
TM4SF5の機能を阻害する抗がん物質のスクリーニング方法及びカルコン系の化合物を含有する抗がん用組成物
本発明は、抗がん物質のスクリーニング方法及び前記方法によりスクリーニングされた抗がん用の化合物に関し、さらに詳しくは、配列番号2のポリペプチドを発現する腫瘍タンパク質であるTM4SF5(transmembrane 4 L6 family member 5)を発現するがん細胞を培養して抗がん候補物質にて処理した後、前記TM4SF5の発現と関連する種々の分子的な機序による諸現象に基づいて腫瘍生成及び転移に対する拮抗作用を示す物質を抗がん物質としてスクリーニングする方法、前記方法によりスクリーニングされたカルコン系の化合物及びこれを有効成分とする抗がん組成物に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は抗がん物質のスクリーニング方法に係り、さらに詳しくは、TM4SF5による腫瘍の形成と発生及び転移を阻害する化合物のスクリーニング方法に関する。また、本発明は、該方法により抗がん物質としてスクリーニングされたカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物に関する。
【背景技術】
【0002】
トランスメンブラン4 L6ファミリーメンバー5(transmembrane 4 L6 family member 5:TM4SF5またはL6H)は腫瘍特異抗原であるTM4SF1(L6)と相同性を有するタンパク質であって、前記TM4SF1(L6)は、トランスメンブラン4 L6ファミリー(transmembrane 4 L6 family)に属する。トランスメンブラン4 L6ファミリーに属するタンパク質は、TM4SF1(L6)に加えて、IL−TMP、L6Dなどがある。腫瘍特異抗原として知られている前記TM4SF1(L6)は、直腸がん、肺がん、乳房がん、さらには卵巣がんなどにおいて高く発現されていることが知られており、TM4SF1(L6)と相同性を有するタンパク質であるTM4SF5(L6H)もまたすい臓がん、胃がん、直腸がん、さらに肝がんなど種々のがん細胞において高く発現されると当業界に報告されている(Muller-Pillasch, F. et al., Gene, 208:25, 1998、 Pascual-Le Tallec, L. et al., J. Clin. Endocrinol. Metab., 87:501, 2002)。また、TM4SF1(L6)に比べてTM4SF5のmRNAのレベルは胃がん組織においては低く発現された場合があるが、肝がん細胞株と胃がん細胞株においては高く発現されたものであるとも報告されている(Kaneko, R. et al., Am. J. Gastroenterol., 96(12):3457, 2001)。
【0003】
これらの研究に基づいて、近年、米国特許US6、350、581B1においてTM4SF5が新規な発がん遺伝子として登録されており(TUANと表記)、本発明者によってCos7繊維芽細胞にTM4SF5を人為的に発現させた結果、アクチン構造の再構成と接着斑の形成、及び接着斑キナーゼ(focal adhesion kinase:FAK)のアミノ酸チロシン925番のリン酸化が、インテグリンα2により引き起こされること、及びこのような現象は細胞成長因子を含む血清により阻害、調節されることを研究結果として報告している(Lee, S.Y. et al., Exp. Cell Res., 312:2983, 2006)。
【0004】
しかしながら、腫瘍形成におけるTM4SF5の分子的な機序はほとんど知られていないのが現状である。すなわち、分子的なレベルにおけるTM4SF5タンパク質の機能及びその発がん機能に係る証拠は生化学的、細胞生物学的に全く知られていない。
【0005】
一方、カルコンは食用の植物に豊富に存在する物質であって、フラボノイドまたはイソフラボノイドの前駆体として知られている。前記のカルコンの誘導体は植物の黄色色素の構成成分であって、植物の色合いに影響するだけではなく、有害な紫外線から植物を保護する役割を果たすこともある(天然物化学研究法、ウ・ウォンシク著、ソウル大学出版部)。カルコン誘導体は菊科植物であるハルシャギク属に多量存在する。代表的なカルコン類には2’、6’−ジヒドロキシ−4’−メトキシカルコン、カルタミンなどがあり、これらは桂皮、紅花、胡椒などの植物に含有されている。ジヒドロカルコンは主としてバラ科及びカラムラサキツツジ科植物に含有されており、りんごに存在するジヒドロカルコンであるフロリジンはりんごの病気に対する抵抗力と関係があると報告されている。
【0006】
このようなカルコン誘導体は、例えば、抗原虫性(Liu, M. et al., J. Med. Chem., 44:4443, 2001)、抗炎症性(Babu, M.A. et al., Bioorg. Med. Chem., 10:4035, 2003)、免疫調節(Barfod, L. et al., int. immunopharmacol., 2:545, 2002)、一酸化窒素阻害(Rojas, J. et al., Bioorg. Med. Chem. Lett., 12:1951, 2002)、抗がん(Kumar, S.K. et al., J. Med. Chem., 46:2813, 2003)、抗HIV(Artico, M. et al., J. Med. Chem., 41:3984, 1998)活性など極めて様々な薬物学的な活性を示すことが知られている。
【0007】
また、大韓民国公開特許10−2003−0036993号公報には、前記カルコン化合物が基底膜成分を分解するマトリックスメタロプロテアーゼ(Matrix metalloproteinase、MMP)の活性を阻害する性質を有していることが記載されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 15:5514, 2004)。
【0008】
カルコン系の化合物の中でも、スルホンアミドまたはスルホネート基が置換されたカルコン系の化合物には自然に存在するカルコン系の化合物が有していない特異な生物学的特性があることが報告されている。特に、本発明者により、スルホンアミドカルコン系の化合物が高いα−グルコシダーゼ阻害活性を有していることが報告されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 15:5514, 2004、大韓民国公開特許10−0751899)。加えて、本発明者により、スルホネートカルコン系の化合物が選択的なカリウムイオンチャンネル阻害活性を有していることも報告されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 2007)。
【0009】
しかしながら、これまで、前記スルホンアミド及びスルホネートカルコン系の化合物が発がん遺伝子であるTM4SF5を阻害する機序を通じて抗がん機能を発現するということは全く解明されていない。
【0010】
そこで、本発明者らは、ヒトのがん細胞において現れるTM4SF5の発がん遺伝子としての機能は、上皮間葉転換(epithelial-mesenchymal transition:EMT)現象の促進、接触阻止現象の消失(loss of contact inhibition)によるものであり、これは、FAKのリン酸化及び細胞質内のp27kip1タンパク質の蓄積によるRhoAタンパク質の活性阻害及び細胞形態の変化につながる一連の過程を通じてなされるという新たな経路を見出し、前記諸現象に基づいてTM4SF5の発現による腫瘍の形成と発生及び転移を阻害する抗がん物質をスクリーニングするために鋭意努力した結果、本発明を完成するに至った。
【発明の詳細な説明】
【0011】
《技術的課題》
本発明の目的は、TM4SF5による腫瘍の形成と発生及び転移を阻害する抗がん物質のスクリーニング方法を提供することにある。
【0012】
本発明の他の目的は、前記方法によりスクリーニングされた抗がん物質、特に、スルホンアミド及びスルホネートカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物を提供することにある。
【技術的解決方法】
【0013】
上記の目的を達成するために、本発明は、下記のステップを含む、抗がん物質のスクリーニング方法を提供する:
(a)配列番号2のポリペプチドを発現するがん細胞を培養した後、抗がん候補物質で処理するステップ、
(b)前記抗がん候補物質で処理されたがん細胞に対して、
(i)接着斑キナーゼ(FAK)を構成するアミノ酸配列のうち577番位のチロシンのリン酸化の確認、(ii)FAKのRho−GTPase activating protein(RhoGAP)への結合、またはFAKのGTPase Regulator Associated with FAK(GRAF)への結合測定、(iii)細胞質内p27Kip1の発現量及び細胞質内安定化測定、(iv)RhoAの活性測定、及び(v)Rac1の活性測定、
の少なくとも1つを検出するステップ、及び
(c)抗がん候補物質で処理しない場合に比べて、
(i)FAKの577番位のチロシンのリン酸化減少、(ii)FAKのRhoGAPへの結合、またはFAKのGRAFへの結合抑制、(iii)細胞質内p27Kip1発現減少及び細胞質内安定化、(iv)RhoA活性増加、または(v)Rac1活性減少
の事象が現れた場合に、抗がん候補物質を抗がん物質として選択するステップ。
【0014】
本発明において、前記ステップ(b)は、細胞の形態確認、細胞接着関連のタンパク質発現確認、細胞間接触パターンまたは接触生長確認、α平滑筋アクチン(α−smooth muscle actin:α−SMA)タンパク質またはビメンチンタンパク質の発現確認、E−カドヘリンタンパク質の発現確認及び上皮間葉転換(epithelial-mesenchymal transition、EMT)確認よりなる群から選ばれる1以上をさらに行い、前記ステップ(c)は、細胞の形態がロッド状から多角形に戻るか、あるいは、細胞接着関連のタンパク質の発現が減少するか、あるいは、細胞−細胞間の接触が維持されるか、あるいは、細胞生長の接触阻止現象が現れるか、あるいは、α−SMAタンパク質またはビメンチンタンパク質の発現が減少するか、あるいは、E−カドヘリンタンパク質の発現が増加するか、あるいは、上皮間葉転換(EMT)が減少する場合をさらに含むことを特徴としてもよい。
【0015】
本発明において、前記ステップ(b)は、細胞外基質(extracellular matrix)または血清の存在下での細胞の移動又は運動性確認、細胞外基質を構成するコラーゲンゲルへの浸潤確認、細胞外基質複合体を構成するマトリゲルへの浸潤確認及びMMP活性測定よりなる群から選ばれる1以上をさらに行い、前記ステップ(c)は、細胞外基質または血清の存在下で細胞の移動又は運動性が減少するか、あるいは、コラーゲンゲル中における浸潤が減少するか、あるいは、マトリゲル中における浸潤が減少するか、あるいは、MMP活性が減少する場合をさらに含むことを特徴としてもよい。
【0016】
本発明は、また、前記方法によりスクリーニングされた、下記の化学式1または化学式2で表わされる抗がん用のカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物、並びに前記化合物の抗がん治療及び予防の用途を提供する。
【0017】
【化1】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【0018】
【化2】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【0019】
本発明の他の特徴及び具現例は、下記の詳細な説明及び特許請求の範囲からなお一層明らかになる。
【図面の簡単な説明】
【0020】
【図1】図1は、SNU449p、SNU449Cp、Tp、T3、T7、T16細胞の写真及びウェスタンブロットの結果である。
【図2】図2は、SNU449CpとSNU449Tpに対してローダミン標識ファロイジンで染色したアクチン写真である。
【図3】図3は、表記のタンパク質に対するウェスタンブロットの結果である。
【図4】図4は、RhoAとRac1に対するin vitroプルダウンアッセイによる活性分析結果である。
【図5】図5は、SNU449p、SNU449Cp及びSNU449Tp細胞に対するウェスタンブロット分析結果である。
【図6】図6は、図5のいくつかの細胞のRhoA活性分析結果である。
【図7】図7は、SNU449Tp細胞にpEGFPとshTM4SF5を一緒に導入して24時間培養後、ローダミン標識ファロイジン及びDAPIを蛍光染色した写真である。
【図8】図8は、FAK−RhoA間の信号伝達に対する模式図である。
【図9】図9は、SNU449Tp細胞におけるp190RhoGAP、GRAF、FAK抗体を用いたウェスタンブロット分析結果である。
【図10】図10は、SNU449Tp細胞に(HA)3−tagged FAK野生型タンパク質とY577F突然変異タンパク質を過剰発現した後のウェスタンブロット分析結果である。
【図11】図11は、対照群細胞にFLAG−tagged GRAFタンパク質を過剰発現した後の蛍光免疫染色写真である。
【図12】図12は、SNU449CpとSNU449Tp細胞に対する蛍光免疫染色写真である。
【図13】図13は、核と細胞質を分画化した後、p27kip1タンパク質に対する抗体とα−チューブリンに対する抗体を用いたウェスタンブロット分析結果である。
【図14】図14は、p27kip1タンパク質発現レベルとSer10リン酸化に対する、表記の抗体を用いたウェスタンブロット分析結果である。
【図15】図15は、p27KiP1のセリン10番のアミノ酸残基のリン酸化と関連するSNU449CpとSNU449Tp細胞に対する蛍光免疫染色写真である。
【図16】図16は、p27kip1またはGAPDHの遺伝子のmRNAへの転写を確認するためのRT−PCR分析結果及びウェスタンブロットの結果である。
【図17】図17は、SNU449Tp細胞をpEGFPとshTM4SF5を一緒に導入した後、p27kip1タンパク質とDAPIで細胞核を免疫蛍光染色法により染色した写真である。
【図18】図18は、図示の抗体を用いてウェスタンブロット分析した結果である。
【図19】図19は、SNU449Tp細胞に(HA)−tagged FAK野生型タンパク質とY577F突然変異体を過剰発現させた後のウェスタンブロット分析結果である。
【図20】図20は、肝がん細胞株Huh7に対して様々なshRNAを導入した後のウェスタンブロット分析結果である。
【図21】図21は、免疫蛍光染色法により染色した写真である。
【図22】図22は、SNU449Cp細胞にFLAG−tagged p27kip1タンパク質を過剰発現させた後の免疫蛍光染色法による共焦点顕微鏡写真である。
【図23】図23は、SNU449Tp細胞においてp27kip1の発現を阻害した後の免疫蛍光染色写真である。
【図24】図24は、図示の抗体を用いたウェスタンブロット分析結果である。
【図25】図25は、SNU449Tp細胞に活性型RhoA63L突然変異ベクターを導入した後のアクチン構造を免疫染色した写真である。
【図26】図26は、SNU449CpとSNU449Tp細胞にLPAを処理した後の免疫染色写真である。
【図27】図27は、SNU449Tp細胞にmDia活性型の突然変異体、活性型のRac1(Rac1 61L)及び活性型のPAK1(PAK1−caax)を導入した後に免疫蛍光染色分析を行った写真である。
【図28】図28は、細胞接触と関連するタンパク質に対するウェスタンブロット分析結果である。
【図29】図29は、SNU449Cp及びSNU449Tp細胞に対するE−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色分析結果である。
【図30】図30は、SNU449Tp細胞においてp27kip1発現を阻害した場合、E−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色分析結果である。
【図31】図31は、shRNAとshTM4SF5を発現するHuh7細胞におけるウェスタンブロット分析結果である。
【図32】図32は、前記樹立された細胞株にHGF(100ng/ml)を24時間処理した後、ZO1の位置を免疫染色した写真である。
【図33】図33は、SNU449CpとSNU449Tp細胞に対してDAPIで細胞核を染色した写真(矢印:細胞分裂と核が重なり合った部分)及び12時間おきに1回ずつ測定した細胞数のグラフである。
【図34】図34は、SNU449Tp細胞が重なり合って細胞分裂を示す染色写真である(矢印:細胞分裂中にある核)。
【図35】図35は、SNU449Cp細胞とTM4SF5が発現されるT7、T16、及びT3の細胞周期のS期分布を調べてまとめたグラフである。
【図36】図36は、SNU449Cp細胞及びSNU449Tp細胞にshRNAとshTM4SF5を導入した後、フローサイトメトリーを用いてS細胞周期を分析したグラフである。
【図37】図37は、SNU449CpとSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図38】図38は、LPAを処理したSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図39】図39は、p27kip1shRNAアデノウィルスを感染させたSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図40】図40は、SNU449T16細胞にE−カドヘリンを再発現させたSNU449T16m細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図41】図41は、SNU449Cp(Cp)、SNU449T16(T16)、及びSNU449T16m(T16m)細胞に対する、表記の分子の発現レベルに対する調査結果である。
【図42】図42は、SNU449T16(T16)及びSNU449T16m(T16m)細胞に対してβ−カテニン、ZO1、またはE−カドヘリンを免疫染色した写真である。
【図43】図43は、SNU449Cp細胞とSNU449Tp細胞に対してDMSOまたはTSAHC(4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン)を表記の濃度にてそれぞれ処理した後、生存細胞の数を数えて記録したグラフである。
【図44】図44は、SNU449Tp細胞にDMSOまたは20μMのTSAHCを処理し、BrdUを24時間かけて浸透させた後、蛍光染色法により確認した写真である。
【図45】図45は、SNU449Tp細胞に対してDMSOまたは20μM TSAHCを処理した後のE−カドヘリンまたはβ−カテニンの免疫染色写真である。
【図46】図46は、表記のタンパク質のウェスタンブロット分析結果である。
【図47】図47は、p27kip1の蛍光免疫染色分析結果である。
【図48】図48は、in vitro RhoA活性分析結果である。
【図49】図49は、DNAとアクチンの免疫染色写真である。
【図50】図50は、SNU449CpとSNU449Tp細胞に対してPNGaseF及びEndoH酵素を処理した後のウェスタンブロットの結果である。
【図51】図51は、TM4SF5を発現する細胞及び発現しない細胞に対するTSAHCの処理による生長/増殖の変化を示すグラフである。
【図52】図52は、SNU449細胞株のソフトアガーアッセイを行った結果である。
【図53】図53は、SNU449Cp、SNU449Tp、及びSNU449T16m細胞による腫瘍形成の結果とTSAHCによる腫瘍生成の低下の度合いを観察した結果である。
【図54】図54は、腫瘍組織に対するH&E(ヘマトキシリン・エオシン)染色結果及び免疫組織化学染色法により分析した結果である。
【図55】図55は、TM4SF5により形成された腫瘍組織におけるp27kip1、pS10p27kip1及びTM4SF5タンパク質に対してウェスタンブロットを行った結果である。
【図56】図56は、TSAHCの腹腔注射による腫瘍の体積及び動物の体重を測定した結果グラフである。
【図57】図57は、TM4SF5による接触阻止消失現象の模式図である。
【図58】図58は、SNU449Cp及びSNU449Tp細胞において、傷が満たされる度合いに対する光学顕微鏡観察写真である。
【図59】図59は、SNU449Cp及びTp細胞において、細胞の移動の度合いを示すクリスタルバイオレット染色後、光学顕微鏡写真及びその染色を溶かし出して260nmの波長において吸光度を測定した結果グラフである。
【図60】図60は、SNU449Tp細胞のコラーゲンゲル上に浸潤する現象を光学顕微鏡にて観察した写真である。
【図61】図61は、SNU449Tp細胞のマトリゲル上に浸潤した細胞を光学顕微鏡にて観察した写真及びその細胞数のグラフである。
【図62】図62は、SNU449Tp、T3、T7、及びT16細胞におけるMMPs酵素活性関係のゼラチンジモグラフィの結果及びMMP9とα−チューブリンタンパク質の発現の度合いを免疫ブロットにて確認した結果である。
【図63】図63は、SNU449P細胞において、MMP−2ルシフェラーゼ及びMMP−9ルシフェラーゼの分析を用いて各遺伝子のプロモーターの転写機能活性を比較したグラフである。
【図64】図64は、TM4SF5が発現される各種の細胞をマウスの尾静脈に注入後、肺組織に形成された腫瘤写真及び転移がんの存在を示すH&E染色写真である。
【図65】図65は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、移動した細胞の染色写真及び細胞数に対するグラフである。
【図66】図66は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、コラーゲンゲルにおける浸潤性現象に対する写真である。
【図67】図67は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、マトリゲルにおける浸潤性に対するグラフである。
【図68】図68は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時または未処理時におけるMMP2及び9ジモグラフィの遂行結果である。
【図69】図69は、SNU449Cp及びSNU449Tp細胞に対して図示の化合物を処理した後の前記細胞の形態変化に関する光学顕微鏡写真である。
【発明を実施するための最良の形態】
【0021】
本発明は、一観点において、腫瘍誘導因子として作用するTM4SF5(transmembrane 4L 6 family member 5)発現と関連する様々な細胞生物学的及び生化学的な現象に基づく抗がん物質のスクリーニング方法に関する。
【0022】
本発明の配列番号1で表わされるポリヌクレオチドから発現される配列番号2で表わされるポリペプチドにより形成される腫瘍タンパク質であるTM4SF5(transmembrane 4L 6 family member 5)は腫瘍特異的な抗原であるL6と相同性を有するタンパク質である。前記TM4SF5は非水溶性のタンパク質であって、細胞膜を通過する4個の領域と、細胞外に存在する2個の環構造、及び細胞質に存在する2個の末端構造を有しており、すい臓がん、胃がん、直腸がん、肝がんなど種々のがん細胞において高く発現することが知られている。
【0023】
TM4SFタンパク質はインテグリンなどの細胞付着分子と複合体を形成して巨大なテトラスパニンウェブをなして、細胞の付着、増殖及び移動などの様々な生物学的機能に寄与すると推定されている。単層よりなる上皮組織は隣り合う細胞間の細胞−細胞接触によるものであって、E−カドヘリンなどの細胞接触に預かる分子間の強い相互結合により、あるいは、インテグリン膜水溶体が細胞外基質となるタンパク質と結合することにより細胞基底部に強く付着して存在する。強く連結された単層組織の崩壊は上皮細胞としての機能を失い、1次的な腫瘍細胞体から腫瘍細胞が離脱してしまう。
【0024】
インテグリンは細胞付着に関連する受容体のファミリーであって、細胞付着がインテグリンと細胞外基質との結合により起こる場合、様々な細胞内信号伝達物質が活性化されてその機能を行い、アクチンフィラメントの再構成にも寄与する。インテグリンによる細胞付着と関連する信号伝達物質としてFAK及びRhoA-GTPase(Rho−GTPase activating protein)などが存在する。
【0025】
本発明者らは、SNU449(KCLB No.00449)肝がん上皮細胞においてTM4SF5はインテグリンによる信号伝達と密接に関連していることを確認し、この関連性を下記の観察により推定した:
(1)TM4SF5の発現はpY397FAKではなく、pY577FAKを増加させる。
(2)Y577FFAK突然変異はTM4SF5によるFAKとRhoGAPとの結合を阻害する。
(3)TM4SF5の発現はFAK−p190RhoGAPまたはFAK−GRAF間の相互作用による結合を促すことによりRhoAの活性を阻害する。
(4)前記機序とは別に、TM4SF5の発現は細胞質内に存在するp27kip1の発現増加及び安定化に寄与し、これはRhoAの活性阻害につながる。
【0026】
すなわち、本発明者らは、前記観察結果に基づいて鋭意努力した結果、TM4SF5の分子的なレベルにおける前記TM4SF5タンパク質の機能及びその発がん機能を生化学的及び細胞生物学的に究明した。
【0027】
Rhoファミリータンパク質の特性
細胞の付着及び離脱と関連するGTP−結合タンパク質であるRhoファミリータンパク質の代表例としてRhoA及びRac1があるが、これらは、GTPが結合されると生理的な活性を示し、GDPが結合されると非活性となってその機能を中止する。一般に、前記Rhoファミリータンパク質は低いGTPase活性を有しており、細胞の付着時に現れる接着斑(focal adhesion、FA)及びインテグリンの集合と解離に重要な役割を果たす。RhoAGTPaseはRhoA、Rac1及びCdc42よりなり、その下位信号伝達媒介分子であるLIMK、PAK1、MLCK、mDia、ROCKなどのタンパク質により、アクチン重合体の形成及びマイオシン軽鎖リン酸化を通じてアクチンの再構成に重要な役割を果たす。
【0028】
FAへの信号伝達成分の集合はFAKの活性に依存するが、これは、FAKによるタンパク質のリン酸化により他のタンパク質が結合可能な位置が露出されるためである。すなわち、FAKが欠如した細胞においてはFAの形成が増加する。すなわち、FAK内チロシンのリン酸化はFA形成を促す。また、FAKは細胞の運動性にも預かるが、FAKが過剰発現されると運動性が増加し、浸透能が増加する。
【0029】
このようなFAKの活性はRhoAタンパク質と密接に関連しているが、FAKが欠如した細胞においてはRhoA活性が増加されている。これは、FAKのC−末端に結合しているRhoGAP(Rho−GTPase activating protein)及びGRAF(GTPase Regulator Associated with FAK)がFAKのリン酸化によりRhoGAPを刺激してRhoAの活性を低下させるためである。
【0030】
しかしながら、Rhoファミリータンパク質のうちRac1は前記RhoAの作用とは反対である。すなわち、Rac1が活性化されるとFAの形成が減少し、細胞の移動を増進させる。細胞の移動を阻害するFA構造物が解離されると、FAの破壊を誘導する成長因子の刺激による細胞移動が促されるためである。
【0031】
TM4SF5の発現によるFAKのリン酸化及びRhoAの活性低下
本発明者らは、TM4SF5の発現はpY397FAKではないpY577FAKを増加させ、FAK−p190RhoGAPまたはFAK−GRAF間の相互作用による結合を促すということを見出した。
【0032】
pY577FAKはFAKタンパク質を構成するアミノ酸のうち577番位にあるチロシンをリン酸化させたものである。このため、TM4SF5の発現によるpY577FAKの増加はFAKのリン酸化増加を意味し、これは、RhoGAPタンパク質を刺激してRhoGAP−FAKまたはGRAF−FAK間結合を促し、これは、RhoAの活性の低下につながる。しかしながら、Rac1の活性は増加する。この明細書における図8に、FAK−RhoA間の信号伝達はRhoGAPと関連するという模式図を示している。
【0033】
FAKタンパク質の577番のアミノ酸であるチロシンをリン酸化(pY577FAK)した場合及びc−Srcタンパク質の416番のアミノ酸であるチロシンをリン酸化(pY416cSrc)した場合に、Rac1とRhoAの活性を調べてみた結果、TM4SF5が発現される細胞においてRac1の活性はいずれも増加するのに対し、RhoAの活性はいずれも減少することを確認した(図3及び図5)。このようなTM4SF5によるpY577FAKの増加とRhoA活性阻害の現象は、TM4SF5に対するsiRNAを用いてTM4SF5の発現を阻害するとさらに反転することが分かる(図4及び図6)。
【0034】
さらに、TM4SF5が発現される細胞において、そうではない母細胞に比べてFAK−p190RhoGAP間の結合、若しくはFAK−GRAF間の結合が強く結合することが分かり(図9)、FAKタンパク質の577番のアミノ酸チロシンをフェニルアラニンに置換した突然変異タンパク質を発現させた場合にはTM4SF5によるFAK−p190RhoGAP間の結合、またはFAK−GRAF間の結合が減少する結果を示している(図10)。
【0035】
これにより、TM4SF5が発現される細胞においてはFAKからRhoAにつながる一連の信号伝達過程においてpY577FAKが重要な役割を果たすということが推察でき、前記結果を踏まえて、pY577FAK発現が減少したり、FAK−RhoGAP間の結合またはFAK−GRAF間の結合が抑制されたり、Rac1の活性が減少したり、RhoA活性が促される場合には前記腫瘍形成タンパク質であるTM4SF5の発現が抑制されることが分かる。
【0036】
TM4SF5の発現による細胞質内p27kip1タンパク質の蓄積及びRhoAの活性低下
本発明者らは、TM4SF5が発現される細胞においてp27kip1タンパク質が顕著に高い濃度にて存在し、これは、細胞質内におけるp27kip1mRNA及びp27kip1タンパク質の高い安定化に起因することを見出した(図12)。これに対し、TM4SF5が発現されない細胞においてp27kip1は、その量も少ないが、存在するほとんどが核に位置している(図13)。
【0037】
p27kip1は核中におけるサイクリン依存性キナーゼ(cyclin-dependent kinase:CDK)の阻害因子であって、腫瘍抑制機能をすることが知られている。前記p27kip1の細胞質への移動は、細胞周期のG0からG1に転移される間に10番のセリン残基がリン酸化されることにより行われる。前記p27kip1Ser10のリン酸化は、キナーゼのスタスミンとの相互作用(Kinase interacting with Stathmin:KiS)及びPKB/Aktタンパク質によりなされるか、あるいは、成長が止まっている細胞またはin vitro環境においてMirk/dyrk1B及びErk1/2によりそれぞれなされる。
【0038】
このようなSer10位のリン酸化は、Thr187またはThr157位よりもTM4SF5の発現とさらに有意に関連しており、p27kip1タンパク質の細胞質内への位置移動及び安定化に寄与する。すなわち、pT157p27kip1とpT187p27kip1はTM4SF5の発現による諸現象との関連性がない。このため、TM4SF5が発現される肝がん細胞などにおいてshRNAなどを用いてTM4SF5の発現を阻害すると、p27kip1の発現及びセリン10番の残基のリン酸化が減少し、さらに、細胞質内に存在するp27kip1タンパク質もまた減少する(図19及び図20)。これは、最近報告されているp27kip1S10A突然変異タンパク質が発現される形質転換マウスにおいて発がん遺伝子Rasによるp27kip1の細胞質への移動が阻害されるという結果と一致する。
【0039】
一方、最近の報告によれば、細胞質内に存在するp27kip1タンパク質は、直接的に結合してRhoAを活性化可能なGTP置換タンパク質(GEF)のRhoAへの結合を抑えることにより、RhoA信号伝達活性を阻害することが知られている。このような細胞質内のp27kip1蓄積によるRhoA信号伝達間の関係を究明するために、RhoGTPaseを活性化させたときのp27kip1タンパク質の細胞内位置変化を調べてみた。RhoAを活性化させたり、その下位伝達媒介因子であるmDiaを活性化させたり、RhoAの活性因子であるLPAを処理した場合、細胞質内のp27kip1タンパク質発現レベルもさらに減少する結果を示している(図27)。これは、RhoA信号伝達が活性化されると、p27kip1タンパク質がそれ以上安定化できず、細胞質ではない核中に位置してしまうためである。すなわち、細胞質内のp27kip1とRhoGTPaseとの間には相互調節の連結環が存在する。このため、TM4SF5の発現はp27kip1の発現増加及び細胞質における安定化に寄与し、これは、RhoAタンパク質の活性阻害につながる。
【0040】
しかしながら、TM4SF5によるp27kip1蓄積に基づくRhoA活性阻害現象と、TM4SF5によるFAK/RhoGAPsタンパク質間の結合に基づくRhoA活性阻害現象は別々に起こる信号伝達経路である。これは、Y577F FAK突然変異タンパク質(チロシン577番がフェニルアラニンに変わった変異体)を過剰発現させると、p27kip1の発現とセリン10番の残基のリン酸化にはいかなる影響も与えないためである(図18)。
【0041】
また、p27kip1の細胞質内の蓄積は、前記RhoA活性とは対照的にRac1タンパク質は活性化させる。すなわち、活性化されたRac1及びその下位伝達媒介体であるPAK1の発現はp27kip1タンパク質の細胞質内蓄積を引き起こす(図27の下)。
【0042】
TM4SF5によるRhoAの活性低下に基づく細胞形態の変化
TM4SF5が発現されない母細胞である肝がん細胞(SNU449)のアクチンは強い繊維状の構造に広がった多角形の細胞形態を維持しており、TM4SF5が発現された細胞(SNU449Tp)におけるアクチンは非正常的な束が長いロッド細胞形態に沿って形成されている(図2)。
【0043】
このような細胞形態の変化に預かる信号伝達を媒介するRac1の活性及びRhoAの活性について、TM4SF5が発現される細胞においてRac1の活性は増加するのに対し、RhoAの活性は減少する。これは、TM4SF5に対するsiRNAを用いてTM4SF5の発現を阻害することにより再び反転される。すなわち、前記siRNAが導入された細胞は母細胞と同様に強い繊維状のアクチン構造を有する多角形の細胞形態に復元されるが、そうではない細胞は依然としてTM4SF5発現の影響により非正常的な束が形成された細胞形態を構成する(図7)。
【0044】
また、TM4SF5が発現される細胞はp27kip1が高く発現されるという特徴があるため、p27kip1タンパク質の変化もまた細胞形態の変化に影響する。TM4SF5が発現されない母細胞にp27kip1を過剰発現させると、細胞の形態が長くなり(図22)、これとは逆に、TM4SF5が発現される細胞においてp27kip1の発現阻害またはセリン残基のリン酸化を阻害する突然変異であるp27kip1S10Aを処理すると、細胞の形態も多角形の母細胞と同じ形態に変化する(図23)。
【0045】
要するに、前記細胞質のp27kip1による細胞形態の変化とRho信号伝達との間の関係と関連して、TM4SF5が発現された細胞においてRhoGTPaseが活性化(RhoA活性化、mDia活性化またはLPA処理)されると、RhoA信号伝達の活性化によりp27kip1タンパク質がそれ以上安定化できずに核中に位置してしまい、細胞質におけるp27kip1タンパク質の発現レベルが減少し、長尺状の細胞形態が母細胞と同じ多角形に戻る。これに対し、活性化されたRac1及びその下位伝達媒介体であるPAK1の発現はTM4SF5が発現されない母細胞の形態を長く変化させ、p27kip1タンパク質の細胞質内蓄積を引き起こす(図27)。
【0046】
このため、結論的に、細胞質内のp27kip1とRhoGTPaseとの間には相互調節の連結環が存在し、細胞質のp27kip1によるRhoGTPaseの活性調節はTM4SF5が発現される細胞形態の変化を誘導するということである。このような細胞形態の変化は後述する現象とも関連がある。
【0047】
TM4SF5の発現によるEMT現象及び細胞間接触
(1)細胞間接触現象の減少及び上皮間葉転換の(EMT、epithelial-mesenchymal transition)誘導
本発明者らは、TM4SF5発現による細胞質へのp27kip1の蓄積は細胞−細胞間接触消失及びEMT現象を誘導することを見出した。
【0048】
細胞間の接触におけるRhoAタンパク質の活性変化は細胞の種類とその信号伝達の特徴に依存するが、上述したTM4SF5によるp27kip1タンパク質の蓄積によるRhoAの活性阻害現象は細胞中のアクチン構造の変化をもたらし、これは、カドヘリン/カテニンタンパク質複合体を通じての細胞−細胞接触とも関連する。
【0049】
TM4SF5が発現されない母細胞においては、E−カドヘリン、ZO1、デスモプラキンなどの細胞接触タンパク質が多量発現し、細胞接触部位によく整列して存在する。これに対し、TM4SF5が発現される細胞においては、前記細胞接触タンパク質の発現は減少されているだけではなく、細胞間接触が上手になされず、散発的な発現パターンを示す(図29)。しかしながら、E−カドヘリンの発現抑制転写因子であるsnail1もまた同時発現減少されることにより、snail1に非依存的で、且つ、TM4SF5に依存的なE−カドヘリンの発現減少を示す(図29)。
【0050】
これに対し、TM4SF5が発現される細胞には、上皮間葉転換(EMT、epithelial-mesenchymal transition)に預かるSMAタンパク質(図28)及びビメンチンタンパク質が高く発現される。このような上皮細胞の移動は細胞の形態形成及び傷治癒などの生理的な過程に重要な役割を果たし、慢性炎症及び腫瘍転移過程に預かる。
【0051】
TM4SF5によるp27kip1タンパク質の蓄積によるRhoAの活性阻害現象はアクチンフィラメントの非正常的な再構成を誘導するため、細胞の形態が長くなり、このような細胞形態の変化は細胞−細胞間の吸着が崩壊するEMT現象を誘導する。要するに、TM4SF5によるSMAタンパク質の発現とEMT現象はRhoAの活性阻害により起こり、細胞形態の長さの変化につながると言える。
【0052】
このため、TM4SF5が発現された細胞においてp27kip1タンパク質の発現を阻害すると、EMT現象が阻止されながら細胞−細胞接触が維持される。これは、p27kip1の発現が阻害されることにより、p27kip1が細胞質ではない核内にさらに多く蓄積されることにより、長く伸びていた細胞の形態が母細胞と同じ多角形の細胞形態に変わるためである。このように、p27kip1タンパク質の細胞内における位置、すなわち、核と細胞質における位置調節は細胞の形態と細胞との間の接触形成に寄与する。
【0053】
加えて、内生性のTM4SF5を有するHuh7肝がん細胞株に対してshRNAを用いてその発現を阻害すると、HGFによる細胞間の接触消失現象が阻害されるため、前記TM4SF5はある信号伝達体系下において本質的にEMTを誘導すると考えられる。
【0054】
(2)接触阻止現象の消失
本発明者らはTM4SF5が発現される細胞においては前記EMT現象により細胞間に高密度にて互いに接触される環境においても隣り合う細胞の存在を認知できずに持続的に細胞分裂及び増殖を維持する、すなわち、細胞接触による生長阻止現象が失われることを確認した。
【0055】
生長の接触阻止現象は、細胞が隣り合う他の細胞と接触したときにそれ以上分裂せず生長が止まるような現象を意味する。既存の報告によれば、E−カドヘリンによる細胞−細胞接触は細胞の増殖と腫瘍生成を抑制することが知られており、最近の報告文献によれば、接触阻止が起こる間に細胞の広がり現象が減少し、成長が止まるということである。このため、EMT現象などにより細胞が互いに接触されていないならば、細胞は隣り合う他の細胞を認識することができずに継続的な分裂をする結果、接触阻止現象が失われるであろう。
【0056】
TM4SF5が発現されない母細胞は多角形の細胞形態を帯びるため、増殖していて、経時的に接触阻止現象によりそれ以上生長しなくなるという飽和曲線を描くが、TM4SF5が発現される長い形態の細胞は細胞間接触消失を誘導するために核が重なり合って存在する程度に細胞が密集しているにも拘わらず、継続的な細胞分裂が起こる(図33)。
【0057】
また、TM4SF5が発現される細胞においては母細胞に比べてS期に存在する細胞集団がさらに多く(図35)、shRNAを用いてTM4SF5の発現を阻害すると、S期に存在するこのような細胞集団が減る(図36)。
【0058】
すなわち、TM4SF5の発現による細胞間接触現象減少及びEMT現象は細胞生長の接触阻止現象を消失させる。図57は、これに対する模式図を示している。図57によれば、発がん遺伝子であると推定されるTM4SF5の過剰発現は細胞質内のp27kip1の発現及び安定化を引き起こし、これは、hKiSまたはPKB/Aktによるセリン10番の残基のリン酸化によるものであり、TM4SF5によるpY577FAKとFAK/RhoGAPsとの結合によりRhoAの活性が阻害される。Rac1の活性はRhoAの不活性化と相反する関係をもって存在するが、RhoAとRac1の下位信号伝達媒介体であるmDiaとPAKはアクチンの構造変化と細胞内の収縮性を調節してしまい、これは、細胞形状の変化につながるため、このような細胞形態の変化は細胞間接触消失現象を誘導して、隣り合う細胞を認識できずに継続的に生長してしまい、接触阻止現象を引き起こしてしまうのである。
【0059】
TM4SF5による細胞移動、浸潤及びがん転移
上述したように、TM4SF5の発現と関連があるFAKが過剰発現されたり、Rac1が活性化されると、細胞の運動性が増加し、Rho活性及びEMT現象による上皮細胞の移動は細胞の形態形成及び傷治癒などの生理的な過程に重要な役割を果たし、さらに、慢性炎症及び腫瘍転移過程に必須である。
【0060】
このため、TM4SF5が発現される細胞株の場合の方が、発現されない細胞株に比べて移動性が大きい(図58及び図59)。このような細胞の移動に伴い、細胞は付着部位から分離されて周りの細胞外基質内に浸潤される。TM4SF5が発現される細胞は細胞外基質を構成するコラーゲンゲルにTM4SF5を発現しない細胞よりもさらに効率よく浸潤するだけではなく(図60)、様々な細胞外基質複合体であるマトリゲルに効率よく浸潤する(図61)。
【0061】
前記作用は部分的にMMPs(matrix metalloproteinase)のタンパク質分解作用によりなされる。MMPsはZn2+依存的なペプチド分解酵素であって、細胞の基質を分解する能力を有している。これらのうちMMP−2とMMP−9はゼラチンとフィブロネクチン基質を分解するが、これらは、TM4SF5が発現される細胞(SNU449Tp、T3、T7及びT16など)内においてその活性が高い(図62)。特に、MMP−9は酵素活性だけではなく、遺伝子発現活性も高い(図62の途中)。
【0062】
ヌードマウスがん転移モデルを用いた実施例9から明らかになったように、TM4SF5を発現するがん細胞において前記細胞移動性及び浸潤性由来のがん転移性はin vivoにおいても現れる。
【0063】
総合的にみたとき、TM4SF5の腫瘍誘導因子としての機能はin vitro細胞とin vivo動物においていずれも細胞の成長が適切に調節されずに継続的に分裂させるため、肝がん細胞などにおいてTM4SF5の発現はFAKのリン酸化増加及び細胞質内のp27kip1タンパク質の蓄積によるRhoAタンパク質の活性阻害を引き起こして、持続的な細胞成長、顕著な細胞周期S期への進入、細胞形態の変化及び付着−非依存的な生長を引き起こす。そして、このTM4SF5の発現細胞株をヌードマウス(in vivo動物)に注入すると腫瘍が形成される。
【0064】
そこで、本発明者らは、腫瘍誘導因子として働くTM4SF5(transmembrane 4L 6 family member 5)発現と関連する上記の様々な細胞生物学的及び生化学的現象に基づいて抗がん物質のスクリーニング方法を発明した。
【0065】
本発明において使用する「抗がん」という用語は、がんの成長を抑制または予防するものを意味する。「がんの成長を抑制または予防する」とは、治療または処理しなかったときと比較時にがんの成長及びがん転移を減少させることを含む概念である。前記「がん転移」は腫瘍(がん)細胞が身体の遠く離れた部分に拡散される過程を意味する。この明細書において前記用語は転移過程により発生されるがんも含む。
【0066】
本発明の方法においては配列番号2で表わされるポリペプチドを発現するがん細胞を培養する。前記ポリペプチドを発現する代表的な場合として、配列番号1で表わされるポリヌクレオチドが挙げられる。
【0067】
配列番号2で表わされるポリペプチドを発現するがん細胞はTM4SF5タンパク質を発現する細胞株であって、一具体例として、上皮細胞においてTM4SF5が持続的に発現されるSNU449肝がん細胞株(KCLB No.00449)を製作し、これを培地において培養することにより行うことができる。前記SNU449細胞の代わりにSNU398細胞(KCLB No.00398)を使用してもよい。
【0068】
本発明において、配列番号1で表わされるヌクレオチドからポリペプチドを発現するがん細胞としては、例えば、すい臓がん細胞、胃がん細胞、肝がん細胞、大腸がん細胞、脳がん細胞、または肺がん細胞または人為的に製造したがん細胞などが挙げられるが、TM4SF5腫瘍タンパク質を発現する細胞であれば非制限的に使用可能である。上記の「人為的に製造したがん細胞」としては、例えば、遺伝子操作を含むクローン技術によりTM4SF5腫瘍タンパク質を発現する細胞が挙げられる。
【0069】
本発明の方法は、次いで、前記工程において培養された細胞を抗がん候補物質にて処理するステップを含む。好ましくは、TM4SF5が細胞内において特異的に誘導する現象を調節すると予想される化合物を処理する。
【0070】
最後に、抗がん候補物質にて処理しない場合と比較して、前記処理した化合物が後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定する。
【0071】
本発明においては、前記物質の抗がん機能を、上述したTM4SF5発現と関連する様々な細胞生物学的及び生化学的現象を基に以下のように定義する:
(i)FAKの577番のチロシン位置のリン酸化減少
(ii)FAK−RhoGAP、もしくはFAK−GRAF間の結合抑制
(iii)細胞質内p27Kip1タンパク質発現減少
(iv)RhoAの活性増加
(v)Rac1の活性減少
【0072】
前記方法において、TM4SF5発現による細胞の形態及び細胞接触パターンと関連する上述した特徴に基づいてさらに後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定する:
(vi)細胞の形態がロッド状から多角形に戻る
(vii)細胞接触関連タンパク質の発現減少
(viii)細胞−細胞間接触が維持される
(ix)細胞生長の接触阻止現象が現れる
(x)SMA(α−smooth muscle actin)タンパク質またはビメンチンタンパク質の発現減少、またはE−カドヘリンタンパク質の発現増加
(xi)上皮間葉転換(EMT)減少
【0073】
このとき、(vii)における前記細胞接触関連タンパク質はE−カドヘリン、ZO1(zonula occludens−1)、β−カテニン及びデスモプラキンよりなる群から選ばれ、(ix)における前記細胞生長の接触阻止の現象は、細胞数の減少、細胞周期のS期に存在する細胞集団の減少または多層増殖の抑制を意味する。すなわち、多層増殖が抑制されて細胞が接触されると単層にのみ伸び、それ以上増殖がなされないことを言う。また、前記細胞数の減少または細胞周期のS期に存在する細胞集団の減少は、前記TM4SF5及びそれと相互作用する膜タンパク質のN結合型グリコシレーションの不活性によりなされる。
【0074】
加えて、上記の方法において、TM4SF5による細胞の移動性、浸潤性増加と関連する上述した特徴に基づいてさらに後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定することもできる:
(xii)細胞外基質若しくは血清の存在下における細胞の移動又は運動性減少
(xiii)細胞外基質を構成するコラーゲンゲル中における浸潤性抑制
(xiv)細胞外基質複合体であるマトリゲル中における浸潤性抑制
(xv)MMP活性減少(発現減少)
【0075】
このとき、前記MMPの好適な例として、MMP−2及びMMP−9が挙げられる。
【0076】
すなわち、腫瘍誘導因子、若しくは、EMT誘導因子TM4SF5の発現の抑制は、FAKのリン酸化または活性変化阻害、細胞質内のp27Kip1発現蓄積によるRhoA活性低下阻害、Rac1及びRhoAを含むRhoGTPaseの活性変化による細胞形態の変化阻害、上皮間葉転換[分化、EMT現象(Epithelial-Mesenchymal Transition)または変換分化]の誘導抑制、S期への進行抑制、接触抑制の消失により、結果的に多層増殖の誘導抑制、吸着−非依存的な増殖またはソフトアガー内における増殖抑制、細胞移動性及び浸潤性阻害、ヌードマウスにおけるがん遺伝子発現がん細胞の注入による腫瘍形成及び転移抑制などの諸現象により確認することができる。
【0077】
本発明は、他の観点において、前記方法によりスクリーニングされた、TM4SF5に対する抗がん機能を有する抗がん物質に関する。「TM4SF5に対する抗がん機能」とは上述した通りである。好ましくは、下記の化学式1または2で表わされるカルコン系の化合物を抗がん物質として決定する。
【0078】
【化3】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールであり、好ましくは、メチル、ベンジル、p−トルイル、p−ニトロフェニルまたはp−フルオロフェニルである。
【0079】
【化4】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールであり、好ましくは、メチル、ベンジル、p−トルイル、p−ニトロフェニルまたはp−フルオロフェニルである。
【0080】
本発明の抗がん物質は、さらに好ましくは、下記表1に記載のカルコン系の化合物である。表1の化合物のうち化合物1で示すTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン]が代表的な化合物である。
【0081】
【表1】
【0082】
前記表1に記載の化合物1から97はそれぞれ以下の通りである。
4′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホネート)−4−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(メタンスルホネート)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンベンゼンスルホニルアミノ)−2−クロロ−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン
【0083】
大韓民国公開特許10−2003−0036993にはカルコン系の化合物が基底膜成分を分解するマトリックスメタロプロテアーゼ(MMP)の活性を阻害する性質を有していることを記載しているが、カルコン系の化合物の抗がん機能については全く言及していないだけではなく、本発明の前記化学式1または2で表わされるカルコン系の化合物はその構造上スルホン基(SO3−)またはスルホンアミド基(SO2NH−)を含有していることを特徴としている。すなわち、カルコン系の化合物のTM4SF5に対する抗がん活性(機能)は前記スルホン基(SO3−)由来のものである。
【0084】
特に、この明細書の実施例12において、カルコンのA−リング(左リング)にOH、NH2、またはOCH3を置換基として有している場合には、TM4SF5の機能を通じて多層構造に伸びる現象を阻害する抗がん性が観察されないが、本発明のカルコン系の化合物でのようにスルホニル基が導入されたスルホンアミドカルコンまたはスルホネートカルコンにおいてはTM4SF5機能阻害による抗がん活性が観察されることを示している。すなわち、カルコン化合物において、「スルホン基(スルホニル)」の導入は現在知られているカルコン誘導体とは全く異なる独特な抗がん活性を発現させる。
【0085】
本発明の実施例においては、これらのうち代表的な化合物であるTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン](表1の1番化合物)を用いて抗がん機能を確認した。
【0086】
前記カルコン系の化合物はTM4SF5による現象を特異的に阻害する拮抗剤として機能する。TM4SF5が発現される細胞株に、前記TSAHCを含むカルコン系の化合物のうち1以上を処理すると、継続的な生長曲線がカルコン系の化合物の濃度、処理時間により依存的に減少し(図43の右)、細胞周期S期への進入(図44)、及び細胞間の接触が復活される(図45)。これに対し、本発明のカルコン系の化合物薬物の効果はTM4SF5が発現されない細胞においては全く効果がないことが判明された(図43の左)。
【0087】
さらに、TM4SF5によるp27kip1、pS10p27kip1、pY577FAKの発現の度合い及びSMAまたはビメンチンの発現においても、前記カルコン系の化合物により効果的に減少し(図46)、TM4SF5が発現されたSNU449細胞株であるSNU449Tp細胞株に本発明のカルコン系の化合物を処理すると、細胞質に存在するp27kip1がそれ以上安定化できずに減少する(図47)。
【0088】
本発明のカルコン系の化合物、特にTSAHCは腫瘍誘導因子TM4SF5及びそれと相互作用する膜タンパク質のN結合型グリコシレーション活性に影響を与えて、TM4SF5が細胞膜において(若しくは、テトラスパニン−ウェブ)インテグリンなどを含む他のタンパク質との結合を通じて発がん機能を行う上で妨害若しくは抑制効果を来たす。
【0089】
このような側面は、TSAHCを処理時にTM4SF5のN結合型グリコシレーション部分のFNGaseFに対する感受性の向上により確認することができる(図50)。このため、本発明のTSAHCなどのカルコン系の化合物はTM4SF5そのものを攻撃してN結合型グリコシレーション作用の構造的な面に変化を与えてTM4SF5のタンパク質−タンパク質結合(テトラスパニン−ウェブ内における結合)に影響を与えることにより、TM4SF5による発がん機能を抑制すると類推することができる。
【0090】
加えて、本発明のカルコン系の化合物はTM4SF5が発現される細胞におけるRhoAの活性阻害現象を修復させ(図48)、長く変化されていた細胞の形態から多角形の形態に復帰させる(図49)。これは、TM4SF5が発現による諸現象を抑制していることを示す結果であって、TM4SF5に対して拮抗剤として働く本発明のカルコン系の化合物に対してTM4SF5による腫瘍形成の治療薬物としての効能を予測可能にする。
【0091】
また、TM4SF5による細胞の移動、浸潤現象もまた本発明のカルコン系の化合物、例えば、TSAHCにより効率よく阻害される。すなわち、TM4SF5が発現される細胞株はTSAHCを処理することにより細胞の移動性(図65)、コラーゲンゲルにおける浸潤性(図66)及びマトリゲルにおける浸潤性(図67)がいずれも有効に阻害される。これは、本発明のカルコン系の化合物が細胞の移動性に預かるMMPsの酵素活性を阻害することを意味し(図68)、これは、TSAHCがTM4SF5による腫瘍誘導だけではなく、腫瘍細胞の転移過程をも有効に阻害可能であることを示唆している。
【0092】
本発明は、さらに他の観点において、前記カルコン系の化合物のうち、好ましくは、前記表1に記載の化合物のうちいずれか1種若しくはそれ以上を有効成分として含有する抗がん用組成物を含む。この明細書の実施例8においては、表1に記載の化合物のうち1種(TSAHC)のみを処理しても抗がん機能を示すことを確認した。
【0093】
本発明による前記カルコン系の化合物は、薬学的に許容可能な塩の形態であるカルコン誘導体の形態で使用することができる。
【0094】
前記塩としては、薬学的に許容可能な遊離酸により形成された酸付加塩が有用である。すなわち、前記カルコン誘導体は当該技術分野における通常の方法により薬学的に許容される酸付加塩から形成可能である。遊離酸としては、無機酸と有機酸を使用することができ、無機酸としては、塩酸、臭酸、硫黄酸、リン酸などを使用することができ、有機酸としては、クエン酸、酢酸、乳酸、マレイン酸、フマル酸、グルコン酸、メタンスルホン酸、コハク酸、酒石酸、4−トルエンスルホン酸、エムボン酸、グルタミン酸またはアスパラギン酸などを使用することができる。好ましくは、無機酸としては塩酸、有機酸としてはメタンスルホン酸を使用することができる。
【0095】
また、本発明による前記カルコン誘導体は、薬学的に許容可能な塩だけではなく、通常の方法により製造可能なあらゆる塩、水和物、溶媒和物をいずれも含むことができる。
【0096】
臨床的な使用のために、本発明による前記カルコン系の化合物またはその塩形態などは単独で投与可能であるが、賦形剤、結合剤、滑沢剤、崩解剤、コーティング物質、乳化剤、懸濁剤、溶媒、安定化剤、吸収促進剤及び/または軟膏基材を混合することにより特定の使用及び好適な目的に適するように剤型化された薬剤学的な混合物の形態で一般的に投与可能である。前記混合物は、経口用、注射用、直腸用または外用投与用として使用可能である。
【0097】
さらに詳しくは、上述したように、前記カルコン系の化合物またはその塩を含有する前記薬剤学的な抗がん用組成物は経口的に投与可能であり、例えば、錠剤、コーティング錠剤、糖衣錠、硬質または軟質ゼラチンカプセル剤、液剤、乳化剤または懸濁剤などの剤型であってもよい。また、投与は、直腸に投与、例えば、座剤を用いて投与可能であり、局所的または経皮的に投与、例えば、軟膏、クリーム、ゲルまたは液剤を用いて投与可能であり、または、非経口的に投与、例えば、注射用の溶液を用いて投与可能である。
【0098】
錠剤、コーティング錠剤、糖衣錠、硬質または軟質ゼラチンカプセル剤の製造のために、本発明のカルコン系の化合物は薬剤学的に不活性な無機または有機賦形剤(薬剤学的に許容される担体)と一緒に混合可能である。錠剤、コーティング錠剤、糖衣錠、硬質ゼラチンカプセル剤に適当な賦形剤の例には、ラクトース、メーズ澱粉またはその誘導体、タルクまたはステアリン酸またはその塩が含まれる。軟質ゼラチンカプセル剤に用いられて好適な賦形剤には、例えば、植物性オイル、ワックス、脂肪、半固形または液体ポリオールなどが含まれる。しかしながら、活性成分の性質により、軟質ゼラチンカプセル剤にいかなる賦形剤も不要になる場合がありうる。液剤及びシロップ剤の製造のために使用可能な賦形剤には、例えば、水、ポリオール、スクロース、転化糖及びグルコースが含まれる。注射用の溶液の製造のために使用可能な賦形剤には、例えば、水、アルコール、ポリオール、グリセリン及び植物性オイルが含まれる。座剤及び局所または経皮適用用の製造のために使用可能な賦形剤には、例えば、天然オイルまたは硬化油、ワックス、脂肪及び半固形または液体ポリオールが含まれる。
【0099】
さらに、前記薬剤学的な組成物は、保存剤、溶解剤、安定化剤、湿潤剤、乳化剤、甘味剤、色素、芳香剤、浸透圧調節用の塩、緩衝剤、コーティング剤または抗酸化剤を含むことができ、それらは治療学的に価値のある他の薬剤を含むこともできる。
【0100】
結果的に、経口投与用の薬剤学的な剤型は顆粒剤、錠剤、糖コーティング錠剤、カプセル剤、丸剤、懸濁剤または乳化剤であってもよく、非経口用の剤型、例えば、靜脈内、筋肉内または皮下、剤型用としては滅菌水溶液の剤型が使用可能であり、これは、等張性溶液を作成するために他の物質、例えば、塩またはグルコースを含むことができる。抗腫瘍剤は、また、座剤またはペッサリーの剤型にて投与可能であり、または、ローション、溶液、クリーム、軟膏または散布剤の形態で外用的に適用可能である。
【0101】
本発明のカルコン系の化合物の1日容量レベルは経口または非経口用に投与されるときに5〜2000mgである。1回投与または2回以上の分割投与が可能である。しかしながら、実際の投与量は投与経路、患者の年齢、性別及び体重、及び疾患の重症度などの種々の関連因子に照らして決定する必要がある。このため、前記投与量はいかなる方法によっても本発明の範囲を限定するものではない。
【0102】
実施例
以下、実施例を挙げて本発明をより詳細に説明する。但し、これらの実施例は単に本発明を一層詳しく説明するためのものであり、本発明の要旨により本発明の範囲がこれらの実施例に限定されるものではないということは、当業界における通常の知識を有する者にとって明らかである。
【0103】
特に、下記の実施例においては肝がん細胞株だけを使用しているが、この他にも、すい臓がん細胞、胃がん細胞、肝がん細胞、大腸がん細胞、脳がん細胞、乳房がん細胞、前立腺がん細胞、子宮がん細胞、食道がん、膀胱がん、または肺がん細胞などのヒト由来の様々な種類のがん組織及びがん細胞株が使用可能であるということは当業界における通常の知識を有する者にとって自明である。
【0104】
実施例1:TM4SF5によるがん細胞培養
(1)細胞株樹立
TM4SF5(GenBank No.:NM−003963)が持続的で且つ安定的に発現される細胞株を樹立するために、ヒトの肝がん細胞であるSNU449細胞(KCLB No.00449)に対照群ベクターであるpLNCX(Clontech Laboratories inc.、PaloAlto、CA)とTM4SF5入りpLNCXを有するレトロウィルスを感染させた。
【0105】
TM4SF5発現レトロウィルスを作成するために、プラスミドpLNCXのHind III/ClaI制限酵素サイトに、Huh7(JCRB No.0403)細胞のcDNAプールからPCR方法により得たcDNAを挿入した後、PT67細胞(ATCC CRL−12284)に導入して安定的にTM4SF5を発現する細胞を得て培養することにより、TM4SF5を発現するレトロウィルスを製作した(Experimental Cell Research, 312:2983, 2006)。以下、前記対照群ベクターを挿入したSNU449細胞はSNU449Cp(またはCp)で表記し、TM4SF5が発現させてなるSNU449細胞はSNU449Tp(またはTp)で表記する。
【0106】
TM4SF5発現レトロウィルスをSNU449細胞に感染させた後、形成された細胞集団を全て集めた細胞株(SNU449Cp及びSNU449Tp)と単一細胞集団を分離して樹立した細胞株(TM4SF5が発現されるT3、T7、T11及びT16細胞)を樹立した。
【0107】
そして、TM4SF5の発現が抑制されるHuh7細胞株を作成するために、TM4SF5に対するshRNAを導入した後、G418(500μg/ml)を用いてshTM4SF5が発現される細胞だけを選択的に分離した。この細胞株に対して10%FBS、0.25μg/mlゲンタマイシンを添加した後、200μg/mlのG418入りまたは200μg/mlのG418未添加のRPMi−1640培地において5%CO2及び37℃の培養機により培養して使用した。
【0108】
(2)材料及び方法
1.TM4SF5抗体の製作
pGEX−5X2ベクター(Phamacia Biotech.)にTM4SF5のc−末端部位(アミノ酸229番から594番までを制限酵素EcoR1を用いて切断)を挿入製作してその塩基配列を確認した。DH5αバクテリアからIPTGに誘導された組換えタンパク質を得るために0.3%SDSとタンパク質分解酵素阻害剤が含まれているPBSを用いた。タンパク質抽出後、電気泳動されたSDSゲルから抗原を抽出してマウスに免疫した。合計で3回の免疫反応を誘導した後、マウスから血清を得て組換えタンパク質と動物細胞抽出物に対する免疫反応性を検査した。
【0109】
2.タンパク質分離及びウェスタンブロット分析
様々な実験条件において得られた細胞溶解物を、種々の条件下の細胞をPBS溶液により洗い取った後にRIPA緩衝溶液を用いて用意した。特定の場合、明示された発現ベクターを導入したり、TM4SF5に対するshRNAを発現させたり、p27kip1に対するsiRNAを発現するアデノウィルスを感染させる条件の細胞が用いられた。
【0110】
加えて、10μMのシクロヘキシミドを明示した時間中に処理した細胞も用いられた。このような溶解物のタンパク質は定量化して同量使用され、ホスホ−Y577FAK、ホスホ−Y416Src、c−Src、RhoA、p120ctn、β−カテニン、pS10p27kip1、FAK、E−カドヘリン、ZO1、Rac1、p190RhoGAP、p27kip1、GRAF、hKiS、デスモプラキン、α−チューブリン、α−カテニン、さらにα−smooth muscle actin(SMA)に対する抗体を用いてウェスタンブロット分析を行った。
【0111】
3.免疫蛍光染色法分析
細胞内のタンパク質を蛍光免疫染色するために、細胞を10%FBS/RPMI−1640により予めコーティングしておいたカバーガラスの上に載せ、細胞の形態が揃って完全に付着するまで37℃において一日中培養した。細胞に上述の特定の遺伝子を導入させたり様々なウィルスを感染させる場合には、上記した時間中に培養した後に使用したり、10mMリゾホスファチジン酸(LPA)を処理した細胞はカバーガラスに付着させた後、1時間かけて処理した後に使用した。蛍光免疫染色法は当該タンパク質の1次抗体を反応させた後、余剰の抗体を洗い取った後、蛍光物質が付着された2次抗体と反応させて再び洗い取って蛍光分析可能なスライドを作成し、アクチンの染色法はローダミンが付着されたファロイジンを反応させて行われた。
【0112】
使用された抗体はE−カドヘリン、β−カテニン、FLAG、DAPI(4’−6−ジアミジノ−2−フェニルインドール)、BrdU(5−ブロモ−2’−デオキシ−ウリジン)、p27kip1、さらにZO1(zonula occludens−1)などが使用された。蛍光免疫染色がなされたカバーガラスは観察のためにスライドに載せて蛍光顕微鏡または共焦点レーザー顕微鏡により観察した。
【0113】
4.RT−PCR
P27kip1のmRNAレベルを調べるためにRT−PCRをTM4SF5非発現若しくは発現細胞株にアクチノマイコンDを処理して様々な時間帯に細胞からトータルRNAを抽出してそれからmRNAを得た後、p27Kip1に対するプライマーを用いてPCR方法により行った。使用されたプライマーはセンス(5’−taa ccc ggg act tgg aga ag−3’:配列番号3)とアンチセンス(5’−gct tct tgg gcg tct gct c−3’:配列番号4)であった。
【0114】
5.p27kip1の細胞質タンパク質層と核タンパク質層の分画化
TM4SF5が発現されるSNU449Tp細胞とT16細胞におけるP27kip1の細胞質発現と核内発現量を調べるためにLiangら(Nature Medicine, 8:1153-1160, 2002)が提示した方法により分画化が行われた。
【0115】
6.ソフトアガーアッセイ
60mm培養皿に0.7%アガーと10%FBSが含まれた培養液を予め敷いて固めた後、その上に106個の細胞を0.3%アガーと10%FBSが含まれた細胞培養液と一緒に投入した。これらの培養皿は5%CO2が与えられる37℃培養機において25日間培養され、2日おきに200μg/mlG418と10%FBSが含まれた培養液に交替した。形成された細胞集団はデジタルカメラ付き顕微鏡により観察、記録した。
【0116】
7.統計の分析
データの意味のある差を得るために平均値の比較をスチューダントt−tsetにより行った。p値が0.05以下である場合にその差は意味あると言える。
【0117】
実施例2:TM4SF5によるpY577FAKタンパク質発現及びRhoAの活性阻害
(1)TM4SF5が発現される細胞の形態学的な特性
実施例1において樹立されたTM4SF5が発現されないSNU449母細胞(SNU449pまたはp)、対照群ベクターが持続的に発現されるSNU449Cp細胞及びTM4SF5を発現する細胞株(SNU449Tp、T3、T7、及びT16)に対する各細胞の形態を光学顕微鏡を用いて確認し、さらにTM4SF5に対する抗体及びα−チューブリンに対する抗体を用いてウェスタンブロットを行った。
【0118】
そして、TM4SF5の発現有無による細胞の様々なアクチン構造を調べるために、対照群細胞(SNU449Cp)及びTM4SF5が発現される細胞(SNU449Tp)をガラスカバースリップの上に生長させた後、ファロイジンが付着されたローダミンにより染色した。
【0119】
その結果、クローン由来のT3、T7、T16及びTpはTM4SF5が発現されない母細胞であるSNU449p細胞と比較したとき、細胞形態の側面においてその長さの伸張が起こっていることが確認された(図1)。そして、SNU449細胞のアクチンを蛍光染色法により染色して観察した結果、強い繊維状の構造に広がった多角形の細胞形態を維持しており、TM4SF5が発現されるSNU449Tp細胞においてアクチンは非正常的な束が長い細胞形態に沿って形成されていた(図2)。
【0120】
(2)pY577FAK発現による効果
上述した細胞形態の変化に預かる信号伝達を調べるために、実施例1において樹立されたTM4SF5を発現する細胞株(SNU449Tp、T3、T7、及びT16)において、FAKタンパク質の577番のアミノ酸であるチロシンがリン酸化されたpY577FAKタンパク質発現の度合い、c−Srcタンパク質の416番のアミノ酸であるチロシンがリン酸化されたpY416cSrcタンパク質発現の度合い、Rac1活性及びRhoA活性を調べてみた。前記全体の細胞溶解物を集めて、図3及び図5に表記のタンパク質に対する抗体を用いてウェスタンブロット分析を行ったり試験管内においてRhoAとRac1をプルダウンしてその活性を分析した。
【0121】
また、TM4SF5に対するshRNA(shTM4SF5)を用いてTM4SF5の発現を阻害する場合の変化を観察した。母細胞であるSNU449p、対照群細胞であるSNU449Cp及びTM4SF5が発現される各種の細胞において、緑蛍光タンパク質(GFP)に対するshRNA、ランダムな塩基配列を有するshRNA及びTM4SF5に対するshRNAを導入した後に細胞を回収してウェスタンブロット分析とRhoA活性分析を行った。
【0122】
その結果、TM4SF5が発現される前記細胞においてはpY577FAKタンパク質とpY416cSrcタンパク質発現及びRac1活性は増加されていたが、RhoA活性は減少されていることを確認した(図3及び図5)。そして、TM4SF5による前記pY577FAKの増加とRhoA活性阻害現象はTM4SF5の発現を阻害することにより再び反転することが分かる(図4及び図6)。
【0123】
そして、SNU449Tp細胞にpEGFPとshTM4SF5を一緒に導入して24時間後にカバースリップの上に載せ、24時間の培養時間が経過した後、ファロイジンが付着されたローダミンとDNA染色のためのDAPIを一緒に蛍光染色した結果、前記shTM4SF5が導入された細胞は対照区に属するSNU449細胞と同様に強い繊維状のアクチン構造を有する多角形の細胞形態に修復されるが、そうではない隣り合う細胞は依然としてTM4SF5発現の影響により細胞形態の変化が現れないことを確認した(図7)。
【0124】
(3)RhoA活性阻害
併せて、RhoGAPタンパク質を通じてのRhoAの不活性化過程におけるpY577FAKの役割を調べてみた。Cp細胞とTp細胞から得られた細胞溶解物に対してFAKに対する抗体を用いて免疫沈降法を行い、免疫沈降された混合物はp190RhoGAP、GRAF、FAK抗体を用いてウェスタンブロット分析を行った。また、前記SNU449Tp細胞に(HA)3−tagged FAK野生型タンパク質とY577F突然変異タンパク質を一時的に過剰発現させて、2日後に細胞溶解物を免疫沈降法を行って図10において表記している抗体を用いてウェスタンブロットを行った。
【0125】
その結果、FAK−p190RhoGAP間の結合、若しくはFAK−GRAF間の結合が、TM4SF5が発現される細胞において、そうではないSNU449Cp細胞に比べて強く結合することが分かる(図9)。そして、このようなTM4SF5によるFAK−p190RhoGAP間の結合若しくはFAK−GRAF間の結合はFAKタンパク質の577番のアミノ酸チロシンをフェニルアラニンに変えた突然変異タンパク質を発現させることにより減少される結果を示し(図10)、TM4SF5が発現される細胞からRhoAにつながる一連の信号伝達過程においてpY577FAKが重要な役割を果たすということを推察することができた。
【0126】
また、FAKと関連するGTPase調節因子であるGRAFに対して、対照群細胞にFLAG−tagged GRAFを一時的に過剰発現して24時間後に上述した方法に従いカバースリップの上に載せ、一日中に培養した後、アクチンとFLAG抗体を用いて蛍光免疫染色法を行った結果、TM4SF5が発現されない対照群細胞におけるGRAFまたはp190RhoGAPの一時的な発現は細胞形態の変化を引き起こすことを確認した(図11)。
【0127】
このため、結論的にSNU449細胞において発現されるTM4SF5はRhoAの活性を阻害して細胞長さの伸長を誘導し、これは、FAK−p190RhoGAP若しくはFAK−GRAFタンパク質間の相互作用によるものであることが分かる。
【0128】
実施例3:TM4SF5による細胞質内のp27kip1の蓄積効果
(1)TM4SF5の発現によるp27kip1の発現増加
TM4SF5によるRhoA活性阻害現象がp27kip1による効果であるかどうかを調べるために、SNU449CpとSNU449Tp細胞をp27kip1に対する抗体とDAPIを用いて蛍光免疫染色した。そして、前記細胞の核と細胞質を実施例1−(2)−5)における方法に従い分画化してp27kip1に対する抗体とa−チューブリンに対する抗体を用いてウェスタンブロット分析を行った。
【0129】
その結果、TM4SF5が発現される細胞においてp27kip1タンパク質が顕著に高濃度にて細胞質に位置することが分かる(図12)。これに対し、TM4SF5が発現されない対照群細胞においては少量ではあるがほとんどが核に位置していた(図13)。
【0130】
TM4SF5によるp27kip1発現レベルとSer10リン酸化の関係を調べるために、実施例1において樹立した様々な細胞株溶解物を得て図14に表記の抗体を用いてウェスタンブロット分析を行った結果、このようなp27kip1タンパク質の細胞質における存在はhKIS(human Kinase-interantigwith Stathmin)タンパク質と活性化されたPKB/Aktタンパク質によりアミノ酸10番のセリンのリン酸化を通じてなされることを確認した(図14)。
【0131】
また、アクチノマイシンD(10mg/ml、ActD)が処理された細胞からRNAを分離してp27kip1及びGAPDHをRT−PCR分析し、シクロヘキサミド(10mM、CHX)を処理した細胞の溶解物を得てp27kip1に対する抗体を用いてウェスタンブロット分析を行った後、その結果として得られるバンドの強度を測定して対照群となるバンドの強度に対する増減をグラフにて示した(図16)。
【0132】
その結果、TM4SF5が発現される細胞においてp27kip1タンパク質発現が顕著に増加されることはp27kip1mRNAとタンパク質の高い安定化のためであることが確認され(図15)、このような現象はTM4SF5に対するsiRNAを用いてその発現を阻害したとき、細胞形態の変化がTM4SF5が発現されないSNU449細胞の形態に戻り、細胞質においてp27kip1タンパク質発現レベルも減少されると共にセリン残基のリン酸化もまた起こらない結果からなお一層明らかになった(図17)。
【0133】
さらに、TM4SF5が細胞質のp27kip1タンパク質の安定化に預かるという仮説はTM4SF5が内生的に発現される様々な肝がん細胞においても有効であることが分かる。
【0134】
内生的にTM4SF5を発現している肝がん細胞株に対して、GFPのshRNA、ランダムな塩基配列(Scr)を有するshRNA及びTM4SF5に対するshRNAを導入して、48時間後に得た全体の細胞溶解物及びSNU449CpとSNU449Tp細胞の溶解物に図18に表記の抗体を用いてウェスタンブロット分析を行った結果、内生的に発現されるTM4SF5の発現の阻害はp27kip1発現及びSer10のリン酸化レベルを減少させ(図18)、SNU449TpTp細胞に(HA)−tagged FAK野生型タンパク質とY577F突然変異を過剰発現させた後、表示された抗体によりウェスタンブロット分析を行った結果、Y577FFAK突然変異タンパク質はp27kip1の発現レベルやSer10のリン酸化には影響しなかった(図19)。また、Huh7肝がん細胞株にScrまたはshTM4SF5と一緒にpEGFPを導入してp27kip1とDAPIを免疫蛍光染色法により染色した結果、内生的に発現されるTM4SF5の阻害は細胞質に存在するp27kip1を減少させることを確認した(図21)。
【0135】
この結果はTM4SF5の発現がp27kip1の発現増加及び細胞質における安定化に寄与し、これは、RhoAタンパク質の活性阻害につながることを証明している。特に、図19の結果はp27kip1の発現とFAKタンパク質のリン酸化は互いに関連性がないことを示唆し、これは、TM4SF5によるp27kip1タンパク質及びRhoA活性阻害現象とTM4SF5によるFAK/RhoGAPs結合によるRhoA活性阻害現象は別個の信号伝達機序において寄与されたものであることが分かる。
【0136】
(2)細胞質のp27kip1蓄積による細胞形態変化
実施例2から明らかなように、TM4SF5が発現される細胞は細胞形態が長く、前記実施例3−(1)から明らかなように、TM4SF5が発現される細胞はp27kip1が高く発現されるという特徴があるため、このようなp27kip1タンパク質の変化が細胞形態の変化にいかなる影響を与えるかを調べてみた。
【0137】
実施例1において樹立したSNU449Cp細胞にFLAG−tagged p27kip1野生型タンパク質を過剰発現させ、48時間後に前記細胞をDAPI、ファロイジンが付着されたローダミン、及びFLAGに対して免疫蛍光染色を行って共焦点顕微鏡により観察した結果、TM4SF5が発現されない母細胞にp27kip1を過剰発現させたときに細胞の形態が長くなることを確認した(図22)。
【0138】
そして、前記SNU449Tp細胞をsiRNAp27kip1アデノウィルスを用いて24時間かけて感染させ、FLAG−tagged p27kip1S10Aを導入して24時間かけて培養した後、免疫蛍光染色を行ない、得られた細胞溶解物を図24に明示された抗体を用いてウェスタンブロットを行った。
【0139】
その結果、このようにTM4SF5が発現される細胞においてp27kip1の発現阻害と一緒にセリン10番がアラニンに変わってその残基のリン酸化が起こらない突然変異体であるp27kip1S10Aを一緒に導入して発現すると、細胞質への移動が阻害され、細胞の形態も多角形のSNU449若しくはSNU449Cp細胞と同じ形態に変化していることを確認することができた(図23及び図24)。
【0140】
前記SNU449Tp細胞にpEGFPと活性化された形態のRhoA63L突然変異ベクターを一緒に導入してアクチンの構造を免疫染色した結果、活性化されたRhoAにより細胞形態変化の反転が起こり、多角形をなしていた(図25)。
【0141】
細胞質のp27kip1による細胞形態の変化とRhoA信号伝達間の関係を究明するために、RhoGTPaseを活性化させたときの細胞形態の変化とp27kip1タンパク質の細胞内位置変化を調べてみた。
【0142】
先ず、SNU449CpとSNU449Tp細胞をカバースリップの上に培養した後、RhoGTPaseを活性化させる因子であるLPA(リゾホスファチジン酸)10mMを1時間かけて処理してDAPIとp27kip1を免疫染色した。その結果、SNU449Tp細胞においてLPA(リゾホスファチジン酸)は細胞質のp27kip1を減少させた(図26)。
【0143】
また、SNU449Tp細胞にpEGFPとmDia活性型突然変異を導入し、且つ、SNU449Cp細胞にpEGFPと活性型のRac1(Rac161L)またはPAK1(PAK1−caax)を導入して免疫蛍光染色分析を行った。その結果、驚くことには、RhoAの下位伝達媒介因子であるmDiaの活性化が細胞の形態をSNU449若しくはSNU449Cp細胞と同じ多角形に戻る結果を示し、細胞質内のp27kip1タンパク質発現レベルも再び減少される結果を示していた(図27)。
【0144】
これは、RhoA信号伝達の活性化によりp27kip1タンパク質がそれ以上安定化できず、核中に位置するためであると推察することができる(図26、図27の上)。これに対し、活性化されたRac1とその下位伝達媒介体であるPAK1の発現はTM4SF5が発現されない細胞であるにも拘わらず、細胞の形態を長く変化させてp27kip1タンパク質の細胞質内蓄積を引き起こした(図27の下)。
【0145】
この結果からみたとき、細胞質内のp27kip1とRhoA及びRac1を含むRhoGTPaseとの間には相互調節の連結環が存在し、細胞質のp27kip1によるRhoGTPaseの活性調節はTM4SF5が発現される細胞形態の変化を誘導することが分かる。
【0146】
実施例4:TM4SF5によるEMT現象
TM4SF5が細胞−細胞間接触の調節にも影響するかどうかを調べてみた。先ず、TM4SF5が発現される細胞とそうではない対照群細胞の溶解物により細胞−細胞接触形成に預かるタンパク質の発現レベルを免疫ブロット方法を用いて確認した。
【0147】
その結果、TM4SF5非発現細胞株においてはE−カドヘリン、ZO1、デスモプラキンなどの細胞接触タンパク質の発現が大幅に増加されていたが、TM4SF5が発現される細胞においてはその発現が減少されていた。しかしながら、上皮間葉転換(EMT)に預かるSMAタンパク質は高く発現されていた(図28)。
【0148】
さらに、高密度にて培養したSNU449Cp細胞とSNU449Tp細胞をカバースリップの上に載せた後、E−カドヘリン、β−カテニン、ZO1タンパク質の発現パターンを免疫染色法を用いて調べてみた結果、TM4SF5を発現しないSNU449細胞とSNU449Cp細胞においては細胞接触部位によく整列されて存在していたE−カドヘリン、β−カテニン、ZO1などが、TM4SF5が発現される細胞においては細胞間接触が上手になされず、散発的に分布する発現パターンを示していた(図29)。
【0149】
SNU449Tp細胞にp27kip1に対するshRNAアデノウィルスを実施例1において説明した通りに感染させた後、E−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色した結果、p27kip1の発現が阻害されたら、E−カドヘリン、β−カテニン、ZO1タンパク質が規則的な形態に再配列され(図30)、SMA発現が減少されることを確認した。
【0150】
また、内生的に発現されるTM4SF5がEMT現象の調節にいかなる役割を果たすかを究明するために、shRNA及びshTM4SF5を発現するHuh7細胞に対してTM4SF5とa−チューブリンの発現を調べ(図31)、HGF(100ng/ml)を24時間かけて処理した後、ZO1を免疫染色した結果を観察した。
【0151】
Huh7細胞株は、TM4SF5発現有無とは無関係に、コロニーをなして成長する特性があるため、細胞間接触パターンが相当良好である。このため、HGF(hepatocyte growth factor)を用いてEMT現象を誘導させた後、TM4SF5が細胞−細胞間接触の形成に及ぼす影響を観察した。その結果、HGFによるHuh7細胞の分散現象(若しくは、接触崩壊)がTM4SF5が阻害されたHuh7細胞において低減することを観察することができた。このため、TM4SF5が細胞−細胞間接触の消失を引き起こしていることが分かる(図32)。
【0152】
実施例5:TM4SF5による細胞生長の接触阻止現象の消失
SNU449CpとSNU449Tp細胞をカバースリップの上において培養した後、DNAとアクチン構造をDAPIにより染色した結果を図33及び図34に示す。
【0153】
矢印にて示す部分は細胞分裂と核が重なり合う部分を示し、細胞がそれ以上成長する空間がないにも拘わらず継続的に分裂し続けていることを示す。また、これらの細胞の生長速度を測定するために6ウェル培養皿に5X104個の細胞をはじめとして12時間おきにその数を数えて記録した結果をグラフにて示す。実際にTM4SF5が発現される449Tp細胞株の生長曲線は継続的に増加するパターンを示し、対照群細胞である449Cp細胞株は経時的に増殖していて、接触阻止現象によりそれ以上成長しなくなる飽和曲線を描いていた。このような特徴は細胞が低密度にて存在する状況においても現れる。すなわち、低密度の細胞のうちTM4SF5発現する細胞は核が重なり合って細胞分裂などをしながら増殖をするが、TM4SF5を発現しないSNU449細胞とSNU449Cp細胞は高密度にて互いに接触される環境においてはそれ以上生長しないという特徴を示している(図34)。
【0154】
また、TM4SF5が発現される様々な細胞株(T7、T16、及びT3)の細胞周期のS期分布を調べてみた結果、TM4SF5を発現しないSNU449細胞と、SNU449Cp細胞細胞に比べてTM4SF5が発現される細胞においてS期に存在する細胞集団がさらに多量であると分かる(図35)。SNU449Cp細胞とSNU449Tp細胞に対照群shRNAとshTM4SF5を導入した後、プロピジウムヨード法によりDNAを染色して流速細胞分析機を用いて細胞周期を分析した結果、shRNAを用いてTM4SF5の発現を阻害したとき、S期に存在する細胞集団が低減することを確認した(図36)。
【0155】
BrdU浸透免疫染色法を用いた調査においても、TM4SF5が発現される細胞はそうではない細胞に比べてDNA合成中のS基の細胞分裂が顕著に高く存在することが分かる(図37)。このような現象は、LPAによるRhoAの活性化や(図38)、p27kip1の発現阻害(図39)、さらにはE−カドヘリンの再発現(図40)などを通じて特異的にDNA合成中のS基の細胞分裂が阻害されると現れた。
【0156】
特に、E−カドヘリンタンパク質の発現を再び高めるSNU449T16m細胞(SNU449T16細胞由来であるため、SNU449T16mと称する。)は、TM4SF5が発現されるにも拘わらず、細胞間の接触がTM4SF5非発現細胞と同じパターンを示し(図42)、SMAタンパク質の発現が減少する結果を示している(図41)。
【0157】
このため、上記の結果からみたとき、TM4SF5が発現される細胞にE−カドヘリンの発現を再び増加することにより、TM4SF5はEMT現象を通じて細胞生長の接触阻止現象を消失させ、これは、細胞間の接触と接触による生長阻止現象との間において緊密な機能の関連性があることを示唆している。
【0158】
実施例6:TM4SF5により形成された腫瘍組織の特徴
TM4SF5による腫瘍発生効果を2種類の方法により検証した。先ず、TM4SF5が発現されたり発現されない様々な細胞を付着−非依存的な生長環境下に放置した。27日間ソフトアガーにおいて培養した後に観察した。実施例1の6)において言及した方法によりSNU449細胞株をもってソフトアガーアッセイを行い、ソフトアガー培養皿に細胞を培養してから27日目に位相差顕微鏡により細胞コロニーの様子を観察・記録した。
【0159】
その結果、TM4SF5が発現される細胞はそうではない細胞に比べて細胞コロニーが顕著に多量生存していることが分かる。しかしながら、TM4SF5が発現される細胞にE−カドヘリンを再発現させた449T3m、T16m細胞においては細胞コロニーの形成が顕著に低減していた。このため、TM4SF5によるEMT現象は細胞付着−非依存的な生長と関連するということを推定することができた(図52)。
【0160】
次に、TM4SF5を発現したり発現しない様々なSNU449細胞株をヌードマウスに注入した。ヌードマウスはBALB/cAnNCrjBgi−nu(Orient.Co.Ltd)を基本種とし、全て生後4〜5週齢、体重が約20gの雌性だけを使用した。滅菌消毒されたねずみかごにおいて滅菌された水と飼料を供給し、層板気流を持続的に維持したクリーンベンチ中に放置して清浄な環境を維持し、全ての術式もクリーンベンチ中において行った。
【0161】
ヌードマウスの昼夜間生物学的な周期のために室内灯を12時間は点灯し、且つ、12時間は消灯した。SNU細胞株を基本細胞株として、SNU449Cp、SNU449Tp、SNU449T16、SNU449T16m細胞の浮遊液を遠心分離して細胞を濃縮し、血球計により細胞数を1X107cells/200μlに算定して、用意された細胞を26ゲージの注射針を用いて1cc注射器によりヌードマウスの脇腹に皮下注射接種した。注射後、実験動物はソウル大学前臨床実験室動物飼育法により維持され、SNU449Tp細胞による腫瘍形成を観察、記録した。
【0162】
TM4SF5が発現される細胞株は腫瘍形成に優れていたが、母細胞(449p)と対照群細胞(449Cp)、449T3m、T16m細胞はこれといった腫瘍が形成されていなかった(図53)。
【0163】
そして、SNU449CpとSNU449Tp細胞を注射したマウスから得られた腫瘍組織をH&E(ヘマトキシリン・エオシン)染色法により染色してその組織形態を観察し、p27kip1とpS10p27kip1発現パターンを免疫組織化学染色法により分析した。免疫化学組織染色法によりTM4SF5が誘導した腫瘍組織の中央部位は死滅した細胞集団が大幅に現れ、細胞質に存在するp27kip1とセリン10番の残基のリン酸化も増加されて現れた(図54)。さらに、前記組織から得られたタンパク質溶解物を用いてウェスタンブロット分析を行った結果、TM4SF5による腫瘍組織は、そうではない細胞による組織と比較したとき、p27kip1の発現レベルとリン酸化レベルが高く現れた(図55)。
【0164】
実施例7:TM4SF5による細胞の移動性増加
(1)細胞の移動性分析
Boyden Transwell chamber 24 well plates(Costar, Cambridge, MA)を使用した。ポリカーボネートフィルター(8μm porosity)上のチャンバーにSNU449Cp及びSNU449Tp細胞を5x104セル/ウェルずつ1日中に37℃、5%のCO2培養機において無血清RPMI(Roswell Park Memorial Institute)培地にて培養した後、24時間後にインサートチャンバー上方のメンブレンを綿棒によりよく拭き取り、70%メタノールに約7分間浸してメンブレン下方にある細胞を固定した。
【0165】
このようにSNU449Cp、SNU449Tp細胞を高密度になるように培養した後、適当な厚さのワウンドを形成した後、経時的にワウンドが満たされる度合いを観察し、光学顕微鏡により記録した。その結果、TM4SF5が発現される細胞株の場合、そうではない細胞株に比べて傷回復速度が増加されることを確認した(図58)。
【0166】
また、前記SNU449Cp、Tp細胞株をトランスウェルのインサートウェルにおき、無血清培地において培養し、インサートの下チャンバーには10%の血清または無血清培地を入れて細胞の経時移動の度合いを観察するために、0.005%クリスタルバイオレットにより1分間染色した後、PBSにより3回洗浄して光学顕微鏡により前記染色された細胞を観察した(図59の上の写真)。前記染色薬を溶解して260nmの波長において吸光度を測定して図表化した(図59の下グラフ)。その結果、TM4SF5が発現される細胞株の場合、そうではない細胞株に比べて細胞移動性が増加されることを確認した(図59)。
【0167】
(2)ジモグラフィ分析
分泌されたMMPsの基質分解活性を測定するためにゼラチンジモグラフィを行った。図62に示す細胞株を60mm培養皿に放置した後、18時間後に血清を除去した培養液に交替して24時間さらに培養した。分泌されたMMPsタンパク質を得るために培養上清だけを取って濃縮した。MMPsの基質となるゼラチンをSDS−PAGEの分離ゲル(ランニングゲル)に添加し、減圧条件下において電気泳動した後、1%トリトンX−100緩衝溶液において再変成させ、次いで、酵素反応緩衝溶液(10mM CaCl2、0.15M NaClと50mMトリス−HCL、pH7.5)により37℃において18時間反応させた。この後、0.5%クマシーブリリアントブルー(Coomassie brilliant blue)R250により3時間染色させた後、10%酢酸、30%メタノールにおいて透明なバンドが現れるまで脱色した。
【0168】
その結果、MMP−2と9はゼラチンとフィブロネクチン基質を分解するが、TM4SF5が発現されるSNU449Tp、T3、T7、及びT16細胞においてその活性が高くなっていることを確認した。すなわち、TM4SF5によりMMPs酵素活性が増加されることを確認した(図62)。
【0169】
そして、SNU449P細胞にMMP−2ルシフェラーゼとMMP−9ルシフェラーゼを様々な濃度のpcDNA3.1−MycHis−TM4SF5と一緒に導入した後、36時間後にルシフェラーゼ活性を測定して、各MMP遺伝子プロモーターの転写活性を比較した結果、MMP−9は、酵素活性だけではなく、遺伝子発現活性も高いことを確認した(図63)。これより、TM4SF5はMMPs遺伝子発現活性にも影響することが分かる。
【0170】
(3)3Dコラゲンを用いた細胞の浸潤性分析
細胞の浸潤性を分析するためにCytodex-3マイクロキャリアを用いた。SNU449CpとSNU449Tp細胞を細胞をマイクロキャリアビーズに付着して培養し、血清と培地成分入りのコラーゲンゲルに混ぜて37℃において培養した後、経時的に現れる現象を観察した。また、TM4SF5が発現されるSNU449Tp細胞がコラーゲンゲル中に浸潤する現象をも光学顕微鏡により観察した(図60)。
【0171】
また、上記のSNU449Tp細胞の浸潤性を分析するために、実施例7−(1)において言及したBoyden Transwell chamber 24 well platesのインサートにマトリゲルを入れて37℃において固めた後、上述した方法に従い細胞を培養して24時間後にインサートの上方のメンブレンを綿棒によりよく拭き取り、36時間後にメンブレンの下方に浸潤された細胞を染色して観察した。その結果、上記のSNU449Tp細胞が様々な細胞外基質複合体であるマトリゲルを有効に浸潤することを確認した(図61)。
【0172】
実施例8:TM4SF5機能を阻害する抗がん物質(拮抗剤)の選択
TM4SF5が細胞内において特異的に誘導する現象を調節すると予想される化合物のうち、前記表1に示すカルコン系の化合物を本発明の方法によりスクリーニングした。図69に記載のカルコン化合物もまた抗がん候補物質として使用した。
【0173】
上述した抗がん機能を示す候補物質を本発明の抗がん物質として決定したが、スクリーニングされた代表的な化合物を前記表1に示す。
【0174】
前記抗がん物質の抗がん機能をより確実に調べるために、表1の化合物のうちTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン]を使用した。
【0175】
TM4SF5が発現される細胞株にTSAHCを処理したとき、継続的な生長曲線がTSAHCの濃度、処理時間により依存的に減少することを観察し(図43の右)、細胞周期S期への進入(図44)、そして細胞間の接触が復活されることを確認した(図45)。
【0176】
これに対し、TSAHC薬物の効果については、TM4SF5が発現されない細胞においては全く効果がないことが分かる(図43の左)。さらに、TM4SF5によるp27kip1、pS10p27kip1、pY577FAKの高い発現や度合い、さらにSMAの発現はTSAHCにより有効に減少されることが分かる(図46)。また、449Tp細胞株にTSAHCを処理した結果、細胞質に存在するp27kip1が減少することから(図47)、TSAHCがTM4SF5に対する拮抗作用をすることによりp27kip1タンパク質がそれ以上安定化できないことが分かる。加えて、TM4SF5が発現される細胞におけるRhoAの活性阻害現象を修復させ(図48)、長く変化されていた細胞の形態が、多角形のTM4SF5が発現されない対照群細胞株と同じ形態に戻ることを確認することができた(図49)。
【0177】
また、TSAHCの処理時にTM4SF5のN結合型グリコシレーション部分のFNGaseFに対する感受性が向上することを確認した(図50)。この結果から、TSAHCはTM4SF5そのものを攻撃してN結合型グリコシレーションの構造的な側面に変化を与えてTM4SF5のタンパク質−タンパク質結合(テトラスパニン−ウェブ内における)に影響することにより、TM4SF5による機能を抑制することができると推定可能である。
【0178】
さらに、TSAHCの処理は、内生的にTM4SF5を発現する細胞については細胞の生長若しくは増殖を抑制したが、発現しない細胞株は根本的にいかなる影響を及ぼさなかった(図51)。このため、これらの結果からみて、TM4SF5に対して拮抗剤として作用するカルコン系の化合物がTM4SF5による腫瘍形成の予防及び治療に効果があることを確認した。
【0179】
カルコン系の化合物が示すTM4SF5阻害率(iC50)を表2に示す。
【0180】
【表2】
【0181】
実施例9:本発明のカルコン誘導体の人体がん細胞株に対する試験管内細胞毒性
本発明の化合物カルコン誘導体の人体がん細胞株に対する試験管内細胞毒性を調べるために、1989年アメリカの国立がん研究所(NCi:National Cancer institute)において開発したスルホロダミンB(SRB)方法を使用した。
【0182】
この実験に使用した人体がん細胞株として、HCT15(colon adenocarcinoma、結腸がん;ATCC CCL−225)、PC−3(prostate adenocarcinoma、前立腺がん;ATCC CRL−1435)、A−549(lung carcinoma、肺がん;ATCC CCL−185)が挙げられ、胃がん細胞はいずれも生命工学研究所において継代培養中のものを使用した。
【0183】
前記人体がん細胞株は10%子牛血清が含まれているRPMi1640培地(ギブコ社カタログno.31800)を用いて、37℃の温度条件下、5%の二酸化炭素培養機において培養し、継代は1週間につき2回行った。
【0184】
細胞を付着面から分離するときには、0.25%トリプシン(シグマ社製)及び3mMCDTA(1、2−シクロヘキサンジアミンテトラアセト酸、シグマ社製)をリン酸塩緩衝溶液(シグマ社製)に溶解させた溶液を使用した。
【0185】
検索用の試料(表1に記載のカルコン化合物誘導体)は少量のDMSO(ジメチルスルホキシド、シグマ社製)に溶かして実験に必要となる濃度まで実験用の培地において希釈して最終的なDMSOの濃度が0.5%以下になるようにし、数段階(最終濃度:30、10、3、1、0.3μg/mL)に亘って希釈された試料溶液はがん細胞に加える前に0.22μmマイクロフィルター(ミリポア社製)によりろ過して無菌状態にした。次いで、通常のSRB法により人体がん細胞株に対する細胞毒性を測定した。
【0186】
本発明の化合物カルコン誘導体の人体がん細胞株に対する細胞毒性(ED50、μg/mL)を下記表3に示す。
【0187】
【表3】
【0188】
上記の結果は、本発明のカルコン系の化合物が人体がん細胞株に対する細胞毒性を示すために抗がん剤として使用可能であることを示唆する。
【0189】
実施例10:TSAHCの腫瘍形成抑制効果
(1)TSAHC投与
前記実施例6に従い用意したヌードマウスに対して、形成された腫瘍の長さと幅をキャリパーにより2日おきに1回ずつ測定して下記の公式(Volume (mm3) = Length × Width2 × 1/2、Length:長径、Width:短径)により腫瘍の体積を算出した。
【0190】
ヌードマウスに誘発された腫瘍の体積が200mm3のサイズにて着床された後、任意に2群に分けて(5mg/kgと50mg/kg)生理食塩水にTSAHCを懸濁させて2日おきに1回ずつ3週間26ゲージの注射針を用いて腹腔内注射する方法により投与した。
【0191】
(2)腫瘍組織の免疫組織化学検査とウェスタンブロット分析
ヌードマウスは、SNU449Tp細胞を注入してから6週目に、またはTSAHCを投薬してから3週目に頸椎脱臼の方法により犠牲し、免疫組織化学検査のために腫瘍組織を切開し、ホルムアルデヒドにより固定した後にパラフィン標本にした。組織のタンパク質発現レベルをウェスタンブロットにより分析するために抽出した腫瘍組織は直ちに液体窒素を用いて凍結標本にして0.1%SDS入りのRIPA緩衝溶液に溶解させてタンパク質混合物を得て分析した。用意された6μm厚さのパラフィン切片に対しては、H&E(ヘマトキシリン・エオシン)染色と免疫組織化学検査を行った。
【0192】
その結果、TM4SF5が発現される細胞を移植したマウスの腫瘍(個体数=6)は持続的に大きくなるのに対し、TSAHCを5mg/Kg、若しくは50mg/Kg濃度にて腹腔注射したマウスの腫瘍はそれぞれ約45%、88%だけそのサイズが減少することを確認した(図56)。
【0193】
実施例11:TSAHCのがん転移抑制効果
(1)がん転移実験動物モデル樹立
がん転移実験動物モデルの準備は、上述した腫瘍誘導モデルと同じ方法により行われた(実施例6参照)。SNU449Cp、SNU449Tp細胞の浮遊液を遠心分離して細胞を濃縮し、血球計により細胞数を1X107と算定して用意された細胞を1cc注射器及び29ゲージのインシュリン注射針を用いてヌードマウス尾靜脈に血管注射して接種した。3週間放置後、マウスを犠牲して肺組織を摘出した後、ホルマリンにより固定してH&E(ヘマトキシリン・エオシン)染色を行った。
【0194】
その結果、TM4SF5が発現される細胞の場合、肺組織に腫瘤が形成されていることを確認した(図64)。図64における矢印の個所が腫瘤部位であって、細胞が密集している個所である。
【0195】
(2)TSAHCのTM4SF5による細胞の移動性及び浸潤性抑制効果
SNU449Cp細胞とSNU449Tp細胞を6−ウェル培養皿におき、24時間後にTSAHC20μMを処理して18時間培養した後、トランスウェルチャンバーのインサートウェルに移した。18時間後に移動した細胞を固定、染色して光学顕微鏡により観察し、その数を数えてグラフ化した(図65)。その結果、TM4SF5による細胞の移動性がTSAHCにより阻害されることを確認した。
【0196】
また、SNU449Cp細胞とSNU449Tp細胞をマイクロキャリアビーズに付着してTSAHC20μMと一緒に18時間かけて培養した。ビーズ−細胞を血清と培地成分入りのコラーゲンゲルに混ぜ、培養を表記の時間だけ行いながら観察、記録した結果、TM4SF5による細胞の浸潤性がTSAHCにより阻害されることを確認した(図66)。
【0197】
上記の如き方法によりマトリゲルが厚くコーティングされたトランスウェルチャンバーを用いて24時間かけて培養した後、マトリゲルを通過した浸潤性の細胞数を数えてグラフ化した結果、マトリゲルにおける浸潤性もまた有効に阻害されることを確認した(図67)。
【0198】
そして、Cp細胞とTp細胞にTSAHC20μMを24時間かけて処理した。24時間後、血清を除去した培地に交換した後、18時間かけて培養した。培養液を取って濃縮した後、上述した方法に従いジモグラフィを行った結果、TM4SF5によるMMPs酵素活性がTSAHCにより阻害されることを確認した(図68)。
【0199】
この結果は、TSAHCがTM4SF5による腫瘍誘導だけではなく、腫瘍細胞の転移過程もまた有効に阻害可能であることを示唆している。
【0200】
実施例12:TSAHC類似体との効果比較
最後に、TM4SF5が預かる現象においてTSAHCと構造的に類似する誘導体の効果を観察した。このとき、使用したDMSO(ジメチルスルホキシド)及びカルコン類似体の化学構造は下記の通りである。
【化5】
【0201】
SNU449Cp細胞とSNU449Tp細胞を6−ウェル培養皿に敷き、前記図に示す薬物を20μMずつ24時間かけて処理した後、光学顕微鏡を用いて細胞の形態変化を観察、記録した。
【0202】
その結果、図69に示すように、TSAHCと構造がほとんど同様であるものの、異なるR基で置換された薬物は、同じ濃度条件(20μMにて24時間処理)下においてTSAHCは有効にTM4SF5が発現される細胞の成長を多層に成長せずに単層に成長させ、同時に細胞の形態をTM4SF5が発現しないSNU449対照群細胞のように変化させることを確認した。これにより、TM4SF5による細胞成長阻害効果はTSAHC化合物の特定のR基により調節されることを類推することができる。
【0203】
前記TSAHC類似体は、前記化学構造式に示すように、カルコンのA−リング(左リング)にOH、NH2、またはOCH3を置換基として有しているが、この場合には、TM4SF5の機能を通じて多層に成長する現象を阻害する抗がん性が観察されなかったが、TSAHCのようにスルホニル基が導入されたスルホンアミドカルコンにおいては、TM4SF5機能阻害を通じての抗がん活性が観察された。すなわち、スルホニルの導入により現在知られているカルコン誘導体とは全く異なる独特な抗がん活性を観察することができた。
【産業上の利用可能性】
【0204】
以上述べたように、本発明は、腫瘍形成におけるTM4SF5の発現と関連する分子的な機序に基づく抗がん物質のスクリーニング方法であって、腫瘍生成及び転移に対する拮抗作用をする治療及び予防目的の物質をスクリーニングする上で有用である。また、前記方法によりスクリーニングされたカルコン系の化合物を含有する抗がん用組成物は、体重の減少、肝及び脾臓という臓器に対して外傷的な異常を来たすことなく、しかも、毒性反応無しに抗がん効果を示すことから、抗がん用の薬剤として有効に使用可能であるというメリットがある。
【0205】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を有する者にとって、このような具体的な記述は単なる好適な実施様態に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかであろう。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定まると言えるであろう。
【技術分野】
【0001】
本発明は抗がん物質のスクリーニング方法に係り、さらに詳しくは、TM4SF5による腫瘍の形成と発生及び転移を阻害する化合物のスクリーニング方法に関する。また、本発明は、該方法により抗がん物質としてスクリーニングされたカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物に関する。
【背景技術】
【0002】
トランスメンブラン4 L6ファミリーメンバー5(transmembrane 4 L6 family member 5:TM4SF5またはL6H)は腫瘍特異抗原であるTM4SF1(L6)と相同性を有するタンパク質であって、前記TM4SF1(L6)は、トランスメンブラン4 L6ファミリー(transmembrane 4 L6 family)に属する。トランスメンブラン4 L6ファミリーに属するタンパク質は、TM4SF1(L6)に加えて、IL−TMP、L6Dなどがある。腫瘍特異抗原として知られている前記TM4SF1(L6)は、直腸がん、肺がん、乳房がん、さらには卵巣がんなどにおいて高く発現されていることが知られており、TM4SF1(L6)と相同性を有するタンパク質であるTM4SF5(L6H)もまたすい臓がん、胃がん、直腸がん、さらに肝がんなど種々のがん細胞において高く発現されると当業界に報告されている(Muller-Pillasch, F. et al., Gene, 208:25, 1998、 Pascual-Le Tallec, L. et al., J. Clin. Endocrinol. Metab., 87:501, 2002)。また、TM4SF1(L6)に比べてTM4SF5のmRNAのレベルは胃がん組織においては低く発現された場合があるが、肝がん細胞株と胃がん細胞株においては高く発現されたものであるとも報告されている(Kaneko, R. et al., Am. J. Gastroenterol., 96(12):3457, 2001)。
【0003】
これらの研究に基づいて、近年、米国特許US6、350、581B1においてTM4SF5が新規な発がん遺伝子として登録されており(TUANと表記)、本発明者によってCos7繊維芽細胞にTM4SF5を人為的に発現させた結果、アクチン構造の再構成と接着斑の形成、及び接着斑キナーゼ(focal adhesion kinase:FAK)のアミノ酸チロシン925番のリン酸化が、インテグリンα2により引き起こされること、及びこのような現象は細胞成長因子を含む血清により阻害、調節されることを研究結果として報告している(Lee, S.Y. et al., Exp. Cell Res., 312:2983, 2006)。
【0004】
しかしながら、腫瘍形成におけるTM4SF5の分子的な機序はほとんど知られていないのが現状である。すなわち、分子的なレベルにおけるTM4SF5タンパク質の機能及びその発がん機能に係る証拠は生化学的、細胞生物学的に全く知られていない。
【0005】
一方、カルコンは食用の植物に豊富に存在する物質であって、フラボノイドまたはイソフラボノイドの前駆体として知られている。前記のカルコンの誘導体は植物の黄色色素の構成成分であって、植物の色合いに影響するだけではなく、有害な紫外線から植物を保護する役割を果たすこともある(天然物化学研究法、ウ・ウォンシク著、ソウル大学出版部)。カルコン誘導体は菊科植物であるハルシャギク属に多量存在する。代表的なカルコン類には2’、6’−ジヒドロキシ−4’−メトキシカルコン、カルタミンなどがあり、これらは桂皮、紅花、胡椒などの植物に含有されている。ジヒドロカルコンは主としてバラ科及びカラムラサキツツジ科植物に含有されており、りんごに存在するジヒドロカルコンであるフロリジンはりんごの病気に対する抵抗力と関係があると報告されている。
【0006】
このようなカルコン誘導体は、例えば、抗原虫性(Liu, M. et al., J. Med. Chem., 44:4443, 2001)、抗炎症性(Babu, M.A. et al., Bioorg. Med. Chem., 10:4035, 2003)、免疫調節(Barfod, L. et al., int. immunopharmacol., 2:545, 2002)、一酸化窒素阻害(Rojas, J. et al., Bioorg. Med. Chem. Lett., 12:1951, 2002)、抗がん(Kumar, S.K. et al., J. Med. Chem., 46:2813, 2003)、抗HIV(Artico, M. et al., J. Med. Chem., 41:3984, 1998)活性など極めて様々な薬物学的な活性を示すことが知られている。
【0007】
また、大韓民国公開特許10−2003−0036993号公報には、前記カルコン化合物が基底膜成分を分解するマトリックスメタロプロテアーゼ(Matrix metalloproteinase、MMP)の活性を阻害する性質を有していることが記載されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 15:5514, 2004)。
【0008】
カルコン系の化合物の中でも、スルホンアミドまたはスルホネート基が置換されたカルコン系の化合物には自然に存在するカルコン系の化合物が有していない特異な生物学的特性があることが報告されている。特に、本発明者により、スルホンアミドカルコン系の化合物が高いα−グルコシダーゼ阻害活性を有していることが報告されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 15:5514, 2004、大韓民国公開特許10−0751899)。加えて、本発明者により、スルホネートカルコン系の化合物が選択的なカリウムイオンチャンネル阻害活性を有していることも報告されている(Park, K.H. et al., Bioorg. Med. Chem. Lett., 2007)。
【0009】
しかしながら、これまで、前記スルホンアミド及びスルホネートカルコン系の化合物が発がん遺伝子であるTM4SF5を阻害する機序を通じて抗がん機能を発現するということは全く解明されていない。
【0010】
そこで、本発明者らは、ヒトのがん細胞において現れるTM4SF5の発がん遺伝子としての機能は、上皮間葉転換(epithelial-mesenchymal transition:EMT)現象の促進、接触阻止現象の消失(loss of contact inhibition)によるものであり、これは、FAKのリン酸化及び細胞質内のp27kip1タンパク質の蓄積によるRhoAタンパク質の活性阻害及び細胞形態の変化につながる一連の過程を通じてなされるという新たな経路を見出し、前記諸現象に基づいてTM4SF5の発現による腫瘍の形成と発生及び転移を阻害する抗がん物質をスクリーニングするために鋭意努力した結果、本発明を完成するに至った。
【発明の詳細な説明】
【0011】
《技術的課題》
本発明の目的は、TM4SF5による腫瘍の形成と発生及び転移を阻害する抗がん物質のスクリーニング方法を提供することにある。
【0012】
本発明の他の目的は、前記方法によりスクリーニングされた抗がん物質、特に、スルホンアミド及びスルホネートカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物を提供することにある。
【技術的解決方法】
【0013】
上記の目的を達成するために、本発明は、下記のステップを含む、抗がん物質のスクリーニング方法を提供する:
(a)配列番号2のポリペプチドを発現するがん細胞を培養した後、抗がん候補物質で処理するステップ、
(b)前記抗がん候補物質で処理されたがん細胞に対して、
(i)接着斑キナーゼ(FAK)を構成するアミノ酸配列のうち577番位のチロシンのリン酸化の確認、(ii)FAKのRho−GTPase activating protein(RhoGAP)への結合、またはFAKのGTPase Regulator Associated with FAK(GRAF)への結合測定、(iii)細胞質内p27Kip1の発現量及び細胞質内安定化測定、(iv)RhoAの活性測定、及び(v)Rac1の活性測定、
の少なくとも1つを検出するステップ、及び
(c)抗がん候補物質で処理しない場合に比べて、
(i)FAKの577番位のチロシンのリン酸化減少、(ii)FAKのRhoGAPへの結合、またはFAKのGRAFへの結合抑制、(iii)細胞質内p27Kip1発現減少及び細胞質内安定化、(iv)RhoA活性増加、または(v)Rac1活性減少
の事象が現れた場合に、抗がん候補物質を抗がん物質として選択するステップ。
【0014】
本発明において、前記ステップ(b)は、細胞の形態確認、細胞接着関連のタンパク質発現確認、細胞間接触パターンまたは接触生長確認、α平滑筋アクチン(α−smooth muscle actin:α−SMA)タンパク質またはビメンチンタンパク質の発現確認、E−カドヘリンタンパク質の発現確認及び上皮間葉転換(epithelial-mesenchymal transition、EMT)確認よりなる群から選ばれる1以上をさらに行い、前記ステップ(c)は、細胞の形態がロッド状から多角形に戻るか、あるいは、細胞接着関連のタンパク質の発現が減少するか、あるいは、細胞−細胞間の接触が維持されるか、あるいは、細胞生長の接触阻止現象が現れるか、あるいは、α−SMAタンパク質またはビメンチンタンパク質の発現が減少するか、あるいは、E−カドヘリンタンパク質の発現が増加するか、あるいは、上皮間葉転換(EMT)が減少する場合をさらに含むことを特徴としてもよい。
【0015】
本発明において、前記ステップ(b)は、細胞外基質(extracellular matrix)または血清の存在下での細胞の移動又は運動性確認、細胞外基質を構成するコラーゲンゲルへの浸潤確認、細胞外基質複合体を構成するマトリゲルへの浸潤確認及びMMP活性測定よりなる群から選ばれる1以上をさらに行い、前記ステップ(c)は、細胞外基質または血清の存在下で細胞の移動又は運動性が減少するか、あるいは、コラーゲンゲル中における浸潤が減少するか、あるいは、マトリゲル中における浸潤が減少するか、あるいは、MMP活性が減少する場合をさらに含むことを特徴としてもよい。
【0016】
本発明は、また、前記方法によりスクリーニングされた、下記の化学式1または化学式2で表わされる抗がん用のカルコン系の化合物及びこれを有効成分として含有する抗がん用組成物、並びに前記化合物の抗がん治療及び予防の用途を提供する。
【0017】
【化1】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【0018】
【化2】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【0019】
本発明の他の特徴及び具現例は、下記の詳細な説明及び特許請求の範囲からなお一層明らかになる。
【図面の簡単な説明】
【0020】
【図1】図1は、SNU449p、SNU449Cp、Tp、T3、T7、T16細胞の写真及びウェスタンブロットの結果である。
【図2】図2は、SNU449CpとSNU449Tpに対してローダミン標識ファロイジンで染色したアクチン写真である。
【図3】図3は、表記のタンパク質に対するウェスタンブロットの結果である。
【図4】図4は、RhoAとRac1に対するin vitroプルダウンアッセイによる活性分析結果である。
【図5】図5は、SNU449p、SNU449Cp及びSNU449Tp細胞に対するウェスタンブロット分析結果である。
【図6】図6は、図5のいくつかの細胞のRhoA活性分析結果である。
【図7】図7は、SNU449Tp細胞にpEGFPとshTM4SF5を一緒に導入して24時間培養後、ローダミン標識ファロイジン及びDAPIを蛍光染色した写真である。
【図8】図8は、FAK−RhoA間の信号伝達に対する模式図である。
【図9】図9は、SNU449Tp細胞におけるp190RhoGAP、GRAF、FAK抗体を用いたウェスタンブロット分析結果である。
【図10】図10は、SNU449Tp細胞に(HA)3−tagged FAK野生型タンパク質とY577F突然変異タンパク質を過剰発現した後のウェスタンブロット分析結果である。
【図11】図11は、対照群細胞にFLAG−tagged GRAFタンパク質を過剰発現した後の蛍光免疫染色写真である。
【図12】図12は、SNU449CpとSNU449Tp細胞に対する蛍光免疫染色写真である。
【図13】図13は、核と細胞質を分画化した後、p27kip1タンパク質に対する抗体とα−チューブリンに対する抗体を用いたウェスタンブロット分析結果である。
【図14】図14は、p27kip1タンパク質発現レベルとSer10リン酸化に対する、表記の抗体を用いたウェスタンブロット分析結果である。
【図15】図15は、p27KiP1のセリン10番のアミノ酸残基のリン酸化と関連するSNU449CpとSNU449Tp細胞に対する蛍光免疫染色写真である。
【図16】図16は、p27kip1またはGAPDHの遺伝子のmRNAへの転写を確認するためのRT−PCR分析結果及びウェスタンブロットの結果である。
【図17】図17は、SNU449Tp細胞をpEGFPとshTM4SF5を一緒に導入した後、p27kip1タンパク質とDAPIで細胞核を免疫蛍光染色法により染色した写真である。
【図18】図18は、図示の抗体を用いてウェスタンブロット分析した結果である。
【図19】図19は、SNU449Tp細胞に(HA)−tagged FAK野生型タンパク質とY577F突然変異体を過剰発現させた後のウェスタンブロット分析結果である。
【図20】図20は、肝がん細胞株Huh7に対して様々なshRNAを導入した後のウェスタンブロット分析結果である。
【図21】図21は、免疫蛍光染色法により染色した写真である。
【図22】図22は、SNU449Cp細胞にFLAG−tagged p27kip1タンパク質を過剰発現させた後の免疫蛍光染色法による共焦点顕微鏡写真である。
【図23】図23は、SNU449Tp細胞においてp27kip1の発現を阻害した後の免疫蛍光染色写真である。
【図24】図24は、図示の抗体を用いたウェスタンブロット分析結果である。
【図25】図25は、SNU449Tp細胞に活性型RhoA63L突然変異ベクターを導入した後のアクチン構造を免疫染色した写真である。
【図26】図26は、SNU449CpとSNU449Tp細胞にLPAを処理した後の免疫染色写真である。
【図27】図27は、SNU449Tp細胞にmDia活性型の突然変異体、活性型のRac1(Rac1 61L)及び活性型のPAK1(PAK1−caax)を導入した後に免疫蛍光染色分析を行った写真である。
【図28】図28は、細胞接触と関連するタンパク質に対するウェスタンブロット分析結果である。
【図29】図29は、SNU449Cp及びSNU449Tp細胞に対するE−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色分析結果である。
【図30】図30は、SNU449Tp細胞においてp27kip1発現を阻害した場合、E−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色分析結果である。
【図31】図31は、shRNAとshTM4SF5を発現するHuh7細胞におけるウェスタンブロット分析結果である。
【図32】図32は、前記樹立された細胞株にHGF(100ng/ml)を24時間処理した後、ZO1の位置を免疫染色した写真である。
【図33】図33は、SNU449CpとSNU449Tp細胞に対してDAPIで細胞核を染色した写真(矢印:細胞分裂と核が重なり合った部分)及び12時間おきに1回ずつ測定した細胞数のグラフである。
【図34】図34は、SNU449Tp細胞が重なり合って細胞分裂を示す染色写真である(矢印:細胞分裂中にある核)。
【図35】図35は、SNU449Cp細胞とTM4SF5が発現されるT7、T16、及びT3の細胞周期のS期分布を調べてまとめたグラフである。
【図36】図36は、SNU449Cp細胞及びSNU449Tp細胞にshRNAとshTM4SF5を導入した後、フローサイトメトリーを用いてS細胞周期を分析したグラフである。
【図37】図37は、SNU449CpとSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図38】図38は、LPAを処理したSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図39】図39は、p27kip1shRNAアデノウィルスを感染させたSNU449Tp細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図40】図40は、SNU449T16細胞にE−カドヘリンを再発現させたSNU449T16m細胞に対してBrdU浸透免疫染色法を用いてS期に入った細胞の分布を分析した結果である。
【図41】図41は、SNU449Cp(Cp)、SNU449T16(T16)、及びSNU449T16m(T16m)細胞に対する、表記の分子の発現レベルに対する調査結果である。
【図42】図42は、SNU449T16(T16)及びSNU449T16m(T16m)細胞に対してβ−カテニン、ZO1、またはE−カドヘリンを免疫染色した写真である。
【図43】図43は、SNU449Cp細胞とSNU449Tp細胞に対してDMSOまたはTSAHC(4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン)を表記の濃度にてそれぞれ処理した後、生存細胞の数を数えて記録したグラフである。
【図44】図44は、SNU449Tp細胞にDMSOまたは20μMのTSAHCを処理し、BrdUを24時間かけて浸透させた後、蛍光染色法により確認した写真である。
【図45】図45は、SNU449Tp細胞に対してDMSOまたは20μM TSAHCを処理した後のE−カドヘリンまたはβ−カテニンの免疫染色写真である。
【図46】図46は、表記のタンパク質のウェスタンブロット分析結果である。
【図47】図47は、p27kip1の蛍光免疫染色分析結果である。
【図48】図48は、in vitro RhoA活性分析結果である。
【図49】図49は、DNAとアクチンの免疫染色写真である。
【図50】図50は、SNU449CpとSNU449Tp細胞に対してPNGaseF及びEndoH酵素を処理した後のウェスタンブロットの結果である。
【図51】図51は、TM4SF5を発現する細胞及び発現しない細胞に対するTSAHCの処理による生長/増殖の変化を示すグラフである。
【図52】図52は、SNU449細胞株のソフトアガーアッセイを行った結果である。
【図53】図53は、SNU449Cp、SNU449Tp、及びSNU449T16m細胞による腫瘍形成の結果とTSAHCによる腫瘍生成の低下の度合いを観察した結果である。
【図54】図54は、腫瘍組織に対するH&E(ヘマトキシリン・エオシン)染色結果及び免疫組織化学染色法により分析した結果である。
【図55】図55は、TM4SF5により形成された腫瘍組織におけるp27kip1、pS10p27kip1及びTM4SF5タンパク質に対してウェスタンブロットを行った結果である。
【図56】図56は、TSAHCの腹腔注射による腫瘍の体積及び動物の体重を測定した結果グラフである。
【図57】図57は、TM4SF5による接触阻止消失現象の模式図である。
【図58】図58は、SNU449Cp及びSNU449Tp細胞において、傷が満たされる度合いに対する光学顕微鏡観察写真である。
【図59】図59は、SNU449Cp及びTp細胞において、細胞の移動の度合いを示すクリスタルバイオレット染色後、光学顕微鏡写真及びその染色を溶かし出して260nmの波長において吸光度を測定した結果グラフである。
【図60】図60は、SNU449Tp細胞のコラーゲンゲル上に浸潤する現象を光学顕微鏡にて観察した写真である。
【図61】図61は、SNU449Tp細胞のマトリゲル上に浸潤した細胞を光学顕微鏡にて観察した写真及びその細胞数のグラフである。
【図62】図62は、SNU449Tp、T3、T7、及びT16細胞におけるMMPs酵素活性関係のゼラチンジモグラフィの結果及びMMP9とα−チューブリンタンパク質の発現の度合いを免疫ブロットにて確認した結果である。
【図63】図63は、SNU449P細胞において、MMP−2ルシフェラーゼ及びMMP−9ルシフェラーゼの分析を用いて各遺伝子のプロモーターの転写機能活性を比較したグラフである。
【図64】図64は、TM4SF5が発現される各種の細胞をマウスの尾静脈に注入後、肺組織に形成された腫瘤写真及び転移がんの存在を示すH&E染色写真である。
【図65】図65は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、移動した細胞の染色写真及び細胞数に対するグラフである。
【図66】図66は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、コラーゲンゲルにおける浸潤性現象に対する写真である。
【図67】図67は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時またはTSAHC未処理時における、マトリゲルにおける浸潤性に対するグラフである。
【図68】図68は、SNU449Cp及びSNU449Tp細胞に対するTSAHC処理時または未処理時におけるMMP2及び9ジモグラフィの遂行結果である。
【図69】図69は、SNU449Cp及びSNU449Tp細胞に対して図示の化合物を処理した後の前記細胞の形態変化に関する光学顕微鏡写真である。
【発明を実施するための最良の形態】
【0021】
本発明は、一観点において、腫瘍誘導因子として作用するTM4SF5(transmembrane 4L 6 family member 5)発現と関連する様々な細胞生物学的及び生化学的な現象に基づく抗がん物質のスクリーニング方法に関する。
【0022】
本発明の配列番号1で表わされるポリヌクレオチドから発現される配列番号2で表わされるポリペプチドにより形成される腫瘍タンパク質であるTM4SF5(transmembrane 4L 6 family member 5)は腫瘍特異的な抗原であるL6と相同性を有するタンパク質である。前記TM4SF5は非水溶性のタンパク質であって、細胞膜を通過する4個の領域と、細胞外に存在する2個の環構造、及び細胞質に存在する2個の末端構造を有しており、すい臓がん、胃がん、直腸がん、肝がんなど種々のがん細胞において高く発現することが知られている。
【0023】
TM4SFタンパク質はインテグリンなどの細胞付着分子と複合体を形成して巨大なテトラスパニンウェブをなして、細胞の付着、増殖及び移動などの様々な生物学的機能に寄与すると推定されている。単層よりなる上皮組織は隣り合う細胞間の細胞−細胞接触によるものであって、E−カドヘリンなどの細胞接触に預かる分子間の強い相互結合により、あるいは、インテグリン膜水溶体が細胞外基質となるタンパク質と結合することにより細胞基底部に強く付着して存在する。強く連結された単層組織の崩壊は上皮細胞としての機能を失い、1次的な腫瘍細胞体から腫瘍細胞が離脱してしまう。
【0024】
インテグリンは細胞付着に関連する受容体のファミリーであって、細胞付着がインテグリンと細胞外基質との結合により起こる場合、様々な細胞内信号伝達物質が活性化されてその機能を行い、アクチンフィラメントの再構成にも寄与する。インテグリンによる細胞付着と関連する信号伝達物質としてFAK及びRhoA-GTPase(Rho−GTPase activating protein)などが存在する。
【0025】
本発明者らは、SNU449(KCLB No.00449)肝がん上皮細胞においてTM4SF5はインテグリンによる信号伝達と密接に関連していることを確認し、この関連性を下記の観察により推定した:
(1)TM4SF5の発現はpY397FAKではなく、pY577FAKを増加させる。
(2)Y577FFAK突然変異はTM4SF5によるFAKとRhoGAPとの結合を阻害する。
(3)TM4SF5の発現はFAK−p190RhoGAPまたはFAK−GRAF間の相互作用による結合を促すことによりRhoAの活性を阻害する。
(4)前記機序とは別に、TM4SF5の発現は細胞質内に存在するp27kip1の発現増加及び安定化に寄与し、これはRhoAの活性阻害につながる。
【0026】
すなわち、本発明者らは、前記観察結果に基づいて鋭意努力した結果、TM4SF5の分子的なレベルにおける前記TM4SF5タンパク質の機能及びその発がん機能を生化学的及び細胞生物学的に究明した。
【0027】
Rhoファミリータンパク質の特性
細胞の付着及び離脱と関連するGTP−結合タンパク質であるRhoファミリータンパク質の代表例としてRhoA及びRac1があるが、これらは、GTPが結合されると生理的な活性を示し、GDPが結合されると非活性となってその機能を中止する。一般に、前記Rhoファミリータンパク質は低いGTPase活性を有しており、細胞の付着時に現れる接着斑(focal adhesion、FA)及びインテグリンの集合と解離に重要な役割を果たす。RhoAGTPaseはRhoA、Rac1及びCdc42よりなり、その下位信号伝達媒介分子であるLIMK、PAK1、MLCK、mDia、ROCKなどのタンパク質により、アクチン重合体の形成及びマイオシン軽鎖リン酸化を通じてアクチンの再構成に重要な役割を果たす。
【0028】
FAへの信号伝達成分の集合はFAKの活性に依存するが、これは、FAKによるタンパク質のリン酸化により他のタンパク質が結合可能な位置が露出されるためである。すなわち、FAKが欠如した細胞においてはFAの形成が増加する。すなわち、FAK内チロシンのリン酸化はFA形成を促す。また、FAKは細胞の運動性にも預かるが、FAKが過剰発現されると運動性が増加し、浸透能が増加する。
【0029】
このようなFAKの活性はRhoAタンパク質と密接に関連しているが、FAKが欠如した細胞においてはRhoA活性が増加されている。これは、FAKのC−末端に結合しているRhoGAP(Rho−GTPase activating protein)及びGRAF(GTPase Regulator Associated with FAK)がFAKのリン酸化によりRhoGAPを刺激してRhoAの活性を低下させるためである。
【0030】
しかしながら、Rhoファミリータンパク質のうちRac1は前記RhoAの作用とは反対である。すなわち、Rac1が活性化されるとFAの形成が減少し、細胞の移動を増進させる。細胞の移動を阻害するFA構造物が解離されると、FAの破壊を誘導する成長因子の刺激による細胞移動が促されるためである。
【0031】
TM4SF5の発現によるFAKのリン酸化及びRhoAの活性低下
本発明者らは、TM4SF5の発現はpY397FAKではないpY577FAKを増加させ、FAK−p190RhoGAPまたはFAK−GRAF間の相互作用による結合を促すということを見出した。
【0032】
pY577FAKはFAKタンパク質を構成するアミノ酸のうち577番位にあるチロシンをリン酸化させたものである。このため、TM4SF5の発現によるpY577FAKの増加はFAKのリン酸化増加を意味し、これは、RhoGAPタンパク質を刺激してRhoGAP−FAKまたはGRAF−FAK間結合を促し、これは、RhoAの活性の低下につながる。しかしながら、Rac1の活性は増加する。この明細書における図8に、FAK−RhoA間の信号伝達はRhoGAPと関連するという模式図を示している。
【0033】
FAKタンパク質の577番のアミノ酸であるチロシンをリン酸化(pY577FAK)した場合及びc−Srcタンパク質の416番のアミノ酸であるチロシンをリン酸化(pY416cSrc)した場合に、Rac1とRhoAの活性を調べてみた結果、TM4SF5が発現される細胞においてRac1の活性はいずれも増加するのに対し、RhoAの活性はいずれも減少することを確認した(図3及び図5)。このようなTM4SF5によるpY577FAKの増加とRhoA活性阻害の現象は、TM4SF5に対するsiRNAを用いてTM4SF5の発現を阻害するとさらに反転することが分かる(図4及び図6)。
【0034】
さらに、TM4SF5が発現される細胞において、そうではない母細胞に比べてFAK−p190RhoGAP間の結合、若しくはFAK−GRAF間の結合が強く結合することが分かり(図9)、FAKタンパク質の577番のアミノ酸チロシンをフェニルアラニンに置換した突然変異タンパク質を発現させた場合にはTM4SF5によるFAK−p190RhoGAP間の結合、またはFAK−GRAF間の結合が減少する結果を示している(図10)。
【0035】
これにより、TM4SF5が発現される細胞においてはFAKからRhoAにつながる一連の信号伝達過程においてpY577FAKが重要な役割を果たすということが推察でき、前記結果を踏まえて、pY577FAK発現が減少したり、FAK−RhoGAP間の結合またはFAK−GRAF間の結合が抑制されたり、Rac1の活性が減少したり、RhoA活性が促される場合には前記腫瘍形成タンパク質であるTM4SF5の発現が抑制されることが分かる。
【0036】
TM4SF5の発現による細胞質内p27kip1タンパク質の蓄積及びRhoAの活性低下
本発明者らは、TM4SF5が発現される細胞においてp27kip1タンパク質が顕著に高い濃度にて存在し、これは、細胞質内におけるp27kip1mRNA及びp27kip1タンパク質の高い安定化に起因することを見出した(図12)。これに対し、TM4SF5が発現されない細胞においてp27kip1は、その量も少ないが、存在するほとんどが核に位置している(図13)。
【0037】
p27kip1は核中におけるサイクリン依存性キナーゼ(cyclin-dependent kinase:CDK)の阻害因子であって、腫瘍抑制機能をすることが知られている。前記p27kip1の細胞質への移動は、細胞周期のG0からG1に転移される間に10番のセリン残基がリン酸化されることにより行われる。前記p27kip1Ser10のリン酸化は、キナーゼのスタスミンとの相互作用(Kinase interacting with Stathmin:KiS)及びPKB/Aktタンパク質によりなされるか、あるいは、成長が止まっている細胞またはin vitro環境においてMirk/dyrk1B及びErk1/2によりそれぞれなされる。
【0038】
このようなSer10位のリン酸化は、Thr187またはThr157位よりもTM4SF5の発現とさらに有意に関連しており、p27kip1タンパク質の細胞質内への位置移動及び安定化に寄与する。すなわち、pT157p27kip1とpT187p27kip1はTM4SF5の発現による諸現象との関連性がない。このため、TM4SF5が発現される肝がん細胞などにおいてshRNAなどを用いてTM4SF5の発現を阻害すると、p27kip1の発現及びセリン10番の残基のリン酸化が減少し、さらに、細胞質内に存在するp27kip1タンパク質もまた減少する(図19及び図20)。これは、最近報告されているp27kip1S10A突然変異タンパク質が発現される形質転換マウスにおいて発がん遺伝子Rasによるp27kip1の細胞質への移動が阻害されるという結果と一致する。
【0039】
一方、最近の報告によれば、細胞質内に存在するp27kip1タンパク質は、直接的に結合してRhoAを活性化可能なGTP置換タンパク質(GEF)のRhoAへの結合を抑えることにより、RhoA信号伝達活性を阻害することが知られている。このような細胞質内のp27kip1蓄積によるRhoA信号伝達間の関係を究明するために、RhoGTPaseを活性化させたときのp27kip1タンパク質の細胞内位置変化を調べてみた。RhoAを活性化させたり、その下位伝達媒介因子であるmDiaを活性化させたり、RhoAの活性因子であるLPAを処理した場合、細胞質内のp27kip1タンパク質発現レベルもさらに減少する結果を示している(図27)。これは、RhoA信号伝達が活性化されると、p27kip1タンパク質がそれ以上安定化できず、細胞質ではない核中に位置してしまうためである。すなわち、細胞質内のp27kip1とRhoGTPaseとの間には相互調節の連結環が存在する。このため、TM4SF5の発現はp27kip1の発現増加及び細胞質における安定化に寄与し、これは、RhoAタンパク質の活性阻害につながる。
【0040】
しかしながら、TM4SF5によるp27kip1蓄積に基づくRhoA活性阻害現象と、TM4SF5によるFAK/RhoGAPsタンパク質間の結合に基づくRhoA活性阻害現象は別々に起こる信号伝達経路である。これは、Y577F FAK突然変異タンパク質(チロシン577番がフェニルアラニンに変わった変異体)を過剰発現させると、p27kip1の発現とセリン10番の残基のリン酸化にはいかなる影響も与えないためである(図18)。
【0041】
また、p27kip1の細胞質内の蓄積は、前記RhoA活性とは対照的にRac1タンパク質は活性化させる。すなわち、活性化されたRac1及びその下位伝達媒介体であるPAK1の発現はp27kip1タンパク質の細胞質内蓄積を引き起こす(図27の下)。
【0042】
TM4SF5によるRhoAの活性低下に基づく細胞形態の変化
TM4SF5が発現されない母細胞である肝がん細胞(SNU449)のアクチンは強い繊維状の構造に広がった多角形の細胞形態を維持しており、TM4SF5が発現された細胞(SNU449Tp)におけるアクチンは非正常的な束が長いロッド細胞形態に沿って形成されている(図2)。
【0043】
このような細胞形態の変化に預かる信号伝達を媒介するRac1の活性及びRhoAの活性について、TM4SF5が発現される細胞においてRac1の活性は増加するのに対し、RhoAの活性は減少する。これは、TM4SF5に対するsiRNAを用いてTM4SF5の発現を阻害することにより再び反転される。すなわち、前記siRNAが導入された細胞は母細胞と同様に強い繊維状のアクチン構造を有する多角形の細胞形態に復元されるが、そうではない細胞は依然としてTM4SF5発現の影響により非正常的な束が形成された細胞形態を構成する(図7)。
【0044】
また、TM4SF5が発現される細胞はp27kip1が高く発現されるという特徴があるため、p27kip1タンパク質の変化もまた細胞形態の変化に影響する。TM4SF5が発現されない母細胞にp27kip1を過剰発現させると、細胞の形態が長くなり(図22)、これとは逆に、TM4SF5が発現される細胞においてp27kip1の発現阻害またはセリン残基のリン酸化を阻害する突然変異であるp27kip1S10Aを処理すると、細胞の形態も多角形の母細胞と同じ形態に変化する(図23)。
【0045】
要するに、前記細胞質のp27kip1による細胞形態の変化とRho信号伝達との間の関係と関連して、TM4SF5が発現された細胞においてRhoGTPaseが活性化(RhoA活性化、mDia活性化またはLPA処理)されると、RhoA信号伝達の活性化によりp27kip1タンパク質がそれ以上安定化できずに核中に位置してしまい、細胞質におけるp27kip1タンパク質の発現レベルが減少し、長尺状の細胞形態が母細胞と同じ多角形に戻る。これに対し、活性化されたRac1及びその下位伝達媒介体であるPAK1の発現はTM4SF5が発現されない母細胞の形態を長く変化させ、p27kip1タンパク質の細胞質内蓄積を引き起こす(図27)。
【0046】
このため、結論的に、細胞質内のp27kip1とRhoGTPaseとの間には相互調節の連結環が存在し、細胞質のp27kip1によるRhoGTPaseの活性調節はTM4SF5が発現される細胞形態の変化を誘導するということである。このような細胞形態の変化は後述する現象とも関連がある。
【0047】
TM4SF5の発現によるEMT現象及び細胞間接触
(1)細胞間接触現象の減少及び上皮間葉転換の(EMT、epithelial-mesenchymal transition)誘導
本発明者らは、TM4SF5発現による細胞質へのp27kip1の蓄積は細胞−細胞間接触消失及びEMT現象を誘導することを見出した。
【0048】
細胞間の接触におけるRhoAタンパク質の活性変化は細胞の種類とその信号伝達の特徴に依存するが、上述したTM4SF5によるp27kip1タンパク質の蓄積によるRhoAの活性阻害現象は細胞中のアクチン構造の変化をもたらし、これは、カドヘリン/カテニンタンパク質複合体を通じての細胞−細胞接触とも関連する。
【0049】
TM4SF5が発現されない母細胞においては、E−カドヘリン、ZO1、デスモプラキンなどの細胞接触タンパク質が多量発現し、細胞接触部位によく整列して存在する。これに対し、TM4SF5が発現される細胞においては、前記細胞接触タンパク質の発現は減少されているだけではなく、細胞間接触が上手になされず、散発的な発現パターンを示す(図29)。しかしながら、E−カドヘリンの発現抑制転写因子であるsnail1もまた同時発現減少されることにより、snail1に非依存的で、且つ、TM4SF5に依存的なE−カドヘリンの発現減少を示す(図29)。
【0050】
これに対し、TM4SF5が発現される細胞には、上皮間葉転換(EMT、epithelial-mesenchymal transition)に預かるSMAタンパク質(図28)及びビメンチンタンパク質が高く発現される。このような上皮細胞の移動は細胞の形態形成及び傷治癒などの生理的な過程に重要な役割を果たし、慢性炎症及び腫瘍転移過程に預かる。
【0051】
TM4SF5によるp27kip1タンパク質の蓄積によるRhoAの活性阻害現象はアクチンフィラメントの非正常的な再構成を誘導するため、細胞の形態が長くなり、このような細胞形態の変化は細胞−細胞間の吸着が崩壊するEMT現象を誘導する。要するに、TM4SF5によるSMAタンパク質の発現とEMT現象はRhoAの活性阻害により起こり、細胞形態の長さの変化につながると言える。
【0052】
このため、TM4SF5が発現された細胞においてp27kip1タンパク質の発現を阻害すると、EMT現象が阻止されながら細胞−細胞接触が維持される。これは、p27kip1の発現が阻害されることにより、p27kip1が細胞質ではない核内にさらに多く蓄積されることにより、長く伸びていた細胞の形態が母細胞と同じ多角形の細胞形態に変わるためである。このように、p27kip1タンパク質の細胞内における位置、すなわち、核と細胞質における位置調節は細胞の形態と細胞との間の接触形成に寄与する。
【0053】
加えて、内生性のTM4SF5を有するHuh7肝がん細胞株に対してshRNAを用いてその発現を阻害すると、HGFによる細胞間の接触消失現象が阻害されるため、前記TM4SF5はある信号伝達体系下において本質的にEMTを誘導すると考えられる。
【0054】
(2)接触阻止現象の消失
本発明者らはTM4SF5が発現される細胞においては前記EMT現象により細胞間に高密度にて互いに接触される環境においても隣り合う細胞の存在を認知できずに持続的に細胞分裂及び増殖を維持する、すなわち、細胞接触による生長阻止現象が失われることを確認した。
【0055】
生長の接触阻止現象は、細胞が隣り合う他の細胞と接触したときにそれ以上分裂せず生長が止まるような現象を意味する。既存の報告によれば、E−カドヘリンによる細胞−細胞接触は細胞の増殖と腫瘍生成を抑制することが知られており、最近の報告文献によれば、接触阻止が起こる間に細胞の広がり現象が減少し、成長が止まるということである。このため、EMT現象などにより細胞が互いに接触されていないならば、細胞は隣り合う他の細胞を認識することができずに継続的な分裂をする結果、接触阻止現象が失われるであろう。
【0056】
TM4SF5が発現されない母細胞は多角形の細胞形態を帯びるため、増殖していて、経時的に接触阻止現象によりそれ以上生長しなくなるという飽和曲線を描くが、TM4SF5が発現される長い形態の細胞は細胞間接触消失を誘導するために核が重なり合って存在する程度に細胞が密集しているにも拘わらず、継続的な細胞分裂が起こる(図33)。
【0057】
また、TM4SF5が発現される細胞においては母細胞に比べてS期に存在する細胞集団がさらに多く(図35)、shRNAを用いてTM4SF5の発現を阻害すると、S期に存在するこのような細胞集団が減る(図36)。
【0058】
すなわち、TM4SF5の発現による細胞間接触現象減少及びEMT現象は細胞生長の接触阻止現象を消失させる。図57は、これに対する模式図を示している。図57によれば、発がん遺伝子であると推定されるTM4SF5の過剰発現は細胞質内のp27kip1の発現及び安定化を引き起こし、これは、hKiSまたはPKB/Aktによるセリン10番の残基のリン酸化によるものであり、TM4SF5によるpY577FAKとFAK/RhoGAPsとの結合によりRhoAの活性が阻害される。Rac1の活性はRhoAの不活性化と相反する関係をもって存在するが、RhoAとRac1の下位信号伝達媒介体であるmDiaとPAKはアクチンの構造変化と細胞内の収縮性を調節してしまい、これは、細胞形状の変化につながるため、このような細胞形態の変化は細胞間接触消失現象を誘導して、隣り合う細胞を認識できずに継続的に生長してしまい、接触阻止現象を引き起こしてしまうのである。
【0059】
TM4SF5による細胞移動、浸潤及びがん転移
上述したように、TM4SF5の発現と関連があるFAKが過剰発現されたり、Rac1が活性化されると、細胞の運動性が増加し、Rho活性及びEMT現象による上皮細胞の移動は細胞の形態形成及び傷治癒などの生理的な過程に重要な役割を果たし、さらに、慢性炎症及び腫瘍転移過程に必須である。
【0060】
このため、TM4SF5が発現される細胞株の場合の方が、発現されない細胞株に比べて移動性が大きい(図58及び図59)。このような細胞の移動に伴い、細胞は付着部位から分離されて周りの細胞外基質内に浸潤される。TM4SF5が発現される細胞は細胞外基質を構成するコラーゲンゲルにTM4SF5を発現しない細胞よりもさらに効率よく浸潤するだけではなく(図60)、様々な細胞外基質複合体であるマトリゲルに効率よく浸潤する(図61)。
【0061】
前記作用は部分的にMMPs(matrix metalloproteinase)のタンパク質分解作用によりなされる。MMPsはZn2+依存的なペプチド分解酵素であって、細胞の基質を分解する能力を有している。これらのうちMMP−2とMMP−9はゼラチンとフィブロネクチン基質を分解するが、これらは、TM4SF5が発現される細胞(SNU449Tp、T3、T7及びT16など)内においてその活性が高い(図62)。特に、MMP−9は酵素活性だけではなく、遺伝子発現活性も高い(図62の途中)。
【0062】
ヌードマウスがん転移モデルを用いた実施例9から明らかになったように、TM4SF5を発現するがん細胞において前記細胞移動性及び浸潤性由来のがん転移性はin vivoにおいても現れる。
【0063】
総合的にみたとき、TM4SF5の腫瘍誘導因子としての機能はin vitro細胞とin vivo動物においていずれも細胞の成長が適切に調節されずに継続的に分裂させるため、肝がん細胞などにおいてTM4SF5の発現はFAKのリン酸化増加及び細胞質内のp27kip1タンパク質の蓄積によるRhoAタンパク質の活性阻害を引き起こして、持続的な細胞成長、顕著な細胞周期S期への進入、細胞形態の変化及び付着−非依存的な生長を引き起こす。そして、このTM4SF5の発現細胞株をヌードマウス(in vivo動物)に注入すると腫瘍が形成される。
【0064】
そこで、本発明者らは、腫瘍誘導因子として働くTM4SF5(transmembrane 4L 6 family member 5)発現と関連する上記の様々な細胞生物学的及び生化学的現象に基づいて抗がん物質のスクリーニング方法を発明した。
【0065】
本発明において使用する「抗がん」という用語は、がんの成長を抑制または予防するものを意味する。「がんの成長を抑制または予防する」とは、治療または処理しなかったときと比較時にがんの成長及びがん転移を減少させることを含む概念である。前記「がん転移」は腫瘍(がん)細胞が身体の遠く離れた部分に拡散される過程を意味する。この明細書において前記用語は転移過程により発生されるがんも含む。
【0066】
本発明の方法においては配列番号2で表わされるポリペプチドを発現するがん細胞を培養する。前記ポリペプチドを発現する代表的な場合として、配列番号1で表わされるポリヌクレオチドが挙げられる。
【0067】
配列番号2で表わされるポリペプチドを発現するがん細胞はTM4SF5タンパク質を発現する細胞株であって、一具体例として、上皮細胞においてTM4SF5が持続的に発現されるSNU449肝がん細胞株(KCLB No.00449)を製作し、これを培地において培養することにより行うことができる。前記SNU449細胞の代わりにSNU398細胞(KCLB No.00398)を使用してもよい。
【0068】
本発明において、配列番号1で表わされるヌクレオチドからポリペプチドを発現するがん細胞としては、例えば、すい臓がん細胞、胃がん細胞、肝がん細胞、大腸がん細胞、脳がん細胞、または肺がん細胞または人為的に製造したがん細胞などが挙げられるが、TM4SF5腫瘍タンパク質を発現する細胞であれば非制限的に使用可能である。上記の「人為的に製造したがん細胞」としては、例えば、遺伝子操作を含むクローン技術によりTM4SF5腫瘍タンパク質を発現する細胞が挙げられる。
【0069】
本発明の方法は、次いで、前記工程において培養された細胞を抗がん候補物質にて処理するステップを含む。好ましくは、TM4SF5が細胞内において特異的に誘導する現象を調節すると予想される化合物を処理する。
【0070】
最後に、抗がん候補物質にて処理しない場合と比較して、前記処理した化合物が後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定する。
【0071】
本発明においては、前記物質の抗がん機能を、上述したTM4SF5発現と関連する様々な細胞生物学的及び生化学的現象を基に以下のように定義する:
(i)FAKの577番のチロシン位置のリン酸化減少
(ii)FAK−RhoGAP、もしくはFAK−GRAF間の結合抑制
(iii)細胞質内p27Kip1タンパク質発現減少
(iv)RhoAの活性増加
(v)Rac1の活性減少
【0072】
前記方法において、TM4SF5発現による細胞の形態及び細胞接触パターンと関連する上述した特徴に基づいてさらに後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定する:
(vi)細胞の形態がロッド状から多角形に戻る
(vii)細胞接触関連タンパク質の発現減少
(viii)細胞−細胞間接触が維持される
(ix)細胞生長の接触阻止現象が現れる
(x)SMA(α−smooth muscle actin)タンパク質またはビメンチンタンパク質の発現減少、またはE−カドヘリンタンパク質の発現増加
(xi)上皮間葉転換(EMT)減少
【0073】
このとき、(vii)における前記細胞接触関連タンパク質はE−カドヘリン、ZO1(zonula occludens−1)、β−カテニン及びデスモプラキンよりなる群から選ばれ、(ix)における前記細胞生長の接触阻止の現象は、細胞数の減少、細胞周期のS期に存在する細胞集団の減少または多層増殖の抑制を意味する。すなわち、多層増殖が抑制されて細胞が接触されると単層にのみ伸び、それ以上増殖がなされないことを言う。また、前記細胞数の減少または細胞周期のS期に存在する細胞集団の減少は、前記TM4SF5及びそれと相互作用する膜タンパク質のN結合型グリコシレーションの不活性によりなされる。
【0074】
加えて、上記の方法において、TM4SF5による細胞の移動性、浸潤性増加と関連する上述した特徴に基づいてさらに後述する抗がん機能のうち1以上の機能を示す物質を抗がん物質として決定することもできる:
(xii)細胞外基質若しくは血清の存在下における細胞の移動又は運動性減少
(xiii)細胞外基質を構成するコラーゲンゲル中における浸潤性抑制
(xiv)細胞外基質複合体であるマトリゲル中における浸潤性抑制
(xv)MMP活性減少(発現減少)
【0075】
このとき、前記MMPの好適な例として、MMP−2及びMMP−9が挙げられる。
【0076】
すなわち、腫瘍誘導因子、若しくは、EMT誘導因子TM4SF5の発現の抑制は、FAKのリン酸化または活性変化阻害、細胞質内のp27Kip1発現蓄積によるRhoA活性低下阻害、Rac1及びRhoAを含むRhoGTPaseの活性変化による細胞形態の変化阻害、上皮間葉転換[分化、EMT現象(Epithelial-Mesenchymal Transition)または変換分化]の誘導抑制、S期への進行抑制、接触抑制の消失により、結果的に多層増殖の誘導抑制、吸着−非依存的な増殖またはソフトアガー内における増殖抑制、細胞移動性及び浸潤性阻害、ヌードマウスにおけるがん遺伝子発現がん細胞の注入による腫瘍形成及び転移抑制などの諸現象により確認することができる。
【0077】
本発明は、他の観点において、前記方法によりスクリーニングされた、TM4SF5に対する抗がん機能を有する抗がん物質に関する。「TM4SF5に対する抗がん機能」とは上述した通りである。好ましくは、下記の化学式1または2で表わされるカルコン系の化合物を抗がん物質として決定する。
【0078】
【化3】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールであり、好ましくは、メチル、ベンジル、p−トルイル、p−ニトロフェニルまたはp−フルオロフェニルである。
【0079】
【化4】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールであり、好ましくは、メチル、ベンジル、p−トルイル、p−ニトロフェニルまたはp−フルオロフェニルである。
【0080】
本発明の抗がん物質は、さらに好ましくは、下記表1に記載のカルコン系の化合物である。表1の化合物のうち化合物1で示すTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン]が代表的な化合物である。
【0081】
【表1】
【0082】
前記表1に記載の化合物1から97はそれぞれ以下の通りである。
4′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホネート)−4−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(メタンスルホネート)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンベンゼンスルホニルアミノ)−2−クロロ−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン
【0083】
大韓民国公開特許10−2003−0036993にはカルコン系の化合物が基底膜成分を分解するマトリックスメタロプロテアーゼ(MMP)の活性を阻害する性質を有していることを記載しているが、カルコン系の化合物の抗がん機能については全く言及していないだけではなく、本発明の前記化学式1または2で表わされるカルコン系の化合物はその構造上スルホン基(SO3−)またはスルホンアミド基(SO2NH−)を含有していることを特徴としている。すなわち、カルコン系の化合物のTM4SF5に対する抗がん活性(機能)は前記スルホン基(SO3−)由来のものである。
【0084】
特に、この明細書の実施例12において、カルコンのA−リング(左リング)にOH、NH2、またはOCH3を置換基として有している場合には、TM4SF5の機能を通じて多層構造に伸びる現象を阻害する抗がん性が観察されないが、本発明のカルコン系の化合物でのようにスルホニル基が導入されたスルホンアミドカルコンまたはスルホネートカルコンにおいてはTM4SF5機能阻害による抗がん活性が観察されることを示している。すなわち、カルコン化合物において、「スルホン基(スルホニル)」の導入は現在知られているカルコン誘導体とは全く異なる独特な抗がん活性を発現させる。
【0085】
本発明の実施例においては、これらのうち代表的な化合物であるTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン](表1の1番化合物)を用いて抗がん機能を確認した。
【0086】
前記カルコン系の化合物はTM4SF5による現象を特異的に阻害する拮抗剤として機能する。TM4SF5が発現される細胞株に、前記TSAHCを含むカルコン系の化合物のうち1以上を処理すると、継続的な生長曲線がカルコン系の化合物の濃度、処理時間により依存的に減少し(図43の右)、細胞周期S期への進入(図44)、及び細胞間の接触が復活される(図45)。これに対し、本発明のカルコン系の化合物薬物の効果はTM4SF5が発現されない細胞においては全く効果がないことが判明された(図43の左)。
【0087】
さらに、TM4SF5によるp27kip1、pS10p27kip1、pY577FAKの発現の度合い及びSMAまたはビメンチンの発現においても、前記カルコン系の化合物により効果的に減少し(図46)、TM4SF5が発現されたSNU449細胞株であるSNU449Tp細胞株に本発明のカルコン系の化合物を処理すると、細胞質に存在するp27kip1がそれ以上安定化できずに減少する(図47)。
【0088】
本発明のカルコン系の化合物、特にTSAHCは腫瘍誘導因子TM4SF5及びそれと相互作用する膜タンパク質のN結合型グリコシレーション活性に影響を与えて、TM4SF5が細胞膜において(若しくは、テトラスパニン−ウェブ)インテグリンなどを含む他のタンパク質との結合を通じて発がん機能を行う上で妨害若しくは抑制効果を来たす。
【0089】
このような側面は、TSAHCを処理時にTM4SF5のN結合型グリコシレーション部分のFNGaseFに対する感受性の向上により確認することができる(図50)。このため、本発明のTSAHCなどのカルコン系の化合物はTM4SF5そのものを攻撃してN結合型グリコシレーション作用の構造的な面に変化を与えてTM4SF5のタンパク質−タンパク質結合(テトラスパニン−ウェブ内における結合)に影響を与えることにより、TM4SF5による発がん機能を抑制すると類推することができる。
【0090】
加えて、本発明のカルコン系の化合物はTM4SF5が発現される細胞におけるRhoAの活性阻害現象を修復させ(図48)、長く変化されていた細胞の形態から多角形の形態に復帰させる(図49)。これは、TM4SF5が発現による諸現象を抑制していることを示す結果であって、TM4SF5に対して拮抗剤として働く本発明のカルコン系の化合物に対してTM4SF5による腫瘍形成の治療薬物としての効能を予測可能にする。
【0091】
また、TM4SF5による細胞の移動、浸潤現象もまた本発明のカルコン系の化合物、例えば、TSAHCにより効率よく阻害される。すなわち、TM4SF5が発現される細胞株はTSAHCを処理することにより細胞の移動性(図65)、コラーゲンゲルにおける浸潤性(図66)及びマトリゲルにおける浸潤性(図67)がいずれも有効に阻害される。これは、本発明のカルコン系の化合物が細胞の移動性に預かるMMPsの酵素活性を阻害することを意味し(図68)、これは、TSAHCがTM4SF5による腫瘍誘導だけではなく、腫瘍細胞の転移過程をも有効に阻害可能であることを示唆している。
【0092】
本発明は、さらに他の観点において、前記カルコン系の化合物のうち、好ましくは、前記表1に記載の化合物のうちいずれか1種若しくはそれ以上を有効成分として含有する抗がん用組成物を含む。この明細書の実施例8においては、表1に記載の化合物のうち1種(TSAHC)のみを処理しても抗がん機能を示すことを確認した。
【0093】
本発明による前記カルコン系の化合物は、薬学的に許容可能な塩の形態であるカルコン誘導体の形態で使用することができる。
【0094】
前記塩としては、薬学的に許容可能な遊離酸により形成された酸付加塩が有用である。すなわち、前記カルコン誘導体は当該技術分野における通常の方法により薬学的に許容される酸付加塩から形成可能である。遊離酸としては、無機酸と有機酸を使用することができ、無機酸としては、塩酸、臭酸、硫黄酸、リン酸などを使用することができ、有機酸としては、クエン酸、酢酸、乳酸、マレイン酸、フマル酸、グルコン酸、メタンスルホン酸、コハク酸、酒石酸、4−トルエンスルホン酸、エムボン酸、グルタミン酸またはアスパラギン酸などを使用することができる。好ましくは、無機酸としては塩酸、有機酸としてはメタンスルホン酸を使用することができる。
【0095】
また、本発明による前記カルコン誘導体は、薬学的に許容可能な塩だけではなく、通常の方法により製造可能なあらゆる塩、水和物、溶媒和物をいずれも含むことができる。
【0096】
臨床的な使用のために、本発明による前記カルコン系の化合物またはその塩形態などは単独で投与可能であるが、賦形剤、結合剤、滑沢剤、崩解剤、コーティング物質、乳化剤、懸濁剤、溶媒、安定化剤、吸収促進剤及び/または軟膏基材を混合することにより特定の使用及び好適な目的に適するように剤型化された薬剤学的な混合物の形態で一般的に投与可能である。前記混合物は、経口用、注射用、直腸用または外用投与用として使用可能である。
【0097】
さらに詳しくは、上述したように、前記カルコン系の化合物またはその塩を含有する前記薬剤学的な抗がん用組成物は経口的に投与可能であり、例えば、錠剤、コーティング錠剤、糖衣錠、硬質または軟質ゼラチンカプセル剤、液剤、乳化剤または懸濁剤などの剤型であってもよい。また、投与は、直腸に投与、例えば、座剤を用いて投与可能であり、局所的または経皮的に投与、例えば、軟膏、クリーム、ゲルまたは液剤を用いて投与可能であり、または、非経口的に投与、例えば、注射用の溶液を用いて投与可能である。
【0098】
錠剤、コーティング錠剤、糖衣錠、硬質または軟質ゼラチンカプセル剤の製造のために、本発明のカルコン系の化合物は薬剤学的に不活性な無機または有機賦形剤(薬剤学的に許容される担体)と一緒に混合可能である。錠剤、コーティング錠剤、糖衣錠、硬質ゼラチンカプセル剤に適当な賦形剤の例には、ラクトース、メーズ澱粉またはその誘導体、タルクまたはステアリン酸またはその塩が含まれる。軟質ゼラチンカプセル剤に用いられて好適な賦形剤には、例えば、植物性オイル、ワックス、脂肪、半固形または液体ポリオールなどが含まれる。しかしながら、活性成分の性質により、軟質ゼラチンカプセル剤にいかなる賦形剤も不要になる場合がありうる。液剤及びシロップ剤の製造のために使用可能な賦形剤には、例えば、水、ポリオール、スクロース、転化糖及びグルコースが含まれる。注射用の溶液の製造のために使用可能な賦形剤には、例えば、水、アルコール、ポリオール、グリセリン及び植物性オイルが含まれる。座剤及び局所または経皮適用用の製造のために使用可能な賦形剤には、例えば、天然オイルまたは硬化油、ワックス、脂肪及び半固形または液体ポリオールが含まれる。
【0099】
さらに、前記薬剤学的な組成物は、保存剤、溶解剤、安定化剤、湿潤剤、乳化剤、甘味剤、色素、芳香剤、浸透圧調節用の塩、緩衝剤、コーティング剤または抗酸化剤を含むことができ、それらは治療学的に価値のある他の薬剤を含むこともできる。
【0100】
結果的に、経口投与用の薬剤学的な剤型は顆粒剤、錠剤、糖コーティング錠剤、カプセル剤、丸剤、懸濁剤または乳化剤であってもよく、非経口用の剤型、例えば、靜脈内、筋肉内または皮下、剤型用としては滅菌水溶液の剤型が使用可能であり、これは、等張性溶液を作成するために他の物質、例えば、塩またはグルコースを含むことができる。抗腫瘍剤は、また、座剤またはペッサリーの剤型にて投与可能であり、または、ローション、溶液、クリーム、軟膏または散布剤の形態で外用的に適用可能である。
【0101】
本発明のカルコン系の化合物の1日容量レベルは経口または非経口用に投与されるときに5〜2000mgである。1回投与または2回以上の分割投与が可能である。しかしながら、実際の投与量は投与経路、患者の年齢、性別及び体重、及び疾患の重症度などの種々の関連因子に照らして決定する必要がある。このため、前記投与量はいかなる方法によっても本発明の範囲を限定するものではない。
【0102】
実施例
以下、実施例を挙げて本発明をより詳細に説明する。但し、これらの実施例は単に本発明を一層詳しく説明するためのものであり、本発明の要旨により本発明の範囲がこれらの実施例に限定されるものではないということは、当業界における通常の知識を有する者にとって明らかである。
【0103】
特に、下記の実施例においては肝がん細胞株だけを使用しているが、この他にも、すい臓がん細胞、胃がん細胞、肝がん細胞、大腸がん細胞、脳がん細胞、乳房がん細胞、前立腺がん細胞、子宮がん細胞、食道がん、膀胱がん、または肺がん細胞などのヒト由来の様々な種類のがん組織及びがん細胞株が使用可能であるということは当業界における通常の知識を有する者にとって自明である。
【0104】
実施例1:TM4SF5によるがん細胞培養
(1)細胞株樹立
TM4SF5(GenBank No.:NM−003963)が持続的で且つ安定的に発現される細胞株を樹立するために、ヒトの肝がん細胞であるSNU449細胞(KCLB No.00449)に対照群ベクターであるpLNCX(Clontech Laboratories inc.、PaloAlto、CA)とTM4SF5入りpLNCXを有するレトロウィルスを感染させた。
【0105】
TM4SF5発現レトロウィルスを作成するために、プラスミドpLNCXのHind III/ClaI制限酵素サイトに、Huh7(JCRB No.0403)細胞のcDNAプールからPCR方法により得たcDNAを挿入した後、PT67細胞(ATCC CRL−12284)に導入して安定的にTM4SF5を発現する細胞を得て培養することにより、TM4SF5を発現するレトロウィルスを製作した(Experimental Cell Research, 312:2983, 2006)。以下、前記対照群ベクターを挿入したSNU449細胞はSNU449Cp(またはCp)で表記し、TM4SF5が発現させてなるSNU449細胞はSNU449Tp(またはTp)で表記する。
【0106】
TM4SF5発現レトロウィルスをSNU449細胞に感染させた後、形成された細胞集団を全て集めた細胞株(SNU449Cp及びSNU449Tp)と単一細胞集団を分離して樹立した細胞株(TM4SF5が発現されるT3、T7、T11及びT16細胞)を樹立した。
【0107】
そして、TM4SF5の発現が抑制されるHuh7細胞株を作成するために、TM4SF5に対するshRNAを導入した後、G418(500μg/ml)を用いてshTM4SF5が発現される細胞だけを選択的に分離した。この細胞株に対して10%FBS、0.25μg/mlゲンタマイシンを添加した後、200μg/mlのG418入りまたは200μg/mlのG418未添加のRPMi−1640培地において5%CO2及び37℃の培養機により培養して使用した。
【0108】
(2)材料及び方法
1.TM4SF5抗体の製作
pGEX−5X2ベクター(Phamacia Biotech.)にTM4SF5のc−末端部位(アミノ酸229番から594番までを制限酵素EcoR1を用いて切断)を挿入製作してその塩基配列を確認した。DH5αバクテリアからIPTGに誘導された組換えタンパク質を得るために0.3%SDSとタンパク質分解酵素阻害剤が含まれているPBSを用いた。タンパク質抽出後、電気泳動されたSDSゲルから抗原を抽出してマウスに免疫した。合計で3回の免疫反応を誘導した後、マウスから血清を得て組換えタンパク質と動物細胞抽出物に対する免疫反応性を検査した。
【0109】
2.タンパク質分離及びウェスタンブロット分析
様々な実験条件において得られた細胞溶解物を、種々の条件下の細胞をPBS溶液により洗い取った後にRIPA緩衝溶液を用いて用意した。特定の場合、明示された発現ベクターを導入したり、TM4SF5に対するshRNAを発現させたり、p27kip1に対するsiRNAを発現するアデノウィルスを感染させる条件の細胞が用いられた。
【0110】
加えて、10μMのシクロヘキシミドを明示した時間中に処理した細胞も用いられた。このような溶解物のタンパク質は定量化して同量使用され、ホスホ−Y577FAK、ホスホ−Y416Src、c−Src、RhoA、p120ctn、β−カテニン、pS10p27kip1、FAK、E−カドヘリン、ZO1、Rac1、p190RhoGAP、p27kip1、GRAF、hKiS、デスモプラキン、α−チューブリン、α−カテニン、さらにα−smooth muscle actin(SMA)に対する抗体を用いてウェスタンブロット分析を行った。
【0111】
3.免疫蛍光染色法分析
細胞内のタンパク質を蛍光免疫染色するために、細胞を10%FBS/RPMI−1640により予めコーティングしておいたカバーガラスの上に載せ、細胞の形態が揃って完全に付着するまで37℃において一日中培養した。細胞に上述の特定の遺伝子を導入させたり様々なウィルスを感染させる場合には、上記した時間中に培養した後に使用したり、10mMリゾホスファチジン酸(LPA)を処理した細胞はカバーガラスに付着させた後、1時間かけて処理した後に使用した。蛍光免疫染色法は当該タンパク質の1次抗体を反応させた後、余剰の抗体を洗い取った後、蛍光物質が付着された2次抗体と反応させて再び洗い取って蛍光分析可能なスライドを作成し、アクチンの染色法はローダミンが付着されたファロイジンを反応させて行われた。
【0112】
使用された抗体はE−カドヘリン、β−カテニン、FLAG、DAPI(4’−6−ジアミジノ−2−フェニルインドール)、BrdU(5−ブロモ−2’−デオキシ−ウリジン)、p27kip1、さらにZO1(zonula occludens−1)などが使用された。蛍光免疫染色がなされたカバーガラスは観察のためにスライドに載せて蛍光顕微鏡または共焦点レーザー顕微鏡により観察した。
【0113】
4.RT−PCR
P27kip1のmRNAレベルを調べるためにRT−PCRをTM4SF5非発現若しくは発現細胞株にアクチノマイコンDを処理して様々な時間帯に細胞からトータルRNAを抽出してそれからmRNAを得た後、p27Kip1に対するプライマーを用いてPCR方法により行った。使用されたプライマーはセンス(5’−taa ccc ggg act tgg aga ag−3’:配列番号3)とアンチセンス(5’−gct tct tgg gcg tct gct c−3’:配列番号4)であった。
【0114】
5.p27kip1の細胞質タンパク質層と核タンパク質層の分画化
TM4SF5が発現されるSNU449Tp細胞とT16細胞におけるP27kip1の細胞質発現と核内発現量を調べるためにLiangら(Nature Medicine, 8:1153-1160, 2002)が提示した方法により分画化が行われた。
【0115】
6.ソフトアガーアッセイ
60mm培養皿に0.7%アガーと10%FBSが含まれた培養液を予め敷いて固めた後、その上に106個の細胞を0.3%アガーと10%FBSが含まれた細胞培養液と一緒に投入した。これらの培養皿は5%CO2が与えられる37℃培養機において25日間培養され、2日おきに200μg/mlG418と10%FBSが含まれた培養液に交替した。形成された細胞集団はデジタルカメラ付き顕微鏡により観察、記録した。
【0116】
7.統計の分析
データの意味のある差を得るために平均値の比較をスチューダントt−tsetにより行った。p値が0.05以下である場合にその差は意味あると言える。
【0117】
実施例2:TM4SF5によるpY577FAKタンパク質発現及びRhoAの活性阻害
(1)TM4SF5が発現される細胞の形態学的な特性
実施例1において樹立されたTM4SF5が発現されないSNU449母細胞(SNU449pまたはp)、対照群ベクターが持続的に発現されるSNU449Cp細胞及びTM4SF5を発現する細胞株(SNU449Tp、T3、T7、及びT16)に対する各細胞の形態を光学顕微鏡を用いて確認し、さらにTM4SF5に対する抗体及びα−チューブリンに対する抗体を用いてウェスタンブロットを行った。
【0118】
そして、TM4SF5の発現有無による細胞の様々なアクチン構造を調べるために、対照群細胞(SNU449Cp)及びTM4SF5が発現される細胞(SNU449Tp)をガラスカバースリップの上に生長させた後、ファロイジンが付着されたローダミンにより染色した。
【0119】
その結果、クローン由来のT3、T7、T16及びTpはTM4SF5が発現されない母細胞であるSNU449p細胞と比較したとき、細胞形態の側面においてその長さの伸張が起こっていることが確認された(図1)。そして、SNU449細胞のアクチンを蛍光染色法により染色して観察した結果、強い繊維状の構造に広がった多角形の細胞形態を維持しており、TM4SF5が発現されるSNU449Tp細胞においてアクチンは非正常的な束が長い細胞形態に沿って形成されていた(図2)。
【0120】
(2)pY577FAK発現による効果
上述した細胞形態の変化に預かる信号伝達を調べるために、実施例1において樹立されたTM4SF5を発現する細胞株(SNU449Tp、T3、T7、及びT16)において、FAKタンパク質の577番のアミノ酸であるチロシンがリン酸化されたpY577FAKタンパク質発現の度合い、c−Srcタンパク質の416番のアミノ酸であるチロシンがリン酸化されたpY416cSrcタンパク質発現の度合い、Rac1活性及びRhoA活性を調べてみた。前記全体の細胞溶解物を集めて、図3及び図5に表記のタンパク質に対する抗体を用いてウェスタンブロット分析を行ったり試験管内においてRhoAとRac1をプルダウンしてその活性を分析した。
【0121】
また、TM4SF5に対するshRNA(shTM4SF5)を用いてTM4SF5の発現を阻害する場合の変化を観察した。母細胞であるSNU449p、対照群細胞であるSNU449Cp及びTM4SF5が発現される各種の細胞において、緑蛍光タンパク質(GFP)に対するshRNA、ランダムな塩基配列を有するshRNA及びTM4SF5に対するshRNAを導入した後に細胞を回収してウェスタンブロット分析とRhoA活性分析を行った。
【0122】
その結果、TM4SF5が発現される前記細胞においてはpY577FAKタンパク質とpY416cSrcタンパク質発現及びRac1活性は増加されていたが、RhoA活性は減少されていることを確認した(図3及び図5)。そして、TM4SF5による前記pY577FAKの増加とRhoA活性阻害現象はTM4SF5の発現を阻害することにより再び反転することが分かる(図4及び図6)。
【0123】
そして、SNU449Tp細胞にpEGFPとshTM4SF5を一緒に導入して24時間後にカバースリップの上に載せ、24時間の培養時間が経過した後、ファロイジンが付着されたローダミンとDNA染色のためのDAPIを一緒に蛍光染色した結果、前記shTM4SF5が導入された細胞は対照区に属するSNU449細胞と同様に強い繊維状のアクチン構造を有する多角形の細胞形態に修復されるが、そうではない隣り合う細胞は依然としてTM4SF5発現の影響により細胞形態の変化が現れないことを確認した(図7)。
【0124】
(3)RhoA活性阻害
併せて、RhoGAPタンパク質を通じてのRhoAの不活性化過程におけるpY577FAKの役割を調べてみた。Cp細胞とTp細胞から得られた細胞溶解物に対してFAKに対する抗体を用いて免疫沈降法を行い、免疫沈降された混合物はp190RhoGAP、GRAF、FAK抗体を用いてウェスタンブロット分析を行った。また、前記SNU449Tp細胞に(HA)3−tagged FAK野生型タンパク質とY577F突然変異タンパク質を一時的に過剰発現させて、2日後に細胞溶解物を免疫沈降法を行って図10において表記している抗体を用いてウェスタンブロットを行った。
【0125】
その結果、FAK−p190RhoGAP間の結合、若しくはFAK−GRAF間の結合が、TM4SF5が発現される細胞において、そうではないSNU449Cp細胞に比べて強く結合することが分かる(図9)。そして、このようなTM4SF5によるFAK−p190RhoGAP間の結合若しくはFAK−GRAF間の結合はFAKタンパク質の577番のアミノ酸チロシンをフェニルアラニンに変えた突然変異タンパク質を発現させることにより減少される結果を示し(図10)、TM4SF5が発現される細胞からRhoAにつながる一連の信号伝達過程においてpY577FAKが重要な役割を果たすということを推察することができた。
【0126】
また、FAKと関連するGTPase調節因子であるGRAFに対して、対照群細胞にFLAG−tagged GRAFを一時的に過剰発現して24時間後に上述した方法に従いカバースリップの上に載せ、一日中に培養した後、アクチンとFLAG抗体を用いて蛍光免疫染色法を行った結果、TM4SF5が発現されない対照群細胞におけるGRAFまたはp190RhoGAPの一時的な発現は細胞形態の変化を引き起こすことを確認した(図11)。
【0127】
このため、結論的にSNU449細胞において発現されるTM4SF5はRhoAの活性を阻害して細胞長さの伸長を誘導し、これは、FAK−p190RhoGAP若しくはFAK−GRAFタンパク質間の相互作用によるものであることが分かる。
【0128】
実施例3:TM4SF5による細胞質内のp27kip1の蓄積効果
(1)TM4SF5の発現によるp27kip1の発現増加
TM4SF5によるRhoA活性阻害現象がp27kip1による効果であるかどうかを調べるために、SNU449CpとSNU449Tp細胞をp27kip1に対する抗体とDAPIを用いて蛍光免疫染色した。そして、前記細胞の核と細胞質を実施例1−(2)−5)における方法に従い分画化してp27kip1に対する抗体とa−チューブリンに対する抗体を用いてウェスタンブロット分析を行った。
【0129】
その結果、TM4SF5が発現される細胞においてp27kip1タンパク質が顕著に高濃度にて細胞質に位置することが分かる(図12)。これに対し、TM4SF5が発現されない対照群細胞においては少量ではあるがほとんどが核に位置していた(図13)。
【0130】
TM4SF5によるp27kip1発現レベルとSer10リン酸化の関係を調べるために、実施例1において樹立した様々な細胞株溶解物を得て図14に表記の抗体を用いてウェスタンブロット分析を行った結果、このようなp27kip1タンパク質の細胞質における存在はhKIS(human Kinase-interantigwith Stathmin)タンパク質と活性化されたPKB/Aktタンパク質によりアミノ酸10番のセリンのリン酸化を通じてなされることを確認した(図14)。
【0131】
また、アクチノマイシンD(10mg/ml、ActD)が処理された細胞からRNAを分離してp27kip1及びGAPDHをRT−PCR分析し、シクロヘキサミド(10mM、CHX)を処理した細胞の溶解物を得てp27kip1に対する抗体を用いてウェスタンブロット分析を行った後、その結果として得られるバンドの強度を測定して対照群となるバンドの強度に対する増減をグラフにて示した(図16)。
【0132】
その結果、TM4SF5が発現される細胞においてp27kip1タンパク質発現が顕著に増加されることはp27kip1mRNAとタンパク質の高い安定化のためであることが確認され(図15)、このような現象はTM4SF5に対するsiRNAを用いてその発現を阻害したとき、細胞形態の変化がTM4SF5が発現されないSNU449細胞の形態に戻り、細胞質においてp27kip1タンパク質発現レベルも減少されると共にセリン残基のリン酸化もまた起こらない結果からなお一層明らかになった(図17)。
【0133】
さらに、TM4SF5が細胞質のp27kip1タンパク質の安定化に預かるという仮説はTM4SF5が内生的に発現される様々な肝がん細胞においても有効であることが分かる。
【0134】
内生的にTM4SF5を発現している肝がん細胞株に対して、GFPのshRNA、ランダムな塩基配列(Scr)を有するshRNA及びTM4SF5に対するshRNAを導入して、48時間後に得た全体の細胞溶解物及びSNU449CpとSNU449Tp細胞の溶解物に図18に表記の抗体を用いてウェスタンブロット分析を行った結果、内生的に発現されるTM4SF5の発現の阻害はp27kip1発現及びSer10のリン酸化レベルを減少させ(図18)、SNU449TpTp細胞に(HA)−tagged FAK野生型タンパク質とY577F突然変異を過剰発現させた後、表示された抗体によりウェスタンブロット分析を行った結果、Y577FFAK突然変異タンパク質はp27kip1の発現レベルやSer10のリン酸化には影響しなかった(図19)。また、Huh7肝がん細胞株にScrまたはshTM4SF5と一緒にpEGFPを導入してp27kip1とDAPIを免疫蛍光染色法により染色した結果、内生的に発現されるTM4SF5の阻害は細胞質に存在するp27kip1を減少させることを確認した(図21)。
【0135】
この結果はTM4SF5の発現がp27kip1の発現増加及び細胞質における安定化に寄与し、これは、RhoAタンパク質の活性阻害につながることを証明している。特に、図19の結果はp27kip1の発現とFAKタンパク質のリン酸化は互いに関連性がないことを示唆し、これは、TM4SF5によるp27kip1タンパク質及びRhoA活性阻害現象とTM4SF5によるFAK/RhoGAPs結合によるRhoA活性阻害現象は別個の信号伝達機序において寄与されたものであることが分かる。
【0136】
(2)細胞質のp27kip1蓄積による細胞形態変化
実施例2から明らかなように、TM4SF5が発現される細胞は細胞形態が長く、前記実施例3−(1)から明らかなように、TM4SF5が発現される細胞はp27kip1が高く発現されるという特徴があるため、このようなp27kip1タンパク質の変化が細胞形態の変化にいかなる影響を与えるかを調べてみた。
【0137】
実施例1において樹立したSNU449Cp細胞にFLAG−tagged p27kip1野生型タンパク質を過剰発現させ、48時間後に前記細胞をDAPI、ファロイジンが付着されたローダミン、及びFLAGに対して免疫蛍光染色を行って共焦点顕微鏡により観察した結果、TM4SF5が発現されない母細胞にp27kip1を過剰発現させたときに細胞の形態が長くなることを確認した(図22)。
【0138】
そして、前記SNU449Tp細胞をsiRNAp27kip1アデノウィルスを用いて24時間かけて感染させ、FLAG−tagged p27kip1S10Aを導入して24時間かけて培養した後、免疫蛍光染色を行ない、得られた細胞溶解物を図24に明示された抗体を用いてウェスタンブロットを行った。
【0139】
その結果、このようにTM4SF5が発現される細胞においてp27kip1の発現阻害と一緒にセリン10番がアラニンに変わってその残基のリン酸化が起こらない突然変異体であるp27kip1S10Aを一緒に導入して発現すると、細胞質への移動が阻害され、細胞の形態も多角形のSNU449若しくはSNU449Cp細胞と同じ形態に変化していることを確認することができた(図23及び図24)。
【0140】
前記SNU449Tp細胞にpEGFPと活性化された形態のRhoA63L突然変異ベクターを一緒に導入してアクチンの構造を免疫染色した結果、活性化されたRhoAにより細胞形態変化の反転が起こり、多角形をなしていた(図25)。
【0141】
細胞質のp27kip1による細胞形態の変化とRhoA信号伝達間の関係を究明するために、RhoGTPaseを活性化させたときの細胞形態の変化とp27kip1タンパク質の細胞内位置変化を調べてみた。
【0142】
先ず、SNU449CpとSNU449Tp細胞をカバースリップの上に培養した後、RhoGTPaseを活性化させる因子であるLPA(リゾホスファチジン酸)10mMを1時間かけて処理してDAPIとp27kip1を免疫染色した。その結果、SNU449Tp細胞においてLPA(リゾホスファチジン酸)は細胞質のp27kip1を減少させた(図26)。
【0143】
また、SNU449Tp細胞にpEGFPとmDia活性型突然変異を導入し、且つ、SNU449Cp細胞にpEGFPと活性型のRac1(Rac161L)またはPAK1(PAK1−caax)を導入して免疫蛍光染色分析を行った。その結果、驚くことには、RhoAの下位伝達媒介因子であるmDiaの活性化が細胞の形態をSNU449若しくはSNU449Cp細胞と同じ多角形に戻る結果を示し、細胞質内のp27kip1タンパク質発現レベルも再び減少される結果を示していた(図27)。
【0144】
これは、RhoA信号伝達の活性化によりp27kip1タンパク質がそれ以上安定化できず、核中に位置するためであると推察することができる(図26、図27の上)。これに対し、活性化されたRac1とその下位伝達媒介体であるPAK1の発現はTM4SF5が発現されない細胞であるにも拘わらず、細胞の形態を長く変化させてp27kip1タンパク質の細胞質内蓄積を引き起こした(図27の下)。
【0145】
この結果からみたとき、細胞質内のp27kip1とRhoA及びRac1を含むRhoGTPaseとの間には相互調節の連結環が存在し、細胞質のp27kip1によるRhoGTPaseの活性調節はTM4SF5が発現される細胞形態の変化を誘導することが分かる。
【0146】
実施例4:TM4SF5によるEMT現象
TM4SF5が細胞−細胞間接触の調節にも影響するかどうかを調べてみた。先ず、TM4SF5が発現される細胞とそうではない対照群細胞の溶解物により細胞−細胞接触形成に預かるタンパク質の発現レベルを免疫ブロット方法を用いて確認した。
【0147】
その結果、TM4SF5非発現細胞株においてはE−カドヘリン、ZO1、デスモプラキンなどの細胞接触タンパク質の発現が大幅に増加されていたが、TM4SF5が発現される細胞においてはその発現が減少されていた。しかしながら、上皮間葉転換(EMT)に預かるSMAタンパク質は高く発現されていた(図28)。
【0148】
さらに、高密度にて培養したSNU449Cp細胞とSNU449Tp細胞をカバースリップの上に載せた後、E−カドヘリン、β−カテニン、ZO1タンパク質の発現パターンを免疫染色法を用いて調べてみた結果、TM4SF5を発現しないSNU449細胞とSNU449Cp細胞においては細胞接触部位によく整列されて存在していたE−カドヘリン、β−カテニン、ZO1などが、TM4SF5が発現される細胞においては細胞間接触が上手になされず、散発的に分布する発現パターンを示していた(図29)。
【0149】
SNU449Tp細胞にp27kip1に対するshRNAアデノウィルスを実施例1において説明した通りに感染させた後、E−カドヘリン、β−カテニン、ZO1タンパク質の免疫染色した結果、p27kip1の発現が阻害されたら、E−カドヘリン、β−カテニン、ZO1タンパク質が規則的な形態に再配列され(図30)、SMA発現が減少されることを確認した。
【0150】
また、内生的に発現されるTM4SF5がEMT現象の調節にいかなる役割を果たすかを究明するために、shRNA及びshTM4SF5を発現するHuh7細胞に対してTM4SF5とa−チューブリンの発現を調べ(図31)、HGF(100ng/ml)を24時間かけて処理した後、ZO1を免疫染色した結果を観察した。
【0151】
Huh7細胞株は、TM4SF5発現有無とは無関係に、コロニーをなして成長する特性があるため、細胞間接触パターンが相当良好である。このため、HGF(hepatocyte growth factor)を用いてEMT現象を誘導させた後、TM4SF5が細胞−細胞間接触の形成に及ぼす影響を観察した。その結果、HGFによるHuh7細胞の分散現象(若しくは、接触崩壊)がTM4SF5が阻害されたHuh7細胞において低減することを観察することができた。このため、TM4SF5が細胞−細胞間接触の消失を引き起こしていることが分かる(図32)。
【0152】
実施例5:TM4SF5による細胞生長の接触阻止現象の消失
SNU449CpとSNU449Tp細胞をカバースリップの上において培養した後、DNAとアクチン構造をDAPIにより染色した結果を図33及び図34に示す。
【0153】
矢印にて示す部分は細胞分裂と核が重なり合う部分を示し、細胞がそれ以上成長する空間がないにも拘わらず継続的に分裂し続けていることを示す。また、これらの細胞の生長速度を測定するために6ウェル培養皿に5X104個の細胞をはじめとして12時間おきにその数を数えて記録した結果をグラフにて示す。実際にTM4SF5が発現される449Tp細胞株の生長曲線は継続的に増加するパターンを示し、対照群細胞である449Cp細胞株は経時的に増殖していて、接触阻止現象によりそれ以上成長しなくなる飽和曲線を描いていた。このような特徴は細胞が低密度にて存在する状況においても現れる。すなわち、低密度の細胞のうちTM4SF5発現する細胞は核が重なり合って細胞分裂などをしながら増殖をするが、TM4SF5を発現しないSNU449細胞とSNU449Cp細胞は高密度にて互いに接触される環境においてはそれ以上生長しないという特徴を示している(図34)。
【0154】
また、TM4SF5が発現される様々な細胞株(T7、T16、及びT3)の細胞周期のS期分布を調べてみた結果、TM4SF5を発現しないSNU449細胞と、SNU449Cp細胞細胞に比べてTM4SF5が発現される細胞においてS期に存在する細胞集団がさらに多量であると分かる(図35)。SNU449Cp細胞とSNU449Tp細胞に対照群shRNAとshTM4SF5を導入した後、プロピジウムヨード法によりDNAを染色して流速細胞分析機を用いて細胞周期を分析した結果、shRNAを用いてTM4SF5の発現を阻害したとき、S期に存在する細胞集団が低減することを確認した(図36)。
【0155】
BrdU浸透免疫染色法を用いた調査においても、TM4SF5が発現される細胞はそうではない細胞に比べてDNA合成中のS基の細胞分裂が顕著に高く存在することが分かる(図37)。このような現象は、LPAによるRhoAの活性化や(図38)、p27kip1の発現阻害(図39)、さらにはE−カドヘリンの再発現(図40)などを通じて特異的にDNA合成中のS基の細胞分裂が阻害されると現れた。
【0156】
特に、E−カドヘリンタンパク質の発現を再び高めるSNU449T16m細胞(SNU449T16細胞由来であるため、SNU449T16mと称する。)は、TM4SF5が発現されるにも拘わらず、細胞間の接触がTM4SF5非発現細胞と同じパターンを示し(図42)、SMAタンパク質の発現が減少する結果を示している(図41)。
【0157】
このため、上記の結果からみたとき、TM4SF5が発現される細胞にE−カドヘリンの発現を再び増加することにより、TM4SF5はEMT現象を通じて細胞生長の接触阻止現象を消失させ、これは、細胞間の接触と接触による生長阻止現象との間において緊密な機能の関連性があることを示唆している。
【0158】
実施例6:TM4SF5により形成された腫瘍組織の特徴
TM4SF5による腫瘍発生効果を2種類の方法により検証した。先ず、TM4SF5が発現されたり発現されない様々な細胞を付着−非依存的な生長環境下に放置した。27日間ソフトアガーにおいて培養した後に観察した。実施例1の6)において言及した方法によりSNU449細胞株をもってソフトアガーアッセイを行い、ソフトアガー培養皿に細胞を培養してから27日目に位相差顕微鏡により細胞コロニーの様子を観察・記録した。
【0159】
その結果、TM4SF5が発現される細胞はそうではない細胞に比べて細胞コロニーが顕著に多量生存していることが分かる。しかしながら、TM4SF5が発現される細胞にE−カドヘリンを再発現させた449T3m、T16m細胞においては細胞コロニーの形成が顕著に低減していた。このため、TM4SF5によるEMT現象は細胞付着−非依存的な生長と関連するということを推定することができた(図52)。
【0160】
次に、TM4SF5を発現したり発現しない様々なSNU449細胞株をヌードマウスに注入した。ヌードマウスはBALB/cAnNCrjBgi−nu(Orient.Co.Ltd)を基本種とし、全て生後4〜5週齢、体重が約20gの雌性だけを使用した。滅菌消毒されたねずみかごにおいて滅菌された水と飼料を供給し、層板気流を持続的に維持したクリーンベンチ中に放置して清浄な環境を維持し、全ての術式もクリーンベンチ中において行った。
【0161】
ヌードマウスの昼夜間生物学的な周期のために室内灯を12時間は点灯し、且つ、12時間は消灯した。SNU細胞株を基本細胞株として、SNU449Cp、SNU449Tp、SNU449T16、SNU449T16m細胞の浮遊液を遠心分離して細胞を濃縮し、血球計により細胞数を1X107cells/200μlに算定して、用意された細胞を26ゲージの注射針を用いて1cc注射器によりヌードマウスの脇腹に皮下注射接種した。注射後、実験動物はソウル大学前臨床実験室動物飼育法により維持され、SNU449Tp細胞による腫瘍形成を観察、記録した。
【0162】
TM4SF5が発現される細胞株は腫瘍形成に優れていたが、母細胞(449p)と対照群細胞(449Cp)、449T3m、T16m細胞はこれといった腫瘍が形成されていなかった(図53)。
【0163】
そして、SNU449CpとSNU449Tp細胞を注射したマウスから得られた腫瘍組織をH&E(ヘマトキシリン・エオシン)染色法により染色してその組織形態を観察し、p27kip1とpS10p27kip1発現パターンを免疫組織化学染色法により分析した。免疫化学組織染色法によりTM4SF5が誘導した腫瘍組織の中央部位は死滅した細胞集団が大幅に現れ、細胞質に存在するp27kip1とセリン10番の残基のリン酸化も増加されて現れた(図54)。さらに、前記組織から得られたタンパク質溶解物を用いてウェスタンブロット分析を行った結果、TM4SF5による腫瘍組織は、そうではない細胞による組織と比較したとき、p27kip1の発現レベルとリン酸化レベルが高く現れた(図55)。
【0164】
実施例7:TM4SF5による細胞の移動性増加
(1)細胞の移動性分析
Boyden Transwell chamber 24 well plates(Costar, Cambridge, MA)を使用した。ポリカーボネートフィルター(8μm porosity)上のチャンバーにSNU449Cp及びSNU449Tp細胞を5x104セル/ウェルずつ1日中に37℃、5%のCO2培養機において無血清RPMI(Roswell Park Memorial Institute)培地にて培養した後、24時間後にインサートチャンバー上方のメンブレンを綿棒によりよく拭き取り、70%メタノールに約7分間浸してメンブレン下方にある細胞を固定した。
【0165】
このようにSNU449Cp、SNU449Tp細胞を高密度になるように培養した後、適当な厚さのワウンドを形成した後、経時的にワウンドが満たされる度合いを観察し、光学顕微鏡により記録した。その結果、TM4SF5が発現される細胞株の場合、そうではない細胞株に比べて傷回復速度が増加されることを確認した(図58)。
【0166】
また、前記SNU449Cp、Tp細胞株をトランスウェルのインサートウェルにおき、無血清培地において培養し、インサートの下チャンバーには10%の血清または無血清培地を入れて細胞の経時移動の度合いを観察するために、0.005%クリスタルバイオレットにより1分間染色した後、PBSにより3回洗浄して光学顕微鏡により前記染色された細胞を観察した(図59の上の写真)。前記染色薬を溶解して260nmの波長において吸光度を測定して図表化した(図59の下グラフ)。その結果、TM4SF5が発現される細胞株の場合、そうではない細胞株に比べて細胞移動性が増加されることを確認した(図59)。
【0167】
(2)ジモグラフィ分析
分泌されたMMPsの基質分解活性を測定するためにゼラチンジモグラフィを行った。図62に示す細胞株を60mm培養皿に放置した後、18時間後に血清を除去した培養液に交替して24時間さらに培養した。分泌されたMMPsタンパク質を得るために培養上清だけを取って濃縮した。MMPsの基質となるゼラチンをSDS−PAGEの分離ゲル(ランニングゲル)に添加し、減圧条件下において電気泳動した後、1%トリトンX−100緩衝溶液において再変成させ、次いで、酵素反応緩衝溶液(10mM CaCl2、0.15M NaClと50mMトリス−HCL、pH7.5)により37℃において18時間反応させた。この後、0.5%クマシーブリリアントブルー(Coomassie brilliant blue)R250により3時間染色させた後、10%酢酸、30%メタノールにおいて透明なバンドが現れるまで脱色した。
【0168】
その結果、MMP−2と9はゼラチンとフィブロネクチン基質を分解するが、TM4SF5が発現されるSNU449Tp、T3、T7、及びT16細胞においてその活性が高くなっていることを確認した。すなわち、TM4SF5によりMMPs酵素活性が増加されることを確認した(図62)。
【0169】
そして、SNU449P細胞にMMP−2ルシフェラーゼとMMP−9ルシフェラーゼを様々な濃度のpcDNA3.1−MycHis−TM4SF5と一緒に導入した後、36時間後にルシフェラーゼ活性を測定して、各MMP遺伝子プロモーターの転写活性を比較した結果、MMP−9は、酵素活性だけではなく、遺伝子発現活性も高いことを確認した(図63)。これより、TM4SF5はMMPs遺伝子発現活性にも影響することが分かる。
【0170】
(3)3Dコラゲンを用いた細胞の浸潤性分析
細胞の浸潤性を分析するためにCytodex-3マイクロキャリアを用いた。SNU449CpとSNU449Tp細胞を細胞をマイクロキャリアビーズに付着して培養し、血清と培地成分入りのコラーゲンゲルに混ぜて37℃において培養した後、経時的に現れる現象を観察した。また、TM4SF5が発現されるSNU449Tp細胞がコラーゲンゲル中に浸潤する現象をも光学顕微鏡により観察した(図60)。
【0171】
また、上記のSNU449Tp細胞の浸潤性を分析するために、実施例7−(1)において言及したBoyden Transwell chamber 24 well platesのインサートにマトリゲルを入れて37℃において固めた後、上述した方法に従い細胞を培養して24時間後にインサートの上方のメンブレンを綿棒によりよく拭き取り、36時間後にメンブレンの下方に浸潤された細胞を染色して観察した。その結果、上記のSNU449Tp細胞が様々な細胞外基質複合体であるマトリゲルを有効に浸潤することを確認した(図61)。
【0172】
実施例8:TM4SF5機能を阻害する抗がん物質(拮抗剤)の選択
TM4SF5が細胞内において特異的に誘導する現象を調節すると予想される化合物のうち、前記表1に示すカルコン系の化合物を本発明の方法によりスクリーニングした。図69に記載のカルコン化合物もまた抗がん候補物質として使用した。
【0173】
上述した抗がん機能を示す候補物質を本発明の抗がん物質として決定したが、スクリーニングされた代表的な化合物を前記表1に示す。
【0174】
前記抗がん物質の抗がん機能をより確実に調べるために、表1の化合物のうちTSAHC[4’−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン]を使用した。
【0175】
TM4SF5が発現される細胞株にTSAHCを処理したとき、継続的な生長曲線がTSAHCの濃度、処理時間により依存的に減少することを観察し(図43の右)、細胞周期S期への進入(図44)、そして細胞間の接触が復活されることを確認した(図45)。
【0176】
これに対し、TSAHC薬物の効果については、TM4SF5が発現されない細胞においては全く効果がないことが分かる(図43の左)。さらに、TM4SF5によるp27kip1、pS10p27kip1、pY577FAKの高い発現や度合い、さらにSMAの発現はTSAHCにより有効に減少されることが分かる(図46)。また、449Tp細胞株にTSAHCを処理した結果、細胞質に存在するp27kip1が減少することから(図47)、TSAHCがTM4SF5に対する拮抗作用をすることによりp27kip1タンパク質がそれ以上安定化できないことが分かる。加えて、TM4SF5が発現される細胞におけるRhoAの活性阻害現象を修復させ(図48)、長く変化されていた細胞の形態が、多角形のTM4SF5が発現されない対照群細胞株と同じ形態に戻ることを確認することができた(図49)。
【0177】
また、TSAHCの処理時にTM4SF5のN結合型グリコシレーション部分のFNGaseFに対する感受性が向上することを確認した(図50)。この結果から、TSAHCはTM4SF5そのものを攻撃してN結合型グリコシレーションの構造的な側面に変化を与えてTM4SF5のタンパク質−タンパク質結合(テトラスパニン−ウェブ内における)に影響することにより、TM4SF5による機能を抑制することができると推定可能である。
【0178】
さらに、TSAHCの処理は、内生的にTM4SF5を発現する細胞については細胞の生長若しくは増殖を抑制したが、発現しない細胞株は根本的にいかなる影響を及ぼさなかった(図51)。このため、これらの結果からみて、TM4SF5に対して拮抗剤として作用するカルコン系の化合物がTM4SF5による腫瘍形成の予防及び治療に効果があることを確認した。
【0179】
カルコン系の化合物が示すTM4SF5阻害率(iC50)を表2に示す。
【0180】
【表2】
【0181】
実施例9:本発明のカルコン誘導体の人体がん細胞株に対する試験管内細胞毒性
本発明の化合物カルコン誘導体の人体がん細胞株に対する試験管内細胞毒性を調べるために、1989年アメリカの国立がん研究所(NCi:National Cancer institute)において開発したスルホロダミンB(SRB)方法を使用した。
【0182】
この実験に使用した人体がん細胞株として、HCT15(colon adenocarcinoma、結腸がん;ATCC CCL−225)、PC−3(prostate adenocarcinoma、前立腺がん;ATCC CRL−1435)、A−549(lung carcinoma、肺がん;ATCC CCL−185)が挙げられ、胃がん細胞はいずれも生命工学研究所において継代培養中のものを使用した。
【0183】
前記人体がん細胞株は10%子牛血清が含まれているRPMi1640培地(ギブコ社カタログno.31800)を用いて、37℃の温度条件下、5%の二酸化炭素培養機において培養し、継代は1週間につき2回行った。
【0184】
細胞を付着面から分離するときには、0.25%トリプシン(シグマ社製)及び3mMCDTA(1、2−シクロヘキサンジアミンテトラアセト酸、シグマ社製)をリン酸塩緩衝溶液(シグマ社製)に溶解させた溶液を使用した。
【0185】
検索用の試料(表1に記載のカルコン化合物誘導体)は少量のDMSO(ジメチルスルホキシド、シグマ社製)に溶かして実験に必要となる濃度まで実験用の培地において希釈して最終的なDMSOの濃度が0.5%以下になるようにし、数段階(最終濃度:30、10、3、1、0.3μg/mL)に亘って希釈された試料溶液はがん細胞に加える前に0.22μmマイクロフィルター(ミリポア社製)によりろ過して無菌状態にした。次いで、通常のSRB法により人体がん細胞株に対する細胞毒性を測定した。
【0186】
本発明の化合物カルコン誘導体の人体がん細胞株に対する細胞毒性(ED50、μg/mL)を下記表3に示す。
【0187】
【表3】
【0188】
上記の結果は、本発明のカルコン系の化合物が人体がん細胞株に対する細胞毒性を示すために抗がん剤として使用可能であることを示唆する。
【0189】
実施例10:TSAHCの腫瘍形成抑制効果
(1)TSAHC投与
前記実施例6に従い用意したヌードマウスに対して、形成された腫瘍の長さと幅をキャリパーにより2日おきに1回ずつ測定して下記の公式(Volume (mm3) = Length × Width2 × 1/2、Length:長径、Width:短径)により腫瘍の体積を算出した。
【0190】
ヌードマウスに誘発された腫瘍の体積が200mm3のサイズにて着床された後、任意に2群に分けて(5mg/kgと50mg/kg)生理食塩水にTSAHCを懸濁させて2日おきに1回ずつ3週間26ゲージの注射針を用いて腹腔内注射する方法により投与した。
【0191】
(2)腫瘍組織の免疫組織化学検査とウェスタンブロット分析
ヌードマウスは、SNU449Tp細胞を注入してから6週目に、またはTSAHCを投薬してから3週目に頸椎脱臼の方法により犠牲し、免疫組織化学検査のために腫瘍組織を切開し、ホルムアルデヒドにより固定した後にパラフィン標本にした。組織のタンパク質発現レベルをウェスタンブロットにより分析するために抽出した腫瘍組織は直ちに液体窒素を用いて凍結標本にして0.1%SDS入りのRIPA緩衝溶液に溶解させてタンパク質混合物を得て分析した。用意された6μm厚さのパラフィン切片に対しては、H&E(ヘマトキシリン・エオシン)染色と免疫組織化学検査を行った。
【0192】
その結果、TM4SF5が発現される細胞を移植したマウスの腫瘍(個体数=6)は持続的に大きくなるのに対し、TSAHCを5mg/Kg、若しくは50mg/Kg濃度にて腹腔注射したマウスの腫瘍はそれぞれ約45%、88%だけそのサイズが減少することを確認した(図56)。
【0193】
実施例11:TSAHCのがん転移抑制効果
(1)がん転移実験動物モデル樹立
がん転移実験動物モデルの準備は、上述した腫瘍誘導モデルと同じ方法により行われた(実施例6参照)。SNU449Cp、SNU449Tp細胞の浮遊液を遠心分離して細胞を濃縮し、血球計により細胞数を1X107と算定して用意された細胞を1cc注射器及び29ゲージのインシュリン注射針を用いてヌードマウス尾靜脈に血管注射して接種した。3週間放置後、マウスを犠牲して肺組織を摘出した後、ホルマリンにより固定してH&E(ヘマトキシリン・エオシン)染色を行った。
【0194】
その結果、TM4SF5が発現される細胞の場合、肺組織に腫瘤が形成されていることを確認した(図64)。図64における矢印の個所が腫瘤部位であって、細胞が密集している個所である。
【0195】
(2)TSAHCのTM4SF5による細胞の移動性及び浸潤性抑制効果
SNU449Cp細胞とSNU449Tp細胞を6−ウェル培養皿におき、24時間後にTSAHC20μMを処理して18時間培養した後、トランスウェルチャンバーのインサートウェルに移した。18時間後に移動した細胞を固定、染色して光学顕微鏡により観察し、その数を数えてグラフ化した(図65)。その結果、TM4SF5による細胞の移動性がTSAHCにより阻害されることを確認した。
【0196】
また、SNU449Cp細胞とSNU449Tp細胞をマイクロキャリアビーズに付着してTSAHC20μMと一緒に18時間かけて培養した。ビーズ−細胞を血清と培地成分入りのコラーゲンゲルに混ぜ、培養を表記の時間だけ行いながら観察、記録した結果、TM4SF5による細胞の浸潤性がTSAHCにより阻害されることを確認した(図66)。
【0197】
上記の如き方法によりマトリゲルが厚くコーティングされたトランスウェルチャンバーを用いて24時間かけて培養した後、マトリゲルを通過した浸潤性の細胞数を数えてグラフ化した結果、マトリゲルにおける浸潤性もまた有効に阻害されることを確認した(図67)。
【0198】
そして、Cp細胞とTp細胞にTSAHC20μMを24時間かけて処理した。24時間後、血清を除去した培地に交換した後、18時間かけて培養した。培養液を取って濃縮した後、上述した方法に従いジモグラフィを行った結果、TM4SF5によるMMPs酵素活性がTSAHCにより阻害されることを確認した(図68)。
【0199】
この結果は、TSAHCがTM4SF5による腫瘍誘導だけではなく、腫瘍細胞の転移過程もまた有効に阻害可能であることを示唆している。
【0200】
実施例12:TSAHC類似体との効果比較
最後に、TM4SF5が預かる現象においてTSAHCと構造的に類似する誘導体の効果を観察した。このとき、使用したDMSO(ジメチルスルホキシド)及びカルコン類似体の化学構造は下記の通りである。
【化5】
【0201】
SNU449Cp細胞とSNU449Tp細胞を6−ウェル培養皿に敷き、前記図に示す薬物を20μMずつ24時間かけて処理した後、光学顕微鏡を用いて細胞の形態変化を観察、記録した。
【0202】
その結果、図69に示すように、TSAHCと構造がほとんど同様であるものの、異なるR基で置換された薬物は、同じ濃度条件(20μMにて24時間処理)下においてTSAHCは有効にTM4SF5が発現される細胞の成長を多層に成長せずに単層に成長させ、同時に細胞の形態をTM4SF5が発現しないSNU449対照群細胞のように変化させることを確認した。これにより、TM4SF5による細胞成長阻害効果はTSAHC化合物の特定のR基により調節されることを類推することができる。
【0203】
前記TSAHC類似体は、前記化学構造式に示すように、カルコンのA−リング(左リング)にOH、NH2、またはOCH3を置換基として有しているが、この場合には、TM4SF5の機能を通じて多層に成長する現象を阻害する抗がん性が観察されなかったが、TSAHCのようにスルホニル基が導入されたスルホンアミドカルコンにおいては、TM4SF5機能阻害を通じての抗がん活性が観察された。すなわち、スルホニルの導入により現在知られているカルコン誘導体とは全く異なる独特な抗がん活性を観察することができた。
【産業上の利用可能性】
【0204】
以上述べたように、本発明は、腫瘍形成におけるTM4SF5の発現と関連する分子的な機序に基づく抗がん物質のスクリーニング方法であって、腫瘍生成及び転移に対する拮抗作用をする治療及び予防目的の物質をスクリーニングする上で有用である。また、前記方法によりスクリーニングされたカルコン系の化合物を含有する抗がん用組成物は、体重の減少、肝及び脾臓という臓器に対して外傷的な異常を来たすことなく、しかも、毒性反応無しに抗がん効果を示すことから、抗がん用の薬剤として有効に使用可能であるというメリットがある。
【0205】
以上、本発明の内容の特定の部分を詳述したが、当業界における通常の知識を有する者にとって、このような具体的な記述は単なる好適な実施様態に過ぎず、これにより本発明の範囲が制限されることはないという点は明らかであろう。よって、本発明の実質的な範囲は特許請求の範囲とこれらの等価物により定まると言えるであろう。
【特許請求の範囲】
【請求項1】
下記のステップを含む、抗がん物質のスクリーニング方法:
(a)配列番号2のポリペプチドを発現するがん細胞を培養した後、抗がん候補物質で処理するステップ、
(b)前記抗がん候補物質で処理されたがん細胞に対して、:
(i)接着斑キナーゼ(FAK)を構成するアミノ酸配列のうち577番位のチロシンのリン酸化の確認、
(ii)FAKのRho−GTPase activating protein(RhoGAP)への結合、またはFAKのGTPase Regulator Associated with FAK(GRAF)への結合測定、
(iii)細胞質内p27Kip1の発現量及び安定性測定、
(iv)RhoAの活性測定、及び
(v)Rac1の活性測定、の少なくとも1つを検出するステップ、及び
(c)抗がん候補物質で処理しない場合に比べて、
(i)FAKの577番位のチロシンのリン酸化減少、
(ii)FAKのRhoGAPへの結合、またはFAKのGRAFへの結合抑制、
(iii)細胞質内p27Kip1発現及び安定性減少、
(iv)RhoA活性増加、または
(v)Rac1活性減少の事象が現れた場合に、抗がん候補物質を抗がん物質として選択するステップ。
【請求項2】
前記細胞質内p27KiP1は、前記p27KiP1を構成するアミノ酸配列の中10番位のセリン残基がリン酸化されていることを特徴とする請求項1に記載の方法。
【請求項3】
前記ステップ(b)は、細胞の形態確認、細胞接着関連のタンパク質発現確認、細胞間接触パターンまたは接触生長確認、α平滑筋アクチン(α−SMA)タンパク質またはビメンチンタンパク質の発現確認、E−カドヘリンタンパク質の発現確認及び上皮間葉転換(EMT)確認、よりなる群から選ばれる1以上をさらに行い、
前記ステップ(c)は、細胞の形態がロッド状から多角形に戻るか、あるいは、細胞接着関連のタンパク質の発現が減少するか、あるいは、細胞−細胞間の接触が維持されるか、あるいは、細胞生長の接触阻止現象が現れるか、あるいは、α−SMAタンパク質またはビメンチンタンパク質の発現が減少するか、あるいは、E−カドヘリンタンパク質の発現が増加するか、あるいは、上皮間葉転換(EMT)が減少する場合をさらに含むことを特徴とする請求項1に記載の方法。
【請求項4】
前記細胞接着関連タンパク質はE−カドヘリン、zonula occludens−1(ZO1)、β−カテニン及びデスモプラキンよりなる群から選ばれることを特徴とする請求項3に記載の方法。
【請求項5】
前記細胞生長の接触阻止現象は、細胞数の減少、細胞周期のS期に存在する細胞集団の減少または多層増殖の抑制であることを特徴とする請求項3に記載の方法。
【請求項6】
前記細胞数の減少または細胞周期のS期に存在する細胞集団の減少は、細胞膜タンパク質のN結合型グリコシレーションの不活性によることを特徴とする請求項5に記載の方法。
【請求項7】
前記ステップ(b)は、細胞外基質または血清の存在下での細胞の移動又は運動性確認、細胞外基質を構成するコラーゲンゲルへの浸潤確認、細胞外基質複合体を構成するマトリゲルへの浸潤確認及びマトリックスメタロプロテアーゼ(MMP)活性測定よりなる群から選ばれる1以上をさらに行い、
前記ステップ(c)は、細胞外基質または血清の存在下で細胞の移動又は運動性が減少するか、あるいは、コラーゲンゲル中における浸潤が減少するか、あるいは、マトリゲル中における浸潤が減少するか、あるいは、MMP活性が減少する場合をさらに含むことを特徴とする請求項1または請求項3に記載の方法。
【請求項8】
前記MMPはMMP−2またはMMP−9であることを特徴とする請求項7に記載の方法。
【請求項9】
前記がん細胞はすい臓がん、胃がん、肝がん、大腸がん、脳がん、乳房がん、甲状腺がん、膀胱がん、食道がん、子宮がん及び肺がんからなる群より選択されたことを特徴とする請求項1に記載の方法。
【請求項10】
下記の化学式1または化学式2で表わされる抗がん用のカルコン系の化合物:
【化1】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである;
【化2】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【請求項11】
次の化合物からなる群より選択される1以上の抗がん用のカルコン系の化合物:
4′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホネート)−4−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(メタンスルホネート)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンベンゼンスルホニルアミノ)−2−クロロ−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン及び
4′−(m−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン。
【請求項12】
請求項10または請求項11のカルコン系の化合物を有効成分として含有する抗がん用組成物。
【請求項1】
下記のステップを含む、抗がん物質のスクリーニング方法:
(a)配列番号2のポリペプチドを発現するがん細胞を培養した後、抗がん候補物質で処理するステップ、
(b)前記抗がん候補物質で処理されたがん細胞に対して、:
(i)接着斑キナーゼ(FAK)を構成するアミノ酸配列のうち577番位のチロシンのリン酸化の確認、
(ii)FAKのRho−GTPase activating protein(RhoGAP)への結合、またはFAKのGTPase Regulator Associated with FAK(GRAF)への結合測定、
(iii)細胞質内p27Kip1の発現量及び安定性測定、
(iv)RhoAの活性測定、及び
(v)Rac1の活性測定、の少なくとも1つを検出するステップ、及び
(c)抗がん候補物質で処理しない場合に比べて、
(i)FAKの577番位のチロシンのリン酸化減少、
(ii)FAKのRhoGAPへの結合、またはFAKのGRAFへの結合抑制、
(iii)細胞質内p27Kip1発現及び安定性減少、
(iv)RhoA活性増加、または
(v)Rac1活性減少の事象が現れた場合に、抗がん候補物質を抗がん物質として選択するステップ。
【請求項2】
前記細胞質内p27KiP1は、前記p27KiP1を構成するアミノ酸配列の中10番位のセリン残基がリン酸化されていることを特徴とする請求項1に記載の方法。
【請求項3】
前記ステップ(b)は、細胞の形態確認、細胞接着関連のタンパク質発現確認、細胞間接触パターンまたは接触生長確認、α平滑筋アクチン(α−SMA)タンパク質またはビメンチンタンパク質の発現確認、E−カドヘリンタンパク質の発現確認及び上皮間葉転換(EMT)確認、よりなる群から選ばれる1以上をさらに行い、
前記ステップ(c)は、細胞の形態がロッド状から多角形に戻るか、あるいは、細胞接着関連のタンパク質の発現が減少するか、あるいは、細胞−細胞間の接触が維持されるか、あるいは、細胞生長の接触阻止現象が現れるか、あるいは、α−SMAタンパク質またはビメンチンタンパク質の発現が減少するか、あるいは、E−カドヘリンタンパク質の発現が増加するか、あるいは、上皮間葉転換(EMT)が減少する場合をさらに含むことを特徴とする請求項1に記載の方法。
【請求項4】
前記細胞接着関連タンパク質はE−カドヘリン、zonula occludens−1(ZO1)、β−カテニン及びデスモプラキンよりなる群から選ばれることを特徴とする請求項3に記載の方法。
【請求項5】
前記細胞生長の接触阻止現象は、細胞数の減少、細胞周期のS期に存在する細胞集団の減少または多層増殖の抑制であることを特徴とする請求項3に記載の方法。
【請求項6】
前記細胞数の減少または細胞周期のS期に存在する細胞集団の減少は、細胞膜タンパク質のN結合型グリコシレーションの不活性によることを特徴とする請求項5に記載の方法。
【請求項7】
前記ステップ(b)は、細胞外基質または血清の存在下での細胞の移動又は運動性確認、細胞外基質を構成するコラーゲンゲルへの浸潤確認、細胞外基質複合体を構成するマトリゲルへの浸潤確認及びマトリックスメタロプロテアーゼ(MMP)活性測定よりなる群から選ばれる1以上をさらに行い、
前記ステップ(c)は、細胞外基質または血清の存在下で細胞の移動又は運動性が減少するか、あるいは、コラーゲンゲル中における浸潤が減少するか、あるいは、マトリゲル中における浸潤が減少するか、あるいは、MMP活性が減少する場合をさらに含むことを特徴とする請求項1または請求項3に記載の方法。
【請求項8】
前記MMPはMMP−2またはMMP−9であることを特徴とする請求項7に記載の方法。
【請求項9】
前記がん細胞はすい臓がん、胃がん、肝がん、大腸がん、脳がん、乳房がん、甲状腺がん、膀胱がん、食道がん、子宮がん及び肺がんからなる群より選択されたことを特徴とする請求項1に記載の方法。
【請求項10】
下記の化学式1または化学式2で表わされる抗がん用のカルコン系の化合物:
【化1】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである;
【化2】
式中、R1はR4SO2−であり、R2及びR3はそれぞれ独立して水素またはヒドロキシ基であり、前記R4はC1〜C5のアルキルまたは水素、ハロゲン、ニトロ及びC1〜C5のアルキルよりなる群から選ばれる1以上の置換基を有するC6〜C10のアリールである。
【請求項11】
次の化合物からなる群より選択される1以上の抗がん用のカルコン系の化合物:
4′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3−ヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホネート)−4−ヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−アミノベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(ベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(メタンスルホネート)−4−ヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−トルエンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−トルエンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(p−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
3′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、3−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、4−ジヒドロキシカルコン、
2′−(m−ヒドロキシベンゼンスルホニルアミノ)−2、5−ジヒドロキシカルコン、
4′−(p−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−フルオロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−ニトロベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(m−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(o−アミノベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(ベンゼンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(メタンスルホニルアミノ)−3、4−ジヒドロキシカルコン、
4′−(p−トルエンベンゼンスルホニルアミノ)−2−クロロ−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−4−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3−ヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2−ヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン、
4′−(p−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−3、4−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、3−ジヒドロキシカルコン、
4′−(m−ヒドロキシベンゼンスルホネート)−2、4−ジヒドロキシカルコン及び
4′−(m−ヒドロキシベンゼンスルホネート)−2、5−ジヒドロキシカルコン。
【請求項12】
請求項10または請求項11のカルコン系の化合物を有効成分として含有する抗がん用組成物。
【図1】

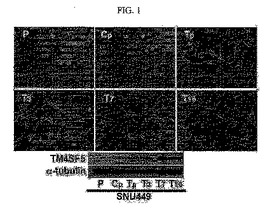
【図2】
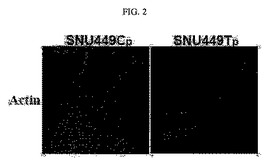
【図3】

【図4】

【図5】
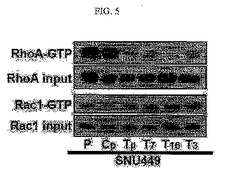
【図6】
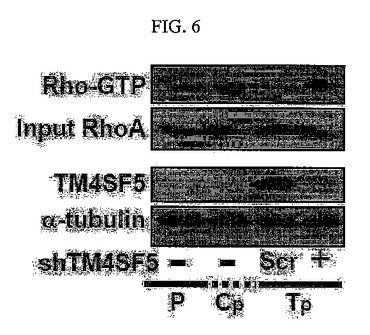
【図7】

【図8】

【図9】
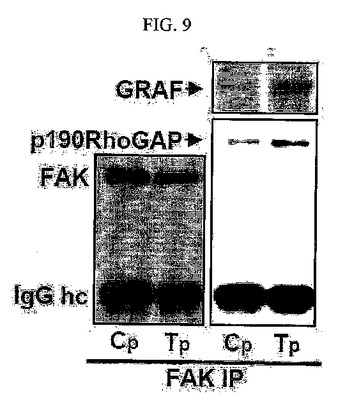
【図10】

【図11】

【図12】

【図13】

【図14】
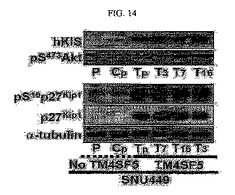
【図15】
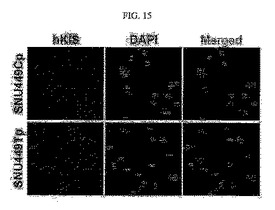
【図16】
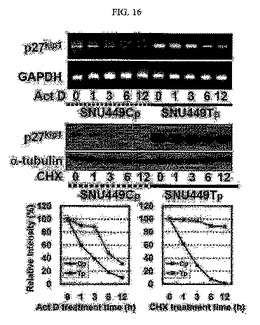
【図17】

【図18】
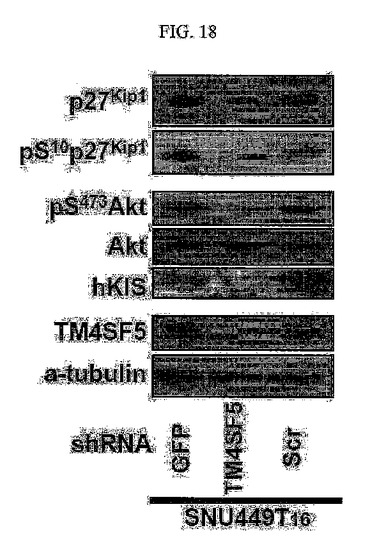
【図19】

【図20】

【図21】

【図22】
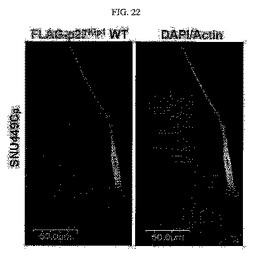
【図23】
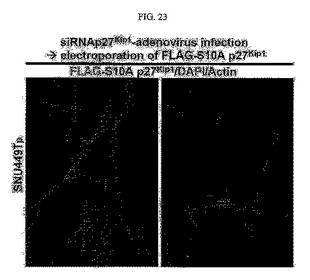
【図24】
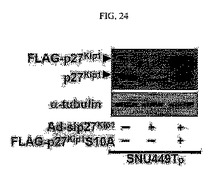
【図25】
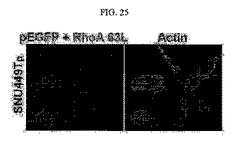
【図26】
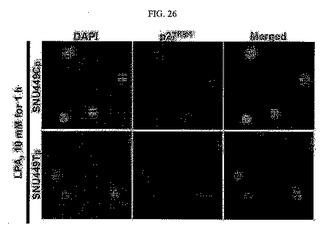
【図27】
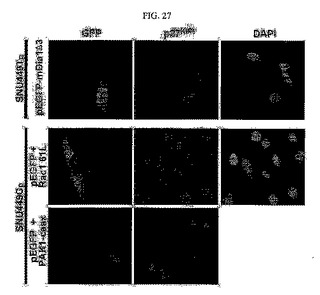
【図28】

【図29】

【図30】
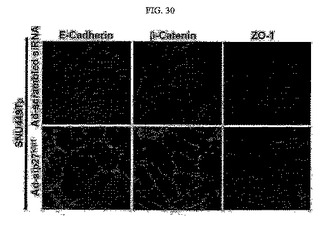
【図31】
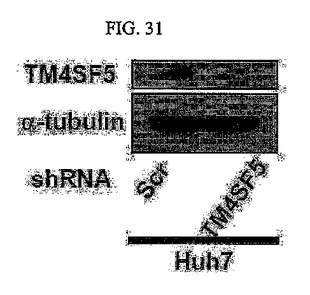
【図32】
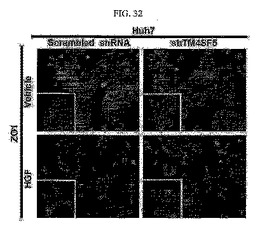
【図33】

【図34】

【図35】
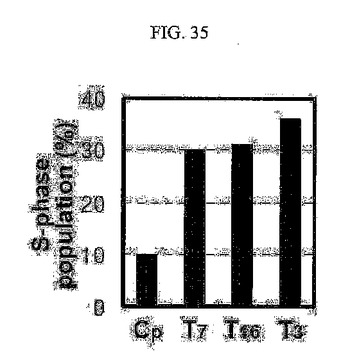
【図36】
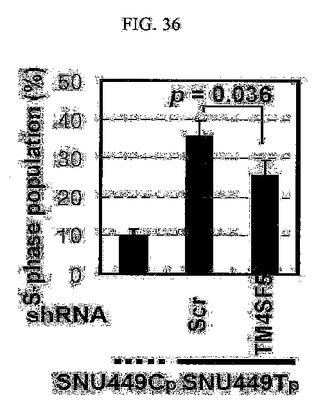
【図37】

【図38】
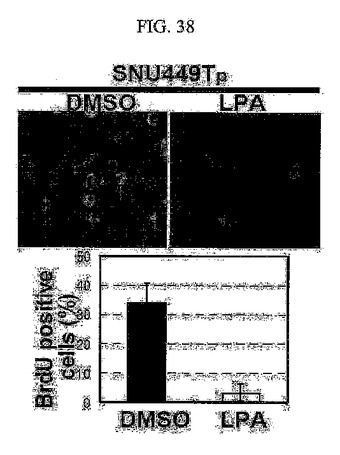
【図39】
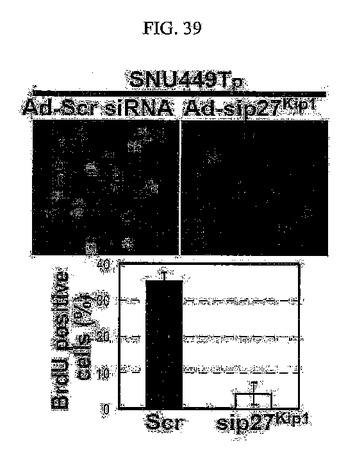
【図40】
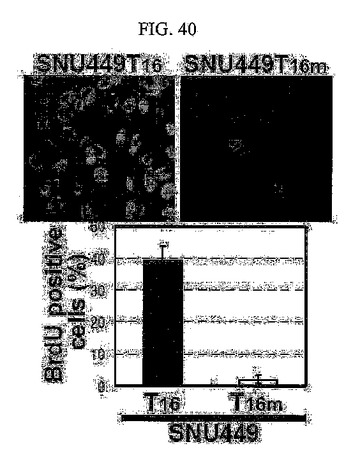
【図41】

【図42】

【図43】

【図44】

【図45】

【図46】

【図47】
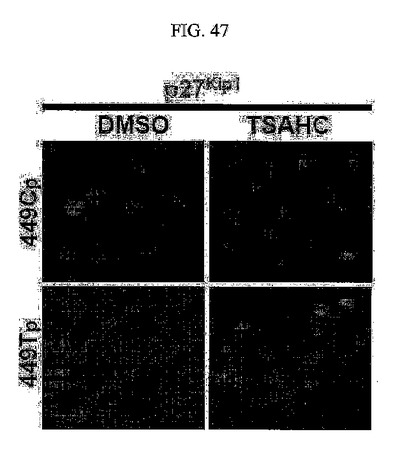
【図48】

【図49】

【図50】

【図51】
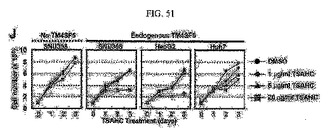
【図52】
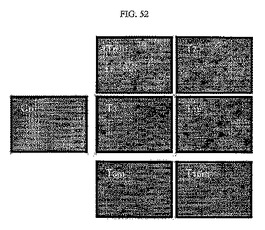
【図53】
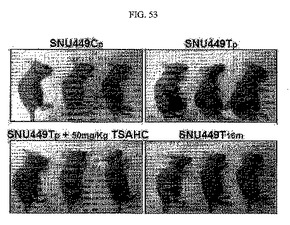
【図54】
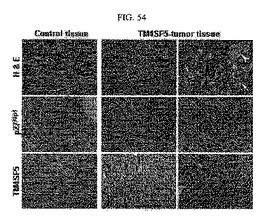
【図55】

【図56】

【図57】
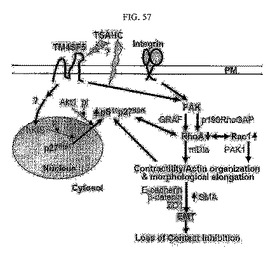
【図58】
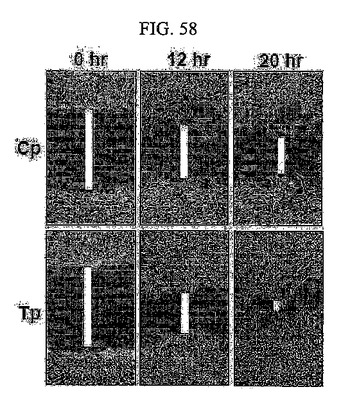
【図59】
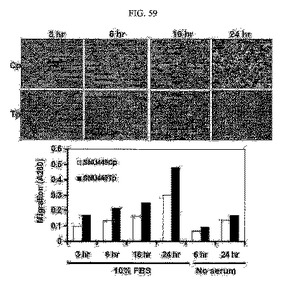
【図60】

【図61】
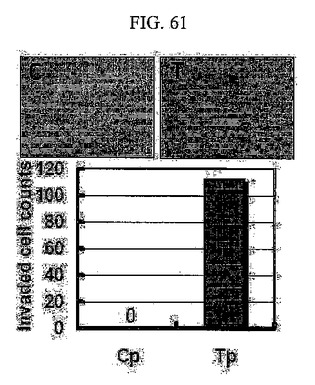
【図62】
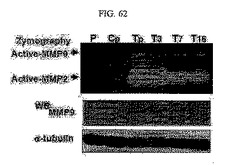
【図63】
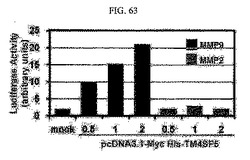
【図64】
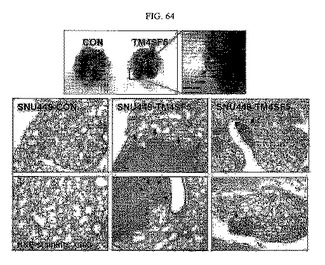
【図65】
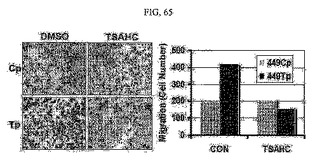
【図66】

【図67】
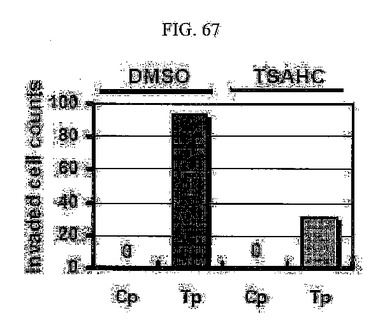
【図68】

【図69】
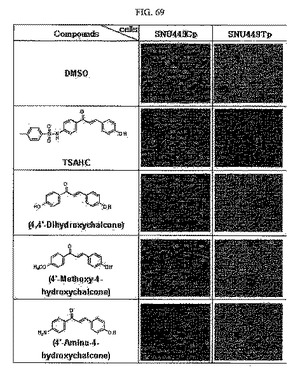
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図42】
【図43】
【図44】
【図45】
【図46】
【図47】
【図48】
【図49】
【図50】
【図51】
【図52】
【図53】
【図54】
【図55】
【図56】
【図57】
【図58】
【図59】
【図60】
【図61】
【図62】
【図63】
【図64】
【図65】
【図66】
【図67】
【図68】
【図69】
【公表番号】特表2010−511892(P2010−511892A)
【公表日】平成22年4月15日(2010.4.15)
【国際特許分類】
【出願番号】特願2009−540168(P2009−540168)
【出願日】平成19年12月7日(2007.12.7)
【国際出願番号】PCT/KR2007/006337
【国際公開番号】WO2008/069608
【国際公開日】平成20年6月12日(2008.6.12)
【出願人】(505326520)ソウル ナショナル ユニバーシティ インダストリー ファウンデーション (10)
【出願人】(509109545)インダストリー−アカデミック コーポレーション ファウンデーション キョンサン ナショナル ユニバーシティ (2)
【Fターム(参考)】
【公表日】平成22年4月15日(2010.4.15)
【国際特許分類】
【出願日】平成19年12月7日(2007.12.7)
【国際出願番号】PCT/KR2007/006337
【国際公開番号】WO2008/069608
【国際公開日】平成20年6月12日(2008.6.12)
【出願人】(505326520)ソウル ナショナル ユニバーシティ インダストリー ファウンデーション (10)
【出願人】(509109545)インダストリー−アカデミック コーポレーション ファウンデーション キョンサン ナショナル ユニバーシティ (2)
【Fターム(参考)】
[ Back to top ]