
RNA中のイノシン化部位の検出方法
【課題】 ノイズやSNPとの判別が容易であり、解析サンプルとしてRNAのみで解析を行うことが可能であり、少量のサンプルでも解析可能であり、さらにバックグラウンドを低く抑えることが可能であり、イノシンの存在を高感度に検出することが可能である、RNA中のイノシン化部位を検出する方法を提供すること。
【解決手段】 RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法
【解決手段】 RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、RNA中のイノシン化部位の検出方法に関する。より詳細には、本発明は、RNA中のイノシン化部位を化学修飾することによって、該イノシン化部位を検出する方法に関する。
【背景技術】
【0002】
脱アミノ化(デアミネーション)による修飾は、RNAエディティングと呼ばれている。RNAエディティングにより、A からIへの変化が生じ、mRNA上のコドンが変化することでアミノ酸配列の変化を引き起こす。二本鎖RNAアデノシンデアミネース(ADAR;adenosine deaminase acting on RNA)はRNAの二本鎖部分を特異的に認識してアデノシン残基をイノシンに変換する酵素である。哺乳動物には、ADAR1、ADAR2、ADAR3の3種類のファミリーが存在する。グルタミン酸受容体のサブユニットGluR-BのmRNAは、ADAR2の基質であり、エキソン11番中のアデノシンがイノシンへとエディティングされ、アミノ酸配列がグルタミンからアルギニンへと変化する。ADAR2のノックアウトマウスはこのエディティングが生じないため、グルタミン酸受容体のカルシウム透過性をコントロールできず、てんかん症状を起こし、早期に死に至ることが知られている。ADARは二本鎖RNAに対し幅広い基質認識能を示すことが知られ、GluR-B以外にも、セロトニンレセプター(5-HT2c)、カリウムチャンネル(Kv1.1)などでAからIへのエディティングが見つかっている。また、興味深いことにADAR2は自身のmRNAのイントロン部位をAからIにエディティングすることで、可変的スプライシングを誘導し、ADAR2の発現量をフィードバック制御していることが知られている。
【0003】
ADARが高発現している脳mRNAには17000塩基に一度の頻度でイノシンが含まれているとの見積もりがあり、ヒトmRNAには未知のエディティング部位が多量に存在することが予測されていた。最近、バイオインフォマティクス的手法を用いた解析により、ヒトmRNAのUTRにおけるAlu因子の反復配列にAからIへのエディティング候補部位が大量に報告された。実験的に実証された例はこのうちのまだわずかであるが、1600遺伝子の12000箇所以上の部位にイノシンが存在するという見積もりは、AからIへのエディティングが遺伝子のグローバルな発現制御に大きな役割を果たしていることを示唆している。Alu因子を持っているのは霊長類だけであり、ADARは脳で高発現していることを考え合わせると、AからIへのエディティングによるトランスクリプトームの複雑性増加は、高度に進化した脳の神経回路の複雑性の獲得と関連している可能性がある。また、RNAエディティング異常に起因する疾患も報告されている。悪性グリオーマ(Malignant glioma)や筋萎縮性側索硬化症(Amyotrophic lateral sclerosis)ではグルタミン酸受容体のサブユニットタンパク質(GluR2)のAからIへのエディティングが顕著に減少している。このように、mRNA中のイノシンはRNAに質的な変化を与えるものであり、高次生命現象と深く関わっている可能性が高い。
【0004】
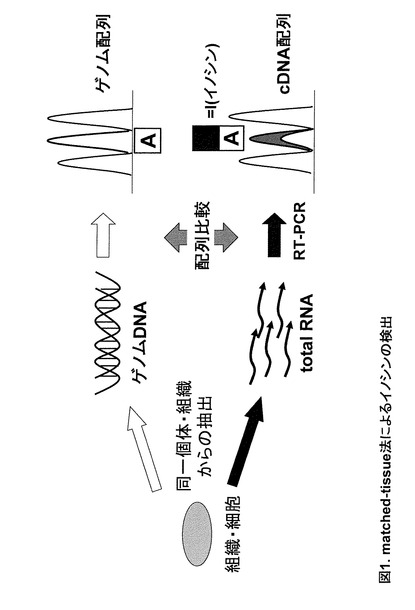
AからIへの RNA エディティング部位(=Iの存在部位)の検出法としてこれまで最も一般的である手法がmatched-tissue 法である(図1)。この方法は同一個体の同一組織由来のRNA(cDNAとして増幅)とゲノムで相当する領域の配列を比較する方法である。IはCと塩基対を形成するため、mRNAから逆転写・PCR増幅後のcDNA中のI相当部位にはGが取り込まれる。よってエディティング部位ではゲノム上の塩基がAであるにも関わらず、cDNA上ではGもしくはA/Gの混在となっている。しかしこの方法ではPCRの非特異増幅・ゲノム混入・シークエンスエラー・ノイズ・対立遺伝子間のSNP (Allele SNP)・偽遺伝子・遺伝子コピー・スプライスバリアント等を原因とする偽シグナルによるGの混在と、目的とするイノシン由来のGの混在を判別することが難しい。また、エキソン境界領域や偽遺伝子の存在する領域の場合、RNAに対応するゲノム領域を判別することが困難であるためこの方法自体用いることができない。さらにこの方法で特定されたイノシン部位は配列上Gとして検出されるため、厳密にはイノシンの存在を証明してはいない。また、同一個体由来のゲノムとRNAの両方を準備する必要があるため、解析可能なサンプルも制限されてしまうという難点も持つ。
【0005】
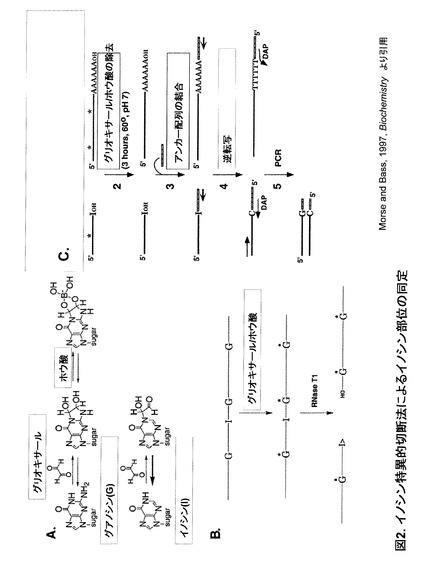
また、これまでに分子生物学分野におけるイノシンの化学修飾法としては2例の報告がある。1つは生体内RNA中のイノシン部位同定を目的としたイノシン特異的切断法である(非特許文献1、2及び3)(図2)。この手法は以下の3ステップからなる。
【0006】
(1)化学修飾試薬としてグリオキサールを用いて、RNA鎖中のグアノシン(G)とイノシン(I)の塩基修飾を行う。Gの場合、グリオキサールは1位及びN'2位に付加してcis-ジオールとなり、さらにホウ酸と化合物(図中G*)される。一方イノシンの場合、2位のアミノ基が存在しないためグリオキサールは1位にのみ付加する。さらに付加した構造は不安定であるため、容易に逆反応を起こして元のイノシンに戻る。
(2)グリオキサール/ホウ酸処理後のRNAをGとイノシンに特異的なRNase T1により切断する。このときGがG*へと修飾されているためにG部位では切断は起らない。一方イノシンは修飾を受けていない(元に戻っている)ためイノシン部位での切断が起る。結果としてイノシン部位特異的な切断が可能となる。
(3)切断後のRNA中のG*から付加物を取り除き、イノシンを含む側のRNAフラグメントにアンカー配列を取り付けてこの部位から逆転写・PCRを行い、その配列を解析する。
【0007】
この方法でMorseらは数例のmRNA中のイノシン部位を同定しているが(非特許文献3)、その後の報告はない。この解析法の成功の鍵はグリオキサール/ホウ酸処理によるGのRNase T1非感受性修飾処理である。RNAを構成する塩基の1/4を占めるGを全て修飾するのは困難であり、未修飾のGが残存することは避けられない。さらに数千〜数万塩基からなる一本のmRNA中にイノシンは0〜30塩基程度しか存在しないほど微量であり、未修飾のG部位の切断によるバックグラウンドが目的のイノシン部位における切断の検出を非常に困難なものにしている。そのため仮に候補部位が得られた場合でも、matched tissue 法による確認が必要である。
【0008】
また、イノシン特異的化学修飾はYoshidaらにより報告されている(非特許文献4)。この解析法では酵母の転位RNA(transfer RNA; tRNA)のアンチコドン部位(遺伝暗号を規定するmRNA上のコドンと対号する)に存在するイノシンの機能解明を目的とし、イノシンの1位をアクリロニトリルを用いてシアノエチル化してコドン−アンチコドン対形成を阻害してその効果を解析している。また反応効率は低いものの、シュードウリジン(□)もシアノエチル化され(非特許文献5)、Mengel-Jorgensenらはシアノエチル化によるシュードウリジンの分子量の変化を質量分析法により解析し、tRNA上の位置を特定する手法を報告している(非特許文献6)。これらの手法は比較的豊富に存在するtRNAを解析対象とし、アンチコドンの不活性化の与える影響もしくはシアノエチル化による質量の変化を解析するものである。
【0009】
【非特許文献1】Morse, D. P. (2004). Identification of substrates for adenosine deaminases that act on RNA. Methods Mol Biol 265, 199-218.
【非特許文献2】Morse, D. P., and Bass, B. L. (1997). Detection of inosine in messenger RNA by inosine-specific cleavage. Biochemistry 36, 8429-8434.
【非特許文献3】Morse, D. P., and Bass, B. L. (1999). Long RNA hairpins that contain inosine are present in Caenorhabditis elegans poly(A)+ RNA. Proc Natl Acad Sci U S A 96, 6048-6053.
【非特許文献4】Yoshida, M., Furuichi, Y., Ukita, T., and Kaziro, Y. (1967). The effect of cyanoethylation on codon recognition of yeast tRNA containing inosine. Biochim Biophys Acta 149, 308-310.
【非特許文献5】Yoshida, M., and Ukita, T. (1968). Modification of nucleosides and nucleotides. 8. The reaction rates of pseudouridine residues with acrylonitrile and its relation to the secondary structure of transfer ribonucleic acid. Biochim Biophys Acta 157, 466-475.
【非特許文献6】Mengel-Jorgensen, J., and Kirpekar, F. (2002). Detection of pseudouridine and other modifications in tRNA by cyanoethylation and MALDI mass spectrometry. Nucleic Acids Res 30, e135.
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、ノイズやSNPとの判別が容易であり、解析サンプルとしてRNAのみで解析を行うことが可能であり、少量のサンプルでも解析可能であり、さらにバックグラウンドを低く抑えることが可能であり、イノシンの存在を高感度に検出することが可能である、RNA中のイノシン化部位を検出する方法を提供することを解決すべき課題とした。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意検討した結果、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾することによって、RNA中のイノシン化部位を検出できることを見出し、本発明を完成するに至った。
【0012】
即ち、本発明によれば、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法が提供される。
【0013】
好ましくは、本発明の方法は、さらに、化学修飾されたRNAを逆転写反応に供してcDNAを合成する工程、及び合成されたcDNAに基づいてイノシン化部位を検出する工程を含む。
好ましくは、イノシン化部位の検出は、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定である。
【0014】
好ましくは、α,β−不飽和結合と電子吸引性基とを有する化合物は、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物である。
好ましくは、電子吸引性基は、CN、NO2、SO3H、CONH2、COCH3、COOCH3、COOC2H5、COCH3、COC2H5、又はCOC6H5である。
【0015】
好ましくは、化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行う。
好ましくは、対照として化学修飾されていないRNAを逆転写反応に供して合成されたcDNAと化学修飾されたRNAから合成されたcDNAとを比較することにより、イノシン化部位を検出する。
好ましくは、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシンを検出する。
好ましくは、化学修飾されたRNAに由来するcDNAの長さを検出することによって、化学修飾されたイノシンを検出する。
【0016】
好ましくは、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによって、化学修飾されたイノシンを検出する。
好ましくは、化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いて検出することによって、化学修飾されたイノシンを検出する。
【0017】
本発明の別の側面によれば、α,β−不飽和結合と電子吸引性基とを有する化合物を含む、RNA中のイノシン化部位修飾剤が提供される。
本発明のさらに別の側面によれば、上記した本発明の修飾剤を含む、上記した本発明の方法を行うための試薬キットが提供される。
【発明の効果】
【0018】
12000箇所以上とも言われるイノシン化部位は、現状では、その大部分が同定されていない。イノシンは脳のmRNAに大量に存在することを考えると、精神疾患(統合失調症、パニック症候群、双極性障害、自閉症など)の異常と関連があると考えられる。これらの疾患と明確に関連するイノシン化部位を本発明の方法によって同定することができれば、疾患の発症機構の解明や治療法を考える上で非常に重要な知見を得ることができる。また、個体ごとにイノシン化部位の変動解析を行うことで、SNPのように生活習慣病や様々な疾患の発症リスクを見積もることが可能になる。
【発明を実施するための最良の形態】
【0019】
以下、本発明の実施の形態についてさらに詳細に説明する。
本発明によるイノシン化部位の検出方法は、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含むことを特徴とする。本発明においてRNAの種類は特に限定されず、mRNA、rRNA又はtRNAの何れでもよい。
【0020】
本発明の方法では、イノシン特異的な修飾・逆転写伸長阻害を検出することにより、cDNA中のイノシンの存在を証明することが可能であり、ノイズやSNPとの判別も容易である。さらに解析サンプルとしてゲノムは必要なくRNAのみで解析が行えるため、少量の貴重なサンプルでも解析可能である。この理由によりスプライス部位などの遺伝子構造や、ゲノム上のコード領域が不明な場合でもイノシン部位を特定することが可能である。これらの点において、本発明の方法は、従来のmatched-tissu法より優れている。また、本発明の方法ではイノシン特異的な修飾を施すため、バックグラウンドを非常に低く抑えることが可能であり、イノシンの存在を高感度に検出・証明することができる利点を持つ。さらに本発明の方法における検出をマイクロアレイで行うことにより、生体内RNA中のイノシン部位を網羅的に特定することができる。さらに、本発明の方法は、全RNA種を対象とし、イノシン特異的な化学修飾による逆転写鎖の伸長阻害を利用するという点において、従来報告されているイノシン特異的化学修飾(Yoshida, M., Furuichi, Y., Ukita, T., and Kaziro, Y. (1967). The effect of cyanoethylation on codon recognition of yeast tRNA containing inosine. Biochim Biophys Acta 149, 308-310)とは区別されるものである。
【0021】
以下、本発明の方法を具体的に説明する。
【0022】
(1)イノシン特異的化学修飾
本発明による方法では先ず、イノシン特異的な化学修飾をRNAに施す。試薬としてはα, β−不飽和−電子求引性基化合物を用いることができる。
【0023】
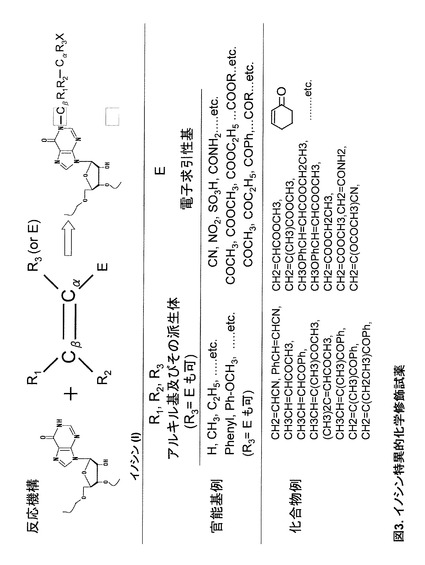
反応機構は基本的にはMichael付加と呼ばれる機構(図3A)で進む。まず、修飾試薬の電子求引性基の影響でβ位の炭素原子が正電荷を帯びる。この炭素原子が、イノシンの活性アミンである1位の窒素原子に求電子付加することにより、イノシンへの付加修飾が進む。修飾試薬の選択によっては、付加官能基特異的にイノシンを含むRNA分子を単離することも可能である。修飾試薬により適切な溶媒を選択して、反応を行うことができる。以下に記載の実施例ではアクリロニトリル(CH2=CHCN)を使用している。アクリロニトリルの場合、同様にシュードウリジンも反応することが知られているが、これは反応時間を短縮することでほぼイノシン特異的な反応条件とすることが可能である。
【0024】
α,β−不飽和結合と電子吸引性基とを有する化合物としては、例えば、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物を挙げることができる。上記化合物中の官能基の例、並びに上記化合物の具体例は、図3B及び図3Cにそれぞれ示す。
【0025】
上記化合物の具体例としては、CH2=CHCN、C6H5CH=CHCN、CH3CH=CHCOCH3、CH3CH=CHCOC6H5、CH3CH=C(CH3)COCH3、(CH3)2C=CHCOCH3、CH3CH=C(CH3)COC6H5、CH2=C(CH3)COC6H5、CH2=C(CH2CH3)COC6H5、CH2=CHCOOCH3、CH2=C(CH3)COOCH3、CH3OC6H5CH=CHCOOCH2CH3、CH3OC6H5CH=CHCOOCH3、CH2=COOCH2CH3、CH2=COOCH3、CH2=CONH2、CH2=C(OCOCH3)CN、及び
【0026】
【化1】

【0027】
などを挙げることができる。
【0028】
(2)逆転写反応(必要に応じて増幅)
本発明の方法では、イノシン特異的に化学修飾を施したRNAに対して逆転写反応を行うことができる。化学修飾をしていない場合、逆転写酵素によりイノシン部位に対してシチジン(C)が取り込まれる。一方、化学修飾をほどこした場合、この化学修飾によりCの取り込みが阻害されて逆転写鎖の伸長はその手前で停止する。
【0029】
逆転写反応は、プライマー、化学修飾を施したRNA(鋳型)、4種のdNTP、逆転写酵素などを用いて常法により行うことができる。即ち、逆転写反応は、上記の試薬を適当な反応液(例えば、適当な塩を含む緩衝液)中で混合して、所定の温度で一定時間インキュベートすることにより行うことができる。
【0030】
本発明においては、化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行うことが好ましい。この場合、逆転写反応と増幅反応は、RT−PCR法により一連の操作により行うことができる。あるいは逆転写後のcDNAを鋳型とした転写増幅により行うことも可能である。
【0031】
また、逆転写反応では、化学修飾されていないRNAを逆転写反応に供してcDNAを合成し、これを以下の検出工程において対照用として用いることもできる。
【0032】
(3)検出
検出においては、質量分析によりイノシンによる化学修飾の有無を検出してもよいし、あるいは、伸長阻害された逆転写鎖を検出し、イノシン化学修飾の有無で比較することによりRNA中のイノシン部位を検出してもよい。この際、検出対象情報としては、その長さ、塩基配列、又は量などが挙げられる。
【0033】
検出の第一の態様としては、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシン化部位を検出することができる。質量分析を行う場合、イノシンの有無及び総量を解析するには、化学修飾後のRNAをサンプルとしてもよいが、好ましくは化学修飾されたRNAをRNaseとフォスファターゼでヌクレオシドまで分解した後に行う。ここで用いるRNaseはヌクレオシドまで分解可能ならば特に限定はなく、例えば、NucleaseP1などを用いることができる。フォスファターゼも特に限定はなく、バクテリアなどの由来する生物種も限定はない。例えば、バクテリAlkaline Phospataseなどを用いることができる。
【0034】
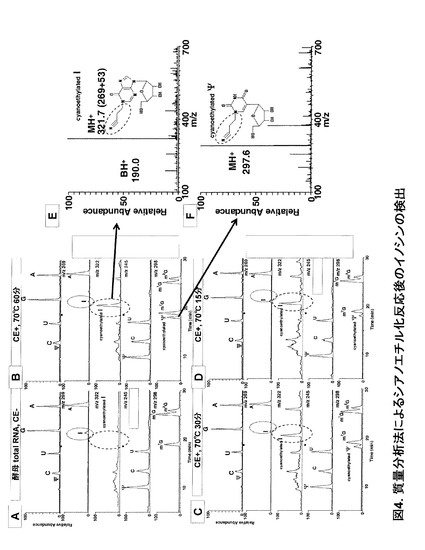
例えば、アクリロニトリルをイノシン化部位の修飾剤として用いてRNAを処理し、その後ヌクレオシドまで分解する。このサンプルを非化学修飾RNA由来のヌクレオチドサンプルと比較して、液体クロマトグラフィー/質量分析法(LC/MS)におけるイノシンのピーク(m/z 269)の減少またはシアノエチル化イノシンのピーク(m/z 322)の出現またはその増加によりイノシン化部位の検出を行うことができる。
【0035】
また、イノシンを含む領域の同定には、化学修飾後のRNAをサンプルとしても良いが、好ましくは化学修飾されたRNAを適当なRNaseにより断片化した後に行う。ここで用いるRNaseはRNAを数塩基から100塩基程度の長さに断片化可能ならば特に限定はなく、例えばG特異的RNase T1などを用いることができる。
【0036】
例えば、アクリロニトリルをイノシン化部位の修飾剤として用いてRNAを処理し、その後RNA断片に分解する。このサンプルを非化学修飾RNA由来のRNA断片化サンプルと比較して、液体クロマトグラフィー/質量分析法(LC/MS)におけるイノシンを含む断片のピークの減少またはシアノエチル化イノシンを含む断片のピーク(イノシン一箇所当たり53の分子量増加)の出現またはその増加によりイノシンを含む領域の同定を行うことができる。
【0037】
この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域の同定が可能である。
【0038】
検出の第二の態様としては、化学修飾されたRNAの逆転写産物であるcDNAあるいはその増幅産物の長さを検出することによってα,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。cDNAの長さは電気泳動など当業者に公知の常法により検出することができる。上述したように化学修飾(例えば、シアノエチル化など)により逆転写鎖の伸長をイノシン化部位特異的に停止させ、逆転写に用いたプライマーの3’末端から伸長した長さを電気泳動の移動度等からイノシン部位を検出することが可能である。この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定が可能である。
【0039】
また、検出の第三の態様としては、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによってα,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。上述のように化学修飾されたイノシンが存在すると、逆転写反応はイノシン部位の手前で停止し、短いcDNAが生成sれる。このcDNAの塩基配列を決定することにより、cDNAの伸長が停止したイノシン部位を検出することができる。すなわち、この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定が可能である。
【0040】
さらに、検出の第四の態様としては、化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いてハイブリダイゼーションにより検出することによって、α,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。例えば、mRNAを化学修飾した後にポリTのプライマーを用いて3'末端から伸長させると、逆転写産物には伸長反応がイノシン部位で停止し、mRNAの上流配列に相補する配列を欠いた短いcDNAが生成される。対照として、非化学修飾RNAに由来するcDNAを用いると、イノシンを含む化学修飾RNAに由来するcDNAでは、RNAのイノシン化部位の上流のみの配列を含むプローブと反応しないが、非化学修飾RNAに由来するcDNAでは該プローブと反応したシグナルを検出することができる。このようにシグナルの有無もしくは減少により、イノシン部位を検出することができる。また、プローブのデザインによっては、イノシン部位を含む領域さらには位置を同定することも可能である。この方法は、マイクロアレイと組み合わせて行ってもよい。上記のように、この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域の同定が可能である。
【0041】
本発明は、RNA中のイノシンを修飾する修飾剤、さらにはこれを含むイノシン部位検出用試薬キットに関する。
【0042】
RNA中のイノシンを修飾する修飾剤は、α,β−不飽和結合と電子吸引性基とを有する化合物を含有する。「α,β−不飽和結合と電子吸引性基とを有する化合物」については上記方法に使用し得る化合物と同様であることからその説明を省略する。
【0043】
イノシン部位の検出用試薬キットは、検出手法に応じて、設計することができる。たとえば、上記第一の検出手法のための試薬キットの場合には、上記イノシン修飾剤に加えて、Rnaseやフォスファターゼを組合せることできる。また、第二の検出手法のための試薬キットとしては、上記イノシン修飾剤に加えて逆転写産物を生成するための逆転写酵素や必要に応じて逆転写反応用緩衝液などを組合せることができる。第三の検出手法のための試薬キットの場合には、上記イノシン修飾剤に加えて、塩基配列決定用の試薬一式を組み合わせることができる。塩基配列決定用の試薬一式としては、例えば、ポリメラーゼ、dNTP、ddNTP、反応用緩衝液などを一例としてあげることができる。第四の検出手法のための試薬としては、上記イノシン修飾剤に加えて、検出したいRNAの上流配列に基づくプローブ(プローブセット)と、ハイブリダイゼーション用の緩衝液、洗浄液などを組み合わせるができる。プローブセットについては、マイクロアレイに固定化して提供してもよい。
【0044】
以下の実施例により本発明をさらに具体的に説明するが、本発明は実施例によって限定されるものではない。
【実施例】
【0045】
実施例1:イノシン特異的化学修飾(1-シアノエチル化)反応
解析対象とするRNA画分10μgを30μlのCEバッファー (41%エタノール, 1.1M TEA-酢酸 (pH8.6))に溶解させた後、15.2M のアクリロニトリル(東京化成) 4μlを加え遮光で70℃ 15分又は30分反応させた。またコントロール(CE-)としてアクリロニトリルを非添加条件でインキュベートしたサンプルも用意した。反応後のサンプルを氷上で急冷し、RNeasy MinElute Cleanup kit (QIAGEN)により精製した。さらに溶出液中のRNAをエタノール沈殿・70%エタノールでリンスした後、凍結乾燥させた。
【0046】
実施例2:
特異的にシアノエチル化されたイノシンを液体クロマトグラフィー/質量分析法 (LC/MS法)により確認した。サンプルには酵母から抽出したtotal RNAを使用し、基本的に実施例1に記載した方法に従い反応した。ただし反応時間を70℃ 0分、15分、30分、又は60分とし、反応後のRNAはRNeasy MinElute Cleanup kitを使用せずそのままエタノール沈殿・70%エタノールでリンスした後、凍結乾燥させた。続いて乾燥後のRNAを、Sakurai, M., Ohtsuki, T., Suzuki, T., and Watanabe, K. (2005). Unusual usage of wobble modifications in mitochondrial tRNAs of the nematode Ascaris suum. FEBS Lett 579, 2767-2772に従ってNuclease P1とbacterial alkaline phosphatase を用いてヌクレオシドまで分解後、LC/MS解析を行った。
【0047】
その結果、CE-(Cyanoethyl化, 図4A)で観察されたイノシンのピーク(m/z 269)がシアノエチル化の反応時間にともない減少し、代わりにCE-では検出されなかったシアノエチル化イノシン(CE-I)(m/z 322)のピークが観察された(図4B、C、D、E)。また、シアノエチル化シュードウリジン(CE-Ψ)のピーク (m/z 298)もシアノエチル化時間に伴い増加していったがその反応速度は遅く(図4B、C、D、F)、60分反応後も6割程度の未反応シュードウリジン(m/z 245)が検出された。これ以外の塩基については変化が検出されなかったことから、このシアノエチル化反応はイノシン特異的であるといえる。
【0048】
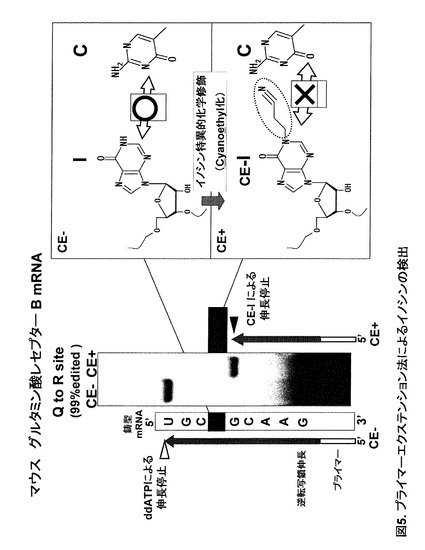
実施例3:プライマーエクステンション法によるイノシンの検出
実施例1に記載された反応手順によりマウス脳total RNA に対してイノシン特異的化学修飾反応を行った。反応後のRNA25μgと、5’末端を32P標識したDNAプライマー0.4pmolを5μl に溶解し、65℃3分インキュベート後、室温まで冷やした。ここで用いたDNAプライマーはマウスグルタミン酸レセプターB mRNA におけるQ /R A-to-I RNA エディティング部位 (Aの99%がイノシンへとエディティングを受けている)に対して下流に設計したものであり、その塩基配列は、5'-GATCTTGGCGAAATATCGCATC-3' (配列番号1)である。室温まで冷却したサンプルを150μM dGTP, dCTP, dTTPと同濃度のddATP, 0.5mM DTT, 3mM MgCl2, 175mM KCl, 50mM Tris-HCl (pH8.3), 100U SuperScript III reverse transcriptase (Invitrogen) を含む10μl 溶液中で50℃60分インキュベートした。
【0049】
反応後 2xLoading solution (7M urea, 0.05%bromophenol blue, 0.05%xylene cyanol, 1x TBE) を10μl, 100%formamideを20μl 加え、95℃ 5分ボイルした後、10μl を泳動に使用した。泳動は15%polyacrylamide, 7M urea, 1xTBEのゲルで行った。
【0050】
未処理(CE-)のtotal RNAの場合、イノシン部位に対して逆転写鎖にCがとりこまれ、滞りなく伸長が進んだ[確認のため最初のU登場部位でddATP(ダイデオキシATP)により伸長を止めている]。一方、シアノエチル化した場合(CE+)、図5に示したようにシアノエチルイノシン(CE-I)の1−シアノエチル基によりCの取り込みが阻害されるため、逆転写鎖の伸長はイノシンの一塩基手前で停止した。この結果から、シアノエチル化により逆転写鎖の伸長をイノシン部位特異的に停止させ、その伸長した長さによりイノシン部位を同定することが可能であるといえる。
【0051】
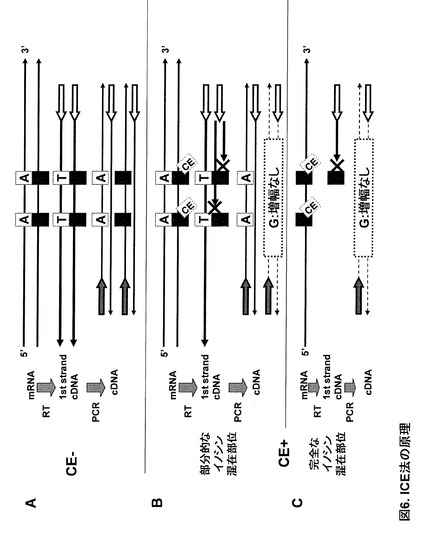
実施例4:ダイレクトシークエンシングによるイノシンの検出(Inosine Chemical Erasing法; ICE法)
実施例1に記載された反応手順によりマウス脳total RNA に対してイノシン特異的化学修飾反応を行った。一方、マウス セロトニンレセプター 2c (5HT2cR) mRNAにおける5箇所のA-to-I RNA エディティング部位を中心とした領域250ntを増幅するようなフォワードプライマーとリバースプライマーを設計した。ここで用いたDNAプライマーの塩基配列は、フォワードプライマー;5'-ATGGAGAAGAAACTGCACAATG-3'(配列番号2), リバースプライマー; 5'-ATGATGGCCTTAGTCCGCGAAT-3' (配列番号3)である。両方のプライマーをそれぞれ2.5pmolと、反応後のマウス脳total RNA を混合した。ここで加えたtotal RNA量は、それぞれ、CE-(アクリロニトリル無しの条件でのコントロール)条件;10ng, CE+ 15min条件; 50ng, CE+ 30min; 50ngである。混合したサンプルを 12.5μl のスケールでSuperScript III One-Step RT-PCR system with Platinum Taq DNA polymerase (Invitrogen)によりRT-PCRを行った。反応は逆転写; 55℃30分, 熱変性; 94℃2分後、94℃15秒, 60℃30秒, 68℃30秒を38サイクル行った。反応溶液5μlをExoSAP-IT (Usb)により処理して鋳型を精製した。続いてBig Dye Terminator v1.1 Cycle sequencing kit (Applied Biosystems)を用いてシークエンス反応を行い、ABI PRISM 3700 DNA Analyzer (Applied Biosystems)により解析した。また、比較のためゲノムに対するプライマーを設計し、genomic PCRを行い同様にシークエンシングした。
【0052】
この手法の原理は次のようである。まず未処理(CE-)のtotal RNAの場合、リバースプライマーから伸長した逆転写鎖はイノシン部位に対してCをとりこみ、フォワードプライマー相補部位まで到達する。この1st strand cDNAのPCR増幅産物センス鎖のイノシン相当部位にはイノシンを反映したGが存在する(図6A)。しかしシアノエチル化を施すとイノシンを含むmRNAに対する逆転写鎖の伸長はイノシン部位の手前で止まり、フォワードプライマー相補部位まで到達せず、その後のPCRで増幅されない。よってイノシンが部分的に混在している場合はPCR産物センス鎖の相当部位からイノシンを反映したGは消失する(図6B)。また、完全にイノシンへの置換が起きている場合は、PCRによる増幅全体が見られなくなる(図6C)。
【0053】
実施例において(図7)、マウス脳 セロトニンレセプター 2c mRNAのゲノム配列とcDNA CE-配列を比較すると、5箇所のA-to-I RNA エディティング部位においてIを反映したGが混在している。しかしこの混在しているGのピークはシアノエチル化反応の進行に伴い特異的に消失し、A単独のピークとなった。よってCE-とCE+の配列を比較することにより、微量RNA中のイノシンを検出することが可能である。この化学修飾によりイノシン由来のGを特異的に消失させる手法を、Inosine Chemical Erasing( ICE法)と称する。
【0054】
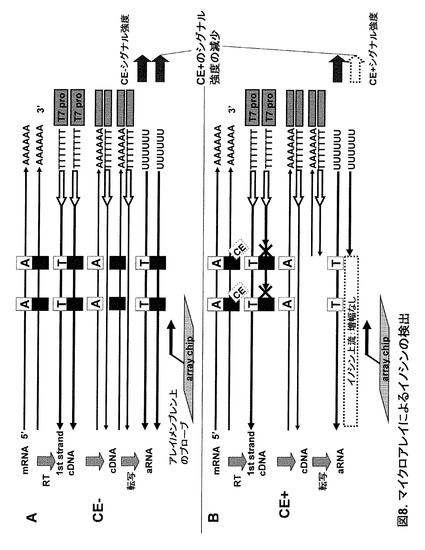
実施例5:マイクロアレイによるイノシンの検出(ICE-マイクロアレイ法)
実施例1で調整したHuman brain total RNA (CE-)と(CE+ 15min)を用いてマイクロアレイ解析を行った。T7プロモーター配列を付加したオリゴdTプライマー [T7(dT)24 cDNA primer (Ambion) ] を用いた逆転写反応から先は日立ソフト・DNAチップ研究所に依頼し、同社のAceGene Human Olig Chip 30K 1Chip versionを用いて行った。同社では逆転写からcDNA合成、アミノアリルラベルaRNA(amplified RNA)転写増幅についてはAmino Allyl MessageAmp aRNA kit (Ambion)を使用している。
【0055】
この手法の原理は以下の通りである。設定はイノシン部位の下流(3')側に逆転写用のプライマーが位置し、イノシン部位の上流(5')側にアレイ上のプローブが設計されている場合である。まず未処理(CE-)のtotal RNAの場合、リバースプライマーから伸長した逆転写鎖はイノシン部位に対してCをとりこみ伸長する。その後二本鎖cDNA化・T7 RNA polymeraseによるaRNAの転写増幅・標識を経てアレイ上のプローブとハイブリダイズさせ、このアレイ上のaRNAの標識強度(シグナル強度)を定量する(図8A)。一方CE+の場合、イノシンを含むmRNAに対する逆転写鎖の伸長はイノシン部位の手前で停止するため、短い逆転写鎖が合成される。合成された逆転写鎖の長さはそのまま転写増幅を経てaRNAの長さに反映される(図8B)。よってイノシン特異的に停止した逆転写鎖に起因する短いaRNAは、アレイ上のプローブとの相補領域を含まず、プローブとハイブリダイズできない。結果的にプローブ上のaRNAのシグナル強度は、イノシン部位の手前で逆転写鎖が停止した量を反映して減少すると考えられる。すなわち、CE-のシグナル強度と比較して、CE+のシグナル強度が減少した場合、そのアレイ上のプローブと逆転写プライマーの間の領域にイノシンが存在するといえる。
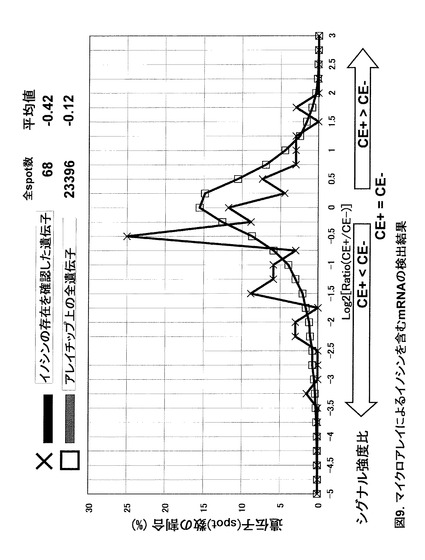
【0056】
実施例においては上記アレイ上のプローブ設計位置の制約から、日立ソフトのAce Geneを用いた。プローブ設計位置と、本発明者らがICE法により確認したA-to-I RNA エディティング部位の情報から、アレイ上のプローブ位置とmRNA末端との間にイノシン部位が存在するmRNA 68種を抽出した(図9, ×印―黒線)。この68種とアレイ上の全遺伝子(図9, □印-灰色線)について、CE+のCE−に対するシグナル強度比 [Log2(CE+/CE-), グラフ中では横軸において負の方向に進むほどCE+のシグナルが減少したことを意味する] を解析したところ、RNAエディティングを受けるmRNA 68種の分布が負の方向に偏り、その平均値は-0.42とアレイ全遺伝子の平均値-0.12に対して優位なシグナル減少が得られた。よって本手法によりmRNA中のイノシンの存在が検出可能であることが示された。
【図面の簡単な説明】
【0057】
【図1】図1は、matched-tissue法によるイノシンの検出を示す。
【図2】図2は、イノシン特異的切断法によるイノシン部位の同定を示す。
【図3】図3は、イノシン特異的化学修飾試薬を示す。
【図4】図4は、質量分析法によるシアノエチル化反応後のイノシンの検出を示す。
【図5】図5は、プライマーエクステンション法によるイノシンの検出を示す。
【図6】図6は、ICE法の原理を示す。
【図7】図7は、ICE法によるイノシンの検出を示す。
【図8】図8は、マイクロアレイによるイノシンの検出を示す。
【図9】図9は、マイクロアレイによるイノシンを含むmRNAの検出結果を示す。
【配列表フリーテキスト】
【0058】
SEQUENCE LISTING
<110> The University of Tokyo
<120> A method for detecting inosine site in RNA
<130> A51628A
<160> 3
<210> 1
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 1
gatcttggcg aaatatcgca tc 22
<210> 2
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 2
atggagaaga aactgcacaa tg 22
<210> 3
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 3
atgatggcct tagtccgcga at 22
【技術分野】
【0001】
本発明は、RNA中のイノシン化部位の検出方法に関する。より詳細には、本発明は、RNA中のイノシン化部位を化学修飾することによって、該イノシン化部位を検出する方法に関する。
【背景技術】
【0002】
脱アミノ化(デアミネーション)による修飾は、RNAエディティングと呼ばれている。RNAエディティングにより、A からIへの変化が生じ、mRNA上のコドンが変化することでアミノ酸配列の変化を引き起こす。二本鎖RNAアデノシンデアミネース(ADAR;adenosine deaminase acting on RNA)はRNAの二本鎖部分を特異的に認識してアデノシン残基をイノシンに変換する酵素である。哺乳動物には、ADAR1、ADAR2、ADAR3の3種類のファミリーが存在する。グルタミン酸受容体のサブユニットGluR-BのmRNAは、ADAR2の基質であり、エキソン11番中のアデノシンがイノシンへとエディティングされ、アミノ酸配列がグルタミンからアルギニンへと変化する。ADAR2のノックアウトマウスはこのエディティングが生じないため、グルタミン酸受容体のカルシウム透過性をコントロールできず、てんかん症状を起こし、早期に死に至ることが知られている。ADARは二本鎖RNAに対し幅広い基質認識能を示すことが知られ、GluR-B以外にも、セロトニンレセプター(5-HT2c)、カリウムチャンネル(Kv1.1)などでAからIへのエディティングが見つかっている。また、興味深いことにADAR2は自身のmRNAのイントロン部位をAからIにエディティングすることで、可変的スプライシングを誘導し、ADAR2の発現量をフィードバック制御していることが知られている。
【0003】
ADARが高発現している脳mRNAには17000塩基に一度の頻度でイノシンが含まれているとの見積もりがあり、ヒトmRNAには未知のエディティング部位が多量に存在することが予測されていた。最近、バイオインフォマティクス的手法を用いた解析により、ヒトmRNAのUTRにおけるAlu因子の反復配列にAからIへのエディティング候補部位が大量に報告された。実験的に実証された例はこのうちのまだわずかであるが、1600遺伝子の12000箇所以上の部位にイノシンが存在するという見積もりは、AからIへのエディティングが遺伝子のグローバルな発現制御に大きな役割を果たしていることを示唆している。Alu因子を持っているのは霊長類だけであり、ADARは脳で高発現していることを考え合わせると、AからIへのエディティングによるトランスクリプトームの複雑性増加は、高度に進化した脳の神経回路の複雑性の獲得と関連している可能性がある。また、RNAエディティング異常に起因する疾患も報告されている。悪性グリオーマ(Malignant glioma)や筋萎縮性側索硬化症(Amyotrophic lateral sclerosis)ではグルタミン酸受容体のサブユニットタンパク質(GluR2)のAからIへのエディティングが顕著に減少している。このように、mRNA中のイノシンはRNAに質的な変化を与えるものであり、高次生命現象と深く関わっている可能性が高い。
【0004】
AからIへの RNA エディティング部位(=Iの存在部位)の検出法としてこれまで最も一般的である手法がmatched-tissue 法である(図1)。この方法は同一個体の同一組織由来のRNA(cDNAとして増幅)とゲノムで相当する領域の配列を比較する方法である。IはCと塩基対を形成するため、mRNAから逆転写・PCR増幅後のcDNA中のI相当部位にはGが取り込まれる。よってエディティング部位ではゲノム上の塩基がAであるにも関わらず、cDNA上ではGもしくはA/Gの混在となっている。しかしこの方法ではPCRの非特異増幅・ゲノム混入・シークエンスエラー・ノイズ・対立遺伝子間のSNP (Allele SNP)・偽遺伝子・遺伝子コピー・スプライスバリアント等を原因とする偽シグナルによるGの混在と、目的とするイノシン由来のGの混在を判別することが難しい。また、エキソン境界領域や偽遺伝子の存在する領域の場合、RNAに対応するゲノム領域を判別することが困難であるためこの方法自体用いることができない。さらにこの方法で特定されたイノシン部位は配列上Gとして検出されるため、厳密にはイノシンの存在を証明してはいない。また、同一個体由来のゲノムとRNAの両方を準備する必要があるため、解析可能なサンプルも制限されてしまうという難点も持つ。
【0005】
また、これまでに分子生物学分野におけるイノシンの化学修飾法としては2例の報告がある。1つは生体内RNA中のイノシン部位同定を目的としたイノシン特異的切断法である(非特許文献1、2及び3)(図2)。この手法は以下の3ステップからなる。
【0006】
(1)化学修飾試薬としてグリオキサールを用いて、RNA鎖中のグアノシン(G)とイノシン(I)の塩基修飾を行う。Gの場合、グリオキサールは1位及びN'2位に付加してcis-ジオールとなり、さらにホウ酸と化合物(図中G*)される。一方イノシンの場合、2位のアミノ基が存在しないためグリオキサールは1位にのみ付加する。さらに付加した構造は不安定であるため、容易に逆反応を起こして元のイノシンに戻る。
(2)グリオキサール/ホウ酸処理後のRNAをGとイノシンに特異的なRNase T1により切断する。このときGがG*へと修飾されているためにG部位では切断は起らない。一方イノシンは修飾を受けていない(元に戻っている)ためイノシン部位での切断が起る。結果としてイノシン部位特異的な切断が可能となる。
(3)切断後のRNA中のG*から付加物を取り除き、イノシンを含む側のRNAフラグメントにアンカー配列を取り付けてこの部位から逆転写・PCRを行い、その配列を解析する。
【0007】
この方法でMorseらは数例のmRNA中のイノシン部位を同定しているが(非特許文献3)、その後の報告はない。この解析法の成功の鍵はグリオキサール/ホウ酸処理によるGのRNase T1非感受性修飾処理である。RNAを構成する塩基の1/4を占めるGを全て修飾するのは困難であり、未修飾のGが残存することは避けられない。さらに数千〜数万塩基からなる一本のmRNA中にイノシンは0〜30塩基程度しか存在しないほど微量であり、未修飾のG部位の切断によるバックグラウンドが目的のイノシン部位における切断の検出を非常に困難なものにしている。そのため仮に候補部位が得られた場合でも、matched tissue 法による確認が必要である。
【0008】
また、イノシン特異的化学修飾はYoshidaらにより報告されている(非特許文献4)。この解析法では酵母の転位RNA(transfer RNA; tRNA)のアンチコドン部位(遺伝暗号を規定するmRNA上のコドンと対号する)に存在するイノシンの機能解明を目的とし、イノシンの1位をアクリロニトリルを用いてシアノエチル化してコドン−アンチコドン対形成を阻害してその効果を解析している。また反応効率は低いものの、シュードウリジン(□)もシアノエチル化され(非特許文献5)、Mengel-Jorgensenらはシアノエチル化によるシュードウリジンの分子量の変化を質量分析法により解析し、tRNA上の位置を特定する手法を報告している(非特許文献6)。これらの手法は比較的豊富に存在するtRNAを解析対象とし、アンチコドンの不活性化の与える影響もしくはシアノエチル化による質量の変化を解析するものである。
【0009】
【非特許文献1】Morse, D. P. (2004). Identification of substrates for adenosine deaminases that act on RNA. Methods Mol Biol 265, 199-218.
【非特許文献2】Morse, D. P., and Bass, B. L. (1997). Detection of inosine in messenger RNA by inosine-specific cleavage. Biochemistry 36, 8429-8434.
【非特許文献3】Morse, D. P., and Bass, B. L. (1999). Long RNA hairpins that contain inosine are present in Caenorhabditis elegans poly(A)+ RNA. Proc Natl Acad Sci U S A 96, 6048-6053.
【非特許文献4】Yoshida, M., Furuichi, Y., Ukita, T., and Kaziro, Y. (1967). The effect of cyanoethylation on codon recognition of yeast tRNA containing inosine. Biochim Biophys Acta 149, 308-310.
【非特許文献5】Yoshida, M., and Ukita, T. (1968). Modification of nucleosides and nucleotides. 8. The reaction rates of pseudouridine residues with acrylonitrile and its relation to the secondary structure of transfer ribonucleic acid. Biochim Biophys Acta 157, 466-475.
【非特許文献6】Mengel-Jorgensen, J., and Kirpekar, F. (2002). Detection of pseudouridine and other modifications in tRNA by cyanoethylation and MALDI mass spectrometry. Nucleic Acids Res 30, e135.
【発明の開示】
【発明が解決しようとする課題】
【0010】
本発明は、ノイズやSNPとの判別が容易であり、解析サンプルとしてRNAのみで解析を行うことが可能であり、少量のサンプルでも解析可能であり、さらにバックグラウンドを低く抑えることが可能であり、イノシンの存在を高感度に検出することが可能である、RNA中のイノシン化部位を検出する方法を提供することを解決すべき課題とした。
【課題を解決するための手段】
【0011】
本発明者らは、上記課題を解決するために鋭意検討した結果、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾することによって、RNA中のイノシン化部位を検出できることを見出し、本発明を完成するに至った。
【0012】
即ち、本発明によれば、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法が提供される。
【0013】
好ましくは、本発明の方法は、さらに、化学修飾されたRNAを逆転写反応に供してcDNAを合成する工程、及び合成されたcDNAに基づいてイノシン化部位を検出する工程を含む。
好ましくは、イノシン化部位の検出は、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定である。
【0014】
好ましくは、α,β−不飽和結合と電子吸引性基とを有する化合物は、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物である。
好ましくは、電子吸引性基は、CN、NO2、SO3H、CONH2、COCH3、COOCH3、COOC2H5、COCH3、COC2H5、又はCOC6H5である。
【0015】
好ましくは、化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行う。
好ましくは、対照として化学修飾されていないRNAを逆転写反応に供して合成されたcDNAと化学修飾されたRNAから合成されたcDNAとを比較することにより、イノシン化部位を検出する。
好ましくは、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシンを検出する。
好ましくは、化学修飾されたRNAに由来するcDNAの長さを検出することによって、化学修飾されたイノシンを検出する。
【0016】
好ましくは、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによって、化学修飾されたイノシンを検出する。
好ましくは、化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いて検出することによって、化学修飾されたイノシンを検出する。
【0017】
本発明の別の側面によれば、α,β−不飽和結合と電子吸引性基とを有する化合物を含む、RNA中のイノシン化部位修飾剤が提供される。
本発明のさらに別の側面によれば、上記した本発明の修飾剤を含む、上記した本発明の方法を行うための試薬キットが提供される。
【発明の効果】
【0018】
12000箇所以上とも言われるイノシン化部位は、現状では、その大部分が同定されていない。イノシンは脳のmRNAに大量に存在することを考えると、精神疾患(統合失調症、パニック症候群、双極性障害、自閉症など)の異常と関連があると考えられる。これらの疾患と明確に関連するイノシン化部位を本発明の方法によって同定することができれば、疾患の発症機構の解明や治療法を考える上で非常に重要な知見を得ることができる。また、個体ごとにイノシン化部位の変動解析を行うことで、SNPのように生活習慣病や様々な疾患の発症リスクを見積もることが可能になる。
【発明を実施するための最良の形態】
【0019】
以下、本発明の実施の形態についてさらに詳細に説明する。
本発明によるイノシン化部位の検出方法は、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含むことを特徴とする。本発明においてRNAの種類は特に限定されず、mRNA、rRNA又はtRNAの何れでもよい。
【0020】
本発明の方法では、イノシン特異的な修飾・逆転写伸長阻害を検出することにより、cDNA中のイノシンの存在を証明することが可能であり、ノイズやSNPとの判別も容易である。さらに解析サンプルとしてゲノムは必要なくRNAのみで解析が行えるため、少量の貴重なサンプルでも解析可能である。この理由によりスプライス部位などの遺伝子構造や、ゲノム上のコード領域が不明な場合でもイノシン部位を特定することが可能である。これらの点において、本発明の方法は、従来のmatched-tissu法より優れている。また、本発明の方法ではイノシン特異的な修飾を施すため、バックグラウンドを非常に低く抑えることが可能であり、イノシンの存在を高感度に検出・証明することができる利点を持つ。さらに本発明の方法における検出をマイクロアレイで行うことにより、生体内RNA中のイノシン部位を網羅的に特定することができる。さらに、本発明の方法は、全RNA種を対象とし、イノシン特異的な化学修飾による逆転写鎖の伸長阻害を利用するという点において、従来報告されているイノシン特異的化学修飾(Yoshida, M., Furuichi, Y., Ukita, T., and Kaziro, Y. (1967). The effect of cyanoethylation on codon recognition of yeast tRNA containing inosine. Biochim Biophys Acta 149, 308-310)とは区別されるものである。
【0021】
以下、本発明の方法を具体的に説明する。
【0022】
(1)イノシン特異的化学修飾
本発明による方法では先ず、イノシン特異的な化学修飾をRNAに施す。試薬としてはα, β−不飽和−電子求引性基化合物を用いることができる。
【0023】
反応機構は基本的にはMichael付加と呼ばれる機構(図3A)で進む。まず、修飾試薬の電子求引性基の影響でβ位の炭素原子が正電荷を帯びる。この炭素原子が、イノシンの活性アミンである1位の窒素原子に求電子付加することにより、イノシンへの付加修飾が進む。修飾試薬の選択によっては、付加官能基特異的にイノシンを含むRNA分子を単離することも可能である。修飾試薬により適切な溶媒を選択して、反応を行うことができる。以下に記載の実施例ではアクリロニトリル(CH2=CHCN)を使用している。アクリロニトリルの場合、同様にシュードウリジンも反応することが知られているが、これは反応時間を短縮することでほぼイノシン特異的な反応条件とすることが可能である。
【0024】
α,β−不飽和結合と電子吸引性基とを有する化合物としては、例えば、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物を挙げることができる。上記化合物中の官能基の例、並びに上記化合物の具体例は、図3B及び図3Cにそれぞれ示す。
【0025】
上記化合物の具体例としては、CH2=CHCN、C6H5CH=CHCN、CH3CH=CHCOCH3、CH3CH=CHCOC6H5、CH3CH=C(CH3)COCH3、(CH3)2C=CHCOCH3、CH3CH=C(CH3)COC6H5、CH2=C(CH3)COC6H5、CH2=C(CH2CH3)COC6H5、CH2=CHCOOCH3、CH2=C(CH3)COOCH3、CH3OC6H5CH=CHCOOCH2CH3、CH3OC6H5CH=CHCOOCH3、CH2=COOCH2CH3、CH2=COOCH3、CH2=CONH2、CH2=C(OCOCH3)CN、及び
【0026】
【化1】
【0027】
などを挙げることができる。
【0028】
(2)逆転写反応(必要に応じて増幅)
本発明の方法では、イノシン特異的に化学修飾を施したRNAに対して逆転写反応を行うことができる。化学修飾をしていない場合、逆転写酵素によりイノシン部位に対してシチジン(C)が取り込まれる。一方、化学修飾をほどこした場合、この化学修飾によりCの取り込みが阻害されて逆転写鎖の伸長はその手前で停止する。
【0029】
逆転写反応は、プライマー、化学修飾を施したRNA(鋳型)、4種のdNTP、逆転写酵素などを用いて常法により行うことができる。即ち、逆転写反応は、上記の試薬を適当な反応液(例えば、適当な塩を含む緩衝液)中で混合して、所定の温度で一定時間インキュベートすることにより行うことができる。
【0030】
本発明においては、化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行うことが好ましい。この場合、逆転写反応と増幅反応は、RT−PCR法により一連の操作により行うことができる。あるいは逆転写後のcDNAを鋳型とした転写増幅により行うことも可能である。
【0031】
また、逆転写反応では、化学修飾されていないRNAを逆転写反応に供してcDNAを合成し、これを以下の検出工程において対照用として用いることもできる。
【0032】
(3)検出
検出においては、質量分析によりイノシンによる化学修飾の有無を検出してもよいし、あるいは、伸長阻害された逆転写鎖を検出し、イノシン化学修飾の有無で比較することによりRNA中のイノシン部位を検出してもよい。この際、検出対象情報としては、その長さ、塩基配列、又は量などが挙げられる。
【0033】
検出の第一の態様としては、RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシン化部位を検出することができる。質量分析を行う場合、イノシンの有無及び総量を解析するには、化学修飾後のRNAをサンプルとしてもよいが、好ましくは化学修飾されたRNAをRNaseとフォスファターゼでヌクレオシドまで分解した後に行う。ここで用いるRNaseはヌクレオシドまで分解可能ならば特に限定はなく、例えば、NucleaseP1などを用いることができる。フォスファターゼも特に限定はなく、バクテリアなどの由来する生物種も限定はない。例えば、バクテリAlkaline Phospataseなどを用いることができる。
【0034】
例えば、アクリロニトリルをイノシン化部位の修飾剤として用いてRNAを処理し、その後ヌクレオシドまで分解する。このサンプルを非化学修飾RNA由来のヌクレオチドサンプルと比較して、液体クロマトグラフィー/質量分析法(LC/MS)におけるイノシンのピーク(m/z 269)の減少またはシアノエチル化イノシンのピーク(m/z 322)の出現またはその増加によりイノシン化部位の検出を行うことができる。
【0035】
また、イノシンを含む領域の同定には、化学修飾後のRNAをサンプルとしても良いが、好ましくは化学修飾されたRNAを適当なRNaseにより断片化した後に行う。ここで用いるRNaseはRNAを数塩基から100塩基程度の長さに断片化可能ならば特に限定はなく、例えばG特異的RNase T1などを用いることができる。
【0036】
例えば、アクリロニトリルをイノシン化部位の修飾剤として用いてRNAを処理し、その後RNA断片に分解する。このサンプルを非化学修飾RNA由来のRNA断片化サンプルと比較して、液体クロマトグラフィー/質量分析法(LC/MS)におけるイノシンを含む断片のピークの減少またはシアノエチル化イノシンを含む断片のピーク(イノシン一箇所当たり53の分子量増加)の出現またはその増加によりイノシンを含む領域の同定を行うことができる。
【0037】
この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域の同定が可能である。
【0038】
検出の第二の態様としては、化学修飾されたRNAの逆転写産物であるcDNAあるいはその増幅産物の長さを検出することによってα,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。cDNAの長さは電気泳動など当業者に公知の常法により検出することができる。上述したように化学修飾(例えば、シアノエチル化など)により逆転写鎖の伸長をイノシン化部位特異的に停止させ、逆転写に用いたプライマーの3’末端から伸長した長さを電気泳動の移動度等からイノシン部位を検出することが可能である。この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定が可能である。
【0039】
また、検出の第三の態様としては、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによってα,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。上述のように化学修飾されたイノシンが存在すると、逆転写反応はイノシン部位の手前で停止し、短いcDNAが生成sれる。このcDNAの塩基配列を決定することにより、cDNAの伸長が停止したイノシン部位を検出することができる。すなわち、この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定が可能である。
【0040】
さらに、検出の第四の態様としては、化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いてハイブリダイゼーションにより検出することによって、α,β−不飽和結合と電子吸引性基とを有する化合物によって化学修飾されたイノシンを検出することができる。例えば、mRNAを化学修飾した後にポリTのプライマーを用いて3'末端から伸長させると、逆転写産物には伸長反応がイノシン部位で停止し、mRNAの上流配列に相補する配列を欠いた短いcDNAが生成される。対照として、非化学修飾RNAに由来するcDNAを用いると、イノシンを含む化学修飾RNAに由来するcDNAでは、RNAのイノシン化部位の上流のみの配列を含むプローブと反応しないが、非化学修飾RNAに由来するcDNAでは該プローブと反応したシグナルを検出することができる。このようにシグナルの有無もしくは減少により、イノシン部位を検出することができる。また、プローブのデザインによっては、イノシン部位を含む領域さらには位置を同定することも可能である。この方法は、マイクロアレイと組み合わせて行ってもよい。上記のように、この方法によれば、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域の同定が可能である。
【0041】
本発明は、RNA中のイノシンを修飾する修飾剤、さらにはこれを含むイノシン部位検出用試薬キットに関する。
【0042】
RNA中のイノシンを修飾する修飾剤は、α,β−不飽和結合と電子吸引性基とを有する化合物を含有する。「α,β−不飽和結合と電子吸引性基とを有する化合物」については上記方法に使用し得る化合物と同様であることからその説明を省略する。
【0043】
イノシン部位の検出用試薬キットは、検出手法に応じて、設計することができる。たとえば、上記第一の検出手法のための試薬キットの場合には、上記イノシン修飾剤に加えて、Rnaseやフォスファターゼを組合せることできる。また、第二の検出手法のための試薬キットとしては、上記イノシン修飾剤に加えて逆転写産物を生成するための逆転写酵素や必要に応じて逆転写反応用緩衝液などを組合せることができる。第三の検出手法のための試薬キットの場合には、上記イノシン修飾剤に加えて、塩基配列決定用の試薬一式を組み合わせることができる。塩基配列決定用の試薬一式としては、例えば、ポリメラーゼ、dNTP、ddNTP、反応用緩衝液などを一例としてあげることができる。第四の検出手法のための試薬としては、上記イノシン修飾剤に加えて、検出したいRNAの上流配列に基づくプローブ(プローブセット)と、ハイブリダイゼーション用の緩衝液、洗浄液などを組み合わせるができる。プローブセットについては、マイクロアレイに固定化して提供してもよい。
【0044】
以下の実施例により本発明をさらに具体的に説明するが、本発明は実施例によって限定されるものではない。
【実施例】
【0045】
実施例1:イノシン特異的化学修飾(1-シアノエチル化)反応
解析対象とするRNA画分10μgを30μlのCEバッファー (41%エタノール, 1.1M TEA-酢酸 (pH8.6))に溶解させた後、15.2M のアクリロニトリル(東京化成) 4μlを加え遮光で70℃ 15分又は30分反応させた。またコントロール(CE-)としてアクリロニトリルを非添加条件でインキュベートしたサンプルも用意した。反応後のサンプルを氷上で急冷し、RNeasy MinElute Cleanup kit (QIAGEN)により精製した。さらに溶出液中のRNAをエタノール沈殿・70%エタノールでリンスした後、凍結乾燥させた。
【0046】
実施例2:
特異的にシアノエチル化されたイノシンを液体クロマトグラフィー/質量分析法 (LC/MS法)により確認した。サンプルには酵母から抽出したtotal RNAを使用し、基本的に実施例1に記載した方法に従い反応した。ただし反応時間を70℃ 0分、15分、30分、又は60分とし、反応後のRNAはRNeasy MinElute Cleanup kitを使用せずそのままエタノール沈殿・70%エタノールでリンスした後、凍結乾燥させた。続いて乾燥後のRNAを、Sakurai, M., Ohtsuki, T., Suzuki, T., and Watanabe, K. (2005). Unusual usage of wobble modifications in mitochondrial tRNAs of the nematode Ascaris suum. FEBS Lett 579, 2767-2772に従ってNuclease P1とbacterial alkaline phosphatase を用いてヌクレオシドまで分解後、LC/MS解析を行った。
【0047】
その結果、CE-(Cyanoethyl化, 図4A)で観察されたイノシンのピーク(m/z 269)がシアノエチル化の反応時間にともない減少し、代わりにCE-では検出されなかったシアノエチル化イノシン(CE-I)(m/z 322)のピークが観察された(図4B、C、D、E)。また、シアノエチル化シュードウリジン(CE-Ψ)のピーク (m/z 298)もシアノエチル化時間に伴い増加していったがその反応速度は遅く(図4B、C、D、F)、60分反応後も6割程度の未反応シュードウリジン(m/z 245)が検出された。これ以外の塩基については変化が検出されなかったことから、このシアノエチル化反応はイノシン特異的であるといえる。
【0048】
実施例3:プライマーエクステンション法によるイノシンの検出
実施例1に記載された反応手順によりマウス脳total RNA に対してイノシン特異的化学修飾反応を行った。反応後のRNA25μgと、5’末端を32P標識したDNAプライマー0.4pmolを5μl に溶解し、65℃3分インキュベート後、室温まで冷やした。ここで用いたDNAプライマーはマウスグルタミン酸レセプターB mRNA におけるQ /R A-to-I RNA エディティング部位 (Aの99%がイノシンへとエディティングを受けている)に対して下流に設計したものであり、その塩基配列は、5'-GATCTTGGCGAAATATCGCATC-3' (配列番号1)である。室温まで冷却したサンプルを150μM dGTP, dCTP, dTTPと同濃度のddATP, 0.5mM DTT, 3mM MgCl2, 175mM KCl, 50mM Tris-HCl (pH8.3), 100U SuperScript III reverse transcriptase (Invitrogen) を含む10μl 溶液中で50℃60分インキュベートした。
【0049】
反応後 2xLoading solution (7M urea, 0.05%bromophenol blue, 0.05%xylene cyanol, 1x TBE) を10μl, 100%formamideを20μl 加え、95℃ 5分ボイルした後、10μl を泳動に使用した。泳動は15%polyacrylamide, 7M urea, 1xTBEのゲルで行った。
【0050】
未処理(CE-)のtotal RNAの場合、イノシン部位に対して逆転写鎖にCがとりこまれ、滞りなく伸長が進んだ[確認のため最初のU登場部位でddATP(ダイデオキシATP)により伸長を止めている]。一方、シアノエチル化した場合(CE+)、図5に示したようにシアノエチルイノシン(CE-I)の1−シアノエチル基によりCの取り込みが阻害されるため、逆転写鎖の伸長はイノシンの一塩基手前で停止した。この結果から、シアノエチル化により逆転写鎖の伸長をイノシン部位特異的に停止させ、その伸長した長さによりイノシン部位を同定することが可能であるといえる。
【0051】
実施例4:ダイレクトシークエンシングによるイノシンの検出(Inosine Chemical Erasing法; ICE法)
実施例1に記載された反応手順によりマウス脳total RNA に対してイノシン特異的化学修飾反応を行った。一方、マウス セロトニンレセプター 2c (5HT2cR) mRNAにおける5箇所のA-to-I RNA エディティング部位を中心とした領域250ntを増幅するようなフォワードプライマーとリバースプライマーを設計した。ここで用いたDNAプライマーの塩基配列は、フォワードプライマー;5'-ATGGAGAAGAAACTGCACAATG-3'(配列番号2), リバースプライマー; 5'-ATGATGGCCTTAGTCCGCGAAT-3' (配列番号3)である。両方のプライマーをそれぞれ2.5pmolと、反応後のマウス脳total RNA を混合した。ここで加えたtotal RNA量は、それぞれ、CE-(アクリロニトリル無しの条件でのコントロール)条件;10ng, CE+ 15min条件; 50ng, CE+ 30min; 50ngである。混合したサンプルを 12.5μl のスケールでSuperScript III One-Step RT-PCR system with Platinum Taq DNA polymerase (Invitrogen)によりRT-PCRを行った。反応は逆転写; 55℃30分, 熱変性; 94℃2分後、94℃15秒, 60℃30秒, 68℃30秒を38サイクル行った。反応溶液5μlをExoSAP-IT (Usb)により処理して鋳型を精製した。続いてBig Dye Terminator v1.1 Cycle sequencing kit (Applied Biosystems)を用いてシークエンス反応を行い、ABI PRISM 3700 DNA Analyzer (Applied Biosystems)により解析した。また、比較のためゲノムに対するプライマーを設計し、genomic PCRを行い同様にシークエンシングした。
【0052】
この手法の原理は次のようである。まず未処理(CE-)のtotal RNAの場合、リバースプライマーから伸長した逆転写鎖はイノシン部位に対してCをとりこみ、フォワードプライマー相補部位まで到達する。この1st strand cDNAのPCR増幅産物センス鎖のイノシン相当部位にはイノシンを反映したGが存在する(図6A)。しかしシアノエチル化を施すとイノシンを含むmRNAに対する逆転写鎖の伸長はイノシン部位の手前で止まり、フォワードプライマー相補部位まで到達せず、その後のPCRで増幅されない。よってイノシンが部分的に混在している場合はPCR産物センス鎖の相当部位からイノシンを反映したGは消失する(図6B)。また、完全にイノシンへの置換が起きている場合は、PCRによる増幅全体が見られなくなる(図6C)。
【0053】
実施例において(図7)、マウス脳 セロトニンレセプター 2c mRNAのゲノム配列とcDNA CE-配列を比較すると、5箇所のA-to-I RNA エディティング部位においてIを反映したGが混在している。しかしこの混在しているGのピークはシアノエチル化反応の進行に伴い特異的に消失し、A単独のピークとなった。よってCE-とCE+の配列を比較することにより、微量RNA中のイノシンを検出することが可能である。この化学修飾によりイノシン由来のGを特異的に消失させる手法を、Inosine Chemical Erasing( ICE法)と称する。
【0054】
実施例5:マイクロアレイによるイノシンの検出(ICE-マイクロアレイ法)
実施例1で調整したHuman brain total RNA (CE-)と(CE+ 15min)を用いてマイクロアレイ解析を行った。T7プロモーター配列を付加したオリゴdTプライマー [T7(dT)24 cDNA primer (Ambion) ] を用いた逆転写反応から先は日立ソフト・DNAチップ研究所に依頼し、同社のAceGene Human Olig Chip 30K 1Chip versionを用いて行った。同社では逆転写からcDNA合成、アミノアリルラベルaRNA(amplified RNA)転写増幅についてはAmino Allyl MessageAmp aRNA kit (Ambion)を使用している。
【0055】
この手法の原理は以下の通りである。設定はイノシン部位の下流(3')側に逆転写用のプライマーが位置し、イノシン部位の上流(5')側にアレイ上のプローブが設計されている場合である。まず未処理(CE-)のtotal RNAの場合、リバースプライマーから伸長した逆転写鎖はイノシン部位に対してCをとりこみ伸長する。その後二本鎖cDNA化・T7 RNA polymeraseによるaRNAの転写増幅・標識を経てアレイ上のプローブとハイブリダイズさせ、このアレイ上のaRNAの標識強度(シグナル強度)を定量する(図8A)。一方CE+の場合、イノシンを含むmRNAに対する逆転写鎖の伸長はイノシン部位の手前で停止するため、短い逆転写鎖が合成される。合成された逆転写鎖の長さはそのまま転写増幅を経てaRNAの長さに反映される(図8B)。よってイノシン特異的に停止した逆転写鎖に起因する短いaRNAは、アレイ上のプローブとの相補領域を含まず、プローブとハイブリダイズできない。結果的にプローブ上のaRNAのシグナル強度は、イノシン部位の手前で逆転写鎖が停止した量を反映して減少すると考えられる。すなわち、CE-のシグナル強度と比較して、CE+のシグナル強度が減少した場合、そのアレイ上のプローブと逆転写プライマーの間の領域にイノシンが存在するといえる。
【0056】
実施例においては上記アレイ上のプローブ設計位置の制約から、日立ソフトのAce Geneを用いた。プローブ設計位置と、本発明者らがICE法により確認したA-to-I RNA エディティング部位の情報から、アレイ上のプローブ位置とmRNA末端との間にイノシン部位が存在するmRNA 68種を抽出した(図9, ×印―黒線)。この68種とアレイ上の全遺伝子(図9, □印-灰色線)について、CE+のCE−に対するシグナル強度比 [Log2(CE+/CE-), グラフ中では横軸において負の方向に進むほどCE+のシグナルが減少したことを意味する] を解析したところ、RNAエディティングを受けるmRNA 68種の分布が負の方向に偏り、その平均値は-0.42とアレイ全遺伝子の平均値-0.12に対して優位なシグナル減少が得られた。よって本手法によりmRNA中のイノシンの存在が検出可能であることが示された。
【図面の簡単な説明】
【0057】
【図1】図1は、matched-tissue法によるイノシンの検出を示す。
【図2】図2は、イノシン特異的切断法によるイノシン部位の同定を示す。
【図3】図3は、イノシン特異的化学修飾試薬を示す。
【図4】図4は、質量分析法によるシアノエチル化反応後のイノシンの検出を示す。
【図5】図5は、プライマーエクステンション法によるイノシンの検出を示す。
【図6】図6は、ICE法の原理を示す。
【図7】図7は、ICE法によるイノシンの検出を示す。
【図8】図8は、マイクロアレイによるイノシンの検出を示す。
【図9】図9は、マイクロアレイによるイノシンを含むmRNAの検出結果を示す。
【配列表フリーテキスト】
【0058】
SEQUENCE LISTING
<110> The University of Tokyo
<120> A method for detecting inosine site in RNA
<130> A51628A
<160> 3
<210> 1
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 1
gatcttggcg aaatatcgca tc 22
<210> 2
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 2
atggagaaga aactgcacaa tg 22
<210> 3
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic DNA
<400> 3
atgatggcct tagtccgcga at 22
【特許請求の範囲】
【請求項1】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法。
【請求項2】
さらに、化学修飾されたRNAを逆転写反応に供してcDNAを合成する工程、及び合成されたcDNAに基づいてイノシン化部位を検出する工程を含む、請求項1に記載の方法。
【請求項3】
イノシン化部位の検出が、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定である、請求項1又は2に記載の方法。
【請求項4】
α,β−不飽和結合と電子吸引性基とを有する化合物が、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物である、請求項1から3の何れかに記載の方法。
【請求項5】
電子吸引性基が、CN、NO2、SO3H、CONH2、COCH3、COOCH3、COOC2H5、COCH3、COC2H5、又はCOC6H5である、請求項4に記載の方法。
【請求項6】
化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行う、請求項1から5の何れかに記載の方法。
【請求項7】
対照として化学修飾されていないRNAを逆転写反応に供して合成されたcDNAと化学修飾されたRNAから合成されたcDNAとを比較することにより、イノシン化部位を検出する、請求項2から6の何れかに記載の方法。
【請求項8】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシンを検出する、請求項1、3、4または5の何れかに記載の方法。
【請求項9】
化学修飾されたRNAに由来するcDNAの長さを検出することによって、化学修飾されたイノシンを検出する、請求項2から7の何れかに記載の方法。
【請求項10】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによって、化学修飾されたイノシンを検出する、請求項1から7の何れかに記載の方法。
【請求項11】
化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いて検出することによって、化学修飾されたイノシンを検出する、請求項2から7の何れかに記載の方法。
【請求項12】
α,β−不飽和結合と電子吸引性基とを有する化合物を含む、RNA中のイノシン化部位修飾剤。
【請求項13】
請求項12記載の修飾剤を含む、請求項1から11の何れかに記載の方法を行うための試薬キット。
【請求項1】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾する工程を含む、イノシン化部位の検出方法。
【請求項2】
さらに、化学修飾されたRNAを逆転写反応に供してcDNAを合成する工程、及び合成されたcDNAに基づいてイノシン化部位を検出する工程を含む、請求項1に記載の方法。
【請求項3】
イノシン化部位の検出が、イノシン化部位の有無の検出、イノシンの量の定量、及び/又はイノシンを含む領域又はイノシン化部位の同定である、請求項1又は2に記載の方法。
【請求項4】
α,β−不飽和結合と電子吸引性基とを有する化合物が、
式:C(R1)R2=C(R3)E
(式中、R1及びR2はそれぞれ独立に水素原子、炭素数1から6のアルキル基、フェニル基、又は炭素数1から6のアルコキシル基を有するフェニル基を示し、またR1及びR2の何れか一方はEと結合して環を形成してもよい。R3は、水素原子、炭素数1から6のアルキル基、フェニル基、炭素数1から6のアルコキシル基を有するフェニル基、又は電子吸引性基を示す。Eは電子吸引性基を示す。)
で表される化合物である、請求項1から3の何れかに記載の方法。
【請求項5】
電子吸引性基が、CN、NO2、SO3H、CONH2、COCH3、COOCH3、COOC2H5、COCH3、COC2H5、又はCOC6H5である、請求項4に記載の方法。
【請求項6】
化学修飾されたRNAを逆転写反応に供してcDNAを合成した後に、cDNAの増幅反応を行う、請求項1から5の何れかに記載の方法。
【請求項7】
対照として化学修飾されていないRNAを逆転写反応に供して合成されたcDNAと化学修飾されたRNAから合成されたcDNAとを比較することにより、イノシン化部位を検出する、請求項2から6の何れかに記載の方法。
【請求項8】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNAを質量分析に供することによって化学修飾されたイノシンを検出する、請求項1、3、4または5の何れかに記載の方法。
【請求項9】
化学修飾されたRNAに由来するcDNAの長さを検出することによって、化学修飾されたイノシンを検出する、請求項2から7の何れかに記載の方法。
【請求項10】
RNAをα,β−不飽和結合と電子吸引性基とを有する化合物で処理することによってイノシン化部位を化学修飾した後に、化学修飾されたRNA又はそれに由来するcDNAの塩基配列を決定することによって、化学修飾されたイノシンを検出する、請求項1から7の何れかに記載の方法。
【請求項11】
化学修飾されたRNAに由来するcDNAを、RNAのイノシン化部位の上流のみの配列を含むプローブを用いて検出することによって、化学修飾されたイノシンを検出する、請求項2から7の何れかに記載の方法。
【請求項12】
α,β−不飽和結合と電子吸引性基とを有する化合物を含む、RNA中のイノシン化部位修飾剤。
【請求項13】
請求項12記載の修飾剤を含む、請求項1から11の何れかに記載の方法を行うための試薬キット。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2008−259425(P2008−259425A)
【公開日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願番号】特願2005−229335(P2005−229335)
【出願日】平成17年8月8日(2005.8.8)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成17年度文部科学省「科学技術試験研究」、産業活力再生特別措置法第30条の適用を受けるもの
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
【公開日】平成20年10月30日(2008.10.30)
【国際特許分類】
【出願日】平成17年8月8日(2005.8.8)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成17年度文部科学省「科学技術試験研究」、産業活力再生特別措置法第30条の適用を受けるもの
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【Fターム(参考)】
[ Back to top ]