
SYKキナーゼ発現の阻害
【課題】小さな干渉RNA(siRNA)を使用してSykキナーゼの発現を阻害する方法を提供する
【解決手段】細胞内に存在する標的SykキナーゼmRNA配列の切断を導く約20〜23のヌクレオチド長である特定の配列を有するsiRNA分子。siRNA分子をコードするヌクレオチド配列を前記細胞に導入し、前記siRNA分子を細胞内に生成させる。
【解決手段】細胞内に存在する標的SykキナーゼmRNA配列の切断を導く約20〜23のヌクレオチド長である特定の配列を有するsiRNA分子。siRNA分子をコードするヌクレオチド配列を前記細胞に導入し、前記siRNA分子を細胞内に生成させる。
【発明の詳細な説明】
【技術分野】
【0001】
この出願は、2003年7月3日に出願された仮出願第60/484,299号からの優先権を主張する。前記出願の全内容は参照によって本明細書中に組み込まれる。
【0002】
本発明は、一般にSykキナーゼに関し、特に小さな干渉RNA(siRNA)を使用してSykキナーゼの発現を阻害する方法に関する。
【背景技術】
【0003】
二本鎖RNAは、線虫およびショウジョウバエ、ならびに植物を含む数多くの生物の遺伝子発現に干渉する強力な物質であることが明らかになってきた(Bernstein et al, RNA 7: 1509-2151 (2001), McManus et al, Nat. Rev. Genet. 3: 737-747 (2992), Hutvagner et al, Curr. Opin. Genet. Dev. 12: 225-232 (2002), Zamore, Nat. Struct. Biol.8 : 746-750 (2001)Tuschl et al, Genes Dev. 13: 3191-3197(1999))。二本鎖RNAで哺乳類の遺伝子を沈黙させる当初の課題は、侵入するRNAウイルスに対する防御手段として進化した哺乳類の免疫系が、インターフェロン反応といった機構を介して二本鎖RNAを含む細胞を破壊してしまうことであった(Clarke et al, RNA 1: 7-20 (1995))。しかしながら、小さな干渉RNA(siRNA)と命名された非常に短いRNA断片(例えば、20〜23ヌクレオチド長)が免疫反応を免れ得ることが実証されてきた。すなわち、導入されたsiRNAが哺乳類細胞内の物質を沈黙させる遺伝子として十分に機能し得る(Elbashir et al, Nature 411: 494-498 (2001), Elbashir et al, Genes Dev. 15: 188-200 (2001), Paddison etal, Genes Dev. 16: 948-958 (2002), Wianny et al, Nat. Cell Biol. 2: 70-75(2000))。
【0004】
現在解っていることとして、RNAiは多段階のプロセスを含む。二本鎖RNAは、エンドヌクレアーゼであるダイサー(Dicer)によって切断され、21〜23のヌクレオチド破片(siRNA)を生成する。siRNAの二重鎖は、2つの単一鎖のRNAに分解され、一つの鎖が標的RNAの切断に導くガイドRNAとして機能するタンパク質含有複合体に組み込まれる(Schwarz et al, Mol. Cell. 10: 537-548 (2002), Zamore et al, Cell 101: 25-33 (2000))。これによって、特定の遺伝子情報を沈黙させる(Zeng et al, Proc. Natl. Acad. Sci. 100: 9779 (2003)も参照)。
【0005】
また、抗センスDNAが、遺伝子発現を阻害するために広く使用されている(Roth et al, Annu. Rev. Biomed. Eng. 1: 265-297 (1999))。いったん細胞に入ると、抗センスオリゴヌクレオチド(ASO)は相補的mRNAを認識して強く結合し、mRNAが細胞のタンパク質翻訳機構と相互作用することを妨げる。
【0006】
SykキナーゼmRNA ASOによるSykキナーゼ発現の阻害は、Fcγ受容体シグナルを劇的に減少させ(Matsuda et al, Molec. Biol. of the Cell 7: 1095-1106(1996))、エアロゾルによってラットの肺に導入されたSykキナーゼmRNA ASOは、Fcγ受容体誘導型の肺の炎症を防ぐことが実証されている(Stenton et al, J. Immunol. 169: 1028-1036 (2002))。
【0007】
少なくともある系において、siRNAは、遺伝子発現の阻害剤としてASOよりも強力で信頼性がある。本発明は、Sykキナーゼ発現の阻害剤としてのsiRNAの効力を試験するために計画された研究によってもたらされる。
【発明の概要】
【0008】
本発明は、一般的にはSykキナーゼに関する。好ましい実施形態において、本発明は、小さな干渉RNA(siRNA)を使用してSykキナーゼ発現を阻害する方法に関し、前記方法に基づいた治療学的方法に関する。
【0009】
本発明の対象および利点は、以下の詳細な説明から明らかになるであろう。
【発明の詳細な説明】
【0010】
本発明は、SykキナーゼmRNAを標的にするRNA分子に関する。例えば、本発明は、SykキナーゼmRNAの切断および/または分解を誘導する約19、20または21〜約23のヌクレオチド長のRNA分子に関する。
【0011】
好ましい実施形態において、本発明は、siRNA分子、典型的には2つの20〜23のヌクレオチド(nt)鎖を含む2本鎖RNA分子の使用に関する。本発明の使用に適したsiRNAは、任意の種々の手法を使用して生成され得る。siRNAはインビトロで調製され得、その後に直接的に細胞に導入される(例えば、トランスフェクションによって)。
【0012】
代わりに、細胞内発現は、細胞内でsiRNAを発現する構築物(例えば、DNAに基づいたベクターまたはカセット)を細胞内にトランスフェクトすることによって達成され得る。
【0013】
より具体的には、本発明の使用に適したsiRNAは、例えば化学的合成、インビトロでの転写、ダイサーまたはRNaseIIIといったRNaseIII酵素を使用する、より長いdsRNAの酵素的消化を介して調製され得、また、siRNA発現プラスミドまたはウイルスベクター由来の細胞内発現、PCRで誘導されたsiRNA発現カセット由来の細胞内発現を介して調製され得る。これらの様々な手法の詳細な説明は容易に理解可能であり、例えば、http://www.ambion.com/techlib/tn/103/2.html, www.bdbiosciences.com, www.oligoengine.com, www.genetherapysestems.com, www.dharmacon.com,
http://www. mpibpc.gwdg.de/abteilungen/100/105/sirna.htmlに見出すことができ、および/または本明細書中で引用した参照文献中にも見出すことができる(これらの参照文献はまた、参照によって本明細書中に組み込まれる)(Sui et al, Proc Natl Acad Sci USA 99: 5515-20 (2002), Brummelkamp et al, Science 296: 550-3 (2002), Paul et al, Nature Biotechnology 20: 505-8 (2002), Lee et al, Nature Biotechnology 20: 500-5 (2002), Castanotto et al, RNA8 : 1454-60 (2002) および US Appln. 20030108923も参照)。
【0014】
様々な手法を利用して本発明のRNAの安定性を増強することができる(例えば、米国特許出願第20020086356, 20020177570 および 20020055162, ならびに米国特許第6,197,944, 6,590,093, 6,399,307, 6,057,134, 5,939,262, および5,256, 555、ならびに本明細書中で引用された参照文献を参照)。
【0015】
上記に示したように、本発明で使用するのに適したsiRNAを化学的に調製することができる。
【0016】
有利には、2’ヒドロキシルを、例えば、酸不安定なオルトエステル保護基を使用して合成プロセス中の分解から保護する{Scaringe et al, J. Am. Chem. Soc. 120: 11820(1998)およびwww.dharmacon.com(例えば、明細書中に記載されたACE技術)を参照}。RNAオリゴマーを、使用する前に2’を脱保護化してアニールさせることができる。
【0017】
化学的に合成されたsiRNAにおいて、2本鎖分子のうちの少なくとも1本鎖は、約1〜6ヌクレオチド(例えばピリミジンおよび/またはプリンヌクレオチド)長の3’オーバーハングをもつことができる。好ましくは、3’のオーバーハングは、約1〜5のヌクレオチド(例えば、チミジンまたはウリジン)、より好ましくは約1〜4のヌクレオチドおよび最も好ましくは2または3のヌクレオチド長である。
【0018】
有利には、各鎖はオーバーハングを有する。オーバーハングの長さは、各鎖の間で同じかまたは異なるものとすることができる。典型的には、両方の鎖は同じ長さのオーバーハングを有する。特定の態様において、本発明のRNAは、RNA鎖の両側の3’末端に約1〜3、特に約2のヌクレオチドのオーバーハングをもつ対になった21または22のヌクレオチドを含む。
【0019】
上記に示したように、本発明の使用に適したsiRNAは、RNaseIII型酵素(例えば、ダイサー)を使用してより長いdsRNAの酵素的消化によって調製され得る(上記に列挙した参考文献およびウェブサイトを参照)。例えば、完全長の標的遺伝子から大量のsiRNAを生成できる商業的に利用可能なダイサーsiRNAの生成キットを使用することができる(Gene Therapy Systems, Inc, MV062603)。siRNAは、PCRに基づいたクローニングを使用して標的DNAおよびT7 RNAポリメラーゼのプロモーター配列から生成され得る。標的配列からのRNA転写の後に、組換え体Dicerは、転写されたRNAiを22bpのsiRNAに切断することができる。
【0020】
また、上記に示したように、本発明における使用に適したsiRNA分子はまた、当該技術において既知の方法を使用して組換え的に生成され得る(上記に列挙した参考文献およびウェブサイトを参照)。組換え技術は、哺乳類細胞内でsiRNAのインビボでの転写を可能にする。この手法に従って、例えば、RNAポリメラーゼIIIまたはU6プロモーター配列を含むベクターを使用することができる。このようなベクター{ウイルスベクターおよびプラスミドベクター(例えばpSIREN)を含む}を、ウイルス系(例えば、アデノウイルスまたはレトロウイルス系)と組み合わせて発現ベクターまたはシャトルベクターとして使用し、siRNAを哺乳類細胞に導入することができる。インビボでアニールして機能的なsiRNAを生成するsiRNAのセンス鎖およびアンチセンス鎖を発現するようにベクターを加工することができる。
【0021】
代わりに、ヘアピン構造をとるRNAは、標的のセンス鎖(例えば、約20 nt)をベクターに挿入し、続いて短いスペーサー(例えば、約4〜約10 nt)を挿入し、その後に標的のアンチセンス鎖(例えば、約20 nt)と、例えば約5〜6のTを転写終結区として挿入することによって発現され得る。得られたRNA転写産物は折り畳まれて、3’末端に2〜3のUをもつ、例えば約20 bpのステムおよび約10 ntのループを含むステムループ構造を形成する(Paddison et al, Proc. Natl. Acad. Sci. 99: 1443-1448 (2002)も参照)。インビボでの生成を達成する使用に適した構成(ベクターおよびプロモーターの選択を含む)は、当業者によって容易に設計され得、例えば、細胞/組織のターゲットおよび要求される効果に応じて変化するであろう。
【0022】
dsRNAは、標的化されたSykキナーゼmRNAに対する十分な相同性を有するという条件で、本発明の方法において使用され得る。siRNA二重鎖は、例えばSykキナーゼcDNAを標的モチーフ「AA(N)19」(式中、Nは任意のヌクレオチドであって、約30%〜70%のG/C含量が好ましく、約50%のG/C含量がより好ましい。)について検索することによって設計され得る。siRNA二重鎖のセンス鎖は、選択されたAA(N)19モチーフのうち3〜21のヌクレオチドと一致することができる。siRNA二重鎖のセンス鎖は、選択されたAA(N)19モチーフのうち1〜21のヌクレオチドと相補的な配列を有し得る。さらなる設計の詳細は、http://www. mpibpc. gwdg. de/abteilungen/100/105/sirna. html.に与えられる。
【0023】
好ましい標的配列は、SykキナーゼmRNAに特有の配列を含む。例えば、標的配列は、Sykキナーゼの2つのSH2ドメイン間の配列、あるいは第二のSH2ドメインとキナーゼドメインとの間の配列から選択され得る。特定のDNA標的配列については、以下の制限のない例において説明する。さらなる標的として、以下のものを含むがこれらに限定されない。
【化1】

【0024】
本明細書中に記載されたsiRNAは、種々の方法において使用され得る。例えば、siRNA分子は、細胞内または生物体内の標的SykキナーゼmRNAに対して使用され得る。特定の態様において、細胞または個体内の細胞においてRNAiを引き起こすために、siRNAがヒト細胞またはヒト体内に導入され得、Sykキナーゼ発現と関連した疾病または望ましくない状態(例えば、肺、関節、眼または膀胱の炎症)を予防または治療する。siRNAはまた、血液細胞の免疫破壊の治療、例えば、自己免疫性溶血性貧血における赤血球、および免疫性血小板減少性紫斑病(ITP)における血小板の治療において使用され得る(例えば、マクロファージおよび脾臓および肝臓細胞内のSykキナーゼmRNAを標的にすることによって)。本発明の方法によれば、Sykキナーゼ遺伝子が標的化され、対応するmRNA(標的化されたSykキナーゼ遺伝子の転写産物)がRNAiによって分解される。肺細胞が標的になるとき、siRNA含有組成物はエアロゾル化して例えば吸入によって投与され得る。関節に対する投与は、siRNA含有溶液の注射によって達成され得る。眼に対する投与は、例えばキャリア内にsiRNAを含む液滴の注入または滴下によって達成され得る。膀胱等に対する投与は、siRNAを含む組成物で標的組織を洗浄または灌注することによって達成され得る。皮膚に対する投与は、局所的な投与を介してなし得る(例えば、液体、クリームまたはゲルとして)。
【0025】
本発明によれば、個体の細胞(例えば、血液単核細胞、好塩基球またはマスト細胞)をex vivoで処理し、SykキナーゼmRNAの分解を達成することができる。処理すべき細胞は、既知の方法を使用して個体から得ることができ、対応するSykキナーゼmRNAの分解を引き起こすsiRNAを細胞内に導入することができ、その後、その細胞を個体に再び戻すことができる。
【0026】
特定の実施形態において、本発明は、Fce受容体を有する細胞、例えばマスト細胞からのメディエータ(例えば、ヒスタミン)の放出を阻害する上述したsiRNAの使用に関する。ヒスタミン(マスト細胞メディエータ)放出の阻害は、例えば喘息の治療における治療学的重要性がある。
【0027】
本発明のsiRNA(またはsiRNAの細胞内生成をもたらす使用に適した構築物)を全身的に(例えばIVを介して)投与することができ、あるいは標的組織に直接的に(例えば、肺に対するエアロゾル投与を介して)投与することができる。送達は、本明細書中に記載された技術を使用して達成され得る(リポソーム処方を含む)。リポソーム処方に加えて、ポリマー処方が使用され得る。ポリエチレンイミン(PEI)は、好適な陽イオン性ポリマーの例である。直鎖22kDaおよび枝分れ鎖25kDa PEIを含む様々なサイズのPEIが使用され得る(他のサイズ、修飾および未修飾、ならびに生分解性バージョンが使用され得る)。送達はまた、例えば非毒性ウイルス送達系(例えば、アデノ関連ウイルス送達系)を使用して達成され得る。最適投与量は患者、siRNA、投与方法、および必要とされる効果に依存する。最適な条件は、過度の実験を行うことなく当業者によって確立され得る。
【0028】
本発明のある側面は、以下の非限定的な例におけるより詳細な説明で説明され得る(また、米国特許20030084471、20030108923、および20020086356も参照)。
【0029】
例1
実験の詳細
試薬
Lipofectamine 2000およびOpti-MemをInvitrogen(San Diego, CA)から購入した。EagleのMEM(EMEM)、FCS、ペニシリン、およびストレプトマイシンをLife Technologies(Grand Island, NY)から購入した。ウサギ抗マウスSykキナーゼポリクローナル抗体(Ab)および抗アクチンAbをSanta Cruz Biotechnology(Santa Cruz, CA)から購入し、F(ab’)2ヤギ抗ウサギAbが、Jackson Laboratory(Bar Harbor, ME)によって供給された。化学ルミネセンス試薬を、DuPont NEN(Boston, MA)から購入した。
【0030】
細胞および細胞系統
ペンシルベニア大学で健康ボランティアから得た末梢血中単球を上述したように単離した(Matsuda et al, Molec. Biol. of the Cell 7: 1095-1106 (1996))。簡単に言うと、ヘパリン処理を施した血液をFicoll-Hypaqueで遠心分離し(Lymphocyte Separation Medium; Organon Teknika, Durham, NC)、RPMI1640(GIBCO BRL, Grand Island, NY)、10%熱不活性化ウシ胎仔血清(FCS)(Intergen,Purchase, NY)および2mM L-グルタミンを含む完全培地中で境界面の細胞を再懸濁した。細胞は、FCSで予めコートされた組織培養フラスコに37℃、30分間で付着した。45〜90分後、付着していない細胞をハンクス液で強く洗浄して取り除いた。細胞を強く撹拌することによって収集した。単球は、トリパンブルー排除法によって判定したときに常に98%を超える生存可能性を示した。単離された単球を、L-グルタミン(2mM)および10%熱不活性化FCSを追加したRPMI1640中に37℃、5%CO2で維持した。
【0031】
ラット好塩基細胞(RBL-2H3)を、17%FBS、100Uペニシリン、100μg/mlストレプトマイシン、および4mMグルタミンを追加したEMEM中で37℃、5%CO2で生育させた。
【0032】
siRNA二重鎖の構築
SykキナーゼについてのsiRNAを、Dharmacon Research Inc. (Lafayette, Co)によって調製した。製造業者によって提供されたガイドラインに基づいてsiRNAを設計し、ヒトSykキナーゼRNAにおける潜在的なsiRNA標的(AA対直後の19ヌクレオチド下流)を最初に特定した。その後、これらの部位をラットおよびマウスのSykキナーゼRNAの配列においてスキャンし、これらの種における共通のSykキナーゼRNA標的配列を同定した。2つの適切な位置を同定し、各々19の相補的ヌクレオチド鎖とチミジンの3’末端非相補的二量体(Elbashir et al, Nature 411: 494-498(2001))からなる2つの21-mer RNAを構築した。siRNAのセンス鎖は、末端チミジン二量体を除いて標的mRNA配列と同一の配列である。siRNAのアンチセンス鎖は、標的配列の逆の相補的配列である。
【0033】
1)siRNA-1:ヒト、bp 296〜bp 316; マウスおよびラット、bp 307〜bp 327。
【0034】
センス 5’- gaagcccuucaaccggccc dTdT 3’
アンチセンス 3’-dTdT cuucgggaaguuggccggg 5’
【0035】
2)siRNA-2:ヒト、bp 364〜bp 382; マウスおよびラット、bp 375〜bp 393
センス 5’- ccucaucagggaauaucug dTdT 3’
アンチセンス 3’-dTdT ggaguagucccuuauagac 5’
【0036】
トランスフェクション
siRNAをトランスフェクションによってRBL-2H3細胞および単球に導入した。トランスフェクションのために、5×104のRBL-2H3細胞または1×105の単球を24ウェルプレートの各ウェルに播種した。24時間後、完全培地を血清および抗生物質を欠く400μlの新鮮培地に置き換え、siRNA/Lipofectamine 2000複合物を各ウェルに加えた。RBL細胞については、siRNA/Lipofectamine 2000複合物は、製造業者のプロトコルに従って血清または抗生物質を加えずに、3μlのsiRNA二重鎖(20μM)と3μlのLipofectamine 2000を、100μlのOpti-memに添加することによって生成した。単球については、siRNA/Lipofectamine 2000複合物は、血清または抗生物質を加えずに、3μlのsiRNA二重鎖(20μM)と1μlのLipofectamine 2000を、100μlのOpti-memに添加することによって生成した。ウェスタンブロットによってキナーゼタンパク質の発現を試験する48時間前から細胞を37℃でインキュベートした。
【0037】
Sykキナーゼタンパク質のウェスタンブロット分析
Laemmliサンプルバッファー(2% SDS, 10% グリセロール, 100 mM DTT, and 60 mM トリス(pH 6.8))中で細胞を5分間にわたって煮沸することによって溶解物を調製した。溶解物中のタンパク質を、SDS-PAGE(10%ポリアクリルアミド)によって分離し、サンプルバッファー(25mMトリス、190mMグリシン、および20%メタノール)中のニトロセルロース膜に転写した。ヤギ抗ウサギHRPでインキュベート(室温で1.5時間)する前に、1μg/mlのウサギ抗マウスSykキナーゼポリクローナルAbでニトロセルロース膜を4℃で終夜インキュベートした。膜上のタンパク質のバンドを、製造業者のプロトコルに従って化学発光試薬で視覚化した。Sykキナーゼタンパク質を検出した後、時折撹拌しながら100 mM 2-ME、2% SDSおよび62.5 mM Tris-HCL (pH 6.7)を含む剥離バッファー中で膜を50℃で30分間にわたりインキュベートすることによって抗SykキナーゼAbを取り除いた。その後、膜を抗アクチンAbで再プローブ化し、膜上のバンドを化学発光試薬で視覚化した。Sykキナーゼのタンパク質のレベルを濃度測定によって数量化した(Personal Densitometer, Molecular Dynamics)。
【0038】
結果
Sykキナーゼの発現についてのsiRNAの効果
ラット好塩基球(RBL-2H3細胞)およびヒト単球に、ラット、マウスおよびヒトのSykキナーゼRNAで共通の配列をもつsiRNAをトランスフェクトした。Sykキナーゼタンパク質の発現を、抗SykキナーゼAbを使用してウェスタンブロットによって分析した(図2および3)。siRNAで処理されたRBL細胞内におけるSykキナーゼ発現の阻害を図2に示し、アクチンタンパク質のレベルで標準化されたSykキナーゼタンパク質のレベルを表1に示した。アクチンは、タンパク質の発現を調べるために基準として使用される共通のタンパク質である。siRNAで処理されたRBL細胞におけるSykキナーゼタンパク質の発現は、未処理のRBL細胞(レーン1)と比較して45〜51%まで阻害された(図2、レーン2および3)。より大きなSykキナーゼ阻害が、インビトロで培養された増殖性細胞と比較して単球といった非増殖性の細胞において達成され得ることがSyk キナーゼmRNA ASOでの実験で示されていることから、培養細胞中のsiRNAによるSykキナーゼ遺伝子発現の阻害の大きさは奨励される。また、培養液中に維持された単球内のsiRNAによるSykキナーゼ発現の阻害が観察された(図3)。siRNAで処理された単球内でのSykキナーゼの発現が阻害された(図3、レーン2)。さらに、siRNAで処理されたU937細胞とsiRNAで処理されたTHP-1細胞中のSykキナーゼの発現が阻害された(U937とTHP-1はマクロファージ様の細胞系統である)。従って、データは、SykキナーゼRNAに対するsiRNAの有効性を実証し、この遺伝子の発現を抑制し、SykキナーゼRNAに対するsiRNAが、炎症に効く強力な治療学的ツールとして役に立つことを示す。
【表1】
【0039】
考察
これらの研究において使用されたsiRNAは化学的に合成されるが(Dharmicon Research Inc., Lafayette, CO)、siRNAは組換え技術によって調製することもできる。siRNA二重鎖は、21ヌクレオチドのセンス鎖と21ヌクレオチドのアンチセンス鎖を、2-ヌクレオチド(dT)3’オーバーハングを有するようにペアにして含み得る。
【0040】
siRNAのための標的化された領域は、開始コドンの50〜100ヌクレオチド下流から始まるように設計されたcDNAから選択された配列AA(N19)(N、任意のヌクレオチド)とすることができる。約50%のG/C含量が好ましい。pol IIIプロモーターからのRNAの発現は、最初に転写されたヌクレオチドがプリンであるときのみ効率的なので、センス鎖およびアンチセンス鎖のsiRNAは、これらが標的部位での変化を伴わずにpol III発現ベクターから発現され得るように、プリンヌクレオチドから始まるのが好ましい。
【0041】
上述した研究において、ヒトSykキナーゼmRNA中の潜在的なsiRNAの標的を選択し、次に、ラットおよびマウスSykキナーゼmRNA中のその配列を調べ、これらの生物種における共通のSykキナーゼmRNA標的配列を同定した。2つの適切な標的配列が以下のように同定された。
【0042】
標的領域(1)(cDNA):5’ aagaagcccttcaaccggccc;
標的領域(2)(cDNA):5’ aacctcatcagggaatatctg。
【0043】
標的配列およびSykキナーゼsiRNAを図1に示す。二本鎖RNAは、ヌクレオチドの分解に対する高い感受性をもたず、デオキシヌクレオチド(ウリジン(U)ではなくてチミジン(T))の使用は、3’オーバーハングの安定性に影響を及ぼし得る。
【0044】
例2
HS-24細胞(2×105)を、24〜48時間にわたって12ウェルプレート中でLipofectamine 2000とともに3μlのsiRNA-1(対照)またはsiRNA-2またはSykアンチセンス鎖で前処理した。そして、終夜にわたって10 ng/mlのTNFによって前記細胞を刺激した。
【0045】
siRNA-1
標的DNA: 5’ AAGAAGCCCTTCAACCGGCCC
センスsiRNA 5’- gaagcccuucaaccggccc dTdT 3’
アンチセンスsiRNA 5’-dTdT cuucgggaaguuggccggg 5’
【0046】
siRNA-2
標的DNA: 5’ AACCTCATCAGGGAATATCTG
センス 5’- ccucaucagggaauaucug dTdT 3’
アンチセンス 3’-dTdT ggaguagucccuuauagac 5’
【0047】
読出し: 培養上清中のIL-6(ELISA)およびICAM-1の細胞表面における発現(フローサイトメトリー)。ウェスタンブロット分析(図4を参照)によって示された、処理の48時間後にSykタンパク質発現の減少を引き起こしたsiRNA-2。
【0048】
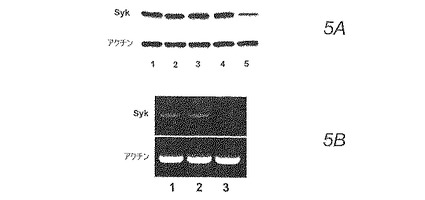
より具体的には、HS-24細胞にsiRNA-2またはsiRNA-1(対照)を一過性的にトランスフェクトした。次に、ICAM-1の細胞表面の発現、およびIL-6の放出について試験した。siRNA-1ではなくsiRNA-2でのHS-24細胞の48時間の処理では、Sykタンパク質(図5A)とmRNA(図5B)発現の両方が著しく抑制された。
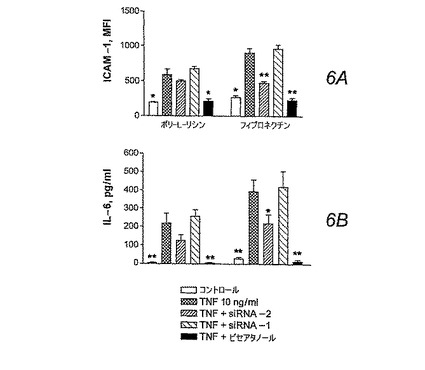
【0049】
HS-24細胞は、siRNA-2の処理による影響を受けずに、恒常的に低レベルのICAM-1(図6A)を発現した(図示せず)。終夜培養における10ng/ml TNFでのHS-24細胞の刺激は、休眠細胞(ポリ-L-リシン上にプレート)およびフィブロネクチンに付着した細胞の両方において、ICAM-1発現の著しい増加を引き起こした(図6A)。フィブロネクチンに付着した細胞は、ポリ-L-リシンに付着した細胞と比較して、TNFで刺激した後にICAM-1のより高い発現を示した(P<0.05)。siRNAでのトランスフェクションは、フィブロネクチンでプレートされたHS-24細胞中のTNFで誘導されたICAM-1の発現を下方制御したが、ポリ-L-リシンでプレートされた細胞中のICAM-1については顕著な効果を示さなかった。薬理学的なSyk阻害剤(ピセアタノール(piceatannol)10μM)での終夜にわたる処理は、ポリ-L-リシンおよびフィブロネクチン付着状態の両方においてTNFで刺激された細胞内のICAM-1の著しい下方制御が引き起こされた(図6A)。トリパンブルー染料除外法によって測定したとき、siRNAまたはピセアタノールでの細胞の処理は、生存度についての顕著な効果が無かった(全ての実験において、生存度は>96%)。
【0050】
TNF刺激を伴わないHS-24細胞によるIL-6の放出は最小であったが、フィブロネクチンに対する細胞付着の培養上清中のIL-6のレベルはより高くなる傾向にあった(図6B)。予期した通り、IL-6レベルの大幅な上昇が両方の培養条件におけるTNF刺激の後に観察され、フィブロネクチン付着細胞においてより高いレベルであった(P<0.05)。siRNA-2の処理は、フィブロネクチン付着培養において統計学的有意性に到達したIL-6放出の下方制御を引き起こした(55〜58%)。ピセアタノールはTNFで誘導されたIL-6の放出を殆ど完全に阻害した(図6B)。
【0051】
従って、Sykキナーゼの阻害は、TNFで誘導されたICAM-1およびIL-6放出の発現を下方制御し、炎症の特徴は気管上皮に現れる。その効果はフィブロネクチンに付着した細胞において顕著であり、これらの炎症前現象におけるSyk関与が少なくとも部分的にβ1インテグリン依存性であることを示した。
【0052】
例3
Sykキナーゼに対するsiRNAの効果が、卵白アルブミン(OA)で誘導された喘息のBrown Norway型ラットにおいてインビボで研究された。
【0053】
Brown NorwayラットをOA腹腔内注射で感作させ(Laberge et al, Am. J. Respir. Crit. Care Med. 151:822(1995)に記載)、感作後21日目に使用した。
【0054】
この例において使用されたsiRNAは以下の通りである:
【0055】
標的DNA: 5’ AACCTCATCAGGGAATATCTG
センス 5’- ccucaucagggaauaucug uu 3’
アンチセンス 3’- uu ggaguagucccuuauagac 5’
【0056】
図7および8で使用されたsiRNA2Mは、追加の安定性を与えるDharmaconによって修飾された上記配列である。図7および8で使用されたsiRNA-2は、未修飾形態における上記配列である。
【0057】
siRNAをリポソーム無しで単独で使用するかまたはsiRNA/リポソーム複合体の形成後に使用した。1,2 ジオレオイル-3-トリメチルアンモニウム-プロパン(DOTAP)/ジオレオイル-ホスファチジル-エタノール-アミン(DOPE)リポソームを以下の文献等に基づいて調製した(Stenton et al, J. Immunol. 169: 1028 (2002)および本明細書中で列挙した参考文献を参照)。陽イオン性DOTAP:DOPEリポソームを、2.5:1のリポソーム対siRNAの比でインキュベートし、125μgのsiRNA(リポソーム内包または非内包)を、噴霧後にエアロゾルによって投与した。
【0058】
Sykキナーゼで標的化されたsiRNAのエアロゾル化投与は、Stenton et al, J. Immunol. 164: 3790 (2000)に記載されたとおりとした。Stenton et al, J. Immunol. 169:1028 (2002)に記載されたSidestream噴霧器を使用して、Nine milliterの生理食塩水、siRNAまたはsiRNA/リポソームを、45分間にわたる噴霧によってラットに投与した。24時間後にこの手順を繰り返し、48時間後に第三の処理を行った。第三の処理直後に、5分間にわたって生理食塩水中でエアロゾル化された生理食塩水または5%OAでラットを抗原刺激した。抗原刺激の24時間後にこの動物を屠殺した。
【0059】
気管支肺胞洗浄(BAL)を、Stenton et al, J. Immunol. 169:1028(2002)によって記載されたとおりに行った。
【0060】
単離されたBAL細胞をカウントし、細胞の塗沫標本をCytospinによって調製した。細胞の違いをHEMA-3試薬(Biochemical Sciences, Swedesboro, NJ)で染色した後に先入観をもたずにカウントした。
【0061】
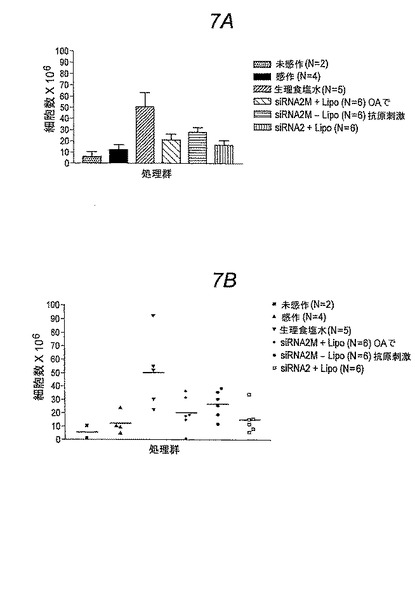
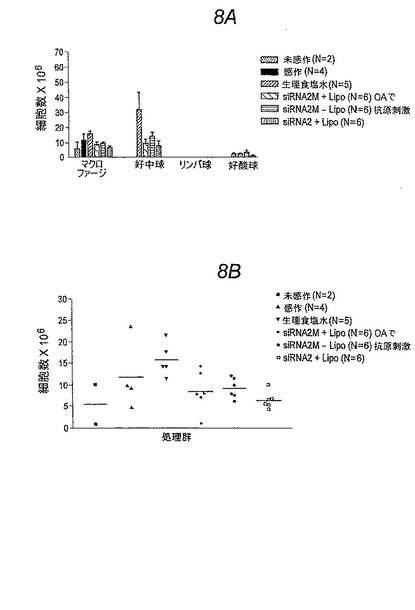
要約すると、OAで誘導されたアレルギー喘息および肺性炎症のBrown Norwayラットモデルを上記に記載された研究において使用した。BAL液に細胞を補充して測定したとき、リポソーム内包および非内包のSykキナーゼ標的化siRNAによって肺の炎症が劇的に抑制された。
【0062】
上記に列挙した全ての文章および他の炎症源は、参照によってその全内容が本明細書中に組み込まれる。
【図面の簡単な説明】
【0063】
【図1】各SykキナーゼsiRNAのセンス鎖は、最初の鋳型アデニン二量体および末端オーバーハングチミジン二量体を除いて、標的配列と同じ配列である。siRNAのアンチセンス鎖は、標的配列の逆の相補的配列である。
【図2】SykキナーゼmRNAに対するsiRNAをトランスフェクトしたRBL-2H3細胞中のSykキナーゼの発現。RBL-2H3細胞にsiRNA-1(レーン2)、siRNA(レーン3)をトランスフェクトし、リポフェクタミンをトランスフェクションしてコントロールとした(レーン1)。細胞溶解液中のタンパク質をSDS-PAGEによって分離し、ニトロセルロース膜に転写した。上のパネルはSykキナーゼのイムノブロットであり;下のパネルはアクチンのイムノブロットである。
【図3】SykキナーゼmRNAに対するsiRNAでトランスフェクトされたヒト単球内におけるSykキナーゼの発現。単球をsiRNA(レーン2)で処理し、リポフェクタミンのトランスフェクションをコントロールとした(レーン1)。細胞溶解物中のタンパク質をSDS-PAGEによって分離し、ニトロセルロース膜に転写し、抗Sykキナーゼ抗体でイムノブロットした。
【図4】ウェスタンブロット分析:HS-24細胞中のSykタンパク質の発現。
【図5】図5Aおよび5B。 (図5A)siRNA処理後のHS-24細胞を溶解し、HS-24細胞溶解物中の総タンパク質等量を10%SDSゲル電気泳動によって溶解し、Sykまたはアクチンに対するモノクローナル抗体を使用してウェスタンブロットによって分析を行った。レーン1-未処理、レーン2-siRNA-1(対照)で24時間処理、レーン3-siRNA-1で48時間処理、レーン4-siRNA-2で24時間処理、レーン5-siRNA-2で48時間処理。 (図5B)RNAを単離し、Sykおよびβ-アクチンのためにRT-PCRを行った。レーン1-未処理、レーン2-siRNA-1(対照)で48時間処理、レーン3-siRNA-2で48時間処理。
【図6】図6Aおよび6B。 ポリリシンでコートされたプレート上に播種されたHS-24細胞(刺激を与えられていない休眠細胞)と、フィブロネクチンでコートされたプレート上に播種されたHS-24細胞(刺激を与えられた細胞)とを、siRNA-2、またはsiRNA-1(対照)、またはピセアタノールで処理した。終夜培養にわたって細胞を10ng/mlのTNFで処理した。(図6A) siRNA(48時間)またはピセアタノール(16時間)処理の後、細胞を取り出し、抗CD54(ICAM-1)で免疫染色してフローサイトメトリーによって分析を行った。 (図6B) IL-6 ELISAキットを使用して細胞培養上清をIL-6の放出について分析した。TNFで刺激された未処理の細胞(例えば、siRNAで処理されていない細胞)と比較したときの*P<0.05、**P<0.005。結果は、3〜5の独立した実験系の代表的な値である。データは、siRNA-2によるSykの発現の阻害は、炎症反応において重要なTNFで誘導されたICAM-1の発現とIL-6の放出を下方制御する。
【図7】図7Aおよび7B。 3回の処理の後、卵白アルブミン(OA)で感作および抗原刺激されたBrown Norwayラットの気管支肺胞洗浄(BAL)液中の総細胞数にエアロゾルを介して送達されたSykキナーゼに対するsiRNAの効果。図7Aは棒グラフとしてデータを提供し、図7Bは個々のデータポイントを示す(個々の動物)。
【図8−1】図8Aおよび8B。 3回の処理の後、OAで感作および抗原刺激されたBrown NorwayラットのBAL液中の多数のマクロファージ、好中球、リンパ球および好酸球にエアロゾルを介して送達されたSykキナーゼに対するsiRNAの効果。(図8Aは棒グラフとしてのデータを提供する。図8Bはマクロファージ数についての個々のデータポイントを示す(個々の動物)。
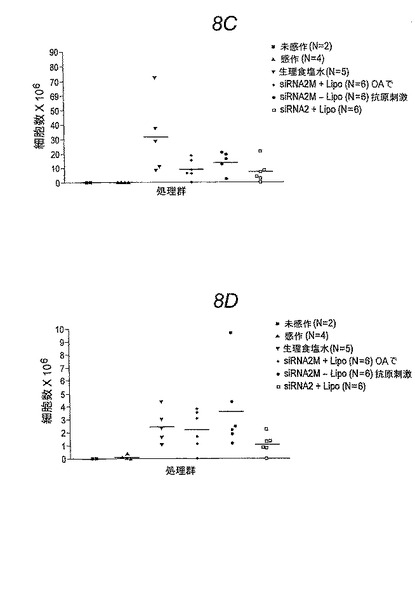
【図8−2】図8Cおよび8D。 図8Cは好中球数についての個々のデータポイントを示し、図8Dは好酸球数についての個々のデータポイントを示す。
【技術分野】
【0001】
この出願は、2003年7月3日に出願された仮出願第60/484,299号からの優先権を主張する。前記出願の全内容は参照によって本明細書中に組み込まれる。
【0002】
本発明は、一般にSykキナーゼに関し、特に小さな干渉RNA(siRNA)を使用してSykキナーゼの発現を阻害する方法に関する。
【背景技術】
【0003】
二本鎖RNAは、線虫およびショウジョウバエ、ならびに植物を含む数多くの生物の遺伝子発現に干渉する強力な物質であることが明らかになってきた(Bernstein et al, RNA 7: 1509-2151 (2001), McManus et al, Nat. Rev. Genet. 3: 737-747 (2992), Hutvagner et al, Curr. Opin. Genet. Dev. 12: 225-232 (2002), Zamore, Nat. Struct. Biol.8 : 746-750 (2001)Tuschl et al, Genes Dev. 13: 3191-3197(1999))。二本鎖RNAで哺乳類の遺伝子を沈黙させる当初の課題は、侵入するRNAウイルスに対する防御手段として進化した哺乳類の免疫系が、インターフェロン反応といった機構を介して二本鎖RNAを含む細胞を破壊してしまうことであった(Clarke et al, RNA 1: 7-20 (1995))。しかしながら、小さな干渉RNA(siRNA)と命名された非常に短いRNA断片(例えば、20〜23ヌクレオチド長)が免疫反応を免れ得ることが実証されてきた。すなわち、導入されたsiRNAが哺乳類細胞内の物質を沈黙させる遺伝子として十分に機能し得る(Elbashir et al, Nature 411: 494-498 (2001), Elbashir et al, Genes Dev. 15: 188-200 (2001), Paddison etal, Genes Dev. 16: 948-958 (2002), Wianny et al, Nat. Cell Biol. 2: 70-75(2000))。
【0004】
現在解っていることとして、RNAiは多段階のプロセスを含む。二本鎖RNAは、エンドヌクレアーゼであるダイサー(Dicer)によって切断され、21〜23のヌクレオチド破片(siRNA)を生成する。siRNAの二重鎖は、2つの単一鎖のRNAに分解され、一つの鎖が標的RNAの切断に導くガイドRNAとして機能するタンパク質含有複合体に組み込まれる(Schwarz et al, Mol. Cell. 10: 537-548 (2002), Zamore et al, Cell 101: 25-33 (2000))。これによって、特定の遺伝子情報を沈黙させる(Zeng et al, Proc. Natl. Acad. Sci. 100: 9779 (2003)も参照)。
【0005】
また、抗センスDNAが、遺伝子発現を阻害するために広く使用されている(Roth et al, Annu. Rev. Biomed. Eng. 1: 265-297 (1999))。いったん細胞に入ると、抗センスオリゴヌクレオチド(ASO)は相補的mRNAを認識して強く結合し、mRNAが細胞のタンパク質翻訳機構と相互作用することを妨げる。
【0006】
SykキナーゼmRNA ASOによるSykキナーゼ発現の阻害は、Fcγ受容体シグナルを劇的に減少させ(Matsuda et al, Molec. Biol. of the Cell 7: 1095-1106(1996))、エアロゾルによってラットの肺に導入されたSykキナーゼmRNA ASOは、Fcγ受容体誘導型の肺の炎症を防ぐことが実証されている(Stenton et al, J. Immunol. 169: 1028-1036 (2002))。
【0007】
少なくともある系において、siRNAは、遺伝子発現の阻害剤としてASOよりも強力で信頼性がある。本発明は、Sykキナーゼ発現の阻害剤としてのsiRNAの効力を試験するために計画された研究によってもたらされる。
【発明の概要】
【0008】
本発明は、一般的にはSykキナーゼに関する。好ましい実施形態において、本発明は、小さな干渉RNA(siRNA)を使用してSykキナーゼ発現を阻害する方法に関し、前記方法に基づいた治療学的方法に関する。
【0009】
本発明の対象および利点は、以下の詳細な説明から明らかになるであろう。
【発明の詳細な説明】
【0010】
本発明は、SykキナーゼmRNAを標的にするRNA分子に関する。例えば、本発明は、SykキナーゼmRNAの切断および/または分解を誘導する約19、20または21〜約23のヌクレオチド長のRNA分子に関する。
【0011】
好ましい実施形態において、本発明は、siRNA分子、典型的には2つの20〜23のヌクレオチド(nt)鎖を含む2本鎖RNA分子の使用に関する。本発明の使用に適したsiRNAは、任意の種々の手法を使用して生成され得る。siRNAはインビトロで調製され得、その後に直接的に細胞に導入される(例えば、トランスフェクションによって)。
【0012】
代わりに、細胞内発現は、細胞内でsiRNAを発現する構築物(例えば、DNAに基づいたベクターまたはカセット)を細胞内にトランスフェクトすることによって達成され得る。
【0013】
より具体的には、本発明の使用に適したsiRNAは、例えば化学的合成、インビトロでの転写、ダイサーまたはRNaseIIIといったRNaseIII酵素を使用する、より長いdsRNAの酵素的消化を介して調製され得、また、siRNA発現プラスミドまたはウイルスベクター由来の細胞内発現、PCRで誘導されたsiRNA発現カセット由来の細胞内発現を介して調製され得る。これらの様々な手法の詳細な説明は容易に理解可能であり、例えば、http://www.ambion.com/techlib/tn/103/2.html, www.bdbiosciences.com, www.oligoengine.com, www.genetherapysestems.com, www.dharmacon.com,
http://www. mpibpc.gwdg.de/abteilungen/100/105/sirna.htmlに見出すことができ、および/または本明細書中で引用した参照文献中にも見出すことができる(これらの参照文献はまた、参照によって本明細書中に組み込まれる)(Sui et al, Proc Natl Acad Sci USA 99: 5515-20 (2002), Brummelkamp et al, Science 296: 550-3 (2002), Paul et al, Nature Biotechnology 20: 505-8 (2002), Lee et al, Nature Biotechnology 20: 500-5 (2002), Castanotto et al, RNA8 : 1454-60 (2002) および US Appln. 20030108923も参照)。
【0014】
様々な手法を利用して本発明のRNAの安定性を増強することができる(例えば、米国特許出願第20020086356, 20020177570 および 20020055162, ならびに米国特許第6,197,944, 6,590,093, 6,399,307, 6,057,134, 5,939,262, および5,256, 555、ならびに本明細書中で引用された参照文献を参照)。
【0015】
上記に示したように、本発明で使用するのに適したsiRNAを化学的に調製することができる。
【0016】
有利には、2’ヒドロキシルを、例えば、酸不安定なオルトエステル保護基を使用して合成プロセス中の分解から保護する{Scaringe et al, J. Am. Chem. Soc. 120: 11820(1998)およびwww.dharmacon.com(例えば、明細書中に記載されたACE技術)を参照}。RNAオリゴマーを、使用する前に2’を脱保護化してアニールさせることができる。
【0017】
化学的に合成されたsiRNAにおいて、2本鎖分子のうちの少なくとも1本鎖は、約1〜6ヌクレオチド(例えばピリミジンおよび/またはプリンヌクレオチド)長の3’オーバーハングをもつことができる。好ましくは、3’のオーバーハングは、約1〜5のヌクレオチド(例えば、チミジンまたはウリジン)、より好ましくは約1〜4のヌクレオチドおよび最も好ましくは2または3のヌクレオチド長である。
【0018】
有利には、各鎖はオーバーハングを有する。オーバーハングの長さは、各鎖の間で同じかまたは異なるものとすることができる。典型的には、両方の鎖は同じ長さのオーバーハングを有する。特定の態様において、本発明のRNAは、RNA鎖の両側の3’末端に約1〜3、特に約2のヌクレオチドのオーバーハングをもつ対になった21または22のヌクレオチドを含む。
【0019】
上記に示したように、本発明の使用に適したsiRNAは、RNaseIII型酵素(例えば、ダイサー)を使用してより長いdsRNAの酵素的消化によって調製され得る(上記に列挙した参考文献およびウェブサイトを参照)。例えば、完全長の標的遺伝子から大量のsiRNAを生成できる商業的に利用可能なダイサーsiRNAの生成キットを使用することができる(Gene Therapy Systems, Inc, MV062603)。siRNAは、PCRに基づいたクローニングを使用して標的DNAおよびT7 RNAポリメラーゼのプロモーター配列から生成され得る。標的配列からのRNA転写の後に、組換え体Dicerは、転写されたRNAiを22bpのsiRNAに切断することができる。
【0020】
また、上記に示したように、本発明における使用に適したsiRNA分子はまた、当該技術において既知の方法を使用して組換え的に生成され得る(上記に列挙した参考文献およびウェブサイトを参照)。組換え技術は、哺乳類細胞内でsiRNAのインビボでの転写を可能にする。この手法に従って、例えば、RNAポリメラーゼIIIまたはU6プロモーター配列を含むベクターを使用することができる。このようなベクター{ウイルスベクターおよびプラスミドベクター(例えばpSIREN)を含む}を、ウイルス系(例えば、アデノウイルスまたはレトロウイルス系)と組み合わせて発現ベクターまたはシャトルベクターとして使用し、siRNAを哺乳類細胞に導入することができる。インビボでアニールして機能的なsiRNAを生成するsiRNAのセンス鎖およびアンチセンス鎖を発現するようにベクターを加工することができる。
【0021】
代わりに、ヘアピン構造をとるRNAは、標的のセンス鎖(例えば、約20 nt)をベクターに挿入し、続いて短いスペーサー(例えば、約4〜約10 nt)を挿入し、その後に標的のアンチセンス鎖(例えば、約20 nt)と、例えば約5〜6のTを転写終結区として挿入することによって発現され得る。得られたRNA転写産物は折り畳まれて、3’末端に2〜3のUをもつ、例えば約20 bpのステムおよび約10 ntのループを含むステムループ構造を形成する(Paddison et al, Proc. Natl. Acad. Sci. 99: 1443-1448 (2002)も参照)。インビボでの生成を達成する使用に適した構成(ベクターおよびプロモーターの選択を含む)は、当業者によって容易に設計され得、例えば、細胞/組織のターゲットおよび要求される効果に応じて変化するであろう。
【0022】
dsRNAは、標的化されたSykキナーゼmRNAに対する十分な相同性を有するという条件で、本発明の方法において使用され得る。siRNA二重鎖は、例えばSykキナーゼcDNAを標的モチーフ「AA(N)19」(式中、Nは任意のヌクレオチドであって、約30%〜70%のG/C含量が好ましく、約50%のG/C含量がより好ましい。)について検索することによって設計され得る。siRNA二重鎖のセンス鎖は、選択されたAA(N)19モチーフのうち3〜21のヌクレオチドと一致することができる。siRNA二重鎖のセンス鎖は、選択されたAA(N)19モチーフのうち1〜21のヌクレオチドと相補的な配列を有し得る。さらなる設計の詳細は、http://www. mpibpc. gwdg. de/abteilungen/100/105/sirna. html.に与えられる。
【0023】
好ましい標的配列は、SykキナーゼmRNAに特有の配列を含む。例えば、標的配列は、Sykキナーゼの2つのSH2ドメイン間の配列、あるいは第二のSH2ドメインとキナーゼドメインとの間の配列から選択され得る。特定のDNA標的配列については、以下の制限のない例において説明する。さらなる標的として、以下のものを含むがこれらに限定されない。
【化1】
【0024】
本明細書中に記載されたsiRNAは、種々の方法において使用され得る。例えば、siRNA分子は、細胞内または生物体内の標的SykキナーゼmRNAに対して使用され得る。特定の態様において、細胞または個体内の細胞においてRNAiを引き起こすために、siRNAがヒト細胞またはヒト体内に導入され得、Sykキナーゼ発現と関連した疾病または望ましくない状態(例えば、肺、関節、眼または膀胱の炎症)を予防または治療する。siRNAはまた、血液細胞の免疫破壊の治療、例えば、自己免疫性溶血性貧血における赤血球、および免疫性血小板減少性紫斑病(ITP)における血小板の治療において使用され得る(例えば、マクロファージおよび脾臓および肝臓細胞内のSykキナーゼmRNAを標的にすることによって)。本発明の方法によれば、Sykキナーゼ遺伝子が標的化され、対応するmRNA(標的化されたSykキナーゼ遺伝子の転写産物)がRNAiによって分解される。肺細胞が標的になるとき、siRNA含有組成物はエアロゾル化して例えば吸入によって投与され得る。関節に対する投与は、siRNA含有溶液の注射によって達成され得る。眼に対する投与は、例えばキャリア内にsiRNAを含む液滴の注入または滴下によって達成され得る。膀胱等に対する投与は、siRNAを含む組成物で標的組織を洗浄または灌注することによって達成され得る。皮膚に対する投与は、局所的な投与を介してなし得る(例えば、液体、クリームまたはゲルとして)。
【0025】
本発明によれば、個体の細胞(例えば、血液単核細胞、好塩基球またはマスト細胞)をex vivoで処理し、SykキナーゼmRNAの分解を達成することができる。処理すべき細胞は、既知の方法を使用して個体から得ることができ、対応するSykキナーゼmRNAの分解を引き起こすsiRNAを細胞内に導入することができ、その後、その細胞を個体に再び戻すことができる。
【0026】
特定の実施形態において、本発明は、Fce受容体を有する細胞、例えばマスト細胞からのメディエータ(例えば、ヒスタミン)の放出を阻害する上述したsiRNAの使用に関する。ヒスタミン(マスト細胞メディエータ)放出の阻害は、例えば喘息の治療における治療学的重要性がある。
【0027】
本発明のsiRNA(またはsiRNAの細胞内生成をもたらす使用に適した構築物)を全身的に(例えばIVを介して)投与することができ、あるいは標的組織に直接的に(例えば、肺に対するエアロゾル投与を介して)投与することができる。送達は、本明細書中に記載された技術を使用して達成され得る(リポソーム処方を含む)。リポソーム処方に加えて、ポリマー処方が使用され得る。ポリエチレンイミン(PEI)は、好適な陽イオン性ポリマーの例である。直鎖22kDaおよび枝分れ鎖25kDa PEIを含む様々なサイズのPEIが使用され得る(他のサイズ、修飾および未修飾、ならびに生分解性バージョンが使用され得る)。送達はまた、例えば非毒性ウイルス送達系(例えば、アデノ関連ウイルス送達系)を使用して達成され得る。最適投与量は患者、siRNA、投与方法、および必要とされる効果に依存する。最適な条件は、過度の実験を行うことなく当業者によって確立され得る。
【0028】
本発明のある側面は、以下の非限定的な例におけるより詳細な説明で説明され得る(また、米国特許20030084471、20030108923、および20020086356も参照)。
【0029】
例1
実験の詳細
試薬
Lipofectamine 2000およびOpti-MemをInvitrogen(San Diego, CA)から購入した。EagleのMEM(EMEM)、FCS、ペニシリン、およびストレプトマイシンをLife Technologies(Grand Island, NY)から購入した。ウサギ抗マウスSykキナーゼポリクローナル抗体(Ab)および抗アクチンAbをSanta Cruz Biotechnology(Santa Cruz, CA)から購入し、F(ab’)2ヤギ抗ウサギAbが、Jackson Laboratory(Bar Harbor, ME)によって供給された。化学ルミネセンス試薬を、DuPont NEN(Boston, MA)から購入した。
【0030】
細胞および細胞系統
ペンシルベニア大学で健康ボランティアから得た末梢血中単球を上述したように単離した(Matsuda et al, Molec. Biol. of the Cell 7: 1095-1106 (1996))。簡単に言うと、ヘパリン処理を施した血液をFicoll-Hypaqueで遠心分離し(Lymphocyte Separation Medium; Organon Teknika, Durham, NC)、RPMI1640(GIBCO BRL, Grand Island, NY)、10%熱不活性化ウシ胎仔血清(FCS)(Intergen,Purchase, NY)および2mM L-グルタミンを含む完全培地中で境界面の細胞を再懸濁した。細胞は、FCSで予めコートされた組織培養フラスコに37℃、30分間で付着した。45〜90分後、付着していない細胞をハンクス液で強く洗浄して取り除いた。細胞を強く撹拌することによって収集した。単球は、トリパンブルー排除法によって判定したときに常に98%を超える生存可能性を示した。単離された単球を、L-グルタミン(2mM)および10%熱不活性化FCSを追加したRPMI1640中に37℃、5%CO2で維持した。
【0031】
ラット好塩基細胞(RBL-2H3)を、17%FBS、100Uペニシリン、100μg/mlストレプトマイシン、および4mMグルタミンを追加したEMEM中で37℃、5%CO2で生育させた。
【0032】
siRNA二重鎖の構築
SykキナーゼについてのsiRNAを、Dharmacon Research Inc. (Lafayette, Co)によって調製した。製造業者によって提供されたガイドラインに基づいてsiRNAを設計し、ヒトSykキナーゼRNAにおける潜在的なsiRNA標的(AA対直後の19ヌクレオチド下流)を最初に特定した。その後、これらの部位をラットおよびマウスのSykキナーゼRNAの配列においてスキャンし、これらの種における共通のSykキナーゼRNA標的配列を同定した。2つの適切な位置を同定し、各々19の相補的ヌクレオチド鎖とチミジンの3’末端非相補的二量体(Elbashir et al, Nature 411: 494-498(2001))からなる2つの21-mer RNAを構築した。siRNAのセンス鎖は、末端チミジン二量体を除いて標的mRNA配列と同一の配列である。siRNAのアンチセンス鎖は、標的配列の逆の相補的配列である。
【0033】
1)siRNA-1:ヒト、bp 296〜bp 316; マウスおよびラット、bp 307〜bp 327。
【0034】
センス 5’- gaagcccuucaaccggccc dTdT 3’
アンチセンス 3’-dTdT cuucgggaaguuggccggg 5’
【0035】
2)siRNA-2:ヒト、bp 364〜bp 382; マウスおよびラット、bp 375〜bp 393
センス 5’- ccucaucagggaauaucug dTdT 3’
アンチセンス 3’-dTdT ggaguagucccuuauagac 5’
【0036】
トランスフェクション
siRNAをトランスフェクションによってRBL-2H3細胞および単球に導入した。トランスフェクションのために、5×104のRBL-2H3細胞または1×105の単球を24ウェルプレートの各ウェルに播種した。24時間後、完全培地を血清および抗生物質を欠く400μlの新鮮培地に置き換え、siRNA/Lipofectamine 2000複合物を各ウェルに加えた。RBL細胞については、siRNA/Lipofectamine 2000複合物は、製造業者のプロトコルに従って血清または抗生物質を加えずに、3μlのsiRNA二重鎖(20μM)と3μlのLipofectamine 2000を、100μlのOpti-memに添加することによって生成した。単球については、siRNA/Lipofectamine 2000複合物は、血清または抗生物質を加えずに、3μlのsiRNA二重鎖(20μM)と1μlのLipofectamine 2000を、100μlのOpti-memに添加することによって生成した。ウェスタンブロットによってキナーゼタンパク質の発現を試験する48時間前から細胞を37℃でインキュベートした。
【0037】
Sykキナーゼタンパク質のウェスタンブロット分析
Laemmliサンプルバッファー(2% SDS, 10% グリセロール, 100 mM DTT, and 60 mM トリス(pH 6.8))中で細胞を5分間にわたって煮沸することによって溶解物を調製した。溶解物中のタンパク質を、SDS-PAGE(10%ポリアクリルアミド)によって分離し、サンプルバッファー(25mMトリス、190mMグリシン、および20%メタノール)中のニトロセルロース膜に転写した。ヤギ抗ウサギHRPでインキュベート(室温で1.5時間)する前に、1μg/mlのウサギ抗マウスSykキナーゼポリクローナルAbでニトロセルロース膜を4℃で終夜インキュベートした。膜上のタンパク質のバンドを、製造業者のプロトコルに従って化学発光試薬で視覚化した。Sykキナーゼタンパク質を検出した後、時折撹拌しながら100 mM 2-ME、2% SDSおよび62.5 mM Tris-HCL (pH 6.7)を含む剥離バッファー中で膜を50℃で30分間にわたりインキュベートすることによって抗SykキナーゼAbを取り除いた。その後、膜を抗アクチンAbで再プローブ化し、膜上のバンドを化学発光試薬で視覚化した。Sykキナーゼのタンパク質のレベルを濃度測定によって数量化した(Personal Densitometer, Molecular Dynamics)。
【0038】
結果
Sykキナーゼの発現についてのsiRNAの効果
ラット好塩基球(RBL-2H3細胞)およびヒト単球に、ラット、マウスおよびヒトのSykキナーゼRNAで共通の配列をもつsiRNAをトランスフェクトした。Sykキナーゼタンパク質の発現を、抗SykキナーゼAbを使用してウェスタンブロットによって分析した(図2および3)。siRNAで処理されたRBL細胞内におけるSykキナーゼ発現の阻害を図2に示し、アクチンタンパク質のレベルで標準化されたSykキナーゼタンパク質のレベルを表1に示した。アクチンは、タンパク質の発現を調べるために基準として使用される共通のタンパク質である。siRNAで処理されたRBL細胞におけるSykキナーゼタンパク質の発現は、未処理のRBL細胞(レーン1)と比較して45〜51%まで阻害された(図2、レーン2および3)。より大きなSykキナーゼ阻害が、インビトロで培養された増殖性細胞と比較して単球といった非増殖性の細胞において達成され得ることがSyk キナーゼmRNA ASOでの実験で示されていることから、培養細胞中のsiRNAによるSykキナーゼ遺伝子発現の阻害の大きさは奨励される。また、培養液中に維持された単球内のsiRNAによるSykキナーゼ発現の阻害が観察された(図3)。siRNAで処理された単球内でのSykキナーゼの発現が阻害された(図3、レーン2)。さらに、siRNAで処理されたU937細胞とsiRNAで処理されたTHP-1細胞中のSykキナーゼの発現が阻害された(U937とTHP-1はマクロファージ様の細胞系統である)。従って、データは、SykキナーゼRNAに対するsiRNAの有効性を実証し、この遺伝子の発現を抑制し、SykキナーゼRNAに対するsiRNAが、炎症に効く強力な治療学的ツールとして役に立つことを示す。
【表1】
【0039】
考察
これらの研究において使用されたsiRNAは化学的に合成されるが(Dharmicon Research Inc., Lafayette, CO)、siRNAは組換え技術によって調製することもできる。siRNA二重鎖は、21ヌクレオチドのセンス鎖と21ヌクレオチドのアンチセンス鎖を、2-ヌクレオチド(dT)3’オーバーハングを有するようにペアにして含み得る。
【0040】
siRNAのための標的化された領域は、開始コドンの50〜100ヌクレオチド下流から始まるように設計されたcDNAから選択された配列AA(N19)(N、任意のヌクレオチド)とすることができる。約50%のG/C含量が好ましい。pol IIIプロモーターからのRNAの発現は、最初に転写されたヌクレオチドがプリンであるときのみ効率的なので、センス鎖およびアンチセンス鎖のsiRNAは、これらが標的部位での変化を伴わずにpol III発現ベクターから発現され得るように、プリンヌクレオチドから始まるのが好ましい。
【0041】
上述した研究において、ヒトSykキナーゼmRNA中の潜在的なsiRNAの標的を選択し、次に、ラットおよびマウスSykキナーゼmRNA中のその配列を調べ、これらの生物種における共通のSykキナーゼmRNA標的配列を同定した。2つの適切な標的配列が以下のように同定された。
【0042】
標的領域(1)(cDNA):5’ aagaagcccttcaaccggccc;
標的領域(2)(cDNA):5’ aacctcatcagggaatatctg。
【0043】
標的配列およびSykキナーゼsiRNAを図1に示す。二本鎖RNAは、ヌクレオチドの分解に対する高い感受性をもたず、デオキシヌクレオチド(ウリジン(U)ではなくてチミジン(T))の使用は、3’オーバーハングの安定性に影響を及ぼし得る。
【0044】
例2
HS-24細胞(2×105)を、24〜48時間にわたって12ウェルプレート中でLipofectamine 2000とともに3μlのsiRNA-1(対照)またはsiRNA-2またはSykアンチセンス鎖で前処理した。そして、終夜にわたって10 ng/mlのTNFによって前記細胞を刺激した。
【0045】
siRNA-1
標的DNA: 5’ AAGAAGCCCTTCAACCGGCCC
センスsiRNA 5’- gaagcccuucaaccggccc dTdT 3’
アンチセンスsiRNA 5’-dTdT cuucgggaaguuggccggg 5’
【0046】
siRNA-2
標的DNA: 5’ AACCTCATCAGGGAATATCTG
センス 5’- ccucaucagggaauaucug dTdT 3’
アンチセンス 3’-dTdT ggaguagucccuuauagac 5’
【0047】
読出し: 培養上清中のIL-6(ELISA)およびICAM-1の細胞表面における発現(フローサイトメトリー)。ウェスタンブロット分析(図4を参照)によって示された、処理の48時間後にSykタンパク質発現の減少を引き起こしたsiRNA-2。
【0048】
より具体的には、HS-24細胞にsiRNA-2またはsiRNA-1(対照)を一過性的にトランスフェクトした。次に、ICAM-1の細胞表面の発現、およびIL-6の放出について試験した。siRNA-1ではなくsiRNA-2でのHS-24細胞の48時間の処理では、Sykタンパク質(図5A)とmRNA(図5B)発現の両方が著しく抑制された。
【0049】
HS-24細胞は、siRNA-2の処理による影響を受けずに、恒常的に低レベルのICAM-1(図6A)を発現した(図示せず)。終夜培養における10ng/ml TNFでのHS-24細胞の刺激は、休眠細胞(ポリ-L-リシン上にプレート)およびフィブロネクチンに付着した細胞の両方において、ICAM-1発現の著しい増加を引き起こした(図6A)。フィブロネクチンに付着した細胞は、ポリ-L-リシンに付着した細胞と比較して、TNFで刺激した後にICAM-1のより高い発現を示した(P<0.05)。siRNAでのトランスフェクションは、フィブロネクチンでプレートされたHS-24細胞中のTNFで誘導されたICAM-1の発現を下方制御したが、ポリ-L-リシンでプレートされた細胞中のICAM-1については顕著な効果を示さなかった。薬理学的なSyk阻害剤(ピセアタノール(piceatannol)10μM)での終夜にわたる処理は、ポリ-L-リシンおよびフィブロネクチン付着状態の両方においてTNFで刺激された細胞内のICAM-1の著しい下方制御が引き起こされた(図6A)。トリパンブルー染料除外法によって測定したとき、siRNAまたはピセアタノールでの細胞の処理は、生存度についての顕著な効果が無かった(全ての実験において、生存度は>96%)。
【0050】
TNF刺激を伴わないHS-24細胞によるIL-6の放出は最小であったが、フィブロネクチンに対する細胞付着の培養上清中のIL-6のレベルはより高くなる傾向にあった(図6B)。予期した通り、IL-6レベルの大幅な上昇が両方の培養条件におけるTNF刺激の後に観察され、フィブロネクチン付着細胞においてより高いレベルであった(P<0.05)。siRNA-2の処理は、フィブロネクチン付着培養において統計学的有意性に到達したIL-6放出の下方制御を引き起こした(55〜58%)。ピセアタノールはTNFで誘導されたIL-6の放出を殆ど完全に阻害した(図6B)。
【0051】
従って、Sykキナーゼの阻害は、TNFで誘導されたICAM-1およびIL-6放出の発現を下方制御し、炎症の特徴は気管上皮に現れる。その効果はフィブロネクチンに付着した細胞において顕著であり、これらの炎症前現象におけるSyk関与が少なくとも部分的にβ1インテグリン依存性であることを示した。
【0052】
例3
Sykキナーゼに対するsiRNAの効果が、卵白アルブミン(OA)で誘導された喘息のBrown Norway型ラットにおいてインビボで研究された。
【0053】
Brown NorwayラットをOA腹腔内注射で感作させ(Laberge et al, Am. J. Respir. Crit. Care Med. 151:822(1995)に記載)、感作後21日目に使用した。
【0054】
この例において使用されたsiRNAは以下の通りである:
【0055】
標的DNA: 5’ AACCTCATCAGGGAATATCTG
センス 5’- ccucaucagggaauaucug uu 3’
アンチセンス 3’- uu ggaguagucccuuauagac 5’
【0056】
図7および8で使用されたsiRNA2Mは、追加の安定性を与えるDharmaconによって修飾された上記配列である。図7および8で使用されたsiRNA-2は、未修飾形態における上記配列である。
【0057】
siRNAをリポソーム無しで単独で使用するかまたはsiRNA/リポソーム複合体の形成後に使用した。1,2 ジオレオイル-3-トリメチルアンモニウム-プロパン(DOTAP)/ジオレオイル-ホスファチジル-エタノール-アミン(DOPE)リポソームを以下の文献等に基づいて調製した(Stenton et al, J. Immunol. 169: 1028 (2002)および本明細書中で列挙した参考文献を参照)。陽イオン性DOTAP:DOPEリポソームを、2.5:1のリポソーム対siRNAの比でインキュベートし、125μgのsiRNA(リポソーム内包または非内包)を、噴霧後にエアロゾルによって投与した。
【0058】
Sykキナーゼで標的化されたsiRNAのエアロゾル化投与は、Stenton et al, J. Immunol. 164: 3790 (2000)に記載されたとおりとした。Stenton et al, J. Immunol. 169:1028 (2002)に記載されたSidestream噴霧器を使用して、Nine milliterの生理食塩水、siRNAまたはsiRNA/リポソームを、45分間にわたる噴霧によってラットに投与した。24時間後にこの手順を繰り返し、48時間後に第三の処理を行った。第三の処理直後に、5分間にわたって生理食塩水中でエアロゾル化された生理食塩水または5%OAでラットを抗原刺激した。抗原刺激の24時間後にこの動物を屠殺した。
【0059】
気管支肺胞洗浄(BAL)を、Stenton et al, J. Immunol. 169:1028(2002)によって記載されたとおりに行った。
【0060】
単離されたBAL細胞をカウントし、細胞の塗沫標本をCytospinによって調製した。細胞の違いをHEMA-3試薬(Biochemical Sciences, Swedesboro, NJ)で染色した後に先入観をもたずにカウントした。
【0061】
要約すると、OAで誘導されたアレルギー喘息および肺性炎症のBrown Norwayラットモデルを上記に記載された研究において使用した。BAL液に細胞を補充して測定したとき、リポソーム内包および非内包のSykキナーゼ標的化siRNAによって肺の炎症が劇的に抑制された。
【0062】
上記に列挙した全ての文章および他の炎症源は、参照によってその全内容が本明細書中に組み込まれる。
【図面の簡単な説明】
【0063】
【図1】各SykキナーゼsiRNAのセンス鎖は、最初の鋳型アデニン二量体および末端オーバーハングチミジン二量体を除いて、標的配列と同じ配列である。siRNAのアンチセンス鎖は、標的配列の逆の相補的配列である。
【図2】SykキナーゼmRNAに対するsiRNAをトランスフェクトしたRBL-2H3細胞中のSykキナーゼの発現。RBL-2H3細胞にsiRNA-1(レーン2)、siRNA(レーン3)をトランスフェクトし、リポフェクタミンをトランスフェクションしてコントロールとした(レーン1)。細胞溶解液中のタンパク質をSDS-PAGEによって分離し、ニトロセルロース膜に転写した。上のパネルはSykキナーゼのイムノブロットであり;下のパネルはアクチンのイムノブロットである。
【図3】SykキナーゼmRNAに対するsiRNAでトランスフェクトされたヒト単球内におけるSykキナーゼの発現。単球をsiRNA(レーン2)で処理し、リポフェクタミンのトランスフェクションをコントロールとした(レーン1)。細胞溶解物中のタンパク質をSDS-PAGEによって分離し、ニトロセルロース膜に転写し、抗Sykキナーゼ抗体でイムノブロットした。
【図4】ウェスタンブロット分析:HS-24細胞中のSykタンパク質の発現。
【図5】図5Aおよび5B。 (図5A)siRNA処理後のHS-24細胞を溶解し、HS-24細胞溶解物中の総タンパク質等量を10%SDSゲル電気泳動によって溶解し、Sykまたはアクチンに対するモノクローナル抗体を使用してウェスタンブロットによって分析を行った。レーン1-未処理、レーン2-siRNA-1(対照)で24時間処理、レーン3-siRNA-1で48時間処理、レーン4-siRNA-2で24時間処理、レーン5-siRNA-2で48時間処理。 (図5B)RNAを単離し、Sykおよびβ-アクチンのためにRT-PCRを行った。レーン1-未処理、レーン2-siRNA-1(対照)で48時間処理、レーン3-siRNA-2で48時間処理。
【図6】図6Aおよび6B。 ポリリシンでコートされたプレート上に播種されたHS-24細胞(刺激を与えられていない休眠細胞)と、フィブロネクチンでコートされたプレート上に播種されたHS-24細胞(刺激を与えられた細胞)とを、siRNA-2、またはsiRNA-1(対照)、またはピセアタノールで処理した。終夜培養にわたって細胞を10ng/mlのTNFで処理した。(図6A) siRNA(48時間)またはピセアタノール(16時間)処理の後、細胞を取り出し、抗CD54(ICAM-1)で免疫染色してフローサイトメトリーによって分析を行った。 (図6B) IL-6 ELISAキットを使用して細胞培養上清をIL-6の放出について分析した。TNFで刺激された未処理の細胞(例えば、siRNAで処理されていない細胞)と比較したときの*P<0.05、**P<0.005。結果は、3〜5の独立した実験系の代表的な値である。データは、siRNA-2によるSykの発現の阻害は、炎症反応において重要なTNFで誘導されたICAM-1の発現とIL-6の放出を下方制御する。
【図7】図7Aおよび7B。 3回の処理の後、卵白アルブミン(OA)で感作および抗原刺激されたBrown Norwayラットの気管支肺胞洗浄(BAL)液中の総細胞数にエアロゾルを介して送達されたSykキナーゼに対するsiRNAの効果。図7Aは棒グラフとしてデータを提供し、図7Bは個々のデータポイントを示す(個々の動物)。
【図8−1】図8Aおよび8B。 3回の処理の後、OAで感作および抗原刺激されたBrown NorwayラットのBAL液中の多数のマクロファージ、好中球、リンパ球および好酸球にエアロゾルを介して送達されたSykキナーゼに対するsiRNAの効果。(図8Aは棒グラフとしてのデータを提供する。図8Bはマクロファージ数についての個々のデータポイントを示す(個々の動物)。
【図8−2】図8Cおよび8D。 図8Cは好中球数についての個々のデータポイントを示し、図8Dは好酸球数についての個々のデータポイントを示す。
【特許請求の範囲】
【請求項1】
細胞内のSykキナーゼの発現を阻害する方法であって、前記細胞内に存在する標的SykキナーゼmRNA配列の切断を導く小さな干渉RNA(siRNA)分子を前記細胞内に導入し、前記阻害を引き起こす方法。
【請求項2】
前記siRNA分子が、約20〜23のヌクレオチド長である請求項1に記載の方法。
【請求項3】
前記siRNA分子が、前記細胞内に直接的に導入される請求項1に記載の方法。
【請求項4】
前記siRNA分子をコードするヌクレオチド配列を前記細胞に導入し、前記siRNA分子を細胞内に生成させる請求項1に記載の方法。
【請求項5】
前記siRNA分子が2つの鎖を含み、前記siRNA分子の少なくとも1つ鎖が約1〜6ヌクレオチド長の3’オーバーハングを有する請求項1に記載の方法。
【請求項6】
前記siRNA分子の両鎖が、約2〜3ヌクレオチド長の3’オーバーハングを有する請求項5に記載の方法。
【請求項7】
前記3’オーバーハングが、ウリジンまたはチミジンを含む請求項6に記載の方法。
【請求項8】
前記siRNA分子が、ステムループ構造を有する請求項1に記載の方法。
【請求項9】
前記ステムループ構造の3’末端が、約1〜6ヌクレオチド長の3’オーバーハングを有する請求項8に記載の方法。
【請求項10】
前記3’オーバーハングが、約2〜3ヌクレオチド長である請求項9に記載の方法。
【請求項11】
前記標的配列が、SykキナーゼmRNAに特有の配列である請求項1に記載の方法。
【請求項12】
前記標的配列(cDNAとして発現)が、以下の配列からなる群から選択されたヌクレオチド配列を含む請求項11に記載の方法。
【化1】
【請求項13】
前記細胞が、哺乳類細胞である請求項1に記載の方法。
【請求項14】
前記細胞が、ヒト細胞である請求項13に記載の方法。
【請求項15】
前記細胞が、ヒトの体内に存在する請求項14に記載の方法。
【請求項16】
前記ヒトが炎症状態にある患者であり、かつ前記siRNA分子が前記状態を治療するために十分な量で投与される請求項15に記載の方法。
【請求項17】
前記炎症状態が、前記患者の気管支、肺、眼、膀胱または皮膚の状態である請求項16に記載の方法。
【請求項18】
前記炎症状態が気管支または肺の状態であり、かつ前記siRNA分子が吸入によって気管支または肺細胞に導入される請求項17に記載の方法。
【請求項19】
以下の配列からなる群から選択される配列に相補的な配列を含むsiRNA分子。
【化2】
【請求項20】
請求項19に記載のsiRNA分子と担体とを含む組成物。
【請求項21】
請求項19に記載のsiRNA分子およびリポソームまたはポリマーを含む組成物。
【請求項1】
細胞内のSykキナーゼの発現を阻害する方法であって、前記細胞内に存在する標的SykキナーゼmRNA配列の切断を導く小さな干渉RNA(siRNA)分子を前記細胞内に導入し、前記阻害を引き起こす方法。
【請求項2】
前記siRNA分子が、約20〜23のヌクレオチド長である請求項1に記載の方法。
【請求項3】
前記siRNA分子が、前記細胞内に直接的に導入される請求項1に記載の方法。
【請求項4】
前記siRNA分子をコードするヌクレオチド配列を前記細胞に導入し、前記siRNA分子を細胞内に生成させる請求項1に記載の方法。
【請求項5】
前記siRNA分子が2つの鎖を含み、前記siRNA分子の少なくとも1つ鎖が約1〜6ヌクレオチド長の3’オーバーハングを有する請求項1に記載の方法。
【請求項6】
前記siRNA分子の両鎖が、約2〜3ヌクレオチド長の3’オーバーハングを有する請求項5に記載の方法。
【請求項7】
前記3’オーバーハングが、ウリジンまたはチミジンを含む請求項6に記載の方法。
【請求項8】
前記siRNA分子が、ステムループ構造を有する請求項1に記載の方法。
【請求項9】
前記ステムループ構造の3’末端が、約1〜6ヌクレオチド長の3’オーバーハングを有する請求項8に記載の方法。
【請求項10】
前記3’オーバーハングが、約2〜3ヌクレオチド長である請求項9に記載の方法。
【請求項11】
前記標的配列が、SykキナーゼmRNAに特有の配列である請求項1に記載の方法。
【請求項12】
前記標的配列(cDNAとして発現)が、以下の配列からなる群から選択されたヌクレオチド配列を含む請求項11に記載の方法。
【化1】
【請求項13】
前記細胞が、哺乳類細胞である請求項1に記載の方法。
【請求項14】
前記細胞が、ヒト細胞である請求項13に記載の方法。
【請求項15】
前記細胞が、ヒトの体内に存在する請求項14に記載の方法。
【請求項16】
前記ヒトが炎症状態にある患者であり、かつ前記siRNA分子が前記状態を治療するために十分な量で投与される請求項15に記載の方法。
【請求項17】
前記炎症状態が、前記患者の気管支、肺、眼、膀胱または皮膚の状態である請求項16に記載の方法。
【請求項18】
前記炎症状態が気管支または肺の状態であり、かつ前記siRNA分子が吸入によって気管支または肺細胞に導入される請求項17に記載の方法。
【請求項19】
以下の配列からなる群から選択される配列に相補的な配列を含むsiRNA分子。
【化2】
【請求項20】
請求項19に記載のsiRNA分子と担体とを含む組成物。
【請求項21】
請求項19に記載のsiRNA分子およびリポソームまたはポリマーを含む組成物。
【図1】


【図2】

【図3】

【図4】

【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【公開番号】特開2011−200238(P2011−200238A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−106343(P2011−106343)
【出願日】平成23年5月11日(2011.5.11)
【分割の表示】特願2006−517793(P2006−517793)の分割
【原出願日】平成16年7月1日(2004.7.1)
【出願人】(500429103)ザ・トラスティーズ・オブ・ザ・ユニバーシティ・オブ・ペンシルバニア (102)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2011−106343(P2011−106343)
【出願日】平成23年5月11日(2011.5.11)
【分割の表示】特願2006−517793(P2006−517793)の分割
【原出願日】平成16年7月1日(2004.7.1)
【出願人】(500429103)ザ・トラスティーズ・オブ・ザ・ユニバーシティ・オブ・ペンシルバニア (102)
【Fターム(参考)】
[ Back to top ]