
T細胞レセプターβ鎖遺伝子及びα鎖遺伝子
【課題】本発明は、メラノーマ特異的ペプチドであるMAGE1 135−143(NYKHCFPEI)、MAGE3 195−203(IMPKAGLLI)、MART1 27−35(AAGIGILTV)、gp100 209−217(IMDQVPFSV)、CMVpp65 341−349(QYDPVAALF)、又はCMVpp65 495−503(NLVPMVATV)を特異的に認識するT細胞レセプターβ鎖やα鎖及びそれらの遺伝子を提供することを目的とする。
【解決手段】樹状細胞に基づく上記ペプチド特異的なCTT細胞の誘導方法を構築し、該CTL細胞をAuto−MACS法にて純化し、純化した細胞からRNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いTCRレパトワ解析により、発現TCRの遺伝子分類を行い、解析結果に基づいてTCR遺伝子をクローニングした。クローニングした断片のDNAシークエンスを解析し、上記ペプチドを特異的に認識する新規なT細胞レセプターβ鎖やα鎖を同定した。
【解決手段】樹状細胞に基づく上記ペプチド特異的なCTT細胞の誘導方法を構築し、該CTL細胞をAuto−MACS法にて純化し、純化した細胞からRNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いTCRレパトワ解析により、発現TCRの遺伝子分類を行い、解析結果に基づいてTCR遺伝子をクローニングした。クローニングした断片のDNAシークエンスを解析し、上記ペプチドを特異的に認識する新規なT細胞レセプターβ鎖やα鎖を同定した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、T細胞レセプターβ鎖タンパク質、並びにT細胞レセプターβ鎖遺伝子、該遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質発現ベクター、及び該発現ベクターが導入された形質転換体に関する。また、本発明は、T細胞レセプターα鎖タンパク質、並びにT細胞レセプターα鎖遺伝子、該遺伝子が組み込まれたT細胞レセプターα鎖タンパク質発現ベクター、及び該発現ベクターが導入された形質転換体に関する。
【背景技術】
【0002】
悪性黒色腫は、インターロイキン2などを用いた免疫療法が比較的有効な癌である(例えば、非特許文献1参照)が、生体内における抗腫瘍効果には、腫瘍細胞に反応する細胞傷害性T細胞(Cytotoxic T lymphocyte:CTL)が重要であることが示されている(例えば、非特許文献2参照)。免疫療法後、腫瘍反応性T細胞が活性化され、T細胞上のT細胞レセプター(T cell receptor:TCR)が、HLA(ヒト白血球抗原)分子によって腫瘍細胞表面に提示される腫瘍細胞タンパク質が分解されてできたペプチド(腫瘍抗原)を認識し、腫瘍細胞を傷害する。更に各種サイトカインを分泌し、マクロファージ等の他の免疫細胞をも誘導活性化し、生体内で腫瘍拒絶を起こすと考えられている。腫瘍反応性T細胞には自己腫瘍細胞のみならず、HLAを共有する他の患者からの腫瘍細胞にも反応するような共通腫瘍抗原を認識する場合と、自己腫瘍細胞しか反応しない固有抗原を認識する場合がある。
【0003】
ヒト腫瘍細胞に対する免疫応答機構を解明して、新しい免疫療法を開発するためには、これらの腫瘍反応性T細胞が認識する腫瘍抗原を同定することが重要である。腫瘍反応性T細胞が樹立されている場合には、その反応性を利用した機能的cDNA発現クローニング法を用い、悪性黒色腫抗原の単離が可能である。例えば、転移メラノーマ患者からの腫瘍反応性細胞傷害性T細胞と、HLA−A2を発現する腫瘍細胞株とを用いて、メラノーマの予防、診断及び治療に用いることができる悪性黒色腫抗原を単離することが可能であり(例えば、特許文献1参照)、その一つMART1は同様な方法によって単離、同定されたものである。現在までに代表的なものとして、上記MART1やgp100のような色素細胞(メラノサイト)組織特異的タンパク質、各種腫瘍と正常組織では精巣に発現が認められるMAGE、BAGE、GAGE、NY−ESO−I等のCT抗原(Cancer-Testis抗原)、腫瘍細胞の遺伝子異常により産生されるβカテニン、CDK4(cyclin dependent kinase 4)、p15、GnT−V等の変異ペプチド抗原が単離されている(例えば、非特許文献3参照)。一方、MAGE抗原は、メラノーマのみならず肺がんや消化器がんなど多くの固形がんでの高い発現が報告されている。
【0004】
また、cytomegalovirus(CMV)感染症は、通常の免疫状態である健常人ではあまり問題にはならないが、特に臓器移植後など免疫低下状態では、重篤な間質性肺炎などの合併症を起こしうる深刻な疾患である。現在、抗ウイルス剤の投与によりCMV感染症の臨床症状は緩和されるが、根治対策となり得る抗ウイルス剤は現在のところまだ知られていない。これまでに主にCMVpp65タンパク由来のHLA−A2拘束性のエピトープに関する研究が行われており、CMVpp65 495−503(NLVPMVATV)ペプチドを用いた特異的CTL細胞の誘導やテトラマー試薬を用いたCTLクローンの評価が行われている。しかし、進行がん患者でのCMVに対するCTL細胞の誘導反応に関して、それほど多くの知見は得られておらず、CMV感染症に特異的なCTL細胞のT細胞レセプターβ鎖遺伝子については何ら知られていない。特に日本人に多いHLA−A24拘束性のCMVエピトープに関する研究は、まだ始まったばかりである。
【0005】
T細胞はT細胞レセプターによりMHC分子/抗原ペプチド複合体を認識して、抗原提示細胞(APC)などの標的細胞に結合するが、CTLはMHCクラスI分子/抗原ペプチドの複合体のみに結合し(MHCクラスI拘束性)、ヘルパーT(Th)細胞はMHCクラスII分子/抗原ペプチドの複合体のみしか結合できない(MHCクラスII拘束性)とされている。また、MHCクラスI分子はほとんどの有核細胞に発現しているが、MHCクラスII分子は限られた一部の細胞しか発現していない。このためTh細胞はMHCクラスII分子を発現した樹状細胞やB細胞、活性化T細胞などとは結合できるが、これら以外の腫瘍細胞や感染細胞などには直接結合することができない。しかし遺伝子操作によってMHCクラスI拘束性とされるCTL由来のT細胞レセプター遺伝子を導入したMHCクラスII拘束性とされるCD4陽性CD8陰性T細胞が、対応抗原をパルスしたAPCとCD8非依存的に反応して活性化されることができ、対応抗原に結合できるT細胞レセプターを発現していることが示された。また、非特異的な抗腫瘍活性をもつ末梢血リンパ球に腫瘍抗原特異的CTL由来のT細胞レセプター遺伝子を導入することにより、特異的に腫瘍を傷害することができることが報告されている(例えば、非特許文献4参照)。
【0006】
他方、T細胞表面に存在する膜表面タンパク質であり、抗原認識を担うT細胞レセプターには、α鎖・β鎖から成るヘテロ二量体とγ鎖・δ鎖から成るヘテロ二量体の二種類が同定されている。T細胞レセプター遺伝子は免疫グロブリン遺伝子と同様に、可変領域(V:variable)、多様性領域(D:diversity)、結合領域(J:joining)からなり分子の多様性を生じるVドメインと、細胞外定常領域、膜貫通領域、細胞内領域を含む定常領域(C:constant)からなるCドメインにより構成されている。これらのアミノ酸配列はT細胞ごとに異なっていることから、T細胞レセプターは抗原認識分子であると同時に個々のT細胞の目印にもなっている。このT細胞レセプターの多様性は後天的なT細胞レセプター遺伝子の再構成によって生みだされており、T細胞レセプターβ,δ鎖はV−D−Jを、T細胞レセプターα,γ鎖はV−Jを再構成することが知られている。
【0007】
T細胞の初期分化は、B細胞と同様にT細胞レセプターのα鎖及びβ鎖遺伝子の再構成と密接に関わっている。T細胞レセプターβ鎖遺伝子の再構成はDN−T細胞(double negative:CD4−8−)の段階で開始され、β鎖遺伝子の再構成に成功した細胞はβ鎖タンパク質と代替α鎖(preTα)の複合体、つまりプレT細胞レセプターを細胞表面に発現する。そして、CD4及びCD8分子の発現が誘導され、DP−T細胞(double positive:CD4+8+)へと移行し、DPとなったT細胞は、次にT細胞レセプターのα鎖遺伝子の再構成に成功した細胞のみが、T細胞レセプターを表面に発現できることが知られている。
【0008】
T細胞レセプターβ鎖の遺伝子構成に関しては、ヒトゲノム遺伝子配列情報データベースによれば、T細胞レセプターβ鎖の遺伝子は約30種類の亜族(遺伝子自体は約70種)からなるV遺伝子、2種類のD遺伝子、14種類のJ遺伝子、2種類のC遺伝子によって構成され、Vβ中に2つの可変領域(CDR1、CDR2)が存在することも知られている。これらの中からVβ、Dβ、Jβが各1つずつ選ばれて結合し、T細胞レセプターβ鎖をコードするDNAが構成されている。この際、Vβ−Dβ−Jβの結合部にはランダムな塩基の挿入や欠失が起こり、無限に近い多様性が生みだされている。このVβ−Dβ−Jβの結合部位(連結部分)は、CDR3(相補性決定領域3:third complementarity determining region)、結合部(junctional)領域と呼ばれ、T細胞レセプターの中で最も多型性に富む部分であり、塩基の挿入・欠失が起こるため、各遺伝子構成が同じであっても連結部分も含めて同一配列になることは極めて稀である。
【0009】
また、T細胞レセプターα鎖の遺伝子構成に関しては、ヒトゲノム遺伝子配列情報データベースによれば、T細胞α鎖の遺伝子は約40種類の亜族(遺伝子自体は約50種)からなるV遺伝子、約60種のJ遺伝子、1種類のC遺伝子によって構成される。T細胞の分化の過程で起きる遺伝子組み換えによって、前述の中からVα、Jα、Cαが各1種ずつ選ばれて結合し、染色体上でT細胞レセプターα鎖をコードするDNAが構成される。染色体上で構成されたT細胞レセプターα鎖遺伝子は発現してそのタンパク質が細胞表面に表出する。T細胞α鎖のVα−Jαの結合部位(連結部分)は、多型性に富む部分であり、塩基の挿入・欠失が起こるため、各遺伝子構成が同じであっても、連結部分を含めて同一配列になることは極めて稀である。
【0010】
【特許文献1】特表平10−505481号公報
【非特許文献1】JAMA 271, 907, 1994
【非特許文献2】Microbiol.Immunol. 42, 803, 1998
【非特許文献3】Immunol Res 16, 313, 1997
【非特許文献4】J.Immunol.,171:3287-3295, 2003
【発明の開示】
【発明が解決しようとする課題】
【0011】
T細胞レパートリー(T細胞レパトワ)の多様性は、T細胞間での多様な抗原認識能と密接に関係していることから、T細胞の機能,免疫応答,免疫機能を考える上でT細胞レセプターレパートリーの機能解明を行うことは極めて重要である。最近、癌,自己免疫疾患,アレルギー疾患等において、患部局所に存在するT細胞のT細胞レセプターレパートリー解析の研究が進められ、疾患特異的なT細胞が解明されつつある。しかし、そのT細胞上のT細胞レセプター分子が、どのような働きをしているか、ということは、T細胞レセプター分子の単離が困難であること等の問題があり、まだその詳細が明らかにされるに至っていない。本発明の課題は、メラノーマ特異的ペプチドであるMAGE1 135−143(NYKHCFPEI:以下、MAGE1−A24ペプチドと称する場合がある)、MAGE3 195−203(IMPKAGLLI:以下、MAGE3−A24ペプチドと称する場合がある)、MART1 27−35(AAGIGILTV:以下、MART1−A2ペプチドと称する場合がある)、gp100 209−217(IMDQVPFSV:以下、gp100−A2ペプチドと称する場合がある)、CMVpp65 341−349(QYDPVAALF:以下、CMVpp65−A24ペプチドと称する場合がある)、又はCMVpp65 495−503(NLVPMVATV:以下、CMVpp65−A2ペプチドと称する場合がある)を特異的に認識するT細胞レセプターβ鎖及びその遺伝子を提供することにある。
また、本発明の課題は、CTL細胞膜上で上記T細胞レセプターβ鎖とヘテロ二量体を構成したときに、がん細胞由来の抗原ペプチドに特異性が高いT細胞レセプターα鎖及びその遺伝子を提供することにもある。
【課題を解決するための手段】
【0012】
T細胞レセプター分子が、T細胞を介する免疫応答および免疫機構に重要な役割を果たしていることから、これら免疫応答および免疫機構の解析には、個々のT細胞レセプター分子を得ることが必要である。しかし、膜表面タンパク質を細胞から精製すると、1010個の細胞からせいぜい10μg程度しか得られない。しかも、T細胞レセプターの多様性により、一種類のT細胞レセプターを分取するためには、それぞれのT細胞レセプターを持つT細胞株を樹立し、その各々からT細胞レセプターを単離することが必須となる。しかし、T細胞のT細胞レセプターレパートリーは1010種類にも及んでおり、1010種のT細胞株を樹立することは、実質的に極めて困難である。
【0013】
MAGE1、MAGE3、MART1及びgp100はメラノーマに発現する癌抗原であり、ペプチドワクチンによる治療を受けている転移性のメラノーマ患者において、抗原抗体反応を強く誘導するものである。また、CMVpp65は、CMV感染症に発現するCMV抗原であり、CMV感染症患者において、抗原抗体反応を誘導するものである。MAGE1やMAGE3はHLA−A24限局的な抗原決定部位を持っており(MAGE1 135−143:NYKHCFPEI; MAGE3 195−203:IMPKAGLLI)、MART1やgp100はHLA−A2限局的な抗原決定部位を持っている(MART1 27−35:AAGIGILTV; gp100 209−217:IMDQVPFSV)。また、CMVpp65はHLA−A24限局的な抗原決定部位(CMVpp65 341−349:QYDPVAALF)と、HLA−A2限局的な抗原決定部位(CMVpp65 495−503:NLVPMVATV)を持っている。
【0014】
本発明者らは、樹状細胞に基づく上記ペプチド特異的なCTL細胞の誘導方法を構築し、該CTL細胞をAuto−MACS法にて純化し、純化した細胞からRNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いTCRレパトワ解析により、発現TCRの遺伝子分類を行い、解析結果に基づいてTCR遺伝子をクローニングした。クローニングした断片のDNAシークエンスを解析し、上記ペプチドを特異的に認識する新規なT細胞レセプターβ鎖及びそれとヘテロ二量体を形成するα鎖を同定した。本発明は、上記知見に基づき完成するに至ったものである。
【0015】
すなわち本発明は、(1)配列番号7、9、11、13、15、17、19、21若しくは23で表される塩基配列、又は、配列番号8、10、12、14、16、18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子や、(2)上記(1)に記載のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターや、(3)上記(2)に記載のT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが導入された形質転換体に関する。
【0016】
また本発明は、(4)配列番号32、34、36、38、40、42、44、46、48、50、52、54、56、58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子や、(5)上記(4)に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターや、(6)上記(5)に記載のT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体や、(7)上記(1)に記載のT細胞レセプターβ鎖遺伝子及び上記(4)に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターや、(8)上記(7)に記載のT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体や、(9)組換えベクターが導入されたリンパ球である上記(3)、(6)又は(8)に記載の形質転換体に関する。
【0017】
さらに本発明は、(10)配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質や、(11)配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質に特異的に結合する抗体に関する。
【0018】
またさらに本発明は、(12)配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質や、(13)配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質に特異的に結合する抗体に関する。
【0019】
また本発明は、(14)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(15)配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(16)配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物に関する。
【0020】
さらに本発明は、(17)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(18)配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(19)配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物や、(20)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子及びT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(21)配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(22)配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物に関する。
【発明の効果】
【0021】
本発明によると、MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の特定の疾患におけるT細胞の機能、免疫応答、免疫機能を解明する上で有用である。また、本発明のT細胞レセプターβ鎖遺伝子やα鎖遺伝子を利用した遺伝子治療や養子免疫療法により、患者の細胞性免疫を疾患特異的に向上させ、MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の特定の疾患を治療しうる可能性がある。
【発明を実施するための最良の形態】
【0022】
本発明のT細胞レセプターβ鎖遺伝子(TCRβ鎖遺伝子ともいう)は、疾患特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子である限り特に制限されず、例えばメラノーマ特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子や、CMV感染症特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子等を好ましく例示することができる。
【0023】
前記メラノーマ特異的抗原ペプチドとしては、MAGE1、MAGE3、MART1、gp100等が知られており、中でも、MAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)、MAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)、MART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)、gp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)を好ましく例示することができ、上記CMV感染症特異的抗原ペプチドとしては、CMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)、CMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)を好ましく例示することができる。
【0024】
前記本発明のT細胞レセプターβ鎖遺伝子として具体的には、配列番号7や配列番号9で表される塩基配列(MAGE1−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号11で表される塩基配列(MAGE3−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号13で表される塩基配列(MART1−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号15で表される塩基配列(gp100−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号17や配列番号19や配列番号21で表される塩基配列(CMVpp65−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号23で表される塩基配列(CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号8や配列番号10で表されるアミノ酸配列をコードする塩基配列(MAGE1−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号12で表されるアミノ酸配列をコードする塩基配列(MAGE3−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号14で表されるアミノ酸配列をコードする塩基配列(MART1−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号16で表されるアミノ酸配列をコードする塩基配列(gp100−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号18や配列番号20や配列番号22で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号24で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子)からなるT細胞レセプターβ鎖遺伝子を好ましく例示することができる。
【0025】
本発明のT細胞レセプターα鎖遺伝子(TCRα鎖遺伝子ともいう)は、疾患特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子である限り特に制限されず、例えば前述のメラノーマ特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子や、前述のCMV感染症特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子等を好ましく例示することができる。
【0026】
前記本発明のT細胞レセプターα鎖遺伝子として具体的には、配列番号32や配列番号34や配列番号36や配列番号38や配列番号40や配列番号42で表される塩基配列(MART1−A2ペプチド特異的TCRα鎖遺伝子)、配列番号44で表される塩基配列(gp100−A2ペプチド特異的TCRα鎖遺伝子)、配列番号46や配列番号48や配列番号50や配列番号52で表される塩基配列(MAGE1−A24ペプチド特異的TCRα鎖遺伝子)、配列番号54や配列番号56で表される塩基配列(MAGE3−A24ペプチド特異的TCRα鎖遺伝子)、配列番号58や配列番号61で表される塩基配列(CMVpp65−A2ペプチド特異的TCRα鎖遺伝子)、配列番号63や配列番号65や配列番号67や配列番号69や配列番号71や配列番号74や配列番号76で表される塩基配列(CMVpp65−A24ペプチド特異的TCRα鎖遺伝子)、配列番号33や配列番号35や配列番号37や配列番号39や配列番号41や配列番号43で表されるアミノ酸配列をコードする塩基配列(MART1−A2ペプチド特異的TCRα鎖遺伝子)、配列番号45で表されるアミノ酸配列をコードする塩基配列(gp100−A2ペプチド特異的TCRα鎖遺伝子)、配列番号47や配列番号49や配列番号51や配列番号53で表されるアミノ酸配列をコードする塩基配列(MAGE1−A24ペプチド特異的TCRα鎖遺伝子)、配列番号55や配列番号57で表されるアミノ酸配列をコードする塩基配列(MAGE3−A24ペプチド特異的TCRα鎖遺伝子)、配列番号59や配列番号62で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A2ペプチド特異的TCRα鎖遺伝子)、配列番号64や配列番号66や配列番号68や配列番号70や配列番号72や配列番号75や配列番号77で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A24ペプチド特異的TCRα鎖遺伝子)からなるT細胞レセプターα鎖遺伝子を好ましく例示することができる。
【0027】
本発明の遺伝子の取得方法や調製方法は特に限定されるものでなく、本明細書中に開示した前記塩基配列や前記アミノ酸配列の各情報に基づいて適当なプローブやプライマーを調製し、それらを用いて当該遺伝子が存在することが予測されるヒトcDNAライブラリーをスクリーニングすることにより目的の遺伝子を単離したり、常法に従って化学合成により調製することができる。例えば、細胞又は組織からの全RNAの分離、mRNAの分離や精製、cDNAの取得とそのクローニングなどはいずれも常法に従って実施することができる。本発明の遺伝子をcDNAライブラリーからスクリーニングする方法は、例えば、Molecular Cloning: A laboratory Manual, 2nd Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY., 1989に記載の方法等、当業者により常用される方法を挙げることができる。
【0028】
本発明のT細胞レセプターβ鎖タンパク質は、配列番号8や配列番号10で表されるアミノ酸配列からなるタンパク質(MAGE1−A24ペプチド特異的TCRβ鎖タンパク質)、配列番号12で表されるアミノ酸配列からなるタンパク質(MAGE3−A24ペプチド特異的TCRβ鎖タンパク質)、配列番号14で表されるアミノ酸配列からなるタンパク質(MART1−A2ペプチド特異的TCRβ鎖タンパク質)、配列番号16で表されるアミノ酸配列からなるタンパク質(gp100−A2ペプチド特異的TCRβ鎖タンパク質)、配列番号18や配列番号20や配列番号22で表されるアミノ酸配列からなるタンパク質(CMVpp65−A24ペプチド特異的TCRβ鎖タンパク質)、又は配列番号24で表されるアミノ酸配列からなるタンパク質(CMVpp65−A2ペプチド特異的TCRβ鎖タンパク質)である。
【0029】
また、本発明のT細胞レセプターα鎖タンパク質は、配列番号33や配列番号35や配列番号37や配列番号39や配列番号41や配列番号43で表されるアミノ酸配列からなるタンパク質(MART1−A2ペプチド特異的TCRα鎖タンパク質)、配列番号45で表されるアミノ酸配列からなるタンパク質(gp100−A2ペプチド特異的TCRα鎖タンパク質)、配列番号47や配列番号49や配列番号51や配列番号53で表されるアミノ酸配列からなるタンパク質(MAGE1−A24ペプチド特異的TCRα鎖タンパク質)、配列番号55や配列番号57で表されるアミノ酸配列からなるタンパク質(MAGE3−A24ペプチド特異的TCRα鎖タンパク質)、配列番号59や配列番号62で表されるアミノ酸配列からなるタンパク質(CMVpp65−A2ペプチド特異的TCRα鎖タンパク質)、配列番号64や配列番号66や配列番号68や配列番号70や配列番号72や配列番号75や配列番号77で表されるアミノ酸配列からなるタンパク質(CMVpp65−A24ペプチド特異的TCRα鎖タンパク質)である。
【0030】
本発明のタンパク質の取得・調製方法は特に限定されず、天然由来のタンパク質でも、化学合成したタンパク質でも、遺伝子組換え技術により作製した組み換えタンパク質の何れでもよい。天然由来のタンパク質を取得する場合には、かかるタンパク質を発現しているT細胞からタンパク質の単離・精製方法を適宜組み合わせることにより、本発明のタンパク質を取得することができる。化学合成によりタンパク質を調製する場合には、例えば、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法に従って本発明のタンパク質を合成することができる。また、各種の市販のペプチド合成機を利用して本発明のタンパク質を合成することもできる。遺伝子組換え技術によりタンパク質を調製する場合には、該タンパク質をコードする塩基配列からなるDNAを好適な発現系に導入することにより本発明のタンパク質を調製することができる。これらの中でも、比較的容易な操作でかつ大量に調製することが可能な遺伝子組換え技術による調製が好ましい。
【0031】
例えば、遺伝子組換え技術によって、本発明のタンパク質を調製する場合、かかるタンパク質を細胞培養物から回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ハイドロキシアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法、好ましくは、高速液体クロマトグラフィーが用いられる。特に、アフィニティークロマトグラフィーに用いるカラムとしては、例えば、本発明のタンパク質に対するモノクローナル抗体等の抗体を結合させたカラムや、上記本発明のタンパク質に通常のペプチドタグを付加した場合は、このペプチドタグに親和性のある物質を結合したカラムを用いることにより、これらのタンパク質の精製物を得ることができる。また、本発明のタンパク質が細胞膜に発現している場合は、細胞膜分解酵素を作用させた後、上記の精製処理を行うことにより精製標品を得ることができる。
【0032】
また、本発明のタンパク質をマーカータンパク質やペプチドタグと結合させることもできる。マーカータンパク質としては、従来知られているマーカータンパク質であれば特に制限されるものではなく、例えば、アルカリフォスファターゼ、HRP等の酵素、抗体のFc領域、GFP等の蛍光物質などを具体的に挙げることができ、また本発明におけるペプチドタグとしては、HA、FLAG、Myc等のエピトープタグや、GST、マルトース結合タンパク質、ビオチン化ペプチド、オリゴヒスチジン等の親和性タグなどの従来知られているペプチドタグを具体的に例示することができる。かかる融合ペプチドは、常法により作製することができ、Ni−NTAとHisタグの親和性を利用した本発明のタンパク質の精製や、本発明のタンパク質の検出や、本発明のタンパク質に対する抗体の定量や、その他当該分野の研究用試薬としても有用である。
【0033】
本発明の組換えベクターとしては、前記本発明のT細胞レセプターβ鎖遺伝子及び/又はT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及び/又はT細胞レセプターα鎖タンパク質を発現しうる組換えベクターである限り特に制限されないが、動物細胞において発現しうるものが好ましい。本発明の組換えベクターは、本発明の遺伝子を発現ベクターに適切にインテグレイトすることにより構築することができる。かかる発現ベクターとしては、宿主細胞において自立複製可能であるものや、あるいは宿主細胞の染色体中へ組込み可能であるものが好ましく、また、本発明の遺伝子を発現できる位置にプロモーター、エンハンサー、ターミネーター等の制御配列を含有しているものを好適に使用することができる。
【0034】
動物細胞用の発現ベクターとして、例えば、pEGFP-C1(Clontech社製)、pGBT−9(Clontech社製)、pcDNAI(フナコシ社製)、pcDM8(フナコシ社製)、pAGE107(Cytotechnology, 3, 133, 1990)、pCDM8(Nature, 329, 840, 1987)、pcDNAI/AmP(Invitrogen社製)、pREP4(Invitrogen社製)、pAGE103(J.Blochem., 101, 1307, 1987)、pAGE210等を例示することができる。動物細胞用のプロモーターとしては、例えば、サイトメガロウイルス(ヒトCMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーター、レトロウイルスのプロモーター、メタロチオネインプロモーター、ヒートショックプロモーター、SRαプロモーター等を挙げることができる。
【0035】
本発明の形質転換体としては、上記本発明の組換えベクターが導入された宿主細胞であれば特に制限されないが、リンパ球が好ましく、中でもナイーブT細胞が好ましい。中でも、上記本発明の組換えベクターがインビトロで導入されたリンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができる疾患(該組換えベクターに組み込まれたT細胞レセプターβ鎖遺伝子やT細胞レセプターα鎖遺伝子にコードされるT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質が特異性を示す抗原ペプチドを特異的に発現する疾患)に対する治療用組成物として有用である。特に、該組換えベクターに組み込まれた本発明のT細胞レセプターβ鎖遺伝子の塩基配列が、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15で表される塩基配列や、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16で表されるアミノ酸配列をコードする塩基配列からなる場合や、前記組換えベクターに組み込まれた本発明のT細胞レセプターα鎖遺伝子の塩基配列が、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56で表される塩基配列や、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57で表されるアミノ酸配列をコードする塩基配列からなる場合は、前記リンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができるMAGE1,3陽性の固形がん又はメラノーマ治療用組成物として有用であり、該組換えベクターに組み込まれた本発明のT細胞レセプターβ鎖遺伝子の塩基配列が、配列番号17、配列番号19、配列番号21、配列番号23で表される塩基配列や、配列番号18、配列番号20、配列番号22、配列番号24で表されるアミノ酸配列をコードする塩基配列からなる場合や、前記組換えベクターに組み込まれた本発明のT細胞レセプターα鎖遺伝子の塩基配列が、配列番号58、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号74、配列番号76で表される塩基配列や、配列番号59、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号75、配列番号77で表されるアミノ酸配列をコードする塩基配列からなる場合は、前記リンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができるCMV感染症治療用組成物として有用である。
なお、本発明における「MAGE1,3陽性の固形がん又はメラノーマ治療用組成物」とは、メラノーマ; MAGE1陽性の固形がん(但しメラノーマを除く); 及び、MAGE3陽性の固形がん(但しメラノーマを除く);から選ばれる1つ又は2つ以上の疾患に対して治療効果を奏する組成物を意味する。
【0036】
養子免疫療法等免疫治療剤に用いる場合は、形質転換T細胞クローンを精製することが好ましく、その手法としては、リン酸緩衝液等に混合して遠心する方法、分離剤を用いた比重遠心法などを用いることができる。MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の疾患の治療用組成物の投与量は、1回あたり形質転換リンパ球として106〜1012個の細胞が好ましい。投与形態としては、注射剤、点滴剤等の液体が好ましく、形質転換リンパ球をヒト血清アルブミンを0.01〜5%となるように添加した生理食塩液に分散した注射剤又は点滴剤がより好ましい。投与方法としては、静脈への点滴又は静脈、動脈、局所等への注射が好ましい。投与する液量は、投与方法、投与する場所等に異なるが、50〜500ccとするのが好ましく、この液量に前記の形質転換リンパ球が含まれるようにするのが好ましい。投与頻度は1回/日〜1回/月とするのが好ましく、投与回数は少なくとも1回、好ましくは5回以上とすることができる。
【0037】
本発明の抗体は、上記本発明のT細胞レセプターβ鎖タンパク質又はT細胞レセプターα鎖タンパク質に特異的に結合する抗体である限り特に制限されず、モノクローナル抗体、ポリクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体等の免疫特異的な抗体を例示することができる。本発明のこれらの抗体は上記本発明のT細胞レセプターβ鎖タンパク質を抗原として用いて常法により作製することができるが、その中でもモノクローナル抗体がその特異性に優れている点でより好ましい。かかるモノクローナル抗体等の本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に特異的に結合する抗体は、例えば、メラノーマやCMV感染症等の疾患の診断や本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質の分子機構を明らかにする上で有用である。
【0038】
本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に対する抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)に本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質又はこれらのタンパク質を膜表面に発現した細胞を投与することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975)、トリオーマ法、ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc., 1985)など任意の方法を用いることができる。以下に本発明のT細胞レセプタータンパク質として、ヒトT細胞由来の本発明のT細胞レセプターβ鎖タンパク質を例に挙げてマウス由来の本発明のT細胞レセプターβ鎖タンパク質に対して特異的に結合するモノクローナル抗体、すなわち抗hT細胞レセプターβ鎖マウスモノクローナル抗体の作製方法を説明する。
【0039】
上記抗hT細胞レセプターβ鎖モノクローナル抗体は、抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマをインビボ又はインビトロで常法により培養することにより生産することができる。例えば、インビボ系においては、齧歯動物、好ましくはマウス又はラットの腹腔内で培養することにより、またインビトロ系においては、動物細胞培養用培地で培養することにより得ることができる。インビトロ系でハイブリドーマを培養するための培地としては、ストレプトマイシンやペニシリン等の抗生物質を含むRPMI1640又はMEM等の細胞培養培地を例示することができる。
【0040】
抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマは、例えば、本発明のT細胞レセプターβ鎖タンパク質又は該タンパク質を膜表面に発現したT細胞を用いてBALB/cマウスを免疫し、免疫されたマウスの脾臓細胞とマウスNS−1細胞(ATCC TIB−18)とを、常法により細胞融合させ、免疫蛍光染色パターンによりスクリーニングすることにより、抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマを作出することができる。また、かかるモノクローナル抗体の分離・精製方法としては、タンパク質の精製に一般的に用いられる方法であればどのような方法でもよく、アフィニティークロマトグラフィー等の液体クロマトグラフィーを具体的に例示することができる。
【0041】
また、本発明の上記本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に対する一本鎖抗体をつくるためには、一本鎖抗体の調製法(米国特許第4,946,778号)を適用することができる。また、ヒト化抗体を発現させるために、トランスジェニックマウス又は他の哺乳動物等を利用したり、上記抗体を用いて、本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質を発現するクローンを単離・同定したり、アフィニティークロマトグラフィーでそのポリペプチドを精製することもできる。
【0042】
また上記本発明の抗hT細胞レセプターβ鎖モノクローナル抗体等の抗体に、例えば、FITC(フルオレセインイソシアネート)又はテトラメチルローダミンイソシアネート等の蛍光物質や、125I、32P、14C、35S又は3H等のラジオアイソトープや、アルカリホスファターゼ、ペルオキシダーゼ、β−ガラクトシダーゼ又はフィコエリトリン等の酵素で標識したものや、グリーン蛍光タンパク質(GFP)等の蛍光発光タンパク質などを融合させた融合タンパク質を用いることによって、上記本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質の機能解析を行うことができる。また免疫学的測定方法としては、RIA法、ELISA法、蛍光抗体法、プラーク法、スポット法、血球凝集反応法、オクタロニー法等の方法を挙げることができる。
【0043】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。なお、実施例1における実験概要は、樹状細胞ワクチン投与を受けたメラノーマ患者(HLA−A24陽性)リンパ球の入手→メラノーマ抗原ペプチドMAGE1(A24拘束性)にて処理した樹状細胞の作製→樹状細胞及びT2−A24細胞刺激によるペプチド特異的CTL細胞の誘導・増幅→増幅したCTL細胞のマーカー解析(FACS)と定量(テトラマー染色)及び機能解析(刺激されたCTL細胞からのIFN−γ産生解析)→AutoMACS法による、テトラマー又はレパトワ抗体陽性CTL細胞の純化→RNA抽出→逆転写反応→PCRによるTCR遺伝子(β鎖)の増幅→DNA Analyzer(ABI3100)を用いたDNA断片解析(TCRβ鎖遺伝子の同定)→PCRによるタンパク質コード領域の増幅とクローニング→ABI3100による塩基配列の決定である。その他の実施例3、5、7及び9における実験概要は、用いるペプチドが異なる点を除いて、ほぼ同様である。また、実施例10における実験概要は、用いるペプチドの他、増幅や同定する対象がTCRβ鎖遺伝子ではなく、TCRα鎖遺伝子である点を除いて、ほぼ同様である。
【実施例1】
【0044】
[MAGE1−A24ペプチド特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行った。今回リンパ球を使用したMEL−001の患者では、MAGE1 135−143にて処理した樹状細胞の投与を行ったが、約4cm大の肺転移巣が著明に縮小を認め、明確な臨床的効果が得られている(図1)。また免疫反応のモニタリング検査であるELISPOTアッセイでは、末梢血中にMAGE1−A24ペプチドに対する特異的なCTL細胞の陽性反応(Δ)が確認され、臨床効果と相関した結果が得られている(図2)。
【0045】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(MAGE1−A24ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたMAGE1−A24テトラマー陽性CTL細胞について定量的な評価を行った。その結果を図3に示す。図3に示されているとおり、対照コントロールのテトラマーであるHIV−A24テトラマーでは、ほとんど陽性細胞は認められない(0.04%)が、MAGE1−A24テトラマー染色では18.4%と高率に陽性CTL細胞の増幅が認められた。
【0046】
また、MAGE1−A24ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社製)を用いて、有意に増加したレパトワの同定を行った。その結果を図4に示す。図4に示されているとおり、MAGE1−A24テトラマー陽性CTL細胞の増幅を認めたMEL−001症例では、レパトワ抗体パネルのスクリーニングにて2種類のレパトワ(TCRVβ5.1及びTCRVβ7.1)の増加が検出された。レパトワ陽性細胞の比率は、TCRVβ5.1が12.8%、TCRVβ7.1が27.1%であった。
【0047】
MAGE1−A24ペプチド特異的CTL細胞を、MAGE1−A24テトラマー及び、TCRVβ5.1又はTCRVβ7.1にて染色し、陽性細胞について定量的な評価を行った。その結果を図5に示す。図5に示されているとおり、MAGE1−A24ペプチド特異的CTL細胞の増幅を認めたMEL−001症例では、MAGE1−A24テトラマー及びTCRVβ7.1の両方が陽性であった細胞が10.6%、MAGE1−A24テトラマー及びTCRVβ5.1の両方が陽性であった細胞が0.84%であり、MAGE1−A24テトラマー陽性のCTL細胞の大半は、TCRVβ7.1が陽性であった。一方、コントロール用のテトラマーであるHIV−A24テトラマーは、ほとんど染色されなかった。
【0048】
MAGE1−A24ペプチドにて処理を行ったLCL(EB virusにて不死化したヒトB細胞培養株)であるTISI細胞にて、前述のMAGE1−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図6に示す。図6に示されているとおり、MAGE1−A24テトラマー及びIFN−γが共に陽性の細胞比率は9.1%であり、TCRVβ7.1及びIFN−γが共に陽性の細胞比率は9.9%であり、TCRVβ5.1及びIFN−γが共に陽性の細胞比率は0.71%であった。これらの結果より、MAGE1−A24テトラマー陽性CTL細胞において、上述の刺激によってIFN−γを産生しうる細胞は、ほとんどがTCRVβ7.1陽性の細胞であることが明らかとなった。
【0049】
上記図5及び図6の実験の結果を考慮して、TCRVβ7.1陽性CTL細胞を、HLA−A24テトラマー又は特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は97%であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0050】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、前述のMAGE1−A24ペプチド特異的TCRβ鎖タンパク質をコードする2クローンの遺伝子(TCRβ鎖遺伝子)にそれぞれ対応するプライマーを用いてPCR増幅を行い、各増幅断片の塩基配列を解析した。具体的には、前述のTCRβ鎖遺伝子(clone1)に対応するセンスプライマー(gctagcatgggctgcaggctgctctgc:配列番号25)とアンチセンスプライマー(tcagaaatcctttctcttgaccatggc:配列番号26)を用いて、本TCRβ鎖遺伝子(clone1)のタンパク質コード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子(clone1)の塩基配列を配列番号7に、そのアミノ酸配列を配列番号8に示す。また、前述のもう一方のTCRβ鎖遺伝子(clone2)についてもそれに対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号26)を用いて、本TCRβ鎖遺伝子(clone2)のタンパク質コード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子(clone2)の塩基配列を配列番号9に、そのアミノ酸配列を配列番号10に示す。クローニングした2つの配列を解析した結果、これらのTCRβ鎖遺伝子は同じ遺伝子ではあるが、異なるクローンであることが判明した。
【0051】
本実施例及び後述の実施例に使用したプライマーの配列は、TCRレパトワ解析ではJeffry R. Currier and Mary Ann Robinsonが成書Current Protocols in Immunology※3 に記載したものを使用し、タンパク質コード領域のクローニングでは、公開データベース(NCBI)に収載されるゲノム配列にもとづいた。
※1 ImMunoGeneTics:1125985409921_0
※2 National Center for Biotechnology Information:http://www.ncbi.nlm.nih.gov/
※3 Current Protocols in Immunology, John E. Coligan et al. ed. (2000) 10.28.1-10.28.24. John Wiley & Sons, Inc.
【実施例2】
【0052】
実施例1で得られたMAGE1−A24ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。ここで、ナイーブT細胞とは、胸腺での分化後、抗原にまだ遭遇していないT細胞のことである。
【0053】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0054】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、mock(4μg)、GFP(2μg)又はTCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、mockとして使用したベクターはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターからGFP遺伝子を切り出したものを用い、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図7に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。また、MAGE1−A24特異的TCRβ鎖遺伝子は、クローン1については29.4%の細胞で、クローン2については32.4%の細胞で発現が確認され、良好な発現効率が認められた。
【0055】
また、MAGE1−A24ペプチドにて処理したTISI細胞に対する、MAGE1−A24ペプチド特異的なTCRβ鎖を発現したHLA−A*2402陽性ナイーブT細胞の細胞障害活性を評価した。TCRβ鎖遺伝子(clone1又はclone2)を導入発現したHLA−A*2402陽性ナイーブT細胞としては、前記エレクトロポレーションによりTCRβ鎖遺伝子のDNA(4μg)又はGFP(2μg)を導入した細胞を使用した。また、TISI細胞をPBS(Mg, Ca含有、Cambrex社製)+1%ヒト血清アルブミン溶液に懸濁し、MAGE1−A24ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該MAGE1−A24ペプチド処理したTISI細胞1×105個を、同数の前記TCRβ鎖遺伝子を導入発現したHLA−A*2402陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、MAGE1−A24ペプチド処理したTISI細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図8に示す。図8の結果から分かるように、TCRβ鎖遺伝子(clone2)を導入したリンパ球のみIFN−γが刺激され、本TCRβ鎖遺伝子がコードするTCRβ鎖を発現したT細胞が、腫瘍特異的な細胞障害活性を有することが示唆された。
【実施例3】
【0056】
[MAGE3−A24ペプチド特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行った。今回リンパ球を使用したMEL−007の患者では、MAGE3 195−203にて処理した樹状細胞の投与を行ったが、約2cm大の頚部リンパ節転移巣が著明に縮小を認め、明確な臨床的効果が得られている(図9)。また免疫反応のモニタリング検査であるELISPOTアッセイでは、末梢血中にMAGE3−A24ペプチドに対する特異的なCTL細胞の陽性反応(◇)が確認され、臨床効果と相関した結果が得られている(図10)。
【0057】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(MAGE3−A24ペプチド特異的CTL細胞)について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark, Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果を図11に示す。図11に示されているとおり、ほとんどのレパトワ抗体では、比率が5%以下であったが、TCRVβ3.0(TCRVβ3)とTCRVβ13.1に対する陽性CTL細胞の比率の増加が確認された。より具体的には、図12に示されているとおり、TCRVβ3.0陽性細胞の比率は24.7%、TCRVβ13.1陽性細胞の比率は11.0%であった。
【0058】
MAGE3−A24ペプチドにて処理を行ったT2−A24細胞(T2細胞にHLAーA−*2402遺伝子を導入後、A24陽性細胞をクローニングした細胞)にて、前述のMAGE3−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図13に示す。図13に示されているとおり、上述の刺激により全体の6.8%の比率のCTL細胞にIFN−γの産生が認められた。一方、MAGE3−A24ペプチドによる処理を行わなかったT2−A24細胞による刺激を行った場合(コントロール)は、IFN−γの産生は見られなかった。
【0059】
上記図12及び図13の実験の結果を考慮して、最終的に増幅効率が高かったTCRVβ3.0陽性のCTL細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は97%であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0060】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、前述のMAGE3−A24ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。具体的には、前述のTCRβ鎖遺伝子に対応するセンスプライマー(gcagccatgggaatcaggctcctctgt:配列番号27)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号11に、そのアミノ酸配列を配列番号12に示す。このTCRβ鎖遺伝子の構成はTRBV28*01―TRBD1*01―TRBJ1−1*01―TRBC1であった。また、V−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果を図14に示す。また、TCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(337−372、36塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【実施例4】
【0061】
実施例3で得られたMAGE3−A24ペプチド特異的CTL由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0062】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0063】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、mock(4μg)、GFP(4μg)又はTCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、mockとして使用したベクターはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターからGFP遺伝子を切り出したものを用い、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、TCRβ遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図15に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。また、MAGE3−A24特異的TCRβ鎖遺伝子の発現は33.9%の細胞に確認され、良好な発現効率が認められた。
【0064】
なお、MAGE3−A24ペプチド特異的TCRβ鎖遺伝子発現リンパ球の細胞障害活性などの機能解析実験は今後行う予定である。
【実施例5】
【0065】
[MART1−A2ペプチド又はgp100−A2特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A2陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)又はgp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)にて処理した樹状細胞及びT2細胞を用いて4回刺激を行った。
【0066】
ex vivoにて前述のHLA−A2陽性の患者由来のリンパ球から誘導・増幅されたA2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なCTL細胞をテトラマーにて染色し、増幅されたA2ペプチド特異的テトラマー陽性CTL細胞(MART1−A2ペプチド特異的CTL細胞やgp100−A2ペプチド特異的CTL細胞)について定量的な評価を行った。その結果を図16に示す。図16に示されているとおり、対照コントロールのテトラマーであるFlu(Influenza)−MPテトラマーでは、ほとんど陽性細胞は認められない(0.04%)が、MART1−A2テトラマー染色やgp100−A2テトラマー染色では、それぞれ78.8%、44.3%と高率に陽性CTL細胞の増幅が認められた。
【0067】
MART1−A2ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果、MART1−A2テトラマー陽性CTL細胞の増幅を認めたMEL−006症例(case1)では、レパトワ抗体パネルのスクリーニングにて4種類のレパトワ(TCRVβ1、TCRVβ2、TCRVβ7.1及びTCRVβ14)の増加が検出された。
【0068】
MART1−A2ペプチドにて処理を行ったT2細胞にて、前述のMART1−A2ペプチド特異的CTL細胞又はgp100−A2ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図17に示す。図17に示されているとおり、TCRVβ1陽性のCTL細胞群において、IFN−γの産生が最も強く認められた。そこで、TCRVβ1陽性のCTL細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は95%以上であった。これらの純化したCTL細胞を、後述のTCRβ鎖の遺伝子解析に用いた。
【0069】
また、gp100−A2テトラマー陽性CTL細胞の増幅を認めたMEL−008症例(case2)において、HLA−A2テトラマーに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、98%以上の純度でgp100−A2テトラマー陽性細胞の集団が得られた(図18左)。また、得られたgp100−A2テトラマー陽性細胞を、レパトワ抗体で染色したところ、テトラマーsortingされたほとんどすべての細胞は、TCRVβ8を有していることが確認された(図18右)。そこで、前述の陽性細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0070】
上述の純化した細胞(MART1−A2ペプチド特異的CTL細胞又はgp100−A2ペプチド特異的CTL細胞)からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。具体的には、MART1−A2ペプチド特異的タンパク質遺伝子(TCRβ鎖遺伝子)に対応するセンスプライマー(cctgccatgggcttcaggctcctctgc:配列番号28)とアンチセンスプライマー(ctagcctctggaatcctttctcttgacc:配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号13に、そのアミノ酸配列を配列番号14に示す。また、gp100−A2ペプチド特異的タンパク質遺伝子(TCRβ鎖遺伝子)についても、それに対応するセンスプライマー(tctgccatggactcctggaccctctgc:配列番号30)とアンチセンスプライマー(配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号15に、そのアミノ酸配列を配列番号16に示す。
【実施例6】
【0071】
実施例5で得られたMART1−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子、又は、gp100−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*0201陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0072】
実施例5で得られたTCRβ鎖遺伝子をクローニングした患者とは異なるHLA−A*0201陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0073】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、TCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図19に示す。図19に示されているとおり、TCRVβ1遺伝子(MART1−A2ペプチド特異的TCRβ鎖遺伝子)及びTCRVβ8遺伝子(gp100−A2ペプチド特異的TCRβ鎖遺伝子)の導入効率は、それぞれ23.9%、13.3%であった。
【0074】
また、実施例6の上記遺伝子導入実験において、TCRβ鎖遺伝子のDNAに代えてGFP遺伝子(4μg)を用いて同様の実験を行った。その結果を図20に示す。図20に示されているとおり、GFP遺伝子のエレクトロポレーション法(EP法)によるナイーブT細胞への導入効率は43.3%であり、良好であった。
【0075】
また、MART1−A2ペプチドにて処理したT2細胞に対する、MART1−A2ペプチド特異的なTCRβ鎖を発現したHLA−A*0201陽性ナイーブT細胞の細胞障害活性を評価した。TCRβ鎖遺伝子を導入発現したHLA−A*0201陽性ナイーブT細胞としては、前記エレクトロポレーションによりTCRβ鎖遺伝子のDNA(4μg)、mock(4μg)、又はGFP(2μg)を導入した細胞を使用した。また、T2細胞をPBS+1%ヒト血清アルブミン溶液に懸濁し、MART1−A2ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該MART1−A2ペプチド処理したT2細胞1×105個を、同数の前記TCRβ鎖遺伝子を導入発現したHLA−A*0201陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、MART1−A2ペプチド処理したT2細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図21に示す。また、gp100−A2ペプチド特異的なTCRβ鎖を発現したHLA−A*0201陽性ナイーブT細胞の細胞障害活性についても同様に評価した。その結果を図21に示す。図21の結果から分かるように、MART1−A2及びgp100−A2ペプチド特異的なTCRβ鎖遺伝子を導入されたナイーブT細胞は、それぞれ標的細胞を処理したペプチド特異的に刺激され、有意なIFN−γの産生が確認された。これにより、これらTCRβ鎖遺伝子が細胞障害活性を有するタンパク質をコードする遺伝子であることが確認された。
【実施例7】
【0076】
[CMVpp65−A24ペプチド特異的TCRβ鎖について]
本研究では、末期がんであるHLA−A24陽性の転移性メラノーマ患者6例(M3、M7、M9、M13、M14、M18)由来のリンパ球を、メラノーマ特異的ペプチドであるCMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行い、CMVpp65−A24ペプチドに特異的なCTL lineの作製を試みた(図22)。CTL lineの作製は具体的には以下のようなプロトコールで行った。
【0077】
まず最初に、患者由来のリンパ球を2.5〜5×106cell/mlの濃度で、6−ウェルプレートに播種した後、非接着細胞の回収を行い、一時凍結保存した。一方、接着細胞については、GM−CSF、IL−4及びTNF−αの存在下にて培養し、成熟樹状細胞(dendritic cell:DC)を得た。播種から7日後に、樹状細胞をCMVpp65−A24ペプチド及びβ2−ミクログロブリンにて処理後、放射線照射した。放射線照射した成熟樹状細胞(DC)と、凍結保存しておいた前述の非接着細胞(T細胞)とで共培養を行った。(DC:T=1:10〜1:100(細胞数比))。共培養中の培養液にIL−7、IL−2を必要量添加した。播種から14日後に、同様の刺激をCMVpp65−A24ペプチド特異的なCTL細胞に対して再度行った。さらに、播種から21日後及び28日後に、CMVpp65−A24ペプチドにて処理したT2−A24細胞により、CTL細胞を刺激増幅した(T2−A24:CTL=1:10(細胞数比))。播種から35日後に、増幅されたCMVpp65−A24ペプチド特異的CTL細胞を、後述のIFN−γの産生刺激などの機能的アッセイやMACSによるソーテイングの実験に利用した。
【0078】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(CMVpp65−A24ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたCMVpp65−A24テトラマー陽性CTL細胞(CMVpp65 A24テトラマー+CD8+陽性CTL)について定量的な評価を行った。その結果を図23に示す。図23左に示されているように、6例中4例(M3、M7、M9、M18)で、CMVpp65−A24テトラマー陽性CTL細胞の比率が、刺激前の1%以下から、最大平均33.3%[65.9%(M3)11.2%(M7)、35%(M9)、21%(M18)]にまで増幅した。特にM3症例では、樹状細胞2回のみの刺激で著明な増幅効果が見られた。しかし、CMVpp65−A24テトラマー陽性CTL細胞の細胞数については、明らかな増加を確認できたのは3例(M3、M7、M18)であった(図23右)。また、CMVpp65−A24テトラマー陽性CTL細胞の細胞数の増幅の程度を、刺激前の細胞数を100とし、刺激後の細胞数をそれに対する比率で示した(図24)。刺激増幅後、細胞数の明らかな増加が認められた3例(M3、M7、M18)では、経過中に2.1〜207倍の範囲内で増加が認められ、最大で平均80.9倍の増幅効果が確認された。特に、最も増幅効果の高かったM3症例では、最大207倍にまでCMVpp65−A24ペプチド特異的CTL細胞を増加させることができた。
【0079】
なお、図23でテトラマー陽性細胞比率の増加が認められた各症例(M3、M7、M9、M18)における、増幅前後のフローサイトメトリーのデータをそれぞれ図25、図26、図27、図28に示す。これらの図から、テトラマー染色のコントロールとして用いたHLA−A24HIVテトラマーによる染色では、前述の陽性細胞は確認されなかったことが示された。
【0080】
また、CMVpp65−A24ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark, Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果、CMVpp65−A24テトラマー陽性CTL細胞の増幅を認めたM3症例では、レパトワ抗体パネルのスクリーニングにてTCRVβ7.1の増加が検出され、M7症例ではTCRVβ17の増加が検出され、M9症例ではTCRVβ7.1の増加が検出された。
【0081】
CMVpp65−A24ペプチド特異的CTL細胞を、各症例について増加が検出されたレパトワにて染色し、陽性細胞について定量的な評価を行った。その結果を図29に示す。図29に示されているとおり、CMVpp65−A24ペプチド特異的CTL細胞の増幅を認めたM3症例では、TCRVβ7.1が陽性であった細胞が94.9%、M7症例では、TCRVβ17が陽性であった細胞が62.5%、M9症例では、TCRVβ7.1が陽性であった細胞が54.3%であった。
【0082】
CMVpp65−A24ペプチドにて処理を行ったLCL(EB virusにて不死化したヒトB細胞培養株)であるTISI細胞にて、前述のCMVpp65−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図30に示す。図30に示されているとおり、すべての症例のCTL細胞において、CMVpp65−A24ペプチド処理したTISI細胞にて刺激した場合にのみ、大量のIFN−γが産生された。また、M9症例のIFN−γの産生量は、他の2症例と比較して低い傾向があった。
【0083】
また、CMVpp65−A24テトラマー又はTCRレパトワ特異的に細胞障害活性が誘導されるかを確認する目的で、標的細胞にて刺激したM3症例由来のCTL細胞のIFN−γ抗体を用いた細胞内染色を行った。その結果を図31に示す。図31に示されているとおり、コントロールペプチドであるCEA A24ペプチドによる刺激では、IFN−γの産生は見られなかったが、CMV A24ペプチドによる刺激の場合、テトラマー陽性細胞の61.5%に、TCRVβ7.1抗体陽性細胞の70.1%にIFN−γの産生が確認された。
【0084】
上記図29の実験の結果を考慮して、M3症例からはTCRVβ7.1陽性CTL細胞を、M7症例からはTCRVβ17陽性CTL細胞を、M9症例からはTCRVβ7.1細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、各陽性細胞は95%以上であった。これらの純化した各CTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0085】
上述の純化した3種の細胞からそれぞれtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいて3種のTCRタンパク質コード領域遺伝子(M3TCRβ鎖遺伝子、M7TCRβ鎖遺伝子、M9TCRβ鎖遺伝子)をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
【0086】
TCRレパトワ解析の結果に基づいて、前述のCMVpp65−A24ペプチド特異的TCRβ鎖タンパク質をコードする3種の遺伝子(M3TCRβ鎖遺伝子、M7TCRβ鎖遺伝子、M9TCRβ鎖遺伝子)に対応するプライマーをそれぞれ用いてPCR増幅を行い、3種の増幅断片の塩基配列を解析した。M3TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号17に、そのアミノ酸配列を配列番号18に示す。また、M7TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(tgcactatgagcaaccaggtgctctgc:配列番号31)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号19に、そのアミノ酸配列を配列番号20に示す。また、M9TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号21に、そのアミノ酸配列を配列番号22に示す。これら3クローンのTCRβ鎖遺伝子の塩基配列を決定したところ、M3TCRβ鎖遺伝子の構成はTRBV4−1*01―TRBD1*01―TRBJ1−2*01―TRBC1であり、M7TCRβ鎖遺伝子の構成は、TRBV19*01―TRBD1*01―TRBJ1−2*01―TRBC1であり、M9TCRβ鎖遺伝子の構成はTRBV4−1*01―TRBD2*01―TRBJ1−2*01―TRBC1であった。また、これら3クローンのTCRβ鎖遺伝子のV−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果をそれぞれ図32〜34に示す。また、これら3クローンのTCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(M3: 337−378、42塩基)(M7:337−372、36塩基)(M9:337−378、42塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【実施例8】
【0087】
実施例7で得られたCMVpp65−A24ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0088】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0089】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、GFP(2μg)又はM3TCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、M3TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をM3TCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びM3TCRβ鎖遺伝子の導入効率を検討した。その結果を図35に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。一方、CMVpp65−A24特異的TCRβ鎖遺伝子(M3TCRβ鎖遺伝子)は、38.3%の細胞で発現が確認され、良好な発現効率が認められた。
【0090】
また、CMVpp65−A24ペプチドにて処理したTISI細胞に対する、CMVpp65−A24ペプチド特異的なTCRβ鎖を発現したHLA−A*2402陽性ナイーブT細胞の細胞障害活性を評価した。前述のTCRβ鎖遺伝子(M3TCRβ鎖遺伝子)を導入発現したHLA−A*2402陽性ナイーブT細胞としては、前記エレクトロポレーションによりM3TCRβ鎖遺伝子のDNA(4μg)又はGFP(2μg)を導入した細胞を使用した。また、TISI細胞をPBS+1%ヒト血清アルブミン溶液に懸濁し、CMVpp65−A24ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該CMVpp65−A24ペプチド処理したTISI細胞1×105個を、同数の前記M3TCRβ鎖遺伝子を導入発現したHLA−A*2402陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、CMVpp65−A24ペプチド処理したTISI細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図36に示す。図36の結果から分かるように、CMVpp65−A24特異的TCRβ鎖遺伝子(M3TCRβ鎖遺伝子)を導入したリンパ球のみIFN−γが刺激され、本TCRβ鎖遺伝子がコードするTCRβ鎖を発現したT細胞が、CMV特異的な細胞障害活性を有することが示唆された。
【実施例9】
【0091】
[CMVpp65−A2ペプチド特異的TCRβ鎖について]
本研究では、末期がんであるHLA−A2陽性の転移性メラノーマ患者4例(M8、M10、M19、M5−SCC)由来のリンパ球を、メラノーマ特異的ペプチドであるCMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)にて処理した樹状細胞及びT2細胞を用いて4回刺激を行い、CMVpp65−A2ペプチドに特異的なCTL lineの作製を試みた(図37)。CTL lineの作製は具体的には以下のようなプロトコールで行った。
【0092】
まず最初に、患者由来のリンパ球を2.5〜5×106cell/mlの濃度で、6−ウェルプレートに播種した後、非接着細胞の回収を行い、一時凍結保存した。一方、接着細胞については、GM−CSF、IL−4及びTNF−αの存在下にて培養し、成熟樹状細胞(dendritic cell:DC)を得た。播種から7日後に、樹状細胞をCMVpp65−A2ペプチド及びβ2−ミクログロブリンにて処理後、放射線照射した。放射線照射した成熟樹状細胞(DC)と、凍結保存しておいた前述の非接着細胞(T細胞)とで共培養を行った。(DC:T=1:10〜1:100(細胞数比))。共培養中の培養液にIL−7、IL−2を必要量添加した。播種から14日後に、同様の刺激をCMVpp65−A24ペプチド特異的なCTL細胞に対して再度行った。さらに、播種から21日後及び28日後に、CMVpp65−A2ペプチドにて処理したT2細胞により、CTL細胞を刺激増幅した(T2:CTL=1:10(細胞数比))。播種から35日後に、増幅されたCMVpp65−A2ペプチド特異的CTL細胞を、後述のIFN−γの産生刺激などの機能的アッセイやMACSによるソーテイングの実験に利用した。
【0093】
ex vivoにて前述のHLA−A2陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(CMVpp65−A2ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたCMVpp65−A2テトラマー陽性CTL細胞(CMVpp65−A2テトラマー+CD8+陽性CTL)について定量的な評価を行った。その結果を図38上に示す。図38上に示されているように、4例中1例(M8)で、CMVpp65−A2テトラマー陽性CTL細胞の比率が、刺激前の1%以下から、最大20%以上にまで増幅した。一方、CMVpp65−A2テトラマー陽性CTL細胞の細胞数については、3例(M8、M10、M5−SSC)で増加傾向が見られ、特に、M8症例では、刺激前の448倍と、最も効率的に特異的CTL細胞の誘導が見られた(図38下)。
【0094】
なお、M8における、増幅前後のフローサイトメトリーのデータをそれぞれ図39に示す。増幅前のテトラマー陽性細胞比率は1%以下(0.27%)であったが、DC2回刺激後は2.14%に、さらにT2細胞で2回刺激後は24.8%まで増加した。一方、テトラマー染色のコントロールとして用いたHLA−A2 Flu−MPテトラマーによる染色では、陽性細胞の増幅は確認されなかった。
【0095】
また、CMVpp65−A2ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果を図40に示す。図40に示されているように、CMVpp65−A2テトラマー陽性CTL細胞の増幅を認めたM8症例では、レパトワ抗体パネルのスクリーニングにてTCRVβ8及びTCRVβ14の増加が検出された。すなわち、TCRVβ8が陽性であった細胞が14.8%、TCRVβ14が陽性であった細胞が7.3%であった。
【0096】
CMVpp65−A2ペプチドにて処理を行ったT2細胞にて、前述のCMVpp65−A2ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図41に示す。図41に示されているとおり、CD8及びIFN−γの両方が陽性のCTL細胞の比率は21.2%であり、そのうち、IFN−γ陽性のCTL細胞のほとんどすべてがTCRVβ8陽性のCTL細胞であった(22.7%)。この結果から、細胞障害活性を有するCTL細胞のTCRレパトワは、TCRVβ8であることが確認された。
【0097】
後述のTCRβ鎖遺伝子の解析に利用するために、M8由来のTCRVβ8陽性CTL細胞を以下の方法で選別した。
【0098】
M8症例由来のリンパ球をIn vitroにてDCで2回、T2細胞で3回刺激をして得られた増幅されたCMVA2テトラマー陽性細胞を、テトラマーsortingしたところ、増幅後のCMVpp65−A2テトラマー及びCD8陽性CTL細胞(CMVpp65−A2テトラマー+CD8+陽性CTL)並びにCMVpp65−A2テトラマー及びTCRVβ8陽性CTL細胞(CMVpp65−A2テトラマー+TCRVβ8+陽性CTL)の細胞の比率は、それぞれ43.1%及び44.6%であった(図42)。PE−CMVA2テトラマー試薬と抗PE抗体−マイクロビーズを用いてCTL細胞を染色し、MACS法にてテトラマー陽性細胞を選別したところ、該陽性細胞の純度は95.6%であった(図42)。選別された前記陽性細胞のうち、ほとんどすべてがTCRVβ8陽性であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0099】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子(M8TCRβ鎖遺伝子)をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
【0100】
TCRレパトワ解析の結果に基づいて、前述のCMVpp65−A2ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(M8TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。M8TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号30)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号23に、そのアミノ酸配列を配列番号24に示す。このM8TCRβ鎖遺伝子の塩基配列を決定したところ、M8TCRβ鎖遺伝子の構成はTRBV12−4*01―TRBD1*01―TRBJ1−2*01―TRBC1であった。また、このM8TCRβ鎖遺伝子のV−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果をそれぞれ図43に示す。また、このM8TCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(340−369、30塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【0101】
なお、CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子発現リンパ球の細胞障害活性などの機能解析実験は今後行う予定である。
【実施例10】
【0102】
[MART1−A2ペプチド特異的TCRα鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、前述のHLA−A2またはA24の患者(M1、M3、M6、M7、M8、M9)を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)、gp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)、MAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)若しくはMAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)、又は、サイトメガロウイルスに特異的なペプチドであるCMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)若しくはCMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)にて処理した樹状細胞を用いて4回刺激を行った。この刺激によって誘導された、前記の各種ペプチドに特異的なCTL細胞を、PEで標識した各種テトラマーにて染色し、テトラマー染色陽性のCTL細胞を、磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化した。純化して得られた各種のテトラマー染色陽性CTL細胞を、次のTCRα鎖のレパトワ解析に用いた。
【実施例11】
【0103】
[TCRα鎖のレパトワ解析]
上記実施例10で得られた各種のテトラマー染色陽性CTL細胞のうち、まずM8症例MART1−A2ペプチド特異的CTL(MART−A2テトラマー陽性)についてTCRα鎖のレパトワ解析を行った。すなわち、このCTLからtotal RNAを抽出し、抽出したtotal RNA2μgを用いた逆転写反応にてcDNAを合成し、合成により得られたcDNA全量に対して1/40量のcDNAとTCRα鎖cDNAレパトワ解析用プライマーとを用いたPCRにより得られたPCR産物の解析を試みた。TCRα鎖cDNAレパトワ解析用プライマーとしては、33種類のセンスプライマー(HAV1Aセンスプライマー:TCTGGTATGTGCAATACCCCAACC:配列番号79; HAV1Bセンスプライマー:CTGAGGAAACCCTCTGTGCA:配列番号80; HAV2センスプライマー:GATGGAAGGTTTACAGCACAGCTC:配列番号81; HAV3センスプライマー:CACAGTGGAAGATTAAGAGTCACGC:配列番号82; HAV4Aセンスプライマー:AACAGAATGGCCTCTCTGGC:配列番号83; HAV4Bセンスプライマー:GGATTGCGCTGAAGGAAGAG:配列番号84; HAV5センスプライマー:TGAAGGTCACCTTTGATACCACCC:配列番号85; HAV6センスプライマー:AATCCGCCAACCTTGTCATCTCCG:配列番号86; HAV7センスプライマー:AACTGCACGTACCAGACATC:配列番号87; HAV8センスプライマー:ACCCTGAGTGTCCAGGAGGG:配列番号88; HAV9センスプライマー:CACTGCTGACCTTAACAAAGGCG:配列番号89; HAV10センスプライマー:TCCTGGTGACAGTAGTTACG:配列番号90; HAV11センスプライマー:AGGCTCAAAGCCTTCTCAGCAGGG:配列番号91; HAV12センスプライマー:TCCACCAGTTCCTTCAACTTCACC:配列番号92; HAV13センスプライマー:TTCATCAAAACCCTTGGGGACAGC:配列番号93; HAV14センスプライマー:CCCAGCAGGCAGATGATTCTCGTT:配列番号94; HAV15センスプライマー:GGATAAACATCTGTCTCTGCG:配列番号95; HAV16センスプライマー:GATAGCCATACGTCCAGATG:配列番号96; HAV18センスプライマー:TGCCACTCTTAATACCAAGGAGGG:配列番号97; HAV19センスプライマー:ACACTGGCTGCAACAGCATC:配列番号98; HAV20センスプライマー:TTACAAACGAAGTGGCCTCC:配列番号99; HAV21センスプライマー:ACCCTGCTGAAGGTCCTACATTCC:配列番号100; HAV22センスプライマー:CTTGGAGAAAGGCTCAGTTC:配列番号101; HAV23センスプライマー:TGCCTCGCTGGATAAATCATCAGG:配列番号102; HAV24センスプライマー:TCCCAGCTCAGCGATTCAGCCTCC:配列番号103; HAV25センスプライマー:GTCCTGTCCTCTTGATAGCC:配列番号104; HAV26センスプライマー:AGCCCAGCCATGCAGGCATCTACC:配列番号105; HAV27センスプライマー:TTGATACCAAAGCCCGTCTC:配列番号106; HAV28センスプライマー:GAACATCACAGCCACCCAGACCGG:配列番号107; HAV29センスプライマー:GCAAAGCTCCCTGTACCTTACGG:配列番号108; HAV30センスプライマー:TTTCTGCACAGCACAGCCCC:配列番号109; HAV31センスプライマー:AGCAAAAACTTCGGAGGCGG:配列番号110; HAV32センスプライマー:AAGGAGAGGACTTCACCACG:配列番号111)からなる群から選ばれる1種類のセンスプライマーと、アンチセンスプライマー(HTCA3アンチセンスプライマー:GTTGCTCTTGAAGTCCATAGACC:配列番号112)とを用いた。
【0104】
上記のPCRにより得られたPCR産物をアガロースゲルで電気泳動した結果を図44(A)に示す。図44(A)において、両脇のマーカーのレーンを除くと、上段の左側のレーンから順に、センスプライマーとして上記HAV1AからHAV14までの16種類をそれぞれ用いたPCR産物を電気泳動し、下段の左側のレーンから順に、センスプライマーとして上記HAV15からHAV32までの17種類をそれぞれ用いたPCR産物を電気泳動した。図44(A)から分かるように、HAV2, 6, 9, 24, 26及び32の各レーンにおいて、PCRの増幅産物のバンドが確認された。これらのバンドをゲルから回収し、TAクローニング法(TOPO TA Cloning Kit、インビトロジェン社製)によりベクターへ組み込んだ。
このベクターに組み込まれたバンド部分(PCR産物)の塩基配列を決定したところ、HAV2, 9及び32のPCR産物はTCRα鎖遺伝子由来のDNA断片であるが、その他のPCR産物は非特異的反応による増幅産物であることが判明した。HAV2, 9及び32のDNA断片が含まれる遺伝子は、NCBIにより使用される新規名称TRAV12-2, 16及び25にそれぞれ相当する遺伝子であった。これら3種類のTCRα遺伝子レパトワについて、全タンパク質コード領域の遺伝子取得を目的として、上記のcDNA断片を鋳型に各レパトワ特異的な16種類のセンスプライマー(TRAV1-2センスプライマー:CAGCAGATGTGGGGAGTTTTCCTTCTT:配列番号113; TRAV4センスプライマー:TAGAATATGAGGCAAGTGGCGAGAGTG:配列番号114; TRAV8-2センスプライマー:TCAGCCATGCTCCTGCTGCTCGTCCCA:配列番号115; TRAV8-3センスプライマー:TCAGCCATGCTCCTGGAGCTTATCCCA:配列番号116; TRAV8-4センスプライマー:TCAGCCATGCTCCTGCTGCTCGTCCCA:配列番号117; TRAV12-1センスプライマー:AAAAGAATGATGATATCCTTGAGAGTT:配列番号118; TRAV12-2センスプライマー:AGAAGAATGATGAAATCCTTGAGAGTT:配列番号119; TRAV16センスプライマー:TCAGACATGAAGCCCACCCTCATCTCA:配列番号120; TRAV17センスプライマー:AGAAGAATGGAAACTCTCCTGGGAGTG:配列番号121; TRAV21センスプライマー:GAAGGAATGGAGACCCTCTTGGGCCTG:配列番号122; TRAV22センスプライマー:AGCATCATGAAGAGGATATTGGGAGCT:配列番号123; TRAV24センスプライマー:GGAAACATGGAGAAGAATCCTTTGGCA:配列番号124; TRAV25センスプライマー:AGGGAGATGCTACTCATCACATCAATG:配列番号125; TRAV13-1センスプライマー:ACAAGGATGACATCCATTCGAGCTGTA:配列番号126; TRAV35センスプライマー:ATAAGGATGCTCCTTGAACATTTATTA:配列番号127; TRAV38-2センスプライマー:GTGAGCATGGCATGCCCTGGCTTCCTG:配列番号128)からなる群から選ばれる1種類のセンスプライマーと、終止コドンを含む定常領域アンチセンスプライマー(TRACアンチセンスプライマー:TCAGCTGGACCACAGCCGCAGCGTCAT:配列番号129)とを用いて遺伝子増幅を行った。増幅したPCR産物(各遺伝子のタンパク質コード領域のDNA断片)をアガロースゲル電気泳動した結果を図44(B)に示す。いずれのPCR産物についても、想定された位置にバンドが検出された。
【0105】
次いで、上記のPCR産物をベクターにクローニングした後、これら3クローンのPCR産物(TCRα鎖遺伝子)の塩基配列を決定したところ、TRAV12-2(HAV2)、TRAV16(HAV9)、TRAV25(HAV32)のTCRα鎖遺伝子の構成は、それぞれ「TRAV12-2*01―TRAJ10*01」(図51:配列番号38の塩基配列; 図52:配列番号39のアミノ酸配列)、「TRAV16*01―TRAJ38*01」(図53:配列番号40の塩基配列; 図54:配列番号41のアミノ酸配列)、「TRAV25*01―TRAJ31*01」(図55:配列番号42の塩基配列; 図56:配列番号43のアミノ酸配列)であることが分かった。
【0106】
また、上記実施例10で得られた各種のテトラマー染色陽性CTL細胞のうち、M8症例MART1−A2ペプチド特異的CTL(MART−A2テトラマー陽性)以外のCTL(M6症例MART1−A2ペプチド特異的CTL、M8症例gp100−A2ペプチド特異的CTL、M1症例MAGE1−A24ペプチド特異的CTL、M7症例MAGE3−A24ペプチド特異的CTL、M8症例CMVpp65−A2ペプチド特異的CTL、M3症例CMVpp65−A24ペプチド特異的CTL、M7症例CMVpp65−A24ペプチド特異的CTL、及び、M9症例CMVpp65−A24ペプチド特異的CTL)についても、同様の手法によってTCRα鎖のレパトワ解析を行った。その結果、M6症例MART1−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV12-2*01―TRAJ40*01」(図45:配列番号32の塩基配列; 図46:配列番号33のアミノ酸配列)、「TRAV12-2*01―TRAJ50*01」(図47:配列番号34の塩基配列; 図48:配列番号35のアミノ酸配列)、「TRAV25*01―TRAJ21*01」(図49:配列番号36の塩基配列; 図50:配列番号37のアミノ酸配列)であり、M8症例gp100−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-4*01―TRAJ9*01」(図57:配列番号44の塩基配列; 図58:配列番号45のアミノ酸配列)であり、M1症例MAGE1−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV1-2*01―TRAJ10*01」(図59:配列番号46の塩基配列; 図60:配列番号47のアミノ酸配列)、「TRAV12-1*01―TRAJ49*01」(図61:配列番号48の塩基配列; 図62:配列番号49のアミノ酸配列)、「TRAV17*01―TRAJ23*01」(図63:配列番号50の塩基配列; 図64:配列番号51のアミノ酸配列)、「TRAV22*01―TRAJ40*01」(図65:配列番号52の塩基配列; 図66:配列番号53のアミノ酸配列)であり、M7症例MAGE3−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV13-1*01―TRAJ45*01」(図67:配列番号54の塩基配列; 図68:配列番号55のアミノ酸配列)、「TRAV38-2/DV8*01―TRAJ57*01」(図69:配列番号56の塩基配列; 図70:配列番号57のアミノ酸配列)であり、M8症例CMVpp65−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-2*01―TRAJ23*01」(図71:配列番号58の塩基配列; 図72:配列番号59のアミノ酸配列)、「TRAV26―stop codon」(図73:配列番号60の塩基配列)、「TRAV35*02―TRAJ50*01」(図74:配列番号61の塩基配列; 図75:配列番号62のアミノ酸配列)であり、M3症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-3*01―TRAJ39*01」(図76:配列番号63の塩基配列; 図77:配列番号64のアミノ酸配列)、「TRAV24*01―TRAJ54*01」(図78:配列番号65の塩基配列; 図79:配列番号66のアミノ酸配列)、「TRAV38-2/DV8*01―TRAJ49*01」(図80:配列番号67の塩基配列; 図81:配列番号68のアミノ酸配列)であり、M7症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV21*02―TRAJ28*01」(図82:配列番号69の塩基配列; 図83:配列番号70のアミノ酸配列)、「TRAV24*01―TRAJ47*02」(図84:配列番号71の塩基配列; 図85:配列番号72のアミノ酸配列)、「TRAV35―stop codon」(図86:配列番号73の塩基配列)であり、M9症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV4*01―TRAJ44*01」(図87:配列番号74の塩基配列; 図88:配列番号75のアミノ酸配列)、「TRAV16*01―TRAJ49*01」(図89:配列番号76の塩基配列; 図90:配列番号77のアミノ酸配列)、「TRAV17―stop codon」(図91:配列番号78の塩基配列)であった。
【実施例12】
【0107】
[TCRα鎖/β鎖遺伝子の導入発現]
上記の腫瘍関連ペプチドに特異的なTCRβ鎖遺伝子とα鎖遺伝子とを同時にmutant Jurkat細胞(ATCC TIB−153)へ導入した場合に、TCRβ鎖タンパク質及びα鎖タンパク質が発現し、細胞障害活性を誘導し得るか否かにつき検討した。
すなわち、実施例5で得られたM8症例MART1−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子と、実施例11で得られたM8症例MART1−A2ペプチド特異的CTL細胞由来のTCRα鎖遺伝子とを、エレクトロポレーション法(Amaxa社のNucleofector装置を使用)にて同時にmutant Jurkat細胞(ATCC TIB−153)に導入し、それらTCRα鎖やβ鎖の発現の解析を行った。なお、Mutant Jurkat細胞は、Jurkat細胞のE6-1クローンの変異型で、TCRのβ鎖を欠失しており、さらに、膜上にTCRα鎖及びβ鎖からなるheterodimer並びにCD3タンパク質を発現しないことが知られているため、TCRβ鎖遺伝子の導入実験によく使用される。
【0108】
まずpmax-GFP発現ベクターをmutant Jurkat細胞へ導入し、pmax-GFP/mutant Jurkat細胞を作製した。その際、mutant Jurkat細胞への遺伝子導入の至適条件を設定した。pmax-GFP発現ベクターの導入から48時間後に、フローサイトメトリー(FACSCalibur(登録商標):ベクトン・ディッキンソン社製)を用いて、pmax-GFP/mutant Jurkat細胞中のGFP遺伝子及びCD3タンパク質の発現効率を測定した。その結果を、図92に示す。図92から分かるように、全細胞の61.8%にGFP遺伝子の発現が確認されたが、CD3タンパク質の発現はほとんど認められなかった。
【0109】
また、TCRVβ3.0(NCBIにより使用される新規名称はTRBV28)を乗せたpmaxベクター8μgと、各TCRα鎖遺伝子(TRAV12、TRAV16又はTRAV25)を乗せたpmaxベクター8μgとを、同時に2×106個のmutant Jurkat細胞に導入し、pmax-TCRVβ3.0・pmax-TCRα/mutant Jurkat細胞を作製した。また、コントロールとして、TCRVβ3.0を乗せたpmaxベクターのみを導入したmutant Jurkat細胞(pmax-TCRVβ3.0/mutant Jurkat細胞)を作製した。
各発現ベクターの導入から48時間後に、フローサイトメトリー(FACSCalibur:ベクトン・ディッキンソン社製)を用いて、pmax-TCRVβ3.0・pmax-TCRα/mutant Jurkat細胞中のTCRVβ3.0 (TRBV28)遺伝子、及び、CD3タンパク質の発現効率を測定した。その結果を、図93に示す。図93から分かるように、TCRVβ3.0 (TRBV28)の発現は、TCRα鎖遺伝子を同時導入した場合の方が高く、TRAV12やTRAV16遺伝子を同時導入した場合はそれぞれ11.5%、10.3%と発現の増加を認めた(TCRVβ3.0単独では、1.7%)。また同様の実験におけるCD3タンパク質の発現も、TRAV12やTRAV16遺伝子を同時導入した場合は、19.8%、21.8%と発現の増加を認めた(TCRVβ3.0単独では、8.7%)(図94)。
これらの結果から、上記のTCRβ鎖とTCRα鎖の組み合わせによって、CD3の発現が向上し、細胞障害活性が誘導されることが示された。
【図面の簡単な説明】
【0110】
【図1】MAGE1−A24ペプチド処理した樹状細胞の、肺転移巣に対する治療効果を示す図である。
【図2】末梢血中のMAGE1−A24ペプチド等に対する特異的なCTL細胞の比率を、ELISPOTアッセイにより測定した結果を示す図である。
【図3】MAGE1−A24ペプチド特異的CTL細胞を、テトラマーにて染色した結果を示す図である。
【図4】MAGE1−A24ペプチド特異的CTL細胞についての2種類のTCRレパトワに対する解析結果を示す図である。
【図5】MAGE1−A24ペプチド特異的CTL細胞を、テトラマー及びTCRVβにて染色した結果を示す図である。
【図6】MAGE1−A24ペプチド処理したTISI細胞で刺激されたMAGE1−A24ペプチド特異的CTL細胞を、テトラマー又はIFN−γ等にて染色した結果を示す図である。
【図7】MAGE1−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図8】MAGE1−A24ペプチド特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図9】MAGE3−A24ペプチド処理した樹状細胞の、頚部リンパ節転移巣に対する治療効果を示す図である。
【図10】末梢血中のMAGE3−A24ペプチド等に対する特異的なCTL細胞の比率を、ELISPOTアッセイにより測定した結果を示す図である。
【図11】MAGE3−A24ペプチド特異的CTL細胞についてのTCRレパトワ解析の結果を示す図である。
【図12】MAGE3−A24ペプチド特異的CTL細胞についての2種類のTCRレパトワに対する解析結果を示す図である。
【図13】MAGE3−A24ペプチド処理したT2−A24細胞等で刺激されたMAGE3−A24ペプチド特異的CTL細胞を、IFN−γにて染色した結果を示す図である。
【図14】本発明の新規なTCRβ鎖遺伝子の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図15】MAGE3−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図16】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なCTL細胞を、テトラマーにて染色した結果を示す図である。
【図17】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)処理したT2細胞で刺激されたA2ペプチド特異的CTL細胞を、テトラマー及びIFN−γにて染色した結果を示す図である。
【図18】gp100−A2ペプチド特異的CTL細胞について、テトラマー(左図)及びTCRレパトワ(右図)に対する解析結果を示す図である。
【図19】MART1−A2ペプチド又はgp100−A2ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図20】GFP遺伝子を導入したナイーブT細胞等を、該GFPにて染色した結果を示す図である。
【図21】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図22】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の刺激増幅プロトコールを示す図である。
【図23】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の増幅効果を示す図である。
【図24】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の増幅効果を示す図である。
【図25】CMVpp65−A24ペプチド特異的CTL細胞(M3症例)をテトラマーにて染色した結果を示す図である。
【図26】CMVpp65−A24ペプチド特異的CTL細胞(M7症例)をテトラマーにて染色した結果を示す図である。
【図27】CMVpp65−A24ペプチド特異的CTL細胞(M9症例)をテトラマーにて染色した結果を示す図である。
【図28】CMVpp65−A24ペプチド特異的CTL細胞(M18症例)をテトラマーにて染色した結果を示す図である。
【図29】CMVpp65−A24ペプチド特異的CTL細胞を、特異的レパトワにて染色した結果を示す図である。
【図30】CMVpp65−A24ペプチド処理したTISI細胞等で刺激されたCMVpp65−A24ペプチド特異的CTL細胞における、IFN−γ量を示す図である。
【図31】CMVpp65−A24ペプチド処理したTISI細胞等で刺激されたCMVpp65−A24ペプチド特異的CTL細胞等を、IFN−γ等にて染色した結果を示す図である。
【図32】本発明の新規なTCRβ鎖遺伝子(M3症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図33】本発明の新規なTCRβ鎖遺伝子(M7症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図34】本発明の新規なTCRβ鎖遺伝子(M9症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図35】CMVpp65−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図36】CMVpp65−A24ペプチド特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図37】In vitroにおけるCMVpp65−A2ペプチド特異的CTL細胞の刺激増幅プロトコールを示す図である。
【図38】In vitroにおけるCMVpp65−A2ペプチド特異的CTL細胞の増幅効果を示す図である。
【図39】CMVpp65−A2ペプチド特異的CTL細胞をテトラマーにて染色した結果を示す図である。
【図40】CMVpp65−A2ペプチド特異的CTL細胞についてのTCRレパトワ解析の結果を示す図である。
【図41】CMVpp65−A2ペプチド処理したT2細胞等で刺激されたCMVpp65−A2ペプチド特異的CTL細胞等を、IFN−γ等にて染色した結果を示す図である。
【図42】In vitroにおいて刺激増幅されたCMVpp65−A2ペプチド特異的CTL細胞をテトラマーにて染色した結果を示す図である。
【図43】本発明の新規なTCRβ鎖遺伝子(M8症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図44】M8症例MART1−A2ペプチド特異的CTL由来のTCRα鎖のレパトワ解析の結果(PCR産物のアガロースゲルでの電気泳動結果)を示す図である。
【図45】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図46】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図47】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図48】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図49】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図50】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図51】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図52】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図53】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図54】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図55】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図56】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図57】M8症例gp100−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図58】M8症例gp100−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図59】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図60】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図61】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図62】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図63】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図64】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図65】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図66】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図67】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図68】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図69】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図70】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図71】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図72】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図73】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図74】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図75】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図76】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図77】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図78】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図79】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図80】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図81】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図82】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図83】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図84】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図85】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図86】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図87】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図88】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図89】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図90】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図91】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図92】GFP遺伝子を導入したmutant Jurkat細胞を、GFPにて染色した結果を示す図(左図)、及び、該細胞をCD3にて染色した結果を示す図(右図)である。
【図93】TCRβ鎖遺伝子とα鎖遺伝子とを同時に導入したmutant Jurkat細胞(右上図、左下図、右下図)やTCRβ鎖遺伝子のみを導入したmutant Jurkat細胞(左上図)を、TCRβ鎖にて染色した結果を示す図である。
【図94】TCRβ鎖遺伝子とα鎖遺伝子とを同時に導入したmutant Jurkat細胞(右上図、左下図、右下図)やTCRβ鎖遺伝子のみを導入したmutant Jurkat細胞(左上図)を、CD3にて染色した結果を示す図である。
【技術分野】
【0001】
本発明は、T細胞レセプターβ鎖タンパク質、並びにT細胞レセプターβ鎖遺伝子、該遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質発現ベクター、及び該発現ベクターが導入された形質転換体に関する。また、本発明は、T細胞レセプターα鎖タンパク質、並びにT細胞レセプターα鎖遺伝子、該遺伝子が組み込まれたT細胞レセプターα鎖タンパク質発現ベクター、及び該発現ベクターが導入された形質転換体に関する。
【背景技術】
【0002】
悪性黒色腫は、インターロイキン2などを用いた免疫療法が比較的有効な癌である(例えば、非特許文献1参照)が、生体内における抗腫瘍効果には、腫瘍細胞に反応する細胞傷害性T細胞(Cytotoxic T lymphocyte:CTL)が重要であることが示されている(例えば、非特許文献2参照)。免疫療法後、腫瘍反応性T細胞が活性化され、T細胞上のT細胞レセプター(T cell receptor:TCR)が、HLA(ヒト白血球抗原)分子によって腫瘍細胞表面に提示される腫瘍細胞タンパク質が分解されてできたペプチド(腫瘍抗原)を認識し、腫瘍細胞を傷害する。更に各種サイトカインを分泌し、マクロファージ等の他の免疫細胞をも誘導活性化し、生体内で腫瘍拒絶を起こすと考えられている。腫瘍反応性T細胞には自己腫瘍細胞のみならず、HLAを共有する他の患者からの腫瘍細胞にも反応するような共通腫瘍抗原を認識する場合と、自己腫瘍細胞しか反応しない固有抗原を認識する場合がある。
【0003】
ヒト腫瘍細胞に対する免疫応答機構を解明して、新しい免疫療法を開発するためには、これらの腫瘍反応性T細胞が認識する腫瘍抗原を同定することが重要である。腫瘍反応性T細胞が樹立されている場合には、その反応性を利用した機能的cDNA発現クローニング法を用い、悪性黒色腫抗原の単離が可能である。例えば、転移メラノーマ患者からの腫瘍反応性細胞傷害性T細胞と、HLA−A2を発現する腫瘍細胞株とを用いて、メラノーマの予防、診断及び治療に用いることができる悪性黒色腫抗原を単離することが可能であり(例えば、特許文献1参照)、その一つMART1は同様な方法によって単離、同定されたものである。現在までに代表的なものとして、上記MART1やgp100のような色素細胞(メラノサイト)組織特異的タンパク質、各種腫瘍と正常組織では精巣に発現が認められるMAGE、BAGE、GAGE、NY−ESO−I等のCT抗原(Cancer-Testis抗原)、腫瘍細胞の遺伝子異常により産生されるβカテニン、CDK4(cyclin dependent kinase 4)、p15、GnT−V等の変異ペプチド抗原が単離されている(例えば、非特許文献3参照)。一方、MAGE抗原は、メラノーマのみならず肺がんや消化器がんなど多くの固形がんでの高い発現が報告されている。
【0004】
また、cytomegalovirus(CMV)感染症は、通常の免疫状態である健常人ではあまり問題にはならないが、特に臓器移植後など免疫低下状態では、重篤な間質性肺炎などの合併症を起こしうる深刻な疾患である。現在、抗ウイルス剤の投与によりCMV感染症の臨床症状は緩和されるが、根治対策となり得る抗ウイルス剤は現在のところまだ知られていない。これまでに主にCMVpp65タンパク由来のHLA−A2拘束性のエピトープに関する研究が行われており、CMVpp65 495−503(NLVPMVATV)ペプチドを用いた特異的CTL細胞の誘導やテトラマー試薬を用いたCTLクローンの評価が行われている。しかし、進行がん患者でのCMVに対するCTL細胞の誘導反応に関して、それほど多くの知見は得られておらず、CMV感染症に特異的なCTL細胞のT細胞レセプターβ鎖遺伝子については何ら知られていない。特に日本人に多いHLA−A24拘束性のCMVエピトープに関する研究は、まだ始まったばかりである。
【0005】
T細胞はT細胞レセプターによりMHC分子/抗原ペプチド複合体を認識して、抗原提示細胞(APC)などの標的細胞に結合するが、CTLはMHCクラスI分子/抗原ペプチドの複合体のみに結合し(MHCクラスI拘束性)、ヘルパーT(Th)細胞はMHCクラスII分子/抗原ペプチドの複合体のみしか結合できない(MHCクラスII拘束性)とされている。また、MHCクラスI分子はほとんどの有核細胞に発現しているが、MHCクラスII分子は限られた一部の細胞しか発現していない。このためTh細胞はMHCクラスII分子を発現した樹状細胞やB細胞、活性化T細胞などとは結合できるが、これら以外の腫瘍細胞や感染細胞などには直接結合することができない。しかし遺伝子操作によってMHCクラスI拘束性とされるCTL由来のT細胞レセプター遺伝子を導入したMHCクラスII拘束性とされるCD4陽性CD8陰性T細胞が、対応抗原をパルスしたAPCとCD8非依存的に反応して活性化されることができ、対応抗原に結合できるT細胞レセプターを発現していることが示された。また、非特異的な抗腫瘍活性をもつ末梢血リンパ球に腫瘍抗原特異的CTL由来のT細胞レセプター遺伝子を導入することにより、特異的に腫瘍を傷害することができることが報告されている(例えば、非特許文献4参照)。
【0006】
他方、T細胞表面に存在する膜表面タンパク質であり、抗原認識を担うT細胞レセプターには、α鎖・β鎖から成るヘテロ二量体とγ鎖・δ鎖から成るヘテロ二量体の二種類が同定されている。T細胞レセプター遺伝子は免疫グロブリン遺伝子と同様に、可変領域(V:variable)、多様性領域(D:diversity)、結合領域(J:joining)からなり分子の多様性を生じるVドメインと、細胞外定常領域、膜貫通領域、細胞内領域を含む定常領域(C:constant)からなるCドメインにより構成されている。これらのアミノ酸配列はT細胞ごとに異なっていることから、T細胞レセプターは抗原認識分子であると同時に個々のT細胞の目印にもなっている。このT細胞レセプターの多様性は後天的なT細胞レセプター遺伝子の再構成によって生みだされており、T細胞レセプターβ,δ鎖はV−D−Jを、T細胞レセプターα,γ鎖はV−Jを再構成することが知られている。
【0007】
T細胞の初期分化は、B細胞と同様にT細胞レセプターのα鎖及びβ鎖遺伝子の再構成と密接に関わっている。T細胞レセプターβ鎖遺伝子の再構成はDN−T細胞(double negative:CD4−8−)の段階で開始され、β鎖遺伝子の再構成に成功した細胞はβ鎖タンパク質と代替α鎖(preTα)の複合体、つまりプレT細胞レセプターを細胞表面に発現する。そして、CD4及びCD8分子の発現が誘導され、DP−T細胞(double positive:CD4+8+)へと移行し、DPとなったT細胞は、次にT細胞レセプターのα鎖遺伝子の再構成に成功した細胞のみが、T細胞レセプターを表面に発現できることが知られている。
【0008】
T細胞レセプターβ鎖の遺伝子構成に関しては、ヒトゲノム遺伝子配列情報データベースによれば、T細胞レセプターβ鎖の遺伝子は約30種類の亜族(遺伝子自体は約70種)からなるV遺伝子、2種類のD遺伝子、14種類のJ遺伝子、2種類のC遺伝子によって構成され、Vβ中に2つの可変領域(CDR1、CDR2)が存在することも知られている。これらの中からVβ、Dβ、Jβが各1つずつ選ばれて結合し、T細胞レセプターβ鎖をコードするDNAが構成されている。この際、Vβ−Dβ−Jβの結合部にはランダムな塩基の挿入や欠失が起こり、無限に近い多様性が生みだされている。このVβ−Dβ−Jβの結合部位(連結部分)は、CDR3(相補性決定領域3:third complementarity determining region)、結合部(junctional)領域と呼ばれ、T細胞レセプターの中で最も多型性に富む部分であり、塩基の挿入・欠失が起こるため、各遺伝子構成が同じであっても連結部分も含めて同一配列になることは極めて稀である。
【0009】
また、T細胞レセプターα鎖の遺伝子構成に関しては、ヒトゲノム遺伝子配列情報データベースによれば、T細胞α鎖の遺伝子は約40種類の亜族(遺伝子自体は約50種)からなるV遺伝子、約60種のJ遺伝子、1種類のC遺伝子によって構成される。T細胞の分化の過程で起きる遺伝子組み換えによって、前述の中からVα、Jα、Cαが各1種ずつ選ばれて結合し、染色体上でT細胞レセプターα鎖をコードするDNAが構成される。染色体上で構成されたT細胞レセプターα鎖遺伝子は発現してそのタンパク質が細胞表面に表出する。T細胞α鎖のVα−Jαの結合部位(連結部分)は、多型性に富む部分であり、塩基の挿入・欠失が起こるため、各遺伝子構成が同じであっても、連結部分を含めて同一配列になることは極めて稀である。
【0010】
【特許文献1】特表平10−505481号公報
【非特許文献1】JAMA 271, 907, 1994
【非特許文献2】Microbiol.Immunol. 42, 803, 1998
【非特許文献3】Immunol Res 16, 313, 1997
【非特許文献4】J.Immunol.,171:3287-3295, 2003
【発明の開示】
【発明が解決しようとする課題】
【0011】
T細胞レパートリー(T細胞レパトワ)の多様性は、T細胞間での多様な抗原認識能と密接に関係していることから、T細胞の機能,免疫応答,免疫機能を考える上でT細胞レセプターレパートリーの機能解明を行うことは極めて重要である。最近、癌,自己免疫疾患,アレルギー疾患等において、患部局所に存在するT細胞のT細胞レセプターレパートリー解析の研究が進められ、疾患特異的なT細胞が解明されつつある。しかし、そのT細胞上のT細胞レセプター分子が、どのような働きをしているか、ということは、T細胞レセプター分子の単離が困難であること等の問題があり、まだその詳細が明らかにされるに至っていない。本発明の課題は、メラノーマ特異的ペプチドであるMAGE1 135−143(NYKHCFPEI:以下、MAGE1−A24ペプチドと称する場合がある)、MAGE3 195−203(IMPKAGLLI:以下、MAGE3−A24ペプチドと称する場合がある)、MART1 27−35(AAGIGILTV:以下、MART1−A2ペプチドと称する場合がある)、gp100 209−217(IMDQVPFSV:以下、gp100−A2ペプチドと称する場合がある)、CMVpp65 341−349(QYDPVAALF:以下、CMVpp65−A24ペプチドと称する場合がある)、又はCMVpp65 495−503(NLVPMVATV:以下、CMVpp65−A2ペプチドと称する場合がある)を特異的に認識するT細胞レセプターβ鎖及びその遺伝子を提供することにある。
また、本発明の課題は、CTL細胞膜上で上記T細胞レセプターβ鎖とヘテロ二量体を構成したときに、がん細胞由来の抗原ペプチドに特異性が高いT細胞レセプターα鎖及びその遺伝子を提供することにもある。
【課題を解決するための手段】
【0012】
T細胞レセプター分子が、T細胞を介する免疫応答および免疫機構に重要な役割を果たしていることから、これら免疫応答および免疫機構の解析には、個々のT細胞レセプター分子を得ることが必要である。しかし、膜表面タンパク質を細胞から精製すると、1010個の細胞からせいぜい10μg程度しか得られない。しかも、T細胞レセプターの多様性により、一種類のT細胞レセプターを分取するためには、それぞれのT細胞レセプターを持つT細胞株を樹立し、その各々からT細胞レセプターを単離することが必須となる。しかし、T細胞のT細胞レセプターレパートリーは1010種類にも及んでおり、1010種のT細胞株を樹立することは、実質的に極めて困難である。
【0013】
MAGE1、MAGE3、MART1及びgp100はメラノーマに発現する癌抗原であり、ペプチドワクチンによる治療を受けている転移性のメラノーマ患者において、抗原抗体反応を強く誘導するものである。また、CMVpp65は、CMV感染症に発現するCMV抗原であり、CMV感染症患者において、抗原抗体反応を誘導するものである。MAGE1やMAGE3はHLA−A24限局的な抗原決定部位を持っており(MAGE1 135−143:NYKHCFPEI; MAGE3 195−203:IMPKAGLLI)、MART1やgp100はHLA−A2限局的な抗原決定部位を持っている(MART1 27−35:AAGIGILTV; gp100 209−217:IMDQVPFSV)。また、CMVpp65はHLA−A24限局的な抗原決定部位(CMVpp65 341−349:QYDPVAALF)と、HLA−A2限局的な抗原決定部位(CMVpp65 495−503:NLVPMVATV)を持っている。
【0014】
本発明者らは、樹状細胞に基づく上記ペプチド特異的なCTL細胞の誘導方法を構築し、該CTL細胞をAuto−MACS法にて純化し、純化した細胞からRNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いTCRレパトワ解析により、発現TCRの遺伝子分類を行い、解析結果に基づいてTCR遺伝子をクローニングした。クローニングした断片のDNAシークエンスを解析し、上記ペプチドを特異的に認識する新規なT細胞レセプターβ鎖及びそれとヘテロ二量体を形成するα鎖を同定した。本発明は、上記知見に基づき完成するに至ったものである。
【0015】
すなわち本発明は、(1)配列番号7、9、11、13、15、17、19、21若しくは23で表される塩基配列、又は、配列番号8、10、12、14、16、18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子や、(2)上記(1)に記載のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターや、(3)上記(2)に記載のT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが導入された形質転換体に関する。
【0016】
また本発明は、(4)配列番号32、34、36、38、40、42、44、46、48、50、52、54、56、58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子や、(5)上記(4)に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターや、(6)上記(5)に記載のT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体や、(7)上記(1)に記載のT細胞レセプターβ鎖遺伝子及び上記(4)に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターや、(8)上記(7)に記載のT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体や、(9)組換えベクターが導入されたリンパ球である上記(3)、(6)又は(8)に記載の形質転換体に関する。
【0017】
さらに本発明は、(10)配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質や、(11)配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質に特異的に結合する抗体に関する。
【0018】
またさらに本発明は、(12)配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質や、(13)配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質に特異的に結合する抗体に関する。
【0019】
また本発明は、(14)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(15)配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(16)配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物に関する。
【0020】
さらに本発明は、(17)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(18)配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(19)配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物や、(20)疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子及びT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物や、(21)配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物や、(22)配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物に関する。
【発明の効果】
【0021】
本発明によると、MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の特定の疾患におけるT細胞の機能、免疫応答、免疫機能を解明する上で有用である。また、本発明のT細胞レセプターβ鎖遺伝子やα鎖遺伝子を利用した遺伝子治療や養子免疫療法により、患者の細胞性免疫を疾患特異的に向上させ、MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の特定の疾患を治療しうる可能性がある。
【発明を実施するための最良の形態】
【0022】
本発明のT細胞レセプターβ鎖遺伝子(TCRβ鎖遺伝子ともいう)は、疾患特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子である限り特に制限されず、例えばメラノーマ特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子や、CMV感染症特異的抗原ペプチドに特異的なCTL細胞のTCRβ鎖遺伝子等を好ましく例示することができる。
【0023】
前記メラノーマ特異的抗原ペプチドとしては、MAGE1、MAGE3、MART1、gp100等が知られており、中でも、MAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)、MAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)、MART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)、gp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)を好ましく例示することができ、上記CMV感染症特異的抗原ペプチドとしては、CMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)、CMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)を好ましく例示することができる。
【0024】
前記本発明のT細胞レセプターβ鎖遺伝子として具体的には、配列番号7や配列番号9で表される塩基配列(MAGE1−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号11で表される塩基配列(MAGE3−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号13で表される塩基配列(MART1−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号15で表される塩基配列(gp100−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号17や配列番号19や配列番号21で表される塩基配列(CMVpp65−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号23で表される塩基配列(CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号8や配列番号10で表されるアミノ酸配列をコードする塩基配列(MAGE1−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号12で表されるアミノ酸配列をコードする塩基配列(MAGE3−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号14で表されるアミノ酸配列をコードする塩基配列(MART1−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号16で表されるアミノ酸配列をコードする塩基配列(gp100−A2ペプチド特異的TCRβ鎖遺伝子)、配列番号18や配列番号20や配列番号22で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A24ペプチド特異的TCRβ鎖遺伝子)、配列番号24で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子)からなるT細胞レセプターβ鎖遺伝子を好ましく例示することができる。
【0025】
本発明のT細胞レセプターα鎖遺伝子(TCRα鎖遺伝子ともいう)は、疾患特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子である限り特に制限されず、例えば前述のメラノーマ特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子や、前述のCMV感染症特異的抗原ペプチドに特異的なCTL細胞のTCRα鎖遺伝子等を好ましく例示することができる。
【0026】
前記本発明のT細胞レセプターα鎖遺伝子として具体的には、配列番号32や配列番号34や配列番号36や配列番号38や配列番号40や配列番号42で表される塩基配列(MART1−A2ペプチド特異的TCRα鎖遺伝子)、配列番号44で表される塩基配列(gp100−A2ペプチド特異的TCRα鎖遺伝子)、配列番号46や配列番号48や配列番号50や配列番号52で表される塩基配列(MAGE1−A24ペプチド特異的TCRα鎖遺伝子)、配列番号54や配列番号56で表される塩基配列(MAGE3−A24ペプチド特異的TCRα鎖遺伝子)、配列番号58や配列番号61で表される塩基配列(CMVpp65−A2ペプチド特異的TCRα鎖遺伝子)、配列番号63や配列番号65や配列番号67や配列番号69や配列番号71や配列番号74や配列番号76で表される塩基配列(CMVpp65−A24ペプチド特異的TCRα鎖遺伝子)、配列番号33や配列番号35や配列番号37や配列番号39や配列番号41や配列番号43で表されるアミノ酸配列をコードする塩基配列(MART1−A2ペプチド特異的TCRα鎖遺伝子)、配列番号45で表されるアミノ酸配列をコードする塩基配列(gp100−A2ペプチド特異的TCRα鎖遺伝子)、配列番号47や配列番号49や配列番号51や配列番号53で表されるアミノ酸配列をコードする塩基配列(MAGE1−A24ペプチド特異的TCRα鎖遺伝子)、配列番号55や配列番号57で表されるアミノ酸配列をコードする塩基配列(MAGE3−A24ペプチド特異的TCRα鎖遺伝子)、配列番号59や配列番号62で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A2ペプチド特異的TCRα鎖遺伝子)、配列番号64や配列番号66や配列番号68や配列番号70や配列番号72や配列番号75や配列番号77で表されるアミノ酸配列をコードする塩基配列(CMVpp65−A24ペプチド特異的TCRα鎖遺伝子)からなるT細胞レセプターα鎖遺伝子を好ましく例示することができる。
【0027】
本発明の遺伝子の取得方法や調製方法は特に限定されるものでなく、本明細書中に開示した前記塩基配列や前記アミノ酸配列の各情報に基づいて適当なプローブやプライマーを調製し、それらを用いて当該遺伝子が存在することが予測されるヒトcDNAライブラリーをスクリーニングすることにより目的の遺伝子を単離したり、常法に従って化学合成により調製することができる。例えば、細胞又は組織からの全RNAの分離、mRNAの分離や精製、cDNAの取得とそのクローニングなどはいずれも常法に従って実施することができる。本発明の遺伝子をcDNAライブラリーからスクリーニングする方法は、例えば、Molecular Cloning: A laboratory Manual, 2nd Ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY., 1989に記載の方法等、当業者により常用される方法を挙げることができる。
【0028】
本発明のT細胞レセプターβ鎖タンパク質は、配列番号8や配列番号10で表されるアミノ酸配列からなるタンパク質(MAGE1−A24ペプチド特異的TCRβ鎖タンパク質)、配列番号12で表されるアミノ酸配列からなるタンパク質(MAGE3−A24ペプチド特異的TCRβ鎖タンパク質)、配列番号14で表されるアミノ酸配列からなるタンパク質(MART1−A2ペプチド特異的TCRβ鎖タンパク質)、配列番号16で表されるアミノ酸配列からなるタンパク質(gp100−A2ペプチド特異的TCRβ鎖タンパク質)、配列番号18や配列番号20や配列番号22で表されるアミノ酸配列からなるタンパク質(CMVpp65−A24ペプチド特異的TCRβ鎖タンパク質)、又は配列番号24で表されるアミノ酸配列からなるタンパク質(CMVpp65−A2ペプチド特異的TCRβ鎖タンパク質)である。
【0029】
また、本発明のT細胞レセプターα鎖タンパク質は、配列番号33や配列番号35や配列番号37や配列番号39や配列番号41や配列番号43で表されるアミノ酸配列からなるタンパク質(MART1−A2ペプチド特異的TCRα鎖タンパク質)、配列番号45で表されるアミノ酸配列からなるタンパク質(gp100−A2ペプチド特異的TCRα鎖タンパク質)、配列番号47や配列番号49や配列番号51や配列番号53で表されるアミノ酸配列からなるタンパク質(MAGE1−A24ペプチド特異的TCRα鎖タンパク質)、配列番号55や配列番号57で表されるアミノ酸配列からなるタンパク質(MAGE3−A24ペプチド特異的TCRα鎖タンパク質)、配列番号59や配列番号62で表されるアミノ酸配列からなるタンパク質(CMVpp65−A2ペプチド特異的TCRα鎖タンパク質)、配列番号64や配列番号66や配列番号68や配列番号70や配列番号72や配列番号75や配列番号77で表されるアミノ酸配列からなるタンパク質(CMVpp65−A24ペプチド特異的TCRα鎖タンパク質)である。
【0030】
本発明のタンパク質の取得・調製方法は特に限定されず、天然由来のタンパク質でも、化学合成したタンパク質でも、遺伝子組換え技術により作製した組み換えタンパク質の何れでもよい。天然由来のタンパク質を取得する場合には、かかるタンパク質を発現しているT細胞からタンパク質の単離・精製方法を適宜組み合わせることにより、本発明のタンパク質を取得することができる。化学合成によりタンパク質を調製する場合には、例えば、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法に従って本発明のタンパク質を合成することができる。また、各種の市販のペプチド合成機を利用して本発明のタンパク質を合成することもできる。遺伝子組換え技術によりタンパク質を調製する場合には、該タンパク質をコードする塩基配列からなるDNAを好適な発現系に導入することにより本発明のタンパク質を調製することができる。これらの中でも、比較的容易な操作でかつ大量に調製することが可能な遺伝子組換え技術による調製が好ましい。
【0031】
例えば、遺伝子組換え技術によって、本発明のタンパク質を調製する場合、かかるタンパク質を細胞培養物から回収し精製するには、硫酸アンモニウムまたはエタノール沈殿、酸抽出、アニオンまたはカチオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ハイドロキシアパタイトクロマトグラフィーおよびレクチンクロマトグラフィーを含めた公知の方法、好ましくは、高速液体クロマトグラフィーが用いられる。特に、アフィニティークロマトグラフィーに用いるカラムとしては、例えば、本発明のタンパク質に対するモノクローナル抗体等の抗体を結合させたカラムや、上記本発明のタンパク質に通常のペプチドタグを付加した場合は、このペプチドタグに親和性のある物質を結合したカラムを用いることにより、これらのタンパク質の精製物を得ることができる。また、本発明のタンパク質が細胞膜に発現している場合は、細胞膜分解酵素を作用させた後、上記の精製処理を行うことにより精製標品を得ることができる。
【0032】
また、本発明のタンパク質をマーカータンパク質やペプチドタグと結合させることもできる。マーカータンパク質としては、従来知られているマーカータンパク質であれば特に制限されるものではなく、例えば、アルカリフォスファターゼ、HRP等の酵素、抗体のFc領域、GFP等の蛍光物質などを具体的に挙げることができ、また本発明におけるペプチドタグとしては、HA、FLAG、Myc等のエピトープタグや、GST、マルトース結合タンパク質、ビオチン化ペプチド、オリゴヒスチジン等の親和性タグなどの従来知られているペプチドタグを具体的に例示することができる。かかる融合ペプチドは、常法により作製することができ、Ni−NTAとHisタグの親和性を利用した本発明のタンパク質の精製や、本発明のタンパク質の検出や、本発明のタンパク質に対する抗体の定量や、その他当該分野の研究用試薬としても有用である。
【0033】
本発明の組換えベクターとしては、前記本発明のT細胞レセプターβ鎖遺伝子及び/又はT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及び/又はT細胞レセプターα鎖タンパク質を発現しうる組換えベクターである限り特に制限されないが、動物細胞において発現しうるものが好ましい。本発明の組換えベクターは、本発明の遺伝子を発現ベクターに適切にインテグレイトすることにより構築することができる。かかる発現ベクターとしては、宿主細胞において自立複製可能であるものや、あるいは宿主細胞の染色体中へ組込み可能であるものが好ましく、また、本発明の遺伝子を発現できる位置にプロモーター、エンハンサー、ターミネーター等の制御配列を含有しているものを好適に使用することができる。
【0034】
動物細胞用の発現ベクターとして、例えば、pEGFP-C1(Clontech社製)、pGBT−9(Clontech社製)、pcDNAI(フナコシ社製)、pcDM8(フナコシ社製)、pAGE107(Cytotechnology, 3, 133, 1990)、pCDM8(Nature, 329, 840, 1987)、pcDNAI/AmP(Invitrogen社製)、pREP4(Invitrogen社製)、pAGE103(J.Blochem., 101, 1307, 1987)、pAGE210等を例示することができる。動物細胞用のプロモーターとしては、例えば、サイトメガロウイルス(ヒトCMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーター、レトロウイルスのプロモーター、メタロチオネインプロモーター、ヒートショックプロモーター、SRαプロモーター等を挙げることができる。
【0035】
本発明の形質転換体としては、上記本発明の組換えベクターが導入された宿主細胞であれば特に制限されないが、リンパ球が好ましく、中でもナイーブT細胞が好ましい。中でも、上記本発明の組換えベクターがインビトロで導入されたリンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができる疾患(該組換えベクターに組み込まれたT細胞レセプターβ鎖遺伝子やT細胞レセプターα鎖遺伝子にコードされるT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質が特異性を示す抗原ペプチドを特異的に発現する疾患)に対する治療用組成物として有用である。特に、該組換えベクターに組み込まれた本発明のT細胞レセプターβ鎖遺伝子の塩基配列が、配列番号7、配列番号9、配列番号11、配列番号13、配列番号15で表される塩基配列や、配列番号8、配列番号10、配列番号12、配列番号14、配列番号16で表されるアミノ酸配列をコードする塩基配列からなる場合や、前記組換えベクターに組み込まれた本発明のT細胞レセプターα鎖遺伝子の塩基配列が、配列番号32、配列番号34、配列番号36、配列番号38、配列番号40、配列番号42、配列番号44、配列番号46、配列番号48、配列番号50、配列番号52、配列番号54、配列番号56で表される塩基配列や、配列番号33、配列番号35、配列番号37、配列番号39、配列番号41、配列番号43、配列番号45、配列番号47、配列番号49、配列番号51、配列番号53、配列番号55、配列番号57で表されるアミノ酸配列をコードする塩基配列からなる場合は、前記リンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができるMAGE1,3陽性の固形がん又はメラノーマ治療用組成物として有用であり、該組換えベクターに組み込まれた本発明のT細胞レセプターβ鎖遺伝子の塩基配列が、配列番号17、配列番号19、配列番号21、配列番号23で表される塩基配列や、配列番号18、配列番号20、配列番号22、配列番号24で表されるアミノ酸配列をコードする塩基配列からなる場合や、前記組換えベクターに組み込まれた本発明のT細胞レセプターα鎖遺伝子の塩基配列が、配列番号58、配列番号61、配列番号63、配列番号65、配列番号67、配列番号69、配列番号71、配列番号74、配列番号76で表される塩基配列や、配列番号59、配列番号62、配列番号64、配列番号66、配列番号68、配列番号70、配列番号72、配列番号75、配列番号77で表されるアミノ酸配列をコードする塩基配列からなる場合は、前記リンパ球、好ましくは、ナイーブT細胞は、養子免疫療法等に好適に用いることができるCMV感染症治療用組成物として有用である。
なお、本発明における「MAGE1,3陽性の固形がん又はメラノーマ治療用組成物」とは、メラノーマ; MAGE1陽性の固形がん(但しメラノーマを除く); 及び、MAGE3陽性の固形がん(但しメラノーマを除く);から選ばれる1つ又は2つ以上の疾患に対して治療効果を奏する組成物を意味する。
【0036】
養子免疫療法等免疫治療剤に用いる場合は、形質転換T細胞クローンを精製することが好ましく、その手法としては、リン酸緩衝液等に混合して遠心する方法、分離剤を用いた比重遠心法などを用いることができる。MAGE1,3陽性の固形がんやメラノーマやCMV感染症等の疾患の治療用組成物の投与量は、1回あたり形質転換リンパ球として106〜1012個の細胞が好ましい。投与形態としては、注射剤、点滴剤等の液体が好ましく、形質転換リンパ球をヒト血清アルブミンを0.01〜5%となるように添加した生理食塩液に分散した注射剤又は点滴剤がより好ましい。投与方法としては、静脈への点滴又は静脈、動脈、局所等への注射が好ましい。投与する液量は、投与方法、投与する場所等に異なるが、50〜500ccとするのが好ましく、この液量に前記の形質転換リンパ球が含まれるようにするのが好ましい。投与頻度は1回/日〜1回/月とするのが好ましく、投与回数は少なくとも1回、好ましくは5回以上とすることができる。
【0037】
本発明の抗体は、上記本発明のT細胞レセプターβ鎖タンパク質又はT細胞レセプターα鎖タンパク質に特異的に結合する抗体である限り特に制限されず、モノクローナル抗体、ポリクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体等の免疫特異的な抗体を例示することができる。本発明のこれらの抗体は上記本発明のT細胞レセプターβ鎖タンパク質を抗原として用いて常法により作製することができるが、その中でもモノクローナル抗体がその特異性に優れている点でより好ましい。かかるモノクローナル抗体等の本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に特異的に結合する抗体は、例えば、メラノーマやCMV感染症等の疾患の診断や本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質の分子機構を明らかにする上で有用である。
【0038】
本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に対する抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)に本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質又はこれらのタンパク質を膜表面に発現した細胞を投与することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975)、トリオーマ法、ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc., 1985)など任意の方法を用いることができる。以下に本発明のT細胞レセプタータンパク質として、ヒトT細胞由来の本発明のT細胞レセプターβ鎖タンパク質を例に挙げてマウス由来の本発明のT細胞レセプターβ鎖タンパク質に対して特異的に結合するモノクローナル抗体、すなわち抗hT細胞レセプターβ鎖マウスモノクローナル抗体の作製方法を説明する。
【0039】
上記抗hT細胞レセプターβ鎖モノクローナル抗体は、抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマをインビボ又はインビトロで常法により培養することにより生産することができる。例えば、インビボ系においては、齧歯動物、好ましくはマウス又はラットの腹腔内で培養することにより、またインビトロ系においては、動物細胞培養用培地で培養することにより得ることができる。インビトロ系でハイブリドーマを培養するための培地としては、ストレプトマイシンやペニシリン等の抗生物質を含むRPMI1640又はMEM等の細胞培養培地を例示することができる。
【0040】
抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマは、例えば、本発明のT細胞レセプターβ鎖タンパク質又は該タンパク質を膜表面に発現したT細胞を用いてBALB/cマウスを免疫し、免疫されたマウスの脾臓細胞とマウスNS−1細胞(ATCC TIB−18)とを、常法により細胞融合させ、免疫蛍光染色パターンによりスクリーニングすることにより、抗hT細胞レセプターβ鎖モノクローナル抗体産生ハイブリドーマを作出することができる。また、かかるモノクローナル抗体の分離・精製方法としては、タンパク質の精製に一般的に用いられる方法であればどのような方法でもよく、アフィニティークロマトグラフィー等の液体クロマトグラフィーを具体的に例示することができる。
【0041】
また、本発明の上記本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質に対する一本鎖抗体をつくるためには、一本鎖抗体の調製法(米国特許第4,946,778号)を適用することができる。また、ヒト化抗体を発現させるために、トランスジェニックマウス又は他の哺乳動物等を利用したり、上記抗体を用いて、本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質を発現するクローンを単離・同定したり、アフィニティークロマトグラフィーでそのポリペプチドを精製することもできる。
【0042】
また上記本発明の抗hT細胞レセプターβ鎖モノクローナル抗体等の抗体に、例えば、FITC(フルオレセインイソシアネート)又はテトラメチルローダミンイソシアネート等の蛍光物質や、125I、32P、14C、35S又は3H等のラジオアイソトープや、アルカリホスファターゼ、ペルオキシダーゼ、β−ガラクトシダーゼ又はフィコエリトリン等の酵素で標識したものや、グリーン蛍光タンパク質(GFP)等の蛍光発光タンパク質などを融合させた融合タンパク質を用いることによって、上記本発明のT細胞レセプターβ鎖タンパク質やT細胞レセプターα鎖タンパク質の機能解析を行うことができる。また免疫学的測定方法としては、RIA法、ELISA法、蛍光抗体法、プラーク法、スポット法、血球凝集反応法、オクタロニー法等の方法を挙げることができる。
【0043】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。なお、実施例1における実験概要は、樹状細胞ワクチン投与を受けたメラノーマ患者(HLA−A24陽性)リンパ球の入手→メラノーマ抗原ペプチドMAGE1(A24拘束性)にて処理した樹状細胞の作製→樹状細胞及びT2−A24細胞刺激によるペプチド特異的CTL細胞の誘導・増幅→増幅したCTL細胞のマーカー解析(FACS)と定量(テトラマー染色)及び機能解析(刺激されたCTL細胞からのIFN−γ産生解析)→AutoMACS法による、テトラマー又はレパトワ抗体陽性CTL細胞の純化→RNA抽出→逆転写反応→PCRによるTCR遺伝子(β鎖)の増幅→DNA Analyzer(ABI3100)を用いたDNA断片解析(TCRβ鎖遺伝子の同定)→PCRによるタンパク質コード領域の増幅とクローニング→ABI3100による塩基配列の決定である。その他の実施例3、5、7及び9における実験概要は、用いるペプチドが異なる点を除いて、ほぼ同様である。また、実施例10における実験概要は、用いるペプチドの他、増幅や同定する対象がTCRβ鎖遺伝子ではなく、TCRα鎖遺伝子である点を除いて、ほぼ同様である。
【実施例1】
【0044】
[MAGE1−A24ペプチド特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行った。今回リンパ球を使用したMEL−001の患者では、MAGE1 135−143にて処理した樹状細胞の投与を行ったが、約4cm大の肺転移巣が著明に縮小を認め、明確な臨床的効果が得られている(図1)。また免疫反応のモニタリング検査であるELISPOTアッセイでは、末梢血中にMAGE1−A24ペプチドに対する特異的なCTL細胞の陽性反応(Δ)が確認され、臨床効果と相関した結果が得られている(図2)。
【0045】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(MAGE1−A24ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたMAGE1−A24テトラマー陽性CTL細胞について定量的な評価を行った。その結果を図3に示す。図3に示されているとおり、対照コントロールのテトラマーであるHIV−A24テトラマーでは、ほとんど陽性細胞は認められない(0.04%)が、MAGE1−A24テトラマー染色では18.4%と高率に陽性CTL細胞の増幅が認められた。
【0046】
また、MAGE1−A24ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社製)を用いて、有意に増加したレパトワの同定を行った。その結果を図4に示す。図4に示されているとおり、MAGE1−A24テトラマー陽性CTL細胞の増幅を認めたMEL−001症例では、レパトワ抗体パネルのスクリーニングにて2種類のレパトワ(TCRVβ5.1及びTCRVβ7.1)の増加が検出された。レパトワ陽性細胞の比率は、TCRVβ5.1が12.8%、TCRVβ7.1が27.1%であった。
【0047】
MAGE1−A24ペプチド特異的CTL細胞を、MAGE1−A24テトラマー及び、TCRVβ5.1又はTCRVβ7.1にて染色し、陽性細胞について定量的な評価を行った。その結果を図5に示す。図5に示されているとおり、MAGE1−A24ペプチド特異的CTL細胞の増幅を認めたMEL−001症例では、MAGE1−A24テトラマー及びTCRVβ7.1の両方が陽性であった細胞が10.6%、MAGE1−A24テトラマー及びTCRVβ5.1の両方が陽性であった細胞が0.84%であり、MAGE1−A24テトラマー陽性のCTL細胞の大半は、TCRVβ7.1が陽性であった。一方、コントロール用のテトラマーであるHIV−A24テトラマーは、ほとんど染色されなかった。
【0048】
MAGE1−A24ペプチドにて処理を行ったLCL(EB virusにて不死化したヒトB細胞培養株)であるTISI細胞にて、前述のMAGE1−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図6に示す。図6に示されているとおり、MAGE1−A24テトラマー及びIFN−γが共に陽性の細胞比率は9.1%であり、TCRVβ7.1及びIFN−γが共に陽性の細胞比率は9.9%であり、TCRVβ5.1及びIFN−γが共に陽性の細胞比率は0.71%であった。これらの結果より、MAGE1−A24テトラマー陽性CTL細胞において、上述の刺激によってIFN−γを産生しうる細胞は、ほとんどがTCRVβ7.1陽性の細胞であることが明らかとなった。
【0049】
上記図5及び図6の実験の結果を考慮して、TCRVβ7.1陽性CTL細胞を、HLA−A24テトラマー又は特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は97%であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0050】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、前述のMAGE1−A24ペプチド特異的TCRβ鎖タンパク質をコードする2クローンの遺伝子(TCRβ鎖遺伝子)にそれぞれ対応するプライマーを用いてPCR増幅を行い、各増幅断片の塩基配列を解析した。具体的には、前述のTCRβ鎖遺伝子(clone1)に対応するセンスプライマー(gctagcatgggctgcaggctgctctgc:配列番号25)とアンチセンスプライマー(tcagaaatcctttctcttgaccatggc:配列番号26)を用いて、本TCRβ鎖遺伝子(clone1)のタンパク質コード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子(clone1)の塩基配列を配列番号7に、そのアミノ酸配列を配列番号8に示す。また、前述のもう一方のTCRβ鎖遺伝子(clone2)についてもそれに対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号26)を用いて、本TCRβ鎖遺伝子(clone2)のタンパク質コード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子(clone2)の塩基配列を配列番号9に、そのアミノ酸配列を配列番号10に示す。クローニングした2つの配列を解析した結果、これらのTCRβ鎖遺伝子は同じ遺伝子ではあるが、異なるクローンであることが判明した。
【0051】
本実施例及び後述の実施例に使用したプライマーの配列は、TCRレパトワ解析ではJeffry R. Currier and Mary Ann Robinsonが成書Current Protocols in Immunology※3 に記載したものを使用し、タンパク質コード領域のクローニングでは、公開データベース(NCBI)に収載されるゲノム配列にもとづいた。
※1 ImMunoGeneTics:1125985409921_0
※2 National Center for Biotechnology Information:http://www.ncbi.nlm.nih.gov/
※3 Current Protocols in Immunology, John E. Coligan et al. ed. (2000) 10.28.1-10.28.24. John Wiley & Sons, Inc.
【実施例2】
【0052】
実施例1で得られたMAGE1−A24ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。ここで、ナイーブT細胞とは、胸腺での分化後、抗原にまだ遭遇していないT細胞のことである。
【0053】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0054】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、mock(4μg)、GFP(2μg)又はTCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、mockとして使用したベクターはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターからGFP遺伝子を切り出したものを用い、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図7に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。また、MAGE1−A24特異的TCRβ鎖遺伝子は、クローン1については29.4%の細胞で、クローン2については32.4%の細胞で発現が確認され、良好な発現効率が認められた。
【0055】
また、MAGE1−A24ペプチドにて処理したTISI細胞に対する、MAGE1−A24ペプチド特異的なTCRβ鎖を発現したHLA−A*2402陽性ナイーブT細胞の細胞障害活性を評価した。TCRβ鎖遺伝子(clone1又はclone2)を導入発現したHLA−A*2402陽性ナイーブT細胞としては、前記エレクトロポレーションによりTCRβ鎖遺伝子のDNA(4μg)又はGFP(2μg)を導入した細胞を使用した。また、TISI細胞をPBS(Mg, Ca含有、Cambrex社製)+1%ヒト血清アルブミン溶液に懸濁し、MAGE1−A24ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該MAGE1−A24ペプチド処理したTISI細胞1×105個を、同数の前記TCRβ鎖遺伝子を導入発現したHLA−A*2402陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、MAGE1−A24ペプチド処理したTISI細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図8に示す。図8の結果から分かるように、TCRβ鎖遺伝子(clone2)を導入したリンパ球のみIFN−γが刺激され、本TCRβ鎖遺伝子がコードするTCRβ鎖を発現したT細胞が、腫瘍特異的な細胞障害活性を有することが示唆された。
【実施例3】
【0056】
[MAGE3−A24ペプチド特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行った。今回リンパ球を使用したMEL−007の患者では、MAGE3 195−203にて処理した樹状細胞の投与を行ったが、約2cm大の頚部リンパ節転移巣が著明に縮小を認め、明確な臨床的効果が得られている(図9)。また免疫反応のモニタリング検査であるELISPOTアッセイでは、末梢血中にMAGE3−A24ペプチドに対する特異的なCTL細胞の陽性反応(◇)が確認され、臨床効果と相関した結果が得られている(図10)。
【0057】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(MAGE3−A24ペプチド特異的CTL細胞)について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark, Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果を図11に示す。図11に示されているとおり、ほとんどのレパトワ抗体では、比率が5%以下であったが、TCRVβ3.0(TCRVβ3)とTCRVβ13.1に対する陽性CTL細胞の比率の増加が確認された。より具体的には、図12に示されているとおり、TCRVβ3.0陽性細胞の比率は24.7%、TCRVβ13.1陽性細胞の比率は11.0%であった。
【0058】
MAGE3−A24ペプチドにて処理を行ったT2−A24細胞(T2細胞にHLAーA−*2402遺伝子を導入後、A24陽性細胞をクローニングした細胞)にて、前述のMAGE3−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図13に示す。図13に示されているとおり、上述の刺激により全体の6.8%の比率のCTL細胞にIFN−γの産生が認められた。一方、MAGE3−A24ペプチドによる処理を行わなかったT2−A24細胞による刺激を行った場合(コントロール)は、IFN−γの産生は見られなかった。
【0059】
上記図12及び図13の実験の結果を考慮して、最終的に増幅効率が高かったTCRVβ3.0陽性のCTL細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は97%であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0060】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、前述のMAGE3−A24ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。具体的には、前述のTCRβ鎖遺伝子に対応するセンスプライマー(gcagccatgggaatcaggctcctctgt:配列番号27)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号11に、そのアミノ酸配列を配列番号12に示す。このTCRβ鎖遺伝子の構成はTRBV28*01―TRBD1*01―TRBJ1−1*01―TRBC1であった。また、V−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果を図14に示す。また、TCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(337−372、36塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【実施例4】
【0061】
実施例3で得られたMAGE3−A24ペプチド特異的CTL由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0062】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0063】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、mock(4μg)、GFP(4μg)又はTCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、mockとして使用したベクターはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターからGFP遺伝子を切り出したものを用い、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、TCRβ遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図15に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。また、MAGE3−A24特異的TCRβ鎖遺伝子の発現は33.9%の細胞に確認され、良好な発現効率が認められた。
【0064】
なお、MAGE3−A24ペプチド特異的TCRβ鎖遺伝子発現リンパ球の細胞障害活性などの機能解析実験は今後行う予定である。
【実施例5】
【0065】
[MART1−A2ペプチド又はgp100−A2特異的TCRβ鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、HLA−A2及びA24の患者を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A2陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)又はgp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)にて処理した樹状細胞及びT2細胞を用いて4回刺激を行った。
【0066】
ex vivoにて前述のHLA−A2陽性の患者由来のリンパ球から誘導・増幅されたA2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なCTL細胞をテトラマーにて染色し、増幅されたA2ペプチド特異的テトラマー陽性CTL細胞(MART1−A2ペプチド特異的CTL細胞やgp100−A2ペプチド特異的CTL細胞)について定量的な評価を行った。その結果を図16に示す。図16に示されているとおり、対照コントロールのテトラマーであるFlu(Influenza)−MPテトラマーでは、ほとんど陽性細胞は認められない(0.04%)が、MART1−A2テトラマー染色やgp100−A2テトラマー染色では、それぞれ78.8%、44.3%と高率に陽性CTL細胞の増幅が認められた。
【0067】
MART1−A2ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果、MART1−A2テトラマー陽性CTL細胞の増幅を認めたMEL−006症例(case1)では、レパトワ抗体パネルのスクリーニングにて4種類のレパトワ(TCRVβ1、TCRVβ2、TCRVβ7.1及びTCRVβ14)の増加が検出された。
【0068】
MART1−A2ペプチドにて処理を行ったT2細胞にて、前述のMART1−A2ペプチド特異的CTL細胞又はgp100−A2ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図17に示す。図17に示されているとおり、TCRVβ1陽性のCTL細胞群において、IFN−γの産生が最も強く認められた。そこで、TCRVβ1陽性のCTL細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、陽性細胞は95%以上であった。これらの純化したCTL細胞を、後述のTCRβ鎖の遺伝子解析に用いた。
【0069】
また、gp100−A2テトラマー陽性CTL細胞の増幅を認めたMEL−008症例(case2)において、HLA−A2テトラマーに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、98%以上の純度でgp100−A2テトラマー陽性細胞の集団が得られた(図18左)。また、得られたgp100−A2テトラマー陽性細胞を、レパトワ抗体で染色したところ、テトラマーsortingされたほとんどすべての細胞は、TCRVβ8を有していることが確認された(図18右)。そこで、前述の陽性細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0070】
上述の純化した細胞(MART1−A2ペプチド特異的CTL細胞又はgp100−A2ペプチド特異的CTL細胞)からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
TCRレパトワ解析の結果に基づいて、ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。具体的には、MART1−A2ペプチド特異的タンパク質遺伝子(TCRβ鎖遺伝子)に対応するセンスプライマー(cctgccatgggcttcaggctcctctgc:配列番号28)とアンチセンスプライマー(ctagcctctggaatcctttctcttgacc:配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号13に、そのアミノ酸配列を配列番号14に示す。また、gp100−A2ペプチド特異的タンパク質遺伝子(TCRβ鎖遺伝子)についても、それに対応するセンスプライマー(tctgccatggactcctggaccctctgc:配列番号30)とアンチセンスプライマー(配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号15に、そのアミノ酸配列を配列番号16に示す。
【実施例6】
【0071】
実施例5で得られたMART1−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子、又は、gp100−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*0201陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0072】
実施例5で得られたTCRβ鎖遺伝子をクローニングした患者とは異なるHLA−A*0201陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0073】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、TCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をTCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びTCRβ鎖遺伝子の導入効率を検討した。その結果を図19に示す。図19に示されているとおり、TCRVβ1遺伝子(MART1−A2ペプチド特異的TCRβ鎖遺伝子)及びTCRVβ8遺伝子(gp100−A2ペプチド特異的TCRβ鎖遺伝子)の導入効率は、それぞれ23.9%、13.3%であった。
【0074】
また、実施例6の上記遺伝子導入実験において、TCRβ鎖遺伝子のDNAに代えてGFP遺伝子(4μg)を用いて同様の実験を行った。その結果を図20に示す。図20に示されているとおり、GFP遺伝子のエレクトロポレーション法(EP法)によるナイーブT細胞への導入効率は43.3%であり、良好であった。
【0075】
また、MART1−A2ペプチドにて処理したT2細胞に対する、MART1−A2ペプチド特異的なTCRβ鎖を発現したHLA−A*0201陽性ナイーブT細胞の細胞障害活性を評価した。TCRβ鎖遺伝子を導入発現したHLA−A*0201陽性ナイーブT細胞としては、前記エレクトロポレーションによりTCRβ鎖遺伝子のDNA(4μg)、mock(4μg)、又はGFP(2μg)を導入した細胞を使用した。また、T2細胞をPBS+1%ヒト血清アルブミン溶液に懸濁し、MART1−A2ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該MART1−A2ペプチド処理したT2細胞1×105個を、同数の前記TCRβ鎖遺伝子を導入発現したHLA−A*0201陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、MART1−A2ペプチド処理したT2細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図21に示す。また、gp100−A2ペプチド特異的なTCRβ鎖を発現したHLA−A*0201陽性ナイーブT細胞の細胞障害活性についても同様に評価した。その結果を図21に示す。図21の結果から分かるように、MART1−A2及びgp100−A2ペプチド特異的なTCRβ鎖遺伝子を導入されたナイーブT細胞は、それぞれ標的細胞を処理したペプチド特異的に刺激され、有意なIFN−γの産生が確認された。これにより、これらTCRβ鎖遺伝子が細胞障害活性を有するタンパク質をコードする遺伝子であることが確認された。
【実施例7】
【0076】
[CMVpp65−A24ペプチド特異的TCRβ鎖について]
本研究では、末期がんであるHLA−A24陽性の転移性メラノーマ患者6例(M3、M7、M9、M13、M14、M18)由来のリンパ球を、メラノーマ特異的ペプチドであるCMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)にて処理した樹状細胞及びT2−A24細胞(T2細胞にHLA−A*2402遺伝子を導入し発現させたもの)を用いて4回刺激を行い、CMVpp65−A24ペプチドに特異的なCTL lineの作製を試みた(図22)。CTL lineの作製は具体的には以下のようなプロトコールで行った。
【0077】
まず最初に、患者由来のリンパ球を2.5〜5×106cell/mlの濃度で、6−ウェルプレートに播種した後、非接着細胞の回収を行い、一時凍結保存した。一方、接着細胞については、GM−CSF、IL−4及びTNF−αの存在下にて培養し、成熟樹状細胞(dendritic cell:DC)を得た。播種から7日後に、樹状細胞をCMVpp65−A24ペプチド及びβ2−ミクログロブリンにて処理後、放射線照射した。放射線照射した成熟樹状細胞(DC)と、凍結保存しておいた前述の非接着細胞(T細胞)とで共培養を行った。(DC:T=1:10〜1:100(細胞数比))。共培養中の培養液にIL−7、IL−2を必要量添加した。播種から14日後に、同様の刺激をCMVpp65−A24ペプチド特異的なCTL細胞に対して再度行った。さらに、播種から21日後及び28日後に、CMVpp65−A24ペプチドにて処理したT2−A24細胞により、CTL細胞を刺激増幅した(T2−A24:CTL=1:10(細胞数比))。播種から35日後に、増幅されたCMVpp65−A24ペプチド特異的CTL細胞を、後述のIFN−γの産生刺激などの機能的アッセイやMACSによるソーテイングの実験に利用した。
【0078】
ex vivoにて前述のHLA−A24陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(CMVpp65−A24ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたCMVpp65−A24テトラマー陽性CTL細胞(CMVpp65 A24テトラマー+CD8+陽性CTL)について定量的な評価を行った。その結果を図23に示す。図23左に示されているように、6例中4例(M3、M7、M9、M18)で、CMVpp65−A24テトラマー陽性CTL細胞の比率が、刺激前の1%以下から、最大平均33.3%[65.9%(M3)11.2%(M7)、35%(M9)、21%(M18)]にまで増幅した。特にM3症例では、樹状細胞2回のみの刺激で著明な増幅効果が見られた。しかし、CMVpp65−A24テトラマー陽性CTL細胞の細胞数については、明らかな増加を確認できたのは3例(M3、M7、M18)であった(図23右)。また、CMVpp65−A24テトラマー陽性CTL細胞の細胞数の増幅の程度を、刺激前の細胞数を100とし、刺激後の細胞数をそれに対する比率で示した(図24)。刺激増幅後、細胞数の明らかな増加が認められた3例(M3、M7、M18)では、経過中に2.1〜207倍の範囲内で増加が認められ、最大で平均80.9倍の増幅効果が確認された。特に、最も増幅効果の高かったM3症例では、最大207倍にまでCMVpp65−A24ペプチド特異的CTL細胞を増加させることができた。
【0079】
なお、図23でテトラマー陽性細胞比率の増加が認められた各症例(M3、M7、M9、M18)における、増幅前後のフローサイトメトリーのデータをそれぞれ図25、図26、図27、図28に示す。これらの図から、テトラマー染色のコントロールとして用いたHLA−A24HIVテトラマーによる染色では、前述の陽性細胞は確認されなかったことが示された。
【0080】
また、CMVpp65−A24ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark, Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果、CMVpp65−A24テトラマー陽性CTL細胞の増幅を認めたM3症例では、レパトワ抗体パネルのスクリーニングにてTCRVβ7.1の増加が検出され、M7症例ではTCRVβ17の増加が検出され、M9症例ではTCRVβ7.1の増加が検出された。
【0081】
CMVpp65−A24ペプチド特異的CTL細胞を、各症例について増加が検出されたレパトワにて染色し、陽性細胞について定量的な評価を行った。その結果を図29に示す。図29に示されているとおり、CMVpp65−A24ペプチド特異的CTL細胞の増幅を認めたM3症例では、TCRVβ7.1が陽性であった細胞が94.9%、M7症例では、TCRVβ17が陽性であった細胞が62.5%、M9症例では、TCRVβ7.1が陽性であった細胞が54.3%であった。
【0082】
CMVpp65−A24ペプチドにて処理を行ったLCL(EB virusにて不死化したヒトB細胞培養株)であるTISI細胞にて、前述のCMVpp65−A24ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図30に示す。図30に示されているとおり、すべての症例のCTL細胞において、CMVpp65−A24ペプチド処理したTISI細胞にて刺激した場合にのみ、大量のIFN−γが産生された。また、M9症例のIFN−γの産生量は、他の2症例と比較して低い傾向があった。
【0083】
また、CMVpp65−A24テトラマー又はTCRレパトワ特異的に細胞障害活性が誘導されるかを確認する目的で、標的細胞にて刺激したM3症例由来のCTL細胞のIFN−γ抗体を用いた細胞内染色を行った。その結果を図31に示す。図31に示されているとおり、コントロールペプチドであるCEA A24ペプチドによる刺激では、IFN−γの産生は見られなかったが、CMV A24ペプチドによる刺激の場合、テトラマー陽性細胞の61.5%に、TCRVβ7.1抗体陽性細胞の70.1%にIFN−γの産生が確認された。
【0084】
上記図29の実験の結果を考慮して、M3症例からはTCRVβ7.1陽性CTL細胞を、M7症例からはTCRVβ17陽性CTL細胞を、M9症例からはTCRVβ7.1細胞を、特異的なTCRレパトワに対するモノクローナル抗体及び磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化したところ、各陽性細胞は95%以上であった。これらの純化した各CTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0085】
上述の純化した3種の細胞からそれぞれtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいて3種のTCRタンパク質コード領域遺伝子(M3TCRβ鎖遺伝子、M7TCRβ鎖遺伝子、M9TCRβ鎖遺伝子)をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
【0086】
TCRレパトワ解析の結果に基づいて、前述のCMVpp65−A24ペプチド特異的TCRβ鎖タンパク質をコードする3種の遺伝子(M3TCRβ鎖遺伝子、M7TCRβ鎖遺伝子、M9TCRβ鎖遺伝子)に対応するプライマーをそれぞれ用いてPCR増幅を行い、3種の増幅断片の塩基配列を解析した。M3TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号17に、そのアミノ酸配列を配列番号18に示す。また、M7TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(tgcactatgagcaaccaggtgctctgc:配列番号31)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号19に、そのアミノ酸配列を配列番号20に示す。また、M9TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号25)とアンチセンスプライマー(配列番号29)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号21に、そのアミノ酸配列を配列番号22に示す。これら3クローンのTCRβ鎖遺伝子の塩基配列を決定したところ、M3TCRβ鎖遺伝子の構成はTRBV4−1*01―TRBD1*01―TRBJ1−2*01―TRBC1であり、M7TCRβ鎖遺伝子の構成は、TRBV19*01―TRBD1*01―TRBJ1−2*01―TRBC1であり、M9TCRβ鎖遺伝子の構成はTRBV4−1*01―TRBD2*01―TRBJ1−2*01―TRBC1であった。また、これら3クローンのTCRβ鎖遺伝子のV−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果をそれぞれ図32〜34に示す。また、これら3クローンのTCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(M3: 337−378、42塩基)(M7:337−372、36塩基)(M9:337−378、42塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【実施例8】
【0087】
実施例7で得られたCMVpp65−A24ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子がHLA−A*2402陽性ナイーブT細胞へ導入された場合に、該遺伝子産物である受容体タンパク質が発現し、腫瘍特異的な細胞障害活性を誘導しうるか否かにつき検討した。
【0088】
HLA−A*2402陽性のメラノーマ患者由来のナイーブリンパ球細胞をGT−T503(タカラバイオ社製)+5%ヒト血清(Cambrex社製)培地に懸濁した。あらかじめ抗ヒトCD3抗体(2.5μg/ml,ヤンセンファーマ社製)と抗ヒトCD28抗体(1μg/ml,BD Pharmingen社製)にて2時間コーテイングを行った75cm2フラスコに、上記培地に懸濁したナイーブリンパ球を2×106cell/mlの濃度で播種し、5日間培養を行った。上記の抗ヒトCD3抗体と抗ヒトCD28抗体との併用により増殖刺激されたリンパ球を回収し、Human T cell Nucleofector kit (Amaxa社製)を用いて、取扱説明書に従い当該TCRβ鎖遺伝子のエレクトロポレーション法(Nucleofector装置, Amaxa社製)による遺伝子導入を行った。エレクトロポレーション法による遺伝子導入は、具体的には、以下のように行った。
【0089】
まず5×106個の刺激後のリンパ球をそれぞれ100μlのエレクトロポレーション用bufferに懸濁した後、GFP(2μg)又はM3TCRβ鎖遺伝子のDNA(4μg)を加え、キュベットに移し変えてNucleofector装置にサンプルをセットし、プログラムNo.23でエレクトロポレーションを実施した。その際、GFP遺伝子としてはAmaxa社のHuman T cell Nucleofector kitに付属のpmaxGFPベクターを用い、M3TCRβ鎖遺伝子としては、pmaxGFPベクターからGFP遺伝子をM3TCRβ鎖遺伝子に乗せ変えたものを作製し、使用した。細胞を回収後、12−ウェルプレートに移してGT−T−503+5%ヒト血清培地にて48時間培養を行った後、フローサイトメトリー(FACSCalibur, BD社製)にてGFP及びM3TCRβ鎖遺伝子の導入効率を検討した。その結果を図35に示す。発現用のコントロール遺伝子であるGFP遺伝子は、全リンパ球の59.3%に導入発現されており(Tリンパ球全体の約62%)、エレクトロポレーション法によるヒトリンパ球細胞への遺伝子導入実験自体は、成功したものと考えられた。一方、CMVpp65−A24特異的TCRβ鎖遺伝子(M3TCRβ鎖遺伝子)は、38.3%の細胞で発現が確認され、良好な発現効率が認められた。
【0090】
また、CMVpp65−A24ペプチドにて処理したTISI細胞に対する、CMVpp65−A24ペプチド特異的なTCRβ鎖を発現したHLA−A*2402陽性ナイーブT細胞の細胞障害活性を評価した。前述のTCRβ鎖遺伝子(M3TCRβ鎖遺伝子)を導入発現したHLA−A*2402陽性ナイーブT細胞としては、前記エレクトロポレーションによりM3TCRβ鎖遺伝子のDNA(4μg)又はGFP(2μg)を導入した細胞を使用した。また、TISI細胞をPBS+1%ヒト血清アルブミン溶液に懸濁し、CMVpp65−A24ペプチドを最終濃度20μg/mlとなるように加え、37℃で2時間インキュベートした。該CMVpp65−A24ペプチド処理したTISI細胞1×105個を、同数の前記M3TCRβ鎖遺伝子を導入発現したHLA−A*2402陽性ナイーブT細胞と共に混合培養した。24時間刺激後、培養上清を回収し、刺激により産生されたIFN−γを定量し、CMVpp65−A24ペプチド処理したTISI細胞に対する、前記ナイーブT細胞の細胞障害活性の指標とした。その結果を図36に示す。図36の結果から分かるように、CMVpp65−A24特異的TCRβ鎖遺伝子(M3TCRβ鎖遺伝子)を導入したリンパ球のみIFN−γが刺激され、本TCRβ鎖遺伝子がコードするTCRβ鎖を発現したT細胞が、CMV特異的な細胞障害活性を有することが示唆された。
【実施例9】
【0091】
[CMVpp65−A2ペプチド特異的TCRβ鎖について]
本研究では、末期がんであるHLA−A2陽性の転移性メラノーマ患者4例(M8、M10、M19、M5−SCC)由来のリンパ球を、メラノーマ特異的ペプチドであるCMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)にて処理した樹状細胞及びT2細胞を用いて4回刺激を行い、CMVpp65−A2ペプチドに特異的なCTL lineの作製を試みた(図37)。CTL lineの作製は具体的には以下のようなプロトコールで行った。
【0092】
まず最初に、患者由来のリンパ球を2.5〜5×106cell/mlの濃度で、6−ウェルプレートに播種した後、非接着細胞の回収を行い、一時凍結保存した。一方、接着細胞については、GM−CSF、IL−4及びTNF−αの存在下にて培養し、成熟樹状細胞(dendritic cell:DC)を得た。播種から7日後に、樹状細胞をCMVpp65−A2ペプチド及びβ2−ミクログロブリンにて処理後、放射線照射した。放射線照射した成熟樹状細胞(DC)と、凍結保存しておいた前述の非接着細胞(T細胞)とで共培養を行った。(DC:T=1:10〜1:100(細胞数比))。共培養中の培養液にIL−7、IL−2を必要量添加した。播種から14日後に、同様の刺激をCMVpp65−A24ペプチド特異的なCTL細胞に対して再度行った。さらに、播種から21日後及び28日後に、CMVpp65−A2ペプチドにて処理したT2細胞により、CTL細胞を刺激増幅した(T2:CTL=1:10(細胞数比))。播種から35日後に、増幅されたCMVpp65−A2ペプチド特異的CTL細胞を、後述のIFN−γの産生刺激などの機能的アッセイやMACSによるソーテイングの実験に利用した。
【0093】
ex vivoにて前述のHLA−A2陽性の患者由来のリンパ球から誘導・増幅されたペプチド特異的なCTL細胞(CMVpp65−A2ペプチド特異的CTL細胞)をテトラマーにて染色し、増幅されたCMVpp65−A2テトラマー陽性CTL細胞(CMVpp65−A2テトラマー+CD8+陽性CTL)について定量的な評価を行った。その結果を図38上に示す。図38上に示されているように、4例中1例(M8)で、CMVpp65−A2テトラマー陽性CTL細胞の比率が、刺激前の1%以下から、最大20%以上にまで増幅した。一方、CMVpp65−A2テトラマー陽性CTL細胞の細胞数については、3例(M8、M10、M5−SSC)で増加傾向が見られ、特に、M8症例では、刺激前の448倍と、最も効率的に特異的CTL細胞の誘導が見られた(図38下)。
【0094】
なお、M8における、増幅前後のフローサイトメトリーのデータをそれぞれ図39に示す。増幅前のテトラマー陽性細胞比率は1%以下(0.27%)であったが、DC2回刺激後は2.14%に、さらにT2細胞で2回刺激後は24.8%まで増加した。一方、テトラマー染色のコントロールとして用いたHLA−A2 Flu−MPテトラマーによる染色では、陽性細胞の増幅は確認されなかった。
【0095】
また、CMVpp65−A2ペプチド特異的CTL細胞について、TCRレパトワに対するモノクローナル抗体パネル(IOTestR Beta Mark,Beckman Coulter社)を用いて、有意に増加したレパトワの同定を行った。その結果を図40に示す。図40に示されているように、CMVpp65−A2テトラマー陽性CTL細胞の増幅を認めたM8症例では、レパトワ抗体パネルのスクリーニングにてTCRVβ8及びTCRVβ14の増加が検出された。すなわち、TCRVβ8が陽性であった細胞が14.8%、TCRVβ14が陽性であった細胞が7.3%であった。
【0096】
CMVpp65−A2ペプチドにて処理を行ったT2細胞にて、前述のCMVpp65−A2ペプチド特異的CTL細胞を刺激し、その刺激により細胞内に産生されたIFN−γを細胞内染色してフローサイトメトリーで検出した。その結果を図41に示す。図41に示されているとおり、CD8及びIFN−γの両方が陽性のCTL細胞の比率は21.2%であり、そのうち、IFN−γ陽性のCTL細胞のほとんどすべてがTCRVβ8陽性のCTL細胞であった(22.7%)。この結果から、細胞障害活性を有するCTL細胞のTCRレパトワは、TCRVβ8であることが確認された。
【0097】
後述のTCRβ鎖遺伝子の解析に利用するために、M8由来のTCRVβ8陽性CTL細胞を以下の方法で選別した。
【0098】
M8症例由来のリンパ球をIn vitroにてDCで2回、T2細胞で3回刺激をして得られた増幅されたCMVA2テトラマー陽性細胞を、テトラマーsortingしたところ、増幅後のCMVpp65−A2テトラマー及びCD8陽性CTL細胞(CMVpp65−A2テトラマー+CD8+陽性CTL)並びにCMVpp65−A2テトラマー及びTCRVβ8陽性CTL細胞(CMVpp65−A2テトラマー+TCRVβ8+陽性CTL)の細胞の比率は、それぞれ43.1%及び44.6%であった(図42)。PE−CMVA2テトラマー試薬と抗PE抗体−マイクロビーズを用いてCTL細胞を染色し、MACS法にてテトラマー陽性細胞を選別したところ、該陽性細胞の純度は95.6%であった(図42)。選別された前記陽性細胞のうち、ほとんどすべてがTCRVβ8陽性であった。これらの純化したCTL細胞を、次のTCRβ鎖の遺伝子解析に用いた。
【0099】
上述の純化した細胞からtotal RNAを抽出し、逆転写反応にてcDNA合成後、PCR法を用いたTCRレパトワ解析により発現TCRの遺伝子分類を行い、その解析結果に基づいてTCRタンパク質コード領域遺伝子(M8TCRβ鎖遺伝子)をクローニングした。CTLは、α鎖とβ鎖からなるヘテロ二量体のTCRタンパク質を発現する。今回我々の決定した遺伝子配列はβ鎖に対応するものである。
【0100】
TCRレパトワ解析の結果に基づいて、前述のCMVpp65−A2ペプチド特異的TCRβ鎖タンパク質をコードする遺伝子(M8TCRβ鎖遺伝子)に対応するプライマーを用いてPCR増幅を行い、増幅断片の塩基配列を解析した。M8TCRβ鎖遺伝子について具体的には、該TCRβ鎖遺伝子に対応するセンスプライマー(配列番号30)とアンチセンスプライマー(配列番号26)を用いて、該TCRβ鎖遺伝子のコード領域全域の遺伝子増幅を行い、これをクローニングした。このTCRβ鎖遺伝子の塩基配列を配列番号23に、そのアミノ酸配列を配列番号24に示す。このM8TCRβ鎖遺伝子の塩基配列を決定したところ、M8TCRβ鎖遺伝子の構成はTRBV12−4*01―TRBD1*01―TRBJ1−2*01―TRBC1であった。また、このM8TCRβ鎖遺伝子のV−D−Jの接合部分の塩基配列につき、前述のIMGTの公開ソフト(JunctionAnalysis)を用いた解析結果をそれぞれ図43に示す。また、このM8TCRβ鎖遺伝子の塩基配列及びアミノ酸配列についてNCBIの相同性検索プログラム(BLAST)による解析を行った。その結果、全長及びV−D−J−Cの接合領域を含む部分断片(340−369、30塩基)の塩基配列については、完全に一致するものはなかった。全長及びV−D−J−Cの接合部を中心とした部分断片(60アミノ酸)のアミノ酸配列について検索した場合でも、相同性が高いものはあるが、抗原特異性を決定する接合領域に関して完全に一致する登録はなかった。
【0101】
なお、CMVpp65−A2ペプチド特異的TCRβ鎖遺伝子発現リンパ球の細胞障害活性などの機能解析実験は今後行う予定である。
【実施例10】
【0102】
[MART1−A2ペプチド特異的TCRα鎖について]
国立がんセンター倫理審査委員会の承認を受けて実施された臨床研究(悪性黒色腫に対する樹状細胞を用いた腫瘍特異的免疫療法)で得られた転移性メラノーマ患者由来のリンパ球を用いた。この臨床研究は、前述のHLA−A2またはA24の患者(M1、M3、M6、M7、M8、M9)を対象に腫瘍関連抗原ペプチドにて処理した樹状細胞を患者皮下に投与し、腫瘍免疫を誘導する臨床試験である。本臨床研究では、樹状細胞ワクチン投与を受けたHLA−A24陽性の患者由来のリンパ球を、メラノーマ特異的ペプチドであるMART1 27−35(MART1−A2ペプチド:AAGIGILTV;配列番号3)、gp100 209−217(gp100−A2ペプチド:IMDQVPFSV;配列番号4)、MAGE1 135−143(MAGE1−A24ペプチド:NYKHCFPEI;配列番号1)若しくはMAGE3 195−203(MAGE3−A24ペプチド:IMPKAGLLI;配列番号2)、又は、サイトメガロウイルスに特異的なペプチドであるCMVpp65 341−349(CMVpp65−A24ペプチド:QYDPVAALF;配列番号5)若しくはCMVpp65 495−503(CMVpp65−A2ペプチド:NLVPMVATV;配列番号6)にて処理した樹状細胞を用いて4回刺激を行った。この刺激によって誘導された、前記の各種ペプチドに特異的なCTL細胞を、PEで標識した各種テトラマーにて染色し、テトラマー染色陽性のCTL細胞を、磁気ビーズ細胞分離装置(Auto−magnet cell sorting system)を利用して純化した。純化して得られた各種のテトラマー染色陽性CTL細胞を、次のTCRα鎖のレパトワ解析に用いた。
【実施例11】
【0103】
[TCRα鎖のレパトワ解析]
上記実施例10で得られた各種のテトラマー染色陽性CTL細胞のうち、まずM8症例MART1−A2ペプチド特異的CTL(MART−A2テトラマー陽性)についてTCRα鎖のレパトワ解析を行った。すなわち、このCTLからtotal RNAを抽出し、抽出したtotal RNA2μgを用いた逆転写反応にてcDNAを合成し、合成により得られたcDNA全量に対して1/40量のcDNAとTCRα鎖cDNAレパトワ解析用プライマーとを用いたPCRにより得られたPCR産物の解析を試みた。TCRα鎖cDNAレパトワ解析用プライマーとしては、33種類のセンスプライマー(HAV1Aセンスプライマー:TCTGGTATGTGCAATACCCCAACC:配列番号79; HAV1Bセンスプライマー:CTGAGGAAACCCTCTGTGCA:配列番号80; HAV2センスプライマー:GATGGAAGGTTTACAGCACAGCTC:配列番号81; HAV3センスプライマー:CACAGTGGAAGATTAAGAGTCACGC:配列番号82; HAV4Aセンスプライマー:AACAGAATGGCCTCTCTGGC:配列番号83; HAV4Bセンスプライマー:GGATTGCGCTGAAGGAAGAG:配列番号84; HAV5センスプライマー:TGAAGGTCACCTTTGATACCACCC:配列番号85; HAV6センスプライマー:AATCCGCCAACCTTGTCATCTCCG:配列番号86; HAV7センスプライマー:AACTGCACGTACCAGACATC:配列番号87; HAV8センスプライマー:ACCCTGAGTGTCCAGGAGGG:配列番号88; HAV9センスプライマー:CACTGCTGACCTTAACAAAGGCG:配列番号89; HAV10センスプライマー:TCCTGGTGACAGTAGTTACG:配列番号90; HAV11センスプライマー:AGGCTCAAAGCCTTCTCAGCAGGG:配列番号91; HAV12センスプライマー:TCCACCAGTTCCTTCAACTTCACC:配列番号92; HAV13センスプライマー:TTCATCAAAACCCTTGGGGACAGC:配列番号93; HAV14センスプライマー:CCCAGCAGGCAGATGATTCTCGTT:配列番号94; HAV15センスプライマー:GGATAAACATCTGTCTCTGCG:配列番号95; HAV16センスプライマー:GATAGCCATACGTCCAGATG:配列番号96; HAV18センスプライマー:TGCCACTCTTAATACCAAGGAGGG:配列番号97; HAV19センスプライマー:ACACTGGCTGCAACAGCATC:配列番号98; HAV20センスプライマー:TTACAAACGAAGTGGCCTCC:配列番号99; HAV21センスプライマー:ACCCTGCTGAAGGTCCTACATTCC:配列番号100; HAV22センスプライマー:CTTGGAGAAAGGCTCAGTTC:配列番号101; HAV23センスプライマー:TGCCTCGCTGGATAAATCATCAGG:配列番号102; HAV24センスプライマー:TCCCAGCTCAGCGATTCAGCCTCC:配列番号103; HAV25センスプライマー:GTCCTGTCCTCTTGATAGCC:配列番号104; HAV26センスプライマー:AGCCCAGCCATGCAGGCATCTACC:配列番号105; HAV27センスプライマー:TTGATACCAAAGCCCGTCTC:配列番号106; HAV28センスプライマー:GAACATCACAGCCACCCAGACCGG:配列番号107; HAV29センスプライマー:GCAAAGCTCCCTGTACCTTACGG:配列番号108; HAV30センスプライマー:TTTCTGCACAGCACAGCCCC:配列番号109; HAV31センスプライマー:AGCAAAAACTTCGGAGGCGG:配列番号110; HAV32センスプライマー:AAGGAGAGGACTTCACCACG:配列番号111)からなる群から選ばれる1種類のセンスプライマーと、アンチセンスプライマー(HTCA3アンチセンスプライマー:GTTGCTCTTGAAGTCCATAGACC:配列番号112)とを用いた。
【0104】
上記のPCRにより得られたPCR産物をアガロースゲルで電気泳動した結果を図44(A)に示す。図44(A)において、両脇のマーカーのレーンを除くと、上段の左側のレーンから順に、センスプライマーとして上記HAV1AからHAV14までの16種類をそれぞれ用いたPCR産物を電気泳動し、下段の左側のレーンから順に、センスプライマーとして上記HAV15からHAV32までの17種類をそれぞれ用いたPCR産物を電気泳動した。図44(A)から分かるように、HAV2, 6, 9, 24, 26及び32の各レーンにおいて、PCRの増幅産物のバンドが確認された。これらのバンドをゲルから回収し、TAクローニング法(TOPO TA Cloning Kit、インビトロジェン社製)によりベクターへ組み込んだ。
このベクターに組み込まれたバンド部分(PCR産物)の塩基配列を決定したところ、HAV2, 9及び32のPCR産物はTCRα鎖遺伝子由来のDNA断片であるが、その他のPCR産物は非特異的反応による増幅産物であることが判明した。HAV2, 9及び32のDNA断片が含まれる遺伝子は、NCBIにより使用される新規名称TRAV12-2, 16及び25にそれぞれ相当する遺伝子であった。これら3種類のTCRα遺伝子レパトワについて、全タンパク質コード領域の遺伝子取得を目的として、上記のcDNA断片を鋳型に各レパトワ特異的な16種類のセンスプライマー(TRAV1-2センスプライマー:CAGCAGATGTGGGGAGTTTTCCTTCTT:配列番号113; TRAV4センスプライマー:TAGAATATGAGGCAAGTGGCGAGAGTG:配列番号114; TRAV8-2センスプライマー:TCAGCCATGCTCCTGCTGCTCGTCCCA:配列番号115; TRAV8-3センスプライマー:TCAGCCATGCTCCTGGAGCTTATCCCA:配列番号116; TRAV8-4センスプライマー:TCAGCCATGCTCCTGCTGCTCGTCCCA:配列番号117; TRAV12-1センスプライマー:AAAAGAATGATGATATCCTTGAGAGTT:配列番号118; TRAV12-2センスプライマー:AGAAGAATGATGAAATCCTTGAGAGTT:配列番号119; TRAV16センスプライマー:TCAGACATGAAGCCCACCCTCATCTCA:配列番号120; TRAV17センスプライマー:AGAAGAATGGAAACTCTCCTGGGAGTG:配列番号121; TRAV21センスプライマー:GAAGGAATGGAGACCCTCTTGGGCCTG:配列番号122; TRAV22センスプライマー:AGCATCATGAAGAGGATATTGGGAGCT:配列番号123; TRAV24センスプライマー:GGAAACATGGAGAAGAATCCTTTGGCA:配列番号124; TRAV25センスプライマー:AGGGAGATGCTACTCATCACATCAATG:配列番号125; TRAV13-1センスプライマー:ACAAGGATGACATCCATTCGAGCTGTA:配列番号126; TRAV35センスプライマー:ATAAGGATGCTCCTTGAACATTTATTA:配列番号127; TRAV38-2センスプライマー:GTGAGCATGGCATGCCCTGGCTTCCTG:配列番号128)からなる群から選ばれる1種類のセンスプライマーと、終止コドンを含む定常領域アンチセンスプライマー(TRACアンチセンスプライマー:TCAGCTGGACCACAGCCGCAGCGTCAT:配列番号129)とを用いて遺伝子増幅を行った。増幅したPCR産物(各遺伝子のタンパク質コード領域のDNA断片)をアガロースゲル電気泳動した結果を図44(B)に示す。いずれのPCR産物についても、想定された位置にバンドが検出された。
【0105】
次いで、上記のPCR産物をベクターにクローニングした後、これら3クローンのPCR産物(TCRα鎖遺伝子)の塩基配列を決定したところ、TRAV12-2(HAV2)、TRAV16(HAV9)、TRAV25(HAV32)のTCRα鎖遺伝子の構成は、それぞれ「TRAV12-2*01―TRAJ10*01」(図51:配列番号38の塩基配列; 図52:配列番号39のアミノ酸配列)、「TRAV16*01―TRAJ38*01」(図53:配列番号40の塩基配列; 図54:配列番号41のアミノ酸配列)、「TRAV25*01―TRAJ31*01」(図55:配列番号42の塩基配列; 図56:配列番号43のアミノ酸配列)であることが分かった。
【0106】
また、上記実施例10で得られた各種のテトラマー染色陽性CTL細胞のうち、M8症例MART1−A2ペプチド特異的CTL(MART−A2テトラマー陽性)以外のCTL(M6症例MART1−A2ペプチド特異的CTL、M8症例gp100−A2ペプチド特異的CTL、M1症例MAGE1−A24ペプチド特異的CTL、M7症例MAGE3−A24ペプチド特異的CTL、M8症例CMVpp65−A2ペプチド特異的CTL、M3症例CMVpp65−A24ペプチド特異的CTL、M7症例CMVpp65−A24ペプチド特異的CTL、及び、M9症例CMVpp65−A24ペプチド特異的CTL)についても、同様の手法によってTCRα鎖のレパトワ解析を行った。その結果、M6症例MART1−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV12-2*01―TRAJ40*01」(図45:配列番号32の塩基配列; 図46:配列番号33のアミノ酸配列)、「TRAV12-2*01―TRAJ50*01」(図47:配列番号34の塩基配列; 図48:配列番号35のアミノ酸配列)、「TRAV25*01―TRAJ21*01」(図49:配列番号36の塩基配列; 図50:配列番号37のアミノ酸配列)であり、M8症例gp100−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-4*01―TRAJ9*01」(図57:配列番号44の塩基配列; 図58:配列番号45のアミノ酸配列)であり、M1症例MAGE1−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV1-2*01―TRAJ10*01」(図59:配列番号46の塩基配列; 図60:配列番号47のアミノ酸配列)、「TRAV12-1*01―TRAJ49*01」(図61:配列番号48の塩基配列; 図62:配列番号49のアミノ酸配列)、「TRAV17*01―TRAJ23*01」(図63:配列番号50の塩基配列; 図64:配列番号51のアミノ酸配列)、「TRAV22*01―TRAJ40*01」(図65:配列番号52の塩基配列; 図66:配列番号53のアミノ酸配列)であり、M7症例MAGE3−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV13-1*01―TRAJ45*01」(図67:配列番号54の塩基配列; 図68:配列番号55のアミノ酸配列)、「TRAV38-2/DV8*01―TRAJ57*01」(図69:配列番号56の塩基配列; 図70:配列番号57のアミノ酸配列)であり、M8症例CMVpp65−A2ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-2*01―TRAJ23*01」(図71:配列番号58の塩基配列; 図72:配列番号59のアミノ酸配列)、「TRAV26―stop codon」(図73:配列番号60の塩基配列)、「TRAV35*02―TRAJ50*01」(図74:配列番号61の塩基配列; 図75:配列番号62のアミノ酸配列)であり、M3症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV8-3*01―TRAJ39*01」(図76:配列番号63の塩基配列; 図77:配列番号64のアミノ酸配列)、「TRAV24*01―TRAJ54*01」(図78:配列番号65の塩基配列; 図79:配列番号66のアミノ酸配列)、「TRAV38-2/DV8*01―TRAJ49*01」(図80:配列番号67の塩基配列; 図81:配列番号68のアミノ酸配列)であり、M7症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV21*02―TRAJ28*01」(図82:配列番号69の塩基配列; 図83:配列番号70のアミノ酸配列)、「TRAV24*01―TRAJ47*02」(図84:配列番号71の塩基配列; 図85:配列番号72のアミノ酸配列)、「TRAV35―stop codon」(図86:配列番号73の塩基配列)であり、M9症例CMVpp65−A24ペプチド特異的CTL由来のTCRα鎖遺伝子の構成は、「TRAV4*01―TRAJ44*01」(図87:配列番号74の塩基配列; 図88:配列番号75のアミノ酸配列)、「TRAV16*01―TRAJ49*01」(図89:配列番号76の塩基配列; 図90:配列番号77のアミノ酸配列)、「TRAV17―stop codon」(図91:配列番号78の塩基配列)であった。
【実施例12】
【0107】
[TCRα鎖/β鎖遺伝子の導入発現]
上記の腫瘍関連ペプチドに特異的なTCRβ鎖遺伝子とα鎖遺伝子とを同時にmutant Jurkat細胞(ATCC TIB−153)へ導入した場合に、TCRβ鎖タンパク質及びα鎖タンパク質が発現し、細胞障害活性を誘導し得るか否かにつき検討した。
すなわち、実施例5で得られたM8症例MART1−A2ペプチド特異的CTL細胞由来のTCRβ鎖遺伝子と、実施例11で得られたM8症例MART1−A2ペプチド特異的CTL細胞由来のTCRα鎖遺伝子とを、エレクトロポレーション法(Amaxa社のNucleofector装置を使用)にて同時にmutant Jurkat細胞(ATCC TIB−153)に導入し、それらTCRα鎖やβ鎖の発現の解析を行った。なお、Mutant Jurkat細胞は、Jurkat細胞のE6-1クローンの変異型で、TCRのβ鎖を欠失しており、さらに、膜上にTCRα鎖及びβ鎖からなるheterodimer並びにCD3タンパク質を発現しないことが知られているため、TCRβ鎖遺伝子の導入実験によく使用される。
【0108】
まずpmax-GFP発現ベクターをmutant Jurkat細胞へ導入し、pmax-GFP/mutant Jurkat細胞を作製した。その際、mutant Jurkat細胞への遺伝子導入の至適条件を設定した。pmax-GFP発現ベクターの導入から48時間後に、フローサイトメトリー(FACSCalibur(登録商標):ベクトン・ディッキンソン社製)を用いて、pmax-GFP/mutant Jurkat細胞中のGFP遺伝子及びCD3タンパク質の発現効率を測定した。その結果を、図92に示す。図92から分かるように、全細胞の61.8%にGFP遺伝子の発現が確認されたが、CD3タンパク質の発現はほとんど認められなかった。
【0109】
また、TCRVβ3.0(NCBIにより使用される新規名称はTRBV28)を乗せたpmaxベクター8μgと、各TCRα鎖遺伝子(TRAV12、TRAV16又はTRAV25)を乗せたpmaxベクター8μgとを、同時に2×106個のmutant Jurkat細胞に導入し、pmax-TCRVβ3.0・pmax-TCRα/mutant Jurkat細胞を作製した。また、コントロールとして、TCRVβ3.0を乗せたpmaxベクターのみを導入したmutant Jurkat細胞(pmax-TCRVβ3.0/mutant Jurkat細胞)を作製した。
各発現ベクターの導入から48時間後に、フローサイトメトリー(FACSCalibur:ベクトン・ディッキンソン社製)を用いて、pmax-TCRVβ3.0・pmax-TCRα/mutant Jurkat細胞中のTCRVβ3.0 (TRBV28)遺伝子、及び、CD3タンパク質の発現効率を測定した。その結果を、図93に示す。図93から分かるように、TCRVβ3.0 (TRBV28)の発現は、TCRα鎖遺伝子を同時導入した場合の方が高く、TRAV12やTRAV16遺伝子を同時導入した場合はそれぞれ11.5%、10.3%と発現の増加を認めた(TCRVβ3.0単独では、1.7%)。また同様の実験におけるCD3タンパク質の発現も、TRAV12やTRAV16遺伝子を同時導入した場合は、19.8%、21.8%と発現の増加を認めた(TCRVβ3.0単独では、8.7%)(図94)。
これらの結果から、上記のTCRβ鎖とTCRα鎖の組み合わせによって、CD3の発現が向上し、細胞障害活性が誘導されることが示された。
【図面の簡単な説明】
【0110】
【図1】MAGE1−A24ペプチド処理した樹状細胞の、肺転移巣に対する治療効果を示す図である。
【図2】末梢血中のMAGE1−A24ペプチド等に対する特異的なCTL細胞の比率を、ELISPOTアッセイにより測定した結果を示す図である。
【図3】MAGE1−A24ペプチド特異的CTL細胞を、テトラマーにて染色した結果を示す図である。
【図4】MAGE1−A24ペプチド特異的CTL細胞についての2種類のTCRレパトワに対する解析結果を示す図である。
【図5】MAGE1−A24ペプチド特異的CTL細胞を、テトラマー及びTCRVβにて染色した結果を示す図である。
【図6】MAGE1−A24ペプチド処理したTISI細胞で刺激されたMAGE1−A24ペプチド特異的CTL細胞を、テトラマー又はIFN−γ等にて染色した結果を示す図である。
【図7】MAGE1−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図8】MAGE1−A24ペプチド特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図9】MAGE3−A24ペプチド処理した樹状細胞の、頚部リンパ節転移巣に対する治療効果を示す図である。
【図10】末梢血中のMAGE3−A24ペプチド等に対する特異的なCTL細胞の比率を、ELISPOTアッセイにより測定した結果を示す図である。
【図11】MAGE3−A24ペプチド特異的CTL細胞についてのTCRレパトワ解析の結果を示す図である。
【図12】MAGE3−A24ペプチド特異的CTL細胞についての2種類のTCRレパトワに対する解析結果を示す図である。
【図13】MAGE3−A24ペプチド処理したT2−A24細胞等で刺激されたMAGE3−A24ペプチド特異的CTL細胞を、IFN−γにて染色した結果を示す図である。
【図14】本発明の新規なTCRβ鎖遺伝子の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図15】MAGE3−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図16】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なCTL細胞を、テトラマーにて染色した結果を示す図である。
【図17】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)処理したT2細胞で刺激されたA2ペプチド特異的CTL細胞を、テトラマー及びIFN−γにて染色した結果を示す図である。
【図18】gp100−A2ペプチド特異的CTL細胞について、テトラマー(左図)及びTCRレパトワ(右図)に対する解析結果を示す図である。
【図19】MART1−A2ペプチド又はgp100−A2ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図20】GFP遺伝子を導入したナイーブT細胞等を、該GFPにて染色した結果を示す図である。
【図21】A2ペプチド(MART1−A2ペプチド又はgp100−A2ペプチド)特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図22】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の刺激増幅プロトコールを示す図である。
【図23】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の増幅効果を示す図である。
【図24】In vitroにおけるCMVpp65−A24ペプチド特異的CTL細胞の増幅効果を示す図である。
【図25】CMVpp65−A24ペプチド特異的CTL細胞(M3症例)をテトラマーにて染色した結果を示す図である。
【図26】CMVpp65−A24ペプチド特異的CTL細胞(M7症例)をテトラマーにて染色した結果を示す図である。
【図27】CMVpp65−A24ペプチド特異的CTL細胞(M9症例)をテトラマーにて染色した結果を示す図である。
【図28】CMVpp65−A24ペプチド特異的CTL細胞(M18症例)をテトラマーにて染色した結果を示す図である。
【図29】CMVpp65−A24ペプチド特異的CTL細胞を、特異的レパトワにて染色した結果を示す図である。
【図30】CMVpp65−A24ペプチド処理したTISI細胞等で刺激されたCMVpp65−A24ペプチド特異的CTL細胞における、IFN−γ量を示す図である。
【図31】CMVpp65−A24ペプチド処理したTISI細胞等で刺激されたCMVpp65−A24ペプチド特異的CTL細胞等を、IFN−γ等にて染色した結果を示す図である。
【図32】本発明の新規なTCRβ鎖遺伝子(M3症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図33】本発明の新規なTCRβ鎖遺伝子(M7症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図34】本発明の新規なTCRβ鎖遺伝子(M9症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図35】CMVpp65−A24ペプチド特異的なTCRβ遺伝子を導入したナイーブT細胞等を、該TCRβ鎖等にて染色した結果を示す図である。
【図36】CMVpp65−A24ペプチド特異的なTCRβ遺伝子等を導入したナイーブT細胞における、IFN−γ量を示す図である。
【図37】In vitroにおけるCMVpp65−A2ペプチド特異的CTL細胞の刺激増幅プロトコールを示す図である。
【図38】In vitroにおけるCMVpp65−A2ペプチド特異的CTL細胞の増幅効果を示す図である。
【図39】CMVpp65−A2ペプチド特異的CTL細胞をテトラマーにて染色した結果を示す図である。
【図40】CMVpp65−A2ペプチド特異的CTL細胞についてのTCRレパトワ解析の結果を示す図である。
【図41】CMVpp65−A2ペプチド処理したT2細胞等で刺激されたCMVpp65−A2ペプチド特異的CTL細胞等を、IFN−γ等にて染色した結果を示す図である。
【図42】In vitroにおいて刺激増幅されたCMVpp65−A2ペプチド特異的CTL細胞をテトラマーにて染色した結果を示す図である。
【図43】本発明の新規なTCRβ鎖遺伝子(M8症例由来)の塩基配列、推定アミノ酸配列、並びに、V−D−J接合領域の配列及びレパトワを示す図である。
【図44】M8症例MART1−A2ペプチド特異的CTL由来のTCRα鎖のレパトワ解析の結果(PCR産物のアガロースゲルでの電気泳動結果)を示す図である。
【図45】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図46】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図47】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図48】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図49】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図50】M6症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図51】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図52】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図53】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図54】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図55】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図56】M8症例MART1−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図57】M8症例gp100−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図58】M8症例gp100−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図59】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図60】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図61】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図62】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図63】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図64】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図65】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図66】M1症例MAGE1−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図67】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図68】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図69】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図70】M7症例MAGE3−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図71】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図72】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図73】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図74】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図75】M8症例CMVpp65−A2ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図76】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図77】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図78】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図79】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図80】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図81】M3症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図82】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図83】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図84】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図85】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図86】M7症例CMVpp65−A24ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図87】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図88】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図89】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の塩基配列、及び、レパトワを示す図である。
【図90】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来の本発明の新規なTCRα鎖遺伝子の推定アミノ酸配列、及び、レパトワを示す図である。
【図91】M9症例CMVpp65−A24ペプチド特異的CTL細胞由来のTCRα鎖遺伝子様の塩基配列、及び、レパトワを示す図である。
【図92】GFP遺伝子を導入したmutant Jurkat細胞を、GFPにて染色した結果を示す図(左図)、及び、該細胞をCD3にて染色した結果を示す図(右図)である。
【図93】TCRβ鎖遺伝子とα鎖遺伝子とを同時に導入したmutant Jurkat細胞(右上図、左下図、右下図)やTCRβ鎖遺伝子のみを導入したmutant Jurkat細胞(左上図)を、TCRβ鎖にて染色した結果を示す図である。
【図94】TCRβ鎖遺伝子とα鎖遺伝子とを同時に導入したmutant Jurkat細胞(右上図、左下図、右下図)やTCRβ鎖遺伝子のみを導入したmutant Jurkat細胞(左上図)を、CD3にて染色した結果を示す図である。
【特許請求の範囲】
【請求項1】
配列番号7、9、11、13、15、17、19、21若しくは23で表される塩基配列、又は、配列番号8、10、12、14、16、18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子。
【請求項2】
請求項1に記載のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクター。
【請求項3】
請求項2に記載のT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項4】
配列番号32、34、36、38、40、42、44、46、48、50、52、54、56、58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子。
【請求項5】
請求項4に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクター。
【請求項6】
請求項5に記載のT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項7】
請求項1に記載のT細胞レセプターβ鎖遺伝子及び請求項4に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクター。
【請求項8】
請求項7に記載のT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項9】
組換えベクターが導入されたリンパ球である請求項3、6又は8に記載の形質転換体。
【請求項10】
配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質。
【請求項11】
配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質に特異的に結合する抗体。
【請求項12】
配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質。
【請求項13】
配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質に特異的に結合する抗体。
【請求項14】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項15】
配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項16】
配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【請求項17】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項18】
配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項19】
配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【請求項20】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子及びT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項21】
配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項22】
配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【請求項1】
配列番号7、9、11、13、15、17、19、21若しくは23で表される塩基配列、又は、配列番号8、10、12、14、16、18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子。
【請求項2】
請求項1に記載のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクター。
【請求項3】
請求項2に記載のT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項4】
配列番号32、34、36、38、40、42、44、46、48、50、52、54、56、58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子。
【請求項5】
請求項4に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクター。
【請求項6】
請求項5に記載のT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項7】
請求項1に記載のT細胞レセプターβ鎖遺伝子及び請求項4に記載のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクター。
【請求項8】
請求項7に記載のT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが導入された形質転換体。
【請求項9】
組換えベクターが導入されたリンパ球である請求項3、6又は8に記載の形質転換体。
【請求項10】
配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質。
【請求項11】
配列番号8、10、12、14、16、18、20、22又は24で表されるアミノ酸配列からなるT細胞レセプターβ鎖タンパク質に特異的に結合する抗体。
【請求項12】
配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質。
【請求項13】
配列番号33、35、37、39、41、43、45、47、49、51、53、55、57、59、62、64、66、68、70、72、75又は77で表されるアミノ酸配列からなるT細胞レセプターα鎖タンパク質に特異的に結合する抗体。
【請求項14】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項15】
配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項16】
配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子が組み込まれたT細胞レセプターβ鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【請求項17】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項18】
配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項19】
配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子が組み込まれたT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【請求項20】
疾患特異的抗原ペプチドに特異的なCTL細胞のT細胞レセプターβ鎖遺伝子及びT細胞レセプターα鎖遺伝子が組み込まれた、T細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターが、インビトロで導入されたリンパ球を有効成分とする疾患治療用組成物。
【請求項21】
配列番号7、9、11、13若しくは15で表される塩基配列、又は、配列番号8、10、12、14若しくは16で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号32、34、36、38、40、42、44、46、48、50、52、54若しくは56で表される塩基配列、又は、配列番号33、35、37、39、41、43、45、47、49、51、53、55若しくは57で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするMAGE1,3陽性の固形がん又はメラノーマ治療用組成物。
【請求項22】
配列番号17、19、21若しくは23で表される塩基配列、又は、配列番号18、20、22若しくは24で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターβ鎖遺伝子と、配列番号58、61、63、65、67、69、71、74若しくは76で表される塩基配列、又は、配列番号59、62、64、66、68、70、72、75若しくは77で表されるアミノ酸配列をコードする塩基配列からなるT細胞レセプターα鎖遺伝子とが組み込まれたT細胞レセプターβ鎖タンパク質及びT細胞レセプターα鎖タンパク質を発現しうる組換えベクターがインビトロで導入されたリンパ球を有効成分とするサイトメガロウイルス(CMV)感染症治療用組成物。
【図23】



【図38】
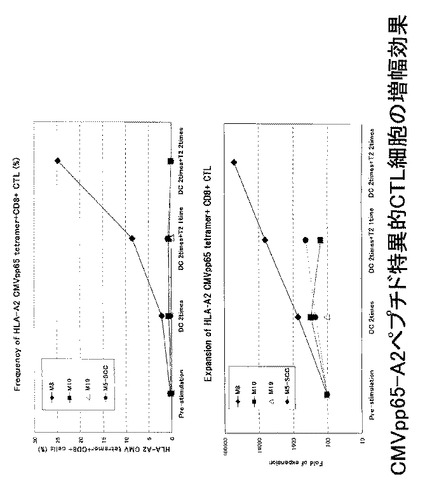
【図1】
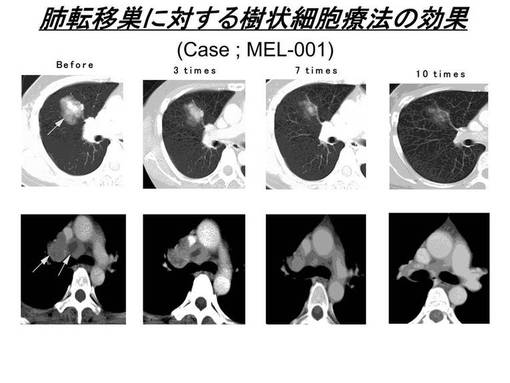
【図2】
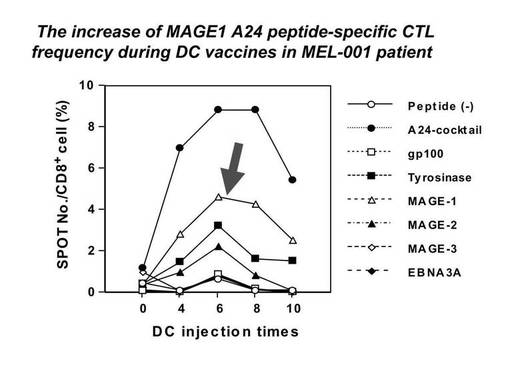
【図3】

【図4】

【図5】
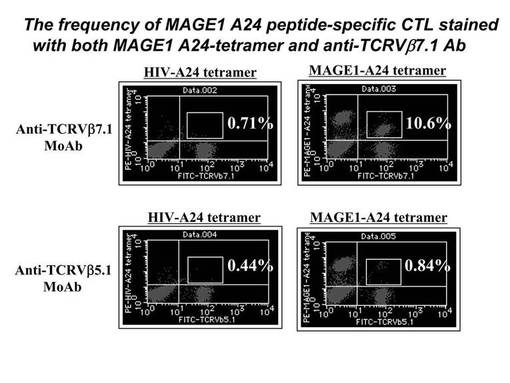
【図6】
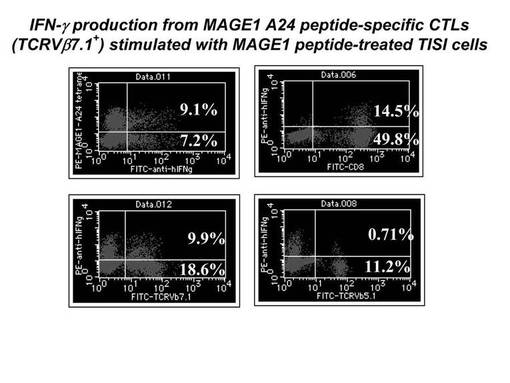
【図7】
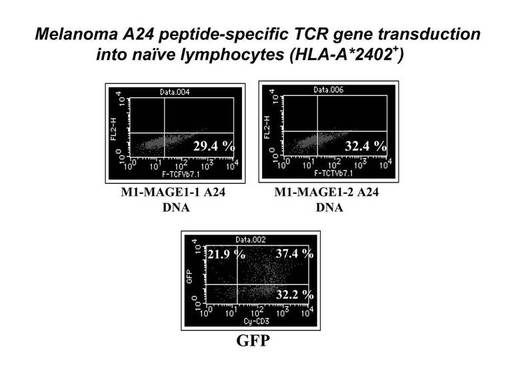
【図8】
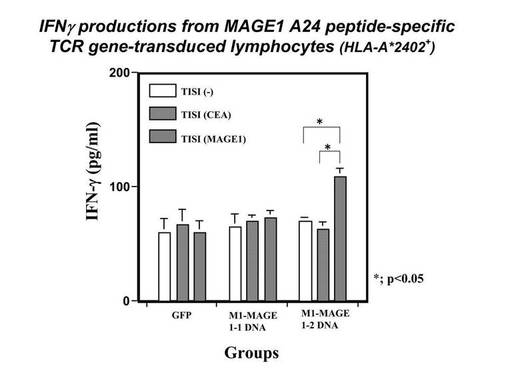
【図9】
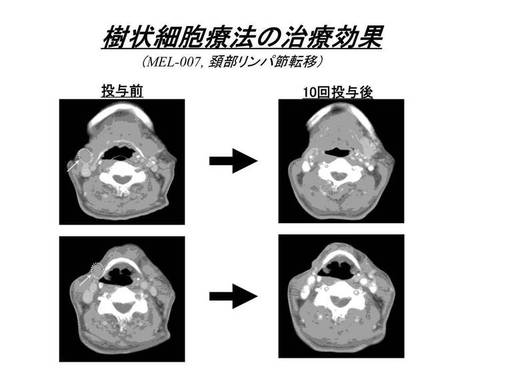
【図10】

【図11】
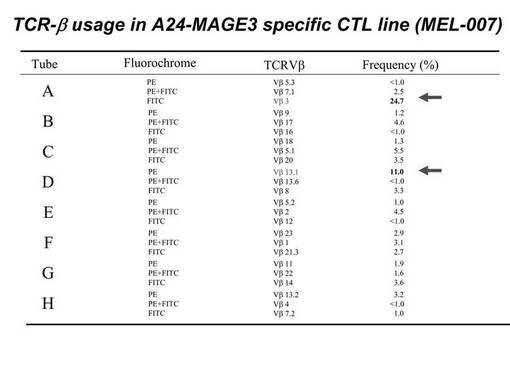
【図12】
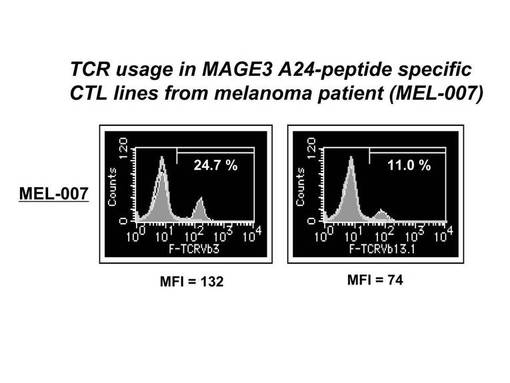
【図13】
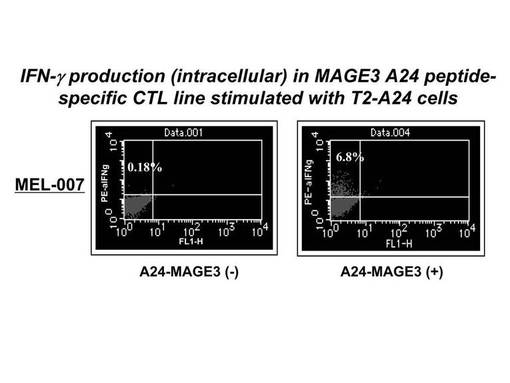
【図14】

【図15】
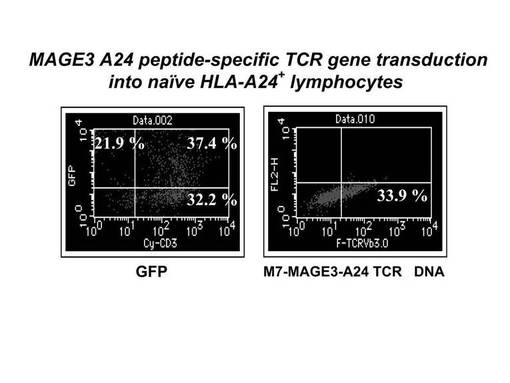
【図16】
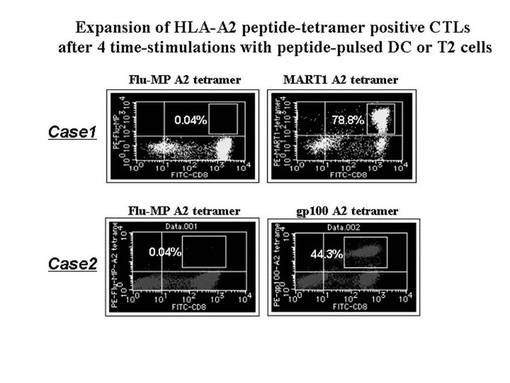
【図17】
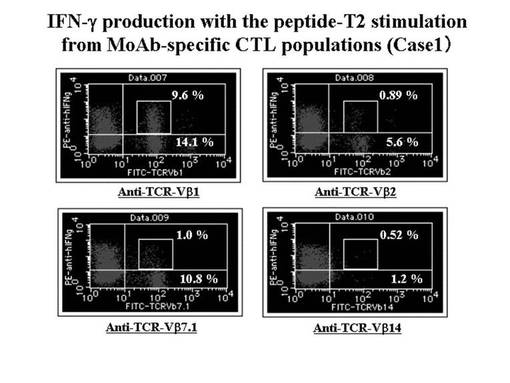
【図18】
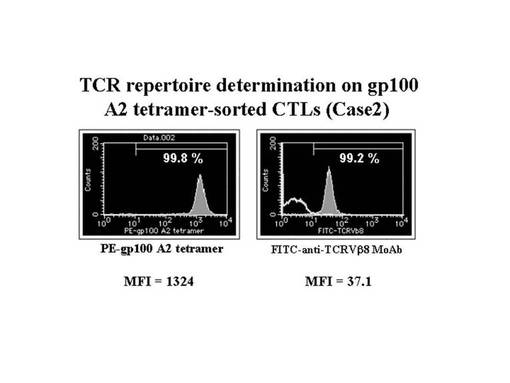
【図19】
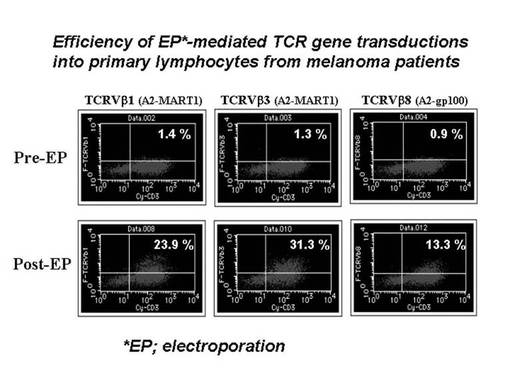
【図20】
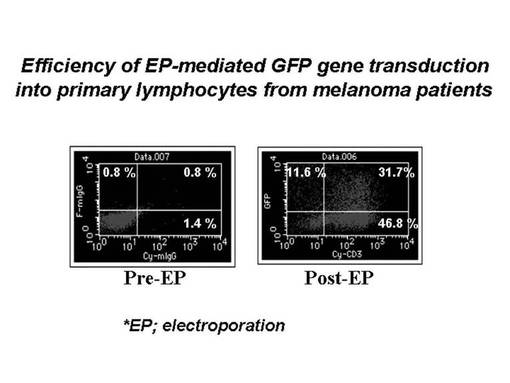
【図21】
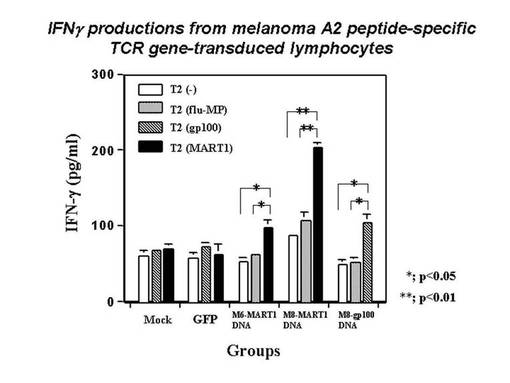
【図22】

【図24】
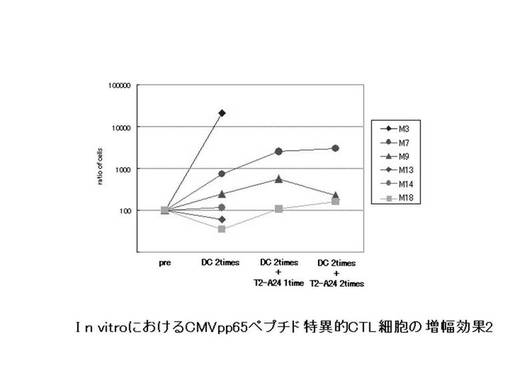
【図25】
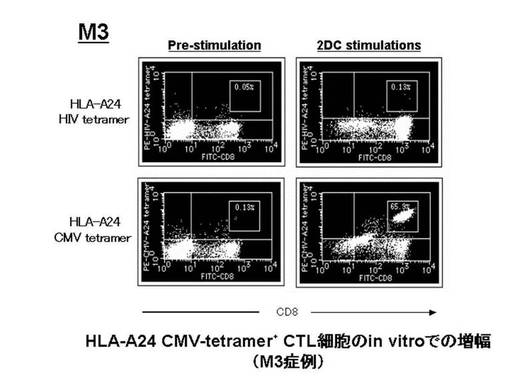
【図26】

【図27】
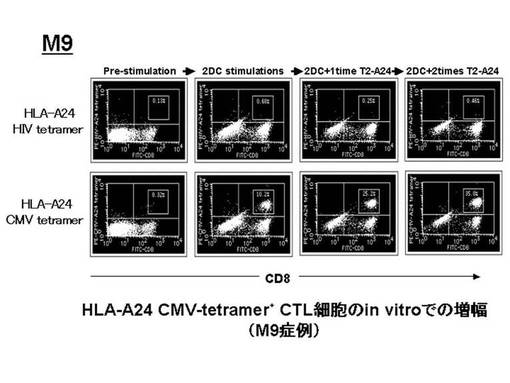
【図28】
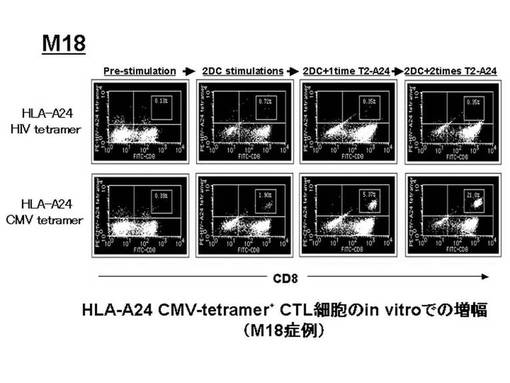
【図29】
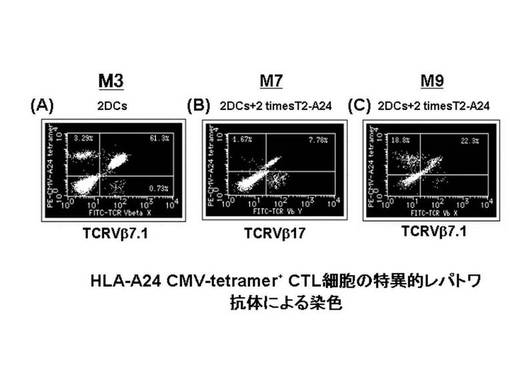
【図30】
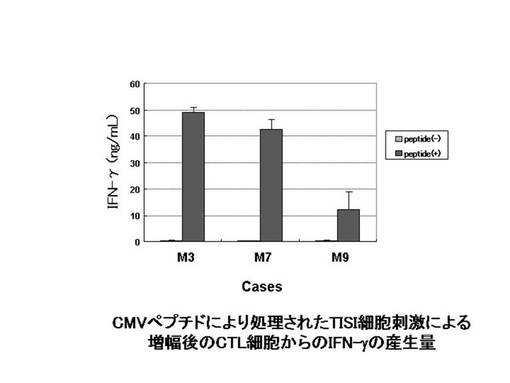
【図31】

【図32】

【図33】

【図34】

【図35】

【図36】
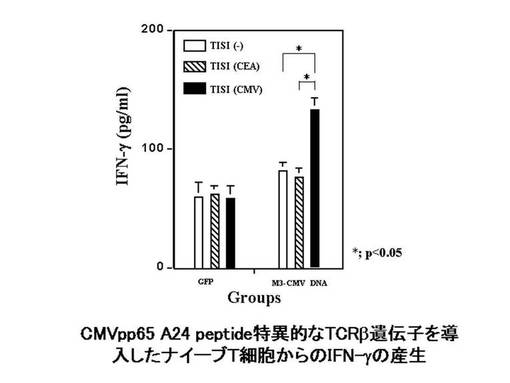
【図37】

【図39】
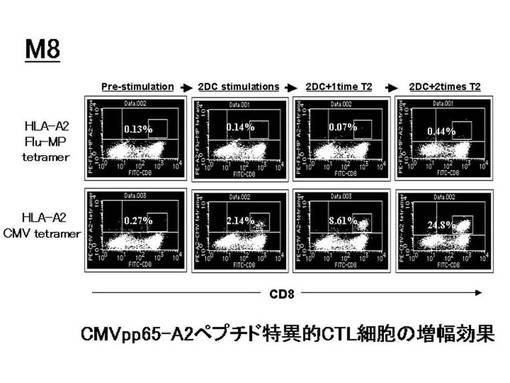
【図40】
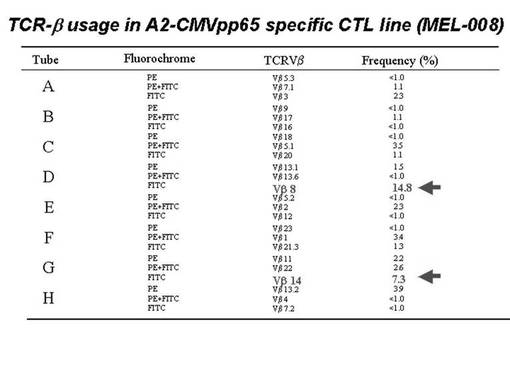
【図41】
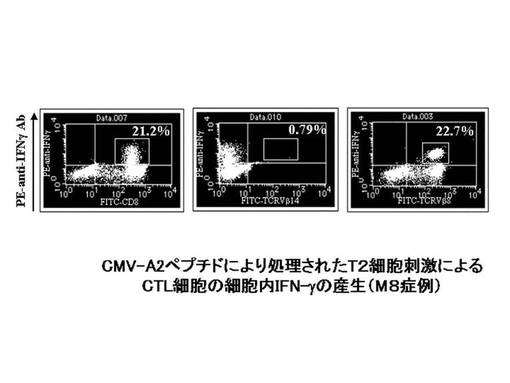
【図42】

【図43】

【図44】

【図45】
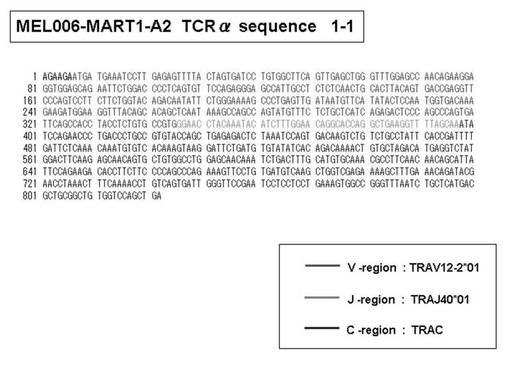
【図46】

【図47】
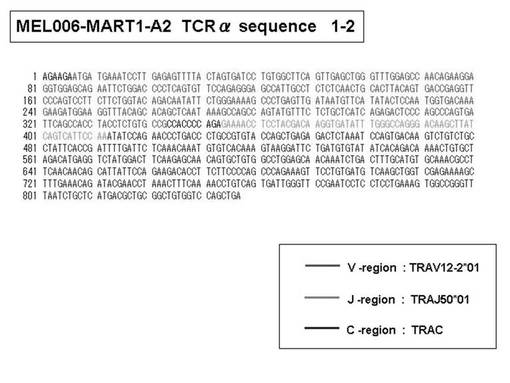
【図48】

【図49】
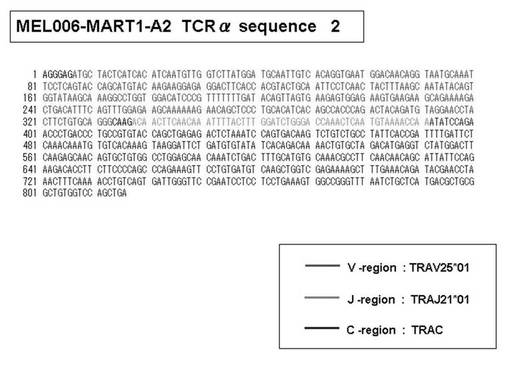
【図50】

【図51】

【図52】

【図53】
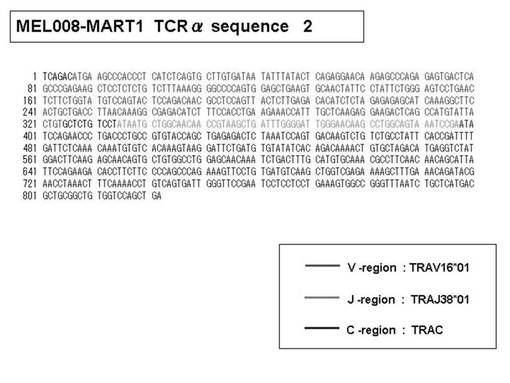
【図54】

【図55】

【図56】

【図57】
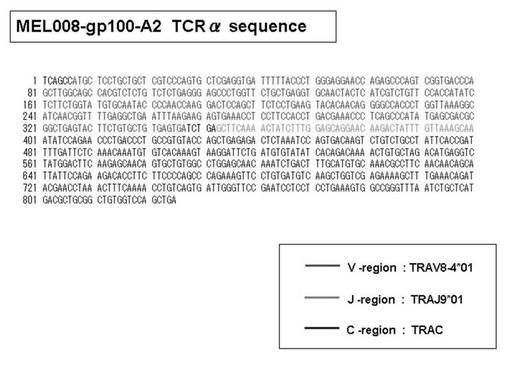
【図58】

【図59】
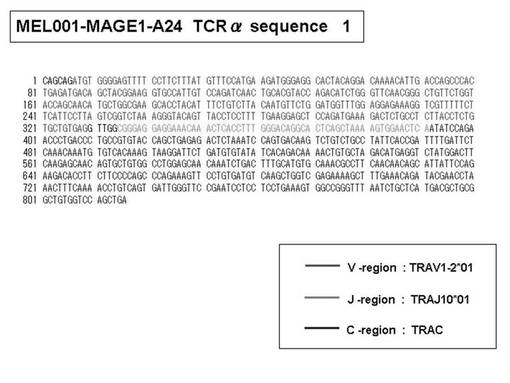
【図60】

【図61】
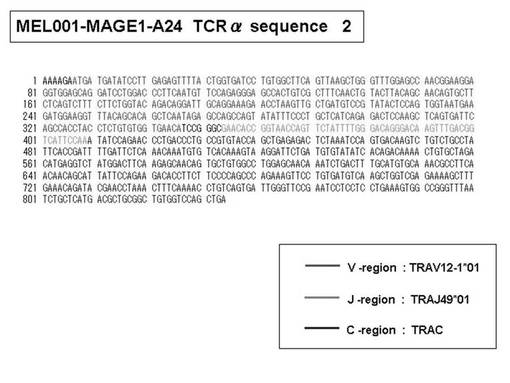
【図62】

【図63】
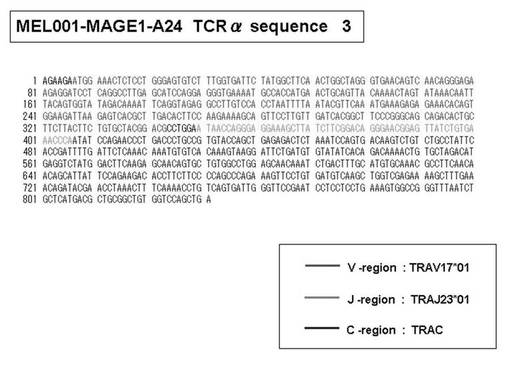
【図64】

【図65】
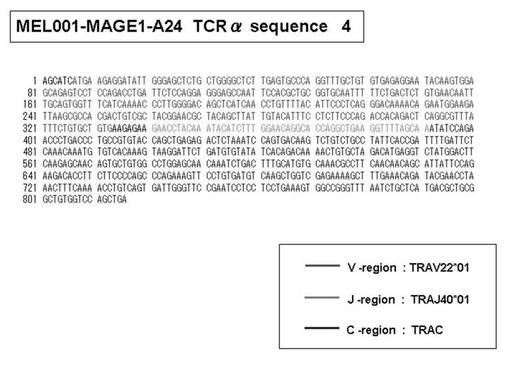
【図66】

【図67】
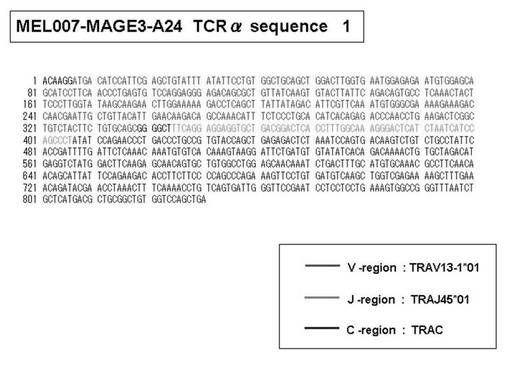
【図68】

【図69】

【図70】

【図71】
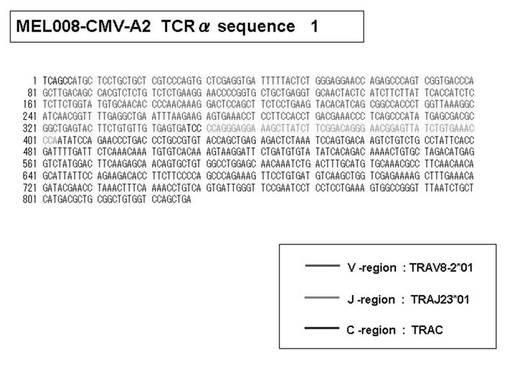
【図72】

【図73】
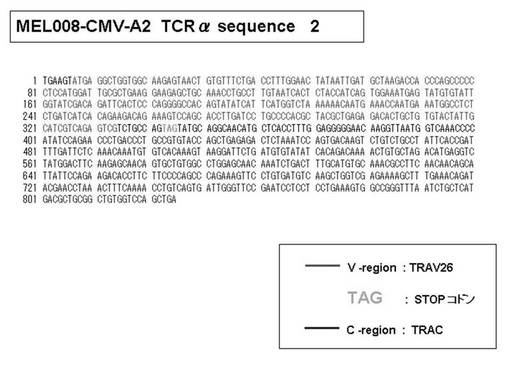
【図74】
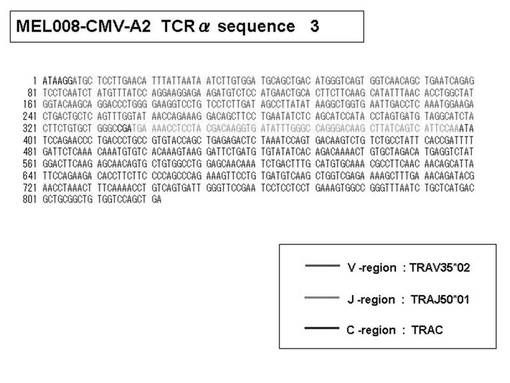
【図75】

【図76】
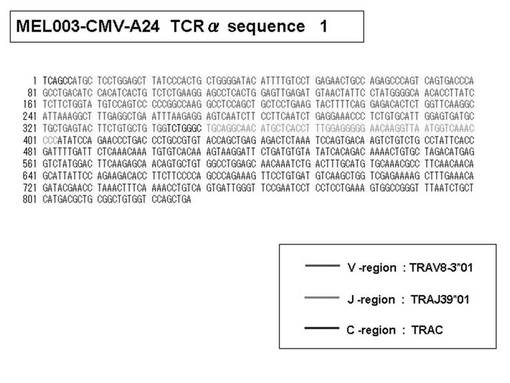
【図77】

【図78】
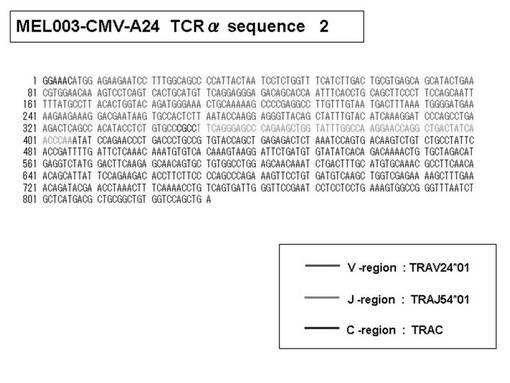
【図79】

【図80】
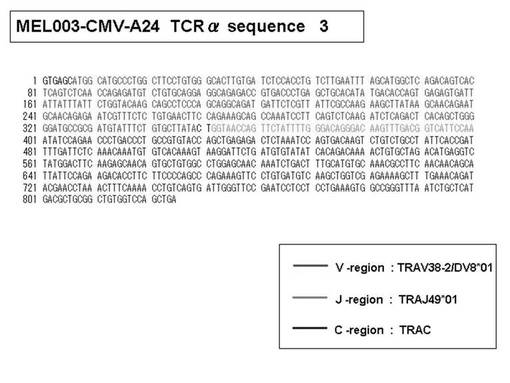
【図81】

【図82】
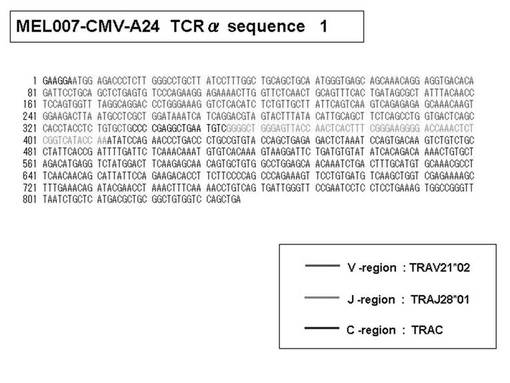
【図83】

【図84】
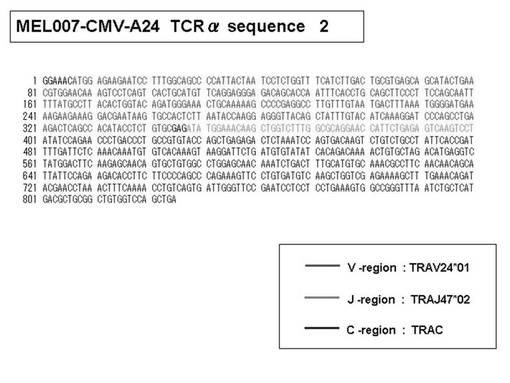
【図85】

【図86】
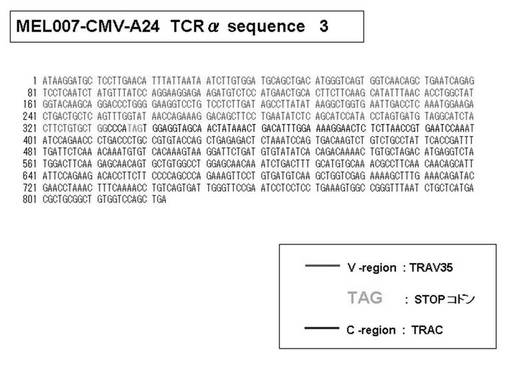
【図87】

【図88】

【図89】
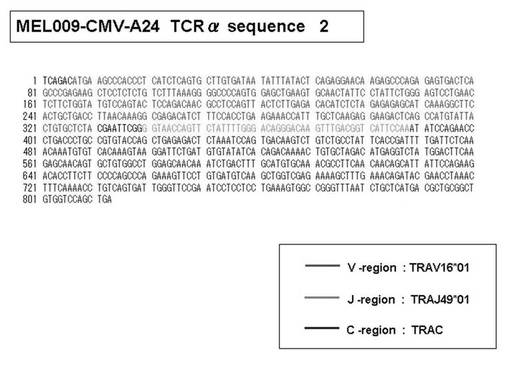
【図90】

【図91】
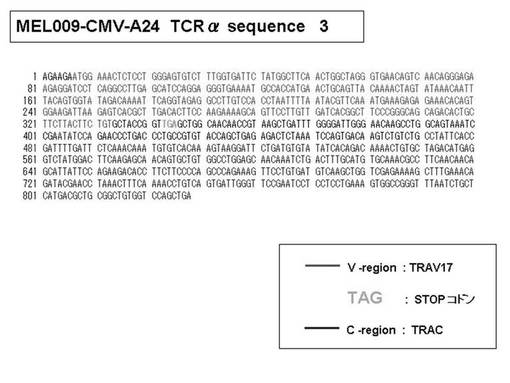
【図92】
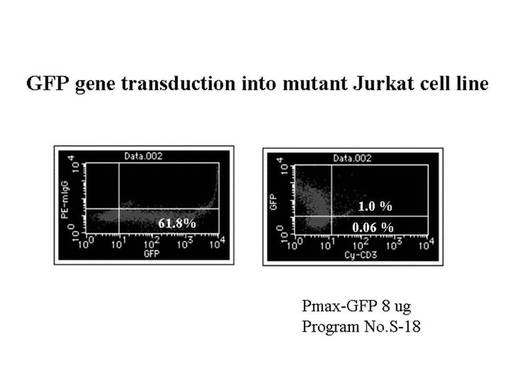
【図93】
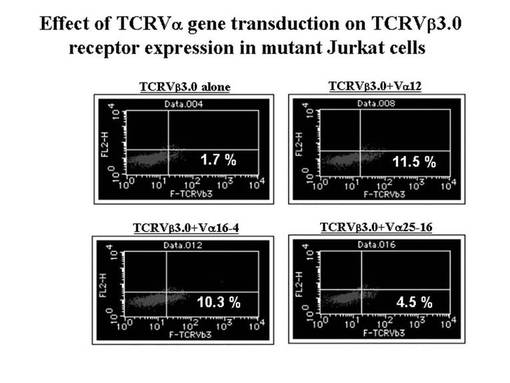
【図94】

【図38】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【図58】
【図59】
【図60】
【図61】
【図62】
【図63】
【図64】
【図65】
【図66】
【図67】
【図68】
【図69】
【図70】
【図71】
【図72】
【図73】
【図74】
【図75】
【図76】
【図77】
【図78】
【図79】
【図80】
【図81】
【図82】
【図83】
【図84】
【図85】
【図86】
【図87】
【図88】
【図89】
【図90】
【図91】
【図92】
【図93】
【図94】
【公開番号】特開2008−263950(P2008−263950A)
【公開日】平成20年11月6日(2008.11.6)
【国際特許分類】
【出願番号】特願2008−8755(P2008−8755)
【出願日】平成20年1月18日(2008.1.18)
【出願人】(590002389)静岡県 (173)
【Fターム(参考)】
【公開日】平成20年11月6日(2008.11.6)
【国際特許分類】
【出願日】平成20年1月18日(2008.1.18)
【出願人】(590002389)静岡県 (173)
【Fターム(参考)】
[ Back to top ]