
RrhJ1I修飾酵素およびその遺伝子
【課題】遺伝子工学の分野で有用なGCCGGCを認識する新規修飾酵素及び該酵素タンパク質をコードする遺伝子の提供。
【解決手段】ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株から得られる修飾酵素。該特定のアミノ酸配列または特定のアミノ酸配列を含むタンパク質、特定の塩基配列または特定の塩基配列からなるDNAを含む遺伝子。また該遺伝子組換えベクターを含む形質転換体によるRrhj1I修飾酵素の製造方法。
【解決手段】ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株から得られる修飾酵素。該特定のアミノ酸配列または特定のアミノ酸配列を含むタンパク質、特定の塩基配列または特定の塩基配列からなるDNAを含む遺伝子。また該遺伝子組換えベクターを含む形質転換体によるRrhj1I修飾酵素の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、遺伝子工学試薬として有用な制限酵素および修飾酵素、該酵素タンパク質をコードする遺伝子、並びに該酵素の製造方法に関する。
【背景技術】
【0002】
制限酵素は、ある特定のDNA塩基配列を特異的に認識し、切断することのできるエンド型ヌクレアーゼであり、これまでに数多くの制限酵素が見出されている。
分子遺伝学、または生化学等の発展により、DNAが遺伝を司る本体であることが明らかになって以来、制限酵素は遺伝病の解明や、遺伝子操作等において現在幅広く用いられている有用な酵素である。このうち、特にDNA塩基配列を特異的に認識し、特定のDNAを切断するII型制限酵素が重要かつ必要なものとして用いられている。
【0003】
これまでにII型制限酵素は300種以上が精製されている。それらの中で、GCCGGCを認識する制限酵素としては40種近くのイソチゾマー(isoschizomer)が存在することが知られており、例えば、NaeI(タカラバイオ他)、NgoMIV(New England Biolabs、Promega)、MroNI(SibEnzyme)、PdiI(Fermentas)が市販されている。ここで、イソチゾマーは、認識配列が互いに一致している酵素のペアを意味する。
【0004】
また、特異的なDNA配列をメチル化する修飾酵素が数多く見出されている。
これまでに種々の微生物において制限酵素および修飾酵素の存在が確認されており、これらの酵素は、宿主内で制限・修飾系に関与していることが知られている。
【0005】
例えば、NaeI制限酵素および修飾酵素は、Lechevalieria aerocolonigenes ATCC23870(Nocardia aerocolonigenes)に存在することが知られている。しかし、この細菌中に含まれる制限酵素および修飾酵素の含量は少ないため、両酵素を大量に得ることは困難であった。
これまでに、染色体中に組み込まれたMspIメチラーゼ遺伝子を有する大腸菌K802株を用いてNaeI制限酵素の生産を行った結果、この大腸菌は、細胞1μg当たり、野生株(Lechevalieria aerocolonigenes ATCC23870)の50倍の活性(37℃、1時間の反応で1μgのDNAを分解できる酵素量)を有していたことが報告されている(特許文献1)。
【0006】
ところで、ロドコッカス属(Rhodococcus)に属する細菌において、制限酵素は15例が報告されているに過ぎない。例えば、6塩基配列を認識する制限酵素としてBamHI(GGATCC)、SalI(GTCGAC)、BspHI(TCATGA)、SphI(GCATCG)のイソチゾマーや、4塩基配列を認識する制限酵素としてMboI(GATC)やNlaIV(GGNNCC)のイソチゾマーが見出されている。しかし、GCCGGCを認識する制限酵素および修飾酵素はこれまでに見出されていなかった。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開平6−98779号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、遺伝子工学の分野で有用なGCCGGCを認識する制限酵素および修飾酵素、さらには該酵素タンパク質をコードする制限酵素遺伝子および修飾酵素遺伝子を提供することである。また、本発明の目的は、該制限酵素および修飾酵素の製造方法を提供することである。
【課題を解決するための手段】
【0009】
本発明者は上記課題を解決するために鋭意研究を行った結果、ロドコッカス属に属する細菌において新規の制限酵素および修飾酵素を見出した。詳しくは、本発明者は、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株から、制限酵素を精製した。このJ1菌由来の制限酵素のDNA切断パターンはNaeIのものと一致したことから、得られたJ1菌制限酵素はNaeIのイソチゾマーであることが示された。また、ロドコッカス・ロドクロウスJ-1株から修飾酵素を単離した。そして、得られたJ1菌修飾酵素の塩基配列情報から、J1菌制限酵素を単離した。このように、本発明者は、ロドコッカス属に属する細菌における新規の制限酵素および修飾酵素を見出し、本発明を完成した。
【0010】
すなわち、本発明は、以下の発明を包含する。
(1)以下の(A)または(B)のタンパク質。
(A)配列番号2記載のアミノ酸配列を含むタンパク質
(B)配列番号2記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質
(2)(1)記載のタンパク質をコードする遺伝子。
(3)以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号1記載の塩基配列からなるDNA
(b)配列番号1記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I制限酵素活性を有するタンパク質をコードするDNA
(4)以下の(A)または(B)のタンパク質。
(A)配列番号4記載のアミノ酸配列を含むタンパク質
(B)配列番号4記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
(5)(4)記載のタンパク質をコードする遺伝子。
(6)以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号3記載の塩基配列からなるDNA
(b)配列番号3記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I修飾酵素活性を有するタンパク質をコードするDNA
(7)(2)、(3)、(5)、または(6)のいずれか1項に記載の遺伝子を含む組換えベクター。
(8)(7)記載の組換えベクターを含む形質転換体。
(9)(8)記載の形質転換体の培養物からRrhJ1I制限酵素活性を有するタンパク質を採取することを含む、RrhJ1I制限酵素の製造方法。
(10)(8)記載の形質転換体の培養物からRrhJ1I修飾酵素活性を有するタンパク質を採取することを含む、RrhJ1I修飾酵素の製造方法。
【発明の効果】
【0011】
本発明により、遺伝子工学の分野で有用なGCCGGCを認識する制限酵素および修飾酵素、さらには該酵素タンパク質をコードする制限酵素遺伝子および修飾酵素遺伝子が提供される。また、本発明の遺伝子を用いて、該制限酵素および修飾酵素の製造が可能となるため、該制限酵素および修飾酵素を大量に生産することができる。
【0012】
また、本発明の制限酵素および修飾酵素は、制限修飾系を構成しているため、本発明の酵素および本発明の遺伝子は、例えば、制限修飾系の研究試薬として、非常に有用である。
【図面の簡単な説明】
【0013】
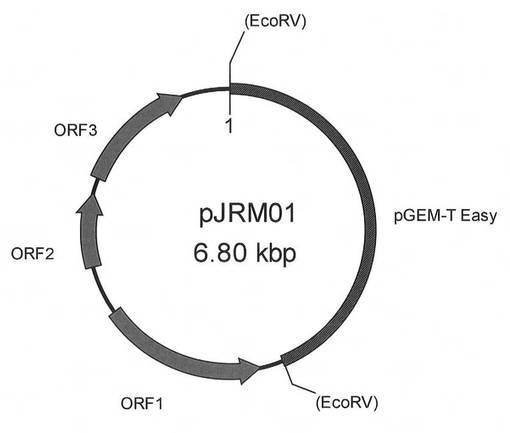
【図1】pJRM01の構造を示す模式図である。
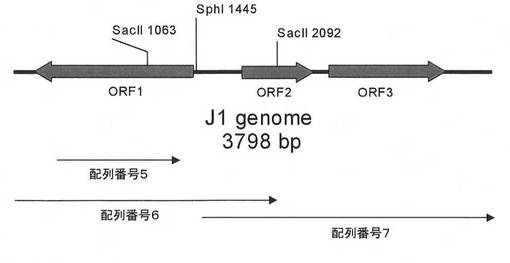
【図2】制限・修飾酵素遺伝子の制限酵素地図を示す図である。
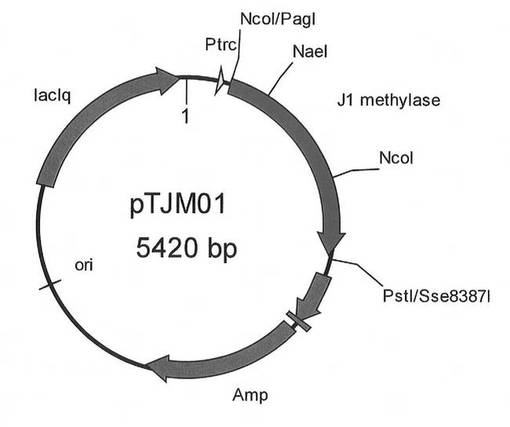
【図3】pTJM01の構築図である。
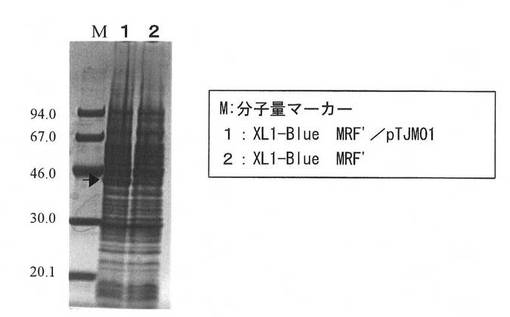
【図4】組換え体の超音波破砕上清を、SDS-PAGEした結果を示す図である。
【図5】pMCLMの構造を示す模式図である。
【図6】pGErR10の構造を示す模式図である。
【図7】無細胞抽出液を使用した切断結果を示す図である。
【発明を実施するための形態】
【0014】
以下、本発明の実施の形態について詳細に説明するが、本実施の形態は、本発明を説明するための例示であり、本発明をこの実施の形態のみに限定させるものではない。
【0015】
本発明は、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株(FERM BP-1478)由来の制限酵素(RrhJ1I制限酵素)および修飾酵素(RrhJ1I修飾酵素)に関するものである。ロドコッカス・ロドクロウスJ-1株(以下、「J1菌」ともいう)は、「Rhodococcus rhodochrous J-1」(FERM BP-1478)として独立行政法人産業技術総合研究所 特許生物寄託センター(茨城県つくば市東1丁目1番地1 中央第6)に寄託されている。
【0016】
本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素は、ロドコッカス属に属する細菌の自己防御系としての制限修飾系を構成していると考えられる。
【0017】
細菌の多くは、制限修飾系によって、外来のDNAから自己のDNAを守っている。つまり、修飾酵素により自己のDNAを修飾(メチル化)することで、自己のDNAを修飾型の異なる外来のDNAと区別している。そして、外来のDNAは、しかるべき位置にメチル基の無いことが制限酵素に認識され、制限酵素により切断される。
【0018】
制限修飾系を保持する細菌が安定な増殖を維持するためには、両酵素活性のバランスが適切に保たれる必要がある。すなわち、制限酵素が発現し、機能する前に、修飾酵素によって染色体遺伝子がメチル化され、制限酵素の切断から保護される必要がある。そのため、制限酵素遺伝子と修飾酵素遺伝子の転写および翻訳は、高度に制御されている必要がある。したがって、両遺伝子はゲノム上において近傍に位置することによって、オペロンを形成したり、特異的な転写制御因子による制御を受けられるように進化したと考えられる。このように、制限酵素と対応する修飾酵素とは密接な関係にあるため、両者を同時に提供する本発明は、非常に有用である。
【0019】
本明細書において、「制限酵素」は、ある特定の塩基配列を特異的に認識し、特定位置でDNAを切断することのできる酵素を意味する。本発明において、制限酵素は、好ましくは、エンド型ヌクレアーゼ(II型制限酵素)である。
【0020】
本明細書において、「RrhJ1I制限酵素」(「J1菌制限酵素」ともいう)は、ロドコッカス・ロドクロウスJ-1株で発現する制限酵素であり、GCCGGCの6塩基配列を認識し、DNAを切断する活性(「RrhJ1I制限酵素活性」ともいう)を有するタンパク質を意味する。本発明のRrhJ1I制限酵素による認識部位と切断部位とは、同一でもよく、または異なっていてもよい。切断されるDNAは、1本鎖DNAまたは2本鎖DNAのいずれでもよい。また、切断後のDNA末端は、例えば、5'突出末端、3'突出末端または平滑末端のいずれでもよい。
【0021】
本明細書において、RrhJ1I制限酵素活性は、RrhJ1I制限酵素をDNAと接触させ、接触後のDNAの分子量またはDNA断片数を測定することにより評価することができる。当業者であれば、基質DNA、接触時の酵素量、温度、溶液組成または接触時間などの条件を設定することができる。DNAの分子量は、例えば、アガロース電気泳動によって測定することができる。接触前のDNAの分子量と接触後のDNAの分子量、または接触前のDNAの断片数と接触後DNAの断片数とを比較することで、RrhJ1I制限酵素活性を評価することができる。また、GCCGGCを認識する既知の制限酵素を対照として使用してもよい。
【0022】
本明細書において、「修飾酵素」は、対応する制限酵素による切断からDNAを保護するようにDNAに修飾することのできる酵素を意味する。この場合、対応する修飾酵素および制限酵素は、同一の配列を特異的に認識する。本明細書において、同一の微生物に由来する酵素であって、同一の塩基配列を認識する制限酵素と修飾酵素との関係を、「対応する」と称する場合がある。本発明において、修飾酵素は、好ましくは、DNAをメチル化するメチラーゼである。
【0023】
本明細書において、「RrhJ1I修飾酵素」(「J1菌修飾酵素」ともいう)は、ロドコッカス・ロドクロウスJ-1株で発現する修飾酵素であり、RrhJ1I制限酵素による切断からDNAを保護する活性(「RrhJ1I修飾酵素活性」ともいう)を有するタンパク質を意味する。本発明のRrhJ1I修飾酵素による認識部位と修飾部位とは、同一でもよく、または異なっていてもよい。発明のRrhJ1I修飾酵素により修飾される塩基およびその数は特に限定されない。例えば、RrhJ1I修飾酵素は、認識配列中の1〜3個のシトシンをメチル化することができる。
【0024】
本明細書において、RrhJ1I修飾酵素活性は、RrhJ1I修飾酵素を接触させたDNAにRrhJ1I制限酵素またはそのイソチゾマーを接触させ、制限酵素接触後のDNAの分子量またはDNAの断片数を測定することにより評価することができる。当業者であれば、基質DNA、修飾酵素または制限酵素による接触時の酵素量、温度、溶液組成または接触時間などの条件を設定することができる。修飾酵素に接触させた後もしくは制限酵素に接触前のDNAの分子量と制限酵素に接触後のDNAの分子量、または修飾酵素に接触させた後もしくは制限酵素に接触前のDNA断片数と制限酵素に接触後のDNA断片数とを比較することで、RrhJ1I修飾酵素活性を評価することができる。RrhJ1I修飾酵素活性を評価する際、GCCGGCを認識する制限酵素に対応する既知の修飾酵素を対照として使用してもよい。
【0025】
(1)RrhJ1I制限・修飾系酵素
本発明のRrhJ1I制限酵素は、例えば、配列番号2に記載のアミノ酸配列を含むものである。配列番号2に記載のアミノ酸配列を含むタンパク質は、例えば、配列番号2に記載のアミノ酸配列からなるタンパク質である。
【0026】
また、本発明のRrhJ1I制限酵素は、前記のものに限定されることはなく、配列番号2に記載のアミノ酸配列の全部または一部を含むタンパク質であって、かつRrhJ1I制限酵素活性を有するタンパク質を含むものである。
また、本発明のRrhJ1I制限酵素には、配列番号2記載のアミノ酸配列と約50%以上、好ましくは約60%以上、より好ましくは約70%以上、さらに好ましくは約80%以上、特に好ましくは約90%以上、さらに特に好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有するアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質も含まれる。
さらに、配列番号2記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質も本発明のRrhJ1I制限酵素に含まれる。
【0027】
本発明のRrhJ1I修飾酵素は、例えば、配列番号4に記載のアミノ酸配列を含むものでである。配列番号4に記載のアミノ酸配列を含むタンパク質は、例えば、配列番号4に記載のアミノ酸配列からなるタンパク質である。
また、本発明のRrhJ1I修飾酵素は、前記のものに限定されることはなく、配列番号4記載のアミノ酸配列の全部または一部を含むタンパク質であって、かつRrhJ1I修飾酵素活性を有するタンパク質を含むものである。
また、本発明のRrhJ1I修飾酵素には、配列番号4記載のアミノ酸配列と約70%以上、好ましくは約80%以上、より好ましくは約90%以上、さらに好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有するアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質も含まれる。
さらに、配列番号4記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質も本発明のRrhJ1I修飾酵素に含まれる。
【0028】
前記の配列番号2または4記載のアミノ酸配列において、1または数個のアミノ酸に欠失、置換または付加等の変異が生じたアミノ酸配列としては、例えば(i)配列番号2または4記載のアミノ酸配列において、1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が欠失したアミノ酸配列、(ii)配列番号2または4記載のアミノ酸配列の1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、(iii)配列番号2または4記載のアミノ酸配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が付加したアミノ酸配列、(iv)配列番号2または4記載のアミノ酸配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が挿入されたアミノ酸配列、(v)上記(i)〜(iv)を組み合わせたアミノ酸配列が挙げられる。
【0029】
(2)RrhJ1I制限酵素遺伝子およびRrhJ1I修飾酵素遺伝子
本発明のRrhJ1I 制限酵素遺伝子は、本発明のRrhJ1I制限酵素をコードする遺伝子である。本発明のRrhJ1I制限酵素は前述のとおりである。本発明のRrhJ1I制限酵素遺伝子は、例えば、配列番号1に記載の塩基配列からなるDNAを含むものが挙げられるが、これに限定されるものではない。
本発明のRrhJ1I 制限酵素遺伝子には、配列番号1記載の塩基配列と約50%以上、好ましくは約60%以上、より好ましくは約70%以上、さらに好ましくは約80%以上、特に好ましくは約90%以上、さらに特に好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有する塩基配列であって、RrhJ1I制限酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I制限酵素遺伝子には、上記のDNAのほか、配列番号1に記載の塩基配列において、1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列であって、RrhJ1I制限酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I制限酵素遺伝子には、配列番号1に記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I 制限酵素活性を有するタンパク質をコードするDNAが含まれる。これらの遺伝子などは本発明のRrhJ1I制限酵素遺伝子に含まれる。ストリンジェントな条件については後述する。
【0030】
本発明のRrhJ1I 修飾酵素遺伝子は、本発明のRrhJ1I修飾酵素をコードする遺伝子である。本発明のRrhJ1I修飾酵素は前述のとおりである。本発明のRrhJ1I修飾酵素遺伝子は、例えば、配列番号3に記載の塩基配列からなるDNAを含むものが挙げられるが、これに限定されるものではない。
本発明のRrhJ1I 修飾酵素遺伝子には、配列番号3記載の塩基配列と約70%以上、好ましくは約80%以上、より好ましくは約90%以上、さらに好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有する塩基配列であって、RrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I修飾酵素遺伝子には、上記のDNAのほか、配列番号3に記載の塩基配列において、1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列であって、RrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I修飾酵素遺伝子には、配列番号3に記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I 修飾酵素活性を有するタンパク質をコードするDNAが含まれる。これらの遺伝子も本発明のRrhJ1I修飾酵素遺伝子に含まれる。
【0031】
前記のストリンジェントな条件としては、例えば、DNAを固定したナイロン膜を、6×SSC(1×SSCは塩化ナトリウム8.76g 、クエン酸ナトリウム4.41g を1リットルの水に溶かしたもの)、1% SDS、100μg/mlサケ精子DNA、0.1% ウシ血清アルブミン、0.1% ポリビニルピロリドン、0.1% フィコールを含む溶液中で65℃にて20時間プローブとともに保温してハイブリダイゼーションを行う条件をあげることができるが、これに限定されるわけではない。当業者であれば、このようなバッファーの塩濃度、温度等の条件に加えて、その他のプローブ濃度、プローブの長さ、反応時間等の諸条件を加味し、ハイブリダイゼーションの条件を設定することができる。
【0032】
ハイブリダイゼーション法の詳細な手順については、Molecular Cloning, A Laboratory Manual 2nd ed.(Cold Spring Harbor Laboratory Press (1989))等を参照することができる。
以下に、ハイブリダイゼーションによりRrhJ1I制限酵素遺伝子またはRrhJ1I修飾酵素遺伝子を得る方法の一例を示すが、これに限定されるわけではない。
【0033】
まず、適当な遺伝子源から得たDNAを常法に従ってプラスミドやファージベクターに接続してDNAライブラリーを作製する。このライブラリーを適当な宿主に導入して得られる形質転換体をプレート上で培養し、生育したコロニーまたはプラークをニトロセルロースやナイロンの膜に移し取り、変性処理の後にDNAを膜に固定する。この膜をあらかじめ32P等で標識したプローブを含む上記の組成の溶液中、上記のストリンジェントな条件で保温し、ハイブリダイゼーションを行う。プローブとしては、配列番号2または配列番号4に記載したアミノ酸配列の全部または一部をコードするポリヌクレオチドを含有していればよく、たとえば配列番号1または配列番号3に記載の塩基配列の全部または一部からなるポリヌクレオチドまたはそれを含むポリヌクレオチドを使用することができる。
ハイブリダイゼーションの終了後、非特異的に吸着したプローブを洗い流し、オートラジオグラフィー等によりプローブとハイブリッドを形成したクローンを同定する。この操作をハイブリッド形成クローンを単離できるまで繰り返す。
こうして得られたクローンの中には、目的の酵素活性を有するタンパク質をコードする遺伝子が保持されている。そして、クローンから目的の遺伝子を得るには、アルカリ法などの公知のポリヌクレオチド抽出方法を使用する。
【0034】
また、前記の配列番号1または3に記載の塩基配列において1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列としては、例えば(i)配列番号1または3に記載の塩基配列において、1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が欠失した塩基配列、(ii)配列番号1または3に記載の塩基配列の1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が他の塩基で置換された塩基配列、(iii)配列番号1または3に記載の塩基配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が付加した塩基配列、(iv)配列番号1または3に記載の塩基配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が挿入された塩基配列、(v)上記(i)〜(iv)を組み合わせた塩基配列が挙げられる。
【0035】
このように種々のDNAが本発明のRrhJ1I制限酵素遺伝子または本発明のRrhJ1I修飾酵素遺伝子の範囲内に含まれるのは、以下の理由によるものである。即ち、遺伝子上でアミノ酸を指定するコドン(3つの塩基の組み合わせ)は、アミノ酸の種類ごとに1〜6種類ずつが存在することが知られている。したがって、あるアミノ酸配列をコードする遺伝子は多数存在することができる。
遺伝子は自然界において決して安定に存在しているものではなく、その塩基配列に変異が起こることはまれではない。遺伝子上に起こった変異によっては、コードされるアミノ酸配列に変化を与えない変異(サイレント変異と呼ばれる)もあり、この場合には同じアミノ酸配列をコードする異なる遺伝子が生じたといえる。したがって、ある特定のアミノ酸配列をコードする遺伝子が単離されても、それを含有する生物が継代されていくうちに同じアミノ酸配列をコードする多種類の遺伝子ができていく可能性は否定できない。
【0036】
さらに、同じアミノ酸配列をコードする多種類の遺伝子を人為的に作製することは、種々の遺伝子工学的手法を用いれば困難なことではない。
例えば、遺伝子工学的なタンパク質の生産において、目的のタンパク質をコードする本来の遺伝子上で使用されているコドンが宿主中では使用頻度の低いものであった場合には、タンパク質の発現量が低いことがある。このような場合にはコードされているアミノ酸配列に変化を与えることなく、コドンを宿主で繁用されているものに人為的に変換することにより、目的タンパク質の高発現を図ることが行われている。
【0037】
このように、特定のアミノ酸配列をコードする多種類の遺伝子は人為的に作製可能なことは言うまでもなく、自然界においても生成されうるものである。したがって、本発明中に開示された塩基配列と同一の遺伝子ではなくても、RrhJ1I 制限酵素活性または修飾酵素活性を示すタンパク質をコードするDNAである限り、本発明のRrhJ1I制限酵素遺伝子またはRrhJ1I修飾酵素遺伝子に含まれるものである。
【0038】
また、本発明は、RrhJ1I制限酵素遺伝子およびRrhJ1I修飾酵素遺伝子の両方を含む遺伝子も含有する。制限酵素をコードする遺伝子および該制限酵素に対応する修飾酵素をコードする遺伝子は、ゲノム上で互いに近傍に位置することが知られている。
本明細書において、「RrhJ1I 制限・修飾系酵素の遺伝子」には、RrhJ1I 制限酵素をコードする遺伝子およびRrhJ1I 制限酵素による切断よりDNAを保護するRrhJ1I修飾酵素をコードする遺伝子が含まれる。
【0039】
本発明のRrhJ1I制限酵素遺伝子、RrhJ1I修飾酵素遺伝子およびRrhJ1I 制限・修飾系酵素の遺伝子は、これらの酵素を発現する微生物から単離することができる。例えば、ロドコッカス・ロドクロウスJ-1株由来のゲノムDNAを鋳型として、既知のアミノ酸配列情報から遺伝子の縮重を考慮して設計したプライマーもしくはプローブまたは既知の塩基配列情報に基づいて設計したプライマーもしくはプローブを用いたPCRまたはハイブリダイゼーション法により、遺伝子を単離することができる。単離した遺伝子から、本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素を製造することができる。
【0040】
また、遺伝子に変異を導入することによって、1個または数個のアミノ酸に欠失、置換または付加等の変異を導入することもできる。アミノ酸配列と遺伝子に変異を導入するには、Kunkel法や Gapped duplex法等の公知手法により、部位特異的突然変異誘発法を利用した変異導入用キット、例えばQuikChangeTM Site-Directed Mutagenesis Kit(ストラタジーン社)、GeneTailorTM Site-Directed Mutagenesis System(インビトロジェン社)、TaKaRa Site-Directed Mutagenesis System(Mutan-K、Mutan-Super Express Km等:タカラバイオ社)等を用いることができる。
【0041】
本発明において、DNAの塩基配列の確認は、慣用の方法により配列決定することにより行うことができる。例えば、ジデオキシヌクレオチドチェーンターミネーション法(Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463)等により行うことができる。また、適当なDNAシークエンサーを利用して配列を解析することも可能である。
塩基配列の決定は、プラスミドベクターを用いて作製された形質転換体の場合、宿主がエシェリヒア・コリであれば試験管等で培養を行い、常法に従ってプラスミドを調製する。得られたプラスミドをそのまま鋳型とするか、あるいは挿入断片を取り出してM13ファージベクター等にサブクローニングした後に、ジデオキシ法により塩基配列を決定する。ファージベクターで作製された形質転換体の場合も基本的に同様な操作により塩基配列を決定することができる。これら培養から塩基配列決定までの基本的な実験法については、例えば、前述のT.ManiatisらのMolecular Cloning, A Laboratory Manual等に記載されている。
【0042】
得られた遺伝子が目的のRrhJ1I 制限・修飾系酵素をコードする遺伝子であるかどうかの確認は、決定された塩基配列を配列番号1または配列番号3に記載の塩基配列と比較して行うことができる。あるいは決定された塩基配列より推定されるアミノ酸配列を配列番号2または配列番号4に記載のアミノ酸配列と比較して行うことができる。
【0043】
(3)本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素の製造方法
本発明のRrhJ1I 制限酵素を製造するには、まず前記のようなRrhJ1I 制限酵素をコードする遺伝子と、RrhJ1I修飾酵素をコードする遺伝子とを組み込んだ組換えベクターを作製する。そして、この組換えベクターで形質転換された形質転換体を作製し、これを培養して、培養物中よりRrhJ1I 制限酵素活性を有するタンパク質を採取することによりRrhJ1I 制限酵素を製造することができる。
この場合、形質転換体はRrhJ1I 制限酵素をコードする遺伝子を含む組換えベクターと、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターの両方の組換えベクターで形質転換された形質転換体を用いてもよい。
また、RrhJ1I修飾酵素をコードする遺伝子の代わりに、RrhJ1I修飾酵素が修飾する塩基配列と同じ塩基配列を修飾する酵素(例としてはNaeI修飾酵素)をコードする遺伝子を用いてもよい。
【0044】
また、本発明のRrhJ1I 修飾酵素を製造するには、まず前記のようなRrhJ1I 制限酵素をコードする遺伝子と、RrhJ1I 修飾酵素をコードする遺伝子とを組み込んだ組換えベクターを作製する。そして、この組換えベクターで形質転換された形質転換体を作製し、これを培養して、培養物中よりRrhJ1I 修飾酵素活性を有するタンパク質を採取することによりRrhJ1I 修飾酵素を製造することができる。
この場合、形質転換体はRrhJ1I 制限酵素をコードする遺伝子を含む組換えベクターと、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターの両方の組換えベクターで形質転換された形質転換体を用いてもよい。
あるいは、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターのみで形質転換された形質転換体を用いてもよい。
【0045】
本発明のRrhJ1I制限酵素遺伝子、本発明のRrhJ1I修飾酵素遺伝子または本発明のRrhJ1I制限・修飾酵素系の遺伝子を含む組換えベクターは、本発明の範囲に含まれる。
【0046】
本発明の組換えベクターは、上記酵素をコードする遺伝子の上流に転写プロモーター、場合によっては下流にターミネーターを挿入して発現カセットを構築し、このカセットを発現ベクターに挿入することにより作製することができる。あるいは、発現ベクターに転写プロモーターおよび/またはターミネーターがすでに存在する場合には、発現カセットを構築することなく、ベクター中のプロモーターおよび/またはターミネーターを利用して、その間に当該酵素をコードする遺伝子を挿入すればよい。
【0047】
本明細書において、プロモーターは、例えば、trcプロモーター、lacプロモーターなどをあげることができるが、これに限定されるわけではない。
本明細書において、ターミネーターは、例えば、trpオペロンターミネータをあげることができるが、これに限定されるわけではない。
【0048】
ベクターに本発明の遺伝子を挿入するには、制限酵素を用いる方法、トポイソメラーゼを用いる方法等を利用することができる。また、挿入の際に必要であれば、適当なリンカーを付加してもよい。また、アミノ酸への翻訳にとって重要な塩基配列として、SD配列やKozak配列などのリボソーム結合配列が知られており、これらの配列を遺伝子の上流に挿入することもできる。挿入にともない、遺伝子がコードするアミノ酸配列の一部を置換してもよい。
【0049】
本発明において使用されるベクターは、本発明の遺伝子を保持するものであれば特に限定されず、それぞれの宿主に適したベクターを用いることができる。ベクターとしては、例えば、プラスミドDNA、バクテリオファージDNA、レトロトランスポゾンDNA、人工染色体DNAなどが挙げられる。例えば、大腸菌を宿主とする場合には、pTrc99A(GEヘルスケアバイオサイエンス)、pACYC184(ニッポンジーン)、pMW118(ニッポンジーン)などを挙げることができる。また、必要に応じて、これらのベクターを改変したものを用いることもできる。
【0050】
本発明の組換えベクターを宿主に導入することで、本発明の形質転換体を作製することができる。当該形質転換体も本発明の範囲に含まれる。
本発明において使用する宿主は、上記組換えベクターが導入された後、目的の制限酵素または修飾酵素を発現することができる限り、特に限定されるものではない。宿主としては、例えば、大腸菌(エシェリヒア・コリ)、枯草菌(バチルス・ズブチリス(Bacillus subtilis))、ロドコッカス菌(Rhodococcus)、放線菌などの細菌、酵母(サッカロミセス・セレビシエ(Saccharomyces cerevisiae))、カビ、動物細胞、植物細胞、昆虫細胞などが挙げられる。本発明において、宿主は好ましくは大腸菌である。
本発明において、大腸菌は、例えば、大腸菌K12株やB株、あるいはそれらの野生株由来の派生株であるJM109株、XL1-Blue株(例えば、XL1-Blue MRF')、K802株、C600株などを挙げることができる。
宿主への組換えベクターの導入方法としては、宿主に適した方法であれば特に限定されるものではなく、当業者であれば公知技術から適宜選択することができる。このような方法としては、例えば、エレクトロポレーション法、カルシウムイオンを用いる方法、スフェロプラスト法、酢酸リチウム法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0051】
本発明の形質転換体を培養して、培養物からRrhJ1I制限酵素活性を有するタンパク質またはRrhJ1I修飾酵素活性を有するタンパク質を採取することにより、RrhJ1I制限酵素またはRrhJ1I修飾酵素を製造することができる。
【0052】
本発明において「培養物」とは、菌体、培養液、無細胞抽出液、細胞膜などの培養により得られるものを意味する。無細胞抽出液は、培養後の菌体を、例えばリン酸ナトリウム緩衝液を加えてホモジナイザーなどで物理的に破砕した後、遠心(15,000rpm, 10min, 4℃)し、破砕できない菌体(細胞)が存在しないように上清を回収して得ることができる。細胞膜は、上記遠心で得られたペレットとして得ることができ、また、ペレットを溶解バッファーで懸濁することにより得ることもできる。
【0053】
本発明の制限酵素または修飾酵素は、培養物をそのまま用いてもよいし、透析や硫安沈殿などの公知の方法、あるいはゲルろ過、イオン交換、アフィニティー等の各種クロマトグラフィーなどの公知の方法を単独または適宜組み合わせることによって、培養物から濃縮、精製したものを用いてもよい。この場合、酵素活性、分子量、等電点などを指標に濃縮、精製することができる。
【0054】
また、本発明においては、本発明の遺伝子または本発明のベクターから、本発明の酵素を製造することも可能である。すなわち、本発明においては、いわゆる無細胞タンパク質合成系を採用して、本発明の制限酵素または修飾酵素を産生することが可能である。
無細胞タンパク質合成系とは、細胞抽出液を用いて試験管などの人工容器内でタンパク質を合成する系である。なお、本発明において使用される無細胞タンパク質合成系には、DNAを鋳型としてRNAを合成する無細胞転写系も含まれる。
ここで、上記細胞抽出液は、真核細胞由来または原核細胞由来の抽出液、例えば、小麦胚芽、大腸菌などの抽出液を使用することができる。なお、これらの細胞抽出液は濃縮されたものであっても濃縮されていないものであってもよい。
【0055】
細胞抽出液は、例えば限外濾過、透析、ポリエチレングリコール(PEG)沈殿等によって得ることができる。さらに本発明において、無細胞タンパク質合成は、市販のキットを用いて行うこともできる。そのようなキットとしては、例えば試薬キットPROTEIOSTM(東洋紡)、TNTTM System(プロメガ)、合成装置のPG-MateTM(東洋紡)、RTS(ロシュ・ダイアグノスティクス)などが挙げられる。
上記のように無細胞タンパク質合成によって得られる本発明の制限酵素および修飾酵素は、前述のように適宜クロマトグラフィーを選択して、濃縮、精製することができる。
【0056】
以下に、本発明の具体的な態様として、本発明のRrhJ1I 制限酵素遺伝子および修飾酵素遺伝子をクローニングし、該遺伝子を発現させるための手順を例示する。
【0057】
(i)J1菌を培養し、得られる菌体より制限酵素を精製し、制限酵素認識配列を決定する工程
J1菌の制限酵素活性を調べるために、まずJ1株を培養し、培養した菌体を集めて超音波処理にて破砕した後、超遠心分離を行って上清を集め、これを活性測定用の試料とする。この試料の適当量を、基質であるλファージDNA(λ−DNA)とともに37℃でインキュベートした後、基質DNAの分解をアガロースゲル電気泳動により確認する。
当該方法を用いてJ1菌の制限酵素活性を調べると、酵素活性は、λ−DNAを13Kbの大きさに切断する活性として検出される。
【0058】
この工程において、種々のプラスミドを基質DNAとして使用してJ1菌制限酵素による切断パターンを解析すると、J1菌制限酵素の認識配列をGCCGGCに決定することができる。
【0059】
(ii)J1菌制限酵素と同じ配列を切断するイソチゾマーに対応する修飾酵素遺伝子からプライマーを設計し、J1株の染色体DNAを鋳型にPCRを行う工程
【0060】
J1菌制限酵素の複数のイソチゾマーに対応する各修飾酵素の間には共通配列部分が存在する場合がある。例えば、GCCGGCを切断するJ1菌制限酵素のイソチゾマーとしてNaeIやNgoMIVがあり、NaeI修飾酵素およびMgoMIV修飾酵素のアミノ酸配列には49%のホモロジーが認められる。次に、これらのアミノ酸配列から設計したDNAをプライマーに用いると、GCCGGCを切断する制限酵素に対応する修飾酵素の遺伝子をクローニングすることができる。例えば、当該方法を用いてJ1株の染色体DNAにおいて修飾酵素遺伝子をPCRで増幅すると、0.9 Kbの断片が増幅し、そのDNA配列を決定することができる。得られるDNA断片は、NaeI修飾酵素またはMgoMIV修飾酵素とのホモロジーが認められる。
【0061】
本工程によって、J1菌制限酵素のイソチゾマーに対応する修飾酵素とホモロジーを有する遺伝子断片を得ることができる。
【0062】
(iii)PCRの増幅産物のDNA配列を確認し、インバースPCR等で修飾酵素遺伝子と制限酵素遺伝子を決定する工程
この工程において、まず、修飾酵素遺伝子および制限酵素遺伝子をクローニングする。クローニングの方法としては、J1菌染色体ライブラリーからのサザンハイブリダイゼーション法やインバースPCR法を用いることができる。得られるDNA断片の塩基配列は通常の方法、たとえばジデオキシ法によって決定することができる。さらに、得られた塩基配列を解析することにより、その塩基配列中のタンパク質をコードしうる領域(オープンリーディングフレーム、ORF)の存在を推定することができる。
【0063】
例えば、修飾酵素遺伝子をクローニングするために工程(ii)で決定したDNA配列を利用し、インバースPCRを行うことができる。先ず、J1菌染色体DNAを制限酵素SacIIで切断し、ライゲーション反応を行って、環状DNAを作製する。次に、工程(ii)で決定したDNA配列の両末端付近のDNA配列をプライマーとしてPCR反応を行い、未知の配列を含むDNA断片を増幅する。最後に、増幅したDNA断片のDNA配列を解析し、修飾酵素遺伝子を決定する。このように得られた配列中には1つのORFが存在する(以下、「ORF1」(修飾酵素)ともいう)。ORF1の塩基配列から推定されるアミノ酸配列を配列番号4に示す。配列番号4のアミノ酸配列のホモロジー検索を行った結果、NaeIメチラーゼのアミノ酸配列に対して67%の相同性が認められる。
【0064】
通常、制限酵素遺伝子は修飾酵素遺伝子の近傍に存在しており、修飾酵素遺伝子が取得できれば制限酵素遺伝子の取得は可能であると考えられる。具体的には、J1菌染色体DNAを制限酵素SphIで切断し、ライゲーション反応によって環状化したDNAを作製する。次に、修飾酵素遺伝子または修飾酵素遺伝子近傍のDNA配列をプライマーとして環状DNAに対してインバースPCRを行ない、約6KbのDNA断片を増幅する。この断片のDNA配列を解析し、制限酵素遺伝子を決定することができる。このように得られた塩基配列中には2つのORFが存在しており、5'側より順に「ORF2」(HNHエンドヌクレアーゼ)、「ORF3」(制限酵素)とする。ORF3の塩基配列から推定されるアミノ酸配列を配列番号2に、ORF2の塩基配列(配列番号8)から推定されるアミノ酸配列を配列番号9に示す。配列番号2に記載のアミノ酸配列のホモロジー検索を行った結果、II型制限酵素のNaeIと47%の相同性が認められ、配列番号9に記載のアミノ酸配列のホモロジー検索を行った結果、Arthrobacter sp. FB24由来のHNHエンドヌクレアーゼと33%の相同性が認められる。
【0065】
本工程により、RrhJ1I制限酵素をコードすると推定されるORF(ORF3)、およびRrhJ1I修飾酵素をコードすると推定されるORF(ORF1)を得ることができる。
【0066】
(iv)RrhJ1I制限酵素および/またはRrhJ1I制限酵素を発現する形質転換体を作製する工程
本工程では、工程(iii)で得られたORF1および/またはORF3の塩基配列を基に、J1菌染色体DNAからPCRでRrhJ1I修飾酵素遺伝子および/またはRrhJ1I制限酵素遺伝子を含むDNA断片を増幅する。次に、増幅したDNA断片を適当なベクターに結合して大腸菌を形質転換し、RrhJ1I修飾酵素および/またはRrhJ1I制限酵素が発現する形質転換体を作製する。
【0067】
J1菌染色体DNAからPCRでRrhJ1I制限・修飾系の遺伝子を増幅する方法は、例えば工程(ii)の方法と同様に行うことができる。
制限酵素および/または修飾酵素を発現する形質転換体を作製するには、まず、得られたRrhJ1I制限・修飾系酵素の遺伝子を適当な宿主細胞、たとえばエシェリヒア・コリ、バチルス・ズブチリス(Bacillus subtilis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ロドコッカス菌(Rhodococcus)、放線菌、動物細胞、昆虫細胞、植物細胞等において発現できるような発現ベクターに常法に従い接続する。そして、得られた組換えベクターを宿主細胞に導入し、形質転換体を作製すればよい。
【0068】
本発明に使用する宿主は、メチル化シトシンに関わる制限系遺伝子(mcrA,B,C)、またはメチル化アデニンに関わる制限系遺伝子(mrr)の欠損株を使用するのが好ましい。大腸菌の一例としては、XL1-Blue MRF'、K802などを用いることができる。この形質転換体を培養することにより、本発明のRrhJ1I 制限酵素または本発明のRrhJ1I修飾酵素タンパク質を発現させることができる。
【0069】
なお、RrhJ1I制限酵素を発現させるための形質転換体は制限・修飾両酵素の遺伝子を保持していることが望ましい。この両遺伝子は同じベクター上に位置した状態で形質転換体に保持されていてもよいし、あるいは別々のベクターに組み込まれた状態で形質転換体に保持されていてもよい。RrhJ1I制限酵素を発現させるための形質転換体としては、例えば、RrhJ1I修飾酵素遺伝子を含むプラスミド(pMCLM、実施例7)およびRrhJ1I制限酵素遺伝子を含むプラスミド(pGErR10、実施例7)を含有する形質転換体XL1-Blue MRF'/pMCLM+pGErR10(実施例8)を挙げることができる。
一方、RrhJ1I 修飾酵素の発現には上記の形質転換体の他、修飾酵素遺伝子のみを保持する形質転換体を用いることもできる。RrhJ1I修飾酵素を発現させるための形質転換体としては、例えば、RrhJ1I修飾酵素を含むプラスミドpTJM01(実施例6)を含有する形質転換体XL1-Blue MRF'/pTJM01を挙げることができる。
【0070】
形質転換体の培養物のRrhJ1I修飾酵素活性は、RrhJ1I認識配列を含む培養物中のDNAが、RrhJ1I 制限酵素、あるいはそのイソチゾマー(例えば、NaeI制限酵素)によって切断されないことにより確認することができる。本工程で得られる形質転換体は、RrhJ1I 修飾酵素遺伝子を含む約1.2 kbのDNA断片を含むプラスミドを保持している。
【0071】
形質転換体の培養物のRrhJ1I制限酵素活性は、以下のようなイン・ビトロの方法により調べることができる。すなわち、試験する形質転換体を培養し、その培養物より菌体を集めて超音波処理にて破砕した後、超遠心分離を行って上清を集め、これを活性測定用の試料とする。この適当量を基質であるλファージDNA(λ−DNA)とともに37℃でインキュベートした後、基質DNAの分解をアガロースゲル電気泳動により確認することができる。当該方法を用いて大腸菌形質転換体のRrhJ1I 制限酵素活性を調べると、RrhJ1Iによりλ−DNAを切断したときの断片の予想サイズと同じ大きさのフラグメントに切断する活性を検出できる。
【0072】
(v)工程(iv)で修飾酵素および制限酵素活性が検出された形質転換体を培養し、該培養物からRrhJ1I修飾酵素および制限酵素を採取する工程
本工程では、工程(iv)で得られた形質転換体から制限酵素、または修飾酵素を精製する。
【0073】
形質転換体の培養物からRrhJ1I 制限酵素および修飾酵素を採取するにあたっては、たとえば培養物より菌体を集菌後、超音波破砕、超遠心分離等により酵素を抽出し、ついで除核酸法、塩析法、アフィニティクロマトグラフィー法、ゲル濾過法、イオン交換クロマトグラフィー法等を組み合わせて精製すればよい。この方法により、RrhJ1I 制限酵素および修飾酵素を大量に得ることができる。
【0074】
用いる発現系によっては、形質転換体中で発現された酵素タンパク質が不溶物[封入体(inclusionbody)]として蓄積される場合がある。この場合にはこの不溶物を回収し、穏和な変性条件、たとえば尿素等の変性剤存在下で可溶化した後に変性剤を除くことによって活性型のタンパク質を得ることができる。さらに上記のようなクロマトグラフィー操作を行って目的とする酵素タンパク質を精製することができる。
【0075】
また、上記で得られた遺伝子をプローブとしてストリンジェントな条件下でハイブリダイゼーションを行えば、得られた遺伝子(配列番号1、配列番号3)と配列は少し異なるが、同様な酵素活性を持つタンパク質をコードする遺伝子を得ることができ、これも本発明の範囲内のものである。
得られた遺伝子が目的の制限・修飾系酵素をコードする領域の全てを含まない場合には、得られた遺伝子の塩基配列をもとにしてプライマーを合成し、これを用いたPCRによって不足する領域を増幅したり、あるいは得られた遺伝子の断片をプローブとしてDNAライブラリーのスクリーニングを繰り返すことにより、全コード領域を得ることができる。
【0076】
こうして得られたRrhJ1I 制限・修飾系酵素をコードする遺伝子を含有する形質転換体を作製して該遺伝子にコードされる酵素タンパク質を発現させ、さらに発現させた酵素タンパク質を精製することができる。形質転換体の作製、酵素タンパク質の発現および精製はいずれも上記と同様の方法で行うことができる。こうして得られた酵素タンパク質はRrhJ1I 制限酵素活性、あるいはRrhJ1I 修飾酵素活性を保持しており、これらは本発明に含まれる。
【0077】
実施例
以下、実施例を用いて本発明をより詳細に説明する。以下は本発明の例示であって本発明を限定する趣旨ではない。
【実施例1】
【0078】
J1菌からの制限酵素精製
J1菌からの制限酵素の精製は以下の方法で行った。
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株をMYKG培地(0.5%ポリペプトン、0.3%バクトイーストエキス、0.3%バクトモルトエキス、1% グルコース、0.2% K2HPO4、0.2% KH2PO4、pH7.0)で、30℃にて72時間振盪培養した。
菌体を破砕用緩衝液(10 mM Tris-HCl (pH 8.0)、10 mM 2-mercaptoethanol、1 mM EDTA)に懸濁したのち、4℃で超音波破砕を行った。遠心分離(12,000 rpm、4℃、30 min)で得られた上清を無細胞抽出液とした。次に、核酸を除去するため、ストレプトマイシン硫酸塩を終濃度4%になるように無細胞抽出液に添加して、氷上で30分間静置後、遠心分離によって沈殿を除去した。得られた上清を酵素精製緩衝液(10 mM Potassium phosphate buffer(pH 7.5)、10 mM 2-mercaptoethanol、5% glycerol)で一晩透析を行った後、DEAE-Sepharose(GEヘルスケア バイオサイエンス社)、HiTrap Heparin(GEヘルスケア バイオサイエンス社)、HiTrap Q(GEヘルスケア バイオサイエンス社)、P-cellulose(ワットマン)、HiPrep Sephacryl S-200 HR(GEヘルスケア バイオサイエンス社)を用いて精製を行い、J1菌制限酵素を得た。全ての精製操作は4℃で行った。
【0079】
J1菌から得られた制限酵素の酵素活性の測定は、以下の方法で行った。市販の制限酵素反応用buffer(LまたはH、タカラバイオ)を使用した。λ-DNAを基質DNAとして0.5μg添加した10μlの反応液を用いて、37℃で反応を行った。反応終了後、反応液をアガロースゲル電気泳動に供し、エチジウムブロマイドで染色を行って切断パターンを解析した。λ-DNAを基質とした時、約13kbの大きさに切断されたバンドが確認された。
【0080】
このJ1菌制限酵素の認識配列の決定は、以下のように行った。基質DNAとして複合プラスミドpK4を使用した。プラスミドpK4はATCC1267/pK4として独立行政法人産業技術総合研究所特許微生物寄託センターに寄託されている(FERM BP-3731)(特開平5-64589 号および同5-68566 号公報参照)。J1菌制限酵素でpK4を切断した結果、約3.4Kb、約0.7Kb、約0.5Kbの断片が生じた。この切断パターンと、pK4を市販酵素で切断したパターンとを比較すると、NaeI(タカラバイオ)の切断パターンと一致することが認められた。
【0081】
次に、プラスミドpK4をJ1菌に導入したJ1組換え体(J1/pK4)から回収したpK4を、上記から得られたJ1菌制限酵素とNaeIでそれぞれ切断したところ、いずれの酵素によっても切断されなかった。よって、J1菌の有する制限酵素はGCCGGCを切断するNaeIのイソチゾマーであることがわかり、認識配列はGCCGGCであることが明らかになった。
【実施例2】
【0082】
J1菌染色体DNAの調製
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株を100mlのMYKG培地中、30℃にて72時間振盪培養した。
培養後、集菌し、集菌された菌体をSaline-EDTA溶液(0.1M EDTA、0.15M NaCl (pH8.0))4mlに懸濁した。懸濁液にリゾチーム40mgを加えて37℃で1〜2時間振盪した後、-20℃で凍結した。
【0083】
次に、10mlのTris-SDS液(1%SDS、0.1M NaCl、0.1M Tris-HCl (pH9.0))を穏やかに振盪しながら加え、さらにプロテイナーゼK(メルク社)(10 mg/ml)を10μl加えて37℃で1時間振盪した。
次に、等量のTE (10mM Tris-HCl、1mM EDTA (pH8.0))飽和フェノールを加え、撹拌後、遠心した。上層を採取し、2倍量のエタノールを加えた後、ガラス棒でDNAを巻きとり、90%、80%、70%のエタノールで順次フェノールを取り除いた。
【0084】
次に、DNAを3mlのTE緩衝液に溶解させ、リボヌクレアーゼA溶液(100℃、15分間の加熱処理済)を10μg/mlになるよう加え、37℃で30分間振盪した。さらに、プロテイナーゼKを加え37℃で30分間振盪した後、等量のTE飽和フェノールを加えて遠心し、上層と下層に分離させた。
上層についてこの操作を2回繰り返した後、同量のクロロホルム(4%イソアミルアルコール含有)を加え、同様の抽出操作を繰り返した。その後、上層に2倍量のエタノールを加え、ガラス棒でDNAを巻きとり回収し、染色体DNA標品を得た。
【実施例3】
【0085】
修飾酵素遺伝子のPCR
J1菌修飾酵素遺伝子をPCRで増幅するためのプライマーを以下の方法で設計した。
NaeIとNgoMIVはGCCGGCを切断する制限酵素である。先ず、これらの制限酵素に対応する修飾酵素のアミノ酸ホモロジー解析を行った。その結果、両酵素のN末端付近のGCQALGL(配列番号22)と、C末端付近のGNAFPPP(配列番号23)が一致していることから、このアミノ酸配列に対応するオリゴヌクレオチドJM-01(配列番号10)、JM-04(配列番号11)を合成し、プライマーに用いた。PCRは以下の条件で実施し、約0.9Kbのバンドの増幅が確認された。
【0086】
反応液組成
鋳型DNA(J1菌染色体DNA、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-01(配列番号10) 1μl
プライマーJM-04(配列番号11) 1μl
2.5mM dNTP 8μl
DMSO 10μl
滅菌水 68μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:1分の反応を30サイクル
プライマー
JM-01:GGN GGN CAR GCN CTN GGN CT(配列番号10)
JM-04:GGN GGN GGR AAN GCR TTN CC(配列番号11)
【0087】
次に、増幅された配列のダイレクトシークエンシングをPCRで使用したプライマーを用いて実施した。その結果、配列番号5に示す配列が得られ、ホモロジー検索の結果、NaeIメチラーゼとの相同性が認められた。
【実施例4】
【0088】
J1菌修飾酵素遺伝子のクローニング
J1菌修飾酵素遺伝子全長のクローニングは以下の方法で行った。
J1菌ゲノムDNAを制限酵素SacIIで分解し、切断したゲノムDNAをGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いて回収し、DNA Ligation Kit(タカラバイオ)を用いて環化した。次に、環化したゲノムDNAを鋳型に使用し、配列番号12、13のオリゴヌクレオチドをプライマーに用いて以下の条件でPCRを行い、約1Kbの断片を増幅した。
【0089】
反応液組成
鋳型DNA(環化J1ゲノムDNA) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-05(配列番号12) 1μl
プライマーJM-06(配列番号13) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:10分の反応を30サイクル
プライマー
JM-05:TTTCCAGACCTAGTGCCTGA(配列番号12)
JM-06:GGCAGTAGAAGTGGCCGGAC(配列番号13)
【0090】
また、プライマーとして、配列番号14、および15のオリゴヌクレオチド用いた以外は、上記と同様の条件でPCRを行い、約2Kbの断片を増幅した。
JM-07:ACGTAACCGAACTCGGTCAG(配列番号14)
JM-08:GTATCGGCAGATCGGCAATG(配列番号15)
【0091】
PCR産物を確認した後、それぞれの反応液からGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を使用してPCR産物を回収した。得られたPCR産物は、pGEM-T Easyベクター(プロメガ)に結合して、大腸菌XL1-Blue MRF'(Stratagene)を形質転換した。得られた組換えコロニーからプラスミドを抽出し、DNA配列の確認を行った。DNA配列を解析した結果、配列番号6に示される塩基配列が得られた。配列番号6に示される塩基配列中に、配列番号3に示す1264bpのオープンリーディングフレーム(ORF1)を見出した。このORF1のコードするアミノ酸配列は、NaeI修飾酵素に対して67%の相同性を持っていることから、ORF1は、GCCGGCを認識配列とするJ1菌修飾酵素をコードしていることが推定された。
【実施例5】
【0092】
J1菌制限酵素遺伝子のクローニング
J1菌制限酵素遺伝子のクローニングを実施例4と同様の方法で行った。
J1菌ゲノムDNAを制限酵素SphIで分解し、切断したゲノムDNAをGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いて回収し、DNA Ligation Kit(タカラバイオ)を用いて環化した。次に、環化したゲノムDNAを鋳型に使用し、配列番号16、17のオリゴヌクレオチドをプライマーに用いて実施例4と同様の条件でPCRを行い、約6Kbの断片を増幅した。
JK-01:CCGGCGCGATCAAACGGGTG(配列番号16)
JK-02:TGCTGACCATCGGGCACCTG(配列番号17)
【0093】
PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物を、pGEM-T Easyベクター(プロメガ)に結合して、得られたベクターで大腸菌XL1-Blue MRF'を形質転換した。得られた組換えコロニーからプラスミドを抽出し、DNA配列の確認を行った。DNA配列を解析した結果、配列番号7に示される塩基配列が得られた。配列番号7に示される塩基配列中には、配列番号2に示されるアミノ酸配列をコードするオープンリーディングフレーム(ORF3:配列番号1)が見出された。得られたORF3のコードするアミノ酸配列は、NaeI制限酵素に対して47%の相同性を持っていることから、ORF3は、GCCGGCを認識配列とするII型のJ1菌制限酵素をコードしていることが推定された。また、HNHエンドヌクレーアゼと33%の相同性を持つアミノ酸をコードするオープンリーディングフレーム(ORF2:配列番号8)も見出された。
【0094】
最後に、J1菌制限酵素とJ1菌修飾酵素の遺伝子を含む約3.8Kbの断片を、J1菌染色体DNAから配列番号18、19のオリゴヌクレオチドをプライマーに用いて実施例3と同様の条件で温度サイクルを:94℃:30秒、65℃:30秒および72℃:3分として30サイクル行いPCR増幅した。得られたPCR産物を、pGEM-T Easyベクター(プロメガ)に結合してpJRM01を作製した。図1はpJRM01を示す模式図であり、約3.8KbのPCR断片にORF1(J1菌修飾酵素遺伝子)、ORF2、ORF3(J1菌制限酵素遺伝子)が含まれている(図2)。
【0095】
JM-32:GGtctagaGTTGGCGATTCCTCATCGCG(配列番号18)
JM-33:CCaagcttATCGCTCGCGGGGGTGCTCCG(配列番号19)
【実施例6】
【0096】
大腸菌組換え体によるJ1菌修飾酵素の生産
J1菌修飾酵素を得るために、実施例5で得られたプラスミドpJRM01を鋳型として使用して、以下に示す反応液組成およびプライマーを用いてPCRを行った。この際、修飾酵素をPtrcプロモーター下流のNcoIサイトに結合させるため、J1菌修飾酵素の2番目のアミノ酸をセリン(TCG)からスレオニン(ACG)に置換した。
【0097】
反応液組成
鋳型DNA(組換えプラスミドpJRM01) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-18(配列番号20) 1μl
プライマーJM-11(配列番号21) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:1分の反応を30サイクル
プライマー
JM-18:GGGTCATGACGCGGTCCAGCTACGAG(配列番号20)
JM-11:CTCCctgCAGGCGGCGTGGAAGCCTGG(配列番号21)
【0098】
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1.3kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物を、制限酵素PagIとSse8387Iで切断した。制限酵素処理を行ったPCR産物を、0.7%アガロースゲルにおける電気泳動に供し、約1.3Kb付近のバンドを回収した。回収したPCR産物を、Ligation Kit(タカラバイオ)を用いてベクターpTrc99AのNcoI-PstI部位に連結し、プラスミドを作製した。得られたプラスミドを、pTJM01と名付けた。図3は、プラスミドpTJM01の構造を示す模式図である。
【0099】
このプラスミドでXL1-Blue MRF'(Strategene)を形質転換し、得られた組換えコロニーをLB培地(1%トリプトン、0.5%バクトイーストエキス、0.5% NaCl、100μg/mlアンピシリン、1mM IPTG)で37℃一晩培養した。培養した菌からQIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製し、NaeIで処理した。その結果、NaeIで切断されなかったことから、修飾酵素によってプラスミドが修飾されていることを確認できた。
【0100】
さらに、培養した大腸菌組換え体(XL1-Blue MRF'/pTJM01)の超音波破砕した上清をSDS-ポリアクリルアミドゲル電気泳動(SDS-PAGE)に供したところ、J1菌修飾酵素の分子量に相当する45KDaにバンドが確認できたことから、J1菌修飾酵素が大量に発現していることが確認された(図4)。図4中、Mは、分子量マーカーを示し、レーン1はXL1-Blue MRF'/pTJM01の超音波破砕液上清のSDS-PAGEの結果を示し、レーン2はXL1-Blue MRF'の超音波破砕液上清のSDS-PAGEの結果を示す。
本実施例により、RrhJ1I修飾酵素遺伝子を含むベクターを含有した本発明の大腸菌組換え体は、大量のRrhJ1I修飾酵素を発現することが明らかになった。
【実施例7】
【0101】
J1菌修飾酵素発現プラスミド、およびJ1菌制限酵素発現プラスミドの構築
(1)J1菌修飾酵素発現プラスミドpMCLMの構築
実施例2で調製したJ1菌染色体DNAを鋳型にして、以下の条件でPCRを行った。
反応液組成
鋳型DNA(J1菌染色体、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJ1-M5(配列番号24) 1μl
プライマーJ1-M6(配列番号25) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、59℃:30秒および72℃:2分の反応を30サイクル
プライマー
J1-M5:GGGAGGTATGATCGATCCGTTC(配列番号24)
J1-M6:CGGCCAGGCGTGGAAG(配列番号25)
【0102】
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1.4kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物をpGEM-T Easy vector(プロメガ)に連結し、プラスミドを作製した。得られたプラスミドはDNA配列の確認を行い、pGTEM2と名付けた。
【0103】
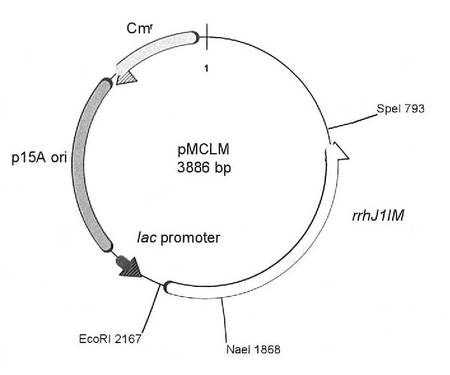
次に、pGTEM2に組み込まれたPCR産物をp15A複製オリジンを持つプラスミドであるpMCL200(Gene 162(1)1995, Pages 157-158)のLacプロモーター下流に挿入した。すなわち、pGTEM2とpMCL200をSpeIとEcoRIで切断して0.7%アガロースゲル電気泳動に供し、GFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いてそれぞれ約1.4Kb(pGTEM2から)、約2.5Kb(pMCL200から)の断片を回収した。回収したDNAをLigation Kit(タカラバイオ)を用いて連結し、プラスミドを作製した。得られたプラスミドを、pMCLMと名付けた。図5は、プラスミドpMCLMの構造を示す模式図である。図5中、「rrhJ1IM」は、J1菌修飾酵素遺伝子を示す。
【0104】
プラスミドpMCLMがJ1菌修飾酵素を発現しているか確認するため、以下の実験を行った。pMCLMでXL1-Blue MRF'(Strategene)を形質転換し、得られた組換えコロニーをLB培地(25μg/ml クロラムフェニコール含有)で37℃一晩培養した。培養した菌からQIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製し、NaeIで処理した。その結果、NaeIで切断されなかったことから、修飾酵素によってプラスミドが修飾されていることを確認できた。
【0105】
(2)J1菌制限酵素発現プラスミドpGErR10の構築
実施例2で調製したJ1菌染色体DNAを鋳型にして、以下の条件でPCRを行った。
反応液組成
鋳型DNA(J1菌染色体、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJ1-R17(配列番号26) 1μl
プライマーJ1-R18(配列番号27) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、66℃:30秒および72℃:1分の反応を30サイクル
プライマー
J1-R17:GATCCACCCGCCCACACCAC(配列番号26)
J1-R18:GGCTTCCGGCCACCGGGAAAG(配列番号27)
【0106】
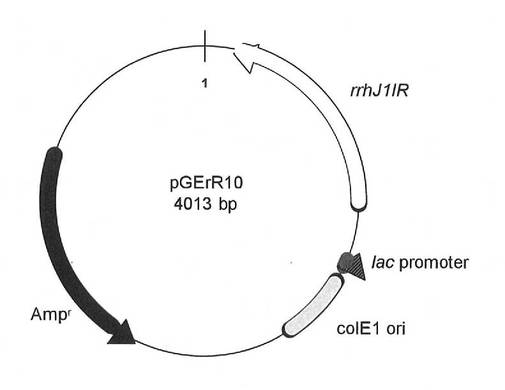
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物をpGEM-T Easy vector(プロメガ)に連結し、(1)で調製したpMCLMと共にXL1-Blue MRF'を形質転換した。得られた形質転換体をLB培地(100μg/ml アンピシリン、25μg/ml クロラムフェニコール含有)で37℃一晩培養し、QIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製した。得られたプラスミドはDNA配列を確認し、pGErR10と名付けた。図6は、プラスミドpGErR10の構造を示す模式図である。図6中、「rrhJ1IR」は、J1菌制限酵素遺伝子を示す。
【実施例8】
【0107】
大腸菌組換え体によるJ1菌制限酵素の生産
(1)無細胞抽出液の調製
実施例7(2)で作製したJ1菌制限酵素発現ベクターを含む大腸菌組換え体(XL1-Blue MRF'/pMCLM+pGErR10)をLB培地(100μg/ml アンピシリン、25μg/ml クロラムフェニコール含有)で37℃で16時間培養した。培養液を遠心分離によって回収し、破砕用緩衝液に懸濁したのち、4℃で10分間、超音波による破砕を行った。破砕液は遠心分離(12,000 rpm、4℃、30 min)し、得られた上清を無細胞抽出液とした。
また比較例として、以下の実験を行った。J1菌を実施例1の方法で培養し、実施例8(1)と同様の手法で調製したサンプルをJ1菌の無細胞抽出液とした。また、それぞれの無細胞抽出液のタンパク質濃度はBioRad Protein Assay Kit(バイオラド)を用いて定量した。
【0108】
(2)制限酵素活性の比較
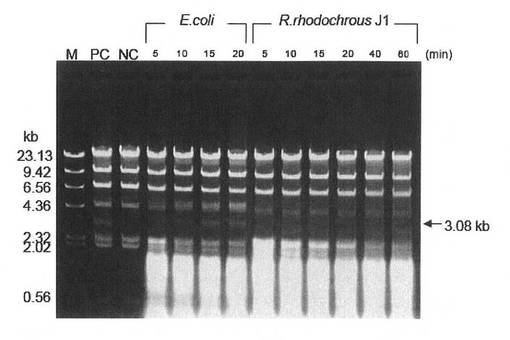
(1)で調製した無細胞抽出液を用いて、RrhJ1I制限酵素活性を調べた。
基質としてλ−DNAをHindIIIで切断したDNA(TOYOBO:λ/HindIII digest)0.5μgを使用し、大腸菌組換え体またはJ1菌の無細胞抽出液(それぞれ40μgタンパク質)を制限酵素用L buffer(タカラバイオ)と共に加え、37℃でサンプリングしながら1時間インキュベートした。サンプリングした反応液はフェノール処理を行って、電気泳動に共した。用いた基質DNAは、RrhJ1I制限酵素で切断されると約3.0Kbのバンドが出現する。
【0109】
結果を図7に示す。図7中、「M」は分子量マーカーを示し、「PC」は実施例1で調製した精製酵素を用いた切断結果を示し、「NC」は精製酵素を添加しないで行った実験結果を示す。
電気泳動の結果より、大腸菌組換え体の無細胞抽出液(図7「E.coli」)は反応20分で約3kbのバンドが出現したが、J1菌の無細胞抽出液(図7「R.rhodochrous J1」)は反応1時間後でもバンドは出現しなかった。以上の結果より、RrhJ1I制限酵素遺伝子を含むベクターを含有した本発明の大腸菌組換え体は、J1菌より大量の制限酵素を生産していることを確認した。
【配列表フリーテキスト】
【0110】
配列番号1:RrhJ1I制限酵素の塩基配列
配列番号2:RrhJ1I制限酵素のアミノ酸配列
配列番号3:RrhJ1I修飾酵素の塩基配列
配列番号4:RrhJ1I修飾酵素のアミノ酸配列
配列番号5:実施例3で得られたDNAの塩基配列
配列番号6:実施例4で得られたDNAの塩基配列
配列番号7:実施例5で得られたDNAの塩基配列
配列番号8:ORF2の塩基配列
配列番号9:配列番号8から推定されるアミノ酸配列
配列番号10:プライマー
配列番号11:プライマー
配列番号12:プライマー
配列番号13:プライマー
配列番号14:プライマー
配列番号15:プライマー
配列番号16:プライマー
配列番号17:プライマー
配列番号18:プライマー
配列番号19:プライマー
配列番号20:プライマー
配列番号21:プライマー
配列番号22:NaeI対応修飾酵素とNgoMIV対応修飾酵素のN末端相同アミノ酸配列
配列番号23:NaeI対応修飾酵素とNgoMIV対応修飾酵素のC末端相同アミノ酸配列
配列番号24:プライマー
配列番号25:プライマー
配列番号26:プライマー
配列番号27:プライマー
配列表中、nはa、t、gまたはcを示す。
配列表中、rはgまたはaを示す。
【技術分野】
【0001】
本発明は、遺伝子工学試薬として有用な制限酵素および修飾酵素、該酵素タンパク質をコードする遺伝子、並びに該酵素の製造方法に関する。
【背景技術】
【0002】
制限酵素は、ある特定のDNA塩基配列を特異的に認識し、切断することのできるエンド型ヌクレアーゼであり、これまでに数多くの制限酵素が見出されている。
分子遺伝学、または生化学等の発展により、DNAが遺伝を司る本体であることが明らかになって以来、制限酵素は遺伝病の解明や、遺伝子操作等において現在幅広く用いられている有用な酵素である。このうち、特にDNA塩基配列を特異的に認識し、特定のDNAを切断するII型制限酵素が重要かつ必要なものとして用いられている。
【0003】
これまでにII型制限酵素は300種以上が精製されている。それらの中で、GCCGGCを認識する制限酵素としては40種近くのイソチゾマー(isoschizomer)が存在することが知られており、例えば、NaeI(タカラバイオ他)、NgoMIV(New England Biolabs、Promega)、MroNI(SibEnzyme)、PdiI(Fermentas)が市販されている。ここで、イソチゾマーは、認識配列が互いに一致している酵素のペアを意味する。
【0004】
また、特異的なDNA配列をメチル化する修飾酵素が数多く見出されている。
これまでに種々の微生物において制限酵素および修飾酵素の存在が確認されており、これらの酵素は、宿主内で制限・修飾系に関与していることが知られている。
【0005】
例えば、NaeI制限酵素および修飾酵素は、Lechevalieria aerocolonigenes ATCC23870(Nocardia aerocolonigenes)に存在することが知られている。しかし、この細菌中に含まれる制限酵素および修飾酵素の含量は少ないため、両酵素を大量に得ることは困難であった。
これまでに、染色体中に組み込まれたMspIメチラーゼ遺伝子を有する大腸菌K802株を用いてNaeI制限酵素の生産を行った結果、この大腸菌は、細胞1μg当たり、野生株(Lechevalieria aerocolonigenes ATCC23870)の50倍の活性(37℃、1時間の反応で1μgのDNAを分解できる酵素量)を有していたことが報告されている(特許文献1)。
【0006】
ところで、ロドコッカス属(Rhodococcus)に属する細菌において、制限酵素は15例が報告されているに過ぎない。例えば、6塩基配列を認識する制限酵素としてBamHI(GGATCC)、SalI(GTCGAC)、BspHI(TCATGA)、SphI(GCATCG)のイソチゾマーや、4塩基配列を認識する制限酵素としてMboI(GATC)やNlaIV(GGNNCC)のイソチゾマーが見出されている。しかし、GCCGGCを認識する制限酵素および修飾酵素はこれまでに見出されていなかった。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開平6−98779号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の目的は、遺伝子工学の分野で有用なGCCGGCを認識する制限酵素および修飾酵素、さらには該酵素タンパク質をコードする制限酵素遺伝子および修飾酵素遺伝子を提供することである。また、本発明の目的は、該制限酵素および修飾酵素の製造方法を提供することである。
【課題を解決するための手段】
【0009】
本発明者は上記課題を解決するために鋭意研究を行った結果、ロドコッカス属に属する細菌において新規の制限酵素および修飾酵素を見出した。詳しくは、本発明者は、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株から、制限酵素を精製した。このJ1菌由来の制限酵素のDNA切断パターンはNaeIのものと一致したことから、得られたJ1菌制限酵素はNaeIのイソチゾマーであることが示された。また、ロドコッカス・ロドクロウスJ-1株から修飾酵素を単離した。そして、得られたJ1菌修飾酵素の塩基配列情報から、J1菌制限酵素を単離した。このように、本発明者は、ロドコッカス属に属する細菌における新規の制限酵素および修飾酵素を見出し、本発明を完成した。
【0010】
すなわち、本発明は、以下の発明を包含する。
(1)以下の(A)または(B)のタンパク質。
(A)配列番号2記載のアミノ酸配列を含むタンパク質
(B)配列番号2記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質
(2)(1)記載のタンパク質をコードする遺伝子。
(3)以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号1記載の塩基配列からなるDNA
(b)配列番号1記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I制限酵素活性を有するタンパク質をコードするDNA
(4)以下の(A)または(B)のタンパク質。
(A)配列番号4記載のアミノ酸配列を含むタンパク質
(B)配列番号4記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
(5)(4)記載のタンパク質をコードする遺伝子。
(6)以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号3記載の塩基配列からなるDNA
(b)配列番号3記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I修飾酵素活性を有するタンパク質をコードするDNA
(7)(2)、(3)、(5)、または(6)のいずれか1項に記載の遺伝子を含む組換えベクター。
(8)(7)記載の組換えベクターを含む形質転換体。
(9)(8)記載の形質転換体の培養物からRrhJ1I制限酵素活性を有するタンパク質を採取することを含む、RrhJ1I制限酵素の製造方法。
(10)(8)記載の形質転換体の培養物からRrhJ1I修飾酵素活性を有するタンパク質を採取することを含む、RrhJ1I修飾酵素の製造方法。
【発明の効果】
【0011】
本発明により、遺伝子工学の分野で有用なGCCGGCを認識する制限酵素および修飾酵素、さらには該酵素タンパク質をコードする制限酵素遺伝子および修飾酵素遺伝子が提供される。また、本発明の遺伝子を用いて、該制限酵素および修飾酵素の製造が可能となるため、該制限酵素および修飾酵素を大量に生産することができる。
【0012】
また、本発明の制限酵素および修飾酵素は、制限修飾系を構成しているため、本発明の酵素および本発明の遺伝子は、例えば、制限修飾系の研究試薬として、非常に有用である。
【図面の簡単な説明】
【0013】
【図1】pJRM01の構造を示す模式図である。
【図2】制限・修飾酵素遺伝子の制限酵素地図を示す図である。
【図3】pTJM01の構築図である。
【図4】組換え体の超音波破砕上清を、SDS-PAGEした結果を示す図である。
【図5】pMCLMの構造を示す模式図である。
【図6】pGErR10の構造を示す模式図である。
【図7】無細胞抽出液を使用した切断結果を示す図である。
【発明を実施するための形態】
【0014】
以下、本発明の実施の形態について詳細に説明するが、本実施の形態は、本発明を説明するための例示であり、本発明をこの実施の形態のみに限定させるものではない。
【0015】
本発明は、ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株(FERM BP-1478)由来の制限酵素(RrhJ1I制限酵素)および修飾酵素(RrhJ1I修飾酵素)に関するものである。ロドコッカス・ロドクロウスJ-1株(以下、「J1菌」ともいう)は、「Rhodococcus rhodochrous J-1」(FERM BP-1478)として独立行政法人産業技術総合研究所 特許生物寄託センター(茨城県つくば市東1丁目1番地1 中央第6)に寄託されている。
【0016】
本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素は、ロドコッカス属に属する細菌の自己防御系としての制限修飾系を構成していると考えられる。
【0017】
細菌の多くは、制限修飾系によって、外来のDNAから自己のDNAを守っている。つまり、修飾酵素により自己のDNAを修飾(メチル化)することで、自己のDNAを修飾型の異なる外来のDNAと区別している。そして、外来のDNAは、しかるべき位置にメチル基の無いことが制限酵素に認識され、制限酵素により切断される。
【0018】
制限修飾系を保持する細菌が安定な増殖を維持するためには、両酵素活性のバランスが適切に保たれる必要がある。すなわち、制限酵素が発現し、機能する前に、修飾酵素によって染色体遺伝子がメチル化され、制限酵素の切断から保護される必要がある。そのため、制限酵素遺伝子と修飾酵素遺伝子の転写および翻訳は、高度に制御されている必要がある。したがって、両遺伝子はゲノム上において近傍に位置することによって、オペロンを形成したり、特異的な転写制御因子による制御を受けられるように進化したと考えられる。このように、制限酵素と対応する修飾酵素とは密接な関係にあるため、両者を同時に提供する本発明は、非常に有用である。
【0019】
本明細書において、「制限酵素」は、ある特定の塩基配列を特異的に認識し、特定位置でDNAを切断することのできる酵素を意味する。本発明において、制限酵素は、好ましくは、エンド型ヌクレアーゼ(II型制限酵素)である。
【0020】
本明細書において、「RrhJ1I制限酵素」(「J1菌制限酵素」ともいう)は、ロドコッカス・ロドクロウスJ-1株で発現する制限酵素であり、GCCGGCの6塩基配列を認識し、DNAを切断する活性(「RrhJ1I制限酵素活性」ともいう)を有するタンパク質を意味する。本発明のRrhJ1I制限酵素による認識部位と切断部位とは、同一でもよく、または異なっていてもよい。切断されるDNAは、1本鎖DNAまたは2本鎖DNAのいずれでもよい。また、切断後のDNA末端は、例えば、5'突出末端、3'突出末端または平滑末端のいずれでもよい。
【0021】
本明細書において、RrhJ1I制限酵素活性は、RrhJ1I制限酵素をDNAと接触させ、接触後のDNAの分子量またはDNA断片数を測定することにより評価することができる。当業者であれば、基質DNA、接触時の酵素量、温度、溶液組成または接触時間などの条件を設定することができる。DNAの分子量は、例えば、アガロース電気泳動によって測定することができる。接触前のDNAの分子量と接触後のDNAの分子量、または接触前のDNAの断片数と接触後DNAの断片数とを比較することで、RrhJ1I制限酵素活性を評価することができる。また、GCCGGCを認識する既知の制限酵素を対照として使用してもよい。
【0022】
本明細書において、「修飾酵素」は、対応する制限酵素による切断からDNAを保護するようにDNAに修飾することのできる酵素を意味する。この場合、対応する修飾酵素および制限酵素は、同一の配列を特異的に認識する。本明細書において、同一の微生物に由来する酵素であって、同一の塩基配列を認識する制限酵素と修飾酵素との関係を、「対応する」と称する場合がある。本発明において、修飾酵素は、好ましくは、DNAをメチル化するメチラーゼである。
【0023】
本明細書において、「RrhJ1I修飾酵素」(「J1菌修飾酵素」ともいう)は、ロドコッカス・ロドクロウスJ-1株で発現する修飾酵素であり、RrhJ1I制限酵素による切断からDNAを保護する活性(「RrhJ1I修飾酵素活性」ともいう)を有するタンパク質を意味する。本発明のRrhJ1I修飾酵素による認識部位と修飾部位とは、同一でもよく、または異なっていてもよい。発明のRrhJ1I修飾酵素により修飾される塩基およびその数は特に限定されない。例えば、RrhJ1I修飾酵素は、認識配列中の1〜3個のシトシンをメチル化することができる。
【0024】
本明細書において、RrhJ1I修飾酵素活性は、RrhJ1I修飾酵素を接触させたDNAにRrhJ1I制限酵素またはそのイソチゾマーを接触させ、制限酵素接触後のDNAの分子量またはDNAの断片数を測定することにより評価することができる。当業者であれば、基質DNA、修飾酵素または制限酵素による接触時の酵素量、温度、溶液組成または接触時間などの条件を設定することができる。修飾酵素に接触させた後もしくは制限酵素に接触前のDNAの分子量と制限酵素に接触後のDNAの分子量、または修飾酵素に接触させた後もしくは制限酵素に接触前のDNA断片数と制限酵素に接触後のDNA断片数とを比較することで、RrhJ1I修飾酵素活性を評価することができる。RrhJ1I修飾酵素活性を評価する際、GCCGGCを認識する制限酵素に対応する既知の修飾酵素を対照として使用してもよい。
【0025】
(1)RrhJ1I制限・修飾系酵素
本発明のRrhJ1I制限酵素は、例えば、配列番号2に記載のアミノ酸配列を含むものである。配列番号2に記載のアミノ酸配列を含むタンパク質は、例えば、配列番号2に記載のアミノ酸配列からなるタンパク質である。
【0026】
また、本発明のRrhJ1I制限酵素は、前記のものに限定されることはなく、配列番号2に記載のアミノ酸配列の全部または一部を含むタンパク質であって、かつRrhJ1I制限酵素活性を有するタンパク質を含むものである。
また、本発明のRrhJ1I制限酵素には、配列番号2記載のアミノ酸配列と約50%以上、好ましくは約60%以上、より好ましくは約70%以上、さらに好ましくは約80%以上、特に好ましくは約90%以上、さらに特に好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有するアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質も含まれる。
さらに、配列番号2記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列を含み、かつRrhJ1I制限酵素活性を有するタンパク質も本発明のRrhJ1I制限酵素に含まれる。
【0027】
本発明のRrhJ1I修飾酵素は、例えば、配列番号4に記載のアミノ酸配列を含むものでである。配列番号4に記載のアミノ酸配列を含むタンパク質は、例えば、配列番号4に記載のアミノ酸配列からなるタンパク質である。
また、本発明のRrhJ1I修飾酵素は、前記のものに限定されることはなく、配列番号4記載のアミノ酸配列の全部または一部を含むタンパク質であって、かつRrhJ1I修飾酵素活性を有するタンパク質を含むものである。
また、本発明のRrhJ1I修飾酵素には、配列番号4記載のアミノ酸配列と約70%以上、好ましくは約80%以上、より好ましくは約90%以上、さらに好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有するアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質も含まれる。
さらに、配列番号4記載のアミノ酸配列において、1または数個のアミノ酸が欠失、置換または付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質も本発明のRrhJ1I修飾酵素に含まれる。
【0028】
前記の配列番号2または4記載のアミノ酸配列において、1または数個のアミノ酸に欠失、置換または付加等の変異が生じたアミノ酸配列としては、例えば(i)配列番号2または4記載のアミノ酸配列において、1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が欠失したアミノ酸配列、(ii)配列番号2または4記載のアミノ酸配列の1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が他のアミノ酸で置換されたアミノ酸配列、(iii)配列番号2または4記載のアミノ酸配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が付加したアミノ酸配列、(iv)配列番号2または4記載のアミノ酸配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)のアミノ酸が挿入されたアミノ酸配列、(v)上記(i)〜(iv)を組み合わせたアミノ酸配列が挙げられる。
【0029】
(2)RrhJ1I制限酵素遺伝子およびRrhJ1I修飾酵素遺伝子
本発明のRrhJ1I 制限酵素遺伝子は、本発明のRrhJ1I制限酵素をコードする遺伝子である。本発明のRrhJ1I制限酵素は前述のとおりである。本発明のRrhJ1I制限酵素遺伝子は、例えば、配列番号1に記載の塩基配列からなるDNAを含むものが挙げられるが、これに限定されるものではない。
本発明のRrhJ1I 制限酵素遺伝子には、配列番号1記載の塩基配列と約50%以上、好ましくは約60%以上、より好ましくは約70%以上、さらに好ましくは約80%以上、特に好ましくは約90%以上、さらに特に好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有する塩基配列であって、RrhJ1I制限酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I制限酵素遺伝子には、上記のDNAのほか、配列番号1に記載の塩基配列において、1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列であって、RrhJ1I制限酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I制限酵素遺伝子には、配列番号1に記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I 制限酵素活性を有するタンパク質をコードするDNAが含まれる。これらの遺伝子などは本発明のRrhJ1I制限酵素遺伝子に含まれる。ストリンジェントな条件については後述する。
【0030】
本発明のRrhJ1I 修飾酵素遺伝子は、本発明のRrhJ1I修飾酵素をコードする遺伝子である。本発明のRrhJ1I修飾酵素は前述のとおりである。本発明のRrhJ1I修飾酵素遺伝子は、例えば、配列番号3に記載の塩基配列からなるDNAを含むものが挙げられるが、これに限定されるものではない。
本発明のRrhJ1I 修飾酵素遺伝子には、配列番号3記載の塩基配列と約70%以上、好ましくは約80%以上、より好ましくは約90%以上、さらに好ましくは約95%以上、最も好ましくは約98%以上の相同性(同一性)を有する塩基配列であって、RrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I修飾酵素遺伝子には、上記のDNAのほか、配列番号3に記載の塩基配列において、1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列であって、RrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNAが含まれる。
また、本発明のRrhJ1I修飾酵素遺伝子には、配列番号3に記載の塩基配列からなるDNAと相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつRrhJ1I 修飾酵素活性を有するタンパク質をコードするDNAが含まれる。これらの遺伝子も本発明のRrhJ1I修飾酵素遺伝子に含まれる。
【0031】
前記のストリンジェントな条件としては、例えば、DNAを固定したナイロン膜を、6×SSC(1×SSCは塩化ナトリウム8.76g 、クエン酸ナトリウム4.41g を1リットルの水に溶かしたもの)、1% SDS、100μg/mlサケ精子DNA、0.1% ウシ血清アルブミン、0.1% ポリビニルピロリドン、0.1% フィコールを含む溶液中で65℃にて20時間プローブとともに保温してハイブリダイゼーションを行う条件をあげることができるが、これに限定されるわけではない。当業者であれば、このようなバッファーの塩濃度、温度等の条件に加えて、その他のプローブ濃度、プローブの長さ、反応時間等の諸条件を加味し、ハイブリダイゼーションの条件を設定することができる。
【0032】
ハイブリダイゼーション法の詳細な手順については、Molecular Cloning, A Laboratory Manual 2nd ed.(Cold Spring Harbor Laboratory Press (1989))等を参照することができる。
以下に、ハイブリダイゼーションによりRrhJ1I制限酵素遺伝子またはRrhJ1I修飾酵素遺伝子を得る方法の一例を示すが、これに限定されるわけではない。
【0033】
まず、適当な遺伝子源から得たDNAを常法に従ってプラスミドやファージベクターに接続してDNAライブラリーを作製する。このライブラリーを適当な宿主に導入して得られる形質転換体をプレート上で培養し、生育したコロニーまたはプラークをニトロセルロースやナイロンの膜に移し取り、変性処理の後にDNAを膜に固定する。この膜をあらかじめ32P等で標識したプローブを含む上記の組成の溶液中、上記のストリンジェントな条件で保温し、ハイブリダイゼーションを行う。プローブとしては、配列番号2または配列番号4に記載したアミノ酸配列の全部または一部をコードするポリヌクレオチドを含有していればよく、たとえば配列番号1または配列番号3に記載の塩基配列の全部または一部からなるポリヌクレオチドまたはそれを含むポリヌクレオチドを使用することができる。
ハイブリダイゼーションの終了後、非特異的に吸着したプローブを洗い流し、オートラジオグラフィー等によりプローブとハイブリッドを形成したクローンを同定する。この操作をハイブリッド形成クローンを単離できるまで繰り返す。
こうして得られたクローンの中には、目的の酵素活性を有するタンパク質をコードする遺伝子が保持されている。そして、クローンから目的の遺伝子を得るには、アルカリ法などの公知のポリヌクレオチド抽出方法を使用する。
【0034】
また、前記の配列番号1または3に記載の塩基配列において1個または数個の塩基に欠失、置換または付加等の変異が生じた塩基配列としては、例えば(i)配列番号1または3に記載の塩基配列において、1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が欠失した塩基配列、(ii)配列番号1または3に記載の塩基配列の1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が他の塩基で置換された塩基配列、(iii)配列番号1または3に記載の塩基配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が付加した塩基配列、(iv)配列番号1または3に記載の塩基配列に1〜20個(例えば1〜10個、好ましくは1〜5個、さらに好ましくは1〜2個)の塩基が挿入された塩基配列、(v)上記(i)〜(iv)を組み合わせた塩基配列が挙げられる。
【0035】
このように種々のDNAが本発明のRrhJ1I制限酵素遺伝子または本発明のRrhJ1I修飾酵素遺伝子の範囲内に含まれるのは、以下の理由によるものである。即ち、遺伝子上でアミノ酸を指定するコドン(3つの塩基の組み合わせ)は、アミノ酸の種類ごとに1〜6種類ずつが存在することが知られている。したがって、あるアミノ酸配列をコードする遺伝子は多数存在することができる。
遺伝子は自然界において決して安定に存在しているものではなく、その塩基配列に変異が起こることはまれではない。遺伝子上に起こった変異によっては、コードされるアミノ酸配列に変化を与えない変異(サイレント変異と呼ばれる)もあり、この場合には同じアミノ酸配列をコードする異なる遺伝子が生じたといえる。したがって、ある特定のアミノ酸配列をコードする遺伝子が単離されても、それを含有する生物が継代されていくうちに同じアミノ酸配列をコードする多種類の遺伝子ができていく可能性は否定できない。
【0036】
さらに、同じアミノ酸配列をコードする多種類の遺伝子を人為的に作製することは、種々の遺伝子工学的手法を用いれば困難なことではない。
例えば、遺伝子工学的なタンパク質の生産において、目的のタンパク質をコードする本来の遺伝子上で使用されているコドンが宿主中では使用頻度の低いものであった場合には、タンパク質の発現量が低いことがある。このような場合にはコードされているアミノ酸配列に変化を与えることなく、コドンを宿主で繁用されているものに人為的に変換することにより、目的タンパク質の高発現を図ることが行われている。
【0037】
このように、特定のアミノ酸配列をコードする多種類の遺伝子は人為的に作製可能なことは言うまでもなく、自然界においても生成されうるものである。したがって、本発明中に開示された塩基配列と同一の遺伝子ではなくても、RrhJ1I 制限酵素活性または修飾酵素活性を示すタンパク質をコードするDNAである限り、本発明のRrhJ1I制限酵素遺伝子またはRrhJ1I修飾酵素遺伝子に含まれるものである。
【0038】
また、本発明は、RrhJ1I制限酵素遺伝子およびRrhJ1I修飾酵素遺伝子の両方を含む遺伝子も含有する。制限酵素をコードする遺伝子および該制限酵素に対応する修飾酵素をコードする遺伝子は、ゲノム上で互いに近傍に位置することが知られている。
本明細書において、「RrhJ1I 制限・修飾系酵素の遺伝子」には、RrhJ1I 制限酵素をコードする遺伝子およびRrhJ1I 制限酵素による切断よりDNAを保護するRrhJ1I修飾酵素をコードする遺伝子が含まれる。
【0039】
本発明のRrhJ1I制限酵素遺伝子、RrhJ1I修飾酵素遺伝子およびRrhJ1I 制限・修飾系酵素の遺伝子は、これらの酵素を発現する微生物から単離することができる。例えば、ロドコッカス・ロドクロウスJ-1株由来のゲノムDNAを鋳型として、既知のアミノ酸配列情報から遺伝子の縮重を考慮して設計したプライマーもしくはプローブまたは既知の塩基配列情報に基づいて設計したプライマーもしくはプローブを用いたPCRまたはハイブリダイゼーション法により、遺伝子を単離することができる。単離した遺伝子から、本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素を製造することができる。
【0040】
また、遺伝子に変異を導入することによって、1個または数個のアミノ酸に欠失、置換または付加等の変異を導入することもできる。アミノ酸配列と遺伝子に変異を導入するには、Kunkel法や Gapped duplex法等の公知手法により、部位特異的突然変異誘発法を利用した変異導入用キット、例えばQuikChangeTM Site-Directed Mutagenesis Kit(ストラタジーン社)、GeneTailorTM Site-Directed Mutagenesis System(インビトロジェン社)、TaKaRa Site-Directed Mutagenesis System(Mutan-K、Mutan-Super Express Km等:タカラバイオ社)等を用いることができる。
【0041】
本発明において、DNAの塩基配列の確認は、慣用の方法により配列決定することにより行うことができる。例えば、ジデオキシヌクレオチドチェーンターミネーション法(Sanger et al. (1977) Proc. Natl. Acad. Sci. USA 74: 5463)等により行うことができる。また、適当なDNAシークエンサーを利用して配列を解析することも可能である。
塩基配列の決定は、プラスミドベクターを用いて作製された形質転換体の場合、宿主がエシェリヒア・コリであれば試験管等で培養を行い、常法に従ってプラスミドを調製する。得られたプラスミドをそのまま鋳型とするか、あるいは挿入断片を取り出してM13ファージベクター等にサブクローニングした後に、ジデオキシ法により塩基配列を決定する。ファージベクターで作製された形質転換体の場合も基本的に同様な操作により塩基配列を決定することができる。これら培養から塩基配列決定までの基本的な実験法については、例えば、前述のT.ManiatisらのMolecular Cloning, A Laboratory Manual等に記載されている。
【0042】
得られた遺伝子が目的のRrhJ1I 制限・修飾系酵素をコードする遺伝子であるかどうかの確認は、決定された塩基配列を配列番号1または配列番号3に記載の塩基配列と比較して行うことができる。あるいは決定された塩基配列より推定されるアミノ酸配列を配列番号2または配列番号4に記載のアミノ酸配列と比較して行うことができる。
【0043】
(3)本発明のRrhJ1I制限酵素およびRrhJ1I修飾酵素の製造方法
本発明のRrhJ1I 制限酵素を製造するには、まず前記のようなRrhJ1I 制限酵素をコードする遺伝子と、RrhJ1I修飾酵素をコードする遺伝子とを組み込んだ組換えベクターを作製する。そして、この組換えベクターで形質転換された形質転換体を作製し、これを培養して、培養物中よりRrhJ1I 制限酵素活性を有するタンパク質を採取することによりRrhJ1I 制限酵素を製造することができる。
この場合、形質転換体はRrhJ1I 制限酵素をコードする遺伝子を含む組換えベクターと、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターの両方の組換えベクターで形質転換された形質転換体を用いてもよい。
また、RrhJ1I修飾酵素をコードする遺伝子の代わりに、RrhJ1I修飾酵素が修飾する塩基配列と同じ塩基配列を修飾する酵素(例としてはNaeI修飾酵素)をコードする遺伝子を用いてもよい。
【0044】
また、本発明のRrhJ1I 修飾酵素を製造するには、まず前記のようなRrhJ1I 制限酵素をコードする遺伝子と、RrhJ1I 修飾酵素をコードする遺伝子とを組み込んだ組換えベクターを作製する。そして、この組換えベクターで形質転換された形質転換体を作製し、これを培養して、培養物中よりRrhJ1I 修飾酵素活性を有するタンパク質を採取することによりRrhJ1I 修飾酵素を製造することができる。
この場合、形質転換体はRrhJ1I 制限酵素をコードする遺伝子を含む組換えベクターと、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターの両方の組換えベクターで形質転換された形質転換体を用いてもよい。
あるいは、RrhJ1I 修飾酵素をコードする遺伝子を含む組換えベクターのみで形質転換された形質転換体を用いてもよい。
【0045】
本発明のRrhJ1I制限酵素遺伝子、本発明のRrhJ1I修飾酵素遺伝子または本発明のRrhJ1I制限・修飾酵素系の遺伝子を含む組換えベクターは、本発明の範囲に含まれる。
【0046】
本発明の組換えベクターは、上記酵素をコードする遺伝子の上流に転写プロモーター、場合によっては下流にターミネーターを挿入して発現カセットを構築し、このカセットを発現ベクターに挿入することにより作製することができる。あるいは、発現ベクターに転写プロモーターおよび/またはターミネーターがすでに存在する場合には、発現カセットを構築することなく、ベクター中のプロモーターおよび/またはターミネーターを利用して、その間に当該酵素をコードする遺伝子を挿入すればよい。
【0047】
本明細書において、プロモーターは、例えば、trcプロモーター、lacプロモーターなどをあげることができるが、これに限定されるわけではない。
本明細書において、ターミネーターは、例えば、trpオペロンターミネータをあげることができるが、これに限定されるわけではない。
【0048】
ベクターに本発明の遺伝子を挿入するには、制限酵素を用いる方法、トポイソメラーゼを用いる方法等を利用することができる。また、挿入の際に必要であれば、適当なリンカーを付加してもよい。また、アミノ酸への翻訳にとって重要な塩基配列として、SD配列やKozak配列などのリボソーム結合配列が知られており、これらの配列を遺伝子の上流に挿入することもできる。挿入にともない、遺伝子がコードするアミノ酸配列の一部を置換してもよい。
【0049】
本発明において使用されるベクターは、本発明の遺伝子を保持するものであれば特に限定されず、それぞれの宿主に適したベクターを用いることができる。ベクターとしては、例えば、プラスミドDNA、バクテリオファージDNA、レトロトランスポゾンDNA、人工染色体DNAなどが挙げられる。例えば、大腸菌を宿主とする場合には、pTrc99A(GEヘルスケアバイオサイエンス)、pACYC184(ニッポンジーン)、pMW118(ニッポンジーン)などを挙げることができる。また、必要に応じて、これらのベクターを改変したものを用いることもできる。
【0050】
本発明の組換えベクターを宿主に導入することで、本発明の形質転換体を作製することができる。当該形質転換体も本発明の範囲に含まれる。
本発明において使用する宿主は、上記組換えベクターが導入された後、目的の制限酵素または修飾酵素を発現することができる限り、特に限定されるものではない。宿主としては、例えば、大腸菌(エシェリヒア・コリ)、枯草菌(バチルス・ズブチリス(Bacillus subtilis))、ロドコッカス菌(Rhodococcus)、放線菌などの細菌、酵母(サッカロミセス・セレビシエ(Saccharomyces cerevisiae))、カビ、動物細胞、植物細胞、昆虫細胞などが挙げられる。本発明において、宿主は好ましくは大腸菌である。
本発明において、大腸菌は、例えば、大腸菌K12株やB株、あるいはそれらの野生株由来の派生株であるJM109株、XL1-Blue株(例えば、XL1-Blue MRF')、K802株、C600株などを挙げることができる。
宿主への組換えベクターの導入方法としては、宿主に適した方法であれば特に限定されるものではなく、当業者であれば公知技術から適宜選択することができる。このような方法としては、例えば、エレクトロポレーション法、カルシウムイオンを用いる方法、スフェロプラスト法、酢酸リチウム法、リン酸カルシウム法、リポフェクション法等が挙げられる。
【0051】
本発明の形質転換体を培養して、培養物からRrhJ1I制限酵素活性を有するタンパク質またはRrhJ1I修飾酵素活性を有するタンパク質を採取することにより、RrhJ1I制限酵素またはRrhJ1I修飾酵素を製造することができる。
【0052】
本発明において「培養物」とは、菌体、培養液、無細胞抽出液、細胞膜などの培養により得られるものを意味する。無細胞抽出液は、培養後の菌体を、例えばリン酸ナトリウム緩衝液を加えてホモジナイザーなどで物理的に破砕した後、遠心(15,000rpm, 10min, 4℃)し、破砕できない菌体(細胞)が存在しないように上清を回収して得ることができる。細胞膜は、上記遠心で得られたペレットとして得ることができ、また、ペレットを溶解バッファーで懸濁することにより得ることもできる。
【0053】
本発明の制限酵素または修飾酵素は、培養物をそのまま用いてもよいし、透析や硫安沈殿などの公知の方法、あるいはゲルろ過、イオン交換、アフィニティー等の各種クロマトグラフィーなどの公知の方法を単独または適宜組み合わせることによって、培養物から濃縮、精製したものを用いてもよい。この場合、酵素活性、分子量、等電点などを指標に濃縮、精製することができる。
【0054】
また、本発明においては、本発明の遺伝子または本発明のベクターから、本発明の酵素を製造することも可能である。すなわち、本発明においては、いわゆる無細胞タンパク質合成系を採用して、本発明の制限酵素または修飾酵素を産生することが可能である。
無細胞タンパク質合成系とは、細胞抽出液を用いて試験管などの人工容器内でタンパク質を合成する系である。なお、本発明において使用される無細胞タンパク質合成系には、DNAを鋳型としてRNAを合成する無細胞転写系も含まれる。
ここで、上記細胞抽出液は、真核細胞由来または原核細胞由来の抽出液、例えば、小麦胚芽、大腸菌などの抽出液を使用することができる。なお、これらの細胞抽出液は濃縮されたものであっても濃縮されていないものであってもよい。
【0055】
細胞抽出液は、例えば限外濾過、透析、ポリエチレングリコール(PEG)沈殿等によって得ることができる。さらに本発明において、無細胞タンパク質合成は、市販のキットを用いて行うこともできる。そのようなキットとしては、例えば試薬キットPROTEIOSTM(東洋紡)、TNTTM System(プロメガ)、合成装置のPG-MateTM(東洋紡)、RTS(ロシュ・ダイアグノスティクス)などが挙げられる。
上記のように無細胞タンパク質合成によって得られる本発明の制限酵素および修飾酵素は、前述のように適宜クロマトグラフィーを選択して、濃縮、精製することができる。
【0056】
以下に、本発明の具体的な態様として、本発明のRrhJ1I 制限酵素遺伝子および修飾酵素遺伝子をクローニングし、該遺伝子を発現させるための手順を例示する。
【0057】
(i)J1菌を培養し、得られる菌体より制限酵素を精製し、制限酵素認識配列を決定する工程
J1菌の制限酵素活性を調べるために、まずJ1株を培養し、培養した菌体を集めて超音波処理にて破砕した後、超遠心分離を行って上清を集め、これを活性測定用の試料とする。この試料の適当量を、基質であるλファージDNA(λ−DNA)とともに37℃でインキュベートした後、基質DNAの分解をアガロースゲル電気泳動により確認する。
当該方法を用いてJ1菌の制限酵素活性を調べると、酵素活性は、λ−DNAを13Kbの大きさに切断する活性として検出される。
【0058】
この工程において、種々のプラスミドを基質DNAとして使用してJ1菌制限酵素による切断パターンを解析すると、J1菌制限酵素の認識配列をGCCGGCに決定することができる。
【0059】
(ii)J1菌制限酵素と同じ配列を切断するイソチゾマーに対応する修飾酵素遺伝子からプライマーを設計し、J1株の染色体DNAを鋳型にPCRを行う工程
【0060】
J1菌制限酵素の複数のイソチゾマーに対応する各修飾酵素の間には共通配列部分が存在する場合がある。例えば、GCCGGCを切断するJ1菌制限酵素のイソチゾマーとしてNaeIやNgoMIVがあり、NaeI修飾酵素およびMgoMIV修飾酵素のアミノ酸配列には49%のホモロジーが認められる。次に、これらのアミノ酸配列から設計したDNAをプライマーに用いると、GCCGGCを切断する制限酵素に対応する修飾酵素の遺伝子をクローニングすることができる。例えば、当該方法を用いてJ1株の染色体DNAにおいて修飾酵素遺伝子をPCRで増幅すると、0.9 Kbの断片が増幅し、そのDNA配列を決定することができる。得られるDNA断片は、NaeI修飾酵素またはMgoMIV修飾酵素とのホモロジーが認められる。
【0061】
本工程によって、J1菌制限酵素のイソチゾマーに対応する修飾酵素とホモロジーを有する遺伝子断片を得ることができる。
【0062】
(iii)PCRの増幅産物のDNA配列を確認し、インバースPCR等で修飾酵素遺伝子と制限酵素遺伝子を決定する工程
この工程において、まず、修飾酵素遺伝子および制限酵素遺伝子をクローニングする。クローニングの方法としては、J1菌染色体ライブラリーからのサザンハイブリダイゼーション法やインバースPCR法を用いることができる。得られるDNA断片の塩基配列は通常の方法、たとえばジデオキシ法によって決定することができる。さらに、得られた塩基配列を解析することにより、その塩基配列中のタンパク質をコードしうる領域(オープンリーディングフレーム、ORF)の存在を推定することができる。
【0063】
例えば、修飾酵素遺伝子をクローニングするために工程(ii)で決定したDNA配列を利用し、インバースPCRを行うことができる。先ず、J1菌染色体DNAを制限酵素SacIIで切断し、ライゲーション反応を行って、環状DNAを作製する。次に、工程(ii)で決定したDNA配列の両末端付近のDNA配列をプライマーとしてPCR反応を行い、未知の配列を含むDNA断片を増幅する。最後に、増幅したDNA断片のDNA配列を解析し、修飾酵素遺伝子を決定する。このように得られた配列中には1つのORFが存在する(以下、「ORF1」(修飾酵素)ともいう)。ORF1の塩基配列から推定されるアミノ酸配列を配列番号4に示す。配列番号4のアミノ酸配列のホモロジー検索を行った結果、NaeIメチラーゼのアミノ酸配列に対して67%の相同性が認められる。
【0064】
通常、制限酵素遺伝子は修飾酵素遺伝子の近傍に存在しており、修飾酵素遺伝子が取得できれば制限酵素遺伝子の取得は可能であると考えられる。具体的には、J1菌染色体DNAを制限酵素SphIで切断し、ライゲーション反応によって環状化したDNAを作製する。次に、修飾酵素遺伝子または修飾酵素遺伝子近傍のDNA配列をプライマーとして環状DNAに対してインバースPCRを行ない、約6KbのDNA断片を増幅する。この断片のDNA配列を解析し、制限酵素遺伝子を決定することができる。このように得られた塩基配列中には2つのORFが存在しており、5'側より順に「ORF2」(HNHエンドヌクレアーゼ)、「ORF3」(制限酵素)とする。ORF3の塩基配列から推定されるアミノ酸配列を配列番号2に、ORF2の塩基配列(配列番号8)から推定されるアミノ酸配列を配列番号9に示す。配列番号2に記載のアミノ酸配列のホモロジー検索を行った結果、II型制限酵素のNaeIと47%の相同性が認められ、配列番号9に記載のアミノ酸配列のホモロジー検索を行った結果、Arthrobacter sp. FB24由来のHNHエンドヌクレアーゼと33%の相同性が認められる。
【0065】
本工程により、RrhJ1I制限酵素をコードすると推定されるORF(ORF3)、およびRrhJ1I修飾酵素をコードすると推定されるORF(ORF1)を得ることができる。
【0066】
(iv)RrhJ1I制限酵素および/またはRrhJ1I制限酵素を発現する形質転換体を作製する工程
本工程では、工程(iii)で得られたORF1および/またはORF3の塩基配列を基に、J1菌染色体DNAからPCRでRrhJ1I修飾酵素遺伝子および/またはRrhJ1I制限酵素遺伝子を含むDNA断片を増幅する。次に、増幅したDNA断片を適当なベクターに結合して大腸菌を形質転換し、RrhJ1I修飾酵素および/またはRrhJ1I制限酵素が発現する形質転換体を作製する。
【0067】
J1菌染色体DNAからPCRでRrhJ1I制限・修飾系の遺伝子を増幅する方法は、例えば工程(ii)の方法と同様に行うことができる。
制限酵素および/または修飾酵素を発現する形質転換体を作製するには、まず、得られたRrhJ1I制限・修飾系酵素の遺伝子を適当な宿主細胞、たとえばエシェリヒア・コリ、バチルス・ズブチリス(Bacillus subtilis)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、ロドコッカス菌(Rhodococcus)、放線菌、動物細胞、昆虫細胞、植物細胞等において発現できるような発現ベクターに常法に従い接続する。そして、得られた組換えベクターを宿主細胞に導入し、形質転換体を作製すればよい。
【0068】
本発明に使用する宿主は、メチル化シトシンに関わる制限系遺伝子(mcrA,B,C)、またはメチル化アデニンに関わる制限系遺伝子(mrr)の欠損株を使用するのが好ましい。大腸菌の一例としては、XL1-Blue MRF'、K802などを用いることができる。この形質転換体を培養することにより、本発明のRrhJ1I 制限酵素または本発明のRrhJ1I修飾酵素タンパク質を発現させることができる。
【0069】
なお、RrhJ1I制限酵素を発現させるための形質転換体は制限・修飾両酵素の遺伝子を保持していることが望ましい。この両遺伝子は同じベクター上に位置した状態で形質転換体に保持されていてもよいし、あるいは別々のベクターに組み込まれた状態で形質転換体に保持されていてもよい。RrhJ1I制限酵素を発現させるための形質転換体としては、例えば、RrhJ1I修飾酵素遺伝子を含むプラスミド(pMCLM、実施例7)およびRrhJ1I制限酵素遺伝子を含むプラスミド(pGErR10、実施例7)を含有する形質転換体XL1-Blue MRF'/pMCLM+pGErR10(実施例8)を挙げることができる。
一方、RrhJ1I 修飾酵素の発現には上記の形質転換体の他、修飾酵素遺伝子のみを保持する形質転換体を用いることもできる。RrhJ1I修飾酵素を発現させるための形質転換体としては、例えば、RrhJ1I修飾酵素を含むプラスミドpTJM01(実施例6)を含有する形質転換体XL1-Blue MRF'/pTJM01を挙げることができる。
【0070】
形質転換体の培養物のRrhJ1I修飾酵素活性は、RrhJ1I認識配列を含む培養物中のDNAが、RrhJ1I 制限酵素、あるいはそのイソチゾマー(例えば、NaeI制限酵素)によって切断されないことにより確認することができる。本工程で得られる形質転換体は、RrhJ1I 修飾酵素遺伝子を含む約1.2 kbのDNA断片を含むプラスミドを保持している。
【0071】
形質転換体の培養物のRrhJ1I制限酵素活性は、以下のようなイン・ビトロの方法により調べることができる。すなわち、試験する形質転換体を培養し、その培養物より菌体を集めて超音波処理にて破砕した後、超遠心分離を行って上清を集め、これを活性測定用の試料とする。この適当量を基質であるλファージDNA(λ−DNA)とともに37℃でインキュベートした後、基質DNAの分解をアガロースゲル電気泳動により確認することができる。当該方法を用いて大腸菌形質転換体のRrhJ1I 制限酵素活性を調べると、RrhJ1Iによりλ−DNAを切断したときの断片の予想サイズと同じ大きさのフラグメントに切断する活性を検出できる。
【0072】
(v)工程(iv)で修飾酵素および制限酵素活性が検出された形質転換体を培養し、該培養物からRrhJ1I修飾酵素および制限酵素を採取する工程
本工程では、工程(iv)で得られた形質転換体から制限酵素、または修飾酵素を精製する。
【0073】
形質転換体の培養物からRrhJ1I 制限酵素および修飾酵素を採取するにあたっては、たとえば培養物より菌体を集菌後、超音波破砕、超遠心分離等により酵素を抽出し、ついで除核酸法、塩析法、アフィニティクロマトグラフィー法、ゲル濾過法、イオン交換クロマトグラフィー法等を組み合わせて精製すればよい。この方法により、RrhJ1I 制限酵素および修飾酵素を大量に得ることができる。
【0074】
用いる発現系によっては、形質転換体中で発現された酵素タンパク質が不溶物[封入体(inclusionbody)]として蓄積される場合がある。この場合にはこの不溶物を回収し、穏和な変性条件、たとえば尿素等の変性剤存在下で可溶化した後に変性剤を除くことによって活性型のタンパク質を得ることができる。さらに上記のようなクロマトグラフィー操作を行って目的とする酵素タンパク質を精製することができる。
【0075】
また、上記で得られた遺伝子をプローブとしてストリンジェントな条件下でハイブリダイゼーションを行えば、得られた遺伝子(配列番号1、配列番号3)と配列は少し異なるが、同様な酵素活性を持つタンパク質をコードする遺伝子を得ることができ、これも本発明の範囲内のものである。
得られた遺伝子が目的の制限・修飾系酵素をコードする領域の全てを含まない場合には、得られた遺伝子の塩基配列をもとにしてプライマーを合成し、これを用いたPCRによって不足する領域を増幅したり、あるいは得られた遺伝子の断片をプローブとしてDNAライブラリーのスクリーニングを繰り返すことにより、全コード領域を得ることができる。
【0076】
こうして得られたRrhJ1I 制限・修飾系酵素をコードする遺伝子を含有する形質転換体を作製して該遺伝子にコードされる酵素タンパク質を発現させ、さらに発現させた酵素タンパク質を精製することができる。形質転換体の作製、酵素タンパク質の発現および精製はいずれも上記と同様の方法で行うことができる。こうして得られた酵素タンパク質はRrhJ1I 制限酵素活性、あるいはRrhJ1I 修飾酵素活性を保持しており、これらは本発明に含まれる。
【0077】
実施例
以下、実施例を用いて本発明をより詳細に説明する。以下は本発明の例示であって本発明を限定する趣旨ではない。
【実施例1】
【0078】
J1菌からの制限酵素精製
J1菌からの制限酵素の精製は以下の方法で行った。
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株をMYKG培地(0.5%ポリペプトン、0.3%バクトイーストエキス、0.3%バクトモルトエキス、1% グルコース、0.2% K2HPO4、0.2% KH2PO4、pH7.0)で、30℃にて72時間振盪培養した。
菌体を破砕用緩衝液(10 mM Tris-HCl (pH 8.0)、10 mM 2-mercaptoethanol、1 mM EDTA)に懸濁したのち、4℃で超音波破砕を行った。遠心分離(12,000 rpm、4℃、30 min)で得られた上清を無細胞抽出液とした。次に、核酸を除去するため、ストレプトマイシン硫酸塩を終濃度4%になるように無細胞抽出液に添加して、氷上で30分間静置後、遠心分離によって沈殿を除去した。得られた上清を酵素精製緩衝液(10 mM Potassium phosphate buffer(pH 7.5)、10 mM 2-mercaptoethanol、5% glycerol)で一晩透析を行った後、DEAE-Sepharose(GEヘルスケア バイオサイエンス社)、HiTrap Heparin(GEヘルスケア バイオサイエンス社)、HiTrap Q(GEヘルスケア バイオサイエンス社)、P-cellulose(ワットマン)、HiPrep Sephacryl S-200 HR(GEヘルスケア バイオサイエンス社)を用いて精製を行い、J1菌制限酵素を得た。全ての精製操作は4℃で行った。
【0079】
J1菌から得られた制限酵素の酵素活性の測定は、以下の方法で行った。市販の制限酵素反応用buffer(LまたはH、タカラバイオ)を使用した。λ-DNAを基質DNAとして0.5μg添加した10μlの反応液を用いて、37℃で反応を行った。反応終了後、反応液をアガロースゲル電気泳動に供し、エチジウムブロマイドで染色を行って切断パターンを解析した。λ-DNAを基質とした時、約13kbの大きさに切断されたバンドが確認された。
【0080】
このJ1菌制限酵素の認識配列の決定は、以下のように行った。基質DNAとして複合プラスミドpK4を使用した。プラスミドpK4はATCC1267/pK4として独立行政法人産業技術総合研究所特許微生物寄託センターに寄託されている(FERM BP-3731)(特開平5-64589 号および同5-68566 号公報参照)。J1菌制限酵素でpK4を切断した結果、約3.4Kb、約0.7Kb、約0.5Kbの断片が生じた。この切断パターンと、pK4を市販酵素で切断したパターンとを比較すると、NaeI(タカラバイオ)の切断パターンと一致することが認められた。
【0081】
次に、プラスミドpK4をJ1菌に導入したJ1組換え体(J1/pK4)から回収したpK4を、上記から得られたJ1菌制限酵素とNaeIでそれぞれ切断したところ、いずれの酵素によっても切断されなかった。よって、J1菌の有する制限酵素はGCCGGCを切断するNaeIのイソチゾマーであることがわかり、認識配列はGCCGGCであることが明らかになった。
【実施例2】
【0082】
J1菌染色体DNAの調製
ロドコッカス・ロドクロウス(Rhodococcus rhodochrous)J-1株を100mlのMYKG培地中、30℃にて72時間振盪培養した。
培養後、集菌し、集菌された菌体をSaline-EDTA溶液(0.1M EDTA、0.15M NaCl (pH8.0))4mlに懸濁した。懸濁液にリゾチーム40mgを加えて37℃で1〜2時間振盪した後、-20℃で凍結した。
【0083】
次に、10mlのTris-SDS液(1%SDS、0.1M NaCl、0.1M Tris-HCl (pH9.0))を穏やかに振盪しながら加え、さらにプロテイナーゼK(メルク社)(10 mg/ml)を10μl加えて37℃で1時間振盪した。
次に、等量のTE (10mM Tris-HCl、1mM EDTA (pH8.0))飽和フェノールを加え、撹拌後、遠心した。上層を採取し、2倍量のエタノールを加えた後、ガラス棒でDNAを巻きとり、90%、80%、70%のエタノールで順次フェノールを取り除いた。
【0084】
次に、DNAを3mlのTE緩衝液に溶解させ、リボヌクレアーゼA溶液(100℃、15分間の加熱処理済)を10μg/mlになるよう加え、37℃で30分間振盪した。さらに、プロテイナーゼKを加え37℃で30分間振盪した後、等量のTE飽和フェノールを加えて遠心し、上層と下層に分離させた。
上層についてこの操作を2回繰り返した後、同量のクロロホルム(4%イソアミルアルコール含有)を加え、同様の抽出操作を繰り返した。その後、上層に2倍量のエタノールを加え、ガラス棒でDNAを巻きとり回収し、染色体DNA標品を得た。
【実施例3】
【0085】
修飾酵素遺伝子のPCR
J1菌修飾酵素遺伝子をPCRで増幅するためのプライマーを以下の方法で設計した。
NaeIとNgoMIVはGCCGGCを切断する制限酵素である。先ず、これらの制限酵素に対応する修飾酵素のアミノ酸ホモロジー解析を行った。その結果、両酵素のN末端付近のGCQALGL(配列番号22)と、C末端付近のGNAFPPP(配列番号23)が一致していることから、このアミノ酸配列に対応するオリゴヌクレオチドJM-01(配列番号10)、JM-04(配列番号11)を合成し、プライマーに用いた。PCRは以下の条件で実施し、約0.9Kbのバンドの増幅が確認された。
【0086】
反応液組成
鋳型DNA(J1菌染色体DNA、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-01(配列番号10) 1μl
プライマーJM-04(配列番号11) 1μl
2.5mM dNTP 8μl
DMSO 10μl
滅菌水 68μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:1分の反応を30サイクル
プライマー
JM-01:GGN GGN CAR GCN CTN GGN CT(配列番号10)
JM-04:GGN GGN GGR AAN GCR TTN CC(配列番号11)
【0087】
次に、増幅された配列のダイレクトシークエンシングをPCRで使用したプライマーを用いて実施した。その結果、配列番号5に示す配列が得られ、ホモロジー検索の結果、NaeIメチラーゼとの相同性が認められた。
【実施例4】
【0088】
J1菌修飾酵素遺伝子のクローニング
J1菌修飾酵素遺伝子全長のクローニングは以下の方法で行った。
J1菌ゲノムDNAを制限酵素SacIIで分解し、切断したゲノムDNAをGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いて回収し、DNA Ligation Kit(タカラバイオ)を用いて環化した。次に、環化したゲノムDNAを鋳型に使用し、配列番号12、13のオリゴヌクレオチドをプライマーに用いて以下の条件でPCRを行い、約1Kbの断片を増幅した。
【0089】
反応液組成
鋳型DNA(環化J1ゲノムDNA) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-05(配列番号12) 1μl
プライマーJM-06(配列番号13) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:10分の反応を30サイクル
プライマー
JM-05:TTTCCAGACCTAGTGCCTGA(配列番号12)
JM-06:GGCAGTAGAAGTGGCCGGAC(配列番号13)
【0090】
また、プライマーとして、配列番号14、および15のオリゴヌクレオチド用いた以外は、上記と同様の条件でPCRを行い、約2Kbの断片を増幅した。
JM-07:ACGTAACCGAACTCGGTCAG(配列番号14)
JM-08:GTATCGGCAGATCGGCAATG(配列番号15)
【0091】
PCR産物を確認した後、それぞれの反応液からGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を使用してPCR産物を回収した。得られたPCR産物は、pGEM-T Easyベクター(プロメガ)に結合して、大腸菌XL1-Blue MRF'(Stratagene)を形質転換した。得られた組換えコロニーからプラスミドを抽出し、DNA配列の確認を行った。DNA配列を解析した結果、配列番号6に示される塩基配列が得られた。配列番号6に示される塩基配列中に、配列番号3に示す1264bpのオープンリーディングフレーム(ORF1)を見出した。このORF1のコードするアミノ酸配列は、NaeI修飾酵素に対して67%の相同性を持っていることから、ORF1は、GCCGGCを認識配列とするJ1菌修飾酵素をコードしていることが推定された。
【実施例5】
【0092】
J1菌制限酵素遺伝子のクローニング
J1菌制限酵素遺伝子のクローニングを実施例4と同様の方法で行った。
J1菌ゲノムDNAを制限酵素SphIで分解し、切断したゲノムDNAをGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いて回収し、DNA Ligation Kit(タカラバイオ)を用いて環化した。次に、環化したゲノムDNAを鋳型に使用し、配列番号16、17のオリゴヌクレオチドをプライマーに用いて実施例4と同様の条件でPCRを行い、約6Kbの断片を増幅した。
JK-01:CCGGCGCGATCAAACGGGTG(配列番号16)
JK-02:TGCTGACCATCGGGCACCTG(配列番号17)
【0093】
PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物を、pGEM-T Easyベクター(プロメガ)に結合して、得られたベクターで大腸菌XL1-Blue MRF'を形質転換した。得られた組換えコロニーからプラスミドを抽出し、DNA配列の確認を行った。DNA配列を解析した結果、配列番号7に示される塩基配列が得られた。配列番号7に示される塩基配列中には、配列番号2に示されるアミノ酸配列をコードするオープンリーディングフレーム(ORF3:配列番号1)が見出された。得られたORF3のコードするアミノ酸配列は、NaeI制限酵素に対して47%の相同性を持っていることから、ORF3は、GCCGGCを認識配列とするII型のJ1菌制限酵素をコードしていることが推定された。また、HNHエンドヌクレーアゼと33%の相同性を持つアミノ酸をコードするオープンリーディングフレーム(ORF2:配列番号8)も見出された。
【0094】
最後に、J1菌制限酵素とJ1菌修飾酵素の遺伝子を含む約3.8Kbの断片を、J1菌染色体DNAから配列番号18、19のオリゴヌクレオチドをプライマーに用いて実施例3と同様の条件で温度サイクルを:94℃:30秒、65℃:30秒および72℃:3分として30サイクル行いPCR増幅した。得られたPCR産物を、pGEM-T Easyベクター(プロメガ)に結合してpJRM01を作製した。図1はpJRM01を示す模式図であり、約3.8KbのPCR断片にORF1(J1菌修飾酵素遺伝子)、ORF2、ORF3(J1菌制限酵素遺伝子)が含まれている(図2)。
【0095】
JM-32:GGtctagaGTTGGCGATTCCTCATCGCG(配列番号18)
JM-33:CCaagcttATCGCTCGCGGGGGTGCTCCG(配列番号19)
【実施例6】
【0096】
大腸菌組換え体によるJ1菌修飾酵素の生産
J1菌修飾酵素を得るために、実施例5で得られたプラスミドpJRM01を鋳型として使用して、以下に示す反応液組成およびプライマーを用いてPCRを行った。この際、修飾酵素をPtrcプロモーター下流のNcoIサイトに結合させるため、J1菌修飾酵素の2番目のアミノ酸をセリン(TCG)からスレオニン(ACG)に置換した。
【0097】
反応液組成
鋳型DNA(組換えプラスミドpJRM01) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJM-18(配列番号20) 1μl
プライマーJM-11(配列番号21) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、65℃:30秒および72℃:1分の反応を30サイクル
プライマー
JM-18:GGGTCATGACGCGGTCCAGCTACGAG(配列番号20)
JM-11:CTCCctgCAGGCGGCGTGGAAGCCTGG(配列番号21)
【0098】
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1.3kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物を、制限酵素PagIとSse8387Iで切断した。制限酵素処理を行ったPCR産物を、0.7%アガロースゲルにおける電気泳動に供し、約1.3Kb付近のバンドを回収した。回収したPCR産物を、Ligation Kit(タカラバイオ)を用いてベクターpTrc99AのNcoI-PstI部位に連結し、プラスミドを作製した。得られたプラスミドを、pTJM01と名付けた。図3は、プラスミドpTJM01の構造を示す模式図である。
【0099】
このプラスミドでXL1-Blue MRF'(Strategene)を形質転換し、得られた組換えコロニーをLB培地(1%トリプトン、0.5%バクトイーストエキス、0.5% NaCl、100μg/mlアンピシリン、1mM IPTG)で37℃一晩培養した。培養した菌からQIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製し、NaeIで処理した。その結果、NaeIで切断されなかったことから、修飾酵素によってプラスミドが修飾されていることを確認できた。
【0100】
さらに、培養した大腸菌組換え体(XL1-Blue MRF'/pTJM01)の超音波破砕した上清をSDS-ポリアクリルアミドゲル電気泳動(SDS-PAGE)に供したところ、J1菌修飾酵素の分子量に相当する45KDaにバンドが確認できたことから、J1菌修飾酵素が大量に発現していることが確認された(図4)。図4中、Mは、分子量マーカーを示し、レーン1はXL1-Blue MRF'/pTJM01の超音波破砕液上清のSDS-PAGEの結果を示し、レーン2はXL1-Blue MRF'の超音波破砕液上清のSDS-PAGEの結果を示す。
本実施例により、RrhJ1I修飾酵素遺伝子を含むベクターを含有した本発明の大腸菌組換え体は、大量のRrhJ1I修飾酵素を発現することが明らかになった。
【実施例7】
【0101】
J1菌修飾酵素発現プラスミド、およびJ1菌制限酵素発現プラスミドの構築
(1)J1菌修飾酵素発現プラスミドpMCLMの構築
実施例2で調製したJ1菌染色体DNAを鋳型にして、以下の条件でPCRを行った。
反応液組成
鋳型DNA(J1菌染色体、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJ1-M5(配列番号24) 1μl
プライマーJ1-M6(配列番号25) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、59℃:30秒および72℃:2分の反応を30サイクル
プライマー
J1-M5:GGGAGGTATGATCGATCCGTTC(配列番号24)
J1-M6:CGGCCAGGCGTGGAAG(配列番号25)
【0102】
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1.4kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物をpGEM-T Easy vector(プロメガ)に連結し、プラスミドを作製した。得られたプラスミドはDNA配列の確認を行い、pGTEM2と名付けた。
【0103】
次に、pGTEM2に組み込まれたPCR産物をp15A複製オリジンを持つプラスミドであるpMCL200(Gene 162(1)1995, Pages 157-158)のLacプロモーター下流に挿入した。すなわち、pGTEM2とpMCL200をSpeIとEcoRIで切断して0.7%アガロースゲル電気泳動に供し、GFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)を用いてそれぞれ約1.4Kb(pGTEM2から)、約2.5Kb(pMCL200から)の断片を回収した。回収したDNAをLigation Kit(タカラバイオ)を用いて連結し、プラスミドを作製した。得られたプラスミドを、pMCLMと名付けた。図5は、プラスミドpMCLMの構造を示す模式図である。図5中、「rrhJ1IM」は、J1菌修飾酵素遺伝子を示す。
【0104】
プラスミドpMCLMがJ1菌修飾酵素を発現しているか確認するため、以下の実験を行った。pMCLMでXL1-Blue MRF'(Strategene)を形質転換し、得られた組換えコロニーをLB培地(25μg/ml クロラムフェニコール含有)で37℃一晩培養した。培養した菌からQIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製し、NaeIで処理した。その結果、NaeIで切断されなかったことから、修飾酵素によってプラスミドが修飾されていることを確認できた。
【0105】
(2)J1菌制限酵素発現プラスミドpGErR10の構築
実施例2で調製したJ1菌染色体DNAを鋳型にして、以下の条件でPCRを行った。
反応液組成
鋳型DNA(J1菌染色体、実施例2) 1μl
10×Ex Buffer(タカラバイオ) 10μl
プライマーJ1-R17(配列番号26) 1μl
プライマーJ1-R18(配列番号27) 1μl
2.5mM dNTP 8μl
滅菌水 78μl
ExTaq DNAポリメラーゼ(タカラバイオ) 1μl
総量 100μl
温度サイクル:94℃:30秒、66℃:30秒および72℃:1分の反応を30サイクル
プライマー
J1-R17:GATCCACCCGCCCACACCAC(配列番号26)
J1-R18:GGCTTCCGGCCACCGGGAAAG(配列番号27)
【0106】
PCR終了後、反応液5μlを0.7%アガロースゲルにおける電気泳動に供し、約1kbのPCR産物の検出を行った。PCR産物を確認した後、反応液からPCR産物をGFX PCR DNA band and GelBand Purification kit(GEヘルスケア バイオサイエンス社)で精製した。得られたPCR産物をpGEM-T Easy vector(プロメガ)に連結し、(1)で調製したpMCLMと共にXL1-Blue MRF'を形質転換した。得られた形質転換体をLB培地(100μg/ml アンピシリン、25μg/ml クロラムフェニコール含有)で37℃一晩培養し、QIAprep Spin Miniprep kit(キアゲン)を使用してプラスミドを調製した。得られたプラスミドはDNA配列を確認し、pGErR10と名付けた。図6は、プラスミドpGErR10の構造を示す模式図である。図6中、「rrhJ1IR」は、J1菌制限酵素遺伝子を示す。
【実施例8】
【0107】
大腸菌組換え体によるJ1菌制限酵素の生産
(1)無細胞抽出液の調製
実施例7(2)で作製したJ1菌制限酵素発現ベクターを含む大腸菌組換え体(XL1-Blue MRF'/pMCLM+pGErR10)をLB培地(100μg/ml アンピシリン、25μg/ml クロラムフェニコール含有)で37℃で16時間培養した。培養液を遠心分離によって回収し、破砕用緩衝液に懸濁したのち、4℃で10分間、超音波による破砕を行った。破砕液は遠心分離(12,000 rpm、4℃、30 min)し、得られた上清を無細胞抽出液とした。
また比較例として、以下の実験を行った。J1菌を実施例1の方法で培養し、実施例8(1)と同様の手法で調製したサンプルをJ1菌の無細胞抽出液とした。また、それぞれの無細胞抽出液のタンパク質濃度はBioRad Protein Assay Kit(バイオラド)を用いて定量した。
【0108】
(2)制限酵素活性の比較
(1)で調製した無細胞抽出液を用いて、RrhJ1I制限酵素活性を調べた。
基質としてλ−DNAをHindIIIで切断したDNA(TOYOBO:λ/HindIII digest)0.5μgを使用し、大腸菌組換え体またはJ1菌の無細胞抽出液(それぞれ40μgタンパク質)を制限酵素用L buffer(タカラバイオ)と共に加え、37℃でサンプリングしながら1時間インキュベートした。サンプリングした反応液はフェノール処理を行って、電気泳動に共した。用いた基質DNAは、RrhJ1I制限酵素で切断されると約3.0Kbのバンドが出現する。
【0109】
結果を図7に示す。図7中、「M」は分子量マーカーを示し、「PC」は実施例1で調製した精製酵素を用いた切断結果を示し、「NC」は精製酵素を添加しないで行った実験結果を示す。
電気泳動の結果より、大腸菌組換え体の無細胞抽出液(図7「E.coli」)は反応20分で約3kbのバンドが出現したが、J1菌の無細胞抽出液(図7「R.rhodochrous J1」)は反応1時間後でもバンドは出現しなかった。以上の結果より、RrhJ1I制限酵素遺伝子を含むベクターを含有した本発明の大腸菌組換え体は、J1菌より大量の制限酵素を生産していることを確認した。
【配列表フリーテキスト】
【0110】
配列番号1:RrhJ1I制限酵素の塩基配列
配列番号2:RrhJ1I制限酵素のアミノ酸配列
配列番号3:RrhJ1I修飾酵素の塩基配列
配列番号4:RrhJ1I修飾酵素のアミノ酸配列
配列番号5:実施例3で得られたDNAの塩基配列
配列番号6:実施例4で得られたDNAの塩基配列
配列番号7:実施例5で得られたDNAの塩基配列
配列番号8:ORF2の塩基配列
配列番号9:配列番号8から推定されるアミノ酸配列
配列番号10:プライマー
配列番号11:プライマー
配列番号12:プライマー
配列番号13:プライマー
配列番号14:プライマー
配列番号15:プライマー
配列番号16:プライマー
配列番号17:プライマー
配列番号18:プライマー
配列番号19:プライマー
配列番号20:プライマー
配列番号21:プライマー
配列番号22:NaeI対応修飾酵素とNgoMIV対応修飾酵素のN末端相同アミノ酸配列
配列番号23:NaeI対応修飾酵素とNgoMIV対応修飾酵素のC末端相同アミノ酸配列
配列番号24:プライマー
配列番号25:プライマー
配列番号26:プライマー
配列番号27:プライマー
配列表中、nはa、t、gまたはcを示す。
配列表中、rはgまたはaを示す。
【特許請求の範囲】
【請求項1】
以下の(A)、(B)、または(C)のタンパク質。
(A)配列番号4記載のアミノ酸配列を含むタンパク質
(B)配列番号4記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
(C)配列番号4記載のアミノ酸配列と相同性が90%以上のアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
【請求項2】
請求項1記載のタンパク質をコードする遺伝子。
【請求項3】
以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号3記載の塩基配列からなるDNA
(b)配列番号3記載の塩基配列からなるDNAと同一性が90%以上で、かつRrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNA
【請求項4】
請求項2または3に記載の遺伝子を含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項5記載の形質転換体の培養物からRrhJ1I修飾酵素活性を有するタンパク質を採取することを含む、RrhJ1I修飾酵素の製造方法。
【請求項1】
以下の(A)、(B)、または(C)のタンパク質。
(A)配列番号4記載のアミノ酸配列を含むタンパク質
(B)配列番号4記載のアミノ酸配列において、1もしくは数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
(C)配列番号4記載のアミノ酸配列と相同性が90%以上のアミノ酸配列を含み、かつRrhJ1I修飾酵素活性を有するタンパク質
【請求項2】
請求項1記載のタンパク質をコードする遺伝子。
【請求項3】
以下の(a)または(b)のDNAを含む遺伝子。
(a)配列番号3記載の塩基配列からなるDNA
(b)配列番号3記載の塩基配列からなるDNAと同一性が90%以上で、かつRrhJ1I修飾酵素活性を有するタンパク質をコードする塩基配列からなるDNA
【請求項4】
請求項2または3に記載の遺伝子を含む組換えベクター。
【請求項5】
請求項4記載の組換えベクターを含む形質転換体。
【請求項6】
請求項5記載の形質転換体の培養物からRrhJ1I修飾酵素活性を有するタンパク質を採取することを含む、RrhJ1I修飾酵素の製造方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2012−157372(P2012−157372A)
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2012−123840(P2012−123840)
【出願日】平成24年5月31日(2012.5.31)
【分割の表示】特願2007−52924(P2007−52924)の分割
【原出願日】平成19年3月2日(2007.3.2)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【Fターム(参考)】
【公開日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成24年5月31日(2012.5.31)
【分割の表示】特願2007−52924(P2007−52924)の分割
【原出願日】平成19年3月2日(2007.3.2)
【出願人】(000006035)三菱レイヨン株式会社 (2,875)
【Fターム(参考)】
[ Back to top ]