
RTK−過剰機能−誘導障害、特に癌の治療のための阻害剤の使用
【課題】増加した受容体チロシンキナーゼ活性の結果である病気、特に癌の治療および/または予防のための阻害剤の提供。
【解決手段】受容体チロシンキナーゼの改変された活性の変化は、特にFGFR−4の突然変異によってトリガーされ、ここに、前記突然変異は特にFGFR−4の膜貫通ドメイン中の点突然変異であり、疎水性アミノ酸の親水性アミノ酸への交換に導く。さらに、癌の治療および/または予防のためのFGFR−4に対する阻害剤の使用に関する。さらに、細胞において過剰発現および/または改変された活性を引き起こす突然変異FGFR−4に対する関する。最後に、突然変異FGFR−4分子のDNAおよびRNA配列、前記の阻害剤を含有する医薬組成物および、さらに、診断およびスクリーニング手法に関する。
【解決手段】受容体チロシンキナーゼの改変された活性の変化は、特にFGFR−4の突然変異によってトリガーされ、ここに、前記突然変異は特にFGFR−4の膜貫通ドメイン中の点突然変異であり、疎水性アミノ酸の親水性アミノ酸への交換に導く。さらに、癌の治療および/または予防のためのFGFR−4に対する阻害剤の使用に関する。さらに、細胞において過剰発現および/または改変された活性を引き起こす突然変異FGFR−4に対する関する。最後に、突然変異FGFR−4分子のDNAおよびRNA配列、前記の阻害剤を含有する医薬組成物および、さらに、診断およびスクリーニング手法に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[発明の属する技術分野]
【0002】
本発明は増大した受容体チロシンキナーゼ活性の結果である病気、特に癌の治療および/または予防のための阻害剤の使用に関する。該使用は、特に、受容体チロシンキナーゼ(RTK)の過剰発現および/または改変された活性の阻害あるいは低下に指向される。特に、受容体チロシンキナーゼのこの改変された活性はFGFR−4の突然変異によってトリガーすることができ、ここに、この突然変異は特にFGFR−4の膜貫通ドメイン中の点突然変異であり、疎水性アミノ酸を親水性アミノ酸への交換に導く。本発明は、さらに、癌の治療および/または予防のためのFGFRキナーゼの阻害剤の使用に関する。さらに、本発明は突然変異FGFR−4に関し、これは細胞において過剰発現および/または改変された活性に導く。最後に、本発明は突然変異FGFR−4分子のDNAおよびRNA配列に関する。最後に、加えて、本発明は前記の阻害剤を含有する医薬組成物および、さらに、診断およびスクリーニング手法に関する。
【従来の技術】
【0003】
細胞の増殖は生物の特別の要求に依存した注意深く調節されるプロセスである。若い生物において、細胞分裂速度は細胞の死滅速度を超え、これは生物のサイズの増加に導く。成体生物においては、細胞の新しい形成および細胞の死滅は「定常状態」が生起するようにバランスしている。しかしながら、稀な場合には、細胞増殖の制御が破壊され、細胞は増殖し分裂を始めるが、このタイプの非常に多数の細胞に対して特別の要求は生物では存在しない。この制御されない細胞増殖は癌の原因である。時々転移形成と関連する制御されない細胞増殖を誘導することができる因子はしばしば化学的性質であるが、例えば放射線照射のごとき物理的性質のものでもあり得る。癌をトリガーするもう1つの原因はある種の生物における遺伝的特別性あるいは突然変異であり、これは多かれ少なかれ細胞変性に導く。
【0004】
今日まで、例えば乳房において正常増殖および分化を制御するプロセスを解明するのは満足すべき程度には依然として可能ではなかった。ホルモン制御に加えて、哺乳動物細胞の発生に介入する異なる局所的に生じた成長因子の複雑なネットワークもある。哺乳動物細胞における癌の細胞の発生の正確な原因、他の細胞にもあてはまるように、それらが発散するほどに不明確かつ知られていない。オンコジイン(癌遺伝子)および腫瘍サプレッサー遺伝子における改変は乳癌の癌発生において重要な役割を演じるようである。加えて、遺伝的に改変された細胞で生起する調節因子による補強された刺激は細胞増殖の増大した進行に導き得る。
【0005】
現在、実質的に2つの代替法が癌の治療で利用できる。外科的介入によって完全に病気生物から癌細胞が首尾よく除去されるか、あるいは例えば、医薬の投与によって(化学療法)あるいは照射のごとき物理的治療手法によって生物中の変性細胞を無害なものとするような試みが成されるかのいずれかである。
【0006】
化学療法においては、ある形態においてはDNAに介入し、迅速に増殖する細胞に損傷を与え、ゆっくりと分裂するか全く分裂しない細胞よりも強くより高いDNA代謝能力を生じさせる必要がある医薬がしばしば使用される。しかしながら、多くの化学療法剤の著しく不利な点は、健康な細胞も化学療法の間に損傷される結果として、使用される活性物質の低い特異性である。活性物質のこの低い特異性は、さらに、その用量が各場合において癌細胞を同時に殺しつつ、損傷される健康な細胞ができるだけ少なくなるようなものでなければならないことを必要とする。これはしばしば可能ではなく、さらに癌が拡大し、最終段階では非常に重要な機能の不全を引き起こすので癌患者は死亡する。
【0007】
ある種の成長因子受容体の過剰発現および/または改変された活性は乳癌を含めた多くの新生物の激化した増殖に寄与すると推定されている。例えば、乳癌におけるEGFR、すなわち上皮因子受容体、あるいはERB B−2受容体の過剰発現は貧弱な予後に関連付けられてきた。FGF(歴史的には:繊維芽細胞成長因子)蛋白質も乳腺中の癌のあるいは他の癌の発生に関与し得た。しかしながら、この点についての結果は矛盾しているか、あるいは結論が出ていない。
【0008】
FGFは、そのうち9つのメンバーがこれまでに知られているペプチド調節因子の大きなファミリーを構成する。これらのうち8つはヒトにおいてよく特徴付けられている(BasilicoおよびMoscatelli, 1992;Coulier, 1993)。FGFは高親和性チロシンキナーゼ受容体を介して働き、これは少なくとも4つの異なる遺伝子によってコードされる。さらに、FGFは腫瘍形成に対して効果を有するのみならず、心臓血管病、組織損傷後の復元、神経生物学および胚発育でも主要な役割を演じ得るマルチ機能の調節ペプチドである。酸性および塩基性FGF(aFGFおよびbFGF)が最初のものであり、該ファミリーの最良に特徴づけられているメンバーである。インビボでは、例えば、胚発生における中胚葉誘導(Slackら、 1987;Kimelmanら、 1988)および血管形成(Thomasら、1985;Thompsonら、 1989; FolkmannおよびKlagsbrun、1987)に関与することが示されている。
【0009】
対応する受容体(FGFR)では、それらをコードする4つの同様の遺伝子が同定されている。これらの遺伝子は、3つの免疫グロブリンループおよび酸性部分、疎水性膜貫通ドメインおよび細胞内ドメインよりなり、チロシンキナーゼ活性を取り込む細胞外ドメインを持つ構造的に関連した蛋白質をコードする。これらの遺伝子のうち2つ、FGFR−1およびFGFR−2では、それらはオルターナティブスプライシングによって生起する複数の転写体を有することを示すことができた(GivolおよびYayon, 1992およびJohnsonおよびWilliams, 1993)。これらの遺伝子から生起するスプライシング変異体は、代替エクソンから生起し得る、受容体の細胞外領域および第3の免疫グロブリンドメインの第2の半分についての配列中の免疫グロブリン様ドメインの数に関して異なる。加えて、膜貫通および膜近接の欠点あるいは欠失が起こりかねず、これは分泌されたあるいはキナーゼ不活性蛋白質産物を生じ得る。
【0010】
FGFR−3では、代替転写体および対応するイソ型を見いだすことができたが、FGFR−4では、ただ1つの知られた蛋白質産物があるにすぎない。非常に多数のFGFR遺伝子および転写体および規定されたFGFに対する特異性の多くの蛋白質産物に対する欠如のため、特異的受容体に対する特異的リガンドの作用を測定するのは困難である。従って、特異的FGF受容体および規定された病気の間の相関は、規定された受容体の作用の特別のメカニズムと病気との相関は別として、大きな困難性を伴って確立できたに過ぎない。従って、FGFRを利用して、複雑な病像の癌を治療するのはかなり困難である。
【0011】
[発明が解決しようとする課題]
従って、本発明の目的は、受容体チロシンキナーゼ(RPK)がその発生に関与する体細胞障害、特に癌の可能な治療および/または予防を特定することにある。特に、本発明の目的は、受容体チロシンキナーゼの過剰発現および/または改変された、例えば構成的活性を阻害しおよび/または低下させることにある。
【0012】
本発明のさらなる目的は、突然変異FGFR−4の受容体チロシンキナーゼの改変された活性を阻害しおよび/または低下させることにある。
【0013】
本発明のさらなる目的は、癌発生および/または転移形成に関与するさらなるRTKを特定することにある。さらに、本発明の目的は、RTKのDNA配列あるいは対応するRNA配列を特定することにある。
【0014】
本発明のさらなる目的は、改良された診断あるいは種々の診断およびスクリーニング手法を特定することにある。
【0015】
最後に、本発明の目的は、特に癌がそれで治療することができる医薬組成物を特定することにある。
【0016】
これらの目的は独立の請求項の目的によって達成される。従属の請求項は本発明の好ましい開発を特定する。
【発明の実施の形態】
【0017】
本発明をよく理解するために、本明細書中で用いる用語を詳細に説明する。
【0018】
「阻害剤」とはRTKを阻害し、あるいはその活性を低下させるいずれもの物質と理解される。これはRTK、キナーゼ−不活性受容体あるいは抗受容体抗体に対して向けられた低分子量物質であり得る。
【0019】
「キナーゼ−不活性受容体」とは、もはやいずれのチロシンキナーゼ活性も有しないいずれもの受容体と理解される。
【0020】
「受容体チロシンキナーゼ」[Sic]とは、チロシンキナーゼ活性を有するいずれもの受容体と理解される。該発現はチロシンキナーゼ活性を有する成長因子受容体、およびHER2あるいはmet−受容体を含む。
【0021】
「RTK−過剰機能」とは過剰発現(後記参照)および/または改変された活性(後記参照)と理解される。
【0022】
「欠陥シグナル伝達活性」とは、突然変異受容体がこの欠陥シグナル発生がもはやリガンド例えば成長因子の存在に依存しないという意味で、突然変異受容体がもはや細胞外増殖シグナルあるいはもう1つのシグナルを細胞内シグナルに変換できないことを意味する。
【0023】
「成長因子」とは、正常および/または形質転換哺乳動物細胞によってとりわけ分泌され、細胞の増殖およびその活性の維持の刺激において、細胞増殖の調節において重要な役割を演じるいずれの細胞分裂促進性化学物質、通常はポリペプチドも意味する。用語「成長因子」とは例えば、上皮成長因子(EGF)、血小板由来成長因子(PDGF)および神経成長因子(NGF)およびFGF、すなわち繊維芽細胞成長因子を含む。
【0024】
「突然変異受容体チロシンキナーゼ」とは、該受容体が野生型受容体からの異なる、もはや調節できないチロシンキナーゼ活性を有するように、野生型受容体と比較して、構造改変を含有する受容体チロシンキナーゼと理解される。突然変異の1つのクラスはRTKの改変された活性に導く。
【0025】
「野生型成長因子受容体」あるいは「野生型」受容体とは、天然に生じる成長因子受容体あるいは非突然変異アミノ酸配列を保有する受容体と理解される。「野生型」は集団で最も通常に起こる受容体に対応する。
【0026】
成長因子受容体あるいは受容体の「細胞外ドメイン」とは、通常細胞から細胞外環境に突き出た受容体の部分と理解される。細胞外ドメインは、例えば、それにもう1つの分子の成長因子が結合する受容体の一部を含む。
【0027】
成長因子受容体あるいは受容体の「膜貫通領域」とは、通常、受容体を発現する細胞の細胞膜に位置する受容体の疎水性部分と理解される。
【0028】
成長因子受容体あるいは受容体の「チロシンキナーゼドメイン」あるいは「細胞質ドメイン」とは、通常、細胞の内部に位置し、チロシン残基のトランスリン酸化をもたらす受容体の部分であると理解される。
【0029】
「有効量」とは、所望の治療効果を達成できる本発明による組成物の量と理解される。
【0030】
「繊維芽細胞成長因子(FGF)」とは、細胞、とりわけ繊維芽細胞の増殖および他の特性に影響する細胞分裂促進性ポリペプチドと理解される。
【0031】
「過剰発現」とは、野生型と比較した細胞によるRTK蛋白質の増大した生産と理解される。これは、たとえばRTK遺伝子の遺伝子の増幅によって、トリガーされ、拡張した、コントロールされていない細胞分列活性を引き起こしうる。
【0032】
「改変された活性」とは、成長因子受容体によって媒介される単一移動経路の永久的活性と理解される。かくして、改変されたRTKをもって、キナーゼ活性がリガンド存在しない場合には存在する。
【0033】
本発明に従えば、突然変異FGFR−4は細胞において対応する受容体チロシンキナーゼの過剰発現および/または改変された活性に至ることができ、よって、癌に至り得る。
【0034】
成長因子受容体はヒト癌細胞の発達および増殖において決定的な役割を演じる。健康な細胞において、成長因子受容体は、とりわけ、細胞増殖の制御のみならず分化、細胞移動に関与する。細胞分裂についての現実のシグナルは、生物の要求に応じて形成される成長因子である。受容体はシグナル伝達の機能を取り、すなわち、それは細胞外増殖シグナルの細胞の内側における分裂活性への変換に関与する。多くの成長因子受容体でもって、成長因子が細胞外ドメインに結合した後、蛋白質におけるチロシン残基へのリン酸を移動させるその能力は決定的な役割を演じる。また、これらの受容体は受容体チロシンキナーゼとして記載される。受容体チロシンキナーゼの総説はYarden Y およびUllichA, REV. Biochem. 1988, 57, 443−78に見いだされる。成長因子の結合後におけるこれらの成長因子受容体の二量体化はシグナル伝達のプロセスにおけるさらなる重要な事象である。細胞外シグナルの、チロシンキナーゼ活性を持つ成長因子受容体によって媒介される細胞内シグナルへの変換は以下の5つの工程に分けることができる:
1.受容体の細胞外ドメインでの(リガンドとも記載される)成長因子の結合は立体配座の変化を誘導し、これは、
2.改変された立体配座の持つ受容体の二量体化を引き起こし、同時に
3.キナーゼ活性の同時の誘導、
4.一旦再度活性化された受容体立体配座を生じ、それを安定化させると、受容体ダイマー中のチロシン残基のトランスリン酸化、および
5.ポリペプチド基質のリン酸化および細胞因子との相互作用を引き起こす。
【0035】
例えば、受容体の過剰発現あるいは改変された活性によるこのシグナル伝達鎖の制御されない過剰機能は、とりわけ、関連細胞の増大した分裂活性および極端な場合には変性癌細胞に至り得る。成長因子受容体および細胞外から細胞内媒体へのシグナル伝達におけるその機能、ならびに癌発生に対する異常に発現された受容体の可能な影響に関する総説は、Ullich A,およびSchlessinger J(1990)Cell 61 203−212に提供されている。
【0036】
今回、驚くべきことに、前記で説明した5段階シグナル伝達鎖において突然変異FGFR−4の結果、突然変異RTKの改変された活性がその発生に決定的に関与するシグナル伝達活性が増大することが見出された。
【0037】
よって、本発明の請求項1によると、受容体チロシンキナーゼの少なくとも1つの阻害剤がRTK−過剰機能−誘導障害、特に癌の治療および/または予防で用いられる。さらに、本発明によると、増大したシグナル伝達に帰すことができる組織の過剰増殖および/または組織の増大した侵入性の結果である病気あるいは体細胞障害を排除し、あるいは軽減することができる。
【0038】
阻害剤ならびに低分子量物質として、例えば、少なくとも1つのキナーゼ不活性受容体を使用することができる。阻害剤の、例えばキナーゼ−不活性受容体の使用を介して、受容体チロシンキナーゼの改変された活性を阻害しおよび/または低下させることができる。すでに記載したごとく、成長因子受容体の過剰発現および/または改変された活性は癌をトリガーし、あるいは進行させるのに重要な因子である。乳癌におけるEGFRあるいはErb B−2受容体の過剰発現は、例えば、貧弱な予後と関連付けられてきた(前記参照)。よって、この過剰発現および/または改変された活性の阻害は癌の治療および/または予防において重要な要素である。FGFR−4は胚形成の間に組織特異的にスイッチを切られる。しかしながら、それは乳癌患者の30%に存在し、またそれは健康な被験者の組織では検出できない。受容体チロシンキナーゼの阻害剤の使用は、過剰発現および/または改変された活性の低下あるいは完全な阻害に至る。同様に、キナーゼ−不活性受容体の使用は、受容体チロシンキナーゼの活性の低下および/または完全な阻害に至る。というのは、ヘテロダイマーのキナーゼ機能はもはやシグナル伝達できないからである。キナーゼ−不活性受容体の作用は、非機能的ヘテロダイマーが形成される(希釈効果)という事実に基づく。シグナルが細胞の生物学的応答に変換されるのが妨げられる結果として、シグナル伝達の欠如は過剰発現および/または改変された活性シグナルの伝達の妨げに至る。その結果、受容体チロシンキナーゼのこの阻害を介して、あるいはこれらのキナーゼ−不活性受容体を介して、効果的かつ積極的に癌の治療および/または予防に介入することは可能である。
【0039】
驚くべきことに、FGFR−4突然変異は健康な個人の生殖細胞系でも起こることが判明した。生殖細胞系突然変異は遺伝的素因に至り、これは個人を種々の病気の活性にかかりやすくすると推定される。癌形成との関係で、腫瘍組織における突然変異受容体の増大発現は癌形成に関与すると推定される。生殖細胞系突然変異は、さらに、とりわけ、以下の病気、すなわち動脈硬化症、白血病、リンパ腫、肝細胞癌腫および胆管癌についての素因と見なされる。
【0040】
その結果、本発明は利用できるさらなる遺伝子マーカーを作成し、これは種々の病気およびこれらに対する罹患性の診断および初期認識において非常に助けとなることが判明する。
【0041】
従って、本発明は本件物質であるFGFR−4をコードする核酸の検出方法に関し、それにより、特に受容体コーディング核酸の突然変異が検出される。これは例えばオリゴヌクレオチドプローブとのハイブリダイゼーションによって行うことができ、これは具体的には突然変異、特に点突然変異の存在あるいは不存在を示すことができる。これにおいて、例えば、突然変異核酸およびオリゴヌクレオチドの間の「ミスマッチ」を、もし「ミスマッチ」が存在すると、ハイブリダイゼーションが起こらず、従ってシグナルがないように利用される。別法として、特異的FGFR−4 PCRプライマーでの核酸の増幅および適当な制限エンドヌクレアーゼでの引き続いての切断によって突然変異を検出することができる。もし、例えば、突然変異認識配列がもはや制限エンドヌクレアーゼによって切断部位として認識されないように制限エンドヌクレアーゼの認識配列に影響するならば、これは、非突然変異野生型におけるよりも異なる制限断片に至る。PCRによって、制限断片は特異的に検出することができ、従って、述べた場合において、例えば、より大きな制限断片が野生型で比較して突然変異体に存在する。しかしながら、別法として、適当な酵素での切断後に「野生型断片」が突然変異体中でより小さくなる結果として、突然変異は新しい制限切断部位の創製に至り得る。野生型におけるGlyの代わりに突然変異体中のArgへの交換に至る、EMBL Gene Bank/DDBJにX57205下で寄託された、配列の位置388におけるFGFR−4の膜貫通ドメイン中の突然変異は、制限エンドヌクレアーゼBstN1の認識配列GGWCCに関する。その結果、80および29bpの2つの新しい制限断片が形成され、これはとりわけ制限分析によって検出できる。
【0042】
本発明によると、さらに、RTKの過剰発現および特に改変された活性は増大された侵入性、すなわち、強化された転移形成に至ることが示され得る。転移形成は癌の腫瘍な問題の1つであるので、これは、過剰発現および/または改変された活性の阻害あるいは低下が、癌と闘うにおいて効果的な薬剤をもたらし、それにより特に転移形成が阻害される。
【0043】
可能な阻害剤は、例えば、Mohammadiら(1997)に記載されている。
【0044】
好ましくは、本発明によると、受容体チロシンキナーゼの過剰発現および/または改変された活性に介入され、これはFGFR−4の突然変異によってトリガーされる。この突然変異は1個あるいは数個の点突然変異であり得る。特に、突然変異/複数の突然変異が、特に疎水性アミノ酸が親水性アミノ酸へと交換される結果として、FGFR−4の膜貫通ドメインで起こる。
【0045】
FGFR−3における疎水性アミノ酸を親水性アミノ酸に代える交換に至る、突然変異はある種の病気と関連することが知られている。かくして、例えば、膜貫通ドメイン中の点突然変異による繊維芽細胞成長因子受容体3の改変された活性が軟骨形成不全で見いだされている。小人症の最も普通に起こる遺伝形態である軟骨形成はある種の骨の成熟過程における血管に実質的に基づく常染色体優性障害である。FGFR−3の膜貫通ドメインにおけるGlyからArgへの置換によって軟骨形成不全がトリガーされることを示すことができた。さらに、FGFR−3におけるArg突然変異はダイマー受容体のキナーゼ機能を活性化することを示すことができた。また、Arg点突然変異はFGFR−3それ自体のチロシンキナーゼ活性のリガンド−非依存性および受容体上のホスホチロシンのかなり増大した改変レベルに至る。これらの結果は、軟骨形成異常の分子的基礎がFGFR−3による調節されからグナル移動であることを示唆する。
【0046】
FGFR−3の膜貫通ドメインにおけるさらなる突然変異は、アラニンをグルタミンへの交換である。このアミノ酸交換はもう1つの病気、すなわち黒色表皮症を持つクルーゾン病に至る。
【0047】
本発明によると、FGFR−4における突然変異、特に、疎水性アミノ酸を親水性アミノ酸への交換に至る膜貫通ドメイン中の点突然変異は癌についてのトリガリングおよび良好でない予後に関し、そのためには受容体チロシンキナーゼの阻害あるいはキナーゼ不活性受容体の使用が癌の治療および/または予防に適し、ここに、受容体チロシンキナーゼは突然変異のため改変された方法にて過剰発現されるかあるいは活性である。
【0048】
特に、グリシンのアルギニンへの交換に至る位置388における点突然変異では、この結果、受容体チロシンキナーゼが改変された方法で活性となり、このホモ−およびヘテロ接合の結果、リガンド刺激なくしてシグナル伝達が起こり、その結果細胞の制御されない増殖がトリガーされ得る。最悪の場合、この制御されない増殖は癌に至る。次いで、膜貫通ドメインは配列(配列番号1):
RYTDIILYASGSLALAVLLLLARLYを有し、
他方、非突然変異ドメインは以下の配列(配列番号2):
RYTDIILYASGSLALAVLLLLAGLYを有する。
【0049】
理論に拘束されるつもりはないが、前記突然変異の1つ、特にグリシンのアルギニンへの交換に至る位置388での点突然変異を保有する受容体チロシンキナーゼの活性化は、それを通じて膜貫通ドメインにおける交換が可能とされる相互作用ため起こる、ダイマー立体配座における受容体の安定化に基づくと推定される。リガンド−非依存性ダイマーの強化された形成は、増大した受容体チロシンキナーゼ活性および細胞形質転換に至る。癌のトリガリングに対する点突然変異の効果についての1つの可能性は、例えば、突然変異がFGFR−4によるシグナル伝達に作用する点に、膜を通じての受容体移動が防止され、それ自体とのあるいは他のFGFRとの二量体化が乱される点に、あるいは受容体のチロシンキナーゼ活性が影響される点に基礎を有し得る。
【0050】
本発明によると、乳癌を持つSt.Petersburgからの患者の56%が(バイオプシーによる検査)が位置388で突然変異を担い、これはグリシンのアルギニンへの交換に結び付けることができた。これらのうち、45%がヘテロ接合であり、11%がホモ接合である。これは、これは、高い割合で、位置388における点突然変異と乳癌の発生との間のつながりを示唆する。
【0051】
乳癌を持つドイツ人患者を調べたさらなる検査において、患者の43%のみが位置388で点突然変異を示した。癌患者の正常組織および正常個体の組織からのDNAでの実験から、突然変異は生殖細胞系突然変異であると帰結することができる。
【0052】
さらに、細胞系からのゲノムDNAおよびcDNAも実験して、位置388における点突然変異の割合を決定した。乳癌、比較としての正常乳房上皮細胞系、偏平細胞癌腫、神経膠芽細胞腫、神経芽細胞腫および子宮癌に由来する細胞系を実験した。正常乳房上皮細胞系を除いて全ての細胞系で、FGFR−4分子における位置388での点突然変異のかなりの割合を見いだすことができた。よって、阻害剤あるいはキナーゼ−不活性受容体の前記使用は癌腫の治療で特に適する。ここに、神経芽細胞腫、子宮癌および膵臓癌の治療は特に有望なようであるが、他のタイプの癌はそうではない。
【0053】
特に好ましいのは、特に膜貫通ドメインにおいて位置388において突然変異GlyからArgへの突然変異を持つ、突然変異FGFR−4を阻害する阻害剤の使用である。
【0054】
さらに、本発明は、細胞において受容体の過剰発現および/または改変された活性に至る突然変異FGFR−4に関する。好ましくは、この突然変異FGFR−4は、野生型受容体における疎水性アミノ酸が、突然変異受容体において親水性アミノ酸に交換されていることに特徴がある。特に好ましくは、点突然変異であり、膜貫通ドメインにおいて起こる、突然変異である。さらにより好ましくは、点突然変異が位置388で起こり、その結果、好ましくは、グリシンがアルギニンによって置換される。
【0055】
従来、専門家に間では、突然変異されていない唯1つのFGFR−4が起こると推定された。突然変異FGFR−4は知られていなかった。従って、本発明により、突然変異FGFR−4が存在することを示すことができたのは驚くべきことであった。特に、本発明により、突然変異、特に位置388における点突然変異と、癌の発生との間の関係を示すことができた。さらに、健康な個人における生殖細胞系突然変異は、とりわけ、動脈硬化症の発生についての遺伝素因と関係付けられた。
【0056】
本発明は、さらに、突然変異FGFR−4をコードする配列を含有するDNA分子に関する。また、本発明は、突然変異FGFR−4をコードするRNA配列を含有するRNA分子を含む。前記配列は癌の診断に使用することができる。これにおいて、配列はFGFR−4における突然変異を特異的に認識することができる。FGFR−4における突然変異の存在は、癌の治療のために貧弱な予後と関係付けられる。この理由は対応する腫瘍の攻撃的増殖挙動であり得る。
【0057】
これとは別に、本発明は、乳癌の差異的診断のための手法に関し、ここで、患者の核酸を前記したDNAおよび/またはRNAのうちの一方と接触させ、従って、突然変異FGFR−4の存在および/または不存在を示すシグナルが得られる。最後に、本発明は、前記阻害剤あるいはキナーゼ−不活性受容体を含有する医薬組成物に関する。これとは別に、チロシンキナーゼ活性の阻害剤の同定のためのスクリーニング手法に関し、ここに、本発明の受容体は可能な阻害剤と接触され、阻害剤の存在下および/または不存在下におけるチロシンキナーゼ活性が測定される。
【0058】
さらに、既に前記にて詳細に記載されたごとく、突然変異の存在の検出がPCR、続いての制限酵素切断によって行うことができる。
【0059】
最後に、他の分子生物学的診断手法も可能である。
【0060】
さらに、本発明の目的は、本発明による突然変異FGFR−4と特異的に反応する抗体である。本発明の意味における「特異的」とは、本発明による抗体は特異的受容体に結合するが、非特異的受容体には結合しないということである。
【0061】
以下、図面および実施例によって本発明を詳細に記載する。
【実施例】
【0062】
細胞培養。ヒト細胞系MDA−MB−453、ZR 75−1、K562およびSKBr3はATCCから得た。個々の供給源は最後の表に見いだすことができる。MDA−MB−453、K562およびZR 75−1は、10%の胎児ウシ血清(Sigma, Taufkirchen)を含有するRPMI(Gibco, Eggenstein)で培養した。SKBR3は15%胎児ウシ血清を含有するMcCoy5a(Gibco,Eggenstein)で培養した。全ての細胞培養培地はペニシリン/ストレプトマイシン(Sigma,Taufkirchen)を含有していた。細胞を水−蒸気飽和雰囲気および8%CO2 中にて37℃でインキュベートした。
【0063】
FGFR−4388Arg/wtのクローニング。K562およびMDA−MB−453細胞からのRNAの調製では、3×107細胞をグアニジニウムイソシアネートで溶解させ、CsClグラジエントでの超遠心によって精製した。cDNA合成は、製造業者の指示に従って、各場合において、逆転写酵素(Boehringer, Mannheim)および10pmolの「ランダムオリゴヌクレオチド」を用いて行った。引き続いてのPCR反応では0.5μlを用いた。
【0064】
FGFR−4388ArgおよびFGFR−4wtをPCR反応によって増幅した。このために、以下のプライマーを用いた:センス−GCTCAGAGGGCGGGCGGGGGTGCCGGCCG、アンチセンスCCGCTCGAGTGCCTGCACAGCCTTGAGCCTTGC。PCR反応では、以下のものを用いた:製造業者の指示に従って1.5U/25μlのExpand−Polymerase(Boehringer, Mannheim)および反応緩衝液:200μM dNTP、0.01% v/vTritonX100;10% v/vDMSO、および各々0.2pmolのセンスおよびα−センスプライマー。以下の反応工程を行った:35サイクル、94℃ 1分、64℃ 1分、72℃ 2.5分。FGFR−4388ArgのクローニングではMDA−MB−453cDNAを用い、FGFR−4wtのクローニングではK562 cDNAを用いた。PCR産物はpcDNA3ベクター(Invitrogen)にクローニングした。このようにして、G388Rを有するFGFR−4および野生型FGFRをさらなる工程のために得ることができた。
【0065】
FGFR−4の膜貫通ドメインの増幅。以下のプライマーを用いた:センス−GACCGCAGCAGCGCCCGAGGCCAG;アンチセンスAGAGGGAAGAGGGAGAGCTTCTG。PCT反応では以下のものを用いた。製造業者の指示に従った1.5U/25μl Taq−Polymerase(Boehringer, Mannheim)および反応緩衝液:200μMdTNP;各々0.2pmolのセンスおよびα−センスプライマー、0.5αl cDNAあるいは腫瘍バイオプシーおよび細胞系からのゲノムDNA。以下の反応工程を用いた:35サイクル、95℃ 45秒、72℃ 45秒。
【0066】
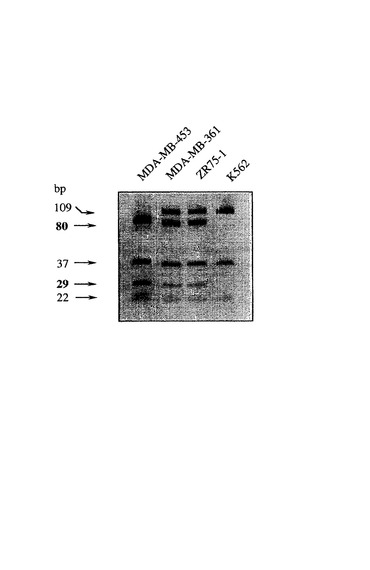
制限酵素消化による分析。ゲノムあるいはcDNAからのFGFR−4の膜貫通ドメインを前記したごとくに増幅した。制限酵素消化を用いて、G1217A突然変異につきバイオプシーおよび細胞系をテストするために、PCR産物を、5U/25μlのBstB1(NEB、Schwalbach/Taunus)と、60℃で1時間インキュベートした。制限酵素消化からのDNA断片を20%ポリアクリルアミドゲルで分離し、臭化エチジウムで染色した。野生型受容体の分析は109、37および22塩基対サイズの断片を生じる(トラック4)。他方、突然変異G1217Aの結果として、BstB1についてのさらなる制限酵素切断部位が形成される。突然変異受容体はさらなる80および29塩基対サイズの断片を示し、他方、109塩基対サイズの断片は消失する(トラック:ホモ接合体;トラック2および3:ヘテロ接合体)(図2参照)。
【0067】
制限酵素消化によるゲノムDNAの遺伝子型
初代腫瘍の組織試料からのゲノムDNAを標準的な方法によって単離した(Current Protocols in Molecular Biology, John Wiley and Sons, Inc., 1995)。ゲノムDNAの遺伝子型分析ができるためには、FGFR−4遺伝子における膜貫通領域をPCR反応において以下のプライマーで増幅した:
5’−GACCGCAGCAGCGCCCGAGGCCAG−3’(bp1275−1142)、および
5’−AGAGGGAAGAGGGAGAGCTTCTG−3’(bp1275−1297)。PCR反応では、Ready−to−Go PCR Beats(Pharmacia, Uppsala, スウェーデン)を用いた。以下のPCRサイクルを用いた:95℃における3分、94℃における45秒、72℃における45秒、および72℃における5分。合計35サイクルを行った。PCR産物を、5U/25μlのBstB1(NEB、Schwalbach/Taunus)と、60℃で1時間インキュベートした。制限酵素消化からのDNA断片を20%ポリアクリルアミドゲルで分離し、臭化エチジウムで染色した。388Argアレレ(対立遺伝子)は80および29bpサイズの2つの断片によって特徴付けられ、他方、388Glyアレレは単一の109bpサイズの断片によって示される。各遺伝子型分析は3回反復された。
【0068】
PCR産物のDNA配列決定。FGFR−4の膜貫通ドメインの配列分析では、PCR産物をBluescriptにクローンニングした。このために、PCR反応を既に記載したごとくに行った。以下のプライマーを用いた:センス−GGGAATTGACCGCAGCAGCGCCCGAGG;α−センス−GCTCTAGAAGAGGGAAGAGGGAGAG。FGFR−4Arg388/wtのクローニングのPCR産物はベクターpcDNA3中で、直接配列決定することができた。T/−プライマーのプラスミドDNAへのアニーリングの後、配列決定反応をT/−DNAポリメラーゼ(Pharmacia,Freisburg)を用いて行った。次いで、配列決定反応の産物を変性5%ポリアクリルアミドゲル(7.5M尿素、1×TBE)で分離し、乾燥した後にX線フイルムに暴露した(図3参照)。これより、野生型および突然変異のDNA配列が得られた。
【0069】
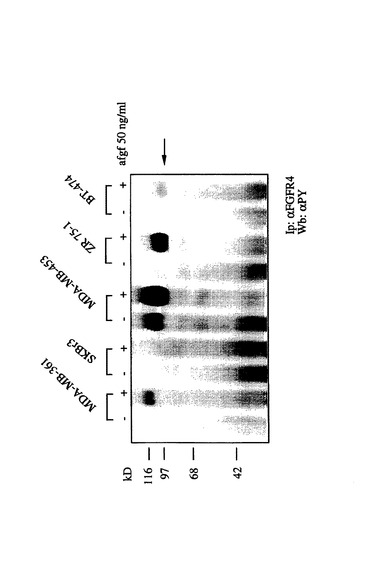
免疫沈殿およびウェスタンブロット分析。2.2×106細胞を10cmのペトリ皿上に広げ、一晩インキュベートした。細胞培地を、胎児ウシ血清を含まない培地によって置換し、さらに24時間インキュベートした。刺激には、細胞を50ngのsFGF/mlで10分間インキュベートし、冷PBSで2回洗浄し、氷上に置いた。細胞を300μlの冷溶解緩衝液(1% w/w NP−40、1.25% w/vデオキシコール酸ナトリウム、0.1% w/vSDS、0.15M NaCl、0.01Mリン酸ナトリウム、pH7.2、2mM EDTA、10mMフッ化ナトリウム、1mM PMSF、20μg/mlアプロチニン、1mMオルトバナデート、10mMピロリン酸ナトリウム)と共に4℃で15分間インキュベートし、溶解物を4℃の遠心(13,000RPM)によって清澄化した。蛋白質値決定では、Micro−BCA Protein Assay(Pierce)を製造業者の指示に従って用いた。免疫沈殿では、細胞溶解物を等しい蛋白質含有量に調整し、次いで、0.5μgの抗−FGFR−4(Santa Cruz)にて、回転するホイール上のプロテイン−A−Sepharose (Pharmacia, Freisburg)にて4℃で18時間インキュベートした。免疫複合体を冷HNTG(20mM HEPES pH7.5、150mM NaCl、0.1%TritonX100、10%グリセリン、10mMピロリン酸ナトリウム)で4回洗浄した。試料の調製では、免疫複合体を50μl3×Laemmli緩衝液で処理し、99℃で5分間インキュベートした。沈殿した蛋白質を7.5%SDS−PAGE上で分離した(図1参照)。
【0070】
ウェスタンブロットでは、SDS−PAGEによって分離した蛋白質をニトロセルロースに移した。膜上の非特異的蛋白質結合部位をTBS−T/0.25%ゼラチン(10mM Tris/HCl pH8.0、0.15M NaCl、0.05%Tween20)と共に室温で2時間のインキュベーションによってブロックした。初代抗体でのインキュベーションは傾いたシェーカー上にて4℃で6時間行った。次いで、非特異的に結合した抗体をTBS−T/0.25%ゼラチンで4回洗浄することによって除去した。二次抗体の結合は室温にて1時間行った。非特異的に結合した二次抗体はさらなる洗浄工程によって除去した。免疫複合体は製造業者の指示に従ってECLTMキット(Amersham, Braunschweig)で可視化した。
【0071】
統計学的方法。統計学的計算は統計学プログラムMedCalc(MedCalc SOftware, ベルギー)およびEpillnfo 6.04(CDC、アトランタ、ジョージア)の助けを借りて行った。異なる群における遺伝子型および臨床的領域との間の相関を決定するために、優劣比、信頼区間(CI)および統計的有意性(P値)を計算した。少数の388Argホモ結合患者のため、この群を統計的計算では388Argヘテロ接合患者の群と組合せた。
【0072】
腫瘍細胞系におけるFGFR−4の検出。表IはRTKの発現および乳癌の間の相関関係を示す。RTKの発現は、明瞭に、乳癌よりも細胞系でしばしば起こり、他方、正常上皮細胞の細胞系では検出できない。
【0073】
【表1】

【0074】
表2から、G388R突然変異は他の癌タイプの細胞系でも起こり、これらと相関性があることが明らかである。健康な上皮細胞系では、前記突然変異は検出できない。
【0075】
【表2−1】
【表2−2】
【0076】
バイオプシーにおけるFGFR−4突然変異G388Rの検出。表3は調べた乳癌を持つSt Petersburgからの61人の女性患者のうち、56%がG388R突然変異(そのうち45%がヘテロ接合であり、11%がホモ接合である)を保有していたことを示す。調べたミュンヘンからの69人の女性乳癌患者のうち、43%がG388R突然変異(そのうち32%がヘテロ的であり、11%がホモ接合的である)を保有していた。St Petersburgおよびミュンヘンからの女性患者における突然変異の全パーセンテージの割合は異なる。これは、G388R突然変異が生殖細胞系突然変異であることを示唆する。
【0077】
【表3−1】
【表3−2】
【表3−3】
【0078】
FGFR−4−G388R突然変異およびFGFR−4発現の検出の間の相関関係。以下の表4から、G388R突然変異(ゲノムDNAおよびcDNA)は、発現および/または強化された発現が起こる場合のみ起こることが明らかである。突然変異は、RTK発現が見いだされない、正常乳房上皮細胞系でも乳癌細胞系でも検出できない。
【0079】
そのRTK発現が特に顕著である細胞系MDA−MB 453はホモ接合G388R突然変異を示す。
【0080】
【表4】
【0081】
FGFR−4突然変異G388Rおよびリンパ節転移状態、あるいは再発なしの生存時間の間の相関関係の研究
表5は、乳癌の腫瘍形成におけるG388R突然変異の役割の研究に参画する全ての患者の臨床的パラメーターを示す。G388R突然変異を持つ患者はG388R突然変異を持たない患者よりも不良な長期予後を有することが見いだされた。
表5に対する記号
Her2:Her2受容体の発現レベル、0=発現なし、〜3=過剰発現
OPDAT:手術の日付
M/R:転移形成/再発、0=無し/1=有り
Vers:死亡;0=無し/1=有り
BERRE:再発なくしての生存時間(月数で)
グレード:腫瘍の分化グレード;1=強力な分化/3=低い分化段階:初代腫瘍のサイズ
E−Rec:エストロゲン受容体の発現、0=発現無し、〜12=最高発現
GEN:FGFR−4の遺伝子型;G=野生型アレレ;R=突然変異アレレ
BEDAT:最後の観察の日付
REZDAT:再発診断の日付
TODDAT:死亡の日付
BERLEB:通算しての生存時間
Nod.:リンパ節における転移;0=無し/1=有り
Men:閉経
P−Rec:プロゲステロン受容体の発現:0=発現無し、〜12=最高発現
【0082】
【表5−1】
【表5−2】
【表5−3】
【0083】
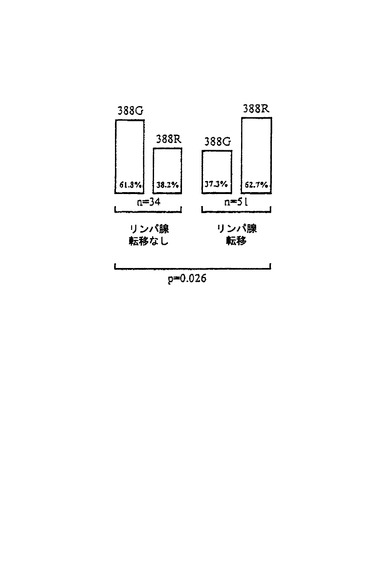
図4から、G388R突然変異は、最初の治療時においてリンパ節に転移を既に有する患者において数が多いことが見いだされる。G388R突然変異を持つ患者のうち、62.7%がリンパ節転移を有し、他方、G388R突然変異を持たない患者のうち、38.2%のみがリンパ節において転移を呈した。リンパ節転移状態は不良な予後を持つ腫瘍および良好な予後を持つ腫瘍のさらなる区別のための重要な予後マーカーであるので、この結果から、調べた85人の患者においてG388Rはよりひどい腫瘍進行に至れしめると結論できる。
【0084】
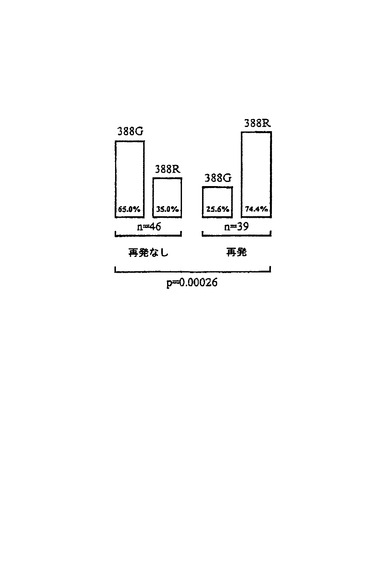
図5より、調べた患者の群において、再発なし生存確率は、G388R突然変異を有しない患者についてよりもG388R突然変異を持つものにつき、非常に低いことが分かる。再発した患者の74.4%が388R遺伝子型を有するが、25.6%のみが388G遺伝子型を有する。これは、G388R突然変異を持つ患者がより速やかに再発し、従って、成功裡に治療できなかったことを示す。
【0085】
要約すると、FGFR−4突然変異G388Rはリンパ節における転移形成の2.7倍(OR=2.7;CI:1.02<OR<7.4)増大した危険性に至らしめ、さらに5.44倍(OR=5.44;CI:1.93<OR<7.4)増大した腫瘍の再発の危険性に至らしめると云うことができる。かくして、突然変異したFGFR−4アレレ(G388R)を持つ患者は、腫瘍再発に対して素因を有するようであり、よって、より良好でない病気予後を有するようである。
【0086】
材料
アクリルアミド Serva,ハイデルベルグ
寒天 Difco,デトロイト
アガロース BRL,Eggenstein
アンピシリン Boehringer,マイハイム
アプロチニン Sigma,Taufkirchen
N,N’−ビスアクリルアミド Roth,Karisruhe
塩化セシウム BRL,Eggenstein
デスオキシヌクレオチド Pharmacia,Freisburg
臭化エチジウム Sigma,Taufkirchen
ゼラチン Sigma,Taufkirchen
グアニジウムイソチオシアネート Fluka,スイス国
HEPES Serva,ハイデルベルグ
フッ化ナトリウム Sigma,Taufkirchen
PMSF Sigma,Taufkirchen
SDS Roth,Karlsruhe
Tris Riedel de Haen, Seeze
TritonX100 Serva,ハイデルベルグ
Tween20 Serva,Taufkirchen
リストされていない全ての物質は、会社Sigma(Taufkirchen), Serva(ハイデルベルグ), Riedel Deursche Patentamt[sic]HaenあるいはMerck(Darmstadt)会社から入手し、最高に可能な純度グレードを用いた。
【0087】
装置
DNAの電気泳動 Workshop, MPI for Biochemistry, Martinsried
蛋白質の電気泳動 Atto,日本冷凍遠心
Biofuge17S Heraeus,Hanau
蛋白質移動 Semidryブロット装置, Workshop, MPI for Biochemistry, Martinstried
滅菌ワークベンチ Biogard, The Baker Company, USA
細胞培養 インキュベーター B5060 EK/Co2 , Hanau
細胞計数 コールターカウンター, Counter Electronics, グラスゴー
【0088】
文献
バシリコ・シイおよびマスカテリィ・デイ:ザ・エフジイエフ・ファミリー・オブ・グロウス・ファクターズ・アンド・オンコジーン アドバーンスト・キャンサー・リサーチズ,59,115−164(1992)
クーリエ・エフ, デ・ラペイリエール・オーおよびビルムバウム・デイ:コンプレクシティー・オブ・ザ・エフジイエフ・ファミリー:ザ・プルーフ・ナイン/メディ/サイ 9,1113−1115(1993)
フォークマン・ジェイおよびクラグスブルン・エム:サイエンス235, 442−447(1987)
ジバル・デイおよびヤヨン・エイ:コンプレクシティー・オブ・エフジイエフ・レセプターズ:ジェネティックス・ベーシス・フォー・ストラクチュラル・デリバリー・アンド・ファンクショナル・スペシフィシティー、エフエイエスイービイ・ジャーナル 6:3362−3369(1992)
ジョンソン・デイ・イーおよびウィリアムズ・エル・ティー:ストラクチュラル・アンド・ファンクショナル・ダイバーシティー・イン・ザ・エフジーエフ・レセプター・マルチジーン・ファミリー, アドバーンスト・キャンサー・リサーチズ 60:1−41(1993)
キメイマン・デイ, アブラハム・ジェイ・エイ, ハーパランタ・テイ,パルシィー・テイ・エムおよびキルシュナー・エム・ダブリュー:サイエンス,242, 1053−1058(1988)
モハマディー・エムら, サイエンス776,955−959(1997)
スラック・ジェイ・エム・ダブリュー, ダーリントン・ジイ・ジイ, ヘルス・ジェイ・ケイおよびゴッドサーブ・エス・エフ:ネイチャー326, 197−200(1987)
トーマス・ケイ・エイ,リオス−カンデロール・エム, グリメズ−ガレゴ・ジイ, ジサルボ・ジェイ, ベルネット・シイ, ロドケイ・ジェイおよびフィッツパトリック・エス:プロシーディングズ・ナショナル・アカデミー・オブ・サイエンシズ・ユーエスエイ, 82, 6409−6413(1985)
トンプソン・ジェイ・エイ, ハウデンシルト・シイ・シイ, アンダーソン・ケイ・デイ, ジピエテロ・ジェイ・ウム, アンダーソン・ダブリ・エフおよびマシアーグ・テイ(1989):プロシーディングズ・ナショナル・アカデミー・オブ・サイエンシズ・ユーエスエイ,86, 7928−7932(1989)
【0089】
【表6】
【図面の簡単な説明】
【0090】
【図1】FGFR−4(リン酸化されたFGR受容体−4は、矢印で示されている)の免疫沈殿物のSDSPAGEを示す。
【図2】突然変異FGFR−4のポリアクリルアミドゲルである。
【図3】FGFR−4の膜貫通ドメインの配列分析である。
【図4】FGFR−4突然変異G388Rとリンパ節転移形成の状態(n:患者数、p:P値)との相間関係である。
【図5】FGFR−4突然変異G388Rと再発なしでの生存時間(n:患者数、p:P値)との相間関係である。
【発明の詳細な説明】
【0001】
[発明の属する技術分野]
【0002】
本発明は増大した受容体チロシンキナーゼ活性の結果である病気、特に癌の治療および/または予防のための阻害剤の使用に関する。該使用は、特に、受容体チロシンキナーゼ(RTK)の過剰発現および/または改変された活性の阻害あるいは低下に指向される。特に、受容体チロシンキナーゼのこの改変された活性はFGFR−4の突然変異によってトリガーすることができ、ここに、この突然変異は特にFGFR−4の膜貫通ドメイン中の点突然変異であり、疎水性アミノ酸を親水性アミノ酸への交換に導く。本発明は、さらに、癌の治療および/または予防のためのFGFRキナーゼの阻害剤の使用に関する。さらに、本発明は突然変異FGFR−4に関し、これは細胞において過剰発現および/または改変された活性に導く。最後に、本発明は突然変異FGFR−4分子のDNAおよびRNA配列に関する。最後に、加えて、本発明は前記の阻害剤を含有する医薬組成物および、さらに、診断およびスクリーニング手法に関する。
【従来の技術】
【0003】
細胞の増殖は生物の特別の要求に依存した注意深く調節されるプロセスである。若い生物において、細胞分裂速度は細胞の死滅速度を超え、これは生物のサイズの増加に導く。成体生物においては、細胞の新しい形成および細胞の死滅は「定常状態」が生起するようにバランスしている。しかしながら、稀な場合には、細胞増殖の制御が破壊され、細胞は増殖し分裂を始めるが、このタイプの非常に多数の細胞に対して特別の要求は生物では存在しない。この制御されない細胞増殖は癌の原因である。時々転移形成と関連する制御されない細胞増殖を誘導することができる因子はしばしば化学的性質であるが、例えば放射線照射のごとき物理的性質のものでもあり得る。癌をトリガーするもう1つの原因はある種の生物における遺伝的特別性あるいは突然変異であり、これは多かれ少なかれ細胞変性に導く。
【0004】
今日まで、例えば乳房において正常増殖および分化を制御するプロセスを解明するのは満足すべき程度には依然として可能ではなかった。ホルモン制御に加えて、哺乳動物細胞の発生に介入する異なる局所的に生じた成長因子の複雑なネットワークもある。哺乳動物細胞における癌の細胞の発生の正確な原因、他の細胞にもあてはまるように、それらが発散するほどに不明確かつ知られていない。オンコジイン(癌遺伝子)および腫瘍サプレッサー遺伝子における改変は乳癌の癌発生において重要な役割を演じるようである。加えて、遺伝的に改変された細胞で生起する調節因子による補強された刺激は細胞増殖の増大した進行に導き得る。
【0005】
現在、実質的に2つの代替法が癌の治療で利用できる。外科的介入によって完全に病気生物から癌細胞が首尾よく除去されるか、あるいは例えば、医薬の投与によって(化学療法)あるいは照射のごとき物理的治療手法によって生物中の変性細胞を無害なものとするような試みが成されるかのいずれかである。
【0006】
化学療法においては、ある形態においてはDNAに介入し、迅速に増殖する細胞に損傷を与え、ゆっくりと分裂するか全く分裂しない細胞よりも強くより高いDNA代謝能力を生じさせる必要がある医薬がしばしば使用される。しかしながら、多くの化学療法剤の著しく不利な点は、健康な細胞も化学療法の間に損傷される結果として、使用される活性物質の低い特異性である。活性物質のこの低い特異性は、さらに、その用量が各場合において癌細胞を同時に殺しつつ、損傷される健康な細胞ができるだけ少なくなるようなものでなければならないことを必要とする。これはしばしば可能ではなく、さらに癌が拡大し、最終段階では非常に重要な機能の不全を引き起こすので癌患者は死亡する。
【0007】
ある種の成長因子受容体の過剰発現および/または改変された活性は乳癌を含めた多くの新生物の激化した増殖に寄与すると推定されている。例えば、乳癌におけるEGFR、すなわち上皮因子受容体、あるいはERB B−2受容体の過剰発現は貧弱な予後に関連付けられてきた。FGF(歴史的には:繊維芽細胞成長因子)蛋白質も乳腺中の癌のあるいは他の癌の発生に関与し得た。しかしながら、この点についての結果は矛盾しているか、あるいは結論が出ていない。
【0008】
FGFは、そのうち9つのメンバーがこれまでに知られているペプチド調節因子の大きなファミリーを構成する。これらのうち8つはヒトにおいてよく特徴付けられている(BasilicoおよびMoscatelli, 1992;Coulier, 1993)。FGFは高親和性チロシンキナーゼ受容体を介して働き、これは少なくとも4つの異なる遺伝子によってコードされる。さらに、FGFは腫瘍形成に対して効果を有するのみならず、心臓血管病、組織損傷後の復元、神経生物学および胚発育でも主要な役割を演じ得るマルチ機能の調節ペプチドである。酸性および塩基性FGF(aFGFおよびbFGF)が最初のものであり、該ファミリーの最良に特徴づけられているメンバーである。インビボでは、例えば、胚発生における中胚葉誘導(Slackら、 1987;Kimelmanら、 1988)および血管形成(Thomasら、1985;Thompsonら、 1989; FolkmannおよびKlagsbrun、1987)に関与することが示されている。
【0009】
対応する受容体(FGFR)では、それらをコードする4つの同様の遺伝子が同定されている。これらの遺伝子は、3つの免疫グロブリンループおよび酸性部分、疎水性膜貫通ドメインおよび細胞内ドメインよりなり、チロシンキナーゼ活性を取り込む細胞外ドメインを持つ構造的に関連した蛋白質をコードする。これらの遺伝子のうち2つ、FGFR−1およびFGFR−2では、それらはオルターナティブスプライシングによって生起する複数の転写体を有することを示すことができた(GivolおよびYayon, 1992およびJohnsonおよびWilliams, 1993)。これらの遺伝子から生起するスプライシング変異体は、代替エクソンから生起し得る、受容体の細胞外領域および第3の免疫グロブリンドメインの第2の半分についての配列中の免疫グロブリン様ドメインの数に関して異なる。加えて、膜貫通および膜近接の欠点あるいは欠失が起こりかねず、これは分泌されたあるいはキナーゼ不活性蛋白質産物を生じ得る。
【0010】
FGFR−3では、代替転写体および対応するイソ型を見いだすことができたが、FGFR−4では、ただ1つの知られた蛋白質産物があるにすぎない。非常に多数のFGFR遺伝子および転写体および規定されたFGFに対する特異性の多くの蛋白質産物に対する欠如のため、特異的受容体に対する特異的リガンドの作用を測定するのは困難である。従って、特異的FGF受容体および規定された病気の間の相関は、規定された受容体の作用の特別のメカニズムと病気との相関は別として、大きな困難性を伴って確立できたに過ぎない。従って、FGFRを利用して、複雑な病像の癌を治療するのはかなり困難である。
【0011】
[発明が解決しようとする課題]
従って、本発明の目的は、受容体チロシンキナーゼ(RPK)がその発生に関与する体細胞障害、特に癌の可能な治療および/または予防を特定することにある。特に、本発明の目的は、受容体チロシンキナーゼの過剰発現および/または改変された、例えば構成的活性を阻害しおよび/または低下させることにある。
【0012】
本発明のさらなる目的は、突然変異FGFR−4の受容体チロシンキナーゼの改変された活性を阻害しおよび/または低下させることにある。
【0013】
本発明のさらなる目的は、癌発生および/または転移形成に関与するさらなるRTKを特定することにある。さらに、本発明の目的は、RTKのDNA配列あるいは対応するRNA配列を特定することにある。
【0014】
本発明のさらなる目的は、改良された診断あるいは種々の診断およびスクリーニング手法を特定することにある。
【0015】
最後に、本発明の目的は、特に癌がそれで治療することができる医薬組成物を特定することにある。
【0016】
これらの目的は独立の請求項の目的によって達成される。従属の請求項は本発明の好ましい開発を特定する。
【発明の実施の形態】
【0017】
本発明をよく理解するために、本明細書中で用いる用語を詳細に説明する。
【0018】
「阻害剤」とはRTKを阻害し、あるいはその活性を低下させるいずれもの物質と理解される。これはRTK、キナーゼ−不活性受容体あるいは抗受容体抗体に対して向けられた低分子量物質であり得る。
【0019】
「キナーゼ−不活性受容体」とは、もはやいずれのチロシンキナーゼ活性も有しないいずれもの受容体と理解される。
【0020】
「受容体チロシンキナーゼ」[Sic]とは、チロシンキナーゼ活性を有するいずれもの受容体と理解される。該発現はチロシンキナーゼ活性を有する成長因子受容体、およびHER2あるいはmet−受容体を含む。
【0021】
「RTK−過剰機能」とは過剰発現(後記参照)および/または改変された活性(後記参照)と理解される。
【0022】
「欠陥シグナル伝達活性」とは、突然変異受容体がこの欠陥シグナル発生がもはやリガンド例えば成長因子の存在に依存しないという意味で、突然変異受容体がもはや細胞外増殖シグナルあるいはもう1つのシグナルを細胞内シグナルに変換できないことを意味する。
【0023】
「成長因子」とは、正常および/または形質転換哺乳動物細胞によってとりわけ分泌され、細胞の増殖およびその活性の維持の刺激において、細胞増殖の調節において重要な役割を演じるいずれの細胞分裂促進性化学物質、通常はポリペプチドも意味する。用語「成長因子」とは例えば、上皮成長因子(EGF)、血小板由来成長因子(PDGF)および神経成長因子(NGF)およびFGF、すなわち繊維芽細胞成長因子を含む。
【0024】
「突然変異受容体チロシンキナーゼ」とは、該受容体が野生型受容体からの異なる、もはや調節できないチロシンキナーゼ活性を有するように、野生型受容体と比較して、構造改変を含有する受容体チロシンキナーゼと理解される。突然変異の1つのクラスはRTKの改変された活性に導く。
【0025】
「野生型成長因子受容体」あるいは「野生型」受容体とは、天然に生じる成長因子受容体あるいは非突然変異アミノ酸配列を保有する受容体と理解される。「野生型」は集団で最も通常に起こる受容体に対応する。
【0026】
成長因子受容体あるいは受容体の「細胞外ドメイン」とは、通常細胞から細胞外環境に突き出た受容体の部分と理解される。細胞外ドメインは、例えば、それにもう1つの分子の成長因子が結合する受容体の一部を含む。
【0027】
成長因子受容体あるいは受容体の「膜貫通領域」とは、通常、受容体を発現する細胞の細胞膜に位置する受容体の疎水性部分と理解される。
【0028】
成長因子受容体あるいは受容体の「チロシンキナーゼドメイン」あるいは「細胞質ドメイン」とは、通常、細胞の内部に位置し、チロシン残基のトランスリン酸化をもたらす受容体の部分であると理解される。
【0029】
「有効量」とは、所望の治療効果を達成できる本発明による組成物の量と理解される。
【0030】
「繊維芽細胞成長因子(FGF)」とは、細胞、とりわけ繊維芽細胞の増殖および他の特性に影響する細胞分裂促進性ポリペプチドと理解される。
【0031】
「過剰発現」とは、野生型と比較した細胞によるRTK蛋白質の増大した生産と理解される。これは、たとえばRTK遺伝子の遺伝子の増幅によって、トリガーされ、拡張した、コントロールされていない細胞分列活性を引き起こしうる。
【0032】
「改変された活性」とは、成長因子受容体によって媒介される単一移動経路の永久的活性と理解される。かくして、改変されたRTKをもって、キナーゼ活性がリガンド存在しない場合には存在する。
【0033】
本発明に従えば、突然変異FGFR−4は細胞において対応する受容体チロシンキナーゼの過剰発現および/または改変された活性に至ることができ、よって、癌に至り得る。
【0034】
成長因子受容体はヒト癌細胞の発達および増殖において決定的な役割を演じる。健康な細胞において、成長因子受容体は、とりわけ、細胞増殖の制御のみならず分化、細胞移動に関与する。細胞分裂についての現実のシグナルは、生物の要求に応じて形成される成長因子である。受容体はシグナル伝達の機能を取り、すなわち、それは細胞外増殖シグナルの細胞の内側における分裂活性への変換に関与する。多くの成長因子受容体でもって、成長因子が細胞外ドメインに結合した後、蛋白質におけるチロシン残基へのリン酸を移動させるその能力は決定的な役割を演じる。また、これらの受容体は受容体チロシンキナーゼとして記載される。受容体チロシンキナーゼの総説はYarden Y およびUllichA, REV. Biochem. 1988, 57, 443−78に見いだされる。成長因子の結合後におけるこれらの成長因子受容体の二量体化はシグナル伝達のプロセスにおけるさらなる重要な事象である。細胞外シグナルの、チロシンキナーゼ活性を持つ成長因子受容体によって媒介される細胞内シグナルへの変換は以下の5つの工程に分けることができる:
1.受容体の細胞外ドメインでの(リガンドとも記載される)成長因子の結合は立体配座の変化を誘導し、これは、
2.改変された立体配座の持つ受容体の二量体化を引き起こし、同時に
3.キナーゼ活性の同時の誘導、
4.一旦再度活性化された受容体立体配座を生じ、それを安定化させると、受容体ダイマー中のチロシン残基のトランスリン酸化、および
5.ポリペプチド基質のリン酸化および細胞因子との相互作用を引き起こす。
【0035】
例えば、受容体の過剰発現あるいは改変された活性によるこのシグナル伝達鎖の制御されない過剰機能は、とりわけ、関連細胞の増大した分裂活性および極端な場合には変性癌細胞に至り得る。成長因子受容体および細胞外から細胞内媒体へのシグナル伝達におけるその機能、ならびに癌発生に対する異常に発現された受容体の可能な影響に関する総説は、Ullich A,およびSchlessinger J(1990)Cell 61 203−212に提供されている。
【0036】
今回、驚くべきことに、前記で説明した5段階シグナル伝達鎖において突然変異FGFR−4の結果、突然変異RTKの改変された活性がその発生に決定的に関与するシグナル伝達活性が増大することが見出された。
【0037】
よって、本発明の請求項1によると、受容体チロシンキナーゼの少なくとも1つの阻害剤がRTK−過剰機能−誘導障害、特に癌の治療および/または予防で用いられる。さらに、本発明によると、増大したシグナル伝達に帰すことができる組織の過剰増殖および/または組織の増大した侵入性の結果である病気あるいは体細胞障害を排除し、あるいは軽減することができる。
【0038】
阻害剤ならびに低分子量物質として、例えば、少なくとも1つのキナーゼ不活性受容体を使用することができる。阻害剤の、例えばキナーゼ−不活性受容体の使用を介して、受容体チロシンキナーゼの改変された活性を阻害しおよび/または低下させることができる。すでに記載したごとく、成長因子受容体の過剰発現および/または改変された活性は癌をトリガーし、あるいは進行させるのに重要な因子である。乳癌におけるEGFRあるいはErb B−2受容体の過剰発現は、例えば、貧弱な予後と関連付けられてきた(前記参照)。よって、この過剰発現および/または改変された活性の阻害は癌の治療および/または予防において重要な要素である。FGFR−4は胚形成の間に組織特異的にスイッチを切られる。しかしながら、それは乳癌患者の30%に存在し、またそれは健康な被験者の組織では検出できない。受容体チロシンキナーゼの阻害剤の使用は、過剰発現および/または改変された活性の低下あるいは完全な阻害に至る。同様に、キナーゼ−不活性受容体の使用は、受容体チロシンキナーゼの活性の低下および/または完全な阻害に至る。というのは、ヘテロダイマーのキナーゼ機能はもはやシグナル伝達できないからである。キナーゼ−不活性受容体の作用は、非機能的ヘテロダイマーが形成される(希釈効果)という事実に基づく。シグナルが細胞の生物学的応答に変換されるのが妨げられる結果として、シグナル伝達の欠如は過剰発現および/または改変された活性シグナルの伝達の妨げに至る。その結果、受容体チロシンキナーゼのこの阻害を介して、あるいはこれらのキナーゼ−不活性受容体を介して、効果的かつ積極的に癌の治療および/または予防に介入することは可能である。
【0039】
驚くべきことに、FGFR−4突然変異は健康な個人の生殖細胞系でも起こることが判明した。生殖細胞系突然変異は遺伝的素因に至り、これは個人を種々の病気の活性にかかりやすくすると推定される。癌形成との関係で、腫瘍組織における突然変異受容体の増大発現は癌形成に関与すると推定される。生殖細胞系突然変異は、さらに、とりわけ、以下の病気、すなわち動脈硬化症、白血病、リンパ腫、肝細胞癌腫および胆管癌についての素因と見なされる。
【0040】
その結果、本発明は利用できるさらなる遺伝子マーカーを作成し、これは種々の病気およびこれらに対する罹患性の診断および初期認識において非常に助けとなることが判明する。
【0041】
従って、本発明は本件物質であるFGFR−4をコードする核酸の検出方法に関し、それにより、特に受容体コーディング核酸の突然変異が検出される。これは例えばオリゴヌクレオチドプローブとのハイブリダイゼーションによって行うことができ、これは具体的には突然変異、特に点突然変異の存在あるいは不存在を示すことができる。これにおいて、例えば、突然変異核酸およびオリゴヌクレオチドの間の「ミスマッチ」を、もし「ミスマッチ」が存在すると、ハイブリダイゼーションが起こらず、従ってシグナルがないように利用される。別法として、特異的FGFR−4 PCRプライマーでの核酸の増幅および適当な制限エンドヌクレアーゼでの引き続いての切断によって突然変異を検出することができる。もし、例えば、突然変異認識配列がもはや制限エンドヌクレアーゼによって切断部位として認識されないように制限エンドヌクレアーゼの認識配列に影響するならば、これは、非突然変異野生型におけるよりも異なる制限断片に至る。PCRによって、制限断片は特異的に検出することができ、従って、述べた場合において、例えば、より大きな制限断片が野生型で比較して突然変異体に存在する。しかしながら、別法として、適当な酵素での切断後に「野生型断片」が突然変異体中でより小さくなる結果として、突然変異は新しい制限切断部位の創製に至り得る。野生型におけるGlyの代わりに突然変異体中のArgへの交換に至る、EMBL Gene Bank/DDBJにX57205下で寄託された、配列の位置388におけるFGFR−4の膜貫通ドメイン中の突然変異は、制限エンドヌクレアーゼBstN1の認識配列GGWCCに関する。その結果、80および29bpの2つの新しい制限断片が形成され、これはとりわけ制限分析によって検出できる。
【0042】
本発明によると、さらに、RTKの過剰発現および特に改変された活性は増大された侵入性、すなわち、強化された転移形成に至ることが示され得る。転移形成は癌の腫瘍な問題の1つであるので、これは、過剰発現および/または改変された活性の阻害あるいは低下が、癌と闘うにおいて効果的な薬剤をもたらし、それにより特に転移形成が阻害される。
【0043】
可能な阻害剤は、例えば、Mohammadiら(1997)に記載されている。
【0044】
好ましくは、本発明によると、受容体チロシンキナーゼの過剰発現および/または改変された活性に介入され、これはFGFR−4の突然変異によってトリガーされる。この突然変異は1個あるいは数個の点突然変異であり得る。特に、突然変異/複数の突然変異が、特に疎水性アミノ酸が親水性アミノ酸へと交換される結果として、FGFR−4の膜貫通ドメインで起こる。
【0045】
FGFR−3における疎水性アミノ酸を親水性アミノ酸に代える交換に至る、突然変異はある種の病気と関連することが知られている。かくして、例えば、膜貫通ドメイン中の点突然変異による繊維芽細胞成長因子受容体3の改変された活性が軟骨形成不全で見いだされている。小人症の最も普通に起こる遺伝形態である軟骨形成はある種の骨の成熟過程における血管に実質的に基づく常染色体優性障害である。FGFR−3の膜貫通ドメインにおけるGlyからArgへの置換によって軟骨形成不全がトリガーされることを示すことができた。さらに、FGFR−3におけるArg突然変異はダイマー受容体のキナーゼ機能を活性化することを示すことができた。また、Arg点突然変異はFGFR−3それ自体のチロシンキナーゼ活性のリガンド−非依存性および受容体上のホスホチロシンのかなり増大した改変レベルに至る。これらの結果は、軟骨形成異常の分子的基礎がFGFR−3による調節されからグナル移動であることを示唆する。
【0046】
FGFR−3の膜貫通ドメインにおけるさらなる突然変異は、アラニンをグルタミンへの交換である。このアミノ酸交換はもう1つの病気、すなわち黒色表皮症を持つクルーゾン病に至る。
【0047】
本発明によると、FGFR−4における突然変異、特に、疎水性アミノ酸を親水性アミノ酸への交換に至る膜貫通ドメイン中の点突然変異は癌についてのトリガリングおよび良好でない予後に関し、そのためには受容体チロシンキナーゼの阻害あるいはキナーゼ不活性受容体の使用が癌の治療および/または予防に適し、ここに、受容体チロシンキナーゼは突然変異のため改変された方法にて過剰発現されるかあるいは活性である。
【0048】
特に、グリシンのアルギニンへの交換に至る位置388における点突然変異では、この結果、受容体チロシンキナーゼが改変された方法で活性となり、このホモ−およびヘテロ接合の結果、リガンド刺激なくしてシグナル伝達が起こり、その結果細胞の制御されない増殖がトリガーされ得る。最悪の場合、この制御されない増殖は癌に至る。次いで、膜貫通ドメインは配列(配列番号1):
RYTDIILYASGSLALAVLLLLARLYを有し、
他方、非突然変異ドメインは以下の配列(配列番号2):
RYTDIILYASGSLALAVLLLLAGLYを有する。
【0049】
理論に拘束されるつもりはないが、前記突然変異の1つ、特にグリシンのアルギニンへの交換に至る位置388での点突然変異を保有する受容体チロシンキナーゼの活性化は、それを通じて膜貫通ドメインにおける交換が可能とされる相互作用ため起こる、ダイマー立体配座における受容体の安定化に基づくと推定される。リガンド−非依存性ダイマーの強化された形成は、増大した受容体チロシンキナーゼ活性および細胞形質転換に至る。癌のトリガリングに対する点突然変異の効果についての1つの可能性は、例えば、突然変異がFGFR−4によるシグナル伝達に作用する点に、膜を通じての受容体移動が防止され、それ自体とのあるいは他のFGFRとの二量体化が乱される点に、あるいは受容体のチロシンキナーゼ活性が影響される点に基礎を有し得る。
【0050】
本発明によると、乳癌を持つSt.Petersburgからの患者の56%が(バイオプシーによる検査)が位置388で突然変異を担い、これはグリシンのアルギニンへの交換に結び付けることができた。これらのうち、45%がヘテロ接合であり、11%がホモ接合である。これは、これは、高い割合で、位置388における点突然変異と乳癌の発生との間のつながりを示唆する。
【0051】
乳癌を持つドイツ人患者を調べたさらなる検査において、患者の43%のみが位置388で点突然変異を示した。癌患者の正常組織および正常個体の組織からのDNAでの実験から、突然変異は生殖細胞系突然変異であると帰結することができる。
【0052】
さらに、細胞系からのゲノムDNAおよびcDNAも実験して、位置388における点突然変異の割合を決定した。乳癌、比較としての正常乳房上皮細胞系、偏平細胞癌腫、神経膠芽細胞腫、神経芽細胞腫および子宮癌に由来する細胞系を実験した。正常乳房上皮細胞系を除いて全ての細胞系で、FGFR−4分子における位置388での点突然変異のかなりの割合を見いだすことができた。よって、阻害剤あるいはキナーゼ−不活性受容体の前記使用は癌腫の治療で特に適する。ここに、神経芽細胞腫、子宮癌および膵臓癌の治療は特に有望なようであるが、他のタイプの癌はそうではない。
【0053】
特に好ましいのは、特に膜貫通ドメインにおいて位置388において突然変異GlyからArgへの突然変異を持つ、突然変異FGFR−4を阻害する阻害剤の使用である。
【0054】
さらに、本発明は、細胞において受容体の過剰発現および/または改変された活性に至る突然変異FGFR−4に関する。好ましくは、この突然変異FGFR−4は、野生型受容体における疎水性アミノ酸が、突然変異受容体において親水性アミノ酸に交換されていることに特徴がある。特に好ましくは、点突然変異であり、膜貫通ドメインにおいて起こる、突然変異である。さらにより好ましくは、点突然変異が位置388で起こり、その結果、好ましくは、グリシンがアルギニンによって置換される。
【0055】
従来、専門家に間では、突然変異されていない唯1つのFGFR−4が起こると推定された。突然変異FGFR−4は知られていなかった。従って、本発明により、突然変異FGFR−4が存在することを示すことができたのは驚くべきことであった。特に、本発明により、突然変異、特に位置388における点突然変異と、癌の発生との間の関係を示すことができた。さらに、健康な個人における生殖細胞系突然変異は、とりわけ、動脈硬化症の発生についての遺伝素因と関係付けられた。
【0056】
本発明は、さらに、突然変異FGFR−4をコードする配列を含有するDNA分子に関する。また、本発明は、突然変異FGFR−4をコードするRNA配列を含有するRNA分子を含む。前記配列は癌の診断に使用することができる。これにおいて、配列はFGFR−4における突然変異を特異的に認識することができる。FGFR−4における突然変異の存在は、癌の治療のために貧弱な予後と関係付けられる。この理由は対応する腫瘍の攻撃的増殖挙動であり得る。
【0057】
これとは別に、本発明は、乳癌の差異的診断のための手法に関し、ここで、患者の核酸を前記したDNAおよび/またはRNAのうちの一方と接触させ、従って、突然変異FGFR−4の存在および/または不存在を示すシグナルが得られる。最後に、本発明は、前記阻害剤あるいはキナーゼ−不活性受容体を含有する医薬組成物に関する。これとは別に、チロシンキナーゼ活性の阻害剤の同定のためのスクリーニング手法に関し、ここに、本発明の受容体は可能な阻害剤と接触され、阻害剤の存在下および/または不存在下におけるチロシンキナーゼ活性が測定される。
【0058】
さらに、既に前記にて詳細に記載されたごとく、突然変異の存在の検出がPCR、続いての制限酵素切断によって行うことができる。
【0059】
最後に、他の分子生物学的診断手法も可能である。
【0060】
さらに、本発明の目的は、本発明による突然変異FGFR−4と特異的に反応する抗体である。本発明の意味における「特異的」とは、本発明による抗体は特異的受容体に結合するが、非特異的受容体には結合しないということである。
【0061】
以下、図面および実施例によって本発明を詳細に記載する。
【実施例】
【0062】
細胞培養。ヒト細胞系MDA−MB−453、ZR 75−1、K562およびSKBr3はATCCから得た。個々の供給源は最後の表に見いだすことができる。MDA−MB−453、K562およびZR 75−1は、10%の胎児ウシ血清(Sigma, Taufkirchen)を含有するRPMI(Gibco, Eggenstein)で培養した。SKBR3は15%胎児ウシ血清を含有するMcCoy5a(Gibco,Eggenstein)で培養した。全ての細胞培養培地はペニシリン/ストレプトマイシン(Sigma,Taufkirchen)を含有していた。細胞を水−蒸気飽和雰囲気および8%CO2 中にて37℃でインキュベートした。
【0063】
FGFR−4388Arg/wtのクローニング。K562およびMDA−MB−453細胞からのRNAの調製では、3×107細胞をグアニジニウムイソシアネートで溶解させ、CsClグラジエントでの超遠心によって精製した。cDNA合成は、製造業者の指示に従って、各場合において、逆転写酵素(Boehringer, Mannheim)および10pmolの「ランダムオリゴヌクレオチド」を用いて行った。引き続いてのPCR反応では0.5μlを用いた。
【0064】
FGFR−4388ArgおよびFGFR−4wtをPCR反応によって増幅した。このために、以下のプライマーを用いた:センス−GCTCAGAGGGCGGGCGGGGGTGCCGGCCG、アンチセンスCCGCTCGAGTGCCTGCACAGCCTTGAGCCTTGC。PCR反応では、以下のものを用いた:製造業者の指示に従って1.5U/25μlのExpand−Polymerase(Boehringer, Mannheim)および反応緩衝液:200μM dNTP、0.01% v/vTritonX100;10% v/vDMSO、および各々0.2pmolのセンスおよびα−センスプライマー。以下の反応工程を行った:35サイクル、94℃ 1分、64℃ 1分、72℃ 2.5分。FGFR−4388ArgのクローニングではMDA−MB−453cDNAを用い、FGFR−4wtのクローニングではK562 cDNAを用いた。PCR産物はpcDNA3ベクター(Invitrogen)にクローニングした。このようにして、G388Rを有するFGFR−4および野生型FGFRをさらなる工程のために得ることができた。
【0065】
FGFR−4の膜貫通ドメインの増幅。以下のプライマーを用いた:センス−GACCGCAGCAGCGCCCGAGGCCAG;アンチセンスAGAGGGAAGAGGGAGAGCTTCTG。PCT反応では以下のものを用いた。製造業者の指示に従った1.5U/25μl Taq−Polymerase(Boehringer, Mannheim)および反応緩衝液:200μMdTNP;各々0.2pmolのセンスおよびα−センスプライマー、0.5αl cDNAあるいは腫瘍バイオプシーおよび細胞系からのゲノムDNA。以下の反応工程を用いた:35サイクル、95℃ 45秒、72℃ 45秒。
【0066】
制限酵素消化による分析。ゲノムあるいはcDNAからのFGFR−4の膜貫通ドメインを前記したごとくに増幅した。制限酵素消化を用いて、G1217A突然変異につきバイオプシーおよび細胞系をテストするために、PCR産物を、5U/25μlのBstB1(NEB、Schwalbach/Taunus)と、60℃で1時間インキュベートした。制限酵素消化からのDNA断片を20%ポリアクリルアミドゲルで分離し、臭化エチジウムで染色した。野生型受容体の分析は109、37および22塩基対サイズの断片を生じる(トラック4)。他方、突然変異G1217Aの結果として、BstB1についてのさらなる制限酵素切断部位が形成される。突然変異受容体はさらなる80および29塩基対サイズの断片を示し、他方、109塩基対サイズの断片は消失する(トラック:ホモ接合体;トラック2および3:ヘテロ接合体)(図2参照)。
【0067】
制限酵素消化によるゲノムDNAの遺伝子型
初代腫瘍の組織試料からのゲノムDNAを標準的な方法によって単離した(Current Protocols in Molecular Biology, John Wiley and Sons, Inc., 1995)。ゲノムDNAの遺伝子型分析ができるためには、FGFR−4遺伝子における膜貫通領域をPCR反応において以下のプライマーで増幅した:
5’−GACCGCAGCAGCGCCCGAGGCCAG−3’(bp1275−1142)、および
5’−AGAGGGAAGAGGGAGAGCTTCTG−3’(bp1275−1297)。PCR反応では、Ready−to−Go PCR Beats(Pharmacia, Uppsala, スウェーデン)を用いた。以下のPCRサイクルを用いた:95℃における3分、94℃における45秒、72℃における45秒、および72℃における5分。合計35サイクルを行った。PCR産物を、5U/25μlのBstB1(NEB、Schwalbach/Taunus)と、60℃で1時間インキュベートした。制限酵素消化からのDNA断片を20%ポリアクリルアミドゲルで分離し、臭化エチジウムで染色した。388Argアレレ(対立遺伝子)は80および29bpサイズの2つの断片によって特徴付けられ、他方、388Glyアレレは単一の109bpサイズの断片によって示される。各遺伝子型分析は3回反復された。
【0068】
PCR産物のDNA配列決定。FGFR−4の膜貫通ドメインの配列分析では、PCR産物をBluescriptにクローンニングした。このために、PCR反応を既に記載したごとくに行った。以下のプライマーを用いた:センス−GGGAATTGACCGCAGCAGCGCCCGAGG;α−センス−GCTCTAGAAGAGGGAAGAGGGAGAG。FGFR−4Arg388/wtのクローニングのPCR産物はベクターpcDNA3中で、直接配列決定することができた。T/−プライマーのプラスミドDNAへのアニーリングの後、配列決定反応をT/−DNAポリメラーゼ(Pharmacia,Freisburg)を用いて行った。次いで、配列決定反応の産物を変性5%ポリアクリルアミドゲル(7.5M尿素、1×TBE)で分離し、乾燥した後にX線フイルムに暴露した(図3参照)。これより、野生型および突然変異のDNA配列が得られた。
【0069】
免疫沈殿およびウェスタンブロット分析。2.2×106細胞を10cmのペトリ皿上に広げ、一晩インキュベートした。細胞培地を、胎児ウシ血清を含まない培地によって置換し、さらに24時間インキュベートした。刺激には、細胞を50ngのsFGF/mlで10分間インキュベートし、冷PBSで2回洗浄し、氷上に置いた。細胞を300μlの冷溶解緩衝液(1% w/w NP−40、1.25% w/vデオキシコール酸ナトリウム、0.1% w/vSDS、0.15M NaCl、0.01Mリン酸ナトリウム、pH7.2、2mM EDTA、10mMフッ化ナトリウム、1mM PMSF、20μg/mlアプロチニン、1mMオルトバナデート、10mMピロリン酸ナトリウム)と共に4℃で15分間インキュベートし、溶解物を4℃の遠心(13,000RPM)によって清澄化した。蛋白質値決定では、Micro−BCA Protein Assay(Pierce)を製造業者の指示に従って用いた。免疫沈殿では、細胞溶解物を等しい蛋白質含有量に調整し、次いで、0.5μgの抗−FGFR−4(Santa Cruz)にて、回転するホイール上のプロテイン−A−Sepharose (Pharmacia, Freisburg)にて4℃で18時間インキュベートした。免疫複合体を冷HNTG(20mM HEPES pH7.5、150mM NaCl、0.1%TritonX100、10%グリセリン、10mMピロリン酸ナトリウム)で4回洗浄した。試料の調製では、免疫複合体を50μl3×Laemmli緩衝液で処理し、99℃で5分間インキュベートした。沈殿した蛋白質を7.5%SDS−PAGE上で分離した(図1参照)。
【0070】
ウェスタンブロットでは、SDS−PAGEによって分離した蛋白質をニトロセルロースに移した。膜上の非特異的蛋白質結合部位をTBS−T/0.25%ゼラチン(10mM Tris/HCl pH8.0、0.15M NaCl、0.05%Tween20)と共に室温で2時間のインキュベーションによってブロックした。初代抗体でのインキュベーションは傾いたシェーカー上にて4℃で6時間行った。次いで、非特異的に結合した抗体をTBS−T/0.25%ゼラチンで4回洗浄することによって除去した。二次抗体の結合は室温にて1時間行った。非特異的に結合した二次抗体はさらなる洗浄工程によって除去した。免疫複合体は製造業者の指示に従ってECLTMキット(Amersham, Braunschweig)で可視化した。
【0071】
統計学的方法。統計学的計算は統計学プログラムMedCalc(MedCalc SOftware, ベルギー)およびEpillnfo 6.04(CDC、アトランタ、ジョージア)の助けを借りて行った。異なる群における遺伝子型および臨床的領域との間の相関を決定するために、優劣比、信頼区間(CI)および統計的有意性(P値)を計算した。少数の388Argホモ結合患者のため、この群を統計的計算では388Argヘテロ接合患者の群と組合せた。
【0072】
腫瘍細胞系におけるFGFR−4の検出。表IはRTKの発現および乳癌の間の相関関係を示す。RTKの発現は、明瞭に、乳癌よりも細胞系でしばしば起こり、他方、正常上皮細胞の細胞系では検出できない。
【0073】
【表1】
【0074】
表2から、G388R突然変異は他の癌タイプの細胞系でも起こり、これらと相関性があることが明らかである。健康な上皮細胞系では、前記突然変異は検出できない。
【0075】
【表2−1】
【表2−2】
【0076】
バイオプシーにおけるFGFR−4突然変異G388Rの検出。表3は調べた乳癌を持つSt Petersburgからの61人の女性患者のうち、56%がG388R突然変異(そのうち45%がヘテロ接合であり、11%がホモ接合である)を保有していたことを示す。調べたミュンヘンからの69人の女性乳癌患者のうち、43%がG388R突然変異(そのうち32%がヘテロ的であり、11%がホモ接合的である)を保有していた。St Petersburgおよびミュンヘンからの女性患者における突然変異の全パーセンテージの割合は異なる。これは、G388R突然変異が生殖細胞系突然変異であることを示唆する。
【0077】
【表3−1】
【表3−2】
【表3−3】
【0078】
FGFR−4−G388R突然変異およびFGFR−4発現の検出の間の相関関係。以下の表4から、G388R突然変異(ゲノムDNAおよびcDNA)は、発現および/または強化された発現が起こる場合のみ起こることが明らかである。突然変異は、RTK発現が見いだされない、正常乳房上皮細胞系でも乳癌細胞系でも検出できない。
【0079】
そのRTK発現が特に顕著である細胞系MDA−MB 453はホモ接合G388R突然変異を示す。
【0080】
【表4】
【0081】
FGFR−4突然変異G388Rおよびリンパ節転移状態、あるいは再発なしの生存時間の間の相関関係の研究
表5は、乳癌の腫瘍形成におけるG388R突然変異の役割の研究に参画する全ての患者の臨床的パラメーターを示す。G388R突然変異を持つ患者はG388R突然変異を持たない患者よりも不良な長期予後を有することが見いだされた。
表5に対する記号
Her2:Her2受容体の発現レベル、0=発現なし、〜3=過剰発現
OPDAT:手術の日付
M/R:転移形成/再発、0=無し/1=有り
Vers:死亡;0=無し/1=有り
BERRE:再発なくしての生存時間(月数で)
グレード:腫瘍の分化グレード;1=強力な分化/3=低い分化段階:初代腫瘍のサイズ
E−Rec:エストロゲン受容体の発現、0=発現無し、〜12=最高発現
GEN:FGFR−4の遺伝子型;G=野生型アレレ;R=突然変異アレレ
BEDAT:最後の観察の日付
REZDAT:再発診断の日付
TODDAT:死亡の日付
BERLEB:通算しての生存時間
Nod.:リンパ節における転移;0=無し/1=有り
Men:閉経
P−Rec:プロゲステロン受容体の発現:0=発現無し、〜12=最高発現
【0082】
【表5−1】
【表5−2】
【表5−3】
【0083】
図4から、G388R突然変異は、最初の治療時においてリンパ節に転移を既に有する患者において数が多いことが見いだされる。G388R突然変異を持つ患者のうち、62.7%がリンパ節転移を有し、他方、G388R突然変異を持たない患者のうち、38.2%のみがリンパ節において転移を呈した。リンパ節転移状態は不良な予後を持つ腫瘍および良好な予後を持つ腫瘍のさらなる区別のための重要な予後マーカーであるので、この結果から、調べた85人の患者においてG388Rはよりひどい腫瘍進行に至れしめると結論できる。
【0084】
図5より、調べた患者の群において、再発なし生存確率は、G388R突然変異を有しない患者についてよりもG388R突然変異を持つものにつき、非常に低いことが分かる。再発した患者の74.4%が388R遺伝子型を有するが、25.6%のみが388G遺伝子型を有する。これは、G388R突然変異を持つ患者がより速やかに再発し、従って、成功裡に治療できなかったことを示す。
【0085】
要約すると、FGFR−4突然変異G388Rはリンパ節における転移形成の2.7倍(OR=2.7;CI:1.02<OR<7.4)増大した危険性に至らしめ、さらに5.44倍(OR=5.44;CI:1.93<OR<7.4)増大した腫瘍の再発の危険性に至らしめると云うことができる。かくして、突然変異したFGFR−4アレレ(G388R)を持つ患者は、腫瘍再発に対して素因を有するようであり、よって、より良好でない病気予後を有するようである。
【0086】
材料
アクリルアミド Serva,ハイデルベルグ
寒天 Difco,デトロイト
アガロース BRL,Eggenstein
アンピシリン Boehringer,マイハイム
アプロチニン Sigma,Taufkirchen
N,N’−ビスアクリルアミド Roth,Karisruhe
塩化セシウム BRL,Eggenstein
デスオキシヌクレオチド Pharmacia,Freisburg
臭化エチジウム Sigma,Taufkirchen
ゼラチン Sigma,Taufkirchen
グアニジウムイソチオシアネート Fluka,スイス国
HEPES Serva,ハイデルベルグ
フッ化ナトリウム Sigma,Taufkirchen
PMSF Sigma,Taufkirchen
SDS Roth,Karlsruhe
Tris Riedel de Haen, Seeze
TritonX100 Serva,ハイデルベルグ
Tween20 Serva,Taufkirchen
リストされていない全ての物質は、会社Sigma(Taufkirchen), Serva(ハイデルベルグ), Riedel Deursche Patentamt[sic]HaenあるいはMerck(Darmstadt)会社から入手し、最高に可能な純度グレードを用いた。
【0087】
装置
DNAの電気泳動 Workshop, MPI for Biochemistry, Martinsried
蛋白質の電気泳動 Atto,日本冷凍遠心
Biofuge17S Heraeus,Hanau
蛋白質移動 Semidryブロット装置, Workshop, MPI for Biochemistry, Martinstried
滅菌ワークベンチ Biogard, The Baker Company, USA
細胞培養 インキュベーター B5060 EK/Co2 , Hanau
細胞計数 コールターカウンター, Counter Electronics, グラスゴー
【0088】
文献
バシリコ・シイおよびマスカテリィ・デイ:ザ・エフジイエフ・ファミリー・オブ・グロウス・ファクターズ・アンド・オンコジーン アドバーンスト・キャンサー・リサーチズ,59,115−164(1992)
クーリエ・エフ, デ・ラペイリエール・オーおよびビルムバウム・デイ:コンプレクシティー・オブ・ザ・エフジイエフ・ファミリー:ザ・プルーフ・ナイン/メディ/サイ 9,1113−1115(1993)
フォークマン・ジェイおよびクラグスブルン・エム:サイエンス235, 442−447(1987)
ジバル・デイおよびヤヨン・エイ:コンプレクシティー・オブ・エフジイエフ・レセプターズ:ジェネティックス・ベーシス・フォー・ストラクチュラル・デリバリー・アンド・ファンクショナル・スペシフィシティー、エフエイエスイービイ・ジャーナル 6:3362−3369(1992)
ジョンソン・デイ・イーおよびウィリアムズ・エル・ティー:ストラクチュラル・アンド・ファンクショナル・ダイバーシティー・イン・ザ・エフジーエフ・レセプター・マルチジーン・ファミリー, アドバーンスト・キャンサー・リサーチズ 60:1−41(1993)
キメイマン・デイ, アブラハム・ジェイ・エイ, ハーパランタ・テイ,パルシィー・テイ・エムおよびキルシュナー・エム・ダブリュー:サイエンス,242, 1053−1058(1988)
モハマディー・エムら, サイエンス776,955−959(1997)
スラック・ジェイ・エム・ダブリュー, ダーリントン・ジイ・ジイ, ヘルス・ジェイ・ケイおよびゴッドサーブ・エス・エフ:ネイチャー326, 197−200(1987)
トーマス・ケイ・エイ,リオス−カンデロール・エム, グリメズ−ガレゴ・ジイ, ジサルボ・ジェイ, ベルネット・シイ, ロドケイ・ジェイおよびフィッツパトリック・エス:プロシーディングズ・ナショナル・アカデミー・オブ・サイエンシズ・ユーエスエイ, 82, 6409−6413(1985)
トンプソン・ジェイ・エイ, ハウデンシルト・シイ・シイ, アンダーソン・ケイ・デイ, ジピエテロ・ジェイ・ウム, アンダーソン・ダブリ・エフおよびマシアーグ・テイ(1989):プロシーディングズ・ナショナル・アカデミー・オブ・サイエンシズ・ユーエスエイ,86, 7928−7932(1989)
【0089】
【表6】
【図面の簡単な説明】
【0090】
【図1】FGFR−4(リン酸化されたFGR受容体−4は、矢印で示されている)の免疫沈殿物のSDSPAGEを示す。
【図2】突然変異FGFR−4のポリアクリルアミドゲルである。
【図3】FGFR−4の膜貫通ドメインの配列分析である。
【図4】FGFR−4突然変異G388Rとリンパ節転移形成の状態(n:患者数、p:P値)との相間関係である。
【図5】FGFR−4突然変異G388Rと再発なしでの生存時間(n:患者数、p:P値)との相間関係である。
【特許請求の範囲】
【請求項1】
癌の治療および/または予防のための受容体チロシンキナーゼ(RTK)の少なくとも1つの阻害剤の使用。
【請求項2】
前記阻害剤がキナーゼ−不活性受容体、低分子量物質及び抗FGFR−4抗体から選択されることを特徴とする、請求項1に記載の使用。
【請求項3】
前記阻害剤がFGFR−4の過剰発現および/または改変された活性を低下しおよび/または阻害することを特徴とする、請求項1に記載の使用。
【請求項4】
前記過剰発現および/または前記改変された活性がFGFR−4の突然変異によってトリガーされることを特徴とする、請求項3に記載の使用。
【請求項5】
前記突然変異が1個あるいは数個の点突然変異であることを特徴とする、請求項4に記載の使用。
【請求項6】
前記突然変異がFGFR−4の膜貫通ドメインでおこることを特徴とする、請求項5に記載の使用。
【請求項7】
前記突然変異が疎水性アミノ酸を親水性アミノ酸への交換に導く請求項5または6に記載の使用。
【請求項8】
前記点突然変異がFGFR−4分子中のAA位置388で起こることを特徴とする、請求項5〜7のいずれか一項に記載の使用。
【請求項9】
前記AA位置388における点突然変異がグリシンに代えてアルギニンへの交換に導く(G388R)ことを特徴とする、請求項5〜8のいずれか一項に記載の使用。
【請求項10】
前記突然変異が生殖細胞系突然変異であることを特徴とする請求項4〜9のいずれか一項に記載の使用。
【請求項11】
特に転移形成の阻害によって過剰増殖および/または侵入、特に癌種に帰せられる癌および/または病気の治療のための、請求項1〜10のいずれか一項に記載の使用。
【請求項12】
乳癌の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項13】
偏平細胞癌腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項14】
神経膠芽細胞腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項15】
神経芽細胞腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項16】
子宮癌の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項17】
阻害剤がFGFR−4、好ましくは突然変異FGFR−4を阻害することを特徴とする、請求項1〜16のいずれか一項に記載の使用。
【請求項18】
細胞中での受容体の過剰発現および/または改変された活性に導く突然変異FGFR−4。
【請求項19】
野生型受容体における疎水性アミノ酸が突然変異受容体中の親水性アミノ酸に交換されたことを特徴とする、請求項18に記載の突然変異FGFR−4。
【請求項20】
前記突然変異が点突然変異であり、好ましくは膜貫通ドメインで起こることを特徴とする、請求項18または19に記載の突然変異FGFR−4。
【請求項21】
点突然変異が位置388で起こり、好ましくはグリシンがアルギニンによって置換されることを特徴とする、請求項18に記載の突然変異FGFR−4。
【請求項22】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4をコードする配列を含有するDNA分子。
【請求項23】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4をコードする配列を含有するRNA分子。
【請求項24】
病気、特に癌の診断における請求項22および23のいずれか一項に記載の配列の使用。
【請求項25】
前記配列がFGFR−4の位置388における点突然変異を特異的に検出することができることを特徴とする、請求項24に記載の使用。
【請求項26】
症例試料中の、突然変異FGFR−4蛋白質またはそれをコードする核酸の検出の工程を含む、特に癌の差異的診断のための診断手法。
【請求項27】
突然変異FGFR−4の存在および/または不存在を示すシグナルが得られるように患者の核酸をDNAおよび/またはRNAと接触させ、および/または患者の核酸をPCRによって増幅し、引き続いて、その認識配列が突然変異によって影響される制限酵素で切断し、および/または患者の蛋白質を突然変異蛋白質につき特異的な抗体と接触させることを特徴とする、請求項26に記載の診断手法。
【請求項28】
症例試料中の、突然変異FGFR−4蛋白質またはそれをコードする核酸、あるいは増幅されたFGFR−4核酸を検出する工程を含む、癌および/または他の病気の発生についての遺伝的素因の存在につきスクリーニングする手法。
【請求項29】
請求項1〜17のいずれか一項に記載された阻害剤を含む医薬組成物。
【請求項30】
チロシンキナーゼ活性の阻害剤の同定のためのスクリーニング手法であって、請求項18〜21のいずれか一項に記載の受容体を、潜在的な阻害剤と接触させ、該阻害剤の存在および/または不存在においてチロシンキナーゼ活性を測定する前記手法。
【請求項31】
癌治療における標的としての、好ましくは請求項18〜21のいずれか一項に記載の突然変異FGFR−4。
【請求項32】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4蛋白質と特異的に反応する抗体。
【請求項1】
癌の治療および/または予防のための受容体チロシンキナーゼ(RTK)の少なくとも1つの阻害剤の使用。
【請求項2】
前記阻害剤がキナーゼ−不活性受容体、低分子量物質及び抗FGFR−4抗体から選択されることを特徴とする、請求項1に記載の使用。
【請求項3】
前記阻害剤がFGFR−4の過剰発現および/または改変された活性を低下しおよび/または阻害することを特徴とする、請求項1に記載の使用。
【請求項4】
前記過剰発現および/または前記改変された活性がFGFR−4の突然変異によってトリガーされることを特徴とする、請求項3に記載の使用。
【請求項5】
前記突然変異が1個あるいは数個の点突然変異であることを特徴とする、請求項4に記載の使用。
【請求項6】
前記突然変異がFGFR−4の膜貫通ドメインでおこることを特徴とする、請求項5に記載の使用。
【請求項7】
前記突然変異が疎水性アミノ酸を親水性アミノ酸への交換に導く請求項5または6に記載の使用。
【請求項8】
前記点突然変異がFGFR−4分子中のAA位置388で起こることを特徴とする、請求項5〜7のいずれか一項に記載の使用。
【請求項9】
前記AA位置388における点突然変異がグリシンに代えてアルギニンへの交換に導く(G388R)ことを特徴とする、請求項5〜8のいずれか一項に記載の使用。
【請求項10】
前記突然変異が生殖細胞系突然変異であることを特徴とする請求項4〜9のいずれか一項に記載の使用。
【請求項11】
特に転移形成の阻害によって過剰増殖および/または侵入、特に癌種に帰せられる癌および/または病気の治療のための、請求項1〜10のいずれか一項に記載の使用。
【請求項12】
乳癌の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項13】
偏平細胞癌腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項14】
神経膠芽細胞腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項15】
神経芽細胞腫の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項16】
子宮癌の治療のための請求項1〜11のいずれか一項に記載の使用。
【請求項17】
阻害剤がFGFR−4、好ましくは突然変異FGFR−4を阻害することを特徴とする、請求項1〜16のいずれか一項に記載の使用。
【請求項18】
細胞中での受容体の過剰発現および/または改変された活性に導く突然変異FGFR−4。
【請求項19】
野生型受容体における疎水性アミノ酸が突然変異受容体中の親水性アミノ酸に交換されたことを特徴とする、請求項18に記載の突然変異FGFR−4。
【請求項20】
前記突然変異が点突然変異であり、好ましくは膜貫通ドメインで起こることを特徴とする、請求項18または19に記載の突然変異FGFR−4。
【請求項21】
点突然変異が位置388で起こり、好ましくはグリシンがアルギニンによって置換されることを特徴とする、請求項18に記載の突然変異FGFR−4。
【請求項22】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4をコードする配列を含有するDNA分子。
【請求項23】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4をコードする配列を含有するRNA分子。
【請求項24】
病気、特に癌の診断における請求項22および23のいずれか一項に記載の配列の使用。
【請求項25】
前記配列がFGFR−4の位置388における点突然変異を特異的に検出することができることを特徴とする、請求項24に記載の使用。
【請求項26】
症例試料中の、突然変異FGFR−4蛋白質またはそれをコードする核酸の検出の工程を含む、特に癌の差異的診断のための診断手法。
【請求項27】
突然変異FGFR−4の存在および/または不存在を示すシグナルが得られるように患者の核酸をDNAおよび/またはRNAと接触させ、および/または患者の核酸をPCRによって増幅し、引き続いて、その認識配列が突然変異によって影響される制限酵素で切断し、および/または患者の蛋白質を突然変異蛋白質につき特異的な抗体と接触させることを特徴とする、請求項26に記載の診断手法。
【請求項28】
症例試料中の、突然変異FGFR−4蛋白質またはそれをコードする核酸、あるいは増幅されたFGFR−4核酸を検出する工程を含む、癌および/または他の病気の発生についての遺伝的素因の存在につきスクリーニングする手法。
【請求項29】
請求項1〜17のいずれか一項に記載された阻害剤を含む医薬組成物。
【請求項30】
チロシンキナーゼ活性の阻害剤の同定のためのスクリーニング手法であって、請求項18〜21のいずれか一項に記載の受容体を、潜在的な阻害剤と接触させ、該阻害剤の存在および/または不存在においてチロシンキナーゼ活性を測定する前記手法。
【請求項31】
癌治療における標的としての、好ましくは請求項18〜21のいずれか一項に記載の突然変異FGFR−4。
【請求項32】
請求項18〜21のいずれか一項に記載の突然変異FGFR−4蛋白質と特異的に反応する抗体。
【図1】

【図2】
【図3】

【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2009−1549(P2009−1549A)
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願番号】特願2008−127466(P2008−127466)
【出願日】平成20年5月14日(2008.5.14)
【分割の表示】特願2000−528281(P2000−528281)の分割
【原出願日】平成11年1月22日(1999.1.22)
【出願人】(596070445)マックス−プランク−ゲゼルシャフト ツール フォーデルング デル ヴィッセンシャフテン エー.ヴェー. (13)
【Fターム(参考)】
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願日】平成20年5月14日(2008.5.14)
【分割の表示】特願2000−528281(P2000−528281)の分割
【原出願日】平成11年1月22日(1999.1.22)
【出願人】(596070445)マックス−プランク−ゲゼルシャフト ツール フォーデルング デル ヴィッセンシャフテン エー.ヴェー. (13)
【Fターム(参考)】
[ Back to top ]