
Y群髄膜炎菌ワクチン及びそれらの髄膜炎菌組合せワクチン
本発明は、改変髄膜炎菌Y多糖体(GYMP)、改変多糖体と担体を含む複合体、Y群髄膜炎菌に対するヒトを含めた温血動物の免疫用ワクチンに関し、これらの改変多糖体、複合体及びワクチンを製造する方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、改変髄膜炎菌Y多糖体(GYMP)、改変多糖体と担体を含む複合体、Y群髄膜炎菌に対するヒトを含めた温血動物の免疫用ワクチンに関し、これらの改変多糖体、複合体及びワクチンを製造する方法に関する。
【背景技術】
【0002】
細菌性髄膜炎は、世界規模での健康に対する重大な脅威である。髄膜炎菌は、細菌性髄膜炎及び敗血症の主要な原因である。髄膜炎菌は、多糖きょう膜で被包されている。髄膜炎菌単離体は、その化学的及び抗原性的に異なる多糖きょう膜に基づいて12の群に分類することができる。これらの群のうちA、B、C、Y及びW135の5種類は、ヒトにおける細菌性髄膜炎及び敗血症の実質的にすべての症例の原因である。
【0003】
Y群髄膜炎菌によって引き起こされる細菌性髄膜炎の発生率は増加している。米国のいくつかの郡において1989年から1995年に実施された活動的な研究室によるサーベイランスによれば、血清型Y髄膜炎菌性疾患の比率は1989年の0%から1995年の32.5%に急激に増加した。この期間中、髄膜炎菌性疾患の全発生率は一定していた(Anonymous 1996, Serogroup Y meningococcal disease - Illinois, Connecticut, and selected areas, USA 1989-1996, Morb. Mortal. Rep. 45, 1010-1013)。
【0004】
Y群髄膜炎菌によって引き起こされる細菌性髄膜炎は、子供や若年成人においても起こり得るが、高齢者において発症し易い傾向にあり、細菌性髄膜炎の他の系統よりも肺炎を引き起こす可能性が高い。
【0005】
多糖体ワクチンは、いくつかの血清型の髄膜炎菌によって引き起こされる疾患を予防するために使用されてきた。しかし、これらのワクチンは、一部の人種においては髄膜炎菌によって引き起こされる疾患の予防に有効ではない。
【0006】
多糖体はT非依存性(胸腺非依存性)抗原である。多糖体は、タンパク質に対する免疫応答を誘導するときに遭遇しないいくつかの免疫学的諸特性を有する。これらの諸特性としては、免疫応答を誘導するT細胞が存在する明白な必要性がないこと、IgMの優勢、免疫後の記憶誘導(memory induction)や親和性成熟がないこと、乳児、高齢者及び免疫無防備状態の人における低免疫原性などが挙げられる。その結果、純粋な多糖体ワクチンは、これらの患者に対して有効に使用することができない。
【0007】
血清型Y多糖体は、等モルのN−アセチルノイラミン酸とD−グルコースからなり、部分的にO−アセチル化されていることが、1970年代にJenningsによって報告された(Bhattacharjee et al,. Can. J. Biochem. 54: 1-8, 1976)。
【0008】
ほとんどのY群髄膜炎菌単離体は、O−アセチル基がもっぱらそのシアル酸残基のC−7とC−9の間に分布するO−アセチル陽性(OA)多糖体を産生することが見出された。O−アセチル基分布の位置及び濃度におけるこの不均一性は、多糖複合体の処方を複雑にしている。
【非特許文献1】Anonymous 1996, Serogroup Y meningococcal disease - Illinois, Connecticut, and selected areas, USA 1989-1996, Morb. Mortal. Rep. 45, 1010-1013
【非特許文献2】Bhattacharjee et al,. Can. J. Biochem. 54: 1-8, 1976
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の一目的は、Y群髄膜炎菌に対する免疫化への使用に適切なワクチンを提供することである。
【課題を解決するための手段】
【0010】
本発明は、少なくとも1個の改変髄膜炎菌Y多糖体又は改変髄膜炎菌Y多糖体の断片又は髄膜炎菌Y多糖体の断片を含むワクチンを提供する。以下の文において、「髄膜炎菌Y多糖体」という用語及びその語形(forms)は、改変髄膜炎菌Y多糖体、改変髄膜炎菌Y多糖体の断片及び髄膜炎菌Y多糖体の断片を含む。これらの多糖体は単独で使用して、T非依存性応答を誘発する抗原組成物を産生することができる。
【0011】
改変髄膜炎菌Y多糖体は、一般に、T依存性免疫応答を誘発するために担体タンパク質と複合化される。したがって、本発明は、改変髄膜炎菌Y多糖体と担体タンパク質を含む複合材料及びその複合材料を製造する方法も提供する。
【0012】
本発明は、改変髄膜炎菌Y多糖体を製造する方法も提供する。髄膜炎菌Y多糖体は、そのシアル酸残基のC−7とC−9の間にもっぱら分布するO−アセチル基を含有するので、脱O−アセチル化によって改変することができる。したがって、本発明は、髄膜炎菌Y多糖体の脱O−アセチル化方法も提供する。一実施形態においては、塩基加水分解を使用してこの多糖体を脱O−アセチル化することができる。
【0013】
本発明は、脱O−アセチル化髄膜炎菌Y多糖体又はO−アセチル化髄膜炎菌Y多糖体を断片化する方法も提供する。一実施形態においては、弱酸加水分解を使用して多糖体を断片化する。多糖体のグリコシド結合を切断する他の方法としては、オゾン分解、超音波処理、塩基加水分解などが挙げられる。
【発明を実施するための最良の形態】
【0014】
本発明によれば、髄膜炎菌Y多糖体は、塩基加水分解によって脱O−アセチル化することができる。塩基加水分解は、多糖体の溶液を塩基と一緒に加熱することによって実施することができる。
【0015】
適切な塩基としては、アルカリ金属水酸化物、アルカリ金属アルキル酸化物などの強塩基が挙げられる。適切な塩基の具体例としては、NaOH、KOH、LiOH、NaH、NaOMe及びKOtBuが挙げられる。
【0016】
使用される塩基の濃度は、塩基の解離定数などの考慮事項を含めて、使用される塩基の性質によって決まることはいうまでもない。当業者は、適切な塩基濃度を容易に決定することができるはずである。塩基がNaOHである場合、多糖体と混合されたときの適切な塩基濃度は0.1から10Nである。様々な例示的実施形態において、適切な濃度は2N、3N、4Nなどの1から5Nである。
【0017】
適切な反応温度は、使用される塩基及び選択される濃度によって決まる。一般に、50℃から100℃の反応温度が適切である。特に、約80℃などの75℃から85℃の温度が適切である。
【0018】
反応時間は、使用される塩基、塩基濃度、反応温度などいくつかの要因によって決まる。一般に、反応時間は塩基濃度又は温度の低下とともに長くなる。適切な反応時間は一般に10から25時間であり、より典型的には15から20時間、例えば、16、17、18又は19時間である。
【0019】
炭水化物化学の当業者は、特定のパラメータを最適化し、収率を最適化するために上記パラメータを操作できることを理解されたい。そうすることによって、化学薬品消費(塩基)、温度、時間などの商業的に関連する変数を特定のシステムにおける製造に対して最適化することができる。例えば、脱O−アセチル化Y髄膜炎菌多糖体を製造するのに必要な時間を最小限に抑え、かくして塩基濃度及び温度の好ましい範囲の上部を使用することを選択したい場合がある。本明細書に提供される例は、これらの多糖体の比較的小規模な製造を説明するものであり、より大規模な製造システムに適合させるために、上記パラメータを示された範囲内で変えることができる。
【0020】
この方法によって達成される脱O−アセチル化度は、H−NMR分光法によって測定して完全である。脱O−アセチル化多糖体は酸加水分解に場合によっては供されてもよい。このステップは、大きい分子多糖体をより小さなMwの断片に分解し、出発材料1グラムにつきより多くの個体Y群髄膜炎菌多糖体エピトープ提示分子を提供するために使用される。
【0021】
酸加水分解は、多糖体溶液を酸又は緩衝剤形成化合物と一緒に撹拌することによって実施することができる。適切な酸又は緩衝剤形成化合物としては、約3〜6の酸性pHにわたって緩衝する、酢酸ナトリウムなどのアルカリ金属アセタートなどが挙げられる。化学の当業者は、他の弱酸塩を使用してこのpH範囲の適切な緩衝剤を調製できることを理解されたい。緩衝剤は、水素イオン(H+)源を全反応期間にわたって比較的一定した濃度に維持するので、本発明の方法において特に有用である。
【0022】
使用される酸又は緩衝剤形成化合物の濃度が、使用される酸又は緩衝剤形成化合物の性質によって決まることはいうまでもない。当業者は、適切な酸及び緩衝剤形成化合物濃度を容易に決定することができるはずである。酢酸ナトリウムが多糖体と混合されるときの適切な濃度は、0.1N酢酸ナトリウムなどの0.01から1N、例えば0.05から0.5Nである。
【0023】
適切な反応温度も使用される酸又は緩衝剤形成化合物によって決まる。一般に、50℃から100℃、より典型的には約70℃など約65℃から80℃の反応温度が適切である。
【0024】
本発明の様々な実施形態においては、反応混合物を撹拌することができる。撹拌速度は、使用される酸又は緩衝剤形成化合物、酸又は緩衝剤形成化合物の濃度、使用される撹拌機のタイプ、反応温度などいくつかの要因によって決まる。反応器が回転振とうされる場合には、適切な撹拌速度は一般に50から100rpm、例えば、65rpmである。
【0025】
脱O−アセチル化多糖体は、酸加水分解に供されたかどうかにかかわらず、再N−アセチル化(re−N−acetylation)を場合によっては起こすことができる。このステップによって、塩基加水分解中に生成され得る遊離第一級アミンがすべて確実に再N−アセチル化される。
【0026】
再N−アセチル化は、無水酢酸を用いて実施することができる。一般に、このステップは中性又は塩基性溶液中で起こり、例えば、溶液のpHは7から9などの7から13、例えば8とすることができる。他の再N−アセチル化用化学薬品としては、塩化アセチル、酢酸ペンタフルオロフェニル、酢酸4−ニトロフェニルなどが挙げられる。これらの薬剤の使用は化学分野において公知である。
【0027】
適切な反応温度は、反応混合物のpH、使用される試薬の性質などの要因によって決まる。一般に、反応は15℃から35℃などの室温付近で起こり、より典型的には20℃から25℃で起こる。
【0028】
本発明の一実施形態においては、脱O−アセチル化髄膜炎菌Y多糖体断片を調製する方法は、
a)少なくともある程度精製された髄膜炎菌Y多糖体を用意するステップと、
b)前記多糖体の塩基加水分解ステップと、
c)ステップ(a)の生成物の酸加水分解ステップと
を含む。
【0029】
ステップ(b)の生成物は場合によっては再N−アセチル化してもよい。
【0030】
多糖体を精製するための強塩基処理の使用は、参照により全体を本明細書に援用する米国特許第6248570号に記載されている。
【0031】
O−アセチル化髄膜炎菌Y多糖体は、上記酸加水分解プロセスによって断片化することができる。
【0032】
脱O−アセチル化多糖体は、酸加水分解に供されたかどうかにかかわらず、上記再N−アセチル化プロセスを場合によっては受けてもよい。
【0033】
上記方法に使用される髄膜炎菌Y多糖体は、使用前に髄膜炎菌性細菌から単離することができる。当分野で公知の任意の単離方法を使用することができる(米国特許第6248570号参照)。適切な単離方法としては、遠心分離とそれに続く限外ろ過などが挙げられる。単離された多糖体の本質の確認及び定量は、H−NMR分光法、糖成分のGC−MS分析、タイピング抗血清を用いたELISA、又は当業者に公知の他の方法によって実施することができる。
【0034】
酸加水分解ステップを含む上記方法によって改変された多糖体は一般に低分子量である。本明細書では、「低分子量」という用語は、多角度レーザー光散乱に接続されたサイズ排除クロマトグラフィー(SEC−MALLS)によって測定して5から50kDaなどの100kDa未満、例えば10から20kDaの分子量を意味する。
【0035】
脱O−アセチル化多糖体とO−アセチル化多糖体の両方は、酸化により還元性基を生成させることによって活性化することができる。還元性基の適切な生成方法としては、アルデヒド末端を生成する過ヨウ素酸塩(又はパラ過ヨウ素酸、メタ過ヨウ素酸ナトリウム、メタ過ヨウ素酸カリウムなどの関係する試薬)による限定的酸化切断(limited oxidative cleavage)などが挙げられる。
【0036】
特定の理論に拘泥するものではないが、出願人らは、アルデヒド基がC−8において多糖体のシアル酸環外側鎖に選択的に導入されることを提唱する。すなわち、酸化はC−8位とC−9位の間で起こると考えられる。
【0037】
活性化された多糖体群は、適切な担体タンパク質との複合に適切である。したがって、本発明は、適切な担体タンパク質と複合化された脱O−アセチル化Y髄膜炎菌多糖体又はその断片を含む複合産物(conjugated product)も提供する。O−アセチル化Y髄膜炎菌多糖体は活性化することができるので、開示された方法は、適切な担体タンパク質と複合化されたO−アセチル化Y髄膜炎菌多糖体又はその断片も提供する。
【0038】
任意の適切なタンパク質を担体タンパク質として使用することができる。本発明における使用に適切であるために、担体タンパク質は、治療すべき哺乳動物、例えば乳児に投与するのに安全でなければならず、免疫学的に有効でなければならない。安全性要件としては、主要な毒性がないこと、アレルギー反応のリスクが最小であることなどが挙げられる。
【0039】
適切な担体タンパク質としては、細菌毒素、トキソイドなどが挙げられる。適切な細菌毒素及びトキソイドの例としては、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、(米国特許第5439808号に記載の)rPorBなどのPorB及びすべての他のそれらの誘導体、百日咳菌、並びに大腸菌を含めた腸毒素原性細菌の毒素又はトキソイドが挙げられるが、これらだけに限定されない。全毒素及びトキソイドに加えて、適切な免疫賦活性効果を示すタンパク質トキソイドの断片又は一部も使用することができる。例えば、破傷風トキソイドの断片Cを担体として使用することができる。使用される毒素又はトキソイドは、その自然の出所(例えば、百日咳トキソイドの場合は百日咳菌)に由来することができ、又は組換えによって産生することができる。
【0040】
適切な担体タンパク質を選択することによって、「担体効果」効果が得られる。「担体効果」によって、Y髄膜炎菌多糖体は、Y髄膜炎菌多糖体が単独で存在する場合よりも担体付着のためにより免疫原性になる。
【0041】
免疫される哺乳動物が担体のみで以前に免疫されている場合には、より大きな応答が得られることがある。乳児は、慣例的に破傷風及びジフテリアトキソイドの予防接種を受ける。したがって、Y髄膜炎菌に対して乳児を免疫するように意図されたワクチン中にこれらのトキソイドを使用することによって、効果が増大すると予想することができる。「担体効果」が得られたかどうかは、多糖体−担体タンパク質複合体による暴露に対する動物モデル又はヒトの免疫応答を監視することによって確認することができる。
【0042】
破傷風毒素、ジフテリア毒素などの一部の細菌毒素は、2個以上のタンパク質で構成される。これらのタンパク質の1個は、哺乳動物細胞表面への結合に対して強い親和性を有する。やはり理論に拘泥するものではないが、この強い結合能力を有するタンパク質を使用することによって、免疫系からの応答をより効果的に惹起することが可能である。
【0043】
担体タンパク質は、未変性毒素又は無毒化毒素(トキソイド)とすることができる。或いは、毒素に抗原性的に類似しているが無毒であるタンパク質を与える突然変異技術によって遺伝子改変されたタンパク質を使用することができる。このようにして改変されたタンパク質は「交差反応性材料」又はCRMとして知られる。本発明に使用することができるかかる材料の1つはCRM197である。CRM197は未変性ジフテリア毒素に基づく。CRM197は単一のアミノ酸変化を有し、すなわち野生型毒素中のグリシン52がCRM197ではグルタミン酸で置換され、未変性ジフテリア毒素から免疫学的に識別不能である。
【0044】
本発明の一態様においては、細菌毒素又はトキソイドは破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドである。
【0045】
したがって、本発明は、破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドに複合化された脱O−アセチル化Y髄膜炎菌多糖体若しくはO−アセチル化Y髄膜炎菌多糖体を含む複合産物を提供する。
【0046】
未変性毒素が使用されるときには、Y髄膜炎菌多糖体との複合化によって未変性毒素の毒性は低下することがある。しかし、残留毒性レベルは高すぎる恐れがある。したがって、さらに解毒が必要になることがある。当分野で公知の任意の適切な方法を使用してこのタンパク質を解毒することができる。従来の方法としては、タンパク質上の遊離アミノ基と反応するホルマリンの使用などが挙げられる。
【0047】
或いは、未変性毒素は、Y髄膜炎菌多糖体と複合化する前に、例えばホルマリンで解毒してトキソイドを生成することができる。
【0048】
Y髄膜炎菌多糖体と適切な担体タンパク質の複合体は、活性化多糖体を適切な担体と接触させることによって得ることができる。通常、タンパク質担体よりも過剰の多糖体が使用され、一般に重量で2から3倍過剰の多糖体が使用される。当業者は、多糖体とタンパク質担体の比を変更して、複合体1個当たり異なる数の抗原多糖体成分を得ることができる。複合体1個当たりの多糖体成分の比がより高い場合には、担体タンパク質1個につきより多量の多糖体から出発する。より低い比を得るには、担体タンパク質1個につきより少量の多糖体から出発する。
【0049】
還元的アミノ化は、参照によりその全体を本明細書に援用する米国特許第4356170号に記載のように好ましいカップリング形式である。したがって、活性化された多糖体と担体を還元性カップリング剤の存在下で接触させることが好ましい。適切なカップリング剤としては、シアノ水素化ホウ素イオンなどの還元剤又はその等価物などが挙げられ、シアノ水素化ホウ素ナトリウムなどの適切な塩の形で得ることができる。目的とする還元末端を還元せず、担体タンパク質にも多糖体にも悪影響を及ぼさない任意の還元剤をカップリング剤として使用することができる。例えば、ピリジンボランは、この目的に適切な別の選択的還元剤である。他の適切な還元剤も当業者には明白なはずである。
【0050】
選択的還元性カップリング剤は、多糖体のカルボニル基と担体タンパク質のアミノ基の間で形成されるシッフ塩基中間体の穏和な選択的還元剤として作用すると考えられる。これらのイオンは、複合化された後に多糖体上に残留する任意の活性なアルデヒド基の還元を遅らせる副次的効果を有すると考えられる。
【0051】
カップリング剤は、通常は、複合産物の一部を形成せず、カップリング反応中に消費される。次いで、複合体を精製して残留カップリング剤及び反応副生物を除去することができる。精製された複合体は、炭水化物をタンパク質に複合化するために以前に使用された、アジピン酸ジヒドラジド、p−ニトロ−フェニル−エチルアミンなどの潜在的毒性連結剤を含まないことが好ましい。
【0052】
活性化多糖体と担体を接触させる条件は担体の性質によって決まる。例えば、担体が破傷風トキソイドであるときには、活性化多糖体と担体は、約7.4などのほぼ中性のpHで30℃から40℃などの約20℃から50℃、例えば37℃の温度で接触させることができる。
【0053】
カップリング反応が終了した後に、多糖体上のあらゆる残留アルデヒド基をキャップすることができる。これによって、活性化ステップ中に生成されたが、多糖体を担体タンパク質に複合化するのに利用されなかった髄膜炎菌多糖鎖上のあらゆる残留アルデヒド基が還元される。適切なキャッピング剤としては、NaBH4、残留アルデヒド基をアルコール基に還元することができる当業者に公知の他の還元剤などが挙げられる。
【0054】
本発明の方法によって生成される脱O−アセチル化Y髄膜炎菌多糖体は、ワクチン調製物中の抗原として利用することができるが、この調製物はT依存性免疫応答をもたらさない。Y髄膜炎菌多糖体と適切な担体タンパク質の複合体は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとして又はワクチン組成物中でより好ましく使用される。したがって、一部の好ましい実施形態においては、本発明のワクチンは、脱O−アセチル化多糖体を含む複合体を含む。実施例のデータに示されたとおり、脱O−アセチル化多糖複合体は、O−アセチル化多糖体と交差反応性の抗体を産生する動物において免疫応答を誘発することができる。
【0055】
通常の製剤操作によれば、多糖体又は複合体は安定化され、ワクチンとして処方される前又は処方後に発熱原性がないことが確認される。一態様においては、本発明は、破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドと複合化された脱O−アセチル化Y髄膜炎菌多糖体を含むワクチンを提供する。このワクチンは、ワクチン組成物中に従来含まれる成分を含有することができる。例えば、このワクチンは、適切な担体媒体、賦形剤、希釈剤又はアジュバントの1種類若しくは複数を含むことができる。
【0056】
適切な担体としては、生理的リン酸ナトリウム緩衝食塩水(pH7.4)などが挙げられる。アジュバントを含む担体の例は、pH6のリン酸ナトリウム緩衝食塩水に懸濁された0.125Mリン酸アルミニウムゲルである。ワクチンでの使用に適切な他の薬剤として許容される担体は当分野で公知であり、本発明に使用することができる。適切なアジュバントとしては、水酸化アルミニウム、当業者に公知の他のアジュバント、例えば概して免疫原性である核酸、ペプチド化合物などが挙げられる。
【0057】
本発明のワクチンは任意の適切な方法によって投与することができる。例えば、本発明のワクチンは注射、例えば、筋肉内又は皮下注射によって投与することができる。適切な担体を用いた無針経皮、鼻腔内又は粘膜投与を利用することもできる。
【0058】
一般に、本発明のワクチンは5から100μg、好ましくは10から50μgの複合材料を含む。正確な投与量は、通常の用量/応答実験によって決定することができる。ワクチンは単一投与量又は2回以上、例えば3回のより少ない投与量で投与することができる。
【0059】
本発明の複合産物は、1系統を超える髄膜炎菌に対する免疫化に使用されるワクチンに使用することもできる。例えば、本発明の複合産物は、髄膜炎菌Yに加えて髄膜炎菌A、C及びW135の1種類又は複数に対して免疫化が意図されたワクチンに使用することができる。複合産物は、髄膜炎菌に加えて他の疾患に対して免疫化が意図されたワクチンに使用することもできる。例えば、本発明の複合体をヘモフィルスインフルエンザ複合体と併用して組合せHib−髄膜炎菌性ワクチンを製造することができる。
【0060】
本発明の複合産物をワクチンとして使用すると「担体効果」が得られる。Y髄膜炎菌多糖体は、Y髄膜炎菌多糖体が単独で存在する場合よりも、担体としてより強力な抗原に付着するためにより免疫原性になる。特に、本発明のワクチンは、ヒトを含めた若年哺乳動物において有効なレベルの抗髄膜炎菌Y抗体形成を誘発するために使用することができ、非複合多糖を含むワクチンよりも年齢依存性が極めて低い。
【0061】
担体タンパク質が、本発明のワクチンのレシピエントがワクチン接種された疾患、例えば、破傷風又はジフテリアの細菌毒素又はトキソイドであるときには、髄膜炎菌Yに対する免疫と同時に担体毒素又はトキソイドに対する所望の免疫を得ることができる。
【0062】
或いは、本発明の複合産物を含有するワクチンのレシピエントが、担体毒素又はトキソイドに関連する疾患に対して以前にワクチン接種を受けた場合には、「追加免疫」効果によって髄膜炎菌Yに対する抗体の産生を増大させることができる。
【0063】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンの製造における、上述された脱O−アセチル化Y髄膜炎菌多糖体及び/又はO−アセチル化Y髄膜炎菌多糖体の使用も提供する。
【0064】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンの製造における、上述された脱O−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物及び/或いはO−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物の使用も提供する。
【0065】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとしての、上述された脱O−アセチル化Y髄膜炎菌多糖体及び/又はO−アセチル化Y髄膜炎菌多糖体の使用も提供する。
【0066】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとしての、上述された脱O−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物及び/或いはO−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物の使用も提供する。
【0067】
以下の非限定的な実施例によって本発明を説明する。
【実施例1】
【0068】
様々な髄膜炎菌性Y系統に由来する多糖体のO−アセチル体の評価
未変性多糖体の調製:
髄膜炎菌性Y系統S1975、S225、S3536及びS3790はCarl Frasch博士(CBER/FDA、Bethesda、MD)の厚意によって提供された。Yスラテラス(Slaterus)系統は、Francoise Collins博士(LCDC、Ontario、Canada)によって提供された。これらの系統は、振とうフラスコ中、撹拌下、グルコース及び酵母抽出物を含む培地中で37℃で増殖された。培養物を8000rpmの遠心分離によって収集し、上清を回収し、0.22μmフィルターユニットによってろ過滅菌した。
【0069】
マイクロろ過された培養上清は、Biomax 300kDa Pellicon膜(0.5m2)(Millipore Corp.、Bedford、Mass.USA)を備えたフィルター装置によって限外ろ過されて濃縮された。濃縮された保持液を1M NaClで12回、次いで脱イオン(DI)水で10回透析ろ過し、凍結乾燥させた。高分子量精製「未変性」多糖体は、GC−MSによって糖組成が分析され、H−NMR分光法によって500MHzでO−アセチル含量及び位置が分析された。
【0070】
表1にGC−MS及びH−NMR分析結果を要約する。
【0071】
【表1】

【0072】
この表からわかるように、それらのきょう膜多糖体のO−アセチル含量並びに存在する場合のこれらのO−アセチル基の位置に関連して系統間の不均一性がかなりある。例えば、系統S3790とスラテラスはOAc含量がほぼ同じであるが、それらのシアル酸残基上のこれらの基(C−7対C−9)の百分率はまったく異なる。系統S3790由来の多糖体はそのシアル酸のほぼ1個の位置(C−7)にのみOAc基を有するのに対して、スラテラス由来の多糖体はそれらを主にC−9上に有する。
【実施例2】
【0073】
Y群髄膜炎菌多糖体(GYMP)の精製
dOA GYMPの調製:
300kDa MWCO膜を用いたUFによる多糖体の捕捉:
細胞を含まないマイクロろ過発酵透過液約13Lを、Pellicon Biomax 300kDa膜(0.5m2)(Millipore Corp.、Bedford、Mass.USA)を備えたフィルター装置を用いて限外ろ過によって約1リットルに濃縮した。濃縮された保持液を1M NaClで12回、次いでDI水で10回透析ろ過した。それをさらに約0.2Lに濃縮し、回収した。
【0074】
多糖体の塩基加水分解:
300kDa保持溶液(約5mg PS/mL)を最終濃度2N NaOHに調節し、80℃に設定された乾燥器に16〜18時間置いた。反応混合物を50℃未満に冷却した後にDI水10L中に希釈した。30kDa MWCO Pellicon膜を通して濃縮した後に、濃縮された保持液を1M NaClで12回、次いでDI水で10回透析ろ過した。それをさらに約0.2Lに濃縮し、回収した。
【0075】
dOA GYMPの酸加水分解:
保持溶液をテフロン反応器に移し、酢酸ナトリウム(NaOAc)を最終濃度0.1Nまで添加した。反応混合物を6N HClを用いてpH5に調節し、70℃に設定された水浴中に置いた。多糖体がSuperose 12(Pharmacia)カラムを用いたSEC−MALLS(多角度レーザー光散乱に接続されたサイズ排除クロマトグラフィー)によって測定して20,000ダルトンの目標MWに達するまで65rpmで振とうした。
【0076】
断片化されたdOA多糖体の再N−アセチル化:
溶液のpHを6N HCl溶液を用いて8に調節し、次いで無水酢酸を0.8M無水酢酸の最終濃度まで室温で滴下した。5N NaOHを用いて反応混合物をpH7から9に維持した。反応終了後、反応混合物のpHを13に上昇させ、混合物をさらに1.5時間撹拌した。次いで、6N HCl溶液を用いて反応pHをpH8に調節した。反応混合物を1M NaCl 4Lに注ぎ、Pellicon Biomax 100kDa膜(0.5m2)を備えた装置を用いて約1Lに濃縮し、透過液を収集した。100K最終透過液を、Biomax 5K Pellicon膜(0.5m2)を用いてUPによって約1リットルに濃縮した。濃縮保持液をDI水で10回透析ろ過し、次いで約0.2Lに濃縮し、収集した。
【0077】
OA GYMPの調製:
OA GYMPの精製プロセスは、プロセスから省略されたアルカリ分解(2N NaOH)ステップ以外、dOA多糖体に対して上述されたものと同様であった。このプロセスから得られたGYMPは、それらのシアル酸のC7位とC9位の間に一様に分布する繰返し単位1個につきO−アセチル基約0.6M(又はシアル酸のN−アセチル基に対して60%)を有した。
【0078】
複合化前のGYMPの活性化:
低分子量(LMW)OA及びdOAc多糖体(10〜20kDa)をメタ過ヨウ素酸ナトリウムを用いて酸化して、アルデヒド基をC−8においてそれらの多糖体のシアル酸環外側鎖に選択的に導入した(C−9とC−8の間の酸化)。
【0079】
Y群髄膜炎菌多糖(GYMP)複合体の調製
破傷風トキソイドへの髄膜炎菌Y多糖体の複合化:
過ヨウ素酸塩酸化型LMW GYMP多糖体(25mg)及び破傷風トキソイド(TT)(10mg)(Serum Statens Institute、Copenhagen、Denmark)を0.2Mホスフェート緩衝剤(pH7.4)0.5mlに溶解した。シアノ水素化ホウ素ナトリウム(5mg)を添加し、その混合物を37℃で1日インキュベートした。複合化が終了した後に、溶液のpHを1M酢酸を用いて7から9に維持しながら新たに調製された1mM NaOH中の100mg/mL NaBH4溶液40〜60μlを氷浴中で極めてゆっくり添加することによって残留アルデヒドのキャッピングを実施した。
【0080】
生成した複合体は、100kDa MWCO膜及びタンジェンシャルフローろ過(TFF)システム(Millipore)を用いた限外ろ過によって1M NaClで、次いでダイアフィルトレーション法によって食塩水で精製された。
【0081】
バルク複合体は、処方される前に2℃〜4℃で貯蔵された。
【実施例3】
【0082】
マウスにおけるGYMP複合体の効力
免疫化:
4〜6週齢のメスのSwiss Websterマウスに水酸化アルミニウム(Alhydrogel、Superfos、Denmark)に吸着した結合型ワクチンを皮下注射した。各マウスに複合多糖2μgを0、28及び42日の3回投与した。0、28、38及び52日目にマウスから採血した。
【0083】
ELISAによるGYMP特異抗体:
GYMP特異的IgG力価は、被覆抗原としてヒト血清アルブミンに連結されたLMW GYMP(OAc又はdOA)を用いてELISAによって推定された。
【0084】
1回目及び2回目の注射後にGYMP−TT複合体によって誘発されるmenY PS特異的IgG力価(相乗平均)を図1に示す。これらは被覆抗原としてdOA GYMP−HSAを用いたELISAによって測定された。図1からわかるとおり、両方のタイプのGYMP複合体(OA及びdOA)はほぼ同じレベルのGYMP特異的IgGを産生し、PS上のOA基が免疫原性にさほど重要ではないことを示唆している。図2及び図3に、それぞれOA及びdOA GYMP−HSA被覆プレート上のOA及びdOA GYMP−TT抗血清のELISA結合(38日目)を示す。両方の抗原被覆プレート上で、抗dOA GYMP−TT抗血清は、対応するOA複合抗血清よりも反応性がわずかに高く、PS上のOAが免疫原性に必ずしも必要でないことを示している。
【0085】
血清殺菌活性(SBA):
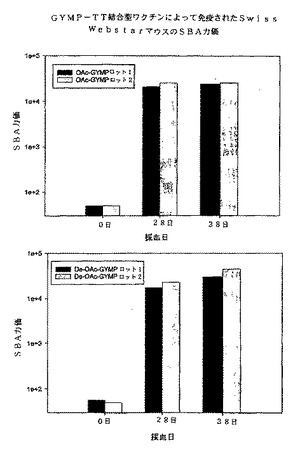
血清の血清殺菌活性は、様々なレベルのOAを有する多糖体を発現する表1に記載されたGYM系統を用いて試験された。図4に、ウサギ補体並びにOA及びdOA GYMP−TT複合体を用いて誘導されたマウス抗血清のOA menY系統3790を用いて測定されたSBA力価を示す。図4に示されたとおり、dOA GYMP複合体は、(OAきょう膜PSを発現する)menY系統3790に対して、対応するOA複合体と類似したSBAレベルを誘導し、PS上のOA基が免疫原性及び効力にさほど重要でないことを示している。
【0086】
また、dOA GYMP複合体によって誘導された抗体は、様々なOA率のきょう膜PSを発現するGYM系統を死滅させることができる(表2)。
【0087】
【表2】
【実施例4】
【0088】
結合型ワクチンによって誘導される抗体の特異性、多糖体上のOA基の役割
結合型ワクチン誘導抗体の結合の特異性:
OA GYMP HAS複合体又はdOA GYMP−HSA複合体に対するワクチン誘導抗体(OA GYMP複合体又はdOA GYMP複合体を含むワクチン)のELISA結合の競合阻害は、主としてC9(GYMスラテラス)又はC7(GYM S3790)にシアル酸OAを含む、又はOAを全く含まない(GYM S1975)高分子量(HMW)GYMP阻害剤を用いて実施された。
【0089】
図5に、dOA GYMP、主としてC7のOA GYMP及び主としてC9のOA GYMPに対するdOA GYMP複合抗血清の特異性を示す。全3種類の多糖体阻害剤は、OA GYMP HSA被覆プレートへのこれらの抗血清の結合を完全に阻害することができる。OA GYMP(C9)は最も有効ではなく、dOA GYMP複合体に対して産生された抗体がOA多糖体とdOA多糖体で実際に識別されないことを示している。
【0090】
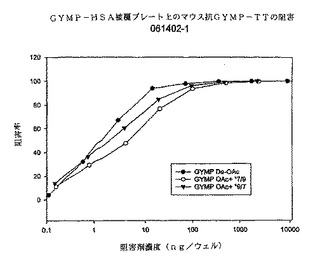
図6に、上記同じ多糖体阻害剤に対するdOA GYMP複合抗血清の特異性を示す。ただし、今回は、dOA GYMP HSA被覆プレートに対する結合である。ここでもやはり、多糖体阻害剤は結合を完全に阻害することができ、dOA多糖体が最良の阻害剤である。
【0091】
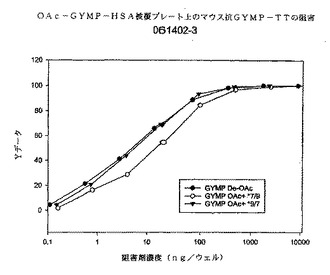
図7に、OA GYMP HSA被覆プレートを用いたOA GYMP複合抗血清の特異性を示す。3種類の多糖体阻害剤は被覆プレートへの抗体の結合を十分に阻害することができるが、この場合は、C9のOA GYMPが最良の阻害剤であり、続いてdOA GYMP、次いでC7のOA GYMPと考えられ、OA複合体はエピトープ特異性がC9のGYMP OAに向かってわずかに増加する抗体を産生したことがおそらくは示唆される。しかし、これらの抗体は、dOAエピトープを極めて明確に認識する。
【0092】
図8は、dOA GYMP HSA被覆プレートへの抗OA GYMP複合抗血清結合の特異性を検討したものである。3種類の多糖体阻害剤は、被覆プレートへのこの抗血清の結合を十分に阻害することができる。dOA GYMPとC7のOA GYMPが最良であり効力が等しい阻害剤である。
【0093】
まとめると、これらのデータによれば、マウスにおいてはGYMP複合体のdOA体はOA GYM多糖体とdOA GYM多糖体(それぞれ図5及び図6)の両方を同様に認識する抗体を誘導する。
【0094】
抗GYMP機能性抗体の特異性、OAの役割:
シアル酸上のOAを主としてC7又はC9に有する可溶性GYMP阻害剤、或いはシアル酸上のOAをまったく持たない可溶性GYMP阻害剤を用いたOA GYM系統(3790)に対する(ウサギ補体を用いた)SBAの競合阻害によって、OA又はdOA GYMP複合体によって誘導される機能性抗体の特異性を検討した。
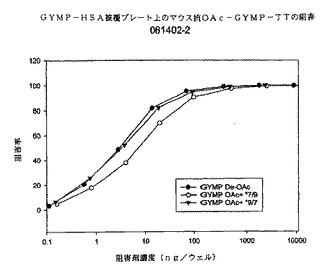
【0095】
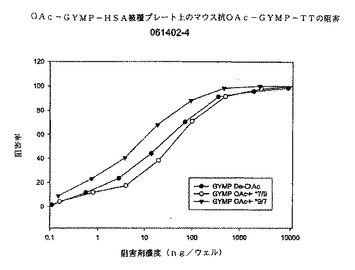
図9に、マウス抗dOA GYMP複合抗血清を用いたかかる阻害の結果を示す。3種類の多糖体阻害剤は系統3790の死滅を完全に阻害することができる。dOA PSは(50%において)両方のOA多糖体阻害剤のほぼ5倍有効である。両方のOA多糖体阻害剤は互いに同じように阻害するように見える。これらのデータによれば、OA多糖体を発現する生きたGYM生物の表面では、dOA GYMPに対する抗体が、dOAシアル酸(多糖体上約10%)を含む多糖体エピトープを認識し、或いは産生される抗体の親和性が高いことを示している。
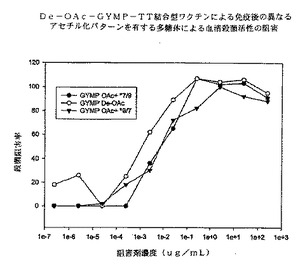
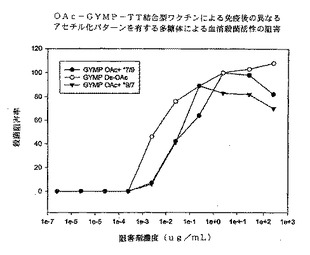
【0096】
図10に、マウス抗OA GYMP複合抗血清を用いたSBAの競合阻害の結果を示す。dOA GYMP複合抗血清に関しては、dOA GYMPがGYM 3790生物に対してSBAを阻害する最良の多糖体であり、他の2種類のOA GYMPがそれに続く。これらのデータによれば、多糖体上のOA基は機能性抗体によって認識される防御エピトープにさほど重要ではないことが強く示唆される。
【実施例5】
【0097】
GYMPエピトープのサイズの決定
dOA GYMPからの長さの異なる(1個から9個の繰返し)オリゴ糖阻害剤の生成:
dOA GYMPを0.1N酢酸ナトリウム緩衝剤pH3を用いて80℃で2時間部分加水分解した。次いで、生成した長さの異なる(1個から9個の繰返し単位)オリゴ糖をサイズ排除クロマトグラフィーによってSuperdex G−30(Pharmacia)カラムにかけて分離した。オリゴ糖の溶出を214nmにおけるUV検出によって監視した。長さの異なる個々のオリゴ糖を含む画分を混合し、凍結乾燥させた。かくして得られた阻害剤の純度及びサイズをH−NMR分光法によって600MHzで実施した。各GYMオリゴ糖阻害剤は、その還元末端に還元性ベータシアル酸を有し、もう一方の非還元性末端にα−D−グルコースを有する。
【0098】
オリゴ糖阻害剤を用いたELISAによる競合阻害:
GYMPエピトープのサイズは、長さが漸増するGYMオリゴ糖によるGYMP特異的ウサギ抗体とGYMP−HSA被覆プレートの結合の競合阻害(ELISA)によって決定された。
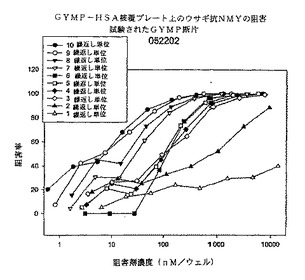
【0099】
図11は、dOA GYMP−HSA被覆プレートへのウサギ抗全GYM抗体(DIFCO、Maryland)結合のELISA阻害である。dOA GYMオリゴ糖は1個から10個の繰返し単位の範囲にある。この図からわかるように、1個及び2個の繰返し単位は不十分な結合阻害剤であり、3個の繰返し単位オリゴ糖(6個のグリコシル残基)は結合を完全に阻害することができる。これらのデータによれば、6番目の残基の還元性シアル酸はエクアトリアル位のカルボキシラート基とベータコンホメーションの関係にあるので、抗体の結合部位はおそらく5個の糖残基と接触する。オリゴ糖のサイズが増加すると(4から10RU)、モル/モル基準での阻害が増大し、長さが漸増する抗体によって認識されるエピトープコンホメーションが安定化することが示唆される。
【0100】
図12は、同じ阻害剤によるOA GYMP−HSA被覆プレート上の同じ抗体の結合阻害である。阻害パターンは図11において認められた阻害パターンと類似しており、3RUオリゴ糖構造を超えると結合が増加する。これらのデータはGYMP複合体を設計する上で意味がある。というのは、担体タンパク質と複合化されるGYMPハプテンの最小サイズが10個の繰返し単位であることをこれらのデータは示唆しているからである。
【実施例6】
【0101】
組み合わせられた髄膜炎菌性Y結合型ワクチンの効力
dOA GCMP−TTとdOA GWMP rPorB結合型ワクチンと組み合わせられたdOA GYMP−TTの組合せの効力をSwiss Websterマウスにおいて試験した。各多糖体複合抗原2μgを組み合わせて、又は単独で0日及び28日にマウスに皮下投与した。0、28及び38日に採血し、OA GYMPを発現するGYM系統3790に対するSBAについて血清を分析した。
【0102】
図13にかかる実験の結果を示す。組合せ複合抗血清におけるGYMに対するSBAは、単独GYMP複合抗血清において測定されたSBAと大きく異ならず、3種類の複合体間に免疫学的干渉が認められないことが示された。同様に、組合せ複合体でも単独複合体でも、GCMP複合体成分とGWMP複合体成分から生じるSBAに免疫学的干渉は認められなかった。
【図面の簡単な説明】
【0103】
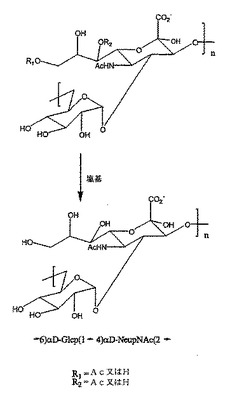
【図1】繰返し単位脱O−アセチル化Y群(「deOAc−Y」)髄膜炎菌多糖体の構造、下図及び未変性O−アセチル化Y群(「OAc−Y」)髄膜炎菌多糖体、上図からのその調製を示す図である。
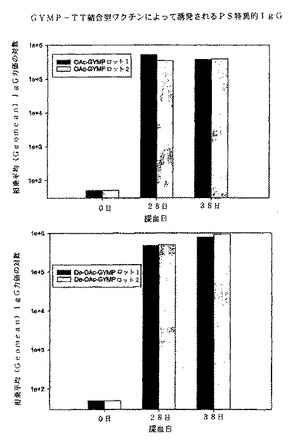
【図2】メスのSwiss Websterマウスにおいて1回又は2回の注射後に実施例3に記載されたように測定された、OAc−Y(上図)及びdeOAc−Y(下図)髄膜炎菌多糖体−破傷風トキソイド複合体によって誘発されたY髄膜炎菌多糖体特異的IgG力価の相乗平均を示すグラフである。
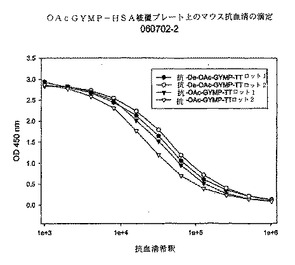
【図3】実施例3において測定された、OAc−Y髄膜炎菌多糖体−HSA被覆プレート上のOAc−Y及びdeOAc−Y髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されるマウス抗血清の38日目におけるELISA滴定を示すグラフである。
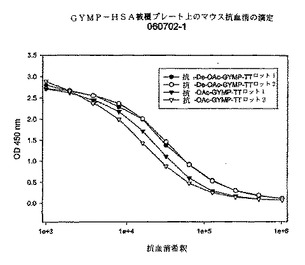
【図4】実施例3において測定された、deOAc−Y髄膜炎菌多糖体−HSA被覆プレート上のOAc−Y及びdeOAc−Y髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されるマウス抗血清の38日目におけるELISA滴定を示すグラフである。
【図5】実施例3において測定された、ウサギ補体と、OAc−Y(上図)及びdeOAc−Y(下図)髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されたマウス抗血清のOAc−Y髄膜炎菌系統3790とを用いて測定された血清の殺菌活性力価を示す図である。
【図6】実施例4において測定された、炭素9(*9/7)又は炭素7(*7/9)におけるO−アセチル化量の多いOAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、OAc−GYMP−HSA複合体で被覆されたマイクロタイタープレート上でのdeOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図7】実施例4において測定された、OAc及びdeOAc−Y髄膜炎菌多糖体に対する、deOAc−GYMP−HSA複合体で被覆されたマイクロタイタープレート上でのdeOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図8】実施例4において測定された、OAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、OAc−Y髄膜炎菌多糖体−HSA複合体で被覆されたマイクロタイタープレート上でのOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図9】実施例4において測定された、OAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、deOAc−Y髄膜炎菌多糖体−HSA複合体で被覆されたマイクロタイタープレート上でのOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図10】実施例4において測定され、マウス抗dOAc−Y髄膜炎菌多糖体複合抗血清を用いて可溶性Y髄膜炎菌多糖体阻害剤によるOAc−Y髄膜炎菌多糖体系統(3790)に対する(ウサギ補体を用いた)SBAの競合阻害によって測定された、OAc−Y又はdeOAc−Y髄膜炎菌多糖体複合体によって誘導される機能性抗体の特異性を示すグラフである。
【図11】実施例4において測定された、マウス抗OAc−Y髄膜炎菌多糖体複合抗血清を用いたSBAの競合阻害の結果を示すグラフである。
【図12】実施例5において測定された、1個から10個の繰返し単位のdeOAc−Y髄膜炎菌オリゴ糖によるdeOAc−Y髄膜炎菌多糖体−HSA被覆プレートに対するウサギ抗ホールセルOAc−Y髄膜炎菌多糖体抗体の結合のELISA阻害を示すグラフである。
【図13】実施例5において測定された、1個から10個の繰返し単位のdeOAc−Y髄膜炎菌オリゴ糖によるOAc−Y髄膜炎菌多糖体−HSA被覆プレートに対するウサギ抗ホールセルOAc−Y髄膜炎菌多糖体抗体の結合のELISA阻害を示すグラフである。
【図14】実施例6において測定された、Swiss WebsterマウスにおいてdeOAc−C髄膜炎菌多糖体及び破傷風トキソイド複合体及びdeOAc−W髄膜炎菌多糖体及びrPorB複合体と組み合わせられたdeOAc−Y髄膜炎菌多糖体と破傷風トキソイドの複合体の効力に関する試験結果を示すグラフである。
【技術分野】
【0001】
本発明は、改変髄膜炎菌Y多糖体(GYMP)、改変多糖体と担体を含む複合体、Y群髄膜炎菌に対するヒトを含めた温血動物の免疫用ワクチンに関し、これらの改変多糖体、複合体及びワクチンを製造する方法に関する。
【背景技術】
【0002】
細菌性髄膜炎は、世界規模での健康に対する重大な脅威である。髄膜炎菌は、細菌性髄膜炎及び敗血症の主要な原因である。髄膜炎菌は、多糖きょう膜で被包されている。髄膜炎菌単離体は、その化学的及び抗原性的に異なる多糖きょう膜に基づいて12の群に分類することができる。これらの群のうちA、B、C、Y及びW135の5種類は、ヒトにおける細菌性髄膜炎及び敗血症の実質的にすべての症例の原因である。
【0003】
Y群髄膜炎菌によって引き起こされる細菌性髄膜炎の発生率は増加している。米国のいくつかの郡において1989年から1995年に実施された活動的な研究室によるサーベイランスによれば、血清型Y髄膜炎菌性疾患の比率は1989年の0%から1995年の32.5%に急激に増加した。この期間中、髄膜炎菌性疾患の全発生率は一定していた(Anonymous 1996, Serogroup Y meningococcal disease - Illinois, Connecticut, and selected areas, USA 1989-1996, Morb. Mortal. Rep. 45, 1010-1013)。
【0004】
Y群髄膜炎菌によって引き起こされる細菌性髄膜炎は、子供や若年成人においても起こり得るが、高齢者において発症し易い傾向にあり、細菌性髄膜炎の他の系統よりも肺炎を引き起こす可能性が高い。
【0005】
多糖体ワクチンは、いくつかの血清型の髄膜炎菌によって引き起こされる疾患を予防するために使用されてきた。しかし、これらのワクチンは、一部の人種においては髄膜炎菌によって引き起こされる疾患の予防に有効ではない。
【0006】
多糖体はT非依存性(胸腺非依存性)抗原である。多糖体は、タンパク質に対する免疫応答を誘導するときに遭遇しないいくつかの免疫学的諸特性を有する。これらの諸特性としては、免疫応答を誘導するT細胞が存在する明白な必要性がないこと、IgMの優勢、免疫後の記憶誘導(memory induction)や親和性成熟がないこと、乳児、高齢者及び免疫無防備状態の人における低免疫原性などが挙げられる。その結果、純粋な多糖体ワクチンは、これらの患者に対して有効に使用することができない。
【0007】
血清型Y多糖体は、等モルのN−アセチルノイラミン酸とD−グルコースからなり、部分的にO−アセチル化されていることが、1970年代にJenningsによって報告された(Bhattacharjee et al,. Can. J. Biochem. 54: 1-8, 1976)。
【0008】
ほとんどのY群髄膜炎菌単離体は、O−アセチル基がもっぱらそのシアル酸残基のC−7とC−9の間に分布するO−アセチル陽性(OA)多糖体を産生することが見出された。O−アセチル基分布の位置及び濃度におけるこの不均一性は、多糖複合体の処方を複雑にしている。
【非特許文献1】Anonymous 1996, Serogroup Y meningococcal disease - Illinois, Connecticut, and selected areas, USA 1989-1996, Morb. Mortal. Rep. 45, 1010-1013
【非特許文献2】Bhattacharjee et al,. Can. J. Biochem. 54: 1-8, 1976
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の一目的は、Y群髄膜炎菌に対する免疫化への使用に適切なワクチンを提供することである。
【課題を解決するための手段】
【0010】
本発明は、少なくとも1個の改変髄膜炎菌Y多糖体又は改変髄膜炎菌Y多糖体の断片又は髄膜炎菌Y多糖体の断片を含むワクチンを提供する。以下の文において、「髄膜炎菌Y多糖体」という用語及びその語形(forms)は、改変髄膜炎菌Y多糖体、改変髄膜炎菌Y多糖体の断片及び髄膜炎菌Y多糖体の断片を含む。これらの多糖体は単独で使用して、T非依存性応答を誘発する抗原組成物を産生することができる。
【0011】
改変髄膜炎菌Y多糖体は、一般に、T依存性免疫応答を誘発するために担体タンパク質と複合化される。したがって、本発明は、改変髄膜炎菌Y多糖体と担体タンパク質を含む複合材料及びその複合材料を製造する方法も提供する。
【0012】
本発明は、改変髄膜炎菌Y多糖体を製造する方法も提供する。髄膜炎菌Y多糖体は、そのシアル酸残基のC−7とC−9の間にもっぱら分布するO−アセチル基を含有するので、脱O−アセチル化によって改変することができる。したがって、本発明は、髄膜炎菌Y多糖体の脱O−アセチル化方法も提供する。一実施形態においては、塩基加水分解を使用してこの多糖体を脱O−アセチル化することができる。
【0013】
本発明は、脱O−アセチル化髄膜炎菌Y多糖体又はO−アセチル化髄膜炎菌Y多糖体を断片化する方法も提供する。一実施形態においては、弱酸加水分解を使用して多糖体を断片化する。多糖体のグリコシド結合を切断する他の方法としては、オゾン分解、超音波処理、塩基加水分解などが挙げられる。
【発明を実施するための最良の形態】
【0014】
本発明によれば、髄膜炎菌Y多糖体は、塩基加水分解によって脱O−アセチル化することができる。塩基加水分解は、多糖体の溶液を塩基と一緒に加熱することによって実施することができる。
【0015】
適切な塩基としては、アルカリ金属水酸化物、アルカリ金属アルキル酸化物などの強塩基が挙げられる。適切な塩基の具体例としては、NaOH、KOH、LiOH、NaH、NaOMe及びKOtBuが挙げられる。
【0016】
使用される塩基の濃度は、塩基の解離定数などの考慮事項を含めて、使用される塩基の性質によって決まることはいうまでもない。当業者は、適切な塩基濃度を容易に決定することができるはずである。塩基がNaOHである場合、多糖体と混合されたときの適切な塩基濃度は0.1から10Nである。様々な例示的実施形態において、適切な濃度は2N、3N、4Nなどの1から5Nである。
【0017】
適切な反応温度は、使用される塩基及び選択される濃度によって決まる。一般に、50℃から100℃の反応温度が適切である。特に、約80℃などの75℃から85℃の温度が適切である。
【0018】
反応時間は、使用される塩基、塩基濃度、反応温度などいくつかの要因によって決まる。一般に、反応時間は塩基濃度又は温度の低下とともに長くなる。適切な反応時間は一般に10から25時間であり、より典型的には15から20時間、例えば、16、17、18又は19時間である。
【0019】
炭水化物化学の当業者は、特定のパラメータを最適化し、収率を最適化するために上記パラメータを操作できることを理解されたい。そうすることによって、化学薬品消費(塩基)、温度、時間などの商業的に関連する変数を特定のシステムにおける製造に対して最適化することができる。例えば、脱O−アセチル化Y髄膜炎菌多糖体を製造するのに必要な時間を最小限に抑え、かくして塩基濃度及び温度の好ましい範囲の上部を使用することを選択したい場合がある。本明細書に提供される例は、これらの多糖体の比較的小規模な製造を説明するものであり、より大規模な製造システムに適合させるために、上記パラメータを示された範囲内で変えることができる。
【0020】
この方法によって達成される脱O−アセチル化度は、H−NMR分光法によって測定して完全である。脱O−アセチル化多糖体は酸加水分解に場合によっては供されてもよい。このステップは、大きい分子多糖体をより小さなMwの断片に分解し、出発材料1グラムにつきより多くの個体Y群髄膜炎菌多糖体エピトープ提示分子を提供するために使用される。
【0021】
酸加水分解は、多糖体溶液を酸又は緩衝剤形成化合物と一緒に撹拌することによって実施することができる。適切な酸又は緩衝剤形成化合物としては、約3〜6の酸性pHにわたって緩衝する、酢酸ナトリウムなどのアルカリ金属アセタートなどが挙げられる。化学の当業者は、他の弱酸塩を使用してこのpH範囲の適切な緩衝剤を調製できることを理解されたい。緩衝剤は、水素イオン(H+)源を全反応期間にわたって比較的一定した濃度に維持するので、本発明の方法において特に有用である。
【0022】
使用される酸又は緩衝剤形成化合物の濃度が、使用される酸又は緩衝剤形成化合物の性質によって決まることはいうまでもない。当業者は、適切な酸及び緩衝剤形成化合物濃度を容易に決定することができるはずである。酢酸ナトリウムが多糖体と混合されるときの適切な濃度は、0.1N酢酸ナトリウムなどの0.01から1N、例えば0.05から0.5Nである。
【0023】
適切な反応温度も使用される酸又は緩衝剤形成化合物によって決まる。一般に、50℃から100℃、より典型的には約70℃など約65℃から80℃の反応温度が適切である。
【0024】
本発明の様々な実施形態においては、反応混合物を撹拌することができる。撹拌速度は、使用される酸又は緩衝剤形成化合物、酸又は緩衝剤形成化合物の濃度、使用される撹拌機のタイプ、反応温度などいくつかの要因によって決まる。反応器が回転振とうされる場合には、適切な撹拌速度は一般に50から100rpm、例えば、65rpmである。
【0025】
脱O−アセチル化多糖体は、酸加水分解に供されたかどうかにかかわらず、再N−アセチル化(re−N−acetylation)を場合によっては起こすことができる。このステップによって、塩基加水分解中に生成され得る遊離第一級アミンがすべて確実に再N−アセチル化される。
【0026】
再N−アセチル化は、無水酢酸を用いて実施することができる。一般に、このステップは中性又は塩基性溶液中で起こり、例えば、溶液のpHは7から9などの7から13、例えば8とすることができる。他の再N−アセチル化用化学薬品としては、塩化アセチル、酢酸ペンタフルオロフェニル、酢酸4−ニトロフェニルなどが挙げられる。これらの薬剤の使用は化学分野において公知である。
【0027】
適切な反応温度は、反応混合物のpH、使用される試薬の性質などの要因によって決まる。一般に、反応は15℃から35℃などの室温付近で起こり、より典型的には20℃から25℃で起こる。
【0028】
本発明の一実施形態においては、脱O−アセチル化髄膜炎菌Y多糖体断片を調製する方法は、
a)少なくともある程度精製された髄膜炎菌Y多糖体を用意するステップと、
b)前記多糖体の塩基加水分解ステップと、
c)ステップ(a)の生成物の酸加水分解ステップと
を含む。
【0029】
ステップ(b)の生成物は場合によっては再N−アセチル化してもよい。
【0030】
多糖体を精製するための強塩基処理の使用は、参照により全体を本明細書に援用する米国特許第6248570号に記載されている。
【0031】
O−アセチル化髄膜炎菌Y多糖体は、上記酸加水分解プロセスによって断片化することができる。
【0032】
脱O−アセチル化多糖体は、酸加水分解に供されたかどうかにかかわらず、上記再N−アセチル化プロセスを場合によっては受けてもよい。
【0033】
上記方法に使用される髄膜炎菌Y多糖体は、使用前に髄膜炎菌性細菌から単離することができる。当分野で公知の任意の単離方法を使用することができる(米国特許第6248570号参照)。適切な単離方法としては、遠心分離とそれに続く限外ろ過などが挙げられる。単離された多糖体の本質の確認及び定量は、H−NMR分光法、糖成分のGC−MS分析、タイピング抗血清を用いたELISA、又は当業者に公知の他の方法によって実施することができる。
【0034】
酸加水分解ステップを含む上記方法によって改変された多糖体は一般に低分子量である。本明細書では、「低分子量」という用語は、多角度レーザー光散乱に接続されたサイズ排除クロマトグラフィー(SEC−MALLS)によって測定して5から50kDaなどの100kDa未満、例えば10から20kDaの分子量を意味する。
【0035】
脱O−アセチル化多糖体とO−アセチル化多糖体の両方は、酸化により還元性基を生成させることによって活性化することができる。還元性基の適切な生成方法としては、アルデヒド末端を生成する過ヨウ素酸塩(又はパラ過ヨウ素酸、メタ過ヨウ素酸ナトリウム、メタ過ヨウ素酸カリウムなどの関係する試薬)による限定的酸化切断(limited oxidative cleavage)などが挙げられる。
【0036】
特定の理論に拘泥するものではないが、出願人らは、アルデヒド基がC−8において多糖体のシアル酸環外側鎖に選択的に導入されることを提唱する。すなわち、酸化はC−8位とC−9位の間で起こると考えられる。
【0037】
活性化された多糖体群は、適切な担体タンパク質との複合に適切である。したがって、本発明は、適切な担体タンパク質と複合化された脱O−アセチル化Y髄膜炎菌多糖体又はその断片を含む複合産物(conjugated product)も提供する。O−アセチル化Y髄膜炎菌多糖体は活性化することができるので、開示された方法は、適切な担体タンパク質と複合化されたO−アセチル化Y髄膜炎菌多糖体又はその断片も提供する。
【0038】
任意の適切なタンパク質を担体タンパク質として使用することができる。本発明における使用に適切であるために、担体タンパク質は、治療すべき哺乳動物、例えば乳児に投与するのに安全でなければならず、免疫学的に有効でなければならない。安全性要件としては、主要な毒性がないこと、アレルギー反応のリスクが最小であることなどが挙げられる。
【0039】
適切な担体タンパク質としては、細菌毒素、トキソイドなどが挙げられる。適切な細菌毒素及びトキソイドの例としては、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、(米国特許第5439808号に記載の)rPorBなどのPorB及びすべての他のそれらの誘導体、百日咳菌、並びに大腸菌を含めた腸毒素原性細菌の毒素又はトキソイドが挙げられるが、これらだけに限定されない。全毒素及びトキソイドに加えて、適切な免疫賦活性効果を示すタンパク質トキソイドの断片又は一部も使用することができる。例えば、破傷風トキソイドの断片Cを担体として使用することができる。使用される毒素又はトキソイドは、その自然の出所(例えば、百日咳トキソイドの場合は百日咳菌)に由来することができ、又は組換えによって産生することができる。
【0040】
適切な担体タンパク質を選択することによって、「担体効果」効果が得られる。「担体効果」によって、Y髄膜炎菌多糖体は、Y髄膜炎菌多糖体が単独で存在する場合よりも担体付着のためにより免疫原性になる。
【0041】
免疫される哺乳動物が担体のみで以前に免疫されている場合には、より大きな応答が得られることがある。乳児は、慣例的に破傷風及びジフテリアトキソイドの予防接種を受ける。したがって、Y髄膜炎菌に対して乳児を免疫するように意図されたワクチン中にこれらのトキソイドを使用することによって、効果が増大すると予想することができる。「担体効果」が得られたかどうかは、多糖体−担体タンパク質複合体による暴露に対する動物モデル又はヒトの免疫応答を監視することによって確認することができる。
【0042】
破傷風毒素、ジフテリア毒素などの一部の細菌毒素は、2個以上のタンパク質で構成される。これらのタンパク質の1個は、哺乳動物細胞表面への結合に対して強い親和性を有する。やはり理論に拘泥するものではないが、この強い結合能力を有するタンパク質を使用することによって、免疫系からの応答をより効果的に惹起することが可能である。
【0043】
担体タンパク質は、未変性毒素又は無毒化毒素(トキソイド)とすることができる。或いは、毒素に抗原性的に類似しているが無毒であるタンパク質を与える突然変異技術によって遺伝子改変されたタンパク質を使用することができる。このようにして改変されたタンパク質は「交差反応性材料」又はCRMとして知られる。本発明に使用することができるかかる材料の1つはCRM197である。CRM197は未変性ジフテリア毒素に基づく。CRM197は単一のアミノ酸変化を有し、すなわち野生型毒素中のグリシン52がCRM197ではグルタミン酸で置換され、未変性ジフテリア毒素から免疫学的に識別不能である。
【0044】
本発明の一態様においては、細菌毒素又はトキソイドは破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドである。
【0045】
したがって、本発明は、破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドに複合化された脱O−アセチル化Y髄膜炎菌多糖体若しくはO−アセチル化Y髄膜炎菌多糖体を含む複合産物を提供する。
【0046】
未変性毒素が使用されるときには、Y髄膜炎菌多糖体との複合化によって未変性毒素の毒性は低下することがある。しかし、残留毒性レベルは高すぎる恐れがある。したがって、さらに解毒が必要になることがある。当分野で公知の任意の適切な方法を使用してこのタンパク質を解毒することができる。従来の方法としては、タンパク質上の遊離アミノ基と反応するホルマリンの使用などが挙げられる。
【0047】
或いは、未変性毒素は、Y髄膜炎菌多糖体と複合化する前に、例えばホルマリンで解毒してトキソイドを生成することができる。
【0048】
Y髄膜炎菌多糖体と適切な担体タンパク質の複合体は、活性化多糖体を適切な担体と接触させることによって得ることができる。通常、タンパク質担体よりも過剰の多糖体が使用され、一般に重量で2から3倍過剰の多糖体が使用される。当業者は、多糖体とタンパク質担体の比を変更して、複合体1個当たり異なる数の抗原多糖体成分を得ることができる。複合体1個当たりの多糖体成分の比がより高い場合には、担体タンパク質1個につきより多量の多糖体から出発する。より低い比を得るには、担体タンパク質1個につきより少量の多糖体から出発する。
【0049】
還元的アミノ化は、参照によりその全体を本明細書に援用する米国特許第4356170号に記載のように好ましいカップリング形式である。したがって、活性化された多糖体と担体を還元性カップリング剤の存在下で接触させることが好ましい。適切なカップリング剤としては、シアノ水素化ホウ素イオンなどの還元剤又はその等価物などが挙げられ、シアノ水素化ホウ素ナトリウムなどの適切な塩の形で得ることができる。目的とする還元末端を還元せず、担体タンパク質にも多糖体にも悪影響を及ぼさない任意の還元剤をカップリング剤として使用することができる。例えば、ピリジンボランは、この目的に適切な別の選択的還元剤である。他の適切な還元剤も当業者には明白なはずである。
【0050】
選択的還元性カップリング剤は、多糖体のカルボニル基と担体タンパク質のアミノ基の間で形成されるシッフ塩基中間体の穏和な選択的還元剤として作用すると考えられる。これらのイオンは、複合化された後に多糖体上に残留する任意の活性なアルデヒド基の還元を遅らせる副次的効果を有すると考えられる。
【0051】
カップリング剤は、通常は、複合産物の一部を形成せず、カップリング反応中に消費される。次いで、複合体を精製して残留カップリング剤及び反応副生物を除去することができる。精製された複合体は、炭水化物をタンパク質に複合化するために以前に使用された、アジピン酸ジヒドラジド、p−ニトロ−フェニル−エチルアミンなどの潜在的毒性連結剤を含まないことが好ましい。
【0052】
活性化多糖体と担体を接触させる条件は担体の性質によって決まる。例えば、担体が破傷風トキソイドであるときには、活性化多糖体と担体は、約7.4などのほぼ中性のpHで30℃から40℃などの約20℃から50℃、例えば37℃の温度で接触させることができる。
【0053】
カップリング反応が終了した後に、多糖体上のあらゆる残留アルデヒド基をキャップすることができる。これによって、活性化ステップ中に生成されたが、多糖体を担体タンパク質に複合化するのに利用されなかった髄膜炎菌多糖鎖上のあらゆる残留アルデヒド基が還元される。適切なキャッピング剤としては、NaBH4、残留アルデヒド基をアルコール基に還元することができる当業者に公知の他の還元剤などが挙げられる。
【0054】
本発明の方法によって生成される脱O−アセチル化Y髄膜炎菌多糖体は、ワクチン調製物中の抗原として利用することができるが、この調製物はT依存性免疫応答をもたらさない。Y髄膜炎菌多糖体と適切な担体タンパク質の複合体は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとして又はワクチン組成物中でより好ましく使用される。したがって、一部の好ましい実施形態においては、本発明のワクチンは、脱O−アセチル化多糖体を含む複合体を含む。実施例のデータに示されたとおり、脱O−アセチル化多糖複合体は、O−アセチル化多糖体と交差反応性の抗体を産生する動物において免疫応答を誘発することができる。
【0055】
通常の製剤操作によれば、多糖体又は複合体は安定化され、ワクチンとして処方される前又は処方後に発熱原性がないことが確認される。一態様においては、本発明は、破傷風毒素又はトキソイド或いはジフテリア毒素又はトキソイドと複合化された脱O−アセチル化Y髄膜炎菌多糖体を含むワクチンを提供する。このワクチンは、ワクチン組成物中に従来含まれる成分を含有することができる。例えば、このワクチンは、適切な担体媒体、賦形剤、希釈剤又はアジュバントの1種類若しくは複数を含むことができる。
【0056】
適切な担体としては、生理的リン酸ナトリウム緩衝食塩水(pH7.4)などが挙げられる。アジュバントを含む担体の例は、pH6のリン酸ナトリウム緩衝食塩水に懸濁された0.125Mリン酸アルミニウムゲルである。ワクチンでの使用に適切な他の薬剤として許容される担体は当分野で公知であり、本発明に使用することができる。適切なアジュバントとしては、水酸化アルミニウム、当業者に公知の他のアジュバント、例えば概して免疫原性である核酸、ペプチド化合物などが挙げられる。
【0057】
本発明のワクチンは任意の適切な方法によって投与することができる。例えば、本発明のワクチンは注射、例えば、筋肉内又は皮下注射によって投与することができる。適切な担体を用いた無針経皮、鼻腔内又は粘膜投与を利用することもできる。
【0058】
一般に、本発明のワクチンは5から100μg、好ましくは10から50μgの複合材料を含む。正確な投与量は、通常の用量/応答実験によって決定することができる。ワクチンは単一投与量又は2回以上、例えば3回のより少ない投与量で投与することができる。
【0059】
本発明の複合産物は、1系統を超える髄膜炎菌に対する免疫化に使用されるワクチンに使用することもできる。例えば、本発明の複合産物は、髄膜炎菌Yに加えて髄膜炎菌A、C及びW135の1種類又は複数に対して免疫化が意図されたワクチンに使用することができる。複合産物は、髄膜炎菌に加えて他の疾患に対して免疫化が意図されたワクチンに使用することもできる。例えば、本発明の複合体をヘモフィルスインフルエンザ複合体と併用して組合せHib−髄膜炎菌性ワクチンを製造することができる。
【0060】
本発明の複合産物をワクチンとして使用すると「担体効果」が得られる。Y髄膜炎菌多糖体は、Y髄膜炎菌多糖体が単独で存在する場合よりも、担体としてより強力な抗原に付着するためにより免疫原性になる。特に、本発明のワクチンは、ヒトを含めた若年哺乳動物において有効なレベルの抗髄膜炎菌Y抗体形成を誘発するために使用することができ、非複合多糖を含むワクチンよりも年齢依存性が極めて低い。
【0061】
担体タンパク質が、本発明のワクチンのレシピエントがワクチン接種された疾患、例えば、破傷風又はジフテリアの細菌毒素又はトキソイドであるときには、髄膜炎菌Yに対する免疫と同時に担体毒素又はトキソイドに対する所望の免疫を得ることができる。
【0062】
或いは、本発明の複合産物を含有するワクチンのレシピエントが、担体毒素又はトキソイドに関連する疾患に対して以前にワクチン接種を受けた場合には、「追加免疫」効果によって髄膜炎菌Yに対する抗体の産生を増大させることができる。
【0063】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンの製造における、上述された脱O−アセチル化Y髄膜炎菌多糖体及び/又はO−アセチル化Y髄膜炎菌多糖体の使用も提供する。
【0064】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンの製造における、上述された脱O−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物及び/或いはO−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物の使用も提供する。
【0065】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとしての、上述された脱O−アセチル化Y髄膜炎菌多糖体及び/又はO−アセチル化Y髄膜炎菌多糖体の使用も提供する。
【0066】
本発明は、Y群髄膜炎菌に対する免疫化に使用されるワクチンとしての、上述された脱O−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物及び/或いはO−アセチル化Y髄膜炎菌多糖体と破傷風毒素又はトキソイド、ジフテリア毒素又はトキソイドなどの適切な担体とを含む複合産物の使用も提供する。
【0067】
以下の非限定的な実施例によって本発明を説明する。
【実施例1】
【0068】
様々な髄膜炎菌性Y系統に由来する多糖体のO−アセチル体の評価
未変性多糖体の調製:
髄膜炎菌性Y系統S1975、S225、S3536及びS3790はCarl Frasch博士(CBER/FDA、Bethesda、MD)の厚意によって提供された。Yスラテラス(Slaterus)系統は、Francoise Collins博士(LCDC、Ontario、Canada)によって提供された。これらの系統は、振とうフラスコ中、撹拌下、グルコース及び酵母抽出物を含む培地中で37℃で増殖された。培養物を8000rpmの遠心分離によって収集し、上清を回収し、0.22μmフィルターユニットによってろ過滅菌した。
【0069】
マイクロろ過された培養上清は、Biomax 300kDa Pellicon膜(0.5m2)(Millipore Corp.、Bedford、Mass.USA)を備えたフィルター装置によって限外ろ過されて濃縮された。濃縮された保持液を1M NaClで12回、次いで脱イオン(DI)水で10回透析ろ過し、凍結乾燥させた。高分子量精製「未変性」多糖体は、GC−MSによって糖組成が分析され、H−NMR分光法によって500MHzでO−アセチル含量及び位置が分析された。
【0070】
表1にGC−MS及びH−NMR分析結果を要約する。
【0071】
【表1】
【0072】
この表からわかるように、それらのきょう膜多糖体のO−アセチル含量並びに存在する場合のこれらのO−アセチル基の位置に関連して系統間の不均一性がかなりある。例えば、系統S3790とスラテラスはOAc含量がほぼ同じであるが、それらのシアル酸残基上のこれらの基(C−7対C−9)の百分率はまったく異なる。系統S3790由来の多糖体はそのシアル酸のほぼ1個の位置(C−7)にのみOAc基を有するのに対して、スラテラス由来の多糖体はそれらを主にC−9上に有する。
【実施例2】
【0073】
Y群髄膜炎菌多糖体(GYMP)の精製
dOA GYMPの調製:
300kDa MWCO膜を用いたUFによる多糖体の捕捉:
細胞を含まないマイクロろ過発酵透過液約13Lを、Pellicon Biomax 300kDa膜(0.5m2)(Millipore Corp.、Bedford、Mass.USA)を備えたフィルター装置を用いて限外ろ過によって約1リットルに濃縮した。濃縮された保持液を1M NaClで12回、次いでDI水で10回透析ろ過した。それをさらに約0.2Lに濃縮し、回収した。
【0074】
多糖体の塩基加水分解:
300kDa保持溶液(約5mg PS/mL)を最終濃度2N NaOHに調節し、80℃に設定された乾燥器に16〜18時間置いた。反応混合物を50℃未満に冷却した後にDI水10L中に希釈した。30kDa MWCO Pellicon膜を通して濃縮した後に、濃縮された保持液を1M NaClで12回、次いでDI水で10回透析ろ過した。それをさらに約0.2Lに濃縮し、回収した。
【0075】
dOA GYMPの酸加水分解:
保持溶液をテフロン反応器に移し、酢酸ナトリウム(NaOAc)を最終濃度0.1Nまで添加した。反応混合物を6N HClを用いてpH5に調節し、70℃に設定された水浴中に置いた。多糖体がSuperose 12(Pharmacia)カラムを用いたSEC−MALLS(多角度レーザー光散乱に接続されたサイズ排除クロマトグラフィー)によって測定して20,000ダルトンの目標MWに達するまで65rpmで振とうした。
【0076】
断片化されたdOA多糖体の再N−アセチル化:
溶液のpHを6N HCl溶液を用いて8に調節し、次いで無水酢酸を0.8M無水酢酸の最終濃度まで室温で滴下した。5N NaOHを用いて反応混合物をpH7から9に維持した。反応終了後、反応混合物のpHを13に上昇させ、混合物をさらに1.5時間撹拌した。次いで、6N HCl溶液を用いて反応pHをpH8に調節した。反応混合物を1M NaCl 4Lに注ぎ、Pellicon Biomax 100kDa膜(0.5m2)を備えた装置を用いて約1Lに濃縮し、透過液を収集した。100K最終透過液を、Biomax 5K Pellicon膜(0.5m2)を用いてUPによって約1リットルに濃縮した。濃縮保持液をDI水で10回透析ろ過し、次いで約0.2Lに濃縮し、収集した。
【0077】
OA GYMPの調製:
OA GYMPの精製プロセスは、プロセスから省略されたアルカリ分解(2N NaOH)ステップ以外、dOA多糖体に対して上述されたものと同様であった。このプロセスから得られたGYMPは、それらのシアル酸のC7位とC9位の間に一様に分布する繰返し単位1個につきO−アセチル基約0.6M(又はシアル酸のN−アセチル基に対して60%)を有した。
【0078】
複合化前のGYMPの活性化:
低分子量(LMW)OA及びdOAc多糖体(10〜20kDa)をメタ過ヨウ素酸ナトリウムを用いて酸化して、アルデヒド基をC−8においてそれらの多糖体のシアル酸環外側鎖に選択的に導入した(C−9とC−8の間の酸化)。
【0079】
Y群髄膜炎菌多糖(GYMP)複合体の調製
破傷風トキソイドへの髄膜炎菌Y多糖体の複合化:
過ヨウ素酸塩酸化型LMW GYMP多糖体(25mg)及び破傷風トキソイド(TT)(10mg)(Serum Statens Institute、Copenhagen、Denmark)を0.2Mホスフェート緩衝剤(pH7.4)0.5mlに溶解した。シアノ水素化ホウ素ナトリウム(5mg)を添加し、その混合物を37℃で1日インキュベートした。複合化が終了した後に、溶液のpHを1M酢酸を用いて7から9に維持しながら新たに調製された1mM NaOH中の100mg/mL NaBH4溶液40〜60μlを氷浴中で極めてゆっくり添加することによって残留アルデヒドのキャッピングを実施した。
【0080】
生成した複合体は、100kDa MWCO膜及びタンジェンシャルフローろ過(TFF)システム(Millipore)を用いた限外ろ過によって1M NaClで、次いでダイアフィルトレーション法によって食塩水で精製された。
【0081】
バルク複合体は、処方される前に2℃〜4℃で貯蔵された。
【実施例3】
【0082】
マウスにおけるGYMP複合体の効力
免疫化:
4〜6週齢のメスのSwiss Websterマウスに水酸化アルミニウム(Alhydrogel、Superfos、Denmark)に吸着した結合型ワクチンを皮下注射した。各マウスに複合多糖2μgを0、28及び42日の3回投与した。0、28、38及び52日目にマウスから採血した。
【0083】
ELISAによるGYMP特異抗体:
GYMP特異的IgG力価は、被覆抗原としてヒト血清アルブミンに連結されたLMW GYMP(OAc又はdOA)を用いてELISAによって推定された。
【0084】
1回目及び2回目の注射後にGYMP−TT複合体によって誘発されるmenY PS特異的IgG力価(相乗平均)を図1に示す。これらは被覆抗原としてdOA GYMP−HSAを用いたELISAによって測定された。図1からわかるとおり、両方のタイプのGYMP複合体(OA及びdOA)はほぼ同じレベルのGYMP特異的IgGを産生し、PS上のOA基が免疫原性にさほど重要ではないことを示唆している。図2及び図3に、それぞれOA及びdOA GYMP−HSA被覆プレート上のOA及びdOA GYMP−TT抗血清のELISA結合(38日目)を示す。両方の抗原被覆プレート上で、抗dOA GYMP−TT抗血清は、対応するOA複合抗血清よりも反応性がわずかに高く、PS上のOAが免疫原性に必ずしも必要でないことを示している。
【0085】
血清殺菌活性(SBA):
血清の血清殺菌活性は、様々なレベルのOAを有する多糖体を発現する表1に記載されたGYM系統を用いて試験された。図4に、ウサギ補体並びにOA及びdOA GYMP−TT複合体を用いて誘導されたマウス抗血清のOA menY系統3790を用いて測定されたSBA力価を示す。図4に示されたとおり、dOA GYMP複合体は、(OAきょう膜PSを発現する)menY系統3790に対して、対応するOA複合体と類似したSBAレベルを誘導し、PS上のOA基が免疫原性及び効力にさほど重要でないことを示している。
【0086】
また、dOA GYMP複合体によって誘導された抗体は、様々なOA率のきょう膜PSを発現するGYM系統を死滅させることができる(表2)。
【0087】
【表2】
【実施例4】
【0088】
結合型ワクチンによって誘導される抗体の特異性、多糖体上のOA基の役割
結合型ワクチン誘導抗体の結合の特異性:
OA GYMP HAS複合体又はdOA GYMP−HSA複合体に対するワクチン誘導抗体(OA GYMP複合体又はdOA GYMP複合体を含むワクチン)のELISA結合の競合阻害は、主としてC9(GYMスラテラス)又はC7(GYM S3790)にシアル酸OAを含む、又はOAを全く含まない(GYM S1975)高分子量(HMW)GYMP阻害剤を用いて実施された。
【0089】
図5に、dOA GYMP、主としてC7のOA GYMP及び主としてC9のOA GYMPに対するdOA GYMP複合抗血清の特異性を示す。全3種類の多糖体阻害剤は、OA GYMP HSA被覆プレートへのこれらの抗血清の結合を完全に阻害することができる。OA GYMP(C9)は最も有効ではなく、dOA GYMP複合体に対して産生された抗体がOA多糖体とdOA多糖体で実際に識別されないことを示している。
【0090】
図6に、上記同じ多糖体阻害剤に対するdOA GYMP複合抗血清の特異性を示す。ただし、今回は、dOA GYMP HSA被覆プレートに対する結合である。ここでもやはり、多糖体阻害剤は結合を完全に阻害することができ、dOA多糖体が最良の阻害剤である。
【0091】
図7に、OA GYMP HSA被覆プレートを用いたOA GYMP複合抗血清の特異性を示す。3種類の多糖体阻害剤は被覆プレートへの抗体の結合を十分に阻害することができるが、この場合は、C9のOA GYMPが最良の阻害剤であり、続いてdOA GYMP、次いでC7のOA GYMPと考えられ、OA複合体はエピトープ特異性がC9のGYMP OAに向かってわずかに増加する抗体を産生したことがおそらくは示唆される。しかし、これらの抗体は、dOAエピトープを極めて明確に認識する。
【0092】
図8は、dOA GYMP HSA被覆プレートへの抗OA GYMP複合抗血清結合の特異性を検討したものである。3種類の多糖体阻害剤は、被覆プレートへのこの抗血清の結合を十分に阻害することができる。dOA GYMPとC7のOA GYMPが最良であり効力が等しい阻害剤である。
【0093】
まとめると、これらのデータによれば、マウスにおいてはGYMP複合体のdOA体はOA GYM多糖体とdOA GYM多糖体(それぞれ図5及び図6)の両方を同様に認識する抗体を誘導する。
【0094】
抗GYMP機能性抗体の特異性、OAの役割:
シアル酸上のOAを主としてC7又はC9に有する可溶性GYMP阻害剤、或いはシアル酸上のOAをまったく持たない可溶性GYMP阻害剤を用いたOA GYM系統(3790)に対する(ウサギ補体を用いた)SBAの競合阻害によって、OA又はdOA GYMP複合体によって誘導される機能性抗体の特異性を検討した。
【0095】
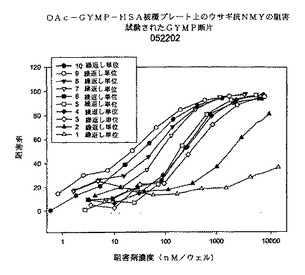
図9に、マウス抗dOA GYMP複合抗血清を用いたかかる阻害の結果を示す。3種類の多糖体阻害剤は系統3790の死滅を完全に阻害することができる。dOA PSは(50%において)両方のOA多糖体阻害剤のほぼ5倍有効である。両方のOA多糖体阻害剤は互いに同じように阻害するように見える。これらのデータによれば、OA多糖体を発現する生きたGYM生物の表面では、dOA GYMPに対する抗体が、dOAシアル酸(多糖体上約10%)を含む多糖体エピトープを認識し、或いは産生される抗体の親和性が高いことを示している。
【0096】
図10に、マウス抗OA GYMP複合抗血清を用いたSBAの競合阻害の結果を示す。dOA GYMP複合抗血清に関しては、dOA GYMPがGYM 3790生物に対してSBAを阻害する最良の多糖体であり、他の2種類のOA GYMPがそれに続く。これらのデータによれば、多糖体上のOA基は機能性抗体によって認識される防御エピトープにさほど重要ではないことが強く示唆される。
【実施例5】
【0097】
GYMPエピトープのサイズの決定
dOA GYMPからの長さの異なる(1個から9個の繰返し)オリゴ糖阻害剤の生成:
dOA GYMPを0.1N酢酸ナトリウム緩衝剤pH3を用いて80℃で2時間部分加水分解した。次いで、生成した長さの異なる(1個から9個の繰返し単位)オリゴ糖をサイズ排除クロマトグラフィーによってSuperdex G−30(Pharmacia)カラムにかけて分離した。オリゴ糖の溶出を214nmにおけるUV検出によって監視した。長さの異なる個々のオリゴ糖を含む画分を混合し、凍結乾燥させた。かくして得られた阻害剤の純度及びサイズをH−NMR分光法によって600MHzで実施した。各GYMオリゴ糖阻害剤は、その還元末端に還元性ベータシアル酸を有し、もう一方の非還元性末端にα−D−グルコースを有する。
【0098】
オリゴ糖阻害剤を用いたELISAによる競合阻害:
GYMPエピトープのサイズは、長さが漸増するGYMオリゴ糖によるGYMP特異的ウサギ抗体とGYMP−HSA被覆プレートの結合の競合阻害(ELISA)によって決定された。
【0099】
図11は、dOA GYMP−HSA被覆プレートへのウサギ抗全GYM抗体(DIFCO、Maryland)結合のELISA阻害である。dOA GYMオリゴ糖は1個から10個の繰返し単位の範囲にある。この図からわかるように、1個及び2個の繰返し単位は不十分な結合阻害剤であり、3個の繰返し単位オリゴ糖(6個のグリコシル残基)は結合を完全に阻害することができる。これらのデータによれば、6番目の残基の還元性シアル酸はエクアトリアル位のカルボキシラート基とベータコンホメーションの関係にあるので、抗体の結合部位はおそらく5個の糖残基と接触する。オリゴ糖のサイズが増加すると(4から10RU)、モル/モル基準での阻害が増大し、長さが漸増する抗体によって認識されるエピトープコンホメーションが安定化することが示唆される。
【0100】
図12は、同じ阻害剤によるOA GYMP−HSA被覆プレート上の同じ抗体の結合阻害である。阻害パターンは図11において認められた阻害パターンと類似しており、3RUオリゴ糖構造を超えると結合が増加する。これらのデータはGYMP複合体を設計する上で意味がある。というのは、担体タンパク質と複合化されるGYMPハプテンの最小サイズが10個の繰返し単位であることをこれらのデータは示唆しているからである。
【実施例6】
【0101】
組み合わせられた髄膜炎菌性Y結合型ワクチンの効力
dOA GCMP−TTとdOA GWMP rPorB結合型ワクチンと組み合わせられたdOA GYMP−TTの組合せの効力をSwiss Websterマウスにおいて試験した。各多糖体複合抗原2μgを組み合わせて、又は単独で0日及び28日にマウスに皮下投与した。0、28及び38日に採血し、OA GYMPを発現するGYM系統3790に対するSBAについて血清を分析した。
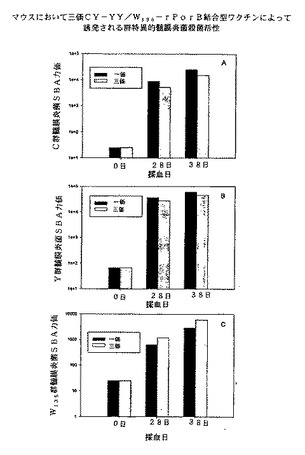
【0102】
図13にかかる実験の結果を示す。組合せ複合抗血清におけるGYMに対するSBAは、単独GYMP複合抗血清において測定されたSBAと大きく異ならず、3種類の複合体間に免疫学的干渉が認められないことが示された。同様に、組合せ複合体でも単独複合体でも、GCMP複合体成分とGWMP複合体成分から生じるSBAに免疫学的干渉は認められなかった。
【図面の簡単な説明】
【0103】
【図1】繰返し単位脱O−アセチル化Y群(「deOAc−Y」)髄膜炎菌多糖体の構造、下図及び未変性O−アセチル化Y群(「OAc−Y」)髄膜炎菌多糖体、上図からのその調製を示す図である。
【図2】メスのSwiss Websterマウスにおいて1回又は2回の注射後に実施例3に記載されたように測定された、OAc−Y(上図)及びdeOAc−Y(下図)髄膜炎菌多糖体−破傷風トキソイド複合体によって誘発されたY髄膜炎菌多糖体特異的IgG力価の相乗平均を示すグラフである。
【図3】実施例3において測定された、OAc−Y髄膜炎菌多糖体−HSA被覆プレート上のOAc−Y及びdeOAc−Y髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されるマウス抗血清の38日目におけるELISA滴定を示すグラフである。
【図4】実施例3において測定された、deOAc−Y髄膜炎菌多糖体−HSA被覆プレート上のOAc−Y及びdeOAc−Y髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されるマウス抗血清の38日目におけるELISA滴定を示すグラフである。
【図5】実施例3において測定された、ウサギ補体と、OAc−Y(上図)及びdeOAc−Y(下図)髄膜炎菌多糖体−破傷風トキソイド複合体によって誘導されたマウス抗血清のOAc−Y髄膜炎菌系統3790とを用いて測定された血清の殺菌活性力価を示す図である。
【図6】実施例4において測定された、炭素9(*9/7)又は炭素7(*7/9)におけるO−アセチル化量の多いOAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、OAc−GYMP−HSA複合体で被覆されたマイクロタイタープレート上でのdeOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図7】実施例4において測定された、OAc及びdeOAc−Y髄膜炎菌多糖体に対する、deOAc−GYMP−HSA複合体で被覆されたマイクロタイタープレート上でのdeOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図8】実施例4において測定された、OAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、OAc−Y髄膜炎菌多糖体−HSA複合体で被覆されたマイクロタイタープレート上でのOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図9】実施例4において測定された、OAc−Y及びdeOAc−Y髄膜炎菌多糖体に対する、deOAc−Y髄膜炎菌多糖体−HSA複合体で被覆されたマイクロタイタープレート上でのOAc−Y髄膜炎菌多糖体複合抗血清の結合阻害を示すグラフである。
【図10】実施例4において測定され、マウス抗dOAc−Y髄膜炎菌多糖体複合抗血清を用いて可溶性Y髄膜炎菌多糖体阻害剤によるOAc−Y髄膜炎菌多糖体系統(3790)に対する(ウサギ補体を用いた)SBAの競合阻害によって測定された、OAc−Y又はdeOAc−Y髄膜炎菌多糖体複合体によって誘導される機能性抗体の特異性を示すグラフである。
【図11】実施例4において測定された、マウス抗OAc−Y髄膜炎菌多糖体複合抗血清を用いたSBAの競合阻害の結果を示すグラフである。
【図12】実施例5において測定された、1個から10個の繰返し単位のdeOAc−Y髄膜炎菌オリゴ糖によるdeOAc−Y髄膜炎菌多糖体−HSA被覆プレートに対するウサギ抗ホールセルOAc−Y髄膜炎菌多糖体抗体の結合のELISA阻害を示すグラフである。
【図13】実施例5において測定された、1個から10個の繰返し単位のdeOAc−Y髄膜炎菌オリゴ糖によるOAc−Y髄膜炎菌多糖体−HSA被覆プレートに対するウサギ抗ホールセルOAc−Y髄膜炎菌多糖体抗体の結合のELISA阻害を示すグラフである。
【図14】実施例6において測定された、Swiss WebsterマウスにおいてdeOAc−C髄膜炎菌多糖体及び破傷風トキソイド複合体及びdeOAc−W髄膜炎菌多糖体及びrPorB複合体と組み合わせられたdeOAc−Y髄膜炎菌多糖体と破傷風トキソイドの複合体の効力に関する試験結果を示すグラフである。
【特許請求の範囲】
【請求項1】
髄膜炎菌感染に対するワクチンとして使用される、O−脱アセチル化されたO−アセチル陽性Y群髄膜炎菌多糖体又はその断片を含み、重合体担体に共有結合されたY群髄膜炎菌多糖体を含有する免疫原性複合体であって、脱O−アセチル化度が80%を超える免疫原性複合体。
【請求項2】
脱O−アセチル化度が100%であることを特徴とする、請求項1に記載の免疫原性複合体。
【請求項3】
分子量平均が10kDa、50kDa及び150kDaからなる群から選択される、請求項1に記載の多糖体。
【請求項4】
断片化され、前記断片サイズが5個の繰返し単位(約2.5kDa)から200個の繰返し単位(約100kDa)を含む、請求項1又は請求項2に記載の多糖体。
【請求項5】
断片化され、前記断片サイズが20個の繰返し単位(約10kDa)から40個の繰返し単位(約20kDa)を含む、請求項1又は請求項2に記載の多糖体。
【請求項6】
担体タンパク質に複合化された脱O−アセチル化髄膜炎菌Y多糖体を含む複合産物(conjugate product)。
【請求項7】
前記担体タンパク質が細菌毒素又はトキソイドである、請求項4に記載の複合産物。
【請求項8】
前記細菌毒素又はトキソイドが、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、百日咳及び大腸菌毒素又はトキソイドからなる群から選択される、請求項5に記載の複合産物。
【請求項9】
前記細菌毒素又はトキソイドが破傷風毒素又はトキソイドである、請求項6に記載の複合産物。
【請求項10】
改変髄膜炎菌Y多糖体が請求項2に記載されたものである、複合産物。
【請求項11】
請求項4に記載の複合産物を含むワクチン。
【請求項12】
前記細菌毒素又はトキソイドが、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、髄膜炎菌性ポリンB、百日咳及び大腸菌毒素又はトキソイドからなる群から選択される、請求項9に記載のワクチン。
【請求項13】
前記細菌毒素又はトキソイドが破傷風毒素又はトキソイドである、請求項10に記載のワクチン。
【請求項14】
アジュバントを含む、請求項1に記載のワクチン。
【請求項15】
前記アジュバントが水酸化アルミニウムである、請求項12に記載のワクチン。
【請求項16】
注射投与用の請求項9に記載のワクチン。
【請求項17】
前記複合材料が請求項2に記載の多糖体を含む、請求項9に記載のワクチン。
【請求項18】
髄膜炎において使用されるY群髄膜炎菌用ワクチンの製造における、請求項1に記載の改変多糖体の使用。
【請求項19】
髄膜炎において使用されるY群髄膜炎菌用ワクチンの製造における、請求項4に記載の複合材料の使用。
【請求項20】
Y群髄膜炎菌に対する免疫化に使用されるワクチンを製造する方法であって、請求項1に記載の改変多糖体を用意するステップを含み、薬剤として許容される担体媒体、希釈剤又はアジュバントの1個若しくは複数とその改変多糖体を混合するステップを場合によっては含んでもよい方法。
【請求項21】
Y群髄膜炎菌に対する免疫化に使用されるワクチンを製造する方法であって、請求項4に記載の複合材料を用意するステップを含み、薬剤として許容される担体媒体、希釈剤又はアジュバントの1個若しくは複数とその複合材料を混合するステップを場合によっては含んでもよい方法。
【請求項22】
髄膜炎に対する請求項9に記載のY群髄膜炎菌用ワクチンの使用。
【請求項23】
Y群髄膜炎菌の免疫になるように温血動物に予防接種をする方法であって、請求項9に記載のワクチンを前記動物に投与するステップを含む方法。
【請求項24】
改変髄膜炎菌Y多糖体を調製する方法であって、髄膜炎菌Y多糖体が少なくとも部分的に脱O−アセチル化されるように前記髄膜炎菌Y多糖体を塩基加水分解に供するステップを含む方法。
【請求項25】
改変髄膜炎菌Y多糖体を調製する方法であって、髄膜炎菌Y多糖体が断片化されるように前記髄膜炎菌Y多糖体を酸加水分解に供するステップを含む方法。
【請求項26】
分子量が10から20kDaである改変髄膜炎菌Y多糖体断片を調製する方法であって、
(a)少なくともある程度精製された髄膜炎菌Y多糖体を用意するステップと、
(b)前記多糖体の塩基加水分解ステップと、
(c)ステップ(a)の生成物の酸加水分解ステップと
を含み、場合によっては
(d)ステップ(b)の生成物を再N−アセチル化するステップ
を含んでもよい方法。
【請求項27】
請求項4に記載の複合産物(conjugated product)を製造する方法であって、場合によってはカップリング剤の存在下で、改変髄膜炎菌Y多糖体を担体タンパク質と接触させるステップを含む方法。
【請求項28】
髄膜炎菌性Y、C及びW135疾患を予防するためのY群、C群及びW135群髄膜炎菌多糖体の各de−OAc体を含む、組合せ髄膜炎菌性結合型ワクチン。
【請求項1】
髄膜炎菌感染に対するワクチンとして使用される、O−脱アセチル化されたO−アセチル陽性Y群髄膜炎菌多糖体又はその断片を含み、重合体担体に共有結合されたY群髄膜炎菌多糖体を含有する免疫原性複合体であって、脱O−アセチル化度が80%を超える免疫原性複合体。
【請求項2】
脱O−アセチル化度が100%であることを特徴とする、請求項1に記載の免疫原性複合体。
【請求項3】
分子量平均が10kDa、50kDa及び150kDaからなる群から選択される、請求項1に記載の多糖体。
【請求項4】
断片化され、前記断片サイズが5個の繰返し単位(約2.5kDa)から200個の繰返し単位(約100kDa)を含む、請求項1又は請求項2に記載の多糖体。
【請求項5】
断片化され、前記断片サイズが20個の繰返し単位(約10kDa)から40個の繰返し単位(約20kDa)を含む、請求項1又は請求項2に記載の多糖体。
【請求項6】
担体タンパク質に複合化された脱O−アセチル化髄膜炎菌Y多糖体を含む複合産物(conjugate product)。
【請求項7】
前記担体タンパク質が細菌毒素又はトキソイドである、請求項4に記載の複合産物。
【請求項8】
前記細菌毒素又はトキソイドが、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、百日咳及び大腸菌毒素又はトキソイドからなる群から選択される、請求項5に記載の複合産物。
【請求項9】
前記細菌毒素又はトキソイドが破傷風毒素又はトキソイドである、請求項6に記載の複合産物。
【請求項10】
改変髄膜炎菌Y多糖体が請求項2に記載されたものである、複合産物。
【請求項11】
請求項4に記載の複合産物を含むワクチン。
【請求項12】
前記細菌毒素又はトキソイドが、ジフテリア、破傷風、シュードモナス、スタフィロコッカス、ストレプトコッカス、髄膜炎菌性ポリンB、百日咳及び大腸菌毒素又はトキソイドからなる群から選択される、請求項9に記載のワクチン。
【請求項13】
前記細菌毒素又はトキソイドが破傷風毒素又はトキソイドである、請求項10に記載のワクチン。
【請求項14】
アジュバントを含む、請求項1に記載のワクチン。
【請求項15】
前記アジュバントが水酸化アルミニウムである、請求項12に記載のワクチン。
【請求項16】
注射投与用の請求項9に記載のワクチン。
【請求項17】
前記複合材料が請求項2に記載の多糖体を含む、請求項9に記載のワクチン。
【請求項18】
髄膜炎において使用されるY群髄膜炎菌用ワクチンの製造における、請求項1に記載の改変多糖体の使用。
【請求項19】
髄膜炎において使用されるY群髄膜炎菌用ワクチンの製造における、請求項4に記載の複合材料の使用。
【請求項20】
Y群髄膜炎菌に対する免疫化に使用されるワクチンを製造する方法であって、請求項1に記載の改変多糖体を用意するステップを含み、薬剤として許容される担体媒体、希釈剤又はアジュバントの1個若しくは複数とその改変多糖体を混合するステップを場合によっては含んでもよい方法。
【請求項21】
Y群髄膜炎菌に対する免疫化に使用されるワクチンを製造する方法であって、請求項4に記載の複合材料を用意するステップを含み、薬剤として許容される担体媒体、希釈剤又はアジュバントの1個若しくは複数とその複合材料を混合するステップを場合によっては含んでもよい方法。
【請求項22】
髄膜炎に対する請求項9に記載のY群髄膜炎菌用ワクチンの使用。
【請求項23】
Y群髄膜炎菌の免疫になるように温血動物に予防接種をする方法であって、請求項9に記載のワクチンを前記動物に投与するステップを含む方法。
【請求項24】
改変髄膜炎菌Y多糖体を調製する方法であって、髄膜炎菌Y多糖体が少なくとも部分的に脱O−アセチル化されるように前記髄膜炎菌Y多糖体を塩基加水分解に供するステップを含む方法。
【請求項25】
改変髄膜炎菌Y多糖体を調製する方法であって、髄膜炎菌Y多糖体が断片化されるように前記髄膜炎菌Y多糖体を酸加水分解に供するステップを含む方法。
【請求項26】
分子量が10から20kDaである改変髄膜炎菌Y多糖体断片を調製する方法であって、
(a)少なくともある程度精製された髄膜炎菌Y多糖体を用意するステップと、
(b)前記多糖体の塩基加水分解ステップと、
(c)ステップ(a)の生成物の酸加水分解ステップと
を含み、場合によっては
(d)ステップ(b)の生成物を再N−アセチル化するステップ
を含んでもよい方法。
【請求項27】
請求項4に記載の複合産物(conjugated product)を製造する方法であって、場合によってはカップリング剤の存在下で、改変髄膜炎菌Y多糖体を担体タンパク質と接触させるステップを含む方法。
【請求項28】
髄膜炎菌性Y、C及びW135疾患を予防するためのY群、C群及びW135群髄膜炎菌多糖体の各de−OAc体を含む、組合せ髄膜炎菌性結合型ワクチン。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2007−524623(P2007−524623A)
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願番号】特願2006−517558(P2006−517558)
【出願日】平成16年6月23日(2004.6.23)
【国際出願番号】PCT/US2004/020048
【国際公開番号】WO2005/000347
【国際公開日】平成17年1月6日(2005.1.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(591013229)バクスター・インターナショナル・インコーポレイテッド (448)
【氏名又は名称原語表記】BAXTER INTERNATIONAL INCORP0RATED
【出願人】(505474924)バクスター ヘルスケア エス.エイ. (2)
【Fターム(参考)】
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願日】平成16年6月23日(2004.6.23)
【国際出願番号】PCT/US2004/020048
【国際公開番号】WO2005/000347
【国際公開日】平成17年1月6日(2005.1.6)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.テフロン
【出願人】(591013229)バクスター・インターナショナル・インコーポレイテッド (448)
【氏名又は名称原語表記】BAXTER INTERNATIONAL INCORP0RATED
【出願人】(505474924)バクスター ヘルスケア エス.エイ. (2)
【Fターム(参考)】
[ Back to top ]