
invitro再構成タンパク質合成系による膜タンパク質合成方法
【課題】アポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子に膜タンパク質を、invitro再構成タンパク質合成系を用いて、再構成する。
【解決手段】(1)厳密に成分とその量を制御できるinvitro再構成タンパク質合成系を用いることにより、(2)リン脂質-アポリポタンパク質複合体粒子に、膜タンパク質を再構成すること。
【解決手段】(1)厳密に成分とその量を制御できるinvitro再構成タンパク質合成系を用いることにより、(2)リン脂質-アポリポタンパク質複合体粒子に、膜タンパク質を再構成すること。
【発明の詳細な説明】
【技術分野】
【0001】
本願発明は、無細胞タンパク質合成系を用いた膜タンパク質の合成に関する。
より具体的には、存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬を含むin vitro再構成タンパク質合成系による膜タンパク質合成方法に関する。
【背景技術】
【0002】
医薬品や試薬として利用可能なタンパク質を遺伝子組換え技術を用いて生産する試みが広く行われている。遺伝子組換え技術では、取扱いの簡便さや効率の面から、現在のところ大腸菌、枯草菌、カビ、又は酵母等の微生物、蚕等の昆虫、ウシ等の哺乳動物、或は培養可能な植物細胞、昆虫細胞、動物細胞が好んで用いられている。遺伝子組換え技術を用いてタンパク質を生産させる方法は汎用されているが、目的のタンパク質の発現量が少ない、発現できても活性を持たない、或は凝集体を形成する等の問題があり、培養条件、生育条件、及び誘導条件の検討や、種々の発現系を試す等の試行錯誤が必要である。そのような種々の条件検討を行っても、生産することが難しいタンパク質も数多く報告されている。
【0003】
一方、このような生物や細胞を使わない、無細胞タンパク質合成と呼ばれるタンパク質合成法が知られている。無細胞タンパク質合成系は、in vitroタンパク質合成系やin vitro転写・翻訳系とも呼ばれ、大腸菌、ウサギ網状赤血球、又はコムギ胚芽細胞等から調製した抽出液又はこれらの粗画分を利用することによって、鋳型遺伝子が転写・翻訳されタンパク質が合成される。無細胞タンパク質合成系においては、生物や細胞の機能を乱すようなタンパク質も合成できる可能性が高いこと、96穴や384穴のフォーマットに適応させ多品種なタンパク質を合成したり、多様な合成反応条件を一度にたくさん試すことが可能である等、生物や細胞を用いることによる種々の制限を解除できるという特徴がある。
【0004】
ところで、タンパク質の内、膜タンパク質は、全タンパク質の20−30%を占め、種々の情報伝達や酵素反応など、生物機能に重要な役割を担っているといわれている。例えば、細胞質膜上の膜タンパク質には、細胞外でのリガンドとの結合により細胞内に必要な情報を伝達する役割をもつ膜タンパク質が含まれる。このように、膜タンパク質であれば、主にリン脂質からなる生体膜に固定された状態で、膜により隔てられた2つの部位で異なる機能を果たす場合も多い。
【0005】
ところが、膜タンパク質は、水溶液中ではその疎水性の高さから凝集などを起こして機能を示さず、脂質膜に結合又は付着した構造ではじめて活性を示すために、膜タンパク質を通常の無細胞系で遺伝子工学的に製造しても機能しないという問題があった。そこで、膜タンパク質の機能解析に、膜タンパク質を脂質二重膜に埋め込む手法が種々検討されている。例えば、リポソームに膜タンパク質を取り込む研究なども行われてきた。しかしながら、膜タンパク質自体の調製が難しいうえ、調製した膜タンパク質をリポソームに正しい方向性を持って導入することが困難で、解析が難しいという問題が残っている。
【0006】
ところで、従来から、血清中でコレステロール輸送に関与しているアポリポタンパク質(APO)は、リン脂質と混合することにより、アポリポタンパク質が脂質二重層の周りを取り囲む高密度リポ蛋白質様の円盤状の小粒子に再構成できることが知られていた(非特許文献1)。最近になり、この円盤状の小粒子の、膜タンパク質の機能解析への利用が提唱され、このような人工的な円盤状の小粒子は、リン脂質−アポリポタンパク質複合体粒子、ナノリポタンパク質、ナノディスクなどと呼ばれている。すなわち、リン脂質−アポリポタンパク質複合体粒子を再構成させる際に、膜タンパク質を共存させると、再構成されたリン脂質−アポリポタンパク質複合体粒子に膜タンパク質が含まれることが明らかとなり、膜タンパク質の機能性質の研究への応用が提唱されている(特許文献1、非特許文献2,3)。また、上記無細胞タンパク質合成系を用いて、ナノディスク(リン脂質−アポリポタンパク質複合体粒子)に膜タンパク質を埋め込んで合成する方法が発表されている(特許文献2)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4303468号
【特許文献2】特表2009-521209号
【非特許文献】
【0008】
【非特許文献1】Methods in Enzymology (1986) Vol.128, p.553-582
【非特許文献2】Protein Science (2003) vol.12, p.2476-2481
【非特許文献3】Journal of Biological Chemistry (2007) Vol.282, p.28309-28320
【発明の概要】
【発明が解決しようとする課題】
【0009】
本願発明は、アポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子に、in vitro 再構成タンパク質合成系を用いて、膜タンパク質を再構成することを第1の課題とする。
【0010】
更に本願発明は、in vitro 再構成タンパク質合成系の成分やその成分量を変化させることにより、膜タンパク質を前記リン脂質-アポリポタンパク質複合体粒子に再構成する最適な条件を検討することを第2の課題とする。
【課題を解決するための手段】
【0011】
本願発明者らは、(1)厳密に成分とその量を制御できるin vitro再構成タンパク質合成系を用いることにより、(2)アポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子に、膜タンパク質を挿入することに成功して、本願発明を完成させた。
【0012】
さらに本願発明者は、厳密に成分とその量を制御できるin vitro再構成タンパク質合成系を用いることにより、前記in vitro 再構成タンパク質合成系中のリボソームの量とリン脂質-アポリポタンパク質複合体粒子の量とを一定割合範囲内とすることにより、効率的に膜タンパク質を前記リン脂質-アポリポタンパク質複合体粒子に再構成できることを見出した。
【0013】
更に、リン脂質の組成を種々に変化させたり、更に/或いは、アポリポタンパク質の種類を変えてリン脂質-アポリポタンパク質複合体粒子を調製することにより、膜タンパク質の活性の高い状態で、前記リン脂質-アポリポタンパク質複合体粒子に膜タンパク質を再構成する方法を提供する。
【発明の効果】
【0014】
従来の無細胞タンパク質合成系を用いては、機能を保持した形で製造することが困難であった膜タンパク質を、成分が特定されているin vitro 再構成タンパク質合成系を用いることにより、定量的に活性を保つ形態で、製造することが可能となった。さらに本願発明者は、厳密に成分とその量を制御できるin vitro 再構成タンパク質合成系を用いることにより、前記in vitro 再構成タンパク質合成系中のリボソームとリン脂質-アポリポタンパク質複合体粒子とのモル比を一定割合、具体的には、1:0.01〜1:100、より好適には、1:0.1〜1:10とすることにより、膜タンパク質の前記リン脂質-アポリポタンパク質複合体粒子への効率的な再構成を可能とするという優れた効果を奏するものである。
【0015】
また、本願発明は、リン脂質及び/又はアポリポタンパク質の組成を種々に変化させてアポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子を調製することにより、膜タンパク質を、機能できる高次構造を保って前記リン脂質-アポリポタンパク質複合体粒子に再構成することができるという優れた効果を奏するものである。
【図面の簡単な説明】
【0016】
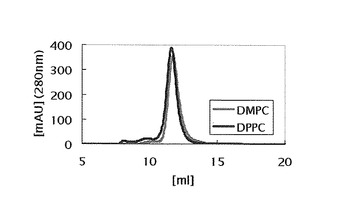
【図1】調製したDPPCもしくは、DMPCを含むリン脂質-アポリポタンパク質複合体粒子のゲル濾過クロマトグラム。
【図2】アポリポタンパク質としてMSP1-E1、脂質としてDMPCを含むリン脂質-アポリポタンパク質複合体粒子存在下(+)、および非存在下(-)で合成したバクテリオロドプシンを検出したゲル。Tは合成画分、Sは遠心上清画分を示す。
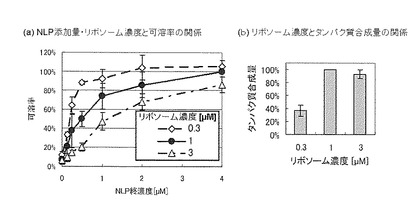
【図3】(a)は、0.3 μM、1 μM、又は3 μMのリボソームを含むPUREシステムに、リン脂質-アポリポタンパク質複合体粒子を0から4 μMまで変化させて添加した合成反応液を使用してバクテリオロドプシンを合成したときの可溶率の変化を示すグラフ。(b)は、0.3、1、3 μMのリボソーム濃度のPUREシステムでのバクテリオロドプシンの合成量を示すグラフ。リボソーム濃度が1 μMのときの合成量を100とした相対値で示す。
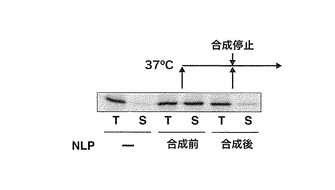
【図4】バクテリオロドプシン合成後に、リン脂質-アポリポタンパク質複合体粒子を反応液に添加しても可溶性は増大しないことを示すゲル。PUREシステムでバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた後、リン脂質-アポリポタンパク質複合体粒子を添加し、さらに1時間反応を続けた反応液、およびその遠心上清画分をSDS-PAGEで分離後、オートラジオグラフィーによりバンドを検出した。ネガティブコントロールとして、リン脂質-アポリポタンパク質複合体粒子を加えていない場合、ポジティブコントロールとして合成反応前にリン脂質-アポリポタンパク質複合体粒子を添加しておいた場合を示す。また、Tは合成画分、Sは遠心上清画分を示す。
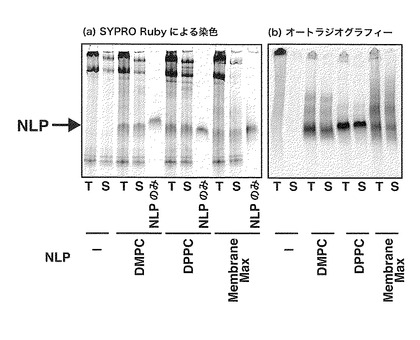
【図5】DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含むPUREシステムを用いてバクテリオロドプシンを合成し、合成画分(T)、遠心上清画分(S)をNativePAGEに供した結果を示すゲル。(a)は、リン脂質-アポリポタンパク質複合体粒子中のアポリポタンパク質をSYPRO Rubyで染色したゲルを、(b)は合成したバクテリオロドプシンをオートラジオグラフィーで検出したゲルを示す。
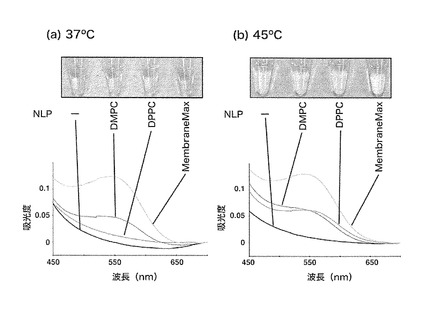
【図6】DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含むPUREシステムを用いて、37℃でバクテリオロドプシンを合成し、クロラムフェニコール添加により合成反応を停止させた後、レチナールを添加して色の変化を、(a) 37℃、(b) 45℃で観察した結果で、チューブの写真、および波長550nm付近における溶液の吸光度を測定し、それぞれ波長700nmの値を0としたときの値を示す。
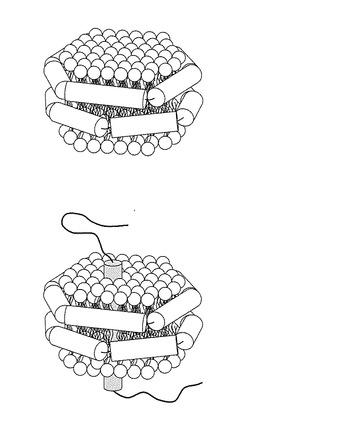
【図7】リン脂質−アポリポタンパク質複合体粒子を示す概念図である。
【発明を実施するための形態】
【0017】
1.はじめに
本願発明者らは、本願発明者らが既に開発したin vitro再構成タンパク質合成系(PUREシステムとも呼ばれる)と、リン脂質−アポリポタンパク質複合体粒子を組み合わせることで、膜タンパク質をコードする核酸を鋳型として合成された膜タンパク質をモデル膜であるリン脂質−アポリポタンパク質複合体粒子に再構成できるシステムを開発したものである。
【0018】
特に本願発明者等が開発したin vitro再構成タンパク質合成系を用いることにより、タンパク質合成系の種々の成分を厳密に制御できることから、本願発明は種々の応用ができるものである。
【0019】
例えば、本願発明は、存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、前記複数の成分の存在量及びリン脂質−アポリポタンパク質複合体粒子の存在量を変化させて、膜タンパク質の合成の最適な条件を検討する方法並びにその最適条件を用いた前記膜タンパク質を合成する方法を包含する。
【0020】
また、本願発明は、上記の膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子の種類及び/又は濃度及び/又は添加条件を変化させて膜タンパク質を合成し、当該膜タンパク質の可溶性、および/又は活性を測定することにより当該膜タンパク質の活性と、使用したリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件との相関を検証する方法を包含する。
【0021】
更に、本願発明は、存在量および純度がそれぞれ特定されている成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、及び存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子を含む、in vitro再構成タンパク質合成キットを包含する。
【0022】
2.膜タンパク質合成方法の概要
本願発明では、膜タンパク質は、a)目的とする膜タンパク質をコードする1または複数の鋳型核酸、b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、及びc) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子を適宜な溶媒に添加することで製造できる。溶媒としては、通常の無細胞タンパク質合成系で使用される緩衝液であれば、いずれのものでもよい。
【0023】
リン脂質−アポリポタンパク質複合体粒子は、in vitro再構成タンパク質合成系に共存して、タンパク質が合成されることが望ましく、或いは、必須である。in vitro再構成タンパク質合成系で膜タンパク質をその鋳型核酸から翻訳して調製した後に、リン脂質−アポリポタンパク質複合体粒子を添加しても、リン脂質−アポリポタンパク質複合体粒子に膜タンパク質が再構成されないことがある。
【0024】
本願発明において、膜タンパク質がリン脂質−アポリポタンパク質複合体粒子に再構成されるとは、膜タンパク質がリン脂質−アポリポタンパク質複合体粒子中の脂質二重層に挿入され、埋め込まれ、或いは貫通していることを意味している。
以下、まず上記a)、b)、c)の構成要素について、それぞれ以下に説明する。
【0025】
2−1.
膜タンパク質をコードする核酸
膜タンパク質は、いずれの膜タンパク質であっても膜タンパク質である限り、どのような生物種の由来であって、いずれのものも利用できる。例えば、膜タンパク質は、係留膜タンパク質、包埋膜タンパク質、または内在性膜タンパク質などに分類できるが、そのいずれでもよい。係留膜タンパク質の例としては、NADPH−シトクロムP450レダクターゼ、シトクロムb5およびヒト組織因子が挙げられる。包埋膜タンパク質の例としては、肝臓ミクロソーム由来のチトクロムP450が挙げられる。内在性膜タンパク質は、7回膜貫通タンパク質によって例示され、これには、バクテリオロドプシン、やGタンパク質共役タンパク質レセプターが挙げられる。
【0026】
核酸としては、DNAやmRNAなど下記のタンパク合成試薬で、翻訳、又は転写及び翻訳できるものであればよい。
【0027】
2−2タンパク質合成基本試薬
2−2−1
タンパク質合成基本試薬は、その成分として、リボソーム、開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、メチオニルtRNAトランスフォルミラーゼ類、tRNA類、アミノ酸類、リボヌクレオシド3リン酸類、10-フォルミル5,6,7,8-テトラヒドロ葉酸(FD)、塩類及び水をそれぞれ予め決められた純度の成分を決められた量だけ含むことを特徴とするが、全ての成分が必要でなく、成分は適宜選択することができる。
【0028】
系を構成するこれらの成分は、細胞抽出液又はその粗画分を用いないことはもちろんであるが、タンパク質のジスルフィド結合形成に影響を与える物質濃度を計算可能であることが望ましい。
【0029】
本発明のin vitro再構成タンパク質合成系は、系を構成する全ての成分を再構成するため、このような成分を特定し、その含有量を計算することは容易である。
【0030】
本発明におけるタンパク質合成反応基本試薬は、DNAからの転写・翻訳、或は、RNAの翻訳を行わせるタンパク質合成のための反応系として使用できる。本発明に言うタンパク質とは、2個以上のアミノ酸がペプチド結合によって結合したものを言い、ペプチド、オリゴペプチド、ポリペプチドを含む。本発明に言うRNAは化学合成されたRNA及びmRNAを含み、DNAは、合成されたDNA、DNAベクター、ゲノムDNA、PCR産物及びcDNAを含む。
【0031】
本発明におけるタンパク質合成反応基本試薬において、存在量および純度がそれぞれ特定されている成分を含むとは、それぞれの成分について、個々に精製され、その純度が測定可能であり、かつ定量可能であることを意味している。本発明では、存在量および純度がそれぞれ特定された複数の成分とは、予め塩析、クロマトグラフィー、電気泳動、溶解度の差、再結晶、遠心等の物質の精製法により、それぞれが精製された物質であって、クロマトグラフィー、電気泳動、質量分析、遠心等の分析方法により、それぞれの純度がおおむね80%以上、好適には90%以上である物質を言う。例えば、タンパク質であれば、主にクロマトグラフィーにより精製され、SDS−ポリアクリルアミドゲル電気泳動(SDS-PAGE)により純度が決定され、リボソームであれば、主に超遠心法により精製され、超遠心分離による沈降分析により純度が決定される。リボソームは複数のRNA分子(原核生物では、23S、5S、及び16Sの3つ、真核生物では28S、5.8S、5S、18Sの4つのRNA分子)と複数のリボソームタンパク質(原核生物では約50個、真核生物では約80個のタンパク質)とからなる分子量数百万の集合体分子であるが、沈降分析によって集合体分子として分子を同定し純度を測定することが可能である。tRNA類はほとんどが74から94ヌクレオチドから成り、異なる塩基配列をもつ分子であるが、電気泳動などにより分子を分離・同定し、260 nm及び280 nmの吸光度測定により純度を測定することが可能である。その他、アミノ酸や塩等の低分子の物質はいずれもクロマトグラフィー、融点測定、元素分析、質量分析等の常法により、物質を同定し純度を測定することが可能である。
【0032】
タンパク質合成基本試薬としての転写/翻訳のための因子・酵素としては、大腸菌等の原核細胞由来のものに限らず、真核細胞由来のものも使用でき、(1)RNAからの翻訳の場合は、リボソーム、開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、tRNA類、アデノシン3リン酸(ATP),グアノシン3リン酸(GTP),アミノ酸類、10-フォルミル5,6,7,8-テトラヒドロ葉酸(FD)、塩類及び水であり、大腸菌等の原核細胞由来の反応系である場合は更にメチオニルtRNAトランスフォルミラーゼ類を含む;(2)DNAからの転写/翻訳の場合は、(1)に加えウリジン3リン酸(UTP),シチジン3リン酸(CTP)及びRNAポリメラーゼ類、例えばT7RNAポリメラーゼを含む。
【0033】
本発明の反応系を構成する各種因子・酵素は、大腸菌、カビ、酵母、及び培養細胞等全ての生物が本来備えているものであるから、これをそれぞれ高度に精製し、成分として用いることもできるが、各タンパク質が多量に得られ、未知の不要又は阻害成分が反応系内に持ち込まれる可能性が低くなることから、組換え生産されたものを用いることがより好ましい。
【0034】
具体的には開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、メチオニルtRNAトランスフォルミラーゼ類、又はRNAポリメラーゼ類をコードする遺伝子を適切なベクターにつなぎ、大腸菌、枯草菌、カビ、又は酵母等に形質転換し、発現誘導を行い、該タンパク質を精製し、本発明の反応系を構成する成分とすることができる。組換え体により、各種因子・酵素を生産する場合、インタクトな状態でそのタンパク質を発現させても良いが、融合タンパク質として発現しても良い。そのような融合タンパク質として、ヒスチジンタグ(以下His-Tag)、ストレプトタグ、GSTタグ、及びFLAGタグ等を例示することができる。(ApplMicrobiol Biotechnol. 60(5):523-533(2003))
【0035】
一例として、His-Tagとニッケルカラムを利用した、His-Tagを付したタンパク質成分の精製方法の概略を示せば次の通りである。これ以外にも様々なバリエーションが知られており、適宜選択して使用できる。
1.遺伝子工学的手法により、目的タンパク質のN末にHis-Tag(6個のHisよりなる)を結合させた融合タンパク質を得る。
2.タグをつけたタンパク質が発現している細胞を氷中で超音波処理し、ローディングバッファー(300 mM NaCl, 50 mM NaH2PO4, pH 8.0)に懸濁させる。
3.細胞の溶解物を遠心分離する(30,000 g、4℃で30分間)。
4.上記で得られた上清に、氷で冷やしたローディングバッファーの中 で平衡化した50%のNi2+-NTA slurry (Qiagen社製)を加える。4℃で1時間撹拌する。
【0036】
5.樹脂をカラムにロードし、カラム容量の20倍のローディングバッファーで、4℃でカラムの洗浄を行う。
6.カラム容量の20倍のローディングバッファー(10 mMimidazole, pH 8.0を含む)で、4℃でカラムの洗浄を行う。
7.カラム容量の20倍のローディングバッファーを用いて、imidazoleの濃度勾配を10から250 mMになるように設定し、カラムから目的タンパク質の溶出を行わせ、1 mlずつフラクションを集める。SDS-PAGEで目的タンパク質を確認する。
【0037】
また、反応系を構成する各種因子・酵素として、直接タンパク質の合成反応には関わらないが、クレアチンキナーゼ類、ミオキナーゼ類又はヌクレオシドジフォスフェートキナーゼ類といったエネルギー再生に関わる酵素、及び無機ピロフォスファターゼ類といった、転写・翻訳反応で生じる無機ピロリン酸の分解のための酵素類も、同様な方法で該タンパク質を精製し、本発明の反応系に加えることがより好ましい。
【0038】
塩類としては、転写・翻訳に必須な陽イオン・陰イオンを含むことが必須であり、グルタミン酸カリウム、塩化アンモニウム、酢酸マグネシウム、塩化カルシウム等が通常使用される。なお、上記以外にも適宜選択して使用できることは言うまでもない。水はイオンや微生物類、酵素類を含まないもので、例えばミリポア社製のミリQ水製造装置によって製造される水や市販の純水を挙げることができる。
【0039】
リボソームは、ペプチド合成の場であり、mRNAと結合し、アミノアシルtRNAをA部位に、フォルミルメチオニルtRNA又はペプチジルtRNAをP部位にそれぞれ配位してペプチド結合を形成させる反応を行う(Science289:920-930(2000))。本発明においては、かかる機能を有するものであれば、由来を問わず使用することが可能である。例えば、大腸菌由来のリボソームが使用されるが、真核細胞由来のものも使用できる。本発明において用いられるリボソームの好ましい例は、大腸菌由来のものであり、例えば大腸菌A19株、MRE600株から得られるものを挙げることができる。
【0040】
リボソームの精製方法として、特許公開2007-121361号に記載の方法があげられるが、その他、従来使用されているスクロース密度勾配遠心を利用した方法なども使用できる。
【0041】
本発明のin vitro再構成タンパク質合成系で使用される開始因子類には、翻訳開始複合体の形成に必須であるか、又は、これを著しく促進する因子であり、大腸菌由来のものとして、IF1、IF2及びIF3が知られている(Biochemistry29:5881-5889(1990))。開始因子IF3は、翻訳の開始に必要な段階である、70Sリボソームの30Sサブユニットと50Sサブユニットへの解離を促進し、また、翻訳開始複合体の形成の際に、フォルミルメチオニルtRNA以外のtRNAのP部位への挿入を阻害する。開始因子IF2は、フォルミルメチオニルtRNAと結合し、30SリボソームサブユニットのP部位へフォルミルメチオニルtRNAを運び、翻訳開始複合体を形成する。開始因子IF1は開始因子IF2, IF3の機能を促進する。本発明において用いられる開始因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0042】
延長因子EF-Tuは、GTP型とGDP型の2種類があり、GTP型はアミノアシルtRNAと結合してこれをリボソームのA部位へ運ぶ。EF-Tuがリボソームから離れる際にGTPが加水分解され、GDP型へ転換する。(EMBOJ.17:7490-7497(1998))。延長因子EF-Tsは、EF-Tu(GDP型)に結合し、GTP型への転換を促進する(ArchivesofBiochemistryandBiophysics348:157-162(1997))。延長因子EF-Gは、ペプチド鎖伸長過程において、ペプチド結合形成反応の後の転位(translocation)反応を促進する(Nature Structure Biology 6:643-647(1999),FEMS Microbiology Reviews 23:317-333(1999))。本発明において用いられる延長因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0043】
解離因子は、終結因子とも呼ばれ、タンパク質合成の終結、翻訳されたペプチド鎖の解離、更に次のmRNAの翻訳開始へのリボソームの再生に必須である。解離因子RF1及びRF2は、リボソームのA部位に終止コドン(UAA,UAG,UGA)が来た時、A部位に入ってペプチジルtRNA(P部位にある)からのペプチド鎖の解離を促進する。RF1は終止コドンのうちUAA,UAGを認識し、RF2はUAA,UGAを認識する。解離因子RF3は、RF1,RF2によるペプチド鎖の解離反応後の、RF1,RF2のリボソームからの解離を促進する。リボソーム再生因子(RRF)は、タンパク質合成の停止後、P部位に残っているtRNAの脱離と、次のタンパク質合成へのリボソームの再生を促進する。本発明においては、RRFも解離因子類の一つとして取扱うことにする。なお、解離因子RF1,RF2,RF3及びRRFの機能については、EMBOJ.16:4126-4133(1997)、EMBOJ.16:4134-4141(1997)に解説されている。解離因子を含まない反応系でタンパク合成を行った場合は、終始コドンの手前で反応が止まり、リボソーム・ペプチド・mRNAの安定な3者複合体の形成が容易に行われる(ポリソームディスプレイ法、リボソームディスプレイ法)。またペプチド鎖への非天然アミノ酸の導入は、RF1及び/又はRF2を反応系から省くことにより行われる。即ち、RF1を省いた場合はUAGコドン、RF2を省いた場合はUGAコドンへの非天然アミノ酸の導入が高い効率で行われる。本発明において用いられる解離因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0044】
アミノアシルtRNA合成酵素は、ATPの存在下でアミノ酸とtRNAを共有結合させ、アミノアシルtRNAを合成する酵素である(RNA 3:954-960(1997),タンパク質核酸酵素39:1215-1225(1994))。本発明において用いられるアミノアシルtRNA合成酵素の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。また、非天然アミノ酸を認識する人工アミノアシルtRNA合成酵素(特許2668701号)を用いることもできる。
【0045】
メチオニルtRNAトランスフォルミラーゼ(MTF)は原核生物におけるタンパク質合成においてメチオニルtRNAのアミノ基にフォルミル基がついたN-フォルミルメチオニル(fMet)tRNAを合成する酵素である。即ち、メチオニルtRNAトランスフォルミラ―ゼは、N10‐フォルミルテトラヒドロ葉酸のフォルミル基を、開始コドンに対応するメチオニルtRNAのN末端に転移させ、fMet-tRNAにする(Proc.Natl.Acad.Sci.USA96:875-880(1999))。付加されたフォルミル基は開始因子IF2により認識され、タンパク質合成の開始シグナルとして作用する。真核生物の細胞質における合成系にはMTFはないが、真核生物のミトコンドリア及び葉緑体における合成系には存在する。本発明において用いられるMTFの好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものである。
【0046】
RNAポリメラーゼは、DNA配列をRNAに転写する酵素であり、様々な生物に存在することが知られている。その一例として、T7ファージ由来の、T7RNAポリメラーゼを挙げることができ、このポリメラーゼはT7プロモーターと呼ばれる特異的なDNA配列に結合してその下流のDNA配列をRNAに転写する酵素である。本発明者等は、T7RNAポリメラーゼのN末端にヒスタグを付加して、融合タンパク質として大腸菌BL21株において大量発現を行い、ニッケルカラムを用いるアフィニティクロマトグラフィーにより精製を行った。本発明においては、T7RNAポリメラーゼ以外にも種々のRNAポリメラーゼを用いることができる。例えば、T3RNAポリメラーゼやSP6RNAポリメラーゼが市販されており、これ等を利用することもできる。
【0047】
アミノ酸類としては、天然型アミノ酸に加え、非天然型アミノ酸も用いることができる。これらのアミノ酸は、in vitro再構成タンパク質合成系を構成するアミノアシルtRNA合成酵素の作用によってtRNAに保持される。あるいは、予めアミノ酸をtRNAにチャージしてin vitro再構成タンパク質合成系に加えることができる。本発明において、tRNAへのアミノ酸のチャージとは、tRNAにアミノ酸を結合させ、リボソームにおける翻訳反応に利用される状態にすることを言う。非天然アミノ酸を認識する人工アミノアシル合成酵素存在下で非天然アミノ酸を添加したり、非天然アミノ酸でチャージされたtRNAを用いたりすることで、蛋白質の特定のコドンの部位に非天然アミノ酸を導入することが可能となる。
【0048】
tRNA類としては、大腸菌、酵母等の細胞から精製したtRNAを用いることができる。またアンチコドンやその他の塩基を任意に変更した人工tRNAも用いることができる(Hohsaka, T et al. (1996) J. Am. Chem. Soc., vol.121, p.34-40, Hirao I et al (2002) Nat. Biotechnol., vol.20, p.177-182)。例えば、CUAをアンチコドンとして持つtRNAに非天然のアミノ酸をチャージすることで、本来終止コドンであるUAGコドンを非天然アミノ酸に翻訳することが可能である。また,4塩基コドンをアンチコドンとして持つtRNAに非天然アミノ酸をチャージした人工アミノアシルtRNAを用いることにより、天然には存在しない4塩基コドンを非天然アミノ酸に翻訳することが可能である(Hohsaka et al. (1999) J.Am.Chem.Soc., vol.121, p.12194-12195)。このような人工tRNAを作製する方法としては,RNAを用いる方法も使用できる(特表2003-514572)。これらの方法により部位特異的に非天然アミノ酸を導入した蛋白質を合成することができる。
【0049】
上記in vitro再構成タンパク質合成系を構成する各因子は、転写や翻訳に好適なpH7-8を維持する緩衝液に加えることによって、in vitro再構成タンパク質合成系とすることができる。本発明に用いられる緩衝液としては、リン酸カリウム緩衝液(pH 7.3)、Hepes-KOH(pH 7.6)などをあげることができる。
【0050】
上記in vitro再構成タンパク質合成系には、酵素の基質として、および/もしくは、活性の向上、維持を目的として、その他の低分子化合物を添加できる。具体的には、ヌクレオシド三リン酸(ATP, GTP, CTP, UTPなど)などの基質、プトレシン(putrescine)、スペルミジン(spermidine)などのポリアミン類、およびクレアチンリン酸などのエネルギー再生のための基質などをin vitro再構成タンパク質合成系に加えることができる。
【0051】
上記in vitro再構成タンパク質合成系では、例えば分子シャペロンと呼ばれる一群のタンパク質に代表される、タンパク質の凝集を抑制する因子は含んでいない。すなわち、本願発明においては、前記分子シャペロンを含まないin vitro再構成タンパク質合成系を用いても、膜タンパク質をリン脂質−アポリポタンパク質複合体粒子に挿入することができる。しかしながら、膜タンパク質の合成を行なう際に、これらの因子の添加することにより、凝集を抑制しながら合成することも可能である。
【0052】
分子シャペロンとしては、具体的には、例えば、Hsp70ファミリー、Hsp60ファミリー、Hsp90ファミリー、Hsp100ファミリー、低分子量Hspファミリー、イソメラーゼ類、および、これらの補助因子をあげることができる。分子シャペロン類も上記のタンパク質合成に関与する因子類と同様、上記のような精製法により、精製し、本発明の反応系に加えることがより好ましい。
【0053】
2−2−2 膜タンパク質がS-S結合を含む場合
更に、本願タンパク質合成基本試薬は、膜タンパク質が、S-S結合を含む場合、上記2−2−1記載のタンパク質合成基本試薬に加えて、ジスルフィド及びチオール間の酸化還元平衡状態の人為的調整をするための(イ)酸化還元を触媒する酵素、及び/又は(ロ)酸化還元状態を調節する試薬を含むことができる。
【0054】
2−2−2(1) ジスルフィド結合形成に影響を与える物質
ジスルフィド結合形成に影響を与える物質の例として、ジスルフィド結合の酸化還元を触媒する酵素である酸化還元酵素及び/又はジスルフィド結合の酸化還元状態を調節する試薬である酸化還元試薬が挙げられる。より具体的には、ジスルフィド結合形成に影響を与える酵素及び又は試薬として、(イ)酸化還元を触媒する酵素:グルタチオンレダクターゼ類、チオレドキシンレダクターゼ類、プロテインジスルフィドイソメラーゼ類、ジスルフィドインターチェインジタンパク質類、チオレドキシン様タンパク質等のタンパク質及び/又は(ロ)酸化還元状態を調節する試薬:還元型グルタチオン、酸化型グルタチオン、ジチオスレイトール(DTT)、2-メルカプトエタノール及びチオレドキシン等の低分子化合物を例示することができる。本発明における酸化還元試薬とはジスルフィドを還元しチオールにしたり、逆にチオールを酸化しジスルフィドを形成させることのできる試薬を言う。
【0055】
ジスルフィド結合形成に影響を与える物質の例としての、(イ)酸化還元酵素(酸化還元を触媒する酵素)及び/又は(ロ)酸化還元試薬(酸化還元状態を調節する試薬)は、タンパク質のジスルフィド結合の架橋を促進・調整する酵素類及び/又はその基質を用いることが望ましい。なお、(イ)酸化還元を触媒する酵素及び/又は(ロ)酸化還元状態を調節する試薬は、必ずしも翻訳反応中に加える必要はなく、翻訳終了後に加えても良い。翻訳終了後に添加する場合には、添加後更に数十分から1時間程度37℃に静置するのが好ましい。
【0056】
酸化還元状態を調節する試薬として、DTTを用いる場合は濃度が0〜5 mM、好適には0.001 〜2 mMであることが好ましい。更に好適には、0.060〜1 mMであることが好ましい。
【0057】
酸化還元状態を調節する試薬として、酸化型グルタチオンを用いる場合は濃度が0〜8mM、好適には、0.1〜4mMであることが好ましい。更に好適には1 〜4mMであることが好ましい。
【0058】
前記タンパク質の合成方法おいて、ジスルフィドインターチェインジタンパク質類はDsbA及び/又はDsbCであることが好ましい。
【0059】
プロテインジスルフィドイソメラーゼ(EC 5.3.4.1.)は真核生物の小胞体内膜に存在する約55kDaのジスルフィド結合の形成や異性化・還元反応を触媒する酵素であり、シャペロン様の活性も持つと言われている。このタンパク質は前述の通り、生物から精製し、成分として用いることもできるが、組換え生産されたものを用いてもよい。ウシの肝臓から精製したプロテインジスルフィドイソメラーゼ(PIR データベース アクセッション番号ISBOSS)、酵母のプロテインジスルフィドイソメラーゼ遺伝子を大腸菌に組換え発現させ、精製したタンパク質等を用いることができる。
【0060】
その他にプロテインジスルフィドイソメラーゼ類として下記のタンパク質知られており、本発明のプロテインジスルフィドイソメラーゼ質類として使用可能である。
ヒトのprotein disulfide-isomerase PIRデータベース アクセッションナンバー ISHUSS
ヒトのprotein disulfide isomerase related protein GenBank アクセッションナンバー4758304
酵母のprotein disulfide isomerase homolog PIR データベース A44483
【0061】
ジスルフィドインターチェインジタンパク質類は、ジスルフィドインターチェインジタンパク質A、B、C、D(DsbA, B , C, D)の4種のタンパク質が大腸菌等で知られている。DsbAは、21kDaのチオレドキシン様フォールド構造を持つ、ジスルフィド結合の形成を触媒すると考えられている酵素である。DsbBは4個の膜貫通部位と、2個のペリプラズムドメインを持つ20kDaのタンパク質で、DsbAを酸化型に維持すると言われている。DsbCはホモ2量体を形成するペリプラズムタンパク質であり、チオレドキシン様のフォールドを持ち、主にジスルフィド結合の異性化を担う酵素であると言われているが、他にシャペロンとしての働きもあると考えられている。DsbDは分子量59kDaのタンパク質で2つのペリプラズムドメイン、8個の膜貫通領域からなるタンパク質で、DsbCの活性中心のシステインを還元型に維持する働きを持っていると考えられる。
【0062】
ジスルフィドインターチェインジタンパク質類として下記のタンパク質が知られており、本発明のジスルフィドインターチェインジタンパク質類として使用可能である。
E. coliのDsbA SWISS-PROTprotein database Accession P24991
E. coli のDsbC SWISS-PROTprotein database Accession P21892
Salmonella typhimuriumのprotein disulfide-isomerase dsbA homolog PIRデータベース アクセッションナンバーS32895
Neisseria meningitidisのthiol-disulfide interchange protein dsbA homolog NMB0278
PIRデータベース アクセッションナンバーC81217
Caenorhabditis elegansのprotein disulphide isomerase isoform I Gene Bank アクセッションナンバー AAB94647
Datisca glomerataのprotein disulfide isomerase homolog Gene Bank アクセッションナンバーAAD28260
【0063】
これらジスルフィドインタージェインジタンパク質類は前述の通り、生物から精製し、成分として用いることもできるが、組換え生産されたものを用いてもよい。
【0064】
酸化還元酵素としてプロテインジスルフィドイソメラーゼ(PDI)を用いる場合、濃度が0 μM〜10 μMであることが好ましい。好適には0.001〜5 μMであることが好ましい。更に好適には0.001 μM〜2 μMであることが好ましい。酸化還元酵素としてジスルフィドインターチェインジタンパク質を用いた場合、0 μM〜10 μMであることが好ましい。好適には0.01から10 μMであることが好ましい。更に好適には0.1 μM〜10 μMであることが好ましい。
【0065】
なお、グルタチオンレダクターゼ類、チオレドキシンレダクターゼ類については、できるだけ含まないことが望ましく、タンパク質合成系を構成する成分におけるチオレドキシンレダクターゼ及び/又はグルタチオンレダクターゼの含量が100 ng/ml以下であることが好ましい。
【0066】
プロテインジスルフィドイソメラーゼ類、ジスルフィドインターチェインジタンパク質類も、上記のタンパク質合成に関与する因子類と同様、上記のような精製法により、精製し、本発明の反応系に加えることがより好ましい。
【0067】
2−3.リン脂質−アポリポタンパク質複合体粒子
本願発明で言う、リン脂質−アポリポタンパク質複合体粒子は、ナノリポタンパク質、(nanolipoprotein)、ナノディスク(Nanodisc)、ナノリポタンパク質粒子(nanolipoproteinparticles)、リン脂質−アポリポタンパク質粒子と称されることもある。当該粒子はリン脂質を含んでなり、アポリポタンパク質により二重層ディスクとして構成される。その構造の概略を図7に示す。
【0068】
本願発明明細書中では、アポリポタンパク質という用語には、天然のアポリポタンパク質又はその変異体であって、リン脂質−アポリポタンパク質粒子を形成できるものはすべて包含する。
【0069】
本願発明で言う、リン脂質−アポリポタンパク質複合体粒子は、例えば、アポリポタンパク質及びリン脂質を、コール酸若しくはコール酸塩を含む適宜な溶媒中で混合した後、混合液をコール酸及びコール酸塩を含まない溶媒に対して透析することにより得られる。
【0070】
従来技術において記載のように、リン脂質−アポリポタンパク質粒子は、単離されたアポリポタンパク質及びリン脂質を混合し、リン脂質−アポリポタンパク質として凝集させて調製してもよい。
【0071】
リン脂質−アポリポタンパク質複合体粒子の調製方法は従来技術において公知であり、例えば、Jonasら、(1984)J.Biol.Chem.259:6369−6375、Jonasら、(1989)J.Biol.Chem.264:4818−4824、Jonasら、(1993)J.Biol.Chem.268:1596−1602、米国特許第7048949号、米国特許出願公開第2005/0182243号A1、第2005/0152984号A1、第2004/0053384号A1及び第2006/0088524号A1、特許公報4303468号に記載されている。
【0072】
調製した前記リン脂質−アポリポタンパク質複合体粒子は、例えば、ゲル濾過クロマトグラフィーによって精製され、純度が決定される。
【0073】
本発明でいうアポリポタンパク質は、1つ以上の両親媒性のヘリックス状ドメインを有する。本願発明で言うアポリポタンパク質には、例えば、アポリポタンパク質A−I、アポリポタンパク質A−II、アポリポタンパク質A−IV、アポリポタンパク質A−V、アポリポタンパク質B−100、アポリポタンパク質B−48、アポリポタンパク質C−I、アポリポタンパク質C−II、アポリポタンパク質C−III、アポリポタンパク質D、アポリポタンパク質E、アポリポタンパク質H、リポタンパク質(a)、アポリポフォリンI(アポリポフォリンII)、及びアポリポフォリンIIIが挙げられる。また上述したように、本発明でいうアポリポタンパク質には、前記アポリポタンパク質のアミノ酸配列に対して1以上のアミノ酸が置換、欠失及び/又は付加した変異体であって、リン脂質と複合粒子を形成する能力を保持するもの、又は他のタンパク質との融合タンパク質であってリン脂質と複合粒子を形成する能力を保持するもの、その他誘導体も包含される。また、本発明でいうアポリポタンパク質には、たとえば。特表2007−525490号や特開2008−44958号に示される、MSP1、MSP2、MSP1E1、MSP1E2、MSP1E3およびMSP2などの膜スカホールドタンパク質(MSP)も包含される。
【0074】
本発明で用いるアポリポタンパク質としては、好適には、ApoA−I、ApoA−IV、ApoA−V、ApoC−I、ApoC−II、ApoC−III、Apo−E又はアポリポフォリンIII又はこれらのいずれかの変異体が挙げられる。
【0075】
リン脂質−アポリポタンパク質複合体粒子に用いるリン脂質は、グリセロール又はスフィンゴ脂質ベースであってもよく、例えば、2分子の6〜20の炭素原子の飽和脂肪酸、及び一般的に用いられるヘッド基(例えばホスファチジルコリン、ホスファチジルエタノールアミン及びホスファチジルセリン)を含んでなってもよい。ヘッド基は、荷電しなくともよく、正に荷電してもよく、負に荷電してもよく、また双性イオンであってもよい。リン脂質は、天然物)若しくは合成物、又は天然物と合成物の混合物であってもよい。リン脂質の非限定的な例としては、PC(ホスファチジルコリン)、PE(ホスファチジルエタノールアミン)、PI(ホスファチジルイノシトール)、DPPC(ジパルミトイル−ホスファチジルコリン)、DMPC(ジミリストイルホスファチジルコリン)、POPC(1−パルミトイル−2−オレオイル−ホスファチジルコリン)、DHPC(ジヘキサノイルホスファチジルコリン)、ジパルミトイルホスファチジルエタノールアミン、ジパルミトイルホスファチジルイノシトール、ジミリストイルホスファチジルエタノールアミン、ジミリストイルホスファチジルイノシトール、ジヘキサノイルホスファチジルエタノールアミン、ジヘキサノイルホスファチジルイノシトール、1−パルミトイル−2−オレオイル−ホスファチジルエタノールアミン、又は1−パルミトイル−2−オレオイル−ホスファチジル基イノシトールが挙げられる。
好適には、DMPCが挙げられる。
【0076】
3.本願発明のin vitro再構成タンパク質合成系による膜タンパク質合成方法でリン脂質-アポリポタンパク質複合体粒子に再構成された膜タンパク質の評価又は検証
3−1.膜タンパク質が「リン脂質-アポリポタンパク質複合体粒子に再構成」しているか否かの評価
本発明の方法により合成された膜タンパク質が、リン脂質−アポリポタンパク質複合体粒子に再構成されている、つまり、合成された膜タンパク質がリン脂質−アポリポタンパク質複合体粒子に挿入され、付着し、埋め込まれ、又は貫通等により再構成しているかは、目的の膜タンパク質がリン脂質−アポリポタンパク質複合体粒子と共に溶媒上清に存在するか、つまり可溶化しているか、それとも沈殿している、つまり可溶化していないかを計測することにより評価することができる。
【0077】
更に目的とする膜タンパク質の天然のリガンドや膜タンパク質と会合する因子が、リン脂質−アポリポタンパク質複合体粒子に再構成された膜タンパク質と会合するか否かを調べることにより確認することができる。
【0078】
例えば、バクテリオロドプシンがレチナールと結合するとレチナールが呈色反応をすることを利用することができる。
【0079】
3−2.リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能の評価・検証
目的とする膜タンパク質に応じた周知の機能測定方法により、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がその機能を有しているか、評価、検証することができる。
【0080】
4.膜タンパク質合成方法におけるタンパク質合成基本試薬中の各成分の量及びリン脂質−アポリポタンパク質複合体粒子の量の検討
本願発明のin vitro再構成タンパク質合成系による膜タンパク質合成方法は、「b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬」を用いるので、個々の成分の純度及び量を変化させて膜タンパク質合成することができ、更に、「c)リン脂質−アポリポタンパク質複合体粒子」についても、リン脂質−アポリポタンパク質複合体粒子の量を測定し、又はその量を変えて、或いは、濃度を変えて、膜タンパク質の合成を行うことができる。そこで、本願発明者らは、タンパク質合成反応基本試薬の種々の成分の量及び濃度並びにc)リン脂質−アポリポタンパク質複合体粒子の量及び濃度について検討を加えた。
【0081】
本願発明者は、様々な試行錯誤の上、タンパク質基本試薬の種々の成分の内、特にリボソーム量と、リン脂質−アポリポタンパク質複合体粒子の量の比率及びそれぞれの濃度が、膜タンパク質の合成に大きな影響を与えることを見出した。
【0082】
具体的には、in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.01〜1:100が望ましく、より好適には、in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.1〜1:10とすることがよいことをみいだした。
【0083】
さらに、リボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、上記比率とする時、in vitro再構成タンパク質合成系におけるリボソームの濃度が、0.1〜3 μM、リン脂質−アポリポタンパク質複合体粒子の濃度が0.01〜10μMであることが望ましいことを見出した。
【0084】
5.膜タンパク質の活性発現に最適なリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検証する方法
膜タンパク質の活性発現に最適な条件を求めるために、リン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検討し、検証することができる。
【0085】
5−1.膜タンパク質合成方法におけるリン脂質−アポリポタンパク質複合体粒子中のリン脂質の検討
リン脂質−アポリポタンパク質複合体粒子中のリン脂質を変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がより適切に構造的に再構成されているかを上記3.記載の評価方法により評価し、適切なリン脂質を選択することができる。
【0086】
また、リン脂質−アポリポタンパク質複合体粒子中のリン脂質を変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、適切なリン脂質を選択することもできる。
【0087】
たとえば、リン脂質としてDMPCを採用することにより、リン脂質−アポリポタンパク質複合体粒子に再構成した膜タンパク質は、より適切な高次構造をとり、膜タンパク質の機能を発揮しやすい。
【0088】
5−2.膜タンパク質合成方法におけるリン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の検討、並びにアポリポタンパク質及びリン脂質の組み合わせの検討
リン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の種類を例えば、上記2−3に記載されるアポリポタンパク質の内から選ばれるものに変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がより適切に構造的に再構成されているかを上記3.記載の評価方法により評価し、適切なアポリポタンパク質を選択することができる。
【0089】
また、同様にリン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の種類を変更し、リン脂質の種類も変更することにより、アポリポタンパク質及びリン脂質の組み合わせを変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定し、適切なアポリポタンパク質及びリン脂質の組み合わせを選択することもできる。
【0090】
5−3.膜タンパク質合成における、リン脂質−アポリポタンパク質複合体粒子中の膜タンパク質合成系への添加時期及び添加時点の温度の検討
存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加する時期を変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加する適切な時期を選択することもできる。
【0091】
また、存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加するときの温度を変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加するときの適切な温度を選択することもできる。
【実施例】
【0092】
<実施例1>リン脂質-アポリポタンパク質複合体の調製
特許第4303468号公報に従い、以下のようにリン脂質-アポリポタンパク質複合体粒子を調製した。アポリポタンパク質としては、ApoA-Iの改変体であるMSP1E1 (Denisov, I. G., et al. (2004) J. Am. Chem. Soc., vol. 126, p. 3477-3487) を文献に従って精製して使用した。ジパルミトイルホスファチジルコリン(DPPC)もしくは、ジミリストイルホスファチジルコリン(DMPC)のクロロホルム溶液(Avanti Polar Lipids製)を乾燥し、最終脂質濃度が50 mMになるように0.1 M NaClおよび100 mMコール酸ナトリウムを含む10 mM Tris-HClpH7.4の緩衝液に懸濁し、50℃に加熱しながらボルテックスすることで完全に溶解した。各リン脂質溶液を、モル比で、MSP1E1:リン脂質=2:200になるようにMSP1E1溶液と混合した。この混合溶液を、DMPCを含む混合溶液は室温(25℃)で、DPPCの場合は37℃で一晩インキュベートし、次いでコール酸ナトリウムを含まない1000倍容量の緩衝液に対して、緩衝液を4回交換しながら2日かけて透析した。透析後の混合溶液を、Superdex 200 10/300 GL (GE Healthcare)を用いたゲル濾過クロマトグラフィーにより分画し、図1に示すようなクロマトグラムを得た。ピーク画分を回収することにより、リン脂質-アポリポタンパク質複合体粒子を調製した。
【0093】
<実施例2>タンパク質合成反応試薬
タンパク質合成反応試薬の基本組成は、リボソーム濃度以外は、Shimizu et al. (2005) Methods, vol.36, p.299-304に従った。すなわち、50 mM HEPES-KOH pH7.6、2 mM ATP、2 mMGTP、1 mM CTP、1 mM UTP、20 mM クレアチンリン酸(Creatine phosphate)、56 A260 units/ml 大腸菌tRNA混合液、0.01 μg/μl 10-formyl-5,6,7,8-tetrahydrofolic acid(FD)、0.3 mM 各アミノ酸、13 mM 酢酸マグネシウム、100 mMグルタミン酸カリウム、2 mM スペルミジン(spermidine)、1 mM ジチオスレイトール(DTT),1 μM大腸菌リボソーム,0.02 μg/μl IF1,0.04 μg/μl IF2,0.015 μg/μl IF3,0.02 μg/μl EF-G,0.04 μg/μl EF-Tu,0.02 μg/μl EF-Ts,0.01 μg/μl RF1,0.01 μg/μl RF2,0.01 μg/μl RF3,0.01 μg/μl RRF,0.6-6 units/μl各アミノアシルtRNA合成酵素(ARS)及びメチオニルtRNAトランスフォルミラーゼ(MTF),0.004 μg/μl クレアチンキナーゼ(CK; creatine kinase),0.003 μg/μlミオキナーゼ(MK; myokinase),0.001 μg/μl ヌクレオシド二リン酸キナーゼ(NDK; nucleoside diphosphate kinase),0.0356 units/μl ピロフォスファターゼ(PPiase; Pyrophosphatase)及び0.01 μg/μl T7 RNAポリメラーゼを基本組成とした。通常のタンパク質合成では、上記基本組成のタンパク質合成反応試薬に、鋳型DNAを最終濃度が0.02-0.1 μMになるように添加して37℃で反応を行なった。なお、タンパク質合成の目的に応じて、上記基本組成に追加および、もしくは除去した反応試薬を使用した。上記組成のうち、リボソームは、Ohashi et al. (2007) BBRC, vol.352, p.270-276、及びタンパク質因子は、Shimizu et al. (2001) Nat. Biotechnol., vol.19, p.751-755に従って調製され、純度が測定されたものを、その他の成分は市販の精製試薬を使用した。
【0094】
<実施例3>リン脂質-アポリポタンパク質複合体粒子存在下でのバクテリオロドプシンの合成
EF-Tu、EF-Ts、EF-Gを3倍量含む上記基本試薬に、アポリポタンパク質としてMSP1-E1、脂質としてDMPCを含むリン脂質-アポリポタンパク質複合体粒子を最終濃度が4 μMになるように添加した膜タンパク質合成用PUREシステムを調製した。膜タンパク質合成用PUREシステムに、[35S]メチオニン、およびバクテリオロドプシンをコードする鋳型DNAを加えて15 μlに調製し、37℃で1時間反応させた。反応終了後、15 μlのミリQ水を加え、10 μlの合成画分を分取した。残りの反応液を20,000xgで30分間遠心し、上清画分を得た。合成画分、および遠心上清画分をSDS-PAGEにかけ、オートラジオグラフィーにより、合成されたバクテリオロドプシンの可溶性を評価した。図2に示すように、リン脂質-アポリポタンパク質複合体粒子非存在下で合成されたバクテリオロドプシンは、上清画分にほとんど回収されていないが、リン脂質-アポリポタンパク質複合体粒子存在下で合成した場合は、ほぼ全量が上清画分に回収された。このことは、細胞抽出液のように、分子シャペロンなど膜タンパク質の膜挿入を助ける因子を全く含まないPUREシステムにおいても、リン脂質-アポリポタンパク質複合体粒子存在下では合成膜タンパク質が可溶性に回収できることを示している。
【0095】
<実施例4>膜タンパク質合成におけるリン脂質-アポリポタンパク質複合体粒子濃度依存性
上記膜タンパク質合成用PUREシステムにおいて、リボソーム濃度を、0.3、1、3 μM、添加するリン脂質-アポリポタンパク質複合体粒子濃度を、0.25-4 μMのように変化させて、バクテリオロドプシンを合成した。合成画分、上清画分に含まれるバクテリオロドプシンを定量し、可溶率(%)=上清画分/合成画分×100として、各合成条件における可溶率を算出した。図3に示すように、リボソーム濃度を0.3 μMに減少させると、合成量も低下するため、必要となるリン脂質-アポリポタンパク質複合体粒子も低下した。一方、リボソーム濃度を増加させた場合、合成量は増加しなかったが、可溶率が低下した。この結果により、膜タンパク質合成におけるリボソーム濃度とリン脂質-アポリポタンパク質複合体粒子濃度には最適な条件があることが明らかになった。すなわち、1 μMリボソーム、4 μM リン脂質-アポリポタンパク質複合体粒子が最適であることが分かった。
【0096】
<実施例5>膜タンパク質合成におけるリン脂質-アポリポタンパク質複合体粒子添加時期依存性
上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた後、4 μMになるようにリン脂質-アポリポタンパク質複合体粒子を添加し、さらに1時間反応を続けた。合成反応画分を分取後、上記の遠心行ない、上清画分を回収した。一方で、上記のように合成前にリン脂質-アポリポタンパク質複合体粒子を添加した膜タンパク質合成用PUREシステムを用いて合成を行なった。合成画分、および遠心上清画分をSDS-PAGEで分離後、オートラジオグラフィーによりバンドを検出した(図4)。この結果、効率よく可溶性の膜タンパク質を合成するためには、あらかじめリン脂質-アポリポタンパク質複合体粒子を添加したPUREシステムを使用する必要があることが明らかとなった。
【0097】
<実施例6>膜タンパク質とリン脂質-アポリポタンパク質複合体粒子の複合体形成
DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含む上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンを合成し、上記と同様、合成画分、遠心上清画分を得た。それぞれNative-PAGEに供し、リン脂質-アポリポタンパク質複合体粒子をSYPRO Ruby(Invitrogen社製)で染色した(図5(a))。一方、合成された[35S]メチオニンを含むバクテリオロドプシンをオートラジオグラフィーで検出した(図5(b))。リン脂質-アポリポタンパク質複合体粒子存在下で合成した場合、いずれのリン脂質-アポリポタンパク質複合体粒子を用いても、リン脂質-アポリポタンパク質複合体粒子とほぼ同じ移動度のバンドを生じた。これは、バクテリオロドプシンがリン脂質-アポリポタンパク質複合体粒子と複合体を形成していることを示している。一方、リン脂質-アポリポタンパク質複合体粒子を添加せずに合成した場合、合成されたバクテリオロドプシンは、ゲルに入らないことが確認された。これは、リン脂質-アポリポタンパク質複合体粒子非存在下の合成では電気泳動されない凝集体を形成していることを示している。
【0098】
<実施例7>膜タンパク質の高次構造形成
DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含む上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた。合成反応液にレチナールを添加し、37℃、45℃でさらに1時間反応した。1時間後、反応液の色の変化を記録し、波長550nm付近の吸光度を測定した(図6)。DMPCを含むリン脂質-アポリポタンパク質複合体粒子、およびMembraneMaxを使用した場合は、反応温度に関わらず、溶液の色が黄色から紫色に変化することを確認した。レチナールは、正しい構造を形成したバクテリオロドプシンに結合し、紫色に変化することが知られている。すなわち、上記のリン脂質-アポリポタンパク質複合体粒子を使用した場合、正しい立体構造を形成したバクテリオロドプシンが合成されていることを示している。一方、DPPCを含むリン脂質-アポリポタンパク質複合体粒子を使用した場合、合成温度である37℃でレチナールを反応させた場合には色の変化が観察されなかったが、45℃で観察された。DMPC、DPPCの転移温度はそれぞれ約29℃、44℃であり、レチナールの結合には相転移温度以上が必要であることが分かった。実施例6において、合成されたバクテリオロドプシンは、37℃でDPPCを含むリン脂質-アポリポタンパク質複合体粒子と複合体を形成し、可溶性になることが示されている。しかしながら、レチナールとの結合には相転移温度以上が必要であることは、合成された膜タンパク質の評価には、可溶性の上昇だけでは不十分であることを示している。
【産業上の利用可能性】
【0099】
本願発明は、膜タンパク質の合成に利用できるほか、膜タンパク質の機能の測定、更には、ハイスループットに種々の膜タンパク質を合成し、その機能や、リガンド結合性などによりスクリーニングすることもできる。
【技術分野】
【0001】
本願発明は、無細胞タンパク質合成系を用いた膜タンパク質の合成に関する。
より具体的には、存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬を含むin vitro再構成タンパク質合成系による膜タンパク質合成方法に関する。
【背景技術】
【0002】
医薬品や試薬として利用可能なタンパク質を遺伝子組換え技術を用いて生産する試みが広く行われている。遺伝子組換え技術では、取扱いの簡便さや効率の面から、現在のところ大腸菌、枯草菌、カビ、又は酵母等の微生物、蚕等の昆虫、ウシ等の哺乳動物、或は培養可能な植物細胞、昆虫細胞、動物細胞が好んで用いられている。遺伝子組換え技術を用いてタンパク質を生産させる方法は汎用されているが、目的のタンパク質の発現量が少ない、発現できても活性を持たない、或は凝集体を形成する等の問題があり、培養条件、生育条件、及び誘導条件の検討や、種々の発現系を試す等の試行錯誤が必要である。そのような種々の条件検討を行っても、生産することが難しいタンパク質も数多く報告されている。
【0003】
一方、このような生物や細胞を使わない、無細胞タンパク質合成と呼ばれるタンパク質合成法が知られている。無細胞タンパク質合成系は、in vitroタンパク質合成系やin vitro転写・翻訳系とも呼ばれ、大腸菌、ウサギ網状赤血球、又はコムギ胚芽細胞等から調製した抽出液又はこれらの粗画分を利用することによって、鋳型遺伝子が転写・翻訳されタンパク質が合成される。無細胞タンパク質合成系においては、生物や細胞の機能を乱すようなタンパク質も合成できる可能性が高いこと、96穴や384穴のフォーマットに適応させ多品種なタンパク質を合成したり、多様な合成反応条件を一度にたくさん試すことが可能である等、生物や細胞を用いることによる種々の制限を解除できるという特徴がある。
【0004】
ところで、タンパク質の内、膜タンパク質は、全タンパク質の20−30%を占め、種々の情報伝達や酵素反応など、生物機能に重要な役割を担っているといわれている。例えば、細胞質膜上の膜タンパク質には、細胞外でのリガンドとの結合により細胞内に必要な情報を伝達する役割をもつ膜タンパク質が含まれる。このように、膜タンパク質であれば、主にリン脂質からなる生体膜に固定された状態で、膜により隔てられた2つの部位で異なる機能を果たす場合も多い。
【0005】
ところが、膜タンパク質は、水溶液中ではその疎水性の高さから凝集などを起こして機能を示さず、脂質膜に結合又は付着した構造ではじめて活性を示すために、膜タンパク質を通常の無細胞系で遺伝子工学的に製造しても機能しないという問題があった。そこで、膜タンパク質の機能解析に、膜タンパク質を脂質二重膜に埋め込む手法が種々検討されている。例えば、リポソームに膜タンパク質を取り込む研究なども行われてきた。しかしながら、膜タンパク質自体の調製が難しいうえ、調製した膜タンパク質をリポソームに正しい方向性を持って導入することが困難で、解析が難しいという問題が残っている。
【0006】
ところで、従来から、血清中でコレステロール輸送に関与しているアポリポタンパク質(APO)は、リン脂質と混合することにより、アポリポタンパク質が脂質二重層の周りを取り囲む高密度リポ蛋白質様の円盤状の小粒子に再構成できることが知られていた(非特許文献1)。最近になり、この円盤状の小粒子の、膜タンパク質の機能解析への利用が提唱され、このような人工的な円盤状の小粒子は、リン脂質−アポリポタンパク質複合体粒子、ナノリポタンパク質、ナノディスクなどと呼ばれている。すなわち、リン脂質−アポリポタンパク質複合体粒子を再構成させる際に、膜タンパク質を共存させると、再構成されたリン脂質−アポリポタンパク質複合体粒子に膜タンパク質が含まれることが明らかとなり、膜タンパク質の機能性質の研究への応用が提唱されている(特許文献1、非特許文献2,3)。また、上記無細胞タンパク質合成系を用いて、ナノディスク(リン脂質−アポリポタンパク質複合体粒子)に膜タンパク質を埋め込んで合成する方法が発表されている(特許文献2)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特許第4303468号
【特許文献2】特表2009-521209号
【非特許文献】
【0008】
【非特許文献1】Methods in Enzymology (1986) Vol.128, p.553-582
【非特許文献2】Protein Science (2003) vol.12, p.2476-2481
【非特許文献3】Journal of Biological Chemistry (2007) Vol.282, p.28309-28320
【発明の概要】
【発明が解決しようとする課題】
【0009】
本願発明は、アポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子に、in vitro 再構成タンパク質合成系を用いて、膜タンパク質を再構成することを第1の課題とする。
【0010】
更に本願発明は、in vitro 再構成タンパク質合成系の成分やその成分量を変化させることにより、膜タンパク質を前記リン脂質-アポリポタンパク質複合体粒子に再構成する最適な条件を検討することを第2の課題とする。
【課題を解決するための手段】
【0011】
本願発明者らは、(1)厳密に成分とその量を制御できるin vitro再構成タンパク質合成系を用いることにより、(2)アポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子に、膜タンパク質を挿入することに成功して、本願発明を完成させた。
【0012】
さらに本願発明者は、厳密に成分とその量を制御できるin vitro再構成タンパク質合成系を用いることにより、前記in vitro 再構成タンパク質合成系中のリボソームの量とリン脂質-アポリポタンパク質複合体粒子の量とを一定割合範囲内とすることにより、効率的に膜タンパク質を前記リン脂質-アポリポタンパク質複合体粒子に再構成できることを見出した。
【0013】
更に、リン脂質の組成を種々に変化させたり、更に/或いは、アポリポタンパク質の種類を変えてリン脂質-アポリポタンパク質複合体粒子を調製することにより、膜タンパク質の活性の高い状態で、前記リン脂質-アポリポタンパク質複合体粒子に膜タンパク質を再構成する方法を提供する。
【発明の効果】
【0014】
従来の無細胞タンパク質合成系を用いては、機能を保持した形で製造することが困難であった膜タンパク質を、成分が特定されているin vitro 再構成タンパク質合成系を用いることにより、定量的に活性を保つ形態で、製造することが可能となった。さらに本願発明者は、厳密に成分とその量を制御できるin vitro 再構成タンパク質合成系を用いることにより、前記in vitro 再構成タンパク質合成系中のリボソームとリン脂質-アポリポタンパク質複合体粒子とのモル比を一定割合、具体的には、1:0.01〜1:100、より好適には、1:0.1〜1:10とすることにより、膜タンパク質の前記リン脂質-アポリポタンパク質複合体粒子への効率的な再構成を可能とするという優れた効果を奏するものである。
【0015】
また、本願発明は、リン脂質及び/又はアポリポタンパク質の組成を種々に変化させてアポリポタンパク質とリン脂質を含むリン脂質-アポリポタンパク質複合体粒子を調製することにより、膜タンパク質を、機能できる高次構造を保って前記リン脂質-アポリポタンパク質複合体粒子に再構成することができるという優れた効果を奏するものである。
【図面の簡単な説明】
【0016】
【図1】調製したDPPCもしくは、DMPCを含むリン脂質-アポリポタンパク質複合体粒子のゲル濾過クロマトグラム。
【図2】アポリポタンパク質としてMSP1-E1、脂質としてDMPCを含むリン脂質-アポリポタンパク質複合体粒子存在下(+)、および非存在下(-)で合成したバクテリオロドプシンを検出したゲル。Tは合成画分、Sは遠心上清画分を示す。
【図3】(a)は、0.3 μM、1 μM、又は3 μMのリボソームを含むPUREシステムに、リン脂質-アポリポタンパク質複合体粒子を0から4 μMまで変化させて添加した合成反応液を使用してバクテリオロドプシンを合成したときの可溶率の変化を示すグラフ。(b)は、0.3、1、3 μMのリボソーム濃度のPUREシステムでのバクテリオロドプシンの合成量を示すグラフ。リボソーム濃度が1 μMのときの合成量を100とした相対値で示す。
【図4】バクテリオロドプシン合成後に、リン脂質-アポリポタンパク質複合体粒子を反応液に添加しても可溶性は増大しないことを示すゲル。PUREシステムでバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた後、リン脂質-アポリポタンパク質複合体粒子を添加し、さらに1時間反応を続けた反応液、およびその遠心上清画分をSDS-PAGEで分離後、オートラジオグラフィーによりバンドを検出した。ネガティブコントロールとして、リン脂質-アポリポタンパク質複合体粒子を加えていない場合、ポジティブコントロールとして合成反応前にリン脂質-アポリポタンパク質複合体粒子を添加しておいた場合を示す。また、Tは合成画分、Sは遠心上清画分を示す。
【図5】DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含むPUREシステムを用いてバクテリオロドプシンを合成し、合成画分(T)、遠心上清画分(S)をNativePAGEに供した結果を示すゲル。(a)は、リン脂質-アポリポタンパク質複合体粒子中のアポリポタンパク質をSYPRO Rubyで染色したゲルを、(b)は合成したバクテリオロドプシンをオートラジオグラフィーで検出したゲルを示す。
【図6】DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含むPUREシステムを用いて、37℃でバクテリオロドプシンを合成し、クロラムフェニコール添加により合成反応を停止させた後、レチナールを添加して色の変化を、(a) 37℃、(b) 45℃で観察した結果で、チューブの写真、および波長550nm付近における溶液の吸光度を測定し、それぞれ波長700nmの値を0としたときの値を示す。
【図7】リン脂質−アポリポタンパク質複合体粒子を示す概念図である。
【発明を実施するための形態】
【0017】
1.はじめに
本願発明者らは、本願発明者らが既に開発したin vitro再構成タンパク質合成系(PUREシステムとも呼ばれる)と、リン脂質−アポリポタンパク質複合体粒子を組み合わせることで、膜タンパク質をコードする核酸を鋳型として合成された膜タンパク質をモデル膜であるリン脂質−アポリポタンパク質複合体粒子に再構成できるシステムを開発したものである。
【0018】
特に本願発明者等が開発したin vitro再構成タンパク質合成系を用いることにより、タンパク質合成系の種々の成分を厳密に制御できることから、本願発明は種々の応用ができるものである。
【0019】
例えば、本願発明は、存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、前記複数の成分の存在量及びリン脂質−アポリポタンパク質複合体粒子の存在量を変化させて、膜タンパク質の合成の最適な条件を検討する方法並びにその最適条件を用いた前記膜タンパク質を合成する方法を包含する。
【0020】
また、本願発明は、上記の膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子の種類及び/又は濃度及び/又は添加条件を変化させて膜タンパク質を合成し、当該膜タンパク質の可溶性、および/又は活性を測定することにより当該膜タンパク質の活性と、使用したリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件との相関を検証する方法を包含する。
【0021】
更に、本願発明は、存在量および純度がそれぞれ特定されている成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、及び存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子を含む、in vitro再構成タンパク質合成キットを包含する。
【0022】
2.膜タンパク質合成方法の概要
本願発明では、膜タンパク質は、a)目的とする膜タンパク質をコードする1または複数の鋳型核酸、b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、及びc) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子を適宜な溶媒に添加することで製造できる。溶媒としては、通常の無細胞タンパク質合成系で使用される緩衝液であれば、いずれのものでもよい。
【0023】
リン脂質−アポリポタンパク質複合体粒子は、in vitro再構成タンパク質合成系に共存して、タンパク質が合成されることが望ましく、或いは、必須である。in vitro再構成タンパク質合成系で膜タンパク質をその鋳型核酸から翻訳して調製した後に、リン脂質−アポリポタンパク質複合体粒子を添加しても、リン脂質−アポリポタンパク質複合体粒子に膜タンパク質が再構成されないことがある。
【0024】
本願発明において、膜タンパク質がリン脂質−アポリポタンパク質複合体粒子に再構成されるとは、膜タンパク質がリン脂質−アポリポタンパク質複合体粒子中の脂質二重層に挿入され、埋め込まれ、或いは貫通していることを意味している。
以下、まず上記a)、b)、c)の構成要素について、それぞれ以下に説明する。
【0025】
2−1.
膜タンパク質をコードする核酸
膜タンパク質は、いずれの膜タンパク質であっても膜タンパク質である限り、どのような生物種の由来であって、いずれのものも利用できる。例えば、膜タンパク質は、係留膜タンパク質、包埋膜タンパク質、または内在性膜タンパク質などに分類できるが、そのいずれでもよい。係留膜タンパク質の例としては、NADPH−シトクロムP450レダクターゼ、シトクロムb5およびヒト組織因子が挙げられる。包埋膜タンパク質の例としては、肝臓ミクロソーム由来のチトクロムP450が挙げられる。内在性膜タンパク質は、7回膜貫通タンパク質によって例示され、これには、バクテリオロドプシン、やGタンパク質共役タンパク質レセプターが挙げられる。
【0026】
核酸としては、DNAやmRNAなど下記のタンパク合成試薬で、翻訳、又は転写及び翻訳できるものであればよい。
【0027】
2−2タンパク質合成基本試薬
2−2−1
タンパク質合成基本試薬は、その成分として、リボソーム、開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、メチオニルtRNAトランスフォルミラーゼ類、tRNA類、アミノ酸類、リボヌクレオシド3リン酸類、10-フォルミル5,6,7,8-テトラヒドロ葉酸(FD)、塩類及び水をそれぞれ予め決められた純度の成分を決められた量だけ含むことを特徴とするが、全ての成分が必要でなく、成分は適宜選択することができる。
【0028】
系を構成するこれらの成分は、細胞抽出液又はその粗画分を用いないことはもちろんであるが、タンパク質のジスルフィド結合形成に影響を与える物質濃度を計算可能であることが望ましい。
【0029】
本発明のin vitro再構成タンパク質合成系は、系を構成する全ての成分を再構成するため、このような成分を特定し、その含有量を計算することは容易である。
【0030】
本発明におけるタンパク質合成反応基本試薬は、DNAからの転写・翻訳、或は、RNAの翻訳を行わせるタンパク質合成のための反応系として使用できる。本発明に言うタンパク質とは、2個以上のアミノ酸がペプチド結合によって結合したものを言い、ペプチド、オリゴペプチド、ポリペプチドを含む。本発明に言うRNAは化学合成されたRNA及びmRNAを含み、DNAは、合成されたDNA、DNAベクター、ゲノムDNA、PCR産物及びcDNAを含む。
【0031】
本発明におけるタンパク質合成反応基本試薬において、存在量および純度がそれぞれ特定されている成分を含むとは、それぞれの成分について、個々に精製され、その純度が測定可能であり、かつ定量可能であることを意味している。本発明では、存在量および純度がそれぞれ特定された複数の成分とは、予め塩析、クロマトグラフィー、電気泳動、溶解度の差、再結晶、遠心等の物質の精製法により、それぞれが精製された物質であって、クロマトグラフィー、電気泳動、質量分析、遠心等の分析方法により、それぞれの純度がおおむね80%以上、好適には90%以上である物質を言う。例えば、タンパク質であれば、主にクロマトグラフィーにより精製され、SDS−ポリアクリルアミドゲル電気泳動(SDS-PAGE)により純度が決定され、リボソームであれば、主に超遠心法により精製され、超遠心分離による沈降分析により純度が決定される。リボソームは複数のRNA分子(原核生物では、23S、5S、及び16Sの3つ、真核生物では28S、5.8S、5S、18Sの4つのRNA分子)と複数のリボソームタンパク質(原核生物では約50個、真核生物では約80個のタンパク質)とからなる分子量数百万の集合体分子であるが、沈降分析によって集合体分子として分子を同定し純度を測定することが可能である。tRNA類はほとんどが74から94ヌクレオチドから成り、異なる塩基配列をもつ分子であるが、電気泳動などにより分子を分離・同定し、260 nm及び280 nmの吸光度測定により純度を測定することが可能である。その他、アミノ酸や塩等の低分子の物質はいずれもクロマトグラフィー、融点測定、元素分析、質量分析等の常法により、物質を同定し純度を測定することが可能である。
【0032】
タンパク質合成基本試薬としての転写/翻訳のための因子・酵素としては、大腸菌等の原核細胞由来のものに限らず、真核細胞由来のものも使用でき、(1)RNAからの翻訳の場合は、リボソーム、開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、tRNA類、アデノシン3リン酸(ATP),グアノシン3リン酸(GTP),アミノ酸類、10-フォルミル5,6,7,8-テトラヒドロ葉酸(FD)、塩類及び水であり、大腸菌等の原核細胞由来の反応系である場合は更にメチオニルtRNAトランスフォルミラーゼ類を含む;(2)DNAからの転写/翻訳の場合は、(1)に加えウリジン3リン酸(UTP),シチジン3リン酸(CTP)及びRNAポリメラーゼ類、例えばT7RNAポリメラーゼを含む。
【0033】
本発明の反応系を構成する各種因子・酵素は、大腸菌、カビ、酵母、及び培養細胞等全ての生物が本来備えているものであるから、これをそれぞれ高度に精製し、成分として用いることもできるが、各タンパク質が多量に得られ、未知の不要又は阻害成分が反応系内に持ち込まれる可能性が低くなることから、組換え生産されたものを用いることがより好ましい。
【0034】
具体的には開始因子類、延長因子類、解離因子類、アミノアシルtRNA合成酵素類、メチオニルtRNAトランスフォルミラーゼ類、又はRNAポリメラーゼ類をコードする遺伝子を適切なベクターにつなぎ、大腸菌、枯草菌、カビ、又は酵母等に形質転換し、発現誘導を行い、該タンパク質を精製し、本発明の反応系を構成する成分とすることができる。組換え体により、各種因子・酵素を生産する場合、インタクトな状態でそのタンパク質を発現させても良いが、融合タンパク質として発現しても良い。そのような融合タンパク質として、ヒスチジンタグ(以下His-Tag)、ストレプトタグ、GSTタグ、及びFLAGタグ等を例示することができる。(ApplMicrobiol Biotechnol. 60(5):523-533(2003))
【0035】
一例として、His-Tagとニッケルカラムを利用した、His-Tagを付したタンパク質成分の精製方法の概略を示せば次の通りである。これ以外にも様々なバリエーションが知られており、適宜選択して使用できる。
1.遺伝子工学的手法により、目的タンパク質のN末にHis-Tag(6個のHisよりなる)を結合させた融合タンパク質を得る。
2.タグをつけたタンパク質が発現している細胞を氷中で超音波処理し、ローディングバッファー(300 mM NaCl, 50 mM NaH2PO4, pH 8.0)に懸濁させる。
3.細胞の溶解物を遠心分離する(30,000 g、4℃で30分間)。
4.上記で得られた上清に、氷で冷やしたローディングバッファーの中 で平衡化した50%のNi2+-NTA slurry (Qiagen社製)を加える。4℃で1時間撹拌する。
【0036】
5.樹脂をカラムにロードし、カラム容量の20倍のローディングバッファーで、4℃でカラムの洗浄を行う。
6.カラム容量の20倍のローディングバッファー(10 mMimidazole, pH 8.0を含む)で、4℃でカラムの洗浄を行う。
7.カラム容量の20倍のローディングバッファーを用いて、imidazoleの濃度勾配を10から250 mMになるように設定し、カラムから目的タンパク質の溶出を行わせ、1 mlずつフラクションを集める。SDS-PAGEで目的タンパク質を確認する。
【0037】
また、反応系を構成する各種因子・酵素として、直接タンパク質の合成反応には関わらないが、クレアチンキナーゼ類、ミオキナーゼ類又はヌクレオシドジフォスフェートキナーゼ類といったエネルギー再生に関わる酵素、及び無機ピロフォスファターゼ類といった、転写・翻訳反応で生じる無機ピロリン酸の分解のための酵素類も、同様な方法で該タンパク質を精製し、本発明の反応系に加えることがより好ましい。
【0038】
塩類としては、転写・翻訳に必須な陽イオン・陰イオンを含むことが必須であり、グルタミン酸カリウム、塩化アンモニウム、酢酸マグネシウム、塩化カルシウム等が通常使用される。なお、上記以外にも適宜選択して使用できることは言うまでもない。水はイオンや微生物類、酵素類を含まないもので、例えばミリポア社製のミリQ水製造装置によって製造される水や市販の純水を挙げることができる。
【0039】
リボソームは、ペプチド合成の場であり、mRNAと結合し、アミノアシルtRNAをA部位に、フォルミルメチオニルtRNA又はペプチジルtRNAをP部位にそれぞれ配位してペプチド結合を形成させる反応を行う(Science289:920-930(2000))。本発明においては、かかる機能を有するものであれば、由来を問わず使用することが可能である。例えば、大腸菌由来のリボソームが使用されるが、真核細胞由来のものも使用できる。本発明において用いられるリボソームの好ましい例は、大腸菌由来のものであり、例えば大腸菌A19株、MRE600株から得られるものを挙げることができる。
【0040】
リボソームの精製方法として、特許公開2007-121361号に記載の方法があげられるが、その他、従来使用されているスクロース密度勾配遠心を利用した方法なども使用できる。
【0041】
本発明のin vitro再構成タンパク質合成系で使用される開始因子類には、翻訳開始複合体の形成に必須であるか、又は、これを著しく促進する因子であり、大腸菌由来のものとして、IF1、IF2及びIF3が知られている(Biochemistry29:5881-5889(1990))。開始因子IF3は、翻訳の開始に必要な段階である、70Sリボソームの30Sサブユニットと50Sサブユニットへの解離を促進し、また、翻訳開始複合体の形成の際に、フォルミルメチオニルtRNA以外のtRNAのP部位への挿入を阻害する。開始因子IF2は、フォルミルメチオニルtRNAと結合し、30SリボソームサブユニットのP部位へフォルミルメチオニルtRNAを運び、翻訳開始複合体を形成する。開始因子IF1は開始因子IF2, IF3の機能を促進する。本発明において用いられる開始因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0042】
延長因子EF-Tuは、GTP型とGDP型の2種類があり、GTP型はアミノアシルtRNAと結合してこれをリボソームのA部位へ運ぶ。EF-Tuがリボソームから離れる際にGTPが加水分解され、GDP型へ転換する。(EMBOJ.17:7490-7497(1998))。延長因子EF-Tsは、EF-Tu(GDP型)に結合し、GTP型への転換を促進する(ArchivesofBiochemistryandBiophysics348:157-162(1997))。延長因子EF-Gは、ペプチド鎖伸長過程において、ペプチド結合形成反応の後の転位(translocation)反応を促進する(Nature Structure Biology 6:643-647(1999),FEMS Microbiology Reviews 23:317-333(1999))。本発明において用いられる延長因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0043】
解離因子は、終結因子とも呼ばれ、タンパク質合成の終結、翻訳されたペプチド鎖の解離、更に次のmRNAの翻訳開始へのリボソームの再生に必須である。解離因子RF1及びRF2は、リボソームのA部位に終止コドン(UAA,UAG,UGA)が来た時、A部位に入ってペプチジルtRNA(P部位にある)からのペプチド鎖の解離を促進する。RF1は終止コドンのうちUAA,UAGを認識し、RF2はUAA,UGAを認識する。解離因子RF3は、RF1,RF2によるペプチド鎖の解離反応後の、RF1,RF2のリボソームからの解離を促進する。リボソーム再生因子(RRF)は、タンパク質合成の停止後、P部位に残っているtRNAの脱離と、次のタンパク質合成へのリボソームの再生を促進する。本発明においては、RRFも解離因子類の一つとして取扱うことにする。なお、解離因子RF1,RF2,RF3及びRRFの機能については、EMBOJ.16:4126-4133(1997)、EMBOJ.16:4134-4141(1997)に解説されている。解離因子を含まない反応系でタンパク合成を行った場合は、終始コドンの手前で反応が止まり、リボソーム・ペプチド・mRNAの安定な3者複合体の形成が容易に行われる(ポリソームディスプレイ法、リボソームディスプレイ法)。またペプチド鎖への非天然アミノ酸の導入は、RF1及び/又はRF2を反応系から省くことにより行われる。即ち、RF1を省いた場合はUAGコドン、RF2を省いた場合はUGAコドンへの非天然アミノ酸の導入が高い効率で行われる。本発明において用いられる解離因子の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。
【0044】
アミノアシルtRNA合成酵素は、ATPの存在下でアミノ酸とtRNAを共有結合させ、アミノアシルtRNAを合成する酵素である(RNA 3:954-960(1997),タンパク質核酸酵素39:1215-1225(1994))。本発明において用いられるアミノアシルtRNA合成酵素の好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものを挙げることができるが、真核細胞由来のものも使用できる。また、非天然アミノ酸を認識する人工アミノアシルtRNA合成酵素(特許2668701号)を用いることもできる。
【0045】
メチオニルtRNAトランスフォルミラーゼ(MTF)は原核生物におけるタンパク質合成においてメチオニルtRNAのアミノ基にフォルミル基がついたN-フォルミルメチオニル(fMet)tRNAを合成する酵素である。即ち、メチオニルtRNAトランスフォルミラ―ゼは、N10‐フォルミルテトラヒドロ葉酸のフォルミル基を、開始コドンに対応するメチオニルtRNAのN末端に転移させ、fMet-tRNAにする(Proc.Natl.Acad.Sci.USA96:875-880(1999))。付加されたフォルミル基は開始因子IF2により認識され、タンパク質合成の開始シグナルとして作用する。真核生物の細胞質における合成系にはMTFはないが、真核生物のミトコンドリア及び葉緑体における合成系には存在する。本発明において用いられるMTFの好ましい例は、大腸菌由来のものであり、例えば大腸菌K12株から得られるものである。
【0046】
RNAポリメラーゼは、DNA配列をRNAに転写する酵素であり、様々な生物に存在することが知られている。その一例として、T7ファージ由来の、T7RNAポリメラーゼを挙げることができ、このポリメラーゼはT7プロモーターと呼ばれる特異的なDNA配列に結合してその下流のDNA配列をRNAに転写する酵素である。本発明者等は、T7RNAポリメラーゼのN末端にヒスタグを付加して、融合タンパク質として大腸菌BL21株において大量発現を行い、ニッケルカラムを用いるアフィニティクロマトグラフィーにより精製を行った。本発明においては、T7RNAポリメラーゼ以外にも種々のRNAポリメラーゼを用いることができる。例えば、T3RNAポリメラーゼやSP6RNAポリメラーゼが市販されており、これ等を利用することもできる。
【0047】
アミノ酸類としては、天然型アミノ酸に加え、非天然型アミノ酸も用いることができる。これらのアミノ酸は、in vitro再構成タンパク質合成系を構成するアミノアシルtRNA合成酵素の作用によってtRNAに保持される。あるいは、予めアミノ酸をtRNAにチャージしてin vitro再構成タンパク質合成系に加えることができる。本発明において、tRNAへのアミノ酸のチャージとは、tRNAにアミノ酸を結合させ、リボソームにおける翻訳反応に利用される状態にすることを言う。非天然アミノ酸を認識する人工アミノアシル合成酵素存在下で非天然アミノ酸を添加したり、非天然アミノ酸でチャージされたtRNAを用いたりすることで、蛋白質の特定のコドンの部位に非天然アミノ酸を導入することが可能となる。
【0048】
tRNA類としては、大腸菌、酵母等の細胞から精製したtRNAを用いることができる。またアンチコドンやその他の塩基を任意に変更した人工tRNAも用いることができる(Hohsaka, T et al. (1996) J. Am. Chem. Soc., vol.121, p.34-40, Hirao I et al (2002) Nat. Biotechnol., vol.20, p.177-182)。例えば、CUAをアンチコドンとして持つtRNAに非天然のアミノ酸をチャージすることで、本来終止コドンであるUAGコドンを非天然アミノ酸に翻訳することが可能である。また,4塩基コドンをアンチコドンとして持つtRNAに非天然アミノ酸をチャージした人工アミノアシルtRNAを用いることにより、天然には存在しない4塩基コドンを非天然アミノ酸に翻訳することが可能である(Hohsaka et al. (1999) J.Am.Chem.Soc., vol.121, p.12194-12195)。このような人工tRNAを作製する方法としては,RNAを用いる方法も使用できる(特表2003-514572)。これらの方法により部位特異的に非天然アミノ酸を導入した蛋白質を合成することができる。
【0049】
上記in vitro再構成タンパク質合成系を構成する各因子は、転写や翻訳に好適なpH7-8を維持する緩衝液に加えることによって、in vitro再構成タンパク質合成系とすることができる。本発明に用いられる緩衝液としては、リン酸カリウム緩衝液(pH 7.3)、Hepes-KOH(pH 7.6)などをあげることができる。
【0050】
上記in vitro再構成タンパク質合成系には、酵素の基質として、および/もしくは、活性の向上、維持を目的として、その他の低分子化合物を添加できる。具体的には、ヌクレオシド三リン酸(ATP, GTP, CTP, UTPなど)などの基質、プトレシン(putrescine)、スペルミジン(spermidine)などのポリアミン類、およびクレアチンリン酸などのエネルギー再生のための基質などをin vitro再構成タンパク質合成系に加えることができる。
【0051】
上記in vitro再構成タンパク質合成系では、例えば分子シャペロンと呼ばれる一群のタンパク質に代表される、タンパク質の凝集を抑制する因子は含んでいない。すなわち、本願発明においては、前記分子シャペロンを含まないin vitro再構成タンパク質合成系を用いても、膜タンパク質をリン脂質−アポリポタンパク質複合体粒子に挿入することができる。しかしながら、膜タンパク質の合成を行なう際に、これらの因子の添加することにより、凝集を抑制しながら合成することも可能である。
【0052】
分子シャペロンとしては、具体的には、例えば、Hsp70ファミリー、Hsp60ファミリー、Hsp90ファミリー、Hsp100ファミリー、低分子量Hspファミリー、イソメラーゼ類、および、これらの補助因子をあげることができる。分子シャペロン類も上記のタンパク質合成に関与する因子類と同様、上記のような精製法により、精製し、本発明の反応系に加えることがより好ましい。
【0053】
2−2−2 膜タンパク質がS-S結合を含む場合
更に、本願タンパク質合成基本試薬は、膜タンパク質が、S-S結合を含む場合、上記2−2−1記載のタンパク質合成基本試薬に加えて、ジスルフィド及びチオール間の酸化還元平衡状態の人為的調整をするための(イ)酸化還元を触媒する酵素、及び/又は(ロ)酸化還元状態を調節する試薬を含むことができる。
【0054】
2−2−2(1) ジスルフィド結合形成に影響を与える物質
ジスルフィド結合形成に影響を与える物質の例として、ジスルフィド結合の酸化還元を触媒する酵素である酸化還元酵素及び/又はジスルフィド結合の酸化還元状態を調節する試薬である酸化還元試薬が挙げられる。より具体的には、ジスルフィド結合形成に影響を与える酵素及び又は試薬として、(イ)酸化還元を触媒する酵素:グルタチオンレダクターゼ類、チオレドキシンレダクターゼ類、プロテインジスルフィドイソメラーゼ類、ジスルフィドインターチェインジタンパク質類、チオレドキシン様タンパク質等のタンパク質及び/又は(ロ)酸化還元状態を調節する試薬:還元型グルタチオン、酸化型グルタチオン、ジチオスレイトール(DTT)、2-メルカプトエタノール及びチオレドキシン等の低分子化合物を例示することができる。本発明における酸化還元試薬とはジスルフィドを還元しチオールにしたり、逆にチオールを酸化しジスルフィドを形成させることのできる試薬を言う。
【0055】
ジスルフィド結合形成に影響を与える物質の例としての、(イ)酸化還元酵素(酸化還元を触媒する酵素)及び/又は(ロ)酸化還元試薬(酸化還元状態を調節する試薬)は、タンパク質のジスルフィド結合の架橋を促進・調整する酵素類及び/又はその基質を用いることが望ましい。なお、(イ)酸化還元を触媒する酵素及び/又は(ロ)酸化還元状態を調節する試薬は、必ずしも翻訳反応中に加える必要はなく、翻訳終了後に加えても良い。翻訳終了後に添加する場合には、添加後更に数十分から1時間程度37℃に静置するのが好ましい。
【0056】
酸化還元状態を調節する試薬として、DTTを用いる場合は濃度が0〜5 mM、好適には0.001 〜2 mMであることが好ましい。更に好適には、0.060〜1 mMであることが好ましい。
【0057】
酸化還元状態を調節する試薬として、酸化型グルタチオンを用いる場合は濃度が0〜8mM、好適には、0.1〜4mMであることが好ましい。更に好適には1 〜4mMであることが好ましい。
【0058】
前記タンパク質の合成方法おいて、ジスルフィドインターチェインジタンパク質類はDsbA及び/又はDsbCであることが好ましい。
【0059】
プロテインジスルフィドイソメラーゼ(EC 5.3.4.1.)は真核生物の小胞体内膜に存在する約55kDaのジスルフィド結合の形成や異性化・還元反応を触媒する酵素であり、シャペロン様の活性も持つと言われている。このタンパク質は前述の通り、生物から精製し、成分として用いることもできるが、組換え生産されたものを用いてもよい。ウシの肝臓から精製したプロテインジスルフィドイソメラーゼ(PIR データベース アクセッション番号ISBOSS)、酵母のプロテインジスルフィドイソメラーゼ遺伝子を大腸菌に組換え発現させ、精製したタンパク質等を用いることができる。
【0060】
その他にプロテインジスルフィドイソメラーゼ類として下記のタンパク質知られており、本発明のプロテインジスルフィドイソメラーゼ質類として使用可能である。
ヒトのprotein disulfide-isomerase PIRデータベース アクセッションナンバー ISHUSS
ヒトのprotein disulfide isomerase related protein GenBank アクセッションナンバー4758304
酵母のprotein disulfide isomerase homolog PIR データベース A44483
【0061】
ジスルフィドインターチェインジタンパク質類は、ジスルフィドインターチェインジタンパク質A、B、C、D(DsbA, B , C, D)の4種のタンパク質が大腸菌等で知られている。DsbAは、21kDaのチオレドキシン様フォールド構造を持つ、ジスルフィド結合の形成を触媒すると考えられている酵素である。DsbBは4個の膜貫通部位と、2個のペリプラズムドメインを持つ20kDaのタンパク質で、DsbAを酸化型に維持すると言われている。DsbCはホモ2量体を形成するペリプラズムタンパク質であり、チオレドキシン様のフォールドを持ち、主にジスルフィド結合の異性化を担う酵素であると言われているが、他にシャペロンとしての働きもあると考えられている。DsbDは分子量59kDaのタンパク質で2つのペリプラズムドメイン、8個の膜貫通領域からなるタンパク質で、DsbCの活性中心のシステインを還元型に維持する働きを持っていると考えられる。
【0062】
ジスルフィドインターチェインジタンパク質類として下記のタンパク質が知られており、本発明のジスルフィドインターチェインジタンパク質類として使用可能である。
E. coliのDsbA SWISS-PROTprotein database Accession P24991
E. coli のDsbC SWISS-PROTprotein database Accession P21892
Salmonella typhimuriumのprotein disulfide-isomerase dsbA homolog PIRデータベース アクセッションナンバーS32895
Neisseria meningitidisのthiol-disulfide interchange protein dsbA homolog NMB0278
PIRデータベース アクセッションナンバーC81217
Caenorhabditis elegansのprotein disulphide isomerase isoform I Gene Bank アクセッションナンバー AAB94647
Datisca glomerataのprotein disulfide isomerase homolog Gene Bank アクセッションナンバーAAD28260
【0063】
これらジスルフィドインタージェインジタンパク質類は前述の通り、生物から精製し、成分として用いることもできるが、組換え生産されたものを用いてもよい。
【0064】
酸化還元酵素としてプロテインジスルフィドイソメラーゼ(PDI)を用いる場合、濃度が0 μM〜10 μMであることが好ましい。好適には0.001〜5 μMであることが好ましい。更に好適には0.001 μM〜2 μMであることが好ましい。酸化還元酵素としてジスルフィドインターチェインジタンパク質を用いた場合、0 μM〜10 μMであることが好ましい。好適には0.01から10 μMであることが好ましい。更に好適には0.1 μM〜10 μMであることが好ましい。
【0065】
なお、グルタチオンレダクターゼ類、チオレドキシンレダクターゼ類については、できるだけ含まないことが望ましく、タンパク質合成系を構成する成分におけるチオレドキシンレダクターゼ及び/又はグルタチオンレダクターゼの含量が100 ng/ml以下であることが好ましい。
【0066】
プロテインジスルフィドイソメラーゼ類、ジスルフィドインターチェインジタンパク質類も、上記のタンパク質合成に関与する因子類と同様、上記のような精製法により、精製し、本発明の反応系に加えることがより好ましい。
【0067】
2−3.リン脂質−アポリポタンパク質複合体粒子
本願発明で言う、リン脂質−アポリポタンパク質複合体粒子は、ナノリポタンパク質、(nanolipoprotein)、ナノディスク(Nanodisc)、ナノリポタンパク質粒子(nanolipoproteinparticles)、リン脂質−アポリポタンパク質粒子と称されることもある。当該粒子はリン脂質を含んでなり、アポリポタンパク質により二重層ディスクとして構成される。その構造の概略を図7に示す。
【0068】
本願発明明細書中では、アポリポタンパク質という用語には、天然のアポリポタンパク質又はその変異体であって、リン脂質−アポリポタンパク質粒子を形成できるものはすべて包含する。
【0069】
本願発明で言う、リン脂質−アポリポタンパク質複合体粒子は、例えば、アポリポタンパク質及びリン脂質を、コール酸若しくはコール酸塩を含む適宜な溶媒中で混合した後、混合液をコール酸及びコール酸塩を含まない溶媒に対して透析することにより得られる。
【0070】
従来技術において記載のように、リン脂質−アポリポタンパク質粒子は、単離されたアポリポタンパク質及びリン脂質を混合し、リン脂質−アポリポタンパク質として凝集させて調製してもよい。
【0071】
リン脂質−アポリポタンパク質複合体粒子の調製方法は従来技術において公知であり、例えば、Jonasら、(1984)J.Biol.Chem.259:6369−6375、Jonasら、(1989)J.Biol.Chem.264:4818−4824、Jonasら、(1993)J.Biol.Chem.268:1596−1602、米国特許第7048949号、米国特許出願公開第2005/0182243号A1、第2005/0152984号A1、第2004/0053384号A1及び第2006/0088524号A1、特許公報4303468号に記載されている。
【0072】
調製した前記リン脂質−アポリポタンパク質複合体粒子は、例えば、ゲル濾過クロマトグラフィーによって精製され、純度が決定される。
【0073】
本発明でいうアポリポタンパク質は、1つ以上の両親媒性のヘリックス状ドメインを有する。本願発明で言うアポリポタンパク質には、例えば、アポリポタンパク質A−I、アポリポタンパク質A−II、アポリポタンパク質A−IV、アポリポタンパク質A−V、アポリポタンパク質B−100、アポリポタンパク質B−48、アポリポタンパク質C−I、アポリポタンパク質C−II、アポリポタンパク質C−III、アポリポタンパク質D、アポリポタンパク質E、アポリポタンパク質H、リポタンパク質(a)、アポリポフォリンI(アポリポフォリンII)、及びアポリポフォリンIIIが挙げられる。また上述したように、本発明でいうアポリポタンパク質には、前記アポリポタンパク質のアミノ酸配列に対して1以上のアミノ酸が置換、欠失及び/又は付加した変異体であって、リン脂質と複合粒子を形成する能力を保持するもの、又は他のタンパク質との融合タンパク質であってリン脂質と複合粒子を形成する能力を保持するもの、その他誘導体も包含される。また、本発明でいうアポリポタンパク質には、たとえば。特表2007−525490号や特開2008−44958号に示される、MSP1、MSP2、MSP1E1、MSP1E2、MSP1E3およびMSP2などの膜スカホールドタンパク質(MSP)も包含される。
【0074】
本発明で用いるアポリポタンパク質としては、好適には、ApoA−I、ApoA−IV、ApoA−V、ApoC−I、ApoC−II、ApoC−III、Apo−E又はアポリポフォリンIII又はこれらのいずれかの変異体が挙げられる。
【0075】
リン脂質−アポリポタンパク質複合体粒子に用いるリン脂質は、グリセロール又はスフィンゴ脂質ベースであってもよく、例えば、2分子の6〜20の炭素原子の飽和脂肪酸、及び一般的に用いられるヘッド基(例えばホスファチジルコリン、ホスファチジルエタノールアミン及びホスファチジルセリン)を含んでなってもよい。ヘッド基は、荷電しなくともよく、正に荷電してもよく、負に荷電してもよく、また双性イオンであってもよい。リン脂質は、天然物)若しくは合成物、又は天然物と合成物の混合物であってもよい。リン脂質の非限定的な例としては、PC(ホスファチジルコリン)、PE(ホスファチジルエタノールアミン)、PI(ホスファチジルイノシトール)、DPPC(ジパルミトイル−ホスファチジルコリン)、DMPC(ジミリストイルホスファチジルコリン)、POPC(1−パルミトイル−2−オレオイル−ホスファチジルコリン)、DHPC(ジヘキサノイルホスファチジルコリン)、ジパルミトイルホスファチジルエタノールアミン、ジパルミトイルホスファチジルイノシトール、ジミリストイルホスファチジルエタノールアミン、ジミリストイルホスファチジルイノシトール、ジヘキサノイルホスファチジルエタノールアミン、ジヘキサノイルホスファチジルイノシトール、1−パルミトイル−2−オレオイル−ホスファチジルエタノールアミン、又は1−パルミトイル−2−オレオイル−ホスファチジル基イノシトールが挙げられる。
好適には、DMPCが挙げられる。
【0076】
3.本願発明のin vitro再構成タンパク質合成系による膜タンパク質合成方法でリン脂質-アポリポタンパク質複合体粒子に再構成された膜タンパク質の評価又は検証
3−1.膜タンパク質が「リン脂質-アポリポタンパク質複合体粒子に再構成」しているか否かの評価
本発明の方法により合成された膜タンパク質が、リン脂質−アポリポタンパク質複合体粒子に再構成されている、つまり、合成された膜タンパク質がリン脂質−アポリポタンパク質複合体粒子に挿入され、付着し、埋め込まれ、又は貫通等により再構成しているかは、目的の膜タンパク質がリン脂質−アポリポタンパク質複合体粒子と共に溶媒上清に存在するか、つまり可溶化しているか、それとも沈殿している、つまり可溶化していないかを計測することにより評価することができる。
【0077】
更に目的とする膜タンパク質の天然のリガンドや膜タンパク質と会合する因子が、リン脂質−アポリポタンパク質複合体粒子に再構成された膜タンパク質と会合するか否かを調べることにより確認することができる。
【0078】
例えば、バクテリオロドプシンがレチナールと結合するとレチナールが呈色反応をすることを利用することができる。
【0079】
3−2.リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能の評価・検証
目的とする膜タンパク質に応じた周知の機能測定方法により、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がその機能を有しているか、評価、検証することができる。
【0080】
4.膜タンパク質合成方法におけるタンパク質合成基本試薬中の各成分の量及びリン脂質−アポリポタンパク質複合体粒子の量の検討
本願発明のin vitro再構成タンパク質合成系による膜タンパク質合成方法は、「b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬」を用いるので、個々の成分の純度及び量を変化させて膜タンパク質合成することができ、更に、「c)リン脂質−アポリポタンパク質複合体粒子」についても、リン脂質−アポリポタンパク質複合体粒子の量を測定し、又はその量を変えて、或いは、濃度を変えて、膜タンパク質の合成を行うことができる。そこで、本願発明者らは、タンパク質合成反応基本試薬の種々の成分の量及び濃度並びにc)リン脂質−アポリポタンパク質複合体粒子の量及び濃度について検討を加えた。
【0081】
本願発明者は、様々な試行錯誤の上、タンパク質基本試薬の種々の成分の内、特にリボソーム量と、リン脂質−アポリポタンパク質複合体粒子の量の比率及びそれぞれの濃度が、膜タンパク質の合成に大きな影響を与えることを見出した。
【0082】
具体的には、in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.01〜1:100が望ましく、より好適には、in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.1〜1:10とすることがよいことをみいだした。
【0083】
さらに、リボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、上記比率とする時、in vitro再構成タンパク質合成系におけるリボソームの濃度が、0.1〜3 μM、リン脂質−アポリポタンパク質複合体粒子の濃度が0.01〜10μMであることが望ましいことを見出した。
【0084】
5.膜タンパク質の活性発現に最適なリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検証する方法
膜タンパク質の活性発現に最適な条件を求めるために、リン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検討し、検証することができる。
【0085】
5−1.膜タンパク質合成方法におけるリン脂質−アポリポタンパク質複合体粒子中のリン脂質の検討
リン脂質−アポリポタンパク質複合体粒子中のリン脂質を変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がより適切に構造的に再構成されているかを上記3.記載の評価方法により評価し、適切なリン脂質を選択することができる。
【0086】
また、リン脂質−アポリポタンパク質複合体粒子中のリン脂質を変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、適切なリン脂質を選択することもできる。
【0087】
たとえば、リン脂質としてDMPCを採用することにより、リン脂質−アポリポタンパク質複合体粒子に再構成した膜タンパク質は、より適切な高次構造をとり、膜タンパク質の機能を発揮しやすい。
【0088】
5−2.膜タンパク質合成方法におけるリン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の検討、並びにアポリポタンパク質及びリン脂質の組み合わせの検討
リン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の種類を例えば、上記2−3に記載されるアポリポタンパク質の内から選ばれるものに変更することにより、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質がより適切に構造的に再構成されているかを上記3.記載の評価方法により評価し、適切なアポリポタンパク質を選択することができる。
【0089】
また、同様にリン脂質−アポリポタンパク質複合体粒子中のアポリポタンパク質の種類を変更し、リン脂質の種類も変更することにより、アポリポタンパク質及びリン脂質の組み合わせを変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定し、適切なアポリポタンパク質及びリン脂質の組み合わせを選択することもできる。
【0090】
5−3.膜タンパク質合成における、リン脂質−アポリポタンパク質複合体粒子中の膜タンパク質合成系への添加時期及び添加時点の温度の検討
存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加する時期を変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加する適切な時期を選択することもできる。
【0091】
また、存在量及び純度がそれぞれ特定されている複数の成分からなるin vitro再構成タンパク質合成系と、リン脂質−アポリポタンパク質複合体粒子を用いて、膜タンパク質を合成する方法において、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加するときの温度を変化させて、リン脂質-アポリポタンパク質複合体に再構成された膜タンパク質の機能を上記3.記載の方法により測定して、リン脂質−アポリポタンパク質複合体粒子をin vitro再構成タンパク質合成系に添加するときの適切な温度を選択することもできる。
【実施例】
【0092】
<実施例1>リン脂質-アポリポタンパク質複合体の調製
特許第4303468号公報に従い、以下のようにリン脂質-アポリポタンパク質複合体粒子を調製した。アポリポタンパク質としては、ApoA-Iの改変体であるMSP1E1 (Denisov, I. G., et al. (2004) J. Am. Chem. Soc., vol. 126, p. 3477-3487) を文献に従って精製して使用した。ジパルミトイルホスファチジルコリン(DPPC)もしくは、ジミリストイルホスファチジルコリン(DMPC)のクロロホルム溶液(Avanti Polar Lipids製)を乾燥し、最終脂質濃度が50 mMになるように0.1 M NaClおよび100 mMコール酸ナトリウムを含む10 mM Tris-HClpH7.4の緩衝液に懸濁し、50℃に加熱しながらボルテックスすることで完全に溶解した。各リン脂質溶液を、モル比で、MSP1E1:リン脂質=2:200になるようにMSP1E1溶液と混合した。この混合溶液を、DMPCを含む混合溶液は室温(25℃)で、DPPCの場合は37℃で一晩インキュベートし、次いでコール酸ナトリウムを含まない1000倍容量の緩衝液に対して、緩衝液を4回交換しながら2日かけて透析した。透析後の混合溶液を、Superdex 200 10/300 GL (GE Healthcare)を用いたゲル濾過クロマトグラフィーにより分画し、図1に示すようなクロマトグラムを得た。ピーク画分を回収することにより、リン脂質-アポリポタンパク質複合体粒子を調製した。
【0093】
<実施例2>タンパク質合成反応試薬
タンパク質合成反応試薬の基本組成は、リボソーム濃度以外は、Shimizu et al. (2005) Methods, vol.36, p.299-304に従った。すなわち、50 mM HEPES-KOH pH7.6、2 mM ATP、2 mMGTP、1 mM CTP、1 mM UTP、20 mM クレアチンリン酸(Creatine phosphate)、56 A260 units/ml 大腸菌tRNA混合液、0.01 μg/μl 10-formyl-5,6,7,8-tetrahydrofolic acid(FD)、0.3 mM 各アミノ酸、13 mM 酢酸マグネシウム、100 mMグルタミン酸カリウム、2 mM スペルミジン(spermidine)、1 mM ジチオスレイトール(DTT),1 μM大腸菌リボソーム,0.02 μg/μl IF1,0.04 μg/μl IF2,0.015 μg/μl IF3,0.02 μg/μl EF-G,0.04 μg/μl EF-Tu,0.02 μg/μl EF-Ts,0.01 μg/μl RF1,0.01 μg/μl RF2,0.01 μg/μl RF3,0.01 μg/μl RRF,0.6-6 units/μl各アミノアシルtRNA合成酵素(ARS)及びメチオニルtRNAトランスフォルミラーゼ(MTF),0.004 μg/μl クレアチンキナーゼ(CK; creatine kinase),0.003 μg/μlミオキナーゼ(MK; myokinase),0.001 μg/μl ヌクレオシド二リン酸キナーゼ(NDK; nucleoside diphosphate kinase),0.0356 units/μl ピロフォスファターゼ(PPiase; Pyrophosphatase)及び0.01 μg/μl T7 RNAポリメラーゼを基本組成とした。通常のタンパク質合成では、上記基本組成のタンパク質合成反応試薬に、鋳型DNAを最終濃度が0.02-0.1 μMになるように添加して37℃で反応を行なった。なお、タンパク質合成の目的に応じて、上記基本組成に追加および、もしくは除去した反応試薬を使用した。上記組成のうち、リボソームは、Ohashi et al. (2007) BBRC, vol.352, p.270-276、及びタンパク質因子は、Shimizu et al. (2001) Nat. Biotechnol., vol.19, p.751-755に従って調製され、純度が測定されたものを、その他の成分は市販の精製試薬を使用した。
【0094】
<実施例3>リン脂質-アポリポタンパク質複合体粒子存在下でのバクテリオロドプシンの合成
EF-Tu、EF-Ts、EF-Gを3倍量含む上記基本試薬に、アポリポタンパク質としてMSP1-E1、脂質としてDMPCを含むリン脂質-アポリポタンパク質複合体粒子を最終濃度が4 μMになるように添加した膜タンパク質合成用PUREシステムを調製した。膜タンパク質合成用PUREシステムに、[35S]メチオニン、およびバクテリオロドプシンをコードする鋳型DNAを加えて15 μlに調製し、37℃で1時間反応させた。反応終了後、15 μlのミリQ水を加え、10 μlの合成画分を分取した。残りの反応液を20,000xgで30分間遠心し、上清画分を得た。合成画分、および遠心上清画分をSDS-PAGEにかけ、オートラジオグラフィーにより、合成されたバクテリオロドプシンの可溶性を評価した。図2に示すように、リン脂質-アポリポタンパク質複合体粒子非存在下で合成されたバクテリオロドプシンは、上清画分にほとんど回収されていないが、リン脂質-アポリポタンパク質複合体粒子存在下で合成した場合は、ほぼ全量が上清画分に回収された。このことは、細胞抽出液のように、分子シャペロンなど膜タンパク質の膜挿入を助ける因子を全く含まないPUREシステムにおいても、リン脂質-アポリポタンパク質複合体粒子存在下では合成膜タンパク質が可溶性に回収できることを示している。
【0095】
<実施例4>膜タンパク質合成におけるリン脂質-アポリポタンパク質複合体粒子濃度依存性
上記膜タンパク質合成用PUREシステムにおいて、リボソーム濃度を、0.3、1、3 μM、添加するリン脂質-アポリポタンパク質複合体粒子濃度を、0.25-4 μMのように変化させて、バクテリオロドプシンを合成した。合成画分、上清画分に含まれるバクテリオロドプシンを定量し、可溶率(%)=上清画分/合成画分×100として、各合成条件における可溶率を算出した。図3に示すように、リボソーム濃度を0.3 μMに減少させると、合成量も低下するため、必要となるリン脂質-アポリポタンパク質複合体粒子も低下した。一方、リボソーム濃度を増加させた場合、合成量は増加しなかったが、可溶率が低下した。この結果により、膜タンパク質合成におけるリボソーム濃度とリン脂質-アポリポタンパク質複合体粒子濃度には最適な条件があることが明らかになった。すなわち、1 μMリボソーム、4 μM リン脂質-アポリポタンパク質複合体粒子が最適であることが分かった。
【0096】
<実施例5>膜タンパク質合成におけるリン脂質-アポリポタンパク質複合体粒子添加時期依存性
上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた後、4 μMになるようにリン脂質-アポリポタンパク質複合体粒子を添加し、さらに1時間反応を続けた。合成反応画分を分取後、上記の遠心行ない、上清画分を回収した。一方で、上記のように合成前にリン脂質-アポリポタンパク質複合体粒子を添加した膜タンパク質合成用PUREシステムを用いて合成を行なった。合成画分、および遠心上清画分をSDS-PAGEで分離後、オートラジオグラフィーによりバンドを検出した(図4)。この結果、効率よく可溶性の膜タンパク質を合成するためには、あらかじめリン脂質-アポリポタンパク質複合体粒子を添加したPUREシステムを使用する必要があることが明らかとなった。
【0097】
<実施例6>膜タンパク質とリン脂質-アポリポタンパク質複合体粒子の複合体形成
DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含む上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンを合成し、上記と同様、合成画分、遠心上清画分を得た。それぞれNative-PAGEに供し、リン脂質-アポリポタンパク質複合体粒子をSYPRO Ruby(Invitrogen社製)で染色した(図5(a))。一方、合成された[35S]メチオニンを含むバクテリオロドプシンをオートラジオグラフィーで検出した(図5(b))。リン脂質-アポリポタンパク質複合体粒子存在下で合成した場合、いずれのリン脂質-アポリポタンパク質複合体粒子を用いても、リン脂質-アポリポタンパク質複合体粒子とほぼ同じ移動度のバンドを生じた。これは、バクテリオロドプシンがリン脂質-アポリポタンパク質複合体粒子と複合体を形成していることを示している。一方、リン脂質-アポリポタンパク質複合体粒子を添加せずに合成した場合、合成されたバクテリオロドプシンは、ゲルに入らないことが確認された。これは、リン脂質-アポリポタンパク質複合体粒子非存在下の合成では電気泳動されない凝集体を形成していることを示している。
【0098】
<実施例7>膜タンパク質の高次構造形成
DMPC、またはDPPCを含むリン脂質-アポリポタンパク質複合体粒子、またはMembraneMax(Invitrogen社製)を含む上記膜タンパク質合成用PUREシステムを用いてバクテリオロドプシンの合成を37℃で1時間行ない、クロラムフェニコール添加により合成反応を停止させた。合成反応液にレチナールを添加し、37℃、45℃でさらに1時間反応した。1時間後、反応液の色の変化を記録し、波長550nm付近の吸光度を測定した(図6)。DMPCを含むリン脂質-アポリポタンパク質複合体粒子、およびMembraneMaxを使用した場合は、反応温度に関わらず、溶液の色が黄色から紫色に変化することを確認した。レチナールは、正しい構造を形成したバクテリオロドプシンに結合し、紫色に変化することが知られている。すなわち、上記のリン脂質-アポリポタンパク質複合体粒子を使用した場合、正しい立体構造を形成したバクテリオロドプシンが合成されていることを示している。一方、DPPCを含むリン脂質-アポリポタンパク質複合体粒子を使用した場合、合成温度である37℃でレチナールを反応させた場合には色の変化が観察されなかったが、45℃で観察された。DMPC、DPPCの転移温度はそれぞれ約29℃、44℃であり、レチナールの結合には相転移温度以上が必要であることが分かった。実施例6において、合成されたバクテリオロドプシンは、37℃でDPPCを含むリン脂質-アポリポタンパク質複合体粒子と複合体を形成し、可溶性になることが示されている。しかしながら、レチナールとの結合には相転移温度以上が必要であることは、合成された膜タンパク質の評価には、可溶性の上昇だけでは不十分であることを示している。
【産業上の利用可能性】
【0099】
本願発明は、膜タンパク質の合成に利用できるほか、膜タンパク質の機能の測定、更には、ハイスループットに種々の膜タンパク質を合成し、その機能や、リガンド結合性などによりスクリーニングすることもできる。
【特許請求の範囲】
【請求項1】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系による膜タンパク質合成方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項2】
タンパク質合成基本試薬が、濃度及び純度がそれぞれ特定されているリボソーム、開始因子、延長因子、解離因子、アミノアシルtRNA合成酵素、およびメチオニルtRNAトランスフォルミラーゼからなる群から選択される、少なくとも1つの成分を含むことを特徴とする請求項1に記載の膜タンパク質合成方法。
【請求項3】
リン脂質−アポリポタンパク質複合体粒子が、合成された膜タンパク質の可溶性を増大させることを特徴とする請求項1又は2に記載の膜タンパク質合成方法。
【請求項4】
リン脂質−アポリポタンパク質複合体粒子が、アポリポタンパク質及びリン脂質を含む請求項1又は2に記載の膜タンパク質合成方法。
【請求項5】
リン脂質がDMPCである請求項4記載の膜タンパク質合成方法
【請求項6】
in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.01〜1:100である請求項1から5のいずれかに記載の方法。
【請求項7】
in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.1〜1:10である請求項6に記載の方法。
【請求項8】
in vitro再構成タンパク質合成系におけるリボソームの濃度が、0.1〜3 μMである請求項6又は7に記載の方法。
【請求項9】
in vitro再構成タンパク質合成系におけるリン脂質−アポリポタンパク質複合体粒子の濃度が、0.01〜10μMである請求項6又は7に記載の方法。
【請求項10】
タンパク質合成反応基本試薬には分子シャペロンが含まれないことを特徴とする請求項1〜9のいずれか1項に記載の膜タンパク質合成方法。
【請求項11】
タンパク質合成反応基本試薬に含まれるタンパク質が、膜タンパク質合成終了後に反応系から金属アフィニティ樹脂で除去できる請求項1〜10のいずれか1項に記載の膜タンパク質合成方法。
【請求項12】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系において、リン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を変化させて膜タンパク質を合成し、当該膜タンパク質の可溶性、および/もしくは活性を測定することにより、当該膜タンパク質の活性発現に最適なリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検証する方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項13】
タンパク質合成基本試薬が、濃度及び純度がそれぞれ特定されているリボソーム、開始因子、延長因子、解離因子、アミノアシルtRNA合成酵素、およびメチオニルtRNAトランスフォルミラーゼからなる群から選択される、少なくとも1つの成分を含むことを特徴とする請求項12に記載の検証方法。
【請求項14】
リン脂質−アポリポタンパク質複合体粒子が、合成された膜タンパク質の可溶性を増大させることを特徴とする請求項12又は13に記載の検証方法。
【請求項15】
リン脂質−アポリポタンパク質複合体粒子が、アポリポタンパク質及びリン脂質を含む請求項12又は13に記載の検証方法。
【請求項16】
タンパク質合成反応基本試薬には分子シャペロンが含まれないことを特徴とする請求項12〜15のいずれか1項に記載の検証方法。
【請求項17】
下記a)及びb)を含むin vitro再構成タンパク質合成キット。
a) 存在量および純度がそれぞれ特定されている成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
b) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項18】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系において、タンパク基本試薬中の成分とリン脂質−アポリポタンパク質複合体粒子の比率を変化させることを含む、膜タンパク質合成方法における最適化評価方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項1】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系による膜タンパク質合成方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項2】
タンパク質合成基本試薬が、濃度及び純度がそれぞれ特定されているリボソーム、開始因子、延長因子、解離因子、アミノアシルtRNA合成酵素、およびメチオニルtRNAトランスフォルミラーゼからなる群から選択される、少なくとも1つの成分を含むことを特徴とする請求項1に記載の膜タンパク質合成方法。
【請求項3】
リン脂質−アポリポタンパク質複合体粒子が、合成された膜タンパク質の可溶性を増大させることを特徴とする請求項1又は2に記載の膜タンパク質合成方法。
【請求項4】
リン脂質−アポリポタンパク質複合体粒子が、アポリポタンパク質及びリン脂質を含む請求項1又は2に記載の膜タンパク質合成方法。
【請求項5】
リン脂質がDMPCである請求項4記載の膜タンパク質合成方法
【請求項6】
in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.01〜1:100である請求項1から5のいずれかに記載の方法。
【請求項7】
in vitro再構成タンパク質合成系を構成するリボソームとリン脂質−アポリポタンパク質複合体粒子のモル比が、1:0.1〜1:10である請求項6に記載の方法。
【請求項8】
in vitro再構成タンパク質合成系におけるリボソームの濃度が、0.1〜3 μMである請求項6又は7に記載の方法。
【請求項9】
in vitro再構成タンパク質合成系におけるリン脂質−アポリポタンパク質複合体粒子の濃度が、0.01〜10μMである請求項6又は7に記載の方法。
【請求項10】
タンパク質合成反応基本試薬には分子シャペロンが含まれないことを特徴とする請求項1〜9のいずれか1項に記載の膜タンパク質合成方法。
【請求項11】
タンパク質合成反応基本試薬に含まれるタンパク質が、膜タンパク質合成終了後に反応系から金属アフィニティ樹脂で除去できる請求項1〜10のいずれか1項に記載の膜タンパク質合成方法。
【請求項12】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系において、リン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を変化させて膜タンパク質を合成し、当該膜タンパク質の可溶性、および/もしくは活性を測定することにより、当該膜タンパク質の活性発現に最適なリン脂質−アポリポタンパク質複合体粒子の種類及び/または濃度及び/または添加条件を検証する方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項13】
タンパク質合成基本試薬が、濃度及び純度がそれぞれ特定されているリボソーム、開始因子、延長因子、解離因子、アミノアシルtRNA合成酵素、およびメチオニルtRNAトランスフォルミラーゼからなる群から選択される、少なくとも1つの成分を含むことを特徴とする請求項12に記載の検証方法。
【請求項14】
リン脂質−アポリポタンパク質複合体粒子が、合成された膜タンパク質の可溶性を増大させることを特徴とする請求項12又は13に記載の検証方法。
【請求項15】
リン脂質−アポリポタンパク質複合体粒子が、アポリポタンパク質及びリン脂質を含む請求項12又は13に記載の検証方法。
【請求項16】
タンパク質合成反応基本試薬には分子シャペロンが含まれないことを特徴とする請求項12〜15のいずれか1項に記載の検証方法。
【請求項17】
下記a)及びb)を含むin vitro再構成タンパク質合成キット。
a) 存在量および純度がそれぞれ特定されている成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
b) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【請求項18】
下記a)及びb)及びc)からなるin vitro再構成タンパク質合成系において、タンパク基本試薬中の成分とリン脂質−アポリポタンパク質複合体粒子の比率を変化させることを含む、膜タンパク質合成方法における最適化評価方法。
a) 目的とするタンパク質をコードする1または複数の鋳型核酸、
b) 存在量及び純度がそれぞれ特定されている複数の成分からなり、鋳型核酸を添加することにより鋳型核酸がコードするタンパク質の合成反応が生じるタンパク質合成反応基本試薬、
c) 存在量及び純度が特定されているリン脂質−アポリポタンパク質複合体粒子。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2011−188776(P2011−188776A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2010−56454(P2010−56454)
【出願日】平成22年3月12日(2010.3.12)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、文部科学省、地域科学技術振興事業委託事業、産業技術力強化法第19条の適用をうける特許出願
【出願人】(507371294)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成22年3月12日(2010.3.12)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度、文部科学省、地域科学技術振興事業委託事業、産業技術力強化法第19条の適用をうける特許出願
【出願人】(507371294)
【Fターム(参考)】
[ Back to top ]