
SARSコロナウイルスSタンパク質およびその使用
本発明は、SARSコロナウイルスの成熟グリコシル化スパイク(S)タンパク質の使用、Sタンパク質のフラグメント、該フラグメントの産生方法、該フラグメントのSARS感染の検出における使用、並びにSARSに罹患している患者の治療のための該フラグメントの使用または該フラグメントの相応する抗体の使用に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
(技術分野)
本発明は、SARSコロナウイルスの成熟グリコシル化スパイク(S)タンパク質の使用、Sタンパク質のフラグメント、該フラグメントの産生方法、該フラグメントのSARS感染の検出における使用、並びにSARSに罹患している患者のワクチン接種または治療のための該フラグメントの使用または該フラグメントの相応する抗体の使用に関する。
(技術背景)
2002年〜2003年において、新型肺炎、即ち、SARSと称する疾病を発症させる新たなウイルスが、中国南部から発生し、少なくとも30カ国の人々に感染した。2003年7月の発生終了時には、報告された患者およびSARSによる死亡者の世界保健機構集計数は、それぞれ、約8500人および800人以上であった。この発生は、多大な社会的および医療的費用を感染国にもたらしただけでなく、地域および国際経済に計り知れない影響を与えた。
新規のコロナウイルスは、SARSの病原因子として確立され(文献7、Drosten等、2003年;文献39、Ksiazek等、2003年)、その後、SARSコロナウイルスまたはSARS CoVと命名された。その29.6kbのゲノムにより、他のコロナウイルスに類似するレプリカーゼ、スパイク、膜、エンベロープおよびヌクレオカプシド(N)並びに幾つかの他の特異的なタンパク質をコード化する14個の読み取り枠(orf)が明らかになった(文献40、Marra等、2003年;文献41、Rota等、2003年)。SARS CoVスパイクタンパク質における困難性は、グリコールタンパク質であるという困難性であり、抗体、ワクチン並びに他の治療用、診断用および予防用手段の製造に適するであろうヒト化形の十分な該タンパク質を産生させることは困難である。ヒト化形のタンパク質は、人体内のタンパク質形に類似するタンパク質である。
【0002】
(発明の開示)
この問題に対する1つの解決法は、グリコシル化されてヒト化されるようなSARS CoVスパイクタンパク質を産生し得る系を見出すことであろう。
もう1つの解決法は、中和抗体を誘発し得るSARS CoVスパイクタンパク質のフラグメントを見出すことである。
本発明においては、本発明者等は、細胞外ドメイン全体(48〜1192のアミノ酸)に及ぶSの5種のフラグメントを大腸菌中のGST融合タンパク質として発現させ、その精製タンパク質を使用して、ウサギ中で抗体を産生させた。ウェスタンブロット分析および免疫沈降試験により、本発明者等は、これら抗体の全てが、それらのターゲット領域に対し特異性であり且つ高感受性であることを証明した。固定させたが浸透化はしていない細胞において行なった間接免疫蛍光法は、これら抗体が、細胞表面に発現するSの細胞外ドメイン上の線状エピトープを認識し得ることを示した。これらの抗体をVero E6培養物中でSARS Cov複製を抑制するそれらの能力についで試験したとき、S SEQ ID NO.4の1029〜1192のアミノ酸にターゲッティングさせた抗S 10抗体は、強い中和活性を有し、SARS CoV中和ドメインを含有するSのこの領域がウイルス進入および/または複製において極めて重要であることを示唆していた。
【0003】
本発明の1つの局面によれば、コロナウイルスの成熟グリコシル化スパイクタンパク質を提供する。
本発明のもう1つの局面によれば、下記の工程を含む、コロナウイルスの成熟グリコシル化スパイクタンパク質の産生方法を提供する:
細胞にコロナウイルスのスパイクタンパク質またはその一部をコード化する核酸を移入する工程;上記細胞内で上記スパイクタンパク質を発現させる工程;および、上記スパイクタンパク質を分離する工程。
本発明のさらなる局面は、下記の工程を含む、コロナウイルスの成熟グリコシル化スパイクタンパク質のスクリーニング方法を提供する:
スパイクタンパク質を分離する工程;Endo-Hを有する上記分離スパイクタンパク質を免疫沈降させる工程;および、成熟グリコシル化スパイクタンパク質である残存スパイクタンパク質を検出する工程。
本発明のもう1つの局面は、コロナウイルスの成熟グリコシル化スパイクタンパク質またはその一部に対する抗体を提供する。
以下の説明は、本発明の上記局面に関連する。
1つの実施態様においては、好ましくは、上記コロナウイルスは、SARSコロナウイルスである。好ましくは、上記コロナウイルスはSARSコロナウイルス株、2774である。
もう1つの実施態様においては、好ましくは、上記成熟グリコシル化スパイクタンパク質は、膜貫通ドメイン(TMD)を含有する。
もう1つの実施態様においては、好ましくは、上記成熟グリコシル化スパイクタンパク質は、210KDaタンパク質である。
もう1つの実施態様においては、好ましくは、上記細胞は、肺細胞系A549である。
もう1つの実施態様においては、好ましくは、上記抗体は、SARSコロナウイルス感染の免疫検出において使用する。
もう1つの実施態様においては、好ましくは、上記スパイクタンパク質または上記抗体は、ワクチンの製造において使用する。
【0004】
本発明のさらなる局面によれば、本発明は、SARSコロナウイルスのSタンパク質(SEQ ID NO.2)のペプチドまたはタンパク質フラグメントを提供し、該フラグメントは、SARSコロナウイルスのS遺伝子由来のアミノ酸数1055〜1192の配列(SEQ ID NO.5)、或いは上記S遺伝子のアミノ酸数1029〜1192の配列(SEQ ID NO.4)を含む。好ましくは、上記ペプチドまたはタンパク質は、コロナウイルスSタンパク質のHR2ヘプタッド領域を有する。上記ペプチドまたはタンパク質は、後述するようなS 10(SEQ ID NO.4)、S 11(SEQ ID NO.6)、S 17(SEQ ID NO.7)、S 18(SEQ ID NO.8)、S 19(SEQ ID NO.9)またはS 20(SEQ ID NO.10)であり得る。
また、本発明は、下記の工程を含む、コロナウイルスのSタンパク質のフラグメントの産生方法を提供する:
a) 細胞に、SARSコロナウイルスのS遺伝子由来のアミノ酸数1055〜1192の配列(SEQ ID NO.5)を含むコロナウイルスのSタンパク質のフラグメントをコード化する核酸を移入する工程(該核酸は細胞内でのその発現を誘導し得る調節配列と操作的に関連する);
b) 上記タンパク質フラグメントを上記細胞内で発現させる工程;および、
c) 上記タンパク質フラグメントを分離する工程。
さらに、本発明は、SARSコロナウイルスのSEQ ID NO.2のペプチドまたはタンパク質フラグメントに対する抗体を提供し、該フラグメントは、SEQ ID NO.5、好ましくはSEQ ID NO.4を含む。好ましくは、上記ペプチドまたはタンパク質は、コロナウイルスSタンパク質のHR2ヘプタッド領域を含む。さらに詳細には、本発明は、ペプチドまたはタンパク質S 10(SEQ ID NO.4)、S 11(SEQ ID NO.6)、S 17(SEQ ID NO.7)、S 18(SEQ ID NO.8)、S 19(SEQ ID NO.9)またはS 20(SEQ ID NO.10)に対する抗体を提供する。
上記抗体は、上記抗体を患者から収集した細胞の少なくとも一部に適用する工程を含む、患者のSARSコロナウイルス感染の検出方法において使用する。上記抗体を含有するSARSコロナウイルス検出用の関連キットも、本発明によって提供される。
最後に、本発明は、患者に上述したようなペプチドまたはタンパク質或いはそのようなペプチドまたはタンパク質の抗体を投与することを含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法を提供する。例えば、有効量の上記ペプチドまたはタンパク質或いはそのようなペプチドまたはタンパク質の抗体を含むワクチンが本発明によって提供される。
【0005】
(発明を実施するための最良の形態)
コロナウイルスのSタンパク質は、Sタンパク質が宿主細胞上の細胞レセプターに結合し、さらにウイルスと細胞膜との融合に不可欠であることから、組織親和性の重要な決定因子である。SARS-CosVにおいては、S単独に対する液性応答は、SARS-CoV感染を防御するのに十分であるようである(文献14)。
上記SARSコロナウイルス株、2774のSタンパク質を、サル腎臓細胞Vero E6およびCos-7中、並びにヒト腎臓293T、肺細胞A549およびMRC-5中で、ワクチニア-T7発現系において発現させた。Sタンパク質は、ウサギ、ウマおよびマウス内で産生させたSに対するポリ-およびモノ-クロナール抗体を使用したときに、免疫沈降法(IP)、ウェスタンブロット法(WB)、免疫蛍光法(IF)によって検出した。これらの抗体は、Sの細胞外ドメイン全体に及ぶ種々の領域を認識する。本発明者等は、パルスチェイス試験において、Sの200kDのコアグリコシル化形が210kDの完全グリコシル化Endo-H抵抗性形にプロセシングされることを見出した。さらにまた、1群のC-末端切断および内部欠失構築物を発現させたとき、膜貫通ドメイン(TMD)を有していないSポリペプチドが、TMDを有するSポリペプチドとは対照的に、Endo-H感受性であり、トランスゴルジに輸送されず、細胞表面上で発現しないことを見出した。また、本発明者等は、2種の宿主タンパク質(ぞれぞれ、62kDおよび64kD)が、試験したいずれの細胞系においても、動物内で産生させた抗体および回復患者由来の抗体によってSと共免疫沈降することも見出した。Sのアミノ酸残基31〜200は、この領域の欠失が特異結合を完全に消滅させるので、これら2種のタンパク質と相互作用するドメインであると同定した。これら2種のタンパク質は、糖タンパク質である;何故ならば、これらタンパク質がEndo-H処理に感受性であり且つこれらタンパク質がSARSコロナウイルスのSタンパク質と相互作用するが鳥類の群3のコロナウイルスの一員であるIBVとは相互作用しない。興味あることに、Sタンパク質は、WB試験において検出した移入細胞中で少なくとも2個の片として開裂するものの、通常の開裂部位は、配列比較によっては予測されなかった。ウイルス感染細胞においても、同様の開裂産生物が観察された。Sタンパク質は、感染細胞の上清中で検出することができ、およそその半分が開裂していた。本発明者等は、最も効率的な発現および成熟が、並行移入試験における移入Cos-7およびA549細胞中で生じていたことを見出した。該プロセシングは、Sタンパク質の大多数が成熟した完全グリコシル化210kD形であり、ウイルス感染細胞の上清中の天然形のSタンパク質と同時移行するので、Cos-7細胞におけるよりもA549細胞においてより完全である。
【0006】
また、本発明においては、本発明者等は、Sタンパク質のフラグメントも取得し、細胞外ドメイン全体(48〜1192アミノ酸)に及ぶ5種の細菌発現Sフラグメントに対するウサギポリクロナール抗体を産生させた。これらのタンパク質を大腸菌内で発現させたとき、これらの抗体は、グリコシル化部位のようなS上のある種の翻訳後修飾を認識し得ないであろう。また、本発明者等の分析は、上記タンパク質をSDS変性ゲルから抽出し、SDSの存在下に溶出させたので、線状エピトープに限られているようであるが、本発明者等は、上記タンパク質がある程度の折り畳み(folding)を維持し得ることを除外できない。興味あることに、本発明者等は、上記抗体が、ウェスタンブロット分析において変性Sタンパク質(哺乳類細胞において発現させた)を認識するのみならず、細胞溶解物からの天然Sタンパク質を免疫沈降させるのにも使用し得ることを見出した(図1および2)。その結果は、これらの抗体が、折り畳みS中に存在する線状および露出エピトープおよび/またはSの構造中に見出される回旋のような可撓性領域に結合することを示唆している。重要なことに、免疫沈降試験は、これらの抗体が、非グリコシル化(140kDa)およびコアグリコシル化全長S(200kDa)の双方に、さらにまた、Endo-H抵抗性完全グリコシル化S(210kDa)に結合し得ることを示した。また、このことは、これらの抗体が細胞表面上に発現するSタンパク質を認識し得ることを示している間接免疫蛍光試験によっても裏付けされた。コアグリコシル化Sタンパク質が、成熟に達する前に、ゴルジ体に移送されて、200kDa〜210kDaの見掛け分子量の増大を生じなければならず、その後、細胞表面に輸送されることは既知である(文献11、15)。また、Sの210kDaの完全グリコシル化形は、後でビリオンに取込まれ、SARS-Cov感染Vero E6培養物の倍地中に分泌することも知られている。
【0007】
中和アッセイは、GST-S 10に対し産生させた抗体が、200 TCID50において1:364までの力価でVero E6細胞中のSARS-CoV複製を中和し得ることを示しており、この力価は、回復期患者において得られた値に匹敵し得る。累積的免疫化後に採血した血清の分析は中和力価の定常的な増大を示しており、免疫化ウサギがGST-S 10に対する特異的免疫応答を示すことを示唆していた。他の抗体のうちで抗体中和を誘発し得るようなものはなく、このことは、Sタンパク質の48〜1055アミノ酸(aa)中に中和エピトープが存在しないことを示唆し得る。しかしながら、この現象は、261〜672aaに対するヒト抗体が中和特性を有することが証明されているので、ほぼありそうもない(レセプター結合性ドメインについての下記の説明を参照されたい)。より可能性ある説明としては、この領域内の中和エピトープが濃密にグリコシル化されているかおよび/または堅固な三次構造を含有することである。従って、本発明者等の結果は、Sタンパク質の48〜1055aaが、ヒト細胞中でのグリコシル化過程を擬態し得ない非哺乳類系において発現させたペプチド系またはタンパク質をベースとするワクチンの開発において使用するにはあまり適切ではないことを示した。S 9は798〜1055aa由来であり、S 10は1029〜1192aa由来であるので、Sタンパク質SEQ ID NO.5の1055〜1192aa領域が、サブユニットワクチン開発において適し得る線状中和エピトープ(1個以上)を含有することは明らかである。
【0008】
コロナウイルス感染の第1段階では、宿主細胞へのビリオンの付着であり、SARS-CoVの場合は、ACE-2が、SARS-CoV Sタンパク質に結合する細胞レセプターとして同定されている(文献9)。Sタンパク質のN-末端中のドメイン、即ち、およそ300〜510のアミノ酸(aa)は、レセプター結合性ドメインである(文献16)。その後、ウイルスエンベロープの脂質二分子層の宿主細胞膜との融合が生じ、この過程もSタンパク質が介在する(文献4)。コロナウイルスSタンパク質は、群Iのウイルス融合タンパク質であり、2つのヘプタッド繰返し領域(HR1およびHR2)を含有し、S2ドメインまたはC-末端ドメインにおいて見出される。これらのドメインは、Sのオリゴマー構造を決定する重要な役割を果たし、従って、ウイルスと細胞膜間の融合を介在することを前提とする(文献4)。SARS-CoVにおいては、HR2は膜貫通アンカー(1148〜1193aa)近くに位置し、HR1はHR2 (900〜1005aa)の約140aa上流にある(文献14)。興味あることに、S 10 (1029〜1192aa)は、HR2領域にまで及ぶ。
生化学研究においては、SARS-CoV Sタンパク質のHR1およびHR2に相応するペプチドが、他の既知の群I融合タンパク質の典型的な構造的特徴を有する逆平行6螺旋束中に結合し得ることが証明されており、SARS-CoVが果たす膜融合および細胞侵入メカニズムが、MHVのような他のコロナウイルスにおけるメカニズムに類似することを示唆している(文献1)。全長Sタンパク質においては、上記HR1-HR2構造は、HR1のN-末端近くにあると予想される融合ペプチドを膜貫通ドメインの極めて近くに接近させ、このことが、ウイルスと細胞膜の融合を容易にし、ウイルスの細胞への侵入を可能にしている。高親和性でもってHR2ドメインに結合することにより、抗S 10抗体が、HR1とHR2と間の相互作用を妨げ、従って、SARS-CoVと宿主細胞との融合を阻止し得ることは極めて可能性のあることであろう。
【0009】
細菌発現させたタンパク質は、大規模に製造するのに容易で且つ費用効率的であろうから、本研究において同定したS 10フラグメント(1029〜1192aa) SEQ ID NO.4は、SARS-CoV用の理想的なワクチン候補であり得る。将来の研究においては、抗S 10抗体が動物モデルにおけるSARS-CoV感染を防御し得るかどうか決定すること並びにこのS領域の膜融合に対する正確な寄与を描き出すことが重要であろう。後者については、本発明者等は、現在、GST-S 10タンパク質で免疫化したマウスから得られたモノクロナール抗体をマッピングし、その結合および中和特性を判定しているところである。最後に、組換えS 10タンパク質および抗S 10抗体を抗ウイルス治療用に開発することが潜在的には可能であろう。
コロナウイルスはプラス鎖RNAウイルスであり、そのビリオンは、当該種の全メンバーに共通している3種の膜タンパク質、即ち、スパイク(S)、膜(M)およびエンベロープ(E)を含有するエンベロープによって囲まれているヌクレオカプシドコアからなる(参考として、文献8、13を参照されたい)。Sタンパク質は、ビリオン表面上に形態学的に特徴のある突起を形成しており、宿主レセプターへの結合および膜融合を介在する。上記MおよびEタンパク質はウイルス集合にとって重要であり、一方、NはウイルスRNAパッケージングにとって重要である。
コロナウイルスのSタンパク質は、宿主免疫応答および抗体によるウイルス中和を誘発させることに関与している(文献6、14)。SARS-CoVにおいては、以前の感染がマウスモデルにおいて保護免疫をもたらし、また、新生マウスへの中和抗体の受動移行により、新生マウスが感染から保護されることが可能であろう。このことは、ネコ伝染性腹膜炎ウイルス(文献10)における場合とは異なり、再感染時または免疫血清投与後のマウスにおいてはSARS-CoV感染の増進を伴わないであろうし、従って、SARS-CoVに対してワクチン接種をすることは安全であろう。Sタンパク質をコード化するDNAワクチンは、単独で、T細胞および中和抗体応答を誘発し、マウスをSARS-CoV感染から保護し得、Sタンパク質が実際にSARS-CoV感染におけるウイルス中和に対する初期ターゲットであることを示唆している。この知見は、代理/保菌者ウイルスを使用してSを霊長類に発現させている別の研究(文献5)によっても裏付けられた。これらの研究から、Sに対する体液応答のみで、SARS-CoV感染を防御するのに十分なようである。
【0010】
不活化全菌SARS-CoVはワクチン接種において見込みのある結果を示し得るものの、エピトープ系ワクチンがあり得る病原性復帰の可能性を回避しまた製造するのに安全で且つ安価であろうことから、中和抗体を誘発し得るS中のエピトープ(1個以上)を同定することが重要である。本試験において、本発明者等は、SARS-CoV感染に対するワクチンおよび治療薬の開発に使用すべく、SARS-CoVのSタンパク質中の中和エピトープを同定した。本発明者等は、Sの種々の領域をGST融合タンパク質として発現させ、これらを使用してウサギ中で抗体を産生させた。その後、これらのポリクロナール抗体を、Sタンパク質に対するこれら抗体の特異性についてウェスタンブロット分析、免疫沈降法および免疫蛍光分析において試験し、さらにまた、Vero E6培養物中でSARS-CoV複製を抑制するこれら抗体の能力について試験した。
コロナウイルス群のメンバーについては、以前の研究により、殆どの場合、Sタンパク質は、おそらくは細胞フリン(furin)様プロテアーゼによって、2つのサブユニット、即ち、N-末端S1とC-末端S2に開裂することが証明されている。タンパク質分解開裂は、膜融合および感染性を増強し得るが、必須条件ではあり得ない(文献20〜23参照)。開裂と膜融合(または感染性)との関係は、使用する個々の株および細胞に依存しているようである(文献24)。おそらく、このことは、Sの融合活性の複数の決定因子および試験する細胞中の酵素の発現レベルの差異を反映している。SARS-CoVのSタンパク質のタンパク質分解開裂は、今日まで解明されていない。従って、SARS-CoVのSタンパク質がS1およびS2として開裂するかどうかを研究することは興味あることであろう。このタンパク質も開裂するとすれば、その場合、融合および感染性に対するその効果をこの新たに出現したコロナウイルスにおいて試験することができる。本発明者等は、SARS-CoVのSタンパク質が110kDaのN-末端S1および90kDaのC-末端S2として開裂することを、これらのN-末端S1およびC-末端S2を、感染Vero E6細胞の培地中および精製ビリオン中で、SのNおよびC-末端に対して特異性の抗体を使用して検出したときに見出した。また、200kDaの全長Sタンパク質もビリオン中に検出されたので、本発明者等は、SARS-CoVのSタンパク質は部分的に開裂すると結論付けた。相対的に豊富な開裂産生物は、S1およびS2に対する各抗体の親和性が異なるため、各ブロットを濃度計により直接スキャンすることによっては正確に算定することができなかった。これらの抗体は、それぞれ、精製ビリオンおよび大腸菌発現Sフラグメントによって産生させた。
【0011】
他のコロナウイルスにおける典型的なS1/S2開裂シグナルがSARS CoVのS配列中に存在しないという事実は、上記の知見をより興味あるものとしている。以前の研究は、RRARRまたはRRAHRモチーフはコロナウイルスSタンパク質の開裂部位の直ぐ上流に位置してことを示唆している。複数の塩基性残基の幾つかの点変異は、開裂を阻止または停止させる可能性がある(文献23、25)。おそらくは、このモチーフは、他のウイルス融合タンパク質の開裂に関与していることが証明されている哺乳類細胞のフリン様プロテアーゼに対して必要とする(文献26)。本発明者等は、哺乳類細胞系における単一Sタンパク質の発現系を開発し、次いで、各変異体を使用してSARS CoVのSの代替開裂モチーフを特定することを決定した。感染細胞中で同定したS2と同様なサイズを有する90-kDaタンパク質を、S遺伝子SEQ ID 1を移入したCos-7細胞中で検出した。この系は、Sの内部欠失変異体を使用することによって開裂シグナルを決定することを可能にする。本発明者等は、551から570の残基を除去したとき、90-kDaタンパク質は検出されなかったことを見出した。上記変異体において欠失した20個の残基の配列は、FGRDVSDFTDSVRDPKTSEIであった。上記90-kDaタンパク質は、上記S遺伝子を移入した細胞中の開裂タンパク質S2であった。上記20個のアミノ酸残基は、代替翻訳開始コドンを含有しておらず、従って、上記90-kDaタンパク質は、翻訳の内部開始に基づき得なかった。上記90-kDaタンパク質は、C-末端に対する抗体によってのみ検出可能であり、N-末端に対する抗体によっては検出し得ず、従って、上記90-kDaタンパク質は、翻訳の早期終結に由来する産生物ではあり得なかった。興味あることに、これらの20個の残基は、フリン様プロテアーゼが必要とする複数または対の塩基性残基のモチーフを含有していなかった。従って、SARS CoVのSタンパク質は、他のコロナウイルスにおいて使用される同じ酵素によってではなく異なる細胞エンドプロテアーゼによって開裂し得る。結果は、昆虫細胞中でのマウスフリンによるSの共発現がSARS CoVのS開裂を生じさせなかったという第三者による観察と一致している。また、インフルエンザAおよびセンダイ(Sendai)ウイルスの融合タンパク質のコンセンサス開裂モチーフである血液凝固因子Xa様(またはトリプシン様)プロテアーゼにおける配列モチーフGln(Glu)-X-Arg (Xは、Gly、SerまたはThrである)は存在していなかった。タンパク質分解開裂は、これらウイルスの融合タンパク質を活性化していた。
【0012】
マウス肝炎ウイルスのSタンパク質の早期の研究によれば、該Sタンパク質は、小胞体内でN-結合グルコシル化され、三量体化される(文献10)。高マンノース側鎖は、ゴルジ体への輸送中にトリミングされ、さらに修飾される。開裂は、ゴルジ体内またはゴルジ体後に生じる。SARS CoVのSタンパク質は、1255個の残基を含有し、23個の潜在的N-結合グリコシル化部位を担持する。開裂が残基551〜570において生じる場合、S1およびS2は、それぞれ、12個および11個の部位を有するであろう。S1がS2よりも約20-kDa大きいので、S1における潜在的部位の全てではないが殆どがグルコシル化において使用されるが、S2における潜在的部位の全てが使用されないと想定するのが合理的であった。この試験において、本発明者等は、細胞溶解物中で検出した開裂S1およびS2のサイズが上清および精製ビリオン中で検出した開裂S1およびS2のサイズよりも僅かに小さいことも観察した。このことは、(1) さらなる修飾が、Sの解裂後おそらくはビリオンの集合前に生じていること、および(2) Sタンパク質は、宿主細胞プロテアーゼにより細胞内で開裂し得ることを示唆し得る。
単一S遺伝子を移入したCos-7細胞においては、SARS CoVを感染させたVero E6細胞におけるのと異なり、サイズ55-および38-kDaの小N-末端産生物は検出されたが110-kDa S1は検出されなかった。これらは、S1の開裂/劣化産生物または早期終結産生物であり得る。興味あることに、64/62-kDaの他の2つのタンパク質が移入および感染細胞の双方において同定され、これら2つのN-末端タンパク質の産生は移入細胞において人工的ではないことを示唆していた。
Sの開裂とその機能(膜融合、感染性および病原性)との関係は、矛盾したままである。Sプロセシングに関する問題に対処することが依然として求められている。例えば、何故、Sの開裂は、この開裂がSの機能と関連しない場合に、多くのコロナウイルスにおいて進化的に保存されるのか? 最近の報告は、SARS CoVの取込みSタンパク質による偽ビリオン類 HIV類のトリプシン活性化が細胞-細胞膜において必要であるが、この活性化は感染性を増進しないこと示唆している(文献32)。
【0013】
材料および方法
材料
本試験におけるCos7およびVero E6は、American Type Culture Collection (米国バージニア州マナッサス)から購入した。Cos7細胞は、1gグルコース/リットル、2mM L-グルタミン、1.5g重炭酸ナトリウム/リットル、0.1mM 非不可欠アミノ酸類、0.1mgストレプトマイシン/ml、100Uのペニシリンおよび5%のウシ胎仔血清を含有するダルベッコ(Dulbecco)変性イーグル培地(HyClone社、米国ユタ州)中で、5%CO2インキュベーター内で37℃にて培養した。Vero E6細胞は、2mM L-グルタミンおよびL-アミノ酸を含有する培地199 (HyClone社、米国ユタ州)中で、5%CO2インキュベーター内で37℃にて培養した。
Sarsコロナウイルスのシンガポール株 SARS CoV 2003VA2774 (“2774”)をTan Tock Seng病院において分離し、シンガポールのEnvironmental Health Institute (EHI)の研究室において、Vero E6細胞中で増殖するよう順応化させた。Vero E6細胞中の継代3を、直接RNA抽出、逆転写およびポリメラーゼ連鎖反応(RT-PCR)、および配列分析において使用した。組換えワクチニア/T7ウイルス(VT3)を増殖させ、Vero細胞上で滴定した(該ウイルスは、鳥類伝染性気管支炎ウイルス、IBVの増殖用サブクローンである) (文献54、ShenおよびLiu、2003年)。
DNA構築物のクローニング
(a) 大腸菌中での発現用:全長S (1〜1255アミノ酸(aa))を含有するシンガポール分離物2774を使用して増幅およびRT-PCRを実施し、RT-PCRからのcDNAをS構築物のクローニング用テンプレートとして使用した。5種の構築物 S 1、S 2、S 3、S 9、およびS 10 (表1)を、表2に示すプライマーによるPCRによって取得した。使用したプライマーは、全てResearch Biolabs社またはProligo Pte社(シンガポール)から購入した。PCR産生物をBamHI/XhoIにより消化し、BamHI/XhoI切断pGEX4T1ベクター(Amersham Pharmacia Biotech社、スウェーデン国ウプサラ)中に連結反応させて、グルタチオンS-トランスフェラーゼ(GST)融合タンパク質の発現用プラスミドpGEX-S 1, pGEX-S 2, pGEX-S 3, pGEX-S 9 and pGEX-S 10を得た。
(b) 哺乳類細胞での発現用:T7ワクチニアウイルス系と一緒に使用して哺乳類細胞中でSを効率的に発現させ得る、全長Sを含有するpKT-Sの構築。プラスミドpKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16 (図1)を下記のようにしてクローニングした。
【0014】
表1:本試験において使用するプラスミド

【0015】
表2:本試験において使用するオリゴヌクレオチドプライマー
【0016】
プラスミド類の構築:特異性正および逆プライマーをシンガポール株2774のS遺伝子を増幅するように設計した。各PCR産生物をBamHIおよびStuIにより消化し、BamHI/EcoRV切断pKT0中に連結反応させてT7プロモーターの制御下にプラスミドpKT-Sを得た。各特異性プライマーを、それぞれ、ヌクレオチド位置21476〜25171、〜25066、〜24934、〜24415、〜24157、および〜23866からの株2774配列を増幅するように設計した。6種のRT-PCR産生物をBamHIにより消化し、T7プロモーターの制御下にBamHI/EcoRV切断pKT0中に連結反応させて、それぞれ、プラスミドpKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15、pKT-S 16およびpKT-S 22を産生させた。これらのS構築物がコード化するタンパク質のサイズは、図1aに示している。2回のPCRを、各特異性プライマーを使用して実施して、内部欠失を有するSフラグメントを産生させた。各PCRフラグメントをpKT0中にクローニングして、プラスミドpKT-S 17、pKT-S 18、pKT-S 19、pKT-S 32、pKT-S 33およびpKT-S 34を産生させた。これらの変異体は、図1aに示す位置で200または20個のアミノ酸残基の欠失を有するSタンパク質をコード化する。
感染Vero E6細胞中でのSタンパク質の分析:融合性細胞を株2774により感染多重度(m.o.i) 1で感染させ、37℃で12〜15時間インキュベートした。培地中の細胞屑片を低速遠心分離により除去した。細胞をPBSで洗浄し、PBS中に再懸濁させた。1容量の5×標準タンパク質サンプル緩衝液を4容量の細胞懸濁液即ち培養培地に添加した。各サンプルを5分間100℃に加熱し、ウェスタンブロット分析前に−20℃に保った。ウイルス精製においては、 -プロピオラクトンを感染細胞培養物に0.05%の最終濃度まで添加して、感染性を不活化させた。不活化をVero E6細胞中での処理サンプルの滴定によって試験した。ウイルスを3回の凍結/解凍により採集し、細胞屑片を10分間の5,000rpmでの遠心分離によって除去した。限外濾過を行なってウイルスを濃縮した(300,000 NMWL、Millipore社)。濃縮サンプルを、Sepharose 4B急速流カラム(Pharmacia社)に、製造者の使用説明書に従い塗布した。溶出画分を透過型電子顕微鏡によって検査した。ウイルス粒子を含有する画分をウェスタンブロットによるSタンパク質分析において使用した。
【0017】
移入Cos-7細胞中でのSタンパク質の分析:60mmのペトリ皿内の細胞の50%の融合性単分子層を、バクテリオファージT7 RNAポリメラーゼを発現する組換えワクチニアウイルスvTF7-3によりm.o.i 1で感染させた。1時間の吸着後、細胞に、製造業者の使用説明書に従いEffectene Reagents (QIAGEN社)を使用して2〜5μgのプラスミドを移入した。移入細胞を37℃で1夜インキュベートし、細胞溶解物を、1×ウェスタンブロット分析用タンパク質サンプル緩衝液中に細胞ペレットを再懸濁させることによって調製した。
大腸菌中で発現させた組換えSタンパク質の精製:プラスミドpGEX-S 1、pGEX-S 2、pGEX-S 3、pGEX-S 9およびpGEX-S 10を、個々に、BL21 (DE3)細胞中に形質転換させた。各プレートからの単一コロニーを、アンピシリン (100 g/ml)を含有するLB-寒天プレート中で37℃にて1夜増殖させた。5ミリリットルの得られた各培養物を、2リットルのアンピシリン (100 g/ml)を含有するLB培地中に接種し、37℃のシェーカー内でOD600が0.6に達するまでインキュベートした。タンパク質の発現を、1mM IPTGを使用して誘発させた。細胞を、誘導後2時間で、4℃で10分間の5,000gでの遠心分離により採集した。得られた細胞ペレットをPBS-1mM PMSF-20 g/ml DNase I中に再懸濁させ、フレンチプレスへの2回通過により溶解させた。溶解物を22,000gで30分間遠心分離した。ペレット中の不溶性タンパク質を3回洗浄し、1% Triton X-100を含有するPBS中に再懸濁させた。タンパク質を10% PAGE-SDSゲル中で分離した。GST融合タンパク質を含有するゲルストリップを切断し、タンパク質を、Mini Trans-Blot細胞(BIORAD社、米国カリフォルニア州ハーキュリーズ)を使用して溶出させた。得られた融合タンパク質を、ウェスタンブロットにおいてマウス抗-GST抗体(Santa Cruz Biotechnology者、米国カリフォルニア州サンタクルズ)を使用して検出し、その濃度をクーマシーブリリアントブルーR-250染色SDS-PAGEゲル中のBSA標準と比較することによって推定した。
【0018】
各種S構築物に対する抗体の産生:1ミリグラムの種々の抗原の各々を等容量の完全フロイントアジュバント(Sigma社、米国ミズーリ州セントルイス)と混合し、ニュージーランドシロウサギの免疫化に使用した。2羽のウサギを使用して各それぞれの抗原に対する抗体を産生させた。初期免疫化の2週間後、各ウサギに3週間間隔でブースター注入を行なった。不完全フロイントアジュバント(Sigma社)をその後のブースター注入において使用した。10mlの血液を、各ウサギから、4回目、6回目、8回目、12回目、14回目および16回目の注入後の各時点で採血した。実験動物の使用についての手順は、全てAnimal Research Ethics Committee (AREC)の規定およびガイドラインに従って実施した。
哺乳類細胞中での組換えSフラグメントの発現:Cos7細胞を、ウェスタンブロット分析、免疫沈降法および免疫蛍光法のための哺乳類発現系として使用した。60mm皿中で増殖させたCos7細胞の単分子層をT7ワクチニアウイルス感染に感染多重度(m.o.i) 1で1時間供した。pKT-S、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16プラスミドによる細胞の一時的移入を、Effectene移入試薬(Qiagen社、米国カリフォルニア州バレンシア)を使用し、製造業者のプロトコールに従って実施した。
対照試験においては、細胞をT7ワクチニアウイルスにより感染させ、空ベクター、pKT0によりmock移入した。
Sタンパク質のエンドグリコシダーゼHおよびN-グリコシダーゼF処理:細胞を組換えワクチニア-T7ウイルスで感染させ、上述のようにして、プラスミドpKT-Sを移入しまたは空プラスミドをmock移入した。細胞をPBSで洗浄し、製造業者(Roche社)が推奨する溶解緩衝液中に再懸濁させた。細胞屑片を12,000rpmでの遠心分離により除去した。10μlの上清を、Endo-H (0.5単位/20μl、Roche社)を含むまたは含まない製造業者(Roche社)が推奨する10μlの緩衝液と混合し、37℃で2〜3時間インキュベートした。使用した溶解緩衝液は、製造業者(Roche社)が推奨する濃度のプロテアーゼインヒビターCompleteRカクテル錠剤および0.2mM PMSFを含有する。
【0019】
ウェスタンブロット分析:細胞溶解物を還元条件(0.2M DTTを含む1%SDS)下に1×SDS負荷緩衝液中で調製し、10%PAGEゲル中で分解させ、ニトロセルロース膜に移した。各ブロットを0.05% Tween-20を含むPBS中5%無油脂乳中でブロッキングし、ウサギ抗血清によって検証し(1:20,000希釈)、各種S融合タンパク質に対して4℃で1夜産生させた。上記膜をヤギ抗-ウサギホースラディシュペルオキシダーゼ(HRP)接合二次抗体と一緒に1:2000希釈で室温にて1時間インキュベートし、増強化学発光試薬(Pierce社、米国イリノイ州ロックフォード)により展開した。
放射能標識免疫沈降法:細胞をワクチニア-T7ウイルスで感染させ、上述のようにして、pKT-S或いはSのC-末端欠失変異体を発現するプラスミド、即ち、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15またはpKT-S16で移入処理した。pKT0でmock移入した細胞は、対照として調製した。細胞を、35S-metで1時間半標識化する前に、30分間飢餓させ、2時間追跡した。経時試験においては、細胞採集前の追跡時間は、それぞれ、0時間、1/2時間、1時間、2時間、4時間および6時間であった。細胞を、50mM Tris、1mM PMSF、1% NP40を含有する溶解緩衝液(pH 7.4)を使用して溶解させ、10分間16,000gで遠心分離した。300 lの上清を5 lのウサギ抗-GTS-S 1、2、3、9または10と一緒に1/2時間インキュベートし、その後、プロテイン-Aセファロースビーズ(Roche Diagnostics社)と一緒に1時間インキュベートした。ビーズを溶解緩衝液で3回洗浄した。20 lの1×SDS負荷緩衝液(0.2M DTTを含有する)をビーズに添加し、100℃で10分間煮沸した。各サンプルをエンドグリコシダーゼH (EndoH)酵素((Roche Diagnostics社)で処理し、該EndoH試験において37℃で2時間インキュベートする。各サンプルを7.5% SDS-PAGEゲル中で分離し、オートラヂオグラフィーによって展開する。
【0020】
間接免疫蛍光法:Permanoxスライド(Nalge Nunc International社、米国イリノイ州ネーパービル)上で増殖させたCos-7細胞を、上述のようにして、感染させ移入処理した。移入後8時間で、細胞を、4%パラホルムアルデヒドにより室温で10分間固定し、PBS+1%ウシ血清アルブミン(BSA)により30分間ブロッキングし、次いで、一次抗体(1:200)と一緒に1.5時間インキュベートし、洗浄し、その後、フルオレセインイソチオシアネート(FITC)接合二次抗体(1:200、Santa Cruz社)と一緒に1時間インキュベートした。インキュベーションおよび洗浄は、全て室温で実施した。各スライドをFluorescence Mounting Medium (DakoCytomation社)によって取付け、AxioVision Fluorescence Light Microscope (Carl Zeiss社、ドイツ)上で分析した。
中和アッセイ:ウサギ血清中の中和抗体を測定するために、本発明者等は、96ウェルプレートを使用して、血清の連続希釈によるアッセイを実施した。2×104 Vero E6細胞を96ウェルプレートの各ウェル内の200 lのMedium 199中で増殖させ、37℃でインキュベートした。ウサギ血清の培地による比1:10〜1:1280の連続希釈液を調製した。0.1mlの希釈ウサギ抗血清を、それぞれのウェルに添加する前に、0.1mlのSARS CoVと200 TCID50で室温にて1時間混合した。96ウェルプレートをCO2インキュベーター内で3〜5日間インキュベートして細胞変性効果(CPE)を観察した。CPEを有する細胞のパーセントを、各ウェルから10 lの再懸濁細胞を採取し、顕微鏡で計数することによって測定した。中和力価を算出する式は、Log (試験血清の50%中和力価) = Log (CPE%の50%よりも高い血清希釈)−Log (希釈計数)×(50%よりも高いCPE%−50%のCPE%)/(50%よりも高いCPE%−50%よりも低いCPE%)である。上記計算における逆対数は、試験血清の50%中和力価として判定する。試験は、全て2回実施する。
【0021】
結果
ワクチニア-T7発現系でのSのプロセシング:移入細胞中での発現およびプロセシングを分析するために、S遺伝子をT7プロモーターの制御下にベクター(pKT0)中にクローニングした。Cos-7細胞をワクチニア-T7組換えウイルスで感染させ、引続き、S遺伝子を含有するプラスミドを移入した。Sタンパク質発現プロフィールを、死滅させ精製したウイルス粒子によって産生させたウマ- -SARS抗体を使用して、ウェスタンブロットにより分析した。図1bに示しているように、200-、140-、110-、90-、64/62-、55-および38-kDaタンパク質を検出した(レーン2)。これらの種は、S-特異性であり、そのようなタンパク質は陰性対照細胞においては検出されなかったので(レーン1)、Sの発現およびプロセシングに基づいているようであった。200-および140-kDaタンパク質は、全長Sタンパク質(グリコシル化および未グリコシル化)に相応しており、他の小さい産生物は、おそらく開裂タンパク質および/または早期翻訳産生物であった。結果は、SARS CoVのSタンパク質が本生体外発現系において開裂し得、SARS CoV Sタンパク質のプロセシングの分析において使用し得ることを示唆していた。
SARS CoV感染細胞中での開裂産生物の検出:Sタンパク質がウイルス感染Vero E6細胞中でもプロセシングされるかどうかを調べるために、ウェスタンブロットを、細胞溶解物中およびウイルス感染培養物の培地中の双方においてS由来産生物を検出ために実施した。Sタンパク質がN-末端S1およびC-末端S2として開裂するならば、これら産生物は、細胞溶解物中で検出されるべきである;さらに、2つの開裂タンパク質がビリオン中に集合するのであれば、これらタンパク質は、感染細胞の培地中で検出されるべきである。この目的において、N-末端領域およびC-末端領域のそれぞれに対して産生させた2種のウサギ抗-S抗体を使用した。ウサギ- -S 1はアミノ酸残基48から358の領域を認識し、ウサギ- -S 10は1029から1192の領域を認識した(図1a参照)。図2aに示すように、ウサギ- -S 1を使用した場合、200-および110-kDaの2種の特異性タンパク質を細胞溶融物中(レーン2)および培地中(レーン4)の双方で検出した。ウサギ- -S 10を使用したときは、200-および90-kDaの2種のタンパク質を細胞溶融物中(レーン6)および培地中(レーン8)の双方で検出した。結果は、(1) 110-および90-kDa種は、ウイルス感染細胞中のSのそれぞれN-およびC-末端開裂産生物S1およびS2を示し得ること、および(2) Sタンパク質は、SARS CoVにおいて完全ではないが部分的に開裂することを示唆していた。培地中の200-、110-および90-kDaタンパク質は、細胞溶解物中のそれらの対応物よりも僅かに大きいことを観察した。これらの産生物は、開裂後でビリオン中への集合前にさらなる変性を受け得るようである。
【0022】
上記の観察を確認するために、精製ウイルス粒子を、上記の同じ抗体を使用するウェスタンブロットにおいて使用した。図2bに示すように、S1サブユニットはウサギ- -S 1を使用して検出し(レーン1)、S2サブユニットはウサギ- -S 10を使用して検出した(レーン2)。S1およびS2サブユニットの双方は、ウマ- -SARS抗体によって検出した(レーン3)。また、全長Sタンパク質は、使用した3種の抗体の全てによって検出した(レーン1、2および3)。S、S1およびS2のサイズは、感染細胞の培地中で検出した成熟形と同じであった。結果は、開裂産生物S1およびS2並びに全長Sがビリオン中に集合していることを明らかに示していた。
移入細胞中でも検出された62/64-kDaタンパク質(図1b参照)は、感染細胞の溶解物中で同定されたが(図2a、レーン2)、培養培地(レーン4)および精製ビリオン(図2b、レーン1および3)においては同定されなかった。移入細胞中で検出された140-、55−および38kDaタンパク質(図1bに示す)は、感染細胞および精製ウイルス粒子中では見出せず、これらタンパク質は、ビリオン中には集合しないことを示唆していた。140-kDaタンパク質は、Sの未グリコシル化形であり得る。140-、55-および38-kDaタンパク質は、S移入細胞においてのみ産生していたようであった。
移入細胞中でのSの開裂産生物の検出:コロナウイルスにおける開裂部位の直ぐ上流の典型的モチーフ(RRAR/HR)をSARS CoVのS中に見出せなかったので、S1およびS2サブユニット間に代替的開裂部位を有する配列を実験的に判定する必要があった。この目的において、6種のC-末端切断S構築物(S 11〜S 16、図1a参照)をCos-7細胞中で発現させた。Sタンパク質が開裂するとすれば、C-末端産生物は、C-末端欠失を有する変異体を使用するとき、より小さくなるであろう。一方、N-末端産生物のサイズは、これらの変異体を使用するとき、同じままであろう。種々の領域およびドメインに対して産生させたさらなるポリクローナルおよびモノクローナル抗体を、全ての潜在的開裂および/または他のS由来産生物を検出するために使用した。ウサギ- -S 2、ウサギ- -S 3およびウサギ- -S 9は、残基362〜790 (S 2)、168〜461 (S 3)および798〜1055 (S 9)内の領域を認識した(図1a参照)。モノクローナル抗体MAb12-6およびMAbC1は、それぞれ、Sの残基281〜300および631〜650を含有するドメインを認識した。
【0023】
(1) C-末端産生物の検出:ウェスタンブロット分析を、全長SおよびS 11〜S 16構築物を移入したCos-7細胞中のSタンパク質を検出するために実施した。図3aに示すように、200-、140-および90-kDaバンドを、全長Sタンパク質を発現する細胞において、ウサギ -S 2、-S 9、-S 10およびMAbC1抗体によって精査したときに検出した(レーン1および9、上方および下方パネル)。200-および140-kDaバンドよりも次第に小さくなる産生物をC-末端欠失変異体(S 11〜S 16)を発現する細胞において上記と同じ4種の抗体によって検出した(図3a、レーン2〜7および10〜15)。これらのうち、ウサギ- -S 2およびモノクローナルMAbC1は、欠失構築物各々からの相応する産生物を検出し得ていた(図3a、上方パネル、レーン2〜7および10〜15)。ウサギ- -S 9およびウサギ- -S 10のみは、予測したとおりのより大きい切断変異体に由来する産生物を認識していた(図3a、下方パネル) (また、図1aも参照されたい)。これら産生物の見掛け分子量は、これら産生物の算出分子量およびグリコシル化後のこれら産生物の予測分子量変化と一致しており、これら産生物が、各構築物によってコード化された産生物のそれぞれの全長グリコシル化および未グリコシル化形であることを示唆していた。
【0024】
同様に、90-kDaタンパク質よりも次第に小さくなる産生物を、C-末端欠失構築物(S 11〜 16)から、同じC-末端特異性抗体によって検出した(図3a、レーン1〜7および9〜15、上方パネルのレーン1〜7における矢印によって示されているように)。これら産生物の移行パターンおよびこれら産生物の見掛け分子量は、これら産生物がSタンパク質のC-末端開裂産生物を示すことを強力に示唆していた。S 16タンパク質はN-末端797アミノ酸残基を含有し、そのC-末端開裂産生物はサイズ的に30-kDaである(レーン7および15、上方パネル)。モノクローナル抗体MAbC1を使用したとき、S 16に由来する30-kDa産生物を検出しており(レーン15、上方パネル)、開裂部位がこの抗体の結合配列(残基631〜650)の上流に存在することを示していた。
55-kDaタンパク質(レーン1〜7、上方パネル)が、大きいS1産生物の代りに、ウサギ- -S 2によって検出されたことが注目された。該55-kDaタンパク質は、モノクローナル抗体MAbC1によっては検出されなかった(レーン9〜15、上方パネル)。また、もう1群の次第に小さくなり比較的弱い産生物が、ウサギ- -S 2およびMAbC1を使用したときに検出されたことも注目された(レーン1〜7および9〜15、上方パネル、上方パネルのレーン9〜15における矢印によって示されているように)。これら産生物は、中間産生物であり得、S1は、本試験において使用する条件下において、移入細胞中で分解または開裂し得ることを示唆している。このことは、110-kDa S1が、残基362〜790に及び且つS1とS2の双方を認識する可能性を有するウサギ- -S 2によって何故検出されなかったかを説明している。
【0025】
(2) N-末端産生物の検出:ウサギ- -S 1、ウサギ- -S 3およびモノクローナル抗体MAb12-6を、上述した同じ構築物を移入した細胞中のSのN-末端産生物を検出するために使用した。全長構築物(図3b、レーン1および9)においては、200-、140-、64-/62-、55-および38-kDaバンドが上記3種の抗体を使用したときに観察された(矢印で示している)。140-kDaタンパク質(および200-kDaタンパク質)は、これらのN-末端特異性抗体によっても検出されたことを観察し、該タンパク質がSの全長未グリコシル化形であることを確認した。C-末端切断S構築物S 11〜 16においては、各レーンの上部の2つのバンドは、各切断構築物によってコード化されたグリコシル化または未グリコシル化の全長タンパク質であった(図3b、レーン2〜7および9〜15)。64-/62-、55-および38-kDaタンパク質は、全ての構築物を発現する細胞において、これらのN-末端特異性抗体によって検出された(図3b、レーン1〜7および9〜15)。64-/62-、55-および38-kDaタンパク質のサイズは、C-末端切断変異体を使用したとき同じままであり、これらのタンパク質がSのN-末端産生物であったことを確認した。
上記ウマ-α-SARS抗体を使用して上記同じ細胞溶解物中のタンパク質を検出した。この抗体は、N-およびC-末端産生物の双方を検出し得る。上述したN-およびC-末端タンパク質の全てが検出されており(図3b、レーン9〜15、下方パネル)、上述したタンパク質の全てが特異的にS由来であったことを確認した。
110-kDa S1は、使用した条件下においては、移入細胞中で検出されなかった(或いは、ウマ -SARSにより弱いバンドとして検出された、図1b参照)。代りに、より小さいN-末端産生物が検出された。結果は、S1がこの発現系においては急速に分解されることを強力に示唆していた。また、このことは、64/62-kDaタンパク質が感染細胞においても検出されているので、ウイルス感染細胞においても生じ得る(図2a、レーン2参照)。とは言え、この発現およびプロセシング系は、SARS CoV Sタンパク質における開裂部位配列をマッピングすることを可能にしている。
【0026】
開裂部位を有するアミノ酸配列のマッピング:全長Sタンパク質においては、C-末端開裂産生物は、サイズとして90-kDaであった。最も短い切断構築物S 16 (残基1〜797をコード化している)においては、開裂産生物は、サイズとして30-kDaであった。モノクローナル抗体MAbC1は、631から650までの残基を認識した。従って、S1およびS2の推定開裂部位は、アミノ酸残基600辺りの領域に位置し得る。Sの開裂に関与する配列を判定するために、S遺伝子の数種の内部欠失変異体を構築した。欠失配列が開裂部位を含有するならば、90-kDaは、産生されないであろう。先ず、残基601〜800 (S 17)、401〜600 (S 18)および201〜400 (S 19)からの欠失を担持する3種の変異体を発現させた。これらS特異性タンパク質を、ウサギ- -S 10を使用して、ウェスタンブロットにより分析した。図4に示すように、S 17を移入した細胞においては、90-kDaよりも200残基短いタンパク質が観察され(レーン2)、開裂部位は、残基600の上流にあり得ることを示していた。残基401〜600の欠失(S 18)は、90-kDa開裂産生物を実際に消滅させていた(レーン3)。残基201〜400 (S 18)を欠失させた場合、90-kDa産生物は、予測通り依然として産生していた(レーン4)。事実、90-kDa S2よりも僅かに大きい産生物を、S-18を移入した細胞中で検出し、S1がおそらくはS1の中央でエンドプロテアーゼによって開裂されるさらなる証拠を提供していた。このことは、C-末端に対する抗体が1群の中間開裂産生物を検出したという観察と一致していた(図3a参照、上方パネルのレーン8〜15の矢印で示している)。
【0027】
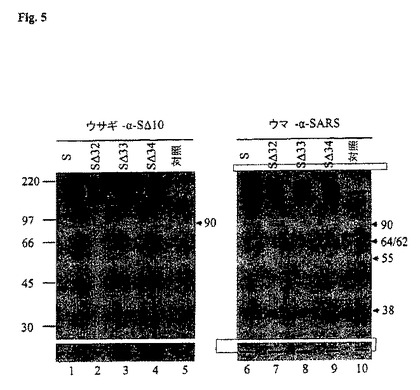
開裂部位を有する配列をさらに明確にするため、3種のさらなる欠失変異体を発現させ、分析した。これらの構築物は、残基531〜550 (S 32)、551〜570 (S 33)および571〜590 (S 34)からの欠失を有するSタンパク質をコード化していた。ウサギ- -S 10を使用してのウェスタンブロット分析(図4b)は、残基551〜579 (S 33)を除去したとき、90-kDaタンパク質の産生は消滅していたことを示していた(レーン3)。531〜550 (S 32)および571〜590 (S 34)の欠失は、90-kDa開裂産生物の検出に影響していなかった(レーン2および4)。このことを、ウマ- -SARSを使用して同じ細胞溶解物を分析したときに確認した(図5、レーン6〜10)。90-kDaおよび55-kDaタンパク質の双方がS 33を発現させたときのみ検出されず、55-kDaタンパク質はS1の解裂または分解産生物であることを示していることが注目された。S 33中で欠失させた20残基の配列は、FGRDVSDFTDSVRDPKTSEIであった。SARS CoV Sのアミノ酸配列に対するBLAST調査によれば、残基551〜570は、現在までに公表された分離株のうちで十分に保存されていることが証明された(データは示していない)。他のコロナウイルスのSタンパク質と異なり、RRARRまたはRRAHR (または、対の塩基性残基でさえもの)モチーフは、この小領域内には存在しない。結果は、SARS CoV Sタンパク質の開裂部位が他のコロナウイルスの開裂部位と異なっていることおよびフリン様プロテアーゼよりもむしろ他の細胞プロテアーゼが含まれていることを強力に示唆している。
【0028】
エンドグリコシダーゼ-H (Endo-H)およびN−グリコシダーゼF (N-gly-F)処理によるSタンパク質のさらなるグリコシル化試験:図6に示すように、移入細胞中のSタンパク質をN-gly-F (レーン1〜4)およびEndo-H (レーン5〜8)によって処理した。結果は、200-kDaタンパク質が、これらの2つの酵素による処理に対して感受性であったので、Sのグリコシル化形であることを示していた。140-kDaタンパク質は、これら酵素による処理が該タンパク質に対して効果を有してなかったので、未グリコシル化の全長Sであった(図6、レーン1、2、5および6)。興味があるのは、200-kDaタンパク質よりも僅かに大きい上部バンドがEndo-H抵抗性であることを見出したことであった(レーン5および6)。結果は、200-kDaタンパク質が、感染細胞培養物において示した(本試験の早期で説明した)ように、おそらくはビリオン中に取込まれる前に、さらに変性されていることを裏付けている。また、結果は、64/62-および38-kDaタンパク質もN-末端産生物のグリコシル化形であることも証明していた。90-および55-kDaタンパク質がグリコシル化形またはEndo-H抵抗性形であるかどうかを検証することは興味あることであろう。しかしながら、開裂の効率は移入細胞においてはるかに低いので、これらのタンパク質は、本試験において使用した条件下では検出し得なかった。
【0029】
Sタンパク質上の種々の領域に対するウサギ抗体の産生:哺乳類発現天然S構築物を使用した以前の研究によれば、中和すべきS1領域内のSの270〜510、548〜567および607〜627(文献11、30)、並びにS2領域内の803〜828をターゲットする抗体も中和特性を有することを知見していることが判明している(文献13)。しかしながら、細菌発現系における、グリコシル化のような翻訳後修飾の欠落は、中和抗体を発現する種々の潜在的部位を示し得る。細菌発現SARS CoV Sタンパク質上の中和領域を試験するために、本発明者等は、細胞外ドメイン全体に及ぶSの5種のDNA構築物をクローニングした(図1)。本発明者等は、S1領域をカバーする2種のフラグメント(S 1、S 3)、S2領域をカバーする2種のフラグメント(S 9、S 10)およびS1とS2の双方の部分をカバーする1種のフラグメント(S 2)をクローニングした。構築物S 9およびS 10は、それぞれ、ヘプタッド繰返し1 (HR1)およびヘプタッド繰返し2 (HR2)領域を含有する。構築物S 1、S 2、S 3、S 9およびS 10用の抗原を、細菌発現系を使用して産生させた。これらの抗原をウサギに注入してそれらそれぞれのターゲッティング領域に対するポリクローナル抗体を産生させた。2羽のウサギを使用してそれぞれの抗原に対する抗体を産生させた。初期免疫化の2週間後、各ウサギに3週間間隔でブースター注入をおこなった。10mlの血液を、各ウサギから、4回目、6回目、8回目、12回目、14回目および16回目の注入後の各時点で採血した。これらのポリクローナル抗体を、ウェスタンブロット分析、免疫沈降法および免疫蛍光法によって特性決定した。
ウェスタンブロットにおいてのSARS CoV Sタンパク質に対するウサギ抗体の特異性:哺乳類細胞中で発現させた全長Sタンパク質に対するウサギ抗体の特異性を、ウェスタンブロット分析により測定する。想定したように、SARS CoV感染から回復した患者(P6)の血清由来の抗-S抗体(文献26は)は、全長Sタンパク質の2つの主要バンド、即ち、140kDa未グリコシル化形および200kDaグリコシル化形を、pKT-Sを移入したCos-7細胞の溶解物中で検出し得た(図2a)。これらのバンドは、陰性対照においては検出されなかったので、特異性Sバンドであった。本発明者等は、5 S組換えタンパク質に対して産生させた抗体を使用して、同様な結果を観察した(図2b〜f)。このことは、各種S構築物に対して産生させた全ての抗体が、線状化Sタンパク質の種々の領域をターゲッティングすることによって、変性条件においてSに特異的に結合し得たことを示している。従って、これらの抗体は、変性全長SARS CoV Sタンパク質に対して特異性である。
【0030】
免疫沈降法においての天然形のSARS CoVの検出:天然Sタンパク質に対する抗体の特異性およびこれら抗体のそれぞれのSタンパク質上のターゲット領域を判定するために、本発明者等は、種々の抗体を使用して、免疫沈降試験を実施した。T7得ワクチニアウイルスを感染させ、pKT-SおよびC-末端欠失変異体、即ち、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16を移入したCos-7細胞の溶解物を、P6血清並びにS 1、S 2、S 3、S 9およびS 10に対して産生させた5種の抗体によって免疫沈降させた。SARS CoV Sタンパク質およびC-末端欠失組換えタンパク質は、P6血清による免疫沈降によって検出し得る(図3f)。コアグリコシル化Sタンパク質(200kDa)および完全グリコシル化Sタンパク質(210kDa)も、各種組換えSタンパク質に対して産生させた抗体を使用したときに明らかに検出した(図3a〜e、レーン1)。領域S 1、S 2およびS 3をターゲットする抗体は、それぞれ、全長Sおよび全てのC-欠失変異体を検出し得る(図3a〜c)。領域S 9ををターゲットする抗体は、全長SおよびpKT-S 11〜pKT-S 15を検出し得る。pKT-S 16レーンにおいては、pKT-S 16がS 9領域を含んでいなかったので、特異性バンドは検出されなかった(図3d)。S 10対してターゲットする抗体は、pKT-S 11〜pKT-S 13によって発現させたタンパク質バンドを検出するだけである(図3e)。pKT-S 14〜pKT-S 16によって発現させた領域は、S 10領域を含んでなく、従って、ウサギ抗-S 10は、これらのレーンにおいていずれのバンドも検出せず、これらの抗体がこれら抗体のターゲット領域に対し高度に特異性であることを示していた。Sタンパク質の生来の構造は免疫沈降における溶解緩衝液中で保持されており、結果は、天然タンパク質が、上記5種の組換えタンパク質に対して産生させた5種の抗体全部が認識し得るエピトープ(1個以上)を有することを示していた。
【0031】
200kDa Sタンパク質の210kDa EndoH抵抗性形への成熟:2つのS特異性バンドを免疫沈降試験において観察した。上記2つのバンド間の関係を解明するために、本発明者等は、経時試験を実施した。pKT-Sを移入し、35S-metで標識し、さらにウサギ抗-S 10により免疫沈降させたCos-7細胞を、1/2時間、1時間、2時間、4時間および6時間で採集した。結果は、200kDaバンドの漸次的減少を伴っての210kDaバンドの次第の増大を示した(図4、レーン1〜8)。EndoH試験を行なって各バンドがEndoH処理に感受性であったどうかを解明する。Cos-7細胞をT7ワクチニアウイルス感染に供し、pKT-Sを移入し、EndoH酵素で処理したが、対照細胞はEndoHで処理しなかった。結果は、210kDaバンドがEndoH抵抗性であり、200kDaバンドがEndoH感受性であることを示していた(図4、レーン9、10)。従って、結果は、200kDaバンドの210kDaバンドへの成熟を実証していた。
Cos-7細胞表面上でのSARS CoV Sタンパク質の検出:産生させた各抗体が天然形のSタンパク質を認識し得るさらなる証拠を提供するために、本発明者等は、免疫蛍光試験を行なった。間接免疫蛍光試験において使用したCos-7細胞は、非浸透性であった。上述したようなT7ワクチニアウイルスによる感染およびpKT-Sによる移入後に、十分な時間(移入後8時間)で、Sタンパク質を発現させ、細胞表面に移送させたところ、これらのタンパク質は、各種Sタンパク質構築物に対して産生させた抗体のいずれを使用しても明らかに検出し得た(図5)。緑色蛍光は、Cos-7細胞に結合している各種一次抗体の位置を示していた。Sタンパク質は、Cos-7細胞により、それらの生来の構造において発現し、プロセシングされていた。S 1、S 2、S 3、S 9およびS 10に対する抗体の細胞表面上のSタンパク質への結合は、これらの抗体がSARS CoV Sタンパク質の生来構造に対して特異性であり感受性であるというさらなる証拠を提供している。
S2内の領域は、中和活性を顕在化させる:各種Sタンパク質を注入したウサギ由来の血清の全てを、各採血後の中和活性について試験した。pGEX-S 1、S 2、S 3およびS 9を注入したウサギにおける結果は、陰性応答を示した。pGEX-S 10を注入したウサギ由来の血清は、4回目注入後に中和活性を示した。SARS CoVを200 TCID50で使用した初期試験は、8回目の注入後の血清採血から始まるウサギ抗-S 10採血の全てにおいて高力価(1:364)の中和抗体を示した(表3)。16回目の注入は最後のブースター注入であり、ウサギは、この段階で死亡させた。上記の結果を確認するため、pGEX-S 10による4回目、6回目、8回目、12回目、14回目および16回目の注入後の血清サンプルを、SARS CoVを1000 TCID50で使用する中和試験に送った(表4)。中和活性は、1000 TCID50において1:189.2ほど高く、SARS患者において検出した力価よりも高いとはいえないまでも、それに匹敵していることが判明した。SARS患者における100 TCID50での抗体応答は、210日間に亘って1:150〜1:475の範囲である(文献35)。この結果は、S 10がSARS CoVに対する中和抗体の産生を刺激し得る領域である強力な証拠を提供している。
【0032】
表3:200 TCID50での中和試験
表4:1000 TCID50での中和試験
【0033】
SEQ ID NO.1:SARS CoV、2774株のクローン12のスパイク(S)遺伝子の全長ヌクレオチド配列。核酸1から3765までの線状のRNA。
生物体:SARSコロナウイルス2774株ウイルス;ssRNAプラス鎖ウイルス、DNA段階無し;ニドウイルス目;コロナウイルス科;コロナウイルス。
SEQ ID NO.2:SARS-CoV、2774株のクローン12のスパイク(S)遺伝子の全長アミノ酸配列。アミノ酸1〜1255。
SEQ ID NO.3:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 10フラグメントのヌクレオチド配列。核酸3087〜3581。
SEQ ID NO.4:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 10フラグメントのアミノ酸配列。アミノ酸1029〜1192。
SEQ ID NO.5:SARS-CoV、2774株のクローン12のスパイク(S)遺伝子の中和フラグメントのアミノ酸配列。アミノ酸1055〜1192。
SEQ ID NO.6:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 11フラグメントのアミノ酸配列。アミノ酸1〜1232。
SEQ ID NO.7:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 17フラグメントのアミノ酸配列。601から800までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.8:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 18フラグメントのアミノ酸配列。401から600までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.9:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 19フラグメントのアミノ酸配列。201から400までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.10:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 20フラグメントのアミノ酸配列。30から200までの欠失を有するアミノ酸1〜1255。
【0034】
2003年12月10日に出願された米国仮特許出願第60/528,596号を含む本明細書において引用した文献は、すべて、その全体を参考として本明細書に合体させる。
【0035】
参考文献
【0036】
【0037】
【0038】
【0039】
【0040】
【図面の簡単な説明】
【0041】
【図1】Sタンパク質検出のためのウェスタンブロット分析:Cos7細胞由来の溶解物に、プラスミドpKT-Sを移入する(レーン1、3、5、7、9、11)。Cos7細胞由来の溶解物に、陰性対照としての挿入物を含まないプラスミドを移入する(レーン2、4、6、8、10、12)。BenchMark Prestained Protein Ladder(Invitrogen社)を右側のマーカーとして使用する。200kDaおよび140kDaバンドは、特異性Sタンパク質バンドである。
【図2】放射線標識免疫沈降法によって測定した各抗体の特異性:Cos7細胞の溶解物に、それぞれS、S 11、S 12、S 13、S 14、S 15およびS 16を移入する(レーン1、2、3、4、5、6および7)。Cos7細胞の溶解物に、陰性対照としての挿入物を含まないプラスミドを移入する(レーン8)。High-Range Rainbow Molecular Weight Markers(Amersham社)を、左側のマーカーとして使用する。
【図3】Sタンパク質のグリコシル化の経時変化:pKT-Sを移入したCos7細胞の溶解物を、それぞれ、0時間、0.5時間、1時間、2時間、4時間および6時間で採集する(レーン1、2、3、4、5、6および7)。挿入物を含まないプラスミドを移入したCos7細胞の溶解物を、陰性対照として、6時間で採集する(レーン8)。Cos7細胞の溶解物にpKT-Sを移入し、Endo-H処理する(移入後6時間)。pKT-Sを移入し、陰性対照として移入後6時間で採集したCos7細胞の溶解物。ウサギ抗-S10をSタンパク質の検出において使用した。
【図4】一時的に移入したCos7細胞の表面上で発現させたSタンパク質:Cos7細胞の表面上で発現させたSタンパク質を、それぞれ、緑色蛍光によって示すように、Rb -S 1、Rb -S 2、Rb -S 3、Rb -S 9およびRb -S 10により検出した。上記細胞の位置は、白色灯を使用して得た。蛍光は、対照試験物においては検出されなかった。
【図5】S1/S2の開裂部位を含む配列のマッピング:全長S (レーン1および6)、内部欠失突然変異体S 32 (レーン2および7)、S 33 (レーン3および8)および残基531〜550、551〜570および571〜590を欠失するS 34 (レーン4および9)を、それぞれ、Cos-7細胞中で発現させた。ウェスタンブロット分析を、ウサギ- -S 10(左側パネル)およびウマ- -SARS(右側パネル)を使用して実施した。各タンパク質を、10%PAGEゲル上で分離した。特異性タンパク質の分子量を右側に、マーカーの分子量左側にキロダルトンで示している。上記各ゲルをストリッピングし、負荷対照としてのマウス抗- -アクチンで再精査した。
【図6】N-グリコシダーゼFおよびエンドグリコシダーゼHによるS由来タンパク質の処理:全長SをCos-7細胞中で発現させた。細胞を溶解緩衝液中に再懸濁させた。各サンプルを、(a)N-グリコシダーゼF (N-gly-F)および(b)エンドグリコシダーゼH (Endo-H)で処理(+)またはmock処理(−)した。タンパク質を10%PAGEゲル上で分離し、ウェスタンブロットをウサギ- -S1で実施した。mock移入細胞由来の溶解物を陰性対照として使用した(レーン3、4、7および8)。特異性タンパク質の分子量を右側に、マーカーの分子量を左側にキロダルトンで示している。
【図1a】
【図1b】
【発明の詳細な説明】
【0001】
(技術分野)
本発明は、SARSコロナウイルスの成熟グリコシル化スパイク(S)タンパク質の使用、Sタンパク質のフラグメント、該フラグメントの産生方法、該フラグメントのSARS感染の検出における使用、並びにSARSに罹患している患者のワクチン接種または治療のための該フラグメントの使用または該フラグメントの相応する抗体の使用に関する。
(技術背景)
2002年〜2003年において、新型肺炎、即ち、SARSと称する疾病を発症させる新たなウイルスが、中国南部から発生し、少なくとも30カ国の人々に感染した。2003年7月の発生終了時には、報告された患者およびSARSによる死亡者の世界保健機構集計数は、それぞれ、約8500人および800人以上であった。この発生は、多大な社会的および医療的費用を感染国にもたらしただけでなく、地域および国際経済に計り知れない影響を与えた。
新規のコロナウイルスは、SARSの病原因子として確立され(文献7、Drosten等、2003年;文献39、Ksiazek等、2003年)、その後、SARSコロナウイルスまたはSARS CoVと命名された。その29.6kbのゲノムにより、他のコロナウイルスに類似するレプリカーゼ、スパイク、膜、エンベロープおよびヌクレオカプシド(N)並びに幾つかの他の特異的なタンパク質をコード化する14個の読み取り枠(orf)が明らかになった(文献40、Marra等、2003年;文献41、Rota等、2003年)。SARS CoVスパイクタンパク質における困難性は、グリコールタンパク質であるという困難性であり、抗体、ワクチン並びに他の治療用、診断用および予防用手段の製造に適するであろうヒト化形の十分な該タンパク質を産生させることは困難である。ヒト化形のタンパク質は、人体内のタンパク質形に類似するタンパク質である。
【0002】
(発明の開示)
この問題に対する1つの解決法は、グリコシル化されてヒト化されるようなSARS CoVスパイクタンパク質を産生し得る系を見出すことであろう。
もう1つの解決法は、中和抗体を誘発し得るSARS CoVスパイクタンパク質のフラグメントを見出すことである。
本発明においては、本発明者等は、細胞外ドメイン全体(48〜1192のアミノ酸)に及ぶSの5種のフラグメントを大腸菌中のGST融合タンパク質として発現させ、その精製タンパク質を使用して、ウサギ中で抗体を産生させた。ウェスタンブロット分析および免疫沈降試験により、本発明者等は、これら抗体の全てが、それらのターゲット領域に対し特異性であり且つ高感受性であることを証明した。固定させたが浸透化はしていない細胞において行なった間接免疫蛍光法は、これら抗体が、細胞表面に発現するSの細胞外ドメイン上の線状エピトープを認識し得ることを示した。これらの抗体をVero E6培養物中でSARS Cov複製を抑制するそれらの能力についで試験したとき、S SEQ ID NO.4の1029〜1192のアミノ酸にターゲッティングさせた抗S 10抗体は、強い中和活性を有し、SARS CoV中和ドメインを含有するSのこの領域がウイルス進入および/または複製において極めて重要であることを示唆していた。
【0003】
本発明の1つの局面によれば、コロナウイルスの成熟グリコシル化スパイクタンパク質を提供する。
本発明のもう1つの局面によれば、下記の工程を含む、コロナウイルスの成熟グリコシル化スパイクタンパク質の産生方法を提供する:
細胞にコロナウイルスのスパイクタンパク質またはその一部をコード化する核酸を移入する工程;上記細胞内で上記スパイクタンパク質を発現させる工程;および、上記スパイクタンパク質を分離する工程。
本発明のさらなる局面は、下記の工程を含む、コロナウイルスの成熟グリコシル化スパイクタンパク質のスクリーニング方法を提供する:
スパイクタンパク質を分離する工程;Endo-Hを有する上記分離スパイクタンパク質を免疫沈降させる工程;および、成熟グリコシル化スパイクタンパク質である残存スパイクタンパク質を検出する工程。
本発明のもう1つの局面は、コロナウイルスの成熟グリコシル化スパイクタンパク質またはその一部に対する抗体を提供する。
以下の説明は、本発明の上記局面に関連する。
1つの実施態様においては、好ましくは、上記コロナウイルスは、SARSコロナウイルスである。好ましくは、上記コロナウイルスはSARSコロナウイルス株、2774である。
もう1つの実施態様においては、好ましくは、上記成熟グリコシル化スパイクタンパク質は、膜貫通ドメイン(TMD)を含有する。
もう1つの実施態様においては、好ましくは、上記成熟グリコシル化スパイクタンパク質は、210KDaタンパク質である。
もう1つの実施態様においては、好ましくは、上記細胞は、肺細胞系A549である。
もう1つの実施態様においては、好ましくは、上記抗体は、SARSコロナウイルス感染の免疫検出において使用する。
もう1つの実施態様においては、好ましくは、上記スパイクタンパク質または上記抗体は、ワクチンの製造において使用する。
【0004】
本発明のさらなる局面によれば、本発明は、SARSコロナウイルスのSタンパク質(SEQ ID NO.2)のペプチドまたはタンパク質フラグメントを提供し、該フラグメントは、SARSコロナウイルスのS遺伝子由来のアミノ酸数1055〜1192の配列(SEQ ID NO.5)、或いは上記S遺伝子のアミノ酸数1029〜1192の配列(SEQ ID NO.4)を含む。好ましくは、上記ペプチドまたはタンパク質は、コロナウイルスSタンパク質のHR2ヘプタッド領域を有する。上記ペプチドまたはタンパク質は、後述するようなS 10(SEQ ID NO.4)、S 11(SEQ ID NO.6)、S 17(SEQ ID NO.7)、S 18(SEQ ID NO.8)、S 19(SEQ ID NO.9)またはS 20(SEQ ID NO.10)であり得る。
また、本発明は、下記の工程を含む、コロナウイルスのSタンパク質のフラグメントの産生方法を提供する:
a) 細胞に、SARSコロナウイルスのS遺伝子由来のアミノ酸数1055〜1192の配列(SEQ ID NO.5)を含むコロナウイルスのSタンパク質のフラグメントをコード化する核酸を移入する工程(該核酸は細胞内でのその発現を誘導し得る調節配列と操作的に関連する);
b) 上記タンパク質フラグメントを上記細胞内で発現させる工程;および、
c) 上記タンパク質フラグメントを分離する工程。
さらに、本発明は、SARSコロナウイルスのSEQ ID NO.2のペプチドまたはタンパク質フラグメントに対する抗体を提供し、該フラグメントは、SEQ ID NO.5、好ましくはSEQ ID NO.4を含む。好ましくは、上記ペプチドまたはタンパク質は、コロナウイルスSタンパク質のHR2ヘプタッド領域を含む。さらに詳細には、本発明は、ペプチドまたはタンパク質S 10(SEQ ID NO.4)、S 11(SEQ ID NO.6)、S 17(SEQ ID NO.7)、S 18(SEQ ID NO.8)、S 19(SEQ ID NO.9)またはS 20(SEQ ID NO.10)に対する抗体を提供する。
上記抗体は、上記抗体を患者から収集した細胞の少なくとも一部に適用する工程を含む、患者のSARSコロナウイルス感染の検出方法において使用する。上記抗体を含有するSARSコロナウイルス検出用の関連キットも、本発明によって提供される。
最後に、本発明は、患者に上述したようなペプチドまたはタンパク質或いはそのようなペプチドまたはタンパク質の抗体を投与することを含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法を提供する。例えば、有効量の上記ペプチドまたはタンパク質或いはそのようなペプチドまたはタンパク質の抗体を含むワクチンが本発明によって提供される。
【0005】
(発明を実施するための最良の形態)
コロナウイルスのSタンパク質は、Sタンパク質が宿主細胞上の細胞レセプターに結合し、さらにウイルスと細胞膜との融合に不可欠であることから、組織親和性の重要な決定因子である。SARS-CosVにおいては、S単独に対する液性応答は、SARS-CoV感染を防御するのに十分であるようである(文献14)。
上記SARSコロナウイルス株、2774のSタンパク質を、サル腎臓細胞Vero E6およびCos-7中、並びにヒト腎臓293T、肺細胞A549およびMRC-5中で、ワクチニア-T7発現系において発現させた。Sタンパク質は、ウサギ、ウマおよびマウス内で産生させたSに対するポリ-およびモノ-クロナール抗体を使用したときに、免疫沈降法(IP)、ウェスタンブロット法(WB)、免疫蛍光法(IF)によって検出した。これらの抗体は、Sの細胞外ドメイン全体に及ぶ種々の領域を認識する。本発明者等は、パルスチェイス試験において、Sの200kDのコアグリコシル化形が210kDの完全グリコシル化Endo-H抵抗性形にプロセシングされることを見出した。さらにまた、1群のC-末端切断および内部欠失構築物を発現させたとき、膜貫通ドメイン(TMD)を有していないSポリペプチドが、TMDを有するSポリペプチドとは対照的に、Endo-H感受性であり、トランスゴルジに輸送されず、細胞表面上で発現しないことを見出した。また、本発明者等は、2種の宿主タンパク質(ぞれぞれ、62kDおよび64kD)が、試験したいずれの細胞系においても、動物内で産生させた抗体および回復患者由来の抗体によってSと共免疫沈降することも見出した。Sのアミノ酸残基31〜200は、この領域の欠失が特異結合を完全に消滅させるので、これら2種のタンパク質と相互作用するドメインであると同定した。これら2種のタンパク質は、糖タンパク質である;何故ならば、これらタンパク質がEndo-H処理に感受性であり且つこれらタンパク質がSARSコロナウイルスのSタンパク質と相互作用するが鳥類の群3のコロナウイルスの一員であるIBVとは相互作用しない。興味あることに、Sタンパク質は、WB試験において検出した移入細胞中で少なくとも2個の片として開裂するものの、通常の開裂部位は、配列比較によっては予測されなかった。ウイルス感染細胞においても、同様の開裂産生物が観察された。Sタンパク質は、感染細胞の上清中で検出することができ、およそその半分が開裂していた。本発明者等は、最も効率的な発現および成熟が、並行移入試験における移入Cos-7およびA549細胞中で生じていたことを見出した。該プロセシングは、Sタンパク質の大多数が成熟した完全グリコシル化210kD形であり、ウイルス感染細胞の上清中の天然形のSタンパク質と同時移行するので、Cos-7細胞におけるよりもA549細胞においてより完全である。
【0006】
また、本発明においては、本発明者等は、Sタンパク質のフラグメントも取得し、細胞外ドメイン全体(48〜1192アミノ酸)に及ぶ5種の細菌発現Sフラグメントに対するウサギポリクロナール抗体を産生させた。これらのタンパク質を大腸菌内で発現させたとき、これらの抗体は、グリコシル化部位のようなS上のある種の翻訳後修飾を認識し得ないであろう。また、本発明者等の分析は、上記タンパク質をSDS変性ゲルから抽出し、SDSの存在下に溶出させたので、線状エピトープに限られているようであるが、本発明者等は、上記タンパク質がある程度の折り畳み(folding)を維持し得ることを除外できない。興味あることに、本発明者等は、上記抗体が、ウェスタンブロット分析において変性Sタンパク質(哺乳類細胞において発現させた)を認識するのみならず、細胞溶解物からの天然Sタンパク質を免疫沈降させるのにも使用し得ることを見出した(図1および2)。その結果は、これらの抗体が、折り畳みS中に存在する線状および露出エピトープおよび/またはSの構造中に見出される回旋のような可撓性領域に結合することを示唆している。重要なことに、免疫沈降試験は、これらの抗体が、非グリコシル化(140kDa)およびコアグリコシル化全長S(200kDa)の双方に、さらにまた、Endo-H抵抗性完全グリコシル化S(210kDa)に結合し得ることを示した。また、このことは、これらの抗体が細胞表面上に発現するSタンパク質を認識し得ることを示している間接免疫蛍光試験によっても裏付けされた。コアグリコシル化Sタンパク質が、成熟に達する前に、ゴルジ体に移送されて、200kDa〜210kDaの見掛け分子量の増大を生じなければならず、その後、細胞表面に輸送されることは既知である(文献11、15)。また、Sの210kDaの完全グリコシル化形は、後でビリオンに取込まれ、SARS-Cov感染Vero E6培養物の倍地中に分泌することも知られている。
【0007】
中和アッセイは、GST-S 10に対し産生させた抗体が、200 TCID50において1:364までの力価でVero E6細胞中のSARS-CoV複製を中和し得ることを示しており、この力価は、回復期患者において得られた値に匹敵し得る。累積的免疫化後に採血した血清の分析は中和力価の定常的な増大を示しており、免疫化ウサギがGST-S 10に対する特異的免疫応答を示すことを示唆していた。他の抗体のうちで抗体中和を誘発し得るようなものはなく、このことは、Sタンパク質の48〜1055アミノ酸(aa)中に中和エピトープが存在しないことを示唆し得る。しかしながら、この現象は、261〜672aaに対するヒト抗体が中和特性を有することが証明されているので、ほぼありそうもない(レセプター結合性ドメインについての下記の説明を参照されたい)。より可能性ある説明としては、この領域内の中和エピトープが濃密にグリコシル化されているかおよび/または堅固な三次構造を含有することである。従って、本発明者等の結果は、Sタンパク質の48〜1055aaが、ヒト細胞中でのグリコシル化過程を擬態し得ない非哺乳類系において発現させたペプチド系またはタンパク質をベースとするワクチンの開発において使用するにはあまり適切ではないことを示した。S 9は798〜1055aa由来であり、S 10は1029〜1192aa由来であるので、Sタンパク質SEQ ID NO.5の1055〜1192aa領域が、サブユニットワクチン開発において適し得る線状中和エピトープ(1個以上)を含有することは明らかである。
【0008】
コロナウイルス感染の第1段階では、宿主細胞へのビリオンの付着であり、SARS-CoVの場合は、ACE-2が、SARS-CoV Sタンパク質に結合する細胞レセプターとして同定されている(文献9)。Sタンパク質のN-末端中のドメイン、即ち、およそ300〜510のアミノ酸(aa)は、レセプター結合性ドメインである(文献16)。その後、ウイルスエンベロープの脂質二分子層の宿主細胞膜との融合が生じ、この過程もSタンパク質が介在する(文献4)。コロナウイルスSタンパク質は、群Iのウイルス融合タンパク質であり、2つのヘプタッド繰返し領域(HR1およびHR2)を含有し、S2ドメインまたはC-末端ドメインにおいて見出される。これらのドメインは、Sのオリゴマー構造を決定する重要な役割を果たし、従って、ウイルスと細胞膜間の融合を介在することを前提とする(文献4)。SARS-CoVにおいては、HR2は膜貫通アンカー(1148〜1193aa)近くに位置し、HR1はHR2 (900〜1005aa)の約140aa上流にある(文献14)。興味あることに、S 10 (1029〜1192aa)は、HR2領域にまで及ぶ。
生化学研究においては、SARS-CoV Sタンパク質のHR1およびHR2に相応するペプチドが、他の既知の群I融合タンパク質の典型的な構造的特徴を有する逆平行6螺旋束中に結合し得ることが証明されており、SARS-CoVが果たす膜融合および細胞侵入メカニズムが、MHVのような他のコロナウイルスにおけるメカニズムに類似することを示唆している(文献1)。全長Sタンパク質においては、上記HR1-HR2構造は、HR1のN-末端近くにあると予想される融合ペプチドを膜貫通ドメインの極めて近くに接近させ、このことが、ウイルスと細胞膜の融合を容易にし、ウイルスの細胞への侵入を可能にしている。高親和性でもってHR2ドメインに結合することにより、抗S 10抗体が、HR1とHR2と間の相互作用を妨げ、従って、SARS-CoVと宿主細胞との融合を阻止し得ることは極めて可能性のあることであろう。
【0009】
細菌発現させたタンパク質は、大規模に製造するのに容易で且つ費用効率的であろうから、本研究において同定したS 10フラグメント(1029〜1192aa) SEQ ID NO.4は、SARS-CoV用の理想的なワクチン候補であり得る。将来の研究においては、抗S 10抗体が動物モデルにおけるSARS-CoV感染を防御し得るかどうか決定すること並びにこのS領域の膜融合に対する正確な寄与を描き出すことが重要であろう。後者については、本発明者等は、現在、GST-S 10タンパク質で免疫化したマウスから得られたモノクロナール抗体をマッピングし、その結合および中和特性を判定しているところである。最後に、組換えS 10タンパク質および抗S 10抗体を抗ウイルス治療用に開発することが潜在的には可能であろう。
コロナウイルスはプラス鎖RNAウイルスであり、そのビリオンは、当該種の全メンバーに共通している3種の膜タンパク質、即ち、スパイク(S)、膜(M)およびエンベロープ(E)を含有するエンベロープによって囲まれているヌクレオカプシドコアからなる(参考として、文献8、13を参照されたい)。Sタンパク質は、ビリオン表面上に形態学的に特徴のある突起を形成しており、宿主レセプターへの結合および膜融合を介在する。上記MおよびEタンパク質はウイルス集合にとって重要であり、一方、NはウイルスRNAパッケージングにとって重要である。
コロナウイルスのSタンパク質は、宿主免疫応答および抗体によるウイルス中和を誘発させることに関与している(文献6、14)。SARS-CoVにおいては、以前の感染がマウスモデルにおいて保護免疫をもたらし、また、新生マウスへの中和抗体の受動移行により、新生マウスが感染から保護されることが可能であろう。このことは、ネコ伝染性腹膜炎ウイルス(文献10)における場合とは異なり、再感染時または免疫血清投与後のマウスにおいてはSARS-CoV感染の増進を伴わないであろうし、従って、SARS-CoVに対してワクチン接種をすることは安全であろう。Sタンパク質をコード化するDNAワクチンは、単独で、T細胞および中和抗体応答を誘発し、マウスをSARS-CoV感染から保護し得、Sタンパク質が実際にSARS-CoV感染におけるウイルス中和に対する初期ターゲットであることを示唆している。この知見は、代理/保菌者ウイルスを使用してSを霊長類に発現させている別の研究(文献5)によっても裏付けられた。これらの研究から、Sに対する体液応答のみで、SARS-CoV感染を防御するのに十分なようである。
【0010】
不活化全菌SARS-CoVはワクチン接種において見込みのある結果を示し得るものの、エピトープ系ワクチンがあり得る病原性復帰の可能性を回避しまた製造するのに安全で且つ安価であろうことから、中和抗体を誘発し得るS中のエピトープ(1個以上)を同定することが重要である。本試験において、本発明者等は、SARS-CoV感染に対するワクチンおよび治療薬の開発に使用すべく、SARS-CoVのSタンパク質中の中和エピトープを同定した。本発明者等は、Sの種々の領域をGST融合タンパク質として発現させ、これらを使用してウサギ中で抗体を産生させた。その後、これらのポリクロナール抗体を、Sタンパク質に対するこれら抗体の特異性についてウェスタンブロット分析、免疫沈降法および免疫蛍光分析において試験し、さらにまた、Vero E6培養物中でSARS-CoV複製を抑制するこれら抗体の能力について試験した。
コロナウイルス群のメンバーについては、以前の研究により、殆どの場合、Sタンパク質は、おそらくは細胞フリン(furin)様プロテアーゼによって、2つのサブユニット、即ち、N-末端S1とC-末端S2に開裂することが証明されている。タンパク質分解開裂は、膜融合および感染性を増強し得るが、必須条件ではあり得ない(文献20〜23参照)。開裂と膜融合(または感染性)との関係は、使用する個々の株および細胞に依存しているようである(文献24)。おそらく、このことは、Sの融合活性の複数の決定因子および試験する細胞中の酵素の発現レベルの差異を反映している。SARS-CoVのSタンパク質のタンパク質分解開裂は、今日まで解明されていない。従って、SARS-CoVのSタンパク質がS1およびS2として開裂するかどうかを研究することは興味あることであろう。このタンパク質も開裂するとすれば、その場合、融合および感染性に対するその効果をこの新たに出現したコロナウイルスにおいて試験することができる。本発明者等は、SARS-CoVのSタンパク質が110kDaのN-末端S1および90kDaのC-末端S2として開裂することを、これらのN-末端S1およびC-末端S2を、感染Vero E6細胞の培地中および精製ビリオン中で、SのNおよびC-末端に対して特異性の抗体を使用して検出したときに見出した。また、200kDaの全長Sタンパク質もビリオン中に検出されたので、本発明者等は、SARS-CoVのSタンパク質は部分的に開裂すると結論付けた。相対的に豊富な開裂産生物は、S1およびS2に対する各抗体の親和性が異なるため、各ブロットを濃度計により直接スキャンすることによっては正確に算定することができなかった。これらの抗体は、それぞれ、精製ビリオンおよび大腸菌発現Sフラグメントによって産生させた。
【0011】
他のコロナウイルスにおける典型的なS1/S2開裂シグナルがSARS CoVのS配列中に存在しないという事実は、上記の知見をより興味あるものとしている。以前の研究は、RRARRまたはRRAHRモチーフはコロナウイルスSタンパク質の開裂部位の直ぐ上流に位置してことを示唆している。複数の塩基性残基の幾つかの点変異は、開裂を阻止または停止させる可能性がある(文献23、25)。おそらくは、このモチーフは、他のウイルス融合タンパク質の開裂に関与していることが証明されている哺乳類細胞のフリン様プロテアーゼに対して必要とする(文献26)。本発明者等は、哺乳類細胞系における単一Sタンパク質の発現系を開発し、次いで、各変異体を使用してSARS CoVのSの代替開裂モチーフを特定することを決定した。感染細胞中で同定したS2と同様なサイズを有する90-kDaタンパク質を、S遺伝子SEQ ID 1を移入したCos-7細胞中で検出した。この系は、Sの内部欠失変異体を使用することによって開裂シグナルを決定することを可能にする。本発明者等は、551から570の残基を除去したとき、90-kDaタンパク質は検出されなかったことを見出した。上記変異体において欠失した20個の残基の配列は、FGRDVSDFTDSVRDPKTSEIであった。上記90-kDaタンパク質は、上記S遺伝子を移入した細胞中の開裂タンパク質S2であった。上記20個のアミノ酸残基は、代替翻訳開始コドンを含有しておらず、従って、上記90-kDaタンパク質は、翻訳の内部開始に基づき得なかった。上記90-kDaタンパク質は、C-末端に対する抗体によってのみ検出可能であり、N-末端に対する抗体によっては検出し得ず、従って、上記90-kDaタンパク質は、翻訳の早期終結に由来する産生物ではあり得なかった。興味あることに、これらの20個の残基は、フリン様プロテアーゼが必要とする複数または対の塩基性残基のモチーフを含有していなかった。従って、SARS CoVのSタンパク質は、他のコロナウイルスにおいて使用される同じ酵素によってではなく異なる細胞エンドプロテアーゼによって開裂し得る。結果は、昆虫細胞中でのマウスフリンによるSの共発現がSARS CoVのS開裂を生じさせなかったという第三者による観察と一致している。また、インフルエンザAおよびセンダイ(Sendai)ウイルスの融合タンパク質のコンセンサス開裂モチーフである血液凝固因子Xa様(またはトリプシン様)プロテアーゼにおける配列モチーフGln(Glu)-X-Arg (Xは、Gly、SerまたはThrである)は存在していなかった。タンパク質分解開裂は、これらウイルスの融合タンパク質を活性化していた。
【0012】
マウス肝炎ウイルスのSタンパク質の早期の研究によれば、該Sタンパク質は、小胞体内でN-結合グルコシル化され、三量体化される(文献10)。高マンノース側鎖は、ゴルジ体への輸送中にトリミングされ、さらに修飾される。開裂は、ゴルジ体内またはゴルジ体後に生じる。SARS CoVのSタンパク質は、1255個の残基を含有し、23個の潜在的N-結合グリコシル化部位を担持する。開裂が残基551〜570において生じる場合、S1およびS2は、それぞれ、12個および11個の部位を有するであろう。S1がS2よりも約20-kDa大きいので、S1における潜在的部位の全てではないが殆どがグルコシル化において使用されるが、S2における潜在的部位の全てが使用されないと想定するのが合理的であった。この試験において、本発明者等は、細胞溶解物中で検出した開裂S1およびS2のサイズが上清および精製ビリオン中で検出した開裂S1およびS2のサイズよりも僅かに小さいことも観察した。このことは、(1) さらなる修飾が、Sの解裂後おそらくはビリオンの集合前に生じていること、および(2) Sタンパク質は、宿主細胞プロテアーゼにより細胞内で開裂し得ることを示唆し得る。
単一S遺伝子を移入したCos-7細胞においては、SARS CoVを感染させたVero E6細胞におけるのと異なり、サイズ55-および38-kDaの小N-末端産生物は検出されたが110-kDa S1は検出されなかった。これらは、S1の開裂/劣化産生物または早期終結産生物であり得る。興味あることに、64/62-kDaの他の2つのタンパク質が移入および感染細胞の双方において同定され、これら2つのN-末端タンパク質の産生は移入細胞において人工的ではないことを示唆していた。
Sの開裂とその機能(膜融合、感染性および病原性)との関係は、矛盾したままである。Sプロセシングに関する問題に対処することが依然として求められている。例えば、何故、Sの開裂は、この開裂がSの機能と関連しない場合に、多くのコロナウイルスにおいて進化的に保存されるのか? 最近の報告は、SARS CoVの取込みSタンパク質による偽ビリオン類 HIV類のトリプシン活性化が細胞-細胞膜において必要であるが、この活性化は感染性を増進しないこと示唆している(文献32)。
【0013】
材料および方法
材料
本試験におけるCos7およびVero E6は、American Type Culture Collection (米国バージニア州マナッサス)から購入した。Cos7細胞は、1gグルコース/リットル、2mM L-グルタミン、1.5g重炭酸ナトリウム/リットル、0.1mM 非不可欠アミノ酸類、0.1mgストレプトマイシン/ml、100Uのペニシリンおよび5%のウシ胎仔血清を含有するダルベッコ(Dulbecco)変性イーグル培地(HyClone社、米国ユタ州)中で、5%CO2インキュベーター内で37℃にて培養した。Vero E6細胞は、2mM L-グルタミンおよびL-アミノ酸を含有する培地199 (HyClone社、米国ユタ州)中で、5%CO2インキュベーター内で37℃にて培養した。
Sarsコロナウイルスのシンガポール株 SARS CoV 2003VA2774 (“2774”)をTan Tock Seng病院において分離し、シンガポールのEnvironmental Health Institute (EHI)の研究室において、Vero E6細胞中で増殖するよう順応化させた。Vero E6細胞中の継代3を、直接RNA抽出、逆転写およびポリメラーゼ連鎖反応(RT-PCR)、および配列分析において使用した。組換えワクチニア/T7ウイルス(VT3)を増殖させ、Vero細胞上で滴定した(該ウイルスは、鳥類伝染性気管支炎ウイルス、IBVの増殖用サブクローンである) (文献54、ShenおよびLiu、2003年)。
DNA構築物のクローニング
(a) 大腸菌中での発現用:全長S (1〜1255アミノ酸(aa))を含有するシンガポール分離物2774を使用して増幅およびRT-PCRを実施し、RT-PCRからのcDNAをS構築物のクローニング用テンプレートとして使用した。5種の構築物 S 1、S 2、S 3、S 9、およびS 10 (表1)を、表2に示すプライマーによるPCRによって取得した。使用したプライマーは、全てResearch Biolabs社またはProligo Pte社(シンガポール)から購入した。PCR産生物をBamHI/XhoIにより消化し、BamHI/XhoI切断pGEX4T1ベクター(Amersham Pharmacia Biotech社、スウェーデン国ウプサラ)中に連結反応させて、グルタチオンS-トランスフェラーゼ(GST)融合タンパク質の発現用プラスミドpGEX-S 1, pGEX-S 2, pGEX-S 3, pGEX-S 9 and pGEX-S 10を得た。
(b) 哺乳類細胞での発現用:T7ワクチニアウイルス系と一緒に使用して哺乳類細胞中でSを効率的に発現させ得る、全長Sを含有するpKT-Sの構築。プラスミドpKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16 (図1)を下記のようにしてクローニングした。
【0014】
表1:本試験において使用するプラスミド
【0015】
表2:本試験において使用するオリゴヌクレオチドプライマー
【0016】
プラスミド類の構築:特異性正および逆プライマーをシンガポール株2774のS遺伝子を増幅するように設計した。各PCR産生物をBamHIおよびStuIにより消化し、BamHI/EcoRV切断pKT0中に連結反応させてT7プロモーターの制御下にプラスミドpKT-Sを得た。各特異性プライマーを、それぞれ、ヌクレオチド位置21476〜25171、〜25066、〜24934、〜24415、〜24157、および〜23866からの株2774配列を増幅するように設計した。6種のRT-PCR産生物をBamHIにより消化し、T7プロモーターの制御下にBamHI/EcoRV切断pKT0中に連結反応させて、それぞれ、プラスミドpKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15、pKT-S 16およびpKT-S 22を産生させた。これらのS構築物がコード化するタンパク質のサイズは、図1aに示している。2回のPCRを、各特異性プライマーを使用して実施して、内部欠失を有するSフラグメントを産生させた。各PCRフラグメントをpKT0中にクローニングして、プラスミドpKT-S 17、pKT-S 18、pKT-S 19、pKT-S 32、pKT-S 33およびpKT-S 34を産生させた。これらの変異体は、図1aに示す位置で200または20個のアミノ酸残基の欠失を有するSタンパク質をコード化する。
感染Vero E6細胞中でのSタンパク質の分析:融合性細胞を株2774により感染多重度(m.o.i) 1で感染させ、37℃で12〜15時間インキュベートした。培地中の細胞屑片を低速遠心分離により除去した。細胞をPBSで洗浄し、PBS中に再懸濁させた。1容量の5×標準タンパク質サンプル緩衝液を4容量の細胞懸濁液即ち培養培地に添加した。各サンプルを5分間100℃に加熱し、ウェスタンブロット分析前に−20℃に保った。ウイルス精製においては、 -プロピオラクトンを感染細胞培養物に0.05%の最終濃度まで添加して、感染性を不活化させた。不活化をVero E6細胞中での処理サンプルの滴定によって試験した。ウイルスを3回の凍結/解凍により採集し、細胞屑片を10分間の5,000rpmでの遠心分離によって除去した。限外濾過を行なってウイルスを濃縮した(300,000 NMWL、Millipore社)。濃縮サンプルを、Sepharose 4B急速流カラム(Pharmacia社)に、製造者の使用説明書に従い塗布した。溶出画分を透過型電子顕微鏡によって検査した。ウイルス粒子を含有する画分をウェスタンブロットによるSタンパク質分析において使用した。
【0017】
移入Cos-7細胞中でのSタンパク質の分析:60mmのペトリ皿内の細胞の50%の融合性単分子層を、バクテリオファージT7 RNAポリメラーゼを発現する組換えワクチニアウイルスvTF7-3によりm.o.i 1で感染させた。1時間の吸着後、細胞に、製造業者の使用説明書に従いEffectene Reagents (QIAGEN社)を使用して2〜5μgのプラスミドを移入した。移入細胞を37℃で1夜インキュベートし、細胞溶解物を、1×ウェスタンブロット分析用タンパク質サンプル緩衝液中に細胞ペレットを再懸濁させることによって調製した。
大腸菌中で発現させた組換えSタンパク質の精製:プラスミドpGEX-S 1、pGEX-S 2、pGEX-S 3、pGEX-S 9およびpGEX-S 10を、個々に、BL21 (DE3)細胞中に形質転換させた。各プレートからの単一コロニーを、アンピシリン (100 g/ml)を含有するLB-寒天プレート中で37℃にて1夜増殖させた。5ミリリットルの得られた各培養物を、2リットルのアンピシリン (100 g/ml)を含有するLB培地中に接種し、37℃のシェーカー内でOD600が0.6に達するまでインキュベートした。タンパク質の発現を、1mM IPTGを使用して誘発させた。細胞を、誘導後2時間で、4℃で10分間の5,000gでの遠心分離により採集した。得られた細胞ペレットをPBS-1mM PMSF-20 g/ml DNase I中に再懸濁させ、フレンチプレスへの2回通過により溶解させた。溶解物を22,000gで30分間遠心分離した。ペレット中の不溶性タンパク質を3回洗浄し、1% Triton X-100を含有するPBS中に再懸濁させた。タンパク質を10% PAGE-SDSゲル中で分離した。GST融合タンパク質を含有するゲルストリップを切断し、タンパク質を、Mini Trans-Blot細胞(BIORAD社、米国カリフォルニア州ハーキュリーズ)を使用して溶出させた。得られた融合タンパク質を、ウェスタンブロットにおいてマウス抗-GST抗体(Santa Cruz Biotechnology者、米国カリフォルニア州サンタクルズ)を使用して検出し、その濃度をクーマシーブリリアントブルーR-250染色SDS-PAGEゲル中のBSA標準と比較することによって推定した。
【0018】
各種S構築物に対する抗体の産生:1ミリグラムの種々の抗原の各々を等容量の完全フロイントアジュバント(Sigma社、米国ミズーリ州セントルイス)と混合し、ニュージーランドシロウサギの免疫化に使用した。2羽のウサギを使用して各それぞれの抗原に対する抗体を産生させた。初期免疫化の2週間後、各ウサギに3週間間隔でブースター注入を行なった。不完全フロイントアジュバント(Sigma社)をその後のブースター注入において使用した。10mlの血液を、各ウサギから、4回目、6回目、8回目、12回目、14回目および16回目の注入後の各時点で採血した。実験動物の使用についての手順は、全てAnimal Research Ethics Committee (AREC)の規定およびガイドラインに従って実施した。
哺乳類細胞中での組換えSフラグメントの発現:Cos7細胞を、ウェスタンブロット分析、免疫沈降法および免疫蛍光法のための哺乳類発現系として使用した。60mm皿中で増殖させたCos7細胞の単分子層をT7ワクチニアウイルス感染に感染多重度(m.o.i) 1で1時間供した。pKT-S、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16プラスミドによる細胞の一時的移入を、Effectene移入試薬(Qiagen社、米国カリフォルニア州バレンシア)を使用し、製造業者のプロトコールに従って実施した。
対照試験においては、細胞をT7ワクチニアウイルスにより感染させ、空ベクター、pKT0によりmock移入した。
Sタンパク質のエンドグリコシダーゼHおよびN-グリコシダーゼF処理:細胞を組換えワクチニア-T7ウイルスで感染させ、上述のようにして、プラスミドpKT-Sを移入しまたは空プラスミドをmock移入した。細胞をPBSで洗浄し、製造業者(Roche社)が推奨する溶解緩衝液中に再懸濁させた。細胞屑片を12,000rpmでの遠心分離により除去した。10μlの上清を、Endo-H (0.5単位/20μl、Roche社)を含むまたは含まない製造業者(Roche社)が推奨する10μlの緩衝液と混合し、37℃で2〜3時間インキュベートした。使用した溶解緩衝液は、製造業者(Roche社)が推奨する濃度のプロテアーゼインヒビターCompleteRカクテル錠剤および0.2mM PMSFを含有する。
【0019】
ウェスタンブロット分析:細胞溶解物を還元条件(0.2M DTTを含む1%SDS)下に1×SDS負荷緩衝液中で調製し、10%PAGEゲル中で分解させ、ニトロセルロース膜に移した。各ブロットを0.05% Tween-20を含むPBS中5%無油脂乳中でブロッキングし、ウサギ抗血清によって検証し(1:20,000希釈)、各種S融合タンパク質に対して4℃で1夜産生させた。上記膜をヤギ抗-ウサギホースラディシュペルオキシダーゼ(HRP)接合二次抗体と一緒に1:2000希釈で室温にて1時間インキュベートし、増強化学発光試薬(Pierce社、米国イリノイ州ロックフォード)により展開した。
放射能標識免疫沈降法:細胞をワクチニア-T7ウイルスで感染させ、上述のようにして、pKT-S或いはSのC-末端欠失変異体を発現するプラスミド、即ち、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15またはpKT-S16で移入処理した。pKT0でmock移入した細胞は、対照として調製した。細胞を、35S-metで1時間半標識化する前に、30分間飢餓させ、2時間追跡した。経時試験においては、細胞採集前の追跡時間は、それぞれ、0時間、1/2時間、1時間、2時間、4時間および6時間であった。細胞を、50mM Tris、1mM PMSF、1% NP40を含有する溶解緩衝液(pH 7.4)を使用して溶解させ、10分間16,000gで遠心分離した。300 lの上清を5 lのウサギ抗-GTS-S 1、2、3、9または10と一緒に1/2時間インキュベートし、その後、プロテイン-Aセファロースビーズ(Roche Diagnostics社)と一緒に1時間インキュベートした。ビーズを溶解緩衝液で3回洗浄した。20 lの1×SDS負荷緩衝液(0.2M DTTを含有する)をビーズに添加し、100℃で10分間煮沸した。各サンプルをエンドグリコシダーゼH (EndoH)酵素((Roche Diagnostics社)で処理し、該EndoH試験において37℃で2時間インキュベートする。各サンプルを7.5% SDS-PAGEゲル中で分離し、オートラヂオグラフィーによって展開する。
【0020】
間接免疫蛍光法:Permanoxスライド(Nalge Nunc International社、米国イリノイ州ネーパービル)上で増殖させたCos-7細胞を、上述のようにして、感染させ移入処理した。移入後8時間で、細胞を、4%パラホルムアルデヒドにより室温で10分間固定し、PBS+1%ウシ血清アルブミン(BSA)により30分間ブロッキングし、次いで、一次抗体(1:200)と一緒に1.5時間インキュベートし、洗浄し、その後、フルオレセインイソチオシアネート(FITC)接合二次抗体(1:200、Santa Cruz社)と一緒に1時間インキュベートした。インキュベーションおよび洗浄は、全て室温で実施した。各スライドをFluorescence Mounting Medium (DakoCytomation社)によって取付け、AxioVision Fluorescence Light Microscope (Carl Zeiss社、ドイツ)上で分析した。
中和アッセイ:ウサギ血清中の中和抗体を測定するために、本発明者等は、96ウェルプレートを使用して、血清の連続希釈によるアッセイを実施した。2×104 Vero E6細胞を96ウェルプレートの各ウェル内の200 lのMedium 199中で増殖させ、37℃でインキュベートした。ウサギ血清の培地による比1:10〜1:1280の連続希釈液を調製した。0.1mlの希釈ウサギ抗血清を、それぞれのウェルに添加する前に、0.1mlのSARS CoVと200 TCID50で室温にて1時間混合した。96ウェルプレートをCO2インキュベーター内で3〜5日間インキュベートして細胞変性効果(CPE)を観察した。CPEを有する細胞のパーセントを、各ウェルから10 lの再懸濁細胞を採取し、顕微鏡で計数することによって測定した。中和力価を算出する式は、Log (試験血清の50%中和力価) = Log (CPE%の50%よりも高い血清希釈)−Log (希釈計数)×(50%よりも高いCPE%−50%のCPE%)/(50%よりも高いCPE%−50%よりも低いCPE%)である。上記計算における逆対数は、試験血清の50%中和力価として判定する。試験は、全て2回実施する。
【0021】
結果
ワクチニア-T7発現系でのSのプロセシング:移入細胞中での発現およびプロセシングを分析するために、S遺伝子をT7プロモーターの制御下にベクター(pKT0)中にクローニングした。Cos-7細胞をワクチニア-T7組換えウイルスで感染させ、引続き、S遺伝子を含有するプラスミドを移入した。Sタンパク質発現プロフィールを、死滅させ精製したウイルス粒子によって産生させたウマ- -SARS抗体を使用して、ウェスタンブロットにより分析した。図1bに示しているように、200-、140-、110-、90-、64/62-、55-および38-kDaタンパク質を検出した(レーン2)。これらの種は、S-特異性であり、そのようなタンパク質は陰性対照細胞においては検出されなかったので(レーン1)、Sの発現およびプロセシングに基づいているようであった。200-および140-kDaタンパク質は、全長Sタンパク質(グリコシル化および未グリコシル化)に相応しており、他の小さい産生物は、おそらく開裂タンパク質および/または早期翻訳産生物であった。結果は、SARS CoVのSタンパク質が本生体外発現系において開裂し得、SARS CoV Sタンパク質のプロセシングの分析において使用し得ることを示唆していた。
SARS CoV感染細胞中での開裂産生物の検出:Sタンパク質がウイルス感染Vero E6細胞中でもプロセシングされるかどうかを調べるために、ウェスタンブロットを、細胞溶解物中およびウイルス感染培養物の培地中の双方においてS由来産生物を検出ために実施した。Sタンパク質がN-末端S1およびC-末端S2として開裂するならば、これら産生物は、細胞溶解物中で検出されるべきである;さらに、2つの開裂タンパク質がビリオン中に集合するのであれば、これらタンパク質は、感染細胞の培地中で検出されるべきである。この目的において、N-末端領域およびC-末端領域のそれぞれに対して産生させた2種のウサギ抗-S抗体を使用した。ウサギ- -S 1はアミノ酸残基48から358の領域を認識し、ウサギ- -S 10は1029から1192の領域を認識した(図1a参照)。図2aに示すように、ウサギ- -S 1を使用した場合、200-および110-kDaの2種の特異性タンパク質を細胞溶融物中(レーン2)および培地中(レーン4)の双方で検出した。ウサギ- -S 10を使用したときは、200-および90-kDaの2種のタンパク質を細胞溶融物中(レーン6)および培地中(レーン8)の双方で検出した。結果は、(1) 110-および90-kDa種は、ウイルス感染細胞中のSのそれぞれN-およびC-末端開裂産生物S1およびS2を示し得ること、および(2) Sタンパク質は、SARS CoVにおいて完全ではないが部分的に開裂することを示唆していた。培地中の200-、110-および90-kDaタンパク質は、細胞溶解物中のそれらの対応物よりも僅かに大きいことを観察した。これらの産生物は、開裂後でビリオン中への集合前にさらなる変性を受け得るようである。
【0022】
上記の観察を確認するために、精製ウイルス粒子を、上記の同じ抗体を使用するウェスタンブロットにおいて使用した。図2bに示すように、S1サブユニットはウサギ- -S 1を使用して検出し(レーン1)、S2サブユニットはウサギ- -S 10を使用して検出した(レーン2)。S1およびS2サブユニットの双方は、ウマ- -SARS抗体によって検出した(レーン3)。また、全長Sタンパク質は、使用した3種の抗体の全てによって検出した(レーン1、2および3)。S、S1およびS2のサイズは、感染細胞の培地中で検出した成熟形と同じであった。結果は、開裂産生物S1およびS2並びに全長Sがビリオン中に集合していることを明らかに示していた。
移入細胞中でも検出された62/64-kDaタンパク質(図1b参照)は、感染細胞の溶解物中で同定されたが(図2a、レーン2)、培養培地(レーン4)および精製ビリオン(図2b、レーン1および3)においては同定されなかった。移入細胞中で検出された140-、55−および38kDaタンパク質(図1bに示す)は、感染細胞および精製ウイルス粒子中では見出せず、これらタンパク質は、ビリオン中には集合しないことを示唆していた。140-kDaタンパク質は、Sの未グリコシル化形であり得る。140-、55-および38-kDaタンパク質は、S移入細胞においてのみ産生していたようであった。
移入細胞中でのSの開裂産生物の検出:コロナウイルスにおける開裂部位の直ぐ上流の典型的モチーフ(RRAR/HR)をSARS CoVのS中に見出せなかったので、S1およびS2サブユニット間に代替的開裂部位を有する配列を実験的に判定する必要があった。この目的において、6種のC-末端切断S構築物(S 11〜S 16、図1a参照)をCos-7細胞中で発現させた。Sタンパク質が開裂するとすれば、C-末端産生物は、C-末端欠失を有する変異体を使用するとき、より小さくなるであろう。一方、N-末端産生物のサイズは、これらの変異体を使用するとき、同じままであろう。種々の領域およびドメインに対して産生させたさらなるポリクローナルおよびモノクローナル抗体を、全ての潜在的開裂および/または他のS由来産生物を検出するために使用した。ウサギ- -S 2、ウサギ- -S 3およびウサギ- -S 9は、残基362〜790 (S 2)、168〜461 (S 3)および798〜1055 (S 9)内の領域を認識した(図1a参照)。モノクローナル抗体MAb12-6およびMAbC1は、それぞれ、Sの残基281〜300および631〜650を含有するドメインを認識した。
【0023】
(1) C-末端産生物の検出:ウェスタンブロット分析を、全長SおよびS 11〜S 16構築物を移入したCos-7細胞中のSタンパク質を検出するために実施した。図3aに示すように、200-、140-および90-kDaバンドを、全長Sタンパク質を発現する細胞において、ウサギ -S 2、-S 9、-S 10およびMAbC1抗体によって精査したときに検出した(レーン1および9、上方および下方パネル)。200-および140-kDaバンドよりも次第に小さくなる産生物をC-末端欠失変異体(S 11〜S 16)を発現する細胞において上記と同じ4種の抗体によって検出した(図3a、レーン2〜7および10〜15)。これらのうち、ウサギ- -S 2およびモノクローナルMAbC1は、欠失構築物各々からの相応する産生物を検出し得ていた(図3a、上方パネル、レーン2〜7および10〜15)。ウサギ- -S 9およびウサギ- -S 10のみは、予測したとおりのより大きい切断変異体に由来する産生物を認識していた(図3a、下方パネル) (また、図1aも参照されたい)。これら産生物の見掛け分子量は、これら産生物の算出分子量およびグリコシル化後のこれら産生物の予測分子量変化と一致しており、これら産生物が、各構築物によってコード化された産生物のそれぞれの全長グリコシル化および未グリコシル化形であることを示唆していた。
【0024】
同様に、90-kDaタンパク質よりも次第に小さくなる産生物を、C-末端欠失構築物(S 11〜 16)から、同じC-末端特異性抗体によって検出した(図3a、レーン1〜7および9〜15、上方パネルのレーン1〜7における矢印によって示されているように)。これら産生物の移行パターンおよびこれら産生物の見掛け分子量は、これら産生物がSタンパク質のC-末端開裂産生物を示すことを強力に示唆していた。S 16タンパク質はN-末端797アミノ酸残基を含有し、そのC-末端開裂産生物はサイズ的に30-kDaである(レーン7および15、上方パネル)。モノクローナル抗体MAbC1を使用したとき、S 16に由来する30-kDa産生物を検出しており(レーン15、上方パネル)、開裂部位がこの抗体の結合配列(残基631〜650)の上流に存在することを示していた。
55-kDaタンパク質(レーン1〜7、上方パネル)が、大きいS1産生物の代りに、ウサギ- -S 2によって検出されたことが注目された。該55-kDaタンパク質は、モノクローナル抗体MAbC1によっては検出されなかった(レーン9〜15、上方パネル)。また、もう1群の次第に小さくなり比較的弱い産生物が、ウサギ- -S 2およびMAbC1を使用したときに検出されたことも注目された(レーン1〜7および9〜15、上方パネル、上方パネルのレーン9〜15における矢印によって示されているように)。これら産生物は、中間産生物であり得、S1は、本試験において使用する条件下において、移入細胞中で分解または開裂し得ることを示唆している。このことは、110-kDa S1が、残基362〜790に及び且つS1とS2の双方を認識する可能性を有するウサギ- -S 2によって何故検出されなかったかを説明している。
【0025】
(2) N-末端産生物の検出:ウサギ- -S 1、ウサギ- -S 3およびモノクローナル抗体MAb12-6を、上述した同じ構築物を移入した細胞中のSのN-末端産生物を検出するために使用した。全長構築物(図3b、レーン1および9)においては、200-、140-、64-/62-、55-および38-kDaバンドが上記3種の抗体を使用したときに観察された(矢印で示している)。140-kDaタンパク質(および200-kDaタンパク質)は、これらのN-末端特異性抗体によっても検出されたことを観察し、該タンパク質がSの全長未グリコシル化形であることを確認した。C-末端切断S構築物S 11〜 16においては、各レーンの上部の2つのバンドは、各切断構築物によってコード化されたグリコシル化または未グリコシル化の全長タンパク質であった(図3b、レーン2〜7および9〜15)。64-/62-、55-および38-kDaタンパク質は、全ての構築物を発現する細胞において、これらのN-末端特異性抗体によって検出された(図3b、レーン1〜7および9〜15)。64-/62-、55-および38-kDaタンパク質のサイズは、C-末端切断変異体を使用したとき同じままであり、これらのタンパク質がSのN-末端産生物であったことを確認した。
上記ウマ-α-SARS抗体を使用して上記同じ細胞溶解物中のタンパク質を検出した。この抗体は、N-およびC-末端産生物の双方を検出し得る。上述したN-およびC-末端タンパク質の全てが検出されており(図3b、レーン9〜15、下方パネル)、上述したタンパク質の全てが特異的にS由来であったことを確認した。
110-kDa S1は、使用した条件下においては、移入細胞中で検出されなかった(或いは、ウマ -SARSにより弱いバンドとして検出された、図1b参照)。代りに、より小さいN-末端産生物が検出された。結果は、S1がこの発現系においては急速に分解されることを強力に示唆していた。また、このことは、64/62-kDaタンパク質が感染細胞においても検出されているので、ウイルス感染細胞においても生じ得る(図2a、レーン2参照)。とは言え、この発現およびプロセシング系は、SARS CoV Sタンパク質における開裂部位配列をマッピングすることを可能にしている。
【0026】
開裂部位を有するアミノ酸配列のマッピング:全長Sタンパク質においては、C-末端開裂産生物は、サイズとして90-kDaであった。最も短い切断構築物S 16 (残基1〜797をコード化している)においては、開裂産生物は、サイズとして30-kDaであった。モノクローナル抗体MAbC1は、631から650までの残基を認識した。従って、S1およびS2の推定開裂部位は、アミノ酸残基600辺りの領域に位置し得る。Sの開裂に関与する配列を判定するために、S遺伝子の数種の内部欠失変異体を構築した。欠失配列が開裂部位を含有するならば、90-kDaは、産生されないであろう。先ず、残基601〜800 (S 17)、401〜600 (S 18)および201〜400 (S 19)からの欠失を担持する3種の変異体を発現させた。これらS特異性タンパク質を、ウサギ- -S 10を使用して、ウェスタンブロットにより分析した。図4に示すように、S 17を移入した細胞においては、90-kDaよりも200残基短いタンパク質が観察され(レーン2)、開裂部位は、残基600の上流にあり得ることを示していた。残基401〜600の欠失(S 18)は、90-kDa開裂産生物を実際に消滅させていた(レーン3)。残基201〜400 (S 18)を欠失させた場合、90-kDa産生物は、予測通り依然として産生していた(レーン4)。事実、90-kDa S2よりも僅かに大きい産生物を、S-18を移入した細胞中で検出し、S1がおそらくはS1の中央でエンドプロテアーゼによって開裂されるさらなる証拠を提供していた。このことは、C-末端に対する抗体が1群の中間開裂産生物を検出したという観察と一致していた(図3a参照、上方パネルのレーン8〜15の矢印で示している)。
【0027】
開裂部位を有する配列をさらに明確にするため、3種のさらなる欠失変異体を発現させ、分析した。これらの構築物は、残基531〜550 (S 32)、551〜570 (S 33)および571〜590 (S 34)からの欠失を有するSタンパク質をコード化していた。ウサギ- -S 10を使用してのウェスタンブロット分析(図4b)は、残基551〜579 (S 33)を除去したとき、90-kDaタンパク質の産生は消滅していたことを示していた(レーン3)。531〜550 (S 32)および571〜590 (S 34)の欠失は、90-kDa開裂産生物の検出に影響していなかった(レーン2および4)。このことを、ウマ- -SARSを使用して同じ細胞溶解物を分析したときに確認した(図5、レーン6〜10)。90-kDaおよび55-kDaタンパク質の双方がS 33を発現させたときのみ検出されず、55-kDaタンパク質はS1の解裂または分解産生物であることを示していることが注目された。S 33中で欠失させた20残基の配列は、FGRDVSDFTDSVRDPKTSEIであった。SARS CoV Sのアミノ酸配列に対するBLAST調査によれば、残基551〜570は、現在までに公表された分離株のうちで十分に保存されていることが証明された(データは示していない)。他のコロナウイルスのSタンパク質と異なり、RRARRまたはRRAHR (または、対の塩基性残基でさえもの)モチーフは、この小領域内には存在しない。結果は、SARS CoV Sタンパク質の開裂部位が他のコロナウイルスの開裂部位と異なっていることおよびフリン様プロテアーゼよりもむしろ他の細胞プロテアーゼが含まれていることを強力に示唆している。
【0028】
エンドグリコシダーゼ-H (Endo-H)およびN−グリコシダーゼF (N-gly-F)処理によるSタンパク質のさらなるグリコシル化試験:図6に示すように、移入細胞中のSタンパク質をN-gly-F (レーン1〜4)およびEndo-H (レーン5〜8)によって処理した。結果は、200-kDaタンパク質が、これらの2つの酵素による処理に対して感受性であったので、Sのグリコシル化形であることを示していた。140-kDaタンパク質は、これら酵素による処理が該タンパク質に対して効果を有してなかったので、未グリコシル化の全長Sであった(図6、レーン1、2、5および6)。興味があるのは、200-kDaタンパク質よりも僅かに大きい上部バンドがEndo-H抵抗性であることを見出したことであった(レーン5および6)。結果は、200-kDaタンパク質が、感染細胞培養物において示した(本試験の早期で説明した)ように、おそらくはビリオン中に取込まれる前に、さらに変性されていることを裏付けている。また、結果は、64/62-および38-kDaタンパク質もN-末端産生物のグリコシル化形であることも証明していた。90-および55-kDaタンパク質がグリコシル化形またはEndo-H抵抗性形であるかどうかを検証することは興味あることであろう。しかしながら、開裂の効率は移入細胞においてはるかに低いので、これらのタンパク質は、本試験において使用した条件下では検出し得なかった。
【0029】
Sタンパク質上の種々の領域に対するウサギ抗体の産生:哺乳類発現天然S構築物を使用した以前の研究によれば、中和すべきS1領域内のSの270〜510、548〜567および607〜627(文献11、30)、並びにS2領域内の803〜828をターゲットする抗体も中和特性を有することを知見していることが判明している(文献13)。しかしながら、細菌発現系における、グリコシル化のような翻訳後修飾の欠落は、中和抗体を発現する種々の潜在的部位を示し得る。細菌発現SARS CoV Sタンパク質上の中和領域を試験するために、本発明者等は、細胞外ドメイン全体に及ぶSの5種のDNA構築物をクローニングした(図1)。本発明者等は、S1領域をカバーする2種のフラグメント(S 1、S 3)、S2領域をカバーする2種のフラグメント(S 9、S 10)およびS1とS2の双方の部分をカバーする1種のフラグメント(S 2)をクローニングした。構築物S 9およびS 10は、それぞれ、ヘプタッド繰返し1 (HR1)およびヘプタッド繰返し2 (HR2)領域を含有する。構築物S 1、S 2、S 3、S 9およびS 10用の抗原を、細菌発現系を使用して産生させた。これらの抗原をウサギに注入してそれらそれぞれのターゲッティング領域に対するポリクローナル抗体を産生させた。2羽のウサギを使用してそれぞれの抗原に対する抗体を産生させた。初期免疫化の2週間後、各ウサギに3週間間隔でブースター注入をおこなった。10mlの血液を、各ウサギから、4回目、6回目、8回目、12回目、14回目および16回目の注入後の各時点で採血した。これらのポリクローナル抗体を、ウェスタンブロット分析、免疫沈降法および免疫蛍光法によって特性決定した。
ウェスタンブロットにおいてのSARS CoV Sタンパク質に対するウサギ抗体の特異性:哺乳類細胞中で発現させた全長Sタンパク質に対するウサギ抗体の特異性を、ウェスタンブロット分析により測定する。想定したように、SARS CoV感染から回復した患者(P6)の血清由来の抗-S抗体(文献26は)は、全長Sタンパク質の2つの主要バンド、即ち、140kDa未グリコシル化形および200kDaグリコシル化形を、pKT-Sを移入したCos-7細胞の溶解物中で検出し得た(図2a)。これらのバンドは、陰性対照においては検出されなかったので、特異性Sバンドであった。本発明者等は、5 S組換えタンパク質に対して産生させた抗体を使用して、同様な結果を観察した(図2b〜f)。このことは、各種S構築物に対して産生させた全ての抗体が、線状化Sタンパク質の種々の領域をターゲッティングすることによって、変性条件においてSに特異的に結合し得たことを示している。従って、これらの抗体は、変性全長SARS CoV Sタンパク質に対して特異性である。
【0030】
免疫沈降法においての天然形のSARS CoVの検出:天然Sタンパク質に対する抗体の特異性およびこれら抗体のそれぞれのSタンパク質上のターゲット領域を判定するために、本発明者等は、種々の抗体を使用して、免疫沈降試験を実施した。T7得ワクチニアウイルスを感染させ、pKT-SおよびC-末端欠失変異体、即ち、pKT-S 11、pKT-S 12、pKT-S 13、pKT-S 14、pKT-S 15およびpKT-S 16を移入したCos-7細胞の溶解物を、P6血清並びにS 1、S 2、S 3、S 9およびS 10に対して産生させた5種の抗体によって免疫沈降させた。SARS CoV Sタンパク質およびC-末端欠失組換えタンパク質は、P6血清による免疫沈降によって検出し得る(図3f)。コアグリコシル化Sタンパク質(200kDa)および完全グリコシル化Sタンパク質(210kDa)も、各種組換えSタンパク質に対して産生させた抗体を使用したときに明らかに検出した(図3a〜e、レーン1)。領域S 1、S 2およびS 3をターゲットする抗体は、それぞれ、全長Sおよび全てのC-欠失変異体を検出し得る(図3a〜c)。領域S 9ををターゲットする抗体は、全長SおよびpKT-S 11〜pKT-S 15を検出し得る。pKT-S 16レーンにおいては、pKT-S 16がS 9領域を含んでいなかったので、特異性バンドは検出されなかった(図3d)。S 10対してターゲットする抗体は、pKT-S 11〜pKT-S 13によって発現させたタンパク質バンドを検出するだけである(図3e)。pKT-S 14〜pKT-S 16によって発現させた領域は、S 10領域を含んでなく、従って、ウサギ抗-S 10は、これらのレーンにおいていずれのバンドも検出せず、これらの抗体がこれら抗体のターゲット領域に対し高度に特異性であることを示していた。Sタンパク質の生来の構造は免疫沈降における溶解緩衝液中で保持されており、結果は、天然タンパク質が、上記5種の組換えタンパク質に対して産生させた5種の抗体全部が認識し得るエピトープ(1個以上)を有することを示していた。
【0031】
200kDa Sタンパク質の210kDa EndoH抵抗性形への成熟:2つのS特異性バンドを免疫沈降試験において観察した。上記2つのバンド間の関係を解明するために、本発明者等は、経時試験を実施した。pKT-Sを移入し、35S-metで標識し、さらにウサギ抗-S 10により免疫沈降させたCos-7細胞を、1/2時間、1時間、2時間、4時間および6時間で採集した。結果は、200kDaバンドの漸次的減少を伴っての210kDaバンドの次第の増大を示した(図4、レーン1〜8)。EndoH試験を行なって各バンドがEndoH処理に感受性であったどうかを解明する。Cos-7細胞をT7ワクチニアウイルス感染に供し、pKT-Sを移入し、EndoH酵素で処理したが、対照細胞はEndoHで処理しなかった。結果は、210kDaバンドがEndoH抵抗性であり、200kDaバンドがEndoH感受性であることを示していた(図4、レーン9、10)。従って、結果は、200kDaバンドの210kDaバンドへの成熟を実証していた。
Cos-7細胞表面上でのSARS CoV Sタンパク質の検出:産生させた各抗体が天然形のSタンパク質を認識し得るさらなる証拠を提供するために、本発明者等は、免疫蛍光試験を行なった。間接免疫蛍光試験において使用したCos-7細胞は、非浸透性であった。上述したようなT7ワクチニアウイルスによる感染およびpKT-Sによる移入後に、十分な時間(移入後8時間)で、Sタンパク質を発現させ、細胞表面に移送させたところ、これらのタンパク質は、各種Sタンパク質構築物に対して産生させた抗体のいずれを使用しても明らかに検出し得た(図5)。緑色蛍光は、Cos-7細胞に結合している各種一次抗体の位置を示していた。Sタンパク質は、Cos-7細胞により、それらの生来の構造において発現し、プロセシングされていた。S 1、S 2、S 3、S 9およびS 10に対する抗体の細胞表面上のSタンパク質への結合は、これらの抗体がSARS CoV Sタンパク質の生来構造に対して特異性であり感受性であるというさらなる証拠を提供している。
S2内の領域は、中和活性を顕在化させる:各種Sタンパク質を注入したウサギ由来の血清の全てを、各採血後の中和活性について試験した。pGEX-S 1、S 2、S 3およびS 9を注入したウサギにおける結果は、陰性応答を示した。pGEX-S 10を注入したウサギ由来の血清は、4回目注入後に中和活性を示した。SARS CoVを200 TCID50で使用した初期試験は、8回目の注入後の血清採血から始まるウサギ抗-S 10採血の全てにおいて高力価(1:364)の中和抗体を示した(表3)。16回目の注入は最後のブースター注入であり、ウサギは、この段階で死亡させた。上記の結果を確認するため、pGEX-S 10による4回目、6回目、8回目、12回目、14回目および16回目の注入後の血清サンプルを、SARS CoVを1000 TCID50で使用する中和試験に送った(表4)。中和活性は、1000 TCID50において1:189.2ほど高く、SARS患者において検出した力価よりも高いとはいえないまでも、それに匹敵していることが判明した。SARS患者における100 TCID50での抗体応答は、210日間に亘って1:150〜1:475の範囲である(文献35)。この結果は、S 10がSARS CoVに対する中和抗体の産生を刺激し得る領域である強力な証拠を提供している。
【0032】
表3:200 TCID50での中和試験
表4:1000 TCID50での中和試験
【0033】
SEQ ID NO.1:SARS CoV、2774株のクローン12のスパイク(S)遺伝子の全長ヌクレオチド配列。核酸1から3765までの線状のRNA。
生物体:SARSコロナウイルス2774株ウイルス;ssRNAプラス鎖ウイルス、DNA段階無し;ニドウイルス目;コロナウイルス科;コロナウイルス。
SEQ ID NO.2:SARS-CoV、2774株のクローン12のスパイク(S)遺伝子の全長アミノ酸配列。アミノ酸1〜1255。
SEQ ID NO.3:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 10フラグメントのヌクレオチド配列。核酸3087〜3581。
SEQ ID NO.4:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 10フラグメントのアミノ酸配列。アミノ酸1029〜1192。
SEQ ID NO.5:SARS-CoV、2774株のクローン12のスパイク(S)遺伝子の中和フラグメントのアミノ酸配列。アミノ酸1055〜1192。
SEQ ID NO.6:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 11フラグメントのアミノ酸配列。アミノ酸1〜1232。
SEQ ID NO.7:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 17フラグメントのアミノ酸配列。601から800までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.8:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 18フラグメントのアミノ酸配列。401から600までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.9:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 19フラグメントのアミノ酸配列。201から400までの欠失を有するアミノ酸1〜1255。
SEQ ID NO.10:SARS CoV、2774株のクローン12のスパイク(S)遺伝子のS 20フラグメントのアミノ酸配列。30から200までの欠失を有するアミノ酸1〜1255。
【0034】
2003年12月10日に出願された米国仮特許出願第60/528,596号を含む本明細書において引用した文献は、すべて、その全体を参考として本明細書に合体させる。
【0035】
参考文献
【0036】
【0037】
【0038】
【0039】
【0040】
【図面の簡単な説明】
【0041】
【図1】Sタンパク質検出のためのウェスタンブロット分析:Cos7細胞由来の溶解物に、プラスミドpKT-Sを移入する(レーン1、3、5、7、9、11)。Cos7細胞由来の溶解物に、陰性対照としての挿入物を含まないプラスミドを移入する(レーン2、4、6、8、10、12)。BenchMark Prestained Protein Ladder(Invitrogen社)を右側のマーカーとして使用する。200kDaおよび140kDaバンドは、特異性Sタンパク質バンドである。
【図2】放射線標識免疫沈降法によって測定した各抗体の特異性:Cos7細胞の溶解物に、それぞれS、S 11、S 12、S 13、S 14、S 15およびS 16を移入する(レーン1、2、3、4、5、6および7)。Cos7細胞の溶解物に、陰性対照としての挿入物を含まないプラスミドを移入する(レーン8)。High-Range Rainbow Molecular Weight Markers(Amersham社)を、左側のマーカーとして使用する。
【図3】Sタンパク質のグリコシル化の経時変化:pKT-Sを移入したCos7細胞の溶解物を、それぞれ、0時間、0.5時間、1時間、2時間、4時間および6時間で採集する(レーン1、2、3、4、5、6および7)。挿入物を含まないプラスミドを移入したCos7細胞の溶解物を、陰性対照として、6時間で採集する(レーン8)。Cos7細胞の溶解物にpKT-Sを移入し、Endo-H処理する(移入後6時間)。pKT-Sを移入し、陰性対照として移入後6時間で採集したCos7細胞の溶解物。ウサギ抗-S10をSタンパク質の検出において使用した。
【図4】一時的に移入したCos7細胞の表面上で発現させたSタンパク質:Cos7細胞の表面上で発現させたSタンパク質を、それぞれ、緑色蛍光によって示すように、Rb -S 1、Rb -S 2、Rb -S 3、Rb -S 9およびRb -S 10により検出した。上記細胞の位置は、白色灯を使用して得た。蛍光は、対照試験物においては検出されなかった。
【図5】S1/S2の開裂部位を含む配列のマッピング:全長S (レーン1および6)、内部欠失突然変異体S 32 (レーン2および7)、S 33 (レーン3および8)および残基531〜550、551〜570および571〜590を欠失するS 34 (レーン4および9)を、それぞれ、Cos-7細胞中で発現させた。ウェスタンブロット分析を、ウサギ- -S 10(左側パネル)およびウマ- -SARS(右側パネル)を使用して実施した。各タンパク質を、10%PAGEゲル上で分離した。特異性タンパク質の分子量を右側に、マーカーの分子量左側にキロダルトンで示している。上記各ゲルをストリッピングし、負荷対照としてのマウス抗- -アクチンで再精査した。
【図6】N-グリコシダーゼFおよびエンドグリコシダーゼHによるS由来タンパク質の処理:全長SをCos-7細胞中で発現させた。細胞を溶解緩衝液中に再懸濁させた。各サンプルを、(a)N-グリコシダーゼF (N-gly-F)および(b)エンドグリコシダーゼH (Endo-H)で処理(+)またはmock処理(−)した。タンパク質を10%PAGEゲル上で分離し、ウェスタンブロットをウサギ- -S1で実施した。mock移入細胞由来の溶解物を陰性対照として使用した(レーン3、4、7および8)。特異性タンパク質の分子量を右側に、マーカーの分子量を左側にキロダルトンで示している。
【図1a】
【図1b】
【特許請求の範囲】
【請求項1】
中和ドメインを含むことを特徴とする、SEQ ID NO.1のペプチドまたはタンパク質フラグメント。
【請求項2】
SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10の群から選ばれたアミノ酸配列を含む、請求項1記載のペプチドまたはタンパク質。
【請求項3】
コロナウイルスのSタンパク質のHR2ヘプタッド領域を有する、請求項1記載のペプチドまたはタンパク質。
【請求項4】
SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10から本質的になる群から選ばれた、請求項1記載のペプチドまたはタンパク質。
【請求項5】
前記フラグメントが、SEQ ID NO.4を含む、請求項1記載のペプチドまたはタンパク質。
【請求項6】
前記フラグメントが、SEQ ID NO.5を含む、請求項1記載のペプチドまたはタンパク質。
【請求項7】
下記の工程を含むことを特徴とする、コロナウイルスのSタンパク質のフラグメントの産生方法:
a)細胞を、コロナウイルスのSタンパク質のフラグメントをコード化する核酸により形質転換する工程(前記フラグメントは中和ドメインを含み、前記核酸は細胞内でのその発現を誘導し得る調節配列と操作的に関連する);
b)前記タンパク質フラグメントを前記細胞内で発現させる工程;および、
c)前記タンパク質フラグメントを分離する工程。
【請求項8】
SEQ ID NO.1のペプチドまたはタンパク質フラグメントに対する抗体であって、前記フラグメントがSEQ ID NO.5を含むことを特徴とする抗体。
【請求項9】
前記フラグメントが、SEQ ID NO.4を含む、請求項8記載の抗体。
【請求項10】
前記ペプチドまたはタンパク質が、コロナウイルスSタンパク質のHR2ヘプタッド領域を含む、請求項8記載の抗体。
【請求項11】
前記ペプチドまたはタンパク質が、SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10からなる群から選ばれる、請求項8記載の抗体。
【請求項12】
患者から収集した細胞の少なくとも1部に請求項8記載の抗体を適用する工程を含むことを特徴とする、患者のSARSコロナウイルス感染の検出方法。
【請求項13】
請求項8記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項14】
請求項1記載のペプチドまたはタンパク質を患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法。
【請求項15】
請求項8記載の抗体を患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法。
【請求項16】
有効量の請求項1記載のペプチドまたはタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防するワクチン。
【請求項17】
有効量の請求項8記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防するワクチン。
【請求項18】
S 1、S 2、S 3およびS 9からなる群から選ばれたSタンパク質フラグメントに対する抗体。
【請求項19】
患者から収集した細胞の少なくとも一部に、請求項18記載の抗体を適用する工程を含む、患者のSARSコロナウイルス感染の検出方法。
【請求項20】
請求項18記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項21】
コロナウイルスの成熟グリコシル化スパイクタンパク質。
【請求項22】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項23】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項24】
前記コロナウイルスが、SARSコロナウイルスである、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項25】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項26】
下記の工程を含むことを特徴とする、コロナウイルスの成熟グリコシル化スパイクタンパク質の産生方法:
a)細胞を、コロナウイルスのスパイクタンパク質またはその一部をコード化する核酸により形質転換する工程;
b)前記スパイクタンパク質を前記細胞中で発現させる工程;および、
c)前記コロナウイルスの成熟グリコシル化スパイクタンパク質を分離する工程。
【請求項27】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項26記載の方法。
【請求項28】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項26記載の方法。
【請求項29】
前記コロナウイルスが、SARSコロナウイルスである、請求項26記載の方法。
【請求項30】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項26記載の方法。
【請求項31】
前記細胞を、ヒト肺細胞およびcos-7細胞からなる群から選択する、請求項26記載の方法。
【請求項32】
下記の工程を含むことを特徴とする、コロナウイルスの成熟グリコシル化スパイクタンパク質のスクリーニング方法:
a) スパイクタンパク質を分離する工程;
b) 分離したスパイクタンパク質をEndo-Hにより免疫沈降させる工程;および、
c) 成熟グリコシル化スパイクタンパク質である残存スパイクタンパク質を検出する工程。
【請求項33】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項32記載の方法。
【請求項34】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項32記載の方法。
【請求項35】
前記コロナウイルスが、SARSコロナウイルスである、請求項32記載の方法。
【請求項36】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項32記載の方法。
【請求項37】
コロナウイルスの成熟グリコシル化スパイクタンパク質またはその一部に対する抗体。
【請求項38】
前記成熟グリコシル化スパイクタンパク質が、抗膜貫通ドメイン(TMD)を含有する、請求項37記載の抗体。
【請求項39】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項37記載の抗体。
【請求項40】
前記コロナウイルスが、SARSコロナウイルスである、請求項37記載の抗体。
【請求項41】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項37記載の抗体。
【請求項42】
患者から収集した細胞の少なくとも一部に、請求項37記載の抗体を適用する工程を含む、患者のSARSコロナウイルス感染の検出方法。
【請求項43】
請求項37記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項44】
請求項37記載の抗体を必要とする患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の進行を遅延させる方法。
【請求項45】
請求項21記載の成熟グリコシル化スパイクタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項46】
請求項37記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項47】
請求項22記載の成熟グリコシル化スパイクタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項48】
請求項38記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項1】
中和ドメインを含むことを特徴とする、SEQ ID NO.1のペプチドまたはタンパク質フラグメント。
【請求項2】
SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10の群から選ばれたアミノ酸配列を含む、請求項1記載のペプチドまたはタンパク質。
【請求項3】
コロナウイルスのSタンパク質のHR2ヘプタッド領域を有する、請求項1記載のペプチドまたはタンパク質。
【請求項4】
SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10から本質的になる群から選ばれた、請求項1記載のペプチドまたはタンパク質。
【請求項5】
前記フラグメントが、SEQ ID NO.4を含む、請求項1記載のペプチドまたはタンパク質。
【請求項6】
前記フラグメントが、SEQ ID NO.5を含む、請求項1記載のペプチドまたはタンパク質。
【請求項7】
下記の工程を含むことを特徴とする、コロナウイルスのSタンパク質のフラグメントの産生方法:
a)細胞を、コロナウイルスのSタンパク質のフラグメントをコード化する核酸により形質転換する工程(前記フラグメントは中和ドメインを含み、前記核酸は細胞内でのその発現を誘導し得る調節配列と操作的に関連する);
b)前記タンパク質フラグメントを前記細胞内で発現させる工程;および、
c)前記タンパク質フラグメントを分離する工程。
【請求項8】
SEQ ID NO.1のペプチドまたはタンパク質フラグメントに対する抗体であって、前記フラグメントがSEQ ID NO.5を含むことを特徴とする抗体。
【請求項9】
前記フラグメントが、SEQ ID NO.4を含む、請求項8記載の抗体。
【請求項10】
前記ペプチドまたはタンパク質が、コロナウイルスSタンパク質のHR2ヘプタッド領域を含む、請求項8記載の抗体。
【請求項11】
前記ペプチドまたはタンパク質が、SEQ ID NO.4、SEQ ID NO.5、SEQ ID NO.6、SEQ ID NO.7、SEQ ID NO.8、SEQ ID NO.9またはSEQ ID NO.10からなる群から選ばれる、請求項8記載の抗体。
【請求項12】
患者から収集した細胞の少なくとも1部に請求項8記載の抗体を適用する工程を含むことを特徴とする、患者のSARSコロナウイルス感染の検出方法。
【請求項13】
請求項8記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項14】
請求項1記載のペプチドまたはタンパク質を患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法。
【請求項15】
請求項8記載の抗体を患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防する方法。
【請求項16】
有効量の請求項1記載のペプチドまたはタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防するワクチン。
【請求項17】
有効量の請求項8記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたはその発症を予防するワクチン。
【請求項18】
S 1、S 2、S 3およびS 9からなる群から選ばれたSタンパク質フラグメントに対する抗体。
【請求項19】
患者から収集した細胞の少なくとも一部に、請求項18記載の抗体を適用する工程を含む、患者のSARSコロナウイルス感染の検出方法。
【請求項20】
請求項18記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項21】
コロナウイルスの成熟グリコシル化スパイクタンパク質。
【請求項22】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項23】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項24】
前記コロナウイルスが、SARSコロナウイルスである、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項25】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項21記載の成熟グリコシル化スパイクタンパク質。
【請求項26】
下記の工程を含むことを特徴とする、コロナウイルスの成熟グリコシル化スパイクタンパク質の産生方法:
a)細胞を、コロナウイルスのスパイクタンパク質またはその一部をコード化する核酸により形質転換する工程;
b)前記スパイクタンパク質を前記細胞中で発現させる工程;および、
c)前記コロナウイルスの成熟グリコシル化スパイクタンパク質を分離する工程。
【請求項27】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項26記載の方法。
【請求項28】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項26記載の方法。
【請求項29】
前記コロナウイルスが、SARSコロナウイルスである、請求項26記載の方法。
【請求項30】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項26記載の方法。
【請求項31】
前記細胞を、ヒト肺細胞およびcos-7細胞からなる群から選択する、請求項26記載の方法。
【請求項32】
下記の工程を含むことを特徴とする、コロナウイルスの成熟グリコシル化スパイクタンパク質のスクリーニング方法:
a) スパイクタンパク質を分離する工程;
b) 分離したスパイクタンパク質をEndo-Hにより免疫沈降させる工程;および、
c) 成熟グリコシル化スパイクタンパク質である残存スパイクタンパク質を検出する工程。
【請求項33】
前記成熟グリコシル化スパイクタンパク質が、膜貫通ドメイン(TMD)を含有する、請求項32記載の方法。
【請求項34】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項32記載の方法。
【請求項35】
前記コロナウイルスが、SARSコロナウイルスである、請求項32記載の方法。
【請求項36】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項32記載の方法。
【請求項37】
コロナウイルスの成熟グリコシル化スパイクタンパク質またはその一部に対する抗体。
【請求項38】
前記成熟グリコシル化スパイクタンパク質が、抗膜貫通ドメイン(TMD)を含有する、請求項37記載の抗体。
【請求項39】
前記成熟グリコシル化スパイクタンパク質が、210KDaタンパク質である、請求項37記載の抗体。
【請求項40】
前記コロナウイルスが、SARSコロナウイルスである、請求項37記載の抗体。
【請求項41】
前記コロナウイルスが、SARSコロナウイルス株、2774である、請求項37記載の抗体。
【請求項42】
患者から収集した細胞の少なくとも一部に、請求項37記載の抗体を適用する工程を含む、患者のSARSコロナウイルス感染の検出方法。
【請求項43】
請求項37記載の抗体を含有する、SARSコロナウイルスの検出用キット。
【請求項44】
請求項37記載の抗体を必要とする患者に投与することを特徴とする、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の進行を遅延させる方法。
【請求項45】
請求項21記載の成熟グリコシル化スパイクタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項46】
請求項37記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項47】
請求項22記載の成熟グリコシル化スパイクタンパク質を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【請求項48】
請求項38記載の抗体を含む、重症後天性呼吸器症候群を有する患者を治療するまたは重症後天性呼吸器症候群の発症を予防するワクチン。
【図2】

【図3a】
【図3b】
【図4】
【図5】
【図6】
【図3a】
【図3b】
【図4】
【図5】
【図6】
【公表番号】特表2008−505050(P2008−505050A)
【公表日】平成20年2月21日(2008.2.21)
【国際特許分類】
【出願番号】特願2006−543656(P2006−543656)
【出願日】平成16年12月10日(2004.12.10)
【国際出願番号】PCT/IB2004/004073
【国際公開番号】WO2005/056585
【国際公開日】平成17年6月23日(2005.6.23)
【出願人】(506200337)エイジェンシー フォア サイエンス テクノロジー アンド リサーチ (1)
【Fターム(参考)】
【公表日】平成20年2月21日(2008.2.21)
【国際特許分類】
【出願日】平成16年12月10日(2004.12.10)
【国際出願番号】PCT/IB2004/004073
【国際公開番号】WO2005/056585
【国際公開日】平成17年6月23日(2005.6.23)
【出願人】(506200337)エイジェンシー フォア サイエンス テクノロジー アンド リサーチ (1)
【Fターム(参考)】
[ Back to top ]