
RNA分解酵素活性を有するタンパク質及びその利用
【課題】RNA研究に有用なRNA分解酵素活性を有するタンパク質の提供。
【解決手段】特定のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含み、RNAを切断する活性を有する、タンパク質、該タンパク質の検索方法、該タンパク質をコードする、ポリヌクレオチド、該ポリヌクレオチドを含む形質転換体、該RNA分解酵素を用いてRNAを切断する工程を備える、核酸の処理方法。
【解決手段】特定のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含み、RNAを切断する活性を有する、タンパク質、該タンパク質の検索方法、該タンパク質をコードする、ポリヌクレオチド、該ポリヌクレオチドを含む形質転換体、該RNA分解酵素を用いてRNAを切断する工程を備える、核酸の処理方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、RNA分解酵素活性を有するタンパク質に関し、例えば、アデニン(A)塩基を認識してリボヌクレオチド鎖等を切断する活性を有するタンパク質及びその利用に関する。
【背景技術】
【0002】
近年、生物体におけるDNAのみならずRNAの機能が研究されるようになってきている。特に、塩基特異的な切断活性を有するRNA分解酵素(RNase)は、RNA分子の二次構造の決定やタンパク質-RNA相互作用の研究に広く用いられている。現在入手可能なエンド型RNaseのうち、一本鎖RNA分子を切断するRNase Aはピリミジン塩基(U及びC)の3’側、RNase T1はグアニン塩基(G)の3’側を、RNaseT2はAとUの3’側をそれぞれ切断する。RNase V1とRNase IIIは二本鎖RNA分子を特異的に切断し、RNase Hは、RNA/DNA二本鎖のRNA鎖を特異的に分解することが知られている(非特許文献1)。
【非特許文献1】NUCLEASES, Edited by Linn, S. M. and Roberts, R. J., Cold Spring Harbor Laboratory (1982)
【発明の開示】
【発明が解決しようとする課題】
【0003】
RNA分子の二次構造の決定等には、なんらかの選択性を持ってRNAを切断するものであることが必要であるが、これらの他にRNA分子の二次構造決定等に利用可能なRNaseは未だ見い出されていない。切断部位を塩基特異的に認識するRNA分解酵素は未だ見出されていない。また、これら各種のRNaseはいずれもアデニン塩基(A)を高い選択性で認識するものでなかった。
【0004】
そこで、本発明は、RNA研究に有用なRNA分解酵素活性を有するタンパク質を提供することを一つの目的とする。また、本発明は、アデニン塩基を認識してRNA鎖を切断する酵素活性を有するタンパク質を提供することを一つの目的とする。さらに、本発明は、アデニン塩基を高い選択性で認識してRNAを切断する酵素活性を有するタンパク質を提供することを一つの目的とする。また、本発明は、こうしたRNA分解酵素の利用を提供することを他の一つの目的とし、こうした酵素活性を有するタンパク質を発現させるためのポリヌクレオチド、当該ポリヌクレオチドを、前記タンパク質を発現可能に保持する形質転換体を提供することを他の一つの目的とする、さらにまた、本発明は、こうした酵素を用いた核酸の分析キット及び処理方法を提供することを他の一つの目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、植物に普遍的に存在するタンパク質であってRNA結合活性を有するタンパク質について検討していたところ、細胞器官におけるRNAプロセッシングに関与すると考えられるPPRモチーフ(Pentatrico Peptide Repeat:PPR)に連結されることのあるDYWドメイン及び当該DYWに類するドメインが、RNA切断活性、すなわち、RNA分解酵素として機能するという知見を得た。また、本発明者らは、同時に、このドメインがアデニン塩基に選択性の高い切断活性を有するという知見も得た。本発明は、これらの知見に基づきなされたものであり、本発明によれば、以下の手段が提供される。
【0006】
本発明によれば、
以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、RNAを切断する活性を有する、タンパク質が提供される。
【0007】
さらに、本発明のタンパク質は、以下の特徴;
(b)チトクロームCヘム結合モチーフCxxCHを有する。
を有することができる。また、本発明のタンパク質は、以下の特徴;
(c)PPRモチーフを備える天然タンパク質のC末端領域に備えられるアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列である。
を有することもできる。
【0008】
本発明のタンパク質は、前記アミノ酸配列は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれかを問い合わせ配列としてホモロジー検索を行って得られるE値が所定値以下(好ましくは1.0×10-10以下)である配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列とすることができる。前記アミノ酸配列は、好ましくは、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列である。
【0009】
本発明のタンパク質は、単子葉植物及び双子葉植物のいずれかのタンパク質とすることができる。また、前記RNA切断活性はエンド型であることが好ましく、さらに好ましくは、前記RNA切断活性は塩基特異的である。また、前記RNA切断活性は、アデニン塩基を有するリボヌクレオチドの5’側を部位特異的に切断する活性であることが好ましい。
【0010】
また、本発明によれば、RNA分解酵素のスクリーニング方法であって、配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、を備える、方法も提供される。前記タンパク質の準備工程は、配列番号:1、配列番号:4及び配列番号:6のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程とすることができる。
【0011】
さらに、本発明によれば、RNA分解酵素の製造方法であって、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列とチトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することを特徴とする、方法も提供される。
【0012】
本発明によれば、上記いずれかに記載のタンパク質を含む、核酸分析キットも提供され、さらに、本発明によれば、上記いずれかに記載のタンパク質をコードするポリヌクレオチドも提供され、本発明のポリヌクレオチドを、上記いずれかのタンパク質を発現可能に保持する形質転換体も提供され、上記いずれかのタンパク質を用いてRNAを切断する工程を備える、核酸の処理方法も提供される。この処理方法は、核酸を分離精製する工程、RNAを抽出する工程及びRNA断片を検出する工程のいずれかあるいは2工程以上を備えることもできる。
【発明を実施するための最良の形態】
【0013】
本発明のタンパク質は、以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、RNAを切断する活性を有することができる。
【0014】
本発明のタンパク質(以下、単に本タンパク質という。)は、上記アミノ酸配列を有することにより、RNA切断活性を有する酵素として機能することができる。また、前記アミノ酸配列は、PPRをモチーフを有しRNAプロセッシングに関与すると考えられる天然タンパク質のC末端に備えられるアミノ酸配列であることから、RNAの機能解析等の研究に有用である。特に、アデニン塩基特異的にRNAを切断することができるため、RNA等における塩基配列の正確な決定に有用である。また、本発明のタンパク質は、PPRタンパク質のC末端側の配列であるDYWモチーフに関連付けられるアミノ酸配列を含んでかつRNA切断活性を有するタンパク質であるということもできる。
【0015】
また、本発明の核酸分析キット(以下、単に、本分析キットという。)は、本タンパク質を含むことができる。本分析キットは、RNAやDNA等の抽出分離、RNAの配列や機能解析に有用である。
【0016】
また、本発明の形質転換体(以下、単に、本形質転換体という。)は、本タンパク質をコードするポリヌクレオチドを、本タンパク質を発現可能に保持することができる。本形質転換体によれば、RNA分解酵素活性を有するタンパク質を供給するのに都合がよい。さらに、本タンパク質をコードする本発明のポリヌクレオチド(以下、単に本ポリヌクレオチドという。)によれば、有用なRNA分解酵素を効率的にあるいは任意の宿主に生産させることができるコンストラクトや本タンパク質を発現可能な形質転換体を提供することができる。さらにまた、本発明の核酸の処理方法によれば、本タンパク質をRNA分解酵素として利用することでRNAを分解して従来にない核酸の分離精製やRNA代謝の研究等が可能となる。
【0017】
以下、本発明の実施の実施形態であるタンパク質、核酸分析キット、核酸処理方法、ポリヌクレオチド及び形質転換体等について詳細に説明する。
【0018】
(タンパク質)
本タンパク質は、1又は2以上の特徴を有するアミノ酸配列を備えることができる。本タンパク質が備えることができる一つのアミノ酸配列は、配列番号:1に記載の配列とすることができる。配列番号:1に記載のアミノ酸配列は、シロイヌナズナ(Arabidopsis thaliana、ecotype columbia)の機能未知タンパク質遺伝子at2g02980(MATDB:http://mips.gsf.de/proj/thal/db/index.html)がコードするPPRモチーフを有する603アミノ酸からなるタンパク質(PPRタンパク質)(配列番号:3)の一部からなる配列である。具体的には、配列番号:3に記載のアミノ酸配列のC末端側の第495位〜第603位の109個のアミノ酸からなる配列である。PPRタンパク質は、RNAプロセッシングに関連していると考えられているが、そのC末端領域がRNase活性を有することは本出願前において確認されていない。
【0019】
また、本タンパク質が備えることのできる他の一つのアミノ酸配列は、配列番号:4に記載の配列である。配列番号:4に記載のアミノ酸配列(Os5g30710(OS_M23)の一部、データベース:TIGR Rice Genome Annotation (http://www.tigr.org/tdb/e2k1/osa1/)は、イネ(Oryza sativa)由来の天然タンパク質(Os5g30710(OS_M23)に含まれるアミノ酸配列である。このアミノ酸配列が含まれるタンパク質も、PPRモチーフを有するPPRタンパク質である。
【0020】
本タンパク質は、これらの2種のアミノ酸配列に限定されないで、さらに他のアミノ酸配列を備えることができる。すなわち、本タンパク質は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を備えることもできる。すなわち、これらの2種のアミノ酸配列に対してアミノ酸の置換、欠失、挿入及び付加から選択される1又は2以上のアミノ酸の変異を有するアミノ酸配列(以下、改変配列ともいう。)である。アミノ酸変異の数は、特に限定しないが、好ましくは20個以下であり、より好ましくは10個以下である。より好ましくは5個以下である。
【0021】
アミノ酸配列への部位特異的な変異の導入は当業者において公知の手法によって可能である。アミノ酸配列が改変されたタンパク質をコードするDNAを調製するための当業者によく知られた方法としては、例えば、site-directed mutagenesis法(Kramer W & Fritz H-J: Methods Enzymol 154: 350,
1987)が挙げられる。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得る。
【0022】
また、こうした改変配列の他の一つとしては、もとのアミノ酸配列をコードする塩基配列又はその一部からなるDNA(その相補鎖であってもよい。)とハイブリダイズするDNAによってコードされるアミノ酸配列が挙げられる。例えば、配列番号:1に記載のアミノ酸配列は、配列番号:2に記載の塩基配列によってコードされている。また、配列番号:4に記載のアミノ酸配列は、配列番号:5に記載の塩基配列によってコードされている。これらの塩基配列に基づくDNAを利用することができる。
【0023】
このような改変配列は、ハイブリダイゼーション技術(Southern EM: J Mol Biol 98: 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki
RK, et al: Science 230: 1350, 1985、Saiki
RK,et al: Science 239: 487, 1988)を利用する方法が挙げられる。すなわち、配列番号:2に記載されるDNA若しくはその一部又はこれらの相補鎖をプローブとして、あるいは、こうしたタンパク質のゲノムDNAの塩基配列に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、シロイヌナズナや他の植物から、ハイブリダイゼーションによってDNAを単離し、該タンパク質を取得することは当業者において通常行い得ることである。
【0024】
なお、こうしたタンパク質をコードするDNAは、好ましくは、ストリンジェントな条件下でハイブリダイゼーション反応を行うことによって得ることができる。本発明においてストリンジェントなハイブリダイゼーション条件とは、6M 尿素、0.4%
SDS、0.5×SSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を指す。よりストリンジェンシーの高い条件、例えば、6M 尿素、0.4%
SDS、0.1×SSCの条件下では、より相同性の高いDNAを単離できると期待される。なお、高い相同性とは、アミノ酸配列全体で少なくとも50%以上、好ましくは70%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を意味するものとする。なお、アミノ酸配列や塩基配列の同一性は、BLAST等を用いて決定できる。
【0025】
配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列においてアミノ酸変異を導入した改変配列は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列として任意のデータベースをホモロジー検索して得られるE値(Expected value, E-value;期待値)が所定値以下のアミノ酸配列とすることができる。E値は、一般的に、あるいは用いるホモロジー検索プログラムにおいて定義されている。本発明におけるE値については、ホモロジー検索手法における一般的な定義あるいは用いる特定のホモロジー検索手法において個別に定義されていれば、その定義に従うものとする
【0026】
E値が小さければ小さいほど配列番号1:及び配列番号:4のいずれかに記載の配列と類似性が高い。E値は好ましくは、1.0×10-10以下である。一般にE値が1.0×10-10以下であれば、配列番号:1等に記載のアミノ酸配列に類似することが偶然には起きにくいということができる。こうしたアミノ酸配列であれば、機能的にも配列番号:1に記載のアミノ酸配列を有するタンパク質と同質あるいは類似の活性を有していると考えられる。好ましくは、E値は1.0×10-15以下である。よりE値が小さいアミノ酸配列を備えることで配列番号:1等に記載のアミノ酸配列からなるタンパク質と同質で同等の活性を得られやすくなる。
【0027】
ホモロジー検索は、問い合わせ配列に対するダイナミックプログラミングによるローカルペアワイズアライメントによるものとすることができる。この種のホモロジー検索プログラムは、インターネット等を通じて公開又は提供されているものから適宜選択して用いることができる。例えば、SSEARCH(http://www.ddbj.nig.ac.jp/search/ssearch-j.html等);BLAST(Proc.
Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc
Natl Acad Sci USA 90: 5873, 1993,http://www.ncbi.nlm.nih.gov/BLAST/)、PSI-BLAST(http://www.ncbi.nlm.nih.gov/BLAST/等);FASTA(http://www.ebi.ac.uk/fasta33/)及び等を用いることができる。なかでも、BLAST及びPSI-BLASTを好ましく用いることができる。
【0028】
検索対象とするデータベースは、特に限定しない。例えば、ホモロジー検索プログラムを公開するサイトにおいて準備されているものから適宜選択して用いることができる。また、プログラムとは別個に公開されている又は非公開状態のデータベースを用いることもできる。本タンパク質は、真核生物、特に植物のゲノム又はタンパク質あるいはこれらの一部(モチーフ)を含むデータベースを利用することで多くの類似配列を抽出することができる。本タンパク質のアミノ酸配列は、PPRモチーフを有するPPRタンパク質のC末端領域に相当するかあるいは類似することが多く、PPRタンパク質は植物に多く含まれているからである。PPRタンパク質は、シロイヌナズナなどの双子葉植物及びイネ(Oryza sativa)のような単子葉植物の双方に含まれており、こうした植物のタンパク質のデータベースを検索対象とすることが好ましい。
【0029】
なお、ホモロジー検索にあたっては、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列とするほか、当該アミノ酸配列をコードする塩基配列を問い合わせ配列として塩基配列やアミノ酸配列のデータベースを検索することにより、所定値以下のE値を有するアミノ酸配列を取得することも包含される。この場合、例えば、必要に応じてBLAST等に包含される各種ホモロジー検索手法を用いればよい。
【0030】
例えば、配列番号:1を問い合わせ配列としたホモロジー検索(BLAST)によって抽出されるアミノ酸配列としては、配列番号:4に記載のアミノ酸配列が挙げられる(Os5g30710(OS_M23)、データベース:TIGR Rice Genome Annotation (http://www.tigr.org/tdb/e2k1/osa1/)、E値3×10-27)。
【0031】
また、本タンパク質は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索をしたときに所定値以下のE値であるアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含んでいてもよい。すなわち、抽出されたアミノ酸配列の改変配列を含んでいてもよい。こうしたアミノ酸変異の数は、特に限定しないが、好ましくは20個以下であり、より好ましくは10個以下である。より好ましくは5個以下である。
【0032】
本タンパク質が有することのできるアミノ酸配列は、また、PPRタンパク質ファミリーのマルチプルアライメントによって得られるコンセンサス配列に基づくものであってもよい。コンセンサス配列を得るためのマルチプルアライメントに用いる手法やコンセンサス配列を取得する手法は特に限定されない。例えば、ClustalW: http://align.genome.jp/; HMMER(隠れマルコフモデル): http://hmmer.wustl.edu/;
MultiAlin:http://prodes.toulouse.inra.fr/multalin/multalin.html; mkdom/xdom:
http://prodes.toulouse.inra.fr/prodom/xdom/等各種の手法を採用することができる。コンセンサス配列が得られたなら、コンセンサス配列において高度に保存されているアミノ酸を維持するような改変配列や予測される活性部位や結合部位におけるアミノ酸をこれらの部位における相互作用を強めるように変異させた改変配列を得ることができる。例えば、こうしたコンセンサス配列の一例としては、HMMERマトリックスを利用してシロイヌナズナ85種のPPRタンパク質ファミリーのマルチプルアライメントから得られたもの(C末端側のDYWモチーフ)が開示されている(図1及び配列番号:6)(The Plant Cell,
Vol. 16, 2089-2103, August 2004、Fig.2)。図1において、保存されているアミノ酸は大文字で示され、特によく保存されているアミノ酸は太字で示されている。
【0033】
本タンパク質が備えることのできるアミノ酸配列の他の一つの特徴としては、チトクロームCヘム結合モチーフCxxCH(C:システィン、H:ヒスチジン、x:任意のアミノ酸)を有していることが挙げられる。このモチーフは、金属配位モチーフとして知られている。このモチーフを保持することがRNA分解酵素活性及びそのアデニン塩基特異性に好ましい。このモチーフは、例えば、配列番号:1において第69位〜第72位に相当する配列に位置することができる。当該モチーフは、C末端側から約40アミノ酸程度の位置、例えば、C末端側から35〜45アミノ酸に位置することができる。当該モチーフの有無は、例えば、PROSITE(http://au.expasy.org/prosite/)など各種公知のモチーフデータベースにて確認することができる。既に例示した配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列においてもこのモチーフを備えている。
【0034】
本タンパク質が備えることができるアミノ酸配列のさらに他の一つの特徴としては、PPRモチーフを備える天然タンパク質(PPRタンパク質)のC末端領域に位置するアミノ酸配列又はその改変配列が挙げられる。PPR(pentatricopepetide repeat)モチーフは、35アミノ酸の保存配列がリピートしたものであり、植物に多く存在し、なかでも葉緑体やミトコンドリアに局在することが知られている(Biochem Soc Trans. 2004 Aug;32(Pt 4):571-4.)。PPRモチーフの有無は、例えば、PFAM(http://www.sanger.ac.uk/Software/Pfam/)などのドメインデータベースにより知ることができる(Accession No.PF01535)。35アミノ酸のコンセンサス配列を、図2及び配列番号:7に示す。図2において、よく保存されているアミノ酸は大文字で示されている。配列番号:1及び配列番号:4に記載のアミノ酸配列はいずれもこうしたPPRタンパク質のC末端領域に位置される配列である。なお、PPRタンパク質のC末端側領域に位置されるアミノ酸配列の改変配列における改変配列の意味は、既に説明した意味がここでも適用される。
【0035】
こうしたタンパク質は、PPRタンパク質が植物の葉緑体やミトコンドリアに多く存在することから、単子葉植物及び双子葉植物の天然タンパク質中のアミノ酸配列及びこの改変配列であるアミノ酸配列を有することができる。植物種は特に限定されないが、例えば、単子葉植物としては、イネ、ムギ、キビ等が挙げられ、双子葉植物としては、シロイヌナズナ、ダイズ、ピーナッツ、ごま、ナタネ、綿実、ヒマワリ、サフラワー、、バレイショ、サツマイモ等が挙げられる。
【0036】
本タンパク質は、RNA切断活性を有することができる。本タンパク質のRNA切断活性は、RNAを切断することができる限りどのような態様であってもよい。本タンパク質のRNA切断活性は、RNA特異的であることが好ましい。RNA特異的であるとDNAに作用しないのでDNA等と容易に分離することができるし、DNA配列を維持させることができる。また、エキソ型でもエンド型でもよいが、好ましくはエンド型である。エンド型であることで、RNAの機能解析等に有用である。なお、本明細書において、RNA切断活性とは、切断部位又はその近傍にRNA鎖を備えるオリゴヌクレオチド又はポリヌクレオチドを切断するものであればよい。したがって、DNA/RNAハイブリッドやDNA-RNAキメラ、修飾塩基やその他の修飾や付加を含むヌクレオチド鎖を切断対象として含めることができる。
【0037】
本タンパク質は、塩基特異的なRNA切断活性を有していることが好ましい。本明細書において。塩基特異的なRNA切断活性を有しているとは、1又は2以上の特定塩基を認識して認識した特定塩基に対して所定の位置にある結合を切断する活性を有していることを意味している。本発明においては、好ましくはアデニン塩基を認識する。さらに、アデニン塩基を認識してその5’側においてRNAを切断することが好ましい。従来、アデニン塩基を高い特異性を持って認識して切断するRNaseは見出されておらず、研究及びその他の各種用途に有用である。
【0038】
本タンパク質は、EDTAが所定濃度範囲にあるとき、換言すれば、所定濃度の2価金属イオンの存在下でRNA切断活性を発揮することができることが好ましい。こうした2価金属イオン濃度依存性を備えることで2価金属イオン濃度やEDTA等のキレート剤の添加濃度でその活性を制御することができる。好ましくは、本タンパク質はそのRNA切断活性を発揮するのにあたって、2価金属イオン濃度について至適範囲を有している。例えば、配列番号:1に記載のアミノ酸配列を含むタンパク質は、EDTA等のキレート剤の不存在下及び高濃度存在下ではRNA切断活性を有していないが、20mM以上、好ましくは25mM以上の濃度で、また、70mM以下、好ましくは、62.5mM以下の濃度で高いRNA切断活性を発揮することができる。また、例えば、マグネシウムイオンなどの2価金属イオン濃度としては、1mM以上10mM以下程度が好ましく、より好ましくは、下限が2mMであり、上限が5mMである。
【0039】
RNA切断活性は、公知の方法によって測定することができる。一般的には、適当な媒体下で基質RNAとタンパク質とを接触させるようにすればよい。基質RNAには放射性元素などの適当な標識を付して置くことができる。適当な温度下で一定時間反応させた後、RNAを抽出し尿素変性ポリアクリルアミドゲルなどで電気泳動することによりRNAの分解程度を検知することができる。なお、部位特異的な切断活性を測定する場合には、分解反応産物に対してコンプレメンタリーDNAを調製するとともに、基質RNAの鋳型であるDNAからDNAラダーを調製し、これらをそれぞれ尿素変性ポリアクリルアミドゲル電気泳動などで分離させることによって切断部位を特定することができる。
【0040】
なお、本タンパク質は、RNA切断活性を発揮できる限り、さらに他のアミノ酸配列を備えていてもよい。製造上の都合及び使用用途等に応じて各種のアミノ酸配列を付加することもできる。
【0041】
(本タンパク質の製造方法)
本タンパク質は、天然タンパク質又はその一部であるときには、自然界において保持される植物などの生物体又はそのタンパク質が局在するオルガネラ画分から、タンパク質を抽出し、分離精製等することによっても得ることができる。また、本タンパク質が天然タンパク質の全部又は一部であっても、非天然のタンパク質であっても、化学的又は遺伝子工学的に得ることもできる。遺伝子工学的手法によって得る場合には、本タンパク質をコードするポリヌクレオチドを用いて無細胞的にあるいはこのポリヌクレオチドを適当な宿主に導入して形質転換体を作出し、当該形質転換体の宿主内又は宿主外で発現させるようにすることができる。
【0042】
(RNA分解酵素のスクリーニング方法及び製造方法)
本発明のRNA分解酵素のスクリーニング方法は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、を備えることができる。このスクリーニング方法によれば、RNA切断活性を有するタンパク質(RNA分解酵素)を効率的に得ることができる。また、前記タンパク質の準備工程は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの改変配列を備えるタンパク質を準備する工程とすることもできる。配列番号:1及び配列番号:4に記載のアミノ酸配列は、酵素活性が確認されたDYW配列であるため、これらの配列を問い合わせ配列とすることで同等活性のあるアミノ酸配列を効率的に抽出することができる。さらに、本発明のRNA分解酵素のスクリーニング方法においては、配列番号:6に記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って所定値以下のE値を有するアミノ酸配列を抽出するようにしてもよい。配列番号:6に記載の配列は、DYWモチーフのコンセンサス配列であるから、この配列と所定値以下のE値を有するアミノ酸配列を有するタンパク質をRNA切断活性のスクリーニング対象とすることが可能である。さらに、チトクロームCヘム結合モチーフを有するアミノ酸配列を準備するようにするとより一層効果的にRNA分解酵素をスクリーニングすることができる。
【0043】
タンパク質の準備工程では、例えば、所定のホモロジー検索において所定値以下のE値を有するアミノ酸配列を有するタンパク質を抽出し、当該タンパク質をコードするポリヌクレオチドなどを用いて当該タンパク質を遺伝子工学的に生産することもできる。
【0044】
なお、所定値以下のE値を有するアミノ酸配列を備えるタンパク質を準備するにあたっては、既に説明した各種のホモロジー検索プログラムを用いることができる。また、E値については、好ましくは、1.0×10-10以下であり、より好ましくは、1.0×10-15以下である。また、改変配列についても既に説明した各種の態様をこの方法で準備するタンパク質に適用することができる。RNA切断活性の有無は、既に説明した方法等、当業者において周知の方法を用いて測定することができる。
【0045】
こうしたRNA分解酵素のスクリーニングによってRNA切断活性を有することが検出されたタンパク質は、既に説明した本タンパク質を製造する場合と同様、天然の生物体から取得することもできるし、人工的に取得することもできる。RNA分解酵素は、また、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの改変配列を備えるとともに、チトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することによっても得ることができる。こうしたタンパク質は、高い確率でRNA切断活性を発揮できるからである。
【0046】
(核酸分析キット)
本分析キットは、本タンパク質をキット構成に含めることができる。キットに含められるタンパク質は、どういった形態で含められていてもよいが、例えば、保存性等を考慮して凍結乾燥体として含められていることが好ましい。また、本分析キットには、その他のRNase、DNase等、核酸の分離精製、分析等に必要な酵素や試薬を含められていてもよい。
【0047】
(ポリヌクレオチド)
本ポリヌクレオチドは、本タンパク質をコードする領域を有することができる。本タンパク質をコードするポリヌクレオチドの塩基配列としては、例えば、配列番号:2及び配列番号:5が挙げられる。ポリヌクレオチドにおける塩基配列は、本タンパク質をコードしている以上コドン用法は特に限定されない。
【0048】
本ポリヌクレオチドは、例えば、本ポリヌクレオチドを発現可能にプロモーター等の調節領域を備える発現カセット、ベクター、プラスミド、人工染色体及び感染可能なアグロバクテリウム等のウイルスの態様であってもよい。
【0049】
(形質転換体)
本形質転換体は、本ポリヌクレオチドを、本タンパク質を発現可能に保持する形質転換体とすることができる。本形質転換体は、大腸菌や乳酸菌などの原核細胞のほか、酵母、植物培養細胞及び植物体を宿主とすることができる。植物体としては、植物個体、その植物個体を構成しうる全ての種類、形態の細胞、植物固体の一部である組織や器官ならびに生殖細胞を含んでいる。また、植物個体の一部としては、その繁殖媒体(種子、根茎、果実、切穂等)も包含する。
【0050】
本形質転換体が、本タンパク質の発現可能な本ポリヌクレオチドを保持するには、本ポリヌクレオチドは適当なプロモーターの制御下に連結されることが好ましい。また、適当なターミネータも連結されることが好ましい。その他、発現の増強に有効なエレメントを適宜備えることができる。
【0051】
こうした形質転換体は、本タンパク質を大量に生産するほか、RNA/DNA代謝研究に有用である。
【0052】
また、本形質転換体は、本タンパク質の発現を抑制可能に構築されていてもよい。例えば、植物等に内在する、配列番号:1に記載のアミノ酸配列又は配列番号:4に記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って所定値以下のE値を有するアミノ酸配列又は当該アミノ酸配列を含むタンパク質あるいはさらにチトクロームCヘム結合モチーフを有するタンパク質の発現を抑制可能に構築されていてもよい。こうした形質転換体は、RNA/DNA代謝研究に有用である。本タンパク質の発現を抑制する態様としては、内在する本タンパク質をコードする遺伝子の破壊や当該遺伝子の転写や翻訳を抑制するほか、変異等を導入して不完全なタンパク質等として発現させてその作用を低下又は失活させたり、あるいは本タンパク質と相互作用して本タンパク質の作用を低下させる物質を発現させることが挙げられる。
【0053】
例えば、特定の遺伝子の発現を抑制する方法としては、特定遺伝子のノックアウトのほか、、アンチセンス法、コサプレッション法、RNA干渉法及びリボザイム法等を用いて、mRNAを対象にした発現抑制可能な核酸コンストラクトを導入し保持させることが挙げられる。
【0054】
なお、形質転換体は、当業者の公知の手法を採用して作出することができる。形質転換体が植物体の場合、例えば、本タンパク質を発現可能に保持する形質転換体を作成するには、植物細胞内で本タンパク質を発現可能なコンストラクトを構築し、当該コンストラクトを植物細胞に導入し、該植物細胞から植物体を再生させればよい。ベクターは特異に限定されない。また、植物細胞内で恒常的に遺伝子を発現させるためのプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクターや、外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることもできる。コンストラクトが導入される細胞としては、植物体に再生可能なあらゆる種類の形態の植物細胞を含めることができる。例えば、培養細胞、プロトプラスト、苗条原基、多芽体、毛状根、カルス、葉など特定器官の切片等が挙げられる。植物の形質転体は、所定の再生工程を実施することで細胞を植物体に変換することができる。再生の方法は、植物の種類によって異なるが、各種公知の方法を使用できる。植物細胞への核酸コンストラクトの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。また、パーティクルガン法においては、例えば、バイオラッド社のものを用いることが可能である。形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。また、再生した植物体から有性生殖または無性生殖により子孫を得ることができる。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなど)を得て、それらを基に該植物体を量産することも可能である。
【0055】
(核酸の処理方法)
本処理方法は、タンパク質を用いてRNAを切断する工程を備えることができる。この処理方法は、RNAを切断し、RNAを断片化することができる。この切断工程によれば、例えば、試験管内においてRNA断片を生成させて、当該RNAの機能探索のための材料を得ることができる。また、塩基特異的にRNAを切断するため、生じたRNA断片を分析することで切断対象となったRNAの構造等を推測することもできる。
【0056】
本処理方法は、さらに、DNAなどの核酸を分離精製する工程を備えていてもよいし、RNAを抽出する工程を備えていてもよいし、さらに、RNA切断後に当該切断したRNA断片を電気泳動やMALDI等で検出する工程を備えることもできる。
【0057】
核酸の分離精製工程において、分離精製対象は、RNAとDNAとが混在する可能性のある試料であることが好ましい。こうした試料としては、生体から抽出された試料のほか、試験内でDNA合成した際の反応試料等であってもよい。例えば、RNAとDNAが混在する場合には、本タンパク質は、DNAには作用しないため共存するRNAから未切断断片としてDNAを容易に精製することができる。一方、本タンパク質はRNAに作用するため、RNAを切断断片として分離することができる。こうした分離精製工程においては、本タンパク質を適当な固相に固定化しておくと都合がよい。また、こうした固相を充填したカラムを用いてもよい。核酸の分離精製工程はRNAの切断工程後に備えることが好ましい。
【0058】
RNAを抽出する工程において、抽出対象は生体試料であることが好ましい。こうしたRNAを抽出して、その後、本タンパク質で切断することで、生体内におけるRNAの代謝研究が可能となる。RNA抽出工程は、RNA切断工程に先立って備えることが好ましい。
【0059】
さらに、RNA切断後に生じたRNA断片を検出する工程は、RNA断片自体、あるいはコンプレメンタリーDNAを調製した上で、電気泳動、マイクロアレイ及びMALDI等で分析するようにしてもよい。
【0060】
以下、本発明に関して、実施例を挙げて具体的に説明するが、本発明は以下の実施例の限定されるものではない。
【実施例1】
【0061】
(実施例1: 新規RNA分解酵素、T-DYWの調製とRNA分解活性の測定)
(シロイヌナズナからのゲノムDNAの調製)
シロイヌナズナ(Arabidopsis thaliana, ecotype Columbia)をムラシゲ・スクーク培地(2%ショ糖、0.5% Gellangamを含む)で3週間培養した。培養した植物の緑葉(0.5 g)をフェノール/クロロホルム抽出した後、エタノールを加えてDNAを不溶化した。回収したDNAを100 μlのTE液(10 mM トリス・塩酸 (pH 8.0)、1mM EDTA)に溶解し、10ユニットのRNase A(DNase-free、タカラバイオ社)を加えて、37℃で30分反応させた。その後、反応液を再度フェノール/クロロホルム抽出した後、エタノール沈殿によりDNAを回収した。10 μgのDNAが得られた。
【0062】
(DYWモチーフをコードする遺伝子のクローニング)
シロイヌナズナからのゲノムDNAの調製は上記の実施例1に記載されている方法により行った。シロイヌナズナゲノム情報データベース(MATDB:
http://mips.gsf.de/proj/thal/db/index.html)に掲載されている配列情報を参照し、DYWモチーフを持つタンパク質遺伝子at2g02980の当該DYWモチーフに相当するDNA配列を増幅するためのオリゴヌクレオチドプライマー(02980DYW-F、02980_m-R;配列番号:8、9に記載)を調製した。オリゴヌクレオチドプライマーのフォワードプライマーである02980DYW-F、リバースプライマーである02980_m-Rの5'側にはそれぞれEco
RI、SalI配列を付加した。EcoRIとSalI配列は得られたクローンを制限酵素処理で挿入配列を切り出すのに利用できるように組み入れた。
【0063】
at2g02980を含むDNA断片は、100 ngのゲノムDNAと上記プライマーを含む50 μlの反応液を95℃ 30秒、60℃ 30秒、72℃ 30秒の25サイクルでPrime Star (タカラバイオ社)
をDNA 伸長酵素として用い、PCRすることによって増幅した。その結果、339塩基対のDNA断片の増幅が確認できた。得られたDNA断片は、pBAD/Thio-TOPO ベクター(Invitrogen社)を用いて、クローニングした。クローニングするために必要なDNA断片の3'末端突出アデノシンは、DNA断片を0.1
ユニット TaKaRa Ex-Taq(タカラバイオ社)と2.5 mM ATPで70℃、10分間反応することで付加した。クローン化したDYWモチーフをコードするDNA配列を決定し、上記データベース上においてDYWモチーフに相当するDNA配列と相同な配列(配列番号:2)であることを確認し、プラスミドpTrx_DYWと命名した。
【0064】
(T-DYWタンパク質の調製)
得られたプラスミドpTrx_DYWをEscherichia coli LMG194株(Invitrogen社)に形質転換した。この大腸菌をアンピシリンが100 μg/mlの濃度で存在する0.2%グルコースを含むRM培地4,000 ml(1リットル培地を含む2リットル三角フラスコ、計4本)中で、37℃で培養した。培養液の濁度が波長600 nmでの吸光度が0.5に達した時に、誘導物質であるL-アラビノースを最終濃度が0.2%になるように添加し、さらに4時間培養を行った。遠心による集菌後、菌体を1 mg/mlのリゾチームを含む200 mlのバッファーA(50 mM トリス・塩酸 pH 8.0、500 mM KCl、2
mM imidazole、10 mM MgCl2、0.5% Triton X100、10% グリセロール)に懸濁し、超音波破砕と凍結溶解により菌体を破壊した。15,000 x g、20分間の遠心分離後に、上清を粗抽出液として回収した。この粗抽出液をバッファーAで平衡化したニッケルカラム樹脂(ProBond A、Invitrogen社)を充填したカラムに供した。
【0065】
カラムクロマトグラフィーは、20
mM imidazoleを含むバッファーAで十分に洗浄した後、200
mM imidazoleを含むバッファーAで目的タンパク質を溶出する二段階濃度勾配により行った。得られた組み換えタンパク質をSDSポリアクリルアミドゲル電気泳動により確認したところ、30 kDaのタンパク質として検出された。これをT-DYWタンパク質と命名した(配列番号:10)。なお、このタンパク質は、配列番号:1に記載のアミノ酸配列を備えるとともに、N末端側に溶解性を高めるためのチオレドキンのアミノ酸配列、C末端側にヒスチジンタグ配列を備える融合タンパク質である。RM培地1リットルあたり、600 μgのT-DYWタンパク質が得られた。T-DYWタンパク質を含む精製画分1 mlを1リットルのバッファー E(20 mM トリス・塩酸 pH 7.9、60
mM KCl、12.5 mM MgCl2、0.1 mM EDTA、17% グリセロール、2 mM DTT)で透析した後、T-DYWタンパク質の精製標品とした。
【0066】
(基質RNAの調製)
基質RNAとして、シロイヌナズナ葉緑体ndhB遺伝子の開始コドンから500塩基のRNAを用いた。基質RNAとして用いるRNAをNB500と命名した(配列番号:11)。NB500を合成するためのDNA断片は、オリゴヌクレオチドプライマーndhB-FとndhB-R(配列番号12、13に記載)を用いて、上記のシロイヌナズナゲノムDNA 10
ngを鋳型DNAとして含む50 μlの反応液を95℃ 30秒、60℃ 30秒、72℃ 30秒の25サイクルでPrime
Star (タカラバイオ 社) をDNA 伸長酵素として用い、PCRすることによって増幅した。ndhB-Fプライマーの5'末端側には基質RNAを試験管内で合成するためのT7 プロモータ配列を、ndhB-Rプライマーの5'末端側には3'→5'エクソヌクレアーゼによる非特異的な分解を防ぐためのステムループ構造を形成する塩基配列をそれぞれ付加した。得られたDNA断片は、アガロースゲルで展開後、ゲルから切り出すことによって精製した。精製DNA断片を鋳型にNTP mix(10 nmol GTP、CTP、ATP、0.5 nmol UTP)、4 μl [32P] α-UTP (GE ヘルスケア社、3000 Ci/mmol)、T7 RNA polymerase(タカラバイオ社)を含む20μlの反応液を37℃ 60分間反応させることで、基質RNAを合成した。基質RNAはフェノール/クロロホルム抽出、エタノール沈殿後、全量を6 M 尿素を含む変性6%ポリアクリルアミドゲル電気泳動で展開し、X線フィルムで60秒間感光させることによって、32P標識RNAを検出した。32P標識RNAをゲルから切り出し、200 μlのゲル溶出液(0.3 M 酢酸ナトリウム、2.5 mM EDTA、0.01% SDS)中に、4℃で12時間浸し、RNAをゲルから溶出した。溶出したRNAのうち、1 μlの放射活性を測定し、合成したRNAの総量を算出した。エタノール沈殿後、8,250 cpm/μl (1 fmol/μl)になるように、RNAを超純水に溶解した。この調製方法で通常、8,250 cpm/μlのRNAが約50 μl得られる。
【0067】
(T-DYWタンパク質のRNA分解活性)
T-DYWタンパク質存在下もしくは非存在下でのRNAの分解活性を調べた。50 mM EDTA を含む反応液(10 mM トリス・塩酸 pH 7.9、30
mM KCl、6 mM MgCl2、2
mM DTT、8% グリセロール、0.0067% of
Triton X-100)20 μl中に上記の1 fmolの基質RNA(NB500)と50 ng〜1.5 μgのT-DYWタンパク質を混合し、25℃で30分間反応した。反応後にフェノール/クロロホルム抽出、エタノール沈殿により、NB500 RNAを抽出した。RNAは6 M尿素を含む変性6%ポリアクリルアミドゲルで展開し、電気泳動後にゲルを乾燥させた。ゲル中のRNAの放射活性をバイオイメージングアナライザーBAS2000(フジフィルム社)で測定した。その結果を以下の図3に示す。
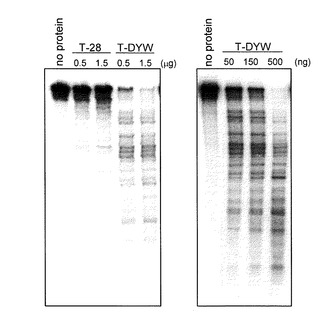
【0068】
図3より明らかなように、T-DYWタンパク質とRNAを混合するとタンパク質濃度依存的に基質RNAの分解活性が観察された。対照実験として、RNA結合タンパク質であるcp28をT-DYWと同様のベクター、発現系で調製した組み換えタンパク質をT-28として(配列番号:14)、同様の実験を行ったところ、RNAの分解活性は観察されなかった。タンパク質非存在下(no protein)でもRNAの分解は観察されなかった。
【0069】
(T-DYWタンパク質のRNA分解活性におけるEDTAの効果)
多くのRNA分解酵素は、そのRNA分解活性に二価金属イオンを必要とする。T-DYW精製標本中に含まれる二価金属イオンのRNA分解活性における影響を検証するために、EDTAを添加して、T-DYWタンパク質のRNA分解活性を測定した。0〜200
mM EDTAを含む反応溶液20 μl中に1 fmolのNB500 RNAと50
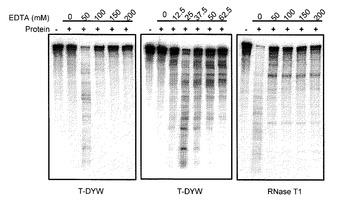
ngのT-DYWタンパク質を混合し、25℃で30分間反応した。反応後のRNA抽出、電気泳動、検出は実施例1に記載したのと同様の方法で行った。結果を図4に示す。
【0070】
図4に示すように、T-DYWタンパク質のRNA分解活性は50 mM EDTA存在下のみで確認された(図4左)。至適EDTA濃度を検証するために、EDTA濃度をさらに細かく変化させたところ、この反応条件では12.5mM以上62.5mM以下の濃度でRNA分解活性が検出され、25 mM EDTA存在下でもっとも効率のよいRNA分解活性が検出された(図4中)。反応液中の二価金属イオンを十分にキレートする高濃度のEDTAではT-DYWタンパク質のRNA分解活性は観察されなかったことから、T-DYWタンパク質のRNA分解活性には二価金属イオンが影響を与えることが示唆された。対照実験として、既知RNA分解酵素である100 ユニットのRNase
T1(Ambion 社)を用いて、同様の実験を行った。その結果、RNase T1はEDTA未添加で最大のRNA分解活性を示し、高濃度のEDTA存在下でもRNA分解活性を示した(図4右)。また、T-DYWとRNase T1では、RNA分解中間産物のパターンが異なることから、両タンパク質のRNA分解における認識塩基は異なることが明らかである。RNase T1はRNA中のシトシン残基の3'側を切断することが既に明らかになっている。
【0071】
(T-DYWタンパク質の酵素活性の核酸特異性)
T-DYWタンパク質の核酸切断活性をRNA、一本鎖DNA、二本鎖DNAを基質に用いて測定した。32P 標識二本鎖DNAは、プライマーndhB-F、ndhB-Rと鋳型DNAであるNB500 DNAを用い、dNTP mix(10 nmol dATP、dGTP、TTP、0.5 nmol dCTP)、4 μl
[32P] α-dCTP (GE ヘルスケア社、3000 Ci/mmol)、Ex-Taq(タカラバイオ社)を含む溶液でPCR反応を行うことにより調製した。PCR反応は実施例1と同様に行った。得られたDNA断片は変性6%ポリアクリルアミドゲル電気泳動による展開後、ゲルからDNAを溶出させることによって精製した。一本鎖DNAは二本鎖DNAを90℃で3分間処理した後に、急冷することによって調製した。T-DYWタンパク質とNB500 RNA (RNA)、一本鎖NB500 DNA (ssDNA)、二本鎖NB500 DNA(dsDNA)を混合し、実施例1 と同様の条件で、それぞれの基質核酸への切断活性を調べた。結果を図5に示す。
【0072】
図5に示したように、T-DYWタンパク質はRNAにのみ切断活性を示した。対照実験として、既知RNaseである100ユニットのRNase T1(Ambion社)、既知DNaseである10ユニットのDNase
I(RNase-free、タカラバイオ社)を用いて、酵素活性の核酸特異性を調べた。その結果、RNase T1はRNA、DNase IはDNAのみへの切断活性を呈したことから、T-DYWタンパク質はRNA特異的な核酸分解活性を呈することが明らかである。
【0073】
(T-DYWのRNA切断部位)
T-DYWタンパク質によって分解されたRNAの分解中間産物は、明瞭なバンドとして検出できる(図5)。これは既知RNaseであるRNase T1、RNaseAなどのRNAを内部で切断するエンドヌクレアーゼで観察される現象である。そこで、T-DYWタンパク質によって切断されるRNA残基の特性を検証するために、切断されたRNAの5'末端をプライマー伸長法により決定した。10 fmolの未標識ndhB RNAと1 μgのT-DYWタンパク質を用いて、実施例1 と同様の条件で反応した後、反応液からRNAを抽出した。
【0074】
次に5'末端を[32P]で標識したndh-Rプライマー0.4 pmolと回収したRNAを混合し、逆転写酵素(ReverTraAce、TOYOBO社)を用いて、コンプリメンタリーDNAを合成した。同時に、切断された塩基を特定するためのDNAラダーを調製した。DNAラダーは5'末端を[32P]で標識したndh-Rプライマーと鋳型DNAであるNB500を用いて、Thermo Sequenase Primer Cycle Sequencing kit (GEヘルスケア社)により調製した。得られたコンプリメンタリーDNAとDNAラダーを6M 尿素を含む6%ポリアクリルアミドゲルで電気泳動した。電気泳動後にゲルを乾燥させた。ゲル中のRNAの放射活性をBAS2000(Fuji
Film社)で測定した。その結果を以下の図6に示す。
【0075】
図6に示したように、T-DYWタンパク質によって切断されたRNAの5'末端がいくつか検出された。得られた5'末端の位置(NB500の5'末端を1とする)と塩基を示した。得られた5'末端は全てアデノシン残基であったことから、T-DYWはRNA中のアデノシン残基の5'側を選択的に切断するRNA分解酵素であることがわかった。
【実施例2】
【0076】
(イネ由来のDYWタンパク質の調製と分解活性)
まず、イネ (Oryza
sativa, Nipponbare)からのDNA調製を実施例1と同様に行った。次に、調製したイネのゲノムDNAを鋳型に、イネゲノム情報データベース(http://www.tigr.org/tdb/e2k1/osa1/overview.shtml)に掲載されている配列情報を参照し、シロイヌナズナのタンパク質 (AT2980(At2g02980)中のDYWモチーフの配列(配列番号:1)と高い相同性を示すモチーフを持つタンパク質遺伝子Os5g30710のDNA配列を増幅するためのオリゴヌクレオチドプライマー(Os5g30710-F、Os5g30710-R; 配列番号:15,16に記載)を調製した。Os5g30710を含むDNA断片の増幅(配列番号:17)、pBAD/Thio-TOPOベクター(Invitrogen社)へのクローニングは、実施例1に記載したT-DYWタンパク質遺伝子のクローニングと同様に行い、得られたプラスミドをpTrx-osDYWと命名した。このプラスミドpTrx-osDYWを用い、イネ由来のタンパク質T-osDYWを調製した(配列番号:18)。組み換えタンパク質の調製は、実施例1に記載した方法と同様に行った。
【0077】
このT-osDYWタンパク質のRNA分解活性は、50ngのT-osDYWタンパク質を用いて、実施例1 に記載したT-DYWタンパク質のRNA分解活性の測定と同様に行った。その結果を図7に示す。
【0078】
図7より明らかなように、イネ由来T-osDYWタンパク質においても、ナズナ由来T-DYWと同様なRNA分解活性が測定された。コントロールとして、タンパク質非存在下(レーン、no protein)、インサートのないpBAD-Thio-TOPOベクターから調製したチオレドキン及びヒスチジンタグのみからなるタンパク質(50 ng、control、配列番号:19)のみでもRNA分解活性を測定したが、RNAの分解は観察されなかった。
【実施例3】
【0079】
(変異DYWタンパク質の調製とRNA分解活性)
T-DYW、T-osDYWタンパク質は、そのアミノ酸配列中に金属配位モチーフ、CxxCH、を持つ(C、システイン;H、ヒスチジン;x、任意のアミノ酸)。このモチーフのRNA分解における働きを検証するために、以下の方法で、変異T-DYWタンパク質、T-DYW_Mタンパク質、を調製した。変異はT-DYWタンパク質中のアミノ酸配列CxxCHを、GxxGH(G、グリシン)に置換することによって行った。以下に詳細な調製法を示す。
【0080】
実施例1に記載したpTrx-DYWを鋳型に、まず2つのDNA断片、D1とD2をそれぞれ、オリゴヌクレオチドプライマー02980DYW-F(配列番号:8)と02980M2-R(配列番号:21)、02980M2-F(配列番号:20)と02980_m-R(配列番号:9)を用いて、実施例1 と同様の条件でPCRすることによって調製した。次に、得られたD1およびD2のDNA断片を混合したものを鋳型に、オリゴヌクレオチドプライマー02980DYW-F(配列番号:8)と02980_m-R(配列番号:9)を用いて、PCRすることによって、変異DYWタンパク質をコードするDNA断片を得た(配列番号:22)。DNA断片のpBAD-Thio/TOPOベクターへのクローニングと、組み換えタンパク質T-DYW_M(配列番号:23)の調製は、前述の実施例1 と同様の方法で行った。
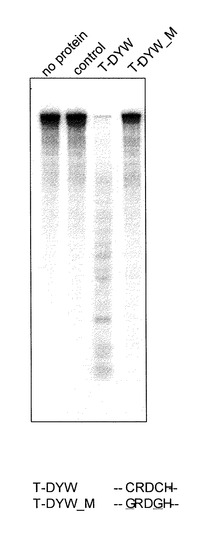
【0081】
得られたT-DYW_Mタンパク質のRNA分解活性は、実施例1に記載したT-DYWタンパク質のRNA分解活性の測定と同様の方法で行った。得られた結果を図8に示す。
【0082】
図8より明らかなように、T-DYWタンパク質(50 ng)では、RNA分解活性が観察できたが、T-DYWタンパク質中の2つのシステイン残基に変異を導入したT-DYW_Mタンパク質(50 ng)では、RNA分解活性は観察されなかった。コントロールとして、タンパク質非存在下(レーン、-protein)、pBAD/Thio-TOPOベクターのタグタンパク質のみ(50 ngインサートなし、配列番号:19)でもRNA分解活性を測定したが、RNAの分解は観察されなかった。このことは、T-DYWタンパク質のRNA分解活性には、金属配位モチーフであるCxxCHのアミノ酸配列が重要な働きを担うことを示している。
【配列表フリーテキスト】
【0083】
配列番号:8,9,12,13,15,16,20,21:プライマー
配列番号:10,14,18,19,23:融合タンパク質
配列番号:17,22:融合タンパク質をコードするDNA
【図面の簡単な説明】
【0084】
【図1】PPRタンパク質において見出されているDYWモチーフのコンセンサス配列の一例を示す図。
【図2】PPRタンパク質における35アミノ酸モチーフの一例を示す図。
【図3】実施例1で調製したT-DYWタンパク質のRNA切断活性を示す図。
【図4】実施例1で調製したT-DYWタンパク質のRNA切断活性に対するEDTAの効果を示す図。
【図5】実施例1で調製したT-TYWタンパク質のRNA切断活性の核酸特異性を示す図。
【図6】実施例1で調製したT-TYWタンパク質のRNA切断活性が塩基特異的であることを示す図。
【図7】実施例2で調製したイネDYWタンパク質のRNA切断活性を示す図。
【図8】実施例3で調製した変異を導入したDYWタンパク質のRNA切断活性を示す図。
【技術分野】
【0001】
本発明は、RNA分解酵素活性を有するタンパク質に関し、例えば、アデニン(A)塩基を認識してリボヌクレオチド鎖等を切断する活性を有するタンパク質及びその利用に関する。
【背景技術】
【0002】
近年、生物体におけるDNAのみならずRNAの機能が研究されるようになってきている。特に、塩基特異的な切断活性を有するRNA分解酵素(RNase)は、RNA分子の二次構造の決定やタンパク質-RNA相互作用の研究に広く用いられている。現在入手可能なエンド型RNaseのうち、一本鎖RNA分子を切断するRNase Aはピリミジン塩基(U及びC)の3’側、RNase T1はグアニン塩基(G)の3’側を、RNaseT2はAとUの3’側をそれぞれ切断する。RNase V1とRNase IIIは二本鎖RNA分子を特異的に切断し、RNase Hは、RNA/DNA二本鎖のRNA鎖を特異的に分解することが知られている(非特許文献1)。
【非特許文献1】NUCLEASES, Edited by Linn, S. M. and Roberts, R. J., Cold Spring Harbor Laboratory (1982)
【発明の開示】
【発明が解決しようとする課題】
【0003】
RNA分子の二次構造の決定等には、なんらかの選択性を持ってRNAを切断するものであることが必要であるが、これらの他にRNA分子の二次構造決定等に利用可能なRNaseは未だ見い出されていない。切断部位を塩基特異的に認識するRNA分解酵素は未だ見出されていない。また、これら各種のRNaseはいずれもアデニン塩基(A)を高い選択性で認識するものでなかった。
【0004】
そこで、本発明は、RNA研究に有用なRNA分解酵素活性を有するタンパク質を提供することを一つの目的とする。また、本発明は、アデニン塩基を認識してRNA鎖を切断する酵素活性を有するタンパク質を提供することを一つの目的とする。さらに、本発明は、アデニン塩基を高い選択性で認識してRNAを切断する酵素活性を有するタンパク質を提供することを一つの目的とする。また、本発明は、こうしたRNA分解酵素の利用を提供することを他の一つの目的とし、こうした酵素活性を有するタンパク質を発現させるためのポリヌクレオチド、当該ポリヌクレオチドを、前記タンパク質を発現可能に保持する形質転換体を提供することを他の一つの目的とする、さらにまた、本発明は、こうした酵素を用いた核酸の分析キット及び処理方法を提供することを他の一つの目的とする。
【課題を解決するための手段】
【0005】
本発明者らは、植物に普遍的に存在するタンパク質であってRNA結合活性を有するタンパク質について検討していたところ、細胞器官におけるRNAプロセッシングに関与すると考えられるPPRモチーフ(Pentatrico Peptide Repeat:PPR)に連結されることのあるDYWドメイン及び当該DYWに類するドメインが、RNA切断活性、すなわち、RNA分解酵素として機能するという知見を得た。また、本発明者らは、同時に、このドメインがアデニン塩基に選択性の高い切断活性を有するという知見も得た。本発明は、これらの知見に基づきなされたものであり、本発明によれば、以下の手段が提供される。
【0006】
本発明によれば、
以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、RNAを切断する活性を有する、タンパク質が提供される。
【0007】
さらに、本発明のタンパク質は、以下の特徴;
(b)チトクロームCヘム結合モチーフCxxCHを有する。
を有することができる。また、本発明のタンパク質は、以下の特徴;
(c)PPRモチーフを備える天然タンパク質のC末端領域に備えられるアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列である。
を有することもできる。
【0008】
本発明のタンパク質は、前記アミノ酸配列は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれかを問い合わせ配列としてホモロジー検索を行って得られるE値が所定値以下(好ましくは1.0×10-10以下)である配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列とすることができる。前記アミノ酸配列は、好ましくは、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列である。
【0009】
本発明のタンパク質は、単子葉植物及び双子葉植物のいずれかのタンパク質とすることができる。また、前記RNA切断活性はエンド型であることが好ましく、さらに好ましくは、前記RNA切断活性は塩基特異的である。また、前記RNA切断活性は、アデニン塩基を有するリボヌクレオチドの5’側を部位特異的に切断する活性であることが好ましい。
【0010】
また、本発明によれば、RNA分解酵素のスクリーニング方法であって、配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、を備える、方法も提供される。前記タンパク質の準備工程は、配列番号:1、配列番号:4及び配列番号:6のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程とすることができる。
【0011】
さらに、本発明によれば、RNA分解酵素の製造方法であって、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加、および/または挿入されたアミノ酸配列とチトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することを特徴とする、方法も提供される。
【0012】
本発明によれば、上記いずれかに記載のタンパク質を含む、核酸分析キットも提供され、さらに、本発明によれば、上記いずれかに記載のタンパク質をコードするポリヌクレオチドも提供され、本発明のポリヌクレオチドを、上記いずれかのタンパク質を発現可能に保持する形質転換体も提供され、上記いずれかのタンパク質を用いてRNAを切断する工程を備える、核酸の処理方法も提供される。この処理方法は、核酸を分離精製する工程、RNAを抽出する工程及びRNA断片を検出する工程のいずれかあるいは2工程以上を備えることもできる。
【発明を実施するための最良の形態】
【0013】
本発明のタンパク質は、以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、RNAを切断する活性を有することができる。
【0014】
本発明のタンパク質(以下、単に本タンパク質という。)は、上記アミノ酸配列を有することにより、RNA切断活性を有する酵素として機能することができる。また、前記アミノ酸配列は、PPRをモチーフを有しRNAプロセッシングに関与すると考えられる天然タンパク質のC末端に備えられるアミノ酸配列であることから、RNAの機能解析等の研究に有用である。特に、アデニン塩基特異的にRNAを切断することができるため、RNA等における塩基配列の正確な決定に有用である。また、本発明のタンパク質は、PPRタンパク質のC末端側の配列であるDYWモチーフに関連付けられるアミノ酸配列を含んでかつRNA切断活性を有するタンパク質であるということもできる。
【0015】
また、本発明の核酸分析キット(以下、単に、本分析キットという。)は、本タンパク質を含むことができる。本分析キットは、RNAやDNA等の抽出分離、RNAの配列や機能解析に有用である。
【0016】
また、本発明の形質転換体(以下、単に、本形質転換体という。)は、本タンパク質をコードするポリヌクレオチドを、本タンパク質を発現可能に保持することができる。本形質転換体によれば、RNA分解酵素活性を有するタンパク質を供給するのに都合がよい。さらに、本タンパク質をコードする本発明のポリヌクレオチド(以下、単に本ポリヌクレオチドという。)によれば、有用なRNA分解酵素を効率的にあるいは任意の宿主に生産させることができるコンストラクトや本タンパク質を発現可能な形質転換体を提供することができる。さらにまた、本発明の核酸の処理方法によれば、本タンパク質をRNA分解酵素として利用することでRNAを分解して従来にない核酸の分離精製やRNA代謝の研究等が可能となる。
【0017】
以下、本発明の実施の実施形態であるタンパク質、核酸分析キット、核酸処理方法、ポリヌクレオチド及び形質転換体等について詳細に説明する。
【0018】
(タンパク質)
本タンパク質は、1又は2以上の特徴を有するアミノ酸配列を備えることができる。本タンパク質が備えることができる一つのアミノ酸配列は、配列番号:1に記載の配列とすることができる。配列番号:1に記載のアミノ酸配列は、シロイヌナズナ(Arabidopsis thaliana、ecotype columbia)の機能未知タンパク質遺伝子at2g02980(MATDB:http://mips.gsf.de/proj/thal/db/index.html)がコードするPPRモチーフを有する603アミノ酸からなるタンパク質(PPRタンパク質)(配列番号:3)の一部からなる配列である。具体的には、配列番号:3に記載のアミノ酸配列のC末端側の第495位〜第603位の109個のアミノ酸からなる配列である。PPRタンパク質は、RNAプロセッシングに関連していると考えられているが、そのC末端領域がRNase活性を有することは本出願前において確認されていない。
【0019】
また、本タンパク質が備えることのできる他の一つのアミノ酸配列は、配列番号:4に記載の配列である。配列番号:4に記載のアミノ酸配列(Os5g30710(OS_M23)の一部、データベース:TIGR Rice Genome Annotation (http://www.tigr.org/tdb/e2k1/osa1/)は、イネ(Oryza sativa)由来の天然タンパク質(Os5g30710(OS_M23)に含まれるアミノ酸配列である。このアミノ酸配列が含まれるタンパク質も、PPRモチーフを有するPPRタンパク質である。
【0020】
本タンパク質は、これらの2種のアミノ酸配列に限定されないで、さらに他のアミノ酸配列を備えることができる。すなわち、本タンパク質は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を備えることもできる。すなわち、これらの2種のアミノ酸配列に対してアミノ酸の置換、欠失、挿入及び付加から選択される1又は2以上のアミノ酸の変異を有するアミノ酸配列(以下、改変配列ともいう。)である。アミノ酸変異の数は、特に限定しないが、好ましくは20個以下であり、より好ましくは10個以下である。より好ましくは5個以下である。
【0021】
アミノ酸配列への部位特異的な変異の導入は当業者において公知の手法によって可能である。アミノ酸配列が改変されたタンパク質をコードするDNAを調製するための当業者によく知られた方法としては、例えば、site-directed mutagenesis法(Kramer W & Fritz H-J: Methods Enzymol 154: 350,
1987)が挙げられる。また、自然界においても、塩基配列の変異によりコードするタンパク質のアミノ酸配列が変異することは起こり得る。
【0022】
また、こうした改変配列の他の一つとしては、もとのアミノ酸配列をコードする塩基配列又はその一部からなるDNA(その相補鎖であってもよい。)とハイブリダイズするDNAによってコードされるアミノ酸配列が挙げられる。例えば、配列番号:1に記載のアミノ酸配列は、配列番号:2に記載の塩基配列によってコードされている。また、配列番号:4に記載のアミノ酸配列は、配列番号:5に記載の塩基配列によってコードされている。これらの塩基配列に基づくDNAを利用することができる。
【0023】
このような改変配列は、ハイブリダイゼーション技術(Southern EM: J Mol Biol 98: 503, 1975)やポリメラーゼ連鎖反応(PCR)技術(Saiki
RK, et al: Science 230: 1350, 1985、Saiki
RK,et al: Science 239: 487, 1988)を利用する方法が挙げられる。すなわち、配列番号:2に記載されるDNA若しくはその一部又はこれらの相補鎖をプローブとして、あるいは、こうしたタンパク質のゲノムDNAの塩基配列に特異的にハイブリダイズするオリゴヌクレオチドをプライマーとして、シロイヌナズナや他の植物から、ハイブリダイゼーションによってDNAを単離し、該タンパク質を取得することは当業者において通常行い得ることである。
【0024】
なお、こうしたタンパク質をコードするDNAは、好ましくは、ストリンジェントな条件下でハイブリダイゼーション反応を行うことによって得ることができる。本発明においてストリンジェントなハイブリダイゼーション条件とは、6M 尿素、0.4%
SDS、0.5×SSCの条件またはこれと同等のストリンジェンシーのハイブリダイゼーション条件を指す。よりストリンジェンシーの高い条件、例えば、6M 尿素、0.4%
SDS、0.1×SSCの条件下では、より相同性の高いDNAを単離できると期待される。なお、高い相同性とは、アミノ酸配列全体で少なくとも50%以上、好ましくは70%以上、さらに好ましくは90%以上、最も好ましくは95%以上の配列の同一性を意味するものとする。なお、アミノ酸配列や塩基配列の同一性は、BLAST等を用いて決定できる。
【0025】
配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列においてアミノ酸変異を導入した改変配列は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列として任意のデータベースをホモロジー検索して得られるE値(Expected value, E-value;期待値)が所定値以下のアミノ酸配列とすることができる。E値は、一般的に、あるいは用いるホモロジー検索プログラムにおいて定義されている。本発明におけるE値については、ホモロジー検索手法における一般的な定義あるいは用いる特定のホモロジー検索手法において個別に定義されていれば、その定義に従うものとする
【0026】
E値が小さければ小さいほど配列番号1:及び配列番号:4のいずれかに記載の配列と類似性が高い。E値は好ましくは、1.0×10-10以下である。一般にE値が1.0×10-10以下であれば、配列番号:1等に記載のアミノ酸配列に類似することが偶然には起きにくいということができる。こうしたアミノ酸配列であれば、機能的にも配列番号:1に記載のアミノ酸配列を有するタンパク質と同質あるいは類似の活性を有していると考えられる。好ましくは、E値は1.0×10-15以下である。よりE値が小さいアミノ酸配列を備えることで配列番号:1等に記載のアミノ酸配列からなるタンパク質と同質で同等の活性を得られやすくなる。
【0027】
ホモロジー検索は、問い合わせ配列に対するダイナミックプログラミングによるローカルペアワイズアライメントによるものとすることができる。この種のホモロジー検索プログラムは、インターネット等を通じて公開又は提供されているものから適宜選択して用いることができる。例えば、SSEARCH(http://www.ddbj.nig.ac.jp/search/ssearch-j.html等);BLAST(Proc.
Natl. Acad. Sci. USA 87:2264-2268, 1990、Proc
Natl Acad Sci USA 90: 5873, 1993,http://www.ncbi.nlm.nih.gov/BLAST/)、PSI-BLAST(http://www.ncbi.nlm.nih.gov/BLAST/等);FASTA(http://www.ebi.ac.uk/fasta33/)及び等を用いることができる。なかでも、BLAST及びPSI-BLASTを好ましく用いることができる。
【0028】
検索対象とするデータベースは、特に限定しない。例えば、ホモロジー検索プログラムを公開するサイトにおいて準備されているものから適宜選択して用いることができる。また、プログラムとは別個に公開されている又は非公開状態のデータベースを用いることもできる。本タンパク質は、真核生物、特に植物のゲノム又はタンパク質あるいはこれらの一部(モチーフ)を含むデータベースを利用することで多くの類似配列を抽出することができる。本タンパク質のアミノ酸配列は、PPRモチーフを有するPPRタンパク質のC末端領域に相当するかあるいは類似することが多く、PPRタンパク質は植物に多く含まれているからである。PPRタンパク質は、シロイヌナズナなどの双子葉植物及びイネ(Oryza sativa)のような単子葉植物の双方に含まれており、こうした植物のタンパク質のデータベースを検索対象とすることが好ましい。
【0029】
なお、ホモロジー検索にあたっては、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列とするほか、当該アミノ酸配列をコードする塩基配列を問い合わせ配列として塩基配列やアミノ酸配列のデータベースを検索することにより、所定値以下のE値を有するアミノ酸配列を取得することも包含される。この場合、例えば、必要に応じてBLAST等に包含される各種ホモロジー検索手法を用いればよい。
【0030】
例えば、配列番号:1を問い合わせ配列としたホモロジー検索(BLAST)によって抽出されるアミノ酸配列としては、配列番号:4に記載のアミノ酸配列が挙げられる(Os5g30710(OS_M23)、データベース:TIGR Rice Genome Annotation (http://www.tigr.org/tdb/e2k1/osa1/)、E値3×10-27)。
【0031】
また、本タンパク質は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索をしたときに所定値以下のE値であるアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含んでいてもよい。すなわち、抽出されたアミノ酸配列の改変配列を含んでいてもよい。こうしたアミノ酸変異の数は、特に限定しないが、好ましくは20個以下であり、より好ましくは10個以下である。より好ましくは5個以下である。
【0032】
本タンパク質が有することのできるアミノ酸配列は、また、PPRタンパク質ファミリーのマルチプルアライメントによって得られるコンセンサス配列に基づくものであってもよい。コンセンサス配列を得るためのマルチプルアライメントに用いる手法やコンセンサス配列を取得する手法は特に限定されない。例えば、ClustalW: http://align.genome.jp/; HMMER(隠れマルコフモデル): http://hmmer.wustl.edu/;
MultiAlin:http://prodes.toulouse.inra.fr/multalin/multalin.html; mkdom/xdom:
http://prodes.toulouse.inra.fr/prodom/xdom/等各種の手法を採用することができる。コンセンサス配列が得られたなら、コンセンサス配列において高度に保存されているアミノ酸を維持するような改変配列や予測される活性部位や結合部位におけるアミノ酸をこれらの部位における相互作用を強めるように変異させた改変配列を得ることができる。例えば、こうしたコンセンサス配列の一例としては、HMMERマトリックスを利用してシロイヌナズナ85種のPPRタンパク質ファミリーのマルチプルアライメントから得られたもの(C末端側のDYWモチーフ)が開示されている(図1及び配列番号:6)(The Plant Cell,
Vol. 16, 2089-2103, August 2004、Fig.2)。図1において、保存されているアミノ酸は大文字で示され、特によく保存されているアミノ酸は太字で示されている。
【0033】
本タンパク質が備えることのできるアミノ酸配列の他の一つの特徴としては、チトクロームCヘム結合モチーフCxxCH(C:システィン、H:ヒスチジン、x:任意のアミノ酸)を有していることが挙げられる。このモチーフは、金属配位モチーフとして知られている。このモチーフを保持することがRNA分解酵素活性及びそのアデニン塩基特異性に好ましい。このモチーフは、例えば、配列番号:1において第69位〜第72位に相当する配列に位置することができる。当該モチーフは、C末端側から約40アミノ酸程度の位置、例えば、C末端側から35〜45アミノ酸に位置することができる。当該モチーフの有無は、例えば、PROSITE(http://au.expasy.org/prosite/)など各種公知のモチーフデータベースにて確認することができる。既に例示した配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列においてもこのモチーフを備えている。
【0034】
本タンパク質が備えることができるアミノ酸配列のさらに他の一つの特徴としては、PPRモチーフを備える天然タンパク質(PPRタンパク質)のC末端領域に位置するアミノ酸配列又はその改変配列が挙げられる。PPR(pentatricopepetide repeat)モチーフは、35アミノ酸の保存配列がリピートしたものであり、植物に多く存在し、なかでも葉緑体やミトコンドリアに局在することが知られている(Biochem Soc Trans. 2004 Aug;32(Pt 4):571-4.)。PPRモチーフの有無は、例えば、PFAM(http://www.sanger.ac.uk/Software/Pfam/)などのドメインデータベースにより知ることができる(Accession No.PF01535)。35アミノ酸のコンセンサス配列を、図2及び配列番号:7に示す。図2において、よく保存されているアミノ酸は大文字で示されている。配列番号:1及び配列番号:4に記載のアミノ酸配列はいずれもこうしたPPRタンパク質のC末端領域に位置される配列である。なお、PPRタンパク質のC末端側領域に位置されるアミノ酸配列の改変配列における改変配列の意味は、既に説明した意味がここでも適用される。
【0035】
こうしたタンパク質は、PPRタンパク質が植物の葉緑体やミトコンドリアに多く存在することから、単子葉植物及び双子葉植物の天然タンパク質中のアミノ酸配列及びこの改変配列であるアミノ酸配列を有することができる。植物種は特に限定されないが、例えば、単子葉植物としては、イネ、ムギ、キビ等が挙げられ、双子葉植物としては、シロイヌナズナ、ダイズ、ピーナッツ、ごま、ナタネ、綿実、ヒマワリ、サフラワー、、バレイショ、サツマイモ等が挙げられる。
【0036】
本タンパク質は、RNA切断活性を有することができる。本タンパク質のRNA切断活性は、RNAを切断することができる限りどのような態様であってもよい。本タンパク質のRNA切断活性は、RNA特異的であることが好ましい。RNA特異的であるとDNAに作用しないのでDNA等と容易に分離することができるし、DNA配列を維持させることができる。また、エキソ型でもエンド型でもよいが、好ましくはエンド型である。エンド型であることで、RNAの機能解析等に有用である。なお、本明細書において、RNA切断活性とは、切断部位又はその近傍にRNA鎖を備えるオリゴヌクレオチド又はポリヌクレオチドを切断するものであればよい。したがって、DNA/RNAハイブリッドやDNA-RNAキメラ、修飾塩基やその他の修飾や付加を含むヌクレオチド鎖を切断対象として含めることができる。
【0037】
本タンパク質は、塩基特異的なRNA切断活性を有していることが好ましい。本明細書において。塩基特異的なRNA切断活性を有しているとは、1又は2以上の特定塩基を認識して認識した特定塩基に対して所定の位置にある結合を切断する活性を有していることを意味している。本発明においては、好ましくはアデニン塩基を認識する。さらに、アデニン塩基を認識してその5’側においてRNAを切断することが好ましい。従来、アデニン塩基を高い特異性を持って認識して切断するRNaseは見出されておらず、研究及びその他の各種用途に有用である。
【0038】
本タンパク質は、EDTAが所定濃度範囲にあるとき、換言すれば、所定濃度の2価金属イオンの存在下でRNA切断活性を発揮することができることが好ましい。こうした2価金属イオン濃度依存性を備えることで2価金属イオン濃度やEDTA等のキレート剤の添加濃度でその活性を制御することができる。好ましくは、本タンパク質はそのRNA切断活性を発揮するのにあたって、2価金属イオン濃度について至適範囲を有している。例えば、配列番号:1に記載のアミノ酸配列を含むタンパク質は、EDTA等のキレート剤の不存在下及び高濃度存在下ではRNA切断活性を有していないが、20mM以上、好ましくは25mM以上の濃度で、また、70mM以下、好ましくは、62.5mM以下の濃度で高いRNA切断活性を発揮することができる。また、例えば、マグネシウムイオンなどの2価金属イオン濃度としては、1mM以上10mM以下程度が好ましく、より好ましくは、下限が2mMであり、上限が5mMである。
【0039】
RNA切断活性は、公知の方法によって測定することができる。一般的には、適当な媒体下で基質RNAとタンパク質とを接触させるようにすればよい。基質RNAには放射性元素などの適当な標識を付して置くことができる。適当な温度下で一定時間反応させた後、RNAを抽出し尿素変性ポリアクリルアミドゲルなどで電気泳動することによりRNAの分解程度を検知することができる。なお、部位特異的な切断活性を測定する場合には、分解反応産物に対してコンプレメンタリーDNAを調製するとともに、基質RNAの鋳型であるDNAからDNAラダーを調製し、これらをそれぞれ尿素変性ポリアクリルアミドゲル電気泳動などで分離させることによって切断部位を特定することができる。
【0040】
なお、本タンパク質は、RNA切断活性を発揮できる限り、さらに他のアミノ酸配列を備えていてもよい。製造上の都合及び使用用途等に応じて各種のアミノ酸配列を付加することもできる。
【0041】
(本タンパク質の製造方法)
本タンパク質は、天然タンパク質又はその一部であるときには、自然界において保持される植物などの生物体又はそのタンパク質が局在するオルガネラ画分から、タンパク質を抽出し、分離精製等することによっても得ることができる。また、本タンパク質が天然タンパク質の全部又は一部であっても、非天然のタンパク質であっても、化学的又は遺伝子工学的に得ることもできる。遺伝子工学的手法によって得る場合には、本タンパク質をコードするポリヌクレオチドを用いて無細胞的にあるいはこのポリヌクレオチドを適当な宿主に導入して形質転換体を作出し、当該形質転換体の宿主内又は宿主外で発現させるようにすることができる。
【0042】
(RNA分解酵素のスクリーニング方法及び製造方法)
本発明のRNA分解酵素のスクリーニング方法は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、を備えることができる。このスクリーニング方法によれば、RNA切断活性を有するタンパク質(RNA分解酵素)を効率的に得ることができる。また、前記タンパク質の準備工程は、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの改変配列を備えるタンパク質を準備する工程とすることもできる。配列番号:1及び配列番号:4に記載のアミノ酸配列は、酵素活性が確認されたDYW配列であるため、これらの配列を問い合わせ配列とすることで同等活性のあるアミノ酸配列を効率的に抽出することができる。さらに、本発明のRNA分解酵素のスクリーニング方法においては、配列番号:6に記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って所定値以下のE値を有するアミノ酸配列を抽出するようにしてもよい。配列番号:6に記載の配列は、DYWモチーフのコンセンサス配列であるから、この配列と所定値以下のE値を有するアミノ酸配列を有するタンパク質をRNA切断活性のスクリーニング対象とすることが可能である。さらに、チトクロームCヘム結合モチーフを有するアミノ酸配列を準備するようにするとより一層効果的にRNA分解酵素をスクリーニングすることができる。
【0043】
タンパク質の準備工程では、例えば、所定のホモロジー検索において所定値以下のE値を有するアミノ酸配列を有するタンパク質を抽出し、当該タンパク質をコードするポリヌクレオチドなどを用いて当該タンパク質を遺伝子工学的に生産することもできる。
【0044】
なお、所定値以下のE値を有するアミノ酸配列を備えるタンパク質を準備するにあたっては、既に説明した各種のホモロジー検索プログラムを用いることができる。また、E値については、好ましくは、1.0×10-10以下であり、より好ましくは、1.0×10-15以下である。また、改変配列についても既に説明した各種の態様をこの方法で準備するタンパク質に適用することができる。RNA切断活性の有無は、既に説明した方法等、当業者において周知の方法を用いて測定することができる。
【0045】
こうしたRNA分解酵素のスクリーニングによってRNA切断活性を有することが検出されたタンパク質は、既に説明した本タンパク質を製造する場合と同様、天然の生物体から取得することもできるし、人工的に取得することもできる。RNA分解酵素は、また、配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの改変配列を備えるとともに、チトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することによっても得ることができる。こうしたタンパク質は、高い確率でRNA切断活性を発揮できるからである。
【0046】
(核酸分析キット)
本分析キットは、本タンパク質をキット構成に含めることができる。キットに含められるタンパク質は、どういった形態で含められていてもよいが、例えば、保存性等を考慮して凍結乾燥体として含められていることが好ましい。また、本分析キットには、その他のRNase、DNase等、核酸の分離精製、分析等に必要な酵素や試薬を含められていてもよい。
【0047】
(ポリヌクレオチド)
本ポリヌクレオチドは、本タンパク質をコードする領域を有することができる。本タンパク質をコードするポリヌクレオチドの塩基配列としては、例えば、配列番号:2及び配列番号:5が挙げられる。ポリヌクレオチドにおける塩基配列は、本タンパク質をコードしている以上コドン用法は特に限定されない。
【0048】
本ポリヌクレオチドは、例えば、本ポリヌクレオチドを発現可能にプロモーター等の調節領域を備える発現カセット、ベクター、プラスミド、人工染色体及び感染可能なアグロバクテリウム等のウイルスの態様であってもよい。
【0049】
(形質転換体)
本形質転換体は、本ポリヌクレオチドを、本タンパク質を発現可能に保持する形質転換体とすることができる。本形質転換体は、大腸菌や乳酸菌などの原核細胞のほか、酵母、植物培養細胞及び植物体を宿主とすることができる。植物体としては、植物個体、その植物個体を構成しうる全ての種類、形態の細胞、植物固体の一部である組織や器官ならびに生殖細胞を含んでいる。また、植物個体の一部としては、その繁殖媒体(種子、根茎、果実、切穂等)も包含する。
【0050】
本形質転換体が、本タンパク質の発現可能な本ポリヌクレオチドを保持するには、本ポリヌクレオチドは適当なプロモーターの制御下に連結されることが好ましい。また、適当なターミネータも連結されることが好ましい。その他、発現の増強に有効なエレメントを適宜備えることができる。
【0051】
こうした形質転換体は、本タンパク質を大量に生産するほか、RNA/DNA代謝研究に有用である。
【0052】
また、本形質転換体は、本タンパク質の発現を抑制可能に構築されていてもよい。例えば、植物等に内在する、配列番号:1に記載のアミノ酸配列又は配列番号:4に記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って所定値以下のE値を有するアミノ酸配列又は当該アミノ酸配列を含むタンパク質あるいはさらにチトクロームCヘム結合モチーフを有するタンパク質の発現を抑制可能に構築されていてもよい。こうした形質転換体は、RNA/DNA代謝研究に有用である。本タンパク質の発現を抑制する態様としては、内在する本タンパク質をコードする遺伝子の破壊や当該遺伝子の転写や翻訳を抑制するほか、変異等を導入して不完全なタンパク質等として発現させてその作用を低下又は失活させたり、あるいは本タンパク質と相互作用して本タンパク質の作用を低下させる物質を発現させることが挙げられる。
【0053】
例えば、特定の遺伝子の発現を抑制する方法としては、特定遺伝子のノックアウトのほか、、アンチセンス法、コサプレッション法、RNA干渉法及びリボザイム法等を用いて、mRNAを対象にした発現抑制可能な核酸コンストラクトを導入し保持させることが挙げられる。
【0054】
なお、形質転換体は、当業者の公知の手法を採用して作出することができる。形質転換体が植物体の場合、例えば、本タンパク質を発現可能に保持する形質転換体を作成するには、植物細胞内で本タンパク質を発現可能なコンストラクトを構築し、当該コンストラクトを植物細胞に導入し、該植物細胞から植物体を再生させればよい。ベクターは特異に限定されない。また、植物細胞内で恒常的に遺伝子を発現させるためのプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクターや、外的な刺激により誘導的に活性化されるプロモーターを有するベクターを用いることもできる。コンストラクトが導入される細胞としては、植物体に再生可能なあらゆる種類の形態の植物細胞を含めることができる。例えば、培養細胞、プロトプラスト、苗条原基、多芽体、毛状根、カルス、葉など特定器官の切片等が挙げられる。植物の形質転体は、所定の再生工程を実施することで細胞を植物体に変換することができる。再生の方法は、植物の種類によって異なるが、各種公知の方法を使用できる。植物細胞への核酸コンストラクトの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。また、パーティクルガン法においては、例えば、バイオラッド社のものを用いることが可能である。形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。また、再生した植物体から有性生殖または無性生殖により子孫を得ることができる。また、該植物体やその子孫あるいはクローンから繁殖材料(例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなど)を得て、それらを基に該植物体を量産することも可能である。
【0055】
(核酸の処理方法)
本処理方法は、タンパク質を用いてRNAを切断する工程を備えることができる。この処理方法は、RNAを切断し、RNAを断片化することができる。この切断工程によれば、例えば、試験管内においてRNA断片を生成させて、当該RNAの機能探索のための材料を得ることができる。また、塩基特異的にRNAを切断するため、生じたRNA断片を分析することで切断対象となったRNAの構造等を推測することもできる。
【0056】
本処理方法は、さらに、DNAなどの核酸を分離精製する工程を備えていてもよいし、RNAを抽出する工程を備えていてもよいし、さらに、RNA切断後に当該切断したRNA断片を電気泳動やMALDI等で検出する工程を備えることもできる。
【0057】
核酸の分離精製工程において、分離精製対象は、RNAとDNAとが混在する可能性のある試料であることが好ましい。こうした試料としては、生体から抽出された試料のほか、試験内でDNA合成した際の反応試料等であってもよい。例えば、RNAとDNAが混在する場合には、本タンパク質は、DNAには作用しないため共存するRNAから未切断断片としてDNAを容易に精製することができる。一方、本タンパク質はRNAに作用するため、RNAを切断断片として分離することができる。こうした分離精製工程においては、本タンパク質を適当な固相に固定化しておくと都合がよい。また、こうした固相を充填したカラムを用いてもよい。核酸の分離精製工程はRNAの切断工程後に備えることが好ましい。
【0058】
RNAを抽出する工程において、抽出対象は生体試料であることが好ましい。こうしたRNAを抽出して、その後、本タンパク質で切断することで、生体内におけるRNAの代謝研究が可能となる。RNA抽出工程は、RNA切断工程に先立って備えることが好ましい。
【0059】
さらに、RNA切断後に生じたRNA断片を検出する工程は、RNA断片自体、あるいはコンプレメンタリーDNAを調製した上で、電気泳動、マイクロアレイ及びMALDI等で分析するようにしてもよい。
【0060】
以下、本発明に関して、実施例を挙げて具体的に説明するが、本発明は以下の実施例の限定されるものではない。
【実施例1】
【0061】
(実施例1: 新規RNA分解酵素、T-DYWの調製とRNA分解活性の測定)
(シロイヌナズナからのゲノムDNAの調製)
シロイヌナズナ(Arabidopsis thaliana, ecotype Columbia)をムラシゲ・スクーク培地(2%ショ糖、0.5% Gellangamを含む)で3週間培養した。培養した植物の緑葉(0.5 g)をフェノール/クロロホルム抽出した後、エタノールを加えてDNAを不溶化した。回収したDNAを100 μlのTE液(10 mM トリス・塩酸 (pH 8.0)、1mM EDTA)に溶解し、10ユニットのRNase A(DNase-free、タカラバイオ社)を加えて、37℃で30分反応させた。その後、反応液を再度フェノール/クロロホルム抽出した後、エタノール沈殿によりDNAを回収した。10 μgのDNAが得られた。
【0062】
(DYWモチーフをコードする遺伝子のクローニング)
シロイヌナズナからのゲノムDNAの調製は上記の実施例1に記載されている方法により行った。シロイヌナズナゲノム情報データベース(MATDB:
http://mips.gsf.de/proj/thal/db/index.html)に掲載されている配列情報を参照し、DYWモチーフを持つタンパク質遺伝子at2g02980の当該DYWモチーフに相当するDNA配列を増幅するためのオリゴヌクレオチドプライマー(02980DYW-F、02980_m-R;配列番号:8、9に記載)を調製した。オリゴヌクレオチドプライマーのフォワードプライマーである02980DYW-F、リバースプライマーである02980_m-Rの5'側にはそれぞれEco
RI、SalI配列を付加した。EcoRIとSalI配列は得られたクローンを制限酵素処理で挿入配列を切り出すのに利用できるように組み入れた。
【0063】
at2g02980を含むDNA断片は、100 ngのゲノムDNAと上記プライマーを含む50 μlの反応液を95℃ 30秒、60℃ 30秒、72℃ 30秒の25サイクルでPrime Star (タカラバイオ社)
をDNA 伸長酵素として用い、PCRすることによって増幅した。その結果、339塩基対のDNA断片の増幅が確認できた。得られたDNA断片は、pBAD/Thio-TOPO ベクター(Invitrogen社)を用いて、クローニングした。クローニングするために必要なDNA断片の3'末端突出アデノシンは、DNA断片を0.1
ユニット TaKaRa Ex-Taq(タカラバイオ社)と2.5 mM ATPで70℃、10分間反応することで付加した。クローン化したDYWモチーフをコードするDNA配列を決定し、上記データベース上においてDYWモチーフに相当するDNA配列と相同な配列(配列番号:2)であることを確認し、プラスミドpTrx_DYWと命名した。
【0064】
(T-DYWタンパク質の調製)
得られたプラスミドpTrx_DYWをEscherichia coli LMG194株(Invitrogen社)に形質転換した。この大腸菌をアンピシリンが100 μg/mlの濃度で存在する0.2%グルコースを含むRM培地4,000 ml(1リットル培地を含む2リットル三角フラスコ、計4本)中で、37℃で培養した。培養液の濁度が波長600 nmでの吸光度が0.5に達した時に、誘導物質であるL-アラビノースを最終濃度が0.2%になるように添加し、さらに4時間培養を行った。遠心による集菌後、菌体を1 mg/mlのリゾチームを含む200 mlのバッファーA(50 mM トリス・塩酸 pH 8.0、500 mM KCl、2
mM imidazole、10 mM MgCl2、0.5% Triton X100、10% グリセロール)に懸濁し、超音波破砕と凍結溶解により菌体を破壊した。15,000 x g、20分間の遠心分離後に、上清を粗抽出液として回収した。この粗抽出液をバッファーAで平衡化したニッケルカラム樹脂(ProBond A、Invitrogen社)を充填したカラムに供した。
【0065】
カラムクロマトグラフィーは、20
mM imidazoleを含むバッファーAで十分に洗浄した後、200
mM imidazoleを含むバッファーAで目的タンパク質を溶出する二段階濃度勾配により行った。得られた組み換えタンパク質をSDSポリアクリルアミドゲル電気泳動により確認したところ、30 kDaのタンパク質として検出された。これをT-DYWタンパク質と命名した(配列番号:10)。なお、このタンパク質は、配列番号:1に記載のアミノ酸配列を備えるとともに、N末端側に溶解性を高めるためのチオレドキンのアミノ酸配列、C末端側にヒスチジンタグ配列を備える融合タンパク質である。RM培地1リットルあたり、600 μgのT-DYWタンパク質が得られた。T-DYWタンパク質を含む精製画分1 mlを1リットルのバッファー E(20 mM トリス・塩酸 pH 7.9、60
mM KCl、12.5 mM MgCl2、0.1 mM EDTA、17% グリセロール、2 mM DTT)で透析した後、T-DYWタンパク質の精製標品とした。
【0066】
(基質RNAの調製)
基質RNAとして、シロイヌナズナ葉緑体ndhB遺伝子の開始コドンから500塩基のRNAを用いた。基質RNAとして用いるRNAをNB500と命名した(配列番号:11)。NB500を合成するためのDNA断片は、オリゴヌクレオチドプライマーndhB-FとndhB-R(配列番号12、13に記載)を用いて、上記のシロイヌナズナゲノムDNA 10
ngを鋳型DNAとして含む50 μlの反応液を95℃ 30秒、60℃ 30秒、72℃ 30秒の25サイクルでPrime
Star (タカラバイオ 社) をDNA 伸長酵素として用い、PCRすることによって増幅した。ndhB-Fプライマーの5'末端側には基質RNAを試験管内で合成するためのT7 プロモータ配列を、ndhB-Rプライマーの5'末端側には3'→5'エクソヌクレアーゼによる非特異的な分解を防ぐためのステムループ構造を形成する塩基配列をそれぞれ付加した。得られたDNA断片は、アガロースゲルで展開後、ゲルから切り出すことによって精製した。精製DNA断片を鋳型にNTP mix(10 nmol GTP、CTP、ATP、0.5 nmol UTP)、4 μl [32P] α-UTP (GE ヘルスケア社、3000 Ci/mmol)、T7 RNA polymerase(タカラバイオ社)を含む20μlの反応液を37℃ 60分間反応させることで、基質RNAを合成した。基質RNAはフェノール/クロロホルム抽出、エタノール沈殿後、全量を6 M 尿素を含む変性6%ポリアクリルアミドゲル電気泳動で展開し、X線フィルムで60秒間感光させることによって、32P標識RNAを検出した。32P標識RNAをゲルから切り出し、200 μlのゲル溶出液(0.3 M 酢酸ナトリウム、2.5 mM EDTA、0.01% SDS)中に、4℃で12時間浸し、RNAをゲルから溶出した。溶出したRNAのうち、1 μlの放射活性を測定し、合成したRNAの総量を算出した。エタノール沈殿後、8,250 cpm/μl (1 fmol/μl)になるように、RNAを超純水に溶解した。この調製方法で通常、8,250 cpm/μlのRNAが約50 μl得られる。
【0067】
(T-DYWタンパク質のRNA分解活性)
T-DYWタンパク質存在下もしくは非存在下でのRNAの分解活性を調べた。50 mM EDTA を含む反応液(10 mM トリス・塩酸 pH 7.9、30
mM KCl、6 mM MgCl2、2
mM DTT、8% グリセロール、0.0067% of
Triton X-100)20 μl中に上記の1 fmolの基質RNA(NB500)と50 ng〜1.5 μgのT-DYWタンパク質を混合し、25℃で30分間反応した。反応後にフェノール/クロロホルム抽出、エタノール沈殿により、NB500 RNAを抽出した。RNAは6 M尿素を含む変性6%ポリアクリルアミドゲルで展開し、電気泳動後にゲルを乾燥させた。ゲル中のRNAの放射活性をバイオイメージングアナライザーBAS2000(フジフィルム社)で測定した。その結果を以下の図3に示す。
【0068】
図3より明らかなように、T-DYWタンパク質とRNAを混合するとタンパク質濃度依存的に基質RNAの分解活性が観察された。対照実験として、RNA結合タンパク質であるcp28をT-DYWと同様のベクター、発現系で調製した組み換えタンパク質をT-28として(配列番号:14)、同様の実験を行ったところ、RNAの分解活性は観察されなかった。タンパク質非存在下(no protein)でもRNAの分解は観察されなかった。
【0069】
(T-DYWタンパク質のRNA分解活性におけるEDTAの効果)
多くのRNA分解酵素は、そのRNA分解活性に二価金属イオンを必要とする。T-DYW精製標本中に含まれる二価金属イオンのRNA分解活性における影響を検証するために、EDTAを添加して、T-DYWタンパク質のRNA分解活性を測定した。0〜200
mM EDTAを含む反応溶液20 μl中に1 fmolのNB500 RNAと50
ngのT-DYWタンパク質を混合し、25℃で30分間反応した。反応後のRNA抽出、電気泳動、検出は実施例1に記載したのと同様の方法で行った。結果を図4に示す。
【0070】
図4に示すように、T-DYWタンパク質のRNA分解活性は50 mM EDTA存在下のみで確認された(図4左)。至適EDTA濃度を検証するために、EDTA濃度をさらに細かく変化させたところ、この反応条件では12.5mM以上62.5mM以下の濃度でRNA分解活性が検出され、25 mM EDTA存在下でもっとも効率のよいRNA分解活性が検出された(図4中)。反応液中の二価金属イオンを十分にキレートする高濃度のEDTAではT-DYWタンパク質のRNA分解活性は観察されなかったことから、T-DYWタンパク質のRNA分解活性には二価金属イオンが影響を与えることが示唆された。対照実験として、既知RNA分解酵素である100 ユニットのRNase
T1(Ambion 社)を用いて、同様の実験を行った。その結果、RNase T1はEDTA未添加で最大のRNA分解活性を示し、高濃度のEDTA存在下でもRNA分解活性を示した(図4右)。また、T-DYWとRNase T1では、RNA分解中間産物のパターンが異なることから、両タンパク質のRNA分解における認識塩基は異なることが明らかである。RNase T1はRNA中のシトシン残基の3'側を切断することが既に明らかになっている。
【0071】
(T-DYWタンパク質の酵素活性の核酸特異性)
T-DYWタンパク質の核酸切断活性をRNA、一本鎖DNA、二本鎖DNAを基質に用いて測定した。32P 標識二本鎖DNAは、プライマーndhB-F、ndhB-Rと鋳型DNAであるNB500 DNAを用い、dNTP mix(10 nmol dATP、dGTP、TTP、0.5 nmol dCTP)、4 μl
[32P] α-dCTP (GE ヘルスケア社、3000 Ci/mmol)、Ex-Taq(タカラバイオ社)を含む溶液でPCR反応を行うことにより調製した。PCR反応は実施例1と同様に行った。得られたDNA断片は変性6%ポリアクリルアミドゲル電気泳動による展開後、ゲルからDNAを溶出させることによって精製した。一本鎖DNAは二本鎖DNAを90℃で3分間処理した後に、急冷することによって調製した。T-DYWタンパク質とNB500 RNA (RNA)、一本鎖NB500 DNA (ssDNA)、二本鎖NB500 DNA(dsDNA)を混合し、実施例1 と同様の条件で、それぞれの基質核酸への切断活性を調べた。結果を図5に示す。
【0072】
図5に示したように、T-DYWタンパク質はRNAにのみ切断活性を示した。対照実験として、既知RNaseである100ユニットのRNase T1(Ambion社)、既知DNaseである10ユニットのDNase
I(RNase-free、タカラバイオ社)を用いて、酵素活性の核酸特異性を調べた。その結果、RNase T1はRNA、DNase IはDNAのみへの切断活性を呈したことから、T-DYWタンパク質はRNA特異的な核酸分解活性を呈することが明らかである。
【0073】
(T-DYWのRNA切断部位)
T-DYWタンパク質によって分解されたRNAの分解中間産物は、明瞭なバンドとして検出できる(図5)。これは既知RNaseであるRNase T1、RNaseAなどのRNAを内部で切断するエンドヌクレアーゼで観察される現象である。そこで、T-DYWタンパク質によって切断されるRNA残基の特性を検証するために、切断されたRNAの5'末端をプライマー伸長法により決定した。10 fmolの未標識ndhB RNAと1 μgのT-DYWタンパク質を用いて、実施例1 と同様の条件で反応した後、反応液からRNAを抽出した。
【0074】
次に5'末端を[32P]で標識したndh-Rプライマー0.4 pmolと回収したRNAを混合し、逆転写酵素(ReverTraAce、TOYOBO社)を用いて、コンプリメンタリーDNAを合成した。同時に、切断された塩基を特定するためのDNAラダーを調製した。DNAラダーは5'末端を[32P]で標識したndh-Rプライマーと鋳型DNAであるNB500を用いて、Thermo Sequenase Primer Cycle Sequencing kit (GEヘルスケア社)により調製した。得られたコンプリメンタリーDNAとDNAラダーを6M 尿素を含む6%ポリアクリルアミドゲルで電気泳動した。電気泳動後にゲルを乾燥させた。ゲル中のRNAの放射活性をBAS2000(Fuji
Film社)で測定した。その結果を以下の図6に示す。
【0075】
図6に示したように、T-DYWタンパク質によって切断されたRNAの5'末端がいくつか検出された。得られた5'末端の位置(NB500の5'末端を1とする)と塩基を示した。得られた5'末端は全てアデノシン残基であったことから、T-DYWはRNA中のアデノシン残基の5'側を選択的に切断するRNA分解酵素であることがわかった。
【実施例2】
【0076】
(イネ由来のDYWタンパク質の調製と分解活性)
まず、イネ (Oryza
sativa, Nipponbare)からのDNA調製を実施例1と同様に行った。次に、調製したイネのゲノムDNAを鋳型に、イネゲノム情報データベース(http://www.tigr.org/tdb/e2k1/osa1/overview.shtml)に掲載されている配列情報を参照し、シロイヌナズナのタンパク質 (AT2980(At2g02980)中のDYWモチーフの配列(配列番号:1)と高い相同性を示すモチーフを持つタンパク質遺伝子Os5g30710のDNA配列を増幅するためのオリゴヌクレオチドプライマー(Os5g30710-F、Os5g30710-R; 配列番号:15,16に記載)を調製した。Os5g30710を含むDNA断片の増幅(配列番号:17)、pBAD/Thio-TOPOベクター(Invitrogen社)へのクローニングは、実施例1に記載したT-DYWタンパク質遺伝子のクローニングと同様に行い、得られたプラスミドをpTrx-osDYWと命名した。このプラスミドpTrx-osDYWを用い、イネ由来のタンパク質T-osDYWを調製した(配列番号:18)。組み換えタンパク質の調製は、実施例1に記載した方法と同様に行った。
【0077】
このT-osDYWタンパク質のRNA分解活性は、50ngのT-osDYWタンパク質を用いて、実施例1 に記載したT-DYWタンパク質のRNA分解活性の測定と同様に行った。その結果を図7に示す。
【0078】
図7より明らかなように、イネ由来T-osDYWタンパク質においても、ナズナ由来T-DYWと同様なRNA分解活性が測定された。コントロールとして、タンパク質非存在下(レーン、no protein)、インサートのないpBAD-Thio-TOPOベクターから調製したチオレドキン及びヒスチジンタグのみからなるタンパク質(50 ng、control、配列番号:19)のみでもRNA分解活性を測定したが、RNAの分解は観察されなかった。
【実施例3】
【0079】
(変異DYWタンパク質の調製とRNA分解活性)
T-DYW、T-osDYWタンパク質は、そのアミノ酸配列中に金属配位モチーフ、CxxCH、を持つ(C、システイン;H、ヒスチジン;x、任意のアミノ酸)。このモチーフのRNA分解における働きを検証するために、以下の方法で、変異T-DYWタンパク質、T-DYW_Mタンパク質、を調製した。変異はT-DYWタンパク質中のアミノ酸配列CxxCHを、GxxGH(G、グリシン)に置換することによって行った。以下に詳細な調製法を示す。
【0080】
実施例1に記載したpTrx-DYWを鋳型に、まず2つのDNA断片、D1とD2をそれぞれ、オリゴヌクレオチドプライマー02980DYW-F(配列番号:8)と02980M2-R(配列番号:21)、02980M2-F(配列番号:20)と02980_m-R(配列番号:9)を用いて、実施例1 と同様の条件でPCRすることによって調製した。次に、得られたD1およびD2のDNA断片を混合したものを鋳型に、オリゴヌクレオチドプライマー02980DYW-F(配列番号:8)と02980_m-R(配列番号:9)を用いて、PCRすることによって、変異DYWタンパク質をコードするDNA断片を得た(配列番号:22)。DNA断片のpBAD-Thio/TOPOベクターへのクローニングと、組み換えタンパク質T-DYW_M(配列番号:23)の調製は、前述の実施例1 と同様の方法で行った。
【0081】
得られたT-DYW_Mタンパク質のRNA分解活性は、実施例1に記載したT-DYWタンパク質のRNA分解活性の測定と同様の方法で行った。得られた結果を図8に示す。
【0082】
図8より明らかなように、T-DYWタンパク質(50 ng)では、RNA分解活性が観察できたが、T-DYWタンパク質中の2つのシステイン残基に変異を導入したT-DYW_Mタンパク質(50 ng)では、RNA分解活性は観察されなかった。コントロールとして、タンパク質非存在下(レーン、-protein)、pBAD/Thio-TOPOベクターのタグタンパク質のみ(50 ngインサートなし、配列番号:19)でもRNA分解活性を測定したが、RNAの分解は観察されなかった。このことは、T-DYWタンパク質のRNA分解活性には、金属配位モチーフであるCxxCHのアミノ酸配列が重要な働きを担うことを示している。
【配列表フリーテキスト】
【0083】
配列番号:8,9,12,13,15,16,20,21:プライマー
配列番号:10,14,18,19,23:融合タンパク質
配列番号:17,22:融合タンパク質をコードするDNA
【図面の簡単な説明】
【0084】
【図1】PPRタンパク質において見出されているDYWモチーフのコンセンサス配列の一例を示す図。
【図2】PPRタンパク質における35アミノ酸モチーフの一例を示す図。
【図3】実施例1で調製したT-DYWタンパク質のRNA切断活性を示す図。
【図4】実施例1で調製したT-DYWタンパク質のRNA切断活性に対するEDTAの効果を示す図。
【図5】実施例1で調製したT-TYWタンパク質のRNA切断活性の核酸特異性を示す図。
【図6】実施例1で調製したT-TYWタンパク質のRNA切断活性が塩基特異的であることを示す図。
【図7】実施例2で調製したイネDYWタンパク質のRNA切断活性を示す図。
【図8】実施例3で調製した変異を導入したDYWタンパク質のRNA切断活性を示す図。
【特許請求の範囲】
【請求項1】
以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、
RNAを切断する活性を有する、タンパク質。
【請求項2】
さらに、以下の特徴;
(b)チトクロームCヘム結合モチーフCxxCHを含む。
を有する、請求項1に記載のタンパク質。
【請求項3】
さらに、以下の特徴;
(c)PPRモチーフを備える天然タンパク質のC末端領域に備えられるアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有する、請求項1又は2に記載のタンパク質。
【請求項4】
前記アミノ酸配列は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれかを問い合わせ配列としてホモロジー検索を行って得られるE値が1.0×10-10以下である配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列である、請求項1〜3のいずれかに記載のタンパク質。
【請求項5】
前記天然タンパク質は、単子葉植物及び双子葉植物のいずれかのタンパク質である、請求項1〜4のいずれかに記載のタンパク質。
【請求項6】
前記RNA切断活性はエンド型である、請求項1〜5のいずれかに記載のタンパク質。
【請求項7】
前記RNA切断活性は塩基特異的である、請求項1〜6のいずれかに記載のタンパク質。
【請求項8】
前記RNA切断活性は、アデニン塩基を有するリボヌクレオチドの5’側を部位特異的に切断する活性である、請求項1〜7のいずれかに記載のタンパク質。
【請求項9】
RNA分解酵素のスクリーニング方法であって、
配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、
前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、
を備える、方法。
【請求項10】
前記タンパク質の準備工程は、配列番号:1、配列番号:4及び配列番号:6のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質タンパク質を準備する工程である、請求項9に記載の方法。
【請求項11】
RNA分解酵素の製造方法であって、
配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列とチトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することを特徴とする、方法。
【請求項12】
請求項1〜8のいずれかに記載のタンパク質を含む、核酸分析キット。
【請求項13】
請求項1〜8のいずれかに記載のタンパク質をコードする、ポリヌクレオチド。
【請求項14】
請求項13に記載のポリヌクレオチドを、請求項1〜9のいずれかに記載のタンパク質を発現可能に保持する、形質転換体。
【請求項15】
請求項1〜8のいずれかに記載のタンパク質を用いてRNAを切断する工程を備える、核酸の処理方法。
【請求項16】
核酸を分離精製する工程を備える、請求項15に記載の方法。
【請求項17】
RNAを抽出する工程を備える、請求項15に記載の方法。
【請求項18】
RNA切断後のRNA断片を検出する工程を備える、請求項15に記載の方法。
【請求項1】
以下の特徴;
(a)配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有し、
RNAを切断する活性を有する、タンパク質。
【請求項2】
さらに、以下の特徴;
(b)チトクロームCヘム結合モチーフCxxCHを含む。
を有する、請求項1に記載のタンパク質。
【請求項3】
さらに、以下の特徴;
(c)PPRモチーフを備える天然タンパク質のC末端領域に備えられるアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を含む。
を有する、請求項1又は2に記載のタンパク質。
【請求項4】
前記アミノ酸配列は、配列番号:1及び配列番号:4に記載のアミノ酸配列のいずれかを問い合わせ配列としてホモロジー検索を行って得られるE値が1.0×10-10以下である配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列である、請求項1〜3のいずれかに記載のタンパク質。
【請求項5】
前記天然タンパク質は、単子葉植物及び双子葉植物のいずれかのタンパク質である、請求項1〜4のいずれかに記載のタンパク質。
【請求項6】
前記RNA切断活性はエンド型である、請求項1〜5のいずれかに記載のタンパク質。
【請求項7】
前記RNA切断活性は塩基特異的である、請求項1〜6のいずれかに記載のタンパク質。
【請求項8】
前記RNA切断活性は、アデニン塩基を有するリボヌクレオチドの5’側を部位特異的に切断する活性である、請求項1〜7のいずれかに記載のタンパク質。
【請求項9】
RNA分解酵素のスクリーニング方法であって、
配列番号:1、配列番号:4及び配列番号:6に記載のアミノ酸配列のいずれか又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質を準備する工程と、
前記準備したタンパク質についてRNA切断活性の有無を検出する工程と、
を備える、方法。
【請求項10】
前記タンパク質の準備工程は、配列番号:1、配列番号:4及び配列番号:6のいずれかに記載のアミノ酸配列を問い合わせ配列としてホモロジー検索を行って得られる所定値以下のE値を有するアミノ酸配列又はこれらの配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列を有するタンパク質タンパク質を準備する工程である、請求項9に記載の方法。
【請求項11】
RNA分解酵素の製造方法であって、
配列番号:1及び配列番号:4のいずれかに記載のアミノ酸配列又はこれらのアミノ酸配列において1若しくは複数個のアミノ酸が置換、欠失、付加及び/又は挿入されたアミノ酸配列とチトクロームCヘム結合モチーフCxxCHを備えるタンパク質を製造することを特徴とする、方法。
【請求項12】
請求項1〜8のいずれかに記載のタンパク質を含む、核酸分析キット。
【請求項13】
請求項1〜8のいずれかに記載のタンパク質をコードする、ポリヌクレオチド。
【請求項14】
請求項13に記載のポリヌクレオチドを、請求項1〜9のいずれかに記載のタンパク質を発現可能に保持する、形質転換体。
【請求項15】
請求項1〜8のいずれかに記載のタンパク質を用いてRNAを切断する工程を備える、核酸の処理方法。
【請求項16】
核酸を分離精製する工程を備える、請求項15に記載の方法。
【請求項17】
RNAを抽出する工程を備える、請求項15に記載の方法。
【請求項18】
RNA切断後のRNA断片を検出する工程を備える、請求項15に記載の方法。
【図1】



【図2】

【図3】
【図4】
【図5】

【図6】

【図8】
【図7】

【図2】
【図3】
【図4】
【図5】
【図6】
【図8】
【図7】
【公開番号】特開2008−141960(P2008−141960A)
【公開日】平成20年6月26日(2008.6.26)
【国際特許分類】
【出願番号】特願2006−328869(P2006−328869)
【出願日】平成18年12月5日(2006.12.5)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
【公開日】平成20年6月26日(2008.6.26)
【国際特許分類】
【出願日】平成18年12月5日(2006.12.5)
【出願人】(504139662)国立大学法人名古屋大学 (996)
【Fターム(参考)】
[ Back to top ]