
SIRT1およびLXRのコレステロール制御複合体ならびに使用法
SIRT1とLXRとのコレステロール調節複合体および使用方法が開示されている。SIRT1は、LXR要素に結合しているLXRと複合体を形成する。複合体を形成する方法、複合体の形成を調節する作用剤を同定する方法、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法、哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法、正常未満のSIRT1活性レベルを有すると見なされる対象を処置する方法、候補物質がLXR依存性プロセスを調節するか否かを評価する方法、および候補物質がLXRのSIRT1依存性作用を調節するか否かを評価する方法が、開示されている。

【発明の詳細な説明】
【背景技術】
【0001】
コレステロールの恒常性は、食事の摂取、デノボ合成、輸送、代謝および排泄の間の均衡により維持されている。低濃度の高密度リポタンパク質(HDL)および高濃度の低密度リポタンパク質(LDL)は、高コレステロール血症、高グリセリド血症、ならびに中年および更に高年齢の人々の主要な死因である加齢性疾患の心血管疾患1およびアルツハイマー病2−6のリスク増加と関係している。遺伝因子、環境の双方が、心血管疾患およびアルツハイマー病の進行の原因となり、こうした疾患のリスクは年齢と共に増加する。しかし、遺伝因子が環境を感知してこうした加齢性疾患を媒介する根本的な機構の理解は、不十分である。
【0002】
Sir2(silent information regulator 2)は、環境変化に応答する生涯の決定的な調節因子である。Sir2遺伝子は、酵母、C.elegans7およびDrosophila8,9における寿命決定因子である。酵母10,11、Drosophila8,9のいずれでも、Sir2活性は、カロリー制限で得られる寿命延長にも必要である。生化学的には、Sir2およびその同種物質(サーチュイン)は、高度に保存されたNAD+依存性タンパク質デアセチラーゼの一群である12−14。NAD+を必要とすることにより、サーチュインは、細胞代謝をモニターし、加齢に影響する細胞過程を調節することができる。哺乳類において、Sir2タンパク質の哺乳類オーソログ(orthologue)であるSIRT1は、多数のタンパク質基質を有し、細胞周期、アポトーシス、酸化的ストレス応答、神経変性などの加齢過程を調節することができる15−20。こうした過程において、SIRT1は、細胞死と細胞生存との均衡を変化させ、それによりストレス耐性を生じることができる21。
【0003】
コレステロールおよび脂肪の代謝は、通常の多くの細胞因子および環境因子により調節される。脂質感知核内受容体の2種のサブクラスである、ペルオキシソーム増殖剤応答性受容体(PPAR)および肝X受容体(LXR)は、脂質および炭水化物の代謝に決定的な役割を果たしている。最近の研究によれば、白色脂肪組織では、SIRT1は、核内受容体コリプレッサー(N−CoR)を介して核内受容体PPARγと相互作用し、食物欠乏時に脂肪代謝を促進することが示された22(非特許文献1)。SIRT1は、PPARγコアクチベーターのPGC−1αと相互作用し、それを修飾することにより、肝グルコースの恒常性を調節することもできる23,24(非特許文献2、非特許文献3)。特許文献1では、ヒト対象の高密度リポタンパク質(HDL)の血中濃度は、対象にトランスレスベラトロールを投与することにより増加させることができ、この物質は、対象の低密度リポタンパク質(LDL)の血中濃度を低下させることが報告されている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】米国特許第6,048,903号明細書
【非特許文献】
【0005】
【非特許文献1】Picard,F.ら、Nature(2004)429、771−6
【非特許文献2】Rodgers,J.T.ら、Nature(2005)434、113−8
【非特許文献3】Nemoto,S.ら、J Biol Chem(2005)280,16456−60
【発明の概要】
【発明が解決しようとする課題】
【0006】
脂肪代謝およびグルコース恒常性におけるSIRT1の関与によって、このタンパク質が哺乳類においてコレステロール恒常性も調節し得るという可能性が得られる。本発明は、コレステロール恒常性に関するこうした新規な機構を取り扱う。
【課題を解決するための手段】
【0007】
発明の要旨
本発明は、全般的に、コレステロール恒常性およびコレステロール逆輸送の分野で有用な組成物および方法に関する。
【0008】
第1の態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する、単離した複合体を提供する。ある種の実施形態では、この複合体は、LXR応答エレメントを更に含有する。
【0009】
関連する態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体を形成する方法を提供する。この方法には、哺乳類SIRT1タンパク質、哺乳類LXRタンパク質、およびLXR応答エレメントを含む細胞内核酸の断片を含有する組成物を組み合わせることが含まれる。
【0010】
その上更なる態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を調節する、作用剤を同定する方法を提供する。この方法は、
哺乳類SIRT1タンパク質、哺乳類LXRタンパク質、およびLXR応答エレメントを含む細胞内核酸の断片を含有する組成物を組み合わせ、それにより複合体組成物を提供する工程、
更に、哺乳類SIRT1タンパク質を含有する組成物、哺乳類LXRタンパク質を含有する組成物、もしくは細胞内核酸の該断片を含有する組成物の1つを、組合せ工程の前に該作用剤を含む第4の組成物と接触させる、または複合体組成物を組合せ工程の後に該作用剤を含む第4の組成物と接触させるステップ、ならびに
複合体の形成が、該作用剤によって調節されているか否かを決定するステップ
を包含する。
この方法のある種の実施形態では、該作用剤は複合体の形成を増加させる。追加の実施形態では、判定ステップは、該作用剤を含有していない対照組成物との比較により行われる。
【0011】
更に追加の態様では、本発明は、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法であって、SIRT1活性を刺激する作用剤をその哺乳動物に投与することを含む方法を提供する。有利な実施形態では、該作用剤はT0901317を含む。
【0012】
その上更なる態様では、本発明は、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法であって、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体の形成を促進する作用剤をその哺乳動物に投与することを含む方法を提供する。この方法の有利な実施形態では、該作用剤は22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む。
【0013】
更に追加の態様では、本発明は、哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含有する核酸をその細胞に導入することを含む方法を提供する。この方法の幾つかの実施形態では、タンパク質デアセチラーゼは、真核生物Sir2または哺乳類SIRT1である。この方法の更に他の実施形態は、T0901317などのSIRT1活性を刺激する作用剤と該細胞を更に接触させるステップを含む。この方法の他の実施形態では、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体の形成を促進する作用剤と、該細胞を更に接触させる。ある種の実施形態では、複合体促進性作用剤は、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む。
【0014】
本発明の更なる態様は、SIRT1活性のレベルが正常未満と見なされる対象を処置する方法を提供する。このような対象は、正常未満と見なされる、HDL−コレステロール濃度およびHDL−コレステロール対LDL−コレステロールの比率を示す。この方法は、種特異的なSIRT1をコードする核酸をその対象に投与することを含み、その核酸は、対象の細胞内にSIRT1の治療有効量を発現するのに有効である。
【0015】
更なる態様では、本発明は、候補物質がLXR依存性プロセスを調節するか否かを評価する方法を提供する。この方法は、LXREに作動可能に駆動されるレポーター遺伝子を保持するプラスミドを細胞に移入し、その細胞を候補物質と接触させ、更に、候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定することによって、候補物質の有無により検出される差異が、候補物質がLXR依存性プロセスを調節することを示すことを含む。
【0016】
追加の態様では、本発明は、候補物質がLXRのSIRT1依存性作用を調節するか否かを評価する方法を提供する。この方法は、SIRT1遺伝子を保持するプラスミドを細胞に移入し、LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するプラスミドを、その細胞に更に移入し、その細胞を候補物質と接触させ、更に、候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定し、それにより、候補物質の有無により検出される差異が、候補物質がLXRのSIRT1依存性作用を調節することを示すことを含む。
【0017】
更なる態様では、本発明は、候補物質がSIRT1のLXRとの相互作用を調節するか否かを評価する方法を提供する。この方法は、エピトープタグ付きLXRをコードする配列を保持するベクターで細胞をトランスフェクトし、その細胞を候補物質と接触させ、その細胞を溶解し、エピトープタグに特異的な抗体を細胞溶解物と接触させ、更に、SIRT1とLXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収し、SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行し、それにより、候補物質の不在下と比較した、候補物質の存在下で検出した複合体形成の差異が、候補物質がSIRT1のLXRとの相互作用を調節することを示すことを含む。
【図面の簡単な説明】
【0018】
【図1】SIRT1の機能喪失がコレステロール恒常性の変化を起こすことを示す図である。(a〜c)野生型(塗潰し棒)、SIRT1+/−(斜線付き棒)およびSIRT1−/−(白抜き棒)の各マウスからの血漿試料を、(a)総コレステロール、(b)HDLコレステロール、(c)LDLコレステロールについて分析した。(d)野生型(塗潰し菱形)およびSIRT1−/−(白抜き菱形)の各マウスにおけるFPLC血漿リポタンパク質のコレステロール像。(e)野生型(塗潰し棒)およびSIRT1−/−(白抜き棒)の各マウスのHDL画分における相対脂質含量。(f)野生型対照(塗潰し棒)および同腹子SIRT1−/−(白抜き棒)の精巣および肝臓からの総コレステロール。(g)ニコチンアミドによる前処理有りまたは無しの、マウス初代胚性線維芽細胞(MEF)(左パネル)またはTHP1ヒト単球(右パネル)からの[3H]コレステロールのApoAI媒介排出。
【図2】ABCA1の発現およびLXR核内受容体との相互作用を示す図である。(a)肝臓(左パネル、定量的リアルタイムPCRで決定)、ならびに精巣および卵巣(右パネル、野生型(+/+)およびSIRT1−/−(−/−)動物のノーザンブロットで決定)におけるABCA1 mRNAの発現。(b)SIRT1活性の抑制はABCA1プロモーター活性を低下させる。(c)MEFにおけるSIRT1欠損のABCA1プロモーター活性に対する効果。(d)SIRT1とABCA1プロモーターのLXREとの相互作用。(e)SIRT1はLXREαおよびLXREβと相互作用する。
【図3】SIRT1欠損が、インビボでのLXRアゴニストに対する正常な応答を損なうことを示す図である。(a)SIRT1−/−マウスの肝臓におけるLXR標的mRNA。(b)LXRアゴニストT0901317で処理したマウスの肝臓におけるLXR標的mRNAの相対量。(c)T0901317を有する、または有していない、野生型およびSIRT1−/−マウスにおけるABCA1タンパク質濃度。(d)T0901317投与後のトリグリセリド濃度。SIRT1−/−マウスでは、血漿および肝臓におけるトリグリセリド蓄積量が低下する。(e)T0901317供給後の血漿におけるコレステロールの誘導。(f)T0901317供給の前(塗潰した四角)および後(白抜きの四角)の野生型およびSIRT1−/−マウスにおける、代表的なFPLC血漿リポタンパク質像。
【図4】SIRT1が、LXRタンパク質の脱アセチル化によりLXRの安定性および活性を調節することを示す図である。(a)LXRタンパク質発現SIRT1に対するSIRT1の効果。(b)RNA干渉によるSIRT1のノックダウンがLXR濃度に及ぼす効果。(c)LXRのタンパク質発現に対する阻害剤の効果。(d)SIRT1はLXRのユビキチン化を促進する。(e)インビボでのLXRのアセチル化、および22(R)−HCおよび9−シスRAによるLXRの活性化。(f)SIRT1はHA−LXRβを脱アセチル化する。
【図5】SIRT1またはプロテアソーム媒介LXR分解の抑制が、ABCA1プロモーター上のLXRの転写活性に及ぼす効果の概要を表示する図である。
【発明を実施するための形態】
【0019】
発明の詳細な説明
配列の表
表1.本開示における配列の表
【0020】
【化1】
本明細書で使用する場合、HDL(高密度リポタンパク質)は、アポリポタンパク質A、C、DおよびEを含めた、高いタンパク質比率を有する一部類の血漿リポタンパク質に関する。HDLは、血漿中で遊離しているかエステル化されているかに関係なく、コレステロールをHDL−コレステロール複合体として取り込み、輸送する。HDLという用語は、HDL粒子に結合しているコレステロールを指定するために、文脈に応じて使用し得る。
【0021】
本明細書で使用する場合、LDL(低密度リポタンパク質)は、コレステロール、コレステロールエステルおよびトリグリセリドを含んだ、高い脂質比率を有する一部類の血漿リポタンパク質に関する。その中には、主にアポリポタンパク質B−100およびアポリポタンパク質Eが含まれる。LDLは、血漿中でコレステロールを取り込み、輸送する。LDLという用語は、LDL粒子に結合しているコレステロールを指定するために、文脈に応じて使用し得る。
【0022】
本明細書で使用する場合、用語「単離された」および同類用語は、自然状態で、ある物質の出現に付随する少なくとも1種の成分の除去により、自然出現状態から精製された前記物質に関する。用語「精製された」は、「単離された」と同義に使用し得る。これらの用語は、該物質が何らかの所定の純度を有することを必要としないことが理解される。
【0023】
本明細書で使用する場合、用語「複合体」および同類用語は、単離手順を受け、そのような結合体として特性決定されるのに十分に、相互に安定に結合している2種以上の物質の結合体に関する。複合体の各成分が相互に直接結合してもよく、またはその結合体は、同定した各成分が相互に直接には接触しないように、介在促進成分を含んでもよい。複合体は、各成分を結合する非共役結合性相互作用、または各成分間の共役結合によって形成し得る。
【0024】
本明細書で使用する場合、用語「コレステロール」とは、コレステロール、コレステリルエステル双方としてのコレステロールの出現全体を指す。当業者には明らかと思われる特定の文脈では、「コレステロール」は、遊離コレステロールだけを指すために、本明細書では使用することもある。
【0025】
本明細書で使用する場合、用語「処置」ならびに同類の用語および語句は、疾患もしくは病状、症状または疾患素因を有する対象に対する治療剤の適用または投与、あるいはその対象由来の単離した組織または細胞系に対する治療剤の適用または投与に関する。処置は、その治療もしくは治癒を促進すること、または、疾患、症状もしくは疾患素因を軽減、緩和、改変、治癒、改善、改良する、もしくはそれに影響を及ぼすことを意図している。
【0026】
本明細書で使用する場合、「治療有効量」および「予防有効量」という語句は、疾患の処置において治療上有益となる量、またはそれぞれ、疾患を予防するもしくはその重度を低減する効果を指す。治療上有効な特定の量は、治療される対象における応答の評価を用いて、普通の医者により容易に決定することができ、疾患の特質、対象の病歴および年齢、疾患の段階、ならびに他の治療剤の投与などの、当技術分野で公知の要因に応じて変化し得る。
【0027】
本明細書で使用する場合、「医薬組成物」は、薬理有効量のポリヌクレオチドおよび医薬として許容可能な担体を含む組成物に関する。本明細書で使用する場合、「薬理有効量」、「治療有効量」または単に「有効量」とは、意図した薬理的、治療的または予防的結果を生じるのに有効な阻害性ポリヌクレオチドの量を指す。例えば、疾患または障害に関連する臨床パラメーターに測定し得る最小限の変化が少なくともあるときに、所与の臨床処置が有効であると見なす場合、その疾患または障害を処置するための薬物の治療有効量は、そのパラメーターに少なくとも変化度を起こすのに必要な量である。
【0028】
用語「医薬として許容可能な担体」とは、少なくとも、生理学的に許容可能であり、規制当局により認可され得る、治療剤投与用の組成物を指す。
SIRT1ポリペプチド
本明細書で使用する場合、「SIRT1ポリペプチド」、「SIRT1タンパク質」ならびに関連する用語および語句は、野生型SIRT1、突然変異体SIRT1、変異体SIRT1、ならびに生物活性なその断片および成熟形態に関する。本発明で使用すべき重要なSIRT1タンパク質は、ヒトSIRT1である。ヒトSIRT1のアミノ酸配列は、GenBank寄託番号NP_036370に示され、アミノ酸残基747個からなるものと開示されており、従来の1文字アミノ酸記号を用いて表2に示される(International Union Of Biochemistry And Molecular Biology,Recommendations on Biochemical & Organic Nomenclature, Symbols &Terminology etc., Part 1, Section A: Amino-Acid Nomenclature, Section 3AA-1,Names Of Common Alpha-Amino Acids, http://www.chem.qmul.ac.uk/iubmb/ およびJ.Biol. Chem., 1985年、260号、14〜42頁)。
【0029】
表2.ヒトSIRT1のアミノ酸配列
【0030】
【化2】
マウスSIRT1のアミノ酸配列は、GenBank寄託番号NP_062786に示され、アミノ酸残基737個からなるものと開示されており、表3に示される。
【0031】
表3.マウスSIRT1のアミノ酸配列
【0032】
【化3】
ヒト肝X受容体αのアミノ酸配列は、GenBank寄託番号NP_062786に示され、アミノ酸残基447個からなるものと開示されており、表4に示される。
【0033】
表4.ヒト肝X受容体αのアミノ酸配列
【0034】
【化4】
ヒト肝X受容体βのアミノ酸配列は、GenBank寄託番号NP_009052.3に示され、アミノ酸残基461個からなるものと開示されており、表5に示される。
【0035】
表5.ヒト肝X受容体βのアミノ酸配列
【0036】
【化5】
マウス肝X受容体αのアミノ酸配列は、GenBank寄託番号NP_038867に示され、アミノ酸残基445個からなるものと開示されており、表6に示される。
【0037】
表6.マウス肝X受容体αのアミノ酸配列
【0038】
【化6】
マウス肝X受容体βのアミノ酸配列は、GenBank寄託番号NP_033499.1に示され、アミノ酸残基446個からなるものと開示されており、表7に示される。
【0039】
表7.マウス肝X受容体βのアミノ酸配列
【0040】
【化7】
一般に、本発明の方法および組成物に使用されるような「SIRT1ポリペプチド」および多様な「LXRポリペプチド」には、表2〜7に示すようなヒト野生型のSIRT1およびLXRポリペプチド、またはマウス野生型のSIRT1およびマウスLXRポリペプチド、ならびに脊椎動物野生型のそれらのオーソログと、それらのドメイン、モチーフおよび断片とが含まれる。それに加え、これらの用語には、組換え変異体のポリペプチド、ドメイン、モチーフおよび断片も含まれ、そうしたものでは、少なくとも1個のアミノ酸残基が異なるアミノ酸残基に変化してしまい、または1個もしくは複数の残基が欠失していることもあり、または1個もしくは複数の残基が、元の配列における隣接残基の間に挿入されていることもある。突然変異体または変異体のSIRT1ポリペプチドでは、アミノ酸残基1個から残基の1%までが変化し、または残基の2%まで、もしくは5%まで、もしくは8%まで、もしくは10%まで、もしくは15%まで、もしくは20%まで、もしくはそれよりやや高い比率まで、野生型または基準配列から変化していることもある。SIRT1の突然変異体または変異体の組換えポリペプチド、ドメイン、モチーフおよび断片は、野生型のSIRT1およびLXRの少なくとも1種の生物活性または機能を示すことが立証し得る限り、本発明の方法および組成物に使用される。生物活性または機能の保有は、非限定的な例として分子生物学、細胞生物学、病理学、臨床医学などを含む、本発明に関連する分野の技術者により判定し得る。本発明の各分野のこのような技術者は、少なくとも本発明の実施例に記載の方法により、SIRT1およびLXRの組換え変異体ポリペプチド、ドメイン、モチーフおよび断片をアッセイしてもよい。
【0041】
SIRT1またはLXRポリペプチドのアミノ酸配列が、そのタンパク質の構造または機能にさしたる作用をせずに変化させ得ることは、当技術分野では認識されていよう。配列のこのような差異を考察する場合、該タンパク質上には、その活性にとって重要な特定の領域が存在すると見込まれることを想起されたい。一般に、類似機能をもたらす残基を使用することを前提とすれば、三次構造を形成する残基を置き換えることは可能である。他の例では、当該変化がタンパク質の非重要領域で起こる場合、残基の種類が全く重要でないこともある。
【0042】
核酸
本明細書で使用する場合、用語「SIRT1ポリヌクレオチド」もしくは「SIRT1核酸」、または「LXRポリヌクレオチド」もしくは「LXR核酸」、あるいは関連する用語および語句は、本明細書に記載の任意のSIRT1ポリペプチドをコードする、任意のポリヌクレオチドに関する。一般に、前記のSIRT1ポリペプチドをコードする任意のヌクレオチド配列は、本発明内に包含される。幾つかの実施形態では、表2に示すヒトSIRT1のアミノ酸配列を有するポリペプチドをコードする核酸は、表8に示すGenBank寄託番号NM_012238に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表8では、そのコード配列が54位から2297位まで拡がっている。
【0043】
表8
【0044】
【化8−1】
【0045】
【化8−2】
幾つかの実施形態では、表3に示すマウスSIRT1のアミノ酸配列を有するポリペプチドをコードする核酸は、表9に示すGenBank寄託番号NM_019812に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表9では、そのコード配列が48位から2261位まで拡がっている。
【0046】
表9
【0047】
【化9−1】
【0048】
【化9−2】
幾つかの実施形態では、表4に示すヒト肝X受容体αのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_005693に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表10では、そのコード配列が36位から1379位まで拡がっている。
【0049】
表10.ヒト肝X受容体αをコードするポリヌクレオチド配列
【0050】
【化10】
幾つかの実施形態では、表5に示すヒト肝X受容体βのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_007121に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表11では、そのコード配列が259位から1644位まで拡がっている。
【0051】
表11.ヒト肝X受容体βをコードするポリヌクレオチド配列
【0052】
【化11】
幾つかの実施形態では、表6に示すマウス肝X受容体αのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_013839に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表12では、そのコード配列が190位から1527位まで拡がっている。
【0053】
表12.マウス肝X受容体αをコードするポリヌクレオチド配列
【0054】
【化12】
幾つかの実施形態では、表7に示すマウス肝X受容体βのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_009473に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表13では、そのコード配列が271位から1611位まで拡がっている。
【0055】
表13.マウス肝X受容体βをコードするポリヌクレオチド配列
【0056】
【化13】
それに加え、本発明には、表8〜13に示す配列の突然変異体もしくは変異体核酸、またはその断片であって、その塩基のいずれかが開示された配列から変化しているが、同時に、SIRT1タンパク質様の活性および生理機能を維持するポリペプチドを依然としてコードし得るポリヌクレオチドも含まれる。SIRT1突然変異体または変異体のポリヌクレオチドは、突然変異体または変異体のSIRT1またはLXRポリペプチドであって、アミノ酸残基1個から残基の1%までが変化し、または残基の2%まで、もしくは5%まで、もしくは8%まで、もしくは10%まで、もしくは15%まで、もしくは20%まで、もしくはそれよりやや高い比率まで、野生型または基準配列から変化していることもあるポリペプチドをコードする。「核酸」または「ポリヌクレオチド」とは、DNA、RNA、1個もしくは複数の修飾ヌクレオチドもしくは修飾ペントースリン酸骨格構造を含んだDNAまたはRNA、ポリペプチド−核酸、および構築体中に含まれる塩基の配列のコード特性を保持する類似の構築体を意味する。本発明には更に、その断片、誘導体、類縁体および相同体を含めた、任意のSIRT1またはLXRコード配列の核酸配列の相補体も含まれる。それに加え、本発明には、構造中に化学修飾を含んだ核酸もしくは核酸断片、またはその相補体も含まれる。
【0057】
また、核酸断片も含まれる。核酸断片は、SIRT1またはLXRポリペプチドの断片をコードすることもある。それに加え、SIRT1またはLXR核酸断片は、SIRT1またはLXRタンパク質コード核酸(例えば、SIRT1またはLXR mRNA)を同定するためのハイブリッド形成用プローブ、およびSIRT1またはLXR核酸分子を増幅または変異させるためのポリメラーゼ連鎖反応(PCR)用プライマーに用いる断片として使用し得る。本明細書で使用する場合、用語「核酸分子」とは、DNA分子(例えば、cDNAまたはゲノムDNA)、RNA分子(例えばmRNA)、ヌクレオチド類縁体を用いて生成したDNAまたはRNAの類縁体、ならびにそれらの誘導体、断片および相補体を包含することを意図している。核酸分子は、1本鎖、2本鎖のいずれでもよいが、好ましくは2本鎖DNAである。
【0058】
核酸
本明細書で使用する場合、「核酸」または「ポリヌクレオチド」、ならびに類似の用語および語句は、天然のヌクレオチドからなるポリマー、ならびに合成または修飾ヌクレオチドからなるポリマーに関する。したがって、本明細書で使用する場合、RNAであるポリヌクレオチド、またはDNAであるポリヌクレオチド、またはデオキシリボヌクレオチド、リボヌクレオチドの両方を含有するポリヌクレオチドは、天然の塩基およびリボース環もしくはデオキシリボース環などの天然の部分を含んでもよく、あるいは以下に記載するような合成または修飾部分で構成されてもよい。本発明に使用するポリヌクレオチドは、1本鎖でもよく、または塩基対2本鎖構造でもよく、または塩基対3本鎖構造でさえもよい。
【0059】
核酸およびポリヌクレオチドは、長さがヌクレオチド20個以上、または長さがヌクレオチド30個以上、または長さがヌクレオチド50個以上、または長さが100個以上、または100個以上、または1000個以上、または数万個以上、または数十万個以上の場合もある。本明細書で使用する場合、「オリゴヌクレオチド」およびこれを基にした類似の用語は、直前の段落に記載したような、天然のヌクレオチドからなる短いポリマー、ならびに合成または修飾ヌクレオチドからなるポリマーに関する。オリゴヌクレオチドは、長さがヌクレオチド10個以上、または長さがヌクレオチド15個もしくは16個もしくは17個もしくは18個もしくは19個もしくは20個以上、または長さがヌクレオチド21個もしくは22個もしくは23個もしくは24個以上、または長さがヌクレオチド25個もしくは26個もしくは27個もしくは28個もしくは29個もしくは30個以上、または長さがヌクレオチド35個以上、40個以上、45個以上、約50個までの場合もある。オリゴヌクレオチドは、化学的に合成してもよく、siRNA、PCRプライマー、またはプローブとして使用し得る。
【0060】
前段落に示したサイズ範囲が重複するために、用語「ポリヌクレオチド」および「オリゴヌクレオチド」は、本明細書では同義に使用し得る。
【0061】
本明細書で使用する場合、「ヌクレオチド配列」、「オリゴヌクレオチド配列」または「ポリヌクレオチド配列」および類似の用語は、互換的に、オリゴヌクレオチドまたはポリヌクレオチドが有する塩基の配列にも、その配列を保持するオリゴヌクレオチドまたはポリヌクレオチドの構造にも関する。更に、ヌクレオチド配列またはポリヌクレオチド配列は、塩基の配列が、当分野で従来より使用されているような、塩基を指定する文字の特定の配列を記載または記述することにより規定される、任意の天然または合成のポリヌクレオチドまたはオリゴヌクレオチドにも関する。
【0062】
「ヌクレオシド」は、本発明の分野に関係する生化学、分子生物学、ゲノム学および類似分野などの分野における技術者によって、プリンまたはピリミジン塩基とグリコシド結合で連結した単糖を含むものと従来より理解され、「ヌクレオチド」は、通常その糖の(ペントースの)3’または5’位に付加するが、その糖の他の位置にあってもよい、少なくとも1個のリン酸基を有するヌクレオシドからなる。ヌクレオチド残基は、オリゴヌクレオチドまたはポリヌクレオチドにおいて連番位置を占める。ヌクレオチドの修飾体または誘導体は、オリゴヌクレオチドまたはポリヌクレオチドにおいて任意の連番位置に出現してもよい。全ての修飾または誘導体化されたオリゴヌクレオチドおよびポリヌクレオチドは、本発明内に包含され、特許請求の範囲内に入る。修飾体または誘導体は、リン酸基、単糖または塩基中に出現することができる。
【0063】
非限定的な例として、以下の記述では、本発明のポリヌクレオチドの範囲内に全て入る修飾または誘導体化された特定のヌクレオチドが示される。その単糖は、例えば、リボースまたはデオキシリボース以外のペントースまたはヘキソースとすることにより、修飾してもよい。該単糖は、ヒドロキシ基をヒドロ基またはアミノ基で置換すること、追加のヒドロキシ基をアルキル化またはエステル化することなどによっても、修飾してもよい。2’−O−メチル、2’−O−エチル、2’−O−プロピル、2’−O−アリール、2’−O−アミノアルキルまたは2’−デオキシ−2’−フルオロ基などの2’位における置換基は、オリゴヌクレオチドに強化されたハイブリッド形成特性を付与する。
【0064】
オリゴヌクレオチドおよびポリヌクレオチド中の塩基は、「非修飾」または「天然」塩基でもよく、プリン塩基のアデニン(A)およびグアニン(G)、ならびにピリミジン塩基のチミン(T)、シトシン(C)およびウラシル(U)を包含する。それに加え、該塩基は修飾または置換がある塩基でもよい。修飾塩基の非限定的例には、ヒポキサンチン、キサンチン、4−アセチルシトシン、5−(カルボキシヒドロキシメチル)ウラシル、5−カルボキシメチルアミノメチル−2−チオウリジン、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、β−D−ガラクトシルケオシン(queosine)、イノシン、N6−イソペンテニルアデニン、1−メチルグアニン、1−メチルイノシン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−アデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、β−D−マンノシルケオシン、5’−メトキシカルボキシメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸(v)、ワイブトキソシン(wybutoxosine)、プソイドウラシル、ケオシン、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸(v)、5−メチル−2−チオウラシル、3−(3−アミノ−3−N−2−カルボキシプロピル)ウラシル、(acp3)wおよび2,6−ジアミノプリン、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチルおよび他のアルキル誘導体、ならびにアデニンおよびグアニンの2−プロピルおよび他のアルキル誘導体などの、他の合成および天然の塩基が含まれる。
【0065】
ヌクレオチド間の結合は、通常は3’−5’リン酸結合であり、それは天然のホスホジエステル結合、ホスホチオエステル結合、および更に他の合成結合でもよい。ホスホロチオエート骨格を含有するオリゴヌクレオチドは、強化されたヌクレアーゼ安定性を有する。修飾された骨格の非限定的例には、ホスホロチオエート、キラルなホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、3’−アルキレンホスホネート、5’−アルキレンホスホネートおよびキラルなホスホネートを含むメチルおよび他のアルキルホスホネート、ホスフィネート、3’−アミノホスホロアミデートおよびアミノアルキルホスホロアミデートを含むホスホロアミデート、チオノホスホロアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートならびにボラノホスフェートが含まれる。
【0066】
上記に例示した修飾を含むいずれの修飾も、本発明のポリヌクレオチド中に容易に組み入れることができ、その範囲中に含まれる。任意の修飾ヌクレオチドの使用は、当業者には理解されるように、同じ塩基対形成特性を有する天然ヌクレオチドの使用と同等である。全ての同等の修飾ヌクレオチドは、本明細書に開示し、主張するように、本発明の範囲内に入る。
【0067】
「単離」された核酸分子は、その核酸の天然源中に存在する他の核酸分子から分離された核酸分子である。単離された核酸分子の例には、それだけに限らないが、ベクターに含有される組換えDNA分子、異種宿主細胞中に保持される組換えDNA分子、部分的または実質的に精製された核酸分子、および合成DNAまたはRNA分子が含まれる。
【0068】
本明細書に開示する核酸分子、例えば、表8〜13のヌクレオチド配列を有する核酸分子、またはその相補体は、標準的な分子生物学技法およびここに示した配列情報を用いて単離することができる。表8〜13の核酸配列の全体または一部をハイブリッド形成プローブとして用い、SIRT1またはLXRの核酸配列を標準的なハイブリッド形成法およびクローニング法を用いて単離することができる(例えば、Sambrookら編、 MOLECULAR CLONING: A LABORATORY MANUAL 第2版、Cold Spring HarborLaboratory Press、Cold Spring Harbor、NY、1989年およびAusubelら編者、CURRENT PROTOCOLS INMOLECULAR BIOLOGY、John Wiley & Sons、New York、NY、1993年に記載のような技法)。
【0069】
本明細書に開示する核酸は、鋳型としてcDNA、mRNA、あるいはゲノムDNAと、標準的なPCR増幅法に従った適当なオリゴヌクレオチドプライマーとを用いて増幅することができる。このように増幅した核酸は、適当なベクター中にクローニングし、DNA配列分析により特性を決定することができる。更に、SIRT1またはLXRのヌクレオチド配列に対応したオリゴヌクレオチドが、標準的な合成法、例えば自動DNAシンセサイザーを用いて調製することができる。
【0070】
ポリヌクレオチドの合成
オリゴヌクレオチドおよびポリヌクレオチドは、標準的な合成法、例えば自動DNAシンセサイザーを用いて調製することができる。オリゴヌクレオチドを合成する方法には、それだけに限らないが、表面誘導体化粒子上へのヌクレオチドホスホロアミダイトの逐次付加を含めた周知の化学プロセスが含まれ、その逐次付加は、Oligonucleotides and Analogues A Practical Approach、F. Eckstein編、OxfordUniversity Press、Oxford、1〜24頁(1991年)におけるT. Brown およびDorcas J. S. Brownが記載し、参照により本明細書に組み込まれる通りである。
【0071】
合成手順の一例では、Expedite RNAホスホロアミダイトおよびチミジンホスホロアミダイト(Proligo、ドイツ)が使用される。合成オリゴヌクレオチドは、脱保護され、ゲル精製された(Elbashirら(2001年)Genes & Dev. 15巻、188〜200頁)後、Sep−Pak C18カートリッジ(Waters、Milford、Mass.、米国)で精製される(Tuschlら(1993年)Biochemistry、32巻、11658〜11668頁)。相補的なssRNAは、90℃、1分間アニーリング緩衝液(100mM酢酸カリウム、pH7.4の30mM HEPES−KOH、2mM酢酸マグネシウム)中で、その後37℃、1時間インキュベートして、対応するdsポリヌクレオチドへハイブリダイズさせる。
【0072】
オリゴヌクレオチド合成の他の方法には、それだけに限らないが、ホスホトリエステル法およびホスホジエステル法による固相オリゴヌクレオチド合成(Narangら、(1979年)Meth. Enzymol.、68巻、90頁)、ならびにH−ホスホネート法(とりわけ、Garegg,P.J.ら、(1985年)「Formation of internucleotidic bonds via phosphonate intermediates」、Chem.Scripta 25巻、280〜282頁;およびFroehler, B.C.ら、(1986年a)「Synthesis of DNA viadeoxynucleoside H-phosphonate intermediates」、Nucleic Acid Res.、14巻、5399〜5407頁)および支持体上での合成(Beaucageら(1981年)TetrahedronLetters 22巻、1859〜1862頁)、更に、ホスホロアミデート法(Caruthers, M. H.ら、「Methods in Enzymology」、154巻、287〜314頁(1988年)、米国特許第5153319号、第5132418号、第4500707号、第4458066号、第4973679号、第4668777号および第4415732号、ならびに「Synthesisand Applications of DNA and RNA」、S. A. Narang編、Academic Press、New York、1987年に記載の他の技法、およびその中に含まれる参考文献)、および非ホスホロアミダイト法が含まれる。固相合成は、不純物および過剰試薬からのオリゴヌクレオチドの単離を促進する。固体支持体から切断されると、オリゴヌクレオチドは公知の方法により更に単離し得る。
【0073】
ベクター
本発明は、本発明の1種または複数のポリヌクレオチドを含有する多種のベクターを提供する。有利には、本発明において開示されるいずれのベクターも、該ヌクレオチド配列に作動可能に連結されたプロモーター、エンハンサーまたはその両方を含む。
【0074】
本発明において開示されるベクターを調製する方法は、分子生物学、細胞生物学、腫瘍学の分野、および本発明に関連する他の分野において広く知られている。ベクターを調製する有用な方法は、非限定的な例として、Molecular Cloning: ALaboratory Manual(第3版)(Sambrook, Jら(2001年)Cold Spring Harbor Laboratory Press、ColdSpring Harbor、NY)、およびShort protocols in molecular biology(第5版)(Ausubel FMら(2002年)JohnWiley & Sons、New York City)に記載されている。
【0075】
抗体
本明細書で使用する場合の用語「抗体」とは、免疫グロブリン分子および免疫グロブリン(Ig)分子の免疫活性な部分、即ち、抗原と特異的に結合する(免疫反応を起こす)抗原結合部位を含有する分子を指す。このような抗体には、それだけに限らないが、ポリクローナル、モノクローナル、キメラ、単鎖、Fab、Fab’およびF(ab’)2断片、ならびにFab発現ライブラリーが含まれる。一般に、ヒトから得た抗体分子は、分子中に存在する重鎖の特質が相互に異なるクラスIgG、IgM、IgA、IgEおよびIgDのいずれかに関する。特定のクラスは、IgG1、IgG2、その他などのサブクラスも有する。更に、ヒトでは、軽鎖がκ鎖またはλ鎖の場合がある。本明細書における抗体への言及には、ヒト抗体種のこのような全てのクラス、サブクラスおよび型への言及を包含する。本明細書に開示するいずれの抗体も、そのコグネイト抗原に「免疫特異的に」結合する。免疫特異的な結合とは、特定の免疫原を宿主に接種することにより生成した抗体が、その免疫原性部分を含んだ、抗原などの分子と高い親和性で結合し、免疫原非含有分子とは弱い親和性でしか、または全く結合しないことを意味する。この定義で使用する場合、高親和性とは、1×10−6M未満の解離定数を有することを意味し、弱親和性とは、約1×10−6Mより高い解離定数を有することを意味する。
【0076】
抗原またはその一部分もしくは断片として作用させることを意図した、本発明の単離タンパク質またはそのようなタンパク質を含有する複合体は、ポリクローナルおよびモノクローナル抗体調製の標準的技法を用いて、その抗原に免疫特異的に結合する抗体を生成するための免疫原として使用することができる。その全長タンパク質を使用することができ、あるいは本発明は、免疫原として使用するために、該抗原の抗原性ペプチド断片を提供する。抗原性ペプチド断片は、表2〜7に示したアミノ酸配列などの、全長タンパク質のアミノ酸配列のアミノ酸残基少なくとも6個を含み、そのペプチドに対して生成した抗体が、全長タンパク質、またはエピトープを含有する任意の断片と特異的な免疫複合体を形成するように、全長タンパク質の該エピトープを包含する。好ましくは、抗原性ペプチドは、アミノ酸残基少なくとも10個、またはアミノ酸残基少なくとも15個、またはアミノ酸残基少なくとも20個、またはアミノ酸残基少なくとも30個を含む。抗原性ペプチドが包含する好ましいエピトープは、そのタンパク質の表面上に位置した領域であり、こうした領域は、通常、親水性領域である。
【0077】
本発明のある種の実施形態では、抗原性ペプチド断片が包含する少なくとも1個のエピトープは、SIRT1またはLXRタンパク質の表面上に位置した、そのタンパク質の一領域、例えば親水性領域である。ヒトのSIRT1またはLXRタンパク質配列の疎水性分析により、成長促進性ポリペプチドのいずれの領域が、特に親水性であり、したがって抗体産生を目標とするのに有用な表面残基をコードしそうであるかが示されよう。抗体産生を目標とする手段として、親水性および疎水性の領域を示す水親疎性プロットを、例えば、フーリエ変換を伴う、または伴わないKyte Doolittle法またはHopp Woods法を含めた、当技術分野で周知の任意の方法により作成し得る。例えば、その全体が参照により本明細書に各々組み込まれるHopp and Woods、1981年、Proc. Nat. Acad. Sci. USA 78巻、3824〜3828頁; Kyteand Doolittle 1982年、J. Mol. Biol. 157巻、105〜142頁を参照されたい。抗原性タンパク質、またはその誘導体、断片、類縁体もしくは相同体内にある、1つまたは複数のドメインに特異的な抗体も、本明細書に示されている。
【0078】
本発明のタンパク質、またはその誘導体、断片、類縁体、相同体もしくはオーソログは、こうしたタンパク質成分に免疫特異的に結合する抗体の生成において、免疫原として利用し得る。
【0079】
本発明のタンパク質、またはその誘導体、断片、類縁体、相同体もしくはオーソログに対する、ポリクローナルまたはモノクローナル抗体の産生のために、当技術分野内で公知の多様な手順を使用し得る(例えば、参照により本明細書に組み込まれるAntibodies: A Laboratory Manual、Harlow E, and Lane D、1988年、ColdSpring Harbor Laboratory Press、Cold Spring Harbor、NYを参照されたい)。こうした抗体の一部を以下に考察する。
【0080】
ポリクローナル抗体
ポリクローナル抗体を産生するために、適切な多種の宿主動物(例えば、ウサギ、ヤギ、マウスまたは他の動物)を、自然のタンパク質、その合成変異体、または前記タンパク質の誘導体による1回または複数回の注射で免疫し得る。適当な免疫原調製物は、例えば、天然の免疫原タンパク質、その免疫原タンパク質に相当する化学合成ポリペプチド、または組換え発現した免疫原タンパク質を含有することができる。更に、該タンパク質は、免疫される哺乳動物中で免疫原性であることが知られている第2のタンパク質にコンジュゲートしてもよい。このような免疫原タンパク質の例には、それだけに限らないが、キーホールリンペットヘモシニアン、血清アルブミン、ウシサイログロブリンおよび大豆トリプシンインヒビターが挙げられる。この調製物は、アジュバントを更に含むことができる。
【0081】
免疫原タンパク質に対するポリクローナル抗体分子は、当該哺乳動物から(例えば、その血液から)単離し、プロテインAもしくはプロテインGを用いる、または抗原のカラム上でのアフィニティークロマトグラフィーなどの周知の技法によって、更に精製することができる。免疫グロブリンの精製は、例えばD.Wilkinson(The Scientist, Inc., Philadelphia PA出版のThe Scientist、14巻、8号(2000年4月17日)、25〜28頁)により考察されている。
【0082】
モノクローナル抗体
モノクローナル抗体は、Kohler and Milstein、Nature、256巻、495頁(1975年)に記載のものなどのハイブリドーマ法を用いて調製することができる。ハイブリドーマ法では、マウス、ハムスターまたは適当な他の宿主動物を通常、免疫剤で免疫することにより、免疫剤に特異的に結合すると見込まれる抗体を産生する、または産生できるリンパ球を誘導する。あるいは、リンパ球をインビトロで免疫することができる。
【0083】
免疫剤には、通常、タンパク質抗原、その断片、またはその融合タンパク質が含まれることになろう。次いで、ポリエチレングリコールなどの適切な融合剤を用いて、リンパ球を不死化細胞系と融合し、ハイブリドーマ細胞を形成する[Goding、Monoclonal Antibodies: Principles and Practice、Academic Press、(1986年)59〜103頁]。
【0084】
次いで、ハイブリドーマ細胞を培養した培地を、抗原に対するモノクローナル抗体の有無についてアッセイすることができる。所望のハイブリドーマ細胞を特定した後、限界希釈手順によりクローンをサブクローニングし、標準的方法(Goding、1986年)により増殖させることができる。この目的のための適切な培地には、例えば、ダルベッコの改良イーグル培地およびRPMI−1640培地が挙げられる。あるいは、哺乳動物中の腹水としてハイブリドーマ細胞をインビボで増殖させることができる。
【0085】
サブクローンが分泌するモノクローナル抗体は、例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィーなどの従来からの免疫グロブリン精製手順により、培地または腹水から単離または精製することができる。
【0086】
組換えベクターおよび宿主細胞
本発明のある種の態様では、哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含有する核酸を細胞中に導入することを含む方法が開示される。哺乳類細胞はヒト細胞でもよい。タンパク質デアセチラーゼは、多様に、真核生物Sir2タンパク質または哺乳類SIRT1タンパク質でもよい。哺乳類細胞に意図した核酸配列を移入する技法は、本発明の分野の技術者に広く知られている。このような方法の特定の非限定的な例を以下に詳細に説明する。
【0087】
本発明で開示する一態様は、あるタンパク質をコードする核酸、またはその誘導体、断片、類縁体もしくは相同体を含有するベクター、好ましくは発現ベクターに関する。本明細書で使用する場合、用語「ベクター」とは、結合している別の核酸を輸送できる核酸分子を指す。1種のベクターは「プラスミド」であり、追加のDNAセグメントをその中に連結できる環状の2本鎖DNAループを指す。別種のベクターはウィルスベクターであり、そこでは追加のDNAセグメントをウィルスゲノム中に連結できる。ある種のベクターは、導入先の宿主細胞中で自律複製をすることができる(例えば、細菌性複製起点を有する細菌ベクターおよび哺乳類エピソームベクター)。他のベクター(例えば、哺乳類非エピソームベクター)は、宿主細胞中に導入した際、宿主細胞のゲノム内に組み込まれ、それにより宿主ゲノムと共に複製される。その上、ある種のベクターは、作動的に連結されている遺伝子の発現を指示することができる。このようなベクターは、本明細書では「発現ベクター」と称する。一般に、組換えDNA技法で有用な発現ベクターは、プラスミドの形態を取ることが多い。本明細書では、プラスミドがベクターの最も汎用される形態であるので、「プラスミド」および「ベクター」は、互換的に使用することができる。しかし、本発明は、同等な機能を発揮するような、ウィルスベクター(例えば、複製欠陥レトロウィルス、アデノウィルスおよびアデノ随伴ウィルス)などの他形態の発現ベクターも包含することを意図している。
【0088】
本発明に開示する組換え発現ベクターは、本発明に開示する核酸を宿主細胞中で核酸を発現するのに適切な形態で含み、そのことは、組換え発現ベクターが、発現に用いる宿主細胞に基づいて選択されている、即ち、発現しようとする核酸配列に作動的に連結されている1個または複数の調節配列を含むことを意味している。組換え発現ベクター内で、「作動可能に連結されている」とは、対象とするヌクレオチド配列が、そのヌクレオチド配列の発現(例えば、インビトロでの転写/翻訳系において、または宿主細胞中にベクターを導入した場合は宿主細胞において)を可能とするように調節配列(複数も)に連結されていることを意味することが、その意図するところである。用語「調節配列」は、プロモーター、エンハンサーおよび他の発現制御要素(例えば、ポリアデニル化シグナル)を包含することを意図している。このような調節配列は、例えば、Goeddel、GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185、AcademicPress、San Diego、Calif.(1990年)に記載されている。調節配列には、多種の宿主細胞においてヌクレオチド配列の構成的発現を指示するもの、および特定の宿主細胞においてのみヌクレオチド配列の発現を指示するもの(例えば、組織特異的調節配列)が含まれる。当業者であれば、発現ベクターの設計が、形質転換しようとする宿主細胞の選択、所望するタンパク質の発現量などの要因に依存し得ることは認識されよう。本発明に開示する発現ベクターは、宿主細胞中に導入することにより、融合タンパク質またはペプチドを含め、本明細書に記載するような核酸がコードするタンパク質またはペプチドを産生することができる(例えば、SIRT1またはLXRタンパク質、SIRT1またはLXRタンパク質の変異型、融合タンパク質など)。
【0089】
本発明に開示する組換え発現ベクターは、原核または真核細胞中にSIRT1またはLXRタンパク質を発現するように設計することができる。例えば、SIRT1またはLXRタンパク質は、大腸菌(E.coli)などの細菌細胞、昆虫細胞(バキュロウィルス発現ベクターを用いて)、酵母細胞、または哺乳類細胞中に発現させることができる。適切な宿主細胞は、Goeddel、GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185、AcademicPress、San Diego、Calif.(1990年)で更に考察されている。あるいは、組換え発現ベクターは、例えば、T7プロモーター調節配列およびT7ポリメラーゼを用いて、インビトロで転写し、翻訳することができる。
【0090】
原核生物中でのタンパク質の発現は、融合、非融合いずれかのタンパク質の発現を指示する構成または誘導プロモーターを含有するベクターを用いて、大腸菌中で実施することが最も多い。融合ベクターは、その中にコードされるタンパク質に幾つものアミノ酸を、普通その組換えタンパク質のアミノ末端へ付加する。このような融合ベクターは、通常3つの目的:(1)組換えタンパク質の発現を増加させること、(2)組換えタンパク質の溶解性を増加させること、および(3)アフィニティー精製におけるリガンドとして作用することにより、組換えタンパク質の精製を補助することを実現する。典型的な融合発現ベクターには、それぞれグルタチオンS−トランスフェラーゼ(GST)、マルトースE結合タンパク質またはプロテインAを標的組換えタンパク質に融合する、pGEX(Pharmacia Biotech Inc;Smith and Johnson(1988年)Gene 67巻、31〜40頁)、pMAL(New England Biolabs,Beverly,Mass.)およびpRIT5(Pharmacia,Piscataway,N.J.)が含まれる。
【0091】
適切な誘導性の非融合大腸菌発現ベクターには、pTrc(Amrannら、(1988年)Gene69巻、301〜315頁)およびpET11d(Studierら、GENE EXPRESSION TECHNOLOGY: METHODS INENZYMOLOGY 185、Academic Press、San Diego、Calif.(1990年)60〜89頁)が含まれる。
【0092】
別の実施形態では、本発明に開示するタンパク質は、酵母発現ベクターを用いて発現される。酵母S.セレビシエ(S.cerevisiae)に発現するためのベクターの例には、pYepSec1(Baldariら、(1987年)EMBO J 6巻、229〜234頁)、pMFa(Kurjan and Herskowitz、(1982年)Cell30巻、933〜943頁)、pJRY88(Schultzら、(1987年)Gene 54巻、113〜123頁)、pYES2(Invitrogen Corporation,San Diego,Calif.)、およびpicZ(Invitrogen Corp.San Diego,Calif.)が挙げられる。
【0093】
あるいは、本発明の複合体中に開示されるタンパク質は、バキュロウィルス発現ベクターを用いて昆虫細胞中に発現することができる。培養する昆虫細胞(例えばSF9細胞)中にタンパク質を発現するために利用可能なバキュロウィルスベクターには、pAc系列(Smithら(1983年)Mol Cell Biol 3巻、2156〜2165頁)およびpVL系列(Lucklow andSummers(1989年)Virology 170巻、31〜39頁)が挙げられる。
【0094】
更に別の実施形態では、本発明に開示する核酸は、哺乳類発現ベクターを用いて哺乳類細胞中に発現される。哺乳類発現ベクターの例には、pCDM8(Seed(1987年)Nature 329巻、840頁)およびpMT2PC(Kaufmanら(1987年)EMBO J 6巻、187〜195頁)が挙げられる。哺乳類細胞中で使用する場合、発現ベクターの制御機能は、ウィルス性調節要素から得ることが多い。例えば、汎用されるプロモーターは、ポリオーマ、アデノウィルス2、サイトメガロウィルスおよびシミアンウィルス40に由来する。原核、真核双方の細胞用に適切な他の発現系については、例えば、Sambrookら、MOLECULAR CLONING: A LABORATORY MANUAL 第2版、Cold Spring Harbor Laboratory、ColdSpring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1989年を参照されたい。
【0095】
別の実施形態では、組換え哺乳類発現ベクターは、特定の細胞型中で選択的に核酸を発現するように指示することができる(例えば、核酸を発現するために、組織特異的な調節要素が使用される)。適切な組織特異的プロモーターの非限定的な例には、アルブミンプロモーター(肝特異的;Pinkertら(1987年)Genes Dev 1巻、268〜277頁)、リンパ系特異的プロモーター(Calame and Eaton(1988年)AdvImmunol 43巻、235〜275頁)、特にT細胞受容体(Winoto and Baltimore(1989年)EMBO J 8巻、729〜733頁)および免疫グロブリン(Banerjiら(1983年)Cell33巻、729〜740頁;Queen and Baltimore(1983年)Cell 33巻、741〜748頁)のプロモーター、ニューロン特異的プロモーター(例えばニューロフィラメントプロモーター;Byrneand Ruddle(1989年)PNAS 86巻、5473〜5477頁)、膵特異的プロモーター(Edlundら(1985年)Science 230巻、912〜916頁)、および乳腺特異的プロモーター(例えば、乳清プロモーター;米国特許第4873316号および欧州出願公開第264166号)が挙げられる。発生時調節されるプロモーター、例えば、マウスホックス(hox)プロモーター(Kesseland Gruss(1990年)Science 249巻、374〜379頁)およびα−フェトプロテインプロモーター(Campes and Tilghman(1989年)GenesDev 3巻、537〜546頁)も包含される。
【0096】
本発明に開示する別の態様は、本発明に開示する組換え発現ベクターを導入した宿主細胞に関する。用語「宿主細胞」および「組換え宿主細胞」は、本明細書では互換的に使用される。このような用語は、特定の対象細胞だけでなく、このような細胞の子孫または潜在的子孫も指すことが理解される。ある種の改変が、突然変異、環境影響のいずれかにより後継世代に起こり得るので、このような子孫は、実際には親細胞と同一でないこともあるが、それでも本明細書で使用する場合の該用語の範囲内に含められる。
【0097】
宿主細胞は、任意の原核または真核細胞でもよい。例えば、SIRT1またはLXRタンパク質は、大腸菌などの細菌細胞、昆虫細胞、酵母または哺乳類細胞(チャイニーズハムスター卵巣細胞(CHO)もしくはCOS細胞など)中に発現することができる。他の適切な宿主細胞は、当業者に公知である。
【0098】
ベクターDNAは、従来の形質転換またはトランスフェクション技法を介して原核または真核細胞中に導入することができる。本明細書で使用する場合、用語「形質転換」および「トランスフェクション」は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE−デキストラン媒介トランスフェクション、リポフェクションまたは電気穿孔を含む、宿主細胞中に外来核酸(例えばDNA)を導入するための当分野認知の多様な技法を指すことを意図している。宿主細胞を形質転換またはトランスフェクションする適切な方法は、Sambrookら(2001年)、Ausubelら(2002年)および他の実験手引に見出すことができる。
【0099】
哺乳類細胞の安定なトランスフェクションのために、使用する発現ベクターおよびトランスフェクション法に応じて、ほんの小さな分率の細胞が、外来DNAをそのゲノム中に組み込み得ることが知られている。こうした組込み体を特定し、選択するために、選択マーカー(例えば、抗生物質に対する耐性)をコードする遺伝子が、一般に、対象の遺伝子と共に宿主細胞中に導入される。多種の選択マーカーには、G418、ハイグロマイシン、メソトレキセートなどの薬物に対して、耐性を付与するものが含まれる。選択マーカーをコードする核酸は、成長促進物質をコードする同じベクター上で宿主細胞中に導入することができ、または別のベクター上で導入することができる。導入される核酸を安定に移入した細胞は、薬物選択によって特定することができる(例えば、選択マーカー遺伝子を組み込んだ細胞は生存することになろうが、その他の細胞は死滅する)。
【0100】
培養物中の原核または真核宿主細胞などの本発明の宿主細胞は、SIRT1またはLXRタンパク質を産生(即ち、発現)するために、使用することができる。したがって、本発明は、本発明の宿主細胞を用いてSIRT1またはLXRタンパク質を産生する方法を更に提供する。一実施形態では、この方法は、SIRT1またはLXRタンパク質が産生するように、適切な培地中で本発明の宿主細胞(SIRT1またはLXRタンパク質をコードする組換え発現ベクターを導入した後の)を培養することを含む。別の実施形態では、この方法は、培地または宿主細胞からSIRT1またはLXRタンパク質を単離することを含む。
【0101】
脊椎動物細胞のトランスフェクションは、細胞中にDNAを導入するリポソームなどの他の粒子以外に、それだけに限らないが、アデノウィルス、アデノ随伴ウィルス、レトロウィルスの各ベクターを含めた、組換えベクターを用いて更に実現することができる。上記のものなどの技法は、SIRT1またはLXRポリペプチドをコードする任意のヌクレオチド配列を脊椎動物細胞中に導入するために、利用することができる。例えば、哺乳類細胞のトランスフェクションのために、幾種ものウィルス依拠発現系を利用し得る。発現ベクターとしてアデノウィルスを使用する場合、対象とするSIRT1またはLXRヌクレオチド配列を、アデノウィルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三部分(tripartite)リーダー配列に連結してもよい。次いで、このキメラ遺伝子は、インビトロまたはインビボでの組換えにより、アデノウィルスゲノム中に挿入されることがある。ウィルスゲノムの非必須領域(例えば、E1またはE3領域)に挿入されると、感染宿主中で生存でき、SIRT1またはLXR産物を発現できる組換えウィルスが生じることになろう(例えば、Logan & Shenk、1984年、Proc. Natl. Acad. Sci. USA 81巻、3655〜3659頁を参照されたい)。SIRT1またはLXRコード配列の一部分だけが挿入される場合には、多分ATG開始コドンを含んだ外因性翻訳制御シグナルを供給しなければならない。こうした外因性翻訳制御シグナルおよび開始コドンは、天然、合成いずれの多様な起源のものでもよい。発現の効率は、適当な転写エンハンサー要素、転写ターミネーターなどを含めることにより、高めることができる(Bitterら、1987年、Methodsin Enzymol. 153巻、516〜544頁を参照されたい)。
【0102】
治療的処置
ある種の病状および病的状態は、対象の細胞における異種SIRT1もしくはLXR、または異種LXRαもしくはLXRβの発現に有利に応答すると考えられている。したがって、本発明は、対象における病状、疾患または病的状態を処置する方法であって、該病状が、SIRT1またはLXR、LXRαまたはLXRβのポリペプチドに応答する方法を開示する。この方法は、病状の軽減または改善に有効な量で、SIRT1またはLXR、LXRαまたはLXRβのポリペプチドをコードする核酸を対象に投与することを含む。病状の軽減とは、症状の悪化傾向が、より緩やかまたはより穏やかな悪化傾向に弱まることを意味する。病状の改善とは、病状の兆候および症候が弱まり、患者がより良好な健康へと改善するような、患者の実際の改善を意味する。この方法の重要な実施において、病状は、心筋梗塞、脳血管発作、腎疾患、アルツハイマー病を含む神経疾患などから選定される。病状を処置する方法の有利な実施形態では、対象はヒトである。
【0103】
本明細書に記載の処置法の多様な実施形態では、SIRT1もしくはLXR、LXRαもしくはLXRβのポリペプチド、その変異体またはその断片をコードする核酸を、対象の細胞中への送達を含む、その核酸配列の細胞中への効率的送達を保証する各種組成物のいずれか中で、対象へ投与し得る。
【0104】
SIRT1またはLXR核酸配列による対象の処置は、SIRT1またはLXR核酸配列の脊椎動物細胞中への導入に有効である、適切な核酸、プラスミド、ベクター、ウィルスベクター、リポソームまたは類似組成物を投与することにより、実現することができる。核酸のトランスフェクションは、陽イオン両親媒性物質の使用で補助し得る(米国特許第6503945号およびその中に開示されている文献)。エクスビボでのレトロウィルス遺伝子治療は、例えばHacein-Bey-Abinaら(2003年、Science 302巻、415〜419頁)により記載されている。対象中へのトランス遺伝子の治療的導入法は、「GeneTransfer Methods: Introducing DNA Into Living Cells and Organisms」、P.A. Nortonand L.F. Steel、Eaton Publishing、2000年において考察されている。細胞および生物中へのトランス遺伝子の治療的導入手法は、「GeneTherapy Protocols」、Paul D. Robbins(編)、Humana Press(1997年)に示されている。
【0105】
ポリヌクレオチドを含む医薬組成物
治療用途のための医薬組成物には、1種または複数のポリヌクレオチドおよび担体が含まれる。1種または複数のポリヌクレオチドを含む医薬組成物は、標的遺伝子の発現または活性と関連する疾患または障害の処置に有用である。担体には、それだけに限らないが、塩水、緩衝塩水、デキストロース、水、グリセロール、エタノール、およびそれらの組合せが挙げられる。経口投与される薬物について、医薬として許容可能な担体には、それだけに限らないが、不活性希釈剤、崩壊剤、結合剤、潤滑剤、甘味剤、香味剤、着色剤、防腐剤などの医薬として許容可能な賦形剤が挙げられる。
【0106】
1日につき受容者の体重1kg当たりポリヌクレオチド5mgの最大投与量は、標的遺伝子の発現を阻害する、または完全に抑制するのに十分である。一般に、適切なポリヌクレオチド用量は、1日につき受容者の体重1kg当たり0.01〜5.0mgの範囲、好ましくは1日につき体重1kg当たり0.1〜200μg(μg/kg)の範囲、より好ましくは1日につき0.1〜100μg/kgの範囲、より一層好ましくは1日につき1.0〜50μg/kgの範囲、最も好ましくは1日につき1.0〜25μg/kgの範囲に入ることとなろう。医薬組成物は、毎日1回投与してもよく、またはポリヌクレオチドは、1日を通して適当な間隔の分割用量として、2回、3回、4回、5回、6回もしくはそれより多い回数で投与してもよい。その場合、各分割用量中に含有するポリヌクレオチドは、1日総投与量を実現するために、それ相当に減量させねばならない。投与単位は、例えば、数日の期間に亘ってポリヌクレオチドを持続放出する従来の製剤を用いて、数日に亘る送達用の持続放出製剤として調合することもできる。持続放出製剤は、当技術分野で周知のものである。この実施形態では、投与単位は、1日用量の何倍かに相当する量を含有する。
【0107】
当業者であれば、それだけに限らないが、疾患または障害の重度、以前の処置、対象の健康全般および/または年齢、ならびに罹っている他の疾患を含めた特定の要因が、対象の有効な処置に必要な投与量および適時に影響し得ることは、認識されていよう。その上、治療有効量の組成物による対象の処置は、単回処置または一連の処置を包含することができる。本発明により包含される個々のポリヌクレオチドに関する有効投与量および体内半減期の見積りは、従来手法を用いて、または適当な動物モデルを用いたインビボ試験に基づいて行うことができ、また、適当な用量−応答特性を決定するための確立した規準に従って、処置中に調整することができる。
【0108】
本発明により包含される医薬組成物は、それだけに限らないが、経口経路、または静脈内、筋肉内、腹腔内、皮下、経皮、気道(エアゾル)、直腸、経膣および局所(口腔および舌下を含む)投与を含めた非経口経路を含む、当技術分野で公知の任意の手段によって投与してもよい。ある種の実施形態では、医薬組成物は、静脈内または非経口内注入または注射により投与され、追加の一般的実施形態では、ポリヌクレオチドを含む医薬組成物は、腹腔鏡および類似のマイクロ手術の手順を用いて、その場の腫瘍、癌または前癌増殖物に直接投与してもよい。
【0109】
筋肉内、腹腔内、皮下および静脈内使用のために、本発明の医薬組成物は、一般に、適当なpHおよび等張性に緩衝させた、滅菌、水性の溶液または懸濁液として供給されよう。適切な水性媒体には、リンゲル液および等張塩化ナトリウムが挙げられる。好ましい実施形態では、担体は専ら緩衝水溶液らなる。これに関しては、「専ら」とは、標的遺伝子を発現する細胞におけるポリヌクレオチドの取込みに影響し得る、またはそれを媒介し得る、助剤または封入物質が全く存在しないことを意味する。このような物質には、例えば、リポソームやキャプシドなどの以下に説明するようなミセル構造が挙げられる。驚くべきことに、本発明者らは、裸ポリヌクレオチドおよび生理的に許容可能な溶媒だけを含有する組成物が、細胞により取り込まれ、そのポリヌクレオチドが、標的遺伝子の発現を有効に抑制することを発見した。マイクロインジェクション、リポフェクション、ウィルス、ウィロイド、キャプシド、キャプソイドまたは他の助剤が、細胞培養物中へのポリヌクレオチドの導入には必要とされるが、驚くべきことに、こうした方法および助剤が、インビボでのポリヌクレオチドの取込みには必要ではない。本発明による水性懸濁液は、セルロース誘導体、アルギン酸ナトリウム、ポリビニルピロリドン、トラガカントガムなどの懸濁剤、およびレシチンなどの湿潤剤を含んでもよい。水性懸濁液用の適切な防腐剤には、p−ヒドロキシ安息香酸エチルおよびn−プロピルが挙げられる。
【0110】
本発明に従って有用な医薬組成物は、埋め込み材およびマイクロカプセル化送達系を含めた制御放出製剤などの、体内からの急速消失からポリヌクレオチドを保護するためのカプセル化製剤も包含する。エチレン酢酸ビニル、ポリ酸無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、ポリ乳酸などの生分解性で生体適合性のポリマーを使用することができる。このような製剤の調製法は、当業者には明らかであろう。その各材料は、Alza CorporationおよびNova Pharmaceuticals,Inc.から商業的に入手することもできる。リポソーム懸濁液(ウィルス抗原に対するモノクローナル抗体を有する、感染細胞を標的としたリポソームを含める)も、医薬として許容可能な担体として使用することができる。こうしたものは、例えば、参照により本明細書に組み込まれる米国特許第4522811号、PCT公開WO第91/06309号および欧州特許公開第EP−A−43075号に記載のように、当業者に公知の方法に従って調製することができる。
【0111】
ある種の実施形態では、カプセル化製剤はウィルスコートタンパク質を含む。この実施形態では、ポリヌクレオチドは、少なくとも1つのウィルスコートタンパク質と結合し、会合し、またはそれに囲い込まれてもよい。ウィルスコートタンパク質は、ポリオーマウィルスなどのウィルスに由来するか、それと会合していてもよく、または、部分的もしくは完全に人工的なものでもよい。例えば、コートタンパク質は、ポリオーマウィルスのウィルスプロテイン1および/またはウィルスプロテイン2、あるいはその誘導体でもよい。
【0112】
このような化合物の毒性および治療効果は、例えば、LD50(その集団の50%に致死的な用量)およびED50(その集団の50%に治療有効な用量)を決定するための、細胞培養物または実験動物における標準的な薬学手順によって決定することができる。毒性効果と治療効果との用量比が、治療指数であり、比率LD50/ED50として表現することができる。高い治療指数を示す化合物が好ましい。
【0113】
細胞培養アッセイおよび動物試験から得られるデータは、ヒトに使用する投与量の範囲を処方する際に使用することができる。本発明の組成物の投与量は、毒性を殆どまたは全く示さずに、ED50を含む循環濃度の範囲内にあるのが好ましい。その投与量は、使用する剤形および利用する投与経路に応じてこの範囲内で変化させてもよい。本発明の方法に使用するいずれの化合物についても、細胞培養物において決定した場合のIC50(即ち、症状の最大抑制の半量を実現する試験化合物の濃度)を含む、その化合物の循環血漿濃度範囲となるように、治療有効用量を、細胞培養アッセイおよび動物モデルから当初に推定することができる。このような情報は、ヒトにおいて有用な用量をより正確に決定するために、使用することができる。
【0114】
以上に考察したように、単独または複数として投与する以外に、本発明に従って有用なポリヌクレオチドは、疾患の処置に有効な他の既知の作用剤と組み合わせて投与することもできる。いずれにせよ、投薬担当内科医は、当技術分野で知られているか、または本明細書に記載した効力の標準尺度を用いた観察結果に基づいて、ポリヌクレオチド投与の量および適時を調節することができる。
【0115】
医薬組成物
本発明に開示するポリヌクレオチドは、本明細書では「活性化合物」または「治療剤」と呼称される。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0116】
本発明は、SIRT1活性を刺激する作用剤を投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法、ならびに、SIRT1活性を刺激する作用剤を哺乳動物に投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、該作用剤の使用を含む。このような作用剤は、本明細書では「活性化合物」または「治療剤」と呼称される。このような作用剤の一例はT0901317である。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0117】
本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤を投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法も含む。それに加え、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤を哺乳動物に投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、該作用剤の使用を含む。このような作用剤は、本明細書では「活性化合物」または「治療剤」と呼称される。このような作用剤の例には、22(R)−ヒドロキシコレステロール、9−シスレチノイン酸のいずれか、または22(R)−ヒドロキシコレステロール、9−シスレチノイン酸の両者の組合せが挙げられる。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0118】
本明細書で使用する場合、「医薬として許容可能な担体」は、医薬投与に適合できる、ありとあらゆる溶媒、分散媒、コーティング、抗細菌剤および抗真菌剤、等張剤および吸収遅延剤などを包含することを意図している。適切な担体は、参照により本明細書に組み込まれるRemington's Pharmaceutical Sciences、Gennaro AR(編)20版(2000年)Williams& Wilkins PA、米国、およびWilson and Gisvold's Textbook of Organic Medicinal andPharmaceutical Chemistry、Delgado and Remers著、Lippincott-Ravenなどの教科書に記載されている。このような担体または希釈剤に使用し得る成分の好ましい例には、それだけに限らないが、水、塩水、リン酸塩、カルボン酸塩、アミノ酸溶液、リンゲル液、デキストロース(グルコースの同義語)溶液、および5%ヒト血清アルブミンが挙げられる。非限定的な例として、デキストロースは5%または10%水溶液として使用してもよい。リポソーム、および不揮発性油などの非水性媒体も使用し得る。医薬活性物質用のこのような媒体および作用剤の使用は、当技術分野で周知のことである。補助的な活性化合物も、該組成物中に組み入れることができる。
【0119】
本発明の医薬組成物は、意図する投与経路に適合するように処方される。投与経路の例には、非経口経路、例えば、静脈内、皮内、皮下、経口、経鼻、吸入、経皮(局所)、経粘膜および直腸投与が挙げられる。非経口、静脈内、皮内または皮下投与に使用される溶液または懸濁液は、以下の成分:注射水、塩水溶液、不揮発性油、ポリエチレングリコール、グリセリン、プロピレングリコールもしくは他の合成溶媒などの希釈剤;ベンジルアルコールもしくはメチルパラベンなどの抗細菌剤;アスコルビン酸もしくは重亜硫酸ナトリウムなどの抗酸化剤;エチレンジアミン四酢酸などのキレート剤;酢酸塩、クエン酸塩もしくはリン酸塩などの緩衝剤、および塩化ナトリウムもしくはデキストロースなどの等張化剤を含むことができる。
【0120】
吸入による投与のために、該化合物は、適切な高圧ガス、例えば二酸化炭素などのガスを含有する加圧容器もしくはディスペンサー、または噴霧器からのエアゾールスプレーの形態で送達される。
【0121】
一実施形態では、活性化合物は、埋め込み材およびマイクロカプセル化送達系を含めた制御放出製剤などの、体内からの急速消失から該化合物を保護すると見込まれる担体と共に調製される。持続放出製剤の適切な例には、抗体を含有する固形疎水性ポリマーの半透性マトリックスであって、成形品、例えばフィルムまたはマイクロカプセルの形態をしたマトリックスが挙げられる。持続放出性マトリックスの例には、ポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチルメタクリレート)またはポリ(ビニルアルコール))、ポリラクチド(米国特許第3773919号)、L−グルタミン酸およびγエチル−L−グルタミン酸のコポリマー、非分解性エチレン−酢酸ビニル、LUPRON DEPOT(商標)(乳酸−グリコール酸コポリマーおよび酢酸ロイプロリドからなる注射用ミクロスフェア)などの分解性乳酸−グリコール酸コポリマー、およびポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。エチレン−酢酸ビニル、乳酸−グリコール酸などのポリマーは、分子の放出を100日超の間可能とするが、ある種のヒドロゲルでは、医薬活性剤を放出する期間がより短い。有利なポリマーは、生分解性または生体適合性である。リポソーム懸濁液(ウィルス抗原に対するモノクローナル抗体を有する、感染細胞を標的としたリポソームを含める)も、医薬として許容可能な担体として使用することができる。こうしたものは、例えば、米国特許第4522811号に記載のように、当業者に公知の方法に従って調製することができる。ミクロスフェアなどの有利な形態を有する持続放出製剤は、上記の材料などから調製することができる。
【0122】
本発明に開示するポリヌクレオチドは、ベクター中に挿入し、遺伝子治療用ベクターとして使用することができる。遺伝子治療用ベクターは、例えば、米国特許第5703055号に記載のように、幾つもの経路のいずれかにより対象へ送達することができる。したがって、送達には、例えば、静脈注射、局所投与(米国特許第5328470号を参照されたい)、または定位注射(例えば、Chenら(1994年)Proc. Natl. Acad. Sci. USA 91巻、3054〜3057頁を参照されたい)も挙げることができる。遺伝子治療用ベクターの医薬製剤は、許容可能な希釈剤中に遺伝子治療用ベクターを含むことができ、または遺伝子送達媒体を埋め込んだ徐放性マトリックスを含むことができる。あるいは、完全な遺伝子送達ベクター、例えばレトロウィルスベクターを組換え細胞から無傷で産生することができる場合、該医薬製剤は、遺伝子送達系を産生する1個または複数の細胞を含むことができる。
【0123】
医薬組成物は、キット中、例えば容器、包み、またはディスペンサー中に投与説明書と共に含めることができる。
【0124】
対象の呼吸器ウィルス感染症を処置する医薬組成物または薬剤の製造における治療剤の使用も、本発明に入る。
【0125】
幾つかの実施形態では、本発明に開示するポリヌクレオチドは、リポソーム媒介トランスフェクションにより、例えば、市販の試薬または技法、例えばOligofectamine(商標)、LipofectAmine(商標)試薬、LipofectAmine2000(商標)(Invitrogen)の使用により、ならびに電気穿孔および類似の技法により、送達される。それに加え、ポリヌクレオチドは、呼吸器中への吸入および点滴注入により、げっ歯類または非ヒト霊長類などの動物モデルへ送達される。動物モデルに用いる付加的経路には、静脈内(IV)、皮下(SC)、および関連投与経路が挙げられる。医薬組成物は、ポリヌクレオチドの安定性を保護し、その寿命を延長し、その機能を増強し、または特定の組織/細胞をその標的とさせる、追加の成分を含む。これらの成分には、多種の生分解性ポリマー、陽イオン性ポリマー(ポリエチレンイミンなど)、ヒスチジン−リシン(HK)ポリペプチドなどの陽イオン性コポリペプチド(例えば、MixonらのPCT公開WO第01/47496号、BiomerieuxのWO第02/096941号、およびMassachusetts Institute of TechnologyのWO第99/42091号を参照されたい)、PEG化陽イオン性ポリペプチドおよびリガンド導入ポリマーなど、陽荷電ポリペプチド、PolyTranポリマー(スクレログルカンの名でも知られる天然多糖;Pillsbury Co.,Minneapolis,Minn)、界面活性剤(Infasurf;Forest Laboratories,Inc.;ONY Inc.)、ならびに陽イオン性ポリマー(ポリエチレンイミンなど)が挙げられる。Infasurf(登録商標)(カーファクタント(calfactant))は、子ウシの肺から単離し、気管内点滴注入に使用する天然の肺界面活性剤であり、リン脂質、中性脂質、ならびに疎水性の界面活性剤関連タンパク質BおよびCを含有する。該ポリマーは、一次元、多次元のいずれでもよく、直径が20ミクロン未満、20〜100ミクロンの間、または100ミクロン超のマイクロ粒子またはナノ粒子でもよかろう。前記ポリマーは、特殊な組織または細胞の受容体または分子に特異的なリガンド分子を搬送し、したがってポリヌクレオチドの標的向け送達に使用することができよう。ポリヌクレオチドは、DOTAP、DOTAP/コレステロール(Qbiogen,Inc.)などの陽イオン性リポソーム系担体、および他種の脂質水溶液によっても送達される。その上、低率(5〜10%)グルコース水溶液およびInfasurfは、ポリヌクレオチドの気道送達用の有効な担体である。
【0126】
遺伝子治療
現在では、遺伝子治療は、治療および予防(例えば免疫用接種)環境で使用するための高度な開発状態にある。新規な送達法が利用でき、多様な遺伝子治療様式に関する幾つかの臨床試験が進行中である。「Direct DNA delivery to bone cells」という名称の米国特許出願公開第20060115462号は、裸ポリヌクレオチドまたは遺伝子発現ベクターなどのポリヌクレオチドの哺乳類四肢骨部の細胞に対するインビボ送達を可能とする方法を開示している。
【0127】
「Replication-competent adenoviral vectors」という名称の米国特許出願公開第20060115456号は、アデノウィルス2型、4型、5型および7型を含み、治療および予防用途向けに核酸およびポリヌクレオチドを送達する際に使用するための、改良された複製能のあるアデノウィルスベクターを開示している。
【0128】
「Methods of Tranducing genes into T cells」という名称の米国特許出願公開第20060104950号は、パラミクソウィルスベクターを用いた活性化T細胞中への遺伝子の形質導入を開示している。
【0129】
「Methods of making viral particles havinga modified cell binding activity and uses thereof」という名称の米国特許出願公開第20060110361号は、その物品の表面上にある1種または複数のペブチドがパッケージング細胞に由来するように、ウィルス粒子をパッケージする方法を開示している。この出願は、このような系が、例えば遺伝子治療処置に有用であると明言している。
【0130】
「Vector-mediated delivery ofpolynucleotides encoding soluble VEGF receptors」という名称の米国特許出願公開第20060110364号は、可溶型VEGFR3を発現するためのベクター組成物、および癌の処置におけるその使用方法を開示している。
【0131】
「Multifunctional molecular complexes forgene transfer to cells」という名称の米国特許出願公開第20050239204号は、核酸組成物を標的細胞へ移送するための非ウィルス多機能分子複合体を開示している。
【0132】
VandenDriesscheらは、ヒトVIII因子遺伝子をレトロウィルスベクターによりVIII因子欠損マウス中に導入するインビボ遺伝子治療によって、A型血友病を有効に処置できることを示した79。遺伝子導入は、少なくとも肝臓、脾臓および肺臓において行われた。A型血友病の重症患者における非ウィルス体細胞遺伝子治療系の安全性が、Rothらにより試験されてきた80。
【0133】
安定な有糸分裂後の状態のために、骨格筋は、組込み型、非組込み型双方のウィルスおよび非ウィルスベクターの使用による遺伝子操作の重要な標的である。非肥満性糖尿病(NOD)マウスにおける自己免疫性糖尿病を予防するための筋肉誘導遺伝子治療が、Goudyらにより報告された81。彼らは、免疫調節サイトカインのIL4またはIL10のためのマウスcDNAを含有する、組換えアデノ付随ウィルスベクターを開発した。IL4ではなく、IL10による雌性NODマウスの骨格筋形質導入によって、糖尿病が完全に消失した。
【0134】
Wells82によれば、アデノ付随ウィルスベクターおよび裸プラスミドDNAは、現在、筋肉中に遺伝子導入をするための優れたベクターである。ベクター送達系、特に血管経路を用いる系における主要な躍進は、ここ2年ほど以内になされており、その結果、筋ジストロフィーの遺伝子治療、および分泌タンパク質の産生用プラットホームとしての筋肉の使用が、臨床的可能性となった。
【0135】
癌溶解治療は、アデノウィルス(Ad)などの弱毒化溶解性ウィルスに基づく新規な抗癌処置である83。これらのウィルスは、溶解によって宿主細胞の破壊を誘発し、したがって古典的な遺伝子治療用ウィルスとは異なる。癌溶解性Adは、一般に、癌細胞のみで複製するように遺伝子操作を受けている。3種の癌溶解性Adが、癌患者において第1相および第2相臨床試験を完了した。これらのウィルスは、追加の治療遺伝子を備えるように、更に操作されているところである83。
【0136】
Takahashiらは、多剤耐性(MDR1)遺伝子治療の臨床研究を開始した。現在まで処置された患者2名は、完全に寛解しており、MDR1遺伝子導入による明らかな有害作用を受けなかった84。
【0137】
Broedlらは、様々な高脂肪血症の処置が無効な現状にあって、体細胞遺伝子治療は、有力な治療手法と考えられると述べている。多くの症例で、原理証明前臨床試験が既に行われており、ホモ接合型の家族性高コレステロール血症の症例では、臨床試験が行われた85。
【0138】
クラスタリンを対象とするアンチセンスオリゴデオキシヌクレオチドが、前立腺癌の処置のために第1相臨床試験において使用された86。この処置により、前立腺癌におけるクラスタリンが90%までも抑制された。Miyakeらは、第2相試験が2005年の早期に開始する予定であったと明言している86。
【0139】
Satohらは、単純疱疹ウィルスチミジンキナーゼ遺伝子のウィルス媒介形質導入と、その後のガンシクロビルプロドラッグによる一連の治療を含め、前立腺癌遺伝子治療の現状を総説している87。その報告によれば、人体前立腺癌の遺伝子治療臨床試験では、安全性、臨床的効力、および抗腫瘍活性の生物学的効果が実証された。Satohらによれば、多数の好ましい前臨床試験の後、インターロイキン−2、インターロイキン−12、B7−1(CD80)、B7−2(CD86)、顆粒球−マクロファージコロニー刺激因子などの免疫調節性サイトカインを用いる前立腺癌の遺伝子治療に対して、幾つかの臨床試験が承認された87。
【0140】
潜在的活性剤を同定するためのアッセイ法
多様な態様では、本発明は、候補化合物または候補物質が、本発明において対象とする活性を調節するか否かを決定することを意図したアッセイ法を開示する。このような方法には以下のものが挙げられる。
【0141】
本発明は、候補物質がLXR依存性プロセスを調節するか否かを評価する方法であって、
a)LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターで細胞をトランスフェクトし、
b)その細胞を候補物質と接触させ、更に、
c)候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定する
ことを含み、それによって、候補物質の存在と不在との間で検出されるLXR依存性プロセスの発生の程度における差異が、候補物質がLXR依存性プロセスを調節することを示す
方法を開示する。
【0142】
本発明は、更に、候補物質がLXRのSIRT1依存性作用を調節するか否かを評価する方法であって、
a)SIRT1遺伝子を保持するベクターで細胞をトランスフェクトし、
b)LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するベクターで該細胞をさらにトランスフェクトし、
c)その細胞を候補物質と接触させ、更に、
d)候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定する
ことを含み、それにより、候補物質の存在と不在との間で検出されるLXRのSIRT1依存性作用の程度における差異が、候補物質がLXRのSIRT1依存性作用を調節することを示す
方法を開示する。
【0143】
本発明は、更に、候補物質が、特異的結合対(specific binding pair)のメンバーのSIRT1およびLXRを含む特異的結合対の形成を調節するか否かを評価する方法であって、
a)特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトし、
b)特異的結合対の第2メンバーをコードする配列を保持するベクターで該細胞をさらにトランスフェクトし、
c)その細胞を候補物質と接触させ、
d)その細胞を溶解し、エピトープタグに特異的な抗体を細胞溶解物と接触させ、更に、SIRT1とLXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収し、
e)SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行する
ことを含み、それにより、候補物質の不在下と比較した、候補物質の存在下で検出した複合体形成の差異が、候補物質がSIRT1のLXRとの相互作用を調節することを示す方法を開示する。
【0144】
こうしたアッセイ法では、参照により本明細書に組み込まれる、上述した技法などにより、ベクターのトランスフェクションを実施し得る。本発明の分野における技術者に公知であり、本明細書に具体的に記載していない任意の同等な技法も、トランスフェクションを実現するために同様に使用してもよい。
【0145】
多様な該アッセイ法は、更に、LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターのトランスフェクションを含む。レポーター遺伝子は、レポーター遺伝子を発現するためにLXREに特異的に結合する細胞成分により、活性化されることになろう。レポーター遺伝子産物は、アッセイを行うために工夫した実験系において検出できる。一般に、レポーター遺伝子は、化学的または物理的検出法によって、発現した際に容易に検出できるタンパク質、因子、または酵素活性をコードする。レポーター遺伝子の非限定的な例には、西洋ワサビペルオキシダーゼまたはホタルルシフェラーゼなどの酵素、発色タンパク質または緑色蛍光色素などの蛍光タンパク質などをコードする遺伝子が挙げられる。本発明の分野における技術者に公知であり、本明細書に具体的に記載していない任意の同等なレポーター遺伝子も、細胞に移入することにより、潜在的な活性剤を同定するアッセイを実施するために、同様に使用してもよい。
【0146】
該アッセイ法の多様な付加的実施形態は、SIRT1およびLXRを含む特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトすることを含む。この目的に使用できるエピトープには、特異抗体が結合する任意のエピトープが挙げられる。エピトープの非限定的な一例は、FLAGエピトープであり、その他の例は、本発明の分野における技術者に周知のものである。本明細書に具体的に記載していない任意の同等なエピトープも、潜在的な活性剤を同定するアッセイを実施するために、特異的結合対の一メンバーとのキメラとして使用してもよい。
【0147】
潜在的な活性剤を求めるアッセイは、高度反復並行アッセイ装置におけるハイスループットスクリーンを用いて実施することが多い。こうしたシステムでは、多数のアッセイを同時に実施するために、多重ウェルプレートを使用してもよい。潜在的な活性剤を検出するアッセイに使用される候補化合物は、一般に、一群の関連化合物における多様な部分および/または置換基を系統的に変化させた、化合物のコンビナトリアルライブラリーを化学合成した結果として取得される。類似の化学品ライブラリーは、通常、個人または組織が利用できる保管化合物として既に入手が可能である。更なる類似の化学品ライブラリーには、異種の天然源から得られる化学品、ならびにコンビナトリアルケミストリーの方法を用いて更に修飾した、このような天然化合物の誘導体が含まれる。一般に、化合物のライブラリーは、本発明の分野における技術者に広く知られている。候補化合物を調製する、または候補化合物のライブラリーを調製する、本発明の分野における技術者に公知の同等な任意の方法を、潜在的な活性剤を同定するアッセイを実施する際に使用してもよい。
【実施例】
【0148】
方法
プラスミド
プラスミドのpBabe−SIRT1およびpCMV−FLAG−HsSIRT1は、既に記載されている15,16。
動物および血漿リポタンパク質分析
129/sv−CD1を混合バックグランドとするSIRT1+/+、SIRT1+/−およびSIRT1−/−マウスは、7a.m.〜7p.m.の昼光サイクルの温度制御(22℃)施設に収容し、食物(0.02%コレステロール入り)および水を自由に摂取させた。
【0149】
細胞、レトロウィルス感染およびトランスフェクション
SIRT1+/+、SIRT1+/−およびSIRT1−/−マウスの胚性線維芽細胞(MEF)を以前に記載のようにE14.5の胚から分離し73、10%FBSおよび抗生物質を含むダルベッコ改良イーグル培地(DMEM)中で培養した。不死化MEF(MEFI)は、初代MEFにpRS−SV40Tプラスミドを移入することにより生成した。HEK293TおよびPhoenix細胞(ATCC,Rockville,MD)は、10%FBSおよび抗生物質を含むDMEM中で培養した。THP−1細胞(ATCC,Rockville,MD)は、10%FBSおよび0.5mM β−メルカプトエタノールを含むRPMI1640中で培養した。
【0150】
全てのトランスフェクションは、製造業者の説明書(Invitrogen,Carlsbad,Ca)に従ってリポフェクタミン2000plus試薬を用いて行った。レトロウィルス感染については、Phoenix細胞にpBabeまたはpBabe−SIRT1を移入した。レトロウィルスを含有する培地は、48時間後に収集し、ろ過し、ポリブレン(1mg/ml)で処理し、MEF上に移した。次いで、1μg/mlのピューロマイシンを含有する通常培地で感染細胞を培養した。
【0151】
(実施例1)
野生型およびSIRT1ノックアウトマウスにおける血漿コレステロール
各群における野生型10匹、SIRT1+/−13匹、および同腹子SIRT1−/−13匹の雄性マウスを分析した。動物を昼光サイクルの開始から4時間絶食させ、次いで血液を採集し、K3−EDTA処理により血漿を取得した。総コレステロール、HDLおよびLDLの血漿濃度を、酵素呈色アッセイキット(Wako Diagnostics,Richmond,Va)によって測定した。SIRT1−/−4匹、および同腹子SIRT1+/+4匹の雄性マウスから得た保管血漿100μlを、高速液体クロマトグラフィー(FPLC)のカラム(Superose 6Bカラム、Amersham−Pharmacia Biotech,Piscataway,NJ)2本を用いてサイズ分画した。個別の3実験からの代表的プロファイルを示してある。次いで、FPLCから得た画分のコレステロール含量をWako製酵素呈色アッセイキットで分析した。実験は、合計してSIRT1−/−12匹および同腹子SIRT1+/+12匹の雄を用いて3回繰り返した。結果は、スチューデントのt検定を用いて比較した。
【0152】
図1aは、SIRT1−/−マウスが、週齢および性別が対応する野生型マウスと比較して、僅かに低い総血漿コレステロール濃度を有したことを示す。図1のパネルb(直接酵素アッセイ)およびパネルc(サイズ排除クロマトグラフィー/酵素アッセイ)を点検すると、マウス血漿中の主要な富コレステロールリポタンパク質(全体の80〜90%)である、HDL中に保持されるコレステロールが約40%低下した(*、p<0.001)ために、そうなることが分かる。血漿LDLコレステロール濃度は、SIRT1−/−および対照マウスにおいて類似していた(図1cおよびd)。その結果、SIRT1−/−マウスの総コレステロール/HDL比は、異常に高く(野生型対照の1.36±0.19に対して2.57±0.98、p<0.001)、HDL/LDL比は異常に低かった(3.56±1.24に対して1.92±0.94、p<0.001)。SIRT1−/−マウスにおける多面発現効果のために、野生型と表現型が類似しているSIRT1+/−ヘテロ接合型も調べた。HDL濃度は、SIRT1+/−ヘテロ接合型でも有意に低下していることが判明した(図1b(*、p<0.001))。
【0153】
総コレステロール(TC)、遊離コレステロール(FC)、コレステロールエステル(CE)およびリン脂質(PL)の含量を、12対の動物からのHDL画分(25〜38番画分)において測定しており、それを図1パネルeに示す。血漿HDL中の総コレステロール、遊離コレステロール、コレステロールエステルおよびリン脂質の量のSIRT1−/−マウスにおける相対的低下は、類似しており、約25〜30%であった(図1e;**、p<0.001)。HDLのサイズ分布は、SIRT1−/−マウスにおいて正常であった(図1d)が、それは、HDLの低下が、粒子の組成または構造が変化したためより、粒子数が減少した結果であったことを示唆している。
【0154】
(実施例2)
野生型およびSIRT1ノックアウトマウスの組織におけるコレステロール
マウス組織中の総コレステロールを決定するために、SIRT1−/−マウスおよび対照の同腹子を昼光サイクルの開始から4時間絶食させ、その後犠牲にした。次いで、組織を採集し、秤量した。既述のように70,71、肝臓および精巣の総コレステロールを抽出し、GCにより測定した。トリグリセリドを含めた総脂質も、60%ブタノール、13%メタノールおよび27% Triton X−100を含有する溶液中に溶解し、Wako製酵素呈色アッセイキットで測定した。
【0155】
SIRT1−/−マウスにおいて、異常に低い血漿HDL(実施例1)は、HDLが外因性コレステロール源として重要な以下の2種の組織内で、コレステロールの蓄積量が増加したことと関係していたが、そうした組織とは、ステロール保存およびステロイド合成25のためにHDLコレステロールを使用し、相対量最高のSIRT1タンパク質を有する器官26,27である精巣、ならびにコレステロールおよびHDLの恒常性に中心的役割を果たす28肝臓である(図1f;n=8;***、p<0.01)。
【0156】
SIRT1−/−マウスにおける血漿HDLコレステロールの減少、ならびに肝臓および精巣中の組織コレステロールの蓄積量増加は、リポタンパク質コレステロールの取込み増加、または細胞コレステロールのリポタンパク質への排出量(移出量)減少のために起こり得よう。組織コレステロールの増加は、局所的なコレステロール合成の増加によることもあろう。しかし、発明者らは、肝臓および精巣において、コレステロール生合成における律速酵素のHMG−CoAレダクターゼをコードする高感受性コレステロール調節遺伝子に対応するmRNAの量が、本質的に正常であることを観察した(データは示していない)。また、SIRT1−/−マウス胚性線維芽細胞において(MEF、データは示していない)、HDLコレステロールの細胞取込みを媒介する主要なHDL受容体SR−BI29のタンパク質量の明らかな増加が、こうした組織中で見られず(データは示していない)、HDLからの脂質取込み速度の増加も見られなかった。更に、他の主要な2種のリポタンパク質受容体であるLDL受容体(LDLR)およびVLDL/キロミクロン受容体LRPのmRNA量が、肝臓および精巣では増加するのでなく、実質的に減少していた(データは示していない)。新規コレステロール合成の増加でも受容体媒介移入でも、SIRT1−/−マウスにおける組織内増加および血漿コレステロール減少を説明できないようなので、細胞のコレステロール流出量の低下が、こうした観察を説明し得るのではないかと思われた。
【0157】
(実施例3)
野生型およびSIRT1−/−細胞からのコレステロールの排出
コレステロール逆輸送は、末梢組織中の過剰コレステロールを、体内から除去するために肝臓へ輸送する過程である30,31。この過程の第1段階は、細胞からリポタンパク質、特にHDLへのコレステロールの排出である。数種の細胞表面コレステロール輸送タンパク質が、コレステロール流出を媒介することができ、そうしたものには、SR−BI32−35、ABCGI36,37、およびこれらのうちで特性決定が最も良くなされたもの、即ち、非エステル化コレステロールおよびリン脂質を貧脂質アポリポタンパク質(主にアポA−I)へ移し、HDL粒子を形成する、ABCA1と称するATP結合カセット(ABC)輸送体30,38−44が含まれる。この過程に対するSIRT1の効果を試験するために、アポA−I媒介コレステロール流出を、[3H]コレステロールで細胞を標識した異なる2種の培養細胞系で測定し、細胞外培地中にあるアポA−Iへの標識コレステロールの排出をモニターした。
【0158】
MEFについては、コレステロール流出アッセイを、改良を加えたが記載してあるように72行った。初代SIRT1−/−および同腹子対照SIRT1+/+またはSIRT1+/−のMEFを、12ウェルプレート中に集密度50%で播種し、終夜培養した。次いで、細胞をRPMI1640/0.2%ウシ血清アルブミン(BSA)中に24時間移し、LDL50μg/mlおよび[3H]コレステロール1μCi/ml(NEN Life Science Products,Boston,Ma.)と更に24時間インキュベーションすることにより、細胞にコレステロールを担持させた。細胞をPBSで洗浄し、RPMI1640/0.2%BSA中で4時間平衡化し、次いでRPMI1640/0.2%BSA中、15μg/mlアポA−Iと共に、またはそれ無しで24時間インキュベートした。次いで培地を採集し、細胞を0.2N NaOHで溶解した。培地および細胞溶解物中に回収された放射活性を測定し、アポAI媒介コレステロール流出を、アポAI非含有培地中の非特異的排出量を差し引いた全放射活性に対する、培地中に回収した放射活性の比率(%)として計算した。コレステロール流出アッセイは、SIRT1−/−および同腹子3匹からの対照SIRT1+/+またはSIRT1+/−のMEFを7対用いて、2重に行った。
【0159】
初代MEFを用いて、[3H]コレステロールを担持した、野生型対照細胞およびSIRT1−/−細胞を、15μg/mlアポAIを含む、または含まないRPMI1640培地と共にインキュベートした。細胞および培地中の[3H]コレステロール量を24時間後に測定し、アポAI媒介コレステロール流出を計算した。図1g(左パネル)は、対照SIRT1+/+の初代MEFからの排出(塗潰し棒)が、SIRT1−/−のMEFからの排出(白抜き棒)よりほぼ3倍高かったことを示している(n=7;****、p<0.001)。
【0160】
ヒト単球細胞系THP−1細胞については、細胞をRPMI1640/10%FBS中、10mMニコチンアミドで48時間処理した後、RPMI1640/1%FBS/0.2%BSA培地中、LDL50μg/mlおよび[3H]コレステロール1μCi/mlで24時間、細胞にコレステロールを担持させた。細胞をPBSで洗浄し、RPMI1640/0.2%BSA中、15μg/mlアポA−Iと共に、またはそれ無しで24時間インキュベートした。次いで、培地と細胞とを分離し、細胞を0.2N NaOHで溶解した。培地および細胞溶解物中に回収された放射活性を測定し、アポAI媒介コレステロール流出を上記の通り計算した。
【0161】
SIRT1阻害剤のニコチンアミド(Nico)45によるTHP−1の処理も、[3H]コレステロール流出を大まかに1/3に抑制した(図1g、右パネル)。この実験は、三重に行い、2回繰り返した(p<0.01)。したがって、MEF中でのSIRT1の喪失、および単球中でのSIRT1活性の阻害は、アポAI媒介コレステロール流出を低下させる。こうした結果は、コレステロール流出の低下が、SIRT1−/−マウスにおいて、血漿HDLコレステロールの低下および組織コレステロールの増加を起こすのではないかという可能性を支持している。
【0162】
(実施例4)
野生型およびSIRT1ノックアウト組織におけるABCA1 mRNAの量
コレステロール流出の低下が、SIRT1−/−マウスにおいて、血漿HDLコレステロールの低下および組織コレステロールの増加を起こし得るか否かを調べるために、肝臓、精巣および卵巣におけるABCA1 mRNAの量を比較した。
【0163】
Qiagen RNeasyミニキット(Qiagen Inc.,Calencia,Ca)により、マウスの肝臓、精巣、チオグリコール酸賦活腹腔マクロファージおよび卵巣から総RNAを分離した。リアルタイムPCR分析のために、ランダムプライマーと共にSuperScript III逆転写酵素(Invitrogen,Carlsbad,Ca)によってcDNAを合成した。次いで、生成cDNAに、Cybergreen(Qiagen Inc.,Calencia,Ca)の存在下、遺伝子特異的プライマーを用いてPCR分析を行った。mRNAの相対的存在量を、シクロフィリン量へ規格化することによって得た。ノーザンハイブリッド形成のために、標準的手順に従って、RNA(10μg/レーン)をホルムアルデヒド−アガロースゲル(1.0%)上で分離し、GeneScreen Plus膜(NEN Life Science Products,Boston,Ma.)に移し、ハイブリッド形成した。ABCA1のmRNA量をホスホイメージ装置(phosphoimager)で定量し、相当するアクチン量に対して規格化した。
【0164】
ABCA1 mRNA発現量が、野生型と比較して、SIRT1−/−マウスでは3種の試験組織において40〜50%低下した(図2a)。この低下が、血漿HDL量の低下の原因になったのではないかという発想と合致して、ABCA1+/−ヘテロ接合型マウスでは、HDLコレステロールが、SIRT1−/−マウスにおける低下同様に、ABCA1+/+対照の約60%に低下している。
【0165】
(実施例5)
ABCA1プロモーターの活性
ABCA1の転写は、細胞コレステロールの増加によって誘発され、LXR核内受容体転写因子がこの応答を媒介する47−49。LXR/レチノイドX受容体(RXR)ヘテロ二量体は、オキシステロールにより活性化され50、全身コレステロールの恒常性、脂質生合成、炎症反応、炭水化物代謝および肥満の調節に重要な役割を果たす51。ABCA1の発現に対するSIRT1の効果が、LXRにより媒介されるか否かを決定するために、ホタルルシフェラーゼ遺伝子の発現を、マウスABCA1プロモーター、またはヌクレオチド8個の欠失でLXR応答エレメント(LXRE)を除去した対照変異体プロモーターのいずれかで推進する、レポーター構築体を生成した。以前の研究によれば、22(R)−ヒドロキシコレステロール(22(R)−HC、LXRアゴニスト)および9−シスレチノイン酸(9−シスRA、RXRアゴニスト)の併用で、ABCA1プロモーター中のLXREの活性が劇的に刺激されることが示されている47。
【0166】
ルシフェラーゼレポータープラスミドのpGL3−ABCA1を、pGL3ベクター(Promega,Madison,Wi)のKpnI−XhoI部位中にマウスABCA1プロモーター領域の−847〜+244を挿入することにより、創製した。pGL3−ABCA1のLXRE変異体型であるpGL3−ABCA1−LXREを、マウスABCA1の野生型プロモーターからLXREを欠失させることにより、創製した。
【0167】
ルシフェラーゼアッセイについては、24ウェルプレート中に播種したMEFIまたはHEK293T細胞に、pGL3−ABCA1またはpGL3−ABCA1−LXREを100ng、およびpRL−TK(ウミシイタケ(Renilla)ルシフェラーゼ、Promega)を10ng移入した。野生型(WT)またはLXRE変異体(−LXRE)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを移入した細胞を、クラスIおよびIIデアセチラーゼの阻害剤であるTrichostatin A(TSA)1μM、もしくはニコチンアミド(Nico)10mM、または両者のニコチンアミド10mM+TSA1μMで処理した。LXRE変異体ベクターを移入した細胞を、10%FBS入りDMEM中、22(R)−HCを10μMおよび9−シスRAを1μMの不在下(塗潰し棒)、または存在下(白抜き棒)でインキュベートした(図2b)。次いで、Dual−Luciferase Reporter Assay System(Promega)を用いて、6時間または24時間後にルシフェラーゼ活性を測定した。最終的なGL(ホタルルシフェラーゼ)活性は、同時発現したRL(ウミシイタケルシフェラーゼ)活性を用いて規格化した。実験は、二重に行い、3回繰り返した。
【0168】
野生型および変異体レポーター構築体をHEK293T細胞中に発現させた(図2b、WT(左パネル)または−LXRE(右パネル))。野生型からのルシフェラーゼ発現は、22(R)−HC+9−シスRAでの処理により8倍賦活された(図2b、左パネル)。その上、この誘導発現は、SIRT1(a)などのクラスIIIデアセチラーゼを阻害する阻害剤ニコチンアミドにより、50%抑制されたが、TSAでは抑制されなかった。LXREがない構築体は、LXRアゴニストに応答せず、低い基準活性を示した(図2b、右パネル)。纏めると、こうした知見は、他クラスのデアセチラーゼではなく、SIRT1が、注目すべきことに正の調節剤として、ABCA1プロモーターにおけるLXR活性を調節することを示唆している。
【0169】
それに加え、こうした実験に使用される系が、候補物質によるLXR依存性プロセスの調節を評価する方法を与えることが分かる。
【0170】
(実施例6)
ABCA1プロモーターに対するSIRT1活性の効果
ABCA1プロモーターに対するSIRT1の影響を直接決定するために、これらのプロモーターレポーター構築体を、SIRT1タンパク質を発現するようにpBabe−mT1ベクターを感染させた、または感染させていない、不死化したSIRT1+/+およびSIRT1−/−MEF中に導入した(図2c)。SIRT1+/+およびSIRT1−/−MEFを生成し、それらに、実施例3に記載のように、pBabe−SIRT1(+)またはpBabe対照ベクター(−)を感染させた。
【0171】
図2c、右パネルの差込図は、これら4種の細胞系におけるSIRT1の発現量に関する免疫ブロットの結果を示し、チューブリンをローディング対照として使用した。このベクターからのSIRT1の発現は、SIRT1+/+MEFにおける内因性SIRT1よりやや低かった。
【0172】
次いで、MEFに、野生型(WT)またはLXRE変異体(−LXRE)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを、22(R)−HCを10μMおよび9−シスRAを1μMの不在下(塗潰し棒)、または存在下(白抜き棒)で移入した。ルシフェラーゼ活性を24時間後に測定した。野生型プロモーターを有する全ての細胞は、LXR/RXR活性化剤で処理した際、ルシフェラーゼ発現の顕著な賦活を示した(図2c、左パネル)が、対照変異体(−LXRE)レポーターを有する細胞の低い基準活性は、これらの活性化剤で賦活されなかった(図2c、右パネル)。最も重要なことは、野生型プロモーター構築体のLXRE依存性活性が、SIRT1欠損性の細胞(SIRT1−/−MEF)では、SIRT1+/+細胞またはSIRT1で再構成された細胞(pBabe−T1)中より、40%低かった(p<0.02)ことである。こうした知見は、SIRT1が、LXREにおいて作用するLXR/RXRを介してABCA1プロモーターを刺激するというモデルを支持している。
【0173】
この実施例における実験系は、LXRのSIRT1依存性作用に対する候補物質の調節作用を評価する方法を与える。
【0174】
(実施例7)
SIRT1とABCA1プロモーターのLXREとの相互作用
SIRT1が、ABCA1プロモーター中のLXREに直接結合するか否かを決定するために、発明者らは、クロマチン免疫沈降(ChiP)アッセイを用い、マウスABCA1プロモーターを様々なプライマーで捜し求めた。
【0175】
ChiP分析は、改良を加えたが記載してあるように76行った。手短に言えば、細胞を培地中の1%パラホルムアルデヒドで15分間、室温で架橋し、0.125Mグリシンで架橋を終結させた。次いで、細胞をHEPES緩衝液(50mM HEPES−NaOH、pH7.9、140mM NaCl、1mM EDTA、10%グリセロール、0.5% NP40、0.25% Triton X−100、およびComplete(商標)プロテアーゼインヒビター混合物(Roche,Indianapolis,In))中に採集し、核を精製した。精製核をTE緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、およびComplete(商標)プロテアーゼインヒビター混合物)中に再懸濁し、氷上で超音波処理して平均長1kbにした。超音波処理上清をIP緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、10%グリセロール、1% Triton X−100、0.1%デオキシコール酸Na、およびComplete(商標)プロテアーゼインヒビター混合物)中に希釈し、プロテインAビーズで2回予備浄化した。次いで、予備浄化クロマチン試料を抗SIRT1ウサギポリクローナル抗体(Upstate,Charlottesville,Va)と4℃で終夜インキュベートし、プロテインAビーズを2時間添加することにより、免疫複合体を回収した。次いで、回収した免疫沈降物をIP緩衝液で2回、500mM NaCl含有IP緩衝液で2回、RIPA緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、250mM NaCl、0.5% NP40、0.5%デオキシコール酸Na、およびComplete(商標)プロテアーゼインヒビター混合物)で2回洗浄した。次いで、免疫沈降物中のクロマチンを10×TE/1%SDSで65℃、15分間溶出した。溶出クロマチンを65℃で少なくとも6時間インキュベートして、架橋を無効にし、プロテイナーゼKおよびRNアーゼAで処理し、フェノール/クロロホルムおよびクロロホルムで抽出した。最終的なDNA断片をLiCl/グリコーゲン/エタノール沈降により回収し、ペレットを水中に再懸濁し、PCRでアッセイした。
【0176】
マウスABCA1プロモーターのLXREについては、下記のプライマーを用いてPCR反応を行った。
【0177】
【化14】
前記ChiPアッセイの陰性対照として、ABCA1プロモーター上のLXREの上流にある6.2kb断片も、下記のプライマーで増幅した。
【0178】
【化15】
ヒトプロモーターのLXREについては、下記のプライマーを用いてPCR反応を行った。
【0179】
【化16】
陰性対照として、LXREの上流にある7.5kb断片を下記のプライマーで増幅した。
【0180】
【化17】
実験は少なくとも3回行ったが、その代表的データを示してある。
【0181】
図2d上部は、SIRT1に対する抗体で免疫沈降する前(入力)または後(αSIRT1)で、LXRE特異的または上流特異的なPCRプローブで探った結果を示す。実験処理において、(αSIRT1)抗体処理により、SIRT1+/+MEF(左レーン、暗灰色バンドを示す上部像)からのLXREを含有するクロマチンの断片は、沈降したが、SIRT1−/−MEF対照(右レーン、地黒を示す上部像)では沈降せず、LXREの上流にある6.2kbDNAを含有する断片は、いずれの細胞系でも沈降しなかった(地黒を示す両レーンの下部像)。FLAGエピトープタグ付きヒトSIRT1の発現ベクターまたは陰性対照ベクターを移入した、HEK293T(図2d、下部)およびTHP−1(データは示していない)ヒト細胞においても、同様な結果が見出された。したがって、SIRT1は、ABCA1プロモーターと、LXREと直ぐの近接部で結合する。
【0182】
(実施例8)
SIRT1のLXRαおよびLXRβとの相互作用
SIRT1は、DNAと直接には結合しない26ため、ABCA1プロモーター上のその存在には、DNAと会合するLXRなどのタンパク質に結合することが必要である。LXR転写因子ファミリーには2種のメンバー、LXRαおよびLXRβがある50,52。LXRαの発現は組織特異的であるが、LXRβの方は遍在的である。SIRT1とLXRとの会合があるか否かを共免疫沈降により決定するために、LXRαおよびLXRβのマウスcDNAをHAタグ付き発現ベクター中にクローニングした。次いで、SIRT1発現ベクターpBabe−SIRT1を、そのHAタグ付きLXR用ベクターと共に、またはそれ無しでHEK293T細胞中に移入した。
【0183】
この実施例では、SIRT1とLXRαまたはLXRβとの相互作用の可能性を調べた。NH2末端HAタグ付きLXRα(HA−LXRα)およびLXRβ(HA−LXRβ)を発現するプラスミドを、修飾pcDNA3−NHAベクター中のHAエピトープの下流に(それとインフレームで)、LXRαまたはLXRβの全長マウスcDNAをクローニングすることにより、創製した。マウスSIRT1を過剰発現するHEK293T細胞(MmSIRT1)およびHA−LXRα、またはMmSIRT1およびHA−LXRβを、NP40緩衝液(10mM Tris−HCl、pH8.0、150mM NaCl、0.5% NP40、およびComplete(商標)プロテアーゼインヒビター混合物)中に採集し、18ゲージシリンジ針の中を10回通過させることにより、溶解した。4℃で1時間揺動させた後、溶解物を16000×gで15分間遠心分離した。次いで、生成上清を1%BSA入りNP40緩衝液中に希釈し、抗HAモノクローナル抗体(Santa Cruz Biotechnology)3μgの添加により、免疫沈降を開始した。次いで、混合物を4℃で2時間から終夜の間、揺動させながらインキュベートした。更に1時間プロテインAビーズの添加により、免疫複合体を回収し、免疫沈降物をNP40緩衝液1mlで5回洗浄した。次いで、全溶解物および対応する免疫沈降試料をSDS−PAGEで分離し、抗SIRT1ポリクローナル抗体および抗mycまたは抗HA抗体を用いて免疫ブロットした。
【0184】
モノクローナルマウス抗LXRα抗体は、R&D system製であり、ポリクローナルヤギ抗LXRβ抗体は、Santa Cruz Biotechnology製であった。指示したタンパク質を発現するヒトHEK293T細胞を溶解し、上記のように抗HAモノクローナル抗体で免疫沈降(IP)させた(図2e)。次いで、全溶解物(レーン1〜4)および抗HA免疫沈降試料(レーン5〜8)を、SDS−PAGEで分離し、抗SIRT1ポリクローナル抗体(上端像)および抗HA−LXRα抗体(上部パネル)または抗HA−LXRβ抗体(下部パネル)を用いて探査した。図2eに示すように、移入したマウスSIRT1タンパク質(mT1)は、タグ付きLXRαまたはLXRβが細胞中に同時発現した場合だけ、抗HA抗体により交差沈降した(レーン8、上部および下部パネル)。HA−LXRは、ゲル中でやや低い移動度を示す内因性ヒトSIRT1タンパク質(hT1)も共沈降させた。したがって、SIRT1のLXRとの直接または間接結合が、恐らく、未処置細胞中でのABCA1プロモーターのLXREとSIRT1が会合する原因である。
【0185】
更に、この実施例に記載した免疫沈降実験は、SIRT1のLXRαまたはLXRβとの相互作用を試験物質が調節する能力のための感度良いアッセイを与える。
【0186】
(実施例9)
SIRT1−/−マウスの肝臓におけるLXR標的mRNA
LXRとSIRT1との相互作用のために、SIRT1が機能喪失すると、食事コレステロールに応答するLXR媒介脂質恒常性が損なわれ得るという可能性が持ち上がる。この仮説を試験するために、ABCA1を含めた一連のLXR標的遺伝子のmRNA量を、SIRT1−/−マウスの肝臓(図3a)において、野生型対照動物(塗潰し棒)およびSIRT1−/−動物(白抜き棒)の肝臓からの総mRNAを用いて分析した。分析は、リアルタイムPCRで行った。LXRのmRNA量が、SIRT1−/−肝臓においてたとえ正常であっても、SREBP1、ABCA1、ABCG1、ABCG5、LDLRなどの多くのLXR標的は、SIRT1+/+対照に比較して減少していた(n=3、*、P<0.05)。LXR標的ではないことが知られている対照遺伝子53,54のmRNA量は、SIRT−/−マウスにおいて有意に変化しなかった。
【0187】
(実施例10)
LXRアゴニストT0901317で処理したマウスの肝臓における代謝物質の相対量
インビボでのLXR機能に対するSIRT1の効果を更に洞察するために、野生型およびSIRT1−/−動物を、LXRアゴニストT0901317(N−(2,2,2−トリフルオロエチル)−N−[4−(2,2,2−トリフルオロ−1−ヒドロキシ−1−トリフルオロメチルエチル)フェニル]ベンゼンスルホンアミド)53の8日経口投与に対する応答について、分析した。
【0188】
【化18】
マウス組織中のABCA1タンパク質を検出するために、SIRT1+/+またはSIRT1−/−肝臓をRIPA緩衝液中で均質化した。次いで、肝臓抽出物をSDS試料緩衝液と混合し、37℃で20分間変性することにより、高温でのABCA1の凝集を防止した。次いで、試料を7.5%SDS−PAGE中に分離し、抗ABCA1ポリクローナル抗体(R80抗体、M.FitzgeraldおよびM.Freeman両博士からの贈呈品)を用いてブロットした。
【0189】
SIRT1−/−および対象の2〜6月齢雄に、経口による強制補給でT0901317(10mg/kg)を供給し、血漿を供給の前後に採集して、血漿脂質分析用とした。組織を供給後に採集して、更なる分析用とした。T0901317(10mg/kg)を供給していない(黒色塗潰し棒)、または供給した(黒色斜線付き棒)野生型動物、およびT0901317を供給していない(白抜き棒)、または供給した(灰色斜線付き棒)SIRT1−/−マウスの肝臓からの総RNAを、リアルタイムPCRで分析した。野生型動物へのT0901317供給は、数種のLXR標的:SREBP−1c、LPLおよびABCA1のRNA段階での発現を有意に誘発した(図3b)(n=4、**、p<0.05)。しかし、SIRT1−/−マウスにおけるこれらの標的の誘発は、鈍らせられた。総タンパク質溶解物を指示したマウスの肝臓から取得し、免疫ブロットによりそのABCA1タンパク質量を分析した(図3c)(以前に観察したように、ABCA1タンパク質は、ウェスタンブロットで多重のバンドを含む78)。
【0190】
図3のパネルd、eおよびfにおける血漿の結果は、T0901317を8日投与した前後の同じマウスを表わす。図3のパネルdにおける肝臓の結果は、図3の上のパネルb(実施例9)およびc(本実施例)の場合のように、T0901317を供給していない、または供給した同腹子を比較している。T0901317を供給していない(黒色塗潰し棒)、または供給した(左斜線付き棒)野生型マウス、およびT0901317を供給していない(白抜き棒)、または供給した(右斜線付き棒)SIRT1−/−動物の、血漿(図3d、左パネル)および肝臓(図3d、右パネル)からのトリグリセリドを分析した。血漿および肝臓のトリグリセリド蓄積のLXRアゴニストT0901317による誘発53に対する、SIRT1−/−動物における応答の減少が認められた(図3d;n=5、***、p<0.001)。処置マウスにおける血漿コレステロール濃度の低下(図3e;n=5、****、p<0.02)も、FPLCリポタンパク質画分のコレステロールアッセイで決定した場合の特にHDL粒子において見られた(図3f;n=5)。T0901317は、SREBP−1cの誘導により血漿および肝臓のトリグリセリドを増加させる53ので、SIRT1−/−マウスにおけるSREBP−1c mRNAの少ない量(図3a)が、T0901317が誘発するトリグリセリドの低下を説明し得る。したがって、LXRに依存するレポーターアッセイにおける、ABCA1プロモーターに対するSIRT1の効果だけでなく、LXRおよびそのアゴニストのインビボでの機能も、SIRT1に依存していた。
【0191】
(実施例11)
LXRタンパク質発現に対するSIRT1の効果
SIRT1は、ヒストン12、p53 15〜17、FOXO転写因子18、19、およびPGC−1α 23、24などの多くの標的タンパク質と直接、相互作用し、それらを脱アセチル化する。SIRT1によるこうした標的の脱アセチル化は、その転写活性を抑制(一部のプロモーターにおけるp53およびFOXO)または活性化(他のプロモーターにおけるPGC−1αおよびFOXO)する。SIRT1が、LXRに対してどのように正の調節をするのか調べるために、SIRT1+/+およびSIRT1−/−マウスの肝臓内の内因性LXRタンパク質量を最初に決定した。LXRの標的活性化能が、野生型と比較して低下していた(実施例9;図3a)とはいえ、SIRT1−/−動物においてLXRタンパク質量が激増していた(図4a)。
【0192】
外因性CMVプロモーターで推進されるHA−LXRβを発現するHEK293T細胞に、pSuperベクター、pSuper−SIRT1 RNAi構築体のいずれかを移入した。トランフェクションから3日後、細胞を採集し、HAおよびSIRT1に対する抗体を用いたウェスタンブロットで、HA−LXRβおよびSIRT1の量を分析した。SIRT1発現のRNAi媒介ノックダウン(図4b、レーン3および4)、ニコチンアミドによるSIRT1活性の抑制−しかし、TSAによる他のデアセチラーゼの阻害はなかった−(図4c)の双方が、HA−LXRβタンパク質量を増加させた。SIRT1−/−マウスではLXR mRNAの低下が検出されなかった(図3a)ので、SIRT1は転写後にLXRタンパク質量を調節する。
【0193】
HA−LXRβを発現するHEK293T細胞を、対照培地(図3c、レーン1)、または1μM TSA(レーン2)、10mMニコチンアミド(レーン3)、25μM MG132(カルボベンジルオキシ−Leu−Leu−Leu−al)(レーン4)、もしくは10mMニコチンアミド+25μM MG132(レーン5)を含む培地のいずれかで、6時間処理した。次いで、細胞を採集し、HAおよびSIRT1に対する抗体を用いたウェスタンブロットで、HA−LXRβ(上部像)およびSIRT1(中央像)の量を分析した。ニコチンアミドによるSIRT1活性の抑制(レーン3)−しかし、TSAによる他のデアセチラーゼの阻害はなかった(レーン2)−によって、HA−LXRβタンパク質量が増加した(上部像)。SIRT1−/−マウスではLXR mRNAの低下が検出されなかった(実施例9、図3aを参照されたい)ので、SIRT1は転写後にLXRタンパク質量を調節する。ニコチンアミドによるSIRT1活性の抑制は、プロテアソーム媒介LXR分解を妨害することにより、HA−LXRのタンパク質量を増加させると考えられる。
【0194】
次に、LXR活性がタンパク質量と逆相関しているという知見の根拠を調べた。リシン残基のアセチル化はユビキチン化を遮断することができ、SIRT1はデアセチラーゼであるので、SIRT1の活性喪失は、LXRアセチル化の定常状態レベルを増加させ、したがってユビキチン/プロテアソーム分解経路を抑制するのではないかとも思われる。これが事実であるとすれば、MG132によるプロテアソーム活性の抑制は、SIRT1が活性である場合はLXRの定常状態レベルを増加させ、SIRT1がニコチンアミドにより阻害される場合はそのレベルを増加させないはずである。事実、これは、HAタグ付きLXRβを発現するHEK293T細胞において認められた(図4c、レーン4対レーン5)。
【0195】
HEK293T細胞に、空pBabeベクター、HA−LXRβ、pBabe−SIRT1、およびpBapbe−hSIRT1HY(不活性デアセチラーゼであるSIRT1変異体(Vaziriら、2001年を参照されたい))を発現する各構築体を同時移入した。40時間後、細胞を25μM MG132と共に1時間インキュベートし、採集した。次いで、HAに対する抗体を用いて、HA−LXRβを免疫沈降させた後、ユビキチン、HAおよびSIRT1に対する抗体を用いたウェスタンブロットを施した。その結果から、不活性変異体のhSIRT1HYではなく、活性SIRT1トランス遺伝子(hSIRT1)の発現が、移入されたHEK293T細胞中でユビキチン結合LXRの量を増加させたことが示される(図4d、レーン3対レーン4)。
【0196】
(実施例12)
インビボでのLXRのアセチル化
インビボでのLXRのアセチル化、およびSIRT1によるLXRのデアセチル化を検出するために、HEK293T細胞に、HA−LXRβと、pBabe、pBabe−HsSIRT1、pBabe−HsSIRT1H355Yのいずれかとを同時移入し、改良を加えたが記載してあるように77、[3H]酢酸ナトリウムを含有する培地中で培養した。手短に言うと、トランスフェクションから40時間後、2mCi/ml[3H]酢酸ナトリウム、プロテアソームインヒビターMG132(25μM)、22(R)−HC(10μM)、および9−シス−RA(1μM)を培養物に1時間添加した。次いで、細胞を冷リン酸緩衝塩水で2回洗浄し、NP40緩衝液中で溶解した。溶解物を16000×gで4℃、15分間遠心分離した。次いで、HA−LXRβを抗HA抗体で免疫沈降させ、SDS−PAGEで分離した。[3H]酢酸標識HA−LXRβを含有するゲルを、クーマシーブリリャントブルーで先ず染色し、次いで市販のフルオログラフィー増強溶液(Amplify,Amersham Biosciences)で30分間浸漬することにより、増強した。乾燥したゲルを−70℃、3〜7日間オートラジオグラフィーに掛けた。同じ試料に抗SIRT1抗体で免疫ブロットも行った。
【0197】
その結果を図4eに示してある。HA−LXRβは、通常の培養条件下でアセチル化された(レーン1)。更に、22(R)−HC+9−シスRAでの処理によるLXRβの活性化が、脱アセチル化を賦活した(および/または、多分アセチル化を抑制した。レーン2)。図4fに示すように、[3H]酢酸標識HA−LXRの量が、空ベクターを移入した対照細胞(pBabe;レーン1)、または酵素不活性SIRT1(pBabe−hSIRT1HY;レーン3)より、hSIRT1を発現する細胞(pBabe−hSIRT1;レーン2)において実質的に少なかったので、SIRT1は、少なくとも部分的にはLXRの脱アセチル化の原因に多分なっているようである。こうした知見は、SIRT1によるLXRβの脱アセチル化が、その代謝回転率を促進し、その活性も増加させることを示唆している。
【0198】
(実施例13)
SIRT1媒介またはプロテアソーム媒介LXR分解の抑制が、ABCA1プロモーター上のLXRの転写活性に及ぼす効果
HEK293T細胞に、野生型(WT;図4g、左パネル)またはLXRE変異体(−LXRE;図4g、右パネル)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを移入し、10μM 22(R)−HCおよび1μM 9−シスRAの不在下(塗潰し棒)または存在下(白抜き棒)、10mMニコチンアミド(Nico)、25μM MG132、10mMニコチンアミド+25μM MG132の両方のいずれかで処理した。実施例5の場合のように、ルシフェラーゼ活性を測定し、規格化した。
【0199】
ニコチンアミドは、ABCA1プロモーターに推進されるルシフェラーゼレポーターの誘導を有意に鈍らせることが判明した(図4g、左パネル)。その上、プロテアソームインヒビターのMG132も、ニコチンアミドと同程度に該レポーターを阻害したが、ニコチンアミドとMG132とを併用しても更に阻害することはなかった。LXREを失っている対照レポーターは、ニコチンアミドまたはMG132で変化しない基準活性を示した(図4g、右パネル)。こうした知見は、以下に考察するように、SIRT1によるLXRβの脱アセチル化、ならびにプロテアソームによる、後続するそのユビキチン化および分解が、ABCA1プロモーターの活性化を補助することを示唆している。
【0200】
以上の実施例は、SIRT1がLXR転写因子を正に調節し、したがってコレステロール恒常性の調節に重要な役割を果たしていることを示す。LXRおよびSIRT1は、ABCA1輸送体をコードする遺伝子の転写を活性化し、その輸送体が、コレステロール逆輸送およびHDL合成を媒介する。遺伝子および疫学研究によれば、血漿HDL濃度は、心血管疾患30および恐らくはアルツハイマー病5,6の危険性と逆関連することが示されているので、こうした知見はヒトの医療に直接関係し得る。コレステロールのHDL媒介逆輸送は、動脈細胞から過剰のコレステロールを除去することにより、アテローム性動脈硬化症から防御し55,56、脳内の富コレステロールラフト形成を減少させることにより、アルツハイマー病から防御し得る57,58。LXRの活性化によって、SIRT1は、老化に付随する心血管疾患および恐らくはアルツハイマー病の危険性を減少させ得る。
【0201】
SIRT1とLXRとの物理的および機能的相互作用は、SIRT1が、LXRの調節によって、インビボでその効果の少なくとも幾つかを発揮することを示唆している。実際に、SIRT1欠乏マウスは、HDLのコレステロール量(図1)およびトリグリセリド量(図3d)の減少などの多くの代謝欠陥を、LXR欠乏マウスと共有している53,54,63。また、肝LXRアゴニストにより抑制できる64糖新生の律速酵素であるPEPCKは、SIRT1−/−マウスにおいて誘導される19。LXR依存性の遺伝子発現およびエネルギー恒常性は、微生物に対する先天性免疫応答にとって重要であることが最近示されており、LXRを失っているマウスは、細菌感染を非常に受け易い65。したがって、肺感染症26および眼の感染症(未公表の観察)に対する、SIRT1ノックアウト動物の観察された感受性は、こうした動物における鈍ったLXR活性に因ることも考えられる。
【0202】
本知見は、SIRT1が、LXRの脱アセチル化およびその代謝回転の生起によって、LXR活性を促進することを示唆する(図5)。SIRT1によるLXRの調節により、このサーチュインがコレステロール恒常性経路の中心に位置している。
【0203】
参考文献
【0204】
【化19】
【0205】
【化20】
【0206】
【化21】
【0207】
【化22】
【0208】
【化23】
【0209】
【化24】
【0210】
【化25】
【背景技術】
【0001】
コレステロールの恒常性は、食事の摂取、デノボ合成、輸送、代謝および排泄の間の均衡により維持されている。低濃度の高密度リポタンパク質(HDL)および高濃度の低密度リポタンパク質(LDL)は、高コレステロール血症、高グリセリド血症、ならびに中年および更に高年齢の人々の主要な死因である加齢性疾患の心血管疾患1およびアルツハイマー病2−6のリスク増加と関係している。遺伝因子、環境の双方が、心血管疾患およびアルツハイマー病の進行の原因となり、こうした疾患のリスクは年齢と共に増加する。しかし、遺伝因子が環境を感知してこうした加齢性疾患を媒介する根本的な機構の理解は、不十分である。
【0002】
Sir2(silent information regulator 2)は、環境変化に応答する生涯の決定的な調節因子である。Sir2遺伝子は、酵母、C.elegans7およびDrosophila8,9における寿命決定因子である。酵母10,11、Drosophila8,9のいずれでも、Sir2活性は、カロリー制限で得られる寿命延長にも必要である。生化学的には、Sir2およびその同種物質(サーチュイン)は、高度に保存されたNAD+依存性タンパク質デアセチラーゼの一群である12−14。NAD+を必要とすることにより、サーチュインは、細胞代謝をモニターし、加齢に影響する細胞過程を調節することができる。哺乳類において、Sir2タンパク質の哺乳類オーソログ(orthologue)であるSIRT1は、多数のタンパク質基質を有し、細胞周期、アポトーシス、酸化的ストレス応答、神経変性などの加齢過程を調節することができる15−20。こうした過程において、SIRT1は、細胞死と細胞生存との均衡を変化させ、それによりストレス耐性を生じることができる21。
【0003】
コレステロールおよび脂肪の代謝は、通常の多くの細胞因子および環境因子により調節される。脂質感知核内受容体の2種のサブクラスである、ペルオキシソーム増殖剤応答性受容体(PPAR)および肝X受容体(LXR)は、脂質および炭水化物の代謝に決定的な役割を果たしている。最近の研究によれば、白色脂肪組織では、SIRT1は、核内受容体コリプレッサー(N−CoR)を介して核内受容体PPARγと相互作用し、食物欠乏時に脂肪代謝を促進することが示された22(非特許文献1)。SIRT1は、PPARγコアクチベーターのPGC−1αと相互作用し、それを修飾することにより、肝グルコースの恒常性を調節することもできる23,24(非特許文献2、非特許文献3)。特許文献1では、ヒト対象の高密度リポタンパク質(HDL)の血中濃度は、対象にトランスレスベラトロールを投与することにより増加させることができ、この物質は、対象の低密度リポタンパク質(LDL)の血中濃度を低下させることが報告されている。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】米国特許第6,048,903号明細書
【非特許文献】
【0005】
【非特許文献1】Picard,F.ら、Nature(2004)429、771−6
【非特許文献2】Rodgers,J.T.ら、Nature(2005)434、113−8
【非特許文献3】Nemoto,S.ら、J Biol Chem(2005)280,16456−60
【発明の概要】
【発明が解決しようとする課題】
【0006】
脂肪代謝およびグルコース恒常性におけるSIRT1の関与によって、このタンパク質が哺乳類においてコレステロール恒常性も調節し得るという可能性が得られる。本発明は、コレステロール恒常性に関するこうした新規な機構を取り扱う。
【課題を解決するための手段】
【0007】
発明の要旨
本発明は、全般的に、コレステロール恒常性およびコレステロール逆輸送の分野で有用な組成物および方法に関する。
【0008】
第1の態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する、単離した複合体を提供する。ある種の実施形態では、この複合体は、LXR応答エレメントを更に含有する。
【0009】
関連する態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体を形成する方法を提供する。この方法には、哺乳類SIRT1タンパク質、哺乳類LXRタンパク質、およびLXR応答エレメントを含む細胞内核酸の断片を含有する組成物を組み合わせることが含まれる。
【0010】
その上更なる態様では、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を調節する、作用剤を同定する方法を提供する。この方法は、
哺乳類SIRT1タンパク質、哺乳類LXRタンパク質、およびLXR応答エレメントを含む細胞内核酸の断片を含有する組成物を組み合わせ、それにより複合体組成物を提供する工程、
更に、哺乳類SIRT1タンパク質を含有する組成物、哺乳類LXRタンパク質を含有する組成物、もしくは細胞内核酸の該断片を含有する組成物の1つを、組合せ工程の前に該作用剤を含む第4の組成物と接触させる、または複合体組成物を組合せ工程の後に該作用剤を含む第4の組成物と接触させるステップ、ならびに
複合体の形成が、該作用剤によって調節されているか否かを決定するステップ
を包含する。
この方法のある種の実施形態では、該作用剤は複合体の形成を増加させる。追加の実施形態では、判定ステップは、該作用剤を含有していない対照組成物との比較により行われる。
【0011】
更に追加の態様では、本発明は、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法であって、SIRT1活性を刺激する作用剤をその哺乳動物に投与することを含む方法を提供する。有利な実施形態では、該作用剤はT0901317を含む。
【0012】
その上更なる態様では、本発明は、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法であって、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体の形成を促進する作用剤をその哺乳動物に投与することを含む方法を提供する。この方法の有利な実施形態では、該作用剤は22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む。
【0013】
更に追加の態様では、本発明は、哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含有する核酸をその細胞に導入することを含む方法を提供する。この方法の幾つかの実施形態では、タンパク質デアセチラーゼは、真核生物Sir2または哺乳類SIRT1である。この方法の更に他の実施形態は、T0901317などのSIRT1活性を刺激する作用剤と該細胞を更に接触させるステップを含む。この方法の他の実施形態では、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含有する複合体の形成を促進する作用剤と、該細胞を更に接触させる。ある種の実施形態では、複合体促進性作用剤は、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む。
【0014】
本発明の更なる態様は、SIRT1活性のレベルが正常未満と見なされる対象を処置する方法を提供する。このような対象は、正常未満と見なされる、HDL−コレステロール濃度およびHDL−コレステロール対LDL−コレステロールの比率を示す。この方法は、種特異的なSIRT1をコードする核酸をその対象に投与することを含み、その核酸は、対象の細胞内にSIRT1の治療有効量を発現するのに有効である。
【0015】
更なる態様では、本発明は、候補物質がLXR依存性プロセスを調節するか否かを評価する方法を提供する。この方法は、LXREに作動可能に駆動されるレポーター遺伝子を保持するプラスミドを細胞に移入し、その細胞を候補物質と接触させ、更に、候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定することによって、候補物質の有無により検出される差異が、候補物質がLXR依存性プロセスを調節することを示すことを含む。
【0016】
追加の態様では、本発明は、候補物質がLXRのSIRT1依存性作用を調節するか否かを評価する方法を提供する。この方法は、SIRT1遺伝子を保持するプラスミドを細胞に移入し、LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するプラスミドを、その細胞に更に移入し、その細胞を候補物質と接触させ、更に、候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定し、それにより、候補物質の有無により検出される差異が、候補物質がLXRのSIRT1依存性作用を調節することを示すことを含む。
【0017】
更なる態様では、本発明は、候補物質がSIRT1のLXRとの相互作用を調節するか否かを評価する方法を提供する。この方法は、エピトープタグ付きLXRをコードする配列を保持するベクターで細胞をトランスフェクトし、その細胞を候補物質と接触させ、その細胞を溶解し、エピトープタグに特異的な抗体を細胞溶解物と接触させ、更に、SIRT1とLXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収し、SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行し、それにより、候補物質の不在下と比較した、候補物質の存在下で検出した複合体形成の差異が、候補物質がSIRT1のLXRとの相互作用を調節することを示すことを含む。
【図面の簡単な説明】
【0018】
【図1】SIRT1の機能喪失がコレステロール恒常性の変化を起こすことを示す図である。(a〜c)野生型(塗潰し棒)、SIRT1+/−(斜線付き棒)およびSIRT1−/−(白抜き棒)の各マウスからの血漿試料を、(a)総コレステロール、(b)HDLコレステロール、(c)LDLコレステロールについて分析した。(d)野生型(塗潰し菱形)およびSIRT1−/−(白抜き菱形)の各マウスにおけるFPLC血漿リポタンパク質のコレステロール像。(e)野生型(塗潰し棒)およびSIRT1−/−(白抜き棒)の各マウスのHDL画分における相対脂質含量。(f)野生型対照(塗潰し棒)および同腹子SIRT1−/−(白抜き棒)の精巣および肝臓からの総コレステロール。(g)ニコチンアミドによる前処理有りまたは無しの、マウス初代胚性線維芽細胞(MEF)(左パネル)またはTHP1ヒト単球(右パネル)からの[3H]コレステロールのApoAI媒介排出。
【図2】ABCA1の発現およびLXR核内受容体との相互作用を示す図である。(a)肝臓(左パネル、定量的リアルタイムPCRで決定)、ならびに精巣および卵巣(右パネル、野生型(+/+)およびSIRT1−/−(−/−)動物のノーザンブロットで決定)におけるABCA1 mRNAの発現。(b)SIRT1活性の抑制はABCA1プロモーター活性を低下させる。(c)MEFにおけるSIRT1欠損のABCA1プロモーター活性に対する効果。(d)SIRT1とABCA1プロモーターのLXREとの相互作用。(e)SIRT1はLXREαおよびLXREβと相互作用する。
【図3】SIRT1欠損が、インビボでのLXRアゴニストに対する正常な応答を損なうことを示す図である。(a)SIRT1−/−マウスの肝臓におけるLXR標的mRNA。(b)LXRアゴニストT0901317で処理したマウスの肝臓におけるLXR標的mRNAの相対量。(c)T0901317を有する、または有していない、野生型およびSIRT1−/−マウスにおけるABCA1タンパク質濃度。(d)T0901317投与後のトリグリセリド濃度。SIRT1−/−マウスでは、血漿および肝臓におけるトリグリセリド蓄積量が低下する。(e)T0901317供給後の血漿におけるコレステロールの誘導。(f)T0901317供給の前(塗潰した四角)および後(白抜きの四角)の野生型およびSIRT1−/−マウスにおける、代表的なFPLC血漿リポタンパク質像。
【図4】SIRT1が、LXRタンパク質の脱アセチル化によりLXRの安定性および活性を調節することを示す図である。(a)LXRタンパク質発現SIRT1に対するSIRT1の効果。(b)RNA干渉によるSIRT1のノックダウンがLXR濃度に及ぼす効果。(c)LXRのタンパク質発現に対する阻害剤の効果。(d)SIRT1はLXRのユビキチン化を促進する。(e)インビボでのLXRのアセチル化、および22(R)−HCおよび9−シスRAによるLXRの活性化。(f)SIRT1はHA−LXRβを脱アセチル化する。
【図5】SIRT1またはプロテアソーム媒介LXR分解の抑制が、ABCA1プロモーター上のLXRの転写活性に及ぼす効果の概要を表示する図である。
【発明を実施するための形態】
【0019】
発明の詳細な説明
配列の表
表1.本開示における配列の表
【0020】
【化1】
本明細書で使用する場合、HDL(高密度リポタンパク質)は、アポリポタンパク質A、C、DおよびEを含めた、高いタンパク質比率を有する一部類の血漿リポタンパク質に関する。HDLは、血漿中で遊離しているかエステル化されているかに関係なく、コレステロールをHDL−コレステロール複合体として取り込み、輸送する。HDLという用語は、HDL粒子に結合しているコレステロールを指定するために、文脈に応じて使用し得る。
【0021】
本明細書で使用する場合、LDL(低密度リポタンパク質)は、コレステロール、コレステロールエステルおよびトリグリセリドを含んだ、高い脂質比率を有する一部類の血漿リポタンパク質に関する。その中には、主にアポリポタンパク質B−100およびアポリポタンパク質Eが含まれる。LDLは、血漿中でコレステロールを取り込み、輸送する。LDLという用語は、LDL粒子に結合しているコレステロールを指定するために、文脈に応じて使用し得る。
【0022】
本明細書で使用する場合、用語「単離された」および同類用語は、自然状態で、ある物質の出現に付随する少なくとも1種の成分の除去により、自然出現状態から精製された前記物質に関する。用語「精製された」は、「単離された」と同義に使用し得る。これらの用語は、該物質が何らかの所定の純度を有することを必要としないことが理解される。
【0023】
本明細書で使用する場合、用語「複合体」および同類用語は、単離手順を受け、そのような結合体として特性決定されるのに十分に、相互に安定に結合している2種以上の物質の結合体に関する。複合体の各成分が相互に直接結合してもよく、またはその結合体は、同定した各成分が相互に直接には接触しないように、介在促進成分を含んでもよい。複合体は、各成分を結合する非共役結合性相互作用、または各成分間の共役結合によって形成し得る。
【0024】
本明細書で使用する場合、用語「コレステロール」とは、コレステロール、コレステリルエステル双方としてのコレステロールの出現全体を指す。当業者には明らかと思われる特定の文脈では、「コレステロール」は、遊離コレステロールだけを指すために、本明細書では使用することもある。
【0025】
本明細書で使用する場合、用語「処置」ならびに同類の用語および語句は、疾患もしくは病状、症状または疾患素因を有する対象に対する治療剤の適用または投与、あるいはその対象由来の単離した組織または細胞系に対する治療剤の適用または投与に関する。処置は、その治療もしくは治癒を促進すること、または、疾患、症状もしくは疾患素因を軽減、緩和、改変、治癒、改善、改良する、もしくはそれに影響を及ぼすことを意図している。
【0026】
本明細書で使用する場合、「治療有効量」および「予防有効量」という語句は、疾患の処置において治療上有益となる量、またはそれぞれ、疾患を予防するもしくはその重度を低減する効果を指す。治療上有効な特定の量は、治療される対象における応答の評価を用いて、普通の医者により容易に決定することができ、疾患の特質、対象の病歴および年齢、疾患の段階、ならびに他の治療剤の投与などの、当技術分野で公知の要因に応じて変化し得る。
【0027】
本明細書で使用する場合、「医薬組成物」は、薬理有効量のポリヌクレオチドおよび医薬として許容可能な担体を含む組成物に関する。本明細書で使用する場合、「薬理有効量」、「治療有効量」または単に「有効量」とは、意図した薬理的、治療的または予防的結果を生じるのに有効な阻害性ポリヌクレオチドの量を指す。例えば、疾患または障害に関連する臨床パラメーターに測定し得る最小限の変化が少なくともあるときに、所与の臨床処置が有効であると見なす場合、その疾患または障害を処置するための薬物の治療有効量は、そのパラメーターに少なくとも変化度を起こすのに必要な量である。
【0028】
用語「医薬として許容可能な担体」とは、少なくとも、生理学的に許容可能であり、規制当局により認可され得る、治療剤投与用の組成物を指す。
SIRT1ポリペプチド
本明細書で使用する場合、「SIRT1ポリペプチド」、「SIRT1タンパク質」ならびに関連する用語および語句は、野生型SIRT1、突然変異体SIRT1、変異体SIRT1、ならびに生物活性なその断片および成熟形態に関する。本発明で使用すべき重要なSIRT1タンパク質は、ヒトSIRT1である。ヒトSIRT1のアミノ酸配列は、GenBank寄託番号NP_036370に示され、アミノ酸残基747個からなるものと開示されており、従来の1文字アミノ酸記号を用いて表2に示される(International Union Of Biochemistry And Molecular Biology,Recommendations on Biochemical & Organic Nomenclature, Symbols &Terminology etc., Part 1, Section A: Amino-Acid Nomenclature, Section 3AA-1,Names Of Common Alpha-Amino Acids, http://www.chem.qmul.ac.uk/iubmb/ およびJ.Biol. Chem., 1985年、260号、14〜42頁)。
【0029】
表2.ヒトSIRT1のアミノ酸配列
【0030】
【化2】
マウスSIRT1のアミノ酸配列は、GenBank寄託番号NP_062786に示され、アミノ酸残基737個からなるものと開示されており、表3に示される。
【0031】
表3.マウスSIRT1のアミノ酸配列
【0032】
【化3】
ヒト肝X受容体αのアミノ酸配列は、GenBank寄託番号NP_062786に示され、アミノ酸残基447個からなるものと開示されており、表4に示される。
【0033】
表4.ヒト肝X受容体αのアミノ酸配列
【0034】
【化4】
ヒト肝X受容体βのアミノ酸配列は、GenBank寄託番号NP_009052.3に示され、アミノ酸残基461個からなるものと開示されており、表5に示される。
【0035】
表5.ヒト肝X受容体βのアミノ酸配列
【0036】
【化5】
マウス肝X受容体αのアミノ酸配列は、GenBank寄託番号NP_038867に示され、アミノ酸残基445個からなるものと開示されており、表6に示される。
【0037】
表6.マウス肝X受容体αのアミノ酸配列
【0038】
【化6】
マウス肝X受容体βのアミノ酸配列は、GenBank寄託番号NP_033499.1に示され、アミノ酸残基446個からなるものと開示されており、表7に示される。
【0039】
表7.マウス肝X受容体βのアミノ酸配列
【0040】
【化7】
一般に、本発明の方法および組成物に使用されるような「SIRT1ポリペプチド」および多様な「LXRポリペプチド」には、表2〜7に示すようなヒト野生型のSIRT1およびLXRポリペプチド、またはマウス野生型のSIRT1およびマウスLXRポリペプチド、ならびに脊椎動物野生型のそれらのオーソログと、それらのドメイン、モチーフおよび断片とが含まれる。それに加え、これらの用語には、組換え変異体のポリペプチド、ドメイン、モチーフおよび断片も含まれ、そうしたものでは、少なくとも1個のアミノ酸残基が異なるアミノ酸残基に変化してしまい、または1個もしくは複数の残基が欠失していることもあり、または1個もしくは複数の残基が、元の配列における隣接残基の間に挿入されていることもある。突然変異体または変異体のSIRT1ポリペプチドでは、アミノ酸残基1個から残基の1%までが変化し、または残基の2%まで、もしくは5%まで、もしくは8%まで、もしくは10%まで、もしくは15%まで、もしくは20%まで、もしくはそれよりやや高い比率まで、野生型または基準配列から変化していることもある。SIRT1の突然変異体または変異体の組換えポリペプチド、ドメイン、モチーフおよび断片は、野生型のSIRT1およびLXRの少なくとも1種の生物活性または機能を示すことが立証し得る限り、本発明の方法および組成物に使用される。生物活性または機能の保有は、非限定的な例として分子生物学、細胞生物学、病理学、臨床医学などを含む、本発明に関連する分野の技術者により判定し得る。本発明の各分野のこのような技術者は、少なくとも本発明の実施例に記載の方法により、SIRT1およびLXRの組換え変異体ポリペプチド、ドメイン、モチーフおよび断片をアッセイしてもよい。
【0041】
SIRT1またはLXRポリペプチドのアミノ酸配列が、そのタンパク質の構造または機能にさしたる作用をせずに変化させ得ることは、当技術分野では認識されていよう。配列のこのような差異を考察する場合、該タンパク質上には、その活性にとって重要な特定の領域が存在すると見込まれることを想起されたい。一般に、類似機能をもたらす残基を使用することを前提とすれば、三次構造を形成する残基を置き換えることは可能である。他の例では、当該変化がタンパク質の非重要領域で起こる場合、残基の種類が全く重要でないこともある。
【0042】
核酸
本明細書で使用する場合、用語「SIRT1ポリヌクレオチド」もしくは「SIRT1核酸」、または「LXRポリヌクレオチド」もしくは「LXR核酸」、あるいは関連する用語および語句は、本明細書に記載の任意のSIRT1ポリペプチドをコードする、任意のポリヌクレオチドに関する。一般に、前記のSIRT1ポリペプチドをコードする任意のヌクレオチド配列は、本発明内に包含される。幾つかの実施形態では、表2に示すヒトSIRT1のアミノ酸配列を有するポリペプチドをコードする核酸は、表8に示すGenBank寄託番号NM_012238に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表8では、そのコード配列が54位から2297位まで拡がっている。
【0043】
表8
【0044】
【化8−1】
【0045】
【化8−2】
幾つかの実施形態では、表3に示すマウスSIRT1のアミノ酸配列を有するポリペプチドをコードする核酸は、表9に示すGenBank寄託番号NM_019812に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表9では、そのコード配列が48位から2261位まで拡がっている。
【0046】
表9
【0047】
【化9−1】
【0048】
【化9−2】
幾つかの実施形態では、表4に示すヒト肝X受容体αのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_005693に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表10では、そのコード配列が36位から1379位まで拡がっている。
【0049】
表10.ヒト肝X受容体αをコードするポリヌクレオチド配列
【0050】
【化10】
幾つかの実施形態では、表5に示すヒト肝X受容体βのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_007121に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表11では、そのコード配列が259位から1644位まで拡がっている。
【0051】
表11.ヒト肝X受容体βをコードするポリヌクレオチド配列
【0052】
【化11】
幾つかの実施形態では、表6に示すマウス肝X受容体αのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_013839に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表12では、そのコード配列が190位から1527位まで拡がっている。
【0053】
表12.マウス肝X受容体αをコードするポリヌクレオチド配列
【0054】
【化12】
幾つかの実施形態では、表7に示すマウス肝X受容体βのアミノ酸配列を有するポリペプチドをコードする核酸は、GenBank寄託番号NM_009473に開示されたmRNA核酸配列、またはその断片のコード配列を含む。表13では、そのコード配列が271位から1611位まで拡がっている。
【0055】
表13.マウス肝X受容体βをコードするポリヌクレオチド配列
【0056】
【化13】
それに加え、本発明には、表8〜13に示す配列の突然変異体もしくは変異体核酸、またはその断片であって、その塩基のいずれかが開示された配列から変化しているが、同時に、SIRT1タンパク質様の活性および生理機能を維持するポリペプチドを依然としてコードし得るポリヌクレオチドも含まれる。SIRT1突然変異体または変異体のポリヌクレオチドは、突然変異体または変異体のSIRT1またはLXRポリペプチドであって、アミノ酸残基1個から残基の1%までが変化し、または残基の2%まで、もしくは5%まで、もしくは8%まで、もしくは10%まで、もしくは15%まで、もしくは20%まで、もしくはそれよりやや高い比率まで、野生型または基準配列から変化していることもあるポリペプチドをコードする。「核酸」または「ポリヌクレオチド」とは、DNA、RNA、1個もしくは複数の修飾ヌクレオチドもしくは修飾ペントースリン酸骨格構造を含んだDNAまたはRNA、ポリペプチド−核酸、および構築体中に含まれる塩基の配列のコード特性を保持する類似の構築体を意味する。本発明には更に、その断片、誘導体、類縁体および相同体を含めた、任意のSIRT1またはLXRコード配列の核酸配列の相補体も含まれる。それに加え、本発明には、構造中に化学修飾を含んだ核酸もしくは核酸断片、またはその相補体も含まれる。
【0057】
また、核酸断片も含まれる。核酸断片は、SIRT1またはLXRポリペプチドの断片をコードすることもある。それに加え、SIRT1またはLXR核酸断片は、SIRT1またはLXRタンパク質コード核酸(例えば、SIRT1またはLXR mRNA)を同定するためのハイブリッド形成用プローブ、およびSIRT1またはLXR核酸分子を増幅または変異させるためのポリメラーゼ連鎖反応(PCR)用プライマーに用いる断片として使用し得る。本明細書で使用する場合、用語「核酸分子」とは、DNA分子(例えば、cDNAまたはゲノムDNA)、RNA分子(例えばmRNA)、ヌクレオチド類縁体を用いて生成したDNAまたはRNAの類縁体、ならびにそれらの誘導体、断片および相補体を包含することを意図している。核酸分子は、1本鎖、2本鎖のいずれでもよいが、好ましくは2本鎖DNAである。
【0058】
核酸
本明細書で使用する場合、「核酸」または「ポリヌクレオチド」、ならびに類似の用語および語句は、天然のヌクレオチドからなるポリマー、ならびに合成または修飾ヌクレオチドからなるポリマーに関する。したがって、本明細書で使用する場合、RNAであるポリヌクレオチド、またはDNAであるポリヌクレオチド、またはデオキシリボヌクレオチド、リボヌクレオチドの両方を含有するポリヌクレオチドは、天然の塩基およびリボース環もしくはデオキシリボース環などの天然の部分を含んでもよく、あるいは以下に記載するような合成または修飾部分で構成されてもよい。本発明に使用するポリヌクレオチドは、1本鎖でもよく、または塩基対2本鎖構造でもよく、または塩基対3本鎖構造でさえもよい。
【0059】
核酸およびポリヌクレオチドは、長さがヌクレオチド20個以上、または長さがヌクレオチド30個以上、または長さがヌクレオチド50個以上、または長さが100個以上、または100個以上、または1000個以上、または数万個以上、または数十万個以上の場合もある。本明細書で使用する場合、「オリゴヌクレオチド」およびこれを基にした類似の用語は、直前の段落に記載したような、天然のヌクレオチドからなる短いポリマー、ならびに合成または修飾ヌクレオチドからなるポリマーに関する。オリゴヌクレオチドは、長さがヌクレオチド10個以上、または長さがヌクレオチド15個もしくは16個もしくは17個もしくは18個もしくは19個もしくは20個以上、または長さがヌクレオチド21個もしくは22個もしくは23個もしくは24個以上、または長さがヌクレオチド25個もしくは26個もしくは27個もしくは28個もしくは29個もしくは30個以上、または長さがヌクレオチド35個以上、40個以上、45個以上、約50個までの場合もある。オリゴヌクレオチドは、化学的に合成してもよく、siRNA、PCRプライマー、またはプローブとして使用し得る。
【0060】
前段落に示したサイズ範囲が重複するために、用語「ポリヌクレオチド」および「オリゴヌクレオチド」は、本明細書では同義に使用し得る。
【0061】
本明細書で使用する場合、「ヌクレオチド配列」、「オリゴヌクレオチド配列」または「ポリヌクレオチド配列」および類似の用語は、互換的に、オリゴヌクレオチドまたはポリヌクレオチドが有する塩基の配列にも、その配列を保持するオリゴヌクレオチドまたはポリヌクレオチドの構造にも関する。更に、ヌクレオチド配列またはポリヌクレオチド配列は、塩基の配列が、当分野で従来より使用されているような、塩基を指定する文字の特定の配列を記載または記述することにより規定される、任意の天然または合成のポリヌクレオチドまたはオリゴヌクレオチドにも関する。
【0062】
「ヌクレオシド」は、本発明の分野に関係する生化学、分子生物学、ゲノム学および類似分野などの分野における技術者によって、プリンまたはピリミジン塩基とグリコシド結合で連結した単糖を含むものと従来より理解され、「ヌクレオチド」は、通常その糖の(ペントースの)3’または5’位に付加するが、その糖の他の位置にあってもよい、少なくとも1個のリン酸基を有するヌクレオシドからなる。ヌクレオチド残基は、オリゴヌクレオチドまたはポリヌクレオチドにおいて連番位置を占める。ヌクレオチドの修飾体または誘導体は、オリゴヌクレオチドまたはポリヌクレオチドにおいて任意の連番位置に出現してもよい。全ての修飾または誘導体化されたオリゴヌクレオチドおよびポリヌクレオチドは、本発明内に包含され、特許請求の範囲内に入る。修飾体または誘導体は、リン酸基、単糖または塩基中に出現することができる。
【0063】
非限定的な例として、以下の記述では、本発明のポリヌクレオチドの範囲内に全て入る修飾または誘導体化された特定のヌクレオチドが示される。その単糖は、例えば、リボースまたはデオキシリボース以外のペントースまたはヘキソースとすることにより、修飾してもよい。該単糖は、ヒドロキシ基をヒドロ基またはアミノ基で置換すること、追加のヒドロキシ基をアルキル化またはエステル化することなどによっても、修飾してもよい。2’−O−メチル、2’−O−エチル、2’−O−プロピル、2’−O−アリール、2’−O−アミノアルキルまたは2’−デオキシ−2’−フルオロ基などの2’位における置換基は、オリゴヌクレオチドに強化されたハイブリッド形成特性を付与する。
【0064】
オリゴヌクレオチドおよびポリヌクレオチド中の塩基は、「非修飾」または「天然」塩基でもよく、プリン塩基のアデニン(A)およびグアニン(G)、ならびにピリミジン塩基のチミン(T)、シトシン(C)およびウラシル(U)を包含する。それに加え、該塩基は修飾または置換がある塩基でもよい。修飾塩基の非限定的例には、ヒポキサンチン、キサンチン、4−アセチルシトシン、5−(カルボキシヒドロキシメチル)ウラシル、5−カルボキシメチルアミノメチル−2−チオウリジン、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、β−D−ガラクトシルケオシン(queosine)、イノシン、N6−イソペンテニルアデニン、1−メチルグアニン、1−メチルイノシン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−アデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、β−D−マンノシルケオシン、5’−メトキシカルボキシメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸(v)、ワイブトキソシン(wybutoxosine)、プソイドウラシル、ケオシン、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸(v)、5−メチル−2−チオウラシル、3−(3−アミノ−3−N−2−カルボキシプロピル)ウラシル、(acp3)wおよび2,6−ジアミノプリン、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチルおよび他のアルキル誘導体、ならびにアデニンおよびグアニンの2−プロピルおよび他のアルキル誘導体などの、他の合成および天然の塩基が含まれる。
【0065】
ヌクレオチド間の結合は、通常は3’−5’リン酸結合であり、それは天然のホスホジエステル結合、ホスホチオエステル結合、および更に他の合成結合でもよい。ホスホロチオエート骨格を含有するオリゴヌクレオチドは、強化されたヌクレアーゼ安定性を有する。修飾された骨格の非限定的例には、ホスホロチオエート、キラルなホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、3’−アルキレンホスホネート、5’−アルキレンホスホネートおよびキラルなホスホネートを含むメチルおよび他のアルキルホスホネート、ホスフィネート、3’−アミノホスホロアミデートおよびアミノアルキルホスホロアミデートを含むホスホロアミデート、チオノホスホロアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートならびにボラノホスフェートが含まれる。
【0066】
上記に例示した修飾を含むいずれの修飾も、本発明のポリヌクレオチド中に容易に組み入れることができ、その範囲中に含まれる。任意の修飾ヌクレオチドの使用は、当業者には理解されるように、同じ塩基対形成特性を有する天然ヌクレオチドの使用と同等である。全ての同等の修飾ヌクレオチドは、本明細書に開示し、主張するように、本発明の範囲内に入る。
【0067】
「単離」された核酸分子は、その核酸の天然源中に存在する他の核酸分子から分離された核酸分子である。単離された核酸分子の例には、それだけに限らないが、ベクターに含有される組換えDNA分子、異種宿主細胞中に保持される組換えDNA分子、部分的または実質的に精製された核酸分子、および合成DNAまたはRNA分子が含まれる。
【0068】
本明細書に開示する核酸分子、例えば、表8〜13のヌクレオチド配列を有する核酸分子、またはその相補体は、標準的な分子生物学技法およびここに示した配列情報を用いて単離することができる。表8〜13の核酸配列の全体または一部をハイブリッド形成プローブとして用い、SIRT1またはLXRの核酸配列を標準的なハイブリッド形成法およびクローニング法を用いて単離することができる(例えば、Sambrookら編、 MOLECULAR CLONING: A LABORATORY MANUAL 第2版、Cold Spring HarborLaboratory Press、Cold Spring Harbor、NY、1989年およびAusubelら編者、CURRENT PROTOCOLS INMOLECULAR BIOLOGY、John Wiley & Sons、New York、NY、1993年に記載のような技法)。
【0069】
本明細書に開示する核酸は、鋳型としてcDNA、mRNA、あるいはゲノムDNAと、標準的なPCR増幅法に従った適当なオリゴヌクレオチドプライマーとを用いて増幅することができる。このように増幅した核酸は、適当なベクター中にクローニングし、DNA配列分析により特性を決定することができる。更に、SIRT1またはLXRのヌクレオチド配列に対応したオリゴヌクレオチドが、標準的な合成法、例えば自動DNAシンセサイザーを用いて調製することができる。
【0070】
ポリヌクレオチドの合成
オリゴヌクレオチドおよびポリヌクレオチドは、標準的な合成法、例えば自動DNAシンセサイザーを用いて調製することができる。オリゴヌクレオチドを合成する方法には、それだけに限らないが、表面誘導体化粒子上へのヌクレオチドホスホロアミダイトの逐次付加を含めた周知の化学プロセスが含まれ、その逐次付加は、Oligonucleotides and Analogues A Practical Approach、F. Eckstein編、OxfordUniversity Press、Oxford、1〜24頁(1991年)におけるT. Brown およびDorcas J. S. Brownが記載し、参照により本明細書に組み込まれる通りである。
【0071】
合成手順の一例では、Expedite RNAホスホロアミダイトおよびチミジンホスホロアミダイト(Proligo、ドイツ)が使用される。合成オリゴヌクレオチドは、脱保護され、ゲル精製された(Elbashirら(2001年)Genes & Dev. 15巻、188〜200頁)後、Sep−Pak C18カートリッジ(Waters、Milford、Mass.、米国)で精製される(Tuschlら(1993年)Biochemistry、32巻、11658〜11668頁)。相補的なssRNAは、90℃、1分間アニーリング緩衝液(100mM酢酸カリウム、pH7.4の30mM HEPES−KOH、2mM酢酸マグネシウム)中で、その後37℃、1時間インキュベートして、対応するdsポリヌクレオチドへハイブリダイズさせる。
【0072】
オリゴヌクレオチド合成の他の方法には、それだけに限らないが、ホスホトリエステル法およびホスホジエステル法による固相オリゴヌクレオチド合成(Narangら、(1979年)Meth. Enzymol.、68巻、90頁)、ならびにH−ホスホネート法(とりわけ、Garegg,P.J.ら、(1985年)「Formation of internucleotidic bonds via phosphonate intermediates」、Chem.Scripta 25巻、280〜282頁;およびFroehler, B.C.ら、(1986年a)「Synthesis of DNA viadeoxynucleoside H-phosphonate intermediates」、Nucleic Acid Res.、14巻、5399〜5407頁)および支持体上での合成(Beaucageら(1981年)TetrahedronLetters 22巻、1859〜1862頁)、更に、ホスホロアミデート法(Caruthers, M. H.ら、「Methods in Enzymology」、154巻、287〜314頁(1988年)、米国特許第5153319号、第5132418号、第4500707号、第4458066号、第4973679号、第4668777号および第4415732号、ならびに「Synthesisand Applications of DNA and RNA」、S. A. Narang編、Academic Press、New York、1987年に記載の他の技法、およびその中に含まれる参考文献)、および非ホスホロアミダイト法が含まれる。固相合成は、不純物および過剰試薬からのオリゴヌクレオチドの単離を促進する。固体支持体から切断されると、オリゴヌクレオチドは公知の方法により更に単離し得る。
【0073】
ベクター
本発明は、本発明の1種または複数のポリヌクレオチドを含有する多種のベクターを提供する。有利には、本発明において開示されるいずれのベクターも、該ヌクレオチド配列に作動可能に連結されたプロモーター、エンハンサーまたはその両方を含む。
【0074】
本発明において開示されるベクターを調製する方法は、分子生物学、細胞生物学、腫瘍学の分野、および本発明に関連する他の分野において広く知られている。ベクターを調製する有用な方法は、非限定的な例として、Molecular Cloning: ALaboratory Manual(第3版)(Sambrook, Jら(2001年)Cold Spring Harbor Laboratory Press、ColdSpring Harbor、NY)、およびShort protocols in molecular biology(第5版)(Ausubel FMら(2002年)JohnWiley & Sons、New York City)に記載されている。
【0075】
抗体
本明細書で使用する場合の用語「抗体」とは、免疫グロブリン分子および免疫グロブリン(Ig)分子の免疫活性な部分、即ち、抗原と特異的に結合する(免疫反応を起こす)抗原結合部位を含有する分子を指す。このような抗体には、それだけに限らないが、ポリクローナル、モノクローナル、キメラ、単鎖、Fab、Fab’およびF(ab’)2断片、ならびにFab発現ライブラリーが含まれる。一般に、ヒトから得た抗体分子は、分子中に存在する重鎖の特質が相互に異なるクラスIgG、IgM、IgA、IgEおよびIgDのいずれかに関する。特定のクラスは、IgG1、IgG2、その他などのサブクラスも有する。更に、ヒトでは、軽鎖がκ鎖またはλ鎖の場合がある。本明細書における抗体への言及には、ヒト抗体種のこのような全てのクラス、サブクラスおよび型への言及を包含する。本明細書に開示するいずれの抗体も、そのコグネイト抗原に「免疫特異的に」結合する。免疫特異的な結合とは、特定の免疫原を宿主に接種することにより生成した抗体が、その免疫原性部分を含んだ、抗原などの分子と高い親和性で結合し、免疫原非含有分子とは弱い親和性でしか、または全く結合しないことを意味する。この定義で使用する場合、高親和性とは、1×10−6M未満の解離定数を有することを意味し、弱親和性とは、約1×10−6Mより高い解離定数を有することを意味する。
【0076】
抗原またはその一部分もしくは断片として作用させることを意図した、本発明の単離タンパク質またはそのようなタンパク質を含有する複合体は、ポリクローナルおよびモノクローナル抗体調製の標準的技法を用いて、その抗原に免疫特異的に結合する抗体を生成するための免疫原として使用することができる。その全長タンパク質を使用することができ、あるいは本発明は、免疫原として使用するために、該抗原の抗原性ペプチド断片を提供する。抗原性ペプチド断片は、表2〜7に示したアミノ酸配列などの、全長タンパク質のアミノ酸配列のアミノ酸残基少なくとも6個を含み、そのペプチドに対して生成した抗体が、全長タンパク質、またはエピトープを含有する任意の断片と特異的な免疫複合体を形成するように、全長タンパク質の該エピトープを包含する。好ましくは、抗原性ペプチドは、アミノ酸残基少なくとも10個、またはアミノ酸残基少なくとも15個、またはアミノ酸残基少なくとも20個、またはアミノ酸残基少なくとも30個を含む。抗原性ペプチドが包含する好ましいエピトープは、そのタンパク質の表面上に位置した領域であり、こうした領域は、通常、親水性領域である。
【0077】
本発明のある種の実施形態では、抗原性ペプチド断片が包含する少なくとも1個のエピトープは、SIRT1またはLXRタンパク質の表面上に位置した、そのタンパク質の一領域、例えば親水性領域である。ヒトのSIRT1またはLXRタンパク質配列の疎水性分析により、成長促進性ポリペプチドのいずれの領域が、特に親水性であり、したがって抗体産生を目標とするのに有用な表面残基をコードしそうであるかが示されよう。抗体産生を目標とする手段として、親水性および疎水性の領域を示す水親疎性プロットを、例えば、フーリエ変換を伴う、または伴わないKyte Doolittle法またはHopp Woods法を含めた、当技術分野で周知の任意の方法により作成し得る。例えば、その全体が参照により本明細書に各々組み込まれるHopp and Woods、1981年、Proc. Nat. Acad. Sci. USA 78巻、3824〜3828頁; Kyteand Doolittle 1982年、J. Mol. Biol. 157巻、105〜142頁を参照されたい。抗原性タンパク質、またはその誘導体、断片、類縁体もしくは相同体内にある、1つまたは複数のドメインに特異的な抗体も、本明細書に示されている。
【0078】
本発明のタンパク質、またはその誘導体、断片、類縁体、相同体もしくはオーソログは、こうしたタンパク質成分に免疫特異的に結合する抗体の生成において、免疫原として利用し得る。
【0079】
本発明のタンパク質、またはその誘導体、断片、類縁体、相同体もしくはオーソログに対する、ポリクローナルまたはモノクローナル抗体の産生のために、当技術分野内で公知の多様な手順を使用し得る(例えば、参照により本明細書に組み込まれるAntibodies: A Laboratory Manual、Harlow E, and Lane D、1988年、ColdSpring Harbor Laboratory Press、Cold Spring Harbor、NYを参照されたい)。こうした抗体の一部を以下に考察する。
【0080】
ポリクローナル抗体
ポリクローナル抗体を産生するために、適切な多種の宿主動物(例えば、ウサギ、ヤギ、マウスまたは他の動物)を、自然のタンパク質、その合成変異体、または前記タンパク質の誘導体による1回または複数回の注射で免疫し得る。適当な免疫原調製物は、例えば、天然の免疫原タンパク質、その免疫原タンパク質に相当する化学合成ポリペプチド、または組換え発現した免疫原タンパク質を含有することができる。更に、該タンパク質は、免疫される哺乳動物中で免疫原性であることが知られている第2のタンパク質にコンジュゲートしてもよい。このような免疫原タンパク質の例には、それだけに限らないが、キーホールリンペットヘモシニアン、血清アルブミン、ウシサイログロブリンおよび大豆トリプシンインヒビターが挙げられる。この調製物は、アジュバントを更に含むことができる。
【0081】
免疫原タンパク質に対するポリクローナル抗体分子は、当該哺乳動物から(例えば、その血液から)単離し、プロテインAもしくはプロテインGを用いる、または抗原のカラム上でのアフィニティークロマトグラフィーなどの周知の技法によって、更に精製することができる。免疫グロブリンの精製は、例えばD.Wilkinson(The Scientist, Inc., Philadelphia PA出版のThe Scientist、14巻、8号(2000年4月17日)、25〜28頁)により考察されている。
【0082】
モノクローナル抗体
モノクローナル抗体は、Kohler and Milstein、Nature、256巻、495頁(1975年)に記載のものなどのハイブリドーマ法を用いて調製することができる。ハイブリドーマ法では、マウス、ハムスターまたは適当な他の宿主動物を通常、免疫剤で免疫することにより、免疫剤に特異的に結合すると見込まれる抗体を産生する、または産生できるリンパ球を誘導する。あるいは、リンパ球をインビトロで免疫することができる。
【0083】
免疫剤には、通常、タンパク質抗原、その断片、またはその融合タンパク質が含まれることになろう。次いで、ポリエチレングリコールなどの適切な融合剤を用いて、リンパ球を不死化細胞系と融合し、ハイブリドーマ細胞を形成する[Goding、Monoclonal Antibodies: Principles and Practice、Academic Press、(1986年)59〜103頁]。
【0084】
次いで、ハイブリドーマ細胞を培養した培地を、抗原に対するモノクローナル抗体の有無についてアッセイすることができる。所望のハイブリドーマ細胞を特定した後、限界希釈手順によりクローンをサブクローニングし、標準的方法(Goding、1986年)により増殖させることができる。この目的のための適切な培地には、例えば、ダルベッコの改良イーグル培地およびRPMI−1640培地が挙げられる。あるいは、哺乳動物中の腹水としてハイブリドーマ細胞をインビボで増殖させることができる。
【0085】
サブクローンが分泌するモノクローナル抗体は、例えば、プロテインA−セファロース、ヒドロキシアパタイトクロマトグラフィー、ゲル電気泳動、透析またはアフィニティークロマトグラフィーなどの従来からの免疫グロブリン精製手順により、培地または腹水から単離または精製することができる。
【0086】
組換えベクターおよび宿主細胞
本発明のある種の態様では、哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含有する核酸を細胞中に導入することを含む方法が開示される。哺乳類細胞はヒト細胞でもよい。タンパク質デアセチラーゼは、多様に、真核生物Sir2タンパク質または哺乳類SIRT1タンパク質でもよい。哺乳類細胞に意図した核酸配列を移入する技法は、本発明の分野の技術者に広く知られている。このような方法の特定の非限定的な例を以下に詳細に説明する。
【0087】
本発明で開示する一態様は、あるタンパク質をコードする核酸、またはその誘導体、断片、類縁体もしくは相同体を含有するベクター、好ましくは発現ベクターに関する。本明細書で使用する場合、用語「ベクター」とは、結合している別の核酸を輸送できる核酸分子を指す。1種のベクターは「プラスミド」であり、追加のDNAセグメントをその中に連結できる環状の2本鎖DNAループを指す。別種のベクターはウィルスベクターであり、そこでは追加のDNAセグメントをウィルスゲノム中に連結できる。ある種のベクターは、導入先の宿主細胞中で自律複製をすることができる(例えば、細菌性複製起点を有する細菌ベクターおよび哺乳類エピソームベクター)。他のベクター(例えば、哺乳類非エピソームベクター)は、宿主細胞中に導入した際、宿主細胞のゲノム内に組み込まれ、それにより宿主ゲノムと共に複製される。その上、ある種のベクターは、作動的に連結されている遺伝子の発現を指示することができる。このようなベクターは、本明細書では「発現ベクター」と称する。一般に、組換えDNA技法で有用な発現ベクターは、プラスミドの形態を取ることが多い。本明細書では、プラスミドがベクターの最も汎用される形態であるので、「プラスミド」および「ベクター」は、互換的に使用することができる。しかし、本発明は、同等な機能を発揮するような、ウィルスベクター(例えば、複製欠陥レトロウィルス、アデノウィルスおよびアデノ随伴ウィルス)などの他形態の発現ベクターも包含することを意図している。
【0088】
本発明に開示する組換え発現ベクターは、本発明に開示する核酸を宿主細胞中で核酸を発現するのに適切な形態で含み、そのことは、組換え発現ベクターが、発現に用いる宿主細胞に基づいて選択されている、即ち、発現しようとする核酸配列に作動的に連結されている1個または複数の調節配列を含むことを意味している。組換え発現ベクター内で、「作動可能に連結されている」とは、対象とするヌクレオチド配列が、そのヌクレオチド配列の発現(例えば、インビトロでの転写/翻訳系において、または宿主細胞中にベクターを導入した場合は宿主細胞において)を可能とするように調節配列(複数も)に連結されていることを意味することが、その意図するところである。用語「調節配列」は、プロモーター、エンハンサーおよび他の発現制御要素(例えば、ポリアデニル化シグナル)を包含することを意図している。このような調節配列は、例えば、Goeddel、GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185、AcademicPress、San Diego、Calif.(1990年)に記載されている。調節配列には、多種の宿主細胞においてヌクレオチド配列の構成的発現を指示するもの、および特定の宿主細胞においてのみヌクレオチド配列の発現を指示するもの(例えば、組織特異的調節配列)が含まれる。当業者であれば、発現ベクターの設計が、形質転換しようとする宿主細胞の選択、所望するタンパク質の発現量などの要因に依存し得ることは認識されよう。本発明に開示する発現ベクターは、宿主細胞中に導入することにより、融合タンパク質またはペプチドを含め、本明細書に記載するような核酸がコードするタンパク質またはペプチドを産生することができる(例えば、SIRT1またはLXRタンパク質、SIRT1またはLXRタンパク質の変異型、融合タンパク質など)。
【0089】
本発明に開示する組換え発現ベクターは、原核または真核細胞中にSIRT1またはLXRタンパク質を発現するように設計することができる。例えば、SIRT1またはLXRタンパク質は、大腸菌(E.coli)などの細菌細胞、昆虫細胞(バキュロウィルス発現ベクターを用いて)、酵母細胞、または哺乳類細胞中に発現させることができる。適切な宿主細胞は、Goeddel、GENE EXPRESSION TECHNOLOGY: METHODS IN ENZYMOLOGY 185、AcademicPress、San Diego、Calif.(1990年)で更に考察されている。あるいは、組換え発現ベクターは、例えば、T7プロモーター調節配列およびT7ポリメラーゼを用いて、インビトロで転写し、翻訳することができる。
【0090】
原核生物中でのタンパク質の発現は、融合、非融合いずれかのタンパク質の発現を指示する構成または誘導プロモーターを含有するベクターを用いて、大腸菌中で実施することが最も多い。融合ベクターは、その中にコードされるタンパク質に幾つものアミノ酸を、普通その組換えタンパク質のアミノ末端へ付加する。このような融合ベクターは、通常3つの目的:(1)組換えタンパク質の発現を増加させること、(2)組換えタンパク質の溶解性を増加させること、および(3)アフィニティー精製におけるリガンドとして作用することにより、組換えタンパク質の精製を補助することを実現する。典型的な融合発現ベクターには、それぞれグルタチオンS−トランスフェラーゼ(GST)、マルトースE結合タンパク質またはプロテインAを標的組換えタンパク質に融合する、pGEX(Pharmacia Biotech Inc;Smith and Johnson(1988年)Gene 67巻、31〜40頁)、pMAL(New England Biolabs,Beverly,Mass.)およびpRIT5(Pharmacia,Piscataway,N.J.)が含まれる。
【0091】
適切な誘導性の非融合大腸菌発現ベクターには、pTrc(Amrannら、(1988年)Gene69巻、301〜315頁)およびpET11d(Studierら、GENE EXPRESSION TECHNOLOGY: METHODS INENZYMOLOGY 185、Academic Press、San Diego、Calif.(1990年)60〜89頁)が含まれる。
【0092】
別の実施形態では、本発明に開示するタンパク質は、酵母発現ベクターを用いて発現される。酵母S.セレビシエ(S.cerevisiae)に発現するためのベクターの例には、pYepSec1(Baldariら、(1987年)EMBO J 6巻、229〜234頁)、pMFa(Kurjan and Herskowitz、(1982年)Cell30巻、933〜943頁)、pJRY88(Schultzら、(1987年)Gene 54巻、113〜123頁)、pYES2(Invitrogen Corporation,San Diego,Calif.)、およびpicZ(Invitrogen Corp.San Diego,Calif.)が挙げられる。
【0093】
あるいは、本発明の複合体中に開示されるタンパク質は、バキュロウィルス発現ベクターを用いて昆虫細胞中に発現することができる。培養する昆虫細胞(例えばSF9細胞)中にタンパク質を発現するために利用可能なバキュロウィルスベクターには、pAc系列(Smithら(1983年)Mol Cell Biol 3巻、2156〜2165頁)およびpVL系列(Lucklow andSummers(1989年)Virology 170巻、31〜39頁)が挙げられる。
【0094】
更に別の実施形態では、本発明に開示する核酸は、哺乳類発現ベクターを用いて哺乳類細胞中に発現される。哺乳類発現ベクターの例には、pCDM8(Seed(1987年)Nature 329巻、840頁)およびpMT2PC(Kaufmanら(1987年)EMBO J 6巻、187〜195頁)が挙げられる。哺乳類細胞中で使用する場合、発現ベクターの制御機能は、ウィルス性調節要素から得ることが多い。例えば、汎用されるプロモーターは、ポリオーマ、アデノウィルス2、サイトメガロウィルスおよびシミアンウィルス40に由来する。原核、真核双方の細胞用に適切な他の発現系については、例えば、Sambrookら、MOLECULAR CLONING: A LABORATORY MANUAL 第2版、Cold Spring Harbor Laboratory、ColdSpring Harbor Laboratory Press、Cold Spring Harbor、N.Y.、1989年を参照されたい。
【0095】
別の実施形態では、組換え哺乳類発現ベクターは、特定の細胞型中で選択的に核酸を発現するように指示することができる(例えば、核酸を発現するために、組織特異的な調節要素が使用される)。適切な組織特異的プロモーターの非限定的な例には、アルブミンプロモーター(肝特異的;Pinkertら(1987年)Genes Dev 1巻、268〜277頁)、リンパ系特異的プロモーター(Calame and Eaton(1988年)AdvImmunol 43巻、235〜275頁)、特にT細胞受容体(Winoto and Baltimore(1989年)EMBO J 8巻、729〜733頁)および免疫グロブリン(Banerjiら(1983年)Cell33巻、729〜740頁;Queen and Baltimore(1983年)Cell 33巻、741〜748頁)のプロモーター、ニューロン特異的プロモーター(例えばニューロフィラメントプロモーター;Byrneand Ruddle(1989年)PNAS 86巻、5473〜5477頁)、膵特異的プロモーター(Edlundら(1985年)Science 230巻、912〜916頁)、および乳腺特異的プロモーター(例えば、乳清プロモーター;米国特許第4873316号および欧州出願公開第264166号)が挙げられる。発生時調節されるプロモーター、例えば、マウスホックス(hox)プロモーター(Kesseland Gruss(1990年)Science 249巻、374〜379頁)およびα−フェトプロテインプロモーター(Campes and Tilghman(1989年)GenesDev 3巻、537〜546頁)も包含される。
【0096】
本発明に開示する別の態様は、本発明に開示する組換え発現ベクターを導入した宿主細胞に関する。用語「宿主細胞」および「組換え宿主細胞」は、本明細書では互換的に使用される。このような用語は、特定の対象細胞だけでなく、このような細胞の子孫または潜在的子孫も指すことが理解される。ある種の改変が、突然変異、環境影響のいずれかにより後継世代に起こり得るので、このような子孫は、実際には親細胞と同一でないこともあるが、それでも本明細書で使用する場合の該用語の範囲内に含められる。
【0097】
宿主細胞は、任意の原核または真核細胞でもよい。例えば、SIRT1またはLXRタンパク質は、大腸菌などの細菌細胞、昆虫細胞、酵母または哺乳類細胞(チャイニーズハムスター卵巣細胞(CHO)もしくはCOS細胞など)中に発現することができる。他の適切な宿主細胞は、当業者に公知である。
【0098】
ベクターDNAは、従来の形質転換またはトランスフェクション技法を介して原核または真核細胞中に導入することができる。本明細書で使用する場合、用語「形質転換」および「トランスフェクション」は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE−デキストラン媒介トランスフェクション、リポフェクションまたは電気穿孔を含む、宿主細胞中に外来核酸(例えばDNA)を導入するための当分野認知の多様な技法を指すことを意図している。宿主細胞を形質転換またはトランスフェクションする適切な方法は、Sambrookら(2001年)、Ausubelら(2002年)および他の実験手引に見出すことができる。
【0099】
哺乳類細胞の安定なトランスフェクションのために、使用する発現ベクターおよびトランスフェクション法に応じて、ほんの小さな分率の細胞が、外来DNAをそのゲノム中に組み込み得ることが知られている。こうした組込み体を特定し、選択するために、選択マーカー(例えば、抗生物質に対する耐性)をコードする遺伝子が、一般に、対象の遺伝子と共に宿主細胞中に導入される。多種の選択マーカーには、G418、ハイグロマイシン、メソトレキセートなどの薬物に対して、耐性を付与するものが含まれる。選択マーカーをコードする核酸は、成長促進物質をコードする同じベクター上で宿主細胞中に導入することができ、または別のベクター上で導入することができる。導入される核酸を安定に移入した細胞は、薬物選択によって特定することができる(例えば、選択マーカー遺伝子を組み込んだ細胞は生存することになろうが、その他の細胞は死滅する)。
【0100】
培養物中の原核または真核宿主細胞などの本発明の宿主細胞は、SIRT1またはLXRタンパク質を産生(即ち、発現)するために、使用することができる。したがって、本発明は、本発明の宿主細胞を用いてSIRT1またはLXRタンパク質を産生する方法を更に提供する。一実施形態では、この方法は、SIRT1またはLXRタンパク質が産生するように、適切な培地中で本発明の宿主細胞(SIRT1またはLXRタンパク質をコードする組換え発現ベクターを導入した後の)を培養することを含む。別の実施形態では、この方法は、培地または宿主細胞からSIRT1またはLXRタンパク質を単離することを含む。
【0101】
脊椎動物細胞のトランスフェクションは、細胞中にDNAを導入するリポソームなどの他の粒子以外に、それだけに限らないが、アデノウィルス、アデノ随伴ウィルス、レトロウィルスの各ベクターを含めた、組換えベクターを用いて更に実現することができる。上記のものなどの技法は、SIRT1またはLXRポリペプチドをコードする任意のヌクレオチド配列を脊椎動物細胞中に導入するために、利用することができる。例えば、哺乳類細胞のトランスフェクションのために、幾種ものウィルス依拠発現系を利用し得る。発現ベクターとしてアデノウィルスを使用する場合、対象とするSIRT1またはLXRヌクレオチド配列を、アデノウィルス転写/翻訳制御複合体、例えば、後期プロモーターおよび三部分(tripartite)リーダー配列に連結してもよい。次いで、このキメラ遺伝子は、インビトロまたはインビボでの組換えにより、アデノウィルスゲノム中に挿入されることがある。ウィルスゲノムの非必須領域(例えば、E1またはE3領域)に挿入されると、感染宿主中で生存でき、SIRT1またはLXR産物を発現できる組換えウィルスが生じることになろう(例えば、Logan & Shenk、1984年、Proc. Natl. Acad. Sci. USA 81巻、3655〜3659頁を参照されたい)。SIRT1またはLXRコード配列の一部分だけが挿入される場合には、多分ATG開始コドンを含んだ外因性翻訳制御シグナルを供給しなければならない。こうした外因性翻訳制御シグナルおよび開始コドンは、天然、合成いずれの多様な起源のものでもよい。発現の効率は、適当な転写エンハンサー要素、転写ターミネーターなどを含めることにより、高めることができる(Bitterら、1987年、Methodsin Enzymol. 153巻、516〜544頁を参照されたい)。
【0102】
治療的処置
ある種の病状および病的状態は、対象の細胞における異種SIRT1もしくはLXR、または異種LXRαもしくはLXRβの発現に有利に応答すると考えられている。したがって、本発明は、対象における病状、疾患または病的状態を処置する方法であって、該病状が、SIRT1またはLXR、LXRαまたはLXRβのポリペプチドに応答する方法を開示する。この方法は、病状の軽減または改善に有効な量で、SIRT1またはLXR、LXRαまたはLXRβのポリペプチドをコードする核酸を対象に投与することを含む。病状の軽減とは、症状の悪化傾向が、より緩やかまたはより穏やかな悪化傾向に弱まることを意味する。病状の改善とは、病状の兆候および症候が弱まり、患者がより良好な健康へと改善するような、患者の実際の改善を意味する。この方法の重要な実施において、病状は、心筋梗塞、脳血管発作、腎疾患、アルツハイマー病を含む神経疾患などから選定される。病状を処置する方法の有利な実施形態では、対象はヒトである。
【0103】
本明細書に記載の処置法の多様な実施形態では、SIRT1もしくはLXR、LXRαもしくはLXRβのポリペプチド、その変異体またはその断片をコードする核酸を、対象の細胞中への送達を含む、その核酸配列の細胞中への効率的送達を保証する各種組成物のいずれか中で、対象へ投与し得る。
【0104】
SIRT1またはLXR核酸配列による対象の処置は、SIRT1またはLXR核酸配列の脊椎動物細胞中への導入に有効である、適切な核酸、プラスミド、ベクター、ウィルスベクター、リポソームまたは類似組成物を投与することにより、実現することができる。核酸のトランスフェクションは、陽イオン両親媒性物質の使用で補助し得る(米国特許第6503945号およびその中に開示されている文献)。エクスビボでのレトロウィルス遺伝子治療は、例えばHacein-Bey-Abinaら(2003年、Science 302巻、415〜419頁)により記載されている。対象中へのトランス遺伝子の治療的導入法は、「GeneTransfer Methods: Introducing DNA Into Living Cells and Organisms」、P.A. Nortonand L.F. Steel、Eaton Publishing、2000年において考察されている。細胞および生物中へのトランス遺伝子の治療的導入手法は、「GeneTherapy Protocols」、Paul D. Robbins(編)、Humana Press(1997年)に示されている。
【0105】
ポリヌクレオチドを含む医薬組成物
治療用途のための医薬組成物には、1種または複数のポリヌクレオチドおよび担体が含まれる。1種または複数のポリヌクレオチドを含む医薬組成物は、標的遺伝子の発現または活性と関連する疾患または障害の処置に有用である。担体には、それだけに限らないが、塩水、緩衝塩水、デキストロース、水、グリセロール、エタノール、およびそれらの組合せが挙げられる。経口投与される薬物について、医薬として許容可能な担体には、それだけに限らないが、不活性希釈剤、崩壊剤、結合剤、潤滑剤、甘味剤、香味剤、着色剤、防腐剤などの医薬として許容可能な賦形剤が挙げられる。
【0106】
1日につき受容者の体重1kg当たりポリヌクレオチド5mgの最大投与量は、標的遺伝子の発現を阻害する、または完全に抑制するのに十分である。一般に、適切なポリヌクレオチド用量は、1日につき受容者の体重1kg当たり0.01〜5.0mgの範囲、好ましくは1日につき体重1kg当たり0.1〜200μg(μg/kg)の範囲、より好ましくは1日につき0.1〜100μg/kgの範囲、より一層好ましくは1日につき1.0〜50μg/kgの範囲、最も好ましくは1日につき1.0〜25μg/kgの範囲に入ることとなろう。医薬組成物は、毎日1回投与してもよく、またはポリヌクレオチドは、1日を通して適当な間隔の分割用量として、2回、3回、4回、5回、6回もしくはそれより多い回数で投与してもよい。その場合、各分割用量中に含有するポリヌクレオチドは、1日総投与量を実現するために、それ相当に減量させねばならない。投与単位は、例えば、数日の期間に亘ってポリヌクレオチドを持続放出する従来の製剤を用いて、数日に亘る送達用の持続放出製剤として調合することもできる。持続放出製剤は、当技術分野で周知のものである。この実施形態では、投与単位は、1日用量の何倍かに相当する量を含有する。
【0107】
当業者であれば、それだけに限らないが、疾患または障害の重度、以前の処置、対象の健康全般および/または年齢、ならびに罹っている他の疾患を含めた特定の要因が、対象の有効な処置に必要な投与量および適時に影響し得ることは、認識されていよう。その上、治療有効量の組成物による対象の処置は、単回処置または一連の処置を包含することができる。本発明により包含される個々のポリヌクレオチドに関する有効投与量および体内半減期の見積りは、従来手法を用いて、または適当な動物モデルを用いたインビボ試験に基づいて行うことができ、また、適当な用量−応答特性を決定するための確立した規準に従って、処置中に調整することができる。
【0108】
本発明により包含される医薬組成物は、それだけに限らないが、経口経路、または静脈内、筋肉内、腹腔内、皮下、経皮、気道(エアゾル)、直腸、経膣および局所(口腔および舌下を含む)投与を含めた非経口経路を含む、当技術分野で公知の任意の手段によって投与してもよい。ある種の実施形態では、医薬組成物は、静脈内または非経口内注入または注射により投与され、追加の一般的実施形態では、ポリヌクレオチドを含む医薬組成物は、腹腔鏡および類似のマイクロ手術の手順を用いて、その場の腫瘍、癌または前癌増殖物に直接投与してもよい。
【0109】
筋肉内、腹腔内、皮下および静脈内使用のために、本発明の医薬組成物は、一般に、適当なpHおよび等張性に緩衝させた、滅菌、水性の溶液または懸濁液として供給されよう。適切な水性媒体には、リンゲル液および等張塩化ナトリウムが挙げられる。好ましい実施形態では、担体は専ら緩衝水溶液らなる。これに関しては、「専ら」とは、標的遺伝子を発現する細胞におけるポリヌクレオチドの取込みに影響し得る、またはそれを媒介し得る、助剤または封入物質が全く存在しないことを意味する。このような物質には、例えば、リポソームやキャプシドなどの以下に説明するようなミセル構造が挙げられる。驚くべきことに、本発明者らは、裸ポリヌクレオチドおよび生理的に許容可能な溶媒だけを含有する組成物が、細胞により取り込まれ、そのポリヌクレオチドが、標的遺伝子の発現を有効に抑制することを発見した。マイクロインジェクション、リポフェクション、ウィルス、ウィロイド、キャプシド、キャプソイドまたは他の助剤が、細胞培養物中へのポリヌクレオチドの導入には必要とされるが、驚くべきことに、こうした方法および助剤が、インビボでのポリヌクレオチドの取込みには必要ではない。本発明による水性懸濁液は、セルロース誘導体、アルギン酸ナトリウム、ポリビニルピロリドン、トラガカントガムなどの懸濁剤、およびレシチンなどの湿潤剤を含んでもよい。水性懸濁液用の適切な防腐剤には、p−ヒドロキシ安息香酸エチルおよびn−プロピルが挙げられる。
【0110】
本発明に従って有用な医薬組成物は、埋め込み材およびマイクロカプセル化送達系を含めた制御放出製剤などの、体内からの急速消失からポリヌクレオチドを保護するためのカプセル化製剤も包含する。エチレン酢酸ビニル、ポリ酸無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、ポリ乳酸などの生分解性で生体適合性のポリマーを使用することができる。このような製剤の調製法は、当業者には明らかであろう。その各材料は、Alza CorporationおよびNova Pharmaceuticals,Inc.から商業的に入手することもできる。リポソーム懸濁液(ウィルス抗原に対するモノクローナル抗体を有する、感染細胞を標的としたリポソームを含める)も、医薬として許容可能な担体として使用することができる。こうしたものは、例えば、参照により本明細書に組み込まれる米国特許第4522811号、PCT公開WO第91/06309号および欧州特許公開第EP−A−43075号に記載のように、当業者に公知の方法に従って調製することができる。
【0111】
ある種の実施形態では、カプセル化製剤はウィルスコートタンパク質を含む。この実施形態では、ポリヌクレオチドは、少なくとも1つのウィルスコートタンパク質と結合し、会合し、またはそれに囲い込まれてもよい。ウィルスコートタンパク質は、ポリオーマウィルスなどのウィルスに由来するか、それと会合していてもよく、または、部分的もしくは完全に人工的なものでもよい。例えば、コートタンパク質は、ポリオーマウィルスのウィルスプロテイン1および/またはウィルスプロテイン2、あるいはその誘導体でもよい。
【0112】
このような化合物の毒性および治療効果は、例えば、LD50(その集団の50%に致死的な用量)およびED50(その集団の50%に治療有効な用量)を決定するための、細胞培養物または実験動物における標準的な薬学手順によって決定することができる。毒性効果と治療効果との用量比が、治療指数であり、比率LD50/ED50として表現することができる。高い治療指数を示す化合物が好ましい。
【0113】
細胞培養アッセイおよび動物試験から得られるデータは、ヒトに使用する投与量の範囲を処方する際に使用することができる。本発明の組成物の投与量は、毒性を殆どまたは全く示さずに、ED50を含む循環濃度の範囲内にあるのが好ましい。その投与量は、使用する剤形および利用する投与経路に応じてこの範囲内で変化させてもよい。本発明の方法に使用するいずれの化合物についても、細胞培養物において決定した場合のIC50(即ち、症状の最大抑制の半量を実現する試験化合物の濃度)を含む、その化合物の循環血漿濃度範囲となるように、治療有効用量を、細胞培養アッセイおよび動物モデルから当初に推定することができる。このような情報は、ヒトにおいて有用な用量をより正確に決定するために、使用することができる。
【0114】
以上に考察したように、単独または複数として投与する以外に、本発明に従って有用なポリヌクレオチドは、疾患の処置に有効な他の既知の作用剤と組み合わせて投与することもできる。いずれにせよ、投薬担当内科医は、当技術分野で知られているか、または本明細書に記載した効力の標準尺度を用いた観察結果に基づいて、ポリヌクレオチド投与の量および適時を調節することができる。
【0115】
医薬組成物
本発明に開示するポリヌクレオチドは、本明細書では「活性化合物」または「治療剤」と呼称される。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0116】
本発明は、SIRT1活性を刺激する作用剤を投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法、ならびに、SIRT1活性を刺激する作用剤を哺乳動物に投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、該作用剤の使用を含む。このような作用剤は、本明細書では「活性化合物」または「治療剤」と呼称される。このような作用剤の一例はT0901317である。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0117】
本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤を投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる方法も含む。それに加え、本発明は、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤を哺乳動物に投与することにより、哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、該作用剤の使用を含む。このような作用剤は、本明細書では「活性化合物」または「治療剤」と呼称される。このような作用剤の例には、22(R)−ヒドロキシコレステロール、9−シスレチノイン酸のいずれか、または22(R)−ヒドロキシコレステロール、9−シスレチノイン酸の両者の組合せが挙げられる。こうした治療剤は、対象への投与に適した医薬組成物中に組み入れることができる。
【0118】
本明細書で使用する場合、「医薬として許容可能な担体」は、医薬投与に適合できる、ありとあらゆる溶媒、分散媒、コーティング、抗細菌剤および抗真菌剤、等張剤および吸収遅延剤などを包含することを意図している。適切な担体は、参照により本明細書に組み込まれるRemington's Pharmaceutical Sciences、Gennaro AR(編)20版(2000年)Williams& Wilkins PA、米国、およびWilson and Gisvold's Textbook of Organic Medicinal andPharmaceutical Chemistry、Delgado and Remers著、Lippincott-Ravenなどの教科書に記載されている。このような担体または希釈剤に使用し得る成分の好ましい例には、それだけに限らないが、水、塩水、リン酸塩、カルボン酸塩、アミノ酸溶液、リンゲル液、デキストロース(グルコースの同義語)溶液、および5%ヒト血清アルブミンが挙げられる。非限定的な例として、デキストロースは5%または10%水溶液として使用してもよい。リポソーム、および不揮発性油などの非水性媒体も使用し得る。医薬活性物質用のこのような媒体および作用剤の使用は、当技術分野で周知のことである。補助的な活性化合物も、該組成物中に組み入れることができる。
【0119】
本発明の医薬組成物は、意図する投与経路に適合するように処方される。投与経路の例には、非経口経路、例えば、静脈内、皮内、皮下、経口、経鼻、吸入、経皮(局所)、経粘膜および直腸投与が挙げられる。非経口、静脈内、皮内または皮下投与に使用される溶液または懸濁液は、以下の成分:注射水、塩水溶液、不揮発性油、ポリエチレングリコール、グリセリン、プロピレングリコールもしくは他の合成溶媒などの希釈剤;ベンジルアルコールもしくはメチルパラベンなどの抗細菌剤;アスコルビン酸もしくは重亜硫酸ナトリウムなどの抗酸化剤;エチレンジアミン四酢酸などのキレート剤;酢酸塩、クエン酸塩もしくはリン酸塩などの緩衝剤、および塩化ナトリウムもしくはデキストロースなどの等張化剤を含むことができる。
【0120】
吸入による投与のために、該化合物は、適切な高圧ガス、例えば二酸化炭素などのガスを含有する加圧容器もしくはディスペンサー、または噴霧器からのエアゾールスプレーの形態で送達される。
【0121】
一実施形態では、活性化合物は、埋め込み材およびマイクロカプセル化送達系を含めた制御放出製剤などの、体内からの急速消失から該化合物を保護すると見込まれる担体と共に調製される。持続放出製剤の適切な例には、抗体を含有する固形疎水性ポリマーの半透性マトリックスであって、成形品、例えばフィルムまたはマイクロカプセルの形態をしたマトリックスが挙げられる。持続放出性マトリックスの例には、ポリエステル、ヒドロゲル(例えば、ポリ(2−ヒドロキシエチルメタクリレート)またはポリ(ビニルアルコール))、ポリラクチド(米国特許第3773919号)、L−グルタミン酸およびγエチル−L−グルタミン酸のコポリマー、非分解性エチレン−酢酸ビニル、LUPRON DEPOT(商標)(乳酸−グリコール酸コポリマーおよび酢酸ロイプロリドからなる注射用ミクロスフェア)などの分解性乳酸−グリコール酸コポリマー、およびポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。エチレン−酢酸ビニル、乳酸−グリコール酸などのポリマーは、分子の放出を100日超の間可能とするが、ある種のヒドロゲルでは、医薬活性剤を放出する期間がより短い。有利なポリマーは、生分解性または生体適合性である。リポソーム懸濁液(ウィルス抗原に対するモノクローナル抗体を有する、感染細胞を標的としたリポソームを含める)も、医薬として許容可能な担体として使用することができる。こうしたものは、例えば、米国特許第4522811号に記載のように、当業者に公知の方法に従って調製することができる。ミクロスフェアなどの有利な形態を有する持続放出製剤は、上記の材料などから調製することができる。
【0122】
本発明に開示するポリヌクレオチドは、ベクター中に挿入し、遺伝子治療用ベクターとして使用することができる。遺伝子治療用ベクターは、例えば、米国特許第5703055号に記載のように、幾つもの経路のいずれかにより対象へ送達することができる。したがって、送達には、例えば、静脈注射、局所投与(米国特許第5328470号を参照されたい)、または定位注射(例えば、Chenら(1994年)Proc. Natl. Acad. Sci. USA 91巻、3054〜3057頁を参照されたい)も挙げることができる。遺伝子治療用ベクターの医薬製剤は、許容可能な希釈剤中に遺伝子治療用ベクターを含むことができ、または遺伝子送達媒体を埋め込んだ徐放性マトリックスを含むことができる。あるいは、完全な遺伝子送達ベクター、例えばレトロウィルスベクターを組換え細胞から無傷で産生することができる場合、該医薬製剤は、遺伝子送達系を産生する1個または複数の細胞を含むことができる。
【0123】
医薬組成物は、キット中、例えば容器、包み、またはディスペンサー中に投与説明書と共に含めることができる。
【0124】
対象の呼吸器ウィルス感染症を処置する医薬組成物または薬剤の製造における治療剤の使用も、本発明に入る。
【0125】
幾つかの実施形態では、本発明に開示するポリヌクレオチドは、リポソーム媒介トランスフェクションにより、例えば、市販の試薬または技法、例えばOligofectamine(商標)、LipofectAmine(商標)試薬、LipofectAmine2000(商標)(Invitrogen)の使用により、ならびに電気穿孔および類似の技法により、送達される。それに加え、ポリヌクレオチドは、呼吸器中への吸入および点滴注入により、げっ歯類または非ヒト霊長類などの動物モデルへ送達される。動物モデルに用いる付加的経路には、静脈内(IV)、皮下(SC)、および関連投与経路が挙げられる。医薬組成物は、ポリヌクレオチドの安定性を保護し、その寿命を延長し、その機能を増強し、または特定の組織/細胞をその標的とさせる、追加の成分を含む。これらの成分には、多種の生分解性ポリマー、陽イオン性ポリマー(ポリエチレンイミンなど)、ヒスチジン−リシン(HK)ポリペプチドなどの陽イオン性コポリペプチド(例えば、MixonらのPCT公開WO第01/47496号、BiomerieuxのWO第02/096941号、およびMassachusetts Institute of TechnologyのWO第99/42091号を参照されたい)、PEG化陽イオン性ポリペプチドおよびリガンド導入ポリマーなど、陽荷電ポリペプチド、PolyTranポリマー(スクレログルカンの名でも知られる天然多糖;Pillsbury Co.,Minneapolis,Minn)、界面活性剤(Infasurf;Forest Laboratories,Inc.;ONY Inc.)、ならびに陽イオン性ポリマー(ポリエチレンイミンなど)が挙げられる。Infasurf(登録商標)(カーファクタント(calfactant))は、子ウシの肺から単離し、気管内点滴注入に使用する天然の肺界面活性剤であり、リン脂質、中性脂質、ならびに疎水性の界面活性剤関連タンパク質BおよびCを含有する。該ポリマーは、一次元、多次元のいずれでもよく、直径が20ミクロン未満、20〜100ミクロンの間、または100ミクロン超のマイクロ粒子またはナノ粒子でもよかろう。前記ポリマーは、特殊な組織または細胞の受容体または分子に特異的なリガンド分子を搬送し、したがってポリヌクレオチドの標的向け送達に使用することができよう。ポリヌクレオチドは、DOTAP、DOTAP/コレステロール(Qbiogen,Inc.)などの陽イオン性リポソーム系担体、および他種の脂質水溶液によっても送達される。その上、低率(5〜10%)グルコース水溶液およびInfasurfは、ポリヌクレオチドの気道送達用の有効な担体である。
【0126】
遺伝子治療
現在では、遺伝子治療は、治療および予防(例えば免疫用接種)環境で使用するための高度な開発状態にある。新規な送達法が利用でき、多様な遺伝子治療様式に関する幾つかの臨床試験が進行中である。「Direct DNA delivery to bone cells」という名称の米国特許出願公開第20060115462号は、裸ポリヌクレオチドまたは遺伝子発現ベクターなどのポリヌクレオチドの哺乳類四肢骨部の細胞に対するインビボ送達を可能とする方法を開示している。
【0127】
「Replication-competent adenoviral vectors」という名称の米国特許出願公開第20060115456号は、アデノウィルス2型、4型、5型および7型を含み、治療および予防用途向けに核酸およびポリヌクレオチドを送達する際に使用するための、改良された複製能のあるアデノウィルスベクターを開示している。
【0128】
「Methods of Tranducing genes into T cells」という名称の米国特許出願公開第20060104950号は、パラミクソウィルスベクターを用いた活性化T細胞中への遺伝子の形質導入を開示している。
【0129】
「Methods of making viral particles havinga modified cell binding activity and uses thereof」という名称の米国特許出願公開第20060110361号は、その物品の表面上にある1種または複数のペブチドがパッケージング細胞に由来するように、ウィルス粒子をパッケージする方法を開示している。この出願は、このような系が、例えば遺伝子治療処置に有用であると明言している。
【0130】
「Vector-mediated delivery ofpolynucleotides encoding soluble VEGF receptors」という名称の米国特許出願公開第20060110364号は、可溶型VEGFR3を発現するためのベクター組成物、および癌の処置におけるその使用方法を開示している。
【0131】
「Multifunctional molecular complexes forgene transfer to cells」という名称の米国特許出願公開第20050239204号は、核酸組成物を標的細胞へ移送するための非ウィルス多機能分子複合体を開示している。
【0132】
VandenDriesscheらは、ヒトVIII因子遺伝子をレトロウィルスベクターによりVIII因子欠損マウス中に導入するインビボ遺伝子治療によって、A型血友病を有効に処置できることを示した79。遺伝子導入は、少なくとも肝臓、脾臓および肺臓において行われた。A型血友病の重症患者における非ウィルス体細胞遺伝子治療系の安全性が、Rothらにより試験されてきた80。
【0133】
安定な有糸分裂後の状態のために、骨格筋は、組込み型、非組込み型双方のウィルスおよび非ウィルスベクターの使用による遺伝子操作の重要な標的である。非肥満性糖尿病(NOD)マウスにおける自己免疫性糖尿病を予防するための筋肉誘導遺伝子治療が、Goudyらにより報告された81。彼らは、免疫調節サイトカインのIL4またはIL10のためのマウスcDNAを含有する、組換えアデノ付随ウィルスベクターを開発した。IL4ではなく、IL10による雌性NODマウスの骨格筋形質導入によって、糖尿病が完全に消失した。
【0134】
Wells82によれば、アデノ付随ウィルスベクターおよび裸プラスミドDNAは、現在、筋肉中に遺伝子導入をするための優れたベクターである。ベクター送達系、特に血管経路を用いる系における主要な躍進は、ここ2年ほど以内になされており、その結果、筋ジストロフィーの遺伝子治療、および分泌タンパク質の産生用プラットホームとしての筋肉の使用が、臨床的可能性となった。
【0135】
癌溶解治療は、アデノウィルス(Ad)などの弱毒化溶解性ウィルスに基づく新規な抗癌処置である83。これらのウィルスは、溶解によって宿主細胞の破壊を誘発し、したがって古典的な遺伝子治療用ウィルスとは異なる。癌溶解性Adは、一般に、癌細胞のみで複製するように遺伝子操作を受けている。3種の癌溶解性Adが、癌患者において第1相および第2相臨床試験を完了した。これらのウィルスは、追加の治療遺伝子を備えるように、更に操作されているところである83。
【0136】
Takahashiらは、多剤耐性(MDR1)遺伝子治療の臨床研究を開始した。現在まで処置された患者2名は、完全に寛解しており、MDR1遺伝子導入による明らかな有害作用を受けなかった84。
【0137】
Broedlらは、様々な高脂肪血症の処置が無効な現状にあって、体細胞遺伝子治療は、有力な治療手法と考えられると述べている。多くの症例で、原理証明前臨床試験が既に行われており、ホモ接合型の家族性高コレステロール血症の症例では、臨床試験が行われた85。
【0138】
クラスタリンを対象とするアンチセンスオリゴデオキシヌクレオチドが、前立腺癌の処置のために第1相臨床試験において使用された86。この処置により、前立腺癌におけるクラスタリンが90%までも抑制された。Miyakeらは、第2相試験が2005年の早期に開始する予定であったと明言している86。
【0139】
Satohらは、単純疱疹ウィルスチミジンキナーゼ遺伝子のウィルス媒介形質導入と、その後のガンシクロビルプロドラッグによる一連の治療を含め、前立腺癌遺伝子治療の現状を総説している87。その報告によれば、人体前立腺癌の遺伝子治療臨床試験では、安全性、臨床的効力、および抗腫瘍活性の生物学的効果が実証された。Satohらによれば、多数の好ましい前臨床試験の後、インターロイキン−2、インターロイキン−12、B7−1(CD80)、B7−2(CD86)、顆粒球−マクロファージコロニー刺激因子などの免疫調節性サイトカインを用いる前立腺癌の遺伝子治療に対して、幾つかの臨床試験が承認された87。
【0140】
潜在的活性剤を同定するためのアッセイ法
多様な態様では、本発明は、候補化合物または候補物質が、本発明において対象とする活性を調節するか否かを決定することを意図したアッセイ法を開示する。このような方法には以下のものが挙げられる。
【0141】
本発明は、候補物質がLXR依存性プロセスを調節するか否かを評価する方法であって、
a)LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターで細胞をトランスフェクトし、
b)その細胞を候補物質と接触させ、更に、
c)候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定する
ことを含み、それによって、候補物質の存在と不在との間で検出されるLXR依存性プロセスの発生の程度における差異が、候補物質がLXR依存性プロセスを調節することを示す
方法を開示する。
【0142】
本発明は、更に、候補物質がLXRのSIRT1依存性作用を調節するか否かを評価する方法であって、
a)SIRT1遺伝子を保持するベクターで細胞をトランスフェクトし、
b)LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するベクターで該細胞をさらにトランスフェクトし、
c)その細胞を候補物質と接触させ、更に、
d)候補物質と接触していない細胞と比較して、候補物質がレポーター遺伝子の発現を調節するか否かを決定する
ことを含み、それにより、候補物質の存在と不在との間で検出されるLXRのSIRT1依存性作用の程度における差異が、候補物質がLXRのSIRT1依存性作用を調節することを示す
方法を開示する。
【0143】
本発明は、更に、候補物質が、特異的結合対(specific binding pair)のメンバーのSIRT1およびLXRを含む特異的結合対の形成を調節するか否かを評価する方法であって、
a)特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトし、
b)特異的結合対の第2メンバーをコードする配列を保持するベクターで該細胞をさらにトランスフェクトし、
c)その細胞を候補物質と接触させ、
d)その細胞を溶解し、エピトープタグに特異的な抗体を細胞溶解物と接触させ、更に、SIRT1とLXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収し、
e)SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行する
ことを含み、それにより、候補物質の不在下と比較した、候補物質の存在下で検出した複合体形成の差異が、候補物質がSIRT1のLXRとの相互作用を調節することを示す方法を開示する。
【0144】
こうしたアッセイ法では、参照により本明細書に組み込まれる、上述した技法などにより、ベクターのトランスフェクションを実施し得る。本発明の分野における技術者に公知であり、本明細書に具体的に記載していない任意の同等な技法も、トランスフェクションを実現するために同様に使用してもよい。
【0145】
多様な該アッセイ法は、更に、LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターのトランスフェクションを含む。レポーター遺伝子は、レポーター遺伝子を発現するためにLXREに特異的に結合する細胞成分により、活性化されることになろう。レポーター遺伝子産物は、アッセイを行うために工夫した実験系において検出できる。一般に、レポーター遺伝子は、化学的または物理的検出法によって、発現した際に容易に検出できるタンパク質、因子、または酵素活性をコードする。レポーター遺伝子の非限定的な例には、西洋ワサビペルオキシダーゼまたはホタルルシフェラーゼなどの酵素、発色タンパク質または緑色蛍光色素などの蛍光タンパク質などをコードする遺伝子が挙げられる。本発明の分野における技術者に公知であり、本明細書に具体的に記載していない任意の同等なレポーター遺伝子も、細胞に移入することにより、潜在的な活性剤を同定するアッセイを実施するために、同様に使用してもよい。
【0146】
該アッセイ法の多様な付加的実施形態は、SIRT1およびLXRを含む特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトすることを含む。この目的に使用できるエピトープには、特異抗体が結合する任意のエピトープが挙げられる。エピトープの非限定的な一例は、FLAGエピトープであり、その他の例は、本発明の分野における技術者に周知のものである。本明細書に具体的に記載していない任意の同等なエピトープも、潜在的な活性剤を同定するアッセイを実施するために、特異的結合対の一メンバーとのキメラとして使用してもよい。
【0147】
潜在的な活性剤を求めるアッセイは、高度反復並行アッセイ装置におけるハイスループットスクリーンを用いて実施することが多い。こうしたシステムでは、多数のアッセイを同時に実施するために、多重ウェルプレートを使用してもよい。潜在的な活性剤を検出するアッセイに使用される候補化合物は、一般に、一群の関連化合物における多様な部分および/または置換基を系統的に変化させた、化合物のコンビナトリアルライブラリーを化学合成した結果として取得される。類似の化学品ライブラリーは、通常、個人または組織が利用できる保管化合物として既に入手が可能である。更なる類似の化学品ライブラリーには、異種の天然源から得られる化学品、ならびにコンビナトリアルケミストリーの方法を用いて更に修飾した、このような天然化合物の誘導体が含まれる。一般に、化合物のライブラリーは、本発明の分野における技術者に広く知られている。候補化合物を調製する、または候補化合物のライブラリーを調製する、本発明の分野における技術者に公知の同等な任意の方法を、潜在的な活性剤を同定するアッセイを実施する際に使用してもよい。
【実施例】
【0148】
方法
プラスミド
プラスミドのpBabe−SIRT1およびpCMV−FLAG−HsSIRT1は、既に記載されている15,16。
動物および血漿リポタンパク質分析
129/sv−CD1を混合バックグランドとするSIRT1+/+、SIRT1+/−およびSIRT1−/−マウスは、7a.m.〜7p.m.の昼光サイクルの温度制御(22℃)施設に収容し、食物(0.02%コレステロール入り)および水を自由に摂取させた。
【0149】
細胞、レトロウィルス感染およびトランスフェクション
SIRT1+/+、SIRT1+/−およびSIRT1−/−マウスの胚性線維芽細胞(MEF)を以前に記載のようにE14.5の胚から分離し73、10%FBSおよび抗生物質を含むダルベッコ改良イーグル培地(DMEM)中で培養した。不死化MEF(MEFI)は、初代MEFにpRS−SV40Tプラスミドを移入することにより生成した。HEK293TおよびPhoenix細胞(ATCC,Rockville,MD)は、10%FBSおよび抗生物質を含むDMEM中で培養した。THP−1細胞(ATCC,Rockville,MD)は、10%FBSおよび0.5mM β−メルカプトエタノールを含むRPMI1640中で培養した。
【0150】
全てのトランスフェクションは、製造業者の説明書(Invitrogen,Carlsbad,Ca)に従ってリポフェクタミン2000plus試薬を用いて行った。レトロウィルス感染については、Phoenix細胞にpBabeまたはpBabe−SIRT1を移入した。レトロウィルスを含有する培地は、48時間後に収集し、ろ過し、ポリブレン(1mg/ml)で処理し、MEF上に移した。次いで、1μg/mlのピューロマイシンを含有する通常培地で感染細胞を培養した。
【0151】
(実施例1)
野生型およびSIRT1ノックアウトマウスにおける血漿コレステロール
各群における野生型10匹、SIRT1+/−13匹、および同腹子SIRT1−/−13匹の雄性マウスを分析した。動物を昼光サイクルの開始から4時間絶食させ、次いで血液を採集し、K3−EDTA処理により血漿を取得した。総コレステロール、HDLおよびLDLの血漿濃度を、酵素呈色アッセイキット(Wako Diagnostics,Richmond,Va)によって測定した。SIRT1−/−4匹、および同腹子SIRT1+/+4匹の雄性マウスから得た保管血漿100μlを、高速液体クロマトグラフィー(FPLC)のカラム(Superose 6Bカラム、Amersham−Pharmacia Biotech,Piscataway,NJ)2本を用いてサイズ分画した。個別の3実験からの代表的プロファイルを示してある。次いで、FPLCから得た画分のコレステロール含量をWako製酵素呈色アッセイキットで分析した。実験は、合計してSIRT1−/−12匹および同腹子SIRT1+/+12匹の雄を用いて3回繰り返した。結果は、スチューデントのt検定を用いて比較した。
【0152】
図1aは、SIRT1−/−マウスが、週齢および性別が対応する野生型マウスと比較して、僅かに低い総血漿コレステロール濃度を有したことを示す。図1のパネルb(直接酵素アッセイ)およびパネルc(サイズ排除クロマトグラフィー/酵素アッセイ)を点検すると、マウス血漿中の主要な富コレステロールリポタンパク質(全体の80〜90%)である、HDL中に保持されるコレステロールが約40%低下した(*、p<0.001)ために、そうなることが分かる。血漿LDLコレステロール濃度は、SIRT1−/−および対照マウスにおいて類似していた(図1cおよびd)。その結果、SIRT1−/−マウスの総コレステロール/HDL比は、異常に高く(野生型対照の1.36±0.19に対して2.57±0.98、p<0.001)、HDL/LDL比は異常に低かった(3.56±1.24に対して1.92±0.94、p<0.001)。SIRT1−/−マウスにおける多面発現効果のために、野生型と表現型が類似しているSIRT1+/−ヘテロ接合型も調べた。HDL濃度は、SIRT1+/−ヘテロ接合型でも有意に低下していることが判明した(図1b(*、p<0.001))。
【0153】
総コレステロール(TC)、遊離コレステロール(FC)、コレステロールエステル(CE)およびリン脂質(PL)の含量を、12対の動物からのHDL画分(25〜38番画分)において測定しており、それを図1パネルeに示す。血漿HDL中の総コレステロール、遊離コレステロール、コレステロールエステルおよびリン脂質の量のSIRT1−/−マウスにおける相対的低下は、類似しており、約25〜30%であった(図1e;**、p<0.001)。HDLのサイズ分布は、SIRT1−/−マウスにおいて正常であった(図1d)が、それは、HDLの低下が、粒子の組成または構造が変化したためより、粒子数が減少した結果であったことを示唆している。
【0154】
(実施例2)
野生型およびSIRT1ノックアウトマウスの組織におけるコレステロール
マウス組織中の総コレステロールを決定するために、SIRT1−/−マウスおよび対照の同腹子を昼光サイクルの開始から4時間絶食させ、その後犠牲にした。次いで、組織を採集し、秤量した。既述のように70,71、肝臓および精巣の総コレステロールを抽出し、GCにより測定した。トリグリセリドを含めた総脂質も、60%ブタノール、13%メタノールおよび27% Triton X−100を含有する溶液中に溶解し、Wako製酵素呈色アッセイキットで測定した。
【0155】
SIRT1−/−マウスにおいて、異常に低い血漿HDL(実施例1)は、HDLが外因性コレステロール源として重要な以下の2種の組織内で、コレステロールの蓄積量が増加したことと関係していたが、そうした組織とは、ステロール保存およびステロイド合成25のためにHDLコレステロールを使用し、相対量最高のSIRT1タンパク質を有する器官26,27である精巣、ならびにコレステロールおよびHDLの恒常性に中心的役割を果たす28肝臓である(図1f;n=8;***、p<0.01)。
【0156】
SIRT1−/−マウスにおける血漿HDLコレステロールの減少、ならびに肝臓および精巣中の組織コレステロールの蓄積量増加は、リポタンパク質コレステロールの取込み増加、または細胞コレステロールのリポタンパク質への排出量(移出量)減少のために起こり得よう。組織コレステロールの増加は、局所的なコレステロール合成の増加によることもあろう。しかし、発明者らは、肝臓および精巣において、コレステロール生合成における律速酵素のHMG−CoAレダクターゼをコードする高感受性コレステロール調節遺伝子に対応するmRNAの量が、本質的に正常であることを観察した(データは示していない)。また、SIRT1−/−マウス胚性線維芽細胞において(MEF、データは示していない)、HDLコレステロールの細胞取込みを媒介する主要なHDL受容体SR−BI29のタンパク質量の明らかな増加が、こうした組織中で見られず(データは示していない)、HDLからの脂質取込み速度の増加も見られなかった。更に、他の主要な2種のリポタンパク質受容体であるLDL受容体(LDLR)およびVLDL/キロミクロン受容体LRPのmRNA量が、肝臓および精巣では増加するのでなく、実質的に減少していた(データは示していない)。新規コレステロール合成の増加でも受容体媒介移入でも、SIRT1−/−マウスにおける組織内増加および血漿コレステロール減少を説明できないようなので、細胞のコレステロール流出量の低下が、こうした観察を説明し得るのではないかと思われた。
【0157】
(実施例3)
野生型およびSIRT1−/−細胞からのコレステロールの排出
コレステロール逆輸送は、末梢組織中の過剰コレステロールを、体内から除去するために肝臓へ輸送する過程である30,31。この過程の第1段階は、細胞からリポタンパク質、特にHDLへのコレステロールの排出である。数種の細胞表面コレステロール輸送タンパク質が、コレステロール流出を媒介することができ、そうしたものには、SR−BI32−35、ABCGI36,37、およびこれらのうちで特性決定が最も良くなされたもの、即ち、非エステル化コレステロールおよびリン脂質を貧脂質アポリポタンパク質(主にアポA−I)へ移し、HDL粒子を形成する、ABCA1と称するATP結合カセット(ABC)輸送体30,38−44が含まれる。この過程に対するSIRT1の効果を試験するために、アポA−I媒介コレステロール流出を、[3H]コレステロールで細胞を標識した異なる2種の培養細胞系で測定し、細胞外培地中にあるアポA−Iへの標識コレステロールの排出をモニターした。
【0158】
MEFについては、コレステロール流出アッセイを、改良を加えたが記載してあるように72行った。初代SIRT1−/−および同腹子対照SIRT1+/+またはSIRT1+/−のMEFを、12ウェルプレート中に集密度50%で播種し、終夜培養した。次いで、細胞をRPMI1640/0.2%ウシ血清アルブミン(BSA)中に24時間移し、LDL50μg/mlおよび[3H]コレステロール1μCi/ml(NEN Life Science Products,Boston,Ma.)と更に24時間インキュベーションすることにより、細胞にコレステロールを担持させた。細胞をPBSで洗浄し、RPMI1640/0.2%BSA中で4時間平衡化し、次いでRPMI1640/0.2%BSA中、15μg/mlアポA−Iと共に、またはそれ無しで24時間インキュベートした。次いで培地を採集し、細胞を0.2N NaOHで溶解した。培地および細胞溶解物中に回収された放射活性を測定し、アポAI媒介コレステロール流出を、アポAI非含有培地中の非特異的排出量を差し引いた全放射活性に対する、培地中に回収した放射活性の比率(%)として計算した。コレステロール流出アッセイは、SIRT1−/−および同腹子3匹からの対照SIRT1+/+またはSIRT1+/−のMEFを7対用いて、2重に行った。
【0159】
初代MEFを用いて、[3H]コレステロールを担持した、野生型対照細胞およびSIRT1−/−細胞を、15μg/mlアポAIを含む、または含まないRPMI1640培地と共にインキュベートした。細胞および培地中の[3H]コレステロール量を24時間後に測定し、アポAI媒介コレステロール流出を計算した。図1g(左パネル)は、対照SIRT1+/+の初代MEFからの排出(塗潰し棒)が、SIRT1−/−のMEFからの排出(白抜き棒)よりほぼ3倍高かったことを示している(n=7;****、p<0.001)。
【0160】
ヒト単球細胞系THP−1細胞については、細胞をRPMI1640/10%FBS中、10mMニコチンアミドで48時間処理した後、RPMI1640/1%FBS/0.2%BSA培地中、LDL50μg/mlおよび[3H]コレステロール1μCi/mlで24時間、細胞にコレステロールを担持させた。細胞をPBSで洗浄し、RPMI1640/0.2%BSA中、15μg/mlアポA−Iと共に、またはそれ無しで24時間インキュベートした。次いで、培地と細胞とを分離し、細胞を0.2N NaOHで溶解した。培地および細胞溶解物中に回収された放射活性を測定し、アポAI媒介コレステロール流出を上記の通り計算した。
【0161】
SIRT1阻害剤のニコチンアミド(Nico)45によるTHP−1の処理も、[3H]コレステロール流出を大まかに1/3に抑制した(図1g、右パネル)。この実験は、三重に行い、2回繰り返した(p<0.01)。したがって、MEF中でのSIRT1の喪失、および単球中でのSIRT1活性の阻害は、アポAI媒介コレステロール流出を低下させる。こうした結果は、コレステロール流出の低下が、SIRT1−/−マウスにおいて、血漿HDLコレステロールの低下および組織コレステロールの増加を起こすのではないかという可能性を支持している。
【0162】
(実施例4)
野生型およびSIRT1ノックアウト組織におけるABCA1 mRNAの量
コレステロール流出の低下が、SIRT1−/−マウスにおいて、血漿HDLコレステロールの低下および組織コレステロールの増加を起こし得るか否かを調べるために、肝臓、精巣および卵巣におけるABCA1 mRNAの量を比較した。
【0163】
Qiagen RNeasyミニキット(Qiagen Inc.,Calencia,Ca)により、マウスの肝臓、精巣、チオグリコール酸賦活腹腔マクロファージおよび卵巣から総RNAを分離した。リアルタイムPCR分析のために、ランダムプライマーと共にSuperScript III逆転写酵素(Invitrogen,Carlsbad,Ca)によってcDNAを合成した。次いで、生成cDNAに、Cybergreen(Qiagen Inc.,Calencia,Ca)の存在下、遺伝子特異的プライマーを用いてPCR分析を行った。mRNAの相対的存在量を、シクロフィリン量へ規格化することによって得た。ノーザンハイブリッド形成のために、標準的手順に従って、RNA(10μg/レーン)をホルムアルデヒド−アガロースゲル(1.0%)上で分離し、GeneScreen Plus膜(NEN Life Science Products,Boston,Ma.)に移し、ハイブリッド形成した。ABCA1のmRNA量をホスホイメージ装置(phosphoimager)で定量し、相当するアクチン量に対して規格化した。
【0164】
ABCA1 mRNA発現量が、野生型と比較して、SIRT1−/−マウスでは3種の試験組織において40〜50%低下した(図2a)。この低下が、血漿HDL量の低下の原因になったのではないかという発想と合致して、ABCA1+/−ヘテロ接合型マウスでは、HDLコレステロールが、SIRT1−/−マウスにおける低下同様に、ABCA1+/+対照の約60%に低下している。
【0165】
(実施例5)
ABCA1プロモーターの活性
ABCA1の転写は、細胞コレステロールの増加によって誘発され、LXR核内受容体転写因子がこの応答を媒介する47−49。LXR/レチノイドX受容体(RXR)ヘテロ二量体は、オキシステロールにより活性化され50、全身コレステロールの恒常性、脂質生合成、炎症反応、炭水化物代謝および肥満の調節に重要な役割を果たす51。ABCA1の発現に対するSIRT1の効果が、LXRにより媒介されるか否かを決定するために、ホタルルシフェラーゼ遺伝子の発現を、マウスABCA1プロモーター、またはヌクレオチド8個の欠失でLXR応答エレメント(LXRE)を除去した対照変異体プロモーターのいずれかで推進する、レポーター構築体を生成した。以前の研究によれば、22(R)−ヒドロキシコレステロール(22(R)−HC、LXRアゴニスト)および9−シスレチノイン酸(9−シスRA、RXRアゴニスト)の併用で、ABCA1プロモーター中のLXREの活性が劇的に刺激されることが示されている47。
【0166】
ルシフェラーゼレポータープラスミドのpGL3−ABCA1を、pGL3ベクター(Promega,Madison,Wi)のKpnI−XhoI部位中にマウスABCA1プロモーター領域の−847〜+244を挿入することにより、創製した。pGL3−ABCA1のLXRE変異体型であるpGL3−ABCA1−LXREを、マウスABCA1の野生型プロモーターからLXREを欠失させることにより、創製した。
【0167】
ルシフェラーゼアッセイについては、24ウェルプレート中に播種したMEFIまたはHEK293T細胞に、pGL3−ABCA1またはpGL3−ABCA1−LXREを100ng、およびpRL−TK(ウミシイタケ(Renilla)ルシフェラーゼ、Promega)を10ng移入した。野生型(WT)またはLXRE変異体(−LXRE)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを移入した細胞を、クラスIおよびIIデアセチラーゼの阻害剤であるTrichostatin A(TSA)1μM、もしくはニコチンアミド(Nico)10mM、または両者のニコチンアミド10mM+TSA1μMで処理した。LXRE変異体ベクターを移入した細胞を、10%FBS入りDMEM中、22(R)−HCを10μMおよび9−シスRAを1μMの不在下(塗潰し棒)、または存在下(白抜き棒)でインキュベートした(図2b)。次いで、Dual−Luciferase Reporter Assay System(Promega)を用いて、6時間または24時間後にルシフェラーゼ活性を測定した。最終的なGL(ホタルルシフェラーゼ)活性は、同時発現したRL(ウミシイタケルシフェラーゼ)活性を用いて規格化した。実験は、二重に行い、3回繰り返した。
【0168】
野生型および変異体レポーター構築体をHEK293T細胞中に発現させた(図2b、WT(左パネル)または−LXRE(右パネル))。野生型からのルシフェラーゼ発現は、22(R)−HC+9−シスRAでの処理により8倍賦活された(図2b、左パネル)。その上、この誘導発現は、SIRT1(a)などのクラスIIIデアセチラーゼを阻害する阻害剤ニコチンアミドにより、50%抑制されたが、TSAでは抑制されなかった。LXREがない構築体は、LXRアゴニストに応答せず、低い基準活性を示した(図2b、右パネル)。纏めると、こうした知見は、他クラスのデアセチラーゼではなく、SIRT1が、注目すべきことに正の調節剤として、ABCA1プロモーターにおけるLXR活性を調節することを示唆している。
【0169】
それに加え、こうした実験に使用される系が、候補物質によるLXR依存性プロセスの調節を評価する方法を与えることが分かる。
【0170】
(実施例6)
ABCA1プロモーターに対するSIRT1活性の効果
ABCA1プロモーターに対するSIRT1の影響を直接決定するために、これらのプロモーターレポーター構築体を、SIRT1タンパク質を発現するようにpBabe−mT1ベクターを感染させた、または感染させていない、不死化したSIRT1+/+およびSIRT1−/−MEF中に導入した(図2c)。SIRT1+/+およびSIRT1−/−MEFを生成し、それらに、実施例3に記載のように、pBabe−SIRT1(+)またはpBabe対照ベクター(−)を感染させた。
【0171】
図2c、右パネルの差込図は、これら4種の細胞系におけるSIRT1の発現量に関する免疫ブロットの結果を示し、チューブリンをローディング対照として使用した。このベクターからのSIRT1の発現は、SIRT1+/+MEFにおける内因性SIRT1よりやや低かった。
【0172】
次いで、MEFに、野生型(WT)またはLXRE変異体(−LXRE)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを、22(R)−HCを10μMおよび9−シスRAを1μMの不在下(塗潰し棒)、または存在下(白抜き棒)で移入した。ルシフェラーゼ活性を24時間後に測定した。野生型プロモーターを有する全ての細胞は、LXR/RXR活性化剤で処理した際、ルシフェラーゼ発現の顕著な賦活を示した(図2c、左パネル)が、対照変異体(−LXRE)レポーターを有する細胞の低い基準活性は、これらの活性化剤で賦活されなかった(図2c、右パネル)。最も重要なことは、野生型プロモーター構築体のLXRE依存性活性が、SIRT1欠損性の細胞(SIRT1−/−MEF)では、SIRT1+/+細胞またはSIRT1で再構成された細胞(pBabe−T1)中より、40%低かった(p<0.02)ことである。こうした知見は、SIRT1が、LXREにおいて作用するLXR/RXRを介してABCA1プロモーターを刺激するというモデルを支持している。
【0173】
この実施例における実験系は、LXRのSIRT1依存性作用に対する候補物質の調節作用を評価する方法を与える。
【0174】
(実施例7)
SIRT1とABCA1プロモーターのLXREとの相互作用
SIRT1が、ABCA1プロモーター中のLXREに直接結合するか否かを決定するために、発明者らは、クロマチン免疫沈降(ChiP)アッセイを用い、マウスABCA1プロモーターを様々なプライマーで捜し求めた。
【0175】
ChiP分析は、改良を加えたが記載してあるように76行った。手短に言えば、細胞を培地中の1%パラホルムアルデヒドで15分間、室温で架橋し、0.125Mグリシンで架橋を終結させた。次いで、細胞をHEPES緩衝液(50mM HEPES−NaOH、pH7.9、140mM NaCl、1mM EDTA、10%グリセロール、0.5% NP40、0.25% Triton X−100、およびComplete(商標)プロテアーゼインヒビター混合物(Roche,Indianapolis,In))中に採集し、核を精製した。精製核をTE緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、およびComplete(商標)プロテアーゼインヒビター混合物)中に再懸濁し、氷上で超音波処理して平均長1kbにした。超音波処理上清をIP緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、10%グリセロール、1% Triton X−100、0.1%デオキシコール酸Na、およびComplete(商標)プロテアーゼインヒビター混合物)中に希釈し、プロテインAビーズで2回予備浄化した。次いで、予備浄化クロマチン試料を抗SIRT1ウサギポリクローナル抗体(Upstate,Charlottesville,Va)と4℃で終夜インキュベートし、プロテインAビーズを2時間添加することにより、免疫複合体を回収した。次いで、回収した免疫沈降物をIP緩衝液で2回、500mM NaCl含有IP緩衝液で2回、RIPA緩衝液(10mM Tris−HCl、pH8.0、1mM EDTA、250mM NaCl、0.5% NP40、0.5%デオキシコール酸Na、およびComplete(商標)プロテアーゼインヒビター混合物)で2回洗浄した。次いで、免疫沈降物中のクロマチンを10×TE/1%SDSで65℃、15分間溶出した。溶出クロマチンを65℃で少なくとも6時間インキュベートして、架橋を無効にし、プロテイナーゼKおよびRNアーゼAで処理し、フェノール/クロロホルムおよびクロロホルムで抽出した。最終的なDNA断片をLiCl/グリコーゲン/エタノール沈降により回収し、ペレットを水中に再懸濁し、PCRでアッセイした。
【0176】
マウスABCA1プロモーターのLXREについては、下記のプライマーを用いてPCR反応を行った。
【0177】
【化14】
前記ChiPアッセイの陰性対照として、ABCA1プロモーター上のLXREの上流にある6.2kb断片も、下記のプライマーで増幅した。
【0178】
【化15】
ヒトプロモーターのLXREについては、下記のプライマーを用いてPCR反応を行った。
【0179】
【化16】
陰性対照として、LXREの上流にある7.5kb断片を下記のプライマーで増幅した。
【0180】
【化17】
実験は少なくとも3回行ったが、その代表的データを示してある。
【0181】
図2d上部は、SIRT1に対する抗体で免疫沈降する前(入力)または後(αSIRT1)で、LXRE特異的または上流特異的なPCRプローブで探った結果を示す。実験処理において、(αSIRT1)抗体処理により、SIRT1+/+MEF(左レーン、暗灰色バンドを示す上部像)からのLXREを含有するクロマチンの断片は、沈降したが、SIRT1−/−MEF対照(右レーン、地黒を示す上部像)では沈降せず、LXREの上流にある6.2kbDNAを含有する断片は、いずれの細胞系でも沈降しなかった(地黒を示す両レーンの下部像)。FLAGエピトープタグ付きヒトSIRT1の発現ベクターまたは陰性対照ベクターを移入した、HEK293T(図2d、下部)およびTHP−1(データは示していない)ヒト細胞においても、同様な結果が見出された。したがって、SIRT1は、ABCA1プロモーターと、LXREと直ぐの近接部で結合する。
【0182】
(実施例8)
SIRT1のLXRαおよびLXRβとの相互作用
SIRT1は、DNAと直接には結合しない26ため、ABCA1プロモーター上のその存在には、DNAと会合するLXRなどのタンパク質に結合することが必要である。LXR転写因子ファミリーには2種のメンバー、LXRαおよびLXRβがある50,52。LXRαの発現は組織特異的であるが、LXRβの方は遍在的である。SIRT1とLXRとの会合があるか否かを共免疫沈降により決定するために、LXRαおよびLXRβのマウスcDNAをHAタグ付き発現ベクター中にクローニングした。次いで、SIRT1発現ベクターpBabe−SIRT1を、そのHAタグ付きLXR用ベクターと共に、またはそれ無しでHEK293T細胞中に移入した。
【0183】
この実施例では、SIRT1とLXRαまたはLXRβとの相互作用の可能性を調べた。NH2末端HAタグ付きLXRα(HA−LXRα)およびLXRβ(HA−LXRβ)を発現するプラスミドを、修飾pcDNA3−NHAベクター中のHAエピトープの下流に(それとインフレームで)、LXRαまたはLXRβの全長マウスcDNAをクローニングすることにより、創製した。マウスSIRT1を過剰発現するHEK293T細胞(MmSIRT1)およびHA−LXRα、またはMmSIRT1およびHA−LXRβを、NP40緩衝液(10mM Tris−HCl、pH8.0、150mM NaCl、0.5% NP40、およびComplete(商標)プロテアーゼインヒビター混合物)中に採集し、18ゲージシリンジ針の中を10回通過させることにより、溶解した。4℃で1時間揺動させた後、溶解物を16000×gで15分間遠心分離した。次いで、生成上清を1%BSA入りNP40緩衝液中に希釈し、抗HAモノクローナル抗体(Santa Cruz Biotechnology)3μgの添加により、免疫沈降を開始した。次いで、混合物を4℃で2時間から終夜の間、揺動させながらインキュベートした。更に1時間プロテインAビーズの添加により、免疫複合体を回収し、免疫沈降物をNP40緩衝液1mlで5回洗浄した。次いで、全溶解物および対応する免疫沈降試料をSDS−PAGEで分離し、抗SIRT1ポリクローナル抗体および抗mycまたは抗HA抗体を用いて免疫ブロットした。
【0184】
モノクローナルマウス抗LXRα抗体は、R&D system製であり、ポリクローナルヤギ抗LXRβ抗体は、Santa Cruz Biotechnology製であった。指示したタンパク質を発現するヒトHEK293T細胞を溶解し、上記のように抗HAモノクローナル抗体で免疫沈降(IP)させた(図2e)。次いで、全溶解物(レーン1〜4)および抗HA免疫沈降試料(レーン5〜8)を、SDS−PAGEで分離し、抗SIRT1ポリクローナル抗体(上端像)および抗HA−LXRα抗体(上部パネル)または抗HA−LXRβ抗体(下部パネル)を用いて探査した。図2eに示すように、移入したマウスSIRT1タンパク質(mT1)は、タグ付きLXRαまたはLXRβが細胞中に同時発現した場合だけ、抗HA抗体により交差沈降した(レーン8、上部および下部パネル)。HA−LXRは、ゲル中でやや低い移動度を示す内因性ヒトSIRT1タンパク質(hT1)も共沈降させた。したがって、SIRT1のLXRとの直接または間接結合が、恐らく、未処置細胞中でのABCA1プロモーターのLXREとSIRT1が会合する原因である。
【0185】
更に、この実施例に記載した免疫沈降実験は、SIRT1のLXRαまたはLXRβとの相互作用を試験物質が調節する能力のための感度良いアッセイを与える。
【0186】
(実施例9)
SIRT1−/−マウスの肝臓におけるLXR標的mRNA
LXRとSIRT1との相互作用のために、SIRT1が機能喪失すると、食事コレステロールに応答するLXR媒介脂質恒常性が損なわれ得るという可能性が持ち上がる。この仮説を試験するために、ABCA1を含めた一連のLXR標的遺伝子のmRNA量を、SIRT1−/−マウスの肝臓(図3a)において、野生型対照動物(塗潰し棒)およびSIRT1−/−動物(白抜き棒)の肝臓からの総mRNAを用いて分析した。分析は、リアルタイムPCRで行った。LXRのmRNA量が、SIRT1−/−肝臓においてたとえ正常であっても、SREBP1、ABCA1、ABCG1、ABCG5、LDLRなどの多くのLXR標的は、SIRT1+/+対照に比較して減少していた(n=3、*、P<0.05)。LXR標的ではないことが知られている対照遺伝子53,54のmRNA量は、SIRT−/−マウスにおいて有意に変化しなかった。
【0187】
(実施例10)
LXRアゴニストT0901317で処理したマウスの肝臓における代謝物質の相対量
インビボでのLXR機能に対するSIRT1の効果を更に洞察するために、野生型およびSIRT1−/−動物を、LXRアゴニストT0901317(N−(2,2,2−トリフルオロエチル)−N−[4−(2,2,2−トリフルオロ−1−ヒドロキシ−1−トリフルオロメチルエチル)フェニル]ベンゼンスルホンアミド)53の8日経口投与に対する応答について、分析した。
【0188】
【化18】
マウス組織中のABCA1タンパク質を検出するために、SIRT1+/+またはSIRT1−/−肝臓をRIPA緩衝液中で均質化した。次いで、肝臓抽出物をSDS試料緩衝液と混合し、37℃で20分間変性することにより、高温でのABCA1の凝集を防止した。次いで、試料を7.5%SDS−PAGE中に分離し、抗ABCA1ポリクローナル抗体(R80抗体、M.FitzgeraldおよびM.Freeman両博士からの贈呈品)を用いてブロットした。
【0189】
SIRT1−/−および対象の2〜6月齢雄に、経口による強制補給でT0901317(10mg/kg)を供給し、血漿を供給の前後に採集して、血漿脂質分析用とした。組織を供給後に採集して、更なる分析用とした。T0901317(10mg/kg)を供給していない(黒色塗潰し棒)、または供給した(黒色斜線付き棒)野生型動物、およびT0901317を供給していない(白抜き棒)、または供給した(灰色斜線付き棒)SIRT1−/−マウスの肝臓からの総RNAを、リアルタイムPCRで分析した。野生型動物へのT0901317供給は、数種のLXR標的:SREBP−1c、LPLおよびABCA1のRNA段階での発現を有意に誘発した(図3b)(n=4、**、p<0.05)。しかし、SIRT1−/−マウスにおけるこれらの標的の誘発は、鈍らせられた。総タンパク質溶解物を指示したマウスの肝臓から取得し、免疫ブロットによりそのABCA1タンパク質量を分析した(図3c)(以前に観察したように、ABCA1タンパク質は、ウェスタンブロットで多重のバンドを含む78)。
【0190】
図3のパネルd、eおよびfにおける血漿の結果は、T0901317を8日投与した前後の同じマウスを表わす。図3のパネルdにおける肝臓の結果は、図3の上のパネルb(実施例9)およびc(本実施例)の場合のように、T0901317を供給していない、または供給した同腹子を比較している。T0901317を供給していない(黒色塗潰し棒)、または供給した(左斜線付き棒)野生型マウス、およびT0901317を供給していない(白抜き棒)、または供給した(右斜線付き棒)SIRT1−/−動物の、血漿(図3d、左パネル)および肝臓(図3d、右パネル)からのトリグリセリドを分析した。血漿および肝臓のトリグリセリド蓄積のLXRアゴニストT0901317による誘発53に対する、SIRT1−/−動物における応答の減少が認められた(図3d;n=5、***、p<0.001)。処置マウスにおける血漿コレステロール濃度の低下(図3e;n=5、****、p<0.02)も、FPLCリポタンパク質画分のコレステロールアッセイで決定した場合の特にHDL粒子において見られた(図3f;n=5)。T0901317は、SREBP−1cの誘導により血漿および肝臓のトリグリセリドを増加させる53ので、SIRT1−/−マウスにおけるSREBP−1c mRNAの少ない量(図3a)が、T0901317が誘発するトリグリセリドの低下を説明し得る。したがって、LXRに依存するレポーターアッセイにおける、ABCA1プロモーターに対するSIRT1の効果だけでなく、LXRおよびそのアゴニストのインビボでの機能も、SIRT1に依存していた。
【0191】
(実施例11)
LXRタンパク質発現に対するSIRT1の効果
SIRT1は、ヒストン12、p53 15〜17、FOXO転写因子18、19、およびPGC−1α 23、24などの多くの標的タンパク質と直接、相互作用し、それらを脱アセチル化する。SIRT1によるこうした標的の脱アセチル化は、その転写活性を抑制(一部のプロモーターにおけるp53およびFOXO)または活性化(他のプロモーターにおけるPGC−1αおよびFOXO)する。SIRT1が、LXRに対してどのように正の調節をするのか調べるために、SIRT1+/+およびSIRT1−/−マウスの肝臓内の内因性LXRタンパク質量を最初に決定した。LXRの標的活性化能が、野生型と比較して低下していた(実施例9;図3a)とはいえ、SIRT1−/−動物においてLXRタンパク質量が激増していた(図4a)。
【0192】
外因性CMVプロモーターで推進されるHA−LXRβを発現するHEK293T細胞に、pSuperベクター、pSuper−SIRT1 RNAi構築体のいずれかを移入した。トランフェクションから3日後、細胞を採集し、HAおよびSIRT1に対する抗体を用いたウェスタンブロットで、HA−LXRβおよびSIRT1の量を分析した。SIRT1発現のRNAi媒介ノックダウン(図4b、レーン3および4)、ニコチンアミドによるSIRT1活性の抑制−しかし、TSAによる他のデアセチラーゼの阻害はなかった−(図4c)の双方が、HA−LXRβタンパク質量を増加させた。SIRT1−/−マウスではLXR mRNAの低下が検出されなかった(図3a)ので、SIRT1は転写後にLXRタンパク質量を調節する。
【0193】
HA−LXRβを発現するHEK293T細胞を、対照培地(図3c、レーン1)、または1μM TSA(レーン2)、10mMニコチンアミド(レーン3)、25μM MG132(カルボベンジルオキシ−Leu−Leu−Leu−al)(レーン4)、もしくは10mMニコチンアミド+25μM MG132(レーン5)を含む培地のいずれかで、6時間処理した。次いで、細胞を採集し、HAおよびSIRT1に対する抗体を用いたウェスタンブロットで、HA−LXRβ(上部像)およびSIRT1(中央像)の量を分析した。ニコチンアミドによるSIRT1活性の抑制(レーン3)−しかし、TSAによる他のデアセチラーゼの阻害はなかった(レーン2)−によって、HA−LXRβタンパク質量が増加した(上部像)。SIRT1−/−マウスではLXR mRNAの低下が検出されなかった(実施例9、図3aを参照されたい)ので、SIRT1は転写後にLXRタンパク質量を調節する。ニコチンアミドによるSIRT1活性の抑制は、プロテアソーム媒介LXR分解を妨害することにより、HA−LXRのタンパク質量を増加させると考えられる。
【0194】
次に、LXR活性がタンパク質量と逆相関しているという知見の根拠を調べた。リシン残基のアセチル化はユビキチン化を遮断することができ、SIRT1はデアセチラーゼであるので、SIRT1の活性喪失は、LXRアセチル化の定常状態レベルを増加させ、したがってユビキチン/プロテアソーム分解経路を抑制するのではないかとも思われる。これが事実であるとすれば、MG132によるプロテアソーム活性の抑制は、SIRT1が活性である場合はLXRの定常状態レベルを増加させ、SIRT1がニコチンアミドにより阻害される場合はそのレベルを増加させないはずである。事実、これは、HAタグ付きLXRβを発現するHEK293T細胞において認められた(図4c、レーン4対レーン5)。
【0195】
HEK293T細胞に、空pBabeベクター、HA−LXRβ、pBabe−SIRT1、およびpBapbe−hSIRT1HY(不活性デアセチラーゼであるSIRT1変異体(Vaziriら、2001年を参照されたい))を発現する各構築体を同時移入した。40時間後、細胞を25μM MG132と共に1時間インキュベートし、採集した。次いで、HAに対する抗体を用いて、HA−LXRβを免疫沈降させた後、ユビキチン、HAおよびSIRT1に対する抗体を用いたウェスタンブロットを施した。その結果から、不活性変異体のhSIRT1HYではなく、活性SIRT1トランス遺伝子(hSIRT1)の発現が、移入されたHEK293T細胞中でユビキチン結合LXRの量を増加させたことが示される(図4d、レーン3対レーン4)。
【0196】
(実施例12)
インビボでのLXRのアセチル化
インビボでのLXRのアセチル化、およびSIRT1によるLXRのデアセチル化を検出するために、HEK293T細胞に、HA−LXRβと、pBabe、pBabe−HsSIRT1、pBabe−HsSIRT1H355Yのいずれかとを同時移入し、改良を加えたが記載してあるように77、[3H]酢酸ナトリウムを含有する培地中で培養した。手短に言うと、トランスフェクションから40時間後、2mCi/ml[3H]酢酸ナトリウム、プロテアソームインヒビターMG132(25μM)、22(R)−HC(10μM)、および9−シス−RA(1μM)を培養物に1時間添加した。次いで、細胞を冷リン酸緩衝塩水で2回洗浄し、NP40緩衝液中で溶解した。溶解物を16000×gで4℃、15分間遠心分離した。次いで、HA−LXRβを抗HA抗体で免疫沈降させ、SDS−PAGEで分離した。[3H]酢酸標識HA−LXRβを含有するゲルを、クーマシーブリリャントブルーで先ず染色し、次いで市販のフルオログラフィー増強溶液(Amplify,Amersham Biosciences)で30分間浸漬することにより、増強した。乾燥したゲルを−70℃、3〜7日間オートラジオグラフィーに掛けた。同じ試料に抗SIRT1抗体で免疫ブロットも行った。
【0197】
その結果を図4eに示してある。HA−LXRβは、通常の培養条件下でアセチル化された(レーン1)。更に、22(R)−HC+9−シスRAでの処理によるLXRβの活性化が、脱アセチル化を賦活した(および/または、多分アセチル化を抑制した。レーン2)。図4fに示すように、[3H]酢酸標識HA−LXRの量が、空ベクターを移入した対照細胞(pBabe;レーン1)、または酵素不活性SIRT1(pBabe−hSIRT1HY;レーン3)より、hSIRT1を発現する細胞(pBabe−hSIRT1;レーン2)において実質的に少なかったので、SIRT1は、少なくとも部分的にはLXRの脱アセチル化の原因に多分なっているようである。こうした知見は、SIRT1によるLXRβの脱アセチル化が、その代謝回転率を促進し、その活性も増加させることを示唆している。
【0198】
(実施例13)
SIRT1媒介またはプロテアソーム媒介LXR分解の抑制が、ABCA1プロモーター上のLXRの転写活性に及ぼす効果
HEK293T細胞に、野生型(WT;図4g、左パネル)またはLXRE変異体(−LXRE;図4g、右パネル)マウスABCA1プロモーターに推進されるルシフェラーゼレポーターベクターを移入し、10μM 22(R)−HCおよび1μM 9−シスRAの不在下(塗潰し棒)または存在下(白抜き棒)、10mMニコチンアミド(Nico)、25μM MG132、10mMニコチンアミド+25μM MG132の両方のいずれかで処理した。実施例5の場合のように、ルシフェラーゼ活性を測定し、規格化した。
【0199】
ニコチンアミドは、ABCA1プロモーターに推進されるルシフェラーゼレポーターの誘導を有意に鈍らせることが判明した(図4g、左パネル)。その上、プロテアソームインヒビターのMG132も、ニコチンアミドと同程度に該レポーターを阻害したが、ニコチンアミドとMG132とを併用しても更に阻害することはなかった。LXREを失っている対照レポーターは、ニコチンアミドまたはMG132で変化しない基準活性を示した(図4g、右パネル)。こうした知見は、以下に考察するように、SIRT1によるLXRβの脱アセチル化、ならびにプロテアソームによる、後続するそのユビキチン化および分解が、ABCA1プロモーターの活性化を補助することを示唆している。
【0200】
以上の実施例は、SIRT1がLXR転写因子を正に調節し、したがってコレステロール恒常性の調節に重要な役割を果たしていることを示す。LXRおよびSIRT1は、ABCA1輸送体をコードする遺伝子の転写を活性化し、その輸送体が、コレステロール逆輸送およびHDL合成を媒介する。遺伝子および疫学研究によれば、血漿HDL濃度は、心血管疾患30および恐らくはアルツハイマー病5,6の危険性と逆関連することが示されているので、こうした知見はヒトの医療に直接関係し得る。コレステロールのHDL媒介逆輸送は、動脈細胞から過剰のコレステロールを除去することにより、アテローム性動脈硬化症から防御し55,56、脳内の富コレステロールラフト形成を減少させることにより、アルツハイマー病から防御し得る57,58。LXRの活性化によって、SIRT1は、老化に付随する心血管疾患および恐らくはアルツハイマー病の危険性を減少させ得る。
【0201】
SIRT1とLXRとの物理的および機能的相互作用は、SIRT1が、LXRの調節によって、インビボでその効果の少なくとも幾つかを発揮することを示唆している。実際に、SIRT1欠乏マウスは、HDLのコレステロール量(図1)およびトリグリセリド量(図3d)の減少などの多くの代謝欠陥を、LXR欠乏マウスと共有している53,54,63。また、肝LXRアゴニストにより抑制できる64糖新生の律速酵素であるPEPCKは、SIRT1−/−マウスにおいて誘導される19。LXR依存性の遺伝子発現およびエネルギー恒常性は、微生物に対する先天性免疫応答にとって重要であることが最近示されており、LXRを失っているマウスは、細菌感染を非常に受け易い65。したがって、肺感染症26および眼の感染症(未公表の観察)に対する、SIRT1ノックアウト動物の観察された感受性は、こうした動物における鈍ったLXR活性に因ることも考えられる。
【0202】
本知見は、SIRT1が、LXRの脱アセチル化およびその代謝回転の生起によって、LXR活性を促進することを示唆する(図5)。SIRT1によるLXRの調節により、このサーチュインがコレステロール恒常性経路の中心に位置している。
【0203】
参考文献
【0204】
【化19】
【0205】
【化20】
【0206】
【化21】
【0207】
【化22】
【0208】
【化23】
【0209】
【化24】
【0210】
【化25】
【特許請求の範囲】
【請求項1】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む、単離された複合体。
【請求項2】
LXR応答エレメントを更に含む、請求項1に記載の複合体。
【請求項3】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体を形成する方法であって、該方法は、哺乳類SIRT1タンパク質を含む第1の組成物と、哺乳類LXRタンパク質を含む第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む第3の組成物とを組み合わせる工程を含む、方法。
【請求項4】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を調節する、作用剤を同定する方法であって、該方法は:
a)哺乳類SIRT1タンパク質を含む第1の組成物と、哺乳類LXRタンパク質を含む第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む第3の組成物とを組み合わせて、複合体組成物を提供する工程、
b)更に、
i)該組合せ工程の前に、該第1の組成物、該第2の組成物、もしくは該第3の組成物を、該作用剤を含む第4の組成物と接触させるか、または
ii)該組合せ工程の後に、該複合体組成物を、該作用剤を含む該第4の組成物と接触させ、
それにより試験組成物を提供する工程、および
c)該複合体の形成が該作用剤により調節されるか否かを決定する工程
を含む方法。
【請求項5】
前記作用剤が前記複合体の形成を増加させる、請求項4に記載の方法。
【請求項6】
前記決定する工程が、
d)哺乳類SIRT1タンパク質を含む前記第1の組成物と、哺乳類LXRタンパク質を含む前記第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む前記第3の組成物とを組み合わせて、前記作用剤を含まない対照組成物を提供する工程、および
e)該試験組成物における前記複合体の形成が、該対照組成物における該複合体の形成と比較して、調節されるか否かを決定する工程
を更に含む、請求項4に記載の方法。
【請求項7】
作用剤を哺乳動物に投与することにより、該哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、SIRT1活性を刺激する該作用剤の使用。
【請求項8】
前記哺乳動物がヒトである、請求項7に記載の使用。
【請求項9】
前記作用剤がT0901317を含む、請求項7に記載の使用。
【請求項10】
作用剤を哺乳動物に投与することにより、該哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤の使用。
【請求項11】
前記哺乳動物がヒトである、請求項10に記載の使用。
【請求項12】
前記作用剤が、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む、請求項10に記載の使用。
【請求項13】
哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含んだ核酸を該細胞に導入する工程を含む、方法。
【請求項14】
前記哺乳類細胞がヒト細胞である、請求項13に記載の方法。
【請求項15】
前記タンパク質デアセチラーゼが真核生物Sir2である、請求項13に記載の方法。
【請求項16】
前記タンパク質デアセチラーゼが哺乳類SIRT1である、請求項13に記載の方法。
【請求項17】
SIRT1活性を刺激する作用剤と、前記細胞を更に接触させる、請求項13に記載の方法。
【請求項18】
前記作用剤がT0901317を含む、請求項17に記載の方法。
【請求項19】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤と、前記細胞を更に接触させる、請求項13に記載の方法。
【請求項20】
前記作用剤が、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む、請求項19に記載の方法。
【請求項21】
候補物質がLXR依存性プロセスを調節するか否かを評価するための方法であって、該方法は、
a)LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターで細胞をトランスフェクトする工程、
b)該細胞を該候補物質と接触させる工程、および
c)該候補物質と接触していない細胞と比較して、該候補物質が該レポーター遺伝子の発現を調節するか否かを決定する工程
を含み、それによって、該候補の存在と不在との間で検出される該LXR依存性プロセスの発生の程度における差異は、該候補物質が該LXR依存性プロセスを調節することを示す、
方法。
【請求項22】
候補物質がLXRのSIRT1依存性作用を調節するか否かを評価するための方法であって、該方法は、
a)SIRT1遺伝子を保持するベクターで細胞をトランスフェクトする工程、
b)LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するベクターで該細胞をさらにトランスフェクトする工程、
c)該細胞を該候補と接触させる工程、および
d)該候補と接触していない細胞と比較して、該候補が該レポーター遺伝子の発現を調節するか否かを決定する工程、
を含み、それにより、該候補の存在と不在との間で検出される該LXRのSIRT1依存性作用の程度における差異は、該候補がLXRの該SIRT1依存性作用を調節することを示す、
方法。
【請求項23】
候補物質が、特異的結合対のメンバーのSIRT1およびLXRを含む特異的結合対の形成を調節するか否かを評価するための方法であって、該方法は、
a)該特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトする工程、
b)該特異的結合対の第2メンバーをコードする配列を保持するベクターで該細胞をさらにトランスフェクトする工程、
c)該細胞を該候補物質と接触させる工程、
d)該細胞を溶解し、該エピトープタグに特異的な抗体と細胞溶解物を接触させ、そして、該SIRT1と該LXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収する工程、
e)SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行する工程
を含み、それにより、該候補の不在下と比較した、該候補の存在下で検出した複合体形成の差異は、該候補が該SIRT1の該LXRとの相互作用を調節することを示す、方法。
【請求項1】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む、単離された複合体。
【請求項2】
LXR応答エレメントを更に含む、請求項1に記載の複合体。
【請求項3】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体を形成する方法であって、該方法は、哺乳類SIRT1タンパク質を含む第1の組成物と、哺乳類LXRタンパク質を含む第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む第3の組成物とを組み合わせる工程を含む、方法。
【請求項4】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を調節する、作用剤を同定する方法であって、該方法は:
a)哺乳類SIRT1タンパク質を含む第1の組成物と、哺乳類LXRタンパク質を含む第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む第3の組成物とを組み合わせて、複合体組成物を提供する工程、
b)更に、
i)該組合せ工程の前に、該第1の組成物、該第2の組成物、もしくは該第3の組成物を、該作用剤を含む第4の組成物と接触させるか、または
ii)該組合せ工程の後に、該複合体組成物を、該作用剤を含む該第4の組成物と接触させ、
それにより試験組成物を提供する工程、および
c)該複合体の形成が該作用剤により調節されるか否かを決定する工程
を含む方法。
【請求項5】
前記作用剤が前記複合体の形成を増加させる、請求項4に記載の方法。
【請求項6】
前記決定する工程が、
d)哺乳類SIRT1タンパク質を含む前記第1の組成物と、哺乳類LXRタンパク質を含む前記第2の組成物と、LXR応答エレメントを含む細胞の核酸の断片を含む前記第3の組成物とを組み合わせて、前記作用剤を含まない対照組成物を提供する工程、および
e)該試験組成物における前記複合体の形成が、該対照組成物における該複合体の形成と比較して、調節されるか否かを決定する工程
を更に含む、請求項4に記載の方法。
【請求項7】
作用剤を哺乳動物に投与することにより、該哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、SIRT1活性を刺激する該作用剤の使用。
【請求項8】
前記哺乳動物がヒトである、請求項7に記載の使用。
【請求項9】
前記作用剤がT0901317を含む、請求項7に記載の使用。
【請求項10】
作用剤を哺乳動物に投与することにより、該哺乳動物の血漿において、高密度リポタンパク質(HDL)に結合するコレステロールの総コレステロールに対する比率を増加させる薬剤の製造における、哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤の使用。
【請求項11】
前記哺乳動物がヒトである、請求項10に記載の使用。
【請求項12】
前記作用剤が、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む、請求項10に記載の使用。
【請求項13】
哺乳類細胞からのABCA1媒介コレステロール流出を促進する方法であって、タンパク質デアセチラーゼをコードする配列を含んだ核酸を該細胞に導入する工程を含む、方法。
【請求項14】
前記哺乳類細胞がヒト細胞である、請求項13に記載の方法。
【請求項15】
前記タンパク質デアセチラーゼが真核生物Sir2である、請求項13に記載の方法。
【請求項16】
前記タンパク質デアセチラーゼが哺乳類SIRT1である、請求項13に記載の方法。
【請求項17】
SIRT1活性を刺激する作用剤と、前記細胞を更に接触させる、請求項13に記載の方法。
【請求項18】
前記作用剤がT0901317を含む、請求項17に記載の方法。
【請求項19】
哺乳類SIRT1タンパク質および哺乳類LXRタンパク質を含む複合体の形成を促進する作用剤と、前記細胞を更に接触させる、請求項13に記載の方法。
【請求項20】
前記作用剤が、22(R)−ヒドロキシコレステロールもしくは9−シスレチノイン酸、またはその両方を含む、請求項19に記載の方法。
【請求項21】
候補物質がLXR依存性プロセスを調節するか否かを評価するための方法であって、該方法は、
a)LXREに作動可能に駆動されるレポーター遺伝子を保持するベクターで細胞をトランスフェクトする工程、
b)該細胞を該候補物質と接触させる工程、および
c)該候補物質と接触していない細胞と比較して、該候補物質が該レポーター遺伝子の発現を調節するか否かを決定する工程
を含み、それによって、該候補の存在と不在との間で検出される該LXR依存性プロセスの発生の程度における差異は、該候補物質が該LXR依存性プロセスを調節することを示す、
方法。
【請求項22】
候補物質がLXRのSIRT1依存性作用を調節するか否かを評価するための方法であって、該方法は、
a)SIRT1遺伝子を保持するベクターで細胞をトランスフェクトする工程、
b)LXREプロモーターに作動可能に駆動されるレポーター遺伝子を保持するベクターで該細胞をさらにトランスフェクトする工程、
c)該細胞を該候補と接触させる工程、および
d)該候補と接触していない細胞と比較して、該候補が該レポーター遺伝子の発現を調節するか否かを決定する工程、
を含み、それにより、該候補の存在と不在との間で検出される該LXRのSIRT1依存性作用の程度における差異は、該候補がLXRの該SIRT1依存性作用を調節することを示す、
方法。
【請求項23】
候補物質が、特異的結合対のメンバーのSIRT1およびLXRを含む特異的結合対の形成を調節するか否かを評価するための方法であって、該方法は、
a)該特異的結合対のエピトープタグ付き第1メンバーをコードする配列を保持するベクターで細胞をトランスフェクトする工程、
b)該特異的結合対の第2メンバーをコードする配列を保持するベクターで該細胞をさらにトランスフェクトする工程、
c)該細胞を該候補物質と接触させる工程、
d)該細胞を溶解し、該エピトープタグに特異的な抗体と細胞溶解物を接触させ、そして、該SIRT1と該LXRとの複合体を含む免疫沈降物を抗体特異的吸着剤で回収する工程、
e)SIRT1およびLXRに特異的な抗体を用いてウェスタンブロット手順を実行する工程
を含み、それにより、該候補の不在下と比較した、該候補の存在下で検出した複合体形成の差異は、該候補が該SIRT1の該LXRとの相互作用を調節することを示す、方法。
【図1】

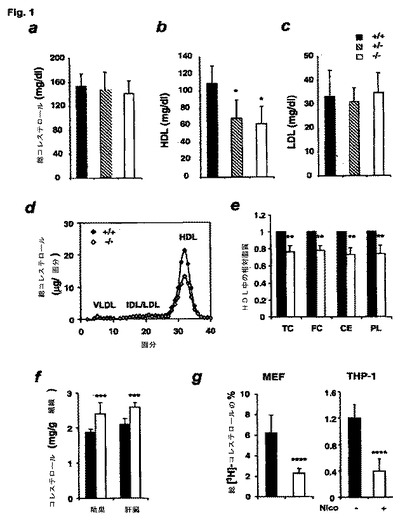
【図2】
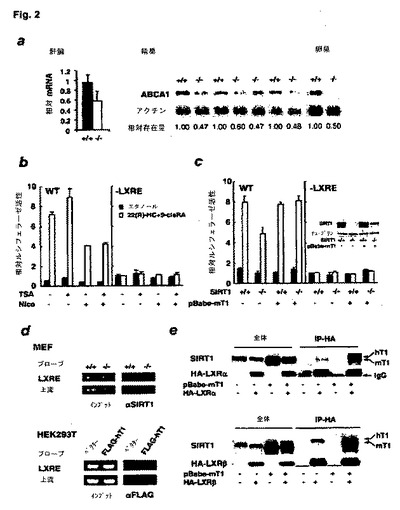
【図3】
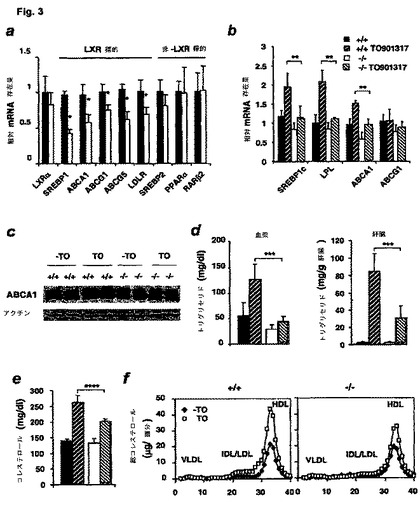
【図4】
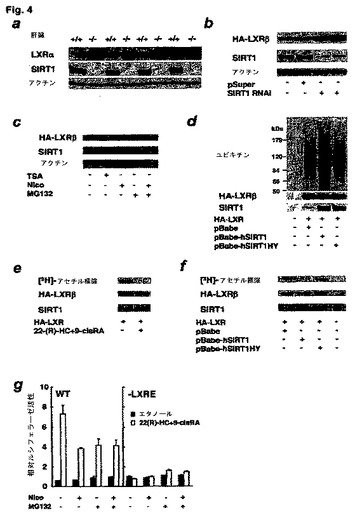
【図5】
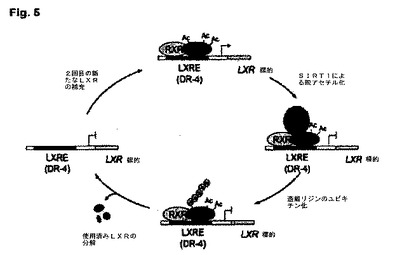
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2009−539844(P2009−539844A)
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願番号】特願2009−514364(P2009−514364)
【出願日】平成19年6月6日(2007.6.6)
【国際出願番号】PCT/US2007/013364
【国際公開番号】WO2007/146023
【国際公開日】平成19年12月21日(2007.12.21)
【出願人】(596060697)マサチューセッツ・インスティテュート・オブ・テクノロジー (233)
【Fターム(参考)】
【公表日】平成21年11月19日(2009.11.19)
【国際特許分類】
【出願日】平成19年6月6日(2007.6.6)
【国際出願番号】PCT/US2007/013364
【国際公開番号】WO2007/146023
【国際公開日】平成19年12月21日(2007.12.21)
【出願人】(596060697)マサチューセッツ・インスティテュート・オブ・テクノロジー (233)
【Fターム(参考)】
[ Back to top ]