
SMYD3によるVEGFR1のメチル化調整因子を同定する方法
本発明は、ポリペプチドのメチルトランスフェラーゼ活性を決定する方法及びメチルトランスフェラーゼ活性の調整剤、特に、SMYD3によるVEGFR1のメチル化の調整剤のスクリーニング方法に関する。本発明は、さらに、このように同定された調整剤を用いる結腸直腸癌、肝細胞癌、膀胱癌及び/または乳癌を治療及び予防するための方法及び薬学的組成物を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、転写制御、より具体的には、メチルトランスフェラーゼ活性を調整する因子、例えば、SMYD3(“ZNFN3A1”としても知られる)によるVEGFR1のメチル化を調整する因子の同定に関する。SMYD3が多くの癌種において上方制御されるので、そのように同定されるSMYD3モジュレーターは、例えば、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌を含む癌の治療に有用なことが判明する可能性がある。
【背景技術】
【0002】
ヒストン末端の修飾は、転写制御、テロメア維持、DNA複製及び染色体分離に重要な役割を果たす(Kouzarides T, Cell 2007, 128(4): 693-705)。このような修飾の例としては、アセチル化、リン酸化、メチル化、及び/又はユビキチン化が挙げられる。共有結合修飾は特に、クロマチン構造だけでなく、クロマチン結合タンパク質との相互作用も制御する(Kouzarides T, Cell 2007, 128(4): 693-705.; Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80)。さらに、H3K4、H3K36及びH3K79のメチル化は、ユークロマチン構造に関連し、一方、H3K9、H3K27及びH4K20のメチル化はヘテロクロマチン構造に関連する(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80)。
【0003】
クロマチンの構造は、転写の重要レギュレーターの一つである;非転写遺伝子はヘテロクロマチン内に小型化される一方、転写遺伝子はユークロマチン内に存在し、ユークロマチンでは、転写複合体は標的DNAにアクセス可能である(Li B et al., Cell 2007, 128(4): 707-19)。さらに、ヒストン残基の修飾は、その結合タンパク質との相互作用を促進し、他のヒストン末端へのその後の修飾に影響する(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80.; Li B et al., Cell 2007, 128(4): 707-19.; Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。例えば、H3セリン10(H3S10)のリン酸化は、H3K9のメチル化を抑制する。逆に、H3K9のメチル化は、H3S10のリン酸化に拮抗する。H3S10のリン酸化は、GCN5によるH3K14のアセチル化を促進する(Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。
【0004】
H3K9のメチル化は、HP1の輸送に関与して、染色体領域を明確化し、これは、一方で、ヘテロクロマチンのドメインの確立及び維持に不可欠である(Nakayama J et al., Science 2001, 292(5514): 110-3.; Lachner M et al., Nature 2001, 410(6824): 116-20.; Bannister AJ et al., Nature 2001, 410(6824): 120-4)。HP1タンパク質のゲノム特定部位への動員は、クロマチンの多成分との相互作用に関与する(Nielsen SJ et al., Nature 2001, 412(6846): 561-5)。この動員は、転写活性に不可欠なタンパク質であるH3K4のメチル化を抑制すると考えられている。これらのデータは、異なる修飾間の相互作用により制御されるヒストン修飾の複雑な性質を示している。実際、これらのタンパク質の増大が、そのヒストンメチルトランスフェラーゼ活性を通じて腫瘍形成を促進又は阻害することが証明されてきている(Varambally S et al., Nature 2002, 419(6907): 624-9;Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40;Gibbons RJ, Human molecular genetics 2005, 14 Spec No 1: R85-92)。
【0005】
ヒストン末端のメチル化は熱心に研究が進められてきたものの、非ヒストンタンパク質のメチル化は、不明なままである。最近の研究では、ヒストンH3K4メチルトランスフェラーゼ(MTase)であるSET7/9が、TAF10及びp53を基質として触媒することが報告されている(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82;Chuikov S et al., Nature 2004, 432(7015): 353-60)。血管内皮成長因子受容体−1(VEGFR1)(Accession No.:NM_002019)は、受容体二量体化及びそのリガンドとの相互作用の点で、生理的及び病理学的な血管形成において機能する、受容体チロシンキナーゼ(RTK)である(Shibuya M et al., Oncogene 1990, 5(4): 519-24.; Rahimi N, Experimental eye research 2006, 83(5): 1005-16)。
【0006】
VEGFR1は、細胞外ドメイン、7つの免疫グロブリン(Ig)様配列、及び、長いキナーゼインサートを有する細胞質チロシンキナーゼドメインを含有する、FMS/KIT/PDGFRファミリーと構造的類似性を共有する。VEGFR1は、全長のチロシンキナーゼ受容体として及び細胞外ドメインのみ保持する可溶性の形態として、2つの形態で発現される。VEGFR2及びVEGFR3等の他のRTKとのホモ二量体化又はヘテロ二量体化を通じて、VEGFR1の全長形態は積極的にそのリガンドとの結合のシグナル伝達を媒介する。しかし、VEGFR1の可溶性形態は、リガンドの捕捉を通じて阻害剤として働き、血管形成を抑制する(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。これらの相反する機能の制御機序は、不明なままである。
【0007】
VEGFRリガンドの1つ、血管内皮成長因子A(VEGFA)は、VEGFR1及びVEGFR2の双方と会合するが、VEGFAとVEGFR1との結合性は、少なくともVEGFR2に対するものよりも一桁超高い(Sawano A et al., Cell Growth Differ 1996, 7(2): 213-21)。他方、VEGFR1の内在性チロシンキナーゼ活性は、VEGFR2のものと比較してはるかに低い。VEGFAとの結合の際、これらの受容体は、その自己リン酸化レベルを劇的に増大させ、内皮細胞(EC)の成長、EC前駆体の動員、ナチュラルキラー細胞のECへの固着、及び、単球の遊走を誘導する(Shibuya M et al., Oncogene 1990, 5(4): 519-24.; Rahimi N, Experimental eye research 2006, 83(5): 1005-16.; Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。いくつかのVEGFR1チロシンリン酸化部位及びそれらの可能な結合相手は、異なる発現モデルで記述されているが(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)、下流のシグナル伝達現象は、いまだに明らかにされておらず、それはひとえに、この受容体の低い生物学的活性による。
【0008】
本発明者らは、SMYD3(Accession No.:AB057595)が、ヒストンH3(H3−K4)のリジン4上にジ-メチルトランスフェラーゼ活性及びトリメチルトランスフェラーゼ活性を有することを先に報告している(WO2005/071102(その全体は本明細書中に参照により引用される)参照)。さらに、先の報告では、SMYD3の発現増大が結腸直腸癌(CRC)及び肝細胞癌(HCC)細胞の増殖に重大な役割を果たすことを実証している(WO2003/027143(その全体は本明細書中に参照により引用される)、並びにHamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004))及びHamamoto R et al., Cancer Sci 2006, 97(2): 113-8参照)。特に、SMYD3の過剰発現は、NIH3T3細胞の成長促進をもたらすことが示され、いくつかの癌細胞での内在性SMYD3発現のノックダウンはこれらの細胞の成長阻害及びアポトーシスを誘導することが示された。本発明者らはまた、網膜芽腫タンパク質(RB1)がSMYD3のSETドメインとの相互作用を通じてメチル化され、このようなメチル化は、インビトロ及びインビボでのCDK2/サイクリンE又はCDK6/サイクリンD3複合体により、スレオニン821/826及びセリン807/811でのRB1のリン酸化を促進することを示してきた(WO2007/004526(その全体は本明細書中に参照により引用される))。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】WO2003/027143 (JP 2005-511023)
【特許文献2】WO2004/076623 (JP 2006-519009)
【特許文献3】WO2005/071102 (JP 2007-519391)
【特許文献4】WO2006/092958, A1
【特許文献5】WO2007/004526, A2
【非特許文献】
【0010】
【非特許文献1】Kunizaki, et al. Cancer Res. 2007 Nov 15;67(22):10759-65
【発明の概要】
【0011】
SMYD3メチルトランスフェラーゼの他の基質の探索において、本発明者らは、SMYD3がインビボ及びインビトロでVEGFR1のリジン831をメチル化すること、及び、かかるVEGFR1のメチル化はそのキナーゼ活性を強めることを見出した。これらの知見は、VEGFR1及びヒト腫瘍形成の制御機序のよりよい理解をもたらすであろう。
【0012】
従って、本発明は、SMYD3によるリジン831のメチル化を介した、VEGFR1制御の新規な機序の発見に少なくとも一部基づく。“ZNFN3A1”の遺伝子名でも知られるSMYD3は、結腸直腸癌及び肝細胞癌(例えば、上記WO2003/027143を参照)並びに膀胱癌及び乳癌(例えば、上記WO2006/085684及びWO2006/092958)の大部分で上昇制御されるヒストンH3メチルトランスフェラーゼである(文献はそれぞれその全体が本明細書中に参照により引用される)。
【0013】
本明細書中で示されるように、VEGFR1はSMYD3のSETドメインと相互作用する。この相互作用は、VEGFR1のメチル化をもたらし、これが本明細書中で示されるように、インビトロで強いキナーゼ活性をもたらす。これらの知見は、SMYD3の生物学的役割だけでなく、VEGFR1の制御機序についてのより深い理解を提供する。さらに、該発見は、腫瘍形成、より具体的には、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌の腫瘍形成のよりよい理解に寄与し、それゆえ、これらの腫瘍の新規な治療戦略の発展に寄与する。
【0014】
したがって、本発明の目的は、SMYD3によるVEGFR1のメチル化を調整する因子を同定する方法を提供することにあり、該方法は:
(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、試験因子の存在下、VEGFR1ペプチドのメチル化に適した条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)で検出したメチル化レベルを前記因子の非存在下で検出した対照レベルと比較する工程を含み、ここで、
対照レベルと比較したメチル化レベルの減少又は増大は、前記因子がSMYD3によるVEGFR1のメチル化を調整することを示す。
【0015】
本発明の更なる目的は、VEGFR1のメチル化を制御する試験化合物の能力を検出するキットを提供することにあり、かかるキットは、(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド、(b)SMYD3ポリペプチドによりメチル化されうるVEGFR1ペプチド、及び(c)VEGFR1ペプチドのメチル化のコファクター、を含む。更なる態様において、該キットは、S−アデノシルホモシステインヒドロラーゼ(SAHH)を任意に含んでもよい。
【0016】
本発明は、さらに、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌等の、癌治療用化合物のスクリーニング方法を提供するものであり、かかる方法は、(a)上述の方法に従ってメチル化を調整する試験化合物を同定する工程、及び、(b)試験化合物の非存在下で検出した対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程、を含む。
【0017】
本発明は、さらに、ポリペプチドのメチルトランスフェラーゼ活性を測定する方法を提供するものであり、該方法は、
(a)以下からなる群から選択されるポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化が可能な条件下で接触させる工程;
(i)配列番号2(SMYD3)のアミノ酸配列を有するポリペプチド;
(ii)配列番号2のアミノ酸配列を有するポリペプチドであって、1または2以上のアミノ酸が置換、欠失、又は挿入され、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価な生物学的活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を有するアミノ酸配列を有するポリペプチド;
(vi)配列番号1のヌクレオチド配列を有するポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされたポリペプチドであって、配列番号2のアミノ酸配列に対応するポリペプチドと等価な生物学的活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルとメチルトランスフェラーゼ活性とを関連させて、メチルトランスフェラーゼ活性を測定する工程
を含む。
【0018】
さらに、本発明はまた、SMYD3のメチルトランスフェラーゼ活性を測定するためのキットを提供するものであり、該キットは、以下の要素:
(a)SMYD3によりメチル化されうるVEGFR1ペプチド;
(b)VEGFR1ペプチドのメチル化のコファクター;及び
(c)リジン831のメチル化を検出するための検出試薬
を含む。
【0019】
本発明の上記及び他の目的、特徴及び利点は、以下の詳細な説明を添付の図面及び実施例、並びに本明細書に添付の特許請求の範囲を読んだとき、より十分に明らかとなるであろう。
【0020】
他に定義しない限り、本明細書中で用いられる全ての技術的及び科学的用語は、本発明の属する技術の分野における通常の技術、知識を有する者により一般に理解されるものと同じ意味を有する。また、本明細書中で用いられる「1つの(a, an)」及び「該(the)」は、他に特に示さない限り、「少なくとも1つ」を意味する。
【0021】
本明細書中に記載されたものと同様または等価な方法と材料は本発明の実施又は試験に用いることができるが、適切な方法と材料を本明細書において以下に示す。
【0022】
本明細書において言及される全ての出版物、特許出願、特許、およびその他の参考文献は、その全体が参照により引用される。矛盾する場合、定義を含む本明細書によって統制される。さらに、材料、方法、および実施例は、例示的なものに過ぎず、本発明を制限するためのものではない。
【図面の簡単な説明】
【0023】
【図1】図1はインビトロでのVEGFR1の細胞質領域のメチル化を示す。(a)は、基質として、VEGFR1の異なる細胞質領域をそれぞれ含む組換えタンパク質VEGFR1#1、#2及び#3を用いたSMYD3のインビトロでのメチルトランスフェラーゼ(MTase)アッセイの結果を示す。「Ig−LD」は免疫グロブリン様ドメインに、「TM」は膜貫通ドメインに、「TK」はチロシンキナーゼドメインに対応する。組換え体VEGFR1#1、#2及び#3タンパク質は、3HでラベルしたSAM(メチルドナーである)とともに、免疫沈降したFlagタグ化SMYD3の存在下(■)又は非存在下でインキュベートした。メチル化基質は、液体シンチレーションカウンターで定量化した。モックプラスミドをトランスフェクトした細胞からの免疫沈降物を対照に用いた(□)。独立した実験を異なる免疫沈降物について3回行った。(b)は、フルオログラムによるメチル化VEGFR1の検出を示す。組換えSMYD3は、S−(5’−アデノシル)−L−ホモシステイン ヒドロラーゼ(SAHH)の存在下又は非存在下で、組換えVEGFR1とともにインキュベートした。基質は、抗GST抗体を用いてイムノブロット分析により定量化した(下パネル)。(c)はVEGFR1#1のメチル化領域の決定を示す。
【図2】図2は、インビトロ及びインビボ両方でのVEGFR1のリジン831のメチル化を示す。(a)は、ヒストンメチルトランスフェラーゼによりメチル化されたリジン、及び、VEGFR1中の候補リジン(*)の隣接配列のアラインメントを示す。(b)は、VEGFR1−N1の野生型、及び、変異体(K819A及びK831A)のインビトロメチル化を示す(上パネル)。(c)は、VEGFR1−K831の異なる置換を含むVEGFR1変異体のインビトロメチル化を示す。(d)は、抗メチル化K831(K831me2)特異的抗体を用いたイムノブロット分析により、VEGFR1のインビトロメチル化を検出したことを示す(中パネル)。メチル化は、フルオログラムによる異なる実験により確認した(上パネル)。VEGFR1は、抗GST抗体を用いたイムノブロット分析で定量化した(下パネル)。(e)は、HAタグ化VEGFR1を発現する293−SMYD3細胞及び293−モック細胞内でのVEGFR1のメチル化を示す。細胞は、タンパク質合成阻害剤の存在下で、L−[メチル−3H]メチオニンで処理した。VEGFR1は抗HA抗体で免疫沈降し、フルオログラムで調べた(上パネル)。(f)は、抗メチル化K831(K831me2)特異的抗体を用いたイムノブロット分析で検出したVEGFR1のメチル化を示す。(g)は、SMYD3のノックダウンによるK831メチル化の抑制を示す。293−SMYD3細胞からの全ての細胞抽出物は、SMYD3特異的siRNA又は対照siRNAで処理し、イムノブロット分析に用いた。
【図3】図3は、SMYD3及びVEGFR1の間の相互作用を示す。(a)は、HAタグ化SMYD3及びFlagタグ化VEGFR1の異なる領域(VEGFR1#1、#2及び#3)を発現するHEK293細胞からの抽出物を用いた免疫共沈降アッセイにより調べた、SMYD3及びVEGFR1の間の相互作用を示す。(b)は、内在性SMYD3とVEGFR1との相互作用を示す。いずれもSMYD3及びVEGFR1を発現する、SNU423、SW480及びMCF7細胞からの全ての細胞抽出物を、抗SMYD3抗体で沈降させた。沈降物は、抗VEGFR1抗体(上パネル)、又は、抗SMYD3抗体(下パネル)でイムノブロットした。(c)は、VEGFR1とSMYD3のSETドメインとの会合を示す。Flagタグ化SMYD3の野生型又は欠失型(N−20、Δ2又はΔ3)を、HEK293細胞においてHAタグ化VEGFR1と共発現させた。IP(免疫沈降)を抗HA又は抗Flag抗体のいずれかで行い、沈降物について、ウェスタンブロット分析を、抗Flag抗体(上パネル)又は抗HA抗体(下パネル)を用いて行った。(d)は、VEGFR1の細胞質領域とSMYD3のSETドメインとのインビボでの相互作用を示す。Flagタグ化野生型又はSMYD3のSETドメインを、HEK293細胞においてHAタグ化VEGFR1と共発現させた。細胞抽出物は、抗Flag抗体(上パネル)又は抗HA抗体(下パネル)を用いて沈降させ、沈降物を、抗HA抗体を用いてウェスタンブロット分析により分析した。
【図4】図4に示される結果は、VEGFR1のキナーゼ活性がそのメチル化によって強められることを示す。(a)は、メチル化又は非メチル化VEGFR1を用いて行ったインビトロキナーゼアッセイの結果を示す。組換えGST融合VEGFR1は、SMYD3で処理し(■)、又は、処理しなかった(□)。そのキナーゼ活性について、そのペプチド基質を用いて引き続き分析した。リン酸化基質は、抗リン酸化チロシン抗体で検出し、特異的二次抗体を用いた蛍光免疫アッセイにより定量化した。(b)に示される結果は、組換えVEGFR1の自己リン酸化がSMYD3により強められることを示す。VEGFR1はPAGE−SDSで分離し、その後、ニトロセルロース膜に移した。抗リン酸化チロシン又は抗VEGFR1抗体を用いてイムノブロット分析を行った。
【発明を実施するための形態】
【0024】
SMYD3のcDNAは、配列番号1に示されるように、1284個のヌクレオチドのオープンリーディングフレームを含む、1622個のヌクレオチドから構成される。オープンリーディングフレームは、配列番号2に示されるように、ジンクフィンガーモチーフ及びSETドメインを有する428アミノ酸タンパク質をコードする。ジンクフィンガードメイン(MYND)は、アミノ酸49からアミノ酸87にわたり、SET(Su3−9、Enhancer−of−zeste、Trihorrax)ドメインは、アミノ酸117からアミノ酸246にわたる。
【0025】
SMYD3タンパク質の細胞内局在は、細胞周期進行の間に変わり、また、培養細胞の密度によっても変わる。SMYD3タンパク質は、細胞が、S期の中間〜後半にあるか、低密度状態で培養されるときに、核に蓄積される。しかし、SMYD3タンパク質は、細胞が細胞周期の他の期、又は、密集状態で成長するとき、細胞質内及び核内に局在化する。
【0026】
本発明は、VEGFR1基質のメチル化のためのSMYD3のメチルトランスフェラーゼ活性を決定する方法を提供する。該方法は、SMYD3ポリペプチド又はメチルトランスフェラーゼ活性を有するその機能的等価物をVEGFR1タンパク質と接触させ、接触させたSMYD3又はその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることにより実施されてもよい。この態様(context)において、SMYD3のメチルトランスフェラーゼ活性は、VEGFR1のメチル化の程度に対応する。したがって、メチルトランスフェラーゼ活性はVEGFR1基質のメチル化レベルを検出することにより測定可能である。より具体的には、本発明の態様において、測定されるメチルトランスフェラーゼ活性は、参照サンプルとの関連付けを介してVEGFR1ペプチドのメチル化レベルに較正されてもよい。本発明のこの態様において、既知のメチルトランスフェラーゼ活性を有するいかなる生物学的サンプルも、参照サンプルとして用いることが可能である。例えば、必要な較正曲線は、精製SMYD3ペプチドの段階希釈を介して得ることができる。
【0027】
さらに、メチルトランスフェラーゼ活性調整剤のスクリーニング方法もまた提供される。本発明は、したがって、SMYD3メチルトランスフェラーゼ活性を調整する因子をスクリーニングする方法を提供する。該方法は、SMYD3ポリペプチド又はメチルトランスフェラーゼ活性を有するその機能的等価物をVEGFR1タンパク質と接触させ、接触させたSMYD3又はその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることにより実施されてもよい。SMYD3又はその機能的等価物のメチルトランスフェラーゼ活性を調整する因子は、これにより同定可能である。
【0028】
本発明の態様において、「機能的に等価」との語は、対象タンパク質又はポリペプチドが、SMYD3と同一の又は実質的に同一のメチルトランスフェラーゼ活性を有することを意味する。特に、該タンパク質は、VEGFR1タンパク質、又は、リジン831を含むVEGFR1タンパク質の断片のメチル化を触媒する。対象タンパク質が標的活性を有するかどうかは、本発明により常に決定できる。すなわち、メチルトランスフェラーゼ活性は、(a)ポリペプチドを基質(例えば、VEGFR1タンパク質又はリジン831を含むその断片)及びコファクター(例えば、S−アデノシル−L−メチオニン)と、該基質のメチル化に適した条件下で接触させ、(b)該基質のメチル化レベルを検出することにより、決定可能である。
【0029】
本明細書中で用いられるとき、「VEGFR1ペプチド」の語は、全長VEGFR1タンパク質(例えば、配列番号4)、及び、その機能的変異体及び断片を指す。機能的断片の例には、配列番号4のアミノ酸800から841で構成される断片のようなC末端断片が含まれるが、これに限られない。好適な断片は、位置831のリジン残基を含む。機能的変異体の例には、全長VEGFR1たんぱく質のメチル化可能性を保持する以下のVEGFR1変異体:K819A、K819E及びK819R、が含まれるが、これに限られない。
【0030】
特定のタンパク質の機能的等価物であるタンパク質を調製する方法は、当業者にはよく知られており、タンパク質に変異を導入する一般的な方法が含まれる。例えば、当業者は、部位特異的変異導入法を用いてヒトSMYD3タンパク質のアミノ酸配列中に適当な変異を導入することにより、ヒトSMYD3タンパク質に機能的に等価なタンパク質を調製できる(例えば、Hashimoto-Gotoh, T. et al. (1995), Gene 152, 271-275;Zoller, MJ, and Smith, M. (1983), Methods Enzymol. 100, 468-500;Kramer, W. et al. (1984), Nucleic Acids Res. 12, 9441-9456;Kramer W, and Fritz HJ. (1987) Methods. Enzymol. 154, 350-367;Kunkel, TA (1985), Proc. Natl. Acad. Sci. USA. 82, 488-492)。アミノ酸変異は、自然にも発生しうる。本発明の態様において有用なSMYD3ポリペプチドには、生じる変異タンパク質がヒトSMYD3タンパク質の機能的等価物、より具体的には、ヒトSMYD3タンパク質のメチルトランスフェラーゼ活性を保持する、1又は2以上のアミノ酸が変異したヒトSMYD3タンパク質のアミノ酸配列を有するタンパク質が含まれる。かかる変異体において変異されるアミノ酸の数は、通常、20アミノ酸以下、典型的には10アミノ酸以下、好ましくは6アミノ酸以下、より好ましくは3アミノ酸以下である。メチルトランスフェラーゼ活性を維持するため、SETドメイン“NHSCXXN”及び“GEELXXXY”が好ましくは変異タンパク質のアミノ酸配列に保存される(“X”は任意のアミノ酸を示す)。したがって、本発明の態様において、SMYD3ポリペプチドは、配列番号2のアミノ酸配列を含み、ここで、1又は2以上のアミノ酸は置換、欠失、又は挿入され、さらにここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドのメチルトランスフェラーゼ活性と等価な活性を有し、さらにここで、SETドメイン“NHSCXXN”及び“GEELXXXY”は保存されている。より詳細には、配列番号2の117位のアミノ酸から246位までのアミノ酸が好ましくは保存される。
【0031】
変異又は修飾されたタンパク質、すなわち、特定アミノ酸配列の1又は2以上のアミノ酸残基を欠失、付加及び/又は置換することにより修飾されたアミノ酸配列を有するタンパク質は、元のタンパク質の生物学的活性を保持することが知られている(Mark, D. F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666;Zoller, M. J. & Smith, M., Nucleic Acids Research (1982) 10, 6487-6500;Wang, A. et al., Science (1984) 224, 1431-1433;Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413)。例えば、SMYD3ポリペプチドは、SETドメインを保持する限り、そのメチルトランスフェラーゼ活性を保持することが予測される。したがって、本発明に係る方法は、配列番号2のアミノ酸配列の117〜246の位置のアミノ酸配列を含むポリペプチド、例えば、実施例に示される配列番号2のアミノ酸配列の110〜250の位置のアミノ酸配列を含むポリペプチドを用いて実施可能である。
【0032】
変異されるアミノ酸残基は、アミノ酸側鎖の性質の保存が可能な、異なるアミノ酸に好適に変異される(保存的アミノ酸置換として知られるプロセス)。アミノ酸側鎖の性質の例は、以下の群に含まれる。
疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、
親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)。
他に、以下の官能基又は特性を共通に有する側鎖:
脂肪族側鎖(G、A、V、L、I、P);
水酸基を含有する側鎖(S、T、Y);
硫黄原子を含有する側鎖(C、M);
カルボン酸及びアミドを含有する側鎖(D、N、E、Q);
塩基を含有する側鎖(R、K、H);及び
芳香族を含有する側鎖(F、H、Y、W)。
さらに、例えば、アミノ酸の分類は、変異マトリックスとして周知である(Taylor 1986, J, Theor. Biol. 119, 205-218;Sambrook, J. et al., Molecular Cloning 3rd ed. A7.6-A7.9, Cold Spring Harbor Lab. Press, 2001)。このような分類は、以下のように要約できる:
脂肪族アミノ酸:L、I、V
芳香族アミノ酸:H、W、Y、F
荷電アミノ酸:D、E、R、K、H
陽荷電アミノ酸:R、K、H
陰荷電アミノ酸:D、E
疎水性アミノ酸:H、W、Y、F、M、L、I、V、C、A、G、T、K
極性アミノ酸:T、S、N、D、E、Q、R、K、H、W、Y
小アミノ酸:P、V、C、A、G、T、S、N、D
極小アミノ酸:A、G、S
大(非小)アミノ酸:Q、E、R、K、H、W、Y、F、M、L、I
ここで、挿入文字は、アミノ酸の1文字コードを示す。
【0033】
ヒトSMYD3タンパク質(配列番号2)のアミノ酸配列に1又は2以上のアミノ酸残基が付加されたタンパク質の例は、ヒトSMYD3タンパク質を含有する融合タンパク質である。融合タンパク質には、ヒトSMYD3タンパク質及び他のペプチド又はタンパク質の融合が含まれ、本発明において用いられる。融合タンパク質は、当業者には周知の技術、たとえば、本発明に係るヒトSMYD3タンパク質をコードするDNAを他のペプチド又はタンパク質をコードするDNAとフレームが一致するように連結し、融合DNAを発現ベクターに挿入して、宿主内で発現させることにより作成可能である。本発明のタンパク質に融合されるペプチド又はタンパク質について何ら制約はない。
【0034】
SMYD3タンパク質に融合されるペプチドとして使用可能な既知のペプチドには、例えば、FLAG(Hopp, T. P. et al., Biotechnology (1988) 6, 1204-1210)、6つのHis(ヒスチジン)残基を含有する6xHis、10xHis、インフルエンザアグルチニン(HA)、ヒトc−myc断片、VSP−GP断片、p18HIV断片、T7−タグ、HSV−タグ、E−タグ、SV40T抗原断片、lckタグ、αチューブリン断片、B−タグ、プロテインC断片などが含まれる。本発明に係るタンパク質に融合可能なタンパク質の例には、GST(グルタチオン−S−トランスフェラーゼ)、インフルエンザアグルチニン(HA)、免疫グロブリン定常領域、βガラクトシダーゼ、MBP(マルトース結合タンパク質)などが含まれる。
【0035】
融合タンパク質は、上述の融合ペプチド又はタンパク質をコードする、商業的に入手可能なDNAを、本発明に係るタンパク質をコードするDNAと融合し、調製した融合DNAを発現することより調製できる。
【0036】
機能的に等価なタンパク質を単離する当業者に既知の他の方法では、相同配列を同定するハイブリダイズ技術を用いる(Sambrook, J. et al., Molecular Cloning 2nd ed. 9.47-9.58, Cold Spring Harbor Lab. Press, 1989)。当業者であれば容易にヒトSMYD3タンパク質をコードするSMYD3 DNA配列(例えば、配列番号1)の全部又は一部と高い相同性を有するDNAを単離し、単離したDNAからヒトSMYD3タンパク質と機能的に等価なタンパク質を単離することができる。本発明に使用されるタンパク質には、ヒトSMYD3タンパク質をコードするDNA配列の全部又は一部とハイブリダイズするDNAによりコードされるもの、及び、ヒトSMYD3タンパク質と機能的に等価なものが含まれる。これらのタンパク質には、ヒト又はマウス由来のタンパク質に対応する哺乳類の相同体が含まれる(例えば、サル、ラット、ウサギ及びウシ遺伝子によりコードされるタンパク質)。動物からヒトSMYD3タンパク質をコードするDNAに高い相同性を示すcDNAを単離する場合、骨格筋、精巣、HCC又は結腸直腸腫瘍由来の組織を用いることが特に好ましい。
【0037】
ヒトSMYD3タンパク質の機能的等価物をコードするDNAを単離するためのハイブリダイズ条件は、当業者により通常通りに選択可能である。例えば、“Rapid−hyb buffer”(Amersham LIFE SCIENCE)を用いて68℃で30分又はそれ以上プリハイブリダイゼーションを行い、ラベルしたプローブを添加し、68℃で1時間又はそれ以上温めることにより実施してもよい。その後の洗浄ステップは、例えば、低ストリンジェント条件で行うことができる。低ストリンジェント条件とは、例えば、42℃、2xSSC、0.1%SDS、又は、50℃、2xSSC、0.1%SDSである。より好ましくは、高ストリンジェント条件が用いられる。本発明の態様において、高ストリンジェント条件には、例えば、2xSSC、0.01%SDS、室温で20分間の洗浄を3回行い、次いで、1xSSC、0.1%SDS、37℃、20分間の洗浄を3回行い、次いで、1xSSC、0.1%SDS、50℃、20分間の洗浄を2回行うことが含まれる。しかし、温度及び塩濃度などのいくつかの要因がハイブリダイゼーションのストリンジェンシーに影響を与える可能性があり、当業者は要求されるストリンジェンシーを達成するためにこれらの要因を適切に選択できる。
【0038】
ハイブリダイゼーションに代えて、遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法を利用し、ヒトSMYD3タンパク質(配列番号2)をコードするDNA(配列番号1)の配列情報に基づいて合成されたプライマーを用いて、ヒトSMYD3タンパク質に機能的に等価なタンパク質をコードするDNAを単離することができる。
【0039】
上記ハイブリダイゼーション技術又は遺伝子増幅技術により単離されたDNAがコードする、ヒトSMYD3タンパク質に機能的に等価なタンパク質は、通常、ヒトSMYD3タンパク質のアミノ酸配列と高い相同性を有する。「高い相同性」(「高い同一性」ともいう)とは、通常、2つの任意に整列された配列(ポリペプチド又はポリヌクレオチド配列のいずれか)間の同一性の程度をいう。典型的には、高い相同性又は同一性とは、40%以上、好ましくは60%以上、より好ましくは80%以上、さらにより好ましくは85%、90%、95%、98%、99%以上をいう。2つのポリペプチド又はポリヌクレオチド配列間の相同性又は同一性の程度は、「Wilbur, W. J. and Lipman, D. J. Proc. Natl. Acad. Sci. USA (1983) 80, 726-730」中のアルゴリズムに従って決定可能である。
【0040】
本発明の態様において有用なタンパク質は、その製造に用いる細胞又は宿主或いは用いる精製方法によって、アミノ酸配列、分子量、等電点、糖鎖の有無、又は、形態においてバリエーションをとることが可能である。にもかかわらず、ヒトSMYD3タンパク質(配列番号2)の機能的等価物である限り、本発明に有用である。
【0041】
本発明の態様において有用なタンパク質は、当業者には周知の方法により、組換えタンパク質又は天然タンパク質として調製可能である。組換えタンパク質は、本発明のタンパク質をコードするDNA(例えば、配列番号1のヌクレオチド配列に対応するDNA)を適切な発現ベクターに挿入し、該ベクターを適切な宿主細胞に導入し、抽出物を得て、該抽出物をクロマトグラフィー、例えば、イオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲルろ過、又は本発明に係るタンパク質に対する抗体が固定されるカラムを利用するアフィニティクロマトグラフィーにかけて、或いは、1以上の上記カラムを組み合わせて、タンパク質を精製することにより調製可能である。
【0042】
さらに、本発明の態様において有用なタンパク質が宿主細胞内(例えば、動物細胞及び大腸菌)でグルタチオン−S−トランスフェラーゼとの融合タンパク質として、又は、多数のヒスチジンが付加された組換えタンパク質として発現される場合には、発現された組換えタンパク質はグルタチオンカラム又はニッケルカラムを用いて精製できる。
【0043】
融合タンパク質の精製後、必要に応じて、トロンビン又は活性化第Xa因子で切断して、標的タンパク質以外の領域を除くこともできる。
【0044】
天然タンパク質は、当業者に既知の方法により、例えば、以下に記載するSMYD3タンパク質に結合する抗体が結合されたアフィニティカラムを、本発明に係るタンパク質を発現する組織又は細胞抽出物に接触させることにより単離可能である。抗体は、ポリクローナル抗体又はモノクローナル抗体が可能である。
【0045】
本発明において、SMYD3ポリペプチドのメチルトランスフェラーゼ活性は当業者には既知の方法により決定できる。例えば、SMYD3ポリペプチド及びVEGFR1ペプチド基質を、ラベルしたメチルドナーとともに適切なアッセイ条件でインキュベートしてもよい。好ましいメチルドナーの例としては、S−アデノシル−[メチル−14C]−L−メチオニン及びS−アデノシル−[メチル−3H]−L−メチオニンが挙げられるが、これに限られない。VEGFR1ペプチドへの放射ラベルの転移は、例えば、SDS−PAGE電気泳動及びフルオログラフィーにより検出できる。また、反応後、VEGFR1ペプチドはろ過によりメチルドナーから分離でき、フィルターに残った放射ラベルの量はシンチレーションカウントにより定量化できる。色素生産性及び蛍光ラベルのようなメチルドナーに付加できる他の適したラベル、及び、これらのラベルのVEGFR1ペプチドへの転移を検出する方法は、当業者には既知である。
【0046】
また、SMYD3のメチルトランスフェラーゼ活性は、非ラベル化メチルドナー(例えば、S−アデノシル−L−メチオニン)及びメチル化VEGFR1ペプチドを選択的に認識する試薬を用いて決定できる。例えば、SMYD3、メチル化される基質、及びメチルドナーを基質のメチル化に適した条件下でインキュベートした後、メチル化された基質は通常の免疫学的方法を用いて検出できる。メチル化基質を認識する抗体を用いるどのような免疫学的技術も検出に用いることができる。例えば、メチル化VEGFR1 K831(例えば、抗K831me2)に対する抗体を抗体として使用できる。VEGFR1のメチル化、特に、VEGFR1のK831でのメチル化を認識するこのような抗体も、本発明により提供される。メチル化VEGFR1 K831を認識する抗体を用いるELISA又はイムノブロット分析は、本発明に適している場合がある。
【0047】
本明細書中で示されるように、Lys831でのVEGFR1のメチル化は、VEGFR1のリン酸化を強めた。したがって、他の実施形態において、VEGFR1のメチル化レベルはVEGFR1のリン酸化レベルを介して評価可能である。VEGFR1のリン酸化は、放射ラベル化リン酸ドナーを用いて検出してもよい。また、VEGFR1のリン酸化部位を認識する抗体をVEGFR1のリン酸化レベルの評価に用いてもよい。
【0048】
本発明によれば、SMYD3によりリン酸化されるVEGFR1ペプチドの上記性質を、SMYD3及び他のH3K4メチルトランスフェラーゼ活性の測定に利用してもよい。このようなペプチドのメチルトランスフェラーゼ活性の測定方法は:
(a)H3K4メチルトランスフェラーゼを、メチル化されるVEGFR1ペプチド及びコファクターとVEGFR1ペプチドのメチル化が可能な条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルをメチルトランスフェラーゼ活性と関連させてメチルトランスフェラーゼ活性を測定する工程
を含む。
【0049】
本明細書中において、H3K4メチルトランスフェラーゼは、VEGFR1ペプチド上のメチル基を転移させる能力のある限り、いかなるポリペプチドであってもよく、このようなポリペプチドには、上記SMYD3ポリペプチドが含まれるが、これに限られない。本方法の工程は、同じVEGFR1ペプチド、反応条件(コファクターの付加、強化因子等)、検出手段及びSMYD3によるVEGFR1のメチル化を調整する化合物の上記同定方法に類するものを用いて行ってもよい。
【0050】
本発明は、SMYD3を過剰発現する癌を治療するための化合物のスクリーニング方法をさらに提供し、該方法は、上記方法を用いるメチル化を調整する試験化合物を同定する工程、及び、試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程を含む。
【0051】
該スクリーニングは、(a)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片及びSMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片を、VEGFR1ペプチド及びSMYD3ポリペプチドの結合を可能とする条件下、かつ、試験化合物の存在下で接触させる工程、並びに(b)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片、及び、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片の間の結合を阻害する試験化合物を癌治療のための候補化合物として選択する工程を含む。本発明において、SMYD3によるVEGFR1のメチル化、又は、VEGFR1とSMYD3との間の結合を抑制することにより、細胞の成長が低減することが明らかとなった。従って、SMYD3によるVEGFR1の結合又はメチル化を阻害する試験化合物をスクリーニングすることにより、癌を治療又は予防する可能性のある候補化合物を同定することができる。癌を治療又は予防するこれらの候補化合物の可能性は、癌のための治療用因子を同定する第2の及び/又はさらなるスクリーニングにより評価してもよい。
本発明の態様において、SMYD3を過剰発現する癌には、SMYD3のその発現レベルが癌の同じ器官の正常領域と比較して高い癌が含まれる。例えば、本発明者らは、種々の癌、例えば、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌におけるSMYD3の過剰発現を明らかにしてきた(WO2003/027143、WO2006/085684及びWO2006/092958)。従って、本発明の好ましい態様において、SMYD3を過剰発現する癌は、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択されてもよい。
【0052】
試験化合物がVEGFR1ペプチドとSMYD3ポリペプチドとの結合を阻害するかは、VEGFR1ペプチドとSMYD3ポリペプチドとの結合レベルを、該化合物の非存在下で検出した結合レベルと比較し、VEGFR1ポリペプチドとSMYD3との結合レベルを減少させる化合物を選択することにより決定できる。その機能的等価物を含む任意のVEGFR1ペプチド及びSMYD3ポリペプチドは、互いの結合能力が保持される限りこのスクリーニングに用いてもよい。例えば、VEGFR1ペプチドのSMYD3結合領域を含む断片を、VEGFR1に等価なペプチドとしてこのスクリーニングに用いてもよい。本発明において、VEGFR1に等価なかかるペプチドは、配列番号4のアミノ酸配列の800〜1000の位置のアミノ酸配列を好ましくは含みうる。同様に、SMYD3ペプチドのVEGFR1結合領域を含む断片を、SMYD3に等価なペプチドとして用いてもよい。例えば、かかるSMYD3のペプチド等価物は、配列番号2のアミノ酸配列の100〜250の位置のアミノ酸配列を含んでもよい。
本発明の態様において、2つのタンパク質間の「結合を阻害する」とは、該タンパク質間の結合を少なくとも減少させることをいう。従って、ある場合には、サンプル中の結合ペアの割合は、適当な(例えば、試験化合物で処理されていない、或いは、非癌サンプルからの、又は癌サンプルからの)対照と比較して減少しているであろう。結合タンパク質量の減少は、例えば、対照サンプル中の結合ペアよりも、90%、80%、70%、60%、50%、40%、25%、10%、5%、1%未満(例えば0%)でありうる。
【0053】
一般に、かかる会合に干渉する試験化合物の能力を決定するいかなる方法も、本発明での使用に適している。例えば、ELISAフォーマットでの競合的及び非競合的阻害アッセイも利用可能である。対照実験は、系の最大結合能力を決定するために実行されるべきである(例えば、結合VEGFR1をSMYD3ポリペプチドと接触させ、VEGFR1に結合したタンパク質の量を決定する、或いは、その逆)。本発明に係る結合を阻害する化合物を同定する方法として、当業者に周知の方法の多くを用いることができる。かかる同定は、インビトロアッセイ系、例えば、細胞系として実行可能である。より詳細には、まず、VEGFR1ペプチド又はSMYD3ポリペプチドパートナーのいずれか一方を支持体に結合させ、他方のタンパク質を試験化合物とともにそこに接触させる。次いで、混合物をインキュベートし、洗浄し、支持体に結合した他方のタンパク質を検出及び/又は測定する。
【0054】
タンパク質の結合に使用可能な支持体の例には、不溶性多糖、例えば、アガロース、セルロース及びデキストランが含まれ;そして、合成樹脂、例えば、ポリアクリルアミド、ポリスチレン及びシリコン;好ましくは、上記材料から調製された、商業的に入手可能なビーズ及びプレート(例えば、マルチウェルプレート、バイオセンサーチップ等)を用いることができる。ビーズを用いる場合には、これらはカラムに充填してもよい。また、磁性ビーズの使用もまた当業者には知られており、磁性によりビーズ上に結合されたタンパク質の容易な単離が可能とする。
【0055】
タンパク質の支持体への結合は、通常の方法、例えば、化学結合及び物理吸着により行うことができる。また、タンパク質は、該タンパク質を特異的に認識する抗体を介して支持体に結合させてもよい。さらに、タンパク質の支持体への結合はまた、相互作用する分子、例えば、アビジン及びビオチンにより行われてもよい。
【0056】
タンパク質間の結合は、バッファーがタンパク質間の結合を阻害しない限り、バッファー、例えば、リン酸バッファー及びTrisバッファー(これらに限られない)中で行われる。
【0057】
本発明において、表面プラズモン共鳴現象を用いるバイオセンサーを、結合タンパク質を検出又は定量化するための手段として用いてもよい。かかるバイオセンサーを用いる場合、タンパク質間の相互作用は表面プラズモン共鳴信号としてリアルタイムで、ごく少量のポリペプチドを用いてラベル化無しで観察可能である(例えば、BIAcore、Pharmacia)。従って、BIAcoreのようなバイオセンサーを用いて、VEGFR1ペプチドとSMYD3ポリペプチドとの間の結合を評価することができる。
【0058】
また、VEGFR1ペプチド又はSMYD3ポリペプチドのいずれかをラベルしてもよく、結合タンパク質のラベルを該結合タンパク質の検出又は測定に用いてもよい。特に、タンパク質の一方をプレラベル化した後、ラベル化タンパク質を他方のタンパク質と試験化合物の存在下で接触させ、次いで、結合タンパク質を洗浄後ラベルに従って検出又は測定する。
【0059】
放射性同位体(例えば、3H、14C、32P、33P、35S、125I、131I)、酵素(例えば、アルカリフォスファターゼ、西洋わさびペルオキシダーゼ、βガラクトシダーゼ、βグルコシダーゼ)、蛍光化合物(例えば、フルオレセインイソチオシアネート(FITC)、フルオレセイン、テキサスレッド、緑色蛍光タンパク質、ローダミン)、磁性ビーズ(例えば、DYNABEADSTM)、熱量測定ラベル(例えば、コロイド金又は着色グラス或いはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックス等)ビーズ)、及びビオチン/アビジン、等のラベル化合物を本発明の方法におけるタンパク質のラベル化に用いることができる。かかるラベルの使用を教示する特許として、米国特許番号3,817,837;3,850,752;3,939,350;3,996,345;4,277,437;4,275,149;及び4,366,241が挙げられる。しかし、本発明は、これらに限られず、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的又は化学的手段により検出可能ないかなるラベルも使用可能である。
【0060】
タンパク質を放射性同位体でラベルする場合、検出又は測定は、液体シンチレーションにより行うことができる。また、酵素でラベルしたタンパク質は、酵素の基質を付加して基質の酵素的変化、例えば、呈色を吸光光度計で検出することにより検出又は測定できる。さらに、蛍光基質をラベルとして用いる場合には、結合タンパク質は蛍光光度計を用いて検出又は測定することができる。
【0061】
さらに、本発明に係るスクリーニング方法における結合は、VEGFR1ペプチド又はSMYD3ポリペプチドに対する抗体を用いて検出又は測定することもできる。例えば、支持体に固定したVEGFR1ペプチドを試験化合物及びSMYD3ポリペプチドと接触させた後、混合物をインキュベート、洗浄し、検出又は測定をSMYD3ポリペプチドに対する抗体を用いて行うことができる。また、SMYD3ポリペプチドを支持体に固定してもよく、VEGFR1ペプチドに対する抗体をその抗体として用いてもよい。
【0062】
本発明に係るスクリーニングに抗体を用いる場合、抗体は、上記ラベル化物質の1つで好適にラベル化され、ラベル化物質に基づいて検出又は測定される。また、SMYD3ポリペプチド又はVEGFR1ペプチドに対する抗体は、ラベル化物質でラベル化された二次抗体で検出される一次抗体として使用されてもよい。さらに、本発明に係るスクリーニングにおいてタンパク質に結合した抗体は、プロテインG又はプロテインAカラムを用いて検出又は測定できる。
【0063】
また、本発明の同定方法の他の態様において、細胞を用いるツーハイブリッドシステムを用いることができる(“MATCHMAKER Two−Hybrid system”、“Mammalian MATCHMAKER Two−Hybrid Assay Kit”、“MATCHMAKER one−Hybrid system”(Clontech);“HybriZAP Two−Hybrid Vector System”(Stratagene);参考文献Dalton and Treisman, Cell 1992, 68: 597-612、Fields and Sternglanz, Trends Genet 1994, 10: 286-92)。ツーハイブリッドシステムにおいて、例えば、SMYD3ポリペプチドは、SRF結合領域又はGAL4結合領域に融合され、酵母菌で発現される。VEGFR1ペプチドは、VP16又はGAL4転写活性領域に融合され、同様に試験化合物の存在下で酵母菌細胞内に発現される。また、VEGFR1ペプチドは、SRF結合領域又はGAL4結合領域に融合されてもよく、SMYD3ポリペプチドはVP16又はGAL4転写活性領域に融合されてもよい。試験化合物がSMYD3ポリペプチドとVEGFR1ペプチドの間の結合を阻害しない場合、2つの結合は、レポーター遺伝子を活性化し、陽性のクローンを検出可能にする。レポーター遺伝子として、例えば、Ade2遺伝子、lacZ遺伝子、CAT遺伝子、ルシフェラーゼ遺伝子などを、HIS3遺伝子の代わりに用いることができる。
【0064】
ここで、SMYD3ポリペプチドとVEGFR1ペプチドとの間の結合レベルは、SMYD3ポリペプチドとVEGFR1ペプチドの結合後に発生する任意の変化として測定することもできる。特に、かかるスクリーニングは試験化合物をSMYD3ポリペプチド及びVEGFR1ペプチドを発現する細胞と接触させることにより行うこともできる。例えば、細胞増殖の抑制を検出して、SMYD3ポリペプチドとVEGFR1ペプチドとの結合への試験化合物の影響を決定することもできる。
1.競合的アッセイフォーマット
競合的アッセイを本発明に係る試験化合物のスクリーニングに用いることができる。例によれば、競合的ELISAアッセイフォーマットとして、固相支持体に結合されたSMYD3ポリペプチド(又はVEGFR1ペプチド)が挙げられる。結合されたSMYD3ポリペプチド(又はVEGFR1ペプチド)を、VEGFR1ペプチド(又はSMYD3ポリペプチド)及び試験化合物とともにインキュベートする。試験化合物及び/又はVEGFR1ペプチド(又はSMYD3ポリペプチド)をSMYD3ポリペプチド(又はVEGFR1ペプチド)と結合させるのに十分な時間の後、基質を洗浄して結合していない物質を除く。SMYD3ポリペプチドに結合したVEGFR1ペプチドの量はその後決定される。これは当業者に既知のあらゆる種々の方法において、例えば、検出可能なラベルでタグ化されたVEGFR1ペプチド(又はSMYD3ポリペプチド)種を用いることにより、或いは、洗浄した基質を、VEGFR1ペプチド(又はSMYD3ポリペプチド)に対する、ラベルした抗体と接触させることにより、達成してもよい。SMYD3ポリペプチド(又はVEGFR1ペプチド)に結合したVEGFR1ペプチド(又はSMYD3ポリペプチド)の量は、試験化合物のVEGFR1ペプチドとSMYD3ポリペプチドとの会合を阻害する能力に反比例することとなる。タンパク質、例えば、これに限られないが、抗体のラベル化は、Harlow & Lane, Antibodies, A Laboratory Manual (1988)に記載されている。
【0065】
そのほかでは、VEGFR1ペプチド(又はSMYD3ポリペプチド)を、アフィニティータグでラベルする。ラベルしたVEGFR1(又はSMYD3ポリペプチド)を、次いで、試験化合物及びSMYD3ポリペプチド(又はVEGFR1ペプチド)とともにインキュベートし、さらに免疫沈降する。免疫沈降に引き続いて、SMYD3ポリペプチド(又はVEGFR1ペプチド)に対する抗体を用いたウェスタンブロットを行う。上述の競合的アッセイフォーマットのように、VEGFR1ペプチド(又はSMYD3ポリペプチド)に会合して検出されるSMYD3ポリペプチド(VEGFR1ペプチド)の量は、試験化合物のVEGFR1ペプチド及びSMYD3ポリペプチドの会合を阻害する能力に反比例する。
【0066】
2.非競合的アッセイ
非競合的結合アッセイもまた、本明細書中に記載されているような競合的アッセイを用いたスクリーニングを容易に適用できないフォーマットで構築された試験化合物のライブラリの初期スクリーニングとして、有用性が認められ得る(例えば、Barret et al., Anal Biochem 1992, 204: 357-64を参照)。
【0067】
ファージライブラリーは、多数の異なる組換えペプチドの実用的な量を高速に産生する点で有用である。ファージライブラリーは、本発明に係る競合的アッセイに活用できないが、非競合的フォーマットで効率的にスクリーニングして、どの組換えペプチド試験化合物がVEGFR1ペプチド又はSMYD3ポリペプチドに結合するかを決定することができる。結合するものとして同定される試験化合物をその後産生し、競合的アッセイフォーマットを用いてスクリーニング可能である。ファージ及び細胞表示ライブラリーの産生及びスクリーニングは当業者には周知であり、例えば、Ladner et al., WO 88/06630; Fuchs et al., Biotechnology 1991, 9: 1369-72; Goward et al., TIBS 1993, 18: 136-40; Charbit et al., EMBO J 1986, 5: 3029-37; Cull et al., PNAS USA 1992, 89: 1865-9; Cwirla et al., PNAS USA 1990, 87: 6378-82にて論じられている。
【0068】
例示的な非競合的アッセイの後には、一方の構成成分(VEGFR1ペプチド又はSMYD3ポリペプチド)の添加の無い、競合的アッセイについて記載したものと類似の処理が行われる。しかし、非競合的アッセイがVEGFR1ペプチド又はSMYD3ポリペプチドと結合する試験化合物を決定する際、試験化合物のVEGFR1ペプチドとSMYD3ポリペプチドの両方と結合する能力は、各候補について決定しなければならない。従って、例によれば、固定されたVEGFR1ペプチドへの試験化合物の結合は、結合していない試験化合物を洗浄除去し、結合した試験化合物を支持体から溶出させ、次いで、溶出物を、例えば、質量分析、タンパク質決定(Bradford又はLowryアッセイ、或いは280nmの吸光分析)で分析することにより、決定できる。あるいは、溶出工程を無くし、試験化合物の結合を、支持体表面での有機層の分光学的性質の変化をモニタリングすることにより決定してもよい。表面の分光学的性質をモニタリングする方法は、吸光度、反射率、透過率、複屈折、屈折率、回折、表面プラズモン共鳴、偏光解析法、共振ミラー技術、格子結合波導技術(grating coupled waveguide technique)、多極共振解析(multipolar resonance spectroscopy)が挙げられるが、これに限られず、これらの全ては当業者には既知である。アッセイではラベルした試験化合物を用い、溶出工程を省くこともできる。この場合、非結合物質を洗浄除去後、支持体に会合したラベルの量が、試験化合物の結合に正比例する。
【0069】
多数の周知のロボットシステムが、液相化学において開発されてきている。これらのシステムは、武田薬品工業株式会社(大阪、日本)により開発された自動化合成装置のような自動化されたワークステーション、及び、化学者によりなされる手動の合成操作を模倣したロボットアーム(Zymate II、Zymark Corporation、Hopkinton、Mass.;Orca、Hewlett Packard、Palo Alto、Calif.)を利用する多くのロボット合成を含む。上記装置のいずれも本発明での使用に適している。本明細書中で論じたように操作できるようこれらの装置に修正を行う性質及び実行は(もしあれば)、関連技術分野の当業者には明白であろう。さらに、非常に多くのコンビナトリアルライブラリーはそれ自体商業的に入手可能である(例えば、ComGenex、Princeton、N.J.;Asinex、Moscow、Ru;Tripos、Inc.、St. Louis、MO;ChemStar、Ltd、Moscow、RU;3D Pharmaceuticals、Exton、PA、Martek Biosciences、Columbia、MD、など)。
【0070】
種々の低スループット又は高スループット酵素アッセイフォーマットが当業者に知られており、SMYD3のメチルトランスフェラーゼ活性の検出又は測定に容易に適用可能である。高スループットアッセイについては、VEGFR1ペプチド基質は通常のように固体支持体、例えばマルチウェルプレート、スライド又はチップに固定化できる。反応後、メチル化産生物は上述の方法により個体支持体上で検出できる。また、メチルトランスフェラーゼ反応は、溶液中でも起こり、VEGFR1ペプチドが固体支持体上に固定化できた後、メチル化産生物を検出できる。このようなアッセイを実施するため、固体支持体にストレプトアビジン、及びビオチンでラベルしたVEGFR1を塗布してもよく、或いは、固体支持体に抗VEGFR1抗体を塗布してもよい。当業者であれば、所望のスクリーニングのスループット能力によって、適したアッセイフォーマットを決定できる。
【0071】
本発明は、また、本発明に係るタンパク質の部分ペプチドの使用も含む。部分ペプチドはSMYD3タンパク質に特異的なアミノ酸配列を有し、好ましくは、約400アミノ酸未満、通常は約200アミノ酸未満、場合により約100アミノ酸未満、そして、少なくとも約7アミノ酸、好ましくは約8アミノ酸以上、より好ましくは約9アミノ酸以上のアミノ酸配列からなる。部分ペプチドは、例えば、SMYD3タンパク質に結合する因子又は化合物のスクリーニング、及び、SMYD3と例えばSAMのようなそのコファクターとの間の結合の阻害剤のスクリーニングに用いることができる。かかるスクリーニング方法の態様において、SETドメインを含有する部分ペプチドが好ましい。
【0072】
本発明の態様において有用な部分ペプチドは、遺伝子工学により、既知のペプチド合成方法により、又は、本発明にかかるタンパク質を適当なペプチダーゼで分解することにより産生できる。ペプチド合成のため、例えば、固相合成又は液相合成を用いることができる。
【0073】
SETドメインの変異を有するSMYD3変異体は、細胞増殖に抑制効果を示す。従って、SMYD3の部分ペプチドは、好ましくは、SETドメイン“NHSCXXN”及び/又は“GEELXXXY”を含む。
【0074】
任意の試験化合物を用いることができる。例として、細胞抽出物、細胞培養上清、微生物の発酵産生物、海洋生命体の抽出物、植物の抽出物、精製又は粗タンパク質、ペプチド、非ペプチド化合物、合成微小化合物及び天然化合物が挙げられるが、これに限られない。本発明に係る試験因子又は化合物は、技術分野で既知のコンビナトリアルライブラリー方法を用いて得ることもでき、これには、(1)生物学的ライブラリー、(2)空間的にアドレス可能な平行(spatially addressable parallel)固相又は液相ライブラリー、(3)逆重畳積分を必要とする合成ライブラリー方法、(4)「1ビーズ1化合物(one-bead one-compound)」ライブラリー方法、及び、(5)アフィニティークロマトグラフィーによる選択を用いる合成ライブラリー方法が挙げられるが、これに限られない。アフィニティークロマトグラフィーによる選択を用いる生物学的ライブラリー方法はペプチドライブラリーに限られるが、他の4つのアプローチはペプチド、非ペプチドオリゴマー又は化合物の小分子ライブラリーに適用できる(Lam, Anticancer Drug Des. 12: 145-67 (1997))。分子ライブラリーの合成方法の例には、当該技術分野において見出すことができる(DeWitt et al., Proc Natl Acad Sci U S A. 1993 Aug 1;90(15):6909-13.; Erb et al., Proc. Natl. Acad. Sci. USA 91: 11422-6 (1994); Zuckermann et al., J. Med. Chem. 37: 2678-85 (1994); Cho et al., Science 261: 1303-5 (1993); Carell et al., Angew. Chem. Int. Ed. Engl. 33: 2059 (1994); Carell et al., Angew. Chem. Int. Ed. Engl. 33: 2061 (1994); Gallop et al., J. Med. Chem. 37: 1233-51 (1994))。化合物のライブラリーは溶液(Houghten, Bio/Techniques 13: 412-21 (1992)を参照)、ビーズ(Lam, Nature 354: 82-4 (1991))、チップ(Fodor, Nature 364: 555-6 (1993))、バクテリア(米国特許番号第5,223,409)、胞子(米国特許番号第5,571,698;5,403,484及び5,223,409)、プラスミド(Cull et al., Proc. Natl. Acad. Sci. USA 89: 1865-9 (1992))、又はファージ(Scott and Smith, Science 249: 386-90 (1990); Devlin, Science 249: 404-6 (1990); Cwirla et al., Proc. Natl. Acad. Sci. USA 87: 6378-82 (1990); Felici, J. Mol. Biol. 222: 301-10 (1991); 米国特許出願公開20020103360)に提示されてもよい。本発明に係る試験化合物は、単一化合物又は組み合わせ化合物でよい。本発明のスクリーニング方法で組み合わせ化合物が用いられた場合、化合物は連続的に又は同時に接触させることができる。
【0075】
本明細書中に記載されるアッセイに有用な試験因子又は化合物は、SMYD3、又は、メチルトランスフェラーゼ活性を失った部分SMYD3ペプチドに特異的に結合する抗体の形態をとってもよい。例えば、抗体(例えば、モノクローナル抗体)は、SMYD3及びそのVEGFR1基質の間の結合を阻害する能力について試験可能である。
【0076】
本発明のスクリーニング方法により単離される因子又は化合物は、SMYD3のメチルトランスフェラーゼ活性を阻害する候補薬剤であり、従って、肝細胞癌、直腸結腸癌、膀胱癌及び/又は乳癌の治療または予防に適用されうる。
【0077】
さらに、SMYD3のメチルトランスフェラーゼ活性を阻害する因子又は化合物の部分構造が付加、欠失、及び/又は置換により転換された因子又は化合物も、本発明に係るスクリーニング方法により得られ得る因子又は化合物に含まれる。
【0078】
上述のように、SMYD3のメチルトランスフェラーゼ活性を阻害する因子又は化合物は、SMYD3のメチルトランスフェラーゼ活性を失った部分ペプチドであってもよく、又は、SMYD3に対する抗体であってもよい。本明細書中で用いられる場合、「抗体」の語は、抗体の合成に用いられた抗原とのみ、又は、これに非常に関連する抗原と相互作用(すなわち、結合)する特異構造を有する免疫グロブリン分子をさす。さらに、抗体は、SMYD3遺伝子でコードされるタンパク質に結合する限り、抗体の断片又は修飾された抗体であってよい。例えば、抗体断片は、Fab、F(ab’)2、Fv又は単鎖Fv(scFv)であってよく、ここで、H及びL鎖からのFv断片は、適当なリンカーにより接合される(Huston J. S. et al. Proc. Natl. Acad. Sci. U.S.A. 85:5879-5883 (1988))。より詳細には、抗体断片はパパイン又はペプシンのような酵素で抗体を処理することにより生成してもよい。また、抗体断片をコードする遺伝子を構築し、発現ベクターに挿入し、適当な宿主細胞中で発現させてもよい(例えば、Co M. S. et al. J. Immunol. 152:2968-2976 (1994); Better M. and Horwitz A. H. Methods Enzymol. 178:476-496 (1989); Pluckthun A. and Skerra A. Methods Enzymol. 178:497-515 (1989); Lamoyi E. Methods Enzymol. 121:652-663 (1986); Rousseaux J. et al. Methods Enzymol. 121:663-669 (1986); Bird R. E.及びWalker B. W. Trends Biotechnol. 9:132-137 (1991)を参照)。
【0079】
抗体は、例えば、ポリエチレングリコール(PEG)のような種々の分子との接合により修飾されてもよい。本発明は、かかる修飾抗体を提供する。修飾抗体は、抗体を化学的に修飾することにより得ることができる。そのような修飾方法は、当該分野において一般的である。また、抗体は、非ヒト抗体由来の可変領域及びヒト抗体由来の定常領域を有するキメラ抗体、或いは、非ヒト抗体由来の相補性決定領域(CDR)、ヒト抗体由来のフレームワーク領域(FR)及び定常領域からなるヒト化抗体の形態をとってもよい。このような抗体は、通常の技術を用いて調製可能である。ヒト化はげっ歯類のCDR又はCDR配列をヒト抗体の対応する配列と置換することにより行うことができる(例えば、Verhoeyen et al., Science 239:1534-1536 (1988)参照)。従って、かかるヒト化抗体はキメラ抗体であって、実質的には無処理のヒト可変ドメインが、非ヒト種由来の対応する配列により置換されているだけにすぎない抗体である。
【0080】
ヒトの可変領域並びにヒトのフレームワーク及び定常領域よりなる完全なヒト抗体も使用可能である。かかる抗体は当業者には既知の種々の技術を用いて産生可能である。例えば、バクテリオファージ上に表されるヒト抗体断片の組換えライブラリーの使用を含むインビトロ方法が用いられ得る(例えば、Hoogenboom & Winter, J. Mol. Biol. 227:381 (1991))。同様に、ヒト抗体は、ヒト免疫グロブリン座位を、遺伝子組換え動物、例えば、内在性免疫グロブリン遺伝子が部分的に又は完全に不活性化されているマウスに導入することにより作成できる。このアプローチは、例えば、米国特許番号第6,150,584、5,545,807、5,545,806、5,569,825、5,625,126、5,633,425、5,661,016に記載されている。
【0081】
本発明に係る方法により単離された因子又は化合物をヒト及び他の哺乳類、例えば、マウス、ラット、モルモット、ウサギ、ネコ、イヌ、ヒツジ、ブタ、ウシ、サル、ヒヒ及びチンパンジー、の医薬品として投与する場合、単離された因子又は化合物は直接投与してもよく、或いは、既知の医薬品調製方法を用いる投薬形態へ製剤化されてもよい。例えば、必要により、薬剤は、糖衣タブレット、カプセル、エリキシル剤及びマイクロカプセルとして経口で、或いは、滅菌溶液又は懸濁液を水又は薬学的に許容可能な液体とともに注射する形態で非経口で与えられてもよい。さらに、因子又は化合物は薬学的に許容可能なキャリア又は媒体、特に、滅菌水、生理食塩水、植物オイル、乳化剤、懸濁剤、界面活性剤、安定剤、芳香剤、賦形剤、ビヒクル、保存料、結合剤などと、一般的に許容された薬剤付与に必要とされる単位用量形態に混合することができる。これらの調合剤中の活性成分の量は、取得できる指示範囲内の適量な投与量となる。
【0082】
タブレット及びカプセルに混合されうる添加剤の例としては、ゼラチン、コーンスターチ、トラガントゴム及びアラビアゴムなどの結合剤;結晶セルロースなどの賦形剤;コーンスターチ、ゼラチン、アルギニン酸などの膨張剤;ステアリン酸マグネシウムなどの潤滑剤;スクロース、ラクトース又はサッカリンなどの甘味剤;及び、ペパーミント、アカモノ(Gaultheria adenothrix)オイル及びチェリーなどの芳香剤が挙げられる。単位用量形態がカプセルである場合、オイルなどの液体キャリアも上記成分に含まれうる。注射用滅菌組成物は、注射に用いられる滅菌水などのビヒクルを用いる通常の薬剤付与に従って製剤化することができる。
【0083】
生理食塩水、グルコース及び、D-ソルビトール、D−マンノース、D−マンニトール及び食塩などのアジュバントを含む他の等張液を注射用水性液として用いることができる。これらはアルコール、特にエタノールなどの適切な溶解剤、プロピレングリコール及びポリエチレングリコールなどのポリアルコール、Polysorbate80(商標)及びHCO−50などの界面活性剤とあわせて用いることができる。
【0084】
ゴマ油又は大豆オイルは油性の液体として使用でき、溶解剤としての安息香酸ベンジル又はベンジルアルコールと併せて使用でき、リン酸バッファー及び酢酸ナトリウムバッファーなどのバッファー;塩酸プロカインなどの鎮痛剤;ベンジルアルコール及びフェノールなどの安定剤;及び抗酸化剤とともに製剤化されてもよい。調製した注射は、適切なアンプルに充填されてもよい。
【0085】
当業者に既知の方法を、例えば動脈、静脈又は経皮注射として、そしてまた、鼻腔内、筋肉内又は経口投与として、本発明に係る医薬品組成物を患者に投与するために用いることができる。用量及び投与方法は患者の体重及び年齢並びに投与方法によって変わる;しかし、当業者は適した投与方法を慣例的に選択することができる。さらに、当該因子又は化合物がDNAによりコード可能である場合、DNAを遺伝子治療のためのベクターに挿入することができ、該ベクターを、治療を行うために患者に投与することができる。投与の用量及び方法は、患者の体重、年齢及び症状によって変わるが、当業者は適切に選択することができる。
【0086】
例えば、SMYD3に結合し、その活性を制御する因子又は化合物の用量は、症状に依存するが、通常の成人(体重60kg)に経口投与される場合、典型的な用量は、1日当たり約0.1mg〜約100mg、好ましくは1日当たり約1.0mg〜約50mg、より好ましくは1日当たり約1.0mg〜約20mgの範囲である。
【0087】
非経口的に投与する場合、通常の成人(体重60kg)への注射形態では、患者、標的器官、症状及び投与方法に応じていくつか違いがあるものの、1日当たり約0.01mg〜約30mg、好ましくは1日当たり約0.1mg〜約20mg、より好ましくは1日当たり約0.1mg〜約10mgの用量での静脈注射が都合がよい。さらに、他の動物の場合にも、体重60kgに変換した量を投与することができる。
【0088】
本発明はさらに被験体(subject)の癌、例えば、肝細胞癌、直腸結腸癌、膀胱癌及び乳癌、の治療のための方法を提供する。投与は、SMYD3のメチルトランスフェラーゼ活性の異常に関連する障害の危険がある(又は疑いのある)、又は、障害を有する被験体に対し、予防的又は治療的でありうる。該方法は、適した癌細胞でのSMYD3の機能を減少させることを含む。機能は、本発明に係る方法のスクリーニングにより得られる因子又は化合物の投与を介して阻害されうる。
【0089】
他の形態において、本発明は、本明細書中に記載された1又は2以上の治療的因子又は化合物を含有する薬学的、又は治療的組成物を含む。さらに、本発明はまた、本明細書中に記載された1又は2以上の治療的因子又は化合物の、癌、より具体的には、肝細胞癌、直腸結腸癌、膀胱癌及び乳癌の治療及び/又は予防のための、医薬的又は治療的組成物の製造のための使用を提供する。薬学的製剤は、経口、直腸、鼻腔、外用(頬側及び舌下を含む)、腟内又は非経口的(筋肉内、皮下及び静脈)投与、或いは、吸入又はガス吸入による投与に適したものを含んでもよい。前記製剤は、適当な場合、分離した用量単位で簡便に与えられてもよく、薬学業界では既知のいかなる方法により調製されてもよい。全てのかかる薬学的方法は、活性化合物を液体キャリア又は微細に分割された固体キャリア或いは必要に応じて両方とを会合させ、次いで、必要な場合には、生成物を所望の製剤に形作る工程を含む。
【0090】
経口投与に適した薬学的製剤は、それぞれが所定量の活性成分を含有する、カプセル、カシェ剤、又はタブレットなどの分離した用量として;粉末又は顆粒として;或いは溶液、懸濁液又はエマルションとして、簡便に与えられてもよい。活性成分は、巨丸剤又は糊状剤として与えられてもよく、純粋な形態、すなわち、キャリア無しで与えられてもよい。経口投与のためのタブレット及びカプセルは、一般的な賦形剤、例えば、結合剤、充填剤、潤滑剤、崩壊剤、潤湿剤を含有してもよい。タブレットは、圧縮又は成型により、場合により、1又は2以上の製剤成分とともに作成してもよい。圧縮タブレットは適切な機械中で粉末又は顆粒などの流通形態で活性成分を、場合により結合剤、潤滑剤、不活希釈剤、界面活性又は分散剤とともに混合して、圧縮することにより調製できる。成型タブレットは、不活性液体希釈剤で濡らした粉末化化合物の混合物を適した機械中で成型することにより作成できる。タブレットは、当業者に周知の方法によりコートしてもよい。経口液体調製物は、例えば、水性又は油性の懸濁液、溶液、エマルション、シロップ又はエリキシル剤の形態であってよく、又は、使用前に水又は他の適したビヒクルでの構成を目的とする乾燥生成物として与えられてもよい。かかる液体調製物は、通常の添加剤、例えば、懸濁剤、乳化剤、非水性ビヒクル(食用オイルを含んでもよい)、又は保存剤を含んでもよい。タブレットは、場合により、中の活性成分がゆっくりと又は制御されて放出されるように製剤化されていてもよい。
【0091】
非経口投与のための製剤は、製剤を目的レシピエントの血液と等張に保持する抗酸化剤、バッファー、静菌剤および溶質を含んでもよい水性及び非水性の滅菌注射溶液;並びに、懸濁剤及び増粘剤を含んでもよい水性又は非水性の滅菌懸濁液を含んでもよい。製剤は、単位用量又は多用量容器、例えば、密閉アンプル及びバイアルに入れて与えられてもよく、使用直前に滅菌液体キャリア、例えば、生理食塩水、注射水の添加のみが求められるフリーズドライ(凍結乾燥)状態で貯蔵されてもよい。また、製剤は連続的点滴用に与えられてもよい。即席の注射溶液及び懸濁液は、上述した類の滅菌粉末、顆粒及びタブレットから調製することができる。
【0092】
直腸投与のための製剤は、ココアバター又はポリエチレングリコールなどの通常のキャリアを伴う座薬として与えられてもよい。例えば、頬側又は舌下の、口中への局所投与のための製剤には、スクロース及びアラビアゴム又はトラガントなどの風味付けされた基剤に活性成分を含む薬用キャンディー、及び、ゼラチン及びグリセリン又はスクロース又はアラビアゴムなどの基剤に活性成分を含むトローチが含まれる。鼻腔内投与のため、本発明により得られる化合物は液体スプレー又は分散可能な粉末として或いは液滴の形態で使用されてもよい。液滴は、水性又は非水性基剤とともに製剤化され、1又は2以上の分散剤、溶解剤又は懸濁剤も含んでよい。液体スプレーは、圧縮パックから簡便に送達される。
【0093】
吸入による投与のため、化合物は吸入器、噴霧器、圧縮パック又はエアロゾルスプレーを送達する他の簡便な手段から、簡便に送達される。圧縮パックは、ジクロロジフルオロメタン、トリクロロフルオロメタン、ジクロロテトラフルオロエタン、二酸化炭素又は他の適したガスなどの適した噴霧剤を含んでもよい。圧縮エアロゾルの場合、投与単位はある計量量を送達する弁を設けることにより決定可能である。
【0094】
また、吸入又はガス注入による投与のため、化合物は乾燥粉末組成物、例えば、化合物とラクトース又はスターチなどの適した粉末基剤との粉末混合物の形態とされてもよい。粉末組成物は、粉末が吸入又はガス注入の助けで投与され得る単位用量形態、例えば、カプセル、カートリッジ、ゼラチン又はブリスターで与えられてもよい。
【0095】
所望の場合には、活性成分を徐放するよう適合された上述の製剤を用いてもよい。薬学的組成物は、抗菌剤、免疫抑制剤又は保存剤などの他の活性成分を含んでもよい。
【0096】
特に上述した成分に加えて、本発明の製剤は、問題の製剤の種類に関して、当業者には一般的な他の因子を含んでよいこと、例えば、経口投与に適したものが香味料を含んでよいことは理解されるべきである。
【0097】
好適な単位投与製剤は、活性成分の効果用量(後述)又はその適当な画分を含有するものである。
【0098】
上記条件のそれぞれについて、組成物は経口又は注射により、1日当たり約0.1〜約250mg/kgの用量で投与されてもよい。成人の用量範囲は、一般に1日当たり約5mg〜約17.5g、好ましくは1日当たり約5mg〜約10g、最も好ましくは1日当たり約100mg〜約3gである。タブレット又は分散単位で与えられる他の単位用量形態には、そのような用量で効果的である量又はこれと同じ用量の数倍として、例えば、約5mg〜約500mg、通常約100mg〜約500mgを、簡便に含んでもよい。
【0099】
薬学的組成物は、好ましくは、経口又は注射(静脈又は皮下)により投与され、被験体に投与される正確な量は付き添いの医師の責任となる。しかし、用いられる用量は、被験体の年齢及び性別、治療される正確な障害及びその深刻度を含む数々の要因による。投与の経路もまた、状態及びその深刻度により変化しうる。
【0100】
本発明はさらに、試験化合物のVEGFR1のメチル化を制御する能力を検出するキットを提供し、該キットは、以下の構成要素:
(a)以下からなる群から選択されるメチルトランスフェラーゼ活性を有するSMYD3ポリペプチド;
(i)配列番号2のアミノ酸配列を有するポリペプチド;
(ii)配列番号2のアミノ酸配列を有するポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失又は挿入され、さらにここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性であるアミノ酸配列を有するポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(iv)配列番号1のヌクレオチドを有するポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(b)(a)のポリペプチドによりメチル化され得るVEGFR1ペプチド;及び
(c)VEGFR1ペプチドのメチル化のコファクター
を含む。
【0101】
本発明において、好ましいVEGFR1ペプチドは、配列番号4のアミノ酸配列から構成されるポリペプチドまたはその機能的変異体もしくは機能的断片を含む。好ましい断片には、配列番号4のアミノ酸配列の位置800〜841のアミノ酸配列を含むか又は、それらから構成されるポリペプチド、或いは、VEGFR1のリジン831を含むポリペプチドを含むが、これに限られない。さらに、好ましいコファクターは、S−アデノシルホモシステインヒドロラーゼ(SAHH)である。本発明に係るキットは、メチル化されたVEGFR1ペプチドを検出する試薬をさらに含んでもよい。例えば、メチル化VEGFR1ペプチドを認識する抗体は検出試薬として使用可能である。アッセイを行うための説明書(例えば、書類、テープ、VCR、CD−ROM等)がキットに含まれてもよい。キットのアッセイフォーマットは、当業者には既知のメチルトランスフェラーゼアッセイ又は結合アッセイである。
【0102】
以下の例は、単なる例示であり、本発明の範囲を限定することを意図するものではない。本発明の態様が以下の実施例に記載されるものの、当業者は、本明細書中に記載されたものと同様の又は等価な他の方法及び材料が本発明の実施又は試験に用いることができることを認識するであろう。
【0103】
実施例
材料と方法
試薬及びウェスタンブロット分析
タンパク質の抽出、IP(免疫沈降)、及びウェスタンブロット分析は、文献に記載の方法に従って行った(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)。イムノブロット分析は抗SMYD3抗体(Abcam、Cambridge、MA)、抗HA抗体(Sigma、St Louis、MO)、抗Flag抗体(Sigma)、抗GST抗体(BD Pharmingen、San Diego、CA)、抗βアクチン抗体(Sigma)、又は抗リン酸化チロシン抗体(PY20、Sigma)を用いて行った。HRP接合二次抗体を、ECL検出システム(Amersham Biosciences、Piscataway、NJ)に用いた。組換えヒストンH3及びVEGFR1(3108)は、Upstate Biotechnology(Lake Placid、NY)及びCalbiochem(San Diego、CA)からそれぞれ購入した。メチル化リジン831特異的抗体(抗K831me2)は、ジメチル化リジン831を含む合成ペプチドであるRERLKLGKm2SLGRGAで免疫したウサギの血清から精製した。
【0104】
細胞株及び組織標本:
ヒト胎児腎細胞株(HEK293及び293T)及びヒト結腸癌細胞株(SW480)は、American Type Culture Collection(ATCC、Manassas、VA)から入手した。SNU423HCC細胞及びMCF7乳癌細胞はKorea cell−line bank及び財団法人癌研究会からそれぞれ好意に基づいて提供された。全ての細胞株は、適切な培養液中、単層で成長させた。
【0105】
プラスミドの調製
SMYD3を発現するプラスミドをすでに文献に記載されているように調製した(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)。本発明者らは、また、VEGFR1の野生型又は欠失形態のいずれかを含む種々のRT-PCR産生物を、pCMV−HA(Clutch、Palo Alto、CA)、p3xFlag−CMV14(sigma)又はpGEX6P−3ベクター(Amersham Biosciences)の適切な部位にクローニングすることにより、HAタグ化又は3xFlagタグ化VEGFR1を発現するプラスミドを調製した。RT-PCR実験は、プライマーの組(表1)を用いて行った。アミノ酸配列の置換を含む変異体VEGFR1プラスミドは、Quickchange II XL Site-directed Mutagenesis kitを用いてサプライヤーのプロトコルに従って生成した(Strata gene、La Jolla、CA)。
【0106】
本発明者らは、FuGENE(商標)6試薬を用いてサプライヤーのプロトコル(Roche、Indianapolis、IN)に従って培養細胞に哺乳類のプラスミドをトランスフェクトした。組換えGST融合VEGFR1タンパク質を大腸菌、BL21バクテリア細胞からグルタチオンセファロース4B(Amersham Biosciences)を用いて精製した。本実施例で用いたプライマーを以下に示す。
【0107】
(表1)プライマー配列リスト

【0108】
インビトロメチルトランスフェラーゼ及びキナーゼアッセイ
293T細胞に、Flagタグ化野生型SMYD3を発現するプラスミド(p3XFLAG−CMV−SMYD3)をトランスフェクトした。タグ化SMYD3タンパク質は、抗Flag抗体を用いる免疫沈降により精製した。組換えSMYD3タンパク質は、バキュロウイルスシステム(Invitrogen、Carlsbad、CA)を用いてSf9細胞中で調製した。インビトロメチルトランスフェラーゼアッセイは、わずかに修正を施して他の文献に記載されたように(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)行った。
【0109】
簡単にいえば、免疫沈降した又は組換えSMYD3タンパク質を、1μgの組換えヒストンH3又はVEGFR1タンパク質と、メチルドナーとしての2μCiの[メチル−3H]ラベル化S−アデノシル−L−メチオニン(SAM、Amersham Biosciences)の存在下、S−(5’−アデノシル)−L−ホモシステインヒドロラーゼ(SAHH、Sigma)の存在下又は非存在下、メチルトランスフェラーゼバッファー(50mM Tris−HCl pH8.5、100mM NaCl、10mM DTT)中で混合し、30℃で1時間インキュベートした。ラベル化タンパク質は液体シンチレーションカウンターで測定し、又は、SDS−PAGE後、フルオログラフィーで検出した。インビトロキナーゼアッセイをGST融合VEGFR1及びHTScan(商標)VEGFR1キナーゼアッセイキットを用い、サプライヤーのプロトコルに従って行った(Cell Signaling、Danvers、MA)。基質のリン酸化は、Time−Resolved Plate Reader(Perkin−Elmer、Wellesley、MA)を用いて615nmの蛍光発光により定量した。
【0110】
インビボメチル化アッセイ
VEGFR1のインビボメチル化は、Liu及びDreyfuss(Liu Q and Dreyfuss G., Molecular and cellular biology 1995, 15(5): 2800-8)に記載された方法に従い、わずかに修正を施して分析した。簡単に言えば、HAタグ化VEGFR1を発現するHEK293細胞を、100μg/mlのシクロヘキシミドとともに、37℃で30分間インキュベートし、その後、10μCi/mlのL−[メチル−3H]メチオニンを含む培養液中にそのまま3時間保持した。VEGFR1は、細胞抽出物から抗HA抗体で沈降し、SDS−PAGEにより分離し、続いて、BASイメージングシステム(BAS−TR2040、FUJI)により、又は、イムノブロット分析により分析した。
【0111】
RNA干渉
RNAi実験は、Dharmacon(Chicago、IL)から購入した、SMYD3に特異的な又は対照siRNAの二本鎖オリゴヌクレオチドで行った。1x105のHEK293−SMYD3細胞を、オリゴフェクタミンを用い、各siRNAで最終濃度100nMでトランスフェクトした。トランスフェクトから48時間後、タンパク質を細胞から抽出し、ウェスタンブロット分析にかけた。
【0112】
統計分析
全ての分析は、Statviewソフトウェアを用いて行った(SAS Institute、Cary、NC)。
【0113】
実施例1:SMYD3はVEGFR1をインビトロでメチル化する
最近の研究では、VEGFRがヒトの発癌に関与していることが示唆されている。従って、ここで、VEGFR1について、インビトロメチルトランスフェラーゼアッセイを用い、SMYD3の候補メチル化標的として調べた。本発明者らは、基質に関し、VEGFR1の3つの細胞質領域の組換えタンパク質、VEGFR1#1(コドン800−1000)、VEGFR1#2(コドン1000−1200)及びVEGFR1#3(コドン1200−1338)を調製した。SMYD3又は対照タンパク質を、Flagタグ化SMYD3又はモックプラスミドでそれぞれトランスフェクトしたHEK293細胞から抗Flag抗体で免疫沈降した。結果として、VEGFR1#1は、SMYD3によりインビトロで対照タンパク質と比較して有意にメチル化された(図1a)。しかし、VEGFR1#2又はVEGFR1#3において、メチル化は観察されず、VEGFR1#1がメチル化残基を含有することが示唆された。インビトロの組換えヒストンH3と比較して約3倍高いVEGFR1#1のメチル化が検出された(データ示さず)。
【0114】
インビトロメチルトランスフェラーゼアッセイをさらに、昆虫細胞から精製した組換えSMYD3タンパク質について行い、その結果は、VEGFR1#1のメチル化を示したが、VEGFR1#2もVEGFR1#3もメチル化を示さなかった(図1b)。メチル化残基を決定するため、本発明者らは、VEGFR1#1構築物の異なる断片を含む他のVEGFR1タンパク質を調製した(図1c)。インビトロメチルトランスフェラーゼアッセイは、コドン800−841を含むVEGFR1−N1、及びコドン800−878を含むVEGFR1−N7が、SMYD3によりメチル化されたことを示した。一方で、コドン842−878、879−920、842−900、879−900、842−900又は842−1000をそれぞれ含むVEGFR1−N2、N3、N4、N5、N6又はN8に、メチル化バンドは観察されなかった(図1c)。
【0115】
このデータは、800と841の間の領域がもっともメチル化残基を含む可能性が高いことを示唆する。興味深いことに、VEGFR2には、その細胞質領域については基質としても分析したが、メチル化バンドは検出されなかった(データ示さず)。まとめると、このデータはさらにSMYD3が、SET7/9及びSMYD2と同様に、非ヒストンタンパク質のメチル化に関し、基質特異性及び配列特異性の両方を有することを示唆する。
【0116】
実施例2:VEGFR1リジン831がSMYD3によりメチル化される
SMYD3を含む、SET含有メチルトランスフェラーゼのほとんどは、リジンを修飾する。従って、この領域内のリジンを調査した。リジン819、828、831及び840は他種のVEGFR1オルソログの中で保存されていることが決定された(データ示さず)。先の報告では、K/R−S/T/A−KがヒストンメチルトランスフェラーゼSET7/9の共通メチル化モチーフであることが示されているが(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)、他のヒストンメチルトランスフェラーゼの標的リジン、例えば、p53K370、ヒストンH3K4及びH3K27は、K/R−S/T/A−Kに隣接していないが、S/T−K−X又はX−K−Sには隣接している(図2a)。従って、これらのモチーフに一致し、他種に保存されたリジン819及び831は、本アッセイの中心であった。
【0117】
インビトロメチルトランスフェラーゼアッセイを、野生型及びリジンのアラニンへの置換を含む変異体VEGFR1−N1タンパク質を用いて行った(図2b)。アッセイにより、野生型及びK819A変異体のメチル化バンドが示されたが、K831A変異体は、いかなるメチル化バンドも示さなかった(図2b)。VEGFR1 K831が他のアミノ酸に置換された他の変異タンパク質も同様にメチル化を示さなかった(図2c)。リジン831のインビトロメチル化を確認するため、リジンに対するメチル化特異的抗体(抗K831me2)を調製した。インビトロメチルトランスフェラーゼアッセイは、フルオログラム並びに抗K831me2抗体を用いたウェスタンブロット分析により、組換えVEGFR1のメチル化されたリジン831を確認した(図2d)。
【0118】
実施例3:リジン831のインビボメチル化
VEGFR1のインビボメチル化を調べるため、インビボメチル化アッセイを、Flagタグ化SMYD3(293−SMYD3)を安定に発現するHEK293細胞、及び、対照細胞(293−モック)を用いて行った。細胞を、HAタグ化VEGFR1でトランスフェクトし、続いて、タンパク質合成阻害剤の存在下でL−[メチル−3H]メチオニンとともにインキュベートした。抗HA抗体での細胞の免疫沈降物を、SDS−PAGEにより分離し、蛍光映像装置(fluoroimager)によりメチル化されたVEGFR1を調べた。予想したように、293−モック細胞からのものと比較して、VEGFR1の強められたメチル化が293−SMYD3細胞からの沈降物中に検出された(図2e)。これと一致して、抗K831me2抗体を用いた沈降物のイムノブロット分析では、対照細胞と比較して強められたリジン831のメチル化が確認された(図2f)。
【0119】
SMYD3に依存したメチル化を確認すべく、293−SMYD3細胞をSMYD3−siRNA又は対照siRNAで処理した。抗SMYD3抗体を用いたイムノブロット分析は、SMYD3−siRNAが、対照siRNAと比較してSMYD3タンパク質の発現を効果的に減少させたことを示した(図2g、上パネル)。SMYD3のノックダウンと関連して、SMYD3−siRNAは、対照siRNAと比較して細胞中のK831のメチル化を減少させたが(図2g、第2パネル)、VEGFR1の発現はこれらのsiRNA間でほとんど変わらなかった(図2g、第3パネル)。これらのデータは、リジン831がSMYD3によってインビボでメチル化されることを示唆する。
【0120】
実施例4:SMYD3とVEGFR1との間の相互作用
VEGFR1はSMYD3によりメチル化されるため、本発明者らは、SMYD3がVEGFR1と相互作用するか否か調べた。本発明者らは、VEGFR1のFlagタグ化細胞質領域をHAタグ化SMYD3とともにHEK293細胞内で発現させた。抗HA抗体での免疫沈降及びその後の抗Flag抗体でのイムノブロット分析は、SMYD3がVEGFR1#1(コドン800−1000)とは会合するが、VEGFR1#2ともVEGFR1#3とも会合しないことを示した(図3a)。これと一致して、抗Flag抗体での免疫沈降及びその後の抗HA抗体でのイムノブロット分析は、SMYD3とVEGFR1#1との相互作用を実証した(図3a)。内在性SMYD3とVEGFR1との相互作用を調べるため、本発明者らは、さらに、SMYD3及びVEGFR1の両方を発現するSNU423、SW480又はMCF7細胞からの抽出物を用いた、抗SMYD3抗体での免疫沈降を行った。結果として、本発明者らは、抗SMYD3抗体での免疫沈降により、内在性SMYD3はVEGFR1と共免疫沈降することを見出し(図3b)、これは、SMYD3とVEGFR1との間のインビボ相互作用を示唆する。VEGFR1に対する抗体は、免疫沈降には適用できないため、本発明者らは抗VEGFR1抗体で免疫沈降は行わなかった。
【0121】
VEGFR1#1はSMYD3によりメチル化されるリジン831を含むので、これらのデータはVEGFR1のメチル化に一致するものである。本発明者らは、また、SMYD3の野生型及び3つの欠失型:SMYD3−N20(コドン20−428)、SMYD3−Δ2(コドン100−428)及びSMYD3−Δ3(コドン250−428)を発現するプラスミドを用い、結合に関与するSMYD3の領域を調べた。野生型SMYD3、SMYD3−N20及びSMYD3−Δ2はVEGFR1と相互作用するが、SETドメインの無いSMYD3−Δ3はしなかった(図3c)。SETドメイン(100−250)を発現する他のプラスミドは、VEGFR1の細胞質ドメインと会合を示した(図3d)。このデータはSETドメインが相互作用に関与することを示唆する。
【0122】
実施例5:SMYD3によるメチル化を介したVEGFR1のキナーゼ活性増大
リジン831はキナーゼドメインに位置する。そこで、VEGFR1メチル化のそのキナーゼ活性への影響をさらに調べた。キナーゼ活性は、基質のリン酸化チロシンを認識するVEGFR1キナーゼアッセイキットを用いて調べた。VEGFR1の細胞質領域の組換えタンパク質を3Hラベル化SAMで、SMYD3の存在下又は非存在下で処理し、その後、キナーゼ活性を調べた。予想したように、SMYD3によるVEGFR1のメチル化が、3H−BASシステムにより検出された(データ示さず)。重要なことに、SMYD3で処理したVEGFR1は、対照と比較してそのキナーゼ活性の有意な増大を示した(図4a)。VEGFR1無しのSMYD3単独によるリン酸化は観察されなかった(データ示さず)。抗リン酸化チロシン抗体でのウェスタンブロット分析はまた、SMYD3非存在下と比較して、SMYD3存在下では、組換えヒトVEGF165タンパク質を伴うFc領域に融合された組換えヒトVEGFR1タンパク質のリン酸化増大をも明らかとした(図4b)。興味深いことに、VEGFR1の組換え細胞質領域(コドン784−1338)をインビトロキナーゼアッセイに用いた場合には、VEGFR1の自己リン酸化は観察されなかった(データ示さず)。従って、VEGFR1のメチル化は、VEGFR1標的分子のリン酸化に影響し、そのシグナル伝達を強めることとなる。
【0123】
考察
本明細書中で示したように、VEGFR1はSMYD3ヒストンメチルトランスフェラーゼの、新規な非ヒストン標的である。ヒストン末端の修飾は、クロマチン構造の一部変質を介して、転写、DNA修復、テロメア維持、DNA複製及び染色体分離において重要な役割を果たす。最近の分子研究はヒストン末端のリジン修飾、及び、ヒストンメチルトランスフェラーゼ及びデメチラーゼによるその動的制御の重要性を明らかにしている(Volkel P and Angrand PO, Biochimie 2007, 89(1): 1-20)。17を超えるヒストンメチルトランスフェラーゼがこれまで同定されており、これらのメチルトランスフェラーゼは、それぞれの基質に特異性を有している。H3K4は、SET7/9、Set1、MLL1、MLL2、MLL3、MLL4、MLL5、ASH1及びSMYD3によりモノ、ジ、又はトリメチル化される(Kouzarides T, Cell 2007, 128(4): 693-705.; Volkel P and Angrand PO, Biochimie 2007, 89(1): 1-20)。
【0124】
重要なことに、これらのメチルトランスフェラーゼの中で、SET7/9及びSMYD2は、非ヒストンタンパク質をメチル化することが知られている(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82;Chuikov S et al., Nature 2004, 432(7015): 353-60;Huang J et al., Nature 2006, 444(7119): 629-32)。SET7/9はインビトロ及びインビボでp53中のリジン372をメチル化し、p53の安定性を上昇させた(Chuikov S et al., Nature 2004, 432(7015): 353-60)。SET7/9は、TAF10のリジン189(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82)及びTAF7のリジン5(ヒトTATAボックス結合タンパク質関連因子)でのメチルトランスフェラーゼ活性も示した(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)。
【0125】
さらに、p53 K370のメチル化は、SMYD3(SET及びMYNDドメイン)ファミリーのもう1つのメンバーであるSMYD2により示されている(Huang J et al., Nature 2006, 444(7119): 629-32)。本明細書中において、VEGFR1が、細胞質及び核に局在する、H3K4のヒストンメチルトランスフェラーゼの新規な非ヒストン標的であることが確認される。他のH3K4メチルトランスフェラーゼ、例えば、Set1、MLL1、MLL2、MLL3、MLL4、MLL5及びASH,1は、同じヒストンリジン修飾を認識するはずであり、基質がどのように異なるメチルトランスフェラーゼにより修飾されるかを見ることは関心あることであろう。ヒストンメチル化は動的に制御されるので、非ヒストンタンパク質のメチル化もまたヒストンデメチラーゼにより修飾されうる。従って、VEGFRのメチル化されたリジン831は、LSD1(Shi Y et al., Cell 2004, 119(7): 941-53)やJARID1(Seward DJ et al., Nat Struct Mol Biol 2007, 14(3): 240-2)のようなH3K4デメチラーゼにより触媒されるであろう。
【0126】
VEGFRは、細胞外リガンド結合ドメイン、膜貫通ドメイン、キナーゼドメイン及びカルボキシル末端領域からなる受容体チロシンキナーゼ(RTK)である。VEGFR1を含む全てのRTKは、GXGXXG(ATP結合部位)、HRDLA(触媒に不可欠なモチーフ)及び1又は2のチロシン自己リン酸化部位を含む、進化的に保存されたキナーゼドメインを含む(Rahimi N, Experimental eye research 2006, 83(5): 1005-16.; Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。
【0127】
本明細書中で示したように、SMYD3が誘導するリジン831のメチル化は、インビトロでのVEGFR1のキナーゼ活性増大をもたらす。理論に縛られることは望まないものの、この結果はリジン残基が、キナーゼドメインの“GXGXXXG”モチーフからN末端に3アミノ酸に位置するという事実に由来すると考えられる。ゆえに、リジン831のメチル化はキナーゼドメインの立体構造を変化させうる。一方で、VEGFR1の細胞質ドメインのC末端領域がキナーゼ活性に対して阻害的役割を有するため、VEGFR1のメチル化は阻害的ドメインを抑制してキナーゼ活性を増大する(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。
【0128】
ヒストン末端についての最近の研究は、その修飾残基が特異的分子により認識されることを示している;リン酸化されたH3S10はGCN5、PVAF及びp300を増加させ、メチル化されたH3K9はヘテロクロマチンタンパク質1(HP1)を増加させ、かつメチル化されたH3K4はCHD1、SNF2L/ISWI、WDR5、BPTF/NURF301及びING2を増加させる。従って、メチル化リジン831は、VEGFR1のキナーゼ活性に影響を与え得るメチル化特異的タンパク質複合体を増加させる。ヒストン末端の修飾は、他のヒストン末端の修飾に関連し、これは共有結合的修飾間の相互作用を示す(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5;Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80;Li B et al., Cell 2007, 128(4): 707-19;Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。
【0129】
H3S10のリン酸化は、GCN5を媒介としたH3K14のアセチル化を促進するが、SUV39H1によるH3K9のメチル化を阻害する。反対に、H3K9のメチル化は、H3S10のリン酸化を抑制する。同様に、H4K3のメチル化はH4K8及びH4K12のアセチル化を促進する(Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。共有結合ヒストン修飾間の相互作用を見るに、本発明者らは、VEGFR1のメチル化が、その強められたキナーゼ活性による自己リン酸化、並びに、その後の分子間リン酸化を介したVEGFR2のリン酸化を変質しうると推測するよう誘導される。メチル化されたリジン831はTry1169、1213、1242、1327及び1333(VEGFR1のリン酸化されたチロシン)(Olsson AK et al., Nature reviews 2006, 7(5): 359-71)から離れているように見えるが、これらの残基はリジン831に構造的に近い位置を示し得る。さらに、異なるリガンドによるVEGFシグナル伝達についての今後の研究は、VEGFR1メチル化の役割についてのよりよい理解をもたらす可能性がある。
【0130】
認識モチーフ(K/R−S/T/A−K)がSET7/9メチルトランスフェラーゼに対する共通認識部位であることがこれまでの研究により報告されている(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)。しかし、他のヒストンメチルトランスフェラーゼによるメチル化リジンの隣接配列は、必ずしもこのモチーフと適合しない。ヒストンH3K9、H3K27及びp53のリジン370のアミノ酸配列の比較により、X−K−Sがヒストンリジンメチルトランスフェラーゼの別の保存されたモチーフであることが見出された。重要なことに、VEGFR1のメチル化されたリジン831は、この知見によく適合する。VEGFR2のキナーゼドメインはVEGFR1のものと70.1%のアミノ酸類似性を有するが、VEGFR2はSMYD3によりメチル化されない。VEGFR2のリジン831に対応するリジンは共通配列に隣接しないので、この結果は保存されたモチーフに矛盾しない。VEGFR1と比べて高いキナーゼ活性を示すVEGFR2がSMYD3によりメチル化されなかったことは興味深い。
【0131】
SMYD3はVEGFR2のメチル化に影響を与えない場合がある一方で、SMYD3はVEGFR1のリン酸化を介したVEGFR2のリン酸化には影響を与える場合があるが、これはこれら2つの分子がヘテロ2量体を形成するからである(Rahimi N, Experimental eye research 2006, 83(5): 1005-16;Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。1つの受容体のリン酸化が、分子内リン酸化により他の受容体のリン酸化に影響を与えることは周知である(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。酵母Rkm1(SETドメイン含有酵素)がRpl23a及びRpl23bに対するメチルトランスフェラーゼ活性を示したことはさらに留意すべきである(Porras-Yakushi TR et al., J Biol Chem 2007, 282(17):12368-76)。メチル化リジンの周辺の標的配列の比較により、N/P−P−Kがメチル化の標的でありうることが示唆された。従って、リジンの隣接配列は、複数のメチルトランスフェラーゼの間で異なる可能性がある。
【0132】
VEGFR1は血管表皮細胞上で排他的に発現すると言われてきたが、最近の研究では、非EC細胞内でのVEGFR1の発現が示されている。VEGFR1は大腸癌(Lesslie DP et al., Br J Cancer 2006, 94(11): 1710-7;Fan F et al., Oncogene 2005, 24(16): 2647-53;Duff SE et al., Eur J Cancer 2006, 42(1): 112-7;Andre T et al., International journal of cancer 2000, 86(2): 174-81)、乳癌(Li YS et al., Pathology international 2006, 56(5): 256-61;Wu Y et al., International journal of cancer 2006, 119(7): 1519-29)、膵臓癌(Chung GG et al., Cancer 2006, 106(8): 1677-84.; Yang AD et al., Cancer Res 2006, 66(1): 46-51)、前立腺癌(Jackson MW et a., Cancer Res 2002, 62(3): 854-9)、腎癌(Jacobsen J et al., BJU Int 2006, 97(5): 1102-8;Ljungberg BJ et al., BJU Int 2006, 98(3): 661-7)及び卵巣癌(Wang FQ et al., International journal of cancer 2006, 118(4): 879-88;Chen H et al., Gynecol Oncol 2004, 94(3): 630-5)の組織及び癌細胞株(Fan F et al., Oncogene 2005, 24(16): 2647-53;Duff SE et al., Eur J Cancer 2006, 42(1): 112-7;Wu Y et al., International journal of cancer 2006, 119(7): 1519-29;Jackson MW et a., Cancer Res 2002, 62(3): 854-9;Soker S et al., Am J Pathol 2001, 159(2): 651-9;Wu W et al., Oncogene 2003, 22(22): 3361-70)を含むヒト組織の広い範囲で発現されている。報告によれば、VEGFR1の発現は初期大腸癌標本ではかすかであったが、その発現は肝転移では明確に検出された(Fan F et al., Oncogene 2005, 24(16): 2647-53)。
【0133】
さらに、VEGFR1は腫瘍の成長及び進行に関連していることが報告された:VEGFR1の外因性発現は膵臓癌の侵襲と転移を強めた。すなわち、強められたSMYD3の発現は侵襲性の及び/又は転移する性質を癌細胞に付与する可能性がある。この場合、VEGFR1メチル化を標的とする治療アプローチは、癌細胞の侵襲と転移を阻害することにより患者を利するであろう。SMYD3のメチルトランスフェラーゼ活性の抑制は、VEGFR1を媒介とする癌の進行の阻害の助けとなりうる。
【0134】
本明細書中に示されるように、SMYD3はVEGFR1のリジン831をインビトロ及びインビボでメチル化し、メチル化されたVEGFR1はそのキナーゼ活性を非メチル化VEGFR1と比べて強められた。従って、多量のSMYD3タンパク質を発現する癌細胞は、VEGFR1を媒介とした強められたシグナル伝達経路を示し得る。このデータはVEGFR1の新規な制御メカニズム、及び、ヒトの発癌に関与する、調節解除されたVEGFR1シグナル伝達に光を当てる可能性がある。
【産業上の利用可能性】
【0135】
本発明は、直腸結腸癌、肝細胞癌、膀胱癌及び乳癌を含む種々の癌の予防、診断及び治療のためのさらなる分子標的の同定に有用である。さらに、本明細書中に報告されたデータは、癌の包括的な理解を向上させ、新規な診断戦略の開発を容易とし、治療的薬剤及び予防薬剤のための分子標的の同定のカギを提供する。かかる情報は、腫瘍発生のより深い理解に寄与し、癌の診断、治療及び最終的には予防の新規な戦略の開発のための指標を提供する。
【0136】
本発明は、詳細に、その特定の実施形態をもって記載されたが、上記記載は本質的に例示的かつ説明的なものであり、本発明及びその好ましい実施形態を例示することを意図することは理解されるべきである。通常の実験を通して、当業者は、種々の変化及び修正が本発明の精神及び範囲から逸脱することなくなされうることを容易に理解するであろう。従って、本発明は、上記記載によって定義されず、以下の特許請求の範囲及びその均等物により定義されることが意図される。
【技術分野】
【0001】
本発明は、転写制御、より具体的には、メチルトランスフェラーゼ活性を調整する因子、例えば、SMYD3(“ZNFN3A1”としても知られる)によるVEGFR1のメチル化を調整する因子の同定に関する。SMYD3が多くの癌種において上方制御されるので、そのように同定されるSMYD3モジュレーターは、例えば、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌を含む癌の治療に有用なことが判明する可能性がある。
【背景技術】
【0002】
ヒストン末端の修飾は、転写制御、テロメア維持、DNA複製及び染色体分離に重要な役割を果たす(Kouzarides T, Cell 2007, 128(4): 693-705)。このような修飾の例としては、アセチル化、リン酸化、メチル化、及び/又はユビキチン化が挙げられる。共有結合修飾は特に、クロマチン構造だけでなく、クロマチン結合タンパク質との相互作用も制御する(Kouzarides T, Cell 2007, 128(4): 693-705.; Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80)。さらに、H3K4、H3K36及びH3K79のメチル化は、ユークロマチン構造に関連し、一方、H3K9、H3K27及びH4K20のメチル化はヘテロクロマチン構造に関連する(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80)。
【0003】
クロマチンの構造は、転写の重要レギュレーターの一つである;非転写遺伝子はヘテロクロマチン内に小型化される一方、転写遺伝子はユークロマチン内に存在し、ユークロマチンでは、転写複合体は標的DNAにアクセス可能である(Li B et al., Cell 2007, 128(4): 707-19)。さらに、ヒストン残基の修飾は、その結合タンパク質との相互作用を促進し、他のヒストン末端へのその後の修飾に影響する(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5.; Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80.; Li B et al., Cell 2007, 128(4): 707-19.; Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。例えば、H3セリン10(H3S10)のリン酸化は、H3K9のメチル化を抑制する。逆に、H3K9のメチル化は、H3S10のリン酸化に拮抗する。H3S10のリン酸化は、GCN5によるH3K14のアセチル化を促進する(Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。
【0004】
H3K9のメチル化は、HP1の輸送に関与して、染色体領域を明確化し、これは、一方で、ヘテロクロマチンのドメインの確立及び維持に不可欠である(Nakayama J et al., Science 2001, 292(5514): 110-3.; Lachner M et al., Nature 2001, 410(6824): 116-20.; Bannister AJ et al., Nature 2001, 410(6824): 120-4)。HP1タンパク質のゲノム特定部位への動員は、クロマチンの多成分との相互作用に関与する(Nielsen SJ et al., Nature 2001, 412(6846): 561-5)。この動員は、転写活性に不可欠なタンパク質であるH3K4のメチル化を抑制すると考えられている。これらのデータは、異なる修飾間の相互作用により制御されるヒストン修飾の複雑な性質を示している。実際、これらのタンパク質の増大が、そのヒストンメチルトランスフェラーゼ活性を通じて腫瘍形成を促進又は阻害することが証明されてきている(Varambally S et al., Nature 2002, 419(6907): 624-9;Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40;Gibbons RJ, Human molecular genetics 2005, 14 Spec No 1: R85-92)。
【0005】
ヒストン末端のメチル化は熱心に研究が進められてきたものの、非ヒストンタンパク質のメチル化は、不明なままである。最近の研究では、ヒストンH3K4メチルトランスフェラーゼ(MTase)であるSET7/9が、TAF10及びp53を基質として触媒することが報告されている(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82;Chuikov S et al., Nature 2004, 432(7015): 353-60)。血管内皮成長因子受容体−1(VEGFR1)(Accession No.:NM_002019)は、受容体二量体化及びそのリガンドとの相互作用の点で、生理的及び病理学的な血管形成において機能する、受容体チロシンキナーゼ(RTK)である(Shibuya M et al., Oncogene 1990, 5(4): 519-24.; Rahimi N, Experimental eye research 2006, 83(5): 1005-16)。
【0006】
VEGFR1は、細胞外ドメイン、7つの免疫グロブリン(Ig)様配列、及び、長いキナーゼインサートを有する細胞質チロシンキナーゼドメインを含有する、FMS/KIT/PDGFRファミリーと構造的類似性を共有する。VEGFR1は、全長のチロシンキナーゼ受容体として及び細胞外ドメインのみ保持する可溶性の形態として、2つの形態で発現される。VEGFR2及びVEGFR3等の他のRTKとのホモ二量体化又はヘテロ二量体化を通じて、VEGFR1の全長形態は積極的にそのリガンドとの結合のシグナル伝達を媒介する。しかし、VEGFR1の可溶性形態は、リガンドの捕捉を通じて阻害剤として働き、血管形成を抑制する(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。これらの相反する機能の制御機序は、不明なままである。
【0007】
VEGFRリガンドの1つ、血管内皮成長因子A(VEGFA)は、VEGFR1及びVEGFR2の双方と会合するが、VEGFAとVEGFR1との結合性は、少なくともVEGFR2に対するものよりも一桁超高い(Sawano A et al., Cell Growth Differ 1996, 7(2): 213-21)。他方、VEGFR1の内在性チロシンキナーゼ活性は、VEGFR2のものと比較してはるかに低い。VEGFAとの結合の際、これらの受容体は、その自己リン酸化レベルを劇的に増大させ、内皮細胞(EC)の成長、EC前駆体の動員、ナチュラルキラー細胞のECへの固着、及び、単球の遊走を誘導する(Shibuya M et al., Oncogene 1990, 5(4): 519-24.; Rahimi N, Experimental eye research 2006, 83(5): 1005-16.; Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。いくつかのVEGFR1チロシンリン酸化部位及びそれらの可能な結合相手は、異なる発現モデルで記述されているが(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)、下流のシグナル伝達現象は、いまだに明らかにされておらず、それはひとえに、この受容体の低い生物学的活性による。
【0008】
本発明者らは、SMYD3(Accession No.:AB057595)が、ヒストンH3(H3−K4)のリジン4上にジ-メチルトランスフェラーゼ活性及びトリメチルトランスフェラーゼ活性を有することを先に報告している(WO2005/071102(その全体は本明細書中に参照により引用される)参照)。さらに、先の報告では、SMYD3の発現増大が結腸直腸癌(CRC)及び肝細胞癌(HCC)細胞の増殖に重大な役割を果たすことを実証している(WO2003/027143(その全体は本明細書中に参照により引用される)、並びにHamamoto, R. et al., Nat Cell Biol 6, 731-40 (2004))及びHamamoto R et al., Cancer Sci 2006, 97(2): 113-8参照)。特に、SMYD3の過剰発現は、NIH3T3細胞の成長促進をもたらすことが示され、いくつかの癌細胞での内在性SMYD3発現のノックダウンはこれらの細胞の成長阻害及びアポトーシスを誘導することが示された。本発明者らはまた、網膜芽腫タンパク質(RB1)がSMYD3のSETドメインとの相互作用を通じてメチル化され、このようなメチル化は、インビトロ及びインビボでのCDK2/サイクリンE又はCDK6/サイクリンD3複合体により、スレオニン821/826及びセリン807/811でのRB1のリン酸化を促進することを示してきた(WO2007/004526(その全体は本明細書中に参照により引用される))。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】WO2003/027143 (JP 2005-511023)
【特許文献2】WO2004/076623 (JP 2006-519009)
【特許文献3】WO2005/071102 (JP 2007-519391)
【特許文献4】WO2006/092958, A1
【特許文献5】WO2007/004526, A2
【非特許文献】
【0010】
【非特許文献1】Kunizaki, et al. Cancer Res. 2007 Nov 15;67(22):10759-65
【発明の概要】
【0011】
SMYD3メチルトランスフェラーゼの他の基質の探索において、本発明者らは、SMYD3がインビボ及びインビトロでVEGFR1のリジン831をメチル化すること、及び、かかるVEGFR1のメチル化はそのキナーゼ活性を強めることを見出した。これらの知見は、VEGFR1及びヒト腫瘍形成の制御機序のよりよい理解をもたらすであろう。
【0012】
従って、本発明は、SMYD3によるリジン831のメチル化を介した、VEGFR1制御の新規な機序の発見に少なくとも一部基づく。“ZNFN3A1”の遺伝子名でも知られるSMYD3は、結腸直腸癌及び肝細胞癌(例えば、上記WO2003/027143を参照)並びに膀胱癌及び乳癌(例えば、上記WO2006/085684及びWO2006/092958)の大部分で上昇制御されるヒストンH3メチルトランスフェラーゼである(文献はそれぞれその全体が本明細書中に参照により引用される)。
【0013】
本明細書中で示されるように、VEGFR1はSMYD3のSETドメインと相互作用する。この相互作用は、VEGFR1のメチル化をもたらし、これが本明細書中で示されるように、インビトロで強いキナーゼ活性をもたらす。これらの知見は、SMYD3の生物学的役割だけでなく、VEGFR1の制御機序についてのより深い理解を提供する。さらに、該発見は、腫瘍形成、より具体的には、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌の腫瘍形成のよりよい理解に寄与し、それゆえ、これらの腫瘍の新規な治療戦略の発展に寄与する。
【0014】
したがって、本発明の目的は、SMYD3によるVEGFR1のメチル化を調整する因子を同定する方法を提供することにあり、該方法は:
(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、試験因子の存在下、VEGFR1ペプチドのメチル化に適した条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)で検出したメチル化レベルを前記因子の非存在下で検出した対照レベルと比較する工程を含み、ここで、
対照レベルと比較したメチル化レベルの減少又は増大は、前記因子がSMYD3によるVEGFR1のメチル化を調整することを示す。
【0015】
本発明の更なる目的は、VEGFR1のメチル化を制御する試験化合物の能力を検出するキットを提供することにあり、かかるキットは、(a)メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド、(b)SMYD3ポリペプチドによりメチル化されうるVEGFR1ペプチド、及び(c)VEGFR1ペプチドのメチル化のコファクター、を含む。更なる態様において、該キットは、S−アデノシルホモシステインヒドロラーゼ(SAHH)を任意に含んでもよい。
【0016】
本発明は、さらに、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌等の、癌治療用化合物のスクリーニング方法を提供するものであり、かかる方法は、(a)上述の方法に従ってメチル化を調整する試験化合物を同定する工程、及び、(b)試験化合物の非存在下で検出した対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程、を含む。
【0017】
本発明は、さらに、ポリペプチドのメチルトランスフェラーゼ活性を測定する方法を提供するものであり、該方法は、
(a)以下からなる群から選択されるポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化が可能な条件下で接触させる工程;
(i)配列番号2(SMYD3)のアミノ酸配列を有するポリペプチド;
(ii)配列番号2のアミノ酸配列を有するポリペプチドであって、1または2以上のアミノ酸が置換、欠失、又は挿入され、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価な生物学的活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を有するアミノ酸配列を有するポリペプチド;
(vi)配列番号1のヌクレオチド配列を有するポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされたポリペプチドであって、配列番号2のアミノ酸配列に対応するポリペプチドと等価な生物学的活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルとメチルトランスフェラーゼ活性とを関連させて、メチルトランスフェラーゼ活性を測定する工程
を含む。
【0018】
さらに、本発明はまた、SMYD3のメチルトランスフェラーゼ活性を測定するためのキットを提供するものであり、該キットは、以下の要素:
(a)SMYD3によりメチル化されうるVEGFR1ペプチド;
(b)VEGFR1ペプチドのメチル化のコファクター;及び
(c)リジン831のメチル化を検出するための検出試薬
を含む。
【0019】
本発明の上記及び他の目的、特徴及び利点は、以下の詳細な説明を添付の図面及び実施例、並びに本明細書に添付の特許請求の範囲を読んだとき、より十分に明らかとなるであろう。
【0020】
他に定義しない限り、本明細書中で用いられる全ての技術的及び科学的用語は、本発明の属する技術の分野における通常の技術、知識を有する者により一般に理解されるものと同じ意味を有する。また、本明細書中で用いられる「1つの(a, an)」及び「該(the)」は、他に特に示さない限り、「少なくとも1つ」を意味する。
【0021】
本明細書中に記載されたものと同様または等価な方法と材料は本発明の実施又は試験に用いることができるが、適切な方法と材料を本明細書において以下に示す。
【0022】
本明細書において言及される全ての出版物、特許出願、特許、およびその他の参考文献は、その全体が参照により引用される。矛盾する場合、定義を含む本明細書によって統制される。さらに、材料、方法、および実施例は、例示的なものに過ぎず、本発明を制限するためのものではない。
【図面の簡単な説明】
【0023】
【図1】図1はインビトロでのVEGFR1の細胞質領域のメチル化を示す。(a)は、基質として、VEGFR1の異なる細胞質領域をそれぞれ含む組換えタンパク質VEGFR1#1、#2及び#3を用いたSMYD3のインビトロでのメチルトランスフェラーゼ(MTase)アッセイの結果を示す。「Ig−LD」は免疫グロブリン様ドメインに、「TM」は膜貫通ドメインに、「TK」はチロシンキナーゼドメインに対応する。組換え体VEGFR1#1、#2及び#3タンパク質は、3HでラベルしたSAM(メチルドナーである)とともに、免疫沈降したFlagタグ化SMYD3の存在下(■)又は非存在下でインキュベートした。メチル化基質は、液体シンチレーションカウンターで定量化した。モックプラスミドをトランスフェクトした細胞からの免疫沈降物を対照に用いた(□)。独立した実験を異なる免疫沈降物について3回行った。(b)は、フルオログラムによるメチル化VEGFR1の検出を示す。組換えSMYD3は、S−(5’−アデノシル)−L−ホモシステイン ヒドロラーゼ(SAHH)の存在下又は非存在下で、組換えVEGFR1とともにインキュベートした。基質は、抗GST抗体を用いてイムノブロット分析により定量化した(下パネル)。(c)はVEGFR1#1のメチル化領域の決定を示す。
【図2】図2は、インビトロ及びインビボ両方でのVEGFR1のリジン831のメチル化を示す。(a)は、ヒストンメチルトランスフェラーゼによりメチル化されたリジン、及び、VEGFR1中の候補リジン(*)の隣接配列のアラインメントを示す。(b)は、VEGFR1−N1の野生型、及び、変異体(K819A及びK831A)のインビトロメチル化を示す(上パネル)。(c)は、VEGFR1−K831の異なる置換を含むVEGFR1変異体のインビトロメチル化を示す。(d)は、抗メチル化K831(K831me2)特異的抗体を用いたイムノブロット分析により、VEGFR1のインビトロメチル化を検出したことを示す(中パネル)。メチル化は、フルオログラムによる異なる実験により確認した(上パネル)。VEGFR1は、抗GST抗体を用いたイムノブロット分析で定量化した(下パネル)。(e)は、HAタグ化VEGFR1を発現する293−SMYD3細胞及び293−モック細胞内でのVEGFR1のメチル化を示す。細胞は、タンパク質合成阻害剤の存在下で、L−[メチル−3H]メチオニンで処理した。VEGFR1は抗HA抗体で免疫沈降し、フルオログラムで調べた(上パネル)。(f)は、抗メチル化K831(K831me2)特異的抗体を用いたイムノブロット分析で検出したVEGFR1のメチル化を示す。(g)は、SMYD3のノックダウンによるK831メチル化の抑制を示す。293−SMYD3細胞からの全ての細胞抽出物は、SMYD3特異的siRNA又は対照siRNAで処理し、イムノブロット分析に用いた。
【図3】図3は、SMYD3及びVEGFR1の間の相互作用を示す。(a)は、HAタグ化SMYD3及びFlagタグ化VEGFR1の異なる領域(VEGFR1#1、#2及び#3)を発現するHEK293細胞からの抽出物を用いた免疫共沈降アッセイにより調べた、SMYD3及びVEGFR1の間の相互作用を示す。(b)は、内在性SMYD3とVEGFR1との相互作用を示す。いずれもSMYD3及びVEGFR1を発現する、SNU423、SW480及びMCF7細胞からの全ての細胞抽出物を、抗SMYD3抗体で沈降させた。沈降物は、抗VEGFR1抗体(上パネル)、又は、抗SMYD3抗体(下パネル)でイムノブロットした。(c)は、VEGFR1とSMYD3のSETドメインとの会合を示す。Flagタグ化SMYD3の野生型又は欠失型(N−20、Δ2又はΔ3)を、HEK293細胞においてHAタグ化VEGFR1と共発現させた。IP(免疫沈降)を抗HA又は抗Flag抗体のいずれかで行い、沈降物について、ウェスタンブロット分析を、抗Flag抗体(上パネル)又は抗HA抗体(下パネル)を用いて行った。(d)は、VEGFR1の細胞質領域とSMYD3のSETドメインとのインビボでの相互作用を示す。Flagタグ化野生型又はSMYD3のSETドメインを、HEK293細胞においてHAタグ化VEGFR1と共発現させた。細胞抽出物は、抗Flag抗体(上パネル)又は抗HA抗体(下パネル)を用いて沈降させ、沈降物を、抗HA抗体を用いてウェスタンブロット分析により分析した。
【図4】図4に示される結果は、VEGFR1のキナーゼ活性がそのメチル化によって強められることを示す。(a)は、メチル化又は非メチル化VEGFR1を用いて行ったインビトロキナーゼアッセイの結果を示す。組換えGST融合VEGFR1は、SMYD3で処理し(■)、又は、処理しなかった(□)。そのキナーゼ活性について、そのペプチド基質を用いて引き続き分析した。リン酸化基質は、抗リン酸化チロシン抗体で検出し、特異的二次抗体を用いた蛍光免疫アッセイにより定量化した。(b)に示される結果は、組換えVEGFR1の自己リン酸化がSMYD3により強められることを示す。VEGFR1はPAGE−SDSで分離し、その後、ニトロセルロース膜に移した。抗リン酸化チロシン又は抗VEGFR1抗体を用いてイムノブロット分析を行った。
【発明を実施するための形態】
【0024】
SMYD3のcDNAは、配列番号1に示されるように、1284個のヌクレオチドのオープンリーディングフレームを含む、1622個のヌクレオチドから構成される。オープンリーディングフレームは、配列番号2に示されるように、ジンクフィンガーモチーフ及びSETドメインを有する428アミノ酸タンパク質をコードする。ジンクフィンガードメイン(MYND)は、アミノ酸49からアミノ酸87にわたり、SET(Su3−9、Enhancer−of−zeste、Trihorrax)ドメインは、アミノ酸117からアミノ酸246にわたる。
【0025】
SMYD3タンパク質の細胞内局在は、細胞周期進行の間に変わり、また、培養細胞の密度によっても変わる。SMYD3タンパク質は、細胞が、S期の中間〜後半にあるか、低密度状態で培養されるときに、核に蓄積される。しかし、SMYD3タンパク質は、細胞が細胞周期の他の期、又は、密集状態で成長するとき、細胞質内及び核内に局在化する。
【0026】
本発明は、VEGFR1基質のメチル化のためのSMYD3のメチルトランスフェラーゼ活性を決定する方法を提供する。該方法は、SMYD3ポリペプチド又はメチルトランスフェラーゼ活性を有するその機能的等価物をVEGFR1タンパク質と接触させ、接触させたSMYD3又はその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることにより実施されてもよい。この態様(context)において、SMYD3のメチルトランスフェラーゼ活性は、VEGFR1のメチル化の程度に対応する。したがって、メチルトランスフェラーゼ活性はVEGFR1基質のメチル化レベルを検出することにより測定可能である。より具体的には、本発明の態様において、測定されるメチルトランスフェラーゼ活性は、参照サンプルとの関連付けを介してVEGFR1ペプチドのメチル化レベルに較正されてもよい。本発明のこの態様において、既知のメチルトランスフェラーゼ活性を有するいかなる生物学的サンプルも、参照サンプルとして用いることが可能である。例えば、必要な較正曲線は、精製SMYD3ペプチドの段階希釈を介して得ることができる。
【0027】
さらに、メチルトランスフェラーゼ活性調整剤のスクリーニング方法もまた提供される。本発明は、したがって、SMYD3メチルトランスフェラーゼ活性を調整する因子をスクリーニングする方法を提供する。該方法は、SMYD3ポリペプチド又はメチルトランスフェラーゼ活性を有するその機能的等価物をVEGFR1タンパク質と接触させ、接触させたSMYD3又はその機能的等価物のメチルトランスフェラーゼ活性をアッセイすることにより実施されてもよい。SMYD3又はその機能的等価物のメチルトランスフェラーゼ活性を調整する因子は、これにより同定可能である。
【0028】
本発明の態様において、「機能的に等価」との語は、対象タンパク質又はポリペプチドが、SMYD3と同一の又は実質的に同一のメチルトランスフェラーゼ活性を有することを意味する。特に、該タンパク質は、VEGFR1タンパク質、又は、リジン831を含むVEGFR1タンパク質の断片のメチル化を触媒する。対象タンパク質が標的活性を有するかどうかは、本発明により常に決定できる。すなわち、メチルトランスフェラーゼ活性は、(a)ポリペプチドを基質(例えば、VEGFR1タンパク質又はリジン831を含むその断片)及びコファクター(例えば、S−アデノシル−L−メチオニン)と、該基質のメチル化に適した条件下で接触させ、(b)該基質のメチル化レベルを検出することにより、決定可能である。
【0029】
本明細書中で用いられるとき、「VEGFR1ペプチド」の語は、全長VEGFR1タンパク質(例えば、配列番号4)、及び、その機能的変異体及び断片を指す。機能的断片の例には、配列番号4のアミノ酸800から841で構成される断片のようなC末端断片が含まれるが、これに限られない。好適な断片は、位置831のリジン残基を含む。機能的変異体の例には、全長VEGFR1たんぱく質のメチル化可能性を保持する以下のVEGFR1変異体:K819A、K819E及びK819R、が含まれるが、これに限られない。
【0030】
特定のタンパク質の機能的等価物であるタンパク質を調製する方法は、当業者にはよく知られており、タンパク質に変異を導入する一般的な方法が含まれる。例えば、当業者は、部位特異的変異導入法を用いてヒトSMYD3タンパク質のアミノ酸配列中に適当な変異を導入することにより、ヒトSMYD3タンパク質に機能的に等価なタンパク質を調製できる(例えば、Hashimoto-Gotoh, T. et al. (1995), Gene 152, 271-275;Zoller, MJ, and Smith, M. (1983), Methods Enzymol. 100, 468-500;Kramer, W. et al. (1984), Nucleic Acids Res. 12, 9441-9456;Kramer W, and Fritz HJ. (1987) Methods. Enzymol. 154, 350-367;Kunkel, TA (1985), Proc. Natl. Acad. Sci. USA. 82, 488-492)。アミノ酸変異は、自然にも発生しうる。本発明の態様において有用なSMYD3ポリペプチドには、生じる変異タンパク質がヒトSMYD3タンパク質の機能的等価物、より具体的には、ヒトSMYD3タンパク質のメチルトランスフェラーゼ活性を保持する、1又は2以上のアミノ酸が変異したヒトSMYD3タンパク質のアミノ酸配列を有するタンパク質が含まれる。かかる変異体において変異されるアミノ酸の数は、通常、20アミノ酸以下、典型的には10アミノ酸以下、好ましくは6アミノ酸以下、より好ましくは3アミノ酸以下である。メチルトランスフェラーゼ活性を維持するため、SETドメイン“NHSCXXN”及び“GEELXXXY”が好ましくは変異タンパク質のアミノ酸配列に保存される(“X”は任意のアミノ酸を示す)。したがって、本発明の態様において、SMYD3ポリペプチドは、配列番号2のアミノ酸配列を含み、ここで、1又は2以上のアミノ酸は置換、欠失、又は挿入され、さらにここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドのメチルトランスフェラーゼ活性と等価な活性を有し、さらにここで、SETドメイン“NHSCXXN”及び“GEELXXXY”は保存されている。より詳細には、配列番号2の117位のアミノ酸から246位までのアミノ酸が好ましくは保存される。
【0031】
変異又は修飾されたタンパク質、すなわち、特定アミノ酸配列の1又は2以上のアミノ酸残基を欠失、付加及び/又は置換することにより修飾されたアミノ酸配列を有するタンパク質は、元のタンパク質の生物学的活性を保持することが知られている(Mark, D. F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666;Zoller, M. J. & Smith, M., Nucleic Acids Research (1982) 10, 6487-6500;Wang, A. et al., Science (1984) 224, 1431-1433;Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413)。例えば、SMYD3ポリペプチドは、SETドメインを保持する限り、そのメチルトランスフェラーゼ活性を保持することが予測される。したがって、本発明に係る方法は、配列番号2のアミノ酸配列の117〜246の位置のアミノ酸配列を含むポリペプチド、例えば、実施例に示される配列番号2のアミノ酸配列の110〜250の位置のアミノ酸配列を含むポリペプチドを用いて実施可能である。
【0032】
変異されるアミノ酸残基は、アミノ酸側鎖の性質の保存が可能な、異なるアミノ酸に好適に変異される(保存的アミノ酸置換として知られるプロセス)。アミノ酸側鎖の性質の例は、以下の群に含まれる。
疎水性アミノ酸(A、I、L、M、F、P、W、Y、V)、
親水性アミノ酸(R、D、N、C、E、Q、G、H、K、S、T)。
他に、以下の官能基又は特性を共通に有する側鎖:
脂肪族側鎖(G、A、V、L、I、P);
水酸基を含有する側鎖(S、T、Y);
硫黄原子を含有する側鎖(C、M);
カルボン酸及びアミドを含有する側鎖(D、N、E、Q);
塩基を含有する側鎖(R、K、H);及び
芳香族を含有する側鎖(F、H、Y、W)。
さらに、例えば、アミノ酸の分類は、変異マトリックスとして周知である(Taylor 1986, J, Theor. Biol. 119, 205-218;Sambrook, J. et al., Molecular Cloning 3rd ed. A7.6-A7.9, Cold Spring Harbor Lab. Press, 2001)。このような分類は、以下のように要約できる:
脂肪族アミノ酸:L、I、V
芳香族アミノ酸:H、W、Y、F
荷電アミノ酸:D、E、R、K、H
陽荷電アミノ酸:R、K、H
陰荷電アミノ酸:D、E
疎水性アミノ酸:H、W、Y、F、M、L、I、V、C、A、G、T、K
極性アミノ酸:T、S、N、D、E、Q、R、K、H、W、Y
小アミノ酸:P、V、C、A、G、T、S、N、D
極小アミノ酸:A、G、S
大(非小)アミノ酸:Q、E、R、K、H、W、Y、F、M、L、I
ここで、挿入文字は、アミノ酸の1文字コードを示す。
【0033】
ヒトSMYD3タンパク質(配列番号2)のアミノ酸配列に1又は2以上のアミノ酸残基が付加されたタンパク質の例は、ヒトSMYD3タンパク質を含有する融合タンパク質である。融合タンパク質には、ヒトSMYD3タンパク質及び他のペプチド又はタンパク質の融合が含まれ、本発明において用いられる。融合タンパク質は、当業者には周知の技術、たとえば、本発明に係るヒトSMYD3タンパク質をコードするDNAを他のペプチド又はタンパク質をコードするDNAとフレームが一致するように連結し、融合DNAを発現ベクターに挿入して、宿主内で発現させることにより作成可能である。本発明のタンパク質に融合されるペプチド又はタンパク質について何ら制約はない。
【0034】
SMYD3タンパク質に融合されるペプチドとして使用可能な既知のペプチドには、例えば、FLAG(Hopp, T. P. et al., Biotechnology (1988) 6, 1204-1210)、6つのHis(ヒスチジン)残基を含有する6xHis、10xHis、インフルエンザアグルチニン(HA)、ヒトc−myc断片、VSP−GP断片、p18HIV断片、T7−タグ、HSV−タグ、E−タグ、SV40T抗原断片、lckタグ、αチューブリン断片、B−タグ、プロテインC断片などが含まれる。本発明に係るタンパク質に融合可能なタンパク質の例には、GST(グルタチオン−S−トランスフェラーゼ)、インフルエンザアグルチニン(HA)、免疫グロブリン定常領域、βガラクトシダーゼ、MBP(マルトース結合タンパク質)などが含まれる。
【0035】
融合タンパク質は、上述の融合ペプチド又はタンパク質をコードする、商業的に入手可能なDNAを、本発明に係るタンパク質をコードするDNAと融合し、調製した融合DNAを発現することより調製できる。
【0036】
機能的に等価なタンパク質を単離する当業者に既知の他の方法では、相同配列を同定するハイブリダイズ技術を用いる(Sambrook, J. et al., Molecular Cloning 2nd ed. 9.47-9.58, Cold Spring Harbor Lab. Press, 1989)。当業者であれば容易にヒトSMYD3タンパク質をコードするSMYD3 DNA配列(例えば、配列番号1)の全部又は一部と高い相同性を有するDNAを単離し、単離したDNAからヒトSMYD3タンパク質と機能的に等価なタンパク質を単離することができる。本発明に使用されるタンパク質には、ヒトSMYD3タンパク質をコードするDNA配列の全部又は一部とハイブリダイズするDNAによりコードされるもの、及び、ヒトSMYD3タンパク質と機能的に等価なものが含まれる。これらのタンパク質には、ヒト又はマウス由来のタンパク質に対応する哺乳類の相同体が含まれる(例えば、サル、ラット、ウサギ及びウシ遺伝子によりコードされるタンパク質)。動物からヒトSMYD3タンパク質をコードするDNAに高い相同性を示すcDNAを単離する場合、骨格筋、精巣、HCC又は結腸直腸腫瘍由来の組織を用いることが特に好ましい。
【0037】
ヒトSMYD3タンパク質の機能的等価物をコードするDNAを単離するためのハイブリダイズ条件は、当業者により通常通りに選択可能である。例えば、“Rapid−hyb buffer”(Amersham LIFE SCIENCE)を用いて68℃で30分又はそれ以上プリハイブリダイゼーションを行い、ラベルしたプローブを添加し、68℃で1時間又はそれ以上温めることにより実施してもよい。その後の洗浄ステップは、例えば、低ストリンジェント条件で行うことができる。低ストリンジェント条件とは、例えば、42℃、2xSSC、0.1%SDS、又は、50℃、2xSSC、0.1%SDSである。より好ましくは、高ストリンジェント条件が用いられる。本発明の態様において、高ストリンジェント条件には、例えば、2xSSC、0.01%SDS、室温で20分間の洗浄を3回行い、次いで、1xSSC、0.1%SDS、37℃、20分間の洗浄を3回行い、次いで、1xSSC、0.1%SDS、50℃、20分間の洗浄を2回行うことが含まれる。しかし、温度及び塩濃度などのいくつかの要因がハイブリダイゼーションのストリンジェンシーに影響を与える可能性があり、当業者は要求されるストリンジェンシーを達成するためにこれらの要因を適切に選択できる。
【0038】
ハイブリダイゼーションに代えて、遺伝子増幅法、例えば、ポリメラーゼ連鎖反応(PCR)法を利用し、ヒトSMYD3タンパク質(配列番号2)をコードするDNA(配列番号1)の配列情報に基づいて合成されたプライマーを用いて、ヒトSMYD3タンパク質に機能的に等価なタンパク質をコードするDNAを単離することができる。
【0039】
上記ハイブリダイゼーション技術又は遺伝子増幅技術により単離されたDNAがコードする、ヒトSMYD3タンパク質に機能的に等価なタンパク質は、通常、ヒトSMYD3タンパク質のアミノ酸配列と高い相同性を有する。「高い相同性」(「高い同一性」ともいう)とは、通常、2つの任意に整列された配列(ポリペプチド又はポリヌクレオチド配列のいずれか)間の同一性の程度をいう。典型的には、高い相同性又は同一性とは、40%以上、好ましくは60%以上、より好ましくは80%以上、さらにより好ましくは85%、90%、95%、98%、99%以上をいう。2つのポリペプチド又はポリヌクレオチド配列間の相同性又は同一性の程度は、「Wilbur, W. J. and Lipman, D. J. Proc. Natl. Acad. Sci. USA (1983) 80, 726-730」中のアルゴリズムに従って決定可能である。
【0040】
本発明の態様において有用なタンパク質は、その製造に用いる細胞又は宿主或いは用いる精製方法によって、アミノ酸配列、分子量、等電点、糖鎖の有無、又は、形態においてバリエーションをとることが可能である。にもかかわらず、ヒトSMYD3タンパク質(配列番号2)の機能的等価物である限り、本発明に有用である。
【0041】
本発明の態様において有用なタンパク質は、当業者には周知の方法により、組換えタンパク質又は天然タンパク質として調製可能である。組換えタンパク質は、本発明のタンパク質をコードするDNA(例えば、配列番号1のヌクレオチド配列に対応するDNA)を適切な発現ベクターに挿入し、該ベクターを適切な宿主細胞に導入し、抽出物を得て、該抽出物をクロマトグラフィー、例えば、イオン交換クロマトグラフィー、逆相クロマトグラフィー、ゲルろ過、又は本発明に係るタンパク質に対する抗体が固定されるカラムを利用するアフィニティクロマトグラフィーにかけて、或いは、1以上の上記カラムを組み合わせて、タンパク質を精製することにより調製可能である。
【0042】
さらに、本発明の態様において有用なタンパク質が宿主細胞内(例えば、動物細胞及び大腸菌)でグルタチオン−S−トランスフェラーゼとの融合タンパク質として、又は、多数のヒスチジンが付加された組換えタンパク質として発現される場合には、発現された組換えタンパク質はグルタチオンカラム又はニッケルカラムを用いて精製できる。
【0043】
融合タンパク質の精製後、必要に応じて、トロンビン又は活性化第Xa因子で切断して、標的タンパク質以外の領域を除くこともできる。
【0044】
天然タンパク質は、当業者に既知の方法により、例えば、以下に記載するSMYD3タンパク質に結合する抗体が結合されたアフィニティカラムを、本発明に係るタンパク質を発現する組織又は細胞抽出物に接触させることにより単離可能である。抗体は、ポリクローナル抗体又はモノクローナル抗体が可能である。
【0045】
本発明において、SMYD3ポリペプチドのメチルトランスフェラーゼ活性は当業者には既知の方法により決定できる。例えば、SMYD3ポリペプチド及びVEGFR1ペプチド基質を、ラベルしたメチルドナーとともに適切なアッセイ条件でインキュベートしてもよい。好ましいメチルドナーの例としては、S−アデノシル−[メチル−14C]−L−メチオニン及びS−アデノシル−[メチル−3H]−L−メチオニンが挙げられるが、これに限られない。VEGFR1ペプチドへの放射ラベルの転移は、例えば、SDS−PAGE電気泳動及びフルオログラフィーにより検出できる。また、反応後、VEGFR1ペプチドはろ過によりメチルドナーから分離でき、フィルターに残った放射ラベルの量はシンチレーションカウントにより定量化できる。色素生産性及び蛍光ラベルのようなメチルドナーに付加できる他の適したラベル、及び、これらのラベルのVEGFR1ペプチドへの転移を検出する方法は、当業者には既知である。
【0046】
また、SMYD3のメチルトランスフェラーゼ活性は、非ラベル化メチルドナー(例えば、S−アデノシル−L−メチオニン)及びメチル化VEGFR1ペプチドを選択的に認識する試薬を用いて決定できる。例えば、SMYD3、メチル化される基質、及びメチルドナーを基質のメチル化に適した条件下でインキュベートした後、メチル化された基質は通常の免疫学的方法を用いて検出できる。メチル化基質を認識する抗体を用いるどのような免疫学的技術も検出に用いることができる。例えば、メチル化VEGFR1 K831(例えば、抗K831me2)に対する抗体を抗体として使用できる。VEGFR1のメチル化、特に、VEGFR1のK831でのメチル化を認識するこのような抗体も、本発明により提供される。メチル化VEGFR1 K831を認識する抗体を用いるELISA又はイムノブロット分析は、本発明に適している場合がある。
【0047】
本明細書中で示されるように、Lys831でのVEGFR1のメチル化は、VEGFR1のリン酸化を強めた。したがって、他の実施形態において、VEGFR1のメチル化レベルはVEGFR1のリン酸化レベルを介して評価可能である。VEGFR1のリン酸化は、放射ラベル化リン酸ドナーを用いて検出してもよい。また、VEGFR1のリン酸化部位を認識する抗体をVEGFR1のリン酸化レベルの評価に用いてもよい。
【0048】
本発明によれば、SMYD3によりリン酸化されるVEGFR1ペプチドの上記性質を、SMYD3及び他のH3K4メチルトランスフェラーゼ活性の測定に利用してもよい。このようなペプチドのメチルトランスフェラーゼ活性の測定方法は:
(a)H3K4メチルトランスフェラーゼを、メチル化されるVEGFR1ペプチド及びコファクターとVEGFR1ペプチドのメチル化が可能な条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルをメチルトランスフェラーゼ活性と関連させてメチルトランスフェラーゼ活性を測定する工程
を含む。
【0049】
本明細書中において、H3K4メチルトランスフェラーゼは、VEGFR1ペプチド上のメチル基を転移させる能力のある限り、いかなるポリペプチドであってもよく、このようなポリペプチドには、上記SMYD3ポリペプチドが含まれるが、これに限られない。本方法の工程は、同じVEGFR1ペプチド、反応条件(コファクターの付加、強化因子等)、検出手段及びSMYD3によるVEGFR1のメチル化を調整する化合物の上記同定方法に類するものを用いて行ってもよい。
【0050】
本発明は、SMYD3を過剰発現する癌を治療するための化合物のスクリーニング方法をさらに提供し、該方法は、上記方法を用いるメチル化を調整する試験化合物を同定する工程、及び、試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程を含む。
【0051】
該スクリーニングは、(a)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片及びSMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片を、VEGFR1ペプチド及びSMYD3ポリペプチドの結合を可能とする条件下、かつ、試験化合物の存在下で接触させる工程、並びに(b)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片、及び、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片の間の結合を阻害する試験化合物を癌治療のための候補化合物として選択する工程を含む。本発明において、SMYD3によるVEGFR1のメチル化、又は、VEGFR1とSMYD3との間の結合を抑制することにより、細胞の成長が低減することが明らかとなった。従って、SMYD3によるVEGFR1の結合又はメチル化を阻害する試験化合物をスクリーニングすることにより、癌を治療又は予防する可能性のある候補化合物を同定することができる。癌を治療又は予防するこれらの候補化合物の可能性は、癌のための治療用因子を同定する第2の及び/又はさらなるスクリーニングにより評価してもよい。
本発明の態様において、SMYD3を過剰発現する癌には、SMYD3のその発現レベルが癌の同じ器官の正常領域と比較して高い癌が含まれる。例えば、本発明者らは、種々の癌、例えば、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌におけるSMYD3の過剰発現を明らかにしてきた(WO2003/027143、WO2006/085684及びWO2006/092958)。従って、本発明の好ましい態様において、SMYD3を過剰発現する癌は、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択されてもよい。
【0052】
試験化合物がVEGFR1ペプチドとSMYD3ポリペプチドとの結合を阻害するかは、VEGFR1ペプチドとSMYD3ポリペプチドとの結合レベルを、該化合物の非存在下で検出した結合レベルと比較し、VEGFR1ポリペプチドとSMYD3との結合レベルを減少させる化合物を選択することにより決定できる。その機能的等価物を含む任意のVEGFR1ペプチド及びSMYD3ポリペプチドは、互いの結合能力が保持される限りこのスクリーニングに用いてもよい。例えば、VEGFR1ペプチドのSMYD3結合領域を含む断片を、VEGFR1に等価なペプチドとしてこのスクリーニングに用いてもよい。本発明において、VEGFR1に等価なかかるペプチドは、配列番号4のアミノ酸配列の800〜1000の位置のアミノ酸配列を好ましくは含みうる。同様に、SMYD3ペプチドのVEGFR1結合領域を含む断片を、SMYD3に等価なペプチドとして用いてもよい。例えば、かかるSMYD3のペプチド等価物は、配列番号2のアミノ酸配列の100〜250の位置のアミノ酸配列を含んでもよい。
本発明の態様において、2つのタンパク質間の「結合を阻害する」とは、該タンパク質間の結合を少なくとも減少させることをいう。従って、ある場合には、サンプル中の結合ペアの割合は、適当な(例えば、試験化合物で処理されていない、或いは、非癌サンプルからの、又は癌サンプルからの)対照と比較して減少しているであろう。結合タンパク質量の減少は、例えば、対照サンプル中の結合ペアよりも、90%、80%、70%、60%、50%、40%、25%、10%、5%、1%未満(例えば0%)でありうる。
【0053】
一般に、かかる会合に干渉する試験化合物の能力を決定するいかなる方法も、本発明での使用に適している。例えば、ELISAフォーマットでの競合的及び非競合的阻害アッセイも利用可能である。対照実験は、系の最大結合能力を決定するために実行されるべきである(例えば、結合VEGFR1をSMYD3ポリペプチドと接触させ、VEGFR1に結合したタンパク質の量を決定する、或いは、その逆)。本発明に係る結合を阻害する化合物を同定する方法として、当業者に周知の方法の多くを用いることができる。かかる同定は、インビトロアッセイ系、例えば、細胞系として実行可能である。より詳細には、まず、VEGFR1ペプチド又はSMYD3ポリペプチドパートナーのいずれか一方を支持体に結合させ、他方のタンパク質を試験化合物とともにそこに接触させる。次いで、混合物をインキュベートし、洗浄し、支持体に結合した他方のタンパク質を検出及び/又は測定する。
【0054】
タンパク質の結合に使用可能な支持体の例には、不溶性多糖、例えば、アガロース、セルロース及びデキストランが含まれ;そして、合成樹脂、例えば、ポリアクリルアミド、ポリスチレン及びシリコン;好ましくは、上記材料から調製された、商業的に入手可能なビーズ及びプレート(例えば、マルチウェルプレート、バイオセンサーチップ等)を用いることができる。ビーズを用いる場合には、これらはカラムに充填してもよい。また、磁性ビーズの使用もまた当業者には知られており、磁性によりビーズ上に結合されたタンパク質の容易な単離が可能とする。
【0055】
タンパク質の支持体への結合は、通常の方法、例えば、化学結合及び物理吸着により行うことができる。また、タンパク質は、該タンパク質を特異的に認識する抗体を介して支持体に結合させてもよい。さらに、タンパク質の支持体への結合はまた、相互作用する分子、例えば、アビジン及びビオチンにより行われてもよい。
【0056】
タンパク質間の結合は、バッファーがタンパク質間の結合を阻害しない限り、バッファー、例えば、リン酸バッファー及びTrisバッファー(これらに限られない)中で行われる。
【0057】
本発明において、表面プラズモン共鳴現象を用いるバイオセンサーを、結合タンパク質を検出又は定量化するための手段として用いてもよい。かかるバイオセンサーを用いる場合、タンパク質間の相互作用は表面プラズモン共鳴信号としてリアルタイムで、ごく少量のポリペプチドを用いてラベル化無しで観察可能である(例えば、BIAcore、Pharmacia)。従って、BIAcoreのようなバイオセンサーを用いて、VEGFR1ペプチドとSMYD3ポリペプチドとの間の結合を評価することができる。
【0058】
また、VEGFR1ペプチド又はSMYD3ポリペプチドのいずれかをラベルしてもよく、結合タンパク質のラベルを該結合タンパク質の検出又は測定に用いてもよい。特に、タンパク質の一方をプレラベル化した後、ラベル化タンパク質を他方のタンパク質と試験化合物の存在下で接触させ、次いで、結合タンパク質を洗浄後ラベルに従って検出又は測定する。
【0059】
放射性同位体(例えば、3H、14C、32P、33P、35S、125I、131I)、酵素(例えば、アルカリフォスファターゼ、西洋わさびペルオキシダーゼ、βガラクトシダーゼ、βグルコシダーゼ)、蛍光化合物(例えば、フルオレセインイソチオシアネート(FITC)、フルオレセイン、テキサスレッド、緑色蛍光タンパク質、ローダミン)、磁性ビーズ(例えば、DYNABEADSTM)、熱量測定ラベル(例えば、コロイド金又は着色グラス或いはプラスチック(例えば、ポリスチレン、ポリプロピレン、ラテックス等)ビーズ)、及びビオチン/アビジン、等のラベル化合物を本発明の方法におけるタンパク質のラベル化に用いることができる。かかるラベルの使用を教示する特許として、米国特許番号3,817,837;3,850,752;3,939,350;3,996,345;4,277,437;4,275,149;及び4,366,241が挙げられる。しかし、本発明は、これらに限られず、分光学的、光化学的、生化学的、免疫化学的、電気的、光学的又は化学的手段により検出可能ないかなるラベルも使用可能である。
【0060】
タンパク質を放射性同位体でラベルする場合、検出又は測定は、液体シンチレーションにより行うことができる。また、酵素でラベルしたタンパク質は、酵素の基質を付加して基質の酵素的変化、例えば、呈色を吸光光度計で検出することにより検出又は測定できる。さらに、蛍光基質をラベルとして用いる場合には、結合タンパク質は蛍光光度計を用いて検出又は測定することができる。
【0061】
さらに、本発明に係るスクリーニング方法における結合は、VEGFR1ペプチド又はSMYD3ポリペプチドに対する抗体を用いて検出又は測定することもできる。例えば、支持体に固定したVEGFR1ペプチドを試験化合物及びSMYD3ポリペプチドと接触させた後、混合物をインキュベート、洗浄し、検出又は測定をSMYD3ポリペプチドに対する抗体を用いて行うことができる。また、SMYD3ポリペプチドを支持体に固定してもよく、VEGFR1ペプチドに対する抗体をその抗体として用いてもよい。
【0062】
本発明に係るスクリーニングに抗体を用いる場合、抗体は、上記ラベル化物質の1つで好適にラベル化され、ラベル化物質に基づいて検出又は測定される。また、SMYD3ポリペプチド又はVEGFR1ペプチドに対する抗体は、ラベル化物質でラベル化された二次抗体で検出される一次抗体として使用されてもよい。さらに、本発明に係るスクリーニングにおいてタンパク質に結合した抗体は、プロテインG又はプロテインAカラムを用いて検出又は測定できる。
【0063】
また、本発明の同定方法の他の態様において、細胞を用いるツーハイブリッドシステムを用いることができる(“MATCHMAKER Two−Hybrid system”、“Mammalian MATCHMAKER Two−Hybrid Assay Kit”、“MATCHMAKER one−Hybrid system”(Clontech);“HybriZAP Two−Hybrid Vector System”(Stratagene);参考文献Dalton and Treisman, Cell 1992, 68: 597-612、Fields and Sternglanz, Trends Genet 1994, 10: 286-92)。ツーハイブリッドシステムにおいて、例えば、SMYD3ポリペプチドは、SRF結合領域又はGAL4結合領域に融合され、酵母菌で発現される。VEGFR1ペプチドは、VP16又はGAL4転写活性領域に融合され、同様に試験化合物の存在下で酵母菌細胞内に発現される。また、VEGFR1ペプチドは、SRF結合領域又はGAL4結合領域に融合されてもよく、SMYD3ポリペプチドはVP16又はGAL4転写活性領域に融合されてもよい。試験化合物がSMYD3ポリペプチドとVEGFR1ペプチドの間の結合を阻害しない場合、2つの結合は、レポーター遺伝子を活性化し、陽性のクローンを検出可能にする。レポーター遺伝子として、例えば、Ade2遺伝子、lacZ遺伝子、CAT遺伝子、ルシフェラーゼ遺伝子などを、HIS3遺伝子の代わりに用いることができる。
【0064】
ここで、SMYD3ポリペプチドとVEGFR1ペプチドとの間の結合レベルは、SMYD3ポリペプチドとVEGFR1ペプチドの結合後に発生する任意の変化として測定することもできる。特に、かかるスクリーニングは試験化合物をSMYD3ポリペプチド及びVEGFR1ペプチドを発現する細胞と接触させることにより行うこともできる。例えば、細胞増殖の抑制を検出して、SMYD3ポリペプチドとVEGFR1ペプチドとの結合への試験化合物の影響を決定することもできる。
1.競合的アッセイフォーマット
競合的アッセイを本発明に係る試験化合物のスクリーニングに用いることができる。例によれば、競合的ELISAアッセイフォーマットとして、固相支持体に結合されたSMYD3ポリペプチド(又はVEGFR1ペプチド)が挙げられる。結合されたSMYD3ポリペプチド(又はVEGFR1ペプチド)を、VEGFR1ペプチド(又はSMYD3ポリペプチド)及び試験化合物とともにインキュベートする。試験化合物及び/又はVEGFR1ペプチド(又はSMYD3ポリペプチド)をSMYD3ポリペプチド(又はVEGFR1ペプチド)と結合させるのに十分な時間の後、基質を洗浄して結合していない物質を除く。SMYD3ポリペプチドに結合したVEGFR1ペプチドの量はその後決定される。これは当業者に既知のあらゆる種々の方法において、例えば、検出可能なラベルでタグ化されたVEGFR1ペプチド(又はSMYD3ポリペプチド)種を用いることにより、或いは、洗浄した基質を、VEGFR1ペプチド(又はSMYD3ポリペプチド)に対する、ラベルした抗体と接触させることにより、達成してもよい。SMYD3ポリペプチド(又はVEGFR1ペプチド)に結合したVEGFR1ペプチド(又はSMYD3ポリペプチド)の量は、試験化合物のVEGFR1ペプチドとSMYD3ポリペプチドとの会合を阻害する能力に反比例することとなる。タンパク質、例えば、これに限られないが、抗体のラベル化は、Harlow & Lane, Antibodies, A Laboratory Manual (1988)に記載されている。
【0065】
そのほかでは、VEGFR1ペプチド(又はSMYD3ポリペプチド)を、アフィニティータグでラベルする。ラベルしたVEGFR1(又はSMYD3ポリペプチド)を、次いで、試験化合物及びSMYD3ポリペプチド(又はVEGFR1ペプチド)とともにインキュベートし、さらに免疫沈降する。免疫沈降に引き続いて、SMYD3ポリペプチド(又はVEGFR1ペプチド)に対する抗体を用いたウェスタンブロットを行う。上述の競合的アッセイフォーマットのように、VEGFR1ペプチド(又はSMYD3ポリペプチド)に会合して検出されるSMYD3ポリペプチド(VEGFR1ペプチド)の量は、試験化合物のVEGFR1ペプチド及びSMYD3ポリペプチドの会合を阻害する能力に反比例する。
【0066】
2.非競合的アッセイ
非競合的結合アッセイもまた、本明細書中に記載されているような競合的アッセイを用いたスクリーニングを容易に適用できないフォーマットで構築された試験化合物のライブラリの初期スクリーニングとして、有用性が認められ得る(例えば、Barret et al., Anal Biochem 1992, 204: 357-64を参照)。
【0067】
ファージライブラリーは、多数の異なる組換えペプチドの実用的な量を高速に産生する点で有用である。ファージライブラリーは、本発明に係る競合的アッセイに活用できないが、非競合的フォーマットで効率的にスクリーニングして、どの組換えペプチド試験化合物がVEGFR1ペプチド又はSMYD3ポリペプチドに結合するかを決定することができる。結合するものとして同定される試験化合物をその後産生し、競合的アッセイフォーマットを用いてスクリーニング可能である。ファージ及び細胞表示ライブラリーの産生及びスクリーニングは当業者には周知であり、例えば、Ladner et al., WO 88/06630; Fuchs et al., Biotechnology 1991, 9: 1369-72; Goward et al., TIBS 1993, 18: 136-40; Charbit et al., EMBO J 1986, 5: 3029-37; Cull et al., PNAS USA 1992, 89: 1865-9; Cwirla et al., PNAS USA 1990, 87: 6378-82にて論じられている。
【0068】
例示的な非競合的アッセイの後には、一方の構成成分(VEGFR1ペプチド又はSMYD3ポリペプチド)の添加の無い、競合的アッセイについて記載したものと類似の処理が行われる。しかし、非競合的アッセイがVEGFR1ペプチド又はSMYD3ポリペプチドと結合する試験化合物を決定する際、試験化合物のVEGFR1ペプチドとSMYD3ポリペプチドの両方と結合する能力は、各候補について決定しなければならない。従って、例によれば、固定されたVEGFR1ペプチドへの試験化合物の結合は、結合していない試験化合物を洗浄除去し、結合した試験化合物を支持体から溶出させ、次いで、溶出物を、例えば、質量分析、タンパク質決定(Bradford又はLowryアッセイ、或いは280nmの吸光分析)で分析することにより、決定できる。あるいは、溶出工程を無くし、試験化合物の結合を、支持体表面での有機層の分光学的性質の変化をモニタリングすることにより決定してもよい。表面の分光学的性質をモニタリングする方法は、吸光度、反射率、透過率、複屈折、屈折率、回折、表面プラズモン共鳴、偏光解析法、共振ミラー技術、格子結合波導技術(grating coupled waveguide technique)、多極共振解析(multipolar resonance spectroscopy)が挙げられるが、これに限られず、これらの全ては当業者には既知である。アッセイではラベルした試験化合物を用い、溶出工程を省くこともできる。この場合、非結合物質を洗浄除去後、支持体に会合したラベルの量が、試験化合物の結合に正比例する。
【0069】
多数の周知のロボットシステムが、液相化学において開発されてきている。これらのシステムは、武田薬品工業株式会社(大阪、日本)により開発された自動化合成装置のような自動化されたワークステーション、及び、化学者によりなされる手動の合成操作を模倣したロボットアーム(Zymate II、Zymark Corporation、Hopkinton、Mass.;Orca、Hewlett Packard、Palo Alto、Calif.)を利用する多くのロボット合成を含む。上記装置のいずれも本発明での使用に適している。本明細書中で論じたように操作できるようこれらの装置に修正を行う性質及び実行は(もしあれば)、関連技術分野の当業者には明白であろう。さらに、非常に多くのコンビナトリアルライブラリーはそれ自体商業的に入手可能である(例えば、ComGenex、Princeton、N.J.;Asinex、Moscow、Ru;Tripos、Inc.、St. Louis、MO;ChemStar、Ltd、Moscow、RU;3D Pharmaceuticals、Exton、PA、Martek Biosciences、Columbia、MD、など)。
【0070】
種々の低スループット又は高スループット酵素アッセイフォーマットが当業者に知られており、SMYD3のメチルトランスフェラーゼ活性の検出又は測定に容易に適用可能である。高スループットアッセイについては、VEGFR1ペプチド基質は通常のように固体支持体、例えばマルチウェルプレート、スライド又はチップに固定化できる。反応後、メチル化産生物は上述の方法により個体支持体上で検出できる。また、メチルトランスフェラーゼ反応は、溶液中でも起こり、VEGFR1ペプチドが固体支持体上に固定化できた後、メチル化産生物を検出できる。このようなアッセイを実施するため、固体支持体にストレプトアビジン、及びビオチンでラベルしたVEGFR1を塗布してもよく、或いは、固体支持体に抗VEGFR1抗体を塗布してもよい。当業者であれば、所望のスクリーニングのスループット能力によって、適したアッセイフォーマットを決定できる。
【0071】
本発明は、また、本発明に係るタンパク質の部分ペプチドの使用も含む。部分ペプチドはSMYD3タンパク質に特異的なアミノ酸配列を有し、好ましくは、約400アミノ酸未満、通常は約200アミノ酸未満、場合により約100アミノ酸未満、そして、少なくとも約7アミノ酸、好ましくは約8アミノ酸以上、より好ましくは約9アミノ酸以上のアミノ酸配列からなる。部分ペプチドは、例えば、SMYD3タンパク質に結合する因子又は化合物のスクリーニング、及び、SMYD3と例えばSAMのようなそのコファクターとの間の結合の阻害剤のスクリーニングに用いることができる。かかるスクリーニング方法の態様において、SETドメインを含有する部分ペプチドが好ましい。
【0072】
本発明の態様において有用な部分ペプチドは、遺伝子工学により、既知のペプチド合成方法により、又は、本発明にかかるタンパク質を適当なペプチダーゼで分解することにより産生できる。ペプチド合成のため、例えば、固相合成又は液相合成を用いることができる。
【0073】
SETドメインの変異を有するSMYD3変異体は、細胞増殖に抑制効果を示す。従って、SMYD3の部分ペプチドは、好ましくは、SETドメイン“NHSCXXN”及び/又は“GEELXXXY”を含む。
【0074】
任意の試験化合物を用いることができる。例として、細胞抽出物、細胞培養上清、微生物の発酵産生物、海洋生命体の抽出物、植物の抽出物、精製又は粗タンパク質、ペプチド、非ペプチド化合物、合成微小化合物及び天然化合物が挙げられるが、これに限られない。本発明に係る試験因子又は化合物は、技術分野で既知のコンビナトリアルライブラリー方法を用いて得ることもでき、これには、(1)生物学的ライブラリー、(2)空間的にアドレス可能な平行(spatially addressable parallel)固相又は液相ライブラリー、(3)逆重畳積分を必要とする合成ライブラリー方法、(4)「1ビーズ1化合物(one-bead one-compound)」ライブラリー方法、及び、(5)アフィニティークロマトグラフィーによる選択を用いる合成ライブラリー方法が挙げられるが、これに限られない。アフィニティークロマトグラフィーによる選択を用いる生物学的ライブラリー方法はペプチドライブラリーに限られるが、他の4つのアプローチはペプチド、非ペプチドオリゴマー又は化合物の小分子ライブラリーに適用できる(Lam, Anticancer Drug Des. 12: 145-67 (1997))。分子ライブラリーの合成方法の例には、当該技術分野において見出すことができる(DeWitt et al., Proc Natl Acad Sci U S A. 1993 Aug 1;90(15):6909-13.; Erb et al., Proc. Natl. Acad. Sci. USA 91: 11422-6 (1994); Zuckermann et al., J. Med. Chem. 37: 2678-85 (1994); Cho et al., Science 261: 1303-5 (1993); Carell et al., Angew. Chem. Int. Ed. Engl. 33: 2059 (1994); Carell et al., Angew. Chem. Int. Ed. Engl. 33: 2061 (1994); Gallop et al., J. Med. Chem. 37: 1233-51 (1994))。化合物のライブラリーは溶液(Houghten, Bio/Techniques 13: 412-21 (1992)を参照)、ビーズ(Lam, Nature 354: 82-4 (1991))、チップ(Fodor, Nature 364: 555-6 (1993))、バクテリア(米国特許番号第5,223,409)、胞子(米国特許番号第5,571,698;5,403,484及び5,223,409)、プラスミド(Cull et al., Proc. Natl. Acad. Sci. USA 89: 1865-9 (1992))、又はファージ(Scott and Smith, Science 249: 386-90 (1990); Devlin, Science 249: 404-6 (1990); Cwirla et al., Proc. Natl. Acad. Sci. USA 87: 6378-82 (1990); Felici, J. Mol. Biol. 222: 301-10 (1991); 米国特許出願公開20020103360)に提示されてもよい。本発明に係る試験化合物は、単一化合物又は組み合わせ化合物でよい。本発明のスクリーニング方法で組み合わせ化合物が用いられた場合、化合物は連続的に又は同時に接触させることができる。
【0075】
本明細書中に記載されるアッセイに有用な試験因子又は化合物は、SMYD3、又は、メチルトランスフェラーゼ活性を失った部分SMYD3ペプチドに特異的に結合する抗体の形態をとってもよい。例えば、抗体(例えば、モノクローナル抗体)は、SMYD3及びそのVEGFR1基質の間の結合を阻害する能力について試験可能である。
【0076】
本発明のスクリーニング方法により単離される因子又は化合物は、SMYD3のメチルトランスフェラーゼ活性を阻害する候補薬剤であり、従って、肝細胞癌、直腸結腸癌、膀胱癌及び/又は乳癌の治療または予防に適用されうる。
【0077】
さらに、SMYD3のメチルトランスフェラーゼ活性を阻害する因子又は化合物の部分構造が付加、欠失、及び/又は置換により転換された因子又は化合物も、本発明に係るスクリーニング方法により得られ得る因子又は化合物に含まれる。
【0078】
上述のように、SMYD3のメチルトランスフェラーゼ活性を阻害する因子又は化合物は、SMYD3のメチルトランスフェラーゼ活性を失った部分ペプチドであってもよく、又は、SMYD3に対する抗体であってもよい。本明細書中で用いられる場合、「抗体」の語は、抗体の合成に用いられた抗原とのみ、又は、これに非常に関連する抗原と相互作用(すなわち、結合)する特異構造を有する免疫グロブリン分子をさす。さらに、抗体は、SMYD3遺伝子でコードされるタンパク質に結合する限り、抗体の断片又は修飾された抗体であってよい。例えば、抗体断片は、Fab、F(ab’)2、Fv又は単鎖Fv(scFv)であってよく、ここで、H及びL鎖からのFv断片は、適当なリンカーにより接合される(Huston J. S. et al. Proc. Natl. Acad. Sci. U.S.A. 85:5879-5883 (1988))。より詳細には、抗体断片はパパイン又はペプシンのような酵素で抗体を処理することにより生成してもよい。また、抗体断片をコードする遺伝子を構築し、発現ベクターに挿入し、適当な宿主細胞中で発現させてもよい(例えば、Co M. S. et al. J. Immunol. 152:2968-2976 (1994); Better M. and Horwitz A. H. Methods Enzymol. 178:476-496 (1989); Pluckthun A. and Skerra A. Methods Enzymol. 178:497-515 (1989); Lamoyi E. Methods Enzymol. 121:652-663 (1986); Rousseaux J. et al. Methods Enzymol. 121:663-669 (1986); Bird R. E.及びWalker B. W. Trends Biotechnol. 9:132-137 (1991)を参照)。
【0079】
抗体は、例えば、ポリエチレングリコール(PEG)のような種々の分子との接合により修飾されてもよい。本発明は、かかる修飾抗体を提供する。修飾抗体は、抗体を化学的に修飾することにより得ることができる。そのような修飾方法は、当該分野において一般的である。また、抗体は、非ヒト抗体由来の可変領域及びヒト抗体由来の定常領域を有するキメラ抗体、或いは、非ヒト抗体由来の相補性決定領域(CDR)、ヒト抗体由来のフレームワーク領域(FR)及び定常領域からなるヒト化抗体の形態をとってもよい。このような抗体は、通常の技術を用いて調製可能である。ヒト化はげっ歯類のCDR又はCDR配列をヒト抗体の対応する配列と置換することにより行うことができる(例えば、Verhoeyen et al., Science 239:1534-1536 (1988)参照)。従って、かかるヒト化抗体はキメラ抗体であって、実質的には無処理のヒト可変ドメインが、非ヒト種由来の対応する配列により置換されているだけにすぎない抗体である。
【0080】
ヒトの可変領域並びにヒトのフレームワーク及び定常領域よりなる完全なヒト抗体も使用可能である。かかる抗体は当業者には既知の種々の技術を用いて産生可能である。例えば、バクテリオファージ上に表されるヒト抗体断片の組換えライブラリーの使用を含むインビトロ方法が用いられ得る(例えば、Hoogenboom & Winter, J. Mol. Biol. 227:381 (1991))。同様に、ヒト抗体は、ヒト免疫グロブリン座位を、遺伝子組換え動物、例えば、内在性免疫グロブリン遺伝子が部分的に又は完全に不活性化されているマウスに導入することにより作成できる。このアプローチは、例えば、米国特許番号第6,150,584、5,545,807、5,545,806、5,569,825、5,625,126、5,633,425、5,661,016に記載されている。
【0081】
本発明に係る方法により単離された因子又は化合物をヒト及び他の哺乳類、例えば、マウス、ラット、モルモット、ウサギ、ネコ、イヌ、ヒツジ、ブタ、ウシ、サル、ヒヒ及びチンパンジー、の医薬品として投与する場合、単離された因子又は化合物は直接投与してもよく、或いは、既知の医薬品調製方法を用いる投薬形態へ製剤化されてもよい。例えば、必要により、薬剤は、糖衣タブレット、カプセル、エリキシル剤及びマイクロカプセルとして経口で、或いは、滅菌溶液又は懸濁液を水又は薬学的に許容可能な液体とともに注射する形態で非経口で与えられてもよい。さらに、因子又は化合物は薬学的に許容可能なキャリア又は媒体、特に、滅菌水、生理食塩水、植物オイル、乳化剤、懸濁剤、界面活性剤、安定剤、芳香剤、賦形剤、ビヒクル、保存料、結合剤などと、一般的に許容された薬剤付与に必要とされる単位用量形態に混合することができる。これらの調合剤中の活性成分の量は、取得できる指示範囲内の適量な投与量となる。
【0082】
タブレット及びカプセルに混合されうる添加剤の例としては、ゼラチン、コーンスターチ、トラガントゴム及びアラビアゴムなどの結合剤;結晶セルロースなどの賦形剤;コーンスターチ、ゼラチン、アルギニン酸などの膨張剤;ステアリン酸マグネシウムなどの潤滑剤;スクロース、ラクトース又はサッカリンなどの甘味剤;及び、ペパーミント、アカモノ(Gaultheria adenothrix)オイル及びチェリーなどの芳香剤が挙げられる。単位用量形態がカプセルである場合、オイルなどの液体キャリアも上記成分に含まれうる。注射用滅菌組成物は、注射に用いられる滅菌水などのビヒクルを用いる通常の薬剤付与に従って製剤化することができる。
【0083】
生理食塩水、グルコース及び、D-ソルビトール、D−マンノース、D−マンニトール及び食塩などのアジュバントを含む他の等張液を注射用水性液として用いることができる。これらはアルコール、特にエタノールなどの適切な溶解剤、プロピレングリコール及びポリエチレングリコールなどのポリアルコール、Polysorbate80(商標)及びHCO−50などの界面活性剤とあわせて用いることができる。
【0084】
ゴマ油又は大豆オイルは油性の液体として使用でき、溶解剤としての安息香酸ベンジル又はベンジルアルコールと併せて使用でき、リン酸バッファー及び酢酸ナトリウムバッファーなどのバッファー;塩酸プロカインなどの鎮痛剤;ベンジルアルコール及びフェノールなどの安定剤;及び抗酸化剤とともに製剤化されてもよい。調製した注射は、適切なアンプルに充填されてもよい。
【0085】
当業者に既知の方法を、例えば動脈、静脈又は経皮注射として、そしてまた、鼻腔内、筋肉内又は経口投与として、本発明に係る医薬品組成物を患者に投与するために用いることができる。用量及び投与方法は患者の体重及び年齢並びに投与方法によって変わる;しかし、当業者は適した投与方法を慣例的に選択することができる。さらに、当該因子又は化合物がDNAによりコード可能である場合、DNAを遺伝子治療のためのベクターに挿入することができ、該ベクターを、治療を行うために患者に投与することができる。投与の用量及び方法は、患者の体重、年齢及び症状によって変わるが、当業者は適切に選択することができる。
【0086】
例えば、SMYD3に結合し、その活性を制御する因子又は化合物の用量は、症状に依存するが、通常の成人(体重60kg)に経口投与される場合、典型的な用量は、1日当たり約0.1mg〜約100mg、好ましくは1日当たり約1.0mg〜約50mg、より好ましくは1日当たり約1.0mg〜約20mgの範囲である。
【0087】
非経口的に投与する場合、通常の成人(体重60kg)への注射形態では、患者、標的器官、症状及び投与方法に応じていくつか違いがあるものの、1日当たり約0.01mg〜約30mg、好ましくは1日当たり約0.1mg〜約20mg、より好ましくは1日当たり約0.1mg〜約10mgの用量での静脈注射が都合がよい。さらに、他の動物の場合にも、体重60kgに変換した量を投与することができる。
【0088】
本発明はさらに被験体(subject)の癌、例えば、肝細胞癌、直腸結腸癌、膀胱癌及び乳癌、の治療のための方法を提供する。投与は、SMYD3のメチルトランスフェラーゼ活性の異常に関連する障害の危険がある(又は疑いのある)、又は、障害を有する被験体に対し、予防的又は治療的でありうる。該方法は、適した癌細胞でのSMYD3の機能を減少させることを含む。機能は、本発明に係る方法のスクリーニングにより得られる因子又は化合物の投与を介して阻害されうる。
【0089】
他の形態において、本発明は、本明細書中に記載された1又は2以上の治療的因子又は化合物を含有する薬学的、又は治療的組成物を含む。さらに、本発明はまた、本明細書中に記載された1又は2以上の治療的因子又は化合物の、癌、より具体的には、肝細胞癌、直腸結腸癌、膀胱癌及び乳癌の治療及び/又は予防のための、医薬的又は治療的組成物の製造のための使用を提供する。薬学的製剤は、経口、直腸、鼻腔、外用(頬側及び舌下を含む)、腟内又は非経口的(筋肉内、皮下及び静脈)投与、或いは、吸入又はガス吸入による投与に適したものを含んでもよい。前記製剤は、適当な場合、分離した用量単位で簡便に与えられてもよく、薬学業界では既知のいかなる方法により調製されてもよい。全てのかかる薬学的方法は、活性化合物を液体キャリア又は微細に分割された固体キャリア或いは必要に応じて両方とを会合させ、次いで、必要な場合には、生成物を所望の製剤に形作る工程を含む。
【0090】
経口投与に適した薬学的製剤は、それぞれが所定量の活性成分を含有する、カプセル、カシェ剤、又はタブレットなどの分離した用量として;粉末又は顆粒として;或いは溶液、懸濁液又はエマルションとして、簡便に与えられてもよい。活性成分は、巨丸剤又は糊状剤として与えられてもよく、純粋な形態、すなわち、キャリア無しで与えられてもよい。経口投与のためのタブレット及びカプセルは、一般的な賦形剤、例えば、結合剤、充填剤、潤滑剤、崩壊剤、潤湿剤を含有してもよい。タブレットは、圧縮又は成型により、場合により、1又は2以上の製剤成分とともに作成してもよい。圧縮タブレットは適切な機械中で粉末又は顆粒などの流通形態で活性成分を、場合により結合剤、潤滑剤、不活希釈剤、界面活性又は分散剤とともに混合して、圧縮することにより調製できる。成型タブレットは、不活性液体希釈剤で濡らした粉末化化合物の混合物を適した機械中で成型することにより作成できる。タブレットは、当業者に周知の方法によりコートしてもよい。経口液体調製物は、例えば、水性又は油性の懸濁液、溶液、エマルション、シロップ又はエリキシル剤の形態であってよく、又は、使用前に水又は他の適したビヒクルでの構成を目的とする乾燥生成物として与えられてもよい。かかる液体調製物は、通常の添加剤、例えば、懸濁剤、乳化剤、非水性ビヒクル(食用オイルを含んでもよい)、又は保存剤を含んでもよい。タブレットは、場合により、中の活性成分がゆっくりと又は制御されて放出されるように製剤化されていてもよい。
【0091】
非経口投与のための製剤は、製剤を目的レシピエントの血液と等張に保持する抗酸化剤、バッファー、静菌剤および溶質を含んでもよい水性及び非水性の滅菌注射溶液;並びに、懸濁剤及び増粘剤を含んでもよい水性又は非水性の滅菌懸濁液を含んでもよい。製剤は、単位用量又は多用量容器、例えば、密閉アンプル及びバイアルに入れて与えられてもよく、使用直前に滅菌液体キャリア、例えば、生理食塩水、注射水の添加のみが求められるフリーズドライ(凍結乾燥)状態で貯蔵されてもよい。また、製剤は連続的点滴用に与えられてもよい。即席の注射溶液及び懸濁液は、上述した類の滅菌粉末、顆粒及びタブレットから調製することができる。
【0092】
直腸投与のための製剤は、ココアバター又はポリエチレングリコールなどの通常のキャリアを伴う座薬として与えられてもよい。例えば、頬側又は舌下の、口中への局所投与のための製剤には、スクロース及びアラビアゴム又はトラガントなどの風味付けされた基剤に活性成分を含む薬用キャンディー、及び、ゼラチン及びグリセリン又はスクロース又はアラビアゴムなどの基剤に活性成分を含むトローチが含まれる。鼻腔内投与のため、本発明により得られる化合物は液体スプレー又は分散可能な粉末として或いは液滴の形態で使用されてもよい。液滴は、水性又は非水性基剤とともに製剤化され、1又は2以上の分散剤、溶解剤又は懸濁剤も含んでよい。液体スプレーは、圧縮パックから簡便に送達される。
【0093】
吸入による投与のため、化合物は吸入器、噴霧器、圧縮パック又はエアロゾルスプレーを送達する他の簡便な手段から、簡便に送達される。圧縮パックは、ジクロロジフルオロメタン、トリクロロフルオロメタン、ジクロロテトラフルオロエタン、二酸化炭素又は他の適したガスなどの適した噴霧剤を含んでもよい。圧縮エアロゾルの場合、投与単位はある計量量を送達する弁を設けることにより決定可能である。
【0094】
また、吸入又はガス注入による投与のため、化合物は乾燥粉末組成物、例えば、化合物とラクトース又はスターチなどの適した粉末基剤との粉末混合物の形態とされてもよい。粉末組成物は、粉末が吸入又はガス注入の助けで投与され得る単位用量形態、例えば、カプセル、カートリッジ、ゼラチン又はブリスターで与えられてもよい。
【0095】
所望の場合には、活性成分を徐放するよう適合された上述の製剤を用いてもよい。薬学的組成物は、抗菌剤、免疫抑制剤又は保存剤などの他の活性成分を含んでもよい。
【0096】
特に上述した成分に加えて、本発明の製剤は、問題の製剤の種類に関して、当業者には一般的な他の因子を含んでよいこと、例えば、経口投与に適したものが香味料を含んでよいことは理解されるべきである。
【0097】
好適な単位投与製剤は、活性成分の効果用量(後述)又はその適当な画分を含有するものである。
【0098】
上記条件のそれぞれについて、組成物は経口又は注射により、1日当たり約0.1〜約250mg/kgの用量で投与されてもよい。成人の用量範囲は、一般に1日当たり約5mg〜約17.5g、好ましくは1日当たり約5mg〜約10g、最も好ましくは1日当たり約100mg〜約3gである。タブレット又は分散単位で与えられる他の単位用量形態には、そのような用量で効果的である量又はこれと同じ用量の数倍として、例えば、約5mg〜約500mg、通常約100mg〜約500mgを、簡便に含んでもよい。
【0099】
薬学的組成物は、好ましくは、経口又は注射(静脈又は皮下)により投与され、被験体に投与される正確な量は付き添いの医師の責任となる。しかし、用いられる用量は、被験体の年齢及び性別、治療される正確な障害及びその深刻度を含む数々の要因による。投与の経路もまた、状態及びその深刻度により変化しうる。
【0100】
本発明はさらに、試験化合物のVEGFR1のメチル化を制御する能力を検出するキットを提供し、該キットは、以下の構成要素:
(a)以下からなる群から選択されるメチルトランスフェラーゼ活性を有するSMYD3ポリペプチド;
(i)配列番号2のアミノ酸配列を有するポリペプチド;
(ii)配列番号2のアミノ酸配列を有するポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失又は挿入され、さらにここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性であるアミノ酸配列を有するポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(iv)配列番号1のヌクレオチドを有するポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、該ポリペプチドは配列番号2のアミノ酸配列を有するポリペプチドと等価なメチルトランスフェラーゼ活性を有する、ポリペプチド;
(b)(a)のポリペプチドによりメチル化され得るVEGFR1ペプチド;及び
(c)VEGFR1ペプチドのメチル化のコファクター
を含む。
【0101】
本発明において、好ましいVEGFR1ペプチドは、配列番号4のアミノ酸配列から構成されるポリペプチドまたはその機能的変異体もしくは機能的断片を含む。好ましい断片には、配列番号4のアミノ酸配列の位置800〜841のアミノ酸配列を含むか又は、それらから構成されるポリペプチド、或いは、VEGFR1のリジン831を含むポリペプチドを含むが、これに限られない。さらに、好ましいコファクターは、S−アデノシルホモシステインヒドロラーゼ(SAHH)である。本発明に係るキットは、メチル化されたVEGFR1ペプチドを検出する試薬をさらに含んでもよい。例えば、メチル化VEGFR1ペプチドを認識する抗体は検出試薬として使用可能である。アッセイを行うための説明書(例えば、書類、テープ、VCR、CD−ROM等)がキットに含まれてもよい。キットのアッセイフォーマットは、当業者には既知のメチルトランスフェラーゼアッセイ又は結合アッセイである。
【0102】
以下の例は、単なる例示であり、本発明の範囲を限定することを意図するものではない。本発明の態様が以下の実施例に記載されるものの、当業者は、本明細書中に記載されたものと同様の又は等価な他の方法及び材料が本発明の実施又は試験に用いることができることを認識するであろう。
【0103】
実施例
材料と方法
試薬及びウェスタンブロット分析
タンパク質の抽出、IP(免疫沈降)、及びウェスタンブロット分析は、文献に記載の方法に従って行った(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)。イムノブロット分析は抗SMYD3抗体(Abcam、Cambridge、MA)、抗HA抗体(Sigma、St Louis、MO)、抗Flag抗体(Sigma)、抗GST抗体(BD Pharmingen、San Diego、CA)、抗βアクチン抗体(Sigma)、又は抗リン酸化チロシン抗体(PY20、Sigma)を用いて行った。HRP接合二次抗体を、ECL検出システム(Amersham Biosciences、Piscataway、NJ)に用いた。組換えヒストンH3及びVEGFR1(3108)は、Upstate Biotechnology(Lake Placid、NY)及びCalbiochem(San Diego、CA)からそれぞれ購入した。メチル化リジン831特異的抗体(抗K831me2)は、ジメチル化リジン831を含む合成ペプチドであるRERLKLGKm2SLGRGAで免疫したウサギの血清から精製した。
【0104】
細胞株及び組織標本:
ヒト胎児腎細胞株(HEK293及び293T)及びヒト結腸癌細胞株(SW480)は、American Type Culture Collection(ATCC、Manassas、VA)から入手した。SNU423HCC細胞及びMCF7乳癌細胞はKorea cell−line bank及び財団法人癌研究会からそれぞれ好意に基づいて提供された。全ての細胞株は、適切な培養液中、単層で成長させた。
【0105】
プラスミドの調製
SMYD3を発現するプラスミドをすでに文献に記載されているように調製した(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)。本発明者らは、また、VEGFR1の野生型又は欠失形態のいずれかを含む種々のRT-PCR産生物を、pCMV−HA(Clutch、Palo Alto、CA)、p3xFlag−CMV14(sigma)又はpGEX6P−3ベクター(Amersham Biosciences)の適切な部位にクローニングすることにより、HAタグ化又は3xFlagタグ化VEGFR1を発現するプラスミドを調製した。RT-PCR実験は、プライマーの組(表1)を用いて行った。アミノ酸配列の置換を含む変異体VEGFR1プラスミドは、Quickchange II XL Site-directed Mutagenesis kitを用いてサプライヤーのプロトコルに従って生成した(Strata gene、La Jolla、CA)。
【0106】
本発明者らは、FuGENE(商標)6試薬を用いてサプライヤーのプロトコル(Roche、Indianapolis、IN)に従って培養細胞に哺乳類のプラスミドをトランスフェクトした。組換えGST融合VEGFR1タンパク質を大腸菌、BL21バクテリア細胞からグルタチオンセファロース4B(Amersham Biosciences)を用いて精製した。本実施例で用いたプライマーを以下に示す。
【0107】
(表1)プライマー配列リスト
【0108】
インビトロメチルトランスフェラーゼ及びキナーゼアッセイ
293T細胞に、Flagタグ化野生型SMYD3を発現するプラスミド(p3XFLAG−CMV−SMYD3)をトランスフェクトした。タグ化SMYD3タンパク質は、抗Flag抗体を用いる免疫沈降により精製した。組換えSMYD3タンパク質は、バキュロウイルスシステム(Invitrogen、Carlsbad、CA)を用いてSf9細胞中で調製した。インビトロメチルトランスフェラーゼアッセイは、わずかに修正を施して他の文献に記載されたように(Hamamoto R et al., S Nature cell biology 2004, 6(8): 731-40)行った。
【0109】
簡単にいえば、免疫沈降した又は組換えSMYD3タンパク質を、1μgの組換えヒストンH3又はVEGFR1タンパク質と、メチルドナーとしての2μCiの[メチル−3H]ラベル化S−アデノシル−L−メチオニン(SAM、Amersham Biosciences)の存在下、S−(5’−アデノシル)−L−ホモシステインヒドロラーゼ(SAHH、Sigma)の存在下又は非存在下、メチルトランスフェラーゼバッファー(50mM Tris−HCl pH8.5、100mM NaCl、10mM DTT)中で混合し、30℃で1時間インキュベートした。ラベル化タンパク質は液体シンチレーションカウンターで測定し、又は、SDS−PAGE後、フルオログラフィーで検出した。インビトロキナーゼアッセイをGST融合VEGFR1及びHTScan(商標)VEGFR1キナーゼアッセイキットを用い、サプライヤーのプロトコルに従って行った(Cell Signaling、Danvers、MA)。基質のリン酸化は、Time−Resolved Plate Reader(Perkin−Elmer、Wellesley、MA)を用いて615nmの蛍光発光により定量した。
【0110】
インビボメチル化アッセイ
VEGFR1のインビボメチル化は、Liu及びDreyfuss(Liu Q and Dreyfuss G., Molecular and cellular biology 1995, 15(5): 2800-8)に記載された方法に従い、わずかに修正を施して分析した。簡単に言えば、HAタグ化VEGFR1を発現するHEK293細胞を、100μg/mlのシクロヘキシミドとともに、37℃で30分間インキュベートし、その後、10μCi/mlのL−[メチル−3H]メチオニンを含む培養液中にそのまま3時間保持した。VEGFR1は、細胞抽出物から抗HA抗体で沈降し、SDS−PAGEにより分離し、続いて、BASイメージングシステム(BAS−TR2040、FUJI)により、又は、イムノブロット分析により分析した。
【0111】
RNA干渉
RNAi実験は、Dharmacon(Chicago、IL)から購入した、SMYD3に特異的な又は対照siRNAの二本鎖オリゴヌクレオチドで行った。1x105のHEK293−SMYD3細胞を、オリゴフェクタミンを用い、各siRNAで最終濃度100nMでトランスフェクトした。トランスフェクトから48時間後、タンパク質を細胞から抽出し、ウェスタンブロット分析にかけた。
【0112】
統計分析
全ての分析は、Statviewソフトウェアを用いて行った(SAS Institute、Cary、NC)。
【0113】
実施例1:SMYD3はVEGFR1をインビトロでメチル化する
最近の研究では、VEGFRがヒトの発癌に関与していることが示唆されている。従って、ここで、VEGFR1について、インビトロメチルトランスフェラーゼアッセイを用い、SMYD3の候補メチル化標的として調べた。本発明者らは、基質に関し、VEGFR1の3つの細胞質領域の組換えタンパク質、VEGFR1#1(コドン800−1000)、VEGFR1#2(コドン1000−1200)及びVEGFR1#3(コドン1200−1338)を調製した。SMYD3又は対照タンパク質を、Flagタグ化SMYD3又はモックプラスミドでそれぞれトランスフェクトしたHEK293細胞から抗Flag抗体で免疫沈降した。結果として、VEGFR1#1は、SMYD3によりインビトロで対照タンパク質と比較して有意にメチル化された(図1a)。しかし、VEGFR1#2又はVEGFR1#3において、メチル化は観察されず、VEGFR1#1がメチル化残基を含有することが示唆された。インビトロの組換えヒストンH3と比較して約3倍高いVEGFR1#1のメチル化が検出された(データ示さず)。
【0114】
インビトロメチルトランスフェラーゼアッセイをさらに、昆虫細胞から精製した組換えSMYD3タンパク質について行い、その結果は、VEGFR1#1のメチル化を示したが、VEGFR1#2もVEGFR1#3もメチル化を示さなかった(図1b)。メチル化残基を決定するため、本発明者らは、VEGFR1#1構築物の異なる断片を含む他のVEGFR1タンパク質を調製した(図1c)。インビトロメチルトランスフェラーゼアッセイは、コドン800−841を含むVEGFR1−N1、及びコドン800−878を含むVEGFR1−N7が、SMYD3によりメチル化されたことを示した。一方で、コドン842−878、879−920、842−900、879−900、842−900又は842−1000をそれぞれ含むVEGFR1−N2、N3、N4、N5、N6又はN8に、メチル化バンドは観察されなかった(図1c)。
【0115】
このデータは、800と841の間の領域がもっともメチル化残基を含む可能性が高いことを示唆する。興味深いことに、VEGFR2には、その細胞質領域については基質としても分析したが、メチル化バンドは検出されなかった(データ示さず)。まとめると、このデータはさらにSMYD3が、SET7/9及びSMYD2と同様に、非ヒストンタンパク質のメチル化に関し、基質特異性及び配列特異性の両方を有することを示唆する。
【0116】
実施例2:VEGFR1リジン831がSMYD3によりメチル化される
SMYD3を含む、SET含有メチルトランスフェラーゼのほとんどは、リジンを修飾する。従って、この領域内のリジンを調査した。リジン819、828、831及び840は他種のVEGFR1オルソログの中で保存されていることが決定された(データ示さず)。先の報告では、K/R−S/T/A−KがヒストンメチルトランスフェラーゼSET7/9の共通メチル化モチーフであることが示されているが(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)、他のヒストンメチルトランスフェラーゼの標的リジン、例えば、p53K370、ヒストンH3K4及びH3K27は、K/R−S/T/A−Kに隣接していないが、S/T−K−X又はX−K−Sには隣接している(図2a)。従って、これらのモチーフに一致し、他種に保存されたリジン819及び831は、本アッセイの中心であった。
【0117】
インビトロメチルトランスフェラーゼアッセイを、野生型及びリジンのアラニンへの置換を含む変異体VEGFR1−N1タンパク質を用いて行った(図2b)。アッセイにより、野生型及びK819A変異体のメチル化バンドが示されたが、K831A変異体は、いかなるメチル化バンドも示さなかった(図2b)。VEGFR1 K831が他のアミノ酸に置換された他の変異タンパク質も同様にメチル化を示さなかった(図2c)。リジン831のインビトロメチル化を確認するため、リジンに対するメチル化特異的抗体(抗K831me2)を調製した。インビトロメチルトランスフェラーゼアッセイは、フルオログラム並びに抗K831me2抗体を用いたウェスタンブロット分析により、組換えVEGFR1のメチル化されたリジン831を確認した(図2d)。
【0118】
実施例3:リジン831のインビボメチル化
VEGFR1のインビボメチル化を調べるため、インビボメチル化アッセイを、Flagタグ化SMYD3(293−SMYD3)を安定に発現するHEK293細胞、及び、対照細胞(293−モック)を用いて行った。細胞を、HAタグ化VEGFR1でトランスフェクトし、続いて、タンパク質合成阻害剤の存在下でL−[メチル−3H]メチオニンとともにインキュベートした。抗HA抗体での細胞の免疫沈降物を、SDS−PAGEにより分離し、蛍光映像装置(fluoroimager)によりメチル化されたVEGFR1を調べた。予想したように、293−モック細胞からのものと比較して、VEGFR1の強められたメチル化が293−SMYD3細胞からの沈降物中に検出された(図2e)。これと一致して、抗K831me2抗体を用いた沈降物のイムノブロット分析では、対照細胞と比較して強められたリジン831のメチル化が確認された(図2f)。
【0119】
SMYD3に依存したメチル化を確認すべく、293−SMYD3細胞をSMYD3−siRNA又は対照siRNAで処理した。抗SMYD3抗体を用いたイムノブロット分析は、SMYD3−siRNAが、対照siRNAと比較してSMYD3タンパク質の発現を効果的に減少させたことを示した(図2g、上パネル)。SMYD3のノックダウンと関連して、SMYD3−siRNAは、対照siRNAと比較して細胞中のK831のメチル化を減少させたが(図2g、第2パネル)、VEGFR1の発現はこれらのsiRNA間でほとんど変わらなかった(図2g、第3パネル)。これらのデータは、リジン831がSMYD3によってインビボでメチル化されることを示唆する。
【0120】
実施例4:SMYD3とVEGFR1との間の相互作用
VEGFR1はSMYD3によりメチル化されるため、本発明者らは、SMYD3がVEGFR1と相互作用するか否か調べた。本発明者らは、VEGFR1のFlagタグ化細胞質領域をHAタグ化SMYD3とともにHEK293細胞内で発現させた。抗HA抗体での免疫沈降及びその後の抗Flag抗体でのイムノブロット分析は、SMYD3がVEGFR1#1(コドン800−1000)とは会合するが、VEGFR1#2ともVEGFR1#3とも会合しないことを示した(図3a)。これと一致して、抗Flag抗体での免疫沈降及びその後の抗HA抗体でのイムノブロット分析は、SMYD3とVEGFR1#1との相互作用を実証した(図3a)。内在性SMYD3とVEGFR1との相互作用を調べるため、本発明者らは、さらに、SMYD3及びVEGFR1の両方を発現するSNU423、SW480又はMCF7細胞からの抽出物を用いた、抗SMYD3抗体での免疫沈降を行った。結果として、本発明者らは、抗SMYD3抗体での免疫沈降により、内在性SMYD3はVEGFR1と共免疫沈降することを見出し(図3b)、これは、SMYD3とVEGFR1との間のインビボ相互作用を示唆する。VEGFR1に対する抗体は、免疫沈降には適用できないため、本発明者らは抗VEGFR1抗体で免疫沈降は行わなかった。
【0121】
VEGFR1#1はSMYD3によりメチル化されるリジン831を含むので、これらのデータはVEGFR1のメチル化に一致するものである。本発明者らは、また、SMYD3の野生型及び3つの欠失型:SMYD3−N20(コドン20−428)、SMYD3−Δ2(コドン100−428)及びSMYD3−Δ3(コドン250−428)を発現するプラスミドを用い、結合に関与するSMYD3の領域を調べた。野生型SMYD3、SMYD3−N20及びSMYD3−Δ2はVEGFR1と相互作用するが、SETドメインの無いSMYD3−Δ3はしなかった(図3c)。SETドメイン(100−250)を発現する他のプラスミドは、VEGFR1の細胞質ドメインと会合を示した(図3d)。このデータはSETドメインが相互作用に関与することを示唆する。
【0122】
実施例5:SMYD3によるメチル化を介したVEGFR1のキナーゼ活性増大
リジン831はキナーゼドメインに位置する。そこで、VEGFR1メチル化のそのキナーゼ活性への影響をさらに調べた。キナーゼ活性は、基質のリン酸化チロシンを認識するVEGFR1キナーゼアッセイキットを用いて調べた。VEGFR1の細胞質領域の組換えタンパク質を3Hラベル化SAMで、SMYD3の存在下又は非存在下で処理し、その後、キナーゼ活性を調べた。予想したように、SMYD3によるVEGFR1のメチル化が、3H−BASシステムにより検出された(データ示さず)。重要なことに、SMYD3で処理したVEGFR1は、対照と比較してそのキナーゼ活性の有意な増大を示した(図4a)。VEGFR1無しのSMYD3単独によるリン酸化は観察されなかった(データ示さず)。抗リン酸化チロシン抗体でのウェスタンブロット分析はまた、SMYD3非存在下と比較して、SMYD3存在下では、組換えヒトVEGF165タンパク質を伴うFc領域に融合された組換えヒトVEGFR1タンパク質のリン酸化増大をも明らかとした(図4b)。興味深いことに、VEGFR1の組換え細胞質領域(コドン784−1338)をインビトロキナーゼアッセイに用いた場合には、VEGFR1の自己リン酸化は観察されなかった(データ示さず)。従って、VEGFR1のメチル化は、VEGFR1標的分子のリン酸化に影響し、そのシグナル伝達を強めることとなる。
【0123】
考察
本明細書中で示したように、VEGFR1はSMYD3ヒストンメチルトランスフェラーゼの、新規な非ヒストン標的である。ヒストン末端の修飾は、クロマチン構造の一部変質を介して、転写、DNA修復、テロメア維持、DNA複製及び染色体分離において重要な役割を果たす。最近の分子研究はヒストン末端のリジン修飾、及び、ヒストンメチルトランスフェラーゼ及びデメチラーゼによるその動的制御の重要性を明らかにしている(Volkel P and Angrand PO, Biochimie 2007, 89(1): 1-20)。17を超えるヒストンメチルトランスフェラーゼがこれまで同定されており、これらのメチルトランスフェラーゼは、それぞれの基質に特異性を有している。H3K4は、SET7/9、Set1、MLL1、MLL2、MLL3、MLL4、MLL5、ASH1及びSMYD3によりモノ、ジ、又はトリメチル化される(Kouzarides T, Cell 2007, 128(4): 693-705.; Volkel P and Angrand PO, Biochimie 2007, 89(1): 1-20)。
【0124】
重要なことに、これらのメチルトランスフェラーゼの中で、SET7/9及びSMYD2は、非ヒストンタンパク質をメチル化することが知られている(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82;Chuikov S et al., Nature 2004, 432(7015): 353-60;Huang J et al., Nature 2006, 444(7119): 629-32)。SET7/9はインビトロ及びインビボでp53中のリジン372をメチル化し、p53の安定性を上昇させた(Chuikov S et al., Nature 2004, 432(7015): 353-60)。SET7/9は、TAF10のリジン189(Kouskouti A et al., Mol Cell 2004, 14(2): 175-82)及びTAF7のリジン5(ヒトTATAボックス結合タンパク質関連因子)でのメチルトランスフェラーゼ活性も示した(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)。
【0125】
さらに、p53 K370のメチル化は、SMYD3(SET及びMYNDドメイン)ファミリーのもう1つのメンバーであるSMYD2により示されている(Huang J et al., Nature 2006, 444(7119): 629-32)。本明細書中において、VEGFR1が、細胞質及び核に局在する、H3K4のヒストンメチルトランスフェラーゼの新規な非ヒストン標的であることが確認される。他のH3K4メチルトランスフェラーゼ、例えば、Set1、MLL1、MLL2、MLL3、MLL4、MLL5及びASH,1は、同じヒストンリジン修飾を認識するはずであり、基質がどのように異なるメチルトランスフェラーゼにより修飾されるかを見ることは関心あることであろう。ヒストンメチル化は動的に制御されるので、非ヒストンタンパク質のメチル化もまたヒストンデメチラーゼにより修飾されうる。従って、VEGFRのメチル化されたリジン831は、LSD1(Shi Y et al., Cell 2004, 119(7): 941-53)やJARID1(Seward DJ et al., Nat Struct Mol Biol 2007, 14(3): 240-2)のようなH3K4デメチラーゼにより触媒されるであろう。
【0126】
VEGFRは、細胞外リガンド結合ドメイン、膜貫通ドメイン、キナーゼドメイン及びカルボキシル末端領域からなる受容体チロシンキナーゼ(RTK)である。VEGFR1を含む全てのRTKは、GXGXXG(ATP結合部位)、HRDLA(触媒に不可欠なモチーフ)及び1又は2のチロシン自己リン酸化部位を含む、進化的に保存されたキナーゼドメインを含む(Rahimi N, Experimental eye research 2006, 83(5): 1005-16.; Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。
【0127】
本明細書中で示したように、SMYD3が誘導するリジン831のメチル化は、インビトロでのVEGFR1のキナーゼ活性増大をもたらす。理論に縛られることは望まないものの、この結果はリジン残基が、キナーゼドメインの“GXGXXXG”モチーフからN末端に3アミノ酸に位置するという事実に由来すると考えられる。ゆえに、リジン831のメチル化はキナーゼドメインの立体構造を変化させうる。一方で、VEGFR1の細胞質ドメインのC末端領域がキナーゼ活性に対して阻害的役割を有するため、VEGFR1のメチル化は阻害的ドメインを抑制してキナーゼ活性を増大する(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。
【0128】
ヒストン末端についての最近の研究は、その修飾残基が特異的分子により認識されることを示している;リン酸化されたH3S10はGCN5、PVAF及びp300を増加させ、メチル化されたH3K9はヘテロクロマチンタンパク質1(HP1)を増加させ、かつメチル化されたH3K4はCHD1、SNF2L/ISWI、WDR5、BPTF/NURF301及びING2を増加させる。従って、メチル化リジン831は、VEGFR1のキナーゼ活性に影響を与え得るメチル化特異的タンパク質複合体を増加させる。ヒストン末端の修飾は、他のヒストン末端の修飾に関連し、これは共有結合的修飾間の相互作用を示す(Strahl BD and Allis CD, Nature 2000, 403(6765): 41-5;Jenuwein T and Allis CD, Science 2001, 293(5532): 1074-80;Li B et al., Cell 2007, 128(4): 707-19;Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。
【0129】
H3S10のリン酸化は、GCN5を媒介としたH3K14のアセチル化を促進するが、SUV39H1によるH3K9のメチル化を阻害する。反対に、H3K9のメチル化は、H3S10のリン酸化を抑制する。同様に、H4K3のメチル化はH4K8及びH4K12のアセチル化を促進する(Zhang Y and Reinberg D, Genes & development 2001, 15(18): 2343-60)。共有結合ヒストン修飾間の相互作用を見るに、本発明者らは、VEGFR1のメチル化が、その強められたキナーゼ活性による自己リン酸化、並びに、その後の分子間リン酸化を介したVEGFR2のリン酸化を変質しうると推測するよう誘導される。メチル化されたリジン831はTry1169、1213、1242、1327及び1333(VEGFR1のリン酸化されたチロシン)(Olsson AK et al., Nature reviews 2006, 7(5): 359-71)から離れているように見えるが、これらの残基はリジン831に構造的に近い位置を示し得る。さらに、異なるリガンドによるVEGFシグナル伝達についての今後の研究は、VEGFR1メチル化の役割についてのよりよい理解をもたらす可能性がある。
【0130】
認識モチーフ(K/R−S/T/A−K)がSET7/9メチルトランスフェラーゼに対する共通認識部位であることがこれまでの研究により報告されている(Couture JF et al., Nat Struct Mol Biol 2006, 13(2): 140-6)。しかし、他のヒストンメチルトランスフェラーゼによるメチル化リジンの隣接配列は、必ずしもこのモチーフと適合しない。ヒストンH3K9、H3K27及びp53のリジン370のアミノ酸配列の比較により、X−K−Sがヒストンリジンメチルトランスフェラーゼの別の保存されたモチーフであることが見出された。重要なことに、VEGFR1のメチル化されたリジン831は、この知見によく適合する。VEGFR2のキナーゼドメインはVEGFR1のものと70.1%のアミノ酸類似性を有するが、VEGFR2はSMYD3によりメチル化されない。VEGFR2のリジン831に対応するリジンは共通配列に隣接しないので、この結果は保存されたモチーフに矛盾しない。VEGFR1と比べて高いキナーゼ活性を示すVEGFR2がSMYD3によりメチル化されなかったことは興味深い。
【0131】
SMYD3はVEGFR2のメチル化に影響を与えない場合がある一方で、SMYD3はVEGFR1のリン酸化を介したVEGFR2のリン酸化には影響を与える場合があるが、これはこれら2つの分子がヘテロ2量体を形成するからである(Rahimi N, Experimental eye research 2006, 83(5): 1005-16;Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。1つの受容体のリン酸化が、分子内リン酸化により他の受容体のリン酸化に影響を与えることは周知である(Shibuya M, Journal of biochemistry and molecular biology 2006, 39(5): 469-78)。酵母Rkm1(SETドメイン含有酵素)がRpl23a及びRpl23bに対するメチルトランスフェラーゼ活性を示したことはさらに留意すべきである(Porras-Yakushi TR et al., J Biol Chem 2007, 282(17):12368-76)。メチル化リジンの周辺の標的配列の比較により、N/P−P−Kがメチル化の標的でありうることが示唆された。従って、リジンの隣接配列は、複数のメチルトランスフェラーゼの間で異なる可能性がある。
【0132】
VEGFR1は血管表皮細胞上で排他的に発現すると言われてきたが、最近の研究では、非EC細胞内でのVEGFR1の発現が示されている。VEGFR1は大腸癌(Lesslie DP et al., Br J Cancer 2006, 94(11): 1710-7;Fan F et al., Oncogene 2005, 24(16): 2647-53;Duff SE et al., Eur J Cancer 2006, 42(1): 112-7;Andre T et al., International journal of cancer 2000, 86(2): 174-81)、乳癌(Li YS et al., Pathology international 2006, 56(5): 256-61;Wu Y et al., International journal of cancer 2006, 119(7): 1519-29)、膵臓癌(Chung GG et al., Cancer 2006, 106(8): 1677-84.; Yang AD et al., Cancer Res 2006, 66(1): 46-51)、前立腺癌(Jackson MW et a., Cancer Res 2002, 62(3): 854-9)、腎癌(Jacobsen J et al., BJU Int 2006, 97(5): 1102-8;Ljungberg BJ et al., BJU Int 2006, 98(3): 661-7)及び卵巣癌(Wang FQ et al., International journal of cancer 2006, 118(4): 879-88;Chen H et al., Gynecol Oncol 2004, 94(3): 630-5)の組織及び癌細胞株(Fan F et al., Oncogene 2005, 24(16): 2647-53;Duff SE et al., Eur J Cancer 2006, 42(1): 112-7;Wu Y et al., International journal of cancer 2006, 119(7): 1519-29;Jackson MW et a., Cancer Res 2002, 62(3): 854-9;Soker S et al., Am J Pathol 2001, 159(2): 651-9;Wu W et al., Oncogene 2003, 22(22): 3361-70)を含むヒト組織の広い範囲で発現されている。報告によれば、VEGFR1の発現は初期大腸癌標本ではかすかであったが、その発現は肝転移では明確に検出された(Fan F et al., Oncogene 2005, 24(16): 2647-53)。
【0133】
さらに、VEGFR1は腫瘍の成長及び進行に関連していることが報告された:VEGFR1の外因性発現は膵臓癌の侵襲と転移を強めた。すなわち、強められたSMYD3の発現は侵襲性の及び/又は転移する性質を癌細胞に付与する可能性がある。この場合、VEGFR1メチル化を標的とする治療アプローチは、癌細胞の侵襲と転移を阻害することにより患者を利するであろう。SMYD3のメチルトランスフェラーゼ活性の抑制は、VEGFR1を媒介とする癌の進行の阻害の助けとなりうる。
【0134】
本明細書中に示されるように、SMYD3はVEGFR1のリジン831をインビトロ及びインビボでメチル化し、メチル化されたVEGFR1はそのキナーゼ活性を非メチル化VEGFR1と比べて強められた。従って、多量のSMYD3タンパク質を発現する癌細胞は、VEGFR1を媒介とした強められたシグナル伝達経路を示し得る。このデータはVEGFR1の新規な制御メカニズム、及び、ヒトの発癌に関与する、調節解除されたVEGFR1シグナル伝達に光を当てる可能性がある。
【産業上の利用可能性】
【0135】
本発明は、直腸結腸癌、肝細胞癌、膀胱癌及び乳癌を含む種々の癌の予防、診断及び治療のためのさらなる分子標的の同定に有用である。さらに、本明細書中に報告されたデータは、癌の包括的な理解を向上させ、新規な診断戦略の開発を容易とし、治療的薬剤及び予防薬剤のための分子標的の同定のカギを提供する。かかる情報は、腫瘍発生のより深い理解に寄与し、癌の診断、治療及び最終的には予防の新規な戦略の開発のための指標を提供する。
【0136】
本発明は、詳細に、その特定の実施形態をもって記載されたが、上記記載は本質的に例示的かつ説明的なものであり、本発明及びその好ましい実施形態を例示することを意図することは理解されるべきである。通常の実験を通して、当業者は、種々の変化及び修正が本発明の精神及び範囲から逸脱することなくなされうることを容易に理解するであろう。従って、本発明は、上記記載によって定義されず、以下の特許請求の範囲及びその均等物により定義されることが意図される。
【特許請求の範囲】
【請求項1】
SMYD3によるVEGFR1のメチル化を調整する因子を同定する方法であって、該方法が、
(a)以下から構成される群から選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化に適した条件下で、前記因子の存在下で接触させる工程;
(i)配列番号2のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を含むアミノ酸配列を有するポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルを前記因子の非存在下で検出した対照レベルと比較する工程を含み、
前記対照レベルと比較したメチル化レベルの増大又は減少が、前記因子がSMYD3によるVEGFR1のメチル化を調整することを示す、方法。
【請求項2】
前記コファクターが、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項1に記載の方法。
【請求項3】
前記(ii)に定義されるポリペプチドが、20以下の保存的アミノ酸置換を含む配列番号2のアミノ酸配列を含む、請求項1に記載の方法。
【請求項4】
前記(iii)に定義されるポリペプチドが、配列番号2と少なくとも約95%の相同性を有するアミノ酸配列を含む、請求項1に記載の方法。
【請求項5】
前記(iv)に定義されるポリペプチドが、配列番号1のヌクレオチド配列からなるポリペプチドに、高いストリンジェント条件下で特異的にハイブリダイズする、請求項1に記載の方法。
【請求項6】
前記(v)に定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項1に記載の方法。
【請求項7】
前記VEGFR1ペプチドが、配列番号4のアミノ酸配列を含むポリペプチド、または、その機能的変異体もしくは機能的断片である、請求項1に記載の方法。
【請求項8】
前記機能的VEGFR1断片が、配列番号4のアミノ酸配列の位置800〜1000のアミノ酸配列を含む、請求項7に記載の方法。
【請求項9】
前記機能的VEGFR1断片が、配列番号4のアミノ酸配列の位置800〜841のアミノ酸配列を含む、請求項7に記載の方法。
【請求項10】
前記機能的VEGFR1変異体が、以下の変異:K819A、K819E及びK819Rの1又は2以上を含む、配列番号4のアミノ酸配列を含む、請求項7に記載の方法。
【請求項11】
前記メチル化レベルが、VEGFR1のリジン831で検出される、請求項1に記載の方法。
【請求項12】
SMYD3を過剰発現する癌治療のための化合物のスクリーニング方法であって、該方法が、
(a)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片を、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片と、VEGFR1ペプチドとSMYD3ポリペプチドとの結合を可能とする条件下及び試験化合物の存在下で接触させる工程;及び
(b)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片と、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片との間の結合を阻害する試験化合物を、癌治療のための候補化合物として選択する工程
を含む、方法。
【請求項13】
前記VEGFR1ペプチドのSMYD3結合領域を含む断片が、配列番号4のアミノ酸配列の位置800〜1000のアミノ酸配列を含む、請求項12に記載の方法。
【請求項14】
前記SMYD3ペプチドのVEGFR1結合領域を含む断片が、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項12に記載の方法。
【請求項15】
前記癌が、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択される、請求項12に記載の方法。
【請求項16】
SMYD3を過剰発現する癌の治療のための化合物をスクリーニングする方法であって、該方法が、
(a)請求項1に記載の方法を用いてメチル化を調整する試験化合物を同定する工程;及び
(b)前記試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程
を含む、方法。
【請求項17】
前記癌が、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択される、請求項16に記載の方法。
【請求項18】
試験化合物のVEGFR1のメチル化調整能力を検出するキットであって、該キットが、
(a)以下から構成される群から選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド;
(i)配列番号2のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を含むアミノ酸配列を有するポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)前記(a)のポリペプチドによりメチル化され得るVEGFR1ペプチド;及び
(c)VEGFR1ペプチドのメチル化のためのコファクター
を含む、キット。
【請求項19】
前記VEGFR1ペプチドが、配列番号4のアミノ酸配列を含むポリペプチド、または、その機能的変異体もしくは機能的断片である、請求項18に記載の方法。
【請求項20】
前記キットが、以下の:
(d)S−アデノシルホモシステインヒドロラーゼ(SAHH)
をさらに含む、請求項18に記載のキット。
【請求項21】
前記(v)に定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項18に記載のキット。
【請求項22】
ポリペプチドのメチルトランスフェラーゼ活性を測定する方法であって、該方法が、
(a)H3K4メチルトランスフェラーゼを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化を可能とする条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化とメチルトランスフェラーゼ活性とを関連付けることにより、メチルトランスフェラーゼ活性を測定する工程
を含む、方法。
【請求項23】
前記VEGFR1ペプチドが、少なくともK831を含むVEGFR1断片である、請求項22に記載の方法。
【請求項24】
前記コファクターが、S−アデノシル−L−メチオニンである、請求項22に記載の方法。
【請求項25】
前記ポリペプチドを、VEGFR1ペプチド及びコファクターと、メチル化増強因子の存在下で接触させる、請求項22に記載の方法。
【請求項26】
前記メチル化増強因子が、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項25に記載の方法。
【請求項27】
K831が、VEGFR1のメチル化されたリジン831を認識する抗体で検出される、請求項23に記載の方法。
【請求項28】
前記H3K4メチルトランスフェラーゼが、
(i)配列番号2(SMYD3)のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価な生物学的活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を有するアミノ酸配列を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価な生物学的活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
から構成される群から選択される、請求項22に記載の方法。
【請求項29】
前記(v)で定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を有する、請求項28に記載の方法。
【請求項30】
SMYD3のメチルトランスフェラーゼ活性を測定するキットであって、該キットが、
(a)SMYD3によりメチル化され得るVEGFR1ペプチド;
(b)VEGFR1ペプチドのメチル化のためのコファクター;及び
(c)リジン831のメチル化を検出するための検出試薬
を含む、キット。
【請求項31】
前記コファクターが、S−アデノシル−L−メチオニンである、請求項30に記載のキット。
【請求項32】
前記検出試薬が、VEGFR1のメチル化されたリジン831を認識する抗体である、請求項30に記載のキット。
【請求項33】
前記キットが、SMYD3によるVEGFR1のメチル化を増強する因子をさらに含む、請求項30に記載の方法。
【請求項34】
前記メチル化増強因子が、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項33に記載のキット。
【請求項35】
VEGFR1のメチル化を認識する抗体。
【請求項36】
VEGFR1のリジン831のメチル化を認識する、請求項35に記載の抗体。
【請求項1】
SMYD3によるVEGFR1のメチル化を調整する因子を同定する方法であって、該方法が、
(a)以下から構成される群から選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチドを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化に適した条件下で、前記因子の存在下で接触させる工程;
(i)配列番号2のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を含むアミノ酸配列を有するポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化レベルを前記因子の非存在下で検出した対照レベルと比較する工程を含み、
前記対照レベルと比較したメチル化レベルの増大又は減少が、前記因子がSMYD3によるVEGFR1のメチル化を調整することを示す、方法。
【請求項2】
前記コファクターが、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項1に記載の方法。
【請求項3】
前記(ii)に定義されるポリペプチドが、20以下の保存的アミノ酸置換を含む配列番号2のアミノ酸配列を含む、請求項1に記載の方法。
【請求項4】
前記(iii)に定義されるポリペプチドが、配列番号2と少なくとも約95%の相同性を有するアミノ酸配列を含む、請求項1に記載の方法。
【請求項5】
前記(iv)に定義されるポリペプチドが、配列番号1のヌクレオチド配列からなるポリペプチドに、高いストリンジェント条件下で特異的にハイブリダイズする、請求項1に記載の方法。
【請求項6】
前記(v)に定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項1に記載の方法。
【請求項7】
前記VEGFR1ペプチドが、配列番号4のアミノ酸配列を含むポリペプチド、または、その機能的変異体もしくは機能的断片である、請求項1に記載の方法。
【請求項8】
前記機能的VEGFR1断片が、配列番号4のアミノ酸配列の位置800〜1000のアミノ酸配列を含む、請求項7に記載の方法。
【請求項9】
前記機能的VEGFR1断片が、配列番号4のアミノ酸配列の位置800〜841のアミノ酸配列を含む、請求項7に記載の方法。
【請求項10】
前記機能的VEGFR1変異体が、以下の変異:K819A、K819E及びK819Rの1又は2以上を含む、配列番号4のアミノ酸配列を含む、請求項7に記載の方法。
【請求項11】
前記メチル化レベルが、VEGFR1のリジン831で検出される、請求項1に記載の方法。
【請求項12】
SMYD3を過剰発現する癌治療のための化合物のスクリーニング方法であって、該方法が、
(a)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片を、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片と、VEGFR1ペプチドとSMYD3ポリペプチドとの結合を可能とする条件下及び試験化合物の存在下で接触させる工程;及び
(b)VEGFR1ペプチド又はそのSMYD3結合領域を含む断片と、SMYD3ポリペプチド又はそのVEGFR1結合領域を含む断片との間の結合を阻害する試験化合物を、癌治療のための候補化合物として選択する工程
を含む、方法。
【請求項13】
前記VEGFR1ペプチドのSMYD3結合領域を含む断片が、配列番号4のアミノ酸配列の位置800〜1000のアミノ酸配列を含む、請求項12に記載の方法。
【請求項14】
前記SMYD3ペプチドのVEGFR1結合領域を含む断片が、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項12に記載の方法。
【請求項15】
前記癌が、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択される、請求項12に記載の方法。
【請求項16】
SMYD3を過剰発現する癌の治療のための化合物をスクリーニングする方法であって、該方法が、
(a)請求項1に記載の方法を用いてメチル化を調整する試験化合物を同定する工程;及び
(b)前記試験化合物の非存在下で検出される対照メチル化レベルと比較して、メチル化される基質のメチル化レベルを減少させる試験化合物を選択する工程
を含む、方法。
【請求項17】
前記癌が、結腸直腸癌、肝細胞癌、膀胱癌及び乳癌からなる群から選択される、請求項16に記載の方法。
【請求項18】
試験化合物のVEGFR1のメチル化調整能力を検出するキットであって、該キットが、
(a)以下から構成される群から選択される、メチルトランスフェラーゼ活性を有するSMYD3ポリペプチド;
(i)配列番号2のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を含むアミノ酸配列を有するポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
(b)前記(a)のポリペプチドによりメチル化され得るVEGFR1ペプチド;及び
(c)VEGFR1ペプチドのメチル化のためのコファクター
を含む、キット。
【請求項19】
前記VEGFR1ペプチドが、配列番号4のアミノ酸配列を含むポリペプチド、または、その機能的変異体もしくは機能的断片である、請求項18に記載の方法。
【請求項20】
前記キットが、以下の:
(d)S−アデノシルホモシステインヒドロラーゼ(SAHH)
をさらに含む、請求項18に記載のキット。
【請求項21】
前記(v)に定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を含む、請求項18に記載のキット。
【請求項22】
ポリペプチドのメチルトランスフェラーゼ活性を測定する方法であって、該方法が、
(a)H3K4メチルトランスフェラーゼを、メチル化されるVEGFR1ペプチド及びコファクターと、VEGFR1ペプチドのメチル化を可能とする条件下で接触させる工程;
(b)VEGFR1ペプチドのメチル化レベルを検出する工程;及び
(c)工程(b)のメチル化とメチルトランスフェラーゼ活性とを関連付けることにより、メチルトランスフェラーゼ活性を測定する工程
を含む、方法。
【請求項23】
前記VEGFR1ペプチドが、少なくともK831を含むVEGFR1断片である、請求項22に記載の方法。
【請求項24】
前記コファクターが、S−アデノシル−L−メチオニンである、請求項22に記載の方法。
【請求項25】
前記ポリペプチドを、VEGFR1ペプチド及びコファクターと、メチル化増強因子の存在下で接触させる、請求項22に記載の方法。
【請求項26】
前記メチル化増強因子が、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項25に記載の方法。
【請求項27】
K831が、VEGFR1のメチル化されたリジン831を認識する抗体で検出される、請求項23に記載の方法。
【請求項28】
前記H3K4メチルトランスフェラーゼが、
(i)配列番号2(SMYD3)のアミノ酸配列を含むポリペプチド;
(ii)配列番号2のアミノ酸配列を含むポリペプチドであって、ここで、1又は2以上のアミノ酸が置換、欠失、又は挿入され、さらにここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価な生物学的活性を有するポリペプチド;
(iii)配列番号2と少なくとも約80%の相同性を有するアミノ酸配列を有するポリペプチド;
(iv)配列番号1のヌクレオチド配列からなるポリヌクレオチドにストリンジェントな条件下でハイブリダイズするポリヌクレオチドによりコードされるポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価な生物学的活性を有するポリペプチド;及び
(v)配列番号2のアミノ酸配列の位置117〜246のアミノ酸配列を含むポリペプチドであって、ここで、前記ポリペプチドが配列番号2のアミノ酸配列からなるポリペプチドと等価なメチルトランスフェラーゼ活性を有するポリペプチド;
から構成される群から選択される、請求項22に記載の方法。
【請求項29】
前記(v)で定義されるポリペプチドが、配列番号2のアミノ酸配列の位置100〜250のアミノ酸配列を有する、請求項28に記載の方法。
【請求項30】
SMYD3のメチルトランスフェラーゼ活性を測定するキットであって、該キットが、
(a)SMYD3によりメチル化され得るVEGFR1ペプチド;
(b)VEGFR1ペプチドのメチル化のためのコファクター;及び
(c)リジン831のメチル化を検出するための検出試薬
を含む、キット。
【請求項31】
前記コファクターが、S−アデノシル−L−メチオニンである、請求項30に記載のキット。
【請求項32】
前記検出試薬が、VEGFR1のメチル化されたリジン831を認識する抗体である、請求項30に記載のキット。
【請求項33】
前記キットが、SMYD3によるVEGFR1のメチル化を増強する因子をさらに含む、請求項30に記載の方法。
【請求項34】
前記メチル化増強因子が、S−アデノシルホモシステインヒドロラーゼ(SAHH)である、請求項33に記載のキット。
【請求項35】
VEGFR1のメチル化を認識する抗体。
【請求項36】
VEGFR1のリジン831のメチル化を認識する、請求項35に記載の抗体。
【図1】

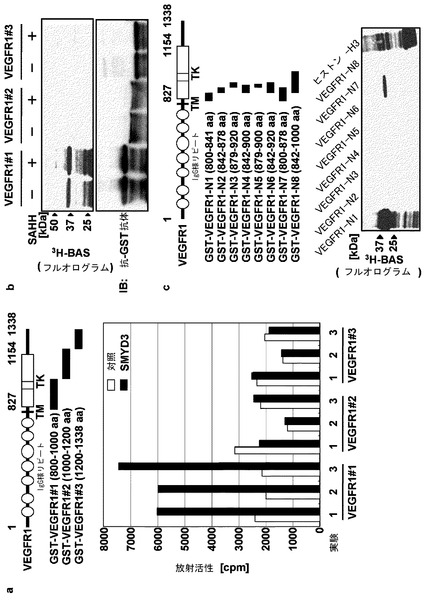
【図2】
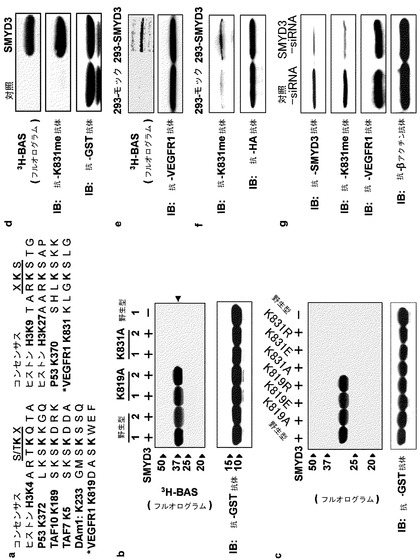
【図3】
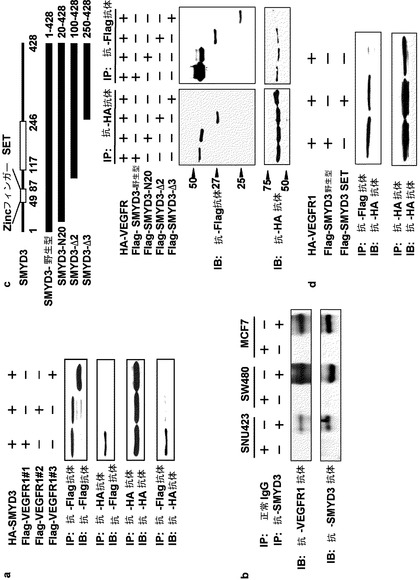
【図4】
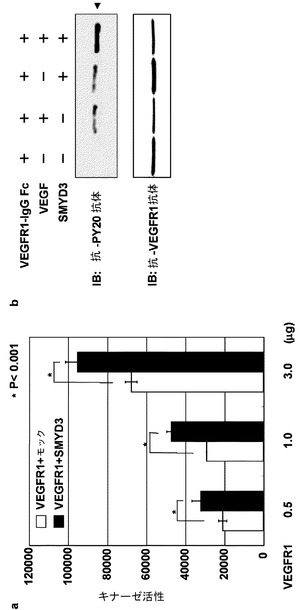
【図2】
【図3】
【図4】
【公表番号】特表2010−528584(P2010−528584A)
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願番号】特願2009−550134(P2009−550134)
【出願日】平成20年6月13日(2008.6.13)
【国際出願番号】PCT/JP2008/001516
【国際公開番号】WO2008/152816
【国際公開日】平成20年12月18日(2008.12.18)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願日】平成20年6月13日(2008.6.13)
【国際出願番号】PCT/JP2008/001516
【国際公開番号】WO2008/152816
【国際公開日】平成20年12月18日(2008.12.18)
【出願人】(502240113)オンコセラピー・サイエンス株式会社 (142)
【Fターム(参考)】
[ Back to top ]