
SODおよびプロラミン系ペプチドフラグメントを含む医薬組成
【課題】医薬投与用に適合された医薬組成を提供する。この医薬組成は、少なくとも1つのスーパーオキシドジスムターゼ(SOD)および少なくとも1つのプロラミン系ペプチドフラグメントの機能的に活性な組み合わせを含む。
【解決手段】本発明は、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントを含む医薬投与用に適合された医薬組成に関する。本発明はまた、特定のスーパーオキシドジスムターゼおよびプロラミン系ペプチドフラグメントに関する。
【解決手段】本発明は、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントを含む医薬投与用に適合された医薬組成に関する。本発明はまた、特定のスーパーオキシドジスムターゼおよびプロラミン系ペプチドフラグメントに関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、スーパーオキシドジスムターゼを含む医薬組成に関し、特に、他の化合物と関連する、スーパーオキシドジスムターゼを含む医薬組成に関する。
【背景技術】
【0002】
スーパーオキシドジスムターゼ(E.C.l.15.1.1.)またはSODという名称は、スーパーオキシド(O2〜°)アニオンの不均化において関与する普遍的金属酵素の仲間である。ラジカル種の蓄積または過度量のラジカル種の生成は、ほとんどの生物にとって有害である。この状態は、酸化ストレスとして未だ知られており、多様な病理学的過程(例えば、発癌現象、アテローム性動脈硬化、加齢および炎症疾患(例えば、セリアック病(グルテン不耐性としても知られる))において発生する多数の代謝異常と関連する。
【0003】
その発展または細胞局在の程度と別に、SODは、主に3つの形態で存在する。これら3つの形態は、分子内に含まれる金属イオン(すなわち、銅亜鉛またはCuZn−SOD、マンガンまたはMn〜SODおよび鉄またはFe〜SOD)によって区別される。CuZn−SODおよびMn〜SODはまた、より特殊な細胞組織(例えば、ペルオキシソームまたは植物中の葉緑体)においても見られ、一方哺乳類において、細胞外CuZn−SODまたはEc−SODが、細胞外区画において特異的に発見されている。より最近では、その他の公知のSODに対して弱い相同性しか持たないNi〜SODが、ストレプトミセスにおいて特定されている。
【0004】
多数の実験において、全SODが同一抗ラジカル活性を持っていても、その有効生物活性は、所与の治療指標および標的生物に対して同一ではないことが示されている。詳細には、ラット中のカラギーナンまたはアドリアマイシンによって誘発される脚部浮腫中の相同SODの抗炎症活動は、異種由来のSODのものと比較すると、非機能性である。異種SODの有効性は、活性部位に存在する金属の種類または酵素の全体的分子量よりも、わずかであっても酵素のアミノ酸の変動により高く依存するように見える。
【0005】
それにもかかわらず、用いられる異種文脈に応じて、異種SODと免疫原性SODとの間の境界をそれぞれについて決定する必要がある。実際、ラットにおける研究により、以下が示唆されている。
(a)相同ラットCuZn−SODまたは特定の異種SOD(例えば、ヒトMn〜SOD)は抗炎症活動を持たず、
(b)特定の異種SOD(例えば、ウシCuZn−SOD)はこのような抗炎症活動を持ち、さらに他のもの(とりわけイーストCuZn−SOD)は、炎症性活動も持ち得る。
【0006】
実施された多様な異なる研究から、臨床用量であっても、相同型酵素の有効性が低いため、ヒトSODは、ヒトにおける特定の抗炎症適応用の異種SODよりも有効性が極めて低いことが分かっている。特にこれらの理由のため、ウシCuZn−SODは、ウシ海綿状脳症(BSE)発病前に投与した場合に臨床試験において有利な効果を示すことが分かっている。
【0007】
ウシCuZn−SODとヒトCuZn−SODとの間には18.3%の相違が存在し、また多数回の注入が必要なことにも関わらず、アレルギーまたはアナフィラキシーショックの例はほとんど観察されていない。後になっても継続されているいくつかの初期研究によれば、抗SOD抗体循環率を増加させることにより、異種SODの薬理活動を漸進的に抑制させることが可能であることが提案されている。他のより最近の報告では、異種SODの提示および内在化を容易化する過程においてこれらの循環抗体が関与することがより多く示されている。
【0008】
SODを経口投与した場合も、胃腸管中において急速劣化することが多いため、バイオアベイラビリティおよび有効性の低減に繋がる点において、問題である。この問題は、SODの有効性が最も高い特定の細胞位置までSODを到達させることが困難であることにより、悪化する。
【発明の開示】
【発明が解決しようとする課題】
【0009】
出願人は、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントを含む医薬組成は、特に上記問題を解消し、また、この結合により、細胞遺伝子座に対するSODの細胞ターゲティングを容易化して有効性を最適化することができる、ことを発見した。
【0010】
従って、本発明の目的は、医薬投与用に適合された医薬組成を提供することである。この医薬組成は、少なくとも1つのスーパーオキシドジスムターゼ(SOD)および少なくとも1つのプロラミン系ペプチドフラグメントの機能的に活性な組み合わせを含む。「機能的に活性な組み合わせ」という用語により、上述した2つの成分(これらの成分は、いずれかが結び付けられているかまたは相互に物理的または化学的に結合している)は成分の蓄積効果を越える相乗効果を生成するように共に作用することが、理解される。詳細には、本発明において、SODとプロラミン系ペプチドフラグメントとの結合は、SODの存在可能時間を向上させると同時に、プロラミン系ペプチドフラグメントによって場合によって発生する炎症反応を顕著に低減させることが分かっている。
【0011】
本発明の他の目的は、本記載全体から明らかになるが、特に好適な目的としては、上記医薬組成中で採用されるアミノ酸分子、そのためのまたはハイブリダイゼーションを通じて得られる任意の核酸配列コーディング、このようなアミノ酸分子の薬剤、抗細胞ストレス剤としての使用、または炎症病理の治療における薬剤の調合のためのこのようなアミノ酸分子の使用がある。本発明は、特定のプロラミン系ペプチドフラグメントの存在あるいはその少なくとも1つのスーパーオキシドジスムターゼとの化学的または物理的組み合わせを検出するために用いられる特定の抗体も網羅する。請求項または記載中で「キメラ」という用語について言及がある場合、この用語は、当該分子が2つ以上の別個の遺伝源から由来する成分を含むことを意味する。
【課題を解決するための手段】
【0012】
[適切なプロラミン]
好適には、上記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンのフラグメントあるいは誘導体、類似体、塩またはその代謝体である。さらに好適には、上記プロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である。最も好適な実施形態において、上記プロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である。
【0013】
本発明の別の好適な実施形態において、上記少なくとも1つのプロラミン系ペプチドフラグメントは、完全に加水分解されたか、実質的に加水分解されたかまたは若干加水分解されたプロラミン系ペプチドフラグメントからなる群から選択される。しかし、一般的には、上記少なくとも1つのプロラミン系ペプチドフラグメントは、胃腸加水分解過程を模倣するPTC(パンクレアチン、トリプシン、キモトリプシン)加水分解されたプロラミンから得られたフラグメントからなる群から選択される。
【0014】
代替的であるが好適な実施形態において、少なくとも1つのプロラミン系ペプチドフラグメントは、腸管内においてターゲティング信号として機能する範囲において、加水分解される。
【0015】
[適切なSOD]
スーパーオキシドジスムターゼは、当業者に一般的に公知のものから選択でき、ヒトスーパーオキシドジスムターゼ、動物スーパーオキシドジスムターゼ、バクテリアスーパーオキシドジスムターゼ、イーストスーパーオキシドジスムターゼおよび植物スーパーオキシドジスムターゼからなる群から選択され得る。しかし、好適には、上記少なくとも1つのスーパーオキシドジスムターゼは、CuZnスーパーオキシドジスムターゼ、Mnスーパーオキシドジスムターゼ、細胞外スーパーオキシドジスムターゼ、NiスーパーオキシドジスムターゼおよびFeスーパーオキシドジスムターゼからなる群から選択される。本発明の1つの好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは相同スーパーオキシドジスムターゼである。本発明の別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは異種スーパーオキシドジスムターゼである。「相同」という用語は、当該分子の任意の所与の標的宿主中の未変性分子と同一由来のSODを指し、「異種」という用語は、当該分子の任意の所与の標的宿主中の未変性分子と異なる由来のSODを指す。最も好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは異種CuZnスーパーオキシドジスムターゼである。
【0016】
上述したように、上記少なくとも1つのスーパーオキシドジスムターゼは好適には、植物スーパーオキシドジスムターゼ、さらにより好適には異種CuZn植物スーパーオキシドジスムターゼである。このようなスーパーオキシドジスムターゼは、多くの任意の異なる様式で入手または生成可能である。例えば、上記少なくとも1つのスーパーオキシドジスムターゼは、植物から抽出可能である。上記少なくとも1つのスーパーオキシドジスムターゼの抽出および生成に植物を用いる場合、これらの植物は、ウリ科のものでよく、好適にはメロンからなる群から選択され、あるいは、ナス科からなる群から選択され、好適にはトマトからなる群から選択される。
【0017】
利用可能な多様な植物スーパーオキシドジスムターゼのうち、本発明によるスーパーオキシドジスムターゼは好適には、ペルオキソームスーパーオキシドジスムターゼ、葉緑体スーパーオキシドジスムターゼおよび細胞質スーパーオキシドジスムターゼからなる植物スーパーオキシドジスムターゼの群から選択される。
【0018】
別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、組換えスーパーオキシドジスムターゼである。上記組換えスーパーオキシドジスムターゼは、上記SODの核酸コーディングで形質転換された宿主細胞中の遺伝工学および発現を通じて、得ることができる。詳細には、出願人らは、上記少なくとも1つのスーパーオキシドジスムターゼが修飾葉緑体、ペルオキソームまたは細胞質CuZn組換えスーパーオキシドジスムターゼである場合に特に好ましいことを発見した。さらに別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、雑種異種/相同スーパーオキシドジスムターゼ(好適には、雑種植物/ヒトスーパーオキシドジスムターゼ)である。
【0019】
一般的な組換えタンパク質の発現と同様に、異質または未変性遺伝子を宿主細胞ゲノム中に導入する技術も当業者に周知である。従って、本発明の1つの好適な実施形態において、上記修飾葉緑体スーパーオキシドジスムターゼ、ペルオキソームスーパーオキシドジスムターゼまたは細胞質スーパーオキシドジスムターゼは、宿主細胞中で発現された修飾核酸配列によってコードされる。これらの修飾コーディング配列を得るために必要なPCR増幅遺伝子技術も当業者に公知であり、ここでの説明は不要である。
【0020】
本発明の最も好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、以下によってコードされる組換えスーパーオキシドジスムターゼである。
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意の1つによる核酸配列
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうち任意のものに対して厳密な条件下でハイブリダイズする核酸配列
SEQ.ID24〜SEQ.ID33として特定される配列のうちの任意の1つと少なくとも70%BLAST同一の相同性を有する核酸配列
【0021】
厳密な条件下でハイブリダイジングすることにより、当該核酸が、以下の条件65℃におけるSSPE×0.2下で述べられた配列にハイブリダイズすることが理解される。
【0022】
最後に、本発明の別の目的は、酸化および炎症病理または疾患の治療のための薬剤(特に、任意のほとんどの病気(グルテン不耐性、および好適にはセリアック病)に伴う病理または疾患のための薬剤)の製造のための、上述したような医薬組成の使用に関する。
【0023】
[異種植物SOD]
出願人は、植物由来のSODは本発明において極めて有利であることを発見した。CuZn−SODは、高等植物の光合成細胞のいくつかの形態において存在する。これらは、主に葉緑体中そしてより低い程度において細胞質およびペルオキシソーム中で見受けられる。これらは、アミノ酸組成、分子量、等電点およびその重合レベルにおいて、相互に異なる。
【0024】
これらのうち多くは、生化学的にまたは遺伝子的に特徴付けられる。葉緑体またはペルオキソーム植物SODが、先ずタンパク質のN末端においてリーダー配列または信号ペプチドを含む前駆体として、生成される。この前駆体は、葉緑体またはペルオキシソーム中への転位後、余分なN末端部分の削除において行われる機能性成熟を経験する。
【0025】
この特異性に加えて、上記ペルオキソームSOD(特に、メロンのもの)または葉緑体SOD(特にトマトのもの)は、(a)細胞質SOD中の70%保存領域に対して葉緑体およびペルオキソームSOD中の約90%保存領域において、細胞質SODよりもこれらの間においてより良好に保存され、(b)他の植物CuZn−SODよりも熱および過酸化水素不活性化に対して高い耐性を持つのにもかかわらず、経腸または非経口投与のいずれにおける治療用途候補として考えられてこられなかった。
【0026】
しかし、出願人の配列アライメントにより、葉緑体またはペルオキソームSODは、その等価な細胞質ゾル由来植物SODよりも2〜4%少なくヒトまたはウシSODと異なることが明らかになった。
【0027】
本発明の好適な実施形態のうちの1つにおいて、例えばメロンからの葉緑体CuZn−SODの選択は、出願人によって保持された。なぜならば、細胞質形態に豊富な植物由来の特定のコドンがバクテリアにおいて稀であることが特定され、また、この理由のため、発現レベルと、これらを豊富にまたは活性部位において含む生合成酵素の特定活動とが有意に変化する。
【0028】
出願人は好適には、バクテリアシステムを生合成SODの組換え発現のための宿主細胞として用いることを選択し、これにより、これら2つの局面が確認される。
【0029】
出願人のバクテリアシステムにおいてヒトCuZn−SODは発現され、機能性であった。これは上記稀なコドンのうち少なくとも2つを含むものの、そこから形成された2つのアルギニン残留物は、N末端領域において発見可能であり、ヒトSODの酵素的機能に関与しているように見えない。
【0030】
一方、細胞質トマトSODにおいて、上記稀なコドンから得られた2つのアルギニン残留物は、上記酵素のC末端部分中に配置され、そのうちの1つは、スーパーオキシドアニオンの誘引に関する酵素的機能に関与する。
【0031】
この場合、このシステム中で発現される生合成トマト細胞質SODは、活性がずっと低い。葉緑体トマトSOD上について他の研究が行われたが、このような稀なコドンが存在せず、バクテリア中でのその発現により完全な機能性の生合成SODが得られ、このSODが完全に活性であることが示されている。
【0032】
このバクテリアシステム中でクローニングおよび発現された第1の2個のメロンSODは、得られる精製レベルにおける細胞質トマトと同様の特定の活動を持つことが分かっている。この場合、上記稀なコドンは、細胞質トマトSOD中で特定されたのと同じ位置を網羅し、上記アルギニンのうち1つは、上記スーパーオキシドの誘引機能を持つ。
【0033】
よって、現在用いられている宿主株を、Ecoli(すなわち、アルギニンAGGまたはAGA)において稀に用いられるコドン向けのさらなるtRNAsを含む株と代替すると、有利であった。この特定の株は、バクテリア中の機能的に活性な植物タンパク質の発現を向上させるように、設計される。
【0034】
従って、植物由来の成熟SODは、その抽出された形態においておよび好適には組換え酵素として、経口投与と組み合わされると、異種活性および免疫耐性双方を調和させることができ、そのため、病理学的炎症過程の治療におけるウシSODの代替物として機能可能である。
【0035】
[SODの経口投与および細胞ターゲティング]
治療的に受容可能なSODの活性を増加させるために、いくつかの異なる処方が考えられた(すなわち、リポソームカプセル封入、アルブミン共役、またはポリエチレングリコールとの結合、またはヘパリン親和性ペプチドとの雑種としての結合)。
【0036】
リポソームまたはセラミドを用いた経口投与された異種ウシCuZn−SODの抗炎症活性は、先ず、カラギーナンによって誘発されるラット下肢浮腫の低減において示された。また、セラミドをグリアジンのような植物プロラミンと代替し、非組換えウシSODを非組換え植物SODと代替した場合、注入可能なウシSODと比較可能な治療活動により、ラット中で誘発される認知疾患の発病を遅延させることが分かった。
【0037】
[グリアジン使用における副作用]
しかし、この種の経口投与の場合、限られてはいるが無視できない数の人数に有意の多くの副作用が発生している。上記修飾処方中で用いられた特定の物質(特に、グリアジンのような植物プロラミン)は、胃腸管中でのペプシン−トリプシン加水分解後に耐性の低い生成物を発生させ得、その結果、グルテンアレルギーまたはさらにはセリアック病の場合のような毒性となり得ることが周知である。このヒトのみに発生する疾病は、ヒト腸細胞の刷毛状縁部の特定容量にリンクし、これにより、HLAクラスII分子が発現する。いくつかのハプロタイプ(例えば、HLA−DR、HLA−DP、HLA−DQ)は、グリアジン〜特異T細胞の活性化に密接に関連することが分かっている。グリアジンペプチドのこの有毒処理は、以下の3つの連続工程が関与すると考えられている。
a)グリアジンの有毒フラグメントがHLA受容体に結合し、腸管微絨毛縁部上の腸細胞へ輸送される。
b)これらが固有層中のCD4+型上皮内細胞へ提示された場合、上記腸細胞側底レベルへ輸送される。
c)最後に、無傷のまたは部分加水分解されたポリペプチドが、上記腸細胞側底膜から上記固有層中のAPC細胞(抗原提示細胞)へと解放される。
【0038】
小麦胚芽油中に存在するグリアジンペプチドおよび胃腸管中での加水分解によって生成されるものにより、負の反応(すなわち、炎症反応)が発生し、一方、酵素的に活性のSODと結び付けられると、このようなペプチドの負の効果は大幅に低減される。
【0039】
加えて、上記負の副作用を除いて小麦胚芽油中に存在するものと同様の挙動を示す二粒系コムギ中に存在するグリアジンの結合が、酵素的に活性のSODと結び付けられた場合に、酸化ストレスに関連する欠乏の治療にとって最も好適であることが分かっている。
【0040】
生理的条件下において、上皮細胞によって提示される抗原に対する反応をTH2型T細胞によって仲介する。これらの細胞は、IgA生成に起因し、とりわけ、食物抗原に関する免疫耐性に起因する。グルテン不耐性またはそのより深刻な形態であるセリアック病の場合、グリアジン由来のペプチドは、すでに酸化促進性であるためグリアジンに対して一層感受性の高い酸化ストレスの状態を生成することにより、腸細胞の酸化代謝を変化させることができる。
【0041】
この炎症状態は、サイトカインおよび一酸化窒素(NO)の生成における重要な変化と関連する、THl細胞型に対する細胞反応の転換により、特徴付けられる。これらの条件下において、脂質過酸化反応の増加および酸化および低減した形態のグルタチオン(GSSG/GSH)の比の増加は、抗酸化物質をグリアジンと融合させるとこの治療指標において有利な効果が得られるはずであるという出願人の仮説を補強した。
【0042】
結果として、そのグリアジンとの組み合わせから得られる経口経路によるSOD抗炎症活性の増強が、細胞膜の親和性増加および関連する代謝経路の基礎制御に起因して、現れることとなる。
【0043】
よって、これらの効果を実証するために、酵素的に活性の異種SODを先ず小麦胚芽油プロラミンの加水分解物の誘導体(詳細にはVulグリアジン)と結合させ、そのアミノ酸配列がNter−QQPYPQPQPF−Cter(SEQ.IDN°01)であることを確認した。その後、二粒系コムギプロラミンの加水分解物のその無毒類似性誘導体(詳細にはDurグリアジン)と結合させ、そのアミノ酸配列がNter−QQPQDAVQPF−Cter(SEQ.IDN°02)であることを確認した。この類似体は、グリアジンの有毒誘導体の競合的抑制物質であり、腸細胞との相互作用サイトのターゲティングが可能である。
【0044】
プロラミン(詳細にはグリアジン)は、胃腸管移行時において自身が包囲している活性成分を保護するという積極的役割を果たし、その加水分解された形態において、腸管粘膜内の細胞における活性成分のターゲティングおよび連続的放出を助ける。これらの誘導体をタンパク質N末端部分に追加された無毒類似体(例えば、異種SOD)と代替した場合、上記誘導体ペプチドフラグメントの腸細胞ターゲッターとしての役割を維持すると同時に、プロラミンの抗原誘導体の有害な効果を回避することができる。
【0045】
いくつかの治療指標における注入可能なウシSODによって実証される薬理有効性と、BSEおよびCFJDの出現によって発生した公的安全の問題とを鑑みて、上記した本発明の目的は、以下の利点を持つことであることが分かった。
1.レシピエントの遺伝的バックグラウンドに関係なく、異種非免疫原性SODまたは免疫耐性の誘発のみを行うもの。
2.植物由来の(有利には食物由来の)SOD。
3.このようなSODの場合に従来では有効ではなかった投与経路を通じて異種SODを薬理的に活性させる処方。
4.この異種SODを無毒腸管ターゲティングペプチドと結合させることで、経口投与されたこれらのSODの異種活性を増強させ、治療法が未だない適応(詳細にはオーファンドラッグ適応)における代替的治療を示すこと。
【0046】
本発明のこれらのおよび他の実施形態について、以下詳細に説明する。
【発明を実施するための最良の形態】
【0047】
[例]
(例1)
銅亜鉛ペルオキソームまたはメロン「キューカミスメロL」から得られた葉緑体スーパーオキシドジスムターゼについてコーディングするメッセンジャーRNAのクローニングと、その完全な核酸配列(コーディングおよび非翻訳配列双方)の決定とを、逐次RT−PCR増幅によって行った。図2中に示すような偽細胞質という名称の第1のタンパク質変異体を得るために、コーディング配列のみを含む第1のクローンを遺伝工学によって改変した。
【0048】
この第1のタンパク質変異体は、メロン葉緑体SODの遺伝子改変の一般図を示す。この第1の変異体は、未変性信号ペプチドに対応するN末端における第1の65個のアミノ酸の置換を示し、上記アミノ酸対メチオニン−バリンは、細胞質植物SODのN末端の特性である。この技術によって改変された過程により、植物細胞中においてSODが経験する葉緑体成熟工程が得られ、その後、図示の2つのアミノ酸が付加される。第2および第3の変異体は、このメチオニン−バリンアミノ酸対に加えて、胃腸管ターゲティングのためのグリアジンペプチド配列(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ)をそれぞれ含む。その由来は小麦胚芽油であり、そのペプチド配列により、感作反応を生成することなく、以下のような、腸細胞と相互作用するグリアジンペプチドの再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ)。その由来は二粒系コムギである。
【0049】
(例2)
コーディング配列全体の直接的RT−PCR増幅により、トマト葉アカナス(PubMedBlast受入番号X14040)から、銅亜鉛SOD細胞質スーパーオキシドジスムターゼをコードするメッセンジャーRNAのクローニングが得られ、これにより、細胞質という名称の第1のトマト変異体が得られた。第2の変異体および第3の変異体も、胃腸管ターゲティング(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ))のためのグリアジンペプチド配列の付加により、遺伝子操作された。その由来は小麦胚芽油であり、上記ペプチド配列により、感作反応を生成することなく、以下のような腸細胞上でのグリアジンペプチド相互作用の再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ)。その由来は二粒系コムギである。
【0050】
(例3)
コーディング配列全体の直接的RT−PCR増幅により、ヒト血液細胞ホモサピエンス(PubMed/Blast受入番号K00065)から得られた銅亜鉛SOD細胞質スーパーオキシドジスムターゼについてコードするメッセンジャーRNAのクローニングを得、これにより、細胞質という名称の第1のヒト変異体を得た。第2変異体および第3の変異体についても、胃腸管ターゲティング(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ))のためのグリアジンペプチド配列の付加により、遺伝子操作を行った。上記ペプチド配列により、感作反応を生成することなく、以下のような腸細胞上でのグリアジンペプチド相互作用の再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ))。
【0051】
(例4)
各種類の構成の第1の変異体の発現物を用いて、得られた多様な生合成SODの抗ラジカル活性を検証した。これらの活性について、未変性PAGE上のNBTの還元により、確認した。その結果、上記組換え生合成タンパク質の活性が、現在最も用いられているバクテリア宿主株中に稀に存在するコドンの植物cDNA中の存在に密接にリンクすることが分かった。
【0052】
現在用いられている株中に発現する3つのcDNAからの低い発現率または活性率については、バクテリア株(とりわけコドンAGA、AGG(アルギニン)およびより低い範囲ではコドンCCC(プロリン)、GGA(グリシン)CTA(ロイシン)、およびATA(イソロイシン))によって稀に用いられるコドンの存在によって説明することができる。この問題を回避するために、上記cDNAをpET30a+(NOVAGENから市販)として特定される別の発現ベクターシステムに移行させ、これを用いて、これらの稀なアミノ酸用の適切なtRNAを含むRosettaDE3pLysS(NOVAGENから市販)として特定される許容バクテリア株を形質転換した。
【0053】
[本発明の詳細なプロトコル]
(キューカミスメロLSODのクローニングプロトコルおよび遺伝子改変)
変種キューカミスメロL(VILMORIN−GNIS−2251029として登録されている市販の変種)または上記変種キューカミスメロLC(ASL−NCIMS−40310として登録されている変種)のいずれかから、真核細胞に使用されるものと同一の技術[TRIzol−TMキット、Life Technologies、フランス]を用いて窒素凍結保存されたキューカミスメロLメロン葉から全RNAを抽出した。
【0054】
(メロンSODの3’RACE−PCR)
25μlの反応緩衝液中の修飾オリゴdTプライマーAAGCAGTGGTATCAACGCAGAGTACTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTVN(SEQ.IDN°03)[Clontech、Smart cDNA ref:k1051]から、メッセンジャーRNAの単鎖DNAマトリックス中への転写を実施した:トリスHCl20mM pH8.3、KCl50mM、MgCl22.5mM、10mM DTTおよび400μMの各dNTPを200単位のSuperscriptII逆転写酵素(Gibco−BRL)の存在下で42℃で1時間。
【0055】
補体としていくつかの単鎖プライマー対(例えば、EndプライマーAAGCAGTGGTATCAACGCAGAGT(SEQ.IDN°04)を修飾オリゴdTプライマーならびにメロンコーディング領域のためのMelcyt5F1 GGTGAYACMACMAATGGYTG(SEQ.IDN°05)およびMelcyt5S2 CATGCKGGKGAYCTDGG(SEQ.IDN°06)それぞれと組み合わせることにより、SODのためのメッセンジャーRNAコーディングの3’末端の増幅を実施した。アンプライマー配列は、いくつかの植物種のペルオキソーム、葉緑体および細胞質CuZn−SODの保存領域に基づくものであるため、本質的に極めて縮重するものである。
【0056】
(メロンSODの5’RACE−PCR)
バクテリアベクターpGEM−T(Promegaから市販)中へのクローニング後、7個の独立クローンを配列し、コンセンサス配列を用いて、3つの新規の特定のプライマーVed3R3 ACAAAGGCTCTTCCAACTACAG(SEQ.IDN°07)、Ved3R2 GCCGCTAAGAGGAATCTG(SEQ.IDN°08)およびVed3R1 TGGTTGCCTCTGCTACTCCATC(SEQ.IDN°09)を構築した。この特定のプライマーVed3R3を用いて、メッセンジャーRNAをdCTPホモポリマー伸長(5’RACEシステム、Life Technologies、ref:18274−058)によって終端された単鎖DNAマトリックス中に転写した。このマトリックスの増幅は、プライマーAAPP GGCCAGGCGTCGACTA GTACGGGGGGGGGGGGGGGG(SEQ.IDN°10)およびプレ増幅工程のためのVed3R2およびAAP GGCCAGGCGTCGACTAGTAC(SEQ.IDN°11)および特定のメッセンジャーRNAの5’末端の最終増幅のためのVed3R1の連続的組み合わせにより、得た。
【0057】
(3つのSOD変異体の増幅および遺伝子改変)
各種(ヒト、トマトおよびメロン)からの異なるSOD変異体の増幅を、3つの単鎖5’アンプライマーを以下のような各種のためのユニークな単鎖3’アンプライマーと組み合わせることにより、可能にした。
【0058】
名称 サイト 配列
Hum5Cyt NcoI atcggatccATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°12)
Hum5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°13)
Hum5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°14)
Hum3Rfw HindIII ctcgagaaqcttTTATTGGGCGATCCCAATTAC(SEQ.IDN°15)
【0059】
名称 サイト 配列
Tom5Cyt NcoI atcggatccATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°16)
Tom5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°17)
Tom5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°18)
Tom3Rfw HindIII ctcgagaagcttTAACCCTGGAGGCCAATAAT(SEQ.IDN°19)
【0060】
名称 サイト 配列
Mel5pCyt NcoI atcggatccATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°20)
Mel5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°21)
Mel5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°22)
Mel3Rfw HindIII ctcgagaagcttCACACAGGAGTCAGACCGAC(SEQ.IDN°23)
【0061】
ここで、Cytは、オリジナルの65アミノ酸配列の代わりにさらなる2個のアミノ酸を含むメロン偽細胞質形態を指定し、ヒトおよびトマト中の細胞質形態については、参照符号DurおよびVulがそれぞれ以下を示す。
【0062】
DURは、二粒系コムギから得られた胃腸管ターゲティング信号ペプチドを含む細胞質形態を示す。
【0063】
VULは、小麦胚芽油から得られた胃腸管ターゲティング信号ペプチドを含む細胞質形態を示す。
【0064】
これらのプライマーは、5’BamHIまたはNcolサイトおよび3’Hindlllサイトのいずれかにおける同相でのバクテリア発現ベクターpET30a+(NOVAGENから市販)中へのその導入を可能にする必要な合成配列を含むように、構築された。これらの増幅および遺伝子改変のためのグローバルプロシージャを図1中に報告する。
【0065】
バクテリア発現システムにより、クローンcDNAのハイブリドタンパク質としての発現が可能となり、SODまたはタンパク質変異体がそのN末端を介してHis−Tag配列へとリンクされる。
【0066】
全ての増幅は、TAPS(pH9.3)を25mM、KClを50mM、MgCI2を1.5mM、各dNTPを200μM、各プライマーを10μM、プラスミドDNAを10ng、ゴールドスターDNAポリメラーゼ(Eurogentec)を2.0単位を含む50μlの反応体積中において行った。各使用サーモサイクリングプログラムを以下の表に示し、GeneAmpPCRサーモサイクルシステム9700(Applied Biosystem)上で実施した。
【0067】
[RT−PCR増幅サイクルの表]
(メロン3’RACE)
95℃、3分
95℃、25秒
55℃、30秒
72℃、40秒
95℃、25秒
58℃、30秒
72℃、40秒
(メロン5’RACE)
95℃、3分
95℃、30秒
55℃、30秒
72℃、60秒
95℃、30秒
61℃、40秒
72℃、50秒
(メロンクロロ)
95℃、3分
95℃、40秒
57℃、40秒
72℃、40秒
95℃、40秒
60℃、40秒
72℃、50秒
(メロン偽Cyt)
95℃、3分
95℃、40秒
57℃、40秒
72℃、40秒
95℃、40秒
60℃、40秒
72℃、50秒
【0068】
ヒト、トマトおよびメロンSODについてエンコーディングする3つのタンパク質変異体「Cyt、Dur、およびVul」の増幅を5’オリゴヌクレオチドプライマーのサイズ差を考慮した同一プロトコルに従って行った。先ず、反応物を94℃で5分間加熱した後、以下のような5回の予備サイクル(94℃での変性を0.45分、56℃でのアニーリングを0.55分および72℃での伸展を0.55分)に暴露した後、さらなる25サイクル(94℃での変性を0.45分、59℃でのアニーリングを0.55分および72°Cでの伸展を0.55分)に暴露した。
【0069】
事前クローンされたCu−ZnSOD CDNAのコーディング領域をPCRによって増幅し、増幅された459bpおよび495bp生成物をその後適切な制限エンドヌクレアーゼNcol〜5’端部およびHindlll〜3’端部によって消化し、その結果、同相でのバクテリア発現pET30a+(NOVAGENから市販)中へのその導入を可能とした。
【0070】
各種(ヒト、トマト、およびメロン)のための3つのクローンのヌクレオチド配列を決定し、事前取得された配列と同一であることが分かり、以下に示す。
【0071】
[葉緑体またはペルオキソームCuZn−SOD形態の場合(SEQ.IDN°24)]

【0072】
[メロン偽細胞質Cu−ZnSOD形態の場合(SEQ.IDN°25)]
【0073】
[二粒系コムギペプチドによるメロン偽細胞質Cu−ZnSOD形態(SEQ.IDN°26)]
【0074】
[小麦胚芽油ペプチドによるメロン偽細胞質Cu−ZnSOD形態の場合(SEQ.IDN°27)]
【0075】
[トマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°28)]
【0076】
[二粒系コムギペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°29)]
【0077】
小麦胚芽油ペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°30)
【0078】
[ヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°31)]
【0079】
二粒系コムギペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°32)
【0080】
[小麦胚芽油ペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°33)]
【0081】
公知の植物スーパーオキシドジスムターゼによるORFおよび配列アライメントの決定のためのコンピュータソフトウェアを用いた核酸配列の分析により、要求に匹敵するペプチド配列の決定と、上記した遺伝子改変から得られた変異体のペプチド配列の決定とが可能となった。
【0082】
[メロン葉緑体CuZn−SOD形態の場合(SEQ.IDN°34)]
【0083】
[メロン偽細胞質CuZnSOD形態の場合(SEQ.IDN°35)]
【0084】
[メロン偽細胞質CuZnSOD形態+二粒系コムギペプチドの場合(SEQ.IDN°36)]
【0085】
[メロン偽細胞質CuZnSOD形態+小麦胚芽油ペプチドの場合(SEQ.IDN°37)]
【0086】
[トマト細胞質Cu−ZnSOD形態の場合の場合(SEQ.IDN°38)]
【0087】
[二粒系コムギペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°39)]
【0088】
[小麦胚芽油ペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°40)]
【0089】
[ヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°41)]
【0090】
[二粒系コムギペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°42)]
【0091】
[小麦胚芽油ペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°43)]
【0092】
バクテリア発現システムpET30aにより、クローンCDNAをハイブリッドタンパク質として発現させることが可能となり、Rosetta(DE3)pLysSバクテリア株(Novagen)中への発現後、SODが切断可能なN末端His−Tag配列(His−Tag)が融合される。
【0093】
OD600nmの値が0.6まで上昇するまで、生成されたCu−ZnSODの変異体形態それぞれについて入手および選択された組換えクローンを、適切な抗生物質(30μg/mlのクロラムフェニコールおよび/または34μg/mlのカナマイシンが補充された1リットルのリッチメディア(トリプトンを10g/l、イースト抽出物を5g/l、NaClを5g/l)中で再懸濁した。その後、37°Cでのインキュベーションを少なくとも3時間行い、この時点において、タンパク質の発現を制御するラックプロモーター誘導物質であるイソプロピルチオガラクトピラノサイド(IPTG)を追加して最終濃度1mMを得、さらに4〜6時間インキュベートした。
【0094】
各調製のための2個のバクテリアペレットを(1)50mlの寒冷PBS緩衝液または(2)50mlの添加液(Novagenから市販)中で再懸濁し、その後、4℃で各30秒の4連続サイクル(Vibracell40W〜20KHz)において、超音波処理緩衝液(トリスHCl pH7.4を10mM、EDTAを1mM、NaClを150mM)に暴露した。遠心分離を13000gで20分間行うことにより、細胞質内タンパク質抽出物を単離した。
【0095】
粗タンパク質抽出物から開始して、His−Bindクイックカートリッジ(NOVAGENからref70155として市販)上で、多様なHis−TagSOD融合タンパク質を単離した。その後、組換えエンテロキナーゼ(ref69066、NOVAGEN)でのハイブリドタンパク質の加水分解後、多様なSODおよびその特定の変異体を新規精製工程によってHis−Bindクイックカートリッジ上で精製した。
【0096】
(例5)
[メロン抗recSOD抗体]
組換えメロンSODのアミノ酸配列を得た後、スイスプロットアライメントを用いて他の植物SODとの比較分析を実施し、メロン特異ポリクローナル抗体(Eurogentec、ベルギー)を得るためにウサギの免疫化を行うために、2個のペプチド配列14−27(EP1668−GVVTLTQEDDGPTS、SEQ.IDN°44))および117−131(EP1669−HELADDLGKGGHELS、SEQ.IDN°45)を選択した。抗体EP1669をイムノアブソープションによる親和性カラム上で精製しウエスタンブロット分析中で1/2000において用いて、生成されたメロン組換えSODに対する抗体の特異性を確認した。
【0097】
SDS−PAGE分析のために、タンパク質抽出物を95℃で5分間加熱した後、電気泳動を12%のSDSポリアクリルアミドゲル上で実施した。分離されたタンパク質を免疫ブロットPVDF膜(BIORAD)上に移行させた。上記抗体の任意のポテンシャルクロス特異性が、上記増幅されたOpti4CNヤギ抗ウサギ検出キット(BIORAD)により、明らかとなった。
【0098】
EP1669抗体は、免疫化ペプチド由来の完全な組換えメロンSODタンパク質に対して良好な特異性を示した。EP1669抗体はまた、組換えヒトおよびトマトSODならびに天然ウシSODまたはヒトSODに対してクロス特異性を示した。従って、この抗体は、本発明の組換えSODタンパク質の薬理活動の検証に必要な実験のための検出ツールとして有用であった。
【0099】
(例6)
[抗ラジカル活性]
メロン偽細胞質Cu−ZnSODおよびヒトまたはトマト細胞質Cu−ZnSODならびにその2つの異なるグリアジンペプチドを支持する等価物をその抗ラジカル活性について分析した。これらの活性は、NATIVE−PAGE電気泳動上でのNBT光還元に従って確認される(BeauchampsおよびFridovitch、1971)。
【0100】
(例7)
[細胞ターゲティング]
標的薬剤として用いられるグリアジンペプチドが先に示したようにHLA−DQ2と相互作用してヒト腸細胞中に進入可能なだけでなく、それと物理的にまたは化学的に結合するタンパク質の併用ターゲティングを容易化することも可能であることを実証するために、ピルビン酸ナトリウム、グルタミン、非必須アミノ酸およびペニシリン/ストレプトマイシンが補充されたダルベッコ変法イーグル培地(DME)中の10%胎児ウシ血清(FBS)中で、ヒト大腸癌細胞株HT−29(ATCC HTB−38)およびCaco−2(ATCCHTB−37)を成長させる。いくつかの場合において、細胞を100lU/mlのIFN―γ(R&Dシステムズ)で48時間インキュベートし、HLAクラスIl発現を誘発する。
【0101】
[グリアジンペプチド提示分析およびフローサイトメトリ]
およそ2×105個の細胞を、37℃で組換えSODで1mg/mlまでの濃度で1〜20時間インキュベートする。0.2μmミリポア膜での濾過により、試験されたSOD化合物を殺菌する。これらの細胞を回収し、L243mAb(抗HLA DR、ATCC HB−55)を用いてHLAクラスII発現用のフローサイトメトリによって分析し、その後、FITC結合型ヤギ抗マウスlgG(ICN−Cappel)を行う。
【0102】
細胞表面におけるSOD化合物を検出するために、これらの細胞をEP−1669抗体またはEP−1668抗体(ISO細胞)でインキュベートし、FITC結合型ヤギ抗ウサギlgGで明らかにする。4°Cで60分間インキュベーションを行い、その後、FBSを10%およびアジ化ナトリウムを0.1%含むPBS中で、これらの細胞を3回洗浄する。その後、これらの細胞をFACScanフローサイトメーター(Becton Dickinson)上で分析する。
【0103】
(例8)
[アネキシンV陽性細胞の検出]
この部分中で報告される実験全てにおいて、48時間の暴露時間が用いられた。アポトーシス接着細胞の検出は、フルオレセインイソチオシアネートでラベルされたアネキシンV〜PS(Martinら、1995)の結合を用いたフローサイトメトリにより、行った。インキュベーション期間後、上記したようなトリプシンおよびEDTAを含むPBS溶液で細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、1×106個の細胞を遠心分離によって収集して、接着細胞中のアポトーシスを評価した。アネキシンV−FITC陽性細胞の評価は、ApoAlertアネキシンVアポトーシスキット(Clontech Laboratories、Palo Alto、CA、米国)を用いて行った。暗所において200μlの結合緩衝液中で10μlのアネキシンV−FITC(最終濃度:0.5μg/ml)および0.5μg/mlのヨウ化プロピジウムによって細胞を再懸濁させ、15分間インキュベートして、壊死細胞を排除した。488nmでチューニングされたアルゴンイオンレーザでCoulter Epics Elite ESP細胞分取器(Miami、FL)上で、二色サイトメトリ(蛍光標示式細胞分取(FACS))分析を行った(Koopmanら、1994)。
【0104】
(例9)
[細胞浸透]
上記したように処理された第2のフラクションの細胞を用いて、EP1669抗体およびEP−1668抗体によるウエスタンブロット分析を用いて試験された異なる細胞区画(すなわち、膜、細胞質)のタンパク質抽出物を調製し、これにより、ターゲティング信号ペプチドによりこれらが上記細胞中へ(詳細には上記細胞区画中へ)転位置していることを示す。
【0105】
(例10)
[細胞内レドックス状態上で異なった挙動を示すSOD化合物]
制御および処理された細胞(5×105個)を回収し、スーパーオキシドアニオン、過酸化水素およびGSHそれぞれの検出のために、ハイドロエチジン(HE;分子プローブ)、ジヒドロローダミン123(DHR123;分子プローブ)または5−クロロメチル−2’、7’−ジクロロ−ジヒドロフルオレセインジアセタート(CM−H2DCFDA、分子プローブ)で、495μlのハンクス平衡塩溶液(pH7.4)中でインキュベートする。37℃15分間後、上述したような血球計算器上でサンプルを分析する。
【0106】
(例11)
[細胞活性]
SODはその高分子量に起因して細胞膜に浸透しなかったと長く考えられてきたが、24時間後に外因的に付加された有意な量の無担体のCuZnSODが細胞膜をマーク付けすることができたことを実証する試みはわずかしか行われていない(Edeas MAら、Cell Mol Biol.1996、vol.42、p1137)。
【0107】
それにもかかわらず、細胞外的に付加された時に上記SOD酵素によって細胞内ROSの解毒をより効率的に行うために、細胞に浸透可能な組換えSODタンパク質が生成されている。ヒトCuZn−スーパーオキシドジスムターゼ(CuZn−SOD)遺伝子を、HIV−1の9アミノ酸Tatタンパク質伝達ドメイン(RKKRRQRRR)をエンコードする遺伝子フラグメントと融合した。このTat−SOD融合タンパク質は、外因的に培地中に付加された際に時間(10〜60分)内に用量依存的(0.2〜2μM)にHeLa細胞中に進入することが分かった。一旦上記細胞中に入ると、伝達したTat−SODタンパク質は酵素的に活性であり、かつ、24時間安定であった。パラコートで処理されたHeLa細胞(細胞内スーパーオキシドアニオン発生装置)の細胞バイアビリティーは、伝達したTat−SODによって増加する(Kwon HYら、FEBS Lett.2000、vol485、p163)。しかし、この場合、HIV−1Tat伝達タンパク質が多様な細胞型におけるMnSOD発現の低減の原因であることが分かっことは、注目すべきである(MareckiJCら、2004 Free Radic Biol Med、vol37、p869/PorntadavitySら、DNAcell Biol、2005、vol24、p299)。
【0108】
上記ヒトCuZn〜スーパーオキシドジスムターゼ(CuZn−SOD)遺伝子をリジンに富んだペプチド(KKKKKKKKK)の9アミノ酸をエンコードする遺伝子フラグメントと融合させた場合、9Lys−SODの伝達有効性は、Tat−SODよりも高効率であった(Park Jら、Mol Cells、2002、vol13、p202)。
【0109】
多様なグリアジン鎖由来のタンパク質ペプチドは、a)その相対的消化性(タンパク質抵抗)、b)その相対的サイズまたは配列(高プロリン領域、c)細胞培養物中でのその相対的濃度(凝集またはアポトーシス)、d)HLAクラスII分子によるその相対的認識(HLA DQ2またはDR3)、e)ゾヌリン誘導のその相対的可能性に応じて、腸管細胞の有意な修飾および/または変更を生成する能力を有すると認識されている。
【0110】
少なくとも1つ(ただし、これに1つに限定されない)特定のグリアジン由来ペプチド(19〜20アミノ酸残留物−LGQQQQPFPPQQPYPQPQPF−)の場合、HLA DR3分子と直面すると、HLADQ2分子液滴に効率的に結合可能であることが分かっている(Shidrawi.RGら、1998、Clin Exp Immunol vol11、p158)。グリアジン鎖から単離された他の多くのペプチドが、HLA DQ2またはDR3に対して同様の能力を有する(Qiao.SWら、2004、J.Immunol、vol173、p1757)が、正常クラスII HLAとのグリアジンペプチド相互作用については、ほとんど知られていない。
【0111】
グリアジンが腸管バリアを通過することを可能にする早期工程は、先ずラット上皮腸管細胞IEC−6上で確立される。グリアジンによるIEC−6細胞のインキュベーションにより、細胞内アクチンフィラメントの再構築が起こるが、この再構築は、グリアジンインキュベーションのわずか15分後に視認可能となり、60分後にピークとなり、2.5時間後にベースライン値に戻った。+4℃の温度ではグリアジンによって誘発されたFアクチン変化を抑制できず、その結果、アクチン重合がグリアジンエンドサイトーシスと関連するという可能性が除外される(KwiatkowskaKら、Bioassays、1999、vol21p422)。
【0112】
グリアジンは、腸細胞からのゾヌリン放出を含む別の経路を用い得る。ゾヌリン生成は、グリアジン濃度に依存し、少なくとも0.1mg/mlのグリアジンでのインキュベーション後15分経過後に早くも細胞培養物上清中で検出可能であった。その後、30分においてピークに達し、60分後にベースラインに戻った。Ussingチャンバ中に取り付けられたウサギ腸管粘膜にグリアジンを付加すると、Rt(抵抗力)が低減し、この抵抗力低減は、インキュベーション30分経過後に有意となった。他の著者は、グリアジンペプチド誘導体ゾヌリン放出および腸管浸透性の同時増加の強力な誘導物質であると推測している(ClementeMGら、Gut.2003、vol52、p218)。
【0113】
腸管上皮とのグリアジン相互作用によりゾヌリン放出を通じて腸管浸透性が上昇し、その結果、グリアジンの傍細胞転位およびその腸管粘膜下層内でのマクロファージとの後続相互作用が可能となり、その結果、炎症性反応およびフリーラジカル生成が発生する(ThomasKEら、2006、J Immunol、vol176、p2512)ことが後に実証された。
【0114】
上述したような組換えメロンCuZnスーパーオキシドジスムターゼ遺伝子が、Aグリアジンの10アミノ酸ペプチドをエンコードする遺伝子フラグメントと融合された(QQPYPQPQPF、SEQ.IDN°01、以下ペプチド982と示す)。Gli−SOD融合タンパク質が発現し、エシェリキアコリ中で精製された。
【0115】
[バクテリアクローン培養物からのタグ付けされた組換えキューカミスメロタンパク質(SODおよびGli−SOD)の生成および精製]
バクテリア発現システムpET30aにより、クローンされたcDNAのハイブリドタンパク質としての発現が可能となった。このハイブリドタンパク質において、組換えメロンSODまたはGli−SOD(QQPYPQPQPF−SOD)は、Rosetta(DE3)pLysSバクテリア株(Novagen)中での発現後、切断可能なN末端His−Tag配列(ポリペプチド−His6−エンテロキナーゼ開裂配列−5kDa)と融合された。
【0116】
[発酵過程および誘導過程]
吸光度A600が1単位の値に達するまで、50μg/mlのカナマイシンおよび35μg/mlのクロラムフェニコールが補充された2リットルのYES培地中で37℃で発酵を行う。その後、IPTGを誘発タンパク質発現に最終濃度0.5mMまで付加した。一晩成長させた後、遠心分離によって細胞を収集して、高圧において細胞切断を実施した(French Press、2サイクル:P>800バール)。
【0117】
[変性工程]
その後、尿素の連続的付加によりバルクを変性させ、その結果、トリス20mMを有する8Mの緩衝液の最終濃度がpH8.0となる。完全溶解後、室温における完全なタンパク質可溶化にβメルカプトエタノールを付加(最終濃度10mM)する。17000gにおける遠心分離により15分間行い、可溶性タンパク質を回収した。
【0118】
[精製工程]
金属固定化アフィニティークロマトグラフィー(NiCl20.25M、Ni2+イオンで活性化されたキレート化セファロース)によるアフィニティー精製により、組換えタグ付けされたタンパク質を単離し、先ず平衡緩衝液中のカラム中にロードした:トリス20mM、尿素8M、βメルカプトエタノール10mM(pH8.0)。
【0119】
[インカラムリフォールディング工程]
固定化組換えタンパク質を大量に洗浄し、尿素量増加により部分的にリフォールディングし、カラム尿素8M〜1.5Mの勾配(緩衝液中、トリス20mM、βメルカプトエタノール10mM、pH8.0)に付加した。
【0120】
その抗酸化物質活性を取り出すため、スーパーオキシドジスムターゼは、再生工程時、その天然補助因子を回復しなければならない。これは、NaAc50mM、尿素1.5M、CuCl20.1mM、ZnCl20.1mMおよびβメルカプトエタノール10mM(pH5.0)からなる緩衝液中のCu/Znの連続的固定により、行われる。
【0121】
組換えSODフォールディングの安定化は、トリス20mM、尿素1.5M、CuCl20.1mM、ZnCl20.1mM、GSH1mM、GSSG4mMおよびpH8.0からなる緩衝液中のシステインアミノ酸間の分子内ジスルフィド架橋の形成により、有利に行われる。
【0122】
[溶出工程]
トリス20mM、尿素1.5M、イミダゾール0.5MおよびpH8.0からなる緩衝液により、上記親和性カラムからの濃縮されたタグ付けされた組換えタンパク質の溶出を得る。その後、これらをトリス20mM、CuCl20.1mM、ZnCl20.1mMおよびpH8.0中で一晩透析した後、0.22μmフィルタによって最終濾過を行って、消化時の微生物成長を回避する。タンパク質内容物推定は、μBCA方法による上清中で評価した。
【0123】
[タグ開裂]
組換えタンパク質をハイブリドタンパク質として発現させる。このハイブリドタンパク質中、SODまたはGli−SODがそのN末端部分において少量の切断可能なリンカーを介してHis−Tag配列(His−Tag)を含む少量のタンパク質に融合している。
【0124】
この開裂は、2.0%エンテロキナーゼでのインキュベーションを室温において44時間行うことにより、達成される。
【0125】
[フォールディングされかつ無菌の組換えSODの精製]
放出後、最終精製工程により、トリス−HCl20mM、pH8.0緩衝液で平衡化されかつトリス−HCl20mM、NaCl0.5M、pH8.0の溶出緩衝液で溶出されたMonoQカラム上で、少量のタギングタンパク質を組換えSODから除去した。
【0126】
消化時の微生物成長を回避するための0.22μmでの濾過を行った。タンパク質内容物推定は、μBCA方法により上清中で評価した。
【0127】
上記精製から得られた結果を図5中に示す。
【0128】
[組換えリフォールディングされたキューカミスSODまたはGli−SODの抗酸化物質活動]
多工程変性/再生プロトコルに従って、キューカミス組換えSODおよびグリアジンペプチド(Gli−SOD)を支持するその等価物を生成および精製した。SDS−PAGE分析により、有意な程度の精製が明らかとなり、その抗酸化物質活動の回復に基づいて、上記リフォールディング過程の有効性を評価する。組換えタンパク質(キューカミスメロSODおよびGli−SOD)双方のNATIVE−PAGE電気泳動上でのNBTを低減させる可能性(NBT光還元、BeauchampsおよびFridovitch、1971)を評価し、天然キューカミスメロ抽出物の可能性と比較した。
【0129】
NATIVE−PAGE電気泳動上で得られた遊走プロファイルおよびNBT低減は、組換えタンパク質、SODまたはGli−SOD(4000〜10000UNBT/mgタンパク質)双方の正確かつ高効率のリフォールディングを裏付けており、これは、その抗酸化物質活動を示すために必須のイベントである。
【0130】
ここから得られた結果を図6中に示す。
【0131】
[メロン抗recSOD抗体]
上記組換えメロンSODのアミノ酸配列を得た後、Swiss Plotアライメントを用いて他の植物SODとの比較分析を行って、2つの相対的に保存されたペプチド配列14−27(EP1668−GVVTLTQEDDGPTS)および117−131(EP1669−HELADDLGKGGHELS)を選択した。これらのペプチド双方を合成し、ウサギの免疫化のための適切なキャリア(KLH)と結合させて、植物特異ポリクローナル抗体を得た(Eurogentec、ベルギー)。抗体EP1669を免疫吸着によって親和性カラム上で精製し、ウエスタンブロット分析中で1/2000で用いて、天然およびメロン組換えSODならびに精製ウシSODに対するこの抗体の特異性を確認した。
【0132】
SDS−PAGE分析のため、これらのタンパク質を95℃まで5分加熱した後、電気泳動をSDS−PAGEポリアクリルアミドゲル上で12%で行った。分離されたタンパク質を免疫ブロットPVDF膜(BIORAD)上に移行させた。抗体の任意のポテンシャルクロス特異性が、増幅Opti4CNヤギ抗ウサギ検出キット(BIORAD)によって明らかとなった。
【0133】
EP1669抗体は、上記天然メロン抽出物中に含まれる2個のSODとのクロス特異性そして精製ウシSODとのクロス特異性を示した。
【0134】
さらに、EP1669抗体は、上記免疫化ペプチドの由来元である完全な組換えメロンSODタンパク質との良好な特異性を示した。よって、抗体は、上記天然抽出物中に存在する反応性SODのうちの少なくとも1つとの相同性だけでなく、多様な種間のSOD科内に存在し得る相同性(完全な相同性)も示すことが確認された。この抗体は、本発明の組換えSODタンパク質の薬理活動の検証に必要な検出実験において有用なツールである。
【0135】
上記抗体結合試験の結果を図7に示す。
【0136】
[細胞培養物]
ヒト大腸癌細胞株HT−29(ATCC HTB−38)を、GlutaMAX−1、4.5g/L Dグルコース、ピルビン酸ナトリウムおよび100単位/mlのペニシリン/ストレプトマイシンが補充されたダルベッコ変法イーグル培地(D−MEM)中の10%胎児ウシ血清(FBS)中で、成長させる。各実験(n=4)において、およそ2×104個の細胞/ウェルを5%CO2雰囲気中で37℃で24時間インキュベートした。任意の実験の前に、既存の培地を新規培地と交換した。上記インキュベーション期間後、0.05%トリプシンおよび0.5mM EDTAを含むPBS溶液で細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、遠心分離によって回収した。その後、GUAVA PC Viacount Flexキットを用いて血球計算器中で細胞をカウントし、細胞数/mlで報告して、細胞増殖を推定した。
【0137】
[グリアジンペプチドの無毒濃度の効果]
グリアジン治療は、濃度に依存する様式で、細胞成長に多様な効果を及ぼすことが以前から分かっている。50〜200μg/mlの濃度は細胞成長を抑制する一方、500μg/mlまでの濃度は深刻な細胞破壊の原因となり得る。グリアジンペプチドは、先ずヒト腸管細胞の酸化バランスを変更することにより、その効果を開始し得ることが知られている(Dolfiniら、2002、Toxicol in Vitro、vol16、p331)。
【0138】
現在の実験において、72時間にわたって培養物を定期的にサンプリングして、多様な成分への暴露を行った場合および行わなかった場合のHT−29細胞の増殖率およびバイアビリティーを評価した。(a)10μMのグリアジンペプチド982(小麦胚芽油、QQPYPOPQPF.SEQ.IDN°01)は、有意な腸細胞凝集(De Vicenzi.Mら、1997、Toxicology、vol120、p207)または腸細胞アポトーシス(Giovanniniら、2000、Toxicology、vol145、p63)を誘発するように事前に関連付けられた濃度よりも少なくとも100倍小さい。
【0139】
我々が試験したこのような低い濃度において、グリアジンペプチドがHT−29細胞株増殖に主な効果を発揮する能力を持つが、この能力を持つのは、培養物の24時間においてのみである。この効果は、48時間後に急速に消え去る。なぜならば、この48時間経過後において、増殖がその基底レベルに戻り、おそらくはペプチド内部移行に起因してペプチドの存在から完全に独立するように見えるからである。グリアジンペプチドは、濃度閾値を越えかつ/またはヒト腸管細胞培養物上の十分な濃度を維持して、凝集またはさらなるアポトーシスをもたらさなければならないように見える。これは、高濃度のグリアジンペプチドで腸管細胞を24時間暴露した後、有意なO20−およびH2O2生成の増加および抗酸化物質酵素の低減が観察された実験によって裏付けられ、上記グリアジンペプチド誘発アポトーシスに先行する(Giovannini.Cら、2003、FEBS Letters、vol540、p117)。
【0140】
上記した試験の結果を図8の図解中に示す。
【0141】
[組換えSODの最小活性用量]
組換えタンパク質用量反応実験のため、組換えSODまたはGli−SOD(0〜50NBT単位)の濃度をさらに48時間増加させて、HT−29細胞をインキュベートした。
【0142】
この実験において、2つの異なる平行な挙動について、低用量または高用量を付加して、48時間でのHT−29成長における組換えSODおよびGli−SOD双方について観察した。
【0143】
HT−29細胞株の成長は、10単位用量の組換えSODまたは5単位用量の組換えGli−SODのいずれかにより変化するが、これらの値未満の値の場合、HT−29の成長は若干減速する。組換えSODおよび組換えGli−SODの用量を増加させると、HT−29成長誘導のその相対レベルは同一レベルに留まり、これは、どんなに高用量であっても、誘導閾値が達成可能であることを示している。
【0144】
これらの結果に従って、最適な用量が、組換えGli−SODの場合に5〜10単位および組換えSODの場合に10〜25単位で区切られ得る。
【0145】
上記実験の結果を図9に示す。
【0146】
[本発明の組換えSODは、「ペプチド効果」の補正を操作する]
培養物を定期的に72時間サンプリングして、多様な成分への暴露を行ったかまたは行わなかった以下の場合において、HT−29細胞の増殖率およびバイアビリティーを評価した。(a)10単位の組換えSODまたは(b)10μMのペプチド982と結合された10単位の組換えSODまたは最後に(c)10単位の組換えGli−SOD(グリアジンペプチド982に物理的にリンクしたSOD)
【0147】
この濃度において、SODまたはGli−SODのいずれも任意の有毒効果を発揮しない。
【0148】
なぜならば、これらの増殖曲線は厳密に基底曲線に適合しかつ平行に構成されているからである。さらに、組換えGli−SODのN末端部分におけるペプチド982の存在にもかかわらず、期待されたような「ペプチド効果」は24時間の培養物において発生しない。我々は、グリアジンペプチドが組換えSOD(Gli−SOD)と関連付けられるかまたは結合し、その結果、グリアジンペプチドによって仲介されるフリーラジカル生成および/または抗酸化物質酵素の障害の一時的増加と相互作用することができる場合、上記にて証明されたグリアジンペプチドのHT−29増殖に対する効果が、24時間の培養物において存在しないことに気がついた(RivabeneRら、1999、Biochem Biophys Acat、vol1453、p152)。
【0149】
この驚くべき相乗効果は、上記グリアジンペプチドおよび上記SODの結合あるいは物理的または化学的結合があろうとなかろうと、機能性組み合わせを含む治療組成および治療的組成において極めて有用な結果を持つ。なぜならば、これは、炎症ペプチドが投与形態中に意図的に存在する場合でも、期待された炎症反応を軽減さらには解消することができることを示すからである。
【0150】
この実験の結果の概要を図10中に示す。
【0151】
[細胞アポトーシス]
2つの基礎的な経路が、アポトーシスを実行し得る。第1の経路において、デスリガンドはその受容体(Fas/FasLigand)を活性化させ、その結果、デスコンプレックスが形成される。その後、このデスコンプレックスは、カスパーゼ8を活性化させる。
【0152】
第2の経路において、細胞ストレスに起因して、細胞質中へのチトクロムCの放出によってミトコンドリア機能障害が発生し、その結果、アポトーシスタンパク質因子−1およびカスパーゼ9との活性化複合体が形成される。その後、これらの複合体は、下流効果またはカスパーゼ3、6および7を活性化させ、その結果、最終アポトーシス変化が実行される。
【0153】
コムギグリアジンからの消化されたペプチドはCD95/Fasアポトーシス経路を介して腸管上皮細胞のアポトーシスを誘発することが知られているが(GiovanniniCら、2003、FEBS Lett.Vol540、p117)、キサンチン/キサンチンオキシダーゼの混合物によって生成されたROS(活性酸素種)は、チトクロムCの細胞質中への放出と共に、カスパーゼ−3を活性化させる(HiguchiMら、1998、oncogene、vol17、p2753)。
【0154】
第1にニューロペプチドとして特徴付けられて、オレキシンだけでなくその受容体も、胃腸管(VoisinTら2000、J Pharmacol Exp Ther、vol292、p638)を含むいくつかの末梢組織中に発現することが分かった(Voisin Tら、2003、Cell Mol Life Sci、vol60、p72)。オレキシンは、ミトコンドリアおよびカスパーゼ活性化からのチトクロムC放出を通じてアポトーシスを促進することにより、24時間後にヒト大腸癌HT−29細胞の成長を全体的に抑制する能力を有する(Rouet-Benzineb Pら、2004、J Biol Chem、vol279、P46875)。
【0155】
本発明の組換えSODまたはGli−SODから期待される細胞ストレスおよびミトコンドリア機能障害と戦う特定の活性により、このような誘発性システムにおけるその強力な抗アポトーシス効率を確認するために、実験を行った。上記組換えSODまたはGli−SODのために用いられた濃度は、増殖曲線に従って規定した。基底アポトーシス過程との相互作用に十分と思われる10単位の組換えSODまたはGli−SODのみを用い、一方、オレキシンまたはキサンチン/キサンチンオキシダーゼによって(X/XO)誘発されたHT−29細胞上でのアポトーシスには、50単位の組換えSODまたはGli−SODがより適しているように見える。
【0156】
[組換えSODは、オレキシン誘発アポトーシスを抑制した]
アポトーシス細胞と非アポトーシス細胞とを区別するGUAVA Nexinキットを用いて、HT−29細胞株中においてアポトーシスを分析した。この方法では、アネキシンVフィコエリトリンを用いて、アポトーシス細胞の外膜上のphopshatidylserine(PS)を検出する(Martinら、1995、J Exp Med、vol182、p1545)。
【0157】
24時間後、トリプシンを0.05%およびEDTAを0.5mM含むPBS溶液によって細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、アポトーシス細胞染色を行い、GUAVAPCAシステムで分析する。
【0158】
基底アポトーシス抑制(部分A)については、10単位の双方の組換えSOD(NBT単位)を含むかまたは含まない新規培地と培地を交換した。オレキシン誘発アポトーシス抑制(部分B)については、1μMのオレキシンおよび/または50単位の各組換えSOD(NBT単位)を含む新規培地と培地を効果した。その結果をアポトーシスアネキシンV−PE陽性細胞の百分率として表し、それは4点の平均である。
【0159】
通常条件下において、外膜PSのアネキシンVラベル付けによって示すように、アポトーシスは20%(20.5)のHT−29細胞中において同時発生した。
【0160】
この速度は、HT−29細胞の培養時において10単位の組換えGli−SODが存在した場合に10単位の組換えメロンSODが存在して16.8%まで到達した際に(18.1%抑制)、18%まで低下した(12.2%抑制)。
【0161】
第2の実験において、HT−29細胞培地へのオレキシンの補充を介してアポトーシスを誘発した際に、組換えSODまたはGli−SODの強力な抗アポトーシス効果を確認した。この場合において、HT−29細胞がアポトーシスとなる速度は、オレキシンを1μM培地中に付加することにより、基底レベルで評価した20%から40%まで向上され得る。オレキシン誘発アポトーシスの速度は、HT−29細胞の培養時において50単位の組換えGli−SODが存在した際に50単位の組換えメロンSODが存在して最終的に28.7%に達した際に(54.7%抑制)、34.4%まで低下した(26%抑制)。
【0162】
この実験の結果の概要を図12中に示す。
【0163】
[組換えSODは、X/XO誘発アポトーシスを抑制した]
腸管上皮細胞機能は、活性酸素種ROSの生成を通じて、活性マクロファージ(腸疾患)などの局所免疫細胞および炎症細胞によって構成され得る。従って、ROSは、腸管上皮細胞アポトーシスと関連するFasおよびFasL発現を誘発し得る(DenningTLら、2002、Free Radic Biol Med vol33、p1641)。
【0164】
キサンチン存在下のキサンチンオキシダーゼは、主にO2−°を生成する(これは、小腸中の活性酸素の代表であり、膜浸透性移行、カルシウム流入、脂質過酸化反応およびミトコンドリア中の膜流動性変化を誘発し得る)(AnupRら、1999;Indian J biochem Biophys、vol36、p266およびAnupRら、2000、BrJ Surg、vol87、p1094)。
【0165】
この第3の実験において、本発明による組換えSODまたはGli−SODの強力な抗アポトーシス効果の試験を、HT−29細胞培地中でのキサンチン/キサンチンオキシダーゼ誘導を介した可能なFas/FasL経路上で行った。
【0166】
この場合、HT−29細胞がアポトーシスとなる速度は、培地中に10mUのX/XO複合体を付加することにより、基底レベルについて評価した20%から30%まで向上され得る。50単位の組換えメロンSODまたはやはり50単位の組換えGli−SODを培地に付加すると、HT−29細胞中でのX/XO誘発アポトーシス経路がそれぞれ71%および86%だけ回避された。
【0167】
この実験の結果を図13のグラフに示す。
【0168】
[組換えGli−SODの結合]
各実験(n=4)において、およそ15×103個の細胞/ラメラ層を5%CO2雰囲気中で37℃で24時間インキュベートした。全実験の前に、既存の培地を新規培地と交換した。
【0169】
その後、細胞単層を新規培地中で50単位のSODまたはGli−SODを用いてまたは用いずにさらに24時間インキュベートし、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、その後、免疫組織化学によって分析した。EP−1669を第1の抗体(ウサギlgG抗SOD、希釈1/2000°)を用いて30分間Gli−SODのHT−29細胞への結合を分析した。その後、第2のFITC結合抗体(ヤギ抗ウサギ抗体、希釈1/100)をさらに30分間分析した。
【0170】
フローサイトメトリではなくこの方法を選択した理由は、フローサイトメトリ技術の場合、トリプシン治療によって細胞を回収し、分析前に収集しなければならず、このような治療は、Gli−SOD分子と細胞表面との間の相互作用に悪影響を与える可能性があるためである。
【0171】
上記FITC蛍光実験の結果を図11中に示す。図11において、細胞近隣のSODの存在がSOD双方において明確に視認され、また、Gli−SODの場合にも、より広範囲で明確に視認される。
【0172】
上記実験は、本発明による、そのN末端において小麦胚芽油グリアジンペプチド配列を持つかまたは持たないキューカミスメロSODをエンコードする遺伝子操作されたcDNAsは、エシェリキアコリ中で効率的に発現され得、抗酸化物質活動を持つ活性の分子として生成され得ることを実証した。加えて、消化されたグリアジン−ペプチドのみが腸管細胞凝集または特定状況の細胞アポトーシスを誘発することで知られているが、本発明者らは、これらの負の効果を回避するより短いグリアジンペプチドを選択する。また、本発明のSODを選択されたグリアジンペプチドと物理的に組み合わせるかまたは単に結合させることで、グリアジンペプチドの確実な炎症効果と相互作用が可能になることも示されている。
【図面の簡単な説明】
【0173】
【図1】キューカミスメロからのCuZnスーパーオキシドジスムターゼのためのcDNAコーディングを得るために用いられるクローニング方法の模式図である。
【図2】本発明の組換えおよびキメラSOD分子を生成するために用いられる遺伝工学の一般図の模式図である。
【図3】本発明による多様なメロンCu−ZnSOD変異体のRT−PCR増幅から得られるアガロースゲル電気泳動を示す。詳細には、ランAは組換え葉緑体メロンCu−ZnSODであり、ランBは組換えメロン偽細胞質Cu−ZnSODであり、ランCは、グリアジンペプチドへのN末端融合を支持するランBの組換え偽細胞質Cu−ZnSODである。
【図4】異なる由来(詳細には、トマト細胞質SOD(線1)、ヒトSOD(線2)およびメロン葉緑体SOD(線3))からの3つの異なるCu−Znスーパーオキシドジスムターゼのアミノ酸アライメントを示す。これらの配列間のアミノ酸残留物の同一性は、円を通じて3つのSODの各群の下側に示される。
【図5】組換えSOD(レーン1〜7)または組換え融合Gli〜SOD(レーンA〜F)の精製過程のクマシーブルー染色ゲルである。図5において、 レーン1およびAは、インビトロジェンから市販されているベンチマークタンパク質ラダーとして公知の分子量マーカである。 レーン2およびBは、His−Tagの開裂を示す。 レーン3、4、5、6およびC、D、Eは、連続精製工程時における本発明による組換えSODおよび組換えGli−SODを示す。 レーン7およびFは、MonoQカラム上の治療後に残っているバンドである。
【図6】NBT還元ゲル電気泳動を用いた組換えSODまたはGli−SODそれぞれのフォールディングを示す。 Aは、125μgの天然メロン抽出物(90U/mgに等価)でロードされたレーンAおよびA’と、0.92μgの未変性SODでロードされたレーン1、1’、および1’’’を有するrecSODを示す。Bは、本発明によるrecGli−SODを示し、レーンAおよびA’は、125μgの天然メロン抽出物(90U/mgに等価)でロードされ、レーン1および1’は1.84μgの組換えGli−SODでロードされ、レーン2および2’は、0.92μgの組換えGli−SODでロードされる。
【図7】以下のSDS−PAGEである。 Aは、以下のクーマシー青色顕色を示す。 レーン1中の50μgの天然キューカミスメロ抽出物 レーン2中のタンパク質マーカ(6.6、14.1、21.1、31、45および67KDa) レーン3中の2.5μgのウシSOD(シグマ) Bは、以下のEP1669抗体顕色を示す。 レーン4中の50μgの天然キューカミスメロ抽出物 レーン5中のタンパク質マーカ(6.6、14.1、21.1、31、45、67KDa) レーン6中の2.5μgのウシSOD(シグマ)
【図8】HT29細胞株の増殖曲線上のグリアジンペプチドの効果のグラフであり、ここでN=4である。
【図9】本発明によるrecSODまたはrecGli−SODの最小有効量の図解比較であり、N=4である。
【図10】本発明によるrecSODまたはrecGli−SODのHT29細胞株の増殖曲線上への効果のグラフであり、N=4である。
【図11】EP1669抗SOD抗体を用いたSODまたはGli−SODのHT29細胞上への細胞結合を示す1組のFITC画像であり、Aは負の制御であり、白色エリアは、細胞近隣のSODの存在を示し、BはrecSODでの治療後であり、白色エリアは、細胞近隣のSODの存在を示し、CはrecGli−SODでの治療後であり、白色エリアは、細胞近隣のSODの存在を示す。
【図12】アネキシンV試験を用いた、本発明によるSODの基底(A)またはオレキシンによって誘発された(B)HT−29アポトーシスに対する効果を図解表示したものであり、N=4である。Aは、1OUを用いて得られた結果を示し、Bは、recSODおよびrecGli−SODそれぞれの50Uを用いて得られた結果である。
【図13】ヒポキサンチン/キサンチンオキシダーゼによって誘発されたHT−29アポトーシスにおける組換えSODまたはGli−SODに対する効果のグラフを示す(N=4、アネキシンV試験)。
【0174】
参考文献:
1) Arentz-Hansen EH, Production of a panel of recombinant gliadins for the characterisation of T cell reactivity in coeliac disease. Gut 2000 Jan;46(1):46-51
2) Baillet F, Treatment of radiofibrosis with liposomal superoxide dismutase. Preliminary results of 50 cases. Free Radic Res Commun 1986;l(6):387-94
3) Beauchamp C. Superoxide dismutase improved assays and an assay applicable to acrylamide gels. Anal. Biochem:1971,44(1)276-287
4) Biagi F, A non-toxic analogue of a coeliac-activating gliadin peptide: a basis for immunomodulation? Aliment Pharmacol Ther 1999 Jul;13(7):945-50
5) Beckett CG, Analysis of interleukin-4 and interleukin-10 and their association with the lymphocytic infiltrate in the small intestine of patients with coeliac disease. Gut 1996 Dec;39(96):818-23
6) Bolte G, Peptic-tryptic digests of gliadin: contaminating trypsin but not pepsin interferes with gastrointestinal protein binding characteristics. Clin Chim Acta 1996 Mar 29;247(1-2):59-70
7) Bueno P, Peroxisomal copper, zinc superoxide dismutase. Characterization of the isoenzyme from watermelon cotyledons. Plant Physiol 1995 Jul;108(3):1151-60
8) Cannon RE, Cloning of cDNA for maize superoxide dismutase 2 (SOD2).Proc Natl Acad Sci USA 1987 Jan;84(1):179-83
9) Cerutti PA. Prooxidant states and tumor promotion. Science 1985 Jan 25;227(4685):375-81
10) Chirdo FG, In vitro presentation of gliadin-derived peptides by different cell lines. Clin Chim Acta 2002 Mar;317(1-2):151-8
11) Cornell HJ, In vitro mucosal digestion of synthetic gliadin-derived peptides in celiac disease. J Protein Chem 1995 Jul;14(5):335-9
12) Corominas M, Hypersensitivity reaction after orgotein (superoxide dismutase) administration. Allergol Immunopathol (Madr) 1990 Sep-Oct;18(5):297-9
13) Delanian S, Successful treatment of radiation-induced fibrosis using liposomal Cu/Zn superoxide dismutase: clinical trial. Radiother. Oncol. 1994, 32(1):12-20.
14) Diez-Gomez ML, Anaphylaxis after intra-articular injection of orgotein. Detection of an IgE-mediated mechanism. Allergy 1987 Jan;42 (1):74-6
15) Freeman BA, Liposome-mediated augmentation of superoxide dismutase in endothelial cells prevents oxygen injury J Biol Chem 1983 Oct 25;258(20):12534-42
16) Giovannini C, Induction of apoptosis in caco-2 cells by wheat gliadin peptides. Toxicology 2000 Apr 7;145(1):63-71
17) Halliwell. B, The antioxidants of human extracellular fluids. Arch Biochem Biophys 1990 Jul;280(1):l-8
18) Hidalgo IJ, Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989 Mar;96(3):736-49
19) Inoue. M, Expression of a hybrid Cu/Zn-type superoxide dismutase which has high affinity for heparin-like proteoglycans on vascular endothelial cells. J Biol Chem 1991 Sep 5;266(25):16409-14
20) Inoue M, Targeting SOD by gene and protein engineering and inhibition of free radical injury. Free Radic Res Commun 1991;12-13 Pt 1:391-9
21) Jadot G, Anti-inflammatory activity of superoxide dismutases: studies on adjuvant induced polyarthritis in rats. Free Radic Res Commun 1986;2(1-2):27-42
22) Jadot G, Anti-inflammatory activity of superoxide dismutases: inhibition of adriamycin induced edema in rats. Free Radic Res Commun 1986;2(1-2):19-26
23) Jadot G, Anti-inflammatory activity of superoxide dismutases: inhibition of carrageenan induced edema in rats. Free Radic Res Commun 1986;l(6):395-403
24) Jadot G, Clinical pharmacokinetics and delivery of bovine superoxide dismutase. Clin Pharmacokinet 1995 Jan;28(1):17-25
25) Jadot G, Comparative anti-inflammatory activity of different superoxide dismutases and liposomal SOD in ischemia. Free Radic Res Commun 1987;3(6):389-94
26) Joral A, Adverse reactions to food in adults. : J Investig Allergol Clin Immunol 1995 Jan-Feb;5(1):47-9
27) Joral A, Systemic anaphylaxis following parenteral orgotein administration. J Investig Allergol Clin Immunol 1993 Mar-Apr;3(2):103-4
28) Kaminaka H, Molecular cloning and characterization of a cDNA for plastidic copper/zinc-superoxide dismutase in rice (Oryza sativa L). Plant Cell Physiol 1997 Jan;38(1):65-9
29) Kim EJ, Transcriptional and post-transcriptional regulation by nickel of sodN gene encoding nickel-containing superoxide dismutase from Streptomyces coelicolor Muller Mol Microbiol 1998 Jan;27(1):187-95
30) Kleber-Janke T, Use of modified BL21(DE3) Escherichia coli cells for high-level expression of recombinant peanut allergens affected by poor codon usage. Protein Expr Purif 2000 Aug;19(3):419-24
31) Komar AA , Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation. FEBS Lett 1999 Dec 3;462(3):387-91
32) Koopman G. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84(5)1415-1420.
33) Kwiatowski J, Isolation and characterization of an iron-containing superoxide dismutase from tomato leaves, Lycopersicon esculentum. Eur J Biochem 1985 Jan 15;146(2):459-66
34) Lavy A, Increased susceptibility to undergo lipid peroxidation of chylomicrons and low-density lipoprotein in celiac disease. Ann Nutr Metab 1993;37(2):68-74
35) Loh EY, Polymerase chain reaction with single-sided specificity: analysis of T cell receptor delta chain. Science 1989 Jan 13;243(4888):217-20
36) Lundin KE, Gliadin-specific, HLA-DQ(alpha l*0501,beta 1*0201) restricted T cells isolated from the small intestinal mucosa of celiac disease patients. J. Exp Med 1993 Jul 1;178(1):187-96
37) Martin SJ. Early distribution of plasma membrane phosphatidylserine in a general feature of apoptosis regardless of the initiating stimulus : inhibition by overexpression of BcL2 and Abl. J. Exp. Med. 1995,182(5)1545-1556.
38) Mayrhofer G., Absorption and presentation of antigens by epithelial cells of the small intestine: hypotheses and predictions relating to the pathogenesis of coeliac disease. Immunol Cell Biol 1995 Oct;73(5):433-9
39) Mayrhofer G, Subcellular distribution of class II major histocompatibility antigens in enterocytes of the human and rat small intestine. Immunol Cell Biol 1989 Aug;67(Pt4):251-60
40) Michelson AM, Anti-inflammatory activity of superoxide dismutases: comparison of enzymes from different sources in different models in rats: mechanism of action. Free Radic Res Commun 1986;2(1-2):43-56
41) Palma JM, Purification and properties of cytosolic copper, zinc superoxide dismutase from watermelon (Citrullus vulgaris Schrad.) cotyledons. Free Radic Res 1997 Jan;26(1):83-91
42) Pathania V, Geriforte stimulates antioxidant defense system. Indian J Exp Biol 1998 Apr;36(4):414-7
43) Perdereau B, [Superoxide dismutase (Cu/Zn) in cutaneous application in the treatment of radiation-induced fibrosis] Bull Cancer 1994 Aug;81(8):659-69
44) Regnault C, Pharmacokinetics of superoxide dismutase in rats after oral administration. Biopharm Drug Dispos 1996 Mar;17(2):165-74
45) Regnault C, Effect of encapsulation on the anti-inflammatory properties of superoxide dismutase after oral administration. Clin Chim Acta 1995 Sep 15;240(2):117-27
46) Rivabene R, In vitro cytotoxic effect of wheat gliadin-derived peptides on the Caco-2 intestinal cell line is associated with intracellular oxidative imbalance: implications for coeliac disease. Biochim Biophys Acta 1999 Jan 6;1453(1):152-60
47) Scioli JR, Cloning and characterization of a cDNA encoding the chloroplastic copper/zinc-superoxide dismutase from pea. Proc Natl Acad Sci USA 1988 Oct;85(20):7661-5
48) Takeda Y, Albumin-binding superoxide dismutase with a prolonged half-life reduces reperfusion brain injury. Am J Physiol 1993 May;264(5Pt2):H1708-15
49) Tatham AS, Conformational studies of peptides corresponding to the coeliac-activating regions of wheat alpha-gliadin. Biochem J 1990 Sep l;270(2):313-8
50) Turrens JF , Protection against oxygen toxicity by intravenous injection of liposome-entrapped catalase and superoxide dismutase. J Clin Invest 1984 Jan;73(1):87-95
51) Vaille A, Anti-inflammatory activity of various superoxide dismutases on polyarthritis in the Lewis rat. Biochem Pharmacol 1990 Jan 15;39(2):247-55
52) Varenne S, The maximum rate of gene expression is dependent on the downstream context of unfavourable codons. Biochimie 1989 Nov-Dec;71(1l-12):1221-9
53) Vouldoukis I, Fc-receptor-mediated intracellular delivery of Cu/Zn- superoxide dismutase (SOD1) protects against redox-induced apoptosis through a nitric oxide dependent mechanism. Mol Med 2000 Dec;6(12):1042-53
54) Yoo HY, Heavy metal-mediated activation of the rat Cu/Zn superoxide dismutase gene via a metal-responsive element. Mol Gen Genet 1999 Sep;262(2):310-3
55) Zidenberg-Cherr S, Dietary superoxide dismutase does not affect tissue levels. Am J Clin Nutr 1983 Jan;37(1):5-7
【技術分野】
【0001】
本発明は、スーパーオキシドジスムターゼを含む医薬組成に関し、特に、他の化合物と関連する、スーパーオキシドジスムターゼを含む医薬組成に関する。
【背景技術】
【0002】
スーパーオキシドジスムターゼ(E.C.l.15.1.1.)またはSODという名称は、スーパーオキシド(O2〜°)アニオンの不均化において関与する普遍的金属酵素の仲間である。ラジカル種の蓄積または過度量のラジカル種の生成は、ほとんどの生物にとって有害である。この状態は、酸化ストレスとして未だ知られており、多様な病理学的過程(例えば、発癌現象、アテローム性動脈硬化、加齢および炎症疾患(例えば、セリアック病(グルテン不耐性としても知られる))において発生する多数の代謝異常と関連する。
【0003】
その発展または細胞局在の程度と別に、SODは、主に3つの形態で存在する。これら3つの形態は、分子内に含まれる金属イオン(すなわち、銅亜鉛またはCuZn−SOD、マンガンまたはMn〜SODおよび鉄またはFe〜SOD)によって区別される。CuZn−SODおよびMn〜SODはまた、より特殊な細胞組織(例えば、ペルオキシソームまたは植物中の葉緑体)においても見られ、一方哺乳類において、細胞外CuZn−SODまたはEc−SODが、細胞外区画において特異的に発見されている。より最近では、その他の公知のSODに対して弱い相同性しか持たないNi〜SODが、ストレプトミセスにおいて特定されている。
【0004】
多数の実験において、全SODが同一抗ラジカル活性を持っていても、その有効生物活性は、所与の治療指標および標的生物に対して同一ではないことが示されている。詳細には、ラット中のカラギーナンまたはアドリアマイシンによって誘発される脚部浮腫中の相同SODの抗炎症活動は、異種由来のSODのものと比較すると、非機能性である。異種SODの有効性は、活性部位に存在する金属の種類または酵素の全体的分子量よりも、わずかであっても酵素のアミノ酸の変動により高く依存するように見える。
【0005】
それにもかかわらず、用いられる異種文脈に応じて、異種SODと免疫原性SODとの間の境界をそれぞれについて決定する必要がある。実際、ラットにおける研究により、以下が示唆されている。
(a)相同ラットCuZn−SODまたは特定の異種SOD(例えば、ヒトMn〜SOD)は抗炎症活動を持たず、
(b)特定の異種SOD(例えば、ウシCuZn−SOD)はこのような抗炎症活動を持ち、さらに他のもの(とりわけイーストCuZn−SOD)は、炎症性活動も持ち得る。
【0006】
実施された多様な異なる研究から、臨床用量であっても、相同型酵素の有効性が低いため、ヒトSODは、ヒトにおける特定の抗炎症適応用の異種SODよりも有効性が極めて低いことが分かっている。特にこれらの理由のため、ウシCuZn−SODは、ウシ海綿状脳症(BSE)発病前に投与した場合に臨床試験において有利な効果を示すことが分かっている。
【0007】
ウシCuZn−SODとヒトCuZn−SODとの間には18.3%の相違が存在し、また多数回の注入が必要なことにも関わらず、アレルギーまたはアナフィラキシーショックの例はほとんど観察されていない。後になっても継続されているいくつかの初期研究によれば、抗SOD抗体循環率を増加させることにより、異種SODの薬理活動を漸進的に抑制させることが可能であることが提案されている。他のより最近の報告では、異種SODの提示および内在化を容易化する過程においてこれらの循環抗体が関与することがより多く示されている。
【0008】
SODを経口投与した場合も、胃腸管中において急速劣化することが多いため、バイオアベイラビリティおよび有効性の低減に繋がる点において、問題である。この問題は、SODの有効性が最も高い特定の細胞位置までSODを到達させることが困難であることにより、悪化する。
【発明の開示】
【発明が解決しようとする課題】
【0009】
出願人は、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントを含む医薬組成は、特に上記問題を解消し、また、この結合により、細胞遺伝子座に対するSODの細胞ターゲティングを容易化して有効性を最適化することができる、ことを発見した。
【0010】
従って、本発明の目的は、医薬投与用に適合された医薬組成を提供することである。この医薬組成は、少なくとも1つのスーパーオキシドジスムターゼ(SOD)および少なくとも1つのプロラミン系ペプチドフラグメントの機能的に活性な組み合わせを含む。「機能的に活性な組み合わせ」という用語により、上述した2つの成分(これらの成分は、いずれかが結び付けられているかまたは相互に物理的または化学的に結合している)は成分の蓄積効果を越える相乗効果を生成するように共に作用することが、理解される。詳細には、本発明において、SODとプロラミン系ペプチドフラグメントとの結合は、SODの存在可能時間を向上させると同時に、プロラミン系ペプチドフラグメントによって場合によって発生する炎症反応を顕著に低減させることが分かっている。
【0011】
本発明の他の目的は、本記載全体から明らかになるが、特に好適な目的としては、上記医薬組成中で採用されるアミノ酸分子、そのためのまたはハイブリダイゼーションを通じて得られる任意の核酸配列コーディング、このようなアミノ酸分子の薬剤、抗細胞ストレス剤としての使用、または炎症病理の治療における薬剤の調合のためのこのようなアミノ酸分子の使用がある。本発明は、特定のプロラミン系ペプチドフラグメントの存在あるいはその少なくとも1つのスーパーオキシドジスムターゼとの化学的または物理的組み合わせを検出するために用いられる特定の抗体も網羅する。請求項または記載中で「キメラ」という用語について言及がある場合、この用語は、当該分子が2つ以上の別個の遺伝源から由来する成分を含むことを意味する。
【課題を解決するための手段】
【0012】
[適切なプロラミン]
好適には、上記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンのフラグメントあるいは誘導体、類似体、塩またはその代謝体である。さらに好適には、上記プロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である。最も好適な実施形態において、上記プロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である。
【0013】
本発明の別の好適な実施形態において、上記少なくとも1つのプロラミン系ペプチドフラグメントは、完全に加水分解されたか、実質的に加水分解されたかまたは若干加水分解されたプロラミン系ペプチドフラグメントからなる群から選択される。しかし、一般的には、上記少なくとも1つのプロラミン系ペプチドフラグメントは、胃腸加水分解過程を模倣するPTC(パンクレアチン、トリプシン、キモトリプシン)加水分解されたプロラミンから得られたフラグメントからなる群から選択される。
【0014】
代替的であるが好適な実施形態において、少なくとも1つのプロラミン系ペプチドフラグメントは、腸管内においてターゲティング信号として機能する範囲において、加水分解される。
【0015】
[適切なSOD]
スーパーオキシドジスムターゼは、当業者に一般的に公知のものから選択でき、ヒトスーパーオキシドジスムターゼ、動物スーパーオキシドジスムターゼ、バクテリアスーパーオキシドジスムターゼ、イーストスーパーオキシドジスムターゼおよび植物スーパーオキシドジスムターゼからなる群から選択され得る。しかし、好適には、上記少なくとも1つのスーパーオキシドジスムターゼは、CuZnスーパーオキシドジスムターゼ、Mnスーパーオキシドジスムターゼ、細胞外スーパーオキシドジスムターゼ、NiスーパーオキシドジスムターゼおよびFeスーパーオキシドジスムターゼからなる群から選択される。本発明の1つの好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは相同スーパーオキシドジスムターゼである。本発明の別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは異種スーパーオキシドジスムターゼである。「相同」という用語は、当該分子の任意の所与の標的宿主中の未変性分子と同一由来のSODを指し、「異種」という用語は、当該分子の任意の所与の標的宿主中の未変性分子と異なる由来のSODを指す。最も好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは異種CuZnスーパーオキシドジスムターゼである。
【0016】
上述したように、上記少なくとも1つのスーパーオキシドジスムターゼは好適には、植物スーパーオキシドジスムターゼ、さらにより好適には異種CuZn植物スーパーオキシドジスムターゼである。このようなスーパーオキシドジスムターゼは、多くの任意の異なる様式で入手または生成可能である。例えば、上記少なくとも1つのスーパーオキシドジスムターゼは、植物から抽出可能である。上記少なくとも1つのスーパーオキシドジスムターゼの抽出および生成に植物を用いる場合、これらの植物は、ウリ科のものでよく、好適にはメロンからなる群から選択され、あるいは、ナス科からなる群から選択され、好適にはトマトからなる群から選択される。
【0017】
利用可能な多様な植物スーパーオキシドジスムターゼのうち、本発明によるスーパーオキシドジスムターゼは好適には、ペルオキソームスーパーオキシドジスムターゼ、葉緑体スーパーオキシドジスムターゼおよび細胞質スーパーオキシドジスムターゼからなる植物スーパーオキシドジスムターゼの群から選択される。
【0018】
別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、組換えスーパーオキシドジスムターゼである。上記組換えスーパーオキシドジスムターゼは、上記SODの核酸コーディングで形質転換された宿主細胞中の遺伝工学および発現を通じて、得ることができる。詳細には、出願人らは、上記少なくとも1つのスーパーオキシドジスムターゼが修飾葉緑体、ペルオキソームまたは細胞質CuZn組換えスーパーオキシドジスムターゼである場合に特に好ましいことを発見した。さらに別の好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、雑種異種/相同スーパーオキシドジスムターゼ(好適には、雑種植物/ヒトスーパーオキシドジスムターゼ)である。
【0019】
一般的な組換えタンパク質の発現と同様に、異質または未変性遺伝子を宿主細胞ゲノム中に導入する技術も当業者に周知である。従って、本発明の1つの好適な実施形態において、上記修飾葉緑体スーパーオキシドジスムターゼ、ペルオキソームスーパーオキシドジスムターゼまたは細胞質スーパーオキシドジスムターゼは、宿主細胞中で発現された修飾核酸配列によってコードされる。これらの修飾コーディング配列を得るために必要なPCR増幅遺伝子技術も当業者に公知であり、ここでの説明は不要である。
【0020】
本発明の最も好適な実施形態において、上記少なくとも1つのスーパーオキシドジスムターゼは、以下によってコードされる組換えスーパーオキシドジスムターゼである。
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意の1つによる核酸配列
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうち任意のものに対して厳密な条件下でハイブリダイズする核酸配列
SEQ.ID24〜SEQ.ID33として特定される配列のうちの任意の1つと少なくとも70%BLAST同一の相同性を有する核酸配列
【0021】
厳密な条件下でハイブリダイジングすることにより、当該核酸が、以下の条件65℃におけるSSPE×0.2下で述べられた配列にハイブリダイズすることが理解される。
【0022】
最後に、本発明の別の目的は、酸化および炎症病理または疾患の治療のための薬剤(特に、任意のほとんどの病気(グルテン不耐性、および好適にはセリアック病)に伴う病理または疾患のための薬剤)の製造のための、上述したような医薬組成の使用に関する。
【0023】
[異種植物SOD]
出願人は、植物由来のSODは本発明において極めて有利であることを発見した。CuZn−SODは、高等植物の光合成細胞のいくつかの形態において存在する。これらは、主に葉緑体中そしてより低い程度において細胞質およびペルオキシソーム中で見受けられる。これらは、アミノ酸組成、分子量、等電点およびその重合レベルにおいて、相互に異なる。
【0024】
これらのうち多くは、生化学的にまたは遺伝子的に特徴付けられる。葉緑体またはペルオキソーム植物SODが、先ずタンパク質のN末端においてリーダー配列または信号ペプチドを含む前駆体として、生成される。この前駆体は、葉緑体またはペルオキシソーム中への転位後、余分なN末端部分の削除において行われる機能性成熟を経験する。
【0025】
この特異性に加えて、上記ペルオキソームSOD(特に、メロンのもの)または葉緑体SOD(特にトマトのもの)は、(a)細胞質SOD中の70%保存領域に対して葉緑体およびペルオキソームSOD中の約90%保存領域において、細胞質SODよりもこれらの間においてより良好に保存され、(b)他の植物CuZn−SODよりも熱および過酸化水素不活性化に対して高い耐性を持つのにもかかわらず、経腸または非経口投与のいずれにおける治療用途候補として考えられてこられなかった。
【0026】
しかし、出願人の配列アライメントにより、葉緑体またはペルオキソームSODは、その等価な細胞質ゾル由来植物SODよりも2〜4%少なくヒトまたはウシSODと異なることが明らかになった。
【0027】
本発明の好適な実施形態のうちの1つにおいて、例えばメロンからの葉緑体CuZn−SODの選択は、出願人によって保持された。なぜならば、細胞質形態に豊富な植物由来の特定のコドンがバクテリアにおいて稀であることが特定され、また、この理由のため、発現レベルと、これらを豊富にまたは活性部位において含む生合成酵素の特定活動とが有意に変化する。
【0028】
出願人は好適には、バクテリアシステムを生合成SODの組換え発現のための宿主細胞として用いることを選択し、これにより、これら2つの局面が確認される。
【0029】
出願人のバクテリアシステムにおいてヒトCuZn−SODは発現され、機能性であった。これは上記稀なコドンのうち少なくとも2つを含むものの、そこから形成された2つのアルギニン残留物は、N末端領域において発見可能であり、ヒトSODの酵素的機能に関与しているように見えない。
【0030】
一方、細胞質トマトSODにおいて、上記稀なコドンから得られた2つのアルギニン残留物は、上記酵素のC末端部分中に配置され、そのうちの1つは、スーパーオキシドアニオンの誘引に関する酵素的機能に関与する。
【0031】
この場合、このシステム中で発現される生合成トマト細胞質SODは、活性がずっと低い。葉緑体トマトSOD上について他の研究が行われたが、このような稀なコドンが存在せず、バクテリア中でのその発現により完全な機能性の生合成SODが得られ、このSODが完全に活性であることが示されている。
【0032】
このバクテリアシステム中でクローニングおよび発現された第1の2個のメロンSODは、得られる精製レベルにおける細胞質トマトと同様の特定の活動を持つことが分かっている。この場合、上記稀なコドンは、細胞質トマトSOD中で特定されたのと同じ位置を網羅し、上記アルギニンのうち1つは、上記スーパーオキシドの誘引機能を持つ。
【0033】
よって、現在用いられている宿主株を、Ecoli(すなわち、アルギニンAGGまたはAGA)において稀に用いられるコドン向けのさらなるtRNAsを含む株と代替すると、有利であった。この特定の株は、バクテリア中の機能的に活性な植物タンパク質の発現を向上させるように、設計される。
【0034】
従って、植物由来の成熟SODは、その抽出された形態においておよび好適には組換え酵素として、経口投与と組み合わされると、異種活性および免疫耐性双方を調和させることができ、そのため、病理学的炎症過程の治療におけるウシSODの代替物として機能可能である。
【0035】
[SODの経口投与および細胞ターゲティング]
治療的に受容可能なSODの活性を増加させるために、いくつかの異なる処方が考えられた(すなわち、リポソームカプセル封入、アルブミン共役、またはポリエチレングリコールとの結合、またはヘパリン親和性ペプチドとの雑種としての結合)。
【0036】
リポソームまたはセラミドを用いた経口投与された異種ウシCuZn−SODの抗炎症活性は、先ず、カラギーナンによって誘発されるラット下肢浮腫の低減において示された。また、セラミドをグリアジンのような植物プロラミンと代替し、非組換えウシSODを非組換え植物SODと代替した場合、注入可能なウシSODと比較可能な治療活動により、ラット中で誘発される認知疾患の発病を遅延させることが分かった。
【0037】
[グリアジン使用における副作用]
しかし、この種の経口投与の場合、限られてはいるが無視できない数の人数に有意の多くの副作用が発生している。上記修飾処方中で用いられた特定の物質(特に、グリアジンのような植物プロラミン)は、胃腸管中でのペプシン−トリプシン加水分解後に耐性の低い生成物を発生させ得、その結果、グルテンアレルギーまたはさらにはセリアック病の場合のような毒性となり得ることが周知である。このヒトのみに発生する疾病は、ヒト腸細胞の刷毛状縁部の特定容量にリンクし、これにより、HLAクラスII分子が発現する。いくつかのハプロタイプ(例えば、HLA−DR、HLA−DP、HLA−DQ)は、グリアジン〜特異T細胞の活性化に密接に関連することが分かっている。グリアジンペプチドのこの有毒処理は、以下の3つの連続工程が関与すると考えられている。
a)グリアジンの有毒フラグメントがHLA受容体に結合し、腸管微絨毛縁部上の腸細胞へ輸送される。
b)これらが固有層中のCD4+型上皮内細胞へ提示された場合、上記腸細胞側底レベルへ輸送される。
c)最後に、無傷のまたは部分加水分解されたポリペプチドが、上記腸細胞側底膜から上記固有層中のAPC細胞(抗原提示細胞)へと解放される。
【0038】
小麦胚芽油中に存在するグリアジンペプチドおよび胃腸管中での加水分解によって生成されるものにより、負の反応(すなわち、炎症反応)が発生し、一方、酵素的に活性のSODと結び付けられると、このようなペプチドの負の効果は大幅に低減される。
【0039】
加えて、上記負の副作用を除いて小麦胚芽油中に存在するものと同様の挙動を示す二粒系コムギ中に存在するグリアジンの結合が、酵素的に活性のSODと結び付けられた場合に、酸化ストレスに関連する欠乏の治療にとって最も好適であることが分かっている。
【0040】
生理的条件下において、上皮細胞によって提示される抗原に対する反応をTH2型T細胞によって仲介する。これらの細胞は、IgA生成に起因し、とりわけ、食物抗原に関する免疫耐性に起因する。グルテン不耐性またはそのより深刻な形態であるセリアック病の場合、グリアジン由来のペプチドは、すでに酸化促進性であるためグリアジンに対して一層感受性の高い酸化ストレスの状態を生成することにより、腸細胞の酸化代謝を変化させることができる。
【0041】
この炎症状態は、サイトカインおよび一酸化窒素(NO)の生成における重要な変化と関連する、THl細胞型に対する細胞反応の転換により、特徴付けられる。これらの条件下において、脂質過酸化反応の増加および酸化および低減した形態のグルタチオン(GSSG/GSH)の比の増加は、抗酸化物質をグリアジンと融合させるとこの治療指標において有利な効果が得られるはずであるという出願人の仮説を補強した。
【0042】
結果として、そのグリアジンとの組み合わせから得られる経口経路によるSOD抗炎症活性の増強が、細胞膜の親和性増加および関連する代謝経路の基礎制御に起因して、現れることとなる。
【0043】
よって、これらの効果を実証するために、酵素的に活性の異種SODを先ず小麦胚芽油プロラミンの加水分解物の誘導体(詳細にはVulグリアジン)と結合させ、そのアミノ酸配列がNter−QQPYPQPQPF−Cter(SEQ.IDN°01)であることを確認した。その後、二粒系コムギプロラミンの加水分解物のその無毒類似性誘導体(詳細にはDurグリアジン)と結合させ、そのアミノ酸配列がNter−QQPQDAVQPF−Cter(SEQ.IDN°02)であることを確認した。この類似体は、グリアジンの有毒誘導体の競合的抑制物質であり、腸細胞との相互作用サイトのターゲティングが可能である。
【0044】
プロラミン(詳細にはグリアジン)は、胃腸管移行時において自身が包囲している活性成分を保護するという積極的役割を果たし、その加水分解された形態において、腸管粘膜内の細胞における活性成分のターゲティングおよび連続的放出を助ける。これらの誘導体をタンパク質N末端部分に追加された無毒類似体(例えば、異種SOD)と代替した場合、上記誘導体ペプチドフラグメントの腸細胞ターゲッターとしての役割を維持すると同時に、プロラミンの抗原誘導体の有害な効果を回避することができる。
【0045】
いくつかの治療指標における注入可能なウシSODによって実証される薬理有効性と、BSEおよびCFJDの出現によって発生した公的安全の問題とを鑑みて、上記した本発明の目的は、以下の利点を持つことであることが分かった。
1.レシピエントの遺伝的バックグラウンドに関係なく、異種非免疫原性SODまたは免疫耐性の誘発のみを行うもの。
2.植物由来の(有利には食物由来の)SOD。
3.このようなSODの場合に従来では有効ではなかった投与経路を通じて異種SODを薬理的に活性させる処方。
4.この異種SODを無毒腸管ターゲティングペプチドと結合させることで、経口投与されたこれらのSODの異種活性を増強させ、治療法が未だない適応(詳細にはオーファンドラッグ適応)における代替的治療を示すこと。
【0046】
本発明のこれらのおよび他の実施形態について、以下詳細に説明する。
【発明を実施するための最良の形態】
【0047】
[例]
(例1)
銅亜鉛ペルオキソームまたはメロン「キューカミスメロL」から得られた葉緑体スーパーオキシドジスムターゼについてコーディングするメッセンジャーRNAのクローニングと、その完全な核酸配列(コーディングおよび非翻訳配列双方)の決定とを、逐次RT−PCR増幅によって行った。図2中に示すような偽細胞質という名称の第1のタンパク質変異体を得るために、コーディング配列のみを含む第1のクローンを遺伝工学によって改変した。
【0048】
この第1のタンパク質変異体は、メロン葉緑体SODの遺伝子改変の一般図を示す。この第1の変異体は、未変性信号ペプチドに対応するN末端における第1の65個のアミノ酸の置換を示し、上記アミノ酸対メチオニン−バリンは、細胞質植物SODのN末端の特性である。この技術によって改変された過程により、植物細胞中においてSODが経験する葉緑体成熟工程が得られ、その後、図示の2つのアミノ酸が付加される。第2および第3の変異体は、このメチオニン−バリンアミノ酸対に加えて、胃腸管ターゲティングのためのグリアジンペプチド配列(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ)をそれぞれ含む。その由来は小麦胚芽油であり、そのペプチド配列により、感作反応を生成することなく、以下のような、腸細胞と相互作用するグリアジンペプチドの再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ)。その由来は二粒系コムギである。
【0049】
(例2)
コーディング配列全体の直接的RT−PCR増幅により、トマト葉アカナス(PubMedBlast受入番号X14040)から、銅亜鉛SOD細胞質スーパーオキシドジスムターゼをコードするメッセンジャーRNAのクローニングが得られ、これにより、細胞質という名称の第1のトマト変異体が得られた。第2の変異体および第3の変異体も、胃腸管ターゲティング(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ))のためのグリアジンペプチド配列の付加により、遺伝子操作された。その由来は小麦胚芽油であり、上記ペプチド配列により、感作反応を生成することなく、以下のような腸細胞上でのグリアジンペプチド相互作用の再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ)。その由来は二粒系コムギである。
【0050】
(例3)
コーディング配列全体の直接的RT−PCR増幅により、ヒト血液細胞ホモサピエンス(PubMed/Blast受入番号K00065)から得られた銅亜鉛SOD細胞質スーパーオキシドジスムターゼについてコードするメッセンジャーRNAのクローニングを得、これにより、細胞質という名称の第1のヒト変異体を得た。第2変異体および第3の変異体についても、胃腸管ターゲティング(Nter−QQPYPQPQPF−Cter(SEQ.IDN°01)、VuIと呼ぶ))のためのグリアジンペプチド配列の付加により、遺伝子操作を行った。上記ペプチド配列により、感作反応を生成することなく、以下のような腸細胞上でのグリアジンペプチド相互作用の再生が可能となる(Nter−QQPQDAVQPF−Cter(SEQ.IDN°02)、Durと呼ぶ))。
【0051】
(例4)
各種類の構成の第1の変異体の発現物を用いて、得られた多様な生合成SODの抗ラジカル活性を検証した。これらの活性について、未変性PAGE上のNBTの還元により、確認した。その結果、上記組換え生合成タンパク質の活性が、現在最も用いられているバクテリア宿主株中に稀に存在するコドンの植物cDNA中の存在に密接にリンクすることが分かった。
【0052】
現在用いられている株中に発現する3つのcDNAからの低い発現率または活性率については、バクテリア株(とりわけコドンAGA、AGG(アルギニン)およびより低い範囲ではコドンCCC(プロリン)、GGA(グリシン)CTA(ロイシン)、およびATA(イソロイシン))によって稀に用いられるコドンの存在によって説明することができる。この問題を回避するために、上記cDNAをpET30a+(NOVAGENから市販)として特定される別の発現ベクターシステムに移行させ、これを用いて、これらの稀なアミノ酸用の適切なtRNAを含むRosettaDE3pLysS(NOVAGENから市販)として特定される許容バクテリア株を形質転換した。
【0053】
[本発明の詳細なプロトコル]
(キューカミスメロLSODのクローニングプロトコルおよび遺伝子改変)
変種キューカミスメロL(VILMORIN−GNIS−2251029として登録されている市販の変種)または上記変種キューカミスメロLC(ASL−NCIMS−40310として登録されている変種)のいずれかから、真核細胞に使用されるものと同一の技術[TRIzol−TMキット、Life Technologies、フランス]を用いて窒素凍結保存されたキューカミスメロLメロン葉から全RNAを抽出した。
【0054】
(メロンSODの3’RACE−PCR)
25μlの反応緩衝液中の修飾オリゴdTプライマーAAGCAGTGGTATCAACGCAGAGTACTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTVN(SEQ.IDN°03)[Clontech、Smart cDNA ref:k1051]から、メッセンジャーRNAの単鎖DNAマトリックス中への転写を実施した:トリスHCl20mM pH8.3、KCl50mM、MgCl22.5mM、10mM DTTおよび400μMの各dNTPを200単位のSuperscriptII逆転写酵素(Gibco−BRL)の存在下で42℃で1時間。
【0055】
補体としていくつかの単鎖プライマー対(例えば、EndプライマーAAGCAGTGGTATCAACGCAGAGT(SEQ.IDN°04)を修飾オリゴdTプライマーならびにメロンコーディング領域のためのMelcyt5F1 GGTGAYACMACMAATGGYTG(SEQ.IDN°05)およびMelcyt5S2 CATGCKGGKGAYCTDGG(SEQ.IDN°06)それぞれと組み合わせることにより、SODのためのメッセンジャーRNAコーディングの3’末端の増幅を実施した。アンプライマー配列は、いくつかの植物種のペルオキソーム、葉緑体および細胞質CuZn−SODの保存領域に基づくものであるため、本質的に極めて縮重するものである。
【0056】
(メロンSODの5’RACE−PCR)
バクテリアベクターpGEM−T(Promegaから市販)中へのクローニング後、7個の独立クローンを配列し、コンセンサス配列を用いて、3つの新規の特定のプライマーVed3R3 ACAAAGGCTCTTCCAACTACAG(SEQ.IDN°07)、Ved3R2 GCCGCTAAGAGGAATCTG(SEQ.IDN°08)およびVed3R1 TGGTTGCCTCTGCTACTCCATC(SEQ.IDN°09)を構築した。この特定のプライマーVed3R3を用いて、メッセンジャーRNAをdCTPホモポリマー伸長(5’RACEシステム、Life Technologies、ref:18274−058)によって終端された単鎖DNAマトリックス中に転写した。このマトリックスの増幅は、プライマーAAPP GGCCAGGCGTCGACTA GTACGGGGGGGGGGGGGGGG(SEQ.IDN°10)およびプレ増幅工程のためのVed3R2およびAAP GGCCAGGCGTCGACTAGTAC(SEQ.IDN°11)および特定のメッセンジャーRNAの5’末端の最終増幅のためのVed3R1の連続的組み合わせにより、得た。
【0057】
(3つのSOD変異体の増幅および遺伝子改変)
各種(ヒト、トマトおよびメロン)からの異なるSOD変異体の増幅を、3つの単鎖5’アンプライマーを以下のような各種のためのユニークな単鎖3’アンプライマーと組み合わせることにより、可能にした。
【0058】
名称 サイト 配列
Hum5Cyt NcoI atcggatccATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°12)
Hum5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°13)
Hum5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGCGACGAAGGCCGTGTGCGTG(SEQ.IDN°14)
Hum3Rfw HindIII ctcgagaaqcttTTATTGGGCGATCCCAATTAC(SEQ.IDN°15)
【0059】
名称 サイト 配列
Tom5Cyt NcoI atcggatccATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°16)
Tom5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°17)
Tom5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGTGAAGGCCGTCGCCGTCCTT(SEQ.IDN°18)
Tom3Rfw HindIII ctcgagaagcttTAACCCTGGAGGCCAATAAT(SEQ.IDN°19)
【0060】
名称 サイト 配列
Mel5pCyt NcoI atcggatccATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°20)
Mel5Dur NcoI atcggatccATGGCTCAACAACCACAAGATGCTGTCCAACCATTCATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°21)
Mel5Vul NcoI atcggatccATGGCTCAACAACCATATCCACAACCACAACCATTCATGGTGAAAGCTGTCGCTGTCCTC(SEQ.IDN°22)
Mel3Rfw HindIII ctcgagaagcttCACACAGGAGTCAGACCGAC(SEQ.IDN°23)
【0061】
ここで、Cytは、オリジナルの65アミノ酸配列の代わりにさらなる2個のアミノ酸を含むメロン偽細胞質形態を指定し、ヒトおよびトマト中の細胞質形態については、参照符号DurおよびVulがそれぞれ以下を示す。
【0062】
DURは、二粒系コムギから得られた胃腸管ターゲティング信号ペプチドを含む細胞質形態を示す。
【0063】
VULは、小麦胚芽油から得られた胃腸管ターゲティング信号ペプチドを含む細胞質形態を示す。
【0064】
これらのプライマーは、5’BamHIまたはNcolサイトおよび3’Hindlllサイトのいずれかにおける同相でのバクテリア発現ベクターpET30a+(NOVAGENから市販)中へのその導入を可能にする必要な合成配列を含むように、構築された。これらの増幅および遺伝子改変のためのグローバルプロシージャを図1中に報告する。
【0065】
バクテリア発現システムにより、クローンcDNAのハイブリドタンパク質としての発現が可能となり、SODまたはタンパク質変異体がそのN末端を介してHis−Tag配列へとリンクされる。
【0066】
全ての増幅は、TAPS(pH9.3)を25mM、KClを50mM、MgCI2を1.5mM、各dNTPを200μM、各プライマーを10μM、プラスミドDNAを10ng、ゴールドスターDNAポリメラーゼ(Eurogentec)を2.0単位を含む50μlの反応体積中において行った。各使用サーモサイクリングプログラムを以下の表に示し、GeneAmpPCRサーモサイクルシステム9700(Applied Biosystem)上で実施した。
【0067】
[RT−PCR増幅サイクルの表]
(メロン3’RACE)
95℃、3分
95℃、25秒
55℃、30秒
72℃、40秒
95℃、25秒
58℃、30秒
72℃、40秒
(メロン5’RACE)
95℃、3分
95℃、30秒
55℃、30秒
72℃、60秒
95℃、30秒
61℃、40秒
72℃、50秒
(メロンクロロ)
95℃、3分
95℃、40秒
57℃、40秒
72℃、40秒
95℃、40秒
60℃、40秒
72℃、50秒
(メロン偽Cyt)
95℃、3分
95℃、40秒
57℃、40秒
72℃、40秒
95℃、40秒
60℃、40秒
72℃、50秒
【0068】
ヒト、トマトおよびメロンSODについてエンコーディングする3つのタンパク質変異体「Cyt、Dur、およびVul」の増幅を5’オリゴヌクレオチドプライマーのサイズ差を考慮した同一プロトコルに従って行った。先ず、反応物を94℃で5分間加熱した後、以下のような5回の予備サイクル(94℃での変性を0.45分、56℃でのアニーリングを0.55分および72℃での伸展を0.55分)に暴露した後、さらなる25サイクル(94℃での変性を0.45分、59℃でのアニーリングを0.55分および72°Cでの伸展を0.55分)に暴露した。
【0069】
事前クローンされたCu−ZnSOD CDNAのコーディング領域をPCRによって増幅し、増幅された459bpおよび495bp生成物をその後適切な制限エンドヌクレアーゼNcol〜5’端部およびHindlll〜3’端部によって消化し、その結果、同相でのバクテリア発現pET30a+(NOVAGENから市販)中へのその導入を可能とした。
【0070】
各種(ヒト、トマト、およびメロン)のための3つのクローンのヌクレオチド配列を決定し、事前取得された配列と同一であることが分かり、以下に示す。
【0071】
[葉緑体またはペルオキソームCuZn−SOD形態の場合(SEQ.IDN°24)]
【0072】
[メロン偽細胞質Cu−ZnSOD形態の場合(SEQ.IDN°25)]
【0073】
[二粒系コムギペプチドによるメロン偽細胞質Cu−ZnSOD形態(SEQ.IDN°26)]
【0074】
[小麦胚芽油ペプチドによるメロン偽細胞質Cu−ZnSOD形態の場合(SEQ.IDN°27)]
【0075】
[トマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°28)]
【0076】
[二粒系コムギペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°29)]
【0077】
小麦胚芽油ペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°30)
【0078】
[ヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°31)]
【0079】
二粒系コムギペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°32)
【0080】
[小麦胚芽油ペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°33)]
【0081】
公知の植物スーパーオキシドジスムターゼによるORFおよび配列アライメントの決定のためのコンピュータソフトウェアを用いた核酸配列の分析により、要求に匹敵するペプチド配列の決定と、上記した遺伝子改変から得られた変異体のペプチド配列の決定とが可能となった。
【0082】
[メロン葉緑体CuZn−SOD形態の場合(SEQ.IDN°34)]
【0083】
[メロン偽細胞質CuZnSOD形態の場合(SEQ.IDN°35)]
【0084】
[メロン偽細胞質CuZnSOD形態+二粒系コムギペプチドの場合(SEQ.IDN°36)]
【0085】
[メロン偽細胞質CuZnSOD形態+小麦胚芽油ペプチドの場合(SEQ.IDN°37)]
【0086】
[トマト細胞質Cu−ZnSOD形態の場合の場合(SEQ.IDN°38)]
【0087】
[二粒系コムギペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°39)]
【0088】
[小麦胚芽油ペプチドによるトマト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°40)]
【0089】
[ヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°41)]
【0090】
[二粒系コムギペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°42)]
【0091】
[小麦胚芽油ペプチドによるヒト細胞質Cu−ZnSOD形態の場合(SEQ.IDN°43)]
【0092】
バクテリア発現システムpET30aにより、クローンCDNAをハイブリッドタンパク質として発現させることが可能となり、Rosetta(DE3)pLysSバクテリア株(Novagen)中への発現後、SODが切断可能なN末端His−Tag配列(His−Tag)が融合される。
【0093】
OD600nmの値が0.6まで上昇するまで、生成されたCu−ZnSODの変異体形態それぞれについて入手および選択された組換えクローンを、適切な抗生物質(30μg/mlのクロラムフェニコールおよび/または34μg/mlのカナマイシンが補充された1リットルのリッチメディア(トリプトンを10g/l、イースト抽出物を5g/l、NaClを5g/l)中で再懸濁した。その後、37°Cでのインキュベーションを少なくとも3時間行い、この時点において、タンパク質の発現を制御するラックプロモーター誘導物質であるイソプロピルチオガラクトピラノサイド(IPTG)を追加して最終濃度1mMを得、さらに4〜6時間インキュベートした。
【0094】
各調製のための2個のバクテリアペレットを(1)50mlの寒冷PBS緩衝液または(2)50mlの添加液(Novagenから市販)中で再懸濁し、その後、4℃で各30秒の4連続サイクル(Vibracell40W〜20KHz)において、超音波処理緩衝液(トリスHCl pH7.4を10mM、EDTAを1mM、NaClを150mM)に暴露した。遠心分離を13000gで20分間行うことにより、細胞質内タンパク質抽出物を単離した。
【0095】
粗タンパク質抽出物から開始して、His−Bindクイックカートリッジ(NOVAGENからref70155として市販)上で、多様なHis−TagSOD融合タンパク質を単離した。その後、組換えエンテロキナーゼ(ref69066、NOVAGEN)でのハイブリドタンパク質の加水分解後、多様なSODおよびその特定の変異体を新規精製工程によってHis−Bindクイックカートリッジ上で精製した。
【0096】
(例5)
[メロン抗recSOD抗体]
組換えメロンSODのアミノ酸配列を得た後、スイスプロットアライメントを用いて他の植物SODとの比較分析を実施し、メロン特異ポリクローナル抗体(Eurogentec、ベルギー)を得るためにウサギの免疫化を行うために、2個のペプチド配列14−27(EP1668−GVVTLTQEDDGPTS、SEQ.IDN°44))および117−131(EP1669−HELADDLGKGGHELS、SEQ.IDN°45)を選択した。抗体EP1669をイムノアブソープションによる親和性カラム上で精製しウエスタンブロット分析中で1/2000において用いて、生成されたメロン組換えSODに対する抗体の特異性を確認した。
【0097】
SDS−PAGE分析のために、タンパク質抽出物を95℃で5分間加熱した後、電気泳動を12%のSDSポリアクリルアミドゲル上で実施した。分離されたタンパク質を免疫ブロットPVDF膜(BIORAD)上に移行させた。上記抗体の任意のポテンシャルクロス特異性が、上記増幅されたOpti4CNヤギ抗ウサギ検出キット(BIORAD)により、明らかとなった。
【0098】
EP1669抗体は、免疫化ペプチド由来の完全な組換えメロンSODタンパク質に対して良好な特異性を示した。EP1669抗体はまた、組換えヒトおよびトマトSODならびに天然ウシSODまたはヒトSODに対してクロス特異性を示した。従って、この抗体は、本発明の組換えSODタンパク質の薬理活動の検証に必要な実験のための検出ツールとして有用であった。
【0099】
(例6)
[抗ラジカル活性]
メロン偽細胞質Cu−ZnSODおよびヒトまたはトマト細胞質Cu−ZnSODならびにその2つの異なるグリアジンペプチドを支持する等価物をその抗ラジカル活性について分析した。これらの活性は、NATIVE−PAGE電気泳動上でのNBT光還元に従って確認される(BeauchampsおよびFridovitch、1971)。
【0100】
(例7)
[細胞ターゲティング]
標的薬剤として用いられるグリアジンペプチドが先に示したようにHLA−DQ2と相互作用してヒト腸細胞中に進入可能なだけでなく、それと物理的にまたは化学的に結合するタンパク質の併用ターゲティングを容易化することも可能であることを実証するために、ピルビン酸ナトリウム、グルタミン、非必須アミノ酸およびペニシリン/ストレプトマイシンが補充されたダルベッコ変法イーグル培地(DME)中の10%胎児ウシ血清(FBS)中で、ヒト大腸癌細胞株HT−29(ATCC HTB−38)およびCaco−2(ATCCHTB−37)を成長させる。いくつかの場合において、細胞を100lU/mlのIFN―γ(R&Dシステムズ)で48時間インキュベートし、HLAクラスIl発現を誘発する。
【0101】
[グリアジンペプチド提示分析およびフローサイトメトリ]
およそ2×105個の細胞を、37℃で組換えSODで1mg/mlまでの濃度で1〜20時間インキュベートする。0.2μmミリポア膜での濾過により、試験されたSOD化合物を殺菌する。これらの細胞を回収し、L243mAb(抗HLA DR、ATCC HB−55)を用いてHLAクラスII発現用のフローサイトメトリによって分析し、その後、FITC結合型ヤギ抗マウスlgG(ICN−Cappel)を行う。
【0102】
細胞表面におけるSOD化合物を検出するために、これらの細胞をEP−1669抗体またはEP−1668抗体(ISO細胞)でインキュベートし、FITC結合型ヤギ抗ウサギlgGで明らかにする。4°Cで60分間インキュベーションを行い、その後、FBSを10%およびアジ化ナトリウムを0.1%含むPBS中で、これらの細胞を3回洗浄する。その後、これらの細胞をFACScanフローサイトメーター(Becton Dickinson)上で分析する。
【0103】
(例8)
[アネキシンV陽性細胞の検出]
この部分中で報告される実験全てにおいて、48時間の暴露時間が用いられた。アポトーシス接着細胞の検出は、フルオレセインイソチオシアネートでラベルされたアネキシンV〜PS(Martinら、1995)の結合を用いたフローサイトメトリにより、行った。インキュベーション期間後、上記したようなトリプシンおよびEDTAを含むPBS溶液で細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、1×106個の細胞を遠心分離によって収集して、接着細胞中のアポトーシスを評価した。アネキシンV−FITC陽性細胞の評価は、ApoAlertアネキシンVアポトーシスキット(Clontech Laboratories、Palo Alto、CA、米国)を用いて行った。暗所において200μlの結合緩衝液中で10μlのアネキシンV−FITC(最終濃度:0.5μg/ml)および0.5μg/mlのヨウ化プロピジウムによって細胞を再懸濁させ、15分間インキュベートして、壊死細胞を排除した。488nmでチューニングされたアルゴンイオンレーザでCoulter Epics Elite ESP細胞分取器(Miami、FL)上で、二色サイトメトリ(蛍光標示式細胞分取(FACS))分析を行った(Koopmanら、1994)。
【0104】
(例9)
[細胞浸透]
上記したように処理された第2のフラクションの細胞を用いて、EP1669抗体およびEP−1668抗体によるウエスタンブロット分析を用いて試験された異なる細胞区画(すなわち、膜、細胞質)のタンパク質抽出物を調製し、これにより、ターゲティング信号ペプチドによりこれらが上記細胞中へ(詳細には上記細胞区画中へ)転位置していることを示す。
【0105】
(例10)
[細胞内レドックス状態上で異なった挙動を示すSOD化合物]
制御および処理された細胞(5×105個)を回収し、スーパーオキシドアニオン、過酸化水素およびGSHそれぞれの検出のために、ハイドロエチジン(HE;分子プローブ)、ジヒドロローダミン123(DHR123;分子プローブ)または5−クロロメチル−2’、7’−ジクロロ−ジヒドロフルオレセインジアセタート(CM−H2DCFDA、分子プローブ)で、495μlのハンクス平衡塩溶液(pH7.4)中でインキュベートする。37℃15分間後、上述したような血球計算器上でサンプルを分析する。
【0106】
(例11)
[細胞活性]
SODはその高分子量に起因して細胞膜に浸透しなかったと長く考えられてきたが、24時間後に外因的に付加された有意な量の無担体のCuZnSODが細胞膜をマーク付けすることができたことを実証する試みはわずかしか行われていない(Edeas MAら、Cell Mol Biol.1996、vol.42、p1137)。
【0107】
それにもかかわらず、細胞外的に付加された時に上記SOD酵素によって細胞内ROSの解毒をより効率的に行うために、細胞に浸透可能な組換えSODタンパク質が生成されている。ヒトCuZn−スーパーオキシドジスムターゼ(CuZn−SOD)遺伝子を、HIV−1の9アミノ酸Tatタンパク質伝達ドメイン(RKKRRQRRR)をエンコードする遺伝子フラグメントと融合した。このTat−SOD融合タンパク質は、外因的に培地中に付加された際に時間(10〜60分)内に用量依存的(0.2〜2μM)にHeLa細胞中に進入することが分かった。一旦上記細胞中に入ると、伝達したTat−SODタンパク質は酵素的に活性であり、かつ、24時間安定であった。パラコートで処理されたHeLa細胞(細胞内スーパーオキシドアニオン発生装置)の細胞バイアビリティーは、伝達したTat−SODによって増加する(Kwon HYら、FEBS Lett.2000、vol485、p163)。しかし、この場合、HIV−1Tat伝達タンパク質が多様な細胞型におけるMnSOD発現の低減の原因であることが分かっことは、注目すべきである(MareckiJCら、2004 Free Radic Biol Med、vol37、p869/PorntadavitySら、DNAcell Biol、2005、vol24、p299)。
【0108】
上記ヒトCuZn〜スーパーオキシドジスムターゼ(CuZn−SOD)遺伝子をリジンに富んだペプチド(KKKKKKKKK)の9アミノ酸をエンコードする遺伝子フラグメントと融合させた場合、9Lys−SODの伝達有効性は、Tat−SODよりも高効率であった(Park Jら、Mol Cells、2002、vol13、p202)。
【0109】
多様なグリアジン鎖由来のタンパク質ペプチドは、a)その相対的消化性(タンパク質抵抗)、b)その相対的サイズまたは配列(高プロリン領域、c)細胞培養物中でのその相対的濃度(凝集またはアポトーシス)、d)HLAクラスII分子によるその相対的認識(HLA DQ2またはDR3)、e)ゾヌリン誘導のその相対的可能性に応じて、腸管細胞の有意な修飾および/または変更を生成する能力を有すると認識されている。
【0110】
少なくとも1つ(ただし、これに1つに限定されない)特定のグリアジン由来ペプチド(19〜20アミノ酸残留物−LGQQQQPFPPQQPYPQPQPF−)の場合、HLA DR3分子と直面すると、HLADQ2分子液滴に効率的に結合可能であることが分かっている(Shidrawi.RGら、1998、Clin Exp Immunol vol11、p158)。グリアジン鎖から単離された他の多くのペプチドが、HLA DQ2またはDR3に対して同様の能力を有する(Qiao.SWら、2004、J.Immunol、vol173、p1757)が、正常クラスII HLAとのグリアジンペプチド相互作用については、ほとんど知られていない。
【0111】
グリアジンが腸管バリアを通過することを可能にする早期工程は、先ずラット上皮腸管細胞IEC−6上で確立される。グリアジンによるIEC−6細胞のインキュベーションにより、細胞内アクチンフィラメントの再構築が起こるが、この再構築は、グリアジンインキュベーションのわずか15分後に視認可能となり、60分後にピークとなり、2.5時間後にベースライン値に戻った。+4℃の温度ではグリアジンによって誘発されたFアクチン変化を抑制できず、その結果、アクチン重合がグリアジンエンドサイトーシスと関連するという可能性が除外される(KwiatkowskaKら、Bioassays、1999、vol21p422)。
【0112】
グリアジンは、腸細胞からのゾヌリン放出を含む別の経路を用い得る。ゾヌリン生成は、グリアジン濃度に依存し、少なくとも0.1mg/mlのグリアジンでのインキュベーション後15分経過後に早くも細胞培養物上清中で検出可能であった。その後、30分においてピークに達し、60分後にベースラインに戻った。Ussingチャンバ中に取り付けられたウサギ腸管粘膜にグリアジンを付加すると、Rt(抵抗力)が低減し、この抵抗力低減は、インキュベーション30分経過後に有意となった。他の著者は、グリアジンペプチド誘導体ゾヌリン放出および腸管浸透性の同時増加の強力な誘導物質であると推測している(ClementeMGら、Gut.2003、vol52、p218)。
【0113】
腸管上皮とのグリアジン相互作用によりゾヌリン放出を通じて腸管浸透性が上昇し、その結果、グリアジンの傍細胞転位およびその腸管粘膜下層内でのマクロファージとの後続相互作用が可能となり、その結果、炎症性反応およびフリーラジカル生成が発生する(ThomasKEら、2006、J Immunol、vol176、p2512)ことが後に実証された。
【0114】
上述したような組換えメロンCuZnスーパーオキシドジスムターゼ遺伝子が、Aグリアジンの10アミノ酸ペプチドをエンコードする遺伝子フラグメントと融合された(QQPYPQPQPF、SEQ.IDN°01、以下ペプチド982と示す)。Gli−SOD融合タンパク質が発現し、エシェリキアコリ中で精製された。
【0115】
[バクテリアクローン培養物からのタグ付けされた組換えキューカミスメロタンパク質(SODおよびGli−SOD)の生成および精製]
バクテリア発現システムpET30aにより、クローンされたcDNAのハイブリドタンパク質としての発現が可能となった。このハイブリドタンパク質において、組換えメロンSODまたはGli−SOD(QQPYPQPQPF−SOD)は、Rosetta(DE3)pLysSバクテリア株(Novagen)中での発現後、切断可能なN末端His−Tag配列(ポリペプチド−His6−エンテロキナーゼ開裂配列−5kDa)と融合された。
【0116】
[発酵過程および誘導過程]
吸光度A600が1単位の値に達するまで、50μg/mlのカナマイシンおよび35μg/mlのクロラムフェニコールが補充された2リットルのYES培地中で37℃で発酵を行う。その後、IPTGを誘発タンパク質発現に最終濃度0.5mMまで付加した。一晩成長させた後、遠心分離によって細胞を収集して、高圧において細胞切断を実施した(French Press、2サイクル:P>800バール)。
【0117】
[変性工程]
その後、尿素の連続的付加によりバルクを変性させ、その結果、トリス20mMを有する8Mの緩衝液の最終濃度がpH8.0となる。完全溶解後、室温における完全なタンパク質可溶化にβメルカプトエタノールを付加(最終濃度10mM)する。17000gにおける遠心分離により15分間行い、可溶性タンパク質を回収した。
【0118】
[精製工程]
金属固定化アフィニティークロマトグラフィー(NiCl20.25M、Ni2+イオンで活性化されたキレート化セファロース)によるアフィニティー精製により、組換えタグ付けされたタンパク質を単離し、先ず平衡緩衝液中のカラム中にロードした:トリス20mM、尿素8M、βメルカプトエタノール10mM(pH8.0)。
【0119】
[インカラムリフォールディング工程]
固定化組換えタンパク質を大量に洗浄し、尿素量増加により部分的にリフォールディングし、カラム尿素8M〜1.5Mの勾配(緩衝液中、トリス20mM、βメルカプトエタノール10mM、pH8.0)に付加した。
【0120】
その抗酸化物質活性を取り出すため、スーパーオキシドジスムターゼは、再生工程時、その天然補助因子を回復しなければならない。これは、NaAc50mM、尿素1.5M、CuCl20.1mM、ZnCl20.1mMおよびβメルカプトエタノール10mM(pH5.0)からなる緩衝液中のCu/Znの連続的固定により、行われる。
【0121】
組換えSODフォールディングの安定化は、トリス20mM、尿素1.5M、CuCl20.1mM、ZnCl20.1mM、GSH1mM、GSSG4mMおよびpH8.0からなる緩衝液中のシステインアミノ酸間の分子内ジスルフィド架橋の形成により、有利に行われる。
【0122】
[溶出工程]
トリス20mM、尿素1.5M、イミダゾール0.5MおよびpH8.0からなる緩衝液により、上記親和性カラムからの濃縮されたタグ付けされた組換えタンパク質の溶出を得る。その後、これらをトリス20mM、CuCl20.1mM、ZnCl20.1mMおよびpH8.0中で一晩透析した後、0.22μmフィルタによって最終濾過を行って、消化時の微生物成長を回避する。タンパク質内容物推定は、μBCA方法による上清中で評価した。
【0123】
[タグ開裂]
組換えタンパク質をハイブリドタンパク質として発現させる。このハイブリドタンパク質中、SODまたはGli−SODがそのN末端部分において少量の切断可能なリンカーを介してHis−Tag配列(His−Tag)を含む少量のタンパク質に融合している。
【0124】
この開裂は、2.0%エンテロキナーゼでのインキュベーションを室温において44時間行うことにより、達成される。
【0125】
[フォールディングされかつ無菌の組換えSODの精製]
放出後、最終精製工程により、トリス−HCl20mM、pH8.0緩衝液で平衡化されかつトリス−HCl20mM、NaCl0.5M、pH8.0の溶出緩衝液で溶出されたMonoQカラム上で、少量のタギングタンパク質を組換えSODから除去した。
【0126】
消化時の微生物成長を回避するための0.22μmでの濾過を行った。タンパク質内容物推定は、μBCA方法により上清中で評価した。
【0127】
上記精製から得られた結果を図5中に示す。
【0128】
[組換えリフォールディングされたキューカミスSODまたはGli−SODの抗酸化物質活動]
多工程変性/再生プロトコルに従って、キューカミス組換えSODおよびグリアジンペプチド(Gli−SOD)を支持するその等価物を生成および精製した。SDS−PAGE分析により、有意な程度の精製が明らかとなり、その抗酸化物質活動の回復に基づいて、上記リフォールディング過程の有効性を評価する。組換えタンパク質(キューカミスメロSODおよびGli−SOD)双方のNATIVE−PAGE電気泳動上でのNBTを低減させる可能性(NBT光還元、BeauchampsおよびFridovitch、1971)を評価し、天然キューカミスメロ抽出物の可能性と比較した。
【0129】
NATIVE−PAGE電気泳動上で得られた遊走プロファイルおよびNBT低減は、組換えタンパク質、SODまたはGli−SOD(4000〜10000UNBT/mgタンパク質)双方の正確かつ高効率のリフォールディングを裏付けており、これは、その抗酸化物質活動を示すために必須のイベントである。
【0130】
ここから得られた結果を図6中に示す。
【0131】
[メロン抗recSOD抗体]
上記組換えメロンSODのアミノ酸配列を得た後、Swiss Plotアライメントを用いて他の植物SODとの比較分析を行って、2つの相対的に保存されたペプチド配列14−27(EP1668−GVVTLTQEDDGPTS)および117−131(EP1669−HELADDLGKGGHELS)を選択した。これらのペプチド双方を合成し、ウサギの免疫化のための適切なキャリア(KLH)と結合させて、植物特異ポリクローナル抗体を得た(Eurogentec、ベルギー)。抗体EP1669を免疫吸着によって親和性カラム上で精製し、ウエスタンブロット分析中で1/2000で用いて、天然およびメロン組換えSODならびに精製ウシSODに対するこの抗体の特異性を確認した。
【0132】
SDS−PAGE分析のため、これらのタンパク質を95℃まで5分加熱した後、電気泳動をSDS−PAGEポリアクリルアミドゲル上で12%で行った。分離されたタンパク質を免疫ブロットPVDF膜(BIORAD)上に移行させた。抗体の任意のポテンシャルクロス特異性が、増幅Opti4CNヤギ抗ウサギ検出キット(BIORAD)によって明らかとなった。
【0133】
EP1669抗体は、上記天然メロン抽出物中に含まれる2個のSODとのクロス特異性そして精製ウシSODとのクロス特異性を示した。
【0134】
さらに、EP1669抗体は、上記免疫化ペプチドの由来元である完全な組換えメロンSODタンパク質との良好な特異性を示した。よって、抗体は、上記天然抽出物中に存在する反応性SODのうちの少なくとも1つとの相同性だけでなく、多様な種間のSOD科内に存在し得る相同性(完全な相同性)も示すことが確認された。この抗体は、本発明の組換えSODタンパク質の薬理活動の検証に必要な検出実験において有用なツールである。
【0135】
上記抗体結合試験の結果を図7に示す。
【0136】
[細胞培養物]
ヒト大腸癌細胞株HT−29(ATCC HTB−38)を、GlutaMAX−1、4.5g/L Dグルコース、ピルビン酸ナトリウムおよび100単位/mlのペニシリン/ストレプトマイシンが補充されたダルベッコ変法イーグル培地(D−MEM)中の10%胎児ウシ血清(FBS)中で、成長させる。各実験(n=4)において、およそ2×104個の細胞/ウェルを5%CO2雰囲気中で37℃で24時間インキュベートした。任意の実験の前に、既存の培地を新規培地と交換した。上記インキュベーション期間後、0.05%トリプシンおよび0.5mM EDTAを含むPBS溶液で細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、遠心分離によって回収した。その後、GUAVA PC Viacount Flexキットを用いて血球計算器中で細胞をカウントし、細胞数/mlで報告して、細胞増殖を推定した。
【0137】
[グリアジンペプチドの無毒濃度の効果]
グリアジン治療は、濃度に依存する様式で、細胞成長に多様な効果を及ぼすことが以前から分かっている。50〜200μg/mlの濃度は細胞成長を抑制する一方、500μg/mlまでの濃度は深刻な細胞破壊の原因となり得る。グリアジンペプチドは、先ずヒト腸管細胞の酸化バランスを変更することにより、その効果を開始し得ることが知られている(Dolfiniら、2002、Toxicol in Vitro、vol16、p331)。
【0138】
現在の実験において、72時間にわたって培養物を定期的にサンプリングして、多様な成分への暴露を行った場合および行わなかった場合のHT−29細胞の増殖率およびバイアビリティーを評価した。(a)10μMのグリアジンペプチド982(小麦胚芽油、QQPYPOPQPF.SEQ.IDN°01)は、有意な腸細胞凝集(De Vicenzi.Mら、1997、Toxicology、vol120、p207)または腸細胞アポトーシス(Giovanniniら、2000、Toxicology、vol145、p63)を誘発するように事前に関連付けられた濃度よりも少なくとも100倍小さい。
【0139】
我々が試験したこのような低い濃度において、グリアジンペプチドがHT−29細胞株増殖に主な効果を発揮する能力を持つが、この能力を持つのは、培養物の24時間においてのみである。この効果は、48時間後に急速に消え去る。なぜならば、この48時間経過後において、増殖がその基底レベルに戻り、おそらくはペプチド内部移行に起因してペプチドの存在から完全に独立するように見えるからである。グリアジンペプチドは、濃度閾値を越えかつ/またはヒト腸管細胞培養物上の十分な濃度を維持して、凝集またはさらなるアポトーシスをもたらさなければならないように見える。これは、高濃度のグリアジンペプチドで腸管細胞を24時間暴露した後、有意なO20−およびH2O2生成の増加および抗酸化物質酵素の低減が観察された実験によって裏付けられ、上記グリアジンペプチド誘発アポトーシスに先行する(Giovannini.Cら、2003、FEBS Letters、vol540、p117)。
【0140】
上記した試験の結果を図8の図解中に示す。
【0141】
[組換えSODの最小活性用量]
組換えタンパク質用量反応実験のため、組換えSODまたはGli−SOD(0〜50NBT単位)の濃度をさらに48時間増加させて、HT−29細胞をインキュベートした。
【0142】
この実験において、2つの異なる平行な挙動について、低用量または高用量を付加して、48時間でのHT−29成長における組換えSODおよびGli−SOD双方について観察した。
【0143】
HT−29細胞株の成長は、10単位用量の組換えSODまたは5単位用量の組換えGli−SODのいずれかにより変化するが、これらの値未満の値の場合、HT−29の成長は若干減速する。組換えSODおよび組換えGli−SODの用量を増加させると、HT−29成長誘導のその相対レベルは同一レベルに留まり、これは、どんなに高用量であっても、誘導閾値が達成可能であることを示している。
【0144】
これらの結果に従って、最適な用量が、組換えGli−SODの場合に5〜10単位および組換えSODの場合に10〜25単位で区切られ得る。
【0145】
上記実験の結果を図9に示す。
【0146】
[本発明の組換えSODは、「ペプチド効果」の補正を操作する]
培養物を定期的に72時間サンプリングして、多様な成分への暴露を行ったかまたは行わなかった以下の場合において、HT−29細胞の増殖率およびバイアビリティーを評価した。(a)10単位の組換えSODまたは(b)10μMのペプチド982と結合された10単位の組換えSODまたは最後に(c)10単位の組換えGli−SOD(グリアジンペプチド982に物理的にリンクしたSOD)
【0147】
この濃度において、SODまたはGli−SODのいずれも任意の有毒効果を発揮しない。
【0148】
なぜならば、これらの増殖曲線は厳密に基底曲線に適合しかつ平行に構成されているからである。さらに、組換えGli−SODのN末端部分におけるペプチド982の存在にもかかわらず、期待されたような「ペプチド効果」は24時間の培養物において発生しない。我々は、グリアジンペプチドが組換えSOD(Gli−SOD)と関連付けられるかまたは結合し、その結果、グリアジンペプチドによって仲介されるフリーラジカル生成および/または抗酸化物質酵素の障害の一時的増加と相互作用することができる場合、上記にて証明されたグリアジンペプチドのHT−29増殖に対する効果が、24時間の培養物において存在しないことに気がついた(RivabeneRら、1999、Biochem Biophys Acat、vol1453、p152)。
【0149】
この驚くべき相乗効果は、上記グリアジンペプチドおよび上記SODの結合あるいは物理的または化学的結合があろうとなかろうと、機能性組み合わせを含む治療組成および治療的組成において極めて有用な結果を持つ。なぜならば、これは、炎症ペプチドが投与形態中に意図的に存在する場合でも、期待された炎症反応を軽減さらには解消することができることを示すからである。
【0150】
この実験の結果の概要を図10中に示す。
【0151】
[細胞アポトーシス]
2つの基礎的な経路が、アポトーシスを実行し得る。第1の経路において、デスリガンドはその受容体(Fas/FasLigand)を活性化させ、その結果、デスコンプレックスが形成される。その後、このデスコンプレックスは、カスパーゼ8を活性化させる。
【0152】
第2の経路において、細胞ストレスに起因して、細胞質中へのチトクロムCの放出によってミトコンドリア機能障害が発生し、その結果、アポトーシスタンパク質因子−1およびカスパーゼ9との活性化複合体が形成される。その後、これらの複合体は、下流効果またはカスパーゼ3、6および7を活性化させ、その結果、最終アポトーシス変化が実行される。
【0153】
コムギグリアジンからの消化されたペプチドはCD95/Fasアポトーシス経路を介して腸管上皮細胞のアポトーシスを誘発することが知られているが(GiovanniniCら、2003、FEBS Lett.Vol540、p117)、キサンチン/キサンチンオキシダーゼの混合物によって生成されたROS(活性酸素種)は、チトクロムCの細胞質中への放出と共に、カスパーゼ−3を活性化させる(HiguchiMら、1998、oncogene、vol17、p2753)。
【0154】
第1にニューロペプチドとして特徴付けられて、オレキシンだけでなくその受容体も、胃腸管(VoisinTら2000、J Pharmacol Exp Ther、vol292、p638)を含むいくつかの末梢組織中に発現することが分かった(Voisin Tら、2003、Cell Mol Life Sci、vol60、p72)。オレキシンは、ミトコンドリアおよびカスパーゼ活性化からのチトクロムC放出を通じてアポトーシスを促進することにより、24時間後にヒト大腸癌HT−29細胞の成長を全体的に抑制する能力を有する(Rouet-Benzineb Pら、2004、J Biol Chem、vol279、P46875)。
【0155】
本発明の組換えSODまたはGli−SODから期待される細胞ストレスおよびミトコンドリア機能障害と戦う特定の活性により、このような誘発性システムにおけるその強力な抗アポトーシス効率を確認するために、実験を行った。上記組換えSODまたはGli−SODのために用いられた濃度は、増殖曲線に従って規定した。基底アポトーシス過程との相互作用に十分と思われる10単位の組換えSODまたはGli−SODのみを用い、一方、オレキシンまたはキサンチン/キサンチンオキシダーゼによって(X/XO)誘発されたHT−29細胞上でのアポトーシスには、50単位の組換えSODまたはGli−SODがより適しているように見える。
【0156】
[組換えSODは、オレキシン誘発アポトーシスを抑制した]
アポトーシス細胞と非アポトーシス細胞とを区別するGUAVA Nexinキットを用いて、HT−29細胞株中においてアポトーシスを分析した。この方法では、アネキシンVフィコエリトリンを用いて、アポトーシス細胞の外膜上のphopshatidylserine(PS)を検出する(Martinら、1995、J Exp Med、vol182、p1545)。
【0157】
24時間後、トリプシンを0.05%およびEDTAを0.5mM含むPBS溶液によって細胞単層を脱離させ、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、アポトーシス細胞染色を行い、GUAVAPCAシステムで分析する。
【0158】
基底アポトーシス抑制(部分A)については、10単位の双方の組換えSOD(NBT単位)を含むかまたは含まない新規培地と培地を交換した。オレキシン誘発アポトーシス抑制(部分B)については、1μMのオレキシンおよび/または50単位の各組換えSOD(NBT単位)を含む新規培地と培地を効果した。その結果をアポトーシスアネキシンV−PE陽性細胞の百分率として表し、それは4点の平均である。
【0159】
通常条件下において、外膜PSのアネキシンVラベル付けによって示すように、アポトーシスは20%(20.5)のHT−29細胞中において同時発生した。
【0160】
この速度は、HT−29細胞の培養時において10単位の組換えGli−SODが存在した場合に10単位の組換えメロンSODが存在して16.8%まで到達した際に(18.1%抑制)、18%まで低下した(12.2%抑制)。
【0161】
第2の実験において、HT−29細胞培地へのオレキシンの補充を介してアポトーシスを誘発した際に、組換えSODまたはGli−SODの強力な抗アポトーシス効果を確認した。この場合において、HT−29細胞がアポトーシスとなる速度は、オレキシンを1μM培地中に付加することにより、基底レベルで評価した20%から40%まで向上され得る。オレキシン誘発アポトーシスの速度は、HT−29細胞の培養時において50単位の組換えGli−SODが存在した際に50単位の組換えメロンSODが存在して最終的に28.7%に達した際に(54.7%抑制)、34.4%まで低下した(26%抑制)。
【0162】
この実験の結果の概要を図12中に示す。
【0163】
[組換えSODは、X/XO誘発アポトーシスを抑制した]
腸管上皮細胞機能は、活性酸素種ROSの生成を通じて、活性マクロファージ(腸疾患)などの局所免疫細胞および炎症細胞によって構成され得る。従って、ROSは、腸管上皮細胞アポトーシスと関連するFasおよびFasL発現を誘発し得る(DenningTLら、2002、Free Radic Biol Med vol33、p1641)。
【0164】
キサンチン存在下のキサンチンオキシダーゼは、主にO2−°を生成する(これは、小腸中の活性酸素の代表であり、膜浸透性移行、カルシウム流入、脂質過酸化反応およびミトコンドリア中の膜流動性変化を誘発し得る)(AnupRら、1999;Indian J biochem Biophys、vol36、p266およびAnupRら、2000、BrJ Surg、vol87、p1094)。
【0165】
この第3の実験において、本発明による組換えSODまたはGli−SODの強力な抗アポトーシス効果の試験を、HT−29細胞培地中でのキサンチン/キサンチンオキシダーゼ誘導を介した可能なFas/FasL経路上で行った。
【0166】
この場合、HT−29細胞がアポトーシスとなる速度は、培地中に10mUのX/XO複合体を付加することにより、基底レベルについて評価した20%から30%まで向上され得る。50単位の組換えメロンSODまたはやはり50単位の組換えGli−SODを培地に付加すると、HT−29細胞中でのX/XO誘発アポトーシス経路がそれぞれ71%および86%だけ回避された。
【0167】
この実験の結果を図13のグラフに示す。
【0168】
[組換えGli−SODの結合]
各実験(n=4)において、およそ15×103個の細胞/ラメラ層を5%CO2雰囲気中で37℃で24時間インキュベートした。全実験の前に、既存の培地を新規培地と交換した。
【0169】
その後、細胞単層を新規培地中で50単位のSODまたはGli−SODを用いてまたは用いずにさらに24時間インキュベートし、カルシウムを含まずかつマグネシウムを含まないリン酸緩衝生理食塩水溶液(PBS)中で洗浄し、その後、免疫組織化学によって分析した。EP−1669を第1の抗体(ウサギlgG抗SOD、希釈1/2000°)を用いて30分間Gli−SODのHT−29細胞への結合を分析した。その後、第2のFITC結合抗体(ヤギ抗ウサギ抗体、希釈1/100)をさらに30分間分析した。
【0170】
フローサイトメトリではなくこの方法を選択した理由は、フローサイトメトリ技術の場合、トリプシン治療によって細胞を回収し、分析前に収集しなければならず、このような治療は、Gli−SOD分子と細胞表面との間の相互作用に悪影響を与える可能性があるためである。
【0171】
上記FITC蛍光実験の結果を図11中に示す。図11において、細胞近隣のSODの存在がSOD双方において明確に視認され、また、Gli−SODの場合にも、より広範囲で明確に視認される。
【0172】
上記実験は、本発明による、そのN末端において小麦胚芽油グリアジンペプチド配列を持つかまたは持たないキューカミスメロSODをエンコードする遺伝子操作されたcDNAsは、エシェリキアコリ中で効率的に発現され得、抗酸化物質活動を持つ活性の分子として生成され得ることを実証した。加えて、消化されたグリアジン−ペプチドのみが腸管細胞凝集または特定状況の細胞アポトーシスを誘発することで知られているが、本発明者らは、これらの負の効果を回避するより短いグリアジンペプチドを選択する。また、本発明のSODを選択されたグリアジンペプチドと物理的に組み合わせるかまたは単に結合させることで、グリアジンペプチドの確実な炎症効果と相互作用が可能になることも示されている。
【図面の簡単な説明】
【0173】
【図1】キューカミスメロからのCuZnスーパーオキシドジスムターゼのためのcDNAコーディングを得るために用いられるクローニング方法の模式図である。
【図2】本発明の組換えおよびキメラSOD分子を生成するために用いられる遺伝工学の一般図の模式図である。
【図3】本発明による多様なメロンCu−ZnSOD変異体のRT−PCR増幅から得られるアガロースゲル電気泳動を示す。詳細には、ランAは組換え葉緑体メロンCu−ZnSODであり、ランBは組換えメロン偽細胞質Cu−ZnSODであり、ランCは、グリアジンペプチドへのN末端融合を支持するランBの組換え偽細胞質Cu−ZnSODである。
【図4】異なる由来(詳細には、トマト細胞質SOD(線1)、ヒトSOD(線2)およびメロン葉緑体SOD(線3))からの3つの異なるCu−Znスーパーオキシドジスムターゼのアミノ酸アライメントを示す。これらの配列間のアミノ酸残留物の同一性は、円を通じて3つのSODの各群の下側に示される。
【図5】組換えSOD(レーン1〜7)または組換え融合Gli〜SOD(レーンA〜F)の精製過程のクマシーブルー染色ゲルである。図5において、 レーン1およびAは、インビトロジェンから市販されているベンチマークタンパク質ラダーとして公知の分子量マーカである。 レーン2およびBは、His−Tagの開裂を示す。 レーン3、4、5、6およびC、D、Eは、連続精製工程時における本発明による組換えSODおよび組換えGli−SODを示す。 レーン7およびFは、MonoQカラム上の治療後に残っているバンドである。
【図6】NBT還元ゲル電気泳動を用いた組換えSODまたはGli−SODそれぞれのフォールディングを示す。 Aは、125μgの天然メロン抽出物(90U/mgに等価)でロードされたレーンAおよびA’と、0.92μgの未変性SODでロードされたレーン1、1’、および1’’’を有するrecSODを示す。Bは、本発明によるrecGli−SODを示し、レーンAおよびA’は、125μgの天然メロン抽出物(90U/mgに等価)でロードされ、レーン1および1’は1.84μgの組換えGli−SODでロードされ、レーン2および2’は、0.92μgの組換えGli−SODでロードされる。
【図7】以下のSDS−PAGEである。 Aは、以下のクーマシー青色顕色を示す。 レーン1中の50μgの天然キューカミスメロ抽出物 レーン2中のタンパク質マーカ(6.6、14.1、21.1、31、45および67KDa) レーン3中の2.5μgのウシSOD(シグマ) Bは、以下のEP1669抗体顕色を示す。 レーン4中の50μgの天然キューカミスメロ抽出物 レーン5中のタンパク質マーカ(6.6、14.1、21.1、31、45、67KDa) レーン6中の2.5μgのウシSOD(シグマ)
【図8】HT29細胞株の増殖曲線上のグリアジンペプチドの効果のグラフであり、ここでN=4である。
【図9】本発明によるrecSODまたはrecGli−SODの最小有効量の図解比較であり、N=4である。
【図10】本発明によるrecSODまたはrecGli−SODのHT29細胞株の増殖曲線上への効果のグラフであり、N=4である。
【図11】EP1669抗SOD抗体を用いたSODまたはGli−SODのHT29細胞上への細胞結合を示す1組のFITC画像であり、Aは負の制御であり、白色エリアは、細胞近隣のSODの存在を示し、BはrecSODでの治療後であり、白色エリアは、細胞近隣のSODの存在を示し、CはrecGli−SODでの治療後であり、白色エリアは、細胞近隣のSODの存在を示す。
【図12】アネキシンV試験を用いた、本発明によるSODの基底(A)またはオレキシンによって誘発された(B)HT−29アポトーシスに対する効果を図解表示したものであり、N=4である。Aは、1OUを用いて得られた結果を示し、Bは、recSODおよびrecGli−SODそれぞれの50Uを用いて得られた結果である。
【図13】ヒポキサンチン/キサンチンオキシダーゼによって誘発されたHT−29アポトーシスにおける組換えSODまたはGli−SODに対する効果のグラフを示す(N=4、アネキシンV試験)。
【0174】
参考文献:
1) Arentz-Hansen EH, Production of a panel of recombinant gliadins for the characterisation of T cell reactivity in coeliac disease. Gut 2000 Jan;46(1):46-51
2) Baillet F, Treatment of radiofibrosis with liposomal superoxide dismutase. Preliminary results of 50 cases. Free Radic Res Commun 1986;l(6):387-94
3) Beauchamp C. Superoxide dismutase improved assays and an assay applicable to acrylamide gels. Anal. Biochem:1971,44(1)276-287
4) Biagi F, A non-toxic analogue of a coeliac-activating gliadin peptide: a basis for immunomodulation? Aliment Pharmacol Ther 1999 Jul;13(7):945-50
5) Beckett CG, Analysis of interleukin-4 and interleukin-10 and their association with the lymphocytic infiltrate in the small intestine of patients with coeliac disease. Gut 1996 Dec;39(96):818-23
6) Bolte G, Peptic-tryptic digests of gliadin: contaminating trypsin but not pepsin interferes with gastrointestinal protein binding characteristics. Clin Chim Acta 1996 Mar 29;247(1-2):59-70
7) Bueno P, Peroxisomal copper, zinc superoxide dismutase. Characterization of the isoenzyme from watermelon cotyledons. Plant Physiol 1995 Jul;108(3):1151-60
8) Cannon RE, Cloning of cDNA for maize superoxide dismutase 2 (SOD2).Proc Natl Acad Sci USA 1987 Jan;84(1):179-83
9) Cerutti PA. Prooxidant states and tumor promotion. Science 1985 Jan 25;227(4685):375-81
10) Chirdo FG, In vitro presentation of gliadin-derived peptides by different cell lines. Clin Chim Acta 2002 Mar;317(1-2):151-8
11) Cornell HJ, In vitro mucosal digestion of synthetic gliadin-derived peptides in celiac disease. J Protein Chem 1995 Jul;14(5):335-9
12) Corominas M, Hypersensitivity reaction after orgotein (superoxide dismutase) administration. Allergol Immunopathol (Madr) 1990 Sep-Oct;18(5):297-9
13) Delanian S, Successful treatment of radiation-induced fibrosis using liposomal Cu/Zn superoxide dismutase: clinical trial. Radiother. Oncol. 1994, 32(1):12-20.
14) Diez-Gomez ML, Anaphylaxis after intra-articular injection of orgotein. Detection of an IgE-mediated mechanism. Allergy 1987 Jan;42 (1):74-6
15) Freeman BA, Liposome-mediated augmentation of superoxide dismutase in endothelial cells prevents oxygen injury J Biol Chem 1983 Oct 25;258(20):12534-42
16) Giovannini C, Induction of apoptosis in caco-2 cells by wheat gliadin peptides. Toxicology 2000 Apr 7;145(1):63-71
17) Halliwell. B, The antioxidants of human extracellular fluids. Arch Biochem Biophys 1990 Jul;280(1):l-8
18) Hidalgo IJ, Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989 Mar;96(3):736-49
19) Inoue. M, Expression of a hybrid Cu/Zn-type superoxide dismutase which has high affinity for heparin-like proteoglycans on vascular endothelial cells. J Biol Chem 1991 Sep 5;266(25):16409-14
20) Inoue M, Targeting SOD by gene and protein engineering and inhibition of free radical injury. Free Radic Res Commun 1991;12-13 Pt 1:391-9
21) Jadot G, Anti-inflammatory activity of superoxide dismutases: studies on adjuvant induced polyarthritis in rats. Free Radic Res Commun 1986;2(1-2):27-42
22) Jadot G, Anti-inflammatory activity of superoxide dismutases: inhibition of adriamycin induced edema in rats. Free Radic Res Commun 1986;2(1-2):19-26
23) Jadot G, Anti-inflammatory activity of superoxide dismutases: inhibition of carrageenan induced edema in rats. Free Radic Res Commun 1986;l(6):395-403
24) Jadot G, Clinical pharmacokinetics and delivery of bovine superoxide dismutase. Clin Pharmacokinet 1995 Jan;28(1):17-25
25) Jadot G, Comparative anti-inflammatory activity of different superoxide dismutases and liposomal SOD in ischemia. Free Radic Res Commun 1987;3(6):389-94
26) Joral A, Adverse reactions to food in adults. : J Investig Allergol Clin Immunol 1995 Jan-Feb;5(1):47-9
27) Joral A, Systemic anaphylaxis following parenteral orgotein administration. J Investig Allergol Clin Immunol 1993 Mar-Apr;3(2):103-4
28) Kaminaka H, Molecular cloning and characterization of a cDNA for plastidic copper/zinc-superoxide dismutase in rice (Oryza sativa L). Plant Cell Physiol 1997 Jan;38(1):65-9
29) Kim EJ, Transcriptional and post-transcriptional regulation by nickel of sodN gene encoding nickel-containing superoxide dismutase from Streptomyces coelicolor Muller Mol Microbiol 1998 Jan;27(1):187-95
30) Kleber-Janke T, Use of modified BL21(DE3) Escherichia coli cells for high-level expression of recombinant peanut allergens affected by poor codon usage. Protein Expr Purif 2000 Aug;19(3):419-24
31) Komar AA , Synonymous codon substitutions affect ribosome traffic and protein folding during in vitro translation. FEBS Lett 1999 Dec 3;462(3):387-91
32) Koopman G. Annexin V for flow cytometric detection of phosphatidylserine expression on B cells undergoing apoptosis. Blood 1994, 84(5)1415-1420.
33) Kwiatowski J, Isolation and characterization of an iron-containing superoxide dismutase from tomato leaves, Lycopersicon esculentum. Eur J Biochem 1985 Jan 15;146(2):459-66
34) Lavy A, Increased susceptibility to undergo lipid peroxidation of chylomicrons and low-density lipoprotein in celiac disease. Ann Nutr Metab 1993;37(2):68-74
35) Loh EY, Polymerase chain reaction with single-sided specificity: analysis of T cell receptor delta chain. Science 1989 Jan 13;243(4888):217-20
36) Lundin KE, Gliadin-specific, HLA-DQ(alpha l*0501,beta 1*0201) restricted T cells isolated from the small intestinal mucosa of celiac disease patients. J. Exp Med 1993 Jul 1;178(1):187-96
37) Martin SJ. Early distribution of plasma membrane phosphatidylserine in a general feature of apoptosis regardless of the initiating stimulus : inhibition by overexpression of BcL2 and Abl. J. Exp. Med. 1995,182(5)1545-1556.
38) Mayrhofer G., Absorption and presentation of antigens by epithelial cells of the small intestine: hypotheses and predictions relating to the pathogenesis of coeliac disease. Immunol Cell Biol 1995 Oct;73(5):433-9
39) Mayrhofer G, Subcellular distribution of class II major histocompatibility antigens in enterocytes of the human and rat small intestine. Immunol Cell Biol 1989 Aug;67(Pt4):251-60
40) Michelson AM, Anti-inflammatory activity of superoxide dismutases: comparison of enzymes from different sources in different models in rats: mechanism of action. Free Radic Res Commun 1986;2(1-2):43-56
41) Palma JM, Purification and properties of cytosolic copper, zinc superoxide dismutase from watermelon (Citrullus vulgaris Schrad.) cotyledons. Free Radic Res 1997 Jan;26(1):83-91
42) Pathania V, Geriforte stimulates antioxidant defense system. Indian J Exp Biol 1998 Apr;36(4):414-7
43) Perdereau B, [Superoxide dismutase (Cu/Zn) in cutaneous application in the treatment of radiation-induced fibrosis] Bull Cancer 1994 Aug;81(8):659-69
44) Regnault C, Pharmacokinetics of superoxide dismutase in rats after oral administration. Biopharm Drug Dispos 1996 Mar;17(2):165-74
45) Regnault C, Effect of encapsulation on the anti-inflammatory properties of superoxide dismutase after oral administration. Clin Chim Acta 1995 Sep 15;240(2):117-27
46) Rivabene R, In vitro cytotoxic effect of wheat gliadin-derived peptides on the Caco-2 intestinal cell line is associated with intracellular oxidative imbalance: implications for coeliac disease. Biochim Biophys Acta 1999 Jan 6;1453(1):152-60
47) Scioli JR, Cloning and characterization of a cDNA encoding the chloroplastic copper/zinc-superoxide dismutase from pea. Proc Natl Acad Sci USA 1988 Oct;85(20):7661-5
48) Takeda Y, Albumin-binding superoxide dismutase with a prolonged half-life reduces reperfusion brain injury. Am J Physiol 1993 May;264(5Pt2):H1708-15
49) Tatham AS, Conformational studies of peptides corresponding to the coeliac-activating regions of wheat alpha-gliadin. Biochem J 1990 Sep l;270(2):313-8
50) Turrens JF , Protection against oxygen toxicity by intravenous injection of liposome-entrapped catalase and superoxide dismutase. J Clin Invest 1984 Jan;73(1):87-95
51) Vaille A, Anti-inflammatory activity of various superoxide dismutases on polyarthritis in the Lewis rat. Biochem Pharmacol 1990 Jan 15;39(2):247-55
52) Varenne S, The maximum rate of gene expression is dependent on the downstream context of unfavourable codons. Biochimie 1989 Nov-Dec;71(1l-12):1221-9
53) Vouldoukis I, Fc-receptor-mediated intracellular delivery of Cu/Zn- superoxide dismutase (SOD1) protects against redox-induced apoptosis through a nitric oxide dependent mechanism. Mol Med 2000 Dec;6(12):1042-53
54) Yoo HY, Heavy metal-mediated activation of the rat Cu/Zn superoxide dismutase gene via a metal-responsive element. Mol Gen Genet 1999 Sep;262(2):310-3
55) Zidenberg-Cherr S, Dietary superoxide dismutase does not affect tissue levels. Am J Clin Nutr 1983 Jan;37(1):5-7
【特許請求の範囲】
【請求項1】
医薬投与用に適合された医薬組成であって、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントの機能的に活性な組み合わせを含む、医薬組成。
【請求項2】
医薬的に受容可能な賦形剤も含む、請求項1に記載の医薬組成。
【請求項3】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンまたは誘導体、類似体、塩またはその代謝体のフラグメントである、請求項1に記載の医薬組成。
【請求項4】
前記プロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項5】
前記プロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項6】
前記少なくとも1つのスーパーオキシドジスムターゼは、CuZnスーパーオキシドジスムターゼ、Mnスーパーオキシドジスムターゼ、細胞外スーパーオキシドジスムターゼ、NiスーパーオキシドジスムターゼおよびFeスーパーオキシドジスムターゼからなる群から選択される、請求項1に記載の医薬組成。
【請求項7】
前記少なくとも1つのスーパーオキシドジスムターゼは相同スーパーオキシドジスムターゼである、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項8】
前記少なくとも1つのスーパーオキシドジスムターゼは異種スーパーオキシドジスムターゼである、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項9】
前記少なくとも1つのスーパーオキシドジスムターゼは異種CuZnスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項10】
前記少なくとも1つのスーパーオキシドジスムターゼは、雑種異種/相同スーパーオキシドジスムターゼであり、好適には雑種植物/ヒトスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項11】
前記少なくとも1つのスーパーオキシドジスムターゼは、ヒトスーパーオキシドジスムターゼ、動物スーパーオキシドジスムターゼ、バクテリアスーパーオキシドジスムターゼ、イーストスーパーオキシドジスムターゼおよび植物スーパーオキシドジスムターゼからなる群から選択される、請求項1に記載の医薬組成。
【請求項12】
前記少なくとも1つのスーパーオキシドジスムターゼは植物スーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項13】
前記少なくとも1つのスーパーオキシドジスムターゼは異種CuZn植物スーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項14】
前記少なくとも1つのスーパーオキシドジスムターゼは植物から抽出される、請求項1に記載の医薬組成。
【請求項15】
前記少なくとも1つのスーパーオキシドジスムターゼは、ウリ科に属する植物から抽出され、好適にはメロンから抽出され、ナス科に属する植物から抽出され、トマトから抽出される、請求項1に記載の医薬組成。
【請求項16】
前記少なくとも1つのスーパーオキシドジスムターゼは、ペルオキソームスーパーオキシドジスムターゼ、葉緑体スーパーオキシドジスムターゼおよび細胞質スーパーオキシドジスムターゼからなる植物スーパーオキシドジスムターゼの群から選択される、請求項1に記載の医薬組成。
【請求項17】
前記少なくとも1つのスーパーオキシドジスムターゼは組換えスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項18】
前記少なくとも1つのスーパーオキシドジスムターゼは、修飾葉緑体またはペルオキソームCuZn組換えスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項19】
前記修飾葉緑体またはペルオキソームスーパーオキシドジスムターゼは、遺伝子操作された核酸配列によってコードされ、前記核酸配列によって転換された宿主細胞形質中で発現される、請求項18に記載の医薬組成。
【請求項20】
前記少なくとも1つのスーパーオキシドジスムターゼは組換えスーパーオキシドジスムターゼであり、前記組換えスーパーオキシドジスムターゼは、
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意の1つによる核酸配列と、
厳密な条件下でSEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意のものをハイブリダイジングする核酸と、
SEQ.ID24〜SEQ.ID33として特定される配列のうちの任意の1つと少なくとも70%のBLAST同一相同性を有する核酸と、
によってコードされる、
請求項1に記載の医薬組成。
【請求項21】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、完全に加水分解されたプロラミン系ペプチドフラグメント、実質的に加水分解されたプロラミン系ペプチドフラグメントまたは若干加水分解されたプロラミン系ペプチドフラグメントからなる群から選択される、請求項1に記載の医薬組成。
【請求項22】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記胃腸加水分解過程を模倣するPTC(パンクレアチン、トリプシン、キモトリプシン)加水分解されたプロラミンから得られたフラグメントからなる群から選択される、請求項1に記載の医薬組成。
【請求項23】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記胃腸管のターゲティング信号として機能する範囲まで加水分解される、請求項1に記載の医薬組成。
【請求項24】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項25】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項26】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記アミノ酸配列Nter−QQPYPQPQPF−Cterを有する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項27】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記アミノ酸配列Nter−QQPQDAVQPF−Cterを有する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項28】
前記少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントは、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される単一ペプチドを形成する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項29】
組換えスーパーオキシドジスムターゼであって、
SEQ.IDN°24〜SEQ.IDN°33として特定される核酸配列のうちの任意の1つによる核酸配列と、
厳密な条件下でSEQ.IDN°24〜SEQ.IDN°33として特定される核酸配列のうちの任意にハイブリダイジングする核酸と、
SEQ.IDN°24〜SEQ.IDN°33として特定される配列のうちの任意の1つに対して少なくとも70%BLAST同一相同性を有する核酸と、
のうちの1つによってコードされる、組換えスーパーオキシドジスムターゼ。
【請求項30】
そのN末端においてリンカーアミノ酸配列に融合する植物スーパーオキシドジスムターゼからのアミノ酸配列を含むスーパーオキシドジスムターゼ活性を有するキメラ組換え分子。
【請求項31】
前記リンカーアミノ配列は、前記アミノ酸メチオニンおよびバリンを含む、請求項30に記載のキメラ組換え分子。
【請求項32】
前記キメラ組換え分子は、前記リンカーアミノ酸配列の前記N末端に融合されたプロラミン系ペプチドフラグメントをさらに含む、請求項30または31に記載のキメラ組換え分子。
【請求項33】
前記分子は、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される、請求項30〜32うちのいずれかの請求項に記載のキメラ組換え分子。
【請求項34】
前記組換えスーパーオキシドジスムターゼは、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される、請求項30に記載のキメラ組換え分子。
【請求項35】
SEQ.IDN°24〜SEQ.IDN°33によって特定される核酸配列からなる群から選択される単離核酸。
【請求項36】
請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの薬剤としての使用。
【請求項37】
請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの抗細胞ストレス剤としての使用。
【請求項38】
請求項1〜28のうちの任意の1つに記載の医薬組成、あるいは請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの炎症病理薬の製造への使用。
【請求項39】
SEQ.IDN°44またはSEQ.IDN°45の数字下で特定される配列の群から対応して選択される抗組換えスーパーオキシドジスムターゼ抗体。
【請求項1】
医薬投与用に適合された医薬組成であって、少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントの機能的に活性な組み合わせを含む、医薬組成。
【請求項2】
医薬的に受容可能な賦形剤も含む、請求項1に記載の医薬組成。
【請求項3】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンまたは誘導体、類似体、塩またはその代謝体のフラグメントである、請求項1に記載の医薬組成。
【請求項4】
前記プロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項5】
前記プロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項6】
前記少なくとも1つのスーパーオキシドジスムターゼは、CuZnスーパーオキシドジスムターゼ、Mnスーパーオキシドジスムターゼ、細胞外スーパーオキシドジスムターゼ、NiスーパーオキシドジスムターゼおよびFeスーパーオキシドジスムターゼからなる群から選択される、請求項1に記載の医薬組成。
【請求項7】
前記少なくとも1つのスーパーオキシドジスムターゼは相同スーパーオキシドジスムターゼである、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項8】
前記少なくとも1つのスーパーオキシドジスムターゼは異種スーパーオキシドジスムターゼである、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項9】
前記少なくとも1つのスーパーオキシドジスムターゼは異種CuZnスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項10】
前記少なくとも1つのスーパーオキシドジスムターゼは、雑種異種/相同スーパーオキシドジスムターゼであり、好適には雑種植物/ヒトスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項11】
前記少なくとも1つのスーパーオキシドジスムターゼは、ヒトスーパーオキシドジスムターゼ、動物スーパーオキシドジスムターゼ、バクテリアスーパーオキシドジスムターゼ、イーストスーパーオキシドジスムターゼおよび植物スーパーオキシドジスムターゼからなる群から選択される、請求項1に記載の医薬組成。
【請求項12】
前記少なくとも1つのスーパーオキシドジスムターゼは植物スーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項13】
前記少なくとも1つのスーパーオキシドジスムターゼは異種CuZn植物スーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項14】
前記少なくとも1つのスーパーオキシドジスムターゼは植物から抽出される、請求項1に記載の医薬組成。
【請求項15】
前記少なくとも1つのスーパーオキシドジスムターゼは、ウリ科に属する植物から抽出され、好適にはメロンから抽出され、ナス科に属する植物から抽出され、トマトから抽出される、請求項1に記載の医薬組成。
【請求項16】
前記少なくとも1つのスーパーオキシドジスムターゼは、ペルオキソームスーパーオキシドジスムターゼ、葉緑体スーパーオキシドジスムターゼおよび細胞質スーパーオキシドジスムターゼからなる植物スーパーオキシドジスムターゼの群から選択される、請求項1に記載の医薬組成。
【請求項17】
前記少なくとも1つのスーパーオキシドジスムターゼは組換えスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項18】
前記少なくとも1つのスーパーオキシドジスムターゼは、修飾葉緑体またはペルオキソームCuZn組換えスーパーオキシドジスムターゼである、請求項1に記載の医薬組成。
【請求項19】
前記修飾葉緑体またはペルオキソームスーパーオキシドジスムターゼは、遺伝子操作された核酸配列によってコードされ、前記核酸配列によって転換された宿主細胞形質中で発現される、請求項18に記載の医薬組成。
【請求項20】
前記少なくとも1つのスーパーオキシドジスムターゼは組換えスーパーオキシドジスムターゼであり、前記組換えスーパーオキシドジスムターゼは、
SEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意の1つによる核酸配列と、
厳密な条件下でSEQ.ID24〜SEQ.ID33として特定される核酸配列のうちの任意のものをハイブリダイジングする核酸と、
SEQ.ID24〜SEQ.ID33として特定される配列のうちの任意の1つと少なくとも70%のBLAST同一相同性を有する核酸と、
によってコードされる、
請求項1に記載の医薬組成。
【請求項21】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、完全に加水分解されたプロラミン系ペプチドフラグメント、実質的に加水分解されたプロラミン系ペプチドフラグメントまたは若干加水分解されたプロラミン系ペプチドフラグメントからなる群から選択される、請求項1に記載の医薬組成。
【請求項22】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記胃腸加水分解過程を模倣するPTC(パンクレアチン、トリプシン、キモトリプシン)加水分解されたプロラミンから得られたフラグメントからなる群から選択される、請求項1に記載の医薬組成。
【請求項23】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記胃腸管のターゲティング信号として機能する範囲まで加水分解される、請求項1に記載の医薬組成。
【請求項24】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、グリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項25】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、免疫原性プロラミン系ペプチドに対して競合的抑制活性を有するグリアジンの非免疫原性類似体である、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項26】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記アミノ酸配列Nter−QQPYPQPQPF−Cterを有する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項27】
前記少なくとも1つのプロラミン系ペプチドフラグメントは、前記アミノ酸配列Nter−QQPQDAVQPF−Cterを有する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項28】
前記少なくとも1つのスーパーオキシドジスムターゼおよび少なくとも1つのプロラミン系ペプチドフラグメントは、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される単一ペプチドを形成する、前記請求項のうち任意の請求項に記載の医薬組成。
【請求項29】
組換えスーパーオキシドジスムターゼであって、
SEQ.IDN°24〜SEQ.IDN°33として特定される核酸配列のうちの任意の1つによる核酸配列と、
厳密な条件下でSEQ.IDN°24〜SEQ.IDN°33として特定される核酸配列のうちの任意にハイブリダイジングする核酸と、
SEQ.IDN°24〜SEQ.IDN°33として特定される配列のうちの任意の1つに対して少なくとも70%BLAST同一相同性を有する核酸と、
のうちの1つによってコードされる、組換えスーパーオキシドジスムターゼ。
【請求項30】
そのN末端においてリンカーアミノ酸配列に融合する植物スーパーオキシドジスムターゼからのアミノ酸配列を含むスーパーオキシドジスムターゼ活性を有するキメラ組換え分子。
【請求項31】
前記リンカーアミノ配列は、前記アミノ酸メチオニンおよびバリンを含む、請求項30に記載のキメラ組換え分子。
【請求項32】
前記キメラ組換え分子は、前記リンカーアミノ酸配列の前記N末端に融合されたプロラミン系ペプチドフラグメントをさらに含む、請求項30または31に記載のキメラ組換え分子。
【請求項33】
前記分子は、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される、請求項30〜32うちのいずれかの請求項に記載のキメラ組換え分子。
【請求項34】
前記組換えスーパーオキシドジスムターゼは、SEQ.IDN°34、SEQ.IDN°35、SEQ.IDN°36、SEQ.IDN°37、SEQ.IDN°38、SEQ.IDN°39、SEQ.IDN°40、SEQ.IDN°41、SEQ.IDN°42およびSEQ.IDN°43からなるアミノ酸配列の群から選択される、請求項30に記載のキメラ組換え分子。
【請求項35】
SEQ.IDN°24〜SEQ.IDN°33によって特定される核酸配列からなる群から選択される単離核酸。
【請求項36】
請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの薬剤としての使用。
【請求項37】
請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの抗細胞ストレス剤としての使用。
【請求項38】
請求項1〜28のうちの任意の1つに記載の医薬組成、あるいは請求項29〜34のうちの任意の1つに記載の組換えスーパーオキシドジスムターゼの炎症病理薬の製造への使用。
【請求項39】
SEQ.IDN°44またはSEQ.IDN°45の数字下で特定される配列の群から対応して選択される抗組換えスーパーオキシドジスムターゼ抗体。
【図1】

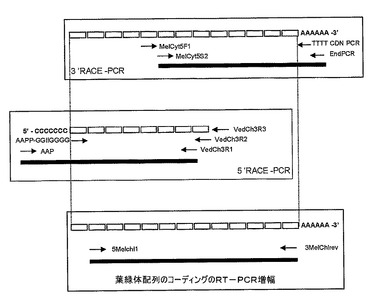
【図2】
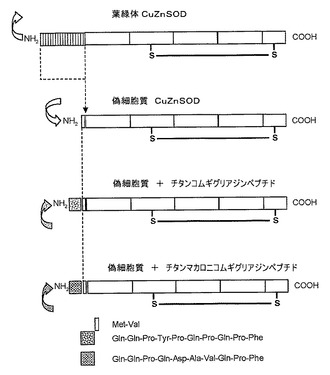
【図3】
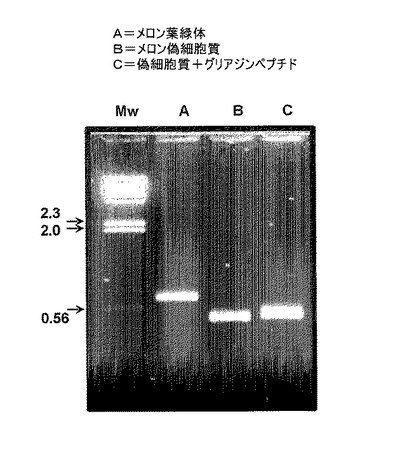
【図4】

【図5】
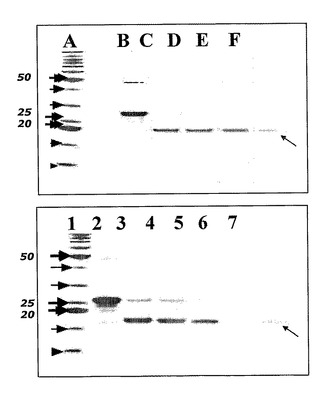
【図6】
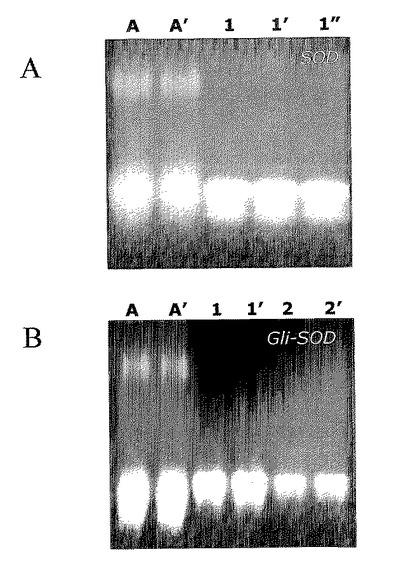
【図7】
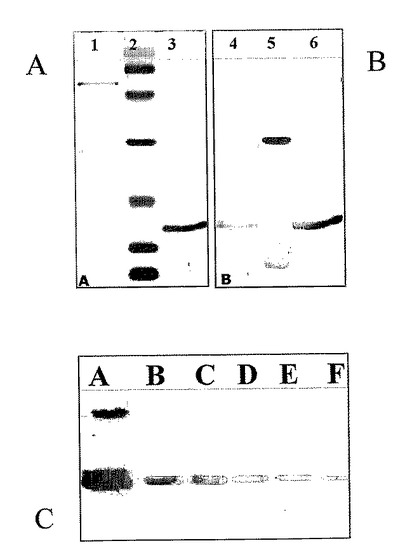
【図8】
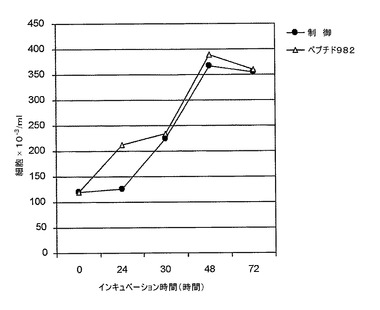
【図9】
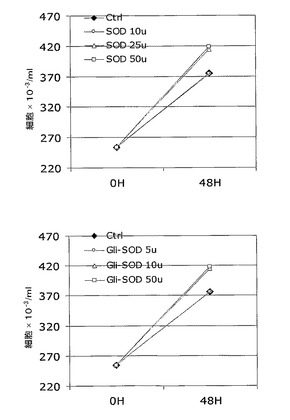
【図10】
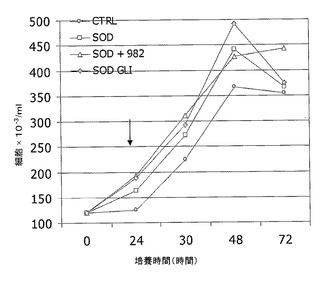
【図11】
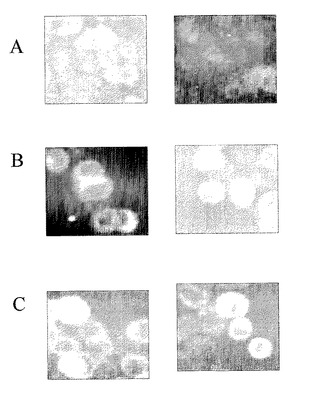
【図12】
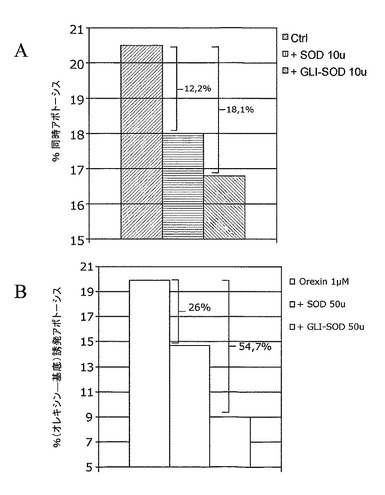
【図13】
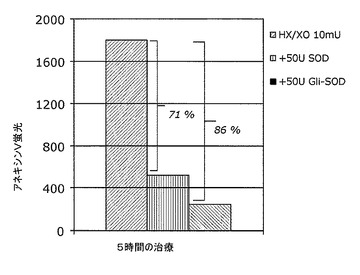
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2009−531311(P2009−531311A)
【公表日】平成21年9月3日(2009.9.3)
【国際特許分類】
【出願番号】特願2008−558917(P2008−558917)
【出願日】平成18年3月15日(2006.3.15)
【国際出願番号】PCT/IB2006/000577
【国際公開番号】WO2007/105024
【国際公開日】平成19年9月20日(2007.9.20)
【出願人】(508268104)
【Fターム(参考)】
【公表日】平成21年9月3日(2009.9.3)
【国際特許分類】
【出願日】平成18年3月15日(2006.3.15)
【国際出願番号】PCT/IB2006/000577
【国際公開番号】WO2007/105024
【国際公開日】平成19年9月20日(2007.9.20)
【出願人】(508268104)
【Fターム(参考)】
[ Back to top ]