
invitroでのタンパク質の連続的な目的に適合した進化のための方法
本発明は、in vitro進化方法におけるタンパク質の変異体の製造方法であって、次の工程:(A)次のものを含有するin vitro発現系の作成工程、(i)変異すべきタンパク質Yをコードする核酸配列S、(ii)タンパク質Y及び/又は少なくとも1種のこの変異体Yに結合することができる目的分子X1、(iii)核酸配列Sを転写することができるRNAポリメラーゼ(Pol)、(iv)核酸配列Sの転写産物を逆転写することができる逆転写酵素(RT)、その際、目的分子XはPolに、そしてタンパク質YはRTにカップリングしているか、又は目的分子XはRTに、そしてタンパク質YはPolにカップリングしている、(B) タンパク質Yの変異体Y及び、これをコードする核酸配列S′の形成下での転写、逆転写及び翻訳を可能にし、かつ、目的分子Xのための改善された結合特性を有する変異体Yの形成を促進する条件下での、(A)からのin vitro発現系のインキュベーション工程、(C)Xに対する結合のための改善された結合特性を有する変異体Yの単離工程及び場合により特性決定工程及び/又はYをコードする核酸配列変異体Sの単離工程を含む、in vitro進化方法におけるタンパク質の変異体の製造方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
発明の詳細な説明
本発明は、in vitro進化方法におけるタンパク質の変異体の製造に関する。
【0002】
発明の背景
医薬、化学産業及び農業経済の範囲におけるバイオテクノロジーの増加する重要性は、そのつどのその使用目的に最適に適合しているタンパク質をますます求めている。まずこれらのタンパク質は主として環境から、大抵はいわゆるメタゲノムスクリーニングの範囲内で単離される。ますます前記タンパク質は将来には様々な方法を介して予定される「人工的な」使用条件に適用される。
【0003】
従って、例えば、天然の変異体よりもより耐熱性であり、他の基質特異性を有するか又はより高い活性を示す酵素が必要とされる。医薬的タンパク質は例えば、より少なく投薬されることができるようにより長い半減期を示すか、又は、目的分子に対する高親和性かつ特異的な結合を介して、疾病に関連した物質代謝経路又は感染経路を阻害することが望ましい。
【0004】
技術水準
この適合は部分的には、合理的タンパク質設計アプローチ(rationale Protein Design Ansaetze)を介して行われる。しかしながらこれまでには、タンパク質の所望の機能−表現型−からその構造を推定するか又はタンパク質の3次元構造から相応する一次配列を導く、極めて限定された手段のみが存在する。それにもかかわらずタンパク質の機能向上を達成すべく、現在のところ、「目的に指向した進化」(Directed Evolution)との概念にまとめられる部分的進化アプローチが使用される。
【0005】
現在の技術水準によれば、Directed Evolutionのこれまでの方法は実質的に、改良すべきタンパク質の多数の変異体(子孫)の産生及び改善された誘導体に応じたその選択に基づく。この際、検査すべき突然変異体の数は、部分的に極めて巨大である可能性があり、しかしながら通常は1011未満である。100個のみのアミノ酸のタンパク質を考慮する場合には、理論的に20100=10130個のこの相違する変異体が存在する。1011の大きさのライブラリーはこれに応じて、可能性のある変異体の極めて少ない部分のみをカバーする。このようなライブラリー中に理論的に最良の変異体が見出されることができる確率はほぼゼロである。
【0006】
目的分子に対して最大の親和性で多数の変異体をスクリーニングするために、既に多数のプロトコールが開発されている(例えば、酵母ツーハイブリッド(Yeast Two-Hybrid)、細菌ディスプレイ(Bacterial Display)、ファージディスプレイ(Phage Display)、リボソームディスプレイ(Ribosomal Display)、mRNAディスプレイ(mRNA Display))。
【0007】
他の特性、例えば酵素活性のスクリーニングのためには、大抵はアッセイフォーマット(Assay-Formate)が必要とされ、これは比較的少ない数の変異体(<106)のみの検査を可能にする。この方法は、突然変異体のライブラリーの産生並びに引き続くその選択に限定されている点で共通している。複製(=最後に行われる選択の「当選者」の突然変異体のすぐ次の世代)は、このプロトコールを用いて確かに手動で可能であるが、しかしながら先行する工程と同様に手間がかかる。従って、相応するアプローチは通例は、1〜2つの世代にわたるのみである。この理由から、述べられたプロトコールは、本来の意味合いにおける進化的アプローチではなく、むしろ存在する変異体プールの排他的選択である。従って、多数の世代を越えた段階的な適合の潜在能力は利用されることができず、これはしかしながら、場合によっては最も活性のある変異体を天文学的に巨大な数の可能性から同定するための必要な前提条件であるかも知れない。
【0008】
その3つの前提条件−複製、突然変異及び選択−を用いた進化の原則は、所定の系のうちでの、単純にそして高度にまで適合された構造の指向された発達を引き起こすことを可能にする。いわゆる「ブラインドウォッチメーカー」のこのモデルは、計画的な助力無しに、かつ、構造データの必要とされる知識無しに、複雑性の作成を可能にする。
【0009】
Bauer et al.は1989年に(PNAS (1989) 86, 7937-7941)、Qβ−レプリカーゼが見出されかつ任意のRNAで末端で接種(animpfen)される細管中で連続的な進化プロセスが生じることを示した。重合フロントがこの細管に沿って生じ、この経過において生じるRNAポリメラーゼはファージ特異的な配列/二次構造へと進化し、というのもこれらはレプリカーゼによりより迅速に複製されるからである。この実験が直接的な実際的な有用性を示さないにもかかわらず、この実験は3つの因子、複製、突然変異及び選択の潜在能力を顕著にカバーする(belegen)。
【0010】
WO 02/22869は、分子ライブラリーのin vitro進化の際の使用方法を記載する。この際、「ツーハイブリッド系(Two Hybrid - System)」系が使用される。ここで、高められたエラー頻度を有するポリメラーゼが単に使用されることができるが、しかし逆転写酵素は使用されない。この刊行物は従って、いわゆる「エラープローンPCR(Error-Prone-PCR)」に関する。
【0011】
WO 2004/024917は、酵素の指向された進化方法を記載し、その際このタンパク質及びそのコードDNAは空間的にカップリングされていて、かつ一区画中に閉じこめられている。このタンパク質はペプチドタグを有する融合コンストラクトとして存在する。出発材料はDNAライブラリーである。しかしながら、リガンド−タンパク質相互作用は選択のために使用されず、かつ、RNAポリメラーゼにより突然変異は導入されない。
【0012】
WO 2005/030957は、融合タンパク質としてコードDNAにカップリングしているタンパク質のin vitro選択を開示する。
【0013】
類似の系は、Bernath, K. et al.によっても記載されている(J,Mol. Biol. (04.02.2005) 345 (5), 1015-1026)。
【0014】
WO 01/51663は、統合された系及び核酸の修飾方法を開示する。特にここでは、Qβ−レプリカーゼの使用下でのNASBA方法が使用される。RNAポリメラーゼによる突然変異の導入に関する示唆はしかしながら開示されていない。
【0015】
逆転写酵素及び他のタンパク質からの融合タンパク質は既に公知であるにもかかわらず、RT−融合タンパク質又はT7−RNAポリメラーゼ−融合タンパク質のin vitro進化における使用についての示唆は存在しない。
【0016】
Qβ−レプリカーゼ系もまた既に詳細に試験されている(McCaskill及びBauer (Proc. Natl. Acad. Sei. USA 1993, 90, 4191-4195)。この刊行物は、Qβ−レプリカーゼ系中での進化波(Evolutionswelle)を記載する。このRNAフロントは、レプリコンの「フィット性」が高いほどより迅速に移動する。本発明による系は、しかしながらこの刊行物から取り出すことができない。
【0017】
WO 2004/108926は、改善された結合能を有するタンパク質の人工的な進化を開示する。前記タンパク質はRNAレプリコン中でコードされ、これは欠点のある複製により準種(Quasi-Spezies)を形成する。これはこの際しかしながら、in vivo発現である。
【0018】
これまでには、タンパク質を進化させることを可能にするin vitro系はいまだ存在しない。突然変異体ライブライリーの手間のかかる作成及びスクリーニングを回避する、改善した特性を有するタンパク質の目的に指向した進化のための系及び方法に関する要求がいまだに存在する。
【0019】
本発明の主題は、次の工程;
(A)次のものを含有するin vitro発現系の作成工程、
(i)変異すべきタンパク質Yをコードする核酸配列S、
(ii)タンパク質Y及び/又は少なくとも1種のこの変異体Y′に結合することができる目的分子X、
(iii)核酸配列Sを転写することができるRNAポリメラーゼ(Pol)、
(iv)核酸配列Sの転写産物を逆転写することができる逆転写酵素(RT)、
その際、目的分子XはPolに、そしてタンパク質YはRTにカップリングしているか、
又は目的分子XはRTに、そしてタンパク質YはPolにカップリングしている、
(B)タンパク質Yの変異体Y′及び、これをコードする核酸配列S′の形成下での転写、逆転写及び翻訳を可能にし、かつ、目的分子Xのための改善された結合特性を有する変異体Y′の形成を促進する条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び場合により特性決定工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含む、タンパク質Yの変異体Y′の製造方法である。
【0020】
特に、本発明は、タンパク質Yの変異体Y′の製造方法であって、次の工程;
(A)次のものを含有するin vitro発現系の作成工程、
(a)次のものをコードする核酸配列S、
(a1)ポリメラーゼ(Pol)によりin transに活性化可能な転写制御配列、
(a2)変異すべきタンパク質Y、及び
(a3)逆転写酵素(RT)、
又は
(a3′)ポリメラーゼ(Pol)、
その際、(a2)及び(a3)又は(a3′)をコードする配列断片は、(a1)によるin transに活性化可能な転写制御配列の制御下で融合タンパク質をコードする、
(b)次のものを含有するタンパク質複合体、
(b1)変異すべきタンパク質Yの変異体少なくとも1種に結合することができる成分X、及び
(b2)(a)からの核酸配列Sの転写のためのRNAポリメラーゼPol、
又は
(b2′)逆転写酵素RT、
(B)タンパク質Yの変異体Y′の形成を可能にする条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び場合により特性決定工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含むタンパク質Yの変異体Y′の製造方法に関する。
【0021】
本発明の内容は従って、一方では所定の変異体ライブラリーの選択を実験室中で可能にし、かつ同時に複製機構を包含する自律系である。この際、この選択は−天然の進化と同様に−、より良好に適合された変異体の有利な複製を介して実現されることが望ましい。
【0022】
本発明は従って、改善された特性、特に改善された結合特性を有するタンパク質の進化を可能にする方法を提供する。この本発明による系は、in vitro転写とin vitro翻訳及び逆転写を組み合わせる。意外にも、in vitro系中においてはこの3つの工程を1つの天然の進化方法へと組み合わせることが可能であることが強調される。この系においては、変異すべきタンパク質をコードする核酸配列のmRNA転写産物が産生され、この転写産物は引き続き逆転写により再度cDNAへと書き換えられ、これは次いで新たに転写されかつ逆転写されることができる。
【0023】
RNAポリメラーゼ及び逆転写酵素の使用下でのこの増幅方法は、PCRの代替品として既に公知である。この既に公知の核酸−増幅方法の1つは、NASBA原則(参照、例えばRomano et al., 1995, J. Virol. Methods, 54 (2-3); 109-119; Romano et al., 1997, Immunol. Invest. 26 (1-2): 15-28)である。
【0024】
有利には、逆転写酵素を用いるこの逆転写工程により、これは公知のとおり、プルーフリーディング機能のエラーのために所定のエラー率を有するが、転写産物から出発してcDNAが産生され、このうち少なくとも若干のものは突然変異によりこの当初のテンプレートとは相違する。このcDNAが繰り返し転写及び逆転写される場合には、核酸レベルで多数の変異体が得られ、これは変異すべきタンパク質の変異体をコードする。
【0025】
本発明によるin vitro系において、この転写産物は、タンパク質及びこの当初のコードされるタンパク質の変異体の形成下で翻訳される。
【0026】
本発明による発現系においては、定義された空間中で、転写、逆転写及び翻訳に必要とされる剤(例えばプライマー、dNTPs、NTPs、tRNAs、アミノ酸その他)が、十分な量で存在する。この定義された空間中に一定の部位で核酸配列S、目的分子X、RNAポリメラーゼ及び逆転写酵素が提供される場合には、例えば接種により提供される場合には、転写、逆転写及び翻訳の経過の際に、この接種部位で相応する剤が調達され、かつ、進行するいわゆる反応フロント(ここで転写産物、タンパク質及び逆転写産物(cDNA)が存在する、(これは最後に形成される))が形成される。
【0027】
使用される複製系は有利には、十分な突然変異率が複製の間に許可されるように構成される。このようにして理論的に試験される、出発コンストラクトの突然変異体の数は、1世代の当選者の子孫の数から、実験の世代の総数の累乗に対して計算される(例えば、1世代につき10個の子孫が400世代では=10400個の可能性のある変異体)。但し、この10400の個々の変異体の各々は明らかに実験において物理的に存在することは不可能であり、しかしながらこの系は自体で、複合体の仮想的な地帯の内部の「小道(Pfad)」を探索し、これは常に、絶対的な最大値へと導く。前記小道に沿った変異体のみが実験の間に存在していて、理論的には前記地帯の全ての点(変異体)が可能である。
【0028】
本発明の発明者は、改善された特性を有するタンパク質変異体をコードする、形成され、突然変異された核酸の特定のものの転写、逆転写及び翻訳が有利に進行するように、このタンパク質変異体の形成を制御するための手段を見出した。これにより、改善されたタンパク質変異体をコードする及びcDNA及び転写産物が、多数存在し、かつ、重合フロントでより迅速に前方に移動する。
【0029】
極めて特定の、改善された変異体の形成に必要とされる進化圧力(Evolutionsdruck)は次のように産生される。
【0030】
変異すべきタンパク質Yは、目的分子Xに結合することができる。前記タンパク質Yを本発明の範囲内で変異させる場合には、変異体Y′が生じ、このうち少なくとも幾つかは目的分子Xに対する結合のための改善された結合特性を示すことができる。このような変異体の産生を促進するため、及び場合によりこのような変異体をまた更に変異させるため、この結合特性を更に改善するために、本発明による方法においてこの条件は、X及び改善された結合特性を有する変異体Y′の間での結合が、改善された結合特性を有する変異体Y′をコードする転写産物を生じるように選択され、有利には逆転写される。これにより、改善された結合特性を有する変異体Y′をコードするcDNAsが生じる。改善された結合特性を有する変異体Y′のための転写産物の逆転写が促進されることにより、変異体の新規の転写及び翻訳もまた定量的に促進される。
【0031】
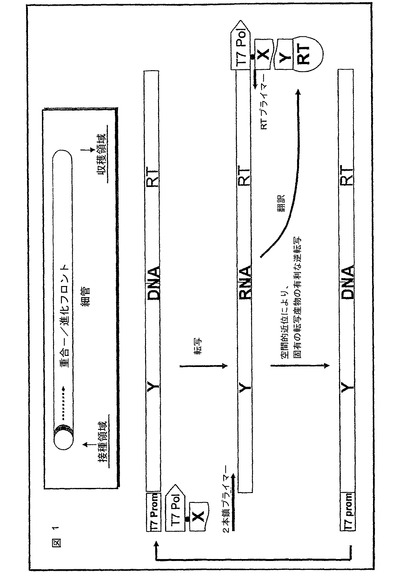
これを達成するために、有利には、RNAポリメラーゼ、逆転写酵素、X及び変異体Y′からなる複合体が形成され、これはこの変異体Y′をコードする転写産物の逆転写を可能にする。Xに対する改善された結合特性を有する変異体Y′をコードする転写産物に対するRTの空間的近位のために、この転写産物は、改善された変異体Y′と複合化されている(又はこれらと融合タンパク質を形成する)RTにより逆に転写される。図1は、本方法の本発明による代替法を図式により示す。
【0032】
有利には本発明による方法において、Yは核酸Sによりタンパク質Pを有する融合タンパク質としてコードされ、その際このタンパク質Pは、転写又は逆転写に関与するタンパク質である。Pは従って、RNAポリメラーゼ(Pol)又は逆転写酵素(RT)であることができ、又はPは、RNAポリメラーゼ又は逆転写酵素と会合(assozieren)しているか又はこれらに結合されることができるタンパク質であることができる。
【0033】
本発明による方法の2つの有利な代替法(1及び2)において、前記タンパク質Yは(1)RTと会合しているか又はRTを有する融合タンパク質としてコードされるか、又は(2)Polと会合するか又はPolを有する融合タンパク質としてコードされる。
【0034】
有利にはYは融合タンパク質として核酸配列Sによりコードされる。これはしかしながら、タンパク質Yとして核酸配列Sによりコードされ、かつ翻訳の後に相応する更なる成分と関連させられることもできる。これは、タンパク質相互作用を介して又は結合分子(例えばビオチン/アビジン、ビオチン ストレプトアビジンその他)を介して達成されることができる。Y又はY′のRT又はPolに対するカップリングの更なる手段は、とりわけ、共有結合的連結を介した化学的カップリング又は架橋性分子、例えばいわゆるリンカー、例えば二官能性クロスリンカーを介した連結をも包含する。この際適した架橋剤は当業者によりこの分野で問題なく選択されることができる。
【0035】
本発明による方法の第1の代替法に対する変法において、タンパク質Y及びRTからなる複合体はまた、2つの異なる核酸配列S1及びS2によっても、そのつど適した転写制御配列の制御下でコードされることができ、その際この場合には、融合タンパク質は生じないが、Y及びRTの間での結合は他の方法で引き起こされることができ、例えばビオチン/アビジン又はストレプトアビジンを介して引き起こされることができる。即ち、核酸配列Sはこの場合に、2つの核酸配列S1及びS2の形で存在する。重要なのは、この翻訳の終了時に、この変異すべきタンパク質Yが、RTタンパク質に結合しているか又はこれと複合化して存在することである。
【0036】
目的分子Xは、前述の代替法に相応して、(1)Polと又は(2)RTと会合しているか又はそのつどの成分を有する融合タンパク質を形成する。
【0037】
目的分子Xは、タンパク質、ペプチド又は核酸であるか、又は他の分子であることができる。従って、前記分子は、核酸(例えば、プラスミドでコードされる)として提供され、かつ本発明による発現系で発現されるか、又は、分子として提供されることができる。
【0038】
第1の代替法においては、変異すべきタンパク質Yは、逆転写酵素を有する融合タンパク質の形で発現カセットによりコードされる。発現カセットは、核酸配列Sの形で提供され、これは本発明による進化方法の経過において転写及び翻訳される。転写及び逆転写の他に、本方法は、核酸Sの転写産物の翻訳を可能にする。これにより、この第1の代替法において、この変異すべきタンパク質Y並びに逆転写酵素RTを含有する融合タンパク質が生じる。
【0039】
この逆転写酵素はしかしながら、第2の代替法に応じて、Xを有する複合体として提供され、同様にRTがその他の様式でXにカップリングしている融合タンパク質として又は複合体として提供されることもできる。第2の代替法によれば、核酸Sによりコードされる融合タンパク質が、RTの代わりにRNAポリメラーゼを含有する。この場合には、反応の開始時に、RNAポリメラーゼが提供されなくてはならない。この方法が次いで、転写及び翻訳を可能にする条件下で実施される場合には、この翻訳により産出される、SによりコードされるポリメラーゼPolは次のサイクルにおいて、出発核酸S並びに場合によりこの変異体S′を転写のためのテンプレートとして使用することができる。
【0040】
転写制御配列は有利には、全ての慣用の、RNA重合反応に適したプロモーターから選択されることができるプロモーターである。有利には、本発明の目的のために、T7−、T3−及びSP6−RNAポリメラーゼプロモーターである。しかしながら他のプロモーターが選択されることもできる。
【0041】
転写の実施のために、相応してRNAポリメラーゼが提供され、有利にはこれはRNAポリメラーゼT7、RNAポリメラーゼT3及びRNAポリメラーゼSP6から選択される。RNAポリメラーゼは、本発明による方法の選択される代替法に応じて、核酸によりコードされて存在することができるか、又はタンパク質(又は融合タンパク質又は複合体)として発現系中に導入されることができる。
【0042】
重要なのは、本発明の目的のためには、X及びY又はX及びY′の間での結合が成功することであり、この結合により逆転写酵素RTはこの転写産物の空間的近傍にもたらされる。即ち、P/X/(Y又はY′)/RTからなる立体複合体が生じる。これは、転写反応及び翻訳反応の経過後にまさに転写された、Y′をコードするmRNA分子が、RNAポリメラーゼPolに対して空間的近傍に存在し、これはタンパク質Xとカップリングしていることを意味する。タンパク質X及びYまたはY′の間の結合が生じる場合には、その際YまたはY′はRTと複合化されて存在するが、有利にはRTタンパク質は転写産物に対する直接的な空間的近傍に見出される。YまたはY′を介したRNA転写産物及びタンパク質RTの間の空間的近位により、Y′のためのこの転写産物の逆転写が促進される。
【0043】
従って、mRNAレベルで核酸配列Sの変異体が産生されることができ、これは次いで、同様に変異体を作出する翻訳産物を生じる。このようにして、変異すべきタンパク質Yの変異体、即ち変異体Y′が生じる。この際、より良好に又はより劣悪に、前記タンパク質Xに結合する変異体が生じる。
【0044】
タンパク質Xに対する結合に関してより良好な特性を有する変異体Y′のための進化の利点は、有利な一実施態様において、転写の直ぐ後に、機能的なRTがその固有の転写産物の直ぐ近傍中で、そして従ってY′について見出され、かつこれが有利には逆転写されることにある。これにより、より迅速に及び/又はより多くの、改善された変異体Y′をコードするcDNAが生じる。
【0045】
選択利点を有するこの変異体は次いで再度、出発核酸配列S′として使用され、これは再度転写、逆転写、かつ翻訳されることができる。このようにして、有利なかつ従って増強した、タンパク質Yの核酸配列変異体S′及び変異体Y′の産生が生じ、これは本方法の経過の後に、適切な間隔の後に単離されることができる。同様に、逆転写酵素の変異体の産生の際のRTのエラー率も利用されることができる。
【0046】
この効果を、ポリメラーゼ及び/又は逆転写酵素及び/又は条件の選択により、当初提供される核酸配列Sの変異体S′及び従って前記核酸によりコードされるタンパク質、特にタンパク質Yの変異体Y′を産生することにより、本発明の系は利用する。
【0047】
有利には、特定のエラー率を有するDNA依存性RNAポリメラーゼが使用され、これにより突然変異を有する転写産物が生じる。この突然変異は例えば点突然変異であることができ、例えばポリメラーゼにより置換、欠失又は挿入が生じる可能性がある。
【0048】
RNAポリメラーゼもRTも、プルーフリーディング機能を有さず、かつ従ってプルーフリーディング機能を有するポリメラーゼと比較してより高い突然変異率を有する。
【0049】
この理由から従って、有利には、相応する野生型ポリメラーゼと比較してより高いエラー率を有するポリメラーゼが使用される。技術水準においては、高められた突然変異率を有するポリメラーゼが既に公知である。有利には、T7ポリメラーゼ及びT3ポリメラーゼ、またしかしSP6ポリメラーゼでもある。T7ポリメラーゼのうち、高められたエラー率を有する変異体が既に存在する(Brakmann及びGrzeszik, 2001, Chem. Biochem. 2, 212-219)。
【0050】
様に、所定の突然変異率を有する転写産物を産生することができる逆転写酵素も使用されることができる。
【0051】
有利には、Pol及びRTポリメラーゼ又は逆転写酵素のために5′−3′−エキソヌクレアーゼ活性無しで使用される。本発明の目的のためには、RNAポリメラーゼのエラー率又は逆転写酵素のいずれかが、又はしかしながら両方もが使用されることができる。
【0052】
転写産物又は/及び逆転写産物の産生のために、高められたエラー率を有する前述のPol分子及びRT分子に代替的に又は補足的に、突然変異率(エラー率)がその他の様式で高められることも可能である。例えば、突然変異作成剤、ヌクレオチド類似体が、Pol及び/又はRTの基質として又は/及びUV線も使用されることができる。当業者はこのような変異原物質を自体で選択することができ、というのもこれらは技術水準において公知であるからである。
【0053】
本発明による方法は、in vitroでの実施に適している。この発現環境はこのために適した全ての発現環境であることができ、有利には細管が使用される。細管の代わりにしかしながら、2次元の発現環境、例えば2つのガラスプレートの間の環境を選択することも可能である。また3次元の発現環境を使用することもできる。
【0054】
細管が使用される場合には、これらは末端でいわゆる接種領域で核酸配列Sでもって接種される。
【0055】
2次元系が使用される場合には、この系を、隅で又は他の部位でも、例えば中間部で、核酸配列Sでもって接種することも可能である。このような2次元系により、小道の自由度及び数は向上し、これは変異すべきタンパク質Yの進化に関連する(einschlagen)。このプロセスは両方の場合において、重合フロントの観察を介して制御可能なままである。これは例えば、マーカー剤、例えばインターカレーション剤により行われることができる。新規の変異体は次いで、より迅速な、線形に細管系中で又は円形に2次元系中で伸展するフロントを形成する。所望の変異体の試料取り出し又は単離は次いで、細管の末端で又は2次元系の周辺で又は任意の他の部位で行われることができる。
【0056】
また、より大きな容量の3次元の発現系を使用することも可能である。このために次いで、反応媒体がより粘性な状態であるように発現環境条件が選択されることが望ましく、というのもこの液体はより大きな容量の系中で毛管力によりあまり安定化されないからである。
【0057】
細管又は2次元又は3次元系に沿った重合フロントの速度は変異体の発生の前進のための、そして従って変異すべきタンパク質Y及びその変異体Y′の結合特性の改善のための指標として用いられることができる。
【0058】
更に、転写及び翻訳のための全ての必要な反応条件は相応して調節される。特にこのために、プライマーとしての相応するオリゴヌクレオチド、及び、ヌクレオチド、特にdNTPs、NTPsその他の作成が属する。翻訳のために相応する酵素及び剤が必要とされ、例えばリボソーム、tRNAs、アミノ酸、エネルギーキャリア(例えばGTP及び類似物)その他が必要とされる。
【0059】
この種の反応条件は、当業者により問題無しに調整されることができ、かつ、この相応するプライマーヌクレオチドは当業者により選択されることもできる。
【0060】
本発明による方法は、提供される系中での自体によるタンパク質Yの変異体Y′の産生を可能にする。従って、変異すべきタンパク質Yを自体で特定の方向に発達させることが可能であり、これは目的分子Xに対する変異体Y′の結合能により制御されることができる。
【0061】
従って、そのつど任意のタンパク質Yを本発明による方法において進化させることが可能である。このために単に、Yに結合することができる相応する目的分子Xが選択され、かつ、Xのための改善された結合特性を有する変異体が手間のかかるスクリーニング無しに得られる。
【0062】
有利には、核酸配列変異体S′(RNA又はDNA又は両方)は、反応フロントで単離される。しかしながら、Y′を単離することもできる。
【0063】
更に、本発明は、改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)RNAポリメラーゼPolによりin transに活性化可能な発現制御配列をコードする核酸配列S、変異すべきタンパク質Y、逆転写酵素RT、
(c)Pol及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができるタンパク質Xを含有する複合体、又は、この種の複合体をコードする核酸配列
を含有する改善された特性を有するタンパク質の製造のためのキットに関する。
【0064】
更に本発明は、改善された特性を有するタンパク質の製造のための代替的なキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(i)RNAポリメラーゼPolによりin transに活性化可能な転写制御配列、
(ii)変異すべきタンパク質Y、
(iii)RNAポリメラーゼPol、
(c)逆転写酵素RT及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができるタンパク質Xを含有する複合体、又は、この種の複合体をコードする核酸配列
を含有する改善された特性を有するタンパク質の製造のための代替的なキットに関する。
【0065】
図の説明
図1は、請求項1による方法を図式により示す。NASBA反応混合物で充填された細管系中で、重合フロント及び進化フロントが進行する。ここでは、空間的に有利には分子が産生され、これは、T7−RNAポリメラーゼに対してカップリングしたタンパク質Xと相互作用できる変異体をコードする。
【0066】
実施例
T7RNAポリメラーゼ(NCBI遺伝子バンク, Acc. No. P00573)及びタンパク質A(Acc. No. CAA43604)からなる融合タンパク質大腸菌(E. coli)中で発現させ、アフィニティクロマトグラフィにより精製し、かつ、5μg/mlの濃度にPBS/10%グリセロール中で調整した。引き続き、抗HIV Env抗体2F5(Hofmann-Lehmann et al., 2001; Ferrantelli et al., 2003)を1:1の比で、キメラ融合タンパク質のこのタンパク質A−ドメインに対して結合させた(=RNA−Pol/2F5複合体)。
【0067】
シラン化した、両末端で開放している、1mmの内径及び10cmの長さを有するガラスキャピラリーを約80μlの次の反応混合物で充填した。
【0068】
大腸菌 in vitro翻訳−反応混合物(Rapid Translation System RTS)、Roche, Penzbergによる、1;1の比で、PBSを用いて
+RNA−Pol/2F5複合体、終濃度0.05μg/mlで、
+10mg/ml PEG4000、粘度向上のため、
+RTプライマー及び2本鎖プライマー、終濃度2pmol/μlで、
+dNTPヌクレオチド、そのつど0.1nmol/μlの終濃度で、
+NTPヌクレオチド、そのつど0.1nmol/μlの終濃度で、
+場合により、エチジウムブロミド、終濃度0.1ng/μlで。
【0069】
この充填された細管を37℃に加熱したチャンバー中に水平に固定し、末端で1μMの、2本鎖DNA分子溶液0.5μlで接種した。このDNA分子は、HIV−Env及びモロニーマウス白血病ウィルス(Acc. No. AAO46154)の逆転写酵素からの融合タンパク質のためのオープンリーディングフレームを含む。
【0070】
この反応チャンバーを閉鎖し、かつ接種されたDNAの複製は重合フロントとして交互に転写産物(RNA)及び逆転写産物(DNA)としてこの細管に沿って、存在する剤の方向に(他の末端へと)伝播することができる。
【0071】
この混合物にエチジウムブロミドを添加する場合には、UVハンドランプを用いて毎時間この目下の重合フロントの位置を確認し、かつこの反応の終点(この細管の末端への到達)を決定した。
【0072】
この細管末端への到達後に、重合フロント(RNA及びDNA)を含有する反応混合物2μlをこの細管から取り出し、かつこの含有されるDNAを、特異的オリゴヌクレオチドを用いたPCR反応を介して増幅した。このPCR産物を適したベクター中でサブクローニングし、かつ大腸菌中で形質転換した。
【0073】
この384クローンの配列評価は、進化したHIV−Env変異体をコードする相違する配列の統計学的分布を生じ、この変異体は抗体2F5に対する高められた親和性を示す。より詳細な検査は、この変異体中の繰り返されるタンパク質モチーフを示し、これは高められた親和性を担う。
【表1】
【図面の簡単な説明】
【0074】
【図1】図1は、請求項1による方法を図式により示す図である。
【技術分野】
【0001】
発明の詳細な説明
本発明は、in vitro進化方法におけるタンパク質の変異体の製造に関する。
【0002】
発明の背景
医薬、化学産業及び農業経済の範囲におけるバイオテクノロジーの増加する重要性は、そのつどのその使用目的に最適に適合しているタンパク質をますます求めている。まずこれらのタンパク質は主として環境から、大抵はいわゆるメタゲノムスクリーニングの範囲内で単離される。ますます前記タンパク質は将来には様々な方法を介して予定される「人工的な」使用条件に適用される。
【0003】
従って、例えば、天然の変異体よりもより耐熱性であり、他の基質特異性を有するか又はより高い活性を示す酵素が必要とされる。医薬的タンパク質は例えば、より少なく投薬されることができるようにより長い半減期を示すか、又は、目的分子に対する高親和性かつ特異的な結合を介して、疾病に関連した物質代謝経路又は感染経路を阻害することが望ましい。
【0004】
技術水準
この適合は部分的には、合理的タンパク質設計アプローチ(rationale Protein Design Ansaetze)を介して行われる。しかしながらこれまでには、タンパク質の所望の機能−表現型−からその構造を推定するか又はタンパク質の3次元構造から相応する一次配列を導く、極めて限定された手段のみが存在する。それにもかかわらずタンパク質の機能向上を達成すべく、現在のところ、「目的に指向した進化」(Directed Evolution)との概念にまとめられる部分的進化アプローチが使用される。
【0005】
現在の技術水準によれば、Directed Evolutionのこれまでの方法は実質的に、改良すべきタンパク質の多数の変異体(子孫)の産生及び改善された誘導体に応じたその選択に基づく。この際、検査すべき突然変異体の数は、部分的に極めて巨大である可能性があり、しかしながら通常は1011未満である。100個のみのアミノ酸のタンパク質を考慮する場合には、理論的に20100=10130個のこの相違する変異体が存在する。1011の大きさのライブラリーはこれに応じて、可能性のある変異体の極めて少ない部分のみをカバーする。このようなライブラリー中に理論的に最良の変異体が見出されることができる確率はほぼゼロである。
【0006】
目的分子に対して最大の親和性で多数の変異体をスクリーニングするために、既に多数のプロトコールが開発されている(例えば、酵母ツーハイブリッド(Yeast Two-Hybrid)、細菌ディスプレイ(Bacterial Display)、ファージディスプレイ(Phage Display)、リボソームディスプレイ(Ribosomal Display)、mRNAディスプレイ(mRNA Display))。
【0007】
他の特性、例えば酵素活性のスクリーニングのためには、大抵はアッセイフォーマット(Assay-Formate)が必要とされ、これは比較的少ない数の変異体(<106)のみの検査を可能にする。この方法は、突然変異体のライブラリーの産生並びに引き続くその選択に限定されている点で共通している。複製(=最後に行われる選択の「当選者」の突然変異体のすぐ次の世代)は、このプロトコールを用いて確かに手動で可能であるが、しかしながら先行する工程と同様に手間がかかる。従って、相応するアプローチは通例は、1〜2つの世代にわたるのみである。この理由から、述べられたプロトコールは、本来の意味合いにおける進化的アプローチではなく、むしろ存在する変異体プールの排他的選択である。従って、多数の世代を越えた段階的な適合の潜在能力は利用されることができず、これはしかしながら、場合によっては最も活性のある変異体を天文学的に巨大な数の可能性から同定するための必要な前提条件であるかも知れない。
【0008】
その3つの前提条件−複製、突然変異及び選択−を用いた進化の原則は、所定の系のうちでの、単純にそして高度にまで適合された構造の指向された発達を引き起こすことを可能にする。いわゆる「ブラインドウォッチメーカー」のこのモデルは、計画的な助力無しに、かつ、構造データの必要とされる知識無しに、複雑性の作成を可能にする。
【0009】
Bauer et al.は1989年に(PNAS (1989) 86, 7937-7941)、Qβ−レプリカーゼが見出されかつ任意のRNAで末端で接種(animpfen)される細管中で連続的な進化プロセスが生じることを示した。重合フロントがこの細管に沿って生じ、この経過において生じるRNAポリメラーゼはファージ特異的な配列/二次構造へと進化し、というのもこれらはレプリカーゼによりより迅速に複製されるからである。この実験が直接的な実際的な有用性を示さないにもかかわらず、この実験は3つの因子、複製、突然変異及び選択の潜在能力を顕著にカバーする(belegen)。
【0010】
WO 02/22869は、分子ライブラリーのin vitro進化の際の使用方法を記載する。この際、「ツーハイブリッド系(Two Hybrid - System)」系が使用される。ここで、高められたエラー頻度を有するポリメラーゼが単に使用されることができるが、しかし逆転写酵素は使用されない。この刊行物は従って、いわゆる「エラープローンPCR(Error-Prone-PCR)」に関する。
【0011】
WO 2004/024917は、酵素の指向された進化方法を記載し、その際このタンパク質及びそのコードDNAは空間的にカップリングされていて、かつ一区画中に閉じこめられている。このタンパク質はペプチドタグを有する融合コンストラクトとして存在する。出発材料はDNAライブラリーである。しかしながら、リガンド−タンパク質相互作用は選択のために使用されず、かつ、RNAポリメラーゼにより突然変異は導入されない。
【0012】
WO 2005/030957は、融合タンパク質としてコードDNAにカップリングしているタンパク質のin vitro選択を開示する。
【0013】
類似の系は、Bernath, K. et al.によっても記載されている(J,Mol. Biol. (04.02.2005) 345 (5), 1015-1026)。
【0014】
WO 01/51663は、統合された系及び核酸の修飾方法を開示する。特にここでは、Qβ−レプリカーゼの使用下でのNASBA方法が使用される。RNAポリメラーゼによる突然変異の導入に関する示唆はしかしながら開示されていない。
【0015】
逆転写酵素及び他のタンパク質からの融合タンパク質は既に公知であるにもかかわらず、RT−融合タンパク質又はT7−RNAポリメラーゼ−融合タンパク質のin vitro進化における使用についての示唆は存在しない。
【0016】
Qβ−レプリカーゼ系もまた既に詳細に試験されている(McCaskill及びBauer (Proc. Natl. Acad. Sei. USA 1993, 90, 4191-4195)。この刊行物は、Qβ−レプリカーゼ系中での進化波(Evolutionswelle)を記載する。このRNAフロントは、レプリコンの「フィット性」が高いほどより迅速に移動する。本発明による系は、しかしながらこの刊行物から取り出すことができない。
【0017】
WO 2004/108926は、改善された結合能を有するタンパク質の人工的な進化を開示する。前記タンパク質はRNAレプリコン中でコードされ、これは欠点のある複製により準種(Quasi-Spezies)を形成する。これはこの際しかしながら、in vivo発現である。
【0018】
これまでには、タンパク質を進化させることを可能にするin vitro系はいまだ存在しない。突然変異体ライブライリーの手間のかかる作成及びスクリーニングを回避する、改善した特性を有するタンパク質の目的に指向した進化のための系及び方法に関する要求がいまだに存在する。
【0019】
本発明の主題は、次の工程;
(A)次のものを含有するin vitro発現系の作成工程、
(i)変異すべきタンパク質Yをコードする核酸配列S、
(ii)タンパク質Y及び/又は少なくとも1種のこの変異体Y′に結合することができる目的分子X、
(iii)核酸配列Sを転写することができるRNAポリメラーゼ(Pol)、
(iv)核酸配列Sの転写産物を逆転写することができる逆転写酵素(RT)、
その際、目的分子XはPolに、そしてタンパク質YはRTにカップリングしているか、
又は目的分子XはRTに、そしてタンパク質YはPolにカップリングしている、
(B)タンパク質Yの変異体Y′及び、これをコードする核酸配列S′の形成下での転写、逆転写及び翻訳を可能にし、かつ、目的分子Xのための改善された結合特性を有する変異体Y′の形成を促進する条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び場合により特性決定工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含む、タンパク質Yの変異体Y′の製造方法である。
【0020】
特に、本発明は、タンパク質Yの変異体Y′の製造方法であって、次の工程;
(A)次のものを含有するin vitro発現系の作成工程、
(a)次のものをコードする核酸配列S、
(a1)ポリメラーゼ(Pol)によりin transに活性化可能な転写制御配列、
(a2)変異すべきタンパク質Y、及び
(a3)逆転写酵素(RT)、
又は
(a3′)ポリメラーゼ(Pol)、
その際、(a2)及び(a3)又は(a3′)をコードする配列断片は、(a1)によるin transに活性化可能な転写制御配列の制御下で融合タンパク質をコードする、
(b)次のものを含有するタンパク質複合体、
(b1)変異すべきタンパク質Yの変異体少なくとも1種に結合することができる成分X、及び
(b2)(a)からの核酸配列Sの転写のためのRNAポリメラーゼPol、
又は
(b2′)逆転写酵素RT、
(B)タンパク質Yの変異体Y′の形成を可能にする条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び場合により特性決定工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含むタンパク質Yの変異体Y′の製造方法に関する。
【0021】
本発明の内容は従って、一方では所定の変異体ライブラリーの選択を実験室中で可能にし、かつ同時に複製機構を包含する自律系である。この際、この選択は−天然の進化と同様に−、より良好に適合された変異体の有利な複製を介して実現されることが望ましい。
【0022】
本発明は従って、改善された特性、特に改善された結合特性を有するタンパク質の進化を可能にする方法を提供する。この本発明による系は、in vitro転写とin vitro翻訳及び逆転写を組み合わせる。意外にも、in vitro系中においてはこの3つの工程を1つの天然の進化方法へと組み合わせることが可能であることが強調される。この系においては、変異すべきタンパク質をコードする核酸配列のmRNA転写産物が産生され、この転写産物は引き続き逆転写により再度cDNAへと書き換えられ、これは次いで新たに転写されかつ逆転写されることができる。
【0023】
RNAポリメラーゼ及び逆転写酵素の使用下でのこの増幅方法は、PCRの代替品として既に公知である。この既に公知の核酸−増幅方法の1つは、NASBA原則(参照、例えばRomano et al., 1995, J. Virol. Methods, 54 (2-3); 109-119; Romano et al., 1997, Immunol. Invest. 26 (1-2): 15-28)である。
【0024】
有利には、逆転写酵素を用いるこの逆転写工程により、これは公知のとおり、プルーフリーディング機能のエラーのために所定のエラー率を有するが、転写産物から出発してcDNAが産生され、このうち少なくとも若干のものは突然変異によりこの当初のテンプレートとは相違する。このcDNAが繰り返し転写及び逆転写される場合には、核酸レベルで多数の変異体が得られ、これは変異すべきタンパク質の変異体をコードする。
【0025】
本発明によるin vitro系において、この転写産物は、タンパク質及びこの当初のコードされるタンパク質の変異体の形成下で翻訳される。
【0026】
本発明による発現系においては、定義された空間中で、転写、逆転写及び翻訳に必要とされる剤(例えばプライマー、dNTPs、NTPs、tRNAs、アミノ酸その他)が、十分な量で存在する。この定義された空間中に一定の部位で核酸配列S、目的分子X、RNAポリメラーゼ及び逆転写酵素が提供される場合には、例えば接種により提供される場合には、転写、逆転写及び翻訳の経過の際に、この接種部位で相応する剤が調達され、かつ、進行するいわゆる反応フロント(ここで転写産物、タンパク質及び逆転写産物(cDNA)が存在する、(これは最後に形成される))が形成される。
【0027】
使用される複製系は有利には、十分な突然変異率が複製の間に許可されるように構成される。このようにして理論的に試験される、出発コンストラクトの突然変異体の数は、1世代の当選者の子孫の数から、実験の世代の総数の累乗に対して計算される(例えば、1世代につき10個の子孫が400世代では=10400個の可能性のある変異体)。但し、この10400の個々の変異体の各々は明らかに実験において物理的に存在することは不可能であり、しかしながらこの系は自体で、複合体の仮想的な地帯の内部の「小道(Pfad)」を探索し、これは常に、絶対的な最大値へと導く。前記小道に沿った変異体のみが実験の間に存在していて、理論的には前記地帯の全ての点(変異体)が可能である。
【0028】
本発明の発明者は、改善された特性を有するタンパク質変異体をコードする、形成され、突然変異された核酸の特定のものの転写、逆転写及び翻訳が有利に進行するように、このタンパク質変異体の形成を制御するための手段を見出した。これにより、改善されたタンパク質変異体をコードする及びcDNA及び転写産物が、多数存在し、かつ、重合フロントでより迅速に前方に移動する。
【0029】
極めて特定の、改善された変異体の形成に必要とされる進化圧力(Evolutionsdruck)は次のように産生される。
【0030】
変異すべきタンパク質Yは、目的分子Xに結合することができる。前記タンパク質Yを本発明の範囲内で変異させる場合には、変異体Y′が生じ、このうち少なくとも幾つかは目的分子Xに対する結合のための改善された結合特性を示すことができる。このような変異体の産生を促進するため、及び場合によりこのような変異体をまた更に変異させるため、この結合特性を更に改善するために、本発明による方法においてこの条件は、X及び改善された結合特性を有する変異体Y′の間での結合が、改善された結合特性を有する変異体Y′をコードする転写産物を生じるように選択され、有利には逆転写される。これにより、改善された結合特性を有する変異体Y′をコードするcDNAsが生じる。改善された結合特性を有する変異体Y′のための転写産物の逆転写が促進されることにより、変異体の新規の転写及び翻訳もまた定量的に促進される。
【0031】
これを達成するために、有利には、RNAポリメラーゼ、逆転写酵素、X及び変異体Y′からなる複合体が形成され、これはこの変異体Y′をコードする転写産物の逆転写を可能にする。Xに対する改善された結合特性を有する変異体Y′をコードする転写産物に対するRTの空間的近位のために、この転写産物は、改善された変異体Y′と複合化されている(又はこれらと融合タンパク質を形成する)RTにより逆に転写される。図1は、本方法の本発明による代替法を図式により示す。
【0032】
有利には本発明による方法において、Yは核酸Sによりタンパク質Pを有する融合タンパク質としてコードされ、その際このタンパク質Pは、転写又は逆転写に関与するタンパク質である。Pは従って、RNAポリメラーゼ(Pol)又は逆転写酵素(RT)であることができ、又はPは、RNAポリメラーゼ又は逆転写酵素と会合(assozieren)しているか又はこれらに結合されることができるタンパク質であることができる。
【0033】
本発明による方法の2つの有利な代替法(1及び2)において、前記タンパク質Yは(1)RTと会合しているか又はRTを有する融合タンパク質としてコードされるか、又は(2)Polと会合するか又はPolを有する融合タンパク質としてコードされる。
【0034】
有利にはYは融合タンパク質として核酸配列Sによりコードされる。これはしかしながら、タンパク質Yとして核酸配列Sによりコードされ、かつ翻訳の後に相応する更なる成分と関連させられることもできる。これは、タンパク質相互作用を介して又は結合分子(例えばビオチン/アビジン、ビオチン ストレプトアビジンその他)を介して達成されることができる。Y又はY′のRT又はPolに対するカップリングの更なる手段は、とりわけ、共有結合的連結を介した化学的カップリング又は架橋性分子、例えばいわゆるリンカー、例えば二官能性クロスリンカーを介した連結をも包含する。この際適した架橋剤は当業者によりこの分野で問題なく選択されることができる。
【0035】
本発明による方法の第1の代替法に対する変法において、タンパク質Y及びRTからなる複合体はまた、2つの異なる核酸配列S1及びS2によっても、そのつど適した転写制御配列の制御下でコードされることができ、その際この場合には、融合タンパク質は生じないが、Y及びRTの間での結合は他の方法で引き起こされることができ、例えばビオチン/アビジン又はストレプトアビジンを介して引き起こされることができる。即ち、核酸配列Sはこの場合に、2つの核酸配列S1及びS2の形で存在する。重要なのは、この翻訳の終了時に、この変異すべきタンパク質Yが、RTタンパク質に結合しているか又はこれと複合化して存在することである。
【0036】
目的分子Xは、前述の代替法に相応して、(1)Polと又は(2)RTと会合しているか又はそのつどの成分を有する融合タンパク質を形成する。
【0037】
目的分子Xは、タンパク質、ペプチド又は核酸であるか、又は他の分子であることができる。従って、前記分子は、核酸(例えば、プラスミドでコードされる)として提供され、かつ本発明による発現系で発現されるか、又は、分子として提供されることができる。
【0038】
第1の代替法においては、変異すべきタンパク質Yは、逆転写酵素を有する融合タンパク質の形で発現カセットによりコードされる。発現カセットは、核酸配列Sの形で提供され、これは本発明による進化方法の経過において転写及び翻訳される。転写及び逆転写の他に、本方法は、核酸Sの転写産物の翻訳を可能にする。これにより、この第1の代替法において、この変異すべきタンパク質Y並びに逆転写酵素RTを含有する融合タンパク質が生じる。
【0039】
この逆転写酵素はしかしながら、第2の代替法に応じて、Xを有する複合体として提供され、同様にRTがその他の様式でXにカップリングしている融合タンパク質として又は複合体として提供されることもできる。第2の代替法によれば、核酸Sによりコードされる融合タンパク質が、RTの代わりにRNAポリメラーゼを含有する。この場合には、反応の開始時に、RNAポリメラーゼが提供されなくてはならない。この方法が次いで、転写及び翻訳を可能にする条件下で実施される場合には、この翻訳により産出される、SによりコードされるポリメラーゼPolは次のサイクルにおいて、出発核酸S並びに場合によりこの変異体S′を転写のためのテンプレートとして使用することができる。
【0040】
転写制御配列は有利には、全ての慣用の、RNA重合反応に適したプロモーターから選択されることができるプロモーターである。有利には、本発明の目的のために、T7−、T3−及びSP6−RNAポリメラーゼプロモーターである。しかしながら他のプロモーターが選択されることもできる。
【0041】
転写の実施のために、相応してRNAポリメラーゼが提供され、有利にはこれはRNAポリメラーゼT7、RNAポリメラーゼT3及びRNAポリメラーゼSP6から選択される。RNAポリメラーゼは、本発明による方法の選択される代替法に応じて、核酸によりコードされて存在することができるか、又はタンパク質(又は融合タンパク質又は複合体)として発現系中に導入されることができる。
【0042】
重要なのは、本発明の目的のためには、X及びY又はX及びY′の間での結合が成功することであり、この結合により逆転写酵素RTはこの転写産物の空間的近傍にもたらされる。即ち、P/X/(Y又はY′)/RTからなる立体複合体が生じる。これは、転写反応及び翻訳反応の経過後にまさに転写された、Y′をコードするmRNA分子が、RNAポリメラーゼPolに対して空間的近傍に存在し、これはタンパク質Xとカップリングしていることを意味する。タンパク質X及びYまたはY′の間の結合が生じる場合には、その際YまたはY′はRTと複合化されて存在するが、有利にはRTタンパク質は転写産物に対する直接的な空間的近傍に見出される。YまたはY′を介したRNA転写産物及びタンパク質RTの間の空間的近位により、Y′のためのこの転写産物の逆転写が促進される。
【0043】
従って、mRNAレベルで核酸配列Sの変異体が産生されることができ、これは次いで、同様に変異体を作出する翻訳産物を生じる。このようにして、変異すべきタンパク質Yの変異体、即ち変異体Y′が生じる。この際、より良好に又はより劣悪に、前記タンパク質Xに結合する変異体が生じる。
【0044】
タンパク質Xに対する結合に関してより良好な特性を有する変異体Y′のための進化の利点は、有利な一実施態様において、転写の直ぐ後に、機能的なRTがその固有の転写産物の直ぐ近傍中で、そして従ってY′について見出され、かつこれが有利には逆転写されることにある。これにより、より迅速に及び/又はより多くの、改善された変異体Y′をコードするcDNAが生じる。
【0045】
選択利点を有するこの変異体は次いで再度、出発核酸配列S′として使用され、これは再度転写、逆転写、かつ翻訳されることができる。このようにして、有利なかつ従って増強した、タンパク質Yの核酸配列変異体S′及び変異体Y′の産生が生じ、これは本方法の経過の後に、適切な間隔の後に単離されることができる。同様に、逆転写酵素の変異体の産生の際のRTのエラー率も利用されることができる。
【0046】
この効果を、ポリメラーゼ及び/又は逆転写酵素及び/又は条件の選択により、当初提供される核酸配列Sの変異体S′及び従って前記核酸によりコードされるタンパク質、特にタンパク質Yの変異体Y′を産生することにより、本発明の系は利用する。
【0047】
有利には、特定のエラー率を有するDNA依存性RNAポリメラーゼが使用され、これにより突然変異を有する転写産物が生じる。この突然変異は例えば点突然変異であることができ、例えばポリメラーゼにより置換、欠失又は挿入が生じる可能性がある。
【0048】
RNAポリメラーゼもRTも、プルーフリーディング機能を有さず、かつ従ってプルーフリーディング機能を有するポリメラーゼと比較してより高い突然変異率を有する。
【0049】
この理由から従って、有利には、相応する野生型ポリメラーゼと比較してより高いエラー率を有するポリメラーゼが使用される。技術水準においては、高められた突然変異率を有するポリメラーゼが既に公知である。有利には、T7ポリメラーゼ及びT3ポリメラーゼ、またしかしSP6ポリメラーゼでもある。T7ポリメラーゼのうち、高められたエラー率を有する変異体が既に存在する(Brakmann及びGrzeszik, 2001, Chem. Biochem. 2, 212-219)。
【0050】
様に、所定の突然変異率を有する転写産物を産生することができる逆転写酵素も使用されることができる。
【0051】
有利には、Pol及びRTポリメラーゼ又は逆転写酵素のために5′−3′−エキソヌクレアーゼ活性無しで使用される。本発明の目的のためには、RNAポリメラーゼのエラー率又は逆転写酵素のいずれかが、又はしかしながら両方もが使用されることができる。
【0052】
転写産物又は/及び逆転写産物の産生のために、高められたエラー率を有する前述のPol分子及びRT分子に代替的に又は補足的に、突然変異率(エラー率)がその他の様式で高められることも可能である。例えば、突然変異作成剤、ヌクレオチド類似体が、Pol及び/又はRTの基質として又は/及びUV線も使用されることができる。当業者はこのような変異原物質を自体で選択することができ、というのもこれらは技術水準において公知であるからである。
【0053】
本発明による方法は、in vitroでの実施に適している。この発現環境はこのために適した全ての発現環境であることができ、有利には細管が使用される。細管の代わりにしかしながら、2次元の発現環境、例えば2つのガラスプレートの間の環境を選択することも可能である。また3次元の発現環境を使用することもできる。
【0054】
細管が使用される場合には、これらは末端でいわゆる接種領域で核酸配列Sでもって接種される。
【0055】
2次元系が使用される場合には、この系を、隅で又は他の部位でも、例えば中間部で、核酸配列Sでもって接種することも可能である。このような2次元系により、小道の自由度及び数は向上し、これは変異すべきタンパク質Yの進化に関連する(einschlagen)。このプロセスは両方の場合において、重合フロントの観察を介して制御可能なままである。これは例えば、マーカー剤、例えばインターカレーション剤により行われることができる。新規の変異体は次いで、より迅速な、線形に細管系中で又は円形に2次元系中で伸展するフロントを形成する。所望の変異体の試料取り出し又は単離は次いで、細管の末端で又は2次元系の周辺で又は任意の他の部位で行われることができる。
【0056】
また、より大きな容量の3次元の発現系を使用することも可能である。このために次いで、反応媒体がより粘性な状態であるように発現環境条件が選択されることが望ましく、というのもこの液体はより大きな容量の系中で毛管力によりあまり安定化されないからである。
【0057】
細管又は2次元又は3次元系に沿った重合フロントの速度は変異体の発生の前進のための、そして従って変異すべきタンパク質Y及びその変異体Y′の結合特性の改善のための指標として用いられることができる。
【0058】
更に、転写及び翻訳のための全ての必要な反応条件は相応して調節される。特にこのために、プライマーとしての相応するオリゴヌクレオチド、及び、ヌクレオチド、特にdNTPs、NTPsその他の作成が属する。翻訳のために相応する酵素及び剤が必要とされ、例えばリボソーム、tRNAs、アミノ酸、エネルギーキャリア(例えばGTP及び類似物)その他が必要とされる。
【0059】
この種の反応条件は、当業者により問題無しに調整されることができ、かつ、この相応するプライマーヌクレオチドは当業者により選択されることもできる。
【0060】
本発明による方法は、提供される系中での自体によるタンパク質Yの変異体Y′の産生を可能にする。従って、変異すべきタンパク質Yを自体で特定の方向に発達させることが可能であり、これは目的分子Xに対する変異体Y′の結合能により制御されることができる。
【0061】
従って、そのつど任意のタンパク質Yを本発明による方法において進化させることが可能である。このために単に、Yに結合することができる相応する目的分子Xが選択され、かつ、Xのための改善された結合特性を有する変異体が手間のかかるスクリーニング無しに得られる。
【0062】
有利には、核酸配列変異体S′(RNA又はDNA又は両方)は、反応フロントで単離される。しかしながら、Y′を単離することもできる。
【0063】
更に、本発明は、改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)RNAポリメラーゼPolによりin transに活性化可能な発現制御配列をコードする核酸配列S、変異すべきタンパク質Y、逆転写酵素RT、
(c)Pol及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができるタンパク質Xを含有する複合体、又は、この種の複合体をコードする核酸配列
を含有する改善された特性を有するタンパク質の製造のためのキットに関する。
【0064】
更に本発明は、改善された特性を有するタンパク質の製造のための代替的なキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(i)RNAポリメラーゼPolによりin transに活性化可能な転写制御配列、
(ii)変異すべきタンパク質Y、
(iii)RNAポリメラーゼPol、
(c)逆転写酵素RT及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができるタンパク質Xを含有する複合体、又は、この種の複合体をコードする核酸配列
を含有する改善された特性を有するタンパク質の製造のための代替的なキットに関する。
【0065】
図の説明
図1は、請求項1による方法を図式により示す。NASBA反応混合物で充填された細管系中で、重合フロント及び進化フロントが進行する。ここでは、空間的に有利には分子が産生され、これは、T7−RNAポリメラーゼに対してカップリングしたタンパク質Xと相互作用できる変異体をコードする。
【0066】
実施例
T7RNAポリメラーゼ(NCBI遺伝子バンク, Acc. No. P00573)及びタンパク質A(Acc. No. CAA43604)からなる融合タンパク質大腸菌(E. coli)中で発現させ、アフィニティクロマトグラフィにより精製し、かつ、5μg/mlの濃度にPBS/10%グリセロール中で調整した。引き続き、抗HIV Env抗体2F5(Hofmann-Lehmann et al., 2001; Ferrantelli et al., 2003)を1:1の比で、キメラ融合タンパク質のこのタンパク質A−ドメインに対して結合させた(=RNA−Pol/2F5複合体)。
【0067】
シラン化した、両末端で開放している、1mmの内径及び10cmの長さを有するガラスキャピラリーを約80μlの次の反応混合物で充填した。
【0068】
大腸菌 in vitro翻訳−反応混合物(Rapid Translation System RTS)、Roche, Penzbergによる、1;1の比で、PBSを用いて
+RNA−Pol/2F5複合体、終濃度0.05μg/mlで、
+10mg/ml PEG4000、粘度向上のため、
+RTプライマー及び2本鎖プライマー、終濃度2pmol/μlで、
+dNTPヌクレオチド、そのつど0.1nmol/μlの終濃度で、
+NTPヌクレオチド、そのつど0.1nmol/μlの終濃度で、
+場合により、エチジウムブロミド、終濃度0.1ng/μlで。
【0069】
この充填された細管を37℃に加熱したチャンバー中に水平に固定し、末端で1μMの、2本鎖DNA分子溶液0.5μlで接種した。このDNA分子は、HIV−Env及びモロニーマウス白血病ウィルス(Acc. No. AAO46154)の逆転写酵素からの融合タンパク質のためのオープンリーディングフレームを含む。
【0070】
この反応チャンバーを閉鎖し、かつ接種されたDNAの複製は重合フロントとして交互に転写産物(RNA)及び逆転写産物(DNA)としてこの細管に沿って、存在する剤の方向に(他の末端へと)伝播することができる。
【0071】
この混合物にエチジウムブロミドを添加する場合には、UVハンドランプを用いて毎時間この目下の重合フロントの位置を確認し、かつこの反応の終点(この細管の末端への到達)を決定した。
【0072】
この細管末端への到達後に、重合フロント(RNA及びDNA)を含有する反応混合物2μlをこの細管から取り出し、かつこの含有されるDNAを、特異的オリゴヌクレオチドを用いたPCR反応を介して増幅した。このPCR産物を適したベクター中でサブクローニングし、かつ大腸菌中で形質転換した。
【0073】
この384クローンの配列評価は、進化したHIV−Env変異体をコードする相違する配列の統計学的分布を生じ、この変異体は抗体2F5に対する高められた親和性を示す。より詳細な検査は、この変異体中の繰り返されるタンパク質モチーフを示し、これは高められた親和性を担う。
【表1】
【図面の簡単な説明】
【0074】
【図1】図1は、請求項1による方法を図式により示す図である。
【特許請求の範囲】
【請求項1】
タンパク質Yの変異体Y′の製造方法であって、次の工程:
(A)次のものを含有するin vitro発現系の作成工程、
(i)変異すべきタンパク質Yをコードする核酸配列S、
(ii)タンパク質Y及び/又は少なくとも1種のこの変異体Y′に結合することができる目的分子X、
(iii)核酸配列Sを転写することができるRNAポリメラーゼ(Pol)、
(iv)核酸配列Sの転写産物を逆転写することができる逆転写酵素(RT)、
その際、目的分子XはPolに、そしてタンパク質YはRTにカップリングしているか、
又は目的分子XはRTに、そしてタンパク質YはPolにカップリングしている、
(B)タンパク質Yの変異体Y′及び、これをコードする核酸配列S′の形成下での転写、逆転写及び翻訳を可能にし、かつ、目的分子Xのための改善された結合特性を有する変異体Y′の形成を促進する条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含む、タンパク質Yの変異体Y′の製造方法。
【請求項2】
次の工程:
(A)次のものを含有するin vitro発現系の作成工程、
(a)以下のものをコードする核酸配列S、
(a1)RNAポリメラーゼPolによりin transに活性化可能な転写制御配列、
(a2)変異すべきタンパク質Y、及び
(a3)逆転写酵素(RT)、
又は
(a3′)RNAポリメラーゼ(Pol)、
その際、(a2)及び(a3)又は(a3′)をコードする配列断片は、(a1)によるin transに活性化可能な転写制御配列の制御下で融合タンパク質をコードする、
(b)次のものを含有するタンパク質複合体、
(b1)タンパク質Y及び/又は変異すべきタンパク質Yの少なくとも1種の変異体Y′に結合することができる成分X、及び
(b2)(a)からの核酸配列Sの転写のためのRNAポリメラーゼ(Pol)、
又は
(b2′)逆転写酵素(RT)、
(B)タンパク質Yの変異体Y′の形成を可能にする条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び/又はY′をコードする核酸配列変異体S′の単離工程
を含む、請求項1記載の方法。
【請求項3】
Pol、RT、X及び変異体Y′から複合体が形成され、これはこの変異体Y′をコードする転写産物の逆転写を可能にする、請求項1又は2記載の方法。
【請求項4】
核酸配列SがY及びRTからなる融合タンパク質又はY及びPolからなる融合タンパク質をコードする、請求項1から3までのいずれか1項記載の方法。
【請求項5】
Pol、RT及び/又はXが核酸によりコードされるか又はタンパク質又は融合タンパク質として提供される、請求項1から4までのいずれか1項記載の方法。
【請求項6】
in vitro発現系として、細管、2次元の発現環境又は3次元の発現環境が使用される、請求項1から5までのいずれか1項記載の方法。
【請求項7】
転写制御配列が、RNA−ポリメラーゼ−T7−プロモーター、RNAポリメラーゼ−T3−プロモーター及びRNA−ポリメラーゼ−SP6−プロモーターから選択される、請求項2から6までのいずれか1項記載の方法。
【請求項8】
PolとしてRNA−ポリメラーゼ−T7、RNA−ポリメラーゼ−T3又はRNAポリメラーゼ−SP6が使用される、請求項1から7までのいずれか1項記載の方法。
【請求項9】
Polとして、相応する野生型ポリメラーゼが示すエラー率と比較して高められたエラー率を示すRNAポリメラーゼが使用される、請求項1から8までのいずれか1項記載の方法。
【請求項10】
改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(b1)RNAポリメラーゼによりin transに活性化可能な発現制御配列、
(b2)変異すべきタンパク質Y、
(b3)逆転写酵素、
(c)RNAポリメラーゼ及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができる目的分子Xを含有する複合体、又は、この種の複合体をコードする核酸配列、
を含有する、改善された特性を有するタンパク質の製造のためのキット。
【請求項11】
改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(b1)タンパク質Pによりin transに活性化可能な転写制御配列、
(b2)変異すべきタンパク質Y、
(b3′)RNAポリメラーゼ、
(c)逆転写酵素及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができる目的分子Xを含有する複合体、又は、この種の複合体をコードする核酸配列、
を含有する、改善された特性を有するタンパク質の製造のためのキット。
【請求項12】
発現環境が、細管、2次元の発現環境及び3次元の発現環境から選択されている、請求項10又は11記載のキット。
【請求項13】
適したプライマー、dNTPs、NTPs、及び/又は緩衝液を更に含有する、請求項10から12までのいずれか1項記載のキット。
【請求項1】
タンパク質Yの変異体Y′の製造方法であって、次の工程:
(A)次のものを含有するin vitro発現系の作成工程、
(i)変異すべきタンパク質Yをコードする核酸配列S、
(ii)タンパク質Y及び/又は少なくとも1種のこの変異体Y′に結合することができる目的分子X、
(iii)核酸配列Sを転写することができるRNAポリメラーゼ(Pol)、
(iv)核酸配列Sの転写産物を逆転写することができる逆転写酵素(RT)、
その際、目的分子XはPolに、そしてタンパク質YはRTにカップリングしているか、
又は目的分子XはRTに、そしてタンパク質YはPolにカップリングしている、
(B)タンパク質Yの変異体Y′及び、これをコードする核酸配列S′の形成下での転写、逆転写及び翻訳を可能にし、かつ、目的分子Xのための改善された結合特性を有する変異体Y′の形成を促進する条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び/又はY′をコードする核酸配列変異体S′の単離工程を含む、タンパク質Yの変異体Y′の製造方法。
【請求項2】
次の工程:
(A)次のものを含有するin vitro発現系の作成工程、
(a)以下のものをコードする核酸配列S、
(a1)RNAポリメラーゼPolによりin transに活性化可能な転写制御配列、
(a2)変異すべきタンパク質Y、及び
(a3)逆転写酵素(RT)、
又は
(a3′)RNAポリメラーゼ(Pol)、
その際、(a2)及び(a3)又は(a3′)をコードする配列断片は、(a1)によるin transに活性化可能な転写制御配列の制御下で融合タンパク質をコードする、
(b)次のものを含有するタンパク質複合体、
(b1)タンパク質Y及び/又は変異すべきタンパク質Yの少なくとも1種の変異体Y′に結合することができる成分X、及び
(b2)(a)からの核酸配列Sの転写のためのRNAポリメラーゼ(Pol)、
又は
(b2′)逆転写酵素(RT)、
(B)タンパク質Yの変異体Y′の形成を可能にする条件下での、(A)からのin vitro発現系のインキュベーション工程、
(C)Xに対する結合のための改善された結合特性を有する変異体Y′の単離工程及び/又はY′をコードする核酸配列変異体S′の単離工程
を含む、請求項1記載の方法。
【請求項3】
Pol、RT、X及び変異体Y′から複合体が形成され、これはこの変異体Y′をコードする転写産物の逆転写を可能にする、請求項1又は2記載の方法。
【請求項4】
核酸配列SがY及びRTからなる融合タンパク質又はY及びPolからなる融合タンパク質をコードする、請求項1から3までのいずれか1項記載の方法。
【請求項5】
Pol、RT及び/又はXが核酸によりコードされるか又はタンパク質又は融合タンパク質として提供される、請求項1から4までのいずれか1項記載の方法。
【請求項6】
in vitro発現系として、細管、2次元の発現環境又は3次元の発現環境が使用される、請求項1から5までのいずれか1項記載の方法。
【請求項7】
転写制御配列が、RNA−ポリメラーゼ−T7−プロモーター、RNAポリメラーゼ−T3−プロモーター及びRNA−ポリメラーゼ−SP6−プロモーターから選択される、請求項2から6までのいずれか1項記載の方法。
【請求項8】
PolとしてRNA−ポリメラーゼ−T7、RNA−ポリメラーゼ−T3又はRNAポリメラーゼ−SP6が使用される、請求項1から7までのいずれか1項記載の方法。
【請求項9】
Polとして、相応する野生型ポリメラーゼが示すエラー率と比較して高められたエラー率を示すRNAポリメラーゼが使用される、請求項1から8までのいずれか1項記載の方法。
【請求項10】
改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(b1)RNAポリメラーゼによりin transに活性化可能な発現制御配列、
(b2)変異すべきタンパク質Y、
(b3)逆転写酵素、
(c)RNAポリメラーゼ及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができる目的分子Xを含有する複合体、又は、この種の複合体をコードする核酸配列、
を含有する、改善された特性を有するタンパク質の製造のためのキット。
【請求項11】
改善された特性を有するタンパク質の製造のためのキットであって、
(a)発現環境、
(b)以下のものをコードする核酸配列S、
(b1)タンパク質Pによりin transに活性化可能な転写制御配列、
(b2)変異すべきタンパク質Y、
(b3′)RNAポリメラーゼ、
(c)逆転写酵素及び変異すべきタンパク質Yの変異体Y′の少なくとも1種に結合することができる目的分子Xを含有する複合体、又は、この種の複合体をコードする核酸配列、
を含有する、改善された特性を有するタンパク質の製造のためのキット。
【請求項12】
発現環境が、細管、2次元の発現環境及び3次元の発現環境から選択されている、請求項10又は11記載のキット。
【請求項13】
適したプライマー、dNTPs、NTPs、及び/又は緩衝液を更に含有する、請求項10から12までのいずれか1項記載のキット。
【図1】

【公表番号】特表2009−504145(P2009−504145A)
【公表日】平成21年2月5日(2009.2.5)
【国際特許分類】
【出願番号】特願2008−525458(P2008−525458)
【出願日】平成18年8月7日(2006.8.7)
【国際出願番号】PCT/EP2006/007798
【国際公開番号】WO2007/017229
【国際公開日】平成19年2月15日(2007.2.15)
【出願人】(508042032)ゲネアルト アクチエンゲゼルシャフト (2)
【氏名又は名称原語表記】GENEART AG
【住所又は居所原語表記】Josef−Engert−Strasse 11, D−93053 Regensburg, Germany
【Fターム(参考)】
【公表日】平成21年2月5日(2009.2.5)
【国際特許分類】
【出願日】平成18年8月7日(2006.8.7)
【国際出願番号】PCT/EP2006/007798
【国際公開番号】WO2007/017229
【国際公開日】平成19年2月15日(2007.2.15)
【出願人】(508042032)ゲネアルト アクチエンゲゼルシャフト (2)
【氏名又は名称原語表記】GENEART AG
【住所又は居所原語表記】Josef−Engert−Strasse 11, D−93053 Regensburg, Germany
【Fターム(参考)】
[ Back to top ]