
UDP−ラムノ−スの製造方法
【課題】 多機能なラムノ-ス付加糖鎖を合成するために必要なUDP-ラムノ-スの効率のよい生産方法を確立し、より大量に供給することにある。
【課題解決手段】
シロイヌナズナ(Arabidopsis thaliana)由来のUDP-glucose4,6-dehydratase-3,5-epimerase/4-keto reductaseをコ-ドする遺伝子(RHM2遺伝子)を宿主細胞内で発現させるか、あるいは該遺伝子中のUDP-glucose 4,6-dehydrataseドメインをコ-ドする遺伝子(RHM2-N)及びUDP-4-keto-6-deoxy-glucose 3,5-epimerase/4-keto reductaseドメインをコ-ドする遺伝子(RHM2-C)を宿主細胞内で共発現させ、細胞内に蓄積したUDP-ラムノ-スを抽出する。もしくは、産生する酵素系によりin vitroでUDP-グルコ-スをUDPラムノ-スに変換する。
【課題解決手段】
シロイヌナズナ(Arabidopsis thaliana)由来のUDP-glucose4,6-dehydratase-3,5-epimerase/4-keto reductaseをコ-ドする遺伝子(RHM2遺伝子)を宿主細胞内で発現させるか、あるいは該遺伝子中のUDP-glucose 4,6-dehydrataseドメインをコ-ドする遺伝子(RHM2-N)及びUDP-4-keto-6-deoxy-glucose 3,5-epimerase/4-keto reductaseドメインをコ-ドする遺伝子(RHM2-C)を宿主細胞内で共発現させ、細胞内に蓄積したUDP-ラムノ-スを抽出する。もしくは、産生する酵素系によりin vitroでUDP-グルコ-スをUDPラムノ-スに変換する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、UDP-ラムノ-スの合成に関与する遺伝子、該遺伝子を有する組換えベクタ-、該組み換えベクタ-により形質転換された形質転換体、ならびに該形質転換体を利用したUDP-ラムノ-スの製造方法に関する。
【背景技術】
【0002】
糖タンパク質などの糖鎖は生体において、非常に重要な働きをしていることが明らかとなっており、このことから糖鎖の構造を自在に変換する糖鎖工学が重要技術となってきている。現在糖鎖を改変する技術としては、化学合成により目的の糖鎖を合成したものをタンパク質に結合させる化学的手法や、細胞が本来持つ内在性の糖鎖合成機能を遺伝子工学の手法を用いて目的の糖鎖に改変する、あるいは糖転移酵素そのものをタンパク質と反応させることで目的の糖タンパク質を得る生物学的手法などがある。化学的手法では、大量合成の手法に道が開けてきてはいるが、糖鎖の複雑性から、いまだに全ての種類の糖鎖を簡単に供給するには至ってはいない、また化学合成した糖鎖を効率良くかつ大量にタンパク質に結合させるのは困難である。一方、生物学的手法は、遺伝子工学の発展により、糖鎖合成関連遺伝子の発現及び機能を制御することが可能となったため、生物の細胞を用いた生体内での糖鎖改変が可能となってきている。また、糖鎖合成酵素を用いた生体外の糖鎖合成も非常に有用であり、均一で、大量の糖鎖の生産が可能となる。一方、生物学的手法を用いた生体内外での糖鎖合成では、糖ヌクレオチドが糖転移酵素の糖供与体として必須となるが、この糖ヌクレオチドは非常に高価であるので、大量生産に用いることは困難となっている。糖ヌクレオチドは、生体内に微量存在するものであり、かつ、高エネルギ-結合で結ばれた非常に反応性に富む不安定な物質であるので、各生物においてその産生量はあまり多くないため大量に生産することは困難であるのが原因である。
【0003】
近年、バクテリアを用いた生産系により、比較的多種類の糖ヌクレオチドの大量生産系が実用的なものへ近づいてきており、その供給は安定する方向へ向かっている。しかしながら、このバクテリアを用いる系では、2種の微生物を混合して、なおかつ、細胞中に含まれる一方の原料を他方の細胞中に送り込むために細胞を破壊した状況で生産を行うことから、ある程度反応過程の長いものでは生産量はそれほど多くなく、新たな手法の開発が求められている。また、バクテリアを用いた生産系によるUDP-ラムノ-スの大量生産系に関する報告は無い。
【0004】
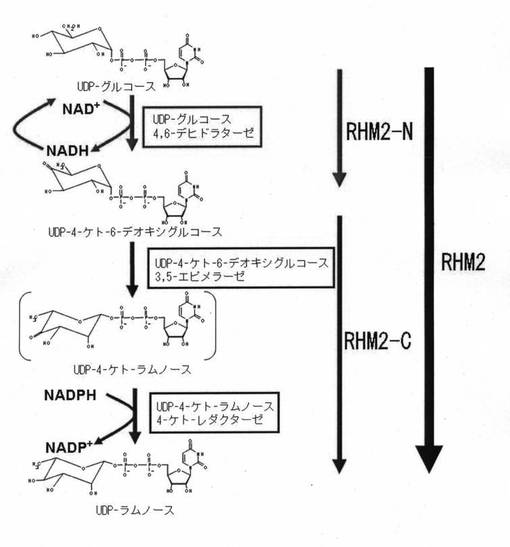
糖ヌクレオチドのうち、UDP-ラムノ-スはラムノ-ス転移酵素の糖供与体としてラムノ-スを含んだ糖鎖合成を行う際には必須のものである。ラムノ-スを含んだ糖鎖にはラムノガラクチュロナンI (RG-I)やラムノガラクチュロナン(RG-II)に代表されるように機能的に重要な役割を果たすことが多いことから、糖供与体の大量で安価な供給が望まれている。また、ラムノ-スを含んだO-結合型糖鎖があり、これらの生合成にもUDP-ラムノ-スが糖供与体として用いられる。よって今後、これらのラムノ-ス含有物質の研究開発が進み重要な機能が明らかになることで新たな需要の拡大が見込まれる。このUDP-ラムノ-スはUDP-グルコ-スから3段階の反応を1つの酵素で触媒し合成される(図1)。この3段階の反応を触媒する酵素は、単一の遺伝子でコ-ドされるが、そのN-末端側ドメインとC-末端側ドメインで機能が異なっている。すなわち、最初の1段階目の反応は、RHM2タンパク質のN-末端側ドメインにコ-ドされるUDP-グルコ-ス4、6-デヒドラタ-ゼによって起こり、UDP-4-ケト-6-デオキシグルコ-スが生成される。続いて2段目、3段目の反応は、RHM2タンパク質のC-末端側でコ-ドされるUDP-4-ケト-6-デオキシグルコ-ス UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼによって起こり、UDP-ラムノ-スに変換される。
これらの酵素はUDP-ラムノ-スを使用する生物であればもっている普遍的な酵素である。ただし、このような生物は合成したUDP-ラムノ-スを使用していくため、細胞内にUDP-ラムノ-スが蓄積していくことはない。このため、生物体からUDP-ラムノ-スを単離する場合、その生産量は極めて低くかつ高価である。また、遺伝子工学的手法を用いてUDP-ラムノ-スを合成した例は見あたらない。
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明の課題は、多機能なラムノ-ス付加糖鎖を合成するために必要なUDP-ラムノ-スの効率のよい生産方法を確立し、より大量に供給することにある。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決すべく鋭意研究を重ねた結果、シロイヌナズナ(Arabidopsis thaliana)のUDP-ラムノ-ス合成を触媒する、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ(以下、UDP-glucose 4,6-dehydratase-3,5-epimerase/4-keto reductaseという場合がある。)をコ-ドする遺伝子を初めて単離し、この遺伝子の構造を明らかにするとともに、この遺伝子を機能的に発現することによって、生体内および生体外で効率よくUDP-ラムノ-スが合成されることを見いだした。また、さらに、上記遺伝子をUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン部分と、 UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメイン部分とに分け、それぞれを共発現させた場合、これら領域を全て含む上記遺伝子を発現させた場合に比較し、UDP-ラムノ-スの生産効率が向上するという驚くべき知見を得て、本発明を完成させるに到ったものである。
【0007】
すなわち、本発明は、以下(1)〜(31)に示されるとおりである。
(1)(a)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列を有するか、あるいは(b)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(2)上記(1)に記載のタンパク質をコ-ドするDNA。
(3)(a)配列番号1、配列番号3もしくは配列番号19で示される塩基配列を有するか、あるいは(b)配列番号1、配列番号3もしくは配列番号・・で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(4)上記(2)又は(3)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(5)上記(4)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(6)(a)配列番号6で示されるアミノ酸配列を有するか、あるいは(b)配列番号6で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質。
(7)上記(6)に記載のタンパク質をコ-ドするDNA。
(8)(a)配列番号5で示される塩基配列を有するか、あるいは(b)配列番号5で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(9)上記(7)又は(8)に記載のDNAにより組み換えられていることを特徴とする、組換えベクタ-。
(10)上記(9)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(11)(a)配列番号8で示されるアミノ酸配列を有するか、あるいは(b)配列番号8で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(12)上記(11)に記載のタンパク質をコ-ドするDNA。
(13)(a)配列番号7で示される塩基配列を有するか、あるいは(b)配列番号7で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(14)上記(12)又は(13)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(15)上記(7)又は(8)に記載のDNA及び上記(12)又は(13)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(16)上記(14)に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
(17)上記(9)及び(14)に記載の組換えベクタ-が、同一宿主細胞に導入されていることを特徴とする形質転換体。
(18)上記(15)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(19)配列番号2、4もしくは20で示されるアミノ酸配列と70%以上の相同性を有するアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(20)イネ(Oryza sativa)またはタバコ(Nicotiana tabacum)由良のタンパク質である、上記(19)に記載のタンパク質。
(21)上記(19)または(20)に記載のタンパク質をコ-ドするDNA。
(22)上記(21)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(23)上記(22)に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
(24)宿主細胞が、UDP-グルコ-スを細胞内に含有していることを特徴とする、上記(5)、(17)、(18)及び(23)のいずれかに記載の形質転換体。
(25)宿主細胞が、UDP-ラムノ-スを消費する酵素系を保持していないことを特徴とする、上記(5)、(17)、(18)及び(23)のいずれかに記載の形質転換体。
(26)宿主細胞が酵母細胞である、上記(5)、(10)、(16)〜(18)及び(23)〜(25)のいずれかに記載の形質転換体。
(27)上記(5)、(17)、(18)及び(23)〜(26)のいずれかに記載の形質転換体を培養し、培養物からUDP-ラムノ-スを採取することを特徴とする、UDP-ラムノ-スの製造方法。
(28)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を培地に培養し、得られた培養物、培養処理物、酵素抽出液または精製酵素をUDP-グルコ-スと接触させることにより、UDP-グルコ-スをUDP-ラムノ-スに変換させることを特徴とする、UDP-ラムノ-スの製造方法。
(29)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を同位体元素により置換されたグルコ-スを炭素源として培養し、培養物から同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
(30)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を培地で培養することにより得られた培養物、培養処理物、酵素抽出液または精製酵素を同位体元素により置換されたUDP-グルコ-スと接触させ、同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
(31)同位体元素がC13またはC14であることを特徴とする、上記29または30に記載の製造方法
【発明の効果】
【0008】
本発明によれば、糖鎖において非常に重要な機能をもつラムノ-スの付加を行うために必須なUDP-ラムノ-スを多量に効率よく生産することができる。現時点において糖タンパク質糖鎖を均一に合成する技術は確立されておらず、最終的には生体外での糖鎖の修飾を行うことで糖鎖の均一な合成を行うことが考えられるが、この際、糖供与体として糖ヌクレオチドは必須なものである。特に、ラムノ-スにおいてはUDP-ラムノ-スが非常に高価なため、生体外での修飾反応を大量に行うことは現在非現実的であるが、本発明により、多量のUDP-ラムノ-スの供給が可能となれば、ラムノ-スが付加された高機能な糖鎖の合成を生体外で行うことができる。したがって、本発明は糖タンパク質の糖鎖の研究において多大な貢献をするものである。
【発明を実施するための最良の形態】
【0009】
本発明における第1の酵素タンパク質は、シロイヌナズナ由来のものであって、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼの3者の活性をともに有するタンパク質であって、配列番号2、配列番号4もしくは配列番号20に示されるアミノ酸配列を有するが(以下、配列番号2に示される酵素タンパク質をRHM1タンパク質、配列番号4で示される酵素タンパク質をRHM2タンパク質、配列番号20で示される酵素タンパク質をRHM3タンパク質とそれぞれいう場合がある。)、これのみでなく、配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ-UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質ものであれば、本発明に含まれる。
本発明の第1の酵素遺伝子は、上記したアミノ酸配列コ-ドするDNAである。具体的な塩基配列は、配列番号1、配列番号3、もしくは配列番号19にそれぞれ示されるが(以下、配列番号1に示される遺伝子をRHM1遺伝子といい、配列番号3に示される遺伝子をRHM2遺伝子、配列番号19で示される遺伝子をRHM3遺伝子とそれぞれいう場合がある。)これのみでなく、配列番号1、配列番号3もしくは配列番号19で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ活性及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性をともに有するタンパク質をコ-ドするものであれば、本発明において使用できる。
本発明においては、上記シロイヌナズナのRHM1、RHM2遺伝子もしくはRHM3遺伝子あるいは上記の変異遺伝子を用いて、遺伝子工学的手法によりUDP-ラムノ-スを効率よく生産可能であるが、この製造法には大別して細胞内で生産する方法(培養法)と、細胞外で生産する方法(酵素変換法)の2つがある。
【0010】
このうち、UDP-ラムノ-スを細胞内で製造するための方法の概要は、例えば、下記の行程よりなる。
(1)シロイヌナズナのRHM1、RHM2もしくはRHM3遺伝子〔UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ遺伝子(UDP-glucose 4,6-dehidratase-3,5-epimerase/4-keto-reductase)〕を取得する;
(2)シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を発現用プラスミド等のベクタ-に導入し、RHM1、RHM2もしくはRHM3遺伝子を発現するベクタ-を構築する;
(3)上記発現ベクタ-を、UDP-グルコ-スを細胞内に持つ宿主細胞に導入し、該宿主細胞を形質転換する;
(4)上記(3)の組換え体を培地に培養し、得られた培養物あるいは細胞破砕物等の培養処理物から抽出によりUDP-ラムノ-スを採取し、さらに単離・精製する。
【0011】
本発明において、UDP-ラムノ-スを細胞外で製造する方法は、下記の工程よりなる。
(1)シロイヌナズナのRHM1、RHM2もしくはRHM3遺伝子(UDP-glucose 4,6-dehidratase-3,5-epimerase/4-keto-reductase遺伝子)を取得する;
(2)シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を発現用プラスミド等のベクタ-に導入し、RHM1、RHM2もしくはRHM3遺伝子を発現するベクタ-を構築する;
(3)上記発現ベクタ-を、上記発現ベクタ-を、UDP-グルコ-スを細胞内に持つ宿主細胞に導入し、該宿主細胞を形質転換する。
(4)上記の形質転換体の培養物、細胞破砕物等の培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素を酵素源とし、UDP-グルコ-スおよびNADPHを添加することによりUDP-グルコ-スをUDP-ラムノ-スに変換し、単離・精製を行う。該UDP-ラムノ-スはラムノ-ス含有糖鎖の合成の際に糖供与体として必須なものであるので、機能的に重要と思われる糖鎖へのラムノ-ス付加の際に有用なものである。
【0012】
また別法として、例えば、シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を含む発現用ベクタ-により形質転換した形質転換体の培養物、培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素と、UDP-グルコ-スを本来的に産生する細胞の培養物、培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素を、各々別途に調整し、これらを順次あるいは同時にUDP-グルコ-スに作用させて、UDP-グルコ-スをUDP-ラムノ-スに変換させることにより、UDP-ラムノ-スを製造してもよい。
上記配列番号4で示されるRHM2タンパク質及びその遺伝子は、その配列中にUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメイン及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインを有する。
本発明の第2の酵素タンパク質は、上記RHM2タンパク質のUDP-グルコ-ス4、6-デヒドラタ-ゼドメインに対応するタンパク質であり、配列番号6に示されるアミノ酸配列を有するか、あるいは該アミノ酸配列において、1又は数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質である(以下、配列番号6に示される酵素タンパク質をRHM2-Nタンパク質という場合がある。)。
また、本発明の第2の遺伝子は、RHM2-Nタンパク質あるいはその上記変異体タンパク質をコ-ドするDNAであり、より具体的には、配列番号5に示されるが(以下、配列番号5に示される遺伝子をRHM2遺伝子という場合がある。)これのみでなく、配列番号5に示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性をコ-ドするものであれば、本発明に含まれる。
本発明の第3の酵素タンパク質は、上記RHM2タンパク質のUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメインとUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインの両者を含むタンパク質であり、配列番号8に示されるアミノ酸配列を有するか、あるいは該アミノ酸配列において、1又は数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有し、かつUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメインとUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼの2つの酵素活性を有するタンパク質である(以下、配列番号8に示される酵素タンパク質をRHM2-Cタンパク質という場合がある。)。
本発明の第3の遺伝子は、RHM2-Cタンパク質あるいはその上記変異体タンパク質をコ-ドするDNAであり、より具体的には、配列番号7に示されるが(以下、配列番号7に示される遺伝子をRHM2-C遺伝子という場合がある。)これのみでなく、配列番号7に示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ活性とUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性の2つの酵素活性を有するタンパク質をコ-ドするものであれば、本発明に含まれる。
これらの第2及び第3の酵素タンパク質は、上記第2及び第3の遺伝子を用いて、通常の遺伝子工学的手法により容易に得ることができる。得られた第2及び第3の酵素タンパク質は、図1に示されるように、UDP-グルコ-スをUDP-4-ケト-デオキシグルコ-スに転換する反応、及びUDP-4-ケト-デオキシグルコ-スをUDP-ラムノ-スに転換する反応をそれぞれ触媒するから、例えば、これら酵素を順次または同時に用いることにより、UDP-グルコ-スからUDP-ラムノ-スを製造することができる。
しかし、本発明におけるUDP-ラムノ-スの生産において、最も生産性が高い方法は、上記第2及び第3の遺伝子を同一宿主細胞において共発現する方法である。この方法による場合、RHM2遺伝子を用いる場合に比べ、UDP-ラムノ-スの生産量は著しく向上する。
以下、本発明をさらに詳細に説明する。本明細書において、アミノ酸配列および塩基配列の略号による表示は、IUPAC-IUBの規定および当該分野における通称または慣行に従うものとする。
【0013】
本発明のRHM1、RHM2もしくはRHM3遺伝子、RHM2-N遺伝子及びRHM2-C遺伝子の取得は、一般的手法に従ってシロイヌナズナから調製したcDNAライブラリを鋳型とし、PCR法により単離することができる。また、RHM2-N遺伝子及びRHM2-C遺伝子については、上記RHM2遺伝子を鋳型として用いてもよい。
【0014】
シロイヌナズナのcDNAライブラリは、通常用いられているプラスミドベクタ-またはλファ-ジ由来のベクタ-を用い、通常の方法に従い作成することができる。
【0015】
PCR法は、生体外でDNAの特殊な領域を、そのセンス・アンチセンスプライマ-、耐熱性のDNAポリメラ-ゼ、DNA増幅システム等の組み合わせを用いて、約2-3時間で10-100万倍に特異的に増幅することができる技術であり、本発明遺伝子およびその断片は、他種の酵素との塩基配列の相同性等をもとに、このPCR法での増幅が可能である。
【0016】
上記遺伝子を組み込むベクタタ-としては、宿主内で複製保持されるものであれば、いずれも使用することができる。例えば大腸菌由来のプラスミドベクタ-pBR322やpUC19などを挙げることができる。
ベクタ-を組み込む方法としては、例えばT.Maniatisらの方法[Molecular Cloning, Cold Spring Harbor Laboratory, p.239 (1982)]に従うことができる。
【0017】
クロ-ン化された遺伝子は、発現に適したベクタ-中のプロモ-タ-の下流に連結して発現ベクタ-を得ることができる。ベクタ-としては、酵母由来のプラスミドYEp352GAP、YEp51、pSH19、などが挙げられる。
【0018】
該遺伝子はその5'末端に翻訳開始コドンとしてのATGを有し、また、3'末端には翻訳終止コドンとしてのTAA、TGAまたはTAGを有してよい。また、5'末端あるいは3'末端には、例えば、タンパク質の単離精製を容易にするため、6 x ヒスチジンやヘマグルチニンタンパク質の一部分である標識抗原遺伝子やGSTタンパク質などの標識タンパク質遺伝子を結合させて発現させてもよい。特に配列番号5に示されるRHM2-N遺伝子のC末端には上記終止コドンを位置させるのが好ましく、また、配列番号7に示されるRHM2-C遺伝子のN末端側には開始コドン(ATG)を位置させるのが好ましいが、得られるタンパク質はメチオニンがなくても当然酵素活性を有する。また、逆に、例えば、N末端側にMet-His tagを付加せしめたタンパク質であってもそれ自体活性を有する。
さらに該遺伝子を発現させるためには、その上流にプロモ-タ-を接続するが、本発明で用いられるプロモ-タ-としては遺伝子の発現に用いる宿主に対応した適切なプロモ-タ-であれば、いかなるものでもよい。
また、形質転換する宿主が酵母である場合、ENO1プロモ-タ-、GAL10プロモ-タ-、GAPDHプロモ-タ-、ADHプロモ-タ-、AOXプロモ-タ-などが挙げられる。
【0019】
このように構築されたRHM1、RHM2もしくはRHM3遺伝子、RHM2-N遺伝子あるいはRHM2-C遺伝子を有する組換えDNAを含むベクタ-を用いて、該ベクタ-を保持する形質転換体を作成する。RHM2-N遺伝子及びRHM2-C遺伝子を共発現させる場合には、例えば、RHM2-N遺伝子を有する組換えベクタ-とRHM2-C遺伝子を有する組換えベクタ-を同一宿主細胞に導入するか、あるいはRHM2-N遺伝子及びRHM2-C遺伝子の両方を同一のベクタ-に導入し、得られた組換えベクタ-を用いて宿主細胞を形質転換する。これらにより得られた形質転換体は、RHM2-N遺伝子を有する組換えベクタ-とRHM2-C遺伝子を有する組換えベクタ-の2種のベクタ-を保持するか、あるいはRHM2-N遺伝子及びRHM2-C遺伝子をともに有する1種類の組換えベクタ-を保持し、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼを産生する。したがって、同形質転換体は、UDP-グルコ-スを原料としてUDP-ラムノ-スを生産することができ、しかも、上記したようにその生産性が高い株である。
宿主としては、UDP-ラムノ-スを生体内で消費しない出芽酵母(Saccharomyces cerevisia)などが挙げられるが、この他UDP-ラムノ-スを消費しない他の酵母(Pichia pastorisなど)などでもよい。また、酵母以外でも、UDP-ラムノ-スの消費する酵素系を持たない宿主であればいかなるものでもよい。
また、細胞破砕物、酵素抽出物等を用い、生体外でUDP-ラムノ-スを生産する場合、細胞質内で該遺伝子産物を生産することが可能な宿主であれば、いかなるものでも用いることができる。
上記形質転換体の作成は、それぞれの宿主について一般的に行われている方法で行う。または一般的ではなくても適用可能な方法ならばよい。例として、宿主が酵母であれば、リチウム法あるいはエレクトロポレ-ション法により組み換えDNAを含むベクタ-を導入する。
【0020】
このようにして、本発明の遺伝子を有する組み換えDNAを含むベクタ-を保持する形質転換体が得られ、該形質転換体を培養することにより、主に該形質転換体の細胞内においてUDP-ラムノ-スとUDP-ラムノ-ス製造に必要な酵素が産生、蓄積する。
【0021】
形質転換体を培養する場合、培養に使用される培地は、それぞれの宿主について一般的に用いられているものを使用する。または、一般的でなくとも適用可能な培地ならばよい。例としては、宿主が酵母であればYPD培地、SD培地などを用いる。培養は、それぞれの宿主について一般的に用いられている条件で行う。また、一般的でなくとも適用可能な条件であればよい。例としては、宿主が酵母であれば約25-37度で、約12時間-5日間行い、必要により通気や撹拌を行うことができる。
【0022】
例えば、上記培養物からUDP-ラムノ-スあるいは酵素を抽出する際は、宿主細胞を遠心分離によって培地と分け、宿主細胞を破砕してUDP-ラムノ-スあるいは酵素を抽出する。宿主が酵母の場合、例えばグラスビ-ズによって細胞を破壊し、遠心分離する。この際、酵素及びUDP-ラムノ-スは、上清画分に存在する。また、1-ブタノ-ルによって飽和させた1Mギ酸溶液中に酵母菌体を懸濁後、30分から3時間程度氷冷し、遠心分離することによってもUDP-ラムノ-スを抽出することが可能である。この場合も、UDP-ラムノ-スは、上清画分に存在する。
上記の遠心分離によって分離された上清画分からUDP-ラムノ-スを分離する場合は、ゲルろ過法等により低分子量の画分を集め、さらにHPLCによる分離を行うことにより、精製単離する。また、イオン交換カラムや逆相カラムを用いて精製単離することも可能である。
また、生体外で酵素変換反応を行い、UDP-ラムノ-スを製造する場合においては、細胞破砕物や上清画分をそのまま酵素源として利用することが可能であるが、上清画分硫酸アンモニウムを75%の濃度に溶かし、これにより沈殿してくるタンパク質画分を集め、透析等により脱塩したものも酵素源として使用することができる。
【0023】
上記酵素源を用いて、基質となるUDP-グルコ-ス、NAD+およびNADPHを加え反応させた後、HPLCでUDP-ラムノ-スを単離精製することにより、UDP-ラムノ-スを得ることができる。
一方、本発明においては、シロイヌナズナ由来の上記配列番号2、配列番号4及び配列番号20に示されるタンパク質(RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質)のみではなく、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質として、RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質に対して相同率70%以上の相同性を有するタンパク質も挙げることができ、本発明においてはこれら相同性を有するタンパク質、及び該タンパク質をコ-ドする塩基配列を有する遺伝子も包含する。これら相同性を有するタンパク質としては、植物由来の、例えばイネ(Oryza Sativa)、タバコ(Nicotiana tabacum)由来のタンパク質が挙げられ、イネにおける相同性を有するタンパク質のアミノ酸配列及びこれをコ-ドする遺伝子の塩基配列はそれぞれ配列番号22、配列番号21に示される。
これら相同性を有するタンパク質は、これらをコ-ドするDNAを用いて、上記RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質について記載したのと同様な遺伝子工学手法を用いて製造することが出来る。すなわち本発明においては、これら相同性を有するタンパク質をコ-ドするDNAにより組み換えられた組換えベクタ-、該組換えベクタ-が導入されている形質転換体、及び該形質転換体を用いた、上記相同性を有する酵素タンパク質の製造方法、及び該形質転換体あるいは該相同酵素タンパク質を用いたUDP-ラムノ-スの製造方法をも包含する。
【0024】
また、本発明の形質転換体を培地に培養するに際し、同位体元素でその構成元素を置換したグルコ-スを炭素源として培地に含有させて培養を行うことにより、培養物から、同位体元素でラベルされたUDP-ラムノ-スを採取することができる。
また別法として、本発明の形質転換体から得られた、酵素タンパク質を含む培養物、培養処理物、酵素抽出液あるいは精製酵素を、同位体元素でその構成元素を置換したUDP-グルコ-スと接触させることにより、同位体元素でラベルされたUDP-ラムノ-スに変換することも可能である。
このような酵素変換法では、上記同位体元素を有するUDP-グルコ-スに対し、上記RHM2-N精製タンパク質あるいは該タンパク質を含有する培養物、培養処理物、酵素抽出液または精製酵素と、同RHM2-C精製タンパク質あるいは該タンパク質を含有する培養物、培養処理物、酵素抽出液を順次または同時に接触させることにより、同位体元素でラベルされたUDP-ラムノ-スを得てもよい。
好ましい同位体元素としては、たとえばC13あるいはC14同位体が挙げられる。このような同位体元素でラベルされたUDP-ラムノ-スは、糖鎖合成系の解明あるいはこの糖鎖合成系に影響を及ぼす物質等の探索等に有用である。
以下、本発明の実施例を示すが、本発明はこの実施例にのみ限定されるものではない。
【実施例】
【0025】
〔実施例1〕RHM1遺伝子(配列番号1)、RHM2遺伝子(配列番号3)およびRHM3遺伝子(配列番号19)の単離
RHM1遺伝子およびRHM2遺伝子に関してはシロイヌナズナのcDNAライブラリを鋳型として用いてPCR法により遺伝子のクロ-ニングを行った。cDNAライブラリはCLONTECH社のQUICK-Clone cDNAを用いた。また、RHM3遺伝子に関しては理化学研究所バイオリソ-スセンタ-よりcDNAクロ-ンを入手し(Resouce Number: pda08705)、鋳型として用いた。プライマ-の設計においては、制限酵素でタンパク質をコ-ドしている部分が容易に切り出せるようにRHM1の場合は、N-末端Sac Iサイト、C-末端部分にKpn Iサイトに位置させ、RHM2およびRHM3の場合は、N-末端Eco RIサイト、C-末端部分にSal Iサイトに位置させた。また、N-末端には、あらかじめ標識抗原であり、ニッケルアガロ-ス結合配列をコ-ドする6 x ヒスチジン配列を付加せしめた。それぞれのプライマ-の配列を以下に示す(配列番号13と14は、RHM1用、同15と16はRHM2用、同23と24はRHM3用)。
AAGAGCTCATGCATCACCATCACCATCACATGGCTTCGTACACTCCCAAGAAC(配列番号13)
AAAGGTACCTCAGGTTTTCTTGTTTGGCCCGTATG(配列番号14)
AGAATTCATGCATCACCATCACCATCACATGGATGATACTACGTATAAGCCAAAGAAC(配列番号15)
AAAAAGTCGACTTAGGTTCTCTTGTTTGGTTCAAAGAC(配列番号16)
AGAATTCATGCATCACCATCACCATCACATGGCTACATATAAGCCTAAGAACATCCTC(配列番号23)
AAAAAGTCGACTTACGTTCTCTTGTTAGGTTCGAAGACG(配列番号24)
PCRの条件は以下の通り。
95度 30秒
57度 30秒
72度 2分15秒
30サイクル
この条件で得られたそれぞれ約2.0 kbpのDNA増幅断片をアガロ-ス電気泳動で分離した後、制限酵素Sac IとKpn IもしくはEco RIとSal Iで切断し、pBluescript II-SK+ベクタ-のEco RIサイトとSal Iサイト部位に挿入した。このクロ-ニングされた遺伝子をダイデオキシ法を用いたシ-クエンスキットにより塩基配列を確認し、配列番号1、3および19に示した塩基配列であることを確認した。
【0026】
〔実施例2〕RHM1、RHM2およびRHM3遺伝子発現ベクタ-の作成およびこのプラスミドを含む酵母形質転換体の作成
pBluescript II-SK+ベクタ-に挿入されているRHM1遺伝子、RHM2遺伝子およびRHM3遺伝子をSac IおよびKpn IもしくはEco RIおよびSal Iで切り出し、酵母の解糖系のプロモ-タ-であるGAPDHとそのタ-ミネ-タ-の間にpUC18のマルチクロ-ニングサイトのうちEco RIからSal Iまでの部分を持ち選択マ-カ-としてURA3遺伝子を持つ発現用ベクタ-YEp352GAP-IIのSacIおよびKpnIサイトもしくはEcoRI およびSal Iサイトに挿入しYEp352-GAP-II-6xHIS-RHM1、YEp352-GAP-II-6xHIS-RHM2およびYEp352-GAP-II-6xHIS-RHM3を構築した。この発現ベクタ-は、酵母のW303-1B株(ura3, lue2, his3, trp1, ade2)に導入し、形質転換体、すなわち、RHM1遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM1株)、RHM2遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2株)およびRHM3遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM3株)を得た。なお、W303-1B株は、ATCC (American Type Culture Collection)から入手可能である(ATCC Number: 201238TM)。
【0027】
〔実施例3〕酵母細胞質タンパク質の抽出
〔実施例2〕で得られた形質転換体より細胞質中のタンパク質を抽出した。まず、〔W303/YEp352-GAP-II-6xHIS-RHM1株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕およびW303株をSD培地にて24時間30度で培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞壁画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。
【0028】
〔実施例4〕RHM1タンパク質、RHM2タンパク質およびRHM3タンパク質の酵母内での発現の確認
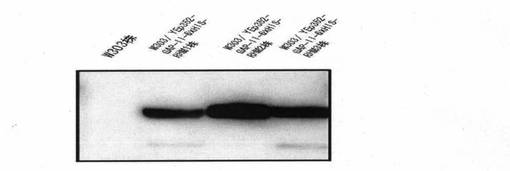
〔実施例2〕で得られた形質転換体について、それぞれの細胞内でタンパク質が発現されていることをウエスタンブロッティング法を用いて確認した。まず、上記2種の形質転換体〔W303/ YEp352-GAP-II-6xHIS-RHM1株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕およびW303株をSD培地にて30℃で24時間培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。このタンパク質を含む細胞質画分を50mM Tris-HCl pH6.8に溶かし、SDS-PAGEを行った後、PVDF膜に電気的に転写し、それぞれ、抗-penta-HIS抗体を用いてタンパク質の発現を確認した(図2)。その結果、RHM1遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM1株〕、RHM2遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕およびRHM3遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕でタンパク質が発現していることが確かめられた。
【0029】
〔実施例5〕UDP-ラムノ-ス合成活性測定
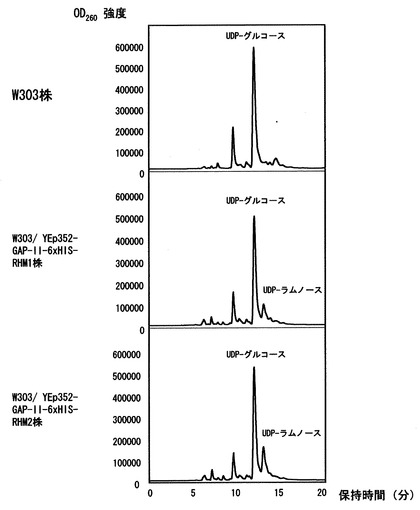
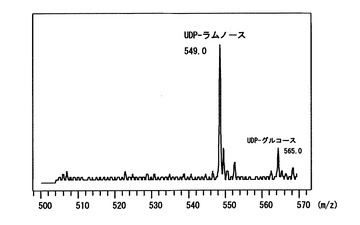
〔実施例3〕で得られた酵母細胞質タンパク質画分のUDP-ラムノ-ス合成活性を検出した。活性測定は基質としてUDP-グルコ-スを、補助因子として1 mM NAD+及び3 mM NADPHを用い、100 mM Tris-HCl、pH 8.0という条件で30度、2時間反応を行い、フェノ-ル:クロロホルム溶液 (24: 1) を加え撹拌した後、遠心分離を行い、上清を回収した。上清を逆相カラムを装備したHPLCに供与しよりUDP-ラムノ-スとUDP-グルコ-スの検出を行った。HPLCはC30カラムを用いて1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い、検出はUV260 nmで行った。その結果、RHM1タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM1株とRHM2タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM2株およびRHM3タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM3株でUDP-ラムノ-ス合成活性を検出した(図3)。さらに、合成したUDP-ラムノ-スについてESI-MS測定を行い分子量を測定した結果、予想される分子量と一致した(図4)。
【0030】
〔実施例6〕RHM2遺伝子導入酵母菌体からのUDP-ラムノ-スの抽出
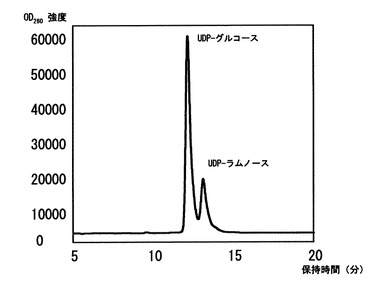
〔実施例3〕で得られた酵母細胞質画分よりUDP-ラムノ-スを分離精製した。まず、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕をSD(-ウラシル)培地にて24時間30度で培養し、得られた酵母細胞O.D.600=10当り1mlの1-ブタノ-ル飽和1 Mギ酸を添加し、十分に撹拌し、氷上にて1時間安置した。この試料より遠心操作を用いて細胞壁画分を取り除き、細胞質画分だけを分離した。上清を凍結乾燥し、純水に再懸濁することによって糖ヌクレオチドを抽出した。抽出した糖ヌクレオチドをC30カラムにより1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い溶出時間からUDP-ラムノ-スを分離した。分離したUDP-ラムノ-スを数度同条件で再分離することによりUDP-ラムノ-スを精製した(図5)。
【0031】
〔実施例7〕RHM2遺伝子のUDP-グルコ-ス4、6-デヒドラタ-ゼドメインの発現とUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインの発現
〔実施例2〕で得られたYEp352-GAP-II-6xHIS-RHM2を用いてPCR法によりRHM2遺伝子中のUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン(RHM2-N)とUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメイン(RHM2-C)のクロ-ニングを行った。プライマ-の設計においては、制限酵素でタンパク質をコ-ドしている部分が容易に切り出せるようにRHM2-N、RHM2-C共に、N-末端Eco RIサイト、C-末端部分にSal Iサイトに位置させた。また、N-末端には、あらかじめ標識抗原であり、ニッケルアガロ-ス結合配列をコ-ドする6 x ヒスチジン配列を付加せしめた。プライマ-は、〔実施例1〕で用いたプライマ-の内、配列番号15と配列番号16を流用した。その他のプライマ-の配列を以下に示す(配列番号15と17は、RHM2-N用、同16と18はRHM2-C用)。
AAAAAGTCGACTTAAGCTTTGTCACCAGAATCACCATT(配列番号17)
AGAATTCATGCATCACCATCACCATCACACACCTAAGAATGGTGATTCTGGTG(配列番号18)
PCRの条件は以下の通り。
95度 30秒
57度 30秒
72度 1分30秒
30サイクル
この条件で得られた約1.2
kb (RHM2-N) と約0.9 kb (RHM2-C)の断片をそれぞれアガロ-ス電気泳動で分離した後、制限酵素Eco RIとSal Iで切断し、YEp352GAP-IIのEcoRI およびSal Iサイトに挿入しYEp352-GAP-II-6xHIS-RHM2-NおよびYEp352-GAP-II-6xHIS-RHM2-Cを構築した。このクロ-ニングされた遺伝子をダイデオキシ法を用いた塩基配列解析により塩基配列を確認し、それぞれ配列番号9及び11に示した塩基配列(対応するタンパク質のアミノ酸配列は10及び12に示す。)であることを確認した。さらに、YEp352-GAP-II-6xHIS-RHM2-Cベクタ-からGAPDHプロモ-タ-、6 x HIS-RHM2-C、GAPDHタ-ミネ-タ-の3つの部分を含む断片を制限酵素Bam HIを用いて切り出し、酵母のプラスミドベクタ-でありLEU2マ-カ-を持つYEp351のBam HIサイトに挿入しYEp351-GAP-II-6xHIS-RHM2-Cを構築した。この発現ベクタ-は、酵母のW303-1B株(ura3, lue2, his3, trp1, ade2)に導入し、形質転換体、すなわち、RHM2-N遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2-N株)とRHM2-C(W303/ YEp352-GAP-II-6xHIS-RHM2-C株)およびRHM2-N遺伝子、RHM2-C遺伝子共発現株を同時に導入したRHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)を得た。
【0032】
〔実施例8〕酵母細胞質タンパク質の抽出
〔実施例7〕で得られた形質転換体(RHM2-N株、RHM2-C株、RHM2-N+RHM2-C株)より〔実施例3〕に示す方法に従って細胞質中のタンパク質を抽出した。まず、形質転換株及びW303株をSD培地にて24時間30度で培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞壁画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。
【0033】
〔実施例9〕RHM2-Nタンパク質、RHM2-Cタンパク質の酵母内での発現の確認
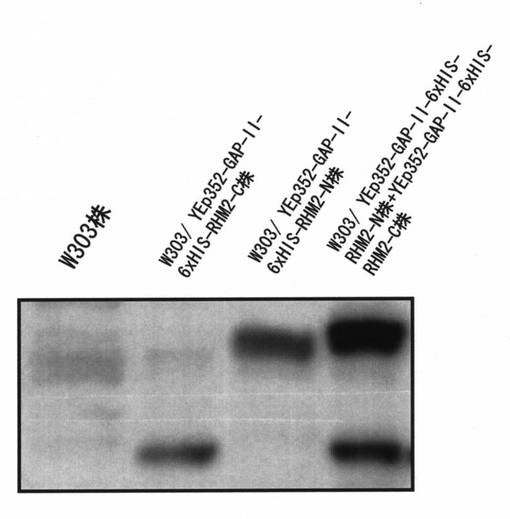
〔実施例7〕で得られた形質転換体について、それぞれの細胞内でタンパク質が発現されていることをウエスタンブロッティング法を用いて確認した。また、ウエスタンブロッティングの試料には〔実施例8〕で抽出した酵母細胞質タンパク質を用いた。このタンパク質を含む細胞質画分をSDS-PAGEを行った後、PVDF膜に電気的に転写し、それぞれ、抗-penta-HIS抗体を用いてタンパク質の発現を確認した(図6)。その結果、RHM2-N遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2-N株)とRHM2-C(W303/ YEp352-GAP-II-6xHIS-RHM2-C株)およびRHM2-N遺伝子、RHM2-C遺伝子共発現株を同時に導入したRHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)でタンパク質が発現していることが確かめられた。
【0034】
〔実施例10〕RHM2-N+C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)におけるUDP-ラムノ-ス合成活性測定
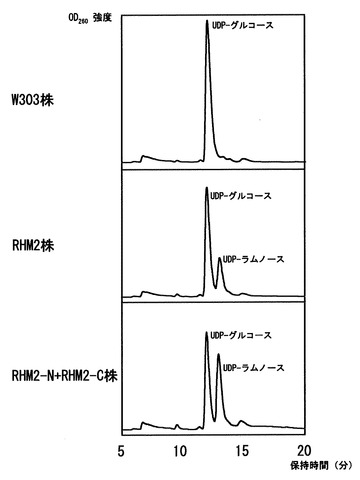
〔実施例3〕もしくは〔実施例8〕で得られた酵母細胞質タンパク質画分のUDP-ラムノ-ス合成活性を検出した。また、同時にW303株から抽出したタンパク質画分とW303/ YEp352-GAP-II-6xHIS-RHM2株から抽出したタンパク質画分についてもUDP-ラムノ-ス合成活性を検出した。活性測定は基質としてUDP-グルコ-スを、補助因子として1 mM NAD+及び3 mM NADPHを用い、100 mM Tris-HCl、pH 7.8という条件で37度、3時間反応を行い、フェノ-ル:クロロホルム溶液 (24: 1) を加え撹拌した後、遠心分離を行い、上清を回収した。上清を逆相カラムを装備したHPLCに供与しよりUDP-ラムノ-スとUDP-グルコ-スの検出を行った。HPLCはC30カラムを用いて1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い、検出はUV260 nmで行った。その結果、RHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)では、W303/ YEp352-GAP-II-6xHIS-RHM2株に比べて高いUDP-ラムノ-ス合成効率を示した(図7)。
【図面の簡単な説明】
【0035】
【図1】UDP-ラムノ-スの合成過程の概略を示す図である。
【図2】〔実施例2〕で得られた各形質転換体がRHM1、RHM2及びRHM3タンパク質を発現していることをウェスタンブロッテイング法により確認した写真である。
【図3】〔実施例5〕で得られた各形質転換体の細胞質画分を用いたUDP-ラムノ-ス合成結果を示す図である。
【図4】上記HPLCにより分離した各分画をESI-MSにより測定した結果を示す図である。
【図5】細胞質中から分離したUDP-ラムノ-スをHPLCにて検出した結果を示す図である。
【図6】〔実施例7〕で得られた各形質転換体がRHM2-NおよびRHM2-Cタンパク質を発現していることをウエスタンブロッテイング法により確認した写真である。
【図7】〔実施例10〕で得られた各形質転換体の細胞質画分を用いたUDP-ラムノ-ス合成結果を示す図である。
【技術分野】
【0001】
本発明は、UDP-ラムノ-スの合成に関与する遺伝子、該遺伝子を有する組換えベクタ-、該組み換えベクタ-により形質転換された形質転換体、ならびに該形質転換体を利用したUDP-ラムノ-スの製造方法に関する。
【背景技術】
【0002】
糖タンパク質などの糖鎖は生体において、非常に重要な働きをしていることが明らかとなっており、このことから糖鎖の構造を自在に変換する糖鎖工学が重要技術となってきている。現在糖鎖を改変する技術としては、化学合成により目的の糖鎖を合成したものをタンパク質に結合させる化学的手法や、細胞が本来持つ内在性の糖鎖合成機能を遺伝子工学の手法を用いて目的の糖鎖に改変する、あるいは糖転移酵素そのものをタンパク質と反応させることで目的の糖タンパク質を得る生物学的手法などがある。化学的手法では、大量合成の手法に道が開けてきてはいるが、糖鎖の複雑性から、いまだに全ての種類の糖鎖を簡単に供給するには至ってはいない、また化学合成した糖鎖を効率良くかつ大量にタンパク質に結合させるのは困難である。一方、生物学的手法は、遺伝子工学の発展により、糖鎖合成関連遺伝子の発現及び機能を制御することが可能となったため、生物の細胞を用いた生体内での糖鎖改変が可能となってきている。また、糖鎖合成酵素を用いた生体外の糖鎖合成も非常に有用であり、均一で、大量の糖鎖の生産が可能となる。一方、生物学的手法を用いた生体内外での糖鎖合成では、糖ヌクレオチドが糖転移酵素の糖供与体として必須となるが、この糖ヌクレオチドは非常に高価であるので、大量生産に用いることは困難となっている。糖ヌクレオチドは、生体内に微量存在するものであり、かつ、高エネルギ-結合で結ばれた非常に反応性に富む不安定な物質であるので、各生物においてその産生量はあまり多くないため大量に生産することは困難であるのが原因である。
【0003】
近年、バクテリアを用いた生産系により、比較的多種類の糖ヌクレオチドの大量生産系が実用的なものへ近づいてきており、その供給は安定する方向へ向かっている。しかしながら、このバクテリアを用いる系では、2種の微生物を混合して、なおかつ、細胞中に含まれる一方の原料を他方の細胞中に送り込むために細胞を破壊した状況で生産を行うことから、ある程度反応過程の長いものでは生産量はそれほど多くなく、新たな手法の開発が求められている。また、バクテリアを用いた生産系によるUDP-ラムノ-スの大量生産系に関する報告は無い。
【0004】
糖ヌクレオチドのうち、UDP-ラムノ-スはラムノ-ス転移酵素の糖供与体としてラムノ-スを含んだ糖鎖合成を行う際には必須のものである。ラムノ-スを含んだ糖鎖にはラムノガラクチュロナンI (RG-I)やラムノガラクチュロナン(RG-II)に代表されるように機能的に重要な役割を果たすことが多いことから、糖供与体の大量で安価な供給が望まれている。また、ラムノ-スを含んだO-結合型糖鎖があり、これらの生合成にもUDP-ラムノ-スが糖供与体として用いられる。よって今後、これらのラムノ-ス含有物質の研究開発が進み重要な機能が明らかになることで新たな需要の拡大が見込まれる。このUDP-ラムノ-スはUDP-グルコ-スから3段階の反応を1つの酵素で触媒し合成される(図1)。この3段階の反応を触媒する酵素は、単一の遺伝子でコ-ドされるが、そのN-末端側ドメインとC-末端側ドメインで機能が異なっている。すなわち、最初の1段階目の反応は、RHM2タンパク質のN-末端側ドメインにコ-ドされるUDP-グルコ-ス4、6-デヒドラタ-ゼによって起こり、UDP-4-ケト-6-デオキシグルコ-スが生成される。続いて2段目、3段目の反応は、RHM2タンパク質のC-末端側でコ-ドされるUDP-4-ケト-6-デオキシグルコ-ス UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼによって起こり、UDP-ラムノ-スに変換される。
これらの酵素はUDP-ラムノ-スを使用する生物であればもっている普遍的な酵素である。ただし、このような生物は合成したUDP-ラムノ-スを使用していくため、細胞内にUDP-ラムノ-スが蓄積していくことはない。このため、生物体からUDP-ラムノ-スを単離する場合、その生産量は極めて低くかつ高価である。また、遺伝子工学的手法を用いてUDP-ラムノ-スを合成した例は見あたらない。
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明の課題は、多機能なラムノ-ス付加糖鎖を合成するために必要なUDP-ラムノ-スの効率のよい生産方法を確立し、より大量に供給することにある。
【課題を解決するための手段】
【0006】
本発明者らは上記課題を解決すべく鋭意研究を重ねた結果、シロイヌナズナ(Arabidopsis thaliana)のUDP-ラムノ-ス合成を触媒する、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ(以下、UDP-glucose 4,6-dehydratase-3,5-epimerase/4-keto reductaseという場合がある。)をコ-ドする遺伝子を初めて単離し、この遺伝子の構造を明らかにするとともに、この遺伝子を機能的に発現することによって、生体内および生体外で効率よくUDP-ラムノ-スが合成されることを見いだした。また、さらに、上記遺伝子をUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン部分と、 UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメイン部分とに分け、それぞれを共発現させた場合、これら領域を全て含む上記遺伝子を発現させた場合に比較し、UDP-ラムノ-スの生産効率が向上するという驚くべき知見を得て、本発明を完成させるに到ったものである。
【0007】
すなわち、本発明は、以下(1)〜(31)に示されるとおりである。
(1)(a)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列を有するか、あるいは(b)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(2)上記(1)に記載のタンパク質をコ-ドするDNA。
(3)(a)配列番号1、配列番号3もしくは配列番号19で示される塩基配列を有するか、あるいは(b)配列番号1、配列番号3もしくは配列番号・・で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(4)上記(2)又は(3)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(5)上記(4)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(6)(a)配列番号6で示されるアミノ酸配列を有するか、あるいは(b)配列番号6で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質。
(7)上記(6)に記載のタンパク質をコ-ドするDNA。
(8)(a)配列番号5で示される塩基配列を有するか、あるいは(b)配列番号5で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(9)上記(7)又は(8)に記載のDNAにより組み換えられていることを特徴とする、組換えベクタ-。
(10)上記(9)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(11)(a)配列番号8で示されるアミノ酸配列を有するか、あるいは(b)配列番号8で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(12)上記(11)に記載のタンパク質をコ-ドするDNA。
(13)(a)配列番号7で示される塩基配列を有するか、あるいは(b)配列番号7で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
(14)上記(12)又は(13)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(15)上記(7)又は(8)に記載のDNA及び上記(12)又は(13)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(16)上記(14)に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
(17)上記(9)及び(14)に記載の組換えベクタ-が、同一宿主細胞に導入されていることを特徴とする形質転換体。
(18)上記(15)に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
(19)配列番号2、4もしくは20で示されるアミノ酸配列と70%以上の相同性を有するアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
(20)イネ(Oryza sativa)またはタバコ(Nicotiana tabacum)由良のタンパク質である、上記(19)に記載のタンパク質。
(21)上記(19)または(20)に記載のタンパク質をコ-ドするDNA。
(22)上記(21)に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
(23)上記(22)に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
(24)宿主細胞が、UDP-グルコ-スを細胞内に含有していることを特徴とする、上記(5)、(17)、(18)及び(23)のいずれかに記載の形質転換体。
(25)宿主細胞が、UDP-ラムノ-スを消費する酵素系を保持していないことを特徴とする、上記(5)、(17)、(18)及び(23)のいずれかに記載の形質転換体。
(26)宿主細胞が酵母細胞である、上記(5)、(10)、(16)〜(18)及び(23)〜(25)のいずれかに記載の形質転換体。
(27)上記(5)、(17)、(18)及び(23)〜(26)のいずれかに記載の形質転換体を培養し、培養物からUDP-ラムノ-スを採取することを特徴とする、UDP-ラムノ-スの製造方法。
(28)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を培地に培養し、得られた培養物、培養処理物、酵素抽出液または精製酵素をUDP-グルコ-スと接触させることにより、UDP-グルコ-スをUDP-ラムノ-スに変換させることを特徴とする、UDP-ラムノ-スの製造方法。
(29)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を同位体元素により置換されたグルコ-スを炭素源として培養し、培養物から同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
(30)上記(5)、(17)、(18)、(23)〜(26)のいずれかに記載の形質転換体を培地で培養することにより得られた培養物、培養処理物、酵素抽出液または精製酵素を同位体元素により置換されたUDP-グルコ-スと接触させ、同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
(31)同位体元素がC13またはC14であることを特徴とする、上記29または30に記載の製造方法
【発明の効果】
【0008】
本発明によれば、糖鎖において非常に重要な機能をもつラムノ-スの付加を行うために必須なUDP-ラムノ-スを多量に効率よく生産することができる。現時点において糖タンパク質糖鎖を均一に合成する技術は確立されておらず、最終的には生体外での糖鎖の修飾を行うことで糖鎖の均一な合成を行うことが考えられるが、この際、糖供与体として糖ヌクレオチドは必須なものである。特に、ラムノ-スにおいてはUDP-ラムノ-スが非常に高価なため、生体外での修飾反応を大量に行うことは現在非現実的であるが、本発明により、多量のUDP-ラムノ-スの供給が可能となれば、ラムノ-スが付加された高機能な糖鎖の合成を生体外で行うことができる。したがって、本発明は糖タンパク質の糖鎖の研究において多大な貢献をするものである。
【発明を実施するための最良の形態】
【0009】
本発明における第1の酵素タンパク質は、シロイヌナズナ由来のものであって、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼの3者の活性をともに有するタンパク質であって、配列番号2、配列番号4もしくは配列番号20に示されるアミノ酸配列を有するが(以下、配列番号2に示される酵素タンパク質をRHM1タンパク質、配列番号4で示される酵素タンパク質をRHM2タンパク質、配列番号20で示される酵素タンパク質をRHM3タンパク質とそれぞれいう場合がある。)、これのみでなく、配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ-UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質ものであれば、本発明に含まれる。
本発明の第1の酵素遺伝子は、上記したアミノ酸配列コ-ドするDNAである。具体的な塩基配列は、配列番号1、配列番号3、もしくは配列番号19にそれぞれ示されるが(以下、配列番号1に示される遺伝子をRHM1遺伝子といい、配列番号3に示される遺伝子をRHM2遺伝子、配列番号19で示される遺伝子をRHM3遺伝子とそれぞれいう場合がある。)これのみでなく、配列番号1、配列番号3もしくは配列番号19で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ活性及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性をともに有するタンパク質をコ-ドするものであれば、本発明において使用できる。
本発明においては、上記シロイヌナズナのRHM1、RHM2遺伝子もしくはRHM3遺伝子あるいは上記の変異遺伝子を用いて、遺伝子工学的手法によりUDP-ラムノ-スを効率よく生産可能であるが、この製造法には大別して細胞内で生産する方法(培養法)と、細胞外で生産する方法(酵素変換法)の2つがある。
【0010】
このうち、UDP-ラムノ-スを細胞内で製造するための方法の概要は、例えば、下記の行程よりなる。
(1)シロイヌナズナのRHM1、RHM2もしくはRHM3遺伝子〔UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ遺伝子(UDP-glucose 4,6-dehidratase-3,5-epimerase/4-keto-reductase)〕を取得する;
(2)シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を発現用プラスミド等のベクタ-に導入し、RHM1、RHM2もしくはRHM3遺伝子を発現するベクタ-を構築する;
(3)上記発現ベクタ-を、UDP-グルコ-スを細胞内に持つ宿主細胞に導入し、該宿主細胞を形質転換する;
(4)上記(3)の組換え体を培地に培養し、得られた培養物あるいは細胞破砕物等の培養処理物から抽出によりUDP-ラムノ-スを採取し、さらに単離・精製する。
【0011】
本発明において、UDP-ラムノ-スを細胞外で製造する方法は、下記の工程よりなる。
(1)シロイヌナズナのRHM1、RHM2もしくはRHM3遺伝子(UDP-glucose 4,6-dehidratase-3,5-epimerase/4-keto-reductase遺伝子)を取得する;
(2)シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を発現用プラスミド等のベクタ-に導入し、RHM1、RHM2もしくはRHM3遺伝子を発現するベクタ-を構築する;
(3)上記発現ベクタ-を、上記発現ベクタ-を、UDP-グルコ-スを細胞内に持つ宿主細胞に導入し、該宿主細胞を形質転換する。
(4)上記の形質転換体の培養物、細胞破砕物等の培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素を酵素源とし、UDP-グルコ-スおよびNADPHを添加することによりUDP-グルコ-スをUDP-ラムノ-スに変換し、単離・精製を行う。該UDP-ラムノ-スはラムノ-ス含有糖鎖の合成の際に糖供与体として必須なものであるので、機能的に重要と思われる糖鎖へのラムノ-ス付加の際に有用なものである。
【0012】
また別法として、例えば、シロイヌナズナRHM1、RHM2もしくはRHM3遺伝子を含む発現用ベクタ-により形質転換した形質転換体の培養物、培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素と、UDP-グルコ-スを本来的に産生する細胞の培養物、培養処理物、細胞内抽出物あるいは該抽出物から単離精製した酵素を、各々別途に調整し、これらを順次あるいは同時にUDP-グルコ-スに作用させて、UDP-グルコ-スをUDP-ラムノ-スに変換させることにより、UDP-ラムノ-スを製造してもよい。
上記配列番号4で示されるRHM2タンパク質及びその遺伝子は、その配列中にUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメイン及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインを有する。
本発明の第2の酵素タンパク質は、上記RHM2タンパク質のUDP-グルコ-ス4、6-デヒドラタ-ゼドメインに対応するタンパク質であり、配列番号6に示されるアミノ酸配列を有するか、あるいは該アミノ酸配列において、1又は数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質である(以下、配列番号6に示される酵素タンパク質をRHM2-Nタンパク質という場合がある。)。
また、本発明の第2の遺伝子は、RHM2-Nタンパク質あるいはその上記変異体タンパク質をコ-ドするDNAであり、より具体的には、配列番号5に示されるが(以下、配列番号5に示される遺伝子をRHM2遺伝子という場合がある。)これのみでなく、配列番号5に示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性をコ-ドするものであれば、本発明に含まれる。
本発明の第3の酵素タンパク質は、上記RHM2タンパク質のUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメインとUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインの両者を含むタンパク質であり、配列番号8に示されるアミノ酸配列を有するか、あるいは該アミノ酸配列において、1又は数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を有し、かつUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼドメインとUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼの2つの酵素活性を有するタンパク質である(以下、配列番号8に示される酵素タンパク質をRHM2-Cタンパク質という場合がある。)。
本発明の第3の遺伝子は、RHM2-Cタンパク質あるいはその上記変異体タンパク質をコ-ドするDNAであり、より具体的には、配列番号7に示されるが(以下、配列番号7に示される遺伝子をRHM2-C遺伝子という場合がある。)これのみでなく、配列番号7に示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有するものであっても、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ活性とUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性の2つの酵素活性を有するタンパク質をコ-ドするものであれば、本発明に含まれる。
これらの第2及び第3の酵素タンパク質は、上記第2及び第3の遺伝子を用いて、通常の遺伝子工学的手法により容易に得ることができる。得られた第2及び第3の酵素タンパク質は、図1に示されるように、UDP-グルコ-スをUDP-4-ケト-デオキシグルコ-スに転換する反応、及びUDP-4-ケト-デオキシグルコ-スをUDP-ラムノ-スに転換する反応をそれぞれ触媒するから、例えば、これら酵素を順次または同時に用いることにより、UDP-グルコ-スからUDP-ラムノ-スを製造することができる。
しかし、本発明におけるUDP-ラムノ-スの生産において、最も生産性が高い方法は、上記第2及び第3の遺伝子を同一宿主細胞において共発現する方法である。この方法による場合、RHM2遺伝子を用いる場合に比べ、UDP-ラムノ-スの生産量は著しく向上する。
以下、本発明をさらに詳細に説明する。本明細書において、アミノ酸配列および塩基配列の略号による表示は、IUPAC-IUBの規定および当該分野における通称または慣行に従うものとする。
【0013】
本発明のRHM1、RHM2もしくはRHM3遺伝子、RHM2-N遺伝子及びRHM2-C遺伝子の取得は、一般的手法に従ってシロイヌナズナから調製したcDNAライブラリを鋳型とし、PCR法により単離することができる。また、RHM2-N遺伝子及びRHM2-C遺伝子については、上記RHM2遺伝子を鋳型として用いてもよい。
【0014】
シロイヌナズナのcDNAライブラリは、通常用いられているプラスミドベクタ-またはλファ-ジ由来のベクタ-を用い、通常の方法に従い作成することができる。
【0015】
PCR法は、生体外でDNAの特殊な領域を、そのセンス・アンチセンスプライマ-、耐熱性のDNAポリメラ-ゼ、DNA増幅システム等の組み合わせを用いて、約2-3時間で10-100万倍に特異的に増幅することができる技術であり、本発明遺伝子およびその断片は、他種の酵素との塩基配列の相同性等をもとに、このPCR法での増幅が可能である。
【0016】
上記遺伝子を組み込むベクタタ-としては、宿主内で複製保持されるものであれば、いずれも使用することができる。例えば大腸菌由来のプラスミドベクタ-pBR322やpUC19などを挙げることができる。
ベクタ-を組み込む方法としては、例えばT.Maniatisらの方法[Molecular Cloning, Cold Spring Harbor Laboratory, p.239 (1982)]に従うことができる。
【0017】
クロ-ン化された遺伝子は、発現に適したベクタ-中のプロモ-タ-の下流に連結して発現ベクタ-を得ることができる。ベクタ-としては、酵母由来のプラスミドYEp352GAP、YEp51、pSH19、などが挙げられる。
【0018】
該遺伝子はその5'末端に翻訳開始コドンとしてのATGを有し、また、3'末端には翻訳終止コドンとしてのTAA、TGAまたはTAGを有してよい。また、5'末端あるいは3'末端には、例えば、タンパク質の単離精製を容易にするため、6 x ヒスチジンやヘマグルチニンタンパク質の一部分である標識抗原遺伝子やGSTタンパク質などの標識タンパク質遺伝子を結合させて発現させてもよい。特に配列番号5に示されるRHM2-N遺伝子のC末端には上記終止コドンを位置させるのが好ましく、また、配列番号7に示されるRHM2-C遺伝子のN末端側には開始コドン(ATG)を位置させるのが好ましいが、得られるタンパク質はメチオニンがなくても当然酵素活性を有する。また、逆に、例えば、N末端側にMet-His tagを付加せしめたタンパク質であってもそれ自体活性を有する。
さらに該遺伝子を発現させるためには、その上流にプロモ-タ-を接続するが、本発明で用いられるプロモ-タ-としては遺伝子の発現に用いる宿主に対応した適切なプロモ-タ-であれば、いかなるものでもよい。
また、形質転換する宿主が酵母である場合、ENO1プロモ-タ-、GAL10プロモ-タ-、GAPDHプロモ-タ-、ADHプロモ-タ-、AOXプロモ-タ-などが挙げられる。
【0019】
このように構築されたRHM1、RHM2もしくはRHM3遺伝子、RHM2-N遺伝子あるいはRHM2-C遺伝子を有する組換えDNAを含むベクタ-を用いて、該ベクタ-を保持する形質転換体を作成する。RHM2-N遺伝子及びRHM2-C遺伝子を共発現させる場合には、例えば、RHM2-N遺伝子を有する組換えベクタ-とRHM2-C遺伝子を有する組換えベクタ-を同一宿主細胞に導入するか、あるいはRHM2-N遺伝子及びRHM2-C遺伝子の両方を同一のベクタ-に導入し、得られた組換えベクタ-を用いて宿主細胞を形質転換する。これらにより得られた形質転換体は、RHM2-N遺伝子を有する組換えベクタ-とRHM2-C遺伝子を有する組換えベクタ-の2種のベクタ-を保持するか、あるいはRHM2-N遺伝子及びRHM2-C遺伝子をともに有する1種類の組換えベクタ-を保持し、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼを産生する。したがって、同形質転換体は、UDP-グルコ-スを原料としてUDP-ラムノ-スを生産することができ、しかも、上記したようにその生産性が高い株である。
宿主としては、UDP-ラムノ-スを生体内で消費しない出芽酵母(Saccharomyces cerevisia)などが挙げられるが、この他UDP-ラムノ-スを消費しない他の酵母(Pichia pastorisなど)などでもよい。また、酵母以外でも、UDP-ラムノ-スの消費する酵素系を持たない宿主であればいかなるものでもよい。
また、細胞破砕物、酵素抽出物等を用い、生体外でUDP-ラムノ-スを生産する場合、細胞質内で該遺伝子産物を生産することが可能な宿主であれば、いかなるものでも用いることができる。
上記形質転換体の作成は、それぞれの宿主について一般的に行われている方法で行う。または一般的ではなくても適用可能な方法ならばよい。例として、宿主が酵母であれば、リチウム法あるいはエレクトロポレ-ション法により組み換えDNAを含むベクタ-を導入する。
【0020】
このようにして、本発明の遺伝子を有する組み換えDNAを含むベクタ-を保持する形質転換体が得られ、該形質転換体を培養することにより、主に該形質転換体の細胞内においてUDP-ラムノ-スとUDP-ラムノ-ス製造に必要な酵素が産生、蓄積する。
【0021】
形質転換体を培養する場合、培養に使用される培地は、それぞれの宿主について一般的に用いられているものを使用する。または、一般的でなくとも適用可能な培地ならばよい。例としては、宿主が酵母であればYPD培地、SD培地などを用いる。培養は、それぞれの宿主について一般的に用いられている条件で行う。また、一般的でなくとも適用可能な条件であればよい。例としては、宿主が酵母であれば約25-37度で、約12時間-5日間行い、必要により通気や撹拌を行うことができる。
【0022】
例えば、上記培養物からUDP-ラムノ-スあるいは酵素を抽出する際は、宿主細胞を遠心分離によって培地と分け、宿主細胞を破砕してUDP-ラムノ-スあるいは酵素を抽出する。宿主が酵母の場合、例えばグラスビ-ズによって細胞を破壊し、遠心分離する。この際、酵素及びUDP-ラムノ-スは、上清画分に存在する。また、1-ブタノ-ルによって飽和させた1Mギ酸溶液中に酵母菌体を懸濁後、30分から3時間程度氷冷し、遠心分離することによってもUDP-ラムノ-スを抽出することが可能である。この場合も、UDP-ラムノ-スは、上清画分に存在する。
上記の遠心分離によって分離された上清画分からUDP-ラムノ-スを分離する場合は、ゲルろ過法等により低分子量の画分を集め、さらにHPLCによる分離を行うことにより、精製単離する。また、イオン交換カラムや逆相カラムを用いて精製単離することも可能である。
また、生体外で酵素変換反応を行い、UDP-ラムノ-スを製造する場合においては、細胞破砕物や上清画分をそのまま酵素源として利用することが可能であるが、上清画分硫酸アンモニウムを75%の濃度に溶かし、これにより沈殿してくるタンパク質画分を集め、透析等により脱塩したものも酵素源として使用することができる。
【0023】
上記酵素源を用いて、基質となるUDP-グルコ-ス、NAD+およびNADPHを加え反応させた後、HPLCでUDP-ラムノ-スを単離精製することにより、UDP-ラムノ-スを得ることができる。
一方、本発明においては、シロイヌナズナ由来の上記配列番号2、配列番号4及び配列番号20に示されるタンパク質(RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質)のみではなく、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質として、RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質に対して相同率70%以上の相同性を有するタンパク質も挙げることができ、本発明においてはこれら相同性を有するタンパク質、及び該タンパク質をコ-ドする塩基配列を有する遺伝子も包含する。これら相同性を有するタンパク質としては、植物由来の、例えばイネ(Oryza Sativa)、タバコ(Nicotiana tabacum)由来のタンパク質が挙げられ、イネにおける相同性を有するタンパク質のアミノ酸配列及びこれをコ-ドする遺伝子の塩基配列はそれぞれ配列番号22、配列番号21に示される。
これら相同性を有するタンパク質は、これらをコ-ドするDNAを用いて、上記RHM1タンパク質、RHM2タンパク質及びRHM3タンパク質について記載したのと同様な遺伝子工学手法を用いて製造することが出来る。すなわち本発明においては、これら相同性を有するタンパク質をコ-ドするDNAにより組み換えられた組換えベクタ-、該組換えベクタ-が導入されている形質転換体、及び該形質転換体を用いた、上記相同性を有する酵素タンパク質の製造方法、及び該形質転換体あるいは該相同酵素タンパク質を用いたUDP-ラムノ-スの製造方法をも包含する。
【0024】
また、本発明の形質転換体を培地に培養するに際し、同位体元素でその構成元素を置換したグルコ-スを炭素源として培地に含有させて培養を行うことにより、培養物から、同位体元素でラベルされたUDP-ラムノ-スを採取することができる。
また別法として、本発明の形質転換体から得られた、酵素タンパク質を含む培養物、培養処理物、酵素抽出液あるいは精製酵素を、同位体元素でその構成元素を置換したUDP-グルコ-スと接触させることにより、同位体元素でラベルされたUDP-ラムノ-スに変換することも可能である。
このような酵素変換法では、上記同位体元素を有するUDP-グルコ-スに対し、上記RHM2-N精製タンパク質あるいは該タンパク質を含有する培養物、培養処理物、酵素抽出液または精製酵素と、同RHM2-C精製タンパク質あるいは該タンパク質を含有する培養物、培養処理物、酵素抽出液を順次または同時に接触させることにより、同位体元素でラベルされたUDP-ラムノ-スを得てもよい。
好ましい同位体元素としては、たとえばC13あるいはC14同位体が挙げられる。このような同位体元素でラベルされたUDP-ラムノ-スは、糖鎖合成系の解明あるいはこの糖鎖合成系に影響を及ぼす物質等の探索等に有用である。
以下、本発明の実施例を示すが、本発明はこの実施例にのみ限定されるものではない。
【実施例】
【0025】
〔実施例1〕RHM1遺伝子(配列番号1)、RHM2遺伝子(配列番号3)およびRHM3遺伝子(配列番号19)の単離
RHM1遺伝子およびRHM2遺伝子に関してはシロイヌナズナのcDNAライブラリを鋳型として用いてPCR法により遺伝子のクロ-ニングを行った。cDNAライブラリはCLONTECH社のQUICK-Clone cDNAを用いた。また、RHM3遺伝子に関しては理化学研究所バイオリソ-スセンタ-よりcDNAクロ-ンを入手し(Resouce Number: pda08705)、鋳型として用いた。プライマ-の設計においては、制限酵素でタンパク質をコ-ドしている部分が容易に切り出せるようにRHM1の場合は、N-末端Sac Iサイト、C-末端部分にKpn Iサイトに位置させ、RHM2およびRHM3の場合は、N-末端Eco RIサイト、C-末端部分にSal Iサイトに位置させた。また、N-末端には、あらかじめ標識抗原であり、ニッケルアガロ-ス結合配列をコ-ドする6 x ヒスチジン配列を付加せしめた。それぞれのプライマ-の配列を以下に示す(配列番号13と14は、RHM1用、同15と16はRHM2用、同23と24はRHM3用)。
AAGAGCTCATGCATCACCATCACCATCACATGGCTTCGTACACTCCCAAGAAC(配列番号13)
AAAGGTACCTCAGGTTTTCTTGTTTGGCCCGTATG(配列番号14)
AGAATTCATGCATCACCATCACCATCACATGGATGATACTACGTATAAGCCAAAGAAC(配列番号15)
AAAAAGTCGACTTAGGTTCTCTTGTTTGGTTCAAAGAC(配列番号16)
AGAATTCATGCATCACCATCACCATCACATGGCTACATATAAGCCTAAGAACATCCTC(配列番号23)
AAAAAGTCGACTTACGTTCTCTTGTTAGGTTCGAAGACG(配列番号24)
PCRの条件は以下の通り。
95度 30秒
57度 30秒
72度 2分15秒
30サイクル
この条件で得られたそれぞれ約2.0 kbpのDNA増幅断片をアガロ-ス電気泳動で分離した後、制限酵素Sac IとKpn IもしくはEco RIとSal Iで切断し、pBluescript II-SK+ベクタ-のEco RIサイトとSal Iサイト部位に挿入した。このクロ-ニングされた遺伝子をダイデオキシ法を用いたシ-クエンスキットにより塩基配列を確認し、配列番号1、3および19に示した塩基配列であることを確認した。
【0026】
〔実施例2〕RHM1、RHM2およびRHM3遺伝子発現ベクタ-の作成およびこのプラスミドを含む酵母形質転換体の作成
pBluescript II-SK+ベクタ-に挿入されているRHM1遺伝子、RHM2遺伝子およびRHM3遺伝子をSac IおよびKpn IもしくはEco RIおよびSal Iで切り出し、酵母の解糖系のプロモ-タ-であるGAPDHとそのタ-ミネ-タ-の間にpUC18のマルチクロ-ニングサイトのうちEco RIからSal Iまでの部分を持ち選択マ-カ-としてURA3遺伝子を持つ発現用ベクタ-YEp352GAP-IIのSacIおよびKpnIサイトもしくはEcoRI およびSal Iサイトに挿入しYEp352-GAP-II-6xHIS-RHM1、YEp352-GAP-II-6xHIS-RHM2およびYEp352-GAP-II-6xHIS-RHM3を構築した。この発現ベクタ-は、酵母のW303-1B株(ura3, lue2, his3, trp1, ade2)に導入し、形質転換体、すなわち、RHM1遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM1株)、RHM2遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2株)およびRHM3遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM3株)を得た。なお、W303-1B株は、ATCC (American Type Culture Collection)から入手可能である(ATCC Number: 201238TM)。
【0027】
〔実施例3〕酵母細胞質タンパク質の抽出
〔実施例2〕で得られた形質転換体より細胞質中のタンパク質を抽出した。まず、〔W303/YEp352-GAP-II-6xHIS-RHM1株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕およびW303株をSD培地にて24時間30度で培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞壁画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。
【0028】
〔実施例4〕RHM1タンパク質、RHM2タンパク質およびRHM3タンパク質の酵母内での発現の確認
〔実施例2〕で得られた形質転換体について、それぞれの細胞内でタンパク質が発現されていることをウエスタンブロッティング法を用いて確認した。まず、上記2種の形質転換体〔W303/ YEp352-GAP-II-6xHIS-RHM1株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕、〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕およびW303株をSD培地にて30℃で24時間培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。このタンパク質を含む細胞質画分を50mM Tris-HCl pH6.8に溶かし、SDS-PAGEを行った後、PVDF膜に電気的に転写し、それぞれ、抗-penta-HIS抗体を用いてタンパク質の発現を確認した(図2)。その結果、RHM1遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM1株〕、RHM2遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕およびRHM3遺伝子導入株〔W303/ YEp352-GAP-II-6xHIS-RHM3株〕でタンパク質が発現していることが確かめられた。
【0029】
〔実施例5〕UDP-ラムノ-ス合成活性測定
〔実施例3〕で得られた酵母細胞質タンパク質画分のUDP-ラムノ-ス合成活性を検出した。活性測定は基質としてUDP-グルコ-スを、補助因子として1 mM NAD+及び3 mM NADPHを用い、100 mM Tris-HCl、pH 8.0という条件で30度、2時間反応を行い、フェノ-ル:クロロホルム溶液 (24: 1) を加え撹拌した後、遠心分離を行い、上清を回収した。上清を逆相カラムを装備したHPLCに供与しよりUDP-ラムノ-スとUDP-グルコ-スの検出を行った。HPLCはC30カラムを用いて1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い、検出はUV260 nmで行った。その結果、RHM1タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM1株とRHM2タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM2株およびRHM3タンパク質を発現させたW303/ YEp352-GAP-II-6xHIS-RHM3株でUDP-ラムノ-ス合成活性を検出した(図3)。さらに、合成したUDP-ラムノ-スについてESI-MS測定を行い分子量を測定した結果、予想される分子量と一致した(図4)。
【0030】
〔実施例6〕RHM2遺伝子導入酵母菌体からのUDP-ラムノ-スの抽出
〔実施例3〕で得られた酵母細胞質画分よりUDP-ラムノ-スを分離精製した。まず、〔W303/ YEp352-GAP-II-6xHIS-RHM2株〕をSD(-ウラシル)培地にて24時間30度で培養し、得られた酵母細胞O.D.600=10当り1mlの1-ブタノ-ル飽和1 Mギ酸を添加し、十分に撹拌し、氷上にて1時間安置した。この試料より遠心操作を用いて細胞壁画分を取り除き、細胞質画分だけを分離した。上清を凍結乾燥し、純水に再懸濁することによって糖ヌクレオチドを抽出した。抽出した糖ヌクレオチドをC30カラムにより1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い溶出時間からUDP-ラムノ-スを分離した。分離したUDP-ラムノ-スを数度同条件で再分離することによりUDP-ラムノ-スを精製した(図5)。
【0031】
〔実施例7〕RHM2遺伝子のUDP-グルコ-ス4、6-デヒドラタ-ゼドメインの発現とUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメインの発現
〔実施例2〕で得られたYEp352-GAP-II-6xHIS-RHM2を用いてPCR法によりRHM2遺伝子中のUDP-グルコ-ス4、6-デヒドラタ-ゼドメイン(RHM2-N)とUDP-4-ケト-6-デオキシグルコ-ス3,5-エピメラ-ゼ/UDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼドメイン(RHM2-C)のクロ-ニングを行った。プライマ-の設計においては、制限酵素でタンパク質をコ-ドしている部分が容易に切り出せるようにRHM2-N、RHM2-C共に、N-末端Eco RIサイト、C-末端部分にSal Iサイトに位置させた。また、N-末端には、あらかじめ標識抗原であり、ニッケルアガロ-ス結合配列をコ-ドする6 x ヒスチジン配列を付加せしめた。プライマ-は、〔実施例1〕で用いたプライマ-の内、配列番号15と配列番号16を流用した。その他のプライマ-の配列を以下に示す(配列番号15と17は、RHM2-N用、同16と18はRHM2-C用)。
AAAAAGTCGACTTAAGCTTTGTCACCAGAATCACCATT(配列番号17)
AGAATTCATGCATCACCATCACCATCACACACCTAAGAATGGTGATTCTGGTG(配列番号18)
PCRの条件は以下の通り。
95度 30秒
57度 30秒
72度 1分30秒
30サイクル
この条件で得られた約1.2
kb (RHM2-N) と約0.9 kb (RHM2-C)の断片をそれぞれアガロ-ス電気泳動で分離した後、制限酵素Eco RIとSal Iで切断し、YEp352GAP-IIのEcoRI およびSal Iサイトに挿入しYEp352-GAP-II-6xHIS-RHM2-NおよびYEp352-GAP-II-6xHIS-RHM2-Cを構築した。このクロ-ニングされた遺伝子をダイデオキシ法を用いた塩基配列解析により塩基配列を確認し、それぞれ配列番号9及び11に示した塩基配列(対応するタンパク質のアミノ酸配列は10及び12に示す。)であることを確認した。さらに、YEp352-GAP-II-6xHIS-RHM2-Cベクタ-からGAPDHプロモ-タ-、6 x HIS-RHM2-C、GAPDHタ-ミネ-タ-の3つの部分を含む断片を制限酵素Bam HIを用いて切り出し、酵母のプラスミドベクタ-でありLEU2マ-カ-を持つYEp351のBam HIサイトに挿入しYEp351-GAP-II-6xHIS-RHM2-Cを構築した。この発現ベクタ-は、酵母のW303-1B株(ura3, lue2, his3, trp1, ade2)に導入し、形質転換体、すなわち、RHM2-N遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2-N株)とRHM2-C(W303/ YEp352-GAP-II-6xHIS-RHM2-C株)およびRHM2-N遺伝子、RHM2-C遺伝子共発現株を同時に導入したRHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)を得た。
【0032】
〔実施例8〕酵母細胞質タンパク質の抽出
〔実施例7〕で得られた形質転換体(RHM2-N株、RHM2-C株、RHM2-N+RHM2-C株)より〔実施例3〕に示す方法に従って細胞質中のタンパク質を抽出した。まず、形質転換株及びW303株をSD培地にて24時間30度で培養し、得られた酵母細胞をガラスビ-ズで破砕した。得られた破砕物より遠心操作を用いて細胞壁画分及びミクロソ-ム画分を取り除き、細胞質画分だけを分離した。
【0033】
〔実施例9〕RHM2-Nタンパク質、RHM2-Cタンパク質の酵母内での発現の確認
〔実施例7〕で得られた形質転換体について、それぞれの細胞内でタンパク質が発現されていることをウエスタンブロッティング法を用いて確認した。また、ウエスタンブロッティングの試料には〔実施例8〕で抽出した酵母細胞質タンパク質を用いた。このタンパク質を含む細胞質画分をSDS-PAGEを行った後、PVDF膜に電気的に転写し、それぞれ、抗-penta-HIS抗体を用いてタンパク質の発現を確認した(図6)。その結果、RHM2-N遺伝子導入株(W303/ YEp352-GAP-II-6xHIS-RHM2-N株)とRHM2-C(W303/ YEp352-GAP-II-6xHIS-RHM2-C株)およびRHM2-N遺伝子、RHM2-C遺伝子共発現株を同時に導入したRHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)でタンパク質が発現していることが確かめられた。
【0034】
〔実施例10〕RHM2-N+C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)におけるUDP-ラムノ-ス合成活性測定
〔実施例3〕もしくは〔実施例8〕で得られた酵母細胞質タンパク質画分のUDP-ラムノ-ス合成活性を検出した。また、同時にW303株から抽出したタンパク質画分とW303/ YEp352-GAP-II-6xHIS-RHM2株から抽出したタンパク質画分についてもUDP-ラムノ-ス合成活性を検出した。活性測定は基質としてUDP-グルコ-スを、補助因子として1 mM NAD+及び3 mM NADPHを用い、100 mM Tris-HCl、pH 7.8という条件で37度、3時間反応を行い、フェノ-ル:クロロホルム溶液 (24: 1) を加え撹拌した後、遠心分離を行い、上清を回収した。上清を逆相カラムを装備したHPLCに供与しよりUDP-ラムノ-スとUDP-グルコ-スの検出を行った。HPLCはC30カラムを用いて1%アセトニトリルを含む20 mMトリエチルアミン(pH 7.0) 溶液を0.7 ml/minで流して分離を行い、検出はUV260 nmで行った。その結果、RHM2-N+RHM2-C株(W303/ YEp352-GAP-II-6xHIS-RHM2, YEp351-GAP-II-6xHIS-RHM2-C株)では、W303/ YEp352-GAP-II-6xHIS-RHM2株に比べて高いUDP-ラムノ-ス合成効率を示した(図7)。
【図面の簡単な説明】
【0035】
【図1】UDP-ラムノ-スの合成過程の概略を示す図である。
【図2】〔実施例2〕で得られた各形質転換体がRHM1、RHM2及びRHM3タンパク質を発現していることをウェスタンブロッテイング法により確認した写真である。
【図3】〔実施例5〕で得られた各形質転換体の細胞質画分を用いたUDP-ラムノ-ス合成結果を示す図である。
【図4】上記HPLCにより分離した各分画をESI-MSにより測定した結果を示す図である。
【図5】細胞質中から分離したUDP-ラムノ-スをHPLCにて検出した結果を示す図である。
【図6】〔実施例7〕で得られた各形質転換体がRHM2-NおよびRHM2-Cタンパク質を発現していることをウエスタンブロッテイング法により確認した写真である。
【図7】〔実施例10〕で得られた各形質転換体の細胞質画分を用いたUDP-ラムノ-ス合成結果を示す図である。
【特許請求の範囲】
【請求項1】
(a)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列を有するか、あるいは(b)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項2】
請求項1に記載のタンパク質をコ-ドするDNA。
【請求項3】
(a)配列番号1、配列番号3もしくは配列番号19で示される塩基配列を有するか、あるいは(b)配列番号1、配列番号3もしくは配列番号19で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項4】
請求項2又は3に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項5】
請求項4に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項6】
(a)配列番号6で示されるアミノ酸配列を有するか、あるいは(b)配列番号6で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質。
【請求項7】
請求項6に記載のタンパク質をコ-ドするDNA。
【請求項8】
(a)配列番号5で示される塩基配列を有するか、あるいは(b)配列番号5で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項9】
請求項7又は8に記載のDNAにより組み換えられていることを特徴とする、組換えベクタ-。
【請求項10】
請求項9に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項11】
(a)配列番号8で示されるアミノ酸配列を有するか、あるいは(b)配列番号8で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項12】
請求項11に記載のタンパク質をコ-ドするDNA。
【請求項13】
(a)配列番号7で示される塩基配列を有するか、あるいは(b)配列番号7で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項14】
請求項12又は13に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項15】
請求項7又は8に記載のDNA及び請求項12又は13に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項16】
請求項14に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項17】
請求項9及び14に記載の組換えベクタ-が、同一宿主細胞に導入されていることを特徴とする形質転換体。
【請求項18】
請求項15に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
【請求項19】
配列番号2、4もしくは20で示されるアミノ酸配列と70%以上の相同性を有するアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項20】
イネ(Oryza sativa)またはタバコ(Nicotiana tabacum)由来のタンパク質である請求項19に記載のタンパク質。
【請求項21】
請求項19または20に記載のタンパク質をコ-ドするDNA。
【請求項22】
請求項21に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項23】
請求項22に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項24】
宿主細胞が、UDP-グルコ-スを細胞内に含有していることを特徴とする、請求項5、17、18及び23のいずれかに記載の形質転換体。
【請求項25】
宿主細胞が、UDP-ラムノ-スを消費する酵素系を保持していないことを特徴とする、請求項5、17、18及び23のいずれかに記載の形質転換体。
【請求項26】
宿主細胞が酵母細胞である、請求項5、10、16〜18及び23〜25のいずれかに記載の形質転換体。
【請求項27】
請求項5、17、18及び23〜26のいずれかに記載の形質転換体を培養し、培養物からUDP-ラムノ-スを採取することを特徴とする、UDP-ラムノ-スの製造方法。
【請求項28】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を培地に培養し、得られた培養物、培養処理物、酵素抽出液または精製酵素をUDP-グルコ-スと接触させることにより、UDP-グルコ-スをUDP-ラムノ-スに変換させることを特徴とする、UDP-ラムノ-スの製造方法。
【請求項29】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を同位体元素により置換されたグルコ-スを炭素源として培養し、培養物から同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
【請求項30】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を培地で培養することにより得られた培養物、培養処理物、酵素抽出液または精製酵素を同位体元素により置換されたUDP-グルコ-スと接触させ、同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
【請求項31】
同位体元素がC13またはC14であることを特徴とする、請求項29または30に記載の製造方法
【請求項1】
(a)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列を有するか、あるいは(b)配列番号2、配列番号4もしくは配列番号20で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項2】
請求項1に記載のタンパク質をコ-ドするDNA。
【請求項3】
(a)配列番号1、配列番号3もしくは配列番号19で示される塩基配列を有するか、あるいは(b)配列番号1、配列番号3もしくは配列番号19で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項4】
請求項2又は3に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項5】
請求項4に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項6】
(a)配列番号6で示されるアミノ酸配列を有するか、あるいは(b)配列番号6で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質。
【請求項7】
請求項6に記載のタンパク質をコ-ドするDNA。
【請求項8】
(a)配列番号5で示される塩基配列を有するか、あるいは(b)配列番号5で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-グルコ-ス4、6-デヒドラタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項9】
請求項7又は8に記載のDNAにより組み換えられていることを特徴とする、組換えベクタ-。
【請求項10】
請求項9に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項11】
(a)配列番号8で示されるアミノ酸配列を有するか、あるいは(b)配列番号8で示されるアミノ酸配列において、1乃至数個のアミノ酸が、欠失、置換若しくは付加されたアミノ酸配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項12】
請求項11に記載のタンパク質をコ-ドするDNA。
【請求項13】
(a)配列番号7で示される塩基配列を有するか、あるいは(b)配列番号7で示される塩基配列において、1乃至数個のヌクレオチドが、欠失、置換若しくは付加された塩基配列を有し、かつ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質をコ-ドするDNA。
【請求項14】
請求項12又は13に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項15】
請求項7又は8に記載のDNA及び請求項12又は13に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項16】
請求項14に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項17】
請求項9及び14に記載の組換えベクタ-が、同一宿主細胞に導入されていることを特徴とする形質転換体。
【請求項18】
請求項15に記載の組換えベクタ-が導入されていることを特徴とする、形質転換体。
【請求項19】
配列番号2、4もしくは20で示されるアミノ酸配列と70%以上の相同性を有するアミノ酸配列を有し、かつUDP-グルコ-ス4、6-デヒドラタ-ゼ、UDP-4-ケト-6-デオキシグルコ-ス3、5-エピメラ-ゼ及びUDP-4-ケト-ラムノ-ス4-ケト-レダクタ-ゼ活性を有するタンパク質。
【請求項20】
イネ(Oryza sativa)またはタバコ(Nicotiana tabacum)由来のタンパク質である請求項19に記載のタンパク質。
【請求項21】
請求項19または20に記載のタンパク質をコ-ドするDNA。
【請求項22】
請求項21に記載のDNAにより組換えられていることを特徴とする、組み換えベクタ-。
【請求項23】
請求項22に記載の組換えベクタ-が宿主細胞に導入されていることを特徴とする、形質転換体。
【請求項24】
宿主細胞が、UDP-グルコ-スを細胞内に含有していることを特徴とする、請求項5、17、18及び23のいずれかに記載の形質転換体。
【請求項25】
宿主細胞が、UDP-ラムノ-スを消費する酵素系を保持していないことを特徴とする、請求項5、17、18及び23のいずれかに記載の形質転換体。
【請求項26】
宿主細胞が酵母細胞である、請求項5、10、16〜18及び23〜25のいずれかに記載の形質転換体。
【請求項27】
請求項5、17、18及び23〜26のいずれかに記載の形質転換体を培養し、培養物からUDP-ラムノ-スを採取することを特徴とする、UDP-ラムノ-スの製造方法。
【請求項28】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を培地に培養し、得られた培養物、培養処理物、酵素抽出液または精製酵素をUDP-グルコ-スと接触させることにより、UDP-グルコ-スをUDP-ラムノ-スに変換させることを特徴とする、UDP-ラムノ-スの製造方法。
【請求項29】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を同位体元素により置換されたグルコ-スを炭素源として培養し、培養物から同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
【請求項30】
請求項5、17、18、23〜26のいずれかに記載の形質転換体を培地で培養することにより得られた培養物、培養処理物、酵素抽出液または精製酵素を同位体元素により置換されたUDP-グルコ-スと接触させ、同位体元素でラベルされたUDP-ラムノ-スを採取することを特徴とする、標識UDP-ラムノ-スの製造方法。
【請求項31】
同位体元素がC13またはC14であることを特徴とする、請求項29または30に記載の製造方法
【図3】


【図4】
【図5】
【図7】
【図1】
【図2】
【図6】
【図4】
【図5】
【図7】
【図1】
【図2】
【図6】
【公開番号】特開2008−124(P2008−124A)
【公開日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願番号】特願2006−204421(P2006−204421)
【出願日】平成18年7月27日(2006.7.27)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成20年1月10日(2008.1.10)
【国際特許分類】
【出願日】平成18年7月27日(2006.7.27)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]