
Tヘルパー細胞エピトープ
【課題】 Tヘルパー細胞エピトープおよび該エピトープを少なくとも1種含む、免疫応答誘導のための組成物を提供する。
【解決手段】 本発明は、SSKTQTHTQQDRPPQPS; QPSTELEETRTSRARHS; RHSTTSAQRSTHYDPRT; PRTSDRPVSYTMNRTRS; TRSRKQTSHRLKNIPVH; SHQYLVIKLIPNASLIE; IGTDNVHYKIMTRPSHQ; YKIMTRPSHQYLVIKLI; KLIPNASLIENCTKAEL; AELGEYEKLLNSVLEPI; KLLNSVLEPINQALTLM; EPINQALTLMTKNVKPL; FAGVVLAGVALGVATAA; GVALGVATAAQITAGIA; TAAQITAGIALHQSNLN; GIALHQSNLNAQAIQSL; NLNAQAIQSLRTSLEQS; QSLRTSLEQSNKAIEEI; EQSNKAIEEIREATQET; TELLSIFGPSLRDPISA; PRYIATNGYLISNFDES; CIRGDTSSCARTLVSGT; DESSCVFVSESAICSQN; TSTIINQSPDKLLTFIA; SPDKLLTFIASDTCPLV及びSGRRQRRFAGVVLAGVAからなる群より選択されるペプチド配列内に含まれているTヘルパー細胞エピトープに関する。
【解決手段】 本発明は、SSKTQTHTQQDRPPQPS; QPSTELEETRTSRARHS; RHSTTSAQRSTHYDPRT; PRTSDRPVSYTMNRTRS; TRSRKQTSHRLKNIPVH; SHQYLVIKLIPNASLIE; IGTDNVHYKIMTRPSHQ; YKIMTRPSHQYLVIKLI; KLIPNASLIENCTKAEL; AELGEYEKLLNSVLEPI; KLLNSVLEPINQALTLM; EPINQALTLMTKNVKPL; FAGVVLAGVALGVATAA; GVALGVATAAQITAGIA; TAAQITAGIALHQSNLN; GIALHQSNLNAQAIQSL; NLNAQAIQSLRTSLEQS; QSLRTSLEQSNKAIEEI; EQSNKAIEEIREATQET; TELLSIFGPSLRDPISA; PRYIATNGYLISNFDES; CIRGDTSSCARTLVSGT; DESSCVFVSESAICSQN; TSTIINQSPDKLLTFIA; SPDKLLTFIASDTCPLV及びSGRRQRRFAGVVLAGVAからなる群より選択されるペプチド配列内に含まれているTヘルパー細胞エピトープに関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イヌジステンパーウイルス(Canine Distemper Virus)(CDV)由来のTヘルパー細胞エピトープに関する。本発明は少なくとも1つのTヘルパー細胞エピトープならびに場合によってはB細胞エピトープおよび/またはCTLエピトープを含む組成物に関する。
【背景技術】
【0002】
有効な抗体応答を誘導できるいずれのペプチドも、免疫系により認識されるエピトープとして知られるアミノ酸の特定の配列を含有しなければならない。特に、抗体応答については、エピトープは、Bリンパ球表面に存在する特異的な免疫グロブリン(Ig)受容体により認識される必要がある。これらの細胞がこのエピトープに対して特異的抗体を産生する能力のある形質細胞に最終的に分化するからである。これらのB細胞エピトープに加えて、免疫原は、抗原提示細胞(APC)がヘルパーTリンパ球上に存在する特異的受容体に対して提示するエピトープも含有しなければならない、というのはヘルパーTリンパ球は、B細胞が抗体産生細胞に分化するために必要とするシグナルを提供する必要があるからである。
【0003】
ウイルス感染症の場合におよび多くの癌の場合に、抗体の再生利用は限られており、その免疫系はウイルス感染細胞または癌細胞を死滅し得る細胞傷害性T細胞(CTL)と応答する。ヘルパーT細胞と同じように、CTLは最初、表面上に提示された特異的なペプチドエピトープをもつAPCとの相互作用により活性化されるが、この時、MHCクラスII分子よりもむしろMHCクラスIに関連する。活性化されると、CTLは、上記ペプチド/クラスI複合体を保持する標的細胞を捕らえて(engage)、溶解を引き起こし得る。このプロセスにおいてヘルパーT細胞がある役割を果たすことも明らかになっており;APCがCTLを活性化できるようになる前に、該APCは最初にヘルパーT細胞から必要な同時刺激性分子(costimulatory molecules)の発現をアップレギュレートするためのシグナルを受けなければならない。
【0004】
ヘルパーT細胞エピトープは、APC表面上に存在して主要組織適合性複合体(MHC)のクラスII遺伝子によりコードされた分子と結合する。次いで、Tヘルパーリンパ球表面の特異的T細胞受容体(TCR)は、クラスII分子とペプチドエピトープとの複合体を認識する。このようにして、MHC分子と関係をもつ抗原エピトープを提示されたT細胞は活性化され、Bリンパ球が分化するために必要なシグナルを提供することができる。伝統的にペプチド免疫原のためのヘルパーT細胞エピトープの供給源はペプチドが共有結合した担体タンパク質であるが、この結合過程は、結合プロセスにおいて抗原決定基を改変することおよび担体に対する抗体誘導にペプチドを指向する抗体を費やすことなどの、他の問題を引き起こしうる(Schutze, M. P., Leclerc, C. Jolivet, M. Audibert, F. Chedid, L. "Carrier-induced epitopic suppression, a major issue for future synthetic vaccines(担体誘導エピトープ抑制、将来の合成ワクチンに対する大きな論点)". J Immunol. 1985, 135, 2319-2322;DiJohn, D., Torrese, J. R. Murillo, J. Herrington, D. A.ら, "Effect of priming with carrier on response to conjugate vaccine(複合ワクチンへの応答に対する担体を用いた初回免疫の効果)". The Lancet. 1989, 2, 1415-1416)。さらに、調製時における無関係なタンパク質の使用は品質管理の議論を起こす。ペプチドワクチンの設計において適当な担体タンパク質を選ぶことは非常に重要であり、それらの選択は毒性および大規模生産の実施可能性などの因子により制限される。この方法には他の制限があり、結合可能なペプチド体のサイズおよび安全に投与できる担体の投与量が挙げられる(Audibert, F. a. C., L. 1984. "Modern approaches to vaccines. Molecular and chemical basis of virus virulence and immunogenicity(ワクチンへの現代的手法 ウイルス毒性および免疫原性の分子的および化学的基礎)"., Cold Spring Harbor Laboratory, New York.)。担体分子は強い免疫応答の誘導を可能にするが、これら担体分子はまた、抗ペプチド抗体応答の抑制のような望まない効果も随伴する(Herzenberg, L. A. and Tokuhisa, T. 1980. "Carrier-priming leads to hapten-specific suppression(担体初回免疫はハプテン特異的な抑制をもたらす)". Nature 285:664;Schutze, M. P., Leclerc, C., Jolivet, M., Audibert, F.およびChedid, L. 1985. "Carrier-induced epitopic suppression, a major issue for futrue synthetic vaccines(担体誘導エピトープ抑制、将来の合成ワクチンの大きな論点)". J Immunol 135:2319;Etlinger, H. M., Felix, A. M., Gillessen, D., Heimer, E.P., Just, M., Pink, J. R., Sinigaglia, F., Sturchler, D., Takacs, Gl, Trzeciak, A.ら, 1988. "Assessment in humans of a synthetic peptide-based vaccine against the sporozoite stage of the human malaria parasite, Plasmodium falciparum(ヒトマラリア寄生虫、Plasmodium falciparumの種虫(sporpzoite)段階に対する合成ペプチドに基づくワクチンのヒトにおける評価)". J Immunol 140:626)。
【0005】
従って、一般的に、免疫原は、細胞傷害性T細胞上に存在する表面Igによりまたは受容体により認識されるエピトープに加えて、ヘルパーT細胞により認識され得るエピトープも含有しなければならない。これらのエピトープのタイプは非常に種々であることを理解されるべきである。B細胞エピトープについては、B細胞受容体は元来の免疫原と直接結合するので、コンフォメーションが重要である。対照的に、T細胞により認識されるエピトープは、エピトープのコンフォメーションの完全さに依存するのでなく、CTLに対してはほぼ9個のアミノ酸の短い配列およびヘルパーT細胞に対しては長さの制限が少ない若干長い配列から構成される。これらのエピトープに対する要件は単に、該エピトープがクラスIまたはクラスII分子の結合クレフト(cleft:裂け目)とそれぞれ適合し得て、かつその複合体が次にT細胞受容体と結合し得ることである。クラスII分子の結合部位は両方末端が開放されていて、結合されるペプチドの長さにおける非常に大きな変化を許容し(Brown, J. H., T. S. Jardetzky, J. C. Gorga, L. J. Stern, R. G. Urban, J. L. Strominger and D. C. Wiley. 1993. "Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1(ヒトのクラスII組織適合性抗原HLA-DR1の3次元構造), Nature 364:33)"、8個という短いアミノ酸残基のエピトープが報じられている(Fahrer, A. M., Geysen, H. M., White, D. O., Jackson, D. C.およびBrown, L. E. "Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-Ed(単一決定基がクラスII制限されたT細胞を認識するための要件の分析は、T細胞応答のかなりの多様性およびI-Edへのペプチド結合の縮重(degeneracy)を表す)". J. Immunol 1995. 155:2849-2857)。
【0006】
イヌジステンパーウイルス(CDV)は、ネガティブ鎖RNAウイルスのパラミキソウイルス(paramyxovirus)ファミリーのモルビリウイルス(morbillivirus)サブグループに属する。このグループのメンバーである他のウイルスは麻疹ウイルス(measles virus)および牛疫ウイルス(rinderpest virus)である。ペプチドベースのワクチンの開発は、タンパク質配列からのBおよびT細胞エピトープの同定に大きな関心を喚起した。CDVのFプロテインのようなタンパク質由来のT細胞エピトープを使う原理は、若いイヌを早期にCDVに対して接種すると、それによって、このタンパク質上に存在するヘルパーT細胞エピトープに対して特異的なヘルパーT細胞を有するようになるだろうということにある。次いで1つ以上のエピトープを含有するワクチンに曝すと、それによって、存在するヘルパーT細胞の補強および従って、増強された免疫応答をもたらすであろう。しかし、このようなヘルパーT細胞エピトープは、初回刺激していない動物に投与して、さらに免疫応答を誘導することもできる。本発明者らは、CDV融合タンパク質の配列からイヌT細胞エピトープを同定し、そして、これらのエピトープを、特にイヌおよび関係種に対する、ペプチドベースのワクチンの設計に使い得るようにすることを目的とした。
【0007】
LHRH(黄体形成ホルモン放出ホルモン)は10個のアミノ酸長のペプチドホルモンであり、その配列は哺乳類に保存されている。該ホルモンは視床下部により分泌され、雄性および雌性両方の生殖生理学を制御する。LHRHベースの免疫避妊ワクチンの開発の原理は、LHRHに対する抗体がホルモンの黄体形成ホルモンの下垂体分泌および卵胞刺激ホルモンに対する作用をブロックし、哺乳類の生殖腺萎縮および不妊に導くという観察に基づく。
【0008】
開発されているほとんどのLHRHワクチンはタンパク質担体と化学的にコンジュゲートしたLHRHからなり、抗LHRH抗体の産生のためのT細胞ヘルプを提供する。LHRT-タンパク質担体コンジュゲートの接種を繰り返すと、「担体に誘導されるエピトープ抑制」として知られる現象により抗LHRH力価は低下することが分かっている。本発明の1つの目的は、ワクチンのタンパク質担体を、定義したTヘルパーエピトープ(THエピトープ)によって置換え、「担体に誘導されるエピトープ抑制」を排除することである。
【発明の概要】
【0009】
本発明者らは、それぞれがTヘルパー細胞エピトープを含有する、多数の17残基のペプチドを同定した。容易に認識されるように、大多数のこれらのペプチドは最小のTヘルパー細胞エピトープではない。典型的には、クラスII分子は8個のアミノ酸のような短いペプチド(Fahrerら, 1995、同書)、しかし通常は12〜19個のアミノ酸のペプチド(Chicz, R. M., Urban, R. G., Gorga, J. C,. Vignali, D. A. A., Lane, W. S.およびStrominger, J. L. 「Specificity and promiscuity among naturally processed peptides bound to HLA-DR alleles(HLA-DR対立遺伝子と結合した天然にプロセシングを受けたペプチド間の特異性および乱交雑性)」. J Exp Med 1993, 178, 27-47;Chicz, R. M., Urban, R. G., Lane, W. S., Gorga, J. C., Stern, L.J., Vignali, D. A. A.,およびStrominger, J. L. 「Predominant naturally processed peptides bound to HLA-DR1 are derived from MHC-related molecules and are heterogeneous in size(HLA-DRIと結合した、天然にプロセシングを受けた主なペプチドはMHC関係分子から誘導され、サイズは不均一である)」. Nature 1992, 358, 764-8)と結合していることが分かっているが、長さで25個までのアミノ酸のペプチドがクラスIIと結合していることも報じられている(Rammensee, H.G. 「Chemistry of peptide associated with class I and class II molecules.(クラスIおよびクラスII分子に関連したペプチドの化学)」 Curr Opin Immunol 1995, 7, 85-95におけるレビュー)。
【0010】
従って、長さが8〜25個の間のアミノ酸残基の範囲にあるペプチドエピトープがクラスII分子と結合し得る。より短いペプチドはより長い配列より低い活性を有し得る「コア」エピトープであり、より長い配列をNまたはC末端で切断して親配列と同じかまたはより高い活性を有するより短い配列を得ようとするのはつまらない試みでしかない。
【図面の簡単な説明】
【0011】
【図1】CDVの融合タンパク質のアミノ酸配列を示す。
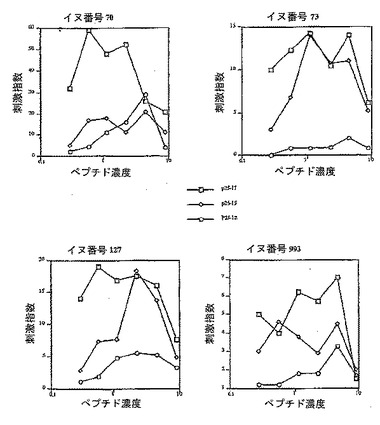
【図2】P25-LHRHで免疫化したイヌからの、Th-エピトープP27およびそのトランケートされた型に対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
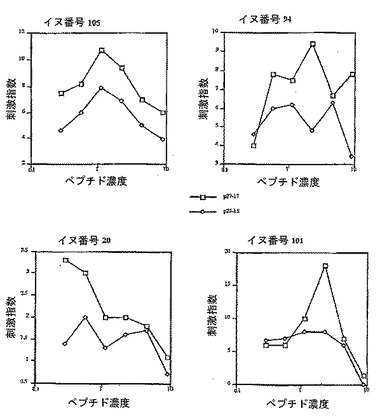
【図3】P27-LHRHで免疫化したイヌからの、Th-エピトープP25およびそのトランケートされた15残基のペプチドに対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
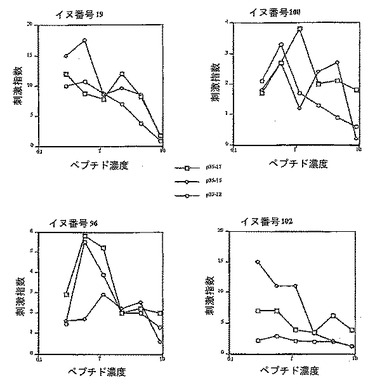
【図4】P35-LHRHで免疫化したイヌからの、Th-エピトープP25およびそのトランケートされた型に対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
【発明を実施するための形態】
【0012】
従って第1の態様においては、本発明はTヘルパー細胞エピトープであって、上記エピトープは、

からなる群から選択されるペプチド配列内に含有されることを特徴とする。
【0013】
第2の態様においては、本発明は、動物に免疫応答を生起させるために使う組成物であって、上記組成物は少なくとも1種のTヘルパー細胞エピトープを含んでなり、少なくとも1種の該Tヘルパー細胞エピトープは、
からなる群から選択されるペプチド配列内に含有されることを特徴とする。
【0014】
本発明の好ましい実施形態においては、該組成物は
からなる群から選択される少なくとも1種のペプチドを含んでなる。
【0015】
該組成物はさらに少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープを含んでなることが、さらに好ましい。
【0016】
また他の好ましい実施形態においては、少なくとも1種の該B細胞エピトープおよび/または少なくとも1種の該CTLエピトープは少なくとも1種のTヘルパー細胞エピトープと連結している。該組成物はまた、各々が少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープを含んでなる複数のエピトープ構築物を含んでなることも好ましい。あるいは、該組成物は、各々が少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のCTLエピトープを含んでなる複数のエピトープ構築物を含んでなっていてもよい。
【0017】
該B細胞エピトープまたは該CTLエピトープはいずれのエピトープであってもよいことは理解されるであろう。本発明で好ましいB細胞エピトープはLHRH B細胞エピトープである。
【0018】
本発明の組成物は複数のTヘルパー細胞エピトープを含んでなってもよい。これらのエピトープは単一であっても一緒に連結して単一のポリペプチドを形成してもよい。エピトープが一緒に単一のポリペプチドに連結される場合、エピトープ同士は近接していてもまたはそれ自身はTヘルパー細胞エピトープの部分でない追加のアミノ酸により隔たれて位置してもよいことは理解されるであろう。
【0019】
ある実施形態においては、上に考察したように、Tヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープおよび/または少なくとも1つのCTLエピトープは、それらのエピトープが連結されている。この連結は、簡単なペプチドの共有結合により実施し得る。他の実施形態においては、エピトープは、最も好ましくはその開示が参照により本明細書に組み入れられるPCT/AU98/00076に記載されたように、重合されている。
【0020】
さらに他の好ましい実施形態においては、該組成物はさらに製薬上許容される賦形剤、好ましくはアジュバントを含んでなる。
【0021】
さらなる態様においては、本発明は動物に免疫応答を誘導する方法であって、動物に本発明の第2の態様の組成物を投与することを含んでなることを特徴とする方法である。
【0022】
製薬上許容される担体または希釈剤は、経口、直腸、鼻、局所(バッカルおよび舌下錠を含む)、膣、非経口(皮下、筋内、静脈内、皮内、くも膜下および硬膜外を含む)投与に適した組成物に使われる担体または希釈剤を含む。これらは受容者に無毒な投与量および濃度にて用いられる。製薬上許容される担体または希釈剤の代表的な例は、限定するものでないが、水、好ましくは生理的pHに緩衝された等張液(リン酸緩衝生理食塩水またはTris緩衝生理食塩水など)を含み、また、マンニトール、ラクトース、トレハロース、デキストロース、グリセロール、エタノールまたはポリペプチド(ヒト血清アルブミンなど)の1以上を含んでもよい。該組成物は単位投与形態で好都合に提示することができ、また製薬業界でよく知られたのいずれかの方法により調製することができる。
【0023】
上記のように、該組成物はアジュバントを含むことが好ましい。理解されるように、「アジュバント」は、ワクチン組成物の免疫原性および効能を増強する1以上の基質を含む組成物を意味する。適当なアジュバントの例は、限定するものでないが、スクアランおよびスクアレン(または他の動物由来の油);ブロックコポリマー;Tween(登録商標)-80のような洗剤;Quil(登録商標) A、DrakeolまたはMarcolのような鉱質油、ピーナッツ油のような植物油;Corynebacterium parvumのようなCorynebacterium由来のアジュバント;Propionibacterium acneのようなPropionibacterium由来のアジュバント;Mycobacterium bovis(Bacille Calmette and GuerinまたはBCG);インターロイキン2およびインターロイキン12のようなインターロイキン;インターロイキン1のようなモノカイン;腫瘍壊死因子;γインターフェロンのようなインターフェロン;サポニン-水酸化アルミニウムまたはQuil-A水酸化アルミニウムのような組合わせ;リポソーム;ISCOMアジュバント;放線菌細胞壁抽出物;ムラミルジペプチドまたは他の誘導体のような合成糖ペプチド;アビリジン;リピドA誘導体;硫酸デキストラン;DEAE-デキストランまたはリン酸アルミニウムを有するもの;Carbopol'EMAのようなカルボキシポリメチレン;Neocryl A640(例えば、米国特許第5,047,238号)のようなアクリルコポリマーエマルジョン;ワクシニアまたは動物ポックスウイルスタンパク質;コレラ毒のようなサブウイルス粒子アジュバント、またはそれらの混合物を含む。
【0024】
当業者には認識されるように、本発明のペプチドは生物学的活性の完全な抑止をすることなしに改変することができる。これらの改変は、付加、欠失および置換、特に保存的置換を含む。Tヘルパー細胞エピトープとしての活性の完全な消失を生じない改変を含むペプチドは本発明の範囲内にあると考えられる。
【0025】
置換の概念は当技術分野では周知であるが、予想される置換の形式を以下に掲げる。
【0026】
予想される他のタイプのペプチドの改変は、限定するものでないが、側鎖の改変、ペプチド合成中の非天然アミノ酸および/またはそれらの誘導体の組込みならびに架橋剤の使用およびペプチドにコンフォメーションの制限を課する他の方法を含む。
【0027】
本発明で考えられる側鎖改変の例は、限定するものでないが、アルデヒドとの反応に続いてNaBH4を用いて還元する還元アルキル化によるようなアミノ基の改変;メチルアセトイミダート(methylacetimidate)によるアミド化;無水酢酸によるアシル化;シアン酸塩によるアミノ基のカルバモイル化;2,4,6-トリニトロベンゼンスルホン酸(TNBS)によるアミノ基のトリニトロベンジル化;無水コハク酸および無水テトラヒドロフタル酸によるアミノ基のアシル化;およびピリドキサール-5'-リン酸によるリシンのピリドキシル化と続いてNaBH4を用いる還元を含む。
【0028】
アルギニン残基のグアニジン基は、2,3-ブタンジオン、フェニルグリオキザールおよびグリオキザールのような試薬を用いて複素環縮合産物の形成により改変することができる。
【0029】
カルボキシル基は、O-アシルイソウレア形成を経由するカルボジイミド活性化とその後、例えば対応するアミドへの続いての誘導体化により改変することができる。
【0030】
トリプトファン残基は、例えばN-ブロモスクシンイミドによる酸化または2-ヒドロキシ-5-ニトロベンジルブロミドまたはハロゲン化スルフェニルによるインドール環のアルキル化により改変することができる。他方、チロシン残基は、テトラニトロメタンによるニトロ化により改変して3-ニトロチロシン誘導体を形成することができる。
【0031】
ヒスチジン残基のイミダゾール環の改変は、ヨード酢酸誘導体によるアルキル化またはジエチルピロカルボン酸によるN-カルベトキシル化により改変することができる。
【0032】
ペプチド合成中に非天然アミノ酸および誘導体を組込む例は、限定するものでないが、ノルロイシン、4-アミノ酪酸、4-アミノ-3-ヒドロキシ-5-フェニルペンタン酸、6-アミノヘキサン酸、t-ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、4-アミノ-3-ヒドロキシ-6-メチルヘプタン酸;2-チエニルアラニンおよび/またはアミノ酸のD-異性体の使用を含む。
【0033】
本発明のペプチドはCDVから誘導することができる。あるいは、ペプチドまたはペプチドエピトープの組合わせは、組換えDNA技術により産生させることができる。しかし、ペプチドは当技術分野で公知の方法を使い合成によって製造するのが好ましい。例えば、ペプチドは、溶液合成または固相合成を使って合成することができる(例えば、Nicholsonにより編集されBlackwell Scientific Publicationsにより出版された標題「Synthetic Vaccines(合成ワクチン)」の出版物に含まれる、AthertonおよびSheppardによる標題「Peptide Synthesis(ペプチド合成)」の第9章に記載)。好ましくは固相支持体を利用し、該固相支持体は、小比率のジビニルベンゼン(例えば1%)により架橋されていてもよいポリスチレンゲルビーズであってもよく、さらにジクロロメタンのような親油性溶媒またはジメチルホルムアミド(DMF)のようなより極性の高い溶媒により膨潤している。該ポリスチレンはクロロメチルまたはアミノメチル基の官能基を付すことができる。あるいは、DMFおよび他の双極性非プロトン性溶媒により高度に溶媒和されかつ膨潤した、架橋されかつ官能基の付されたポリジメチルアクリルアミドゲルを使う。不活性なポリスチレンビーズの表面に通常は移植するかまたは他の方法で結合したポリエチレングリコールを基材にして、他の支持体を利用することができる。好ましい形態では、PAL-PEG-PS、PAC-PEG-PS、KA、KRまたはTGRから選択される市販の固体支持体または樹脂を使用することができる。
【0034】
固相合成において使用するのは可逆性ブロッキング基であって、該可逆性ブロッキング基はα-アミノ、カルボキシまたは側鎖官能基の望まない反応性をマスキングしかつアミノ酸およびペプチドを不活性化するそれらの双極性を破壊する二重の機能を有する。そのような官能基は、RCO-OCMe3-COの構造をもつt-ブチルエステルから選択することができる。RCO-OCH2-C6H6の構造をもつ対応するベンジルエステルならびにベンジルオキシカルボニルまたはZ-誘導体として知られるC6H6CH2-OCO-NHRの構造およびt-ブトキシカルボニルまたはBoc誘導体として知られるMe3-COCO-NHRの構造を有するウレタンも使用することができる。フルオレニルメタノールおよび特にフルオレニル-メトキシカルボニルまたはFmoc基の誘導体も使用することができる。これらのタイプの保護基はそれぞれ、お互いが存在するときに独立して切断することができるのでしばしば使用され、例えばBOC-ベンジルおよびFmoc-三級ブチルの保護方法がある。
【0035】
保護されたアミノ酸またはペプチドのアミノ基およびカルボキシ基と連結する縮合剤に対する対照(reference)も作るべきである。これは、カルボキシ基を活性化して自発的に遊離した一級または二級アミンと反応するようにして行う。この目的のためには、p-ニトロフェノールおよびペンタフルオロフェノールから誘導したような活性化エステルを使うことができる。それらの反応性は、1-ヒドロキシベンゾトリアゾールのような触媒を加えて増加することができる。トリアジンDHBTのエステル(上記のNicholsonの参考文献の215-216頁に記述された)も使うことができる。他のアシル化種は、縮合試薬によるカルボン酸(すなわち、N-α保護したアミノ酸またはペプチド)の処理によりin situで形成させ、直ちにアミノ成分(カルボキシまたはC-保護されたアミノ酸またはペプチド)と反応させる。ジシクロヘキシルカルボジイミド、BOP試薬(Nicholson参考文献の216頁を参照)、O'ベンゾトリアゾール-N,N,N',N'-テトラメチル-ウロニウムヘキサフルオロリン酸(HBTU)およびその類似体のテトラフルオロホウ酸が縮合剤としてしばしば使用される。
【0036】
最初のアミノ酸の固相支持体への結合は、BOC-アミノ酸を使っていずれかの適当な方法で行う。ある方法では、BOCアミノ酸は、そのトリエチルアンモニウム塩を樹脂とともに暖めることによって、クロロメチル樹脂と結合させる。Fmoc-アミノ酸は、同様の方法でp-アルコキシベンジルアルコール樹脂と結合させることができる。あるいは、様々な連結剤または「ハンドル(handle)」を使って、最初のアミノ酸を樹脂に結合することができる。この点については、アミノメチルポリスチレンと連結したp-ヒドロキシメチルフェニル酢酸をこの目的に使うことができる。
【0037】
本明細書を通じて、用語「含んでなる(comprise)」、または「含んでなる(comprises)」または「含んでなる(comprising)」のような変化は、述べられたエレメント、完全体もしくは段階、またはエレメント、完全体または段階の群の包含を意味し、いずれの他のエレメント、完全体もしくは段階またはエレメント、完全体または段階の群の排除を意味するものでないことは理解されるであろう。
【実施例】
【0038】
本発明の性質がより容易に理解されるように、本発明の好ましい型を、以下の限定的ではない実施例を参照しながら記述する。
【0039】
実施例1
Tヘルパー細胞エピトープの同定
方法および結果:
イヌT細胞エピトープ94を同定するために、イヌジステンパーウイルス(CDV)の融合タンパク質の全配列を包含する17残基オーバーラップするペプチドを設計した。同定のために、N-末端から始まるように17merペプチドに順番に番号を付した。Barrettら、1987(Virus Res. 8, 373-386)により決定されたCDVの融合タンパク質の配列を図1に示す。該ペプチドを、1回分のワクチンとして生CDVを含有するCanvacTM 3 (CSL Limited)を用いて免疫化したイヌ由来の末梢血リンパ球(PBMC)を用いるT細胞増殖アッセイに使用した。
【0040】
最初に、4匹のイヌを用いた。これらは、各ワクチン接種の間を4〜6週間あけて、1回分のワクチンとしてCanvacTM 3 を用いて2回追加免疫した。各追加免疫接種の後にイヌから採血し、ペプチドに対してPBMCを試験した。ペプチドに対する有意な増殖は見られなかった。
【0041】
CDVはリンホトロピックウイルスであるとの報告がなされ、前記のワクチンは生CDVからなるため、有意な数の前駆体T細胞の末梢系への侵入を防ぐリンパ性器官中に生CDVが隔離されている可能性があった。末梢血抗CDV T細胞の数を増加させるため、イヌを熱で死滅させたCDV(ウイルス培養培地由来のペレットとして得た。CSL Limited)を用いて追加免疫した。2週間後、イヌから採血し、PBMCをペプチドに対する増殖に関して試験した。再び、該ペプチド抗原に対する増殖は見られなかった。
【0042】
別の方策を用いて、該CDVペプチドを認識する特異的T細胞の前駆体数を増加させた。過剰に免疫化したこれらのイヌから得た新鮮なPBMCを、94のペプチド全てのプールにより、37℃で30分間にわたりin vitroで刺激した。続いて細胞を洗浄して過剰のペプチドの全てを除去し、7日間培養した。次いで、このT細胞の集団を、自己由来APCについて、前記抗原の単一のペプチド全てについて試験した。表1は、有意な増殖レベル(刺激指数>2)の見られたペプチドを示している。
【0043】
この観察を確かめるため、死滅したウイルスの投与を受けた5週間後に、同じ4匹のイヌから再び採血した。PBMCを、94のペプチド全てのプール、またはペプチド21〜40のプール(活性の大部分がこの領域に存在していたため)のいずれかを用いて刺激して7日間培養した後、刺激したT細胞を個々のペプチドについて試験した。全てのペプチドについて有意な刺激指数が得られ、上記結果を裏付けた。1回分のワクチンとして3の1用量のみを受けたさらに4匹のイヌを、in vitro刺激法を用いて試験したところ、4匹のイヌの全てが表2に示したペプチドの大部分に応答した。
【0044】
上記のペプチドを同様に、追加のイヌに由来する細胞についても試験し、表3の結果を得た。ペプチドP64、P74およびP75もまた、CDVで免疫化された、多様な種のイヌ(表4)に由来する末梢血単核細胞と強力に反応することを示しており、それ故に強力なT-ヘルパーエピトープとして同定される。
【表1】
【表2】
【表3】
【表4】
【0045】
再度、同じペプチドと追加のペプチドP32を、追加のイヌに由来する細胞に対して試験した。これらのペプチドもまた同様に、CDVで免疫化された、多様な種のイヌ(表5)に由来する末梢血単核細胞と強力に反応し、それ故に強力なT-ヘルパーエピトープとして同定される。
【0046】
結論として、26種のペプチドが、CDV融合タンパク質中のイヌTヘルパー細胞のエピトープとして同定された。これらの各ペプチドの配列を表6に示す。
【0047】
これらのTヘルパー細胞エピトープは、動物用ワクチン(特にイヌ用ワクチン)の成分として、単に合成ペプチドに基づくワクチンとして、またはより複雑な抗原を含有するワクチンへの添加物としてのいずれかにおいて有用性を有するであろう。
【表5】
【表6】
【0048】
同定されたT細胞エピトープから選択した配列を、結合したB細胞エピトープに対する抗体応答を誘導する能力について試験した。試験は、イヌにおける抗体応答の評価により行った。T細胞エピトープを、B細胞エピトープLHRH(黄体形成ホルモン放出ホルモン)へ、N-末端のT細胞エピトープと、カルボキシル末端に位置させたLHRHと結合させることによって連結した。
【0049】
ペプチドは、Fmoc保護された標準的化学物質を用いて合成した。全てのペプチドを少なくとも80%の純度まで精製し、生成物をマススペクトル法によりチェックした。
【0050】
該ペプチドを、連続したT細胞-B細胞決定基として作成した。ピロGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-GlyのLHRH配列、またはその変異型を、各CDV-Tヘルパーエピトープのカルボキシル末端と結合させた。
【0051】
いくつかのTヘルパーエピトープのin vivoでの評価は、Tヘルパー-LHRH配列でイヌにワクチン接種することにより2回行った。
【0052】
実施例2 (K9-5試験)
全部で14匹の、オスとメスが混在するイヌをこの試験で用いた。全てのイヌは予め生CDVワクチンを接種されており、LHRHに対してもワクチン接種されていた。
【0053】
ワクチン製剤
CDV由来の試験ペプチドP25、P27、P35は、各TヘルパーエピトープのC末端でLHRHを有するよう合成した。使用したLHRH配列は全長が10アミノ酸の天然LHRHであった。各ワクチン構築物は、リピートマラリアB細胞エピトープ(配列を以下の表に示す)に結合したマウスインフルエンザT細胞エピトープを含む対照ペプチドとともに純度80〜90%まで精製した。全てのペプチドを4M尿素に溶解させ、その後適当な容量まで滅菌生理食塩水により希釈し、用量1mLあたり40nmoleとした。IscomatrixTMをアジュバントとしてチオメルサール保存剤(0.01%)とともに終濃度150μg/用量1mLで添加した。
【0054】
ISCOMTM、すなわち免疫刺激性複合体(Barr, SjolanderおよびCox, 1998, Advanced Drug Delivery Systems 32:247-271)は十分に特性決定された種類のアジュバントで、リン脂質、コレステロールおよびサポニンの複合体と、通常は複合対内に取り込まれたタンパク質を含有する。タンパク質抗原の不在下で複合体が形成される場合、この複合体をIscomatrixTMと称する。このアジュバント調製物中で使用されたサポニンはQuil Aであった。
【0055】
ワクチン接種、血液サンプルおよびアッセイ
全てのイヌに1mLの用量で首筋へ送達してワクチン接種した。ワクチン接種は0週と4週に行い、試験期間の間隔で静脈血サンプルを得た。
【0056】
有効なT細胞へのヘルプを、ELISAによってLHRHに対する抗体応答を測定して決定した。ワクチンに基づくペプチドの生物学的有効性を、メスのイヌおける黄体ホルモンおよびオスのイヌにおけるテストステロンのレベルを測定して決定した。
【表7】
【0057】
結果
異なるワクチンを用いて予め免疫化したことにより、全てのイヌにおいて以前から存在している低レベルでのLHRHに対する抗体が見られた。対照群のイヌでは抗体レベルがゆっくり減少した。
【0058】
P25-LHRH、P27-LHRHおよびP35-LHRHで免疫化したイヌは全て、B細胞エピトープ(LHRH)に対する強力な抗体応答を示した。この応答は追加免疫の接種後6週間まで持続した(表8参照)。
【0059】
ワクチンの生物学的効力が、黄体ホルモンとテストステロンのレベルが有意に減少したことで実証された(表9と表10を参照)。
【表8】
【表9】
【表10】
【0060】
CDVのFタンパク質から選択したT細胞エピトープの、イヌにおいて抗体応答を誘導するT細胞ヘルプを提供する際の有効性は、同定した配列が機能性であることを証明する。またこれらの結果は、in vivo活性を有するTヘルパーエピトープ配列の同定のための、in vitroスクリーニング法の科学的アプローチと有用性が妥当であること示す。
【0061】
実施例3 (K9-8試験)
全部で35匹の、オスとメスが混在するイヌをこの試験において用いた。全てのイヌを予め生CDVワクチンでワクチン接種しておいたが、LHRHに対してはワクチン接種をしていなかった。
【0062】
ワクチン製剤
Tヘルパー細胞エピトープを、天然の10アミノ酸配列の、2〜10のアミノ酸を含有するトランケート型のLHRHと以下のように結合させた:
2〜10 LHRH His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly
全てのワクチンは実施例2と同様に配合した、すなわちワクチンの1mL用量あたり40nmoleのペプチド、150μgのIscomatrixTM、および保存剤としてのチオメルサールを含有していた。
【0063】
イヌをペプチドのプールでワクチン接種した場合には、各ペプチドの濃度は等しい濃度となるように調節し、全量で、1mL用量あたり40nmoleのLHRHエピトープとなるように調節した。
【0064】
ワクチン接種、血液サンプルおよびアッセイ
全てのイヌに1mLの用量で首筋へ送達してワクチン接種した。ワクチン接種は0週と4週に行い、試験期間の間隔で静脈血サンプルを得た。
【0065】
有効なT細胞へのヘルプを、ELISAによってLHRHに対する抗体応答を測定して決定した。ワクチンに基づくペプチドの生物学的有効性を、メスのイヌにおける黄体ホルモンおよびオスのイヌにおけるテストステロンのレベルを測定して決定した。
【表11】
【0066】
結果
LHRHに対する強力な抗体応答が、T細胞エピトープP25、P27、P35、P62、P75、およびT細胞エピトープP25、P27およびP35を組み合わせて含むT細胞-LHRHペプチドのプールを有するT細胞LHRH構築物で免疫化したイヌにおいて見られた(表12を参照)。
【0067】
P2-LHRHペプチドおよびP8-LHRHペプチドで免疫化したイヌにおいては、低いかまたは検出不能な抗体応答がみられた(表12を参照)。このことは、これらのT細胞ペプチドはビーグル-フォックスハウンド種のイヌによっては認識されないことを示すと結論付けたが、この結論は、他の種のイヌに由来するPBMCを用いてT細胞ペプチドを同定したことに合致する。ビーグルフォックスハウンドドッグにおける最初のスクリーニングでは、この種のイヌはこれら2種のT細胞エピトープに応答しないことが示された。
【0068】
ペプチドワクチン分野の当業者にはよく理解される通り、個々のペプチドに対する応答は遺伝子によって決定される。クラスII主要組織適合性複合体(MHCII)は多型性である。クラスII分子は細胞表面でT細胞に対する表示のためのペプチドと結合するよう機能する。これはヘルパーT細胞を含むT細胞の活性化プロセスの一部として必要である。MHCクラスIIの対立遺伝子型は別のペプチド抗原のセットと結合し、従って、これらの抗原の応答は遺伝子によって決定される。このように、この結果は、ビーグル-フォックスハウンド種のイヌはP2およびP8に応答するための好適なMHC-II対立遺伝子を有しないが、これらのペプチドを同定するために用いた、プードルシチヅ(shitzu)種など他種のイヌは有することを示すと解釈し得る。
【0069】
対照のイヌにおいては、試験期間中にLHRHに対する抗体レベルに変化は見られず、ホルモンレベルもイヌの年齢、性別に対して正常レベル内であった(表12参照)。
【表12】
【0070】
【0071】
実施例4
ペプチドワクチン中に取り込まれたTh-エピトープの認識を実証するためのin vitro T細胞増殖アッセイ
ペプチド免疫原内のTh-エピトープの認識を実証するために、ペプチドワクチンで免疫化したイヌ(実施例2のイヌ)から得たPBMCを、各Th-エピトープに対して試験した。アッセイは、PBMCを富化することなく行った。フィコールグラジエント精製により得たPBMCを、各Th-エピトープおよびそのトランケート型に対して直接試験した。この研究により、ペプチドワクチンで免疫化したイヌの全てが、取り込まれたTh-エピトープに対して応答することが示され、T細胞活性が各配列中に存在していることが確認された(図2〜4)。各Th-エピトープのトランケート型も同様に試験し、配列内のT細胞活性をより詳細に同定した。P25については、17残基の全配列のほうが、より短い15残基や12残基のペプチド(各ペプチドは配列のN末端から切断)よりも良好であることが観察された(図2)。この結果は、T細胞活性は17残基のペプチドのN末端または中間に対するものであることを示唆する。
【0072】
P27についても同様のことが観察され、長さが17残基の配列のほうが、N末端から切断されたより短い15残基のペプチドよりも良好なシミュレータであった(図3)。この観察もまた、再度、T細胞活性は、全長ペプチドの中間またはN末端に対して存在し得ることを示唆していた。
【0073】
P35とその短い型の場合には、1匹のイヌ(番号102)を除いた他の3匹のイヌは、12残基のペプチドにも全長17残基のペプチドにも同じように応答した(図4)。番号102のイヌにおいては、15残基のペプチドは全長ペプチドよりも刺激性が高かった。この結果から、P35の配列中で最初の2つの残基は必須ではなく、活性はペプチドの中間またはC末端に対するものであることが推定され得る。
【0074】
実施例5
BALB/cマウスにおける試験
実施例3で使用した、CDV-Fから誘導したTh-エピトープとLHRHを有するイヌ用ワクチンを同様に用いてBALB/cマウスを免疫化して、Th-エピトープが異なる動物種において機能性であるか否かを調べた。
【0075】
ワクチン製剤
全てのワクチンは、100μlの用量で2.7nmoleのペプチドと10μgのIscomatrixTMとチオメルソールを保存剤として含有するようにさらに希釈した以外は、実施例3と同じように配合した。
【0076】
ワクチン接種、血液サンプルおよびアッセイ
マウスに、尾の付け根で100μlのワクチンを接種した。ワクチン接種は0週と4週に行い、各ワクチン接種の後に間隔を開けて動物の眼窩後方(retro-orbital)叢から採血した。有効なT細胞ヘルプは、ELISAによりLHRHに対する抗体応答を測定して決定した。
【0077】
結果
P25-LHRHで免疫化したマウスと、P25-LHRH、P27-LHRHPおよびP35-LHRHを含むペプチドプールで免疫化したマウスでは、LHRHに対する高い抗体力価が生じた。ペプチドP35およびP75では生じた抗体力価は低く、一方P2、P8およびP62で免疫化したマウスでは、抗LHRH抗体のレベルは検出限界範囲外であった(表13)。
【0078】
当業者には、広く記載した本発明の意図と範囲から逸脱することなく、具体的な実施例において示された本発明に対して多数の変形および/または改変をなし得ることが理解されよう。従って、本実施形態はあらゆる観点で、例示的であり制限的ではないと解されるべきである。
【表13】
【技術分野】
【0001】
本発明は、イヌジステンパーウイルス(Canine Distemper Virus)(CDV)由来のTヘルパー細胞エピトープに関する。本発明は少なくとも1つのTヘルパー細胞エピトープならびに場合によってはB細胞エピトープおよび/またはCTLエピトープを含む組成物に関する。
【背景技術】
【0002】
有効な抗体応答を誘導できるいずれのペプチドも、免疫系により認識されるエピトープとして知られるアミノ酸の特定の配列を含有しなければならない。特に、抗体応答については、エピトープは、Bリンパ球表面に存在する特異的な免疫グロブリン(Ig)受容体により認識される必要がある。これらの細胞がこのエピトープに対して特異的抗体を産生する能力のある形質細胞に最終的に分化するからである。これらのB細胞エピトープに加えて、免疫原は、抗原提示細胞(APC)がヘルパーTリンパ球上に存在する特異的受容体に対して提示するエピトープも含有しなければならない、というのはヘルパーTリンパ球は、B細胞が抗体産生細胞に分化するために必要とするシグナルを提供する必要があるからである。
【0003】
ウイルス感染症の場合におよび多くの癌の場合に、抗体の再生利用は限られており、その免疫系はウイルス感染細胞または癌細胞を死滅し得る細胞傷害性T細胞(CTL)と応答する。ヘルパーT細胞と同じように、CTLは最初、表面上に提示された特異的なペプチドエピトープをもつAPCとの相互作用により活性化されるが、この時、MHCクラスII分子よりもむしろMHCクラスIに関連する。活性化されると、CTLは、上記ペプチド/クラスI複合体を保持する標的細胞を捕らえて(engage)、溶解を引き起こし得る。このプロセスにおいてヘルパーT細胞がある役割を果たすことも明らかになっており;APCがCTLを活性化できるようになる前に、該APCは最初にヘルパーT細胞から必要な同時刺激性分子(costimulatory molecules)の発現をアップレギュレートするためのシグナルを受けなければならない。
【0004】
ヘルパーT細胞エピトープは、APC表面上に存在して主要組織適合性複合体(MHC)のクラスII遺伝子によりコードされた分子と結合する。次いで、Tヘルパーリンパ球表面の特異的T細胞受容体(TCR)は、クラスII分子とペプチドエピトープとの複合体を認識する。このようにして、MHC分子と関係をもつ抗原エピトープを提示されたT細胞は活性化され、Bリンパ球が分化するために必要なシグナルを提供することができる。伝統的にペプチド免疫原のためのヘルパーT細胞エピトープの供給源はペプチドが共有結合した担体タンパク質であるが、この結合過程は、結合プロセスにおいて抗原決定基を改変することおよび担体に対する抗体誘導にペプチドを指向する抗体を費やすことなどの、他の問題を引き起こしうる(Schutze, M. P., Leclerc, C. Jolivet, M. Audibert, F. Chedid, L. "Carrier-induced epitopic suppression, a major issue for future synthetic vaccines(担体誘導エピトープ抑制、将来の合成ワクチンに対する大きな論点)". J Immunol. 1985, 135, 2319-2322;DiJohn, D., Torrese, J. R. Murillo, J. Herrington, D. A.ら, "Effect of priming with carrier on response to conjugate vaccine(複合ワクチンへの応答に対する担体を用いた初回免疫の効果)". The Lancet. 1989, 2, 1415-1416)。さらに、調製時における無関係なタンパク質の使用は品質管理の議論を起こす。ペプチドワクチンの設計において適当な担体タンパク質を選ぶことは非常に重要であり、それらの選択は毒性および大規模生産の実施可能性などの因子により制限される。この方法には他の制限があり、結合可能なペプチド体のサイズおよび安全に投与できる担体の投与量が挙げられる(Audibert, F. a. C., L. 1984. "Modern approaches to vaccines. Molecular and chemical basis of virus virulence and immunogenicity(ワクチンへの現代的手法 ウイルス毒性および免疫原性の分子的および化学的基礎)"., Cold Spring Harbor Laboratory, New York.)。担体分子は強い免疫応答の誘導を可能にするが、これら担体分子はまた、抗ペプチド抗体応答の抑制のような望まない効果も随伴する(Herzenberg, L. A. and Tokuhisa, T. 1980. "Carrier-priming leads to hapten-specific suppression(担体初回免疫はハプテン特異的な抑制をもたらす)". Nature 285:664;Schutze, M. P., Leclerc, C., Jolivet, M., Audibert, F.およびChedid, L. 1985. "Carrier-induced epitopic suppression, a major issue for futrue synthetic vaccines(担体誘導エピトープ抑制、将来の合成ワクチンの大きな論点)". J Immunol 135:2319;Etlinger, H. M., Felix, A. M., Gillessen, D., Heimer, E.P., Just, M., Pink, J. R., Sinigaglia, F., Sturchler, D., Takacs, Gl, Trzeciak, A.ら, 1988. "Assessment in humans of a synthetic peptide-based vaccine against the sporozoite stage of the human malaria parasite, Plasmodium falciparum(ヒトマラリア寄生虫、Plasmodium falciparumの種虫(sporpzoite)段階に対する合成ペプチドに基づくワクチンのヒトにおける評価)". J Immunol 140:626)。
【0005】
従って、一般的に、免疫原は、細胞傷害性T細胞上に存在する表面Igによりまたは受容体により認識されるエピトープに加えて、ヘルパーT細胞により認識され得るエピトープも含有しなければならない。これらのエピトープのタイプは非常に種々であることを理解されるべきである。B細胞エピトープについては、B細胞受容体は元来の免疫原と直接結合するので、コンフォメーションが重要である。対照的に、T細胞により認識されるエピトープは、エピトープのコンフォメーションの完全さに依存するのでなく、CTLに対してはほぼ9個のアミノ酸の短い配列およびヘルパーT細胞に対しては長さの制限が少ない若干長い配列から構成される。これらのエピトープに対する要件は単に、該エピトープがクラスIまたはクラスII分子の結合クレフト(cleft:裂け目)とそれぞれ適合し得て、かつその複合体が次にT細胞受容体と結合し得ることである。クラスII分子の結合部位は両方末端が開放されていて、結合されるペプチドの長さにおける非常に大きな変化を許容し(Brown, J. H., T. S. Jardetzky, J. C. Gorga, L. J. Stern, R. G. Urban, J. L. Strominger and D. C. Wiley. 1993. "Three-dimensional structure of the human class II histocompatibility antigen HLA-DR1(ヒトのクラスII組織適合性抗原HLA-DR1の3次元構造), Nature 364:33)"、8個という短いアミノ酸残基のエピトープが報じられている(Fahrer, A. M., Geysen, H. M., White, D. O., Jackson, D. C.およびBrown, L. E. "Analysis of the requirements for class II-restricted T-cell recognition of a single determinant reveals considerable diversity in the T-cell response and degeneracy of peptide binding to I-Ed(単一決定基がクラスII制限されたT細胞を認識するための要件の分析は、T細胞応答のかなりの多様性およびI-Edへのペプチド結合の縮重(degeneracy)を表す)". J. Immunol 1995. 155:2849-2857)。
【0006】
イヌジステンパーウイルス(CDV)は、ネガティブ鎖RNAウイルスのパラミキソウイルス(paramyxovirus)ファミリーのモルビリウイルス(morbillivirus)サブグループに属する。このグループのメンバーである他のウイルスは麻疹ウイルス(measles virus)および牛疫ウイルス(rinderpest virus)である。ペプチドベースのワクチンの開発は、タンパク質配列からのBおよびT細胞エピトープの同定に大きな関心を喚起した。CDVのFプロテインのようなタンパク質由来のT細胞エピトープを使う原理は、若いイヌを早期にCDVに対して接種すると、それによって、このタンパク質上に存在するヘルパーT細胞エピトープに対して特異的なヘルパーT細胞を有するようになるだろうということにある。次いで1つ以上のエピトープを含有するワクチンに曝すと、それによって、存在するヘルパーT細胞の補強および従って、増強された免疫応答をもたらすであろう。しかし、このようなヘルパーT細胞エピトープは、初回刺激していない動物に投与して、さらに免疫応答を誘導することもできる。本発明者らは、CDV融合タンパク質の配列からイヌT細胞エピトープを同定し、そして、これらのエピトープを、特にイヌおよび関係種に対する、ペプチドベースのワクチンの設計に使い得るようにすることを目的とした。
【0007】
LHRH(黄体形成ホルモン放出ホルモン)は10個のアミノ酸長のペプチドホルモンであり、その配列は哺乳類に保存されている。該ホルモンは視床下部により分泌され、雄性および雌性両方の生殖生理学を制御する。LHRHベースの免疫避妊ワクチンの開発の原理は、LHRHに対する抗体がホルモンの黄体形成ホルモンの下垂体分泌および卵胞刺激ホルモンに対する作用をブロックし、哺乳類の生殖腺萎縮および不妊に導くという観察に基づく。
【0008】
開発されているほとんどのLHRHワクチンはタンパク質担体と化学的にコンジュゲートしたLHRHからなり、抗LHRH抗体の産生のためのT細胞ヘルプを提供する。LHRT-タンパク質担体コンジュゲートの接種を繰り返すと、「担体に誘導されるエピトープ抑制」として知られる現象により抗LHRH力価は低下することが分かっている。本発明の1つの目的は、ワクチンのタンパク質担体を、定義したTヘルパーエピトープ(THエピトープ)によって置換え、「担体に誘導されるエピトープ抑制」を排除することである。
【発明の概要】
【0009】
本発明者らは、それぞれがTヘルパー細胞エピトープを含有する、多数の17残基のペプチドを同定した。容易に認識されるように、大多数のこれらのペプチドは最小のTヘルパー細胞エピトープではない。典型的には、クラスII分子は8個のアミノ酸のような短いペプチド(Fahrerら, 1995、同書)、しかし通常は12〜19個のアミノ酸のペプチド(Chicz, R. M., Urban, R. G., Gorga, J. C,. Vignali, D. A. A., Lane, W. S.およびStrominger, J. L. 「Specificity and promiscuity among naturally processed peptides bound to HLA-DR alleles(HLA-DR対立遺伝子と結合した天然にプロセシングを受けたペプチド間の特異性および乱交雑性)」. J Exp Med 1993, 178, 27-47;Chicz, R. M., Urban, R. G., Lane, W. S., Gorga, J. C., Stern, L.J., Vignali, D. A. A.,およびStrominger, J. L. 「Predominant naturally processed peptides bound to HLA-DR1 are derived from MHC-related molecules and are heterogeneous in size(HLA-DRIと結合した、天然にプロセシングを受けた主なペプチドはMHC関係分子から誘導され、サイズは不均一である)」. Nature 1992, 358, 764-8)と結合していることが分かっているが、長さで25個までのアミノ酸のペプチドがクラスIIと結合していることも報じられている(Rammensee, H.G. 「Chemistry of peptide associated with class I and class II molecules.(クラスIおよびクラスII分子に関連したペプチドの化学)」 Curr Opin Immunol 1995, 7, 85-95におけるレビュー)。
【0010】
従って、長さが8〜25個の間のアミノ酸残基の範囲にあるペプチドエピトープがクラスII分子と結合し得る。より短いペプチドはより長い配列より低い活性を有し得る「コア」エピトープであり、より長い配列をNまたはC末端で切断して親配列と同じかまたはより高い活性を有するより短い配列を得ようとするのはつまらない試みでしかない。
【図面の簡単な説明】
【0011】
【図1】CDVの融合タンパク質のアミノ酸配列を示す。
【図2】P25-LHRHで免疫化したイヌからの、Th-エピトープP27およびそのトランケートされた型に対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
【図3】P27-LHRHで免疫化したイヌからの、Th-エピトープP25およびそのトランケートされた15残基のペプチドに対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
【図4】P35-LHRHで免疫化したイヌからの、Th-エピトープP25およびそのトランケートされた型に対する刺激指数(X-軸はnmole/ウェルで示したペプチド濃度)を示す。
【発明を実施するための形態】
【0012】
従って第1の態様においては、本発明はTヘルパー細胞エピトープであって、上記エピトープは、
からなる群から選択されるペプチド配列内に含有されることを特徴とする。
【0013】
第2の態様においては、本発明は、動物に免疫応答を生起させるために使う組成物であって、上記組成物は少なくとも1種のTヘルパー細胞エピトープを含んでなり、少なくとも1種の該Tヘルパー細胞エピトープは、
からなる群から選択されるペプチド配列内に含有されることを特徴とする。
【0014】
本発明の好ましい実施形態においては、該組成物は
からなる群から選択される少なくとも1種のペプチドを含んでなる。
【0015】
該組成物はさらに少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープを含んでなることが、さらに好ましい。
【0016】
また他の好ましい実施形態においては、少なくとも1種の該B細胞エピトープおよび/または少なくとも1種の該CTLエピトープは少なくとも1種のTヘルパー細胞エピトープと連結している。該組成物はまた、各々が少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープを含んでなる複数のエピトープ構築物を含んでなることも好ましい。あるいは、該組成物は、各々が少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のCTLエピトープを含んでなる複数のエピトープ構築物を含んでなっていてもよい。
【0017】
該B細胞エピトープまたは該CTLエピトープはいずれのエピトープであってもよいことは理解されるであろう。本発明で好ましいB細胞エピトープはLHRH B細胞エピトープである。
【0018】
本発明の組成物は複数のTヘルパー細胞エピトープを含んでなってもよい。これらのエピトープは単一であっても一緒に連結して単一のポリペプチドを形成してもよい。エピトープが一緒に単一のポリペプチドに連結される場合、エピトープ同士は近接していてもまたはそれ自身はTヘルパー細胞エピトープの部分でない追加のアミノ酸により隔たれて位置してもよいことは理解されるであろう。
【0019】
ある実施形態においては、上に考察したように、Tヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープおよび/または少なくとも1つのCTLエピトープは、それらのエピトープが連結されている。この連結は、簡単なペプチドの共有結合により実施し得る。他の実施形態においては、エピトープは、最も好ましくはその開示が参照により本明細書に組み入れられるPCT/AU98/00076に記載されたように、重合されている。
【0020】
さらに他の好ましい実施形態においては、該組成物はさらに製薬上許容される賦形剤、好ましくはアジュバントを含んでなる。
【0021】
さらなる態様においては、本発明は動物に免疫応答を誘導する方法であって、動物に本発明の第2の態様の組成物を投与することを含んでなることを特徴とする方法である。
【0022】
製薬上許容される担体または希釈剤は、経口、直腸、鼻、局所(バッカルおよび舌下錠を含む)、膣、非経口(皮下、筋内、静脈内、皮内、くも膜下および硬膜外を含む)投与に適した組成物に使われる担体または希釈剤を含む。これらは受容者に無毒な投与量および濃度にて用いられる。製薬上許容される担体または希釈剤の代表的な例は、限定するものでないが、水、好ましくは生理的pHに緩衝された等張液(リン酸緩衝生理食塩水またはTris緩衝生理食塩水など)を含み、また、マンニトール、ラクトース、トレハロース、デキストロース、グリセロール、エタノールまたはポリペプチド(ヒト血清アルブミンなど)の1以上を含んでもよい。該組成物は単位投与形態で好都合に提示することができ、また製薬業界でよく知られたのいずれかの方法により調製することができる。
【0023】
上記のように、該組成物はアジュバントを含むことが好ましい。理解されるように、「アジュバント」は、ワクチン組成物の免疫原性および効能を増強する1以上の基質を含む組成物を意味する。適当なアジュバントの例は、限定するものでないが、スクアランおよびスクアレン(または他の動物由来の油);ブロックコポリマー;Tween(登録商標)-80のような洗剤;Quil(登録商標) A、DrakeolまたはMarcolのような鉱質油、ピーナッツ油のような植物油;Corynebacterium parvumのようなCorynebacterium由来のアジュバント;Propionibacterium acneのようなPropionibacterium由来のアジュバント;Mycobacterium bovis(Bacille Calmette and GuerinまたはBCG);インターロイキン2およびインターロイキン12のようなインターロイキン;インターロイキン1のようなモノカイン;腫瘍壊死因子;γインターフェロンのようなインターフェロン;サポニン-水酸化アルミニウムまたはQuil-A水酸化アルミニウムのような組合わせ;リポソーム;ISCOMアジュバント;放線菌細胞壁抽出物;ムラミルジペプチドまたは他の誘導体のような合成糖ペプチド;アビリジン;リピドA誘導体;硫酸デキストラン;DEAE-デキストランまたはリン酸アルミニウムを有するもの;Carbopol'EMAのようなカルボキシポリメチレン;Neocryl A640(例えば、米国特許第5,047,238号)のようなアクリルコポリマーエマルジョン;ワクシニアまたは動物ポックスウイルスタンパク質;コレラ毒のようなサブウイルス粒子アジュバント、またはそれらの混合物を含む。
【0024】
当業者には認識されるように、本発明のペプチドは生物学的活性の完全な抑止をすることなしに改変することができる。これらの改変は、付加、欠失および置換、特に保存的置換を含む。Tヘルパー細胞エピトープとしての活性の完全な消失を生じない改変を含むペプチドは本発明の範囲内にあると考えられる。
【0025】
置換の概念は当技術分野では周知であるが、予想される置換の形式を以下に掲げる。
【0026】
予想される他のタイプのペプチドの改変は、限定するものでないが、側鎖の改変、ペプチド合成中の非天然アミノ酸および/またはそれらの誘導体の組込みならびに架橋剤の使用およびペプチドにコンフォメーションの制限を課する他の方法を含む。
【0027】
本発明で考えられる側鎖改変の例は、限定するものでないが、アルデヒドとの反応に続いてNaBH4を用いて還元する還元アルキル化によるようなアミノ基の改変;メチルアセトイミダート(methylacetimidate)によるアミド化;無水酢酸によるアシル化;シアン酸塩によるアミノ基のカルバモイル化;2,4,6-トリニトロベンゼンスルホン酸(TNBS)によるアミノ基のトリニトロベンジル化;無水コハク酸および無水テトラヒドロフタル酸によるアミノ基のアシル化;およびピリドキサール-5'-リン酸によるリシンのピリドキシル化と続いてNaBH4を用いる還元を含む。
【0028】
アルギニン残基のグアニジン基は、2,3-ブタンジオン、フェニルグリオキザールおよびグリオキザールのような試薬を用いて複素環縮合産物の形成により改変することができる。
【0029】
カルボキシル基は、O-アシルイソウレア形成を経由するカルボジイミド活性化とその後、例えば対応するアミドへの続いての誘導体化により改変することができる。
【0030】
トリプトファン残基は、例えばN-ブロモスクシンイミドによる酸化または2-ヒドロキシ-5-ニトロベンジルブロミドまたはハロゲン化スルフェニルによるインドール環のアルキル化により改変することができる。他方、チロシン残基は、テトラニトロメタンによるニトロ化により改変して3-ニトロチロシン誘導体を形成することができる。
【0031】
ヒスチジン残基のイミダゾール環の改変は、ヨード酢酸誘導体によるアルキル化またはジエチルピロカルボン酸によるN-カルベトキシル化により改変することができる。
【0032】
ペプチド合成中に非天然アミノ酸および誘導体を組込む例は、限定するものでないが、ノルロイシン、4-アミノ酪酸、4-アミノ-3-ヒドロキシ-5-フェニルペンタン酸、6-アミノヘキサン酸、t-ブチルグリシン、ノルバリン、フェニルグリシン、オルニチン、サルコシン、4-アミノ-3-ヒドロキシ-6-メチルヘプタン酸;2-チエニルアラニンおよび/またはアミノ酸のD-異性体の使用を含む。
【0033】
本発明のペプチドはCDVから誘導することができる。あるいは、ペプチドまたはペプチドエピトープの組合わせは、組換えDNA技術により産生させることができる。しかし、ペプチドは当技術分野で公知の方法を使い合成によって製造するのが好ましい。例えば、ペプチドは、溶液合成または固相合成を使って合成することができる(例えば、Nicholsonにより編集されBlackwell Scientific Publicationsにより出版された標題「Synthetic Vaccines(合成ワクチン)」の出版物に含まれる、AthertonおよびSheppardによる標題「Peptide Synthesis(ペプチド合成)」の第9章に記載)。好ましくは固相支持体を利用し、該固相支持体は、小比率のジビニルベンゼン(例えば1%)により架橋されていてもよいポリスチレンゲルビーズであってもよく、さらにジクロロメタンのような親油性溶媒またはジメチルホルムアミド(DMF)のようなより極性の高い溶媒により膨潤している。該ポリスチレンはクロロメチルまたはアミノメチル基の官能基を付すことができる。あるいは、DMFおよび他の双極性非プロトン性溶媒により高度に溶媒和されかつ膨潤した、架橋されかつ官能基の付されたポリジメチルアクリルアミドゲルを使う。不活性なポリスチレンビーズの表面に通常は移植するかまたは他の方法で結合したポリエチレングリコールを基材にして、他の支持体を利用することができる。好ましい形態では、PAL-PEG-PS、PAC-PEG-PS、KA、KRまたはTGRから選択される市販の固体支持体または樹脂を使用することができる。
【0034】
固相合成において使用するのは可逆性ブロッキング基であって、該可逆性ブロッキング基はα-アミノ、カルボキシまたは側鎖官能基の望まない反応性をマスキングしかつアミノ酸およびペプチドを不活性化するそれらの双極性を破壊する二重の機能を有する。そのような官能基は、RCO-OCMe3-COの構造をもつt-ブチルエステルから選択することができる。RCO-OCH2-C6H6の構造をもつ対応するベンジルエステルならびにベンジルオキシカルボニルまたはZ-誘導体として知られるC6H6CH2-OCO-NHRの構造およびt-ブトキシカルボニルまたはBoc誘導体として知られるMe3-COCO-NHRの構造を有するウレタンも使用することができる。フルオレニルメタノールおよび特にフルオレニル-メトキシカルボニルまたはFmoc基の誘導体も使用することができる。これらのタイプの保護基はそれぞれ、お互いが存在するときに独立して切断することができるのでしばしば使用され、例えばBOC-ベンジルおよびFmoc-三級ブチルの保護方法がある。
【0035】
保護されたアミノ酸またはペプチドのアミノ基およびカルボキシ基と連結する縮合剤に対する対照(reference)も作るべきである。これは、カルボキシ基を活性化して自発的に遊離した一級または二級アミンと反応するようにして行う。この目的のためには、p-ニトロフェノールおよびペンタフルオロフェノールから誘導したような活性化エステルを使うことができる。それらの反応性は、1-ヒドロキシベンゾトリアゾールのような触媒を加えて増加することができる。トリアジンDHBTのエステル(上記のNicholsonの参考文献の215-216頁に記述された)も使うことができる。他のアシル化種は、縮合試薬によるカルボン酸(すなわち、N-α保護したアミノ酸またはペプチド)の処理によりin situで形成させ、直ちにアミノ成分(カルボキシまたはC-保護されたアミノ酸またはペプチド)と反応させる。ジシクロヘキシルカルボジイミド、BOP試薬(Nicholson参考文献の216頁を参照)、O'ベンゾトリアゾール-N,N,N',N'-テトラメチル-ウロニウムヘキサフルオロリン酸(HBTU)およびその類似体のテトラフルオロホウ酸が縮合剤としてしばしば使用される。
【0036】
最初のアミノ酸の固相支持体への結合は、BOC-アミノ酸を使っていずれかの適当な方法で行う。ある方法では、BOCアミノ酸は、そのトリエチルアンモニウム塩を樹脂とともに暖めることによって、クロロメチル樹脂と結合させる。Fmoc-アミノ酸は、同様の方法でp-アルコキシベンジルアルコール樹脂と結合させることができる。あるいは、様々な連結剤または「ハンドル(handle)」を使って、最初のアミノ酸を樹脂に結合することができる。この点については、アミノメチルポリスチレンと連結したp-ヒドロキシメチルフェニル酢酸をこの目的に使うことができる。
【0037】
本明細書を通じて、用語「含んでなる(comprise)」、または「含んでなる(comprises)」または「含んでなる(comprising)」のような変化は、述べられたエレメント、完全体もしくは段階、またはエレメント、完全体または段階の群の包含を意味し、いずれの他のエレメント、完全体もしくは段階またはエレメント、完全体または段階の群の排除を意味するものでないことは理解されるであろう。
【実施例】
【0038】
本発明の性質がより容易に理解されるように、本発明の好ましい型を、以下の限定的ではない実施例を参照しながら記述する。
【0039】
実施例1
Tヘルパー細胞エピトープの同定
方法および結果:
イヌT細胞エピトープ94を同定するために、イヌジステンパーウイルス(CDV)の融合タンパク質の全配列を包含する17残基オーバーラップするペプチドを設計した。同定のために、N-末端から始まるように17merペプチドに順番に番号を付した。Barrettら、1987(Virus Res. 8, 373-386)により決定されたCDVの融合タンパク質の配列を図1に示す。該ペプチドを、1回分のワクチンとして生CDVを含有するCanvacTM 3 (CSL Limited)を用いて免疫化したイヌ由来の末梢血リンパ球(PBMC)を用いるT細胞増殖アッセイに使用した。
【0040】
最初に、4匹のイヌを用いた。これらは、各ワクチン接種の間を4〜6週間あけて、1回分のワクチンとしてCanvacTM 3 を用いて2回追加免疫した。各追加免疫接種の後にイヌから採血し、ペプチドに対してPBMCを試験した。ペプチドに対する有意な増殖は見られなかった。
【0041】
CDVはリンホトロピックウイルスであるとの報告がなされ、前記のワクチンは生CDVからなるため、有意な数の前駆体T細胞の末梢系への侵入を防ぐリンパ性器官中に生CDVが隔離されている可能性があった。末梢血抗CDV T細胞の数を増加させるため、イヌを熱で死滅させたCDV(ウイルス培養培地由来のペレットとして得た。CSL Limited)を用いて追加免疫した。2週間後、イヌから採血し、PBMCをペプチドに対する増殖に関して試験した。再び、該ペプチド抗原に対する増殖は見られなかった。
【0042】
別の方策を用いて、該CDVペプチドを認識する特異的T細胞の前駆体数を増加させた。過剰に免疫化したこれらのイヌから得た新鮮なPBMCを、94のペプチド全てのプールにより、37℃で30分間にわたりin vitroで刺激した。続いて細胞を洗浄して過剰のペプチドの全てを除去し、7日間培養した。次いで、このT細胞の集団を、自己由来APCについて、前記抗原の単一のペプチド全てについて試験した。表1は、有意な増殖レベル(刺激指数>2)の見られたペプチドを示している。
【0043】
この観察を確かめるため、死滅したウイルスの投与を受けた5週間後に、同じ4匹のイヌから再び採血した。PBMCを、94のペプチド全てのプール、またはペプチド21〜40のプール(活性の大部分がこの領域に存在していたため)のいずれかを用いて刺激して7日間培養した後、刺激したT細胞を個々のペプチドについて試験した。全てのペプチドについて有意な刺激指数が得られ、上記結果を裏付けた。1回分のワクチンとして3の1用量のみを受けたさらに4匹のイヌを、in vitro刺激法を用いて試験したところ、4匹のイヌの全てが表2に示したペプチドの大部分に応答した。
【0044】
上記のペプチドを同様に、追加のイヌに由来する細胞についても試験し、表3の結果を得た。ペプチドP64、P74およびP75もまた、CDVで免疫化された、多様な種のイヌ(表4)に由来する末梢血単核細胞と強力に反応することを示しており、それ故に強力なT-ヘルパーエピトープとして同定される。
【表1】
【表2】
【表3】
【表4】
【0045】
再度、同じペプチドと追加のペプチドP32を、追加のイヌに由来する細胞に対して試験した。これらのペプチドもまた同様に、CDVで免疫化された、多様な種のイヌ(表5)に由来する末梢血単核細胞と強力に反応し、それ故に強力なT-ヘルパーエピトープとして同定される。
【0046】
結論として、26種のペプチドが、CDV融合タンパク質中のイヌTヘルパー細胞のエピトープとして同定された。これらの各ペプチドの配列を表6に示す。
【0047】
これらのTヘルパー細胞エピトープは、動物用ワクチン(特にイヌ用ワクチン)の成分として、単に合成ペプチドに基づくワクチンとして、またはより複雑な抗原を含有するワクチンへの添加物としてのいずれかにおいて有用性を有するであろう。
【表5】
【表6】
【0048】
同定されたT細胞エピトープから選択した配列を、結合したB細胞エピトープに対する抗体応答を誘導する能力について試験した。試験は、イヌにおける抗体応答の評価により行った。T細胞エピトープを、B細胞エピトープLHRH(黄体形成ホルモン放出ホルモン)へ、N-末端のT細胞エピトープと、カルボキシル末端に位置させたLHRHと結合させることによって連結した。
【0049】
ペプチドは、Fmoc保護された標準的化学物質を用いて合成した。全てのペプチドを少なくとも80%の純度まで精製し、生成物をマススペクトル法によりチェックした。
【0050】
該ペプチドを、連続したT細胞-B細胞決定基として作成した。ピロGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-GlyのLHRH配列、またはその変異型を、各CDV-Tヘルパーエピトープのカルボキシル末端と結合させた。
【0051】
いくつかのTヘルパーエピトープのin vivoでの評価は、Tヘルパー-LHRH配列でイヌにワクチン接種することにより2回行った。
【0052】
実施例2 (K9-5試験)
全部で14匹の、オスとメスが混在するイヌをこの試験で用いた。全てのイヌは予め生CDVワクチンを接種されており、LHRHに対してもワクチン接種されていた。
【0053】
ワクチン製剤
CDV由来の試験ペプチドP25、P27、P35は、各TヘルパーエピトープのC末端でLHRHを有するよう合成した。使用したLHRH配列は全長が10アミノ酸の天然LHRHであった。各ワクチン構築物は、リピートマラリアB細胞エピトープ(配列を以下の表に示す)に結合したマウスインフルエンザT細胞エピトープを含む対照ペプチドとともに純度80〜90%まで精製した。全てのペプチドを4M尿素に溶解させ、その後適当な容量まで滅菌生理食塩水により希釈し、用量1mLあたり40nmoleとした。IscomatrixTMをアジュバントとしてチオメルサール保存剤(0.01%)とともに終濃度150μg/用量1mLで添加した。
【0054】
ISCOMTM、すなわち免疫刺激性複合体(Barr, SjolanderおよびCox, 1998, Advanced Drug Delivery Systems 32:247-271)は十分に特性決定された種類のアジュバントで、リン脂質、コレステロールおよびサポニンの複合体と、通常は複合対内に取り込まれたタンパク質を含有する。タンパク質抗原の不在下で複合体が形成される場合、この複合体をIscomatrixTMと称する。このアジュバント調製物中で使用されたサポニンはQuil Aであった。
【0055】
ワクチン接種、血液サンプルおよびアッセイ
全てのイヌに1mLの用量で首筋へ送達してワクチン接種した。ワクチン接種は0週と4週に行い、試験期間の間隔で静脈血サンプルを得た。
【0056】
有効なT細胞へのヘルプを、ELISAによってLHRHに対する抗体応答を測定して決定した。ワクチンに基づくペプチドの生物学的有効性を、メスのイヌおける黄体ホルモンおよびオスのイヌにおけるテストステロンのレベルを測定して決定した。
【表7】
【0057】
結果
異なるワクチンを用いて予め免疫化したことにより、全てのイヌにおいて以前から存在している低レベルでのLHRHに対する抗体が見られた。対照群のイヌでは抗体レベルがゆっくり減少した。
【0058】
P25-LHRH、P27-LHRHおよびP35-LHRHで免疫化したイヌは全て、B細胞エピトープ(LHRH)に対する強力な抗体応答を示した。この応答は追加免疫の接種後6週間まで持続した(表8参照)。
【0059】
ワクチンの生物学的効力が、黄体ホルモンとテストステロンのレベルが有意に減少したことで実証された(表9と表10を参照)。
【表8】
【表9】
【表10】
【0060】
CDVのFタンパク質から選択したT細胞エピトープの、イヌにおいて抗体応答を誘導するT細胞ヘルプを提供する際の有効性は、同定した配列が機能性であることを証明する。またこれらの結果は、in vivo活性を有するTヘルパーエピトープ配列の同定のための、in vitroスクリーニング法の科学的アプローチと有用性が妥当であること示す。
【0061】
実施例3 (K9-8試験)
全部で35匹の、オスとメスが混在するイヌをこの試験において用いた。全てのイヌを予め生CDVワクチンでワクチン接種しておいたが、LHRHに対してはワクチン接種をしていなかった。
【0062】
ワクチン製剤
Tヘルパー細胞エピトープを、天然の10アミノ酸配列の、2〜10のアミノ酸を含有するトランケート型のLHRHと以下のように結合させた:
2〜10 LHRH His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly
全てのワクチンは実施例2と同様に配合した、すなわちワクチンの1mL用量あたり40nmoleのペプチド、150μgのIscomatrixTM、および保存剤としてのチオメルサールを含有していた。
【0063】
イヌをペプチドのプールでワクチン接種した場合には、各ペプチドの濃度は等しい濃度となるように調節し、全量で、1mL用量あたり40nmoleのLHRHエピトープとなるように調節した。
【0064】
ワクチン接種、血液サンプルおよびアッセイ
全てのイヌに1mLの用量で首筋へ送達してワクチン接種した。ワクチン接種は0週と4週に行い、試験期間の間隔で静脈血サンプルを得た。
【0065】
有効なT細胞へのヘルプを、ELISAによってLHRHに対する抗体応答を測定して決定した。ワクチンに基づくペプチドの生物学的有効性を、メスのイヌにおける黄体ホルモンおよびオスのイヌにおけるテストステロンのレベルを測定して決定した。
【表11】
【0066】
結果
LHRHに対する強力な抗体応答が、T細胞エピトープP25、P27、P35、P62、P75、およびT細胞エピトープP25、P27およびP35を組み合わせて含むT細胞-LHRHペプチドのプールを有するT細胞LHRH構築物で免疫化したイヌにおいて見られた(表12を参照)。
【0067】
P2-LHRHペプチドおよびP8-LHRHペプチドで免疫化したイヌにおいては、低いかまたは検出不能な抗体応答がみられた(表12を参照)。このことは、これらのT細胞ペプチドはビーグル-フォックスハウンド種のイヌによっては認識されないことを示すと結論付けたが、この結論は、他の種のイヌに由来するPBMCを用いてT細胞ペプチドを同定したことに合致する。ビーグルフォックスハウンドドッグにおける最初のスクリーニングでは、この種のイヌはこれら2種のT細胞エピトープに応答しないことが示された。
【0068】
ペプチドワクチン分野の当業者にはよく理解される通り、個々のペプチドに対する応答は遺伝子によって決定される。クラスII主要組織適合性複合体(MHCII)は多型性である。クラスII分子は細胞表面でT細胞に対する表示のためのペプチドと結合するよう機能する。これはヘルパーT細胞を含むT細胞の活性化プロセスの一部として必要である。MHCクラスIIの対立遺伝子型は別のペプチド抗原のセットと結合し、従って、これらの抗原の応答は遺伝子によって決定される。このように、この結果は、ビーグル-フォックスハウンド種のイヌはP2およびP8に応答するための好適なMHC-II対立遺伝子を有しないが、これらのペプチドを同定するために用いた、プードルシチヅ(shitzu)種など他種のイヌは有することを示すと解釈し得る。
【0069】
対照のイヌにおいては、試験期間中にLHRHに対する抗体レベルに変化は見られず、ホルモンレベルもイヌの年齢、性別に対して正常レベル内であった(表12参照)。
【表12】
【0070】
【0071】
実施例4
ペプチドワクチン中に取り込まれたTh-エピトープの認識を実証するためのin vitro T細胞増殖アッセイ
ペプチド免疫原内のTh-エピトープの認識を実証するために、ペプチドワクチンで免疫化したイヌ(実施例2のイヌ)から得たPBMCを、各Th-エピトープに対して試験した。アッセイは、PBMCを富化することなく行った。フィコールグラジエント精製により得たPBMCを、各Th-エピトープおよびそのトランケート型に対して直接試験した。この研究により、ペプチドワクチンで免疫化したイヌの全てが、取り込まれたTh-エピトープに対して応答することが示され、T細胞活性が各配列中に存在していることが確認された(図2〜4)。各Th-エピトープのトランケート型も同様に試験し、配列内のT細胞活性をより詳細に同定した。P25については、17残基の全配列のほうが、より短い15残基や12残基のペプチド(各ペプチドは配列のN末端から切断)よりも良好であることが観察された(図2)。この結果は、T細胞活性は17残基のペプチドのN末端または中間に対するものであることを示唆する。
【0072】
P27についても同様のことが観察され、長さが17残基の配列のほうが、N末端から切断されたより短い15残基のペプチドよりも良好なシミュレータであった(図3)。この観察もまた、再度、T細胞活性は、全長ペプチドの中間またはN末端に対して存在し得ることを示唆していた。
【0073】
P35とその短い型の場合には、1匹のイヌ(番号102)を除いた他の3匹のイヌは、12残基のペプチドにも全長17残基のペプチドにも同じように応答した(図4)。番号102のイヌにおいては、15残基のペプチドは全長ペプチドよりも刺激性が高かった。この結果から、P35の配列中で最初の2つの残基は必須ではなく、活性はペプチドの中間またはC末端に対するものであることが推定され得る。
【0074】
実施例5
BALB/cマウスにおける試験
実施例3で使用した、CDV-Fから誘導したTh-エピトープとLHRHを有するイヌ用ワクチンを同様に用いてBALB/cマウスを免疫化して、Th-エピトープが異なる動物種において機能性であるか否かを調べた。
【0075】
ワクチン製剤
全てのワクチンは、100μlの用量で2.7nmoleのペプチドと10μgのIscomatrixTMとチオメルソールを保存剤として含有するようにさらに希釈した以外は、実施例3と同じように配合した。
【0076】
ワクチン接種、血液サンプルおよびアッセイ
マウスに、尾の付け根で100μlのワクチンを接種した。ワクチン接種は0週と4週に行い、各ワクチン接種の後に間隔を開けて動物の眼窩後方(retro-orbital)叢から採血した。有効なT細胞ヘルプは、ELISAによりLHRHに対する抗体応答を測定して決定した。
【0077】
結果
P25-LHRHで免疫化したマウスと、P25-LHRH、P27-LHRHPおよびP35-LHRHを含むペプチドプールで免疫化したマウスでは、LHRHに対する高い抗体力価が生じた。ペプチドP35およびP75では生じた抗体力価は低く、一方P2、P8およびP62で免疫化したマウスでは、抗LHRH抗体のレベルは検出限界範囲外であった(表13)。
【0078】
当業者には、広く記載した本発明の意図と範囲から逸脱することなく、具体的な実施例において示された本発明に対して多数の変形および/または改変をなし得ることが理解されよう。従って、本実施形態はあらゆる観点で、例示的であり制限的ではないと解されるべきである。
【表13】
【特許請求の範囲】
【請求項1】
以下の配列:
からなる群より選択されるペプチド配列内に含まれる、Tヘルパー細胞エピトープ。
【請求項2】
以下の配列:
からなる群より選択されるペプチド配列内に含まれるTヘルパー細胞エピトープを少なくとも1種含んでなる、動物において免疫応答を上昇させるために使用する組成物。
【請求項3】
以下の配列:
からなる群より選択されるペプチドを少なくとも1種含んでなる、請求項2に記載の組成物。
【請求項4】
少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープをさらに含んでなる、請求項2または3に記載の組成物。
【請求項5】
少なくとも1種の前記B細胞エピトープおよび/または少なくとも1種の前記CTLエピトープが少なくとも1種の前記Tヘルパー細胞エピトープと結合されている、請求項4に記載の組成物。
【請求項6】
各コンジュゲートが少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープを含んでなる複数のエピトープ構築物を含んでなる、請求項5に記載の組成物。
【請求項7】
各コンジュゲートが少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のCTL細胞エピトープを含んでなる複数のエピトープ構築物を含んでなる、請求項5に記載の組成物。
【請求項8】
1種のLHRH B細胞エピトープを含んでなる、請求項4〜7のいずれか1項に記載の組成物。
【請求項9】
複数のTヘルパー細胞エピトープを含んでなる、請求項2に記載の組成物。
【請求項10】
複数の前記Tヘルパー細胞エピトープが単一のポリペプチドである、請求項9に記載の組成物。
【請求項11】
少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープをさらに含んでなる、請求項9または10に記載の組成物。
【請求項12】
少なくとも1種の前記B細胞エピトープおよび/または少なくとも1種の前記CTLエピトープが複数の前記Tヘルパー細胞エピトープと結合されている、請求項11に記載の組成物。
【請求項13】
1種のLHRH B細胞エピトープを含んでなる、請求項11または12に記載の組成物。
【請求項14】
アジュバントを含んでなる、請求項2〜13のいずれか1項に記載の組成物。
【請求項15】
前記アジュバントがISCOMまたはIscomatrixを含んでなる、請求項14に記載の組成物。
【請求項16】
請求項2〜15のいずれか1項に記載の組成物を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【請求項1】
以下の配列:
からなる群より選択されるペプチド配列内に含まれる、Tヘルパー細胞エピトープ。
【請求項2】
以下の配列:
からなる群より選択されるペプチド配列内に含まれるTヘルパー細胞エピトープを少なくとも1種含んでなる、動物において免疫応答を上昇させるために使用する組成物。
【請求項3】
以下の配列:
からなる群より選択されるペプチドを少なくとも1種含んでなる、請求項2に記載の組成物。
【請求項4】
少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープをさらに含んでなる、請求項2または3に記載の組成物。
【請求項5】
少なくとも1種の前記B細胞エピトープおよび/または少なくとも1種の前記CTLエピトープが少なくとも1種の前記Tヘルパー細胞エピトープと結合されている、請求項4に記載の組成物。
【請求項6】
各コンジュゲートが少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のB細胞エピトープを含んでなる複数のエピトープ構築物を含んでなる、請求項5に記載の組成物。
【請求項7】
各コンジュゲートが少なくとも1種のTヘルパー細胞エピトープおよび少なくとも1種のCTL細胞エピトープを含んでなる複数のエピトープ構築物を含んでなる、請求項5に記載の組成物。
【請求項8】
1種のLHRH B細胞エピトープを含んでなる、請求項4〜7のいずれか1項に記載の組成物。
【請求項9】
複数のTヘルパー細胞エピトープを含んでなる、請求項2に記載の組成物。
【請求項10】
複数の前記Tヘルパー細胞エピトープが単一のポリペプチドである、請求項9に記載の組成物。
【請求項11】
少なくとも1種のB細胞エピトープおよび/または少なくとも1種のCTLエピトープをさらに含んでなる、請求項9または10に記載の組成物。
【請求項12】
少なくとも1種の前記B細胞エピトープおよび/または少なくとも1種の前記CTLエピトープが複数の前記Tヘルパー細胞エピトープと結合されている、請求項11に記載の組成物。
【請求項13】
1種のLHRH B細胞エピトープを含んでなる、請求項11または12に記載の組成物。
【請求項14】
アジュバントを含んでなる、請求項2〜13のいずれか1項に記載の組成物。
【請求項15】
前記アジュバントがISCOMまたはIscomatrixを含んでなる、請求項14に記載の組成物。
【請求項16】
請求項2〜15のいずれか1項に記載の組成物を動物に投与することを含んでなる、動物において免疫応答を誘導するための方法。
【図1】


【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2011−121965(P2011−121965A)
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願番号】特願2011−13646(P2011−13646)
【出願日】平成23年1月26日(2011.1.26)
【分割の表示】特願2000−597449(P2000−597449)の分割
【原出願日】平成12年2月7日(2000.2.7)
【出願人】(501312646)ザ ユニバーシティ オブ メルボーン (1)
【出願人】(500021413)シーエスエル、リミテッド (28)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【出願人】(500057995)ザ カウンシル オブ ザ クイーンズランド インスティテュート オブ メディカル リサーチ (6)
【出願人】(302009017)ウォルター アンド エリザ ホール インスティテュート オブ メディカル リサーチ (3)
【Fターム(参考)】
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願日】平成23年1月26日(2011.1.26)
【分割の表示】特願2000−597449(P2000−597449)の分割
【原出願日】平成12年2月7日(2000.2.7)
【出願人】(501312646)ザ ユニバーシティ オブ メルボーン (1)
【出願人】(500021413)シーエスエル、リミテッド (28)
【出願人】(305039998)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガニゼイション (92)
【出願人】(500057995)ザ カウンシル オブ ザ クイーンズランド インスティテュート オブ メディカル リサーチ (6)
【出願人】(302009017)ウォルター アンド エリザ ホール インスティテュート オブ メディカル リサーチ (3)
【Fターム(参考)】
[ Back to top ]