
WG/WNTシグナル経路の新規成分
本発明は、次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法である。
a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。

a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。
【発明の詳細な説明】
【背景技術】
【0001】
脊椎動物及び無脊椎動物のWnt(ウィント)遺伝子は、システインに富む大きなファミリー分泌タンパク質をコードする。そのタンパク質は、生物的過程の多様性において、細胞間のシグナル分子として重要な役割を果たす(Wodarz及びNusse 1998を広く参照のこと)。最初のWnt遺伝子であるマウスWnt−1は、乳癌におけるマウスの乳癌のウィルスの統合によって活性化された最初の腫瘍形成遺伝子として発見された(Nusse及びVarmus 1982)。その結果、癌におけるWnt(ウィント)シグナル経路の関わりは、大いに研究された。
【0002】
Wnt−1相同染色体としてのショウジョウバエの極性遺伝子ウィングレス(wingless)
の同一性(Cabrera, Alanso et al. 1987; Perrimon and Mahowald 1987; Rijsewijk, Schuermann et al. 1987)で、Wnt遺伝子が重要な発生の調節遺伝子であることが明らかになった。このように、先ず、胚形成と発癌のような、異種の似ていない生物的過程の双方が、同一のシグナル経路を経て細胞コミュニケーションに依存する。
【0003】
標準的なWnt経路の最近のモデルにおいて、分泌されたWntタンパク質は、Frizzle細胞表面の受容体に結合し、細胞質のタンパク質Dishevelled(Dsh)を活性化する。そこで、Dshは、プロテインキナーゼ、Shaggy(Sgg)/GSK3、骨格タンパク質アキシン及びβ−カテニン、脊椎動物のアルマジロ(Armadillo)の相同染色体を含む数種のタンパク質の複合体にシグナルを伝達する。
【0004】
この複合体において、β−カテニンは、Sggによって加リン酸反応が生じた後の、分解のための標的とされる。Wntシグナル及びその結果としてのSgg活性の調節下降の後、β−カテニン(又はそのショウジョウバエ相同染色体アルマジロ(Armadillo))は、分解から逃れ、細胞質内に集まる。遊離細胞質β−カテニンは、未だ知られていないメカニズムによって核へ移動し、転写因子のTcf/Lefファミリーの結合を通じて、遺伝子の転写を調節する(Grosschedl R 1999)。β−カテニンにおける突然変異、APC、及びアキシンは、標準的なWnt経路の本質的な活性化が、ヒトの発癌に寄与することを示唆する、数種のヒトc癌において発見された(Uthoff SM,Eichenberger MR,McAuliffe TL,Hamilton Cl and Galandiuk S.(2001). Mol. Carcinog., 31,56-62)。
【0005】
Wntリガンドのそれらの受容体への結合は、また非標準的な経路の活性化を誘発し、カルシウム融解、c−Jun NH2−末端キナーゼ、及びGタンパク質を通じてシグナルを送ることができる、β−カテニンの独立的なシグナルを送る、Wntシグナルとして言及する。これらの経路は、標準的な、Wntリガンド上昇調節によって特徴づけられる腫瘍における、β−カテニンの依存経路と平行して活性化することができた(Huguet EL, McMahon JA, McMahon AP, Bicknell R and Harris AL(1994). Cancer Res., 54, 2615-2621.; Dale TC, Weber-Hall SJ, Smith K, Huguet EL, Jayatilake H, Gusterson BA, Shuttleworth G, O'Hare M and Harris AL.(1996).Cancer Res., 56, 4320-4323; Vider BZ, Zimber A, Chastre E, Prevot S, Gespach C, Eslein D, Wolloch Y, Tronick SR, Gazit A and Yaniv A.(1996). Oncogene,12, 153-158; Smith K, Bul TD, Poulsom R, Kaklamanis L, Williams G and Harris AL.(1999). Br. J. Cancer,81,496-502)。
【0006】
たとえば、Wnt−2の自由な上昇調節は、ヒトの結腸、直腸ガンや胃ガンにおいて報告された(Katoh M.(2001). Int. J, Oncol., 19, 1003-1007. )。さらに、Holcombe 等(Holcombe RF, Marsh JL, Waterman ML, Lin F, Milovanovic T and Truong T. (2002). Mol. Pathol., 55, 220-226)は、近年において、インサイチューハイブリダイゼーションによって、ヒトの結腸ガンや悪性のメラノーマにおける特異的なWnt遺伝子の発現を分析した。そして、それらの結果は、Wnt−2の過剰な発現が、ヒトの発癌においても関連していることを示唆した(Pham K, Milovanovic T, Barr RJ, Truong T and Holcombe RF. (2003). Mol. Pathol., 56, 280-285.)。
【0007】
発癌における役割に加えて、Wntシグナルは、またスケレトゲネシス(skeletogenesis)、骨形成、フラクチュアリピア(fracture repair)における役割も果たす(Hartmann (2000), Holmen (2005))。たとえばWntタンパク質の上昇調節は、リューマチ関節炎や変形性関節炎の経路生物学に相互に関係するように示された(Sen et al. (2000), Nakamura (2005), Holmen (2005))。
【0008】
近年、β−カテニンの転写活性化を直接的に抑制することによってのみならず、Wntリガンドによって経路の活性化をも抑制することによって、Wnt経路を効果的に抑制する治療薬が知られている。これは、部分的には、その十分な活性化や核移行のために要求される多くの特殊な成分が、未だ知られていないという事実に基づく。従って、これらの高い悪性の疾患に対して有効な薬剤を開発するために、この経路について一層の理解が求められる。
【発明の開示】
【発明が解決しようとする課題】
【0009】
Wg/Wntシグナル経路の活性化のために要求される新しい成分の同定のために、本発明者等は、sev−wg表現型(ショウジョウバエのメラノガスター(melanogaster)の劣性のサプレッサー(抑制遺伝子)をスクリーニングする手法を用いた。
【課題を解決するための手段】
【0010】
この手法において、遺伝子CG6210によってコードされる3L3と称されるタンパク質が見出された。前記遺伝子は、すべての後性動物において相同染色体を有しており、さらに興味深いことには、それぞれの種において、このファミリーの1つの遺伝子のみが十中八九存在している。遺伝学的手段によって、Wg/Wntシグナル経路、特にすべてのWntタンパク質のWg/Wntの分泌経路において、3L3が明確な役割を果たしていることが確かとなった。さらに、3L3が、物理的にWntタンパク質に相互作用することが示された。それ故に、3L3タンパク質は、Wntタンパク分泌物の上昇調節又は下降調節を行い、標準的及び非標準的なWnt経路の双方を抑制するような開発薬剤のための非常に有望な標的である。
【0011】
本発明は、次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法に関するものである。
【0012】
a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。
【0013】
前記d)における誘導体は、任意の分子と接触するa)若しくはb)に記載の核酸分子、若しくはc)に記載の断片であり、前記候補物質は、対応する改変されていない核酸分子若しくは誘導体間の親和性に比べて、多くとも50%増加若しくは減少する、前記誘導体との親和性を示す。
【0014】
次の(ポリ)ペプチドの配列に関する相同率(%)は、配列が直線に整列され、間隙が持ち込まれた後、もし必要であれば、配列表と同一の最大限の百分率を達成するために、そして、配列の同一性の一部としていかなるアミノ酸の保存的な代用を考慮することなく、配列表の配列番号が4、5、6、及び8のアミノ酸残基と同一である候補物質におけるアミノ酸残基の百分率として定義される。このようにして用いられる同一性の数値の%は、(Tatusova TA 1999)から得られたWU−BLAST−2によって発生させることができる。WU−BLAST−2は、ほとんどが不履行の数値にセットされた数種の調査パラメータを用いる。
【0015】
同様の方法により、配列表の配列番号が1、2、3、及び7の核酸分子の相同率(%)は、前記核酸の配列におけるヌクレオチド残基と同一である候補物質におけるヌクレオチド残基の百分率として定義される。このようにして用いられる同一性の数値は、不履行のパラメータにセットされたWU−BLAST−2のBLASTモジュールを用いて発生させることができる。
【0016】
配列表の配列番号1、2、3、及び7の核酸分子によってコードされた、配列表の配列番号が4、5、6、及び8の(ポリ)ペプチドは、それぞれが、たとえばショウジョウバエ、ヒル、ナメクジ、カタツムリ、ミミズ等の無脊椎動物、及び脊椎動物、たとえばヒト、無尾猿(エイプ)、猿(モンキー)、犬、猫、うさぎ、ヤギ、豚、ハムスター、牛、馬、羊、マウス、ラットを含む哺乳類に存在する新規なファミリータンパク質の代表(典型)である。
【0017】
配列表の配列番号が4、5、6、及び8の(ポリ)ペプチドは、3L3又はWLS(Wnt
のない)タンパク質として以下に言及されている。これらのタンパク質は、Wg/Wntシグナル経路において、及びそのようなフォーメーション、空間的な整列の維持、発達途上にある組織の増殖において、及びそのフォーメーション、多くのヒトの腫瘍の成長において、特殊な役割を果たす。
【0018】
配列表の配列番号4の(ポリ)ペプチドは、配列表の配列番号8(3L3−PB)の(ポリ)ペプチドと同様に、ショウジョウバエ(3L3−PA)の3L3タンパク質である。配列表の配列番号5及び6の(ポリ)ペプチドは、それぞれcaenorhabditis(カエノルハブジット)(C.)エレガンス及びヒトの3L3タンパク質である。それらの3L3タンパク質は、ショウジョウバエの3L3タンパク質と構造的及び機能的に相同である。
【0019】
本発明の1つの実施形態において、前記e)工程の下での前記核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同であり、好ましくは少なくとも55%相同であり、より好ましくは少なくとも60%相同であり、さらに好ましくは少なくとも65%相同であり、さらに好ましくは少なくとも70%相同である。
【0020】
他の好ましい実施形態において、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも75%相同であり、好ましくは少なくとも80%相同であり、より好ましくは少なくとも85%相同であり、さらに好ましくは少なくとも86%相同であり、さらに好ましくは少なくとも87%相同である。
【0021】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0022】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも93%相同であり、好ましくは少なくとも94%相同であり、より好ましくは少なくとも95%相同であり、さらに好ましくは少なくとも96%相同であり、さらに好ましくは少なくとも97%相同である。
【0023】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも98%相同であり、好ましくは少なくとも99%相同である。
【0024】
さらに本発明は、次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法に関するものである。
【0025】
a)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドと、候補物質とを、前記(ポリ)ペプチドへの前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部を構成する、(ポリ)ペプチドの断片への候補物質の結合を許容する条件の下で、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの断片と候補物質とを接触させる工程、又は、
c)誘導体への候補物質の結合を許容する条件の下で、a)に記載の(ポリ)ペプチドの誘導体、又はb)に記載の(ポリ)ペプチドの断片の誘導体と候補物質とを接触させる工程、又は、
d)a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチドの断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも50%相同である(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体と候補物質とを接触させる工程、及び
e)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するか、又は、候補物質が、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドとWnt(ウィント)タンパク質間の結合の活性を抑制するか、若しくは活性を促進するかを、検出する。
【0026】
前記c)における誘導体は、N末端若しくはC末端、又はアミノ酸側鎖と接触する任意の分子を持つ、アミノ酸配列の配列表の配列番号4、5、6、若しくは8の(ポリ)ペプチド、又はb)に記載の(ポリ)ペプチドの断片であり、候補物質は、対応する改変されていない(ポリ)ペプチド若しくは(ポリ)ペプチドの誘導体間の親和性に比べて、多くとも50%増加若しくは減少する、前記誘導体との親和性を示す。
【0027】
本発明の1つの実施形態において、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも52%相同であり、好ましくは少なくとも55%相同であり、より好ましくは少なくとも60%相同であり、さらに好ましくは少なくとも65%相同であり、さらに好ましくは少なくとも70%相同である。
【0028】
他の好ましい実施形態において、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも75%相同であり、好ましくは少なくとも80%相同であり、より好ましくは少なくとも85%相同であり、さらに好ましくは少なくとも87%相同である。
【0029】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0030】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0031】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも93%相同であり、好ましくは少なくとも94%相同であり、より好ましくは少なくとも95%相同であり、さらに好ましくは少なくとも96%相同であり、さらに好ましくは少なくとも97%相同である。
【0032】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも98%相同であり、好ましくは少なくとも99%相同である。
【0033】
さらに本発明は、a)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、c)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体、又はアミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドのドメイン、好ましくはWnt(ウィント)タンパク質の分泌物に含有され、若しくはWnt(ウィント)タンパク質と結合する(ポリ)ペプチドのドメインと特異的に結合する抗体に関するものである。
【0034】
さらに本発明は、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子の断片である標的配列、好ましくは配列表の配列番号9又は10の標的配列を有するsiRNAに関するものである。
【0035】
さらに本発明は、前記抗体若しくは前記siRNA、前記スクリーニング方法のa)若しくはb)工程の核酸分子、c)工程の核酸の断片、d)工程の核酸の誘導体、e)工程に記載の核酸分子、断片、若しくは誘導体、前記スクリーニング方法のa)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、c)工程の(ポリ)ペプチドの誘導体、d)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、薬剤としての使用に関するものである。
【0036】
本発明は、前記抗体若しくは前記siRNA、前記スクリーニング方法のa)若しくはb)工程の核酸分子、c)工程の核酸の断片、d)工程の核酸の誘導体、若しくはe)工程の核酸分子、断片、若しくは誘導体、又は前記スクリーニング方法のa)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、若しくはc)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、Wnt(ウィント)シグナルに関連する疾患、たとえば癌、骨若しくは関節の疾患、若しくは発達障害の処置のための医薬準備のための使用に関するものである。
【0037】
好ましい癌の種類は、Wnt依存型の癌、結腸直腸癌、肺癌、咽頭癌、好ましくはWnt−2依存の咽頭癌、小さな腸の悪性腺腫、胃底腺ポリープ、胃癌、胃の腺腫(関連する悪性腺腫のない)、胃腸のカルチノイド腫瘍、esophageal悪性腺腫; Wilmの膵臓(非ductal腺房細胞癌)の少年のnasopharyngeal angiofibromas、黒色腫、pilamatricomas、肺の悪性腺腫、卵巣癌、子宮頚部、子宮のendometrial、胸のfibromatoses、前立腺、甲状腺癌、hepatoblastoma、肝細胞癌、C型肝炎に関連している肝細胞癌、髄芽細胞腫、desmoid腫瘍、腫瘍(腎臓)、pancreatoblastoma、及び滑膜肉腫等である。
【0038】
骨又は関節の疾患の好ましいタイプ(類型)は、それぞれ変形性関節症、又はリューマチ関節炎である。
【0039】
さらに本発明は、前記抗体若しくは前記siRNAからなる医薬組成物に関する。
【0040】
さらに本発明は、増加されたか、減少されたか、3LS表現ではないか、若しくは、変異された3LS表現の(ポリ)ペプチドが少なくとも1つの組織若しくは器官に示される、
好ましくはヒトセルラインシステムであるセルラインシステムか、ショウジョウバエ、マウス、ラット、うさぎ、鶏、蛙、豚、羊、C.エレガント等のワーム、ゼブラフィッシュ等の魚、からなる群から選択される脊椎動物若しくは無脊椎動物等の有機体か、の利用を含む薬剤のスクリーニング、又は改変されたWnt(ウィント)分泌物による誘起された疾患のための検査に関するものである。
【0041】
その検査の好ましい実施形態において、前記器官は、トランス遺伝子として配列表の配列番号1〜3、7に記載された核酸分子の少なくとも1つからなる、3LS遺伝子を表現(発現)するものである。
【0042】
その検査の他の好ましい実施形態において、前記3LS遺伝子が、欠失、点変異、挿入、及び逆位からなる群から選択される変異からなるものである。
【0043】
さらに本発明は、次の工程からなる、細胞のWnt(ウィント)分泌物の改変のための
方法である。細胞を、Wntタンパク質の分泌を抑制し、若しくは促進する物質と接触させる。
【0044】
その方法の好ましい実施形態において、前記物質は、前記抗体若しくは前記siRNA、a)若しくはb)工程の核酸分子、c)工程の断片、d)工程の誘導体、若しくはe)工程の核酸分子、断片、若しくは誘導体、又はa)工程に記載の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、若しくはc)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体である。
【0045】
本発明は、細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、のa)若しくはb)工程に記載の核酸分子、c)工程に記載の断片、d)工程に記載の誘導体、若しくはe)工程に記載の核酸分子、断片、若しくは誘導体、又は抗体若しくはsiRNAの使用に関する。
【0046】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、a)工程に記載の(ポリ)ペプチド、b)工程に記載の(ポリ)ペプチドの断片、c)工程に記載の誘導体、若しくはd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の使用に関する。
【図面の簡単な説明】
【0047】
【図1a】CustalWタンパク質配列は、ショウジョウバエの3L3タンパク質と、C.エレガンスと、ヒトと間の高い相同性を示す。 推定のシグナル配列は、配列上に線によって示され、経膜的なドメインは、下線が付される(TMHMM予測)。
【図1b】図1aと同じである。
【図1c】図1aと同じである。
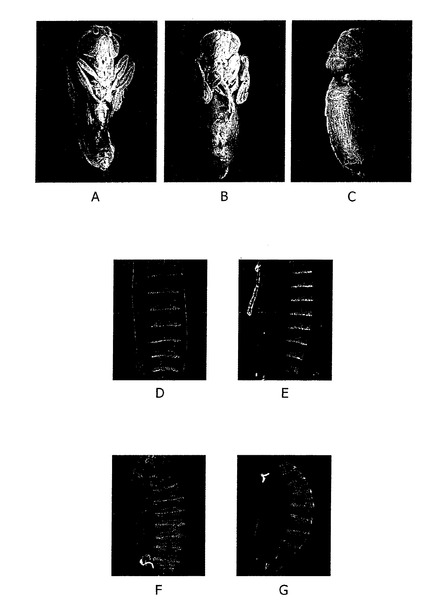
【図2】A) コントロールとしての正常成人pharates(ファラテス)。 B) 知覚力のその触鬚を見せる3L3のための成人pharates homozygous変異体は、すべての3脚の組のrudimental発達である。 C) 知覚力のその触鬚を見せる3L3のための成人pharates homozygous変異体は、すべての3脚の組のrudimental発達と、翼から背板への変化(5-10 %の場合に観測される)。germlineクローン表現型を示す胚。 D)及びE)、: 正常な分割パターンを示す野生型状態。 F)及びG) 及びセグメント(「小歯の芝生」表現型)のWg機能の損失表現型を示す3L3のための胚の変異体。 これらの3L3の突然変異の胚が、セグメントの融合や、典型的なWg又はHhの機能のロス(図2のD−G)を示すことが観察された。この結果からは、3L3が、Wg/Wnt又はHhのシグナル経路の決定的な成分であることが導かれる。
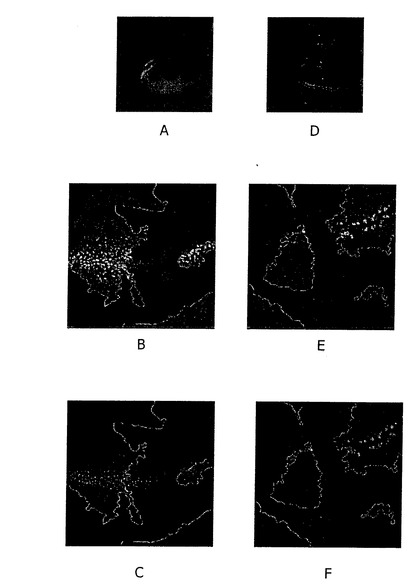
【図3】A) 野生型ディスクの末端の、より少ないLacZ表現。 B) Distalless-LacZ表現は3L3変異体組織で抑えられる。 E) Bと同じであることで、クローンマージンだけが示されます。 D) 野生型ディスクの無意味なタンパク質表現。 C) 無意味なタンパク質表現は3L3変異体で抑圧されます。 F) Bと同じであることで、クローンマージンだけが示される。
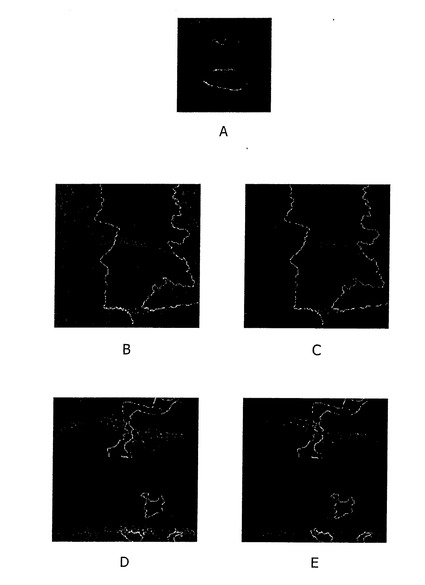
【図4】A) 野生型ディスクのWgタンパク質表現。 B) これらの変異細胞の中では、Wgが蓄積するのを示して、Wgタンパク質表現は3L3変異体組織で増加される。 C) Bと同程度、クローンマージンだけが示される。 D) 細胞外 Wgタンパク質発現はないか、または3L3変異体組織で少し増加されているだけである。B-Cで見られたWgの蓄積が主に細胞内であることが示唆されている。 E) 同様に、Bとして、クローンマージンだけが示される。
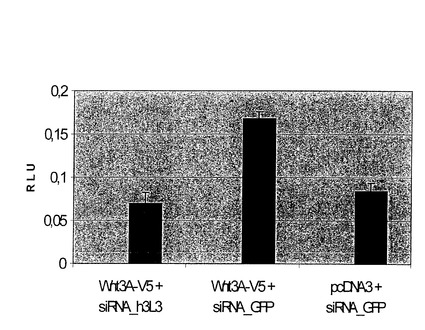
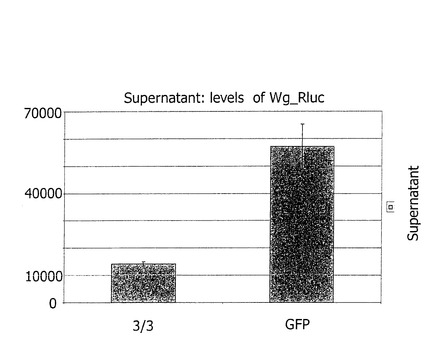
【図5】TOP Flash Luciferaseを含む。 Wnt3A-V5とsiRNA−h3L3 cotransfectedセルは見せかけのコントロールとしてのWntのシグナルの依存するLuciferaseレポーターの遺伝子の誘導を全く示さない。(空のベクトル(pcDNA3)はsiRNA#GFPと共に遺伝子でcotransfectedされた)。 しかしながら、Wnt3A-V5とsiRNA#GFP cotransfectedセルはaを示した。 レポーターの遺伝子の2倍の誘導は見せかけのコントロールと比較された。
【図6】共同免疫沈降反応、IP: α-HA、 α-V5h3L3-HAはWnt3A-V5(レーン1)。 負の制御: CD2-HAとWnt3A-V5(レーン2)の間で相互作用を全く観測することができない。そして、EGFP transfectedセル(レーン3)の中にバンドを全く検出することができない。
【図7】3L3のC..エレガンス相同染色体に対するsiRNA。 A) a wt虫3L3に対するsiRNAの性腺は性腺(B)と、欠陥への末端のチップ細胞移動(C)におけるOocytesの異常なアレンジメントに通ずる。
【図8】TOP Flash Renilla Luciferaseを含む。 GFP RNAiと比べて、WLS RNAiは明確なWgの分泌を示した。
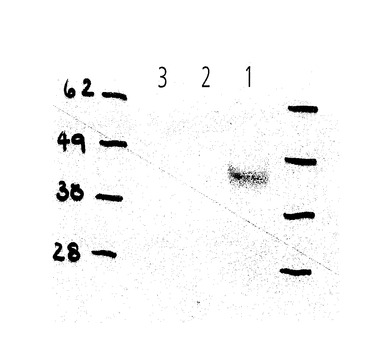
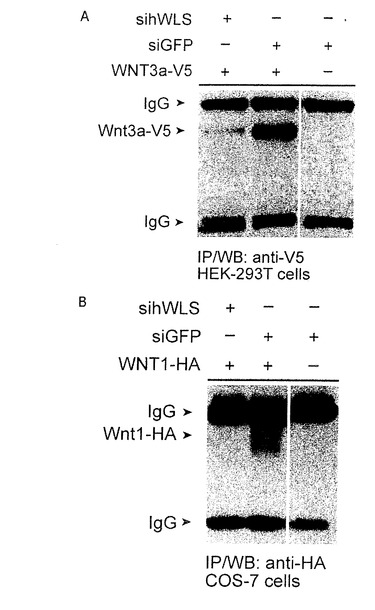
【図9A−B】Wnt3aの分泌はhWLSのノックダウンで損なわれる。 A) HEK-293Tセルについて表面に浮かぶWnt3a-V5のレベルはhWLSの減少で強く減少する。 B) sihWLSはCOS-7セルの媒体の中にWntl-HAの分泌を撤廃する。
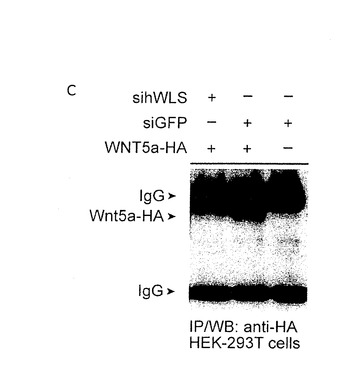
【図9C】siGFPと比べて、sihWLSのアプリケーションはWnt5a-HAの分泌を強く減少させる。
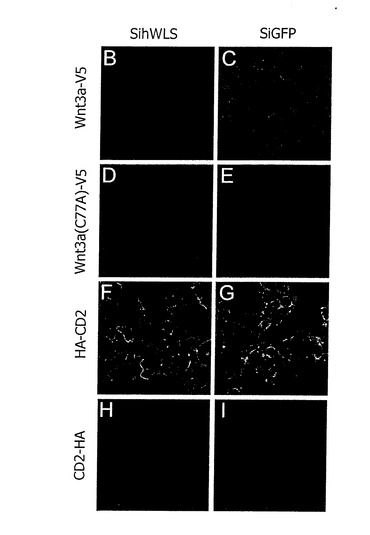
【図10】B-I) HEK-293Tセルのセルの表面stainings。 Wnt3a-V5はsihWLS(B)と共に共同transfectedされたセルのセルの表面に達することができない。siGFPのcotransfectionはセルの表面(C)でWnt3a-V5の検出可能なレベルに通じる。 どちらもsihWLS(D)と共に治療されたセルの表面で検出可能ではない。Wnt3aC77A-V5、または、siGFP(E)。 (F-I) CD2-HAが検出されていない間、HA-CD2はセルの表面(F、G)に検出される(H、I); 従来の汚損(目立たない)で容易にCD2-HAを検出することができる。 HA-CD2の表面レベルはsihWLSによって影響されない。
【図11】C.エレガンス WLS ortholog表現型模写に対してRNAi ABarは母性のオリエンテーション欠陥を紡錘形にする。そして、母性−3変異体肺は左と腹に前方で見せられる。 (A) 野生型肺、ABarスピンドル(黄色)はABprのスピンドルに垂直であって(青い)、すなわち、見せられた光学セクションに垂直な状態で(黄色いドットで、示される)適応する。 (B) RNAi(母性−2)の注入された動物、2個のスピンドルでは、オリエンテーションが平行となる。 (C、D) RNAi(R06B9.6)肺(D)のABarとABprスピンドルと同様に初期の母性−3(or78)変異体段階(C)に、オリエンテーションが平行にある。
【図12】Wnt3aの分泌はhWLSのノックダウンで損なわれる。 A) コントロール(siGFP)と比べて、ShhとsihWLSと共にcotransfectedされたHEK-293Tセルのライセートと培地はShhの生産と分泌における変更を全く示さない。 B) WLSの減少でSSHA-dTNFΔN(TNF-FLAG)の分泌は影響されない。
【発明を実施するための最良の形態】
【0048】
Wnt(ウィント)シグナリングカスケードは、脊椎動物及び無脊椎動物の双方の発生のために重要であり、腫瘍形成に影響を与えてきた。ショウジョウバエのWnt遺伝子は、百以上の遺伝子を含んでいる、Wntタンパクファミリーのなかでも最も特徴的なものの1つである。ショウジョウバエの胚において、Wgは、parasegment boundaries(パラセグメント境界)の形成、及び近接した細胞におけるengrailed(ed)(エングレイルド)の発現の維持に必要である。Wg機能における胚の表皮の欠陥は、不全の分割しか示さない。それは、異常な表皮のパターンに反映される。野性型幼虫の腹の表皮が、裸の領域と交替する歯状ベルトを示している間、Wg変異体幼虫の表皮は歯状物で完全に覆われている。
【0049】
成虫のディスクの発育の間、Wgは、dorso-vental(dorso腹)の位置の情報を制御する。脚のディスクでは、Wgは、腹の運命(Struhl and Basler 1993)の誘導によって、今後の脚をパター(patter)する。Wgの活性が減退している動物においては、腹の脚の半分は、背面のミラーイメージ(mirror image)(ベイカー1988)内で発育する。従って、減退したWgの活性は、notal組織への翼の変化につながり、結果として遺伝子の名前(Sharma and Chopra 1976).につながっている。目のディスクでは、Wgは、頭部の表皮の発育のために、ommatidial分化を抑制し、目の領域を横切って、dorso腹の軸を設立するのにかかわる(Heberlein、Borod et al. 1998)。
【0050】
付加的な遺伝子は、Wgシグナリングの分泌、受容、又は解釈に関係してきた。たとえばショウジョウバエの遺伝学的研究は、Wgシグナリングにおけるfrizzled (Dfz)、 Dishevelled (dsh)、shaggy/zeste-white-3 (sgg/zw-3)、armadillo (arm)、adenomatous polyposis coli (E-apc), axin(アキシン)、及び pangolin (pan)との関わりを示した。これらのトランスデューサー(transducer)の遺伝的な指令は、WgがDshを通じてSggを抑制する作用をし、従って、SggによるArmの抑圧を和らげ、その結果、Armの細胞質での蓄積、及びその核への移行をもたらす。核内では、Armは、標的遺伝子の転写を活性化するために、Panと相互作用をする。脊椎動物の相同染色体は、すべてのこれらの成分のために同定され(最新の論評を参照のこと(Peifer and Polakis 2000))、ショウジョウバエのシグナル経路の、新規な同定されたメンバーが、脊椎動物の相対物を持つかもしれないことを示唆した。
【0051】
Arm、β−カテニンの哺乳類の相同染色体の核への蓄積、及びWg/Wnt経路の重要な活性化に導かれた変異は、直腸癌、乳癌、メラノーマ(黒色腫)、肝細胞癌、卵巣癌、子宮内膜癌、髄芽細胞腫、毛基質細胞腫、及び前立腺癌を含む、多くの種類の癌に観測された。現在では、Wntシグナリングの統制解除が、これらの悪性腫瘍に発生において重要な事象であることが明らかとなっている。しかしながら、Wnt経路を効果的に抑制する治療薬が未だ知られていないばかりか、β−カテニンの転写の活性化を直接的に抑制する治療薬、或いはWntリガンドによる経路の活性化を抑制する治療薬すらも知られていない。これは、十分な活性が要求される多くの特殊な成分が、未だ知られていないという事実に基づくものである。
【0052】
Winglessの活性化のために要求される新しい成分を同定するために、本発明者等はショウジョウバエ遺伝子の手法を用い、ショウジョウバエのメラノガスター(melanogaster)のsev-wg表現型の劣性のサプレッサーのためのスクリーンを行った。この手法において、3L3として下記に引用されているタンパク質が見出された。このタンパク質は、遺伝子CG6210によってコードされている。前記遺伝子は、すべての後生動物において相同物を有し、さらに興味深いことには、それぞれの種において、十中八九は、このファミリーの1つの遺伝子しか存在しない。遺伝学的手段により、本発明者等は、Wg/Wntシグナル経路の決定的な成分としての3L3の役割を確信し、特にWntタンパク質の分泌物における、その重要
性を確信した。さらに、3L3とWntが物理的に相互作用することを、鋭意研究によって見出した。
【0053】
sev-wg表現型の劣性のサプレッサーのためのスクリーンでは、本発明者等は、3L3の近縁種の群から3つの異なる対立遺伝子を見出した。2つの対立遺伝子が、今までのところ、特徴的でない遺伝子CG6210の変異を有している。CG6210は、N末端側がシグナル配列で構成され、推定で7個の経膜的なドメインを有するタンパク質をコードする。3L3の近縁種の群の1つの対立遺伝子であるsu20.53は、シグナル配列の最後のアミノ酸残基と、時期尚早の終止コドンにおけるフレームシフトに導かれる、47bp欠失を含んでいる。他のサプレッサーsu20.54は、最初の経膜的なドメインにおいて、プロリンをセリンに変換するアミノ酸残基に導く点変異を有する。異なる生命情報工学プログラムは、このタンパク質が、知られていない機能のタンパク質のファミリーに属し、また興味深いことに、種ごとには、常にこのファミリーの1つのタンパク質しか存在しないという結論を導いた。さらに、そのタンパク質は、異なる種において非常にうまく保存されており、特に、細胞外での数種の配列や、経膜的で橋架けされたドメイン(図1a−c)の機能が特殊であり、且つ保存されていることを示している。
【0054】
ショウジョウバエの胚は、数種の遺伝子の局在化された活性によって、反復セグメントへ、累進的に再分化される。Wingless(Wg)及びHedgehog(Hh)は、いわゆるセグメント極性遺伝子グループの最も重要な遺伝子であり、A/P軸を、セグメントを未だ形成しておらず将来セグメントとなる、14−15のストライプ(stripes)へ均等に分配する。結果として、Wg又はHhの機能のロスは、胚の腹部の横の反復された外骨格構造のフォーメーションの分裂を生じさせる。これは、セグメント極性遺伝子の活性の、最も明白な表現型の結論である。
【0055】
「germline clones(微生物ラインクローン)」と称される遺伝的な実験は、(3L3の場合のように)興味あるタンパク質が完全に欠乏しているショウジョウバエの突然変異の胚を、通常のものにすることを発明者に許容し、そして分割のパターンを研究することを許容する。特に「germline clones(微生物ラインクローン)」は、多くの遺伝子のために、胚にときどき存在する母性の成分を除去するのに必要である。
【0056】
これらの3L3の突然変異の胚が、セグメントの融合や、典型的なWg又はHhの機能のロス(図2のD−G)を示すことが観察された。この結果からは、3L3が、Wg/Wnt又はHhのシグナル経路の決定的な成分であることが導かれる。
【0057】
また、Wgは、触覚、脚、羽根等の、ショウジョウバエの成虫の付属肢の発育に中枢的な役割を果たす。それ故に、発明者等は、孵化前に飛ぶことを意味する成虫のファラテス(pharates)の3L3の突然変異の表現型を研究した。発育がWgに依存するすべての付属肢が異常であることが観察された(図2A−C)。特に、触覚の端刺が見当たらず、脚が短く、分割の不足が認められ、いわゆる「羽根から胸背板」の5%に変形が観察された。この変形は、Wgの機能のロスのために、病徴的なものである。これらが見出されたことで、
3L3がWg/Wntシグナル経路における決定的な成分であるという明白な結論が得られる。
【0058】
たとえば羽根や脚のような成虫の付属肢は、"imaginal discs(イマジナルディスク、成虫ディスク)"と称されるsack-like 上皮において組織(器官)形成されたimaginal細胞によって幼虫において形成される。特に、羽根のimaginal discは、それが胚の発育過程で形成され、幼虫の時期に75000個の細胞のディスクを一般化するために、3つの幼虫の齢の間、増殖するとき、20個の細胞の周囲に配置されている。これらのimaginal discs(イマジナルディスク)において、Wingless(ウイングレス)は、空間的濃度が変化し、細胞が異なる閾値の濃度において、異なって応答するモルフォゲンとしてふるまう。
【0059】
本発明者等は、たとえばWgによって活性化される遺伝子のような標的遺伝子の抑圧上の3L3突然変異組織の効果の研究するために、羽根のimaginal discs(イマジナルディスク)を実験において用いた。2種の標的遺伝子は、羽根のimaginal discs(イマジナルディスク)において区別される。一方、低いWg濃度でも発現し、それ故にWgの根源から数種の細胞直径距離で発現するDistallessのような「長い範囲の標的遺伝子」がある。他方、
発現のためにモルフォゲンの高いレベルを必要とする(Senseless)のような、「短い範囲の標的遺伝子」と称される標的遺伝子がある。
【0060】
羽根のimaginal disc(イマジナルディスク)において、3L3の突然変異のクローンの形態を含んだ後、長い範囲の標的遺伝子のためのマーカーとしてのDistalless-lacZの発現及び
短い範囲の標的遺伝子(図3A−F)のためのマーカーとしてのSenselessの発現が研究された。この実験において、Distalless-acZ及びSenselessの発現の減少が観察されたが、突然変異の組織が、Wgの産生細胞を含んだ。これらの結果により、3L3が、Wgのモルフォゲンの産生において、十中八九、重要な役割を果たすことが明白である。
【0061】
Wg産生における3L3の重要性を確かめるために、我々は、Wg産生細胞において、3L3のみを選択的に発現する昆虫を釈放することを試みた。動物の休息は、3L3のための相同的な突然変異であった。その結果は、完全な釈放であり、3L3の機能が、Wg産生細胞においてのみ必要であるという結論に言及する。
【0062】
さらに、羽根のimaginal discs(イマジナルディスク)におけるWgタンパク質の発現での3L3突然変異組織の効果が研究された(図4A−E)。3L3突然変異クローンにおいて、Wgの発現量に増加があることが観察された。この結果を良好に特徴づけるために、本発明者等は、類似の実験を遂行し、WgのmRNAの転写のレベルを現すwg-lacZの発現を見た。3L3突然変異組織におけるwg-lacZの発現の増加が全くないという事実は、突然変異組織におけるWgのための、増加するstainingが、Post−転写過程に基づくという推定を正当化する。
【0063】
最後に、Wgの蓄積が細胞内又は細胞外で生じさせるか、区別するために、本発明者等は細胞外タンパク質の検出を許容する異なったプロトコル(protocol)を用いてWgの分配(distribution)を試験した。しかし、3L3突然変異クローンにおける細胞外Wgの蓄積は、検出されなかった。これらの結果は、その機能のロスが、Wg産生細胞の内側でのWgの蓄積を導くので、Wgの分泌における3L3のための役割を示唆する。
【0064】
Wntシグナリングにおいて、3L3のヒト相同染色体が含まれているか否かを試験するために、Wntレポーター(reporter)遺伝子assay(検査)(TOP-flash)が行われた。これは、luciferase(ルシフェラーゼ)遺伝子の前に位置して結合している5TCFを伴うassay(検査)に基礎をおくluciferase(ルシフェラーゼ)である。Wntシグナル経路は、luciferase(ルシフェラーゼ)の蛍光によって測定することができる。以前に(preceding)議論した結果から、ショウジョウバエのWnt産生細胞において3L3が重要であることが知られていた。
【0065】
Wntの産生及び受容細胞を擬態するために、1バッチのヒト293T細胞が、h3L3に対向するmWnt3A−V5及びsiRNAでトランスフェクト(Transfect)された。別のバッチの細胞は、TOP-flash reporter constructでトランスフェクトされた。トランスフェクトがされた後、24時間経過した時、これらの2つの細胞のバッチが混合され、さらに24時間培養した後、luciferase(ルシフェラーゼ)活性が測定された。もし3L3に対向するsiRNAが、「産生」細胞においてWnt3Aでコトランスフェクトされたならば、luciferase(ルシフェラーゼ)活性のレベルは、もし空のベクトル(Wnt3Aの代用の)及びGFPに対向するsiRNAがトランスフェクトされたならば、同程度に低かった。他方、もしGFPに対向するWnt3A及びsiRNAがトランスフェクトされたならば、レポーター遺伝子の明確な誘起が観察された(図5)。これらの結果は、ヒト(h)3L3が、シグナルの受容ではなく、産生において含まれたWnt経路の決定的な成分であることを示している。
【0066】
3L3は、Wnt産生細胞において重要であるので、h3L3−HAがマウス(m)のWnt3A−V5と相互作用すると、調べられた。Co-immunoprecipitation 試験が行われ、その結果は、h3L3−HAがmWnt3A−V5と相互作用することを示唆し、CD2−HAを伴うネガティブコントロールや、他の膜−スパニング(membrane-spanning)タンパク質は、mWnt3A−V5と相互作用しなかった(図6)。
【0067】
ショウジョウバエの羽根ディスク(discs)におけるwntless(WLS)突然変異細胞のクローンにおいて、Wg産生細胞におけるWgの蓄積が観察された。Kc細胞において、我々は、培地へのWgの分泌物を測定するための検査(assay)を開発した。Renilla luciferase
遺伝子は、WgのN末端に融合し、Kc細胞へトランスフェクトした。同時に、その細胞は、WLS又はGFPのdsRNAで扱われた。WLSのRNAiは、GFPのRNAiと比べてWg分泌物の明らかな下降調節を示した(図8)。
【0068】
この分泌物の欠乏が、RNAiによるWLSのヒトの組織培養実験デプレション(depletion)において、Wntシグナルカスケードを活性化することができない結果となった理由であったかどうかを決定するために、我々は、Wnt分泌細胞の培地を分析した。HEK293Tは、siRNAのみならず、Wnt3A−V5、Wnt1−HA、又はWnt5a−HAでもコトランスフェクトした。すべてのこれらのWnt分泌物は、WLSに対するsiRNAがGFPに対するsiRNAと比較してコトランスフェクトされたとき、かなり減少した。これらの結果は、WLSが消耗されたとき、HEK293細胞において、Wnt分泌物が大幅に減少することを示すものである。(図9;2つの異なるsiRNA(siRNAhWLS−A,siRNAhWLS−B)が、HEK293細胞におけるendogenoushWLS遺伝子の発現をノックダウンするのに効果的なことのために、RT−PCRによって有効にさせた。双方の独立したトランスフェクションは、結果として、hWLSの転写の85%減少をさせた(示されていない;siRNAhWLS−Bは、ほとんど用いられた。そして、sihWLSとして言及されている。しかし、siRNAhWLS−Aは、同じ効果を示した。)。sihWLSを持つレスポンダー細胞の取り扱い、又はGFPに対するsiRNAを持つプロデューサー細胞の取り扱いは、Wntシグナリング検査(assay)の結果にさほど影響を与えなかった。
【0069】
WLSが下降調節されるとき、HEK293T細胞において、WNT3aが細胞外表面に到達することができるか否かを試験するために、我々は、siRNA及びWnt3A−V5をコトランスフェクトし、Wnt3aの細胞外分裂のみをステイン(stain)した。コントロールとして、我々は、N末端(細胞外)又はC末端(細胞内)の一方にHA−tagを有するCD2を用いた。我々が用いた細胞外 staining protocolで、我々は、N末端のHA−tagで、CD2のみを欠乏させることができた。WLSに対するRNAiは、細胞外表面にCD2の分泌物の影響を与えなかった。対照的に、Wnt3a−V5の表面のstainingは、WLSに対するsiRNAでコトランスフェクトされた細胞において破壊された。それ故に、我々は、細胞表面へのWNT3a(及びおそらくは他のWntも同様に)の好適な分泌のためにWLSが必要であることを結論づける(図10)。
【0070】
ショウジョウバエの羽根のディスクにおいて、処分されたクローンは、Hhリガンドの明らかな蓄積を示したが、明確な対比で、WLSのクローンにおいては、Hhの蓄積は観察することができなかった。さらに、WLSの突然変異の昆虫は、wg−Ga14::UAS−WLS構造で釈放された。wg産生細胞におけるWLSのみの発現は、致死性を救済し、WLSがWgシグナリングのために特異的(specific)であるという我々の見解を支持するのに十分であった。
【0071】
ヒト組織の培養において、Wntの分泌物のみが影響を受けたが、上述のように、CD2の分泌物は、WLS RNAiによって影響を受けなかった。重要なことは、Shhの分泌物のみならず、培地へのTNFの分泌物も、WLS RNAiによって影響を受けなかった(図12)。これらの結果は、WLSのヒトのorthologueが、Wntシグナリングのために特異的なものであることを示す。
【0072】
我々の結果は、また、Wntの小さなサブセットがWLSの機能のロスに敏感であるばかりでなく、すべてのWntがWLSから独立しているわけではないことを示している。C.エレガンスにおいて、MOM−3は、初期のblastomere polarization (Rocheleau, Thorpe)、VPC specification (Eisenmann and Kim)、及びQ neuroblast migration (Harris)のような、多くの異なる標準的及び非標準的なWnt経路に含まれていることが示されてきた。これらの過程は、3つの異なるWnt(経路)、MOM−2(Rochelau, Thorpe)、LIN−44
(Jiang and Sternberg)及びEGL20(Harris)に依存する。
【0073】
C.エレガンスにおいては、異なる発育過程においてpostembryonically活性である標準的経路と、胚の発育においてのみ観察することができる非標準的経路との、2つのWntシグナル経路が区別されることができる。非標準的Wntシグナリングは、EMSの決定のorientation及びendodermの運命を調節する(EMS細胞は、将来のendoderm及びmesodermのprecursor 細胞のprecursorである)。非標準的Wntシグナリングのロスは、endodermのロスや、mesodermの発育を導く。この非標準的Wntシグナリングの数種の成分は、記載されてきた。それらの1つであるmom−3(more mesoderm-3)は、geneticallyに特徴づけられてきたが、molecularlyには、so far同定されていない(Eisenmann and Kim 2000; Thorpe, Schlesinger et al. 1997).。配列によると、3L3(c3L3)の相同染色体における変異は、異なるmom−3のallelesにおいて見出された。さらに、c3L3に対するRNAiは、
【0074】
distal tip細胞移動における欠乏や、胚のlethality.のような標準的及び非標準的なWnt表現型の典型的なロスを導いた。また、これらの結果は、c3L3がmom−3の表現型のために応答可能であること、及びc3L3に対するRNAiは、mom−3の表現型のロスを擬態することを示している。
【0075】
C.エレガンスにおけるWntシグナリングは、多くの異なる発育過程において含まれている。知られている最初の過程は、mitotic spindlesのMOM−2の発育オリエンテーションであり、胚細胞は4個から8個に分割される。mom−3ワームは、wntless orthologue R06B9.6における突然変異であるので、我々はR06B9.6のdsRNAを、wthermaphroditesの生殖線に注入した。上述のように、分析されたすべての胚には、R06B9.6のdsRNAが注入され、或いはmom−2又はmom−3の突然変異alleleの運搬は、Abar spindle から ABpr spindleへの平行オリエンテーションを示した。対照的に、Wnt胚は、垂直に定位された2つのスピンドル(spindle)を有していた(図11)。
【0076】
要約すると、本発明者等は、遺伝子CG6210によってコードされる3L3と名付けられたリペプチドを見出した。遺伝的手法により、3L3が、Wg/Wntシグナル経路、特に、Wntタンパク質のWg/Wnt分泌経路において、役割を果たしていることが確かめられた。さらに、3L3は、物理的にWntタンパク質と相互作用することが示された。それ故、3L3タンパク質は、Wntリガンドの分泌物を調節することによって、Wnt経路を正又は負に調節する薬剤の開発のために、非常にpromisingな標的である。
【実施例】
【0077】
(実施例1)
[sev−wg表現型の受容サプレッサーのためのスクリーン]
目において、Wgを発現するsev−wgトランス遺伝子は、スクリーンにおいて、表現型マーカー(Brunner et al., 1997)として観察される、ラフな目の表現型に導く。左の腕でFRT80を運び、右の腕で3番目の染色体のsev−wgを運ぶ雄は、12時間の間、21mMのエチルメタンスルフォネート(EMS)で養育された。EMSを適用させた後、24時間経過後に、雄は、X染色体上のeyeless-flp recombinaseである雌と交尾し、3番目の染色体上に、FRT80Mw+を運搬した。ey-flpと両方のFRT染色体でoffspringされた雄は、ラフな目の表現型の抑圧のためにスクリーンされた。異なる対立遺伝子間でのComplementation分析は、2つのallelesを構成するグループ3L3の決定に導いた。
with the ey-flp and both FRT chromosomes were screened for a suppression of the rough eye phenotype. Complementation analysis between different alleles led to the complementation group 3L3 consisting of two alleles.
【0078】
(実施例2)
[すべてのメタゾンにおける相同染色体]
Blast search (blastp)によって、すべての配列のメタゾン有機体において、知られていない機能1171(DUF1171)のドメインを運搬する、この新しいタンパク質ファミリーの1つのコピーのみが存在した。
【0079】
(実施例3)
[ドメインspanningする膜]
TMHMM TOPは、1つのN末端シグナル配列と、CG6210−PBのための7つの推定の経膜的なドメインを予想した。
【0080】
(実施例4)
[Germline クローン]
興味あるタンパク質の完全な不在において、ショウジョウバエの胚の表現型であるメラノガスターの研究に用いられる実験的なprocedureである。
3番目のinstar larvaeをFRTの1つの染色体上に運搬し、推定の変異を同じFRTの相同染色体上及びovoD 変異に運搬する雌は、1.5時間、38℃で熱ショックを受けた。ハッチングの後、雄が同じ推定の変異及びFRTを運搬し、交尾した。
【0081】
雌は、寒天プレート上で、1夜、産卵し、胚が集積された。これを3分間、ブリーチし、タップ(tap)水で洗浄した。胚は、24時間、水に注入された。vitellumを遊離させるために、メタノールとヘプタンを等量ずつ含むbiphasic溶液で浸とうした。胚は、メタノールで4回洗浄され、0.1%のトリトンX−100で4回洗浄された。RTで6時間培養した後、最後に胚をHoyer-lactateでスライド上に集積し、60℃でダイジェストタンパク質に12−24JIKANN培養した。表皮は光学顕微鏡で試験された。
【0082】
(実施例5)
[標的遺伝子の発現に、3L3突然変異組織の効果を研究するための、羽根イマジナルディスク]
イマジナルディスクは、metamorphosisの間、成虫の構造を作る細胞のhollow sacsである。
これらは、胚ectodermにおけるポケットとして上昇し、幼虫がさなぎになるまで体コウの内側に成長し、そのポイントで体壁と付属物を形成するために、「"evaginate"」を内側に戻す。
【0083】
[イマジナルディスクの抗体staining]
幼虫は、氷の上で、リンガー(Ringer)溶液中でdissectedされ、PEM 200μlで200μl管に固定された。5%のホルムアルデヒド 10.7μlと、0.05%のトリトンX−100とを20分間攪拌した。4回洗浄し、1時間追加し、2−3回、PEJTとNa−Azidを加えて培養した。一次抗体(PBTとNa−Azidでdilutedしたもの)で一昼夜、4℃でインキュベートした。Larvaeは、PBT、Na−Azid、1%のHINGSで30分、5回洗浄し、RTに、PBTとNa−Azidでdilutedした二次抗体で、2時間インキュベートした。Larvaeは、2時間で5回PBTとNa−Azidで洗浄し、ディスクはPPDAに集積された。羽根のディスクにおけるStainingは、焦点顕微鏡で分析された。
【0084】
(実施例6)
[羽根のイマジナルディスクにおける3L3突然変異クローンのフォーメーションの誘起、及びDistalless-lacZ (wg-lacZ)の発現の研究]
羽根のイマジナルディスクにおける突然変異クローンは、興味あるタンパク質のための変異である細胞のグループをrepresentする。ショウジョウバエのメラノガスター(melanogaster)において、熱ショックプロモーター(hs- flp)によって操作される、yeast-specific recombinase flippase (flp)を用いて、mitotic recombinationによって遂行される。
【0085】
y w hsp-flp; allele 20.53 FRT80 / TM6b fliesは、y w hsp-flp; πMyc [w+] Minute Pfy+J FRT80 / TM6b,によって交尾され、雌は、2日間、産卵する。
37.0℃で60分間の熱ショックで、24−72時間、AEFが形成され、さらに4時間後に、幼虫が切開された。
【0086】
Distalless-lacZの場合においては、一次抗体にβ−ガラクトシダーゼ(1:2000ラビット、ポリクローナル抗体、Cappel(登録商標))を用いた。
wg-lacZの場合においては、抗体に、α−Wg(4D4)(1:1000マウス、モノクローナル抗体、DSHB)を用いた。
【0087】
(実施例7)
[Wg産生細胞において3L3のみを発現する羽根をrescueするのを試みる手段としての、Wg生産物における3L3の重要性の確立]
トランス遺伝子wg-GAL4 (the transcriptional activator GAL4 は、Wgを発現する細胞内でのみ発現される。) で運搬される3L3heterozygous mutant fliesは、3L3heterozygous mutantであるfliesで交尾され、トランス遺伝子UAS−3L3(GAL4タンパク質のためのUAS結合サイトは、3L3の転写を活性化する。)
【0088】
(実施例8)
[Wg分割の試験]
このプロトコル(protocol)は、一次抗体でのstainingが、PBS中で4℃(endocytosisをブロックするために)で30−60分間行われ、固定が、一次抗体でのstaining後、PBS、4%のホルムアルデヒドで、4℃で20−30分間(Strigini and Cohen, 2000)行われるという事実において、通常のprotocolとは異なる細胞外Wgを見るために用いた。
【0089】
(実施例9)
[TOPフラッシュアッセイ]
細胞は、リン酸カルシウム法によってトランスフェクトした。トランスフェクト後、16時間細胞を洗浄し、24時間混合した。混合後、細胞はPromega からIxPLB でlysed し、分析した。
【0090】
(実施例10)
[Co-immunoprecipitation]
細胞は、リン酸カルシウム法によってトランスフェクトした。トランスフェクト後、36時間、細胞はRIPAバッファーによって、40℃で1時間30分間、lysedした。Lysateは、4℃で30分間、centrifugedされ、その後、タンパク質GSepharoseビーズ及びラビットα−HAで一昼夜インキュベートした。ビーズは、TBSで4回洗浄し、タンパク質は、95℃で10分間、ビーズからSDS−Loadingバッファーでelutedした。マウスα−V5抗体でウエスタンブロットを行った。
【0091】
(コンストラクト)
mWnt3A cDNA配列は、V5−tag及びHis−tagによってpcDNA3にクローンされた。h3LA cDNAは、HA−tagによってpcDNA3にクローンされた。QiagenCD2によって一般化された3L3及びGFPに対するsiRNAは、C末端HA−tagでpcDNA3にクローンされた。
【0092】
(参考文献)
Baker, N. E. (1988). 'Transcription of the segment-polarity gene wingless in the imaginal discs of Drosophila, and the phenotype of a pupal-lethal wg mutation." Development 102(3): 489-97.
Cabrera, C. V., M. C. Alonso, et al. (1987). "Phenocopies induced with antisense RNA identify the wingless gene." Cell 50(4): 659-63.
Eisenmann, D. M., Kim, S. K. (2000). "Protruding vulva mutants identify novel loci and Wnt signaling factors that function during Caenorhabditis elegans vulva development." Genetics 156(3): 1097-116.
Grosschedl R, E. Q. (1999). "Regulation of LEF-ITCF transcription factors by Wnt and other signals." Current Opinion in Cell Biology 11: 233-240.
Harris, J., Honigberg, L., Robinson, N., and Kenyon, C. (1996). Neuronal cell migration in C. elegans: regulation of Hox gene expression and cell position. Development 122, 3117-3131.
Hartmann, C. and Tabin, C. J. (2000). ,,Dual roles of Wnt signaling during chondrogenesis in the chicken limb." Development, 127(14), 3141-3159.
Heberlein, U., E. R. Borod, et al. (1998). "Dorsoventral patterning in the Drosophila retina by wingless." Development 125(4): 567-77.
Holmen, S. L. et al. (2005), ,,Essential role of beta-catenin in postnatal bone acqusition." The Journal of Biological Chemistry, 280 (22), 21162-21168.
Jiang, L. I., and Sternberg, P. W. (1999). Socket cells mediate spicule morphogenesis in Caenorhabditis elegans males. Dev Biol 211, 88-99.
Morin, P. J. (1999). "β-catenin signaling and cancer." Bioessays 21(12): 1021-30.
Nakamura, Y., Nawata, M., Wakitani, S. (2005). "Expression profiles and functional analyses of Wnt-related genes in human joint disorders". American Journal of Pathology, 167(1), 97- 105.
Nusse, R. and H. E. Varmus (1982). "Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome." Cell 31(1): 99- 109.
Perrimon, N. and A. P. Mahowald (1987). "Multiple functions of segment polarity genes in Drosophila." Dev Biol 119(2): 587-600.
Peifer, M. and P. Polakis (2000). "Wnt signaling in oncogenesis and embryogenesis--a look outside the nucleus." Science 287(5458): 1606-9.
Polakis, P., M. Hart, et al. (1999). "Defects in the regulation of β-catenin in colorectal cancer." Adv Exp Med Biol 470: 23-32.
Potter, J. D. (1999). "Colorectal cancer: molecules and populations." Journal of the National Cancer Institute 91(11): 916-32.
Rijsewijk, F., M. Schuermann, et al. (1987). 'The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless." Cell 50(4): 649-57.
Rocheleau, C. E., Downs, W. D., Lin, R., Wittmann, C, Bei, Y., Cha, Y. H., AIi, M., Priess, J. R., and MeIIo, C. C. (1997). Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 90, 707-716.
Roose, J. and H. Clevers (1999). 1TCF transcription factors: molecular switches in carcinogenesis." Biochimica et Biophysica Acta 1424 (2-3): M23-37.
Sen, M., Lauterbach, K., El-Gabalawy, H., Firestein, G. S., Corr, M. Carson, D. A. (2000). ,,Expression and function of wingless and frizzled homologs in rheumatoid arthritis." PNAS, 97 (6), 2791-2796.
Sharma, R. P. and V. L. Chopra (1976). "Effect of the Wingless (wgl) mutation on wing and haltere development in Drosophila melanogaster." Dev Biol 48(2): 461-5.
Strigini, M., and. Cohen, S.. M. (2000). Wingless gradient formation in the Drosophila wing. Curr. Biol. 10, 293-300.
Struhl, G. and K. Basler (1993). "Organizing activity of wingless protein in Drosophila." Cell 72(4): 527-40.
Tatusova TA, M. T. (1999). "Blast 2 sequences-a new tool for comparing protein and nucleotide sequences." FEMS Microbiol Lett. 174: 247-250.
Thorpe, CJ. , Schlesinger, A., et al (1997). "Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm." Cell;90(4): 695-705.
Thorpe, C. 1, Schlesinger, A., and Bowerman, B. (2000). Wnt signalling in Caenorhabditis elegans: regulating repressors and polarizing the cytoskeleton. Trends Cell Biol 10, 10-17.
Waltzer, L. and M. Bienz (1999). 'The control of β-catenin and TCF during embryonic development and cancer." Cancer & Metastasis Reviews 18(2): 231-46.
Wodarz, A. and R. Nusse (1998). "Mechanisms of Wnt signaling in development." Annual Review of Cell & Developmental Biology 14: 59-88.
【背景技術】
【0001】
脊椎動物及び無脊椎動物のWnt(ウィント)遺伝子は、システインに富む大きなファミリー分泌タンパク質をコードする。そのタンパク質は、生物的過程の多様性において、細胞間のシグナル分子として重要な役割を果たす(Wodarz及びNusse 1998を広く参照のこと)。最初のWnt遺伝子であるマウスWnt−1は、乳癌におけるマウスの乳癌のウィルスの統合によって活性化された最初の腫瘍形成遺伝子として発見された(Nusse及びVarmus 1982)。その結果、癌におけるWnt(ウィント)シグナル経路の関わりは、大いに研究された。
【0002】
Wnt−1相同染色体としてのショウジョウバエの極性遺伝子ウィングレス(wingless)
の同一性(Cabrera, Alanso et al. 1987; Perrimon and Mahowald 1987; Rijsewijk, Schuermann et al. 1987)で、Wnt遺伝子が重要な発生の調節遺伝子であることが明らかになった。このように、先ず、胚形成と発癌のような、異種の似ていない生物的過程の双方が、同一のシグナル経路を経て細胞コミュニケーションに依存する。
【0003】
標準的なWnt経路の最近のモデルにおいて、分泌されたWntタンパク質は、Frizzle細胞表面の受容体に結合し、細胞質のタンパク質Dishevelled(Dsh)を活性化する。そこで、Dshは、プロテインキナーゼ、Shaggy(Sgg)/GSK3、骨格タンパク質アキシン及びβ−カテニン、脊椎動物のアルマジロ(Armadillo)の相同染色体を含む数種のタンパク質の複合体にシグナルを伝達する。
【0004】
この複合体において、β−カテニンは、Sggによって加リン酸反応が生じた後の、分解のための標的とされる。Wntシグナル及びその結果としてのSgg活性の調節下降の後、β−カテニン(又はそのショウジョウバエ相同染色体アルマジロ(Armadillo))は、分解から逃れ、細胞質内に集まる。遊離細胞質β−カテニンは、未だ知られていないメカニズムによって核へ移動し、転写因子のTcf/Lefファミリーの結合を通じて、遺伝子の転写を調節する(Grosschedl R 1999)。β−カテニンにおける突然変異、APC、及びアキシンは、標準的なWnt経路の本質的な活性化が、ヒトの発癌に寄与することを示唆する、数種のヒトc癌において発見された(Uthoff SM,Eichenberger MR,McAuliffe TL,Hamilton Cl and Galandiuk S.(2001). Mol. Carcinog., 31,56-62)。
【0005】
Wntリガンドのそれらの受容体への結合は、また非標準的な経路の活性化を誘発し、カルシウム融解、c−Jun NH2−末端キナーゼ、及びGタンパク質を通じてシグナルを送ることができる、β−カテニンの独立的なシグナルを送る、Wntシグナルとして言及する。これらの経路は、標準的な、Wntリガンド上昇調節によって特徴づけられる腫瘍における、β−カテニンの依存経路と平行して活性化することができた(Huguet EL, McMahon JA, McMahon AP, Bicknell R and Harris AL(1994). Cancer Res., 54, 2615-2621.; Dale TC, Weber-Hall SJ, Smith K, Huguet EL, Jayatilake H, Gusterson BA, Shuttleworth G, O'Hare M and Harris AL.(1996).Cancer Res., 56, 4320-4323; Vider BZ, Zimber A, Chastre E, Prevot S, Gespach C, Eslein D, Wolloch Y, Tronick SR, Gazit A and Yaniv A.(1996). Oncogene,12, 153-158; Smith K, Bul TD, Poulsom R, Kaklamanis L, Williams G and Harris AL.(1999). Br. J. Cancer,81,496-502)。
【0006】
たとえば、Wnt−2の自由な上昇調節は、ヒトの結腸、直腸ガンや胃ガンにおいて報告された(Katoh M.(2001). Int. J, Oncol., 19, 1003-1007. )。さらに、Holcombe 等(Holcombe RF, Marsh JL, Waterman ML, Lin F, Milovanovic T and Truong T. (2002). Mol. Pathol., 55, 220-226)は、近年において、インサイチューハイブリダイゼーションによって、ヒトの結腸ガンや悪性のメラノーマにおける特異的なWnt遺伝子の発現を分析した。そして、それらの結果は、Wnt−2の過剰な発現が、ヒトの発癌においても関連していることを示唆した(Pham K, Milovanovic T, Barr RJ, Truong T and Holcombe RF. (2003). Mol. Pathol., 56, 280-285.)。
【0007】
発癌における役割に加えて、Wntシグナルは、またスケレトゲネシス(skeletogenesis)、骨形成、フラクチュアリピア(fracture repair)における役割も果たす(Hartmann (2000), Holmen (2005))。たとえばWntタンパク質の上昇調節は、リューマチ関節炎や変形性関節炎の経路生物学に相互に関係するように示された(Sen et al. (2000), Nakamura (2005), Holmen (2005))。
【0008】
近年、β−カテニンの転写活性化を直接的に抑制することによってのみならず、Wntリガンドによって経路の活性化をも抑制することによって、Wnt経路を効果的に抑制する治療薬が知られている。これは、部分的には、その十分な活性化や核移行のために要求される多くの特殊な成分が、未だ知られていないという事実に基づく。従って、これらの高い悪性の疾患に対して有効な薬剤を開発するために、この経路について一層の理解が求められる。
【発明の開示】
【発明が解決しようとする課題】
【0009】
Wg/Wntシグナル経路の活性化のために要求される新しい成分の同定のために、本発明者等は、sev−wg表現型(ショウジョウバエのメラノガスター(melanogaster)の劣性のサプレッサー(抑制遺伝子)をスクリーニングする手法を用いた。
【課題を解決するための手段】
【0010】
この手法において、遺伝子CG6210によってコードされる3L3と称されるタンパク質が見出された。前記遺伝子は、すべての後性動物において相同染色体を有しており、さらに興味深いことには、それぞれの種において、このファミリーの1つの遺伝子のみが十中八九存在している。遺伝学的手段によって、Wg/Wntシグナル経路、特にすべてのWntタンパク質のWg/Wntの分泌経路において、3L3が明確な役割を果たしていることが確かとなった。さらに、3L3が、物理的にWntタンパク質に相互作用することが示された。それ故に、3L3タンパク質は、Wntタンパク分泌物の上昇調節又は下降調節を行い、標準的及び非標準的なWnt経路の双方を抑制するような開発薬剤のための非常に有望な標的である。
【0011】
本発明は、次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法に関するものである。
【0012】
a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。
【0013】
前記d)における誘導体は、任意の分子と接触するa)若しくはb)に記載の核酸分子、若しくはc)に記載の断片であり、前記候補物質は、対応する改変されていない核酸分子若しくは誘導体間の親和性に比べて、多くとも50%増加若しくは減少する、前記誘導体との親和性を示す。
【0014】
次の(ポリ)ペプチドの配列に関する相同率(%)は、配列が直線に整列され、間隙が持ち込まれた後、もし必要であれば、配列表と同一の最大限の百分率を達成するために、そして、配列の同一性の一部としていかなるアミノ酸の保存的な代用を考慮することなく、配列表の配列番号が4、5、6、及び8のアミノ酸残基と同一である候補物質におけるアミノ酸残基の百分率として定義される。このようにして用いられる同一性の数値の%は、(Tatusova TA 1999)から得られたWU−BLAST−2によって発生させることができる。WU−BLAST−2は、ほとんどが不履行の数値にセットされた数種の調査パラメータを用いる。
【0015】
同様の方法により、配列表の配列番号が1、2、3、及び7の核酸分子の相同率(%)は、前記核酸の配列におけるヌクレオチド残基と同一である候補物質におけるヌクレオチド残基の百分率として定義される。このようにして用いられる同一性の数値は、不履行のパラメータにセットされたWU−BLAST−2のBLASTモジュールを用いて発生させることができる。
【0016】
配列表の配列番号1、2、3、及び7の核酸分子によってコードされた、配列表の配列番号が4、5、6、及び8の(ポリ)ペプチドは、それぞれが、たとえばショウジョウバエ、ヒル、ナメクジ、カタツムリ、ミミズ等の無脊椎動物、及び脊椎動物、たとえばヒト、無尾猿(エイプ)、猿(モンキー)、犬、猫、うさぎ、ヤギ、豚、ハムスター、牛、馬、羊、マウス、ラットを含む哺乳類に存在する新規なファミリータンパク質の代表(典型)である。
【0017】
配列表の配列番号が4、5、6、及び8の(ポリ)ペプチドは、3L3又はWLS(Wnt
のない)タンパク質として以下に言及されている。これらのタンパク質は、Wg/Wntシグナル経路において、及びそのようなフォーメーション、空間的な整列の維持、発達途上にある組織の増殖において、及びそのフォーメーション、多くのヒトの腫瘍の成長において、特殊な役割を果たす。
【0018】
配列表の配列番号4の(ポリ)ペプチドは、配列表の配列番号8(3L3−PB)の(ポリ)ペプチドと同様に、ショウジョウバエ(3L3−PA)の3L3タンパク質である。配列表の配列番号5及び6の(ポリ)ペプチドは、それぞれcaenorhabditis(カエノルハブジット)(C.)エレガンス及びヒトの3L3タンパク質である。それらの3L3タンパク質は、ショウジョウバエの3L3タンパク質と構造的及び機能的に相同である。
【0019】
本発明の1つの実施形態において、前記e)工程の下での前記核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同であり、好ましくは少なくとも55%相同であり、より好ましくは少なくとも60%相同であり、さらに好ましくは少なくとも65%相同であり、さらに好ましくは少なくとも70%相同である。
【0020】
他の好ましい実施形態において、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも75%相同であり、好ましくは少なくとも80%相同であり、より好ましくは少なくとも85%相同であり、さらに好ましくは少なくとも86%相同であり、さらに好ましくは少なくとも87%相同である。
【0021】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0022】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも93%相同であり、好ましくは少なくとも94%相同であり、より好ましくは少なくとも95%相同であり、さらに好ましくは少なくとも96%相同であり、さらに好ましくは少なくとも97%相同である。
【0023】
さらに好ましい実施形態においては、e)工程の下での核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも98%相同であり、好ましくは少なくとも99%相同である。
【0024】
さらに本発明は、次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法に関するものである。
【0025】
a)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドと、候補物質とを、前記(ポリ)ペプチドへの前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部を構成する、(ポリ)ペプチドの断片への候補物質の結合を許容する条件の下で、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの断片と候補物質とを接触させる工程、又は、
c)誘導体への候補物質の結合を許容する条件の下で、a)に記載の(ポリ)ペプチドの誘導体、又はb)に記載の(ポリ)ペプチドの断片の誘導体と候補物質とを接触させる工程、又は、
d)a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチドの断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも50%相同である(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体と候補物質とを接触させる工程、及び
e)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するか、又は、候補物質が、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドとWnt(ウィント)タンパク質間の結合の活性を抑制するか、若しくは活性を促進するかを、検出する。
【0026】
前記c)における誘導体は、N末端若しくはC末端、又はアミノ酸側鎖と接触する任意の分子を持つ、アミノ酸配列の配列表の配列番号4、5、6、若しくは8の(ポリ)ペプチド、又はb)に記載の(ポリ)ペプチドの断片であり、候補物質は、対応する改変されていない(ポリ)ペプチド若しくは(ポリ)ペプチドの誘導体間の親和性に比べて、多くとも50%増加若しくは減少する、前記誘導体との親和性を示す。
【0027】
本発明の1つの実施形態において、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも52%相同であり、好ましくは少なくとも55%相同であり、より好ましくは少なくとも60%相同であり、さらに好ましくは少なくとも65%相同であり、さらに好ましくは少なくとも70%相同である。
【0028】
他の好ましい実施形態において、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも75%相同であり、好ましくは少なくとも80%相同であり、より好ましくは少なくとも85%相同であり、さらに好ましくは少なくとも87%相同である。
【0029】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0030】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同である。
【0031】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも93%相同であり、好ましくは少なくとも94%相同であり、より好ましくは少なくとも95%相同であり、さらに好ましくは少なくとも96%相同であり、さらに好ましくは少なくとも97%相同である。
【0032】
さらに好ましい実施形態においては、d)工程の下での(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも98%相同であり、好ましくは少なくとも99%相同である。
【0033】
さらに本発明は、a)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、c)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体、又はアミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドのドメイン、好ましくはWnt(ウィント)タンパク質の分泌物に含有され、若しくはWnt(ウィント)タンパク質と結合する(ポリ)ペプチドのドメインと特異的に結合する抗体に関するものである。
【0034】
さらに本発明は、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子の断片である標的配列、好ましくは配列表の配列番号9又は10の標的配列を有するsiRNAに関するものである。
【0035】
さらに本発明は、前記抗体若しくは前記siRNA、前記スクリーニング方法のa)若しくはb)工程の核酸分子、c)工程の核酸の断片、d)工程の核酸の誘導体、e)工程に記載の核酸分子、断片、若しくは誘導体、前記スクリーニング方法のa)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、c)工程の(ポリ)ペプチドの誘導体、d)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、薬剤としての使用に関するものである。
【0036】
本発明は、前記抗体若しくは前記siRNA、前記スクリーニング方法のa)若しくはb)工程の核酸分子、c)工程の核酸の断片、d)工程の核酸の誘導体、若しくはe)工程の核酸分子、断片、若しくは誘導体、又は前記スクリーニング方法のa)工程の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、若しくはc)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、Wnt(ウィント)シグナルに関連する疾患、たとえば癌、骨若しくは関節の疾患、若しくは発達障害の処置のための医薬準備のための使用に関するものである。
【0037】
好ましい癌の種類は、Wnt依存型の癌、結腸直腸癌、肺癌、咽頭癌、好ましくはWnt−2依存の咽頭癌、小さな腸の悪性腺腫、胃底腺ポリープ、胃癌、胃の腺腫(関連する悪性腺腫のない)、胃腸のカルチノイド腫瘍、esophageal悪性腺腫; Wilmの膵臓(非ductal腺房細胞癌)の少年のnasopharyngeal angiofibromas、黒色腫、pilamatricomas、肺の悪性腺腫、卵巣癌、子宮頚部、子宮のendometrial、胸のfibromatoses、前立腺、甲状腺癌、hepatoblastoma、肝細胞癌、C型肝炎に関連している肝細胞癌、髄芽細胞腫、desmoid腫瘍、腫瘍(腎臓)、pancreatoblastoma、及び滑膜肉腫等である。
【0038】
骨又は関節の疾患の好ましいタイプ(類型)は、それぞれ変形性関節症、又はリューマチ関節炎である。
【0039】
さらに本発明は、前記抗体若しくは前記siRNAからなる医薬組成物に関する。
【0040】
さらに本発明は、増加されたか、減少されたか、3LS表現ではないか、若しくは、変異された3LS表現の(ポリ)ペプチドが少なくとも1つの組織若しくは器官に示される、
好ましくはヒトセルラインシステムであるセルラインシステムか、ショウジョウバエ、マウス、ラット、うさぎ、鶏、蛙、豚、羊、C.エレガント等のワーム、ゼブラフィッシュ等の魚、からなる群から選択される脊椎動物若しくは無脊椎動物等の有機体か、の利用を含む薬剤のスクリーニング、又は改変されたWnt(ウィント)分泌物による誘起された疾患のための検査に関するものである。
【0041】
その検査の好ましい実施形態において、前記器官は、トランス遺伝子として配列表の配列番号1〜3、7に記載された核酸分子の少なくとも1つからなる、3LS遺伝子を表現(発現)するものである。
【0042】
その検査の他の好ましい実施形態において、前記3LS遺伝子が、欠失、点変異、挿入、及び逆位からなる群から選択される変異からなるものである。
【0043】
さらに本発明は、次の工程からなる、細胞のWnt(ウィント)分泌物の改変のための
方法である。細胞を、Wntタンパク質の分泌を抑制し、若しくは促進する物質と接触させる。
【0044】
その方法の好ましい実施形態において、前記物質は、前記抗体若しくは前記siRNA、a)若しくはb)工程の核酸分子、c)工程の断片、d)工程の誘導体、若しくはe)工程の核酸分子、断片、若しくは誘導体、又はa)工程に記載の(ポリ)ペプチド、b)工程の(ポリ)ペプチドの断片、若しくはc)工程の誘導体、又はd)工程の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体である。
【0045】
本発明は、細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、のa)若しくはb)工程に記載の核酸分子、c)工程に記載の断片、d)工程に記載の誘導体、若しくはe)工程に記載の核酸分子、断片、若しくは誘導体、又は抗体若しくはsiRNAの使用に関する。
【0046】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、a)工程に記載の(ポリ)ペプチド、b)工程に記載の(ポリ)ペプチドの断片、c)工程に記載の誘導体、若しくはd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の使用に関する。
【図面の簡単な説明】
【0047】
【図1a】CustalWタンパク質配列は、ショウジョウバエの3L3タンパク質と、C.エレガンスと、ヒトと間の高い相同性を示す。 推定のシグナル配列は、配列上に線によって示され、経膜的なドメインは、下線が付される(TMHMM予測)。
【図1b】図1aと同じである。
【図1c】図1aと同じである。
【図2】A) コントロールとしての正常成人pharates(ファラテス)。 B) 知覚力のその触鬚を見せる3L3のための成人pharates homozygous変異体は、すべての3脚の組のrudimental発達である。 C) 知覚力のその触鬚を見せる3L3のための成人pharates homozygous変異体は、すべての3脚の組のrudimental発達と、翼から背板への変化(5-10 %の場合に観測される)。germlineクローン表現型を示す胚。 D)及びE)、: 正常な分割パターンを示す野生型状態。 F)及びG) 及びセグメント(「小歯の芝生」表現型)のWg機能の損失表現型を示す3L3のための胚の変異体。 これらの3L3の突然変異の胚が、セグメントの融合や、典型的なWg又はHhの機能のロス(図2のD−G)を示すことが観察された。この結果からは、3L3が、Wg/Wnt又はHhのシグナル経路の決定的な成分であることが導かれる。
【図3】A) 野生型ディスクの末端の、より少ないLacZ表現。 B) Distalless-LacZ表現は3L3変異体組織で抑えられる。 E) Bと同じであることで、クローンマージンだけが示されます。 D) 野生型ディスクの無意味なタンパク質表現。 C) 無意味なタンパク質表現は3L3変異体で抑圧されます。 F) Bと同じであることで、クローンマージンだけが示される。
【図4】A) 野生型ディスクのWgタンパク質表現。 B) これらの変異細胞の中では、Wgが蓄積するのを示して、Wgタンパク質表現は3L3変異体組織で増加される。 C) Bと同程度、クローンマージンだけが示される。 D) 細胞外 Wgタンパク質発現はないか、または3L3変異体組織で少し増加されているだけである。B-Cで見られたWgの蓄積が主に細胞内であることが示唆されている。 E) 同様に、Bとして、クローンマージンだけが示される。
【図5】TOP Flash Luciferaseを含む。 Wnt3A-V5とsiRNA−h3L3 cotransfectedセルは見せかけのコントロールとしてのWntのシグナルの依存するLuciferaseレポーターの遺伝子の誘導を全く示さない。(空のベクトル(pcDNA3)はsiRNA#GFPと共に遺伝子でcotransfectedされた)。 しかしながら、Wnt3A-V5とsiRNA#GFP cotransfectedセルはaを示した。 レポーターの遺伝子の2倍の誘導は見せかけのコントロールと比較された。
【図6】共同免疫沈降反応、IP: α-HA、 α-V5h3L3-HAはWnt3A-V5(レーン1)。 負の制御: CD2-HAとWnt3A-V5(レーン2)の間で相互作用を全く観測することができない。そして、EGFP transfectedセル(レーン3)の中にバンドを全く検出することができない。
【図7】3L3のC..エレガンス相同染色体に対するsiRNA。 A) a wt虫3L3に対するsiRNAの性腺は性腺(B)と、欠陥への末端のチップ細胞移動(C)におけるOocytesの異常なアレンジメントに通ずる。
【図8】TOP Flash Renilla Luciferaseを含む。 GFP RNAiと比べて、WLS RNAiは明確なWgの分泌を示した。
【図9A−B】Wnt3aの分泌はhWLSのノックダウンで損なわれる。 A) HEK-293Tセルについて表面に浮かぶWnt3a-V5のレベルはhWLSの減少で強く減少する。 B) sihWLSはCOS-7セルの媒体の中にWntl-HAの分泌を撤廃する。
【図9C】siGFPと比べて、sihWLSのアプリケーションはWnt5a-HAの分泌を強く減少させる。
【図10】B-I) HEK-293Tセルのセルの表面stainings。 Wnt3a-V5はsihWLS(B)と共に共同transfectedされたセルのセルの表面に達することができない。siGFPのcotransfectionはセルの表面(C)でWnt3a-V5の検出可能なレベルに通じる。 どちらもsihWLS(D)と共に治療されたセルの表面で検出可能ではない。Wnt3aC77A-V5、または、siGFP(E)。 (F-I) CD2-HAが検出されていない間、HA-CD2はセルの表面(F、G)に検出される(H、I); 従来の汚損(目立たない)で容易にCD2-HAを検出することができる。 HA-CD2の表面レベルはsihWLSによって影響されない。
【図11】C.エレガンス WLS ortholog表現型模写に対してRNAi ABarは母性のオリエンテーション欠陥を紡錘形にする。そして、母性−3変異体肺は左と腹に前方で見せられる。 (A) 野生型肺、ABarスピンドル(黄色)はABprのスピンドルに垂直であって(青い)、すなわち、見せられた光学セクションに垂直な状態で(黄色いドットで、示される)適応する。 (B) RNAi(母性−2)の注入された動物、2個のスピンドルでは、オリエンテーションが平行となる。 (C、D) RNAi(R06B9.6)肺(D)のABarとABprスピンドルと同様に初期の母性−3(or78)変異体段階(C)に、オリエンテーションが平行にある。
【図12】Wnt3aの分泌はhWLSのノックダウンで損なわれる。 A) コントロール(siGFP)と比べて、ShhとsihWLSと共にcotransfectedされたHEK-293Tセルのライセートと培地はShhの生産と分泌における変更を全く示さない。 B) WLSの減少でSSHA-dTNFΔN(TNF-FLAG)の分泌は影響されない。
【発明を実施するための最良の形態】
【0048】
Wnt(ウィント)シグナリングカスケードは、脊椎動物及び無脊椎動物の双方の発生のために重要であり、腫瘍形成に影響を与えてきた。ショウジョウバエのWnt遺伝子は、百以上の遺伝子を含んでいる、Wntタンパクファミリーのなかでも最も特徴的なものの1つである。ショウジョウバエの胚において、Wgは、parasegment boundaries(パラセグメント境界)の形成、及び近接した細胞におけるengrailed(ed)(エングレイルド)の発現の維持に必要である。Wg機能における胚の表皮の欠陥は、不全の分割しか示さない。それは、異常な表皮のパターンに反映される。野性型幼虫の腹の表皮が、裸の領域と交替する歯状ベルトを示している間、Wg変異体幼虫の表皮は歯状物で完全に覆われている。
【0049】
成虫のディスクの発育の間、Wgは、dorso-vental(dorso腹)の位置の情報を制御する。脚のディスクでは、Wgは、腹の運命(Struhl and Basler 1993)の誘導によって、今後の脚をパター(patter)する。Wgの活性が減退している動物においては、腹の脚の半分は、背面のミラーイメージ(mirror image)(ベイカー1988)内で発育する。従って、減退したWgの活性は、notal組織への翼の変化につながり、結果として遺伝子の名前(Sharma and Chopra 1976).につながっている。目のディスクでは、Wgは、頭部の表皮の発育のために、ommatidial分化を抑制し、目の領域を横切って、dorso腹の軸を設立するのにかかわる(Heberlein、Borod et al. 1998)。
【0050】
付加的な遺伝子は、Wgシグナリングの分泌、受容、又は解釈に関係してきた。たとえばショウジョウバエの遺伝学的研究は、Wgシグナリングにおけるfrizzled (Dfz)、 Dishevelled (dsh)、shaggy/zeste-white-3 (sgg/zw-3)、armadillo (arm)、adenomatous polyposis coli (E-apc), axin(アキシン)、及び pangolin (pan)との関わりを示した。これらのトランスデューサー(transducer)の遺伝的な指令は、WgがDshを通じてSggを抑制する作用をし、従って、SggによるArmの抑圧を和らげ、その結果、Armの細胞質での蓄積、及びその核への移行をもたらす。核内では、Armは、標的遺伝子の転写を活性化するために、Panと相互作用をする。脊椎動物の相同染色体は、すべてのこれらの成分のために同定され(最新の論評を参照のこと(Peifer and Polakis 2000))、ショウジョウバエのシグナル経路の、新規な同定されたメンバーが、脊椎動物の相対物を持つかもしれないことを示唆した。
【0051】
Arm、β−カテニンの哺乳類の相同染色体の核への蓄積、及びWg/Wnt経路の重要な活性化に導かれた変異は、直腸癌、乳癌、メラノーマ(黒色腫)、肝細胞癌、卵巣癌、子宮内膜癌、髄芽細胞腫、毛基質細胞腫、及び前立腺癌を含む、多くの種類の癌に観測された。現在では、Wntシグナリングの統制解除が、これらの悪性腫瘍に発生において重要な事象であることが明らかとなっている。しかしながら、Wnt経路を効果的に抑制する治療薬が未だ知られていないばかりか、β−カテニンの転写の活性化を直接的に抑制する治療薬、或いはWntリガンドによる経路の活性化を抑制する治療薬すらも知られていない。これは、十分な活性が要求される多くの特殊な成分が、未だ知られていないという事実に基づくものである。
【0052】
Winglessの活性化のために要求される新しい成分を同定するために、本発明者等はショウジョウバエ遺伝子の手法を用い、ショウジョウバエのメラノガスター(melanogaster)のsev-wg表現型の劣性のサプレッサーのためのスクリーンを行った。この手法において、3L3として下記に引用されているタンパク質が見出された。このタンパク質は、遺伝子CG6210によってコードされている。前記遺伝子は、すべての後生動物において相同物を有し、さらに興味深いことには、それぞれの種において、十中八九は、このファミリーの1つの遺伝子しか存在しない。遺伝学的手段により、本発明者等は、Wg/Wntシグナル経路の決定的な成分としての3L3の役割を確信し、特にWntタンパク質の分泌物における、その重要
性を確信した。さらに、3L3とWntが物理的に相互作用することを、鋭意研究によって見出した。
【0053】
sev-wg表現型の劣性のサプレッサーのためのスクリーンでは、本発明者等は、3L3の近縁種の群から3つの異なる対立遺伝子を見出した。2つの対立遺伝子が、今までのところ、特徴的でない遺伝子CG6210の変異を有している。CG6210は、N末端側がシグナル配列で構成され、推定で7個の経膜的なドメインを有するタンパク質をコードする。3L3の近縁種の群の1つの対立遺伝子であるsu20.53は、シグナル配列の最後のアミノ酸残基と、時期尚早の終止コドンにおけるフレームシフトに導かれる、47bp欠失を含んでいる。他のサプレッサーsu20.54は、最初の経膜的なドメインにおいて、プロリンをセリンに変換するアミノ酸残基に導く点変異を有する。異なる生命情報工学プログラムは、このタンパク質が、知られていない機能のタンパク質のファミリーに属し、また興味深いことに、種ごとには、常にこのファミリーの1つのタンパク質しか存在しないという結論を導いた。さらに、そのタンパク質は、異なる種において非常にうまく保存されており、特に、細胞外での数種の配列や、経膜的で橋架けされたドメイン(図1a−c)の機能が特殊であり、且つ保存されていることを示している。
【0054】
ショウジョウバエの胚は、数種の遺伝子の局在化された活性によって、反復セグメントへ、累進的に再分化される。Wingless(Wg)及びHedgehog(Hh)は、いわゆるセグメント極性遺伝子グループの最も重要な遺伝子であり、A/P軸を、セグメントを未だ形成しておらず将来セグメントとなる、14−15のストライプ(stripes)へ均等に分配する。結果として、Wg又はHhの機能のロスは、胚の腹部の横の反復された外骨格構造のフォーメーションの分裂を生じさせる。これは、セグメント極性遺伝子の活性の、最も明白な表現型の結論である。
【0055】
「germline clones(微生物ラインクローン)」と称される遺伝的な実験は、(3L3の場合のように)興味あるタンパク質が完全に欠乏しているショウジョウバエの突然変異の胚を、通常のものにすることを発明者に許容し、そして分割のパターンを研究することを許容する。特に「germline clones(微生物ラインクローン)」は、多くの遺伝子のために、胚にときどき存在する母性の成分を除去するのに必要である。
【0056】
これらの3L3の突然変異の胚が、セグメントの融合や、典型的なWg又はHhの機能のロス(図2のD−G)を示すことが観察された。この結果からは、3L3が、Wg/Wnt又はHhのシグナル経路の決定的な成分であることが導かれる。
【0057】
また、Wgは、触覚、脚、羽根等の、ショウジョウバエの成虫の付属肢の発育に中枢的な役割を果たす。それ故に、発明者等は、孵化前に飛ぶことを意味する成虫のファラテス(pharates)の3L3の突然変異の表現型を研究した。発育がWgに依存するすべての付属肢が異常であることが観察された(図2A−C)。特に、触覚の端刺が見当たらず、脚が短く、分割の不足が認められ、いわゆる「羽根から胸背板」の5%に変形が観察された。この変形は、Wgの機能のロスのために、病徴的なものである。これらが見出されたことで、
3L3がWg/Wntシグナル経路における決定的な成分であるという明白な結論が得られる。
【0058】
たとえば羽根や脚のような成虫の付属肢は、"imaginal discs(イマジナルディスク、成虫ディスク)"と称されるsack-like 上皮において組織(器官)形成されたimaginal細胞によって幼虫において形成される。特に、羽根のimaginal discは、それが胚の発育過程で形成され、幼虫の時期に75000個の細胞のディスクを一般化するために、3つの幼虫の齢の間、増殖するとき、20個の細胞の周囲に配置されている。これらのimaginal discs(イマジナルディスク)において、Wingless(ウイングレス)は、空間的濃度が変化し、細胞が異なる閾値の濃度において、異なって応答するモルフォゲンとしてふるまう。
【0059】
本発明者等は、たとえばWgによって活性化される遺伝子のような標的遺伝子の抑圧上の3L3突然変異組織の効果の研究するために、羽根のimaginal discs(イマジナルディスク)を実験において用いた。2種の標的遺伝子は、羽根のimaginal discs(イマジナルディスク)において区別される。一方、低いWg濃度でも発現し、それ故にWgの根源から数種の細胞直径距離で発現するDistallessのような「長い範囲の標的遺伝子」がある。他方、
発現のためにモルフォゲンの高いレベルを必要とする(Senseless)のような、「短い範囲の標的遺伝子」と称される標的遺伝子がある。
【0060】
羽根のimaginal disc(イマジナルディスク)において、3L3の突然変異のクローンの形態を含んだ後、長い範囲の標的遺伝子のためのマーカーとしてのDistalless-lacZの発現及び
短い範囲の標的遺伝子(図3A−F)のためのマーカーとしてのSenselessの発現が研究された。この実験において、Distalless-acZ及びSenselessの発現の減少が観察されたが、突然変異の組織が、Wgの産生細胞を含んだ。これらの結果により、3L3が、Wgのモルフォゲンの産生において、十中八九、重要な役割を果たすことが明白である。
【0061】
Wg産生における3L3の重要性を確かめるために、我々は、Wg産生細胞において、3L3のみを選択的に発現する昆虫を釈放することを試みた。動物の休息は、3L3のための相同的な突然変異であった。その結果は、完全な釈放であり、3L3の機能が、Wg産生細胞においてのみ必要であるという結論に言及する。
【0062】
さらに、羽根のimaginal discs(イマジナルディスク)におけるWgタンパク質の発現での3L3突然変異組織の効果が研究された(図4A−E)。3L3突然変異クローンにおいて、Wgの発現量に増加があることが観察された。この結果を良好に特徴づけるために、本発明者等は、類似の実験を遂行し、WgのmRNAの転写のレベルを現すwg-lacZの発現を見た。3L3突然変異組織におけるwg-lacZの発現の増加が全くないという事実は、突然変異組織におけるWgのための、増加するstainingが、Post−転写過程に基づくという推定を正当化する。
【0063】
最後に、Wgの蓄積が細胞内又は細胞外で生じさせるか、区別するために、本発明者等は細胞外タンパク質の検出を許容する異なったプロトコル(protocol)を用いてWgの分配(distribution)を試験した。しかし、3L3突然変異クローンにおける細胞外Wgの蓄積は、検出されなかった。これらの結果は、その機能のロスが、Wg産生細胞の内側でのWgの蓄積を導くので、Wgの分泌における3L3のための役割を示唆する。
【0064】
Wntシグナリングにおいて、3L3のヒト相同染色体が含まれているか否かを試験するために、Wntレポーター(reporter)遺伝子assay(検査)(TOP-flash)が行われた。これは、luciferase(ルシフェラーゼ)遺伝子の前に位置して結合している5TCFを伴うassay(検査)に基礎をおくluciferase(ルシフェラーゼ)である。Wntシグナル経路は、luciferase(ルシフェラーゼ)の蛍光によって測定することができる。以前に(preceding)議論した結果から、ショウジョウバエのWnt産生細胞において3L3が重要であることが知られていた。
【0065】
Wntの産生及び受容細胞を擬態するために、1バッチのヒト293T細胞が、h3L3に対向するmWnt3A−V5及びsiRNAでトランスフェクト(Transfect)された。別のバッチの細胞は、TOP-flash reporter constructでトランスフェクトされた。トランスフェクトがされた後、24時間経過した時、これらの2つの細胞のバッチが混合され、さらに24時間培養した後、luciferase(ルシフェラーゼ)活性が測定された。もし3L3に対向するsiRNAが、「産生」細胞においてWnt3Aでコトランスフェクトされたならば、luciferase(ルシフェラーゼ)活性のレベルは、もし空のベクトル(Wnt3Aの代用の)及びGFPに対向するsiRNAがトランスフェクトされたならば、同程度に低かった。他方、もしGFPに対向するWnt3A及びsiRNAがトランスフェクトされたならば、レポーター遺伝子の明確な誘起が観察された(図5)。これらの結果は、ヒト(h)3L3が、シグナルの受容ではなく、産生において含まれたWnt経路の決定的な成分であることを示している。
【0066】
3L3は、Wnt産生細胞において重要であるので、h3L3−HAがマウス(m)のWnt3A−V5と相互作用すると、調べられた。Co-immunoprecipitation 試験が行われ、その結果は、h3L3−HAがmWnt3A−V5と相互作用することを示唆し、CD2−HAを伴うネガティブコントロールや、他の膜−スパニング(membrane-spanning)タンパク質は、mWnt3A−V5と相互作用しなかった(図6)。
【0067】
ショウジョウバエの羽根ディスク(discs)におけるwntless(WLS)突然変異細胞のクローンにおいて、Wg産生細胞におけるWgの蓄積が観察された。Kc細胞において、我々は、培地へのWgの分泌物を測定するための検査(assay)を開発した。Renilla luciferase
遺伝子は、WgのN末端に融合し、Kc細胞へトランスフェクトした。同時に、その細胞は、WLS又はGFPのdsRNAで扱われた。WLSのRNAiは、GFPのRNAiと比べてWg分泌物の明らかな下降調節を示した(図8)。
【0068】
この分泌物の欠乏が、RNAiによるWLSのヒトの組織培養実験デプレション(depletion)において、Wntシグナルカスケードを活性化することができない結果となった理由であったかどうかを決定するために、我々は、Wnt分泌細胞の培地を分析した。HEK293Tは、siRNAのみならず、Wnt3A−V5、Wnt1−HA、又はWnt5a−HAでもコトランスフェクトした。すべてのこれらのWnt分泌物は、WLSに対するsiRNAがGFPに対するsiRNAと比較してコトランスフェクトされたとき、かなり減少した。これらの結果は、WLSが消耗されたとき、HEK293細胞において、Wnt分泌物が大幅に減少することを示すものである。(図9;2つの異なるsiRNA(siRNAhWLS−A,siRNAhWLS−B)が、HEK293細胞におけるendogenoushWLS遺伝子の発現をノックダウンするのに効果的なことのために、RT−PCRによって有効にさせた。双方の独立したトランスフェクションは、結果として、hWLSの転写の85%減少をさせた(示されていない;siRNAhWLS−Bは、ほとんど用いられた。そして、sihWLSとして言及されている。しかし、siRNAhWLS−Aは、同じ効果を示した。)。sihWLSを持つレスポンダー細胞の取り扱い、又はGFPに対するsiRNAを持つプロデューサー細胞の取り扱いは、Wntシグナリング検査(assay)の結果にさほど影響を与えなかった。
【0069】
WLSが下降調節されるとき、HEK293T細胞において、WNT3aが細胞外表面に到達することができるか否かを試験するために、我々は、siRNA及びWnt3A−V5をコトランスフェクトし、Wnt3aの細胞外分裂のみをステイン(stain)した。コントロールとして、我々は、N末端(細胞外)又はC末端(細胞内)の一方にHA−tagを有するCD2を用いた。我々が用いた細胞外 staining protocolで、我々は、N末端のHA−tagで、CD2のみを欠乏させることができた。WLSに対するRNAiは、細胞外表面にCD2の分泌物の影響を与えなかった。対照的に、Wnt3a−V5の表面のstainingは、WLSに対するsiRNAでコトランスフェクトされた細胞において破壊された。それ故に、我々は、細胞表面へのWNT3a(及びおそらくは他のWntも同様に)の好適な分泌のためにWLSが必要であることを結論づける(図10)。
【0070】
ショウジョウバエの羽根のディスクにおいて、処分されたクローンは、Hhリガンドの明らかな蓄積を示したが、明確な対比で、WLSのクローンにおいては、Hhの蓄積は観察することができなかった。さらに、WLSの突然変異の昆虫は、wg−Ga14::UAS−WLS構造で釈放された。wg産生細胞におけるWLSのみの発現は、致死性を救済し、WLSがWgシグナリングのために特異的(specific)であるという我々の見解を支持するのに十分であった。
【0071】
ヒト組織の培養において、Wntの分泌物のみが影響を受けたが、上述のように、CD2の分泌物は、WLS RNAiによって影響を受けなかった。重要なことは、Shhの分泌物のみならず、培地へのTNFの分泌物も、WLS RNAiによって影響を受けなかった(図12)。これらの結果は、WLSのヒトのorthologueが、Wntシグナリングのために特異的なものであることを示す。
【0072】
我々の結果は、また、Wntの小さなサブセットがWLSの機能のロスに敏感であるばかりでなく、すべてのWntがWLSから独立しているわけではないことを示している。C.エレガンスにおいて、MOM−3は、初期のblastomere polarization (Rocheleau, Thorpe)、VPC specification (Eisenmann and Kim)、及びQ neuroblast migration (Harris)のような、多くの異なる標準的及び非標準的なWnt経路に含まれていることが示されてきた。これらの過程は、3つの異なるWnt(経路)、MOM−2(Rochelau, Thorpe)、LIN−44
(Jiang and Sternberg)及びEGL20(Harris)に依存する。
【0073】
C.エレガンスにおいては、異なる発育過程においてpostembryonically活性である標準的経路と、胚の発育においてのみ観察することができる非標準的経路との、2つのWntシグナル経路が区別されることができる。非標準的Wntシグナリングは、EMSの決定のorientation及びendodermの運命を調節する(EMS細胞は、将来のendoderm及びmesodermのprecursor 細胞のprecursorである)。非標準的Wntシグナリングのロスは、endodermのロスや、mesodermの発育を導く。この非標準的Wntシグナリングの数種の成分は、記載されてきた。それらの1つであるmom−3(more mesoderm-3)は、geneticallyに特徴づけられてきたが、molecularlyには、so far同定されていない(Eisenmann and Kim 2000; Thorpe, Schlesinger et al. 1997).。配列によると、3L3(c3L3)の相同染色体における変異は、異なるmom−3のallelesにおいて見出された。さらに、c3L3に対するRNAiは、
【0074】
distal tip細胞移動における欠乏や、胚のlethality.のような標準的及び非標準的なWnt表現型の典型的なロスを導いた。また、これらの結果は、c3L3がmom−3の表現型のために応答可能であること、及びc3L3に対するRNAiは、mom−3の表現型のロスを擬態することを示している。
【0075】
C.エレガンスにおけるWntシグナリングは、多くの異なる発育過程において含まれている。知られている最初の過程は、mitotic spindlesのMOM−2の発育オリエンテーションであり、胚細胞は4個から8個に分割される。mom−3ワームは、wntless orthologue R06B9.6における突然変異であるので、我々はR06B9.6のdsRNAを、wthermaphroditesの生殖線に注入した。上述のように、分析されたすべての胚には、R06B9.6のdsRNAが注入され、或いはmom−2又はmom−3の突然変異alleleの運搬は、Abar spindle から ABpr spindleへの平行オリエンテーションを示した。対照的に、Wnt胚は、垂直に定位された2つのスピンドル(spindle)を有していた(図11)。
【0076】
要約すると、本発明者等は、遺伝子CG6210によってコードされる3L3と名付けられたリペプチドを見出した。遺伝的手法により、3L3が、Wg/Wntシグナル経路、特に、Wntタンパク質のWg/Wnt分泌経路において、役割を果たしていることが確かめられた。さらに、3L3は、物理的にWntタンパク質と相互作用することが示された。それ故、3L3タンパク質は、Wntリガンドの分泌物を調節することによって、Wnt経路を正又は負に調節する薬剤の開発のために、非常にpromisingな標的である。
【実施例】
【0077】
(実施例1)
[sev−wg表現型の受容サプレッサーのためのスクリーン]
目において、Wgを発現するsev−wgトランス遺伝子は、スクリーンにおいて、表現型マーカー(Brunner et al., 1997)として観察される、ラフな目の表現型に導く。左の腕でFRT80を運び、右の腕で3番目の染色体のsev−wgを運ぶ雄は、12時間の間、21mMのエチルメタンスルフォネート(EMS)で養育された。EMSを適用させた後、24時間経過後に、雄は、X染色体上のeyeless-flp recombinaseである雌と交尾し、3番目の染色体上に、FRT80Mw+を運搬した。ey-flpと両方のFRT染色体でoffspringされた雄は、ラフな目の表現型の抑圧のためにスクリーンされた。異なる対立遺伝子間でのComplementation分析は、2つのallelesを構成するグループ3L3の決定に導いた。
with the ey-flp and both FRT chromosomes were screened for a suppression of the rough eye phenotype. Complementation analysis between different alleles led to the complementation group 3L3 consisting of two alleles.
【0078】
(実施例2)
[すべてのメタゾンにおける相同染色体]
Blast search (blastp)によって、すべての配列のメタゾン有機体において、知られていない機能1171(DUF1171)のドメインを運搬する、この新しいタンパク質ファミリーの1つのコピーのみが存在した。
【0079】
(実施例3)
[ドメインspanningする膜]
TMHMM TOPは、1つのN末端シグナル配列と、CG6210−PBのための7つの推定の経膜的なドメインを予想した。
【0080】
(実施例4)
[Germline クローン]
興味あるタンパク質の完全な不在において、ショウジョウバエの胚の表現型であるメラノガスターの研究に用いられる実験的なprocedureである。
3番目のinstar larvaeをFRTの1つの染色体上に運搬し、推定の変異を同じFRTの相同染色体上及びovoD 変異に運搬する雌は、1.5時間、38℃で熱ショックを受けた。ハッチングの後、雄が同じ推定の変異及びFRTを運搬し、交尾した。
【0081】
雌は、寒天プレート上で、1夜、産卵し、胚が集積された。これを3分間、ブリーチし、タップ(tap)水で洗浄した。胚は、24時間、水に注入された。vitellumを遊離させるために、メタノールとヘプタンを等量ずつ含むbiphasic溶液で浸とうした。胚は、メタノールで4回洗浄され、0.1%のトリトンX−100で4回洗浄された。RTで6時間培養した後、最後に胚をHoyer-lactateでスライド上に集積し、60℃でダイジェストタンパク質に12−24JIKANN培養した。表皮は光学顕微鏡で試験された。
【0082】
(実施例5)
[標的遺伝子の発現に、3L3突然変異組織の効果を研究するための、羽根イマジナルディスク]
イマジナルディスクは、metamorphosisの間、成虫の構造を作る細胞のhollow sacsである。
これらは、胚ectodermにおけるポケットとして上昇し、幼虫がさなぎになるまで体コウの内側に成長し、そのポイントで体壁と付属物を形成するために、「"evaginate"」を内側に戻す。
【0083】
[イマジナルディスクの抗体staining]
幼虫は、氷の上で、リンガー(Ringer)溶液中でdissectedされ、PEM 200μlで200μl管に固定された。5%のホルムアルデヒド 10.7μlと、0.05%のトリトンX−100とを20分間攪拌した。4回洗浄し、1時間追加し、2−3回、PEJTとNa−Azidを加えて培養した。一次抗体(PBTとNa−Azidでdilutedしたもの)で一昼夜、4℃でインキュベートした。Larvaeは、PBT、Na−Azid、1%のHINGSで30分、5回洗浄し、RTに、PBTとNa−Azidでdilutedした二次抗体で、2時間インキュベートした。Larvaeは、2時間で5回PBTとNa−Azidで洗浄し、ディスクはPPDAに集積された。羽根のディスクにおけるStainingは、焦点顕微鏡で分析された。
【0084】
(実施例6)
[羽根のイマジナルディスクにおける3L3突然変異クローンのフォーメーションの誘起、及びDistalless-lacZ (wg-lacZ)の発現の研究]
羽根のイマジナルディスクにおける突然変異クローンは、興味あるタンパク質のための変異である細胞のグループをrepresentする。ショウジョウバエのメラノガスター(melanogaster)において、熱ショックプロモーター(hs- flp)によって操作される、yeast-specific recombinase flippase (flp)を用いて、mitotic recombinationによって遂行される。
【0085】
y w hsp-flp; allele 20.53 FRT80 / TM6b fliesは、y w hsp-flp; πMyc [w+] Minute Pfy+J FRT80 / TM6b,によって交尾され、雌は、2日間、産卵する。
37.0℃で60分間の熱ショックで、24−72時間、AEFが形成され、さらに4時間後に、幼虫が切開された。
【0086】
Distalless-lacZの場合においては、一次抗体にβ−ガラクトシダーゼ(1:2000ラビット、ポリクローナル抗体、Cappel(登録商標))を用いた。
wg-lacZの場合においては、抗体に、α−Wg(4D4)(1:1000マウス、モノクローナル抗体、DSHB)を用いた。
【0087】
(実施例7)
[Wg産生細胞において3L3のみを発現する羽根をrescueするのを試みる手段としての、Wg生産物における3L3の重要性の確立]
トランス遺伝子wg-GAL4 (the transcriptional activator GAL4 は、Wgを発現する細胞内でのみ発現される。) で運搬される3L3heterozygous mutant fliesは、3L3heterozygous mutantであるfliesで交尾され、トランス遺伝子UAS−3L3(GAL4タンパク質のためのUAS結合サイトは、3L3の転写を活性化する。)
【0088】
(実施例8)
[Wg分割の試験]
このプロトコル(protocol)は、一次抗体でのstainingが、PBS中で4℃(endocytosisをブロックするために)で30−60分間行われ、固定が、一次抗体でのstaining後、PBS、4%のホルムアルデヒドで、4℃で20−30分間(Strigini and Cohen, 2000)行われるという事実において、通常のprotocolとは異なる細胞外Wgを見るために用いた。
【0089】
(実施例9)
[TOPフラッシュアッセイ]
細胞は、リン酸カルシウム法によってトランスフェクトした。トランスフェクト後、16時間細胞を洗浄し、24時間混合した。混合後、細胞はPromega からIxPLB でlysed し、分析した。
【0090】
(実施例10)
[Co-immunoprecipitation]
細胞は、リン酸カルシウム法によってトランスフェクトした。トランスフェクト後、36時間、細胞はRIPAバッファーによって、40℃で1時間30分間、lysedした。Lysateは、4℃で30分間、centrifugedされ、その後、タンパク質GSepharoseビーズ及びラビットα−HAで一昼夜インキュベートした。ビーズは、TBSで4回洗浄し、タンパク質は、95℃で10分間、ビーズからSDS−Loadingバッファーでelutedした。マウスα−V5抗体でウエスタンブロットを行った。
【0091】
(コンストラクト)
mWnt3A cDNA配列は、V5−tag及びHis−tagによってpcDNA3にクローンされた。h3LA cDNAは、HA−tagによってpcDNA3にクローンされた。QiagenCD2によって一般化された3L3及びGFPに対するsiRNAは、C末端HA−tagでpcDNA3にクローンされた。
【0092】
(参考文献)
Baker, N. E. (1988). 'Transcription of the segment-polarity gene wingless in the imaginal discs of Drosophila, and the phenotype of a pupal-lethal wg mutation." Development 102(3): 489-97.
Cabrera, C. V., M. C. Alonso, et al. (1987). "Phenocopies induced with antisense RNA identify the wingless gene." Cell 50(4): 659-63.
Eisenmann, D. M., Kim, S. K. (2000). "Protruding vulva mutants identify novel loci and Wnt signaling factors that function during Caenorhabditis elegans vulva development." Genetics 156(3): 1097-116.
Grosschedl R, E. Q. (1999). "Regulation of LEF-ITCF transcription factors by Wnt and other signals." Current Opinion in Cell Biology 11: 233-240.
Harris, J., Honigberg, L., Robinson, N., and Kenyon, C. (1996). Neuronal cell migration in C. elegans: regulation of Hox gene expression and cell position. Development 122, 3117-3131.
Hartmann, C. and Tabin, C. J. (2000). ,,Dual roles of Wnt signaling during chondrogenesis in the chicken limb." Development, 127(14), 3141-3159.
Heberlein, U., E. R. Borod, et al. (1998). "Dorsoventral patterning in the Drosophila retina by wingless." Development 125(4): 567-77.
Holmen, S. L. et al. (2005), ,,Essential role of beta-catenin in postnatal bone acqusition." The Journal of Biological Chemistry, 280 (22), 21162-21168.
Jiang, L. I., and Sternberg, P. W. (1999). Socket cells mediate spicule morphogenesis in Caenorhabditis elegans males. Dev Biol 211, 88-99.
Morin, P. J. (1999). "β-catenin signaling and cancer." Bioessays 21(12): 1021-30.
Nakamura, Y., Nawata, M., Wakitani, S. (2005). "Expression profiles and functional analyses of Wnt-related genes in human joint disorders". American Journal of Pathology, 167(1), 97- 105.
Nusse, R. and H. E. Varmus (1982). "Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome." Cell 31(1): 99- 109.
Perrimon, N. and A. P. Mahowald (1987). "Multiple functions of segment polarity genes in Drosophila." Dev Biol 119(2): 587-600.
Peifer, M. and P. Polakis (2000). "Wnt signaling in oncogenesis and embryogenesis--a look outside the nucleus." Science 287(5458): 1606-9.
Polakis, P., M. Hart, et al. (1999). "Defects in the regulation of β-catenin in colorectal cancer." Adv Exp Med Biol 470: 23-32.
Potter, J. D. (1999). "Colorectal cancer: molecules and populations." Journal of the National Cancer Institute 91(11): 916-32.
Rijsewijk, F., M. Schuermann, et al. (1987). 'The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless." Cell 50(4): 649-57.
Rocheleau, C. E., Downs, W. D., Lin, R., Wittmann, C, Bei, Y., Cha, Y. H., AIi, M., Priess, J. R., and MeIIo, C. C. (1997). Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 90, 707-716.
Roose, J. and H. Clevers (1999). 1TCF transcription factors: molecular switches in carcinogenesis." Biochimica et Biophysica Acta 1424 (2-3): M23-37.
Sen, M., Lauterbach, K., El-Gabalawy, H., Firestein, G. S., Corr, M. Carson, D. A. (2000). ,,Expression and function of wingless and frizzled homologs in rheumatoid arthritis." PNAS, 97 (6), 2791-2796.
Sharma, R. P. and V. L. Chopra (1976). "Effect of the Wingless (wgl) mutation on wing and haltere development in Drosophila melanogaster." Dev Biol 48(2): 461-5.
Strigini, M., and. Cohen, S.. M. (2000). Wingless gradient formation in the Drosophila wing. Curr. Biol. 10, 293-300.
Struhl, G. and K. Basler (1993). "Organizing activity of wingless protein in Drosophila." Cell 72(4): 527-40.
Tatusova TA, M. T. (1999). "Blast 2 sequences-a new tool for comparing protein and nucleotide sequences." FEMS Microbiol Lett. 174: 247-250.
Thorpe, CJ. , Schlesinger, A., et al (1997). "Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm." Cell;90(4): 695-705.
Thorpe, C. 1, Schlesinger, A., and Bowerman, B. (2000). Wnt signalling in Caenorhabditis elegans: regulating repressors and polarizing the cytoskeleton. Trends Cell Biol 10, 10-17.
Waltzer, L. and M. Bienz (1999). 'The control of β-catenin and TCF during embryonic development and cancer." Cancer & Metastasis Reviews 18(2): 231-46.
Wodarz, A. and R. Nusse (1998). "Mechanisms of Wnt signaling in development." Annual Review of Cell & Developmental Biology 14: 59-88.
【特許請求の範囲】
【請求項1】
次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法。
a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。
【請求項2】
e)工程の下での前記核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、d)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同であることを特徴とする請求項1記載の方法。
【請求項3】
次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法。
a)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドと、候補物質とを、前記(ポリ)ペプチドへの前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部を構成する、(ポリ)ペプチドの断片への候補物質の結合を許容する条件の下で、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの断片と候補物質とを接触させる工程、又は、
c)誘導体への候補物質の結合を許容する条件の下で、a)に記載の(ポリ)ペプチドの誘導体、又はb)に記載の(ポリ)ペプチドの断片の誘導体と候補物質とを接触させる工程、又は、
d)a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチドの断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも50%相同である(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体と候補物質とを接触させる工程、及び
e)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するか、又は、候補物質が、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドとWnt(ウィント)タンパク質間の結合の活性を抑制するか、若しくは活性を促進するかを、検出する。
【請求項4】
d)工程の下での前記(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同であることを特徴とする請求項3記載の方法。
【請求項5】
請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体、又はアミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドのドメイン、好ましくはWnt(ウィント)タンパク質の分泌物に含有され、若しくはWnt(ウィント)タンパク質と結合する(ポリ)ペプチドのドメインと特異的に結合する抗体。
【請求項6】
アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子の断片である標的配列、好ましくは配列表の配列番号9又は10の標的配列を有するsiRNA。
【請求項7】
薬剤としての使用のための、請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体。
【請求項8】
請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、Wnt(ウィント)シグナルに関連する疾患、たとえば癌、骨若しくは関節の疾患、若しくは発達障害の処置のための医薬準備のための使用。
【請求項9】
請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体を含む医薬組成物。
【請求項10】
増加されたか、減少されたか、3LS表現ではないか、若しくは、変異された3LS表現の(ポリ)ペプチドが少なくとも1つの組織若しくは器官に示される、
好ましくはヒトセルラインシステムであるセルラインシステムか、
ショウジョウバエ、マウス、ラット、うさぎ、鶏、蛙、豚、羊、C.エレガント等のワーム、ゼブラフィッシュ等の魚、からなる群から選択される脊椎動物若しくは無脊椎動物等の有機体か、
の利用を含む薬剤のスクリーニング、
又は
改変されたWnt(ウィント)分泌物による誘起された疾患のための検査。
【請求項11】
前記器官は、トランス遺伝子として配列表の配列番号1〜3、7に記載された核酸分子の少なくとも1つからなる、3LS遺伝子を表現することを特徴とする、請求項10記載の検査。
【請求項12】
前記3LS遺伝子が、欠失、点変異、挿入、及び逆位からなる群から選択される変異を含むことを特徴とする、請求項10記載の検査。
【請求項13】
次の工程からなる、細胞のWnt(ウィント)分泌物の改変のための方法。
細胞と、Wnt(ウィント)タンパクの分泌を抑制し若しくは促進する物質とを接触させる。
【請求項14】
前記物質が、請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体であることを特徴とする、請求項13記載の方法。
【請求項15】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項5記載の抗体若しくは請求項6記載のsiRNAの使用。
【請求項16】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、請求項3のc)工程に記載の誘導体、若しくは請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の使用。
【請求項1】
次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法。
a)塩基配列の配列表の配列番号1、2、3、又は7の核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子と、候補物質とを、前記核酸分子への前記候補物質の結合を許容する条件の下で接触させる工程、又は、
c)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部をコードする断片への候補物質の結合を許容する条件の下で、a)又はb)に記載の核酸分子の断片と候補物質とを接触させる工程、又は、
d)誘導体への候補物質の結合を許容する条件の下で、a)若しくはb)に記載の核酸分子の核酸分子の誘導体、又はc)に記載の断片の誘導体と候補物質とを接触させる工程、又は、
e)a)若しくはb)に記載の核酸分子、c)に記載の断片、若しくはd)に記載の誘導体に対して、それぞれ少なくとも50%相同である核酸分子、断片、若しくは誘導体と候補物質とを接触させる工程、
f)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するかを、検出する。
【請求項2】
e)工程の下での前記核酸分子、断片、若しくは誘導体が、a)若しくはb)に記載の核酸分子、c)に記載の断片、d)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同であることを特徴とする請求項1記載の方法。
【請求項3】
次の工程を含む、Wnt(ウィント)−ファミリータンパク質の分泌を抑制し又は促進する物質のスクリーニング方法。
a)アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドと、候補物質とを、前記(ポリ)ペプチドへの前記候補物質の結合を許容する条件の下で接触させる工程、又は、
b)Wnt(ウィント)タンパク質の分泌を生じさせる、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの一部を構成する、(ポリ)ペプチドの断片への候補物質の結合を許容する条件の下で、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドの断片と候補物質とを接触させる工程、又は、
c)誘導体への候補物質の結合を許容する条件の下で、a)に記載の(ポリ)ペプチドの誘導体、又はb)に記載の(ポリ)ペプチドの断片の誘導体と候補物質とを接触させる工程、又は、
d)a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチドの断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも50%相同である(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体と候補物質とを接触させる工程、及び
e)候補物質が、Wnt(ウィント)タンパク質の分泌の活性を抑制するか、若しくは活性を促進するか、又は、候補物質が、アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドとWnt(ウィント)タンパク質間の結合の活性を抑制するか、若しくは活性を促進するかを、検出する。
【請求項4】
d)工程の下での前記(ポリ)ペプチド、(ポリ)ペプチド断片、若しくは誘導体が、a)に記載の(ポリ)ペプチド、b)に記載の(ポリ)ペプチド断片、若しくはc)に記載の誘導体に対して、それぞれ少なくとも88%相同であり、好ましくは少なくとも89%相同であり、より好ましくは少なくとも90%相同であり、さらに好ましくは少なくとも91%相同であり、さらに好ましくは少なくとも92%相同であることを特徴とする請求項3記載の方法。
【請求項5】
請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体、又はアミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドのドメイン、好ましくはWnt(ウィント)タンパク質の分泌物に含有され、若しくはWnt(ウィント)タンパク質と結合する(ポリ)ペプチドのドメインと特異的に結合する抗体。
【請求項6】
アミノ酸配列の配列表の配列番号4、5、6、又は8の(ポリ)ペプチドをコードする核酸分子の断片である標的配列、好ましくは配列表の配列番号9又は10の標的配列を有するsiRNA。
【請求項7】
薬剤としての使用のための、請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体。
【請求項8】
請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の、Wnt(ウィント)シグナルに関連する疾患、たとえば癌、骨若しくは関節の疾患、若しくは発達障害の処置のための医薬準備のための使用。
【請求項9】
請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体を含む医薬組成物。
【請求項10】
増加されたか、減少されたか、3LS表現ではないか、若しくは、変異された3LS表現の(ポリ)ペプチドが少なくとも1つの組織若しくは器官に示される、
好ましくはヒトセルラインシステムであるセルラインシステムか、
ショウジョウバエ、マウス、ラット、うさぎ、鶏、蛙、豚、羊、C.エレガント等のワーム、ゼブラフィッシュ等の魚、からなる群から選択される脊椎動物若しくは無脊椎動物等の有機体か、
の利用を含む薬剤のスクリーニング、
又は
改変されたWnt(ウィント)分泌物による誘起された疾患のための検査。
【請求項11】
前記器官は、トランス遺伝子として配列表の配列番号1〜3、7に記載された核酸分子の少なくとも1つからなる、3LS遺伝子を表現することを特徴とする、請求項10記載の検査。
【請求項12】
前記3LS遺伝子が、欠失、点変異、挿入、及び逆位からなる群から選択される変異を含むことを特徴とする、請求項10記載の検査。
【請求項13】
次の工程からなる、細胞のWnt(ウィント)分泌物の改変のための方法。
細胞と、Wnt(ウィント)タンパクの分泌を抑制し若しくは促進する物質とを接触させる。
【請求項14】
前記物質が、請求項5記載の抗体若しくは請求項6記載のsiRNA、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、若しくは請求項3のc)工程に記載の誘導体、又は請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体であることを特徴とする、請求項13記載の方法。
【請求項15】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、請求項1のa)若しくはb)工程に記載の核酸分子、請求項1のc)工程に記載の断片、請求項1のd)工程に記載の誘導体、若しくは請求項1のe)工程に記載の核酸分子、断片、若しくは誘導体、又は請求項5記載の抗体若しくは請求項6記載のsiRNAの使用。
【請求項16】
細胞中のWnt(ウィント)シグナル経路の調節、好ましくは細胞の運命の決定を調節し、又は幹細胞の発達を調節するための、請求項3のa)工程に記載の(ポリ)ペプチド、請求項3のb)工程に記載の(ポリ)ペプチドの断片、請求項3のc)工程に記載の誘導体、若しくは請求項3のd)工程に記載の(ポリ)ペプチド、(ポリ)ペプチドの断片、若しくは誘導体の使用。
【図1a】


【図1b】

【図1c】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】
【図9A−B】
【図9C】
【図10】
【図11】

【図12】

【図1b】
【図1c】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9A−B】
【図9C】
【図10】
【図11】
【図12】
【公表番号】特表2008−537484(P2008−537484A)
【公表日】平成20年9月18日(2008.9.18)
【国際特許分類】
【出願番号】特願2008−501241(P2008−501241)
【出願日】平成18年3月20日(2006.3.20)
【国際出願番号】PCT/EP2006/002534
【国際公開番号】WO2006/097336
【国際公開日】平成18年9月21日(2006.9.21)
【出願人】(507310743)ザ ジェネティックス カンパニー,インコーポレイテッド (3)
【氏名又は名称原語表記】THE GENETICS COMPANY,INC.
【住所又は居所原語表記】Wagistrasse 27,CH−8952 Schlieren(CH)
【Fターム(参考)】
【公表日】平成20年9月18日(2008.9.18)
【国際特許分類】
【出願日】平成18年3月20日(2006.3.20)
【国際出願番号】PCT/EP2006/002534
【国際公開番号】WO2006/097336
【国際公開日】平成18年9月21日(2006.9.21)
【出願人】(507310743)ザ ジェネティックス カンパニー,インコーポレイテッド (3)
【氏名又は名称原語表記】THE GENETICS COMPANY,INC.
【住所又は居所原語表記】Wagistrasse 27,CH−8952 Schlieren(CH)
【Fターム(参考)】
[ Back to top ]