
VDCCガンマ−8イオンチャネル
配列番号3、配列番号5、または配列番号7に示されるアミノ酸配列、並びにそれらのホモログ、バリアント、及び誘導体を含むVDCCγ-8ポリペプチドが提供される。VDCCγ-8ポリペプチドをコードし得る核酸、特に配列番号1、配列番号2、配列番号4、または配列番号6に示される核酸配列を含むものも開示される。これらのポリペプチド及びポリヌクレオチドは精神疾患に関与しており、精神疾患を治療するのに有用な治療薬のスクリーニングに有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、イオンチャネル核酸及びポリペプチドに関する。イオンチャネルは、以後「電位依存性カルシウムチャネルガンマ-8」、「VDCCγ-8」、または「γ-8」と称される。本発明はまた、これらの核酸及びポリペプチドの作用を抑制するかまたは活性化することに関するものであり、特に精神疾患の治療のための候補薬物を提供することに関する。
【背景技術】
【0002】
カルシウムチャネルは、排他的ではないが、筋収縮、神経伝達物質放出、ホルモン分泌、遺伝子発現、細胞分裂周期、及び細胞死を介する発達等、広範囲の細胞機能の制御において重要である。細胞内カルシウムは、静止状態において極めて低く、細胞内カルシウムが一過性に上昇するような変化は、第二のメッセンジャーとして作用して、多種多様の範囲の細胞過程を活性化することができる。電位またはリガンド依存性カルシウムチャネルは、細胞内カルシウムの増加を媒介できる。電位依存性カルシウムチャネルは、2つの広範な群:低(またはTタイプ)及び高閾値活性化チャネルに分けることができる。高閾値チャネルは、それらのサブユニット成分によって、L、N、P、Q、及びRチャネルに細分される。
【0003】
電位依存性カルシウムチャネルは、ポア形成アルファ-1サブユニット及びアクセサリサブユニットベータ、及びアルファ-2-デルタを含むヘテロチャネルである。いくつかの状況において、ガンマサブユニットもまた存在する(Caterall、Curr. Opin Neurobiol. 1991年:1;5〜13頁)。ガンマサブユニットタンパク質は、不活性(閉鎖)状態においてカルシウムチャネルを安定化すると考えられる内在性細胞膜内タンパク質である。いくつかのガンマサブユニットタンパク質があり、このVDCCγ-8遺伝子は、2つの同様のガンマサブユニットコード化遺伝子を有するクラスターに配置される、神経カルシウムチャネルガンマサブユニット遺伝子サブファミリーのPMP-22/EMP/MP20ファミリーのメンバーである。非AUG翻訳開始は、VDCCγ-8転写物に対して生じ得ると考えられている。VDCCγ-8サブユニットは、シナプス後肥厚-95、ディスクスラージ、閉鎖帯(PDZ)結合モチーフを含有する。VDCCγ-8は、スタルガジン(stargazin)及びVDCC-γ3と同様に、発達後期に現れ、動物の成熟期に進行的に増加する(胚発育大脳皮質には見られない、Tomitaら、J Cell Biol、2003年:161;805〜816頁を参照)。これは、胎芽期に発現し、動物成熟期に徐々に減少するVDCC-γ4に関連する。
【0004】
スタルガジン、γ-3、及びγ-4とともにγ-8は、AMPA受容体の表面発現(スタルガザー(stargazer)マウスからの小脳細胞におけるGluRの表面発現は〜75%減少するが、一方で、全GluRレベルは10〜20%のみ減少する)を媒介する膜貫通型AMPA制御タンパク質(TARPS)のファミリーに属する。
【0005】
精神疾患は、脳障害、特に感情または情緒不安定性、行動異常、及び/または認知障害を伴うものを定義するために用いられる用語である。精神疾患は、罹患者を衰弱させ、かつかなりの情緒的及び財政的負担を受けながら看護する必要がある。
【0006】
精神病は、重度に障害を受けた思考及び知覚を有する精神状態を定義する用語である。精神病の発症は、衰弱させ、通常、幻覚、偏執症、妄想、思考混乱、及び社会的相互作用の低下を含む。精神病は、一般に、罹患者に対して高度に破壊的となり得る現実との接触の喪失を含む。精神病は、双極性障害、重度の臨床的うつ病及び統合失調症を含む症状に現れる。これらの症状は、十分に理解されておらず、それらを予防及び治療する有効な療法を必要とされている。特に、陰性症状の統合失調症の治療において、かなりの満たされていない医学的ニーズがある。最も一般に用いられる薬物のクロザピン及びオランザピンは、十分効果的になるまでに2〜4週間かかり、統合失調症の陰性症状に対しての効果が比較的弱い。
【0007】
てんかんは、再発性てんかん発作を特徴とする発達障害ともしばしば称される精神病である。この慢性的な神経学的病態は、世界で数百万の人々に影響を及ぼしており、この症状を制御する薬物は存在するが、治療法はない。
【特許文献1】米国特許第5,492,840号
【特許文献2】米国特許第5,482,835号
【特許文献3】WO第84/03564号
【特許文献4】EP-A-239400
【特許文献5】米国特許第5,545,807号
【特許文献6】米国特許第6,075,181号
【特許文献7】米国特許第4,946,779号
【特許文献8】米国特許第4,946,778号
【非特許文献1】Caterall、Curr. Opin Neurobiol. 1991年:1;5〜13頁
【非特許文献2】Tomitaら、J Cell Biol、2003年:161;805〜816頁
【非特許文献3】J. Sambrook、E. F. Fritsch、及びT. Maniatis、1989年、Molecular Cloning:A Laboratory Manual、第二版、Books 1〜3、Cold Spring Harbor Laboratory Press
【非特許文献4】Ausubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州)
【非特許文献5】B. Roe、J. Crabtree、及びA. Kahn、1996年、DNA Isolation and Sequencing:Essential Techniques、John Wiley & Sons
【非特許文献6】J. M. Polak及びJames O'D. McGee、1990年、In Situ Hybridization:Principles and Practice;Oxford University Press
【非特許文献7】M. J. Gait(編集者)、1984年、Oligonucleotide Synthesis:A Practical Approach、Irl Press
【非特許文献8】D. M. J. Lilley及びJ. E. Dahlberg、1992年、Methods of Enzymology:DNA Structure Part A:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Press
【非特許文献9】Edward Harlow、David Lane、Ed HarlowによるUsing Antibodies:A Laboratory Manual:Portable Protocol NO. 1(1999年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-544-7)
【非特許文献10】Ed Harlow(編集者)、David Lane(編集者)によるAntibodies:A Laboratory Manual(1988年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-314-2)、1855
【非特許文献11】Lars-Inge Larsson「Immunocytochemistry:Theory and Practice」、CRC Press Inc.、Baca Raton、フロリダ州、1988年、ISBN 0-8493-6078-1、John D. Pound(編集者)
【非特許文献12】John D. Pound(編集者);「Methods in Molecular Biology」のシリーズにおける「Immunochemical Protocols、80巻」、Humana Press、Totowa、ニュージャージー州、1998年、ISBN 0-89603-493-3
【非特許文献13】Handbook of Drug Screening、Ramakrishna Seethala、Prabhavathi B. Fernandes(2001年、ニューヨーク、ニューヨーク州、Marcel Dekker、ISBN 0-8247-0562-9)
【非特許文献14】Lab Ref:A Handbook of Recipes, Reagents, and Other Refernce Tools for Use at the Bench、Jane Roskams及びLinda Rodgers編集、2002年、Cold Spring Harbor Laboratory、ISBN 0-87969-630-3
【非特許文献15】The Merck Manual of Diagnosis and Therapy(第17版、編集者Beers, M.H.及びBerkow, R、ISBN 0911910107、John Wiley & Sons)
【非特許文献16】http://www.sanger.ac.uk/Software/Pfam/search.shtml
【非特許文献17】Carlsson A. 1988年、The current status of the dopamine theory of schizophrenia. Neuropharmacology 1、179〜186頁
【非特許文献18】Adlardら(2005)J. Neurosci. 25:4217〜21頁
【非特許文献19】Limら(2005)J Neurosci 25:3032〜40頁
【非特許文献20】Proteins-Structure and Molecular Properties、第二版、T. E. Creighton、W. H. Freeman and Company、ニューヨーク、1993年
【非特許文献21】Wold, F.、Posttranslational Protein Modifications:Perspectives and Prospects、Posttranslational Covalent Modification of Proteins の1〜12頁、B. C. Johnson編集、Academic Press、ニューヨーク、1983年
【非特許文献22】Seifterら、「Analysis for protein modifications and nonprotein cofactors」、Meth Enzymol(1990)182:626〜646頁
【非特許文献23】Rattanら、「Protein Synthesis:Posttranslational Modifications and Aging」、Ann NY Acad Sci(1992)663:48〜62頁
【非特許文献24】Devereuxら、1984年、Nucleic Acids Research 12:387頁
【非特許文献25】Atschulら、1990年、J Molec Biol 403〜410頁
【非特許文献26】Ausubelら、1999年 Nucleic Acids Research-18章
【非特許文献27】Ausubelら、1999年、J. Mol. Biol.、7〜58頁から7〜60頁
【非特許文献28】http://www.ncbi.nih.gov/BLAST/blast_help.html
【非特許文献29】Karlin及びAltschul 1990年、Proc. Natl. Acad. Sci. USA 87:2264〜68頁
【非特許文献30】Karlin及びAltschul、1993年、Proc. Natl. Acad. Sci. USA 90:5873〜7頁
【非特許文献31】Altschulら(1994)Nature Genetics 6:119〜129頁
【非特許文献32】http://www.ncbi.nlm.nih.gov/BLAST
【非特許文献33】Coombs J(1994)Dictionary of Biotechnology、Stockton Press、ニューヨーク、ニューヨーク州)
【非特許文献34】Dieffenbach CW及びGS Dveksler(1995年、PCR Primer, a Laboratory Manual、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)
【非特許文献35】Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)
【非特許文献36】Sambrook、上記文献、7章
【非特許文献37】Ausubel, F. M.ら、上記文献、4章及び16章
【非特許文献38】Sambrook, J.ら、(1989年、Molecular Cloning: A Laboratory Manual、4章、8章、及び16〜17章、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)
【非特許文献39】Van Heeks, G.及びS. M. Schuster(1989)J. Biol. Chem. 264:5503〜5509頁
【非特許文献40】Grantら(1987年;Methods Enzymol. 153:516〜544頁)
【非特許文献41】Takamatsu, N.(1987)EMBO J. 6:307〜311頁
【非特許文献42】Coruzzi, G.ら(1984)EMBO J. 3:1671〜1680頁
【非特許文献43】Broglie, R.ら(1984)Science 224:838〜843頁
【非特許文献44】Winter, J.ら(1991)Results Probl. Cell Differ. 17:85〜105頁
【非特許文献45】Hobbs, S.またはMurry, L. E.、McGraw Hill Yearbook of Science and Technology(1992)McGraw Hill、ニューヨーク、ニューヨーク州;191〜196頁
【非特許文献46】Engelhard, E. K.ら(1994)Proc. Nat. Acad. Sci. 91:3224〜3227頁
【非特許文献47】Logan, J.及びT. Shenk(1984)Proc. Natl. Acad. Sci. 81:3655〜3659頁
【非特許文献48】Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125〜162頁
【非特許文献49】Wigler, M.ら(1977)Cell 11:223〜32頁
【非特許文献50】Lowy, I.ら(1980)Cell 22:817〜23頁
【非特許文献51】Wigler, M.ら(1980)Proc. Natl. Acad. Sci. 77:3567〜70頁)
【非特許文献52】Colbere-Garapin, F.ら(1981)J. Mol. Biol. 150:1〜14頁
【非特許文献53】Hartman, S. C.及びR. C. Mulligan(1988)Proc. Natl. Acad. Sci. 85:8047〜51頁
【非特許文献54】Rhodes, C. A.ら(1995)Methods Mol. Biol. 55:121〜131頁
【非特許文献55】Hampton, R.ら(1990年;Serological Methods, a Laboratory Manual, Section IV、APS Press、セントポール、ミネソタ州)
【非特許文献56】Maddox, D. E.ら(1983年;J. Exp. Med. 158:1211〜1216頁)
【非特許文献57】Porath, J.ら(1992)Prot. Exp. Purif. 3:263〜281頁
【非特許文献58】Kroll, D. J.ら(1993年;DNA Cell Biol. 12:441〜453頁)
【非特許文献59】Merrifield J.(1963)J. Am. Chem. Soc. 85:2149〜2154頁
【非特許文献60】Aizawa(1988)、Anal. Chem. Symp. 17:683頁
【非特許文献61】Wattら、J Biol Chem、1998年5月29日;273(22):14053〜8頁
【非特許文献62】Hoffmanら、Proc Natl Acad Sci USA 、2000年10月10日;97(21):11215〜20頁
【非特許文献63】Figlerら、Biochemistry、1997年12月23日;36(51):16288〜99頁
【非特許文献64】Sarrioら、2000年、Mol Cell Biol 2000年7月;20(14):5164〜74頁
【非特許文献65】Coliganら、Current Protocols in Immunology 1(2):5章(1991)
【非特許文献66】Blundell及びJohnson(1976)Protein Crystallography、Academic Press、ニューヨーク
【非特許文献67】Parceら(1989)Science 246:243〜247頁
【非特許文献68】Owickiら(1990)Proc. Nat'l Acad. Sci. USA 87;4007〜4011頁
【非特許文献69】Jaenich, R.(1976)PNAS 73:1260〜1264頁
【非特許文献70】Manipulating the Mouse Embryo、Hogan編集(Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、1986年)
【非特許文献71】Jahnerら(1985)PNAS 82:6927〜6931頁
【非特許文献72】Van der Puttenら(1985)PNAS 82:6148〜6152頁
【非特許文献73】Stewartら(1987)EMBO J. 6:383〜388頁
【非特許文献74】Jahnerら(1982)Nature 298:623〜628頁
【非特許文献75】Evansら(1981)Nature 292:154〜156頁
【非特許文献76】Bradleyら(1984)Nature 309:255〜258頁
【非特許文献77】Gosslerら(1986)PNAS 83:9065〜9069頁
【非特許文献78】Robertsonら(1986)Nature 322:445〜448頁
【非特許文献79】Jaenisch, R.(1988)Science 240:1468〜1474頁
【非特許文献80】Koehler及びMilstein(1975年Nature 256:495〜497頁)
【非特許文献81】Kosborら(1983)Immunol Today 4:72頁
【非特許文献82】Coteら(1983)Proc Natl Acad Sci 80:2026〜2030頁
【非特許文献83】Coleら、Monoclonal Antibodies and Cancer Therapy、77〜96頁、Alan R. Liss社、1985年
【非特許文献84】Morrisonら(1984)Proc Natl Acad Sci 81:6851〜6855頁
【非特許文献85】Neubergerら(1984)Nature 312:604〜608頁
【非特許文献86】Takedaら(1985)Nature 314:452〜454頁
【非特許文献87】Orlandiら(1989年、Proc Natl Acad Sci 86:3833〜3837頁)
【非特許文献88】Winter G及びMilstein C(1991年;Nature 349:293〜299頁)
【非特許文献89】Huse WDら(1989)Science 256:1275〜1281頁
【非特許文献90】Myersら、Science(1985)230:1242頁
【非特許文献91】Cottonら、Proc Natl Acad Sci USA(1985)85:4397〜4401頁
【非特許文献92】M. Cheeら、Science、274巻、610〜613頁(1996)
【非特許文献93】Oritaら(1989)Proc Natl. Acad. Sci USA:86:2766頁
【非特許文献94】Cotton (1993)Mutat Res 285:125〜144頁
【非特許文献95】Hayashi(1992)Genet Anal Tech Appl 9:73〜79頁
【非特許文献96】Keenら(1991)Trends Genet 7:5頁
【非特許文献97】O'Connor、J Neurochem(1991)56:560頁
【非特許文献98】Leeら、Nucleic Acids Res(1979)6:3073頁
【非特許文献99】Cooneyら、Science(1988)241:456頁
【非特許文献100】Dervanら、Science(1991)251:1360頁
【非特許文献101】T. Strachan及びA P Read、Human Molecular Geneticsにおける20章、Gene Therapy and other Molecular Genetic-based Therapeutic Approaches,(及びその中に引用されている文献)、BIOS Scientific Publishers社(1996)
【非特許文献102】Remington's Pharmaceutical Sciences、Mack Publishing社(A. R. Gennaro編集、1985年)
【非特許文献103】Russら、Nature 2000年3月2日;404(6773):95〜99頁
【非特許文献104】Mead及びStephens(2003)J. Neurosci. 23:1041〜1048頁
【非特許文献105】Bakshi VP、Swerdlow NR及びGeyer MA、Clozapine antagonizes phencyclidine-induced deficits in sensorimotor gating of the startle response、J. Pharmcol. Exp. Ther. 19941年;271:787〜794頁
【非特許文献106】McGhie A及びChapman J. Disorder of attention and perception in early schizophrenia、Br. J. Med. Psychol. 1961年;34:103〜116頁
【非特許文献107】Braff DL、Geyer MA及びSwerdlow NR、Human studies of prepulse inhibition of startle:normal subjects, patient groups, nd pharmacological studies、Psychopharmacology、2001年;156:234〜258頁
【発明の開示】
【発明が解決しようとする課題】
【0008】
精神疾患を予防及び治療するための有効な治療法、特に抗精神病及び抗てんかん療法に対する強いニーズが依然としてある。
【課題を解決するための手段】
【0009】
本発明は、VDCCγ-8が精神疾患に関与しており、ゆえに有用な治療標的であるという驚くべき発見に基づいている。特に、VDCCγ-8は、統合失調症及びてんかん等の精神病を含む疾患に関与している。
【0010】
本発明の第一の態様によれば、精神疾患の治療に適した薬剤を同定する方法は、候補薬剤が電位依存性カルシウムチャネルガンマ-8の活性に影響を及ぼすかどうかを判定する工程を含む。
【0011】
[配列表]
配列番号1は、ヒトVDCCγ-8のcDNA配列を示す。
配列番号2は、配列番号1に由来するオープンリーディングフレームを示す。
配列番号3は、配列番号2に由来するヒトVDCCγ-8のアミノ酸配列を示す。
配列番号4は、マウスVDCCγ-8に関するcDNAのオープンリーディングフレームを示す。
配列番号5は、マウスVDCCγ-8のアミノ酸配列を示す。
配列番号6は、ヒトVDCCγ-8に関するcDNAの代替のオープンリーディングフレームを示す。
配列番号7は、配列番号6に由来するヒトVDCCγ-8のアミノ酸配列を示す。
配列番号8〜20は、ノックアウトプラスミドを構築するために用いられた遺伝子型決定プライマーを示す。
配列番号21(図7)は、プライマーの位置及び標的ストラテジーに用いられたVDCCγ-8座領域のゲノム構造を示す。
【発明を実施するための最良の形態】
【0012】
[VDCCγ-8イオンチャネル]
VDCCγ-8イオンチャネル、及びそのホモログ、バリアント、または誘導体、並びにVDCCγ-8関連疾患の治療及び診断におけるその使用を本明細書に記載する。
【0013】
VDCCγ-8は、ヒトVDCCγ-8をコードする増幅cDNA産物を配列決定した結果によって示されるように、イオンチャネルファミリーの他のタンパク質に構造的に関連している。配列番号1のcDNA配列は、2つの可能なオープンリーディングフレーム:425個のアミノ酸のポリペプチド(配列番号3として示される)をコードするヌクレオチド番号104〜1381(配列番号2として示される)と、391個のアミノ酸のポリペプチド(配列番号7として示される)をコードするヌクレオチド番号206〜1381(配列番号6として示される)とを含む。ヒトVDCCγ-8は、ヒト染色体Ch19q13に位置することが分かっている。
【0014】
他に示されない限り、本発明の実施は、通常の当業者の能力の範囲内にある従来の化学、分子生物学、微生物学、組換えDNA、及び免疫学の技術を使用する。このような技術は、文献に説明されている。例えば、J. Sambrook、E. F. Fritsch、及びT. Maniatis、1989年、Molecular Cloning:A Laboratory Manual、第二版、Books 1〜3、Cold Spring Harbor Laboratory Press;Ausubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州);B. Roe、J. Crabtree、及びA. Kahn、1996年、DNA Isolation and Sequencing:Essential Techniques、John Wiley & Sons;J. M. Polak及びJames O'D. McGee、1990年、In Situ Hybridization:Principles and Practice;Oxford University Press;M. J. Gait(編集者)、1984年、Oligonucleotide Synthesis:A Practical Approach、Irl Press;D. M. J. Lilley及びJ. E. Dahlberg、1992年、Methods of Enzymology:DNA Structure Part A:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Press;Edward Harlow、David Lane、Ed HarlowによるUsing Antibodies:A Laboratory Manual:Portable Protocol NO. 1(1999年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-544-7);Ed Harlow(編集者)、David Lane(編集者)によるAntibodies:A Laboratory Manual(1988年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-314-2)、1855;Lars-Inge Larsson「Immunocytochemistry:Theory and Practice」、CRC Press Inc.、Baca Raton、フロリダ州、1988年、ISBN 0-8493-6078-1;John D. Pound(編集者)、「Methods in Molecular Biology」のシリーズにおける「Immunochemical Protocols、80巻」、Humana Press、Totowa、ニュージャージー州、1998年、ISBN 0-89603-493-3;Handbook of Drug Screening、Ramakrishna Seethala、Prabhavathi B. Fernandes(2001年、ニューヨーク、ニューヨーク州、Marcel Dekker、ISBN 0-8247-0562-9);Lab Ref:A Handbook of Recipes, Reagents, and Other Refernce Tools for Use at the Bench、Jane Roskams及びLinda Rodgers編集、2002年、Cold Spring Harbor Laboratory、ISBN 0-87969-630-3;並びにThe Merck Manual of Diagnosis and Therapy (第17版、編集者Beers, M.H.及びBerkow, R、ISBN:0911910107、John Wiley & Sons)を参照されたい。これら各一般テキストは、参照として本明細書に組み込まれている。
【0015】
[VDCCγ-8に対する同一性及び類似性]
pfam(http://www.sanger.ac.uk/Software/Pfam/search.shtml)のHMM構造予測ソフトウェアを用いたVDCCγ-8ポリペプチド(配列番号3)の解析により、VDCCγ-8ペプチドがイオンチャネルであることを確認する。
【0016】
ヒトVDCCγ-8イオンチャネルのマウスホモログをクローニングし、その核酸配列及びアミノ酸配列を、それぞれ配列番号4及び配列番号5として示す。マウスVDCCγ-8イオンチャネルcDNA (配列番号4)は、ヒトVDCCγ-8イオンチャネル(配列番号2及び配列番号6)配列と高度の同一性を示すが、一方、マウスVDCCγ-8イオンチャネルのアミノ酸配列(配列番号5)は、ヒトVDCCγ-8イオンチャネル(配列番号3及び配列番号7)と高度の同一性及び類似性を示す。
【0017】
従って、ヒト及びマウスVDCCγ-8イオンチャネルは、イオンチャネルの大型ファミリーのメンバーである。
【0018】
[VDCCγ-8の発現プロファイル]
VDCCγ-8 cDNAのポリメラーゼ連鎖反応(PCR)増幅により、VDCCγ-8の発現が海馬(+++)、小脳、皮質、線条体、中脳、脳橋(++)、視床下部、視床、前脳、脊髄、下垂体、及び三叉神経節(+)における存在量が変化していることが検出される。
【0019】
VDCCγ-8ノックアウトマウスのLacZ染色により、脳、特に海馬、皮質、扁桃体、嗅覚皮質、前脳、及び視床におけるVDCCγ-8の強い発現が示されている。
【0020】
VDCCγ-8 cDNAを用いて、BLASTNによりヒトまたはマウスESTデータ源をサーチすることによって、cDNAライブラリーにおいて同一性が見られる。このことは、VDCCγ-8が、以下の正常組織または異常組織において発現されていることを示している。
【0021】
登録番号CX223421、DN176885、CX205409:側脳室の側壁
登録番号BE647856、BE864111:マウス脳(小脳、脳幹、嗅球、視床下部、皮質、扁桃体、基底核、松果体、線条体、海馬の混合物由来)
登録番号BB641732:新生児皮質
登録番号BG803929:新生児網膜
【0022】
従って、VDCCγ-8ポリペプチド、核酸、プローブ、抗体、発現ベクター、及びリガンドは、これら及び他の組織におけるVDCCγ-8イオンチャネルの過剰発現、発現不足、及び発現異常に関連する疾患に対する検出、診断、治療、及び他のアッセイに有用である。好ましくは、該疾患は、下記に示されるVDCCγ-8関連疾患を含む。
【0023】
[VDCCγ-8イオンチャネル関連疾患]
本明細書に記載される方法及び組成物によれば、VDCCγ-8イオンチャネルは、様々な疾患を治療及び診断するための治療及び診断に有用である。VDCCγ-8は、下記に検討されるようにVDCCγ-8の活性を変化させる分子をアッセイする際にも有用である。VDCCγ-8を変化させる分子は、VDCCγ-8関連疾患に対して強力な治療薬である。VDCCγ-8が関与する疾患を、便宜上、VDCCγ-8関連疾患と称す。これらの疾患は精神疾患である。本明細書に用いられるように、精神疾患という用語は、感情的または情緒的不安定性、行動異常、及び/または認知機能不全もしくは認知機能障害を含む疾患を含んだ脳機能障害を表す。用語「精神疾患」はまた、発作を示す障害、例えばてんかんを含む。ヒト及び動物(獣医学的)治療及び診断は、本発明の範囲内に入る。
【0024】
ヒトVDCCγ-8は、ヒトの染色体19q13に位置する。従って、特定の実施形態において、Ch19q13イオンチャネルは、この遺伝子座、染色体バンド、領域、アーム、または同じ染色体に位置する疾患を治療または診断するために使用され得る。VDCCγ-8イオンチャネルの染色体遺伝子座(すなわち、ヒトの染色体19q13)と同じ遺伝子座、染色体バンド、領域、アーム、または染色体に結びつけられるものとして決定されている既知の疾患は、熱性発作、貧血、及び糖尿病とともに全般てんかんを含む。
【0025】
VDCCγ-8が欠失しているノックアウトマウスは、実施例に示されているように、様々な表現型を示す。例えば、LABORAS試験において、VDCCγ-8ノックアウトマウスは最初に機能亢進性であり、野生型コントロールマウス(実施例5)と比較して、作業記憶欠損の指標である新規な環境に対するより緩徐な馴化を示唆し、このことは変化したシナプス構成の症状であり得る。従って、我々は、個体におけるシナプス可塑性を変化させる方法を開示し、該方法は、その個体におけるVDCCγ-8のレベルまたは活性を調節することを含む。他で記載されているように、これは、VDCCγ-8の発現を調節することによって、またはVDCCγ-8のモジュレーター(アゴニストまたはアンタゴニスト等)の使用によって達成され得る。
【0026】
VDCCγ-8、及び特にVDCCγ-8のアゴニストを含むVDCCγ-8活性のモジュレーターは、関連する脳領域のシナプス可塑性を特徴とする疾患または症候を治療または緩和させるために使用され得る。このような疾患には、てんかん重積症、全般性発作、及び発作を示すすべての障害、自閉症等の小児性及び発達障害、疾患の自閉症スペクトル、広汎性発達障害、崩壊性障害、アスペルガー症候群、レット症候群、注意欠陥/多動性障害(ADHD)、排他的ではないが、アルツハイマー病、虚血性/血管性認知症、ピック病、広汎性レビー小体認知症、前頭側頭認知症、大脳皮質基底核変性症、ハンチントン病、進行性核上性麻痺、AIDS/HIV認知症、プリオン感染症、脳炎、神経梅毒、血管炎、進行性多病巣性白質脳障害を含む認知障害、並びに統合失調症、全般性不安障害、社会性不安、外傷後ストレス障害、恐怖症、社会的恐怖症、特定恐怖症、パニック障害、強迫性障害、急性ストレス障害、分離障害及びうつ病、大うつ病、気分変調、双極性障害、季節性情動障害、出産後うつ病、急性神経変性疾患、脳卒中、及び外傷性脳傷害を含む精神病が含まれる。
【0027】
好ましい実施形態において、VDCCγ-8関連疾患は、ストレスまたは不安が症状である疾患を含む。
【0028】
さらに好ましい実施形態において、VDCCγ-8関連疾患は、精神病が症状であるか、または症状であり得る疾患を含む。精神病は、一般に、重度の精神疾患の症状であると考えられているが、それ自体は診断ではない。精神病と関連する、または関連し得る任意の精神疾患は、本発明の範囲内にある。精神病に関与する好ましい疾患は、統合失調症、双極性障害(躁うつ病)及び重度の臨床的うつ病である。脳傷害(または他の神経疾患)、薬物中毒及び禁断症状(特にアルコール、バルビツール類及びベンゾジアゼピン類)、狼瘡、高齢者の電解質障害(尿路感染症)、疼痛症候群、睡眠うつ病及び極限ストレス(外傷後ストレス障害等)もまた、精神病の発症を引き起こす可能性があり、本発明の範囲内にある。
【0029】
さらに別の好ましい実施形態において、VDCCγ-8関連疾患は、全般てんかん及びてんかん重積症を含む発作を示す障害である。
【0030】
最も好ましいVDCCγ-8関連疾患は、精神病、統合失調症、ADHD、双極性障害、大うつ病、全般性不安障害、及びてんかん重積症を含むてんかんである。
【0031】
本明細書に詳述された実験データは、統合失調症におけるVDCCγ-8に強く関係する。海馬におけるGluR2受容体の発現の亢進及び低下(実施例5)は、VDCCγ-8KOマウスが、前脳グルタミン酸作動性欠損及び統合失調症の陰性態様に等しい症状を示すことを強く表している。新規な環境の特定マップを作成する能力は、KOマウスにおいて損なわれており(実施例5)、探索行動の時間の増加をもたらす。このことは、Carlsson(Carlsson A. 1988年、The current status of the dopamine theory of schizophrenia. Neuropharmacology 1、179〜186頁)により提案された統合失調症のグルタミン酸作動性仮説と一致して、統合失調症におけるVDCCγ-8の役割を示している。
【0032】
KOマウスはまた、社会的相互作用の欠如を示す(実施例6)。これらのマウスは不安レベルのいずれの変化も示さず、ゆえに、社会的相互作用の欠如は、上記統合失調症の陰性症状と類似した表現型を示唆している。KOマウスはまた、VDCCγ8(TARPγ8)が統合失調症の治療において有用性を有するという極めて強い証拠である、有意なプレパルス抑制破壊を示す(実施例7)。
【0033】
GluR2発現の低下、亢進、社会的相互作用の欠如、及びPPI欠損のすべては、KOマウスが、グルタミン酸作動性NMDA受容体アンタゴニストの投与後、野生型マウスに観察されるものと同様の表現型を有することを含む。これらは、ヒトにおいて精神異常症状を引き起こすことが知られており、学術的研究及び創薬プログラムにおける統合失調症の化学モデルとして常用されている。従って、好ましい実施形態において、VDCCγ-8関連疾患は統合失調症である。
【0034】
上記のとおり、VDCCγ-8イオンチャネルは、本明細書に記載された任意の方法及び組成物を用いることによって、これら特定の疾患のいずれかを診断及び/または治療するために使用され得る。
【0035】
特に、上記に挙げた特定の疾患の治療または診断のために、ホモログ、バリアント、または誘導体を含むVDCCγ-8イオンチャネル核酸、ポリペプチドを含む核酸、ベクターの使用、VDCCγ-8イオンチャネル核酸及び/またはポリペプチドを含む医薬組成物、宿主細胞、及びトランスジェニック動物の使用が意図されている。さらに、上記の特定の疾患の診断または治療において、VDCCγ-8イオンチャネルと相互作用または結合し得る化合物、好ましくはVDCCγ-8のアンタゴニスト、好ましくは該チャネルの伝導性を増大し得る化合物、VDCCγ-8イオンチャネルに対する抗体、並びにこれらを作製し、または同定する方法は本発明の範囲内に入る。特に、特定の疾患の治療または予防のためのワクチンの製造における、これら任意の化合物、組成物、分子等の使用が意図されている。個体の特定における疾患の検出用診断キットもまた、本発明の範囲内にある。
【0036】
VDCCγ-8イオンチャネルの使用によって治療または診断可能な上記のまたはさらなる特定の疾患を同定するための連鎖地図作成の方法は、当該技術分野において既知であり、また本明細書のいずれかに記載されている。
【0037】
[神経興奮、発作、及び認知障害]
神経興奮、発作、及び認知障害、並びに膜貫通型AMPA制御タンパク質(TARP)関連疾患を含むこれらが現れる障害は、当該技術分野において周知である。概要説明は以下のとおりである。
【0038】
神経興奮は、脳細胞の電気的活性化として定義でき、神経興奮は、脳の正常な機能の一部であるか、または機能障害、疾患、または傷害に関連する異常活性の結果でもあり得る。
【0039】
発作は、精神状態の変化、強直性または間代性動作、及び種々の他の症状の中にそれら自体を現す脳機能の一時的変化である。それらは、脳細胞群の一時的な異常神経興奮に起因する。発作は、オンオフ脳侵襲(例えば毒性、衝撃)から、または組織損傷/変性/機能障害の結果としての反復症状として生じ得る。
【0040】
認知は、人が思考または知覚の対象を自覚する精神の働きとして定義でき、それは、知覚、思考、及び記憶のすべての態様を含む。認知障害は、機能障害、疾患、または傷害の結果としての神経興奮の破壊による、上記の働きの破壊である。
【0041】
VDCCγ-8イオンチャネルモジュレーターを、これらの症状のいずれかを治療または軽減するために使用できる。
【0042】
ライフスタイル及び食事療法もまた、神経興奮性、発作、及び/または認知作用を変えるために役立ち得る:1)運動はアルツハイマー病の発症率を減じ得る(アミロイド負荷及び学習欠損の両方に関して、Adlardら(2005)J. Neurosci. 25:4217〜21頁);2)食事のオメガ-3ポリ不飽和脂肪酸、ドコサヘキサエン酸(DHA)は、βアミロイドの産生、蓄積、及び下流毒性の可能性を保護しうるであろう(Limら(2005)J Neurosci 25:3032〜40頁)。我々は、それらの神経興奮性、発作、及び/または認知作用を緩和させるようなライフスタイル処置と組み合わせてVDCCγ-8イオンチャネル及びそのモジュレーターの使用を開示する。
【0043】
レクリエーション及び薬用の両方のある一定の薬物は、薬物からの副作用または禁断症状として神経興奮性、発作、及び/または認知の変化となり得る。このような薬物としては、カフェイン、大麻、アルコール、ニコチン、風邪薬、抗ヒスタミン薬、コカイン、及びアンフェタミンが挙げられる。我々は、それらの神経興奮性、発作、及び/または認知作用を緩和させるような薬物と組み合わせた、VDCCγ-8イオンチャネル及びそのモジュレーターの使用を開示する。
【0044】
[VDCCγ-8イオンチャネルポリペプチド]
本明細書に用いられる用語「VDCCγ-8イオンチャネルポリペプチド」とは、配列番号3、配列番号5、もしくは配列番号7に示されたアミノ酸配列、またはそれらのホモログ、バリアント、または誘導体を含むポリペプチドを指すものとする。該ポリペプチドは、配列番号3に示された配列のホモログ、バリアント、または誘導体を含むか、あるいはそれらであることが好ましい。
【0045】
「ポリペプチド」とは、ペプチド結合または修飾ペプチド結合、すなわちペプチドアイソスターにより互いに結合された2つ以上のアミノ酸を含む任意のペプチドまたはタンパク質のことである。「ポリペプチド」とは、通常、ペプチド、オリゴペプチド、またはオリゴマーと称される短鎖、一般にタンパク質と称される長鎖のことである。ポリペプチドは、20の遺伝子コード化アミノ酸以外のアミノ酸を含有することができる。
【0046】
「ポリペプチド」は、翻訳後処理等の天然プロセシング、または当業者によく知られている化学修飾技術により修飾されたアミノ酸配列を含む。このような修飾は、基礎的テキスト及びより詳細な単行書、並びに多数の研究文献に十分に記載されている。修飾は、ペプチド主鎖、アミノ酸側鎖、及びアミノまたはカルボキシル末端等、ポリペプチドのいずれかの場所で生じ得る。同じタイプの修飾は、所与のポリペプチドのいくつかの部位で同じかまたは変化段階で存在し得ることを認識するであろう。また、所与のポリペプチドは、多くのタイプの修飾を含むことができる。
【0047】
ユビキチン結合の結果、ポリペプチドは分枝でき、それらは、分岐の有り無しで環式化できる。環式、分枝状、及び分枝状環式ポリペプチドは、翻訳後天然プロセシングから生じ得るか、または合成法により作製できる。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスホチジルイノシトールの共有結合、架橋化、環化、ジスルフィド結合形成、脱メチル化、共有架橋形成、シスチン形成、ピログルタメート形成、ホルミル化、ガンマ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨード化、メチル化、ミリストイル化、酸化、タンパク分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニン化等のアミノ酸のタンパク質へのトランスファーRNA媒介付加、及びユビキチン結合が挙げられる。例えば、Proteins-Structure and Molecular Properties、第二版、T. E. Creighton、W. H. Freeman and Company、ニューヨーク、1993年、及びWold, F.、Posttranslational Protein Modifications:Perspectives and Prospects、Posttranslational Covalent Modification of Proteinsの1〜12頁、B. C. Johnson編集、Academic Press、ニューヨーク、1983年;Seifterら、「Analysis for protein modifications and nonprotein cofactors」、Meth Enzymol(1990)182:626〜646頁及びRattanら、「Protein Synthesis:Posttranslational Modifications and Aging」、Ann NY Acad Sci(1992)663:48〜62頁、を参照されたい。
【0048】
本文書に用いられる用語「バリアント」、「ホモログ」、「誘導体」、または「断片」とは、配列からまたは配列への1つ(または複数)のアミノ酸の任意の置換、変化、修飾、置換え、欠失、または付加が挙げられる。他の文脈に認められない限り、「VDCCγ-8」及び「VDCCγ-8イオンチャネル」についての言及は、VDCCγ-8のこのようなバリアント、ホモログ、誘導体、及び断片についての言及を含む。
【0049】
VDCCγ-8に適用される得られたアミノ酸配列は、イオンチャネル活性を有することが好ましく、配列番号3、配列番号5または配列番号7として示されるVDCCγ-8イオンチャネルの少なくとも同じ活性を有することがさらに好ましい。特に、得られたアミノ酸配列がイオンチャネル活性を有するという条件で、用語の「ホモログ」は、構造及び/または機能に関して同一性を包含する。配列同一性(すなわち、類似性)に関して、好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%の配列同一性である。より好ましくは少なくとも95%、より好ましくは少なくとも98%の配列同一性である。これらの用語はまた、VDCCγ-8イオンチャネル核酸配列の対立遺伝子変異であるアミノ酸から誘導されるポリペプチドを包含する。
【0050】
VDCCγ-8イオンチャネル等のイオンチャネルの「チャネル活性」または「生物活性」に言及される場合、これらの用語は、同様の活性または改善された活性もしくは非所望の副作用が減少したこれらの活性等、VDCCγ-8イオンチャネルの代謝機能または生理的機能を指すものとする。VDCCγ-8イオンチャネルの抗原活性及び免疫原性活性もまた含まれる。イオンチャネル活性の例、及びこれらの活性のアッセイ法及び定量化法は当業者に既知である。
【0051】
本明細書に用いられる「欠失」とは、それぞれ1つ以上のヌクレオチド残基またはアミノ酸残基を欠いているヌクレオチド配列またはアミノ酸配列の変化として定義される。本明細書に用いられる「挿入」または「付加」とは、天然物と比較して、それぞれ1つ以上のヌクレオチド残基またはアミノ酸残基の付加が生じたヌクレオチド配列またはアミノ酸配列を変化させることである。本明細書に用いられる「置換」は、それぞれ異なるヌクレオチドまたはアミノ酸による1つ以上のヌクレオチドまたはアミノ酸の置換えから生じる。
【0052】
本明細書に記載されるVDCCγ-8ポリペプチドはまた、サイレント変化を生じ、機能的に等しいアミノ酸配列となるアミノ酸残基の欠失、挿入、または置換を有することができる。計画的なアミノ酸置換は、残基の極性、電荷、溶解性、疎水性、親水性、及び/または両親媒性の類似性に基づいて成すことができる。例えば、負電荷のアミノ酸としては、アスパラギン酸及びグルタミン酸が挙げられ、陽電荷のアミノ酸としては、リシン及びアルギニンが挙げられ、類似の親水性値を有する無電荷極性頭部を有するアミノ酸としては、ロイシン、イソロイシン、バリン、グリシン、アラニン、アスパラギン、グルタミン、セリン、トレオニン、フェニルアラニン、及びチロシンが挙げられる。
【0053】
保存的置換は、例えば、下表に従って作製することができる。第二欄の同じブロック、好ましくは第三欄の同じ行のアミノ酸は互いに置換できる。
【0054】
【表1】

【0055】
VDCCγ-8ポリペプチドは、典型的にN末端またはC末端に、好ましくはN末端に異種のアミノ酸配列をさらに含むことができる。異種の配列としては、細胞内または細胞外タンパク質の標的(リーダー配列等)に影響を及ぼす配列を挙げることができる。異種配列としてはまた、ポリペプチドの免疫原性を増加させ、並びに/あるいはポリペプチドの同定、抽出、及び/または精製を容易にする配列を挙げることができる。特に好ましい別の異種の配列は、好ましくはN末端にあるポリヒスチジン等のポリアミノ酸配列である。少なくとも10のアミノ酸、好ましくは少なくとも17のアミノ酸であるが、50未満のアミノ酸のポリヒスチジン配列が特に好ましい。
【0056】
VDCCγ-8イオンチャネルポリペプチドは、「成熟」タンパク質の形態であり得るか、または融合タンパク質等のより大きなタンパク質の一部であり得る。分泌またはリーダー配列、プロ配列、多ヒスチジン残基等の精製、または組換え産生時の安定性のために、さらなる配列を補助する配列を含有するさらなるアミノ酸配列を含ませることもしばしば有利である。
【0057】
VDCCγ-8ポリペプチドは、既知の技術を用いる組換え手段により有利に作製される。しかしながら、それらは、固相合成等の当業者に周知の技術を用いる合成手段によっても作製できる。本明細書に記載されるポリペプチドは、例えば、抽出及び精製を補助するために、融合タンパク質としても産生できる。融合タンパク質パートナーの例としては、グルタチオン-S-トランスフェラーゼ(GST)、6×His、GAL4(DNA結合及び/または転写活性化ドメイン)、及びβ-ガラクトシダーゼが挙げられる。融合タンパク質配列の除去を可能にするために、トロンビン開裂部位等、融合タンパク質パートナーと対象のタンパク質配列との間のタンパク質開裂部位を含ませることも有利となり得る。融合タンパク質は、対象配列のタンパク質の機能を妨げないことも好ましくなろう。
【0058】
VDCCγ-8ポリペプチドは、実質的に単離形態であり得る。この用語は、天然状態から人の手により変化させることを意味する。「単離」組成物または物質が天然に生じる場合、それを変化させるか、または元の環境から取り出すか、あるいは両方から得る。例えば、生体動物に存在する天然の核酸またはポリペプチドは、「単離」されないが、その天然状態の共存物質から分離される同じポリヌクレオチド、核酸またはポリペプチドは、用語が本明細書に使用されるように「単離」される。
【0059】
しかしながら、VDCCγ-8イオンチャネルタンパク質は、タンパク質の意図された目的を妨害しない担体または希釈剤と混合することができ、さらに実質的に単離されたとみなされ得ると理解されよう。VDCCγ-8ポリペプチドはまた、実質的に精製形態であり得、その場合、それは、調製物中のタンパク質が90%超、例えば、95%、98%または99%がVDCCγ-8ポリペプチドである調製物中のタンパク質を一般に含んでいる。
【0060】
我々はさらに、VDCCγ-8ポリペプチドの一部を含むペプチドを記載する。従って、VDCCγ-8イオンチャネルの断片及びそのホモログ、バリアント、または誘導体が含まれる。該ペプチドは、2〜200の間のアミノ酸長、好ましくは4〜40の間のアミノ酸長であり得る。該ペプチドは、例えば、トリプシン等の好適な酵素による消化により本明細書に開示されたVDCCγ-8ポリペプチドから誘導できる。あるいは、ペプチド、断片等は、組換え手段により作製できるか、または合成的に合成できる。
【0061】
用語の「ペプチド」は、レトロインバーソDペプチド等、当業者に知られている種々の合成ペプチド変異を含む。該ペプチドは、抗原性決定因子及び/またはT細胞エピトープであり得る。該ペプチドはin vivo免疫原性であり得る。好ましくは、該ペプチドは、in vivoで抗体の中和を誘導できる。
【0062】
異なる種からVDCCγ-8イオンチャネル配列を位置合わせを行うことによって、アミノ酸配列の領域が異なる種間で保存されているか(「相同領域」)、領域が異なる種間で異なっているか(「非相同領域」)かを判定することが可能である。
【0063】
従って、VDCCγ-8ポリペプチドは、少なくとも一部の相同領域に相当する配列を含むことができる。相同領域は、少なくとも2種間で高度の相同性を示す。例えば、該相同領域は、上記試験を用いるアミノ酸レベルで少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%、例えば98%または99%の同一性を示すことができる。相同領域に相当する配列を含むペプチドは、さらに下記に詳細に説明されるように治療戦略に使用することができる。あるいは、VDCCγ-8イオンチャネルペプチドは、少なくとも一部の非相同領域に相当する配列を含むことができる。非相同領域は、少なくとも2種間で低度の相同性を示す。
【0064】
[VDCCγ-8イオンチャネルポリヌクレオチド及び核酸]
VDCCγ-8ポリヌクレオチド、VDCCγ-8ヌクレオチド、及びVDCCγ-8核酸、並びにこれら核酸の生産方法及び使用が意図されている。
【0065】
用語の「VDCCγ-8ポリヌクレオチド」、「VDCCγ-8ヌクレオチド」、及び「VDCCγ-8核酸」は、交互に使用することができ、配列番号1、配列番号2、配列番号4、もしくは配列番号6に示される核酸配列、あるいはそれらのホモログ、バリアント、または誘導体を含むポリヌクレオチド/核酸を指すものとする。好ましくは、該ポリヌクレオチド/核酸は、配列番号1、配列番号2、または配列番号6、最も好ましくは配列番号2の核酸配列のホモログ、バリアント、もしくは誘導体を含むか、またはそのものである。
【0066】
これらの用語はまた、ポリペプチド及び/またはペプチド、すなわちVDCCγ-8ポリペプチドをコードし得る核酸配列を含むことが意図されている。従って、VDCCγ-8イオンチャネルポリヌクレオチド及び核酸は、配列番号3、配列番号5、もしくは配列番号7に示されるアミノ酸配列、またはそれらのホモログ、バリアント、もしくは誘導体を含むポリペプチドをコードし得るヌクレオチド配列を含む。VDCCγ-8イオンチャネルポリヌクレオチド及び核酸は、配列番号3に示されるアミノ酸配列またはそのホモログ、バリアント、もしくは誘導体を含むポリペプチドをコードし得るヌクレオチド配列を含むことが好ましい。
【0067】
「ポリヌクレオチド」とは一般に、非修飾RNAまたはDNAもしくは修飾RNAまたはDNAであり得る任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドのことである。「ポリヌクレオチド」としては、限定はしないが、単鎖及び二本鎖DNA、単鎖及び二本鎖領域の混合物であるDNA、単鎖及び二本鎖RNA、並びに単鎖及び二本鎖領域の混合物であるRNA、単鎖、またはより典型的には二本鎖、もしくは単鎖及び二本鎖領域の混合物であり得るDNA及びRNAを含むハイブリッド分子が挙げられる。さらに「ポリヌクレオチド」とは、RNAまたはDNA、あるいはRNA及びDNA両方を含む三本鎖領域のことである。用語のポリヌクレオチドはまた、1つ以上の修飾塩基を含有するDNA及びRNAを含み、安定性または他の理由のために修飾された主鎖を有するDNA及びRNAを含む。「修飾」塩基としては、例えば、トリチル化塩基及びイノシン等の特異塩基が挙げられる。種々の修飾により、DNA及びRNAが作製されており、従って、「ポリヌクレオチド」は、典型的に天然見られるポリヌクレオチドの化学的、酵素的または代謝的に修飾された形態、並びにウイルス及び細胞に特徴的なDNA及びRNAの化学的形態を包含する。ポリヌクレオチドはまた、オリゴヌクレオチドとしばしば称される比較的短鎖のポリヌクレオチドを包含する。
【0068】
多数のヌクレオチド配列は、遺伝子コードの縮退の結果と同一のポリペプチドをコードし得ることが当業者により理解されるであろう。
【0069】
本明細書に用いられる用語「ヌクレオチド配列」とは、ヌクレオチド配列、オリゴヌクレオチド配列、ポリヌクレオチド配列及びそれらのバリアント、ホモログ、断片、及び誘導体(それらの一部等)のことである。ヌクレオチド配列は、センス鎖またはアンチセンス鎖を表そうと、もしくはそれらの組合せを表そうとも、二本鎖または一本鎖であり得るゲノムまたは合成あるいは組換え起源のDNAまたはRNAであり得る。用語のヌクレオチド配列は、組換えDNA技術(例えば、組換えDNA)の使用により調製できる。
【0070】
好ましくは、用語の「ヌクレオチド配列」とは、DNAを意味する。
【0071】
本文書に用いられる用語「バリアント」、「ホモログ」、「誘導体」、または「断片」としては、VDCCγ-8ヌクレオチド配列からまたはその配列に1つ(または複数の)核酸の任意の置換、変化、修飾、置換え、欠失、または付加が挙げられる。他の文脈に認められない限り、「VDCCγ-8」及び「VDCCγ-8イオンチャネル」についての言及は、VDCCγ-8のこのようなバリアント、ホモログ、誘導体、及び断片についての言及を含む。
【0072】
得られたヌクレオチド配列は、イオンチャネル活性を有するポリペプチドをコードすることが好ましく、配列番号3、配列番号5、または配列番号7として示されるイオンチャネルの少なくとも同じ活性を有することが好ましい。用語の「ホモログ」は、得られたヌクレオチド配列がイオンチャネル活性を有するポリペプチドをコードするように、構造及び/または機能に関して同一性を包含するように意図されていることが好ましい。配列同一性(すなわち、類似性)に関して、好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも85%、より好ましくは少なくとも90%の配列同一性である。より好ましくは少なくとも95%、より好ましくは少なくとも98%または99%の配列同一性である。これらの用語はまた、その配列の対立遺伝子変異を包含する。
【0073】
[配列相同性の算出]
本明細書に提出された任意の配列に関する配列同一性は、他の配列が、例えば、該配列(1つ以上)に対して少なくとも70%の配列同一性を有するかどうかを見るために、任意の1つ以上の配列と別の配列との単純な「眼球」比較(すなわち、厳密な比較)により決定され得る。
【0074】
相対的配列同一性はまた、例えば、デフォルトパラメータを用い、同一性を決定するために任意の好適なアルゴリズムを用いて2つ以上の配列間で同一性%を算出できる市販のコンピュータプログラムにより決定され得る。このようなコンピュータプログラムの典型的な例は、CLUSTALである。2つの配列間の同一性及び類似性を決定するための他のコンピュータプログラム法としては、限定はしないが、GCGプログラムパッケージ(Devereuxら、1984年、Nucleic Acids Research 12:387頁)及びFASTA(Atschulら、1990年、J Molec Biol 403〜410頁)が挙げられる。
【0075】
相同性%は、連続配列上で算出できる。すなわち、1つの配列を他の配列と位置合わせし、1つの配列中の各アミノ酸を、他の配列中の対応するアミノ酸と、一度に1つの残基と直接比較する。これは、「アンギャップド」アラインメントと呼ばれる。典型的に、このようなアンギャップドアラインメントは、比較的短い数の残基上で実施される。
【0076】
これは、極めて簡単で一貫した方法であるが、例えば別の同一対の配列において、1つの挿入または欠失により、続くアミノ酸残基がアラインメントから離れる原因となり、従って全域のアラインメントが実施される際に、可能性として相同性%の大きな減少を生じることを考慮できない。その結果、大抵の配列比較法は、総合的相同性スコアを過度に不利にすることなく、可能な挿入及び欠失を考慮する最適のアラインメントを生じるように設計されている。これは、局所的相同性を最大にしようとするために配列アラインメントに「ギャップ」を挿入することにより達成される。
【0077】
しかしながら、これらのより複雑な方法は、アラインメントに生じる各ギャップに「ギャップペナルティー」を与える結果、同じ数のアミノ酸に関して、2つの比較配列間でより高い関連性を反映して、できるだけ少ないギャップによる配列アラインメントは、多数のギャップのものよりも高いスコアを達成するであろう。ギャップの存在のため比較的高いコスト、及びギャップに続く各々の残基に対してより小さなペナルティーを課する「アフィンギャップコスト」が典型的に使用される。これは、最も一般に用いられるギャップスコアリングシステムである。高ギャップペナルティーはもちろん、より少ないギャップにより最適化アラインメントを生じるであろう。大抵のアラインメントプログラムは、ギャップペナルティーを変更させる。しかしながら、配列比較のためにこのようなソフトウェアを使用する場合、デフォルト値を使用することが好ましい。例えば、GCG Wisconsin Bestfitパッケージを用いる場合、アミノ酸配列に対するデフォルトギャップペナルティーは、ギャップに関して-12及び各伸長に関して-4である。
【0078】
従って、最大相同性%の算出は、最初にギャップペナルティーを考慮して最適アラインメントの製作を必要とする。このようなアラインメントを実施するために好適なコンピュータプログラムは、GCG Wisconsin Bestfitパッケージ(ウィスコンシン大学、米国;Devereuxら、1984年、Nucleic Acids Research 12:387頁)である。配列比較を実施できる他のソフトウェアの例としては、限定はしないが、BLASTパッケージ(Ausubelら、1999年 Nucleic Acids Research-18章)、FASTA(Atschulら、1990年J. Mol. Biol.、403〜410頁)及び比較手段のGENEWORKSスイートが挙げられる。BLAST及びFASTAの両方は、オフライン及びオンライン検索で入手できる(Ausubelら、1999年、J. Mol. Biol.、7〜58頁から7〜60頁)。
【0079】
最終相同性%は、同一性によって測定できるが、そのアラインメント処理自体は、典型的には全か無かの対比較に基づかない。その代わり、化学的類似性または進化距離に基づく各対比較に対するスコアを割り当てる基準化類似性スコアマトリックスが一般に用いられる。一般に用いられるこのようなマトリックスの例は、BLOSUM62マトリックス-BLASTスイートのプログラム用のデフォルトマトリックスである。GCG Wisconsinプログラムは一般に、供給される場合、公的デフォルト値または慣習的シンボル比較表を用いる。GCGパッケージに関しては公的デフォルト値、または他のソフトウェアの場合、BLOSUM62等のデフォルトマトリックスを使用することが好ましい。
【0080】
デフォルト値に対してパラメータセットを有するBLASTアルゴリズムを使用することが好都合である。BLASTアルゴリズムは、参照として本明細書に組み込まれている、http://www.ncbi.nih.gov/BLAST/blast_help.htmlに詳細に記載されている。検索パラメータを定義でき、定義されたデフォルトパラメータ上で有利に設定できる。
【0081】
好都合なことに、BLASTにより評価された場合の「実質的な同一性」とは、少なくとも約7、好ましくは少なくとも約9、最も好ましくは10以上のEXPECT値と一致する配列に等しくすることである。BLAST検索においてEXPECTに関するデフォルト閾値は、通常10である。
【0082】
BLAST(Basic Local Alignment Search Tool)は、プログラムblastp、blastn、blastx、tblastn、及びtblastxにより使用される発見的検索アルゴリズムであり、これらのプログラムにより、有意性は、若干増強されたKarlin及びAltschul(Karlin及びAltschul 1990年、Proc. Natl. Acad. Sci. USA 87:2264;68頁Karlin及びAltschul、1993年、Proc. Natl. Acad. Sci. USA 90:5873〜7頁;http://www.ncbi.nih.gov/BLAST/blast_help.htmlを参照)の統計学的方法を用いたそれらの知見に帰する。例えば、問合せの配列に対するホモログを同定するために、BLASTプログラムを配列類似性検索に適合させる。配列データベースの類似性検索における基礎的問題を検討するために、Altschulら(1994)Nature Genetics 6:119〜129頁を参照されたい。
【0083】
配列比較は、http://www.ncbi.nlm.nih.gov/BLASTに提供される簡単なBLAST検索アルゴリズムを用いて実施するのが最も好ましい。いくつかの実施形態において、配列同一性を決定する場合、ギャップペナルティーは使用しない。
【0084】
[ハイブリダイゼーション]
本明細書に提示された配列、またはそれらの任意の断片もしくは誘導体に、あるいは上記の任意の補体にハイブリダイズできるヌクレオチド配列を開示する。
【0085】
ハイブリダイゼーションとは、「核酸の鎖が塩基対を介して相補鎖と接合する処理工程」(Coombs J(1994)Dictionary of Biotechnology、Stockton Press、ニューヨーク、ニューヨーク州)並びにDieffenbach CW及びGS Dveksler(1995年、PCR Primer, a Laboratory Manual、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)に記載されたポリメラーゼ連鎖反応技術において実施される増幅処理工程を意味する。
【0086】
ハイブリダイゼーション条件は、Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)において教示された核酸結合複合体の融解温度(Tm)に基づき、下記に説明されるように規定された「ストリンジェンシー」を付与する。
【0087】
本明細書に提示されたヌクレオチド配列に、またはそれらの補体に選択的にハイブリダイズできるヌクレオチド配列は、少なくとも20、好ましくは少なくとも25または30、例えば、少なくとも40、60、または100以上の連続ヌクレオチドの領域にわたって本明細書に提示された対応するヌクレオチド配列に対して、一般に少なくとも70%、好ましくは少なくとも75%、より好ましくは少なくとも85%または90%、さらにより好ましくは少なくとも95%、98%、または99%相同である。好ましいヌクレオチド配列は、配列番号1、2、4、または6に相同的な領域を含み、1つの配列に対して、好ましくは少なくとも70%、80%、または90%、より好ましくは少なくとも95%相同である。
【0088】
用語の「選択的にハイブリダイズ可能」とは、標的ヌクレオチド配列が有意に上記バックグラウンドのレベルでプローブにハイブリダイズすることが分かっている条件下で、プローブとして用いられるヌクレオチド配列が使用されることを意味する。バックグラウンドハイブリダイゼーションは、例えば、スクリーニングされるcDNAまたはゲノムDNAライブラリーに存在する他のヌクレオチド配列のために生じ得る。この事象において、バックグラウンドとは、プローブと、標的DNAにより見られる特異的相互作用の強度の10倍未満、好ましくは100倍未満であるライブラリーの非特異的DNAメンバーとの間の相互作用により作製されたシグナルレベルを意味する。相互作用の強度は、例えば、プローブを放射標識により、例えば、32Pにより測定できる。
【0089】
中程度から最大のストリンジェンシーの条件下で、本明細書に提示されたヌクレオチド配列にハイブリダイズできるヌクレオチド配列もまた含まれる。ハイブリダイゼーション条件は、Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)において教示された核酸結合複合体の融解温度(Tm)に基づき、下記に説明されるように規定された「ストリンジェンシー」を付与する。
【0090】
最大ストリンジェンシーは、約Tm-5℃(プローブのTmより5℃低い)で;高ストリンジェンシーは、Tmより約5℃から10℃低い;中程度のストリンジェンシーは、Tmより約10℃から20℃低い;低ストリンジェンシーは、Tmより約20℃から25℃低いとき典型的に生じる。当業者により解されるように、最大ストリンジェンシーのハイブリダイゼーションを、同一のヌクレオチド配列を同定または検出するために使用できるが、一方、中程度(または低)ストリンジェンシーのハイブリダイゼーションを、類似のまたは関連ヌクレオチド配列を同定または検出するために使用できる。
【0091】
好ましい実施形態において、ストリンジェント条件下(例えば、65℃及び0.1×SSC{1×SSC = 0.15 M NaCl、0.015 M Na3クエン酸pH 7.0})、1つ以上のVDCCγ-8イオンチャネルヌクレオチド配列にハイブリダイズできるヌクレオチド配列が開示される。ヌクレオチド配列が二本鎖である場合、二重鎖、個々にまたは組合せの両方を包含する。ヌクレオチド配列が一本鎖である場合、そのヌクレオチド配列の相補的配列もまた使用することが理解されよう。
【0092】
本発明の範囲内にあるヌクレオチド配列は、本明細書に提示された配列、あるいはそれらの任意の断片または誘導体に相補的である配列にハイブリダイズできる。同様に、本発明は、配列にハイブリダイズできる配列に相補的であるヌクレオチド配列を包含する。これらのタイプのヌクレオチド配列は、バリアントヌクレオチド配列の例である。この観点から、用語の「バリアント」は、本明細書に提示されたヌクレオチド配列にハイブリダイズできる配列に相補的である配列を包含する。しかしながら、好ましくは、用語の「バリアント」は、ストリンジェント条件下(例えば、65℃及び0.1×SSC{1×SSC = 0.15 M NaCl、0.015 M Na3クエン酸pH 7.0})、本明細書に提示されたヌクレオチド配列にハイブリダイズできる配列に相補的である配列を包含する。
【0093】
[VDCCγ-8イオンチャネル及びホモログのクローニング]
本発明は、本明細書に提示された配列、あるいはそれらの任意の断片または誘導体に相補的であるヌクレオチド配列を包含する。該配列が、その断片に相補的である場合、その配列を、他の生物等における同様のイオンチャネル配列を同定及びクローニングするためのプローブとして使用できる。
【0094】
これは、ヒト及び他の種、例えばマウス、ブタ、ヒツジ等からの、VDCCγ-8イオンチャネル、そのホモログ、及び他の構造的または機能的関連遺伝子のクローニングを可能にする。配列番号1、配列番号2、配列番号4、または配列番号6またはそれらの断片に含まれるヌクレオチド配列と同一か、または十分に同一であるポリヌクレオチドは、適切なライブラリーから部分長または完全長cDNA及びVDCCγ-8イオンチャネルをコードするゲノムクローンを単離するために、cDNA及びゲノムDNA用のハイブリダイゼーションプローブとして使用され得る。このようなプローブはまた、VDCCγ-8イオンチャネル遺伝子に対して配列類似性、好ましくは、高い配列類似性を有する他の遺伝子(ヒト以外の種由来のホモログ及びオルソログをコードする遺伝子を含む)のcDNA及びゲノムクローンを単離するために使用され得る。ハイブリダイゼーションスクリーニング、クローニング、及び配列決定技術は当業者に既知であり、例えばSambrookら(上記文献)に記載されている。
【0095】
プローブとしての使用に好適なヌクレオチド配列は、典型的には、基準のものに対して70%同一、好ましくは80%同一、より好ましくは90%同一、さらにより好ましくは95%同一である。このプローブは一般に、少なくとも15のヌクレオチドを含む。このようなプローブは、少なくとも30のヌクレオチドを有することが好ましく、少なくとも50のヌクレオチドを有することができる。特に好ましいプローブは、150〜500の間の範囲のヌクレオチドであり、さらに特に約300のヌクレオチドである。
【0096】
一実施形態において、ヒト以外の種からホモログ及びオルソログ等、VDCCγ-8ポリペプチドをコードするポリヌクレオチドを得るために、配列番号1、配列番号2、配列番号4、または配列番号6、あるいはそれらの断片を有する標識プローブによりストリンジェントなハイブリダイゼーション下で適切なライブラリーをスクリーニングする工程と、上記ポリヌクレオチド配列を含有する部分長または完全長cDNA及びゲノムクローンを単離する工程とを含む。このようなハイブリダイゼーション技術は、当業者に周知である。ストリンジェントなハイブリダイゼーション条件は、上記に定義されているとおりか、あるいは、50%ホルムアミド、5×SSC(150 mM NaCl、15 mMクエン酸三ナトリウム)、50 mMリン酸ナトリウム(pH 7.6)、5×デンハート液、10%硫酸デキストラン、及び20マイクログラム/ml変性、剪断サケ精巣DNAを含む溶液中、42℃で一晩インキュベーションし、次いで約65℃で0.1×SSC中でろ過洗浄する条件である。
【0097】
[VDCCγ-8イオンチャネルに対する機能的アッセイ]
クローニングされた推定上のVDCCγ-8イオンチャネルポリヌクレオチドは、配列分析または機能的アッセイにより証明され得る。例えば、推定上のVDCCγ-8イオンチャネルまたはホモログは、以下の活性についてアッセイできる。VDCCγ-8受容体cDNAをコードする線形化プラスミドテンプレートからのキャップ付RNA転写物を、標準的手法に従ってin vitroでRNAポリメラーゼにより合成する。in vitro転写物を、最終濃度が0.2 mg/mlで水に懸濁させる。卵巣葉を、成体メスヒキガエルから取り出し、段階Vの濾胞除去卵母細胞を得て、RNA転写物(10 ng/卵母細胞)を注入し、マイクロインジェクション装置を用いて50 nlボーラス中でグルタメート受容体または他のVOCCサブユニットにより共発現させる。2つの電極電圧クランプを、アゴニスト曝露に応答する個々のアフリカツメガエル卵母細胞からの電流を測定するために用いる。ND96溶液媒体中室温で記録する。以下にさらに詳細に記載されるとおり、アフリカツメガエル系はまた、既知のリガンドをスクリーニングし、リガンドを活性化させるための組織/細胞抽出のために使用され得る。代わりの機能的アッセイとしては、全細胞電気生理、蛍光共鳴エネルギー移動(FRET)分析、及びFLIPR分析が挙げられる。
【0098】
[VDCCγ-8イオンチャネルに対する細胞表面発現アッセイ]
段階Vの濾胞除去卵母細胞を得て、RNA転写物(10 ng/卵母細胞)を注入し、グルタメート受容体(GluR1-フリップ等)により共発現させ、マイクロインジェクション装置を用いて50 nlボーラス中、ヘマグルチニンエピトープを細胞外領域に操作した。注入3日後、卵母細胞を、0.25μg/mlのラット抗HA抗体(3F10、Roche)と1時間インキュベートし、次いで西洋ワサビペルオキシダーゼ複合化抗ラット免疫グロブリンと30分インキュベートする。個々の卵母細胞を、50μlのSuperSignal ELISA Femto最大感受性基体(Pierce)に入れ、TD20/20m (Turner Designs)等の好適なルミノメータを用いて化学発光法により定量化する。
【0099】
[VDCCγ-8イオンチャネルに対する発現アッセイ]
VDCCγ-8イオンチャネル関連疾患を治療するのに有用な治療薬を設計するために、VDCCγ-8の発現プロファイル(野生型または特定の変異体かどうか)を測定することは有用である。従って、当業者に既知の方法を、VDCCγ-8が発現される臓器、組織、及び細胞型(並びに発育段階)を決定するために使用できる。例えば、伝統的または「電子」ノーザンを実施できる。逆転写酵素PCR (RT-PCR)もまた、VDCCγ-8遺伝子または変異体の発現をアッセイするために使用できる。VDCCγ-8の発現プロファイルを測定するためにより高感度の方法としては、当業者に既知のRNA分解酵素保護アッセイが挙げられる。
【0100】
ノーザン分析は、遺伝子の転写物の存在を検出するために使用される実験室的技術であり、特定の細胞型または組織からのRNAが結合された膜に対する標識ヌクレオチド配列のハイブリダイゼーションを含む(Sambrook、上記文献、7章及びAusubel, F. M.ら、上記文献、4章及び16章)。BLASTを適用している類似のコンピュータ技術(「電子ノーザン」)を、GenBankまたはLIFESEQデータベース(Incyte Pharmaceuticals)等のヌクレオチドデータベースにおいて同一または関連分子を検索するために使用できる。このタイプの分析は、複数膜ベースのハイブリダイゼーションよりも速く成し得る利点を有する。さらに、コンピュータ検索の感度は、任意の特定のマッチが正確または相同的なものとして分類されるかどうかを判定するために変更され得る。
【0101】
上記のプローブ等、本明細書に記載されたポリヌクレオチド及びポリペプチドを、動物及びヒトの疾患に対する治療法及び診断法の発見のために研究用試薬及び物質として使用できる。
【0102】
[VDCCγ-8イオンチャネルポリペプチドの発現]
当業者に明らかなように、VDCCγ-8イオンチャネルポリペプチド、またはそのホモログ、バリアント、もしくは誘導体をコードする核酸を含む宿主細胞を、好適な条件下(すなわち、VDCCγ-8イオンチャネルポリペプチドが発現される条件)で培養する(一般的に言えば)ことによって、VDCCγ-8ポリペプチドを産生することができる。
【0103】
生物的に活性なVDCCγ-8イオンチャネルを発現させるために、VDCCγ-8イオンチャネルまたはそのホモログ、バリアント、もしくは誘導体をコードするヌクレオチド配列は、適切な発現ベクター、すなわち挿入されたコード化配列の転写及び翻訳に必要な要素を含有するベクターに挿入される。
【0104】
当業者に周知の方法を用いて、VDCCγ-8イオンチャネル、並びに適切な転写及び翻訳調節要素をコードする配列を含有する発現ベクターを構築する。これらの方法としては、in vitro組換えDNA技術、合成技術、及びin vivo遺伝子組換えが挙げられる。このような技術は、Sambrook, J.ら、(1989年、Molecular Cloning:A Laboratory Manual、4章、8章、及び16〜17章、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)及びAusubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州)に記載されている。
【0105】
種々の発現ベクター/宿主系を利用して、VDCCγ-8イオンチャネルをコードする配列を含ませ、発現させることができる。これらとしては、限定はしないが、組換えバクテリオファージ、プラスミド、またはコスミドDNA発現ベクターにより変換された細菌;酵母発現ベクターにより変換された酵母;ウイルス発現ベクター(例えば、バキュロウイルス)により感染させた昆虫細胞系;ウイルス発現ベクター(例えば、カリフラワーモザイクウイルス(CaMV)またはタバコモザイクウイルス(TMV))または細菌発現ベクター(例えば、TiまたはpBR322プラスミド)により変換された植物細胞系;または動物細胞系等の微生物が挙げられる。使用される宿主細胞の性質は重要ではない。
【0106】
「調節要素」または「調節配列」とは、転写及び翻訳を実施するために宿主細胞タンパク質と相互作用するベクターの非翻訳領域(すなわち、エンハンサー、プロモーター、並びに5'及び3'非翻訳領域)のものである。このような要素は、強度及び特異性を変えることができる。利用されるベクター及び宿主系によって、構成的及び誘導的プロモーター等、任意の数の好適な転写及び翻訳要素を使用することができる。例えば、細菌系にクローニングする場合、BLUESCRIPTファージミドのハイブリッドlacZプロモーター(Stratagene、ラジョラ、カリフォルニア州)またはPSPORT1プラスミド(GIBCO/BRL)等の誘導性プロモーターを昆虫細胞に使用できる。バキュロウイルスポリヘドリンプロモーターを、昆虫細胞において使用できる。植物細胞のゲノム(例えば、ヒートショック、PUBISCO、及び保存タンパク質遺伝子)または植物ウイルス(例えば、ウイルスプロモーターまたはリーダー配列)から誘導されたプロモーターまたはエンハンサーをベクター内にクローニングできる。哺乳動物細胞系において、哺乳動物遺伝子または哺乳動物ウイルス由来のプロモーターが好ましい。VDCCγ-8イオンチャネルをコードする配列の複数コピーを含有する細胞系を作製することが必要な場合、SV40またはEBVに基づくベクターを、適切な選択マーカーとともに使用できる。
【0107】
細菌系において、多数の発現ベクターを、VDCCγ-8イオンチャネルのために意図された使用によって選択できる。例えば、大量のVDCCγ-8イオンチャネルが抗体誘導のために必要とされる場合、容易に精製される融合タンパク質の高レベル発現をもたらすベクターを使用できる。このようなベクターとしては、限定はしないが、多機能大腸菌クローニング及びBLUESCRIPT(Stratagene)等の発現ベクターが挙げられ、その中でVDCCγ-8イオンチャネルをコードする配列を、アミノ末端Metに関する配列及び引き続くβ-ガラクトシダーゼの7つの残基を有するフレーム内のベクターに結合でき、その結果、ハイブリッドタンパク質が、pINベクター等を産生する(Van Heeks, G.及びS. M. Schuster(1989)J. Biol. Chem. 264:5503〜5509頁)。pGEXベクター(Promega、マジソン、ウィスコンシン州)を使用して、グルタチオンS-トランスフェラーゼ(GST)との融合タンパク質として異種ポリペプチドを発現させることができる。一般に、このような融合タンパク質は溶解性であり、グルタチオン-アガロースビーズへの吸着、次いで遊離グルタチオンの存在下での溶出により溶解細胞から容易に精製できる。クローニングされた対象のポリペプチドが自由にGST部分から放出できるように、このような系で作製されたタンパク質を、ヘパリン、トロンビン、または因子XAのプロテアーゼ開裂部位を含むように設計できる。
【0108】
酵母菌酵母において、アルファ因子、アルコールオキシダーゼ、及びPGH等の構成的または誘導性プロモーターを含有する多数のベクターを使用できる。レビューに関しては、Ausubel(上記文献)及びGrantら(1987年;Methods Enzymol. 153:516〜544頁)を参照されたい。
【0109】
植物発現ベクターが用いられる場合、VDCCγ-8イオンチャネルをコードする配列の発現は、任意の多数のプロモーターにより誘発され得る。例えば、CaMVの35S及び19Sプロモーター等のウイルスプロモーターを、単独でまたはTMV由来のオメガリーダー配列との組合せで使用できる(Takamatsu, N.(1987)EMBO J. 6:307〜311頁)。あるいは、小型サブユニットのRUBISCOまたはヒートショックプロモーター等の植物プロモーターを使用できる(Coruzzi, G.ら(1984)EMBO J. 3:1671〜1680頁;Broglie, R.ら(1984)Science 224:838〜843頁;及びWinter, J.ら(1991)Results Probl. Cell Differ. 17:85〜105頁)。これらの構築物を、直接のDNA変換または病原体媒介トランスフェクションにより植物細胞に導入できる。このような技術は、多数の一般的に利用できるレビューに記載されている(例えば、Hobbs, S.またはMurry, L. E.、McGraw Hill Yearbook of Science and Technology(1992)McGraw Hill、ニューヨーク、ニューヨーク州;191〜196頁を参照)。
【0110】
昆虫系もまた、VDCCγ-8イオンチャネルを発現させるために使用できる。例えば、1つのこのような系において、オウトグラファカリフォルニア核多汗症ウイルス(AcNPV)は、Spodoptera frugiperda細胞またはTrichoplusia larvaeにおける異種遺伝子を発現させるベクターとして用いられる。VDCCγ-8イオンチャネルをコードする配列を、ポリヘドリン遺伝子等のウイルスの非必須領域にクローニングでき、ポリヘドリンプロモーターの調節下に置くことができる。VDCCγ-8イオンチャネルの好結果の挿入により、ポリヘドリン遺伝子を不活性にし、コートタンパク質を欠く組換えウイルスを生じる。次いで組換えウイルスを、例えば、TrpM8イオンチャネルが発現し得るS. frugiperda細胞またはTrichoplusia larvaeを感染させるために使用できる(Engelhard, E. K.ら(1994)Proc. Nat. Acad. Sci. 91:3224〜3227頁)。
【0111】
哺乳動物宿主細胞において、多くのウイルスベースの発現系を利用できる。アデノウイルスが発現ベクターとして用いられる場合、VDCCγ-8イオンチャネルをコードする配列は、後期プロモーター及び三連リーダー配列からなるアデノウイルス転写/翻訳複合体に結合できる。ウイルスゲノムの非必須E1またはE3領域における挿入を、感染宿主細胞にVDCCγ-8イオンチャネルを発現できる生存可能なウイルスを得るために使用できる(Logan, J.及びT. Shenk(1984)Proc. Natl. Acad. Sci. 81:3655〜3659頁)。さらに、ニワトリ肉腫ウイルス(RSV)エンハンサー等の転写エンハンサーを、哺乳動物宿主細胞における発現を増加させるために使用できる。
【0112】
従って例えば、VDCCγ-8受容体は、ヒト胎生腎細胞293(HEK293)または接着細胞CHOに発現できる。受容体発現を最大にするために、典型的にすべての5'及び3'非翻訳領域(UTR)は、pCDNまたはpCDNA3ベクターへの挿入前に受容体cDNAから除去される。細胞は、リポフェクチンにより個々の受容体cDNAにより形質移入され、400 mg/mlのG418の存在下で選択される。選択3週後、個々のクローンを選択し、さらに分析のために増殖させる。ベクター単独で形質移入されたHEK293またはCHO細胞は、陰性コントロールとして寄与する。個々の受容体を安定に発現する細胞系を単離するために、典型的に約24のクローンを選択し、ノーザンブロット分析により分析する。通常、分析されるG418耐性クローンの約50%において受容体mRNAを検出できる。
【0113】
ヒト人工染色体(HAC)は使用して、プラスミドに含有しかつ発現できるよりも大きな断片のDNAを送達することができる。約6 kb〜10 MbのHACが構築され、治療目的のために従来の送達法(リポソーム、ポリカチオン性アミノポリマー、または小胞)を介して送達される。
【0114】
特定の開始シグナルはまた、VDCCγ-8イオンチャネルをコードする配列のより効率的な翻訳を達成するために使用され得る。このようなシグナルは、ATG開始コドン及び隣接配列を含む。VDCCγ-8イオンチャネル及びその開始コドン、並びに上流配列をコードする配列が、適切な発現ベクターに挿入される場合には、さらなる転写または翻訳調節シグナルを必要としなくてもよい。しかしながら、コード化配列のみ、またはその断片が挿入される場合には、ATG開始コドンを含む外因性翻訳調節シグナルを提供する必要がある。さらに、開始コドンは、全挿入断片の翻訳を確保するために正確なリーディングフレーム内にある必要がある。外因性翻訳要素及び開始コドンは、外因性翻訳要素及び開始コドンは、天然及び合成による種々の起源であり得る。発現効率は、文献(Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125~162頁)に記載されているもの等、使用される特定の細胞系に適切なエンハンサーの包含により増強することができる。文献(Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125〜162頁)に記載されたもの等、使用される特定の細胞系に適切なエンハンサーの包含物により増強できる。
【0115】
さらに、挿入配列の発現を変化させるか、または所望の様式で発現タンパク質を処理する能力に関して、宿主細胞株を選択できる。このようなポリペプチドの修飾としては、限定はしないが、アセチル化、カルボキシル化、グリコシル化、リン酸化、脂質化、及びアシル化が挙げられる。タンパク質の「プレプロ」形態を開裂する翻訳後処理はまた、正しい挿入、折りたたみ、及び/または機能を促進させるために使用され得る。翻訳活性後の特定の細胞機構及び特徴的機構を有する種々の宿主細胞(例えば、CHO、HeLa、MDCK、HEK293、及びWI38)を、American Type Culture Collection (ATCC、ベセスダ、メリーランド州)から入手でき、正しい修飾及び異種タンパク質の処理を確保するために選択できる。
【0116】
長期の組換えタンパク質の高収率産生のために、安定な発現が好ましい。例えば、ウイルスの複製起点及び/または内因性発現要素、並びに選択マーカー遺伝子を、同一または別個のベクター内に含有し得る発現ベクターを用いて、VDCCγ-8イオンチャネルを安定に発現できる細胞系を形質転換できる。該ベクターの導入後、細胞を、選択的培地に切り替える前に富化培地中、約1〜2日間増殖させることができる。選択マーカーの目的とは、耐選択性を付与することであり、その存在は、導入された配列を首尾よく発現する細胞の増殖と回収を可能にする。安定に形質転換された細胞の耐性クローンは、細胞型に対して適切な組織培養技術を用いて増殖できる。
【0117】
任意の数の選択系を、変換細胞系を回収するために使用できる。これらには、限定はしないが、それぞれtk細胞またはapr細胞に使用できる単純ヘルペスウイルスチミジンキナーゼ遺伝子(Wigler, M.ら(1977)Cell 11:223〜32頁)及びアデニンホスホリボシルトランスフェラーゼ遺伝子(Lowy, I.ら(1980)Cell 22:817〜23頁)が挙げられる。また、抗代謝剤、抗生物質、または耐除草剤を、選択基準として使用できる。例えば、dhfrは、耐メトトレキサートを付与し(Wigler, M.ら(1980)Proc. Natl. Acad. Sci. 77:3567〜70頁);nptは、耐アミノグリコシドネオマイシン及びG-418を付与し(Colbere-Garapin, F.ら(1981) J. Mol. Biol. 150: 1〜14頁);及びalsまたはpatは、それぞれ耐クロルスルフロン及び耐ホスフィノトリシンアセチルトランスフェラーゼを付与する(Murry、上記文献)。さらなる選択遺伝子は、例えば、細胞がトリプトファンの代わりにインドールを利用できるtrrB、または細胞がヒスチジンの代わりにヒスチノールを利用できるhisDを記載している(Hartman, S. C.及びR. C. Mulligan(1988)Proc. Natl. Acad. Sci. 85:8047〜51頁)。最近、可視マーカーの使用は、アントシアニン、β-グルクロニダーゼ及びその基質GUS、並びにルシフェラーゼ及びその基質ルシフェリン等のマーカーとして人気を得ている。これらのマーカーを、形質転換体を同定するのみならず、特定のベクター系に起因し得る一過性または安定なタンパク質発現量を定量化するために使用できる(Rhodes, C. A.ら(1995)Methods Mol. Biol. 55:121〜131頁)。
【0118】
マーカー遺伝子発現の存在/非存在は、コントロールの遺伝子もまた存在することを示唆するが、遺伝子の存在及び発現を、確認するために必要であり得る。例えば、VDCCγ-8イオンチャネルをコードする配列が、マーカー遺伝子配列内に挿入される場合、VDCCγ-8イオンチャネルをコードする配列を含有する形質変換細胞は、マーカー遺伝子機能の非存在により同定できる。あるいは、マーカー遺伝子を、単一プロモーターの調節下、VDCCγ-8イオンチャネルをコードする配列と並行して設置することができる。さらに、誘導または選択に応答するマーカー遺伝子の発現は、通常、タンデム遺伝子の発現を示す。
【0119】
あるいは、VDCCγ-8イオンチャネルをコードする核酸配列を含有し、VDCCγ-8イオンチャネルを発現する宿主細胞は、当業者に知られている種々の手法により同定できる。これらの手法としては、限定はしないが、DNA-DNAまたはDNA-RNAハイブリダイゼーション及びタンパク質バイオアッセイまたは免疫アッセイ技術が挙げられ、核酸またはタンパク質配列の検出及び/または定量化に関する技術に基づく膜、溶液、またはチップを含む。
【0120】
VDCCγ-8イオンチャネルをコードするポリヌクレオチド配列の存在は、VDCCγ-8イオンチャネルをコードするプローブまたは断片またはポリヌクレオチドの断片を用いて、DNA-DNAまたはDNA-RNAハイブリダイゼーションあるいは増幅により検出できる。VDCCγ-8イオンチャネルをコードするDNAまたはRNAを含有する形質転換体を検出するために、核酸増幅に基づくアッセイは、VDCCγ-8イオンチャネルをコードする配列に基づくオリゴヌクレオチドまたはオリゴマーの使用を含む。
【0121】
タンパク質に特異的なポリクローナルまたはモノクローナル抗体を用いて、VDCCγ-8イオンチャネルの発現を検出し、測定するための種々のプロトコールは当業者に既知である。このような技術の例としては、酵素結合免疫吸着アッセイ(ELISA)等、放射免疫アッセイ(RIA)等、及び蛍光標示式細胞分取(FACS)が挙げられる。VDCCγ-8イオンチャネルに対する2つの非妨害エピトープに反応性のあるモノクローナル抗体を利用する2部位モノクローナルベースの免疫アッセイが好ましいが、競合結合アッセイを使用できる。これら及び他のアッセイは、当業者、例えば、Hampton, R.ら(1990年;Serological Methods, a Laboratory Manual, Section IV、APS Press、セントポール、ミネソタ州)及びMaddox, D. E.ら(1983年;J. Exp. Med. 158:1211〜1216頁)によく記載されている。
【0122】
多種多様の標識及び複合化技術は、当業者に既知であり、種々の核酸及びアミノ酸アッセイに使用され得る。VDCCγ-8イオンチャネルをコードするポリヌクレオチド関連配列を検出するために標識ハイブリダイゼーションまたはPCRプローブを産生する手段としては、オリゴ標識化、ニック翻訳、エンド標識化、または標識ヌクレオチドを用いるPCR増幅が挙げられる。あるいは、VDCCγ-8イオンチャネルをコードする配列、またはその任意の断片を、mRNAプローブの産生のためにベクターにクローニングできる。このようなベクターは、当業者に既知で市販されており、T7、T3、またはSP6等の適切なRNAポリメラーゼ及び標識ヌクレオチドの添加により、in vitroでRNAプローブを合成するために使用され得る。これらの手法は、Pharmacia & Upjohn(カラマズー、ミシガン州)、GE Healthcare(英国)、及びU. S. Biochemical社(クリーブランド、オハイオ州)により提供されるもの等、種々の市販のキットを用いて実施できる。検出を容易にするために使用できる好適なレポーター分子または標識としては、放射性核種、酵素、蛍光剤、化学発光剤、または色素生成剤、並びに基質、補因子、阻害剤、磁気粒子等が挙げられる。
【0123】
VDCCγ-8サブユニットをコードするヌクレオチド配列により形質転換された宿主細胞を、発現に好適な条件下で培養し、細胞培養からタンパク質を回収できる。形質転換細胞により産生されたタンパク質は、使用される配列及び/またはベクターによって、細胞膜に配置され、分泌され、または細胞内に含ませることができる。当業者により理解されるように、VDCCγ-8サブユニットをコードするポリヌクレオチドを含有する発現ベクターは、原核生物または真核生物細胞膜を介してVDCCγ-8サブユニットの分泌をもたらすシグナル配列を含有するように設計できる。他の構築物を、溶解性タンパク質の精製を促進するポリペプチドドメインをコードするヌクレオチド配列に、VDCCγ-8サブユニットをコードする配列を結合させるために使用できる。このような精製促進ドメインとしては、限定はしないが、固定化金属上で精製を可能にするヒスチジン-トリプトファンモジュール等の金属キレート化ペプチド、固定化免疫グロブリン上で精製を可能にするタンパク質Aドメイン、及びFLAGS拡張/アフィニティー精製システム(Immunex社、シアトル、ワシントン州)に利用されるドメインが挙げられる。精製ドメインと配列をコードするVDCCγ-8サブユニットとの間で、因子XAまたはエンテロキナーゼ(Invitrogen、サンディエゴ、カリフォルニア州)に特異的なもの等の開裂可能なリンカー配列の含有物を、精製を促進させるために使用できる。1つのこのような発現ベクターは、チオレドキシンまたはエンテロキナーゼ開裂部位に先立って6つのヒスチジン残基をコードするVDCCγ-8サブユニット及び核酸を含有する融合タンパク質の発現を提供する。該ヒスチジン残基は、固定化金属イオンアフィニティークロマトグラフィ(IMIAC;Porath, J.ら(1992)Prot. Exp. Purif. 3:263〜281頁に記載)上で精製を促進させるが、一方、エンテロキナーゼ開裂部位は、融合タンパク質からVDCCγ-8サブユニットを精製する手段を提供する。融合タンパク質を含有するベクターの考察は、Kroll, D. J.ら(1993年;DNA Cell Biol. 12:441〜453頁)に提供される。
【0124】
VDCCγ-8サブユニットの断片は、組換え体産生によってだけでなく、固相法を用いた直接的ペプチド合成によって産生され得る(Merrifield J.(1963)J. Am. Chem. Soc. 85:2149〜2154頁)。タンパク質合成は、手動技術または自動操作により実施できる。自動合成は、例えば、Applied Biosystems 431Aペプチドシンセサイザー(Perkin Elmer)を用いて達成できる。VDCCγ-8サブユニットの種々の断片は、別個に合成でき、次いで完全長分子を産生するために結合することができる。
【0125】
[バイオセンサー]
VDCCγ-8ポリペプチド、核酸、プローブ、抗体、発現ベクター、及びリガンドは、バイオセンサー(の産生のため)として有用である。
【0126】
Aizawa(1988)、Anal. Chem. Symp. 17:683頁によれば、バイオセンサーは、分子認識に関する受容体のユニークな組合せ、例えば、VDCCγ-8等の固定化抗体または受容体を有する選択的層、及び測定される値を転送するトランスデューサーとして定義される。このようなバイオセンサーの一群は、受容体と周囲の媒体との相互作用のため、表層の最適性質に生じる変化を検出する。このような技術の中で、特にエリプソメトリー及び表面プラズモン共鳴を挙げることができる。VDCCγ-8を取り込むバイオセンサーを、より好ましくは、上記に定義された疾患に関してVDCCγ-8リガンドの存在またはレベル検出するために使用できる。このようなバイオセンサーの構築は当業者に周知である。
【0127】
従って、VDCCγ-8サブユニットを発現する細胞系を、[3H]イノシトールホスフェートまたは他の第二のメッセンジャーの受容体促進形成を介してATP等のリガンド検出のための受容体系として使用できる(Wattら、1998年、J Biol Chem 5月29日;273(22):14053〜8頁)。受容体リガンドバイオセンサーはまた、Hoffmanら、2000年、Proc Natl Acad Sci USA 10月10日;97(21):11215〜20頁に記載されている。VDCCγ-8を含む光学的及び他のバイオセンサーはまた、例えば、Figlerら、1997年、Biochemistry 12月23日;36(51):16288〜99頁及びSarrioら、2000年、Mol Cell Biol 2000年7月;20(14):5164〜74頁により記載されているG-タンパク質及び他のタンパク質との相互作用のレベルまたは存在を検出するために使用され得る。バイオセンサー用のセンサーユニットは、例えば、米国特許第5,492,840号に記載されている。当業者は、VDCCγ-8のアンタゴニストの価値を等しく認識するであろう。例えば、別の実施形態において、ADHDの治療は、VDCCγ-8アンタゴニストによって達成され得る。
【0128】
[スクリーニングアッセイ]
ホモログ、バリアント、及び誘導体を含むVDCCγ-8ポリペプチドは、天然であろうと組換え体であろうとも、サブユニットを結合し、VDCCγ-8を活性化するか(アゴニスト)、またはその活性を阻害し(アンタゴニスト)、関連チャネルの動態を変えるか、またはその発現を変える化合物に関するスクリーニング法に使用され得る。従って、このようなポリペプチドは、例えば、細胞、無細胞製剤、化学ライブラリー、及び天然物の混合物において、小型分子の基質とリガンドとの結合を評価するためにも使用され得る。これらの基質及びリガンドは、天然の基質及びリガンドであり得るか、または構造的または機能的ミメティックであり得る。Coliganら、Current Protocols in Immunology 1(2):5章(1991)を参照されたい。
【0129】
VDCCγ-8イオンチャネルポリペプチドは、多くの生物学的機能に関与し、上記に掲げた疾患に関連する。従って、一方では、VDCCγ-8イオンチャネルを刺激し、他方では、VDCCγ-8イオンチャネルの機能を阻害できる化合物及び薬物を見出すことが望ましい。一般に、アゴニスト及びアンタゴニストは、興奮毒性及び認知障害のような病態に対する治療及び予防目的のために使用される。VDCCγ-8を欠くマウスが、統合失調症性表現型を示すというデータが本明細書に示されることを当業者は理解するであろう。従って、好ましい実施形態において、VDCCγ-8を活性化する化合物または薬物(すなわち、アゴニスト)は、上記に示されたように統合失調症及び他の関連精神疾患に対する治療薬となる可能性がある。当業者は、VDCCγ-8のアンタゴニストの価値を等しく認識するであろう。例えば、代わりの実施形態において、ADHDの治療は、VDCCγ-8アンタゴニストによって達成され得る。
【0130】
VDCCγ-8イオンチャネルタンパク質と相互作用しやすい候補化合物の合理的な設計は、ポリペプチドの分子形状の構造的研究に基づくことができる。特定の他のタンパク質と相互作用する部位を決定する1つの手段は、物理的構造決定、例えば、X線結晶回折または二次元NMR技術である。これらは、アミノ酸残基が分子接触領域を形成する手引きを提供する。タンパク質構造決定の詳細な説明に関して、例えば、Blundell及びJohnson(1976)Protein Crystallography、Academic Press、ニューヨークを参照されたい。
【0131】
代わりの合理的な設計は、その表面にVDCCγ-8受容体ポリペプチドを発現する適切な細胞を一般に産生することを含むスクリーニング法を用いる。このような細胞としては、動物、酵母、ショウジョウバエ、または大腸菌に由来する細胞が挙げられる。次に、受容体を発現している細胞(または発現受容体を含有する細胞膜)を試験化合物と接触させて、結合、または機能的応答の刺激もしくは阻害を見る。例えば、アフリカツメガエル卵母細胞に、VDCCγ-8 mRNAまたはポリペプチドを注入でき、さらに別途詳細に記載されるように、測定電位クランプの使用により測定される試験化合物への曝露により電流が誘導される。
【0132】
VDCCγ-8イオンチャネルにより個々に各候補化合物を試験する代わりに、候補リガンドのライブラリーまたはバンクが有利には作製され、スクリーニングされ得る。従って、例えば、200を超える推定上の受容体リガンドのバンクを、スクリーニング用に組み立てることができる。このバンクは:イオンチャネルを介して作用することが知られている伝達物質、ホルモン及びケモカイン;イオンチャネル、非哺乳動物、哺乳動物の対応物が未だ同定されていない生物活性ペプチドに対する推定上のアゴニストであり得る天然化合物;及び天然には見られないが、未知の天然リガンドによりイオンチャネルを活性化する化合物、を含む。さらに詳細に別途記載されているように、機能的アッセイ(すなわち、カルシウム、マイクロフィジオメーター(microphysiometer)、FLIPRアッセイ、全細胞電気生理学、卵母細胞電気生理学等、別途参照)並びに結合アッセイの両方を用いて既知のリガンドに関する受容体をスクリーニングするために、このバンクが用いられる。しかしながら、今までのところ、同族でない活性化リガンド(アゴニスト)または不活化リガンド(アンタゴニスト)が残されている多数の哺乳動物の受容体が存在する。このように、これらの受容体に対する活性リガンドは、今日まで同定されたリガンドバンク内に含ませることができない。従って、VDCCγ-8はまた、天然のリガンドを同定するために、組織抽出物に対して機能的にスクリーニング(カルシウム、cAMP、マイクロフィジオメーター(microphysiometer)、卵母細胞電気生理学等を用いて機能的スクリーニング)することができる。陽性の機能的応答を生じる抽出物は、活性化リガンドが単離され、同定されるまで、本明細書に記載されるようにアッセイされるフラクションにより連続的に細分画できる。
【0133】
このような実験において、形質移入されたHEK293細胞またはベクターコントロール細胞における基本のカルシウムレベルは、100〜200 nMの正常な範囲であることが観察される。ホモメリックまたはヘテロメリックVDCCγ-8イオンチャネル、あるいは組換えホモメリックまたはヘテロメリックVDCCγ-8イオンチャネルを発現するHEK293細胞は、fura 2により負荷され、1日に150を超える選択リガンドまたは組織/細胞抽出物は、アゴニスト誘導カルシウム動員について評価される。同様に、VDCCγ-8イオンチャネルまたは組換えVDCCγ-8イオンチャネルを発現するHEK 293細胞は、Ca2+フラックスの増加または減少について評価される。一過性のカルシウムを供するアゴニストは、その応答が受容体を発現する形質移入細胞に特有であるかどうかを判定するために、ベクターコントロール細胞において試験される。
【0134】
別の方法は、VDCCγ-8イオンチャネルの阻害または刺激を判定することにより受容体阻害剤に関するスクリーニングを含む。このような方法は、ホモメリックチャネルを形成するために単独で、または細胞表面に受容体を発現させるヘテロメリックチャネルを形成するために他のTrpチャネルサブユニットとともに、VDCCγ-8サブユニットを真核細胞に形質移入することを含む。次いで、VDCCγ-8受容体の存在下、該細胞を強力なアンタゴニストに曝露させる。電流の電導度または動態の変化を測定するために、全細胞電気生理学を用いて、該細胞を試験することができる。
【0135】
VDCCγ-8のアゴニストまたはアンタゴニストを検出する別の方法は、参照として本明細書に組み込まれている、米国特許第5,482,835号に記載されている酵母ベースの技術である。
【0136】
候補化合物がタンパク質である場合、特定の抗体またはペプチドにおいて、候補化合物のライブラリーを、ファージディスプレイ技術を用いてスクリーニングできる。ファージディスプレイは、組換えバクテリオファージを利用する分子スクリーニングのプロトコールである。この技術は、各々のファージまたはファージミドが特定の候補化合物を発現するように、候補化合物のライブラリーから1つの化合物をコードする遺伝子によりバクテリオファージを形質転換することを含む。形質転換バクテリオファージ(好ましくは固体支持体に係留される)は、適切な候補化合物を発現し、それらのファージコート上に示す。VDCCγ-8ポリペプチドまたはペプチドに結合できる特定の候補化合物は、アフィニティー相互作用に基づく選択様式により富化される。次に好結果の候補薬剤を特性化する。ファージディスプレイは、標準的なアフィニティーリガンドスクリーニング技術よりも利点を有する。ファージ表面は、三次元立体配置で候補薬剤を示し、その天然の立体配座により密接に類似している。これは、スクリーニング目的のために、より特異的かつより高いアフィニティー結合を可能にする。
【0137】
化合物のライブラリーをスクリーニングする別の方法は、化合物のライブラリーを発現する組換えDNA分子により安定に形質転換される真核宿主細胞または原核宿主細胞を利用する。生存形態または固定形態のこのような細胞を、標準的な結合パートナーアッセイに使用できる。また、細胞応答を検出するために高感度法を記載しているParceら(1989)Science 246:243〜247頁;及びOwickiら(1990)Proc. Nat'l Acad. Sci. USA 87;4007〜4011頁を参照されたい。競合アッセイは特に有用であり、化合物のライブラリーを発現している細胞を、125I-抗体等のVDCCγ-8ポリペプチドに結合することが知られている標識抗体、及び結合組成物に対する結合アフィニティーが測定される候補化合物等の試験サンプルと接触させるか、またはインキュベートする。次いでポリペプチドに対する結合及び遊離標識結合パートナーは、結合度を評価するために分離される。結合された試験サンプル量は、ポリペプチドに結合する標識抗体量に反比例する。
【0138】
多くの技術の中で任意の1つを、結合度を評価するために遊離結合パートナーから結合分離するために使用できる。この分離工程は、典型的にフィルターへの接着、フィルターへの接着後の洗浄、プラスチックへの接着後の洗浄、または細胞膜の遠心分離等の手法を典型的に含み得る。
【0139】
さらに別のアプローチは、例えば、変換された真核生物または原核生物宿主細胞から抽出された可溶性、未精製または可溶性の精製ポリペプチドまたはペプチドを使用することである。これは、特異性増加の利点、自動化する能力、及び薬物試験のハイスループットを有して「分子」結合アッセイを可能にする。
【0140】
候補化合物スクリーニングに関する別の技術は、例えば、VDCCγ-8ポリペプチドに対する好適な結合アフィニティーを有する新規な化合物をスクリーニングするハイスループットを提供するアプローチを含み、1984年9月13日に公表された国際特許出願WO第84/03564号(Commonwealth Serum Labs.)に詳細に記載されている。最初に、多数の異なる小型ペプチド試験化合物は、固体支持体、例えば、プラスチックピンまたはいくつかの他の適切な表面上で合成され、Fodorら(1991)を参照されたい。次いで、ピンのすべてを、可溶性VDCCγ-8ポリペプチドと反応させて洗浄する。次の工程は、結合ポリペプチドを検出することを含む。このように、ポリペプチドと特異的に相互作用する化合物が同定される。
【0141】
リガンド結合アッセイは、受容体薬理を確認するための直接的方法を提供し、ハイスループットフォーマットに適応可能である。受容体に対する精製リガンドは、結合研究のために特異的高活性(50〜2000 Ci/mmol)に放射標識できる。次いで、放射標識の処理により、その受容体に対するリガンドの活性を減少させないという決定がなされる。緩衝液、イオン、pH及びヌクレオチド等の他のモジュレーターに関するアッセイ条件は、膜及び全細胞受容体源両方に関して稼動シグナル対ノイズ比を確立するために最適化される。これらのアッセイに関して、特定の受容体結合は、全関連放射活性マイナス過剰の未標識競合リガンドの存在下で測定された放射活性として定義される。可能な場合、2つ以上の競合リガンドは、残存非特異的結合を規定するために使用される。
【0142】
該アッセイは、候補化合物の結合を簡便に試験することができ、その中で、受容体を有する細胞への接着は、候補化合物と直接的または間接的に関連する標識により、または標識競合物との競合を含むアッセイにおいて検出される。さらに、これらのアッセイは、細胞表面に受容体を有する細胞に適切な検出システムを用いて、候補化合物が受容体の活性化により作製されたシグナルを生じるかどうかを試験することができる。活性化阻害剤は、既知のアゴニストの存在下で一般にアッセイされ、候補化合物の存在下、アゴニストによる活性化に対する効果が見られる。
【0143】
さらに、該アッセイは、混合物を形成するためにVDCCγ-8ポリペプチドを含有する溶液と候補化合物とを混合する工程と、混合物中のVDCCγ-8イオンチャネル活性を測定する工程と、標品に対して混合物のVDCCγ-8イオンチャネル活性を比較する工程とを簡便に含ませることができる。
【0144】
VDCCγ-8サブユニットcDNA、タンパク質、及びタンパク質に対する抗体を使用して、細胞中のVDCCγ-8サブユニットmRNA及びタンパク質の産生に対する添加化合物の効果を検出するアッセイを構成することができる。例えばELISAは、当業者に既知の標準的方法によりモノクローナル及びポリクローナル抗体を用いてVDCCγ-8サブユニットタンパク質の分泌または細胞関連レベルを測定するために構成でき、これは、好適に操作された細胞または組織からVDCCγ-8サブユニットの産生を阻害または増強できる薬剤(それぞれアンタゴニストまたはアゴニストとも呼ばれる)を発見するために使用され得る。スクリーニングアッセイを実施するための標準的方法は、当業者に十分に理解される。
【0145】
強力なVDCCγ-8イオンチャネルのアンタゴニスト及びブロッカーの例としては、抗体または、いくつかの場合、プリン及びプリン類縁体等のヌクレオチド及びそれらの類縁体、VDCCγ-8イオンチャネルのリガンド、例えばリガンドの断片、あるいは受容体の活性を抑制するように受容体に結合するが、応答を誘発しない小型分子に密に関連するオリゴヌクレオチドまたはタンパク質が挙げられる。
【0146】
従って、我々はまた、VDCCγ-8ポリペプチド及び/またはペプチドに特異的に結合できる化合物を提供する。
【0147】
用語「化合物」とは、生体高分子(例えば、核酸、タンパク質、非ペプチド、または有機分子)等の化学的化合物(天然または合成)、あるいは細菌、植物、真菌、または動物(特に哺乳動物)細胞もしくは組織等の生体材料から作製された抽出物、あるいは無機要素または分子さえも称する。該化合物は抗体であることが好ましい。
【0148】
実施されるこのようなスクリーニングに必要な材料を、スクリーニング用キット内にパッケージできる。このようなスクリーニング用キットは、VDCCγ-8イオンチャネルポリペプチドの産生を減少させるか、または増強させるVDCCγ-8ポリペプチドまたは化合物に関するアゴニスト、アンタゴニスト、リガンド、受容体、基質、酵素等を同定するために有用である。該スクリーニング用キットは:(a)VDCCγ-8ポリペプチド;(b)VDCCγ-8ポリペプチドを発現する組換え細胞;(c)VDCCγ-8ポリペプチドを発現している細胞膜;または(d)VDCCγ-8ポリペプチドに対する抗体を含む。該スクリーニング用キットは、場合によっては、取扱い説明書を含むことができる。
【0149】
[トランスジェニック動物]
天然もしくは組換えVDCCγ-8イオンチャネル、またはホモログ、バリアントもしくは誘導体を、正常な発現レベルと比較して上昇または減少レベルで発現できるトランスジェニック動物は、本発明の範囲内に入る。VDCCγ-8遺伝子の欠失等1つ以上の機能変異の結果、機能的VDCCγ-8受容体を発現しないトランスジェニック動物(VDCCγ-8ノックアウト動物)が含まれる。このようなトランスジェニック動物は、ブタ、ヒツジ、またはげっ歯類等の非ヒト哺乳動物であることが好ましい。トランスジェニック動物は、マウスまたはラットであることが最も好ましい。このようなトランスジェニック動物をスクリーニング法に使用して、VDCCγ-8イオンチャネルのアゴニスト及び/またはアンタゴニストを同定でき、並びに疾患のin vivo治療としてそれらの有効性を試験することができる。
【0150】
好ましい実施形態において、トランスジェニックVDCCγ-8動物、特にVDCCγ-8ノックアウトは、好ましくは本明細書に設定された試験により測定されるように、実施例に示された表現型を示す。従って、VDCCγ-8動物、特にVDCCγ-8ノックアウトは、好ましくは1つ以上の、以下の:ロータロッド性能の減少、新規環境に対する馴化の減少、認知欠損または発作感受性を示す。
【0151】
非常に好ましい実施形態において、VDCCγ-8動物、特にVDCCγ-8ノックアウトは、対応する野生型マウスと比較して、測定パラメータの少なくとも10%、好ましくは少なくとも20%、より好ましくは少なくとも30%、40%、50%、60%、70%、80%、90%、または100%高いまたは低い(場合によっては)を示す。
【0152】
例えば、VDCCγ-8イオンチャネルの産生を欠くように操作されたトランスジェニック動物を、VDCCγ-8イオンチャネルのアゴニスト及び/またはアンタゴニストを同定するアッセイに使用できる。1つのアッセイは、VDCCγ-8イオンチャネル受容体の非存在下で生理的応答を生じるかどうかを判定するために潜在的薬物(候補リガンドまたは化合物)を評価するように設計される。これは、上記に考察されたようにトランスジェニック動物に薬物を投与し、次いで動物を特定の応答に関してアッセイすることにより達成され得る。任意の生理的パラメータをこのアッセイで測定し得るが、好ましい応答としては、1つ以上の、以下の:耐疾患変化;炎症応答の変化;発作感受性の変化;認知性能の変化;シナプス可塑性の変化;社会的相互作用の変化;が挙げられる。
【0153】
潜在的薬物(候補リガンドまたは化合物)がVDCCγ-8受容体に結合するかどうかを判定するために、VDCCγ-8ノックアウト動物に由来する組織を受容体結合アッセイに使用できる。このようなアッセイは、VDCCγ-8受容体産生を欠くように操作されたトランスジェニック動物からの第一の受容体調製物及び同定された任意のVDCCγ-8リガンドまたは化合物を結合することが知られている供給源からの第二の受容体調製物を得ることにより実施され得る。一般に、第一及び第二の受容体調製物は、それらが得られる供給源を除いてすべての点で類似している。例えば、トランスジェニック動物(上記及び下記等)からの脳組織をアッセイに使用する場合、正常(野生型)動物からの同等の脳組織は、第二の受容体調製物の供給源として使用される。各受容体調製物は、VDCCγ-8受容体に結合することが知られているリガンド単独で、及び候補リガンドまたは化合物の存在下の両方でインキュベートされる。候補リガンドまたは化合物は、いくつかの異なる濃度で調べることが好ましいであろう。
【0154】
既知のリガンドによる結合が、試験化合物により置換される範囲は、第一及び第二の受容体調製物の両方に関して決定される。トランスジェニック動物由来の組織を直接アッセイに使用できるか、または該組織は、それら自体アッセイに使用される膜または膜タンパク質を単離するために処理され得る。好ましいトランスジェニック動物はマウスである。該リガンドは、結合アッセイと適合する任意の手段を用いて標識できる。これには、限定はしないが、放射活性、酵素的、蛍光または化学発光標識(並びにさらに詳細に上記に示される他の標識技術)が挙げられるであろう。
【0155】
さらに、VDCCγ-8イオンチャネル受容体のアンタゴニストは、機能的VDCCγ-8を発現する野生型動物、及びVDCCγ-8受容体機能の発現減少または発現消滅と関連する任意の表現型特徴を示す同定された動物に候補化合物等を投与することにより同定され得る。
【0156】
非ヒトトランスジェニック動物を作製する詳細な方法は、下記にさらに詳細に説明される(実施例1及び2)。トランスジェニック遺伝子構築体を、トランスジェニック哺乳動物を作製するために動物の生殖細胞系に導入できる。例えば、該構築物の1つまたはいくつかのコピーを、標準的なトランスジェニック技術により哺乳動物の胎仔のゲノムに取り込むことができる。
【0157】
代表的な実施形態において、トランスジェニック非ヒト動物は、非ヒト動物の生殖細胞系に導入遺伝子を導入することにより作製される。種々の発達段階における胎仔標的細胞は、導入遺伝子を導入するために使用され得る。胎仔標的細胞の発達段階によって種々の方法が用いられる。任意の動物の特定系は、一般的な良好な健康、良好な胎仔収率、胎仔における良好な前核可視性、及び良好な生殖適合性に関して選択される。さらに、ハプロタイプは重要な因子である。
【0158】
導入遺伝子の胎仔への導入は、例えば、マイクロインジェクション、エレクトロポレーション、またはリポフェクション等、当業者に既知の任意の手段により達成され得る。例えば、該構築体の1つ以上のコピーを発達哺乳動物の細胞に保持させるために哺乳動物の受精卵の前核内に該構築体のマイクロインジェクションにより哺乳動物にVDCCγ-8受容体導入遺伝子を導入できる。受精卵に導入遺伝子構築体の導入後、該卵子は、時間量を変えるためにin vitroでインキュベートできるか、または代理宿主に再移植できるか、あるいはそれらの両方を行うことができる。成熟性に向けてin vitroインキュベーションもまた実施できる。1つの一般的な方法は、種によって約1〜7日間in vitroで胎仔をインキュベートし、次いで胎仔を代理宿主に再移植させることである。
【0159】
遺伝子導入により操作された胎仔の後代を、組織のセグメントのサザンブロット解析により該構築体の存在に関して試験することができる。外因性クローニング構築体の1つ以上のコピーが、このようなトランスジェニック胎仔のゲノムに安定に組込まれたままである場合、遺伝子導入により付加された構築体を担持する永続的トランスジェニック哺乳動物系を確立することが可能である。
【0160】
遺伝子導入により改変された哺乳動物の同腹仔を、子孫のゲノムに該構築体の取込みのために誕生後にアッセイできる。好ましくは、このアッセイは、後代からの染色体材料上の所望の組換えタンパク質産物またはそのセグメントをコードするDNA配列に相当するプローブをハイブリダイズすることによって達成される。それらのゲノムにおける構築体の少なくとも1つのコピーを含有することが分かっているこれらの哺乳動物の後代を、成熟性に向けて成長させる。
【0161】
本文書の目的に関して、受精卵は、本質的に完全生物に成長できる二倍体細胞の形成である。一般に、受精卵は、1つまたは複数の配偶子から2つのハプロイド核の融合によって、天然にまたは人工的に形成された核を含有する卵子を含む。従って、配偶子核は、天然に適合するもの、すなわち、機能的生物への分化及び発達を受けることができる生存受精卵をもたらすものでなければならない。一般に正倍数性受精卵が好ましい。異数性受精卵が得られる場合、染色体数は、どちらかの配偶子が生じた生物の正倍数に関して2つ以上変える必要はない。
【0162】
同様の生物学的考察に加えて、物理的なものは、受精卵核または受精卵核の一部を形成する遺伝物質に加えることができる外因性遺伝物質量(例えば、容量)を調節する。遺伝物質が取り出されない場合、加えることのできる外因性遺伝物質量は、物理的に破壊されることなく吸収される量によって限定される。挿入される外因性遺伝物質の容量は、一般に約10ピコリットルを超えないであろう。添加の物理的効果は、受精卵の生存を物理的に破壊するほど大きなものであってはならない。DNA配列の数及び種類の生物学的限定は、特定の受精卵及び外因性遺伝物質の機能によって変わり、当業者には容易に理解される。というのは、生じた受精卵の外因性遺伝物質等の遺伝物質は、受精卵の機能的生物への分化及び発達を生物学的に開始させ、維持できなければならないからである。
【0163】
受精卵に添加される導入遺伝子構築体のコピー数は、添加される外因性遺伝物質の全量に依存し、遺伝的形質転換をもたらし得る量である。理論的には、1つだけのコピーが必要であり;しかしながら、一般に、1つのコピーが機能的であることを確保するために、多くのコピー、例えば、導入遺伝子構築体の1,000〜20,000のコピーが利用される。しばしば、外因性DNA配列の表現型発現を増強させるために挿入された各外因性DNA配列の2つ以上の機能的コピーを有することが有利である。
【0164】
核遺伝物質への外因性遺伝物質の添加を可能にする任意の技術を、それが、細胞、核膜または他の既存の細胞もしくは遺伝子構造に対して破壊的でない限り利用できる。外因性遺伝物質は、マイクロインジェクションにより核酸遺伝物質に優先的に挿入される。細胞及び細胞構造のマイクロインジェクションは既知であり、当業者に使用される。
【0165】
再移植は、標準的な方法を用いて達成される。通常、代理宿主を麻酔にかけ、胎芽を輸卵管に挿入する。特定の宿主に移植された胎芽数は種によって変わるが、通常、種が天然に生む子孫数に相当する。
【0166】
代理宿主のトランスジェニック子孫は、任意の好適な方法により導入遺伝子の存在及び/または発現についてスクリーニングされ得る。スクリーニングはしばしば、導入遺伝子の少なくとも一部に相補的であるプローブを用いて、サザンブロットまたはノーザンブロット解析により達成される。導入遺伝子によりコード化されたタンパク質に対する抗体を用いるウェスタンブロット解析を、導入遺伝子産物の存在をスクリーニングする代わりのまたはさらなる方法として使用できる。典型的には、DNAは、尾部組織から調製され、導入遺伝子に関するサザン解析またはPCRにより分析される。あるいは、最高レベルで導入遺伝子を発現すると考えられている組織または細胞は、サザン解析またはPCRを用いて導入遺伝子の存在及び発現に関して試験されるが、任意の組織または細胞型を、この解析のために使用できる。
【0167】
導入遺伝子の存在を評価するための代わりのあるいはさらなる方法としては、限定はしないが、酵素及び/または免疫学的アッセイ等の好適な生化学的アッセイ、特定のマーカーまたは酵素活性に関する組織学的染色、フローサイトメトリー分析等が挙げられる。血液分析もまた、血中導入遺伝子産物の存在を検出するために、並びに種々のタイプの血液細胞及び他の血液構成物のレベルに対する導入遺伝子の効果を評価するために有用となり得る。
【0168】
トランスジェニック動物の後代は、トランスジェニック動物と好適なパートナーとの交配、またはトランスジェニック動物から得られた卵子及び/または精子のin vitro受精により得ることができる。パートナーとの交配が実施される場合、パートナーは、トランスジェニック及び/またはノックアウトであってもそうでなくてもよく;パートナーがトランスジェニックである場合、同じかまたは異なる導入遺伝子、あるいは両方を含有できる。あるいは、パートナーは親系であってもよい。in vitro交配が用いられる場合、交配胚芽は、代理宿主に移植してもよく、またはin vitroでインキュベートしてもよく、あるいは両方であってもよい。いずれかの方法を用いる後代は、上記の方法または他の適切な方法を用いて導入遺伝子の存在に関して評価され得る。
【0169】
本明細書に記載された方法に従って作製されたトランスジェニック動物は、外因性遺伝物質を含む。上記に示されたように、一定の実施形態において、外因性遺伝物質は、VDCCγ-8イオンチャネルの産生を生じるDNA配列である。さらに、このような実施形態において、該配列は、転写調節要素、例えば、好ましくは特定タイプの細胞において導入遺伝子産物の発現を可能にするプロモーターに結合する。
【0170】
レトロウイルス感染もまた、導入遺伝子を非ヒト動物に導入するために使用できる。非ヒト胚芽の発達は、in vitroで胚盤胞段階に培養できる。この時期の間、分割球はレトロウイルス感染の標的であり得る(Jaenich, R.(1976)PNAS 73:1260〜1264頁)。分割球の効率的な感染は、透明層を除去するために酵素処理により得られる(Manipulating the Mouse Embryo、Hogan編集(Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、1986年)。導入遺伝子を導入するために用いられるウイルスベクター系は、典型的に導入遺伝子を運搬する複製欠損レトロウイルスである(Jahnerら(1985)PNAS 82:6927〜6931頁;Van der Puttenら(1985)PNAS 82:6148〜6152頁)。形質移入は、ウイルス産生細胞の単層上で分割球を培養することによって容易にかつ効率的に得られる(Van der Putten、上記文献;Stewartら(1987)EMBO J. 6:383〜388頁)。あるいは、感染は後期段階で実施され得る。ウイルスまたはウイルス産生細胞を分割腔に注入できる(Jahnerら(1982)Nature 298:623〜628頁)。トランスジェニック非ヒト動物を形成した細胞のサブセットのみに取込みが生じることから、ファウンダーの大部分は導入遺伝子に関するモザイクである。さらに、ファウンダーは、一般に子孫において分離するゲノムの異なる位置で導入遺伝子の種々のレトロウイルス挿入を含有することができる。さらに、妊娠中期の胎仔の子宮内レトロウイルス感染による生殖細胞系に導入遺伝子を導入することも可能である(Jahnerら(1982)上記文献)。
【0171】
導入遺伝子の導入に関する第三のタイプの標的細胞は、胎芽幹細胞(ES)である。ES細胞は、in vitroで培養され、胎芽と融合された移植前胎芽から得られる(Evansら(1981)Nature 292:154〜156頁;Bradleyら(1984)Nature 309:255〜258頁;Gosslerら(1986)PNAS 83:9065〜9069頁;及びRobertsonら(1986)Nature 322:445〜448頁)。DNA形質移入またはレトロウイルス媒介形質導入により、導入遺伝子をES細胞に効率的に導入できる。このように変換されたES細胞はその後、非ヒト動物から胚盤胞と組み合わせることができる。その後、ES細胞は、胎芽をコロニー化し、生じたキメラ動物の生殖細胞系に寄与する。レビューに関してJaenisch, R.(1988)Science 240:1468〜1474頁を参照されたい。
【0172】
非ヒトトランスジェニック動物が提供され、トランスジェニック動物は、好ましくは上記のとおり、VDCCγ-8イオンチャネル機能のモデルとしてVDCCγ-8遺伝子の変更を有することを特徴とする。遺伝子に対する変更としては、機能変異の欠失または他の喪失、標的化または無作為変異によりヌクレオチド配列を有する外因性遺伝子の導入、別の種からの外因性遺伝子の導入、またはそれらの組合せが挙げられる。トランスジェニック動物は、変更のためにホモ接合性であってもヘテロ接合性であってもよい。それから誘導された動物及び細胞は、VDCCγ-8イオンチャネル機能を調節できる生物活性剤をスクリーニングするために有用である。このスクリーニング法は、疼痛及び癌、特に前立腺癌の潜在的療法の特異性及び作用を判定するために特に有用である。該動物は、脳、心臓、脾臓、及び肝臓等の正常な組織、並びに臓器におけるVDCCγ-8イオンチャネルの役割及びそれらの機能に対する効果を調査するモデルとして有用である。
【0173】
別の態様は、機能的に破壊された内因性VDCCγ-8遺伝子を有するが、また、そのゲノムに運搬し、異種のVDCCγ-8タンパク質(すなわち、別の種からのVDCCγ-8)をコードする導入遺伝子を発現するトランスジェニック非ヒト動物に関する。該動物はマウスであることが好ましく、異種VDCCγ-8はヒトVDCCγ-8である。ヒトVDCCγ-8により再構成されたこのような動物から誘導された動物または細胞系を、in vivo及びin vitroでヒトVDCCγ-8を阻害する薬剤を同定するために使用できる。例えば、ヒトVDCCγ-8を介してシグナル伝達を誘導する刺激剤を、試験される薬剤の存在下及び非存在下で動物または細胞系に投与でき、動物または細胞系の応答を測定できる。in vivo及びin vitroでヒトVDCCγ-8を阻害する薬剤は、薬剤の非存在下の応答と比較して、薬剤の存在下で応答減少に基づいて同定され得る。
【0174】
VDCCγ-8欠損トランスジェニック非ヒト動物(「VDCCγ-8サブユニットノックアウト」)もまた提供される。このような動物は、好ましくは、破壊または削除される内因性VDCCγ-8イオンチャネルゲノム配列の結果、VDCCγ-8イオンチャネルの低い活性を発現する、または活性を発現しないものである。好ましくは、このような動物は、イオンチャネル活性を発現しない。さらに好ましくは、該動物は、配列番号3または配列番号5として示されるVDCCγ-8イオンチャネルの活性を発現しない。VDCCγ-8イオンチャネルノックアウトを、下記にさらに詳細に説明されるように、当業者に既知の種々の手段により作製できる。
【0175】
本発明はまた、宿主細胞においてVDCCγ-8遺伝子を機能的に破壊するための核酸構築体に関する。該核酸構築体は:a)非相同的置換部分;b)非相同的置換部分の上流に位置する第一の相同領域、第一のVDCCγ-8遺伝子配列に対して実質的に同一性を有するヌクレオチド配列を有する第一の相同領域;及びc)非相同的置換部分の下流に位置する第二の相同領域、第二のVDCCγ-8遺伝子配列に対して実質的に同一性を有するヌクレオチド配列を有する第二の相同領域、天然の内因性VDCCγ-8遺伝子において第一のVDCCγ-8遺伝子配列の下流に位置する第二のVDCCγ-8遺伝子配列、を含む。さらに、第一及び第二の相同領域は、核酸分子が宿主細胞に導入される場合、核酸構築体と宿主細胞における内因性VDCCγ-8遺伝子との間の相同的組換えのために十分な長さである。好ましい実施形態において、非相同的置換部分は、好ましくは、調節要素に操作的に連結されたネオマイシンホスホトランスフェラーゼ遺伝子を含み、好ましくは、lacZ及び陽性選択発現カセット等の発現レポーターを含む。
【0176】
好ましくは、第一及び第二のVDCCγ-8遺伝子配列は、配列番号1、配列番号2、配列番号4もしくは配列番号6、またはそれらのホモログ、バリアントもしくは誘導体から誘導される。
【0177】
別の態様は、上記の核酸構築体が取り込まれた組換えベクターに関する。さらに別の態様は、核酸構築体が導入され、それによって核酸構築体と宿主細胞の内因性VDCCγ-8遺伝子との間で相同的組換えを可能にし、内因性VDCCγ-8遺伝子の機能的破壊を生じる宿主細胞に関する。宿主細胞は、肝臓、脳、脾臓または心臓、あるいはマウス胎芽幹細胞等の多能性細胞からVDCCγ-8遺伝子を正常に発現する哺乳動物細胞であり得る。核酸構築体が導入され、内因性VDCCγ-8遺伝子と相同的に組換えられた胎芽幹細胞のさらなる発達によって、胎芽幹細胞からの子孫であり、このようにゲノムにVDCCγ-8遺伝子破壊を担持する細胞を有するトランスジェニック非ヒト動物が作製される。次に生殖細胞系にVDCCγ-8遺伝子破壊を保持する動物は、すべての体細胞及び生殖細胞においてVDCCγ-8遺伝子破壊を有する動物を作製するために選択され、繁殖され得る。次いで、このようなマウスを、VDCCγ-8遺伝子破壊のためのホモ接合性に繁殖できる。
【0178】
[抗体]
本文書の目的に関して、用語の「抗体」とは、それと反対に指定されない限り、ポリクローナル、モノクローナル、キメラ、一本鎖、Fab断片及びFab発現ライブラリーにより生じた断片を含むが、これらに限定されない。このような断片としては、標的物質、Fv、F(ab')及びF(ab')2断片に関する結合活性を保持している全抗体の断片、並びに一本鎖抗体(scFv)、融合タンパク質及び抗体の抗原結合部位を含む他の合成タンパク質の断片が挙げられる。抗体及びその断片は、例えば、EP-A-239400に記載されるようなヒト化抗体であり得る。さらに、例えば、米国特許第5,545,807号及び米国特許第6,075,181号に記載されるような完全ヒト可変領域の抗体(またはそれらの断片)もまた使用できる。中和抗体、すなわち物質のアミノ酸配列の生物学的活性を阻害するものは、診断薬及び治療薬用に特に好ましい。
【0179】
免疫化またはファージディスプレイライブラリーを用いることによる等の標準的な技術により、抗体を作製することができる。
【0180】
ポリペプチドまたはペプチドを、既知の技術によって抗体を産生するために使用できる。このような抗体は、VDCCγ-8イオンチャネルタンパク質またはホモログ、断片等に特異的に結合できる。
【0181】
ポリクローナル抗体が望ましい場合、選択された哺乳動物(例えば、マウス、ウサギ、ヤギ、ウマ等)は、関連ポリペプチドまたはペプチドを含む免疫原性組成物により免疫化され得る。宿主種によって、種々のアジュバントを、免疫学的応答を増加させるために使用できる。このようなアジュバントとしては、限定はしないが、フロイント、水酸化アルミニウム等の無機ゲル、リソレシチン、プルロニックポリオール、ポリアニオン、ペプチド、油乳濁液、スカシ貝ヘモシアニン、及びジニトロフェノール等の表面活性物質が挙げられる。BCG(カルメット・ゲラン杆菌)及びコリネバクテリウムパルブムは、精製物質のアミノ酸配列を、全身防御を刺激する目的で免疫学的に易感染性個体に投与する場合に使用できる潜在的に有用なヒトアジュバントである。
【0182】
免疫化動物からの血清は、既知の方法に従って採取され、処理される。VDCCγ-8ポリペプチドから得ることができるエピトープに対するポリクローナル抗体を含有する血清が他の抗原に対する抗体を含有する場合、ポリクローナル抗体は、免疫アフィニティークロマトグラフィにより精製され得る。ポリクローナル抗血清を作製し、処理する技術は当業者に既知である。このような抗体を作製できるように、我々はまた、動物またはヒトにおける免疫原として使用するために別のアミノ酸配列にハプテン化されたVDCCγ-8のアミノ酸配列またはその断片を提供する。
【0183】
VDCCγ-8イオンチャネルポリペプチドまたはペプチドから得ることができるエピトープに対する特異的なモノクローナル抗体もまた、当業者により容易に作製され得る。ハイブリドーマによるモノクローナル抗体を作製するための一般的な方法論は周知である。不死化抗体産生細胞系は、細胞融合、及び発癌性DNAによるBリンパ球の直接的形質転換、またはエプスタインバルウイルスによる形質移入等の他の技術により作製され得る。軌道(orbit)エピトープに対して作製されたモノクローナル抗体のパネルは、種々の性質、すなわちイソタイプ及びエピトープアフィニティーに関してスクリーニングされ得る。
【0184】
モノクローナル抗体は、連続培養の細胞系により抗体分子の産生を提供する任意の技術を用いて調製され得る。これらとしては、限定はしないが、Koehler及びMilstein(1975年Nature 256:495〜497頁)により原型として記載されたハイブリドーマ技術、トリオーマ技術、ヒトB細胞ハイブリドーマ技術(Kosborら(1983)Immunol Today 4:72頁;Coteら(1983)Proc Natl Acad Sci 80:2026〜2030頁)及びEBVハイブリドーマ技術(Coleら、Monoclonal Antibodies and Cancer Therapy、77〜96頁、Alan R. Liss社、1985年)が挙げられる。
【0185】
さらに、「キメラ抗体」の産生のために開発された技術、すなわち適切な抗原特異性及び生物活性を有する分子を得るためにヒト抗体遺伝子に対するマウス抗体遺伝子のスプライシングを使用できる(Morrisonら(1984)Proc Natl Acad Sci 81:6851〜6855頁;Neubergerら(1984)Nature 312:604〜608頁;Takedaら(1985)Nature 314:452〜454頁)。あるいは、一本鎖抗体の産生のために記載された技術(米国特許第4,946,779号)は、物質特異的一本鎖抗体を産生するために適合できる。
【0186】
VDCCγ-8イオンチャネルポリペプチドまたはペプチドから得ることができるエピトープに対する抗体、モノクローナル及びポリクローナルの両方は、特に診断に有用であり、中和するものは、受動的免疫療法に有用である。特にモノクローナル抗体を、抗イディオタイプ抗体を上昇させるために使用できる。抗イディオタイプ抗体は、保護が望ましい物質及び/または薬剤の「内部像」を担持する免疫グロブリンである。抗イディオタイプ抗体を上昇させる技術は当業者に既知である。これらの抗イディオタイプ抗体もまた治療に有用であり得る。
【0187】
抗体は、リンパ球集団でin vivo産生を誘導することによって、またはOrlandiら(1989年、Proc Natl Acad Sci 86:3833~3837頁) 及びWinter G及びMilstein C(1991年;Nature 349:293~299頁)に開示されているように、抗体がリンパ球集団においてもin vivo産生を誘導することによって、または組換え免疫グロブリンライブラリーもしくは一連の高特異的結合試薬をスクリーニングすることによって産生できる。Orlandiら(1989年、Proc Natl Acad Sci 86:3833〜3837頁)、及びWinter G及びMilstein C(1991年;Nature 349:293〜299頁)によっても産生できる。
【0188】
ポリペプチドまたはペプチドに特異的な結合部位を含有する抗体断片も作製され得る。例えば、このような断片としては、限定はしないが、抗体分子のペプシン消化により産生できるF(ab')2断片及びF(ab')2断片のジスルフィド架橋を還元することにより作製できるFab断片が挙げられる。あるいは、Fab発現ライブラリーは、望ましい特異性を有するモノクローナルFab断片の迅速かつ容易な同定を可能にするために構築できる(Huse WDら(1989)Science 256:1275〜1281頁)。
【0189】
一本鎖抗体の産生技術(米国特許第4,946,778号)はまた、VDCCγ-8イオンチャネルポリペプチドに対する一本鎖抗体を産生するために適合できる。また、トランスジェニックマウス、または哺乳動物を含む他の生物を、ヒト化抗体を発現させるために使用できる。
【0190】
ポリペプチドを発現するクローンを単離または同定するために、あるいはアフィニティークロマトグラフィによりポリペプチドを精製するために、上記抗体を使用できる。
【0191】
VDCCγ-8イオンチャネルポリペプチドに対する抗体を、VDCCγ-8関連疾患を治療するためにも使用できる。
【0192】
[診断アッセイ]
この開示はまた、診断薬としての診断における使用、または遺伝子解析における使用のための、VDCCγ-8イオンチャネルポリヌクレオチド及びポリペプチド(並びにそれらのホモログ、バリアント及び誘導体)の使用に関する。VDCCγ-8イオンチャネル核酸に相補的またはハイブリダイズできる核酸(ホモログ、バリアント及び誘導体を含む)、並びにVDCCγ-8ポリペプチドに対する抗体もまた、このようなアッセイに有用である。
【0193】
好ましい実施形態において、該診断アッセイは、患者から単離されたサンプルに対して実施されるものであり、該アッセイは、ヒトまたは動物の身体に対して実施されるものではない。
【0194】
機能障害と関連するVDCCγ-8イオンチャネル遺伝子の変異形態検出により、VDCCγ-8イオンチャネルの発現不足、過剰発現、または発現の変化から生じる疾患または疾患に対する感受性の診断に加えるか、またはその診断を規定できる診断手段を提供する。VDCCγ-8イオンチャネル遺伝子(コントロール配列を含む)において変異を担持する個体は、種々の技術によるDNAレベルで検出できる。
【0195】
例えば、DNAは患者から単離され、VDCCγ-8のDNA多型パターンを決定できる。この同定されたパターンは、VDCCγ-8の過剰発現、発現不足、または異常発現に関連する疾患に罹患していることが知られている患者のコントロールと比較される。次にVDCCγ-8関連疾患に関連する遺伝子多型パターンを発現する患者を同定できる。VDCCγ-8イオンチャネル遺伝子の遺伝子解析は、当業者に知られた任意の技術により実施できる。例えば、VDCCγ-8対立遺伝子のDNA配列を決定することにより、RFLPまたはSNP解析等により、個体をスクリーニングできる。VDCCγ-8に関する遺伝子配列におけるDNA多型の存在またはその発現を調節する任意の配列を検出することにより、VDCCγ-8の過剰発現、発現不足または異常発現に関連する疾患に関する遺伝子素因を有するものとして、患者を同定できる。
【0196】
そのように同定された患者は、VDCCγ-8関連疾患の発症を防ぐために処置できるか、またはその疾患のさらなる発生または発達を防ぐためにVDCCγ-8関連疾患の早期段階でより積極的に処置できる。
【0197】
VDCCγ-8関連疾患に関連する患者の遺伝子多型パターンの同定用キットもまた提供される。該キットには、DNA採取手段及び遺伝子多型パターンを測定する手段が含まれ、次いでVDCCγ-8関連疾患に対する患者の感受性を判定するためにコントロールサンプルと比較される。VDCCγ-8ポリペプチド及び/またはこのようなポリペプチドに対する抗体(またはその断片)を含むVDCCγ-8関連疾患の診断用キットもまた提供される。
【0198】
診断用核酸は、血液、尿、唾液、組織生検または剖検材料から等、対象の細胞から得ることができる。好ましい実施形態において、DNAは、患者の指穿刺により吸収紙に採取された血液より得られた血液細胞から得られる。さらなる好ましい実施形態において、血液は、AmpliCard(商標)に採取される(University of Sheffield、Department of Medicine and Pharmacology、Royal Hallamshire Hospital、シェフィールド、英国S10 2JF)。
【0199】
DNAは、検出用に直接使用され得るか、または分析前にPCRまたは他の増幅技術を用いることにより酵素的に増幅され得る。対象の遺伝子内の特定の多型DNA領域を標的にするオリゴヌクレオチドDNAプライマーを調製できることによって、標的配列のPCR反応増幅が達成される。RNAまたはcDNAもまた、テンプレートとして同様の方法で使用できる。次にテンプレートDNAからの増幅DNA配列は、制限酵素を用いて分析して増幅配列に存在する遺伝子多型を決定し、それによって患者の遺伝子多型プロファイルを提供できる。制限断片長を、ゲル分析により同定できる。あるいは、または関連してSNP(単一ヌクレオチド多型)解析等の技術を使用できる。
【0200】
欠失及び挿入は、正常な遺伝子型と比較して増幅産物のサイズを変化させて検出できる。点変異は、増幅DNAを標識VDCCγ-8イオンチャネルヌクレオチド配列にハイブリダイズすることにより同定され得る。完全一致配列は、RNアーゼ消化により、または融解温度の相違によりミスマッチ二重鎖と識別され得る。DNA配列の相違はまた、変性剤の有無でゲル中のDNA断片の電気泳動移動度の変化により、または直接的DNA配列決定により検出できる。例えば、Myersら、Science(1985)230:1242頁を参照されたい。特定の位置における配列変化はまた、RNアーゼ及びS1保護等のヌクレアーゼ保護アッセイ、または化学的開裂法により明らかにすることができる。Cottonら、Proc Natl Acad Sci USA(1985)85:4397〜4401頁を参照されたい。別の実施形態において、VDCCγ-8イオンチャネルヌクレオチド配列またはその断片を含むオリゴヌクレオチドプローブのアレイを構築して、例えば、遺伝子変異の効率的なスクリーニングを実施することができる。アレイテクノロジー法は周知であり、一般的な用途があり、遺伝子発現、遺伝子結合、及び遺伝的変異性等、分子遺伝学における種々の問題を扱うために使用され得る(例えば:M. Cheeら、Science、274巻、610〜613頁(1996)を参照)。
【0201】
一本鎖立体配座多型(SSCP)を使用して、変異体と野生型核酸との間の電気泳動移動度の相違を検出することができる(Oritaら(1989)Proc Natl. Acad. Sci USA:86:2766頁;Cotton(1993)Mutat Res 285:125〜144頁;及びHayashi(1992)Genet Anal Tech Appl 9:73〜79頁)。サンプル及びコントロールVDCCγ-8核酸の一本鎖DNA断片を変性でき、再生できる。一本鎖核酸の二次的構造は、配列によって異なり、電気泳動移動度で生じた変化は、単一の塩基変化さえも検出を可能にする。DNA断片は、標識プローブにより標識または検出され得る。アッセイの感度は、RNA(DNAよりも)を用いることにより増強でき、その二次的構造は、配列変化に対してより感度がよい。好ましい実施形態において、主題の方法は、電気泳動移動度の変化に基づいて二本鎖ヘテロ二重鎖分子を分離するためにヘテロ二重鎖分析を利用する(Keenら(1991)Trends Genet 7:5頁)。
【0202】
VDCCγ-8ポリペプチド及び核酸の存在を、サンプル中に検出することができる。このように、上記に掲げた感染症及び疾患は、対象由来のサンプルからVDCCγ-8ポリペプチドまたはVDCCγ-8イオンチャネルmRNAのレベルの異常減少または異常増加を測定することを含む方法により診断され得る。該サンプルは、発現レベルまたは発現パターンの空間的または時間的変化等、VDCCγ-8発現の増減あるいは異常に関連する疾患に罹患しているか、または罹患している疑いのある生物からの細胞または組織のサンプルを含むことができる。このような疾患に罹患している、または罹患している疑いのある生物におけるVDCCγ-8の発現レベルまたは発現パターンは、通常、疾患の診断手段として、正常な生物の発現レベルまたは発現パターンと比較される。
【0203】
従って一般に、サンプル中にVDCCγ-8核酸を含む核酸の存在を検出する方法は、サンプルと、上記核酸に特異的である少なくとも1つの核酸プローブとを接触させること、及び核酸の存在に関して上記サンプルをモニタリングすることを含む。例えば、該核酸プローブは、VDCCγ-8イオンチャネルサブユニット核酸、またはその一部に特異的に結合でき、2つの間の結合を検出でき、複合体自体の存在も検出できる。さらに、我々は、細胞サンプルと、ポリペプチドを結合できる抗体とを接触させること、及びポリペプチドの存在に関して上記サンプルをモニタリングすることによりVDCCγ-8ポリペプチドの存在を検出することを包含する。抗体とポリペプチドとの間で形成された複合体の存在をモニタリングすること、またはポリペプチドと抗体との間の結合をモニタリングすることにより、これを都合よく達成できる。2つの成分の間の結合を検出する方法は当業者に既知であり、FRET(蛍光共鳴エネルギー移動)、表面プラズモン共鳴等を含む。
【0204】
発現の減少または増加は、例えば、PCR、RT-PCR、RNアーゼ保護、ノーザンブロッティング法及び他のハイブリダイゼーション法等のポリヌクレオチドの定量化に関して周知の任意の方法を用いてRNAレベルを測定できる。宿主に由来するサンプル中のVDCCγ-8等のタンパク質のレベルを測定するために使用できるアッセイ技術は、当業者に周知である。このようなアッセイ法としては、放射免疫アッセイ、競合結合アッセイ、ウェスタンブロット解析及びELISAアッセイが挙げられる。
【0205】
疾患用診断キット、または、例えば疼痛及び癌、特に神経障害性痛及び前立腺癌の疾患(感染症を含む)に対する罹病性に関する診断用キットを、我々はさらに開示する。診断用キットは、VDCCγ-8ポリヌクレオチドまたはその断片、相補的ヌクレオチド配列、VDCCγ-8ポリペプチドまたはその断片、あるいはVDCCγ-8ポリペプチドに対する抗体を含む。
【0206】
[染色体アッセイ]
本明細書に記載されたヌクレオチド配列もまた、染色体の同定に役立つ。該配列は特異的に標的とされ、個々のヒト染色体上の特定の位置とハイブリダイズできる。上記のとおり、ヒトVDCCγ-8イオンチャネルは、ヒト染色体19q13にマッピングすることが分かっている。
【0207】
染色体に関連する配列のマッピングは、これらの配列と遺伝子関連疾患とを相関させる上で重要な最初の工程である。配列が、正確な染色体位置にマッピングされたら、染色体上の配列の物理的位置は、遺伝子マップデータと相関づけることができる。このようなデータは、例えば、V. McKusick、ヒトにおけるメンデルヘリタンス(Johns Hopkins University Welch Medical Libraryを介するライン上で入手できる)に見られる。次に、同じ染色体領域にマッピングされた遺伝子と疾患との間の関係を、連鎖解析(物理的に隣接する遺伝子の遺伝的形質)により同定する。
【0208】
cDNAまたはゲノム配列において罹患者と非罹患者との間の相違もまた判定できる。変異が、罹患者の一部またはすべてに見られるが、正常者には見られない場合、変異は疾患の原因物質である可能性が高い。
【0209】
[予防的及び治療的方法]
VDCCγ-8イオンチャネル活性の過剰量及び不足量の両方に関連する異常病態を治療する方法を提供する。
【0210】
VDCCγ-8イオンチャネルの活性が過剰である場合、いくつかのアプローチを利用できる。1つのアプローチは、リガンドのVDCCγ-8イオンチャネルへの結合をブロックすることにより、または第二のシグナルを阻害することにより活性を阻害するのに有効な量で製薬的に許容できる担体とともに、上記の阻害剤化合物(アンタゴニスト)を対象に投与すること、それによって異常病態を緩和することを含む。
【0211】
別のアプローチにおいて、内因性VDCCγ-8イオンチャネルと競合してリガンドをさらに結合できるVDCCγ-8ポリペプチドの溶解形態を投与できる。このような競合物の典型的な実施形態は、VDCCγ-8ポリペプチドの断片を含む。
【0212】
さらに別のアプローチにおいて、内因性VDCCγ-8イオンチャネルをコードする遺伝子発現は、発現ブロッキング技術を用いて阻害できる。既知のこのような技術は、内部で作製されるか、または別個に投与されるアンチセンス配列の使用を含む。例えば、Oligodeoxynucleotides as Antisense Inhibitors of Gene Expression, CRC Press, Boca Raton、フロリダ州(1988)における O'Connor、J Neurochem(1991)56:560頁を参照されたい。あるいは、遺伝子と三重螺旋体を形成するオリゴヌクレオチドを供給できる。例えば、Leeら、Nucleic Acids Res(1979)6:3073頁;Cooneyら、Science(1988)241:456頁;Dervanら、Science(1991)251:1360頁を参照されたい。これらのオリゴマーはそれ自体で投与され得るか、または関連オリゴマーをin vivoで発現させることができる。
【0213】
VDCCγ-8イオンチャネルの発現不足及びその活性に関連する異常病態を治療するために、いくつかのアプローチもまた利用できる。1つのアプローチは、製薬的に許容できる担体と組み合わせてVDCCγ-8イオンチャネルを活性化する化合物、すなわち、上記のアゴニストの治療的有効量を対象に投与することを含み、それによって異常病態を緩和する。あるいは、遺伝子療法を使用して、対象における関連細胞によってVDCCγ-8イオンチャネルの内因性産物をもたらすことができる。例えば、VDCCγ-8ポリヌクレオチドを、上記で考察された複製欠損レトロウイルスベクターにおける発現のために操作することができる。次にレトロウイルス発現構築体を単離でき、VDCCγ-8ポリペプチドをコードするRNAを含有するレトロウイルスプラスミドベクターで形質導入されたパッケージング細胞に導入でき、その結果、該パッケージング細胞は、ここで対象の遺伝子を含有する感染性ウイルス粒子を産生する。これらの産生株細胞を、in vivo細胞操作及びin vivoポリペプチドの発現のために対象に投与できる。遺伝子療法の概要に関しては、T. Strachan及びA P Read、Human Molecular Geneticsにおける20章、Gene Therapy and other Molecular Genetic-based Therapeutic Approaches, (及びその中に引用されている文献)、BIOS Scientific Publishers社(1996)を参照されたい。
【0214】
[製剤及び投与]
VDCCγ-8イオンチャネルポリペプチドの溶解形態等のペプチド、及びアゴニスト並びにアンタゴニストペプチドまたは小型分子を、好適な製薬用担体と組み合わせて製剤化できる。このような製剤は、治療的有効量のポリペプチドまたは化合物、及び製薬的に許容できる担体または賦形剤を含む。このような担体としては、限定はしないが、生理食塩水、緩衝生理食塩水、デキストロース、水、グリセロール、エタノール、及びそれらの組合せが挙げられる。製剤は、投与様式に適合する必要があり、十分当該技術分野の範囲内に入る。本開示は、さらに前述の組成物の1つ以上の成分で満たされた1つ以上の容器を含む製薬用パック及びキットに関する。
【0215】
VDCCγ-8ポリペプチド及び他の化合物を、単独でまたは治療用化合物等の他の化合物と併用して使用できる。
【0216】
製薬組成物の全身投与の好ましい形態としては、典型的には静脈内注射による注射が挙げられる。皮下、筋内、または腹腔内等の他の注射経路を使用できる。全身投与に関する代わりの手段としては、胆汁酸塩またはフシジン酸または他の界面活性剤等の浸透剤を用いて経粘膜及び経皮投与が挙げられる。さらに、腸内またはカプセル化製剤で適切に製剤化される場合、経口投与もまた可能であり得る。これらの化合物の投与はまた、軟膏剤、ペースト剤、ゲル剤等の形態で、局所的及び/または局在化できる。
【0217】
必要とされる投与量範囲は、ペプチドの選択、投与経路、製剤の性質、対象の病態の性質、及び担当医師の判定に依存する。しかしながら、好適な投与量は、0.1〜100μg/kg対象の範囲内である。しかしながら、必要な投与量の広い変動は、利用できる種々の化合物及び種々の投与経路の異なる効率に鑑みて予想することができる。例えば、経口投与は、静脈内注射による投与よりも高投与量を必要とすることが予想されるであろう。これらの投与量レベルの変動は、当業者に十分に解されるように、最適のために標準的な経験的ルーチンを用いて調整できる。
【0218】
治療に用いられるポリペプチドはまた、上記の「遺伝子療法」としばしば称される治療様式の対象において内因的に産生され得る。従って、例えば、対象からの細胞を、例えば、レトロウイルスプラスミドベクターの使用によりex vivoでポリペプチドをコードするためにDNAまたはRNA等のポリヌクレオチドにより操作できる。次いで該細胞を対象に導入する。
【0219】
[製薬組成物]
VDCCγ-8ポリペプチド、ポリヌクレオチド、ペプチド、ベクターまたはそれらの抗体の治療的有効量、及び場合によっては、製薬的に許容できる担体、希釈剤、または賦形剤(それらの組合せ等)を含む製薬組成物を提供する。
【0220】
該製薬組成物は、ヒト及び獣医医療におけるヒトまたは動物使用のためのものであり、1つ以上の製薬的に許容できる任意の希釈剤、担体、または賦形剤を典型的に含む。治療使用のための許容できる担体または希釈剤は、製薬業界に周知であり、例えば、Remington's Pharmaceutical Sciences、Mack Publishing社(A. R. Gennaro編集、1985年)に記載されている。製薬用担体、賦形剤または希釈剤の選択は、目的の投与経路及び標準的な製薬実施に関して選択できる。製薬組成物は、担体としてまたは担体に加えて、賦形剤または希釈剤、任意の好適な結合剤、滑剤、懸濁化剤、コーティング剤、可溶化剤を含むことができる。
【0221】
保存剤、安定化剤、染料及び香味剤さえも、製薬組成物に提供され得る。保存剤の例としては、安息香酸ナトリウム、ソルビン酸及びp-ヒドロキシ安息香酸のエステルが挙げられる。抗酸化剤及び懸濁化剤もまた使用できる。
【0222】
種々の送達系によって種々の組成物/製剤要件があり得る。例として、本明細書に記載された製薬組成物は、ミニポンプを用いるか、または粘膜経路により送達されるように、例えば、吸入用鼻腔スプレーまたはエアロゾルあるいは経口摂取用溶液として製剤化でき、あるいは該製剤は、非経口的に、例えば、静脈内、筋内または皮下経路による送達用の注射剤形態により製剤化される。あるいは、該製剤は、両経路により送達されるように設計され得る。
【0223】
薬剤を、胃腸管粘膜を介して粘膜送達できる場合、胃腸管を介して移行時に安定のままであり得る必要があり、例えば耐タンパク質分解性であり、酸pHに安定で、胆汁酸の耐界面活性作用である必要がある。
【0224】
適切な場合、製薬組成物を、吸入により、座薬またはペッサリーの形態で、局所的にローション、溶液、クリーム、軟膏または粉剤の形態で、皮膚パッチの使用により、経口的に澱粉またはラクトース等の賦形剤を含有する錠剤の形態で、またはカプセルもしくは小卵(ovule)中で単独または賦形剤と混合して、香味剤または着色剤を含有するエリキシル、溶液または懸濁剤の形態で投与でき、またはそれらは、例えば、非経口的に、例えば、静脈内、筋内または皮下に注入できる。非経口投与に関して、該組成物は、他の物質、例えば、血液との等張液を作製するために十分な塩類または単糖類を含有できる滅菌水溶液の形態で最良に使用され得る。口腔または舌下投与に関して、該組成物は、従来の様式で製剤化できる錠剤または舐剤の形態で投与できる。
【0225】
[ワクチン]
別の実施形態は、哺乳類動物にVDCCγ-8イオンチャネルポリペプチドまたはその断片を接種することを含む、哺乳類動物において免疫学的応答を誘導する方法に関するものであり、VDCCγ-8関連疾患から上記動物を保護するために抗体及び/またはT細胞免疫応答を適切に生じさせる。
【0226】
さらに別の実施形態は、疾患から上記動物を保護する抗体を産生するような免疫学的応答を誘導するために、in vivoでVDCCγ-8ポリヌクレオチドの発現をもたらすベクターを介してVDCCγ-8ポリペプチドを送達することを含む、哺乳動物における免疫学的応答を誘導する方法に関する。
【0227】
さらなる実施形態は、哺乳動物宿主に導入される場合、その哺乳動物においてVDCCγ-8ポリペプチドに対する免疫学的応答を誘導する免疫学的/ワクチン製剤(組成物)に関するものであり、該組成物はVDCCγ-8ポリペプチドまたはVDCCγ-8遺伝子を含む。ワクチン製剤は好適な担体をさらに含む。
【0228】
VDCCγ-8ポリペプチドは、胃内で分解されることから、非経口的(皮下、筋内、静脈内、皮膚内等の注射等)に投与されることが好ましい。非経口投与に好適な製剤としては、抗酸化剤、緩衝液、静菌剤及び製剤をレシピエントの血液と等張にさせる溶液を含有できる水性及び非水性滅菌注射液、並びに懸濁化剤または増粘剤を含むことができる水性及び非水性滅菌懸濁液が挙げられる。該製剤は、単位用量または多用量容器、例えば、密封アンプル及びバイアルで提供でき、使用直前に滅菌液担体の添加のみを必要とする凍結乾燥条件で保存することができる。ワクチン製剤は、水中油系及び当業者に既知の他の系等、製剤の免疫原性を増強させるアジュバント系もまた含むことができる。投与量は、ワクチンの特異的活性に依存し、ルーチン的実験により容易に決定できる。
【0229】
ワクチンは、1種以上のVDCCγ-8ポリペプチドまたはペプチドから調製され得る。
【0230】
活性成分として免疫原性ポリペプチドまたはペプチドを含有するワクチンの調製は、当業者に既知である。典型的には、このようなワクチンは、液体溶液または懸濁剤としての注射剤として調製され、注射前の溶液、または懸濁液、液体に好適な固体形態もまた調製され得る。該製剤をまた乳濁化できるか、またはタンパク質はリポソームにカプセル化できる。活性な免疫原性成分は、製薬的に許容でき、活性成分と適合できる賦形剤としばしば混合される。好適な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノール等、及びそれらの組合せである。
【0231】
さらに、所望ならば、ワクチンは、湿潤剤または乳化剤、pH緩衝剤、及び/またはワクチンの効果を増加させるアジュバント等、少量の補助物質を含有できる。効果的となり得るアジュバントの例としては、限定はしないが、水酸化アルミニウム、N-アセチル-ムラミル-L-トレオニル-D-イソグルタミン(thr-MDP)、N-アセチル-ノル-ムラミル-L-アラニル-D-イソグルタミン(CGP 11637、ノル-MDPと称される)、N-アセチルムラミル-L-アラニル-D-イソグルタミル-L-アラニン-2-(1'-2'-ジパルミトイル-sn-グリセロ-3-ヒドロキシホスホリルオキシ)-エチルアミン(CGP 19835A、MTP-PEと称される)、及び細菌から抽出された3つの成分、すなわちモノホスホリル脂質A、トレハロースジミコレート及び2%スクワレン/Tween 80乳濁液中の細胞壁骨格(MPL+TDM+CWS)を含有するRIBIが挙げられる。
【0232】
アジュバント及び他の薬剤のさらなる例としては、水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムカリウム(アルム)、硫酸ベリリウム、シリカ、カオリン、炭素、油中水乳濁液、水中油乳濁液、ムラミルジペプチド、細菌エンドトキシン、脂質X、コリネバクテリウムパルブム(プロピオノバクテリウムアクネス)、百日咳菌、ポリリボヌクレオチド、アルギン酸ナトリウム、ラノリン、リソレシチン、ビタミンA、サポニン、リポソーム、レバミソール、DEAE-デキストラン、ブロック化コポリマーまたは他の合成アジュバントが挙げられる。このようなアジュバントは、種々の供給源、例えば、Merck Adjuvant 65(Merck社、ラーウェイ、ニュージャージー州)またはFreund's Incomplete Adjuvant and Complete Adjuvant(Difco Laboratories、デトロイト、ミシガン州)から市販されている。
【0233】
典型的に、Amphigen(水中油)、Alhydrogel(水酸化アルミニウム)、またはAmphigen及びAlhydrogelの混合物等のアジュバントを使用する。水酸化アルミニウムだけが、ヒトへの使用に承認されている。
【0234】
免疫原とアジュバントとの比率は、両方が有効な量で存在する限り、広範囲にわたって変えることができる。例えば、水酸化ナトリウムは、ワクチン混合物(Al2O3ベース)の約0.5%の量で存在できる。都合のよいことに、ワクチンは、0.2から200μg/ml、好ましくは5から50μg/mlの範囲で、最も好ましくは15μg/mlの免疫原の最終濃度を含有するように製剤化される。
【0235】
製剤化後、ワクチンを滅菌容器に取り込むことができ、次いで密封され、低温で、例えば4℃で保存されるか、または凍結乾燥できる。凍結乾燥は、安定な形態で長期間の保存を可能にする。
【0236】
ワクチンを、非経口的に、注射、例えば、皮下または筋内に投与することが好都合である。他の様式の投与に好適なさらなる製剤としては座薬、いくつかの場合、経口製剤が挙げられる。座薬に関しては、伝統的な結合剤及び担体としては、例えば、ポリアルキレングリコールまたはトリグリセリドを挙げることができ、このような座薬は、0.5%から10%、好ましくは1%から2%の範囲で活性成分を含有する混合物から形成できる。経口製剤は、例えば、製薬グレードのマンニトール、ラクトース、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の正規に使用される賦形剤を含む。これらの組成物は、液剤、懸濁液剤、錠剤、丸剤、カプセル剤、徐放性製剤または散剤の形態をとり、10%から95%、好ましくは25%から70%の活性成分を含有する。ワクチン組成物が凍結乾燥される場合、凍結乾燥物を、投与前、例えば懸濁液として再構成できる。再構成は、緩衝液中で実施されることが好ましい。
【0237】
患者への経口投与用カプセル剤、錠剤及び丸剤は、例えば、Eudragit「S」、Eudragit「L」、セルロースアセテート、セルロースアセテートフタレートまたはヒドロキシプロピルメチルセルロースを含む腸溶性コーティングとともに提供できる。
【0238】
VDCCγ-8ポリペプチドは、中性または塩形態としてワクチンに製剤化され得る。製薬的に許容できる塩類としては、例えば、塩酸またはリン酸等の無機酸、酢酸、シュウ酸、酒石酸及びマレイン酸等の有機酸により形成される酸付加塩(ペプチドの遊離アミノ基と形成される)が挙げられる。遊離カルボキシル基と形成される塩類はまた、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、または水酸化鉄等の無機塩基、及びイソプロピルアミン、トリメチルアミン、2-エチルアミノエタノール、ヒスチジン及びプロカイン等の有機塩基から誘導できる。
【0239】
[投与]
典型的に、医師は、個々の対象に最も好適となる実際の投与量を判定し、特定の患者の年齢、体重及び応答により変わる。下記の投与量は、平均的な症例の代表例である。もちろん、より高いかまたはより低い投与量範囲が個々に利点をもたらす場合もあり得る。
【0240】
本明細書に記載された製薬及びワクチン組成物は、直接注射により投与できる。該組成物は、非経口、粘膜、筋内、静脈内、皮下、眼内または経皮投与用に製剤化できる。典型的に、各タンパク質を、0.01〜30 mg/kg体重、好ましくは、0.1〜10 mg/kg、より好ましくは、0.1〜1 mg/kg体重の用量で投与できる。
【0241】
用語の「投与される」とは、ウイルスまたは非ウイルス技術による送達を含む。ウイルス送達機構としては、限定はしないが、アデノウイルスベクター、アデノ関連ウイルス(AAV)ベクター、ヘルペスウイルスベクター、レトロウイルスベクター、レンチウイルスベクター、及びバキュロウイルスベクターが挙げられる。非ウイルス送達機構としては、脂質媒介形質移入、リポソーム、免疫リポソーム、リポフェクチン、カチオン性面両親媒性物質(CFA)及びそれらの組合せが挙げられる。このような送達機構に対する経路としては、限定はしないが、粘膜、鼻腔、経口、非経口、胃腸管、局所、または舌下経路が挙げられる。
【0242】
用語の「投与される」とは、限定はしないが、例えば、鼻腔用スプレーまたは吸入用エアロゾルとして、または経口摂取用溶液として粘膜経路による送達、送達が、例えば、静脈内、筋内または皮下経路等の注射剤形態による非経口経路による送達が挙げられる。
【0243】
用語の「共投与される」とは、例えば、VDCCγ-8ポリペプチド及びアジュバント等の添加実体の各々の投与部位及び投与時間により、免疫系の必要な調節が達成されるようになることを意味する。従って、ポリペプチド及びアジュバントは、同時に且つ同じ部位で投与され得るが、一方、ポリペプチドを異なる時間で、アジュバントと異なる部位に投与することに利点があり得る。ポリペプチド及びアジュバントを、同じ送達媒体でも送達でき、ポリペプチド及び抗原を結合させても、及び/または結合させなくても、並びに/あるいは遺伝子的に結合させても、及び/または結合させなくてもよい。
【0244】
VDCCγ-8ポリペプチド、ポリヌクレオチド、ペプチド、ヌクレオチド、それらの抗体及びアジュバントを、単位用量または複数用量で宿主対象に別個に投与しても、共投与してもよい。
【0245】
ワクチン組成物及び製薬組成物は、注射(非経口、皮下及び筋内注射を含む)、鼻腔内、粘膜経口、膣内、尿道または眼投与等の多くの異なる経路により投与できる。
【0246】
本明細書に記載されたワクチン及び製薬組成物は、例えば、皮下または筋内注射により、従来からの非経口的に投与できる。他の投与様式に好適であるさらなる製剤としては、座薬及びいくつかの場合、経口用製剤が挙げられる。座薬に関しては、伝統的な結合剤及び担体としては、例えば、ポリアルキレングリコールまたはトリグリセリドを挙げることができ、このような座薬は、0.5〜10%、恐らくは1〜2%の範囲で活性成分を含有する混合物から形成できる。経口製剤は、例えば、製薬グレードのマンニトール、ラクトース、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の正規に使用される賦形剤を含む。これらの組成物は、液剤、懸濁液剤、錠剤、丸剤、カプセル剤、徐放性製剤または散剤の形態をとり、10〜95%、好ましくは25〜70%の活性成分を含有する。ワクチン組成物が凍結乾燥される場合、凍結乾燥物を、投与前、例えば懸濁液として再構成できる。再構成は、緩衝液中で実施されることが好ましい。
【実施例】
【0247】
<実施例1.トランスジェニックVDCCγ-8ノックアウトマウス:VDCCγ-8遺伝子標的ベクターの構築>
VDCCγ-8遺伝子は、ゲノムデータベースの相同検索を用いてバイオ情報科学的に同定された。40kbのマウスゲノムコンティグを、種々のデータベースから集めた。このコンティグは、標的べクター内にクローニングするための相同アームの設計を可能にするのに十分なフランキング配列情報を提供した。
【0248】
マウスVDCCγ-8遺伝子は4つのコード化エクソンを有する。標的ストラテジーは、第一のコード化エクソンの大部分の翻訳部分を除去するように設計される。4.1kbの5'相同アーム及び除去される領域に隣接する2.1kbの3'相同アームは、PCRにより増幅され、断片は、標的ベクターにクローニングされる。アームを増幅するために使用される各オリゴヌクレオチドプライマーの5'末端は、ベクターポリリンカーのクローニング部位と適合し、アーム自体から欠いている希少切断制限酵素に対して異なる認識部位を含有するように合成される。VDCCγ-8の場合、プライマーは、下記のプライマーの表に挙げられるように、NotI/SpeIの5'アームクローニング部位及びAscI/FseIの3'アームクローニング部位(関連制限部位を含む使用される標的ベクターの構造を、図1に示す)を有するように設計される。
【0249】
アームプライマーのペア(5'アームF/5'アームR)及び(3'アームF/3'アームR)に加えて、さらにVDCCγ-8座に特異的なプライマーを以下の目的:単離された推定上の標的クローンにおいて、標的座のサザン解析を可能にするために、各アームの外側にありかつ各アームを越えて延長する非反復ゲノムDNAの2つの短い150〜300bp断片を増幅させるための、5'及び3'プローブプライマーのペア(5'prF/5'prR及び3'prF/3'prR);ベクター特異的プライマー(この場合、asc350)とともに多重PCRに用いられる場合、野生型、ヘテロ接合体、及びホモ接合体のマウス間の区別を可能にするマウス遺伝子型プライマーのペア(hetF及びhetR);最後に、3'アーム領域の末端下流にアニールし、ベクター(TK5IBLMNL)(この場合、asc146)の3'末端に特異的なプライマーとペアにした場合、標的事象特異的2.2kbアンプライマーを産生する標的スクリーニングプライマー(3'scr);のために設計する。このアンプライマーは、所望のゲノム変化が生じ、無作為に組込まれたベクターのコピーを含有するクローンのバックグラウンドから正しく標的化された細胞の同定を可能にする細胞由来のテンプレートDNAからのみ得ることができる。これらプライマーの位置及び標的ストラテジーに用いられるVDCCγ-8座の領域のゲノム構造を、図7に示す(配列番号21)。
【0250】
【表2】
【0251】
相同アームの位置は、VDCCγ-8遺伝子を機能的に破壊するように選択される。除去されるVDCCγ-8領域が、VDCCγ-8遺伝子と同じ配向性で配置された促進ネオマイシンホスホトランスフェラーゼ(neo)遺伝子からなる選択カセットの上流に内因性遺伝子発現レポーター(フレーム独立lacZ遺伝子)からなる非相同配列で置換されている標的ベクターを調製する。
【0252】
5'及び3'相同アームを、標的ベクターTK5IBLMNL内にクローニングすると(図1を参照)、標準的な分子生物学的技術を用いて大規模な高純度DNA調製を行う。20μgの新しく調製されたエンドトキシンなしのDNAを、アンピシリン抵抗性遺伝子と細菌の複製起点との間のベクター骨格の特有な部位に存在する別の希少切断制限酵素PmeIにより制限する。次に線形化DNAを沈殿させ、エレクトロポレーションに備えて、100μlのリン酸緩衝生理食塩水に再懸濁させる。
【0253】
エレクトロポレーションの24時間後、形質移入された細胞を、200μg/mlのネオマイシンを含有する培地中で9日間培養する。クローンを96ウェルプレートに選び取り、PCR(上記のとおり、プライマー3'scr及びasc146を用いて)によるスクリーニング前に、複製及び増殖させ、相同的組換えが、内因性VDCCγ-8遺伝子と標的構築体との間で生じているクローンを同定する。陽性クローンは、1〜5%の割合で同定され得る。これらのクローンを増殖して複製物を凍結し、すべて標準的な手法(Russら、Nature 2000年3月2日;404(6773):95〜99頁)を用いた、上記のように調製された外部5'及び3'プローブを用いることによって、標的事象のサザンブロット確認のために十分に高品質のDNAを調製する。診断用制限酵素によって消化されたDNAのサザンブロットを外部プローブとハイブリダイズする場合、相同的に標的化されたES細胞クローンが、変異体バンド並びに変化していない野生型バンドの存在によって立証される。例えば、5'プローブを用いることによって、EcoRI消化ゲノムDNAから9.3kbの野生型バンド及び6.7kbの標的バンドが得られ、3'プローブを用いることによって、Sspt切断DNAから15kbの野生型バンド及び9.5kbの標的バンドが得られる。
【0254】
<実施例2.トランスジェニックVDCCγ-8ノックアウトマウス:VDCCγ-8イオンチャネル欠損マウスの作製>
C57BL/6のメス及びオスマウスを交尾させ、妊娠3.5日に胚盤胞を単離する。1胚盤胞当り10〜12個の選択されたクローン由来の細胞を注入し、7〜8個の胚盤胞を、偽妊娠F1メスの子宮に移植する。幾匹かの高レベル(100%まで)アグーチオス(アグーチコート色は、標的クローンからの系統を継いだ細胞の寄与を示す)を含んだキメラ同腹仔が誕生する。これらのオスキメラを、メスのMF1及び129マウスと交尾させ、生殖細胞系の伝達を、それぞれアグーチコート色及びPCR遺伝子型決定によって判定する。
【0255】
PCR遺伝子型決定は、第三のベクター特異的プライマー(asc350)とともに、プライマーhetF及びhetRを用いて溶解した尾小端について実施される。この多重PCRにより、プライマーhetF及びhetRから野生型座(存在する場合には)からの増幅が可能となり、216bpバンドが得られる。hetFに対する部位は、ノックアウトマウスでは欠失しており、それゆえ、標的の対立遺伝子からこの増幅は起こらない。しかしながら、asc350プライマーは、3'アームのちょうど内側の領域にアニールするhetRプライマーと組み合わせて、標的座から384 bpバンドを増幅する。従って、この多重PCRにより、以下のとおり同腹仔の遺伝子型が明らかとなる。野生型サンプルは、単一の216 bpバンドを示し、ヘテロ接合体DNAサンプルは、216 bp及び384 bpに2本のバンドを生じ、ホモ接合体サンプルは、標的の特異的384 bpバンドだけを示す。
【0256】
<実施例3.生物学的データ:遺伝子発現パターン>
1)RT-PCR
RT-PCRを用いることによって、海馬(+++)、小脳、皮質、線条体、中脳、脳橋(++)、視床下部、視床、前脳、脊髄、下垂体、及び三叉神経節(+)における該遺伝子発現が示される。
2)Lac Z染色構造のリスト
【0257】
LacZ染色
切開組織のXgal染色を、以下の様式で実施する。
【0258】
代表的な組織薄片は、大きな臓器から作製される。小臓器全体及び管を、固定剤及び染色剤が浸透するように薄切りに開く。組織をPBS(リン酸緩衝生理食塩水)中で十分にすすぎ、血液または腸管内容物を除去する。組織を、固定剤(2%のホルムアルデデヒド、0.2%のグルタルアルデヒド、0.02%のNP40、1mMのMgCl2、0.23mMのデオキシコール酸ナトリウムを含むPBS)中に30〜45分間入れる。PBS中で5分間の洗浄を3回行った後、組織を、30℃で18時間、Xgal染色溶液(PBS中、4mMのKフェロシアニド、4mMのKフェリシアニド、2mMのMgCl2、1mg/mlのX-gal)中で放置する。組織をPBSで3回洗浄し、4%のホルムアルデヒド中で24時間、後染色し、再度PBSで洗浄してから、70%エタノール中に保存する。
【0259】
LacZ染色を用いることによって、VDCCγ-8が、海馬、皮質、扁桃体、嗅覚皮質、前脳、及び視床に発現されることが見出される。
【0260】
<実施例4.生物学的データ:行動:ロータロッド>
全動物を午前7時点灯の12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるため、設定時間に動物を試験する。該試験は、5分間にわたって、一定速度で4rpmから40rpmに加速し、次いで、カットオフ時間(10分)、またはマウスがロータロッドから落下するまで40rpmを保つ加速ロータロッド上での3つの試行を含む。動物がロータロッドから落下する時間を記録する。
【0261】
ノックアウトマウスは、3つのセッションすべてにおいて、野生型マウスと比較して、ロータロッド上での能力低下を示した(図2)。
【0262】
<実施例5.生物学的データ:行動:Laboras>
Laboras(実験動物行動観察記録及び分析システム)システムは、Metris BV(オランダ国)により市販されている自動行動装置である。Laborasは、振動パターンを用いて、行動を自動的に記録し定量化する。全動物を午前7時点灯の12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるために、設定時間に動物を試験する。最初の1時間、マウスをこの装置内に入れ、飼料及び水に自由にアクセスさせる。飼料及び水に自由にアクセスさせたまま、15時間終夜セッションによりこれを引き続き行う。
【0263】
1時間試験において、VDCCγ-8ノックアウトマウスは、セッションを通して、機能亢進(歩行運動頻繁/移動距離/毛づくろい)性である。(図3a)。
【0264】
15時間終夜試験において、VDCCγ-8ノックアウトマウスは、該セッションの最初の3時間は機能亢進性であり、その後、該動物が該装置に慣れるに従い、活動レベルは低下し、野生型マウスの活動レベルと対等になる(図3b)。
【0265】
<実施例5.生物学的データ:免疫組織化学:GluR2免疫組織化学>
GluR抗体に関する標準的免疫組織化学的方法(Mead及びStephens(2003)J. Neurosci. 23:1041〜1048頁)を用いて、Sant Cruz GluR2(N-19、sc-7611)抗体を用いることによって、GluR2 AMPA受容体サブユニットの免疫標識を海馬を含有する脳切片において実施した。野生型マウスと比較して、VDCCγ-8ノックアウトマウスの海馬では、GluR2に対する免疫染色は極めて低かった(図4)。
【0266】
<実施例6.生物学的データ:行動:社会的相互作用>
午前7時点灯の12時間明/12時間暗の明暗サイクル下で、全動物を、食料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるため、設定時間に動物を試験する。
【0267】
動物(野生型コントロールに一致させた対応週齢の各性別の5匹のKOマウス)を、各々9cm径のシリンダーを含有する、1つの中央チャンバー及び2つの「サイド」チャンバーからなる第3のチャンバーの装置に入れる。まず、各試験マウスを、5分間、該装置に入れ、追跡ソフトウェアを用いて活動を記録する。次いで、試験マウスを取り出し、同じ性別の侵入マウスをシリンダー上に置き、次いで、該試験マウスを該中央チャンバー内に戻す。次いで、該活動をさらに5分間記録する。空きチャンバーと比較して、侵入マウスを含有するチャンバーで各試験マウスが費やす時間を記録し、社会的相互作用の測定値として用いる。データは、各チャンバーで費やされた時間もしくは時間パーセントとしてまたは接近回避スコアとして算出され、正のスコアは、社会的相互作用の増加を示し、ゼロのスコアは、社会的相互作用なしを示し、負のスコアは、社会的回避の増加を示す。
【0268】
VDCCγ-8ノックアウトマウスは、この試験において、侵入者チャンバーにおける時間パーセントまたは接近回避スコアとして測定した場合、週齢を一致させた野生型マウスと比較して、社会的相互作用の明らかな欠如を示した(図5a及び5b)。
【0269】
このデータは、統合失調症における有用性を示唆している。
【0270】
<実施例7.生物学的データ:挙動:プレパルス抑制>
プレパルス抑制(PPI)は、大音響(115dB)刺激により喚起される驚愕応答が、驚愕刺激100ms前に供される短時間のそれよりはるかに静かな音響刺激(バックグラウンドより3〜10dB上)によって抑制される観察現象である。このいわゆるプレパルス抑制は、統合失調症患者において破壊されているが、精神運動性の開閉(gating)の欠損を反映している可能性があると考えられる。
【0271】
PPIは、げっ歯類において喚起され得、アンフェタミン、アポモルフィン、フェンシクリジン、及びMK801等の精神薬及び精神作用薬により破壊され得る。この破壊は、ハロペリドール、クロザピン、及びオランザピン等の抗精神病薬によって反転させることができる(Bakshi VP、Swerdlow NR及びGeyer MA、Clozapine antagonizes phencyclidine-induced deficits in sensorimotor gating of the startle response、J. Pharmacol. Exp. Ther. 1941年;271:787〜794頁)。従って、PPIの動物モデルは、構築体及び予測妥当性の両方で存在する。PPIに対する薬物または変異のいずれかの効果は、統合失調症の治療に有用である可能性を強く示している(McGhie A及びChapman J. Disorders of attention and perception in early schizophrenia、Br. J. Med. Psychol. 1961年;34:103〜116頁。Braff DL、Geyer MA及びSwerdlow NR、Human studies of prepulse inhibition of startle:normal subjects, patient groups, and pharmacological studies、Psychopharmacology、2001年;156:234〜258頁)。動物を、午前7時に点灯する12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育し、日周期の作用を避けるために設定時間で試験をする。マウス(対応する週齢に一致させた野生型コントロールとともに各性別の6匹のKOマウス)を、驚愕チャンバー(San Diego Instruments、サンディエゴ、カリフォルニア州、米国)に入れる。各システムは、小型ファンで換気され、天井に据付けられた白熱により照明された音響減衰チャンバーからなった。該チャンバーは、フレームに据付けられた透明のアクリルシリンダー(そこにマウスを入れる)を含んだ。該シリンダー及び該フレームは、フレームの各コーナーの下に配置された4つのスクリューによってPlexiglasベース付近を上昇した。音響刺激は、閉鎖容器の天井に据付けられた小型スピーカーを介して送達された。驚愕応答は、フレームの真下に据付けられた圧電型加速度計によって変換された。アウトプットシグナルをデジタル化し、調整し、San Diego Instruments Startle Reflexソフトウェアを備えたPC上で連続1ミリ秒(ms)の読取りとして記録した。驚愕刺激は、50msの長さで広いスペクトルノイズの115dBパルスであった。プレパルス刺激は、バックグラウンドより3、7、及び10dB上であり、驚愕刺激100ms前に供された。刺激は、擬似ランダム形式で供された。オスKOマウスにおいて、PPIは、週齢を一致させた野生型コントロールと比較して有意に破壊された(対応のあるt検定p=0.01)(図6を参照)。
【0272】
本文書に記載された出願及び特許のそれぞれ、並びに出願及び特許のそれぞれの遂行期間を含む、上記出願及び特許のそれぞれに引用または参照された各文書(「出願引用文書」)、並びに該出願及び特許のそれぞれ、及び任意の出願引用文書に引用または記載された任意の製品に関する任意の製造元の使用説明書またはカタログは、本明細書に参照として組み込まれている。さらに、本文に引用したすべての文書、及び本文に引用した文書に引用または参照したすべての文書、並びに本文に引用または記載された任意の製品に関する任意の製造元の使用説明書またはカタログは、本明細書に参照として組み込まれている。
【0273】
本発明の記載された方法及びシステムの種々の修飾及び変型は、本発明の範囲及び精神を逸脱することなく、当業者に明白であろう。本発明を、特定の好ましい実施形態に関連させて記載したが、当然のことながら、請求された本発明は、そのような特定の実施形態に過度に限定されるべきではなく、それに対する多くの修飾及び付加を、本発明の範囲で行うことができる。実際、分子生物学または関連分野の当業者に明白な、本発明の実施に関して記載された様式の種々の修正形態は本請求項の範囲内にあることとする。さらに、本発明の範囲から逸脱することなく、以下の独立した請求項の特徴と該独立した請求項の特徴との種々の組合せを行うことができる。
【図面の簡単な説明】
【0274】
【図1】ノックアウトベクターを示す図。
【図2】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのロータロッド試験の結果を示すグラフ。
【図3A】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのLABORAS(登録商標)(実験動物行動観察記録及び分析システム:Laboratory Animal Behaviour Observation Registration and Analysis System)における1時間のセッションを示すグラフ。
【図3B】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのLABORAS(登録商標)(実験動物行動観察記録及び分析システム)における15時間の一晩(ON)のセッションを示すグラフ。
【図4】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)の海馬におけるGluR2の免疫組織学的染色を示す図。
【図5a】VDCCγ-8ノックアウトマウスが、野生型マウスと比較して侵入マウスを含むチャンバーにおいて時間減少を示すことを示すグラフ。
【図5b】VDCCγ-8ノックアウトマウス及び野生型マウスに対する10の接近回避スコアを示すグラフ。
【図6】媒体またはMK801を投与された野生型及びノックアウトマウスにおいて観察される、バックグラウンドより上の組合せ音響刺激を用いたプレパルス抑制(PPI)を示す図。
【図7】プライマーの位置及び標的ストラテジーに用いられるVDCCγ-8座領域のゲノム構造を示す図。
【技術分野】
【0001】
本発明は、イオンチャネル核酸及びポリペプチドに関する。イオンチャネルは、以後「電位依存性カルシウムチャネルガンマ-8」、「VDCCγ-8」、または「γ-8」と称される。本発明はまた、これらの核酸及びポリペプチドの作用を抑制するかまたは活性化することに関するものであり、特に精神疾患の治療のための候補薬物を提供することに関する。
【背景技術】
【0002】
カルシウムチャネルは、排他的ではないが、筋収縮、神経伝達物質放出、ホルモン分泌、遺伝子発現、細胞分裂周期、及び細胞死を介する発達等、広範囲の細胞機能の制御において重要である。細胞内カルシウムは、静止状態において極めて低く、細胞内カルシウムが一過性に上昇するような変化は、第二のメッセンジャーとして作用して、多種多様の範囲の細胞過程を活性化することができる。電位またはリガンド依存性カルシウムチャネルは、細胞内カルシウムの増加を媒介できる。電位依存性カルシウムチャネルは、2つの広範な群:低(またはTタイプ)及び高閾値活性化チャネルに分けることができる。高閾値チャネルは、それらのサブユニット成分によって、L、N、P、Q、及びRチャネルに細分される。
【0003】
電位依存性カルシウムチャネルは、ポア形成アルファ-1サブユニット及びアクセサリサブユニットベータ、及びアルファ-2-デルタを含むヘテロチャネルである。いくつかの状況において、ガンマサブユニットもまた存在する(Caterall、Curr. Opin Neurobiol. 1991年:1;5〜13頁)。ガンマサブユニットタンパク質は、不活性(閉鎖)状態においてカルシウムチャネルを安定化すると考えられる内在性細胞膜内タンパク質である。いくつかのガンマサブユニットタンパク質があり、このVDCCγ-8遺伝子は、2つの同様のガンマサブユニットコード化遺伝子を有するクラスターに配置される、神経カルシウムチャネルガンマサブユニット遺伝子サブファミリーのPMP-22/EMP/MP20ファミリーのメンバーである。非AUG翻訳開始は、VDCCγ-8転写物に対して生じ得ると考えられている。VDCCγ-8サブユニットは、シナプス後肥厚-95、ディスクスラージ、閉鎖帯(PDZ)結合モチーフを含有する。VDCCγ-8は、スタルガジン(stargazin)及びVDCC-γ3と同様に、発達後期に現れ、動物の成熟期に進行的に増加する(胚発育大脳皮質には見られない、Tomitaら、J Cell Biol、2003年:161;805〜816頁を参照)。これは、胎芽期に発現し、動物成熟期に徐々に減少するVDCC-γ4に関連する。
【0004】
スタルガジン、γ-3、及びγ-4とともにγ-8は、AMPA受容体の表面発現(スタルガザー(stargazer)マウスからの小脳細胞におけるGluRの表面発現は〜75%減少するが、一方で、全GluRレベルは10〜20%のみ減少する)を媒介する膜貫通型AMPA制御タンパク質(TARPS)のファミリーに属する。
【0005】
精神疾患は、脳障害、特に感情または情緒不安定性、行動異常、及び/または認知障害を伴うものを定義するために用いられる用語である。精神疾患は、罹患者を衰弱させ、かつかなりの情緒的及び財政的負担を受けながら看護する必要がある。
【0006】
精神病は、重度に障害を受けた思考及び知覚を有する精神状態を定義する用語である。精神病の発症は、衰弱させ、通常、幻覚、偏執症、妄想、思考混乱、及び社会的相互作用の低下を含む。精神病は、一般に、罹患者に対して高度に破壊的となり得る現実との接触の喪失を含む。精神病は、双極性障害、重度の臨床的うつ病及び統合失調症を含む症状に現れる。これらの症状は、十分に理解されておらず、それらを予防及び治療する有効な療法を必要とされている。特に、陰性症状の統合失調症の治療において、かなりの満たされていない医学的ニーズがある。最も一般に用いられる薬物のクロザピン及びオランザピンは、十分効果的になるまでに2〜4週間かかり、統合失調症の陰性症状に対しての効果が比較的弱い。
【0007】
てんかんは、再発性てんかん発作を特徴とする発達障害ともしばしば称される精神病である。この慢性的な神経学的病態は、世界で数百万の人々に影響を及ぼしており、この症状を制御する薬物は存在するが、治療法はない。
【特許文献1】米国特許第5,492,840号
【特許文献2】米国特許第5,482,835号
【特許文献3】WO第84/03564号
【特許文献4】EP-A-239400
【特許文献5】米国特許第5,545,807号
【特許文献6】米国特許第6,075,181号
【特許文献7】米国特許第4,946,779号
【特許文献8】米国特許第4,946,778号
【非特許文献1】Caterall、Curr. Opin Neurobiol. 1991年:1;5〜13頁
【非特許文献2】Tomitaら、J Cell Biol、2003年:161;805〜816頁
【非特許文献3】J. Sambrook、E. F. Fritsch、及びT. Maniatis、1989年、Molecular Cloning:A Laboratory Manual、第二版、Books 1〜3、Cold Spring Harbor Laboratory Press
【非特許文献4】Ausubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州)
【非特許文献5】B. Roe、J. Crabtree、及びA. Kahn、1996年、DNA Isolation and Sequencing:Essential Techniques、John Wiley & Sons
【非特許文献6】J. M. Polak及びJames O'D. McGee、1990年、In Situ Hybridization:Principles and Practice;Oxford University Press
【非特許文献7】M. J. Gait(編集者)、1984年、Oligonucleotide Synthesis:A Practical Approach、Irl Press
【非特許文献8】D. M. J. Lilley及びJ. E. Dahlberg、1992年、Methods of Enzymology:DNA Structure Part A:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Press
【非特許文献9】Edward Harlow、David Lane、Ed HarlowによるUsing Antibodies:A Laboratory Manual:Portable Protocol NO. 1(1999年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-544-7)
【非特許文献10】Ed Harlow(編集者)、David Lane(編集者)によるAntibodies:A Laboratory Manual(1988年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-314-2)、1855
【非特許文献11】Lars-Inge Larsson「Immunocytochemistry:Theory and Practice」、CRC Press Inc.、Baca Raton、フロリダ州、1988年、ISBN 0-8493-6078-1、John D. Pound(編集者)
【非特許文献12】John D. Pound(編集者);「Methods in Molecular Biology」のシリーズにおける「Immunochemical Protocols、80巻」、Humana Press、Totowa、ニュージャージー州、1998年、ISBN 0-89603-493-3
【非特許文献13】Handbook of Drug Screening、Ramakrishna Seethala、Prabhavathi B. Fernandes(2001年、ニューヨーク、ニューヨーク州、Marcel Dekker、ISBN 0-8247-0562-9)
【非特許文献14】Lab Ref:A Handbook of Recipes, Reagents, and Other Refernce Tools for Use at the Bench、Jane Roskams及びLinda Rodgers編集、2002年、Cold Spring Harbor Laboratory、ISBN 0-87969-630-3
【非特許文献15】The Merck Manual of Diagnosis and Therapy(第17版、編集者Beers, M.H.及びBerkow, R、ISBN 0911910107、John Wiley & Sons)
【非特許文献16】http://www.sanger.ac.uk/Software/Pfam/search.shtml
【非特許文献17】Carlsson A. 1988年、The current status of the dopamine theory of schizophrenia. Neuropharmacology 1、179〜186頁
【非特許文献18】Adlardら(2005)J. Neurosci. 25:4217〜21頁
【非特許文献19】Limら(2005)J Neurosci 25:3032〜40頁
【非特許文献20】Proteins-Structure and Molecular Properties、第二版、T. E. Creighton、W. H. Freeman and Company、ニューヨーク、1993年
【非特許文献21】Wold, F.、Posttranslational Protein Modifications:Perspectives and Prospects、Posttranslational Covalent Modification of Proteins の1〜12頁、B. C. Johnson編集、Academic Press、ニューヨーク、1983年
【非特許文献22】Seifterら、「Analysis for protein modifications and nonprotein cofactors」、Meth Enzymol(1990)182:626〜646頁
【非特許文献23】Rattanら、「Protein Synthesis:Posttranslational Modifications and Aging」、Ann NY Acad Sci(1992)663:48〜62頁
【非特許文献24】Devereuxら、1984年、Nucleic Acids Research 12:387頁
【非特許文献25】Atschulら、1990年、J Molec Biol 403〜410頁
【非特許文献26】Ausubelら、1999年 Nucleic Acids Research-18章
【非特許文献27】Ausubelら、1999年、J. Mol. Biol.、7〜58頁から7〜60頁
【非特許文献28】http://www.ncbi.nih.gov/BLAST/blast_help.html
【非特許文献29】Karlin及びAltschul 1990年、Proc. Natl. Acad. Sci. USA 87:2264〜68頁
【非特許文献30】Karlin及びAltschul、1993年、Proc. Natl. Acad. Sci. USA 90:5873〜7頁
【非特許文献31】Altschulら(1994)Nature Genetics 6:119〜129頁
【非特許文献32】http://www.ncbi.nlm.nih.gov/BLAST
【非特許文献33】Coombs J(1994)Dictionary of Biotechnology、Stockton Press、ニューヨーク、ニューヨーク州)
【非特許文献34】Dieffenbach CW及びGS Dveksler(1995年、PCR Primer, a Laboratory Manual、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)
【非特許文献35】Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)
【非特許文献36】Sambrook、上記文献、7章
【非特許文献37】Ausubel, F. M.ら、上記文献、4章及び16章
【非特許文献38】Sambrook, J.ら、(1989年、Molecular Cloning: A Laboratory Manual、4章、8章、及び16〜17章、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)
【非特許文献39】Van Heeks, G.及びS. M. Schuster(1989)J. Biol. Chem. 264:5503〜5509頁
【非特許文献40】Grantら(1987年;Methods Enzymol. 153:516〜544頁)
【非特許文献41】Takamatsu, N.(1987)EMBO J. 6:307〜311頁
【非特許文献42】Coruzzi, G.ら(1984)EMBO J. 3:1671〜1680頁
【非特許文献43】Broglie, R.ら(1984)Science 224:838〜843頁
【非特許文献44】Winter, J.ら(1991)Results Probl. Cell Differ. 17:85〜105頁
【非特許文献45】Hobbs, S.またはMurry, L. E.、McGraw Hill Yearbook of Science and Technology(1992)McGraw Hill、ニューヨーク、ニューヨーク州;191〜196頁
【非特許文献46】Engelhard, E. K.ら(1994)Proc. Nat. Acad. Sci. 91:3224〜3227頁
【非特許文献47】Logan, J.及びT. Shenk(1984)Proc. Natl. Acad. Sci. 81:3655〜3659頁
【非特許文献48】Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125〜162頁
【非特許文献49】Wigler, M.ら(1977)Cell 11:223〜32頁
【非特許文献50】Lowy, I.ら(1980)Cell 22:817〜23頁
【非特許文献51】Wigler, M.ら(1980)Proc. Natl. Acad. Sci. 77:3567〜70頁)
【非特許文献52】Colbere-Garapin, F.ら(1981)J. Mol. Biol. 150:1〜14頁
【非特許文献53】Hartman, S. C.及びR. C. Mulligan(1988)Proc. Natl. Acad. Sci. 85:8047〜51頁
【非特許文献54】Rhodes, C. A.ら(1995)Methods Mol. Biol. 55:121〜131頁
【非特許文献55】Hampton, R.ら(1990年;Serological Methods, a Laboratory Manual, Section IV、APS Press、セントポール、ミネソタ州)
【非特許文献56】Maddox, D. E.ら(1983年;J. Exp. Med. 158:1211〜1216頁)
【非特許文献57】Porath, J.ら(1992)Prot. Exp. Purif. 3:263〜281頁
【非特許文献58】Kroll, D. J.ら(1993年;DNA Cell Biol. 12:441〜453頁)
【非特許文献59】Merrifield J.(1963)J. Am. Chem. Soc. 85:2149〜2154頁
【非特許文献60】Aizawa(1988)、Anal. Chem. Symp. 17:683頁
【非特許文献61】Wattら、J Biol Chem、1998年5月29日;273(22):14053〜8頁
【非特許文献62】Hoffmanら、Proc Natl Acad Sci USA 、2000年10月10日;97(21):11215〜20頁
【非特許文献63】Figlerら、Biochemistry、1997年12月23日;36(51):16288〜99頁
【非特許文献64】Sarrioら、2000年、Mol Cell Biol 2000年7月;20(14):5164〜74頁
【非特許文献65】Coliganら、Current Protocols in Immunology 1(2):5章(1991)
【非特許文献66】Blundell及びJohnson(1976)Protein Crystallography、Academic Press、ニューヨーク
【非特許文献67】Parceら(1989)Science 246:243〜247頁
【非特許文献68】Owickiら(1990)Proc. Nat'l Acad. Sci. USA 87;4007〜4011頁
【非特許文献69】Jaenich, R.(1976)PNAS 73:1260〜1264頁
【非特許文献70】Manipulating the Mouse Embryo、Hogan編集(Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、1986年)
【非特許文献71】Jahnerら(1985)PNAS 82:6927〜6931頁
【非特許文献72】Van der Puttenら(1985)PNAS 82:6148〜6152頁
【非特許文献73】Stewartら(1987)EMBO J. 6:383〜388頁
【非特許文献74】Jahnerら(1982)Nature 298:623〜628頁
【非特許文献75】Evansら(1981)Nature 292:154〜156頁
【非特許文献76】Bradleyら(1984)Nature 309:255〜258頁
【非特許文献77】Gosslerら(1986)PNAS 83:9065〜9069頁
【非特許文献78】Robertsonら(1986)Nature 322:445〜448頁
【非特許文献79】Jaenisch, R.(1988)Science 240:1468〜1474頁
【非特許文献80】Koehler及びMilstein(1975年Nature 256:495〜497頁)
【非特許文献81】Kosborら(1983)Immunol Today 4:72頁
【非特許文献82】Coteら(1983)Proc Natl Acad Sci 80:2026〜2030頁
【非特許文献83】Coleら、Monoclonal Antibodies and Cancer Therapy、77〜96頁、Alan R. Liss社、1985年
【非特許文献84】Morrisonら(1984)Proc Natl Acad Sci 81:6851〜6855頁
【非特許文献85】Neubergerら(1984)Nature 312:604〜608頁
【非特許文献86】Takedaら(1985)Nature 314:452〜454頁
【非特許文献87】Orlandiら(1989年、Proc Natl Acad Sci 86:3833〜3837頁)
【非特許文献88】Winter G及びMilstein C(1991年;Nature 349:293〜299頁)
【非特許文献89】Huse WDら(1989)Science 256:1275〜1281頁
【非特許文献90】Myersら、Science(1985)230:1242頁
【非特許文献91】Cottonら、Proc Natl Acad Sci USA(1985)85:4397〜4401頁
【非特許文献92】M. Cheeら、Science、274巻、610〜613頁(1996)
【非特許文献93】Oritaら(1989)Proc Natl. Acad. Sci USA:86:2766頁
【非特許文献94】Cotton (1993)Mutat Res 285:125〜144頁
【非特許文献95】Hayashi(1992)Genet Anal Tech Appl 9:73〜79頁
【非特許文献96】Keenら(1991)Trends Genet 7:5頁
【非特許文献97】O'Connor、J Neurochem(1991)56:560頁
【非特許文献98】Leeら、Nucleic Acids Res(1979)6:3073頁
【非特許文献99】Cooneyら、Science(1988)241:456頁
【非特許文献100】Dervanら、Science(1991)251:1360頁
【非特許文献101】T. Strachan及びA P Read、Human Molecular Geneticsにおける20章、Gene Therapy and other Molecular Genetic-based Therapeutic Approaches,(及びその中に引用されている文献)、BIOS Scientific Publishers社(1996)
【非特許文献102】Remington's Pharmaceutical Sciences、Mack Publishing社(A. R. Gennaro編集、1985年)
【非特許文献103】Russら、Nature 2000年3月2日;404(6773):95〜99頁
【非特許文献104】Mead及びStephens(2003)J. Neurosci. 23:1041〜1048頁
【非特許文献105】Bakshi VP、Swerdlow NR及びGeyer MA、Clozapine antagonizes phencyclidine-induced deficits in sensorimotor gating of the startle response、J. Pharmcol. Exp. Ther. 19941年;271:787〜794頁
【非特許文献106】McGhie A及びChapman J. Disorder of attention and perception in early schizophrenia、Br. J. Med. Psychol. 1961年;34:103〜116頁
【非特許文献107】Braff DL、Geyer MA及びSwerdlow NR、Human studies of prepulse inhibition of startle:normal subjects, patient groups, nd pharmacological studies、Psychopharmacology、2001年;156:234〜258頁
【発明の開示】
【発明が解決しようとする課題】
【0008】
精神疾患を予防及び治療するための有効な治療法、特に抗精神病及び抗てんかん療法に対する強いニーズが依然としてある。
【課題を解決するための手段】
【0009】
本発明は、VDCCγ-8が精神疾患に関与しており、ゆえに有用な治療標的であるという驚くべき発見に基づいている。特に、VDCCγ-8は、統合失調症及びてんかん等の精神病を含む疾患に関与している。
【0010】
本発明の第一の態様によれば、精神疾患の治療に適した薬剤を同定する方法は、候補薬剤が電位依存性カルシウムチャネルガンマ-8の活性に影響を及ぼすかどうかを判定する工程を含む。
【0011】
[配列表]
配列番号1は、ヒトVDCCγ-8のcDNA配列を示す。
配列番号2は、配列番号1に由来するオープンリーディングフレームを示す。
配列番号3は、配列番号2に由来するヒトVDCCγ-8のアミノ酸配列を示す。
配列番号4は、マウスVDCCγ-8に関するcDNAのオープンリーディングフレームを示す。
配列番号5は、マウスVDCCγ-8のアミノ酸配列を示す。
配列番号6は、ヒトVDCCγ-8に関するcDNAの代替のオープンリーディングフレームを示す。
配列番号7は、配列番号6に由来するヒトVDCCγ-8のアミノ酸配列を示す。
配列番号8〜20は、ノックアウトプラスミドを構築するために用いられた遺伝子型決定プライマーを示す。
配列番号21(図7)は、プライマーの位置及び標的ストラテジーに用いられたVDCCγ-8座領域のゲノム構造を示す。
【発明を実施するための最良の形態】
【0012】
[VDCCγ-8イオンチャネル]
VDCCγ-8イオンチャネル、及びそのホモログ、バリアント、または誘導体、並びにVDCCγ-8関連疾患の治療及び診断におけるその使用を本明細書に記載する。
【0013】
VDCCγ-8は、ヒトVDCCγ-8をコードする増幅cDNA産物を配列決定した結果によって示されるように、イオンチャネルファミリーの他のタンパク質に構造的に関連している。配列番号1のcDNA配列は、2つの可能なオープンリーディングフレーム:425個のアミノ酸のポリペプチド(配列番号3として示される)をコードするヌクレオチド番号104〜1381(配列番号2として示される)と、391個のアミノ酸のポリペプチド(配列番号7として示される)をコードするヌクレオチド番号206〜1381(配列番号6として示される)とを含む。ヒトVDCCγ-8は、ヒト染色体Ch19q13に位置することが分かっている。
【0014】
他に示されない限り、本発明の実施は、通常の当業者の能力の範囲内にある従来の化学、分子生物学、微生物学、組換えDNA、及び免疫学の技術を使用する。このような技術は、文献に説明されている。例えば、J. Sambrook、E. F. Fritsch、及びT. Maniatis、1989年、Molecular Cloning:A Laboratory Manual、第二版、Books 1〜3、Cold Spring Harbor Laboratory Press;Ausubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州);B. Roe、J. Crabtree、及びA. Kahn、1996年、DNA Isolation and Sequencing:Essential Techniques、John Wiley & Sons;J. M. Polak及びJames O'D. McGee、1990年、In Situ Hybridization:Principles and Practice;Oxford University Press;M. J. Gait(編集者)、1984年、Oligonucleotide Synthesis:A Practical Approach、Irl Press;D. M. J. Lilley及びJ. E. Dahlberg、1992年、Methods of Enzymology:DNA Structure Part A:Synthesis and Physical Analysis of DNA Methods in Enzymology、Academic Press;Edward Harlow、David Lane、Ed HarlowによるUsing Antibodies:A Laboratory Manual:Portable Protocol NO. 1(1999年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-544-7);Ed Harlow(編集者)、David Lane(編集者)によるAntibodies:A Laboratory Manual(1988年、Cold Spring Harbor Laboratory Press、ISBN、0-87969-314-2)、1855;Lars-Inge Larsson「Immunocytochemistry:Theory and Practice」、CRC Press Inc.、Baca Raton、フロリダ州、1988年、ISBN 0-8493-6078-1;John D. Pound(編集者)、「Methods in Molecular Biology」のシリーズにおける「Immunochemical Protocols、80巻」、Humana Press、Totowa、ニュージャージー州、1998年、ISBN 0-89603-493-3;Handbook of Drug Screening、Ramakrishna Seethala、Prabhavathi B. Fernandes(2001年、ニューヨーク、ニューヨーク州、Marcel Dekker、ISBN 0-8247-0562-9);Lab Ref:A Handbook of Recipes, Reagents, and Other Refernce Tools for Use at the Bench、Jane Roskams及びLinda Rodgers編集、2002年、Cold Spring Harbor Laboratory、ISBN 0-87969-630-3;並びにThe Merck Manual of Diagnosis and Therapy (第17版、編集者Beers, M.H.及びBerkow, R、ISBN:0911910107、John Wiley & Sons)を参照されたい。これら各一般テキストは、参照として本明細書に組み込まれている。
【0015】
[VDCCγ-8に対する同一性及び類似性]
pfam(http://www.sanger.ac.uk/Software/Pfam/search.shtml)のHMM構造予測ソフトウェアを用いたVDCCγ-8ポリペプチド(配列番号3)の解析により、VDCCγ-8ペプチドがイオンチャネルであることを確認する。
【0016】
ヒトVDCCγ-8イオンチャネルのマウスホモログをクローニングし、その核酸配列及びアミノ酸配列を、それぞれ配列番号4及び配列番号5として示す。マウスVDCCγ-8イオンチャネルcDNA (配列番号4)は、ヒトVDCCγ-8イオンチャネル(配列番号2及び配列番号6)配列と高度の同一性を示すが、一方、マウスVDCCγ-8イオンチャネルのアミノ酸配列(配列番号5)は、ヒトVDCCγ-8イオンチャネル(配列番号3及び配列番号7)と高度の同一性及び類似性を示す。
【0017】
従って、ヒト及びマウスVDCCγ-8イオンチャネルは、イオンチャネルの大型ファミリーのメンバーである。
【0018】
[VDCCγ-8の発現プロファイル]
VDCCγ-8 cDNAのポリメラーゼ連鎖反応(PCR)増幅により、VDCCγ-8の発現が海馬(+++)、小脳、皮質、線条体、中脳、脳橋(++)、視床下部、視床、前脳、脊髄、下垂体、及び三叉神経節(+)における存在量が変化していることが検出される。
【0019】
VDCCγ-8ノックアウトマウスのLacZ染色により、脳、特に海馬、皮質、扁桃体、嗅覚皮質、前脳、及び視床におけるVDCCγ-8の強い発現が示されている。
【0020】
VDCCγ-8 cDNAを用いて、BLASTNによりヒトまたはマウスESTデータ源をサーチすることによって、cDNAライブラリーにおいて同一性が見られる。このことは、VDCCγ-8が、以下の正常組織または異常組織において発現されていることを示している。
【0021】
登録番号CX223421、DN176885、CX205409:側脳室の側壁
登録番号BE647856、BE864111:マウス脳(小脳、脳幹、嗅球、視床下部、皮質、扁桃体、基底核、松果体、線条体、海馬の混合物由来)
登録番号BB641732:新生児皮質
登録番号BG803929:新生児網膜
【0022】
従って、VDCCγ-8ポリペプチド、核酸、プローブ、抗体、発現ベクター、及びリガンドは、これら及び他の組織におけるVDCCγ-8イオンチャネルの過剰発現、発現不足、及び発現異常に関連する疾患に対する検出、診断、治療、及び他のアッセイに有用である。好ましくは、該疾患は、下記に示されるVDCCγ-8関連疾患を含む。
【0023】
[VDCCγ-8イオンチャネル関連疾患]
本明細書に記載される方法及び組成物によれば、VDCCγ-8イオンチャネルは、様々な疾患を治療及び診断するための治療及び診断に有用である。VDCCγ-8は、下記に検討されるようにVDCCγ-8の活性を変化させる分子をアッセイする際にも有用である。VDCCγ-8を変化させる分子は、VDCCγ-8関連疾患に対して強力な治療薬である。VDCCγ-8が関与する疾患を、便宜上、VDCCγ-8関連疾患と称す。これらの疾患は精神疾患である。本明細書に用いられるように、精神疾患という用語は、感情的または情緒的不安定性、行動異常、及び/または認知機能不全もしくは認知機能障害を含む疾患を含んだ脳機能障害を表す。用語「精神疾患」はまた、発作を示す障害、例えばてんかんを含む。ヒト及び動物(獣医学的)治療及び診断は、本発明の範囲内に入る。
【0024】
ヒトVDCCγ-8は、ヒトの染色体19q13に位置する。従って、特定の実施形態において、Ch19q13イオンチャネルは、この遺伝子座、染色体バンド、領域、アーム、または同じ染色体に位置する疾患を治療または診断するために使用され得る。VDCCγ-8イオンチャネルの染色体遺伝子座(すなわち、ヒトの染色体19q13)と同じ遺伝子座、染色体バンド、領域、アーム、または染色体に結びつけられるものとして決定されている既知の疾患は、熱性発作、貧血、及び糖尿病とともに全般てんかんを含む。
【0025】
VDCCγ-8が欠失しているノックアウトマウスは、実施例に示されているように、様々な表現型を示す。例えば、LABORAS試験において、VDCCγ-8ノックアウトマウスは最初に機能亢進性であり、野生型コントロールマウス(実施例5)と比較して、作業記憶欠損の指標である新規な環境に対するより緩徐な馴化を示唆し、このことは変化したシナプス構成の症状であり得る。従って、我々は、個体におけるシナプス可塑性を変化させる方法を開示し、該方法は、その個体におけるVDCCγ-8のレベルまたは活性を調節することを含む。他で記載されているように、これは、VDCCγ-8の発現を調節することによって、またはVDCCγ-8のモジュレーター(アゴニストまたはアンタゴニスト等)の使用によって達成され得る。
【0026】
VDCCγ-8、及び特にVDCCγ-8のアゴニストを含むVDCCγ-8活性のモジュレーターは、関連する脳領域のシナプス可塑性を特徴とする疾患または症候を治療または緩和させるために使用され得る。このような疾患には、てんかん重積症、全般性発作、及び発作を示すすべての障害、自閉症等の小児性及び発達障害、疾患の自閉症スペクトル、広汎性発達障害、崩壊性障害、アスペルガー症候群、レット症候群、注意欠陥/多動性障害(ADHD)、排他的ではないが、アルツハイマー病、虚血性/血管性認知症、ピック病、広汎性レビー小体認知症、前頭側頭認知症、大脳皮質基底核変性症、ハンチントン病、進行性核上性麻痺、AIDS/HIV認知症、プリオン感染症、脳炎、神経梅毒、血管炎、進行性多病巣性白質脳障害を含む認知障害、並びに統合失調症、全般性不安障害、社会性不安、外傷後ストレス障害、恐怖症、社会的恐怖症、特定恐怖症、パニック障害、強迫性障害、急性ストレス障害、分離障害及びうつ病、大うつ病、気分変調、双極性障害、季節性情動障害、出産後うつ病、急性神経変性疾患、脳卒中、及び外傷性脳傷害を含む精神病が含まれる。
【0027】
好ましい実施形態において、VDCCγ-8関連疾患は、ストレスまたは不安が症状である疾患を含む。
【0028】
さらに好ましい実施形態において、VDCCγ-8関連疾患は、精神病が症状であるか、または症状であり得る疾患を含む。精神病は、一般に、重度の精神疾患の症状であると考えられているが、それ自体は診断ではない。精神病と関連する、または関連し得る任意の精神疾患は、本発明の範囲内にある。精神病に関与する好ましい疾患は、統合失調症、双極性障害(躁うつ病)及び重度の臨床的うつ病である。脳傷害(または他の神経疾患)、薬物中毒及び禁断症状(特にアルコール、バルビツール類及びベンゾジアゼピン類)、狼瘡、高齢者の電解質障害(尿路感染症)、疼痛症候群、睡眠うつ病及び極限ストレス(外傷後ストレス障害等)もまた、精神病の発症を引き起こす可能性があり、本発明の範囲内にある。
【0029】
さらに別の好ましい実施形態において、VDCCγ-8関連疾患は、全般てんかん及びてんかん重積症を含む発作を示す障害である。
【0030】
最も好ましいVDCCγ-8関連疾患は、精神病、統合失調症、ADHD、双極性障害、大うつ病、全般性不安障害、及びてんかん重積症を含むてんかんである。
【0031】
本明細書に詳述された実験データは、統合失調症におけるVDCCγ-8に強く関係する。海馬におけるGluR2受容体の発現の亢進及び低下(実施例5)は、VDCCγ-8KOマウスが、前脳グルタミン酸作動性欠損及び統合失調症の陰性態様に等しい症状を示すことを強く表している。新規な環境の特定マップを作成する能力は、KOマウスにおいて損なわれており(実施例5)、探索行動の時間の増加をもたらす。このことは、Carlsson(Carlsson A. 1988年、The current status of the dopamine theory of schizophrenia. Neuropharmacology 1、179〜186頁)により提案された統合失調症のグルタミン酸作動性仮説と一致して、統合失調症におけるVDCCγ-8の役割を示している。
【0032】
KOマウスはまた、社会的相互作用の欠如を示す(実施例6)。これらのマウスは不安レベルのいずれの変化も示さず、ゆえに、社会的相互作用の欠如は、上記統合失調症の陰性症状と類似した表現型を示唆している。KOマウスはまた、VDCCγ8(TARPγ8)が統合失調症の治療において有用性を有するという極めて強い証拠である、有意なプレパルス抑制破壊を示す(実施例7)。
【0033】
GluR2発現の低下、亢進、社会的相互作用の欠如、及びPPI欠損のすべては、KOマウスが、グルタミン酸作動性NMDA受容体アンタゴニストの投与後、野生型マウスに観察されるものと同様の表現型を有することを含む。これらは、ヒトにおいて精神異常症状を引き起こすことが知られており、学術的研究及び創薬プログラムにおける統合失調症の化学モデルとして常用されている。従って、好ましい実施形態において、VDCCγ-8関連疾患は統合失調症である。
【0034】
上記のとおり、VDCCγ-8イオンチャネルは、本明細書に記載された任意の方法及び組成物を用いることによって、これら特定の疾患のいずれかを診断及び/または治療するために使用され得る。
【0035】
特に、上記に挙げた特定の疾患の治療または診断のために、ホモログ、バリアント、または誘導体を含むVDCCγ-8イオンチャネル核酸、ポリペプチドを含む核酸、ベクターの使用、VDCCγ-8イオンチャネル核酸及び/またはポリペプチドを含む医薬組成物、宿主細胞、及びトランスジェニック動物の使用が意図されている。さらに、上記の特定の疾患の診断または治療において、VDCCγ-8イオンチャネルと相互作用または結合し得る化合物、好ましくはVDCCγ-8のアンタゴニスト、好ましくは該チャネルの伝導性を増大し得る化合物、VDCCγ-8イオンチャネルに対する抗体、並びにこれらを作製し、または同定する方法は本発明の範囲内に入る。特に、特定の疾患の治療または予防のためのワクチンの製造における、これら任意の化合物、組成物、分子等の使用が意図されている。個体の特定における疾患の検出用診断キットもまた、本発明の範囲内にある。
【0036】
VDCCγ-8イオンチャネルの使用によって治療または診断可能な上記のまたはさらなる特定の疾患を同定するための連鎖地図作成の方法は、当該技術分野において既知であり、また本明細書のいずれかに記載されている。
【0037】
[神経興奮、発作、及び認知障害]
神経興奮、発作、及び認知障害、並びに膜貫通型AMPA制御タンパク質(TARP)関連疾患を含むこれらが現れる障害は、当該技術分野において周知である。概要説明は以下のとおりである。
【0038】
神経興奮は、脳細胞の電気的活性化として定義でき、神経興奮は、脳の正常な機能の一部であるか、または機能障害、疾患、または傷害に関連する異常活性の結果でもあり得る。
【0039】
発作は、精神状態の変化、強直性または間代性動作、及び種々の他の症状の中にそれら自体を現す脳機能の一時的変化である。それらは、脳細胞群の一時的な異常神経興奮に起因する。発作は、オンオフ脳侵襲(例えば毒性、衝撃)から、または組織損傷/変性/機能障害の結果としての反復症状として生じ得る。
【0040】
認知は、人が思考または知覚の対象を自覚する精神の働きとして定義でき、それは、知覚、思考、及び記憶のすべての態様を含む。認知障害は、機能障害、疾患、または傷害の結果としての神経興奮の破壊による、上記の働きの破壊である。
【0041】
VDCCγ-8イオンチャネルモジュレーターを、これらの症状のいずれかを治療または軽減するために使用できる。
【0042】
ライフスタイル及び食事療法もまた、神経興奮性、発作、及び/または認知作用を変えるために役立ち得る:1)運動はアルツハイマー病の発症率を減じ得る(アミロイド負荷及び学習欠損の両方に関して、Adlardら(2005)J. Neurosci. 25:4217〜21頁);2)食事のオメガ-3ポリ不飽和脂肪酸、ドコサヘキサエン酸(DHA)は、βアミロイドの産生、蓄積、及び下流毒性の可能性を保護しうるであろう(Limら(2005)J Neurosci 25:3032〜40頁)。我々は、それらの神経興奮性、発作、及び/または認知作用を緩和させるようなライフスタイル処置と組み合わせてVDCCγ-8イオンチャネル及びそのモジュレーターの使用を開示する。
【0043】
レクリエーション及び薬用の両方のある一定の薬物は、薬物からの副作用または禁断症状として神経興奮性、発作、及び/または認知の変化となり得る。このような薬物としては、カフェイン、大麻、アルコール、ニコチン、風邪薬、抗ヒスタミン薬、コカイン、及びアンフェタミンが挙げられる。我々は、それらの神経興奮性、発作、及び/または認知作用を緩和させるような薬物と組み合わせた、VDCCγ-8イオンチャネル及びそのモジュレーターの使用を開示する。
【0044】
[VDCCγ-8イオンチャネルポリペプチド]
本明細書に用いられる用語「VDCCγ-8イオンチャネルポリペプチド」とは、配列番号3、配列番号5、もしくは配列番号7に示されたアミノ酸配列、またはそれらのホモログ、バリアント、または誘導体を含むポリペプチドを指すものとする。該ポリペプチドは、配列番号3に示された配列のホモログ、バリアント、または誘導体を含むか、あるいはそれらであることが好ましい。
【0045】
「ポリペプチド」とは、ペプチド結合または修飾ペプチド結合、すなわちペプチドアイソスターにより互いに結合された2つ以上のアミノ酸を含む任意のペプチドまたはタンパク質のことである。「ポリペプチド」とは、通常、ペプチド、オリゴペプチド、またはオリゴマーと称される短鎖、一般にタンパク質と称される長鎖のことである。ポリペプチドは、20の遺伝子コード化アミノ酸以外のアミノ酸を含有することができる。
【0046】
「ポリペプチド」は、翻訳後処理等の天然プロセシング、または当業者によく知られている化学修飾技術により修飾されたアミノ酸配列を含む。このような修飾は、基礎的テキスト及びより詳細な単行書、並びに多数の研究文献に十分に記載されている。修飾は、ペプチド主鎖、アミノ酸側鎖、及びアミノまたはカルボキシル末端等、ポリペプチドのいずれかの場所で生じ得る。同じタイプの修飾は、所与のポリペプチドのいくつかの部位で同じかまたは変化段階で存在し得ることを認識するであろう。また、所与のポリペプチドは、多くのタイプの修飾を含むことができる。
【0047】
ユビキチン結合の結果、ポリペプチドは分枝でき、それらは、分岐の有り無しで環式化できる。環式、分枝状、及び分枝状環式ポリペプチドは、翻訳後天然プロセシングから生じ得るか、または合成法により作製できる。修飾としては、アセチル化、アシル化、ADP-リボシル化、アミド化、フラビンの共有結合、ヘム部分の共有結合、ヌクレオチドまたはヌクレオチド誘導体の共有結合、脂質または脂質誘導体の共有結合、ホスホチジルイノシトールの共有結合、架橋化、環化、ジスルフィド結合形成、脱メチル化、共有架橋形成、シスチン形成、ピログルタメート形成、ホルミル化、ガンマ-カルボキシル化、グリコシル化、GPIアンカー形成、ヒドロキシル化、ヨード化、メチル化、ミリストイル化、酸化、タンパク分解処理、リン酸化、プレニル化、ラセミ化、セレノイル化、硫酸化、アルギニン化等のアミノ酸のタンパク質へのトランスファーRNA媒介付加、及びユビキチン結合が挙げられる。例えば、Proteins-Structure and Molecular Properties、第二版、T. E. Creighton、W. H. Freeman and Company、ニューヨーク、1993年、及びWold, F.、Posttranslational Protein Modifications:Perspectives and Prospects、Posttranslational Covalent Modification of Proteinsの1〜12頁、B. C. Johnson編集、Academic Press、ニューヨーク、1983年;Seifterら、「Analysis for protein modifications and nonprotein cofactors」、Meth Enzymol(1990)182:626〜646頁及びRattanら、「Protein Synthesis:Posttranslational Modifications and Aging」、Ann NY Acad Sci(1992)663:48〜62頁、を参照されたい。
【0048】
本文書に用いられる用語「バリアント」、「ホモログ」、「誘導体」、または「断片」とは、配列からまたは配列への1つ(または複数)のアミノ酸の任意の置換、変化、修飾、置換え、欠失、または付加が挙げられる。他の文脈に認められない限り、「VDCCγ-8」及び「VDCCγ-8イオンチャネル」についての言及は、VDCCγ-8のこのようなバリアント、ホモログ、誘導体、及び断片についての言及を含む。
【0049】
VDCCγ-8に適用される得られたアミノ酸配列は、イオンチャネル活性を有することが好ましく、配列番号3、配列番号5または配列番号7として示されるVDCCγ-8イオンチャネルの少なくとも同じ活性を有することがさらに好ましい。特に、得られたアミノ酸配列がイオンチャネル活性を有するという条件で、用語の「ホモログ」は、構造及び/または機能に関して同一性を包含する。配列同一性(すなわち、類似性)に関して、好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも85%、さらにより好ましくは少なくとも90%の配列同一性である。より好ましくは少なくとも95%、より好ましくは少なくとも98%の配列同一性である。これらの用語はまた、VDCCγ-8イオンチャネル核酸配列の対立遺伝子変異であるアミノ酸から誘導されるポリペプチドを包含する。
【0050】
VDCCγ-8イオンチャネル等のイオンチャネルの「チャネル活性」または「生物活性」に言及される場合、これらの用語は、同様の活性または改善された活性もしくは非所望の副作用が減少したこれらの活性等、VDCCγ-8イオンチャネルの代謝機能または生理的機能を指すものとする。VDCCγ-8イオンチャネルの抗原活性及び免疫原性活性もまた含まれる。イオンチャネル活性の例、及びこれらの活性のアッセイ法及び定量化法は当業者に既知である。
【0051】
本明細書に用いられる「欠失」とは、それぞれ1つ以上のヌクレオチド残基またはアミノ酸残基を欠いているヌクレオチド配列またはアミノ酸配列の変化として定義される。本明細書に用いられる「挿入」または「付加」とは、天然物と比較して、それぞれ1つ以上のヌクレオチド残基またはアミノ酸残基の付加が生じたヌクレオチド配列またはアミノ酸配列を変化させることである。本明細書に用いられる「置換」は、それぞれ異なるヌクレオチドまたはアミノ酸による1つ以上のヌクレオチドまたはアミノ酸の置換えから生じる。
【0052】
本明細書に記載されるVDCCγ-8ポリペプチドはまた、サイレント変化を生じ、機能的に等しいアミノ酸配列となるアミノ酸残基の欠失、挿入、または置換を有することができる。計画的なアミノ酸置換は、残基の極性、電荷、溶解性、疎水性、親水性、及び/または両親媒性の類似性に基づいて成すことができる。例えば、負電荷のアミノ酸としては、アスパラギン酸及びグルタミン酸が挙げられ、陽電荷のアミノ酸としては、リシン及びアルギニンが挙げられ、類似の親水性値を有する無電荷極性頭部を有するアミノ酸としては、ロイシン、イソロイシン、バリン、グリシン、アラニン、アスパラギン、グルタミン、セリン、トレオニン、フェニルアラニン、及びチロシンが挙げられる。
【0053】
保存的置換は、例えば、下表に従って作製することができる。第二欄の同じブロック、好ましくは第三欄の同じ行のアミノ酸は互いに置換できる。
【0054】
【表1】
【0055】
VDCCγ-8ポリペプチドは、典型的にN末端またはC末端に、好ましくはN末端に異種のアミノ酸配列をさらに含むことができる。異種の配列としては、細胞内または細胞外タンパク質の標的(リーダー配列等)に影響を及ぼす配列を挙げることができる。異種配列としてはまた、ポリペプチドの免疫原性を増加させ、並びに/あるいはポリペプチドの同定、抽出、及び/または精製を容易にする配列を挙げることができる。特に好ましい別の異種の配列は、好ましくはN末端にあるポリヒスチジン等のポリアミノ酸配列である。少なくとも10のアミノ酸、好ましくは少なくとも17のアミノ酸であるが、50未満のアミノ酸のポリヒスチジン配列が特に好ましい。
【0056】
VDCCγ-8イオンチャネルポリペプチドは、「成熟」タンパク質の形態であり得るか、または融合タンパク質等のより大きなタンパク質の一部であり得る。分泌またはリーダー配列、プロ配列、多ヒスチジン残基等の精製、または組換え産生時の安定性のために、さらなる配列を補助する配列を含有するさらなるアミノ酸配列を含ませることもしばしば有利である。
【0057】
VDCCγ-8ポリペプチドは、既知の技術を用いる組換え手段により有利に作製される。しかしながら、それらは、固相合成等の当業者に周知の技術を用いる合成手段によっても作製できる。本明細書に記載されるポリペプチドは、例えば、抽出及び精製を補助するために、融合タンパク質としても産生できる。融合タンパク質パートナーの例としては、グルタチオン-S-トランスフェラーゼ(GST)、6×His、GAL4(DNA結合及び/または転写活性化ドメイン)、及びβ-ガラクトシダーゼが挙げられる。融合タンパク質配列の除去を可能にするために、トロンビン開裂部位等、融合タンパク質パートナーと対象のタンパク質配列との間のタンパク質開裂部位を含ませることも有利となり得る。融合タンパク質は、対象配列のタンパク質の機能を妨げないことも好ましくなろう。
【0058】
VDCCγ-8ポリペプチドは、実質的に単離形態であり得る。この用語は、天然状態から人の手により変化させることを意味する。「単離」組成物または物質が天然に生じる場合、それを変化させるか、または元の環境から取り出すか、あるいは両方から得る。例えば、生体動物に存在する天然の核酸またはポリペプチドは、「単離」されないが、その天然状態の共存物質から分離される同じポリヌクレオチド、核酸またはポリペプチドは、用語が本明細書に使用されるように「単離」される。
【0059】
しかしながら、VDCCγ-8イオンチャネルタンパク質は、タンパク質の意図された目的を妨害しない担体または希釈剤と混合することができ、さらに実質的に単離されたとみなされ得ると理解されよう。VDCCγ-8ポリペプチドはまた、実質的に精製形態であり得、その場合、それは、調製物中のタンパク質が90%超、例えば、95%、98%または99%がVDCCγ-8ポリペプチドである調製物中のタンパク質を一般に含んでいる。
【0060】
我々はさらに、VDCCγ-8ポリペプチドの一部を含むペプチドを記載する。従って、VDCCγ-8イオンチャネルの断片及びそのホモログ、バリアント、または誘導体が含まれる。該ペプチドは、2〜200の間のアミノ酸長、好ましくは4〜40の間のアミノ酸長であり得る。該ペプチドは、例えば、トリプシン等の好適な酵素による消化により本明細書に開示されたVDCCγ-8ポリペプチドから誘導できる。あるいは、ペプチド、断片等は、組換え手段により作製できるか、または合成的に合成できる。
【0061】
用語の「ペプチド」は、レトロインバーソDペプチド等、当業者に知られている種々の合成ペプチド変異を含む。該ペプチドは、抗原性決定因子及び/またはT細胞エピトープであり得る。該ペプチドはin vivo免疫原性であり得る。好ましくは、該ペプチドは、in vivoで抗体の中和を誘導できる。
【0062】
異なる種からVDCCγ-8イオンチャネル配列を位置合わせを行うことによって、アミノ酸配列の領域が異なる種間で保存されているか(「相同領域」)、領域が異なる種間で異なっているか(「非相同領域」)かを判定することが可能である。
【0063】
従って、VDCCγ-8ポリペプチドは、少なくとも一部の相同領域に相当する配列を含むことができる。相同領域は、少なくとも2種間で高度の相同性を示す。例えば、該相同領域は、上記試験を用いるアミノ酸レベルで少なくとも70%、好ましくは少なくとも80%、より好ましくは少なくとも90%、さらにより好ましくは少なくとも95%、例えば98%または99%の同一性を示すことができる。相同領域に相当する配列を含むペプチドは、さらに下記に詳細に説明されるように治療戦略に使用することができる。あるいは、VDCCγ-8イオンチャネルペプチドは、少なくとも一部の非相同領域に相当する配列を含むことができる。非相同領域は、少なくとも2種間で低度の相同性を示す。
【0064】
[VDCCγ-8イオンチャネルポリヌクレオチド及び核酸]
VDCCγ-8ポリヌクレオチド、VDCCγ-8ヌクレオチド、及びVDCCγ-8核酸、並びにこれら核酸の生産方法及び使用が意図されている。
【0065】
用語の「VDCCγ-8ポリヌクレオチド」、「VDCCγ-8ヌクレオチド」、及び「VDCCγ-8核酸」は、交互に使用することができ、配列番号1、配列番号2、配列番号4、もしくは配列番号6に示される核酸配列、あるいはそれらのホモログ、バリアント、または誘導体を含むポリヌクレオチド/核酸を指すものとする。好ましくは、該ポリヌクレオチド/核酸は、配列番号1、配列番号2、または配列番号6、最も好ましくは配列番号2の核酸配列のホモログ、バリアント、もしくは誘導体を含むか、またはそのものである。
【0066】
これらの用語はまた、ポリペプチド及び/またはペプチド、すなわちVDCCγ-8ポリペプチドをコードし得る核酸配列を含むことが意図されている。従って、VDCCγ-8イオンチャネルポリヌクレオチド及び核酸は、配列番号3、配列番号5、もしくは配列番号7に示されるアミノ酸配列、またはそれらのホモログ、バリアント、もしくは誘導体を含むポリペプチドをコードし得るヌクレオチド配列を含む。VDCCγ-8イオンチャネルポリヌクレオチド及び核酸は、配列番号3に示されるアミノ酸配列またはそのホモログ、バリアント、もしくは誘導体を含むポリペプチドをコードし得るヌクレオチド配列を含むことが好ましい。
【0067】
「ポリヌクレオチド」とは一般に、非修飾RNAまたはDNAもしくは修飾RNAまたはDNAであり得る任意のポリリボヌクレオチドまたはポリデオキシリボヌクレオチドのことである。「ポリヌクレオチド」としては、限定はしないが、単鎖及び二本鎖DNA、単鎖及び二本鎖領域の混合物であるDNA、単鎖及び二本鎖RNA、並びに単鎖及び二本鎖領域の混合物であるRNA、単鎖、またはより典型的には二本鎖、もしくは単鎖及び二本鎖領域の混合物であり得るDNA及びRNAを含むハイブリッド分子が挙げられる。さらに「ポリヌクレオチド」とは、RNAまたはDNA、あるいはRNA及びDNA両方を含む三本鎖領域のことである。用語のポリヌクレオチドはまた、1つ以上の修飾塩基を含有するDNA及びRNAを含み、安定性または他の理由のために修飾された主鎖を有するDNA及びRNAを含む。「修飾」塩基としては、例えば、トリチル化塩基及びイノシン等の特異塩基が挙げられる。種々の修飾により、DNA及びRNAが作製されており、従って、「ポリヌクレオチド」は、典型的に天然見られるポリヌクレオチドの化学的、酵素的または代謝的に修飾された形態、並びにウイルス及び細胞に特徴的なDNA及びRNAの化学的形態を包含する。ポリヌクレオチドはまた、オリゴヌクレオチドとしばしば称される比較的短鎖のポリヌクレオチドを包含する。
【0068】
多数のヌクレオチド配列は、遺伝子コードの縮退の結果と同一のポリペプチドをコードし得ることが当業者により理解されるであろう。
【0069】
本明細書に用いられる用語「ヌクレオチド配列」とは、ヌクレオチド配列、オリゴヌクレオチド配列、ポリヌクレオチド配列及びそれらのバリアント、ホモログ、断片、及び誘導体(それらの一部等)のことである。ヌクレオチド配列は、センス鎖またはアンチセンス鎖を表そうと、もしくはそれらの組合せを表そうとも、二本鎖または一本鎖であり得るゲノムまたは合成あるいは組換え起源のDNAまたはRNAであり得る。用語のヌクレオチド配列は、組換えDNA技術(例えば、組換えDNA)の使用により調製できる。
【0070】
好ましくは、用語の「ヌクレオチド配列」とは、DNAを意味する。
【0071】
本文書に用いられる用語「バリアント」、「ホモログ」、「誘導体」、または「断片」としては、VDCCγ-8ヌクレオチド配列からまたはその配列に1つ(または複数の)核酸の任意の置換、変化、修飾、置換え、欠失、または付加が挙げられる。他の文脈に認められない限り、「VDCCγ-8」及び「VDCCγ-8イオンチャネル」についての言及は、VDCCγ-8のこのようなバリアント、ホモログ、誘導体、及び断片についての言及を含む。
【0072】
得られたヌクレオチド配列は、イオンチャネル活性を有するポリペプチドをコードすることが好ましく、配列番号3、配列番号5、または配列番号7として示されるイオンチャネルの少なくとも同じ活性を有することが好ましい。用語の「ホモログ」は、得られたヌクレオチド配列がイオンチャネル活性を有するポリペプチドをコードするように、構造及び/または機能に関して同一性を包含するように意図されていることが好ましい。配列同一性(すなわち、類似性)に関して、好ましくは少なくとも70%、より好ましくは少なくとも75%、より好ましくは少なくとも85%、より好ましくは少なくとも90%の配列同一性である。より好ましくは少なくとも95%、より好ましくは少なくとも98%または99%の配列同一性である。これらの用語はまた、その配列の対立遺伝子変異を包含する。
【0073】
[配列相同性の算出]
本明細書に提出された任意の配列に関する配列同一性は、他の配列が、例えば、該配列(1つ以上)に対して少なくとも70%の配列同一性を有するかどうかを見るために、任意の1つ以上の配列と別の配列との単純な「眼球」比較(すなわち、厳密な比較)により決定され得る。
【0074】
相対的配列同一性はまた、例えば、デフォルトパラメータを用い、同一性を決定するために任意の好適なアルゴリズムを用いて2つ以上の配列間で同一性%を算出できる市販のコンピュータプログラムにより決定され得る。このようなコンピュータプログラムの典型的な例は、CLUSTALである。2つの配列間の同一性及び類似性を決定するための他のコンピュータプログラム法としては、限定はしないが、GCGプログラムパッケージ(Devereuxら、1984年、Nucleic Acids Research 12:387頁)及びFASTA(Atschulら、1990年、J Molec Biol 403〜410頁)が挙げられる。
【0075】
相同性%は、連続配列上で算出できる。すなわち、1つの配列を他の配列と位置合わせし、1つの配列中の各アミノ酸を、他の配列中の対応するアミノ酸と、一度に1つの残基と直接比較する。これは、「アンギャップド」アラインメントと呼ばれる。典型的に、このようなアンギャップドアラインメントは、比較的短い数の残基上で実施される。
【0076】
これは、極めて簡単で一貫した方法であるが、例えば別の同一対の配列において、1つの挿入または欠失により、続くアミノ酸残基がアラインメントから離れる原因となり、従って全域のアラインメントが実施される際に、可能性として相同性%の大きな減少を生じることを考慮できない。その結果、大抵の配列比較法は、総合的相同性スコアを過度に不利にすることなく、可能な挿入及び欠失を考慮する最適のアラインメントを生じるように設計されている。これは、局所的相同性を最大にしようとするために配列アラインメントに「ギャップ」を挿入することにより達成される。
【0077】
しかしながら、これらのより複雑な方法は、アラインメントに生じる各ギャップに「ギャップペナルティー」を与える結果、同じ数のアミノ酸に関して、2つの比較配列間でより高い関連性を反映して、できるだけ少ないギャップによる配列アラインメントは、多数のギャップのものよりも高いスコアを達成するであろう。ギャップの存在のため比較的高いコスト、及びギャップに続く各々の残基に対してより小さなペナルティーを課する「アフィンギャップコスト」が典型的に使用される。これは、最も一般に用いられるギャップスコアリングシステムである。高ギャップペナルティーはもちろん、より少ないギャップにより最適化アラインメントを生じるであろう。大抵のアラインメントプログラムは、ギャップペナルティーを変更させる。しかしながら、配列比較のためにこのようなソフトウェアを使用する場合、デフォルト値を使用することが好ましい。例えば、GCG Wisconsin Bestfitパッケージを用いる場合、アミノ酸配列に対するデフォルトギャップペナルティーは、ギャップに関して-12及び各伸長に関して-4である。
【0078】
従って、最大相同性%の算出は、最初にギャップペナルティーを考慮して最適アラインメントの製作を必要とする。このようなアラインメントを実施するために好適なコンピュータプログラムは、GCG Wisconsin Bestfitパッケージ(ウィスコンシン大学、米国;Devereuxら、1984年、Nucleic Acids Research 12:387頁)である。配列比較を実施できる他のソフトウェアの例としては、限定はしないが、BLASTパッケージ(Ausubelら、1999年 Nucleic Acids Research-18章)、FASTA(Atschulら、1990年J. Mol. Biol.、403〜410頁)及び比較手段のGENEWORKSスイートが挙げられる。BLAST及びFASTAの両方は、オフライン及びオンライン検索で入手できる(Ausubelら、1999年、J. Mol. Biol.、7〜58頁から7〜60頁)。
【0079】
最終相同性%は、同一性によって測定できるが、そのアラインメント処理自体は、典型的には全か無かの対比較に基づかない。その代わり、化学的類似性または進化距離に基づく各対比較に対するスコアを割り当てる基準化類似性スコアマトリックスが一般に用いられる。一般に用いられるこのようなマトリックスの例は、BLOSUM62マトリックス-BLASTスイートのプログラム用のデフォルトマトリックスである。GCG Wisconsinプログラムは一般に、供給される場合、公的デフォルト値または慣習的シンボル比較表を用いる。GCGパッケージに関しては公的デフォルト値、または他のソフトウェアの場合、BLOSUM62等のデフォルトマトリックスを使用することが好ましい。
【0080】
デフォルト値に対してパラメータセットを有するBLASTアルゴリズムを使用することが好都合である。BLASTアルゴリズムは、参照として本明細書に組み込まれている、http://www.ncbi.nih.gov/BLAST/blast_help.htmlに詳細に記載されている。検索パラメータを定義でき、定義されたデフォルトパラメータ上で有利に設定できる。
【0081】
好都合なことに、BLASTにより評価された場合の「実質的な同一性」とは、少なくとも約7、好ましくは少なくとも約9、最も好ましくは10以上のEXPECT値と一致する配列に等しくすることである。BLAST検索においてEXPECTに関するデフォルト閾値は、通常10である。
【0082】
BLAST(Basic Local Alignment Search Tool)は、プログラムblastp、blastn、blastx、tblastn、及びtblastxにより使用される発見的検索アルゴリズムであり、これらのプログラムにより、有意性は、若干増強されたKarlin及びAltschul(Karlin及びAltschul 1990年、Proc. Natl. Acad. Sci. USA 87:2264;68頁Karlin及びAltschul、1993年、Proc. Natl. Acad. Sci. USA 90:5873〜7頁;http://www.ncbi.nih.gov/BLAST/blast_help.htmlを参照)の統計学的方法を用いたそれらの知見に帰する。例えば、問合せの配列に対するホモログを同定するために、BLASTプログラムを配列類似性検索に適合させる。配列データベースの類似性検索における基礎的問題を検討するために、Altschulら(1994)Nature Genetics 6:119〜129頁を参照されたい。
【0083】
配列比較は、http://www.ncbi.nlm.nih.gov/BLASTに提供される簡単なBLAST検索アルゴリズムを用いて実施するのが最も好ましい。いくつかの実施形態において、配列同一性を決定する場合、ギャップペナルティーは使用しない。
【0084】
[ハイブリダイゼーション]
本明細書に提示された配列、またはそれらの任意の断片もしくは誘導体に、あるいは上記の任意の補体にハイブリダイズできるヌクレオチド配列を開示する。
【0085】
ハイブリダイゼーションとは、「核酸の鎖が塩基対を介して相補鎖と接合する処理工程」(Coombs J(1994)Dictionary of Biotechnology、Stockton Press、ニューヨーク、ニューヨーク州)並びにDieffenbach CW及びGS Dveksler(1995年、PCR Primer, a Laboratory Manual、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)に記載されたポリメラーゼ連鎖反応技術において実施される増幅処理工程を意味する。
【0086】
ハイブリダイゼーション条件は、Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)において教示された核酸結合複合体の融解温度(Tm)に基づき、下記に説明されるように規定された「ストリンジェンシー」を付与する。
【0087】
本明細書に提示されたヌクレオチド配列に、またはそれらの補体に選択的にハイブリダイズできるヌクレオチド配列は、少なくとも20、好ましくは少なくとも25または30、例えば、少なくとも40、60、または100以上の連続ヌクレオチドの領域にわたって本明細書に提示された対応するヌクレオチド配列に対して、一般に少なくとも70%、好ましくは少なくとも75%、より好ましくは少なくとも85%または90%、さらにより好ましくは少なくとも95%、98%、または99%相同である。好ましいヌクレオチド配列は、配列番号1、2、4、または6に相同的な領域を含み、1つの配列に対して、好ましくは少なくとも70%、80%、または90%、より好ましくは少なくとも95%相同である。
【0088】
用語の「選択的にハイブリダイズ可能」とは、標的ヌクレオチド配列が有意に上記バックグラウンドのレベルでプローブにハイブリダイズすることが分かっている条件下で、プローブとして用いられるヌクレオチド配列が使用されることを意味する。バックグラウンドハイブリダイゼーションは、例えば、スクリーニングされるcDNAまたはゲノムDNAライブラリーに存在する他のヌクレオチド配列のために生じ得る。この事象において、バックグラウンドとは、プローブと、標的DNAにより見られる特異的相互作用の強度の10倍未満、好ましくは100倍未満であるライブラリーの非特異的DNAメンバーとの間の相互作用により作製されたシグナルレベルを意味する。相互作用の強度は、例えば、プローブを放射標識により、例えば、32Pにより測定できる。
【0089】
中程度から最大のストリンジェンシーの条件下で、本明細書に提示されたヌクレオチド配列にハイブリダイズできるヌクレオチド配列もまた含まれる。ハイブリダイゼーション条件は、Berger及びKimmel(1987年、Guide to Molecular Cloning Techniques, Methods in Enzymology、152巻、Academic Press、サンディエゴ、カリフォルニア州)において教示された核酸結合複合体の融解温度(Tm)に基づき、下記に説明されるように規定された「ストリンジェンシー」を付与する。
【0090】
最大ストリンジェンシーは、約Tm-5℃(プローブのTmより5℃低い)で;高ストリンジェンシーは、Tmより約5℃から10℃低い;中程度のストリンジェンシーは、Tmより約10℃から20℃低い;低ストリンジェンシーは、Tmより約20℃から25℃低いとき典型的に生じる。当業者により解されるように、最大ストリンジェンシーのハイブリダイゼーションを、同一のヌクレオチド配列を同定または検出するために使用できるが、一方、中程度(または低)ストリンジェンシーのハイブリダイゼーションを、類似のまたは関連ヌクレオチド配列を同定または検出するために使用できる。
【0091】
好ましい実施形態において、ストリンジェント条件下(例えば、65℃及び0.1×SSC{1×SSC = 0.15 M NaCl、0.015 M Na3クエン酸pH 7.0})、1つ以上のVDCCγ-8イオンチャネルヌクレオチド配列にハイブリダイズできるヌクレオチド配列が開示される。ヌクレオチド配列が二本鎖である場合、二重鎖、個々にまたは組合せの両方を包含する。ヌクレオチド配列が一本鎖である場合、そのヌクレオチド配列の相補的配列もまた使用することが理解されよう。
【0092】
本発明の範囲内にあるヌクレオチド配列は、本明細書に提示された配列、あるいはそれらの任意の断片または誘導体に相補的である配列にハイブリダイズできる。同様に、本発明は、配列にハイブリダイズできる配列に相補的であるヌクレオチド配列を包含する。これらのタイプのヌクレオチド配列は、バリアントヌクレオチド配列の例である。この観点から、用語の「バリアント」は、本明細書に提示されたヌクレオチド配列にハイブリダイズできる配列に相補的である配列を包含する。しかしながら、好ましくは、用語の「バリアント」は、ストリンジェント条件下(例えば、65℃及び0.1×SSC{1×SSC = 0.15 M NaCl、0.015 M Na3クエン酸pH 7.0})、本明細書に提示されたヌクレオチド配列にハイブリダイズできる配列に相補的である配列を包含する。
【0093】
[VDCCγ-8イオンチャネル及びホモログのクローニング]
本発明は、本明細書に提示された配列、あるいはそれらの任意の断片または誘導体に相補的であるヌクレオチド配列を包含する。該配列が、その断片に相補的である場合、その配列を、他の生物等における同様のイオンチャネル配列を同定及びクローニングするためのプローブとして使用できる。
【0094】
これは、ヒト及び他の種、例えばマウス、ブタ、ヒツジ等からの、VDCCγ-8イオンチャネル、そのホモログ、及び他の構造的または機能的関連遺伝子のクローニングを可能にする。配列番号1、配列番号2、配列番号4、または配列番号6またはそれらの断片に含まれるヌクレオチド配列と同一か、または十分に同一であるポリヌクレオチドは、適切なライブラリーから部分長または完全長cDNA及びVDCCγ-8イオンチャネルをコードするゲノムクローンを単離するために、cDNA及びゲノムDNA用のハイブリダイゼーションプローブとして使用され得る。このようなプローブはまた、VDCCγ-8イオンチャネル遺伝子に対して配列類似性、好ましくは、高い配列類似性を有する他の遺伝子(ヒト以外の種由来のホモログ及びオルソログをコードする遺伝子を含む)のcDNA及びゲノムクローンを単離するために使用され得る。ハイブリダイゼーションスクリーニング、クローニング、及び配列決定技術は当業者に既知であり、例えばSambrookら(上記文献)に記載されている。
【0095】
プローブとしての使用に好適なヌクレオチド配列は、典型的には、基準のものに対して70%同一、好ましくは80%同一、より好ましくは90%同一、さらにより好ましくは95%同一である。このプローブは一般に、少なくとも15のヌクレオチドを含む。このようなプローブは、少なくとも30のヌクレオチドを有することが好ましく、少なくとも50のヌクレオチドを有することができる。特に好ましいプローブは、150〜500の間の範囲のヌクレオチドであり、さらに特に約300のヌクレオチドである。
【0096】
一実施形態において、ヒト以外の種からホモログ及びオルソログ等、VDCCγ-8ポリペプチドをコードするポリヌクレオチドを得るために、配列番号1、配列番号2、配列番号4、または配列番号6、あるいはそれらの断片を有する標識プローブによりストリンジェントなハイブリダイゼーション下で適切なライブラリーをスクリーニングする工程と、上記ポリヌクレオチド配列を含有する部分長または完全長cDNA及びゲノムクローンを単離する工程とを含む。このようなハイブリダイゼーション技術は、当業者に周知である。ストリンジェントなハイブリダイゼーション条件は、上記に定義されているとおりか、あるいは、50%ホルムアミド、5×SSC(150 mM NaCl、15 mMクエン酸三ナトリウム)、50 mMリン酸ナトリウム(pH 7.6)、5×デンハート液、10%硫酸デキストラン、及び20マイクログラム/ml変性、剪断サケ精巣DNAを含む溶液中、42℃で一晩インキュベーションし、次いで約65℃で0.1×SSC中でろ過洗浄する条件である。
【0097】
[VDCCγ-8イオンチャネルに対する機能的アッセイ]
クローニングされた推定上のVDCCγ-8イオンチャネルポリヌクレオチドは、配列分析または機能的アッセイにより証明され得る。例えば、推定上のVDCCγ-8イオンチャネルまたはホモログは、以下の活性についてアッセイできる。VDCCγ-8受容体cDNAをコードする線形化プラスミドテンプレートからのキャップ付RNA転写物を、標準的手法に従ってin vitroでRNAポリメラーゼにより合成する。in vitro転写物を、最終濃度が0.2 mg/mlで水に懸濁させる。卵巣葉を、成体メスヒキガエルから取り出し、段階Vの濾胞除去卵母細胞を得て、RNA転写物(10 ng/卵母細胞)を注入し、マイクロインジェクション装置を用いて50 nlボーラス中でグルタメート受容体または他のVOCCサブユニットにより共発現させる。2つの電極電圧クランプを、アゴニスト曝露に応答する個々のアフリカツメガエル卵母細胞からの電流を測定するために用いる。ND96溶液媒体中室温で記録する。以下にさらに詳細に記載されるとおり、アフリカツメガエル系はまた、既知のリガンドをスクリーニングし、リガンドを活性化させるための組織/細胞抽出のために使用され得る。代わりの機能的アッセイとしては、全細胞電気生理、蛍光共鳴エネルギー移動(FRET)分析、及びFLIPR分析が挙げられる。
【0098】
[VDCCγ-8イオンチャネルに対する細胞表面発現アッセイ]
段階Vの濾胞除去卵母細胞を得て、RNA転写物(10 ng/卵母細胞)を注入し、グルタメート受容体(GluR1-フリップ等)により共発現させ、マイクロインジェクション装置を用いて50 nlボーラス中、ヘマグルチニンエピトープを細胞外領域に操作した。注入3日後、卵母細胞を、0.25μg/mlのラット抗HA抗体(3F10、Roche)と1時間インキュベートし、次いで西洋ワサビペルオキシダーゼ複合化抗ラット免疫グロブリンと30分インキュベートする。個々の卵母細胞を、50μlのSuperSignal ELISA Femto最大感受性基体(Pierce)に入れ、TD20/20m (Turner Designs)等の好適なルミノメータを用いて化学発光法により定量化する。
【0099】
[VDCCγ-8イオンチャネルに対する発現アッセイ]
VDCCγ-8イオンチャネル関連疾患を治療するのに有用な治療薬を設計するために、VDCCγ-8の発現プロファイル(野生型または特定の変異体かどうか)を測定することは有用である。従って、当業者に既知の方法を、VDCCγ-8が発現される臓器、組織、及び細胞型(並びに発育段階)を決定するために使用できる。例えば、伝統的または「電子」ノーザンを実施できる。逆転写酵素PCR (RT-PCR)もまた、VDCCγ-8遺伝子または変異体の発現をアッセイするために使用できる。VDCCγ-8の発現プロファイルを測定するためにより高感度の方法としては、当業者に既知のRNA分解酵素保護アッセイが挙げられる。
【0100】
ノーザン分析は、遺伝子の転写物の存在を検出するために使用される実験室的技術であり、特定の細胞型または組織からのRNAが結合された膜に対する標識ヌクレオチド配列のハイブリダイゼーションを含む(Sambrook、上記文献、7章及びAusubel, F. M.ら、上記文献、4章及び16章)。BLASTを適用している類似のコンピュータ技術(「電子ノーザン」)を、GenBankまたはLIFESEQデータベース(Incyte Pharmaceuticals)等のヌクレオチドデータベースにおいて同一または関連分子を検索するために使用できる。このタイプの分析は、複数膜ベースのハイブリダイゼーションよりも速く成し得る利点を有する。さらに、コンピュータ検索の感度は、任意の特定のマッチが正確または相同的なものとして分類されるかどうかを判定するために変更され得る。
【0101】
上記のプローブ等、本明細書に記載されたポリヌクレオチド及びポリペプチドを、動物及びヒトの疾患に対する治療法及び診断法の発見のために研究用試薬及び物質として使用できる。
【0102】
[VDCCγ-8イオンチャネルポリペプチドの発現]
当業者に明らかなように、VDCCγ-8イオンチャネルポリペプチド、またはそのホモログ、バリアント、もしくは誘導体をコードする核酸を含む宿主細胞を、好適な条件下(すなわち、VDCCγ-8イオンチャネルポリペプチドが発現される条件)で培養する(一般的に言えば)ことによって、VDCCγ-8ポリペプチドを産生することができる。
【0103】
生物的に活性なVDCCγ-8イオンチャネルを発現させるために、VDCCγ-8イオンチャネルまたはそのホモログ、バリアント、もしくは誘導体をコードするヌクレオチド配列は、適切な発現ベクター、すなわち挿入されたコード化配列の転写及び翻訳に必要な要素を含有するベクターに挿入される。
【0104】
当業者に周知の方法を用いて、VDCCγ-8イオンチャネル、並びに適切な転写及び翻訳調節要素をコードする配列を含有する発現ベクターを構築する。これらの方法としては、in vitro組換えDNA技術、合成技術、及びin vivo遺伝子組換えが挙げられる。このような技術は、Sambrook, J.ら、(1989年、Molecular Cloning:A Laboratory Manual、4章、8章、及び16〜17章、Cold Spring Harbor Press、プレインビュー、ニューヨーク州)及びAusubel, F. M.ら(1995年及び定期的に補遺;Current Protocols in Molecular Biology、9章、13章、及び16章、John Wiley & Sons、ニューヨーク、ニューヨーク州)に記載されている。
【0105】
種々の発現ベクター/宿主系を利用して、VDCCγ-8イオンチャネルをコードする配列を含ませ、発現させることができる。これらとしては、限定はしないが、組換えバクテリオファージ、プラスミド、またはコスミドDNA発現ベクターにより変換された細菌;酵母発現ベクターにより変換された酵母;ウイルス発現ベクター(例えば、バキュロウイルス)により感染させた昆虫細胞系;ウイルス発現ベクター(例えば、カリフラワーモザイクウイルス(CaMV)またはタバコモザイクウイルス(TMV))または細菌発現ベクター(例えば、TiまたはpBR322プラスミド)により変換された植物細胞系;または動物細胞系等の微生物が挙げられる。使用される宿主細胞の性質は重要ではない。
【0106】
「調節要素」または「調節配列」とは、転写及び翻訳を実施するために宿主細胞タンパク質と相互作用するベクターの非翻訳領域(すなわち、エンハンサー、プロモーター、並びに5'及び3'非翻訳領域)のものである。このような要素は、強度及び特異性を変えることができる。利用されるベクター及び宿主系によって、構成的及び誘導的プロモーター等、任意の数の好適な転写及び翻訳要素を使用することができる。例えば、細菌系にクローニングする場合、BLUESCRIPTファージミドのハイブリッドlacZプロモーター(Stratagene、ラジョラ、カリフォルニア州)またはPSPORT1プラスミド(GIBCO/BRL)等の誘導性プロモーターを昆虫細胞に使用できる。バキュロウイルスポリヘドリンプロモーターを、昆虫細胞において使用できる。植物細胞のゲノム(例えば、ヒートショック、PUBISCO、及び保存タンパク質遺伝子)または植物ウイルス(例えば、ウイルスプロモーターまたはリーダー配列)から誘導されたプロモーターまたはエンハンサーをベクター内にクローニングできる。哺乳動物細胞系において、哺乳動物遺伝子または哺乳動物ウイルス由来のプロモーターが好ましい。VDCCγ-8イオンチャネルをコードする配列の複数コピーを含有する細胞系を作製することが必要な場合、SV40またはEBVに基づくベクターを、適切な選択マーカーとともに使用できる。
【0107】
細菌系において、多数の発現ベクターを、VDCCγ-8イオンチャネルのために意図された使用によって選択できる。例えば、大量のVDCCγ-8イオンチャネルが抗体誘導のために必要とされる場合、容易に精製される融合タンパク質の高レベル発現をもたらすベクターを使用できる。このようなベクターとしては、限定はしないが、多機能大腸菌クローニング及びBLUESCRIPT(Stratagene)等の発現ベクターが挙げられ、その中でVDCCγ-8イオンチャネルをコードする配列を、アミノ末端Metに関する配列及び引き続くβ-ガラクトシダーゼの7つの残基を有するフレーム内のベクターに結合でき、その結果、ハイブリッドタンパク質が、pINベクター等を産生する(Van Heeks, G.及びS. M. Schuster(1989)J. Biol. Chem. 264:5503〜5509頁)。pGEXベクター(Promega、マジソン、ウィスコンシン州)を使用して、グルタチオンS-トランスフェラーゼ(GST)との融合タンパク質として異種ポリペプチドを発現させることができる。一般に、このような融合タンパク質は溶解性であり、グルタチオン-アガロースビーズへの吸着、次いで遊離グルタチオンの存在下での溶出により溶解細胞から容易に精製できる。クローニングされた対象のポリペプチドが自由にGST部分から放出できるように、このような系で作製されたタンパク質を、ヘパリン、トロンビン、または因子XAのプロテアーゼ開裂部位を含むように設計できる。
【0108】
酵母菌酵母において、アルファ因子、アルコールオキシダーゼ、及びPGH等の構成的または誘導性プロモーターを含有する多数のベクターを使用できる。レビューに関しては、Ausubel(上記文献)及びGrantら(1987年;Methods Enzymol. 153:516〜544頁)を参照されたい。
【0109】
植物発現ベクターが用いられる場合、VDCCγ-8イオンチャネルをコードする配列の発現は、任意の多数のプロモーターにより誘発され得る。例えば、CaMVの35S及び19Sプロモーター等のウイルスプロモーターを、単独でまたはTMV由来のオメガリーダー配列との組合せで使用できる(Takamatsu, N.(1987)EMBO J. 6:307〜311頁)。あるいは、小型サブユニットのRUBISCOまたはヒートショックプロモーター等の植物プロモーターを使用できる(Coruzzi, G.ら(1984)EMBO J. 3:1671〜1680頁;Broglie, R.ら(1984)Science 224:838〜843頁;及びWinter, J.ら(1991)Results Probl. Cell Differ. 17:85〜105頁)。これらの構築物を、直接のDNA変換または病原体媒介トランスフェクションにより植物細胞に導入できる。このような技術は、多数の一般的に利用できるレビューに記載されている(例えば、Hobbs, S.またはMurry, L. E.、McGraw Hill Yearbook of Science and Technology(1992)McGraw Hill、ニューヨーク、ニューヨーク州;191〜196頁を参照)。
【0110】
昆虫系もまた、VDCCγ-8イオンチャネルを発現させるために使用できる。例えば、1つのこのような系において、オウトグラファカリフォルニア核多汗症ウイルス(AcNPV)は、Spodoptera frugiperda細胞またはTrichoplusia larvaeにおける異種遺伝子を発現させるベクターとして用いられる。VDCCγ-8イオンチャネルをコードする配列を、ポリヘドリン遺伝子等のウイルスの非必須領域にクローニングでき、ポリヘドリンプロモーターの調節下に置くことができる。VDCCγ-8イオンチャネルの好結果の挿入により、ポリヘドリン遺伝子を不活性にし、コートタンパク質を欠く組換えウイルスを生じる。次いで組換えウイルスを、例えば、TrpM8イオンチャネルが発現し得るS. frugiperda細胞またはTrichoplusia larvaeを感染させるために使用できる(Engelhard, E. K.ら(1994)Proc. Nat. Acad. Sci. 91:3224〜3227頁)。
【0111】
哺乳動物宿主細胞において、多くのウイルスベースの発現系を利用できる。アデノウイルスが発現ベクターとして用いられる場合、VDCCγ-8イオンチャネルをコードする配列は、後期プロモーター及び三連リーダー配列からなるアデノウイルス転写/翻訳複合体に結合できる。ウイルスゲノムの非必須E1またはE3領域における挿入を、感染宿主細胞にVDCCγ-8イオンチャネルを発現できる生存可能なウイルスを得るために使用できる(Logan, J.及びT. Shenk(1984)Proc. Natl. Acad. Sci. 81:3655〜3659頁)。さらに、ニワトリ肉腫ウイルス(RSV)エンハンサー等の転写エンハンサーを、哺乳動物宿主細胞における発現を増加させるために使用できる。
【0112】
従って例えば、VDCCγ-8受容体は、ヒト胎生腎細胞293(HEK293)または接着細胞CHOに発現できる。受容体発現を最大にするために、典型的にすべての5'及び3'非翻訳領域(UTR)は、pCDNまたはpCDNA3ベクターへの挿入前に受容体cDNAから除去される。細胞は、リポフェクチンにより個々の受容体cDNAにより形質移入され、400 mg/mlのG418の存在下で選択される。選択3週後、個々のクローンを選択し、さらに分析のために増殖させる。ベクター単独で形質移入されたHEK293またはCHO細胞は、陰性コントロールとして寄与する。個々の受容体を安定に発現する細胞系を単離するために、典型的に約24のクローンを選択し、ノーザンブロット分析により分析する。通常、分析されるG418耐性クローンの約50%において受容体mRNAを検出できる。
【0113】
ヒト人工染色体(HAC)は使用して、プラスミドに含有しかつ発現できるよりも大きな断片のDNAを送達することができる。約6 kb〜10 MbのHACが構築され、治療目的のために従来の送達法(リポソーム、ポリカチオン性アミノポリマー、または小胞)を介して送達される。
【0114】
特定の開始シグナルはまた、VDCCγ-8イオンチャネルをコードする配列のより効率的な翻訳を達成するために使用され得る。このようなシグナルは、ATG開始コドン及び隣接配列を含む。VDCCγ-8イオンチャネル及びその開始コドン、並びに上流配列をコードする配列が、適切な発現ベクターに挿入される場合には、さらなる転写または翻訳調節シグナルを必要としなくてもよい。しかしながら、コード化配列のみ、またはその断片が挿入される場合には、ATG開始コドンを含む外因性翻訳調節シグナルを提供する必要がある。さらに、開始コドンは、全挿入断片の翻訳を確保するために正確なリーディングフレーム内にある必要がある。外因性翻訳要素及び開始コドンは、外因性翻訳要素及び開始コドンは、天然及び合成による種々の起源であり得る。発現効率は、文献(Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125~162頁)に記載されているもの等、使用される特定の細胞系に適切なエンハンサーの包含により増強することができる。文献(Scharf, D.ら(1994)Results Probl. Cell Differ. 20:125〜162頁)に記載されたもの等、使用される特定の細胞系に適切なエンハンサーの包含物により増強できる。
【0115】
さらに、挿入配列の発現を変化させるか、または所望の様式で発現タンパク質を処理する能力に関して、宿主細胞株を選択できる。このようなポリペプチドの修飾としては、限定はしないが、アセチル化、カルボキシル化、グリコシル化、リン酸化、脂質化、及びアシル化が挙げられる。タンパク質の「プレプロ」形態を開裂する翻訳後処理はまた、正しい挿入、折りたたみ、及び/または機能を促進させるために使用され得る。翻訳活性後の特定の細胞機構及び特徴的機構を有する種々の宿主細胞(例えば、CHO、HeLa、MDCK、HEK293、及びWI38)を、American Type Culture Collection (ATCC、ベセスダ、メリーランド州)から入手でき、正しい修飾及び異種タンパク質の処理を確保するために選択できる。
【0116】
長期の組換えタンパク質の高収率産生のために、安定な発現が好ましい。例えば、ウイルスの複製起点及び/または内因性発現要素、並びに選択マーカー遺伝子を、同一または別個のベクター内に含有し得る発現ベクターを用いて、VDCCγ-8イオンチャネルを安定に発現できる細胞系を形質転換できる。該ベクターの導入後、細胞を、選択的培地に切り替える前に富化培地中、約1〜2日間増殖させることができる。選択マーカーの目的とは、耐選択性を付与することであり、その存在は、導入された配列を首尾よく発現する細胞の増殖と回収を可能にする。安定に形質転換された細胞の耐性クローンは、細胞型に対して適切な組織培養技術を用いて増殖できる。
【0117】
任意の数の選択系を、変換細胞系を回収するために使用できる。これらには、限定はしないが、それぞれtk細胞またはapr細胞に使用できる単純ヘルペスウイルスチミジンキナーゼ遺伝子(Wigler, M.ら(1977)Cell 11:223〜32頁)及びアデニンホスホリボシルトランスフェラーゼ遺伝子(Lowy, I.ら(1980)Cell 22:817〜23頁)が挙げられる。また、抗代謝剤、抗生物質、または耐除草剤を、選択基準として使用できる。例えば、dhfrは、耐メトトレキサートを付与し(Wigler, M.ら(1980)Proc. Natl. Acad. Sci. 77:3567〜70頁);nptは、耐アミノグリコシドネオマイシン及びG-418を付与し(Colbere-Garapin, F.ら(1981) J. Mol. Biol. 150: 1〜14頁);及びalsまたはpatは、それぞれ耐クロルスルフロン及び耐ホスフィノトリシンアセチルトランスフェラーゼを付与する(Murry、上記文献)。さらなる選択遺伝子は、例えば、細胞がトリプトファンの代わりにインドールを利用できるtrrB、または細胞がヒスチジンの代わりにヒスチノールを利用できるhisDを記載している(Hartman, S. C.及びR. C. Mulligan(1988)Proc. Natl. Acad. Sci. 85:8047〜51頁)。最近、可視マーカーの使用は、アントシアニン、β-グルクロニダーゼ及びその基質GUS、並びにルシフェラーゼ及びその基質ルシフェリン等のマーカーとして人気を得ている。これらのマーカーを、形質転換体を同定するのみならず、特定のベクター系に起因し得る一過性または安定なタンパク質発現量を定量化するために使用できる(Rhodes, C. A.ら(1995)Methods Mol. Biol. 55:121〜131頁)。
【0118】
マーカー遺伝子発現の存在/非存在は、コントロールの遺伝子もまた存在することを示唆するが、遺伝子の存在及び発現を、確認するために必要であり得る。例えば、VDCCγ-8イオンチャネルをコードする配列が、マーカー遺伝子配列内に挿入される場合、VDCCγ-8イオンチャネルをコードする配列を含有する形質変換細胞は、マーカー遺伝子機能の非存在により同定できる。あるいは、マーカー遺伝子を、単一プロモーターの調節下、VDCCγ-8イオンチャネルをコードする配列と並行して設置することができる。さらに、誘導または選択に応答するマーカー遺伝子の発現は、通常、タンデム遺伝子の発現を示す。
【0119】
あるいは、VDCCγ-8イオンチャネルをコードする核酸配列を含有し、VDCCγ-8イオンチャネルを発現する宿主細胞は、当業者に知られている種々の手法により同定できる。これらの手法としては、限定はしないが、DNA-DNAまたはDNA-RNAハイブリダイゼーション及びタンパク質バイオアッセイまたは免疫アッセイ技術が挙げられ、核酸またはタンパク質配列の検出及び/または定量化に関する技術に基づく膜、溶液、またはチップを含む。
【0120】
VDCCγ-8イオンチャネルをコードするポリヌクレオチド配列の存在は、VDCCγ-8イオンチャネルをコードするプローブまたは断片またはポリヌクレオチドの断片を用いて、DNA-DNAまたはDNA-RNAハイブリダイゼーションあるいは増幅により検出できる。VDCCγ-8イオンチャネルをコードするDNAまたはRNAを含有する形質転換体を検出するために、核酸増幅に基づくアッセイは、VDCCγ-8イオンチャネルをコードする配列に基づくオリゴヌクレオチドまたはオリゴマーの使用を含む。
【0121】
タンパク質に特異的なポリクローナルまたはモノクローナル抗体を用いて、VDCCγ-8イオンチャネルの発現を検出し、測定するための種々のプロトコールは当業者に既知である。このような技術の例としては、酵素結合免疫吸着アッセイ(ELISA)等、放射免疫アッセイ(RIA)等、及び蛍光標示式細胞分取(FACS)が挙げられる。VDCCγ-8イオンチャネルに対する2つの非妨害エピトープに反応性のあるモノクローナル抗体を利用する2部位モノクローナルベースの免疫アッセイが好ましいが、競合結合アッセイを使用できる。これら及び他のアッセイは、当業者、例えば、Hampton, R.ら(1990年;Serological Methods, a Laboratory Manual, Section IV、APS Press、セントポール、ミネソタ州)及びMaddox, D. E.ら(1983年;J. Exp. Med. 158:1211〜1216頁)によく記載されている。
【0122】
多種多様の標識及び複合化技術は、当業者に既知であり、種々の核酸及びアミノ酸アッセイに使用され得る。VDCCγ-8イオンチャネルをコードするポリヌクレオチド関連配列を検出するために標識ハイブリダイゼーションまたはPCRプローブを産生する手段としては、オリゴ標識化、ニック翻訳、エンド標識化、または標識ヌクレオチドを用いるPCR増幅が挙げられる。あるいは、VDCCγ-8イオンチャネルをコードする配列、またはその任意の断片を、mRNAプローブの産生のためにベクターにクローニングできる。このようなベクターは、当業者に既知で市販されており、T7、T3、またはSP6等の適切なRNAポリメラーゼ及び標識ヌクレオチドの添加により、in vitroでRNAプローブを合成するために使用され得る。これらの手法は、Pharmacia & Upjohn(カラマズー、ミシガン州)、GE Healthcare(英国)、及びU. S. Biochemical社(クリーブランド、オハイオ州)により提供されるもの等、種々の市販のキットを用いて実施できる。検出を容易にするために使用できる好適なレポーター分子または標識としては、放射性核種、酵素、蛍光剤、化学発光剤、または色素生成剤、並びに基質、補因子、阻害剤、磁気粒子等が挙げられる。
【0123】
VDCCγ-8サブユニットをコードするヌクレオチド配列により形質転換された宿主細胞を、発現に好適な条件下で培養し、細胞培養からタンパク質を回収できる。形質転換細胞により産生されたタンパク質は、使用される配列及び/またはベクターによって、細胞膜に配置され、分泌され、または細胞内に含ませることができる。当業者により理解されるように、VDCCγ-8サブユニットをコードするポリヌクレオチドを含有する発現ベクターは、原核生物または真核生物細胞膜を介してVDCCγ-8サブユニットの分泌をもたらすシグナル配列を含有するように設計できる。他の構築物を、溶解性タンパク質の精製を促進するポリペプチドドメインをコードするヌクレオチド配列に、VDCCγ-8サブユニットをコードする配列を結合させるために使用できる。このような精製促進ドメインとしては、限定はしないが、固定化金属上で精製を可能にするヒスチジン-トリプトファンモジュール等の金属キレート化ペプチド、固定化免疫グロブリン上で精製を可能にするタンパク質Aドメイン、及びFLAGS拡張/アフィニティー精製システム(Immunex社、シアトル、ワシントン州)に利用されるドメインが挙げられる。精製ドメインと配列をコードするVDCCγ-8サブユニットとの間で、因子XAまたはエンテロキナーゼ(Invitrogen、サンディエゴ、カリフォルニア州)に特異的なもの等の開裂可能なリンカー配列の含有物を、精製を促進させるために使用できる。1つのこのような発現ベクターは、チオレドキシンまたはエンテロキナーゼ開裂部位に先立って6つのヒスチジン残基をコードするVDCCγ-8サブユニット及び核酸を含有する融合タンパク質の発現を提供する。該ヒスチジン残基は、固定化金属イオンアフィニティークロマトグラフィ(IMIAC;Porath, J.ら(1992)Prot. Exp. Purif. 3:263〜281頁に記載)上で精製を促進させるが、一方、エンテロキナーゼ開裂部位は、融合タンパク質からVDCCγ-8サブユニットを精製する手段を提供する。融合タンパク質を含有するベクターの考察は、Kroll, D. J.ら(1993年;DNA Cell Biol. 12:441〜453頁)に提供される。
【0124】
VDCCγ-8サブユニットの断片は、組換え体産生によってだけでなく、固相法を用いた直接的ペプチド合成によって産生され得る(Merrifield J.(1963)J. Am. Chem. Soc. 85:2149〜2154頁)。タンパク質合成は、手動技術または自動操作により実施できる。自動合成は、例えば、Applied Biosystems 431Aペプチドシンセサイザー(Perkin Elmer)を用いて達成できる。VDCCγ-8サブユニットの種々の断片は、別個に合成でき、次いで完全長分子を産生するために結合することができる。
【0125】
[バイオセンサー]
VDCCγ-8ポリペプチド、核酸、プローブ、抗体、発現ベクター、及びリガンドは、バイオセンサー(の産生のため)として有用である。
【0126】
Aizawa(1988)、Anal. Chem. Symp. 17:683頁によれば、バイオセンサーは、分子認識に関する受容体のユニークな組合せ、例えば、VDCCγ-8等の固定化抗体または受容体を有する選択的層、及び測定される値を転送するトランスデューサーとして定義される。このようなバイオセンサーの一群は、受容体と周囲の媒体との相互作用のため、表層の最適性質に生じる変化を検出する。このような技術の中で、特にエリプソメトリー及び表面プラズモン共鳴を挙げることができる。VDCCγ-8を取り込むバイオセンサーを、より好ましくは、上記に定義された疾患に関してVDCCγ-8リガンドの存在またはレベル検出するために使用できる。このようなバイオセンサーの構築は当業者に周知である。
【0127】
従って、VDCCγ-8サブユニットを発現する細胞系を、[3H]イノシトールホスフェートまたは他の第二のメッセンジャーの受容体促進形成を介してATP等のリガンド検出のための受容体系として使用できる(Wattら、1998年、J Biol Chem 5月29日;273(22):14053〜8頁)。受容体リガンドバイオセンサーはまた、Hoffmanら、2000年、Proc Natl Acad Sci USA 10月10日;97(21):11215〜20頁に記載されている。VDCCγ-8を含む光学的及び他のバイオセンサーはまた、例えば、Figlerら、1997年、Biochemistry 12月23日;36(51):16288〜99頁及びSarrioら、2000年、Mol Cell Biol 2000年7月;20(14):5164〜74頁により記載されているG-タンパク質及び他のタンパク質との相互作用のレベルまたは存在を検出するために使用され得る。バイオセンサー用のセンサーユニットは、例えば、米国特許第5,492,840号に記載されている。当業者は、VDCCγ-8のアンタゴニストの価値を等しく認識するであろう。例えば、別の実施形態において、ADHDの治療は、VDCCγ-8アンタゴニストによって達成され得る。
【0128】
[スクリーニングアッセイ]
ホモログ、バリアント、及び誘導体を含むVDCCγ-8ポリペプチドは、天然であろうと組換え体であろうとも、サブユニットを結合し、VDCCγ-8を活性化するか(アゴニスト)、またはその活性を阻害し(アンタゴニスト)、関連チャネルの動態を変えるか、またはその発現を変える化合物に関するスクリーニング法に使用され得る。従って、このようなポリペプチドは、例えば、細胞、無細胞製剤、化学ライブラリー、及び天然物の混合物において、小型分子の基質とリガンドとの結合を評価するためにも使用され得る。これらの基質及びリガンドは、天然の基質及びリガンドであり得るか、または構造的または機能的ミメティックであり得る。Coliganら、Current Protocols in Immunology 1(2):5章(1991)を参照されたい。
【0129】
VDCCγ-8イオンチャネルポリペプチドは、多くの生物学的機能に関与し、上記に掲げた疾患に関連する。従って、一方では、VDCCγ-8イオンチャネルを刺激し、他方では、VDCCγ-8イオンチャネルの機能を阻害できる化合物及び薬物を見出すことが望ましい。一般に、アゴニスト及びアンタゴニストは、興奮毒性及び認知障害のような病態に対する治療及び予防目的のために使用される。VDCCγ-8を欠くマウスが、統合失調症性表現型を示すというデータが本明細書に示されることを当業者は理解するであろう。従って、好ましい実施形態において、VDCCγ-8を活性化する化合物または薬物(すなわち、アゴニスト)は、上記に示されたように統合失調症及び他の関連精神疾患に対する治療薬となる可能性がある。当業者は、VDCCγ-8のアンタゴニストの価値を等しく認識するであろう。例えば、代わりの実施形態において、ADHDの治療は、VDCCγ-8アンタゴニストによって達成され得る。
【0130】
VDCCγ-8イオンチャネルタンパク質と相互作用しやすい候補化合物の合理的な設計は、ポリペプチドの分子形状の構造的研究に基づくことができる。特定の他のタンパク質と相互作用する部位を決定する1つの手段は、物理的構造決定、例えば、X線結晶回折または二次元NMR技術である。これらは、アミノ酸残基が分子接触領域を形成する手引きを提供する。タンパク質構造決定の詳細な説明に関して、例えば、Blundell及びJohnson(1976)Protein Crystallography、Academic Press、ニューヨークを参照されたい。
【0131】
代わりの合理的な設計は、その表面にVDCCγ-8受容体ポリペプチドを発現する適切な細胞を一般に産生することを含むスクリーニング法を用いる。このような細胞としては、動物、酵母、ショウジョウバエ、または大腸菌に由来する細胞が挙げられる。次に、受容体を発現している細胞(または発現受容体を含有する細胞膜)を試験化合物と接触させて、結合、または機能的応答の刺激もしくは阻害を見る。例えば、アフリカツメガエル卵母細胞に、VDCCγ-8 mRNAまたはポリペプチドを注入でき、さらに別途詳細に記載されるように、測定電位クランプの使用により測定される試験化合物への曝露により電流が誘導される。
【0132】
VDCCγ-8イオンチャネルにより個々に各候補化合物を試験する代わりに、候補リガンドのライブラリーまたはバンクが有利には作製され、スクリーニングされ得る。従って、例えば、200を超える推定上の受容体リガンドのバンクを、スクリーニング用に組み立てることができる。このバンクは:イオンチャネルを介して作用することが知られている伝達物質、ホルモン及びケモカイン;イオンチャネル、非哺乳動物、哺乳動物の対応物が未だ同定されていない生物活性ペプチドに対する推定上のアゴニストであり得る天然化合物;及び天然には見られないが、未知の天然リガンドによりイオンチャネルを活性化する化合物、を含む。さらに詳細に別途記載されているように、機能的アッセイ(すなわち、カルシウム、マイクロフィジオメーター(microphysiometer)、FLIPRアッセイ、全細胞電気生理学、卵母細胞電気生理学等、別途参照)並びに結合アッセイの両方を用いて既知のリガンドに関する受容体をスクリーニングするために、このバンクが用いられる。しかしながら、今までのところ、同族でない活性化リガンド(アゴニスト)または不活化リガンド(アンタゴニスト)が残されている多数の哺乳動物の受容体が存在する。このように、これらの受容体に対する活性リガンドは、今日まで同定されたリガンドバンク内に含ませることができない。従って、VDCCγ-8はまた、天然のリガンドを同定するために、組織抽出物に対して機能的にスクリーニング(カルシウム、cAMP、マイクロフィジオメーター(microphysiometer)、卵母細胞電気生理学等を用いて機能的スクリーニング)することができる。陽性の機能的応答を生じる抽出物は、活性化リガンドが単離され、同定されるまで、本明細書に記載されるようにアッセイされるフラクションにより連続的に細分画できる。
【0133】
このような実験において、形質移入されたHEK293細胞またはベクターコントロール細胞における基本のカルシウムレベルは、100〜200 nMの正常な範囲であることが観察される。ホモメリックまたはヘテロメリックVDCCγ-8イオンチャネル、あるいは組換えホモメリックまたはヘテロメリックVDCCγ-8イオンチャネルを発現するHEK293細胞は、fura 2により負荷され、1日に150を超える選択リガンドまたは組織/細胞抽出物は、アゴニスト誘導カルシウム動員について評価される。同様に、VDCCγ-8イオンチャネルまたは組換えVDCCγ-8イオンチャネルを発現するHEK 293細胞は、Ca2+フラックスの増加または減少について評価される。一過性のカルシウムを供するアゴニストは、その応答が受容体を発現する形質移入細胞に特有であるかどうかを判定するために、ベクターコントロール細胞において試験される。
【0134】
別の方法は、VDCCγ-8イオンチャネルの阻害または刺激を判定することにより受容体阻害剤に関するスクリーニングを含む。このような方法は、ホモメリックチャネルを形成するために単独で、または細胞表面に受容体を発現させるヘテロメリックチャネルを形成するために他のTrpチャネルサブユニットとともに、VDCCγ-8サブユニットを真核細胞に形質移入することを含む。次いで、VDCCγ-8受容体の存在下、該細胞を強力なアンタゴニストに曝露させる。電流の電導度または動態の変化を測定するために、全細胞電気生理学を用いて、該細胞を試験することができる。
【0135】
VDCCγ-8のアゴニストまたはアンタゴニストを検出する別の方法は、参照として本明細書に組み込まれている、米国特許第5,482,835号に記載されている酵母ベースの技術である。
【0136】
候補化合物がタンパク質である場合、特定の抗体またはペプチドにおいて、候補化合物のライブラリーを、ファージディスプレイ技術を用いてスクリーニングできる。ファージディスプレイは、組換えバクテリオファージを利用する分子スクリーニングのプロトコールである。この技術は、各々のファージまたはファージミドが特定の候補化合物を発現するように、候補化合物のライブラリーから1つの化合物をコードする遺伝子によりバクテリオファージを形質転換することを含む。形質転換バクテリオファージ(好ましくは固体支持体に係留される)は、適切な候補化合物を発現し、それらのファージコート上に示す。VDCCγ-8ポリペプチドまたはペプチドに結合できる特定の候補化合物は、アフィニティー相互作用に基づく選択様式により富化される。次に好結果の候補薬剤を特性化する。ファージディスプレイは、標準的なアフィニティーリガンドスクリーニング技術よりも利点を有する。ファージ表面は、三次元立体配置で候補薬剤を示し、その天然の立体配座により密接に類似している。これは、スクリーニング目的のために、より特異的かつより高いアフィニティー結合を可能にする。
【0137】
化合物のライブラリーをスクリーニングする別の方法は、化合物のライブラリーを発現する組換えDNA分子により安定に形質転換される真核宿主細胞または原核宿主細胞を利用する。生存形態または固定形態のこのような細胞を、標準的な結合パートナーアッセイに使用できる。また、細胞応答を検出するために高感度法を記載しているParceら(1989)Science 246:243〜247頁;及びOwickiら(1990)Proc. Nat'l Acad. Sci. USA 87;4007〜4011頁を参照されたい。競合アッセイは特に有用であり、化合物のライブラリーを発現している細胞を、125I-抗体等のVDCCγ-8ポリペプチドに結合することが知られている標識抗体、及び結合組成物に対する結合アフィニティーが測定される候補化合物等の試験サンプルと接触させるか、またはインキュベートする。次いでポリペプチドに対する結合及び遊離標識結合パートナーは、結合度を評価するために分離される。結合された試験サンプル量は、ポリペプチドに結合する標識抗体量に反比例する。
【0138】
多くの技術の中で任意の1つを、結合度を評価するために遊離結合パートナーから結合分離するために使用できる。この分離工程は、典型的にフィルターへの接着、フィルターへの接着後の洗浄、プラスチックへの接着後の洗浄、または細胞膜の遠心分離等の手法を典型的に含み得る。
【0139】
さらに別のアプローチは、例えば、変換された真核生物または原核生物宿主細胞から抽出された可溶性、未精製または可溶性の精製ポリペプチドまたはペプチドを使用することである。これは、特異性増加の利点、自動化する能力、及び薬物試験のハイスループットを有して「分子」結合アッセイを可能にする。
【0140】
候補化合物スクリーニングに関する別の技術は、例えば、VDCCγ-8ポリペプチドに対する好適な結合アフィニティーを有する新規な化合物をスクリーニングするハイスループットを提供するアプローチを含み、1984年9月13日に公表された国際特許出願WO第84/03564号(Commonwealth Serum Labs.)に詳細に記載されている。最初に、多数の異なる小型ペプチド試験化合物は、固体支持体、例えば、プラスチックピンまたはいくつかの他の適切な表面上で合成され、Fodorら(1991)を参照されたい。次いで、ピンのすべてを、可溶性VDCCγ-8ポリペプチドと反応させて洗浄する。次の工程は、結合ポリペプチドを検出することを含む。このように、ポリペプチドと特異的に相互作用する化合物が同定される。
【0141】
リガンド結合アッセイは、受容体薬理を確認するための直接的方法を提供し、ハイスループットフォーマットに適応可能である。受容体に対する精製リガンドは、結合研究のために特異的高活性(50〜2000 Ci/mmol)に放射標識できる。次いで、放射標識の処理により、その受容体に対するリガンドの活性を減少させないという決定がなされる。緩衝液、イオン、pH及びヌクレオチド等の他のモジュレーターに関するアッセイ条件は、膜及び全細胞受容体源両方に関して稼動シグナル対ノイズ比を確立するために最適化される。これらのアッセイに関して、特定の受容体結合は、全関連放射活性マイナス過剰の未標識競合リガンドの存在下で測定された放射活性として定義される。可能な場合、2つ以上の競合リガンドは、残存非特異的結合を規定するために使用される。
【0142】
該アッセイは、候補化合物の結合を簡便に試験することができ、その中で、受容体を有する細胞への接着は、候補化合物と直接的または間接的に関連する標識により、または標識競合物との競合を含むアッセイにおいて検出される。さらに、これらのアッセイは、細胞表面に受容体を有する細胞に適切な検出システムを用いて、候補化合物が受容体の活性化により作製されたシグナルを生じるかどうかを試験することができる。活性化阻害剤は、既知のアゴニストの存在下で一般にアッセイされ、候補化合物の存在下、アゴニストによる活性化に対する効果が見られる。
【0143】
さらに、該アッセイは、混合物を形成するためにVDCCγ-8ポリペプチドを含有する溶液と候補化合物とを混合する工程と、混合物中のVDCCγ-8イオンチャネル活性を測定する工程と、標品に対して混合物のVDCCγ-8イオンチャネル活性を比較する工程とを簡便に含ませることができる。
【0144】
VDCCγ-8サブユニットcDNA、タンパク質、及びタンパク質に対する抗体を使用して、細胞中のVDCCγ-8サブユニットmRNA及びタンパク質の産生に対する添加化合物の効果を検出するアッセイを構成することができる。例えばELISAは、当業者に既知の標準的方法によりモノクローナル及びポリクローナル抗体を用いてVDCCγ-8サブユニットタンパク質の分泌または細胞関連レベルを測定するために構成でき、これは、好適に操作された細胞または組織からVDCCγ-8サブユニットの産生を阻害または増強できる薬剤(それぞれアンタゴニストまたはアゴニストとも呼ばれる)を発見するために使用され得る。スクリーニングアッセイを実施するための標準的方法は、当業者に十分に理解される。
【0145】
強力なVDCCγ-8イオンチャネルのアンタゴニスト及びブロッカーの例としては、抗体または、いくつかの場合、プリン及びプリン類縁体等のヌクレオチド及びそれらの類縁体、VDCCγ-8イオンチャネルのリガンド、例えばリガンドの断片、あるいは受容体の活性を抑制するように受容体に結合するが、応答を誘発しない小型分子に密に関連するオリゴヌクレオチドまたはタンパク質が挙げられる。
【0146】
従って、我々はまた、VDCCγ-8ポリペプチド及び/またはペプチドに特異的に結合できる化合物を提供する。
【0147】
用語「化合物」とは、生体高分子(例えば、核酸、タンパク質、非ペプチド、または有機分子)等の化学的化合物(天然または合成)、あるいは細菌、植物、真菌、または動物(特に哺乳動物)細胞もしくは組織等の生体材料から作製された抽出物、あるいは無機要素または分子さえも称する。該化合物は抗体であることが好ましい。
【0148】
実施されるこのようなスクリーニングに必要な材料を、スクリーニング用キット内にパッケージできる。このようなスクリーニング用キットは、VDCCγ-8イオンチャネルポリペプチドの産生を減少させるか、または増強させるVDCCγ-8ポリペプチドまたは化合物に関するアゴニスト、アンタゴニスト、リガンド、受容体、基質、酵素等を同定するために有用である。該スクリーニング用キットは:(a)VDCCγ-8ポリペプチド;(b)VDCCγ-8ポリペプチドを発現する組換え細胞;(c)VDCCγ-8ポリペプチドを発現している細胞膜;または(d)VDCCγ-8ポリペプチドに対する抗体を含む。該スクリーニング用キットは、場合によっては、取扱い説明書を含むことができる。
【0149】
[トランスジェニック動物]
天然もしくは組換えVDCCγ-8イオンチャネル、またはホモログ、バリアントもしくは誘導体を、正常な発現レベルと比較して上昇または減少レベルで発現できるトランスジェニック動物は、本発明の範囲内に入る。VDCCγ-8遺伝子の欠失等1つ以上の機能変異の結果、機能的VDCCγ-8受容体を発現しないトランスジェニック動物(VDCCγ-8ノックアウト動物)が含まれる。このようなトランスジェニック動物は、ブタ、ヒツジ、またはげっ歯類等の非ヒト哺乳動物であることが好ましい。トランスジェニック動物は、マウスまたはラットであることが最も好ましい。このようなトランスジェニック動物をスクリーニング法に使用して、VDCCγ-8イオンチャネルのアゴニスト及び/またはアンタゴニストを同定でき、並びに疾患のin vivo治療としてそれらの有効性を試験することができる。
【0150】
好ましい実施形態において、トランスジェニックVDCCγ-8動物、特にVDCCγ-8ノックアウトは、好ましくは本明細書に設定された試験により測定されるように、実施例に示された表現型を示す。従って、VDCCγ-8動物、特にVDCCγ-8ノックアウトは、好ましくは1つ以上の、以下の:ロータロッド性能の減少、新規環境に対する馴化の減少、認知欠損または発作感受性を示す。
【0151】
非常に好ましい実施形態において、VDCCγ-8動物、特にVDCCγ-8ノックアウトは、対応する野生型マウスと比較して、測定パラメータの少なくとも10%、好ましくは少なくとも20%、より好ましくは少なくとも30%、40%、50%、60%、70%、80%、90%、または100%高いまたは低い(場合によっては)を示す。
【0152】
例えば、VDCCγ-8イオンチャネルの産生を欠くように操作されたトランスジェニック動物を、VDCCγ-8イオンチャネルのアゴニスト及び/またはアンタゴニストを同定するアッセイに使用できる。1つのアッセイは、VDCCγ-8イオンチャネル受容体の非存在下で生理的応答を生じるかどうかを判定するために潜在的薬物(候補リガンドまたは化合物)を評価するように設計される。これは、上記に考察されたようにトランスジェニック動物に薬物を投与し、次いで動物を特定の応答に関してアッセイすることにより達成され得る。任意の生理的パラメータをこのアッセイで測定し得るが、好ましい応答としては、1つ以上の、以下の:耐疾患変化;炎症応答の変化;発作感受性の変化;認知性能の変化;シナプス可塑性の変化;社会的相互作用の変化;が挙げられる。
【0153】
潜在的薬物(候補リガンドまたは化合物)がVDCCγ-8受容体に結合するかどうかを判定するために、VDCCγ-8ノックアウト動物に由来する組織を受容体結合アッセイに使用できる。このようなアッセイは、VDCCγ-8受容体産生を欠くように操作されたトランスジェニック動物からの第一の受容体調製物及び同定された任意のVDCCγ-8リガンドまたは化合物を結合することが知られている供給源からの第二の受容体調製物を得ることにより実施され得る。一般に、第一及び第二の受容体調製物は、それらが得られる供給源を除いてすべての点で類似している。例えば、トランスジェニック動物(上記及び下記等)からの脳組織をアッセイに使用する場合、正常(野生型)動物からの同等の脳組織は、第二の受容体調製物の供給源として使用される。各受容体調製物は、VDCCγ-8受容体に結合することが知られているリガンド単独で、及び候補リガンドまたは化合物の存在下の両方でインキュベートされる。候補リガンドまたは化合物は、いくつかの異なる濃度で調べることが好ましいであろう。
【0154】
既知のリガンドによる結合が、試験化合物により置換される範囲は、第一及び第二の受容体調製物の両方に関して決定される。トランスジェニック動物由来の組織を直接アッセイに使用できるか、または該組織は、それら自体アッセイに使用される膜または膜タンパク質を単離するために処理され得る。好ましいトランスジェニック動物はマウスである。該リガンドは、結合アッセイと適合する任意の手段を用いて標識できる。これには、限定はしないが、放射活性、酵素的、蛍光または化学発光標識(並びにさらに詳細に上記に示される他の標識技術)が挙げられるであろう。
【0155】
さらに、VDCCγ-8イオンチャネル受容体のアンタゴニストは、機能的VDCCγ-8を発現する野生型動物、及びVDCCγ-8受容体機能の発現減少または発現消滅と関連する任意の表現型特徴を示す同定された動物に候補化合物等を投与することにより同定され得る。
【0156】
非ヒトトランスジェニック動物を作製する詳細な方法は、下記にさらに詳細に説明される(実施例1及び2)。トランスジェニック遺伝子構築体を、トランスジェニック哺乳動物を作製するために動物の生殖細胞系に導入できる。例えば、該構築物の1つまたはいくつかのコピーを、標準的なトランスジェニック技術により哺乳動物の胎仔のゲノムに取り込むことができる。
【0157】
代表的な実施形態において、トランスジェニック非ヒト動物は、非ヒト動物の生殖細胞系に導入遺伝子を導入することにより作製される。種々の発達段階における胎仔標的細胞は、導入遺伝子を導入するために使用され得る。胎仔標的細胞の発達段階によって種々の方法が用いられる。任意の動物の特定系は、一般的な良好な健康、良好な胎仔収率、胎仔における良好な前核可視性、及び良好な生殖適合性に関して選択される。さらに、ハプロタイプは重要な因子である。
【0158】
導入遺伝子の胎仔への導入は、例えば、マイクロインジェクション、エレクトロポレーション、またはリポフェクション等、当業者に既知の任意の手段により達成され得る。例えば、該構築体の1つ以上のコピーを発達哺乳動物の細胞に保持させるために哺乳動物の受精卵の前核内に該構築体のマイクロインジェクションにより哺乳動物にVDCCγ-8受容体導入遺伝子を導入できる。受精卵に導入遺伝子構築体の導入後、該卵子は、時間量を変えるためにin vitroでインキュベートできるか、または代理宿主に再移植できるか、あるいはそれらの両方を行うことができる。成熟性に向けてin vitroインキュベーションもまた実施できる。1つの一般的な方法は、種によって約1〜7日間in vitroで胎仔をインキュベートし、次いで胎仔を代理宿主に再移植させることである。
【0159】
遺伝子導入により操作された胎仔の後代を、組織のセグメントのサザンブロット解析により該構築体の存在に関して試験することができる。外因性クローニング構築体の1つ以上のコピーが、このようなトランスジェニック胎仔のゲノムに安定に組込まれたままである場合、遺伝子導入により付加された構築体を担持する永続的トランスジェニック哺乳動物系を確立することが可能である。
【0160】
遺伝子導入により改変された哺乳動物の同腹仔を、子孫のゲノムに該構築体の取込みのために誕生後にアッセイできる。好ましくは、このアッセイは、後代からの染色体材料上の所望の組換えタンパク質産物またはそのセグメントをコードするDNA配列に相当するプローブをハイブリダイズすることによって達成される。それらのゲノムにおける構築体の少なくとも1つのコピーを含有することが分かっているこれらの哺乳動物の後代を、成熟性に向けて成長させる。
【0161】
本文書の目的に関して、受精卵は、本質的に完全生物に成長できる二倍体細胞の形成である。一般に、受精卵は、1つまたは複数の配偶子から2つのハプロイド核の融合によって、天然にまたは人工的に形成された核を含有する卵子を含む。従って、配偶子核は、天然に適合するもの、すなわち、機能的生物への分化及び発達を受けることができる生存受精卵をもたらすものでなければならない。一般に正倍数性受精卵が好ましい。異数性受精卵が得られる場合、染色体数は、どちらかの配偶子が生じた生物の正倍数に関して2つ以上変える必要はない。
【0162】
同様の生物学的考察に加えて、物理的なものは、受精卵核または受精卵核の一部を形成する遺伝物質に加えることができる外因性遺伝物質量(例えば、容量)を調節する。遺伝物質が取り出されない場合、加えることのできる外因性遺伝物質量は、物理的に破壊されることなく吸収される量によって限定される。挿入される外因性遺伝物質の容量は、一般に約10ピコリットルを超えないであろう。添加の物理的効果は、受精卵の生存を物理的に破壊するほど大きなものであってはならない。DNA配列の数及び種類の生物学的限定は、特定の受精卵及び外因性遺伝物質の機能によって変わり、当業者には容易に理解される。というのは、生じた受精卵の外因性遺伝物質等の遺伝物質は、受精卵の機能的生物への分化及び発達を生物学的に開始させ、維持できなければならないからである。
【0163】
受精卵に添加される導入遺伝子構築体のコピー数は、添加される外因性遺伝物質の全量に依存し、遺伝的形質転換をもたらし得る量である。理論的には、1つだけのコピーが必要であり;しかしながら、一般に、1つのコピーが機能的であることを確保するために、多くのコピー、例えば、導入遺伝子構築体の1,000〜20,000のコピーが利用される。しばしば、外因性DNA配列の表現型発現を増強させるために挿入された各外因性DNA配列の2つ以上の機能的コピーを有することが有利である。
【0164】
核遺伝物質への外因性遺伝物質の添加を可能にする任意の技術を、それが、細胞、核膜または他の既存の細胞もしくは遺伝子構造に対して破壊的でない限り利用できる。外因性遺伝物質は、マイクロインジェクションにより核酸遺伝物質に優先的に挿入される。細胞及び細胞構造のマイクロインジェクションは既知であり、当業者に使用される。
【0165】
再移植は、標準的な方法を用いて達成される。通常、代理宿主を麻酔にかけ、胎芽を輸卵管に挿入する。特定の宿主に移植された胎芽数は種によって変わるが、通常、種が天然に生む子孫数に相当する。
【0166】
代理宿主のトランスジェニック子孫は、任意の好適な方法により導入遺伝子の存在及び/または発現についてスクリーニングされ得る。スクリーニングはしばしば、導入遺伝子の少なくとも一部に相補的であるプローブを用いて、サザンブロットまたはノーザンブロット解析により達成される。導入遺伝子によりコード化されたタンパク質に対する抗体を用いるウェスタンブロット解析を、導入遺伝子産物の存在をスクリーニングする代わりのまたはさらなる方法として使用できる。典型的には、DNAは、尾部組織から調製され、導入遺伝子に関するサザン解析またはPCRにより分析される。あるいは、最高レベルで導入遺伝子を発現すると考えられている組織または細胞は、サザン解析またはPCRを用いて導入遺伝子の存在及び発現に関して試験されるが、任意の組織または細胞型を、この解析のために使用できる。
【0167】
導入遺伝子の存在を評価するための代わりのあるいはさらなる方法としては、限定はしないが、酵素及び/または免疫学的アッセイ等の好適な生化学的アッセイ、特定のマーカーまたは酵素活性に関する組織学的染色、フローサイトメトリー分析等が挙げられる。血液分析もまた、血中導入遺伝子産物の存在を検出するために、並びに種々のタイプの血液細胞及び他の血液構成物のレベルに対する導入遺伝子の効果を評価するために有用となり得る。
【0168】
トランスジェニック動物の後代は、トランスジェニック動物と好適なパートナーとの交配、またはトランスジェニック動物から得られた卵子及び/または精子のin vitro受精により得ることができる。パートナーとの交配が実施される場合、パートナーは、トランスジェニック及び/またはノックアウトであってもそうでなくてもよく;パートナーがトランスジェニックである場合、同じかまたは異なる導入遺伝子、あるいは両方を含有できる。あるいは、パートナーは親系であってもよい。in vitro交配が用いられる場合、交配胚芽は、代理宿主に移植してもよく、またはin vitroでインキュベートしてもよく、あるいは両方であってもよい。いずれかの方法を用いる後代は、上記の方法または他の適切な方法を用いて導入遺伝子の存在に関して評価され得る。
【0169】
本明細書に記載された方法に従って作製されたトランスジェニック動物は、外因性遺伝物質を含む。上記に示されたように、一定の実施形態において、外因性遺伝物質は、VDCCγ-8イオンチャネルの産生を生じるDNA配列である。さらに、このような実施形態において、該配列は、転写調節要素、例えば、好ましくは特定タイプの細胞において導入遺伝子産物の発現を可能にするプロモーターに結合する。
【0170】
レトロウイルス感染もまた、導入遺伝子を非ヒト動物に導入するために使用できる。非ヒト胚芽の発達は、in vitroで胚盤胞段階に培養できる。この時期の間、分割球はレトロウイルス感染の標的であり得る(Jaenich, R.(1976)PNAS 73:1260〜1264頁)。分割球の効率的な感染は、透明層を除去するために酵素処理により得られる(Manipulating the Mouse Embryo、Hogan編集(Cold Spring Harbor Laboratory Press、コールドスプリングハーバー、1986年)。導入遺伝子を導入するために用いられるウイルスベクター系は、典型的に導入遺伝子を運搬する複製欠損レトロウイルスである(Jahnerら(1985)PNAS 82:6927〜6931頁;Van der Puttenら(1985)PNAS 82:6148〜6152頁)。形質移入は、ウイルス産生細胞の単層上で分割球を培養することによって容易にかつ効率的に得られる(Van der Putten、上記文献;Stewartら(1987)EMBO J. 6:383〜388頁)。あるいは、感染は後期段階で実施され得る。ウイルスまたはウイルス産生細胞を分割腔に注入できる(Jahnerら(1982)Nature 298:623〜628頁)。トランスジェニック非ヒト動物を形成した細胞のサブセットのみに取込みが生じることから、ファウンダーの大部分は導入遺伝子に関するモザイクである。さらに、ファウンダーは、一般に子孫において分離するゲノムの異なる位置で導入遺伝子の種々のレトロウイルス挿入を含有することができる。さらに、妊娠中期の胎仔の子宮内レトロウイルス感染による生殖細胞系に導入遺伝子を導入することも可能である(Jahnerら(1982)上記文献)。
【0171】
導入遺伝子の導入に関する第三のタイプの標的細胞は、胎芽幹細胞(ES)である。ES細胞は、in vitroで培養され、胎芽と融合された移植前胎芽から得られる(Evansら(1981)Nature 292:154〜156頁;Bradleyら(1984)Nature 309:255〜258頁;Gosslerら(1986)PNAS 83:9065〜9069頁;及びRobertsonら(1986)Nature 322:445〜448頁)。DNA形質移入またはレトロウイルス媒介形質導入により、導入遺伝子をES細胞に効率的に導入できる。このように変換されたES細胞はその後、非ヒト動物から胚盤胞と組み合わせることができる。その後、ES細胞は、胎芽をコロニー化し、生じたキメラ動物の生殖細胞系に寄与する。レビューに関してJaenisch, R.(1988)Science 240:1468〜1474頁を参照されたい。
【0172】
非ヒトトランスジェニック動物が提供され、トランスジェニック動物は、好ましくは上記のとおり、VDCCγ-8イオンチャネル機能のモデルとしてVDCCγ-8遺伝子の変更を有することを特徴とする。遺伝子に対する変更としては、機能変異の欠失または他の喪失、標的化または無作為変異によりヌクレオチド配列を有する外因性遺伝子の導入、別の種からの外因性遺伝子の導入、またはそれらの組合せが挙げられる。トランスジェニック動物は、変更のためにホモ接合性であってもヘテロ接合性であってもよい。それから誘導された動物及び細胞は、VDCCγ-8イオンチャネル機能を調節できる生物活性剤をスクリーニングするために有用である。このスクリーニング法は、疼痛及び癌、特に前立腺癌の潜在的療法の特異性及び作用を判定するために特に有用である。該動物は、脳、心臓、脾臓、及び肝臓等の正常な組織、並びに臓器におけるVDCCγ-8イオンチャネルの役割及びそれらの機能に対する効果を調査するモデルとして有用である。
【0173】
別の態様は、機能的に破壊された内因性VDCCγ-8遺伝子を有するが、また、そのゲノムに運搬し、異種のVDCCγ-8タンパク質(すなわち、別の種からのVDCCγ-8)をコードする導入遺伝子を発現するトランスジェニック非ヒト動物に関する。該動物はマウスであることが好ましく、異種VDCCγ-8はヒトVDCCγ-8である。ヒトVDCCγ-8により再構成されたこのような動物から誘導された動物または細胞系を、in vivo及びin vitroでヒトVDCCγ-8を阻害する薬剤を同定するために使用できる。例えば、ヒトVDCCγ-8を介してシグナル伝達を誘導する刺激剤を、試験される薬剤の存在下及び非存在下で動物または細胞系に投与でき、動物または細胞系の応答を測定できる。in vivo及びin vitroでヒトVDCCγ-8を阻害する薬剤は、薬剤の非存在下の応答と比較して、薬剤の存在下で応答減少に基づいて同定され得る。
【0174】
VDCCγ-8欠損トランスジェニック非ヒト動物(「VDCCγ-8サブユニットノックアウト」)もまた提供される。このような動物は、好ましくは、破壊または削除される内因性VDCCγ-8イオンチャネルゲノム配列の結果、VDCCγ-8イオンチャネルの低い活性を発現する、または活性を発現しないものである。好ましくは、このような動物は、イオンチャネル活性を発現しない。さらに好ましくは、該動物は、配列番号3または配列番号5として示されるVDCCγ-8イオンチャネルの活性を発現しない。VDCCγ-8イオンチャネルノックアウトを、下記にさらに詳細に説明されるように、当業者に既知の種々の手段により作製できる。
【0175】
本発明はまた、宿主細胞においてVDCCγ-8遺伝子を機能的に破壊するための核酸構築体に関する。該核酸構築体は:a)非相同的置換部分;b)非相同的置換部分の上流に位置する第一の相同領域、第一のVDCCγ-8遺伝子配列に対して実質的に同一性を有するヌクレオチド配列を有する第一の相同領域;及びc)非相同的置換部分の下流に位置する第二の相同領域、第二のVDCCγ-8遺伝子配列に対して実質的に同一性を有するヌクレオチド配列を有する第二の相同領域、天然の内因性VDCCγ-8遺伝子において第一のVDCCγ-8遺伝子配列の下流に位置する第二のVDCCγ-8遺伝子配列、を含む。さらに、第一及び第二の相同領域は、核酸分子が宿主細胞に導入される場合、核酸構築体と宿主細胞における内因性VDCCγ-8遺伝子との間の相同的組換えのために十分な長さである。好ましい実施形態において、非相同的置換部分は、好ましくは、調節要素に操作的に連結されたネオマイシンホスホトランスフェラーゼ遺伝子を含み、好ましくは、lacZ及び陽性選択発現カセット等の発現レポーターを含む。
【0176】
好ましくは、第一及び第二のVDCCγ-8遺伝子配列は、配列番号1、配列番号2、配列番号4もしくは配列番号6、またはそれらのホモログ、バリアントもしくは誘導体から誘導される。
【0177】
別の態様は、上記の核酸構築体が取り込まれた組換えベクターに関する。さらに別の態様は、核酸構築体が導入され、それによって核酸構築体と宿主細胞の内因性VDCCγ-8遺伝子との間で相同的組換えを可能にし、内因性VDCCγ-8遺伝子の機能的破壊を生じる宿主細胞に関する。宿主細胞は、肝臓、脳、脾臓または心臓、あるいはマウス胎芽幹細胞等の多能性細胞からVDCCγ-8遺伝子を正常に発現する哺乳動物細胞であり得る。核酸構築体が導入され、内因性VDCCγ-8遺伝子と相同的に組換えられた胎芽幹細胞のさらなる発達によって、胎芽幹細胞からの子孫であり、このようにゲノムにVDCCγ-8遺伝子破壊を担持する細胞を有するトランスジェニック非ヒト動物が作製される。次に生殖細胞系にVDCCγ-8遺伝子破壊を保持する動物は、すべての体細胞及び生殖細胞においてVDCCγ-8遺伝子破壊を有する動物を作製するために選択され、繁殖され得る。次いで、このようなマウスを、VDCCγ-8遺伝子破壊のためのホモ接合性に繁殖できる。
【0178】
[抗体]
本文書の目的に関して、用語の「抗体」とは、それと反対に指定されない限り、ポリクローナル、モノクローナル、キメラ、一本鎖、Fab断片及びFab発現ライブラリーにより生じた断片を含むが、これらに限定されない。このような断片としては、標的物質、Fv、F(ab')及びF(ab')2断片に関する結合活性を保持している全抗体の断片、並びに一本鎖抗体(scFv)、融合タンパク質及び抗体の抗原結合部位を含む他の合成タンパク質の断片が挙げられる。抗体及びその断片は、例えば、EP-A-239400に記載されるようなヒト化抗体であり得る。さらに、例えば、米国特許第5,545,807号及び米国特許第6,075,181号に記載されるような完全ヒト可変領域の抗体(またはそれらの断片)もまた使用できる。中和抗体、すなわち物質のアミノ酸配列の生物学的活性を阻害するものは、診断薬及び治療薬用に特に好ましい。
【0179】
免疫化またはファージディスプレイライブラリーを用いることによる等の標準的な技術により、抗体を作製することができる。
【0180】
ポリペプチドまたはペプチドを、既知の技術によって抗体を産生するために使用できる。このような抗体は、VDCCγ-8イオンチャネルタンパク質またはホモログ、断片等に特異的に結合できる。
【0181】
ポリクローナル抗体が望ましい場合、選択された哺乳動物(例えば、マウス、ウサギ、ヤギ、ウマ等)は、関連ポリペプチドまたはペプチドを含む免疫原性組成物により免疫化され得る。宿主種によって、種々のアジュバントを、免疫学的応答を増加させるために使用できる。このようなアジュバントとしては、限定はしないが、フロイント、水酸化アルミニウム等の無機ゲル、リソレシチン、プルロニックポリオール、ポリアニオン、ペプチド、油乳濁液、スカシ貝ヘモシアニン、及びジニトロフェノール等の表面活性物質が挙げられる。BCG(カルメット・ゲラン杆菌)及びコリネバクテリウムパルブムは、精製物質のアミノ酸配列を、全身防御を刺激する目的で免疫学的に易感染性個体に投与する場合に使用できる潜在的に有用なヒトアジュバントである。
【0182】
免疫化動物からの血清は、既知の方法に従って採取され、処理される。VDCCγ-8ポリペプチドから得ることができるエピトープに対するポリクローナル抗体を含有する血清が他の抗原に対する抗体を含有する場合、ポリクローナル抗体は、免疫アフィニティークロマトグラフィにより精製され得る。ポリクローナル抗血清を作製し、処理する技術は当業者に既知である。このような抗体を作製できるように、我々はまた、動物またはヒトにおける免疫原として使用するために別のアミノ酸配列にハプテン化されたVDCCγ-8のアミノ酸配列またはその断片を提供する。
【0183】
VDCCγ-8イオンチャネルポリペプチドまたはペプチドから得ることができるエピトープに対する特異的なモノクローナル抗体もまた、当業者により容易に作製され得る。ハイブリドーマによるモノクローナル抗体を作製するための一般的な方法論は周知である。不死化抗体産生細胞系は、細胞融合、及び発癌性DNAによるBリンパ球の直接的形質転換、またはエプスタインバルウイルスによる形質移入等の他の技術により作製され得る。軌道(orbit)エピトープに対して作製されたモノクローナル抗体のパネルは、種々の性質、すなわちイソタイプ及びエピトープアフィニティーに関してスクリーニングされ得る。
【0184】
モノクローナル抗体は、連続培養の細胞系により抗体分子の産生を提供する任意の技術を用いて調製され得る。これらとしては、限定はしないが、Koehler及びMilstein(1975年Nature 256:495〜497頁)により原型として記載されたハイブリドーマ技術、トリオーマ技術、ヒトB細胞ハイブリドーマ技術(Kosborら(1983)Immunol Today 4:72頁;Coteら(1983)Proc Natl Acad Sci 80:2026〜2030頁)及びEBVハイブリドーマ技術(Coleら、Monoclonal Antibodies and Cancer Therapy、77〜96頁、Alan R. Liss社、1985年)が挙げられる。
【0185】
さらに、「キメラ抗体」の産生のために開発された技術、すなわち適切な抗原特異性及び生物活性を有する分子を得るためにヒト抗体遺伝子に対するマウス抗体遺伝子のスプライシングを使用できる(Morrisonら(1984)Proc Natl Acad Sci 81:6851〜6855頁;Neubergerら(1984)Nature 312:604〜608頁;Takedaら(1985)Nature 314:452〜454頁)。あるいは、一本鎖抗体の産生のために記載された技術(米国特許第4,946,779号)は、物質特異的一本鎖抗体を産生するために適合できる。
【0186】
VDCCγ-8イオンチャネルポリペプチドまたはペプチドから得ることができるエピトープに対する抗体、モノクローナル及びポリクローナルの両方は、特に診断に有用であり、中和するものは、受動的免疫療法に有用である。特にモノクローナル抗体を、抗イディオタイプ抗体を上昇させるために使用できる。抗イディオタイプ抗体は、保護が望ましい物質及び/または薬剤の「内部像」を担持する免疫グロブリンである。抗イディオタイプ抗体を上昇させる技術は当業者に既知である。これらの抗イディオタイプ抗体もまた治療に有用であり得る。
【0187】
抗体は、リンパ球集団でin vivo産生を誘導することによって、またはOrlandiら(1989年、Proc Natl Acad Sci 86:3833~3837頁) 及びWinter G及びMilstein C(1991年;Nature 349:293~299頁)に開示されているように、抗体がリンパ球集団においてもin vivo産生を誘導することによって、または組換え免疫グロブリンライブラリーもしくは一連の高特異的結合試薬をスクリーニングすることによって産生できる。Orlandiら(1989年、Proc Natl Acad Sci 86:3833〜3837頁)、及びWinter G及びMilstein C(1991年;Nature 349:293〜299頁)によっても産生できる。
【0188】
ポリペプチドまたはペプチドに特異的な結合部位を含有する抗体断片も作製され得る。例えば、このような断片としては、限定はしないが、抗体分子のペプシン消化により産生できるF(ab')2断片及びF(ab')2断片のジスルフィド架橋を還元することにより作製できるFab断片が挙げられる。あるいは、Fab発現ライブラリーは、望ましい特異性を有するモノクローナルFab断片の迅速かつ容易な同定を可能にするために構築できる(Huse WDら(1989)Science 256:1275〜1281頁)。
【0189】
一本鎖抗体の産生技術(米国特許第4,946,778号)はまた、VDCCγ-8イオンチャネルポリペプチドに対する一本鎖抗体を産生するために適合できる。また、トランスジェニックマウス、または哺乳動物を含む他の生物を、ヒト化抗体を発現させるために使用できる。
【0190】
ポリペプチドを発現するクローンを単離または同定するために、あるいはアフィニティークロマトグラフィによりポリペプチドを精製するために、上記抗体を使用できる。
【0191】
VDCCγ-8イオンチャネルポリペプチドに対する抗体を、VDCCγ-8関連疾患を治療するためにも使用できる。
【0192】
[診断アッセイ]
この開示はまた、診断薬としての診断における使用、または遺伝子解析における使用のための、VDCCγ-8イオンチャネルポリヌクレオチド及びポリペプチド(並びにそれらのホモログ、バリアント及び誘導体)の使用に関する。VDCCγ-8イオンチャネル核酸に相補的またはハイブリダイズできる核酸(ホモログ、バリアント及び誘導体を含む)、並びにVDCCγ-8ポリペプチドに対する抗体もまた、このようなアッセイに有用である。
【0193】
好ましい実施形態において、該診断アッセイは、患者から単離されたサンプルに対して実施されるものであり、該アッセイは、ヒトまたは動物の身体に対して実施されるものではない。
【0194】
機能障害と関連するVDCCγ-8イオンチャネル遺伝子の変異形態検出により、VDCCγ-8イオンチャネルの発現不足、過剰発現、または発現の変化から生じる疾患または疾患に対する感受性の診断に加えるか、またはその診断を規定できる診断手段を提供する。VDCCγ-8イオンチャネル遺伝子(コントロール配列を含む)において変異を担持する個体は、種々の技術によるDNAレベルで検出できる。
【0195】
例えば、DNAは患者から単離され、VDCCγ-8のDNA多型パターンを決定できる。この同定されたパターンは、VDCCγ-8の過剰発現、発現不足、または異常発現に関連する疾患に罹患していることが知られている患者のコントロールと比較される。次にVDCCγ-8関連疾患に関連する遺伝子多型パターンを発現する患者を同定できる。VDCCγ-8イオンチャネル遺伝子の遺伝子解析は、当業者に知られた任意の技術により実施できる。例えば、VDCCγ-8対立遺伝子のDNA配列を決定することにより、RFLPまたはSNP解析等により、個体をスクリーニングできる。VDCCγ-8に関する遺伝子配列におけるDNA多型の存在またはその発現を調節する任意の配列を検出することにより、VDCCγ-8の過剰発現、発現不足または異常発現に関連する疾患に関する遺伝子素因を有するものとして、患者を同定できる。
【0196】
そのように同定された患者は、VDCCγ-8関連疾患の発症を防ぐために処置できるか、またはその疾患のさらなる発生または発達を防ぐためにVDCCγ-8関連疾患の早期段階でより積極的に処置できる。
【0197】
VDCCγ-8関連疾患に関連する患者の遺伝子多型パターンの同定用キットもまた提供される。該キットには、DNA採取手段及び遺伝子多型パターンを測定する手段が含まれ、次いでVDCCγ-8関連疾患に対する患者の感受性を判定するためにコントロールサンプルと比較される。VDCCγ-8ポリペプチド及び/またはこのようなポリペプチドに対する抗体(またはその断片)を含むVDCCγ-8関連疾患の診断用キットもまた提供される。
【0198】
診断用核酸は、血液、尿、唾液、組織生検または剖検材料から等、対象の細胞から得ることができる。好ましい実施形態において、DNAは、患者の指穿刺により吸収紙に採取された血液より得られた血液細胞から得られる。さらなる好ましい実施形態において、血液は、AmpliCard(商標)に採取される(University of Sheffield、Department of Medicine and Pharmacology、Royal Hallamshire Hospital、シェフィールド、英国S10 2JF)。
【0199】
DNAは、検出用に直接使用され得るか、または分析前にPCRまたは他の増幅技術を用いることにより酵素的に増幅され得る。対象の遺伝子内の特定の多型DNA領域を標的にするオリゴヌクレオチドDNAプライマーを調製できることによって、標的配列のPCR反応増幅が達成される。RNAまたはcDNAもまた、テンプレートとして同様の方法で使用できる。次にテンプレートDNAからの増幅DNA配列は、制限酵素を用いて分析して増幅配列に存在する遺伝子多型を決定し、それによって患者の遺伝子多型プロファイルを提供できる。制限断片長を、ゲル分析により同定できる。あるいは、または関連してSNP(単一ヌクレオチド多型)解析等の技術を使用できる。
【0200】
欠失及び挿入は、正常な遺伝子型と比較して増幅産物のサイズを変化させて検出できる。点変異は、増幅DNAを標識VDCCγ-8イオンチャネルヌクレオチド配列にハイブリダイズすることにより同定され得る。完全一致配列は、RNアーゼ消化により、または融解温度の相違によりミスマッチ二重鎖と識別され得る。DNA配列の相違はまた、変性剤の有無でゲル中のDNA断片の電気泳動移動度の変化により、または直接的DNA配列決定により検出できる。例えば、Myersら、Science(1985)230:1242頁を参照されたい。特定の位置における配列変化はまた、RNアーゼ及びS1保護等のヌクレアーゼ保護アッセイ、または化学的開裂法により明らかにすることができる。Cottonら、Proc Natl Acad Sci USA(1985)85:4397〜4401頁を参照されたい。別の実施形態において、VDCCγ-8イオンチャネルヌクレオチド配列またはその断片を含むオリゴヌクレオチドプローブのアレイを構築して、例えば、遺伝子変異の効率的なスクリーニングを実施することができる。アレイテクノロジー法は周知であり、一般的な用途があり、遺伝子発現、遺伝子結合、及び遺伝的変異性等、分子遺伝学における種々の問題を扱うために使用され得る(例えば:M. Cheeら、Science、274巻、610〜613頁(1996)を参照)。
【0201】
一本鎖立体配座多型(SSCP)を使用して、変異体と野生型核酸との間の電気泳動移動度の相違を検出することができる(Oritaら(1989)Proc Natl. Acad. Sci USA:86:2766頁;Cotton(1993)Mutat Res 285:125〜144頁;及びHayashi(1992)Genet Anal Tech Appl 9:73〜79頁)。サンプル及びコントロールVDCCγ-8核酸の一本鎖DNA断片を変性でき、再生できる。一本鎖核酸の二次的構造は、配列によって異なり、電気泳動移動度で生じた変化は、単一の塩基変化さえも検出を可能にする。DNA断片は、標識プローブにより標識または検出され得る。アッセイの感度は、RNA(DNAよりも)を用いることにより増強でき、その二次的構造は、配列変化に対してより感度がよい。好ましい実施形態において、主題の方法は、電気泳動移動度の変化に基づいて二本鎖ヘテロ二重鎖分子を分離するためにヘテロ二重鎖分析を利用する(Keenら(1991)Trends Genet 7:5頁)。
【0202】
VDCCγ-8ポリペプチド及び核酸の存在を、サンプル中に検出することができる。このように、上記に掲げた感染症及び疾患は、対象由来のサンプルからVDCCγ-8ポリペプチドまたはVDCCγ-8イオンチャネルmRNAのレベルの異常減少または異常増加を測定することを含む方法により診断され得る。該サンプルは、発現レベルまたは発現パターンの空間的または時間的変化等、VDCCγ-8発現の増減あるいは異常に関連する疾患に罹患しているか、または罹患している疑いのある生物からの細胞または組織のサンプルを含むことができる。このような疾患に罹患している、または罹患している疑いのある生物におけるVDCCγ-8の発現レベルまたは発現パターンは、通常、疾患の診断手段として、正常な生物の発現レベルまたは発現パターンと比較される。
【0203】
従って一般に、サンプル中にVDCCγ-8核酸を含む核酸の存在を検出する方法は、サンプルと、上記核酸に特異的である少なくとも1つの核酸プローブとを接触させること、及び核酸の存在に関して上記サンプルをモニタリングすることを含む。例えば、該核酸プローブは、VDCCγ-8イオンチャネルサブユニット核酸、またはその一部に特異的に結合でき、2つの間の結合を検出でき、複合体自体の存在も検出できる。さらに、我々は、細胞サンプルと、ポリペプチドを結合できる抗体とを接触させること、及びポリペプチドの存在に関して上記サンプルをモニタリングすることによりVDCCγ-8ポリペプチドの存在を検出することを包含する。抗体とポリペプチドとの間で形成された複合体の存在をモニタリングすること、またはポリペプチドと抗体との間の結合をモニタリングすることにより、これを都合よく達成できる。2つの成分の間の結合を検出する方法は当業者に既知であり、FRET(蛍光共鳴エネルギー移動)、表面プラズモン共鳴等を含む。
【0204】
発現の減少または増加は、例えば、PCR、RT-PCR、RNアーゼ保護、ノーザンブロッティング法及び他のハイブリダイゼーション法等のポリヌクレオチドの定量化に関して周知の任意の方法を用いてRNAレベルを測定できる。宿主に由来するサンプル中のVDCCγ-8等のタンパク質のレベルを測定するために使用できるアッセイ技術は、当業者に周知である。このようなアッセイ法としては、放射免疫アッセイ、競合結合アッセイ、ウェスタンブロット解析及びELISAアッセイが挙げられる。
【0205】
疾患用診断キット、または、例えば疼痛及び癌、特に神経障害性痛及び前立腺癌の疾患(感染症を含む)に対する罹病性に関する診断用キットを、我々はさらに開示する。診断用キットは、VDCCγ-8ポリヌクレオチドまたはその断片、相補的ヌクレオチド配列、VDCCγ-8ポリペプチドまたはその断片、あるいはVDCCγ-8ポリペプチドに対する抗体を含む。
【0206】
[染色体アッセイ]
本明細書に記載されたヌクレオチド配列もまた、染色体の同定に役立つ。該配列は特異的に標的とされ、個々のヒト染色体上の特定の位置とハイブリダイズできる。上記のとおり、ヒトVDCCγ-8イオンチャネルは、ヒト染色体19q13にマッピングすることが分かっている。
【0207】
染色体に関連する配列のマッピングは、これらの配列と遺伝子関連疾患とを相関させる上で重要な最初の工程である。配列が、正確な染色体位置にマッピングされたら、染色体上の配列の物理的位置は、遺伝子マップデータと相関づけることができる。このようなデータは、例えば、V. McKusick、ヒトにおけるメンデルヘリタンス(Johns Hopkins University Welch Medical Libraryを介するライン上で入手できる)に見られる。次に、同じ染色体領域にマッピングされた遺伝子と疾患との間の関係を、連鎖解析(物理的に隣接する遺伝子の遺伝的形質)により同定する。
【0208】
cDNAまたはゲノム配列において罹患者と非罹患者との間の相違もまた判定できる。変異が、罹患者の一部またはすべてに見られるが、正常者には見られない場合、変異は疾患の原因物質である可能性が高い。
【0209】
[予防的及び治療的方法]
VDCCγ-8イオンチャネル活性の過剰量及び不足量の両方に関連する異常病態を治療する方法を提供する。
【0210】
VDCCγ-8イオンチャネルの活性が過剰である場合、いくつかのアプローチを利用できる。1つのアプローチは、リガンドのVDCCγ-8イオンチャネルへの結合をブロックすることにより、または第二のシグナルを阻害することにより活性を阻害するのに有効な量で製薬的に許容できる担体とともに、上記の阻害剤化合物(アンタゴニスト)を対象に投与すること、それによって異常病態を緩和することを含む。
【0211】
別のアプローチにおいて、内因性VDCCγ-8イオンチャネルと競合してリガンドをさらに結合できるVDCCγ-8ポリペプチドの溶解形態を投与できる。このような競合物の典型的な実施形態は、VDCCγ-8ポリペプチドの断片を含む。
【0212】
さらに別のアプローチにおいて、内因性VDCCγ-8イオンチャネルをコードする遺伝子発現は、発現ブロッキング技術を用いて阻害できる。既知のこのような技術は、内部で作製されるか、または別個に投与されるアンチセンス配列の使用を含む。例えば、Oligodeoxynucleotides as Antisense Inhibitors of Gene Expression, CRC Press, Boca Raton、フロリダ州(1988)における O'Connor、J Neurochem(1991)56:560頁を参照されたい。あるいは、遺伝子と三重螺旋体を形成するオリゴヌクレオチドを供給できる。例えば、Leeら、Nucleic Acids Res(1979)6:3073頁;Cooneyら、Science(1988)241:456頁;Dervanら、Science(1991)251:1360頁を参照されたい。これらのオリゴマーはそれ自体で投与され得るか、または関連オリゴマーをin vivoで発現させることができる。
【0213】
VDCCγ-8イオンチャネルの発現不足及びその活性に関連する異常病態を治療するために、いくつかのアプローチもまた利用できる。1つのアプローチは、製薬的に許容できる担体と組み合わせてVDCCγ-8イオンチャネルを活性化する化合物、すなわち、上記のアゴニストの治療的有効量を対象に投与することを含み、それによって異常病態を緩和する。あるいは、遺伝子療法を使用して、対象における関連細胞によってVDCCγ-8イオンチャネルの内因性産物をもたらすことができる。例えば、VDCCγ-8ポリヌクレオチドを、上記で考察された複製欠損レトロウイルスベクターにおける発現のために操作することができる。次にレトロウイルス発現構築体を単離でき、VDCCγ-8ポリペプチドをコードするRNAを含有するレトロウイルスプラスミドベクターで形質導入されたパッケージング細胞に導入でき、その結果、該パッケージング細胞は、ここで対象の遺伝子を含有する感染性ウイルス粒子を産生する。これらの産生株細胞を、in vivo細胞操作及びin vivoポリペプチドの発現のために対象に投与できる。遺伝子療法の概要に関しては、T. Strachan及びA P Read、Human Molecular Geneticsにおける20章、Gene Therapy and other Molecular Genetic-based Therapeutic Approaches, (及びその中に引用されている文献)、BIOS Scientific Publishers社(1996)を参照されたい。
【0214】
[製剤及び投与]
VDCCγ-8イオンチャネルポリペプチドの溶解形態等のペプチド、及びアゴニスト並びにアンタゴニストペプチドまたは小型分子を、好適な製薬用担体と組み合わせて製剤化できる。このような製剤は、治療的有効量のポリペプチドまたは化合物、及び製薬的に許容できる担体または賦形剤を含む。このような担体としては、限定はしないが、生理食塩水、緩衝生理食塩水、デキストロース、水、グリセロール、エタノール、及びそれらの組合せが挙げられる。製剤は、投与様式に適合する必要があり、十分当該技術分野の範囲内に入る。本開示は、さらに前述の組成物の1つ以上の成分で満たされた1つ以上の容器を含む製薬用パック及びキットに関する。
【0215】
VDCCγ-8ポリペプチド及び他の化合物を、単独でまたは治療用化合物等の他の化合物と併用して使用できる。
【0216】
製薬組成物の全身投与の好ましい形態としては、典型的には静脈内注射による注射が挙げられる。皮下、筋内、または腹腔内等の他の注射経路を使用できる。全身投与に関する代わりの手段としては、胆汁酸塩またはフシジン酸または他の界面活性剤等の浸透剤を用いて経粘膜及び経皮投与が挙げられる。さらに、腸内またはカプセル化製剤で適切に製剤化される場合、経口投与もまた可能であり得る。これらの化合物の投与はまた、軟膏剤、ペースト剤、ゲル剤等の形態で、局所的及び/または局在化できる。
【0217】
必要とされる投与量範囲は、ペプチドの選択、投与経路、製剤の性質、対象の病態の性質、及び担当医師の判定に依存する。しかしながら、好適な投与量は、0.1〜100μg/kg対象の範囲内である。しかしながら、必要な投与量の広い変動は、利用できる種々の化合物及び種々の投与経路の異なる効率に鑑みて予想することができる。例えば、経口投与は、静脈内注射による投与よりも高投与量を必要とすることが予想されるであろう。これらの投与量レベルの変動は、当業者に十分に解されるように、最適のために標準的な経験的ルーチンを用いて調整できる。
【0218】
治療に用いられるポリペプチドはまた、上記の「遺伝子療法」としばしば称される治療様式の対象において内因的に産生され得る。従って、例えば、対象からの細胞を、例えば、レトロウイルスプラスミドベクターの使用によりex vivoでポリペプチドをコードするためにDNAまたはRNA等のポリヌクレオチドにより操作できる。次いで該細胞を対象に導入する。
【0219】
[製薬組成物]
VDCCγ-8ポリペプチド、ポリヌクレオチド、ペプチド、ベクターまたはそれらの抗体の治療的有効量、及び場合によっては、製薬的に許容できる担体、希釈剤、または賦形剤(それらの組合せ等)を含む製薬組成物を提供する。
【0220】
該製薬組成物は、ヒト及び獣医医療におけるヒトまたは動物使用のためのものであり、1つ以上の製薬的に許容できる任意の希釈剤、担体、または賦形剤を典型的に含む。治療使用のための許容できる担体または希釈剤は、製薬業界に周知であり、例えば、Remington's Pharmaceutical Sciences、Mack Publishing社(A. R. Gennaro編集、1985年)に記載されている。製薬用担体、賦形剤または希釈剤の選択は、目的の投与経路及び標準的な製薬実施に関して選択できる。製薬組成物は、担体としてまたは担体に加えて、賦形剤または希釈剤、任意の好適な結合剤、滑剤、懸濁化剤、コーティング剤、可溶化剤を含むことができる。
【0221】
保存剤、安定化剤、染料及び香味剤さえも、製薬組成物に提供され得る。保存剤の例としては、安息香酸ナトリウム、ソルビン酸及びp-ヒドロキシ安息香酸のエステルが挙げられる。抗酸化剤及び懸濁化剤もまた使用できる。
【0222】
種々の送達系によって種々の組成物/製剤要件があり得る。例として、本明細書に記載された製薬組成物は、ミニポンプを用いるか、または粘膜経路により送達されるように、例えば、吸入用鼻腔スプレーまたはエアロゾルあるいは経口摂取用溶液として製剤化でき、あるいは該製剤は、非経口的に、例えば、静脈内、筋内または皮下経路による送達用の注射剤形態により製剤化される。あるいは、該製剤は、両経路により送達されるように設計され得る。
【0223】
薬剤を、胃腸管粘膜を介して粘膜送達できる場合、胃腸管を介して移行時に安定のままであり得る必要があり、例えば耐タンパク質分解性であり、酸pHに安定で、胆汁酸の耐界面活性作用である必要がある。
【0224】
適切な場合、製薬組成物を、吸入により、座薬またはペッサリーの形態で、局所的にローション、溶液、クリーム、軟膏または粉剤の形態で、皮膚パッチの使用により、経口的に澱粉またはラクトース等の賦形剤を含有する錠剤の形態で、またはカプセルもしくは小卵(ovule)中で単独または賦形剤と混合して、香味剤または着色剤を含有するエリキシル、溶液または懸濁剤の形態で投与でき、またはそれらは、例えば、非経口的に、例えば、静脈内、筋内または皮下に注入できる。非経口投与に関して、該組成物は、他の物質、例えば、血液との等張液を作製するために十分な塩類または単糖類を含有できる滅菌水溶液の形態で最良に使用され得る。口腔または舌下投与に関して、該組成物は、従来の様式で製剤化できる錠剤または舐剤の形態で投与できる。
【0225】
[ワクチン]
別の実施形態は、哺乳類動物にVDCCγ-8イオンチャネルポリペプチドまたはその断片を接種することを含む、哺乳類動物において免疫学的応答を誘導する方法に関するものであり、VDCCγ-8関連疾患から上記動物を保護するために抗体及び/またはT細胞免疫応答を適切に生じさせる。
【0226】
さらに別の実施形態は、疾患から上記動物を保護する抗体を産生するような免疫学的応答を誘導するために、in vivoでVDCCγ-8ポリヌクレオチドの発現をもたらすベクターを介してVDCCγ-8ポリペプチドを送達することを含む、哺乳動物における免疫学的応答を誘導する方法に関する。
【0227】
さらなる実施形態は、哺乳動物宿主に導入される場合、その哺乳動物においてVDCCγ-8ポリペプチドに対する免疫学的応答を誘導する免疫学的/ワクチン製剤(組成物)に関するものであり、該組成物はVDCCγ-8ポリペプチドまたはVDCCγ-8遺伝子を含む。ワクチン製剤は好適な担体をさらに含む。
【0228】
VDCCγ-8ポリペプチドは、胃内で分解されることから、非経口的(皮下、筋内、静脈内、皮膚内等の注射等)に投与されることが好ましい。非経口投与に好適な製剤としては、抗酸化剤、緩衝液、静菌剤及び製剤をレシピエントの血液と等張にさせる溶液を含有できる水性及び非水性滅菌注射液、並びに懸濁化剤または増粘剤を含むことができる水性及び非水性滅菌懸濁液が挙げられる。該製剤は、単位用量または多用量容器、例えば、密封アンプル及びバイアルで提供でき、使用直前に滅菌液担体の添加のみを必要とする凍結乾燥条件で保存することができる。ワクチン製剤は、水中油系及び当業者に既知の他の系等、製剤の免疫原性を増強させるアジュバント系もまた含むことができる。投与量は、ワクチンの特異的活性に依存し、ルーチン的実験により容易に決定できる。
【0229】
ワクチンは、1種以上のVDCCγ-8ポリペプチドまたはペプチドから調製され得る。
【0230】
活性成分として免疫原性ポリペプチドまたはペプチドを含有するワクチンの調製は、当業者に既知である。典型的には、このようなワクチンは、液体溶液または懸濁剤としての注射剤として調製され、注射前の溶液、または懸濁液、液体に好適な固体形態もまた調製され得る。該製剤をまた乳濁化できるか、またはタンパク質はリポソームにカプセル化できる。活性な免疫原性成分は、製薬的に許容でき、活性成分と適合できる賦形剤としばしば混合される。好適な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノール等、及びそれらの組合せである。
【0231】
さらに、所望ならば、ワクチンは、湿潤剤または乳化剤、pH緩衝剤、及び/またはワクチンの効果を増加させるアジュバント等、少量の補助物質を含有できる。効果的となり得るアジュバントの例としては、限定はしないが、水酸化アルミニウム、N-アセチル-ムラミル-L-トレオニル-D-イソグルタミン(thr-MDP)、N-アセチル-ノル-ムラミル-L-アラニル-D-イソグルタミン(CGP 11637、ノル-MDPと称される)、N-アセチルムラミル-L-アラニル-D-イソグルタミル-L-アラニン-2-(1'-2'-ジパルミトイル-sn-グリセロ-3-ヒドロキシホスホリルオキシ)-エチルアミン(CGP 19835A、MTP-PEと称される)、及び細菌から抽出された3つの成分、すなわちモノホスホリル脂質A、トレハロースジミコレート及び2%スクワレン/Tween 80乳濁液中の細胞壁骨格(MPL+TDM+CWS)を含有するRIBIが挙げられる。
【0232】
アジュバント及び他の薬剤のさらなる例としては、水酸化アルミニウム、リン酸アルミニウム、硫酸アルミニウムカリウム(アルム)、硫酸ベリリウム、シリカ、カオリン、炭素、油中水乳濁液、水中油乳濁液、ムラミルジペプチド、細菌エンドトキシン、脂質X、コリネバクテリウムパルブム(プロピオノバクテリウムアクネス)、百日咳菌、ポリリボヌクレオチド、アルギン酸ナトリウム、ラノリン、リソレシチン、ビタミンA、サポニン、リポソーム、レバミソール、DEAE-デキストラン、ブロック化コポリマーまたは他の合成アジュバントが挙げられる。このようなアジュバントは、種々の供給源、例えば、Merck Adjuvant 65(Merck社、ラーウェイ、ニュージャージー州)またはFreund's Incomplete Adjuvant and Complete Adjuvant(Difco Laboratories、デトロイト、ミシガン州)から市販されている。
【0233】
典型的に、Amphigen(水中油)、Alhydrogel(水酸化アルミニウム)、またはAmphigen及びAlhydrogelの混合物等のアジュバントを使用する。水酸化アルミニウムだけが、ヒトへの使用に承認されている。
【0234】
免疫原とアジュバントとの比率は、両方が有効な量で存在する限り、広範囲にわたって変えることができる。例えば、水酸化ナトリウムは、ワクチン混合物(Al2O3ベース)の約0.5%の量で存在できる。都合のよいことに、ワクチンは、0.2から200μg/ml、好ましくは5から50μg/mlの範囲で、最も好ましくは15μg/mlの免疫原の最終濃度を含有するように製剤化される。
【0235】
製剤化後、ワクチンを滅菌容器に取り込むことができ、次いで密封され、低温で、例えば4℃で保存されるか、または凍結乾燥できる。凍結乾燥は、安定な形態で長期間の保存を可能にする。
【0236】
ワクチンを、非経口的に、注射、例えば、皮下または筋内に投与することが好都合である。他の様式の投与に好適なさらなる製剤としては座薬、いくつかの場合、経口製剤が挙げられる。座薬に関しては、伝統的な結合剤及び担体としては、例えば、ポリアルキレングリコールまたはトリグリセリドを挙げることができ、このような座薬は、0.5%から10%、好ましくは1%から2%の範囲で活性成分を含有する混合物から形成できる。経口製剤は、例えば、製薬グレードのマンニトール、ラクトース、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の正規に使用される賦形剤を含む。これらの組成物は、液剤、懸濁液剤、錠剤、丸剤、カプセル剤、徐放性製剤または散剤の形態をとり、10%から95%、好ましくは25%から70%の活性成分を含有する。ワクチン組成物が凍結乾燥される場合、凍結乾燥物を、投与前、例えば懸濁液として再構成できる。再構成は、緩衝液中で実施されることが好ましい。
【0237】
患者への経口投与用カプセル剤、錠剤及び丸剤は、例えば、Eudragit「S」、Eudragit「L」、セルロースアセテート、セルロースアセテートフタレートまたはヒドロキシプロピルメチルセルロースを含む腸溶性コーティングとともに提供できる。
【0238】
VDCCγ-8ポリペプチドは、中性または塩形態としてワクチンに製剤化され得る。製薬的に許容できる塩類としては、例えば、塩酸またはリン酸等の無機酸、酢酸、シュウ酸、酒石酸及びマレイン酸等の有機酸により形成される酸付加塩(ペプチドの遊離アミノ基と形成される)が挙げられる。遊離カルボキシル基と形成される塩類はまた、例えば、ナトリウム、カリウム、アンモニウム、カルシウム、または水酸化鉄等の無機塩基、及びイソプロピルアミン、トリメチルアミン、2-エチルアミノエタノール、ヒスチジン及びプロカイン等の有機塩基から誘導できる。
【0239】
[投与]
典型的に、医師は、個々の対象に最も好適となる実際の投与量を判定し、特定の患者の年齢、体重及び応答により変わる。下記の投与量は、平均的な症例の代表例である。もちろん、より高いかまたはより低い投与量範囲が個々に利点をもたらす場合もあり得る。
【0240】
本明細書に記載された製薬及びワクチン組成物は、直接注射により投与できる。該組成物は、非経口、粘膜、筋内、静脈内、皮下、眼内または経皮投与用に製剤化できる。典型的に、各タンパク質を、0.01〜30 mg/kg体重、好ましくは、0.1〜10 mg/kg、より好ましくは、0.1〜1 mg/kg体重の用量で投与できる。
【0241】
用語の「投与される」とは、ウイルスまたは非ウイルス技術による送達を含む。ウイルス送達機構としては、限定はしないが、アデノウイルスベクター、アデノ関連ウイルス(AAV)ベクター、ヘルペスウイルスベクター、レトロウイルスベクター、レンチウイルスベクター、及びバキュロウイルスベクターが挙げられる。非ウイルス送達機構としては、脂質媒介形質移入、リポソーム、免疫リポソーム、リポフェクチン、カチオン性面両親媒性物質(CFA)及びそれらの組合せが挙げられる。このような送達機構に対する経路としては、限定はしないが、粘膜、鼻腔、経口、非経口、胃腸管、局所、または舌下経路が挙げられる。
【0242】
用語の「投与される」とは、限定はしないが、例えば、鼻腔用スプレーまたは吸入用エアロゾルとして、または経口摂取用溶液として粘膜経路による送達、送達が、例えば、静脈内、筋内または皮下経路等の注射剤形態による非経口経路による送達が挙げられる。
【0243】
用語の「共投与される」とは、例えば、VDCCγ-8ポリペプチド及びアジュバント等の添加実体の各々の投与部位及び投与時間により、免疫系の必要な調節が達成されるようになることを意味する。従って、ポリペプチド及びアジュバントは、同時に且つ同じ部位で投与され得るが、一方、ポリペプチドを異なる時間で、アジュバントと異なる部位に投与することに利点があり得る。ポリペプチド及びアジュバントを、同じ送達媒体でも送達でき、ポリペプチド及び抗原を結合させても、及び/または結合させなくても、並びに/あるいは遺伝子的に結合させても、及び/または結合させなくてもよい。
【0244】
VDCCγ-8ポリペプチド、ポリヌクレオチド、ペプチド、ヌクレオチド、それらの抗体及びアジュバントを、単位用量または複数用量で宿主対象に別個に投与しても、共投与してもよい。
【0245】
ワクチン組成物及び製薬組成物は、注射(非経口、皮下及び筋内注射を含む)、鼻腔内、粘膜経口、膣内、尿道または眼投与等の多くの異なる経路により投与できる。
【0246】
本明細書に記載されたワクチン及び製薬組成物は、例えば、皮下または筋内注射により、従来からの非経口的に投与できる。他の投与様式に好適であるさらなる製剤としては、座薬及びいくつかの場合、経口用製剤が挙げられる。座薬に関しては、伝統的な結合剤及び担体としては、例えば、ポリアルキレングリコールまたはトリグリセリドを挙げることができ、このような座薬は、0.5〜10%、恐らくは1〜2%の範囲で活性成分を含有する混合物から形成できる。経口製剤は、例えば、製薬グレードのマンニトール、ラクトース、澱粉、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウム等の正規に使用される賦形剤を含む。これらの組成物は、液剤、懸濁液剤、錠剤、丸剤、カプセル剤、徐放性製剤または散剤の形態をとり、10〜95%、好ましくは25〜70%の活性成分を含有する。ワクチン組成物が凍結乾燥される場合、凍結乾燥物を、投与前、例えば懸濁液として再構成できる。再構成は、緩衝液中で実施されることが好ましい。
【実施例】
【0247】
<実施例1.トランスジェニックVDCCγ-8ノックアウトマウス:VDCCγ-8遺伝子標的ベクターの構築>
VDCCγ-8遺伝子は、ゲノムデータベースの相同検索を用いてバイオ情報科学的に同定された。40kbのマウスゲノムコンティグを、種々のデータベースから集めた。このコンティグは、標的べクター内にクローニングするための相同アームの設計を可能にするのに十分なフランキング配列情報を提供した。
【0248】
マウスVDCCγ-8遺伝子は4つのコード化エクソンを有する。標的ストラテジーは、第一のコード化エクソンの大部分の翻訳部分を除去するように設計される。4.1kbの5'相同アーム及び除去される領域に隣接する2.1kbの3'相同アームは、PCRにより増幅され、断片は、標的ベクターにクローニングされる。アームを増幅するために使用される各オリゴヌクレオチドプライマーの5'末端は、ベクターポリリンカーのクローニング部位と適合し、アーム自体から欠いている希少切断制限酵素に対して異なる認識部位を含有するように合成される。VDCCγ-8の場合、プライマーは、下記のプライマーの表に挙げられるように、NotI/SpeIの5'アームクローニング部位及びAscI/FseIの3'アームクローニング部位(関連制限部位を含む使用される標的ベクターの構造を、図1に示す)を有するように設計される。
【0249】
アームプライマーのペア(5'アームF/5'アームR)及び(3'アームF/3'アームR)に加えて、さらにVDCCγ-8座に特異的なプライマーを以下の目的:単離された推定上の標的クローンにおいて、標的座のサザン解析を可能にするために、各アームの外側にありかつ各アームを越えて延長する非反復ゲノムDNAの2つの短い150〜300bp断片を増幅させるための、5'及び3'プローブプライマーのペア(5'prF/5'prR及び3'prF/3'prR);ベクター特異的プライマー(この場合、asc350)とともに多重PCRに用いられる場合、野生型、ヘテロ接合体、及びホモ接合体のマウス間の区別を可能にするマウス遺伝子型プライマーのペア(hetF及びhetR);最後に、3'アーム領域の末端下流にアニールし、ベクター(TK5IBLMNL)(この場合、asc146)の3'末端に特異的なプライマーとペアにした場合、標的事象特異的2.2kbアンプライマーを産生する標的スクリーニングプライマー(3'scr);のために設計する。このアンプライマーは、所望のゲノム変化が生じ、無作為に組込まれたベクターのコピーを含有するクローンのバックグラウンドから正しく標的化された細胞の同定を可能にする細胞由来のテンプレートDNAからのみ得ることができる。これらプライマーの位置及び標的ストラテジーに用いられるVDCCγ-8座の領域のゲノム構造を、図7に示す(配列番号21)。
【0250】
【表2】
【0251】
相同アームの位置は、VDCCγ-8遺伝子を機能的に破壊するように選択される。除去されるVDCCγ-8領域が、VDCCγ-8遺伝子と同じ配向性で配置された促進ネオマイシンホスホトランスフェラーゼ(neo)遺伝子からなる選択カセットの上流に内因性遺伝子発現レポーター(フレーム独立lacZ遺伝子)からなる非相同配列で置換されている標的ベクターを調製する。
【0252】
5'及び3'相同アームを、標的ベクターTK5IBLMNL内にクローニングすると(図1を参照)、標準的な分子生物学的技術を用いて大規模な高純度DNA調製を行う。20μgの新しく調製されたエンドトキシンなしのDNAを、アンピシリン抵抗性遺伝子と細菌の複製起点との間のベクター骨格の特有な部位に存在する別の希少切断制限酵素PmeIにより制限する。次に線形化DNAを沈殿させ、エレクトロポレーションに備えて、100μlのリン酸緩衝生理食塩水に再懸濁させる。
【0253】
エレクトロポレーションの24時間後、形質移入された細胞を、200μg/mlのネオマイシンを含有する培地中で9日間培養する。クローンを96ウェルプレートに選び取り、PCR(上記のとおり、プライマー3'scr及びasc146を用いて)によるスクリーニング前に、複製及び増殖させ、相同的組換えが、内因性VDCCγ-8遺伝子と標的構築体との間で生じているクローンを同定する。陽性クローンは、1〜5%の割合で同定され得る。これらのクローンを増殖して複製物を凍結し、すべて標準的な手法(Russら、Nature 2000年3月2日;404(6773):95〜99頁)を用いた、上記のように調製された外部5'及び3'プローブを用いることによって、標的事象のサザンブロット確認のために十分に高品質のDNAを調製する。診断用制限酵素によって消化されたDNAのサザンブロットを外部プローブとハイブリダイズする場合、相同的に標的化されたES細胞クローンが、変異体バンド並びに変化していない野生型バンドの存在によって立証される。例えば、5'プローブを用いることによって、EcoRI消化ゲノムDNAから9.3kbの野生型バンド及び6.7kbの標的バンドが得られ、3'プローブを用いることによって、Sspt切断DNAから15kbの野生型バンド及び9.5kbの標的バンドが得られる。
【0254】
<実施例2.トランスジェニックVDCCγ-8ノックアウトマウス:VDCCγ-8イオンチャネル欠損マウスの作製>
C57BL/6のメス及びオスマウスを交尾させ、妊娠3.5日に胚盤胞を単離する。1胚盤胞当り10〜12個の選択されたクローン由来の細胞を注入し、7〜8個の胚盤胞を、偽妊娠F1メスの子宮に移植する。幾匹かの高レベル(100%まで)アグーチオス(アグーチコート色は、標的クローンからの系統を継いだ細胞の寄与を示す)を含んだキメラ同腹仔が誕生する。これらのオスキメラを、メスのMF1及び129マウスと交尾させ、生殖細胞系の伝達を、それぞれアグーチコート色及びPCR遺伝子型決定によって判定する。
【0255】
PCR遺伝子型決定は、第三のベクター特異的プライマー(asc350)とともに、プライマーhetF及びhetRを用いて溶解した尾小端について実施される。この多重PCRにより、プライマーhetF及びhetRから野生型座(存在する場合には)からの増幅が可能となり、216bpバンドが得られる。hetFに対する部位は、ノックアウトマウスでは欠失しており、それゆえ、標的の対立遺伝子からこの増幅は起こらない。しかしながら、asc350プライマーは、3'アームのちょうど内側の領域にアニールするhetRプライマーと組み合わせて、標的座から384 bpバンドを増幅する。従って、この多重PCRにより、以下のとおり同腹仔の遺伝子型が明らかとなる。野生型サンプルは、単一の216 bpバンドを示し、ヘテロ接合体DNAサンプルは、216 bp及び384 bpに2本のバンドを生じ、ホモ接合体サンプルは、標的の特異的384 bpバンドだけを示す。
【0256】
<実施例3.生物学的データ:遺伝子発現パターン>
1)RT-PCR
RT-PCRを用いることによって、海馬(+++)、小脳、皮質、線条体、中脳、脳橋(++)、視床下部、視床、前脳、脊髄、下垂体、及び三叉神経節(+)における該遺伝子発現が示される。
2)Lac Z染色構造のリスト
【0257】
LacZ染色
切開組織のXgal染色を、以下の様式で実施する。
【0258】
代表的な組織薄片は、大きな臓器から作製される。小臓器全体及び管を、固定剤及び染色剤が浸透するように薄切りに開く。組織をPBS(リン酸緩衝生理食塩水)中で十分にすすぎ、血液または腸管内容物を除去する。組織を、固定剤(2%のホルムアルデデヒド、0.2%のグルタルアルデヒド、0.02%のNP40、1mMのMgCl2、0.23mMのデオキシコール酸ナトリウムを含むPBS)中に30〜45分間入れる。PBS中で5分間の洗浄を3回行った後、組織を、30℃で18時間、Xgal染色溶液(PBS中、4mMのKフェロシアニド、4mMのKフェリシアニド、2mMのMgCl2、1mg/mlのX-gal)中で放置する。組織をPBSで3回洗浄し、4%のホルムアルデヒド中で24時間、後染色し、再度PBSで洗浄してから、70%エタノール中に保存する。
【0259】
LacZ染色を用いることによって、VDCCγ-8が、海馬、皮質、扁桃体、嗅覚皮質、前脳、及び視床に発現されることが見出される。
【0260】
<実施例4.生物学的データ:行動:ロータロッド>
全動物を午前7時点灯の12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるため、設定時間に動物を試験する。該試験は、5分間にわたって、一定速度で4rpmから40rpmに加速し、次いで、カットオフ時間(10分)、またはマウスがロータロッドから落下するまで40rpmを保つ加速ロータロッド上での3つの試行を含む。動物がロータロッドから落下する時間を記録する。
【0261】
ノックアウトマウスは、3つのセッションすべてにおいて、野生型マウスと比較して、ロータロッド上での能力低下を示した(図2)。
【0262】
<実施例5.生物学的データ:行動:Laboras>
Laboras(実験動物行動観察記録及び分析システム)システムは、Metris BV(オランダ国)により市販されている自動行動装置である。Laborasは、振動パターンを用いて、行動を自動的に記録し定量化する。全動物を午前7時点灯の12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるために、設定時間に動物を試験する。最初の1時間、マウスをこの装置内に入れ、飼料及び水に自由にアクセスさせる。飼料及び水に自由にアクセスさせたまま、15時間終夜セッションによりこれを引き続き行う。
【0263】
1時間試験において、VDCCγ-8ノックアウトマウスは、セッションを通して、機能亢進(歩行運動頻繁/移動距離/毛づくろい)性である。(図3a)。
【0264】
15時間終夜試験において、VDCCγ-8ノックアウトマウスは、該セッションの最初の3時間は機能亢進性であり、その後、該動物が該装置に慣れるに従い、活動レベルは低下し、野生型マウスの活動レベルと対等になる(図3b)。
【0265】
<実施例5.生物学的データ:免疫組織化学:GluR2免疫組織化学>
GluR抗体に関する標準的免疫組織化学的方法(Mead及びStephens(2003)J. Neurosci. 23:1041〜1048頁)を用いて、Sant Cruz GluR2(N-19、sc-7611)抗体を用いることによって、GluR2 AMPA受容体サブユニットの免疫標識を海馬を含有する脳切片において実施した。野生型マウスと比較して、VDCCγ-8ノックアウトマウスの海馬では、GluR2に対する免疫染色は極めて低かった(図4)。
【0266】
<実施例6.生物学的データ:行動:社会的相互作用>
午前7時点灯の12時間明/12時間暗の明暗サイクル下で、全動物を、食料及び水に自由にアクセスさせて飼育する。日周期の作用を避けるため、設定時間に動物を試験する。
【0267】
動物(野生型コントロールに一致させた対応週齢の各性別の5匹のKOマウス)を、各々9cm径のシリンダーを含有する、1つの中央チャンバー及び2つの「サイド」チャンバーからなる第3のチャンバーの装置に入れる。まず、各試験マウスを、5分間、該装置に入れ、追跡ソフトウェアを用いて活動を記録する。次いで、試験マウスを取り出し、同じ性別の侵入マウスをシリンダー上に置き、次いで、該試験マウスを該中央チャンバー内に戻す。次いで、該活動をさらに5分間記録する。空きチャンバーと比較して、侵入マウスを含有するチャンバーで各試験マウスが費やす時間を記録し、社会的相互作用の測定値として用いる。データは、各チャンバーで費やされた時間もしくは時間パーセントとしてまたは接近回避スコアとして算出され、正のスコアは、社会的相互作用の増加を示し、ゼロのスコアは、社会的相互作用なしを示し、負のスコアは、社会的回避の増加を示す。
【0268】
VDCCγ-8ノックアウトマウスは、この試験において、侵入者チャンバーにおける時間パーセントまたは接近回避スコアとして測定した場合、週齢を一致させた野生型マウスと比較して、社会的相互作用の明らかな欠如を示した(図5a及び5b)。
【0269】
このデータは、統合失調症における有用性を示唆している。
【0270】
<実施例7.生物学的データ:挙動:プレパルス抑制>
プレパルス抑制(PPI)は、大音響(115dB)刺激により喚起される驚愕応答が、驚愕刺激100ms前に供される短時間のそれよりはるかに静かな音響刺激(バックグラウンドより3〜10dB上)によって抑制される観察現象である。このいわゆるプレパルス抑制は、統合失調症患者において破壊されているが、精神運動性の開閉(gating)の欠損を反映している可能性があると考えられる。
【0271】
PPIは、げっ歯類において喚起され得、アンフェタミン、アポモルフィン、フェンシクリジン、及びMK801等の精神薬及び精神作用薬により破壊され得る。この破壊は、ハロペリドール、クロザピン、及びオランザピン等の抗精神病薬によって反転させることができる(Bakshi VP、Swerdlow NR及びGeyer MA、Clozapine antagonizes phencyclidine-induced deficits in sensorimotor gating of the startle response、J. Pharmacol. Exp. Ther. 1941年;271:787〜794頁)。従って、PPIの動物モデルは、構築体及び予測妥当性の両方で存在する。PPIに対する薬物または変異のいずれかの効果は、統合失調症の治療に有用である可能性を強く示している(McGhie A及びChapman J. Disorders of attention and perception in early schizophrenia、Br. J. Med. Psychol. 1961年;34:103〜116頁。Braff DL、Geyer MA及びSwerdlow NR、Human studies of prepulse inhibition of startle:normal subjects, patient groups, and pharmacological studies、Psychopharmacology、2001年;156:234〜258頁)。動物を、午前7時に点灯する12時間明/12時間暗の明暗サイクル下で、飼料及び水に自由にアクセスさせて飼育し、日周期の作用を避けるために設定時間で試験をする。マウス(対応する週齢に一致させた野生型コントロールとともに各性別の6匹のKOマウス)を、驚愕チャンバー(San Diego Instruments、サンディエゴ、カリフォルニア州、米国)に入れる。各システムは、小型ファンで換気され、天井に据付けられた白熱により照明された音響減衰チャンバーからなった。該チャンバーは、フレームに据付けられた透明のアクリルシリンダー(そこにマウスを入れる)を含んだ。該シリンダー及び該フレームは、フレームの各コーナーの下に配置された4つのスクリューによってPlexiglasベース付近を上昇した。音響刺激は、閉鎖容器の天井に据付けられた小型スピーカーを介して送達された。驚愕応答は、フレームの真下に据付けられた圧電型加速度計によって変換された。アウトプットシグナルをデジタル化し、調整し、San Diego Instruments Startle Reflexソフトウェアを備えたPC上で連続1ミリ秒(ms)の読取りとして記録した。驚愕刺激は、50msの長さで広いスペクトルノイズの115dBパルスであった。プレパルス刺激は、バックグラウンドより3、7、及び10dB上であり、驚愕刺激100ms前に供された。刺激は、擬似ランダム形式で供された。オスKOマウスにおいて、PPIは、週齢を一致させた野生型コントロールと比較して有意に破壊された(対応のあるt検定p=0.01)(図6を参照)。
【0272】
本文書に記載された出願及び特許のそれぞれ、並びに出願及び特許のそれぞれの遂行期間を含む、上記出願及び特許のそれぞれに引用または参照された各文書(「出願引用文書」)、並びに該出願及び特許のそれぞれ、及び任意の出願引用文書に引用または記載された任意の製品に関する任意の製造元の使用説明書またはカタログは、本明細書に参照として組み込まれている。さらに、本文に引用したすべての文書、及び本文に引用した文書に引用または参照したすべての文書、並びに本文に引用または記載された任意の製品に関する任意の製造元の使用説明書またはカタログは、本明細書に参照として組み込まれている。
【0273】
本発明の記載された方法及びシステムの種々の修飾及び変型は、本発明の範囲及び精神を逸脱することなく、当業者に明白であろう。本発明を、特定の好ましい実施形態に関連させて記載したが、当然のことながら、請求された本発明は、そのような特定の実施形態に過度に限定されるべきではなく、それに対する多くの修飾及び付加を、本発明の範囲で行うことができる。実際、分子生物学または関連分野の当業者に明白な、本発明の実施に関して記載された様式の種々の修正形態は本請求項の範囲内にあることとする。さらに、本発明の範囲から逸脱することなく、以下の独立した請求項の特徴と該独立した請求項の特徴との種々の組合せを行うことができる。
【図面の簡単な説明】
【0274】
【図1】ノックアウトベクターを示す図。
【図2】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのロータロッド試験の結果を示すグラフ。
【図3A】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのLABORAS(登録商標)(実験動物行動観察記録及び分析システム:Laboratory Animal Behaviour Observation Registration and Analysis System)における1時間のセッションを示すグラフ。
【図3B】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)についてのLABORAS(登録商標)(実験動物行動観察記録及び分析システム)における15時間の一晩(ON)のセッションを示すグラフ。
【図4】野生型マウス(wt、+/+)と比較した、ノックアウトマウス(変異体、−/−)の海馬におけるGluR2の免疫組織学的染色を示す図。
【図5a】VDCCγ-8ノックアウトマウスが、野生型マウスと比較して侵入マウスを含むチャンバーにおいて時間減少を示すことを示すグラフ。
【図5b】VDCCγ-8ノックアウトマウス及び野生型マウスに対する10の接近回避スコアを示すグラフ。
【図6】媒体またはMK801を投与された野生型及びノックアウトマウスにおいて観察される、バックグラウンドより上の組合せ音響刺激を用いたプレパルス抑制(PPI)を示す図。
【図7】プライマーの位置及び標的ストラテジーに用いられるVDCCγ-8座領域のゲノム構造を示す図。
【特許請求の範囲】
【請求項1】
候補薬剤が電位依存性カルシウムチャネルガンマ-8の活性に影響を及ぼすかどうかを判定する工程を含む、精神疾患の治療に適した薬剤の同定方法。
【請求項2】
上記薬剤が電位依存性カルシウムチャネルガンマ-8の活性を阻害するアンタゴニストである、請求項1に記載の方法。
【請求項3】
上記薬剤が電位依存性カルシウムチャネルガンマ-8の活性を増強するアゴニストである、請求項1に記載の方法。
【請求項4】
上記電位依存性カルシウムチャネルガンマ-8が、配列番号3、配列番号5、もしくは配列番号7として本明細書において特定されているポリペプチド、またはそれらと少なくとも70%の配列同一性を有する配列を含むか、またはそれをコードする、請求項1から3のいずれか一項に記載の方法。
【請求項5】
上記電位依存性カルシウムチャネルガンマ-8が、配列番号3、配列番号5、もしくは配列番号7として本明細書において特定されているポリペプチド、またはそれらと少なくとも70%の同一性を有する配列からなるか、またはそれをコードする、請求項1から4のいずれか一項に記載の方法。
【請求項6】
上記精神疾患が精神病を含む、請求項1から5のいずれか一項に記載の方法。
【請求項7】
上記精神疾患が統合失調症、注意欠陥/多動性障害、双極性障害、または大うつ病である、請求項1から6のいずれか一項に記載の方法。
【請求項8】
上記精神疾患がてんかんである、請求項1から5のいずれか一項に記載の方法。
【請求項9】
上記候補薬剤が、電位依存性カルシウムチャネルガンマ-8ポリペプチドの活性に影響を及ぼすかどうかを判定するために、電位依存性カルシウムチャネルガンマ-8ポリペプチドを上記候補薬剤と接触させる、請求項1から8のいずれか一項に記載の方法。
【請求項10】
上記候補薬剤を、電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している細胞と接触させる、請求項1から9のいずれか一項に記載の方法。
【請求項11】
上記候補薬剤を、電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している非ヒト動物に投与し、上記動物が挙動の変化を示すかどうかを判定する、請求項1から10のいずれか一項に記載の方法。
【請求項12】
上記非ヒト動物が機能的電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している、請求項11に記載の方法。
【請求項13】
上記非ヒト動物が野生型である、請求項11または12に記載の方法。
【請求項14】
上記非ヒト動物がげっ歯類である、請求項11から13のいずれか一項に記載の方法。
【請求項15】
機能的に破壊された内因性電位依存性カルシウムチャネルガンマ-8遺伝子を有する、非ヒトトランスジェニック動物。
【請求項16】
上記動物がヌル変異を有する、請求項15に記載の非ヒトトランスジェニック動物。
【請求項17】
上記動物が上記電位依存性カルシウムチャネルガンマ-8遺伝子に対して−/−である、請求項15または16に記載の非ヒトトランスジェニック動物。
【請求項18】
精神疾患の治療のための電位依存性カルシウムチャネルガンマ-8遺伝子のアゴニストまたはアンタゴニストを同定するための、請求項15から17のいずれか一項に記載の非ヒトトランスジェニック動物またはその単離細胞もしくは組織の使用。
【請求項19】
精神疾患のモデルとしての、請求項15から17のいずれか一項に記載の非ヒトトランスジェニック動物の使用。
【請求項20】
(i)患者から単離されたサンプル中の電位依存性カルシウムチャネルガンマ-8遺伝子の発現レベルを測定する工程と、(ii)コントロールと比較して、上記サンプルが単離された個体が精神疾患を有するか、または精神疾患にかかりやすいかどうかを判定する工程とを含む、個体における精神疾患の存在または感受性の判定方法。
【請求項21】
精神疾患の予防または治療のための医薬の製造における、電位依存性カルシウムチャネルガンマ-8ポリペプチドの活性に影響を及ぼす薬剤の使用。
【請求項22】
精神疾患の治療または予防のための医薬の製造における、外因性電位依存性カルシウムチャネルガンマ-8ポリヌクレオチドの使用。
【請求項23】
精神疾患の治療または予防のための医薬の製造における、電位依存性カルシウムチャネルガンマ-8ポリヌクレオチドの転写または発現を増加または減少させる薬剤の使用。
【請求項24】
上記精神疾患が請求項6から8のいずれか一項に定義されているものである、請求項18から23のいずれか一項に記載の方法または使用。
【請求項1】
候補薬剤が電位依存性カルシウムチャネルガンマ-8の活性に影響を及ぼすかどうかを判定する工程を含む、精神疾患の治療に適した薬剤の同定方法。
【請求項2】
上記薬剤が電位依存性カルシウムチャネルガンマ-8の活性を阻害するアンタゴニストである、請求項1に記載の方法。
【請求項3】
上記薬剤が電位依存性カルシウムチャネルガンマ-8の活性を増強するアゴニストである、請求項1に記載の方法。
【請求項4】
上記電位依存性カルシウムチャネルガンマ-8が、配列番号3、配列番号5、もしくは配列番号7として本明細書において特定されているポリペプチド、またはそれらと少なくとも70%の配列同一性を有する配列を含むか、またはそれをコードする、請求項1から3のいずれか一項に記載の方法。
【請求項5】
上記電位依存性カルシウムチャネルガンマ-8が、配列番号3、配列番号5、もしくは配列番号7として本明細書において特定されているポリペプチド、またはそれらと少なくとも70%の同一性を有する配列からなるか、またはそれをコードする、請求項1から4のいずれか一項に記載の方法。
【請求項6】
上記精神疾患が精神病を含む、請求項1から5のいずれか一項に記載の方法。
【請求項7】
上記精神疾患が統合失調症、注意欠陥/多動性障害、双極性障害、または大うつ病である、請求項1から6のいずれか一項に記載の方法。
【請求項8】
上記精神疾患がてんかんである、請求項1から5のいずれか一項に記載の方法。
【請求項9】
上記候補薬剤が、電位依存性カルシウムチャネルガンマ-8ポリペプチドの活性に影響を及ぼすかどうかを判定するために、電位依存性カルシウムチャネルガンマ-8ポリペプチドを上記候補薬剤と接触させる、請求項1から8のいずれか一項に記載の方法。
【請求項10】
上記候補薬剤を、電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している細胞と接触させる、請求項1から9のいずれか一項に記載の方法。
【請求項11】
上記候補薬剤を、電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している非ヒト動物に投与し、上記動物が挙動の変化を示すかどうかを判定する、請求項1から10のいずれか一項に記載の方法。
【請求項12】
上記非ヒト動物が機能的電位依存性カルシウムチャネルガンマ-8ポリペプチドを発現している、請求項11に記載の方法。
【請求項13】
上記非ヒト動物が野生型である、請求項11または12に記載の方法。
【請求項14】
上記非ヒト動物がげっ歯類である、請求項11から13のいずれか一項に記載の方法。
【請求項15】
機能的に破壊された内因性電位依存性カルシウムチャネルガンマ-8遺伝子を有する、非ヒトトランスジェニック動物。
【請求項16】
上記動物がヌル変異を有する、請求項15に記載の非ヒトトランスジェニック動物。
【請求項17】
上記動物が上記電位依存性カルシウムチャネルガンマ-8遺伝子に対して−/−である、請求項15または16に記載の非ヒトトランスジェニック動物。
【請求項18】
精神疾患の治療のための電位依存性カルシウムチャネルガンマ-8遺伝子のアゴニストまたはアンタゴニストを同定するための、請求項15から17のいずれか一項に記載の非ヒトトランスジェニック動物またはその単離細胞もしくは組織の使用。
【請求項19】
精神疾患のモデルとしての、請求項15から17のいずれか一項に記載の非ヒトトランスジェニック動物の使用。
【請求項20】
(i)患者から単離されたサンプル中の電位依存性カルシウムチャネルガンマ-8遺伝子の発現レベルを測定する工程と、(ii)コントロールと比較して、上記サンプルが単離された個体が精神疾患を有するか、または精神疾患にかかりやすいかどうかを判定する工程とを含む、個体における精神疾患の存在または感受性の判定方法。
【請求項21】
精神疾患の予防または治療のための医薬の製造における、電位依存性カルシウムチャネルガンマ-8ポリペプチドの活性に影響を及ぼす薬剤の使用。
【請求項22】
精神疾患の治療または予防のための医薬の製造における、外因性電位依存性カルシウムチャネルガンマ-8ポリヌクレオチドの使用。
【請求項23】
精神疾患の治療または予防のための医薬の製造における、電位依存性カルシウムチャネルガンマ-8ポリヌクレオチドの転写または発現を増加または減少させる薬剤の使用。
【請求項24】
上記精神疾患が請求項6から8のいずれか一項に定義されているものである、請求項18から23のいずれか一項に記載の方法または使用。
【図1】

【図2】
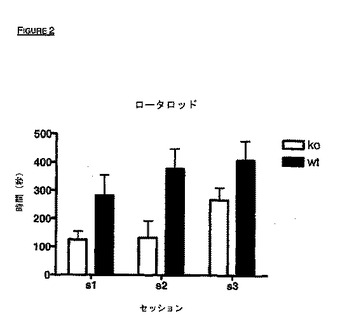
【図3A】
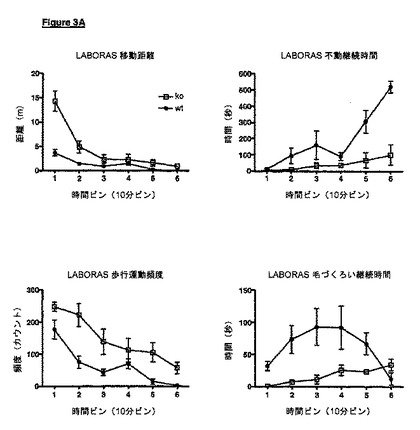
【図3B】
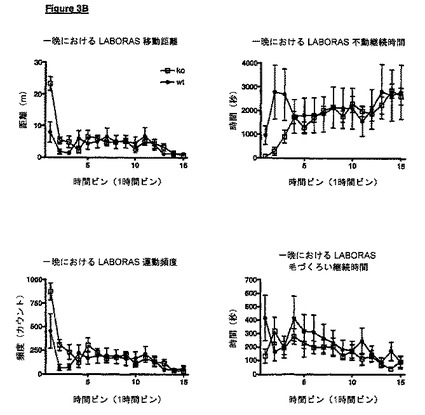
【図4】
【図5a】
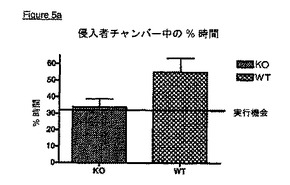
【図5b】
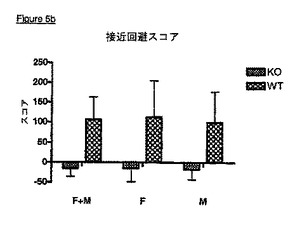
【図6】
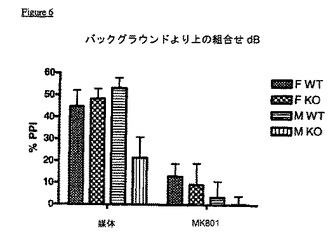
【図7】





【図2】
【図3A】
【図3B】
【図4】
【図5a】
【図5b】
【図6】
【図7】
【公表番号】特表2009−506765(P2009−506765A)
【公表日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願番号】特願2008−528582(P2008−528582)
【出願日】平成18年9月4日(2006.9.4)
【国際出願番号】PCT/GB2006/003263
【国際公開番号】WO2007/026171
【国際公開日】平成19年3月8日(2007.3.8)
【出願人】(507200802)タケダ・ケンブリッジ・リミテッド (6)
【Fターム(参考)】
【公表日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願日】平成18年9月4日(2006.9.4)
【国際出願番号】PCT/GB2006/003263
【国際公開番号】WO2007/026171
【国際公開日】平成19年3月8日(2007.3.8)
【出願人】(507200802)タケダ・ケンブリッジ・リミテッド (6)
【Fターム(参考)】
[ Back to top ]