
RSVFVLPならびにその製造および使用の方法
本発明は、感染因子を含まない、エンベロープウイルスベースのウイルス様粒子(VLP)を単離する分野に関する。好ましい例において、該分野は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、感染因子を不活化する方法を包含する。特定の実施形態では、エンベロープウイルスベースのVLPが、昆虫細胞ベースの発現系内で作製される。本発明は、RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子はエンベロープウイルスコアを含まない、調製物を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、感染因子(infectious agent)を含まない、エンベロープウイルスベースのウイルス様粒子(VLP)を単離する分野に関する。好ましい例において、該分野は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、感染因子を不活化する方法を包含する。特定の実施形態では、エンベロープウイルスベースのVLPを、昆虫細胞ベースの発現系において作製する。
【背景技術】
【0002】
(発明の背景)
RSウイルス(RSV)は、乳児および1歳未満の小児において、細気管支炎および肺炎の主要原因である(CDC National Center for Infectious Diseases(2004年)、Respiratory Syncytial Virus)。RSVはまた、免疫不全である成人および高齢者においても、重要な下気道病原体であり得る。RSVによる天然の感染は、防御的免疫を誘導しないので、個体は、複数回にわたり感染し得る。
【0003】
RSVは、Paramyxoviridae科のPneumovirus属に属する、ネガティブセンスの一本鎖RNAウイルスである。RSVゲノムは、螺旋状のヌクレオカプシドにより取り囲まれ、少なくとも10のタンパク質:3つの膜貫通構造タンパク質(F、G、およびSH)、2つのマトリックスタンパク質(MおよびM2)、3つのヌクレオカプシドタンパク質(N、P、およびL)、および2つの非構造タンパク質(NS1およびNS2)をコードする(Collinsら(1996年)、Respiratory syncytial virus、1313〜1351頁、B.N. Fields(編)、Fields virology. Raven Press、New York、NY)。中和抗体は、Fタンパク質およびGタンパク質によってのみ誘発されると考えられる。RSVが、Gタンパク質に基づいて、AおよびBの亜群に分けられるのに対し、Fは、その亜群間における関連がより密接である。Fタンパク質に対するモノクローナル抗体は、インビトロにおいて中和効果、インビボにおいて予防効果を有することが示されている(例えば、Andersonら、1988年、J. Virol.、62巻:4232〜4238頁; Andersonら、1986年、J. Clin. Micro.、23巻:475〜480頁; Beeler およびCoelingh、1989年、J. Virol.、63巻:2941〜50頁; Garcia−Barrenoら、1989年、J. Virol.、63巻:925〜32頁; Taylorら、1984年、Immunology、52巻:137〜142頁;ならびに特許文献1)。
【0004】
数十年間にわたる研究にもかかわらず、RSVに対する安全で有効なワクチンは存在していない。乳児および小児において試験されたホルマリン不活化ウイルスワクチンは、感染に対して防御せず、野生型のRSVウイルスによるその後の感染において、重篤な症状が生じる危険性の増大と関連していた(Kapikianら、1969年、Am. J. Epidemiol.、89巻:405〜21頁; Chinら、1969年、Am. J. Epidemiol.、89巻:449〜63頁)。生の弱毒化温度感受性変異体の開発に焦点を当てたその後の試みもまた、適切な弱毒化レベルのウイルス候補を同定することができず、また、一部の候補が遺伝子的に不安定であるために失敗した(Hodesら(1974年)、Proc. Soc. Exp. Biol. Med.、145巻、1158〜1164頁; Kimら(1973年)、Pediatrics、52巻、56〜63頁; Wrightら(1976年)、J. Pediatrics、88巻、931〜936頁)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】米国特許第6818216号明細書
【発明の概要】
【発明が解決しようとする課題】
【0006】
ウイルス様粒子(VLP)は、従来のワクチン技術を上回るいくつかの利点を提供する。ワクチンを開発するのに重要なVLPの利点は、それらが、三次元構造との関連で天然ウイルスを模倣するということであり、また、一次エピトープおよび立体配座エピトープの両方に対する中和抗体応答を誘導する能力であり、したがって、他のワクチン処方物より免疫原性であることが示されるはずである。ウイルスベクター法と異なり、VLPは、あらかじめ存在する免疫に対して問題を示さず、したがって、繰り返しの使用を可能とする。RSV抗原を含有するVLPは、昆虫細胞内において、RSV Fタンパク質を、RSV Mタンパク質と、または異種Mタンパク質と共発現させることにより作製されてきた(US2008/0233150)。しかし、US2008/0233150は、RSV Fタンパク質が単独で、互いに凝集しないかまたはRSV Fタンパク質を発現させるのに使用されるウイルスベクターと会合しないVLPを生成できる、哺乳動物細胞系内におけるRSV Fタンパク質単独の実際の発現は教示していない。したがって、RSV Fタンパク質を単独で発現するVLPを生成する方法が必要とされている。
【課題を解決するための手段】
【0007】
(概要)
本発明の好ましい実施形態は、さらなるエンベロープウイルスコア形成ポリペプチドを必要としないRSウイルスFポリペプチドのVLPを生成する方法、およびRSウイルスFポリペプチドのVLP調製物を含む組成物のための、本明細書で開示される各種の方法および組成物を提供することにより、この必要を満たす。好ましい実施形態では、RSウイルスFポリペプチドのVLPは、以下のさらなる特徴のうちの1つまたは複数を有する:該VLPが、エンベロープウイルスコアを含まない;該VLPが、多形性であるか、またはサイズもしくは形状が不均一である;該VLPが、エンベロープウイルスコア形成ポリペプチドを含まない。このような好ましい実施形態は、RSウイルスFポリペプチドが、単独でVLPを形成することが可能であるという、驚くべき観察に基づいている。RSウイルスFポリペプチド単独により形成されるVLPは、エンベロープウイルス、およびエンベロープウイルス成分を用いて形成されるVLPに典型的なタンパク質コアを有さない。
【0008】
本発明の態様は、RSウイルスFポリペプチドを含む、RSウイルスFポリペプチドのウイルス様粒子調製物を包含し、該ウイルス様粒子は、以下の特徴のうちの1つまたは複数を有する:該ウイルス様粒子は、エンベロープウイルスコアを含まない;該ウイルス様粒子は、多形性であるか、サイズが不均一であるか、もしくは形状が不均一である;該ウイルス様粒子は、エンベロープウイルスコア形成ポリペプチドを含まない;および/または該ウイルス様粒子は、哺乳動物グリコシル化を含む。特定の実施形態では、ウイルス様粒子は、他のウイルス様粒子と実質的に凝集しない場合がある。前出の実施形態または態様と組み合わせ得る別の実施形態では、ウイルス様粒子は、ウイルスベクター粒子と実質的に会合しない。
【0009】
前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、調製物はまた、ウイルス様粒子と混合したアジュバントも含む。アジュバントを包含する前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、アジュバントは、ウイルス様粒子の外側に位置する場合もあり、ウイルス様粒子の内側に位置する場合もある。アジュバントを包含する前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、アジュバントをRSウイルスFポリペプチドに共有結合させて、共有結合を形成させ得る。
【0010】
前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、抗RSV−F中和抗体は、RSウイルスFポリペプチドに結合し得る(これにより、該RSV Fポリペプチドが、実質的に天然の立体配座にあることが裏付けられる)。このような中和抗体を包含する、前出の実施形態または態様のうちのいずれかと組み合わせ得る特定の実施形態では、抗RSV−F中和抗体は9C5であり得る。
【0011】
別の態様は、RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法を含み、該方法は、(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと;(b)該発現ベクターを培地中の哺乳動物細胞内へと導入するステップと;(c)該RSウイルスFポリペプチドを発現させて、該RSウイルスFポリペプチドのウイルス様粒子を作製するステップとを含み、該ウイルス様粒子は、以下の特徴のうちの1つまたは複数を有してもよい:該ウイルス様粒子は、エンベロープウイルスコアを含まない;該ウイルス様粒子は、多形性であるか、サイズが不均一であるか、もしくは形状が不均一である;および/または該ウイルス様粒子は、エンベロープウイルスコア形成ポリペプチドを含まない。
【0012】
前出の態様と組み合わせ得る別の実施形態では、該方法は、哺乳動物細胞が培養されている培地から、RSウイルスFポリペプチドのウイルス様粒子を取り出すステップをさらに含む。
【0013】
前出の実施形態および態様と組み合わせ得る別の実施形態では、発現ベクターはウイルスベクターであり得る。前出の実施形態および態様と組み合わせ得る別の実施形態では、ウイルスベクターを、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択することができる。前出の実施形態および態様と組み合わせ得る別の実施形態では、哺乳動物細胞を、BHK細胞、VERO細胞、HT1080細胞、MRC−5細胞、WI 38細胞、MDCK細胞、MDBK細胞、293細胞、293T細胞、RD細胞、COS−7細胞、CHO細胞、Jurkat細胞、HUT細胞、SUPT細胞、C8166細胞、MOLT4/クローン8細胞、MT−2細胞、MT−4細胞、H9細胞、PM1細胞、CEM細胞、黒色腫細胞、SB20細胞、LtK細胞、HeLa細胞、WI−38細胞、L2細胞、CMT−93、およびCEMX174細胞からなる群より選択することができる。
【0014】
前出の実施形態および態様と組み合わせ得る別の実施形態では、抗RSV−F中和抗体は、発現されたRSウイルスFポリペプチドに結合し得る(これにより、該RSウイルスFポリペプチドは、実質的に天然のフォールドであることが裏付けられる)。中和抗体を包含する前出の実施形態および態様と組み合わせ得る別の実施形態では、抗RSV−F中和抗体は9C5である。
【0015】
別の態様は、RSウイルスの感染を処置または予防する方法を含み、該方法は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を被験体に投与するステップを含む。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与は被験体における防御的免疫化反応を誘導する。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与を、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択することができる。
【0016】
別の態様は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を含む医薬組成物を包含する。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、医薬組成物は、薬学的に許容されるキャリアをさらに含む。
【0017】
別の態様は、RSウイルスの感染に対する防御を提供する方法を含み、該方法は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を被験体に投与するステップを含む。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与を、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択することができる。
【0018】
前出の態様およびそれらの実施形態は、本明細書で開示される実施形態のうちのいずれかとさらに組み合わせることができる。前出の実施形態、および/または本明細書で開示されるさらなる実施形態のうちのいずれかと共に包含され得る、本発明のさらなる態様は、本明細書の全体において見出すことができる。
【図面の簡単な説明】
【0019】
【図1】図1は、p3.1−RSVFTのプラスミド地図を示す図である。
【図2】図2は、p3.1−shFv1のプラスミド地図を示す図である。
【図3】図3は、p3.1−shFv2のプラスミド地図を示す図である。
【図4】図4は、p3.1−Gagのプラスミド地図を示す図である。
【図5】図5は、RSV FおよびGag発現ベクターをトランスフェクトされた細胞上のRSV Fの表面発現のサイトメトリー分析を示すグラフである。トランスフェクトされていない細胞およびp3.1−Gagだけをトランスフェクトされた細胞は、バックグラウンド蛍光レベルを示す。p3.1−Gagと共におよび共にではなくRSV F発現ベクターをトランスフェクトされた細胞は、9C5モノクローナル抗体および蛍光2次抗体によるF検出の結果として顕著なレベルの蛍光を示す。
【図6】図14は、Gag遺伝子を共トランスフェクトされておよびされずにRSV F遺伝子をトランスフェクトされた細胞の培地由来の100,000×gペレットにおけるRSV F抗原活性の検出を示すグラフである。
【図7】図7は、p3.1−RSVFTをトランスフェクトされた239−F細胞の培地から収集したVLPの電子顕微鏡像の代表的切片を示す図である。
【図8】図8は、pFB−shFv1のプラスミド地図を示す図である。
【図9】図9は、昆虫細胞中で発現されたshFv1 VLPのショ糖密度勾配を示すグラフである。
【図10】図10は、図9に示すshFv1ショ糖勾配の画分9における凝集したRSV F VLPおよびバキュロウイルス粒子の代表的電子顕微鏡像を示す図である。
【図11】図11は、ヒトカルボキシペプチダーゼM由来のGPIアンカーシグナルに融合したキメラFポリペプチド外部ドメインをコードするp3.1−F−GPIのプラスミド地図を示す図である。
【図12】図12は、p3.1−Gagを共トランスフェクトされておよびされずにp3.1−F−GPIをトランスフェクトされた細胞での改変F−GPIタンパク質の表面発現のフローサイトメトリー分析を示すグラフである。p3.1−Gagだけをトランスフェクトされた細胞は、バックグラウンド蛍光レベルを示した。p3.1−Gagと共におよび共にではなくp3.1−F−GPIをトランスフェクトされた細胞は、9C5モノクローナル抗体および蛍光2次抗体によるF検出の結果として顕著なレベルの蛍光を示した。
【図13】図13は、p3.1−Gag、p3.1−F−GPIおよびp3.1−Gag+p3.1−F−GPIをトランスフェクトされた細胞の培地由来の100,000×gペレットのウエスタンブロット分析を示す図である。レーンは左から右へ:G、p3.1−Gagトランスフェクション由来の試料;F、p3.1−F−GPIトランスフェクション由来の試料;F+G、p3.1−Gag+p3.1−F−GPIトランスフェクション由来の試料である。データは、F発現はGag出芽に干渉するが、Gag発現はF出芽に干渉しないことを示す。
【図14】図14は、Gag遺伝子ベクターを共トランスフェクトされておよびされずにRSV F−GPIベクターをトランスフェクトされた細胞の培地由来の100,000×gペレットにおけるRSV F抗原活性の検出を示すグラフである。生のRSVビリオン粒子を含有しているRSV感染細胞由来の培地の100,000×gペレットを、F−GPI粒子がF抗原に関して生のRSVに抗原的に類似していることを実証する陽性対照として含めた。
【発明を実施するための形態】
【0020】
(好ましい実施形態の詳細な説明)
本発明の好ましい実施形態には、好ましくは、哺乳動物細胞による発現系で作製された、RSウイルスFポリペプチドのVLP調製物と;このようなRSウイルスFポリペプチドのVLP調製物を発現させるかまたは作製する方法と;このような調製物をワクチン組成物へとさらに加工する方法と;このようなワクチン組成物を用いる方法とが含まれるがこれらに限定されない。
【0021】
本発明の特定の態様および実施形態は、適切な宿主細胞内で発現させると、gagポリペプチドまたはマトリックスポリペプチドなど、任意のコアタンパク質またはコア形成タンパク質の不在下で、RSウイルスFポリペプチドは、単独でVLPを生成するのに十分であるという驚くべき発見に基づいている。このようなVLPは、内部におけるタンパク質コアまたはカプシドなしに、脂質またはエンベロープを有する。したがって、このようなVLPは、以下の物理的な特徴のうち1つまたは複数を有する:脂質膜またはエンベロープ内にエンベロープウイルスコア、カプシド、またはヌクレオカプシドを有さない;多形性であるか、またはサイズおよび/もしくは形状が不均一である;該VLPは、互いと凝集しない;該VLPは、それを作製するのに用いられるウイルスベクターと会合しないか、またはそうでなければ凝集しない;ならびに哺乳動物グリコシル化、および/または哺乳動物膜と結合もしくは会合するタンパク質。
【0022】
RSウイルスFポリペプチドを作製するのに好ましい方法は、好ましくは、さらなるポリペプチド抗原の共発現を含めた、哺乳動物細胞内における発現を介する。
【0023】
開示される方法およびプロトコールを実施するには、別段に示さない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を用いる。このような技法は、文献において説明されている。例えば、J. Sambrook、E. F. Fritsch、およびT. Maniatis、1989年、「Molecular Cloning: A Laboratory Manual」、第2版、1〜3巻、Cold Spring Harbor Laboratory Press社; Ausubel, F. M.ら(1995年、および定期的な補遺;「Current Protocols in Molecular Biology」、9、13、および16章、ニューヨーク州、ニューヨーク、John Wiley & Sons社); B. Roe、J. Crabtree、およびA. Kahn、1996年、「DNA Isolation and Sequencing: Essential Techniques」、John Wiley & Sons社; J. M. PolakおよびJames O’D. McGee、1990年、「In Situ Hybridization: Principles and Practice」、Oxford University Press社; M. J. Gait(編)、1984年、「Oligonucleotide Synthesis: A Practical Approach」、Irl Press社;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、「Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology」、Academic Press社を参照されたい。
【0024】
(定義)
本明細書で用いられる「RSウイルスFポリペプチドのVLP」とは、ウイルスカプシド、ヌクレオカプシド、またはコアを形成することが可能な任意のポリペプチドの不在下において発現させることが好ましい、RSウイルスFポリペプチドを用いて形成されるウイルス様粒子を指す。好ましい例には、RSウイルスFポリペプチドの細胞質部分を実質的に欠く、RSウイルスFポリペプチドを用いて生成されるVLP、またはインフルエンザM1ポリペプチドおよび/もしくは赤血球凝集素ポリペプチド(また、場合によってノイラミニダーゼポリペプチド)を用いて生成されるRSウイルスFポリペプチドおよびVLPが含まれるが、これらに限定されない。
【0025】
「哺乳動物グリコシル化」とは、哺乳動物細胞の発現系により生成されるグリコシル化パターンを指す。このようなグリコシル化パターンは、哺乳動物のグリコシル化酵素を包含するように改変された昆虫細胞が、哺乳動物または哺乳動物細胞ベースの発現系により天然にもたらされるグリコシル化パターンではなく「哺乳動物様の」グリコシル化のみをもたらす場合に限り、このような改変昆虫細胞によりもたらされるグリコシル化パターンを包含しない。哺乳動物グリコシル化パターンの好ましい例には、ヒト(例えば、HEK293、HeLa)細胞、チャイニーズハムスター卵巣(CHO)細胞、イヌ(例えば、MDCK)細胞、マウス(例えば、H9)細胞、ラット(例えば、IE)細胞、およびヒト以外の霊長動物(例えば、NCTC)細胞内における発現によりもたらされるグリコシル化が含まれる。
【0026】
本明細書で用いられる「エンベロープウイルスコア形成ポリペプチド」には、エンベロープウイルスのエンベロープ内において、ウイルスカプシド、ヌクレオカプシド、またはコアを形成することが可能な、任意のウイルスポリペプチド、または任意のウイルスポリペプチドの組合せが含まれる。エンベロープウイルスコア形成ポリペプチドは、RSウイルスFポリペプチドのVLPの形成を干渉し得るので、本明細書で記載されるRSウイルスFポリペプチドのVLPの好ましい実施形態は、このようなポリペプチドを除外する。ウイルスカプシド、ヌクレオカプシド、もしくはコアを形成することが不可能な、エンベロープウイルスコア形成ポリペプチドに由来するポリペプチド、またはウイルスカプシド、ヌクレオカプシド、もしくはコアを形成することが不可能な、エンベロープウイルスコア形成ポリペプチドのサブセットは、本明細書で開示されるVLPの形成を干渉しないので、このようなポリペプチドは、本明細書で開示されるVLPに含まれ得る。
【0027】
本明細書で記載されるウイルス様粒子を形成する一因となる、レトロウイルス由来の構造ポリペプチドであるgagポリペプチドは、エンベロープウイルスコア形成ポリペプチドの例である。レトロウイルスのゲノムは、3つの主要な遺伝子産物をコードする:構造タンパク質をコードするgag遺伝子の産物;逆転写酵素および関連のタンパク質分解性ポリペプチド、ヌクレアーゼおよびインテグラーゼに関連する機能をコードするpol遺伝子の産物;envの産物(それによりコードされる糖タンパク質の膜タンパク質が、感染細胞の表面上において、また、放出された成熟ウイルス粒子の表面上においても検出される)。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、レトロウイルスの各群内においても、アミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除くコアタンパク質をもたらす。
【0028】
MLVの場合、Gagの前駆体であるポリタンパク質は、Pr65Gagであり、該前駆体上におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質へと切断される。これらの切断は、ウイルス性プロテアーゼを介し、また、ウイルスに応じて、ウイルス放出の前または後において生じ得る。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態はgPr80Gagから切り出され、gPr80Gagは、非グリコシル化Pr65Gagに対応するAUGコドンの上流に位置する、インフレームの、異なる開始コドンから合成される。グリコシル化Gagを合成しない、MLVの欠失変異体がなおも感染性であり、また、非グリコシル化Gagがなおもウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について、疑問が投げかけられる。ウイルスがコードするプロテアーゼにより、HIV−1 Gagの前駆体であるpr55Gagが翻訳後に切断されることから、N−ミリストイル化されて内部でリン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化されたp24カプシドタンパク質(p24CA)、ならびにヌクレオカプシドタンパク質であるp15(p15NC)がもたらされ、p15は、さらにp9およびp6へと切断される。
【0029】
構造面において、原型的なGagポリタンパク質は、レトロウイルスのgag遺伝子において常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(マトリックスという名称を共有するが、MAとは異なるタンパク質である、インフルエンザマトリックスタンパク質M1と混同しないようにされたい)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。成熟タンパク質へのGagポリタンパク質のプロセシングは、レトロウイルスがコードするプロテアーゼにより触媒され、新たに出芽するウイルス粒子が成熟するときに生じる。機能面において、Gagポリタンパク質は、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメインと;Gagの重合化を促進する相互作用ドメインと;宿主細胞からの新生ビリオンの放出を促進する後期ドメインとに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される任意の切断産物内にもれなく収まる必要はない。これらの重要な機能的エレメントに関する最新技術の進歩は大きい。例えば、
【0030】
【化1】

を参照されたい。
【0031】
エンベロープウイルスコア形成ポリペプチドの範囲内に包含されるgagポリペプチドは、最小限において、ウイルス様粒子を形成するための機能的エレメントを、RSウイルスFポリペプチドのVLPとは独立して包含することになる。RSウイルスFポリペプチドと競合するか、またはこれに干渉する機能的エレメントを除外すれば、このようなgagポリペプチドは、エンベロープウイルスコア形成ポリペプチドではない。
【0032】
Gagポリペプチドについてのレトロウイルスの供給源の例には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)が含まれる。
【0033】
本明細書で用いられる「エンベロープウイルスコア」には、エンベロープウイルスまたはエンベロープウイルスの成分により生成されるウイルス様粒子の脂質膜またはエンベロープの内側で見出される任意のタンパク質性コアが含まれる。タンパク質性コアは、カプシドもしくはヌクレオカプシド、またはマトリックスの全部または一部を含み得る。
【0034】
本明細書で用いられる「脂質ラフト」とは、ウイルス粒子のアセンブリー過程においてgagポリペプチドが濃縮される、細胞膜のマイクロドメインを指す。
【0035】
本明細書で用いられる「脂質ラフト会合ポリペプチド」とは、脂質ラフトと直接的または間接的に会合する任意のポリペプチドを指すが、任意のエンベロープウイルスコア形成ポリペプチドは除外する。本発明で用いられる具体的な脂質ラフト会合ポリペプチドは、VLPの所望の使用、および該脂質ラフト会合ポリペプチドの役割(例えば、RSウイルスFタンパク質の細胞外部分を該VLPへと結合させること、または1つもしくは複数のさらなる抗原もしくはアジュバントを該VLPへと結合させること)に依存する。
【0036】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質もしくは脂質ラフト会合部分;脂質による修飾など、膜との会合を引き起こすタンパク質の修飾により脂質ラフトと直接的に会合するタンパク質もしくはその部分;または脂質ラフト会合ポリペプチドにより脂質ラフトと間接的に会合するポリペプチドであり得る。
【0037】
脂質アンカーを有する多くのタンパク質が、脂質ラフトと会合する。このようなタンパク質の短い断片は、脂質結合に十分であることが多い。このような断片は、天然においてそれ自体では脂質ラフトと会合しないであろう他のタンパク質およびポリペプチドに容易に結合し得るので、該断片は、脂質ラフト会合に理想的となっている。ポリペプチドを脂質ラフトへと連結する脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、および二重アセチル化が含まれる。
【0038】
多くの異なる種類のポリペプチドが、脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスのアセンブリー、また、アセンブルした粒子の出芽を含めた、多くの生物学的活性のためのプラットフォームとして機能し、したがって、これらの過程に関与する各種のポリペプチドと会合する。
【0039】
シグナル伝達カスケードに関与する各種のポリペプチドは、シグナル伝達プラットフォームとして機能する脂質ラフトと会合する。シグナル伝達プラットフォームとして機能する脂質ラフトの1つの種類は、カベオラと呼ばれている。それは、カベオリンファミリーに由来するポリペプチド(例えば、カベオリンおよび/またはフロチリン(flottillin))を含有する、細胞膜のフラスコ型の陥入部である。
【0040】
膜輸送ポリペプチドは、膜輸送プラットフォームとして機能する脂質ラフトと会合する。例には、シンタクシン−1タンパク質、シンタクシン−4タンパク質、シナプシンIタンパク質、アデューシンタンパク質、VAMP2タンパク質、VAMP/シナプトブレビンタンパク質、シナプトブレビンIIタンパク質、SNAREタンパク質、SNAP−25タンパク質、SNAP−23タンパク質、シナプトタグミンIタンパク質、およびシナプトタグミンIIタンパク質など、エンドサイトーシスおよびエクソサイトーシスに関与するタンパク質が含まれる。
【0041】
ウイルス受容体、ウイルス受容体−共受容体複合体、ウイルス侵入過程の調節を補助する他の任意の成分は、ウイルス侵入に特化した膜輸送プラットフォームとして機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体の例には、崩壊促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー膜糖タンパク質;ガングリオシド、Hsc70タンパク質、アルファ2−ベータ1インテグリンおよびアルファ5−ベータ2インテグリンを含めた複数の成分を含有する複合体である、A群ロタウイルスの受容体;HIVウイルス、MLVウイルス、麻疹ウイルス、およびEbolaウイルスなど、複数のエンベロープウイルスの糖タンパク質;ならびにCD5ポリペプチド、CCR5ポリペプチド、およびnefポリペプチドなど、HIVの侵入に関与するポリペプチドが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0042】
ウイルス粒子のアセンブリーに関与するポリペプチドは、ウイルスアセンブリープラットフォームとして機能する脂質ラフトと会合する。ウイルスのヌクレオカプシド、カプシド、またはコアを形成する一因となる部分を除外する限りにおいて、このようなポリペプチドまたはそれらの一部を、脂質ラフト会合ポリペプチドとして用いることができる。このようなポリペプチドの例には、HAインフルエンザエンベロープ糖タンパク質およびNAインフルエンザエンベロープ糖タンパク質、麻疹ウイルスに由来するHタンパク質および成熟F1−F2融合タンパク質、ならびにHIVウイルスに由来するgp160、gp41、およびPr55gagが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. & Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0043】
アセンブルしたウイルスの出芽に関与するポリペプチドは、ウイルス出芽プラットフォームとして機能する脂質ラフトと会合する。宿主細胞からのHIV−1の出芽は、膜ラフト内で生じることを示唆するデータが存在する。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。ウイルスの出芽に関与するポリペプチドについての一般的な情報は、「Fields Virology」(第4版)、2001年において見出すことができる。
【0044】
好ましい脂質ラフト会合ポリペプチドには、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドなどのウイルスポリペプチドが含まれる。これらのポリペプチドの各々は、任意の種類のウイルスに由来し得るが、特定の実施形態は、HIV−1ウイルスに由来するエンベロープタンパク質、RSウイルスまたは麻疹ウイルスに由来する融合タンパク質、RSウイルス、単純ヘルペスウイルス、またはエボラウイルスに由来する糖タンパク質、また、麻疹ウイルスに由来する赤血球凝集素タンパク質を包含する。
【0045】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvumなどが含まれるがこれらに限定されない病原性の原虫、蠕虫、また、他の真核微生物の病原体から得ることができる。このような非ウイルス性脂質ラフト会合ポリペプチドは、それ自体が抗原として作用するので、天然では脂質ラフトと会合しない抗原に連結することなく用いることができる。
【0046】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、赤血球凝集素ポリペプチドである。本明細書で用いられる「赤血球凝集素ポリペプチド」は、感染される細胞に対するウイルスの結合を媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素ポリペプチドはまた、同等の麻疹ウイルスタンパク質にも由来し得る。このタンパク質は、単一の膜貫通ドメインにより、インフルエンザウイルスの表面へとアンカリングすることが見出される抗原性の糖タンパク質である。インフルエンザ赤血球凝集素のうち、H1〜H16と名付けられた、少なくとも16のサブタイプが同定されている。H1、H2、およびH3は、ヒトインフルエンザウイルスにおいて見出される。H5またはH7の赤血球凝集素を有する、高病原性鳥類インフルエンザウイルスがヒトに感染する割合は低いことが判明している。鳥類ウイルス株のH5型赤血球凝集素内における単一のアミノ酸変化がヒト患者において見出されており、これが受容体の特異性を変化させることを可能にし、H5赤血球凝集素が鳥類H5N1ウイルスの受容体特異性を顕著に変化させることによって、これらのウイルスに、ヒト受容体に結合する能力がもたらされると報告されている(109および110)。この知見は、通常ヒトに感染しないH5N1ウイルスが変異し、ヒト細胞に効率的に感染できるようになることを説明する。
【0047】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。天然では、その膜貫通ドメインが、脂質ラフトドメインと会合し、これにより、RSウイルスFポリペプチドと会合することが可能となり、VLP内へと組み込まれる。それは、円筒様の形状をしており、約135Åの長さである。HAを構成する3つの同一の単量体は、中央のコイルドコイルと、VLPの表面上に露出されるシアル酸結合部位を含有する球形の頭状部分とを形成する。HAの単量体は、グリコシル化され、2つのより小型のポリペプチド:HA1サブユニットおよびHA2サブユニットへと切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜へとアンカリングされる、三量体のコイルドコイルを形成し、HA1サブユニットは、球形の頭状部分を形成する。
【0048】
本発明の特定のVLPにおいて脂質ラフト会合ポリペプチドとして用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメインを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない、1または複数のさらなる抗原を包含することが好ましい。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0049】
ウイルス性脂質ラフト会合ポリペプチドの好ましい別の例は、ノイラミニダーゼポリペプチドである。本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼ糖タンパク質は、ウイルス表面上において発現する。ノイラミニダーゼタンパク質は四量体であり、ベータ−ピンホイール構造を有する球形の頭状部分、細いストーク状領域、また、単一の膜貫通ドメインによりウイルス膜内に該タンパク質をアンカリングする小さな疎水性領域からなる共通構造を共有する。シアル酸残基を切断するための活性部位は、すべてのインフルエンザAウイルス内に保存されている、15の荷電アミノ酸により形成される、各サブユニット表面上のポケットを包含する。インフルエンザノイラミニダーゼのうち、N1〜N9と名付けられた、少なくとも9つのサブタイプが同定されている。
【0050】
本発明の特定のVLPにおいて用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメインを包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、1または複数のさらなるポリペプチドのコード配列を赤血球凝集素ポリペプチドにスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない1または複数のさらなる抗原を包含することが好ましい。ノイラミニダーゼポリペプチドコード配列内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0051】
脂質ラフト会合ポリペプチドの好ましい別の例は、ファシクリンI(FasI)と称する、昆虫由来の接着タンパク質である。本明細書で用いられる「ファシクリンIポリペプチド」は、胚の発生に関与する昆虫タンパク質に由来する。この非ウイルス性タンパク質は、昆虫細胞内のバキュロウイルス発現系において発現させることができ、これにより、FasIの脂質ラフト会合がもたらされる(J. Virol.、77巻、6265〜6273頁、2003年)。したがって、ファシクリンIポリペプチドに異種抗原を結合させて、RSウイルスFポリペプチドと共に共発現させると、VLP内にキメラ分子が組み込まれる。本発明のVLPにおいて用いられるファシクリンIポリペプチドは、最小限において、膜アンカードメインを包含するものとする。
【0052】
脂質ラフト会合ポリペプチドの好ましい別の例は、G糖タンパク質と称する、RSVに由来するウイルス性付着タンパク質である。本明細書で用いられる「Gグリコポリペプチド」は、RSV G糖タンパク質に由来する。近年のデータは、脂質ラフトドメインが、インフルエンザウイルスにとって重要であるのと同様に、RSV粒子の出芽にも重要であることを示している(Virol、327巻、175〜185頁、2004年; Arch. Virol.、149巻、199〜210頁、2004年; Virol.、300巻、244〜254頁、2002年)。RSVに由来するG糖タンパク質は、ウイルス性付着タンパク質であり、およびRSV感染に対する防御抗原として機能する、32.5kdの内在性膜タンパク質である。インフルエンザウイルスに由来する赤血球凝集素と同様、その抗原性は、それに付着した、任意の非脂質ラフト抗原の抗原性を増強し得る。非脂質ラフト外来抗原を付着させる形での、Gグリコポリペプチドに対する任意の改変は、結果として、該外来抗原に対して顕著な免疫反応を誘導することが可能なRSウイルスFポリペプチドのVLPをもたらす。
【0053】
VLPが、その文脈により、エンベロープベースのウイルスに基づかないか、または本明細書で開示される特定のエンベロープベースのウイルスの特定の成分に基づくウイルス様粒子を指す場合を除き、用語「RSウイルスFポリペプチドのウイルス様粒子」、「RSウイルスFポリペプチドのVLP」および「VLP」は、本明細書の全体において互換的に用いられる。
【0054】
(抗原)
本発明の特定の態様は、RSウイルスFポリペプチドのVLP調製物と会合するさらなる抗原を包含する。このようなさらなる抗原は、同じ組成物中に組み込むことができ、さらにまた、VLPと共有結合させることもでき、非共有結合させることもできる。好ましい実施形態では、RSウイルスFポリペプチド、および/または他の脂質ラフト会合ポリペプチドが、天然では脂質ラフトと会合しないであろう抗原を含有する、RSウイルスFポリペプチドのVLPを形成するのに容易に適合可能なプラットフォームである。本節では、開示されるVLPと共に用いるのに好ましい抗原について記載する。
【0055】
(抗原と脂質ラフト会合ポリペプチドとの連結)
天然では脂質ラフトと会合しない抗原を含有するVLPを形成する手段として、RSウイルスFポリペプチドおよび/または別の脂質ラフト会合ポリペプチドと、該抗原との連結を形成することができる。単一の抗原または複数の抗原に脂質ラフト会合ポリペプチドを連結することにより、VLPの免疫原性を増大させるか、各種の病原体に免疫原性を付与するか、または、特定の病原体の各種の株に免疫原性を付与することができる。
【0056】
抗原と脂質ラフト会合ポリペプチドとの間の連結は、該抗原を結果としてVLP内へと組み込むのに十分な任意の種類の連結であり得る。結合は、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であり得る。好ましい実施形態では、結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合などの共有結合である。
【0057】
抗原は、脂質ラフト会合ポリペプチドに対する既存の結合を伴なって組換えにより作製することもでき、単離物質として作製し、次いで、後に、脂質ラフト会合ポリペプチドに連結することもできる。
【0058】
(抗原の種類)
本明細書で用いられる抗原は、免疫反応を誘発することが可能であり、また、天然では脂質ラフトと会合しない任意の物質であり得る。抗原には、タンパク質、ポリペプチド(活性タンパク質、また、タンパク質内における個々のポリペプチドエピトープを含めた)、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。天然において、抗原が脂質ラフトと直接的または間接的に会合しない場合、脂質ラフト会合ポリペプチドに連結しない限り、それをVLP内へと組み込むことは期待されない。抗原は、疾患または障害に関与する任意の抗原、例えば、微生物抗原(例えば、ウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、アレルゲンなどであり得る。
【0059】
(抗原の供給源)
本明細書で記載される抗原は、化学的または酵素的に合成することもでき、組換えにより作製することもでき、天然の供給源から単離することもでき、前出の組合せでもあり得る。抗原は、精製することもでき、部分精製することもでき、粗抽出物でもあり得る。
【0060】
ポリペプチド抗原は、液体クロマトグラフィー(例えば、高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ除外クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含めた)、アフィニティークロマトグラフィー、または他の精製法が含まれるがこれらに限定されない、当技術分野で公知の標準的なタンパク質精製法を用いて、天然の供給源から単離することができる。多くの実施形態では、抗原が、例えば、約50%〜約75%純粋、約75%〜約85%純粋、約85%〜約90%純粋、約90%〜約95%純粋、約95%〜約98%純粋、約98%〜約99%純粋、または99%超純粋の精製抗原である。
【0061】
固相ペプチド合成法を用いることができるが、このような技法は、当業者に公知である。Jones、「The Chemical Synthesis of Peptides」(オックスフォード、Clarendon Press社)(1994年)を参照されたい。一般に、このような方法では、活性化させた単量体のユニットを、伸長するペプチド鎖を結合させた固相へと連鎖的に付加することにより、ペプチドを作製する。
【0062】
ポリペプチドを作製するには、十分に確立された組換えDNA法を、脂質ラフト会合ポリペプチドと同じベクター内において用いることもでき、この場合、例えば、ポリペプチドをコードするヌクレオチド配列を含む発現構築物は、適切な宿主細胞(例えば、インビトロの細胞培養物における単細胞実体として増殖させる真核宿主細胞、例えば、酵母細胞、昆虫細胞、哺乳動物細胞など)または、原核細胞(例えば、インビトロの細胞培養物中)中に導入され、これにより遺伝的に改変された宿主細胞を作製し、適切な培養条件下において、該遺伝的に改変された宿主細胞によりタンパク質を作製する。
【0063】
(ウイルス抗原)
適切なウイルス抗原には、以下の群:Retroviridae科(例えば、HIV−1などのヒト免疫不全ウイルス(また、HTLV−III、LAV、もしくはHTLV−III/LAV、またはHIV−IIIとも称する));また、HIV−LPなど、他の単離物; Picomaviridae科(例えば、ポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス); Calciviridae科(例えば、ノーウォークウイルスおよび関連ウイルスを含めた、胃腸炎を引き起こすウイルス株); Togaviridae科(例えば、ウマ脳炎ウイルス、風疹ウイルス); Flaviridae科(例えば、デング熱ウイルス、脳炎ウイルス、黄熱病ウイルス); Coronoviridae科(例えば、コロナウイルス); Rhabdoviradae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Coronaviridae科(例えば、コロナウイルス); Rhabdoviridae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Filoviridae科(例えば、エボラウイルス); Paramyxoviridae科(例えば、パラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、RSウイルス); Orthomyxoviridae科(例えば、インフルエンザウイルス); Bungaviridae科(例えば、ハンタンウイルス、ブンガウイルス、フレボウイルス、およびナイロウイルス); Arena viridae科(出血熱ウイルス); Reoviridae科(例えば、レオウイルス、オルビウイルス、およびロタウイルス); Bimaviridae科; Hepadnaviridae科(B型肝炎ウイルス); Parvovirida科(パルボウイルス); Papovavirida(パピローマウイルス、ポリオーマウイルス); Adenoviridae科(大半のアデノウイルス); Herpesviridae科(1型および2型の単純ヘルペスウイルス(HSV)、水痘帯状疱疹ウイルス、サイトメガロウイルス(CMV)、ヘルペスウイルス); Poxyiridae科(痘瘡ウイルス、ワクシニアウイルス、ポックスウイルス);およびIridoviridae科(例えば、アフリカブタコレラウイルス);ならびに未分類のウイルス(例えば、海綿状脳症の病因因子、デルタ肝炎の因子(B型肝炎ウイルスの欠損サテライトウイルスと思われる)、非A型肝炎、非B型肝炎の因子(クラス1=内部感染による;クラス2=非経口感染による(すなわち、C型肝炎));およびアストロウイルス)のうちの1または複数のウイルスと関連する(例えば、これらにより合成される)抗原が含まれる。
【0064】
(ノロウイルス(Norvirus)抗原)
本明細書で開示されるVLPには、ノロウイルス科に由来する各種の抗原が含まれ得ることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、Caliciviridaeウイルス科内の4つの属のうちの1つを占める。ノロウイルス属内には、遺伝子群Iおよび遺伝子群IIと名付けられた、2つの主要な遺伝子群が存在する。遺伝子群Iのノロウイルス株には、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスが含まれる。遺伝子群IIのノロウイルス株には、ヒューストンウイルス、ハワイウイルス、ローズデールウイルス、グリムズビーウイルス、メキシコウイルス、およびスノーマウンテンエージェントが含まれる(Parker, T.D.ら、J. Virol.(2005年)、79巻(12号):7402〜9頁; Hale, A.D.ら、J Clin. Micro.(2000年)、38巻(4号):1656〜1660頁)。ノーウォークウイルス(NV)は、世界中における、大半の急性ウイルス性胃腸炎の流行的発生の一因となる、ヒトカリチウイルス群の原型株である。ノーウォークウイルスのカプシドタンパク質は、2つのドメイン:殻ドメイン(S)および突出部ドメイン(P)を有する。Pドメイン(アミノ酸226〜530;ノーウォーク株の番号付け)は、2つのサブドメインであるP1およびP2に分けられる。P2ドメインは、P1ドメイン内における127アミノ酸の挿入(アミノ酸279〜405)であり、フォールディングされた単量体の最遠位面に位置する。P2ドメインは、ノロウイルス株のうちで最も保存の程度が低いVP1領域であり、P2内の超可変領域は、受容体の結合および免疫反応性において重要な役割を果たすと考えられている。Pドメインが外部に位置することを踏まえるなら、それは、本明細書で開示されるVLPワクチンのための抗原として用いるのに好ましい抗原、またはポリペプチドエピトープの供給源である。P2ドメインは、遺伝子群Iまたは遺伝子群IIのノロウイルス株に好ましい抗原である。広範にわたるノロウイルス株を通じて保存されるP2ドメインのうちのある領域内に存在するエピトープとして近年になって同定されたmAb 61.21エピトープ、ならびにmAb 54.6エピトープ(Lochridge, V.P.ら、J Gen. Virol.(2005年)、86巻:2799〜2806頁)がさらにより好ましい。
【0065】
(インフルエンザ抗原)
本明細書で開示されるVLPは、赤血球凝集素、ノイラミニダーゼ、またはさらなるインフルエンザ抗原を非制限的に含む、インフルエンザに由来する各種の抗原を包含し得る。さらなる好ましいインフルエンザ抗原は、M2ポリペプチドである。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後において、RNAセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の小型のクラスIII内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであり、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染中または従来のワクチン接種中の免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、インビボにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回の繰り返しコピーを該コアタンパク質のN末端へと融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0066】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされ、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的に結合体化することもでき、マウスおよび他の動物におけるワクチンとして防御的である(92、93)。
【0067】
M2タンパク質は、高度に保存的ではあるが、配列の相違を全く示さないわけではない。一般的な株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的汎流行ワクチンが、ヒトH1単離物およびH3単離物中において現在広まっているM2配列ではなく、病原性の鳥類株に固有のM2配列を反映する必要があることを裏付ける。
【0068】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、RSウイルスFポリペプチドもしくは他の脂質ラフト会合ポリペプチドに連結することにより、VLPワクチン内へと組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、およびNS2が含まれる。これら後者の抗原は、一般に、中和抗体反応の標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【0069】
(他の病原性抗原)
適切な細菌性抗原には、例えば、病原性Pasteurella属種、Staphylococci属種、およびStreptococcus属種などの病原性グラム陽性菌;また、Neisseria属、Escherichia属、Bordetella属、Campylobacter属、Legionella属、Pseudomonas属、Shigella属、Vibrio属、Yersinia属、Salmonella属、Haemophilus属、Brucella属、Francisella属、およびBacterioides属のグラム陰性菌などのグラム陰性菌を含めた、各種の病原性細菌のうちのいずれかと関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。例えば、Schaechter, M、H. Medoff、D. Schlesinger、「Mechanisms of Microbial Disease」、ボルチモア、Williams and Wilkins社(1989年)を参照されたい。
【0070】
感染性の病原性真菌と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Cryptococcus neoformans、Histoplasma capsulatum、Coccidioides immitis、Blastomyces dermatitidis、およびCandida albicans、Candida glabrata、Aspergillus fumigata、Aspergillus flavus、およびSporothrix schenckiiが含まれるがこれらに限定されない感染性真菌と関連する抗原が含まれる。
【0071】
病原性原虫、蠕虫、および他の真核微生物病原体と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvum等が含まれるがこれらに限定されない原虫、蠕虫、および他の真核微生物病原体と関連する抗原が含まれる。
【0072】
適切な抗原には、Helicobacter pyloris、Borelia burgdorferi、Legionella pneumophila、Mycobacteria属種(例えば、M. tuberculosis、M. avium、M. intracellulare、M. kansaii、M. gordonae)、Staphylococcus aureus、Neisseria gonorrhoeae、Neisseria meningitidis、Listeria monocytogenes、Chlamydia trachomatis、Streptococcus pyogenes(Streptococcus属のA群)、Streptococcus agalactiae(Streptococcus属のB群)、Streptococcus属(viridans群)、Streptococcus faecalis、Streptococcus bovis、Streptococcus属(嫌気性の種)、Streptococcus pneumoniae、病原性のCampylobacter属種、Enterococcus属種、Haemophilus influenzae、Bacillus anthracis、Corynebacterium diphtheriae、Corynebacterium(corynebacterium)属種、Erysipelothrix rhusiopathiae、Clostridium perfringens、Clostridium tetani、Enterobacter aerogenes、Klebsiella pneumoniae、Pasturella multocida、Bacteroides属種、Fusobacterium nucleatum、Streptobacillus moniliformis、Treponema pallidium、Treponema pertenue、Leptospira属、Rickettsia属、およびActinomyces israeliなどの病原性微生物と関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。病原性E. coli株の非限定的な例は、ATCC第31618号、同第23505号、同第43886号、同第43892号、同第35401号、同第43896号、同第33985号、同第31619号、および同第31617号である。
【0073】
細胞内病原体と関連する各種のポリペプチドまたは他の抗原のうちのいずれかを、VLP内に組み込むことができる。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープとは、その断片が、MHCクラスI分子と併せて、CD8+リンパ球の表面上におけるT細胞抗原受容体により認識される、例えば、これにより結合されるように、感染細胞の表面上に提示される、細胞内病原体と関連する(例えば、これによりコードされる)任意のポリペプチドである。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープは当技術分野において公知であり、これらには、ヒト免疫不全ウイルス、例えば、HIV gp120またはその抗原性断片と関連する抗原;サイトメガロウイルス抗原; Mycobacterium属抗原(例えば、Mycobacterium avium、Mycobacterium tuberculosisなど);Pneumocystic carinii(PCP)抗原;41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど、Plasmodium falciparumまたは他の任意のマラリア種と関連する抗原が含まれるがこれらに限定されない、マラリア抗原;真菌抗原;酵母抗原(例えば、Candida属種抗原); Toxoplasma gondii、Toxoplasma encephalitis、または他のToxoplasma属種と関連する抗原が含まれるがこれらに限定されないトキソプラズマ抗原;エプスタイン−バーウイルス(EBV)による抗原; Plasmodium属抗原(例えば、gp190/MSP1など)などが含まれるが、これらに限定されない。
【0074】
好ましいVLPワクチンは、Bacillus anthracisに対するものであり得る。Bacillus anthracisは、好気性または通性嫌気性でグラム陽性の非運動性桿菌であり、幅1.0μm、長さ3.0〜5.0μmである。B. anthracisは、悪条件下でも耐性の高い内性胞子を形成し、これは、感染した動物がかつて死亡した場所の土壌で見出すことができる。本明細書で開示されるVLPワクチン内で用いるのに好ましい抗原は、哺乳動物細胞上の受容体に結合し、B. anthracisが疾患を引き起こす能力にとって極めて重要な83kDaのタンパク質である防御抗原(PA)である。より好ましい抗原は、被験体においてBacillus anthracisに対する防御的免疫性を誘導するのに用い得る、Bacillus anthracis PAのC末端における140アミノ酸の断片である。炭疽菌に対するVLPワクチン内において用いられる他の例示的な抗原は、炭疽菌胞子に由来する抗原(例えば、BclA)、該細菌の栄養期に由来する抗原(例えば、細胞壁抗原、被膜抗原(例えば、ポリ−ガンマ−D−グルタミン酸、またはPGA)、分泌抗原(例えば、防御抗原、致死性因子、または浮腫因子などの外毒素))である。VLPワクチンで用いるのに好ましい別の抗原は、B. anthracisに固有のアントロースを含有する四糖である(Daubenspeck J.M.ら、J. Biol. Chem.(2004年)、279巻:30945頁)。四糖は、脂質ラフト会合ポリペプチドに連結することができ、これにより、VLPワクチンとの抗原の会合が可能となる。
【0075】
(腫瘍関連抗原)
公知の各種腫瘍特異抗原または腫瘍関連抗原(TAA)のうちのいずれかを、VLPに組み込むことができる。全TAAを用い得るが、その必要はない。代わりに、TAAの一部、例えば、エピトープを用いることができる。VLP内で用い得る腫瘍関連抗原(またはそれらのエピトープを含有する断片)には、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量黒色腫関連抗原であるMAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原であるOV−TL3およびMOV18、TUAN、アルファ−フェトプロテイン(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原であるG250、EGP−40(EpCAMとしても知られる)、S100(悪性黒色腫関連抗原)、p53、およびp21rasが含まれるがこれらに限定されない。前出のうちのいずれかを含めた、任意のTAA(またはそれらのエピトープ)の合成類似体を用いることができる。さらに、1または複数のTAA(またはそれらのエピトープ)の組合せを、組成物中に組み込むことができる。
【0076】
(アレルゲン)
一態様では、VLPワクチンの一部である抗原が、各種のアレルゲンのうちのいずれかであり得る。アレルゲンベースのワクチンを用い、被験体におけるアレルゲンに対する忍容性を誘導することができる。チロシンとの共沈殿(co−precipitation)を伴うアレルゲンワクチンの例は、米国特許第3,792,159号、同第4,070,455号、および同第6,440,426号において見出すことができる。
【0077】
各種のアレルゲンのうちのいずれかをVLP内に組み込むことができる。アレルゲンには、環境空中アレルゲン;ブタクサ/花粉症などの植物の花粉;雑草花粉アレルゲン;芝草花粉アレルゲン;ジョンソングラス;樹木花粉アレルゲン;ライグラス;イエダニアレルゲン(例えば、Der p I、Der f Iなど)などのクモ形類動物アレルゲン;コナダニ(storage mite)アレルゲン;スギ花粉/花粉症;カビ胞子アレルゲン;動物アレルゲン(例えば、イヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどによるアレルゲン);食物アレルゲン(例えば、甲殻類;ラッカセイなどの堅果;柑橘類によるアレルゲン);昆虫アレルゲン;毒(膜翅目、スズメバチ(yellow jacket)、ミツバチ、カリバチ、スズメバチ(hornet)、ヒアリ);ゴキブリ、ノミ、蚊などによる他の環境昆虫アレルゲン;連鎖球菌抗原などの細菌アレルゲン; Ascaris属抗原などの寄生虫アレルゲン;ウイルス抗原;真菌胞子;薬物アレルゲン;抗生剤;ペニシリンおよび関連化合物;他の抗生剤;ホルモン(インスリン)、酵素(ストレプトキナーゼ)などの全タンパク質;不完全抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝物;ハプテンとして作用し、アレルゲンとして機能することが可能な、産業用化学物質および代謝物(例えば、酸無水物(トリメリト酸無水物など)およびイソシアン酸塩(ジイソシアン酸トルエンなど));小麦粉などの職業アレルゲン(例えば、パン屋喘息(Baker’s asthma)を引き起こすアレルゲン)、トウゴマ、コーヒー豆、また、上記で記載した産業用化学物質;ノミアレルゲン;ヒト以外の動物におけるヒトタンパク質が含まれるが、これらに限定されない。
【0078】
アレルゲンには、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。
【0079】
特定の天然の動物アレルゲンおよび植物アレルゲンの例には、以下の属:Canine属(Canis familiaris); Dermatophagoides属(例えば、Dermatophagoides farinae); Felis属(Felis domesticus); Ambrosia属(Ambrosia artemiisfolia); Lolium属(例えば、Lolium perenneまたはLolium multiflorum); Cryptomeria属(Cryptomeria japonica); Altemaria属(Altemaria altemata); Alder属; Alnus属(Alnus gultinoas); Betula属(Betula verrucosa); Quercus属(Quercus alba); Olea属(Olea europa); Artemisia属(Artemisia vulgaris); Plantago属(例えば、Plantago lanceolata); Parietaria属(例えば、Parietaria officinalisまたはParietaria judaica); Blattella属(例えば、Blattella germanica); Apis属(例えば、Apis multiflorum); Cupressus属(例えば、Cupressus sempervirens、Cupressus arizonica、およびCupressus macrocarpa); Juniperus属(例えば、Juniperus sabinoides、Juniperus virginiana、Juniperus communis、およびJuniperus ashei); Thuya属(例えば、Thuya orientalis); Chamaecyparis属(例えば、Chamaecyparis obtusa); Periplaneta属(例えば、Periplaneta americana); Agropyron属(例えば、Agropyron repens); Secale属(例えば、Secale cereale); Triticum属(例えば、Triticum aestivum); Dactylis属(例えば、Dactylis glomerata); Festuca属(例えば、Festuca elatior); Poa属(例えば、PoapratensisまたはPoa compressa); Avena属(例えば、Avena sativa); Holcus属(例えば、Holcus lanatus); Anthoxanthum属(例えば、Anthoxanthum odoratum); Arrhenatherum属(例えば、Arrhenatherun elatius); Agrostis属(例えば、Agrostis alba); Phleum属(例えば、Phleum pratense); Phalaris属(例えば、Phalaris arundinacea); Paspalum属(例えば、Paspalum notatum); Sorghum属(例えば、Sorghum halepensis); ならびにBromus属(例えば、Bromus inermis)に特異的なタンパク質が含まれるが、これらに限定されない。
【0080】
(哺乳動物細胞からRSV F VLPを作製するのに好ましい方法)
RSウイルスFポリペプチドのVLPは、当業者に適用可能な任意の方法により作製することができる。RSウイルスFポリペプチドのVLPは、VLPを形成する一因となるRSウイルスFポリペプチドを包含するが、任意のエンベロープウイルスコア形成ポリペプチドは除外する。加えて、RSウイルスFポリペプチドのVLPは、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することにより、(VLPを形成する一因となる1または複数のポリペプチドの一部として、天然において、または人工的に存在する抗原以外のさらなる)抗原をもたらすことができる。好ましい実施形態では、ポリペプチドを、任意の適用可能なタンパク質発現系、好ましくは、細胞膜内に脂質ラフトドメインを包含する哺乳動物細胞ベースの系において共発現させることができる。
【0081】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、当技術分野で周知の技法を用いる組換えDNA法により、該ポリペプチドを作製するためのベクターを作製することができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現することによりタンパク質を調製する方法が記載される。当業者に周知の方法を用いて、VLPポリペプチドをコードする配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。例えば、これらの方法には、インビトロにおける組換えDNA法、合成法、また、インビボにおける遺伝子組換えが含まれる。したがって、本発明は、すべてが1または複数のプロモーターに作動可能に連結した、RSウイルスFポリペプチドと、必要に応じて、1または複数のさらなる脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0082】
本明細書で記載されるVLPへとアセンブルされる配列を発現させるのに用い得るベクターの非限定的な例には、ウイルスベースのベクター(例えば、レトロウイルス、アデノウイルス、アデノ関連ウイルス、レンチウイルス)、プラスミドベクター、非ウイルス性ベクター、哺乳動物ベクター、哺乳動物による人工的染色体(例えば、リポソーム、粒子状キャリアなど)、およびこれらの組合せが含まれる。
【0083】
発現ベクター(複数可)は、コード配列と、適切な宿主内において、該コード領域を発現させる発現制御エレメントとを含有することが典型的である。制御エレメントには、一般に、プロモーター、エンハンサー、エクソン、イントロン、スプライシング部位、翻訳開始コドン、ならびに翻訳終結配列および転写終結配列、ならびにベクター内に挿入を導入するための挿入部位が含まれる。翻訳制御エレメントは、M. Kozak(例えば、Kozak, M.、Mamm. Genome、7巻(8号):563〜574頁、1996年; Kozak, M.、Biochimie、76巻(9号):815〜821頁、1994年; Kozak, M.、J Cell Biol、108巻(2号):229〜241頁、1989年; Kozak, M.およびShatkin, A. J.、Methods Enzymol、60巻:360〜375頁、1979年)により総説されている。
【0084】
例えば、哺乳動物細胞による発現に用いられる典型的なプロモーターには、とりわけ、SV40初期プロモーター、CMV前初期プロモーターなどのCMVプロモーター(CMVプロモーターは、イントロンAを包含することができる)、RSV、HIV−LTR、マウス乳癌細胞ウイルスLTRプロモーター(MMLV−LTR)、FIV−LTR、アデノウイルス主要後期プロモーター(Ad MLP)、および単純ヘルペスウイルスプロモーターが含まれる。マウスメタロチオネイン遺伝子に由来するプロモーターなど、他の非ウイルス性プロモーターもまた、哺乳動物による発現に用いられる。翻訳終止コドンに対して3’側に位置する、転写終結配列およびポリアデニル化配列もまた存在することが典型的である。コード配列に対して5’側に位置する、翻訳の開始を最適化するための配列もまた存在することが好ましい。転写ターミネーター/ポリアデニル化シグナルの例には、Sambrookら、前出において記載されている、SV40に由来するシグナル、ならびにウシ成長ホルモンターミネーター配列が含まれる。スプライスドナー部位およびスプライスアクセプター部位を含有するイントロンもまた、本明細書で記載される構築物内にデザインすることができる(Chapmanら、Nuc. Acids Res.(1991年)、19巻:3979〜3986頁)。
【0085】
本明細書では、エンハンサーエレメントもまた、例えば、哺乳動物宿主細胞内における構築物の発現レベルを上昇させるのに用いることができる。例には、Dijkemaら、EMBO J.(1985年)、4巻:761頁において記載されている、SV40初期遺伝子エンハンサー;Gormanら、Proc. Natl. Acad. Sci. USA(1982年b)、79巻:6777頁において記載されている、ラウス肉腫ウイルスの長い末端反復(LTR)に由来するエンハンサー/プロモーター;また、CMVイントロンA配列内に包含されるエレメント(Chapmanら、Nuc. Acids Res.(1991年)、19巻:3979〜3986頁)など、Boshartら、Cell(1985年)、41巻:521頁において記載されている、ヒトCMVに由来するエレメントが含まれる。
【0086】
ベクターが、本明細書で記載される1つまたは複数の配列を含有し得ることは明らかである。例えば、単一のベクターが、VLP内に見出されるすべてのタンパク質をコードする配列を保有し得る。代替的に、複数のベクター(例えば、各々が、単一のポリペプチドコード配列をコードする複数の構築物、または各々が1つもしくは複数のポリペプチドコード配列をコードする複数の構築物)を用いることもできる。単一のベクターが、複数のポリペプチドコード配列を含む実施形態では、該配列を、同じベクター内における、同じであるかまたは異なる転写制御エレメント(例えば、プロモーター)に作動可能に連結することができる。
【0087】
加えて、免疫調節分子(例えば、以下で記載されるアジュバント)、例えば、免疫調節オリゴヌクレオチド(例えば、CpG)、サイトカイン、弱毒化した細菌性毒素などを含む配列、および/またはこれらをコードする配列を非制限的に含む、RSV以外のタンパク質をコードする1つまたは複数の配列も、VLP内で発現させ、また、これに組み込むことができる。
【0088】
従来の技法により、発現ベクターを宿主細胞へと移入することができ、次いで、従来の技法により、トランスフェクトされた細胞を培養して、VLPポリペプチド(複数可)を作製することができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1つまたは複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを作製するのに好ましい実施形態では、以下で詳述する通り、RSウイルスFポリペプチドと、場合によって、抗原またはアジュバントに連結した脂質ラフト会合ポリペプチドとをコードするベクターを、宿主細胞内で(共)発現させて、VLPを作製することができる。
【0089】
トランスフェクション、継代細胞株の確立(当業者に公知の標準的なプロトコールを用いる)、および/または目的のRSV遺伝子を保有するDNA分子での感染後における真核細胞、より好ましくは哺乳動物細胞内において、VLPを作製することが好ましい。VLPを形成するのに必要とされるタンパク質の発現レベルは、選択された遺伝子の転写を駆動する真核細胞プロモーターまたはウイルスプロモーターの配列を最適化することによって最大化する。VLPは、RSウイルスFポリペプチドにより駆動されて培地内へと放出され、そこからVLPを精製し、その後、ワクチンとして処方することができる。VLPは、感染性ワクチンではなく、したがって、ワクチンの不活化は不要である。
【0090】
本明細書で記載される配列から発現するRSウイルスFポリペプチドが、その表面上に抗原タンパク質を提示するVLPへと自己アセンブルする能力は、所望の配列を共導入(co−introduction)することにより、これらのVLPを、任意の宿主細胞内において作製することを可能にする。配列(複数可)(例えば、1つまたは複数の発現ベクター内における)は、多様な組合せで安定的に、および/または一過的に宿主細胞内へと組み込むことができる。
【0091】
適切な宿主細胞には、細菌細胞、哺乳動物細胞、バキュロウイルス/昆虫細胞、酵母細胞、植物細胞、およびXenopus属細胞が含まれるが、これらに限定されない。
【0092】
当技術分野では、例えば、多数の哺乳動物細胞株が公知であり、これらには、BHK、VERO、HT1080、MRC−5、WI 38、MDCK、MDBK、293、293T、RD、COS−7、CHO、Jurkat、HUT、SUPT、C8166、MOLT4/クローン8、MT−2、MT−4、H9、PM1、CEM、黒色腫細胞(例えば、SB20細胞)、LtK、HeLa、WI−38、L2、CMT−93、およびCEMX174(このような細胞株は、例えば、A.T.C.C.から入手可能である)などであるがこれらに限定されない、American Type Culture Collection(A.T.C.C.)から入手可能な初代細胞ならびに不死化細胞株が含まれる。
【0093】
同様に、E.coli、Bacillus subtilis、およびStreptococcus属種などの細菌宿主も、本発現構築物と共に用いられる。
【0094】
本開示において有用な酵母宿主には、とりわけ、Saccharomyces cerevisiae、Candida albicans、Candida maltosa、Hansenula polymorpha、Kluyveromyces fragilis、Kluyveromyces lactis、Pichia guillerimondii、Pichia pastoris、Schizosaccharomyces pombe、およびYarrowia lipolyticaが含まれる。真菌宿主には、例えば、Aspergillus属が含まれる。
【0095】
バキュロウイルス発現ベクターと共に用いられる昆虫細胞には、とりわけ、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusia niが含まれる。LathamおよびGalarza(2001年)、J. Virol.、75巻(13号):6154〜6165頁; Galarzaら(2005年)、Viral. Immunol.、18巻(1号):244〜51頁;ならびに米国特許公開第200550186621号および同第20060263804号を参照されたい。
【0096】
本明細書で示される開示を踏まえれば、VLPのRSウイルスFポリペプチドをコードする1つまたは複数の配列(発現ベクター)を安定的に組み込むことにより、上記で記載した配列のうちの1つまたは複数を発現する細胞株を容易に作製することができる。安定的に組み込まれたRSウイルスFポリペプチド配列(複数可)の発現を制御するプロモーターは、構成的プロモーターの場合もあり、誘導的プロモーターの場合もある。
【0097】
RSウイルスFポリペプチドのVLPを作製する細胞株が由来する親細胞株は、例えば、哺乳動物細胞株、昆虫細胞株、酵母細胞株、細菌細胞株を含めた、上記で記載した任意の細胞から選択することができる。好ましい実施形態では、該細胞株は、哺乳動物細胞株(例えば、293、RD、COS−7、CHO、BHK、VERO、MRC−5、HT1080、および黒色腫細胞)である。哺乳動物細胞を用いてRSV VLPを作製することにより、(i)VLPの形成;(ii)適正なミリストイル化(myristylation)、グリコシル化、および出芽;(iii)非哺乳動物細胞夾雑物の不在;(iv)精製の容易さがもたらされる。
【0098】
細胞株を創出することに加えて、宿主細胞内において、RSVコード配列を一過的に発現させることもできる。適切な組換え発現宿主細胞系には、当技術分野で周知の、細菌発現系、哺乳動物発現系、バキュロウイルス/昆虫発現系、ワクシニア発現系、セムリキ森林熱ウイルス(SFV)発現系、アルファウイルス(シンドビスウイルス、ベネズエラウマ脳炎(VEE)ウイルスなど)発現系、哺乳動物発現系、酵母発現系、およびXenopus属発現系が含まれるが、これらに限定されない。特に好ましい発現系は、哺乳動物細胞株、ワクシニア、シンドビス、昆虫、および酵母系である。
【0099】
例えば、以下:バキュロウイルス発現(Reilly, P. R.ら、BACULOVIRUS EXPRESSION VECTORS: A LABORATORY MANUAL(1992年); Beamesら、Biotechniques、11巻:378頁(1991年);Pharmingen社製;カリフォルニア州、パロアルト、Clontech社製)、ワクシニア発現系(Earl, P. L.ら、「Expression of proteins in mammalian cells using vaccinia」、Current Protocols in Molecular Biology(F. M. Ausubelら編)、Greene Publishing Associates & Wiley Interscience、New York(1991年); 1992年8月4日に発行された、Moss, B.ら、米国特許第5,135,855号)、細菌内における発現(Ausubel, F. M.ら、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY、John Wiley and Sons, Inc.、Media Pa.;Clontech社製)、酵母内における発現(1998年3月17日に発行され、参照により本明細書に組み込まれる、Rosenberg, SおよびTekamp−Olson, P.、米国特許第RE35,749号;1997年5月13日に発行され、参照により本明細書に組み込まれる、Shuster, J. R.、米国特許第5,629,203号; Gellissen, G. ら、Antonie Van Leeuwenhoek、62巻(1〜2号):79〜93頁(1992年); Romanos, M. A. ら、Yeast、8巻(6号):423〜488頁(1992年); Goeddel, D. V.、Methods in Enzymology、185巻(1990年); Guthrie, C.およびG. R. Fink、Methods in Enzymology、194巻(1991年))、動物細胞内における発現(Clontech社製;ニューヨーク州、グラウンドアイランド、Gibco−BRL社製;例えば、チャイニーズハムスター卵巣(CHO)細胞株(Haynes, J. ら、Nuc. Acid. Res.、11巻:687〜706頁(1983年); Lau, Y. F. ら、Mol. Cell. Biol.、4巻:1469〜1475頁(1984年); Kaufman, R. J.、「Selection and coamplification of heterologous genes in mammalian cells」、Methods in Enzymology、185巻、537〜566頁、カリフォルニア州、Academic Press, Inc.、San Diego Calif.(1991年)))、ならびに植物細胞における発現(植物クローニングベクター;カリフォルニア州、パロアルト、Clontech Laboratories社製;およびニュージャージー州、ピスケータウェイ、Pharmacia LKB Biotechnology社製; Hood, E.ら、J. Bacteriol.、168巻:1291〜1301頁(1986年); Nagel, R.ら、FEMS Microbiol. Lett.、67巻:325頁(1990年); Anら、「Binary Vectors」、およびその他、Plant Molecular Biology Manual、A3巻:1〜19頁(1988年); Miki, B. L. A.ら、249〜265頁、およびその他、Plant DNA Infectious Agents(Hohn, T.ら編)、Springer−Verlag、Wien、Austria(1987年);Plant Molecular Biology: Essential Techniques、P. G. JonesおよびJ. M. Sutton、New York、J. Wiley、1997年; Miglani, Gurbachan、Dictionary of Plant Genetics and Molecular Biology、New York、Food Products Press、1998年; Henry, R. J.、Practical Applications of Plant Molecular Biology、New York、Chapman & Hall、1997年)を含めた、多くの適切な発現系が市販されている。
【0100】
VLPを形成するのに必要とされるタンパク質をコードする改変遺伝子を含有する発現ベクターを宿主細胞(複数可)内へと導入し、その後、必要なレベルで発現させると、VLPがアセンブルし、次いで、細胞表面から培地内へと放出される。
【0101】
選択される発現系および宿主に応じて、粒子形成ポリペプチドが発現し、VLPが形成され得る条件下において、発現ベクターにより形質転換された宿主細胞を増殖させることにより、VLPを作製する。適切な増殖条件の選択は、当業者の能力の範囲内にある。
【0102】
次いで、例えば、密度勾配遠心分離、例えば、スクロース勾配、PEG沈殿、ペレット化など(例えば、Kimbauerら、J. Virol.(1993年)、67巻:6929〜6936頁を参照されたい)、ならびに、例えば、イオン交換クロマトグラフィーおよびゲル濾過クロマトグラフィーを含めた標準的な精製法により、その完全性を保存する方法を用いて、粒子を単離する(または実質的に精製する)。
【0103】
(RSウイルスFポリペプチドのVLPのVLP調製物における感染因子を不活化するのに好ましい方法)
電子放射は、RSウイルスFポリペプチドのVLPの免疫原性を実質的に低下させることなく、感染因子を不活化することが可能なので、不活化の好ましい方法は、電子放射を介する方法である。電磁放射で好ましい3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0104】
(光反応性化合物を伴うUV照射)
電磁放射による例示的な不活化法は、光反応性化合物、好ましくは、感染因子中のポリヌクレオチドと反応する光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0105】
好ましい光反応性化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、マイトマイシン、monostral fast blue、norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。好ましい種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖型であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、角度型(angular)であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖型フロクマリンを置換することによりソラーレン誘導体を作製し得る一方、3、4、5、6、4’、または5位において角度型フロクマリンを置換することによりイソソラーレン誘導体を作製し得る。ソラーレンは、二本鎖核酸の塩基対間にインターカレートすることができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.、54巻:1151頁(1985年); Hearstら、Quart. Rev. Biophys.、17巻:1頁(1984頁)を参照されたい。
【0106】
好ましいUV(または場合によって、可視光)放射の波長は、光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が最も有効であり、330〜360nmが最大の有効性を示す。
【0107】
(UV照射単独)
光反応性化合物の存在下におけるUV照射に加え、感染因子は、UV照射単独でも不活化することができる。好ましい実施形態では、放射は、波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの最大吸光度ピーク)であるUVC放射である。UVC放射は、エンベロープを形成する脂質二重層など、本明細書で開示されるRSウイルスFポリペプチドのVLPの成分に対して、安定性および免疫原性の両面で有害性が低い一方で、感染因子を不活化するのに十分なエネルギーを保持するので好ましい。しかし、例えば、UVAおよびUVBなど、他の種類のUV放射もまた用いることができる。
【0108】
(ガンマ照射)
本明細書で開示される方法を実施して組成物を作製するには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。ガンマ照射は、感染因子のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染因子を不活化することもでき、または、該ポリヌクレオチドを攻撃するヒドロキシルラジカルを生成することにより、感染因子を不活化することもできる。
【0109】
(RSウイルスFポリペプチドのVLPを用いる好ましい方法)
(処方物)
本明細書で記載されるRSウイルスFポリペプチドのVLPの好ましい使用は、ワクチン調製物としての使用である。典型的に、このようなワクチンは、液体溶液または懸濁物の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として作製することもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、保湿剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントなどの補助物質を含有し得る。
【0110】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる処方物には坐剤が含まれ、場合によっては、経口処方物、鼻腔内処方物、口腔内処方物、舌下処方物、腹腔内処方物、膣内処方物、経肛門処方物、および頭蓋内処方物も含まれる。坐剤の場合、従来の結合剤およびキャリアには、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが含まれる場合があり、このような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で有効成分を含有する混合物から形成することができる。特定の実施形態では、脂肪酸グリセリドの混合物またはココアバターなどの低温溶融ワックスをまず溶融させ、本明細書で記載されるRSウイルスFポリペプチドのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、好都合なサイズの型に流し込み、冷却して固形化させる。
【0111】
鼻腔内送達に適する処方物には、液体および乾燥粉末が含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、スクロース、トレハロース、およびキトサンなど、通常用いられる賦形剤を包含する。液体処方物または粉末処方物において、キトサンなどの粘膜付着剤を用いて、粘膜絨毛による、鼻腔内投与された処方物のクリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体処方物では安定化剤として用いることができ、また、乾燥粉末処方物では安定化剤およびバルキング剤として用いることができる。加えて、モノホスホリルリピドA(MPL)などのアジュバントを、液体処方物および乾燥粉末処方物の両方において、免疫刺激性アジュバントとして用いることもできる。
【0112】
経口送達に適する処方物には、液体、固体、半固体、ゲル、錠剤、カプセル、ロゼンジなどが含まれる。経口送達に適する処方物には、錠剤、ロゼンジ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のRSウイルスFポリペプチドのVLPワクチン組成物は、溶液、懸濁物、丸薬、持続放出処方物、または粉末の形態を取る場合があり、10〜95%、好ましくは25〜70%の有効成分を含有する場合がある。経口処方物の場合、コレラ毒素が興味深い処方物パートナー(また、可能な結合パートナー)である。
【0113】
膣内投与用に処方する場合、RSウイルスFポリペプチドのVLPワクチンは、ペッサリー、タンポン、クリーム、ジェル、ペースト、フォーム、またはスプレーの形態であり得る。前出の処方物のうちのいずれもが、RSウイルスFポリペプチドのVLPに加えて、キャリアなど、当技術分野で適切であることが公知の薬剤を含有し得る。
【0114】
一部の実施形態では、RSウイルスFポリペプチドのVLPワクチンを、全身送達用に処方する場合もあり、局所送達用に処方する場合もある。このような処方物は、当技術分野において周知である。非経口ビヒクルには、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウムによる溶液、乳酸加リンゲル液、または固定油が含まれる。静脈内ビヒクルには、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロース液に基づく補給剤など)などが含まれる。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが含まれる。
【0115】
RSウイルスFポリペプチドのVLPは、中性処方物または塩ベースの処方物を含めたワクチンへと処方することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が含まれる。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0116】
ワクチンは、投薬処方物に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を起こす能力、また、所望される防御の程度を含め、処置される被験体に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、好ましい範囲は、約0.5μg〜1000μgの範囲、好ましくは1μg〜500μgの範囲、またとりわけ、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)ことが好ましい。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0117】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるヒトの年齢、および抗原処方物により変化する。
【0118】
ワクチン処方物の一部は、それだけでワクチンとして十分に免疫原性であるが、他の処方物では、該ワクチンがアジュバント物質をさらに包含すると、免疫反応が増強される場合もある。
【0119】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達処方物の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤もまた用いることができる。このような化合物の1つである、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬処方物で用いられる(32)。粘膜絨毛によるクリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためのより多くの時間を可能にする能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、それは、接着結合を一過的に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わない、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0120】
キトサンはまた、遺伝的操作により解毒したE.coli易熱性エンテロトキシンの変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に処方することもできる。これは、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果を付加し、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0121】
最後に、キトサン処方物はまた、ワクチンの安定性を改善し、粘膜絨毛によるクリアランスにおいて液体処方物を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができることに注意すべきである(42)。これは、キトサンと共に処方された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子チャレンジ(challenge)に対しても防御的であった(44)。
【0122】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい処方物を表す。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益であり得る。
【0123】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達処方物であるRSウイルスFポリペプチドのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素ポリペプチドを含有するRSウイルスFポリペプチドのVLPがとりわけよく適する場合があり、その結果、HA抗原の結合が増強され、粘膜絨毛によるクリアランスが低減される可能性がある。
【0124】
(アジュバント)
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるRSウイルスFポリペプチドのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons社、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、ニューヨーク、Plenum Press社、ISBN 0−306−45283−9においても詳述されている。
【0125】
一部の実施形態では、RSウイルスFポリペプチドのVLPワクチンが、重量ベースの比率約10:1〜約1010:1のRSウイルスFポリペプチドのVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のRSウイルスFポリペプチドのVLP:アジュバントで、少なくとも1つのアジュバントと混合したRSウイルスFポリペプチドのVLPを包含する。当業者は、アジュバントに関する情報、また、最適の比率を決定する日常的実験により、適切な比率を容易に決定することができる。
【0126】
アジュバントの好ましい例は、RSウイルスFポリペプチドのVLPのポリペプチド成分と共発現させるか、または該ポリペプチド成分と融合させて、キメラポリペプチドを作製することにより、本明細書で記載されるRSウイルスFポリペプチドのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質構成要素である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めている、好ましいアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進(proinflammatory)メディエーターを生成させる(66〜72)。
【0127】
TLR5は、フラジェリン単量体内における保存的構造(これはフラジェリン単量体タンパク質に固有であり、また、鞭毛機能に必要とされる)を認識し、このことは免疫学的圧力に応答したその変異を妨げる(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体へと解体されることが、結合および刺激に必要とされる。
【0128】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応も全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル伝達、また、TLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応がTh2特徴を示すことは、ある意味で驚くべきことである。重要なことに、フラジェリンに対して予め存在する抗体は、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっている。
【0129】
多くの近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝的に解毒されたE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5チャレンジに対する異種サブタイプ的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0130】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン2、インターロイキン7、インターロイキン12、インターフェロン、および他の同様の増殖因子はまた、アジュバントとして用いることができ、これらは
RSウイルスFポリペプチドのVLPのポリペプチド成分と混合または融合させることにより、該ワクチン内に容易に組み込み得るので、これらもまた好ましい。
【0131】
一部の実施形態では、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチン組成物が、5’−TCG−3’配列を含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、イミキモド(R−837)、およびレシキモド(R−848)など、他のTLR7リガンドなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0132】
特定のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア誘導体などの免疫調節アジュバント;油処方物;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群より選択される。
【0133】
アジュバントのさらなる例には、作用物質、例えば、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられるアルミニウム塩、例えば、水酸化物またはリン酸塩(ミョウバン)(例えば、Nicklas(1992年)、Res. Immunol.、143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物が含まれ、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)での再活性化による凝集物、C. parvumなどの細菌細胞またはグラム陰性菌の内毒素もしくはリポ多糖成分との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油ビヒクルにおけるエマルジョン、または保護置換物として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた好ましい。
【0134】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン](poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が含まれる。例えば、アルミニウム塩と併せた、モノホスホリルリピドA、好ましくは、3−脱−O−アシル化モノホスホリルリピドAの組合せを含めた、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントが好ましい。MPL(登録商標)アジュバントは、GlaxoSmithKline社から市販されている(例えば、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0135】
リポソーム処方物もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、RSウイルスFポリペプチドのVLPと合わされる、好ましい例である。
【0136】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であることが示されているため、本発明に従う好ましい選択である。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状処方物は、サポニンが60〜70%w/wを構成することが可能であり、コレステロールおよびリン脂質が10〜15%w/wを構成することが可能であり、タンパク質が10〜15%w/wを構成することが可能な、ISCOM粒子として公知の処方物である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.、3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.、74巻:8〜25頁も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0137】
本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンとアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号;および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で記載される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、また、それらの画分が含まれる。Quil Aの特に好ましい画分は、QS21、QS7、およびQS17である。
【0138】
β−エスシンは、本発明のアジュバント組成物中で用いるのに好ましい、別の溶血性サポニンである。エスシンは、「メルクインデックス」(第12版:エントリー3737)において、ラテン名: Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として記載されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.、4巻、213頁(1953年))による、また、イオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が記載されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月、44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0139】
本発明において用いるのに好ましい別の溶血性サポニンは、ジギトニンである。ジギトニンは、「メルクインデックス」(第12版:エントリー3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において記載される手順により精製されるサポニンとして記載されている。その使用は、コレステロールを判定するための臨床試薬として記載されている。
【0140】
アジュバント効果を達成する別の興味深い(したがって、好ましい)可能性は、Gosselinら、1992年において記載される技法を用いることである。略述すると、本発明の抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)に結合体化することにより増強することができる。とりわけ、抗原と抗FCRIとの結合体は、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、生成後において、または生成の一部として(RSウイルスFポリペプチドのVLPのポリペプチド成分のうちのいずれか1つに対する融合体としての発現によることを含む)、RSウイルスFポリペプチドのVLPに結合体化することができる。
【0141】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0142】
適切なマイコバクテリア誘導体は、ムラミルジペプチド、フロイントの完全アジュバント、RIBI(モンタナ州、ハミルトン、RiBi ImmunoChem Reserch社製)、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群より選択することができる。
【0143】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が含まれる。
【0144】
適切なポリマーアジュバントの例には、炭水化物、例えば、デキストラン、PEG、デンプン、マンナン、およびマンノース;可塑性ポリマー;ならびにラテックスビーズなどのラテックスが含まれる。
【0145】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容されるキャリアおよびビヒクルと併せて)を組み込むことである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、迅速に危険シグナルに応答し、炎症部位へと至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、1998年10月12〜15日、カリフォルニア州、アプトス、シースケープリゾート、「From the Laboratory to the Clinic, Book of Abstracts」において略述されている。
【0146】
RSウイルスFポリペプチドのVLPワクチンと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つの、またはより好ましくは、少なくとも6つ以上のヌクレオチドにより隔てられた2つ以上のジヌクレオチドCpGモチーフを含有することが好ましい。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において記載されている。
【0147】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。好ましい実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはより好ましくは、ホスホロチオエート結合であるが、骨格結合が混合したオリゴヌクレオチドを含め、ホスホジエステル骨格、また、PNAなど他のヌクレオチド骨格も本発明の範囲内にある。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートオリゴヌクレオチドを作製する方法は、米国特許第5,666,153、米国特許第5,278,302号、およびW095/26204において記載されている。
【0148】
好ましいオリゴヌクレオチドの例は、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することが好ましい。
【0149】
【化2】
代替的な好ましいCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。自動合成器を用いてこのようなオリゴヌクレオチドを合成し得ることが好ましい。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0150】
多くの単相または複相のエマルジョン系が記載されている。エマルジョンが、RSウイルスFポリペプチドのVLPの構造を破壊しないように、当業者は、RSウイルスFポリペプチドのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体で、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして記載されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP 0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067;EP0480981B)など、他の油エマルジョンによるアジュバントが記載されている。
【0151】
本明細書で記載されるRSウイルスFポリペプチドのVLPワクチンと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、鉱油性の場合もあり、有機油性の場合もある。鉱油および有機油の例は、当業者に容易に明らかである。
【0152】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含することが好ましい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company社、第25版(1974年))。油は、レシピエントに毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果油(ラッカセイ油など)、種油、穀類油が、植物油の一般的な供給源である。合成油もまた本発明の一部であり、これらには、NEOBEE(登録商標)および他の油など、市販の油が含まれ得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油、および酵母中に低量で見出される不飽和油であり、本発明で用いるのに特に好ましい油である。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、エントリー8619)。
【0153】
特に好ましい油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0154】
加えて、本発明の最も好ましい油エマルジョンによるアジュバントには、抗酸化剤が含まれ、油であるα−トコフェロール(ビタミンE;EP0382271B1)が好ましい。
【0155】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に処方される、スクアレン、α−トコフェロール、TWEEN 80(TM)に基づくエマルジョンアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相へと添加することもできる(WO98/56414)。
【0156】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であることが好ましく、実質的に30〜600nmの範囲内、好ましくは、実質的に直径約30〜500nm、また、最も好ましくは、実質的に直径150〜500nm、また、特に直径約150nmであり得る。この点で、油滴数の80%が好ましい範囲内にあるべきであり、より好ましくは油滴数の90%超、また、最も好ましくは油滴数の95%超が規定のサイズ範囲内にある。本発明の油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内であり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比率は、より安定的なエマルジョンがもたらされるため、1以下であることが好ましい。SPAN 85(TM)もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0157】
水中油エマルジョンを作製する方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを行うステップを包含するが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、少量の液体をホモジナイズするのには適切であることが、当業者には明らかである。同様に、当業者は、Microfluidizer(M110S型マイクロ流体作製器;最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))において乳化過程を適合させることで、より少量または多量のエマルジョンを作製することができる。この適合は、必要とされる直径の油滴を有する調製物が達成されるまでの、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得る。
【0158】
本明細書で開示されるRSウイルスFポリペプチドのVLPによるワクチン調製物は、該ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび針、または固体ワクチンを弾道送達(ballistic delivery)するためにデザインされたデバイス(WO99/27961)、または針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、皮膚に適用することもできる(経皮(transdermalまたはtranscutaneous)送達;WO98/20734;WO98/28037)。したがって、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンは、RSウイルスFポリペプチドのVLPワクチンまたはアジュバント組成物を予め充填した全身投与用の送達デバイスを包含する。したがって、個体、好ましくは哺乳動物または鳥類において免疫反応を誘導する方法であって、本明細書で記載されるRSウイルスFポリペプチドのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/またはキャリアを包含するワクチンを該個体へと投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0159】
本発明のワクチン調製物は、該ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用い得ることが好ましい。代替的な粘膜経路は、膣内経路および直腸内経路である。好ましい経粘膜経路投与は、経鼻経路を介し、鼻腔内ワクチン接種と称する。免疫化される個体の鼻咽頭内へと、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを含め、鼻腔内ワクチン接種の方法は、当技術分野において周知である。したがって、噴霧化またはエアゾール化されたワクチン処方物が、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンの好ましい形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶処方物、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンの処方物である。
【0160】
本明細書で開示される、好ましいRSウイルスFポリペプチドのVLPによるワクチン組成物は、経粘膜ワクチン接種により全身ワクチン接種を代替するヒトにおける適用に適する経粘膜ワクチンのクラスの一例である。
【0161】
RSウイルスFポリペプチドのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性緩衝液、または腸溶性のカプセルもしくはマイクロ顆粒も含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤の他の公知の安定化剤も含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも含まれ得る。
【0162】
代替的に、RSウイルスFポリペプチドのVLPによるワクチン処方物は、キトサン(上記で記載した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチンビヒクルと組み合わせることができる。サポニンはまた、コレステロールの存在下において処方して、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁物中、または小ラメラ(paucilamelar)リポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて処方することもできる。
【0163】
本明細書で記載されるRSウイルスFポリペプチドのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(米国、カリフォルニア州、Chiron社製)、MF−59(Chiron社製、例えば、Granoffら(1997年)、Infect Immun.、65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beecham社製のアジュバントシステム♯2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beecham社製のアジュバントシステム♯4;ミョウバンおよびMPLを含有する)、ベルギー、リクセンサルト、SmithKline Beecham社から市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline社製)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline社製)、ならびに、係属中の米国特許出願第08/853,826号、および同第09/074,720号において記載されるものなどの、他のアミノアルキルグルコサミド4−ホスフェート(AGP)が含まれる。
【0164】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(ジョージア州、ノークロス、CytRx社製);Gerbuアジュバント(ドイツ、ガイベルク、Gerbu Biotechnik GmbH社製);ニトロセルロース(NilssonおよびLarsson(1992年)、Res. Immunol.、143巻:553〜557頁);Seppic ISAシリーズのMontamideアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;フランス、パリ、Seppic社製)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを含めたミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの処方物;およびPROVAX(登録商標)(IDEC Pharmaceuticals社製);OM−174(脂質Aと関連するグルコサミンジサッカリド); Leishmania属の伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント処方物が含まれるがこれらに限定されない。例えば、O’Haganら(2001年)、Biomol Eng.、18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」、D. O’Hagan編(2000年)、Humana Press社を参照されたい。
【0165】
他の好ましいアジュバントには、一般式
HO(CH 2CH2O)n−A−R, (I)
[式中、nは1〜50であり、Aは結合または−−C(O)−−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が含まれる。
【0166】
本発明の一実施形態は、一般式(I)[式中、nは1〜50、好ましくは4〜24、最も好ましくは9であり;R成分はC1〜50、好ましくはC4〜C20アルキル、また最も好ましくはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを含むワクチン処方物からなる。ポリオキシエチレンエーテルの濃度は、0.1〜20%、好ましくは0.1〜10%、また最も好ましくは0.1〜1%の範囲であるべきである。好ましいポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、「メルクインデックス」(第12版;エントリー7717)で記載されている。これらのアジュバント分子は、WO99/52549で記載されている。
【0167】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、好ましいアジュバントの組合せは、上で記載したCpGとの組合せであることが好ましい。
【0168】
本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が含まれる。
【0169】
本発明は、以下の非限定的な実施例を参照することにより、よりよく理解される。本明細書で記載される通り、本発明は、必要に応じて、抗原またはアジュバントに連結したさらなる脂質ラフト会合ポリペプチドを含む、RSウイルスFポリペプチドのVLPを包含する。以下の実施例では、RSウイルスFポリペプチドのVLPを含む、本発明の代表的な実施形態が記載される。
【実施例】
【0170】
(実施例1)
この実施例は、細胞質側末端領域を欠失しているRSV融合(F)糖タンパク質の切断型、またはインフルエンザ赤血球凝集素由来の膜貫通領域および細胞質側末端領域を含有するハイブリッドFタンパク質の哺乳動物細胞発現が、RSV Fを含有するエンベロープウイルス様粒子(VLP)の形成をもたらすことを実証する。
【0171】
この実施例は、マウス白血病ウイルスGag遺伝子産物との共発現で、または共発現ではなくRSV F糖タンパク質が哺乳動物細胞において発現されるように設計された。期待されるのは、RSV Fが細胞から出芽するエンベロープGag VLPに組み込まれることであった。
【0172】
プラスミドp3.1−RSVFTは、細胞質側末端を欠いているが天然の膜貫通ドメインを含有しているF(RSVFT)の切断型をコードした。p3.1−RSVFTの地図を図1に示し、RSVFTのコード配列は以下の通りである。
【0173】
【化3】
天然RSV F遺伝子が、細胞の細胞質で複製するパラミクソウイルス由来であり、したがって細胞の核において適切に発現されないと予測されることから、RSVFTコード配列は、カスタム合成DNA断片として合成し、ウイルスからはクローニングしなかった。コードされたRSVFTのアミノ酸配列(シグナルペプチドおよび膜貫通ドメインを大文字で示す)は以下の通りである。
【0174】
【化4】
プラスミドp3.1−shFv1は、インフルエンザA由来のH5赤血球凝集素の膜貫通ドメインおよび細胞質側末端に融合したRSV Fの外部ドメインからなるハイブリッドタンパク質をコードした。図2は、p3.1−shFv1の地図を示し、shFv1のコード配列は以下の通りである。
【0175】
【化5】
【0176】
【化6】
shFv1のアミノ酸配列(シグナルペプチドコード配列ならびにHA膜貫通および細胞質側末端配列を大文字で示す)は以下の通りである。
【0177】
【化7】
RSVFTと同様にshFv1コード配列は、DNA合成によって作製した。
【0178】
プラスミドp3.1−shFv2は、インフルエンザA由来のH5赤血球凝集素の膜貫通ドメインおよび細胞質側末端に融合したRSV Fの外部ドメインからなるハイブリッドタンパク質をコードした。shFv2は、インフルエンザHA由来膜貫通および末端領域が4アミノ酸長いことにおいてshFv1と異なった。図3は、p3.1−shFv2のプラスミド地図を示す。shFv2のコード配列は以下の通りである。
【0179】
【化8】
shFv2のアミノ酸配列(シグナルペプチドコード配列ならびにHA膜貫通および細胞質側末端配列を大文字で示す)は以下の通りである。
【0180】
【化9】
shFv2コード配列は、DNA合成によって作製した。
【0181】
プラスミドp3.1−Gagは、マウス白血病ウイルス由来のGag遺伝子産物をコードした。図4は、p3.1Gagのプラスミド地図を示す。プラスミドp3.1−Gag由来のGagのコード配列は以下の通りである。
【0182】
【化10】
【0183】
【化11】
MLV Gagのアミノ酸配列は以下の通りである。
【0184】
【化12】
Gag遺伝子は、プラスミドpAMS(ATCC)において見出されたMLVのゲノム由来の天然クローンを表した。
【0185】
10cm2組織培養皿8枚に、10%ウシ胎仔血清を補充したダルベッコ変法イーグル培地で培養した293−F細胞を接種した。細胞がおよそ95%コンフルエンス(confluence)(単層培養)に達したときに、各ウエルに以下のように:
【0186】
【化13】
【0187】
【化14】
プラスミドDNA合計4μgをトランスフェクトした。
【0188】
プラスミドDNAをLIPOFECTAMINE(TM)2000(Invitrogen)を製造者の推奨に従って使用してトランスフェクトした。トランスフェクションの8時間後、トランスフェクション培地をCD293無血清培地で置換し、培養を継続した。トランスフェクションの48時間後、RSV F発現ベクターのいずれかをトランスフェクトした細胞を含むすべての皿は、正しくフォールドしたFの表面発現と一致して有意なレベルの合胞体(融合細胞)を示した。トランスフェクションの48時間後、ピペッティングによって細胞を剥離し、細胞および培地を収集した。細胞の表面でのRSV F抗原活性の発現を、膜に組み込まれた(integrated)状態において正しくフォールドしたF抗原を認識するマウスモノクローナル抗体(9C5)での細胞染色後のフローサイトメトリーによってさらに実証した。細胞をF−陽性細胞の蛍光検出のためにフィコエリトリンに結合体化したヤギ抗マウス2次抗体とさらに反応させた。図5は、Gagベクターと共にまたは共にではなくF発現ベクターのいずれかをトランスフェクトした細胞でのFの有意なレベルの表面発現を実証するフローサイトメトリー実施からのヒストグラムを示す。これらのデータは、トランスフェクトした細胞集団における合胞体の存在と一致し、細胞表面上で正しくフォールドされたF抗原の存在を実証する。
【0189】
トランスフェクトした細胞から収集した増殖培地を合胞体およびすべての細胞細片を除去するために2000rpmで5分間遠心分離し、このステップからの上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×gで1時間遠心分離して、すべてのVLP(Gagおよび/またはF遺伝子発現の結果として培地中に放出された可能性がある)を収集した。このステップからの100,000×gペレットを追加的分析のためにトリス緩衝生理食塩水に再懸濁した。
【0190】
超遠心分離ステップ由来の再懸濁したペレットを、VLP形成の指標としてのGag産物の存在を検出するためにウエスタンブロット分析に供した。予測外に、顕著な量のGag産物をp3.1−Gagベクターだけをトランスフェクトした細胞由来の試料中で検出したが、F遺伝子ベクターのいずれかとの組合せでp3.1−Gagをトランスフェクトした細胞由来のペレットにおいては顕著に減少した量のGagが見出された。結果は、F遺伝子産物の発現がGag産物の出芽機能をある様式で干渉することと一致している。
【0191】
再懸濁した超遠心分離ペレット中のF抗原の存在を次にELISAによって以下の通り検査した:再懸濁した各超遠心分離ペレット試料をNunc MAXISORP(TM)平底96ウエルELISAプレートの列Aの1つのウエルに入れ、各試料をプレートの各カラムを下がりながら連続的に2倍希釈した。4℃、1晩のコーティング後、プレートを0.05%TWEEN20(TM)を含有するPBS(PBS−T)で3回洗浄し、次いでSTARTING BLOCK(TM)(PBS)(Pierce、Biotechnology)でブロックした。ブロッキング後、STARTING BLOCK T20(TM)(Pierce)中に1:1000希釈したモノクローナル抗体9C5 100μlを各ウエルに入れ、プレートを室温で3時間インキュベートした。プレートを再度洗浄し、STARTING BLOCK T20(TM)中に1:1000希釈したヤギ抗マウスHRP結合体化抗体(Southern Biotech)100μlを各ウエルに添加し、室温で1.5時間インキュベートした。PBS−Tでの最終洗浄(3×)に続いて、プレートをABTS試薬(Pierce)100μlの添加によって発色(develop)させ、室温、45分間インキュベートした。
【0192】
図6は、このELISAの結果を示し、顕著なレベルのRSV F抗原が、F遺伝子だけおよびF遺伝子+Gag遺伝子でのトランスフェクションの両方に由来する100,000×gペレットにおいて検出されたことが観察され得る。重要かつ予測外に、より多いRSV F抗原活性がRSV F遺伝子だけをトランスフェクトした細胞由来のペレットにおいて観察された。これらのデータは、F遺伝子単独の発現(Gag発現の非存在下)がF抗原を保有するVLPの形成をもたらすことを実証する。これは、おそらくFによるGag出芽の干渉によって、FとGagとの共発現がGag出芽の抑制を生じさせるという上の観察とも一致する。
【0193】
(実施例2)
この実施例は、p3.1−RSVFTベクターだけをトランスフェクトした哺乳動物細胞の培地中に放出されたRSV F VLPの電子顕微鏡観察を記載する。
【0194】
RSV F遺伝子だけの発現が、単なるRSV F凝集物ではなくVLPの放出を生じさせることを実証するために、10%FBSを補充したDMEM培地中に293−F細胞をおよそ95%コンフルエンスでそれぞれ含むT75組織培養フラスコ5個をp3.1−RSVFT 30μgおよびLIPOFECTAMINE(TM)2000(Invitrogen)75μlで製造者の仕様書に従ってトランスフェクトした。トランスフェクションの8時間後にトランスフェクション培地を除去し、CD293無血清培地で置換した。トランスフェクションの48時間後に顕著なレベルの浮遊合胞体が培地中に観察されたが、これはRSV F遺伝子の発現と一致する。この時点で増殖培地を収集し、合胞体および細胞細片を2000rpm、5分間の遠心分離によって除去した。次いで培地上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×g、1時間、10℃で遠心分離してVLPを収集した。この遠心分離ステップ由来のペレットを、次いで合計300μlのトリス緩衝生理食塩水中に再懸濁した。再懸濁したペレットを次いで低速(およそ5000pm)で短時間遠心分離して、超遠心分離法によって形成されたすべての不可逆的タンパク質沈殿物または凝集物を除去した。再懸濁したVLP調製物を次いで、酢酸ウラニルネガティブ染色を使用する透過型電子顕微鏡によって検査し、代表的な顕微鏡像を図7に示す。顕微鏡像の収集物は、形状およびサイズの両方での多様性を示すエンベロープ粒子または小胞粒子の顕著なアレイを示した。このような多形性の粒子集団は、タンパク質コアを欠いている出芽粒子の形成と一致する。
【0195】
(実施例3)
この実施例は、バキュロウイルス粒子と高度に凝集するRSV F VLPを生じさせた、バキュロウイルス発現系を使用する昆虫細胞でのRSV F VLPの発現を記載する。
【0196】
実施例2に記載した哺乳動物細胞発現系の使用の代替として昆虫細胞バキュロウイルス発現系でRSV F VLPを作製するための試みにおいて、shFv1のコード配列を含有するDNA断片をプラスミドpFastbac1にクローニングし、プラスミドpFB−shFv1を得た。pFB−shFv1の地図を図8に示す。shFv1遺伝子のコード配列およびコードされたアミノ酸配列は上に記載の通りである。プラスミドpFB−shFv1を、高分子量バクミド(bacmid)中にバキュロウイルスゲノムを含有するDH10Bacコンピテント細胞への形質転換によってバキュロウイルスゲノム中に組み換えた。形質転換されたクローンの選択に続いて、高分子量バクミドDNAをこれらの細菌クローンから調製し、shFv1遺伝子を含有する組換えバキュロウイルスを作製するためのSf9昆虫細胞のトランスフェクションに使用した。この系を使用する組換えバキュロウイルス作製に関わる手順の詳細は、website tools.invitrogen.comのfile/content/sfs/manuals/bactobac_man.pdfのPDFドキュメントに見出すことができる。
【0197】
RSV F VLPを作製するために、SF900−II培地中で増殖しているSf9細胞の200mlスピナー培養物3個を27℃で細胞密度が1mlあたり細胞2×106個に達するまでインキュベートした。その時点で2継代shFv1−組換えバキュロウイルス調製物20mlを各スピナーフラスコに添加し、培養物を27℃で細胞生存率が20%未満に下がるまでインキュベートした。次いで培養液を収集し、2k rpmで15分間遠心分離することによって細胞および細片を清澄化した。清澄化した培地を32ml一定分量に分け、100,000×gで1時間、10℃で、トリス緩衝生理食塩水(TBS)、pH7.4および0.05%TWEEN 20(TM)を含有する30%ショ糖クッション4ml上で遠心分離した。ペレットを0.05%TWEEN 20(TM)を含有するTBS合計12mlに再懸濁し、この材料を6ml一定分量2個に分け、TBS/0.05%TWEEN 20(TM)中の20〜60%ショ糖密度勾配上で遠心分離した。勾配を底から分画した。RSV F抗原活性を個々の勾配画分において実施例1に記載のELISA法によって9C5モノクローナル抗体を使用して、以下の変更を伴って検出した:高濃度のショ糖はELISAアッセイを妨げることから、各勾配画分1.5mlの試料100μlをTBSで10倍希釈し、存在したすべてのVLPをショ糖溶液からペレット化させるために100,000×gで40分間遠心分離した。これらの試料をTBSに再懸濁し、実施例1に記載のELISAアッセイに直接使用した。ELISAアッセイの結果を図9にプロットし、これは、それぞれ画分5および9中の2つの可視の薄いバンドと一致するRSV F抗原活性の2つの顕著なピークを示す。凝集したVLPは典型的にはより高密度(より低い画分)にバンド形成することから、画分9中のバンドは凝集していないVLPを含有すると予測された。しかし画分9中の物質の電子顕微鏡検査は、高度に凝集したVLPおよびバキュロウイルス粒子を明らかにした。代表的な電子顕微鏡像を図10に示す。したがって、凝集していないRSV Fを含有するVLPの作製および精製のためには哺乳動物細胞発現系がより適切であることが決定された。
【0198】
(実施例4)
この実施例は、合胞体形成活性を欠いているがVLP出芽活性、Gag阻害活性およびモノクローナル中和抗体9C5との反応性を保持しているRSV FのGPI−アンカー型の発現を実証する。
【0199】
脂質ラフトドメインを標的とするがそれ自身のVLP出芽活性を欠いている改変Fタンパク質の作製をコードする改変RSV F遺伝子コード配列を作製するための試みがなされた。この目的は、ハイブリッドGag−F VLPが、F遺伝子産物が位置する脂質ラフトドメインから出芽するためのGagタンパク質の能力を介して作製され得るように、MLV Gagの出芽活性と適合性があるF遺伝子産物を供給することであった。このために膜貫通および細胞質側末端のコード配列を切断し、それらをGPIアンカーシグナルをコードする配列で置換することによってF遺伝子を改変した。GPIアンカーシグナルは、F糖タンパク質外部ドメインを脂質ラフト位置に方向付けるが、しかしこれらの脂質ラフト位置でFポリペプチド配列の細胞膜貫通は無いことが予測された。この改変が、キメラGag−F VLPを形成するこれらの位置からの出芽のためのGag VLPの能力をもたらすことが予測された。しかし驚くべきことに、RSV FのGPI−アンカー形態は、依然としてVLP出芽活性を示し、このことは、任意の脂質ラフト会合ポリペプチドを使用してRSV Fポリペプチドを脂質ラフトへ方向付けて、RSV Fポリペプチド単独でのVLP出芽を可能にすることができることを実証した。
【0200】
図11は、ヒトカルボキシペプチダーゼMのGPIアンカーシグナルに融合したキメラFタンパク質がコードされた発現ベクターp3.1−F−GPIの地図を示す。プラスミドp3.1−F−GPI由来のF−GPIコード配列のヌクレオチド配列は、以下の通りである。
【0201】
【化15】
ヒトカルボキシペプチダーゼMのGPIアンカーシグナルに融合したRSV Fの外部ドメインからなるF−GPIキメラタンパク質のアミノ酸配列は、(GPIアンカーシグナルを大文字で示す)以下の通りである。
【0202】
【化16】
発現試験を、プラスミドp3.1−Gagもしくはp3.1−F−GPIまたはp3.1−Gagとp3.1−F−GPIとの組合せをトランスフェクトした293F細胞においてRSV F外部ドメインも保有するGag VLPが作製され得ることを実証する目的で実施した。
【0203】
10cm2組織培養皿12枚に、10%ウシ胎仔血清を補充したダルベッコ変法イーグル培地で培養した293−F細胞を接種した。細胞がおよそ95%コンフルエンス(単層培養)に達したときに、各ウエルに以下のように:
【0204】
【化17】
プラスミドDNA合計4μgをトランスフェクトした。
【0205】
プラスミドDNAをLIPOFECTAMINE(TM)2000(Invitrogen)を製造者の推奨に従って使用してトランスフェクトした。トランスフェクションの8時間後、トランスフェクション培地をCD293無血清培地に置換し、培養を継続した。トランスフェクションの48時間後、p3.1−F−GPI発現ベクターをトランスフェクトした細胞を含む皿は、顕著なレベルの合胞体(融合細胞)を示さず、このことは、F糖タンパク質のGPIアンカー改変が細胞融合を促進するF糖タンパク質の能力に干渉したことを示唆していた。
【0206】
トランスフェクションの48時間後、ピペッティングによって細胞を剥離し、細胞および培地を収集した。細胞の表面でのRSV F抗原活性の発現を、膜に組み込まれた状態において正しくフォールドしたF抗原を認識するマウスモノクローナル抗体(9C5)での細胞染色後のフローサイトメトリーによって実証した。細胞をF−陽性細胞の蛍光検出のためにフィコエリトリンに結合体化したヤギ抗マウス2次抗体とさらに反応させた。図12は、Gagベクターと共にまたは共にではなくp3.1−F−GPIベクターをトランスフェクトした細胞上での有意なレベルのFの表面発現を実証するフローサイトメトリー実施からのヒストグラムを示す。これらのデータは、合胞体形成活性はもはや存在しないにもかかわらず、細胞表面上で正しくフォールドされたF抗原の存在を実証した。
【0207】
トランスフェクトした細胞から収集した増殖培地を細胞および細片を除去するために2000rpmで5分間遠心分離し、このステップからの上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×gで1時間遠心分離して、すべてのVLP(Gagおよび/またはF−GPI遺伝子発現の結果として培地中に放出された可能性がある)を収集した。このステップからの100,000×gペレットを追加的分析のためにトリス緩衝生理食塩水に再懸濁した。
【0208】
超遠心分離ステップからの再懸濁ペレットを、VLP形成の指標としてのGag産物の存在を検出するためのウエスタンブロット分析に供した。予測外に、顕著な量のGag産物がp3.1−Gagベクターだけをトランスフェクトした細胞由来の試料中で検出されたが、p3.1−F−GPIベクターと組み合わせてp3.1−Gagをトランスフェクトした細胞由来のペレットにおいてはGagが見出されたとしてもこれは少量であった。これらの結果は、キメラF産物の脂質ラフト標的化機構が変化し、その膜貫通および細胞質側末端領域が除かれているにもかかわらず、F−GPI遺伝子産物の発現がGagタンパク質の出芽機能を依然として干渉することを実証した。これらの結果を図13に示し、これは、GagおよびRSV Fについてのウエスタンブロットを示す。RSVウエスタンブロットは、F抗原活性がF−GPIおよびF−GPI+Gagトランスフェクションの由来のペレットにおいて見出されたことを示し、Gagの有無がF−GPI媒介出芽および粒子形成に影響しないことを示した。
【0209】
再懸濁した超遠心分離ペレット中のF抗原の存在をELISAによっても以下の通り検査した:再懸濁した超遠心分離ペレットの各試料をNunc MAXISORP(TM)平底96ウエルELISAプレートの列Aのウエル1つに入れ、各試料をプレートの各カラムを下がりながら連続的に2倍希釈した。4℃、1晩のコーティング後、プレートを0.05%Tween20を含有するPBS(PBS−T)で3回洗浄し、次いでSTARTING BLOCK(TM)(PBS)(Pierce、Biotechnology)でブロックした。ブロッキング後、STARTING BLOCK(TM)T20(Pierce)中に1:1000希釈したモノクローナル抗体9C5 100μlを各ウエルに入れ、プレートを室温で3時間インキュベートした。プレートを再度洗浄し、STARTING BLOCK(TM)T20中に1:1000希釈したヤギ抗マウスHRP結合体化抗体(Southern Biotech)100μlを各ウエルに添加し、室温で1.5時間インキュベートした。PBS−Tでの最終洗浄(3×)に続いて、プレートをABTS試薬(Pierce)100μlの添加によって発色させ、室温で45分間インキュベートした。
【0210】
図14は、このELISAの結果を示し、顕著なレベルのRSV F抗原活性が、F−GPIだけおよびF−GPI+Gagベクターをトランスフェクトした細胞両方に由来する100,000×gペレットにおいて検出されたことが観察され得る。重要なことに、等量のRSV F抗原活性がF−GPIおよびF−GPI+Gagベクターをトランスフェクトした細胞由来のペレットにおいて観察された。これらのデータは、F−GPI遺伝子単独での発現が(Gag発現の非存在下で)F−GPI抗原を保有するVLPの形成を生じさせるという結論と一致する。これはまた、おそらくF−GPIによるGag出芽の干渉により、F−GPIとGagとの共発現がGag出芽の抑制を生じさせるという上の観察とも一致する。したがってGPIアンカーシグナルの切断および付加によるFポリペプチドの改変は、Fの出芽活性を破壊せず、Gag出芽に対する阻害効果も無効にしない。この実験は、F−GPI出芽粒子が、9C5モノクローナル抗体によって検出されるF抗原活性に関して生のRSVビリオンと同様に挙動することも示す。
【0211】
(さらなる参考文献)
【0212】
【数1】
【0213】
【数2】
【0214】
【数3】
【0215】
【数4】
【0216】
【数5】
【0217】
【数6】
【0218】
【数7】
【0219】
【数8】
【技術分野】
【0001】
(発明の分野)
本発明は、感染因子(infectious agent)を含まない、エンベロープウイルスベースのウイルス様粒子(VLP)を単離する分野に関する。好ましい例において、該分野は、エンベロープウイルスベースのVLPの免疫原性に有害な影響を及ぼさない、感染因子を不活化する方法を包含する。特定の実施形態では、エンベロープウイルスベースのVLPを、昆虫細胞ベースの発現系において作製する。
【背景技術】
【0002】
(発明の背景)
RSウイルス(RSV)は、乳児および1歳未満の小児において、細気管支炎および肺炎の主要原因である(CDC National Center for Infectious Diseases(2004年)、Respiratory Syncytial Virus)。RSVはまた、免疫不全である成人および高齢者においても、重要な下気道病原体であり得る。RSVによる天然の感染は、防御的免疫を誘導しないので、個体は、複数回にわたり感染し得る。
【0003】
RSVは、Paramyxoviridae科のPneumovirus属に属する、ネガティブセンスの一本鎖RNAウイルスである。RSVゲノムは、螺旋状のヌクレオカプシドにより取り囲まれ、少なくとも10のタンパク質:3つの膜貫通構造タンパク質(F、G、およびSH)、2つのマトリックスタンパク質(MおよびM2)、3つのヌクレオカプシドタンパク質(N、P、およびL)、および2つの非構造タンパク質(NS1およびNS2)をコードする(Collinsら(1996年)、Respiratory syncytial virus、1313〜1351頁、B.N. Fields(編)、Fields virology. Raven Press、New York、NY)。中和抗体は、Fタンパク質およびGタンパク質によってのみ誘発されると考えられる。RSVが、Gタンパク質に基づいて、AおよびBの亜群に分けられるのに対し、Fは、その亜群間における関連がより密接である。Fタンパク質に対するモノクローナル抗体は、インビトロにおいて中和効果、インビボにおいて予防効果を有することが示されている(例えば、Andersonら、1988年、J. Virol.、62巻:4232〜4238頁; Andersonら、1986年、J. Clin. Micro.、23巻:475〜480頁; Beeler およびCoelingh、1989年、J. Virol.、63巻:2941〜50頁; Garcia−Barrenoら、1989年、J. Virol.、63巻:925〜32頁; Taylorら、1984年、Immunology、52巻:137〜142頁;ならびに特許文献1)。
【0004】
数十年間にわたる研究にもかかわらず、RSVに対する安全で有効なワクチンは存在していない。乳児および小児において試験されたホルマリン不活化ウイルスワクチンは、感染に対して防御せず、野生型のRSVウイルスによるその後の感染において、重篤な症状が生じる危険性の増大と関連していた(Kapikianら、1969年、Am. J. Epidemiol.、89巻:405〜21頁; Chinら、1969年、Am. J. Epidemiol.、89巻:449〜63頁)。生の弱毒化温度感受性変異体の開発に焦点を当てたその後の試みもまた、適切な弱毒化レベルのウイルス候補を同定することができず、また、一部の候補が遺伝子的に不安定であるために失敗した(Hodesら(1974年)、Proc. Soc. Exp. Biol. Med.、145巻、1158〜1164頁; Kimら(1973年)、Pediatrics、52巻、56〜63頁; Wrightら(1976年)、J. Pediatrics、88巻、931〜936頁)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】米国特許第6818216号明細書
【発明の概要】
【発明が解決しようとする課題】
【0006】
ウイルス様粒子(VLP)は、従来のワクチン技術を上回るいくつかの利点を提供する。ワクチンを開発するのに重要なVLPの利点は、それらが、三次元構造との関連で天然ウイルスを模倣するということであり、また、一次エピトープおよび立体配座エピトープの両方に対する中和抗体応答を誘導する能力であり、したがって、他のワクチン処方物より免疫原性であることが示されるはずである。ウイルスベクター法と異なり、VLPは、あらかじめ存在する免疫に対して問題を示さず、したがって、繰り返しの使用を可能とする。RSV抗原を含有するVLPは、昆虫細胞内において、RSV Fタンパク質を、RSV Mタンパク質と、または異種Mタンパク質と共発現させることにより作製されてきた(US2008/0233150)。しかし、US2008/0233150は、RSV Fタンパク質が単独で、互いに凝集しないかまたはRSV Fタンパク質を発現させるのに使用されるウイルスベクターと会合しないVLPを生成できる、哺乳動物細胞系内におけるRSV Fタンパク質単独の実際の発現は教示していない。したがって、RSV Fタンパク質を単独で発現するVLPを生成する方法が必要とされている。
【課題を解決するための手段】
【0007】
(概要)
本発明の好ましい実施形態は、さらなるエンベロープウイルスコア形成ポリペプチドを必要としないRSウイルスFポリペプチドのVLPを生成する方法、およびRSウイルスFポリペプチドのVLP調製物を含む組成物のための、本明細書で開示される各種の方法および組成物を提供することにより、この必要を満たす。好ましい実施形態では、RSウイルスFポリペプチドのVLPは、以下のさらなる特徴のうちの1つまたは複数を有する:該VLPが、エンベロープウイルスコアを含まない;該VLPが、多形性であるか、またはサイズもしくは形状が不均一である;該VLPが、エンベロープウイルスコア形成ポリペプチドを含まない。このような好ましい実施形態は、RSウイルスFポリペプチドが、単独でVLPを形成することが可能であるという、驚くべき観察に基づいている。RSウイルスFポリペプチド単独により形成されるVLPは、エンベロープウイルス、およびエンベロープウイルス成分を用いて形成されるVLPに典型的なタンパク質コアを有さない。
【0008】
本発明の態様は、RSウイルスFポリペプチドを含む、RSウイルスFポリペプチドのウイルス様粒子調製物を包含し、該ウイルス様粒子は、以下の特徴のうちの1つまたは複数を有する:該ウイルス様粒子は、エンベロープウイルスコアを含まない;該ウイルス様粒子は、多形性であるか、サイズが不均一であるか、もしくは形状が不均一である;該ウイルス様粒子は、エンベロープウイルスコア形成ポリペプチドを含まない;および/または該ウイルス様粒子は、哺乳動物グリコシル化を含む。特定の実施形態では、ウイルス様粒子は、他のウイルス様粒子と実質的に凝集しない場合がある。前出の実施形態または態様と組み合わせ得る別の実施形態では、ウイルス様粒子は、ウイルスベクター粒子と実質的に会合しない。
【0009】
前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、調製物はまた、ウイルス様粒子と混合したアジュバントも含む。アジュバントを包含する前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、アジュバントは、ウイルス様粒子の外側に位置する場合もあり、ウイルス様粒子の内側に位置する場合もある。アジュバントを包含する前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、アジュバントをRSウイルスFポリペプチドに共有結合させて、共有結合を形成させ得る。
【0010】
前出の実施形態または態様のうちのいずれかと組み合わせ得る別の実施形態では、抗RSV−F中和抗体は、RSウイルスFポリペプチドに結合し得る(これにより、該RSV Fポリペプチドが、実質的に天然の立体配座にあることが裏付けられる)。このような中和抗体を包含する、前出の実施形態または態様のうちのいずれかと組み合わせ得る特定の実施形態では、抗RSV−F中和抗体は9C5であり得る。
【0011】
別の態様は、RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法を含み、該方法は、(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと;(b)該発現ベクターを培地中の哺乳動物細胞内へと導入するステップと;(c)該RSウイルスFポリペプチドを発現させて、該RSウイルスFポリペプチドのウイルス様粒子を作製するステップとを含み、該ウイルス様粒子は、以下の特徴のうちの1つまたは複数を有してもよい:該ウイルス様粒子は、エンベロープウイルスコアを含まない;該ウイルス様粒子は、多形性であるか、サイズが不均一であるか、もしくは形状が不均一である;および/または該ウイルス様粒子は、エンベロープウイルスコア形成ポリペプチドを含まない。
【0012】
前出の態様と組み合わせ得る別の実施形態では、該方法は、哺乳動物細胞が培養されている培地から、RSウイルスFポリペプチドのウイルス様粒子を取り出すステップをさらに含む。
【0013】
前出の実施形態および態様と組み合わせ得る別の実施形態では、発現ベクターはウイルスベクターであり得る。前出の実施形態および態様と組み合わせ得る別の実施形態では、ウイルスベクターを、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択することができる。前出の実施形態および態様と組み合わせ得る別の実施形態では、哺乳動物細胞を、BHK細胞、VERO細胞、HT1080細胞、MRC−5細胞、WI 38細胞、MDCK細胞、MDBK細胞、293細胞、293T細胞、RD細胞、COS−7細胞、CHO細胞、Jurkat細胞、HUT細胞、SUPT細胞、C8166細胞、MOLT4/クローン8細胞、MT−2細胞、MT−4細胞、H9細胞、PM1細胞、CEM細胞、黒色腫細胞、SB20細胞、LtK細胞、HeLa細胞、WI−38細胞、L2細胞、CMT−93、およびCEMX174細胞からなる群より選択することができる。
【0014】
前出の実施形態および態様と組み合わせ得る別の実施形態では、抗RSV−F中和抗体は、発現されたRSウイルスFポリペプチドに結合し得る(これにより、該RSウイルスFポリペプチドは、実質的に天然のフォールドであることが裏付けられる)。中和抗体を包含する前出の実施形態および態様と組み合わせ得る別の実施形態では、抗RSV−F中和抗体は9C5である。
【0015】
別の態様は、RSウイルスの感染を処置または予防する方法を含み、該方法は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を被験体に投与するステップを含む。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与は被験体における防御的免疫化反応を誘導する。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与を、皮下(subcutaneous)送達、経皮送達、皮内送達、皮下(subdermal)送達、筋肉内送達、経口(peroral)送達、経口(oral)送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択することができる。
【0016】
別の態様は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を含む医薬組成物を包含する。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、医薬組成物は、薬学的に許容されるキャリアをさらに含む。
【0017】
別の態様は、RSウイルスの感染に対する防御を提供する方法を含み、該方法は、免疫原性量の、本発明のこの態様の前出の実施形態のうちのいずれかによる調製物、または本発明のこの態様の前出の実施形態のうちのいずれかの方法により作製された集団を被験体に投与するステップを含む。前出の態様の実施形態のうちのいずれかと組み合わせ得る別の実施形態では、投与を、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択することができる。
【0018】
前出の態様およびそれらの実施形態は、本明細書で開示される実施形態のうちのいずれかとさらに組み合わせることができる。前出の実施形態、および/または本明細書で開示されるさらなる実施形態のうちのいずれかと共に包含され得る、本発明のさらなる態様は、本明細書の全体において見出すことができる。
【図面の簡単な説明】
【0019】
【図1】図1は、p3.1−RSVFTのプラスミド地図を示す図である。
【図2】図2は、p3.1−shFv1のプラスミド地図を示す図である。
【図3】図3は、p3.1−shFv2のプラスミド地図を示す図である。
【図4】図4は、p3.1−Gagのプラスミド地図を示す図である。
【図5】図5は、RSV FおよびGag発現ベクターをトランスフェクトされた細胞上のRSV Fの表面発現のサイトメトリー分析を示すグラフである。トランスフェクトされていない細胞およびp3.1−Gagだけをトランスフェクトされた細胞は、バックグラウンド蛍光レベルを示す。p3.1−Gagと共におよび共にではなくRSV F発現ベクターをトランスフェクトされた細胞は、9C5モノクローナル抗体および蛍光2次抗体によるF検出の結果として顕著なレベルの蛍光を示す。
【図6】図14は、Gag遺伝子を共トランスフェクトされておよびされずにRSV F遺伝子をトランスフェクトされた細胞の培地由来の100,000×gペレットにおけるRSV F抗原活性の検出を示すグラフである。
【図7】図7は、p3.1−RSVFTをトランスフェクトされた239−F細胞の培地から収集したVLPの電子顕微鏡像の代表的切片を示す図である。
【図8】図8は、pFB−shFv1のプラスミド地図を示す図である。
【図9】図9は、昆虫細胞中で発現されたshFv1 VLPのショ糖密度勾配を示すグラフである。
【図10】図10は、図9に示すshFv1ショ糖勾配の画分9における凝集したRSV F VLPおよびバキュロウイルス粒子の代表的電子顕微鏡像を示す図である。
【図11】図11は、ヒトカルボキシペプチダーゼM由来のGPIアンカーシグナルに融合したキメラFポリペプチド外部ドメインをコードするp3.1−F−GPIのプラスミド地図を示す図である。
【図12】図12は、p3.1−Gagを共トランスフェクトされておよびされずにp3.1−F−GPIをトランスフェクトされた細胞での改変F−GPIタンパク質の表面発現のフローサイトメトリー分析を示すグラフである。p3.1−Gagだけをトランスフェクトされた細胞は、バックグラウンド蛍光レベルを示した。p3.1−Gagと共におよび共にではなくp3.1−F−GPIをトランスフェクトされた細胞は、9C5モノクローナル抗体および蛍光2次抗体によるF検出の結果として顕著なレベルの蛍光を示した。
【図13】図13は、p3.1−Gag、p3.1−F−GPIおよびp3.1−Gag+p3.1−F−GPIをトランスフェクトされた細胞の培地由来の100,000×gペレットのウエスタンブロット分析を示す図である。レーンは左から右へ:G、p3.1−Gagトランスフェクション由来の試料;F、p3.1−F−GPIトランスフェクション由来の試料;F+G、p3.1−Gag+p3.1−F−GPIトランスフェクション由来の試料である。データは、F発現はGag出芽に干渉するが、Gag発現はF出芽に干渉しないことを示す。
【図14】図14は、Gag遺伝子ベクターを共トランスフェクトされておよびされずにRSV F−GPIベクターをトランスフェクトされた細胞の培地由来の100,000×gペレットにおけるRSV F抗原活性の検出を示すグラフである。生のRSVビリオン粒子を含有しているRSV感染細胞由来の培地の100,000×gペレットを、F−GPI粒子がF抗原に関して生のRSVに抗原的に類似していることを実証する陽性対照として含めた。
【発明を実施するための形態】
【0020】
(好ましい実施形態の詳細な説明)
本発明の好ましい実施形態には、好ましくは、哺乳動物細胞による発現系で作製された、RSウイルスFポリペプチドのVLP調製物と;このようなRSウイルスFポリペプチドのVLP調製物を発現させるかまたは作製する方法と;このような調製物をワクチン組成物へとさらに加工する方法と;このようなワクチン組成物を用いる方法とが含まれるがこれらに限定されない。
【0021】
本発明の特定の態様および実施形態は、適切な宿主細胞内で発現させると、gagポリペプチドまたはマトリックスポリペプチドなど、任意のコアタンパク質またはコア形成タンパク質の不在下で、RSウイルスFポリペプチドは、単独でVLPを生成するのに十分であるという驚くべき発見に基づいている。このようなVLPは、内部におけるタンパク質コアまたはカプシドなしに、脂質またはエンベロープを有する。したがって、このようなVLPは、以下の物理的な特徴のうち1つまたは複数を有する:脂質膜またはエンベロープ内にエンベロープウイルスコア、カプシド、またはヌクレオカプシドを有さない;多形性であるか、またはサイズおよび/もしくは形状が不均一である;該VLPは、互いと凝集しない;該VLPは、それを作製するのに用いられるウイルスベクターと会合しないか、またはそうでなければ凝集しない;ならびに哺乳動物グリコシル化、および/または哺乳動物膜と結合もしくは会合するタンパク質。
【0022】
RSウイルスFポリペプチドを作製するのに好ましい方法は、好ましくは、さらなるポリペプチド抗原の共発現を含めた、哺乳動物細胞内における発現を介する。
【0023】
開示される方法およびプロトコールを実施するには、別段に示さない限り、当業者の能力の範囲内にある、化学、分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を用いる。このような技法は、文献において説明されている。例えば、J. Sambrook、E. F. Fritsch、およびT. Maniatis、1989年、「Molecular Cloning: A Laboratory Manual」、第2版、1〜3巻、Cold Spring Harbor Laboratory Press社; Ausubel, F. M.ら(1995年、および定期的な補遺;「Current Protocols in Molecular Biology」、9、13、および16章、ニューヨーク州、ニューヨーク、John Wiley & Sons社); B. Roe、J. Crabtree、およびA. Kahn、1996年、「DNA Isolation and Sequencing: Essential Techniques」、John Wiley & Sons社; J. M. PolakおよびJames O’D. McGee、1990年、「In Situ Hybridization: Principles and Practice」、Oxford University Press社; M. J. Gait(編)、1984年、「Oligonucleotide Synthesis: A Practical Approach」、Irl Press社;ならびにD. M. J. LilleyおよびJ. E. Dahlberg、1992年、「Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology」、Academic Press社を参照されたい。
【0024】
(定義)
本明細書で用いられる「RSウイルスFポリペプチドのVLP」とは、ウイルスカプシド、ヌクレオカプシド、またはコアを形成することが可能な任意のポリペプチドの不在下において発現させることが好ましい、RSウイルスFポリペプチドを用いて形成されるウイルス様粒子を指す。好ましい例には、RSウイルスFポリペプチドの細胞質部分を実質的に欠く、RSウイルスFポリペプチドを用いて生成されるVLP、またはインフルエンザM1ポリペプチドおよび/もしくは赤血球凝集素ポリペプチド(また、場合によってノイラミニダーゼポリペプチド)を用いて生成されるRSウイルスFポリペプチドおよびVLPが含まれるが、これらに限定されない。
【0025】
「哺乳動物グリコシル化」とは、哺乳動物細胞の発現系により生成されるグリコシル化パターンを指す。このようなグリコシル化パターンは、哺乳動物のグリコシル化酵素を包含するように改変された昆虫細胞が、哺乳動物または哺乳動物細胞ベースの発現系により天然にもたらされるグリコシル化パターンではなく「哺乳動物様の」グリコシル化のみをもたらす場合に限り、このような改変昆虫細胞によりもたらされるグリコシル化パターンを包含しない。哺乳動物グリコシル化パターンの好ましい例には、ヒト(例えば、HEK293、HeLa)細胞、チャイニーズハムスター卵巣(CHO)細胞、イヌ(例えば、MDCK)細胞、マウス(例えば、H9)細胞、ラット(例えば、IE)細胞、およびヒト以外の霊長動物(例えば、NCTC)細胞内における発現によりもたらされるグリコシル化が含まれる。
【0026】
本明細書で用いられる「エンベロープウイルスコア形成ポリペプチド」には、エンベロープウイルスのエンベロープ内において、ウイルスカプシド、ヌクレオカプシド、またはコアを形成することが可能な、任意のウイルスポリペプチド、または任意のウイルスポリペプチドの組合せが含まれる。エンベロープウイルスコア形成ポリペプチドは、RSウイルスFポリペプチドのVLPの形成を干渉し得るので、本明細書で記載されるRSウイルスFポリペプチドのVLPの好ましい実施形態は、このようなポリペプチドを除外する。ウイルスカプシド、ヌクレオカプシド、もしくはコアを形成することが不可能な、エンベロープウイルスコア形成ポリペプチドに由来するポリペプチド、またはウイルスカプシド、ヌクレオカプシド、もしくはコアを形成することが不可能な、エンベロープウイルスコア形成ポリペプチドのサブセットは、本明細書で開示されるVLPの形成を干渉しないので、このようなポリペプチドは、本明細書で開示されるVLPに含まれ得る。
【0027】
本明細書で記載されるウイルス様粒子を形成する一因となる、レトロウイルス由来の構造ポリペプチドであるgagポリペプチドは、エンベロープウイルスコア形成ポリペプチドの例である。レトロウイルスのゲノムは、3つの主要な遺伝子産物をコードする:構造タンパク質をコードするgag遺伝子の産物;逆転写酵素および関連のタンパク質分解性ポリペプチド、ヌクレアーゼおよびインテグラーゼに関連する機能をコードするpol遺伝子の産物;envの産物(それによりコードされる糖タンパク質の膜タンパク質が、感染細胞の表面上において、また、放出された成熟ウイルス粒子の表面上においても検出される)。すべてのレトロウイルスのgag遺伝子は、全体的な構造類似性を有し、レトロウイルスの各群内においても、アミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除くコアタンパク質をもたらす。
【0028】
MLVの場合、Gagの前駆体であるポリタンパク質は、Pr65Gagであり、該前駆体上におけるその順序がNH2−p15−pp12−p30−p10−COOHである4つのタンパク質へと切断される。これらの切断は、ウイルス性プロテアーゼを介し、また、ウイルスに応じて、ウイルス放出の前または後において生じ得る。MLV Gagタンパク質は、グリコシル化形態および非グリコシル化形態で存在する。グリコシル化形態はgPr80Gagから切り出され、gPr80Gagは、非グリコシル化Pr65Gagに対応するAUGコドンの上流に位置する、インフレームの、異なる開始コドンから合成される。グリコシル化Gagを合成しない、MLVの欠失変異体がなおも感染性であり、また、非グリコシル化Gagがなおもウイルス様粒子を形成し得ることから、グリコシル化イベントの重要性について、疑問が投げかけられる。ウイルスがコードするプロテアーゼにより、HIV−1 Gagの前駆体であるpr55Gagが翻訳後に切断されることから、N−ミリストイル化されて内部でリン酸化されたp17マトリックスタンパク質(p17MA)、リン酸化されたp24カプシドタンパク質(p24CA)、ならびにヌクレオカプシドタンパク質であるp15(p15NC)がもたらされ、p15は、さらにp9およびp6へと切断される。
【0029】
構造面において、原型的なGagポリタンパク質は、レトロウイルスのgag遺伝子において常に同じ順序で生じる3つの主要なタンパク質:マトリックスタンパク質(MA)(マトリックスという名称を共有するが、MAとは異なるタンパク質である、インフルエンザマトリックスタンパク質M1と混同しないようにされたい)、カプシドタンパク質(CA)、およびヌクレオカプシドタンパク質(NC)に分けられる。成熟タンパク質へのGagポリタンパク質のプロセシングは、レトロウイルスがコードするプロテアーゼにより触媒され、新たに出芽するウイルス粒子が成熟するときに生じる。機能面において、Gagポリタンパク質は、3つのドメイン:細胞膜をGagポリタンパク質の標的とする膜結合ドメインと;Gagの重合化を促進する相互作用ドメインと;宿主細胞からの新生ビリオンの放出を促進する後期ドメインとに分けられる。アセンブリーを媒介するGagタンパク質の形態は、ポリタンパク質である。したがって、アセンブリードメインは、後期に形成される任意の切断産物内にもれなく収まる必要はない。これらの重要な機能的エレメントに関する最新技術の進歩は大きい。例えば、
【0030】
【化1】
を参照されたい。
【0031】
エンベロープウイルスコア形成ポリペプチドの範囲内に包含されるgagポリペプチドは、最小限において、ウイルス様粒子を形成するための機能的エレメントを、RSウイルスFポリペプチドのVLPとは独立して包含することになる。RSウイルスFポリペプチドと競合するか、またはこれに干渉する機能的エレメントを除外すれば、このようなgagポリペプチドは、エンベロープウイルスコア形成ポリペプチドではない。
【0032】
Gagポリペプチドについてのレトロウイルスの供給源の例には、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス、ヤーグジークテヒツジレトロウイルス、およびメーソン−ファイザーサルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトTリンパ向性ウイルス、およびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、またはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ感染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)が含まれる。
【0033】
本明細書で用いられる「エンベロープウイルスコア」には、エンベロープウイルスまたはエンベロープウイルスの成分により生成されるウイルス様粒子の脂質膜またはエンベロープの内側で見出される任意のタンパク質性コアが含まれる。タンパク質性コアは、カプシドもしくはヌクレオカプシド、またはマトリックスの全部または一部を含み得る。
【0034】
本明細書で用いられる「脂質ラフト」とは、ウイルス粒子のアセンブリー過程においてgagポリペプチドが濃縮される、細胞膜のマイクロドメインを指す。
【0035】
本明細書で用いられる「脂質ラフト会合ポリペプチド」とは、脂質ラフトと直接的または間接的に会合する任意のポリペプチドを指すが、任意のエンベロープウイルスコア形成ポリペプチドは除外する。本発明で用いられる具体的な脂質ラフト会合ポリペプチドは、VLPの所望の使用、および該脂質ラフト会合ポリペプチドの役割(例えば、RSウイルスFタンパク質の細胞外部分を該VLPへと結合させること、または1つもしくは複数のさらなる抗原もしくはアジュバントを該VLPへと結合させること)に依存する。
【0036】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質もしくは脂質ラフト会合部分;脂質による修飾など、膜との会合を引き起こすタンパク質の修飾により脂質ラフトと直接的に会合するタンパク質もしくはその部分;または脂質ラフト会合ポリペプチドにより脂質ラフトと間接的に会合するポリペプチドであり得る。
【0037】
脂質アンカーを有する多くのタンパク質が、脂質ラフトと会合する。このようなタンパク質の短い断片は、脂質結合に十分であることが多い。このような断片は、天然においてそれ自体では脂質ラフトと会合しないであろう他のタンパク質およびポリペプチドに容易に結合し得るので、該断片は、脂質ラフト会合に理想的となっている。ポリペプチドを脂質ラフトへと連結する脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、および二重アセチル化が含まれる。
【0038】
多くの異なる種類のポリペプチドが、脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスのアセンブリー、また、アセンブルした粒子の出芽を含めた、多くの生物学的活性のためのプラットフォームとして機能し、したがって、これらの過程に関与する各種のポリペプチドと会合する。
【0039】
シグナル伝達カスケードに関与する各種のポリペプチドは、シグナル伝達プラットフォームとして機能する脂質ラフトと会合する。シグナル伝達プラットフォームとして機能する脂質ラフトの1つの種類は、カベオラと呼ばれている。それは、カベオリンファミリーに由来するポリペプチド(例えば、カベオリンおよび/またはフロチリン(flottillin))を含有する、細胞膜のフラスコ型の陥入部である。
【0040】
膜輸送ポリペプチドは、膜輸送プラットフォームとして機能する脂質ラフトと会合する。例には、シンタクシン−1タンパク質、シンタクシン−4タンパク質、シナプシンIタンパク質、アデューシンタンパク質、VAMP2タンパク質、VAMP/シナプトブレビンタンパク質、シナプトブレビンIIタンパク質、SNAREタンパク質、SNAP−25タンパク質、SNAP−23タンパク質、シナプトタグミンIタンパク質、およびシナプトタグミンIIタンパク質など、エンドサイトーシスおよびエクソサイトーシスに関与するタンパク質が含まれる。
【0041】
ウイルス受容体、ウイルス受容体−共受容体複合体、ウイルス侵入過程の調節を補助する他の任意の成分は、ウイルス侵入に特化した膜輸送プラットフォームとして機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体の例には、崩壊促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー膜糖タンパク質;ガングリオシド、Hsc70タンパク質、アルファ2−ベータ1インテグリンおよびアルファ5−ベータ2インテグリンを含めた複数の成分を含有する複合体である、A群ロタウイルスの受容体;HIVウイルス、MLVウイルス、麻疹ウイルス、およびEbolaウイルスなど、複数のエンベロープウイルスの糖タンパク質;ならびにCD5ポリペプチド、CCR5ポリペプチド、およびnefポリペプチドなど、HIVの侵入に関与するポリペプチドが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0042】
ウイルス粒子のアセンブリーに関与するポリペプチドは、ウイルスアセンブリープラットフォームとして機能する脂質ラフトと会合する。ウイルスのヌクレオカプシド、カプシド、またはコアを形成する一因となる部分を除外する限りにおいて、このようなポリペプチドまたはそれらの一部を、脂質ラフト会合ポリペプチドとして用いることができる。このようなポリペプチドの例には、HAインフルエンザエンベロープ糖タンパク質およびNAインフルエンザエンベロープ糖タンパク質、麻疹ウイルスに由来するHタンパク質および成熟F1−F2融合タンパク質、ならびにHIVウイルスに由来するgp160、gp41、およびPr55gagが含まれる。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. & Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。
【0043】
アセンブルしたウイルスの出芽に関与するポリペプチドは、ウイルス出芽プラットフォームとして機能する脂質ラフトと会合する。宿主細胞からのHIV−1の出芽は、膜ラフト内で生じることを示唆するデータが存在する。ChazalおよびGerlier、2003年、「Virus Entry, Assembly, Budding, and Membrane Rafts」、Microbiol. And Mol. Bio. Rev.、67巻(2号):226〜237頁を参照されたい。ウイルスの出芽に関与するポリペプチドについての一般的な情報は、「Fields Virology」(第4版)、2001年において見出すことができる。
【0044】
好ましい脂質ラフト会合ポリペプチドには、赤血球凝集素ポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質ポリペプチド、糖タンパク質ポリペプチド、およびエンベロープタンパク質ポリペプチドなどのウイルスポリペプチドが含まれる。これらのポリペプチドの各々は、任意の種類のウイルスに由来し得るが、特定の実施形態は、HIV−1ウイルスに由来するエンベロープタンパク質、RSウイルスまたは麻疹ウイルスに由来する融合タンパク質、RSウイルス、単純ヘルペスウイルス、またはエボラウイルスに由来する糖タンパク質、また、麻疹ウイルスに由来する赤血球凝集素タンパク質を包含する。
【0045】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvumなどが含まれるがこれらに限定されない病原性の原虫、蠕虫、また、他の真核微生物の病原体から得ることができる。このような非ウイルス性脂質ラフト会合ポリペプチドは、それ自体が抗原として作用するので、天然では脂質ラフトと会合しない抗原に連結することなく用いることができる。
【0046】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、赤血球凝集素ポリペプチドである。本明細書で用いられる「赤血球凝集素ポリペプチド」は、感染される細胞に対するウイルスの結合を媒介するインフルエンザウイルスタンパク質に由来する。赤血球凝集素ポリペプチドはまた、同等の麻疹ウイルスタンパク質にも由来し得る。このタンパク質は、単一の膜貫通ドメインにより、インフルエンザウイルスの表面へとアンカリングすることが見出される抗原性の糖タンパク質である。インフルエンザ赤血球凝集素のうち、H1〜H16と名付けられた、少なくとも16のサブタイプが同定されている。H1、H2、およびH3は、ヒトインフルエンザウイルスにおいて見出される。H5またはH7の赤血球凝集素を有する、高病原性鳥類インフルエンザウイルスがヒトに感染する割合は低いことが判明している。鳥類ウイルス株のH5型赤血球凝集素内における単一のアミノ酸変化がヒト患者において見出されており、これが受容体の特異性を変化させることを可能にし、H5赤血球凝集素が鳥類H5N1ウイルスの受容体特異性を顕著に変化させることによって、これらのウイルスに、ヒト受容体に結合する能力がもたらされると報告されている(109および110)。この知見は、通常ヒトに感染しないH5N1ウイルスが変異し、ヒト細胞に効率的に感染できるようになることを説明する。
【0047】
赤血球凝集素は、ホモ三量体の内在性膜ポリペプチドである。天然では、その膜貫通ドメインが、脂質ラフトドメインと会合し、これにより、RSウイルスFポリペプチドと会合することが可能となり、VLP内へと組み込まれる。それは、円筒様の形状をしており、約135Åの長さである。HAを構成する3つの同一の単量体は、中央のコイルドコイルと、VLPの表面上に露出されるシアル酸結合部位を含有する球形の頭状部分とを形成する。HAの単量体は、グリコシル化され、2つのより小型のポリペプチド:HA1サブユニットおよびHA2サブユニットへと切断される、単一のポリペプチド前駆体として合成される。HA2サブユニットは、膜へとアンカリングされる、三量体のコイルドコイルを形成し、HA1サブユニットは、球形の頭状部分を形成する。
【0048】
本発明の特定のVLPにおいて脂質ラフト会合ポリペプチドとして用いられる赤血球凝集素ポリペプチドは、最小限において、膜アンカードメインを包含するものとする。赤血球凝集素ポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、H1、H2、H3、H5、H7、およびH9赤血球凝集素に由来することが好ましい。加えて、赤血球凝集素ポリペプチドは、異なるインフルエンザ赤血球凝集素のキメラでもあり得る。赤血球凝集素ポリペプチドは、1または複数のさらなるポリペプチドのコード配列を、赤血球凝集素ポリペプチドのコード配列にスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない、1または複数のさらなる抗原を包含することが好ましい。赤血球凝集素ポリペプチド内へのさらなるポリペプチドの挿入に好ましい部位は、N末端である。
【0049】
ウイルス性脂質ラフト会合ポリペプチドの好ましい別の例は、ノイラミニダーゼポリペプチドである。本明細書で用いられる「ノイラミニダーゼポリペプチド」は、糖タンパク質から末端のシアル酸残基を切断することにより、細胞からのインフルエンザウイルスの放出を媒介する、インフルエンザウイルスタンパク質に由来する。ノイラミニダーゼ糖タンパク質は、ウイルス表面上において発現する。ノイラミニダーゼタンパク質は四量体であり、ベータ−ピンホイール構造を有する球形の頭状部分、細いストーク状領域、また、単一の膜貫通ドメインによりウイルス膜内に該タンパク質をアンカリングする小さな疎水性領域からなる共通構造を共有する。シアル酸残基を切断するための活性部位は、すべてのインフルエンザAウイルス内に保存されている、15の荷電アミノ酸により形成される、各サブユニット表面上のポケットを包含する。インフルエンザノイラミニダーゼのうち、N1〜N9と名付けられた、少なくとも9つのサブタイプが同定されている。
【0050】
本発明の特定のVLPにおいて用いられるノイラミニダーゼポリペプチドは、最小限において、膜アンカードメインを包含するものとする。機能的領域に関する最新技術は極めて高度である。例えば、Vargheseら、Nature、303巻、35〜40頁、1983年; Colmanら、Nature、303巻、41〜44頁、1983年; Lentzら、Biochem、26巻、5321〜5385頁、1987年; Websterら、Virol.、135巻、30〜42頁、1984年を参照されたい。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルス型、インフルエンザウイルス亜型、インフルエンザウイルス株、またはインフルエンザウイルス亜株に由来することが可能であり、N1ノイラミニダーゼおよびN2ノイラミニダーゼに由来することが好ましい。加えて、ノイラミニダーゼポリペプチドは、異なるインフルエンザノイラミニダーゼのキメラでもあり得る。ノイラミニダーゼポリペプチドは、1または複数のさらなるポリペプチドのコード配列を赤血球凝集素ポリペプチドにスプライシングすることにより作製し得る、天然では脂質ラフトと会合しない1または複数のさらなる抗原を包含することが好ましい。ノイラミニダーゼポリペプチドコード配列内へのさらなるポリペプチドの挿入に好ましい部位は、C末端である。
【0051】
脂質ラフト会合ポリペプチドの好ましい別の例は、ファシクリンI(FasI)と称する、昆虫由来の接着タンパク質である。本明細書で用いられる「ファシクリンIポリペプチド」は、胚の発生に関与する昆虫タンパク質に由来する。この非ウイルス性タンパク質は、昆虫細胞内のバキュロウイルス発現系において発現させることができ、これにより、FasIの脂質ラフト会合がもたらされる(J. Virol.、77巻、6265〜6273頁、2003年)。したがって、ファシクリンIポリペプチドに異種抗原を結合させて、RSウイルスFポリペプチドと共に共発現させると、VLP内にキメラ分子が組み込まれる。本発明のVLPにおいて用いられるファシクリンIポリペプチドは、最小限において、膜アンカードメインを包含するものとする。
【0052】
脂質ラフト会合ポリペプチドの好ましい別の例は、G糖タンパク質と称する、RSVに由来するウイルス性付着タンパク質である。本明細書で用いられる「Gグリコポリペプチド」は、RSV G糖タンパク質に由来する。近年のデータは、脂質ラフトドメインが、インフルエンザウイルスにとって重要であるのと同様に、RSV粒子の出芽にも重要であることを示している(Virol、327巻、175〜185頁、2004年; Arch. Virol.、149巻、199〜210頁、2004年; Virol.、300巻、244〜254頁、2002年)。RSVに由来するG糖タンパク質は、ウイルス性付着タンパク質であり、およびRSV感染に対する防御抗原として機能する、32.5kdの内在性膜タンパク質である。インフルエンザウイルスに由来する赤血球凝集素と同様、その抗原性は、それに付着した、任意の非脂質ラフト抗原の抗原性を増強し得る。非脂質ラフト外来抗原を付着させる形での、Gグリコポリペプチドに対する任意の改変は、結果として、該外来抗原に対して顕著な免疫反応を誘導することが可能なRSウイルスFポリペプチドのVLPをもたらす。
【0053】
VLPが、その文脈により、エンベロープベースのウイルスに基づかないか、または本明細書で開示される特定のエンベロープベースのウイルスの特定の成分に基づくウイルス様粒子を指す場合を除き、用語「RSウイルスFポリペプチドのウイルス様粒子」、「RSウイルスFポリペプチドのVLP」および「VLP」は、本明細書の全体において互換的に用いられる。
【0054】
(抗原)
本発明の特定の態様は、RSウイルスFポリペプチドのVLP調製物と会合するさらなる抗原を包含する。このようなさらなる抗原は、同じ組成物中に組み込むことができ、さらにまた、VLPと共有結合させることもでき、非共有結合させることもできる。好ましい実施形態では、RSウイルスFポリペプチド、および/または他の脂質ラフト会合ポリペプチドが、天然では脂質ラフトと会合しないであろう抗原を含有する、RSウイルスFポリペプチドのVLPを形成するのに容易に適合可能なプラットフォームである。本節では、開示されるVLPと共に用いるのに好ましい抗原について記載する。
【0055】
(抗原と脂質ラフト会合ポリペプチドとの連結)
天然では脂質ラフトと会合しない抗原を含有するVLPを形成する手段として、RSウイルスFポリペプチドおよび/または別の脂質ラフト会合ポリペプチドと、該抗原との連結を形成することができる。単一の抗原または複数の抗原に脂質ラフト会合ポリペプチドを連結することにより、VLPの免疫原性を増大させるか、各種の病原体に免疫原性を付与するか、または、特定の病原体の各種の株に免疫原性を付与することができる。
【0056】
抗原と脂質ラフト会合ポリペプチドとの間の連結は、該抗原を結果としてVLP内へと組み込むのに十分な任意の種類の連結であり得る。結合は、共有結合、イオン性相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であり得る。好ましい実施形態では、結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合などの共有結合である。
【0057】
抗原は、脂質ラフト会合ポリペプチドに対する既存の結合を伴なって組換えにより作製することもでき、単離物質として作製し、次いで、後に、脂質ラフト会合ポリペプチドに連結することもできる。
【0058】
(抗原の種類)
本明細書で用いられる抗原は、免疫反応を誘発することが可能であり、また、天然では脂質ラフトと会合しない任意の物質であり得る。抗原には、タンパク質、ポリペプチド(活性タンパク質、また、タンパク質内における個々のポリペプチドエピトープを含めた)、グリコポリペプチド、リポポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。天然において、抗原が脂質ラフトと直接的または間接的に会合しない場合、脂質ラフト会合ポリペプチドに連結しない限り、それをVLP内へと組み込むことは期待されない。抗原は、疾患または障害に関与する任意の抗原、例えば、微生物抗原(例えば、ウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、アレルゲンなどであり得る。
【0059】
(抗原の供給源)
本明細書で記載される抗原は、化学的または酵素的に合成することもでき、組換えにより作製することもでき、天然の供給源から単離することもでき、前出の組合せでもあり得る。抗原は、精製することもでき、部分精製することもでき、粗抽出物でもあり得る。
【0060】
ポリペプチド抗原は、液体クロマトグラフィー(例えば、高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ除外クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含めた)、アフィニティークロマトグラフィー、または他の精製法が含まれるがこれらに限定されない、当技術分野で公知の標準的なタンパク質精製法を用いて、天然の供給源から単離することができる。多くの実施形態では、抗原が、例えば、約50%〜約75%純粋、約75%〜約85%純粋、約85%〜約90%純粋、約90%〜約95%純粋、約95%〜約98%純粋、約98%〜約99%純粋、または99%超純粋の精製抗原である。
【0061】
固相ペプチド合成法を用いることができるが、このような技法は、当業者に公知である。Jones、「The Chemical Synthesis of Peptides」(オックスフォード、Clarendon Press社)(1994年)を参照されたい。一般に、このような方法では、活性化させた単量体のユニットを、伸長するペプチド鎖を結合させた固相へと連鎖的に付加することにより、ペプチドを作製する。
【0062】
ポリペプチドを作製するには、十分に確立された組換えDNA法を、脂質ラフト会合ポリペプチドと同じベクター内において用いることもでき、この場合、例えば、ポリペプチドをコードするヌクレオチド配列を含む発現構築物は、適切な宿主細胞(例えば、インビトロの細胞培養物における単細胞実体として増殖させる真核宿主細胞、例えば、酵母細胞、昆虫細胞、哺乳動物細胞など)または、原核細胞(例えば、インビトロの細胞培養物中)中に導入され、これにより遺伝的に改変された宿主細胞を作製し、適切な培養条件下において、該遺伝的に改変された宿主細胞によりタンパク質を作製する。
【0063】
(ウイルス抗原)
適切なウイルス抗原には、以下の群:Retroviridae科(例えば、HIV−1などのヒト免疫不全ウイルス(また、HTLV−III、LAV、もしくはHTLV−III/LAV、またはHIV−IIIとも称する));また、HIV−LPなど、他の単離物; Picomaviridae科(例えば、ポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス); Calciviridae科(例えば、ノーウォークウイルスおよび関連ウイルスを含めた、胃腸炎を引き起こすウイルス株); Togaviridae科(例えば、ウマ脳炎ウイルス、風疹ウイルス); Flaviridae科(例えば、デング熱ウイルス、脳炎ウイルス、黄熱病ウイルス); Coronoviridae科(例えば、コロナウイルス); Rhabdoviradae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Coronaviridae科(例えば、コロナウイルス); Rhabdoviridae科(例えば、水疱性口内炎ウイルス、狂犬病ウイルス); Filoviridae科(例えば、エボラウイルス); Paramyxoviridae科(例えば、パラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、RSウイルス); Orthomyxoviridae科(例えば、インフルエンザウイルス); Bungaviridae科(例えば、ハンタンウイルス、ブンガウイルス、フレボウイルス、およびナイロウイルス); Arena viridae科(出血熱ウイルス); Reoviridae科(例えば、レオウイルス、オルビウイルス、およびロタウイルス); Bimaviridae科; Hepadnaviridae科(B型肝炎ウイルス); Parvovirida科(パルボウイルス); Papovavirida(パピローマウイルス、ポリオーマウイルス); Adenoviridae科(大半のアデノウイルス); Herpesviridae科(1型および2型の単純ヘルペスウイルス(HSV)、水痘帯状疱疹ウイルス、サイトメガロウイルス(CMV)、ヘルペスウイルス); Poxyiridae科(痘瘡ウイルス、ワクシニアウイルス、ポックスウイルス);およびIridoviridae科(例えば、アフリカブタコレラウイルス);ならびに未分類のウイルス(例えば、海綿状脳症の病因因子、デルタ肝炎の因子(B型肝炎ウイルスの欠損サテライトウイルスと思われる)、非A型肝炎、非B型肝炎の因子(クラス1=内部感染による;クラス2=非経口感染による(すなわち、C型肝炎));およびアストロウイルス)のうちの1または複数のウイルスと関連する(例えば、これらにより合成される)抗原が含まれる。
【0064】
(ノロウイルス(Norvirus)抗原)
本明細書で開示されるVLPには、ノロウイルス科に由来する各種の抗原が含まれ得ることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、Caliciviridaeウイルス科内の4つの属のうちの1つを占める。ノロウイルス属内には、遺伝子群Iおよび遺伝子群IIと名付けられた、2つの主要な遺伝子群が存在する。遺伝子群Iのノロウイルス株には、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスが含まれる。遺伝子群IIのノロウイルス株には、ヒューストンウイルス、ハワイウイルス、ローズデールウイルス、グリムズビーウイルス、メキシコウイルス、およびスノーマウンテンエージェントが含まれる(Parker, T.D.ら、J. Virol.(2005年)、79巻(12号):7402〜9頁; Hale, A.D.ら、J Clin. Micro.(2000年)、38巻(4号):1656〜1660頁)。ノーウォークウイルス(NV)は、世界中における、大半の急性ウイルス性胃腸炎の流行的発生の一因となる、ヒトカリチウイルス群の原型株である。ノーウォークウイルスのカプシドタンパク質は、2つのドメイン:殻ドメイン(S)および突出部ドメイン(P)を有する。Pドメイン(アミノ酸226〜530;ノーウォーク株の番号付け)は、2つのサブドメインであるP1およびP2に分けられる。P2ドメインは、P1ドメイン内における127アミノ酸の挿入(アミノ酸279〜405)であり、フォールディングされた単量体の最遠位面に位置する。P2ドメインは、ノロウイルス株のうちで最も保存の程度が低いVP1領域であり、P2内の超可変領域は、受容体の結合および免疫反応性において重要な役割を果たすと考えられている。Pドメインが外部に位置することを踏まえるなら、それは、本明細書で開示されるVLPワクチンのための抗原として用いるのに好ましい抗原、またはポリペプチドエピトープの供給源である。P2ドメインは、遺伝子群Iまたは遺伝子群IIのノロウイルス株に好ましい抗原である。広範にわたるノロウイルス株を通じて保存されるP2ドメインのうちのある領域内に存在するエピトープとして近年になって同定されたmAb 61.21エピトープ、ならびにmAb 54.6エピトープ(Lochridge, V.P.ら、J Gen. Virol.(2005年)、86巻:2799〜2806頁)がさらにより好ましい。
【0065】
(インフルエンザ抗原)
本明細書で開示されるVLPは、赤血球凝集素、ノイラミニダーゼ、またはさらなるインフルエンザ抗原を非制限的に含む、インフルエンザに由来する各種の抗原を包含し得る。さらなる好ましいインフルエンザ抗原は、M2ポリペプチドである。インフルエンザウイルスのM2ポリペプチドは、スプライシングイベント後において、RNAセグメント7(マトリックスセグメント)によりコードされる、97アミノ酸の小型のクラスIII内在性膜タンパク質である(80、81)。ウイルス粒子上に存在するM2は極めてわずかであり、感染細胞上においてより豊富に見出すことができる。M2は、ウイルスの侵入に必要な、プロトン選択性イオンチャネルとして用いられる(82、83)。感染中または従来のワクチン接種中の免疫原性は最小限であることから、その保存が説明されるが、代替的なフォーマットで存在する場合は、より免疫原性であり防御性である(84〜86)。これは、インビボにおけるM2モノクローナル抗体の受動伝達により、ウイルスクリアランスが加速化され、結果として防御がもたらされるという観察(87)と符合する。M2の外部ドメインエピトープを、融合タンパク質として、HBVのコア粒子に連結すると、非経口接種および鼻腔内接種のいずれによっても、マウスにおいて防御性であり、3回の繰り返しコピーを該コアタンパク質のN末端へと融合させると、最も免疫原性となる(88〜90)。これは、エピトープ密度が増大すると免疫原性が増大することを示す他のキャリア−ハプテンデータ(91)と符合する。
【0066】
M2ワクチンを鼻腔内送達する場合、良好な防御を達成するにはアジュバントが必要とされ、LTR192G(88、90)およびCTA1−DD(89)により良好な結果が達成されている。ペプチドはまた、KLH、またはN. meningitidesの外膜タンパク質複合体、またはヒトパピローマウイルスのVLPなどのキャリアに化学的に結合体化することもでき、マウスおよび他の動物におけるワクチンとして防御的である(92、93)。
【0067】
M2タンパク質は、高度に保存的ではあるが、配列の相違を全く示さないわけではない。一般的な株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除き、今日配列決定されている他のすべてのヒト株と、免疫学的に交差反応性であることが示された(92)。インフルエンザデータベースの配列の検討からも、A/Vietnam/1203/04など、他のより近年の病原性H5N1ヒト単離物のM2配列内において、同様の相違が示されている。この知見は、M2エピトープを組み込む、有効なH5特異的汎流行ワクチンが、ヒトH1単離物およびH3単離物中において現在広まっているM2配列ではなく、病原性の鳥類株に固有のM2配列を反映する必要があることを裏付ける。
【0068】
インフルエンザウイルスに由来するさらなるタンパク質(HA、NA、およびM2以外のタンパク質)も、共発現により、またはさらなる抗原の全部もしくは一部を、RSウイルスFポリペプチドもしくは他の脂質ラフト会合ポリペプチドに連結することにより、VLPワクチン内へと組み込むことができる。これらのさらなる抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、およびNS2が含まれる。これら後者の抗原は、一般に、中和抗体反応の標的ではなく、T細胞により認識される重要なエピトープを含有し得る。このようなエピトープに対してVLPワクチンにより誘導されるT細胞反応が、防御的免疫性を促進するのに有益であることが判明し得る。
【0069】
(他の病原性抗原)
適切な細菌性抗原には、例えば、病原性Pasteurella属種、Staphylococci属種、およびStreptococcus属種などの病原性グラム陽性菌;また、Neisseria属、Escherichia属、Bordetella属、Campylobacter属、Legionella属、Pseudomonas属、Shigella属、Vibrio属、Yersinia属、Salmonella属、Haemophilus属、Brucella属、Francisella属、およびBacterioides属のグラム陰性菌などのグラム陰性菌を含めた、各種の病原性細菌のうちのいずれかと関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。例えば、Schaechter, M、H. Medoff、D. Schlesinger、「Mechanisms of Microbial Disease」、ボルチモア、Williams and Wilkins社(1989年)を参照されたい。
【0070】
感染性の病原性真菌と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Cryptococcus neoformans、Histoplasma capsulatum、Coccidioides immitis、Blastomyces dermatitidis、およびCandida albicans、Candida glabrata、Aspergillus fumigata、Aspergillus flavus、およびSporothrix schenckiiが含まれるがこれらに限定されない感染性真菌と関連する抗原が含まれる。
【0071】
病原性原虫、蠕虫、および他の真核微生物病原体と関連する(例えば、これらにより合成され、また、これらに内因性である)適切な抗原には、Plasmodium falciparum、Plasmodium malariae、Plasmodium ovale、およびPlasmodium vivaxなどのPlasmodium属; Toxoplasma gondii; Trypanosoma brucei、Trypanosoma cruzi; Schistosoma haematobium、Schistosoma mansoni、Schistosoma japonicum; Leishmania donovani; Giardia intestinalis; Cryptosporidium parvum等が含まれるがこれらに限定されない原虫、蠕虫、および他の真核微生物病原体と関連する抗原が含まれる。
【0072】
適切な抗原には、Helicobacter pyloris、Borelia burgdorferi、Legionella pneumophila、Mycobacteria属種(例えば、M. tuberculosis、M. avium、M. intracellulare、M. kansaii、M. gordonae)、Staphylococcus aureus、Neisseria gonorrhoeae、Neisseria meningitidis、Listeria monocytogenes、Chlamydia trachomatis、Streptococcus pyogenes(Streptococcus属のA群)、Streptococcus agalactiae(Streptococcus属のB群)、Streptococcus属(viridans群)、Streptococcus faecalis、Streptococcus bovis、Streptococcus属(嫌気性の種)、Streptococcus pneumoniae、病原性のCampylobacter属種、Enterococcus属種、Haemophilus influenzae、Bacillus anthracis、Corynebacterium diphtheriae、Corynebacterium(corynebacterium)属種、Erysipelothrix rhusiopathiae、Clostridium perfringens、Clostridium tetani、Enterobacter aerogenes、Klebsiella pneumoniae、Pasturella multocida、Bacteroides属種、Fusobacterium nucleatum、Streptobacillus moniliformis、Treponema pallidium、Treponema pertenue、Leptospira属、Rickettsia属、およびActinomyces israeliなどの病原性微生物と関連する(例えば、これらにより合成され、また、これらに内因性である)抗原が含まれる。病原性E. coli株の非限定的な例は、ATCC第31618号、同第23505号、同第43886号、同第43892号、同第35401号、同第43896号、同第33985号、同第31619号、および同第31617号である。
【0073】
細胞内病原体と関連する各種のポリペプチドまたは他の抗原のうちのいずれかを、VLP内に組み込むことができる。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープとは、その断片が、MHCクラスI分子と併せて、CD8+リンパ球の表面上におけるT細胞抗原受容体により認識される、例えば、これにより結合されるように、感染細胞の表面上に提示される、細胞内病原体と関連する(例えば、これによりコードされる)任意のポリペプチドである。細胞内病原体と関連するポリペプチドエピトープおよびペプチドエピトープは当技術分野において公知であり、これらには、ヒト免疫不全ウイルス、例えば、HIV gp120またはその抗原性断片と関連する抗原;サイトメガロウイルス抗原; Mycobacterium属抗原(例えば、Mycobacterium avium、Mycobacterium tuberculosisなど);Pneumocystic carinii(PCP)抗原;41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど、Plasmodium falciparumまたは他の任意のマラリア種と関連する抗原が含まれるがこれらに限定されない、マラリア抗原;真菌抗原;酵母抗原(例えば、Candida属種抗原); Toxoplasma gondii、Toxoplasma encephalitis、または他のToxoplasma属種と関連する抗原が含まれるがこれらに限定されないトキソプラズマ抗原;エプスタイン−バーウイルス(EBV)による抗原; Plasmodium属抗原(例えば、gp190/MSP1など)などが含まれるが、これらに限定されない。
【0074】
好ましいVLPワクチンは、Bacillus anthracisに対するものであり得る。Bacillus anthracisは、好気性または通性嫌気性でグラム陽性の非運動性桿菌であり、幅1.0μm、長さ3.0〜5.0μmである。B. anthracisは、悪条件下でも耐性の高い内性胞子を形成し、これは、感染した動物がかつて死亡した場所の土壌で見出すことができる。本明細書で開示されるVLPワクチン内で用いるのに好ましい抗原は、哺乳動物細胞上の受容体に結合し、B. anthracisが疾患を引き起こす能力にとって極めて重要な83kDaのタンパク質である防御抗原(PA)である。より好ましい抗原は、被験体においてBacillus anthracisに対する防御的免疫性を誘導するのに用い得る、Bacillus anthracis PAのC末端における140アミノ酸の断片である。炭疽菌に対するVLPワクチン内において用いられる他の例示的な抗原は、炭疽菌胞子に由来する抗原(例えば、BclA)、該細菌の栄養期に由来する抗原(例えば、細胞壁抗原、被膜抗原(例えば、ポリ−ガンマ−D−グルタミン酸、またはPGA)、分泌抗原(例えば、防御抗原、致死性因子、または浮腫因子などの外毒素))である。VLPワクチンで用いるのに好ましい別の抗原は、B. anthracisに固有のアントロースを含有する四糖である(Daubenspeck J.M.ら、J. Biol. Chem.(2004年)、279巻:30945頁)。四糖は、脂質ラフト会合ポリペプチドに連結することができ、これにより、VLPワクチンとの抗原の会合が可能となる。
【0075】
(腫瘍関連抗原)
公知の各種腫瘍特異抗原または腫瘍関連抗原(TAA)のうちのいずれかを、VLPに組み込むことができる。全TAAを用い得るが、その必要はない。代わりに、TAAの一部、例えば、エピトープを用いることができる。VLP内で用い得る腫瘍関連抗原(またはそれらのエピトープを含有する断片)には、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量黒色腫関連抗原であるMAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原であるOV−TL3およびMOV18、TUAN、アルファ−フェトプロテイン(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原であるG250、EGP−40(EpCAMとしても知られる)、S100(悪性黒色腫関連抗原)、p53、およびp21rasが含まれるがこれらに限定されない。前出のうちのいずれかを含めた、任意のTAA(またはそれらのエピトープ)の合成類似体を用いることができる。さらに、1または複数のTAA(またはそれらのエピトープ)の組合せを、組成物中に組み込むことができる。
【0076】
(アレルゲン)
一態様では、VLPワクチンの一部である抗原が、各種のアレルゲンのうちのいずれかであり得る。アレルゲンベースのワクチンを用い、被験体におけるアレルゲンに対する忍容性を誘導することができる。チロシンとの共沈殿(co−precipitation)を伴うアレルゲンワクチンの例は、米国特許第3,792,159号、同第4,070,455号、および同第6,440,426号において見出すことができる。
【0077】
各種のアレルゲンのうちのいずれかをVLP内に組み込むことができる。アレルゲンには、環境空中アレルゲン;ブタクサ/花粉症などの植物の花粉;雑草花粉アレルゲン;芝草花粉アレルゲン;ジョンソングラス;樹木花粉アレルゲン;ライグラス;イエダニアレルゲン(例えば、Der p I、Der f Iなど)などのクモ形類動物アレルゲン;コナダニ(storage mite)アレルゲン;スギ花粉/花粉症;カビ胞子アレルゲン;動物アレルゲン(例えば、イヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどによるアレルゲン);食物アレルゲン(例えば、甲殻類;ラッカセイなどの堅果;柑橘類によるアレルゲン);昆虫アレルゲン;毒(膜翅目、スズメバチ(yellow jacket)、ミツバチ、カリバチ、スズメバチ(hornet)、ヒアリ);ゴキブリ、ノミ、蚊などによる他の環境昆虫アレルゲン;連鎖球菌抗原などの細菌アレルゲン; Ascaris属抗原などの寄生虫アレルゲン;ウイルス抗原;真菌胞子;薬物アレルゲン;抗生剤;ペニシリンおよび関連化合物;他の抗生剤;ホルモン(インスリン)、酵素(ストレプトキナーゼ)などの全タンパク質;不完全抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝物;ハプテンとして作用し、アレルゲンとして機能することが可能な、産業用化学物質および代謝物(例えば、酸無水物(トリメリト酸無水物など)およびイソシアン酸塩(ジイソシアン酸トルエンなど));小麦粉などの職業アレルゲン(例えば、パン屋喘息(Baker’s asthma)を引き起こすアレルゲン)、トウゴマ、コーヒー豆、また、上記で記載した産業用化学物質;ノミアレルゲン;ヒト以外の動物におけるヒトタンパク質が含まれるが、これらに限定されない。
【0078】
アレルゲンには、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、多糖、多糖結合体、多糖のペプチド性および非ペプチド性模倣体、ならびに他の分子、低分子、脂質、糖脂質、ならびに炭水化物が含まれるが、これらに限定されない。
【0079】
特定の天然の動物アレルゲンおよび植物アレルゲンの例には、以下の属:Canine属(Canis familiaris); Dermatophagoides属(例えば、Dermatophagoides farinae); Felis属(Felis domesticus); Ambrosia属(Ambrosia artemiisfolia); Lolium属(例えば、Lolium perenneまたはLolium multiflorum); Cryptomeria属(Cryptomeria japonica); Altemaria属(Altemaria altemata); Alder属; Alnus属(Alnus gultinoas); Betula属(Betula verrucosa); Quercus属(Quercus alba); Olea属(Olea europa); Artemisia属(Artemisia vulgaris); Plantago属(例えば、Plantago lanceolata); Parietaria属(例えば、Parietaria officinalisまたはParietaria judaica); Blattella属(例えば、Blattella germanica); Apis属(例えば、Apis multiflorum); Cupressus属(例えば、Cupressus sempervirens、Cupressus arizonica、およびCupressus macrocarpa); Juniperus属(例えば、Juniperus sabinoides、Juniperus virginiana、Juniperus communis、およびJuniperus ashei); Thuya属(例えば、Thuya orientalis); Chamaecyparis属(例えば、Chamaecyparis obtusa); Periplaneta属(例えば、Periplaneta americana); Agropyron属(例えば、Agropyron repens); Secale属(例えば、Secale cereale); Triticum属(例えば、Triticum aestivum); Dactylis属(例えば、Dactylis glomerata); Festuca属(例えば、Festuca elatior); Poa属(例えば、PoapratensisまたはPoa compressa); Avena属(例えば、Avena sativa); Holcus属(例えば、Holcus lanatus); Anthoxanthum属(例えば、Anthoxanthum odoratum); Arrhenatherum属(例えば、Arrhenatherun elatius); Agrostis属(例えば、Agrostis alba); Phleum属(例えば、Phleum pratense); Phalaris属(例えば、Phalaris arundinacea); Paspalum属(例えば、Paspalum notatum); Sorghum属(例えば、Sorghum halepensis); ならびにBromus属(例えば、Bromus inermis)に特異的なタンパク質が含まれるが、これらに限定されない。
【0080】
(哺乳動物細胞からRSV F VLPを作製するのに好ましい方法)
RSウイルスFポリペプチドのVLPは、当業者に適用可能な任意の方法により作製することができる。RSウイルスFポリペプチドのVLPは、VLPを形成する一因となるRSウイルスFポリペプチドを包含するが、任意のエンベロープウイルスコア形成ポリペプチドは除外する。加えて、RSウイルスFポリペプチドのVLPは、膜(脂質ラフトを含めた)会合ポリペプチドなど、1または複数のさらなるポリペプチドを包含することにより、(VLPを形成する一因となる1または複数のポリペプチドの一部として、天然において、または人工的に存在する抗原以外のさらなる)抗原をもたらすことができる。好ましい実施形態では、ポリペプチドを、任意の適用可能なタンパク質発現系、好ましくは、細胞膜内に脂質ラフトドメインを包含する哺乳動物細胞ベースの系において共発現させることができる。
【0081】
組換えによるVLP用ポリペプチドの発現は、該ポリペプチドのうちの1または複数をコードするポリヌクレオチドを含有する発現ベクターを伴う。ポリペプチドのうちの1または複数をコードするポリヌクレオチドが得られたら、当技術分野で周知の技法を用いる組換えDNA法により、該ポリペプチドを作製するためのベクターを作製することができる。このようにして、本明細書では、VLPポリペプチドをコードするヌクレオチド配列のうちのいずれかを含有するポリヌクレオチドを発現することによりタンパク質を調製する方法が記載される。当業者に周知の方法を用いて、VLPポリペプチドをコードする配列と、適切な転写制御シグナルおよび翻訳制御シグナルとを含有する発現ベクターを構築することができる。例えば、これらの方法には、インビトロにおける組換えDNA法、合成法、また、インビボにおける遺伝子組換えが含まれる。したがって、本発明は、すべてが1または複数のプロモーターに作動可能に連結した、RSウイルスFポリペプチドと、必要に応じて、1または複数のさらなる脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含む複製可能なベクターを提供する。
【0082】
本明細書で記載されるVLPへとアセンブルされる配列を発現させるのに用い得るベクターの非限定的な例には、ウイルスベースのベクター(例えば、レトロウイルス、アデノウイルス、アデノ関連ウイルス、レンチウイルス)、プラスミドベクター、非ウイルス性ベクター、哺乳動物ベクター、哺乳動物による人工的染色体(例えば、リポソーム、粒子状キャリアなど)、およびこれらの組合せが含まれる。
【0083】
発現ベクター(複数可)は、コード配列と、適切な宿主内において、該コード領域を発現させる発現制御エレメントとを含有することが典型的である。制御エレメントには、一般に、プロモーター、エンハンサー、エクソン、イントロン、スプライシング部位、翻訳開始コドン、ならびに翻訳終結配列および転写終結配列、ならびにベクター内に挿入を導入するための挿入部位が含まれる。翻訳制御エレメントは、M. Kozak(例えば、Kozak, M.、Mamm. Genome、7巻(8号):563〜574頁、1996年; Kozak, M.、Biochimie、76巻(9号):815〜821頁、1994年; Kozak, M.、J Cell Biol、108巻(2号):229〜241頁、1989年; Kozak, M.およびShatkin, A. J.、Methods Enzymol、60巻:360〜375頁、1979年)により総説されている。
【0084】
例えば、哺乳動物細胞による発現に用いられる典型的なプロモーターには、とりわけ、SV40初期プロモーター、CMV前初期プロモーターなどのCMVプロモーター(CMVプロモーターは、イントロンAを包含することができる)、RSV、HIV−LTR、マウス乳癌細胞ウイルスLTRプロモーター(MMLV−LTR)、FIV−LTR、アデノウイルス主要後期プロモーター(Ad MLP)、および単純ヘルペスウイルスプロモーターが含まれる。マウスメタロチオネイン遺伝子に由来するプロモーターなど、他の非ウイルス性プロモーターもまた、哺乳動物による発現に用いられる。翻訳終止コドンに対して3’側に位置する、転写終結配列およびポリアデニル化配列もまた存在することが典型的である。コード配列に対して5’側に位置する、翻訳の開始を最適化するための配列もまた存在することが好ましい。転写ターミネーター/ポリアデニル化シグナルの例には、Sambrookら、前出において記載されている、SV40に由来するシグナル、ならびにウシ成長ホルモンターミネーター配列が含まれる。スプライスドナー部位およびスプライスアクセプター部位を含有するイントロンもまた、本明細書で記載される構築物内にデザインすることができる(Chapmanら、Nuc. Acids Res.(1991年)、19巻:3979〜3986頁)。
【0085】
本明細書では、エンハンサーエレメントもまた、例えば、哺乳動物宿主細胞内における構築物の発現レベルを上昇させるのに用いることができる。例には、Dijkemaら、EMBO J.(1985年)、4巻:761頁において記載されている、SV40初期遺伝子エンハンサー;Gormanら、Proc. Natl. Acad. Sci. USA(1982年b)、79巻:6777頁において記載されている、ラウス肉腫ウイルスの長い末端反復(LTR)に由来するエンハンサー/プロモーター;また、CMVイントロンA配列内に包含されるエレメント(Chapmanら、Nuc. Acids Res.(1991年)、19巻:3979〜3986頁)など、Boshartら、Cell(1985年)、41巻:521頁において記載されている、ヒトCMVに由来するエレメントが含まれる。
【0086】
ベクターが、本明細書で記載される1つまたは複数の配列を含有し得ることは明らかである。例えば、単一のベクターが、VLP内に見出されるすべてのタンパク質をコードする配列を保有し得る。代替的に、複数のベクター(例えば、各々が、単一のポリペプチドコード配列をコードする複数の構築物、または各々が1つもしくは複数のポリペプチドコード配列をコードする複数の構築物)を用いることもできる。単一のベクターが、複数のポリペプチドコード配列を含む実施形態では、該配列を、同じベクター内における、同じであるかまたは異なる転写制御エレメント(例えば、プロモーター)に作動可能に連結することができる。
【0087】
加えて、免疫調節分子(例えば、以下で記載されるアジュバント)、例えば、免疫調節オリゴヌクレオチド(例えば、CpG)、サイトカイン、弱毒化した細菌性毒素などを含む配列、および/またはこれらをコードする配列を非制限的に含む、RSV以外のタンパク質をコードする1つまたは複数の配列も、VLP内で発現させ、また、これに組み込むことができる。
【0088】
従来の技法により、発現ベクターを宿主細胞へと移入することができ、次いで、従来の技法により、トランスフェクトされた細胞を培養して、VLPポリペプチド(複数可)を作製することができる。したがって、本発明は、異種プロモーターに作動可能に連結したVLPポリペプチドのうちの1つまたは複数をコードするポリヌクレオチドを含有する宿主細胞を包含する。VLPを作製するのに好ましい実施形態では、以下で詳述する通り、RSウイルスFポリペプチドと、場合によって、抗原またはアジュバントに連結した脂質ラフト会合ポリペプチドとをコードするベクターを、宿主細胞内で(共)発現させて、VLPを作製することができる。
【0089】
トランスフェクション、継代細胞株の確立(当業者に公知の標準的なプロトコールを用いる)、および/または目的のRSV遺伝子を保有するDNA分子での感染後における真核細胞、より好ましくは哺乳動物細胞内において、VLPを作製することが好ましい。VLPを形成するのに必要とされるタンパク質の発現レベルは、選択された遺伝子の転写を駆動する真核細胞プロモーターまたはウイルスプロモーターの配列を最適化することによって最大化する。VLPは、RSウイルスFポリペプチドにより駆動されて培地内へと放出され、そこからVLPを精製し、その後、ワクチンとして処方することができる。VLPは、感染性ワクチンではなく、したがって、ワクチンの不活化は不要である。
【0090】
本明細書で記載される配列から発現するRSウイルスFポリペプチドが、その表面上に抗原タンパク質を提示するVLPへと自己アセンブルする能力は、所望の配列を共導入(co−introduction)することにより、これらのVLPを、任意の宿主細胞内において作製することを可能にする。配列(複数可)(例えば、1つまたは複数の発現ベクター内における)は、多様な組合せで安定的に、および/または一過的に宿主細胞内へと組み込むことができる。
【0091】
適切な宿主細胞には、細菌細胞、哺乳動物細胞、バキュロウイルス/昆虫細胞、酵母細胞、植物細胞、およびXenopus属細胞が含まれるが、これらに限定されない。
【0092】
当技術分野では、例えば、多数の哺乳動物細胞株が公知であり、これらには、BHK、VERO、HT1080、MRC−5、WI 38、MDCK、MDBK、293、293T、RD、COS−7、CHO、Jurkat、HUT、SUPT、C8166、MOLT4/クローン8、MT−2、MT−4、H9、PM1、CEM、黒色腫細胞(例えば、SB20細胞)、LtK、HeLa、WI−38、L2、CMT−93、およびCEMX174(このような細胞株は、例えば、A.T.C.C.から入手可能である)などであるがこれらに限定されない、American Type Culture Collection(A.T.C.C.)から入手可能な初代細胞ならびに不死化細胞株が含まれる。
【0093】
同様に、E.coli、Bacillus subtilis、およびStreptococcus属種などの細菌宿主も、本発現構築物と共に用いられる。
【0094】
本開示において有用な酵母宿主には、とりわけ、Saccharomyces cerevisiae、Candida albicans、Candida maltosa、Hansenula polymorpha、Kluyveromyces fragilis、Kluyveromyces lactis、Pichia guillerimondii、Pichia pastoris、Schizosaccharomyces pombe、およびYarrowia lipolyticaが含まれる。真菌宿主には、例えば、Aspergillus属が含まれる。
【0095】
バキュロウイルス発現ベクターと共に用いられる昆虫細胞には、とりわけ、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogaster、Spodoptera frugiperda、およびTrichoplusia niが含まれる。LathamおよびGalarza(2001年)、J. Virol.、75巻(13号):6154〜6165頁; Galarzaら(2005年)、Viral. Immunol.、18巻(1号):244〜51頁;ならびに米国特許公開第200550186621号および同第20060263804号を参照されたい。
【0096】
本明細書で示される開示を踏まえれば、VLPのRSウイルスFポリペプチドをコードする1つまたは複数の配列(発現ベクター)を安定的に組み込むことにより、上記で記載した配列のうちの1つまたは複数を発現する細胞株を容易に作製することができる。安定的に組み込まれたRSウイルスFポリペプチド配列(複数可)の発現を制御するプロモーターは、構成的プロモーターの場合もあり、誘導的プロモーターの場合もある。
【0097】
RSウイルスFポリペプチドのVLPを作製する細胞株が由来する親細胞株は、例えば、哺乳動物細胞株、昆虫細胞株、酵母細胞株、細菌細胞株を含めた、上記で記載した任意の細胞から選択することができる。好ましい実施形態では、該細胞株は、哺乳動物細胞株(例えば、293、RD、COS−7、CHO、BHK、VERO、MRC−5、HT1080、および黒色腫細胞)である。哺乳動物細胞を用いてRSV VLPを作製することにより、(i)VLPの形成;(ii)適正なミリストイル化(myristylation)、グリコシル化、および出芽;(iii)非哺乳動物細胞夾雑物の不在;(iv)精製の容易さがもたらされる。
【0098】
細胞株を創出することに加えて、宿主細胞内において、RSVコード配列を一過的に発現させることもできる。適切な組換え発現宿主細胞系には、当技術分野で周知の、細菌発現系、哺乳動物発現系、バキュロウイルス/昆虫発現系、ワクシニア発現系、セムリキ森林熱ウイルス(SFV)発現系、アルファウイルス(シンドビスウイルス、ベネズエラウマ脳炎(VEE)ウイルスなど)発現系、哺乳動物発現系、酵母発現系、およびXenopus属発現系が含まれるが、これらに限定されない。特に好ましい発現系は、哺乳動物細胞株、ワクシニア、シンドビス、昆虫、および酵母系である。
【0099】
例えば、以下:バキュロウイルス発現(Reilly, P. R.ら、BACULOVIRUS EXPRESSION VECTORS: A LABORATORY MANUAL(1992年); Beamesら、Biotechniques、11巻:378頁(1991年);Pharmingen社製;カリフォルニア州、パロアルト、Clontech社製)、ワクシニア発現系(Earl, P. L.ら、「Expression of proteins in mammalian cells using vaccinia」、Current Protocols in Molecular Biology(F. M. Ausubelら編)、Greene Publishing Associates & Wiley Interscience、New York(1991年); 1992年8月4日に発行された、Moss, B.ら、米国特許第5,135,855号)、細菌内における発現(Ausubel, F. M.ら、CURRENT PROTOCOLS IN MOLECULAR BIOLOGY、John Wiley and Sons, Inc.、Media Pa.;Clontech社製)、酵母内における発現(1998年3月17日に発行され、参照により本明細書に組み込まれる、Rosenberg, SおよびTekamp−Olson, P.、米国特許第RE35,749号;1997年5月13日に発行され、参照により本明細書に組み込まれる、Shuster, J. R.、米国特許第5,629,203号; Gellissen, G. ら、Antonie Van Leeuwenhoek、62巻(1〜2号):79〜93頁(1992年); Romanos, M. A. ら、Yeast、8巻(6号):423〜488頁(1992年); Goeddel, D. V.、Methods in Enzymology、185巻(1990年); Guthrie, C.およびG. R. Fink、Methods in Enzymology、194巻(1991年))、動物細胞内における発現(Clontech社製;ニューヨーク州、グラウンドアイランド、Gibco−BRL社製;例えば、チャイニーズハムスター卵巣(CHO)細胞株(Haynes, J. ら、Nuc. Acid. Res.、11巻:687〜706頁(1983年); Lau, Y. F. ら、Mol. Cell. Biol.、4巻:1469〜1475頁(1984年); Kaufman, R. J.、「Selection and coamplification of heterologous genes in mammalian cells」、Methods in Enzymology、185巻、537〜566頁、カリフォルニア州、Academic Press, Inc.、San Diego Calif.(1991年)))、ならびに植物細胞における発現(植物クローニングベクター;カリフォルニア州、パロアルト、Clontech Laboratories社製;およびニュージャージー州、ピスケータウェイ、Pharmacia LKB Biotechnology社製; Hood, E.ら、J. Bacteriol.、168巻:1291〜1301頁(1986年); Nagel, R.ら、FEMS Microbiol. Lett.、67巻:325頁(1990年); Anら、「Binary Vectors」、およびその他、Plant Molecular Biology Manual、A3巻:1〜19頁(1988年); Miki, B. L. A.ら、249〜265頁、およびその他、Plant DNA Infectious Agents(Hohn, T.ら編)、Springer−Verlag、Wien、Austria(1987年);Plant Molecular Biology: Essential Techniques、P. G. JonesおよびJ. M. Sutton、New York、J. Wiley、1997年; Miglani, Gurbachan、Dictionary of Plant Genetics and Molecular Biology、New York、Food Products Press、1998年; Henry, R. J.、Practical Applications of Plant Molecular Biology、New York、Chapman & Hall、1997年)を含めた、多くの適切な発現系が市販されている。
【0100】
VLPを形成するのに必要とされるタンパク質をコードする改変遺伝子を含有する発現ベクターを宿主細胞(複数可)内へと導入し、その後、必要なレベルで発現させると、VLPがアセンブルし、次いで、細胞表面から培地内へと放出される。
【0101】
選択される発現系および宿主に応じて、粒子形成ポリペプチドが発現し、VLPが形成され得る条件下において、発現ベクターにより形質転換された宿主細胞を増殖させることにより、VLPを作製する。適切な増殖条件の選択は、当業者の能力の範囲内にある。
【0102】
次いで、例えば、密度勾配遠心分離、例えば、スクロース勾配、PEG沈殿、ペレット化など(例えば、Kimbauerら、J. Virol.(1993年)、67巻:6929〜6936頁を参照されたい)、ならびに、例えば、イオン交換クロマトグラフィーおよびゲル濾過クロマトグラフィーを含めた標準的な精製法により、その完全性を保存する方法を用いて、粒子を単離する(または実質的に精製する)。
【0103】
(RSウイルスFポリペプチドのVLPのVLP調製物における感染因子を不活化するのに好ましい方法)
電子放射は、RSウイルスFポリペプチドのVLPの免疫原性を実質的に低下させることなく、感染因子を不活化することが可能なので、不活化の好ましい方法は、電子放射を介する方法である。電磁放射で好ましい3つの方式のすべて(すなわち、光反応性化合物を伴うUV照射、UV照射単独、およびガンマ照射)には、血液、食品、ワクチンなど、多種多様な試料における病原体を不活化するのに用いられてきた長い歴史があるため、不活化用電磁放射を適用するための多種多様な装置が市販されており、これらは、本明細書で開示される方法を実施するのにほとんど〜まったく改変せずに用いることができる。さらに、当技術分野では、波長および線量の最適化がルーチン化しており、したがって、当業者の能力範囲内で容易に実施される。
【0104】
(光反応性化合物を伴うUV照射)
電磁放射による例示的な不活化法は、光反応性化合物、好ましくは、感染因子中のポリヌクレオチドと反応する光反応性化合物の存在下における、UV−A照射などの紫外線照射の組合せである。
【0105】
好ましい光反応性化合物には、アクチノマイシン、アントラサイクリノン、アントラマイシン、ベンゾジピロン、フルオレン、フルオレノン、フロクマリン、マイトマイシン、monostral fast blue、norphillin A、フェナントリジン、フェナザチオニウム塩、フェナジン、フェノチアジン、フェニルアジド、キノリン、およびチオキサンテノン(thiaxanthenones)が含まれる。好ましい種は、2つの主要なカテゴリーのうちの1つに属するフロクマリンである。第1のカテゴリーは、ソラーレン[7H−フロ(3,2−g)−(1)−ベンゾピラン−7−オン、または6−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、直鎖型であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の6位に結合している。第2のカテゴリーは、イソソラーレン[2H−フロ(2,3−h)−(1)−ベンゾピラン−2−オン、または4−ヒドロキシ−5−ベンゾフランアクリル酸のデルタ−ラクトン]であり、これらは、角度型(angular)であり、中央の芳香環部分に付随する2つの酸素残基が1,3の配向性を有し、またさらに、フラン環部分が二環クマリン系の8位に結合している。3、4、5、8、4’、または5’位において直鎖型フロクマリンを置換することによりソラーレン誘導体を作製し得る一方、3、4、5、6、4’、または5位において角度型フロクマリンを置換することによりイソソラーレン誘導体を作製し得る。ソラーレンは、二本鎖核酸の塩基対間にインターカレートすることができ、長波長の紫外光(UVA)を吸収すると、ピリミジン塩基に対する共有結合付加体を形成する。例えば、G. D. Ciminoら、Ann. Rev. Biochem.、54巻:1151頁(1985年); Hearstら、Quart. Rev. Biophys.、17巻:1頁(1984頁)を参照されたい。
【0106】
好ましいUV(または場合によって、可視光)放射の波長は、光付加体が生成される波長に依存し、これは、光反応性化学物質の化学反応に依存する。例示を目的として述べると、多くのソラーレンには、320〜380nmの波長にあるUV放射が最も有効であり、330〜360nmが最大の有効性を示す。
【0107】
(UV照射単独)
光反応性化合物の存在下におけるUV照射に加え、感染因子は、UV照射単独でも不活化することができる。好ましい実施形態では、放射は、波長が、約180〜320nm、または約225〜290nm、または約254nm(すなわち、ポリヌクレオチドの最大吸光度ピーク)であるUVC放射である。UVC放射は、エンベロープを形成する脂質二重層など、本明細書で開示されるRSウイルスFポリペプチドのVLPの成分に対して、安定性および免疫原性の両面で有害性が低い一方で、感染因子を不活化するのに十分なエネルギーを保持するので好ましい。しかし、例えば、UVAおよびUVBなど、他の種類のUV放射もまた用いることができる。
【0108】
(ガンマ照射)
本明細書で開示される方法を実施して組成物を作製するには、ガンマ照射(すなわち、イオン化放射)もまた用いることができる。ガンマ照射は、感染因子のゲノムをコードするポリヌクレオチド内において鎖の切断を導入することにより、感染因子を不活化することもでき、または、該ポリヌクレオチドを攻撃するヒドロキシルラジカルを生成することにより、感染因子を不活化することもできる。
【0109】
(RSウイルスFポリペプチドのVLPを用いる好ましい方法)
(処方物)
本明細書で記載されるRSウイルスFポリペプチドのVLPの好ましい使用は、ワクチン調製物としての使用である。典型的に、このようなワクチンは、液体溶液または懸濁物の注射剤として調製されるが、注射前に液体中に溶解させるか、または懸濁させるのに適する固体形態もまた調製することができる。このような調製物はまた、乳化させることもでき、乾燥粉末として作製することもできる。免疫原性の有効成分は、薬学的に許容され、また、有効成分に適合する、賦形剤と混合することが多い。適切な賦形剤は、例えば、水、生理食塩液、デキストロース、グリセロール、エタノールなどであり、また、これらの組合せである。加えて、所望の場合、ワクチンは、保湿剤もしくは乳化剤、pH緩衝剤、または該ワクチンの有効性を増強するアジュバントなどの補助物質を含有し得る。
【0110】
ワクチンは、注射、例えば、皮下(subcutaneously)注射、皮内注射、皮下(subdermally)注射、または筋肉内注射を介する、従来の非経口投与が可能である。他の投与方式に適する、さらなる処方物には坐剤が含まれ、場合によっては、経口処方物、鼻腔内処方物、口腔内処方物、舌下処方物、腹腔内処方物、膣内処方物、経肛門処方物、および頭蓋内処方物も含まれる。坐剤の場合、従来の結合剤およびキャリアには、例えば、ポリアルキレングリコール(polyalkalene glycol)またはトリグリセリドが含まれる場合があり、このような坐剤は、0.5%〜10%、好ましくは1〜2%の範囲で有効成分を含有する混合物から形成することができる。特定の実施形態では、脂肪酸グリセリドの混合物またはココアバターなどの低温溶融ワックスをまず溶融させ、本明細書で記載されるRSウイルスFポリペプチドのVLPを、例えば、撹拌することにより、均一に分散させる。次いで、溶融した均一の混合物を、好都合なサイズの型に流し込み、冷却して固形化させる。
【0111】
鼻腔内送達に適する処方物には、液体および乾燥粉末が含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、スクロース、トレハロース、およびキトサンなど、通常用いられる賦形剤を包含する。液体処方物または粉末処方物において、キトサンなどの粘膜付着剤を用いて、粘膜絨毛による、鼻腔内投与された処方物のクリアランスを遅延させることができる。マンニトールおよびスクロースなどの糖を、液体処方物では安定化剤として用いることができ、また、乾燥粉末処方物では安定化剤およびバルキング剤として用いることができる。加えて、モノホスホリルリピドA(MPL)などのアジュバントを、液体処方物および乾燥粉末処方物の両方において、免疫刺激性アジュバントとして用いることもできる。
【0112】
経口送達に適する処方物には、液体、固体、半固体、ゲル、錠剤、カプセル、ロゼンジなどが含まれる。経口送達に適する処方物には、錠剤、ロゼンジ、カプセル、ゲル、液体、食品、飲料、栄養補助食品などが含まれる。処方物は、例えば、医薬グレードのマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなど、通常用いられる賦形剤を包含する。他のRSウイルスFポリペプチドのVLPワクチン組成物は、溶液、懸濁物、丸薬、持続放出処方物、または粉末の形態を取る場合があり、10〜95%、好ましくは25〜70%の有効成分を含有する場合がある。経口処方物の場合、コレラ毒素が興味深い処方物パートナー(また、可能な結合パートナー)である。
【0113】
膣内投与用に処方する場合、RSウイルスFポリペプチドのVLPワクチンは、ペッサリー、タンポン、クリーム、ジェル、ペースト、フォーム、またはスプレーの形態であり得る。前出の処方物のうちのいずれもが、RSウイルスFポリペプチドのVLPに加えて、キャリアなど、当技術分野で適切であることが公知の薬剤を含有し得る。
【0114】
一部の実施形態では、RSウイルスFポリペプチドのVLPワクチンを、全身送達用に処方する場合もあり、局所送達用に処方する場合もある。このような処方物は、当技術分野において周知である。非経口ビヒクルには、塩化ナトリウム溶液、リンゲルデキストロース液、デキストロースおよび塩化ナトリウムによる溶液、乳酸加リンゲル液、または固定油が含まれる。静脈内ビヒクルには、流体および栄養物質補給剤、電解質補給剤(リンゲルデキストロース液に基づく補給剤など)などが含まれる。全身投与経路および局所投与経路には、例えば、皮内経路、局所適用経路、静脈内経路、筋肉内経路などが含まれる。
【0115】
RSウイルスFポリペプチドのVLPは、中性処方物または塩ベースの処方物を含めたワクチンへと処方することができる。薬学的に許容される塩には、酸添加塩(ペプチドの遊離アミノ基と共に形成され、また、例えば、塩酸もしくはリン酸などの無機酸、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸と共に形成される)が含まれる。遊離カルボキシル基と共に形成される塩はまた、例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化鉄などの無機塩基、また、イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、プロカインなどの有機塩基に由来し得る。
【0116】
ワクチンは、投薬処方物に適合した形で、また、治療的に有効であり免疫原性となる量で投与することができる。投与量は、例えば、個体の免疫系が免疫反応を起こす能力、また、所望される防御の程度を含め、処置される被験体に依存する。適切な用量範囲は、1回のワクチン接種当たりの有効成分が数百マイクログラムのオーダーであり、好ましい範囲は、約0.5μg〜1000μgの範囲、好ましくは1μg〜500μgの範囲、またとりわけ、約10μg〜100μgの範囲など、約0.1μg〜2000μgの範囲である(1〜10mgの範囲にある高量も意図される)ことが好ましい。初回投与および追加投与に適するレジメンもまた可変的であるが、初回投与にその後の接種または他の投与が続く形が典型的である。
【0117】
適用方式は多様に変化させることができる。従来のワクチン投与方法のうちのいずれもが適用可能である。これらには、固体の生理学的に許容されるベース上における経口適用、または注射など、生理学的に許容される分散剤による非経口適用が含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるヒトの年齢、および抗原処方物により変化する。
【0118】
ワクチン処方物の一部は、それだけでワクチンとして十分に免疫原性であるが、他の処方物では、該ワクチンがアジュバント物質をさらに包含すると、免疫反応が増強される場合もある。
【0119】
とりわけ、鼻腔内ベース、経口ベース、または肺内ベースの送達処方物の場合、送達および免疫原性を改善するには、粘膜付着を改善する送達剤もまた用いることができる。このような化合物の1つである、キチンのN−脱アセチル化形態であるキトサンは、多くの医薬処方物で用いられる(32)。粘膜絨毛によるクリアランスを遅延させ、粘膜による抗原の取込みおよびプロセシングのためのより多くの時間を可能にする能力のために、キトサンは、鼻腔内におけるワクチン送達にとって魅力的な粘膜付着剤である(33、34)。加えて、それは、接着結合を一過的に開口させ、これにより、NALTへの抗原の経上皮輸送が増強され得る。近年のヒト試験では、キトサンを伴うが、さらなるアジュバントは伴わない、三価の不活化インフルエンザワクチンを鼻腔内投与したところ、筋肉内接種後に得られたセロコンバージョンおよびHI力価をごくわずかに下回るセロコンバージョンおよびHI力価がもたらされた(33)。
【0120】
キトサンはまた、遺伝的操作により解毒したE.coli易熱性エンテロトキシンの変異体であるLTK63など、鼻腔内において良好に機能するアジュバントと共に処方することもできる。これは、キトサンにより付与される送達および付着の利益の上に、免疫刺激効果を付加し、その結果、粘膜反応および全身反応の増強をもたらす(35)。
【0121】
最後に、キトサン処方物はまた、ワクチンの安定性を改善し、粘膜絨毛によるクリアランスにおいて液体処方物を上回る遅延を結果としてもたらすことが示されている乾燥粉末フォーマットでも調製することができることに注意すべきである(42)。これは、キトサンと共に処方された、鼻腔内用乾燥粉末によるジフテリア毒素ワクチンを伴う、近年のヒト臨床試験において見られ、ここで、鼻腔内経路は、従来の筋肉内経路と同程度に有効であり、分泌性IgA反応の利益が付加された(43)。該ワクチンはまた、忍容性も極めて良好であった。キトサンおよびMPLを含有する炭疽菌用の鼻腔内乾燥粉末ワクチンは、ウサギにおいて、筋肉内接種より強力な反応を誘導し、また、エアゾールによる胞子チャレンジ(challenge)に対しても防御的であった(44)。
【0122】
鼻腔内ワクチンは、下気道に対してより良好に作用する非経口投与ワクチンに対して、上気道および下気道に作用し得るので、好ましい処方物を表す。これは、アレルゲンベースのワクチンには忍容性を誘導し、また、病原体ベースのワクチンには免疫性を誘導する点で有益であり得る。
【0123】
鼻腔内ワクチンは、上気道および下気道の両方において防御をもたらすのに加え、針による接種の煩雑さを回避し、また、粒子抗原および/または可溶性抗原が、鼻咽頭関連リンパ組織(NALT)と相互作用することを介して、粘膜および全身における体液性反応および細胞性反応の両方を誘導する手段をもたらす(16〜19)。歴史的に、鼻腔内経路は、非経口接種ほど有効ではなかったが、新規の送達処方物であるRSウイルスFポリペプチドのVLPおよびアジュバントを用いることにより、この枠組みが変わりつつある。実際、鼻腔粘膜内にはシアル酸を含有する受容体が豊富であるため、鼻腔内送達には、機能性の赤血球凝集素ポリペプチドを含有するRSウイルスFポリペプチドのVLPがとりわけよく適する場合があり、その結果、HA抗原の結合が増強され、粘膜絨毛によるクリアランスが低減される可能性がある。
【0124】
(アジュバント)
ワクチンに対するアジュバント効果を達成する各種の方法が公知であり、本明細書で開示されるRSウイルスFポリペプチドのVLPと共に用いることができる。一般的な原理および方法は、「The Theory and Practical Application of Adjuvants」、1995年、Duncan E. S. Stewart−Tull(編)、John Wiley & Sons社、ISBN 0−471−95170−6において詳述されており、また、「Vaccines: New Generation Immunological Adjuvants」、1995年、Gregoriadis Gら(編)、ニューヨーク、Plenum Press社、ISBN 0−306−45283−9においても詳述されている。
【0125】
一部の実施形態では、RSウイルスFポリペプチドのVLPワクチンが、重量ベースの比率約10:1〜約1010:1のRSウイルスFポリペプチドのVLP:アジュバント、例えば、約10:1〜約100:1、約100:1〜約103:1、約103:1〜約104:1、約104:1〜約105:1、約105:1〜約106:1、約106:1〜約107:1、約107:1〜約108:1、約108:1〜約109:1、または約109:1〜約1010:1のRSウイルスFポリペプチドのVLP:アジュバントで、少なくとも1つのアジュバントと混合したRSウイルスFポリペプチドのVLPを包含する。当業者は、アジュバントに関する情報、また、最適の比率を決定する日常的実験により、適切な比率を容易に決定することができる。
【0126】
アジュバントの好ましい例は、RSウイルスFポリペプチドのVLPのポリペプチド成分と共発現させるか、または該ポリペプチド成分と融合させて、キメラポリペプチドを作製することにより、本明細書で記載されるRSウイルスFポリペプチドのVLPに容易に添加し得るポリペプチドアジュバントである。鞭毛の主要なタンパク質構成要素である細菌性フラジェリンは、それが、toll様受容体であるTLR5を介する先天性免疫系により認識されるために、アジュバントタンパク質としてますます注目を集めている、好ましいアジュバントである(65)。TLR5を介するフラジェリンシグナル伝達は、DCの成熟化および移動の他、マクロファージ、好中球、および腸上皮細胞の活性化も誘導することにより、先天性免疫機能および後天性免疫機能の両方に影響を及ぼし、その結果、炎症促進(proinflammatory)メディエーターを生成させる(66〜72)。
【0127】
TLR5は、フラジェリン単量体内における保存的構造(これはフラジェリン単量体タンパク質に固有であり、また、鞭毛機能に必要とされる)を認識し、このことは免疫学的圧力に応答したその変異を妨げる(73)。該受容体は、100fMの濃度に対して感受性であるが、完全な線維は認識しない。鞭毛が単量体へと解体されることが、結合および刺激に必要とされる。
【0128】
アジュバントとしてのフラジェリンは、非経口投与または鼻腔内投与された異種抗原に対する防御反応を誘導する強力な活性を有し(66、74〜77)、また、DNAワクチンに対するアジュバント効果もまた報告されている(78)。マウスまたはサルにおいてフラジェリンを用いると、Th2バイアス(これは、インフルエンザなどの呼吸器ウイルスには適切である)は観察されるが、IgE誘導の証拠は観察されていない。加えて、サルにおける鼻腔内投与または全身投与後における、局所炎症反応も全身炎症反応も報告されていない(74)。TLR5を介して、MyD88依存的な形でなされるフラジェリンシグナル伝達、また、TLRを介する、他のすべてのMyD88依存的シグナルは、Th1バイアスを結果としてもたらすことが示されている(67、79)ため、フラジェリンを使用した後で誘発される反応がTh2特徴を示すことは、ある意味で驚くべきことである。重要なことに、フラジェリンに対して予め存在する抗体は、アジュバントの有効性に対してさほどの影響を及ぼさない(74)ことから、フラジェリンは、複数回使用のためのアジュバントとして魅力的となっている。
【0129】
多くの近年における鼻腔内ワクチン試験における共通の主題は、ワクチンの有効性を改善するアジュバントおよび/または送達系の使用である。このような一研究では、遺伝的に解毒されたE. coli易熱性エンテロトキシンアジュバント(LT R192G)を含有するインフルエンザH3ワクチンが、H5チャレンジに対する異種サブタイプ的な防御を結果としてもたらしたが、これは、鼻腔内送達後に限られた。防御は、交差中和抗体の誘導に基づき、これにより、新たなワクチンの開発における鼻腔内経路についての重要な示唆が示された(22)。
【0130】
サイトカイン、コロニー刺激因子(例えば、GM−CSF、CSFなど);腫瘍壊死因子;インターロイキン2、インターロイキン7、インターロイキン12、インターフェロン、および他の同様の増殖因子はまた、アジュバントとして用いることができ、これらは
RSウイルスFポリペプチドのVLPのポリペプチド成分と混合または融合させることにより、該ワクチン内に容易に組み込み得るので、これらもまた好ましい。
【0131】
一部の実施形態では、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチン組成物が、5’−TCG−3’配列を含む、核酸のTLR9リガンド;イミダゾキノリンのTLR7リガンド;置換グアニンのTLR7/8リガンド;ロキソリビン、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、イミキモド(R−837)、およびレシキモド(R−848)など、他のTLR7リガンドなど、Toll様受容体を介して作用する他のアジュバントを包含し得る。
【0132】
特定のアジュバントは、樹状細胞などのAPCによるワクチン分子の取込みを促進し、これらを活性化する。非限定的な例は、免疫標的化アジュバント;毒素、サイトカイン、およびマイコバクテリア誘導体などの免疫調節アジュバント;油処方物;ポリマー;ミセル形成アジュバント;サポニン;免疫刺激複合体マトリックス(ISCOMマトリックス);粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;および封入アジュバントからなる群より選択される。
【0133】
アジュバントのさらなる例には、作用物質、例えば、緩衝生理食塩液中0.05〜0.1パーセントの溶液として一般的に用いられるアルミニウム塩、例えば、水酸化物またはリン酸塩(ミョウバン)(例えば、Nicklas(1992年)、Res. Immunol.、143巻:489〜493頁を参照されたい)、0.25パーセントの溶液として用いられる糖の合成ポリマー(例えば、Carbopol(登録商標))との混合物、それぞれ、70℃〜101℃の範囲の温度で30秒間〜2分間にわたる熱処理によるワクチン中のタンパク質の凝集物が含まれ、また、架橋形成剤による凝集物も可能である。ペプシン処理した、アルブミンに対する抗体(Fab断片)での再活性化による凝集物、C. parvumなどの細菌細胞またはグラム陰性菌の内毒素もしくはリポ多糖成分との混合物、モノオレイン酸マンニド(Aracel A)などの生理学的に許容される油ビヒクルにおけるエマルジョン、または保護置換物として用いられる、ペルフルオロカーボン(Fluosol−DA)の20パーセント溶液によるエマルジョンもまた用いることができる。スクアレンおよびIFAなどの油との混合物もまた好ましい。
【0134】
DDA(ジメチルジオクタデシルアンモニウムブロミド)は、アジュバントの興味深い候補物質であるが、フロイントの完全アジュバントおよびフロイントの不完全アジュバントの他、QuilAおよびQS21などのキラヤサポニンもまた興味深い。さらなる可能性には、モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など、リポ多糖のポリ[ジ(カルボキシレートフェノキシ)ホスファゼン](poly[di(earboxylatophenoxy)phosphazene)(PCPP)誘導体が含まれる。例えば、アルミニウム塩と併せた、モノホスホリルリピドA、好ましくは、3−脱−O−アシル化モノホスホリルリピドAの組合せを含めた、主にTh1型の反応をもたらすリポ多糖ベースのアジュバントが好ましい。MPL(登録商標)アジュバントは、GlaxoSmithKline社から市販されている(例えば、米国特許第4,436,727号;同第4,877,611号;同第4,866,034号;および同第4,912,094号を参照されたい)。
【0135】
リポソーム処方物もまた、アジュバント効果を付与することが知られており、したがって、リポソームアジュバントは、RSウイルスFポリペプチドのVLPと合わされる、好ましい例である。
【0136】
免疫刺激複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、とりわけ、この型のアジュバントが、APCによるMHCクラスII発現を上方制御することが可能であることが示されているため、本発明に従う好ましい選択である。ISCOMマトリックスは、Quillaja saponariaに由来する(場合によって分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質からなる。VLP中のものなどの免疫原性タンパク質と混合する場合、結果として得られる粒子状処方物は、サポニンが60〜70%w/wを構成することが可能であり、コレステロールおよびリン脂質が10〜15%w/wを構成することが可能であり、タンパク質が10〜15%w/wを構成することが可能な、ISCOM粒子として公知の処方物である。免疫刺激複合体の組成および使用に関する詳細は、例えば、アジュバントを扱う上述の教科書中に見出し得るが、また、Morein Bら、1995年、Clin. Immunother.、3巻:461〜475頁の他、Barr I GおよびMitchell G F、1996年、Immunol. and Cell Biol.、74巻:8〜25頁も、完全な免疫刺激複合体を調製するのに有益な教示を提供している。
【0137】
本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンとアジュバントの組合せにおいて用い得るサポニンには、それらがISCOM形態にある場合であれ、そうでない場合であれ、米国特許第5,057,540号;および「Saponins as vaccine adjuvants」、Kensil, C. R.、Crit Rev Ther Drug Carrier Syst、1996年、12巻(1〜2号):1〜55頁;およびEP0362279B1で記載される、Quil Aと称する、Quillaja Saponaria Molinaの樹皮に由来するサポニン、また、それらの画分が含まれる。Quil Aの特に好ましい画分は、QS21、QS7、およびQS17である。
【0138】
β−エスシンは、本発明のアジュバント組成物中で用いるのに好ましい、別の溶血性サポニンである。エスシンは、「メルクインデックス」(第12版:エントリー3737)において、ラテン名: Aesculus hippocastanumであるトチノキの種子内で発生するサポニンの混合物として記載されている。クロマトグラフィーおよび精製(Fiedler、Arzneimittel−Forsch.、4巻、213頁(1953年))による、また、イオン交換樹脂(Erbringら、米国特許第3,238,190号)によるその単離が記載されている。エスシンの画分は精製されており、生物学的に活性であることが示されている(Yoshikawa Mら(Chem Pharm Bull(東京)、1996年8月、44巻(8号):1454〜1464頁))。β−エスシン(escin)はまた、エスシン(aescin)としても公知である。
【0139】
本発明において用いるのに好ましい別の溶血性サポニンは、ジギトニンである。ジギトニンは、「メルクインデックス」(第12版:エントリー3204)において、Digitalis purpureaの種子に由来し、また、Gisvoldら、J.Am.Pharm.Assoc.、1934年、23巻、664頁;およびRuhenstroth−Bauer、Physiol.Chem.、1955年、301巻、621頁において記載される手順により精製されるサポニンとして記載されている。その使用は、コレステロールを判定するための臨床試薬として記載されている。
【0140】
アジュバント効果を達成する別の興味深い(したがって、好ましい)可能性は、Gosselinら、1992年において記載される技法を用いることである。略述すると、本発明の抗原などの関与抗原の提示は、該抗原を、単球/マクロファージ上におけるFC受容体に対する抗体(または抗原に結合する抗体断片)に結合体化することにより増強することができる。とりわけ、抗原と抗FCRIとの結合体は、ワクチン接種のための免疫原性を増強することが示されている。該抗体は、生成後において、または生成の一部として(RSウイルスFポリペプチドのVLPのポリペプチド成分のうちのいずれか1つに対する融合体としての発現によることを含む)、RSウイルスFポリペプチドのVLPに結合体化することができる。
【0141】
他の可能性は、標的化物質および免疫調節物質(すなわち、サイトカイン)の使用を伴う。加えて、ポリI:Cなど、合成のサイトカイン誘導物質もまた用いることができる。
【0142】
適切なマイコバクテリア誘導体は、ムラミルジペプチド、フロイントの完全アジュバント、RIBI(モンタナ州、ハミルトン、RiBi ImmunoChem Reserch社製)、ならびに、TDMおよびTDEなど、トレハロースのジエステルからなる群より選択することができる。
【0143】
適切な免疫標的化アジュバントの例には、CD40リガンドおよびCD40抗体、またはこれらの特異的結合断片(上記の議論を参照されたい)、マンノース、Fab断片、ならびにCTLA−4が含まれる。
【0144】
適切なポリマーアジュバントの例には、炭水化物、例えば、デキストラン、PEG、デンプン、マンナン、およびマンノース;可塑性ポリマー;ならびにラテックスビーズなどのラテックスが含まれる。
【0145】
免疫反応を調節するさらに別の興味深い方法は、「仮想リンパ節」(VLN)(ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017−6501により開発された、特許品の医療デバイス)内に免疫原(場合によって、アジュバント、ならびに薬学的に許容されるキャリアおよびビヒクルと併せて)を組み込むことである。VLN(細長型の管状デバイス)は、リンパ節の構造および機能を模倣する。VLNを皮下に挿入することにより、サイトカインおよびケモカインの急増による無菌的炎症部位が創出される。T細胞およびB細胞の他、APCも、迅速に危険シグナルに応答し、炎症部位へと至り、VLNの多孔性マトリックスの内部に蓄積される。VLNを用いると、抗原に対する免疫反応を誘発するのに必要とされる抗原用量が低下し、VLNを用いるワクチン接種により付与される免疫的防御が、Ribiをアジュバントとして用いる従来の免疫化を凌駕することが示されている。この技法は、Gelber Cら、1998年、「Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node」、1998年10月12〜15日、カリフォルニア州、アプトス、シースケープリゾート、「From the Laboratory to the Clinic, Book of Abstracts」において略述されている。
【0146】
RSウイルスFポリペプチドのVLPワクチンと共に、オリゴヌクレオチドをアジュバントとして用いることができ、少なくとも3つの、またはより好ましくは、少なくとも6つ以上のヌクレオチドにより隔てられた2つ以上のジヌクレオチドCpGモチーフを含有することが好ましい。CpGを含有するオリゴヌクレオチド(ここで、CpGジヌクレオチドは、メチル化されていない)は、主にTh1反応を誘導する。このようなオリゴヌクレオチドは周知であり、例えば、WO96/02555、WO99/33488、ならびに米国特許第6,008,200号および同第5,856,462号において記載されている。
【0147】
このようなオリゴヌクレオチドによるアジュバントは、デオキシヌクレオチドであり得る。好ましい実施形態では、該オリゴヌクレオチド内のヌクレオチド骨格は、ホスホロジチオエート結合であるか、またはより好ましくは、ホスホロチオエート結合であるが、骨格結合が混合したオリゴヌクレオチドを含め、ホスホジエステル骨格、また、PNAなど他のヌクレオチド骨格も本発明の範囲内にある。ホスホロチオエートオリゴヌクレオチドまたはホスホロジチオエートオリゴヌクレオチドを作製する方法は、米国特許第5,666,153、米国特許第5,278,302号、およびW095/26204において記載されている。
【0148】
好ましいオリゴヌクレオチドの例は、以下の配列を有する。配列は、ホスホロチオエート修飾されたヌクレオチド骨格を含有することが好ましい。
【0149】
【化2】
代替的な好ましいCpGオリゴヌクレオチドには、これらに対する重要でない欠失または付加を伴う上記の配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、当技術分野で公知である任意の方法(例えば、EP468520)により合成することができる。自動合成器を用いてこのようなオリゴヌクレオチドを合成し得ることが好ましい。このようなオリゴヌクレオチドによるアジュバントは、10〜50塩基の長さであり得る。別のアジュバント系は、CpGを含有するオリゴヌクレオチドと、サポニン誘導体との組合せを伴い、特に、CpGとQS21との組合せが、WO00/09159において開示されている。
【0150】
多くの単相または複相のエマルジョン系が記載されている。エマルジョンが、RSウイルスFポリペプチドのVLPの構造を破壊しないように、当業者は、RSウイルスFポリペプチドのVLPと共に用いられるこのようなエマルジョン系を容易に適合させることができる。水中油エマルジョンによるアジュバントは、それ自体で、アジュバント組成物として有用であることが示唆されており(EPO399843B)、また、水中油エマルジョンと他の活性薬剤との組合せも、ワクチン用のアジュバントとして記載されている(WO95/17210;WO98/56414;WO99/12565;WO99/11241)。油中水エマルジョン(米国特許第5,422,109号;EP 0480982B2)、および水中油中水エマルジョン(米国特許第5,424,067;EP0480981B)など、他の油エマルジョンによるアジュバントが記載されている。
【0151】
本明細書で記載されるRSウイルスFポリペプチドのVLPワクチンと共に用いられる油エマルジョンによるアジュバントは、天然の場合もあり、合成の場合もあり、また、鉱油性の場合もあり、有機油性の場合もある。鉱油および有機油の例は、当業者に容易に明らかである。
【0152】
任意の水中油組成物が、ヒトへの投与に適するためには、エマルジョン系の油相が、代謝性油を包含することが好ましい。代謝性油という用語の意味は、当技術分野において周知である。代謝性とは、「代謝により変換可能であること」と定義することができる(「Dorland’s Illustrated Medical Dictionary」、W.B. Sanders Company社、第25版(1974年))。油は、レシピエントに毒性でなく、代謝により変換可能な、任意の植物油、魚油、動物油、または合成油であることが可能である。堅果油(ラッカセイ油など)、種油、穀類油が、植物油の一般的な供給源である。合成油もまた本発明の一部であり、これらには、NEOBEE(登録商標)および他の油など、市販の油が含まれ得る。スクアレン(2,6,10,15,19,23−ヘキサメチル−2,6,10,14,18,22−テトラコサヘキサエン)は、サメの肝油中に高量で見出され、また、オリーブ油、コムギ胚種油、コメぬか油、および酵母中に低量で見出される不飽和油であり、本発明で用いるのに特に好ましい油である。スクアレンは、コレステロールの生合成における中間体であるという事実により、代謝性油である(「メルクインデックス」、第10版、エントリー8619)。
【0153】
特に好ましい油エマルジョンは水中油エマルジョンであり、特に、水中スクアレンエマルジョンである。
【0154】
加えて、本発明の最も好ましい油エマルジョンによるアジュバントには、抗酸化剤が含まれ、油であるα−トコフェロール(ビタミンE;EP0382271B1)が好ましい。
【0155】
WO95/17210およびWO99/11241は、場合によって、免疫刺激剤であるQS21および/または3D−MPLと共に処方される、スクアレン、α−トコフェロール、TWEEN 80(TM)に基づくエマルジョンアジュバントを開示している。WO99/12565は、油相へのステロールの添加による、これらのスクアレンエマルジョンに対する改善を開示している。加えて、エマルジョンを安定化させるために、トリカプリリン(C27H50O6)などのトリグリセリドを、油相へと添加することもできる(WO98/56414)。
【0156】
安定的な水中油エマルジョン中で見出される油滴のサイズは、光子相関分光分析により測定したときに、1ミクロン未満であることが好ましく、実質的に30〜600nmの範囲内、好ましくは、実質的に直径約30〜500nm、また、最も好ましくは、実質的に直径150〜500nm、また、特に直径約150nmであり得る。この点で、油滴数の80%が好ましい範囲内にあるべきであり、より好ましくは油滴数の90%超、また、最も好ましくは油滴数の95%超が規定のサイズ範囲内にある。本発明の油エマルジョン中に存在する成分の量は、従来、スクアレンなどの油が2〜10%の範囲内であり;また、存在する場合、アルファトコフェロールが2〜10%であり、ポリオキシエチレンソルビタンモノオレエートなどの界面活性剤が0.3〜3%である。油:アルファトコフェロールの比率は、より安定的なエマルジョンがもたらされるため、1以下であることが好ましい。SPAN 85(TM)もまた、約1%のレベルで存在し得る。一部の場合には、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンが、安定化剤をさらに含有することが有利であり得る。
【0157】
水中油エマルジョンを作製する方法は、当業者に周知である。一般に、該方法は、油相を、PBS/TWEEN80(登録商標)溶液などの界面活性剤と混合し、その後、ホモジナイザーを用いるホモジナイゼーションを行うステップを包含するが、該混合物に、シリンジ針内を2回流過させるステップを含む方法が、少量の液体をホモジナイズするのには適切であることが、当業者には明らかである。同様に、当業者は、Microfluidizer(M110S型マイクロ流体作製器;最大入力圧6バールで2分間にわたり、最大50回の流過(約850バールの出力圧))において乳化過程を適合させることで、より少量または多量のエマルジョンを作製することができる。この適合は、必要とされる直径の油滴を有する調製物が達成されるまでの、結果として得られるエマルジョンの測定を含む日常的な実験により達成し得る。
【0158】
本明細書で開示されるRSウイルスFポリペプチドのVLPによるワクチン調製物は、該ワクチンを、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮投与、静脈内投与、または皮下投与を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用いることができる。ワクチン調製物を全身投与する方法には、従来のシリンジおよび針、または固体ワクチンを弾道送達(ballistic delivery)するためにデザインされたデバイス(WO99/27961)、または針なしの高圧液体ジェットデバイス(米国特許第4,596,556号;米国特許第5,993,412号)、または経皮パッチ(WO97/48440;WO98/28037)が含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、皮膚に適用することもできる(経皮(transdermalまたはtranscutaneous)送達;WO98/20734;WO98/28037)。したがって、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンは、RSウイルスFポリペプチドのVLPワクチンまたはアジュバント組成物を予め充填した全身投与用の送達デバイスを包含する。したがって、個体、好ましくは哺乳動物または鳥類において免疫反応を誘導する方法であって、本明細書で記載されるRSウイルスFポリペプチドのVLP組成物のうちのいずれかを含み、また、場合によって、アジュバントおよび/またはキャリアを包含するワクチンを該個体へと投与するステップを含み、該ワクチンが、非経口経路または全身経路により投与される方法が提供される。
【0159】
本発明のワクチン調製物は、該ワクチンを、経口/経消化管経路、または経鼻経路などの粘膜経路を介して投与することにより、ウイルス感染を受けやすいか、またはこれを患う、哺乳動物または鳥類を保護または処置するのに用い得ることが好ましい。代替的な粘膜経路は、膣内経路および直腸内経路である。好ましい経粘膜経路投与は、経鼻経路を介し、鼻腔内ワクチン接種と称する。免疫化される個体の鼻咽頭内へと、ワクチンの液滴形態、スプレー形態、または乾燥粉末形態を投与することを含め、鼻腔内ワクチン接種の方法は、当技術分野において周知である。したがって、噴霧化またはエアゾール化されたワクチン処方物が、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンの好ましい形態である。経口投与用の胃液耐性のカプセルおよび顆粒などの腸溶処方物、直腸内投与または膣内投与用の坐剤もまた、本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンの処方物である。
【0160】
本明細書で開示される、好ましいRSウイルスFポリペプチドのVLPによるワクチン組成物は、経粘膜ワクチン接種により全身ワクチン接種を代替するヒトにおける適用に適する経粘膜ワクチンのクラスの一例である。
【0161】
RSウイルスFポリペプチドのVLPワクチンはまた、経口経路を介して投与することもできる。このような場合、薬学的に許容される賦形剤にはまた、アルカリ性緩衝液、または腸溶性のカプセルもしくはマイクロ顆粒も含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、膣内経路によっても投与することができる。このような場合、薬学的に許容される賦形剤には、乳化剤、CARBOPOL(登録商標)などのポリマー、また、膣内クリームおよび膣内坐剤の他の公知の安定化剤も含まれ得る。RSウイルスFポリペプチドのVLPワクチンはまた、直腸内経路によっても投与することができる。このような場合、賦形剤には、直腸内坐剤を形成するために当技術分野で公知のワックスおよびポリマーも含まれ得る。
【0162】
代替的に、RSウイルスFポリペプチドのVLPによるワクチン処方物は、キトサン(上記で記載した)または他のポリカチオンポリマー、ポリラクチド粒子およびポリラクチド−コグリコリド粒子、ポリ−N−アセチルグルコサミンベースのポリマーマトリックス、多糖または化学修飾した多糖からなる粒子、リポソームおよび脂質ベースの粒子、グリセロールモノエステルからなる粒子などからなるワクチンビヒクルと組み合わせることができる。サポニンはまた、コレステロールの存在下において処方して、リポソームまたはISCOMなどの粒子構造を形成することもできる。さらに、サポニンは、非粒子状の溶液もしくは懸濁物中、または小ラメラ(paucilamelar)リポソームもしくはISCOMなどの粒子状構造内において、ポリオキシエチレンエーテルまたはポリオキシエチレンエステルと併せて処方することもできる。
【0163】
本明細書で記載されるRSウイルスFポリペプチドのVLPを用いる医薬組成物およびワクチン組成物中で用いられる、さらなる例示的なアジュバントには、SAF(米国、カリフォルニア州、Chiron社製)、MF−59(Chiron社製、例えば、Granoffら(1997年)、Infect Immun.、65巻(5号):1710〜1715頁を参照されたい)、SBASシリーズのアジュバント(例えば、SB−AS2(SmithKline Beecham社製のアジュバントシステム♯2;MPLおよびQS21を含有する水中油エマルジョン);SBAS−4(SmithKline Beecham社製のアジュバントシステム♯4;ミョウバンおよびMPLを含有する)、ベルギー、リクセンサルト、SmithKline Beecham社から市販されている)、Detox(Enhanzyn(登録商標))(GlaxoSmithKline社製)、RC−512、RC−522、RC−527、RC−529、RC−544、およびRC−560(GlaxoSmithKline社製)、ならびに、係属中の米国特許出願第08/853,826号、および同第09/074,720号において記載されるものなどの、他のアミノアルキルグルコサミド4−ホスフェート(AGP)が含まれる。
【0164】
アジュバントの他の例には、HunterによるTiterMax(登録商標)アジュバント(ジョージア州、ノークロス、CytRx社製);Gerbuアジュバント(ドイツ、ガイベルク、Gerbu Biotechnik GmbH社製);ニトロセルロース(NilssonおよびLarsson(1992年)、Res. Immunol.、143巻:553〜557頁);Seppic ISAシリーズのMontamideアジュバント(例えば、ISA−51、ISA−57、ISA−720、ISA−151など;フランス、パリ、Seppic社製)など、鉱油エマルジョン、非鉱油エマルジョン、油中水エマルジョン、または水中油エマルジョンを含めたミョウバン(例えば、水酸化アルミニウム、リン酸アルミニウム)エマルジョンベースの処方物;およびPROVAX(登録商標)(IDEC Pharmaceuticals社製);OM−174(脂質Aと関連するグルコサミンジサッカリド); Leishmania属の伸長因子;CRL 1005など、ミセルを形成する非イオン性のブロックコポリマー;ならびにSyntexアジュバント処方物が含まれるがこれらに限定されない。例えば、O’Haganら(2001年)、Biomol Eng.、18巻(3号):69〜85頁;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」、D. O’Hagan編(2000年)、Humana Press社を参照されたい。
【0165】
他の好ましいアジュバントには、一般式
HO(CH 2CH2O)n−A−R, (I)
[式中、nは1〜50であり、Aは結合または−−C(O)−−であり、RはC1〜50アルキル、またはフェニルC1〜50アルキルである]のアジュバント分子が含まれる。
【0166】
本発明の一実施形態は、一般式(I)[式中、nは1〜50、好ましくは4〜24、最も好ましくは9であり;R成分はC1〜50、好ましくはC4〜C20アルキル、また最も好ましくはC12アルキルであり;また、Aは結合である]のポリオキシエチレンエーテルを含むワクチン処方物からなる。ポリオキシエチレンエーテルの濃度は、0.1〜20%、好ましくは0.1〜10%、また最も好ましくは0.1〜1%の範囲であるべきである。好ましいポリオキシエチレンエーテルは、以下の群:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステアリルエーテル(polyoxyethylene−9−steoryl ether)、ポリオキシエチレン−8−ステアリルエーテル、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテルから選択される。ポリオキシエチレンラウリルエーテルなどのポリオキシエチレンエーテルは、「メルクインデックス」(第12版;エントリー7717)で記載されている。これらのアジュバント分子は、WO99/52549で記載されている。
【0167】
所望の場合、上記の一般式(I)に従うポリオキシエチレンエーテルを、別のアジュバントと組み合わせることができる。例えば、好ましいアジュバントの組合せは、上で記載したCpGとの組合せであることが好ましい。
【0168】
本明細書で開示されるRSウイルスFポリペプチドのVLPワクチンと共に用いるのに適する、薬学的に許容される賦形剤のさらなる例には、水、リン酸緩衝生理食塩液、等張性緩衝液が含まれる。
【0169】
本発明は、以下の非限定的な実施例を参照することにより、よりよく理解される。本明細書で記載される通り、本発明は、必要に応じて、抗原またはアジュバントに連結したさらなる脂質ラフト会合ポリペプチドを含む、RSウイルスFポリペプチドのVLPを包含する。以下の実施例では、RSウイルスFポリペプチドのVLPを含む、本発明の代表的な実施形態が記載される。
【実施例】
【0170】
(実施例1)
この実施例は、細胞質側末端領域を欠失しているRSV融合(F)糖タンパク質の切断型、またはインフルエンザ赤血球凝集素由来の膜貫通領域および細胞質側末端領域を含有するハイブリッドFタンパク質の哺乳動物細胞発現が、RSV Fを含有するエンベロープウイルス様粒子(VLP)の形成をもたらすことを実証する。
【0171】
この実施例は、マウス白血病ウイルスGag遺伝子産物との共発現で、または共発現ではなくRSV F糖タンパク質が哺乳動物細胞において発現されるように設計された。期待されるのは、RSV Fが細胞から出芽するエンベロープGag VLPに組み込まれることであった。
【0172】
プラスミドp3.1−RSVFTは、細胞質側末端を欠いているが天然の膜貫通ドメインを含有しているF(RSVFT)の切断型をコードした。p3.1−RSVFTの地図を図1に示し、RSVFTのコード配列は以下の通りである。
【0173】
【化3】
天然RSV F遺伝子が、細胞の細胞質で複製するパラミクソウイルス由来であり、したがって細胞の核において適切に発現されないと予測されることから、RSVFTコード配列は、カスタム合成DNA断片として合成し、ウイルスからはクローニングしなかった。コードされたRSVFTのアミノ酸配列(シグナルペプチドおよび膜貫通ドメインを大文字で示す)は以下の通りである。
【0174】
【化4】
プラスミドp3.1−shFv1は、インフルエンザA由来のH5赤血球凝集素の膜貫通ドメインおよび細胞質側末端に融合したRSV Fの外部ドメインからなるハイブリッドタンパク質をコードした。図2は、p3.1−shFv1の地図を示し、shFv1のコード配列は以下の通りである。
【0175】
【化5】
【0176】
【化6】
shFv1のアミノ酸配列(シグナルペプチドコード配列ならびにHA膜貫通および細胞質側末端配列を大文字で示す)は以下の通りである。
【0177】
【化7】
RSVFTと同様にshFv1コード配列は、DNA合成によって作製した。
【0178】
プラスミドp3.1−shFv2は、インフルエンザA由来のH5赤血球凝集素の膜貫通ドメインおよび細胞質側末端に融合したRSV Fの外部ドメインからなるハイブリッドタンパク質をコードした。shFv2は、インフルエンザHA由来膜貫通および末端領域が4アミノ酸長いことにおいてshFv1と異なった。図3は、p3.1−shFv2のプラスミド地図を示す。shFv2のコード配列は以下の通りである。
【0179】
【化8】
shFv2のアミノ酸配列(シグナルペプチドコード配列ならびにHA膜貫通および細胞質側末端配列を大文字で示す)は以下の通りである。
【0180】
【化9】
shFv2コード配列は、DNA合成によって作製した。
【0181】
プラスミドp3.1−Gagは、マウス白血病ウイルス由来のGag遺伝子産物をコードした。図4は、p3.1Gagのプラスミド地図を示す。プラスミドp3.1−Gag由来のGagのコード配列は以下の通りである。
【0182】
【化10】
【0183】
【化11】
MLV Gagのアミノ酸配列は以下の通りである。
【0184】
【化12】
Gag遺伝子は、プラスミドpAMS(ATCC)において見出されたMLVのゲノム由来の天然クローンを表した。
【0185】
10cm2組織培養皿8枚に、10%ウシ胎仔血清を補充したダルベッコ変法イーグル培地で培養した293−F細胞を接種した。細胞がおよそ95%コンフルエンス(confluence)(単層培養)に達したときに、各ウエルに以下のように:
【0186】
【化13】
【0187】
【化14】
プラスミドDNA合計4μgをトランスフェクトした。
【0188】
プラスミドDNAをLIPOFECTAMINE(TM)2000(Invitrogen)を製造者の推奨に従って使用してトランスフェクトした。トランスフェクションの8時間後、トランスフェクション培地をCD293無血清培地で置換し、培養を継続した。トランスフェクションの48時間後、RSV F発現ベクターのいずれかをトランスフェクトした細胞を含むすべての皿は、正しくフォールドしたFの表面発現と一致して有意なレベルの合胞体(融合細胞)を示した。トランスフェクションの48時間後、ピペッティングによって細胞を剥離し、細胞および培地を収集した。細胞の表面でのRSV F抗原活性の発現を、膜に組み込まれた(integrated)状態において正しくフォールドしたF抗原を認識するマウスモノクローナル抗体(9C5)での細胞染色後のフローサイトメトリーによってさらに実証した。細胞をF−陽性細胞の蛍光検出のためにフィコエリトリンに結合体化したヤギ抗マウス2次抗体とさらに反応させた。図5は、Gagベクターと共にまたは共にではなくF発現ベクターのいずれかをトランスフェクトした細胞でのFの有意なレベルの表面発現を実証するフローサイトメトリー実施からのヒストグラムを示す。これらのデータは、トランスフェクトした細胞集団における合胞体の存在と一致し、細胞表面上で正しくフォールドされたF抗原の存在を実証する。
【0189】
トランスフェクトした細胞から収集した増殖培地を合胞体およびすべての細胞細片を除去するために2000rpmで5分間遠心分離し、このステップからの上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×gで1時間遠心分離して、すべてのVLP(Gagおよび/またはF遺伝子発現の結果として培地中に放出された可能性がある)を収集した。このステップからの100,000×gペレットを追加的分析のためにトリス緩衝生理食塩水に再懸濁した。
【0190】
超遠心分離ステップ由来の再懸濁したペレットを、VLP形成の指標としてのGag産物の存在を検出するためにウエスタンブロット分析に供した。予測外に、顕著な量のGag産物をp3.1−Gagベクターだけをトランスフェクトした細胞由来の試料中で検出したが、F遺伝子ベクターのいずれかとの組合せでp3.1−Gagをトランスフェクトした細胞由来のペレットにおいては顕著に減少した量のGagが見出された。結果は、F遺伝子産物の発現がGag産物の出芽機能をある様式で干渉することと一致している。
【0191】
再懸濁した超遠心分離ペレット中のF抗原の存在を次にELISAによって以下の通り検査した:再懸濁した各超遠心分離ペレット試料をNunc MAXISORP(TM)平底96ウエルELISAプレートの列Aの1つのウエルに入れ、各試料をプレートの各カラムを下がりながら連続的に2倍希釈した。4℃、1晩のコーティング後、プレートを0.05%TWEEN20(TM)を含有するPBS(PBS−T)で3回洗浄し、次いでSTARTING BLOCK(TM)(PBS)(Pierce、Biotechnology)でブロックした。ブロッキング後、STARTING BLOCK T20(TM)(Pierce)中に1:1000希釈したモノクローナル抗体9C5 100μlを各ウエルに入れ、プレートを室温で3時間インキュベートした。プレートを再度洗浄し、STARTING BLOCK T20(TM)中に1:1000希釈したヤギ抗マウスHRP結合体化抗体(Southern Biotech)100μlを各ウエルに添加し、室温で1.5時間インキュベートした。PBS−Tでの最終洗浄(3×)に続いて、プレートをABTS試薬(Pierce)100μlの添加によって発色(develop)させ、室温、45分間インキュベートした。
【0192】
図6は、このELISAの結果を示し、顕著なレベルのRSV F抗原が、F遺伝子だけおよびF遺伝子+Gag遺伝子でのトランスフェクションの両方に由来する100,000×gペレットにおいて検出されたことが観察され得る。重要かつ予測外に、より多いRSV F抗原活性がRSV F遺伝子だけをトランスフェクトした細胞由来のペレットにおいて観察された。これらのデータは、F遺伝子単独の発現(Gag発現の非存在下)がF抗原を保有するVLPの形成をもたらすことを実証する。これは、おそらくFによるGag出芽の干渉によって、FとGagとの共発現がGag出芽の抑制を生じさせるという上の観察とも一致する。
【0193】
(実施例2)
この実施例は、p3.1−RSVFTベクターだけをトランスフェクトした哺乳動物細胞の培地中に放出されたRSV F VLPの電子顕微鏡観察を記載する。
【0194】
RSV F遺伝子だけの発現が、単なるRSV F凝集物ではなくVLPの放出を生じさせることを実証するために、10%FBSを補充したDMEM培地中に293−F細胞をおよそ95%コンフルエンスでそれぞれ含むT75組織培養フラスコ5個をp3.1−RSVFT 30μgおよびLIPOFECTAMINE(TM)2000(Invitrogen)75μlで製造者の仕様書に従ってトランスフェクトした。トランスフェクションの8時間後にトランスフェクション培地を除去し、CD293無血清培地で置換した。トランスフェクションの48時間後に顕著なレベルの浮遊合胞体が培地中に観察されたが、これはRSV F遺伝子の発現と一致する。この時点で増殖培地を収集し、合胞体および細胞細片を2000rpm、5分間の遠心分離によって除去した。次いで培地上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×g、1時間、10℃で遠心分離してVLPを収集した。この遠心分離ステップ由来のペレットを、次いで合計300μlのトリス緩衝生理食塩水中に再懸濁した。再懸濁したペレットを次いで低速(およそ5000pm)で短時間遠心分離して、超遠心分離法によって形成されたすべての不可逆的タンパク質沈殿物または凝集物を除去した。再懸濁したVLP調製物を次いで、酢酸ウラニルネガティブ染色を使用する透過型電子顕微鏡によって検査し、代表的な顕微鏡像を図7に示す。顕微鏡像の収集物は、形状およびサイズの両方での多様性を示すエンベロープ粒子または小胞粒子の顕著なアレイを示した。このような多形性の粒子集団は、タンパク質コアを欠いている出芽粒子の形成と一致する。
【0195】
(実施例3)
この実施例は、バキュロウイルス粒子と高度に凝集するRSV F VLPを生じさせた、バキュロウイルス発現系を使用する昆虫細胞でのRSV F VLPの発現を記載する。
【0196】
実施例2に記載した哺乳動物細胞発現系の使用の代替として昆虫細胞バキュロウイルス発現系でRSV F VLPを作製するための試みにおいて、shFv1のコード配列を含有するDNA断片をプラスミドpFastbac1にクローニングし、プラスミドpFB−shFv1を得た。pFB−shFv1の地図を図8に示す。shFv1遺伝子のコード配列およびコードされたアミノ酸配列は上に記載の通りである。プラスミドpFB−shFv1を、高分子量バクミド(bacmid)中にバキュロウイルスゲノムを含有するDH10Bacコンピテント細胞への形質転換によってバキュロウイルスゲノム中に組み換えた。形質転換されたクローンの選択に続いて、高分子量バクミドDNAをこれらの細菌クローンから調製し、shFv1遺伝子を含有する組換えバキュロウイルスを作製するためのSf9昆虫細胞のトランスフェクションに使用した。この系を使用する組換えバキュロウイルス作製に関わる手順の詳細は、website tools.invitrogen.comのfile/content/sfs/manuals/bactobac_man.pdfのPDFドキュメントに見出すことができる。
【0197】
RSV F VLPを作製するために、SF900−II培地中で増殖しているSf9細胞の200mlスピナー培養物3個を27℃で細胞密度が1mlあたり細胞2×106個に達するまでインキュベートした。その時点で2継代shFv1−組換えバキュロウイルス調製物20mlを各スピナーフラスコに添加し、培養物を27℃で細胞生存率が20%未満に下がるまでインキュベートした。次いで培養液を収集し、2k rpmで15分間遠心分離することによって細胞および細片を清澄化した。清澄化した培地を32ml一定分量に分け、100,000×gで1時間、10℃で、トリス緩衝生理食塩水(TBS)、pH7.4および0.05%TWEEN 20(TM)を含有する30%ショ糖クッション4ml上で遠心分離した。ペレットを0.05%TWEEN 20(TM)を含有するTBS合計12mlに再懸濁し、この材料を6ml一定分量2個に分け、TBS/0.05%TWEEN 20(TM)中の20〜60%ショ糖密度勾配上で遠心分離した。勾配を底から分画した。RSV F抗原活性を個々の勾配画分において実施例1に記載のELISA法によって9C5モノクローナル抗体を使用して、以下の変更を伴って検出した:高濃度のショ糖はELISAアッセイを妨げることから、各勾配画分1.5mlの試料100μlをTBSで10倍希釈し、存在したすべてのVLPをショ糖溶液からペレット化させるために100,000×gで40分間遠心分離した。これらの試料をTBSに再懸濁し、実施例1に記載のELISAアッセイに直接使用した。ELISAアッセイの結果を図9にプロットし、これは、それぞれ画分5および9中の2つの可視の薄いバンドと一致するRSV F抗原活性の2つの顕著なピークを示す。凝集したVLPは典型的にはより高密度(より低い画分)にバンド形成することから、画分9中のバンドは凝集していないVLPを含有すると予測された。しかし画分9中の物質の電子顕微鏡検査は、高度に凝集したVLPおよびバキュロウイルス粒子を明らかにした。代表的な電子顕微鏡像を図10に示す。したがって、凝集していないRSV Fを含有するVLPの作製および精製のためには哺乳動物細胞発現系がより適切であることが決定された。
【0198】
(実施例4)
この実施例は、合胞体形成活性を欠いているがVLP出芽活性、Gag阻害活性およびモノクローナル中和抗体9C5との反応性を保持しているRSV FのGPI−アンカー型の発現を実証する。
【0199】
脂質ラフトドメインを標的とするがそれ自身のVLP出芽活性を欠いている改変Fタンパク質の作製をコードする改変RSV F遺伝子コード配列を作製するための試みがなされた。この目的は、ハイブリッドGag−F VLPが、F遺伝子産物が位置する脂質ラフトドメインから出芽するためのGagタンパク質の能力を介して作製され得るように、MLV Gagの出芽活性と適合性があるF遺伝子産物を供給することであった。このために膜貫通および細胞質側末端のコード配列を切断し、それらをGPIアンカーシグナルをコードする配列で置換することによってF遺伝子を改変した。GPIアンカーシグナルは、F糖タンパク質外部ドメインを脂質ラフト位置に方向付けるが、しかしこれらの脂質ラフト位置でFポリペプチド配列の細胞膜貫通は無いことが予測された。この改変が、キメラGag−F VLPを形成するこれらの位置からの出芽のためのGag VLPの能力をもたらすことが予測された。しかし驚くべきことに、RSV FのGPI−アンカー形態は、依然としてVLP出芽活性を示し、このことは、任意の脂質ラフト会合ポリペプチドを使用してRSV Fポリペプチドを脂質ラフトへ方向付けて、RSV Fポリペプチド単独でのVLP出芽を可能にすることができることを実証した。
【0200】
図11は、ヒトカルボキシペプチダーゼMのGPIアンカーシグナルに融合したキメラFタンパク質がコードされた発現ベクターp3.1−F−GPIの地図を示す。プラスミドp3.1−F−GPI由来のF−GPIコード配列のヌクレオチド配列は、以下の通りである。
【0201】
【化15】
ヒトカルボキシペプチダーゼMのGPIアンカーシグナルに融合したRSV Fの外部ドメインからなるF−GPIキメラタンパク質のアミノ酸配列は、(GPIアンカーシグナルを大文字で示す)以下の通りである。
【0202】
【化16】
発現試験を、プラスミドp3.1−Gagもしくはp3.1−F−GPIまたはp3.1−Gagとp3.1−F−GPIとの組合せをトランスフェクトした293F細胞においてRSV F外部ドメインも保有するGag VLPが作製され得ることを実証する目的で実施した。
【0203】
10cm2組織培養皿12枚に、10%ウシ胎仔血清を補充したダルベッコ変法イーグル培地で培養した293−F細胞を接種した。細胞がおよそ95%コンフルエンス(単層培養)に達したときに、各ウエルに以下のように:
【0204】
【化17】
プラスミドDNA合計4μgをトランスフェクトした。
【0205】
プラスミドDNAをLIPOFECTAMINE(TM)2000(Invitrogen)を製造者の推奨に従って使用してトランスフェクトした。トランスフェクションの8時間後、トランスフェクション培地をCD293無血清培地に置換し、培養を継続した。トランスフェクションの48時間後、p3.1−F−GPI発現ベクターをトランスフェクトした細胞を含む皿は、顕著なレベルの合胞体(融合細胞)を示さず、このことは、F糖タンパク質のGPIアンカー改変が細胞融合を促進するF糖タンパク質の能力に干渉したことを示唆していた。
【0206】
トランスフェクションの48時間後、ピペッティングによって細胞を剥離し、細胞および培地を収集した。細胞の表面でのRSV F抗原活性の発現を、膜に組み込まれた状態において正しくフォールドしたF抗原を認識するマウスモノクローナル抗体(9C5)での細胞染色後のフローサイトメトリーによって実証した。細胞をF−陽性細胞の蛍光検出のためにフィコエリトリンに結合体化したヤギ抗マウス2次抗体とさらに反応させた。図12は、Gagベクターと共にまたは共にではなくp3.1−F−GPIベクターをトランスフェクトした細胞上での有意なレベルのFの表面発現を実証するフローサイトメトリー実施からのヒストグラムを示す。これらのデータは、合胞体形成活性はもはや存在しないにもかかわらず、細胞表面上で正しくフォールドされたF抗原の存在を実証した。
【0207】
トランスフェクトした細胞から収集した増殖培地を細胞および細片を除去するために2000rpmで5分間遠心分離し、このステップからの上清をトリス緩衝生理食塩水中の30%ショ糖クッション上で100,000×gで1時間遠心分離して、すべてのVLP(Gagおよび/またはF−GPI遺伝子発現の結果として培地中に放出された可能性がある)を収集した。このステップからの100,000×gペレットを追加的分析のためにトリス緩衝生理食塩水に再懸濁した。
【0208】
超遠心分離ステップからの再懸濁ペレットを、VLP形成の指標としてのGag産物の存在を検出するためのウエスタンブロット分析に供した。予測外に、顕著な量のGag産物がp3.1−Gagベクターだけをトランスフェクトした細胞由来の試料中で検出されたが、p3.1−F−GPIベクターと組み合わせてp3.1−Gagをトランスフェクトした細胞由来のペレットにおいてはGagが見出されたとしてもこれは少量であった。これらの結果は、キメラF産物の脂質ラフト標的化機構が変化し、その膜貫通および細胞質側末端領域が除かれているにもかかわらず、F−GPI遺伝子産物の発現がGagタンパク質の出芽機能を依然として干渉することを実証した。これらの結果を図13に示し、これは、GagおよびRSV Fについてのウエスタンブロットを示す。RSVウエスタンブロットは、F抗原活性がF−GPIおよびF−GPI+Gagトランスフェクションの由来のペレットにおいて見出されたことを示し、Gagの有無がF−GPI媒介出芽および粒子形成に影響しないことを示した。
【0209】
再懸濁した超遠心分離ペレット中のF抗原の存在をELISAによっても以下の通り検査した:再懸濁した超遠心分離ペレットの各試料をNunc MAXISORP(TM)平底96ウエルELISAプレートの列Aのウエル1つに入れ、各試料をプレートの各カラムを下がりながら連続的に2倍希釈した。4℃、1晩のコーティング後、プレートを0.05%Tween20を含有するPBS(PBS−T)で3回洗浄し、次いでSTARTING BLOCK(TM)(PBS)(Pierce、Biotechnology)でブロックした。ブロッキング後、STARTING BLOCK(TM)T20(Pierce)中に1:1000希釈したモノクローナル抗体9C5 100μlを各ウエルに入れ、プレートを室温で3時間インキュベートした。プレートを再度洗浄し、STARTING BLOCK(TM)T20中に1:1000希釈したヤギ抗マウスHRP結合体化抗体(Southern Biotech)100μlを各ウエルに添加し、室温で1.5時間インキュベートした。PBS−Tでの最終洗浄(3×)に続いて、プレートをABTS試薬(Pierce)100μlの添加によって発色させ、室温で45分間インキュベートした。
【0210】
図14は、このELISAの結果を示し、顕著なレベルのRSV F抗原活性が、F−GPIだけおよびF−GPI+Gagベクターをトランスフェクトした細胞両方に由来する100,000×gペレットにおいて検出されたことが観察され得る。重要なことに、等量のRSV F抗原活性がF−GPIおよびF−GPI+Gagベクターをトランスフェクトした細胞由来のペレットにおいて観察された。これらのデータは、F−GPI遺伝子単独での発現が(Gag発現の非存在下で)F−GPI抗原を保有するVLPの形成を生じさせるという結論と一致する。これはまた、おそらくF−GPIによるGag出芽の干渉により、F−GPIとGagとの共発現がGag出芽の抑制を生じさせるという上の観察とも一致する。したがってGPIアンカーシグナルの切断および付加によるFポリペプチドの改変は、Fの出芽活性を破壊せず、Gag出芽に対する阻害効果も無効にしない。この実験は、F−GPI出芽粒子が、9C5モノクローナル抗体によって検出されるF抗原活性に関して生のRSVビリオンと同様に挙動することも示す。
【0211】
(さらなる参考文献)
【0212】
【数1】
【0213】
【数2】
【0214】
【数3】
【0215】
【数4】
【0216】
【数5】
【0217】
【数6】
【0218】
【数7】
【0219】
【数8】
【特許請求の範囲】
【請求項1】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子はエンベロープウイルスコアを含まない、調製物。
【請求項2】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子は、多形性であるか、サイズが不均一であるか、または形状が不均一である、調製物。
【請求項3】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子はエンベロープウイルスコア形成ポリペプチドを含まない、調製物。
【請求項4】
前記ウイルス様粒子が哺乳動物グリコシル化を含む、請求項1から3のいずれかに記載の調製物。
【請求項5】
前記ウイルス様粒子が他のウイルス様粒子と実質的に凝集しない、請求項1から4のいずれかに記載の調製物。
【請求項6】
前記ウイルス様粒子がウイルスベクター粒子と実質的に会合しない、請求項1から5のいずれかに記載の調製物。
【請求項7】
前記ウイルス様粒子と混合したアジュバントをさらに含む、請求項1から6のいずれかに記載の調製物。
【請求項8】
前記アジュバントが前記ウイルス様粒子の外側に位置する、請求項7に記載の調製物。
【請求項9】
前記アジュバントが前記ウイルス様粒子の内側に位置する、請求項7に記載の調製物。
【請求項10】
前記アジュバントを前記RSウイルスFポリペプチドに共有結合させて、共有結合を形成させる、請求項7から9のいずれかに記載の調製物。
【請求項11】
抗RSV−F中和抗体が前記RSウイルスFポリペプチドに結合する、請求項1から10のいずれかに記載の調製物。
【請求項12】
前記抗RSV−F中和抗体が9C5である、請求項11に記載の調製物。
【請求項13】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子はエンベロープウイルスコアを含まない、方法。
【請求項14】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子は、多形性であるか、サイズが不均一であるか、または形状が不均一である、方法。
【請求項15】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子はエンベロープウイルスコア形成ポリペプチドを含まない、方法。
【請求項16】
前記哺乳動物細胞が培養されている前記培地から、前記RSウイルスFポリペプチドのウイルス様粒子を取り出すステップをさらに含む、請求項13から15のいずれかに記載の方法。
【請求項17】
前記発現ベクターがウイルスベクターである、請求項13から16のいずれかに記載の方法。
【請求項18】
前記ウイルスベクターが、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択される、請求項17に記載の方法。
【請求項19】
前記哺乳動物細胞が、BHK細胞、VERO細胞、HT1080細胞、MRC−5細胞、WI 38細胞、MDCK細胞、MDBK細胞、293細胞、293T細胞、RD細胞、COS−7細胞、CHO細胞、Jurkat細胞、HUT細胞、SUPT細胞、C8166細胞、MOLT4/クローン8細胞、MT−2細胞、MT−4細胞、H9細胞、PM1細胞、CEM細胞、黒色腫細胞、SB20細胞、LtK細胞、HeLa細胞、WI−38細胞、L2細胞、CMT−93、およびCEMX174細胞からなる群より選択される、請求項13から18のいずれかに記載の方法。
【請求項20】
抗RSV−F中和抗体が、前記発現されたRSウイルスFポリペプチドに結合する、請求項13から19のいずれかに記載の方法。
【請求項21】
前記抗RSV−F中和抗体が9C5である、請求項20に記載の方法。
【請求項22】
RSウイルスの感染を処置または予防する方法であって、免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を被験体に投与するステップを含む、方法。
【請求項23】
前記投与するステップが前記被験体における防御的免疫化反応を誘導する、請求項22に記載の方法。
【請求項24】
前記投与するステップが、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項22から23のいずれかに記載の方法。
【請求項25】
免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を含む、医薬組成物。
【請求項26】
薬学的に許容されるキャリアをさらに含む、請求項25に記載の医薬組成物。
【請求項27】
RSウイルスの感染に対する防御を提供する方法であって、免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を被験体に投与するステップを含む、方法。
【請求項28】
前記投与するステップが、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項27に記載の方法。
【請求項1】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子はエンベロープウイルスコアを含まない、調製物。
【請求項2】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子は、多形性であるか、サイズが不均一であるか、または形状が不均一である、調製物。
【請求項3】
RSウイルスFポリペプチドを含むRSウイルスFポリペプチドのウイルス様粒子調製物であって、前記ウイルス様粒子はエンベロープウイルスコア形成ポリペプチドを含まない、調製物。
【請求項4】
前記ウイルス様粒子が哺乳動物グリコシル化を含む、請求項1から3のいずれかに記載の調製物。
【請求項5】
前記ウイルス様粒子が他のウイルス様粒子と実質的に凝集しない、請求項1から4のいずれかに記載の調製物。
【請求項6】
前記ウイルス様粒子がウイルスベクター粒子と実質的に会合しない、請求項1から5のいずれかに記載の調製物。
【請求項7】
前記ウイルス様粒子と混合したアジュバントをさらに含む、請求項1から6のいずれかに記載の調製物。
【請求項8】
前記アジュバントが前記ウイルス様粒子の外側に位置する、請求項7に記載の調製物。
【請求項9】
前記アジュバントが前記ウイルス様粒子の内側に位置する、請求項7に記載の調製物。
【請求項10】
前記アジュバントを前記RSウイルスFポリペプチドに共有結合させて、共有結合を形成させる、請求項7から9のいずれかに記載の調製物。
【請求項11】
抗RSV−F中和抗体が前記RSウイルスFポリペプチドに結合する、請求項1から10のいずれかに記載の調製物。
【請求項12】
前記抗RSV−F中和抗体が9C5である、請求項11に記載の調製物。
【請求項13】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子はエンベロープウイルスコアを含まない、方法。
【請求項14】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子は、多形性であるか、サイズが不均一であるか、または形状が不均一である、方法。
【請求項15】
RSウイルスFポリペプチドのウイルス様粒子の集団を作製する方法であって、
(a)RSウイルスFポリペプチドを発現する発現ベクターを供給するステップと、
(b)前記発現ベクターを培地中の哺乳動物細胞内へと導入するステップと、
(c)前記RSウイルスFポリペプチドを発現させて、前記RSウイルスFポリペプチドのウイルス様粒子を作製するステップと
を含み、前記ウイルス様粒子はエンベロープウイルスコア形成ポリペプチドを含まない、方法。
【請求項16】
前記哺乳動物細胞が培養されている前記培地から、前記RSウイルスFポリペプチドのウイルス様粒子を取り出すステップをさらに含む、請求項13から15のいずれかに記載の方法。
【請求項17】
前記発現ベクターがウイルスベクターである、請求項13から16のいずれかに記載の方法。
【請求項18】
前記ウイルスベクターが、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスからなる群より選択される、請求項17に記載の方法。
【請求項19】
前記哺乳動物細胞が、BHK細胞、VERO細胞、HT1080細胞、MRC−5細胞、WI 38細胞、MDCK細胞、MDBK細胞、293細胞、293T細胞、RD細胞、COS−7細胞、CHO細胞、Jurkat細胞、HUT細胞、SUPT細胞、C8166細胞、MOLT4/クローン8細胞、MT−2細胞、MT−4細胞、H9細胞、PM1細胞、CEM細胞、黒色腫細胞、SB20細胞、LtK細胞、HeLa細胞、WI−38細胞、L2細胞、CMT−93、およびCEMX174細胞からなる群より選択される、請求項13から18のいずれかに記載の方法。
【請求項20】
抗RSV−F中和抗体が、前記発現されたRSウイルスFポリペプチドに結合する、請求項13から19のいずれかに記載の方法。
【請求項21】
前記抗RSV−F中和抗体が9C5である、請求項20に記載の方法。
【請求項22】
RSウイルスの感染を処置または予防する方法であって、免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を被験体に投与するステップを含む、方法。
【請求項23】
前記投与するステップが前記被験体における防御的免疫化反応を誘導する、請求項22に記載の方法。
【請求項24】
前記投与するステップが、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項22から23のいずれかに記載の方法。
【請求項25】
免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を含む、医薬組成物。
【請求項26】
薬学的に許容されるキャリアをさらに含む、請求項25に記載の医薬組成物。
【請求項27】
RSウイルスの感染に対する防御を提供する方法であって、免疫原性量の、請求項1から12のいずれかに記載の調製物、または請求項13から21のいずれかに記載の方法により作製された集団を被験体に投与するステップを含む、方法。
【請求項28】
前記投与するステップが、皮下送達、経皮送達、皮内送達、皮下送達、筋肉内送達、経口送達、経口送達、鼻腔内送達、口腔内送達、舌下送達、腹腔内送達、膣内送達、経肛門送達、および頭蓋内送達からなる群より選択される、請求項27に記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
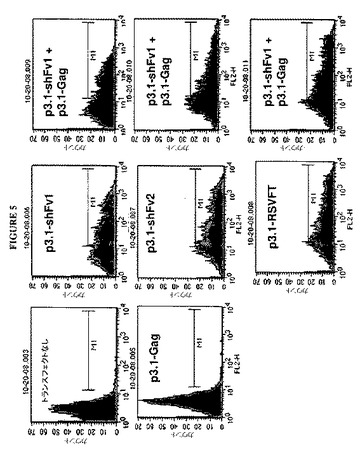
【図6】
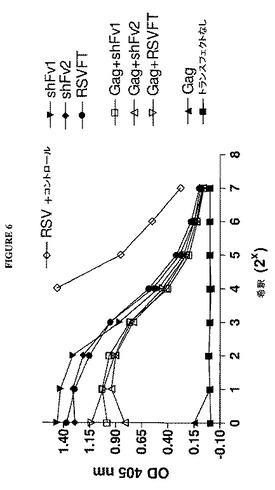
【図7】
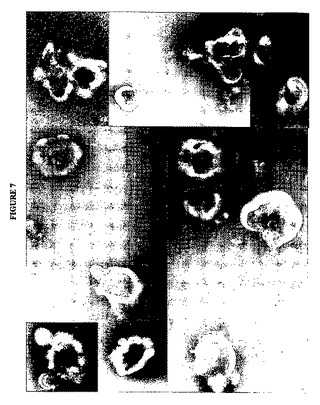
【図8】
【図9】
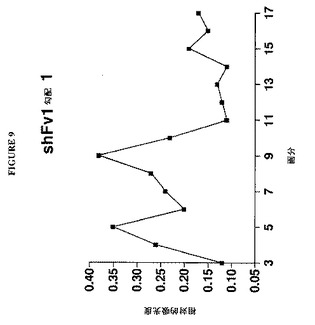
【図10】

【図11】
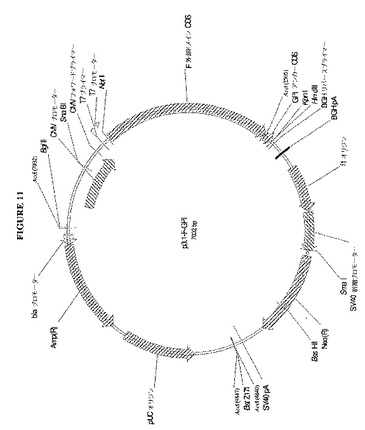
【図12】
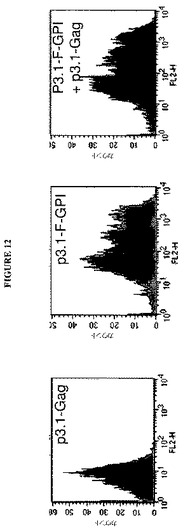
【図13】
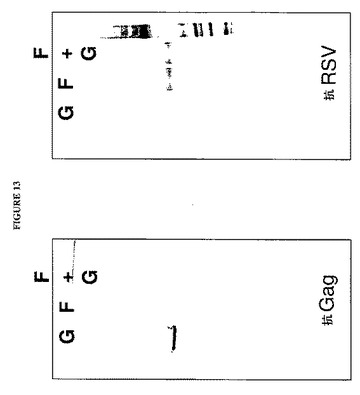
【図14】
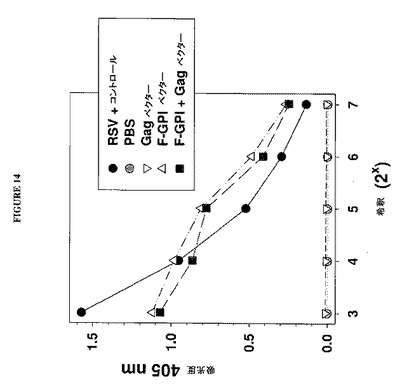
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2012−509280(P2012−509280A)
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願番号】特願2011−536614(P2011−536614)
【出願日】平成21年11月18日(2009.11.18)
【国際出願番号】PCT/US2009/064941
【国際公開番号】WO2010/059689
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
【公表日】平成24年4月19日(2012.4.19)
【国際特許分類】
【出願日】平成21年11月18日(2009.11.18)
【国際出願番号】PCT/US2009/064941
【国際公開番号】WO2010/059689
【国際公開日】平成22年5月27日(2010.5.27)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
[ Back to top ]