
T細胞同時刺激シグナル2(B7/CD28相互作用)を遮断することによる免疫抑制
【課題】異種移植片特異的免疫抑制を促進するための手段を提供すること。
【解決手段】本発明は、レシピエントにおける異種反応性T細胞の活性化を防止するために同時刺激シグナルナル2の送達(B7/CD28相互作用)を遮断することによって、異種移植された臓器のT細胞媒介拒絶を、阻害するための手段および方法を提供する。第1の局面において、同時刺激は、異種ドナー生物由来の可溶性形態のCTLA−4の臓器レシピエントへの投与によって防止される。これは、優先的に、異種移植片上のB7に結合し、そして異種ドナー細胞上のB7とレシピエントT細胞上のCD28との間の相互作用を遮断する。第2の局面において、同時刺激は、異種ドナー細胞上のCTLA−4のリガンドを発現することによってアンタゴナイズされる。このリガンドは、レシピエントの活性化T細胞上のCTLA−4に結合し、シグナル2をアンタゴナイズする。
【解決手段】本発明は、レシピエントにおける異種反応性T細胞の活性化を防止するために同時刺激シグナルナル2の送達(B7/CD28相互作用)を遮断することによって、異種移植された臓器のT細胞媒介拒絶を、阻害するための手段および方法を提供する。第1の局面において、同時刺激は、異種ドナー生物由来の可溶性形態のCTLA−4の臓器レシピエントへの投与によって防止される。これは、優先的に、異種移植片上のB7に結合し、そして異種ドナー細胞上のB7とレシピエントT細胞上のCD28との間の相互作用を遮断する。第2の局面において、同時刺激は、異種ドナー細胞上のCTLA−4のリガンドを発現することによってアンタゴナイズされる。このリガンドは、レシピエントの活性化T細胞上のCTLA−4に結合し、シグナル2をアンタゴナイズする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、異種移植片拒絶の抑制に関する。
【背景技術】
【0002】
(発明の背景)
同種異系の臓器移植の成功は、ここ数十年において確立されているが、ドナー臓器の供給が限定されていることは、多くの患者が、移植される臓器(例えば、腎臓、心臓、または肝臓)を受ける機会がほとんどかまたは全くないことを意味する。これらの人々の非常に多くが、臓器を待っている間に死亡する。1つの潜在的な解決案は、「異種移植」、すなわち、ヒト以外の(「異種」)動物ドナー由来の臓器の使用である。
【0003】
ブタドナー臓器は、適切な候補であると考えられる。なぜなら、ブタは、解剖学的および生理学的にヒトに類似しており、そして供給が豊富であるからである。しかし、ブタの臓器は、超急性拒絶反応(HAR)と呼ばれる体液性のプロセスによって、血管再生の際に迅速に拒絶される。これは、異種移植片の内皮細胞(EC)上の抗原を認識し、そしてそれと交差反応する、レシピエント中に天然に存在する抗体によって引き起こされる。この認識は、補体カスケードを誘発し、これは次いで拒絶を導く。
【0004】
欧州特許第0495852号(Imutran)は、宿主補体の膜結合調節因子が、臓器レシピエント中の補体の完全な活性化を防止するために、異種移植片上で発現されるべきであることを示唆する。このアプローチは、超急性拒絶を生き延びるように設計された臓器を有するトランスジェニック動物を作製するために開発および応用されている(例えば、参考文献1および2)。
【0005】
しかし、HARを生存する臓器は、遅延型異種移植片拒絶(DXR)に供される。これは、レシピエントの炎症性細胞の浸潤および移植片血管の血栓(虚血を導く)によって特徴づけられる。WO98/42850は、異種移植片の表面上での凝固インヒビターの発現が、この型の拒絶の血栓症的局面を阻害し得ることを示す。
【0006】
HARおよびDXRの後には、宿主のTリンパ球媒介応答が続く。T細胞が異種抗原に対して感作され得る、「直接」および「間接」の2つの経路が存在する。直接経路には、異種のドナー細胞上のT細胞とMHC分子との間の相互作用が含まれる。一方、間接経路には、MHCクラスIIの状況における宿主APCによるプロセスされた異種抗原の提示が含まれる。間接T細胞応答は、同種抗原に対してよりも異種抗原に対してより強力であり(3)、これは、直接経路についての知見(4)とは対照的であり、これは、異種抗原に対する直接および間接のヒトT細胞応答の両方が、異種移植が効果的である場合には抑制されなければならないことを示す。
【0007】
抗異種移植片間接T細胞応答の抑制は、異種移植の最大の挑戦の1つであるようである(5,6)。持続的な間接的免疫原性に起因する慢性異種移植片拒絶を予防するのに必要とされる免疫抑制のレベルを維持することは、従来の全身的免疫抑制薬物を使用したのでは、感染および新生物形成の危険性が増大するために、実行することができない(例えば、7)。異種移植が臨床的に成功すべき場合、移植片特異的免疫抑制を促進するための方法が、全身性治療の必要性を減少させるために必要となる。
【0008】
T細胞活性化は、2つの別々のシグナルを必要とする。シグナル1の送達は、単独で、続く抗原への曝露の後のIL−2を産生することができないとして規定される寛容の状態(「アネルギー」)を誘導する。完全な活性化が生じるためには、その細胞は、シグナル2で同時刺激されなければならない。
【0009】
インビボで、シグナル1は、TCR/CD4複合体の、同種異系MHCまたは自己MHCと複合体化した抗原性ペプチドのいずれかとの相互作用によって提供される;シグナル2は、T細胞上の抗原提示細胞(APC)およびCD28上のB7分子(B7.1およびB7.2、それぞれCD80およびCD86としても知られる)間の相互作用によって供給される。
【0010】
モノクローナル抗体(mAb)は、T細胞活性化の研究において重要な役割を果たしてきた。シグナル1は、TCR/CD3複合体に対して指向されるmAbによって供給され得、そしてCD28に対して指向されるmAbは、シグナル2を提供し得る。実際、T細胞は、2つの適切なmAbによって、APCの非存在下でさえ、活性化され得る。活性化はまた、mAbを使用して、提供されるというよりもむしろ、防止され得る。シグナル2は、例えば、B7またはCD28のいずれかを遮断するmAbを使用して遮断され得る。
【0011】
シグナル2はまた、CTLA−4(B7の高親和性リガンド)の改変形態を使用して遮断され得る。CTLA−4は、T細胞活性化の天然の負の調節因子であり、活性化されたT細胞上でのCTLA−4に対するB7結合は、B7/CD28相互作用によって提供される同時刺激シグナルをアンタゴナイズする。抗体の定常ドメインに結合したCTLA−4の細胞外ドメインからなるCTLA−4の可溶性形態は、T細胞活性化を遮断するように構築されている(8,9)。これらの分子(「CTLA4−Ig」または「CTLA4−Fc」)は、抗B7抗体に対して類似の様式で振る舞い、そしてインビトロおよびインビボで、B7の同時刺激機能を予防するため、従って、寛容を促進するために使用されてきた(10)。
【0012】
移植片抗原に対するインビボでのT細胞感作を防止するためにB7/CD28相互作用を標的化することは、移植片生存を増強する有効なストラテジーであることが示されている。CTLA4−Igを使用して、延長された生存が種々の同種移植片モデルにおいて(例えば、11)、およびヒト対マウス島異種移植片モデルにおいて(12)、得られている。異種移植片モデルにおいて、CTLA4−Ig投与は、直接反応性T細胞をアネルギー性にすることによって、異種抗原に対して完全な寛容を引き起こした。
【0013】
従って、本発明の目的は、異種移植片特異的免疫抑制を促進するための手段を提供することである。特に、本発明の目的は、異種移植された臓器のT細胞媒介拒絶を、臓器レシピエントのT細胞が臓器に対する免疫応答を高めることを防止することによって阻害することである。より詳細には、この免疫応答を、異種移植された臓器を認識するレシピエントのT細胞におけるアネルギーを誘導し、結果として異種移植片特異的T細胞寛容を生じることによって防止することが目的である。
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の説明)
本発明は、異種移植された臓器のT細胞媒介拒絶を、レシピエントにおける異種反応性T細胞の活性化を予防するために、同時刺激シグナル2の送達を遮断することによって阻害するための方法および生物学的試薬を提供する。
【0015】
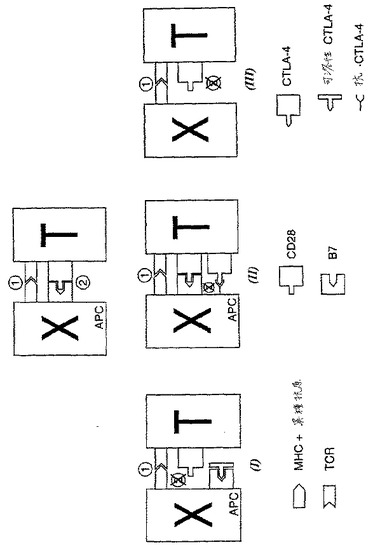
これは、図1に示される3つの局面において具体化される。これらの3つの局面は、単独で、または種々の組み合わせで使用され得る。さらに、従来の免疫抑制技術は、本発明の技術とともに使用され得る。
【0016】
以下は、第8頁から始まる「定義」と題された節と合わせて読まれるべきである。
【0017】
(第1の局面)
第1の局面において、シグナル2による同時刺激は、臓器レシピエントに、異種ドナー臓器からの可溶性形態のCTLA−4を投与することによって防止される。例えば、ブタ臓器(ドナー)がヒト(レシピエント)に移植された場合、可溶性形態のブタCTLA−4(以下を参照のこと)が、ヒトに投与される。
【0018】
ある生物(例えば、ブタ)由来のCTLA−4は、別の生物(例えば、ヒト)由来のB7に結合し得るが、最も高いアビディティーが、同種異系B7に対して見出される。従って、ドナー生物由来の可溶性CTLA−4は、レシピエントB7(正常細胞上の)およびドナーB7(異種移植された細胞上の)の両方に結合し得るが、それは、優先的に異種移植片上のB7に結合する。これは、全身性の免疫抑制を導く傾向があるレシピエント生物からのCTLA−4の投与とは異なり、異種移植片特異的免疫抑制を生じる。異種ドナー細胞上のB7とレシピエントT細胞上のCD28との間の相互作用を遮断することにより、同時刺激シグナル2は、レシピエントのT細胞に送達されない。それゆえ、異種反応性レシピエントT細胞は、アネルギー性にされる。
【0019】
従って、本発明は、臓器レシピエントにおける異種移植片寛容を誘導する方法を提供し、この方法は、そのレシピエントに、異種ドナー生物由来の可溶性形態のCTLA−4タンパク質を投与する工程を包含する。
【0020】
可溶性形態のCTLA−4は、好ましくは、B7に結合する能力を維持しているドナー生物由来のCTLA−4のフラグメントを含む。このフラグメントは、好ましくは、CTLA−4の完全な細胞外ドメインである。
【0021】
好ましくは、可溶性タンパク質は、免疫グロブリンの定常ドメイン(例えば、IgG1のCγ1鎖)をさらに含む。好ましくは、これは、この分子のこの部分に対する免疫応答を予防するために、レシピエント生物由来である。
【0022】
それゆえ、ブタ対ヒト移植のための代表的な実施態様において、可溶性タンパク質は、ヒトCγ1配列に融合したブタCTLA−4の細胞外ドメインを包含し得る。
【0023】
他の生物由来のCTLA−4の可溶性形態は、例えば、参考文献8(ヒトCTLA−4/ヒトIgγ1定常領域)および9(マウスCTLA−4/ヒトIgγ1)に記載される。
【0024】
本発明はまた、臓器レシピエントにおいて異種移植片寛容を誘導するための医薬の調製における異種CTLA−4の可溶性形態の使用を提供する。
【0025】
タンパク質は、異種移植の前、間、または後に投与され得る。前異種移植投与は、レシピエントが異種細胞への曝露を包含する前移植免疫プログラムを実行しているとき、最も有用である。
【0026】
ドナー生物がブタの状況において、本発明は、ブタ細胞からクローニングされたCTLA−4である、配列番号1として図2に示されるアミノ酸配列を含むタンパク質を提供する。これは、本発明における使用のためのCTLA−4の好ましい形態である。このタンパク質の細胞外ドメインはまた、図2に示される。
【0027】
本発明はまた、タンパク質配列番号1(またはそのフラグメント)をコードする核酸を提供する。これは、好ましくは、配列番号2として図3に示されるヌクレオチド配列を含む。
【0028】
さらに、本発明は、本発明の核酸を包含するベクターおよびこのようなベクターで形質転換された細胞を提供する。
【0029】
(第2の局面)
第2の局面において、シグナル2による同時刺激は、異種ドナー細胞上のCTLA−4についてのリガンドを発現することによってアンタゴナイズされる。このリガンドは、レシピエントの活性化されたT細胞上のCTLA−4に結合し、そしてドナーB7とレシピエントCD28との間の相互作用によって提供される同時刺激シグナルをアンタゴナイズする。これは、異種反応性T細胞をアネルギー性にする。
【0030】
従って、本発明は、CTLA−4に結合し得る膜結合タンパク質を提供する。
【0031】
これは、代表的には、CTLA−4結合領域および膜結合領域を含むキメラタンパク質(すなわち、異なるタンパク質の領域を1つのタンパク質へ組み合わせることによって産生されるタンパク質)である。従って、その最も単純な形態において、タンパク質は、このように、融合タンパク質である。
【0032】
「膜結合タンパク質」により、タンパク質は、その細胞外ドメインがCTLA−4に結合し得るように、細胞膜に結合していることを意味する。そのタンパク質を細胞膜に結合させるために、そのタンパク質は、例えば、膜タンパク質からの膜貫通配列、またはGPIアンカーを含み得る。好ましい膜貫通配列は、CD4またはCD8の膜貫通配列である。あるいは、そのタンパク質は、それをそのタンパク質自体が細胞膜に挿入されることなしに、膜タンパク質と細胞外で結合するようにする配列を含み得る。
【0033】
そのタンパク質が、細胞膜に標的化されるように、そのタンパク質がその膜貫通領域(例えば、CD8細胞質ドメイン)と通常結合する細胞質ドメインを含むこともまた望ましいかもしれない。同様に、そのタンパク質が、CTLA−4結合ドメインが細胞膜から分離するために、細胞膜と直に並列する細胞外配列(例えば、CD4ドメイン3および4)を含むことは望ましいかもしれない。グリシンリンカーのような合成リンカーは、同じ目的で使用され得る。
【0034】
タンパク質のCTLA−4結合ドメインは、好ましくは、CTLA−4に対する特異性を有する抗体を含む。これは、好ましくは、1本鎖抗体(sFv)である。それは、好ましくは、レシピエント生物のCTLA−4に対して特異的である。
【0035】
従って、代表的な実施態様において、第2の局面のタンパク質は、リンカーを介してCD8の膜貫通ドメインおよび細胞質ドメインに融合した1本鎖抗体を含み得る。
【0036】
本発明はまた、第2の局面のタンパク質をコードする核酸を提供する。
【0037】
さらに、本発明は、本発明のその核酸を含むベクター、およびそのベクターで形質転換された細胞を提供する。
【0038】
本発明はまた、本発明の第2の局面に従う核酸および/またはベクターを包含する送達システム、およびこの材料を標的細胞に送達する手段を提供する。
【0039】
さらに、本発明は、好ましくは、そのタンパク質が利用可能なCTLA−4に結合し得るように、その表面上に第2の局面のタンパク質を発現する細胞を提供する。
【0040】
細胞がレシピエントT細胞に結合し得るように、その細胞はまた、好ましくは、その表面上にMHC(クラスIまたはクラスII)を発現する。従って、適切には、本発明の細胞は、ドナープロフェッショナルAPCである。しかし、抗CTLA−4タンパク質によって提供されるアンタゴニスト的シグナルのために、これらのプロフェッショナルAPCは、B7ネガティブ細胞として機能的に振る舞う。
【0041】
本発明はまた、このような細胞を含む生物学的組織を提供する。
【0042】
本発明はさらに、第2の局面に従う細胞および/または生物学的組織を含む動物を提供する。
【0043】
本発明はまた、生物学的組織を異種移植に適切にするためのプロセスを提供し、このプロセスは、その生物学的組織を、それがその細胞の表面上の第2の局面に従う1つ以上のタンパク質を発現するように処理する工程を包含する。
【0044】
本発明はまた、ドナー動物(例えば、ブタ)から異種レシピエント動物(例えば、ヒト)へ、本発明に従う生物学的組織を移植する工程を包含する移植の方法を提供する。
【0045】
さらに、本発明の細胞は、前移植投与に適切である。これは、異種移植片自体が移植される前にレシピエントT細胞において誘導される寛容をもたらす。このような前移植レジメンにおいて使用される細胞が、好ましくは、MHCクラスIIを発現するのに対し、その細胞はプロフェッショナルAPCである必要はないことが理解される。
【0046】
さらに、本発明は、医薬として使用するための第2の局面に従うタンパク質または核酸を提供する。
【0047】
本発明はまた、異種移植片レシピエントまたはドナーへの投与のための処方物の製造における、第2の局面に従うタンパク質、核酸、ベクター、または送達システムの使用を提供する。
【0048】
(第3の局面)
第3の局面において、シグナル2による同時刺激は、異種ドナー臓器の細胞の表面上で、レシピエント生物MHCクラスIIを発現することによって防止される。例えば、ブタ臓器(ドナー)が、ヒト(レシピエント)に移植される場合、そのブタ臓器は、ヒトMHCクラスIIを発現する。
【0049】
レシピエントT細胞の直接的活性化が、例えば、上記の本発明の最初の2つの局面の一方または両方を利用することによって回避される場合でも、間接的活性化は、なお生じ得、レシピエントAPCによるMHCクラスIIに対する異種抗原のプロセシングおよび提示を包含する。
【0050】
レシピエントMHCクラスIIを異種ドナーの細胞上に発現することにより、ドナー細胞は、自己MHCクラスIIの状況において異種抗原をレシピエントT細胞に提示する。ドナー細胞がB7を発現しない場合、またはB7が遮断されている場合、異種反応性レシピエントT細胞は、同時刺激シグナル2を受けず、そしてそのレシピエントのAPCが異種抗原自体を提示する機会を有する前に、アネルギー性になる。
【0051】
従って、本発明は、その細胞表面上に、異なる生物のMHCクラスIIを発現する細胞を提供する。好ましくは、これは、その表面上にヒトMHCクラスIIを発現するブタ細胞である。
【0052】
MHCクラスIIは、好ましくは、HLA−DRファミリーのものである。
【0053】
MHCクラスIIは、好ましくは、細胞の表面上に構成的に発現される。
【0054】
同種異系抗MHCクラスII応答を防止するために、MHCクラスIIは、好ましくは、その特定のレシピエントと最大適合性の組織型である。これは、代表的には、例えば、異種ドナー細胞上に発現されるHLA−DRが、特定のレシピエントのHLA−DRと整合するはずであることを確実にすることを包含する。
【0055】
異種抗原が、プロフェッショナルAPC上に見出されるMHCクラスII分子ミラーの溝内に提示することを確実にするために、その細胞は、以下の3つのタンパク質(そのそれぞれが、抗原プロセッシングにおいて重要な役割を有する(不変鎖(invariant chain)、HLA−DRα、およびHLA−DRβ))の1つ以上をまた、発現するはずであることが好ましい。
【0056】
その細胞は、好ましくは、同時刺激分子(例えば、B7)をその表面上に発現しない。従って、代表的には、ドナー細胞は、プロフェッショナルAPCではない。しかし、トランスフェクトされた非免疫原性APC(例えば、B7+であり得る、未成熟樹状細胞)であり得る。
【0057】
本発明はまた、第3の局面に従う細胞を含む生物学的組織を提供する。
【0058】
本発明はさらに、第3の局面に従う細胞および/または生物学的組織を含む動物を提供する。
【0059】
本発明はまた、生物学的組織を異種移植に適切にするためのプロセスを提供し、そのプロセスは、それが異種MHCクラスIIをその細胞の表面上に発現するように、その生物学的組織を処置する工程を包含する。
【0060】
好ましくは、このプロセスは、異種生物由来の非免疫原性細胞(すなわち、同時刺激シグナルを提供し得ない細胞、例えば、B7ネガティブ細胞)を単離する工程、およびこれらの細胞をHLA−DRでトランスフェクトする工程を包含する。HLA−DRは、特定のレシピエントに対して好ましい組織型である。さらに、その細胞はまた、効率的な抗原プロセッシングのために必要とされる他のタンパク質でトランスフェクトされ得る。適切な非免疫原性細胞の例には、B7ネガティブであり、そして同種異系移植の齧歯類モデルにおける寛容を誘導することが示されている腎尿細管上皮細胞が含まれる。
【0061】
本発明はまた、ドナー動物(例えば、ブタ)から異種レシピエント動物(例えば、ヒト)へ、第3の局面に従う生物学的組織を移植する工程を包含する移植の方法を提供する。
【0062】
さらに、本発明の細胞は、前移植投与に適切である。これは、異種移植片自体が移植される前にレシピエントT細胞において誘導される、寛容をもたらす。
【0063】
さらに、本発明は、医薬としての使用のための第3の局面に従う細胞を提供する。
【0064】
本発明はまた、異種移植レシピエントに投与するための処方物の製造における第3の局面に従う細胞または生物学的組織の使用を提供する。
【0065】
本発明はまた、異種移植ドナーに投与するための処方物の調製における異種MHCクラスIIまたは異種MHCクラスIIをコードする核酸の使用を提供する。
【0066】
(定義)
上記のように、用語「核酸」には、DNAおよびRNAの両方が含まれるが、改変された核酸および合成核酸もまた、含まれる。例えば、核酸は、合成(例えば、PNA)であり得るか、または改変されたヌクレオチド間連結(例えば、ホスホロチオエート)を有し得る。さらに、その用語は、センスおよびアンチセンス核酸配列の両方、ならびに二本鎖配列を含む。
【0067】
好ましくは、核酸は、本発明に従うタンパク質の発現を制御するために適切な配列を含む。この発現は、好ましくは、細胞特異的制御、誘導性制御、または時間的制御のように、制御され得る。
【0068】
上記で使用されるように、用語「ベクター」は、核酸を宿主細胞に移入し得る分子を意味し、そして多数の適切なベクターが当該分野で公知である。
【0069】
好ましくは、そのベクターは、トランスジェニック動物の産生について適切である。例えば、トランスジェニックブタの生成のために適切なベクターは、参考文献13、14、15、16、および17に記載される。
【0070】
上記で使用されるように、用語「送達システム」は、遺伝子材料を標的細胞に送達するための手段をいう。
【0071】
上記の特定のベクターはまた、適切な送達システムとして機能し得る。同様に、特定の送達システムはまた、固有にベクターであり得るが、必ずしもそうであるとは限らない。例えば、ウイルスベクターはまた、送達システムとして機能し得るが、リポソーム送達システムは、ベクターではない。送達システムは、ウイルスであってもよく、またはウイルスでなくてもよい。リポソームのような非ウイルスシステムは、大規模産生の費用、発現の持続性の悪さ、安全性に関する懸念のような、ウイルスベースのシステムに関連するいくつかの困難性を回避する。好ましくは、送達システムは、遺伝子療法において使用するのに適している。多数の適切な送達システムが、当該分野で公知である。
【0072】
好ましくは、送達システムは、本発明に従う分子が、異種移植のために適切な細胞、または移植されている細胞によって取り込まれるように、標的化される。より好ましくは、送達システムは、これらの細胞に特異的である。例えば、送達システムは、特定の臓器(例えば、心臓または腎臓)、または特定の細胞型(例えば、内皮細胞またはプロフェッショナルAPC)に標的化され得る。
【0073】
これを達成するために、その送達システムは、例えば、標的細胞上に見出されるレセプターに標的化されたレセプター媒介送達システムであり得る。例えば、送達システムは、心臓細胞上に見出されるレセプター、好ましくは、心臓細胞上に独占的に見出されるレセプターに標的化され得るか、または内皮細胞上に見出されるレセプター、好ましくは、内皮細胞上に独占的に見出されるレセプターに標的化され得る。
【0074】
その送達システムは、好ましくは、トランスジェニック動物の作製に適している。例えば、送達システムは、配偶子、接合子、または胚幹細胞に標的化され得る。
【0075】
本発明のベクターおよび送達システムは、細胞をトランスフェクトして、本発明に従う細胞を生成するために使用され得る。トランスフェクションは、インビボまたはエキソビボで生じ得る。
【0076】
上記で使用される用語「生物学的組織」には、細胞、組織、および臓器の集団(collection)が含まれる。従って、その定義には、例えば、線維芽細胞、角膜、神経組織、心臓、肝臓、または腎臓が含まれる。
【0077】
本発明の第2および第3の局面が「動物」を提供する場合、その動物は、好ましくは、異種移植のための臓器および/または本発明の細胞(例えば、異種移植片レシピエントへの前異種移植片投与のための細胞)の作製に適切である。好ましくは、その動物は哺乳動物であり、そしてより好ましくは、それはトランスジェニックブタまたはトランスジェニックヒツジである。
【0078】
その動物は、それがトランスジェニック生物学的組織を含む(すなわち、遺伝子治療によって処置される)ように、生存している間に処置され得る。好ましくは、生存動物は、トランスジェニック動物を産生するために、本発明に従うベクターでトランスフェクトされる。例えば、本発明に従うベクターは、異種移植のために適切な生物学的組織を産生するためにブタの心臓に特異的に送達され得る。
【0079】
あるいは、動物は、トランスジェニック動物として生まれ得る。このようなトランスジェニック動物を作製するための多くの適切なアプローチが、当該分野で公知である(例えば、参考文献18、19、20)。例えば、DNAのマイクロインジェクションによる接合子または初期胚の直接的操作は、周知であり、胚幹細胞のような多能性の細胞のインビトロ操作も同様である。初期胚のレトロウイルス感染は、種々の種で成功していることが証明されており、そして帯を含まない(zona−free)卵のアデノウイルス感染が報告されている。ヒツジの核移入による遺伝子導入およびクローニングがまた記載されている(例えば、WO97/07668)。
【0080】
本発明が、生物学的組織を異種移植に適切にするためのプロセスを提供する場合、その生物学的組織は、インビボまたはエキソビボのいずれかでそのようにされ得る。例えば、動物の臓器は、本発明のベクターでインビボでトランスフェクトされ得るか、または臓器は、移植前にエキソビボでもしくは移植後にインビボでトランスフェクトされ得る。
本発明は、例えば以下の項目を提供する。
(項目1) レシピエントにおいて異種反応性T細胞の活性化を防止するために同時刺激シグナル2の送達を遮断することによって、異種移植された臓器のT細胞媒介拒絶を阻害し得る、生物学的試薬。
(項目2) 異種移植された臓器のT細胞媒介拒絶を阻害するための方法であって、レシピエントにおける異種反応性T細胞の活性化を防止するために、同時刺激シグナル2の送達を遮断する工程を包含する、方法。
(項目3) 異種ドナー生物由来の可溶性形態のCTLA−4タンパク質を、前記レシピエントに投与することを包含する、請求項2に記載の方法。
(項目4) 前記可溶性タンパク質が、ヒトCγ1配列に融合したブタCTLA−4の細胞外ドメインを含む、請求項3に記載の方法。
(項目5) 医薬として使用するための、可溶性形態の異種CTLA−4。
(項目6) 配列番号1のアミノ酸配列を含む、タンパク質。
(項目7) 請求項6に記載のタンパク質をコードする、核酸。
(項目8) 前記試薬が、CTLA−4に結合し得る膜結合タンパク質である、請求項1に記載の生物学的試薬。
(項目9) CTLA−4に特異的な一本鎖抗体を含む、請求項8に記載のタンパク質。
(項目10) 請求項8または請求項9に記載のタンパク質をコードする、核酸。
(項目11) その表面に請求項8または請求項9に記載のタンパク質を発現する、細胞。
(項目12) 請求項11に記載の細胞を含む、生物学的組織。
(項目13) 請求項11に記載の細胞および/または請求項12に記載の生物学的組織を含む、動物。
(項目14) ドナー動物から異種レシピエント動物へ、請求項12に記載の生物学的組織を移植する工程を包含する、移植法。
(項目15) 生物学的組織を異種移植に適切にするためのプロセスであって、該生物学的組織が請求項8または請求項9に記載のタンパク質をその細胞表面上に発現するように、該生物学的組織を処置する工程を包含する、プロセス。
(項目16) 医薬として使用するための、請求項8もしくは請求項9に記載のタンパク質、または請求項10に記載の核酸。
(項目17) 異種移植片レシピエントまたはドナーに投与するための処方物の調製における、請求項8もしくは請求項9に記載のタンパク質、または請求項10に記載の核酸の、使用。
(項目18) 前記試薬が、異なる生物のMHCクラスIIをその表面上に発現する細胞である、請求項1に記載の生物学的試薬。
(項目19) 前記細胞が、ヒトMHCクラスIIをその表面に発現するブタ細胞である、請求項18に記載の細胞。
(項目20) 前記細胞が、その表面上にB7を発現しない、請求項18または請求項19に記載の細胞。
(項目21) 前記細胞が、トランスフェクトされた未成熟樹状細胞である、請求項18または請求項19に記載の細胞。
(項目22) 請求項18、19、20、または21のいずれか1項に記載の細胞を含む、生物学的組織。
(項目23) 請求項22に記載の生物学的組織を含む、動物。
(項目24) ドナー動物から異種レシピエント動物へ、請求項22に記載の生物学的組織を移植する工程を包含する、移植法。
(項目25) 生物学的組織を異種移植に適切にするためのプロセスであって、該生物学的組織がその細胞の表面上に異種MHCクラスIIを発現するように、該生物学的組織を処置する工程を包含する、プロセス。
(項目26) 医薬として使用するための、請求項18、19、20、または21のいずれか1項に記載の細胞。
(項目27) 異種移植レシピエントに投与するための処方物の製造における、請求項22に記載の生物学的組織の使用。
(項目28) 異種移植ドナーに投与するための処方物の調製における、異種MHCクラスIIまたは異種MHCクラスIIをコードする核酸の、使用。
【図面の簡単な説明】
【0081】
本発明は、添付の図面を参照して記載される。
【図1】図1は、本発明の3つの局面を例示する。「X」は異種(xenogeneic)細胞(または、間接的活性化経路においては、異種抗原(xenoantigen)提示レシピエントAPC)を示し、「T」はレシピエントT細胞を示す。実施態様Iにおいて、同時刺激シグナル2の送達は、CTLA−4の可溶性形態を用いることによって妨げられる。実施態様IIにおいて、抗CTLA−4を使用して、シグナル2を拮抗する。実施態様IIIにおいて、XはレシピエントMHC−IIを発現するが、B7を発現しない。
【図2】図2は、pCTLA−4のアミノ酸配列(配列番号1)を示す。以下の接合部を「*」によって例示する:シグナルペプチド/細胞外ドメイン;細胞外ドメイン/膜貫通ドメイン;膜貫通ドメイン/細胞質ドメイン。ヒト配列とウシ配列とアラインメントもまた示される。pCTLA4との相同性は以下のとおりである:
【化100】

【図3−1】図3は、類似のアラインメントを示すが、ヌクレオチドレベルにおいてである。相同性は以下のとおりである:
【化101】
【図3−2】図3は、類似のアラインメントを示すが、ヌクレオチドレベルにおいてである。相同性は以下のとおりである:
【化102】
【図4】図4は、pCTLA4−Ig構築物のアミノ酸配列を示す。下線を付した配列は、可撓性リンカーGGSGGAAを示し、これはまた、pCTLA4ドメインとIgG1ドメインとの間の接合部を示す。
【図5】図5は、ヒトB7(下の2つの線)またはブタB7(上の2つの線)のいずれかでトランスフェクトしたヒト線維芽細胞に結合するhCTLA4−Ig(白丸および白四角)およびpCTLA4−Ig(白菱形および白三角)のフローサイトメトリー分析の結果を示す。
【図6】図6は、ヒトB7(白四角および白丸)またはブタB7(白菱形および白三角)によって同時刺激した場合の、hCTLA4−Ig(白四角および白菱形)と比較した、pCTLA4−Ig(白丸および白三角)による増殖の選択的阻害を示す。
【図7】図7は、MHC−クラスIIを発現するヒト細胞(7A)またはブタ細胞(7B)を5日間混合した白血球反応における刺激剤として使用した場合の、hCTLA4−Ig(白四角)またはpCTLA4−Ig(白菱形)によるヒトCD4+T細胞増殖の阻害を示す。
【図8】図8は、抗ヒトCTLA−4 sFvのヌクレオチド配列を示す。
【図9】図9は、抗ヒトCTLA−4 sFvの推測タンパク質配列を示す。図9に示したセリン−グリシンリンカーによって重鎖および軽鎖が連結される。
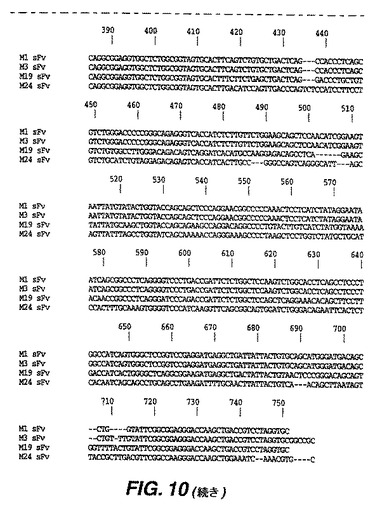
【図10−1】図10は、4つの抗マウスCTLA−4 sFvのヌクレオチド配列を示す。
【図10−2】図10は、4つの抗マウスCTLA−4 sFvのヌクレオチド配列を示す。
【図11】図11は、4つの抗マウスCTLA−4 sFvの推測タンパク質配列を示す。図11に示したセリン−グリシンリンカーによって重鎖および軽鎖が連結される。
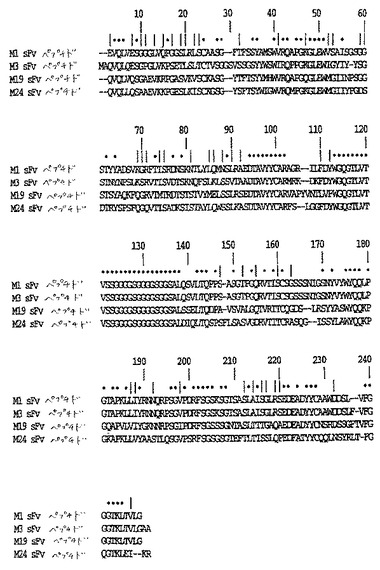
【図12】図12は、CTLA−4特異的sFVの可溶性Ig融合物をコードする構築物を示す。
【図13】図13は、抗hCTLA−4 sFv(白四角)またはコントロールsFv(白丸)のいずれかを発現する細胞によるT細胞増殖の阻害を示す。
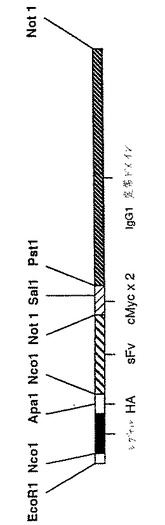
【図14】図14は、抗CTLA−4 sFvの膜結合形態をコードする構築物を示す。
【図15】図15は、ヒトCTLA−4のヌクレオチド配列(A)およびアミノ酸配列(B)を示す。開始コドンに下線を付す。−21位では、その配列はGenBank配列L15006とは異なり、そして110位では、その配列はL15006およびM74363の両方と異なる。
【図16】図16は、クローニングしたヒトCD8αの配列を示す。これは、231位(T→G)、244位(A→G)、266位(T→C)、および437位(T→C)においてGenBank配列と異なる。
【図17】図17は、B7ネガティブまたはB7ポジティブとしてクローンを規定するための、ヒトおよびマウスのCTLA4−IgのIPECへの結合を示す。
【図18】図18は、HLA−DR特異的mAb L243のトランスフェクト細胞に対する結合を示す。
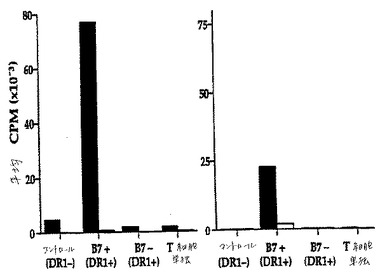
【図19】図19は、HLA−DR−1でトランスフェクトしたIPECに対するヒトT細胞による増殖を示す。
【図20】図20は、HLA−DR−1でトランスフェクトした細胞をインキュベーション下2日後の、ヒトT細胞増殖アッセイの結果を示す。X軸は、増殖アッセイの第2工程において使用した刺激細胞を示す。黒棒は、B7ポジティブトランスフェクタントでプライムしたCD4 T細胞での結果を示す;白棒(見るのは困難である)は、B7ネガティブトランスフェクタントでプライムした後の結果を示す。第1のグラフは、3日で採取した細胞での結果を示す;第2のグラフは、6日目での採取物からの結果を示す。
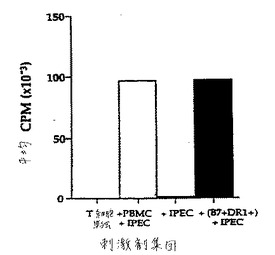
【図21】図21は、IPECに対して惹起された、APC依存性HLA−DR−1拘束T細胞株の増殖を示す。刺激剤集団を、X軸上に示す。
【発明を実施するための形態】
【0082】
(実施態様の説明)
(可溶性ブタCTLA−4)
ブタCTLA−4(「pCTLA4」)を、PHA活性化ブタT細胞からクローニングした。RNAを、標準的な技術を用いて調製し、そしてpCTLA−4を、以下のプライマーを用いてPCRによって増幅した:
【0083】
【数1】
得られた700bpのフラグメントを、EcoRI/HindIII消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を図3に示す。この図はまた、ヒトおよびウシのCTLA−4配列との比較を示す。
【0084】
pCTLA4の予測したアミノ酸配列を、ヒトおよびウシと比較しながら図2に示す。重要なことは、残基97における推定アミノ酸の差異であり、この残基はB7結合において重要であり、保存されたヘキサペプチドモチーフMYPPPYの一部である。pCTLA4において、残基97はロイシンである(LYPPPYとなる)のに対し、(ロイシンもまた、ウシCD28において見出さされているが(21))他の種はメチオニンを有する。この重要なアミノ酸差異は、pCTLA4のヒトおよびブタB7への有利に異なる結合の重要な鍵であると考えられる。
【0085】
pCTLA4の細胞外ドメインを、上記の5’プライマーおよび以下のプライマーを用いて増幅した:
【0086】
【数2】
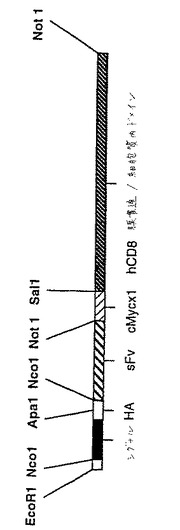
これを484位から増幅し、リンカーGGSGGAA配列(下線を付した)をコードする18塩基対のセグメントを導入し、そしてPstI部位(太字)を導入してヒトIgG1のヒンジ領域へのインフレーム連結を可能にした。得られた500bpのフラグメントを、HindIII/PstI消化したpBluescript−IgG1(ゲノムDNAコードイントロン性配列ならびにPstI/Not1部位の間のヒトIgG1のヒンジ、CH2、CH3および3’非翻訳エキソンを含む)にサブクローニングした。得られた可溶性pCTLA4−Igのアミノ酸配列を図4に示す。
【0087】
(pCTLA4−Igの発現)
キメラpCTLA4−Ig DNA配列を、HindIII/BstXIフラグメントとしてpBluescriptから遊離させ、そして発現ベクターpHOOK−3TM(Invitrogen)にサブクローニングした。これを使用して、標準的なリン酸カルシウム沈殿を用いてDAP.3細胞またはCHO−K1細胞をトランスフェクトした。G418耐性細胞を、CaptureTecTMシステムを用いて分離した。これらのトランスフェクト細胞を、組織培養フラスコ中でコンフルエントになるまで増殖させ、コンフルエントになった時点で、培地を交換し、そして細胞を培養物中でさらに3日間維持した。この段階で、培地を採取し、そしてプロテインGカラムを通して灌流した。pCTLA−4−Igを酸性条件下で溶出させた。この溶出したタンパク質の濃度を、ヒトIgG1に対する抗体および標準的なヒトIgG1骨髄腫タンパク質を用いるELISAを用いて計算した。
【0088】
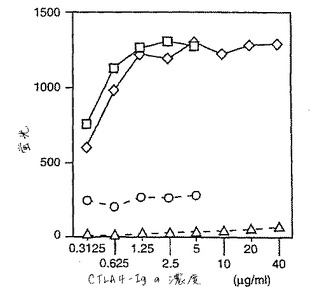
pCTLA4−Igの結合特徴を、ヒトB7−1またはブタB7−2のいずれかでトランスフェクトしたヒト線維芽細胞を用いるフローサイトメトリー分析を使用して、ヒトCTLA4−Igの結合特徴と比較した。これらの実験に関して、ブタおよびヒトのCTLA4−Igの濃度は、ELISAによってアッセイしたとき、等しかった。図5に例示したように、ヒトおよびブタCTLA4−Igは、ブタB7を発現するヒト細胞において類似の結合特徴を有するようであった。しかし、ヒトCTLA4−Igとは異なり、pCTLA4−IgはヒトB7には結合しなかった。このことは、pCTLA4−IgがブタB7に優先的に結合し、そして種特異的試薬として有用であることを意味する。
【0089】
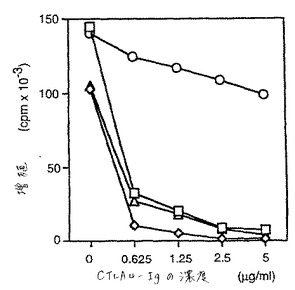
pCTLA4−Igを使用して、種々の刺激剤に対するヒトT細胞増殖応答を阻害した。これらのアッセイにおいて、T細胞応答の同時刺激は、プロフェッショナルAPCにおいてトランスフェクションまたは天然のいずれかによって発現された、ブタまたはヒトのいずれかのB7によって提供された。これらの実験を、図6および7において実証する。
【0090】
トランスフェクトした線維芽細胞刺激剤(HLAクラスIIおよびヒトB7またはブタB7のいずれかを発現する)を用いる実験について、hCTLA4−Igは全ての増殖応答を阻害した(図6、白四角および白菱形)。対照的に、pCTLA4−Igのみが、刺激剤がブタB7を発現した場合の応答を完全に阻害した(白三角);ヒトB7を発現する細胞に対する増殖応答は、最小限に影響されたのみであった(白丸)。
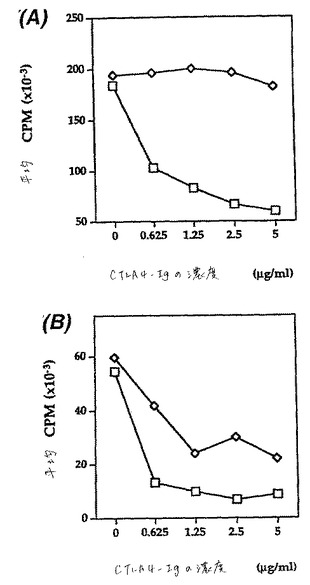
【0091】
類似の実験において、pCTLA4−Igは、MHCクラスIIおよびヒトB7を発現するヒト細胞への増殖応答に対する有意な阻害効果を有しなかったが、ブタ刺激剤に対する応答は阻害した(図7)。
【0092】
これらの結果は、T細胞の同時刺激シグナルがブタB7によって提供された場合のpCTLA4−Igの有効な阻害特性を強調する。同時刺激がヒトB7によって媒介される場合にT細胞増殖を防止できないことはまた、種特異的作用を実証する。pCTAL4−Igは、ブタB7に対して種特異的結合を示し、ブタB7の機能的効果の阻害を示すが、ヒトB7に対しては種特異的結合を示さず、その機能的効果の阻害も示さないと、結論付け得る。
【0093】
(pCTLA−4−Igの特性)
pCTLA4−Igのヒトおよびブタの両方のB7ファミリー分子に対する結合特徴は、例えば、以下の試験を用いて、hCTLA4−Igの結合特徴と比較され得る:
(i)ブタおよびヒトのAPCに対する結合、およびブタまたはヒトのB7を発現するトランスフェクタントに対する結合のフローサイトメトリー分析(上記を参照のこと)
(ii)BiacoreTMを用いる結合の定量的特徴づけ
(iii)ヒト抗ブタおよびヒト同種異系混合したリンパ球培養物に対するCTLA4−Igの効果の機能的分析
(iv)pCTLA4−IgのB6マウスへの移植後のブタ島異種移植生存を延長する能力の機能的評価。
【0094】
(CTLA−4に結合する膜結合型タンパク質)
1012の半合成可変配列を含むファージディスプレイライブラリーを、ヒトまたはマウスのCTLA4−IgおよびコントロールヒトIgG1骨髄腫タンパク質を用いてスクリーニングした。4回のスクリーニング後にヒトCTLA4−Igタンパク質への差示的な結合を提示するファージからsFvを単離し、そして標準的な技術を用いて精製した。そのヌクレオチド配列および推測したアミノ酸配列を、図8、9、10、および11に示す。
【0095】
sFvを、ヌクレオチド配列に基づく特異的プライマーを用いてPCRによって増幅した。プライマーの遠位部分は、pHOOK1内の配列に基づいた。5’プライマーはApaI部位を含み、そして3’プライマーはSalI部位を含んだ。これらは両方とも、独特であると予測された。得られたsFvを、配列決定のためにpBluescriptにサブクローニングして、忠実な増幅を決定した。次いでApaI/SalIフラグメントを、pHOOK1にサブクローニングした。ここでは、ApaI/SalIフラグメントは、上流ではマウスIgκ鎖からのインフレームのシグナル配列および血球凝集素Aエピトープ配列に、そして下流では2つのインフレームのmyc配列およびPDGFレセプターからの膜貫通配列に隣接する。
【0096】
pHOOK1からのmyc配列を、NotI部位(下線を付した)を含む以下の5’プライマー:
【0097】
【数3】
およびPstI部位(下線を付した)を導入した以下の3’プライマー:
【0098】
【数4】
を用いてPCRにより増幅した。得られた113塩基対のフラグメントを、NotI/PstI消化したpBluescriptにサブクローニングした。
【0099】
sFvを、pHOOK1からEcoRI/NotIフラグメントとして遊離させ、そしてEcoRI/PstI消化したpBluescript−IgG1に、NotI/PstI PCR産物とともに連結した(図12)。この構築物は、CTLA−4特異的sFvの可溶性Ig融合物をコードする。真核生物細胞における発現のために、この構築物を、pHOOK3にHindIII/BstXIフラグメントとしてサブクローニングした。
【0100】
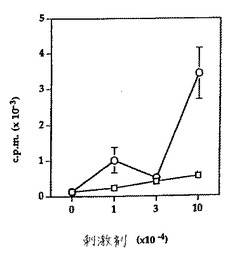
sFvの細胞表面発現を確認するために、pHOOK構築物を、既にHLA−DR分子およびヒトB7を発現している細胞にトランスフェクトした。G418またはミコフェノール酸(使用したベクターに依存する)に耐性である細胞を、培養において増殖させた。その細胞表面上で抗CTLA4−sFv構築物を発現する細胞を、hCTLA4−Igを使用してフローサイトメトリー分析によって同定した。これらの細胞を、限定希釈によってクローニングし、そして5日間の培養におけるT細胞の増殖の刺激剤として使用した。1つの実験の結果を図13に示す。抗hCTLA4 sFvを発現する細胞は、T細胞増殖を刺激しないが(白四角)、一方コントロールsFvを発現する細胞は、正常細胞と同じ方法で増殖を刺激した(白丸)。
【0101】
異なる実験において、図12に示した構築物のEcoRI/SalIフラグメントを、ヒトCD8の膜貫通ドメインおよび細胞質ドメイン(pBluescript−hCD8由来のSalI/BamHIフラグメントとして単離した)とともに、EcoRI/BamHI消化したpBluescriptに同時連結した(図14)。
【0102】
pBluescriptからのEcoRI/BamHIフラグメントを、発現ベクターpHβApr−1−neoまたはシスターベクターpHβApr−1−gptにサブクローニングした。これらを、既にHLA−DR分子およびB7を発現している細胞にトランスフェクトし、そしてpHOOK構築物について上記のように選択した。
【0103】
(膜結合型CTLA−4構築物)
活性化T細胞によるCTLA−4の発現は、一過性であるのみであり、それゆえ、抗CTLA4−sFvの機能的特徴を試験するために、ヒトまたはマウスのCTLA4の細胞外ドメインをコードするDNA配列、ならびにヒトCD8の膜貫通/細胞質配列を含むキメラ構築物を作製した。これらの構築物を発現する細胞を、抗CTLA4−sFvタンパク質の研究のために使用し得る。
【0104】
PHA活性化ヒトT細胞由来のRNAを、標準的な技術を用いて調製した。hCTLA4を、以下のプライマーを用いてPCR増幅した:
HindIII部位を導入する(5’プライマー)
【0105】
【数5】
EcoRI部位を導入する(3’プライマー)
【0106】
【数6】
得られたフラグメントを、HindIII/EcoRI消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を、図15に示す。これは、ヒトCTLA−4についてのGenBankリスト配列とは1つの塩基(439位)が異なる。hCTLA4の推定アミノ酸配列もまた示す。
【0107】
hCTLA−4の細胞外ドメインを、上記の5’プライマーおよび以下のプライマーを用いて増幅した:
【0108】
【数7】
これは、457位から増幅され、そして可撓性GGSGGアミノ酸リンカーをコードする15塩基のセグメント(下線を付した)を、独特のSalI部位(強調表示)とともに含んでいだ。得られたフラグメントを、HindIII/SalI消化したpBluescriptにサブクローニングし、そして配列決定した。
【0109】
hCD8を、得られたT細胞から以下のプライマーを用いてPCR増幅した:
HindIII部位を導入する(5’プライマー)
【0110】
【数8】
EcoRI部位を導入する(3’プライマー)
【0111】
【数9】
得られたフラグメントを、HindIII/EcoRI消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を、図16に示す。このクローンは、GenBankに登録された配列とは4つの位置で異なっていたが、これらはいずれも、続いて増幅した領域内では存在しなかった。
【0112】
hCD8の膜貫通(TM)ドメインおよび細胞質(C)ドメインを、上記の3’プライマーおよび以下の5’プライマーを用いて増幅した:
【0113】
【数10】
これは、532位から増幅され、そして可撓性GGSGGアミノ酸リンカーをコードする15塩基のセグメント(下線を付した)を、独特のSalI部位(強調表示)とともに含んでいた。得られたフラグメントを、HindIII/SalI消化したpBluescriptにサブクローニングし、そしてpBluescript−hCD8と称した。
【0114】
ヒトCTLA−4の細胞外ドメインを、pBluescriptからEcoRI/SalIフラグメントとして切り出し、そしてCD8のTM−ICドメインを、SalI/BamHIフラグメントとして切り出した。これらをともに、EcoRI/BamHI消化したpBluescriptに再び連結した。次いで、全CTLA−4−CD8キメラを単一のEcoRIフラグメントとして取り出し、そしてヒトT細胞白血病株J6への発現のための多数の発現ベクターにサブクローニングした。
【0115】
(細胞表面抗CTLA4タンパク質の特性)
細胞表面抗CTLA−4タンパク質は、以下の機能性試験によってさらに特徴付けられ得る:
(i)膜結合抗CTLA4−sFv−CD8タンパク質を発現する細胞と、可溶性ヒトCTLA4−Igを発現する細胞との間の相互作用のフローサイトメトリー評価
(ii)BiacoreTMを用いる、可溶性抗CTLA4−sFv−Ig融合タンパク質と可溶性ヒトCTLA4−Igとの間の相互作用の定量的評価
(iii)抗CTLA4−sFv−CD8融合タンパク質を発現するJ6トランスフェクタントへの可溶性ヒトCTLA4−Igの結合から生じるシグナル伝達事象についての分析
(iv)同種異系混合リンパ球応答における刺激が、HLA−DRポジティブ、B7ポジティブ、抗CTLA4−sFv−CD8ポジティブ細胞株によって提供される場合のT細胞応答(例えば、増殖、サイトカイン産生、アネルギー誘導)の分析。
【0116】
(マウスMHCクラスIIを発現するB7ネガティブブタ細胞)
50のクローニングした不死化した大動脈内皮細胞(PAEC)を、PAECの単層から、pZipSVU19 DNA(22)を用いる核内マイクロインジェクションにより生成した。不死化細胞(IPEC)から、B7ネガティブクローンを、hCTLA4−IgおよびmCTLA4−Igを用いてフローサイトメトリースクリーニングによって同定した(図17を参照のこと)。次いで、これらを、プラスミド発現ベクターpcExV1−gptおよびpHβApr−1neo中のHLA−DRAおよびDRB1*0101をコードするcDNAでトランスフェクトし、そして細胞を、MXHおよびG418での選択下に置いた。比較のために、B7−ポジティブIPECコントロールを同様に生成した(4)。
【0117】
マウスMHCクラスII分子I−Abを発現するIPECトランスフェクタントの別のシリーズもまた、マウスにおける実験のために生成した。
【0118】
トランスフェクトIPEC細胞におけるMHCクラスIIの表面発現を、モノクローナル抗体L243(HLA−DRに特異的)(図18)またはM5−114(マウスMHCクラスIIに特異的)を用いて検出した。MHCクラスIIポジティブ細胞は、限界希釈によるクローニングの前に数回の蛍光活性化細胞ソーティングを受けた。
【0119】
トランスフェクタントの第2のバッチを、発現ベクターpCMU中のHLA−DMAおよびHLA−DMBおよびp31Ii(不変鎖)をコードするさらなるトランスフェクトcDNAを用いて正確に同じ方法で調製した。
【0120】
(MHCクラスII発現細胞による同種異系T細胞におけるアネルギー誘導)
ヒトT細胞を、標準的なプロトコルを使用して精製した(3)。一次増幅アッセイのために、ガラス繊維フィルターへの採取の16時間前に1マイクロCi3H−チミジンの添加の前に、T細胞を、一定数の照射した刺激細胞(stimulator cell)とともに5日間インキュベートした。このフィルターを、シンチレーションカウンターにおいて読み取った。
【0121】
B7ポジティブIPECは、有意な抗DR1特異的増殖応答を生じ、一方B7ネガティブIPECは任意の増殖応答を開始しなかった(図19)。
【0122】
2工程のアネルギー誘導アッセイを、標準的なプロトコルによって確立した(23)。一次寛容性誘導工程において、B7ポジティブIPECとともにインキュベートしたT細胞を、プライムされた二次免疫応答(3日で最大)の動力学を用いる二次工程における抗DR1増殖応答をマウントした。しかし、一次工程においてB7ネガティブIPECとともにインキュベートしたT細胞は、DR1に対して寛容になり、そしてB7ポジティブIPECにおいて発現されたDR1に対する続く曝露の際に、応答をマウントしなかった(図20)。
【0123】
(DR1を発現するブタ細胞によるDR1拘束T細胞におけるアネルギー誘導)
DR1を発現する個体由来のCD4+T細胞を、標準的な手順に従って精製した。一次増殖アッセイにおいて、それらは、HLA−DR1でトランスフェクトしたB7ポジティブIPECに増殖した。このことは、DR1が、ブタペプチド特異的T細胞についての制限エレメントとして機能し得ることを示す。B7ポジティブおよびB7ネガティブDR1+トランスフェクタントに対する増殖応答を比較するアッセイを行っている。
【0124】
2工程のアネルギー導入アッセイもまた、DR1でトランスフェクトしたB7ネガティブブタ細胞がHLA−DR拘束ヒトT細胞においてアネルギーを誘導することを実証するために行い得る。
【0125】
(HLA−DRにおける提示についてのプロフェッショナルヒトAPCによってプロセスされたブタペプチドと、HLA−DRでトランスフェクトしたIPECのMHCクラスIIにおいて提示されるものとの間の重複)
野生型IPECに対するヒトT細胞株を、ヒトPBMCから得た。この株の増殖応答は、ヒトAPCの存在に依存し、そしてHLA−DRに対する抗体によって阻害可能であり、これらのことは、この株が、ヒトAPCによって提示される、プロセスされたブタ異種抗原(xenoantigen)に対する間接的な特異性を有した。
【0126】
少なくともいくつかのプロセスされたブタペプチドがプロフェッショナルヒトAPCによって間接的に提示されることを意味する、B7ポジティブHLA−DR1でトランスフェクトされたIPECに対して増殖したこの株(図21)もまた、トランスフェクトされたブタ細胞によって提示された。
【0127】
(ブタ島対マウスモデルにおける研究)
インビボでは、ブタ膵島細胞は、ストレプトゾトシン(streptozotocin)処置した糖尿病マウスの腎臓被膜下に移植され得る。島異種移植片(非血管性である)は、T細胞によって単に拒絶される。ブタ島を、末梢麻酔下でブタの膵臓から調製し、そしてレシピエントにおけるその生存を、正常血糖値の維持によって評価する。マウスに、ブタ島の移植前に、B7ネガティブのI−Abを発現するIPECを静脈内注射する。このストラテジーを、本発明の他の局面と組み合わせて、T細胞認識の直接経路を寛容化し、直接経路を介する拒絶が生じないことを確実にする。特定のストラテジーが特異的T細胞寛容性を誘導したか否かを評価するために、島保有腎臓の腎摘出を、同一のブタ島または第三者のブタ島の再移植の前に行う(生存する腎臓の被膜下)。
【0128】
本発明は、例示のみのために上に記載され、そして本発明の範囲および精神内に残る間は改変がなされ得ることが理解される。
【0129】
(参考文献(その内容を完全に援用する)
【0130】
【表1】
【技術分野】
【0001】
本発明は、異種移植片拒絶の抑制に関する。
【背景技術】
【0002】
(発明の背景)
同種異系の臓器移植の成功は、ここ数十年において確立されているが、ドナー臓器の供給が限定されていることは、多くの患者が、移植される臓器(例えば、腎臓、心臓、または肝臓)を受ける機会がほとんどかまたは全くないことを意味する。これらの人々の非常に多くが、臓器を待っている間に死亡する。1つの潜在的な解決案は、「異種移植」、すなわち、ヒト以外の(「異種」)動物ドナー由来の臓器の使用である。
【0003】
ブタドナー臓器は、適切な候補であると考えられる。なぜなら、ブタは、解剖学的および生理学的にヒトに類似しており、そして供給が豊富であるからである。しかし、ブタの臓器は、超急性拒絶反応(HAR)と呼ばれる体液性のプロセスによって、血管再生の際に迅速に拒絶される。これは、異種移植片の内皮細胞(EC)上の抗原を認識し、そしてそれと交差反応する、レシピエント中に天然に存在する抗体によって引き起こされる。この認識は、補体カスケードを誘発し、これは次いで拒絶を導く。
【0004】
欧州特許第0495852号(Imutran)は、宿主補体の膜結合調節因子が、臓器レシピエント中の補体の完全な活性化を防止するために、異種移植片上で発現されるべきであることを示唆する。このアプローチは、超急性拒絶を生き延びるように設計された臓器を有するトランスジェニック動物を作製するために開発および応用されている(例えば、参考文献1および2)。
【0005】
しかし、HARを生存する臓器は、遅延型異種移植片拒絶(DXR)に供される。これは、レシピエントの炎症性細胞の浸潤および移植片血管の血栓(虚血を導く)によって特徴づけられる。WO98/42850は、異種移植片の表面上での凝固インヒビターの発現が、この型の拒絶の血栓症的局面を阻害し得ることを示す。
【0006】
HARおよびDXRの後には、宿主のTリンパ球媒介応答が続く。T細胞が異種抗原に対して感作され得る、「直接」および「間接」の2つの経路が存在する。直接経路には、異種のドナー細胞上のT細胞とMHC分子との間の相互作用が含まれる。一方、間接経路には、MHCクラスIIの状況における宿主APCによるプロセスされた異種抗原の提示が含まれる。間接T細胞応答は、同種抗原に対してよりも異種抗原に対してより強力であり(3)、これは、直接経路についての知見(4)とは対照的であり、これは、異種抗原に対する直接および間接のヒトT細胞応答の両方が、異種移植が効果的である場合には抑制されなければならないことを示す。
【0007】
抗異種移植片間接T細胞応答の抑制は、異種移植の最大の挑戦の1つであるようである(5,6)。持続的な間接的免疫原性に起因する慢性異種移植片拒絶を予防するのに必要とされる免疫抑制のレベルを維持することは、従来の全身的免疫抑制薬物を使用したのでは、感染および新生物形成の危険性が増大するために、実行することができない(例えば、7)。異種移植が臨床的に成功すべき場合、移植片特異的免疫抑制を促進するための方法が、全身性治療の必要性を減少させるために必要となる。
【0008】
T細胞活性化は、2つの別々のシグナルを必要とする。シグナル1の送達は、単独で、続く抗原への曝露の後のIL−2を産生することができないとして規定される寛容の状態(「アネルギー」)を誘導する。完全な活性化が生じるためには、その細胞は、シグナル2で同時刺激されなければならない。
【0009】
インビボで、シグナル1は、TCR/CD4複合体の、同種異系MHCまたは自己MHCと複合体化した抗原性ペプチドのいずれかとの相互作用によって提供される;シグナル2は、T細胞上の抗原提示細胞(APC)およびCD28上のB7分子(B7.1およびB7.2、それぞれCD80およびCD86としても知られる)間の相互作用によって供給される。
【0010】
モノクローナル抗体(mAb)は、T細胞活性化の研究において重要な役割を果たしてきた。シグナル1は、TCR/CD3複合体に対して指向されるmAbによって供給され得、そしてCD28に対して指向されるmAbは、シグナル2を提供し得る。実際、T細胞は、2つの適切なmAbによって、APCの非存在下でさえ、活性化され得る。活性化はまた、mAbを使用して、提供されるというよりもむしろ、防止され得る。シグナル2は、例えば、B7またはCD28のいずれかを遮断するmAbを使用して遮断され得る。
【0011】
シグナル2はまた、CTLA−4(B7の高親和性リガンド)の改変形態を使用して遮断され得る。CTLA−4は、T細胞活性化の天然の負の調節因子であり、活性化されたT細胞上でのCTLA−4に対するB7結合は、B7/CD28相互作用によって提供される同時刺激シグナルをアンタゴナイズする。抗体の定常ドメインに結合したCTLA−4の細胞外ドメインからなるCTLA−4の可溶性形態は、T細胞活性化を遮断するように構築されている(8,9)。これらの分子(「CTLA4−Ig」または「CTLA4−Fc」)は、抗B7抗体に対して類似の様式で振る舞い、そしてインビトロおよびインビボで、B7の同時刺激機能を予防するため、従って、寛容を促進するために使用されてきた(10)。
【0012】
移植片抗原に対するインビボでのT細胞感作を防止するためにB7/CD28相互作用を標的化することは、移植片生存を増強する有効なストラテジーであることが示されている。CTLA4−Igを使用して、延長された生存が種々の同種移植片モデルにおいて(例えば、11)、およびヒト対マウス島異種移植片モデルにおいて(12)、得られている。異種移植片モデルにおいて、CTLA4−Ig投与は、直接反応性T細胞をアネルギー性にすることによって、異種抗原に対して完全な寛容を引き起こした。
【0013】
従って、本発明の目的は、異種移植片特異的免疫抑制を促進するための手段を提供することである。特に、本発明の目的は、異種移植された臓器のT細胞媒介拒絶を、臓器レシピエントのT細胞が臓器に対する免疫応答を高めることを防止することによって阻害することである。より詳細には、この免疫応答を、異種移植された臓器を認識するレシピエントのT細胞におけるアネルギーを誘導し、結果として異種移植片特異的T細胞寛容を生じることによって防止することが目的である。
【発明の概要】
【課題を解決するための手段】
【0014】
(発明の説明)
本発明は、異種移植された臓器のT細胞媒介拒絶を、レシピエントにおける異種反応性T細胞の活性化を予防するために、同時刺激シグナル2の送達を遮断することによって阻害するための方法および生物学的試薬を提供する。
【0015】
これは、図1に示される3つの局面において具体化される。これらの3つの局面は、単独で、または種々の組み合わせで使用され得る。さらに、従来の免疫抑制技術は、本発明の技術とともに使用され得る。
【0016】
以下は、第8頁から始まる「定義」と題された節と合わせて読まれるべきである。
【0017】
(第1の局面)
第1の局面において、シグナル2による同時刺激は、臓器レシピエントに、異種ドナー臓器からの可溶性形態のCTLA−4を投与することによって防止される。例えば、ブタ臓器(ドナー)がヒト(レシピエント)に移植された場合、可溶性形態のブタCTLA−4(以下を参照のこと)が、ヒトに投与される。
【0018】
ある生物(例えば、ブタ)由来のCTLA−4は、別の生物(例えば、ヒト)由来のB7に結合し得るが、最も高いアビディティーが、同種異系B7に対して見出される。従って、ドナー生物由来の可溶性CTLA−4は、レシピエントB7(正常細胞上の)およびドナーB7(異種移植された細胞上の)の両方に結合し得るが、それは、優先的に異種移植片上のB7に結合する。これは、全身性の免疫抑制を導く傾向があるレシピエント生物からのCTLA−4の投与とは異なり、異種移植片特異的免疫抑制を生じる。異種ドナー細胞上のB7とレシピエントT細胞上のCD28との間の相互作用を遮断することにより、同時刺激シグナル2は、レシピエントのT細胞に送達されない。それゆえ、異種反応性レシピエントT細胞は、アネルギー性にされる。
【0019】
従って、本発明は、臓器レシピエントにおける異種移植片寛容を誘導する方法を提供し、この方法は、そのレシピエントに、異種ドナー生物由来の可溶性形態のCTLA−4タンパク質を投与する工程を包含する。
【0020】
可溶性形態のCTLA−4は、好ましくは、B7に結合する能力を維持しているドナー生物由来のCTLA−4のフラグメントを含む。このフラグメントは、好ましくは、CTLA−4の完全な細胞外ドメインである。
【0021】
好ましくは、可溶性タンパク質は、免疫グロブリンの定常ドメイン(例えば、IgG1のCγ1鎖)をさらに含む。好ましくは、これは、この分子のこの部分に対する免疫応答を予防するために、レシピエント生物由来である。
【0022】
それゆえ、ブタ対ヒト移植のための代表的な実施態様において、可溶性タンパク質は、ヒトCγ1配列に融合したブタCTLA−4の細胞外ドメインを包含し得る。
【0023】
他の生物由来のCTLA−4の可溶性形態は、例えば、参考文献8(ヒトCTLA−4/ヒトIgγ1定常領域)および9(マウスCTLA−4/ヒトIgγ1)に記載される。
【0024】
本発明はまた、臓器レシピエントにおいて異種移植片寛容を誘導するための医薬の調製における異種CTLA−4の可溶性形態の使用を提供する。
【0025】
タンパク質は、異種移植の前、間、または後に投与され得る。前異種移植投与は、レシピエントが異種細胞への曝露を包含する前移植免疫プログラムを実行しているとき、最も有用である。
【0026】
ドナー生物がブタの状況において、本発明は、ブタ細胞からクローニングされたCTLA−4である、配列番号1として図2に示されるアミノ酸配列を含むタンパク質を提供する。これは、本発明における使用のためのCTLA−4の好ましい形態である。このタンパク質の細胞外ドメインはまた、図2に示される。
【0027】
本発明はまた、タンパク質配列番号1(またはそのフラグメント)をコードする核酸を提供する。これは、好ましくは、配列番号2として図3に示されるヌクレオチド配列を含む。
【0028】
さらに、本発明は、本発明の核酸を包含するベクターおよびこのようなベクターで形質転換された細胞を提供する。
【0029】
(第2の局面)
第2の局面において、シグナル2による同時刺激は、異種ドナー細胞上のCTLA−4についてのリガンドを発現することによってアンタゴナイズされる。このリガンドは、レシピエントの活性化されたT細胞上のCTLA−4に結合し、そしてドナーB7とレシピエントCD28との間の相互作用によって提供される同時刺激シグナルをアンタゴナイズする。これは、異種反応性T細胞をアネルギー性にする。
【0030】
従って、本発明は、CTLA−4に結合し得る膜結合タンパク質を提供する。
【0031】
これは、代表的には、CTLA−4結合領域および膜結合領域を含むキメラタンパク質(すなわち、異なるタンパク質の領域を1つのタンパク質へ組み合わせることによって産生されるタンパク質)である。従って、その最も単純な形態において、タンパク質は、このように、融合タンパク質である。
【0032】
「膜結合タンパク質」により、タンパク質は、その細胞外ドメインがCTLA−4に結合し得るように、細胞膜に結合していることを意味する。そのタンパク質を細胞膜に結合させるために、そのタンパク質は、例えば、膜タンパク質からの膜貫通配列、またはGPIアンカーを含み得る。好ましい膜貫通配列は、CD4またはCD8の膜貫通配列である。あるいは、そのタンパク質は、それをそのタンパク質自体が細胞膜に挿入されることなしに、膜タンパク質と細胞外で結合するようにする配列を含み得る。
【0033】
そのタンパク質が、細胞膜に標的化されるように、そのタンパク質がその膜貫通領域(例えば、CD8細胞質ドメイン)と通常結合する細胞質ドメインを含むこともまた望ましいかもしれない。同様に、そのタンパク質が、CTLA−4結合ドメインが細胞膜から分離するために、細胞膜と直に並列する細胞外配列(例えば、CD4ドメイン3および4)を含むことは望ましいかもしれない。グリシンリンカーのような合成リンカーは、同じ目的で使用され得る。
【0034】
タンパク質のCTLA−4結合ドメインは、好ましくは、CTLA−4に対する特異性を有する抗体を含む。これは、好ましくは、1本鎖抗体(sFv)である。それは、好ましくは、レシピエント生物のCTLA−4に対して特異的である。
【0035】
従って、代表的な実施態様において、第2の局面のタンパク質は、リンカーを介してCD8の膜貫通ドメインおよび細胞質ドメインに融合した1本鎖抗体を含み得る。
【0036】
本発明はまた、第2の局面のタンパク質をコードする核酸を提供する。
【0037】
さらに、本発明は、本発明のその核酸を含むベクター、およびそのベクターで形質転換された細胞を提供する。
【0038】
本発明はまた、本発明の第2の局面に従う核酸および/またはベクターを包含する送達システム、およびこの材料を標的細胞に送達する手段を提供する。
【0039】
さらに、本発明は、好ましくは、そのタンパク質が利用可能なCTLA−4に結合し得るように、その表面上に第2の局面のタンパク質を発現する細胞を提供する。
【0040】
細胞がレシピエントT細胞に結合し得るように、その細胞はまた、好ましくは、その表面上にMHC(クラスIまたはクラスII)を発現する。従って、適切には、本発明の細胞は、ドナープロフェッショナルAPCである。しかし、抗CTLA−4タンパク質によって提供されるアンタゴニスト的シグナルのために、これらのプロフェッショナルAPCは、B7ネガティブ細胞として機能的に振る舞う。
【0041】
本発明はまた、このような細胞を含む生物学的組織を提供する。
【0042】
本発明はさらに、第2の局面に従う細胞および/または生物学的組織を含む動物を提供する。
【0043】
本発明はまた、生物学的組織を異種移植に適切にするためのプロセスを提供し、このプロセスは、その生物学的組織を、それがその細胞の表面上の第2の局面に従う1つ以上のタンパク質を発現するように処理する工程を包含する。
【0044】
本発明はまた、ドナー動物(例えば、ブタ)から異種レシピエント動物(例えば、ヒト)へ、本発明に従う生物学的組織を移植する工程を包含する移植の方法を提供する。
【0045】
さらに、本発明の細胞は、前移植投与に適切である。これは、異種移植片自体が移植される前にレシピエントT細胞において誘導される寛容をもたらす。このような前移植レジメンにおいて使用される細胞が、好ましくは、MHCクラスIIを発現するのに対し、その細胞はプロフェッショナルAPCである必要はないことが理解される。
【0046】
さらに、本発明は、医薬として使用するための第2の局面に従うタンパク質または核酸を提供する。
【0047】
本発明はまた、異種移植片レシピエントまたはドナーへの投与のための処方物の製造における、第2の局面に従うタンパク質、核酸、ベクター、または送達システムの使用を提供する。
【0048】
(第3の局面)
第3の局面において、シグナル2による同時刺激は、異種ドナー臓器の細胞の表面上で、レシピエント生物MHCクラスIIを発現することによって防止される。例えば、ブタ臓器(ドナー)が、ヒト(レシピエント)に移植される場合、そのブタ臓器は、ヒトMHCクラスIIを発現する。
【0049】
レシピエントT細胞の直接的活性化が、例えば、上記の本発明の最初の2つの局面の一方または両方を利用することによって回避される場合でも、間接的活性化は、なお生じ得、レシピエントAPCによるMHCクラスIIに対する異種抗原のプロセシングおよび提示を包含する。
【0050】
レシピエントMHCクラスIIを異種ドナーの細胞上に発現することにより、ドナー細胞は、自己MHCクラスIIの状況において異種抗原をレシピエントT細胞に提示する。ドナー細胞がB7を発現しない場合、またはB7が遮断されている場合、異種反応性レシピエントT細胞は、同時刺激シグナル2を受けず、そしてそのレシピエントのAPCが異種抗原自体を提示する機会を有する前に、アネルギー性になる。
【0051】
従って、本発明は、その細胞表面上に、異なる生物のMHCクラスIIを発現する細胞を提供する。好ましくは、これは、その表面上にヒトMHCクラスIIを発現するブタ細胞である。
【0052】
MHCクラスIIは、好ましくは、HLA−DRファミリーのものである。
【0053】
MHCクラスIIは、好ましくは、細胞の表面上に構成的に発現される。
【0054】
同種異系抗MHCクラスII応答を防止するために、MHCクラスIIは、好ましくは、その特定のレシピエントと最大適合性の組織型である。これは、代表的には、例えば、異種ドナー細胞上に発現されるHLA−DRが、特定のレシピエントのHLA−DRと整合するはずであることを確実にすることを包含する。
【0055】
異種抗原が、プロフェッショナルAPC上に見出されるMHCクラスII分子ミラーの溝内に提示することを確実にするために、その細胞は、以下の3つのタンパク質(そのそれぞれが、抗原プロセッシングにおいて重要な役割を有する(不変鎖(invariant chain)、HLA−DRα、およびHLA−DRβ))の1つ以上をまた、発現するはずであることが好ましい。
【0056】
その細胞は、好ましくは、同時刺激分子(例えば、B7)をその表面上に発現しない。従って、代表的には、ドナー細胞は、プロフェッショナルAPCではない。しかし、トランスフェクトされた非免疫原性APC(例えば、B7+であり得る、未成熟樹状細胞)であり得る。
【0057】
本発明はまた、第3の局面に従う細胞を含む生物学的組織を提供する。
【0058】
本発明はさらに、第3の局面に従う細胞および/または生物学的組織を含む動物を提供する。
【0059】
本発明はまた、生物学的組織を異種移植に適切にするためのプロセスを提供し、そのプロセスは、それが異種MHCクラスIIをその細胞の表面上に発現するように、その生物学的組織を処置する工程を包含する。
【0060】
好ましくは、このプロセスは、異種生物由来の非免疫原性細胞(すなわち、同時刺激シグナルを提供し得ない細胞、例えば、B7ネガティブ細胞)を単離する工程、およびこれらの細胞をHLA−DRでトランスフェクトする工程を包含する。HLA−DRは、特定のレシピエントに対して好ましい組織型である。さらに、その細胞はまた、効率的な抗原プロセッシングのために必要とされる他のタンパク質でトランスフェクトされ得る。適切な非免疫原性細胞の例には、B7ネガティブであり、そして同種異系移植の齧歯類モデルにおける寛容を誘導することが示されている腎尿細管上皮細胞が含まれる。
【0061】
本発明はまた、ドナー動物(例えば、ブタ)から異種レシピエント動物(例えば、ヒト)へ、第3の局面に従う生物学的組織を移植する工程を包含する移植の方法を提供する。
【0062】
さらに、本発明の細胞は、前移植投与に適切である。これは、異種移植片自体が移植される前にレシピエントT細胞において誘導される、寛容をもたらす。
【0063】
さらに、本発明は、医薬としての使用のための第3の局面に従う細胞を提供する。
【0064】
本発明はまた、異種移植レシピエントに投与するための処方物の製造における第3の局面に従う細胞または生物学的組織の使用を提供する。
【0065】
本発明はまた、異種移植ドナーに投与するための処方物の調製における異種MHCクラスIIまたは異種MHCクラスIIをコードする核酸の使用を提供する。
【0066】
(定義)
上記のように、用語「核酸」には、DNAおよびRNAの両方が含まれるが、改変された核酸および合成核酸もまた、含まれる。例えば、核酸は、合成(例えば、PNA)であり得るか、または改変されたヌクレオチド間連結(例えば、ホスホロチオエート)を有し得る。さらに、その用語は、センスおよびアンチセンス核酸配列の両方、ならびに二本鎖配列を含む。
【0067】
好ましくは、核酸は、本発明に従うタンパク質の発現を制御するために適切な配列を含む。この発現は、好ましくは、細胞特異的制御、誘導性制御、または時間的制御のように、制御され得る。
【0068】
上記で使用されるように、用語「ベクター」は、核酸を宿主細胞に移入し得る分子を意味し、そして多数の適切なベクターが当該分野で公知である。
【0069】
好ましくは、そのベクターは、トランスジェニック動物の産生について適切である。例えば、トランスジェニックブタの生成のために適切なベクターは、参考文献13、14、15、16、および17に記載される。
【0070】
上記で使用されるように、用語「送達システム」は、遺伝子材料を標的細胞に送達するための手段をいう。
【0071】
上記の特定のベクターはまた、適切な送達システムとして機能し得る。同様に、特定の送達システムはまた、固有にベクターであり得るが、必ずしもそうであるとは限らない。例えば、ウイルスベクターはまた、送達システムとして機能し得るが、リポソーム送達システムは、ベクターではない。送達システムは、ウイルスであってもよく、またはウイルスでなくてもよい。リポソームのような非ウイルスシステムは、大規模産生の費用、発現の持続性の悪さ、安全性に関する懸念のような、ウイルスベースのシステムに関連するいくつかの困難性を回避する。好ましくは、送達システムは、遺伝子療法において使用するのに適している。多数の適切な送達システムが、当該分野で公知である。
【0072】
好ましくは、送達システムは、本発明に従う分子が、異種移植のために適切な細胞、または移植されている細胞によって取り込まれるように、標的化される。より好ましくは、送達システムは、これらの細胞に特異的である。例えば、送達システムは、特定の臓器(例えば、心臓または腎臓)、または特定の細胞型(例えば、内皮細胞またはプロフェッショナルAPC)に標的化され得る。
【0073】
これを達成するために、その送達システムは、例えば、標的細胞上に見出されるレセプターに標的化されたレセプター媒介送達システムであり得る。例えば、送達システムは、心臓細胞上に見出されるレセプター、好ましくは、心臓細胞上に独占的に見出されるレセプターに標的化され得るか、または内皮細胞上に見出されるレセプター、好ましくは、内皮細胞上に独占的に見出されるレセプターに標的化され得る。
【0074】
その送達システムは、好ましくは、トランスジェニック動物の作製に適している。例えば、送達システムは、配偶子、接合子、または胚幹細胞に標的化され得る。
【0075】
本発明のベクターおよび送達システムは、細胞をトランスフェクトして、本発明に従う細胞を生成するために使用され得る。トランスフェクションは、インビボまたはエキソビボで生じ得る。
【0076】
上記で使用される用語「生物学的組織」には、細胞、組織、および臓器の集団(collection)が含まれる。従って、その定義には、例えば、線維芽細胞、角膜、神経組織、心臓、肝臓、または腎臓が含まれる。
【0077】
本発明の第2および第3の局面が「動物」を提供する場合、その動物は、好ましくは、異種移植のための臓器および/または本発明の細胞(例えば、異種移植片レシピエントへの前異種移植片投与のための細胞)の作製に適切である。好ましくは、その動物は哺乳動物であり、そしてより好ましくは、それはトランスジェニックブタまたはトランスジェニックヒツジである。
【0078】
その動物は、それがトランスジェニック生物学的組織を含む(すなわち、遺伝子治療によって処置される)ように、生存している間に処置され得る。好ましくは、生存動物は、トランスジェニック動物を産生するために、本発明に従うベクターでトランスフェクトされる。例えば、本発明に従うベクターは、異種移植のために適切な生物学的組織を産生するためにブタの心臓に特異的に送達され得る。
【0079】
あるいは、動物は、トランスジェニック動物として生まれ得る。このようなトランスジェニック動物を作製するための多くの適切なアプローチが、当該分野で公知である(例えば、参考文献18、19、20)。例えば、DNAのマイクロインジェクションによる接合子または初期胚の直接的操作は、周知であり、胚幹細胞のような多能性の細胞のインビトロ操作も同様である。初期胚のレトロウイルス感染は、種々の種で成功していることが証明されており、そして帯を含まない(zona−free)卵のアデノウイルス感染が報告されている。ヒツジの核移入による遺伝子導入およびクローニングがまた記載されている(例えば、WO97/07668)。
【0080】
本発明が、生物学的組織を異種移植に適切にするためのプロセスを提供する場合、その生物学的組織は、インビボまたはエキソビボのいずれかでそのようにされ得る。例えば、動物の臓器は、本発明のベクターでインビボでトランスフェクトされ得るか、または臓器は、移植前にエキソビボでもしくは移植後にインビボでトランスフェクトされ得る。
本発明は、例えば以下の項目を提供する。
(項目1) レシピエントにおいて異種反応性T細胞の活性化を防止するために同時刺激シグナル2の送達を遮断することによって、異種移植された臓器のT細胞媒介拒絶を阻害し得る、生物学的試薬。
(項目2) 異種移植された臓器のT細胞媒介拒絶を阻害するための方法であって、レシピエントにおける異種反応性T細胞の活性化を防止するために、同時刺激シグナル2の送達を遮断する工程を包含する、方法。
(項目3) 異種ドナー生物由来の可溶性形態のCTLA−4タンパク質を、前記レシピエントに投与することを包含する、請求項2に記載の方法。
(項目4) 前記可溶性タンパク質が、ヒトCγ1配列に融合したブタCTLA−4の細胞外ドメインを含む、請求項3に記載の方法。
(項目5) 医薬として使用するための、可溶性形態の異種CTLA−4。
(項目6) 配列番号1のアミノ酸配列を含む、タンパク質。
(項目7) 請求項6に記載のタンパク質をコードする、核酸。
(項目8) 前記試薬が、CTLA−4に結合し得る膜結合タンパク質である、請求項1に記載の生物学的試薬。
(項目9) CTLA−4に特異的な一本鎖抗体を含む、請求項8に記載のタンパク質。
(項目10) 請求項8または請求項9に記載のタンパク質をコードする、核酸。
(項目11) その表面に請求項8または請求項9に記載のタンパク質を発現する、細胞。
(項目12) 請求項11に記載の細胞を含む、生物学的組織。
(項目13) 請求項11に記載の細胞および/または請求項12に記載の生物学的組織を含む、動物。
(項目14) ドナー動物から異種レシピエント動物へ、請求項12に記載の生物学的組織を移植する工程を包含する、移植法。
(項目15) 生物学的組織を異種移植に適切にするためのプロセスであって、該生物学的組織が請求項8または請求項9に記載のタンパク質をその細胞表面上に発現するように、該生物学的組織を処置する工程を包含する、プロセス。
(項目16) 医薬として使用するための、請求項8もしくは請求項9に記載のタンパク質、または請求項10に記載の核酸。
(項目17) 異種移植片レシピエントまたはドナーに投与するための処方物の調製における、請求項8もしくは請求項9に記載のタンパク質、または請求項10に記載の核酸の、使用。
(項目18) 前記試薬が、異なる生物のMHCクラスIIをその表面上に発現する細胞である、請求項1に記載の生物学的試薬。
(項目19) 前記細胞が、ヒトMHCクラスIIをその表面に発現するブタ細胞である、請求項18に記載の細胞。
(項目20) 前記細胞が、その表面上にB7を発現しない、請求項18または請求項19に記載の細胞。
(項目21) 前記細胞が、トランスフェクトされた未成熟樹状細胞である、請求項18または請求項19に記載の細胞。
(項目22) 請求項18、19、20、または21のいずれか1項に記載の細胞を含む、生物学的組織。
(項目23) 請求項22に記載の生物学的組織を含む、動物。
(項目24) ドナー動物から異種レシピエント動物へ、請求項22に記載の生物学的組織を移植する工程を包含する、移植法。
(項目25) 生物学的組織を異種移植に適切にするためのプロセスであって、該生物学的組織がその細胞の表面上に異種MHCクラスIIを発現するように、該生物学的組織を処置する工程を包含する、プロセス。
(項目26) 医薬として使用するための、請求項18、19、20、または21のいずれか1項に記載の細胞。
(項目27) 異種移植レシピエントに投与するための処方物の製造における、請求項22に記載の生物学的組織の使用。
(項目28) 異種移植ドナーに投与するための処方物の調製における、異種MHCクラスIIまたは異種MHCクラスIIをコードする核酸の、使用。
【図面の簡単な説明】
【0081】
本発明は、添付の図面を参照して記載される。
【図1】図1は、本発明の3つの局面を例示する。「X」は異種(xenogeneic)細胞(または、間接的活性化経路においては、異種抗原(xenoantigen)提示レシピエントAPC)を示し、「T」はレシピエントT細胞を示す。実施態様Iにおいて、同時刺激シグナル2の送達は、CTLA−4の可溶性形態を用いることによって妨げられる。実施態様IIにおいて、抗CTLA−4を使用して、シグナル2を拮抗する。実施態様IIIにおいて、XはレシピエントMHC−IIを発現するが、B7を発現しない。
【図2】図2は、pCTLA−4のアミノ酸配列(配列番号1)を示す。以下の接合部を「*」によって例示する:シグナルペプチド/細胞外ドメイン;細胞外ドメイン/膜貫通ドメイン;膜貫通ドメイン/細胞質ドメイン。ヒト配列とウシ配列とアラインメントもまた示される。pCTLA4との相同性は以下のとおりである:
【化100】
【図3−1】図3は、類似のアラインメントを示すが、ヌクレオチドレベルにおいてである。相同性は以下のとおりである:
【化101】
【図3−2】図3は、類似のアラインメントを示すが、ヌクレオチドレベルにおいてである。相同性は以下のとおりである:
【化102】
【図4】図4は、pCTLA4−Ig構築物のアミノ酸配列を示す。下線を付した配列は、可撓性リンカーGGSGGAAを示し、これはまた、pCTLA4ドメインとIgG1ドメインとの間の接合部を示す。
【図5】図5は、ヒトB7(下の2つの線)またはブタB7(上の2つの線)のいずれかでトランスフェクトしたヒト線維芽細胞に結合するhCTLA4−Ig(白丸および白四角)およびpCTLA4−Ig(白菱形および白三角)のフローサイトメトリー分析の結果を示す。
【図6】図6は、ヒトB7(白四角および白丸)またはブタB7(白菱形および白三角)によって同時刺激した場合の、hCTLA4−Ig(白四角および白菱形)と比較した、pCTLA4−Ig(白丸および白三角)による増殖の選択的阻害を示す。
【図7】図7は、MHC−クラスIIを発現するヒト細胞(7A)またはブタ細胞(7B)を5日間混合した白血球反応における刺激剤として使用した場合の、hCTLA4−Ig(白四角)またはpCTLA4−Ig(白菱形)によるヒトCD4+T細胞増殖の阻害を示す。
【図8】図8は、抗ヒトCTLA−4 sFvのヌクレオチド配列を示す。
【図9】図9は、抗ヒトCTLA−4 sFvの推測タンパク質配列を示す。図9に示したセリン−グリシンリンカーによって重鎖および軽鎖が連結される。
【図10−1】図10は、4つの抗マウスCTLA−4 sFvのヌクレオチド配列を示す。
【図10−2】図10は、4つの抗マウスCTLA−4 sFvのヌクレオチド配列を示す。
【図11】図11は、4つの抗マウスCTLA−4 sFvの推測タンパク質配列を示す。図11に示したセリン−グリシンリンカーによって重鎖および軽鎖が連結される。
【図12】図12は、CTLA−4特異的sFVの可溶性Ig融合物をコードする構築物を示す。
【図13】図13は、抗hCTLA−4 sFv(白四角)またはコントロールsFv(白丸)のいずれかを発現する細胞によるT細胞増殖の阻害を示す。
【図14】図14は、抗CTLA−4 sFvの膜結合形態をコードする構築物を示す。
【図15】図15は、ヒトCTLA−4のヌクレオチド配列(A)およびアミノ酸配列(B)を示す。開始コドンに下線を付す。−21位では、その配列はGenBank配列L15006とは異なり、そして110位では、その配列はL15006およびM74363の両方と異なる。
【図16】図16は、クローニングしたヒトCD8αの配列を示す。これは、231位(T→G)、244位(A→G)、266位(T→C)、および437位(T→C)においてGenBank配列と異なる。
【図17】図17は、B7ネガティブまたはB7ポジティブとしてクローンを規定するための、ヒトおよびマウスのCTLA4−IgのIPECへの結合を示す。
【図18】図18は、HLA−DR特異的mAb L243のトランスフェクト細胞に対する結合を示す。
【図19】図19は、HLA−DR−1でトランスフェクトしたIPECに対するヒトT細胞による増殖を示す。
【図20】図20は、HLA−DR−1でトランスフェクトした細胞をインキュベーション下2日後の、ヒトT細胞増殖アッセイの結果を示す。X軸は、増殖アッセイの第2工程において使用した刺激細胞を示す。黒棒は、B7ポジティブトランスフェクタントでプライムしたCD4 T細胞での結果を示す;白棒(見るのは困難である)は、B7ネガティブトランスフェクタントでプライムした後の結果を示す。第1のグラフは、3日で採取した細胞での結果を示す;第2のグラフは、6日目での採取物からの結果を示す。
【図21】図21は、IPECに対して惹起された、APC依存性HLA−DR−1拘束T細胞株の増殖を示す。刺激剤集団を、X軸上に示す。
【発明を実施するための形態】
【0082】
(実施態様の説明)
(可溶性ブタCTLA−4)
ブタCTLA−4(「pCTLA4」)を、PHA活性化ブタT細胞からクローニングした。RNAを、標準的な技術を用いて調製し、そしてpCTLA−4を、以下のプライマーを用いてPCRによって増幅した:
【0083】
【数1】
得られた700bpのフラグメントを、EcoRI/HindIII消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を図3に示す。この図はまた、ヒトおよびウシのCTLA−4配列との比較を示す。
【0084】
pCTLA4の予測したアミノ酸配列を、ヒトおよびウシと比較しながら図2に示す。重要なことは、残基97における推定アミノ酸の差異であり、この残基はB7結合において重要であり、保存されたヘキサペプチドモチーフMYPPPYの一部である。pCTLA4において、残基97はロイシンである(LYPPPYとなる)のに対し、(ロイシンもまた、ウシCD28において見出さされているが(21))他の種はメチオニンを有する。この重要なアミノ酸差異は、pCTLA4のヒトおよびブタB7への有利に異なる結合の重要な鍵であると考えられる。
【0085】
pCTLA4の細胞外ドメインを、上記の5’プライマーおよび以下のプライマーを用いて増幅した:
【0086】
【数2】
これを484位から増幅し、リンカーGGSGGAA配列(下線を付した)をコードする18塩基対のセグメントを導入し、そしてPstI部位(太字)を導入してヒトIgG1のヒンジ領域へのインフレーム連結を可能にした。得られた500bpのフラグメントを、HindIII/PstI消化したpBluescript−IgG1(ゲノムDNAコードイントロン性配列ならびにPstI/Not1部位の間のヒトIgG1のヒンジ、CH2、CH3および3’非翻訳エキソンを含む)にサブクローニングした。得られた可溶性pCTLA4−Igのアミノ酸配列を図4に示す。
【0087】
(pCTLA4−Igの発現)
キメラpCTLA4−Ig DNA配列を、HindIII/BstXIフラグメントとしてpBluescriptから遊離させ、そして発現ベクターpHOOK−3TM(Invitrogen)にサブクローニングした。これを使用して、標準的なリン酸カルシウム沈殿を用いてDAP.3細胞またはCHO−K1細胞をトランスフェクトした。G418耐性細胞を、CaptureTecTMシステムを用いて分離した。これらのトランスフェクト細胞を、組織培養フラスコ中でコンフルエントになるまで増殖させ、コンフルエントになった時点で、培地を交換し、そして細胞を培養物中でさらに3日間維持した。この段階で、培地を採取し、そしてプロテインGカラムを通して灌流した。pCTLA−4−Igを酸性条件下で溶出させた。この溶出したタンパク質の濃度を、ヒトIgG1に対する抗体および標準的なヒトIgG1骨髄腫タンパク質を用いるELISAを用いて計算した。
【0088】
pCTLA4−Igの結合特徴を、ヒトB7−1またはブタB7−2のいずれかでトランスフェクトしたヒト線維芽細胞を用いるフローサイトメトリー分析を使用して、ヒトCTLA4−Igの結合特徴と比較した。これらの実験に関して、ブタおよびヒトのCTLA4−Igの濃度は、ELISAによってアッセイしたとき、等しかった。図5に例示したように、ヒトおよびブタCTLA4−Igは、ブタB7を発現するヒト細胞において類似の結合特徴を有するようであった。しかし、ヒトCTLA4−Igとは異なり、pCTLA4−IgはヒトB7には結合しなかった。このことは、pCTLA4−IgがブタB7に優先的に結合し、そして種特異的試薬として有用であることを意味する。
【0089】
pCTLA4−Igを使用して、種々の刺激剤に対するヒトT細胞増殖応答を阻害した。これらのアッセイにおいて、T細胞応答の同時刺激は、プロフェッショナルAPCにおいてトランスフェクションまたは天然のいずれかによって発現された、ブタまたはヒトのいずれかのB7によって提供された。これらの実験を、図6および7において実証する。
【0090】
トランスフェクトした線維芽細胞刺激剤(HLAクラスIIおよびヒトB7またはブタB7のいずれかを発現する)を用いる実験について、hCTLA4−Igは全ての増殖応答を阻害した(図6、白四角および白菱形)。対照的に、pCTLA4−Igのみが、刺激剤がブタB7を発現した場合の応答を完全に阻害した(白三角);ヒトB7を発現する細胞に対する増殖応答は、最小限に影響されたのみであった(白丸)。
【0091】
類似の実験において、pCTLA4−Igは、MHCクラスIIおよびヒトB7を発現するヒト細胞への増殖応答に対する有意な阻害効果を有しなかったが、ブタ刺激剤に対する応答は阻害した(図7)。
【0092】
これらの結果は、T細胞の同時刺激シグナルがブタB7によって提供された場合のpCTLA4−Igの有効な阻害特性を強調する。同時刺激がヒトB7によって媒介される場合にT細胞増殖を防止できないことはまた、種特異的作用を実証する。pCTAL4−Igは、ブタB7に対して種特異的結合を示し、ブタB7の機能的効果の阻害を示すが、ヒトB7に対しては種特異的結合を示さず、その機能的効果の阻害も示さないと、結論付け得る。
【0093】
(pCTLA−4−Igの特性)
pCTLA4−Igのヒトおよびブタの両方のB7ファミリー分子に対する結合特徴は、例えば、以下の試験を用いて、hCTLA4−Igの結合特徴と比較され得る:
(i)ブタおよびヒトのAPCに対する結合、およびブタまたはヒトのB7を発現するトランスフェクタントに対する結合のフローサイトメトリー分析(上記を参照のこと)
(ii)BiacoreTMを用いる結合の定量的特徴づけ
(iii)ヒト抗ブタおよびヒト同種異系混合したリンパ球培養物に対するCTLA4−Igの効果の機能的分析
(iv)pCTLA4−IgのB6マウスへの移植後のブタ島異種移植生存を延長する能力の機能的評価。
【0094】
(CTLA−4に結合する膜結合型タンパク質)
1012の半合成可変配列を含むファージディスプレイライブラリーを、ヒトまたはマウスのCTLA4−IgおよびコントロールヒトIgG1骨髄腫タンパク質を用いてスクリーニングした。4回のスクリーニング後にヒトCTLA4−Igタンパク質への差示的な結合を提示するファージからsFvを単離し、そして標準的な技術を用いて精製した。そのヌクレオチド配列および推測したアミノ酸配列を、図8、9、10、および11に示す。
【0095】
sFvを、ヌクレオチド配列に基づく特異的プライマーを用いてPCRによって増幅した。プライマーの遠位部分は、pHOOK1内の配列に基づいた。5’プライマーはApaI部位を含み、そして3’プライマーはSalI部位を含んだ。これらは両方とも、独特であると予測された。得られたsFvを、配列決定のためにpBluescriptにサブクローニングして、忠実な増幅を決定した。次いでApaI/SalIフラグメントを、pHOOK1にサブクローニングした。ここでは、ApaI/SalIフラグメントは、上流ではマウスIgκ鎖からのインフレームのシグナル配列および血球凝集素Aエピトープ配列に、そして下流では2つのインフレームのmyc配列およびPDGFレセプターからの膜貫通配列に隣接する。
【0096】
pHOOK1からのmyc配列を、NotI部位(下線を付した)を含む以下の5’プライマー:
【0097】
【数3】
およびPstI部位(下線を付した)を導入した以下の3’プライマー:
【0098】
【数4】
を用いてPCRにより増幅した。得られた113塩基対のフラグメントを、NotI/PstI消化したpBluescriptにサブクローニングした。
【0099】
sFvを、pHOOK1からEcoRI/NotIフラグメントとして遊離させ、そしてEcoRI/PstI消化したpBluescript−IgG1に、NotI/PstI PCR産物とともに連結した(図12)。この構築物は、CTLA−4特異的sFvの可溶性Ig融合物をコードする。真核生物細胞における発現のために、この構築物を、pHOOK3にHindIII/BstXIフラグメントとしてサブクローニングした。
【0100】
sFvの細胞表面発現を確認するために、pHOOK構築物を、既にHLA−DR分子およびヒトB7を発現している細胞にトランスフェクトした。G418またはミコフェノール酸(使用したベクターに依存する)に耐性である細胞を、培養において増殖させた。その細胞表面上で抗CTLA4−sFv構築物を発現する細胞を、hCTLA4−Igを使用してフローサイトメトリー分析によって同定した。これらの細胞を、限定希釈によってクローニングし、そして5日間の培養におけるT細胞の増殖の刺激剤として使用した。1つの実験の結果を図13に示す。抗hCTLA4 sFvを発現する細胞は、T細胞増殖を刺激しないが(白四角)、一方コントロールsFvを発現する細胞は、正常細胞と同じ方法で増殖を刺激した(白丸)。
【0101】
異なる実験において、図12に示した構築物のEcoRI/SalIフラグメントを、ヒトCD8の膜貫通ドメインおよび細胞質ドメイン(pBluescript−hCD8由来のSalI/BamHIフラグメントとして単離した)とともに、EcoRI/BamHI消化したpBluescriptに同時連結した(図14)。
【0102】
pBluescriptからのEcoRI/BamHIフラグメントを、発現ベクターpHβApr−1−neoまたはシスターベクターpHβApr−1−gptにサブクローニングした。これらを、既にHLA−DR分子およびB7を発現している細胞にトランスフェクトし、そしてpHOOK構築物について上記のように選択した。
【0103】
(膜結合型CTLA−4構築物)
活性化T細胞によるCTLA−4の発現は、一過性であるのみであり、それゆえ、抗CTLA4−sFvの機能的特徴を試験するために、ヒトまたはマウスのCTLA4の細胞外ドメインをコードするDNA配列、ならびにヒトCD8の膜貫通/細胞質配列を含むキメラ構築物を作製した。これらの構築物を発現する細胞を、抗CTLA4−sFvタンパク質の研究のために使用し得る。
【0104】
PHA活性化ヒトT細胞由来のRNAを、標準的な技術を用いて調製した。hCTLA4を、以下のプライマーを用いてPCR増幅した:
HindIII部位を導入する(5’プライマー)
【0105】
【数5】
EcoRI部位を導入する(3’プライマー)
【0106】
【数6】
得られたフラグメントを、HindIII/EcoRI消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を、図15に示す。これは、ヒトCTLA−4についてのGenBankリスト配列とは1つの塩基(439位)が異なる。hCTLA4の推定アミノ酸配列もまた示す。
【0107】
hCTLA−4の細胞外ドメインを、上記の5’プライマーおよび以下のプライマーを用いて増幅した:
【0108】
【数7】
これは、457位から増幅され、そして可撓性GGSGGアミノ酸リンカーをコードする15塩基のセグメント(下線を付した)を、独特のSalI部位(強調表示)とともに含んでいだ。得られたフラグメントを、HindIII/SalI消化したpBluescriptにサブクローニングし、そして配列決定した。
【0109】
hCD8を、得られたT細胞から以下のプライマーを用いてPCR増幅した:
HindIII部位を導入する(5’プライマー)
【0110】
【数8】
EcoRI部位を導入する(3’プライマー)
【0111】
【数9】
得られたフラグメントを、HindIII/EcoRI消化したpBluescriptにサブクローニングし、そしてそのヌクレオチド配列を、標準的なT3プライマーおよびT7プライマーを用いて決定した。単一のクローンの配列を、図16に示す。このクローンは、GenBankに登録された配列とは4つの位置で異なっていたが、これらはいずれも、続いて増幅した領域内では存在しなかった。
【0112】
hCD8の膜貫通(TM)ドメインおよび細胞質(C)ドメインを、上記の3’プライマーおよび以下の5’プライマーを用いて増幅した:
【0113】
【数10】
これは、532位から増幅され、そして可撓性GGSGGアミノ酸リンカーをコードする15塩基のセグメント(下線を付した)を、独特のSalI部位(強調表示)とともに含んでいた。得られたフラグメントを、HindIII/SalI消化したpBluescriptにサブクローニングし、そしてpBluescript−hCD8と称した。
【0114】
ヒトCTLA−4の細胞外ドメインを、pBluescriptからEcoRI/SalIフラグメントとして切り出し、そしてCD8のTM−ICドメインを、SalI/BamHIフラグメントとして切り出した。これらをともに、EcoRI/BamHI消化したpBluescriptに再び連結した。次いで、全CTLA−4−CD8キメラを単一のEcoRIフラグメントとして取り出し、そしてヒトT細胞白血病株J6への発現のための多数の発現ベクターにサブクローニングした。
【0115】
(細胞表面抗CTLA4タンパク質の特性)
細胞表面抗CTLA−4タンパク質は、以下の機能性試験によってさらに特徴付けられ得る:
(i)膜結合抗CTLA4−sFv−CD8タンパク質を発現する細胞と、可溶性ヒトCTLA4−Igを発現する細胞との間の相互作用のフローサイトメトリー評価
(ii)BiacoreTMを用いる、可溶性抗CTLA4−sFv−Ig融合タンパク質と可溶性ヒトCTLA4−Igとの間の相互作用の定量的評価
(iii)抗CTLA4−sFv−CD8融合タンパク質を発現するJ6トランスフェクタントへの可溶性ヒトCTLA4−Igの結合から生じるシグナル伝達事象についての分析
(iv)同種異系混合リンパ球応答における刺激が、HLA−DRポジティブ、B7ポジティブ、抗CTLA4−sFv−CD8ポジティブ細胞株によって提供される場合のT細胞応答(例えば、増殖、サイトカイン産生、アネルギー誘導)の分析。
【0116】
(マウスMHCクラスIIを発現するB7ネガティブブタ細胞)
50のクローニングした不死化した大動脈内皮細胞(PAEC)を、PAECの単層から、pZipSVU19 DNA(22)を用いる核内マイクロインジェクションにより生成した。不死化細胞(IPEC)から、B7ネガティブクローンを、hCTLA4−IgおよびmCTLA4−Igを用いてフローサイトメトリースクリーニングによって同定した(図17を参照のこと)。次いで、これらを、プラスミド発現ベクターpcExV1−gptおよびpHβApr−1neo中のHLA−DRAおよびDRB1*0101をコードするcDNAでトランスフェクトし、そして細胞を、MXHおよびG418での選択下に置いた。比較のために、B7−ポジティブIPECコントロールを同様に生成した(4)。
【0117】
マウスMHCクラスII分子I−Abを発現するIPECトランスフェクタントの別のシリーズもまた、マウスにおける実験のために生成した。
【0118】
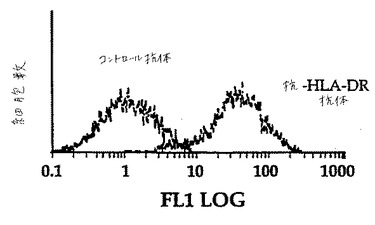
トランスフェクトIPEC細胞におけるMHCクラスIIの表面発現を、モノクローナル抗体L243(HLA−DRに特異的)(図18)またはM5−114(マウスMHCクラスIIに特異的)を用いて検出した。MHCクラスIIポジティブ細胞は、限界希釈によるクローニングの前に数回の蛍光活性化細胞ソーティングを受けた。
【0119】
トランスフェクタントの第2のバッチを、発現ベクターpCMU中のHLA−DMAおよびHLA−DMBおよびp31Ii(不変鎖)をコードするさらなるトランスフェクトcDNAを用いて正確に同じ方法で調製した。
【0120】
(MHCクラスII発現細胞による同種異系T細胞におけるアネルギー誘導)
ヒトT細胞を、標準的なプロトコルを使用して精製した(3)。一次増幅アッセイのために、ガラス繊維フィルターへの採取の16時間前に1マイクロCi3H−チミジンの添加の前に、T細胞を、一定数の照射した刺激細胞(stimulator cell)とともに5日間インキュベートした。このフィルターを、シンチレーションカウンターにおいて読み取った。
【0121】
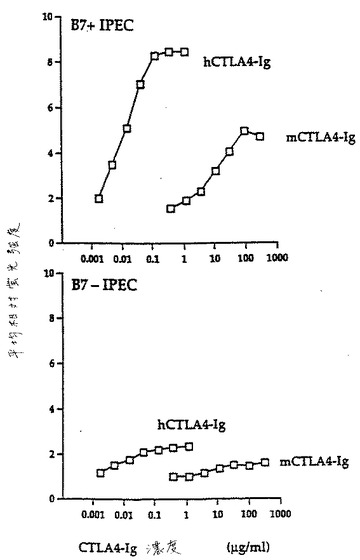
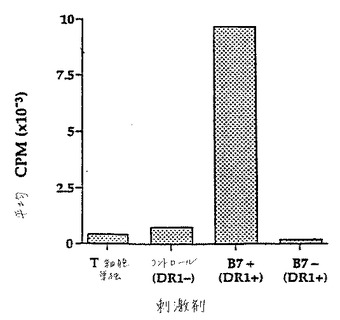
B7ポジティブIPECは、有意な抗DR1特異的増殖応答を生じ、一方B7ネガティブIPECは任意の増殖応答を開始しなかった(図19)。
【0122】
2工程のアネルギー誘導アッセイを、標準的なプロトコルによって確立した(23)。一次寛容性誘導工程において、B7ポジティブIPECとともにインキュベートしたT細胞を、プライムされた二次免疫応答(3日で最大)の動力学を用いる二次工程における抗DR1増殖応答をマウントした。しかし、一次工程においてB7ネガティブIPECとともにインキュベートしたT細胞は、DR1に対して寛容になり、そしてB7ポジティブIPECにおいて発現されたDR1に対する続く曝露の際に、応答をマウントしなかった(図20)。
【0123】
(DR1を発現するブタ細胞によるDR1拘束T細胞におけるアネルギー誘導)
DR1を発現する個体由来のCD4+T細胞を、標準的な手順に従って精製した。一次増殖アッセイにおいて、それらは、HLA−DR1でトランスフェクトしたB7ポジティブIPECに増殖した。このことは、DR1が、ブタペプチド特異的T細胞についての制限エレメントとして機能し得ることを示す。B7ポジティブおよびB7ネガティブDR1+トランスフェクタントに対する増殖応答を比較するアッセイを行っている。
【0124】
2工程のアネルギー導入アッセイもまた、DR1でトランスフェクトしたB7ネガティブブタ細胞がHLA−DR拘束ヒトT細胞においてアネルギーを誘導することを実証するために行い得る。
【0125】
(HLA−DRにおける提示についてのプロフェッショナルヒトAPCによってプロセスされたブタペプチドと、HLA−DRでトランスフェクトしたIPECのMHCクラスIIにおいて提示されるものとの間の重複)
野生型IPECに対するヒトT細胞株を、ヒトPBMCから得た。この株の増殖応答は、ヒトAPCの存在に依存し、そしてHLA−DRに対する抗体によって阻害可能であり、これらのことは、この株が、ヒトAPCによって提示される、プロセスされたブタ異種抗原(xenoantigen)に対する間接的な特異性を有した。
【0126】
少なくともいくつかのプロセスされたブタペプチドがプロフェッショナルヒトAPCによって間接的に提示されることを意味する、B7ポジティブHLA−DR1でトランスフェクトされたIPECに対して増殖したこの株(図21)もまた、トランスフェクトされたブタ細胞によって提示された。
【0127】
(ブタ島対マウスモデルにおける研究)
インビボでは、ブタ膵島細胞は、ストレプトゾトシン(streptozotocin)処置した糖尿病マウスの腎臓被膜下に移植され得る。島異種移植片(非血管性である)は、T細胞によって単に拒絶される。ブタ島を、末梢麻酔下でブタの膵臓から調製し、そしてレシピエントにおけるその生存を、正常血糖値の維持によって評価する。マウスに、ブタ島の移植前に、B7ネガティブのI−Abを発現するIPECを静脈内注射する。このストラテジーを、本発明の他の局面と組み合わせて、T細胞認識の直接経路を寛容化し、直接経路を介する拒絶が生じないことを確実にする。特定のストラテジーが特異的T細胞寛容性を誘導したか否かを評価するために、島保有腎臓の腎摘出を、同一のブタ島または第三者のブタ島の再移植の前に行う(生存する腎臓の被膜下)。
【0128】
本発明は、例示のみのために上に記載され、そして本発明の範囲および精神内に残る間は改変がなされ得ることが理解される。
【0129】
(参考文献(その内容を完全に援用する)
【0130】
【表1】
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】

【図2】

【図3−1】

【図3−2】

【図4】

【図5】
【図6】
【図7】
【図8】

【図9】

【図10−1】

【図10−2】
【図11】
【図12】
【図13】
【図14】
【図15】

【図16】

【図17】
【図18】
【図19】
【図20】
【図21】
【図2】
【図3−1】
【図3−2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10−1】
【図10−2】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公開番号】特開2011−144205(P2011−144205A)
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−96600(P2011−96600)
【出願日】平成23年4月22日(2011.4.22)
【分割の表示】特願2000−547221(P2000−547221)の分割
【原出願日】平成11年4月30日(1999.4.30)
【出願人】(505167543)インペリアル・イノベ−ションズ・リミテッド (23)
【Fターム(参考)】
【公開日】平成23年7月28日(2011.7.28)
【国際特許分類】
【出願番号】特願2011−96600(P2011−96600)
【出願日】平成23年4月22日(2011.4.22)
【分割の表示】特願2000−547221(P2000−547221)の分割
【原出願日】平成11年4月30日(1999.4.30)
【出願人】(505167543)インペリアル・イノベ−ションズ・リミテッド (23)
【Fターム(参考)】
[ Back to top ]