
TALエフェクターに媒介されるDNA修飾
遺伝子標的化(たとえば、転写活性化因子様エフェクターヌクレアーゼ、「TALEN」を用いた遺伝子標的化)に関連する材料および方法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、すべてその全体で本明細書中に参考として組み込まれている、2009年12月10日に出願の米国仮出願第61/285,324号、2010年6月7日に出願の米国仮出願第61/352,108号、および2010年7月22日に出願の米国仮出願第61/366,685号の優先権を主張するものである。
【0002】
政府出資の研究に関する表明
本発明は、国立科学財団(National Science Foundation)によって授与された助成金第0820831号および第0504304号の下の政府支援で行った。政府が本発明の特定の権利を有する。
【0003】
本発明は、遺伝子標的化の方法、特に転写活性化因子様(TAL)エフェクター配列の使用が含まれる方法に関する。
【背景技術】
【0004】
相同組換え(遺伝子標的化)を介して染色体を改変する能力は、生物学者の念願の目標であった。たとえば、植物では、遺伝子標的化は、植物遺伝子の機能の識別を助けて、作物の改善の新しい可能性を開き得る。たとえば、遺伝子標的化を用いて、代謝経路を再編成するために必要な遺伝手術を実施して、変更された油もしくは炭水化物プロフィールを有する種子、増強された栄養品質を有する食品、または疾患およびストレスに対して増加した耐性を有する植物を含めた高価値の作物を作製することが可能である。動物(たとえば哺乳動物)では、遺伝子標的化を疾患の処置に使用し得る。たとえば、遺伝子標的化は、様々な種類の突然変異が原因で欠損性である遺伝子中の補正を設計するために使用し得る。そのような遺伝子標的化の効率的な方法は、達成が困難であった。
【0005】
キサントモナス属中の植物病原細菌のTALエフェクターは、宿主DNAと結合してエフェクターに特異的な宿主遺伝子を活性化することによって、疾患において重要な役割を果たす、または防御を始動する(たとえば、Guら(2005)Nature、435:1122、Yangら(2006)Proc.Natl.Acad.Sci.USA、103:10503、Kayら(2007)Science、318:648、Sugioら(2007)Proc.Natl.Acad.Sci.USA、104:10720、およびRomerら(2007)Science、318:645を参照)。特異性は、エフェクター可変性の、不完全な、典型的には34個のアミノ酸の反復に依存する(Schornackら(2006)J.Plant Physiol.、163:256)。多型は主に反復位置12および13に存在し、本明細書中ではこれを反復可変性二残基(RVD、repeat variable−diresidue)と呼ぶ。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、TALエフェクターのRVDとその標的部位中のヌクレオチドとが一部の縮重を伴って1個のRVD対1個のヌクレオチドの直接的な線形様式で対応しており、明らかなコンテキスト依存性がないことに部分的に基づく。この驚くべき発見は、新しい標的特異的TALエフェクターの標的部位の予測を可能にするタンパク質−DNAの認識の新規機構を表す。本明細書中に記載のように、これらのタンパク質は、ゲノム操作における相同組換え(たとえば、生物燃料または植物におけるバイオ再生可能物質に有用な特色を付加または増強させるため)を促進することができる標的化キメラヌクレアーゼとして、研究および生命工学において有用であり得る。また、これらのタンパク質は、たとえば、非限定的な例として、転写因子および特に病原体(たとえばウイルス)に対する治療剤などの非常に高いレベルの特異性を要する治療的応用としても有用であり得る。
【課題を解決するための手段】
【0007】
一態様では、本発明は、(a)標的DNA配列を含有する細胞を提供するステップと、(b)TALエフェクター−DNA修飾酵素が細胞またはその子孫中の特定のヌクレオチド配列内にあるまたはそれに隣接する標的DNAを修飾するように、細胞内に、(i)二本鎖DNAを修飾することができるDNA修飾酵素ドメイン、および(ii)組み合わせて標的DNA配列中の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメインを含む、転写活性化因子様(TAL)エフェクター−DNA修飾酵素を導入するステップとを含む、細胞の遺伝物質を改変する方法を特長とする。本方法は、標的DNA配列と核酸との間の相同組換えが起こるように、標的DNA配列の少なくとも一部分に相同的な配列を含む核酸を細胞に提供することをさらに含むことができる。細胞は、真核細胞、哺乳動物細胞、植物細胞、または原核細胞であることができる。標的DNAは染色体DNAであることができる。導入ステップは、TALエフェクター−DNA修飾酵素をコードしているベクターを用いて細胞を形質移入すること、TALエフェクター−DNA修飾酵素を細胞内にタンパク質として機械的に注入すること、TALエフェクター−DNA修飾酵素を細胞内にタンパク質として細菌III型分泌系によって送達すること、またはTALエフェクター−DNA修飾酵素を細胞内にタンパク質として電気穿孔によって導入することを含むことができる。DNA修飾酵素は、エンドヌクレアーゼ(たとえばFokIなどのII型制限エンドヌクレアーゼ)であることができる。
【0008】
標的DNA内の特定のヌクレオチド配列と結合するTALエフェクタードメインは、10個以上のDNA結合反復、好ましくは15個以上のDNA結合反復を含むことができる。それぞれのDNA結合反復には、標的DNA中の塩基対の認識配列を決定する反復可変性二残基(RVD)が含まれることができ、それぞれのDNA結合反復は、標的DNA中の1つの塩基対配列の認識を司っており、RVDは、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのHG、Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含む。それぞれのDNA結合反復は、標的DNA中の塩基対の認識を決定するRVD配列を含むことができ、それぞれのDNA結合反復は、標的DNA中の1つの塩基対配列の認識を司っており、RVDは、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのHG、Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、およびTを認識するためのIGのうちの1つまたは複数を含む。
【0009】
別の態様では、本発明は、(1)開始プラスミドをPspXIで直鎖状にするステップであって、開始プラスミドが、選択されたヌクレオチド配列の第1ヌクレオチドに特異的な反復可変性二残基(RVD)を有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含み、第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有するステップと、(2)開始プラスミドのPspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチド(複数可)に特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしている、XhoI粘着末端を有するDNAモジュールをライゲーションするステップと、(3)核酸が選択されたヌクレオチド配列と結合することができるTALエフェクターをコードするまでステップ(1)および(2)を繰り返すステップとを含む、選択されたヌクレオチド配列に特異的なTALエフェクターをコードしている核酸を作製する方法を特長とする。本方法は、ライゲーション後に、PspXI部位中のDNAモジュールの配向を決定することをさらに含むことができる。本方法は、ステップ(1)および(2)を1〜30回繰り返すことを含むことができる。
【0010】
別の態様では、本発明は、(a)細胞のゲノム中の第1のヌクレオチド配列を同定するステップと、(b)(i)組み合わせて第1のユニークなヌクレオチド配列と結合する複数のDNA結合反復、および(ii)第1のヌクレオチド配列内にあるまたはそれに隣接する位置で二本鎖切断を生じるエンドヌクレアーゼを含む、TALENをコードしている核酸を合成するステップであって、それぞれのDNA結合反復が、標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っており、TALENが、以下のRVD、すなわち、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含むステップとを含む、転写活性化因子様エフェクターエンドヌクレアーゼ(TALEN)をコードしている核酸を作製する方法を特長とする。
【0011】
TALENは、以下のRVD、すなわち、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、およびTを認識するためのIGのうちの1つまたは複数を含むことができる。
【0012】
第1のヌクレオチド配列は、以下の基準のうちの少なくとも1つを満たすことができる:i)最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあること、ii)第1(5’)位置にTまたは第2位置にAを有さないこと、iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さないこと、ならびにiv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの基本組成を有すること。
【0013】
本方法は、上述の基準のうちの少なくとも1つを満たしており、15〜18bpによって分離されている、細胞のゲノム中の第1のヌクレオチド配列および第2のヌクレオチド配列を同定することを含むことができる。エンドヌクレアーゼは、第1および第2のヌクレオチド配列の間に二本鎖切断を生じることができる。
【0014】
別の実施形態では、本発明は、DNA結合ドメインが複数のDNA結合反復を含み、それぞれの反復が、標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、エンドヌクレアーゼドメインと標的DNAに特異的なTALエフェクターDNA結合ドメインとを含むTALENであって、以下のRVD、すなわち、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含むTALENを特長とする。TALENは、以下のRVD、すなわち、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、およびTを認識するためのIGのうちの1つまたは複数を含むことができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。
【0015】
さらに別の態様では、本発明は、前記TALENのアミノ酸配列が、配列番号33〜配列番号55、配列番号72、および配列番号73からなる群から選択される、エンドヌクレアーゼドメインおよびTALエフェクタードメインを含むTALENを特長とする。
【0016】
また、本発明は、それ内に遺伝子改変を導入することが所望される標的DNA配列を含む真核細胞を提供するステップと、エンドヌクレアーゼドメインと標的DNA配列と結合するTALエフェクタードメインとを含むTALENを用いて標的DNA配列内に二本鎖切断を生じるステップと、二本鎖切断が起こった細胞またはその子孫から動物を作製するステップとを含む、動物を作製する方法も特長とする。本方法は、標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、細胞内に、外因性核酸と細胞またはその子孫中の標的DNA配列との間の相同組換えが起こることを可能にする条件下で導入するステップと、相同組換えが起こった細胞またはその子孫から動物を作製するステップとをさらに含むことができる。動物は哺乳動物であることができる。遺伝子改変は、置換、挿入、または欠失を含むことができる。
【0017】
さらに別の態様では、本発明は、事前に選択された遺伝子改変をそれ内に導入することが所望される標的DNA配列を含む植物細胞を提供するステップと、エンドヌクレアーゼドメインと標的DNA配列と結合するTALエフェクタードメインとを含むTALENを用いて標的DNA配列内に二本鎖切断を生じるステップと、二本鎖切断が起こった細胞またはその子孫から植物を作製するステップとを含む、植物を作製する方法を特長とする。本方法は、標的DNA配列の少なくとも一部分に相同的な配列を含む外因性核酸を、植物細胞内に、外因性核酸と細胞またはその子孫中の標的DNA配列との間の相同組換えが起こることを可能にする条件下で導入するステップと、相同組換えが起こった細胞またはその子孫から植物を作製するステップとをさらに含むことができる。
【0018】
別の態様では、本発明は、選択されたDNA標的配列を標的としたTALエフェクターエンドヌクレアーゼをコードしている核酸を細胞内に導入するステップと、細胞内でのTALエフェクターエンドヌクレアーゼの発現を誘導するステップと、選択されたDNA標的配列が突然変異を示している細胞を同定するステップとを含む、細胞における標的化遺伝子組換えの方法を特長とする。突然変異は、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択されることができる。本方法は、細胞内にドナーDNAを導入することをさらに含むことができる。細胞は、昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞であることができる。
【0019】
別の態様では、本発明は、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませ、したがって結合部位の−1位でのTの必要性を排除することを含むステップを含む、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法を特長とする。
【0020】
別の態様では、本発明は、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、Gに対して増強された特異性を有するRVDを含有するTALエフェクターDNA結合ドメインをコードしている1つまたは複数の核酸を取り込ませることを含み、前記RVDが、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択され、*がRVDの第2位置でのギャップを示すステップを含む、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法を特長とする。
【0021】
また、本発明は、反復ドメインを含むポリペプチドを合成するステップを含む、標的DNA配列中の少なくとも1つの塩基対を選択的に認識するポリペプチドを生成する方法であって、反復ドメインが、転写活性化因子様(TAL)エフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、標的DNA中の塩基対の認識配列を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法も特長とする。さらに、本発明は、上記方法によって生成されたポリペプチド、および上記方法によって生成されたポリペプチドのコード配列を含むDNAを特長とする。また、上述のDNAと作動可能に連結されたプロモーターを含む発現カセット、および発現カセットを含む非ヒト宿主細胞も特長とされている。別の態様では、本発明は、発現カセットを含む形質転換させた非ヒト生物を特長とする。
【0022】
さらに別の態様では、反復ドメインを含むポリペプチドを構築するステップを含む、ポリペプチドによってDNA配列中の塩基対を選択的に認識する方法であって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法を特長とする。
【0023】
また、本発明は、細胞における標的遺伝子の発現を変調させる方法であって、反復ドメインを含むポリペプチドを含有する細胞が提供され、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法を特長とする。
【0024】
別の態様では、本発明は、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む、反復ドメインを含むポリペプチドを特長とする。また、本発明は、上述のポリペプチドのコード配列を含むDNAも特長とする。
【0025】
別の態様では、本発明は、塩基対が反復ドメインを含むポリペプチドによって特異的に認識されることができるように、標的DNA配列上に位置する塩基対が含まれるように改変されたDNAであって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、塩基対が、超可変領域による選択的かつ決定された認識を受けるために、(a)HDによる認識のためのC/G、(b)NIによる認識のためのA/T、(c)NGによる認識のためのT/A、(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、(e)NNによる認識のためのG/CまたはA/T、(f)IGによる認識のためのT/A、(g)Nによる認識のためのC/GまたはT/A、(h)HGによる認識のためのT/A、(i)Hによる認識のためのT/A、および(j)NKによる認識のためのG/Cからなる群から選択されるDNAを特長とする。また、上述のDNAを含むベクター、DNAを含む非ヒト宿主細胞、およびDNAを含む形質転換させた非ヒト生物も特長とされている。
【0026】
さらに別の態様では、本発明は、反復ドメインを含むポリペプチドによって選択的に認識される標的DNA配列を含むDNAを生成する方法であって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、標的DNA中の塩基対の認識配列を決定する超可変領域を含み、かつ標的DNA中の1つの塩基対配列の認識を司っており、方法が、反復単位によって認識されることができる塩基対を含むDNAを合成するステップを含み、塩基対が、(a)HDによる認識のためのC/G、(b)NIによる認識のためのA/T、(c)NGによる認識のためのT/A、(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、(e)NNによる認識のためのG/CまたはA/T、(f)IGによる認識のためのT/A、(g)Nによる認識のためのC/GまたはT/A、(h)HGによる認識のためのT/A、(i)Hによる認識のためのT/A、および(j)NKによる認識のためのG/Cからなる群から選択される方法を特長とする。
【0027】
別の態様では、本発明は、植物細胞の遺伝物質を改変する方法を特長とする。本方法には、(a)(i)植物細胞中に存在する対応する標的ヌクレオチド配列に関してヌクレオチド配列中の1つまたは複数の修飾を含み、配列特異的TALエフェクターエンドヌクレアーゼ(TALEN)の認識部位をさらに含む、修飾された標的ヌクレオチド配列を含む第1の組換え核酸、および(ii)配列特異的転写活性化因子様(TAL)エフェクターエンドヌクレアーゼをコードしているヌクレオチド配列を含む第2の組換え核酸を、植物細胞内に導入するステップと(b)植物細胞を含有する植物を作製するステップと、(c)植物またはその子孫から得られた細胞、種子、または組織を、標的ヌクレオチド配列での組換えについて分析するステップとが含まれることができる。本方法には、(iii)選択マーカーをコードしているヌクレオチド配列を含む第3の組換え核酸を植物細胞内に導入するステップと、植物またはその子孫が選択マーカーを発現するかどうかを決定するステップとがさらに含まれることができる。本方法には、植物またはその子孫を、選択マーカーの非存在についてスクリーニングするステップがさらに含まれることができる。選択マーカーをコードしているヌクレオチド配列は、片側または両側が植物細胞内在配列(たとえば、第2の配列特異的ヌクレアーゼの切断部位での配列)に類似または同一の配列によって隣接されていても、されていないくてもよい。選択マーカーをコードしているヌクレオチド配列は、両側が配列特異的リコンビナーゼの認識部位によって隣接されていてもよい。本方法には、植物を異系交配させるステップがさらに含まれることができ、異系交配の子孫を選択マーカーの非存在についてスクリーニングするステップを用いるまたは用いない。第1および第2の組換え核酸は、植物細胞内に同時に導入することができる。組換え核酸の一方または両方を、導入ステップの前に直鎖状にすることができる。第1および第2の組換え核酸は、同じ構築体中に存在していてよい。
【0028】
別の態様では、本発明は、細胞の遺伝物質を改変する別の方法を特長とする。本方法には、それ内で相同組換えが起こることが所望される染色体標的DNA配列を含有する初代細胞を提供するステップと、二本鎖DNAを切断することができるエンドヌクレアーゼドメイン、および組み合わせて細胞中の標的DNA内の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメインを含む、TALENを提供するステップと、TALENが細胞中の標的DNA配列内にあるまたはそれに隣接するヌクレオチド配列の両方の鎖を切断するように、標的DNA配列とTALENとを細胞中で接触させるステップとが含まれることができる。本方法には、標的DNA配列と核酸との間の相同組換えが起こるように、標的DNAの少なくとも一部分に相同的な配列を含む核酸を提供することがさらに含まれることができる。標的DNA配列は、細胞に内在性であることができる。細胞は、植物細胞、哺乳動物細胞、魚細胞、昆虫細胞もしくはin vitro培養のためにこれらの生物から誘導した細胞系、または生組織から直接採取し、in vitro培養のために確立させた初代細胞であることができる。接触ステップには、TALENのコード配列を含むベクターで細胞を形質転換させて、TALENタンパク質を細胞中で発現させること、TALENタンパク質を細胞内に機械的に注入すること、TALエフェクターエンドヌクレアーゼタンパク質を細胞内に細菌III型分泌系によって送達すること、またはTALENタンパク質を細胞内に電気穿孔によって導入することが含まれることができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。標的DNA内の特定のヌクレオチド配列と結合するTALエフェクタードメインには、10個以上のDNA結合反復、より好ましくは15個以上のDNA結合反復が含まれることができる。細胞は任意の原核または真核生物からのものであることができる。
【0029】
別の態様では、本発明は、特定の位置でDNAを切断することができる配列特異的TALENを設計する方法を特長とする。本方法には、それ内に二本鎖切断を導入することが所望される第2のヌクレオチド配列に隣接する、第1のユニークな内在染色体ヌクレオチド配列を同定するステップと、(a)組み合わせて第1のユニークな内在染色体ヌクレオチド配列と結合する複数のDNA結合反復ドメイン、および(b)第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含む、配列特異的TALENを設計するステップとが含まれることができる。
【0030】
また、本発明は、エンドヌクレアーゼドメインおよび特定のDNA配列に特異的なTALエフェクターDNA結合ドメインを含むTALENも特長とする。TALENには、精製タグがさらに含まれることができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。
【0031】
別の態様では、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した動物を作製する方法を特長とする。本方法には、それ内に核酸を導入することが所望される内在染色体標的DNA配列を含む初代細胞を提供するステップと、エンドヌクレアーゼドメインおよび内在染色体標的DNA配列と結合するTALエフェクタードメインを含むTALENを用いて、内在染色体標的DNA配列内に二本鎖切断を生じるステップと、内在染色体標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、初代細胞内に、外因性核酸と内在染色体標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった初代細胞から動物を作製するステップとが含まれることができる。動物は哺乳動物であることができる。相同配列は、相同組換え後に遺伝子を破壊するヌクレオチド配列、相同組換え後に遺伝子を置き換えるヌクレオチド配列、相同組換え後に遺伝子内に点突然変異を導入するヌクレオチド配列、および相同組換え後に調節部位を導入するヌクレオチド配列からなる群から選択されるヌクレオチド配列であることができる。
【0032】
さらに別の態様では、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した植物を作製する方法を特長とする。本方法には、それ内に核酸を導入することが所望される内在標的DNA配列を含む植物細胞を提供するステップと、エンドヌクレアーゼドメインおよび内在標的ヌクレオチド配列と結合するTALエフェクタードメインを含むTALENを用いて、内在標的DNA配列内に二本鎖切断を生じるステップと、内在標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、植物細胞内に、外因性核酸と内在標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった植物細胞から植物を作製するステップとが含まれることができる。
【0033】
別の態様では、本発明は、細胞における標的化遺伝子組換えの方法を特長とする。本方法には、選択されたDNA標的配列を標的としたTALENをコードしている核酸分子を細胞内に導入するステップと、細胞内でのTALENの発現を誘導するステップと、選択されたDNA標的配列が突然変異を示している細胞を同定するステップとが含まれることができる。突然変異は、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択されることができる。本方法には、細胞内にドナーDNAを導入することがさらに含まれることができる。細胞は、昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞であることができる。
【0034】
さらに別の態様では、本発明は、(1)選択されたヌクレオチド配列の第1ヌクレオチドに特異的なRVDを有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含む開始プラスミドを選択し、第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有するステップと、(2)開始プラスミドをPspXIで直鎖状にするステップと、(3)PspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチド(複数可)に特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしている、XhoI粘着末端を有するDNAモジュールをライゲーションするステップと、(4)核酸が選択されたヌクレオチド配列と結合することができるTALENをコードするまでステップ(2)および(3)を繰り返すステップとを含む、配列特異的TALENをコードしている核酸を作製する方法を特長とする。一部の事例では、本方法には、ステップ(3)のライゲーションの後に、PspXI部位中のDNAモジュールの配向を確認することがさらに含まれることができる。
【0035】
別段に定義しない限りは、本明細書中で使用したすべての技術用語および科学用語は、本発明が関連する分野の技術者によって一般に理解されているものと同じ意味を有する。本明細書中に記載のものに類似または均等の方法および材料を使用して本発明を実施することができるが、適切な方法および材料が以下に記載されている。本明細書中で言及したすべての出版物、特許出願、特許、および他の参考文献は、その全体で参考として組み込まれている。矛盾する場合は、定義を含めて本明細書が支配する。さらに、材料、方法、および実施例は、例示的なものにすぎず、限定することを意図しない。
【0036】
本発明の1つまたは複数の実施形態の詳細は、添付の図面および以下の説明中に記載されている。本発明の他の特長、目的、および利点は、説明および図面、ならびに特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0037】
【図1】TALエフェクター−DNA認識の暗号を示す。図1Aは、反復領域(白い四角)および代表的な反復配列(配列番号1)を示す一般的なTALエフェクターの図であり、RVDに下線が引いてある。図1Bは、様々なTALエフェクターのRVDおよび標的遺伝子プロモーター配列(配列番号2〜11)の最良のパターン一致(低エントロピーのアラインメント)を示す図である。星印は、残基13での欠失を示す。図1Cは、Bのアラインメント、およびすべてのコメプロモーターを40個の追加のX.oryzaeのTALエフェクターで走査することによって得られ、それぞれのエフェクターについて下流遺伝子が感染中に活性化された最良のアラインメントを保持したさらに10個のアラインメントにおける、RVD−ヌクレオチドの会合を示す図である。図1Dは、20個のTALエフェクター標的部位の隣接ヌクレオチドの頻度を示す図である。位置は標的部位の5’末端に対するものであり、Nは標的部位の長さである。ロゴはWebLogoを使用して作成した。
【図2】OsHen1がXanthomonas oryzae pv.oryzicola株BLS256のTal1cによって活性化されるという証拠を提供する。図2Aは、マーカー交換突然変異体M51、空のコスミドベクター(ev)を保有するM51、tal1a、tal1b、およびtal1cを含有するコスミドpIJF92を保有するM51、ならびに野生型(WT)株における、BLS256を接種した24時間後のコメ葉中の、アクチン遺伝子を参照としたOsHen1の相対的な転写物の存在量を示す半定量的RT−PCR結果の像である。図2Bは、XmalI断片を含有するマーカーの救出および末端配列決定による、M51における単一のマーカー交換突然変異のマッピングに基づいた模式図である。コスミドpIJF92中に含有されていたゲノム領域、救出された断片の座標、およびBLS256ゲノム断片の座標が示されている。
【図3】参照AvrBs3のアミノ酸配列である(配列番号12)。
【図4】参照AvrBs3の核酸配列である(配列番号13)。
【図5】TALヌクレアーゼ発現ベクターのマップである。
【図6】標的レポータープラスミドのマップである。
【図7】TALヌクレアーゼの模式的構造の図である。TALのDNA結合ドメインの認識部位は大文字として表されており、一方でスペーサー配列は小文字で示されている。
【図8】AvrBs3認識ドメインの17回と半分のタンデムの反復のアミノ酸配列である(配列番号16)。位置12および13の超可変アミノ酸が四角で囲まれている。
【図9】TALの有効性を試験するための酵母アッセイのスキームを示す図である。
【図10】AvrBs3 TALヌクレアーゼの酵母アッセイの結果をプロットするグラフである。
【図11】単一、二重、または三重のAsvBs3反復モジュールおよびクローニングベクターの模式図を示す図である。
【図12】ほとんどのTALエフェクター中の反復領域の末端に存在する単一の代表的なTALエフェクター反復(図12A)および代表的な切断された反復(図12B)を示す。ヌクレオチドおよびコードされているアミノ酸配列が示されている。Nは、「XX」と示すRVDをコードしているヌクレオチドを表す。数字はアミノ酸の位置について与えられている。配列はtal1cからとったものである。
【図13】tal1c遺伝子、および反復領域を単一の切断された反復まで縮小してpCS487(やはり示されている)をもたらすプロセスを示す模式図である。MはMscI部位であり、SはSphI部位である。
【図14】pCS487中の元の切断された反復の末端に翻訳的にサイレントな突然変異を導入して、PspXIおよびXhoI部位を生じ、pCS489を得ることを示す模式図である。元の反復のコドン18〜21の配列(配列番号21)および突然変異させた反復(配列番号23)が示されている。コードされているアミノ酸配列(配列番号22)は、突然変異によって変化していなかった。突然変異させたヌクレオチドを斜体にしている。
【図15】GatewayエントリーベクターpENTR−D(Invitrogen、カリフォルニア州Carlsbad)中で反復領域を含まずにtal1cのNおよびC末端のみをコードしているカナマイシン耐性プラスミドである、pCS488のマップである。
【図16】RVD NIを有する反復をコードしているpCS493と命名された単一反復の開始プラスミドのマップである。pCS494、pCS495、およびpCS496と命名された3つの他のプラスミドは、それらがコードするRVD(右に示す)以外は同一であった。
【図17】図17Aは、RVD NIを有する単一反復モジュールのヌクレオチドおよびコードされているアミノ酸配列を示す。5’XhoI適合性の付着末端、MscI部位、および3’PspXI/XhoI適合性の付着末端に下線が引いてある。RVDおよびそれをコードしているヌクレオチドは太字である。HD、NI、およびNGをそれぞれコードしているRVDのコード配列以外は示したものに同一である、3つの他の反復モジュールを構築した。図17Bは、図17Aに示されている反復をコードしている配列を含有するpCS502と命名された単一反復モジュールのプラスミドのマップである。また、pCS503、pCS504、およびpCS505と命名されたプラスミドも作製し、これらは、それらがコードしているRVD(右に示す)以外はpCS502と同一である。
【図18】図18Aは、RVD NIを有する単一反復モジュールのヌクレオチドおよびコードされているアミノ酸配列を示し、ヌクレオチド置換(斜体)が、PspXI/XhoI部位内へのライゲーション後の5’末端でのXhoI部位の再構成を妨げ、内部MscI部位を破壊する。RVDおよびそれがコードしているヌクレオチドは太字である。HD、NI、およびNGをそれぞれコードしているRVDのコード配列以外は同一である、3つの追加の反復モジュールを構築した。図18Bは、追加の反復モジュールを単一反復モジュールのプラスミド内に順次ライゲーションさせることによってアセンブルした3回反復モジュールの模式図である。第1反復中のMscI部位および3’末端のPspXI部位はユニークに保たれ、モジュール全体は2つのXhoI部位によって隣接されている。
【図19】1回、2回、および3回の反復モジュールのプラスミドの完全な組のリストである。
【図20】任意の反復配列をtal1c「主鎖」内にアセンブルして誂えのTALエフェクター遺伝子を作製するために使用できる方法中のステップを示す流れ図である。
【図21】示されているヌクレオチド配列を標的とするTALエンドヌクレアーゼの構築における反復モジュールのアセンブリを示す模式図である。図21Aでは、pCS519、pCS524、pCS537、pCS551、pCS583、およびpCS529と命名されたプラスミドからの反復モジュールを、pCS493と命名された開始プラスミド中の配列に順次付加して、pMAT55、pMAT56、pMAT57、pMAT58、pMAT59、およびpMAT60と命名されたプラスミドがもたらされる。図21Bでは、pCS530、pCS533、pCS522、およびpCS541と命名されたプラスミドからの反復モジュールを、pMAT1と命名されたプラスミド中の配列に順次付加して、pMAT61、pMAT62、pMAT63、およびpMAT64と命名されたプラスミドがもたらされる。
【図22】図22Aは、TALエフェクタータンパク質の模式図である。BamHI断片(Bによって示す)をFokIエンドヌクレアーゼの触媒ドメインと融合させて、TALENを作製した。NはN末端、NLSは核移行シグナル、BはBamHI部位、ADは酸性活性化ドメインである。図22Bは、TALエフェクターAvrBs3およびPthXo1を用いて構築したTALENの活性をプロットするグラフである。Avr−FokIはAvrBs3 TALENであり、Pth−FokIはPthXo1 TALENであり、Avr−FokIおよびPth−FokIは、AvrBs3およびPthXo1とFokIの触媒的に不活性な型との融合体であり(Bitinaiteら(1998)Proc.Natl.Acad.Sci.USA、95:10570〜10575)、ZFNは、Zif268DNA結合ドメインを含有するジンクフィンガーヌクレアーゼである(PorteusおよびBaltimore(2003)Science、300:763)。
【図23】参照PthXo1のアミノ酸配列である(配列番号31)。
【図24】参照PthXo1の核酸配列である(配列番号32)。
【図25】pFZ85ベクターの図である。
【図26】avrBs3_TALENのアミノ酸配列を示す(配列番号33)。
【図27】pthXo1 TALENのアミノ酸配列を示す(配列番号34)。
【図28】図28Aは、様々なスペーサー長を有する標的に対するAvrBs3およびPthXo1 TALENの活性をプロットするグラフである。ZFNはZif268由来のジンクフィンガーヌクレアーゼである。図28Bは、ヘテロ二量体TALENの活性をプロットするグラフである。PthXo1−FokIおよびAvrBs3−FokI発現ベクター、ならびに頭から尾の配向で15bpによって分離されたそれぞれに対する認識部位からなる標的を有するプラスミドを含有する、酵母における活性が示されている(Avr−FokI、Pth−FokI)。また、そのそれぞれの標的に対する、AvrBs3(Avr−FokI)およびPthXo1(Pth−FokI)TALENの個々の活性ならびにZif268(ZFN)の活性も、参照のために示されている。陰性対照として、Avr−FokI、Pth−FokIに対する標的部位プラスミドのみを有する酵母培養物を、LacZ活性についてアッセイした((−)として示す)。
【図29】図29Aは、個々の誂えのTALENのRVD配列およびそのそれぞれのDNA認識配列を示す表である。図29Bは、誂えのTALENの活性をプロットするグラフである。(−)は標的部位プラスミドのみの陰性対照であり、ZFNはジンクフィンガーヌクレアーゼ陽性対照である。
【図30】20個の標的およびTALエフェクターの対の末端のヌクレオチドおよびRVDの頻度の描写である。
【図31】Golden Gateクローニングシステムの模式図である[Englerら(2008)PLoS One、3:e3647、およびEnglerら(2009)PLoS One、4:e5553]。
【図32】本明細書中に記載のGolden Gateクローニング手法を使用した、誂えのTALエフェクター反復をコードしているアレイをアセンブリおよびクローニングするための58個のプラスミドの組を示す。Tetは、プラスミド選択のマーカーであるテトラサイクリン耐性遺伝子であり、specは、プラスミド選択のマーカーであるスペクチノマイシン耐性遺伝子であり、ampは、プラスミド選択のマーカーであるアンピシリン耐性遺伝子である。
【図33】図32に示されているプラスミドの組を使用したGolden Gate手法によって誂えのTALエフェクター反復をコードしているアレイをアセンブリおよびクローニングするための方法の模式図である。例示目的のために、自由裁量による反復アレイのアセンブリが示されている。specは、プラスミド選択のマーカーであるスペクチノマイシン耐性遺伝子であり、ampは、プラスミド選択のマーカーであるアンピシリン耐性遺伝子である。
【図34】本明細書中の実施例9に記載のように作製したTALENのアミノ酸配列を示す。図34Aはテロメラーゼ−TALEN124であり、図34Bはgridlock−TALEN105であり、図34Cはadhl−TALEN58であり、図34Dはadhl−TALEN63であり、図34Eはadhl−TALEN68であり、図34Fはadhl−TALEN73であり、図34Gはadhl−TALEN89であり、図34Hはgridlock−TALEN106であり、図34Iはadhl−TALEN64であり、図34Jはadhl−TALEN69であり、図34Kはadhl−TALEN74であり、図34Lはtt4−TALEN90であり、図34Mはテロメラーゼ−TALEN121であり、図34Nはテロメラーゼ−TALEN126であり、図34Oはgridlock−TALEN107であり、図34Pはgridlock−TALEN117であり、図34Qはテロメラーゼ−TALEN131であり、図34Rはテロメラーゼ−TALEN136であり、図34Sはadhl−TALEN60であり、図34Tはtt4−TALEN85であり、図34Uはgridlock−TALEN102である。
【図35】増加していく長さの誂えのTALEN単量体(9、10、12、13、15、16、17、または18量体)を使用した酵母アッセイによって測定したTALEN活性プロットするグラフである。示したように、TALENはシロイヌナズナおよびゼブラフィッシュの遺伝子に対して標的化した。
【図36】図36Aは、2つのTALEN対によって標的化されているシロイヌナズナADH1遺伝子からの2つの異なるDNA標的配列を示す図である。図36Bは、シロイヌナズナADH1遺伝子を標的とする機能的TALEN対の酵母アッセイデータプロットするグラフである。
【図37】図37Aは、シロイヌナズナのプロトプラスト中のTALEN誘導性突然変異を検出するために使用した制限エンドヌクレアーゼアッセイの模式図である。図37Bは、制限エンドヌクレアーゼアッセイにおける未消化のDNAからの9個のクローンの配列を示す。クローンのうちの6個が、非相同末端結合(NHEJ)によって導入された突然変異を有する。
【図38】図38Aは、いくつかの系統学的に明白に異なるTALエフェクター、すなわち、Xanthomonas gardneriからのAvrHah1、X.campestris pv.vesicatoriaからのAvrBs3、X.oryzae pv.oryzaeからのPthXo1、X.citriからのPthA、およびX.oryzae pv.oryzicolaからのTal1cの第0反復配列を示す。多型位置が四角で囲まれている。図38Bは、PthXo1の第0および第1反復を示す模式図である。「第0」反復は第1反復の直前にあり、35%の同一性を示し、同様の予測された二次構造を有する。第1反復のRVDおよび第0反復の候補の類似残基に下線が引いてある。*はギャップであり、Hはヘリックスであり、Eは伸長である。構造はJPredを使用して予測した(Coleら(2008)Nucl.Acids Res.、36:W197〜W201)。
【図39】示したようにV5でタグ付けしたTALエフェクタータンパク質AvrBs3、PthXo1、およびTal1cをコードしているプラスミドで形質移入したヒト胚性腎臓293T細胞から単離した全タンパク質の、マウス−抗V5抗体を使用した免疫検出後のウエスタンブロットを示す。免疫標識したアクチンがそれぞれのレーンの等価なローディングの対照として示されている。
【図40】図40Aは、TALEN HPRT−3254−17のアミノ酸配列を示し、図40Bは、TALEN HPRT−3286−20rのアミノ酸配列を示す。
【図41】図41Aは、ヒト染色体HPRT遺伝子中のTALEN標的部位を示す模式図である。HPRT−3254−17およびHPRT−3286−20r TALENの結合部位、これらの部位の間のスペーサー中のBpu10I部位、ならびに領域を増幅するためのプライマー部位が示されている。下の座標は、コード配列の第1ヌクレオチドからの距離を塩基対で示している。図41Bは、TALENで処理したおよび処理していない細胞から単離したゲノムDNAを鋳型として使用した、図41Aに示されている領域のPCR増幅産物のBpu10I消化の結果を示す。増幅の前にゲノムDNAをBpu10Iで消化した。DNA断片はアガロースゲル電気泳動によって分離し、臭化エチジウムを使用して可視化した。
【発明を実施するための形態】
【0038】
本特許出願は、TALエフェクターによって媒介される配列特異的DNA認識に関する材料および方法を提供する。本明細書中に記載のように、TALエフェクターの一次アミノ酸配列が、それらが結合するヌクレオチド配列を指示する。本発明者らは、TALエフェクターのアミノ酸配列とそのDNA標的配列との間の関係性は直接的であり、それにより、TALエフェクターの標的部位の予測が可能となり、また、特定のヌクレオチド配列と結合するようにTALエフェクターを誂えることも可能となることを見い出した。そのような予測および誂えは、様々な目的のために利用することができる。一例では、特定のTALエフェクター配列をエンドヌクレアーゼ配列と融合させて、特定のDNA配列に対するエンドヌクレアーゼの標的化、続いて標的配列でまたはその付近でのDNAの切断を可能にすることができる。DNA中の切断(すなわち二本鎖切断)は、相同組換えの頻度を劇的に増加させる可能性がある。したがって、特定の標的DNA配列に対して高い度合の配列類似度を有する配列を保有するDNA構築体と組み合わせて、TALENを使用して、高い精度および効率で複雑なゲノム中の部位特異的突然変異誘発すること、すなわち、遺伝子機能をノックアウトもしくは変更すること、または遺伝子もしくは他の配列を付加することを促進することができる。
【0039】
したがって、本明細書中で提供されている主題には、とりわけ、遺伝子改変した生物(それだけには限定されないが、植物、真菌、ショウジョウバエ(Drosophila)、線虫、ゼブラフィッシュ、マウス、他の哺乳動物およびヒトが含まれる)を作製するための材料および方法が含まれる。そのような方法には、たとえば、細胞をいくつかの組換え核酸で形質移入することが含まれることができる。たとえば、細胞(たとえば真核細胞)を、細胞内に見つかる対応する標的ヌクレオチド配列と比較して変更が含まれるドナーヌクレオチド配列を含有する第1の組換え核酸構築体、およびTALヌクレアーゼをコードしている第2の組換え核酸構築体で形質転換させることができる。一部の実施形態では、細胞を、選択マーカーをコードしている第3の組換え核酸構築体でも形質転換させることができる。ドナー核酸構築体からの核酸配列は、本明細書中に記載の形質転換細胞のゲノム内に取り込まれることができる。たとえば、本明細書中に記載の方法を使用して生成した植物細胞を成長させて、変更されたドナーヌクレオチド配列がそのゲノム内に取り込まれた植物を生成することができる。そのような植物からの種子を使用して、たとえば、改変されていない植物と比較して変更された成長特徴(たとえば、様々な生物的および非生物的なストレスに対する耐性または寛容の増加)、変更された外見(たとえば、変更された色または高さ)、あるいは変更された組成(たとえば、炭素、窒素、油、タンパク質、炭水化物(たとえば糖もしくはデンプン)、アミノ酸、脂肪酸、または二次代謝物のレベルの増加または減少)などの表現型を有する植物を生成することができる。
【0040】
ポリヌクレオチドおよびポリペプチド
単離した核酸およびポリペプチドが本明細書中で提供されている。用語「核酸」および「ポリヌクレオチド」とは、互換性があるように使用され、cDNA、ゲノムDNA、合成(たとえば化学合成)DNA、および核酸類似体を含有するDNA(またはRNA)を含めた、RNAおよびDNAをどちらもいう。ポリヌクレオチドは任意の三次元構造を有することができる。核酸は二本鎖または一本鎖(すなわち、センス鎖またはアンチセンスの一本鎖)であることができる。ポリヌクレオチドの非限定的な例には、遺伝子、遺伝子断片、エクソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝状ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離したDNA、任意の配列の単離したRNA、核酸プローブ、およびプライマー、ならびに核酸類似体が含まれる。
【0041】
本発明のポリペプチド(非限定的な例としてTALエフェクター−DNA修飾酵素など)は、たとえば前記ポリペプチドをコードしているベクターを使用することによってまたはポリペプチド自体として、送達ベクターを超音波穿孔もしくは電気穿孔またはこれらの技法の派生法などの任意の細胞透過処理技法と関連または組み合わせて使用することによって、細胞中に導入することができる。
【0042】
核酸に言及する場合、本明細書中で使用する「単離した」とは、ゲノム中で核酸の片側または両側に通常隣接している核酸を含めた、ゲノム、たとえば植物ゲノム中に存在する他の核酸から分離されている核酸をいう。また、核酸に関して本明細書中で使用する用語「単離した」には、任意の天然に存在しない配列も含まれる。これは、そのような天然に存在しない配列は自然には見つからず、したがって天然に存在するゲノム中のすぐ近くに近接する配列を有さないからである。
【0043】
単離した核酸は、たとえばDNA分子であることができるが、ただし、天然に存在するゲノム中でそのDNA分子のすぐ近くに隣接して通常見つかる核酸配列のうちの1つが除去されているまたは非存在である。したがって、単離した核酸には、それだけには限定されないが、他の配列から独立して別の分子として存在するDNA分子(たとえば、化学合成核酸、またはPCRもしくは制限エンドヌクレアーゼ処理によって生成されたcDNAもしくはゲノムDNA断片)、およびベクター、自己複製プラスミド、ウイルス(たとえば、パラレトロウイルス、レトロウイルス、レンチウイルス、アデノウイルス、もしくはヘルペスウイルス)、または原核生物もしくは真核生物のゲノムDNA内に取り込まれているDNAが含まれる。さらに、単離した核酸には、ハイブリッドまたは融合核酸の一部であるDNA分子などの組換え核酸が含まれることができる。数百から数百万個の他の核酸と共に、たとえば、cDNAライブラリもしくはゲノムライブラリ内、またはゲノムDNA制限消化物を含有するゲルスライス内に存在する核酸は、単離した核酸とみなされるべきでない。
【0044】
核酸は、たとえば化学合成またはポリメラーゼ連鎖反応(PCR)によって作製することができる。PCRとは、標的核酸が増幅される手順または技法をいう。PCRを使用して、全ゲノムDNAまたは全細胞RNAからの配列を含めた、DNAおよびRNAからの特定の配列を増幅することができる。様々なPCR方法は、たとえば、PCRプライマー:実験室の手引き(PCR Primer:A Laboratory Manual)、DieffenbachおよびDveksler編、Cold Spring Harbor Laboratory Press、1995に記載されている。一般に、対象の領域の末端またはそれを越える配列情報を用いて、増幅する鋳型の逆の鎖と配列が同一または類似のオリゴヌクレオチドプライマーを設計する。また、部位特異的なヌクレオチド配列修飾を鋳型核酸内に導入することができる様々なPCR戦略も利用可能である。
【0045】
また、単離した核酸は、突然変異誘発によっても得ることができる。たとえば、ドナー核酸配列を、PCRによるオリゴヌクレオチド特異的突然変異誘発および部位特異的突然変異誘発を含めた標準の技法を使用して突然変異させることができる。分子生物学の手短なプロトコル(Short Protocols in Molecular Biology)、第8章、Green Publishing AssociatesおよびJohn Wiley&Sons、Ausubelら編、1992を参照されたい。
【0046】
本明細書中で使用する用語「ポリペプチド」とは、翻訳後修飾(たとえばリン酸化またはグリコシル化)にかかわらず、2つ以上のサブユニットのアミノ酸の化合物をいう。サブユニットは、ペプチド結合またはたとえばエステルもしくはエーテル結合などの他の結合によって連結され得る。用語「アミノ酸」とは、D/L光学異性体を含めた天然および/または非天然もしくは合成のアミノ酸のいずれかをいう。
【0047】
ポリペプチドに関して「単離した」または「精製した」とは、ポリペプチドが、自然においてそれと共に通常見つかる細胞成分(たとえば、他のポリペプチド、脂質、炭水化物、および核酸)からある程度分離されていることを意味する。精製したポリペプチドは、非還元ポリアクリルアミドゲル上に単一の主バンドを与えることができる。精製したポリペプチドは、少なくとも約75%純粋(たとえば、少なくとも80%、85%、90%、95%、97%、98%、99%、または100%純粋)であることができる。精製したポリペプチドは、たとえば、天然源から抽出することによって、化学合成によって、または宿主細胞もしくはトランスジェニック植物中での組換え産生によって得ることができ、たとえば、アフィニティークロマトグラフィー、免疫沈降、サイズ排除クロマトグラフィー、およびイオン交換クロマトグラフィーを使用して精製することができる。精製の程度は、それだけには限定されないが、カラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、または高性能液体クロマトグラフィーを含めた任意の適切な方法を使用して測定することができる。
【0048】
組換え構築体
また、組換え核酸構築体(たとえばベクター)も本明細書中で提供されている。「ベクター」とは、それ内に別のDNAセグメントを挿入して、挿入したセグメントを複製させ得る、プラスミド、ファージ、またはコスミドなどのレプリコンである。一般に、ベクターは、適当な制御要素と会合している場合に複製が可能である。適切なベクター主鎖には、たとえば、プラスミド、ウイルス、人工染色体、BAC、YAC、またはPACなどの、当分野でルーチン的に使用されているものが含まれる。用語「ベクター」には、クローニングおよび発現ベクター、ならびにウイルスベクターおよび組込みベクターが含まれる。「発現ベクター」とは、1つまたは複数の発現制御配列が含まれるベクターであり、「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。適切な発現ベクターには、それだけには限定されないが、たとえば、バクテリオファージ、バキュロウイルス、タバコモザイクウイルス、ヘルペスウイルス、サイトメガロウイルス、レトロウイルス、ワクシニアウイルス、アデノウイルス、およびアデノ関連ウイルスに由来するプラスミドおよびウイルスベクターが含まれる。数々のベクターおよび発現系がNovagen(ウィスコンシン州Madison)、Clontech(カリフォルニア州Palo Alto)、Stratagene(カリフォルニア州La Jolla)、およびInvitrogen/Life Technologies(カリフォルニア州Carlsbad)などの企業から市販されている。
【0049】
用語「調節領域」、「制御要素」、および「発現制御配列」とは、転写または翻訳の開始および速度、ならびに転写物またはポリペプチド産物の安定性および/または移動度に影響を与えるヌクレオチド配列をいう。調節領域には、それだけには限定されないが、プロモーター配列、エンハンサー配列、応答要素、タンパク質認識部位、誘導性要素、プロモーター制御要素、タンパク質結合配列、5’および3’非翻訳領域(UTR)、転写開始部位、終結配列、ポリアデニル化配列、イントロン、ならびに分泌シグナル、核移行配列(NLS)およびプロテアーゼ切断部位などのコード配列内に存在することができる他の調節領域が含まれる。
【0050】
本明細書中で使用する「作動可能に連結された」とは、発現制御配列が対象のコード配列の発現を有効に制御するように遺伝子構築体内に取り込まれていることを意味する。コード配列は、RNAポリメラーゼがコード配列をRNAへと転写することができる場合に、細胞中の発現制御配列と「作動可能に連結」されているかつその「制御下」にあり、mRNAの場合は、その後に、コード配列によってコードされているタンパク質へと翻訳されることができる。したがって、調節領域は、修飾された標的核酸をそれ内で発現させることが所望される植物細胞、植物、または植物組織中での転写を変調させる、たとえば、調節する、促進するまたは駆動することができる。
【0051】
プロモーターとは、転写が開始される点の典型的には100個以内のヌクレオチド上流(一般にRNAポリメラーゼIIの開始部位付近)の、DNA分子の領域から構成される発現制御配列である。プロモーターは、RNAポリメラーゼおよび他のタンパク質を認識および結合して、転写を開始および変調させることに関与している。コード配列をプロモーターの制御下にするためには、ポリペプチドの翻訳リーディングフレームの翻訳開始部位をプロモーターの1〜約50個のヌクレオチド下流に配置することが典型的には必要である。しかし、プロモーターは、翻訳開始部位の約5,000個と多数のヌクレオチド上流、または転写開始部位の約2,000個のヌクレオチド上流に配置することができる。プロモーターは、典型的には少なくともコア(基底)プロモーターを含む。また、プロモーターには、上流要素などの少なくとも1つの制御要素も含まれ得る。そのような要素には、上流活性化領域(UAR)および任意選択で合成上流要素などのポリヌクレオチドの転写に影響を与える他のDNA配列が含まれる。
【0052】
含めるプロモーターの選択肢は、それだけには限定されないが、効率、選択性、誘導性、所望の発現レベル、および細胞または組織特異性を含めたいくつかの要因に依存する。たとえば、それぞれ特定の組織、器官、および細胞種中でのみまたは優勢に転写を与える組織、器官および細胞に特異的なプロモーターを使用することができる。一部の実施形態では、茎、柔組織、基本分裂組織、維管束、形成層、師部、皮層、茎頂分裂組織、側枝分裂組織、根端分裂組織、側根分裂組織、葉原基、葉肉、または葉表皮などの植物組織に特異的なプロモーターが適切な調節領域である可能性がある。一部の実施形態では、種子に本質的に特異的なプロモーター(「種子に優先的なプロモーター」)が有用な可能性がある。種子に特異的なプロモーターは、種子の発生中に、胚乳および子葉組織中で作動可能に連結された核酸の転写を促進することができる。あるいは、構成的プロモーターは、植物の発生の全体にわたって、植物のほとんどまたはすべての組織中で作動可能に連結された核酸の転写を促進することができる。他のプロモーターのクラスには、それだけには限定されないが、化学薬品、発生刺激、または環境刺激などの外部刺激に応答して転写を与えるプロモーターなどの誘導性プロモーターが含まれる。
【0053】
基底プロモーターとは、転写開始に必要な転写複合体のアセンブリに必要な最小限の配列である。基底プロモーターには、しばしば、転写開始部位から約15〜約35個のヌクレオチド上流に位置し得る「TATAボックス」要素が含まれる。また、基底プロモーターには、転写開始部位から約40〜約200個のヌクレオチド、典型的には約60〜約120個のヌクレオチド上流に位置することができる「CCAATボックス」要素(典型的には配列CCAAT)および/またはGGGCG配列も含まれ得る。
【0054】
本明細書中で提供されている核酸構築体中に含まれることができるプロモーターの非限定的な例には、カリフラワーモザイクウイルス(CaMV)の35S転写開始領域、Agrobacterium tumefaciensのT−DNAに由来する1’または2’プロモーター、Busk((1997)Plant J.、11:1285〜1295)によって記載されているトウモロコシの葉に特異的な遺伝子からのプロモーター、トウモロコシおよび他の種からのknl関連遺伝子、ならびにトウモロコシユビキチン−1プロモーターなどの様々な植物遺伝子からの転写開始領域が含まれる。
【0055】
5’非翻訳領域(UTR)は、転写されるが翻訳されず、転写開始部位と翻訳開始コドンとの間に位置し、+1ヌクレオチドが含まれ得る。3’UTRは、翻訳終結コドンと転写終端との間に位置することができる。UTRは、増加したmRNAメッセージの安定性または減弱した翻訳などの特定の機能を有する場合がある。3’UTRの例には、それだけには限定されないが、ポリアデニル化シグナルおよび転写終結配列が含まれる。また、コード領域の3’末端のポリアデニル化領域もコード配列と作動可能に連結されていることができる。ポリアデニル化領域は、天然遺伝子、様々な他の植物遺伝子、またはAgrobacteriumのT−DNAに由来することができる。
【0056】
また、本明細書中で提供されているベクターには、たとえば、複製起点および/または足場付着領域(SAR)も含まれることができる。さらに、発現ベクターには、発現されたポリペプチドの操作または検出(たとえば精製もしくは局在化)を促進するように設計されたタグ配列が含まれることができる。緑色蛍光タンパク質(GFP)、グルタチオンS−トランスフェラーゼ(GST)、ポリヒスチジン、c−myc、赤血球凝集素、またはFlag(商標)タグ(Kodak、コネチカット州New Haven)配列などのタグ配列は、典型的には、コードされているポリペプチドとの融合体として発現される。そのようなタグは、カルボキシルまたはアミノ末端のどちらかを含めた、ポリペプチド内のどこにでも挿入することができる。
【0057】
「送達ベクター」または「複数の送達ベクター」とは、本発明において、本発明に必要な薬剤/化合物および分子(タンパク質または核酸)を細胞と接触させるまたは細胞内もしくは細胞内区画内に送達するために使用することができる、任意の送達ベクターを意図する。これには、それだけには限定されないが、リポソーム送達ベクター、ウイルス送達ベクター、薬物送達ベクター、化学担体、ポリマー担体、リポプレックス、ポリプレックス、デンドリマー、マイクロバブル(超音波造影剤)、ナノ粒子、乳濁液または他の適切な転移ベクターが含まれる。これらの送達ベクターは、分子、化合物、巨大分子(遺伝子、タンパク質)、またはプラスミド、Diatosによって開発されたペプチドなどの他のベクターの送達を可能にする。これらの事例では、送達ベクターは分子担体である。また、「送達ベクター」または「複数の送達ベクター」とは、形質移入を行うための送達方法も意図する。
【0058】
用語「ベクター」または「複数のベクター」とは、それが連結されている別の核酸を輸送することができる核酸分子をいう。本発明における「ベクター」には、それだけには限定されないが、ウイルスベクター、プラスミド、RNAベクター、または染色体、非染色体、半合成もしくは合成の核酸からなり得る直鎖状もしくは環状のDNAもしくはRNA分子が含まれる。好ましいベクターは、自律複製が可能なもの(エピソームベクター)および/またはそれらが連結されている核酸の発現が可能なもの(発現ベクター)である。多数の適切なベクターが当業者に知られており、市販されている。
【0059】
ウイルスベクターには、レトロウイルス、アデノウイルス、パルボウイルス(たとえばアデノ関連ウイルス)、コロナウイルス、オルトミクソウイルス(たとえばインフルエンザウイルス)などのマイナス鎖RNAウイルス、ラブドウイルス(たとえば狂犬病および水疱性口内炎ウイルス)、パラミクソウイルス(たとえば麻疹およびセンダイ)、ピコルナウイルスおよびアルファウイルスなどのプラス鎖RNAウイルス、ならびに、アデノウイルス、ヘルペスウイルス(たとえば、1および2型単純ヘルペスウイルス、エプスタイン−バーウイルス、サイトメガロウイルス)、およびポックスウイルス(たとえば、ワクシニア、鶏痘、カナリア痘)を含めた二本鎖DNAウイルスが含まれる。他のウイルスには、たとえば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス、および肝炎ウイルスが含まれる。レトロウイルスの例には、トリ白血症肉腫、哺乳動物C型、B型ウイルス、D型ウイルス、HTLV−BLV群、レンチウイルス、スプマウイルスが含まれる(Coffin,J.M.、レトロウイルス科:ウイルスおよびその複製(Retroviridae:The viruses and their replication)、基礎ウイルス学(Fundamental Virology)、第3版、B.N.Fieldsら編、Lippincott−Raven Publishers、Philadelphia、1996)。
【0060】
「レンチウイルスベクター」とは、その比較的大きなパッケージング能力、低下した免疫原性および広い範囲の様々な細胞種を高い効率で安定に形質導入するその能力が理由で遺伝子送達に非常に有望な、HIV系のレンチウイルスベクターを意味する。レンチウイルスベクターは、通常、産生細胞内への3つ(パッケージング、エンベロープおよび転移)以上のプラスミドの一過的な形質移入の後に作製される。HIVと同様、レンチウイルスベクターは、ウイルス表面糖タンパク質と細胞表面上の受容体との相互作用を介して標的細胞に進入する。進入の際、ウイルスRNAは、ウイルス逆転写酵素複合体によって媒介される逆転写を受ける。逆転写の産物は二本鎖の直鎖状ウイルスDNAであり、これは、感染した細胞のDNA中へのウイルス組込みの基質である。前記レンチウイルスベクターは、「非組込み」または「組込み」であることができる。
【0061】
「組込みレンチウイルスベクター(またはLV)」とは、非限定的な例として、標的細胞のゲノムに組み込まれることができるベクターを意味する。
【0062】
逆に、「非組込みレンチウイルスベクター(またはNILV)」とは、ウイルスインテグラーゼの作用によって標的細胞のゲノムに組み込まれない効率的な遺伝子送達ベクターを意味する。
【0063】
好ましいベクターの一種は、エピソーム、すなわち染色体外複製が可能な核酸である。好ましいベクターは、自律複製および/またはそれらが連結されている核酸の発現が可能なものである。それらが作動可能に連結されている遺伝子の発現を指示することができるベクターは、本明細書中で「発現ベクター」と呼ぶ。本発明によるベクターは、それだけには限定されないが、YAC(酵母人工染色体)、BAC(細菌人工)、バキュロウイルスベクター、ファージ、ファージミド、コスミド、ウイルスベクター、プラスミド、RNAベクター、または染色体、非染色体、半合成もしくは合成のDNAからなり得る直鎖状もしくは環状のDNAもしくはRNA分子を含む。一般に、組換えDNA技法において有用な発現ベクターは、多くの場合は「プラスミド」の形態であり、これは一般に、そのベクター形態では染色体と結合していない環状の二本鎖DNAループをいう。多数の適切なベクターが当業者に知られている。ベクターは、選択マーカー、たとえば、真核細胞培養物用のネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸還元酵素、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミン合成酵素、およびヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ、出芽酵母用のTRP1、大腸菌中のテトラサイクリン、リファンピシンまたはアンピシリン耐性を含むことができる。好ましくは、前記ベクターは発現ベクターであり、対象のポリペプチドをコードしている配列は、前記ポリペプチドの産生または合成を可能にするために適切な転写および翻訳制御要素の制御下に置かれる。したがって、前記ポリヌクレオチドは発現カセット中に含まれる。より具体的には、ベクターは、複製起点、前記コードしているポリヌクレオチドと作動可能に連結されたプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを使用する場合)、ポリアデニル化部位および転写終結部位を含む。また、これは、エンハンサーまたはサイレンサー要素も含むことができる。プロモーターの選択は、ポリペプチドを発現させる細胞に依存する。適切なプロモーターには、組織特異的および/または誘導性プロモーターが含まれる。誘導性プロモーターの例は、増加した重金属レベルによって誘導される真核メタロチオニンプロモーター、イソプロピル−β−D−チオガラクト−ピラノシド(IPTG)に応答して誘導される原核lacZプロモーターおよび増加した温度によって誘導される真核熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異抗原(PSA)、α−抗トリプシンプロテアーゼ、ヒト界面活性化因子(SP)AおよびBタンパク質、β−カゼインならびに酸性ホエータンパク質遺伝子である。
【0064】
誘導性プロモーターは、病原体またはストレス、より好ましくは低温、熱、UV光、または高イオン濃度などのストレスによって誘導され得る(Potenzaら(2004)In vitro Cell Dev Biol、40:1〜22に総説)。誘導性プロモーターは化合物によって誘導され得る[Mooreら(2006)、Padidam(2003)、(Wangら(2003)、ならびに(ZuoおよびChua(2000)に総説]。
【0065】
送達ベクターおよびベクターは、超音波穿孔もしくは電気穿孔またはこれらの技法の派生法などの任意の細胞透過処理技法と関連させるまたは組み合わせることができる。
【0066】
複数の調節領域、たとえば、イントロン、エンハンサー、上流活性化領域、および誘導性要素が組換えポリヌクレオチド中に存在し得ることを理解されよう。
【0067】
組換え核酸構築体には、細胞(たとえば植物細胞または動物細胞)の形質転換に適したベクター内に挿入されたポリヌクレオチド配列が含まれることができる。組換えベクターは、たとえば標準の組換えDNA技法を使用して作製することができる(たとえばSambrookら(1989)分子クローニング:実験室の手引き(Molecular Cloning:A Laboratory Manual)、第2版、Cold Spring Harbor Laboratory、ニューヨーク州Cold Spring Harborを参照)。
【0068】
本明細書中に記載の組換え核酸配列は、非正統的な(すなわち、ランダム、非相同的、非部位特異的)組換えによって細胞のゲノム内に組み込まれることができるか、または、本明細書中に記載の組換え核酸配列は、相同組換えによって細胞のゲノム内に組み込まれるように適応させることができる。相同組換えによる組込みのために適応させた核酸配列は、両側が内在標的ヌクレオチド配列に類似または同一の配列で隣接されており、これは、内在標的ヌクレオチド配列を含有するゲノム中の特定の部位(複数可)での組換え核酸の組込みを促進する。また、相同組換えによる組込みのために適応させた核酸配列には、配列特異的ヌクレアーゼの認識部位も含まれることができる。あるいは、配列特異的ヌクレアーゼの認識部位は、形質転換させる細胞のゲノム中に位置することができる。以下に記載のドナー核酸配列は、典型的には、相同組換えによる組込みのために適応せる。
【0069】
また、一部の実施形態では、選択マーカーをコードしている核酸も相同組換えによる組み込のために適応させることができ、したがって、両側が植物ゲノム内の内在配列(たとえば、配列特異的ヌクレアーゼの切断部位での内在配列)と類似または同一の配列で隣接されていることができる。また、一部の事例では、選択マーカーのコード配列を含有する核酸には、配列特異的ヌクレアーゼの認識部位も含まれることができる。これらの実施形態では、配列特異的ヌクレアーゼの認識部位は、ドナー核酸配列内に含有されるものと同じまたは異なることができる(すなわち、ドナー核酸配列と同じヌクレアーゼによって認識される、またはドナー核酸配列とは異なるヌクレアーゼによって認識されることができる)。
【0070】
一部の事例では、組換え核酸配列は、部位特異的な組換えによって細胞のゲノム内に組み込まれるように適応させることができる。本明細書中で使用する「部位特異的な」組換えとは、組換え核酸中の配列とゲノム中の配列との間の相同性によってではなく、特定の核酸配列を認識してこれらの部位間のDNA鎖の相互交換を触媒するリコンビナーゼ酵素の作用によって、核酸配列がゲノム内の特定の部位(複数可)に対して標的化されている場合に起こる組換えをいう。したがって、部位特異的な組換えとは、2つの定義されたヌクレオチド配列の酵素媒介性の切断およびライゲーションをいう。たとえばCre−lox系またはFLP−FRT系を含めた、任意の適切な部位特異的な組換え系を使用することができる。そのような実施形態では、ドナーヌクレオチド配列およびヌクレアーゼをコードしている配列、ならびに一部の事例では選択マーカー配列に加えて、リコンビナーゼ酵素をコードしている核酸を細胞内に導入し得る。たとえば米国特許第4,959,317号を参照されたい。
【0071】
配列特異的エンドヌクレアーゼ
配列特異的ヌクレアーゼおよび配列特異的エンドヌクレアーゼをコードしている組換え核酸が本明細書中で提供されている。配列特異的エンドヌクレアーゼには、TALエフェクターDNA結合ドメインおよびエンドヌクレアーゼドメインが含まれることができる。したがって、そのような配列特異的エンドヌクレアーゼをコードしている核酸には、ヌクレアーゼからのヌクレオチド配列と連結された配列特異的TALエフェクターからのヌクレオチド配列が含まれることができる。
【0072】
TALエフェクターとは、病原体によって植物細胞内に注入され、ここで核へと移動して転写因子として機能して特定の植物遺伝子を活性化する、植物病原細菌のタンパク質である。TALエフェクターの一次アミノ酸配列が、それが結合するヌクレオチド配列指示する。したがって、TALエフェクターの標的部位を予測することができ、また、TALエフェクターは、本明細書中に記載のように、特定のヌクレオチド配列と結合させる目的で操作設計および作製することもできる。
【0073】
TALエフェクターをコードしている核酸配列には、ヌクレアーゼまたはヌクレアーゼの一部分、典型的にはFokIなどのII型制限エンドヌクレアーゼからの非特異的な切断ドメインをコードしている配列が融合している(Kimら(1996)Proc.Natl.Acad.Sci.USA、93:1156〜1160)。他の有用なエンドヌクレアーゼには、たとえば、HhaI、HindIII、NotI、BbvCI、EcoRI、BglI、およびAlwIが含まれ得る。一部のエンドヌクレアーゼ(たとえばFokI)が二量体としてのみ機能することは、TALエフェクターの標的特異性を増強させる際に利用することができる。たとえば、一部の事例では、それぞれのFokI単量体を、異なるDNA標的配列を認識するTALエフェクター配列と融合させることができ、2つの認識部位が近く近接している場合にのみ、不活性な単量体が一体となって機能的な酵素を生じる。ヌクレアーゼを活性化させるためにDNA結合を必要とすることによって、高度に部位特異的な制限酵素を作製することができる。
【0074】
本明細書中で提供されている配列特異的TALENは、細胞中に存在する事前に選択された標的ヌクレオチド配列内の特定の配列を認識することができる。したがって、一部の実施形態では、標的ヌクレオチド配列をヌクレアーゼ認識部位について走査することができ、標的配列に基づいて特定のヌクレアーゼを選択することができる。他の事例では、特定の細胞配列を標的とするようにTALENを操作することができる。所望のTALENをコードしているヌクレオチド配列を任意の適切な発現ベクター内に挿入することができ、1つまたは複数の発現制御配列と連結させることができる。たとえば、ヌクレアーゼのコード配列は、形質転換させる植物種中でエンドヌクレアーゼの構成的発現をもたらすプロモーター配列と作動可能に連結されていることができる。あるいは、エンドヌクレアーゼのコード配列は、条件的発現(たとえば特定の栄養条件下での発現)をもたらすプロモーター配列と作動可能に連結されていることができる。たとえば、カリフラワーモザイクウイルスの35Sプロモーターを構成的発現に使用することができる。他の構成的プロモーターには、それだけには限定されないが、ノパリン合成酵素プロモーター、ユビキチンプロモーター、およびアクチンプロモーターが含まれる。一部の実施形態では、人工エストロゲン誘導性プロモーターを条件的発現に使用することができ、植物がエストロゲンに曝露された際に高レベルの転写を達成することができる。使用することができる他の条件的プロモーターには、たとえば、熱誘導性熱ショック遺伝子プロモーター、およびリブロースビスリン酸カルボキシラーゼの大サブユニットをコードしている遺伝子からのものなどの光調節性プロモーターが含まれる。
【0075】
治療目的には、本発明のTALエフェクター−DNA修飾酵素および薬学的に許容される賦形剤を治療上有効な量で投与する。そのような組合せは、投与した量が生理的に有意である場合に、「治療上有効な量」で投与されたという。薬剤は、その存在がレシピエントの生理学の検出可能な変化をもたらす場合に、生理的に有意である。本コンテキストでは、薬剤は、その存在が標的疾患の1つまたは複数の症状の重篤度の減少および病変または異常のゲノム補正をもたらす場合に、生理的に有意である。標的化DNAおよび/またはTALエフェクター−DNA修飾酵素をコードしている核酸を含むベクターは、様々な方法(たとえば、注入、直接取り込み、噴出性照射、リポソーム、電気穿孔)によって細胞内に導入することができる。TALエフェクター−DNA修飾酵素は、発現ベクターを使用して細胞内で安定にまたは一過的に発現させることができる。真核細胞中での発現の技法は当業者に周知である。(ヒト遺伝学の最新プロトコル(Current Protocols in Human Genetics):第12章「遺伝子治療のベクター(Vectors For Gene Therapy)」および第13章「遺伝子治療の送達系(Delivery Systems for Gene Therapy)」を参照)。
【0076】
本発明のさらなる一態様では、TALエフェクター−DNA修飾酵素は実質的に非免疫原性である、すなわち、有害な免疫学的応答をわずかしかまたはまったく生じない。この種の有害な免疫学的反応を寛解させるまたは排除するための様々な方法を、本発明に従って使用することができる。好ましい実施形態では、TALエフェクター−DNA修飾酵素はN−ホルミルメチオニンを実質的に含まない。望まない免疫学的反応を回避するための別の方法は、TALエフェクター−DNA修飾酵素をポリエチレングリコール(「PEG」)またはポリプロピレングリコール(「PPG」)(好ましくは500〜20,000ダルトンの平均分子量(MW)のもの)とコンジュゲートさせることである。たとえばDavisら(US4,179,337号)によって記載されている、PEGまたはPPGとのコンジュゲーションは、抗ウイルス活性を有する、非免疫原性の生理的に活性のある水溶性のTALエフェクター−DNA修飾酵素のコンジュゲートを提供することができる。やはりポリエチレン−ポリプロピレングリコールのコポリマーを使用した類似の方法が、Saiferら(US5,006,333号)に記載されている。
【0077】
ドナーベクター
また、ドナーヌクレオチド配列を含めた組換え核酸も、本明細書中で提供されている。ドナーヌクレオチド配列には、形質転換させる細胞のゲノム内に内在的に見つかる事前に選択された標的ヌクレオチド配列に関して1つまたは複数の修飾(すなわち、置換、欠失、または挿入)を有する変異体配列が含まれることができる(本明細書中で「修飾された標的ヌクレオチド配列」とも呼ばれる)。ドナー核酸内の変異体配列は、典型的には、両側が細胞内の内在標的ヌクレオチド配列と類似または同一の配列で隣接されている。隣接配列は任意の適切な長さを有することができ、典型的には、少なくとも50個のヌクレオチドの長さである(たとえば、少なくとも50個のヌクレオチド、少なくとも75個のヌクレオチド、少なくとも100個のヌクレオチド、少なくとも200個のヌクレオチド、少なくとも250個のヌクレオチド、少なくとも300個のヌクレオチド、少なくとも500個のヌクレオチド、少なくとも750個のヌクレオチド、少なくとも1000個のヌクレオチド、約50〜約5000個のヌクレオチド、約100〜2500個のヌクレオチド、約100〜約1000個のヌクレオチド、約100〜500個のヌクレオチド、約200〜約500個のヌクレオチド、または約250〜400個のヌクレオチド)。したがって、相同組換えは、生じる細胞のゲノムが、たとえば同じ遺伝子からの内在配列のコンテキスト内の変異体配列を含有するように、組換えドナー核酸構築体と変異体配列の両側の内在標的との間で起こることができる。ドナーヌクレオチド配列は、ゲノム内の任意の適切な配列を標的とするように作製することができる。たとえば、植物では、ドナーヌクレオチド配列は、脂質生合成遺伝子、炭水化物生合成遺伝子、種子貯蔵タンパク質遺伝子、疾患もしくは害虫耐性遺伝子、ストレス寛容遺伝子、乾燥寛容遺伝子、または抗栄養因子を産生する遺伝子を標的とすることができる。さらに、ドナーヌクレオチド配列は、本明細書中に記載の配列特異的ヌクレアーゼの認識部位を含有する。
【0078】
選択マーカー
本明細書中で提供されている方法の一部には、選択可能またはスクリーニング可能なマーカーをコードしている第3の組換え核酸の使用が含まれる。選択可能な特色をもたらすポリペプチドをコードしているヌクレオチド配列を、1つまたは複数の発現制御配列を含有する発現ベクター内に取り込ませることができる。たとえば、発現ベクターには、形質転換させる植物細胞中で構成的発現をもたらすプロモーター配列と作動可能に連結された選択マーカーをコードしている配列が含まれることができる。適切な選択マーカーには、それだけには限定されないが、カナマイシン、G418、ブレオマイシン、アンピシリン、もしくはハイグロマイシンなどの抗生物質、またはグルホシネート、クロロスルフロン(chlorosulfuron)、もしくはホスフィノスリシンなどの除草剤に対する耐性を与えるポリペプチドが含まれることができる。
【0079】
たとえば、植物で使用するための実施形態では、選択マーカーは、イミダゾリノンまたはスルホニル尿素などの成長点または分裂組織を阻害する除草剤に対する耐性を与えることができる。この突然変異ALSおよびAHAS酵素の分類コード中の例示的なポリペプチドは、たとえば、米国特許第5,767,366号および第5,928,937号に記載されている。米国特許第4,761,373号および第5,013,659号は、様々なイミダゾリノンまたはスルホンアミド除草剤に耐性の植物に向けられている。米国特許第4,975,374号は、GSを阻害することが知られている除草剤、たとえばホスフィノスリシンおよびメチオニンスルホキシミンによる阻害に耐性の突然変異グルタミン合成酵素(GS)をコードしている遺伝子を含有する植物細胞および植物に関する。米国特許第5,162,602号は、シクロヘキサンジオンおよびアリールオキシフェノキシプロパン酸除草剤による阻害に耐性の植物を開示している。耐性は、変更されたアセチル補酵素Aカルボキシラーゼ(ACCase)によって与えられる。
【0080】
また、グリフォセート(商品名Roundup(登録商標)の下で販売)に耐性のポリペプチドも、植物における使用に適している。たとえば、米国特許第4,940,835号および第4,769,061号を参照されたい。米国特許第5,554,798号は、トランスジェニックグリフォセート耐性トウモロコシ植物を開示しており、耐性は変更された5−エノールピルビル−3−ホスホシキメート(EPSP)合成酵素によって与えられる。そのようなポリペプチドは、それだけには限定されないが、トリメチルスフホニウム塩、イソプロピルアミン塩、ナトリウム塩、カリウム塩およびアンモニウム塩などのグリフォセート塩を含めた、グリフォセート除草組成物に対する耐性を与えることができる。たとえば、米国特許第6,451,735号および第6,451,732号を参照されたい。
【0081】
また、グルホシネートアンモニウムまたはホスフィノスリシンなどのホスホノ化合物、ならびにピリジノキシまたはフェノキシプロピオン酸およびシクロヘキソンに耐性のポリペプチドも適切である。たとえば、欧州公開第0 242 246号、ならびに米国特許第5,879,903号、第5,276,268号、および第5,561,236号を参照されたい。
【0082】
他の除草剤には、トリアジンおよびベンゾニトリル(ニトリラーゼ)などの光合成を阻害するものが含まれる。たとえば米国特許第4,810,648号を参照されたい。他の除草剤には、2,2−ジクロロプロピオン酸、セトキシジム、ハロキシホップ、イミダゾリノン除草剤、スルホニル尿素除草剤、トリアゾロピリミジン除草剤、s−トリアジン除草剤およびブロモキシニルが含まれる。また、プロトックス(protox)酵素に対する耐性を与える除草剤も適している。たとえば、米国特許公開第20010016956号および米国特許第6,084,155号を参照されたい。
【0083】
一部の実施形態では、選択マーカーをコードしている組換え核酸は、部位特異的な組換えによって細胞(たとえば植物細胞または動物細胞)のゲノム内に組み込ませるために適応させることができる。たとえば、選択マーカーをコードしている配列は、たとえばCreまたはFLPなどのリコンビナーゼの認識配列によって隣接されていることができる。他の実施形態では、選択マーカーをコードしている組換え核酸は、相同組換えによる植物ゲノム内への組込みのために適応させることができる。そのような核酸では、選択マーカーをコードしている配列は、それ内に組換え核酸を導入する植物細胞のゲノム中に見つかる内在ヌクレオチド配列と類似または同一の配列によって隣接されていることができる。内在配列のうちの少なくとも1つが配列特異的ヌクレアーゼの切断部位にあることができる。また、選択マーカーをコードしている核酸は、配列特異的ヌクレアーゼの認識部位も含有することができる。ヌクレアーゼは、ドナーヌクレオチド配列に対して標的化されているものと同じ配列特異的ヌクレアーゼ、またはドナーヌクレオチド配列に対して標的化されているものとは異なる配列特異的ヌクレアーゼであることができる。さらに他の実施形態では、選択マーカーをコードしている組換え核酸は、非正統的な組換えによる植物細胞のゲノム内への組込みのために適応させることができる。そのような核酸は、典型的には、本明細書中に記載の相同的または部位特異的な組換えのために適応させた核酸内に含有される隣接配列およびヌクレアーゼ認識部位を欠く。
【0084】
方法
本明細書中で提供されている構築体のうちの1つまたは複数を使用して、遺伝子改変した生物(たとえば植物または動物)が作製されるように、細胞を形質転換させることができるおよび/またはDNA修飾酵素を細胞内に導入することができる。したがって、本明細書中に記載の核酸および/またはポリペプチド(polypeptdes)を含有する遺伝子改変した生物および細胞も提供される。一部の実施形態では、形質転換細胞は、組換え核酸構築体がそのゲノム内に組み込まれている、すなわち安定に形質転換させることができる。安定に形質転換させた細胞は、典型的には、導入された核酸配列を各細胞分裂で保持する。構築体は、形質転換細胞に内在性のヌクレオチド配列が、内在配列に対応する配列を含有するが内在配列に関して1つまたは複数の修飾を含有する構築体によって置き換えられるように、相同的な様式で組み込まれることができる。そのような修飾された内在配列を含有する植物または動物は本明細書中で「遺伝子改変した生物」(GMO)と呼び得るが、修飾された内在配列は導入遺伝子とみなされないことに留意されたい。また、構築体は、形質転換細胞のゲノム内にランダムに組み込まれるように、非正統的な様式で組み込まれることもできる。
【0085】
あるいは、構築体がそのゲノム内に組み込まれないように、細胞を一過的に形質転換させることができる。たとえば、TALENのコード配列が発現されるが、ベクターがゲノム中に安定に組み込まれていないように、TALENのコード配列を含有するプラスミドベクターを細胞内に導入することができる。一過的に形質転換させた細胞は、典型的には、導入された核酸が十分な回数の細胞分裂の後に娘細胞中で検出できないように、導入された核酸構築体の一部またはすべてを各それぞれの細胞分裂で失う。それにもかかわらず、TALENのコード配列の発現は、ドナー配列と内在標的配列との間の相同組換えを達成するために十分である。一過的に形質転換させたおよび安定に形質転換させた細胞はどちらも本明細書中に記載の方法において有用な可能性がある。
【0086】
特に遺伝子改変した植物細胞に関して、本明細書中に記載の方法で使用する細胞は、全植物の一部または全体を構成することができる。そのような植物は、検討中の種に適した様式で、成長チャンバ、温室、または畑のいずれかで成長させることができる。遺伝子改変した植物は、特定の目的、たとえば、組換え核酸を他の系に導入する、組換え核酸を他の種に転移させる、または他の望ましい特色をさらに選択するために所望されるように、育種することができる。あるいは、遺伝子改変した植物は、そのような技法を受け入れられる種では栄養繁殖させることができる。子孫には特定の植物または植物系の子孫が含まれる。本発明の植物の子孫には、F1、F2、F3、F4、F5、F6および続く世代の植物上で形成された種子、またはBC1、BC2、BC3、および続く世代の植物上で形成された種子、またはF1BC1、F1BC2、F1BC3、および続く世代の植物上で形成された種子が含まれる。遺伝子改変した植物によって生成された種子を成長させ、その後、自家受粉(または異系交配および自家受粉)させて、核酸構築体にホモ接合性の種子を得ることができる。
【0087】
遺伝子改変した細胞(たとえば植物細胞または動物細胞)は、所望する場合は、懸濁培養、または組織もしくは器官培養中で成長させることができる。本明細書中で提供されている方法の目的のために、固体および/または液体の組織培養技法を使用することができる。固体培地を使用する場合、細胞を培地上に直接置くことができるか、またはフィルターフィルム上に置き、その後それを培地と接触させることができる。液体培地を使用する場合、細胞を浮遊装置、たとえば液体培地と接触する多孔性の膜上に置くことができる。固体培地は、典型的には、液体培地から、寒天を加えることによって作製する。たとえば、固体培地は、寒天および適切な濃度のオーキシン、たとえば2,4−ジクロロフェノキシ酢酸(2,4−D)、および適切な濃度のサイトカイニン、たとえばキネチンを含有する、MurashigeおよびSkoog(MS)培地であることができる。
【0088】
細胞は、1個の組換え核酸構築体または複数(たとえば、2、3、4、もしくは5個)の組換え核酸構築体で形質転換させることができる。複数の構築体を利用する場合は、これらは同時にまたは順次形質転換させることができる。多種多様の種を形質転換させる技法が当分野で知られている。本明細書中に記載のポリヌクレオチドおよび/または組換えベクターは、電気穿孔、微量注入、および微粒子銃方法を含めたいくつかの知られている方法のうちの任意のものを使用して、宿主のゲノム内に導入することができる。あるいは、ポリヌクレオチドまたはベクターを適切なT−DNA隣接領域と組み合わせて、慣用のAgrobacterium tumefaciens宿主ベクター内に導入することができる。無害化(disarming)およびバイナリーベクターの使用を含めた、そのようなAgrobacterium tumefaciensに媒介される形質転換技法は当分野で周知である。他の遺伝子移入および形質転換技法には、カルシウムまたはPEGによるプロトプラストの形質転換、電気穿孔に媒介される裸DNAの取り込み、リポソームに媒介される形質移入、電気穿孔、ウイルスベクターに媒介される形質転換、および微粒子銃が含まれる(たとえば、米国特許第5,538,880号、第5,204,253号、第5,591,616号、および第6,329,571号を参照)。植物細胞または組織培養物を形質転換のレシピエント組織として使用する場合は、当業者に知られている技法を使用して、形質転換させた培養物から植物を再生することができる。
【0089】
一部の実施形態では、DNA修飾酵素(たとえばTALEN)を細胞内に直接導入することができる。たとえば、機械的注入、細菌III型分泌系による送達、電気穿孔、またはAgrobacteriumに媒介される転移によって、ポリペプチドを細胞内に導入することができる。たとえば、AgrobacteriumのVirB/D4輸送系、および核タンパク質T複合体の植物細胞への転移を媒介するためのその使用に関する記述には、Vergunstら(2000)Science、290:979〜982を参照されたい。
【0090】
さらに植物に関して、本明細書中に記載のポリヌクレオチド、ベクターおよびポリペプチドは、ベニバナ、アルファルファ、ダイズ、コーヒー、アマランス、ナタネ(高エルカ酸およびアブラナ)、ピーナッツまたはヒマワリなどの双子葉植物、およびアブラヤシ、サトウキビ、バナナ、スーダングラス、トウモロコシ、コムギ、ライムギ、オオムギ、カラスムギ、コメ、アワ、またはソルガムなどの単子葉植物を含めた、いくつかの単子葉および双子葉の植物および植物細胞系内に導入することができる。また、モミおよびマツなどの裸子植物も適切である。
【0091】
したがって、本明細書中に記載の方法は、たとえば、Magniolales、Illiciales、Laurales、Piperales、Aristochiales、Nymphaeales、Ranunculales、Papeverales、Sarraceniaceae、Trochodendrales、Hamamelidales、Eucomiales、Leitneriales、Myricales、Fagales、Casuarinales、Caryophyllales、Batales、Polygonales、Plumbaginales、Dilleniales、Theales、Malvales、Urticales、Lecythidales、Violales、Salicales、Capparales、Ericales、Diapensales、Ebenales、Primulales、Rosales、Fabales、Podostemales、Haloragales、Myrtales、Cornales、Proteales、Santales、Rafflesiales、Celastrales、Euphorbiales、Rhamnales、Sapindales、Juglandales、Geraniales、Polygalales、Umbellales、Gentianales、Polemoniales、Lamiales、Plantaginales、Scrophulariales、Campanulales、Rubiales、Dipsacales、およびAsteralesの目に属する双子葉植物を用いて利用することができる。また、本明細書中に記載の方法は、Alismatales、Hydrocharitales、Najadales、Triuridales、Commelinales、Eriocaulales、Restionales、Poales、Juncales、Cyperales、Typhales、Bromeliales、Zingiberales、Arecales、Cyclanthales、Pandanales、Arales、Lilliales、およびOrchidalesの目に属するものなどの単子葉植物を用いて、または裸子植物類に属する植物、たとえば、Pinales、Ginkgoales、CycadalesおよびGnetalesを用いても利用することができる。
【0092】
本方法は、双子葉植物の属であるAtropa、Alseodaphne、Anacardium、Arachis、Beilschmiedia、Brassica、Carthamus、Cocculus、Croton、Cucumis、Citrus、Citrullus、Capsicum、Catharanthus、Cocos、Coffea、Cucurbita、Daucus、Duguetia、Eschscholzia、Ficus、Fragaria、Glaucium、Glycine、Gossypium、Helianthus、Hevea、Hyoscyamus、Lactuca、Landolphia、Linum、Litsea、Lycopersicon、Lupinus、Manihot、Majorana、Malus、Medicago、Nicotiana、Olea、Parthenium、Papaver、Persea、Phaseolus、Pistacia、Pisum、Pyrus、Prunus、Raphanus、Ricinus、Senecio、Sinomenium、Stephania、Sinapis、Solanum、Theobroma、Trifolium、Trigonella、Vicia、Vinca、Vitis、およびVigna、単子葉植物の属であるAllium、Andropogon、Aragrostis、Asparagus、Avena、Cynodon、Elaeis、Festuca、Festulolium、Heterocallis、Hordeum、Lemna、Lolium、Musa、Oryza、Panicum、Pannesetum、Phleum、Poa、Secale、Sorghum、Triticum、およびZea、または裸子植物の属であるAbies、Cunninghamia、Picea、Pinus、およびPseudotsugaからの種を含めた、幅広い範囲の植物種にわたって使用することができる。
【0093】
形質転換細胞、カルス、組織、または植物は、操作された細胞を、特定の特色または活性、たとえばマーカー遺伝子または抗生物質耐性遺伝子によってコードされているものについて選択またはスクリーニングすることによって、同定および単離することができる。そのようなスクリーニングおよび選択方法は当業者には周知である。さらに、物理的および生化学的方法を使用して形質転換体を同定することができる。これらには、ポリヌクレオチドを検出するためのサザン分析およびPCR増幅、RNA転写物を検出するためのノーザンブロット、S1 RNase保護、プライマー伸長、またはRT−PCR増幅、ポリペプチドおよびポリヌクレオチドの酵素またはリボザイム活性を検出するための酵素アッセイ、ならびにポリペプチドを検出するためのタンパク質ゲル電気泳動、ウエスタンブロット、免疫沈降、および酵素連結免疫アッセイが含まれる。また、in situハイブリダイゼーション、酵素染色、および免疫染色などの他の技法も、ポリペプチドおよび/またはポリヌクレオチドの存在または発現を検出するために使用することができる。言及した技法のすべてを行う方法は周知である。植物細胞内に安定に取り込まれたポリヌクレオチドは、たとえば標準の育種技法を使用して他の植物内に導入することができる。
【0094】
本発明のコンテキストでは、「真核細胞」とは、真菌、酵母、植物もしくは動物細胞、または以下に記載し、in vitro培養のために確立させた生物に由来する細胞系をいう。より好ましくは、真菌は、Aspergillus、Penicillium、Acremonium、Trichoderma、Chrysoporium、Mortierella、KluyveromycesまたはPichiaの属のものであることができる。より好ましくは、真菌は、Aspergillus niger、Aspergillus nidulans、Aspergillus oryzae、Aspergillus terreus、Penicillium chrysogenum、Penicillium citrinum、Acremonium chrysogenum、Trichoderma reesei、Mortierella alpine、Chrysosporium lucknowense、Kluyveromyces lactis、Pichia pastorisまたはPichia ciferriiの種のものであることができる。
【0095】
本発明では、植物は、Arabidospis、Nicotiana、Solanum、Lactuca、Brassica、Oryza、Asparagus、Pisum、Medicago、Zea、Hordeum、Secale、Triticum、Capsicum、Cucumis、Cucurbita、Citrullis、Citrus、またはSorghumの属のものであることができる。より好ましくは、植物は、Arabidospis thaliana、Nicotiana tabaccum、Solanum lycopersicum、Solanum tuberosum、Solanum melongena、Solanum esculentum、Lactuca saliva、Brassica napus、Brassica oleracea、Brassica rapa、Oryza glaberrima、Oryza sativa、Asparagus officinalis、Pisum sativum、Medicago sativa、Zea mays、Hordeum vulgare、Secale cereal、Triticum aestivum、Triticum durum、Capsicum sativus、Cucurbita pepo、Citrullus lanatus、Cucumis melo、Citrus aurantifolia、Citrus maxima、Citrus medica、またはCitrus reticulataの種のものであることができる。
【0096】
本発明では、動物細胞は、Homo、Rattus、Mus、Sus、Bos、Danio、Canis、Felis、Equus、Salmo、Oncorhynchus、Gallus、Meleagris、Drosophila、またはCaenorhabditisの属のものであることができる。より好ましくは、動物細胞は、Homo sapiens、Rattus norvegicus、Mus musculus、Sus scrofa、Bos taurus、Danio rerio、Canis lupus、Felis catus、Equus caballus、Oncorhynchus mykiss、Gallus gallus、またはMeleagris gallopavoの種のものであることができる。動物細胞は、非限定的な例としてSalmo salar、硬骨魚またはブラフィッシュの種からの魚細胞であることができる。また、本発明における動物細胞は、非限定的な例としてキイロショウジョウバエ(Drosophila melanogaster)からの昆虫細胞であることもできる。また、動物細胞は、非限定的な例としてエレガンス線虫(Caenorhabditis elegans)からの寄生虫細胞であることもできる。
【0097】
本発明では、細胞は、植物細胞、哺乳動物細胞、魚細胞、昆虫細胞もしくはin vitro培養のためにこれらの生物から誘導した細胞系、または生組織から直接採取し、in vitro培養のために確立させた初代細胞であることができる。非限定的な例として、細胞系は、CHO−K1細胞、HEK293細胞、Caco2細胞、U2−OS細胞、NIH 3T3細胞、NSO細胞、SP2細胞、CHO−S細胞、DG44細胞、K−562細胞、U−937細胞、MRC5細胞、IMR90細胞、ジャーカット細胞、HepG2細胞、HeLa細胞、HT−1080細胞、HCT−116細胞、Hu−h7細胞、Huvec細胞、Molt 4細胞からなる群から選択されることができる。
【0098】
これらの細胞系はすべて、本発明の方法によって改変して、対象の遺伝子またはタンパク質を産生、発現、定量、検出、研究するための細胞系モデルを提供することができる。また、これらのモデルを使用して、非限定的な例として化学薬品、生物燃料、治療剤および農学などの様々な分野における研究および生成において、対象の生物活性のある分子をスクリーニングすることもできる。
【0099】
また、本発明は、たとえば、細胞内の遺伝物質を変更するため、遺伝子発現を変調させるため、およびたとえば抗ウイルス治療において病原性配列を標的とするために、TALエフェクター内の配列特異的DNA結合ドメインを利用する方法も提供する。たとえば、一部の実施形態では、本発明は、細胞遺伝物質を改変する方法を提供する。一部の実施形態では、本方法には、TALエフェクターDNA結合ドメインを含有するポリペプチド、またはそのようなポリペプチドをコードしている核酸を、細胞内に導入することが含まれる。TALエフェクターDNA結合ドメインは、DNA修飾酵素(たとえばエンドヌクレアーゼ)の全体または一部分と融合させることができる。一部の実施形態では、本方法には、2つ以上の組換え核酸を細胞内に導入することが含まれる。第1の組換え核酸は、細胞中に見つかる対応する事前に選択された標的ヌクレオチド配列に関して1つまたは複数の修飾(すなわち、置換、欠失、または挿入)が含まれる、ドナーヌクレオチド配列を含有する。ドナーヌクレオチド配列は、内在配列またはその一部分がドナー配列またはその一部分で置き換えられているように、内在標的ヌクレオチド配列との相同組換えを受けることができる。標的ヌクレオチド配列には、典型的には配列特異的TALENの認識部位が含まれる。一部の事例では、標的ヌクレオチド配列には、2つ以上の明白に異なるTALENの認識部位(たとえば、明白に異なるDNA配列結合特異性を有するTALENを使用できるように、明白に異なる2つの向かい合った標的配列)が含まれることができる。そのような事例では、DNA切断の特異性は、1つのみの標的配列(または同じ標的配列の複数コピー)を使用する事例と比較して増加している場合がある。
【0100】
第2の組換え核酸は、標的ヌクレオチド配列中の認識部位と結合する配列特異的TALENをコードしているヌクレオチド配列を含有する。一部の事例では、ドナーヌクレオチド配列および配列特異的ヌクレアーゼをコードしているヌクレオチド配列は、同じ核酸構築体中に含有されることができる。あるいは、ドナーヌクレオチド配列およびTALENのコード配列は、別々の構築体中に含有されることができるか、または、TALENポリペプチドを生成して細胞内に直接導入することができる。
【0101】
一部の実施形態では、選択マーカーをコードしているヌクレオチド配列を含有する第3の組換え核酸も使用し得る。第2および第3の組換え核酸は、内在配列との組換えを受け、したがって細胞のゲノム内に組み込まれ得る。これらの組換え事象は非正統的(すなわちランダム)であることができるか、または、これらは相同組換えもしくは部位特異的な組換えによって起こることができる。組換え核酸は、細胞内に同時にまたは順次形質転換させることができ、形質転換の前に直鎖状にすることができる。
【0102】
細胞が植物細胞である場合は、本明細書中で提供されている方法には、形質転換細胞を含有する植物を作製するステップ、植物の子孫を作製するステップ、選択マーカー(含めた場合)を発現する植物を選択またはスクリーニングするステップ、選択された植物の子孫を作製するステップ、および植物(たとえば、組織、種子、前駆細胞、もしくは全植物)または植物の子孫を標的ヌクレオチド配列での組換えについて試験するステップなどのステップがさらに含まれることができる。一部の事例では、本方法には、選択された植物を異系交配させて選択マーカーを除去すること、および/または選択されたもしくは異系交配させた植物を配列特異的ヌクレアーゼの非存在についてスクリーニングすることが含まれることができる。
【0103】
一部の実施形態では、本発明は、細胞、たとえば、原核細胞、動物細胞、または植物細胞の遺伝物質を改変する方法を提供する。本方法には、細胞中に存在する対応する標的ヌクレオチド配列に関してヌクレオチド配列中に1つまたは複数の修飾が含まれる修飾された標的ヌクレオチド配列と、配列特異的TALENの認識部位とを含有する第1の組換え核酸、および配列特異的TALENをコードしているヌクレオチド配列を含有する第2の組換え核酸を、細胞内に導入することが含まれることができる。細胞が植物細胞である場合は、細胞を含有する植物を作製することができ、植物(またはその子孫)から得られた細胞、種子、または組織を、標的ヌクレオチド配列での組換えについて分析することができる。第1および第2の組換え核酸を同時にまたは連続的に細胞内に形質転換させることができ、一方または両方を形質転換の前に直鎖状にし得る。一部の事例では、第1および第2の組換え核酸は同じ構築体中に存在することができる。
【0104】
一部の事例では、本方法には、選択マーカーをコードしているヌクレオチド配列を含有する第3の組換え核酸を細胞内に導入すること、および細胞、細胞から作製した生物、またはその子孫が選択マーカーを発現するかどうかを決定することも含まれることができる。本方法には、さらに、細胞、生物またはその子孫を選択マーカーの非存在についてスクリーニングすることが含まれることができる。選択マーカーをコードしているヌクレオチド配列は、その両側が、第2の配列特異的ヌクレアーゼの切断部位で細胞に内在性のヌクレオチド配列と類似もしくは同一のヌクレオチド配列によって、または配列特異的リコンビナーゼの認識部位によって隣接されていてもいなくてもよい。一部の事例では、本方法には、生物を異系交配させるステップも含まれることができる。異系交配の子孫は、選択マーカーの非存在についてスクリーニングすることができる。
【0105】
また、本発明は、それ内で相同組換えが起こることが所望される標的DNA配列、たとえば、染色体、ミトコンドリア、または葉緑体の配列を含有する細胞を提供するステップと、DNA修飾酵素ドメイン(たとえばエンドヌクレアーゼドメイン)および組み合わせて標的DNA配列内の特定のヌクレオチド配列と結合する複数のTALエフェクター反復を有するTALエフェクタードメインを含有するTALENを提供するステップと、標的DNAの少なくとも一部分に相同的な配列を含有する核酸を提供するステップと、細胞中の標的DNA配列内にあるまたはそれに隣接するヌクレオチド配列の両方の鎖が切断されるように、細胞中の標的DNA配列をTALエンドヌクレアーゼと接触させるステップとを含む、細胞(たとえば植物細胞または動物細胞)の遺伝物質を改変する方法も提供する。そのような切断は、標的DNA配列での相同組換えの頻度を増強させることができる。標的DNA配列は細胞に内在性であることができる。本方法には、TALエンドヌクレアーゼをコードしているcDNAを含有するベクターを細胞内に導入すること、およびTALエンドヌクレアーゼタンパク質を細胞中で発現させることが含まれることができる。一部の事例では、TALエンドヌクレアーゼタンパク質自体を、たとえば、機械的注入、細菌III型分泌系による送達、電気穿孔、またはAgrobacteriumに媒介される転移によって、細胞内に導入することができる。
【0106】
本明細書中に記載の方法は、様々な状況において使用することができる。たとえば、農業では、本明細書中に記載の方法は、以前に組み込んだ導入遺伝子(たとえば除草剤耐性の導入遺伝子)を植物の系、変種、またはハイブリッドから除去するために使用できる、標的部位での相同組換えを促進するために有用である。また、本明細書中に記載の方法を使用して、遺伝子によってコードされている酵素が除草剤耐性を与えるように内在遺伝子を改変することもでき、その例は、改変された酵素がグリフォセート除草剤に対する耐性を与えるような、内在性5−エノールピルビルシキメート−3−リン酸(EPSP)合成酵素遺伝子の改変である。別の例として、本明細書中に記載の方法は、植物または哺乳動物の代謝経路(たとえば脂肪酸生合成)中の1つまたは複数の内在遺伝子の調節領域での相同組換えを、そのような遺伝子の発現が所望の様式で改変されているように促進するために有用である。本明細書中に記載の方法は、動物(たとえばラットまたはマウス)において、非限定的な例として、細胞表面マーカーをコードしているものなどの代謝および内部シグナル伝達経路に関与している1つまたは複数の対象の内在遺伝子、特定の疾患に連関していると同定されている遺伝子、ならびに動物細胞の特定の表現型を司っていることが知られている任意の遺伝子の相同組換えを促進するために有用である。
【0107】
また、本発明は、特定のDNA配列(たとえば、特定の位置でDNAを切断することができるTALEN)と相互作用することができる配列特異的TALエフェクターを設計する方法も提供する。本方法には、TALエフェクターが結合することが所望される標的ヌクレオチド配列(たとえば、内在染色体配列、ミトコンドリアDNA配列、または葉緑体DNA配列)(たとえば、それ内に二本鎖切断を導入することが所望される第2のヌクレオチド配列に隣接する配列)を同定すること、および組み合わせて標的配列と結合する複数のDNA結合反復を含有する配列特異的TALエフェクターを設計することが含まれることができる。本明細書中に記載のように、TALエフェクターには、それらがDNAと相互作用する特異性を決定するいくつかの不完全な反復が含まれる。それぞれの反復は、反復の残基12および13での特定のジアミノ酸配列に応じて、単一の塩基と結合する。したがって、TALエフェクター内の反復を操作することによって(たとえば、標準の技法または本明細書中に記載の技法を使用)、特定のDNA部位を標的化することができる。そのような操作したTALエフェクターは、たとえば、特定のDNA配列を標的とした転写因子として使用することができる。一般的なTALエフェクターの図が図1Aに示されており、反復領域は白い四角によって示されており、代表的な反復配列(配列番号1)中のRVDに下線が引いてある。
【0108】
RVDおよびその対応する標的ヌクレオチドの例が表1Aに示されている(PCT公開WO2010/079430号も参照)。
【表1】

【0109】
他のRVDおよびその対応する標的ヌクレオチドが表1Bに示されている。
【表2】
【0110】
配列特異的なDNA切断が所望される場合は、たとえば、配列特異的TALENを、(a)組み合わせて内在染色体ヌクレオチド配列と結合する複数のDNA結合反復ドメイン、および(b)第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含有するように設計することができる。そのような配列特異的なDNA切断は、本明細書中に記載のように、相同組換えを増強させるために有用な可能性がある。TALENの他の使用には、たとえばウイルスに対する治療剤としての使用が含まれる。TALENは、特定のウイルス配列を標的とする、ウイルスDNAを切断するおよび病原性を低下または消失させるために操作することができる。
【0111】
本明細書中で提供されている材料および方法を使用して、特定の遺伝子の配列を標的化された様式で改変することができる。遺伝子は、操作したTALエフェクターの標的とすることができる複数の配列を含有し得る。しかし、本明細書中に記載のように、特定の標的配列がより有効に標的化され得る。たとえば、実施例9に記載のように、特定の特徴を有する配列は、TALエフェクターによってより有効に標的化され得る。したがって、本明細書中で提供されている方法には、特定の基準を満たしている標的配列を同定することが含まれることができる。これらには、i)15個の塩基の最小の長さかつ5’から3’の配向を有しており、Tが5’末端の部位の直前にある、ii)第1(5’)位置にTまたは第2位置にAを有さない、iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さない、ならびにiv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの基本組成を有する配列が含まれる。
【0112】
本明細書中に記載のTALENは一般に二量体として働くため、本明細書中で提供されている方法の一部の実施形態には、上述の基準のうちの少なくとも1つを満たしており、15〜18bpによって分離されている、細胞中の第1のゲノムヌクレオチド配列および第2のゲノムヌクレオチド配列を同定することが含まれることができる。一部の事例では、1つのTALENポリペプチドがそれぞれのヌクレオチド配列と結合することができ、TALEN中に含有されるエンドヌクレアーゼが15〜18bpのスペーサー内を切断することができる。
【0113】
また、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した動物を作製する方法も提供する。そのような方法には、それ内に核酸を導入することが所望される内在染色体標的DNA配列を含有する細胞を得るステップと、細胞内にTALENを導入して内在染色体標的DNA配列内に二本鎖切断を生じるステップと、内在染色体標的DNAの少なくとも一部分に相同的な配列を含有する外因性核酸を、細胞内に、外因性核酸と内在染色体標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった初代細胞から動物を作製するステップとが含まれることができる。相同的な核酸には、たとえば、相同組換え後に遺伝子を破壊するヌクレオチド配列、相同組換え後に遺伝子を置き換えるヌクレオチド配列、相同組換え後に遺伝子内に点突然変異を導入するヌクレオチド配列、または相同組換え後に調節部位を導入するヌクレオチド配列が含まれることができる。
【0114】
また、本明細書中で提供されている方法を使用して、それ内に所望の核酸が導入されている、遺伝子改変した植物を作製することもできる。そのような方法には、それ内に核酸を導入することが所望される内在標的DNA配列を含有する植物細胞を得るステップと、TALENを導入して内在標的DNA配列内に二本鎖切断を生じるステップと、内在標的DNAの少なくとも一部分に相同的な配列を含有する外因性核酸を、植物細胞内に、外因性核酸と内在標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった植物細胞から植物を作製するステップとが含まれることができる。
【0115】
本明細書中で提供されているTALENに促進される相同組換え方法によって作製された細胞中のDNAは、そのような方法を経ていない細胞と比較して改変されており、改変されたDNAを含有する細胞は「遺伝子改変した」と呼ばれる。しかし、そのような改変は相同組換えを含み、導入遺伝子のランダム組込みを含まないため、そのような細胞を含有する生物は調節目的のGMOとみなされない場合があることに留意されたい。したがって、遺伝子改変を生じるために本明細書中に記載のTALENに促進される方法を使用することは、たとえば、標準の調節手順ならびにその関連する時間および費用を回避し得るという点で、有利であり得る。
【0116】
本明細書中で提供されている標的化遺伝子組換えの他の方法には、選択されたDNA標的配列を標的としたTALENをコードしている核酸分子を細胞(たとえば、植物細胞、昆虫細胞、硬骨魚細胞、または動物細胞)内に導入するステップと、細胞内でのTALENの発現を誘導するステップと、選択されたDNA標的配列が突然変異(たとえば、遺伝物質の欠失、遺伝物質の挿入、または遺伝物質の欠失および挿入両方)を示している組換え細胞を同定するステップとが含まれることができる。また、ドナーDNAを細胞内に導入することもできる。
【0117】
一部の実施形態では、単量体TALENを使用することができる。本明細書中に記載のTALENは、典型的には、2つのTALエフェクタードメインがそれぞれFokI制限酵素の触媒ドメインと融合しており、それぞれの生じるTALENのDNA認識部位がスペーサー配列によって分離されており、それぞれのTALEN単量体と認識部位との結合によりFokIが二量体化してスペーサー内に二本鎖切断を生じることが可能となるように、スペーサーを有する二連認識部位にわたる二量体として機能する(たとえばMoscouおよびBogdanove(2009)Science、326:1501を参照)。単量体TALENも構築することができるが、単一のTALエフェクターと、機能するために二量体化を必要としないヌクレアーゼとが融合しているようにする。たとえば、1つのそのようなヌクレアーゼは、2つの単量体が単一のポリペプチドとして発現される、FokIの単鎖変異体である(Minczukら(2008)Nucleic Acids Res.、36:3926〜3938)。また、他の天然に存在するまたは操作した単量体ヌクレアーゼもこの役割を果たすことができる。単量体TALENに使用されるDNA認識ドメインは、天然に存在するTALエフェクターに由来することができる。あるいは、DNA認識ドメインは、特定のDNA標的を認識するように操作することができる。操作した単鎖TALENは、1つの操作したDNA認識ドメインしか必要としないため、構築および配置がより容易であり得る。
【0118】
一部の実施形態では、二量体DNA配列特異的ヌクレアーゼは、2つの異なるDNA結合ドメイン(たとえば、1個のTALエフェクター結合ドメインおよび別の種類の分子からの1個の結合ドメイン)を使用して作製することができる。上述のように、本明細書中に記載のTALENは、典型的には、スペーサーを有する二連認識部位にわたる二量体として機能する。このヌクレアーゼ構造は、たとえば1個のTALEN単量体および1個のジンクフィンガーヌクレアーゼ単量体から作製した標的特異的なヌクレアーゼにも使用することができる。そのような事例では、TALENおよびジンクフィンガーヌクレアーゼ単量体のDNA認識部位は、適切な長さのスペーサーによって分離されていることができる。2つの単量体の結合により、FokIが二量体化してスペーサー配列内に二本鎖切断を生じることを可能にすることができる。また、ホメオドメイン、myb反復またはロイシンジッパーなどのジンクフィンガー以外のDNA結合ドメインも、FokIと融合させて、機能的ヌクレアーゼを作製するためのTALEN単量体とのパートナーとして役割を果たすことができる。
【0119】
一部の実施形態では、TALエフェクターを使用して、他のタンパク質ドメイン(たとえば非ヌクレアーゼタンパク質ドメイン)を特定のヌクレオチド配列に対して標的化させることができる。たとえば、TALエフェクターは、それだけには限定されないが、DNA相互作用酵素(たとえば、メチラーゼ、トポイソメラーゼ、インテグラーゼ、トランスポサーゼ、もしくはリガーゼ)、転写活性化因子もしくはリプレッサー、またはヒストンなどの他のタンパク質と相互作用するもしくはそれを修飾するタンパク質からのタンパク質ドメインと連結されていることができる。そのようなTALエフェクター融合体の応用には、たとえば、後成的調節要素を作製または改変すること、DNA中に部位特異的な挿入、欠失、または修復を行うこと、遺伝子発現を制御すること、およびクロマチン構造を改変することが含まれる。
【0120】
一部の実施形態では、標的配列のスペーサーは、TALENの特異性および活性を変調させるために選択または変動させることができる。スペーサーを有する二連認識部位にわたる二量体として機能するTALENの、本明細書中に提示した結果は、TALENが様々なスペーサー長にわたって機能することができ、TALENの活性がスペーサー長に伴って変動することを実証している。たとえば以下の実施例6を参照されたい。スペーサー長の柔軟性は、高い特異性で特定の配列(たとえばゲノム中のもの)を標的化するようにスペーサー長を選択できることを示している。さらに、様々なスペーサー長で観察される活性の変動は、所望のレベルのTALEN活性を達成するためにスペーサー長を選択できることを示している。
【0121】
一部の実施形態では、TALEN活性は、DNA結合ドメイン(複数可)内の反復の数および組成を変動させることによって変調させることができる。たとえば、本明細書中の実施例7に記載のように、PthXo1に基づくTALENはAvrBs3に基づくTALENよりも高い活性を示した。PthXo1は、その反復の数およびRVD組成がどちらもAvrBs3とは異なる。さらに、これらのタンパク質の天然に存在するDNA認識部位は、その多様性が、MoscouおよびBogdanove(上記)によって記載されているTALエフェクターDNA暗号に基づいて予測されたそのそれぞれの認識配列とは異なる。さらに、いくつかの同じ長さ(12個のRVD)であるが異なるRVD組成を有する誂えのTALENは、その活性が異なっており、13個のRVDの誂えのTALENは、12個のRVDの誂えのTALENよりも高い活性を有していた。したがって、TALENは、対象のDNA配列を認識するように操作できるだけでなく、(1)活性を変調させるために反復の数を変動させることができる、(2)様々なレベルの活性を達成するために様々な結合部位を選択することができる、ならびに(3)TALEN活性を変調させるためにRVDの組成および標的部位に対するその当てはめ(暗号に従う)を変動させることができる。
【0122】
TALENがヘテロ二量体である場合、たとえば、それぞれTALエフェクタードメインおよびFokIヌクレアーゼ触媒ドメインが含まれる2つの異なる単量体を有する場合、RVDは、2つのTALエフェクタードメインのそれぞれ中で等しい数で見つけることができるか、または、それぞれのドメインは異なる数のRVDを示すことができる。たとえば、合計22個のRVDがDNAを特定のヘテロ二量体TALENと結合させるために使用されている場合は、2つのTALエフェクタードメインのそれぞれ中で11個の反復を見つけることができるか、あるいは、2つのTALエフェクタードメインのうちの一方で10個の反復および他方中で12個を見つけることができる。また、本発明には、単量体として機能するDNA修飾酵素ドメインを有するTALENも包含される。この場合、単量体酵素と融合している単一のTALエフェクタードメイン中ですべてのRVDを見つけることができる。この場合、効率的な結合にするためには、RVDの数が等価な二量体TALEN中で見つかるであろうRVDの合計数に等しくなければならない。たとえば、2つの異なるTALエフェクタードメイン上に10個の反復を有する代わりに(二量体TALENの場合のように)、単一のTALエフェクタードメイン中に20個の反復を有する(単量体TALENの場合のように)。
【0123】
本発明のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも14である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも20である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも24である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも30である。
【0124】
また、本特許出願は、標的DNAに対して増強された標的化能力を有するTALエフェクタータンパク質を作製する方法も提供する。本方法には、たとえば、それぞれのDNA結合反復が標的DNA中の塩基対の認識を決定するRVDを含有し、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを有するTALエフェクターをコードしている核酸を作製することが含まれることができる。以下の実施例12に記載のように、結合部位の−1位でのTの必要性を緩めることで、操作したTALエフェクタータンパク質の標的化能力を増強させ得る。したがって、TALエフェクターをコードしている核酸の作製には、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませ、したがって結合部位の−1位でのTの必要性を排除することが含まれることができる。
【0125】
さらに、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法が本明細書中で提供されている。そのような方法には、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含有する、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製することが含まれることができる。以下の実施例12に記載のように、NN(Gを認識する最も一般的なRVD)の特異性は一般に弱く、コンテキストに伴って変動する場合があるように見えるが、特定のRVDはGに対して増強された特異性を有し得る。したがって、本明細書中で提供されている方法には、Gに対してより頑強な特異性を有し得る代替のRVDを使用することが含まれることができる。たとえば、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択される1つまたは複数のRVDを使用することができ、星印はRVDの第2位置でのギャップを示す。
【0126】
製品
また、本発明は、たとえば、TALENをコードしている核酸分子、TALENポリペプチド、そのような核酸分子もしくはポリペプチドを含有する組成物、またはTALエンドヌクレアーゼを操作した細胞系を含有する製品も提供する。そのような物品は、たとえば、研究ツールとして、または治療的に使用することができる。
【0127】
一部の実施形態では、製品には、本明細書中で提供されている方法を使用して作製した植物からの種子が含まれることができる。種子は、当分野で知られている手段を使用して調整し、当分野で周知の梱包材料を使用して梱包して、製品を調製することができる。種子の梱包は、ラベル、たとえば、梱包材料に固定されたタグもしくはラベル、梱包材料上に印刷されたラベルまたは梱包内に挿入されたラベルを有することができる。ラベルは、梱包内に含有された種子が遺伝子改変した植物の作物を生じることができることを示すことができ、改変していない植物に対して遺伝子改変によって変更されている特色を記載することができる。
【0128】
他の定義
ポリペプチド配列中のアミノ酸残基またはサブユニットは、本明細書中で一文字のコードに従って命名されており、たとえば、QはGlnまたはグルタミン残基を意味し、RはArgまたはアルギニン残基を意味し、DはAspまたはアスパラギン酸残基を意味する。
【0129】
アミノ酸置換とは、1個のアミノ酸残基を別のもので置き換えることを意味し、たとえば、ペプチド配列中のアルギニン残基をグルタミン残基で置き換えることはアミノ酸置換である。
【0130】
ヌクレオチドは以下のように命名されている:一文字のコードがヌクレオシドの塩基を命名するために使用されている:aはアデニンであり、tはチミンであり、cはシトシンであり、gはグアニンである。縮重ヌクレオチドには、rはgまたはa(プリンヌクレオチド)を表し、kはgまたはtを表し、sはgまたはcを表し、wはaまたはtを表し、mはaまたはcを表し、yはtまたはc(ピリミジンヌクレオチド)を表し、dはg、aまたはtを表し、vはg、aまたはcを表し、bはg、tまたはcを表し、hはa、tまたはcを表し、nはg、a、tまたはcを表す。
【0131】
用語「DNA修飾酵素」とは、DNA修飾のレベルが何であれ(切断、共有的相互作用、水媒介性の相互作用・・・)、細胞の遺伝物質を改変することができる任意のタンパク質をいう。DNA相互作用タンパク質(たとえば、メチラーゼ、トポイソメラーゼ、インテグラーゼ、トランスポサーゼ、またはリガーゼ)、転写活性化因子またはリプレッサー、ヒストンなどの他のタンパク質、およびヌクレアーゼが、「DNA修飾酵素」の意味に含まれることを意図する。TALエフェクター−DNA修飾酵素内に含まれている場合、DNA修飾酵素はDNA修飾酵素ドメインと呼ばれる。
【0132】
用語「ヌクレアーゼ」には、エキソヌクレアーゼおよびエンドヌクレアーゼが含まれることを意図する。
【0133】
用語「エンドヌクレアーゼ」とは、DNAまたはRNA分子、好ましくはDNA分子内の核酸間の結合の加水分解(切断)を触媒することができる、任意の野生型または変異体酵素をいう。エンドヌクレアーゼの非限定的な例には、FokI、HhaI、HindIII、NotI、BbvCI、EcoRI、BglI、およびAlwIなどのII型制限エンドヌクレアーゼが含まれる。また、エンドヌクレアーゼは、典型的には約12〜45個の塩基対(bp)の長さ、より好ましくは14〜45bpのポリヌクレオチド認識部位を有する場合は、レアカッターエンドヌクレアーゼも含む。レアカッターエンドヌクレアーゼは、定義された座位でのDNA二本鎖切断(DSB)を誘導することによって、HRを有意に増加させる(Rouet、Smihら、1994、Rouet、Smihら、1994、Choulika、Perrinら、1995、PingoudおよびSilva、2007)。レアカッターエンドヌクレアーゼは、たとえば、ホーミングエンドヌクレアーゼ(PaquesおよびDuchateau、2007)、操作したジンクフィンガードメインとFokIなどの制限酵素の触媒ドメインとの融合から生じるキメラジンクフィンガーヌクレアーゼ(ZFN)(PorteusおよびCarroll、2005)、または化学エンドヌクレアーゼ(Eisenschmidt、Lanioら、2005、Arimondo、Thomasら、2006、Simon、Cannataら、2008)であることができる。化学エンドヌクレアーゼでは、化学またはペプチド切断剤は、特定の標的配列を認識する核酸のポリマーまたは別のDNAのどちらかとコンジュゲートされており、したがって、切断活性を特定の配列を標的とする。また、化学エンドヌクレアーゼには、特定のDNA配列と結合することが知られている、オルトフェナントロリンのコンジュゲートなどの合成ヌクレアーゼ、DNA切断分子、および三重鎖形成オリゴヌクレオチド(TFO)も包含される(KalishおよびGlazer、2005)。そのような化学エンドヌクレアーゼが本発明による用語「エンドヌクレアーゼ」中に含まれる。そのようなエンドヌクレアーゼの例には、I−Sce I、I−Chu I、I−Cre I、I−Csm I、PI−Sce I、PI−Tli I、PI−Mtu I、I−Ceu I、I−Sce II、I−Sce III、HO、PI−Civ I、PI−Ctr I、PI−Aae I、PI−Bsu I、PI−Dha I、PI−Dra I、PI−Mav I、PI−Mch I、PI−Mfu I、PI−Mfl I、PI−Mga I、PI−Mgo I、PI−Min I、PI−Mka I、PI−Mle I、PI−Mma I、PI−Msh I、PI−Msm I、PI−Mth I、PI−Mtu I、PI−Mxe I、PI−Npu I、PI−Pfu I、PI−Rma I、PI−Spb I、PI−Ssp I、PI−Fac I、PI−Mja I、PI−Pho I、PI−Tag I、PI−Thy I、PI−Tko I、PI−Tsp I、I−MsoIが含まれる。
【0134】
本発明によるエンドヌクレアーゼは、転写活性化因子様(TAL)エフェクターエンドヌクレアーゼ(TALEN)の一部であることができる。
【0135】
「TALEN」とは、転写活性化因子様(TAL)エフェクター結合ドメインおよびエンドヌクレアーゼドメインを含むタンパク質を意図し、両方のドメインの融合が「単量体TALEN」をもたらす。一部の単量体TALENはそれ自体で機能的な場合があり、他のものは別の単量体TALENとの二量体化を必要とする。二量体化は、両方の単量体TALENが同一である場合にホモ二量体TALENをもたらす可能性があるか、または、単量体TALENが異なる場合にヘテロ二量体TALENをもたらす可能性がある。2つの単量体TALENは、たとえば、そのRVDの数が異なるおよび/または少なくとも1つのRVDの内容(すなわちアミノ酸配列)が異なる場合に、異なる。「TALエフェクター−DNA修飾酵素」とは、転写活性化因子様エフェクター結合ドメインおよびDNA修飾酵素ドメインを含むタンパク質を意図する。
【0136】
「変異体」とは、「変異体」タンパク質、すなわち、自然には天然に存在せず、遺伝子操作またはランダム突然変異誘発によって得られるタンパク質、すなわち操作したタンパク質を意図する。この変異体タンパク質は、たとえば、野生型の天然に存在するタンパク質のアミノ酸配列中の少なくとも1つの残基を異なるアミノ酸で置換することによって得ることができる。前記置換(複数可)は、たとえば部位特異的突然変異誘発および/またはランダム突然変異誘発によって導入することができる。
【0137】
「細胞」または「複数の細胞」とは、任意の原核または真核の、生細胞、in vitro培養のためにこれらの生物から誘導した細胞系、動物もしくは植物起源の初代細胞を意図する。
【0138】
「初代細胞」または「複数の初代細胞」とは、非常に少ない回数の集団倍加しか受けておらず、したがって、連続的な腫瘍原性細胞または人工的に不死化した細胞系と比較してそれらが由来する組織の主な機能的構成要素および特徴をより表している、生組織から直接採取(すなわち生検材料)し、in vitroでの成長のために確立させた細胞を意図する。したがって、これらの細胞は、それらが指すin vivo状態に対するより価値のあるモデルを表す。
【0139】
「相同的」とは、配列間の相同組換えをもたらすために十分な別の配列との同一性を有する、より具体的には、少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%を有する配列を意図する。
【0140】
「同一性」とは、2つの核酸分子またはポリペプチド間の配列同一性をいう。同一性は、比較の目的のためにアラインメントし得るそれぞれの配列中の位置を比較することによって決定することができる。比較した配列中の位置が同じ塩基によって占有されている場合は、分子はその位置で同一である。核酸またはアミノ酸配列間の類似性または同一性の度合は、核酸配列によって共有される位置での同一または一致するヌクレオチドの数の関数である。たとえば初期設定で使用することができるGCG配列解析パッケージ(ウィスコンシン大学、ウィスコンシン州Madison)の一部として利用可能なFASTA、またはBLASTを含めた、様々なアラインメントアルゴリズムおよび/またはプログラムを使用して、2つの配列間の同一性を計算し得る。
【0141】
「突然変異」とは、ポリヌクレオチド(cDNA、遺伝子)またはポリペプチド配列中の1つまたは複数のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。前記突然変異は、遺伝子またはその調節配列のコード配列に影響を与える場合がある。また、これは、ゲノム配列の構造またはコードされているmRNAの構造/安定性にも影響を与え得る。
【0142】
「遺伝子」とは、染色体に沿って直線的に配置されているDNAのセグメントからなり、特定のタンパク質またはタンパク質のセグメントをコードしている、遺伝の基本単位を意味する。遺伝子には、典型的には、プロモーター、5’非翻訳領域、1つまたは複数のコード配列(エクソン)、任意選択でイントロン、3’非翻訳領域が含まれる。遺伝子は、ターミネーター、エンハンサーおよび/またはサイレンサーをさらに含み得る。
【0143】
用語「対象の遺伝子」とは、既知または推定上の遺伝子産物をコードしている任意のヌクレオチド配列をいう。
【0144】
本明細書中で使用する用語「座位」とは、染色体上のDNA配列(たとえば遺伝子のもの)の特定の物理的位置である。用語「座位」とは、通常、染色体上の標的配列の特定の物理的位置をいう。
【0145】
「融合タンパク質」とは、元は別々のタンパク質をコードしている2つ以上の遺伝子を結合させることからなる当分野の周知のプロセスの結果を意図し、前記「融合遺伝子」の翻訳の結果、元のタンパク質のそれぞれに由来する機能的特性を有する単一のポリペプチドがもたらされる。
【0146】
「触媒ドメイン」とは、酵素の活性部位を含有する、酵素のタンパク質ドメインまたはモジュールを意図する。活性部位とは、基質の触媒作用がそこで起こる、前記酵素の部分を意図する。酵素だけでなくその触媒ドメインは、それらが触媒する反応に従って分類および命名される。酵素番号(EC番号)とは、それらが触媒する化学反応に基づく、酵素の数値分類スキームである(ワールドワイドウェブchem.qmul.ac.uk/iubmb/enzyme/)。本発明の範囲では、任意の触媒ドメインをパートナーとして使用し、TALエフェクタードメインと融合させてキメラ融合タンパク質を作製して、TALエフェクター−DNA修飾酵素をもたらすことができる。そのような触媒ドメインの非限定的な例は、MmeI、EsaSSII、CstMI、NucA、EndA大腸菌、NucM、EndA肺炎連鎖球菌、SNase黄色ブドウ球菌、SNase Staphylococcus hyicus、SNaseフレクスナー赤痢菌、枯草菌yncB、エンドデオキシリボヌクレアーゼI腸内細菌ファージT7、EndoGウシ、ttSmr DNAミスマッチ修復タンパク質mutS、Metnaseの切断ドメインのものであることができる。
【0147】
別段に指定しない限りは、本発明の実施は、細胞生物学、細胞培養物、分子生物学、トランスジェニック生物学、微生物学、組換えDNA、および免疫学の慣用技術を用い、これらは十分に当分野の技術範囲内にある。そのような技法は文献中で完全に説明されている。たとえば、分子生物学の最新プロトコル(Current Protocols in Molecular Biology)(Ausubel、2000、Wiley and son Inc、米国議会図書館、分子クローニング:実験室の手引き(Molecular Cloning:A Laboratory Manual)、第3版、(Sambrookら、2001、ニューヨーク州Cold Spring Harbor:Cold Spring Harbor Laboratory Press)、オリゴヌクレオチド合成(Oligonucleotide Synthesis)(M.J.Gait編、1984)、米国特許第4,683,195号、核酸ハイブリダイゼーション(Nucleic Acid Hybridization)(HarriesおよびHiggins編、1984)、転写および翻訳(Transcription and Translation)(HamesおよびHiggins編、1984)、動物細胞の培養(Culture of Animal Cells)(Freshney、Alan R.Liss,Inc.、1987)、固定細胞および酵素(Immobilized Cells and Enzymes)(IRL Press、1986)、Perbal、分子クローニングの実用的な指針(A Practical Guide to Molecular Cloning)(1984)、シリーズ酵素学の方法(Methods in Enzymology)(AbelsonおよびSimon編集長、Academic Press,Inc.、New York)、特に第154巻および第155巻(Wuら編)ならびに第185巻、「遺伝子発現技術(Gene Expression Technology)」(Goeddel編)、哺乳動物細胞の遺伝子転移ベクター(Gene Transfer Vectors For Mammalian Cells)(MillerおよびCalos編、1987、Cold Spring Harbor Laboratory)、細胞および分子生物学における免疫化学的方法(Immunochemical Methods in Cell and Molecular Biology)(MayerおよびWalker編、Academic Press、London、1987)、実験免疫学のハンドブック(Handbook of Experimental Immunology)、第I〜IV巻(WeirおよびBlackwell編、1986)、ならびにマウス胚の操作(Manipulating the Mouse Embryo)、(Cold Spring Harbor Laboratory Press、ニューヨーク州Cold Spring Harbor、1986)を参照されたい。
【0148】
本発明の上記明細書は、任意の当業者による本発明の作製および使用が可能となるようにそれを作製および使用する様式およびプロセスを提供し、この可能にすることは、元の説明の一部を構成する添付の特許請求の範囲の主題のために特に提供されている。
【0149】
上記で使用するように、語句「からなる群から選択される」、「から選択される」などには、指定した物質の混合物が含まれる。
【0150】
数値の限界または範囲が本明細書中に記述されている場合は、端点が含まれる。また、数値の限界または範囲内のすべての値および部分範囲は、明確に書き出されている場合と同様に具体的に含まれる。
【0151】
上記説明は、当業者が本発明を作製および使用することを可能にするために提供されており、特定の応用およびその要件のコンテキストで提供されている。好ましい実施形態への様々な改変は当業者に容易に明らかであり、本明細書中に定義した一般的な原理は、本発明の精神および範囲から逸脱せずに、他の実施形態および応用に適用し得る。したがって、本発明は、示されている実施形態に限定されず、本明細書中に開示した原理および特長と一貫性のある最も広い範囲に従うことを意図する。
【0152】
本発明を一般に説明したが、さらなる理解は、例示の目的でのみ本明細書中で提供されている特定の具体的な実施例を参照して得ることができ、本発明は、別段に指定しない限りは特許請求の範囲に記載されている本発明の範囲を限定しない、以下の実施例中にさらに記載されている。
【実施例】
【0153】
【実施例1】
【0154】
暗号がTALエフェクター−DNA認識を支配する
TAL標的部位中のRVDと近接したヌクレオチドとの間に1対1の直線的な対応が存在するかどうかを決定するために、10個のTALエフェクターのそれぞれの既知の標的遺伝子の予測されたプロモーター領域(すなわち注釈付けされた翻訳開始部位の直前の1,000bp)を、TALエフェクターのRVD配列を用いて、RVD−ヌクレオチドの会合においてエントロピー(ランダム性)を最小限にしたアラインメントについて走査した。以下の式を使用してエントロピーを定量し、Rは、エフェクターのRVDの組であり、Dは、4個のヌクレオチド(A、C、G、T)の組であり、fi,jは、第i番目のRVDが第j番目のヌクレオチドと会合する観察された頻度を表す:
【化1】
【0155】
複数の低エントロピー部位がそれぞれのプロモーター中に存在していた。しかし、エフェクターAvrBs3では、1つのみが活性化に十分かつ必要であると以前に同定された54bpのupa20プロモーター断片にマッピングされ、これは、AvrBs3によって直接活性化された遺伝子に一般的なUPAボックスと一致した(Kayら、上記)。また、エフェクターPthXo1およびAvrXa27では、1つの部位のみが、そのそれぞれの標的であるOs8N3およびXa27の活性化された対立遺伝子と活性化されなかった対立遺伝子との間の多型にそれぞれ重複していた。これら3つの部位でのアラインメントにわたってRVD−ヌクレオチドの会合は一貫していたため、残りのアラインメントをこれらの会合に基づいて選択し、その結果、1対のTALエフェクター−標的あたり正確に1つの部位がもたらされた(図1Bおよび表2)。それぞれの部位にはTが先行している(図1D)。
【0156】
RVD−ヌクレオチドの会合によって与えられた特異性を評価するために、最初に、10個の最小限のエントロピーのTALエフェクター−標的部位のアラインメントにわたって観察されたすべてのRVD−ヌクレオチドの会合の頻度に基づいて重量マトリックスを作成した(図1B)。その後、重量マトリックスを使用して、コメ、Oryza sativa spp.japonica cv.Nipponbare(Osa1、リリース6.0、rice.plantbiology.msu.edu)におけるそれぞれの非冗長遺伝子モデルの翻訳開始に先行する1,000bpのプロモーター領域を、コメの病原体Xanthomonas oryzaeの5個のTALエフェクター(AvrXa27、PthXo1、PthXo6、PthXo7、およびTal1c)に対する最良の一致について走査した。AvrXa27には、Xa27(GenBank受託AY986492)の上流の配列を含めた。この上流配列はNipponbare中には存在しない。観察された会合頻度は90%で重み付けされ、残りの10%はすべての可能な会合の頻度に均等に分布していた。アラインメントは重量マトリックススコア(y軸)を使用して順位付けし、図1BのRVD−ヌクレオチドの会合頻度に由来する頻度スコアの負の対数としてとった。したがって、スコアが低ければ低いほど、一致はより良好である。PthXo1、PthXo6、PthXo7、およびTal1cでは、実験的に同定した標的遺伝子が最良またはほぼ最良の一致であった。より良好な一致には、Tが先行していなかったか、標的を同定するために使用したマイクロアレイ上に表されていなかったか、またはイントロンおよびESTの証拠を欠いていなかった。既知の標的に対する逆相補プロモーター配列の走査は、順方向の部位よりも良好なスコアのアラインメントを与えなかった。この結果は、TALエフェクターがプラス鎖と結合することを暗示しているのではなく、これらがプラス鎖に対して順方向の配向で機能することを示している。5個目のエフェクターAvrXa27の既知の標的は疾患耐性遺伝子Xa27である(Guら、上記)。この一致の順位(5,368)がより低いことは、較正したまたは最近および最適以下の宿主の適応を反映している可能性がある。より良好なスコアの部位は、病因についてAvrXa27によって標的化されている遺伝子を含む可能性が高い。
【0157】
重量マトリックスを再度使用して、すべてのコメプロモーターを40個の追加のX.oryzaeのTALエフェクターで走査し、公的マイクロアレイデータ(PLEXdb.org、受託OS3)に基づいて感染中に下流遺伝子が活性化された最良のアラインメントのものを保持することによって、10個の追加のアラインメントが得られた(表3)。初期の組と同様、Tがそれぞれの部位に先行しており、どの逆鎖の部位もより良好なスコアではなかった。合計20個のアラインメントのRVD−ヌクレオチドの会合頻度が図1Cに示されている。これらは、驚くべきほど単純な暗号を構成している。
【0158】
20個のTALエフェクターヌクレオチドのアラインメントの拡張組中のRVD−ヌクレオチドの頻度を使用して新しい重量マトリックスを作成し、コンピュータスクリプトをPython v2.5(www.python.org)によって書いた。スクリプトは、観察されたvs.観察されていないRVD−ヌクレオチドの会合についてユーザ定義可能な重量因子を用いて、特定のTALエフェクターとの一致についてDNA配列の任意のコレクションを走査するために使用することができる。MoscouおよびBogdanove(上記)を参照されたい。
【0159】
暗号中にはある程度の縮重が存在する。強力な会合は結合親和性のほとんどを占めるアンカーを表している場合があり、弱い会合は柔軟性の測度を提供する。あるいは、隣接効果が関与している場合がある。後者の可能性は、RVDの両側に条件付けられたすべてのRVDのヌクレオチド会合頻度を決定し、それらを観察された全頻度と比較することによって、言い換えれば、RVD−ヌクレオチドの対合を左または右に隣接するRVDに従って選別し、そのようにして選別されたそれぞれの対の相対的頻度をその対の全体的な頻度と比較することによって調査した。隣接するものによって選別したRVD−ヌクレオチドの会合の頻度は観察された全頻度から顕著に逸脱せず、これは、会合がコンテキスト非依存性であることを示唆している。
【0160】
20個の標的部位に隣接する配列によって−1のT以外の保存的なヌクレオチドは明らかとならなかったが、この部位に続いてCが富み、全体的にGが乏しい傾向にある(図1D)。いくつかの例外を除いて、部位は注釈付けされた転写開始の60bp以内上流に始まり、翻訳開始に87bpより近いものはない(表2および表3)。RVD/ヌクレオチドの会合を支配するさらなる規則は、実施例4および5に記載されている。
【0161】
これらの結果を考慮して、ゲノム中のTALエフェクター標的の予測および標的の新規構築が現在可能である。部位を予測する能力は、疾患に重要な宿主遺伝子の同定をはかどらせる。標的を構築する能力は、保存的または複数のTALエフェクターに応答性の、耐久性のある耐性遺伝子の設計に期待される。また、本明細書中に記載のように、自由裁量による遺伝子の活性化のためにTALエフェクターを誂えること、またはDNA修飾のためにの融合タンパク質を標的化することも可能である。
【表3】
【表4】
【実施例2】
【0162】
TALENは酵母中で機能できる
プラスミドの構築:TALエフェクター、AvrBs3のタンパク質コード配列は、プラスミドをBamHIで消化することから得られた。主に反復ドメインをコードしているDNA断片をSphIで切り出した。AvrBs3のアミノ酸配列はGENBANK受託番号P14727および配列番号12の下(図3)、核酸配列は受託番号X16130配列番号13の下(図4)で見つけることができる。図4中、BamHIおよびSphI部位は太字で下線が引かれている。AvrBs3のBamHIおよびSphI断片を、ヌクレアーゼ発現ベクターpDW1789_TAL(図5)内に、FokIヌクレアーゼドメインをコードしている配列に隣接してクローニングした。AvrBs3標的部位を標的レポータープラスミド内にクローニングするために、18bpのスペーサー配列をその間に含んで逆配向に配置された2つのAvrBs3認識部位を含有する2つの相補的DNAオリゴを、5’および3’末端でそれぞれBglIIおよびSpeIのオーバーハングを有するように合成した。6、9、12および15bpのスペーサー長を用いて、認識部位を有する他のレポータープラスミドを作製した。アニーリングさせたDNAオリゴをレポータープラスミドpCP5内にクローニングし(図6)、これをBglIIおよびSpeIで消化した。
【0163】
酵母アッセイ:標的レポータープラスミドを酵母株YPH499(MAT a株)内に形質転換させ、形質転換体を、トリプトファンを欠く合成完全培地(SC−W)上で選択した。TALEN発現プラスミドをYPH500(MAT α株)内に形質転換させ、形質転換体を、ヒスチジンを欠くSC培地(SC−H)上にプレートした。標的レポータープラスミドを保有する酵母コロニーおよびTALEN発現プラスミドを保有するコロニーを、終夜、30℃で、それぞれ液体SC−WおよびSC−H培地中で培養した。培養物を同じOD600に調節し、それぞれの200μlを200μlのYPD培地に混ぜた。混合物を30℃で4時間インキュベーションして、2種類の酵母株を接合させた。混合培養物を遠心沈殿させ、5mlのSC−W−H培地中に30Cで終夜、またはOD600が0.5〜1の範囲に達成するまで再懸濁させた。細胞を収穫し、定量的β−ガラクトシダーゼアッセイを記載のように行った(Townsendら(2009)Nature、459:442〜445)。
【0164】
結果:TAL−FokI融合体は、TALのDNA認識ドメインおよび非特異的なFokI DNA切断ドメインからなる部位特異的なヌクレアーゼである。TALのDNA認識ドメインは、様々なDNA配列と結合するように操作することができる。本明細書中の実施例1に記載のように、DNA結合ドメインの新規クラスであるTALエフェクターのDNA認識特異性が解読されている。具体的には、TALエフェクターのDNA結合ドメインは、特定のDNA配列を認識および結合することができる様々な数のタンデムの34個のアミノ酸の反復を含有する。反復のアミノ酸配列は、反復の位置12および13での2つの隣接する高度に可変性の残基以外は保存的である。これらの位置は一緒になって、DNA結合部位中の個々のヌクレオチドを1つの反復対1個のヌクレオチドで指定する。TALENの構造は図7に例示されている。TALENは二量体として機能し、それぞれの単量体は、FokIエンドヌクレアーゼからの非特異的な切断ドメインと融合した、操作されたTALのDNA認識反復から構成される。DNA認識反復は、対象のゲノム内の標的DNA配列と結合するように操作することができる。TALヌクレアーゼ単量体は、スペーサー配列によって分離されている2つのDNA半分部位のうちの1つと結合する。この間隔が、FokI単量体が二量体化して、半分部位の間のスペーサー配列中に二本鎖DNAの切断(DSB)を生じることを可能にする。
【0165】
TALエフェクターDNA認識ドメインの潜在性を調査するために、ネイティブTALエフェクターが、FokIヌクレアーゼドメインと融合した場合にヌクレアーゼとして機能できるかどうかを決定するための実験を実施した。TALヌクレアーゼ発現構築体および標的レポーター構築体を使用することによって、酵母に基づくアッセイを実施した。図5に例示されているように、ヌクレアーゼ発現構築体の主鎖は、FokIヌクレアーゼドメインおよび酵母TEF1プロモーターの制御下のN末端核移行シグナル(NLS)を含有する。様々なTALエフェクターのクローニングを促進するために、いくつかの制限部位がFokIヌクレアーゼドメインとNLSモチーフとの間に位置している。図6に示されているように、標的レポーター構築体は、コード配列の125bpの重複を有する破壊されたlacZレポーター遺伝子を有する。重複は、URA3遺伝子およびTALのDNA結合ドメインによって認識される標的配列(2つの半分部位およびスペーサー配列から構成される)に隣接する。TALENが標的部位と結合してDNA二本鎖切断(DSB)を生じる場合、そのような切断は、酵母では、重複するlacZ配列間の相同組換えによって、一本鎖アニーリングによって主に修復される(Haber(1995)Bioessays、17:609)。組換えは機能的lacZ遺伝子の再構成およびURA3の損失(5−フルオロオロト酸耐性を与える)をもたらす。lacZ酵素活性を決定することによってTALENの相対的切断活性を測定した。
【0166】
これらの研究では、配列番号16(図8)に記載のように中央ヌクレアーゼ反復領域を有していたネイティブTALエフェクター、AvrBs3を、ヌクレアーゼ発現ベクター内にクローニングし、18bpのスペーサー配列を有するAvrBs3標的部位(逆配向に配置された2つの結合部位)を標的レポーターベクター内にクローニングした。酵母アッセイは、図9に示されており上述されているスキームを使用して行った。結果は、AvrBs3ヌクレアーゼプラスミドおよび標的レポータープラスミドの両方で形質転換させた酵母細胞からのlacZ活性が、標的レポータープラスミドのみを含有していた対照酵母細胞よりも有意に高かった(15.8倍高い)ことを示した(図10)。反復ドメインを主にコードしているSphI断片のみで作製したヌクレアーゼ融合体では、活性は観察されなかった。これは、DNA結合ドメイン以外の配列がTALEN活性に必要であることを示した。また、6および9bpのスペーサー長を有するレポータープラスミドも活性を示すことに失敗し、これは、2つの結合部位間の間隔がFokIを二量体化させるために重要であることを示している。これらのデータは、AvrBs3 TALヌクレアーゼが、酵母中のその同族標的配列を切断する部位特異的なヌクレアーゼとして機能できることを示している。
【実施例3】
【0167】
誂えたTALENのための、TALエフェクター反復のモジュールアセンブリ
それぞれが異なるヌクレオチドを指定する4つの個々のTALエフェクター反復のそれぞれの、102個の塩基対に対応する相補的オリゴヌクレオチドを合成し、アニーリングし、個々にならびにすべての順列の2回および3回の反復の組合せで高コピーの細菌クローニングベクター内にクローニングして、標準の制限消化およびライゲーション技法を使用して4個の単一、16個の二重、および64個の三重反復モジュールを得た(たとえば図11に例示)。所望のTALエフェクターのコード配列は、適切なモジュールを、特徴的な最後の半反復以外の中央反復領域を欠くtal1c遺伝子の切断された形態を含有するGateway−ready高コピー細菌クローニングベクター内に順次導入することによってアセンブルする。たとえば、18回反復のTALエフェクターのコード配列は、5個の三重モジュールおよび1個の二重モジュールを切断されたtal1cベクター内に順次導入することによってアセンブルすることができる。
【実施例4】
【0168】
TALエフェクター反復のモジュールアセンブリのためのシステム
誂えのTALエフェクターをコードしている遺伝子を作製するためのプラスミドおよび方法を開発した。本明細書中に記載のように、TALエフェクターの機能的特異性は反復中のRVDによって決定され、反復およびタンパク質中の他の箇所中の他の多型は稀であり、機能的特異性に関して重要でない。したがって、自由裁量によるTALエフェクター遺伝子の反復領域を所望のRVDを含有する反復で置き換えることによって、誂えのTALエフェクター遺伝子を作製した。RVDの外の反復配列はコンセンサス配列に一致した(以下を参照)。TALエフェクター反復をコードしているDNA断片を、1回、2回、または3回の反復をコードしているモジュール内に順次アセンブルし、元の反復が除去されたTALエフェクター遺伝子内にモジュールをクローニングした。最後の(半)反復を例外として、それぞれのコードされている反復は、配列LTPDQVVAIASXXGGKQALETVQRLLPVLCQDHGを有していた(配列番号18、図12A)。最後の(半)反復は配列LTPDQVVAIASXXGGKQALESを有していた(配列番号20、図12B)。どちらの配列でも、「XX」はRVDの位置を示す。モジュール反復中で使用したRVDは、それぞれA、C、G、およびTとの結合を指定するNI、HD、NN、およびNGであった。以下に記載の実験では、その反復が除去されたXanthomonas oryzae pv.oryzicola株BLS256のtal1c遺伝子を、誂えのTALエフェクター遺伝子を構築するための「主鎖」として使用した。
【0169】
本明細書中に記載の方法には、(1)単一反復の開始プラスミドの作製、(2)単一反復モジュールのプラスミドの作製、(3)複数反復モジュールの作製、(4)1回、2回、および3回の反復モジュールのプラスミドの完全な組の作製、ならびに(5)誂えのTALエフェクターのコード配列のアセンブリの5個の構成要素が含まれていた。
【0170】
単一反復の開始プラスミドを作製するために、第1反復の最初の部分および最後の切断された反復の最後の部分以外の反復領域全体を除去するためにtal1c遺伝子をMscIで消化し、再度ライゲーションさせて、pCS487と命名されたプラスミドがもたらされた(図13)。生じる遺伝子はRVD NIをコードしており、ほとんどのTALエフェクター遺伝子と同様、反復領域に隣接していた2つのSphI部位を含有していた。この遺伝子はXhoI部位を含有していなかった。
【0171】
次に、翻訳的にサイレントな突然変異をpCS487内に導入して、コドン19および20を中心とするユニークなXhoI部位が包含される、ユニークなPspXI部位を作製した。突然変異は、どちらもアミノ酸配列ALES(配列番号22)をコードしているコドン18〜21の元のおよび変更されたヌクレオチド配列(それぞれ配列番号21および配列番号23)を示す、図14に示されている。生じるプラスミドはpCS489と命名された。
【0172】
さらなる突然変異誘発によって、RVD HD、NN、およびNGを用いて3つのさらなる構築体を作製して、それぞれpCS490、pCS491、およびpCS492と命名されたプラスミドを作製した。修飾された反復領域を包含するSphI断片を、pCS489、pCS490、pCS491、およびpCS492から、GatewayエントリーベクターpENTR−D(Invitrogen、カリフォルニア州Carlsbad)中で反復領域を含まずにtal1cのNおよびC末端部分のみをコードしたpCS488と命名されたカナマイシン耐性プラスミド(図15)に移した。この転移は、それぞれpCS493(図16)、pCS494、pCS495、およびpCS496と命名された単一反復の開始プラスミドをもたらした。切断された反復中のPspXI/XhoI部位はこれらのプラスミド中でユニークに保たれた。pCS488およびその誘導体のそれぞれ中のTALエフェクター遺伝子には、原核生物および真核生物中での効率的な翻訳のためにそれぞれシャイン−ダルガノおよびコザック配列が先行していた。
【0173】
その後、単一反復モジュールのプラスミドを構築した。4つの選択されたRVD(NI、HD、NN、およびNG)のそれぞれについて1個のプラスミドを作製した。それぞれのプラスミドは、PspXI部位内にライゲーションした際にXhoIを再構成したがPspXI部位を再構成しなかった5’適合性の付着末端、ならびにXhoIおよびPspXI部位をどちらも再構成した3’適合性の付着末端を有していた。プラスミドは、アニーリングしたオーバーハングを有する合成の相補的オリゴヌクレオチド(図17A)を、pBluescript SK−のPspXI/XhoI部位内にクローニングすることによって作製し、それぞれpCS502(図17B)、pCS503、pCS504、およびpCS505と命名されたプラスミドがもたらされた。それぞれのプラスミドは、ユニークな再構成されたPspXI部位での単一反復モジュールの3’末端の追加の反復の導入、または再構成されたXhoI部位を使用した反復モジュールの切り出しを可能とした。
【0174】
NI、HD、NN、およびNGについてそれぞれ1個の追加の単一反復モジュールを作製した。それぞれは、PspXI部位内にライゲーションした際にPspXIまたはXhoI部位を再構成しなかった5’適合性の付着末端、XhoIおよびPspXI部位をどちらも再構成した3’適合性の付着末端、ならびに内部MscI部位を破壊した翻訳的にサイレントなヌクレオチド置換を有していた(図18A)。これらのモジュールは、オーバーハングを有する合成の相補的なオリゴヌクレオチドをアニーリングすることによって作製した。これらの追加の単一反復モジュールのうちの任意のものを単一反復モジュールのプラスミド(pCS502、pCS503、pCS504、またはpCS505)のユニークなPspXI/XhoI部位内にライゲーションすることで、5’接合部で新しいXhoI部位はもたらされなかったが、ユニークな3’PspXI/XhoI部位の再建がもたられ、それにより、生じるプラスミドをPspXIで切断することによってさらなる追加の反復を導入するために直鎖状にできる。このプロセスの繰返しにより、複数反復を含有するモジュールがもたらされた(図18B)。さらに、XhoIを使用してそれぞれの複数反復モジュール全体を切り出すことができる。MscI部位は追加の単一反復モジュール中で破壊されていたため、第1反復中のMscI部位はユニークに保たれ、続く複数反復モジュールのサブクローニングの際に配向を確認するために有用であった。
【0175】
追加の単一反復モジュールを単一反復モジュールのプラスミド内に繰り返してクローニングして、単一反復モジュールのプラスミドと共に、すべての可能な1回、2回、および3回の反復モジュールの完全な組を作製し、pCS502〜pCS585と命名された合計84個のプラスミドとした(図19)。3回より多くの反復(たとえば、4、5、6、7、8、9、10、または10回より多くの反復)を含有するモジュールは同じ様式で作製される。
【0176】
その後、誂えのTALエフェクター遺伝子を作製するために任意の反復の配列をtal1c「主鎖」内にアセンブルするための方法が考案された。本方法には以下のステップが含まれ、これは図20にも示されている:
(1)第1の所望の反復を有する単一反復の開始プラスミド(それぞれRVD NI、HD、NN、またはNGをコードしているpCS493、pCS494、pCS495、またはpCS495)を選択するステップ、
(2)プラスミドをPspXIで直鎖状にするステップ、
(3)XhoIを使用して次の反復(複数可)のモジュールを適切なモジュールプラスミド(pCS502〜pCS585)から単離するステップ、
(4)ライゲーションするステップ、
(5)MscIで消化することによって配向を確認し、ベクターに基づくプライマーを使用して配列を3’末端から確認するステップ、および
(6)すべての反復がアセンブルされるまでステップ2〜5を繰り返すステップ。
【実施例5】
【0177】
TALENのモジュールアセンブリ用プラスミドのライブラリ
本明細書中に記載のTALEN反復のアセンブリ(たとえば図20に示したステップを使用)は、増加していく数の反復を含有する数々の中間体プラスミドをもたらす。これらのプラスミドのそれぞれは、TALENのモジュールアセンブリ用プラスミド(pMAT)のライブラリが作製されるように保管されている。たとえば、図21Aおよび21Bは、示されているヌクレオチド配列を標的とするTALエンドヌクレアーゼの構築における反復モジュールのアセンブリを示す。図21Aでは、pCS519、pCS524、pCS537、pCS551、pCS583、およびpCS529と命名されたプラスミドからの反復モジュールを、pCS493と命名された開始プラスミド中の配列に順次付加して、pMAT55、pMAT56、pMAT57、pMAT58、pMAT59、およびpMAT60と命名されたプラスミドがもたらされる。図21Bでは、pCS530、pCS533、pCS522、およびpCS541と命名されたプラスミドからの反復モジュールを、pMAT1と命名されたプラスミド中の配列に順次付加して、pMAT61、pMAT62、pMAT63、およびpMAT64と命名されたプラスミドがもたらされる。
【実施例6】
【0178】
誂えたTALENの作製および試験
実施例4および5に記載のシステムを使用して、TALのDNA認識ドメインを使用して特定のDNA標的を認識および切断するTALENを作製した(図22A)。TALENの機能を評価するために、LacZ活性がDNA切断の指標として役割を果たす酵母アッセイを適応させた(Townsendら、上記)。このアッセイでは、標的プラスミドおよびTALEN発現プラスミドを接合によって同じ細胞中で一緒にした。標的プラスミドは、コード配列の125bpの重複を有するlacZレポーター遺伝子有する。重複は、所定のTALENによって認識される標的部位に隣接する。二本鎖DNAの切断が標的部位で起こる際、これが重複した配列間の一本鎖のアニーリングによって修復され、それにより、機能的なlacZ遺伝子が作製され、その発現は定量可能な読取値を提供する標準のβ−ガラクトシダーゼアッセイによって測定することができる(図22A)。このアッセイは、NHEJによって染色体の突然変異を生じる、または高等真核生物において遺伝子編集のために相同組換えを刺激する、ZFNの能力の良好な予測子であることが実証されている(Townsendら、上記、およびZhangら(2010)Proc.Natl.Acad.Sci.USA、107:12028〜12033)。
【0179】
2つの十分に特徴づけられたTALエフェクター、すなわち、コショウの病原体Xanthomonas campestris pv.vesicatoriaからのAvrBs3およびコメの病原体X.oryzae pv.oryzaeからのPthXo1を使用した(Bonasら(1989)Mol.Gen.Genet.、218:127〜136、およびYangら(2006)Proc.Natl.Acad.Sci.USA、103:10503〜10508)。AvrBs3のアミノ酸配列はGENBANK受託番号P14727および配列番号12の下(図3)、核酸配列は受託番号X16130および配列番号13の下(図4)で見つけることができる。PthXo1のアミノ酸配列はGENBANK受託番号ACD58243および配列番号31の下(図23)、核酸配列は受託番号CP000967、遺伝子ID6305128、および配列番号32の下(図24)で見つけることができる。GENBANK受託番号ACD58243の下のPthXo1のアミノ酸配列は、開始コドンの誤った注釈付けが原因で、N末端で切断されている。完全な配列は図23中に提示されている。
【0180】
AvrBs3およびPthXo1の反復ドメインはどちらも保存的なSphI断片内に完全にコードされている(図4および24)。また、どちらのTALエフェクターコード遺伝子も、反復ドメインのコード配列ならびにその前の287個のアミノ酸およびその後の231個のアミノ酸を包含するBamHI制限断片を有する(図4および24、図22Aも参照)。BamHI断片からはTALエフェクター転写活性化ドメインが欠けている。SphI断片およびBamHI断片はどちらも、ヌクレアーゼ発現ベクターpFZ85中に存在するFokIをコードしているDNA断片と融合させた(図25)。FokIヌクレアーゼとAvrBs3およびPthXo1によってコードされているBamHI断片との間の融合タンパク質を図26および27、配列番号33および34に示す。
【0181】
FokI単量体は切断のために二量体化しなければならないが、2つのDNA認識部位間の適切なスペーサー長は不明確であった。ジンクフィンガーアレイが4〜7個のアミノ酸のリンカーによってFokIから分離されているZFNでは、2つの認識部位の間の典型的なスペーサーは5〜7bpである(Handelら(2009)Mol.Ther.、17:104〜111)。たとえば、本明細書中で使用したBamHI TALEN構築体中では235個のアミノ酸が反復ドメインをFokIから分離しているため、BamHIおよびSphI構築体の両方で様々なスペーサー長(6、9、12、15、および18bp)を使用した。陽性対照としては、マウス転写因子Zif268(PorteusおよびBaltimore(2003)Science、300:763)に由来するDNA結合ドメインを有する良好に特徴づけられているジンクフィンガーヌクレアーゼを使用した。陰性対照としては、TALエフェクタードメインを触媒的に不活性なFokI変異体と融合させたか、または非同族DNA標的に対して試験した。
【0182】
200μlの終夜培養物中の、TALEN発現または標的プラスミドのどちらかを含有する一倍体細胞種を、YPD培地中、30℃で接合させた。4時間後、YPD培地を5mlの選択培地で置き換え、終夜、30℃でインキュベーションした。接合した培養物を溶解し、ONPG基質を加え、96ウェルプレートリーダーを使用して吸光度を415nmで読み取った(Townsendら、上記)。β−ガラクトシダーゼレベルを基質切断速度の関数として計算した。15bpのスペーサーが2つの認識部位を分離している標的レポーター構築体で得られた結果が図22Bに示されている。主に反復アレイをコードしたSphI断片に由来するすべてのヌクレアーゼ発現構築体は、活性を示すことに失敗し、これは、機能には反復アレイ中のものに追加のアミノ酸配列が必要であることを示している(図22B)。しかし、BamHI断片に由来するAvrBs3およびPthXo1 TALENの両方で頑強な活性が観察された(図22B)。PthXo1 TALENの活性はZFN陽性対照のものに近かった。活性は機能的FokIドメインを必要とし、所定のTALENによって認識されるDNA標的に特異的であった。
【0183】
また、FokIが最も効率的に二量体化することを可能にするスペーサー長を同定するために、TALエフェクター結合部位間の様々な距離(12〜30bpの11個の長さの変異体)を試験するための実験も実施した(図28A)。どちらの酵素も、一方は15bp、他方は21bp(AvrBs3)または24bp(PthXo1)のどちらかである、2つのスペーサー長の最適条件を示した。PthXo1では、13bp以上のすべての試験したスペーサー長で活性が観察された。しかし、AvrBs3の一部のスペーサー長では活性を示されず、スペーサー長が特定のTALENに重要であることが示唆された。
【0184】
上記実験は、スペーサーの両側に反対方向で配置した2つの同一の認識配列と結合するホモ二量体TALENの活性を試験した。そのような回文部位はゲノム標的中で自然に起こる可能性が低いため、TALENがヘテロ二量体として機能できるかどうかを試験するための実験を実施した。AvrBs3およびPthXo1の認識部位を頭から尾部の配向で15bpのスペーサーの両側に配置した。そのそれぞれの標的に対するAvrBs3およびPthXo1 TALENの個々の活性ならびにZif268の活性を対照として測定した。陰性対照として、ヘテロ二量体部位の標的部位プラスミドのみを有する酵母培養物をLacZ活性についてアッセイした。生じるヘテロ二量体TALENの活性は、2つのホモ二量体酵素で観察された活性の平均に近かった(図28B)。
【0185】
自由裁量による染色体配列に対するTALENを標的とするように反復ドメインをアセンブルできるかどうかを試験するために、以前にZFNを用いた突然変異誘発の標的とされていた2つの遺伝子、すなわちシロイヌナズナからのADH1およびゼブラフィッシュからのgridlockを選択した(Foleyら(2009)PLoS One、4:e4348、およびZhangら、上記)。5’のTによって先行されており、MoscouおよびBogdanove(上記)によって同定されたTALエフェクター結合部位に類似のヌクレオチド組成を有する、コード領域中の12〜13bpの配列の検索を行った。ADH1およびgridlockでは、そのような部位は平均して7〜9bpごとに存在した。4個の12bpの部位がADH1中で選択され(染色体遺伝子配列の位置360、408、928、および975)、1個の13bpの部位がgridlock中で選択された(染色体遺伝子配列の位置2356、図29A)。ネイティブTALエフェクターからの最も豊富なRVD(AにはNI、CにはHD、GにはNN、およびTにはNG)を使用して、これらの標的を認識するようにTALエフェクター反復ドメインを構築した。誂えのTALENを構築するために、実施例4および5に記載のように、これらのRVDを有する反復を個々に合成し、1回、2回、または3回の反復のモジュール内にアセンブルした。これらのモジュールを、元の反復が除去されたtal1c遺伝子の誘導体内に順次ライゲーションさせ(MoscouおよびBogdanove、上記)、これらの操作したTALエフェクターからのBamHI断片を、pFZ85中のFokIの触媒ドメインをコードしている配列と融合させた(図25)。シロイヌナズナからのADH1およびゼブラフィッシュのgridlock遺伝子を標的とした5個の誂えのTALENを作製した。
【0186】
生じる誂えのTALENは、酵母アッセイにおいてホモ二量体TALENとして(すなわち、同一のDNA結合部位が16〜18bpのスペーサーの両側に逆の配向で重複していた)試験したが、天然に存在するDNA標的での切断を指示するためにはヘテロ二量体TALENを構築しなければならないことに留意されたい。スペーサー長は、次の隣接する(かつ向かい合う)候補部位の3’末端から、15bpに最も近い距離に基づいて選択した。16bpのスペーサーをADH1−360−12、ADH1−408−12rに使用し、18bpのスペーサーをADH1−928−12、ADH1−975−12r、およびgridlock−2356−13rに使用した。酵母アッセイは上述のように行った。
【0187】
頑強なヌクレアーゼ活性がADH1−360−12およびgridlock−2356−13r TALENで観察された(図29B)。ADH1−928−12 TALENは控えめな活性を有していたが、それでも陰性対照より有意に高かった。陽性結果を与えたそれぞれのTALENについて、ヌクレアーゼ活性は同族標的に特異的であった。これらの結果は、新規の機能的なTALENを誂えた反復ドメインのアセンブリによって作製できることを示している。
【実施例7】
【0188】
天然に存在する標的およびTALエフェクターの対はヌクレオチドおよびRVDの組成の全体的および位置的な偏りを示す
MoscouおよびBogdanove(上記)によって分析した20個の対合した標的およびTALエフェクターを、全体的な組成の偏りおよびヌクレオチドまたはRVD頻度に対する位置効果について評価した。部位(プラス鎖上)は一般にAおよびCに富み、Gに乏しいことが観察された。平均パーセントAは31±16%であった(1単位の標準偏差)。平均パーセントCは37±13%であった。平均パーセントGは9±8%であり、平均パーセントTは22±10%であった。アラインメントは長さが変動するため、位置効果の分析はそれぞれの末端上の5個の位置に限定された。驚くべきことに、位置1および3でAが有利およびTが不利であり、位置Nおよび場合によっては位置2でTが有利である、標的配列中の偏りが明らかであった。Gは位置N−1で特に稀であった。この偏りはエフェクター中の一致するRVDによって反映されており、位置1および3にはNIが最も一般的であり、位置1にはNGが存在せず、位置Nにはほぼ必ずNGが存在し、位置N−1にはNNは稀である(図30)。
【実施例8】
【0189】
誂えのTALエフェクター反復アレイを迅速にアセンブリおよびクローニングするための方法および試薬
Golden Gateクローニング方法[Englerら(2008)、上記、およびEnglerら(2009)、上記]は、その認識部位の外側で切断して、複数のDNA断片を同時に規則的にライゲーションさせるための誂えのオーバーハングを作製する、IIS型制限エンドヌクレアーゼ(たとえばBsaI)の能力を用いる。この方法を使用して、単一の反応で、いくつかのDNA断片を特定の順序でアレイへと融合させ、所望のデスティネーションベクター(destination vector)内にクローニングすることができる(図31)。
【0190】
誂えのTALエフェクター反復をコードしているアレイをアセンブルするための方法および試薬をGolden Gateシステムに基づいて開発した。BsaI部位がTALエフェクター反復のコード配列の両側に位置している場合は、切断により4bpのオーバーハングによって隣接される反復断片が放出される。切断部位は配列特異的でないため、互い違いにずらす(staggering)ことによって、規則的な相補的オーバーハング(粘着末端)を有する反復クローンを放出させることができ、複数反復アレイの規則的なアセンブリが可能になる。
【0191】
58個のプラスミドのライブラリ(図32Aおよび32B)を作製して、10個までの反復単位の「サブアレイ」への同時アセンブリ、次いでこれらのサブアレイのうちの1、2、または3個および最終的な切断された反復の、完全な誂えのアレイへの同時アセンブリを可能にした。組中のそれぞれの断片が4つの最も一般的なRVDであるHD、NG、NI、およびNNのうちの異なる1つを有する反復モジュールをコードしている、4個の断片の10個のずらし型(staggered)の組を合成し、テトラサイクリン耐性遺伝子を保有するベクター内にクローニングして、合計40個のプラスミドとした。それぞれの断片が4つの最も一般的なRVDのうちの異なる1つをコードしている、20個のアミノ酸の末端切断されたTALエフェクター反復をコードしたさらに4個の断片を合成し、スペクチノマイシン耐性遺伝子を保有する異なるベクター内にクローニングしてさらに4個のプラスミドを得て、「最終反復プラスミド」と命名した、図32A)。ずらし型の組中のすべての断片は、BsaIでの切断にって適切な順序でのアセンブリを可能にする様々な粘着末端を有する断片が放出されるように、ベクター中でBsaI部位によって隣接されている。すなわち、反復モジュール1の断片の3’末端でのオーバーハングは反復モジュール2の断片の5’末端でのオーバーハングにのみ相補的であり、反復モジュール2の3’末端でのオーバーハングは反復モジュール3の5’末端でのオーバーハングにのみ相補的であり、以下同様である。最終反復プラスミド中の断片は、異なるIIS型制限エンドヌクレアーゼ、Esp3Iの部位によって隣接されている。以下に記載の14個の追加のプラスミドを、アセンブルされたサブアレイを受けるためのデスティネーションベクターとして構築した。
【0192】
第1のデスティネーションベクター、プラスミドpFUS Aは、21回以下の反復(最後の切断された反復を数える)の最終アレイへとアセンブルする、第1の10回反復サブアレイを受けるために構築した。pFUS Aは、BsaIによる切断によって、片側で第1反復モジュールの5’末端でのオーバーハングに相補的なオーバーハングが作製され、反対側で第10反復モジュールの3’末端でのオーバーハングに相補的なオーバーハングが作製されるように構築した。そのような最終アレイへとアセンブルする10回以下の反復の第2のサブアレイを受けるために、デスティネーションベクタープラスミドpFUS_B1、pFUS_B2、pFUS_B3、pFUS_B4、pFUS_B5、pFUS_B6、pFUS_B7、pFUS_B8、pFUS_B9、およびpFUS_B10を構築し、これらは、BsaIによって切断された際に、それぞれ第1反復モジュールの5’末端および対応する符番した位置の反復モジュールの3’末端でのオーバーハングに相補的なオーバーハングを有する(たとえば、サブアレイの3’末端のpFUS_B6のオーバーハングは、位置6の4回反復モジュール断片のオーバーハングに一致する)。pFUS_AおよびpFUS_Bシリーズのプラスミド中にクローニングしたアレイは、ベクター中でEsp3I部位によって隣接されており、Esp3Iを用いた消化によって放出された際、アレイは、反復領域を欠いたTALENをコードしているデスティネーションベクターpTAL内に最終の切断された反復断片と共にそれらが順番にライゲーションされることを可能にする、ユニークな相補的なオーバーハングを有する。Esp3Iを用いた切断が、第1の10回反復サブアレイの5’末端でのオーバーハングに相補的な一端でのオーバーハングおよび最終の切断された反復断片の3’末端でのオーバーハングに相補的な他端でのオーバーハングのおかげで正しい位置および正しい配向での反復アレイの挿入を可能にするように、pTALを構築した(図33)。
【0193】
22〜31回反復の最終アレイへとアセンブルする第1および第2の10回反復サブアレイを受けるために、最終の2つのデスティネーションベクタープラスミド、pFUS_A30AおよびpFUS_A30Bを構築した。アレイをpFUS_Bベクターからの第3のアレイおよびEsp3Iを用いた消化によって同様に放出された最終反復プラスミドからの最終の切断された反復断片と共に順番にpTAL内にライゲーションさせることができるよう、Esp3Iを用いた消化により適切な相補的オーバーハングを有するアレイが放出されるように、pFUS_A30AおよびpFUS_A30Bを構築した(図32)。
【0194】
すべてのデスティネーションベクターはIIS型制限エンドヌクレアーゼ部位間にクローニングされたLacZ遺伝子を有しており、組換えのブルー−ホワイトスクリーニングを可能にする。アンピシリン耐性の遺伝子を保有するpTAL以外は、すべてのデスティネーションベクターがスペクチノマイシン耐性の遺伝子を保有する。
【0195】
これらの試薬を使用して誂えのTALエフェクター反復アレイを迅速に構築するために、以下の方法を確立した。第1のステップでは、10回以下の反復の必要なサブアレイの適切な個々のRVDモジュールプラスミドを、適切なデスティネーションベクターと、1本のチューブ中で混合する。T4 DNAリガーゼおよびBsaIエンドヌクレアーゼを加え、反応を、PCR機械中で、2つの酵素のそれぞれの最適温度である37℃で5分間および16℃で10分間、10サイクルの間インキュベーションする。その後、in vivo組換えによるより短い不完全なアレイのクローニングを防止するために、反応混合物をPLASMID−SAFE(商標)ヌクレアーゼで処理してすべての直鎖状dsDNA断片を加水分解し、その後、混合物を使用して化学的にコンピテントな大腸菌細胞を形質転換させる。生じる組換えプラスミドを単離し、正しい構築体を確認する。その後、第2のステップでは、第1のステップからの確認されたプラスミドを適切な最終反復プラスミドおよびpTALと混合し、消化およびライゲーション反応サイクルを第1のステップと同様に実施する。最後に、反応生成物を大腸菌内に導入し、完全長の最終アレイ構築体を単離および確認する。このプロトコルは、1人の人によって1週間以内に完了することができる。
【0196】
表4A中のTALEN85、102および117、ならびに以下の実施例14に記載のTALEN HPRT−3254−17およびHPRT−3286−20rの発現構築体を、本実施例中に記載の方法および試薬を使用して作製した。
【0197】
pTAL中にクローニングした反復アレイは、反復領域に隣接する保存的なSphI制限エンドヌクレアーゼ部位を使用して、他のTALエフェクター遺伝子のコンテキスト内に容易にサブクローニングされる。
【実施例9】
【0198】
誂えのTALENのデータは「規則」およびRVDの数と活性との間の相関について初期の支持を示す
実施例6は、ユニークなDNA配列を認識できるようにTALENのDNA結合ドメインを操作するために実施した実験を記載している。記載のように、これらの誂えのTALENは、シロイヌナズナADH1およびゼブラフィッシュgridlock遺伝子中の部位を認識した。これらの遺伝子だけでなく、シロイヌナズナからのTT4遺伝子、およびゼブラフィッシュからのテロメラーゼ中の部位を認識するように、追加の誂えのTALエフェクターDNA結合ドメインを操作した(Foleyら、上記、およびZhangら、上記)。これらの誂えのTALENは、実施例3、4および8に記載の方法を使用して作製した。誂えのTALENの操作において、観察された組成的および位置的な偏りは、設計原理または「規則」として適応した。最初に、5’のTによって先行されており、少なくとも15bpの長さであり、上述の平均と一貫性のあるヌクレオチド組成を有するコード領域中の配列の検索を実施した。具体的には、0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTを有する部位のみを選択した。そのような部位は平均して7〜9bpごとに存在した。その後、上述の観察された位置的な偏りに順応した部位を選択した。操作したTALENの結合によりFokIの二量体化が可能となるように、この組から、15〜19bpの長さであり、15〜18bpによって分離されている、それぞれの遺伝子中の結合部位の2つの対を同定した。モジュールアセンブリ方法(実施例3および4)は、部分的な長さの構築体を生じた。
【0199】
それぞれが9回以上の反復のアレイを有する、16個のヌクレオチドの配列を標的とするように設計された合計21個の中間体および完全長のTALEN。これらのTALENのアミノ酸配列は図34A〜34U(配列番号35〜55)に提供されている。これら21個のTALENを、実施例2および6に記載の酵母アッセイを使用して、DNAを切断するその能力について試験した。活性データは図35に示されており、表4Aに要約されている。
【0200】
中間の部分的な長さのTALENの一部は、ヌクレオチド組成および末端Tの規則を破っている標的に対応する。表4Aは、長さ、これら2つの規則への順応、およびZFN268と比較した活性を、それぞれのTALENについて示す。結果は、RVDアレイの長さを増加させることで生じるTALENの活性が増加するという大きな傾向を明らかにしている。これは、DNA標的がin vivoで認識可能となる前に必要な最小数のRVDが存在することを示唆している。さらに、規則の順応が重要であると考えられる。検出可能な活性を示さない6個のTALENのうち、2つが標的の組成の規則に違反しており、2つがNGで終わっておらず、別の1つが両方の規則を破っていた(1つが両方の規則に従っていた)。ZFN268の25%未満の活性を有する8個のTALENのうちの3個が規則の一方に違反しており、ZFN268の25〜50%の活性を有する4個のTALENのうちの1個がNGで終わるRVD配列を有していなかった。ZFN268の50%以上の活性を有するTALENはすべての規則に従っており、同じ長さのTALENでは、規則を破ったものは従順なアレイよりも一般に低い活性を有していたことに留意されたい。長さに関する全体的な傾向に一致して、規則を破らなかった中間体でも、対応する完全長のTALENの方がより高い活性を有していた(表4Aおよび図35)。同じ標的に対するTALEN長の相違が原因であるスペーサー長の変動が、この観察に寄与した可能性があるが、スペーサー長のある程度の範囲は耐容される(Christianら、上記)。
【0201】
データ中のある程度の複雑さが明らかであった。たとえば、活性は同じ長さの従順なTALEN間で変動しており、一部の短いアレイは中等度に高い活性を有しており、一部の長い従順なアレイは活性が少ないまたはなかった(表4B)。それにもかかわらず、提供された結果は、1)一般により多数の反復がより高い活性をもたらす、および2)組成および位置的な偏りの規則の順応が活性に重要であるという結論を支持している。したがって、以下の設計原理を導いた。
【0202】
・TALエフェクター結合部位は、最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあるように設計される。
・部位は第1(5’)位置にTまたは第2位置にAを有し得ない。
・部位はT(3’)で終わらなければならず、最後から2番目の位置にGを有し得ない。
・部位の基本組成は指定した範囲内(平均±2単位の標準偏差)でなければならない:Aは0〜63%、Cは11〜63%、Gは0〜25%、およびTは2〜42%。
【表5】
【表6】
【実施例10】
【0203】
ヘテロ二量体TALEN対は酵母アッセイにおいてその意図する天然に存在する標的配列を切断する
実施例2、6および9のデータは、新規標的DNA配列を認識するように誂えのTALENを操作できることを実証している。ホモ二量体標的部位を認識した個々のTALEN単量体を使用して、誂えのTALENの酵母活性データを集めた。すなわち、TALENの標的配列を15〜18bpのスペーサー両側に逆の配向で重複させた。しかし、内在染色体配列の切断は、一般に、2つの異なる誂えのTALENがスペーサーの両側の2つの異なる配列を認識することを必要とする。実施例6に記載のように、この能力は、一緒にしたAvrBs3およびPthXo1 TALENについて、酵母アッセイにおいて、対応するキメラ標的部位を使用して実証した。本発明者らは、2つの異なる誂えのTALENが天然に存在するDNA配列を認識および切断することができるかどうかを試験した。実施例2に記載の酵母アッセイを使用して、シロイヌナズナADH1遺伝子中の2つの異なる標的配列を切断するように設計された誂えのTALENを、これらの標的に対する活性についてアッセイした。標的部位および対応するTALENのDNA配列は図36Aに示されている。TALENのアミノ酸配列は図34に提供されている。酵母アッセイで得られたベータ−ガラクトシダーゼ活性は、図36Bに示されているグラフ中にプロットされている。TALENのその天然に存在する標的配列に対する活性は陰性対照よりも有意に高く、これは、内在標的DNA配列を認識および切断するためにTALENを設計できることを示している。
【実施例11】
【0204】
TALENはシロイヌナズナ中のネイティブ遺伝子を切断し、不正確な非相同末端結合によって突然変異を導入する
シロイヌナズナADH1遺伝子中の標的配列を認識するように設計された活性TALEN対のうちの1つを、染色体DNAを結合、切断および突然変異させることができるかどうかを決定するために試験した。この対(pTALEN69および74)を含む個々のADH1 TALENのそれぞれを、TALENを構成的な35Sプロモーターの制御下に置く植物発現ベクターpFZ14内にクローニングした(Zhangら、上記)。その後、生じる構築体を、シロイヌナズナのプロトプラスト内に電気穿孔によって導入した。48時間後、ゲノムDNAを単離し、Tth111lで消化した。Tth111l切断部位は2つのTALEN認識部位の間のスペーサー配列中に位置する(図37A)。TALENによる染色体DNAの切断は、不正確な非相同末端結合(NHEJ)によって突然変異を導入することが予想され、これはTth111lによる切断の失敗をもたらす。その後、TALEN認識部位を包含する375bpの断片をPCR増幅した。PCR産物を再度Tth111lで消化して、TALENに媒介されるNHEJによって改変されなかった残りのゲノムDNAのほとんどを除去する。その後、消化産物をアガロースゲル上に流す。未切断のPCR産物が観察され、そのような未切断のPCR産物は、内在標的配列でのヌクレアーゼ活性(この事例ではTALEN活性)の指標である(Zhangら、上記)。未切断のDNAをクローニングし、DNA配列決定によって分析した。9個の独立したクローンの配列決定により、6個がNHEJによって導入された突然変異を保有していたことが明らかとなった(図37B)。したがって、TALENは、内在染色体座位を切断し、DNA二本鎖切断および突然変異を導入する。
【実施例12】
【0205】
標的化能力の増強
TALエフェクターDNA暗号の中核では、4つの最も一般的なRVDは、会合頻度に基づいて4個のヌクレオチドに対して見かけ上1対1の特異性を有する。これは、HD、NG、およびNIで顕著にそうであるが、NNではその度合がより低い(図1C)。NNは最も頻繁にはGと会合するが、ほぼ同じように一般的にAと、時折CまたはTと会合する。13個のRVD配列中の4つの位置にNNを有するランダムにアセンブルされたTALエフェクターでは、人工標的中のすべての対応する位置にGを持つことで最良の活性が得られた(Bochら(2009)Science、326:1509〜1512)。Aは活性を減少させたが消失させず、CおよびTは検出可能な活性を排除した。NNである24個のRVDのエフェクターPthXo1の結合部位中の第1位置のみでGをC、T、またはAで置換した場合に、活性の劇的な損失が観察された(Romerら(2010)New Phytol.、187:1048〜1057)。しかし、これは、はるかにより短いAvrHah1(14個のRVD)がAとアラインメントするNNで始まり、23個のRVDのエフェクターPthXo6が位置4〜6にそれぞれAとアラインメントするNNを連続して3個有するが、これらのタンパク質はどちらの活性が高いという観察と対照的であった(Schornackら(2008)New Phytol.、179:546〜556、およびRomerら、上記を参照)。したがって、Gに対するNNの特異性は一般に弱いと考えられ、コンテキストに伴って変動する場合がある。
【0206】
チミンがTALエフェクター標的部位の直前にあるという観察された不変性は、いくつかのエフェクターの要件である[Bochら、上記、Romerら、上記、およびRomerら(2009)Plant Physiol.、150:1697〜1712]。高度に保存的な、TALエフェクター中の反復領域の直前のアミノ酸配列は(図38A)、アミノ酸配列および予測された二次構造のどちらにおいても、反復と顕著な類似度を共有する(図38BおよびBodganoveら(2010)Curr.Opin.Plant Biol.、13:394〜401)。「第0」反復と呼ばれるこの配列が、結合部位の−1位にTが必要であること根拠であり、RVDに類似の位置中の残基(図38B)がヌクレオチドを指定するという仮説が立てられた。
【0207】
これらの発見に基づいて、Gに対して高い特異性を有する反復を取り込ませることによって、および−1でのTの必要性を緩めることによって、操作したTALエフェクタータンパク質の標的化能力を増強できると仮説が立てられた。NNが表示するよりも頑強なGに対する特異性について新規かつ稀なRVDを試験するため、および第0反復のRVDに類似の残基を一般的なRVDで置き換えるための実験を開始した。
【0208】
Gに対する頑強な特異性のための新規かつ稀なRVD:上記開示したモジュール(たとえば実施例4を参照)では、4つのヌクレオチド塩基(それぞれA、C、G、およびT)との結合を指定するために4つの特定のRVD(NI、HD、NN、およびNG)を使用した。他のRVDを含有する反復も有用で有り得、これらはNI、HD、NN、およびNGと比較して4つの塩基に対して増加した特異性および/または親和性を有し得る。Gに対する特異性を改善させることに向けては、新規かつ稀なRVDをコードしているいくつかの反復を構築した。稀なRVD NK、HN、およびNAはGと会合し、これは、Nが残基のうちの一方または他方として重要であり得ることを示唆している(図1C)。したがって、表5に示されているRVDを有する反復をコードしている幅広い誘導体の組を構築した。左の列は、位置12に極性アミノ酸(R、K、D、E、Q、H、S、T、またはY)および位置13にNを有するRVDを記載している。右の列は、RVDの第1位置のNと第2位置の17個の他のアミノ酸のうちの任意のもの(G、L、V、R、K、D、E、Q、H、T、M、C、P、Y、W、またはF)との組合せを記載している。Nを含まないより高い特異性の可能性を考慮するために、位置12に極性アミノ酸(R、K、D、E、Q、H、S、T、またはY)および位置13にギャップ(*)を用いた反復も作製した(中央の列)。
【0209】
新規人工RVDは、Nicotiana benthamianaにおける、GUSもしくは二重ルシフェラーゼレポーターに基づくAgrobacteriumに媒介される一過性発現アッセイなどの、TALエフェクターの転写活性化活性のための定量的レポーター遺伝子に基づくアッセイ、または上述の出芽酵母におけるlacZレポーターに基づくTALENアッセイにおいて、機能について試験する(たとえば実施例2を参照)。試験するRVDを含有する反復モジュールを、測定可能かつ亜飽和レベルの活性を有するTALエフェクターまたはTALEN内に取り込ませ、生じるタンパク質を、対応する位置での4個すべてのヌクレオチドの統合順列を用いて、DNA標的の組に対する活性の相違について試験する。特に、植物体内および酵母アッセイにおいて最小限活性であり、3個の追加の反復でミスマッチに応答性であるPthXo1変異体(複数可)から始めて、新規かつ稀な反復のそれぞれを含有するTALENを(同価の3つ組で)、対応する位置のそれぞれにGを有する標的に対してin vivoで試験する。増加した活性を示す任意のものについて、これらの位置で他のヌクレオチドに入れ替えた標的を用いてアッセイを繰り返して、特異性を確認する。
【表7】
【0210】
位置−1でのTの特異性を緩めるための、第0反復のRVDに類似の位置の一般的なRVD置換:二次構造の予測および第0反復と反復コンセンサス配列とのアラインメントは、第0反復中のKR*(星印はギャップを示す)によって占有される位置がRVDに類似であり、−1でTを指定する残基であったことを示唆した。KRをHD、NG、NI、およびNNで置換し、別にR*を置換した、PthXo1の変異体を、上述のTal1c「主鎖」構築体中で構築した。植物体内および酵母アッセイにおいて、−1位に対応するヌクレオチド、すなわちそれぞれC、T、A、およびGを有する標的を使用して、これらの変異体の活性を野生型エフェクターと比較する。第0反復の位置11のKがコンセンサス反復配列の位置11の残基のSで置換されている、PthXo1のさらなる変異体を構築する。また、この置換と、コンセンサス反復配列の位置16の残基であるKでの第0反復の位置15のVの置換との組合せを有する他の変異体を構築する(表6)。TALエフェクター活性のための近位のTATAボックスを含め得る。さらに、−1のTがTATAボックスの一部であるように見えるAvrBs3とは異なり、PthXo1結合部位に最も近いTATAボックスは46bp下流であり、−1での修飾によって乱されないため、PthXo1はこの実験に有用である。
【0211】
上記修飾が、Gに対する標的化の増強またはT以外のヌクレオチドによって先行される配列を標的化する能力の増加をもたらさない場合は、より包括的な人工RVDの組をG特異性について試験し、また、一般的なRVD以外の置換を第0反復について試験する。
【表8】
【実施例13】
【0212】
新規の予測されたヌクレオチド特異的RVD
表1Aおよび1Bに記載されたRVDをRVD中の第2アミノ酸残基(すなわち、全体的な反復中の13番目)によって群分けした場合に、RVDの第1位置のアミノ酸とは無関係に、そのアミノ酸とRVDによって指定されるヌクレオチド(複数可)とのほぼ完璧な相関が存在したことが観察された(表7)。したがって、ギャップ(星印によって示す)で終わるRVDはCもしくはT、またはTを指定し、Dで終わるRVDはCを指定し、Gで終わるRVDはTを指定し、Nで終わるRVDはGもしくはA、またはGを指定する。また、RVDの位置1のアミノ酸はH、I、N、S、またはYのいずれかであったことも観察された。これらの観察は、RVDの特異性は、第1位置の残基がH、I、N、S、またはYであるかに依存せずに、第2位置の残基によって決定されることを示唆した。したがって、第2位置で観察された残基を第1位置の残基H、I、S、N、またはYと組み合わせるいくつかの新規(すなわち未だ観察されていない)RVDについて、特異性を予測した。したがって、I*、S*、およびY*はCもしくはT、またはTを指定すると予測され、ID、SD、およびYDはCを指定すると予測され、SGはTを指定すると予測され、INおよびYNはGもしくはA、またはGを指定すると予測された。また、第2位置のKは1つの事例しかなかったが、観察されたNKの特異性に基づいて、HK、IK、SK、およびYKはGを指定すると予測された。
【0213】
これらの新規RVDを、実施例2および11に記載の定量的TALエフェクターおよびTALEN活性アッセイにおいて機能および特異性について試験し、既存のRVDと比較した。
【表9】
【実施例14】
【0214】
誂えのTALENは動物細胞において内在標的を切断し、不正確な非相同末端結合によって突然変異を導入する
TALENを動物細胞における標的化突然変異誘発に使用できるかどうかを試験するために、最初に、TALエフェクターAvrBs3、PthXo1、およびTal1cの発現をヒト胚性腎臓(HEK)293T細胞中で試験した。AvrBs3、PthXo1、およびTal1cをコードしている遺伝子からストップコドンを除去し、遺伝子を、哺乳動物発現ベクターpcDNA3.2/V5−DEST(Invitrogen、カリフォルニア州Carlsbad)内に、タンパク質の免疫検出のためのV5エピトープをコードしているそのベクター中の下流配列とインフレームでサブクローニングした。pcDNA3.2/V5−DESTは、TALエフェクター遺伝子を構成的ヒトサイトメガロウイルス(CMV)プロモーターの制御下に置く。生じるプラスミドを個々に用いて、Lipofectamine 2000(Invitrogen)を使用してHEK293T細胞を形質移入し、24時間後、全タンパク質をそれぞれの形質移入した細胞のバッチから単離し、ポリアクリルアミドゲル電気泳動、ウエスタンブロッティングおよびマウス抗V5抗体を使用した免疫標識に供した。標識されたタンパク質は、ヤギ抗マウス抗体−西洋ワサビペルオキシダーゼのコンジュゲートを用いて、SuperSignal Weat Pico化学発光キット(ThermoScientific,Inc.)を使用して検出した。等価なローディングは、アクチンの免疫標識および検出によって確認した。それぞれのTALエフェクタータンパク質は、明らかな分解なしに、検出可能に発現された(図39)。
【0215】
次に、内在性ヒトHPRT遺伝子中の配列を標的とするために、実施例9に記載のように一対のTALENを設計し、HPRT−3254−17およびHPRT−3286−20rと命名した(図40Aおよび図40B)。Golden Gateクローニングに基づく方法および実施例8に記載の試薬を使用して、HPRT−3254−17をコードしているプラスミドpTALEN141およびHPRT−3286−20rをコードしているプラスミドpTALEN142を構築した。その後、TALEN遺伝子を、それらを構成的CMVプロモーターの制御下に置く哺乳動物発現ベクターpCDNA3.1(−)(Invitrogen,Inc.)内にサブクローニングし、プラスミドpTALEN141MおよびpTALEN142Mが得られた。その後、pTALEN141MおよびpTALEN142Mの両方を一緒に用いて、ならびに別に陰性対照としてpCDNA3.1(−)を用いて、HEK293T細胞を形質移入した。72時間後、ゲノムDNAを単離し、制限エンドヌクレアーゼBpu10Iで消化した。Bpu10I部位は、HPRT中でHPRT−3254−17およびHPRT−3286−20rの結合部位を分離するスペーサー内に存在する(図41A)。Bpu10I消化に次いで、PCRを使用して、TALENで処理した試料および対照試料の両方からのTALEN標的部位にまたがる244bpの断片を増幅した。予想された断片がどちらの試料からも増幅され、ゲノムDNAのBpu10I消化が不完全であったことを示す。しかし、続くBpu10Iを用いたPCR産物の消化は、対照試料から増幅された産物の完全な切断をもたらしたが、TALENで処理した試料からの産物の不完全な切断をもたらした(図41B)。TALENで処理した試料中の切断耐性PCR産物の存在は、HPRT中の意図する標的での、TALENに媒介される二本鎖切断の非相同末端結合による不完全な修復の結果、内在性Bpu10I部位がin vivoで突然変異したことの証拠を提供する。したがって、TALENは、哺乳動物細胞における標的化突然変異誘発に使用することができる。
【0216】
他の実施形態
本発明をその詳細な説明と併せて記載したが、前述の説明は、添付の特許請求の範囲によって定義される本発明の範囲を例示し、限定しないことを意図することを理解されたい。他の態様、利点、および改変が以下の特許請求の範囲内にある。
【技術分野】
【0001】
関連出願の相互参照
本出願は、すべてその全体で本明細書中に参考として組み込まれている、2009年12月10日に出願の米国仮出願第61/285,324号、2010年6月7日に出願の米国仮出願第61/352,108号、および2010年7月22日に出願の米国仮出願第61/366,685号の優先権を主張するものである。
【0002】
政府出資の研究に関する表明
本発明は、国立科学財団(National Science Foundation)によって授与された助成金第0820831号および第0504304号の下の政府支援で行った。政府が本発明の特定の権利を有する。
【0003】
本発明は、遺伝子標的化の方法、特に転写活性化因子様(TAL)エフェクター配列の使用が含まれる方法に関する。
【背景技術】
【0004】
相同組換え(遺伝子標的化)を介して染色体を改変する能力は、生物学者の念願の目標であった。たとえば、植物では、遺伝子標的化は、植物遺伝子の機能の識別を助けて、作物の改善の新しい可能性を開き得る。たとえば、遺伝子標的化を用いて、代謝経路を再編成するために必要な遺伝手術を実施して、変更された油もしくは炭水化物プロフィールを有する種子、増強された栄養品質を有する食品、または疾患およびストレスに対して増加した耐性を有する植物を含めた高価値の作物を作製することが可能である。動物(たとえば哺乳動物)では、遺伝子標的化を疾患の処置に使用し得る。たとえば、遺伝子標的化は、様々な種類の突然変異が原因で欠損性である遺伝子中の補正を設計するために使用し得る。そのような遺伝子標的化の効率的な方法は、達成が困難であった。
【0005】
キサントモナス属中の植物病原細菌のTALエフェクターは、宿主DNAと結合してエフェクターに特異的な宿主遺伝子を活性化することによって、疾患において重要な役割を果たす、または防御を始動する(たとえば、Guら(2005)Nature、435:1122、Yangら(2006)Proc.Natl.Acad.Sci.USA、103:10503、Kayら(2007)Science、318:648、Sugioら(2007)Proc.Natl.Acad.Sci.USA、104:10720、およびRomerら(2007)Science、318:645を参照)。特異性は、エフェクター可変性の、不完全な、典型的には34個のアミノ酸の反復に依存する(Schornackら(2006)J.Plant Physiol.、163:256)。多型は主に反復位置12および13に存在し、本明細書中ではこれを反復可変性二残基(RVD、repeat variable−diresidue)と呼ぶ。
【発明の概要】
【発明が解決しようとする課題】
【0006】
本発明は、TALエフェクターのRVDとその標的部位中のヌクレオチドとが一部の縮重を伴って1個のRVD対1個のヌクレオチドの直接的な線形様式で対応しており、明らかなコンテキスト依存性がないことに部分的に基づく。この驚くべき発見は、新しい標的特異的TALエフェクターの標的部位の予測を可能にするタンパク質−DNAの認識の新規機構を表す。本明細書中に記載のように、これらのタンパク質は、ゲノム操作における相同組換え(たとえば、生物燃料または植物におけるバイオ再生可能物質に有用な特色を付加または増強させるため)を促進することができる標的化キメラヌクレアーゼとして、研究および生命工学において有用であり得る。また、これらのタンパク質は、たとえば、非限定的な例として、転写因子および特に病原体(たとえばウイルス)に対する治療剤などの非常に高いレベルの特異性を要する治療的応用としても有用であり得る。
【課題を解決するための手段】
【0007】
一態様では、本発明は、(a)標的DNA配列を含有する細胞を提供するステップと、(b)TALエフェクター−DNA修飾酵素が細胞またはその子孫中の特定のヌクレオチド配列内にあるまたはそれに隣接する標的DNAを修飾するように、細胞内に、(i)二本鎖DNAを修飾することができるDNA修飾酵素ドメイン、および(ii)組み合わせて標的DNA配列中の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメインを含む、転写活性化因子様(TAL)エフェクター−DNA修飾酵素を導入するステップとを含む、細胞の遺伝物質を改変する方法を特長とする。本方法は、標的DNA配列と核酸との間の相同組換えが起こるように、標的DNA配列の少なくとも一部分に相同的な配列を含む核酸を細胞に提供することをさらに含むことができる。細胞は、真核細胞、哺乳動物細胞、植物細胞、または原核細胞であることができる。標的DNAは染色体DNAであることができる。導入ステップは、TALエフェクター−DNA修飾酵素をコードしているベクターを用いて細胞を形質移入すること、TALエフェクター−DNA修飾酵素を細胞内にタンパク質として機械的に注入すること、TALエフェクター−DNA修飾酵素を細胞内にタンパク質として細菌III型分泌系によって送達すること、またはTALエフェクター−DNA修飾酵素を細胞内にタンパク質として電気穿孔によって導入することを含むことができる。DNA修飾酵素は、エンドヌクレアーゼ(たとえばFokIなどのII型制限エンドヌクレアーゼ)であることができる。
【0008】
標的DNA内の特定のヌクレオチド配列と結合するTALエフェクタードメインは、10個以上のDNA結合反復、好ましくは15個以上のDNA結合反復を含むことができる。それぞれのDNA結合反復には、標的DNA中の塩基対の認識配列を決定する反復可変性二残基(RVD)が含まれることができ、それぞれのDNA結合反復は、標的DNA中の1つの塩基対配列の認識を司っており、RVDは、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのHG、Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含む。それぞれのDNA結合反復は、標的DNA中の塩基対の認識を決定するRVD配列を含むことができ、それぞれのDNA結合反復は、標的DNA中の1つの塩基対配列の認識を司っており、RVDは、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、Tを認識するためのHG、Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、およびTを認識するためのIGのうちの1つまたは複数を含む。
【0009】
別の態様では、本発明は、(1)開始プラスミドをPspXIで直鎖状にするステップであって、開始プラスミドが、選択されたヌクレオチド配列の第1ヌクレオチドに特異的な反復可変性二残基(RVD)を有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含み、第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有するステップと、(2)開始プラスミドのPspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチド(複数可)に特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしている、XhoI粘着末端を有するDNAモジュールをライゲーションするステップと、(3)核酸が選択されたヌクレオチド配列と結合することができるTALエフェクターをコードするまでステップ(1)および(2)を繰り返すステップとを含む、選択されたヌクレオチド配列に特異的なTALエフェクターをコードしている核酸を作製する方法を特長とする。本方法は、ライゲーション後に、PspXI部位中のDNAモジュールの配向を決定することをさらに含むことができる。本方法は、ステップ(1)および(2)を1〜30回繰り返すことを含むことができる。
【0010】
別の態様では、本発明は、(a)細胞のゲノム中の第1のヌクレオチド配列を同定するステップと、(b)(i)組み合わせて第1のユニークなヌクレオチド配列と結合する複数のDNA結合反復、および(ii)第1のヌクレオチド配列内にあるまたはそれに隣接する位置で二本鎖切断を生じるエンドヌクレアーゼを含む、TALENをコードしている核酸を合成するステップであって、それぞれのDNA結合反復が、標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っており、TALENが、以下のRVD、すなわち、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含むステップとを含む、転写活性化因子様エフェクターエンドヌクレアーゼ(TALEN)をコードしている核酸を作製する方法を特長とする。
【0011】
TALENは、以下のRVD、すなわち、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、およびTを認識するためのIGのうちの1つまたは複数を含むことができる。
【0012】
第1のヌクレオチド配列は、以下の基準のうちの少なくとも1つを満たすことができる:i)最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあること、ii)第1(5’)位置にTまたは第2位置にAを有さないこと、iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さないこと、ならびにiv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの基本組成を有すること。
【0013】
本方法は、上述の基準のうちの少なくとも1つを満たしており、15〜18bpによって分離されている、細胞のゲノム中の第1のヌクレオチド配列および第2のヌクレオチド配列を同定することを含むことができる。エンドヌクレアーゼは、第1および第2のヌクレオチド配列の間に二本鎖切断を生じることができる。
【0014】
別の実施形態では、本発明は、DNA結合ドメインが複数のDNA結合反復を含み、それぞれの反復が、標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、エンドヌクレアーゼドメインと標的DNAに特異的なTALエフェクターDNA結合ドメインとを含むTALENであって、以下のRVD、すなわち、Cを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、Tを認識するためのIG、Gを認識するためのNK、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、およびTを認識するためのYGのうちの1つまたは複数を含むTALENを特長とする。TALENは、以下のRVD、すなわち、Cを認識するためのHA、Cを認識するためのND、Cを認識するためのHI、Gを認識するためのHN、Gを認識するためのNA、GまたはAを認識するためのSN、Tを認識するためのYG、およびGを認識するためのNKのうちの1つまたは複数、ならびにCを認識するためのHD、Tを認識するためのNG、Aを認識するためのNI、GまたはAを認識するためのNN、AまたはCまたはGまたはTを認識するためのNS、CまたはTを認識するためのN*、Tを認識するためのHG、Tを認識するためのH*、およびTを認識するためのIGのうちの1つまたは複数を含むことができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。
【0015】
さらに別の態様では、本発明は、前記TALENのアミノ酸配列が、配列番号33〜配列番号55、配列番号72、および配列番号73からなる群から選択される、エンドヌクレアーゼドメインおよびTALエフェクタードメインを含むTALENを特長とする。
【0016】
また、本発明は、それ内に遺伝子改変を導入することが所望される標的DNA配列を含む真核細胞を提供するステップと、エンドヌクレアーゼドメインと標的DNA配列と結合するTALエフェクタードメインとを含むTALENを用いて標的DNA配列内に二本鎖切断を生じるステップと、二本鎖切断が起こった細胞またはその子孫から動物を作製するステップとを含む、動物を作製する方法も特長とする。本方法は、標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、細胞内に、外因性核酸と細胞またはその子孫中の標的DNA配列との間の相同組換えが起こることを可能にする条件下で導入するステップと、相同組換えが起こった細胞またはその子孫から動物を作製するステップとをさらに含むことができる。動物は哺乳動物であることができる。遺伝子改変は、置換、挿入、または欠失を含むことができる。
【0017】
さらに別の態様では、本発明は、事前に選択された遺伝子改変をそれ内に導入することが所望される標的DNA配列を含む植物細胞を提供するステップと、エンドヌクレアーゼドメインと標的DNA配列と結合するTALエフェクタードメインとを含むTALENを用いて標的DNA配列内に二本鎖切断を生じるステップと、二本鎖切断が起こった細胞またはその子孫から植物を作製するステップとを含む、植物を作製する方法を特長とする。本方法は、標的DNA配列の少なくとも一部分に相同的な配列を含む外因性核酸を、植物細胞内に、外因性核酸と細胞またはその子孫中の標的DNA配列との間の相同組換えが起こることを可能にする条件下で導入するステップと、相同組換えが起こった細胞またはその子孫から植物を作製するステップとをさらに含むことができる。
【0018】
別の態様では、本発明は、選択されたDNA標的配列を標的としたTALエフェクターエンドヌクレアーゼをコードしている核酸を細胞内に導入するステップと、細胞内でのTALエフェクターエンドヌクレアーゼの発現を誘導するステップと、選択されたDNA標的配列が突然変異を示している細胞を同定するステップとを含む、細胞における標的化遺伝子組換えの方法を特長とする。突然変異は、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択されることができる。本方法は、細胞内にドナーDNAを導入することをさらに含むことができる。細胞は、昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞であることができる。
【0019】
別の態様では、本発明は、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませ、したがって結合部位の−1位でのTの必要性を排除することを含むステップを含む、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法を特長とする。
【0020】
別の態様では、本発明は、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含み、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、Gに対して増強された特異性を有するRVDを含有するTALエフェクターDNA結合ドメインをコードしている1つまたは複数の核酸を取り込ませることを含み、前記RVDが、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択され、*がRVDの第2位置でのギャップを示すステップを含む、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法を特長とする。
【0021】
また、本発明は、反復ドメインを含むポリペプチドを合成するステップを含む、標的DNA配列中の少なくとも1つの塩基対を選択的に認識するポリペプチドを生成する方法であって、反復ドメインが、転写活性化因子様(TAL)エフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、標的DNA中の塩基対の認識配列を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法も特長とする。さらに、本発明は、上記方法によって生成されたポリペプチド、および上記方法によって生成されたポリペプチドのコード配列を含むDNAを特長とする。また、上述のDNAと作動可能に連結されたプロモーターを含む発現カセット、および発現カセットを含む非ヒト宿主細胞も特長とされている。別の態様では、本発明は、発現カセットを含む形質転換させた非ヒト生物を特長とする。
【0022】
さらに別の態様では、反復ドメインを含むポリペプチドを構築するステップを含む、ポリペプチドによってDNA配列中の塩基対を選択的に認識する方法であって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法を特長とする。
【0023】
また、本発明は、細胞における標的遺伝子の発現を変調させる方法であって、反復ドメインを含むポリペプチドを含有する細胞が提供され、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む方法を特長とする。
【0024】
別の態様では、本発明は、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、超可変領域が、(a)C/Gを認識するためのHD、(b)A/Tを認識するためのNI、(c)T/Aを認識するためのNG、(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、(e)G/CまたはA/Tを認識するためのNN、(f)T/Aを認識するためのIG、(g)C/Gを認識するためのN、(h)C/GまたはT/Aを認識するためのHG、(i)T/Aを認識するためのH、および(j)G/Cを認識するためのNKからなる群から選択されるメンバーを含む、反復ドメインを含むポリペプチドを特長とする。また、本発明は、上述のポリペプチドのコード配列を含むDNAも特長とする。
【0025】
別の態様では、本発明は、塩基対が反復ドメインを含むポリペプチドによって特異的に認識されることができるように、標的DNA配列上に位置する塩基対が含まれるように改変されたDNAであって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、かつDNA配列中の1つの塩基対の認識を司っており、塩基対が、超可変領域による選択的かつ決定された認識を受けるために、(a)HDによる認識のためのC/G、(b)NIによる認識のためのA/T、(c)NGによる認識のためのT/A、(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、(e)NNによる認識のためのG/CまたはA/T、(f)IGによる認識のためのT/A、(g)Nによる認識のためのC/GまたはT/A、(h)HGによる認識のためのT/A、(i)Hによる認識のためのT/A、および(j)NKによる認識のためのG/Cからなる群から選択されるDNAを特長とする。また、上述のDNAを含むベクター、DNAを含む非ヒト宿主細胞、およびDNAを含む形質転換させた非ヒト生物も特長とされている。
【0026】
さらに別の態様では、本発明は、反復ドメインを含むポリペプチドによって選択的に認識される標的DNA配列を含むDNAを生成する方法であって、反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、反復単位が、標的DNA中の塩基対の認識配列を決定する超可変領域を含み、かつ標的DNA中の1つの塩基対配列の認識を司っており、方法が、反復単位によって認識されることができる塩基対を含むDNAを合成するステップを含み、塩基対が、(a)HDによる認識のためのC/G、(b)NIによる認識のためのA/T、(c)NGによる認識のためのT/A、(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、(e)NNによる認識のためのG/CまたはA/T、(f)IGによる認識のためのT/A、(g)Nによる認識のためのC/GまたはT/A、(h)HGによる認識のためのT/A、(i)Hによる認識のためのT/A、および(j)NKによる認識のためのG/Cからなる群から選択される方法を特長とする。
【0027】
別の態様では、本発明は、植物細胞の遺伝物質を改変する方法を特長とする。本方法には、(a)(i)植物細胞中に存在する対応する標的ヌクレオチド配列に関してヌクレオチド配列中の1つまたは複数の修飾を含み、配列特異的TALエフェクターエンドヌクレアーゼ(TALEN)の認識部位をさらに含む、修飾された標的ヌクレオチド配列を含む第1の組換え核酸、および(ii)配列特異的転写活性化因子様(TAL)エフェクターエンドヌクレアーゼをコードしているヌクレオチド配列を含む第2の組換え核酸を、植物細胞内に導入するステップと(b)植物細胞を含有する植物を作製するステップと、(c)植物またはその子孫から得られた細胞、種子、または組織を、標的ヌクレオチド配列での組換えについて分析するステップとが含まれることができる。本方法には、(iii)選択マーカーをコードしているヌクレオチド配列を含む第3の組換え核酸を植物細胞内に導入するステップと、植物またはその子孫が選択マーカーを発現するかどうかを決定するステップとがさらに含まれることができる。本方法には、植物またはその子孫を、選択マーカーの非存在についてスクリーニングするステップがさらに含まれることができる。選択マーカーをコードしているヌクレオチド配列は、片側または両側が植物細胞内在配列(たとえば、第2の配列特異的ヌクレアーゼの切断部位での配列)に類似または同一の配列によって隣接されていても、されていないくてもよい。選択マーカーをコードしているヌクレオチド配列は、両側が配列特異的リコンビナーゼの認識部位によって隣接されていてもよい。本方法には、植物を異系交配させるステップがさらに含まれることができ、異系交配の子孫を選択マーカーの非存在についてスクリーニングするステップを用いるまたは用いない。第1および第2の組換え核酸は、植物細胞内に同時に導入することができる。組換え核酸の一方または両方を、導入ステップの前に直鎖状にすることができる。第1および第2の組換え核酸は、同じ構築体中に存在していてよい。
【0028】
別の態様では、本発明は、細胞の遺伝物質を改変する別の方法を特長とする。本方法には、それ内で相同組換えが起こることが所望される染色体標的DNA配列を含有する初代細胞を提供するステップと、二本鎖DNAを切断することができるエンドヌクレアーゼドメイン、および組み合わせて細胞中の標的DNA内の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメインを含む、TALENを提供するステップと、TALENが細胞中の標的DNA配列内にあるまたはそれに隣接するヌクレオチド配列の両方の鎖を切断するように、標的DNA配列とTALENとを細胞中で接触させるステップとが含まれることができる。本方法には、標的DNA配列と核酸との間の相同組換えが起こるように、標的DNAの少なくとも一部分に相同的な配列を含む核酸を提供することがさらに含まれることができる。標的DNA配列は、細胞に内在性であることができる。細胞は、植物細胞、哺乳動物細胞、魚細胞、昆虫細胞もしくはin vitro培養のためにこれらの生物から誘導した細胞系、または生組織から直接採取し、in vitro培養のために確立させた初代細胞であることができる。接触ステップには、TALENのコード配列を含むベクターで細胞を形質転換させて、TALENタンパク質を細胞中で発現させること、TALENタンパク質を細胞内に機械的に注入すること、TALエフェクターエンドヌクレアーゼタンパク質を細胞内に細菌III型分泌系によって送達すること、またはTALENタンパク質を細胞内に電気穿孔によって導入することが含まれることができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。標的DNA内の特定のヌクレオチド配列と結合するTALエフェクタードメインには、10個以上のDNA結合反復、より好ましくは15個以上のDNA結合反復が含まれることができる。細胞は任意の原核または真核生物からのものであることができる。
【0029】
別の態様では、本発明は、特定の位置でDNAを切断することができる配列特異的TALENを設計する方法を特長とする。本方法には、それ内に二本鎖切断を導入することが所望される第2のヌクレオチド配列に隣接する、第1のユニークな内在染色体ヌクレオチド配列を同定するステップと、(a)組み合わせて第1のユニークな内在染色体ヌクレオチド配列と結合する複数のDNA結合反復ドメイン、および(b)第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含む、配列特異的TALENを設計するステップとが含まれることができる。
【0030】
また、本発明は、エンドヌクレアーゼドメインおよび特定のDNA配列に特異的なTALエフェクターDNA結合ドメインを含むTALENも特長とする。TALENには、精製タグがさらに含まれることができる。エンドヌクレアーゼドメインは、II型制限エンドヌクレアーゼ(たとえばFokI)からのものであることができる。
【0031】
別の態様では、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した動物を作製する方法を特長とする。本方法には、それ内に核酸を導入することが所望される内在染色体標的DNA配列を含む初代細胞を提供するステップと、エンドヌクレアーゼドメインおよび内在染色体標的DNA配列と結合するTALエフェクタードメインを含むTALENを用いて、内在染色体標的DNA配列内に二本鎖切断を生じるステップと、内在染色体標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、初代細胞内に、外因性核酸と内在染色体標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった初代細胞から動物を作製するステップとが含まれることができる。動物は哺乳動物であることができる。相同配列は、相同組換え後に遺伝子を破壊するヌクレオチド配列、相同組換え後に遺伝子を置き換えるヌクレオチド配列、相同組換え後に遺伝子内に点突然変異を導入するヌクレオチド配列、および相同組換え後に調節部位を導入するヌクレオチド配列からなる群から選択されるヌクレオチド配列であることができる。
【0032】
さらに別の態様では、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した植物を作製する方法を特長とする。本方法には、それ内に核酸を導入することが所望される内在標的DNA配列を含む植物細胞を提供するステップと、エンドヌクレアーゼドメインおよび内在標的ヌクレオチド配列と結合するTALエフェクタードメインを含むTALENを用いて、内在標的DNA配列内に二本鎖切断を生じるステップと、内在標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、植物細胞内に、外因性核酸と内在標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった植物細胞から植物を作製するステップとが含まれることができる。
【0033】
別の態様では、本発明は、細胞における標的化遺伝子組換えの方法を特長とする。本方法には、選択されたDNA標的配列を標的としたTALENをコードしている核酸分子を細胞内に導入するステップと、細胞内でのTALENの発現を誘導するステップと、選択されたDNA標的配列が突然変異を示している細胞を同定するステップとが含まれることができる。突然変異は、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択されることができる。本方法には、細胞内にドナーDNAを導入することがさらに含まれることができる。細胞は、昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞であることができる。
【0034】
さらに別の態様では、本発明は、(1)選択されたヌクレオチド配列の第1ヌクレオチドに特異的なRVDを有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含む開始プラスミドを選択し、第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有するステップと、(2)開始プラスミドをPspXIで直鎖状にするステップと、(3)PspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチド(複数可)に特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしている、XhoI粘着末端を有するDNAモジュールをライゲーションするステップと、(4)核酸が選択されたヌクレオチド配列と結合することができるTALENをコードするまでステップ(2)および(3)を繰り返すステップとを含む、配列特異的TALENをコードしている核酸を作製する方法を特長とする。一部の事例では、本方法には、ステップ(3)のライゲーションの後に、PspXI部位中のDNAモジュールの配向を確認することがさらに含まれることができる。
【0035】
別段に定義しない限りは、本明細書中で使用したすべての技術用語および科学用語は、本発明が関連する分野の技術者によって一般に理解されているものと同じ意味を有する。本明細書中に記載のものに類似または均等の方法および材料を使用して本発明を実施することができるが、適切な方法および材料が以下に記載されている。本明細書中で言及したすべての出版物、特許出願、特許、および他の参考文献は、その全体で参考として組み込まれている。矛盾する場合は、定義を含めて本明細書が支配する。さらに、材料、方法、および実施例は、例示的なものにすぎず、限定することを意図しない。
【0036】
本発明の1つまたは複数の実施形態の詳細は、添付の図面および以下の説明中に記載されている。本発明の他の特長、目的、および利点は、説明および図面、ならびに特許請求の範囲から明らかとなるであろう。
【図面の簡単な説明】
【0037】
【図1】TALエフェクター−DNA認識の暗号を示す。図1Aは、反復領域(白い四角)および代表的な反復配列(配列番号1)を示す一般的なTALエフェクターの図であり、RVDに下線が引いてある。図1Bは、様々なTALエフェクターのRVDおよび標的遺伝子プロモーター配列(配列番号2〜11)の最良のパターン一致(低エントロピーのアラインメント)を示す図である。星印は、残基13での欠失を示す。図1Cは、Bのアラインメント、およびすべてのコメプロモーターを40個の追加のX.oryzaeのTALエフェクターで走査することによって得られ、それぞれのエフェクターについて下流遺伝子が感染中に活性化された最良のアラインメントを保持したさらに10個のアラインメントにおける、RVD−ヌクレオチドの会合を示す図である。図1Dは、20個のTALエフェクター標的部位の隣接ヌクレオチドの頻度を示す図である。位置は標的部位の5’末端に対するものであり、Nは標的部位の長さである。ロゴはWebLogoを使用して作成した。
【図2】OsHen1がXanthomonas oryzae pv.oryzicola株BLS256のTal1cによって活性化されるという証拠を提供する。図2Aは、マーカー交換突然変異体M51、空のコスミドベクター(ev)を保有するM51、tal1a、tal1b、およびtal1cを含有するコスミドpIJF92を保有するM51、ならびに野生型(WT)株における、BLS256を接種した24時間後のコメ葉中の、アクチン遺伝子を参照としたOsHen1の相対的な転写物の存在量を示す半定量的RT−PCR結果の像である。図2Bは、XmalI断片を含有するマーカーの救出および末端配列決定による、M51における単一のマーカー交換突然変異のマッピングに基づいた模式図である。コスミドpIJF92中に含有されていたゲノム領域、救出された断片の座標、およびBLS256ゲノム断片の座標が示されている。
【図3】参照AvrBs3のアミノ酸配列である(配列番号12)。
【図4】参照AvrBs3の核酸配列である(配列番号13)。
【図5】TALヌクレアーゼ発現ベクターのマップである。
【図6】標的レポータープラスミドのマップである。
【図7】TALヌクレアーゼの模式的構造の図である。TALのDNA結合ドメインの認識部位は大文字として表されており、一方でスペーサー配列は小文字で示されている。
【図8】AvrBs3認識ドメインの17回と半分のタンデムの反復のアミノ酸配列である(配列番号16)。位置12および13の超可変アミノ酸が四角で囲まれている。
【図9】TALの有効性を試験するための酵母アッセイのスキームを示す図である。
【図10】AvrBs3 TALヌクレアーゼの酵母アッセイの結果をプロットするグラフである。
【図11】単一、二重、または三重のAsvBs3反復モジュールおよびクローニングベクターの模式図を示す図である。
【図12】ほとんどのTALエフェクター中の反復領域の末端に存在する単一の代表的なTALエフェクター反復(図12A)および代表的な切断された反復(図12B)を示す。ヌクレオチドおよびコードされているアミノ酸配列が示されている。Nは、「XX」と示すRVDをコードしているヌクレオチドを表す。数字はアミノ酸の位置について与えられている。配列はtal1cからとったものである。
【図13】tal1c遺伝子、および反復領域を単一の切断された反復まで縮小してpCS487(やはり示されている)をもたらすプロセスを示す模式図である。MはMscI部位であり、SはSphI部位である。
【図14】pCS487中の元の切断された反復の末端に翻訳的にサイレントな突然変異を導入して、PspXIおよびXhoI部位を生じ、pCS489を得ることを示す模式図である。元の反復のコドン18〜21の配列(配列番号21)および突然変異させた反復(配列番号23)が示されている。コードされているアミノ酸配列(配列番号22)は、突然変異によって変化していなかった。突然変異させたヌクレオチドを斜体にしている。
【図15】GatewayエントリーベクターpENTR−D(Invitrogen、カリフォルニア州Carlsbad)中で反復領域を含まずにtal1cのNおよびC末端のみをコードしているカナマイシン耐性プラスミドである、pCS488のマップである。
【図16】RVD NIを有する反復をコードしているpCS493と命名された単一反復の開始プラスミドのマップである。pCS494、pCS495、およびpCS496と命名された3つの他のプラスミドは、それらがコードするRVD(右に示す)以外は同一であった。
【図17】図17Aは、RVD NIを有する単一反復モジュールのヌクレオチドおよびコードされているアミノ酸配列を示す。5’XhoI適合性の付着末端、MscI部位、および3’PspXI/XhoI適合性の付着末端に下線が引いてある。RVDおよびそれをコードしているヌクレオチドは太字である。HD、NI、およびNGをそれぞれコードしているRVDのコード配列以外は示したものに同一である、3つの他の反復モジュールを構築した。図17Bは、図17Aに示されている反復をコードしている配列を含有するpCS502と命名された単一反復モジュールのプラスミドのマップである。また、pCS503、pCS504、およびpCS505と命名されたプラスミドも作製し、これらは、それらがコードしているRVD(右に示す)以外はpCS502と同一である。
【図18】図18Aは、RVD NIを有する単一反復モジュールのヌクレオチドおよびコードされているアミノ酸配列を示し、ヌクレオチド置換(斜体)が、PspXI/XhoI部位内へのライゲーション後の5’末端でのXhoI部位の再構成を妨げ、内部MscI部位を破壊する。RVDおよびそれがコードしているヌクレオチドは太字である。HD、NI、およびNGをそれぞれコードしているRVDのコード配列以外は同一である、3つの追加の反復モジュールを構築した。図18Bは、追加の反復モジュールを単一反復モジュールのプラスミド内に順次ライゲーションさせることによってアセンブルした3回反復モジュールの模式図である。第1反復中のMscI部位および3’末端のPspXI部位はユニークに保たれ、モジュール全体は2つのXhoI部位によって隣接されている。
【図19】1回、2回、および3回の反復モジュールのプラスミドの完全な組のリストである。
【図20】任意の反復配列をtal1c「主鎖」内にアセンブルして誂えのTALエフェクター遺伝子を作製するために使用できる方法中のステップを示す流れ図である。
【図21】示されているヌクレオチド配列を標的とするTALエンドヌクレアーゼの構築における反復モジュールのアセンブリを示す模式図である。図21Aでは、pCS519、pCS524、pCS537、pCS551、pCS583、およびpCS529と命名されたプラスミドからの反復モジュールを、pCS493と命名された開始プラスミド中の配列に順次付加して、pMAT55、pMAT56、pMAT57、pMAT58、pMAT59、およびpMAT60と命名されたプラスミドがもたらされる。図21Bでは、pCS530、pCS533、pCS522、およびpCS541と命名されたプラスミドからの反復モジュールを、pMAT1と命名されたプラスミド中の配列に順次付加して、pMAT61、pMAT62、pMAT63、およびpMAT64と命名されたプラスミドがもたらされる。
【図22】図22Aは、TALエフェクタータンパク質の模式図である。BamHI断片(Bによって示す)をFokIエンドヌクレアーゼの触媒ドメインと融合させて、TALENを作製した。NはN末端、NLSは核移行シグナル、BはBamHI部位、ADは酸性活性化ドメインである。図22Bは、TALエフェクターAvrBs3およびPthXo1を用いて構築したTALENの活性をプロットするグラフである。Avr−FokIはAvrBs3 TALENであり、Pth−FokIはPthXo1 TALENであり、Avr−FokIおよびPth−FokIは、AvrBs3およびPthXo1とFokIの触媒的に不活性な型との融合体であり(Bitinaiteら(1998)Proc.Natl.Acad.Sci.USA、95:10570〜10575)、ZFNは、Zif268DNA結合ドメインを含有するジンクフィンガーヌクレアーゼである(PorteusおよびBaltimore(2003)Science、300:763)。
【図23】参照PthXo1のアミノ酸配列である(配列番号31)。
【図24】参照PthXo1の核酸配列である(配列番号32)。
【図25】pFZ85ベクターの図である。
【図26】avrBs3_TALENのアミノ酸配列を示す(配列番号33)。
【図27】pthXo1 TALENのアミノ酸配列を示す(配列番号34)。
【図28】図28Aは、様々なスペーサー長を有する標的に対するAvrBs3およびPthXo1 TALENの活性をプロットするグラフである。ZFNはZif268由来のジンクフィンガーヌクレアーゼである。図28Bは、ヘテロ二量体TALENの活性をプロットするグラフである。PthXo1−FokIおよびAvrBs3−FokI発現ベクター、ならびに頭から尾の配向で15bpによって分離されたそれぞれに対する認識部位からなる標的を有するプラスミドを含有する、酵母における活性が示されている(Avr−FokI、Pth−FokI)。また、そのそれぞれの標的に対する、AvrBs3(Avr−FokI)およびPthXo1(Pth−FokI)TALENの個々の活性ならびにZif268(ZFN)の活性も、参照のために示されている。陰性対照として、Avr−FokI、Pth−FokIに対する標的部位プラスミドのみを有する酵母培養物を、LacZ活性についてアッセイした((−)として示す)。
【図29】図29Aは、個々の誂えのTALENのRVD配列およびそのそれぞれのDNA認識配列を示す表である。図29Bは、誂えのTALENの活性をプロットするグラフである。(−)は標的部位プラスミドのみの陰性対照であり、ZFNはジンクフィンガーヌクレアーゼ陽性対照である。
【図30】20個の標的およびTALエフェクターの対の末端のヌクレオチドおよびRVDの頻度の描写である。
【図31】Golden Gateクローニングシステムの模式図である[Englerら(2008)PLoS One、3:e3647、およびEnglerら(2009)PLoS One、4:e5553]。
【図32】本明細書中に記載のGolden Gateクローニング手法を使用した、誂えのTALエフェクター反復をコードしているアレイをアセンブリおよびクローニングするための58個のプラスミドの組を示す。Tetは、プラスミド選択のマーカーであるテトラサイクリン耐性遺伝子であり、specは、プラスミド選択のマーカーであるスペクチノマイシン耐性遺伝子であり、ampは、プラスミド選択のマーカーであるアンピシリン耐性遺伝子である。
【図33】図32に示されているプラスミドの組を使用したGolden Gate手法によって誂えのTALエフェクター反復をコードしているアレイをアセンブリおよびクローニングするための方法の模式図である。例示目的のために、自由裁量による反復アレイのアセンブリが示されている。specは、プラスミド選択のマーカーであるスペクチノマイシン耐性遺伝子であり、ampは、プラスミド選択のマーカーであるアンピシリン耐性遺伝子である。
【図34】本明細書中の実施例9に記載のように作製したTALENのアミノ酸配列を示す。図34Aはテロメラーゼ−TALEN124であり、図34Bはgridlock−TALEN105であり、図34Cはadhl−TALEN58であり、図34Dはadhl−TALEN63であり、図34Eはadhl−TALEN68であり、図34Fはadhl−TALEN73であり、図34Gはadhl−TALEN89であり、図34Hはgridlock−TALEN106であり、図34Iはadhl−TALEN64であり、図34Jはadhl−TALEN69であり、図34Kはadhl−TALEN74であり、図34Lはtt4−TALEN90であり、図34Mはテロメラーゼ−TALEN121であり、図34Nはテロメラーゼ−TALEN126であり、図34Oはgridlock−TALEN107であり、図34Pはgridlock−TALEN117であり、図34Qはテロメラーゼ−TALEN131であり、図34Rはテロメラーゼ−TALEN136であり、図34Sはadhl−TALEN60であり、図34Tはtt4−TALEN85であり、図34Uはgridlock−TALEN102である。
【図35】増加していく長さの誂えのTALEN単量体(9、10、12、13、15、16、17、または18量体)を使用した酵母アッセイによって測定したTALEN活性プロットするグラフである。示したように、TALENはシロイヌナズナおよびゼブラフィッシュの遺伝子に対して標的化した。
【図36】図36Aは、2つのTALEN対によって標的化されているシロイヌナズナADH1遺伝子からの2つの異なるDNA標的配列を示す図である。図36Bは、シロイヌナズナADH1遺伝子を標的とする機能的TALEN対の酵母アッセイデータプロットするグラフである。
【図37】図37Aは、シロイヌナズナのプロトプラスト中のTALEN誘導性突然変異を検出するために使用した制限エンドヌクレアーゼアッセイの模式図である。図37Bは、制限エンドヌクレアーゼアッセイにおける未消化のDNAからの9個のクローンの配列を示す。クローンのうちの6個が、非相同末端結合(NHEJ)によって導入された突然変異を有する。
【図38】図38Aは、いくつかの系統学的に明白に異なるTALエフェクター、すなわち、Xanthomonas gardneriからのAvrHah1、X.campestris pv.vesicatoriaからのAvrBs3、X.oryzae pv.oryzaeからのPthXo1、X.citriからのPthA、およびX.oryzae pv.oryzicolaからのTal1cの第0反復配列を示す。多型位置が四角で囲まれている。図38Bは、PthXo1の第0および第1反復を示す模式図である。「第0」反復は第1反復の直前にあり、35%の同一性を示し、同様の予測された二次構造を有する。第1反復のRVDおよび第0反復の候補の類似残基に下線が引いてある。*はギャップであり、Hはヘリックスであり、Eは伸長である。構造はJPredを使用して予測した(Coleら(2008)Nucl.Acids Res.、36:W197〜W201)。
【図39】示したようにV5でタグ付けしたTALエフェクタータンパク質AvrBs3、PthXo1、およびTal1cをコードしているプラスミドで形質移入したヒト胚性腎臓293T細胞から単離した全タンパク質の、マウス−抗V5抗体を使用した免疫検出後のウエスタンブロットを示す。免疫標識したアクチンがそれぞれのレーンの等価なローディングの対照として示されている。
【図40】図40Aは、TALEN HPRT−3254−17のアミノ酸配列を示し、図40Bは、TALEN HPRT−3286−20rのアミノ酸配列を示す。
【図41】図41Aは、ヒト染色体HPRT遺伝子中のTALEN標的部位を示す模式図である。HPRT−3254−17およびHPRT−3286−20r TALENの結合部位、これらの部位の間のスペーサー中のBpu10I部位、ならびに領域を増幅するためのプライマー部位が示されている。下の座標は、コード配列の第1ヌクレオチドからの距離を塩基対で示している。図41Bは、TALENで処理したおよび処理していない細胞から単離したゲノムDNAを鋳型として使用した、図41Aに示されている領域のPCR増幅産物のBpu10I消化の結果を示す。増幅の前にゲノムDNAをBpu10Iで消化した。DNA断片はアガロースゲル電気泳動によって分離し、臭化エチジウムを使用して可視化した。
【発明を実施するための形態】
【0038】
本特許出願は、TALエフェクターによって媒介される配列特異的DNA認識に関する材料および方法を提供する。本明細書中に記載のように、TALエフェクターの一次アミノ酸配列が、それらが結合するヌクレオチド配列を指示する。本発明者らは、TALエフェクターのアミノ酸配列とそのDNA標的配列との間の関係性は直接的であり、それにより、TALエフェクターの標的部位の予測が可能となり、また、特定のヌクレオチド配列と結合するようにTALエフェクターを誂えることも可能となることを見い出した。そのような予測および誂えは、様々な目的のために利用することができる。一例では、特定のTALエフェクター配列をエンドヌクレアーゼ配列と融合させて、特定のDNA配列に対するエンドヌクレアーゼの標的化、続いて標的配列でまたはその付近でのDNAの切断を可能にすることができる。DNA中の切断(すなわち二本鎖切断)は、相同組換えの頻度を劇的に増加させる可能性がある。したがって、特定の標的DNA配列に対して高い度合の配列類似度を有する配列を保有するDNA構築体と組み合わせて、TALENを使用して、高い精度および効率で複雑なゲノム中の部位特異的突然変異誘発すること、すなわち、遺伝子機能をノックアウトもしくは変更すること、または遺伝子もしくは他の配列を付加することを促進することができる。
【0039】
したがって、本明細書中で提供されている主題には、とりわけ、遺伝子改変した生物(それだけには限定されないが、植物、真菌、ショウジョウバエ(Drosophila)、線虫、ゼブラフィッシュ、マウス、他の哺乳動物およびヒトが含まれる)を作製するための材料および方法が含まれる。そのような方法には、たとえば、細胞をいくつかの組換え核酸で形質移入することが含まれることができる。たとえば、細胞(たとえば真核細胞)を、細胞内に見つかる対応する標的ヌクレオチド配列と比較して変更が含まれるドナーヌクレオチド配列を含有する第1の組換え核酸構築体、およびTALヌクレアーゼをコードしている第2の組換え核酸構築体で形質転換させることができる。一部の実施形態では、細胞を、選択マーカーをコードしている第3の組換え核酸構築体でも形質転換させることができる。ドナー核酸構築体からの核酸配列は、本明細書中に記載の形質転換細胞のゲノム内に取り込まれることができる。たとえば、本明細書中に記載の方法を使用して生成した植物細胞を成長させて、変更されたドナーヌクレオチド配列がそのゲノム内に取り込まれた植物を生成することができる。そのような植物からの種子を使用して、たとえば、改変されていない植物と比較して変更された成長特徴(たとえば、様々な生物的および非生物的なストレスに対する耐性または寛容の増加)、変更された外見(たとえば、変更された色または高さ)、あるいは変更された組成(たとえば、炭素、窒素、油、タンパク質、炭水化物(たとえば糖もしくはデンプン)、アミノ酸、脂肪酸、または二次代謝物のレベルの増加または減少)などの表現型を有する植物を生成することができる。
【0040】
ポリヌクレオチドおよびポリペプチド
単離した核酸およびポリペプチドが本明細書中で提供されている。用語「核酸」および「ポリヌクレオチド」とは、互換性があるように使用され、cDNA、ゲノムDNA、合成(たとえば化学合成)DNA、および核酸類似体を含有するDNA(またはRNA)を含めた、RNAおよびDNAをどちらもいう。ポリヌクレオチドは任意の三次元構造を有することができる。核酸は二本鎖または一本鎖(すなわち、センス鎖またはアンチセンスの一本鎖)であることができる。ポリヌクレオチドの非限定的な例には、遺伝子、遺伝子断片、エクソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、リボザイム、cDNA、組換えポリヌクレオチド、分枝状ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離したDNA、任意の配列の単離したRNA、核酸プローブ、およびプライマー、ならびに核酸類似体が含まれる。
【0041】
本発明のポリペプチド(非限定的な例としてTALエフェクター−DNA修飾酵素など)は、たとえば前記ポリペプチドをコードしているベクターを使用することによってまたはポリペプチド自体として、送達ベクターを超音波穿孔もしくは電気穿孔またはこれらの技法の派生法などの任意の細胞透過処理技法と関連または組み合わせて使用することによって、細胞中に導入することができる。
【0042】
核酸に言及する場合、本明細書中で使用する「単離した」とは、ゲノム中で核酸の片側または両側に通常隣接している核酸を含めた、ゲノム、たとえば植物ゲノム中に存在する他の核酸から分離されている核酸をいう。また、核酸に関して本明細書中で使用する用語「単離した」には、任意の天然に存在しない配列も含まれる。これは、そのような天然に存在しない配列は自然には見つからず、したがって天然に存在するゲノム中のすぐ近くに近接する配列を有さないからである。
【0043】
単離した核酸は、たとえばDNA分子であることができるが、ただし、天然に存在するゲノム中でそのDNA分子のすぐ近くに隣接して通常見つかる核酸配列のうちの1つが除去されているまたは非存在である。したがって、単離した核酸には、それだけには限定されないが、他の配列から独立して別の分子として存在するDNA分子(たとえば、化学合成核酸、またはPCRもしくは制限エンドヌクレアーゼ処理によって生成されたcDNAもしくはゲノムDNA断片)、およびベクター、自己複製プラスミド、ウイルス(たとえば、パラレトロウイルス、レトロウイルス、レンチウイルス、アデノウイルス、もしくはヘルペスウイルス)、または原核生物もしくは真核生物のゲノムDNA内に取り込まれているDNAが含まれる。さらに、単離した核酸には、ハイブリッドまたは融合核酸の一部であるDNA分子などの組換え核酸が含まれることができる。数百から数百万個の他の核酸と共に、たとえば、cDNAライブラリもしくはゲノムライブラリ内、またはゲノムDNA制限消化物を含有するゲルスライス内に存在する核酸は、単離した核酸とみなされるべきでない。
【0044】
核酸は、たとえば化学合成またはポリメラーゼ連鎖反応(PCR)によって作製することができる。PCRとは、標的核酸が増幅される手順または技法をいう。PCRを使用して、全ゲノムDNAまたは全細胞RNAからの配列を含めた、DNAおよびRNAからの特定の配列を増幅することができる。様々なPCR方法は、たとえば、PCRプライマー:実験室の手引き(PCR Primer:A Laboratory Manual)、DieffenbachおよびDveksler編、Cold Spring Harbor Laboratory Press、1995に記載されている。一般に、対象の領域の末端またはそれを越える配列情報を用いて、増幅する鋳型の逆の鎖と配列が同一または類似のオリゴヌクレオチドプライマーを設計する。また、部位特異的なヌクレオチド配列修飾を鋳型核酸内に導入することができる様々なPCR戦略も利用可能である。
【0045】
また、単離した核酸は、突然変異誘発によっても得ることができる。たとえば、ドナー核酸配列を、PCRによるオリゴヌクレオチド特異的突然変異誘発および部位特異的突然変異誘発を含めた標準の技法を使用して突然変異させることができる。分子生物学の手短なプロトコル(Short Protocols in Molecular Biology)、第8章、Green Publishing AssociatesおよびJohn Wiley&Sons、Ausubelら編、1992を参照されたい。
【0046】
本明細書中で使用する用語「ポリペプチド」とは、翻訳後修飾(たとえばリン酸化またはグリコシル化)にかかわらず、2つ以上のサブユニットのアミノ酸の化合物をいう。サブユニットは、ペプチド結合またはたとえばエステルもしくはエーテル結合などの他の結合によって連結され得る。用語「アミノ酸」とは、D/L光学異性体を含めた天然および/または非天然もしくは合成のアミノ酸のいずれかをいう。
【0047】
ポリペプチドに関して「単離した」または「精製した」とは、ポリペプチドが、自然においてそれと共に通常見つかる細胞成分(たとえば、他のポリペプチド、脂質、炭水化物、および核酸)からある程度分離されていることを意味する。精製したポリペプチドは、非還元ポリアクリルアミドゲル上に単一の主バンドを与えることができる。精製したポリペプチドは、少なくとも約75%純粋(たとえば、少なくとも80%、85%、90%、95%、97%、98%、99%、または100%純粋)であることができる。精製したポリペプチドは、たとえば、天然源から抽出することによって、化学合成によって、または宿主細胞もしくはトランスジェニック植物中での組換え産生によって得ることができ、たとえば、アフィニティークロマトグラフィー、免疫沈降、サイズ排除クロマトグラフィー、およびイオン交換クロマトグラフィーを使用して精製することができる。精製の程度は、それだけには限定されないが、カラムクロマトグラフィー、ポリアクリルアミドゲル電気泳動、または高性能液体クロマトグラフィーを含めた任意の適切な方法を使用して測定することができる。
【0048】
組換え構築体
また、組換え核酸構築体(たとえばベクター)も本明細書中で提供されている。「ベクター」とは、それ内に別のDNAセグメントを挿入して、挿入したセグメントを複製させ得る、プラスミド、ファージ、またはコスミドなどのレプリコンである。一般に、ベクターは、適当な制御要素と会合している場合に複製が可能である。適切なベクター主鎖には、たとえば、プラスミド、ウイルス、人工染色体、BAC、YAC、またはPACなどの、当分野でルーチン的に使用されているものが含まれる。用語「ベクター」には、クローニングおよび発現ベクター、ならびにウイルスベクターおよび組込みベクターが含まれる。「発現ベクター」とは、1つまたは複数の発現制御配列が含まれるベクターであり、「発現制御配列」とは、別のDNA配列の転写および/または翻訳を制御および調節するDNA配列である。適切な発現ベクターには、それだけには限定されないが、たとえば、バクテリオファージ、バキュロウイルス、タバコモザイクウイルス、ヘルペスウイルス、サイトメガロウイルス、レトロウイルス、ワクシニアウイルス、アデノウイルス、およびアデノ関連ウイルスに由来するプラスミドおよびウイルスベクターが含まれる。数々のベクターおよび発現系がNovagen(ウィスコンシン州Madison)、Clontech(カリフォルニア州Palo Alto)、Stratagene(カリフォルニア州La Jolla)、およびInvitrogen/Life Technologies(カリフォルニア州Carlsbad)などの企業から市販されている。
【0049】
用語「調節領域」、「制御要素」、および「発現制御配列」とは、転写または翻訳の開始および速度、ならびに転写物またはポリペプチド産物の安定性および/または移動度に影響を与えるヌクレオチド配列をいう。調節領域には、それだけには限定されないが、プロモーター配列、エンハンサー配列、応答要素、タンパク質認識部位、誘導性要素、プロモーター制御要素、タンパク質結合配列、5’および3’非翻訳領域(UTR)、転写開始部位、終結配列、ポリアデニル化配列、イントロン、ならびに分泌シグナル、核移行配列(NLS)およびプロテアーゼ切断部位などのコード配列内に存在することができる他の調節領域が含まれる。
【0050】
本明細書中で使用する「作動可能に連結された」とは、発現制御配列が対象のコード配列の発現を有効に制御するように遺伝子構築体内に取り込まれていることを意味する。コード配列は、RNAポリメラーゼがコード配列をRNAへと転写することができる場合に、細胞中の発現制御配列と「作動可能に連結」されているかつその「制御下」にあり、mRNAの場合は、その後に、コード配列によってコードされているタンパク質へと翻訳されることができる。したがって、調節領域は、修飾された標的核酸をそれ内で発現させることが所望される植物細胞、植物、または植物組織中での転写を変調させる、たとえば、調節する、促進するまたは駆動することができる。
【0051】
プロモーターとは、転写が開始される点の典型的には100個以内のヌクレオチド上流(一般にRNAポリメラーゼIIの開始部位付近)の、DNA分子の領域から構成される発現制御配列である。プロモーターは、RNAポリメラーゼおよび他のタンパク質を認識および結合して、転写を開始および変調させることに関与している。コード配列をプロモーターの制御下にするためには、ポリペプチドの翻訳リーディングフレームの翻訳開始部位をプロモーターの1〜約50個のヌクレオチド下流に配置することが典型的には必要である。しかし、プロモーターは、翻訳開始部位の約5,000個と多数のヌクレオチド上流、または転写開始部位の約2,000個のヌクレオチド上流に配置することができる。プロモーターは、典型的には少なくともコア(基底)プロモーターを含む。また、プロモーターには、上流要素などの少なくとも1つの制御要素も含まれ得る。そのような要素には、上流活性化領域(UAR)および任意選択で合成上流要素などのポリヌクレオチドの転写に影響を与える他のDNA配列が含まれる。
【0052】
含めるプロモーターの選択肢は、それだけには限定されないが、効率、選択性、誘導性、所望の発現レベル、および細胞または組織特異性を含めたいくつかの要因に依存する。たとえば、それぞれ特定の組織、器官、および細胞種中でのみまたは優勢に転写を与える組織、器官および細胞に特異的なプロモーターを使用することができる。一部の実施形態では、茎、柔組織、基本分裂組織、維管束、形成層、師部、皮層、茎頂分裂組織、側枝分裂組織、根端分裂組織、側根分裂組織、葉原基、葉肉、または葉表皮などの植物組織に特異的なプロモーターが適切な調節領域である可能性がある。一部の実施形態では、種子に本質的に特異的なプロモーター(「種子に優先的なプロモーター」)が有用な可能性がある。種子に特異的なプロモーターは、種子の発生中に、胚乳および子葉組織中で作動可能に連結された核酸の転写を促進することができる。あるいは、構成的プロモーターは、植物の発生の全体にわたって、植物のほとんどまたはすべての組織中で作動可能に連結された核酸の転写を促進することができる。他のプロモーターのクラスには、それだけには限定されないが、化学薬品、発生刺激、または環境刺激などの外部刺激に応答して転写を与えるプロモーターなどの誘導性プロモーターが含まれる。
【0053】
基底プロモーターとは、転写開始に必要な転写複合体のアセンブリに必要な最小限の配列である。基底プロモーターには、しばしば、転写開始部位から約15〜約35個のヌクレオチド上流に位置し得る「TATAボックス」要素が含まれる。また、基底プロモーターには、転写開始部位から約40〜約200個のヌクレオチド、典型的には約60〜約120個のヌクレオチド上流に位置することができる「CCAATボックス」要素(典型的には配列CCAAT)および/またはGGGCG配列も含まれ得る。
【0054】
本明細書中で提供されている核酸構築体中に含まれることができるプロモーターの非限定的な例には、カリフラワーモザイクウイルス(CaMV)の35S転写開始領域、Agrobacterium tumefaciensのT−DNAに由来する1’または2’プロモーター、Busk((1997)Plant J.、11:1285〜1295)によって記載されているトウモロコシの葉に特異的な遺伝子からのプロモーター、トウモロコシおよび他の種からのknl関連遺伝子、ならびにトウモロコシユビキチン−1プロモーターなどの様々な植物遺伝子からの転写開始領域が含まれる。
【0055】
5’非翻訳領域(UTR)は、転写されるが翻訳されず、転写開始部位と翻訳開始コドンとの間に位置し、+1ヌクレオチドが含まれ得る。3’UTRは、翻訳終結コドンと転写終端との間に位置することができる。UTRは、増加したmRNAメッセージの安定性または減弱した翻訳などの特定の機能を有する場合がある。3’UTRの例には、それだけには限定されないが、ポリアデニル化シグナルおよび転写終結配列が含まれる。また、コード領域の3’末端のポリアデニル化領域もコード配列と作動可能に連結されていることができる。ポリアデニル化領域は、天然遺伝子、様々な他の植物遺伝子、またはAgrobacteriumのT−DNAに由来することができる。
【0056】
また、本明細書中で提供されているベクターには、たとえば、複製起点および/または足場付着領域(SAR)も含まれることができる。さらに、発現ベクターには、発現されたポリペプチドの操作または検出(たとえば精製もしくは局在化)を促進するように設計されたタグ配列が含まれることができる。緑色蛍光タンパク質(GFP)、グルタチオンS−トランスフェラーゼ(GST)、ポリヒスチジン、c−myc、赤血球凝集素、またはFlag(商標)タグ(Kodak、コネチカット州New Haven)配列などのタグ配列は、典型的には、コードされているポリペプチドとの融合体として発現される。そのようなタグは、カルボキシルまたはアミノ末端のどちらかを含めた、ポリペプチド内のどこにでも挿入することができる。
【0057】
「送達ベクター」または「複数の送達ベクター」とは、本発明において、本発明に必要な薬剤/化合物および分子(タンパク質または核酸)を細胞と接触させるまたは細胞内もしくは細胞内区画内に送達するために使用することができる、任意の送達ベクターを意図する。これには、それだけには限定されないが、リポソーム送達ベクター、ウイルス送達ベクター、薬物送達ベクター、化学担体、ポリマー担体、リポプレックス、ポリプレックス、デンドリマー、マイクロバブル(超音波造影剤)、ナノ粒子、乳濁液または他の適切な転移ベクターが含まれる。これらの送達ベクターは、分子、化合物、巨大分子(遺伝子、タンパク質)、またはプラスミド、Diatosによって開発されたペプチドなどの他のベクターの送達を可能にする。これらの事例では、送達ベクターは分子担体である。また、「送達ベクター」または「複数の送達ベクター」とは、形質移入を行うための送達方法も意図する。
【0058】
用語「ベクター」または「複数のベクター」とは、それが連結されている別の核酸を輸送することができる核酸分子をいう。本発明における「ベクター」には、それだけには限定されないが、ウイルスベクター、プラスミド、RNAベクター、または染色体、非染色体、半合成もしくは合成の核酸からなり得る直鎖状もしくは環状のDNAもしくはRNA分子が含まれる。好ましいベクターは、自律複製が可能なもの(エピソームベクター)および/またはそれらが連結されている核酸の発現が可能なもの(発現ベクター)である。多数の適切なベクターが当業者に知られており、市販されている。
【0059】
ウイルスベクターには、レトロウイルス、アデノウイルス、パルボウイルス(たとえばアデノ関連ウイルス)、コロナウイルス、オルトミクソウイルス(たとえばインフルエンザウイルス)などのマイナス鎖RNAウイルス、ラブドウイルス(たとえば狂犬病および水疱性口内炎ウイルス)、パラミクソウイルス(たとえば麻疹およびセンダイ)、ピコルナウイルスおよびアルファウイルスなどのプラス鎖RNAウイルス、ならびに、アデノウイルス、ヘルペスウイルス(たとえば、1および2型単純ヘルペスウイルス、エプスタイン−バーウイルス、サイトメガロウイルス)、およびポックスウイルス(たとえば、ワクシニア、鶏痘、カナリア痘)を含めた二本鎖DNAウイルスが含まれる。他のウイルスには、たとえば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス、および肝炎ウイルスが含まれる。レトロウイルスの例には、トリ白血症肉腫、哺乳動物C型、B型ウイルス、D型ウイルス、HTLV−BLV群、レンチウイルス、スプマウイルスが含まれる(Coffin,J.M.、レトロウイルス科:ウイルスおよびその複製(Retroviridae:The viruses and their replication)、基礎ウイルス学(Fundamental Virology)、第3版、B.N.Fieldsら編、Lippincott−Raven Publishers、Philadelphia、1996)。
【0060】
「レンチウイルスベクター」とは、その比較的大きなパッケージング能力、低下した免疫原性および広い範囲の様々な細胞種を高い効率で安定に形質導入するその能力が理由で遺伝子送達に非常に有望な、HIV系のレンチウイルスベクターを意味する。レンチウイルスベクターは、通常、産生細胞内への3つ(パッケージング、エンベロープおよび転移)以上のプラスミドの一過的な形質移入の後に作製される。HIVと同様、レンチウイルスベクターは、ウイルス表面糖タンパク質と細胞表面上の受容体との相互作用を介して標的細胞に進入する。進入の際、ウイルスRNAは、ウイルス逆転写酵素複合体によって媒介される逆転写を受ける。逆転写の産物は二本鎖の直鎖状ウイルスDNAであり、これは、感染した細胞のDNA中へのウイルス組込みの基質である。前記レンチウイルスベクターは、「非組込み」または「組込み」であることができる。
【0061】
「組込みレンチウイルスベクター(またはLV)」とは、非限定的な例として、標的細胞のゲノムに組み込まれることができるベクターを意味する。
【0062】
逆に、「非組込みレンチウイルスベクター(またはNILV)」とは、ウイルスインテグラーゼの作用によって標的細胞のゲノムに組み込まれない効率的な遺伝子送達ベクターを意味する。
【0063】
好ましいベクターの一種は、エピソーム、すなわち染色体外複製が可能な核酸である。好ましいベクターは、自律複製および/またはそれらが連結されている核酸の発現が可能なものである。それらが作動可能に連結されている遺伝子の発現を指示することができるベクターは、本明細書中で「発現ベクター」と呼ぶ。本発明によるベクターは、それだけには限定されないが、YAC(酵母人工染色体)、BAC(細菌人工)、バキュロウイルスベクター、ファージ、ファージミド、コスミド、ウイルスベクター、プラスミド、RNAベクター、または染色体、非染色体、半合成もしくは合成のDNAからなり得る直鎖状もしくは環状のDNAもしくはRNA分子を含む。一般に、組換えDNA技法において有用な発現ベクターは、多くの場合は「プラスミド」の形態であり、これは一般に、そのベクター形態では染色体と結合していない環状の二本鎖DNAループをいう。多数の適切なベクターが当業者に知られている。ベクターは、選択マーカー、たとえば、真核細胞培養物用のネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸還元酵素、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミン合成酵素、およびヒポキサンチン−グアニンホスホリボシルトランスフェラーゼ、出芽酵母用のTRP1、大腸菌中のテトラサイクリン、リファンピシンまたはアンピシリン耐性を含むことができる。好ましくは、前記ベクターは発現ベクターであり、対象のポリペプチドをコードしている配列は、前記ポリペプチドの産生または合成を可能にするために適切な転写および翻訳制御要素の制御下に置かれる。したがって、前記ポリヌクレオチドは発現カセット中に含まれる。より具体的には、ベクターは、複製起点、前記コードしているポリヌクレオチドと作動可能に連結されたプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを使用する場合)、ポリアデニル化部位および転写終結部位を含む。また、これは、エンハンサーまたはサイレンサー要素も含むことができる。プロモーターの選択は、ポリペプチドを発現させる細胞に依存する。適切なプロモーターには、組織特異的および/または誘導性プロモーターが含まれる。誘導性プロモーターの例は、増加した重金属レベルによって誘導される真核メタロチオニンプロモーター、イソプロピル−β−D−チオガラクト−ピラノシド(IPTG)に応答して誘導される原核lacZプロモーターおよび増加した温度によって誘導される真核熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異抗原(PSA)、α−抗トリプシンプロテアーゼ、ヒト界面活性化因子(SP)AおよびBタンパク質、β−カゼインならびに酸性ホエータンパク質遺伝子である。
【0064】
誘導性プロモーターは、病原体またはストレス、より好ましくは低温、熱、UV光、または高イオン濃度などのストレスによって誘導され得る(Potenzaら(2004)In vitro Cell Dev Biol、40:1〜22に総説)。誘導性プロモーターは化合物によって誘導され得る[Mooreら(2006)、Padidam(2003)、(Wangら(2003)、ならびに(ZuoおよびChua(2000)に総説]。
【0065】
送達ベクターおよびベクターは、超音波穿孔もしくは電気穿孔またはこれらの技法の派生法などの任意の細胞透過処理技法と関連させるまたは組み合わせることができる。
【0066】
複数の調節領域、たとえば、イントロン、エンハンサー、上流活性化領域、および誘導性要素が組換えポリヌクレオチド中に存在し得ることを理解されよう。
【0067】
組換え核酸構築体には、細胞(たとえば植物細胞または動物細胞)の形質転換に適したベクター内に挿入されたポリヌクレオチド配列が含まれることができる。組換えベクターは、たとえば標準の組換えDNA技法を使用して作製することができる(たとえばSambrookら(1989)分子クローニング:実験室の手引き(Molecular Cloning:A Laboratory Manual)、第2版、Cold Spring Harbor Laboratory、ニューヨーク州Cold Spring Harborを参照)。
【0068】
本明細書中に記載の組換え核酸配列は、非正統的な(すなわち、ランダム、非相同的、非部位特異的)組換えによって細胞のゲノム内に組み込まれることができるか、または、本明細書中に記載の組換え核酸配列は、相同組換えによって細胞のゲノム内に組み込まれるように適応させることができる。相同組換えによる組込みのために適応させた核酸配列は、両側が内在標的ヌクレオチド配列に類似または同一の配列で隣接されており、これは、内在標的ヌクレオチド配列を含有するゲノム中の特定の部位(複数可)での組換え核酸の組込みを促進する。また、相同組換えによる組込みのために適応させた核酸配列には、配列特異的ヌクレアーゼの認識部位も含まれることができる。あるいは、配列特異的ヌクレアーゼの認識部位は、形質転換させる細胞のゲノム中に位置することができる。以下に記載のドナー核酸配列は、典型的には、相同組換えによる組込みのために適応せる。
【0069】
また、一部の実施形態では、選択マーカーをコードしている核酸も相同組換えによる組み込のために適応させることができ、したがって、両側が植物ゲノム内の内在配列(たとえば、配列特異的ヌクレアーゼの切断部位での内在配列)と類似または同一の配列で隣接されていることができる。また、一部の事例では、選択マーカーのコード配列を含有する核酸には、配列特異的ヌクレアーゼの認識部位も含まれることができる。これらの実施形態では、配列特異的ヌクレアーゼの認識部位は、ドナー核酸配列内に含有されるものと同じまたは異なることができる(すなわち、ドナー核酸配列と同じヌクレアーゼによって認識される、またはドナー核酸配列とは異なるヌクレアーゼによって認識されることができる)。
【0070】
一部の事例では、組換え核酸配列は、部位特異的な組換えによって細胞のゲノム内に組み込まれるように適応させることができる。本明細書中で使用する「部位特異的な」組換えとは、組換え核酸中の配列とゲノム中の配列との間の相同性によってではなく、特定の核酸配列を認識してこれらの部位間のDNA鎖の相互交換を触媒するリコンビナーゼ酵素の作用によって、核酸配列がゲノム内の特定の部位(複数可)に対して標的化されている場合に起こる組換えをいう。したがって、部位特異的な組換えとは、2つの定義されたヌクレオチド配列の酵素媒介性の切断およびライゲーションをいう。たとえばCre−lox系またはFLP−FRT系を含めた、任意の適切な部位特異的な組換え系を使用することができる。そのような実施形態では、ドナーヌクレオチド配列およびヌクレアーゼをコードしている配列、ならびに一部の事例では選択マーカー配列に加えて、リコンビナーゼ酵素をコードしている核酸を細胞内に導入し得る。たとえば米国特許第4,959,317号を参照されたい。
【0071】
配列特異的エンドヌクレアーゼ
配列特異的ヌクレアーゼおよび配列特異的エンドヌクレアーゼをコードしている組換え核酸が本明細書中で提供されている。配列特異的エンドヌクレアーゼには、TALエフェクターDNA結合ドメインおよびエンドヌクレアーゼドメインが含まれることができる。したがって、そのような配列特異的エンドヌクレアーゼをコードしている核酸には、ヌクレアーゼからのヌクレオチド配列と連結された配列特異的TALエフェクターからのヌクレオチド配列が含まれることができる。
【0072】
TALエフェクターとは、病原体によって植物細胞内に注入され、ここで核へと移動して転写因子として機能して特定の植物遺伝子を活性化する、植物病原細菌のタンパク質である。TALエフェクターの一次アミノ酸配列が、それが結合するヌクレオチド配列指示する。したがって、TALエフェクターの標的部位を予測することができ、また、TALエフェクターは、本明細書中に記載のように、特定のヌクレオチド配列と結合させる目的で操作設計および作製することもできる。
【0073】
TALエフェクターをコードしている核酸配列には、ヌクレアーゼまたはヌクレアーゼの一部分、典型的にはFokIなどのII型制限エンドヌクレアーゼからの非特異的な切断ドメインをコードしている配列が融合している(Kimら(1996)Proc.Natl.Acad.Sci.USA、93:1156〜1160)。他の有用なエンドヌクレアーゼには、たとえば、HhaI、HindIII、NotI、BbvCI、EcoRI、BglI、およびAlwIが含まれ得る。一部のエンドヌクレアーゼ(たとえばFokI)が二量体としてのみ機能することは、TALエフェクターの標的特異性を増強させる際に利用することができる。たとえば、一部の事例では、それぞれのFokI単量体を、異なるDNA標的配列を認識するTALエフェクター配列と融合させることができ、2つの認識部位が近く近接している場合にのみ、不活性な単量体が一体となって機能的な酵素を生じる。ヌクレアーゼを活性化させるためにDNA結合を必要とすることによって、高度に部位特異的な制限酵素を作製することができる。
【0074】
本明細書中で提供されている配列特異的TALENは、細胞中に存在する事前に選択された標的ヌクレオチド配列内の特定の配列を認識することができる。したがって、一部の実施形態では、標的ヌクレオチド配列をヌクレアーゼ認識部位について走査することができ、標的配列に基づいて特定のヌクレアーゼを選択することができる。他の事例では、特定の細胞配列を標的とするようにTALENを操作することができる。所望のTALENをコードしているヌクレオチド配列を任意の適切な発現ベクター内に挿入することができ、1つまたは複数の発現制御配列と連結させることができる。たとえば、ヌクレアーゼのコード配列は、形質転換させる植物種中でエンドヌクレアーゼの構成的発現をもたらすプロモーター配列と作動可能に連結されていることができる。あるいは、エンドヌクレアーゼのコード配列は、条件的発現(たとえば特定の栄養条件下での発現)をもたらすプロモーター配列と作動可能に連結されていることができる。たとえば、カリフラワーモザイクウイルスの35Sプロモーターを構成的発現に使用することができる。他の構成的プロモーターには、それだけには限定されないが、ノパリン合成酵素プロモーター、ユビキチンプロモーター、およびアクチンプロモーターが含まれる。一部の実施形態では、人工エストロゲン誘導性プロモーターを条件的発現に使用することができ、植物がエストロゲンに曝露された際に高レベルの転写を達成することができる。使用することができる他の条件的プロモーターには、たとえば、熱誘導性熱ショック遺伝子プロモーター、およびリブロースビスリン酸カルボキシラーゼの大サブユニットをコードしている遺伝子からのものなどの光調節性プロモーターが含まれる。
【0075】
治療目的には、本発明のTALエフェクター−DNA修飾酵素および薬学的に許容される賦形剤を治療上有効な量で投与する。そのような組合せは、投与した量が生理的に有意である場合に、「治療上有効な量」で投与されたという。薬剤は、その存在がレシピエントの生理学の検出可能な変化をもたらす場合に、生理的に有意である。本コンテキストでは、薬剤は、その存在が標的疾患の1つまたは複数の症状の重篤度の減少および病変または異常のゲノム補正をもたらす場合に、生理的に有意である。標的化DNAおよび/またはTALエフェクター−DNA修飾酵素をコードしている核酸を含むベクターは、様々な方法(たとえば、注入、直接取り込み、噴出性照射、リポソーム、電気穿孔)によって細胞内に導入することができる。TALエフェクター−DNA修飾酵素は、発現ベクターを使用して細胞内で安定にまたは一過的に発現させることができる。真核細胞中での発現の技法は当業者に周知である。(ヒト遺伝学の最新プロトコル(Current Protocols in Human Genetics):第12章「遺伝子治療のベクター(Vectors For Gene Therapy)」および第13章「遺伝子治療の送達系(Delivery Systems for Gene Therapy)」を参照)。
【0076】
本発明のさらなる一態様では、TALエフェクター−DNA修飾酵素は実質的に非免疫原性である、すなわち、有害な免疫学的応答をわずかしかまたはまったく生じない。この種の有害な免疫学的反応を寛解させるまたは排除するための様々な方法を、本発明に従って使用することができる。好ましい実施形態では、TALエフェクター−DNA修飾酵素はN−ホルミルメチオニンを実質的に含まない。望まない免疫学的反応を回避するための別の方法は、TALエフェクター−DNA修飾酵素をポリエチレングリコール(「PEG」)またはポリプロピレングリコール(「PPG」)(好ましくは500〜20,000ダルトンの平均分子量(MW)のもの)とコンジュゲートさせることである。たとえばDavisら(US4,179,337号)によって記載されている、PEGまたはPPGとのコンジュゲーションは、抗ウイルス活性を有する、非免疫原性の生理的に活性のある水溶性のTALエフェクター−DNA修飾酵素のコンジュゲートを提供することができる。やはりポリエチレン−ポリプロピレングリコールのコポリマーを使用した類似の方法が、Saiferら(US5,006,333号)に記載されている。
【0077】
ドナーベクター
また、ドナーヌクレオチド配列を含めた組換え核酸も、本明細書中で提供されている。ドナーヌクレオチド配列には、形質転換させる細胞のゲノム内に内在的に見つかる事前に選択された標的ヌクレオチド配列に関して1つまたは複数の修飾(すなわち、置換、欠失、または挿入)を有する変異体配列が含まれることができる(本明細書中で「修飾された標的ヌクレオチド配列」とも呼ばれる)。ドナー核酸内の変異体配列は、典型的には、両側が細胞内の内在標的ヌクレオチド配列と類似または同一の配列で隣接されている。隣接配列は任意の適切な長さを有することができ、典型的には、少なくとも50個のヌクレオチドの長さである(たとえば、少なくとも50個のヌクレオチド、少なくとも75個のヌクレオチド、少なくとも100個のヌクレオチド、少なくとも200個のヌクレオチド、少なくとも250個のヌクレオチド、少なくとも300個のヌクレオチド、少なくとも500個のヌクレオチド、少なくとも750個のヌクレオチド、少なくとも1000個のヌクレオチド、約50〜約5000個のヌクレオチド、約100〜2500個のヌクレオチド、約100〜約1000個のヌクレオチド、約100〜500個のヌクレオチド、約200〜約500個のヌクレオチド、または約250〜400個のヌクレオチド)。したがって、相同組換えは、生じる細胞のゲノムが、たとえば同じ遺伝子からの内在配列のコンテキスト内の変異体配列を含有するように、組換えドナー核酸構築体と変異体配列の両側の内在標的との間で起こることができる。ドナーヌクレオチド配列は、ゲノム内の任意の適切な配列を標的とするように作製することができる。たとえば、植物では、ドナーヌクレオチド配列は、脂質生合成遺伝子、炭水化物生合成遺伝子、種子貯蔵タンパク質遺伝子、疾患もしくは害虫耐性遺伝子、ストレス寛容遺伝子、乾燥寛容遺伝子、または抗栄養因子を産生する遺伝子を標的とすることができる。さらに、ドナーヌクレオチド配列は、本明細書中に記載の配列特異的ヌクレアーゼの認識部位を含有する。
【0078】
選択マーカー
本明細書中で提供されている方法の一部には、選択可能またはスクリーニング可能なマーカーをコードしている第3の組換え核酸の使用が含まれる。選択可能な特色をもたらすポリペプチドをコードしているヌクレオチド配列を、1つまたは複数の発現制御配列を含有する発現ベクター内に取り込ませることができる。たとえば、発現ベクターには、形質転換させる植物細胞中で構成的発現をもたらすプロモーター配列と作動可能に連結された選択マーカーをコードしている配列が含まれることができる。適切な選択マーカーには、それだけには限定されないが、カナマイシン、G418、ブレオマイシン、アンピシリン、もしくはハイグロマイシンなどの抗生物質、またはグルホシネート、クロロスルフロン(chlorosulfuron)、もしくはホスフィノスリシンなどの除草剤に対する耐性を与えるポリペプチドが含まれることができる。
【0079】
たとえば、植物で使用するための実施形態では、選択マーカーは、イミダゾリノンまたはスルホニル尿素などの成長点または分裂組織を阻害する除草剤に対する耐性を与えることができる。この突然変異ALSおよびAHAS酵素の分類コード中の例示的なポリペプチドは、たとえば、米国特許第5,767,366号および第5,928,937号に記載されている。米国特許第4,761,373号および第5,013,659号は、様々なイミダゾリノンまたはスルホンアミド除草剤に耐性の植物に向けられている。米国特許第4,975,374号は、GSを阻害することが知られている除草剤、たとえばホスフィノスリシンおよびメチオニンスルホキシミンによる阻害に耐性の突然変異グルタミン合成酵素(GS)をコードしている遺伝子を含有する植物細胞および植物に関する。米国特許第5,162,602号は、シクロヘキサンジオンおよびアリールオキシフェノキシプロパン酸除草剤による阻害に耐性の植物を開示している。耐性は、変更されたアセチル補酵素Aカルボキシラーゼ(ACCase)によって与えられる。
【0080】
また、グリフォセート(商品名Roundup(登録商標)の下で販売)に耐性のポリペプチドも、植物における使用に適している。たとえば、米国特許第4,940,835号および第4,769,061号を参照されたい。米国特許第5,554,798号は、トランスジェニックグリフォセート耐性トウモロコシ植物を開示しており、耐性は変更された5−エノールピルビル−3−ホスホシキメート(EPSP)合成酵素によって与えられる。そのようなポリペプチドは、それだけには限定されないが、トリメチルスフホニウム塩、イソプロピルアミン塩、ナトリウム塩、カリウム塩およびアンモニウム塩などのグリフォセート塩を含めた、グリフォセート除草組成物に対する耐性を与えることができる。たとえば、米国特許第6,451,735号および第6,451,732号を参照されたい。
【0081】
また、グルホシネートアンモニウムまたはホスフィノスリシンなどのホスホノ化合物、ならびにピリジノキシまたはフェノキシプロピオン酸およびシクロヘキソンに耐性のポリペプチドも適切である。たとえば、欧州公開第0 242 246号、ならびに米国特許第5,879,903号、第5,276,268号、および第5,561,236号を参照されたい。
【0082】
他の除草剤には、トリアジンおよびベンゾニトリル(ニトリラーゼ)などの光合成を阻害するものが含まれる。たとえば米国特許第4,810,648号を参照されたい。他の除草剤には、2,2−ジクロロプロピオン酸、セトキシジム、ハロキシホップ、イミダゾリノン除草剤、スルホニル尿素除草剤、トリアゾロピリミジン除草剤、s−トリアジン除草剤およびブロモキシニルが含まれる。また、プロトックス(protox)酵素に対する耐性を与える除草剤も適している。たとえば、米国特許公開第20010016956号および米国特許第6,084,155号を参照されたい。
【0083】
一部の実施形態では、選択マーカーをコードしている組換え核酸は、部位特異的な組換えによって細胞(たとえば植物細胞または動物細胞)のゲノム内に組み込ませるために適応させることができる。たとえば、選択マーカーをコードしている配列は、たとえばCreまたはFLPなどのリコンビナーゼの認識配列によって隣接されていることができる。他の実施形態では、選択マーカーをコードしている組換え核酸は、相同組換えによる植物ゲノム内への組込みのために適応させることができる。そのような核酸では、選択マーカーをコードしている配列は、それ内に組換え核酸を導入する植物細胞のゲノム中に見つかる内在ヌクレオチド配列と類似または同一の配列によって隣接されていることができる。内在配列のうちの少なくとも1つが配列特異的ヌクレアーゼの切断部位にあることができる。また、選択マーカーをコードしている核酸は、配列特異的ヌクレアーゼの認識部位も含有することができる。ヌクレアーゼは、ドナーヌクレオチド配列に対して標的化されているものと同じ配列特異的ヌクレアーゼ、またはドナーヌクレオチド配列に対して標的化されているものとは異なる配列特異的ヌクレアーゼであることができる。さらに他の実施形態では、選択マーカーをコードしている組換え核酸は、非正統的な組換えによる植物細胞のゲノム内への組込みのために適応させることができる。そのような核酸は、典型的には、本明細書中に記載の相同的または部位特異的な組換えのために適応させた核酸内に含有される隣接配列およびヌクレアーゼ認識部位を欠く。
【0084】
方法
本明細書中で提供されている構築体のうちの1つまたは複数を使用して、遺伝子改変した生物(たとえば植物または動物)が作製されるように、細胞を形質転換させることができるおよび/またはDNA修飾酵素を細胞内に導入することができる。したがって、本明細書中に記載の核酸および/またはポリペプチド(polypeptdes)を含有する遺伝子改変した生物および細胞も提供される。一部の実施形態では、形質転換細胞は、組換え核酸構築体がそのゲノム内に組み込まれている、すなわち安定に形質転換させることができる。安定に形質転換させた細胞は、典型的には、導入された核酸配列を各細胞分裂で保持する。構築体は、形質転換細胞に内在性のヌクレオチド配列が、内在配列に対応する配列を含有するが内在配列に関して1つまたは複数の修飾を含有する構築体によって置き換えられるように、相同的な様式で組み込まれることができる。そのような修飾された内在配列を含有する植物または動物は本明細書中で「遺伝子改変した生物」(GMO)と呼び得るが、修飾された内在配列は導入遺伝子とみなされないことに留意されたい。また、構築体は、形質転換細胞のゲノム内にランダムに組み込まれるように、非正統的な様式で組み込まれることもできる。
【0085】
あるいは、構築体がそのゲノム内に組み込まれないように、細胞を一過的に形質転換させることができる。たとえば、TALENのコード配列が発現されるが、ベクターがゲノム中に安定に組み込まれていないように、TALENのコード配列を含有するプラスミドベクターを細胞内に導入することができる。一過的に形質転換させた細胞は、典型的には、導入された核酸が十分な回数の細胞分裂の後に娘細胞中で検出できないように、導入された核酸構築体の一部またはすべてを各それぞれの細胞分裂で失う。それにもかかわらず、TALENのコード配列の発現は、ドナー配列と内在標的配列との間の相同組換えを達成するために十分である。一過的に形質転換させたおよび安定に形質転換させた細胞はどちらも本明細書中に記載の方法において有用な可能性がある。
【0086】
特に遺伝子改変した植物細胞に関して、本明細書中に記載の方法で使用する細胞は、全植物の一部または全体を構成することができる。そのような植物は、検討中の種に適した様式で、成長チャンバ、温室、または畑のいずれかで成長させることができる。遺伝子改変した植物は、特定の目的、たとえば、組換え核酸を他の系に導入する、組換え核酸を他の種に転移させる、または他の望ましい特色をさらに選択するために所望されるように、育種することができる。あるいは、遺伝子改変した植物は、そのような技法を受け入れられる種では栄養繁殖させることができる。子孫には特定の植物または植物系の子孫が含まれる。本発明の植物の子孫には、F1、F2、F3、F4、F5、F6および続く世代の植物上で形成された種子、またはBC1、BC2、BC3、および続く世代の植物上で形成された種子、またはF1BC1、F1BC2、F1BC3、および続く世代の植物上で形成された種子が含まれる。遺伝子改変した植物によって生成された種子を成長させ、その後、自家受粉(または異系交配および自家受粉)させて、核酸構築体にホモ接合性の種子を得ることができる。
【0087】
遺伝子改変した細胞(たとえば植物細胞または動物細胞)は、所望する場合は、懸濁培養、または組織もしくは器官培養中で成長させることができる。本明細書中で提供されている方法の目的のために、固体および/または液体の組織培養技法を使用することができる。固体培地を使用する場合、細胞を培地上に直接置くことができるか、またはフィルターフィルム上に置き、その後それを培地と接触させることができる。液体培地を使用する場合、細胞を浮遊装置、たとえば液体培地と接触する多孔性の膜上に置くことができる。固体培地は、典型的には、液体培地から、寒天を加えることによって作製する。たとえば、固体培地は、寒天および適切な濃度のオーキシン、たとえば2,4−ジクロロフェノキシ酢酸(2,4−D)、および適切な濃度のサイトカイニン、たとえばキネチンを含有する、MurashigeおよびSkoog(MS)培地であることができる。
【0088】
細胞は、1個の組換え核酸構築体または複数(たとえば、2、3、4、もしくは5個)の組換え核酸構築体で形質転換させることができる。複数の構築体を利用する場合は、これらは同時にまたは順次形質転換させることができる。多種多様の種を形質転換させる技法が当分野で知られている。本明細書中に記載のポリヌクレオチドおよび/または組換えベクターは、電気穿孔、微量注入、および微粒子銃方法を含めたいくつかの知られている方法のうちの任意のものを使用して、宿主のゲノム内に導入することができる。あるいは、ポリヌクレオチドまたはベクターを適切なT−DNA隣接領域と組み合わせて、慣用のAgrobacterium tumefaciens宿主ベクター内に導入することができる。無害化(disarming)およびバイナリーベクターの使用を含めた、そのようなAgrobacterium tumefaciensに媒介される形質転換技法は当分野で周知である。他の遺伝子移入および形質転換技法には、カルシウムまたはPEGによるプロトプラストの形質転換、電気穿孔に媒介される裸DNAの取り込み、リポソームに媒介される形質移入、電気穿孔、ウイルスベクターに媒介される形質転換、および微粒子銃が含まれる(たとえば、米国特許第5,538,880号、第5,204,253号、第5,591,616号、および第6,329,571号を参照)。植物細胞または組織培養物を形質転換のレシピエント組織として使用する場合は、当業者に知られている技法を使用して、形質転換させた培養物から植物を再生することができる。
【0089】
一部の実施形態では、DNA修飾酵素(たとえばTALEN)を細胞内に直接導入することができる。たとえば、機械的注入、細菌III型分泌系による送達、電気穿孔、またはAgrobacteriumに媒介される転移によって、ポリペプチドを細胞内に導入することができる。たとえば、AgrobacteriumのVirB/D4輸送系、および核タンパク質T複合体の植物細胞への転移を媒介するためのその使用に関する記述には、Vergunstら(2000)Science、290:979〜982を参照されたい。
【0090】
さらに植物に関して、本明細書中に記載のポリヌクレオチド、ベクターおよびポリペプチドは、ベニバナ、アルファルファ、ダイズ、コーヒー、アマランス、ナタネ(高エルカ酸およびアブラナ)、ピーナッツまたはヒマワリなどの双子葉植物、およびアブラヤシ、サトウキビ、バナナ、スーダングラス、トウモロコシ、コムギ、ライムギ、オオムギ、カラスムギ、コメ、アワ、またはソルガムなどの単子葉植物を含めた、いくつかの単子葉および双子葉の植物および植物細胞系内に導入することができる。また、モミおよびマツなどの裸子植物も適切である。
【0091】
したがって、本明細書中に記載の方法は、たとえば、Magniolales、Illiciales、Laurales、Piperales、Aristochiales、Nymphaeales、Ranunculales、Papeverales、Sarraceniaceae、Trochodendrales、Hamamelidales、Eucomiales、Leitneriales、Myricales、Fagales、Casuarinales、Caryophyllales、Batales、Polygonales、Plumbaginales、Dilleniales、Theales、Malvales、Urticales、Lecythidales、Violales、Salicales、Capparales、Ericales、Diapensales、Ebenales、Primulales、Rosales、Fabales、Podostemales、Haloragales、Myrtales、Cornales、Proteales、Santales、Rafflesiales、Celastrales、Euphorbiales、Rhamnales、Sapindales、Juglandales、Geraniales、Polygalales、Umbellales、Gentianales、Polemoniales、Lamiales、Plantaginales、Scrophulariales、Campanulales、Rubiales、Dipsacales、およびAsteralesの目に属する双子葉植物を用いて利用することができる。また、本明細書中に記載の方法は、Alismatales、Hydrocharitales、Najadales、Triuridales、Commelinales、Eriocaulales、Restionales、Poales、Juncales、Cyperales、Typhales、Bromeliales、Zingiberales、Arecales、Cyclanthales、Pandanales、Arales、Lilliales、およびOrchidalesの目に属するものなどの単子葉植物を用いて、または裸子植物類に属する植物、たとえば、Pinales、Ginkgoales、CycadalesおよびGnetalesを用いても利用することができる。
【0092】
本方法は、双子葉植物の属であるAtropa、Alseodaphne、Anacardium、Arachis、Beilschmiedia、Brassica、Carthamus、Cocculus、Croton、Cucumis、Citrus、Citrullus、Capsicum、Catharanthus、Cocos、Coffea、Cucurbita、Daucus、Duguetia、Eschscholzia、Ficus、Fragaria、Glaucium、Glycine、Gossypium、Helianthus、Hevea、Hyoscyamus、Lactuca、Landolphia、Linum、Litsea、Lycopersicon、Lupinus、Manihot、Majorana、Malus、Medicago、Nicotiana、Olea、Parthenium、Papaver、Persea、Phaseolus、Pistacia、Pisum、Pyrus、Prunus、Raphanus、Ricinus、Senecio、Sinomenium、Stephania、Sinapis、Solanum、Theobroma、Trifolium、Trigonella、Vicia、Vinca、Vitis、およびVigna、単子葉植物の属であるAllium、Andropogon、Aragrostis、Asparagus、Avena、Cynodon、Elaeis、Festuca、Festulolium、Heterocallis、Hordeum、Lemna、Lolium、Musa、Oryza、Panicum、Pannesetum、Phleum、Poa、Secale、Sorghum、Triticum、およびZea、または裸子植物の属であるAbies、Cunninghamia、Picea、Pinus、およびPseudotsugaからの種を含めた、幅広い範囲の植物種にわたって使用することができる。
【0093】
形質転換細胞、カルス、組織、または植物は、操作された細胞を、特定の特色または活性、たとえばマーカー遺伝子または抗生物質耐性遺伝子によってコードされているものについて選択またはスクリーニングすることによって、同定および単離することができる。そのようなスクリーニングおよび選択方法は当業者には周知である。さらに、物理的および生化学的方法を使用して形質転換体を同定することができる。これらには、ポリヌクレオチドを検出するためのサザン分析およびPCR増幅、RNA転写物を検出するためのノーザンブロット、S1 RNase保護、プライマー伸長、またはRT−PCR増幅、ポリペプチドおよびポリヌクレオチドの酵素またはリボザイム活性を検出するための酵素アッセイ、ならびにポリペプチドを検出するためのタンパク質ゲル電気泳動、ウエスタンブロット、免疫沈降、および酵素連結免疫アッセイが含まれる。また、in situハイブリダイゼーション、酵素染色、および免疫染色などの他の技法も、ポリペプチドおよび/またはポリヌクレオチドの存在または発現を検出するために使用することができる。言及した技法のすべてを行う方法は周知である。植物細胞内に安定に取り込まれたポリヌクレオチドは、たとえば標準の育種技法を使用して他の植物内に導入することができる。
【0094】
本発明のコンテキストでは、「真核細胞」とは、真菌、酵母、植物もしくは動物細胞、または以下に記載し、in vitro培養のために確立させた生物に由来する細胞系をいう。より好ましくは、真菌は、Aspergillus、Penicillium、Acremonium、Trichoderma、Chrysoporium、Mortierella、KluyveromycesまたはPichiaの属のものであることができる。より好ましくは、真菌は、Aspergillus niger、Aspergillus nidulans、Aspergillus oryzae、Aspergillus terreus、Penicillium chrysogenum、Penicillium citrinum、Acremonium chrysogenum、Trichoderma reesei、Mortierella alpine、Chrysosporium lucknowense、Kluyveromyces lactis、Pichia pastorisまたはPichia ciferriiの種のものであることができる。
【0095】
本発明では、植物は、Arabidospis、Nicotiana、Solanum、Lactuca、Brassica、Oryza、Asparagus、Pisum、Medicago、Zea、Hordeum、Secale、Triticum、Capsicum、Cucumis、Cucurbita、Citrullis、Citrus、またはSorghumの属のものであることができる。より好ましくは、植物は、Arabidospis thaliana、Nicotiana tabaccum、Solanum lycopersicum、Solanum tuberosum、Solanum melongena、Solanum esculentum、Lactuca saliva、Brassica napus、Brassica oleracea、Brassica rapa、Oryza glaberrima、Oryza sativa、Asparagus officinalis、Pisum sativum、Medicago sativa、Zea mays、Hordeum vulgare、Secale cereal、Triticum aestivum、Triticum durum、Capsicum sativus、Cucurbita pepo、Citrullus lanatus、Cucumis melo、Citrus aurantifolia、Citrus maxima、Citrus medica、またはCitrus reticulataの種のものであることができる。
【0096】
本発明では、動物細胞は、Homo、Rattus、Mus、Sus、Bos、Danio、Canis、Felis、Equus、Salmo、Oncorhynchus、Gallus、Meleagris、Drosophila、またはCaenorhabditisの属のものであることができる。より好ましくは、動物細胞は、Homo sapiens、Rattus norvegicus、Mus musculus、Sus scrofa、Bos taurus、Danio rerio、Canis lupus、Felis catus、Equus caballus、Oncorhynchus mykiss、Gallus gallus、またはMeleagris gallopavoの種のものであることができる。動物細胞は、非限定的な例としてSalmo salar、硬骨魚またはブラフィッシュの種からの魚細胞であることができる。また、本発明における動物細胞は、非限定的な例としてキイロショウジョウバエ(Drosophila melanogaster)からの昆虫細胞であることもできる。また、動物細胞は、非限定的な例としてエレガンス線虫(Caenorhabditis elegans)からの寄生虫細胞であることもできる。
【0097】
本発明では、細胞は、植物細胞、哺乳動物細胞、魚細胞、昆虫細胞もしくはin vitro培養のためにこれらの生物から誘導した細胞系、または生組織から直接採取し、in vitro培養のために確立させた初代細胞であることができる。非限定的な例として、細胞系は、CHO−K1細胞、HEK293細胞、Caco2細胞、U2−OS細胞、NIH 3T3細胞、NSO細胞、SP2細胞、CHO−S細胞、DG44細胞、K−562細胞、U−937細胞、MRC5細胞、IMR90細胞、ジャーカット細胞、HepG2細胞、HeLa細胞、HT−1080細胞、HCT−116細胞、Hu−h7細胞、Huvec細胞、Molt 4細胞からなる群から選択されることができる。
【0098】
これらの細胞系はすべて、本発明の方法によって改変して、対象の遺伝子またはタンパク質を産生、発現、定量、検出、研究するための細胞系モデルを提供することができる。また、これらのモデルを使用して、非限定的な例として化学薬品、生物燃料、治療剤および農学などの様々な分野における研究および生成において、対象の生物活性のある分子をスクリーニングすることもできる。
【0099】
また、本発明は、たとえば、細胞内の遺伝物質を変更するため、遺伝子発現を変調させるため、およびたとえば抗ウイルス治療において病原性配列を標的とするために、TALエフェクター内の配列特異的DNA結合ドメインを利用する方法も提供する。たとえば、一部の実施形態では、本発明は、細胞遺伝物質を改変する方法を提供する。一部の実施形態では、本方法には、TALエフェクターDNA結合ドメインを含有するポリペプチド、またはそのようなポリペプチドをコードしている核酸を、細胞内に導入することが含まれる。TALエフェクターDNA結合ドメインは、DNA修飾酵素(たとえばエンドヌクレアーゼ)の全体または一部分と融合させることができる。一部の実施形態では、本方法には、2つ以上の組換え核酸を細胞内に導入することが含まれる。第1の組換え核酸は、細胞中に見つかる対応する事前に選択された標的ヌクレオチド配列に関して1つまたは複数の修飾(すなわち、置換、欠失、または挿入)が含まれる、ドナーヌクレオチド配列を含有する。ドナーヌクレオチド配列は、内在配列またはその一部分がドナー配列またはその一部分で置き換えられているように、内在標的ヌクレオチド配列との相同組換えを受けることができる。標的ヌクレオチド配列には、典型的には配列特異的TALENの認識部位が含まれる。一部の事例では、標的ヌクレオチド配列には、2つ以上の明白に異なるTALENの認識部位(たとえば、明白に異なるDNA配列結合特異性を有するTALENを使用できるように、明白に異なる2つの向かい合った標的配列)が含まれることができる。そのような事例では、DNA切断の特異性は、1つのみの標的配列(または同じ標的配列の複数コピー)を使用する事例と比較して増加している場合がある。
【0100】
第2の組換え核酸は、標的ヌクレオチド配列中の認識部位と結合する配列特異的TALENをコードしているヌクレオチド配列を含有する。一部の事例では、ドナーヌクレオチド配列および配列特異的ヌクレアーゼをコードしているヌクレオチド配列は、同じ核酸構築体中に含有されることができる。あるいは、ドナーヌクレオチド配列およびTALENのコード配列は、別々の構築体中に含有されることができるか、または、TALENポリペプチドを生成して細胞内に直接導入することができる。
【0101】
一部の実施形態では、選択マーカーをコードしているヌクレオチド配列を含有する第3の組換え核酸も使用し得る。第2および第3の組換え核酸は、内在配列との組換えを受け、したがって細胞のゲノム内に組み込まれ得る。これらの組換え事象は非正統的(すなわちランダム)であることができるか、または、これらは相同組換えもしくは部位特異的な組換えによって起こることができる。組換え核酸は、細胞内に同時にまたは順次形質転換させることができ、形質転換の前に直鎖状にすることができる。
【0102】
細胞が植物細胞である場合は、本明細書中で提供されている方法には、形質転換細胞を含有する植物を作製するステップ、植物の子孫を作製するステップ、選択マーカー(含めた場合)を発現する植物を選択またはスクリーニングするステップ、選択された植物の子孫を作製するステップ、および植物(たとえば、組織、種子、前駆細胞、もしくは全植物)または植物の子孫を標的ヌクレオチド配列での組換えについて試験するステップなどのステップがさらに含まれることができる。一部の事例では、本方法には、選択された植物を異系交配させて選択マーカーを除去すること、および/または選択されたもしくは異系交配させた植物を配列特異的ヌクレアーゼの非存在についてスクリーニングすることが含まれることができる。
【0103】
一部の実施形態では、本発明は、細胞、たとえば、原核細胞、動物細胞、または植物細胞の遺伝物質を改変する方法を提供する。本方法には、細胞中に存在する対応する標的ヌクレオチド配列に関してヌクレオチド配列中に1つまたは複数の修飾が含まれる修飾された標的ヌクレオチド配列と、配列特異的TALENの認識部位とを含有する第1の組換え核酸、および配列特異的TALENをコードしているヌクレオチド配列を含有する第2の組換え核酸を、細胞内に導入することが含まれることができる。細胞が植物細胞である場合は、細胞を含有する植物を作製することができ、植物(またはその子孫)から得られた細胞、種子、または組織を、標的ヌクレオチド配列での組換えについて分析することができる。第1および第2の組換え核酸を同時にまたは連続的に細胞内に形質転換させることができ、一方または両方を形質転換の前に直鎖状にし得る。一部の事例では、第1および第2の組換え核酸は同じ構築体中に存在することができる。
【0104】
一部の事例では、本方法には、選択マーカーをコードしているヌクレオチド配列を含有する第3の組換え核酸を細胞内に導入すること、および細胞、細胞から作製した生物、またはその子孫が選択マーカーを発現するかどうかを決定することも含まれることができる。本方法には、さらに、細胞、生物またはその子孫を選択マーカーの非存在についてスクリーニングすることが含まれることができる。選択マーカーをコードしているヌクレオチド配列は、その両側が、第2の配列特異的ヌクレアーゼの切断部位で細胞に内在性のヌクレオチド配列と類似もしくは同一のヌクレオチド配列によって、または配列特異的リコンビナーゼの認識部位によって隣接されていてもいなくてもよい。一部の事例では、本方法には、生物を異系交配させるステップも含まれることができる。異系交配の子孫は、選択マーカーの非存在についてスクリーニングすることができる。
【0105】
また、本発明は、それ内で相同組換えが起こることが所望される標的DNA配列、たとえば、染色体、ミトコンドリア、または葉緑体の配列を含有する細胞を提供するステップと、DNA修飾酵素ドメイン(たとえばエンドヌクレアーゼドメイン)および組み合わせて標的DNA配列内の特定のヌクレオチド配列と結合する複数のTALエフェクター反復を有するTALエフェクタードメインを含有するTALENを提供するステップと、標的DNAの少なくとも一部分に相同的な配列を含有する核酸を提供するステップと、細胞中の標的DNA配列内にあるまたはそれに隣接するヌクレオチド配列の両方の鎖が切断されるように、細胞中の標的DNA配列をTALエンドヌクレアーゼと接触させるステップとを含む、細胞(たとえば植物細胞または動物細胞)の遺伝物質を改変する方法も提供する。そのような切断は、標的DNA配列での相同組換えの頻度を増強させることができる。標的DNA配列は細胞に内在性であることができる。本方法には、TALエンドヌクレアーゼをコードしているcDNAを含有するベクターを細胞内に導入すること、およびTALエンドヌクレアーゼタンパク質を細胞中で発現させることが含まれることができる。一部の事例では、TALエンドヌクレアーゼタンパク質自体を、たとえば、機械的注入、細菌III型分泌系による送達、電気穿孔、またはAgrobacteriumに媒介される転移によって、細胞内に導入することができる。
【0106】
本明細書中に記載の方法は、様々な状況において使用することができる。たとえば、農業では、本明細書中に記載の方法は、以前に組み込んだ導入遺伝子(たとえば除草剤耐性の導入遺伝子)を植物の系、変種、またはハイブリッドから除去するために使用できる、標的部位での相同組換えを促進するために有用である。また、本明細書中に記載の方法を使用して、遺伝子によってコードされている酵素が除草剤耐性を与えるように内在遺伝子を改変することもでき、その例は、改変された酵素がグリフォセート除草剤に対する耐性を与えるような、内在性5−エノールピルビルシキメート−3−リン酸(EPSP)合成酵素遺伝子の改変である。別の例として、本明細書中に記載の方法は、植物または哺乳動物の代謝経路(たとえば脂肪酸生合成)中の1つまたは複数の内在遺伝子の調節領域での相同組換えを、そのような遺伝子の発現が所望の様式で改変されているように促進するために有用である。本明細書中に記載の方法は、動物(たとえばラットまたはマウス)において、非限定的な例として、細胞表面マーカーをコードしているものなどの代謝および内部シグナル伝達経路に関与している1つまたは複数の対象の内在遺伝子、特定の疾患に連関していると同定されている遺伝子、ならびに動物細胞の特定の表現型を司っていることが知られている任意の遺伝子の相同組換えを促進するために有用である。
【0107】
また、本発明は、特定のDNA配列(たとえば、特定の位置でDNAを切断することができるTALEN)と相互作用することができる配列特異的TALエフェクターを設計する方法も提供する。本方法には、TALエフェクターが結合することが所望される標的ヌクレオチド配列(たとえば、内在染色体配列、ミトコンドリアDNA配列、または葉緑体DNA配列)(たとえば、それ内に二本鎖切断を導入することが所望される第2のヌクレオチド配列に隣接する配列)を同定すること、および組み合わせて標的配列と結合する複数のDNA結合反復を含有する配列特異的TALエフェクターを設計することが含まれることができる。本明細書中に記載のように、TALエフェクターには、それらがDNAと相互作用する特異性を決定するいくつかの不完全な反復が含まれる。それぞれの反復は、反復の残基12および13での特定のジアミノ酸配列に応じて、単一の塩基と結合する。したがって、TALエフェクター内の反復を操作することによって(たとえば、標準の技法または本明細書中に記載の技法を使用)、特定のDNA部位を標的化することができる。そのような操作したTALエフェクターは、たとえば、特定のDNA配列を標的とした転写因子として使用することができる。一般的なTALエフェクターの図が図1Aに示されており、反復領域は白い四角によって示されており、代表的な反復配列(配列番号1)中のRVDに下線が引いてある。
【0108】
RVDおよびその対応する標的ヌクレオチドの例が表1Aに示されている(PCT公開WO2010/079430号も参照)。
【表1】
【0109】
他のRVDおよびその対応する標的ヌクレオチドが表1Bに示されている。
【表2】
【0110】
配列特異的なDNA切断が所望される場合は、たとえば、配列特異的TALENを、(a)組み合わせて内在染色体ヌクレオチド配列と結合する複数のDNA結合反復ドメイン、および(b)第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含有するように設計することができる。そのような配列特異的なDNA切断は、本明細書中に記載のように、相同組換えを増強させるために有用な可能性がある。TALENの他の使用には、たとえばウイルスに対する治療剤としての使用が含まれる。TALENは、特定のウイルス配列を標的とする、ウイルスDNAを切断するおよび病原性を低下または消失させるために操作することができる。
【0111】
本明細書中で提供されている材料および方法を使用して、特定の遺伝子の配列を標的化された様式で改変することができる。遺伝子は、操作したTALエフェクターの標的とすることができる複数の配列を含有し得る。しかし、本明細書中に記載のように、特定の標的配列がより有効に標的化され得る。たとえば、実施例9に記載のように、特定の特徴を有する配列は、TALエフェクターによってより有効に標的化され得る。したがって、本明細書中で提供されている方法には、特定の基準を満たしている標的配列を同定することが含まれることができる。これらには、i)15個の塩基の最小の長さかつ5’から3’の配向を有しており、Tが5’末端の部位の直前にある、ii)第1(5’)位置にTまたは第2位置にAを有さない、iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さない、ならびにiv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの基本組成を有する配列が含まれる。
【0112】
本明細書中に記載のTALENは一般に二量体として働くため、本明細書中で提供されている方法の一部の実施形態には、上述の基準のうちの少なくとも1つを満たしており、15〜18bpによって分離されている、細胞中の第1のゲノムヌクレオチド配列および第2のゲノムヌクレオチド配列を同定することが含まれることができる。一部の事例では、1つのTALENポリペプチドがそれぞれのヌクレオチド配列と結合することができ、TALEN中に含有されるエンドヌクレアーゼが15〜18bpのスペーサー内を切断することができる。
【0113】
また、本発明は、それ内に所望の核酸が導入されている、遺伝子改変した動物を作製する方法も提供する。そのような方法には、それ内に核酸を導入することが所望される内在染色体標的DNA配列を含有する細胞を得るステップと、細胞内にTALENを導入して内在染色体標的DNA配列内に二本鎖切断を生じるステップと、内在染色体標的DNAの少なくとも一部分に相同的な配列を含有する外因性核酸を、細胞内に、外因性核酸と内在染色体標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった初代細胞から動物を作製するステップとが含まれることができる。相同的な核酸には、たとえば、相同組換え後に遺伝子を破壊するヌクレオチド配列、相同組換え後に遺伝子を置き換えるヌクレオチド配列、相同組換え後に遺伝子内に点突然変異を導入するヌクレオチド配列、または相同組換え後に調節部位を導入するヌクレオチド配列が含まれることができる。
【0114】
また、本明細書中で提供されている方法を使用して、それ内に所望の核酸が導入されている、遺伝子改変した植物を作製することもできる。そのような方法には、それ内に核酸を導入することが所望される内在標的DNA配列を含有する植物細胞を得るステップと、TALENを導入して内在標的DNA配列内に二本鎖切断を生じるステップと、内在標的DNAの少なくとも一部分に相同的な配列を含有する外因性核酸を、植物細胞内に、外因性核酸と内在標的DNAとの間の相同組換えが起こることを可能にする条件下で導入するステップと、それ内で相同組換えが起こった植物細胞から植物を作製するステップとが含まれることができる。
【0115】
本明細書中で提供されているTALENに促進される相同組換え方法によって作製された細胞中のDNAは、そのような方法を経ていない細胞と比較して改変されており、改変されたDNAを含有する細胞は「遺伝子改変した」と呼ばれる。しかし、そのような改変は相同組換えを含み、導入遺伝子のランダム組込みを含まないため、そのような細胞を含有する生物は調節目的のGMOとみなされない場合があることに留意されたい。したがって、遺伝子改変を生じるために本明細書中に記載のTALENに促進される方法を使用することは、たとえば、標準の調節手順ならびにその関連する時間および費用を回避し得るという点で、有利であり得る。
【0116】
本明細書中で提供されている標的化遺伝子組換えの他の方法には、選択されたDNA標的配列を標的としたTALENをコードしている核酸分子を細胞(たとえば、植物細胞、昆虫細胞、硬骨魚細胞、または動物細胞)内に導入するステップと、細胞内でのTALENの発現を誘導するステップと、選択されたDNA標的配列が突然変異(たとえば、遺伝物質の欠失、遺伝物質の挿入、または遺伝物質の欠失および挿入両方)を示している組換え細胞を同定するステップとが含まれることができる。また、ドナーDNAを細胞内に導入することもできる。
【0117】
一部の実施形態では、単量体TALENを使用することができる。本明細書中に記載のTALENは、典型的には、2つのTALエフェクタードメインがそれぞれFokI制限酵素の触媒ドメインと融合しており、それぞれの生じるTALENのDNA認識部位がスペーサー配列によって分離されており、それぞれのTALEN単量体と認識部位との結合によりFokIが二量体化してスペーサー内に二本鎖切断を生じることが可能となるように、スペーサーを有する二連認識部位にわたる二量体として機能する(たとえばMoscouおよびBogdanove(2009)Science、326:1501を参照)。単量体TALENも構築することができるが、単一のTALエフェクターと、機能するために二量体化を必要としないヌクレアーゼとが融合しているようにする。たとえば、1つのそのようなヌクレアーゼは、2つの単量体が単一のポリペプチドとして発現される、FokIの単鎖変異体である(Minczukら(2008)Nucleic Acids Res.、36:3926〜3938)。また、他の天然に存在するまたは操作した単量体ヌクレアーゼもこの役割を果たすことができる。単量体TALENに使用されるDNA認識ドメインは、天然に存在するTALエフェクターに由来することができる。あるいは、DNA認識ドメインは、特定のDNA標的を認識するように操作することができる。操作した単鎖TALENは、1つの操作したDNA認識ドメインしか必要としないため、構築および配置がより容易であり得る。
【0118】
一部の実施形態では、二量体DNA配列特異的ヌクレアーゼは、2つの異なるDNA結合ドメイン(たとえば、1個のTALエフェクター結合ドメインおよび別の種類の分子からの1個の結合ドメイン)を使用して作製することができる。上述のように、本明細書中に記載のTALENは、典型的には、スペーサーを有する二連認識部位にわたる二量体として機能する。このヌクレアーゼ構造は、たとえば1個のTALEN単量体および1個のジンクフィンガーヌクレアーゼ単量体から作製した標的特異的なヌクレアーゼにも使用することができる。そのような事例では、TALENおよびジンクフィンガーヌクレアーゼ単量体のDNA認識部位は、適切な長さのスペーサーによって分離されていることができる。2つの単量体の結合により、FokIが二量体化してスペーサー配列内に二本鎖切断を生じることを可能にすることができる。また、ホメオドメイン、myb反復またはロイシンジッパーなどのジンクフィンガー以外のDNA結合ドメインも、FokIと融合させて、機能的ヌクレアーゼを作製するためのTALEN単量体とのパートナーとして役割を果たすことができる。
【0119】
一部の実施形態では、TALエフェクターを使用して、他のタンパク質ドメイン(たとえば非ヌクレアーゼタンパク質ドメイン)を特定のヌクレオチド配列に対して標的化させることができる。たとえば、TALエフェクターは、それだけには限定されないが、DNA相互作用酵素(たとえば、メチラーゼ、トポイソメラーゼ、インテグラーゼ、トランスポサーゼ、もしくはリガーゼ)、転写活性化因子もしくはリプレッサー、またはヒストンなどの他のタンパク質と相互作用するもしくはそれを修飾するタンパク質からのタンパク質ドメインと連結されていることができる。そのようなTALエフェクター融合体の応用には、たとえば、後成的調節要素を作製または改変すること、DNA中に部位特異的な挿入、欠失、または修復を行うこと、遺伝子発現を制御すること、およびクロマチン構造を改変することが含まれる。
【0120】
一部の実施形態では、標的配列のスペーサーは、TALENの特異性および活性を変調させるために選択または変動させることができる。スペーサーを有する二連認識部位にわたる二量体として機能するTALENの、本明細書中に提示した結果は、TALENが様々なスペーサー長にわたって機能することができ、TALENの活性がスペーサー長に伴って変動することを実証している。たとえば以下の実施例6を参照されたい。スペーサー長の柔軟性は、高い特異性で特定の配列(たとえばゲノム中のもの)を標的化するようにスペーサー長を選択できることを示している。さらに、様々なスペーサー長で観察される活性の変動は、所望のレベルのTALEN活性を達成するためにスペーサー長を選択できることを示している。
【0121】
一部の実施形態では、TALEN活性は、DNA結合ドメイン(複数可)内の反復の数および組成を変動させることによって変調させることができる。たとえば、本明細書中の実施例7に記載のように、PthXo1に基づくTALENはAvrBs3に基づくTALENよりも高い活性を示した。PthXo1は、その反復の数およびRVD組成がどちらもAvrBs3とは異なる。さらに、これらのタンパク質の天然に存在するDNA認識部位は、その多様性が、MoscouおよびBogdanove(上記)によって記載されているTALエフェクターDNA暗号に基づいて予測されたそのそれぞれの認識配列とは異なる。さらに、いくつかの同じ長さ(12個のRVD)であるが異なるRVD組成を有する誂えのTALENは、その活性が異なっており、13個のRVDの誂えのTALENは、12個のRVDの誂えのTALENよりも高い活性を有していた。したがって、TALENは、対象のDNA配列を認識するように操作できるだけでなく、(1)活性を変調させるために反復の数を変動させることができる、(2)様々なレベルの活性を達成するために様々な結合部位を選択することができる、ならびに(3)TALEN活性を変調させるためにRVDの組成および標的部位に対するその当てはめ(暗号に従う)を変動させることができる。
【0122】
TALENがヘテロ二量体である場合、たとえば、それぞれTALエフェクタードメインおよびFokIヌクレアーゼ触媒ドメインが含まれる2つの異なる単量体を有する場合、RVDは、2つのTALエフェクタードメインのそれぞれ中で等しい数で見つけることができるか、または、それぞれのドメインは異なる数のRVDを示すことができる。たとえば、合計22個のRVDがDNAを特定のヘテロ二量体TALENと結合させるために使用されている場合は、2つのTALエフェクタードメインのそれぞれ中で11個の反復を見つけることができるか、あるいは、2つのTALエフェクタードメインのうちの一方で10個の反復および他方中で12個を見つけることができる。また、本発明には、単量体として機能するDNA修飾酵素ドメインを有するTALENも包含される。この場合、単量体酵素と融合している単一のTALエフェクタードメイン中ですべてのRVDを見つけることができる。この場合、効率的な結合にするためには、RVDの数が等価な二量体TALEN中で見つかるであろうRVDの合計数に等しくなければならない。たとえば、2つの異なるTALエフェクタードメイン上に10個の反復を有する代わりに(二量体TALENの場合のように)、単一のTALエフェクタードメイン中に20個の反復を有する(単量体TALENの場合のように)。
【0123】
本発明のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも14である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも20である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも24である。本発明の別のさらなる態様では、二量体または単量体TALEN内の反復の合計数は少なくとも30である。
【0124】
また、本特許出願は、標的DNAに対して増強された標的化能力を有するTALエフェクタータンパク質を作製する方法も提供する。本方法には、たとえば、それぞれのDNA結合反復が標的DNA中の塩基対の認識を決定するRVDを含有し、かつ標的DNA中の1つの塩基対の認識を司っている、複数のDNA結合反復を有するDNA結合ドメインを有するTALエフェクターをコードしている核酸を作製することが含まれることができる。以下の実施例12に記載のように、結合部位の−1位でのTの必要性を緩めることで、操作したTALエフェクタータンパク質の標的化能力を増強させ得る。したがって、TALエフェクターをコードしている核酸の作製には、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませ、したがって結合部位の−1位でのTの必要性を排除することが含まれることができる。
【0125】
さらに、標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法が本明細書中で提供されている。そのような方法には、それぞれの反復が標的DNA中の塩基対の認識を決定するRVDを含有する、複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製することが含まれることができる。以下の実施例12に記載のように、NN(Gを認識する最も一般的なRVD)の特異性は一般に弱く、コンテキストに伴って変動する場合があるように見えるが、特定のRVDはGに対して増強された特異性を有し得る。したがって、本明細書中で提供されている方法には、Gに対してより頑強な特異性を有し得る代替のRVDを使用することが含まれることができる。たとえば、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択される1つまたは複数のRVDを使用することができ、星印はRVDの第2位置でのギャップを示す。
【0126】
製品
また、本発明は、たとえば、TALENをコードしている核酸分子、TALENポリペプチド、そのような核酸分子もしくはポリペプチドを含有する組成物、またはTALエンドヌクレアーゼを操作した細胞系を含有する製品も提供する。そのような物品は、たとえば、研究ツールとして、または治療的に使用することができる。
【0127】
一部の実施形態では、製品には、本明細書中で提供されている方法を使用して作製した植物からの種子が含まれることができる。種子は、当分野で知られている手段を使用して調整し、当分野で周知の梱包材料を使用して梱包して、製品を調製することができる。種子の梱包は、ラベル、たとえば、梱包材料に固定されたタグもしくはラベル、梱包材料上に印刷されたラベルまたは梱包内に挿入されたラベルを有することができる。ラベルは、梱包内に含有された種子が遺伝子改変した植物の作物を生じることができることを示すことができ、改変していない植物に対して遺伝子改変によって変更されている特色を記載することができる。
【0128】
他の定義
ポリペプチド配列中のアミノ酸残基またはサブユニットは、本明細書中で一文字のコードに従って命名されており、たとえば、QはGlnまたはグルタミン残基を意味し、RはArgまたはアルギニン残基を意味し、DはAspまたはアスパラギン酸残基を意味する。
【0129】
アミノ酸置換とは、1個のアミノ酸残基を別のもので置き換えることを意味し、たとえば、ペプチド配列中のアルギニン残基をグルタミン残基で置き換えることはアミノ酸置換である。
【0130】
ヌクレオチドは以下のように命名されている:一文字のコードがヌクレオシドの塩基を命名するために使用されている:aはアデニンであり、tはチミンであり、cはシトシンであり、gはグアニンである。縮重ヌクレオチドには、rはgまたはa(プリンヌクレオチド)を表し、kはgまたはtを表し、sはgまたはcを表し、wはaまたはtを表し、mはaまたはcを表し、yはtまたはc(ピリミジンヌクレオチド)を表し、dはg、aまたはtを表し、vはg、aまたはcを表し、bはg、tまたはcを表し、hはa、tまたはcを表し、nはg、a、tまたはcを表す。
【0131】
用語「DNA修飾酵素」とは、DNA修飾のレベルが何であれ(切断、共有的相互作用、水媒介性の相互作用・・・)、細胞の遺伝物質を改変することができる任意のタンパク質をいう。DNA相互作用タンパク質(たとえば、メチラーゼ、トポイソメラーゼ、インテグラーゼ、トランスポサーゼ、またはリガーゼ)、転写活性化因子またはリプレッサー、ヒストンなどの他のタンパク質、およびヌクレアーゼが、「DNA修飾酵素」の意味に含まれることを意図する。TALエフェクター−DNA修飾酵素内に含まれている場合、DNA修飾酵素はDNA修飾酵素ドメインと呼ばれる。
【0132】
用語「ヌクレアーゼ」には、エキソヌクレアーゼおよびエンドヌクレアーゼが含まれることを意図する。
【0133】
用語「エンドヌクレアーゼ」とは、DNAまたはRNA分子、好ましくはDNA分子内の核酸間の結合の加水分解(切断)を触媒することができる、任意の野生型または変異体酵素をいう。エンドヌクレアーゼの非限定的な例には、FokI、HhaI、HindIII、NotI、BbvCI、EcoRI、BglI、およびAlwIなどのII型制限エンドヌクレアーゼが含まれる。また、エンドヌクレアーゼは、典型的には約12〜45個の塩基対(bp)の長さ、より好ましくは14〜45bpのポリヌクレオチド認識部位を有する場合は、レアカッターエンドヌクレアーゼも含む。レアカッターエンドヌクレアーゼは、定義された座位でのDNA二本鎖切断(DSB)を誘導することによって、HRを有意に増加させる(Rouet、Smihら、1994、Rouet、Smihら、1994、Choulika、Perrinら、1995、PingoudおよびSilva、2007)。レアカッターエンドヌクレアーゼは、たとえば、ホーミングエンドヌクレアーゼ(PaquesおよびDuchateau、2007)、操作したジンクフィンガードメインとFokIなどの制限酵素の触媒ドメインとの融合から生じるキメラジンクフィンガーヌクレアーゼ(ZFN)(PorteusおよびCarroll、2005)、または化学エンドヌクレアーゼ(Eisenschmidt、Lanioら、2005、Arimondo、Thomasら、2006、Simon、Cannataら、2008)であることができる。化学エンドヌクレアーゼでは、化学またはペプチド切断剤は、特定の標的配列を認識する核酸のポリマーまたは別のDNAのどちらかとコンジュゲートされており、したがって、切断活性を特定の配列を標的とする。また、化学エンドヌクレアーゼには、特定のDNA配列と結合することが知られている、オルトフェナントロリンのコンジュゲートなどの合成ヌクレアーゼ、DNA切断分子、および三重鎖形成オリゴヌクレオチド(TFO)も包含される(KalishおよびGlazer、2005)。そのような化学エンドヌクレアーゼが本発明による用語「エンドヌクレアーゼ」中に含まれる。そのようなエンドヌクレアーゼの例には、I−Sce I、I−Chu I、I−Cre I、I−Csm I、PI−Sce I、PI−Tli I、PI−Mtu I、I−Ceu I、I−Sce II、I−Sce III、HO、PI−Civ I、PI−Ctr I、PI−Aae I、PI−Bsu I、PI−Dha I、PI−Dra I、PI−Mav I、PI−Mch I、PI−Mfu I、PI−Mfl I、PI−Mga I、PI−Mgo I、PI−Min I、PI−Mka I、PI−Mle I、PI−Mma I、PI−Msh I、PI−Msm I、PI−Mth I、PI−Mtu I、PI−Mxe I、PI−Npu I、PI−Pfu I、PI−Rma I、PI−Spb I、PI−Ssp I、PI−Fac I、PI−Mja I、PI−Pho I、PI−Tag I、PI−Thy I、PI−Tko I、PI−Tsp I、I−MsoIが含まれる。
【0134】
本発明によるエンドヌクレアーゼは、転写活性化因子様(TAL)エフェクターエンドヌクレアーゼ(TALEN)の一部であることができる。
【0135】
「TALEN」とは、転写活性化因子様(TAL)エフェクター結合ドメインおよびエンドヌクレアーゼドメインを含むタンパク質を意図し、両方のドメインの融合が「単量体TALEN」をもたらす。一部の単量体TALENはそれ自体で機能的な場合があり、他のものは別の単量体TALENとの二量体化を必要とする。二量体化は、両方の単量体TALENが同一である場合にホモ二量体TALENをもたらす可能性があるか、または、単量体TALENが異なる場合にヘテロ二量体TALENをもたらす可能性がある。2つの単量体TALENは、たとえば、そのRVDの数が異なるおよび/または少なくとも1つのRVDの内容(すなわちアミノ酸配列)が異なる場合に、異なる。「TALエフェクター−DNA修飾酵素」とは、転写活性化因子様エフェクター結合ドメインおよびDNA修飾酵素ドメインを含むタンパク質を意図する。
【0136】
「変異体」とは、「変異体」タンパク質、すなわち、自然には天然に存在せず、遺伝子操作またはランダム突然変異誘発によって得られるタンパク質、すなわち操作したタンパク質を意図する。この変異体タンパク質は、たとえば、野生型の天然に存在するタンパク質のアミノ酸配列中の少なくとも1つの残基を異なるアミノ酸で置換することによって得ることができる。前記置換(複数可)は、たとえば部位特異的突然変異誘発および/またはランダム突然変異誘発によって導入することができる。
【0137】
「細胞」または「複数の細胞」とは、任意の原核または真核の、生細胞、in vitro培養のためにこれらの生物から誘導した細胞系、動物もしくは植物起源の初代細胞を意図する。
【0138】
「初代細胞」または「複数の初代細胞」とは、非常に少ない回数の集団倍加しか受けておらず、したがって、連続的な腫瘍原性細胞または人工的に不死化した細胞系と比較してそれらが由来する組織の主な機能的構成要素および特徴をより表している、生組織から直接採取(すなわち生検材料)し、in vitroでの成長のために確立させた細胞を意図する。したがって、これらの細胞は、それらが指すin vivo状態に対するより価値のあるモデルを表す。
【0139】
「相同的」とは、配列間の相同組換えをもたらすために十分な別の配列との同一性を有する、より具体的には、少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%を有する配列を意図する。
【0140】
「同一性」とは、2つの核酸分子またはポリペプチド間の配列同一性をいう。同一性は、比較の目的のためにアラインメントし得るそれぞれの配列中の位置を比較することによって決定することができる。比較した配列中の位置が同じ塩基によって占有されている場合は、分子はその位置で同一である。核酸またはアミノ酸配列間の類似性または同一性の度合は、核酸配列によって共有される位置での同一または一致するヌクレオチドの数の関数である。たとえば初期設定で使用することができるGCG配列解析パッケージ(ウィスコンシン大学、ウィスコンシン州Madison)の一部として利用可能なFASTA、またはBLASTを含めた、様々なアラインメントアルゴリズムおよび/またはプログラムを使用して、2つの配列間の同一性を計算し得る。
【0141】
「突然変異」とは、ポリヌクレオチド(cDNA、遺伝子)またはポリペプチド配列中の1つまたは複数のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。前記突然変異は、遺伝子またはその調節配列のコード配列に影響を与える場合がある。また、これは、ゲノム配列の構造またはコードされているmRNAの構造/安定性にも影響を与え得る。
【0142】
「遺伝子」とは、染色体に沿って直線的に配置されているDNAのセグメントからなり、特定のタンパク質またはタンパク質のセグメントをコードしている、遺伝の基本単位を意味する。遺伝子には、典型的には、プロモーター、5’非翻訳領域、1つまたは複数のコード配列(エクソン)、任意選択でイントロン、3’非翻訳領域が含まれる。遺伝子は、ターミネーター、エンハンサーおよび/またはサイレンサーをさらに含み得る。
【0143】
用語「対象の遺伝子」とは、既知または推定上の遺伝子産物をコードしている任意のヌクレオチド配列をいう。
【0144】
本明細書中で使用する用語「座位」とは、染色体上のDNA配列(たとえば遺伝子のもの)の特定の物理的位置である。用語「座位」とは、通常、染色体上の標的配列の特定の物理的位置をいう。
【0145】
「融合タンパク質」とは、元は別々のタンパク質をコードしている2つ以上の遺伝子を結合させることからなる当分野の周知のプロセスの結果を意図し、前記「融合遺伝子」の翻訳の結果、元のタンパク質のそれぞれに由来する機能的特性を有する単一のポリペプチドがもたらされる。
【0146】
「触媒ドメイン」とは、酵素の活性部位を含有する、酵素のタンパク質ドメインまたはモジュールを意図する。活性部位とは、基質の触媒作用がそこで起こる、前記酵素の部分を意図する。酵素だけでなくその触媒ドメインは、それらが触媒する反応に従って分類および命名される。酵素番号(EC番号)とは、それらが触媒する化学反応に基づく、酵素の数値分類スキームである(ワールドワイドウェブchem.qmul.ac.uk/iubmb/enzyme/)。本発明の範囲では、任意の触媒ドメインをパートナーとして使用し、TALエフェクタードメインと融合させてキメラ融合タンパク質を作製して、TALエフェクター−DNA修飾酵素をもたらすことができる。そのような触媒ドメインの非限定的な例は、MmeI、EsaSSII、CstMI、NucA、EndA大腸菌、NucM、EndA肺炎連鎖球菌、SNase黄色ブドウ球菌、SNase Staphylococcus hyicus、SNaseフレクスナー赤痢菌、枯草菌yncB、エンドデオキシリボヌクレアーゼI腸内細菌ファージT7、EndoGウシ、ttSmr DNAミスマッチ修復タンパク質mutS、Metnaseの切断ドメインのものであることができる。
【0147】
別段に指定しない限りは、本発明の実施は、細胞生物学、細胞培養物、分子生物学、トランスジェニック生物学、微生物学、組換えDNA、および免疫学の慣用技術を用い、これらは十分に当分野の技術範囲内にある。そのような技法は文献中で完全に説明されている。たとえば、分子生物学の最新プロトコル(Current Protocols in Molecular Biology)(Ausubel、2000、Wiley and son Inc、米国議会図書館、分子クローニング:実験室の手引き(Molecular Cloning:A Laboratory Manual)、第3版、(Sambrookら、2001、ニューヨーク州Cold Spring Harbor:Cold Spring Harbor Laboratory Press)、オリゴヌクレオチド合成(Oligonucleotide Synthesis)(M.J.Gait編、1984)、米国特許第4,683,195号、核酸ハイブリダイゼーション(Nucleic Acid Hybridization)(HarriesおよびHiggins編、1984)、転写および翻訳(Transcription and Translation)(HamesおよびHiggins編、1984)、動物細胞の培養(Culture of Animal Cells)(Freshney、Alan R.Liss,Inc.、1987)、固定細胞および酵素(Immobilized Cells and Enzymes)(IRL Press、1986)、Perbal、分子クローニングの実用的な指針(A Practical Guide to Molecular Cloning)(1984)、シリーズ酵素学の方法(Methods in Enzymology)(AbelsonおよびSimon編集長、Academic Press,Inc.、New York)、特に第154巻および第155巻(Wuら編)ならびに第185巻、「遺伝子発現技術(Gene Expression Technology)」(Goeddel編)、哺乳動物細胞の遺伝子転移ベクター(Gene Transfer Vectors For Mammalian Cells)(MillerおよびCalos編、1987、Cold Spring Harbor Laboratory)、細胞および分子生物学における免疫化学的方法(Immunochemical Methods in Cell and Molecular Biology)(MayerおよびWalker編、Academic Press、London、1987)、実験免疫学のハンドブック(Handbook of Experimental Immunology)、第I〜IV巻(WeirおよびBlackwell編、1986)、ならびにマウス胚の操作(Manipulating the Mouse Embryo)、(Cold Spring Harbor Laboratory Press、ニューヨーク州Cold Spring Harbor、1986)を参照されたい。
【0148】
本発明の上記明細書は、任意の当業者による本発明の作製および使用が可能となるようにそれを作製および使用する様式およびプロセスを提供し、この可能にすることは、元の説明の一部を構成する添付の特許請求の範囲の主題のために特に提供されている。
【0149】
上記で使用するように、語句「からなる群から選択される」、「から選択される」などには、指定した物質の混合物が含まれる。
【0150】
数値の限界または範囲が本明細書中に記述されている場合は、端点が含まれる。また、数値の限界または範囲内のすべての値および部分範囲は、明確に書き出されている場合と同様に具体的に含まれる。
【0151】
上記説明は、当業者が本発明を作製および使用することを可能にするために提供されており、特定の応用およびその要件のコンテキストで提供されている。好ましい実施形態への様々な改変は当業者に容易に明らかであり、本明細書中に定義した一般的な原理は、本発明の精神および範囲から逸脱せずに、他の実施形態および応用に適用し得る。したがって、本発明は、示されている実施形態に限定されず、本明細書中に開示した原理および特長と一貫性のある最も広い範囲に従うことを意図する。
【0152】
本発明を一般に説明したが、さらなる理解は、例示の目的でのみ本明細書中で提供されている特定の具体的な実施例を参照して得ることができ、本発明は、別段に指定しない限りは特許請求の範囲に記載されている本発明の範囲を限定しない、以下の実施例中にさらに記載されている。
【実施例】
【0153】
【実施例1】
【0154】
暗号がTALエフェクター−DNA認識を支配する
TAL標的部位中のRVDと近接したヌクレオチドとの間に1対1の直線的な対応が存在するかどうかを決定するために、10個のTALエフェクターのそれぞれの既知の標的遺伝子の予測されたプロモーター領域(すなわち注釈付けされた翻訳開始部位の直前の1,000bp)を、TALエフェクターのRVD配列を用いて、RVD−ヌクレオチドの会合においてエントロピー(ランダム性)を最小限にしたアラインメントについて走査した。以下の式を使用してエントロピーを定量し、Rは、エフェクターのRVDの組であり、Dは、4個のヌクレオチド(A、C、G、T)の組であり、fi,jは、第i番目のRVDが第j番目のヌクレオチドと会合する観察された頻度を表す:
【化1】
【0155】
複数の低エントロピー部位がそれぞれのプロモーター中に存在していた。しかし、エフェクターAvrBs3では、1つのみが活性化に十分かつ必要であると以前に同定された54bpのupa20プロモーター断片にマッピングされ、これは、AvrBs3によって直接活性化された遺伝子に一般的なUPAボックスと一致した(Kayら、上記)。また、エフェクターPthXo1およびAvrXa27では、1つの部位のみが、そのそれぞれの標的であるOs8N3およびXa27の活性化された対立遺伝子と活性化されなかった対立遺伝子との間の多型にそれぞれ重複していた。これら3つの部位でのアラインメントにわたってRVD−ヌクレオチドの会合は一貫していたため、残りのアラインメントをこれらの会合に基づいて選択し、その結果、1対のTALエフェクター−標的あたり正確に1つの部位がもたらされた(図1Bおよび表2)。それぞれの部位にはTが先行している(図1D)。
【0156】
RVD−ヌクレオチドの会合によって与えられた特異性を評価するために、最初に、10個の最小限のエントロピーのTALエフェクター−標的部位のアラインメントにわたって観察されたすべてのRVD−ヌクレオチドの会合の頻度に基づいて重量マトリックスを作成した(図1B)。その後、重量マトリックスを使用して、コメ、Oryza sativa spp.japonica cv.Nipponbare(Osa1、リリース6.0、rice.plantbiology.msu.edu)におけるそれぞれの非冗長遺伝子モデルの翻訳開始に先行する1,000bpのプロモーター領域を、コメの病原体Xanthomonas oryzaeの5個のTALエフェクター(AvrXa27、PthXo1、PthXo6、PthXo7、およびTal1c)に対する最良の一致について走査した。AvrXa27には、Xa27(GenBank受託AY986492)の上流の配列を含めた。この上流配列はNipponbare中には存在しない。観察された会合頻度は90%で重み付けされ、残りの10%はすべての可能な会合の頻度に均等に分布していた。アラインメントは重量マトリックススコア(y軸)を使用して順位付けし、図1BのRVD−ヌクレオチドの会合頻度に由来する頻度スコアの負の対数としてとった。したがって、スコアが低ければ低いほど、一致はより良好である。PthXo1、PthXo6、PthXo7、およびTal1cでは、実験的に同定した標的遺伝子が最良またはほぼ最良の一致であった。より良好な一致には、Tが先行していなかったか、標的を同定するために使用したマイクロアレイ上に表されていなかったか、またはイントロンおよびESTの証拠を欠いていなかった。既知の標的に対する逆相補プロモーター配列の走査は、順方向の部位よりも良好なスコアのアラインメントを与えなかった。この結果は、TALエフェクターがプラス鎖と結合することを暗示しているのではなく、これらがプラス鎖に対して順方向の配向で機能することを示している。5個目のエフェクターAvrXa27の既知の標的は疾患耐性遺伝子Xa27である(Guら、上記)。この一致の順位(5,368)がより低いことは、較正したまたは最近および最適以下の宿主の適応を反映している可能性がある。より良好なスコアの部位は、病因についてAvrXa27によって標的化されている遺伝子を含む可能性が高い。
【0157】
重量マトリックスを再度使用して、すべてのコメプロモーターを40個の追加のX.oryzaeのTALエフェクターで走査し、公的マイクロアレイデータ(PLEXdb.org、受託OS3)に基づいて感染中に下流遺伝子が活性化された最良のアラインメントのものを保持することによって、10個の追加のアラインメントが得られた(表3)。初期の組と同様、Tがそれぞれの部位に先行しており、どの逆鎖の部位もより良好なスコアではなかった。合計20個のアラインメントのRVD−ヌクレオチドの会合頻度が図1Cに示されている。これらは、驚くべきほど単純な暗号を構成している。
【0158】
20個のTALエフェクターヌクレオチドのアラインメントの拡張組中のRVD−ヌクレオチドの頻度を使用して新しい重量マトリックスを作成し、コンピュータスクリプトをPython v2.5(www.python.org)によって書いた。スクリプトは、観察されたvs.観察されていないRVD−ヌクレオチドの会合についてユーザ定義可能な重量因子を用いて、特定のTALエフェクターとの一致についてDNA配列の任意のコレクションを走査するために使用することができる。MoscouおよびBogdanove(上記)を参照されたい。
【0159】
暗号中にはある程度の縮重が存在する。強力な会合は結合親和性のほとんどを占めるアンカーを表している場合があり、弱い会合は柔軟性の測度を提供する。あるいは、隣接効果が関与している場合がある。後者の可能性は、RVDの両側に条件付けられたすべてのRVDのヌクレオチド会合頻度を決定し、それらを観察された全頻度と比較することによって、言い換えれば、RVD−ヌクレオチドの対合を左または右に隣接するRVDに従って選別し、そのようにして選別されたそれぞれの対の相対的頻度をその対の全体的な頻度と比較することによって調査した。隣接するものによって選別したRVD−ヌクレオチドの会合の頻度は観察された全頻度から顕著に逸脱せず、これは、会合がコンテキスト非依存性であることを示唆している。
【0160】
20個の標的部位に隣接する配列によって−1のT以外の保存的なヌクレオチドは明らかとならなかったが、この部位に続いてCが富み、全体的にGが乏しい傾向にある(図1D)。いくつかの例外を除いて、部位は注釈付けされた転写開始の60bp以内上流に始まり、翻訳開始に87bpより近いものはない(表2および表3)。RVD/ヌクレオチドの会合を支配するさらなる規則は、実施例4および5に記載されている。
【0161】
これらの結果を考慮して、ゲノム中のTALエフェクター標的の予測および標的の新規構築が現在可能である。部位を予測する能力は、疾患に重要な宿主遺伝子の同定をはかどらせる。標的を構築する能力は、保存的または複数のTALエフェクターに応答性の、耐久性のある耐性遺伝子の設計に期待される。また、本明細書中に記載のように、自由裁量による遺伝子の活性化のためにTALエフェクターを誂えること、またはDNA修飾のためにの融合タンパク質を標的化することも可能である。
【表3】
【表4】
【実施例2】
【0162】
TALENは酵母中で機能できる
プラスミドの構築:TALエフェクター、AvrBs3のタンパク質コード配列は、プラスミドをBamHIで消化することから得られた。主に反復ドメインをコードしているDNA断片をSphIで切り出した。AvrBs3のアミノ酸配列はGENBANK受託番号P14727および配列番号12の下(図3)、核酸配列は受託番号X16130配列番号13の下(図4)で見つけることができる。図4中、BamHIおよびSphI部位は太字で下線が引かれている。AvrBs3のBamHIおよびSphI断片を、ヌクレアーゼ発現ベクターpDW1789_TAL(図5)内に、FokIヌクレアーゼドメインをコードしている配列に隣接してクローニングした。AvrBs3標的部位を標的レポータープラスミド内にクローニングするために、18bpのスペーサー配列をその間に含んで逆配向に配置された2つのAvrBs3認識部位を含有する2つの相補的DNAオリゴを、5’および3’末端でそれぞれBglIIおよびSpeIのオーバーハングを有するように合成した。6、9、12および15bpのスペーサー長を用いて、認識部位を有する他のレポータープラスミドを作製した。アニーリングさせたDNAオリゴをレポータープラスミドpCP5内にクローニングし(図6)、これをBglIIおよびSpeIで消化した。
【0163】
酵母アッセイ:標的レポータープラスミドを酵母株YPH499(MAT a株)内に形質転換させ、形質転換体を、トリプトファンを欠く合成完全培地(SC−W)上で選択した。TALEN発現プラスミドをYPH500(MAT α株)内に形質転換させ、形質転換体を、ヒスチジンを欠くSC培地(SC−H)上にプレートした。標的レポータープラスミドを保有する酵母コロニーおよびTALEN発現プラスミドを保有するコロニーを、終夜、30℃で、それぞれ液体SC−WおよびSC−H培地中で培養した。培養物を同じOD600に調節し、それぞれの200μlを200μlのYPD培地に混ぜた。混合物を30℃で4時間インキュベーションして、2種類の酵母株を接合させた。混合培養物を遠心沈殿させ、5mlのSC−W−H培地中に30Cで終夜、またはOD600が0.5〜1の範囲に達成するまで再懸濁させた。細胞を収穫し、定量的β−ガラクトシダーゼアッセイを記載のように行った(Townsendら(2009)Nature、459:442〜445)。
【0164】
結果:TAL−FokI融合体は、TALのDNA認識ドメインおよび非特異的なFokI DNA切断ドメインからなる部位特異的なヌクレアーゼである。TALのDNA認識ドメインは、様々なDNA配列と結合するように操作することができる。本明細書中の実施例1に記載のように、DNA結合ドメインの新規クラスであるTALエフェクターのDNA認識特異性が解読されている。具体的には、TALエフェクターのDNA結合ドメインは、特定のDNA配列を認識および結合することができる様々な数のタンデムの34個のアミノ酸の反復を含有する。反復のアミノ酸配列は、反復の位置12および13での2つの隣接する高度に可変性の残基以外は保存的である。これらの位置は一緒になって、DNA結合部位中の個々のヌクレオチドを1つの反復対1個のヌクレオチドで指定する。TALENの構造は図7に例示されている。TALENは二量体として機能し、それぞれの単量体は、FokIエンドヌクレアーゼからの非特異的な切断ドメインと融合した、操作されたTALのDNA認識反復から構成される。DNA認識反復は、対象のゲノム内の標的DNA配列と結合するように操作することができる。TALヌクレアーゼ単量体は、スペーサー配列によって分離されている2つのDNA半分部位のうちの1つと結合する。この間隔が、FokI単量体が二量体化して、半分部位の間のスペーサー配列中に二本鎖DNAの切断(DSB)を生じることを可能にする。
【0165】
TALエフェクターDNA認識ドメインの潜在性を調査するために、ネイティブTALエフェクターが、FokIヌクレアーゼドメインと融合した場合にヌクレアーゼとして機能できるかどうかを決定するための実験を実施した。TALヌクレアーゼ発現構築体および標的レポーター構築体を使用することによって、酵母に基づくアッセイを実施した。図5に例示されているように、ヌクレアーゼ発現構築体の主鎖は、FokIヌクレアーゼドメインおよび酵母TEF1プロモーターの制御下のN末端核移行シグナル(NLS)を含有する。様々なTALエフェクターのクローニングを促進するために、いくつかの制限部位がFokIヌクレアーゼドメインとNLSモチーフとの間に位置している。図6に示されているように、標的レポーター構築体は、コード配列の125bpの重複を有する破壊されたlacZレポーター遺伝子を有する。重複は、URA3遺伝子およびTALのDNA結合ドメインによって認識される標的配列(2つの半分部位およびスペーサー配列から構成される)に隣接する。TALENが標的部位と結合してDNA二本鎖切断(DSB)を生じる場合、そのような切断は、酵母では、重複するlacZ配列間の相同組換えによって、一本鎖アニーリングによって主に修復される(Haber(1995)Bioessays、17:609)。組換えは機能的lacZ遺伝子の再構成およびURA3の損失(5−フルオロオロト酸耐性を与える)をもたらす。lacZ酵素活性を決定することによってTALENの相対的切断活性を測定した。
【0166】
これらの研究では、配列番号16(図8)に記載のように中央ヌクレアーゼ反復領域を有していたネイティブTALエフェクター、AvrBs3を、ヌクレアーゼ発現ベクター内にクローニングし、18bpのスペーサー配列を有するAvrBs3標的部位(逆配向に配置された2つの結合部位)を標的レポーターベクター内にクローニングした。酵母アッセイは、図9に示されており上述されているスキームを使用して行った。結果は、AvrBs3ヌクレアーゼプラスミドおよび標的レポータープラスミドの両方で形質転換させた酵母細胞からのlacZ活性が、標的レポータープラスミドのみを含有していた対照酵母細胞よりも有意に高かった(15.8倍高い)ことを示した(図10)。反復ドメインを主にコードしているSphI断片のみで作製したヌクレアーゼ融合体では、活性は観察されなかった。これは、DNA結合ドメイン以外の配列がTALEN活性に必要であることを示した。また、6および9bpのスペーサー長を有するレポータープラスミドも活性を示すことに失敗し、これは、2つの結合部位間の間隔がFokIを二量体化させるために重要であることを示している。これらのデータは、AvrBs3 TALヌクレアーゼが、酵母中のその同族標的配列を切断する部位特異的なヌクレアーゼとして機能できることを示している。
【実施例3】
【0167】
誂えたTALENのための、TALエフェクター反復のモジュールアセンブリ
それぞれが異なるヌクレオチドを指定する4つの個々のTALエフェクター反復のそれぞれの、102個の塩基対に対応する相補的オリゴヌクレオチドを合成し、アニーリングし、個々にならびにすべての順列の2回および3回の反復の組合せで高コピーの細菌クローニングベクター内にクローニングして、標準の制限消化およびライゲーション技法を使用して4個の単一、16個の二重、および64個の三重反復モジュールを得た(たとえば図11に例示)。所望のTALエフェクターのコード配列は、適切なモジュールを、特徴的な最後の半反復以外の中央反復領域を欠くtal1c遺伝子の切断された形態を含有するGateway−ready高コピー細菌クローニングベクター内に順次導入することによってアセンブルする。たとえば、18回反復のTALエフェクターのコード配列は、5個の三重モジュールおよび1個の二重モジュールを切断されたtal1cベクター内に順次導入することによってアセンブルすることができる。
【実施例4】
【0168】
TALエフェクター反復のモジュールアセンブリのためのシステム
誂えのTALエフェクターをコードしている遺伝子を作製するためのプラスミドおよび方法を開発した。本明細書中に記載のように、TALエフェクターの機能的特異性は反復中のRVDによって決定され、反復およびタンパク質中の他の箇所中の他の多型は稀であり、機能的特異性に関して重要でない。したがって、自由裁量によるTALエフェクター遺伝子の反復領域を所望のRVDを含有する反復で置き換えることによって、誂えのTALエフェクター遺伝子を作製した。RVDの外の反復配列はコンセンサス配列に一致した(以下を参照)。TALエフェクター反復をコードしているDNA断片を、1回、2回、または3回の反復をコードしているモジュール内に順次アセンブルし、元の反復が除去されたTALエフェクター遺伝子内にモジュールをクローニングした。最後の(半)反復を例外として、それぞれのコードされている反復は、配列LTPDQVVAIASXXGGKQALETVQRLLPVLCQDHGを有していた(配列番号18、図12A)。最後の(半)反復は配列LTPDQVVAIASXXGGKQALESを有していた(配列番号20、図12B)。どちらの配列でも、「XX」はRVDの位置を示す。モジュール反復中で使用したRVDは、それぞれA、C、G、およびTとの結合を指定するNI、HD、NN、およびNGであった。以下に記載の実験では、その反復が除去されたXanthomonas oryzae pv.oryzicola株BLS256のtal1c遺伝子を、誂えのTALエフェクター遺伝子を構築するための「主鎖」として使用した。
【0169】
本明細書中に記載の方法には、(1)単一反復の開始プラスミドの作製、(2)単一反復モジュールのプラスミドの作製、(3)複数反復モジュールの作製、(4)1回、2回、および3回の反復モジュールのプラスミドの完全な組の作製、ならびに(5)誂えのTALエフェクターのコード配列のアセンブリの5個の構成要素が含まれていた。
【0170】
単一反復の開始プラスミドを作製するために、第1反復の最初の部分および最後の切断された反復の最後の部分以外の反復領域全体を除去するためにtal1c遺伝子をMscIで消化し、再度ライゲーションさせて、pCS487と命名されたプラスミドがもたらされた(図13)。生じる遺伝子はRVD NIをコードしており、ほとんどのTALエフェクター遺伝子と同様、反復領域に隣接していた2つのSphI部位を含有していた。この遺伝子はXhoI部位を含有していなかった。
【0171】
次に、翻訳的にサイレントな突然変異をpCS487内に導入して、コドン19および20を中心とするユニークなXhoI部位が包含される、ユニークなPspXI部位を作製した。突然変異は、どちらもアミノ酸配列ALES(配列番号22)をコードしているコドン18〜21の元のおよび変更されたヌクレオチド配列(それぞれ配列番号21および配列番号23)を示す、図14に示されている。生じるプラスミドはpCS489と命名された。
【0172】
さらなる突然変異誘発によって、RVD HD、NN、およびNGを用いて3つのさらなる構築体を作製して、それぞれpCS490、pCS491、およびpCS492と命名されたプラスミドを作製した。修飾された反復領域を包含するSphI断片を、pCS489、pCS490、pCS491、およびpCS492から、GatewayエントリーベクターpENTR−D(Invitrogen、カリフォルニア州Carlsbad)中で反復領域を含まずにtal1cのNおよびC末端部分のみをコードしたpCS488と命名されたカナマイシン耐性プラスミド(図15)に移した。この転移は、それぞれpCS493(図16)、pCS494、pCS495、およびpCS496と命名された単一反復の開始プラスミドをもたらした。切断された反復中のPspXI/XhoI部位はこれらのプラスミド中でユニークに保たれた。pCS488およびその誘導体のそれぞれ中のTALエフェクター遺伝子には、原核生物および真核生物中での効率的な翻訳のためにそれぞれシャイン−ダルガノおよびコザック配列が先行していた。
【0173】
その後、単一反復モジュールのプラスミドを構築した。4つの選択されたRVD(NI、HD、NN、およびNG)のそれぞれについて1個のプラスミドを作製した。それぞれのプラスミドは、PspXI部位内にライゲーションした際にXhoIを再構成したがPspXI部位を再構成しなかった5’適合性の付着末端、ならびにXhoIおよびPspXI部位をどちらも再構成した3’適合性の付着末端を有していた。プラスミドは、アニーリングしたオーバーハングを有する合成の相補的オリゴヌクレオチド(図17A)を、pBluescript SK−のPspXI/XhoI部位内にクローニングすることによって作製し、それぞれpCS502(図17B)、pCS503、pCS504、およびpCS505と命名されたプラスミドがもたらされた。それぞれのプラスミドは、ユニークな再構成されたPspXI部位での単一反復モジュールの3’末端の追加の反復の導入、または再構成されたXhoI部位を使用した反復モジュールの切り出しを可能とした。
【0174】
NI、HD、NN、およびNGについてそれぞれ1個の追加の単一反復モジュールを作製した。それぞれは、PspXI部位内にライゲーションした際にPspXIまたはXhoI部位を再構成しなかった5’適合性の付着末端、XhoIおよびPspXI部位をどちらも再構成した3’適合性の付着末端、ならびに内部MscI部位を破壊した翻訳的にサイレントなヌクレオチド置換を有していた(図18A)。これらのモジュールは、オーバーハングを有する合成の相補的なオリゴヌクレオチドをアニーリングすることによって作製した。これらの追加の単一反復モジュールのうちの任意のものを単一反復モジュールのプラスミド(pCS502、pCS503、pCS504、またはpCS505)のユニークなPspXI/XhoI部位内にライゲーションすることで、5’接合部で新しいXhoI部位はもたらされなかったが、ユニークな3’PspXI/XhoI部位の再建がもたられ、それにより、生じるプラスミドをPspXIで切断することによってさらなる追加の反復を導入するために直鎖状にできる。このプロセスの繰返しにより、複数反復を含有するモジュールがもたらされた(図18B)。さらに、XhoIを使用してそれぞれの複数反復モジュール全体を切り出すことができる。MscI部位は追加の単一反復モジュール中で破壊されていたため、第1反復中のMscI部位はユニークに保たれ、続く複数反復モジュールのサブクローニングの際に配向を確認するために有用であった。
【0175】
追加の単一反復モジュールを単一反復モジュールのプラスミド内に繰り返してクローニングして、単一反復モジュールのプラスミドと共に、すべての可能な1回、2回、および3回の反復モジュールの完全な組を作製し、pCS502〜pCS585と命名された合計84個のプラスミドとした(図19)。3回より多くの反復(たとえば、4、5、6、7、8、9、10、または10回より多くの反復)を含有するモジュールは同じ様式で作製される。
【0176】
その後、誂えのTALエフェクター遺伝子を作製するために任意の反復の配列をtal1c「主鎖」内にアセンブルするための方法が考案された。本方法には以下のステップが含まれ、これは図20にも示されている:
(1)第1の所望の反復を有する単一反復の開始プラスミド(それぞれRVD NI、HD、NN、またはNGをコードしているpCS493、pCS494、pCS495、またはpCS495)を選択するステップ、
(2)プラスミドをPspXIで直鎖状にするステップ、
(3)XhoIを使用して次の反復(複数可)のモジュールを適切なモジュールプラスミド(pCS502〜pCS585)から単離するステップ、
(4)ライゲーションするステップ、
(5)MscIで消化することによって配向を確認し、ベクターに基づくプライマーを使用して配列を3’末端から確認するステップ、および
(6)すべての反復がアセンブルされるまでステップ2〜5を繰り返すステップ。
【実施例5】
【0177】
TALENのモジュールアセンブリ用プラスミドのライブラリ
本明細書中に記載のTALEN反復のアセンブリ(たとえば図20に示したステップを使用)は、増加していく数の反復を含有する数々の中間体プラスミドをもたらす。これらのプラスミドのそれぞれは、TALENのモジュールアセンブリ用プラスミド(pMAT)のライブラリが作製されるように保管されている。たとえば、図21Aおよび21Bは、示されているヌクレオチド配列を標的とするTALエンドヌクレアーゼの構築における反復モジュールのアセンブリを示す。図21Aでは、pCS519、pCS524、pCS537、pCS551、pCS583、およびpCS529と命名されたプラスミドからの反復モジュールを、pCS493と命名された開始プラスミド中の配列に順次付加して、pMAT55、pMAT56、pMAT57、pMAT58、pMAT59、およびpMAT60と命名されたプラスミドがもたらされる。図21Bでは、pCS530、pCS533、pCS522、およびpCS541と命名されたプラスミドからの反復モジュールを、pMAT1と命名されたプラスミド中の配列に順次付加して、pMAT61、pMAT62、pMAT63、およびpMAT64と命名されたプラスミドがもたらされる。
【実施例6】
【0178】
誂えたTALENの作製および試験
実施例4および5に記載のシステムを使用して、TALのDNA認識ドメインを使用して特定のDNA標的を認識および切断するTALENを作製した(図22A)。TALENの機能を評価するために、LacZ活性がDNA切断の指標として役割を果たす酵母アッセイを適応させた(Townsendら、上記)。このアッセイでは、標的プラスミドおよびTALEN発現プラスミドを接合によって同じ細胞中で一緒にした。標的プラスミドは、コード配列の125bpの重複を有するlacZレポーター遺伝子有する。重複は、所定のTALENによって認識される標的部位に隣接する。二本鎖DNAの切断が標的部位で起こる際、これが重複した配列間の一本鎖のアニーリングによって修復され、それにより、機能的なlacZ遺伝子が作製され、その発現は定量可能な読取値を提供する標準のβ−ガラクトシダーゼアッセイによって測定することができる(図22A)。このアッセイは、NHEJによって染色体の突然変異を生じる、または高等真核生物において遺伝子編集のために相同組換えを刺激する、ZFNの能力の良好な予測子であることが実証されている(Townsendら、上記、およびZhangら(2010)Proc.Natl.Acad.Sci.USA、107:12028〜12033)。
【0179】
2つの十分に特徴づけられたTALエフェクター、すなわち、コショウの病原体Xanthomonas campestris pv.vesicatoriaからのAvrBs3およびコメの病原体X.oryzae pv.oryzaeからのPthXo1を使用した(Bonasら(1989)Mol.Gen.Genet.、218:127〜136、およびYangら(2006)Proc.Natl.Acad.Sci.USA、103:10503〜10508)。AvrBs3のアミノ酸配列はGENBANK受託番号P14727および配列番号12の下(図3)、核酸配列は受託番号X16130および配列番号13の下(図4)で見つけることができる。PthXo1のアミノ酸配列はGENBANK受託番号ACD58243および配列番号31の下(図23)、核酸配列は受託番号CP000967、遺伝子ID6305128、および配列番号32の下(図24)で見つけることができる。GENBANK受託番号ACD58243の下のPthXo1のアミノ酸配列は、開始コドンの誤った注釈付けが原因で、N末端で切断されている。完全な配列は図23中に提示されている。
【0180】
AvrBs3およびPthXo1の反復ドメインはどちらも保存的なSphI断片内に完全にコードされている(図4および24)。また、どちらのTALエフェクターコード遺伝子も、反復ドメインのコード配列ならびにその前の287個のアミノ酸およびその後の231個のアミノ酸を包含するBamHI制限断片を有する(図4および24、図22Aも参照)。BamHI断片からはTALエフェクター転写活性化ドメインが欠けている。SphI断片およびBamHI断片はどちらも、ヌクレアーゼ発現ベクターpFZ85中に存在するFokIをコードしているDNA断片と融合させた(図25)。FokIヌクレアーゼとAvrBs3およびPthXo1によってコードされているBamHI断片との間の融合タンパク質を図26および27、配列番号33および34に示す。
【0181】
FokI単量体は切断のために二量体化しなければならないが、2つのDNA認識部位間の適切なスペーサー長は不明確であった。ジンクフィンガーアレイが4〜7個のアミノ酸のリンカーによってFokIから分離されているZFNでは、2つの認識部位の間の典型的なスペーサーは5〜7bpである(Handelら(2009)Mol.Ther.、17:104〜111)。たとえば、本明細書中で使用したBamHI TALEN構築体中では235個のアミノ酸が反復ドメインをFokIから分離しているため、BamHIおよびSphI構築体の両方で様々なスペーサー長(6、9、12、15、および18bp)を使用した。陽性対照としては、マウス転写因子Zif268(PorteusおよびBaltimore(2003)Science、300:763)に由来するDNA結合ドメインを有する良好に特徴づけられているジンクフィンガーヌクレアーゼを使用した。陰性対照としては、TALエフェクタードメインを触媒的に不活性なFokI変異体と融合させたか、または非同族DNA標的に対して試験した。
【0182】
200μlの終夜培養物中の、TALEN発現または標的プラスミドのどちらかを含有する一倍体細胞種を、YPD培地中、30℃で接合させた。4時間後、YPD培地を5mlの選択培地で置き換え、終夜、30℃でインキュベーションした。接合した培養物を溶解し、ONPG基質を加え、96ウェルプレートリーダーを使用して吸光度を415nmで読み取った(Townsendら、上記)。β−ガラクトシダーゼレベルを基質切断速度の関数として計算した。15bpのスペーサーが2つの認識部位を分離している標的レポーター構築体で得られた結果が図22Bに示されている。主に反復アレイをコードしたSphI断片に由来するすべてのヌクレアーゼ発現構築体は、活性を示すことに失敗し、これは、機能には反復アレイ中のものに追加のアミノ酸配列が必要であることを示している(図22B)。しかし、BamHI断片に由来するAvrBs3およびPthXo1 TALENの両方で頑強な活性が観察された(図22B)。PthXo1 TALENの活性はZFN陽性対照のものに近かった。活性は機能的FokIドメインを必要とし、所定のTALENによって認識されるDNA標的に特異的であった。
【0183】
また、FokIが最も効率的に二量体化することを可能にするスペーサー長を同定するために、TALエフェクター結合部位間の様々な距離(12〜30bpの11個の長さの変異体)を試験するための実験も実施した(図28A)。どちらの酵素も、一方は15bp、他方は21bp(AvrBs3)または24bp(PthXo1)のどちらかである、2つのスペーサー長の最適条件を示した。PthXo1では、13bp以上のすべての試験したスペーサー長で活性が観察された。しかし、AvrBs3の一部のスペーサー長では活性を示されず、スペーサー長が特定のTALENに重要であることが示唆された。
【0184】
上記実験は、スペーサーの両側に反対方向で配置した2つの同一の認識配列と結合するホモ二量体TALENの活性を試験した。そのような回文部位はゲノム標的中で自然に起こる可能性が低いため、TALENがヘテロ二量体として機能できるかどうかを試験するための実験を実施した。AvrBs3およびPthXo1の認識部位を頭から尾部の配向で15bpのスペーサーの両側に配置した。そのそれぞれの標的に対するAvrBs3およびPthXo1 TALENの個々の活性ならびにZif268の活性を対照として測定した。陰性対照として、ヘテロ二量体部位の標的部位プラスミドのみを有する酵母培養物をLacZ活性についてアッセイした。生じるヘテロ二量体TALENの活性は、2つのホモ二量体酵素で観察された活性の平均に近かった(図28B)。
【0185】
自由裁量による染色体配列に対するTALENを標的とするように反復ドメインをアセンブルできるかどうかを試験するために、以前にZFNを用いた突然変異誘発の標的とされていた2つの遺伝子、すなわちシロイヌナズナからのADH1およびゼブラフィッシュからのgridlockを選択した(Foleyら(2009)PLoS One、4:e4348、およびZhangら、上記)。5’のTによって先行されており、MoscouおよびBogdanove(上記)によって同定されたTALエフェクター結合部位に類似のヌクレオチド組成を有する、コード領域中の12〜13bpの配列の検索を行った。ADH1およびgridlockでは、そのような部位は平均して7〜9bpごとに存在した。4個の12bpの部位がADH1中で選択され(染色体遺伝子配列の位置360、408、928、および975)、1個の13bpの部位がgridlock中で選択された(染色体遺伝子配列の位置2356、図29A)。ネイティブTALエフェクターからの最も豊富なRVD(AにはNI、CにはHD、GにはNN、およびTにはNG)を使用して、これらの標的を認識するようにTALエフェクター反復ドメインを構築した。誂えのTALENを構築するために、実施例4および5に記載のように、これらのRVDを有する反復を個々に合成し、1回、2回、または3回の反復のモジュール内にアセンブルした。これらのモジュールを、元の反復が除去されたtal1c遺伝子の誘導体内に順次ライゲーションさせ(MoscouおよびBogdanove、上記)、これらの操作したTALエフェクターからのBamHI断片を、pFZ85中のFokIの触媒ドメインをコードしている配列と融合させた(図25)。シロイヌナズナからのADH1およびゼブラフィッシュのgridlock遺伝子を標的とした5個の誂えのTALENを作製した。
【0186】
生じる誂えのTALENは、酵母アッセイにおいてホモ二量体TALENとして(すなわち、同一のDNA結合部位が16〜18bpのスペーサーの両側に逆の配向で重複していた)試験したが、天然に存在するDNA標的での切断を指示するためにはヘテロ二量体TALENを構築しなければならないことに留意されたい。スペーサー長は、次の隣接する(かつ向かい合う)候補部位の3’末端から、15bpに最も近い距離に基づいて選択した。16bpのスペーサーをADH1−360−12、ADH1−408−12rに使用し、18bpのスペーサーをADH1−928−12、ADH1−975−12r、およびgridlock−2356−13rに使用した。酵母アッセイは上述のように行った。
【0187】
頑強なヌクレアーゼ活性がADH1−360−12およびgridlock−2356−13r TALENで観察された(図29B)。ADH1−928−12 TALENは控えめな活性を有していたが、それでも陰性対照より有意に高かった。陽性結果を与えたそれぞれのTALENについて、ヌクレアーゼ活性は同族標的に特異的であった。これらの結果は、新規の機能的なTALENを誂えた反復ドメインのアセンブリによって作製できることを示している。
【実施例7】
【0188】
天然に存在する標的およびTALエフェクターの対はヌクレオチドおよびRVDの組成の全体的および位置的な偏りを示す
MoscouおよびBogdanove(上記)によって分析した20個の対合した標的およびTALエフェクターを、全体的な組成の偏りおよびヌクレオチドまたはRVD頻度に対する位置効果について評価した。部位(プラス鎖上)は一般にAおよびCに富み、Gに乏しいことが観察された。平均パーセントAは31±16%であった(1単位の標準偏差)。平均パーセントCは37±13%であった。平均パーセントGは9±8%であり、平均パーセントTは22±10%であった。アラインメントは長さが変動するため、位置効果の分析はそれぞれの末端上の5個の位置に限定された。驚くべきことに、位置1および3でAが有利およびTが不利であり、位置Nおよび場合によっては位置2でTが有利である、標的配列中の偏りが明らかであった。Gは位置N−1で特に稀であった。この偏りはエフェクター中の一致するRVDによって反映されており、位置1および3にはNIが最も一般的であり、位置1にはNGが存在せず、位置Nにはほぼ必ずNGが存在し、位置N−1にはNNは稀である(図30)。
【実施例8】
【0189】
誂えのTALエフェクター反復アレイを迅速にアセンブリおよびクローニングするための方法および試薬
Golden Gateクローニング方法[Englerら(2008)、上記、およびEnglerら(2009)、上記]は、その認識部位の外側で切断して、複数のDNA断片を同時に規則的にライゲーションさせるための誂えのオーバーハングを作製する、IIS型制限エンドヌクレアーゼ(たとえばBsaI)の能力を用いる。この方法を使用して、単一の反応で、いくつかのDNA断片を特定の順序でアレイへと融合させ、所望のデスティネーションベクター(destination vector)内にクローニングすることができる(図31)。
【0190】
誂えのTALエフェクター反復をコードしているアレイをアセンブルするための方法および試薬をGolden Gateシステムに基づいて開発した。BsaI部位がTALエフェクター反復のコード配列の両側に位置している場合は、切断により4bpのオーバーハングによって隣接される反復断片が放出される。切断部位は配列特異的でないため、互い違いにずらす(staggering)ことによって、規則的な相補的オーバーハング(粘着末端)を有する反復クローンを放出させることができ、複数反復アレイの規則的なアセンブリが可能になる。
【0191】
58個のプラスミドのライブラリ(図32Aおよび32B)を作製して、10個までの反復単位の「サブアレイ」への同時アセンブリ、次いでこれらのサブアレイのうちの1、2、または3個および最終的な切断された反復の、完全な誂えのアレイへの同時アセンブリを可能にした。組中のそれぞれの断片が4つの最も一般的なRVDであるHD、NG、NI、およびNNのうちの異なる1つを有する反復モジュールをコードしている、4個の断片の10個のずらし型(staggered)の組を合成し、テトラサイクリン耐性遺伝子を保有するベクター内にクローニングして、合計40個のプラスミドとした。それぞれの断片が4つの最も一般的なRVDのうちの異なる1つをコードしている、20個のアミノ酸の末端切断されたTALエフェクター反復をコードしたさらに4個の断片を合成し、スペクチノマイシン耐性遺伝子を保有する異なるベクター内にクローニングしてさらに4個のプラスミドを得て、「最終反復プラスミド」と命名した、図32A)。ずらし型の組中のすべての断片は、BsaIでの切断にって適切な順序でのアセンブリを可能にする様々な粘着末端を有する断片が放出されるように、ベクター中でBsaI部位によって隣接されている。すなわち、反復モジュール1の断片の3’末端でのオーバーハングは反復モジュール2の断片の5’末端でのオーバーハングにのみ相補的であり、反復モジュール2の3’末端でのオーバーハングは反復モジュール3の5’末端でのオーバーハングにのみ相補的であり、以下同様である。最終反復プラスミド中の断片は、異なるIIS型制限エンドヌクレアーゼ、Esp3Iの部位によって隣接されている。以下に記載の14個の追加のプラスミドを、アセンブルされたサブアレイを受けるためのデスティネーションベクターとして構築した。
【0192】
第1のデスティネーションベクター、プラスミドpFUS Aは、21回以下の反復(最後の切断された反復を数える)の最終アレイへとアセンブルする、第1の10回反復サブアレイを受けるために構築した。pFUS Aは、BsaIによる切断によって、片側で第1反復モジュールの5’末端でのオーバーハングに相補的なオーバーハングが作製され、反対側で第10反復モジュールの3’末端でのオーバーハングに相補的なオーバーハングが作製されるように構築した。そのような最終アレイへとアセンブルする10回以下の反復の第2のサブアレイを受けるために、デスティネーションベクタープラスミドpFUS_B1、pFUS_B2、pFUS_B3、pFUS_B4、pFUS_B5、pFUS_B6、pFUS_B7、pFUS_B8、pFUS_B9、およびpFUS_B10を構築し、これらは、BsaIによって切断された際に、それぞれ第1反復モジュールの5’末端および対応する符番した位置の反復モジュールの3’末端でのオーバーハングに相補的なオーバーハングを有する(たとえば、サブアレイの3’末端のpFUS_B6のオーバーハングは、位置6の4回反復モジュール断片のオーバーハングに一致する)。pFUS_AおよびpFUS_Bシリーズのプラスミド中にクローニングしたアレイは、ベクター中でEsp3I部位によって隣接されており、Esp3Iを用いた消化によって放出された際、アレイは、反復領域を欠いたTALENをコードしているデスティネーションベクターpTAL内に最終の切断された反復断片と共にそれらが順番にライゲーションされることを可能にする、ユニークな相補的なオーバーハングを有する。Esp3Iを用いた切断が、第1の10回反復サブアレイの5’末端でのオーバーハングに相補的な一端でのオーバーハングおよび最終の切断された反復断片の3’末端でのオーバーハングに相補的な他端でのオーバーハングのおかげで正しい位置および正しい配向での反復アレイの挿入を可能にするように、pTALを構築した(図33)。
【0193】
22〜31回反復の最終アレイへとアセンブルする第1および第2の10回反復サブアレイを受けるために、最終の2つのデスティネーションベクタープラスミド、pFUS_A30AおよびpFUS_A30Bを構築した。アレイをpFUS_Bベクターからの第3のアレイおよびEsp3Iを用いた消化によって同様に放出された最終反復プラスミドからの最終の切断された反復断片と共に順番にpTAL内にライゲーションさせることができるよう、Esp3Iを用いた消化により適切な相補的オーバーハングを有するアレイが放出されるように、pFUS_A30AおよびpFUS_A30Bを構築した(図32)。
【0194】
すべてのデスティネーションベクターはIIS型制限エンドヌクレアーゼ部位間にクローニングされたLacZ遺伝子を有しており、組換えのブルー−ホワイトスクリーニングを可能にする。アンピシリン耐性の遺伝子を保有するpTAL以外は、すべてのデスティネーションベクターがスペクチノマイシン耐性の遺伝子を保有する。
【0195】
これらの試薬を使用して誂えのTALエフェクター反復アレイを迅速に構築するために、以下の方法を確立した。第1のステップでは、10回以下の反復の必要なサブアレイの適切な個々のRVDモジュールプラスミドを、適切なデスティネーションベクターと、1本のチューブ中で混合する。T4 DNAリガーゼおよびBsaIエンドヌクレアーゼを加え、反応を、PCR機械中で、2つの酵素のそれぞれの最適温度である37℃で5分間および16℃で10分間、10サイクルの間インキュベーションする。その後、in vivo組換えによるより短い不完全なアレイのクローニングを防止するために、反応混合物をPLASMID−SAFE(商標)ヌクレアーゼで処理してすべての直鎖状dsDNA断片を加水分解し、その後、混合物を使用して化学的にコンピテントな大腸菌細胞を形質転換させる。生じる組換えプラスミドを単離し、正しい構築体を確認する。その後、第2のステップでは、第1のステップからの確認されたプラスミドを適切な最終反復プラスミドおよびpTALと混合し、消化およびライゲーション反応サイクルを第1のステップと同様に実施する。最後に、反応生成物を大腸菌内に導入し、完全長の最終アレイ構築体を単離および確認する。このプロトコルは、1人の人によって1週間以内に完了することができる。
【0196】
表4A中のTALEN85、102および117、ならびに以下の実施例14に記載のTALEN HPRT−3254−17およびHPRT−3286−20rの発現構築体を、本実施例中に記載の方法および試薬を使用して作製した。
【0197】
pTAL中にクローニングした反復アレイは、反復領域に隣接する保存的なSphI制限エンドヌクレアーゼ部位を使用して、他のTALエフェクター遺伝子のコンテキスト内に容易にサブクローニングされる。
【実施例9】
【0198】
誂えのTALENのデータは「規則」およびRVDの数と活性との間の相関について初期の支持を示す
実施例6は、ユニークなDNA配列を認識できるようにTALENのDNA結合ドメインを操作するために実施した実験を記載している。記載のように、これらの誂えのTALENは、シロイヌナズナADH1およびゼブラフィッシュgridlock遺伝子中の部位を認識した。これらの遺伝子だけでなく、シロイヌナズナからのTT4遺伝子、およびゼブラフィッシュからのテロメラーゼ中の部位を認識するように、追加の誂えのTALエフェクターDNA結合ドメインを操作した(Foleyら、上記、およびZhangら、上記)。これらの誂えのTALENは、実施例3、4および8に記載の方法を使用して作製した。誂えのTALENの操作において、観察された組成的および位置的な偏りは、設計原理または「規則」として適応した。最初に、5’のTによって先行されており、少なくとも15bpの長さであり、上述の平均と一貫性のあるヌクレオチド組成を有するコード領域中の配列の検索を実施した。具体的には、0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTを有する部位のみを選択した。そのような部位は平均して7〜9bpごとに存在した。その後、上述の観察された位置的な偏りに順応した部位を選択した。操作したTALENの結合によりFokIの二量体化が可能となるように、この組から、15〜19bpの長さであり、15〜18bpによって分離されている、それぞれの遺伝子中の結合部位の2つの対を同定した。モジュールアセンブリ方法(実施例3および4)は、部分的な長さの構築体を生じた。
【0199】
それぞれが9回以上の反復のアレイを有する、16個のヌクレオチドの配列を標的とするように設計された合計21個の中間体および完全長のTALEN。これらのTALENのアミノ酸配列は図34A〜34U(配列番号35〜55)に提供されている。これら21個のTALENを、実施例2および6に記載の酵母アッセイを使用して、DNAを切断するその能力について試験した。活性データは図35に示されており、表4Aに要約されている。
【0200】
中間の部分的な長さのTALENの一部は、ヌクレオチド組成および末端Tの規則を破っている標的に対応する。表4Aは、長さ、これら2つの規則への順応、およびZFN268と比較した活性を、それぞれのTALENについて示す。結果は、RVDアレイの長さを増加させることで生じるTALENの活性が増加するという大きな傾向を明らかにしている。これは、DNA標的がin vivoで認識可能となる前に必要な最小数のRVDが存在することを示唆している。さらに、規則の順応が重要であると考えられる。検出可能な活性を示さない6個のTALENのうち、2つが標的の組成の規則に違反しており、2つがNGで終わっておらず、別の1つが両方の規則を破っていた(1つが両方の規則に従っていた)。ZFN268の25%未満の活性を有する8個のTALENのうちの3個が規則の一方に違反しており、ZFN268の25〜50%の活性を有する4個のTALENのうちの1個がNGで終わるRVD配列を有していなかった。ZFN268の50%以上の活性を有するTALENはすべての規則に従っており、同じ長さのTALENでは、規則を破ったものは従順なアレイよりも一般に低い活性を有していたことに留意されたい。長さに関する全体的な傾向に一致して、規則を破らなかった中間体でも、対応する完全長のTALENの方がより高い活性を有していた(表4Aおよび図35)。同じ標的に対するTALEN長の相違が原因であるスペーサー長の変動が、この観察に寄与した可能性があるが、スペーサー長のある程度の範囲は耐容される(Christianら、上記)。
【0201】
データ中のある程度の複雑さが明らかであった。たとえば、活性は同じ長さの従順なTALEN間で変動しており、一部の短いアレイは中等度に高い活性を有しており、一部の長い従順なアレイは活性が少ないまたはなかった(表4B)。それにもかかわらず、提供された結果は、1)一般により多数の反復がより高い活性をもたらす、および2)組成および位置的な偏りの規則の順応が活性に重要であるという結論を支持している。したがって、以下の設計原理を導いた。
【0202】
・TALエフェクター結合部位は、最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあるように設計される。
・部位は第1(5’)位置にTまたは第2位置にAを有し得ない。
・部位はT(3’)で終わらなければならず、最後から2番目の位置にGを有し得ない。
・部位の基本組成は指定した範囲内(平均±2単位の標準偏差)でなければならない:Aは0〜63%、Cは11〜63%、Gは0〜25%、およびTは2〜42%。
【表5】
【表6】
【実施例10】
【0203】
ヘテロ二量体TALEN対は酵母アッセイにおいてその意図する天然に存在する標的配列を切断する
実施例2、6および9のデータは、新規標的DNA配列を認識するように誂えのTALENを操作できることを実証している。ホモ二量体標的部位を認識した個々のTALEN単量体を使用して、誂えのTALENの酵母活性データを集めた。すなわち、TALENの標的配列を15〜18bpのスペーサー両側に逆の配向で重複させた。しかし、内在染色体配列の切断は、一般に、2つの異なる誂えのTALENがスペーサーの両側の2つの異なる配列を認識することを必要とする。実施例6に記載のように、この能力は、一緒にしたAvrBs3およびPthXo1 TALENについて、酵母アッセイにおいて、対応するキメラ標的部位を使用して実証した。本発明者らは、2つの異なる誂えのTALENが天然に存在するDNA配列を認識および切断することができるかどうかを試験した。実施例2に記載の酵母アッセイを使用して、シロイヌナズナADH1遺伝子中の2つの異なる標的配列を切断するように設計された誂えのTALENを、これらの標的に対する活性についてアッセイした。標的部位および対応するTALENのDNA配列は図36Aに示されている。TALENのアミノ酸配列は図34に提供されている。酵母アッセイで得られたベータ−ガラクトシダーゼ活性は、図36Bに示されているグラフ中にプロットされている。TALENのその天然に存在する標的配列に対する活性は陰性対照よりも有意に高く、これは、内在標的DNA配列を認識および切断するためにTALENを設計できることを示している。
【実施例11】
【0204】
TALENはシロイヌナズナ中のネイティブ遺伝子を切断し、不正確な非相同末端結合によって突然変異を導入する
シロイヌナズナADH1遺伝子中の標的配列を認識するように設計された活性TALEN対のうちの1つを、染色体DNAを結合、切断および突然変異させることができるかどうかを決定するために試験した。この対(pTALEN69および74)を含む個々のADH1 TALENのそれぞれを、TALENを構成的な35Sプロモーターの制御下に置く植物発現ベクターpFZ14内にクローニングした(Zhangら、上記)。その後、生じる構築体を、シロイヌナズナのプロトプラスト内に電気穿孔によって導入した。48時間後、ゲノムDNAを単離し、Tth111lで消化した。Tth111l切断部位は2つのTALEN認識部位の間のスペーサー配列中に位置する(図37A)。TALENによる染色体DNAの切断は、不正確な非相同末端結合(NHEJ)によって突然変異を導入することが予想され、これはTth111lによる切断の失敗をもたらす。その後、TALEN認識部位を包含する375bpの断片をPCR増幅した。PCR産物を再度Tth111lで消化して、TALENに媒介されるNHEJによって改変されなかった残りのゲノムDNAのほとんどを除去する。その後、消化産物をアガロースゲル上に流す。未切断のPCR産物が観察され、そのような未切断のPCR産物は、内在標的配列でのヌクレアーゼ活性(この事例ではTALEN活性)の指標である(Zhangら、上記)。未切断のDNAをクローニングし、DNA配列決定によって分析した。9個の独立したクローンの配列決定により、6個がNHEJによって導入された突然変異を保有していたことが明らかとなった(図37B)。したがって、TALENは、内在染色体座位を切断し、DNA二本鎖切断および突然変異を導入する。
【実施例12】
【0205】
標的化能力の増強
TALエフェクターDNA暗号の中核では、4つの最も一般的なRVDは、会合頻度に基づいて4個のヌクレオチドに対して見かけ上1対1の特異性を有する。これは、HD、NG、およびNIで顕著にそうであるが、NNではその度合がより低い(図1C)。NNは最も頻繁にはGと会合するが、ほぼ同じように一般的にAと、時折CまたはTと会合する。13個のRVD配列中の4つの位置にNNを有するランダムにアセンブルされたTALエフェクターでは、人工標的中のすべての対応する位置にGを持つことで最良の活性が得られた(Bochら(2009)Science、326:1509〜1512)。Aは活性を減少させたが消失させず、CおよびTは検出可能な活性を排除した。NNである24個のRVDのエフェクターPthXo1の結合部位中の第1位置のみでGをC、T、またはAで置換した場合に、活性の劇的な損失が観察された(Romerら(2010)New Phytol.、187:1048〜1057)。しかし、これは、はるかにより短いAvrHah1(14個のRVD)がAとアラインメントするNNで始まり、23個のRVDのエフェクターPthXo6が位置4〜6にそれぞれAとアラインメントするNNを連続して3個有するが、これらのタンパク質はどちらの活性が高いという観察と対照的であった(Schornackら(2008)New Phytol.、179:546〜556、およびRomerら、上記を参照)。したがって、Gに対するNNの特異性は一般に弱いと考えられ、コンテキストに伴って変動する場合がある。
【0206】
チミンがTALエフェクター標的部位の直前にあるという観察された不変性は、いくつかのエフェクターの要件である[Bochら、上記、Romerら、上記、およびRomerら(2009)Plant Physiol.、150:1697〜1712]。高度に保存的な、TALエフェクター中の反復領域の直前のアミノ酸配列は(図38A)、アミノ酸配列および予測された二次構造のどちらにおいても、反復と顕著な類似度を共有する(図38BおよびBodganoveら(2010)Curr.Opin.Plant Biol.、13:394〜401)。「第0」反復と呼ばれるこの配列が、結合部位の−1位にTが必要であること根拠であり、RVDに類似の位置中の残基(図38B)がヌクレオチドを指定するという仮説が立てられた。
【0207】
これらの発見に基づいて、Gに対して高い特異性を有する反復を取り込ませることによって、および−1でのTの必要性を緩めることによって、操作したTALエフェクタータンパク質の標的化能力を増強できると仮説が立てられた。NNが表示するよりも頑強なGに対する特異性について新規かつ稀なRVDを試験するため、および第0反復のRVDに類似の残基を一般的なRVDで置き換えるための実験を開始した。
【0208】
Gに対する頑強な特異性のための新規かつ稀なRVD:上記開示したモジュール(たとえば実施例4を参照)では、4つのヌクレオチド塩基(それぞれA、C、G、およびT)との結合を指定するために4つの特定のRVD(NI、HD、NN、およびNG)を使用した。他のRVDを含有する反復も有用で有り得、これらはNI、HD、NN、およびNGと比較して4つの塩基に対して増加した特異性および/または親和性を有し得る。Gに対する特異性を改善させることに向けては、新規かつ稀なRVDをコードしているいくつかの反復を構築した。稀なRVD NK、HN、およびNAはGと会合し、これは、Nが残基のうちの一方または他方として重要であり得ることを示唆している(図1C)。したがって、表5に示されているRVDを有する反復をコードしている幅広い誘導体の組を構築した。左の列は、位置12に極性アミノ酸(R、K、D、E、Q、H、S、T、またはY)および位置13にNを有するRVDを記載している。右の列は、RVDの第1位置のNと第2位置の17個の他のアミノ酸のうちの任意のもの(G、L、V、R、K、D、E、Q、H、T、M、C、P、Y、W、またはF)との組合せを記載している。Nを含まないより高い特異性の可能性を考慮するために、位置12に極性アミノ酸(R、K、D、E、Q、H、S、T、またはY)および位置13にギャップ(*)を用いた反復も作製した(中央の列)。
【0209】
新規人工RVDは、Nicotiana benthamianaにおける、GUSもしくは二重ルシフェラーゼレポーターに基づくAgrobacteriumに媒介される一過性発現アッセイなどの、TALエフェクターの転写活性化活性のための定量的レポーター遺伝子に基づくアッセイ、または上述の出芽酵母におけるlacZレポーターに基づくTALENアッセイにおいて、機能について試験する(たとえば実施例2を参照)。試験するRVDを含有する反復モジュールを、測定可能かつ亜飽和レベルの活性を有するTALエフェクターまたはTALEN内に取り込ませ、生じるタンパク質を、対応する位置での4個すべてのヌクレオチドの統合順列を用いて、DNA標的の組に対する活性の相違について試験する。特に、植物体内および酵母アッセイにおいて最小限活性であり、3個の追加の反復でミスマッチに応答性であるPthXo1変異体(複数可)から始めて、新規かつ稀な反復のそれぞれを含有するTALENを(同価の3つ組で)、対応する位置のそれぞれにGを有する標的に対してin vivoで試験する。増加した活性を示す任意のものについて、これらの位置で他のヌクレオチドに入れ替えた標的を用いてアッセイを繰り返して、特異性を確認する。
【表7】
【0210】
位置−1でのTの特異性を緩めるための、第0反復のRVDに類似の位置の一般的なRVD置換:二次構造の予測および第0反復と反復コンセンサス配列とのアラインメントは、第0反復中のKR*(星印はギャップを示す)によって占有される位置がRVDに類似であり、−1でTを指定する残基であったことを示唆した。KRをHD、NG、NI、およびNNで置換し、別にR*を置換した、PthXo1の変異体を、上述のTal1c「主鎖」構築体中で構築した。植物体内および酵母アッセイにおいて、−1位に対応するヌクレオチド、すなわちそれぞれC、T、A、およびGを有する標的を使用して、これらの変異体の活性を野生型エフェクターと比較する。第0反復の位置11のKがコンセンサス反復配列の位置11の残基のSで置換されている、PthXo1のさらなる変異体を構築する。また、この置換と、コンセンサス反復配列の位置16の残基であるKでの第0反復の位置15のVの置換との組合せを有する他の変異体を構築する(表6)。TALエフェクター活性のための近位のTATAボックスを含め得る。さらに、−1のTがTATAボックスの一部であるように見えるAvrBs3とは異なり、PthXo1結合部位に最も近いTATAボックスは46bp下流であり、−1での修飾によって乱されないため、PthXo1はこの実験に有用である。
【0211】
上記修飾が、Gに対する標的化の増強またはT以外のヌクレオチドによって先行される配列を標的化する能力の増加をもたらさない場合は、より包括的な人工RVDの組をG特異性について試験し、また、一般的なRVD以外の置換を第0反復について試験する。
【表8】
【実施例13】
【0212】
新規の予測されたヌクレオチド特異的RVD
表1Aおよび1Bに記載されたRVDをRVD中の第2アミノ酸残基(すなわち、全体的な反復中の13番目)によって群分けした場合に、RVDの第1位置のアミノ酸とは無関係に、そのアミノ酸とRVDによって指定されるヌクレオチド(複数可)とのほぼ完璧な相関が存在したことが観察された(表7)。したがって、ギャップ(星印によって示す)で終わるRVDはCもしくはT、またはTを指定し、Dで終わるRVDはCを指定し、Gで終わるRVDはTを指定し、Nで終わるRVDはGもしくはA、またはGを指定する。また、RVDの位置1のアミノ酸はH、I、N、S、またはYのいずれかであったことも観察された。これらの観察は、RVDの特異性は、第1位置の残基がH、I、N、S、またはYであるかに依存せずに、第2位置の残基によって決定されることを示唆した。したがって、第2位置で観察された残基を第1位置の残基H、I、S、N、またはYと組み合わせるいくつかの新規(すなわち未だ観察されていない)RVDについて、特異性を予測した。したがって、I*、S*、およびY*はCもしくはT、またはTを指定すると予測され、ID、SD、およびYDはCを指定すると予測され、SGはTを指定すると予測され、INおよびYNはGもしくはA、またはGを指定すると予測された。また、第2位置のKは1つの事例しかなかったが、観察されたNKの特異性に基づいて、HK、IK、SK、およびYKはGを指定すると予測された。
【0213】
これらの新規RVDを、実施例2および11に記載の定量的TALエフェクターおよびTALEN活性アッセイにおいて機能および特異性について試験し、既存のRVDと比較した。
【表9】
【実施例14】
【0214】
誂えのTALENは動物細胞において内在標的を切断し、不正確な非相同末端結合によって突然変異を導入する
TALENを動物細胞における標的化突然変異誘発に使用できるかどうかを試験するために、最初に、TALエフェクターAvrBs3、PthXo1、およびTal1cの発現をヒト胚性腎臓(HEK)293T細胞中で試験した。AvrBs3、PthXo1、およびTal1cをコードしている遺伝子からストップコドンを除去し、遺伝子を、哺乳動物発現ベクターpcDNA3.2/V5−DEST(Invitrogen、カリフォルニア州Carlsbad)内に、タンパク質の免疫検出のためのV5エピトープをコードしているそのベクター中の下流配列とインフレームでサブクローニングした。pcDNA3.2/V5−DESTは、TALエフェクター遺伝子を構成的ヒトサイトメガロウイルス(CMV)プロモーターの制御下に置く。生じるプラスミドを個々に用いて、Lipofectamine 2000(Invitrogen)を使用してHEK293T細胞を形質移入し、24時間後、全タンパク質をそれぞれの形質移入した細胞のバッチから単離し、ポリアクリルアミドゲル電気泳動、ウエスタンブロッティングおよびマウス抗V5抗体を使用した免疫標識に供した。標識されたタンパク質は、ヤギ抗マウス抗体−西洋ワサビペルオキシダーゼのコンジュゲートを用いて、SuperSignal Weat Pico化学発光キット(ThermoScientific,Inc.)を使用して検出した。等価なローディングは、アクチンの免疫標識および検出によって確認した。それぞれのTALエフェクタータンパク質は、明らかな分解なしに、検出可能に発現された(図39)。
【0215】
次に、内在性ヒトHPRT遺伝子中の配列を標的とするために、実施例9に記載のように一対のTALENを設計し、HPRT−3254−17およびHPRT−3286−20rと命名した(図40Aおよび図40B)。Golden Gateクローニングに基づく方法および実施例8に記載の試薬を使用して、HPRT−3254−17をコードしているプラスミドpTALEN141およびHPRT−3286−20rをコードしているプラスミドpTALEN142を構築した。その後、TALEN遺伝子を、それらを構成的CMVプロモーターの制御下に置く哺乳動物発現ベクターpCDNA3.1(−)(Invitrogen,Inc.)内にサブクローニングし、プラスミドpTALEN141MおよびpTALEN142Mが得られた。その後、pTALEN141MおよびpTALEN142Mの両方を一緒に用いて、ならびに別に陰性対照としてpCDNA3.1(−)を用いて、HEK293T細胞を形質移入した。72時間後、ゲノムDNAを単離し、制限エンドヌクレアーゼBpu10Iで消化した。Bpu10I部位は、HPRT中でHPRT−3254−17およびHPRT−3286−20rの結合部位を分離するスペーサー内に存在する(図41A)。Bpu10I消化に次いで、PCRを使用して、TALENで処理した試料および対照試料の両方からのTALEN標的部位にまたがる244bpの断片を増幅した。予想された断片がどちらの試料からも増幅され、ゲノムDNAのBpu10I消化が不完全であったことを示す。しかし、続くBpu10Iを用いたPCR産物の消化は、対照試料から増幅された産物の完全な切断をもたらしたが、TALENで処理した試料からの産物の不完全な切断をもたらした(図41B)。TALENで処理した試料中の切断耐性PCR産物の存在は、HPRT中の意図する標的での、TALENに媒介される二本鎖切断の非相同末端結合による不完全な修復の結果、内在性Bpu10I部位がin vivoで突然変異したことの証拠を提供する。したがって、TALENは、哺乳動物細胞における標的化突然変異誘発に使用することができる。
【0216】
他の実施形態
本発明をその詳細な説明と併せて記載したが、前述の説明は、添付の特許請求の範囲によって定義される本発明の範囲を例示し、限定しないことを意図することを理解されたい。他の態様、利点、および改変が以下の特許請求の範囲内にある。
【特許請求の範囲】
【請求項1】
細胞の遺伝物質を修飾する方法であって、
(a)標的DNA配列を含有する細胞を提供するステップと、
(b)転写活性化因子様(TAL)エフェクター−DNA修飾酵素を前記細胞へ導入するステップであって、
前記TALエフェクター−DNA修飾酵素は、
(i)二本鎖DNAを修飾することができるDNA修飾酵素ドメイン、および
(ii)組み合わせて標的DNA配列中の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメイン
を含むものであり、
前記TALエフェクター−DNA修飾酵素が、前記細胞またはその子孫中の前記特定のヌクレオチド配列内にあるまたはそれに隣接する前記標的DNAを修飾するように、TALエフェクター−DNA修飾酵素を前記細胞へ導入する、ステップと
を含む、方法。
【請求項2】
請求項1に記載の方法であって、
前記標的DNA配列と前記核酸との間で相同組換えが起こるように、前記標的DNA配列の少なくとも一部分に相同的な配列を含む核酸を、前記細胞に提供するステップをさらに含む、方法。
【請求項3】
前記細胞が真核細胞である、請求項1に記載の方法。
【請求項4】
前記細胞が哺乳動物細胞である、請求項1に記載の方法。
【請求項5】
前記細胞が植物細胞である、請求項1に記載の方法。
【請求項6】
前記細胞が原核細胞である、請求項1に記載の方法。
【請求項7】
前記標的DNAが染色体DNAである、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素をコードしているベクターを用いて細胞を形質移入することを含む、方法。
【請求項9】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を前記細胞内にタンパク質として機械的に注入することを含む、方法。
【請求項10】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を、細菌III型分泌系によってタンパク質として前記細胞内に送達することを含む、方法。
【請求項11】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を、電気穿孔によってタンパク質として前記細胞内に導入することを含む、方法。
【請求項12】
前記DNA修飾酵素ドメインがエンドヌクレアーゼである、請求項1に記載の方法。
【請求項13】
前記エンドヌクレアーゼがII型制限エンドヌクレアーゼである、請求項12に記載の方法。
【請求項14】
前記II型制限エンドヌクレアーゼがFokIである、請求項13に記載の方法。
【請求項15】
請求項1に記載の方法であって、
前記標的DNA内の特定のヌクレオチド配列と結合する前記TALエフェクタードメインが、10個以上のDNA結合反復、より好ましくは15個以上のDNA結合反復を含む、方法。
【請求項16】
請求項15に記載の方法であって、
それぞれのDNA結合反復が、前記標的DNA配列中の塩基対の認識を決定する反復可変性二残基(RVD)を含み、
それぞれのDNA結合反復が、前記標的DNA配列中の1つの塩基対の認識を司っており、
前記RVDが、
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのHG、
Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含む、方法。
【請求項17】
請求項16に記載の方法であって、
それぞれのDNA結合反復が、前記標的DNA配列中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA配列中の1つの塩基対の認識を司っており、
前記RVDが、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのHG、
Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、および
Tを認識するためのIG
のうちの1つまたは複数を含む、方法。
【請求項18】
選択されたヌクレオチド配列に特異的なTALエフェクターをコードしている核酸を作製する方法であって、
(1)開始プラスミドをPspXIで直鎖状にするステップであって、
前記開始プラスミドが、選択されたヌクレオチド配列の第1ヌクレオチドに特異的な反復可変性二残基(RVD)を有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含み、
第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有する、ステップと、
(2)前記開始プラスミドのPspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチドに特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしているDNAモジュールをライゲーションするステップであって、
前記DNAモジュールがXhoI粘着末端を有する、ステップと、
(3)前記核酸が、前記選択されたヌクレオチド配列と結合することができるTALエフェクターをコードするまで、ステップ(1)および(2)を繰り返すステップと
を含む、方法。
【請求項19】
、請求項18に記載の方法であって、
ライゲーション後に、前記PspXI部位中の前記DNAモジュールの配向を決定するステップをさらに含む、方法。
【請求項20】
ステップ(1)および(2)を1〜30回繰り返すことを含む、請求項18に記載の方法。
【請求項21】
転写活性化因子様エフェクターエンドヌクレアーゼ(TALEN)をコードしている核酸を作製する方法であって、
(a)細胞のゲノム中の第1のユニークなヌクレオチド配列を同定するステップと、
(b)(i)組み合わせて前記第1のユニークなヌクレオチド配列と結合する複数のDNA結合反復、および(ii)第1のヌクレオチド配列内にあるまたはそれに隣接する第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含む、TALENをコードしている核酸を合成するステップであって、
それぞれのDNA結合反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA中の1つの塩基対の認識を司っており、
前記TALENが、以下のRVD:
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含むステップと
を含む、方法。
【請求項22】
請求項21に記載の方法であって、
前記TALENが、以下のRVD:
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、および
Tを認識するためのIG
のうちの1つまたは複数を含む、方法。
【請求項23】
請求項21に記載の方法であって、
前記第1のヌクレオチド配列が以下の基準:
i)最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあること、
ii)第1(5’)位置にTを有さない、または、第2位置にAを有さないこと、
iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さないこと、ならびに
iv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの塩基組成を有すること
のうちの少なくとも1つを満たす、方法。
【請求項24】
請求項23に記載の方法であって、
前記細胞のゲノム中の第1のヌクレオチド配列および第2のヌクレオチド配列を同定するステップを含み、
前記第1および第2のヌクレオチド配列が、請求項23に記載の基準のうちの少なくとも1つを満たしており、15〜18bpで分離されている、方法。
【請求項25】
前記エンドヌクレアーゼが、前記第1および第2のヌクレオチド配列の間に二本鎖切断を生じる、請求項24に記載の方法。
【請求項26】
エンドヌクレアーゼドメインと標的DNAに特異的なTALエフェクターDNA結合ドメインとを含むTALENであって、
前記DNA結合ドメインが複数のDNA結合反復を含み、
それぞれの反復が前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA中の1つの塩基対の認識を司っており、
前記TALENが、以下のRVD:
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含むTALEN。
【請求項27】
請求項26に記載のTALENであって、
前記TALENが、以下のRVD:
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、および
Tを認識するためのIG
のうちの少なくとも1つまたは複数を含む、TALEN。
【請求項28】
前記エンドヌクレアーゼドメインがII型制限エンドヌクレアーゼ由来のものである、請求項26に記載のTALEN。
【請求項29】
前記II型制限エンドヌクレアーゼがFokIである、請求項28に記載のTALEN。
【請求項30】
エンドヌクレアーゼドメインおよびTALエフェクタードメインを含むTALENであって、
前記TALENのアミノ酸配列が、配列番号33〜配列番号55、配列番号72、および配列番号73からなる群から選択される、TALEN。
【請求項31】
動物を作製する方法であって、
遺伝子修飾を導入することが所望される標的DNA配列を含む真核細胞を提供するステップと、
エンドヌクレアーゼドメインと前記標的DNA配列へ結合するTALエフェクタードメインとを含むTALENを用いて、前記標的DNA配列内で二本鎖切断を生じるステップと、
前記二本鎖切断が起こった細胞またはその子孫から動物を作製するステップと
を含む、方法。
【請求項32】
請求項31に記載の方法であって、
前記標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、前記細胞内に導入するステップであって、
前記導入するステップが、前記外因性核酸と前記細胞またはその子孫中の前記標的DNA配列との間に相同組換えが起こることを可能にする条件下で行われる、ステップと、
前記相同組換えが起こった細胞またはその子孫から動物を作製するステップと
をさらに含む、方法。
【請求項33】
前記動物が哺乳動物である、請求項31に記載の方法。
【請求項34】
前記遺伝子修飾が置換、挿入、または欠失を含む、請求項31に記載の方法。
【請求項35】
植物を作製する方法であって、
事前に選択された遺伝子修飾の導入が所望される標的DNA配列を含む植物細胞を提供するステップと、
エンドヌクレアーゼドメインと前記標的DNA配列へ結合するTALエフェクタードメインとを含むTALENを用いて、前記標的DNA配列内に二本鎖切断を生じるステップと、
前記二本鎖切断が起こった細胞またはその子孫から植物を作製するステップと
を含む、方法。
【請求項36】
請求項35に記載の方法であって、
前記標的DNA配列の少なくとも一部分に相同的な配列を含む外因性核酸を、前記植物細胞内に導入するステップであって、前記導入するステップが、前記外因性核酸と前記細胞またはその子孫中の前記標的DNA配列との間で相同組換えが起こることを可能にする条件下で行われる、ステップと、
前記相同組換えが起こった細胞またはその子孫から植物を作製するステップと
をさらに含む、方法。
【請求項37】
細胞における標的化遺伝子組換えの方法であって、
選択されたDNA標的配列を標的としたTALエフェクターエンドヌクレアーゼをコードしている核酸を前記細胞内に導入するステップと、
前記細胞内での前記TALエフェクターエンドヌクレアーゼの発現を誘導するステップと、
前記選択されたDNA標的配列が突然変異を示している細胞を同定するステップと
を含む、方法。
【請求項38】
前記突然変異が、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択される、請求項37に記載の方法。
【請求項39】
前記細胞内にドナーDNAを導入するステップをさらに含む、請求項37に記載の方法。
【請求項40】
前記細胞が昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞である、請求項37に記載の方法。
【請求項41】
標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法であって、前記方法は、
複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、
それぞれの反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれの反復が、前記標的DNA中の1つの塩基対の認識を司っている、ステップを含み、
前記作製するステップが、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませることを含み、これにより結合部位の−1位でのTの必要性を排除する、方法。
【請求項42】
標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法であって、前記方法は、
複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、
それぞれの反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれの反復が、標的DNA中の1つの塩基対の認識を司っている、ステップ
を含み、
前記作製するステップが、Gに対して増強された特異性を有するRVDを含有するTALエフェクターDNA結合ドメインをコードしている1つまたは複数の核酸を取り込ませることを含み、
前記RVDが、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択され、*がRVDの第2位置でのギャップを示す、方法。
【請求項43】
標的DNA配列中の少なくとも1つの塩基対を選択的に認識するポリペプチドを生成する方法であって、前記方法は、
反復ドメインを含むポリペプチドを合成するステップを含み、
前記反復ドメインが、転写活性化因子様(TAL)エフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記標的DNA中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項44】
請求項43に記載の方法によって生成されたポリペプチド。
【請求項45】
請求項43に記載の方法によって生成されたポリペプチドのコード配列を含むDNA。
【請求項46】
請求項45に記載のDNAと作動可能に連結されたプロモーターを含む発現カセット。
【請求項47】
請求項46に記載の発現カセットを含む非ヒト宿主細胞。
【請求項48】
請求項46に記載の発現カセットを含む形質転換させた非ヒト生物。
【請求項49】
ポリペプチドによってDNA配列中の塩基対を選択的に認識する方法であって、前記方法は、
反復ドメインを含むポリペプチドを構築するステップを含み、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項50】
細胞における標的遺伝子の発現を調節する方法であって、
ポリペプチドを含有する細胞が提供され、
前記ポリペプチドが反復ドメインを含み、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項51】
反復ドメインを含むポリペプチドであって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、ポリペプチド。
【請求項52】
請求項51に記載のポリペプチドのコード配列を含むDNA。
【請求項53】
塩基対が反復ドメインを含むポリペプチドによって特異的に認識されることができるように、標的DNA配列上に位置する塩基対が含まれるように修飾されたDNAであって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記塩基対が、超可変領域による選択的かつ決定された認識を受けるために、
(a)HDによる認識のためのC/G、
(b)NIによる認識のためのA/T、
(c)NGによる認識のためのT/A、
(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、
(e)NNによる認識のためのG/CまたはA/T、
(f)IGによる認識のためのT/A、
(g)Nによる認識のためのC/GまたはT/A、
(h)HGによる認識のためのT/A、
(i)Hによる認識のためのT/A、および
(j)NKによる認識のためのG/C
からなる群から選択される、DNA。
【請求項54】
請求項53に記載のDNAを含むベクター。
【請求項55】
請求項53に記載のDNAを含む非ヒト宿主細胞。
【請求項56】
請求項53に記載のDNAを含む形質転換された非ヒト生物。
【請求項57】
反復ドメインを含むポリペプチドによって選択的に認識される標的DNA配列を含むDNAを生成する方法であって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記標的DNA中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記標的DNA中の1つの塩基対配列の認識を司っており、
前記方法が、前記反復単位によって認識されることができる塩基対を含むDNAを合成するステップを含み、
前記塩基対が、
(a)HDによる認識のためのC/G、
(b)NIによる認識のためのA/T、
(c)NGによる認識のためのT/A、
(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、
(e)NNによる認識のためのG/CまたはA/T、
(f)IGによる認識のためのT/A、
(g)Nによる認識のためのC/GまたはT/A、
(h)HGによる認識のためのT/A、
(i)Hによる認識のためのT/A、および
(j)NKによる認識のためのG/C
からなる群から選択される、方法。
【請求項1】
細胞の遺伝物質を修飾する方法であって、
(a)標的DNA配列を含有する細胞を提供するステップと、
(b)転写活性化因子様(TAL)エフェクター−DNA修飾酵素を前記細胞へ導入するステップであって、
前記TALエフェクター−DNA修飾酵素は、
(i)二本鎖DNAを修飾することができるDNA修飾酵素ドメイン、および
(ii)組み合わせて標的DNA配列中の特定のヌクレオチド配列と結合する複数のTALエフェクター反復配列を含むTALエフェクタードメイン
を含むものであり、
前記TALエフェクター−DNA修飾酵素が、前記細胞またはその子孫中の前記特定のヌクレオチド配列内にあるまたはそれに隣接する前記標的DNAを修飾するように、TALエフェクター−DNA修飾酵素を前記細胞へ導入する、ステップと
を含む、方法。
【請求項2】
請求項1に記載の方法であって、
前記標的DNA配列と前記核酸との間で相同組換えが起こるように、前記標的DNA配列の少なくとも一部分に相同的な配列を含む核酸を、前記細胞に提供するステップをさらに含む、方法。
【請求項3】
前記細胞が真核細胞である、請求項1に記載の方法。
【請求項4】
前記細胞が哺乳動物細胞である、請求項1に記載の方法。
【請求項5】
前記細胞が植物細胞である、請求項1に記載の方法。
【請求項6】
前記細胞が原核細胞である、請求項1に記載の方法。
【請求項7】
前記標的DNAが染色体DNAである、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素をコードしているベクターを用いて細胞を形質移入することを含む、方法。
【請求項9】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を前記細胞内にタンパク質として機械的に注入することを含む、方法。
【請求項10】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を、細菌III型分泌系によってタンパク質として前記細胞内に送達することを含む、方法。
【請求項11】
請求項1に記載の方法であって、
前記導入ステップが、前記TALエフェクター−DNA修飾酵素を、電気穿孔によってタンパク質として前記細胞内に導入することを含む、方法。
【請求項12】
前記DNA修飾酵素ドメインがエンドヌクレアーゼである、請求項1に記載の方法。
【請求項13】
前記エンドヌクレアーゼがII型制限エンドヌクレアーゼである、請求項12に記載の方法。
【請求項14】
前記II型制限エンドヌクレアーゼがFokIである、請求項13に記載の方法。
【請求項15】
請求項1に記載の方法であって、
前記標的DNA内の特定のヌクレオチド配列と結合する前記TALエフェクタードメインが、10個以上のDNA結合反復、より好ましくは15個以上のDNA結合反復を含む、方法。
【請求項16】
請求項15に記載の方法であって、
それぞれのDNA結合反復が、前記標的DNA配列中の塩基対の認識を決定する反復可変性二残基(RVD)を含み、
それぞれのDNA結合反復が、前記標的DNA配列中の1つの塩基対の認識を司っており、
前記RVDが、
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのHG、
Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含む、方法。
【請求項17】
請求項16に記載の方法であって、
それぞれのDNA結合反復が、前記標的DNA配列中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA配列中の1つの塩基対の認識を司っており、
前記RVDが、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*[ただし*はRVDの第2位置中のギャップを表す]、
Tを認識するためのHG、
Tを認識するためのH*[ただし*はRVDの第2位置中のギャップを表す]、および
Tを認識するためのIG
のうちの1つまたは複数を含む、方法。
【請求項18】
選択されたヌクレオチド配列に特異的なTALエフェクターをコードしている核酸を作製する方法であって、
(1)開始プラスミドをPspXIで直鎖状にするステップであって、
前記開始プラスミドが、選択されたヌクレオチド配列の第1ヌクレオチドに特異的な反復可変性二残基(RVD)を有する第1のTALエフェクターDNA結合反復ドメインをコードしているヌクレオチド配列を含み、
第1のTALエフェクターDNA結合反復ドメインが、その3’末端にユニークなPspXI部位を有する、ステップと、
(2)前記開始プラスミドのPspXI部位内に、選択されたヌクレオチド配列の次のヌクレオチドに特異的なRVDを有する1つまたは複数のTALエフェクターDNA結合反復ドメインをコードしているDNAモジュールをライゲーションするステップであって、
前記DNAモジュールがXhoI粘着末端を有する、ステップと、
(3)前記核酸が、前記選択されたヌクレオチド配列と結合することができるTALエフェクターをコードするまで、ステップ(1)および(2)を繰り返すステップと
を含む、方法。
【請求項19】
、請求項18に記載の方法であって、
ライゲーション後に、前記PspXI部位中の前記DNAモジュールの配向を決定するステップをさらに含む、方法。
【請求項20】
ステップ(1)および(2)を1〜30回繰り返すことを含む、請求項18に記載の方法。
【請求項21】
転写活性化因子様エフェクターエンドヌクレアーゼ(TALEN)をコードしている核酸を作製する方法であって、
(a)細胞のゲノム中の第1のユニークなヌクレオチド配列を同定するステップと、
(b)(i)組み合わせて前記第1のユニークなヌクレオチド配列と結合する複数のDNA結合反復、および(ii)第1のヌクレオチド配列内にあるまたはそれに隣接する第2のヌクレオチド配列で二本鎖切断を生じるエンドヌクレアーゼを含む、TALENをコードしている核酸を合成するステップであって、
それぞれのDNA結合反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA中の1つの塩基対の認識を司っており、
前記TALENが、以下のRVD:
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含むステップと
を含む、方法。
【請求項22】
請求項21に記載の方法であって、
前記TALENが、以下のRVD:
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、および
Tを認識するためのIG
のうちの1つまたは複数を含む、方法。
【請求項23】
請求項21に記載の方法であって、
前記第1のヌクレオチド配列が以下の基準:
i)最小で15個の塩基の長さであり、5’から3’に配向されており、Tが5’末端の部位の直前にあること、
ii)第1(5’)位置にTを有さない、または、第2位置にAを有さないこと、
iii)最終(3’)位置がTで終わり、最後から2番目の位置にGを有さないこと、ならびに
iv)0〜63%のA、11〜63%のC、0〜25%のG、および2〜42%のTの塩基組成を有すること
のうちの少なくとも1つを満たす、方法。
【請求項24】
請求項23に記載の方法であって、
前記細胞のゲノム中の第1のヌクレオチド配列および第2のヌクレオチド配列を同定するステップを含み、
前記第1および第2のヌクレオチド配列が、請求項23に記載の基準のうちの少なくとも1つを満たしており、15〜18bpで分離されている、方法。
【請求項25】
前記エンドヌクレアーゼが、前記第1および第2のヌクレオチド配列の間に二本鎖切断を生じる、請求項24に記載の方法。
【請求項26】
エンドヌクレアーゼドメインと標的DNAに特異的なTALエフェクターDNA結合ドメインとを含むTALENであって、
前記DNA結合ドメインが複数のDNA結合反復を含み、
それぞれの反復が前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれのDNA結合反復が、前記標的DNA中の1つの塩基対の認識を司っており、
前記TALENが、以下のRVD:
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、
Tを認識するためのIG、
Gを認識するためのNK、
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、および
Tを認識するためのYG
のうちの1つまたは複数を含むTALEN。
【請求項27】
請求項26に記載のTALENであって、
前記TALENが、以下のRVD:
Cを認識するためのHA、
Cを認識するためのND、
Cを認識するためのHI、
Gを認識するためのHN、
Gを認識するためのNA、
GまたはAを認識するためのSN、
Tを認識するためのYG、および
Gを認識するためのNK、
のうちの1つまたは複数、ならびに
Cを認識するためのHD、
Tを認識するためのNG、
Aを認識するためのNI、
GまたはAを認識するためのNN、
AまたはCまたはGまたはTを認識するためのNS、
CまたはTを認識するためのN*、
Tを認識するためのHG、
Tを認識するためのH*、および
Tを認識するためのIG
のうちの少なくとも1つまたは複数を含む、TALEN。
【請求項28】
前記エンドヌクレアーゼドメインがII型制限エンドヌクレアーゼ由来のものである、請求項26に記載のTALEN。
【請求項29】
前記II型制限エンドヌクレアーゼがFokIである、請求項28に記載のTALEN。
【請求項30】
エンドヌクレアーゼドメインおよびTALエフェクタードメインを含むTALENであって、
前記TALENのアミノ酸配列が、配列番号33〜配列番号55、配列番号72、および配列番号73からなる群から選択される、TALEN。
【請求項31】
動物を作製する方法であって、
遺伝子修飾を導入することが所望される標的DNA配列を含む真核細胞を提供するステップと、
エンドヌクレアーゼドメインと前記標的DNA配列へ結合するTALエフェクタードメインとを含むTALENを用いて、前記標的DNA配列内で二本鎖切断を生じるステップと、
前記二本鎖切断が起こった細胞またはその子孫から動物を作製するステップと
を含む、方法。
【請求項32】
請求項31に記載の方法であって、
前記標的DNAの少なくとも一部分に相同的な配列を含む外因性核酸を、前記細胞内に導入するステップであって、
前記導入するステップが、前記外因性核酸と前記細胞またはその子孫中の前記標的DNA配列との間に相同組換えが起こることを可能にする条件下で行われる、ステップと、
前記相同組換えが起こった細胞またはその子孫から動物を作製するステップと
をさらに含む、方法。
【請求項33】
前記動物が哺乳動物である、請求項31に記載の方法。
【請求項34】
前記遺伝子修飾が置換、挿入、または欠失を含む、請求項31に記載の方法。
【請求項35】
植物を作製する方法であって、
事前に選択された遺伝子修飾の導入が所望される標的DNA配列を含む植物細胞を提供するステップと、
エンドヌクレアーゼドメインと前記標的DNA配列へ結合するTALエフェクタードメインとを含むTALENを用いて、前記標的DNA配列内に二本鎖切断を生じるステップと、
前記二本鎖切断が起こった細胞またはその子孫から植物を作製するステップと
を含む、方法。
【請求項36】
請求項35に記載の方法であって、
前記標的DNA配列の少なくとも一部分に相同的な配列を含む外因性核酸を、前記植物細胞内に導入するステップであって、前記導入するステップが、前記外因性核酸と前記細胞またはその子孫中の前記標的DNA配列との間で相同組換えが起こることを可能にする条件下で行われる、ステップと、
前記相同組換えが起こった細胞またはその子孫から植物を作製するステップと
をさらに含む、方法。
【請求項37】
細胞における標的化遺伝子組換えの方法であって、
選択されたDNA標的配列を標的としたTALエフェクターエンドヌクレアーゼをコードしている核酸を前記細胞内に導入するステップと、
前記細胞内での前記TALエフェクターエンドヌクレアーゼの発現を誘導するステップと、
前記選択されたDNA標的配列が突然変異を示している細胞を同定するステップと
を含む、方法。
【請求項38】
前記突然変異が、遺伝物質の欠失、遺伝物質の挿入、ならびに遺伝物質の欠失および挿入の両方からなる群から選択される、請求項37に記載の方法。
【請求項39】
前記細胞内にドナーDNAを導入するステップをさらに含む、請求項37に記載の方法。
【請求項40】
前記細胞が昆虫細胞、植物細胞、魚細胞、または哺乳動物細胞である、請求項37に記載の方法。
【請求項41】
標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法であって、前記方法は、
複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、
それぞれの反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれの反復が、前記標的DNA中の1つの塩基対の認識を司っている、ステップを含み、
前記作製するステップが、A、C、またはGに対する特異性を有する変異第0DNA結合反復配列をコードしている核酸を取り込ませることを含み、これにより結合部位の−1位でのTの必要性を排除する、方法。
【請求項42】
標的DNAに対して増強された標的化能力を有するTALエフェクターを作製する方法であって、前記方法は、
複数のDNA結合反復を有するDNA結合ドメインを含むTALエフェクターをコードしている核酸を作製するステップであって、
それぞれの反復が、前記標的DNA中の塩基対の認識を決定するRVDを含み、
それぞれの反復が、標的DNA中の1つの塩基対の認識を司っている、ステップ
を含み、
前記作製するステップが、Gに対して増強された特異性を有するRVDを含有するTALエフェクターDNA結合ドメインをコードしている1つまたは複数の核酸を取り込ませることを含み、
前記RVDが、RN、R*、NG、NH、KN、K*、NA、NT、DN、D*、NL、NM、EN、E*、NV、NC、QN、Q*、NR、NP、HN、H*、NK、NY、SN、S*、ND、NW、TN、T*、NE、NF、YN、Y*、およびNQからなる群から選択され、*がRVDの第2位置でのギャップを示す、方法。
【請求項43】
標的DNA配列中の少なくとも1つの塩基対を選択的に認識するポリペプチドを生成する方法であって、前記方法は、
反復ドメインを含むポリペプチドを合成するステップを含み、
前記反復ドメインが、転写活性化因子様(TAL)エフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記標的DNA中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項44】
請求項43に記載の方法によって生成されたポリペプチド。
【請求項45】
請求項43に記載の方法によって生成されたポリペプチドのコード配列を含むDNA。
【請求項46】
請求項45に記載のDNAと作動可能に連結されたプロモーターを含む発現カセット。
【請求項47】
請求項46に記載の発現カセットを含む非ヒト宿主細胞。
【請求項48】
請求項46に記載の発現カセットを含む形質転換させた非ヒト生物。
【請求項49】
ポリペプチドによってDNA配列中の塩基対を選択的に認識する方法であって、前記方法は、
反復ドメインを含むポリペプチドを構築するステップを含み、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項50】
細胞における標的遺伝子の発現を調節する方法であって、
ポリペプチドを含有する細胞が提供され、
前記ポリペプチドが反復ドメインを含み、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、方法。
【請求項51】
反復ドメインを含むポリペプチドであって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、DNA配列中の1つの塩基対の認識を司っており、
前記超可変領域が、
(a)C/Gを認識するためのHD、
(b)A/Tを認識するためのNI、
(c)T/Aを認識するためのNG、
(d)C/GまたはA/TまたはT/AまたはG/Cを認識するためのNS、
(e)G/CまたはA/Tを認識するためのNN、
(f)T/Aを認識するためのIG、
(g)C/Gを認識するためのN、
(h)C/GまたはT/Aを認識するためのHG、
(i)T/Aを認識するためのH、および
(j)G/Cを認識するためのNK
からなる群から選択されるメンバーを含む、ポリペプチド。
【請求項52】
請求項51に記載のポリペプチドのコード配列を含むDNA。
【請求項53】
塩基対が反復ドメインを含むポリペプチドによって特異的に認識されることができるように、標的DNA配列上に位置する塩基対が含まれるように修飾されたDNAであって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記DNA配列中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記DNA配列中の1つの塩基対の認識を司っており、
前記塩基対が、超可変領域による選択的かつ決定された認識を受けるために、
(a)HDによる認識のためのC/G、
(b)NIによる認識のためのA/T、
(c)NGによる認識のためのT/A、
(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、
(e)NNによる認識のためのG/CまたはA/T、
(f)IGによる認識のためのT/A、
(g)Nによる認識のためのC/GまたはT/A、
(h)HGによる認識のためのT/A、
(i)Hによる認識のためのT/A、および
(j)NKによる認識のためのG/C
からなる群から選択される、DNA。
【請求項54】
請求項53に記載のDNAを含むベクター。
【請求項55】
請求項53に記載のDNAを含む非ヒト宿主細胞。
【請求項56】
請求項53に記載のDNAを含む形質転換された非ヒト生物。
【請求項57】
反復ドメインを含むポリペプチドによって選択的に認識される標的DNA配列を含むDNAを生成する方法であって、
前記反復ドメインが、TALエフェクターに由来する少なくとも1つの反復単位を含み、
前記反復単位が、前記標的DNA中の塩基対の認識を決定する超可変領域を含み、
前記反復単位が、前記標的DNA中の1つの塩基対配列の認識を司っており、
前記方法が、前記反復単位によって認識されることができる塩基対を含むDNAを合成するステップを含み、
前記塩基対が、
(a)HDによる認識のためのC/G、
(b)NIによる認識のためのA/T、
(c)NGによる認識のためのT/A、
(d)NSによる認識のためのCTまたはA/TまたはT/AまたはG/C、
(e)NNによる認識のためのG/CまたはA/T、
(f)IGによる認識のためのT/A、
(g)Nによる認識のためのC/GまたはT/A、
(h)HGによる認識のためのT/A、
(i)Hによる認識のためのT/A、および
(j)NKによる認識のためのG/C
からなる群から選択される、方法。
【図1】


【図2】
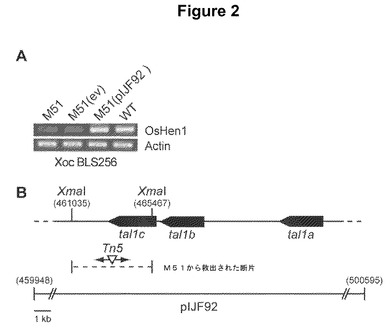
【図3】

【図4】

【図5】
【図6】
【図7】
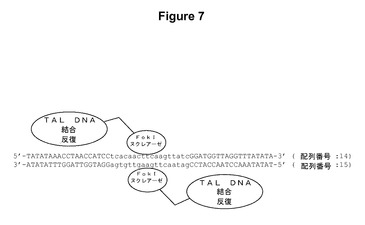
【図8】

【図9】
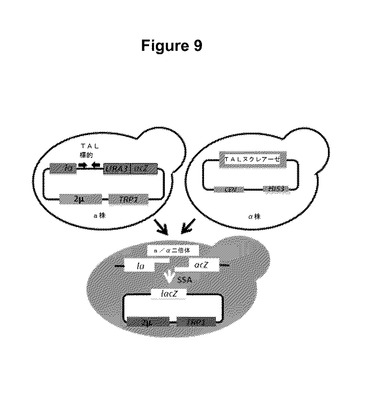
【図10】
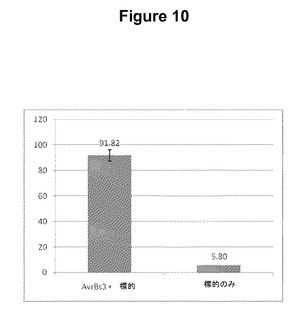
【図11】
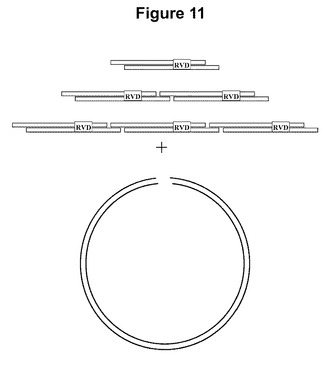
【図12】
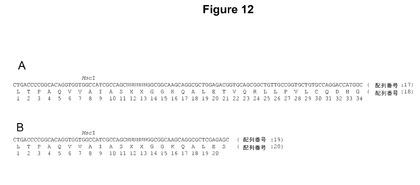
【図13】
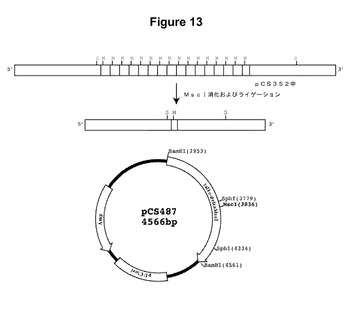
【図14】
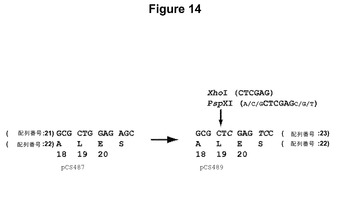
【図15】
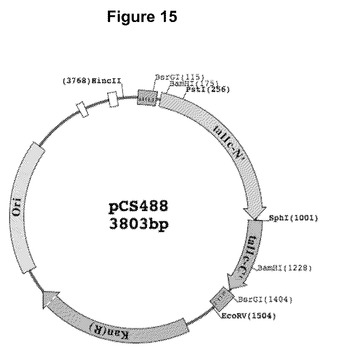
【図16】
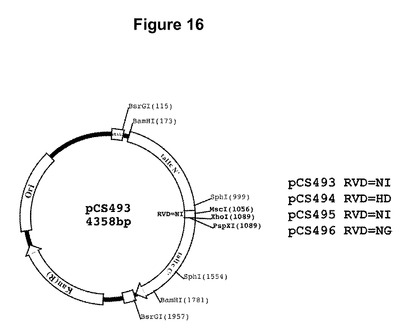
【図17】
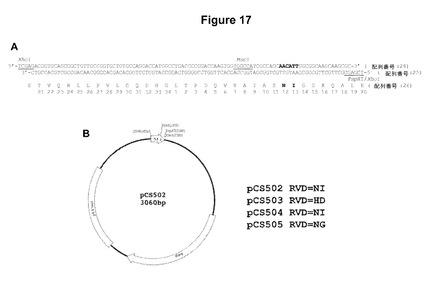
【図18】
【図19】
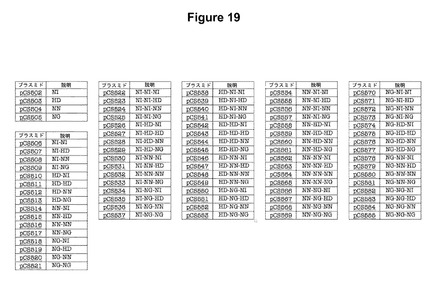
【図20】
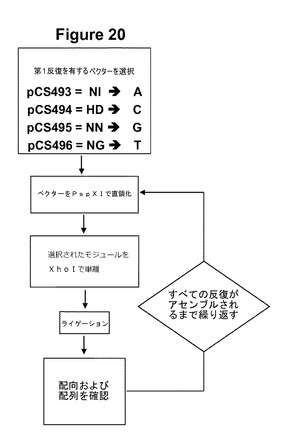
【図21A】
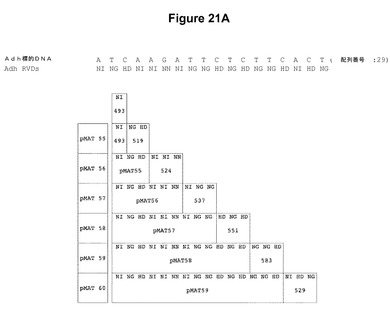
【図21B】
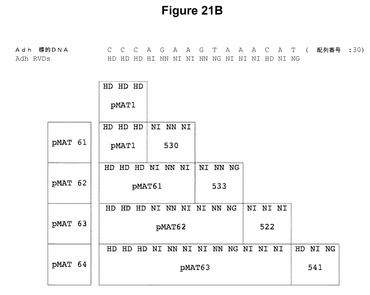
【図22】
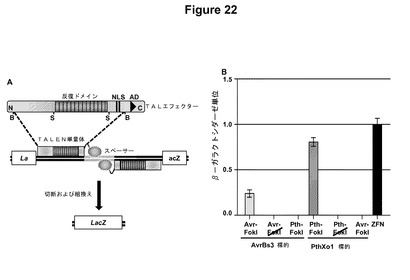
【図23】

【図24】

【図25】
【図26】

【図27】

【図28】
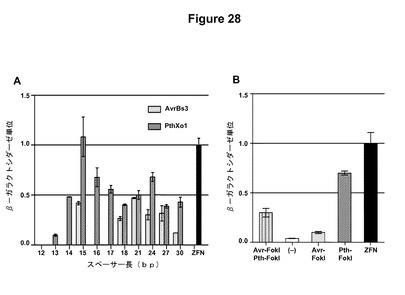
【図29A】
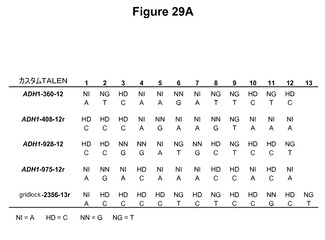
【図29B】
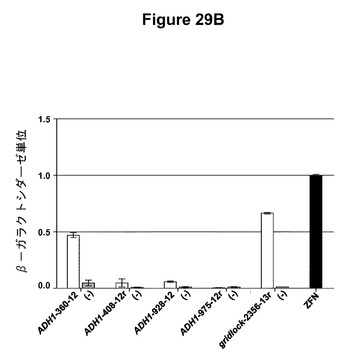
【図30】
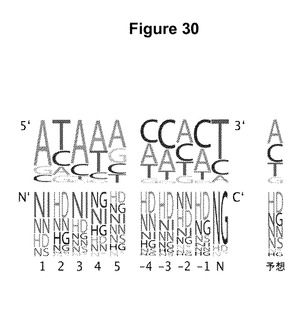
【図31】
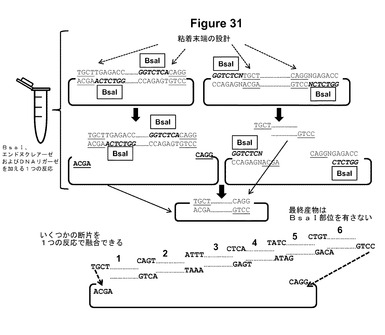
【図32A】
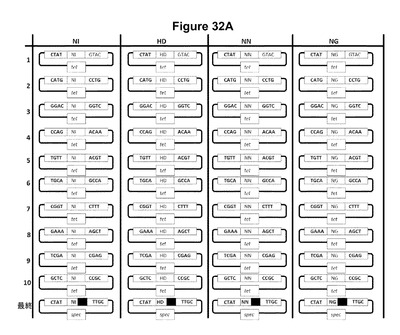
【図32B】
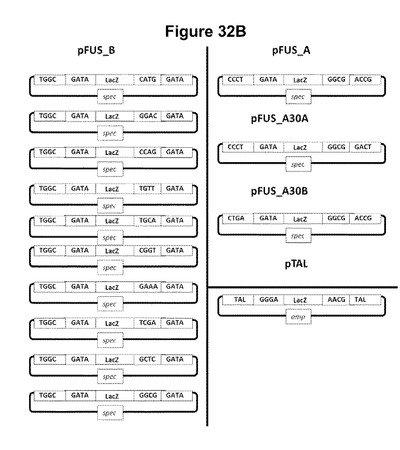
【図33】
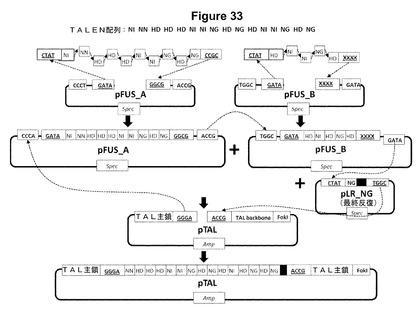
【図34A】

【図34B】

【図34C】

【図34D】

【図34E】

【図34F】

【図34G】

【図34H】

【図34I】

【図34J】

【図34K】

【図34L】

【図34M】

【図34N】

【図34O】

【図34P】

【図34Q】

【図34R】

【図34S】

【図34T】

【図34U】

【図35】
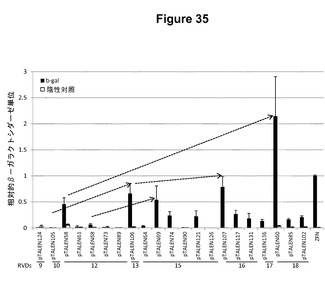
【図36】
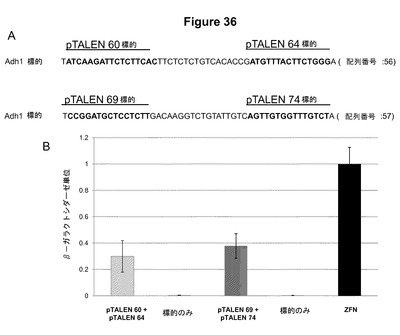
【図37】
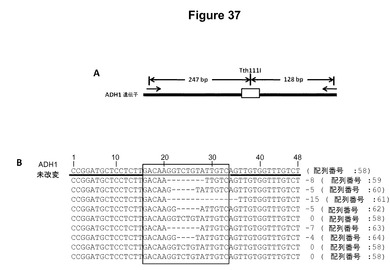
【図38】
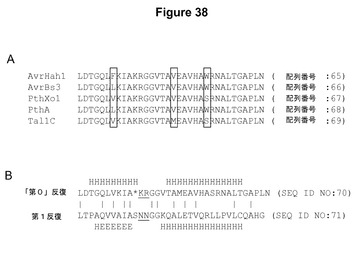
【図39】
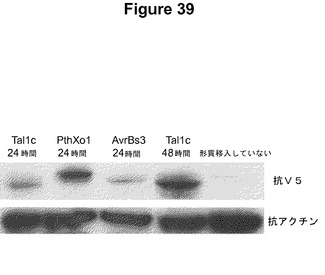
【図40A】

【図40B】

【図41A】
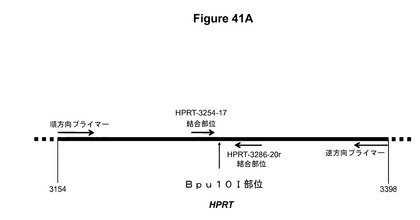
【図41B】
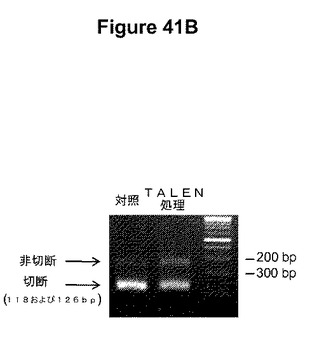
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21A】
【図21B】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29A】
【図29B】
【図30】
【図31】
【図32A】
【図32B】
【図33】
【図34A】
【図34B】
【図34C】
【図34D】
【図34E】
【図34F】
【図34G】
【図34H】
【図34I】
【図34J】
【図34K】
【図34L】
【図34M】
【図34N】
【図34O】
【図34P】
【図34Q】
【図34R】
【図34S】
【図34T】
【図34U】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40A】
【図40B】
【図41A】
【図41B】
【公表番号】特表2013−513389(P2013−513389A)
【公表日】平成25年4月22日(2013.4.22)
【国際特許分類】
【出願番号】特願2012−543313(P2012−543313)
【出願日】平成22年12月10日(2010.12.10)
【国際出願番号】PCT/US2010/059932
【国際公開番号】WO2011/072246
【国際公開日】平成23年6月16日(2011.6.16)
【出願人】(305023366)リージェンツ オブ ザ ユニバーシティ オブ ミネソタ (39)
【出願人】(304057287)アイオワ・ステイト・ユニバーシティ・リサーチ・ファウンデイション・インコーポレイテッド (4)
【氏名又は名称原語表記】Iowa State University Research Foundation, Inc.
【Fターム(参考)】
【公表日】平成25年4月22日(2013.4.22)
【国際特許分類】
【出願日】平成22年12月10日(2010.12.10)
【国際出願番号】PCT/US2010/059932
【国際公開番号】WO2011/072246
【国際公開日】平成23年6月16日(2011.6.16)
【出願人】(305023366)リージェンツ オブ ザ ユニバーシティ オブ ミネソタ (39)
【出願人】(304057287)アイオワ・ステイト・ユニバーシティ・リサーチ・ファウンデイション・インコーポレイテッド (4)
【氏名又は名称原語表記】Iowa State University Research Foundation, Inc.
【Fターム(参考)】
[ Back to top ]