
UVI4−like遺伝子欠損植物
【課題】単一遺伝子の変異によって雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成し、その結果、自殖によって親植物の約2倍のゲノム量を持つ後代種子を得ることができる植物を提供する。
【解決手段】特定のcDNA塩基配列を有するUV14-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失し、雄生側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成して、自殖により約2倍のゲノム量を持つ後代種子が得られる植物。
【解決手段】特定のcDNA塩基配列を有するUV14-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失し、雄生側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成して、自殖により約2倍のゲノム量を持つ後代種子が得られる植物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、UVI4-like遺伝子の機能の低下または欠失によって、雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成し、自殖によって親植物の約2倍のゲノム量を持つ後代種子を得ることができる植物に関するものである。
【背景技術】
【0002】
近年のゲノム解析から、被子植物の進化においては複数回の全ゲノム倍加が起きていることが明らかになった(非特許文献1)。植物で最も小さなゲノムサイズを持つシロイヌナズナ(Arabidopsis)でさえ、少なくとも3回の全ゲノム倍加を経験したと考えられている(非特許文献2;非特許文献3)。このように、今日存在する多くの2倍性植物が実は多倍体起源であると考えられている。
【0003】
これまでの研究から、非還元配偶子の形成が多倍体の形成に大きな役割を果たしており、非還元配偶子との受精によって形成された3倍体を経由して多倍体が形成されたという見解が得られている(非特許文献4;非特許文献5)。非還元配偶子の形成頻度は概して極めて低いが、多くの植物種で確認されている現象である(非特許文献6;非特許文献7)。
【0004】
ジャガイモにおけるいくつかの減数分裂に関する変異体の研究から、非還元配偶子の形成は単一劣性因子の制御下で起き得ることが示されたが、遺伝子は単離されていない(非特許文献8;非特許文献9)。シロイヌナズナにおいては、1遺伝子の変異によって受精能力のある非還元の雌性配偶子が形成されることが報告されている(非特許文献10)。この変異株の自殖後代植物の80%が3倍体であったと報告されているが、非還元配偶子の形成頻度は低く、得られる種子数は1個体あたり10粒以下と極めて少ないものであった。
【0005】
3倍体を経由して4倍体以上の高次倍数体が得られる経路の他にも、非還元の雌性及び雄性配偶子間の受精によって直接4倍体が得られる可能性があるが、これまでに確認されている非還元配偶子の形成頻度が概して低いことならびに雌性及び雄性配偶子における非還元配偶子の形成には相関がみられないことから、非還元の雌性及び雄性配偶子間の受精によって直接4倍体が得られる経路は起こりにくいと考えられていた。さらに、1遺伝子の変異によって直接4倍体の後代植物が得られる現象ならびに雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子の形成につながる遺伝子はこれまでに報告されていない。
【0006】
なお、SEQ ID NO: 2にその配列を記載したUVI4-likeタンパク質は、突然変異によって葉で核内倍加が亢進することが報告されているUVI4(UV-B非感受性4、At2g42260)タンパク質(非特許文献11、特許文献1)とアミノ酸配列の高い類似性を有するが、これまで機能が証明されていなかったタンパク質であり、突然変異によって核内倍加が亢進するという特徴以外に関する機能の推定は不可能である。
【特許文献1】特開2007-228922号
【非特許文献1】Comai (2005) Nature Rev. Genet. 6, 836-846.
【非特許文献2】Vision et al. (2000) Science290, 2114-2117.
【非特許文献3】Adams & Wendel (2005) Curr. Opin. Plant. Biol. 8, 135-141.
【非特許文献4】Ramsey & Schemske (1998) Ann. Rev. Eco. Syst. 29. 467-501.
【非特許文献5】Ramsey & Schemske (2002) Ann. Rev. Eco. Syst. 33. 589-639.
【非特許文献6】Veilleux (1985) Plant Breed. Rev. 3, 253-288.
【非特許文献7】Bretagnolle & Thompson (1995) New Phytologist 129, 1-11.
【非特許文献8】Carputo et al. (2003) Genetics163, 287-294.
【非特許文献9】Peloquin et al. (1999) Genetics 153, 1493-1499.
【非特許文献10】Ravi et al. (2008) Nature 451, 1121-1125.
【非特許文献11】Hase et al. (2006) Plant J. 46, 317-326.
【発明の概要】
【発明が解決しようとする課題】
【0007】
「倍数性育種」という言葉で知られるように、植物の倍数性を高めることは、収量の増加につながる場合がある。
【0008】
本発明は、植物でその機能が低下または欠失することによって雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子が形成される新規遺伝子を特定し、この遺伝子の変異によって雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成し、その結果、自殖によって親植物の約2倍のゲノム量を持つ後代種子を得ることができる植物を提供することを課題としている。
【課題を解決するための手段】
【0009】
本発明者らは、シロイヌナズナ(Arabidopsis)のUVI4-like(UV-B非感受性4類似、At3g57860)遺伝子の破壊株の特性を解析した結果、UVI4-likeタンパク質の機能が低下または欠失した植物では、雄性(花粉)側及び雌性(卵)側の両者において受精能力のある非還元配偶子が形成されることを見いだした。さらに、当該植物の自殖によって当該植物の2倍のゲノム量を持つ後代種子が得られることを見いだし、本発明を完成するに至った。
【0010】
すなわち本発明は、以上のとおりの新規な知見に基づき、SEQ ID NO: 1にそのcDNA塩基配列を記載したUVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物を提供する。
【0011】
本発明の植物は、好ましい態様として、雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成する。さらに好ましい態様として、本発明の植物は、自殖により約2倍のゲノム量を持つ後代種子が得られる。
【0012】
さらにまた、本発明の別の好ましい態様は、アブラナ科の植物であり、さらに詳しくはシロイヌナズナである。
【発明の効果】
【0013】
本発明により、雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成する植物を提供することができる。また、当該植物を自殖することにより、当該植物の2倍のゲノム量を持つ後代種子を生産することができる。
【図面の簡単な説明】
【0014】
【図1】At3g57860と推定ホモログのアミノ酸配列のアラインメント。At3g57860のアミノ酸と同一のものは白抜きで示した。2つの鏃マークはpst11867およびpst15307系統におけるDsトランスポゾンの挿入位置を示す。配列のアラインメントはClustal W program v1.83(http://www.ddbj.nig.ac.jp/)を用いて作製した。
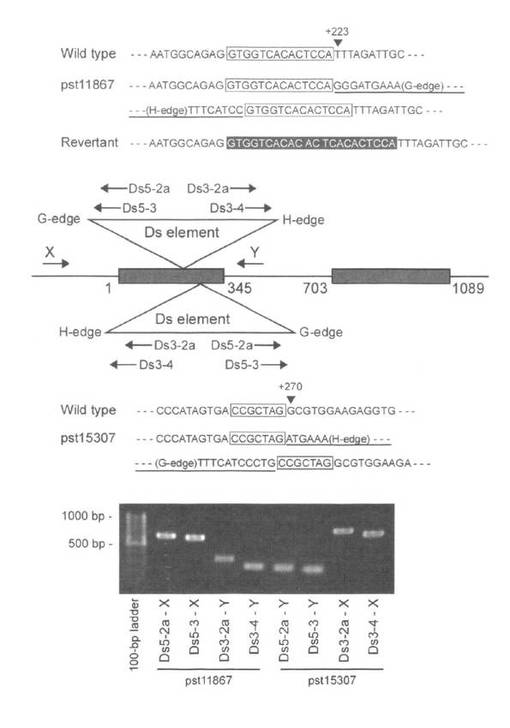
【図2】Dsトランスポゾン挿入部位の模式図とヌクレオチド配列。At3g57860遺伝子のタンパクをコードする領域は灰色の四角で示した。三角形はDsトランスポゾンを示す。矢印は挿入位置を決定するために使用したプライマーを示す。Dsトランスポゾンの端に位置するプライマー(Ds5-2a、Ds5-3、Ds3-2aおよびDs3-4)とゲノムDNA上に設計したプライマー(XおよびY)によって増幅されたPCR断片の長さとゲノム配列は、両系統においてDsトランスポゾンがそれぞれ異なった方向で挿入されたことを示す。PCR断片の配列から、pst11867系統では、開始コドンから矢頭で示した+223のヌクレオチド位置にDsトランスポゾンが挿入され、そして囲み配列で示した14-bpの標的複製を伴っていることが示された。一方、pst15307系統では、+270の位置に挿入され、7-bpの複製を伴っていた。下線を付した配列はDsトランスポゾンの末端の配列を示す。反転表示した配列はDsトランスポゾンの再転移によって生じたフットプリントの塩基配列を示す。
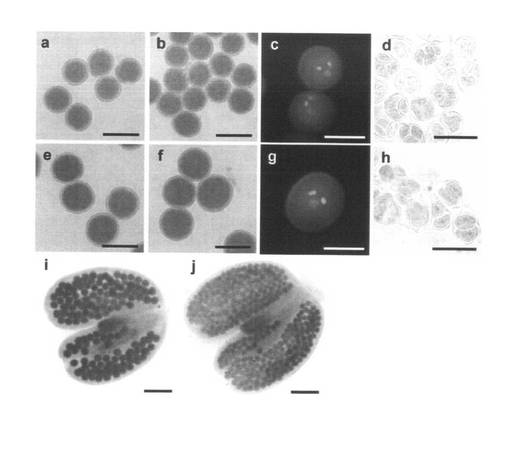
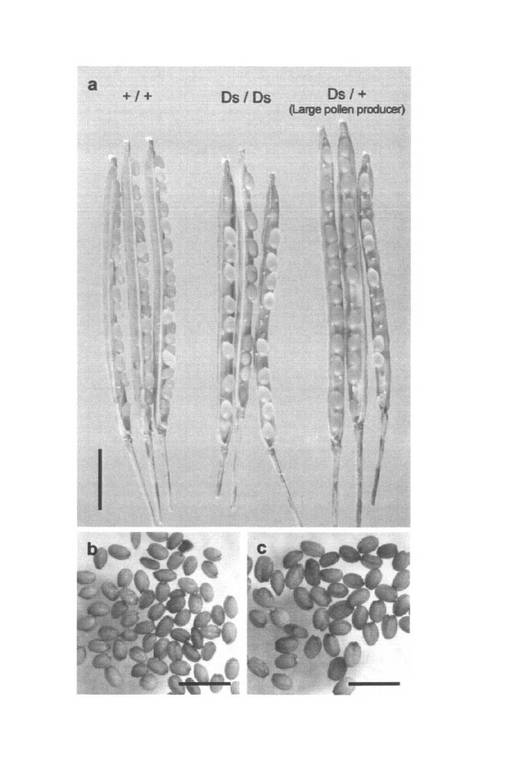
【図3】二分子形成による大型花粉の形成。(a)野生型の花粉。(b、c)ヘテロ接合体の正常サイズの花粉。(d)野生型の四分子。(e)ヘテロ接合体で形成された大型の花粉。(f、g)ホモ接合体で形成された大型の花粉。(h)ホモ接合体で観察された二分子。(i、j)ホモ接合体(i)及び野生型(j)植物体の葯室内の花粉。成熟花粉はアレキサンダー染色液(a、b、e、f、i、j)またはHoechst 33342(c、g)で染色した。四分子(d)および二分子(h)はトルイジンブルーで染色した。スケールバーは(a, b, d - f, h)は30μm 、(c, g)は20μm、(i, j)は100μmの長さを示す。
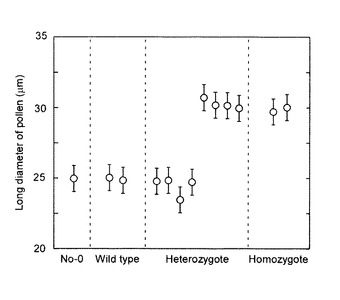
【図4】無作為に抽出した1個体のpst11867系統の2倍性ヘテロ接合体の自殖後代植物体で形成された花粉のサイズ。データは各個体から得られた花粉の長径の平均値±標準偏差を示す。
【図5】葯室内の花粉の比較。開花直前の葯を酢酸カーミンで染色した。ホモ接合体(中央)は、野生型(左)の花粉より大型の非還元花粉を形成する。大型花粉を形成する個体の自殖によって得られた4倍性のホモ接合体(Ds/Ds/Ds/Ds)(右)は、4倍体相当のDNA量を含むと推定される巨大花粉を形成する。
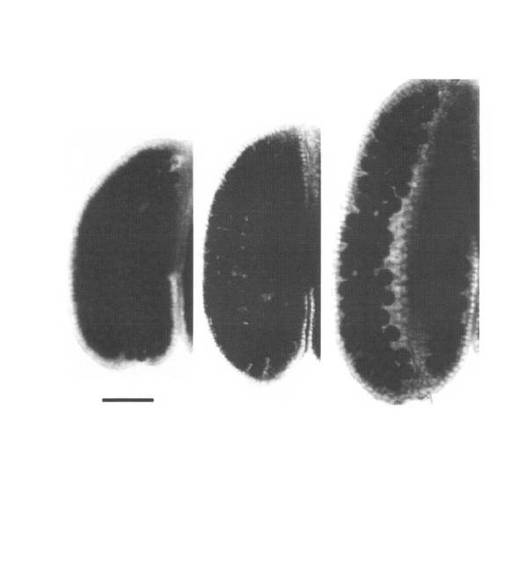
【図6】UVI4-like遺伝子の欠損による倍数性が上昇した種子の形成。(a)野生型(左)、ホモ接合体(中央)および大型花粉を形成するヘテロ接合体(右)の鞘内の種子。(b、c)野生型(b)およびホモ接合体(c)から得られた種子。スケールバーは、(a)が2mm、(b、c)は1mm。
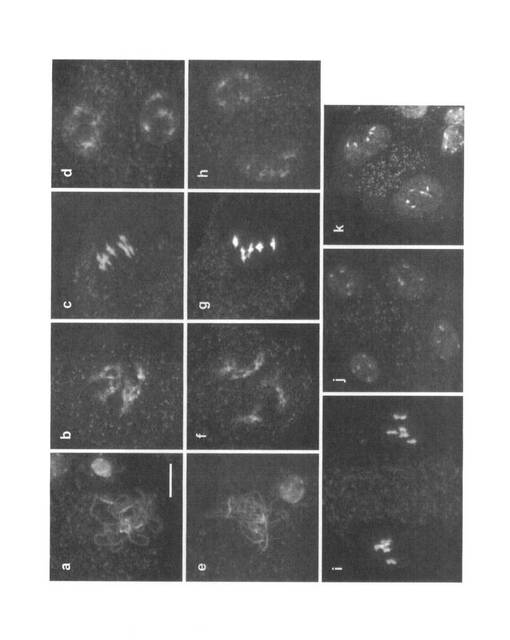
【図7】野生型及びホモ接合体における雄性減数分裂。野生型(a - d, i, j)及びホモ接合体(e - h, k)の染色体伸展標本を示す。(a, e)pachytene期。(b, f) diakinesis期。(c, g)第一減数分裂中期。(d, h)第一減数分裂終期から第二減数分裂前期。(i)第二減数分裂中期。(j)第二減数分裂終期、(k)第一減数分裂終期。スケールバーは10μmの長さを示し、全ての図に共通である。
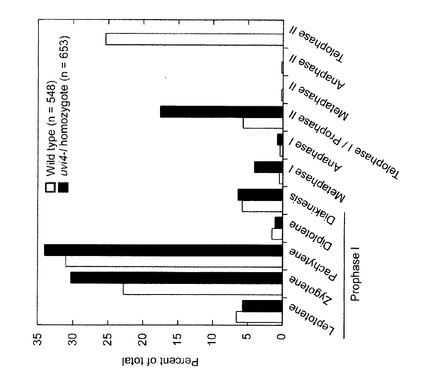
【図8】野生型及びホモ接合体における雄性減数分裂細胞の分布。観察した細胞の総数に対する割合を示す。観察には、野生型及びホモ接合体のそれぞれについて5つの花序を用いた。
【発明を実施するための最良の形態】
【0015】
本発明は、UVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物であるが、「遺伝子の機能が低下または欠失」とは、当該遺伝子の転写の低下または欠失であり、あるいは当該遺伝子のタンパク質への翻訳の低下またはタンパク質の欠失が含まれる。
【0016】
このような遺伝子の機能の低下または形質は、下記実施例で示すように、標的遺伝子(UVI4-like遺伝子または他種ホモログ遺伝子)へのトランスポゾンの挿入によって達成することができる。またそれ以外にも、植物遺伝子の機能低下や欠失に当該技術分野で使用されている方法を適宜に使用することができる。例えば、標的遺伝子と相補的なアンチセンスRNAをコードするDNAを植物体に導入する方法、標的遺伝子のmRNAを特異的に切断するリボザイム(ハンマーヘッド型リボザイムやヘヤピン形リボザイム)をコードするDNAを使用する方法、標的遺伝子のmRNAに相補的二重鎖RNA(dsRNA)をコードするDNAを使用する方法などである。あるいは、標的遺伝子を組込んだウイルスベクターを植物に導入することによるRNAサイレンシング法などを使用することもできる。さらには、ゲノムDNAにおける標的遺伝子を対象としたジーンターゲッティング法による遺伝子ノックアウト法を使用することもできる。すなわち、UVI4-like遺伝子はシロイヌナズナ第3染色体の21437291−21439230塩基配列に位置することが公知であり(GenBank/NC_003074)、このゲノム配列情報に基づいて作製した相同組換えベクターを用いたるジーンターゲッティング法による遺伝子ノックアウトによって、標的遺伝子の機能を欠失させることができる。
【0017】
また、本発明における「UVI4-like遺伝子または他種ホモログ遺伝子」とは、シロイヌナズナのUVI4-like遺伝子、またはシロイヌナズナ以外の植物のUVI4-likeホモログ遺伝子(シロイヌナズナUVI4-like遺伝子と相同の構造および機能を有する遺伝子)である。シロイヌナズナ以外の植物としては、UVI4-like遺伝子と相同性を有する遺伝子が知られているポプラ、ブドウ、イネ(図1)等を対象とすることができるが、これらに限定されない。例えば、シロイヌナズナが属するアブラナ科(キャベツや大根等)の有用植物からUVI4-likeホモログ遺伝子を単離し、これらのアブラナ科植物のUVI4-likeホモログ遺伝子の機能を低下または欠失させることもできる。UVI4-likeホモログ遺伝子を単離するには、例えば、UVI4-like遺伝子の配列情報(SEQ ID NO:1)に基づいて作製したプライマーオリゴヌクレオチドを用いたPCR法、あるいは配列情報に基づいて作製したプローブオリゴヌクレオチドを用いて特定植物のcDNAライブラリーをスクリーニングする方法など、当該技術分野でホモログ遺伝子を特定する方法に準じて行うことができる。
【0018】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例によって限定されるものではない。
【実施例】
【0019】
(1)方法
(1−1)植物材料および育成条件
シロイヌナズナのDsトランスポゾンタグライン(系統名pst11867およびpst15307)は理研バイオリソースセンターから提供されたものである。これらの系統はシロイヌナズナのNo-0エコタイプをバックグラウンドとして作成されたものである。植物体の育成は、市販の培養土(MetroMix350, Scotts-Sierra Horticultural Products Company, Marysville, OH, USA)を用い、23℃、16時間明期、3,000−4,000ルクスの白色蛍光灯を備えた育成室で行った。
(1−2)PCRによる遺伝子型解析
ゲノムDNAはクラボウ製PI-50α自動DNA抽出システム、キアゲン製DNeasy Plant Mini Kit、シグマ製 Extract-N-Amp Plant PCR KitsまたはDNA抽出用溶液(200 mM Tris-HCl (pH7.5)、250 mM NaCl、25 mM EDTA及び0.5% SDSを含む)を用いて抽出した。遺伝子型の調査は、図2中でXおよびYで示したUVI4-like遺伝子に特異的なプライマー配列の組み合わせ
X:5’-TAGACGTTAGGGTGGGGTTTAC-3’(SEQ ID NO:3)および
Y:5’-CAACAGGAAAACACAAAAGAGC-3’(SEQ ID NO:4)、
ならびに、Dsトランスポゾンに特異的なDs5-2a、Ds5-3、Ds3-2a及びDs3-4のプライマー配列(Kuromori et al. (2004) Plant J. 37, 897-905)を用いた。Acエレメントの検出には、Acエレメントの塩基配列に特異的なプライマーの組み合わせ
Ac2656: 5’-TAAAGCCGAGGAGTGGAAGA-3’(SEQ ID NO:5)及び
Ac3244: 5’-TCCCCTCCACCATGATAAAA-3’(SEQ ID NO:6)を用いた。
(1−3)顕微鏡観察
成熟花粉はアレキサンダー染色液(Alexander et al. (1969)Stain Technology 44,117-122)で染色した。葯室中の花粉の観察は、開葯直前の葯を1%酢酸カーミン溶液またはアレキサンダー染色液で一晩染色した後、スライドグラス上でゆるやかに押しつぶすことによって行った。花粉の直径の測定は、成熟花粉を1%酢酸カーミン溶液で染色して行った。花粉の直径はソフトウェアImageJ 1.37vを用いて測定した。各植物体について50粒以上の花粉を測定した。核の大きさの観察は、成熟花粉をHochest33342の0.1%溶液で染色して行った。四分子及び二分子の観察は、新鮮な葯をトルイジンブルー0.02%溶液中に置き、メスを使って内容物をスライドガラス上に取り出して行った。雄性減数分裂細胞の染色体伸展標本は、文献(Ross et al. 1996)に記載の方法で作成し、10 μlの4’,6-diamidino-2-phenylindole(1 μg/ml)で染色した。
(1−4)倍数性解析
2−3週齢の植物体の若いロゼット葉を用い、文献(Haseet al. (2006) Plant J. 46, 317-326)に記載の方法で測定した。最も低いピーク位置のDNA量をPartec製ソフトウェアDPACを用いて測定した。シロイヌナズナの約8.4倍の核DNA量を持つダイズの若い葉を内部標準として同時に測定した。
(2)結果と考察
UVI4-like遺伝子の機能を調査するために、Dsトランスポゾンの挿入によるUVI4-like遺伝子の破壊株を2系統(pst11867およびpst15307)得た。PCRおよび塩基配列解析の結果から、Dsトランスポゾンは、UVI4-like遺伝子の塩基配列の翻訳開始点を+1とした場合、pst11867では+223、pst15307では+270の位置に挿入されていることがわかった(図2)。これらの挿入位置から上記2系統の破壊株は機能損失株であると推測される。PCRを利用した遺伝子型解析によって、調査した11個体のpst11867系統の中には、ノックアウトアリル(トランスポゾンが挿入されることによって機能が失われたUVI4-like対立遺伝子)をホモに持つ個体は無いと考えられた。さらに、種子の大きさが不揃いであったことから核DNA量の測定により倍数性を調査したところ、上記11個体中5個体が4倍性であると考えられた。これらの結果から、調査した11個体のpst11867系統は、2個体の2倍性野生型、4個体の2倍性ヘテロ接合体及び5個体の4倍性ヘテロ接合体であると考えられた。この結果は、UVI4-like遺伝子の破壊によって倍数性が上昇することを示唆する。同様な結果がpst15307系統でも見られたことから、観察された現象はUVI4-like遺伝子の破壊によるものであることを示唆している。
【0020】
無作為に選んだpst11867系統の2倍性のノックアウトアリルのヘテロ接合体から得られた204個体の自殖後代の遺伝子型は、47個体の野生型、141個体のヘテロ接合体及び16個体のホモ接合体であると考えられた。204個体の内、16個体の野生型、27個体のヘテロ接合体及び9個体のホモ接合体を含む計52個体の倍数性を測定した結果、全ての植物体が2倍性であった。ここで発明者らは、全てのホモ接合体ならびに一部のヘテロ接合体(141個体中25個体)が通常よりも大きな花粉を形成することを発見した(表1及び図3e, f)。大きな花粉はシロイヌナズナの花粉に典型的な3つの核、つまり、2つの明るく染色される精核及び1つのぼんやりと染色される栄養核を持っていた(図3g)。しかしながら、核のサイズが図3cに示される野生型のものよりも明らかに大きいことは、倍数性が高いことを示している。ヘテロ接合体は、大きな花粉を形成する個体と野生型と同等サイズの花粉を形成する個体のどちらかに二分され、同一個体で大きな花粉と野生型と同等サイズの花粉の両方を形成する個体は確認されなかった(図4)。ホモ接合体が形成した花粉の長径は29.9±1.1μmであったが野生型の花粉の長径は25.0±1.0μmであった(図4)。野生型の葯室には計420〜440粒の花粉が含まれていたが、大きな花粉を形成する個体では240〜280粒に減少した。大きな花粉を形成する個体では花粉の大きさはほぼ均一であり、形成不全の花粉はほとんど見られなかった(図3i及び図5)。
【0021】
【表1】

【0022】
ホモ接合体及び大きな花粉を形成するヘテロ接合体では、RT-PCRによってUVI4-like遺伝子の転写産物は検出されなかった。これまでの試験結果からpst11867系統はトランスポゾンを転移させるAcエレメントを持っている可能性が示唆された。大きな花粉を形成するヘテロ接合体からAcエレメントに特異的なPCR産物が確認され、さらに、Dsトランスポゾンが再転移した後に残されるフットプリントの塩基配列がUVI4-like遺伝子座に存在することが確認された(図2)。PCRによる遺伝子型の判定は、増幅されたDNAの長さに基づくことから、正常なUVI4-like遺伝子座とDsトランスポゾンが再転移したことによって機能を失ったUVI4-like遺伝子座を区別することができない。以上の結果から、大きな花粉を形成するヘテロ接合体は、Dsトランスポゾンが挿入された遺伝子座とDsトランスポゾンが再転移したことによって機能を失った遺伝子座の両方を持つと考えられた。さらに、pst15307系統とAcエレメントを含まないpst11867系統間の相補試験によって、両系統が同じ遺伝子に変異を持つことが示されたことから、大きな花粉を形成する表現型がUVI4-like遺伝子座の不活化によって生じることが示された。これらの結果から、シロイヌナズナの2倍体では、2つのUVI4-like遺伝子座の両方が不活化された場合、すなわち劣性ホモ接合体においてのみ、大きな花粉を形成する表現型を示すことが示された。
【0023】
大きな花粉を形成する個体の自殖によって生存可能な種子が得られ、それらは野生型で得られる種子よりも明らかに大きかった(図6)。倍数性調査の結果、ホモ接合体から得られた20個体の内19個体(=95%)が4倍体であり、残る1個体は3倍体と考えられた(表2)。4倍体と推定される個体は、大きな花粉を形成するヘテロ接合体の自殖によっても得られた(73個体中63個体=83%)(表2)。これらの結果は、UVI4-like遺伝子の破壊によって生殖能力のある非還元配偶子が雄性(花粉)側及び雌性(卵)側の両者で形成されることを示している。また、3倍体が少数得られたことから、雄性(花粉)側及び雌性(卵)側のいずれか一方においては、正常な半数性の配偶子と非還元配偶子の両者が形成されることが示唆される。
【0024】
【表2】
【0025】
正常なシロイヌナズナの花粉形成においては、減数分裂によって四分子が形成される(図3d)。これと対照的に、大きな花粉を形成する個体では同じ時期に二分子だけが観察された(n = 281)(図3h)。四分子やその他の異常な減数分裂細胞は観察されなかった。この結果はUVI4-like遺伝子の破壊が、雄性(花粉)側の配偶子形成における減数分裂の異常を引き起こし、ほぼ100%に近い頻度で非還元花粉が形成されることを示している。
【0026】
さらに減数分裂の異常について調査するため、野生型との正逆交配を行った。ホモ接合体をを花粉親として野生型と交配することにより得られたF1植物体は、35個体全てが3倍体と考えられた。一方、ホモ接合体を種子親として野生型と交配した場合は、調査した23個体のF1植物体の内、15個体が3倍体、8個体が2倍体であった。これらの結果から、雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成することが改めて示された。さらに、雄性側(花粉)では非還元配偶子の形成頻度がほぼ100%であり、一方、雌性側(卵)では、正常な半数性の配偶子と非還元配偶子の両者が形成されることが示された。なお、対照実験として、野生型と同じサイズの花粉を形成するヘテロ接合体を野生型と交配した場合は、花粉親と種子親のいずれに用いた場合でも、F1植物体は全て2倍性であり、遺伝子型は野生型とヘテロ接合体が1:1に分離することが確かめられた。
【0027】
4倍性のホモ接合体は直径約37μmの巨大花粉を形成した。この個体の自殖によって8倍体と推定される個体が得られた。しかしながら、8倍体は不稔であり、その後代は得られなかった。
【0028】
さらに減数分裂の異常について調査するため、雄性減数分裂における染色体の動態を観察した。野生型での染色体の動態は文献(Ma (2006) The Arabidopsis Book, June 6)等に詳しく記述されているため、ここでは簡潔に述べる。まず、減数第一分裂前期(prophase I)のleptotene期において、やや凝縮した細い線状の染色体が観察される。その後、zygotene期で相同染色体が対合を開始する。pachytene期で相同染色体は完全に対合し、太い糸状の染色体が観察される(図7a)。diplotene期で相同染色体間の対合が解離し始め、相同染色体はさらに凝縮し、diakinesis期で染色体が交叉したキアズマと呼ばれる構造が観察される(図7b)。二価染色体と呼ばれる5対の相同染色体は、第一減数分裂中期(metaphase I)で赤道面に配列する(図7c)。第一減数分裂後期(anaphase I)で相同染色体は二極に分離し、第一減数分裂終期(telophase I)から第二減数分裂前期(prophase II)にかけて染色体はやや脱凝縮する(図7d)。2組の各5本の染色体は、第二減数分裂中期(metaphase II)において異なる赤道面に配列し(図7i)、第二減数分裂の後期(anaphase II)で2回目の染色体の分離が行われる。第二減数分裂終期(telophase II)は減数分裂の最後のステージであり、四分子と呼ばれる4つの半数性の細胞が形成される(図7j)。UVI4-like遺伝子のノックアウトアリルのホモ接合体では、第一減数分裂においては野生型との顕著な差異は見出せなかった。つまり、相同染色体間の対合(図7e)及びキアズマの形成(図7f)は正常と考えられた。また、5対の二価染色体は赤道面に配列した後(図7g)、二極に分離した(図7h)。しかしながら、第二減数分裂に相当する染色体像は観察されなかった。第一減数分裂終期から第二減数分裂前期にかけて、野生型では染色体がやや脱凝縮するが(図7d)、ホモ接合体ではこのような脱凝縮した染色体は稀であり、多くは図7kに示したように凝縮した染色体を持っていた。この凝縮した染色体は、野生型の第二減数分裂終期における染色体の様相(図7j)と類似していることから、ホモ接合体では第二減数分裂を行わずに減数分裂を終えてしまうことが示唆された。
【0029】
減数分裂の進行について調査するため、減数分裂の各ステージに属する細胞の割合を調査した(図8)。野生型ではzygotene期及びpachytene期の細胞の割合が多く、また、第二減数分裂終期の細胞の割合も多かった。第一及び第二減数分裂の中期ならびに後期の細胞の割合は低かった(全体の1%未満)。UVI4-like遺伝子のノックアウトアリルのホモ接合体では、第一減数分裂前期(prophase I)における分布は野生型のものと類似していた。しかしながら、第一減数分裂中期にある細胞の割合(全体の4.1%)は野生型の同ステージ(0.5%)に比べて著しく高かった。第二減数分裂の細胞は観察されなかった。これらの結果から、ホモ接合体では第一減数分裂中期から後期にかけて減数分裂の進行が遅延することが示唆された。
【0030】
細胞分裂の中期から後期への移行は、Anaphase Promoting Complex(APC)と呼ばれるタンパク複合体が重要な役割を果たすことが知られている(Eloy et al. (2006) Cell Cycle 5, 1957-1965)。APCは分裂サイクリンを標的としてこれを分解させる働きがあり、結果として分裂サイクリンーサイクリン依存性キナーゼの複合体を不活化することにより、後期への移行を促進する。また、分裂サイクリンーサイクリン依存性キナーゼ活性の適切な制御が、第一減数分裂から第二減数分裂への移行に重要であると考えられている(Pesin and Orr-Weaver (2008) Annu. Rev. Cell Dev. Biol. 24, 475-499)。以上のことから、UVI4-likeの変異によってAPCの活性が影響を受けている可能性が示唆される。
【技術分野】
【0001】
本発明は、UVI4-like遺伝子の機能の低下または欠失によって、雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成し、自殖によって親植物の約2倍のゲノム量を持つ後代種子を得ることができる植物に関するものである。
【背景技術】
【0002】
近年のゲノム解析から、被子植物の進化においては複数回の全ゲノム倍加が起きていることが明らかになった(非特許文献1)。植物で最も小さなゲノムサイズを持つシロイヌナズナ(Arabidopsis)でさえ、少なくとも3回の全ゲノム倍加を経験したと考えられている(非特許文献2;非特許文献3)。このように、今日存在する多くの2倍性植物が実は多倍体起源であると考えられている。
【0003】
これまでの研究から、非還元配偶子の形成が多倍体の形成に大きな役割を果たしており、非還元配偶子との受精によって形成された3倍体を経由して多倍体が形成されたという見解が得られている(非特許文献4;非特許文献5)。非還元配偶子の形成頻度は概して極めて低いが、多くの植物種で確認されている現象である(非特許文献6;非特許文献7)。
【0004】
ジャガイモにおけるいくつかの減数分裂に関する変異体の研究から、非還元配偶子の形成は単一劣性因子の制御下で起き得ることが示されたが、遺伝子は単離されていない(非特許文献8;非特許文献9)。シロイヌナズナにおいては、1遺伝子の変異によって受精能力のある非還元の雌性配偶子が形成されることが報告されている(非特許文献10)。この変異株の自殖後代植物の80%が3倍体であったと報告されているが、非還元配偶子の形成頻度は低く、得られる種子数は1個体あたり10粒以下と極めて少ないものであった。
【0005】
3倍体を経由して4倍体以上の高次倍数体が得られる経路の他にも、非還元の雌性及び雄性配偶子間の受精によって直接4倍体が得られる可能性があるが、これまでに確認されている非還元配偶子の形成頻度が概して低いことならびに雌性及び雄性配偶子における非還元配偶子の形成には相関がみられないことから、非還元の雌性及び雄性配偶子間の受精によって直接4倍体が得られる経路は起こりにくいと考えられていた。さらに、1遺伝子の変異によって直接4倍体の後代植物が得られる現象ならびに雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子の形成につながる遺伝子はこれまでに報告されていない。
【0006】
なお、SEQ ID NO: 2にその配列を記載したUVI4-likeタンパク質は、突然変異によって葉で核内倍加が亢進することが報告されているUVI4(UV-B非感受性4、At2g42260)タンパク質(非特許文献11、特許文献1)とアミノ酸配列の高い類似性を有するが、これまで機能が証明されていなかったタンパク質であり、突然変異によって核内倍加が亢進するという特徴以外に関する機能の推定は不可能である。
【特許文献1】特開2007-228922号
【非特許文献1】Comai (2005) Nature Rev. Genet. 6, 836-846.
【非特許文献2】Vision et al. (2000) Science290, 2114-2117.
【非特許文献3】Adams & Wendel (2005) Curr. Opin. Plant. Biol. 8, 135-141.
【非特許文献4】Ramsey & Schemske (1998) Ann. Rev. Eco. Syst. 29. 467-501.
【非特許文献5】Ramsey & Schemske (2002) Ann. Rev. Eco. Syst. 33. 589-639.
【非特許文献6】Veilleux (1985) Plant Breed. Rev. 3, 253-288.
【非特許文献7】Bretagnolle & Thompson (1995) New Phytologist 129, 1-11.
【非特許文献8】Carputo et al. (2003) Genetics163, 287-294.
【非特許文献9】Peloquin et al. (1999) Genetics 153, 1493-1499.
【非特許文献10】Ravi et al. (2008) Nature 451, 1121-1125.
【非特許文献11】Hase et al. (2006) Plant J. 46, 317-326.
【発明の概要】
【発明が解決しようとする課題】
【0007】
「倍数性育種」という言葉で知られるように、植物の倍数性を高めることは、収量の増加につながる場合がある。
【0008】
本発明は、植物でその機能が低下または欠失することによって雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子が形成される新規遺伝子を特定し、この遺伝子の変異によって雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成し、その結果、自殖によって親植物の約2倍のゲノム量を持つ後代種子を得ることができる植物を提供することを課題としている。
【課題を解決するための手段】
【0009】
本発明者らは、シロイヌナズナ(Arabidopsis)のUVI4-like(UV-B非感受性4類似、At3g57860)遺伝子の破壊株の特性を解析した結果、UVI4-likeタンパク質の機能が低下または欠失した植物では、雄性(花粉)側及び雌性(卵)側の両者において受精能力のある非還元配偶子が形成されることを見いだした。さらに、当該植物の自殖によって当該植物の2倍のゲノム量を持つ後代種子が得られることを見いだし、本発明を完成するに至った。
【0010】
すなわち本発明は、以上のとおりの新規な知見に基づき、SEQ ID NO: 1にそのcDNA塩基配列を記載したUVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物を提供する。
【0011】
本発明の植物は、好ましい態様として、雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成する。さらに好ましい態様として、本発明の植物は、自殖により約2倍のゲノム量を持つ後代種子が得られる。
【0012】
さらにまた、本発明の別の好ましい態様は、アブラナ科の植物であり、さらに詳しくはシロイヌナズナである。
【発明の効果】
【0013】
本発明により、雄性側(花粉)及び雌性側(卵)の両者において生殖能力のある非還元配偶子を形成する植物を提供することができる。また、当該植物を自殖することにより、当該植物の2倍のゲノム量を持つ後代種子を生産することができる。
【図面の簡単な説明】
【0014】
【図1】At3g57860と推定ホモログのアミノ酸配列のアラインメント。At3g57860のアミノ酸と同一のものは白抜きで示した。2つの鏃マークはpst11867およびpst15307系統におけるDsトランスポゾンの挿入位置を示す。配列のアラインメントはClustal W program v1.83(http://www.ddbj.nig.ac.jp/)を用いて作製した。
【図2】Dsトランスポゾン挿入部位の模式図とヌクレオチド配列。At3g57860遺伝子のタンパクをコードする領域は灰色の四角で示した。三角形はDsトランスポゾンを示す。矢印は挿入位置を決定するために使用したプライマーを示す。Dsトランスポゾンの端に位置するプライマー(Ds5-2a、Ds5-3、Ds3-2aおよびDs3-4)とゲノムDNA上に設計したプライマー(XおよびY)によって増幅されたPCR断片の長さとゲノム配列は、両系統においてDsトランスポゾンがそれぞれ異なった方向で挿入されたことを示す。PCR断片の配列から、pst11867系統では、開始コドンから矢頭で示した+223のヌクレオチド位置にDsトランスポゾンが挿入され、そして囲み配列で示した14-bpの標的複製を伴っていることが示された。一方、pst15307系統では、+270の位置に挿入され、7-bpの複製を伴っていた。下線を付した配列はDsトランスポゾンの末端の配列を示す。反転表示した配列はDsトランスポゾンの再転移によって生じたフットプリントの塩基配列を示す。
【図3】二分子形成による大型花粉の形成。(a)野生型の花粉。(b、c)ヘテロ接合体の正常サイズの花粉。(d)野生型の四分子。(e)ヘテロ接合体で形成された大型の花粉。(f、g)ホモ接合体で形成された大型の花粉。(h)ホモ接合体で観察された二分子。(i、j)ホモ接合体(i)及び野生型(j)植物体の葯室内の花粉。成熟花粉はアレキサンダー染色液(a、b、e、f、i、j)またはHoechst 33342(c、g)で染色した。四分子(d)および二分子(h)はトルイジンブルーで染色した。スケールバーは(a, b, d - f, h)は30μm 、(c, g)は20μm、(i, j)は100μmの長さを示す。
【図4】無作為に抽出した1個体のpst11867系統の2倍性ヘテロ接合体の自殖後代植物体で形成された花粉のサイズ。データは各個体から得られた花粉の長径の平均値±標準偏差を示す。
【図5】葯室内の花粉の比較。開花直前の葯を酢酸カーミンで染色した。ホモ接合体(中央)は、野生型(左)の花粉より大型の非還元花粉を形成する。大型花粉を形成する個体の自殖によって得られた4倍性のホモ接合体(Ds/Ds/Ds/Ds)(右)は、4倍体相当のDNA量を含むと推定される巨大花粉を形成する。
【図6】UVI4-like遺伝子の欠損による倍数性が上昇した種子の形成。(a)野生型(左)、ホモ接合体(中央)および大型花粉を形成するヘテロ接合体(右)の鞘内の種子。(b、c)野生型(b)およびホモ接合体(c)から得られた種子。スケールバーは、(a)が2mm、(b、c)は1mm。
【図7】野生型及びホモ接合体における雄性減数分裂。野生型(a - d, i, j)及びホモ接合体(e - h, k)の染色体伸展標本を示す。(a, e)pachytene期。(b, f) diakinesis期。(c, g)第一減数分裂中期。(d, h)第一減数分裂終期から第二減数分裂前期。(i)第二減数分裂中期。(j)第二減数分裂終期、(k)第一減数分裂終期。スケールバーは10μmの長さを示し、全ての図に共通である。
【図8】野生型及びホモ接合体における雄性減数分裂細胞の分布。観察した細胞の総数に対する割合を示す。観察には、野生型及びホモ接合体のそれぞれについて5つの花序を用いた。
【発明を実施するための最良の形態】
【0015】
本発明は、UVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物であるが、「遺伝子の機能が低下または欠失」とは、当該遺伝子の転写の低下または欠失であり、あるいは当該遺伝子のタンパク質への翻訳の低下またはタンパク質の欠失が含まれる。
【0016】
このような遺伝子の機能の低下または形質は、下記実施例で示すように、標的遺伝子(UVI4-like遺伝子または他種ホモログ遺伝子)へのトランスポゾンの挿入によって達成することができる。またそれ以外にも、植物遺伝子の機能低下や欠失に当該技術分野で使用されている方法を適宜に使用することができる。例えば、標的遺伝子と相補的なアンチセンスRNAをコードするDNAを植物体に導入する方法、標的遺伝子のmRNAを特異的に切断するリボザイム(ハンマーヘッド型リボザイムやヘヤピン形リボザイム)をコードするDNAを使用する方法、標的遺伝子のmRNAに相補的二重鎖RNA(dsRNA)をコードするDNAを使用する方法などである。あるいは、標的遺伝子を組込んだウイルスベクターを植物に導入することによるRNAサイレンシング法などを使用することもできる。さらには、ゲノムDNAにおける標的遺伝子を対象としたジーンターゲッティング法による遺伝子ノックアウト法を使用することもできる。すなわち、UVI4-like遺伝子はシロイヌナズナ第3染色体の21437291−21439230塩基配列に位置することが公知であり(GenBank/NC_003074)、このゲノム配列情報に基づいて作製した相同組換えベクターを用いたるジーンターゲッティング法による遺伝子ノックアウトによって、標的遺伝子の機能を欠失させることができる。
【0017】
また、本発明における「UVI4-like遺伝子または他種ホモログ遺伝子」とは、シロイヌナズナのUVI4-like遺伝子、またはシロイヌナズナ以外の植物のUVI4-likeホモログ遺伝子(シロイヌナズナUVI4-like遺伝子と相同の構造および機能を有する遺伝子)である。シロイヌナズナ以外の植物としては、UVI4-like遺伝子と相同性を有する遺伝子が知られているポプラ、ブドウ、イネ(図1)等を対象とすることができるが、これらに限定されない。例えば、シロイヌナズナが属するアブラナ科(キャベツや大根等)の有用植物からUVI4-likeホモログ遺伝子を単離し、これらのアブラナ科植物のUVI4-likeホモログ遺伝子の機能を低下または欠失させることもできる。UVI4-likeホモログ遺伝子を単離するには、例えば、UVI4-like遺伝子の配列情報(SEQ ID NO:1)に基づいて作製したプライマーオリゴヌクレオチドを用いたPCR法、あるいは配列情報に基づいて作製したプローブオリゴヌクレオチドを用いて特定植物のcDNAライブラリーをスクリーニングする方法など、当該技術分野でホモログ遺伝子を特定する方法に準じて行うことができる。
【0018】
以下、実施例を示して本発明をさらに詳細かつ具体的に説明するが、本発明は以下の例によって限定されるものではない。
【実施例】
【0019】
(1)方法
(1−1)植物材料および育成条件
シロイヌナズナのDsトランスポゾンタグライン(系統名pst11867およびpst15307)は理研バイオリソースセンターから提供されたものである。これらの系統はシロイヌナズナのNo-0エコタイプをバックグラウンドとして作成されたものである。植物体の育成は、市販の培養土(MetroMix350, Scotts-Sierra Horticultural Products Company, Marysville, OH, USA)を用い、23℃、16時間明期、3,000−4,000ルクスの白色蛍光灯を備えた育成室で行った。
(1−2)PCRによる遺伝子型解析
ゲノムDNAはクラボウ製PI-50α自動DNA抽出システム、キアゲン製DNeasy Plant Mini Kit、シグマ製 Extract-N-Amp Plant PCR KitsまたはDNA抽出用溶液(200 mM Tris-HCl (pH7.5)、250 mM NaCl、25 mM EDTA及び0.5% SDSを含む)を用いて抽出した。遺伝子型の調査は、図2中でXおよびYで示したUVI4-like遺伝子に特異的なプライマー配列の組み合わせ
X:5’-TAGACGTTAGGGTGGGGTTTAC-3’(SEQ ID NO:3)および
Y:5’-CAACAGGAAAACACAAAAGAGC-3’(SEQ ID NO:4)、
ならびに、Dsトランスポゾンに特異的なDs5-2a、Ds5-3、Ds3-2a及びDs3-4のプライマー配列(Kuromori et al. (2004) Plant J. 37, 897-905)を用いた。Acエレメントの検出には、Acエレメントの塩基配列に特異的なプライマーの組み合わせ
Ac2656: 5’-TAAAGCCGAGGAGTGGAAGA-3’(SEQ ID NO:5)及び
Ac3244: 5’-TCCCCTCCACCATGATAAAA-3’(SEQ ID NO:6)を用いた。
(1−3)顕微鏡観察
成熟花粉はアレキサンダー染色液(Alexander et al. (1969)Stain Technology 44,117-122)で染色した。葯室中の花粉の観察は、開葯直前の葯を1%酢酸カーミン溶液またはアレキサンダー染色液で一晩染色した後、スライドグラス上でゆるやかに押しつぶすことによって行った。花粉の直径の測定は、成熟花粉を1%酢酸カーミン溶液で染色して行った。花粉の直径はソフトウェアImageJ 1.37vを用いて測定した。各植物体について50粒以上の花粉を測定した。核の大きさの観察は、成熟花粉をHochest33342の0.1%溶液で染色して行った。四分子及び二分子の観察は、新鮮な葯をトルイジンブルー0.02%溶液中に置き、メスを使って内容物をスライドガラス上に取り出して行った。雄性減数分裂細胞の染色体伸展標本は、文献(Ross et al. 1996)に記載の方法で作成し、10 μlの4’,6-diamidino-2-phenylindole(1 μg/ml)で染色した。
(1−4)倍数性解析
2−3週齢の植物体の若いロゼット葉を用い、文献(Haseet al. (2006) Plant J. 46, 317-326)に記載の方法で測定した。最も低いピーク位置のDNA量をPartec製ソフトウェアDPACを用いて測定した。シロイヌナズナの約8.4倍の核DNA量を持つダイズの若い葉を内部標準として同時に測定した。
(2)結果と考察
UVI4-like遺伝子の機能を調査するために、Dsトランスポゾンの挿入によるUVI4-like遺伝子の破壊株を2系統(pst11867およびpst15307)得た。PCRおよび塩基配列解析の結果から、Dsトランスポゾンは、UVI4-like遺伝子の塩基配列の翻訳開始点を+1とした場合、pst11867では+223、pst15307では+270の位置に挿入されていることがわかった(図2)。これらの挿入位置から上記2系統の破壊株は機能損失株であると推測される。PCRを利用した遺伝子型解析によって、調査した11個体のpst11867系統の中には、ノックアウトアリル(トランスポゾンが挿入されることによって機能が失われたUVI4-like対立遺伝子)をホモに持つ個体は無いと考えられた。さらに、種子の大きさが不揃いであったことから核DNA量の測定により倍数性を調査したところ、上記11個体中5個体が4倍性であると考えられた。これらの結果から、調査した11個体のpst11867系統は、2個体の2倍性野生型、4個体の2倍性ヘテロ接合体及び5個体の4倍性ヘテロ接合体であると考えられた。この結果は、UVI4-like遺伝子の破壊によって倍数性が上昇することを示唆する。同様な結果がpst15307系統でも見られたことから、観察された現象はUVI4-like遺伝子の破壊によるものであることを示唆している。
【0020】
無作為に選んだpst11867系統の2倍性のノックアウトアリルのヘテロ接合体から得られた204個体の自殖後代の遺伝子型は、47個体の野生型、141個体のヘテロ接合体及び16個体のホモ接合体であると考えられた。204個体の内、16個体の野生型、27個体のヘテロ接合体及び9個体のホモ接合体を含む計52個体の倍数性を測定した結果、全ての植物体が2倍性であった。ここで発明者らは、全てのホモ接合体ならびに一部のヘテロ接合体(141個体中25個体)が通常よりも大きな花粉を形成することを発見した(表1及び図3e, f)。大きな花粉はシロイヌナズナの花粉に典型的な3つの核、つまり、2つの明るく染色される精核及び1つのぼんやりと染色される栄養核を持っていた(図3g)。しかしながら、核のサイズが図3cに示される野生型のものよりも明らかに大きいことは、倍数性が高いことを示している。ヘテロ接合体は、大きな花粉を形成する個体と野生型と同等サイズの花粉を形成する個体のどちらかに二分され、同一個体で大きな花粉と野生型と同等サイズの花粉の両方を形成する個体は確認されなかった(図4)。ホモ接合体が形成した花粉の長径は29.9±1.1μmであったが野生型の花粉の長径は25.0±1.0μmであった(図4)。野生型の葯室には計420〜440粒の花粉が含まれていたが、大きな花粉を形成する個体では240〜280粒に減少した。大きな花粉を形成する個体では花粉の大きさはほぼ均一であり、形成不全の花粉はほとんど見られなかった(図3i及び図5)。
【0021】
【表1】
【0022】
ホモ接合体及び大きな花粉を形成するヘテロ接合体では、RT-PCRによってUVI4-like遺伝子の転写産物は検出されなかった。これまでの試験結果からpst11867系統はトランスポゾンを転移させるAcエレメントを持っている可能性が示唆された。大きな花粉を形成するヘテロ接合体からAcエレメントに特異的なPCR産物が確認され、さらに、Dsトランスポゾンが再転移した後に残されるフットプリントの塩基配列がUVI4-like遺伝子座に存在することが確認された(図2)。PCRによる遺伝子型の判定は、増幅されたDNAの長さに基づくことから、正常なUVI4-like遺伝子座とDsトランスポゾンが再転移したことによって機能を失ったUVI4-like遺伝子座を区別することができない。以上の結果から、大きな花粉を形成するヘテロ接合体は、Dsトランスポゾンが挿入された遺伝子座とDsトランスポゾンが再転移したことによって機能を失った遺伝子座の両方を持つと考えられた。さらに、pst15307系統とAcエレメントを含まないpst11867系統間の相補試験によって、両系統が同じ遺伝子に変異を持つことが示されたことから、大きな花粉を形成する表現型がUVI4-like遺伝子座の不活化によって生じることが示された。これらの結果から、シロイヌナズナの2倍体では、2つのUVI4-like遺伝子座の両方が不活化された場合、すなわち劣性ホモ接合体においてのみ、大きな花粉を形成する表現型を示すことが示された。
【0023】
大きな花粉を形成する個体の自殖によって生存可能な種子が得られ、それらは野生型で得られる種子よりも明らかに大きかった(図6)。倍数性調査の結果、ホモ接合体から得られた20個体の内19個体(=95%)が4倍体であり、残る1個体は3倍体と考えられた(表2)。4倍体と推定される個体は、大きな花粉を形成するヘテロ接合体の自殖によっても得られた(73個体中63個体=83%)(表2)。これらの結果は、UVI4-like遺伝子の破壊によって生殖能力のある非還元配偶子が雄性(花粉)側及び雌性(卵)側の両者で形成されることを示している。また、3倍体が少数得られたことから、雄性(花粉)側及び雌性(卵)側のいずれか一方においては、正常な半数性の配偶子と非還元配偶子の両者が形成されることが示唆される。
【0024】
【表2】
【0025】
正常なシロイヌナズナの花粉形成においては、減数分裂によって四分子が形成される(図3d)。これと対照的に、大きな花粉を形成する個体では同じ時期に二分子だけが観察された(n = 281)(図3h)。四分子やその他の異常な減数分裂細胞は観察されなかった。この結果はUVI4-like遺伝子の破壊が、雄性(花粉)側の配偶子形成における減数分裂の異常を引き起こし、ほぼ100%に近い頻度で非還元花粉が形成されることを示している。
【0026】
さらに減数分裂の異常について調査するため、野生型との正逆交配を行った。ホモ接合体をを花粉親として野生型と交配することにより得られたF1植物体は、35個体全てが3倍体と考えられた。一方、ホモ接合体を種子親として野生型と交配した場合は、調査した23個体のF1植物体の内、15個体が3倍体、8個体が2倍体であった。これらの結果から、雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成することが改めて示された。さらに、雄性側(花粉)では非還元配偶子の形成頻度がほぼ100%であり、一方、雌性側(卵)では、正常な半数性の配偶子と非還元配偶子の両者が形成されることが示された。なお、対照実験として、野生型と同じサイズの花粉を形成するヘテロ接合体を野生型と交配した場合は、花粉親と種子親のいずれに用いた場合でも、F1植物体は全て2倍性であり、遺伝子型は野生型とヘテロ接合体が1:1に分離することが確かめられた。
【0027】
4倍性のホモ接合体は直径約37μmの巨大花粉を形成した。この個体の自殖によって8倍体と推定される個体が得られた。しかしながら、8倍体は不稔であり、その後代は得られなかった。
【0028】
さらに減数分裂の異常について調査するため、雄性減数分裂における染色体の動態を観察した。野生型での染色体の動態は文献(Ma (2006) The Arabidopsis Book, June 6)等に詳しく記述されているため、ここでは簡潔に述べる。まず、減数第一分裂前期(prophase I)のleptotene期において、やや凝縮した細い線状の染色体が観察される。その後、zygotene期で相同染色体が対合を開始する。pachytene期で相同染色体は完全に対合し、太い糸状の染色体が観察される(図7a)。diplotene期で相同染色体間の対合が解離し始め、相同染色体はさらに凝縮し、diakinesis期で染色体が交叉したキアズマと呼ばれる構造が観察される(図7b)。二価染色体と呼ばれる5対の相同染色体は、第一減数分裂中期(metaphase I)で赤道面に配列する(図7c)。第一減数分裂後期(anaphase I)で相同染色体は二極に分離し、第一減数分裂終期(telophase I)から第二減数分裂前期(prophase II)にかけて染色体はやや脱凝縮する(図7d)。2組の各5本の染色体は、第二減数分裂中期(metaphase II)において異なる赤道面に配列し(図7i)、第二減数分裂の後期(anaphase II)で2回目の染色体の分離が行われる。第二減数分裂終期(telophase II)は減数分裂の最後のステージであり、四分子と呼ばれる4つの半数性の細胞が形成される(図7j)。UVI4-like遺伝子のノックアウトアリルのホモ接合体では、第一減数分裂においては野生型との顕著な差異は見出せなかった。つまり、相同染色体間の対合(図7e)及びキアズマの形成(図7f)は正常と考えられた。また、5対の二価染色体は赤道面に配列した後(図7g)、二極に分離した(図7h)。しかしながら、第二減数分裂に相当する染色体像は観察されなかった。第一減数分裂終期から第二減数分裂前期にかけて、野生型では染色体がやや脱凝縮するが(図7d)、ホモ接合体ではこのような脱凝縮した染色体は稀であり、多くは図7kに示したように凝縮した染色体を持っていた。この凝縮した染色体は、野生型の第二減数分裂終期における染色体の様相(図7j)と類似していることから、ホモ接合体では第二減数分裂を行わずに減数分裂を終えてしまうことが示唆された。
【0029】
減数分裂の進行について調査するため、減数分裂の各ステージに属する細胞の割合を調査した(図8)。野生型ではzygotene期及びpachytene期の細胞の割合が多く、また、第二減数分裂終期の細胞の割合も多かった。第一及び第二減数分裂の中期ならびに後期の細胞の割合は低かった(全体の1%未満)。UVI4-like遺伝子のノックアウトアリルのホモ接合体では、第一減数分裂前期(prophase I)における分布は野生型のものと類似していた。しかしながら、第一減数分裂中期にある細胞の割合(全体の4.1%)は野生型の同ステージ(0.5%)に比べて著しく高かった。第二減数分裂の細胞は観察されなかった。これらの結果から、ホモ接合体では第一減数分裂中期から後期にかけて減数分裂の進行が遅延することが示唆された。
【0030】
細胞分裂の中期から後期への移行は、Anaphase Promoting Complex(APC)と呼ばれるタンパク複合体が重要な役割を果たすことが知られている(Eloy et al. (2006) Cell Cycle 5, 1957-1965)。APCは分裂サイクリンを標的としてこれを分解させる働きがあり、結果として分裂サイクリンーサイクリン依存性キナーゼの複合体を不活化することにより、後期への移行を促進する。また、分裂サイクリンーサイクリン依存性キナーゼ活性の適切な制御が、第一減数分裂から第二減数分裂への移行に重要であると考えられている(Pesin and Orr-Weaver (2008) Annu. Rev. Cell Dev. Biol. 24, 475-499)。以上のことから、UVI4-likeの変異によってAPCの活性が影響を受けている可能性が示唆される。
【特許請求の範囲】
【請求項1】
SEQ ID NO: 1にそのcDNA塩基配列を記載したUVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物。
【請求項2】
雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成する請求項1に記載の植物。
【請求項3】
雄性側での非還元配偶子が100%に近い頻度で形成されることを特徴とする請求項1に記載の植物
【請求項4】
自殖により約2倍のゲノム量を持つ後代種子が得られる請求項1に記載の植物。
【請求項5】
アブラナ科の植物である請求項1に記載の植物。
【請求項6】
シロイヌナズナである請求項5に記載の植物。
【請求項1】
SEQ ID NO: 1にそのcDNA塩基配列を記載したUVI4-like遺伝子または他種ホモログ遺伝子の機能が低下または欠失した植物。
【請求項2】
雄性側(花粉)および雌性側(卵)の両方において生殖能力のある非還元配偶子を形成する請求項1に記載の植物。
【請求項3】
雄性側での非還元配偶子が100%に近い頻度で形成されることを特徴とする請求項1に記載の植物
【請求項4】
自殖により約2倍のゲノム量を持つ後代種子が得られる請求項1に記載の植物。
【請求項5】
アブラナ科の植物である請求項1に記載の植物。
【請求項6】
シロイヌナズナである請求項5に記載の植物。
【図1】


【図4】
【図8】
【図2】
【図3】
【図5】
【図6】
【図7】
【図4】
【図8】
【図2】
【図3】
【図5】
【図6】
【図7】
【公開番号】特開2010−154848(P2010−154848A)
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願番号】特願2009−270452(P2009−270452)
【出願日】平成21年11月27日(2009.11.27)
【出願人】(505374783)独立行政法人 日本原子力研究開発機構 (727)
【Fターム(参考)】
【公開日】平成22年7月15日(2010.7.15)
【国際特許分類】
【出願日】平成21年11月27日(2009.11.27)
【出願人】(505374783)独立行政法人 日本原子力研究開発機構 (727)
【Fターム(参考)】
[ Back to top ]