生され得る。
【0140】
実験の概説
バクテリオファージMS2のウイルス様粒子(VLP)の上でランダム−配列ペプチドライブラリーの構築を容易にする2つのプラスミドベクターを記載する。最初のpDSP1は、PCR−生成されたフラグメント又は他の二本鎖DNAフラグメントを、コートタンパク質一本鎖二量体の下流のコピーのAB−ループ中に都合よくクローニングするために構築された。二番目はpDSP62と呼ばれ、これは、Kunkelら[16]の部位指向性突然変異誘発法によって一本鎖二量体中の事実上任意の位置(通常はAB−ループ)にペプチド配列を導入するために、特異的に構築された。これらのプラスミドの一般的な特徴を下に示す。
【実施例1】
【0141】
pDSP1−都合のよいAB−ループ中の挿入のためのクローニング部位を有する一本鎖二量体を発現するプラスミド
プラスミドpDSP1(図5a及び図7aを参照)は、pET3dのT7転写シグナル、カナマイシン耐性、及びpET9dの複製起点を含む(pET3d及びpET9dに関する情報は、New England Biolabs vectorのデータベース、https://www.lablife.org/ct?f=v&a=listvecinfoで見られる)。これは、1つだけのSalI及びKpnI制限部位を含むように改変された、MS2一本鎖コートタンパク質二量体(6)のコード配列を発現する。これによって、外来の配列をAB−ループに簡易にクローニングすることが容易になる。これらの部位を1つだけにするには、ベクター中及び上流コート配列において、他のSalI及びKpnI部位を破壊することが必要であった。
【0142】
pDSP1のAB−ループ挿入部位の近傍のMS2コート配列を下に示す。SalI及びKpnI部位の存在に注意されたい。
【0143】
【化2】
【0144】
下に示すのは、pDSP1中のランダムな7マーのライブラリーの例であって、ここではランダムな配列が、いわゆる13/16モードで挿入されている。
【0145】
【化3】
【0146】
AB−ループをコードする配列中又はその付近に、1つしかない複数の制限部位が存在することは、それらがその切断によって適合性の「付着末端」を生じる部位に隣接する場合、外来配列を簡易に挿入することが可能になる。しかし、これは時に、図5bに示されるようなPCR法及び組み換えDNA法の組み合わせを用いて外来配列を結合するのにさらに便利である。例えば、下に示される5’PCRプライマーを、BamHIの下流のプラスミドベクター配列にアニールする3’プライマーとともに用いて、アミノ酸13と16との間に挿入されたランダム配列を有するコートタンパク質コード配列のフラグメントを生成してもよい。N=A、C、G又はTであり、S=G又はCである。SalI及びBamHIでの消化後、このフラグメントは、pDSP1のSalIとBamHIとの間に挿入される。下に示すのは、このようなライブラリーを生成するために用いられ得る5’−プライマーの例である:
【0147】
【化4】
【0148】
pDSP1を用いれば、ランダム−配列ペプチドライブラリーは通常、テンプレートとして単量体のコートタンパク質配列を用いて生成されたPCRフラグメントをAB−ループ中にクローニングすることによって構築される(例えば、pMCT)。合成のオリゴヌクレオチド5’−プライマーは、SalI(又はKpnI)部位及びランダムなコドンの配列(例えば、6〜10コピーのNNY)をAB−ループのすぐ上流の部位に結合するように設計される。3’−プライマーは、BamHIのすぐ下流のプラスミドベクター中の配列にアニールする。得られたPCR産物をSalI(又はKpnI)及びBamHIで消化して、pDSP1の対応する部位にクローニングする。これによってAB−ループへのペプチドの挿入が生じるが、挿入の正確な部位は、5’−プライマーの特異的な設計に依存する。ほとんどの挿入に関し、SalI部位の使用は、挿入部位の選択においてKpnIよりも柔軟性を与えるので好ましい。これらの方法によれば、最大で108〜109個の個々のメンバーを有するペプチドVLPライブラリーを比較的直接的に生成する。
【0149】
ペプチドディスプレイ価数の制御のための手段を導入するために、pDSP1の誘導体(pDSP1(am)と呼ばれる)を、一本鎖二量体の半分の間の接合部でのナンセンスコドンの導入によって構築した。pNMsupAによって産生されたようなナンセンスサプレッサーtRNAの存在下で発現される場合、少量の一本鎖二量体が(その外来ペプチドとともに)産生される。しかし、pDSP1(am)から産生されるほとんどのコートタンパク質は、野生型の単位長タンパク質の形態で合成される。2つの形態のコートタンパク質は、平均でわずか約3個のペプチドしか提示しないハイブリッド粒子へとコアセンブル(共集合)する。提示価数の平均レベルは、サプレッサーtRNAの発現レベルを変更することによって、又はより大きいか若しくは小さい抑制効率を示すサプレッサーを使用することによって、大小に調節され得る。
【実施例2】
【0150】
pDSP62−効率的な部位指向性突然変異誘発方法を用いるライブラリー構築のために適切なプラスミド
M13複製起点の導入
pDSP1について上述したようなライブラリー生成のための方法は、大規模にするのが困難である。これは、DNA制限フラグメントを必要な量で精製するのに不便なためである。さらに、ライゲーション反応の間、無用な副産物へ否応なく変換されるDNAがあり、これが所望のプラスミドの収率を下げる。複雑なライブラリーの構築は、代表的なライゲーション反応で見られるよりも正確な組み換えDNAをより高い収率で効率的に生じる方法によって容易にされる。具体的には、部位指向性の突然変異誘発のための古い方法のバリエーションを用いることが好ましく、これは既に他者により、1011個の複雑性の範囲で、繊維状ファージ上でペプチドライブラリーが生成されている(2,6)。この方法を、M13複製起点を含む特定の種類のプラスミド(ファージミドとしても公知)から生成された一本鎖の環状DNAに適用する。プラスミドを含んでいるdut-、ung-株(例えば、BW313)のM13ヘルパーファージ(例えば、M13K07)による感染の結果として、dUTP−置換の一本鎖のDNAの産生が容易になる。現実の突然変異誘発の反応では、ミスマッチのオリゴヌクレオチドプライマーは、一本鎖のDNAテンプレートにアニールされて、DNAポリメラーゼ(例えば、T7ファージのもの)を用いて延長される。DNAを、閉鎖した環状DNAを生成するために連結し、形質転換によってung+株に導入し、ここでこの変異株を優先的に複製する。ペプチド−VLPライブラリーの産生における以前の実験では、典型的には約90%の形質転換体が所望のペプチド挿入物を含むことが示される。プライマー伸長の突然変異誘発反応は、エレクトロポレーションによって1011個程度の個々の組み換え体を容易に生成するのに十分な、比較的大量のDNA(例えば、20μg)で行われることができる。
【0151】
一本鎖のDNAの産生を容易にするため、M13複製起点をpDSP1に導入した。これを行うために、pUC119で見られるM13起点をPCRによって増幅して、pDSP1における唯一のA1wNI部位にクローニングした。pDSP1−IGと呼ばれるこのプラスミドは、pDSP62の前駆体である。これは、pDSP62Iの構築における唯一の中間体であるので、その配列は示していない。
【0152】
合成の「コドンジャグリングされた」コート遺伝子の使用を通じた一本鎖二量体の半分のみへの標的化挿入
効率的なペプチドライブラリー構築物のためにプライマー伸長突然変異誘発を用いるという要望によって、新しい問題がもたらされた。本発明の提示方法は、外来ペプチドを一本鎖二量体の2つのAB−ループの一方にのみ特異的に導入する能力に依拠する。pDSP1に存在する一本鎖二量体配列を用いると、変異原性のプライマーは、両方の半分での配列にアニールして二重の挿入を生じたが、AB−ループの両方における挿入は、タンパク質折り畳みの失敗を高頻度で生じることが既に公知である。さらに、挿入に耐えた場合でも、一本鎖二量体の半分のみを標的とすることができなかった部位指向性の突然変異誘発は、それぞれのVLPで、2つの異なるペプチドの提示をもたらすであろう。
【0153】
これらの理由で、コートタンパク質の「コドンジャグリングされた」ものを合成して、一本鎖二量体の正常な上流の半分と交換した。このコドンジャグリングされた配列は、可能な最大数のサイレントヌクレオチド置換を含み、従って野生型コートタンパク質アミノ酸配列を有するポリペプチドを生じる。しかし、多数の変異体の存在によって、このジャグリングされた配列は、変異原性のオリゴヌクレオチドに対して効率的にアニールできなくなり、従ってこの変異原性のプライマーは特異的に、下流のAB−ループ配列に指向される。プラスミドpDSP62は図6に示され、その配列は図7bで提供される。
【0154】
一本鎖pDSP62産生のためのクロラムフェニコール耐性M13ヘルパーファージ
プラスミドpDSP62は、カナマイシンに対する耐性を付与する。一本鎖ファージミドDNAの産生のために通常用いられるヘルパーファージ(例えば、M13KO7)もカナマイシン耐性を付与し、従ってここで記載されるプラスミドでの使用には不適切である。この理由で、M13KO7のクロラムフェニコール耐性誘導体である、M13CM1を構築した。pACYC184(7)のクロラムフェニコール耐性遺伝子を、XhoI及びSacIに対する認識配列を結合したプライマーを用いてPCRによって増幅し、そのフラグメントを、カナマイシン耐性決定基にほぼ隣接するXhoI及びSacI部位を利用して、カナマイシン耐性遺伝子の代わりにM13KO7に挿入した。カナマイシン(pDSP62維持を選択する)及びクロラムフェニコール(ヘルパーファージを選択する)の存在下で、細胞は、M13 CM1での感染後に大量の一本鎖のプラスミドDNAを産生する。これらの一本鎖のテンプレート及びKunkelら(2)の方法を用いて、[NNS]6、[NNS]7、[NNS]8及び[NNS]10について1010個を超える個々のメンバーを含むランダム配列ペプチドライブラリーが容易に産生された。著しく高い複雑性が、大規模化とともに可能である。
【0155】
ペプチドディスプレイ価数の制御のための手段を導入するために、pDSP62の誘導体(pDSP62(am)と呼ばれる)を、一本鎖二量体の半分の間の接合部におけるナンセンスコドンの導入によって構築した。pNMsupAによって産生されるもののようなナンセンスサプレッサーtRNAの存在下で発現した場合、少量の一本鎖二量体が(その外来ペプチドとともに)産生される。しかし、pDSP1(am)から産生されるほとんどのコートタンパク質は、野生型の単位長のタンパク質の形態で合成される。2つの形態のコートタンパク質は、平均でわずか約3個のペプチドしか提示しないハイブリッド粒子へとコアセンブル(共集合)する。提示価数の平均レベルは、サプレッサーtRNAの発現レベルを変更することによって、又はより大きいか若しくは小さい抑制効率を示すサプレッサーを使用することによって、大小に調節され得る。
【0156】
上述されるとおり、バクテリオファージMS2のウイルス様粒子がペプチドディスプレイのために用いられて、MS2コートタンパク質一本鎖二量体がペプチド挿入に極めて耐性であること、及びそれらがその合成をコードするmRNAを特異的にカプシドで包む正確にアセンブルされたVLPを生成することが確認された[2]。しかし、MS2は、個々のメンバーが同様の分子生物学を共有するウイルスの大きいファミリーの1つのメンバーでしかない。上記のプラスミド及び方法は、本発明者が、MS2コートタンパク質一本鎖二量体の挿入耐性、及びMS2 VLPがその合成を指向するmRNAをカプシドで包む能力を実証した、以前に記載されたMS2 VLPディスプレイシステムに対して改良点を示す[2]。以下の例は、PP7 VLPについて最近得られたもの[1]と同様の結果を実証しており、このことは、PP7一本鎖コートタンパク質二量体の折り畳み及びアセンブリがまた、外来ペプチド挿入に対して高い耐性を示すこと、並びにこのようにして得られたVLPが、mRNAの形態でそれらの合成のための遺伝的情報を含むことを明確に示している。
【0157】
ここで次に、ペプチドディスプレイを目的として、緑膿菌のバクテリオファージである、PP7のVLPの操作を記載する。
【0158】
PP7 VLPは、MS2 VLPを上回るいくつかの潜在的な利点及び改善を提供する。第一に、安定化した内部サブユニットジスルフィド結合が存在するおかげで、この粒子は熱力学的に劇的により安定である(8)。ワクチンを含む多くの実質的な適用のために、安定性の増大は所望の特質である。第二に、PP7 VLPは、MS2のものと免疫学的に交差反応性でない(9)。このことは、VLPの一連の投与が必要となる場合があるワクチン又は標的化薬物送達の適用において、重要であり得る。第三に、PP7 VLPの正確な折り畳み及びアセンブリがペプチド挿入の脱安定化効果に対してさらに耐性であり得ること、又はこれが、MS2 VLPにおいて耐性でないいくつかのペプチドの耐性を少なくとも示し得ることが予想される。
【実施例3】
【0159】
PP7ペプチドディスプレイベクターの設計
大腸菌中でのPP7コートタンパク質の合成のために、2つの一般的な種類のプラスミドを構築した(図9及び図13を参照)。第一は、lacプロモーターからコートタンパク質を発現し、これを使用して(pRZP7と組み合わせて、以下を参照)、翻訳リプレッサー及びVLPアセンブリ試験を用いて、ペプチド挿入のコートタンパク質の耐性について試験する。第二の種類のプラスミドは、T7プロモーター及び転写ターミネーターからタンパク質を発現する。これらのプラスミドは、正確にVLPにアセンブルする大量のコートタンパク質を産生する。それらはまた、VLPへのカプシド形成のために別個の5’及び3’末端を有するコート特異的なmRNAを生じる。
【0160】
ペプチド挿入部位の設計
PP7カプシドの三次元構造は、たとえ2つのタンパク質のアミノ酸配列がわずか約12%の配列同一性しか示さなくても、三次構造がMS2の構造をよく模倣しているコートタンパク質から構成されることを示す(10)[17]。PP7タンパク質は、AB−ループを保有し、その中にペプチドが、MS2について以前に記載されたスキームと同様のスキームに従って挿入され得る[2]。MS2の場合のように、これは、制限エンドヌクレアーゼKpnIの部位を含むようにPP7コート配列を変異させることによって開始し、これによって、pP7Kと呼ばれるプラスミド中の外来配列の挿入が容易になる(図9a)。この改変により、図10に示されるアミノ酸置換(E11T)が生じた。この置換は、変異体コートタンパク質は翻訳を抑制して正確にVLPにアセンブルするので、十分に耐えられた。やはりMS2の実施例によれば、PP7コートタンパク質の一本鎖二量体バージョンの折り畳みは、従来の二量体よりもAB−ループ挿入に対してさらに耐性であると考えられた。その構築は以前に記載されたが(8)、ここでペプチドディスプレイへの使用については初めて記載される。一本鎖二量体は、コード配列の下流コピー中にのみKpnI部位を含むように改変され、p2P7K32を生成した(図9b)。この設計では、ペプチドをアミノ酸11に挿入したが、他の特定の挿入部位が、おそらくAB−ループ内のいずれかで用いられ得ることに注目すべきである。実際には、下記の試験は、2つの別の挿入モードから記載される(図10及び11)。第一は、11番目のアミノ酸が、挿入されたペプチドのN末端側のthr及びC末端側の野生型glu11として2回出現するので、11/11モードと呼ばれる。いわゆる11/12モードでは、挿入物は片側ではthr11に、他の側ではala12に隣接する。
【0161】
AB−ループ挿入物に対するPP7コートタンパク質の一般的な耐性を試験するために、PP7コートタンパク質のAB−ループ中に挿入されるランダム配列ペプチドのライブラリーを、図11に示されるスキームを用いて作成した。このランダム配列は、6、8又は10コピーの配列NNY(ここでNは任意のヌクレオチドであり、Yはピリミジンである)から構成された。このようなライブラリーは、20個の可能なアミノ酸のうち15個を含み、従ってかなり多様性であり得る。しかし、終止コドンの可能性を回避することによって、それに続く分析はかなり容易になる。
【実施例4】
【0162】
コートタンパク質機能の保持のためのランダム−配列ペプチドライブラリーの個々のメンバーを試験するための方法
MS2コートタンパク質と同様に、PP7コートは、翻訳リプレッサーである。大腸菌 lacZ遺伝子に対してPP7翻訳オペレーターを融合するプラスミドであるpRZP7の構築物は以前に記載されており、これは、コートタンパク質の翻訳リプレッサー活性の制御下にβ−ガラクトシダーゼ合成を置く(11)。これは、異なる抗生物質(クロラムフェニコール)に対する耐性を付与するとともに、異なる不適合性群に由来する(すなわち、これはp15A複製起点を用いる)ので、その両方がアンピシリンに対する耐性を付与しかつcolE1起点を用いるpP7K又はp2P7K32のいずれかと同じ大腸菌株で容易に維持され得る。これらのプラスミドの両方とも、pUC119に由来し、lacプロモーターから比較的低レベルでコートタンパク質を発現する。pP7K又はp2P7K32からのPP7コートタンパク質の発現は、pRZP7から発現されるβ−ガラクトシダーゼの翻訳を抑制する。このことは、所定のペプチド挿入物が、コートタンパク質が正確に折り畳む能力を妨害したか否かを決定することを容易にする。それは、不完全なコートタンパク質は、「xgal」として公知のβ−ガラクトシダーゼ発色性基質を含むプレート上に青色のコロニーを生じ、一方では適切に機能するコートタンパク質は白いコロニーを生じるからである。
【0163】
機能の維持のさらに厳密な試験は、ペプチドコートタンパク質組み換え体を発現する細胞の溶解液中のVLPの存在について直接試験することである。これは、超音波処理によって溶解される細胞のアガロースゲル上での電気泳動によって達成される。エチジウムブロマイド染色によってRNA含有VLPを検出し、次にその存在を、抗PP7血清を用いるウエスタンブロット分析によって確認する。
【0164】
次のアイデアは、PP7コートタンパク質のペプチド挿入耐性を試験し、ランダム配列ペプチドライブラリーを作成すること及び翻訳リプレッサー機能を保持する(すなわち、白色コロニーを生成する)クローンの画分を決定することによってアセンブルし、VLPを生成することである。MS2コートタンパク質一本鎖二量体のペプチド挿入耐性の同様の試験は、本願の発明者によって以前に報告されたことに注意されたい[2]。
【実施例5】
【0165】
ほとんどのランダムな6マー、8マー、及び10マーのペプチド挿入物に耐えるPP7コートタンパク質の折り畳み及びアセンブリ
(NNY)8ライブラリーは、pP7K及びp2P7K32中で構築して、形質転換によって大腸菌株CSH41F−/pRZP7中に導入した。形質転換体は、xgalを含む固形培地上にプレートした。挿入フラグメントなしの対照ライゲーションで、1000分の1のコロニーが得られたので、大部分が真正の組み換え体であった。ごく少数のpP7K組み換え体を試験したが、全てがタンパク質の折り畳みに関して欠損しており、かつVLPを作成できなかったことが分かった。これによって、従来の二量体は、通常はAB−ループ中のペプチド挿入によって不安定化されるというMS2での以前の実験[2]からの予想が確認される。しかし、p2P7K32の一本鎖二量体では、ほぼ100%のコロニーが白色であった。それぞれのライブラリー由来の24個の白色コロニーから1mlの培養物を複製した。1つの培養セットから、粗細胞溶解液をVLPのアガロースゲル分析のために調製した。他のセットでプラスミドを単離して制限酵素消化及びゲル電気泳動に供し、全てが予想される長さの挿入物を含むことを確認した。2〜3個の青色コロニーからのプラスミドも単離した。
【0166】
全てが、異常なライゲーション事象から生じるプラスミドを含むことが判明し、一般にはインタクトなコート配列を含まなかった。ほぼ100%のペプチド挿入物が、コートタンパク質の翻訳リプレッサー機能と適合した。すなわち、それらは十分に適切に折り畳まれたコートタンパク質を生じて、野生型タンパク質のような翻訳を抑制する。この結果は、全てのNNYライブラリー(6マー、8マー及び10マー)で、それらが11/11モードでクローニングされたか、又は11/12モードでクローニングされたかには依存せずに得られた。
【0167】
VLPの存在について直接試験するために、粗細胞溶解液をそれぞれの複製培養から調製して、アガロースゲル電気泳動に供した。ゲルをエチジウムブロマイドで染色して、複製をニトロセルロースでブロットして、マウスの抗−PP7血清と西洋ワサビペルオキシダーゼ標識した二次抗体とでプローブした。クローンは、それが染色したゲル及びウエスタンブロットの両方にバンドを含む場合、VLP合成について陽性とみなす。結果を図13に示す。これによって、いくつかは収率の低下を示すが、6マーのクローンのほぼ全てがある程度のレベルでVLPを生成することが示された。このことは、11/11挿入モード及び11/12挿入モードの両方について当てはまった。8マーのクローンの大部分は、容易に同定可能なVLPを生じる。しかし、この効率は、挿入長が10マーまで増大するにつれていくらか低下するようであるが、やはりクローンの明らかに大部分がVLPを生じる。いくつかのペプチドが荷電されたアミノ酸を組み込むことによってVLPの表面荷電を変更するという予想と一致して、個々の粒子の移動度は可変であることに注意されたい。
【実施例6】
【0168】
PP7 VLPは、コート−特異的なmRNAをカプシドで包む。
高レベルの発現及びRNAカプシド形成試験のために、pP7K及びp2P7K32のPP7コート配列を、T7プロモーター及び転写ターミネーターを含んでいるプラスミド中にクローニングして、pETP7K及びpET2P7K32と呼ばれるプラスミドを生成した。大腸菌株BL21(DE3)中のプラスミドは、大量のコートタンパク質を合成した。得られたVLPを、以前に記載(11、12)されたようなセファロースCL4Bでのクロマトグラフィーによって精製した。RNAをフェノール/クロロホルム抽出によってVLPから精製して、ホルムアルデヒドを含んでいる複製アガロースゲル中の電気泳動に供した。一方のゲルをエチジウムブロマイドで染色し、他のゲルはニトロセルロースにブロットして(ノーザンブロット)、ここでPP7配列を、コートのセンス鎖に特異的な標識された合成のオリゴヌクレオチドを用いて検出した。T7のRNAポリメラーゼを用いたpETP7K及びpET2P732のインビトロでの転写によって生成したRNAを、標準として利用した。それぞれのVLPは、その移動度がインビトロ転写生成物の移動度と同一であり、PP7コート特異的なプローブと特異的にハイブリダイズする、優越した種を含む。PP7 VLPがそれらのmRNAをカプシドで包むことが確認され、これによって、アフィニティー選択のために必要な遺伝子型−表現型の連鎖を確立した。MS2コートタンパク質がそのmRNAをカプシドで包む能力の同様の試験が、本願の発明者によって以前に報告されていることに注意されたい。
【実施例7】
【0169】
PP7 VLPに提示されるペプチドは、免疫系に提示され、かつ免疫原性である。
図10及び図15に示される特異的なペプチド配列を提示するPP7 VLPを構築した。これらは、いわゆるFlagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来の配列、HIV−1のgp120のV3ループ、及び炭疽菌の防御抗原を含んだ。これらの挿入されたペプチドがVLPの表面上に実際に提示されることを示すために、HPV16L2に対するモノクローナル抗体(RG−1と呼ばれる)がPP7 L2−VLPに結合する能力をELISAによって評価した。図16に示されるとおり、RG−1は、L2−VLPには結合したが、V3ペプチドを提示したPP7 VLPには結合しなかった。VLPの免疫原性を示すために、マウスを以前に記載されたように[2]、筋肉内注射によってV3−VLPで免疫した。図17に示されるように、マウス由来の血清がELISAによって試験され、V3ペプチドの合成バージョンと特異的に反応する高力価のIgG抗体を有することが示された。MS2 VLPで提示されるペプチドの免疫原性の同様の試験は、本願の発明者によって以前に報告されていることに注意されたい。
【0170】
実施例8。プラスミドpET2P7K32及びpDSP7は、上記のMS2コートタンパク質プロデューサーpDSP1及びpDSP62のPP7アナログである。PP7 VLPでのペプチドディスプレイ価数の制御のために、pET2P7K32(am)及びpDSP7(am)も構築した。それらは、それぞれpDSP1(am)及びpDSP7(am)のアナログである。
【0171】
本明細書に引用される全ての特許、特許出願、刊行物、及び電子的に利用可能なもの(例えばGenBank and RefSeqにおけるヌクレオチド配列寄託、例えばSwissProt、PIR、PRF、PDBにおけるアミノ酸配列寄託、並びにGenBank and RefSeqにおけるアノテーションされたコード領域からの翻訳など)の全体の開示は、参照によって援用される。援用されるものと、オリジナルとして提出された明細書中に示されるものとの間の何らかの不一致は、オリジナルとして提出された明細書を優先して解消されるものとする。前述の詳細な説明及び実施例は、理解の明確化のためにのみ示された。それらから不必要な制限が理解されることはない。本発明は、表示及び記載された厳密な詳細に限定されるものではなく、当業者に明白な変形については、特許請求の範囲に規定される本発明内に含まれる。
【0172】
全ての見出しは、読む者の便利のためであって、そうであると特定されない限り、見出しの下にある文章の意味を限定するために用いられるべきではない。
【0173】
引用文献(参考文献の第一セット)
1.Peabody, DS, Jordan, SK, Caldeira, JC, Manifold-Wheeler, B. Medford, A and Chackerian, B (2007年) Immunogenic display of diverse peptides on virus-like particles of RNA phage MS2. J. Mol. Biol. 380: 252-263.
2.Kunkel TA, Bebenek. McClary J. (1991年) Efficient Site-Directed Mutagenesis Using Uracil-Containing DNA. Methods Enzymol. 204:125-139.
3.Sidhu SS, Lowman HB, Cunningham BC, Wells JA. (2000年) Phage display for selection of novel binding peptides. Methods Enzymol. 328:333-363.
4.Kleina LG, Masson J-M. Normanly J, Abelson J, and Miller JH. (1990年) Construction of Escherichia coli Amber Suppressor tRNA Genes. J. Mol. Biol. 213: 705-717.
5.Lee SK and Deasling JD. (2005年) A propionate-inducible expression system for enteric bacteria. Appl. Env. Microbiol. 71:6856-6862.
6.Peabody. D. S., and Lim, F. (1996年) Nucleic Acids Res 24, 2352-2359.
7.Chang, A. C, and Cohen, S. N. (1978年) J Bacterial 134, 1141-1156.
8.Caldeira, J. C, and Peabody, D. S. (2007年) J Nanobiotechnology 5, 10
9.Olsthoom, R. C, Garde, G., Dayhuff, T., Atkins, J. F., and Van Duin, J. (1995年) Virology 206, 611-625.
10.Tars K, F. K., Bundule M, Liljas L. (2000年) Virology 272. 331-337
11.Lim, F., Downey, T. D., and Peabody, D. S. (2001年) Journal of Biological Chemistry 276, 22507-22512.
12.Peabody, D. S. (1990年) J Biol Chem 265, 5684-5689.
【0174】
引用文献(参考文献の第二セット)
1.Caldeira Jdo C. Medford A, Kines RC. Lino CA, Schiller JT, Chackerian B, Peabody PS: Immunogenic display of diverse peptides, including a broadly cross-type neutralizing human papillomavirus L2 epitope, on virus-like particles of the RNA bacteriophage PP7. Vaccine, 28(27):4384-4393.
2.Peabody PS. Manifold-Wheeler B, Medford A, Jordan SK, do Carmo Caldeira J,
Chackerian B: Immunogenic display of diverse peptides on virus-like particles of RNA phage MS2. J Mol Biol 2008年, 380(1):252-263.
3.Lowman HB, Bass SH, Simpson N, Wells JA: Selecting high-affinity binding proteins by monovalent phage display, Biochemistry 1991, 30(45'): 10832- 10838.
4.Sidhu SS, Lowman HB, Cunningham BC, Wells JA: Phage display for selection of novel binding peptides. Methods in Ezymology 2000年, 328:333-363.
5.Wells JA: Hormone mimicry. Science 1996年, 273(5274):449-450.
6.Peabody PS: Subunit fusion confers tolerance to peptide insertions in a virus coat protein. Arch Biochem Biophvs 1997年, 347(1):85-92.
7.Peabody PS, Chakerian A: Asymmetric contributions to RNA binding by the Thr(45) residues of the MS2 coat protein dimmer. J Biol Chem 1999年, 274(36):25403-25410.
8.Lim F, Peabody PS: RNA recognition site of PP7 coat protein. Nucleic Acids Res 2002年, 30(19):4138-4144.
9.Lim F, Powney TP, Peabody PS: Translational repression and specific RNA binding by the coat protein of the Pseudomonas phage PP7. J Biol Chem 2001年, 276(25):22507-22513.
10.Kunkel TA, Bebenek K, Mcclary J: Efficient Site-Directed Mutagenesis Using Uracil-Containing DNA. Methods in Enzymoloev 1991年, 204:125-139.
11.Tars K FK, Bundule M, Liljas L: The Three-Dimensional Structure of Bacteriophage PP7 from Pseudomonas aeruginosa at 3.7 A Resolution. Virology 2000年, 272:331-337.
12.Kleina LG, Masson JM, Normanly J, Abelson J, Miller JH: Construction of Escherichia coli amber suppressor tRNA genes. II. Synthesis of additional tRNA genes and improvement of suppressor efficiency. J Mol Biol 1990年, 213(4):705-717.
13.Normanly J, Kleina LG, Masson JM, Abelson J, Miller JH: Construction of Escherichia coli amber suppressor tRNA genes. III. Determination of tRNA specificity. J Mol Biol 1990年, 213(4):719-726.
14.Kunkel TA: Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc Natl Acad Sci USA 1985年, 82(2):488-492.
15.Maruyama IN, Maruyama HI, Brenner S: Lambda foo: a lambda phage vector for the expression of foreign proteins, Proc Natl Acad Sci USA 1994年, 91(17):8273-8277.
16.Mikawa YG, Maruyama IN, Brenner S: Surface display of proteins on bacteriophage lambda heads. J Mol Biol 1996年, 262(1):21-30.
17.Dunn IS: Assembly of functional bacteriophage lambda virions incorporating C-terminal peptide or protein fusions with the major tail protein. J Mol Biol 1995, 248(3):497-506.
【技術分野】
【0001】
関連出願及び支援助成金
本出願は、発明の名称「VLPでのペプチドディスプレイ価数の制御(Control of Peptide Display Valency on VLP)」である2009年12月31日出願の米国特許仮出願第61/335,122号、発明の名称「バクテリオファージMS2 VLPでのランダム配列ペプチドライブラリーの容易な構築のためのプラスミドベクター、並びに関連の構築物、ライブラリー及び方法(Plasmid Vectors for Facile Construction of Random Sequence Peptide Libraries on Bacteriophage MS2 VLPs and Related Constructs, Libraries,and Methods」である2009年12月31日出願の米国特許仮出願第61/335,120号、及び発明の名称「バクテリオファージPP7のウイルス様粒子でのペプチドディスプレイ(Peptide Display on Virus−Like Particles of Bacteriophage PP7)」である2009年12月31日出願の米国特許仮出願第61/335,121号から優先権の利益を主張し、それぞれの出願の内容は、その全体が本明細書に援用される。
【0002】
本特許出願は、国立衛生研究所によって助成金番号GM042901及びR01 AI065240により支援された。米国政府は、本発明において一定の権利を有する。
【0003】
本発明の分野
本発明は、RNAバクテリオファージ、特にMS2及びPP7のウイルス様粒子(virus-like particule, VLP)でのペプチドのディスプレイ(提示)のためのシステム及び方法に関する。所望の結合機能を有するペプチドがアフィニティー選択によって単離される、高い複雑性のランダム配列及び抗原フラグメントライブラリーの構築を容易にする方法及びプラスミドベクターが記載されている。ペプチドディスプレイの密度は、アフィニティー選択のストリンジェンシーにおける重要な決定因子であるので、VLPでペプチドディスプレイ価数を制御するための方法及びプラスミドもまた記載される。本発明の方法によって、VLP、特にMS2 VLPでの広い範囲(すなわち、1粒子当たり平均で1未満から90まで)にわたるディスプレイ価数レベルの容易な調節が可能になり、これによって、低価数を示すVLPを含めて、免疫原及びワクチンの特定及び産生が容易になる。このシステムは、主にワクチン開発を視野に入れて開発されてきたが、薬物及び造影剤の細胞又は組織型特異的な標的送達に有用性を有するペプチド−VLPの特定を含め、様々な他の適用において、並びに新規な物質の鋳型合成において、有用性を有する。
【0004】
本出願は、VLPディスプレイシステムの実現を容易にする方法及びプラスミドベクターを記載する。本発明は、ウイルス様粒子上で提示され、VLPの合成及びVLPで提示されるペプチドの合成を指向する特定のRNA(MS2又はPP7のいずれか)をカプシドで包む異種ペプチドの挿入によってそれぞれが改変される、MS2又はPP7のコートポリペプチドから構成されるウイルス様粒子の発現において有用な核酸構築物を提供する。関連のウイルス様粒子、方法及び免疫原性組成物もまた提供される。
【背景技術】
【0005】
発明の背景
ワクチンとしてのVLP。近年の組み換えDNA技術の進歩によって、動物及びヒトの両方において有効な防御免疫を可能にするための十分な量のタンパク質が得られるように、免疫原性タンパク質が特定され、クローニングされ、適切な宿主中で発現されたワクチンの導入がもたらされた。ほとんどの有効なワクチンの多くは、ビリオン表面が中和抗体を誘発する強力な能力に基づく。これらとしては、防御抗体の応答を効果的に誘導する、ポリオ、インフルエンザ及び狂犬病などの認可された不活化ウイルスワクチン又は弱毒化ウイルスワクチンが挙げられる。さらに最近では、ヒトパピローマウイルス(HPV)及びB型肝炎ウイルス(HBV)の構造タンパク質の自己アセンブルに基づくサブユニットワクチンが、食品医薬品局によって承認された。このサブユニットは適切な宿主で発現され、次いで本物のウイルスを構造的に模倣するが、ウイルスのゲノムを欠いているので非感染性である粒子へと自己アセンブルされる。これらのいわゆるウイルス様粒子(VLP)は、個々の粒子それぞれで、ウイルス様粒子を含む構造タンパク質が多数のコピーにおいて存在するので、一般に極めて免疫原性である。この高密度の抗原提示は、それらの粒子が強固な抗体応答を誘発するのを特に有効にする。HBVワクチン及びHPVワクチンは、それぞれのウイルス自体の構造タンパク質からアセンブルされたVLPに基づくが、VLPはまた、異種エピトープの高密度提示のための足場として利用されてもよい。VLPは一般に高度反復性を示し、従って極めて免疫原性の構造であるので、それらは多数の異なるウイルス種に由来し得る。本発明の方法は、ファージディスプレイと類似の方法によって、免疫原性提示及びエピトープ開発の両方のために、RNAバクテリオファージ(特にMS2及びPP7)のVLPを利用することを目的に行われる[1,2]。
【0006】
RNAバクテリオファージ。一本鎖RNAバクテリオファージは、天然に広く分布して見られるウイルスのグループである。いくつかは、ゲノム配列、分子生物学、並びにカプシド構造及び集合に関して詳細に特徴付けられた。MS2はおそらく、最も研究されたグループのメンバーであって、本発明者の研究室で行われたほとんどの研究の焦点であったが、近年の研究はまた、PP7と呼ばれる関連のファージも開拓している。MS2は、4つのタンパク質(マチュラーゼ、コート、溶解(lysis)及びレプリカーゼ)のみをコードする3569ヌクレオチドの一本鎖RNAゲノムを有する。このウイルス粒子は、180個のコートポリペプチド、1分子のマチュラーゼ、及び1コピーのRNAゲノムから構成される。コートタンパク質自体が完全に正二十面体のシェルの形成を担うので、MS2 VLPは、プラスミドから単独の遺伝子の生成物として生成され得る。従って、ペプチドディスプレイのために用いられる他のファージと比較して、RNA VLPは著しく簡易である。ペプチドディスプレイ及びアフィニティー選択のためのMS2及びPP7のVLPの操作は、本願の発明者によって近年示されており[1,2]、また本明細書の後段にも記載される。
【0007】
従来のファージディスプレイによるエピトープ特定。ファージディスプレイは、ランダムなアミノ酸配列の大きいライブラリーの提示を可能にするいくつかの技術のうちの1つであり、それらから特定の専門的な機能(例えば、特定の抗体に結合する能力)を有するペプチドを選択する目的を備える。最も一般的に用いられるファージディスプレイ法は、繊維状ファージ(例えば、M13)に基づく。この基本的な概念は、ウイルス構造タンパク質のうちの1つに対して、ファージのDNAゲノム中に遺伝子的に融合されたランダムな配列のライブラリーを含んでいる組み換えバクテリオファージゲノムを作成することである。このような組み換え体が細菌にトランスフェクトされる時、それぞれがウイルス粒子を生成し、そのウイルス粒子は、その表面上に特定のペプチドを提示し、そのペプチドをコードする同じ組み換えゲノムをパッケージングする。これによって、この方法に必須の遺伝子型及び表現型の連鎖が確立される。任意関数(例えば、レセプターの結合、免疫原性)は、アフィニティー選択の使用と、続く大腸菌での増殖による選択物の増幅とによって、ペプチドディスプレイファージの複雑なライブラリーから選択され得る。ペプチドディスプレイファージの膨大なライブラリーでは、特定のレセプター(例えば、モノクローナル抗体)に結合できるごく少数が、アフィニティー精製されて、次いで大腸菌中での増殖によって増幅され得る。比較的単純な集団を得るには、通常数回の反復回数の選択及び増幅で十分であり、それから所望の活性を有するペプチドを提示する個々のファージをクローニングし、次いで特徴付けすることができる。選択する分子が抗体である場合、このようにして特定されたペプチドは、抗体によって認識されるエピトープであり、そして適切な条件下では、免疫された患者又は動物において、その天然の抗原中でそのエピトープに特異的な抗体応答を誘発し得る。
【0008】
しかし、繊維状ファージディスプレイには不利な点がある。最も重要なことに、繊維状ファージの分子生物学の特異な様式によって、本当に強力な免疫原性について必要な高密度でペプチドを提示することが困難又は不可能となる場合が多い。このことは、免疫原(すなわち、ワクチン)として有用であるべきペプチドエピトープはアフィニティー選択によって特定され得るが、これらのペプチドエピトープは、通常は化学的に合成されて、次いでさらに免疫原性の担体に結合されなければならないということを意味する。不幸にも、ペプチドは、そのアフィニティー選択及び最適化の間にそのペプチドが存在する構造的状況からこのように分離された場合、活性を失う場合が多い。ここで記載したRNAファージVLPディスプレイシステムは、他方では、アフィニティー選択及び免疫原性エピトープの提示を単一のプラットフォーム上で行う能力を生み出す。これは、高密度ペプチドエピトープディスプレイとアフィニティー選択能力とを組み合わせた結果であって、エピトープのアフィニティー選択の間に存在する構造的制約が、免疫過程の間維持され得るということを意味し、エピトープが所望の抗体応答を誘発するのに必要な構造を保持する確率が増大する。
【0009】
RNAファージVLPディスプレイ法の概説。本発明者は、MS2及びPP7を含めて、RNAバクテリオファージのVLPに基づくペプチドディスプレイ及びアフィニティー選択のための技術を以前に記載したが[1,2]、ここでその技術を簡略に説明する。VLPディスプレイ法の開発には、2つの前提条件が満足されることを必要とする:第一に、RNAファージコートタンパク質の形態、及びその中の適切に折り畳んでVLPにアセンブルする能力を破壊することなく外来ペプチドの挿入を許容する部位を特定する必要があった。コートタンパク質の表面上のAB−ループを、ペプチド挿入のための部位として選択した。ここに挿入されたペプチドは、VLPの表面上で顕著に提示される。不幸にも、コートタンパク質の野生型は、AB−ループ中のペプチド挿入に極めて耐性がなく、その圧倒的多数(通常は98%を超える)が折り畳みの失敗に至る。コートタンパク質は、通常は二量体として折り畳まれるが、そのうちの90が正二十面体VLPへとアセンブルされる。本発明者は、新規な形態のコートタンパク質を操作して、それを安定化し、さらなるAB−ループ挿入の耐性を与えた。そうするために、本発明者は、二量体中で2つの同一のポリペプチド鎖のN末端及びC末端の近接性を利用した(図1)。コートタンパク質コード配列を複製すること、次いで2つのコピーを単一のリーディングフレームに融合することによって、本発明者は、いわゆる一本鎖の二量体を生成した。この形態のタンパク質は、劇的にさらに熱力学的に安定であり、その折り畳みは一本鎖二量体の下流のコピーのAB−ループに挿入されたペプチドにかなり耐性である[1,2]。得られたVLPは、1つの二量体当たり1つのペプチド、又は1つのVLP当たり90のペプチドを提示する。ペプチドディスプレイ/アフィニティー選択能力についての第二の前提条件は、アフィニティー選択された配列を増幅するための手段を提供することが必須であることから、遺伝子型に対する表現型の連鎖である。この要件は、本発明者が、RNAファージVLPがその合成を指示するメッセンジャーRNAをカプシドで包むということを示した際に満たされた[1,2]。このことは、アフィニティー選択されたペプチド−VLPの配列が、逆転写及びポリメラーゼ連鎖反応によって増幅され得ることを意味する。選択の標的がモノクローナル抗体である場合、得られたアフィニティー選択されたVLPは、動物又は患者において、その活性が選択抗体の活性を模倣する抗体を引き出すためのワクチン候補物である。
【0010】
ペプチドディスプレイ価数及びその制御に関する考慮。本出願は、RNAファージVLPでの複雑なランダム配列及び抗原フラグメントのライブラリーの構築、並びに特定のモノクローナル抗体(又は他の任意のレセプター)に結合するペプチドのこのようなライブラリーからのアフィニティー選択を容易にするプラスミドベクターを記載する。元々記載されるとおり、RNAファージVLPディスプレイ技術は、それぞれのVLPに90個のペプチドを提示することに注意されたい。これは、ペプチドが一本鎖二量体の1つのAB−ループに挿入され、90個の二量体がVLPを作成するからである[1,2]。これらの粒子の多価数は、MS2 VLPのほとんどの適用について望ましい。例えば、この粒子の高い免疫原性は、提示されるペプチドの高密度に関連し、従ってワクチン中で価値のある特性である。しかし、複数の同時の弱い相互作用という理由のみにより、アフィニティー選択過程の間、多価数によって、選択標的について内因性の高い結合親和性を有するペプチドを提示する粒子を、密接に結合する粒子から識別することが困難になる。この「結合活性対親和性」のジレンマは、繊維状ファージディスプレイを用いての高い親和性のペプチドリガンドの選択において十分立証された問題である[3−5]。本発明は、広い範囲にわたって、すなわち1粒子当たり1未満から90まで、平均のペプチドディスプレイ価数を調節する手段を導入することによって、VLPディスプレイシステムにおいてこの問題に取り組む。これによって、アフィニティー選択過程の間にペプチドディスプレイの密度を変更することが可能になる。選択は数回行われ、初回は通常は多価数の提示を用いて行われ、これによって標的に関してある程度の最小の親和性を有する全てのペプチドを含む比較的複雑な集団を得る。その後の回では、ペプチドディスプレイの価数は減少されることができ、それによって選択のストリンジェンシーが増大し、結果として抗体標的に関して高い親和性を有するペプチドの単離及び好ましいエピトープの良好な分子模倣物が得られる。
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明の目的は、低価数の異種ペプチドを提示するVLPを提供することである。
【0012】
本発明のさらなる目的は、低価数又は高価数の異種ペプチドを提示するVLPなどのVLPを産生するための核酸構築物を提供することである。
【0013】
本発明のさらに別の目的は、低価数又は高価数の異種ペプチドの提示を制御し得るVLPなどのVLPを産生するための核酸構築物を提供することである。
【0014】
本発明のさらに別の目的は、高い親和性の免疫原を特定し、これらの免疫原を治療及び他の製剤又は適用において用いるために、低価数又は高価数の異種ペプチドの提示を制御し得るVLPを産生するための核酸構築物を提供することである。
【0015】
本発明のさらなる目的は、選択された抗体に対して高い親和性を示す免疫原性ペプチドを特定するための方法を提供することである。
【0016】
本発明のさらに別の目的は、高い親和性のペプチドをVLPに組み込むことである。
【0017】
本発明のさらなる目的は、本発明によるVLPを用いて免疫原性の方法及び組成物を提供することである。
【0018】
本発明のこれら及び/又は他の目的のいずれか1つ以上は、以下の本発明の説明から容易に得られる。
【課題を解決するための手段】
【0019】
発明の要旨
本発明は、選択的に免疫原性のペプチドを組み込むVLPを特定及び提供することのさらに効果的な手段を提供するために、高い複雑性のランダム配列及び抗原フラグメントペプチドライブラリーの構築を容易にし、かつMS2 VLPでのペプチドディスプレイの価数(すなわち、密度)を制御することを可能にする、組成物、構造(プラスミドを含む)、システム及び方法に関する。本発明は、その内容全体がここに援用され、上記及び[1,2,6,7]において簡略に記載された米国特許出願公開第2009/0054246号及び出願番号PCT/US2007/018614(WO08/024427として公開)において別に開示された、方法、組成物、粒子、単位及びその他の開示の拡張を示す。
【0020】
本発明は、所定のモノクローナル抗体に最も高い親和性を有するペプチドの選択が、天然の抗原の最良の分子模倣物を提供すること、そしてこれらのペプチドが、関係する抗体応答を提供又は誘導する可能性が最も高いことをもたらす。これらのペプチドは、患者において免疫原性を誘導し、防御応答を提供するために特に適しているものとして提案される。これらのペプチドから調製されるワクチン及び/又はこれらのペプチドを組み込むワクチンは、特にアジュバント無しで投与される本発明によるワクチンを含み、低減された副作用を有し、より効果的である。
【0021】
RNAファージMS2(pDSP1及びpDSP62)のVLP及びRNAファージPP7(pET2P7K32及びpDSP7)由来のVLPにおいて、ランダム配列又は抗原フラグメントペプチドライブラリーの構築を容易にするプラスミドベクターが記載される。これらのベクターは、1011〜1012個を超える個々のメンバーを有するライブラリーの作製を可能にする。しかし、これらのベクターは、高密度で(すなわち、1つのVLP当たり90個)の外来ペプチドを均一に提示するVLPを生じる。上記で説明したとおり、このことは、ペプチドに対して高レベルの免疫原性を与えるが、多価数が選択のストリンジェンシーを低下して、集団の中で最も緊密に結合する種を優先的に選択することを困難にすることでアフィニティー選択の間に問題を生じることが多いので、ワクチン適用のための明白な利点である。
【0022】
本発明は、ペプチドディスプレイ価数制御の問題に対する簡易な解決である、単一のRNAからの少なくとも四(4)アミノ酸長の異種ペプチドを含む大量の野生型及び少量の一本鎖(好ましくは、二量体)のコートタンパク質の産生を可能にするシステムを示す。このアプローチは、pDSP1(am)、pDSP62(am)、pET2P7K32(am)及びpDSP7(am)のようなプラスミドを構築することを含む。これらは、例えば、一本鎖二量体中でコーティングタンパク質の下流の第一のアミノ酸を通常コードするコドン(アラニン)の代わりに、終止コドン(好ましくは、アンバー終止コドン)を含むように改変された、上記のプラスミドの単純な改変体である。これらのプラスミドは、一本鎖二量体の二等分の接合部に終止コドンを有するので(例えば、pDSP1を参照)、それらのプラスミドは正常には、単位長さの野生型コートタンパク質しか産生せず、このタンパク質は当然にVLPにアセンブルする。しかし、本発明のアプローチでは、このプラスミド又は第二のプラスミド(例えば、pNMsupA)は、コート配列を翻訳するわずかな割合のリボソームに終止コドンを読み飛ばしさせ、異種ペプチドを含む一本鎖二量体を生じさせる量でサプレッサーtRNAがもたらされるようにして、プロモーター(例えば、異なる不適合性群からのクロラムフェニコール耐性プラスミド上のlacプロモーター)の制御下で発現されるアラニン挿入サプレッサーtRNA遺伝子などのtRNA遺伝子[8,9]を挿入して改変される。得られたタンパク質は、ゲストの異種ペプチドが、好ましくは、例えば第二のAB−ループ中若しくはカルボキシル末端又は下流サブユニット内の他の位置に挿入されており、同じmRNAから発現された野生型タンパク質とコアセンブル(共集合)して、低い価数を呈するモザイクのVLPを形成する。
【0023】
本発明によれば、VLPは、1つのVLP当たり1未満から九十(90)の異種ペプチド、好ましくは1つのVLP当たり約1〜約十(10)の異種ペプチド、さらに好ましくは1つのVLP当たり約1〜約5の異種ペプチド、さらに好ましくは1つのVLP当たり約1〜約3の異種ペプチド、最も好ましくは1つのVLP当たり約2〜約4の異種ペプチドを提示するように制御された方式で生成され得る。本発明によるペプチド密度の低下は、アフィニティー選択のストリンジェンシーが増大したVLPを生じ、後に高い提示密度の形式に戻った場合に強力に免疫原性となる高い親和性のペプチドの容易な特定が可能になる。アガロースゲル電気泳動及びノーザンブロットによって、本発明によって生じた粒子が関連のRNAをカプシドで包むことを確認する。本発明の方法はさらに、価数(1つのVLP当たりに生じるペプチドの数)が、サプレッサーtRNAの発現レベルを制御することによって、例えばサプレッサーtRNA合成のレベルを調節することによって、広い範囲にわたって調節され得ることになり、これは、例えば誘導因子濃度の関数としてその活性が調節され得るプロモーター(例えば、proB又は他の適切なプロモーター)からtRNAを発現することによって、適宜達成され得る。価数のレベルはまた、異なるサプレッサーtRNA又はその変異体の利用を通じて制御されることができ、より大きい又はより小さい内因性の抑制の効率を有する。
【0024】
当業者は、価数の制御が過剰な野生型コートタンパク質と一緒に組み換えタンパク質を同時発現することによってより簡単に達成されることができ、これによって外来のペプチドの含量が低下されたモザイクカプシドが生じると考えるかもしれない。不幸にも、このアプローチは、価数を適切なレベルまで(例えば、平均で1つのVLP当たり1未満から2〜3のペプチドまで)低下することが、野生型タンパク質が組み換えペプチドよりもかなり過剰に発現されることを必要とするので実用的でない。このような同時発現の方法は、過剰な関連のない(すなわち、非外来ペプチドをコードする)RNAを生じ、これがかなり多量であることによって、ペプチドコードRNAよりも優先してVLP内にパッケージングされる。特異的なカプシド形成は、RNA中のいかなる簡易なパッケージングシグナルの存在からも独立して出現するので(1)、選択的なカプシド形成のために少数派のペプチドコード種を選び出すことは簡単な方法ではないと考えられる。この同時発現アプローチの実行可能な改変型が存在することは想像できるが、これらの改変型は、あまりにも複雑になりがちである。本発明者のシステムは、単一のmRNAから両方の型のタンパク質を生成することによって、この問題を解決する。
【0025】
例として、以下の実施形態は、本発明をさらに実証するために用いられ得る。以下に記載される実施形態のそれぞれでは、MS2コートポリペプチド又はPP7コートポリペプチドが個々の核酸構築物において用いられることができるが、MS2又はPP7コートポリペプチドへの限定は、このような限定が記載の文脈の中で適当でない限り、考慮されるべきではないことに注意されたい。
【0026】
本発明の一実施形態は、次のものを含む核酸構築物(例えば、pDSP1を参照)を提供する:
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーター(例えば、本発明の詳細な説明において後述されるような)であって、このコートポリペプチド二量体のコード配列は、コートポリペプチド二量体のABループを規定する配列のその部分に対して、5’側に配置された第一の制限部位(例えば、SalI又はKpnI)を規定するように改変されている;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位(例えば、BamHI);
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)。
(d)抗生物質耐性遺伝子(例えば、カナマイシン)、及び
(e)原核生物細胞での複製のための複製起点(例えば、プラスミドColE1由来の複製起点)。
【0027】
別の実施形態では、本発明は、次のものを含む核酸構築物(たとえば、pDSP62を参照)を提供する:
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、この一本鎖二量体配列の二分の一が、改変された二量体を生成するために複数のサイレントヌクレオチド置換体で改変されて(すなわち、「コドンジャグリングされて(codon juggled)」)、突然変異のオリゴヌクレオチドプライマーが、この改変された二量体(好ましくは、2つの配列がプライマーのハイブリダイゼーションによって識別されることを可能にするAB−ループのいずれかの側における20ヌクレオチド単位内の十分な数の変異)の改変された半分又は未改変の半分に対して、特異的にアニールされ得る、プロモーターと;
(b)ABループを特定するコート配列のその部分に対して5’側に配置された第一の制限部位(例えば、SalI)と、
(c)コートポリペプチド二量体のコード配列に対して3’側の第二の制限部位(例えば、BamHI)と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と;
(e)第一の抗生物質に対して耐性の遺伝子と;
(f)原核生物細胞での複製のための複製起点と;
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点(例えば、M13又はfd)と;
(h)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージ(例えば、M13、fd)。
【0028】
別の実施形態では、本発明の核酸構築物に関して、バクテリオファージMS2(又はPP7)一本鎖コートポリペプチド二量体のコード配列はさらに、異種ペプチドをコードする核酸配列を含み、この構築物は必要に応じて、第二の制限部位に対して3’側に配置された転写ターミネーターを含む。そのような核酸構築物は、異種ペプチドの挿入によって改変されたMS2(又はPP7)のコートポリペプチドから構成されるウイルス様粒子の発現に有用であり、ここでは異種ペプチドがウイルス様粒子に提示されて、MS2(又はPP7)のmRNAをカプシドで包む。
【0029】
別の実施形態では(例えば、pET2P7K32)、核酸構築物は次のものを含む:
(a)バクテリオファージPP7(又はMS2)一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、このコートポリペプチド二量体のコード配列は、コートポリペプチド二量体のコード配列の下流部分に位置され、かつコートポリペプチド二量体のABループを規定する配列に対して5’側に、又はその中に配置された、第一の制限部位を規定するように改変されている、プロモーターと;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と;
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と、
(d)抗生物質耐性遺伝子と;
(e)原核生物細胞での複製のための複製起点。
【0030】
別の実施形態(例えば、pDSP7)では、核酸構築物は次のものを含む:
(a)バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、この一本鎖二量体配列の半分が、複数のサイレントヌクレオチド置換体で改変されて(すなわち、「コドンジャグリングされて(codon juggled)」)、変異原性のオリゴヌクレオチドプライマーが、一方又はもう一方の半分に対して特異的にアニールされ得る(好ましくは、2つの配列がプライマーのハイブリダイゼーションによって識別されることを可能にするAB−ループのいずれかの側における20ヌクレオチド単位内の十分な数の変異である)、プロモーターと;
(b)コートポリペプチド二量体のコード配列の下流部分に位置し、かつコートポリペプチド二量体のABループを規定する配列に対して5’側に、又はその中に配置された第一の制限部位と;
(c)コートポリペプチド二量体のコード配列に対して3’側に配置された制限部位と;
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマー(関連するPCRプライマーの定義及び列挙は、本発明の詳細な説明に示される)と、
(e)第一の抗生物質に対して耐性の遺伝子と;
(f)原核生物細胞での複製のための複製起点と;
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点(例えば、M13、fd)と;
(h)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージ(例えば、M13、fd)。
【0031】
本発明の別の実施形態では、核酸構築物は次のものを含む:
(a)バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、このコートポリペプチド二量体のコード配列は、(1)コートポリペプチド二量体のコード配列の下流部分に位置され、コートポリペプチド二量体のABループを規定する配列に対して5’側に又はその中に配置された第一の制限部位を規定し、及び(2)ヌクレオチド配列(NNS)xを含み、ここでNは任意のヌクレオチドであり、Sはグアノシンヌクレオチド(G)又はシチジンヌクレオチド(C)であり、xは1〜500の整数である、ように改変されている、プロモーターと;
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と;
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと;
(d)抗生物質耐性遺伝子と;
(e)原核生物細胞での複製のための複製起点。
【0032】
本明細書に記載される本発明の特定の実施形態では、本明細書に記載されるような、上記の特定の構成要素は、選択的ではあるが、構築物内に含まれ得ることが好ましい。
【0033】
他の実施形態(例えば、DSP1(am)、pDSP62(am)、pET2P7K32(am)及びpDSP7(am)と呼ばれる上記のプラスミドの誘導体)では、提示価数の制御は、以下の追加の特徴を含むことによってもたらされる:
(a)一本鎖二量体の上流半分と下流半分との間の接合部のナンセンスコドン(例えば、本明細書で他に記載されるようなアンバーコドン);
(b)上記の(a)に記載されるナンセンスコドンで翻訳終結を部分的に抑制することができるサプレッサーtRNA(例えば、アラニン−挿入アンバー−サプレッサー)を生成するプラスミド(例えば、pNMsupA)。このプラスミドは、第二の不適合性群由来の複製起点を有し、第二の抗生物質に対する耐性を付与し、これによって上記のコートタンパク質産生プラスミドの1つもまた含む細菌中で、そのプラスミドの安定な維持が可能になる。
【0034】
ワクチン候補物のライブラリー構築及びアフィニティー選択の概略
ここではMS2 VLPシステムを例として用いるが、同様の方法が、本出願でやはり示されるPP7システムにもあてはまることが理解されるべきである。
【0035】
1.ライブラリー構築。ランダムな配列ライブラリーは、2つの方法のいずれかによって生成され得る。これらは、後にさらに詳細に記載され、図9b及び図15に図示される。(A)PCR産物は、pDSP1へ、ランダムな配列でalIとBamHIとの間にクローニングされ、このランダムな配列は、AB−ループ中にそれを導入するような方法で結合される。この方法は、比較的低い複雑性のライブラリー(典型的には107〜108個のメンバー)の便利な構築物について適切であるが、より高レベルまで大規模化するには不便である。(B)より複雑性が高いライブラリー(例えば、109〜1011個以上)については、合成オリゴヌクレオチドプライマーは、pDSP62の一本鎖バージョンにアニールされ、DNAポリメラーゼで伸長されて、二本鎖の環状分子を生じ、次にこれがDNAリガーゼの作用によって共有結合的に閉じられる(図12を参照)。いずれかの方法によって産生される組み換えDNA分子は、大腸菌の適切な発現株(例えば、BL21(DE3))に導入され、そこでそれらの大腸菌がVLPを合成する。pDSP1又はpDSP62から産生される粒子は、1粒子当たり90という密度(高密度)で外来ペプチドを提示する。
【0036】
2.アフィニティー選択は、例えば、ある手順(例えば、バイオパニング、又は抗体に高い親和性を示すペプチドを迅速に特定及び分離するための他のアプローチ)にVLPライブラリーを供することによって行われ、ここでは、モノクローナル抗体はマルチウェルのプラスチックプレートの1つ以上のウェルの表面に吸着される。VLPライブラリー溶液を、免疫された抗体とともにインキュベートし、次いで未結合のVLPを洗い流して廃棄し、いずれかの結合したVLPを、通常は溶液のpHを低下することによって溶出する。続いて、溶出したVLPは熱変性され、それが含むRNAは、RNAに対してその3’末端(BamHI部位の十分下流)付近でアニールするオリゴヌクレオチドプライマーを用いて逆転写によってDNAにコピーされる。次いで、得られたcDNAを、第一の制限部位(例えば、SalI)の上流及び第二の制限部位(例えば、BamHI)の下流に特異的にアニールするプライマーを用いてPCRによって増幅する。
【0037】
3.再クローニング(recloning)。上記で得られたPCR産物を、第一及び第二の制限酵素(例えば、SalI及びBamHI)で分解し、次いで適切な発現ベクター中に再クローニングする。2回目の選択回を低ペプチド価数で行うべき場合、PCR産物は、pDSP1(am)又はpDSP62(am)中にクローニングされ、選択された集団全体を、pNMsupAを含む大腸菌の発現株中に導入する(図7を参照)。このように生成されたVLPは、一旦アフィニティー選択されれば、最初のライブラリーに存在したペプチドの集団よりもかなり簡素な集団しか提示しないので、平均して、その外来ペプチドの2〜3のコピーしか提示しない(すなわち、低価数である)。
【0038】
4.アフィニティー選択及び再クローニングのさらなる反復回は、上記のとおり行う。2〜4回又は好ましくは3回若しくは4回(高い価数で1又は2回、続いて低い価数で1又は2回)が典型的には、選択のモノクローナル抗体に緊密に結合するペプチドを提示するVLPの簡素な集団を生じるのには十分である。選択が完了したと考えられる場合、配列をpDSP1又はpDSP62中に再度クローニングし、ここでペプチドは、最大の免疫原性について高密度(1カプシド当たり最大90ペプチドまで)で再度提示される。個々のクローンが得られて、DNA配列分析による特徴付けに供される。選択抗体についてのインビトロにおけるそれらの親和性を評価し、それらが所望の抗体応答を引き出す能力を決定する。十分特徴付けられたモノクローナル抗体標的を使用する実証実験では、本発明者は、その配列が以前に特定されたエピトープの配列を模倣するペプチドを回収した。得られたVLPで免疫された動物は、もとの抗原のエピトープを認識する抗体を生じる。
【0039】
本発明のプラスミド及び他の側面の詳細は、以下の発明の詳細な説明にさらに記載される。
【0040】
本発明による上記の実施形態及び/又は他の実施形態のいずれか1つ以上が、以下の本発明の説明からさらに容易に得られる。
【図面の簡単な説明】
【0041】
【図1】MS2コートタンパク質二量体の構造を図示する。左側上の写真は、2つのサブユニット鎖のN末端及びC末端の近接性を強調する。この特徴によって、外来ペプチド挿入の高い耐性を有する、一本鎖二量体の構築が容易になった。また、図示(左下)されるのは、二量体及びインタクトなVLPの両方における、AB−ループの高い接近性である。垂直方向には、VLP自体の構造が示される。VLPが構成されるコートタンパク質は、準同一によって3つのわずかに異なる配置をとり、赤、青及び緑で示される。AB−ループは黄色で示される。VLP表面上でのそれらの反復性の性質及びそれらの露出に注目されたい。
【図2】VLPのランダム配列ペプチドライブラリーの提示の基礎を示す。ランダム配列ペプチド挿入物のライブラリーは、組み換えDNA方法によって作成され、得られたプラスミド集団は、形質転換によって大腸菌に導入され、VLPのライブラリーが生成される。集団中の個々のVLPのそれぞれは、その表面上に異なるペプチドを提示し、そしてその中に(mRNAの形態で)、その合成のための遺伝情報を含む。
【図3】アフィニティー選択の過程を図示する。ランダム配列ペプチドライブラリーを提示するVLPの集団を、表面で免疫されたモノクローナル抗体とともにインキュベートする。VLPのほとんどは、通常は抗体に結合できず、洗い流されて廃棄される。次いで、ペプチドが抗体との結合を示すいずれかのVLPを特異的に溶出し、それらが含むRNAを逆転写によってDNAにコピーし、PCRによって増幅し、そしてアフィニティー選択されたVLPの産生のために発現プラスミド(例えば、pDSP62又はpDSP62(am))中に再クローニングする。選択は、典型的には反復して(すなわち、2回を超えて)、好ましくは3〜5回行われる。
【図4】ペプチドディスプレイ価数の重要性を示す。多価数のディスプレイによって、本質的に緊密な結合因子を複数のレセプターと同時に相互作用する弱い結合因子と識別することが困難になり得る。通常は、初回のアフィニティー選択は高い価数で行うが、その後の回では、価数は低レベルまで低下され、それによって選択のストリンジェンシーを増大し、選択抗体について最高の親和性を有するペプチドの単離を確実にする。
【図5】AB−ループ中に挿入するために都合のよいクローニング部位を有するプラスミドpDSP1を示す。pDSP1は、例えば1つだけのSalI及びKpnI制限部位を含むように改変された、一本鎖二量体のコード配列を発現する。この二量体は、AB−ループへの外来配列の簡易なクローニングを容易にする。これらの部位を1つだけにするには、ベクター中及び上流のコート配列中で、多数のSalI及びKpnI部位を破壊することが必要であった。
【図6】プラスミドpDSP62を示す。一本鎖のDNAの産生を容易にするために、M13複製起点をプラスミド中に導入し、M13起点の配向に応じてpDSP61及びpDSP62として特定した。このプラスミドはまた、いわゆる「コドンジャグリングされた(codon juggled)」コート配列を、一本鎖二量体の上流半分に含む。コドンジャグリングされた半分は、下流の半分と同じアミノ酸配列をコードするが、可能な最大数のサイレント置換を含むことによってその配列とは異なり、2つの半分をオリゴヌクレオチドのアニールのために識別可能とする。
【図7】単一のRNAから大量の野生型及び少量のAB−ループ組み換えタンパク質を産生するための例示的なシステムを示す。一本鎖二量体においてコートタンパク質の下流のコピーの第一のアミノ酸を通常はコードするアラニンコドンの代わりにアンバー終止コドンを含む、pDSP1の改変体(pDSP1(am))を構築した。さらに、アラニン挿入サプレッサーtRNA遺伝子を合成して、クローニングした。この遺伝子は、コート配列を翻訳する少ない割合のリボソームに終止コドンを読み飛ばしさせて一本鎖二量体を生じさせる量で産生される。得られたタンパク質(とそのゲストペプチド)は、同じmRNAから発現される野生型タンパク質とコアセンブル(共集合)して、本明細書の他で記載されるようなモザイクカプシド及び低減された価数を生じる。
【図8】下に列挙するプラスミドで産生される精製VLPのSDSゲル電気泳動を示す。実験のポイントは、精製VLPにおける、上の図7について及び本出願の実施例の項において記載するような、一本鎖「読み飛ばし」(リードスルー、readthrough)生成物の含量を示すことである。精製されたVLPは、以下のプラスミドから生成された(左から右へ): pDSP1は、コートタンパク質一本鎖二量体のみを含むVLPを生成する。 pDSP1−Flagは、その第二のAB−ループ中に挿入されたFlagエピトープを有する一本鎖二量体のみを含むVLPを生成する。 pDSP1(am)は、一本鎖二量体中のコート配列の接合部にナンセンス変異を有し、これによってpDSP1(am)は、大量の野生型コートタンパク質を生じ、そしてサプレッサー−tRNA(pNMsupAから提供される)の存在下では、少量の一本鎖二量体を生じる。ナンセンス抑制は、この場合低い効率を有するので、ごくわずかなリボソームが、終止コドンを読み飛ばして一本鎖二量体を生成する。この2つの形態のコートタンパク質は、コアセンブルしてモザイク又はハイブリッドのVLPになり、これは大部分の野生型コートタンパク質及び少量の一本鎖二量体(野生型コートタンパク質のレベルの約3%と推定される)からなる。 pDSP1(am)−Flagは、pDSP1(am)のようであるが、一本鎖二量体の第二のAB−ループにフラグエピトープが挿入されている。それは、大部分の野生型コートタンパク質、及びそのAB−ループにフラグペプチドが挿入されている少量の一本鎖二量体を生成する。 図の右の2つの矢印は、pDSP1(am)及びpDSP1(am)−Flagの終止コドンの抑制によって生成される2つのタンパク質の位置を示す。
【図9】図9a及び図9bは、異種ペプチドをコードする核酸配列をそのプラスミド中に挿入するためのpDSP1プラスミド(9a)及び技術(9b)を示す。
【図10】pDSP62プラスミドを表し、一本鎖二量体の下流コピーのAB−ループ中へ配列を特異的に挿入するために、どのように合成オリゴヌクレオチドプライマーが設計され得るかを模式的に示す。オリゴヌクレオチドは、AB−ループに隣接するコート配列に対して完全にアニールするように設計され、それらの間に外来配列を挿入する。
【図11】図11aは、pDSP1プラスミドの核酸配列(配列番号1)を含む。図11bは、pDSP1(am)の核酸配列(配列番号2)を示す。図11cは、pDSP62プラスミドの核酸配列(配列番号3)を示す。図11dは、pDSP62(am)の核酸配列(配列番号4)を示す。pDSP1(am)及びpDSP62(am)のプラスミド配列は、アンバーコドンを導入するヌクレオチド置換によってのみ、pDSP1及びpDSP62と異なる。図11eは、M13K07のクロラムフェニコール耐性誘導体であるM13CM1の核酸配列(配列番号5)である。
【図12】ランダムなペプチド配列ライブラリーを生成するためにpDSP62及びM13CM1を利用する方法を示す。M13CM1は、pDSP62がそのM13起点から複製することを可能にするヘルパーウイルスである。従って、pDSP62を含む大腸菌細胞のM13CM1による感染によって、一本鎖の環状pDSP62の生成が可能になる。dut−、ung−細菌株中で増殖した場合、DNAは、DNA中に通常存在するdTTPのいくつかに代えてdUTPで置換される。ペプチドコード配列の部位特異的な挿入は、DNAポリメラーゼでの合成オリゴヌクレオチドの伸長によって達成され、その環は、DNAリガーゼの作用によって閉じられる。この得られた共有結合的に閉じられた環状DNAを、大腸菌のdut+、ung+株中にエレクトロポレーションによって導入し、そこでdUTP置換鎖が変性され、挿入の高頻度(通常は85〜90%)の組み込みを生じる[10]。
【図13】図13a及び図13bは、pP7Kプラスミド(13a)及びp2P7K32プラスミド(13b)を示す。これらのプラスミドは、翻訳抑制及びVLPアセンブリ試験によってPP7コートタンパク質AB−ループの挿入耐性を試験するためだけに、PP7 VLPプラットフォームの開発の間に利用された(実施例の項を参照)。それらは、後述されたアフィニティー選択のためのVLPライブラリーの構築に用いられる、pET2P7K32又はpDSP7と混同されるべきではない。
【図14】PP7コートタンパク質AB−ループ付近のヌクレオチド及びアミノ酸の配列を示し、また特異的なペプチド挿入物の挿入のために用いたプライマー配列を示す。多数の特異的及びランダムなペプチド配列を挿入して、AB−ループ挿入に対するPP7一本鎖二量体の一般的な耐性を試験し、特定の挿入されたペプチドの免疫原性を決定した。
【図15】PP7一本鎖コートタンパク質のAB−ループの挿入耐性を実証するために、p2P7K32中のランダムな配列ペプチドライブラリーの構築のために用いたスキームを図示する。この方法は、pDSP1のMS2一本鎖二量体中への挿入について図5bで示される方法と同様である。
【図16】本明細書に記載される種々のライブラリーそれぞれからの24個のクローンの細胞溶解液すべてのアガロースゲル電気泳動を示す。得られたクローンのほぼ100%が翻訳抑制をすることができ、このことは、それぞれのタンパク質が適切に折り畳まれた確率を示している。タンパク質が正確に折り畳まれたことをさらに確認するために、それぞれのVLPを形成する能力を、クローンのランダムな選択をこの図に示されるように電気泳動及びウエスタンブロットに供することによって評価した。それぞれのセットの上半分は、エチジウムブロマイド染色したゲルであり、下の半分は、複製のゲルのウエスタンブロットである。それぞれのセットにおける最も左側のレーンは、p2P7K32対照である。それぞれのクローンは、ランダムに生成された異なるペプチド配列を含み、ほぼ全てがVLPを生じるという事実によって、PP7コートタンパク質一本鎖二量体の高レベルの挿入耐性が実証される。
【図17】図17aはpETP7Kプラスミドを示し、図17bはpETP7K32プラスミドを示し、図17cはpDSP7プラスミドを示す。これらのプラスミドは、PP7コートタンパク質の高レベルの過剰発現のために生成された。pDSP7プラスミドは、PP7コートタンパク質の一本鎖二量体を含み、ここでは配列の二分の一が、AB−ループコード配列の近傍に十分な数のヌクレオチド置換を含んで、変異原性のオリゴヌクレオチドをアニールするために二つの半分を識別可能にする。ここで示される実施例では、上流の配列全体が、可能な最大数のサイレント変異を含むように改変される。pET2P7K32及びpDSP7は、MS2コートタンパク質プロデューサー、pDSP1及びpDSP62のPP7アナログとみなされるべきである。
【図18】pETP7K又はpET2P7K32を含む細菌中で生成されたVLPから抽出されたRNAのホルムアルデヒド/アガロースゲル上での電気泳動によって、それらが生成するVLPが、それらの合成を指向するmRNAをカプシドで包むことが示される。これは、逆転写及びPCRによってアフィニティー選択された配列を回収する手段を提供する。別のレーンは、同じプラスミドのインビトロでの転写によって生成されたRNAを含む。左の写真は、エチジウム染色ゲルを示す。右側は、PP7コートタンパク質センス鎖に特異的なオリゴヌクレオチドでプローブされた複製ゲルのブロットである。このプローブは、MS2又はQβコートタンパク質配列(示さず)のインビトロ転写に由来する同様の量のRNAとは反応できない。
【図19】PP7コートタンパク質中にクローニングされたいくつかの特異的なペプチド配列の列挙を示す。
【図20】抗−L2 mAb(RG−1)が、PP7 L2−VLPには結合するが、PP7 V3−VLPには結合しないことを示す。mAb RG−1の希釈物を、500ng/ウェルのL2−VLP又はV3−VLPと反応させた。結合は、西洋ワサビペルオキシダーゼ標識したヤギ抗−マウスIgG二次を用い、続いてABTSでの発色によって検出した。反応性は、405nmでの吸光度(OD405)の測定によって決定した。
【図21】V3ペプチド提示のPP7 VLPが免疫の際に抗−V3 IgG応答を誘導することを図示する。PP7 V3−VLPで、又は対照としてL2−VLPで免疫されたマウスでの抗−V3 IgG抗体応答を示す。マウスを、10μgのVLPと不完全フロイントアジュバントとを用いて3回免疫し、次いで血清を最終の追加免疫の2週後に収集した。別々のマウス7匹から得た希釈血清(6匹はV3−VLPで免疫し、1匹はL2−VLPで免疫した)を、HIVLAI由来のV3ループの一部を提示するペプチドとの反応性についてELISAによって試験した。結合は、西洋ワサビペルオキシダーゼ標識したヤギ抗−マウスIgG二次を用い、続いてABTSでの発色によって検出した。反応性は、405nmでの吸光度(OD405)の測定によって決定した。
【図22】抗Flag M2抗体でのMS2 VLPディスプレイを用いてのアフィニティー選択の結果を示す。抗Flag M2抗体のエピトープ配列は、これ以前に他者により十分に明らかにされている。4回のそれぞれにおけるいくつかのペプチドの配列は、選択の進行を明らかにするために示される。1回及び2回で見い出されるペプチドは、天然のエピトープの認識可能なエレメントを含むが、3回及び4回では類似性は明白であり、配列は野性型エピトープの配列に密接にマッチしている。実際に、4回目の配列は、十分に確立された繊維状ファージディスプレイの方法によって以前に得られた配列よりも密接に天然のエピトープにマッチする(「NEB Transcript,Summer,1006(New England Biolabsのウェブサイト、www.neb.comで入手可能)を参照)。
【図23】図23a〜23dは、pET2P7K32(配列番号6)、pET2P7K32(am)(配列番号7)、pDSP7(配列番号8)、及びpDSP7(am)(配列番号9)のプラスミドのヌクレオチド配列を示す。これらは、図11a〜11dに示される対応するMS2VLPプロデューサーのPP7コートタンパク質−産生アナログである。
【発明を実施するための形態】
【0042】
発明の詳細な説明
本発明によれば、当該分野の技術の範囲内の従来の分子生物学、微生物学及び組み換えDNA技術が使用され得る。このような技術は、文献中で詳細に説明される。例えば、Sambrookら、2001年,「Molecular Cloning:A Laboratory Manual」;Ausubel編集,1994年,「Current Protocols in Molecular Biology」第I〜III巻;Celis編集,1994年,「Cell Biology:A Laboratory Handbook」第I〜III巻;Coligan編集,1994年,「Current Protocols in Immunology」第I〜III巻;Gait編集、1984年,「Oligonucleotide Synthesis」;Hames & Higgins編集、1985年,「Nucleic Acid Hybridization」;Hames & Higgins編集、1984年,「Transcription And Translation」;Freshney編集、1986年,「Animal Cell Culture」;IRL Press,1986年,「Immobilized Cells And Enzymes」;Perbal,1984年,「A Practical Guide To Molecular Cloning」を参照のこと。
【0043】
ある範囲の値が示される場合、その範囲の上限と下限との間の、その状況が他を明確に示すのでない場合における下限の単位の十分の一までのそれぞれのその間の値、及びいずれかの他の言及された範囲における言及された値又はその間の値は、本発明の範囲内に包含されることが理解される。これらのより小さい範囲の上限及び下限は、独立してそのより小さい範囲内に含まれることができ、そしてその言及された範囲でいずれかの特に除外される限界が付された本発明の範囲内にもまた包含される。その言及される範囲が限界の一方又は両方を含む場合、それらの含まれる限界のいずれか又は両方を除外した範囲もまた本発明に含まれる。
【0044】
別段規定しない限り、本明細書において用いられるすべての技術及び科学的用語は、本発明が属する技術分野の当業者によって通常理解されるのと同じ意味を有する。本明細書に記載される方法及び材料と同様又は等価の任意の方法及び材料もまた、本発明の実行又は試験において用いられ得るが、好ましい方法及び材料がここに記載される。
【0045】
本明細書及び添付の特許請求の範囲で用いる場合、単数形(a、an及びthe)は、その状況が明らかに他を示すのでない限り、複数形の言及を包含する。
【0046】
さらに、次の用語は、下記の定義を有するものとする。
【0047】
本明細書において、「ポリヌクレオチド」という用語は、多量体形の任意の長さのヌクレオチド、リボヌクレオチド又はデオキシヌクレオチドのいずれかを指し、そして二本鎖及び一本鎖のDNA及びRNAの両方を包含する。ポリヌクレオチドは、例えばコード領域及び調節性配列(例えば、プロモーター又は転写ターミネーター)などの非コード領域といった、異なる機能を有するヌクレオチド配列を含んでもよい。ポリヌクレオチドは、天然の供給源から直接得られてもよいし、又は組み換え技術、酵素技術若しくは化学的技術の補助によって調製されてもよい。ポリヌクレオチドは、直線状であっても又は位相幾何学における環状であってもよい。ポリヌクレオチドは、例えば、発現ベクター若しくはクローニングベクターなどのプラスミド若しくはバクテリオファージベクターの一部、又はフラグメントであってもよい。
【0048】
制限エンドヌクレアーゼは、明確に規定された配列でDNAを切断する酵素である。それらは、例えば、同じ制限エンドヌクレアーゼでの消化によって生成される他のDNAフラグメントに、DNAリガーゼの作用を通じて容易に連結される特定のDNAフラグメントを生成するために、組み換えDNA技術で用いられる。本出願では、その認識配列がそれぞれGTCGAC、GGTACC、及びGGATCCであるSalI、KpnI、及びBamHIを含め、いくつかの特異的な制限エンドヌクレアーゼに対して言及する。
【0049】
本明細書において、「ポリペプチド」という用語は広義には、ペプチド結合によって一緒に連結される2つ以上のアミノ酸のポリマーを指す。「ポリペプチド」という用語はまた、ジスルフィド結合によって結合された2つ以上のポリペプチドを含む分子、又は多量体(例えば、二量体、四量体)として共有結合若しくは非共有結合的に結合されるポリペプチドの複合体を包含する。したがって、ペプチド、オリゴペプチド及びタンパク質という用語は、すべてがポリペプチドの定義内に包含され、そしてこれらの用語は交換可能に用いられる。これらの用語は、アミノ酸のポリマーの特定の長さを暗示するものでも、ポリペプチドが組み換え技術、化学的若しくは酵素的な合成を用いて生成されるか、又は天然に存在するかを暗示したり識別したりすることを意図するものでもないことが理解されるべきである。
【0050】
「一本鎖二量体」という用語は、通常は二量体のタンパク質であって、その2つのサブユニットが遺伝子的に(化学的に、共有結合を通して)単一のポリペプチド鎖に融合されたタンパク質を指す。具体的には、本発明では、MS2及びPP7コートタンパク質の両方の一本鎖二量体が構築された。これらのタンパク質のそれぞれは、天然には同一のポリペプチド鎖の二量体である。MS2及びPP7コートタンパク質二量体の両方において、1つのサブユニットのN末端は、コンパニオンサブユニットのC末端に対して物理的に近接して存在する(図1を参照)。一本鎖コートタンパク質二量体は、コートタンパク質のDNAコード配列を複製し、次いでそれらを尾−頭方式で互いに融合することによる組み換えDNA方法を用いて生成された。この結果は、単一のポリペプチド鎖であって、ここではコートタンパク質アミノ酸は2回存在し、上流コピーのC末端が、下流コピーのN末端に共有結合されている。通常の(野性型)の2つのサブユニットは、2つの鎖の間の非共有結合的な相互作用を通じてのみ会合される。一本鎖二量体では、これらの非共有結合的な相互作用が維持されるが、2つのサブユニットは、互いにさらに共有結合的に連結された。これは、タンパク質の折り畳み構造をかなり安定化し、そのタンパク質に対して上記のようなペプチド挿入の高い耐性を付与する。
【0051】
本出願は、コートタンパク質の「AB−ループ」に言及することが多い。RNAファージコートタンパク質は、保存された三級構造を保有する。MS2及びPP7コートタンパク質は、例えば、それぞれが図1で示されるMS2コートタンパク質の構造によって例示される構造を有する。それぞれのポリペプチド鎖は、AからGの文字で示される多数のβ鎖に折り畳まれる。β鎖のA及びBは、VLPの表面に露出されるヘアピンの頂部に2つの鎖を接続する3アミノ酸鎖のループを有するヘアピンを形成する。本出願に示されるように、ABループに挿入されるペプチドは、VLPの表面上に露出され、強力な免疫原性である。
【0052】
本明細書に記載されるアミノ酸残基は、「L」異性体型であることが好ましい。しかし、「D」異性体型の残基は、そのポリペプチドによって所望の機能が保持される限り、いずれのLアミノ酸残基とも代えることができる。NH2は、ポリペプチドのアミノ末端に存在する遊離のアミノ基を指す。COOHは、ポリペプチドのカルボキシ末端に存在する遊離のカルボキシル基を指す。
【0053】
「コード配列」の用語は、本明細書では、そのタンパク質生成物のアミノ酸配列を直接特定する核酸配列の部分として規定される。コード配列の境界は、一般に原核生物においては、リボソーム結合(又はシャイン・ダルガノ)部位及び翻訳開始コドン(通常はAUG)によって決定される。真核生物においては、オープンリーディングフレームの開始部位に位置するAUG開始コドン(mRNAの5’末端に通常は近い)、及びオープンリーディングフレームの末端、通常はmRNAの3’末端付近に位置してそれを特定する翻訳終結配列(ナンセンスコドン:UAG、UGA、又はUAAのうちの1つ)によって決定される。限定するものではないが、コード配列としては、DNA、cDNA、及び組み換え核酸配列などを挙げることができる。
【0054】
上記で簡略に注記したとおり、「終止コドン」又は「終結コドン」とは、翻訳の終結をシグナル伝達するメッセンジャーRNA内のヌクレオチドトリプレットである。タンパク質は、アミノ酸の固有の配列であって、メッセンジャーRNAにおけるほとんどのコドンは、成長しているタンパク質鎖へのアミノ酸の付加に相当する−終止コドンは、このプロセスの終結をシグナル伝達し、アミノ酸鎖を遊離する。標準的な遺伝子コードでは、3つの終止コドンがある:UAG(RNA中)/TAG(DNA中)(「アンバー」終止コドンとしても知られる)、UAA/TAA(「オーカー」終止コドンとしても知られる)、及びUGA/TGA(「オパール」又は「アンバー」終止コドンとしても知られる)。この主要な群に対するいくつかの改変が知られている。本発明における終止コドンの使用は、通常はタンパク質合成を終止又は終結させる。しかし、tRNA中には、終止コドンを認識することを可能にさせ、リボソームに終止コドンを読み飛ばさせ、終止コドンの下流のコードされるペプチドの合成を可能にする変異がある[11−13]。例えば、アンバー終止コドンを認識するtRNA中の変異は、翻訳がコドンを「読み飛ばし」て全長タンパク質を生じることを可能にし、それによって、正常な形態のタンパク質を回収し、終止コドンを「抑制する」。ほとんどの場合、終止コドンの抑制は、部分的にしか効果的でない−わずかな割合のリボソームしか終止コドンを読み飛ばすことができないことが多い。しかし、ある場合には、抑制はかなりより効果的であり得る。2〜3のサプレッサーtRNAは、簡単に高度な内因性の抑制効率を保有する。他の場合には、弱いサプレッサーは、単にそれをより高レベルで発現することによってさらに効率的にされ得る。本発明の特定の実施形態では、終止コドンは、その終止コドンの下流の転写単位内にコードされるペプチドの合成を制御するために転写単位内に組み込まれる。終止コドンを認識し、終止コドンの下流のペプチドの合成を可能にするtRNAの制御された合成を提供することによって、そのほとんどが異種ペプチドを含まないコートタンパク質の集団内に、異種ペプチドを含むコートタンパク質が生成され得る。野生型(異種ペプチドが存在しない)コートタンパク質を含む異種ペプチドのこの混合物からアセンブルされるこの得られたVLPでは、異種提示の価数がかなり低くなる。
【0055】
組み換え細胞の「異種」領域とは、より大きい分子と関連して見られることが全く無い、より大きい核酸分子内の核酸の特定可能なセグメントである。「異種」ペプチドとは、より大きいポリペプチドと関連して見られることが全く無い、ポリペプチドの特定可能なセグメントであるペプチドである。
【0056】
VLPの価数とは、粒子に提示される異種ペプチドのコピー数を指す。「低い価数」の異種ペプチド、好ましくは少なくとも4のペプチド単位の免疫原性ペプチドを示すウイルス粒子は、このウイルス粒子を含むコートポリペプチド二量体中で、1未満から最大で約10以上の異種ペプチドを提示する粒子である。低い価数を示すウイルス粒子は、異種ペプチド(好ましくは、野生型コートポリペプチド)を含まない複数のコートポリペプチド二量体、及び異種ペプチドを、好ましくはコートポリペプチドの下流サブユニットのA−Bループ内又は一本鎖二量体コートポリペプチドのカルボキシ末端に含む少数のコートポリペプチド二量体から形成され、従ってモザイクVLPを形成する。
【0057】
文脈内で用いられる「複製起点」とは、通常はDNA複製開始領域を特定することによってDNA合成に関与するDNA配列を指す。必要な因子(DNAポリメラーゼなど)の存在下で、複製起点は、それに会合するDNAを複製させる。例えば、ColE1複製起点(pDSP1などのようなプラスミド中で用いられる)は、細菌の染色体とは独立して複製する能力を、多くの通常用いられるプラスミドクローニングベクターに与える。別の例はp15A複製起点であり、それは本願の他の箇所で記載されるプラスミドpNMsupAで用いらる(図7を参照)。ファージM13由来の追加の複製起点がプラスミド上に存在すること(例えば、pDSP62と同様)によって、大腸菌細胞がいわゆるヘルパーファージ(例えば、本出願中で記載されるM13CM1)で感染される場合にその起点を用いて複製する追加の能力が付与され、これによって必須のタンパク質因子がもたらされる。M13は、細胞内で二本鎖の環状DNAとして複製するが、一本鎖環状型も生じ、これはファージ粒子内にそれをパッケージングする。これらの粒子によって、図12に図示される方法を用いたライブラリー構築のために有用である、pDSP62及びpDSP7(本出願中の他の箇所に記載される)のようなプラスミドのための一本鎖環状DNAの都合のよい供給源が得られる。
【0058】
「プロモーター配列」とは、細胞中でRNAポリメラーゼを結合し、下流(3’方向)のコード配列の転写を開始することができるDNA調節領域である。本発明を規定するためには、このプロモーター配列は、バックグラウンドを超える検出可能なレベルで転写を開始するために必要な最小数の塩基又はエレメントを含む。プロモーター内では、RNAポリメラーゼの結合を担うDNA配列、及び転写開始に必須であるあらゆる関連の因子が見られる。細菌中では、プロモーターは通常、−35及び−10のコンセンサス配列と、多かれ少なかれ特定の転写開始部位とからなる。真核生物プロモーターは、常にではないが、「TATA」ボックス及び「CAT」ボックスを含む場合が多い。細菌の発現ベクター(通常はプラスミド又はファージ)は、一般的には大腸菌のLactose、Arabinose、Tryptophan及びProBオペロン由来のプロモーター、並びにバクテリオファージ供給源由来の他のプロモーターなどの、天然の供給源由来のプロモーターを利用する。例としては、バクテリオファージλ、T7、T3及びSP6由来のプロモーターが挙げられる。
【0059】
細菌では、転写は、一般的に細菌のrho因子の作用をその活性のために必要とするか否かに応じてrho依存性及びrho非依存性(又は内因性)ターミネーターに分類される、特定の転写終結配列で通常は終わる。これらのターミネーターは、RNAポリメラーゼにその転写活性を停止させる部位を特定し、そのためそれらは主としてRNAの3’末端を規定するが、リボヌクレアーゼの続いての作用がRNAをさらに整える場合もある。
【0060】
「抗生物質耐性遺伝子」とは、所定の抗生物質に対する耐性を細菌に付与するタンパク質をコードする遺伝子を指す。例えば、カナマイシン耐性遺伝子は、薬物を改変して不活性化するホスホトランスフェラーゼの合成を指向する。カナマイシン耐性遺伝子がプラスミド(例えば、pDSP1)上に存在すれば、形質転換された細菌内のプラスミドの存在について選択する機序が提供される。同様に、クロラムフェニコール耐性遺伝子は、アセチル化を通じて抗生物質を不活性化するアセチルトランスフェラーゼ酵素を生成することによって、細菌が薬物の存在下で増殖することを可能にする。本出願では、クロラムフェニコール耐性を用いて、pNMsupA及びM13CM1の細菌内での維持を確実にする。
【0061】
「逆転写及びPCR」は、アフィニティー選択されたVLPの核酸配列を増幅する手段として本出願で示される。「逆転写」とは、RNA分子のDNAコピー(又はcDNA)が、逆転写酵素の作用によって生成されるプロセスを指す。本出願では、逆転写を用いて、アフィニティー選択されたVLPにおいてカプシドで包まれたRNA配列のDNAコピーを生成する。逆転写酵素は、プライマーがRNAにアニールされることを必要とする(以下を参照)。
【0062】
「PCR」という用語は、インビトロで特定のDNA配列の増幅のために用いられる技術である、ポリメラーゼ連鎖反応を指す。「PCRプライマー」という用語は、標的DNAにアニールすることを可能にし、それによりDNAポリメラーゼ(例えば、Taq DNAポリメラーゼ)がDNA合成を開始することができるDNA配列(通常は合成オリゴヌクレオチド)を指す。PCRプライマーの対をポリメラーゼ連鎖反応で用いて、DNAの2つの鎖のそれぞれでDNA合成を開始し、これによって、例えば図9b及び図15に図示されるように2つのプライマーの間でDNAセグメントを増幅する。
【0063】
逆転写及びPCRに用いられるプライマーの例をここに示す。
E2:5’ TCA GCG GTG GCA GCA GCC AA 3’−本出願に記載されるVLPによってカプシドで包まれるRNAの3’末端付近にアニールする;逆転写をプライムするために用いられる。
E3.2:5’ CGG GCT TTG TTA GCA GCC GG 3’−E2プライマー部位のすぐ上流の部位において、上述の逆転写によって生成されるcDNAの3’末端付近にアニールする;PCR反応中で3’−プライマーとして機能して、アフィニティー選択された配列を増幅する。
E2及びE3.2プライマーは、pDSP1、pDSP1(am)、pDSP62、pDSP62(am)、pET2P7K32、pET2P7K32(am)、pDSP7、及びpDSP7(am)と共通の配列を含み、従ってこれらの供給源のそれぞれに由来するVLPによってカプシドで包まれるRNAの逆転写及びPCRに有用である。PCRは、下に記載されるプラスミド特異5’プライマーのうちの1つとのE3.2の対合に依存する。簡略のために、pDSP1及びpDSP1(am)についてのプライマーのみを示すが、他のVLPで見られる配列に特異的な類似のプライマーが必要に応じて利用されることも理解されるべきである。
J2:5’ACT CCG GCC TCT ACG GCA AC 3’−pDSP1の一本鎖二量体配列の間の接合部で配列に特異的にアニールするプライマー。E3.2とのPCR反応では、一本鎖二量体の下流の半分を含むDNAセグメントが増幅される(AB−ループ中に挿入されたペプチドの配列を含む)。SalI及びBamHIによるPCR産物の消化によって、関連のプラスミドのSalIとBamHIとの間に挿入され得るフラグメントを生成する(例えば、図9aを参照)。
J2(アンバー):5’ AC TCC GGC ATC TAC TAG AAC TTT AC 3’−このプライマーは、上記J2と全く同じように機能するが、pDSP1(am)から生成される配列に特異的である。
【0064】
「発現制御配列」とは、別のDNA配列の転写及び翻訳を制御及び調節するDNA配列である。コード配列は、RNAポリメラーゼがコード配列をmRNA(これは、次にコード配列によってコードされるタンパク質に翻訳される)に転写する場合、細胞中で転写及び翻訳制御配列の「制御下」にある。転写制御配列とは、宿主細胞中でコード配列の発現をもたらす、例えばプロモーター、エンハンサー、ポリアデニル化シグナル、ターミネーターなどのDNA調節性配列である。翻訳制御配列は、メッセンジャーRNAの翻訳の効率を、通常はリボソーム結合及び翻訳開始の効率を制御することによって決定する。例えば、本出願の他の箇所で考察されるように、RNAファージのコートタンパク質は、周知のファージレプリカーゼの翻訳リプレッサーである。コートタンパク質は、感染した細胞中において十分に高濃度で蓄積することで、ファージレプリカーゼ遺伝子の翻訳開始領域(シャイン・ダルガーノ及びイニシエーターAUG)を含むRNAヘアピンに結合する。これは、複製からウイルスアセンブリまでの移行が起きるウイルスのライフサイクルのある時点で、リボソーム結合を妨げて、レプリカーゼの合成を遮断する。
【0065】
細胞は、外因性又は異種のDNAによって、このようなDNAが細胞の内側に導入された場合に、「形質転換される」。形質転換するDNAは、細胞のゲノムを構成している染色体DNAに組み込まれ(共有結合され)てもよいし、又は組み込まれなくてもよい。例えば原核生物、酵母及び哺乳動物細胞では、形質転換するDNAは、複製起点のプラスミド上に存在するおかげで通常は細菌染色体とは独立に複製する、プラスミドのようなエピソームエレメント上で維持されてもよい。真核生物細胞に関して、安定に形質転換された細胞は、形質転換するDNAが染色体中に組み込まれることで、染色体複製を通じて娘細胞によってDNAが受け継がれる細胞である。この安定性は、形質転換するDNAを含んでいる娘細胞の集団から構成される細胞株又はクローンを樹立する真核生物細胞の能力によって実証される。
【0066】
本明細書に開示される配列と同じアミノ酸配列を有するポリペプチドをコードするが、本明細書に開示される核酸に縮重する、本発明のポリペプチド(単数又は複数)をコードする核酸配列もまた本発明の範囲内であることが理解されるべきである。「縮重する(degenerate to)」とは、異なる3文字コドンを用いて個別のアミノ酸を特定することを意味する。
【0067】
本明細書において、「エピトープ」とは、ポリペプチドの抗原決定基を指す。エピトープは、エピトープに特有の空間的高次構造中に3個のアミノ酸を含み得る。一般的には、エピトープは、少なくとも5個のこのようなアミノ酸からなり、さらに一般的には、少なくとも8〜10個のこのようなアミノ酸からなる。アミノ酸の空間的高次構造を決定する方法は、当該分野で公知であり、例えばX線結晶学及び二次元核磁気共鳴が挙げられる。
【0068】
本明細書において、「ミモトープ(mimotope)」とは、本物の抗原性エピトープを模倣するペプチドである。場合によっては、そのアミノ酸配列は、もとの抗原のエピトープとある程度の類似を示し得るが、場合によっては、配列の類似性はほとんど又は全く存在しない。そのような場合、ミモトープは、異なるアミノ酸配列を用いるエピトープの3D構造を模倣する。糖鎖抗原のエピトープのような、非ペプチドのエピトープさえも模倣するミモトープが確認され得る。
【0069】
本明細書において、「コートタンパク質(単数又は複数)」という用語は、バクテリオファージ又はRNAファージのカプシドアセンブリ内に組み込まれ得る、このバクテリオファージ又はRNA−ファージのタンパク質(単数又は複数)を指す。
【0070】
本明細書において、「コートポリペプチド」とは、コートタンパク質の機能を保有し、さらに全長のコートタンパク質及び/又はその一本鎖改変体を包含する、コートタンパク質のポリペプチドフラグメントとして本明細書において規定される。
【0071】
本明細書において、「免疫応答」という用語は、B−リンパ球、T−リンパ球及び/又は抗原提示細胞の活性化若しくは増殖をもたらす体液性免疫応答及び/又は細胞性免疫応答を指す。しかし、ある場合には、免疫応答は、低い強度であって、本発明による少なくとも1つの物質を用いる場合にのみ検出可能になることがある。「免疫原性」とは、生きている生物体の免疫系を刺激するために用いられる作用物であり、それによりその免疫系の1つ以上の機能が増大されて、免疫原に向けられる。「免疫原性ポリペプチド」とは、細胞性及び/又は体液性の免疫応答を引き出すポリペプチドであり、アジュバントの有無にかかわらず、単独か又は担体に対して連結されている。好ましくは、抗原提示細胞は活性化され得る。
【0072】
本明細書において、「自己抗原」という用語は、宿主のDNAによってコードされるタンパク質、及び宿主のDNAによってコードされるタンパク質又はRNAによって生成される生成物を指す。さらに、2つの若しくはいくつかの自己分子の組み合わせから生じるか又は自己分子の画分に相当するタンパク質、及び高い相同性又は上記のように2つの自己分子を有する(>95%、好ましくは>97%、さらに好ましくは>99%)タンパク質もまた、自己とみなしてもよい。自己抗原の例としては、限定するものではないが、ErbB−2、アミロイド−β、免疫グロブリンE(IgE)、ガストリン、グレリン、血管内皮成長因子(VEGF)、インターロイキン(IL)−17、IL−23、IL−13、CCR5、CXCR4、神経成長因子(NGF)、アンジオテンシンII、TRANCE/RANKL及びMUC−1が挙げられる。
【0073】
本明細書において、「ワクチン」という用語は、本発明の組成物を含みかつ動物に投与され得る形態である処方物を指す。
【0074】
本明細書において、「バクテリオファージのウイルス様粒子」という用語は、バクテリオファージの構造を模倣し、非複製的及び非感染性であり、通常は感染性ウイルスとしてそのバクテリオファージの増殖に必要な1つ以上のウイルス遺伝子を欠いているウイルス様粒子(VLP)を指す。RNAバクテリオファージのVLPは、通常はまた、宿主に対するウイルスの結合又は進入を担う1又は複数のタンパク質をコードする1又は複数の遺伝子を欠いている。
【0075】
この定義はまた、前述の1又は複数の遺伝子が依然として存在しているが不活性であり、従ってやはりバクテリオファージの非複製的及び非感染性のウイルス様粒子ももたらす、バクテリオファージのウイルス様粒子を包含する。
【0076】
「RNAバクテリオファージコートタンパク質のVLP」とは、RNAバクテリオファージコートタンパク質の1つ以上のサブユニットの自己アセンブリから形成されるカプシド構造として規定され、通常はそのコートタンパク質自体のmRNAを含み、さらに任意で宿主RNAを含む。本出願の目的のために、RNAファージVLPは通常、180コピーの野生型コートタンパク質二量体から、若しくは90コピーの一本鎖二量体コートタンパク質から、又は可変数の野生型二量体及び1粒子当たり全部で90個の一本鎖二量体コートタンパク質を含むモザイクVLPとして、アセンブルされる。
【0077】
核酸分子は、発現制御配列が核酸配列の転写及び翻訳を制御及び調節する場合、その発現制御配列に対して「作動可能に連結される」か、又は「作動可能に会合される」。「作動可能に連結される」という用語は、発現されるべき核酸配列の前に適切な開始シグナル(例えば、ATG)を有すること、並びに発現制御配列の制御下で核酸配列の発現及びこの核酸配列によってコードされる所望の生成物の産生を可能にするために正確なリーディングフレームを維持することを包含する。組み換えDNA分子に挿入することが望ましい遺伝子が適切な開始シグナルを含まない場合、このような開始シグナルを遺伝子の前に挿入してもよい。
【0078】
「ストリンジェントなハイブリダイゼーション条件」という用語は、当業者に知られており、Current Protocols in Molecular Biology,John Wiley & Sons.N.Y.(1989年),6.3.1−6.3.6.に見られる。好ましくは、ストリンジェントなハイブリダイゼーション条件の非限定的な例は、約45℃で6×の塩化ナトリウム/クエン酸ナトリウム(SSC)中でのハイブリダイゼーション、続いて0.2.×SSC、0.1%SDSにおいて50℃で、好ましくは55℃で、さらに好ましくは60℃又は65℃での1回以上の洗浄である。
【0079】
ウイルス様粒子の生成
本発明は、ウイルス様ファージ粒子、並びにこれらの粒子をインビボ及びインビトロで生成するための方法に関する。通常、これらの粒子はインビボで生成されるが、これらの粒子の使用はインビトロ状況で適用されてもよく、このアプローチは、本発明によって予想される。本発明によって、実験の複雑性を増大して、相互作用選択に必要な時間を短縮することが可能になる。この方法は典型的には、インビトロでビリオンを産生する工程、及びこのビリオンを回収する工程を包含する。本明細書において、ビリオンを「インビトロで」生成するとは、細胞の外側で、例えば無細胞系で、ビリオンを生成する工程を指し、一方で「インビボで」ビリオンを生成するとは、細胞、例えば大腸菌又は緑膿菌細胞の内側でビリオンを生成する工程を指す。
【0080】
バクテリオファージ
本明細書で想定される系(システム)は、一本鎖RNAバクテリオファージの特性に基づく(The BacteriophagesでのRNA Bacteriophages, Calendar,RL編集.Oxford University Press.2005年を参照)。この群の公知のウイルスは、大腸菌、緑膿菌及びアシネトバクターのなど多様な細菌を攻撃する。それぞれ、極めて類似のゲノム構成、複製ストラテジー及びビリオン構造を保有している。特に、バクテリオファージは、一本鎖の(+)−センスRNAゲノムを含み、マチュラーゼ、コート及びレプリカーゼの遺伝子を含み、かつ小型(300オングストローム未満)の正二重面体のカプシドを有する。これらとしては、限定するものではないが、MS2、Qβ、R17、SP、PP7、GA、M11、MX1、f4、Cb5、Cb12r、Cb23r、7s及びf2 RNAバクテリオファージが挙げられる。
【0081】
PP7は、緑膿菌の一本鎖RNAバクテリオファージであって、MS2及びQβのような大腸菌ファージとは遠縁である。PP7バクテリオファージは、通常は緑膿菌に感染するが、PP7コートタンパク質が大腸菌中で図9及び図13に記載されるプラスミドのようなプラスミドから発現される場合、ウイルス様粒子が容易に生成される。PP7コートタンパク質は、特異的なRNA結合活性を備え、PP7レプリカーゼの翻訳開始領域に融合された配列の翻訳を抑制し得る、特異的なRNA−結合タンパク質であるということが確認された。これは、PP7の翻訳オペレーターを抑制するが、MS2又はQβファージのオペレーターは抑制しないためである。脱凝集したウイルス様粒子からのコートタンパク質の精製及びそのRNA結合活性を再構成するための条件は、確立されている。インビトロにおけるPP7オペレーターRNAの解離定数は、約1nMであることが確認された。コートタンパク質がレプリカーゼ−β−ガラクトシダーゼ融合タンパク質の翻訳を抑制する遺伝子系を用いて、PP7 RNAの結合に重要なアミノ酸残基が特定された。Peabodyら、Translational repression and specific RNA binding by the coat protein of the Pseudomonas phage PP72001,J.Biol.Chem.,Jun 22;276(25):22507−13.Epub 2001年4月16日。
【0082】
いくつかの一本鎖RNAバクテリオファージのコートタンパク質は、公知の翻訳リプレッサーであり、レプリカーゼリボソーム結合部位を含むRNAヘアピンを結合することによってウイルスレプリカーゼ合成を遮断する。RNAファージのX線構造決定によって、コートタンパク質アミノ酸配列の比較から明らかになる相同性が、三次構造において反映されることが示される。リプレッサー及びウイルス粒子の基本的構築ブロックの両方であるコートタンパク質二量体は、一緒になってRNAが結合される大きいβシート表面を形成する2つの結び付けられた単量体からなる。コートタンパク質のそれぞれは、異なるRNAを結合するために共通の構造的なフレームワークを用い、それによって特定のRNA−タンパク質認識の根拠を調査する機会を与える。そのコートタンパク質がMS2のアミノ酸配列に対してわずか13%のアミノ酸配列同一性しか示さない緑膿菌のRNAバクテリオファージである、PP7のコートタンパク質のRNA結合特性を本明細書に記載する。以下の知見もまた示される:(1)PP7のコートタンパク質は、翻訳リプレッサーである;(2)PP7レプリカーゼ翻訳開始部位を含むRNAヘアピンは、インビボ及びインビトロの両方でPP7コートタンパク質によって特異的に結合され、このことはこの構造が翻訳オペレーターであることを示す;並びに(3)RNA結合部位がコートタンパク質β−シートに存在する。この部位のマップは上に示した。
【0083】
例示のために、この群での特によく特徴付けられたメンバー、MS2のゲノムを利用し、これはわずか4個のタンパク質(そのうち2つがビリオンの構造的な成分である)をコードする一本鎖の(+)−センスRNAの3569ヌクレオチド長である。ウイルス粒子は、180コピーのコートタンパク質から形成される正二重面体のカプシド及び1分子のマチュラーゼタンパク質と一緒になった1分子のRNAゲノムから構成される。コートタンパク質はまた、特異的なRNA結合タンパク質である。アセンブリは、コートタンパク質が、レプリカーゼシストロンの5’末端に近いRNAヘアピンである、その特異的な認識標識と会合する場合に開始され得る(その全体が本明細書に援用される、2009年2月26日公開の米国特許出願公開第20090054246の図1B及び配列番号1を参照)。次いでウイルス粒子は、細胞がウイルス溶解タンパク質の影響下で破裂する時に培地中に遊離される。感染性ウイルスの形成は、少なくとも3つの構成要素、すなわちコートタンパク質、マチュラーゼ及びウイルスゲノムRNAを必要とするが、実験では、正二重面体のカプシドのシェルのアセンブリに必要な情報は、コートタンパク質自体の中に完全に含まれることが示される。例えば、精製されたコートタンパク質は、RNAの存在によって刺激されるプロセスにおいてインビトロでカプシドを形成し得る(Beckettら、,1988年,J.Mol Biol 204:939〜47)。さらに、細胞中でプラスミドから発現されるコートタンパク質は、インビボでウイルス様粒子にアセンブルする(Peabody,D.S.,1990年,J Biol Chem 265:5684〜5689)。
【0084】
コートポリペプチド
コード領域によってコードされるコートポリペプチドは、典型的には、少なくとも120、好ましくは少なくとも125アミノ酸長であり、約135アミノ酸長以下、好ましくは130アミノ酸長以下である。本質的にいずれかの一本鎖のRNAバクテリオファージ由来のコートポリペプチドが用いられ得ることが予想される。コートポリペプチドの例としては、限定するものではないが、MS2コートポリペプチド(例えば、米国特許出願公開第20090054246号の配列番号2を参照)、R17コートポリペプチド(例えば、ジーンバンク・アクセッション番号(Genbank Accession No.)P03612を参照)、PRR1コートポリペプチド(例えば、ジーンバンク・アクセッション番号ABH03627を参照)、frファージコートポリペプチド(例えば、ジーンバンク・アクセッション番号NP_039624を参照)、GAコートポリペプチド(例えば、ジーンバンク・アクセッション番号P07234を参照)、Qβコートポリペプチド(例えば、ジーンバンク・アクセッション番号P03615を参照)、SPコートポリペプチド(例えば、ジーンバンク・アクセッション番号P09673を参照)、f4コートポリペプチド(例えば、ジーンバンク・アクセッション番号M37979.1を参照)及びPP7コートポリペプチド(例えば、ジーンバンク・アクセッション番号P0363 0を参照)が挙げられる。
【0085】
PP7コートポリペプチドの例としては、限定するものではないが、RNAヘアピンを有する複合体でのPP7コートタンパク質二量体の種々の鎖が挙げられる(例えば、ジーンバンク・アクセッション番号2QUXR;2QUXO;2QUX_L;2QUX_I;2QUX_F;及び2QUX_C)。また、本明細書の実施例1、及びPeabodyら、RNA recognition site of PP7 coat protein,Nucleic Acids Research.2002年,第30巻,第19号、4138〜4144.[14,15]も参照のこと。
【0086】
本発明において有用なコートポリペプチドとしてはまた、上記で開示されたコートポリペプチドの1つ以上と類似性を有するポリペプチドが挙げられる。この類似性は、構造的類似性と見なされる。構造的類似性は、2つのアミノ酸配列(すなわち、候補アミノ酸配列及びアミノ酸配列)の残基を整列させて、それらの配列の長さに沿った同一のアミノ酸の数を最大限に利用することによって決定され得る;いずれか又は両方の配列におけるギャップは、同一のアミノ酸の数を最大限に利用するために整列させることにおいて許されるが、それぞれの配列におけるアミノ酸は、それにかかわらずその適切な順序のままでなければならない。候補アミノ酸配列とは、例えば米国特許出願公開第2009/0054246号の配列番号2に示されるアミノ酸配列と比較されるアミノ酸配列である。候補アミノ酸配列は、一本鎖RNAウイルスから単離されることも、組み換え技術を用いて産生されることも、又は化学的若しくは酵素的に合成されることもできる。好ましくは、2つのアミノ酸配列は、Tatusovaら(FEMS Microbial Lett 1999年,174:247〜250)に記載されたような、またhttp://www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html.で入手可能である、GCGパッケージ(バージョン10.2,Madison WI)でのBESTFITアルゴリズム、又はBLAST2サーチアルゴリズムのBlastpプログラムを用いて比較される。好ましくは、全てのBLAST 2サーチパラメーターのデフォールト値を用い、これはマトリックス=BLOSUM62;オープンギャップペナルティー=11、エクステンションギャップペナルティー=1、ギャップ xドロップオフ=50、期待値=10、ワードサイズ=3、及び任意でフィルターオンを含む。BLASTサーチアルゴリズムを用いる2つのアミノ酸配列の比較において、構造的類似性は、「同一性」と呼ばれる。
【0087】
また好ましくは、コートポリペプチドとしては、上記で開示されるアミノ酸配列の1つ以上に対して、少なくとも80%のアミノ酸同一性、少なくとも85%のアミノ酸同一性、少なくとも90%のアミノ酸同一性、又は少なくとも95%のアミノ酸同一性を有するアミノ酸配列を有するポリペプチドが挙げられる。好ましくは、コートポリペプチドは活性である。コートポリペプチドが活性であるか否かは、カプシドを形成して一本鎖RNA分子をパッケージングするポリペプチドの能力を評価することによって決定され得る。このような評価は、インビボ又はインビトロの系を用いて行われることができ、このような方法は当該分野及び通常作業において公知である。あるいは、ポリペプチドは、言及されたコートポリペプチド及び/又は機能的活性のように類似の3次元構造を有する場合に、構造的に類似であるとみなされ得る。
【0088】
コートポリペプチド又はポリペプチド中に挿入された異種ペプチド配列は、ランダムなペプチド配列であり得る。特定の実施形態では、ランダム配列は配列Xaanを有し、ここでnは、少なくとも4、少なくとも6、又は少なくとも8及び20以下、18以下、又は16以下であり、そしてそれぞれのXaaは、独立したランダムなアミノ酸である。あるいは、ペプチドフラグメントは、規定された配列を有し、公知の機能(例えば、抗原性、免疫原性)を保有し得る。異種配列は、コートポリペプチドのアミノ末端、コートポリペプチドのカルボキシ末端に存在してもよいし、又はコートポリペプチド内の他の箇所に存在してもよい。好ましくは、異種配列は、挿入配列がカプシドの外面に発現されるようにして、コートポリペプチド中のある位置に存在する。特定の実施形態において、及び本明細書の後述の実施例に記載されるように、ペプチド配列は、上述のコートポリペプチドのABループ領域に挿入されてもよい。このような位置の例としては、例えば、コートポリペプチドのアミノ酸11〜17、又はアミノ酸13〜17の直後のコートポリペプチド中への挿入配列の挿入が挙げられる。最も特殊な実施形態では、異種ペプチドが、MS−2のアミノ酸11〜17、又は特に13〜17に対応する部位に挿入される。
【0089】
本発明による特定の実施形態では、異種ペプチドが、以下に対応する部位に挿入される:
(a)MS−2、R17及びfrコートポリペプチドのアミノ酸11〜17、又は特に13〜17;
(b)GAコートポリペプチドのアミノ酸10〜16
(c)Qβ及びSPコートポリペプチドのアミノ酸10〜17;
(d)PP7コートポリペプチドのアミノ酸8〜11、及び
(e)PRR1コートポリペプチドのアミノ酸9〜17。
【0090】
あるいは、異種ペプチドは、コートポリペプチドのN−末端に挿入されても、又はC−末端に挿入されてもよい。
【0091】
異種ペプチドは、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択できる。
【0092】
この異種ペプチドとしては、限定するものではないが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択されるペプチドが挙げられる。
【0093】
構造的に類似のコートポリペプチドにおいて対応する位置を決定するために、この構造的に類似のコートポリペプチドのアミノ酸配列を、上記で特定されたような名前を挙げられたコートポリペプチドの配列と整列させる。例えば、MS−2コートポリペプチドに対して構造的に類似のコートポリペプチドの対応する位置を、配列番号2(本明細書に援用される米国特許出願公開第2009/0054246号の)と整列させる。この整列から、配列番号1(やはり米国特許出願公開第2009/0054246号の)の所定の位置に対応する他のコートポリペプチドにおける位置を決定し得る。
【0094】
特定の実施形態では、コートポリペプチドは、上流及び下流のサブユニットを含む一本鎖二量体である。それぞれのサブユニットは、機能的なコートポリペプチド配列を含む。異種ペプチドは、本明細書において上述された部位で上流及び/又は下流のサブユニットに挿入されることができ、例えば、好ましくは下流サブユニットのA−Bループ領域に挿入されることができる。特定の実施形態では、このコートポリペプチドは、本明細書に援用される米国特許出願公開第2009/0054246号の配列番号12に示される配列を有し得るMS2コートポリペプチドの一本鎖二量体である。
【0095】
特定の実施形態では、コートポリペプチドは、上流及び下流のサブユニットを含む一本鎖二量体である。それぞれのサブユニットは、機能的なコートポリペプチド配列を含む。異種ペプチドは、本明細書において上述された部位で上流及び/又は下流のサブユニットに挿入されることができ、例えば、下流サブユニットのA−B領域に挿入されることができる。特定の実施形態では、このコートポリペプチドは、PP7コートポリペプチドの一本鎖二量体である。
【0096】
転写単位の調製
本発明の転写単位は、発現調節性領域(例えばプロモーター)、コートポリペプチド
をコードする配列、及び転写ターミネーターを含む。RNAポリヌクレオチドは、任意でコート認識部位(「パッケージングシグナル」、「翻訳オペレーター配列」、「コート認識部位」とも呼ばれる)を含んでもよい。あるいは、転写単位は、翻訳オペレーター配列を含まなくてもよい。プロモーター、コード領域、転写ターミネーター、及び存在する場合はコート認識部位は、一般的に作動可能に連結される。「作動可能に連結される」又は「〜と作動可能に会合される」とは、そのように記載された構成要素が、それらの意図する方法で機能することを可能にする関係にある並列状態を指す。調節性配列は、コード領域の発現が、調節性配列と両立する条件下で達成されるような方法で連結される場合に、そのコード領域と「作動可能に連結される」か、又は「作動可能に会合される」。コート認識部位は、存在する場合、それが意図する方法で機能するならば、RNAポリヌクレオチド内の任意の位置でよい。
【0097】
本発明は、いずれかの特定のプロモーターの使用によって限定されず、広範な種々のプロモーターが公知である。本発明で用いられるプロモーターは、構成的プロモーターであっても、誘導プロモーターであってもよい。好ましいプロモーターは、コートポリペプチドをコードするコード領域によってコードされた高レベルのRNAを駆動することができる。このようなプロモーターの例は、当該分野で公知であり、例えばlacプロモーターT7、T3、及びSP6プロモーターなどが挙げられる。
【0098】
本明細書に記載されるコートポリペプチドをコードするコード領域のヌクレオチド配列は、容易に決定される。本明細書に記載されるコートポリペプチドの1つをコードするヌクレオチド配列の分類の例は、配列番号3(本明細書に援用される米国特許出願公開第2009/0054246の)のヌクレオチド4080〜4470である。これらの分類のヌクレオチド配列は、大型だが有限であり、この分類のそれぞれのメンバーのヌクレオチド配列は、標準的な遺伝子コードを参照して当業者によって容易に決定され得る。
【0099】
さらに、RNAバクテリオファージ一本鎖コートポリペプチドのコード配列は、異種ペプチドの挿入のための部位、及びその異種ペプチドのコード配列自体を含む。特定の実施形態では、その異種ペプチドの挿入のための部位は、制限酵素部位である。
【0100】
特定の実施形態では、コード領域は、コートポリペプチドの一本鎖二量体をコードする。最も特殊な実施形態では、コード領域は、改変された一本鎖コートポリペプチド二量体をコードし、ここでこの改変は、挿入部位に少なくとも4個のアミノ酸のコード配列の挿入を含む。このような転写単位の特定の実施形態の概略図は、米国特許出願公開第2009/0054246号の図3に示される。転写単位は、lacプロモーターのような細菌のプロモーターを含んでもよく、又はT7プロモーターのようなバクテリオファージプロモーター、及び任意でT7転写ターミネーターを含んでもよい。
【0101】
プロモーター及び融合ポリペプチドをコードするコード領域を含むことに加えて、RNAポリヌクレオチドは、通常は転写ターミネーター及び任意でコート認識部位を含む。コート認識部位は、RNAとして存在する場合、ヘアピンを形成するヌクレオチド配列である。これはまた、当該分野では、翻訳オペレーター、パッケージングシグナル、及びRNA結合部位とも呼ばれる。限定する意図はないが、この構造は、翻訳リプレッサー(例えば、コートポリペプチド)によって認識される結合部位として機能し、RNAパッケージングを開始すると考えられる。コート認識部位のヌクレオチド配列は当該分野で公知であり、これには例えば配列番号1(米国特許出願公開第2009/0054246号の図1Bを参照)のヌクレオチドが挙げられる。他のコート認識配列は、一本鎖RNAバクテリオファージR17、GA、Qβ、SP、及びPP7において特徴付けられており、そして当業者にとって容易に利用可能である。本質的に任意の転写ターミネーターが、それがプロモーターとともに機能するならば、RNAポリヌクレオチド中で用いられ得る。転写ターミネーターは、当業者に公知であり、容易に利用可能であり、そしてごく普通に用いられる。
【0102】
合成
後にさらに詳細に記載されるように、本発明のVLPは、特に転写単位が細菌のプロモーターを含むならば、転写単位を細菌に導入することによってインビボで合成され得る。あるいは、VLPは、カップリングされた無細胞の転写/翻訳システムにおいてインビトロで生成されることができる。
【0103】
異種の物質をカプシドで包むVLPのアセンブリ
合成の間、VLPは通常、VLPが翻訳によって生成されるメッセンジャー−RNAと会合する。これは、本出願に記載されるシステムのアフィニティー選択能力にとって重要である。しかし、いくつかの他の適用(例えば、薬物又は造影剤の標的化送達)では、VLPへの他の物質の導入が所望され得る。これらのVLPは、異種物質の存在下でインビトロのVLPアセンブリ反応を行うことによってアセンブルされ得る。具体的には、精製されたコートタンパク質サブユニットは、変性剤(通常は酢酸)で脱凝集されたVLPから得られる。タンパク質サブユニットは、異種の物質と混合される。特定の実施形態では、この物質は、VLPの内部に対してある程度の親和性を有し、好ましくは負に荷電されている。
【0104】
別の方法は、翻訳オペレーターの合成RNAバージョンに対して異種の物質を結合する工程を含む。インビトロのアセンブリ反応の間、RNAは、その認識部位に緊密に結合し、得られたVLP中に効率的に組み込まれ、それと一緒に外来物質を保持する。
【0105】
別の実施形態では、この物質は、VLP表面に自然に存在する細孔を通じてVLP中に受動的に拡散される。特定の実施形態では、この物質はこれらの細孔を通過するのに十分小さく(MSでは、それは直径約10オングストロームである)、かつVLPの内部に高い親和性を有する。
【0106】
VLP集団
上記のとおり、本発明は、VLP集団又はライブラリーに関する。本明細書における「集団」及び「ライブラリー」という用語は、交換可能に用いられ、従って同義であるとみなされる。1つの特定の実施形態では、ライブラリーはランダムなライブラリーであってもよい;別の実施形態では、このライブラリーは、抗原性ポリペプチド由来のフラグメントのライブラリーである、抗原フラグメントライブラリーである。
【0107】
ランダムライブラリー(集団)
ペプチドをコードするオリゴヌクレオチドが調製され得る。1つの特定の実施形態では、特定のアミノ酸をコードするトリプレットは、組成NNS(ここでNはA、G、C又はTであり、SはG又はTである)か、あるいはNNY(ここでNはA、G、C、又はTであり、YはC又はTである)を有する。複数のトリプレットがコートタンパク質遺伝子に挿入されて、タンパク質生成物への対応するペプチドの挿入がもたらされる。終止コドンの存在を最小化するために、ペプチドライブラリーは、天然のアミノ酸組成をより正確に反映するように設計された特注の三ヌクレオチドのホスホラミダイト混合物(Glen Research,Inc.から入手可能)から合成され、終止コドンを完全に欠いているオリゴヌクレオチドを用いて構築されることができる。
【0108】
コートタンパク質へのこのようなランダム配列の挿入によって、VLPの集団(又はライブラリー)の合成がもたらされ、それぞれの粒子は、その表面上で異なるペプチドを提示する。このような集団は、極度に大きくてもよく、数十億の個々のメンバーからなってもよい。実質的に任意のレセプター(例えば、モノクローナル抗体)に特異的なリガンドがこのようなライブラリーに存在することが一般に観察されるが、それらは通常、集団全体のわずかな画分に相当する。アフィニティー選択とその後の増幅(本発明の場合は、カプシドで包まれたRNAの逆転写及びPCRによる)によって、このような希少な種の回収、分析及び開発が可能になる。
【実施例】
【0109】
本発明は、本発明の例示として示される以下の非限定的な例を参照することによってさらによく理解され得る。以下の実施例は、本発明の好ましい実施形態をさらに詳細に例示するために示しているが、決して本発明の範囲を限定するものとして解釈されるべきではない。
【0110】
低い価数の異種ペプチドを示すVLPの生成
本出願は、MS2 VLPのペプチドディスプレイのための技術の開発における近年の進歩を詳述しており、これはランダム配列ペプチドライブラリーの容易な生成のため及びペプチドディスプレイの価数の制御のための新規なプラスミドベクターの説明を含む。これは、米国特許出願公開20090054246号に記載される方法の進歩を表す。
【0111】
本発明は、繊維状ファージディスプレイの能力に類似するアフィニティー選択能力を有するMS2ウイルス様粒子(VLP)プラットフォームの開発に関する。本発明は、部分的には、RNAバクテリオファージのコートタンパク質の、外来のペプチドを提示する能力及びそれらの合成のためにテンプレートとして働く同じmRNAをカプシドで包む能力の両方に基づく。このプロセスは、VLPを形成する細菌細胞中のプラスミドからのコートタンパク質のランダムなペプチド融合物の合成を必要とする。VLPは、細胞から抽出されて、特定の抗体に対する結合のためのアフィニティー選択に供される。最終的に、RNAは、選択されたVLPから抽出されて、カプシドで包まれた配列を回収及び増幅するために逆転写及びPCRに供され、この配列が次にクローニングされて細菌中に再導入され、ここでそれらの配列は、別の回の合成、アセンブリ及び選択のためのテンプレートとして機能する。このプロセスは、必要な回数だけサイクルが繰り返され、最後に、選択された配列を選択されたVLPの高レベルの細菌発現のためにクローニングする。
【0112】
いくつかの他の用途の中でも、ワクチン開発を容易にするために開発されている本発明の方法は、単一のプラットフォームでは一緒に利用できなかった重要な特徴を有する。第一に、本発明の方法は、ウイルス様粒子(VLP)の表面上の高密度反復性アレイとして外来ペプチドを提示することによって高い免疫原性を確実なものにする。これは、外来抗原に対する活発な免疫応答を生じるだけでなく、免疫寛容を克服し、さらに自己抗原に対する抗体を誘導することもできる。第二に、ファージディスプレイと類似のプロセスでは、本発明のプラットフォームは、複雑なランダム配列ライブラリーからアフィニティー選択された配列の回収及び増幅を可能にする。従って、本発明は、単一のプラットフォームにおいて、価数制限及びアフィニティー選択によって関連のエピトープを特定する能力と、その次に免疫系に対してそれらのエピトープをワクチンとして示す能力とを合わせ持つ。エピトープは、抗体標的に対するアフィニティー選択によって特定され、最適化され、次いでプラットフォームを変化することなく、免疫系に対して直接ワクチンとして提示され得る。このペプチドは、それらのもともとの選択の間に存在する構造的連結を変更することなく高密度で提示され、これによって天然のエピトープの忠実な分子模倣物が単離され、それらの最適構造が免疫プロセスの間に維持され、関係する抗体応答が誘導される確率が増大する。
【0113】
MS2 VLPでの高い複雑性のランダム配列ペプチドライブラリー構築物のためのプラスミドベクター
MS2 VLPのランダム配列ペプチドライブラリーを利用する最初の実験は、T7転写単位を有するアンピシリン耐性プラスミドであるpET3dの単純な誘導体で実施された。これは、高い複雑性のライブラリーの作成に便利な最適なシステムよりはいくらか劣ると判明したが、そのため本発明者は、重要であると思われる多数の最適特徴を組み合わせた一連のベクターを作成した。
これらとしては以下が挙げられる:
【0114】
A.AB−ループでの挿入のために従来のクローニング部位を有する一本鎖二量体。
pDSP1(図1)は、AB−ループ−コード配列又はその近位において、例えば1つだけのSalI及びKpnI制限部位を含むように改変された、一本鎖二量体のコード配列を発現する。典型的には、BamHI部位は、コート配列の下流に含まれる。この形態の二量体は、外来配列のAB−ループ中への簡易なクローニングを容易にする。これらの部位を1つだけにするため、ベクター中、上流コート配列中及びプラスミド配列中で、多数のSalI及びKpnI部位を破壊する必要があった。
【0115】
B.カナマイシン耐性。pDSP1は、カナマイシンに対する耐性を付与する。これによって、形質転換されてない細胞の最初の高いバックグラウンドに対して、液体培養物中の形質転換された細菌の選択を可能にすることにより、ライブラリー構築がかなり容易になる。最初の生成プラスミドベクターによって付与されたアンピシリン耐性は、この目的に対して不適当であった。これは、形質転換された大腸菌による抗生物質の急速な分解によって、培養物中で驚くほど短時間後に選択の損失が生じるからである。これは形質転換されていない細胞の過剰増殖を可能にした。当然ながらこの細胞は、効率的な形質転換方法が利用される場合でさえ、通常はこの集団の大部分に相当する。
【0116】
C.M13の複製起点。今日に至るまで、本発明者は、PCRプライマーを利用して一方の末端にランダムな配列が結合されたフラグメントを生成する手順によって、ランダム配列ライブラリーを構築している。次いで、このフラグメントを適切な制限エンドヌクレアーゼによって消化して、上述される1つだけの部位(典型的にはSalIとBamHIとの間、図1を参照)においてpDSP1中にクローニングする。このような方法を用いて、本発明者は、最大109までの個々の組み換え体からなるライブラリーを作成した。しかし、このサイズのライブラリーは、これらの方法を用いて構築するには不便である;かなり大きいライブラリーの構築は、典型的なライゲーション反応で見られるよりもさらに高収率の組み換えDNAを効果的に生じる方法によって容易にされる。具体的には、本発明者は、部位特異的な突然変異誘発(2)のための方法のバリエーションを利用するが、この方法は、1011個の複雑性の範囲で繊維状ファージライブラリーを生成するために他者によって既に用いられている(3)。この方法は、dUTP置換一本鎖環状テンプレート上のミスマッチのプライマーの伸長に依拠する(dUによるdTの置換は、CJ236又はBW313のようなdut−、ung−宿主におけるテンプレートDNAの成長によって達成される)。ランダム配列ペプチドライブラリーを作成するために、プライマーは、AB−ループのいずれかの側でのコート配列に対する相補的な配列が、それぞれの側に配置されたランダムなDNA配列を含む。このプライマーは、DNAポリメラーゼで伸長されて、共有結合的に閉鎖された二本鎖環が、DNAリガーゼの作用によって生成される。ung+株の形質転換(例えば、エレクトロポレーションによる)は、変異鎖の選択的増殖のための強力な選択をもたらし、そして挿入された配列を保有する組み換え体を高収率で生じる。これらの挿入の変異原性反応は、エレクトロポレーションによっておよそ1011個以上の個々の組み換え体を容易に生成するのに十分な比較的大量のDNA(例えば、20ug)で行われることができる。
【0117】
一本鎖DNAの生成を容易にするために、M13の複製起点がプラスミド中に導入され、そしてM13起点の方向に応じて、pDSP61及びpDSP62と呼ばれる(図2)。また、M13CM1と呼ばれるヘルパーファージを構築した。これは、クロラムフェニコールでカナマイシン耐性を置き換えるM13K07の誘導体であり、これによってカナマイシン耐性を付与するM13cm1ヘルパー及びpDSP6の同時の存在が選択可能になる。プラスミドを含むdut-、ung-株(例えば、CJ236)のM13ヘルパーファージによる重複感染によって、dUTP−置換一本鎖DNAの容易な生成がもたらされる。
【0118】
D.合成の「コドンジャグリングされた」コート遺伝子。ライブラリー構築のためにプライマー伸長突然変異を用いる要望によって、新しい問題が導入され、いわゆる「コドンジャグリングされた」コート配列をpDSP6へ導入することが必要になった。本発明のペプチドディスプレイ法は、外来のペプチドを一本鎖二量体の2つのAB−ループのうちの一方にのみ特異的に導入する能力に依拠する。上記のスキームでは、変異原性プライマーは、通常は一本鎖二量体の両方の半分での配列にアニールして、二重挿入が生じる。しかし、AB−ループの両方における同時挿入は、タンパク質折り畳みの失敗を高頻度に生じる。この理由のために、コートタンパク質のコドンジャグリングされたバージョンの合成が完成された。このコドンジャグリングされたバージョンは、可能な最大数のサイレントヌクレオチド置換を一本鎖二量体の上流側半分に導入し、これによって野生型コートタンパク質のアミノ酸配列を有するポリペプチドを生じる。しかし、多くの変異の存在によって、ジャグリングされた配列は、変異原性のオリゴヌクレオチドに対して効率的にアニールできなくなる。この方法では、挿入は、一本鎖二量体の下流側半分を標的にする。
【0119】
それぞれpDSP1(am)及びpDSP62(am)と呼ばれるpDSP1及びpDSP62の改変体は、pDSP1及びPDSP62の特徴の全てを保持するが、部位指向性の突然変異によって改変されて、アラニンコドン(一本鎖二量体中のコートタンパク質の下流のコピーの最初のアミノ酸を特定する)が、UAG(アンバーコドンとして公知の特異的なナンセンスコドン又は終止コドン)に変換される。この終止コドンの抑制によって、単一のmRNAから、野性型及び一本鎖二量体のコートタンパク質の両方の合成が可能になる。これによって、後でさらに詳細に記載されるような平均のペプチドディスプレイ価数を制御する能力が得られる。
【0120】
ナンセンスコドンの読み飛ばしは、通常は抑制されている特定の終止コドンに特異的なサプレッサーtRNAの存在を必要とする。3つの終止コドン全てに活性なサプレッサーtRNAが記載されたが、分子生物学者には周知である。本明細書に記載される研究は、アンバーコドンを利用し、従ってこの場合、アラニンを挿入するために特別に設計されたアンバー−サプレッサーtRNAの使用を必要とする。しかし、他の終止コドン(UAA及びUGA)のそれぞれについて特異的なサプレッサーtRNA、及び種々のアミノ酸を挿入することが同様に利用され得ることが理解されるべきである。
【0121】
細菌細胞中でサプレッサーtRNAを生成するために、pNMsupAと呼ばれるプラスミドを生成した(図3を参照)。これは、pACYC18の誘導体であり、p15Aプラスミド不適合性群由来の複製起点を含む。これはまた、クロラムフェニコールに対する耐性も提供する。このことは、このプラスミドが、ColE1起点を有しかつカナマイシンに対する耐性を付与するpDSP1(am)又はpDSP62(am)をすでに含んでいる細菌細胞では、安定に維持され得るということを意味する。プラスミドpNMsupAはまた、lacプロモーター、及び合成のサプレッサーtRNA遺伝子が挿入されたpUC18のポリリンカー領域を含む。tRNA遺伝子の転写は、lacプロモーターの制御下であり、このことは、サプレッサーtRNAの合成が、lacオペロンの誘導因子(例えば、IPTG)によって制御されるということを意味する。合成tRNA遺伝子の配列は、Kleinaら(4)によって以前に記載された配列を基にしており、下に示される。これには、pNMsupA中でそのクローニングを容易にするEcoRI及びPstI部位が隣接する。
【0122】
【化1】
【0123】
サプレッサーtRNAのレベルを変更することは、ナンセンス抑制のレベルを変更するためにも知られている。pDSP1(am)又はpDSP62(am)からの一本鎖二量体の合成のレベルをこのように変更することによって、1個のVLP当たりに提示されるペプチドの平均数を制御することが可能になり、これによって広い範囲にわたって提示価数の制御が可能になると考えられた。pNMsupAはlacプロモーターを使用するが、サプレッサーtRNA合成の制御のために種々のプロモーターが用いられ得ること、及びいくつか(例えば、プロピオン酸塩オペロン(5)のプロモーター)は、実際に、それらのそれぞれの誘導因子の種々の濃度に対して、適切に管理された、段階的な応答を良好に提供できるということは明らかであろう。
【0124】
III.提示価数の制御
MS2 VLPで提示されたペプチドの数を制御する手段があることが望ましい。多価数のMS2 VLPディスプレイ(1粒子当たり90コピーのエピトープ)によって、抗体について高い内因性の親和性を有するペプチドを提示するVLPを、低い内因性の親和性を有するが、複数の弱い相互作用をする能力のおかげで依然として緊密に結合するものから、区別することが困難になるはずである(すなわち、結合活性対親和性)。これは、繊維状ファージディスプレイについての周知の問題であり、ここでは高い親和性の相互作用の選択は、通常は低い提示価数の使用を必要とする。最高の親和性のペプチドを選択する場合に、適切な分子模倣物の発見はかなり増加される。以下は、選択のストリンジェンシーを改善するために提示価数を減少するための方法の説明である。
【0125】
価数の制御。所定のモノクローナル抗体に対する最高の親和性を有するペプチドの選択が天然の抗原の最良の分子模倣物を提供すること、及びこれらは関係する抗体応答を誘導する可能性が最も高いことが仮定される。理想的には、このアプローチは、多価提示を用いて初回の選択を行い、これにより標的に対していくつかの最少の親和性を有する全てのペプチドを含む比較的複雑な集団を得る。次いで、親和性選択のストリンジェンシーを増大するために、その後の回では提示価数を少なくすることが望ましい。
【0126】
このアプローチでは、単一のRNAから大量の野性型及び少量のAB−ループ組み換えタンパク質の生成を可能にするシステム(系)が提供される。一本鎖二量体中のコートタンパク質の下流のコピーの最初のアミノ酸を普通はコードするアラニンコドンの代わりにアンバー終止コドンを含む、pDSP1の改変体(pDSP1(am))を構築した(図3を参照)。従って、pDSP1(am)は、普通は野性型コートタンパク質のみを産生し、これは当然ながら普通はVLPにアセンブルされる。さらに、本発明者は、アラニン−挿入サプレッサーtRNA遺伝子を合成してクローニングした。異なる不適合性群由来のクロラムフェニコール耐性プラスミド上でのlacプロモーターの制御下で発現されて、サプレッサーtRNAは、コート配列を翻訳するわずかな割合のリボソームにアンバー(終止)コドンを読み飛ばしさせて、一本鎖二量体を産生する量で産生される。得られたタンパク質は(そのゲストペプチドとともに)、同じmRNAから発現される野性型タンパク質とアセンブルして、モザイクカプシドを形成する。精製されたVLPはこのベクターから産生され、それらは平均して、1個のVLP当たり約3個のペプチドを提示すると予測された。SDSゲル電気泳動(図4を参照)によって、精製されたVLP中の「読み飛ばし」生成物の内容を示す。これらを試験して、予想どおりアフィニティー選択のストリンジェンシーの増大を確認した。アガロースゲル電気泳動及びノーザンブロットによって、粒子が関係するRNAをカプシドで包むことが確認される。必要であれば、価数はさらに、サプレッサーtRNAの発現レベルを減少することによって減少されることができる。実際に、サプレッサーtRNA合成のレベルを調節することによって、広い範囲にわたって読み飛ばし生成物の発現(及び提示価数)を制御することが可能である。これは、活性が誘導因子の濃度に応じて正確に調節され得るプロモーター(例えば、proB)からtRNAを発現することによって達成される。
【0127】
抗原フラグメントライブラリー
別の方法では、クローニングされた抗原遺伝子又は表現型ゲノムの存在を利用してランダムな抗原フラグメントライブラリーを作成する。このようなライブラリーの作製のためには、いくつかの方法が存在する。1つは、例えば適切な平均サイズ(例えば、30bp)のフラグメントを生成するためにDNaseIで処理することによる、抗原遺伝子のランダムなフラグメントを要件とする。これらは、コートポリペプチドをコードする遺伝子中の適切な部位に対して平滑末端で連結(ライゲート)される(例えば、一本鎖コートタンパク質二量体のAB−ループ中において)。次いで得られたライブラリーを、アフィニティー選択に供して、抗体によって認識されるペプチドを提示するVLPを回収する。特定の実施形態では、制限部位を、コートポリペプチドのAB−ループ又はN−末端に挿入してもよい。
【0128】
合成
RNAファージVLPは、溶解されてVLPが抽出される生きている大腸菌細胞中のプラスミドから通常は生成される。本発明者によって今まで行われ、本出願の別の箇所で記載された実験では、MS2及びPP7のVLPのランダム配列ペプチドライブラリーは、この方法で正確に生成された。しかし、特定の別の実施形態では、本発明の集団は、当該分野で公知の手順を用いて、結合されたインビトロの転写/翻訳システムにおいて合成されてもよい(例えば、米国特許第7,008,651号;Krarnerら、1999年,Cell−free Coupled Transcription−translation Systems From E.coli.In Protein Expression.A Practical Approach,Higgins及びHames(編集),Oxford University Pressを参照)。特定の実施形態では、バクテリオファージT7の(又は関連の)RNAポリメラーゼを用いて、このようにして生成された大量のRNAを効率的に翻訳するために最適化されたシステム(系)において、T7プロモーターの制御下で、クローニングされた遺伝子の高レベル転写を指向する(例えば、Kimら、1996年,Eur J Biochem 239:881〜886;Jewettら、2004,Biotech及びBioeng 86:19〜26を参照)。
【0129】
VLPがインビボで生成される場合、大腸菌の細胞自体が区画化を提供して、これが所定のコートタンパク質−ペプチド組み換え体の複数のコピーがそのmRNAと特異的にアセンブルすることを確実にする。同様の形態の区画化が提供されないと、特に本発明の集団において、テンプレートの混合物(すなわち、ライブラリー)からインビトロで合成の間に、異なるペプチドに対するそれらの融合によって識別される異なる個々のコートポリペプチドは、恐らく互いのmRNAをパッケージングし、それによって有効なファージディスプレイに必要な遺伝子型/表現型の連鎖を破壊する可能性がある。さらに、それぞれのVLPは、多数のサブユニットからアセンブルされるので、ハイブリッドVLPの形成が生じ得る。従って、1つの好ましい実施形態では、本発明の集団又はライブラリーを調製する場合、1回以上のサイクルの転写/翻訳反応を、水/オイルのエマルジョン中で行う(Tawfikら,1998年,Nat Biotechnol 16:652〜6)。この現在では十分に確立された方法では、個々のテンプレートを、水/オイルのエマルジョンの水の部分に隔離する。適切な条件下で、多数の水性の微小液滴が形成されることができ、それぞれが平均して単一のDNAテンプレート分子及び転写/翻訳の機構を含む。それらはオイルに囲まれるので、これらの部分は互いに通じない。このような液滴中で合成されたコートポリペプチドは、それらをコードする同じmRNAと特異的に会合するはずであり、1つのペプチドのみを提示するVLPへとアセンブルするはずである。合成後に、このエマルジョンは破壊され、VLPは回収されて選択に供されることができる。1つの特定の実施形態では、転写/翻訳反応の全てが水/オイルのエマルジョン中で行われる。1つの特定の実施形態では、1つの液滴当たり1つのテンプレートのみを含む液滴(1つのペプチドのみを提示するVLP)だけが単離される。別の実施形態では、混合VLPを含む液滴が、1回以上のサイクルの転写/翻訳反応で単離され(1つの液滴当たり複数のテンプレート)、その後にペプチドを1つだけ提示するVLP(1つの液滴当たり1個のテンプレート)が単離される。
【0130】
VLP及びVLP集団の使用
本発明のVLP及びVLP集団は、多くの用途の可能性がある。以下にさらに詳細に記載されるように、VLPは、免疫原性組成物、特にワクチンとして、薬物送達デバイスとして、生体医学造影剤として、又は自己アセンブリナノデバイスとして用いられ得る。本発明のVLP集団は、適切なワクチン候補を選択するために用いられ得る。
【0131】
ワクチン候補の選択
本発明のVLP集団又はライブラリーは、ワクチン候補物を選択するために用いられ得る。このライブラリーは、ランダムライブラリーであっても、又は抗原性ライブラリーであってもよい。ランダムあるいは抗原由来ペプチドの配列ライブラリーは、VLPの表面及び特異的な標的エピトープ、又はおそらくミモトープ上に提示され、次いで抗体を用いるアフィニティー選択によって単離される。VLPは、それら自体のmRNAをカプシドで包むので、それら(及びそれらのゲストペプチド)をコードする配列は、逆転写及びPCRによって回収され得る。個々のアフィニティー選択されたVLPは、続いてクローニング、過剰発現及び精製される。
【0132】
ファージディスプレイにおけるアフィニティー選択のための技術は十分開発されており、本発明のVLPディスプレイシステムに直接適用可能である。つまり、抗体(又は抗血清)は、ランダム配列又は抗原フラグメント提示ライブラリー中のVLPのペプチドとの複合体を形成することができる。典型的には、抗体はビオチンで標識され、それによりこの複合体は、ストレプトアビジンでコーティングされた表面、磁気ビーズ又は他の適切な固定媒体に対する結合によって捕獲されることができる。洗浄後、結合されたVLPが溶出され、RNAは、アフィニティー選択された集団から抽出されて、逆転写及びPCRに供されてコートをコードする配列を回収し、この配列が次に再クローニングされて、最良結合の改変体が得られるまで、発現及びアフィニティー選択のさらなる回に供される。VLPからRNAを回収するための多数のスキームが、容易に想定される。1つの魅力的な可能性は、ストレプトアビジンコーティングされたPCRチューブ中でビオチン−mAb−VLP複合体を単に捕獲し、次にVLPを熱的に変性し、それらのRNA内容物を直接RT−PCRに供することである。多くの明らかな代替手段が存在し、種々の固定媒体の結合能力などの考慮に応じて調節が必要な場合もある。一旦、選択された配列がRT−PCRによって回収されれば、それらをクローニングして大腸菌中に再導入することは簡単なことであり、それぞれの段階で注意して必須のライブラリー多様性(これは当然ながら各回の選択で減少する)を保存する。選択が完了したら、それぞれのクローンは、VLPワクチン候補を産生するために過剰発現されることができる。
【0133】
MS2 VLPプラットフォーム上でのアフィニティー選択の効率を確立するため、エピトープが十分特徴付けされたモノクローナル抗体標的を用いて上記の方法によって選択が行われた。M2 抗−Flagモノクローナル抗体、及びライブラリーは、pDSP1中で構築され、アミノ酸13及び16についてのコドンの間に挿入された(すなわち、13/16挿入モード)10個のNNSトリプレットを含む。この特定のライブラリーは、約108個の独立したクローンを含み、かつ外来のペプチドを高い価数で提示した。それ以降、さらに複雑なライブラリーをpDSP62(上記参照)を用いて構築した。しかし、pDSP1ライブラリーが、Flagエピトープに遭遇する合理的な可能性を得るために十分複雑であると考えられた。第一回の選択を、抗体分子に対して10倍過剰であると推測されたVLPを用いて、プラスチックウェルに対する吸着によって固定された250ngの抗体に対して行った。過剰の洗浄後に、結合したVLPを溶出し、次に逆転写及びPCRに供した。PCR生成物をSalI及びBamHIで消化し、2回目で使用されるVLPの産生のためにpDSP62中にクローニングした。ここにおいて、及び全ての引き続く回において、選択物のクローニングによって、少なくとも5×106個の独立したクローンを得た。2回目の選択は、1回目と同じ条件下で行われた。2回目及び3回目の生成物を、pDSP62(am)にクローニングして、VLPを上記のアンバーサプレッサー(pNMsupA)の存在下で生成した。このことは、3回目及び4回目では、ペプチドが低価数で提示されたことを意味する。4回目では、抗体の量は50ngまで減少され、それによりVLPは、抗体に比較して約50倍過剰で存在する。4回目の後、VLPの過剰産生及び精製を期待して、高価数の提示のために生成物をpDSP62中にクローニングした。それぞれの回からの2〜3の選択物の配列を図22に示す。初期の回に得られた配列は、公知のFlagエピトープDYKDDDDKLに対して限られた類似性を示すが、特にYKジペプチドなどの、特定の重要なエレメントが既に明白である。3回目まで、全ての配列は、少なくとも1つの下流DとともにDYKエレメントを示す。4回目までに、1個の配列のみを、配列分析に供された7個のクローンから得た。野生型フラグエピトープの配列に対するその類似性は明白である。Flagエピトープは、従来の繊維状ファージディスプレイ方法を用いて以前にマッピングされており、その結果は、NEB Transcript,1996年 夏で報告された(The NEB Transcriptは、ファージディスプレイライブラリー及びアフィニティー選択キットの商業的供給業者である、New England Biolabsの刊行物である(彼らのウェブサイト、www.Neb.comで入手可能))。図22に報告される配列は、現実にNEBの実験で得られた配列よりも実際のエピトープに対してよく適合しており、このことは、MS2 VLPディスプレイシステムが、アフィニティー選択によるエピトープ特定のための繊維状ファージディスプレイと少なくとも同じ程度に効率的であることを示す。
【0134】
免疫原性組成物
上述したとおり、本発明のスクリーニング手順によって特定されるVLPを用いて、免疫原性組成物、特にワクチンを処方してもよい。ワクチンは、動物に対して投与され得る形態でなければならない。典型的には、ワクチンは、本発明の組成物が懸濁又は溶解されている、従来の生理食塩水又は緩衝化水溶液媒体を含む。この形態では、本発明の組成物は、ある状態又は障害を予防、緩和又は処置するために都合よく用いられ得る。宿主への導入の際、ワクチンは、これらに限定されないが、抗体及び/若しくはサイトカインの産生、細胞傷害性T細胞、抗原提示細胞、ヘルパーT細胞、樹状細胞及び/若しくは他の細胞の活性化などの免疫応答、並びに/又は他の細胞性応答を誘発することができる。
【0135】
任意で、本発明のワクチンはさらに、本発明の化合物に対してわずかな割合でも又は大きな割合でも存在することができるアジュバントを含む。「アジュバント」という用語は、本明細書において、免疫応答の非特異的な刺激因子又は宿主中でデポの生成を可能にする物質であって、本発明のワクチンと組み合された場合、さらにより増強された免疫応答を提供するものを指す。種々のアジュバントが用いられることができる。例としては、完全及び不完全フロイントアジュバント、水酸化アルミニウム及び改変ムラミルジペプチドが挙げられる。
【0136】
任意で、本発明のワクチンは、本発明の化合物に対してわずかな割合でも又は大きな割合でも存在することができるアジュバントをさらに含む。
【0137】
標的化薬物送達
特定の細胞レセプターに結合するペプチドを特定し、それによって標的細胞に結合かつ進入する能力を持った(例えば、エンドサイトーシスによる)粒子を生成するために、アフィニティー選択を用いることができる。MS2 VLPは、20nmの大きさの内径を有する中空の球体である。特定の実施形態では、VLPは、薬物、例えば送達されるべきタンパク質毒素及び任意で細胞型特異的なレセプターに結合するリガンドを含む。このような粒子の内部組成は、例えば、リシンのようなタンパク質毒素をそれに特異的に充填することによって、又は合成の翻訳オペレーター模倣物に対してカップリングすることによって調整され得る。このような粒子の外面に対して細胞特異的なレセプターを結合する能力を付与することによって、選択される細胞型を毒素(又は他の薬物)の送達の標的とすることができる。関連する側面では、コートポリペプチド二量体を含むVLPは、細菌毒素、アジュバント又は免疫刺激性核酸などの異種物質を実際にカプシドで包み得る。
【0138】
生体医学造影剤
薬物が特定の細胞型を標的とし得る同じ方法では、核磁気共鳴画像化のための造影剤が、特定の細胞又は組織に対して送達されることができると考えられ、可能性としては、MRIの診断力を大いに増大する。実際に、MS2粒子は、MRIのコントラストを大きく増大するために、既にガンドリニウムで標識されている(Andersonら,2006年,Nano Letters 6(6),1160〜1164)。従って、特定の実施形態では、このような粒子は、それらの表面上で適切なレセプター特異的ペプチドを提示することによって、特定の部位を標的とすることができるであろう。関連の側面では、コートポリペプチド二量体を含んでいるVLPは、実際に造影剤をカプシドで包み得る。
【0139】
自己アセンブリするナノデバイス
本発明のVLPは、金属結合タンパク質を提示する繊維状ファージ粒子のいずれかの末端に親和性を有するペプチドを含み得る。繊維状ファージ粒子のいずれかの末端に親和性を有するVLPは、繊維状ファージナノワイヤの末端に対してこれらの球体(及びそれらが含むものなら何でも)を連結する可能性を生じる。あるいは、VLPは、金属結合ペプチド(例えば、金及び亜鉛)を提示でき、それにより通常ではない電気的及び光学的な特性を有するアレイが得られる場合がある。あるいは、これらのアレイに自己アセンブリする能力が改善されたVLPは、特定の表面に対する親和性を有するペプチド、又はVLP自体の自己会合の特性を変更するペプチドを提示することによって産生され得る。
【0140】
実験の概説
バクテリオファージMS2のウイルス様粒子(VLP)の上でランダム−配列ペプチドライブラリーの構築を容易にする2つのプラスミドベクターを記載する。最初のpDSP1は、PCR−生成されたフラグメント又は他の二本鎖DNAフラグメントを、コートタンパク質一本鎖二量体の下流のコピーのAB−ループ中に都合よくクローニングするために構築された。二番目はpDSP62と呼ばれ、これは、Kunkelら[16]の部位指向性突然変異誘発法によって一本鎖二量体中の事実上任意の位置(通常はAB−ループ)にペプチド配列を導入するために、特異的に構築された。これらのプラスミドの一般的な特徴を下に示す。
【実施例1】
【0141】
pDSP1−都合のよいAB−ループ中の挿入のためのクローニング部位を有する一本鎖二量体を発現するプラスミド
プラスミドpDSP1(図5a及び図7aを参照)は、pET3dのT7転写シグナル、カナマイシン耐性、及びpET9dの複製起点を含む(pET3d及びpET9dに関する情報は、New England Biolabs vectorのデータベース、https://www.lablife.org/ct?f=v&a=listvecinfoで見られる)。これは、1つだけのSalI及びKpnI制限部位を含むように改変された、MS2一本鎖コートタンパク質二量体(6)のコード配列を発現する。これによって、外来の配列をAB−ループに簡易にクローニングすることが容易になる。これらの部位を1つだけにするには、ベクター中及び上流コート配列において、他のSalI及びKpnI部位を破壊することが必要であった。
【0142】
pDSP1のAB−ループ挿入部位の近傍のMS2コート配列を下に示す。SalI及びKpnI部位の存在に注意されたい。
【0143】
【化2】
【0144】
下に示すのは、pDSP1中のランダムな7マーのライブラリーの例であって、ここではランダムな配列が、いわゆる13/16モードで挿入されている。
【0145】
【化3】
【0146】
AB−ループをコードする配列中又はその付近に、1つしかない複数の制限部位が存在することは、それらがその切断によって適合性の「付着末端」を生じる部位に隣接する場合、外来配列を簡易に挿入することが可能になる。しかし、これは時に、図5bに示されるようなPCR法及び組み換えDNA法の組み合わせを用いて外来配列を結合するのにさらに便利である。例えば、下に示される5’PCRプライマーを、BamHIの下流のプラスミドベクター配列にアニールする3’プライマーとともに用いて、アミノ酸13と16との間に挿入されたランダム配列を有するコートタンパク質コード配列のフラグメントを生成してもよい。N=A、C、G又はTであり、S=G又はCである。SalI及びBamHIでの消化後、このフラグメントは、pDSP1のSalIとBamHIとの間に挿入される。下に示すのは、このようなライブラリーを生成するために用いられ得る5’−プライマーの例である:
【0147】
【化4】
【0148】
pDSP1を用いれば、ランダム−配列ペプチドライブラリーは通常、テンプレートとして単量体のコートタンパク質配列を用いて生成されたPCRフラグメントをAB−ループ中にクローニングすることによって構築される(例えば、pMCT)。合成のオリゴヌクレオチド5’−プライマーは、SalI(又はKpnI)部位及びランダムなコドンの配列(例えば、6〜10コピーのNNY)をAB−ループのすぐ上流の部位に結合するように設計される。3’−プライマーは、BamHIのすぐ下流のプラスミドベクター中の配列にアニールする。得られたPCR産物をSalI(又はKpnI)及びBamHIで消化して、pDSP1の対応する部位にクローニングする。これによってAB−ループへのペプチドの挿入が生じるが、挿入の正確な部位は、5’−プライマーの特異的な設計に依存する。ほとんどの挿入に関し、SalI部位の使用は、挿入部位の選択においてKpnIよりも柔軟性を与えるので好ましい。これらの方法によれば、最大で108〜109個の個々のメンバーを有するペプチドVLPライブラリーを比較的直接的に生成する。
【0149】
ペプチドディスプレイ価数の制御のための手段を導入するために、pDSP1の誘導体(pDSP1(am)と呼ばれる)を、一本鎖二量体の半分の間の接合部でのナンセンスコドンの導入によって構築した。pNMsupAによって産生されたようなナンセンスサプレッサーtRNAの存在下で発現される場合、少量の一本鎖二量体が(その外来ペプチドとともに)産生される。しかし、pDSP1(am)から産生されるほとんどのコートタンパク質は、野生型の単位長タンパク質の形態で合成される。2つの形態のコートタンパク質は、平均でわずか約3個のペプチドしか提示しないハイブリッド粒子へとコアセンブル(共集合)する。提示価数の平均レベルは、サプレッサーtRNAの発現レベルを変更することによって、又はより大きいか若しくは小さい抑制効率を示すサプレッサーを使用することによって、大小に調節され得る。
【実施例2】
【0150】
pDSP62−効率的な部位指向性突然変異誘発方法を用いるライブラリー構築のために適切なプラスミド
M13複製起点の導入
pDSP1について上述したようなライブラリー生成のための方法は、大規模にするのが困難である。これは、DNA制限フラグメントを必要な量で精製するのに不便なためである。さらに、ライゲーション反応の間、無用な副産物へ否応なく変換されるDNAがあり、これが所望のプラスミドの収率を下げる。複雑なライブラリーの構築は、代表的なライゲーション反応で見られるよりも正確な組み換えDNAをより高い収率で効率的に生じる方法によって容易にされる。具体的には、部位指向性の突然変異誘発のための古い方法のバリエーションを用いることが好ましく、これは既に他者により、1011個の複雑性の範囲で、繊維状ファージ上でペプチドライブラリーが生成されている(2,6)。この方法を、M13複製起点を含む特定の種類のプラスミド(ファージミドとしても公知)から生成された一本鎖の環状DNAに適用する。プラスミドを含んでいるdut-、ung-株(例えば、BW313)のM13ヘルパーファージ(例えば、M13K07)による感染の結果として、dUTP−置換の一本鎖のDNAの産生が容易になる。現実の突然変異誘発の反応では、ミスマッチのオリゴヌクレオチドプライマーは、一本鎖のDNAテンプレートにアニールされて、DNAポリメラーゼ(例えば、T7ファージのもの)を用いて延長される。DNAを、閉鎖した環状DNAを生成するために連結し、形質転換によってung+株に導入し、ここでこの変異株を優先的に複製する。ペプチド−VLPライブラリーの産生における以前の実験では、典型的には約90%の形質転換体が所望のペプチド挿入物を含むことが示される。プライマー伸長の突然変異誘発反応は、エレクトロポレーションによって1011個程度の個々の組み換え体を容易に生成するのに十分な、比較的大量のDNA(例えば、20μg)で行われることができる。
【0151】
一本鎖のDNAの産生を容易にするため、M13複製起点をpDSP1に導入した。これを行うために、pUC119で見られるM13起点をPCRによって増幅して、pDSP1における唯一のA1wNI部位にクローニングした。pDSP1−IGと呼ばれるこのプラスミドは、pDSP62の前駆体である。これは、pDSP62Iの構築における唯一の中間体であるので、その配列は示していない。
【0152】
合成の「コドンジャグリングされた」コート遺伝子の使用を通じた一本鎖二量体の半分のみへの標的化挿入
効率的なペプチドライブラリー構築物のためにプライマー伸長突然変異誘発を用いるという要望によって、新しい問題がもたらされた。本発明の提示方法は、外来ペプチドを一本鎖二量体の2つのAB−ループの一方にのみ特異的に導入する能力に依拠する。pDSP1に存在する一本鎖二量体配列を用いると、変異原性のプライマーは、両方の半分での配列にアニールして二重の挿入を生じたが、AB−ループの両方における挿入は、タンパク質折り畳みの失敗を高頻度で生じることが既に公知である。さらに、挿入に耐えた場合でも、一本鎖二量体の半分のみを標的とすることができなかった部位指向性の突然変異誘発は、それぞれのVLPで、2つの異なるペプチドの提示をもたらすであろう。
【0153】
これらの理由で、コートタンパク質の「コドンジャグリングされた」ものを合成して、一本鎖二量体の正常な上流の半分と交換した。このコドンジャグリングされた配列は、可能な最大数のサイレントヌクレオチド置換を含み、従って野生型コートタンパク質アミノ酸配列を有するポリペプチドを生じる。しかし、多数の変異体の存在によって、このジャグリングされた配列は、変異原性のオリゴヌクレオチドに対して効率的にアニールできなくなり、従ってこの変異原性のプライマーは特異的に、下流のAB−ループ配列に指向される。プラスミドpDSP62は図6に示され、その配列は図7bで提供される。
【0154】
一本鎖pDSP62産生のためのクロラムフェニコール耐性M13ヘルパーファージ
プラスミドpDSP62は、カナマイシンに対する耐性を付与する。一本鎖ファージミドDNAの産生のために通常用いられるヘルパーファージ(例えば、M13KO7)もカナマイシン耐性を付与し、従ってここで記載されるプラスミドでの使用には不適切である。この理由で、M13KO7のクロラムフェニコール耐性誘導体である、M13CM1を構築した。pACYC184(7)のクロラムフェニコール耐性遺伝子を、XhoI及びSacIに対する認識配列を結合したプライマーを用いてPCRによって増幅し、そのフラグメントを、カナマイシン耐性決定基にほぼ隣接するXhoI及びSacI部位を利用して、カナマイシン耐性遺伝子の代わりにM13KO7に挿入した。カナマイシン(pDSP62維持を選択する)及びクロラムフェニコール(ヘルパーファージを選択する)の存在下で、細胞は、M13 CM1での感染後に大量の一本鎖のプラスミドDNAを産生する。これらの一本鎖のテンプレート及びKunkelら(2)の方法を用いて、[NNS]6、[NNS]7、[NNS]8及び[NNS]10について1010個を超える個々のメンバーを含むランダム配列ペプチドライブラリーが容易に産生された。著しく高い複雑性が、大規模化とともに可能である。
【0155】
ペプチドディスプレイ価数の制御のための手段を導入するために、pDSP62の誘導体(pDSP62(am)と呼ばれる)を、一本鎖二量体の半分の間の接合部におけるナンセンスコドンの導入によって構築した。pNMsupAによって産生されるもののようなナンセンスサプレッサーtRNAの存在下で発現した場合、少量の一本鎖二量体が(その外来ペプチドとともに)産生される。しかし、pDSP1(am)から産生されるほとんどのコートタンパク質は、野生型の単位長のタンパク質の形態で合成される。2つの形態のコートタンパク質は、平均でわずか約3個のペプチドしか提示しないハイブリッド粒子へとコアセンブル(共集合)する。提示価数の平均レベルは、サプレッサーtRNAの発現レベルを変更することによって、又はより大きいか若しくは小さい抑制効率を示すサプレッサーを使用することによって、大小に調節され得る。
【0156】
上述されるとおり、バクテリオファージMS2のウイルス様粒子がペプチドディスプレイのために用いられて、MS2コートタンパク質一本鎖二量体がペプチド挿入に極めて耐性であること、及びそれらがその合成をコードするmRNAを特異的にカプシドで包む正確にアセンブルされたVLPを生成することが確認された[2]。しかし、MS2は、個々のメンバーが同様の分子生物学を共有するウイルスの大きいファミリーの1つのメンバーでしかない。上記のプラスミド及び方法は、本発明者が、MS2コートタンパク質一本鎖二量体の挿入耐性、及びMS2 VLPがその合成を指向するmRNAをカプシドで包む能力を実証した、以前に記載されたMS2 VLPディスプレイシステムに対して改良点を示す[2]。以下の例は、PP7 VLPについて最近得られたもの[1]と同様の結果を実証しており、このことは、PP7一本鎖コートタンパク質二量体の折り畳み及びアセンブリがまた、外来ペプチド挿入に対して高い耐性を示すこと、並びにこのようにして得られたVLPが、mRNAの形態でそれらの合成のための遺伝的情報を含むことを明確に示している。
【0157】
ここで次に、ペプチドディスプレイを目的として、緑膿菌のバクテリオファージである、PP7のVLPの操作を記載する。
【0158】
PP7 VLPは、MS2 VLPを上回るいくつかの潜在的な利点及び改善を提供する。第一に、安定化した内部サブユニットジスルフィド結合が存在するおかげで、この粒子は熱力学的に劇的により安定である(8)。ワクチンを含む多くの実質的な適用のために、安定性の増大は所望の特質である。第二に、PP7 VLPは、MS2のものと免疫学的に交差反応性でない(9)。このことは、VLPの一連の投与が必要となる場合があるワクチン又は標的化薬物送達の適用において、重要であり得る。第三に、PP7 VLPの正確な折り畳み及びアセンブリがペプチド挿入の脱安定化効果に対してさらに耐性であり得ること、又はこれが、MS2 VLPにおいて耐性でないいくつかのペプチドの耐性を少なくとも示し得ることが予想される。
【実施例3】
【0159】
PP7ペプチドディスプレイベクターの設計
大腸菌中でのPP7コートタンパク質の合成のために、2つの一般的な種類のプラスミドを構築した(図9及び図13を参照)。第一は、lacプロモーターからコートタンパク質を発現し、これを使用して(pRZP7と組み合わせて、以下を参照)、翻訳リプレッサー及びVLPアセンブリ試験を用いて、ペプチド挿入のコートタンパク質の耐性について試験する。第二の種類のプラスミドは、T7プロモーター及び転写ターミネーターからタンパク質を発現する。これらのプラスミドは、正確にVLPにアセンブルする大量のコートタンパク質を産生する。それらはまた、VLPへのカプシド形成のために別個の5’及び3’末端を有するコート特異的なmRNAを生じる。
【0160】
ペプチド挿入部位の設計
PP7カプシドの三次元構造は、たとえ2つのタンパク質のアミノ酸配列がわずか約12%の配列同一性しか示さなくても、三次構造がMS2の構造をよく模倣しているコートタンパク質から構成されることを示す(10)[17]。PP7タンパク質は、AB−ループを保有し、その中にペプチドが、MS2について以前に記載されたスキームと同様のスキームに従って挿入され得る[2]。MS2の場合のように、これは、制限エンドヌクレアーゼKpnIの部位を含むようにPP7コート配列を変異させることによって開始し、これによって、pP7Kと呼ばれるプラスミド中の外来配列の挿入が容易になる(図9a)。この改変により、図10に示されるアミノ酸置換(E11T)が生じた。この置換は、変異体コートタンパク質は翻訳を抑制して正確にVLPにアセンブルするので、十分に耐えられた。やはりMS2の実施例によれば、PP7コートタンパク質の一本鎖二量体バージョンの折り畳みは、従来の二量体よりもAB−ループ挿入に対してさらに耐性であると考えられた。その構築は以前に記載されたが(8)、ここでペプチドディスプレイへの使用については初めて記載される。一本鎖二量体は、コード配列の下流コピー中にのみKpnI部位を含むように改変され、p2P7K32を生成した(図9b)。この設計では、ペプチドをアミノ酸11に挿入したが、他の特定の挿入部位が、おそらくAB−ループ内のいずれかで用いられ得ることに注目すべきである。実際には、下記の試験は、2つの別の挿入モードから記載される(図10及び11)。第一は、11番目のアミノ酸が、挿入されたペプチドのN末端側のthr及びC末端側の野生型glu11として2回出現するので、11/11モードと呼ばれる。いわゆる11/12モードでは、挿入物は片側ではthr11に、他の側ではala12に隣接する。
【0161】
AB−ループ挿入物に対するPP7コートタンパク質の一般的な耐性を試験するために、PP7コートタンパク質のAB−ループ中に挿入されるランダム配列ペプチドのライブラリーを、図11に示されるスキームを用いて作成した。このランダム配列は、6、8又は10コピーの配列NNY(ここでNは任意のヌクレオチドであり、Yはピリミジンである)から構成された。このようなライブラリーは、20個の可能なアミノ酸のうち15個を含み、従ってかなり多様性であり得る。しかし、終止コドンの可能性を回避することによって、それに続く分析はかなり容易になる。
【実施例4】
【0162】
コートタンパク質機能の保持のためのランダム−配列ペプチドライブラリーの個々のメンバーを試験するための方法
MS2コートタンパク質と同様に、PP7コートは、翻訳リプレッサーである。大腸菌 lacZ遺伝子に対してPP7翻訳オペレーターを融合するプラスミドであるpRZP7の構築物は以前に記載されており、これは、コートタンパク質の翻訳リプレッサー活性の制御下にβ−ガラクトシダーゼ合成を置く(11)。これは、異なる抗生物質(クロラムフェニコール)に対する耐性を付与するとともに、異なる不適合性群に由来する(すなわち、これはp15A複製起点を用いる)ので、その両方がアンピシリンに対する耐性を付与しかつcolE1起点を用いるpP7K又はp2P7K32のいずれかと同じ大腸菌株で容易に維持され得る。これらのプラスミドの両方とも、pUC119に由来し、lacプロモーターから比較的低レベルでコートタンパク質を発現する。pP7K又はp2P7K32からのPP7コートタンパク質の発現は、pRZP7から発現されるβ−ガラクトシダーゼの翻訳を抑制する。このことは、所定のペプチド挿入物が、コートタンパク質が正確に折り畳む能力を妨害したか否かを決定することを容易にする。それは、不完全なコートタンパク質は、「xgal」として公知のβ−ガラクトシダーゼ発色性基質を含むプレート上に青色のコロニーを生じ、一方では適切に機能するコートタンパク質は白いコロニーを生じるからである。
【0163】
機能の維持のさらに厳密な試験は、ペプチドコートタンパク質組み換え体を発現する細胞の溶解液中のVLPの存在について直接試験することである。これは、超音波処理によって溶解される細胞のアガロースゲル上での電気泳動によって達成される。エチジウムブロマイド染色によってRNA含有VLPを検出し、次にその存在を、抗PP7血清を用いるウエスタンブロット分析によって確認する。
【0164】
次のアイデアは、PP7コートタンパク質のペプチド挿入耐性を試験し、ランダム配列ペプチドライブラリーを作成すること及び翻訳リプレッサー機能を保持する(すなわち、白色コロニーを生成する)クローンの画分を決定することによってアセンブルし、VLPを生成することである。MS2コートタンパク質一本鎖二量体のペプチド挿入耐性の同様の試験は、本願の発明者によって以前に報告されたことに注意されたい[2]。
【実施例5】
【0165】
ほとんどのランダムな6マー、8マー、及び10マーのペプチド挿入物に耐えるPP7コートタンパク質の折り畳み及びアセンブリ
(NNY)8ライブラリーは、pP7K及びp2P7K32中で構築して、形質転換によって大腸菌株CSH41F−/pRZP7中に導入した。形質転換体は、xgalを含む固形培地上にプレートした。挿入フラグメントなしの対照ライゲーションで、1000分の1のコロニーが得られたので、大部分が真正の組み換え体であった。ごく少数のpP7K組み換え体を試験したが、全てがタンパク質の折り畳みに関して欠損しており、かつVLPを作成できなかったことが分かった。これによって、従来の二量体は、通常はAB−ループ中のペプチド挿入によって不安定化されるというMS2での以前の実験[2]からの予想が確認される。しかし、p2P7K32の一本鎖二量体では、ほぼ100%のコロニーが白色であった。それぞれのライブラリー由来の24個の白色コロニーから1mlの培養物を複製した。1つの培養セットから、粗細胞溶解液をVLPのアガロースゲル分析のために調製した。他のセットでプラスミドを単離して制限酵素消化及びゲル電気泳動に供し、全てが予想される長さの挿入物を含むことを確認した。2〜3個の青色コロニーからのプラスミドも単離した。
【0166】
全てが、異常なライゲーション事象から生じるプラスミドを含むことが判明し、一般にはインタクトなコート配列を含まなかった。ほぼ100%のペプチド挿入物が、コートタンパク質の翻訳リプレッサー機能と適合した。すなわち、それらは十分に適切に折り畳まれたコートタンパク質を生じて、野生型タンパク質のような翻訳を抑制する。この結果は、全てのNNYライブラリー(6マー、8マー及び10マー)で、それらが11/11モードでクローニングされたか、又は11/12モードでクローニングされたかには依存せずに得られた。
【0167】
VLPの存在について直接試験するために、粗細胞溶解液をそれぞれの複製培養から調製して、アガロースゲル電気泳動に供した。ゲルをエチジウムブロマイドで染色して、複製をニトロセルロースでブロットして、マウスの抗−PP7血清と西洋ワサビペルオキシダーゼ標識した二次抗体とでプローブした。クローンは、それが染色したゲル及びウエスタンブロットの両方にバンドを含む場合、VLP合成について陽性とみなす。結果を図13に示す。これによって、いくつかは収率の低下を示すが、6マーのクローンのほぼ全てがある程度のレベルでVLPを生成することが示された。このことは、11/11挿入モード及び11/12挿入モードの両方について当てはまった。8マーのクローンの大部分は、容易に同定可能なVLPを生じる。しかし、この効率は、挿入長が10マーまで増大するにつれていくらか低下するようであるが、やはりクローンの明らかに大部分がVLPを生じる。いくつかのペプチドが荷電されたアミノ酸を組み込むことによってVLPの表面荷電を変更するという予想と一致して、個々の粒子の移動度は可変であることに注意されたい。
【実施例6】
【0168】
PP7 VLPは、コート−特異的なmRNAをカプシドで包む。
高レベルの発現及びRNAカプシド形成試験のために、pP7K及びp2P7K32のPP7コート配列を、T7プロモーター及び転写ターミネーターを含んでいるプラスミド中にクローニングして、pETP7K及びpET2P7K32と呼ばれるプラスミドを生成した。大腸菌株BL21(DE3)中のプラスミドは、大量のコートタンパク質を合成した。得られたVLPを、以前に記載(11、12)されたようなセファロースCL4Bでのクロマトグラフィーによって精製した。RNAをフェノール/クロロホルム抽出によってVLPから精製して、ホルムアルデヒドを含んでいる複製アガロースゲル中の電気泳動に供した。一方のゲルをエチジウムブロマイドで染色し、他のゲルはニトロセルロースにブロットして(ノーザンブロット)、ここでPP7配列を、コートのセンス鎖に特異的な標識された合成のオリゴヌクレオチドを用いて検出した。T7のRNAポリメラーゼを用いたpETP7K及びpET2P732のインビトロでの転写によって生成したRNAを、標準として利用した。それぞれのVLPは、その移動度がインビトロ転写生成物の移動度と同一であり、PP7コート特異的なプローブと特異的にハイブリダイズする、優越した種を含む。PP7 VLPがそれらのmRNAをカプシドで包むことが確認され、これによって、アフィニティー選択のために必要な遺伝子型−表現型の連鎖を確立した。MS2コートタンパク質がそのmRNAをカプシドで包む能力の同様の試験が、本願の発明者によって以前に報告されていることに注意されたい。
【実施例7】
【0169】
PP7 VLPに提示されるペプチドは、免疫系に提示され、かつ免疫原性である。
図10及び図15に示される特異的なペプチド配列を提示するPP7 VLPを構築した。これらは、いわゆるFlagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来の配列、HIV−1のgp120のV3ループ、及び炭疽菌の防御抗原を含んだ。これらの挿入されたペプチドがVLPの表面上に実際に提示されることを示すために、HPV16L2に対するモノクローナル抗体(RG−1と呼ばれる)がPP7 L2−VLPに結合する能力をELISAによって評価した。図16に示されるとおり、RG−1は、L2−VLPには結合したが、V3ペプチドを提示したPP7 VLPには結合しなかった。VLPの免疫原性を示すために、マウスを以前に記載されたように[2]、筋肉内注射によってV3−VLPで免疫した。図17に示されるように、マウス由来の血清がELISAによって試験され、V3ペプチドの合成バージョンと特異的に反応する高力価のIgG抗体を有することが示された。MS2 VLPで提示されるペプチドの免疫原性の同様の試験は、本願の発明者によって以前に報告されていることに注意されたい。
【0170】
実施例8。プラスミドpET2P7K32及びpDSP7は、上記のMS2コートタンパク質プロデューサーpDSP1及びpDSP62のPP7アナログである。PP7 VLPでのペプチドディスプレイ価数の制御のために、pET2P7K32(am)及びpDSP7(am)も構築した。それらは、それぞれpDSP1(am)及びpDSP7(am)のアナログである。
【0171】
本明細書に引用される全ての特許、特許出願、刊行物、及び電子的に利用可能なもの(例えばGenBank and RefSeqにおけるヌクレオチド配列寄託、例えばSwissProt、PIR、PRF、PDBにおけるアミノ酸配列寄託、並びにGenBank and RefSeqにおけるアノテーションされたコード領域からの翻訳など)の全体の開示は、参照によって援用される。援用されるものと、オリジナルとして提出された明細書中に示されるものとの間の何らかの不一致は、オリジナルとして提出された明細書を優先して解消されるものとする。前述の詳細な説明及び実施例は、理解の明確化のためにのみ示された。それらから不必要な制限が理解されることはない。本発明は、表示及び記載された厳密な詳細に限定されるものではなく、当業者に明白な変形については、特許請求の範囲に規定される本発明内に含まれる。
【0172】
全ての見出しは、読む者の便利のためであって、そうであると特定されない限り、見出しの下にある文章の意味を限定するために用いられるべきではない。
【0173】
引用文献(参考文献の第一セット)
1.Peabody, DS, Jordan, SK, Caldeira, JC, Manifold-Wheeler, B. Medford, A and Chackerian, B (2007年) Immunogenic display of diverse peptides on virus-like particles of RNA phage MS2. J. Mol. Biol. 380: 252-263.
2.Kunkel TA, Bebenek. McClary J. (1991年) Efficient Site-Directed Mutagenesis Using Uracil-Containing DNA. Methods Enzymol. 204:125-139.
3.Sidhu SS, Lowman HB, Cunningham BC, Wells JA. (2000年) Phage display for selection of novel binding peptides. Methods Enzymol. 328:333-363.
4.Kleina LG, Masson J-M. Normanly J, Abelson J, and Miller JH. (1990年) Construction of Escherichia coli Amber Suppressor tRNA Genes. J. Mol. Biol. 213: 705-717.
5.Lee SK and Deasling JD. (2005年) A propionate-inducible expression system for enteric bacteria. Appl. Env. Microbiol. 71:6856-6862.
6.Peabody. D. S., and Lim, F. (1996年) Nucleic Acids Res 24, 2352-2359.
7.Chang, A. C, and Cohen, S. N. (1978年) J Bacterial 134, 1141-1156.
8.Caldeira, J. C, and Peabody, D. S. (2007年) J Nanobiotechnology 5, 10
9.Olsthoom, R. C, Garde, G., Dayhuff, T., Atkins, J. F., and Van Duin, J. (1995年) Virology 206, 611-625.
10.Tars K, F. K., Bundule M, Liljas L. (2000年) Virology 272. 331-337
11.Lim, F., Downey, T. D., and Peabody, D. S. (2001年) Journal of Biological Chemistry 276, 22507-22512.
12.Peabody, D. S. (1990年) J Biol Chem 265, 5684-5689.
【0174】
引用文献(参考文献の第二セット)
1.Caldeira Jdo C. Medford A, Kines RC. Lino CA, Schiller JT, Chackerian B, Peabody PS: Immunogenic display of diverse peptides, including a broadly cross-type neutralizing human papillomavirus L2 epitope, on virus-like particles of the RNA bacteriophage PP7. Vaccine, 28(27):4384-4393.
2.Peabody PS. Manifold-Wheeler B, Medford A, Jordan SK, do Carmo Caldeira J,
Chackerian B: Immunogenic display of diverse peptides on virus-like particles of RNA phage MS2. J Mol Biol 2008年, 380(1):252-263.
3.Lowman HB, Bass SH, Simpson N, Wells JA: Selecting high-affinity binding proteins by monovalent phage display, Biochemistry 1991, 30(45'): 10832- 10838.
4.Sidhu SS, Lowman HB, Cunningham BC, Wells JA: Phage display for selection of novel binding peptides. Methods in Ezymology 2000年, 328:333-363.
5.Wells JA: Hormone mimicry. Science 1996年, 273(5274):449-450.
6.Peabody PS: Subunit fusion confers tolerance to peptide insertions in a virus coat protein. Arch Biochem Biophvs 1997年, 347(1):85-92.
7.Peabody PS, Chakerian A: Asymmetric contributions to RNA binding by the Thr(45) residues of the MS2 coat protein dimmer. J Biol Chem 1999年, 274(36):25403-25410.
8.Lim F, Peabody PS: RNA recognition site of PP7 coat protein. Nucleic Acids Res 2002年, 30(19):4138-4144.
9.Lim F, Powney TP, Peabody PS: Translational repression and specific RNA binding by the coat protein of the Pseudomonas phage PP7. J Biol Chem 2001年, 276(25):22507-22513.
10.Kunkel TA, Bebenek K, Mcclary J: Efficient Site-Directed Mutagenesis Using Uracil-Containing DNA. Methods in Enzymoloev 1991年, 204:125-139.
11.Tars K FK, Bundule M, Liljas L: The Three-Dimensional Structure of Bacteriophage PP7 from Pseudomonas aeruginosa at 3.7 A Resolution. Virology 2000年, 272:331-337.
12.Kleina LG, Masson JM, Normanly J, Abelson J, Miller JH: Construction of Escherichia coli amber suppressor tRNA genes. II. Synthesis of additional tRNA genes and improvement of suppressor efficiency. J Mol Biol 1990年, 213(4):705-717.
13.Normanly J, Kleina LG, Masson JM, Abelson J, Miller JH: Construction of Escherichia coli amber suppressor tRNA genes. III. Determination of tRNA specificity. J Mol Biol 1990年, 213(4):719-726.
14.Kunkel TA: Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc Natl Acad Sci USA 1985年, 82(2):488-492.
15.Maruyama IN, Maruyama HI, Brenner S: Lambda foo: a lambda phage vector for the expression of foreign proteins, Proc Natl Acad Sci USA 1994年, 91(17):8273-8277.
16.Mikawa YG, Maruyama IN, Brenner S: Surface display of proteins on bacteriophage lambda heads. J Mol Biol 1996年, 262(1):21-30.
17.Dunn IS: Assembly of functional bacteriophage lambda virions incorporating C-terminal peptide or protein fusions with the major tail protein. J Mol Biol 1995, 248(3):497-506.
【特許請求の範囲】
【請求項1】
ウイルス様粒子(VLP)の集団であって、それぞれのVLPは、(a)RNAバクテリオファージのVLPであり、前記VLPの一部は、(b)異種ペプチドの挿入によって改変された前記RNAバクテリオファージのコートポリペプチドの一本鎖二量体を含み、前記二量体は、上流及び下流のサブユニットを含み、前記異種ペプチドは、前記VLPに提示されかつ(c)前記バクテリオファージのmRNAをカプシドで包み、前記VLPは低い価数を示す、ウイルス様粒子(VLP)の集団。
【請求項2】
前記異種ペプチドが少なくとも4アミノ酸長である、請求項1に記載の集団。
【請求項3】
前記バクテリオファージの複数の前記コートポリペプチドが、野生型コートポリペプチドであり、残りの前記コートポリペプチドが、異種ペプチドを含む前記コートポリペプチドの一本鎖二量体を含み、前記バクテリオファージが、MS2及びPP7からなる群より選択される、請求項1に記載の集団。
【請求項4】
前記異種ペプチドが、前記コートポリペプチドの下流サブユニット中のABループでの部位又は前記RNAバクテリオファージのコートポリペプチドのカルボキシ末端での部位に挿入される、請求項1に記載の集団。
【請求項5】
前記異種ペプチドが、前記RNAバクテリオファージのコートポリペプチドのカルボキシ末端に対応する部位に挿入される、請求項4に記載の集団。
【請求項6】
前記RNAバクテリオファージが、MS2又はPP7、Qβ、R17、SP、PP7、GA、M11、MX1、AP205、PRR1、f4、Cb5、Cb12r、Cb23r、7s及びf2からなる群より選択される、請求項1に記載の集団。
【請求項7】
前記コートポリペプチドが一本鎖二量体であり、前記コートポリペプチドの下流のコピーが、少なくとも4アミノ酸長を有する異種ペプチドの挿入によってABループ中で改変される、請求項1に記載の集団。
【請求項8】
前記VLPで前記異種ペプチドの1コピーから10コピーまでの間のコピーが提示される、請求項1に記載の集団。
【請求項9】
前記VLPで前記異種ペプチドの1コピーから5コピーまでの間のコピーが提示される、請求項1に記載の集団。
【請求項10】
前記VLPで前記異種ペプチドの1コピーより少ないコピーが提示される、請求項1に記載の集団。
【請求項11】
前記異種ペプチドが、前記二量体タンパク質のカルボキシ末端に、又は
(a)MS−2、R−17及びfrコートポリペプチドのアミノ酸11〜17、
(b)GAコートポリペプチドのアミノ酸10〜16、
(c)Qβ及びSPコートポリペプチドのアミノ酸10〜17、
(d)PP7コートポリペプチドのアミノ酸8〜11、
(e)PRR1コートポリペプチドのアミノ酸9〜17
に対応する前記コートタンパク質のAB−ループ中の部位に挿入される、請求項4に記載の集団。
【請求項12】
細菌又はバクテリオファージプロモーターと、
上流サブユニット及び下流サブユニット、前記下流サブユニットに配置される異種ペプチドの挿入のための部位を含むRNAバクテリオファージ一本鎖コートポリペプチド二量体のコード配列と、
前記コートタンパク質の下流サブユニットの第一のアミノ酸をコードするコドンを置き換える終止コドンと、
細菌又はバクテリオファージターミネーターと、
を含む、単離された転写単位。
【請求項13】
アラニン挿入サプレッサーtRNA遺伝子をさらに含む、請求項12に記載の転写単位。
【請求項14】
前記プロモーターがバクテリオファージプロモーターであり、前記ターミネーターがバクテリオファージターミネーターである、請求項12又は13に記載の単離された転写単位。
【請求項15】
前記異種ペプチドの挿入のための部位が制限酵素部位である、請求項12〜14のいずれかに記載の単離された転写単位。
【請求項16】
一本鎖二量体が、改変された一本鎖コートポリペプチド二量体であり、前記改変が、前記挿入部位に挿入されている異種ペプチド配列を少なくとも4アミノ酸含む、請求項12〜15のいずれかに記載の単離された転写単位。
【請求項17】
前記転写単位が、翻訳オペレーター配列を含まない、請求項12に記載の単離された転写単位。
【請求項18】
前記転写単位が、翻訳オペレーター配列を含む、請求項12に記載の単離された転写単位。
【請求項19】
前記終止コドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項12〜18のいずれかに記載の単離された転写単位。
【請求項20】
バクテリオファージプロモーターを含む単離された転写単位であって、RNAバクテリオファージ一本鎖コートポリペプチド二量体のコード配列が、前記二量体の下流半分において前記コード配列中の異種ペプチドの挿入のための部位を有し、前記コード配列が、前記二量体の上流半分におけるコドンジャグリングされた配列と、必要に応じてバクテリオファージターミネーターとを含む、単離された転写単位。
【請求項21】
前記コドンジャグリングされたコード配列が、前記二量体の上流半分に挿入された複数のサイレントヌクレオチド置換を含む、請求項20に記載の単離された転写単位。
【請求項22】
前記一本鎖コートポリペプチド二量体の前記コード配列が、前記コートタンパク質の下流サブユニットの第一のアミノ酸をコードするコドンを置き換える終止コドンを含む、請求項19又は20に記載の単離された転写単位。
【請求項23】
前記終止コドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項21に記載の単離された転写単位。
【請求項24】
低い価数を示すウイルス様粒子のライブラリーを構築するための方法であって、それぞれのウイルス様粒子が、少なくとも1つの一本鎖二量体の下流の一本鎖RNA中に異種ペプチドを含むが、前記ウイルス様粒子において全てがバクテリオファージコートポリペプチド二量体ではない方法であって、
(a)請求項12に記載の複数の転写単位を提供する工程と、
(b)(a)の前記転写単位を制限酵素で処理する工程と、
(c)異種ペプチドのためのコード配列を前記転写単位に挿入して転写単位の集団を得る工程と、
(d)(c)の前記転写単位を発現する工程と、
(e)前記ライブラリーを単離する工程と、
を含む方法。
【請求項25】
前記転写単位が、結合された転写翻訳システムで発現される、請求項24に記載の方法。
【請求項26】
前記異種ペプチドが、少なくとも4ヌクレオチドの長さを有する、請求項24に記載の方法。
【請求項27】
結合された転写翻訳システムにおいて前記転写単位を発現する工程と、区画化された水/オイルエマルジョン中で結合された転写/翻訳の少なくとも1サイクルを行う工程とをさらに含む、請求項24〜26のいずれかに記載の方法。
【請求項28】
目的の特性を有するペプチドを特定するための方法であって、
(a)請求項1の低い価数のVLPの集団を提供する工程と、
(b)目的の特性を有するペプチドを特定するために目的の特性について請求項1に記載の集団中においてVLPで発現された異種ペプチドを試験する工程と、
を含む方法。
【請求項29】
請求項28に記載の方法であって、(a)におけるVLPの前記集団が、バクテリオファージプロモーター及び3’ターミネーターを含む転写単位を発現することによって得られ、RNAバクテリオファージ一本鎖コートポリペプチド二量体の改変コード配列が上流サブユニット及び下流サブユニットを含み、異種ペプチドを挿入するための部位が前記二量体の前記下流サブユニット中で少なくとも4アミノ酸単位の長さであり、終止コドンが、前記コートタンパク質の下流サブユニットの第一のアミノ酸又は前記下流サブユニットのカルボキシ末端後の第一のアミノ酸をコードするコドンを、区画化された水/オイルエマルジョン中の核酸テンプレート由来の結合された転写/翻訳システム中で置き換え、前記水/オイルのエマルジョンから前記集団を回収する方法。
【請求項30】
目的の特性を有する前記ペプチドが免疫原性ペプチドである、請求項28又は29に記載の方法。
【請求項31】
目的の特性を有する前記ペプチドが自己抗原の免疫原性フラグメントである、請求項28〜30のいずれかに記載の方法。
【請求項32】
前記免疫原性フラグメントが、免疫原性HIVペプチドのフラグメントである、請求項30に記載の方法。
【請求項33】
目的の特性を有する前記ペプチドがミモトープである、請求項32に記載の方法。
【請求項34】
免疫原性タンパク質を単離するための方法であって、
(a)請求項24に記載の方法によってVLPの集団から前記免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)前記免疫原性ペプチドを単離する工程と、
を含む方法。
【請求項35】
前記免疫原性ペプチドが、アフィニティー選択によって単離される、請求項34に記載の方法。
【請求項36】
低い価数を示すRNAバクテリオファージの単離されたVLPであって、コートポリペプチドの下流サブユニット中の異種ペプチドの挿入によって改変された前記バクテリオファージの前記コートポリペプチドの一本鎖二量体を含み、前記異種ペプチドは、前記VLPで提示されており、前記異種ペプチドは、HIVペプチド、自己抗原、レセプター及び細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に対して親和性を有するペプチド、金属結合ペプチド又は前記バクテリオファージの表面に親和性を有するペプチドからなる群より選択され、約1〜10個以下の異種ペプチドが前記VLPで提示される、単離されたVLP。
【請求項37】
前記VLPが検出可能標識に結合されている、請求項36に記載の単離されたVLP。
【請求項38】
低い価数を示すRNAバクテリオファージの単離されたVLPであって、異種ペプチドの挿入によって改変された前記ファージのコートポリペプチドの一本鎖二量体を含み、前記異種ペプチドは、前記VLPで提示されており、前記異種ペプチドは、細胞表面レセプターに結合するためのリガンドに結合された薬学的に有効な異種ポリペプチドであり、約1〜10個以下の異種ペプチドが前記VLPで提示される、単離されたVLP。
【請求項39】
請求項36〜38のいずれかに記載のVLPを含む組成物。
【請求項40】
免疫原性組成物であって、低い価数のRNAバクテリオファージの1つ以上のVLPと、前記ファージのコートポリペプチドの一本鎖二量体とを含み、前記コートポリペプチドは、上流サブユニット及び下流サブユニットを含み、前記下流サブユニットは、免疫原性の異種ペプチドの挿入によって改変されており、約1〜10個以下の異種ペプチドが前記VLPで提示される、免疫原性組成物。
【請求項41】
前記異種ペプチドが、カルボキシル末端で前記下流サブユニット中に挿入されている、請求項40に記載の免疫原性組成物。
【請求項42】
前記異種ペプチドが、前記ABループ内の前記下流サブユニット中に挿入されている、請求項40に記載の免疫原性組成物。
【請求項43】
前記異種ペプチドが、自己抗原又はその免疫原性フラグメントである、請求項40〜42のいずれかに記載の免疫原性組成物。
【請求項44】
前記自己抗原が、ErbB−2、アミロイド−β、免疫グロブリンE(IgE)、ガストリン、グレリン、血管内皮成長因子(VEGF)、インターロイキン(IL)−17、IL−23、IL−13、CCR5、CXCR4、神経成長因子(NGF)、アンジオテンシンII、TRANCE/RANKL及びMUC−1からなる群由来のペプチドからなる群から選択される、請求項43に記載の免疫原性組成物。
【請求項45】
前記異種ペプチドがHIV免疫原性ペプチドである、請求項40に記載の免疫原性組成物。
【請求項46】
前記異種ペプチドが、前記コートポリペプチドの前記下流サブユニットのN末端中に挿入される、請求項40に記載の免疫原性組成物。
【請求項47】
核酸構築物であって、
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、前記コートポリペプチド二量体のコード配列が、前記コートポリペプチド二量体のABループを規定する配列の部分に対して5’側に配置された第一の制限部位を規定するように改変されている、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)抗生物質耐性遺伝子と、
(d)原核生物中での複製のための複製起点と、
を含み、前記構築物は、必要に応じて前記第一の制限部位に対する5’側と前記第二の制限部位に対する3’側とに配置されたPCRプライマーと、
を含む核酸構築物。
【請求項48】
前記第一の制限部位に対して5’側に配置された第一のプライマーと、前記第二の制限部位に対して3’側に配置された第二のプライマーとを含む、請求項47に記載の核酸構築物。
【請求項49】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がSalI及びKpnI制限部位のいずれかであり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項47に記載の核酸構築物。
【請求項50】
前記構築物がさらに、前記第二の制限部位に対して5’側に配置された転写ターミネーターを含む、請求項47、48又は49に記載の核酸構築物。
【請求項51】
請求項47、48又は49に記載の核酸構築物であって、
(1)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、
核酸構築物。
【請求項52】
請求項51に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項53】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つが、改変された二量体を生じるために複数のサイレントヌクレオチド置換で改変され、それにより変異原性のオリゴヌクレオチドプライマーが、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された任意の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)第一の抗生物質に対する耐性のための遺伝子と、
(e)原核生物細胞における複製のための複製起点と、
(f)一本鎖DNAバクテリオファージからの第二の複製起点と、
(g)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項54】
コドン配列が、AB−ループのいずれかの側に20ヌクレオチド単位内の十分な数のサイレントなヌクレオチド置換を含む、請求項53に記載の核酸構築物。
【請求項55】
前記第一の抗生物質に対して耐性のリプレッサーがカナマイシン耐性遺伝子であり、ヘルパーファージがクロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されている、請求項53又は54に記載の核酸構築物。
【請求項56】
前記核酸構築物が、第二の制限部位に対して3’側に配置された転写ターミネーターをさらに含む、請求項53又は54に記載の核酸構築物。
【請求項57】
請求項53又は54に記載の核酸構築物であって、前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記RNAバクテリオファージ一本鎖コートポリペプチド二量体が、MS2コートタンパク質一本鎖二量体であり、コドン配列が可能な最大数のサイレントヌクレオチド置換を含み、前記制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記第一の抗生物質に対する耐性遺伝子がカナマイシン耐性遺伝子であり、前記ヘルパーファージの遺伝子が、クロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されており、前記複製起点がcolE1 oriである、核酸構築物。
【請求項58】
請求項53又は54に記載の核酸構築物であって、
(1)前記バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、
核酸構築物。
【請求項59】
請求項53又は54に記載の核酸構築物であって、
(1)前記バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含み、
(3)前記第一の抗生物質に対する耐性遺伝子が、カナマイシン耐性遺伝子であり、かつ前記ヘルパーファージの遺伝子が、クロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されている、
核酸構築物。
【請求項60】
請求項58に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項61】
請求項59に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項62】
配列番号1〜3の核酸構築物。
【請求項63】
請求項47〜62に記載の核酸構築物によって形質転換された原核生物。
【請求項64】
前記原核生物が大腸菌株である、請求項63に記載の原核生物。
【請求項65】
請求項51に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項66】
請求項52に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項67】
請求項58に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項68】
請求項59に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項69】
請求項60に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項70】
請求項61に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項71】
請求項65〜70に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつMS2のmRNAをカプシドで包む、
ウイルス様粒子の集団。
【請求項72】
請求項71に記載のウイルス様粒子の集団であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、ウイルス様粒子の集団。
【請求項73】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項47〜62に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつ前記MS2のmRNAをカプシドで包む、方法。
【請求項74】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項19〜24に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつMS2のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項75】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項73に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項76】
請求項65〜70に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつMS2のmRNAをカプシドで包む、免疫原性組成物。
【請求項77】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のABループを規定する配列の部分に対して5’側に配置されるか又はその中に配置される第一の制限部位を規定するように改変される、プロモーターと、
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)抗生物質耐性遺伝子と、
(d)原核生物細胞中の複製のための複製起点と、
を含み、前記構築物が、必要に応じて第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーを含む、核酸構築物。
【請求項78】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がカナマイシンであり、前記複製起点がcolE1 oriである、請求項77に記載の核酸構築物。
【請求項79】
前記核酸構築物が、さらに前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項77又は78に記載の核酸構築物。
【請求項80】
請求項77又は78に記載の核酸構築物であって、
(1)バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が異種ペプチドをコードする核酸配列をさらに含み、
(2)前記構築物が、必要に応じて前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項81】
請求項80に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項82】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のコード配列の下流部分に配置されかつ前記コートポリペプチド二量体のABループを規定する配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように改変される、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)前記第一の制限部位に対する5’側と前記第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
を含む、核酸構築物。
【請求項83】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項82に記載の核酸構築物。
【請求項84】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項82又は83に記載の核酸構築物。
【請求項85】
請求項82又は83に記載の核酸構築物であって、
(1)バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項86】
請求項85に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項87】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のコード配列の下流部分に配置されかつ前記コートポリペプチド二量体のABループを規定する前記配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように改変される、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
(f)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(g)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項88】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項87に記載の核酸構築物。
【請求項89】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項87又は88に記載の核酸構築物。
【請求項90】
請求項87又は88に記載の核酸構築物であって、
(1)前記バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項91】
請求項90に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項92】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、(1)前記コートポリペプチド二量体のコード配列の下流部分に配置され、前記コートポリペプチド二量体のABループを規定する前記配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように、かつ(2)ヌクレオチド配列(NNS)xを含むように改変され、ここでNは任意のヌクレオチドであり、Sはグアノシンヌクレオチド(G)又はシチジンヌクレオチド(C)であり、xは1〜500の整数である、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
を含む、核酸構築物。
【請求項93】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がアンピシリン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項92に記載の核酸構築物。
【請求項94】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項92又は93に記載の核酸構築物。
【請求項95】
請求項92又は93に記載の核酸構築物であって、
(1)前記バクテリオファージMS7又はPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項96】
請求項95に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項97】
請求項77〜96に記載の核酸構築物によって形質転換された原核生物。
【請求項98】
前記原核生物が大腸菌株である、請求項97に記載の原核生物。
【請求項99】
請求項85に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項100】
請求項86に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項101】
請求項90に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項102】
請求項91に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項103】
請求項95に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項104】
請求項76に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項105】
請求項99〜104に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつPP7のmRNAをカプシドで包む、
ウイルス様粒子の集団。
【請求項106】
一本鎖二量体の上流サブユニットと下流サブユニットとの間の接合部にナンセンスコドンを含む、請求項47〜62及び77〜96のいずれかに記載の構築物。
【請求項107】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成する、請求項106に記載の構築物。
【請求項108】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成するプラスミドと会合されている、請求項106に記載の構築物。
【請求項109】
前記ナンセンスコドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項106〜108のいずれかに記載の構築物。
【請求項109】
前記サプレッサーtRNAがアラニン−挿入アンバー−サプレッサーである、請求項107又は108に記載の構築物。
【請求項110】
前記プラスミドが、第二の不適合性群由来の複製起点を有し、第二の抗生物質に対して耐性を付与する、請求項108に記載の構築物。
【請求項111】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項77〜96に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつPP7のmRNAをカプシドで包む、方法。
【請求項112】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項99〜104に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項113】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項112に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項114】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項112に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて免疫原性ペプチドを単離する工程と、
を含み、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、方法。
【請求項115】
請求項99〜104に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包む、免疫原性組成物。
【請求項115】
請求項99〜104に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが(i)前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包み、(ii)HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、免疫原性組成物。
【請求項116】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つは、改変された二量体を生成するために複数のサイレントヌクレオチドで改変され、それにより変異原性オリゴヌクレオチドプライマーは、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)A−Bループに特異的であるコートポリペプチド配列の部分に対して5’側に配置された第一の制限部位と、
(c)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(e)第一の抗生物質に対して耐性の遺伝子と、
(f)原核生物細胞中の複製のための複製起点と、
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(h)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項117】
前記第二の複製起点がM13由来である、請求項116に記載の核酸構築物。
【請求項118】
前記ヘルパーバクテリオファージがM13である、請求項116又は117に記載の核酸構築物。
【請求項119】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、かつ前記複製起点がcolE1 oriである、請求項116に記載の核酸構築物。
【請求項120】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項116又は117に記載の核酸構築物。
【請求項121】
請求項116又は117に記載の核酸構築物であって、
(1)前記バクテリオファージMS2の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項122】
請求項121に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項123】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つは、改変された二量体を生成するために複数のサイレントヌクレオチドで改変され、それにより変異原性オリゴヌクレオチドプライマーは、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)A−Bループに特異的であるコートポリペプチド配列の部分に対して5’側に配置された第一の制限部位と、
(c)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(e)第一の抗生物質に対して耐性の遺伝子と、
(f)原核生物細胞中の複製のための複製起点と、
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(h)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項125】
前記第二の複製起点がM13由来である、請求項124に記載の核酸構築物。
【請求項126】
前記ヘルパーバクテリオファージがM13である、請求項124又は125に記載の核酸構築物。
【請求項127】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、かつ前記複製起点がcolE1 oriである、請求項124に記載の核酸構築物。
【請求項127】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項124又は125に記載の核酸構築物。
【請求項128】
請求項124又は125に記載の核酸構築物であって、
(1)前記バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項129】
請求項128に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項130】
請求項116〜129に記載の核酸構築物によって形質転換された原核生物。
【請求項131】
前記原核生物が大腸菌株である、請求項130に記載の原核生物。
【請求項132】
請求項116〜122のいずれかに記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項133】
請求項123〜129のいずれか1項に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項134】
請求項132に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつMS2のmRNAをそれぞれカプシドで包む、
ウイルス様粒子の集団。
【請求項135】
請求項133に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつPP7のmRNAをそれぞれカプシドで包む、
ウイルス様粒子の集団。
【請求項140】
一本鎖二量体の上流サブユニットと下流サブユニットとの間の接合部にナンセンスコドンを含む、請求項116〜129のいずれかに記載の構築物。
【請求項141】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成する、請求項140に記載の構築物。
【請求項142】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成するプラスミドと会合されている、請求項140に記載の構築物。
【請求項143】
前記ナンセンスコドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項140〜142のいずれかに記載の構築物。
【請求項144】
前記サプレッサーtRNAが、アラニン−挿入アンバー−サプレッサーである、請求項141又は142に記載の構築物。
【請求項145】
前記プラスミドが、第二の不適合性群由来の複製起点を有し、追加の抗生物質に対して耐性を付与する、請求項116〜144のいずれかに記載の構築物。
【請求項146】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項116〜129に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、方法。
【請求項147】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項132〜133に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項148】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項147に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項149】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項147に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて免疫原性ペプチドを単離する工程と、
を含み、前記異種ペプチドは、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、方法。
【請求項150】
請求項132〜133に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、
免疫原性組成物。
【請求項151】
請求項132〜133に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、(i)前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包み、(ii)HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、
免疫原性組成物。
【請求項1】
ウイルス様粒子(VLP)の集団であって、それぞれのVLPは、(a)RNAバクテリオファージのVLPであり、前記VLPの一部は、(b)異種ペプチドの挿入によって改変された前記RNAバクテリオファージのコートポリペプチドの一本鎖二量体を含み、前記二量体は、上流及び下流のサブユニットを含み、前記異種ペプチドは、前記VLPに提示されかつ(c)前記バクテリオファージのmRNAをカプシドで包み、前記VLPは低い価数を示す、ウイルス様粒子(VLP)の集団。
【請求項2】
前記異種ペプチドが少なくとも4アミノ酸長である、請求項1に記載の集団。
【請求項3】
前記バクテリオファージの複数の前記コートポリペプチドが、野生型コートポリペプチドであり、残りの前記コートポリペプチドが、異種ペプチドを含む前記コートポリペプチドの一本鎖二量体を含み、前記バクテリオファージが、MS2及びPP7からなる群より選択される、請求項1に記載の集団。
【請求項4】
前記異種ペプチドが、前記コートポリペプチドの下流サブユニット中のABループでの部位又は前記RNAバクテリオファージのコートポリペプチドのカルボキシ末端での部位に挿入される、請求項1に記載の集団。
【請求項5】
前記異種ペプチドが、前記RNAバクテリオファージのコートポリペプチドのカルボキシ末端に対応する部位に挿入される、請求項4に記載の集団。
【請求項6】
前記RNAバクテリオファージが、MS2又はPP7、Qβ、R17、SP、PP7、GA、M11、MX1、AP205、PRR1、f4、Cb5、Cb12r、Cb23r、7s及びf2からなる群より選択される、請求項1に記載の集団。
【請求項7】
前記コートポリペプチドが一本鎖二量体であり、前記コートポリペプチドの下流のコピーが、少なくとも4アミノ酸長を有する異種ペプチドの挿入によってABループ中で改変される、請求項1に記載の集団。
【請求項8】
前記VLPで前記異種ペプチドの1コピーから10コピーまでの間のコピーが提示される、請求項1に記載の集団。
【請求項9】
前記VLPで前記異種ペプチドの1コピーから5コピーまでの間のコピーが提示される、請求項1に記載の集団。
【請求項10】
前記VLPで前記異種ペプチドの1コピーより少ないコピーが提示される、請求項1に記載の集団。
【請求項11】
前記異種ペプチドが、前記二量体タンパク質のカルボキシ末端に、又は
(a)MS−2、R−17及びfrコートポリペプチドのアミノ酸11〜17、
(b)GAコートポリペプチドのアミノ酸10〜16、
(c)Qβ及びSPコートポリペプチドのアミノ酸10〜17、
(d)PP7コートポリペプチドのアミノ酸8〜11、
(e)PRR1コートポリペプチドのアミノ酸9〜17
に対応する前記コートタンパク質のAB−ループ中の部位に挿入される、請求項4に記載の集団。
【請求項12】
細菌又はバクテリオファージプロモーターと、
上流サブユニット及び下流サブユニット、前記下流サブユニットに配置される異種ペプチドの挿入のための部位を含むRNAバクテリオファージ一本鎖コートポリペプチド二量体のコード配列と、
前記コートタンパク質の下流サブユニットの第一のアミノ酸をコードするコドンを置き換える終止コドンと、
細菌又はバクテリオファージターミネーターと、
を含む、単離された転写単位。
【請求項13】
アラニン挿入サプレッサーtRNA遺伝子をさらに含む、請求項12に記載の転写単位。
【請求項14】
前記プロモーターがバクテリオファージプロモーターであり、前記ターミネーターがバクテリオファージターミネーターである、請求項12又は13に記載の単離された転写単位。
【請求項15】
前記異種ペプチドの挿入のための部位が制限酵素部位である、請求項12〜14のいずれかに記載の単離された転写単位。
【請求項16】
一本鎖二量体が、改変された一本鎖コートポリペプチド二量体であり、前記改変が、前記挿入部位に挿入されている異種ペプチド配列を少なくとも4アミノ酸含む、請求項12〜15のいずれかに記載の単離された転写単位。
【請求項17】
前記転写単位が、翻訳オペレーター配列を含まない、請求項12に記載の単離された転写単位。
【請求項18】
前記転写単位が、翻訳オペレーター配列を含む、請求項12に記載の単離された転写単位。
【請求項19】
前記終止コドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項12〜18のいずれかに記載の単離された転写単位。
【請求項20】
バクテリオファージプロモーターを含む単離された転写単位であって、RNAバクテリオファージ一本鎖コートポリペプチド二量体のコード配列が、前記二量体の下流半分において前記コード配列中の異種ペプチドの挿入のための部位を有し、前記コード配列が、前記二量体の上流半分におけるコドンジャグリングされた配列と、必要に応じてバクテリオファージターミネーターとを含む、単離された転写単位。
【請求項21】
前記コドンジャグリングされたコード配列が、前記二量体の上流半分に挿入された複数のサイレントヌクレオチド置換を含む、請求項20に記載の単離された転写単位。
【請求項22】
前記一本鎖コートポリペプチド二量体の前記コード配列が、前記コートタンパク質の下流サブユニットの第一のアミノ酸をコードするコドンを置き換える終止コドンを含む、請求項19又は20に記載の単離された転写単位。
【請求項23】
前記終止コドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項21に記載の単離された転写単位。
【請求項24】
低い価数を示すウイルス様粒子のライブラリーを構築するための方法であって、それぞれのウイルス様粒子が、少なくとも1つの一本鎖二量体の下流の一本鎖RNA中に異種ペプチドを含むが、前記ウイルス様粒子において全てがバクテリオファージコートポリペプチド二量体ではない方法であって、
(a)請求項12に記載の複数の転写単位を提供する工程と、
(b)(a)の前記転写単位を制限酵素で処理する工程と、
(c)異種ペプチドのためのコード配列を前記転写単位に挿入して転写単位の集団を得る工程と、
(d)(c)の前記転写単位を発現する工程と、
(e)前記ライブラリーを単離する工程と、
を含む方法。
【請求項25】
前記転写単位が、結合された転写翻訳システムで発現される、請求項24に記載の方法。
【請求項26】
前記異種ペプチドが、少なくとも4ヌクレオチドの長さを有する、請求項24に記載の方法。
【請求項27】
結合された転写翻訳システムにおいて前記転写単位を発現する工程と、区画化された水/オイルエマルジョン中で結合された転写/翻訳の少なくとも1サイクルを行う工程とをさらに含む、請求項24〜26のいずれかに記載の方法。
【請求項28】
目的の特性を有するペプチドを特定するための方法であって、
(a)請求項1の低い価数のVLPの集団を提供する工程と、
(b)目的の特性を有するペプチドを特定するために目的の特性について請求項1に記載の集団中においてVLPで発現された異種ペプチドを試験する工程と、
を含む方法。
【請求項29】
請求項28に記載の方法であって、(a)におけるVLPの前記集団が、バクテリオファージプロモーター及び3’ターミネーターを含む転写単位を発現することによって得られ、RNAバクテリオファージ一本鎖コートポリペプチド二量体の改変コード配列が上流サブユニット及び下流サブユニットを含み、異種ペプチドを挿入するための部位が前記二量体の前記下流サブユニット中で少なくとも4アミノ酸単位の長さであり、終止コドンが、前記コートタンパク質の下流サブユニットの第一のアミノ酸又は前記下流サブユニットのカルボキシ末端後の第一のアミノ酸をコードするコドンを、区画化された水/オイルエマルジョン中の核酸テンプレート由来の結合された転写/翻訳システム中で置き換え、前記水/オイルのエマルジョンから前記集団を回収する方法。
【請求項30】
目的の特性を有する前記ペプチドが免疫原性ペプチドである、請求項28又は29に記載の方法。
【請求項31】
目的の特性を有する前記ペプチドが自己抗原の免疫原性フラグメントである、請求項28〜30のいずれかに記載の方法。
【請求項32】
前記免疫原性フラグメントが、免疫原性HIVペプチドのフラグメントである、請求項30に記載の方法。
【請求項33】
目的の特性を有する前記ペプチドがミモトープである、請求項32に記載の方法。
【請求項34】
免疫原性タンパク質を単離するための方法であって、
(a)請求項24に記載の方法によってVLPの集団から前記免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)前記免疫原性ペプチドを単離する工程と、
を含む方法。
【請求項35】
前記免疫原性ペプチドが、アフィニティー選択によって単離される、請求項34に記載の方法。
【請求項36】
低い価数を示すRNAバクテリオファージの単離されたVLPであって、コートポリペプチドの下流サブユニット中の異種ペプチドの挿入によって改変された前記バクテリオファージの前記コートポリペプチドの一本鎖二量体を含み、前記異種ペプチドは、前記VLPで提示されており、前記異種ペプチドは、HIVペプチド、自己抗原、レセプター及び細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に対して親和性を有するペプチド、金属結合ペプチド又は前記バクテリオファージの表面に親和性を有するペプチドからなる群より選択され、約1〜10個以下の異種ペプチドが前記VLPで提示される、単離されたVLP。
【請求項37】
前記VLPが検出可能標識に結合されている、請求項36に記載の単離されたVLP。
【請求項38】
低い価数を示すRNAバクテリオファージの単離されたVLPであって、異種ペプチドの挿入によって改変された前記ファージのコートポリペプチドの一本鎖二量体を含み、前記異種ペプチドは、前記VLPで提示されており、前記異種ペプチドは、細胞表面レセプターに結合するためのリガンドに結合された薬学的に有効な異種ポリペプチドであり、約1〜10個以下の異種ペプチドが前記VLPで提示される、単離されたVLP。
【請求項39】
請求項36〜38のいずれかに記載のVLPを含む組成物。
【請求項40】
免疫原性組成物であって、低い価数のRNAバクテリオファージの1つ以上のVLPと、前記ファージのコートポリペプチドの一本鎖二量体とを含み、前記コートポリペプチドは、上流サブユニット及び下流サブユニットを含み、前記下流サブユニットは、免疫原性の異種ペプチドの挿入によって改変されており、約1〜10個以下の異種ペプチドが前記VLPで提示される、免疫原性組成物。
【請求項41】
前記異種ペプチドが、カルボキシル末端で前記下流サブユニット中に挿入されている、請求項40に記載の免疫原性組成物。
【請求項42】
前記異種ペプチドが、前記ABループ内の前記下流サブユニット中に挿入されている、請求項40に記載の免疫原性組成物。
【請求項43】
前記異種ペプチドが、自己抗原又はその免疫原性フラグメントである、請求項40〜42のいずれかに記載の免疫原性組成物。
【請求項44】
前記自己抗原が、ErbB−2、アミロイド−β、免疫グロブリンE(IgE)、ガストリン、グレリン、血管内皮成長因子(VEGF)、インターロイキン(IL)−17、IL−23、IL−13、CCR5、CXCR4、神経成長因子(NGF)、アンジオテンシンII、TRANCE/RANKL及びMUC−1からなる群由来のペプチドからなる群から選択される、請求項43に記載の免疫原性組成物。
【請求項45】
前記異種ペプチドがHIV免疫原性ペプチドである、請求項40に記載の免疫原性組成物。
【請求項46】
前記異種ペプチドが、前記コートポリペプチドの前記下流サブユニットのN末端中に挿入される、請求項40に記載の免疫原性組成物。
【請求項47】
核酸構築物であって、
(a)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合されている細菌又はバクテリオファージのプロモーターであって、前記コートポリペプチド二量体のコード配列が、前記コートポリペプチド二量体のABループを規定する配列の部分に対して5’側に配置された第一の制限部位を規定するように改変されている、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)抗生物質耐性遺伝子と、
(d)原核生物中での複製のための複製起点と、
を含み、前記構築物は、必要に応じて前記第一の制限部位に対する5’側と前記第二の制限部位に対する3’側とに配置されたPCRプライマーと、
を含む核酸構築物。
【請求項48】
前記第一の制限部位に対して5’側に配置された第一のプライマーと、前記第二の制限部位に対して3’側に配置された第二のプライマーとを含む、請求項47に記載の核酸構築物。
【請求項49】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がSalI及びKpnI制限部位のいずれかであり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項47に記載の核酸構築物。
【請求項50】
前記構築物がさらに、前記第二の制限部位に対して5’側に配置された転写ターミネーターを含む、請求項47、48又は49に記載の核酸構築物。
【請求項51】
請求項47、48又は49に記載の核酸構築物であって、
(1)バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、
核酸構築物。
【請求項52】
請求項51に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項53】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つが、改変された二量体を生じるために複数のサイレントヌクレオチド置換で改変され、それにより変異原性のオリゴヌクレオチドプライマーが、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された任意の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)第一の抗生物質に対する耐性のための遺伝子と、
(e)原核生物細胞における複製のための複製起点と、
(f)一本鎖DNAバクテリオファージからの第二の複製起点と、
(g)第二の抗生物質に対して耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項54】
コドン配列が、AB−ループのいずれかの側に20ヌクレオチド単位内の十分な数のサイレントなヌクレオチド置換を含む、請求項53に記載の核酸構築物。
【請求項55】
前記第一の抗生物質に対して耐性のリプレッサーがカナマイシン耐性遺伝子であり、ヘルパーファージがクロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されている、請求項53又は54に記載の核酸構築物。
【請求項56】
前記核酸構築物が、第二の制限部位に対して3’側に配置された転写ターミネーターをさらに含む、請求項53又は54に記載の核酸構築物。
【請求項57】
請求項53又は54に記載の核酸構築物であって、前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記RNAバクテリオファージ一本鎖コートポリペプチド二量体が、MS2コートタンパク質一本鎖二量体であり、コドン配列が可能な最大数のサイレントヌクレオチド置換を含み、前記制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記第一の抗生物質に対する耐性遺伝子がカナマイシン耐性遺伝子であり、前記ヘルパーファージの遺伝子が、クロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されており、前記複製起点がcolE1 oriである、核酸構築物。
【請求項58】
請求項53又は54に記載の核酸構築物であって、
(1)前記バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、
核酸構築物。
【請求項59】
請求項53又は54に記載の核酸構築物であって、
(1)前記バクテリオファージMS2一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含み、
(3)前記第一の抗生物質に対する耐性遺伝子が、カナマイシン耐性遺伝子であり、かつ前記ヘルパーファージの遺伝子が、クロラムフェニコールに対する耐性を付与する遺伝子を含むように改変されている、
核酸構築物。
【請求項60】
請求項58に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項61】
請求項59に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項62】
配列番号1〜3の核酸構築物。
【請求項63】
請求項47〜62に記載の核酸構築物によって形質転換された原核生物。
【請求項64】
前記原核生物が大腸菌株である、請求項63に記載の原核生物。
【請求項65】
請求項51に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項66】
請求項52に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項67】
請求項58に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項68】
請求項59に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項69】
請求項60に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項70】
請求項61に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項71】
請求項65〜70に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつMS2のmRNAをカプシドで包む、
ウイルス様粒子の集団。
【請求項72】
請求項71に記載のウイルス様粒子の集団であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はMS2の表面に親和性を有するペプチドからなる群より選択される、ウイルス様粒子の集団。
【請求項73】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項47〜62に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつ前記MS2のmRNAをカプシドで包む、方法。
【請求項74】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項19〜24に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつMS2のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項75】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項73に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項76】
請求項65〜70に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつMS2のmRNAをカプシドで包む、免疫原性組成物。
【請求項77】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のABループを規定する配列の部分に対して5’側に配置されるか又はその中に配置される第一の制限部位を規定するように改変される、プロモーターと、
(b)コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)抗生物質耐性遺伝子と、
(d)原核生物細胞中の複製のための複製起点と、
を含み、前記構築物が、必要に応じて第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーを含む、核酸構築物。
【請求項78】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がカナマイシンであり、前記複製起点がcolE1 oriである、請求項77に記載の核酸構築物。
【請求項79】
前記核酸構築物が、さらに前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項77又は78に記載の核酸構築物。
【請求項80】
請求項77又は78に記載の核酸構築物であって、
(1)バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が異種ペプチドをコードする核酸配列をさらに含み、
(2)前記構築物が、必要に応じて前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項81】
請求項80に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項82】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のコード配列の下流部分に配置されかつ前記コートポリペプチド二量体のABループを規定する配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように改変される、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)前記第一の制限部位に対する5’側と前記第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
を含む、核酸構築物。
【請求項83】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項82に記載の核酸構築物。
【請求項84】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項82又は83に記載の核酸構築物。
【請求項85】
請求項82又は83に記載の核酸構築物であって、
(1)バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項86】
請求項85に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項87】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、前記コートポリペプチド二量体のコード配列の下流部分に配置されかつ前記コートポリペプチド二量体のABループを規定する前記配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように改変される、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
(f)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(g)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項88】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項87に記載の核酸構築物。
【請求項89】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項87又は88に記載の核酸構築物。
【請求項90】
請求項87又は88に記載の核酸構築物であって、
(1)前記バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて前記第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項91】
請求項90に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項92】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2又はPP7の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、前記コートポリペプチド二量体のコード配列は、(1)前記コートポリペプチド二量体のコード配列の下流部分に配置され、前記コートポリペプチド二量体のABループを規定する前記配列に対して5’側に配置されるか又はその中に配置される、第一の制限部位を規定するように、かつ(2)ヌクレオチド配列(NNS)xを含むように改変され、ここでNは任意のヌクレオチドであり、Sはグアノシンヌクレオチド(G)又はシチジンヌクレオチド(C)であり、xは1〜500の整数である、プロモーターと、
(b)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(c)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(d)抗生物質耐性遺伝子と、
(e)原核生物細胞中の複製のための複製起点と、
を含む、核酸構築物。
【請求項93】
前記細菌又はバクテリオファージのプロモーターがlacプロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記抗生物質耐性遺伝子がアンピシリン耐性遺伝子であり、前記複製起点がcolE1 oriである、請求項92に記載の核酸構築物。
【請求項94】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項92又は93に記載の核酸構築物。
【請求項95】
請求項92又は93に記載の核酸構築物であって、
(1)前記バクテリオファージMS7又はPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項96】
請求項95に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項97】
請求項77〜96に記載の核酸構築物によって形質転換された原核生物。
【請求項98】
前記原核生物が大腸菌株である、請求項97に記載の原核生物。
【請求項99】
請求項85に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項100】
請求項86に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項101】
請求項90に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項102】
請求項91に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項103】
請求項95に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項104】
請求項76に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項105】
請求項99〜104に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつPP7のmRNAをカプシドで包む、
ウイルス様粒子の集団。
【請求項106】
一本鎖二量体の上流サブユニットと下流サブユニットとの間の接合部にナンセンスコドンを含む、請求項47〜62及び77〜96のいずれかに記載の構築物。
【請求項107】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成する、請求項106に記載の構築物。
【請求項108】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成するプラスミドと会合されている、請求項106に記載の構築物。
【請求項109】
前記ナンセンスコドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項106〜108のいずれかに記載の構築物。
【請求項109】
前記サプレッサーtRNAがアラニン−挿入アンバー−サプレッサーである、請求項107又は108に記載の構築物。
【請求項110】
前記プラスミドが、第二の不適合性群由来の複製起点を有し、第二の抗生物質に対して耐性を付与する、請求項108に記載の構築物。
【請求項111】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項77〜96に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつPP7のmRNAをカプシドで包む、方法。
【請求項112】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項99〜104に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項113】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項112に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項114】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項112に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて免疫原性ペプチドを単離する工程と、
を含み、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、方法。
【請求項115】
請求項99〜104に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包む、免疫原性組成物。
【請求項115】
請求項99〜104に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが(i)前記ウイルス様粒子上で提示され、かつPP7のmRNAをカプシドで包み、(ii)HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、免疫原性組成物。
【請求項116】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージMS2の一本鎖コートポリペプチド二量体のコード配列と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つは、改変された二量体を生成するために複数のサイレントヌクレオチドで改変され、それにより変異原性オリゴヌクレオチドプライマーは、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)A−Bループに特異的であるコートポリペプチド配列の部分に対して5’側に配置された第一の制限部位と、
(c)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(e)第一の抗生物質に対して耐性の遺伝子と、
(f)原核生物細胞中の複製のための複製起点と、
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(h)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項117】
前記第二の複製起点がM13由来である、請求項116に記載の核酸構築物。
【請求項118】
前記ヘルパーバクテリオファージがM13である、請求項116又は117に記載の核酸構築物。
【請求項119】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、かつ前記複製起点がcolE1 oriである、請求項116に記載の核酸構築物。
【請求項120】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項116又は117に記載の核酸構築物。
【請求項121】
請求項116又は117に記載の核酸構築物であって、
(1)前記バクテリオファージMS2の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項122】
請求項121に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項123】
核酸構築物であって、
(a)細菌又はバクテリオファージのプロモーターであって、バクテリオファージPP7の一本鎖コートポリペプチド二量体と作動可能に会合され、一本鎖二量体配列の二等分のうちの1つは、改変された二量体を生成するために複数のサイレントヌクレオチドで改変され、それにより変異原性オリゴヌクレオチドプライマーは、前記改変された二量体の前記改変された半分又は未改変の半分に対して特異的にアニールされることができる、プロモーターと、
(b)A−Bループに特異的であるコートポリペプチド配列の部分に対して5’側に配置された第一の制限部位と、
(c)前記コートポリペプチド二量体のコード配列に対して3’側に配置された第二の制限部位と、
(d)第一の制限部位に対する5’側と第二の制限部位に対する3’側とに配置されたPCRプライマーと、
(e)第一の抗生物質に対して耐性の遺伝子と、
(f)原核生物細胞中の複製のための複製起点と、
(g)一本鎖DNAバクテリオファージ由来の第二の複製起点と、
(h)第二の抗生物質に対する耐性を付与する遺伝子を含むように改変されたヘルパー一本鎖DNAバクテリオファージと、
を含む、核酸構築物。
【請求項125】
前記第二の複製起点がM13由来である、請求項124に記載の核酸構築物。
【請求項126】
前記ヘルパーバクテリオファージがM13である、請求項124又は125に記載の核酸構築物。
【請求項127】
前記細菌又はバクテリオファージのプロモーターがT7プロモーターであり、前記第一の制限部位がKpnI制限部位であり、前記第二の制限部位がBamHI部位であり、前記PCRプライマーがTP7であり、前記抗生物質耐性遺伝子がカナマイシン耐性遺伝子であり、かつ前記複製起点がcolE1 oriである、請求項124に記載の核酸構築物。
【請求項127】
前記核酸構築物が、さらに第二の制限部位に対して3’側に配置された転写ターミネーターを含む、請求項124又は125に記載の核酸構築物。
【請求項128】
請求項124又は125に記載の核酸構築物であって、
(1)前記バクテリオファージPP7の一本鎖コートポリペプチド二量体のコード配列が、さらに異種ペプチドをコードする核酸配列を含み、
(2)前記構築物が、必要に応じて第二の制限部位に対して3’側に配置された転写ターミネーターを含む、核酸構築物。
【請求項129】
請求項128に記載の核酸構築物であって、前記異種ペプチドが、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、核酸構築物。
【請求項130】
請求項116〜129に記載の核酸構築物によって形質転換された原核生物。
【請求項131】
前記原核生物が大腸菌株である、請求項130に記載の原核生物。
【請求項132】
請求項116〜122のいずれかに記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項133】
請求項123〜129のいずれか1項に記載の核酸構築物によって形質転換された原核生物によって発現されたウイルス様粒子。
【請求項134】
請求項132に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつMS2のmRNAをそれぞれカプシドで包む、
ウイルス様粒子の集団。
【請求項135】
請求項133に記載のウイルス様粒子の集団であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、前記ウイルス様粒子上に提示され、かつPP7のmRNAをそれぞれカプシドで包む、
ウイルス様粒子の集団。
【請求項140】
一本鎖二量体の上流サブユニットと下流サブユニットとの間の接合部にナンセンスコドンを含む、請求項116〜129のいずれかに記載の構築物。
【請求項141】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成する、請求項140に記載の構築物。
【請求項142】
前記ナンセンスコドンで翻訳終結を少なくとも部分的に抑制することができるサプレッサーtRNAを生成するプラスミドと会合されている、請求項140に記載の構築物。
【請求項143】
前記ナンセンスコドンが、アンバー(UAG)、オパール(UGA)又はオーカー(UAA)終止コドンである、請求項140〜142のいずれかに記載の構築物。
【請求項144】
前記サプレッサーtRNAが、アラニン−挿入アンバー−サプレッサーである、請求項141又は142に記載の構築物。
【請求項145】
前記プラスミドが、第二の不適合性群由来の複製起点を有し、追加の抗生物質に対して耐性を付与する、請求項116〜144のいずれかに記載の構築物。
【請求項146】
ウイルス様粒子のライブラリーを構築するための方法であって、前記方法は、
(a)請求項116〜129に記載の複数の核酸構築物を提供する工程と、
(b)前記核酸構築物を制限酵素で処理する工程と、
(c)異種ペプチドのコード配列を前記核酸構築物中に挿入して、転写単位の集団を得る工程と、
(d)前記転写単位を発現する工程及び必要に応じてライブラリーを単離する工程と、
を含み、それぞれの粒子は、前記異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、前記異種ペプチドは、前記ウイルス様粒子上に提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、方法。
【請求項147】
目的の特性を有するペプチドを特定するための方法であって、前記方法は、
(a)請求項132〜133に記載のウイルス様粒子の集団を提供する工程であって、(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、工程と、
(b)目的の特性について前記ウイルス様粒子上に発現される異種ペプチドを試験する工程と、
を含む、方法。
【請求項148】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項147に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて前記免疫原性ペプチドを単離する工程と、
を含む、方法。
【請求項149】
免疫原性タンパク質を単離するための方法であって、前記方法は、
(a)請求項147に記載の方法によってウイルス様粒子の集団から免疫原性ペプチドを特定する工程と、
(b)前記特定された免疫原性ペプチドを増幅する工程と、
(c)必要に応じて免疫原性ペプチドを単離する工程と、
を含み、前記異種ペプチドは、HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、方法。
【請求項150】
請求項132〜133に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包む、
免疫原性組成物。
【請求項151】
請求項132〜133に記載の1つ以上のウイルス様粒子を含む免疫原性組成物であって、
(1)それぞれの粒子が、異種ペプチドの挿入によって改変されたMS2又はPP7のコートポリペプチドを含み、
(2)前記異種ペプチドが、(i)前記ウイルス様粒子上で提示され、かつそれぞれMS2又はPP7のmRNAをカプシドで包み、(ii)HIVペプチド、自己抗原、Flagペプチド、ヒトパピローマウイルス16型(HPV16)のマイナーカプシドタンパク質L2由来のアミノ酸配列、HIV−1のgp120のV3ループ、炭疽菌の防御抗原、レセプター、細胞表面レセプターに結合するリガンド、繊維状ファージ粒子特異的ペプチドのいずれかの末端に親和性を有するペプチド、金属結合ペプチド又はPP7の表面に親和性を有するペプチドからなる群より選択される、
免疫原性組成物。
【図1】

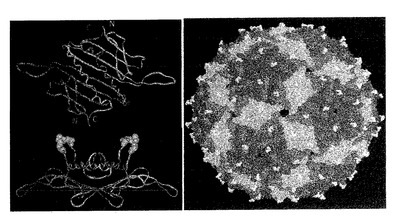
【図2】
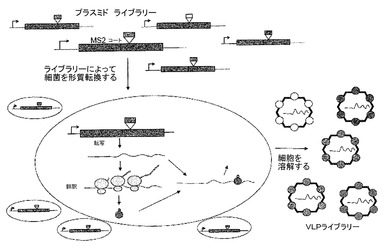
【図3】
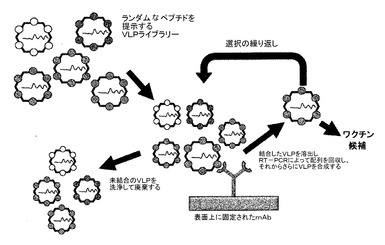
【図4】
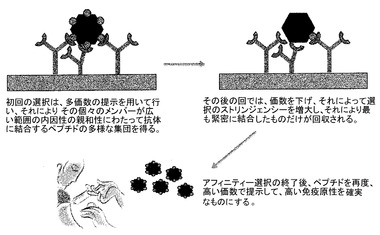
【図5】
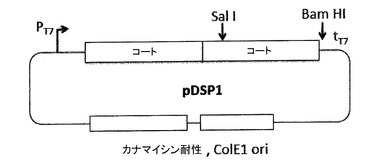
【図6】
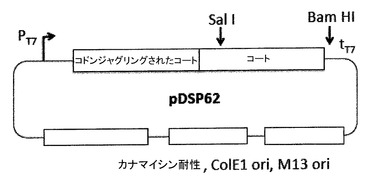
【図7】
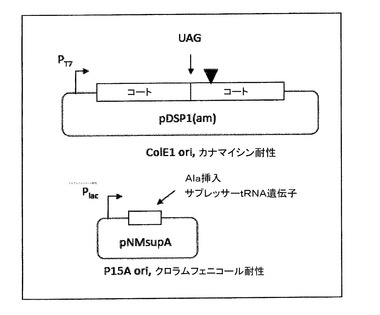
【図8】
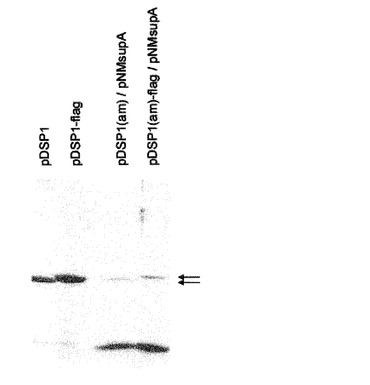
【図9a】
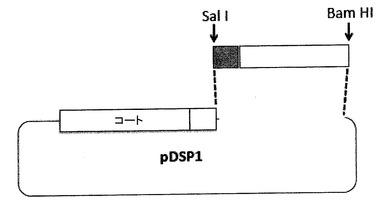
【図9b】
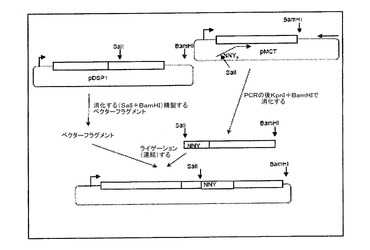
【図10】
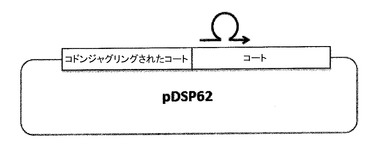
【図11a(1)】

【図11a(2)】

【図11a(3)】

【図11b(1)】

【図11b(2)】

【図11b(3)】

【図11c(1)】

【図11c(2)】

【図11c(3)】

【図11d(1)】

【図11d(2)】

【図11e(1)】

【図11e(2)】

【図11e(3)】

【図11e(4)】

【図12】
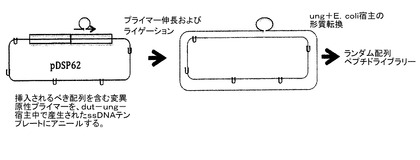
【図13a】
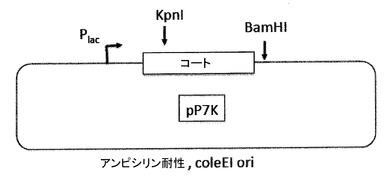
【図13b】
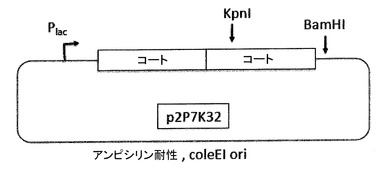
【図14】

【図15】
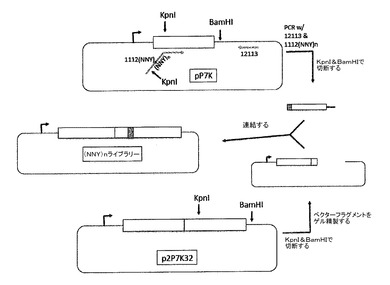
【図16】
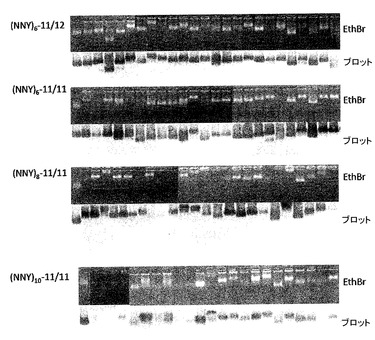
【図17a】
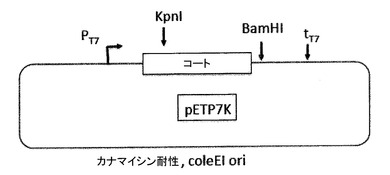
【図17b】
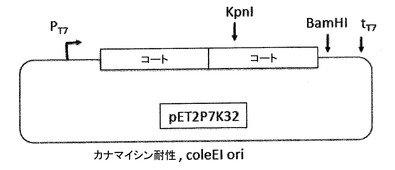
【図17c】
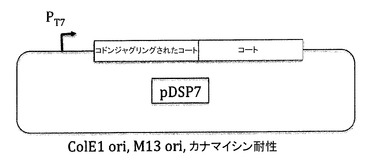
【図18】
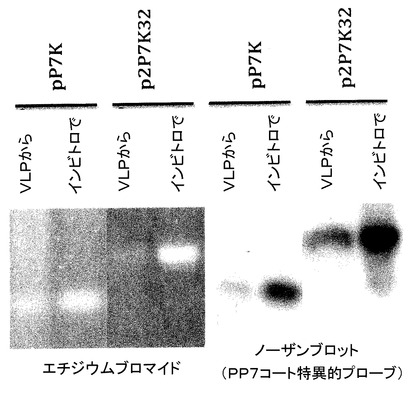
【図19】
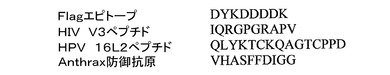
【図20】
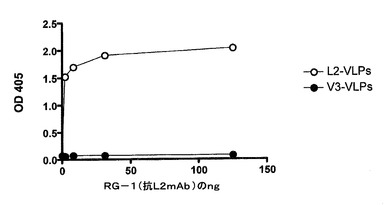
【図21】
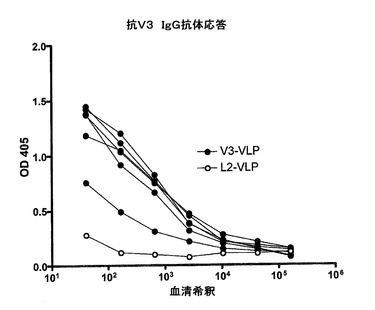
【図22】
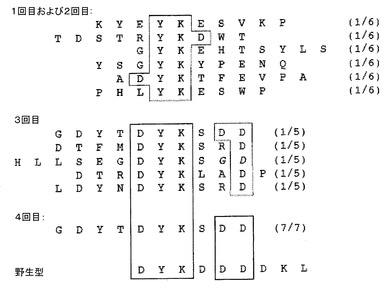
【図23a(1)】

【図23a(2)】

【図23b(1)】

【図23b(2)】

【図23c(1)】

【図23c(2)】

【図23d(1)】

【図23d(2)】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9a】
【図9b】
【図10】
【図11a(1)】
【図11a(2)】
【図11a(3)】
【図11b(1)】
【図11b(2)】
【図11b(3)】
【図11c(1)】
【図11c(2)】
【図11c(3)】
【図11d(1)】
【図11d(2)】
【図11e(1)】
【図11e(2)】
【図11e(3)】
【図11e(4)】
【図12】
【図13a】
【図13b】
【図14】
【図15】
【図16】
【図17a】
【図17b】
【図17c】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23a(1)】
【図23a(2)】
【図23b(1)】
【図23b(2)】
【図23c(1)】
【図23c(2)】
【図23d(1)】
【図23d(2)】
【公表番号】特表2013−516181(P2013−516181A)
【公表日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願番号】特願2012−547321(P2012−547321)
【出願日】平成22年12月31日(2010.12.31)
【国際出願番号】PCT/US2010/062638
【国際公開番号】WO2011/082381
【国際公開日】平成23年7月7日(2011.7.7)
【出願人】(503128320)
【氏名又は名称原語表記】STC.UNM
【Fターム(参考)】
【公表日】平成25年5月13日(2013.5.13)
【国際特許分類】
【出願日】平成22年12月31日(2010.12.31)
【国際出願番号】PCT/US2010/062638
【国際公開番号】WO2011/082381
【国際公開日】平成23年7月7日(2011.7.7)
【出願人】(503128320)
【氏名又は名称原語表記】STC.UNM
【Fターム(参考)】
[ Back to top ]