
下位運動ニューロン疾患の治療方法およびそれを含む組成物
本発明は、(脊髄性筋萎縮症などの)下位運動ニューロン疾患に関する症状の治療、予防、および/または緩和のための方法を提供する。この方法は、アゴニスト抗trkC抗体の投与を含む。組成物およびキットも提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、その全体が参照により本明細書に援用される2005年4月26日出願の米国仮特許出願第60/675393号の優先権の利益を主張するものである。
【0002】
本発明は、下位運動ニューロン疾患の治療および/または予防におけるアゴニスト抗trkC抗体の使用に関する。
【背景技術】
【0003】
運動ニューロン疾患は、運動ニューロンが変性し、死に至る障害である。上位運動ニューロンおよび下位運動ニューロンを含む運動ニューロンは、随意筋に影響を及ぼし、随意筋を刺激して収縮させる。上位運動ニューロンは、大脳皮質に始まり、脳幹および脊髄を経由して線維を送り、下位運動ニューロンの制御に関与する。下位運動ニューロンは、脳幹および脊髄に位置し、線維を筋肉に送る。下位運動ニューロン疾患は、下位運動ニューロンの変性を含む疾患である。下位運動ニューロンが変性すると、通常は活性化する筋線維が連絡切断され、収縮せず、筋力低下および反射の減弱が引き起こされる。どちらの型のニューロンの損失も、脱力、すなわち筋萎縮(消耗)をもたらし、無痛性の脱力は、運動ニューロン疾患の臨床的な特徴である。
【0004】
SMARD1は、第2の一般的な常染色体劣性遺伝疾患である脊髄性筋萎縮症(SMA)の臨床的な変形形態であり、小児期における最も一般的な遺伝的死因である。SMAおよびSMARD1は、進行性の筋肉運動麻痺を随伴する下位運動ニューロンの変性を特徴とする。SMA症例の大多数が、生存運動神経細胞遺伝子(SMN)の突然変異によるものである(1)のに対し、SMARD1は、異なる遺伝子、すなわち免疫グロブリンμ−結合タンパク質2遺伝子(IGHMBP2)の突然変異によって引き起こされる(2〜5)。SMARD1患者は、横隔膜の関与により呼吸器機能の早期の機能障害を患う(4)。一部の予備的な臨床試験で肯定的な結果が出ているにもかかわらず(7)、現在、一般にSMARD1またはSMAの有効な療法は存在しない(6)。
【0005】
nmdマウスは、DNA/RNAヘリカーゼ/ATPaseタンパク質ファミリーの一員であるマウスIghmbp2遺伝子の自然突然変異を有する(8〜10)。この遺伝的欠陥は、イントロン4における単一の突然変異(A→G)からなるため、転写物の80%が異常にスプライスされ、20%が全長である(9)。マウスではIGHMBP2の機能発現が完全に消滅しないので、nmdマウスは、より軽度の形のヒトSMARD1に類似した疾患表現型を示す(9)。筋力低下は、出生から2週間後に発現し始め、四肢の重度の神経性筋萎縮に進行する(8〜11)。
【0006】
神経栄養因子は、運動神経疾患の有望な治療薬であると考えられている。この期待は、培養中の動物胚の運動ニューロンにおけるそれら分子の生存促進特性、すなわち軸索切断後の神経に対するそのプラスの生物学的効果、および神経変性疾患動物モデルにおける病理学的症状の軽減(12〜15)に基づいている。in vitroおよびin vivo研究のほとんどで得られる有望な結果を踏まえて、外来のニューロトロフィンが、アルツハイマー病、筋萎縮性側索硬化症(ALS)、末梢ニューロパチー、パーキン病、およびハンチントン病患者のための臨床試験で使用されている。
【発明の開示】
【発明が解決しようとする課題】
【0007】
神経栄養因子を適用する際の1つの実際的な難点は、これらのタンパク質がすべて、比較的短い半減期を有する一方で、神経変性疾患は慢性的であり、長期の治療が必要であることである。ニューロトロフィン受容体を標的とする治療用のアゴニスト抗体は、特異性が高く半減期が長いために、神経変性疾患のための新規な手法となり得る。
【0008】
特許出願および特許公開を含む、本明細書で引用するすべての参考文献は、その全体が参照により援用される。
【課題を解決するための手段】
【0009】
本発明は、個体における(下位運動ニューロン変性を含む)下位運動ニューロン疾患の治療方法を提供する。下位運動ニューロン疾患の例は、脊髄性筋萎縮症(SMA)および呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)である。この方法は、個体への有効量のアゴニスト抗trkC抗体の投与を含む。
【0010】
一態様では、本発明は、個体における下位運動ニューロン疾患の治療方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。別の態様では、本発明は、個体において下位運動ニューロン疾患に関連する症状の出現を遅らせる方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。別の態様では、本発明は、個体において下位運動ニューロン疾患の症状を緩和させる方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。
【0011】
一部の実施形態では、アゴニスト抗trkC抗体の投与後に個体の筋肉強度が向上する。一部の実施形態では、アゴニスト抗trkC抗体の投与後に個体の筋肉強度の低下を遅らせる。
【0012】
一部の実施形態では、個体は、ヒトなどの哺乳動物である。
【0013】
アゴニスト抗trkC抗体は、当業界で知られている。一部の実施形態では、アゴニスト抗trkC抗体は、モノクローナル抗体である。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトtrkCを結合する。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトtrkCを特異的に結合する。アゴニスト抗trkC抗体は、ヒトおよびげっ歯類trkCを結合するものでよい。アゴニスト抗trkC抗体は、(抗体6.4.1(PCT公開WO01/98361)などの)ヒト抗体でもよいし、または(ヒト化モノクローナル抗体2256を含む)ヒト化抗体でもよい。別の実施形態では、アゴニスト抗trkC抗体は、PCT WO2004/058190および本明細書に記載のヒト化抗体A5である。さらに他の実施形態では、抗trkCアゴニスト抗体は、表1に示す重鎖可変部アミノ酸配列(配列番号1)と表2に示す軽鎖可変部アミノ酸配列(配列番号2)とを含む。他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。CDRの同定は、多分に当分野の技能の範囲内である。一部の実施形態では、CDRはKabatCDRを含む。他の実施形態では、CDRはChothiaCDRである。さらに他の実施形態では、CDRは、KabatおよびChothiaの両方のCDRを含む。一部の実施形態では、抗体は、受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる軽鎖を含む。一部の実施形態では、抗体は、受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖を含む。一部の実施形態では、抗体は、(a)受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる軽鎖と、(b)受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖とを含む。一部の実施形態では、抗体は、(a)受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる1個または複数のCDR、および/または(b)受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖を含む。
【0014】
抗体は、次のもの、すなわち6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256のいずれか1種または複数から選択される抗体と本質的に同じtrkCエピトープを結合するものでも、またはそうした抗体と結合をめぐって競合するものでもよい。PCT公開番号WO01/98361を参照されたい。抗体は、免疫学的に不活性である、例えば補体によって媒介される溶解のきっかけとならず、または抗体依存性の細胞によって媒介される細胞傷害性(ADCC)を刺激しない定常部などの、改変された定常部を含むものでもよい。他の実施形態では、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ、PCT出願番号PCT/GB99/01441、および/または英国特許出願第9809951.8号に記載の通りに定常部を改変する。
【0015】
抗体は、次のもの、すなわち、Fab、Fab’、F(ab’)2、Fv断片、ダイアボディ、抗体断片から生成される単鎖抗体分子および多特異性抗体、ならびに単鎖Fv(scFv)分子のうちの1種または複数から選択される抗体断片などの抗体断片でもよい。抗体はキメラでもよく、二重特異性でもよい。
【0016】
アゴニスト抗trkC抗体の投与は、次の手段、すなわち、静脈内、皮下、吸入、動脈内、筋肉内、心臓内、脳室内、くも膜下腔内、脊髄内、および腹腔内のうちの1種または複数を含めて、当業界で知られている適切な任意の方法でよい。投与は、全身(例えば静脈内)および/または局所でよい。投与は、短時間および/または長期間でよい。投与は、下位運動ニューロン疾患の発症前に行うことができる。
【0017】
別の態様では、本発明は、本発明の方法のいずれかで使用するための、アゴニスト抗trkC抗体を含む組成物およびキットを提供する。これらのキットは、アゴニスト抗trkC抗体を本明細書に記載の方法のいずれかで使用するための説明書をさらに含むこともある。本発明は、本明細書に記載の方法のいずれかで使用するための、アゴニスト抗trkC抗体と薬学的に許容できる担体とを含む医薬組成物も提供する。
【0018】
本発明はまた、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べた組成物およびキットのいずれかを提供する。
【発明を実施するための最良の形態】
【0019】
本発明は、個体における下位運動ニューロン疾患の治療方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。下位運動ニューロン疾患の例には、SMA、およびSMAの臨床的な変形形態であるSMARD1が含まれる。
【0020】
一般技術
本発明の実践では、別段の指定がない限り、当分野の技能の範囲内にある(組換え技術を含む)分子生物学、微生物学、細胞生物学、生化学、および免疫学の従来の技術が使用される。このような技術は、「Molecular Cloning:A Laboratory Manual」、第2版(Sambrookら、1989年)、Cold Spring Harbor Press;「Oligonucleotide Synthesis」(M.J.Gait編、1984年);「Methods in Molecular Biology」、Humana Press;「Cell Biology:A Laboratory Notebook」(J.E.Cellis編、1998年)、Academic Press;「Animal Cell Culture」(R.I.Freshney編、1987年);「Introduction to Cell and Tissue Culture」(J.P.MatherおよびP.E.Roberts、1998年)、Plenum Press;「Cell and Tissue Culture:Laboratory Procedures」(A.Doyle、J.B.Griffiths、およびD.G.Newell編、1993〜8年)、J.Wiley and Sons;「Methods in Enzymology」(Academic Press,Inc.);「Handbook of Experimental Immunology」(D.M.WeirおよびC.C.Blackwell編);「Gene Transfer Vectors for Mammalian Cells」(J.M.MillerおよびM.P.Calos編、1987年);「Current Protocols in Molecular Biology」(F.M.Ausubelら編、1987年);「PCR:The Polymerase Chain Reaction」(Mullisら編、1994年);「Current Protocols in Immunology」(J.E.Coliganら編、1991年);「Short Protocols in Molecular Biology」(Wiley and Sons、1999年);「Immunobiology」(C.A.JanewayおよびP.Travers、1997年);「Antibodies」(P.Finch、1997);「Antibodies:a practical approach」(D.Catty編、IRL Press、1988〜1989年);「Monoclonal antibodies:a practical approach」(P.ShepherdおよびC.Dean編、Oxford University Press、2000年);「Using antibodies:a laboratory manual」(E.HarlowおよびD.Lane(Cold Spring Harbor Laboratory Press、1999年);「The Antibodies」(M.ZanettiおよびJ.D.Capra編、Harwood Academic Publishers、1995年);および「Cancer:Principles and Practice of Oncology」(V.T.DeVitaら編、J.B.Lippincott Company、1993年)などの文献で十分に説明されている。
【0021】
定義
(複数形でも区別なく使用される)「抗体」とは、炭水化物、ポリヌクレオチド、脂質、ポリペプチドなどの標的に、免疫グロブリン分子の可変部に位置する少なくとも1個の抗原認識部位を介して特異的に結合することのできる免疫グロブリン分子である。本明細書では、この用語は、無処置のポリクローナルまたはモノクローナル抗体だけでなく、(Fab、Fab’、F(ab’)2、Fvなどの)これらの断片、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ヒト化抗体、キメラ抗体、ダイアボディ、線形抗体、単鎖抗体、多特異性抗体(例えば二重特異性抗体)、および必要な特異性の抗原認識部位を含む他の任意の改変された形状の免疫グロブリン分子も含む。抗体には、IgG、IgA、IgMなどのどんなクラスの抗体も含まれ、抗体は、任意の特定のクラスのものである必要はない。その重鎖の定常ドメインの抗体アミノ酸配列に応じて、免疫グロブリンを異なるクラスに割り振ることができる。免疫グロブリンの5つの主要なクラスは、IgA、IgG、IgD、IgE、およびIgMであり、これらのいくつかは、さらにサブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA1、およびIgA2に分けられる場合もある。異なるクラスの免疫グロブリンに対応する重鎖定常ドメインは、それぞれα、γ、δ、ε、およびμと呼ばれる。軽鎖にも、κおよびλと呼ばれる2つのクラスがある。異なるクラスの免疫グロブリンのサブユニット構造および三次元立体配置はよく知られている。
【0022】
「モノクローナル抗体」とは、抗原の選択的な結合に関与するアミノ酸(自然に存在するものおよび自然に存在しないもの)からなる同種の抗体集団を指す。モノクローナル抗体の集団は、単一の抗原部位に向けられ、高度に特異的である。用語「モノクローナル抗体」は、無処置のモノクローナル抗体および全長モノクローナル抗体だけでなく、(Fab、Fab’、F(ab’)2、Fvなどの)これらの断片、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ヒト化モノクローナル抗体、キメラモノクローナル抗体、ならびに必要な特異性および抗原結合能の抗原認識部位を含む他の任意の改変された形状の免疫グロブリン分子も含む。抗体の供給源または(例えば、ハイブリドーマ、ファージ選択、組換え発現、トランスジェニック動物などによる)その製法については、限定されないものとする。
【0023】
「ヒト化」抗体とは、実質的にヒトでない種からの免疫グロブリンに由来する抗原結合部位を有し、分子の残りの免疫グロブリン構造がヒト免疫グロブリンの構造および/または配列に基づく分子を指す。抗原結合部位は、定常ドメインに融合した完全な可変ドメインを含んでもよいし、または可変ドメイン中の適切なフレームワーク領域に接ぎ合わされた相補性決定領域(CDR)のみを含んでもよい。抗原結合部位は、野生型でも、または1つまたは複数のアミノ酸置換によって改変されたもの、例えばヒト免疫グロブリンにより近似するように改変されたものでもよい。ヒト化抗体の一部の形態は、すべてのCDR配列を保存している(例えば、マウス抗体からの6個のCDRすべてを含むヒト化マウス抗体)。他の形態のヒト化抗体は、もとの抗体に関して変更されている1個または複数のCDR(1、2、3、4、5、6個)を有する。
【0024】
本明細書では、「ヒト抗体」とは、ヒトによって産生される抗体に相当するアミノ酸配列を有する抗体を意味し、かつ/または当業界で知られ、または本明細書で開示するヒト抗体の製造技術のいずれかを使用して製造されている。このヒト抗体の定義には、少なくとも1本のヒト重鎖ポリペプチドまたは少なくとも1本のヒト軽鎖ポリペプチドを含む抗体が含まれる。そのような1つの例は、ネズミ軽鎖ポリペプチドとヒト重鎖ポリペプチドとを含む抗体である。ヒト抗体は、当業界で知られている様々な技術を使用して生成することができる。一実施形態では、ヒト抗体は、ヒト抗体を発現させるファージライブラリーから選択される(Vaughanら、1996年、Nature Biotechnology、第14巻:309〜314ページ;Sheetsら、1998年、PNAS(米)、第95巻:6157〜6162ページ;HoogenboomおよびWinter、1991年、J.Mol.Biol.、227〜381ページ;Marksら、1991年、J.Mol.Biol.、第222巻:581ページ)。ヒト抗体は、ヒト免疫グロブリン座を、内在性の免疫グロブリン遺伝子が部分的または完全に不活化されているトランスジェニック動物、例えばマウスに導入して作ることもできる。この手法は、米国特許第5545807号、同第5545806号、同第5569825号、同第5625126号、同第5633425号、および同第5661016号に記載されている。別法として、ヒト抗体は、標的抗原に向けられる抗体を産生するヒトBリンパ球を不死化して調製することができる(そのようなBリンパ球は、個体から回収してもよいし、またはin vitroで免疫感作したものでもよい)。例えば、Coleら、「Monoclonal Antibodies and Cancer Therapy」、Alan R.Liss、77ページ(1985年);Boernerら、1991年、J.Immunol.、第147巻(1):86〜95ページ;および米国特許第5750373号を参照されたい。
【0025】
「キメラ抗体」とは、重鎖および軽鎖のアミノ酸配列それぞれの一部分が、ある特定の種に由来し、またはある特定のクラスに属する抗体中の対応する配列と相同であり、鎖の残りの区域が別の抗体中の対応する配列と相同である抗体を指す。通常、これらのキメラ抗体では、軽鎖および重鎖の両方の可変部が、哺乳動物のある種に由来する抗体の可変部を模倣し、定常部分は、別のものに由来する抗体中の配列と相同である。そのようなキメラ形態の1つの明白な利点は、例えば、非ヒト宿主生物から容易に入手可能なハイブリドーマまたはB細胞を使用して、現在知られている供給源から、例えばヒト細胞調製物由来の定常部と組み合わせて、可変部を好都合に得ることができる点である。可変部は、調製が容易であるという利点を有し、特異性はその供給源によって影響を受けないが、ヒトである定常部は、抗体が注射されたときに、ヒトでない供給源からの定常部よりもヒト対象からの免疫応答を誘発しにくい。しかし、この定義は、この特定の例に限定されない。
【0026】
抗体またはポリペプチドに「特異的に結合する」または「優先的に結合する」(本明細書では区別なく使用する)エピトープとは、当業界でよく理解されている用語であり、そのような特異的または優先的な結合を測定する方法も当業界でよく知られている。分子は、ある特定の細胞または物質と、別の細胞または物質より頻繁に、より迅速に、より長期間、かつ/またはより強い親和性で反応または結合するならば、「特異的結合」または「優先的な結合」を示すと言われる。抗体は、他の物質に結合するよりも強い親和性、すなわち結合力で、より容易に、かつ/またはより長い期間で結合するならば、標的に「特異的に結合する」または「優先的に結合する」。例えば、trkCエピトープに特異的または優先的に結合する抗体とは、他のtrkCエピトープまたは非trkCエピトープに結合するのより強い親和性、すなわち結合力で、より容易に、かつ/またはより長い期間、このtrkCエピトープを結合する抗体である。例えば、第1の標的に特異的または優先的に結合する抗体(または部分もしくはエピトープ)が、第2の標的に特異的または優先的に結合してもしなくてもよいことも、この定義を読むことで理解される。「特異的結合」または「優先的な結合」は、それ自体として、必ずしも独占的な結合を(含むことがあるとしても)必要としない。必ずしもそうでないが、一般に、結合への言及は優先的な結合を意味する。
【0027】
「機能Fc領域」は、未変性配列Fc領域の少なくとも1種のエフェクター機能を有する。好例となる「エフェクター機能」には、C1q結合、補体依存性細胞傷害性(CDC)、Fc受容体結合、抗体依存性細胞傷害(ADCC)、食作用、細胞表面受容体(例えばB細胞受容体、BCR)の下方調節などが含まれる。そのようなエフェクター機能には、一般に、Fc領域が結合ドメイン(例えば、抗体可変ドメイン)と合体することが必要であり、エフェクター機能は、そのような抗体エフェクター機能を評価するための当業界で知られている様々なアッセイを使用して評価することができる。
【0028】
「未変性配列Fc領域」は、自然界で見られるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。「変異体Fc領域」は、少なくとも1個のアミノ酸の変更によって未変性配列Fc領域と異なるが、未変性配列Fc領域の少なくとも1種のエフェクター機能を保持するアミノ酸配列を含む。変異体Fc領域は、未変性配列Fc領域または親ポリペプチドFc領域と比べて少なくとも1つのアミノ酸置換を有することが好ましく、例えば、未変性配列Fc領域または親ポリペプチドFc領域中に約1〜約10個のアミノ酸置換、好ましくは約1〜約5個のアミノ酸置換を有する。変異体Fc領域は、本明細書では、未変性配列Fc領域および/または親ポリペプチドFc領域と、少なくとも約80%の配列同一性、最も好ましくは少なくとも約90%の配列同一性、より好ましくは少なくとも約95%の配列同一性を有することが好ましい。
【0029】
本明細書では、「抗体依存性細胞傷害」および「ADCC」とは、Fc受容体(FcR)を発現する非特異的な細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、およびマクロファージ)が、標的細胞上の結合した抗体を認識し、その後標的細胞の溶解を引き起こす細胞性の反応を指す。対象とする分子のADCC活性は、米国特許第5500362号または同第5821337号に記載のものなどのin vitroADCCアッセイを使用して評価することができる。そのようなアッセイに有用なエフェクター細胞には、末梢血単核球(PBMC)およびナチュラルキラー細胞が含まれる。別法としてまたは追加として、対象とする分子のADCC活性は、in vivoで、例えばClynesら、1998年、PNAS(USA)、第95巻:652〜656で開示されているものなどの動物モデルで評価することもできる。
【0030】
「アゴニスト抗trkC抗体」(区別なく「抗trkCアゴニスト抗体」とも呼ぶ)とは、trkC受容体を結合し、および/またはtrkCシグナル伝達機能によって媒介される下流の経路を活性化することのできる抗体を指す。例えば、アゴニスト抗体は、trkC受容体の細胞外ドメインに結合し、それによって受容体の二量体化を引き起こし、細胞内の触媒キナーゼドメインの活性化をもたらすことができる。したがって、アゴニスト抗体は、in vitroおよび/またはin vivoで受容体を発現する細胞の増殖および/または分化を刺激し得る。一部の実施形態では、アゴニスト抗trkC抗体は、trkCに結合し、trkC生物活性を活性化する。一部の実施形態では、本発明の方法で有用なアゴニスト抗体は、trkCのドメインVおよび/またはドメインIVを認識する。Urferら、J.Biol.Chem.第273巻:5829〜5840ページ(1998年)を参照されたい。
【0031】
抗体の「可変部」とは、単独または複合語で、抗体軽鎖の可変部または抗体重鎖の可変部を指す。重鎖および軽鎖の可変部はそれぞれ、超可変領域としても知られている3つの相補性決定領域(CDR)によって連結された4つのフレームワーク領域(FR)からなる。それぞれの鎖のCDRは、FRによってきわめて接近してまとまり、他の鎖のCDRと共に、抗体の抗原結合性部位の形成に寄与する。CDRを決定する技術は、少なくとも2種存在する。すなわち(1)異種間の配列可変性に基づく手法(すなわち、Kabatら、「Sequences of Proteins of Immunological Interest」(第5版、1991年、National Institutes of Health、米メリーランド州ベセズダ))、および(2)抗原−抗体複合体の結晶学的研究に基づく手法(Al−lazikaniら(1997年)、J.Molec.Biol.第273巻:927〜948ページ))。本明細書では、CDRとは、どちらかの手法または両方の手法の組合せによって規定されるCDRを指す場合もある。
【0032】
抗体の「定常部」とは、単独または複合語で抗体軽鎖の定常部または抗体重鎖の定常部を指す。
【0033】
本明細書では、「Fc受容体」および「FcR」とは、抗体のFc領域に結合する受容体について述べるものである。好ましいFcRは、未変性配列ヒトFcRである。さらに、好ましいFcRは、IgG抗体(γ受容体)を結合するものであり、これには、そのアレル変異体および選択的にスプライスされた形態を含めてFcγRI、FcγRII、およびFcγRIIIサブクラスの受容体が含まれる。FcγRII受容体には、主にその細胞質ドメインが異なる同様のアミノ酸配列を有するFcγRIIA(「活性化する受容体」)およびFcγRIIB(「抑制する受容体」)が含まれる。FcRは、RavetchおよびKinet、1991年、Ann.Rev.Immunol.、第9巻:457〜92ページ;Capelら、1994年、Immunomethods、第4巻:25〜34ページ;およびde Haasら、1995年、J.Lab.Clin.Med.、第126巻:330〜41ページで総説されている。「FcR」には、母親のIgGの胎仔への移動を司る新生児受容体、すなわちFcRnも含まれる(Guyerら、1976年、J.Immunol.、第117巻:587ページ;およびKimら、1994年、J.Immunol.、第24巻:249ページ)。
【0034】
「補体依存性細胞傷害性」および「CDC」とは、補体の存在下で標的を溶解させることを指す。補体活性化経路は、補体系(C1q)の第1の構成成分が、同族抗原と複合体を形成した分子(例えば抗体)に結合することで開始される。補体活性化を評価するために、例えば、Gazzano−Santoroら、J.Immunol.Methods、第202巻:163ページ(1996年)に記載されているようにCDCアッセイを実施することができる。
【0035】
本明細書では、「親和性成熟」抗体とは、その1個または複数のCDR中に、抗体の抗原に対する親和性を、変更をもたない親抗体と比べて向上させる1つまたは複数の変更を有する抗体を意味する。一部の実施形態では、親和性成熟抗体は、標的抗原に対してナノモルまたはさらにはピコモルの親和性を有する。親和性成熟抗体は、当業界で知られている手順によって生成される(Marksら、1992年、Bio/Technology、第10巻:779〜783ページ;Barbasら、1994年、Proc Nat.Acad.Sci、USA第91巻:3809〜3813ページ;Schierら、1995年、Gene、第169巻:147〜155ページ;Yeltonら、1995年、J.Immunol.、第155巻:1994〜2004ページ;Jacksonら、1995年、J.Immunol.、第154巻(7):3310〜9ページ;Hawkinsら、1992年、J.Mol.Biol.、第226巻:889〜896ページ)。
【0036】
本明細書では、「trkC」とは、チロシンキナーゼスーパーファミリーの一員であるtrkC受容体ポリペプチドを指す。trkCは、その限りでないが、ヒト、イヌ、ネコ、ウシ、ウマ、霊長類、ならびに(マウスおよびラットを含む)げっ歯類を含めて、任意の哺乳類種の未変性trkC受容体を含む。全長未変性trkCの細胞外ドメインは、様々な他のタンパク質で同定されている相同またはさもなければ同様の構造に即して規定されている。そのドメインは、成熟trkC受容体のN末端から始めて、1)アミノ酸1からアミノ酸48まで伸びる第1のシステインリッチドメイン、2)アミノ酸49からアミノ酸120まで伸びるロイシンリッチドメイン、3)アミノ酸121からアミノ酸177まで伸びる第2のシステインリッチドメイン、4)約アミノ酸196からアミノ酸257まで伸びる第1の免疫グロブリン様ドメイン、および5)約アミノ酸288からアミノ酸351まで伸びる第2の免疫グロブリン様ドメインと呼ばれている。例えば、PCT公開第WO0198361号を参照されたい。ヒトtrkC受容体のドメイン構造は、結晶構造に即して次のようにも呼ばれている。すなわち、アミノ酸1〜アミノ酸47のドメイン1、アミノ酸48〜アミノ酸130のドメイン2、アミノ酸131〜アミノ酸177のドメイン3、アミノ酸178〜アミノ酸165のドメイン4、およびアミノ酸166〜アミノ酸381のドメイン5。例えば、PCT公開WO0198361;Urferら、J.Biol.Chem.第273巻:5829〜5840ページ(1998年)を参照されたい。trkCの変異体も含まれ、その例には、その限りでないが、キナーゼドメインを含まない変異体(Sheltonら.、J.Neurosci.第15巻(1):477〜491ページ(1995年))および改変されたキナーゼドメインを含む変異体(Sheltonら.、J.Neurosci.第15巻(1):477〜491ページ(1995年))が含まれる。
【0037】
「生物活性」とは、本発明のアゴニスト抗trkC抗体と共に使用するとき、一般にtrkC受容体チロシンキナーゼを結合し、および/またはtrkCシグナル伝達機能によって媒介される下流の経路を活性化する能力を有することを指す。本明細書では、「生物活性」は、trkCの未変性リガンドであるNT−3のtrkC発現細胞に対する作用によって誘発されるものと同じような、1種または複数のエフェクター機能を含む。trkCの「生物活性」は、下流のシグナル伝達経路、またはNT−3の作用によって誘発されるものと異なるエフェクター機能も含み得る。限定するものではないが、生物活性には、次のもの、すなわち、trkCを結合し活性化する能力;trkC受容体の二量体化を促進する能力;(損傷を受けた細胞を含む)細胞、特に、末梢(交感神経系、感覚、および腸管)ニューロンおよび中枢(脳および脊髄)ニューロンを含むin vitroまたはin vivoでのニューロン、ならびにニューロンでない細胞、例えば末梢血白血球の発生、生存、機能、維持、および/または再生を促進する能力のいずれか1種または複数が含まれる。特に好ましい生物活性は、下位運動ニューロン疾患の1つまたは複数の症状を(予防を含めて)治療し、および/または下位運動ニューロンの機能を修復および/もしくは改善する能力である。
【0038】
本明細書では、「治療」とは、有益なまたは所望の臨床的な成果を得るための手法である。本発明の目的では、有益なまたは所望の臨床的な成果には、その限りでないが、次のもの、すなわち、下位運動ニューロン疾患に関連する1種または複数の症状の軽減(例えば、下位運動ニューロンの変性、進行性の筋力低下または運動麻痺、呼吸不全、および低下する反射);下位運動ニューロン疾患の進行を遅らせまたは緩慢にすること;下位運動ニューロンの変性を安定化する(すなわち、悪化させない)こと;下位運動ニューロンの変性および/または筋肉の脱力の回復;車いす使用の必要を遅らせること;人工呼吸の必要を遅らせること;ならびに最終的に全体としての生存時間を延長することのうちの1種または複数が含まれる。
【0039】
下位運動ニューロン疾患または下位運動ニューロン疾患の1種または複数の症状を「軽減する」とは、本発明によるアゴニスト抗trkC抗体による治療を受ける個体または個体集団において、疾患の望ましくない臨床病態の程度および/または時間経過を減らすことを意味する。
【0040】
下位運動ニューロン疾患の「症状の重症度を軽減する」または下位運動ニューロン疾患の「症状を緩和させる」とは、下位運動ニューロン疾患の1つまたは複数の症状を和らげ、かつ/またはアゴニスト抗trkC抗体を投与しないものと比べて改善させることを意味する。「重症度を低減する」には、症状の期間の短縮または縮小も含まれる。
【0041】
本明細書では、下位運動ニューロン疾患の出現を「遅らせる」とは、この疾患の出現を先送りし、妨げ、緩慢にし、遅延させ、安定化し、かつ/または延期することを意味する。この遅れは、治療対象となる疾患および/または個体の経歴に応じて様々な長さの時間となり得る。当業者には明らかなように、十分または有意な遅れは、個体が下位運動ニューロンの変性ならびに筋力低下または運動麻痺にかからないという点で、事実上予防を含む。下位運動ニューロン疾患の出現を「遅らせる」方法は、その方法を使用しないのと比べて、所与の時間枠で下位運動ニューロンの変性が出現する確率を低下させ、かつ/または所与の時間枠で下位運動ニューロンの変性の程度を低減する方法である。そのような比較は、通常は、統計学的に有意な数の対象を使用する臨床的な調査に基づく。
【0042】
下位運動ニューロン疾患の「出現」とは、個体内での下位運動ニューロン疾患に関連する症状(例えば、下位運動ニューロン機能の低下ならびに筋力低下および運動麻痺)の発現および/または進行を意味する。出現は、本明細書に記載の標準の臨床的な技術を使用して検出可能となり得る。しかし、出現は、最初に検出不可能な場合もある疾患の進行をも指す。本発明の目的では、進行とは、この場合では、標準の神経学的検査または患者の問診によって決定され、あるいはより専門の量的試験によって決定される場合もある、疾患状態の生物学的な経過を指す。これらのより専門の量的試験には、その限りでないが、微小神経電図検査による冒されたニューロンの伝導速度の測定、専門の平衡性試験、反射試験、専門の固有受容および/または運動感覚の試験、力の試験(例えば、その限りでないが量的な筋力測定、筋電図検査、MRIを含む、筋力の臨床検査)、その限りでないが血圧コントロール試験、様々な生理的および薬理学的な刺激に対する心拍応答試験を含む自律神経機能の試験を含めることができる。例として、SMAでは、試験には、その限りでないが量的な筋力測定、筋電図検査、および/またはMRIを含む筋力の臨床検査などの、運動技能および/または運動力の試験を含めることができる。「出現」には、発生、再発、および発症が含まれる。本明細書では、下位運動ニューロン疾患の「発症」または「発生」には、最初の発症および/または再発が含まれる。
【0043】
本明細書では、「リスクのある」個体とは、下位運動ニューロン疾患が出現するリスクのある個体である。「リスクのある」個体は、検出可能な疾患にかかっていてもいなくてもよく、本明細書に記載の治療方法の前に検出可能な疾患を呈していてもいなくてもよい。「リスクのある」とは、個体が、生存運動神経細胞遺伝子(SMN)または免疫グロブリンμ結合タンパク質2遺伝子(IGHMBP2)の突然変異などの疾患の出現と相互に関係がある測定可能なパラメータである、1種または複数のいわゆるリスクファクターを有することを意味する。これらのリスクファクターの1つまたは複数を有する個体は、これらのリスクファクターなしの個体よりも下位運動ニューロン疾患にかかる確率が高い。
【0044】
「有効量」とは、疾患の発症を遅らせる臨床的な成果を含む、有益なまたは所望の臨床的な成果をもたらすのに十分な量である。有効量は、1回または複数の投与で投与することができる。本発明の目的では、本明細書に記載のアゴニスト抗trkC抗体の有効量は、下位運動ニューロン疾患を緩和させ、安定化し、逆戻りさせ、その進行を緩慢にしかつ/または遅らせ、あるいは予防するのに十分な量である。臨床的な状況の中で理解されるように、薬物、化合物、または医薬組成物の有効量は、別の薬物、化合物、または医薬組成物と共に実現されることもあれば、そうでないこともある。したがって、「有効量」は、1種または複数の治療薬を投与するという状況で考えることができ、1種または複数の他の薬剤と共に望ましい成果が実現される場合がありまたは実現されるならば、単剤が有効量で与えられたと考えることができる。
【0045】
本明細書では、「共に」投与は、同時の投与および/または異なる時点での投与を含む。共に投与は、合剤(co−formulation)としての投与または別個の組成物としての投与も含む。本明細書では、共投与は、アゴニスト抗trkC抗体および別の下位運動ニューロン疾患治療薬またはこれらの組合せが個体に投与され、それが同時および/または別々に起こり得る任意の状況を含む意味である。本明細書でさらに論述するように、アゴニスト抗trkC抗体と他の治療薬を異なる投薬回数または投薬間隔で投与できることは理解される。アゴニスト抗trkC抗体と他の治療薬を、同じ投与経路または異なる投与経路を使用して投与できることも理解される。
【0046】
「個体」は、哺乳動物、より好ましくはヒトである。哺乳動物には、その限りでないが、飼養動物、競技動物、愛玩動物、霊長類、ウマ、乳牛、ネコ、イヌ、および(マウスやラットなどの)げっ歯類も含まれる。
【0047】
本明細書では、単数形の「a」、「an」、および「the」の形は、別段の指摘がない限り、複数形の言及を含む。例えば、「an」antibodyは1種または複数の抗体を含み、「a symptom」は1種または複数の症状を意味する。
【0048】
本明細書では、「ベクター」とは、宿主細胞中で1種または複数の対象とする遺伝子または配列を放出する、好ましくは発現させることのできる構築物を意味する。ベクターの例には、その限りでないが、ウイルスベクター、裸DNAまたはRNA発現ベクター、プラスミド、コスミドまたはファージベクター、カチオン縮合剤と結合したDNAまたはRNA発現ベクター、リポソーム中にカプセル封入されたDNAまたはRNA発現ベクター、ならびに産生細胞などのある種の真核細胞が含まれる。
【0049】
本明細書では、「発現制御配列」とは、核酸の転写を指示する核酸配列を意味する。発現制御配列は、構成的もしくは誘導型プロモーターなどのプロモーター、またはエンハンサーでよい。発現制御配列は、転写される核酸配列に作動可能に連結される。
【0050】
本明細書では、「核酸」または「ポリヌクレオチド」とは、一本鎖または二本鎖の形のデオキシリボヌクレオチドまたはリボヌクレオチドポリマーを指し、別段の制限がない限り、自然に存在するヌクレオチドと同様にして核酸とハイブリッド形成する天然ヌクレオチドの既知の類似体を含む。
【0051】
本明細書では、「薬学的に許容できる担体」には、活性成分と組み合わせたとき、活性成分が生物活性を保持するのを可能にし、対象の免疫系と反応性でない任意の材料が含まれる。例には、その限りでないが、リン酸緩衝溶液、水、水中油型乳濁液などの乳濁液、および様々な種類の湿潤剤などの標準の医薬担体のいずれかが含まれる。エアロゾルまたは非経口投与用の好ましい希釈剤は、リン酸緩衝溶液または通常の(0.9%)生理食塩水である。
【0052】
そのような担体を含む組成物は、よく知られている従来の方法によって製剤される(例えば、「Remington’s Pharmaceutical Sciences」、第18版、A.Gennaro編、Mack Publishing Co.、米ペンシルヴェニア州イーストン、1990年;およびRemington、「The Science and Practice of Pharmacy」、第20版、Mack Publishing、2000年を参照されたい)。
【0053】
本発明の方法
本明細書に記載のすべての方法に関して、アゴニスト抗trkC抗体への言及は、これらの抗体の1種または複数を含む組成物も含む。これらの組成物は、緩衝剤を含めて薬学的に許容できる賦形剤などの、当業界でよく知られている適切な賦形剤をさらに含んでもよい。
【0054】
アゴニスト抗trkC抗体を使用して下位運動ニューロン疾患を治療する方法
本発明は、アゴニスト抗trkC抗体を使用して、下位運動ニューロン疾患を治療し、予防し、その症状の出現を遅らせ、かつ/または軽減する方法を含む。この方法は、その必要のある個体にこれらの抗体を有効量投与するものである(様々な適応症および態様は本明細書に記載する)。有効量のアゴニスト抗trkC抗体は、他の治療薬と共に投与しても、またはそれなしで投与してもよい。個体は、下位運動ニューロン疾患と診断されているものでもよいし、下位運動ニューロン疾患の出現にかかるリスクのあるものでもよい。一部の実施形態では、個体はヒトである。しかし、記載する方法は、獣医学の状況(例えば、イヌ、ネコ、ウシ、ウマなどのヒトでない哺乳動物)にも適用できる。
【0055】
SMAやSMARD1などの下位運動ニューロン疾患を評価する方法およびその治療は、当業界で知られており、本明細書に記載される。
【0056】
アゴニスト抗trkC抗体
本発明の方法は、trkCを活性化する方法でtrkCと相互作用する抗trkC抗体を使用するものである。抗trkCアゴニスト抗体は、以下の特性のいずれか1つまたは複数を示す。すなわち、(a)trkC受容体に結合する、(b)trkC受容体の1つまたは複数のエピトープに結合する、(c)trkC受容体に結合し、trkC生物活性、またはtrkCシグナル伝達機能によって媒介される1つまたは複数の下流経路を活性化する、(d)trkC受容体二量体化を促進し、(e)trkC受容体の活性化を増大させ、(f)好ましい薬物動態および生体利用度特性を示し、(g)細胞の発生、生存、機能、維持、および/または再生を促進する、(h)trkC受容体に結合し、下位運動ニューロン疾患の1つまたは複数の症状を治療し、予防し、逆戻りさせ、または緩和させる。
【0057】
アゴニスト抗trkC抗体は、当業界で知られている。PCT公開WO01/98361;Urferら、J.Biol.Chem.(1998年)第273巻:5829〜5840ページを参照されたい。一部の実施形態では、抗trkCアゴニスト抗体は、抗体「A5」と呼ばれるヒト化マウス抗trkCアゴニスト抗体であり、A330P331→S330S331の突然変異(アミノ酸の番号付けは野生型IgG2a配列に即したKabatの番号付けに基づく;Eur.J.Immunol.(1999年)第29巻:2613〜2624ページを参照されたい)を含むヒト重鎖IgG2a定常部;ヒト軽鎖κ定常部;ならびに表1および2に示す重鎖および軽鎖可変部を含む。
【0058】
表1:A5重鎖可変部。ChothiaCDRは下線を付したイタリック体として示し、KabatCDRは太字として示す。
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYRIHWVRQAPGQGLEWMGEIYPSNARTNYNEKFKSRVTMTRDTSTSTVYMELSSLRSEDTAVYYCARKYYYGNTRRSWYFDVWGQGTTVTVS(配列番号1)
【0059】
表2:A5軽鎖可変部。KabatCDRは下線を付したイタリック体として示し、ChothiaCDRは太字として示す。
【0060】
DIQMTQSPSSLSASVGDRVTITCRASESIDNYGISFLAWYQQKPGKAPKLLIYAASNRGSGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQSKTVPRTFGQGTKLEIKRT(配列番号2)
【0061】
A5の重鎖可変部または軽鎖可変部をコードする以下のポリヌクレオチドは、アメリカ培養細胞系統保存機関、10801 University Boulevard、Manassas、Virginia、USA(ATCC)に寄託した。
【0062】
【表1】

【0063】
ベクターEb.pur.2256.A5は、A5軽鎖可変部をコードするポリヌクレオチドであり、ベクターDb.2256.A5は、A5重鎖可変部をコードするポリヌクレオチドである。
【0064】
他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。CDR領域の決定は、多分に当分野の技能の範囲内である。CDRを決定するいくつかの技術は次の通りである。(1)異種間の配列可変性に基づく手法(すなわち、Kabatら、「Sequences of Proteins of Immunological Interest」(第5版、1991年、National Institutes of Health、米メリーランド州ベセズダ))、(2)抗原−抗体複合体の結晶学的研究に基づく手法(Al−lazikaniら(1997年)、J.Molec.Biol.第273巻:927〜948ページ))。CDRの同定は、多分に当分野の技能の範囲内である。一部の実施形態では、CDRはKabatCDRを含む。他の実施形態では、CDRはChothiaCDRである。さらに他の実施形態では、CDRは、KabatおよびChothia両方のCDRを含む。
【0065】
抗体は、モノクローナル抗体、ポリクローナル抗体、抗体断片(例えば、Fab、Fab’、F(ab’)2、Fv、Fcなど)、キメラ抗体、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ならびに必要な特異性の抗原認識部位を含む他の改変された形状の免疫グロブリン分子を含み得る。抗体は、(ヒト化抗体を含めて)ネズミ、ラット、ヒト、または他の任意の起源のものでよい。したがって、アゴニスト抗trkC抗体は、(抗体6.4.1(PCT公開WO01/98361)などの)ヒト抗体でもよいし、または(ヒト化モノクローナル抗体A5を含む)ヒト化抗体でもよい。
【0066】
アゴニスト抗trkC抗体は、ヒトtrkCを結合するものでよい。アゴニスト抗trkC抗体は、ヒトおよびげっ歯類のtrkCを結合するものでもよい。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトおよびラットのtrkCを結合するものでよい。一部の実施形態では、抗trkC抗体は、ヒトおよびマウスのtrkCを結合するものでよい。一実施形態では、抗体は、ヒトtrkC細胞外ドメイン上の1つまたは複数のエピトープを認識する抗体である。別の実施形態では、抗体は、ヒトtrkC細胞外ドメイン上の1つまたは複数のエピトープを認識するマウスまたはラット抗体である。一部の実施形態では、抗体はヒトtrkCを結合し、別の哺乳類種(一部の実施形態では脊椎動物種)からのtrkCはほとんど結合しない。一部の実施形態では、抗体は、ヒトtrkCならびに別の哺乳類種(一部の実施形態では脊椎動物種)からの1つまたは複数のtrkCを結合する。別の実施形態では、抗体は、霊長類、イヌ、ネコ、ウマ、およびウシのもののうちの1種または複数から選択されるtrkC上の1つまたは複数のエピトープを認識する。一部の実施形態では、抗体は、trkCを結合し、(同類のニューロトロフィン受容体であるtrkAおよび/またはtrkBなどの)他のニューロトロフィン受容体とほとんど交差反応(結合)しない。一部の実施形態では、抗体はtrkCを結合し、さらにtrkAおよび/またはtrkBを結合する。
【0067】
trkCアゴニスト抗体によって認識されるエピトープは、連続的なものでも、連続的でないものでもよい。一部の実施形態では、抗体は、6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256のいずれか1種または複数から選択される抗体と本質的に同じtrkCエピトープを結合する。PCT公開WO01/98361を参照されたい。抗体が向かう先となり得るエピトープの例には、その限りでないが、trkCのドメインVおよび/またはドメインIVが含まれる。別の実施形態では、エピトープには、ヒトtrkCの残基L284、E287、およびN335のうちの1種または複数が含まれる。Urferら、J.Biol.Chem.(1998年)第273巻:5829〜5840ページ)を参照されたい。さらに他の実施形態では、抗体は、免疫学的に不活性な定常部、例えば、補体によって媒介される溶解のきっかけとならず、または抗体依存性細胞によって媒介される細胞傷害性(ADCC)を刺激しない定常部などの、改変された定常部を含む。他の実施形態では、定常部は、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ;PCT出願PCT/GB99/01441;および/または英国特許出願第9809951.8号に記載の通りに改変する。一部の実施形態では、定常部は、A330P331→S330S331の突然変異(野生型IgG2a配列に即したアミノ酸の番号付け;Eur.J.Immunol.(1999年)第29巻:2613〜2624ページを参照されたい)を含むヒト重鎖IgG2a定常部を含む。
【0068】
抗trkCアゴニスト抗体のtrkCに対する結合親和性は、約500nM、400nM、300nM、200nM、100nM、約50nM、約10nM、約1nM、約500pM、約100pM、または約50pMのいずれかから、約2pM、約5pM、約10pM、約15pM、約20pM、または約40pMのいずれかまでである場合がある。一部の実施形態では、結合親和性は、約100nM、約50nM、約10nM、約1nM、約500pM、約100pM、約50pM、または約50pM未満のいずれかである。一部の実施形態では、結合親和性は、約100nM、約50nM、約10nM、約1nM、約500pM、約100pM、または約50pM未満のいずれかである。さらに他の実施形態では、結合親和性は、約2pM、約5pM、約10pM、約15pM、約20pM、約40pM、または約40pM超である。当業界でよく知られているように、結合親和性は、KDまたは解離定数として表すことができ、結合親和性の増大は、KDの低下に一致する。BIAcore分析を使用して評価したマウス抗trkCアゴニストモノクローナル抗体2256のヒトtrkCに対する結合親和性は、約40nMであり、BIAcore分析を使用して評価した(本明細書に記載の)ヒト化抗trkCアゴニスト抗体A5のヒトtrkCに対する結合親和性は、約0.28nMである。
【0069】
抗体のtrkCに対する結合親和性を測定する1つの方法は、抗体の単機能Fab断片の結合親和性を測定することによるものである。単機能Fab断片を得るために、抗体(例えばIgG)をパパインで切断し、または組換えによって発現させることができる。抗体の抗trkC Fab断片の親和性は、表面プラズモン共鳴(BlAcore3000(商標)表面プラズモン共鳴(SPR)システム、BIAcore,INC、米ニュージャージー州Piscaway)によって測定することができる。CM5チップは、供給元の説明書に従って、N−エチル−N’−(3−ジメチルアミノプロピル)−カルボジイニド塩酸塩(EDC)およびN−ヒドロキシスクシンイミド(NHS)で活性化することができる。ヒトtrkC−Fc融合タンパク質(「htrkC」)(またはラットtrkCなどの他の任意のtrkC)を10mMの酢酸ナトリウムpH5.0に希釈し、0.0005mg/mLの濃度で活性化したチップ上に通液(inject)することができる。個々のチップチャネルでの変動する流動時間を使用して、詳細な動力学研究のための200〜400応答単位(RU)とスクリーニングアッセイのための500〜1000RUの2通りの範囲の抗原密度を実現することができる。チップは、エタノールアミンでブロックすることができる。再生研究では、Pierce溶離緩衝液(製品番号21004、Pierce Biotechnology、米イリノイ州ロックフォード)と4M NaCl(2:1)の混合物が、200回を超える通液の間チップ上のhtrkCの活性を保ちながら、結合したFabを効果的に除去することが示されている。HBS−EP緩衝液(0.01MのHEPES、pH7.4、0.15のNaCl、3mMのEDTA、0.005%のSurfactantP20)をBIAcoreアッセイ用の実施緩衝液として使用する。精製したFabサンプルの段階希釈物(推定されるKDの0.1〜10倍)を100μL/分で1分間かけて通液し、最長で2時間の解離時間を与える。Fabタンパク質の濃度は、(アミノ酸分析によって測定した)既知の濃度のFabを標準として使用するELISAおよび/またはSDS−PAGE電気泳動によって測定する。BIAevaluationプログラムを使用してデータを1:1のLangmuir結合モデルに当てはめることによって(Karlsson,R.、Roos,H.、Fagerstam,L.、Petersson,B.(1994年)、Methods Enzymology第6巻:99〜110ページ)、動力学的な結合速度(kon)と解離速度(koff)を同時に得る。平衡解離定数(KD)値をkoff/konとして算出する。
【0070】
別の態様では、ヒトtrkC受容体を活性化することのできる(例えば、ヒト、ヒト化、マウス、キメラの)抗体は、trkCの1つまたは複数の細胞外ドメインを発現する免疫原を使用して生成することもできる。免疫原の一例は、trkCを高度に発現する細胞であり、本明細書に記載の通りに得ることができる。使用することのできる免疫原の別の例は、trkC受容体の細胞外ドメインまたは細胞外ドメインの部分を含む(trkCイムノアドヘシンなどの)可溶性タンパク質である。
【0071】
宿主動物を免疫化する経路およびスケジュールは一般に、本明細書でさらに記載する、抗体の刺激および生成のための確立した技術および従来の技術に合わせることにある。ヒトおよびマウス抗体を生成するための一般技術は、当業界で知られており、本明細書に記載されている。
【0072】
ヒトを含む任意の哺乳類対象またはその抗体産生細胞は、ヒトを含む哺乳類のハイブリドーマ細胞系を生成する基盤として役立つように操作することができると予想される。通常は、宿主動物に、本明細書に記載のものを含むある量の免疫原を腹腔内接種する。
【0073】
ハイブリドーマは、Kohler,B.およびMilstein,C.(1975年)、Nature第256巻:495〜497ページの一般の体細胞ハイブリッド形成技術を使用して、またはBuck,D.W.ら(1982年)、In Vitro、第18巻:377〜381ページによって変更されている通りに、リンパ球および不死化した骨髄腫細胞から調製することができる。その限りでないが、X63−Ag8.653、およびSalk Institute、Cell Distribution Center、米カリフォルニア州サンディエゴからのものを含めて、入手可能な骨髄腫系統をハイブリッド形成で使用することができる。一般に、この技術は、ポリエチレングリコールなどの融合源(fusogen)を使用して、または当業界でよく知られている電気的な手段によって骨髄腫細胞とリンパ球を融合させるものである。融合させた後、細胞を融合培地から分離し、ヒポキサンチン−アミノプテリン−チミジン(HAT)培地などの選択的な増殖培地で増殖させて、ハイブリッド形成していない親細胞を排除する。モノクローナル抗体を分泌するハイブリドーマの培養には、血清を補充し、または補充しない本明細書に記載の培地のいずれかを使用することができる。細胞融合技術の別の選択肢として、EBVによって不死化したB細胞を使用して、本発明の抗trkCモノクローナル抗体を生成することができる。ハイブリドーマを拡張させ、所望ならばサブクローニングし、従来のイムノアッセイ手順(例えば、放射性免疫測定法、酵素免疫測定法、または蛍光免疫測定法)によって抗免疫原活性があるかどうか上清を検定する。
【0074】
抗体供給源として使用することのできるハイブリドーマとは、trkCまたはその部分に特異的なモノクローナル抗体を産生するすべての派生物、すなわち親ハイブリドーマの子孫細胞を含む。
【0075】
そのような抗体を産生するハイブリドーマは、知られている手順を使用して、in vitroまたはin vivoで増殖させることができる。モノクローナル抗体は、所望ならば、硫安塩析、ゲル電気泳動、透析、クロマトグラフィー、限外ろ過などの従来の免疫グロブリン精製手順によって培地または体液から単離してもよい。所望でない活性が存在するならば、例えば、固相に結合している免疫原でできた吸着剤上での調製を実施し、所望の抗体を免疫原から溶離または解放することによって除去できる。ヒトもしくは他の種のtrkC受容体またはヒトもしくは他の種のtrkC受容体の断片、あるいは免疫化する種において免疫原性であるタンパク質、例えば、キーホールリンペットヘモシアニン、血清アルブミン、ウシのサイログロブリン、または大豆トリプシンインヒビターに、二官能性または誘導体化の薬剤、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介して結合)、N−ヒドロキシスクシンイミド(リジン残基を介して)、グルタルアルデヒド、無水コハク酸、SOCl2、またはR1N=C=NR(RおよびR1は異なるアルキル基である)を使用して結合させたターゲットアミノ酸配列を含むヒトまたは他の種のtrkC受容体または断片で宿主動物を免疫化すると、抗体の集団(例えばモノクローナル抗体)を得ることができる。免疫原の別の例は、trkCを高度に発現する細胞であり、これは、組換えの手段によって、あるいは高レベルのtrkCを発現する天然供給源から細胞を単離または濃縮して得ることができる。これらの細胞は、ヒトまたは他の動物に由来するものでよく、直接に単離された免疫原として使用してもよいし、または免疫原性が増大し、または(trkC断片の)trkCの発現が増加しもしくは濃縮されるように処理してもよい。その処理には、その限りでないが、細胞またはその断片を、例えばホルムアルデヒド、グルタルアルデヒド、エタノール、アセトン、および/または様々な酸などの、その安定性または免疫原性を増大させるように設計された薬剤で処理することが含まれる。さらに、このような処理の前または後に、所望の免疫原、この場合ではtrkCまたはその断片を濃縮するため、細胞の処理を行ってもよい。これらの処理ステップには、当業界でよく知られている膜分画法が含まれる。
【0076】
所望ならば、目的の抗trkC抗体(モノクローナルまたはポリクローナル)の配列を決定してもよく、次いでポリヌクレオチド配列をベクターにクローン化して発現または増殖させてもよい。目的の抗体をコードする配列は、宿主細胞中のベクターの中で維持することができ、次いで将来の使用に向けて宿主細胞を拡張させ、凍結することができる。別法として、抗体を「ヒト化」し、または抗体の親和性もしくは他の特性を向上させるために、ポリヌクレオチド配列を遺伝子操作に使用してもよい。例えば、抗体をヒトの臨床試験および治療で使用するならば、定常部をヒト定常部により似るように操作して、免疫応答を回避する場合もある。抗体配列を遺伝子操作して、trkC受容体に対するより強い親和性およびtrkC受容体の活性化におけるより高い有効性を得ることが望ましい場合もある。抗trkC抗体のポリヌクレオチドに1つまたは複数の変更が加えられても、trkC細胞外ドメインまたはtrkCエピトープへのその結合能力が維持され得ることは、当業者には明白であろう。
【0077】
モノクローナル抗体をヒト化するには、大まかな4ステップを踏む。すなわち、(1)出発抗体の軽鎖および重鎖可変ドメインのヌクレオチドおよび推定されるアミノ酸配列を決定するステップ、(2)ヒト化抗体を設計する、すなわち、ヒト化処理の際にどちらの抗体フレームワーク領域を使用するかを決定するステップ、(3)実際のヒト化方法/技術のステップ、および(4)ヒト化抗体の形質移入および発現のステップ。例えば、米国特許第4816567号、同第5807715号、同第5866692号、同第6331415号、同第5530101号、同第5693761号、同第5693762号、同第5585089号、同第6180370号、および同第6548640号を参照されたい。例えば、抗体をヒトの臨床試験および治療で使用するならば、定常部をヒト定常部により似るように操作して、免疫応答を回避する場合もある。例えば、米国特許第5997867号および同第5866692号を参照されたい。
【0078】
組換えヒト化抗体では、Fcγ部分を変更して、Fcγ受容体および補体免疫系との相互作用を回避することができる。この種類の変更は、ケンブリッジ大学Department of PathologyのMike Clark博士によって設計されたものであり、その抗体を調製する技術は、1999年11月18日公開のPCT公開WO99/58572に記載されている。
【0079】
ヒト定常ドメインと融合させたげっ歯類可変部または改変されたげっ歯類可変部ならびにそれに伴う相補性決定領域(CDR)を有するキメラ抗体を含めて、非ヒト免疫グロブリンに由来する抗原結合部位を含むいくつかの「ヒト化」抗体分子が記載されている。例えば、Winterら、Nature第349巻:293〜299ページ(1991年)、Lobuglioら、Proc.Nat.Acad.Sci.USA第86巻:4220〜4224ページ(1989年)、Shawら、J Immunol.第138巻:4534〜4538ページ(1987年)、およびBrownら、Cancer Res.第47巻:3577〜3583ページ(1987年)を参照されたい。他の参考文献は、適切なヒト抗体定常ドメインと融合させる前にヒト支持フレームワーク領域(FR)に接ぎ合わせたげっ歯類CDRについて述べている。例えば、Riechmannら、Nature第332巻:323〜327ページ(1988年)、Verhoeyenら、Science第239巻:1534〜1536ページ(1988年)、およびJonesら、Nature第321巻:522〜525ページ(1986年)を参照されたい。別の参考文献は、組換えによって張り合わせたげっ歯類フレームワーク領域を支持体とするげっ歯類CDRについて述べている。例えば、欧州特許公開第519596号を参照されたい。これらの「ヒト化」分子は、ヒトレシピエントの治療にこれらの部分を適用する期間および有効性を制限する、げっ歯類抗ヒト抗体分子に対する不必要な免疫応答を最小限に抑えるように設計される。抗体定常部は、免疫学的に不活性になるように、例えば、補体によって媒介される溶解のきっかけとならず、抗体依存性の細胞によって媒介される細胞傷害性(ADCC)を刺激しないように操作することができる。他の実施形態では、定常部は、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ;PCT出願PCT/GB99/01441;および/または英国特許出願第9809951.8号に記載の通りに改変する。例えば、PCT/GB99/01441;英国特許出願第9809951.8号を参照されたい。利用することのできる、抗体をヒト化する他の方法は、Daughertyら、Nucl.Acids Res.第19巻:2471〜2476ページ(1991年)および米国特許第6180377号、同第6054297号、同第5997867号、同第5866692号、同第6210671号、同第6350861号、およびPCT公開WO01/27160号で開示されている。
【0080】
さらに別の選択肢では、特異的なヒト免疫グロブリンタンパク質を発現させるように操作された市販のマウスを使用して、完全なヒト抗体を得ることもできる。より望ましい(例えば、完全なヒト抗体)またはより強固な免疫応答を生み出すように設計されたトランスジェニック動物を、ヒト化抗体またはヒト抗体の産生に使用してもよい。そのような技術の例は、Abgenix,Inc.(米カリフォルニア州フリーモント)のXenomouse(商標)ならびにHuMAb−Mouse(登録商標)およびMedarex,Inc.(米ニュージャージー州プリンストン)のTC Mouse(商標)である。
【0081】
別法では、抗体を組換えによって生成し、当業界で知られている任意の方法を使用して発現させることができる。別の選択肢では、抗体をファージディスプレイ技術によって組換えで生成することができる。例えば、米国特許第5565332号、同第5580717号、同第5733743号、および同第6265150号、およびWinterら、Annu.Rev.Immunol.第12巻:433〜455ページ(1994年)を参照されたい。別法として、ファージディスプレイ技術(McCaffertyら、Nature第348巻:552〜553ページ(1990年))を使用すると、未免疫ドナーの免疫グロブリン可変(V)ドメイン遺伝子レパートリーからin vitroでヒト抗体および抗体断片を生成することができる。この技術によれば、抗体Vドメイン遺伝子を、M13やfdなどの糸状のバクテリオファージの主要または非主要外殻タンパク質遺伝子にインフレームでクローン化し、ファージ粒子表面上に機能性の抗体断片として提示する。糸状の粒子は、ファージゲノムの一本鎖DNAコピーを含むので、抗体の機能の特性に基づく選択は、これらの特性を示す抗体をコードする遺伝子の選択ともなる。したがって、ファージは、B細胞の特性の一部を模倣する。ファージディスプレイは、様々な形式で実施することができ、総説については、例えば、Johnson,Kevin S.およびChiswell,David J.、Current Opinion in Structural Biology第3巻、564〜571ページ(1993年)を参照されたい。いくつかのV遺伝子区域供給源をファージディスプレイに使用することができる。Clacksonら、Nature第352巻:624〜628ページ(1991年)は、免疫感作されたマウスの脾臓に由来するV遺伝子の小規模のランダムなコンビナトリアルライブラリーから、多様な並び方の抗オキサゾロン抗体を単離した。本質的にマークら、J.Mol.Biol.第222巻:581〜597ページ(1991年)またはGriffithら、EMBO J.第12巻:725〜734ページ(1993年)に記載の技術に従って、未免疫のヒトドナーからの一レパートリーのV遺伝子を構築することができ、多様なアレイの抗原に対する抗体(自己抗原を含む)を単離することができる。自然の免疫応答では、抗体遺伝子は、高い比率で突然変異を蓄積する(体細胞突然変異)。導入される変化の一部は、より高い親和性を付与することになり、高親和性の表面免疫グロブリンを示すB細胞は、その後の抗原による攻撃の際に優先的に複製され、分化する。この自然の過程は、「鎖シャフリング」として知られている技術を使用して模倣することができる。Marksら、Bio/Technol.第10巻:779〜783ページ(1992年)。この方法では、重鎖および軽鎖可変部遺伝子を、未免疫のドナーから得られたVドメイン遺伝子の(レパートリー)の自然に存在する変異体のレパートリーと順次交換することによって、ファージディスプレイによって得られた「一次」ヒト抗体の親和性を改良することができる。この技術は、pM〜nMの範囲で親和性を有する抗体および抗体断片の生成を可能にする。(「すべてのライブラリーの母なるもの」としても知られている)非常に大きなファージ抗体レパートリーを作製する戦略は、Waterhouseら、Nucl.Acids Res.第21巻:2265〜2266ページ(1993年)に記載されている。遺伝子シャフリングを使用して、げっ歯類抗体から、その出発げっ歯類抗体に対して同様の親和性および特異性を有するヒト抗体を得ることもできる。「エピトープインプリンティング」とも呼ばれるこの方法によれば、ファージディスプレイ技術によって得られたげっ歯類抗体の重鎖または軽鎖Vドメイン遺伝子が、ヒトVドメイン遺伝子のレパートリーと交換され、げっ歯類−ヒトキメラが作り出される。抗原上での選択により、機能性の抗原結合部位を回復させることのできるヒト可変部が単離される、すなわち、エピトープがパートナーの選択を支配(刷り込み)する。この過程を繰り返して残りのげっ歯類Vドメインを交換すると、ヒト抗体が得られる(1993年4月1日公開のPCT公開WO93/06213を参照されたい)。CDR移植によるげっ歯類抗体の伝統的なヒト化とは異なって、この技術は、げっ歯類由来のフレームワークまたはCDR残基をもたない完全にヒトの抗体を提供する。上記論述はヒト化抗体に関係するものであるが、論述した一般原理が、例えばイヌ、ネコ、霊長類、ウマ、およびウシで使用するための抗体のカスタマイズに適用できることは言うまでもない。
【0082】
抗体は、二重特異性抗体、すなわち、少なくとも2種の異なる抗原に対する結合特異性を有するモノクローナル抗体である場合もあり、本明細書で開示する抗体を使用して調製することができる。二重特異性抗体の生成方法は、当業界で知られている(例えば、Sureshら、1986年、Methods in Enzymology第121巻:210ページを参照されたい)。伝統的に、二重特異性抗体の組換え生成は、2種の重鎖が異なる特異性を有する2種の免疫グロブリン重鎖−軽鎖対の同時発現に基づくものであった(MillsteinおよびCuello、1983年、Nature第305巻、537〜539ページ)。
【0083】
二重特異性抗体を生成する1つの手法によれば、所望の結合特異性を有する抗体可変ドメイン(抗体−抗原合体部位)を、免疫グロブリン定常ドメイン配列と融合させる。ヒンジ、CH2、およびCH3領域の少なくとも一部を含む免疫グロブリン重鎖定常ドメインと融合させることが好ましい。第1の重鎖定常部(CH1)が、融合物の少なくとも1つに存在する軽鎖結合に必要な部位を含むことが好ましい。免疫グロブリン重鎖融合物、および所望ならば免疫グロブリン軽鎖をコードするDNAを、別個の発現ベクターに挿入し、適切な宿主生物に同時形質移入する。これは、その構築で使用される3種のポリペプチド鎖の不均等な割合が最適な収率をもたらすとき、実施形態の3種のポリペプチド断片の相互の割合の調整において高い柔軟性をもたらす。しかし、同等の割合の少なくとも2種のポリペプチド鎖の発現が高い収率をもたらすとき、またはその割合が特定の意味をもたないものであるとき、1つの発現ベクター中に2種または3種すべてのポリペプチド鎖のコード配列を挿入することは可能である。
【0084】
1つの手法では、二重特異性抗体は、一方の腕の第1の結合特異性を有するハイブリッド免疫グロブリン重鎖と、他方の腕のハイブリッド免疫グロブリン重鎖−軽鎖対(第2の結合特異性をもたらす)から構成される。二重特異性の半分にしか免疫グロブリン軽鎖がないこの非対称の構造は、望ましくない免疫グロブリン鎖の組合せからの所望の二重特異性化合物の分離を容易にする。この手法は、1994年3月3日公開のPCT公開WO94/04690に記載されている。
【0085】
共有結合によって結合した2種の抗体を含むヘテロコンジュゲート抗体も、本発明の範囲内である。そのような抗体は、免疫系細胞を望ましくない細胞に向けること(米国特許第4676980号)、およびHIV感染の治療(PCT公開WO91/00360およびWO92/200373、ならびにEP03089)に使用されている。ヘテロコンジュゲート抗体は、任意の好都合な架橋結合法を使用して作製することができる。適切な架橋剤および技術は、当業界でよく知られており、米国特許第4676980号に記載されている。
【0086】
抗体は、最初に宿主動物から作られた抗体を単離し、遺伝子配列を得、その遺伝子配列を使用して宿主細胞(例えばCHO細胞)中で組換えによって抗体を発現させることで、組換えで作製することができる。使用することのできる別の方法は、植物(例えばタバコ)、トランスジェニック乳汁、または他の生物において抗体配列を発現させることである。植物または乳汁において抗体を組換え発現させる方法は開示されている。例えば、Peetersら(2001年)、Vaccine第19巻:2756ページ;Lonberg,N.およびD.Huszar(1995年)、Int.Rev.Immunol第13巻:65ページ;およびPollockら(1999年)、J Immunol Methods第231巻:147ページを参照されたい。例えばヒト化、単鎖などの抗体派生物を生成する方法は、当業界で知られている。
【0087】
キメラまたはハイブリッド抗体も、架橋剤を使用するものを含む既知の合成タンパク質化学の方法を使用して、in vitroで調製することができる。例えば、ジスルフィド交換反応を使用して、またはチオエーテル結合を形成して、免疫毒素を構築することができる。この目的に適する試薬の例には、イミノチオラートおよびメチル−4−メルカプトブチルイミダートが含まれる。
【0088】
単鎖Fv断片も、Iliadesら、1997年、FEBS Letters、第409巻:437〜441ページに記載されているように生成することができる。様々なリンカーを使用するそのような単鎖断片の結合は、Korttら、1997年、Protein Engineering、第10巻:423〜433ページに記載されている。組換え産生および抗体の操作のための様々な技術は、当業界でよく知られている。
【0089】
抗体は、1999年11月18日公開のPCT公開WO99/58572号に記載の通りに改変することができる。これらの抗体は、標的分子に向けられる結合ドメインに加え、ヒト免疫グロブリン重鎖の定常ドメインの全部または一部と実質的に相同であるアミノ酸配列を有するエフェクタードメインを含む。これらの抗体は、著しい補体依存性の溶解または細胞性の標的の破壊を誘発することなく、標的分子を結合することができる。エフェクタードメインは、FcRnおよび/またはFcγRIIbを特異的に結合し得ることが好ましい。これらは通常、2種以上のヒト免疫グロブリン重鎖CH2ドメインから得られるキメラドメインに基づくものである。このようにして改変された抗体は、長期にわたる抗体療法で使用して、従来の抗体療法に対する炎症性および他の副作用を回避するのに好ましい。
【0090】
宿主動物の免疫化によってまたは組換えによって生成される抗体は、次の特性のいずれか1つまたは複数を示すべきである。すなわち、(a)trkC受容体に結合する、(b)trkC受容体の1つまたは複数のエピトープに結合する、(c)trkC受容体に結合し、trkCの生物活性、またはtrkCシグナル伝達機能に媒介される1つまたは複数の下流の経路を活性化する、(d)trkC受容体の二量体化を促進する、(e)trkC受容体の活性化を増大させる、(f)好ましい薬物動態および生体利用度特性を示す、(g)細胞の発生、生存、機能、維持、および/または再生を促進する、(h)trkC受容体に結合し、下位運動ニューロン疾患の1つまたは複数の症状を治療し、予防し、逆戻りさせ、または緩和させる。
【0091】
イムノアッセイ、およびfluorescence activated cell sorting(FACS)などのフローサイトメトリー選別技術を使用すると、trkCに特異的な抗体を単離することもできる。
【0092】
抗体は、多くの異なる担体に結合させることができる。担体は、活性のあるものおよび/または不活性なものでよい。よく知られた担体の例には、ポリプロピレン、ポリスチレン、ポリエチレン、デキストラン、ナイロン、アミラーゼ、ガラス、天然および改変型のセルロース、ポリアクリルアミド、アガロース、ならびに磁鉄鉱が含まれる。担体の性質は、本発明の目的では可溶性または不溶性のどちらでもよい。当業者ならば、抗体の結合に適する他の担体について知っており、または常法通りの実験を使用してそれを突き止めることができるであろう。
【0093】
アゴニスト抗trkC抗体をコードするDNAは、当業界で知られているように配列決定することができる。PCT公開WO01/98361を参照されたい。一般に、モノクローナル抗体は、従来の手順を使用して(例えば、モノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを使用して)容易に単離および配列決定がなされる。ハイブリドーマ細胞は、そのようなcDNAの好ましい供給源として役立つ。単離されたなら、DNAは、(PCT公開WO87/04462で開示されている発現ベクターなどの)発現ベクター中に入れ、次いでそれを、別に免疫グロブリンタンパク質を産生しない大腸菌細胞、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、骨髄腫細胞などの宿主細胞に形質移入して、組換え宿主細胞中でモノクローナル抗体の合成を実現することができる。例えば、PCT公開WO87/04462を参照されたい。DNAは、例えば、相同のネズミ配列の代わりにヒト重鎖および軽鎖定常ドメインのコード配列を用いて(Morrisonら、Proc.Nat.Acad.Sci.第81巻:6851ページ(1984年))、または非免疫グロブリンポリペプチドのコード配列の全部もしくは一部に免疫グロブリンコード配列を共有結合によって連結して、改変することもできる。そのようにして、本明細書では、抗trkCモノクローナル抗体の結合特異性を有する「キメラ」または「ハイブリッド」抗体が調製される。(その抗原結合断片などの)アゴニスト抗trkC抗体をコードするDNAは、ここに記載するような、所望の細胞におけるアゴニスト抗trkC抗体の送達および発現に使用することもできる。DNA送達技術については、本明細書でさらに記載する。
【0094】
抗trkC抗体は、当業界でよく知られている方法を使用して特徴付けることができる。例えば、1つの方法は、それが結合するエピトープを同定することであり、例えば、HarlowおよびLane、「Using Antibodies,a Laboratory Manual」、第11章、Cold Spring Harbor Laboratory Press、米ニューヨーク州コールドスプリング港、1999年に記載されているように、抗体−抗原複合体の結晶構造の解明、競合アッセイ、遺伝子断片発現アッセイ、および合成ペプチドに基づくアッセイがこれに含まれる。追加の例では、エピトープマッピングを使用して、抗trkC抗体が結合する配列を決定することができる。エピトープマッピングは、様々な供給元、例えば、Pepscan Systems(Edelhertweg 15、8219 PH Lelystad、The Netherlands)からの市販品を利用することができる。エピトープは、線形のエピトープ、すなわち、一続きのアミノ酸中に含まれるものである場合もあり、または必ずしも一続きの中に含まれなくてもよい、アミノ酸の三次元の相互作用によって形成される高次構造的なエピトープである場合もある。様々な長さ(例えば少なくとも4〜6アミノ酸長)のペプチドを単離し、または(例えば組換えによって)合成し、抗trkC抗体との結合アッセイに使用することができる。別の例では、trkC細胞外配列由来のオーバーラップペプチドを使用し、抗trkC抗体による結合を判定する系統的なスクリーニングにおいて抗trkC抗体が結合するエピトープを決定することができる。遺伝子断片発現アッセイによれば、trkCをコードするオープンリーディングフレームを、無作為に、または特異的な遺伝子構築物によって分断し、発現されるtrkC断片と試験される抗体との反応性を測定する。遺伝子断片は、例えば、PCRによって生成し、次いで、放射性アミノ酸の存在下、in vitroでタンパク質へと転写、翻訳することができる。次いで、放射標識されたtrkC断片への抗体の結合を、免疫沈殿法およびゲル電気泳動によって測定する。ある種のエピトープは、ファージ粒子表面上に示されるランダムなペプチド配列の大規模なライブラリー(ファージライブラリー)を使用して同定することもできる。
【0095】
抗trkC抗体の特徴付けに使用することのできるさらに別の方法は、同じ抗原、すなわちtrkC細胞外ドメインに結合することがわかっている他の抗体との競合アッセイを使用して、抗trkC抗体が他の抗体と同じエピトープに結合するかどうかを判定することである。競合アッセイは、当業者によく知られている。競合アッセイにおいて有用な抗体の例には、抗体6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256が含まれる。PCT公開WO01/98361を参照されたい。
【0096】
ドメインスワップ突然変異体を使用して、PCT公開WO01/98361に記載の通りにエピトープマッピングを実施してもよい。一般に、この手法は、trkAまたはtrkBとほとんど交差反応しない抗trkC抗体に有用である。trkCのドメインスワップ突然変異体は、trkCの細胞外ドメインをtrkBまたはtrkAの対応するドメインと交換して作製することができる。ELISAまたは当業界で知られている他の方法を使用して、各アゴニスト抗trkC抗体の様々なドメインスワップ突然変異体への結合を評価し、その野生型(未変性)trkCへの結合と比較することができる。別の手法では、アラニン走査を実施することができる。抗原、すなわちtrkC受容体の個々の残基を、意図的に別のアミノ酸(通常はアラニン)に突然変異させ、ELISAまたは当業界で知られている他の方法を使用して、その改変trkCの抗体への結合能を試験することによって、その変化の影響を評価する。
【0097】
アゴニスト抗trkC抗体の同定
アゴニスト抗体は、以下の方法の1つまたは複数を含む業界で認められている方法を使用して同定することができる。例えば、米国特許第5766863号および同第5891650号に記載されているキナーゼ受容体活性化(KIRA)アッセイを使用することができる。このELISA型アッセイは、受容体タンパク質チロシンキナーゼ(rPTK、例えばtrk受容体)のキナーゼドメインの自己リン酸化を測定することによるキナーゼ活性化の定性的または定量的な測定、ならびに選択されたrPTKの有望なアゴニストまたはアンタゴニストの同定および特徴付けに適する。アッセイの第1期は、キナーゼ受容体、この場合では真核細胞の細胞膜に存在するtrkC受容体のキナーゼドメインのリン酸化を含む。受容体は、内因性の受容体または受容体をコードする核酸でもよいし、受容体構築物を細胞に入れて形質転換してもよい。通常、第1の固相(例えば、第1のアッセイプレートのウェル)を、そのような細胞(普通は哺乳動物細胞系)の実質的に均一な集団でコートして、細胞が固相に付着するようにする。細胞はしばしば接着性であり、そのために自然に第1の固相に付着する。「受容体構築物」を使用する場合、通常これは、キナーゼ受容体とflagポリペプチドの融合物を含む。flagポリペプチドは、アッセイのELISAの部分で捕捉剤、しばしば捕捉抗体によって認識される。次いで、候補アゴニストなどの分析物を、付着細胞を有するウェルに加えて、チロシンキナーゼ受容体(例えばtrkC受容体)が、分析物にさらされる(または分析物と接触する)ようにする。このアッセイは、目的のチロシンキナーゼ受容体(例えばtrkC)のアゴニストリガンドの同定を可能にするものである。分析物にさらした後、(その中に可溶化洗浄剤を含む)溶解緩衝液を使用して付着細胞を可溶化し、穏やかに攪拌し、それによって細胞可溶化物を放出させ、それを細胞可溶化物の濃縮または清澄の必要なしに直接にアッセイのELISA部分にかけることができる。
【0098】
このように細胞可溶化物を調製し、次いでアッセイのELISAステージにかける準備をする。ELISA段階の第1ステップとして、第2の固相(通常ELISAマイクロタイタープレートのウェル)を、チロシンキナーゼ受容体、または受容体構築物の場合ではflagポリペプチドに特異的に結合する捕捉剤(しばしば捕捉抗体)でコートする。第2の固相のコーティングは、捕捉剤が第2の固相に付着するように実施する。捕捉剤は、一般にモノクローナル抗体であるが、本明細書で実施例に記載するように、ポリクローナル抗体または他の薬剤を使用してもよい。次いで得られる細胞可溶化物を付着性の捕捉剤にさらし、またはそれと接触させて、受容体または受容体構築物が第2の固相に付着する(またはそれに捕捉される)ようにする。次いで、捕捉された受容体または受容体構築物を残して非結合の細胞可溶化物を除去するように洗浄ステップを実施する。次いで、付着しまたは捕捉された受容体または受容体構築物を、チロシンキナーゼ受容体中のリン酸化されたチロシン残基を同定する抗ホスホチロシン抗体にさらし、または接触させる。好ましい実施形態では、抗ホスホチロシン抗体を、非放射性呈色試薬の色調変化を触媒する酵素に(直接または間接的に)結合させる。したがって、受容体のリン酸化は、その後の試薬の色調変化によって測定することができる。酵素は、抗ホスホチロシン抗体に直接に結合することもでき、または結合性の分子(例えばビオチン)を抗ホスホチロシン抗体に結合し、その後酵素をその結合性の分子を介して抗ホスホチロシン抗体に結合することもできる。最後に、捕捉された受容体または受容体構築物への抗ホスホチロシン抗体の結合を、例えば呈色試薬の色調変化によって測定する。
【0099】
最初の同定の後、候補抗体のアゴニスト活性は、標的指向化された生物活性を試験することが知られているバイオアッセイによってさらに確認し、さらに正確にすることができる。例えば、抗trkCモノクローナル抗体がtrkCのアゴニストになる能力を、全長ヒトtrkCを形質移入したPC12細胞を使用するPC12神経突起伸長アッセイで試験することができる(Urferら、Biochem.第36巻:4775〜4781ページ(1997年);Tsoulfasら、Neuron第10巻:975〜990ページ(1993年))。このアッセイは、適切なリガンドによる刺激に応答したラット好チトクロム腫(pheocytochroma)細胞(PC12)による神経突起伸長の過程を測定するものである。これらの細胞は、内因性のtrkAを発現し、したがってNGFに応答する。しかし、これらの細胞は、内因性のtrkCを発現せず、したがって、trkCアゴニストに対する応答を誘発するためにtrkC発現構築物を形質移入する。形質移入された細胞を抗trkC抗体と共にインキュベートした後、神経突起の伸長を測定、例えば、細胞の直径の2倍を超える神経突起を有する細胞をカウントする。形質移入されたPC12細胞において神経突起の伸長を刺激する抗trkC抗体は、trkCアゴニスト活性を実証するものである。
【0100】
trkCの活性化は、胚発生の特定の段階にある様々な特定のニューロンを使用して判定することもできる。適切に選択したニューロンの生存は、trkCの活性化に応じて決まる場合があり、これらのニューロンのin vitroでの生存に従ってtrkCの活性化を測定することが可能である。候補抗体がtrkCを活性化するならば、適切なニューロンの初代培養物に候補抗体を加えると、これらのニューロンが少なくとも数日間生存するようになる。これによって、候補抗体がtrkCを活性化する能力の判定が可能になる。この種類のアッセイの一例では、E11マウス胎仔の三叉神経節を解剖し、分離し、得られるニューロンを低い密度で組織培養皿に播く。次いで、候補抗体を培地およびプレートに加え、24〜48時間インキュベートする。その後、ニューロンの生存を様々な方法のいずれかによって評価する。アゴニスト抗trkC抗体を加えたサンプルは通常、対照抗体を加えたサンプルよりも増大した生存率を示し、これによってアゴニスト抗trkC抗体の存在を判定することができる。例えば、Buchmanら(1993年)、Development第118巻(3):989〜1001ページを参照されたい。
【0101】
アゴニスト抗体は、自然にまたはtrkCをコードするDNAを形質移入した後にtrkCを発現する様々な細胞型において下流のシグナル伝達を活性化するその能力によって同定することができる。そのtrkCは、ヒトまたは(げっ歯類や霊長類などの)他の哺乳類のtrkCでよい。下流のシグナルカスケードは、タンパク質のタンパク質発現またはタンパク質リン酸化のレベルなどの、trkC発現細胞の様々な生化学的または生理学的パラメータへの変化、あるいは(本明細書に記載するようなニューロンの生存および/または神経突起の伸長を含む)細胞の代謝的状態または増殖状態への変化によって検出することができる。関連する生化学的または生理学的パラメータの検出方法は、当業界で知られている。
【0102】
アゴニスト抗trkC抗体の投与
アゴニスト抗trkC抗体の様々な製剤を投与に使用することができる。一部の実施形態では、アゴニスト抗trkC抗体を希釈せずに投与する場合もある。一部の実施形態では、アゴニスト抗trkC抗体は、薬学的に許容できる賦形剤を含む組成物にして投与する。薬学的に許容できる賦形剤は、当業界で知られており、薬理学的に有効な物質の投与を容易にする比較的不活性な物質である。例えば、賦形剤は、形または稠性を与えることができ、または希釈剤として働く。適切な賦形剤には、その限りでないが、安定剤、湿潤剤および乳化剤、様々なモル浸透圧濃度にするための塩、カプセル化剤、緩衝剤、ならびに皮膚浸透性改善剤が含まれる。非経口および非経口でない薬物送達のための賦形剤ならびに製剤は、Remington、「The Science and Practice of Pharmacy」、第20版、Mack Publishing(2000年)に記載されている。
【0103】
アゴニスト抗trkC抗体は、注射(例えば、腹腔内、静脈内、皮下、筋肉内など)による投与向けに製剤することができる。したがって、抗体は、生理食塩水、リンガー液、デキストロース溶液などの薬学的に許容できる媒体と組み合わせることができる。特定の投与計画、すなわち、投与量、タイミング、および繰返しは、特定の個体およびその個体の治療歴に応じて決まることになる。一般に、約1ug/kg体重未満、少なくとも約1ug/kg体重;少なくとも約2ug/kg体重、少なくとも約5ug/kg体重、少なくとも約10ug/kg体重、少なくとも約20ug/kg体重、少なくとも約50ug/kg体重、少なくとも約100ug/kg体重、少なくとも約200ug/kg体重、少なくとも約500ug/kg体重、少なくとも約1mg/kg体重、少なくとも約2mg/kg体重、少なくとも約5mg/kg体重、少なくとも約10mg/kg体重、少なくとも約30mg/kg体重、またはこれより多い量(約50mg/kg、約100mg/kg、約200mg/kg、約500mg/kgなど)の投与量が投与される。
【0104】
半減期などの経験に基づく考慮事項は、一般に投与量決定の一因となる。ヒト化抗体や完全ヒト抗体などのヒト免疫系と適合性のある抗体を使用すると、抗体の半減期を延長し、抗体が宿主の免疫系によって攻撃されないようにすることができる。投与の頻度は、治療の過程で決定し調整することができ、一般に、必ずしもそうでないが、患者におけるアゴニスト抗trkC抗体の有効濃度の維持、ならびに下位運動ニューロン疾患の1つまたは複数の症状の抑制および/または回復および/または遅延に基づくものである。別法として、アゴニスト抗trkC抗体の持続型の連続放出製剤が適切な場合もある。持続放出を実現するための様々な製剤および装置が当業界で知られている。本発明の方法に従うアゴニスト抗trkC抗体の投与は、例えば、投与の目的が治療であろうと予防であろうとレシピエントの生理的な状態、および熟練した従業者に知られている他の要素に応じて、継続的である場合もあれば断続的である場合もある。アゴニスト抗trkC抗体の投与は、予め選択された期間にわたる本質的に継続的なものでもよいし、あるいは例えば、下位運動ニューロン疾患の症状の出現の前、最中、もしくは後のいずれか;下位運動ニューロン疾患の症状の出現の前および最中;前および後;最中および後;および/または前、最中、および後の、間隙を挟んだ一連の投与にしてもよい。
【0105】
一般に、アゴニスト抗trkC抗体の投与では、初期の候補投与量は、約2mg/kgでよい。本発明の目的では、典型的な1日投与量は、上述の要素に応じて約30μg/kg〜100mg/kgまたはそれ以上の範囲となるであろう。数日またはそれ以上にわたって繰り返される投与では、状態に応じて、疾患症状の所望の抑制が起こるまで、または下位運動ニューロン疾患を治療もしくは予防するのに十分な治療レベルが実現されるまで治療を継続する。好例となる投与計画は、約2mg/kgの初回量の後、毎週約1mg/kgの維持量または隔週で約1mg/kgの維持量のtrkCアゴニスト抗体の投与を含む。
【0106】
一実施形態では、抗体の投与量は、trkC受容体を活性化して下位運動ニューロン疾患を治療するアゴニスト抗trkC抗体の1回または複数の投与を受けたことのある個体において経験に基づいて決定する場合もある。個体に、アゴニスト抗trkC抗体の投与量を徐々に増やして与える。アゴニスト抗trkC抗体の有効性を評価するには、本明細書に記載するような下位運動ニューロン疾患状態の指標に従うことができる。
【0107】
他の製剤は、その限りでないがリポソームなどの担体を含めて、当業界で知られている適切な送達形態を含む。例えば、Mahatoら(1997年)、Pharm.Res.第14巻:853〜859ページを参照されたい。リポソーム製剤には、その限りでないが、シトフェクチン(cytofectin)、多層ベシクル、および単層ベシクルが含まれる。
【0108】
in vivo投与に使用する製剤は、無菌でなければならない。これは、例えば無菌濾過膜でのろ過によって容易に実現される。治療用のアゴニスト抗trkC抗体組成物は、一般に、無菌の出入口を有する容器、例えば、静脈内溶液バッグ、または皮下投与針によって突き刺すことのできる栓を有するバイアルに入れられる。
【0109】
アゴニスト抗trkC抗体は、筋肉内、腹腔内、皮下、経口、くも膜下腔内、または局所的な経路によって、例えば大量瞬時投与としてまたは一定期間にわたる継続的な注入による静脈内投与などの既知の方法に従って個体に投与する。アゴニスト抗trkC抗体は、吸入によって投与することもできる。ジェットネブライザーおよび超音波ネブライザーを含む市販の液体製剤用ネブライザーは、投与に有用である。液体製剤をそのまま噴霧することもでき、凍結乾燥された粉末を再形成した後に噴霧することもできる。別法として、アゴニスト抗trkC抗体は、フルオロカーボン製剤および計量投与吸入器を使用してエアロゾル化してもよいし、または凍結乾燥および粉砕した粉末として吸入してもよい。
【0110】
一部の実施形態では、複数種の抗体が存在する場合もある。その抗体は、同じもので場合もあれば、互いに異なる場合もある。一部の実施形態では、少なくとも1種、少なくとも2種、少なくとも3種、少なくとも4種、少なくとも5種の異なるtrkCアゴニスト抗体が存在する。これらの抗体は、互いに不利に影響を及ぼさない相補的な活性を有することが好ましい。
【0111】
(その抗原結合断片などの)アゴニスト抗trkC抗体をコードするポリヌクレオチドは、所望の細胞におけるアゴニスト抗trkC抗体の送達および発現に使用することもできる。発現ベクターを使用してアゴニスト抗trkC抗体の発現を管理できることは言うまでもない。発現ベクターは、腹腔内、静脈内、筋肉内、皮下、くも膜下腔内、脳室内、経口的、経腸的、非経口的、鼻腔内、経皮的に、または吸入によって投与することができる。例えば、発現ベクターの投与には、注射、経口投与、粒子銃、またはカテーテルによる投与を含む局部または全身の投与、ならびに局所投与が含まれる。当業者は、外来タンパク質の発現をin vivoで実現するための発現ベクターの投与に精通している。例えば、米国特許第6436908号、同第6413942号、および同第6376471号を参照されたい。
【0112】
アゴニスト抗trkC抗体をコードするポリヌクレオチドを含む治療用組成物の標的指向化された送達を使用することもできる。受容体を媒介とするDNA送達技術は、例えば、Findeisら、Trends Biotechnol.(1993年)第11巻:202ページ;Chiouら、「Gene Therapeutics:Methods And Applications Of Direct Gene Transfer」(J.A.Wolff編)(1994年);Wuら、J.Biol.Chem.(1988年)第263巻:621ページ;Wuら、J.Biol.Chem.(1994年)第269巻:542ページ;Zenkeら、Proc.Natl.Acad.Sci.(USA)(1990年)第87巻:3655ページ;Wuら、J.Biol.Chem.(1991年)第266巻:338ページに記載されている。遺伝子治療プロトコルでは、ポリヌクレオチドを含む治療用組成物を、局所投与ではDNA約100ng〜約200mgの範囲で投与する。遺伝子治療プロトコルの際に、約500ng〜約50mg、約1μg〜約2mg、約5μg〜約500μg、および約20μg〜約100μgの濃度範囲のDNAを使用することもできる。本発明の治療用ポリヌクレオチドおよびポリペプチドは、遺伝子送達媒体を使用して送達することができる。遺伝子送達媒体は、ウイルス由来のものでも、ウイルス由来でないものでもよい(一般に、Jolly、Cancer Gene Therapy(1994年)第1巻:51ページ;Kimura、Human Gene Therapy(1994年)第5巻:845ページ;Connelly、Human Gene Therapy(1995年)第1巻:185ページ;およびKaplitt、Nature Genetics(1994年)第6巻:148ページ)を参照されたい。そのようなコード配列の発現は、内因性の哺乳類プロモーターまたは異種プロモーターを使用して誘発することができる。コード配列の発現は、構成性または調節性のどちらでもよい。
【0113】
所望のポリヌクレオチドの送達および所望の細胞での発現のためのウイルス系ベクターは、当業界でよく知られている。好例となるウイルス系媒体には、その限りでないが、組換えレトロウイルス(例えば、PCT公開WO90/07936、WO94/03622、WO93/25698、WO93/25234、WO93/11230、WO93/10218、WO91/02805、米国特許第5219740号、同第4777127号、英国特許第2200651号、およびEP0345242を参照されたい)、アルファウイルス系ベクター(例えば、シンドビスウイルスベクター、セムリキ森林熱ウイルス(ATCC VR−67、ATCC VR−1247)、ロスリバーウイルス(ATCC VR−373、ATCC VR−1246)、およびベネズエラウマ脳炎ウイルス(ATCC VR−923、ATCC VR−1250、ATCC VR 1249、ATCC VR−532))、ならびにアデノ随伴ウイルス(AAV)ベクター(例えば、PCT公開WO94/12649、WO93/03769、WO93/19191、WO94/28938、WO95/11984、およびWO95/00655を参照されたい)が含まれる。Curiel、Hum.Gene Ther.(1992年)第3巻:147ページに記載の、死滅させたアデノウイルスに連結されたDNAの投与を使用することもできる。
【0114】
その限りでないが、死滅させたアデノウイルスのみに連結されまたは連結されていないポリカチオン縮合DNA(例えば、Curiel、Hum.Gene Ther.(1992年)第3巻:147ページを参照されたい)、リガンドに連結されたDNA(例えば、Wu、J.Biol.Chem.(1989年)第264巻:16985ページを参照されたい)、真核細胞送達媒体細胞(例えば、米国特許第5814482号、PCT公開WO95/07994、WO96/17072、WO95/30763、およびWO97/42338を参照されたい)、ならびに核電荷中和(nucleic charge neutralization)または細胞膜との融合を含めて、非ウイルスの送達媒体および方法を使用することもできる。裸DNAを用いてもよい。好例となる裸DNA導入方法は、PCT公開WO90/11092および米国特許第5580859号に記載されている。遺伝子送達媒体として働き得るリポソームは、米国特許第5422120号、PCT公開WO95/13796、WO94/23697、WO91/14445、およびEP0524968に記載されている。追加の手法は、Philip、Mol.Cell Biol.(1994年)第14巻:2411ページ、およびWoffendin、Proc.Natl.Acad.Sci.(1994年)第91巻:1581ページに記載されている。
【0115】
アゴニスト抗trkC抗体は、下位運動ニューロン疾患を治療するための(1種または複数のニューロトロフィンなどの)1種または複数の他の薬剤と共に、すなわち、1種または複数の薬剤と組み合わせて、一斉に、または順次投与することができる。例えば、アゴニスト抗trkC抗体は、NT−4と共に投与することができる。共に投与とは、本明細書では、同時投与および/または異なる時点での投与を含む。共に投与は、合剤(すなわち、アゴニスト抗trkC抗体と他の薬剤とが同じ組成物中に存在するもの(組み合わされたもの))としての投与、および/または別個の組成物としての投与も含む。本明細書では、「共投与」は、アゴニスト抗trkC抗体と他の薬剤が有効量で個体に投与される任意の状況を含む意味である。本明細書でさらに論述するように、アゴニスト抗trkC抗体と他の薬剤を、異なる投薬頻度および/または投薬間隔で投与できることは理解される。例えば、アゴニスト抗trkC抗体は、毎週投与することができ、その間他の薬剤(例えばNT−4)をより頻繁に投与することができる。アゴニスト抗trkC抗体と他の薬剤は、同じ投与経路または異なる投与経路を使用して投与することができ、その異なる投与計画を投与の過程で変化させてよいことは理解される。投与は、下位運動ニューロン疾患の発症前でよい。
【0116】
アゴニスト抗trkC抗体をニューロトロフィンと共に投与するものでは、ニューロトロフィン(例えば、CNTF、NT−4)をコードするポリヌクレオチドを、本明細書に記載の発現ベクターを利用する所望の細胞(例えば骨格筋細胞)でのニューロトロフィンの送達および発現に使用してもよい。
【0117】
アゴニスト抗trkC抗体による治療の有効性を評価する方法
治療有効性の評価は、いくつかの異なるレベルで実施することができる。評価は、臨床徴候(例えば、体力試験、電気生理学的な応答、または分子的な変化)をモニターして行うことができる。これらには、標準の神経学的な検査または患者への問診によって決定されるパラメータを含めることができ、あるいは例えば本明細書に記載のより専門の定量的試験によって測定できるものでよい。これらのより専門の定量的試験には、微小神経電図検査などの手段による冒されたニューロンの伝導速度測定、筋電図検査(EMG)、握力などの随意筋力測定、音節の反復、15フィート(4.57メートル)歩くのにかかる時間によって測定される歩行速度、努力性肺活量(FVC)の測定を含む呼吸器の機能試験、呼吸器の能力障害の進行に関連する選択された事象の発生率、聴力、平衡性の試験、固有受容または運動感覚の専門試験、体力試験、筋電図検査;その限りでないが、血圧コントロール試験、様々な生理的および薬理学的な刺激に対する心拍応答試験を含む自律神経機能の試験を含めることができるが、この限りでない。これらの試験には、運動技能または体力の試験を含めることもできる。
【0118】
下位運動ニューロン疾患の治療で使用するための組成物
本発明は、本明細書に記載の方法のいずれかで使用するための組成物も提供する。本発明の方法で使用する組成物は、有効量のアゴニスト抗trkC抗体を含む。そのような組成物の例ならびに製剤方法は、より前の項および以下でも記載する。本発明は、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べた組成物のいずれかも提供する。
【0119】
本発明で使用する組成物は、凍結乾燥製剤または水溶液の形で、薬学的に許容できる担体、賦形剤、または安定剤をさらに含む場合もある(「Remington:The Science and practice of Pharmacy」、第20版(2000年)、Lippincott Williams and Wilkins、K.E.Hoover編)。許容される担体、賦形剤、または安定剤は、その投与量および濃度でレシピエントに対して非毒性であり、リン酸塩、クエン酸塩、他の有機酸などの緩衝剤;アスコルビン酸およびメチオニンを含む抗酸化剤;保存剤(塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメソニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール;ブチルアルコールまたはベンジルアルコール;メチルパラベンやプロピルパラベンなどのアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;m−クレゾールなど);低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、リジンなどのアミノ酸;単糖、二糖、ならびにグルコース、マンノース、もしくはデキストランを含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロース、ソルビトールなどの糖;ナトリウムなどの、塩を生成する対イオン;金属複合体(例えばZn−タンパク質複合体);および/またはTWEEN(商標)、PLURONICS(商標)、ポリエチレングリコール(PEG)などの非イオン性界面活性剤を含む場合もある。薬学的に許容できる賦形剤については本明細書でさらに述べる。
【0120】
一態様では、本発明は、アゴニスト抗trkC抗体を含む組成物を提供する。他の実施形態では、アゴニスト抗trkC抗体はヒトtrkCを認識する。さらに他の実施形態では、アゴニスト抗trkC抗体は、(本明細書に記載の抗体A5などの)ヒト化されたものである。他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。さらに他の実施形態では、抗trkCアゴニスト抗体は、表1に示す重鎖可変部のアミノ酸配列(配列番号1)と表2に示す軽鎖可変部アミノ酸配列(配列番号2)とを含む。さらに他の実施形態では、アゴニスト抗trkC抗体はヒト抗体である。
【0121】
組成物が、複数種のアゴニスト抗trkC抗体(例えば、異なるtrkCエピトープを認識するアゴニスト抗trkC抗体の混合物)を含む場合もあることは理解される。他の好例となる組成物は、同じエピトープを認識する複数種のアゴニスト抗trkC抗体、または異なるtrkCエピトープに結合する異なる種のアゴニスト抗trkC抗体を含む。
【0122】
アゴニスト抗trkC抗体およびその組成物は、アゴニスト抗trkC抗体の有効性を強化および/または補完するのに役立つ他の薬剤と共に使用することもできる。例えば、そのような追加の化合物には、下位運動ニューロン疾患の治療に有用であることがわかっている化合物、(CNTF、NT−3、NT−4、BDNF、およびGDNFを含む)1種または複数のニューロトロフィン、およびアゴニスト抗trkBアゴニストを含めることができる。このような分子は、意図した目的に有効な量で組み合わされて存在することが適切である。trkCアゴニスト抗体およびその組成物は、抗体の有効性を強化および/または補完するのに役立つ他の薬剤と共に使用することもできる。
【0123】
キット
本発明は、この方法で使用するためのキットも提供する。本発明のキットは、抗trkCアゴニスト抗体を含む1個または複数の容器を含み、一部の実施形態では、(下位運動ニューロン疾患の治療方法などの)本明細書に記載の本発明の方法のいずれかに従って使用するための説明書をさらに含む。一部の実施形態では、これらの説明書は、個体が下位運動ニューロン疾患を有しているかどうか、かつ/または下位運動ニューロン疾患にかかるリスクがあるかどうかの確認に基づき治療に適する個体を選択することについての記述を含み、この疾患の治療および/または予防のためのtrkCアゴニスト抗体の投与についてさらに述べる場合もある。本発明はまた、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べたキットのいずれかを提供する。
【0124】
したがって、一実施形態では、本発明は、アゴニスト抗trkC抗体を含むキットを提供する。一部の実施形態では、本発明は、本明細書に記載の方法で使用するための、アゴニスト抗trkC抗体を含むキットを提供する。本発明のキットは、適切な包装が施されている。適切な包装には、その限りでないが、バイアル、瓶、広口瓶、フレキシブルな包装(例えば、密閉されたMylar製またはプラスチック製バッグ)などが含まれる。一部の実施形態では、キットは、容器と、容器に貼られまたは添えられるラベルまたは添付文書とを含む。ラベルまたは添付文書は、組成物が下位運動ニューロン疾患を治療し、予防し、または緩和させるのに有用であることを表示する。説明書は、本明細書に記載の方法のいずれかの実施に向けて用意することができる。容器は、下位運動ニューロン疾患の治療に有効な組成物を保持し、無菌の出入口を有する場合もある(例えば、容器は、静脈内溶液バッグ、または皮下投与針によって突き刺すことのできる栓を有するバイアルでよい)。組成物中の少なくとも1種の活性薬剤は、trkCアゴニスト抗体である。容器は、第2の薬学的に活性のある薬剤をさらに含む場合もある。キットは、緩衝剤や説明的な情報などの追加の構成成分を場合により提供することもある。
【0125】
以下の実施例は、本発明を限定するのでなく例示するためのものである。
【実施例】
【0126】
SMARD1動物モデルに対するアゴニスト抗trkC抗体およびNT−4の効果
呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)は、小児の致命的な常染色体劣性遺伝疾患である。この疾患は、下位運動ニューロンの変性、進行性の筋肉麻痺、および呼吸不全を特徴とし、これらに対して有効な治療は存在しない。nmd(神経筋変性)マウスの表現型は、ヒトSMARD1に酷似している。nmdマウスにおける変異型のマウス遺伝子、すなわちIghmbp2の同定は、SMARD1に罹患したヒトにおける相同遺伝子の突然変異の発見をもたらした。発明者らは、in vivoの電気生理学的な技術を用いてnmdマウスモデルを研究し、Mab2256、すなわちチロシンキナーゼ受容体C(trkC)に対してアゴニスト効果を有するモノクローナル抗体の、nmdマウスにおける疾患の進行に対する有効性を評価した。Mab2256による治療は、nmdマウスにおいて、著しいが一過性の筋力向上をもたらすだけでなく、高頻度の神経刺激の際の神経筋の衰弱を正常化した。これらの結果は、SMARD1などの下位運動ニューロン疾患においてニューロトロフィン受容体に対するモノクローナルアゴニスト抗体を使用することの可能性を示唆するものである。
【0127】
この研究では、SMARD1のnmdマウスモデルの後肢筋および横隔膜筋のin vivoの電気生理学的な特徴付けを実施し、このマウスモデルにおいてtrkC受容体に対するアゴニストのモノクローナル抗体(Mab2256)の潜在的な有効性を評価した。Mab2256治療が筋力の初期の衰えを予防し、電気刺激を繰返し与えた際の衰弱レベルが正常に回復したことからなる筋肉機能の電気生理学的な改善をもたらしたことが示される。しかし、そのような初期の改善は、筋線維の保存または生存の利益には至らず、この治療戦略をさらに最適化することが強調された。
【0128】
結果
trkC抗体Mab2256のアゴニスト特性および薬物動態特性
ヒトtrkC細胞外ドメインに対するモノクローナル抗体を作製し、アゴニスト活性があるかどうか、細胞に基づく受容体チロシンリン酸化アッセイ(16)を使用してスクリーニングした。ネズミIgG1アイソタイプの1つのモノクローナル抗体であるMab2256は、trkAまたはtrkBとの交差反応性なしにtrkC受容体の特異的なバインダーとなることがわかった(データは示さず)。trkCを発現する安定なCHO細胞中で、Mab2256は、最大半有効濃度(EC50)0.87nMでtrkCのチロシンリン酸化を誘発し、一方NT3は、EC50 1.09nMでtrkCのチロシンリン酸化を誘発する(図1A)。そのtrkC受容体活性化能と一致して、Mab2256は、ラット胚三叉神経ニューロン培養物の生存もEC50 2.58nMで用量依存的に援助し、一方NT3のEC50は0.73pMである(図1B)。
【0129】
次に、Mab2256の薬物動態特性を検討した。発明者らは、マウスの腹腔内に投与したとき、Mab2256の血清からの排出相半減期(t1/2)が約199時間であることを見出した。これは、抗体に期待される長く循環する半減期であり、発表されている、内因性trkCアゴニストNT3の約1.28分の血漿半減期と比較すると有利である(17)。
【0130】
nmdマウスの運動能力および生存に対するMab2256の効果
変異マウスは、出生時には野生型と識別不能に見受けられたが、出生の2週間後には摂食および成長が不十分になった。したがって、ホモ変異体は、そのより少ない体重によって野生型およびヘテロ接合体の同腹仔と容易に識別可能であった(図2A)。さらに、変異マウスは、筋肉質量が進行性に損失し、前肢および後肢の握力が顕著に低下した(8〜11)。変異マウスは、重力に対してその身体を支えられず、ケージカバーを掴むことができず、肩および骨盤帯の筋肉質量を徐々に失った。
【0131】
本発明者らは、Mab2256による処置(5mg/体重kg、週2回、出生後20日目、すなわちP20に開始)が、nmdマウスで起こる筋肉機能の進行性の損失を予防し得るかどうかを検討した。Mab2256(n=10)またはPBS(n=5)を6〜8.5週間注射したマウスにおいて、ELISA分析によって血清中のモノクローナル抗体の存在を確認した。血清中のMab2256抗体の平均濃度は、10.6±3.4μg/mLであった(Mab2256注射したマウスn=10)。野生型およびnmdのMab2256処置マウスに副作用は認められなかった。処置したマウスと未処置のマウスとの体重に有意差はなく(図2A、B)、P60では、Mab2256処置した突然変異体の体重は14.5±0.2g(n=16)であり、未処置または媒体対照の体重は14.2±0.83g(n=8)であった。
【0132】
未処置およびMab2256処置した4〜10週齢のマウスにおいて、標準の神経学的検査を実施した。前肢握力を調べるために、マウスをその前肢によって床から10cm上に金属ワイヤーに吊るした。マウスが吊るされたままであった期間を記録した。未処置のnmdマウス(PBS注射したものn=5、注射しなかったものn=9)は、試験全体を通してその前肢のほとんど完全な握力の損失を示した(図2C、塗りつぶされた四角形)。Mab2256処置したnmdマウスは、著しく数週間握力を保持した(図2C、三角形)。PBS注射したnmdマウスが自身をワイヤー上に保つことができた平均期間は、4週齢で0.8±0.6秒であり(n=5)、Mab2256処置したnmdマウスでは同じ年齢で5.1±1.6秒(9匹のマウス、6匹の同腹仔)(p<0.03)、1週間後には4.9±1.4秒(p<0.034)であった。ヘテロ接合体の能力(図2C、灰色四角形、n=8)は、野生型の同胞と同一であった(図2C、白色の四角)。
【0133】
nmdマウスが自身を一定のスピードの回転棒上に保つ能力によって、運動協調性も測定した(試験時間:10秒)。未処置のnmdマウスは、野生型および異型接合性の同腹仔と比べて平衡性が非常に不十分であった(図2D、E)(p<0.001)が、Mab2256処置したnmdマウスは、処置してから1週間後に有意により良好な運動協調性を示した(図2D、F)(p<0.018)。これらの結果は、Mab2256処置がnmdマウスにおいて疾患の進行を緩和させたことを示唆するものであった。
【0134】
Mab2256が生存に影響するかどうかに解答するために、未処置およびMab2256処置したnmdマウスの寿命を離乳から成人期まで記録した。著しい寿命の延長は認められず(マンホイットニーの順位和検定)、Mab2256処置した変異マウスの中央値の寿命は69日間(n=15)であり、未処置nmdマウスでは62日間(n=30)(図2G)であった。
【0135】
nmdマウスにおける電気的神経筋活性
P70に内側腓腹筋(MG)でin vivoEMG測定を実施した。まず最大上振幅の単一の電流パルスによって坐骨神経を刺激し、複合筋活動電位(CMAP)を記録した。nmdマウスにおけるCMAPの平均振幅は、対照値の<50%に縮小した(図3A;それぞれ22.9±5.6mV、n=9、および48.8±4.8mV、n=14;p<0.002)。Mab2256の投与は、P70のnmd(24±3.1mV、n=7)または野生型マウス(61±9.2mV、n=8)においてCMAPの平均振幅に著しい効果を与えず(図3A)、Mab2256による治療プロトコルでは運動線維の損失を止められないことが示唆された。trkBアゴニストがtrkCアゴニストより良好となり得るかどうかを問うため、一群の変異マウスをtrkBアゴニストNT−4/5(5mg/体重kg、週2回、P20に開始)でも処置した。この場合では、CMAPの平均振幅は、NT−4/5処置によっていずれも変化しなかった(P70で26.7±7.1mV、n=3)。
【0136】
Mab2256およびNT−4/5は、高周波によって誘発される神経筋衰弱を正常レベルに回復させる
nmdマウスにおける神経筋の電気的特性ならびにMab2256およびNT−4/5の効果をさらに調査するために、対パルス、および周波数の異なる短い一続きの刺激を使用し、筋電計の応答を調査した。MGでは、nmdマウスにおける対パルス最大上刺激(10秒間隔)に対するCMAP応答の振幅(A1およびA2)は、野生型のものと大いに異なっていた。野生型マウスでは、第2のCMAP(A2)の振幅は、第1の応答(A1)よりかすかに大きいまたは同等であった(図3B)。変異マウスでは、第2の応答の振幅は、第1の応答よりも21.4±0.03%小さかった(n=7)(図3C)(p<0.005)。したがって、CMAP曲線下の面積はその振幅によって変化し、活性化した線維数が実際に変化したものであり、見せかけの低下ではなかったことが示された(データは示さず)。
【0137】
250msの一続きの100Hz刺激では、未処置の野生型MGにおけるCMAPの振幅は、一貫したパターンを示した。すなわち、CMAP振幅は、最初の3〜4回の刺激の間に徐々に増大し、次いで縮小の疑似定常状態値に達するまでの刺激期間にわたって段々に低下した(図3D、上部のトレース;図3E、空の四角形)。このパターンは、その刺激の第1と第2のCMAPの間で振幅が最大に落ち込んだ後、記録期間にわたってさらに縮小することを特徴とする、未処置のnmdマウスで見られるものと非常に異なっていた(図3D、2番目のトレース;図3E、塗りつぶされた四角形)。未処置の突然変異体では、縮小が急速であり、CMAPの標準化された振幅は、刺激の終わりには野生型よりも大いに小さくなり(それぞれ55±6.9%、n=6、および82.5±7.4%、n=6)(p<0.02)、伝達が活動電位発生の閾値を下回っている線維数の増加が示唆された。刺激しない間隔(2〜4分)を挟んだ後同じ刺激パターンを繰り返すと、刺激のそれぞれの実施に対してほとんど同一の応答パターンが得られた。
【0138】
神経筋機能に対するMab2256の有効性を評価するために、モノクローナル抗体で処置した動物の記録を行い、結果を未処置マウスと比較した。繰返しの神経刺激(100Hz)では、疑似定常状態での標準化されたCMAP振幅は、Mab2256処置nmdマウス(74±4.1%、n=6)(図3D、3番目のトレース)において、PBS注射した突然変異体(55±6.9%、n=6)(図3E、上部のグラフ)よりも有意に大きい(p<0.05)。しかし、刺激の終わりの応答の低下量は、Mab2256処置マウスと未処置の野生型マウスで有意には異なっておらず(それぞれ78±9%、n=8、および82.5±7.4%、n=6)(図3E、上部のグラフ)(p<0.73)、モノクローナル抗体による処置が正常な神経筋機能を変化させなかったことが示唆された。
【0139】
NT−4/5処置したnmdマウスでも、最後の定常状態CMAPの標準化された振幅が、100HzでPBS注射した突然変異体より有意に大きかった(79±2.5%、n=3)(図3D、下方のトレース;図3E、下方のグラフの塗りつぶされた円)(p<0.02)。野生型、Mab2256処置、および未処置突然変異体における異なる周波数(10、20、50、および100Hz)での縮小量を図3Fに示す。これらの結果は、Mab2256およびNT−4/5の両方が、高周波によって誘発される神経筋の衰弱をほぼ完全に正常レベルに回復し得たことを示唆している。
【0140】
nmdマウスにおける紡錘からの求心性線維の機能状態
筋紡錘からの求心性線維の機能状態を評価するために、未処置およびMab2256処置nmdマウスにおいて、H波が容易に検出可能である背側の足筋からの記録をとった。H波は、運動線維の活性化によって、単シナプスの固有受容性感覚求心回路(挿入図4A)を通して誘発され、神経運動線維の直接の刺激によって誘発されるM波の後に続く(図4A)。M/H波の比は、野生型(6.1±1.3、n=6)、未処置nmd(6.1±0.9、n=5)、およびMab2256処置したnmdマウス(7.6±2.6、n=4)で異なっておらず、背側の足筋の伸展反射を司る筋紡錘線維が突然変異体で優先的に失われないこと、およびこの処置がこの回路に影響を及ぼさなかったことが示唆された。
【0141】
背側の足筋では、nmdマウスおよび対照マウスにおける異なる刺激周波数でのEMG応答の低下量も調査した。10〜50Hzの周波数で野生型マウスとnmdマウスとにCMAPの平均疑似定常状態低下の有意差はなかった(図4B)。100Hz(図4C)でのみ、低下量がnmdマウス(40±1.6%、n=6)において野生型(32±2.85%、n=6)よりわずかに大きく(P<0.036)、この動物モデルでは背側の足筋がMGほど影響を受けないことが示唆された。
【0142】
MUNE分析
nmdマウスのMGで記録されたCMAP振幅の縮小は、機能運動単位数の減少を示唆するものである。運動単位数推定(MUNE)を使用して、nmdマウスのMGにおける運動ニューロン損失の程度を評価した。機能運動単位数を変異マウス(P215〜P230)の老齢期に測定し、対照同腹仔と比較した。その年齢では、突然変異体においては後肢の筋萎縮症が非常に重篤であったが、ヘテロ接合体では筋肉消耗または体力減退の徴候が認められなかった。連続的な漸増刺激は、対照では運動単位電位の規則的な増加を生じるが(図5A)、nmdマウスでは異常に大きい運動単位電位を誘発した(図5B)。強度が段々と増す10回の「成功した」刺激(すなわち、応答の振幅を増大させた刺激)の後の最終電位サイズは、突然変異体では大きな歩幅の増大(すなわち、巨大な運動単位)が存在するために、対照同胞マウスよりも突然変異体においてはるかに大きかった。2匹の対照(ヘテロ接合体)および2匹の突然変異体同腹仔の電位サイズの定量化を、図5C〜Fのグラフに示す。平均の単一運動単位活動電位(SMUAP)振幅は、nmdマウスでは1.21±0.62mV(n=3)であり、対照マウスでは0.178±0.06mV(n=3)であった。したがって、MUNEは、nmdマウス(平均値は37.5±11.4、n=3)において異型接合性の同腹仔(105.5±12.4、n=3、P<0.0015)と比べて50%超の減少があった。さらに、対照マウスに十分な弱い強度の刺激は、突然変異体では少しの応答も誘発せず、突然変異体の残りの運動単位の大部分で線維活性化の閾値が増大したことが示唆された(データは示さず)。巨大な運動単位の存在は、おそらくは神経筋伝達の重度の損失を少なくとも部分的に代償する、軸索の出芽および除神経された筋線維の神経再支配の徴候である。しかし、突然変異体の力が主として巨大な運動単位に依存するならば、これらの単位の損失が後に筋肉機能の突然の不全をもたらす場合もある。
【0143】
nmdマウスにおける横隔膜の機能
呼吸不全は、早くも生まれてから最初の年にヒトSMARD1に特徴的なものである(4)。nmdマウスにおける呼吸機能およびモノクローナル抗体の潜在的な有効性を評価するために、麻酔条件下でのマウス横隔膜の電気生理学的な特性を検討する。10週齢を超える12匹のマウス、すなわち、対照(野生型およびヘテロ接合体、n=6)、未処置nmdマウス(n=3)、およびMab2256処置したnmdマウス(n=3)がこの研究に含まれた。横隔膜の電気活性は、交互に起こる、自発性の活動電位のバースト(吸息バースト)と呼息と同時に起こる無変化の期間を特徴とする。対照および未処置nmdマウスの横隔膜からの典型となる記録を、図6A、Bの下方のトレースに示す。一般に、対照およびnmdマウスで吸息後の電気活性は、認められなかった。本発明者らは、持続時間および吸息バーストの活性(TI)が少ししか変動しなかったことを発見した。対照マウスの呼吸回数(140.2±15.7bpm、n=6)は、未処置のnmd(141.7±6.2bpm、n=3)およびMab2256処置したnmdマウス(132.9±22.6、n=3)と同様であった。しかし、平均吸息持続時間(TI)は、未処置のnmdマウス(131.6±4.1ms)において対照同腹仔(184.1±11.7ms)と比べて著しく短縮し(28%)(P<0.005)、老年期のnmdマウスが軽度の異常な吸息運動の消失を有することが示唆された。Mab2256処置したnmdマウスでは、TIは146.3±8.2msであり、未処置のnmdマウスよりわずかに長かった。この差は、おそらくはサンプルサイズが小さいために統計学的有意性には到達しなかった(P<0.08)(図6C)。非常に高週齢の突然変異体(38週齢)の横隔神経横断面(図6D)の組織学的検査では、nmd(軸索337±18本、n=2)と対照同腹仔(軸索364±11本、n=2)の有髄軸索数に有意差は示されなかった(図6E)。このことは、nmdマウスにおける呼吸機能不全の根底にあるものが、横隔神経の解剖学的な損失というよりも機能の欠陥であることを示唆している。
【0144】
考察
本発明者らは、nmdマウスの筋電図記録の特性を調査し、疾患の臨床的および電気生理学的な進行に対するtrkC受容体のアゴニストモノクローナル抗体(Mab2256)の有効性を試験した。Mab2256処置が筋力低下を数週間遅らせ、筋肉機能の電気生理学的な改善をもたらしたことを示す。
【0145】
nmdマウスにおける神経筋機能障害
nmd神経筋機能に若干の著しい変化が認められた。主な欠陥には、運動神経線維の重度の損失、およびnmdマウスが繰返しの神経刺激で正常な神経筋伝達を維持できないことが含まれる。通常、繰返しの神経刺激では、CMAPは、いくつかのパルスの後、安定した低下レベルに到達するまで振幅が徐々に縮小し、最大低下度は刺激回数に正比例する。本発明者らは、nmdと野生型のMGでこの生理的応答を比較し、nmdマウスが、その臨床的な欠点と一致した、同腹仔対照よりもはるかに重度のCMAP振幅の縮小を示したことを発見したが、このことは、他の運動ニューロン機能障害マウスモデルでも共通の知見である(18)。nmdマウスにおけるこの現象が、シナプス前終末での欠陥によるものか、シナプス後の効力の低下によるものか、または筋肉線維のより疲労性の表現型への移行によるものかを確認することが将来的に問題となるはずである。
【0146】
CMAP平均振幅は、P70のnmdマウスMGにおいて50%を超えて縮小した。これは、この突然変異体で以前に記載されている5週間での腰椎運動ニューロンの41%までの低下に一致している(11)。疾患のその後の段階(P150〜230)では、MG筋に残存する運動単位数は、対照値の35%に減少したが、12〜14週間の腰髄に28%の運動ニューロンが残存するという以前の推定と離れてはいない(11)。巨大な運動単位電位は、活性化の閾値が高いので、補充が不十分な場合もある。運動単位の重度の損失と共に、繰返しの刺激で有効な伝達を維持できないことは、これらの動物の筋力の減弱を説明するものである。
【0147】
神経栄養因子の治療的な可能性
神経栄養因子は、脊髄運動ニューロンの生存を支え、運動ニューロン疾患の動物モデルにおいて病理学的な症状の軽減にプラスの影響を及ぼすことが示されている(14、19、20)。これらの結果に基づき、組換え型の神経栄養因子は、十年以上の間運動ニューロン疾患有望な治療薬であると考えられてきた。しかし、臨床試験では、不適当な投与量、副作用などの問題に遭遇している(21、22)。ニューロトロフィン受容体のアゴニストモノクローナル抗体は、外来の投与された神経栄養因子に優るいくつかの理論上の利点、例えば、副作用を低減し得る、所与の栄養受容体に対するその特異性、ならびに薬物投与および治療濃度の維持を容易にするその長い循環半減期を有する。それにもかかわらず、これらの分子は、動物モデルで調査し確認する必要がある。例えば、一部のモデルでは、神経栄養因子による運動ニューロンの救出が、一過性の性質のものであることが判明した(23、24)。本発明者らのnmdマウスでの実験では、Mab2256は、疾患の進行を遅らせるが阻止しなかった。
【0148】
繊毛様神経栄養因子(CNTF)およびNT3は、マウス突然変異体pmn(進行性運動神経細胞障害)の寿命を延長し得ることが示されている(14、25、26)。nmdマウスの平均寿命は、54日であることが記載されている(10)。本発明者らの動物施設でも、未処置nmdマウスの生存中央値は同様であった(64日)。出生後第3週〜第11週にnmdマウスをMab2256で処置しても、生存確率は有意に増大しなかった(中央値:69日)。この結果は、nmdマウスでの全長IGHMBP2のニューロン発現もその生存状態を改善し得なかったという事実に従うものである(10)。
【0149】
Mab2256の一過性の効果について考えられる説明は、ニューロトロフィンを用いる他の研究で示されているように、一部の動物が到達する、血漿中で比較的高レベルとなった薬物が、trkC受容体の下方制御を引き起こしたのではないかということである(27、28)。これが事実であるならば、治療的な投与量および投薬回数は、この効果を回避するために調整すべきである。
【0150】
また、Mab2256は、ニューロン生存バイオアッセイにおいてNT−3ほど強力でなく(EC50が約3000倍の差)、有効でもなかった(最大効果が約3倍の差)。したがって、nmdマウスモデルにおいてより長くより高い有効性を実現するために、Mab2256抗体のより高い親和性/活性のものが必要であると思われる。
【0151】
その上、下位運動ニューロンは、trkBおよびtrkCの両方の受容体を発現する。これらおよびことによると他のニューロトロフィン受容体の活性化は、nmdマウスにおける臨床的に有益な成果に必要であるかもしれない。将来的には、この疾患モデルにおいてtrkBおよびtrkCの両方のアゴニストの同時の適用がより大きな治療利益を提供するかどうかを試験することが重要となる。実際に、本発明者らは、Mab2256および内因性のtrkBアゴニストであるNT−4/5が、繰返しの刺激プロトコルにおいてなど、nmdマウス神経筋接合部の電気生理学的特性の側面を回復させ得ることを見出した。NT−3と他の神経栄養因子の組合せが適切な場合もあり、例えば、CNTF遺伝子およびNT−3遺伝子をコードするアデノウイルスベクターのpmnマウス骨格筋細胞への同時注射は、どちらかのベクター単独でよりも軸索の生存時間を大きく増大させる(26、29)。
【0152】
nmdマウスにおいてMab2256でin vivoで見られるプラスの影響の原因である正確な細胞ターゲットはまだ不明であるが、運動ニューロン、筋細胞、グリア細胞、または上記の組合せである可能性がある。運動ニューロンは、NT−3受容体であるtrkCを発現し、NT−3に応答し、生存時間が段々と増す(15、30〜32)。最近の証拠は、NT−3が、運動ニューロンの生存に対するその効果のほかに、神経筋伝達の有効性に影響を及ぼす場合もあることを示唆している(33〜36)。外来のBNDFまたはNT−3は(神経成長因子NGFではないが)、培養物中で神経筋シナプスを発生させる、自発性およびインパルス誘発型のシナプス活性、すなわち、trkC受容体を媒介とし、因子が存在する限り存続するように思われる効果を増強する(37)。その上、単離したニューロンをNT−3で2日間処理すると、新たに形成された神経−筋肉シナプスでの素量的なAChパケットの平均サイズが増大するのに対し、NT−3に対する抗体、またはチロシンキナーゼ受容体の特異的な阻害剤であるK252aで処理すると、存在するシナプスでの素量サイズが縮小するが、これは、NT−3が素量的なパケットの発生および維持を司ることを示唆している(34)。NT−3によるシナプス伝達の増強も成体海馬スライスで報告されている(38)。筋肉によって合成されるNT−3およびNT−4/5は、シナプス前の運動ニューロンに対して逆方向に働き、それによって、例えばAChおよびニューレグリンの合成を増大させることで、途切れないニューロンの機能分化に影響を及ぼす場合がある(39)。これらのシナプス前の効果のほかに、筋肉によって分泌されるニューロトロフィン(BDNF、NT−3、NT−4/5、およびGNDF(15、40〜43)は、筋線維それ自体に対して自己分泌の方法で働く場合がある。NT−3の外からの投与は、クラーレ処理した筋肉において、その処理によって予め変更されたNMJ構造を回復させることが示されている(44)。筋細胞からのNT−3の放出は、NMJでのシナプスの活性によって次々に調節されるようである(36)。
【0153】
アデノウイルスを媒介とする、NT−3の遺伝子導入は、pmnマウスにおいて運動線維の終端の出芽を促進するが、このことは、NT−3が遠位の軸索構造の維持および再生にも関与することを示唆している(26、29)。さらに、神経栄養因子は、運動神経から脊髄への軸索輸送にも影響を及ぼし得る。チューブリンの重合が変更されたために運動の軸索が変性しているpmnマウス(45)において、腓腹筋に注射された、または切断された坐骨神経に直接に適用された蛍光トレーサーの逆方向性の輸送は、CNTF、BDNF、またはNT−3によって改善することができるが、GDNFまたはNGFによって改善することはできない(46)。
【0154】
Mab2256は分子サイズが大きいため、無処置の脳血液関門を通過できないと思われる。しかし、NT−3および他のニューロトロフィン(NT−4/5、BDNF)は、運動ニューロンの軸索に沿って逆行してその細胞体に輸送することができる(31、42、47、48)。一度trkCに結合したこのモノクローナル抗体が、逆行して輸送され、運動ニューロンの細胞体に直接の効果を与え得るかどうかを知ることは大きな関心の的となるはずである。正確な部位およびtrkC抗体によって発揮される作用様式は、疑いなくさらなる研究が待たれる。
【0155】
横隔膜電気活性
SMARD1患者では、横隔膜の麻痺が生後最初の13カ月の間に現れるが(2)、nmdマウスでは、比較的遅くに呼吸異常が現れる(10)。疾患の最終段階(P150〜230)の際の横隔膜の機能状態を評価するために、変異マウスの横隔膜の自発性の電気活性をin vivoで記録した。麻酔下のマウスでは、対照と突然変異体同腹仔との平均呼吸回数に差は見られなかったが、本発明者らは、吸息持続時間(TI)の26%の短縮と、それに付随する、横隔膜強度のある種の低下を生み出す場合もある各吸息バーストの際の活動電位数の減少を見出した。電気的な活性のない帯域と探査電極の位置決め誤りを識別するのが困難であったので、その記録から、横隔膜内に機能していない領域があるかどうかを見分けることはできない(方法を参照されたい)。最近では、nmdマウス横隔膜中の大量の筋障害性変化の存在が記載されている(11)。さらに、これらのマウスは、二次的に呼吸困難の一因となり得るうっ血性心不全および筋ジストロフィー様の表現型も患う(10)。しかし、有髄の軸索数は、非常に高週齢のnmdマウス(38週齢)では減少しなかったが、これは14週齢nmdマウスで見られたものに従うものであり(11)、ある量の全長機能性のIGHMBP2タンパク質の発現に一致するものである(9)。
【0156】
要するに、本発明者らは、nmdマウスへのMab2256処置が、著しいが一過性の筋力の改善、ならびに高頻度の神経刺激の際の神経筋の機能低下量の正常化を生じたことを発見した。この効果の作用機序を解明する別の研究、ならびに神経筋接合部での神経伝達の際のシナプス前およびシナプス後の事象のより完全な特徴付けは、我々がnmdマウスにおける神経筋の欠陥、運動ニューロン中のIGHMBP2の生理的役割を理解し、このひどい疾患の合理的な治療手法を探索する助けとなるはずである。
【0157】
材料および方法
マウスの繁殖および遺伝子型
B6.BLKS−nmd2JマウスをThe Jackson Laboratoryから入手した。nmd2Jについてヘテロ接合体のマウスを交雑し、野生型、ヘテロ接合体、および突然変異体マウスを実験に使用した。マウスを繁殖させ、突然変異体については食物および水をケージの床の高さで利用可能にしたことを除き、標準の条件で維持した。
【0158】
マウスの遺伝子型を記載されている通りに同定した(9)。簡潔に述べると、(その突然変異についてホモ接合性である)nmdマウスの表現型の原因である点突然変異は、野生型マウスには存在しない新しいDdeI制限部位を生じる。罹患していない子孫の中の保因動物(carrier)を特定するPCRアッセイは、突然変異が存在する694bpのPCR産物を増幅する、順方向プライマー:5’−GCTGGAAACGATCACATACCG−3’および逆方向プライマー:5’−AGCTCCTGATGATCCAATGG−3’の2種のオリゴヌクレオチドプライマーを用いて実施した。
【0159】
マウスの処置
両方の性別のコード化された同腹仔マウスのランダムな群に、モノクローナル抗体Mab2256(5mg/体重kg、週2回、20〜21日齢から;Rinat Neuroscience、米カリフォルニア州パロアルト)、ヒト組換え型NT−4/5(5mg/体重kg、週2回、21日齢から;Genentech、米カリフォルニア州サンフランシスコ)、またはPBSのいずれかを腹腔内注射した。すべての動物処置は、制度上の指針および認可に従って実施した。
【0160】
受容体チロシンリン酸化アッセイ
Mab2256のアゴニスト活性は、細胞に基づくtrkC受容体チロシンリン酸化アッセイで、以前に記載されている通りに評価した(16)。Prism Software(GraphPad、米サンディエゴ)を使用する非線形カーブフィッティングによって、最大半有効濃度を推定した。
【0161】
胚三叉神経ニューロン生存アッセイ
E12 Sprague Dawleyラットから、三叉神経ニューロンの分離した培養物を樹立した。解体した神経節をトリプシン処理し、摩砕によって分断した(49)。ニューロンを、96ウェル組織培養プレート中のポリオルニチン/ラミニン基層上の限定された無血清培地に低密度で播いた。プレーティング時に、様々な濃度のNT3およびMab2256抗体を、それぞれ3通りおよび4通りにして培養物に加えた。異なるそれぞれの条件下でのニューロンの生存を定量化するために、プレーティングから48時間後に生存していたニューロン数をカウントした。Prism Software(GraphPad、サンディエゴ)を使用する非線形カーブフィッティングによって、最大半量有効濃度を推定した。
【0162】
薬物動態研究
メスの成体CD−1マウス(n=3)にMab2256を2mg/kgで腹腔内注射した。次いで、注射してから24、48、136、および184時間後に動物の採血を行った。各時点でのMab2256の血清濃度を以下で述べる通りに測定した。
【0163】
モノクローナル抗体濃度の測定
マウスモノクローナル抗体Mab2256の血清レベルは、標準のサンドイッチELISAによって測定した。96ウェルMaxisorpプレート(Nunc)に、HEK293細胞への一過性の形質移入によって発現させた組換え型ヒトtrkC細胞外ドメイン−IgG Fc融合タンパク質(Rinat Neuroscience)をプロテインAカラムで精製したもの0.2mg/mLを一晩かけて4℃で予め吸収させた。次いで、trkCコートされたプレートを、0.5%のウシ血清アルブミンおよび0.05%のTween−20を含むリン酸緩衝溶液(PBS)で25℃で1時間かけてブロックした。プレートを、0.05%のTween−20を含むPBSで3回洗浄した。Mab2256の標準の希釈系列、ならびに適切に希釈した血清サンプルをプレートに入れて25℃で1時間インキュベートした後、3回洗浄した。西洋わさびペルオキシダーゼ(HRP)結合ウサギ抗マウスIgG(Jackson ImmunoResearch)を1:5000で適用し、25℃で1時間インキュベートして、結合したマウスモノクローナル抗体を検出した。最後に、HRPとTMB基質(KPL、デンマーク)の比色反応によってシグナルを検出した。
【0164】
神経学的試験
突然変異体および同腹仔対照マウスに標準の神経学的検査を行い、神経筋の欠陥の発生および拡大を定量化した(SHIRPAプロトコル;www.mgu.har.mrc.ac.uk/mutabase/shirpa_summary.html)(50)。平衡能測定では、マウスを尾から吊り下げて四角い薄い棒切れの上に降ろし、逆立ちさせた。次いで、棒切れを10秒間1秒毎に手で回転させ、マウスが棒切れに留まる能力を秒数で測定した。
【0165】
前肢握力については、マウスを水平な針金の上に保持し、降ろして前肢で針金を握らせるようにした。マウスが前肢によってつながっている能力を10秒間にわたり記録した。
【0166】
筋電図検査(EMG)
以前に記載されている通り(18)に、麻酔下(100mg/Kgケタミン+10mg/Kgキシラジン)のP70の野生型およびnmdマウスからの筋肉内複合活動電位(CMAP)を記録した。簡潔に述べると、記録針電極を、第5指骨の基部に不関電極を添えて背側の足筋中に置き、または腓腹筋の内側部分(MG)中に置いた。アース電極を尾の付け根に置いた。刺激針電極を坐骨の切込みおよび腓骨上部に置いた。過最大電流パルスの刺激プロトコル(持続時間0.05ms、振幅5mA)を単一パルスまたは10、20、50、および100Hzの短い持続時間のパルス列として適用した。刺激パルスは、孤立したパルス刺激装置(A−MSystems、2100モデル)によって作成した。記録された出力を示差的に増幅し(Brownlee Precision、210Aモデル)、20000サンプル/秒でデジタル方式で取得し(ADInstruments、PowerLab/4SP)、後の分析に向けてコンピュータに保存した。
【0167】
MGからの運動単位数推定(MUNE)は、全運動単位の和である最大CMAPを単一の運動単位電位の平均サイズで割ることによって算出した。単一の運動単位電位のサンプリングは、精密に制御された電流を、10回の応答が漸進的に補充されるまで閾値下のレベルから非常に小さな歩幅で適用するものである漸増法(51、52)によって行った。各電流振幅を3回適用し、安定であるとみなされたので、それらが同一であったなら受け入れた。個々の運動単位振幅は、各応答の振幅を以前の応答の振幅から差し引いて得た。個々の値の平均が単一の運動単位活動電位(SMUAP)サイズの推定となった。
【0168】
in vivo横隔膜記録は、針電極を、中線からわずかにそれた剣状突起の後ろでどちらかの部位に挿入して実施した。不関電極を胸の上、アース電極を尾の付け根に置いた。横隔膜は、呼吸と同時に起こるリズミカルなバーストの発生によって容易に確認された。ピーク振幅および記録の積分による面積によって吸息を定量化した。吸息持続時間(TI)も分析した。6連続の呼吸サイクルから平均値を算出した。
【0169】
研究は、コード化されたマウスを用い、どのマウスを試験しているかについて筋電図記録者は盲検化されるように実施した。
【0170】
すべてのデータは平均±SEMとして報告する。統計学的有意性は、スチューデントのt検定を使用して評価した。統計学的有意性判定のための判定基準レベルは、すべての実験についてP<0.05に設定した。
【0171】
組織像
マウスを過量のケタミン/キシラジンで屠殺し、右側横隔神経を、その横隔膜筋への入口近くで採取した。神経を、4%のパラホルムアルデヒドおよび2.5%のグルタルアルデヒドのPBS中固定液で固定し、次いで2%の四酸化オスミウムで固定した後、spurr樹脂中に包埋した(プラスチック包埋)。厚さ2μmの切片をトルイジンブルーで染色し、光学顕微鏡(Axiovert35、Zeiss)で調べた。各神経について有髄の軸索をカウントした。
【0172】
前述の発明は、理解を明確にする目的で図解および例として若干詳細に記載してきたが、ある種の変更および修正を実行してもよいことは当業者には明白となろう。したがって、その記述および例は、添付の請求項によって叙述される本発明の範囲を限定するものと解釈すべきでない。
【0173】
参考文献
1.Lefebvre,S.、Burglen,L.、Reboullet,S.、Clermont,O.、Burlet,P.、Viollet,L.、Benichou,B.、Cruaud,C.、Millasseau,P.、Zeviani,M.ら(1995年)、「Identification and characterization of a spinal muscular atrophy−determining gene」、Cell、第80巻、155〜65ページ
2.Grohmann,K.、Wienker,T.F.、Saar,K.、Rudnik−Schoneborn,S.、Stoltenburg−Didinger,G.、Rossi,R.、Novelli,G.、Nurnberg,G.、Pfeufer,A.、Wirth,B.ら(1999年)、「Diaphragmatic spinal muscular atrophy with respiratory distress is heterogeneous,and one form Is linked to chromosome 11q13−q21」、Am J Hum Genet、第65巻、1459〜62ページ
3.Grohmann,K.、Schuelke,M.、Diers,A.、Hoffmann,K.、Lucke,B.、Adams,C.、Bertini,E.、Leonhardt−Horti,H.、Muntoni,F.、Ouvrier,R.ら(2001年)、「Mutations in the gene encoding immunoglobulin mu−binding protein 2 cause spinal muscular atrophy with respiratory distress type 1」、Nat Genet、第29巻、75〜7ページ
4.Grohmann,K.、Varon,R.、Stolz,P.、Schuelke,M.、Janetzki,C.、Bertini,E.、Bushby,K.、Muntoni,F.、Ouvrier,R.、Van Maldergem,L.ら(2003年)、「Infantile spinal muscular atrophy with respiratory distress type 1(SMARD1)」、Ann Neurol、第54巻、719〜24ページ
5.Pitt,M.、Houlden,H.、Jacobs,J.、Mok,Q.、Harding,B.、Reilly,M.、およびSurtees,R.(2003年)、「Severe infantile neuropathy with diaphragmatic weakness and its relationship to SMARD1」、Brain、第126巻、2682〜92ページ
6.Wirth,B.(2000年)、「An update of the mutation spectrum of the survival motor neuron gene(SMN1)in autosomal recessive spinal muscular atrophy(SMA)」、Hum Mutat、第15巻、228〜37ページ
7.Mercuri,E.、Bertini,E.、Messina,S.、Pelliccioni,M.、D’Amico,A.、Colitto,F.、Mirabella,M.、Tiziano,F.D.、Vitali,T.、Angelozzi,C.ら(2004年)、「Pilot trial of phenylbutyrate in spinal muscular atrophy」、Neuromuscul Disord、第14巻、130〜5ページ
8.Cook,S.A.、Johnson,K.R.、Bronson,R.T.、およびDavisson,M.T.(1995年)、「Neuromuscular degeneration(nmd):a mutation on mouse chromosome 19 that causes motor neuron degeneration」、Mamm Genome、第6巻、187〜91ページ
9.Cox,G.A.、Mahaffey,C.L.、およびFrankel,W.N.(1998年)、「Identification of the mouse neuromuscular degeneration gene and mapping of a second site suppressor allele」、Neuron、第21巻、1327〜37ページ
10.Maddatu,T.P.、Garvey,S.M.、Schroeder,D.G.、Hampton,T.G.、およびCox,G.A.(2004年)、「Transgenic rescue of neurogenic atrophy in the nmd mouse reveals a role for Ighmbp2 in dilated cardiomyopathy」、Hum Mol Genet、第13巻、1105〜15ページ
11.Grohmann,K.、Rossoll,W.、Kobsar,I.、Holtmann,B.、Jablonka,S.、Wessig,C.、Stoltenburg−Didinger,G.、Fischer,U.、Hubner,C.、Martini,R.ら(2004年)、「Characterization of Ighmbp2 in motor neurons and implications for the pathomechanism in a mouse model of human spinal muscular atrophy with respiratory distress type1(SMARD1)」、Hum Mol Genet
12.Arakawa,Y.、Sendtner,M.、およびThoenen,H.(1990年)、「Survival effect of ciliary neurotrophic factor(CNTF)on chick embryonic motoneurons in culture:comparison with other neurotrophic factors and cytokines」、J Neurosci、第10巻、3507〜15ページ
13.Sendtner,M.、Kreutzberg,G.W.、およびThoenen,H.(1990年)、「Ciliary neurotrophic factor prevents the degeneration of motor neurons after axotomy」、Nature、第345巻、440〜1ページ
14.Sendtner,M.、Schmalbruch,H.、Stockli,K.A.、Carroll,P.、Kreutzberg,G.W.、およびThoenen,H.(1992年)、「Ciliary neurotrophic factor prevents degeneration of motor neurons in mouse mutant progressive motor neuronopathy」、Nature、第358巻、502〜4ページ
15.Henderson,C.E.、Camu,W.、Mettling,C.、Gouin,A.、Poulsen,K.、Karihaloo,M.、Rullamas,J.、Evans,T.、McMahon,S.B.、Armanini,M.P.ら(1993年)、「Neurotrophins promote motor neuron survival and are present in embryonic limb bud」、Nature、第363巻、266〜70ページ
16.Sadick,M.D.、Galloway,A.、Shelton,D.、Hale,V.、Weck,S.、Anicetti,V.、およびWong,W.L.(1997年)、「Analysis of neurotrophin/receptor interactions with a gD−flag−modified quantitative kinase receptor activation(gD.KIRA)enzyme−linked immunosorbent assay」、Exp Cell Res、第234巻、354〜61ページ
17.Poduslo,J.F.およびCurran,G.L.(1996年)、「Permeability at the blood−brain and blood−nerve barriers of the neurotrophic factors:NGF,CNTF,NT−3,BDNF」、Brain Res Mol Brain Res、第36巻、280〜6ページ
18.Fernandez−Chacon,R.、Wolfel,M.、Nishimune,H.、Tabares,L.、Schmitz,F.、Castellano−Munoz,M.、Rosenmund,C.、Montesinos,M.L.、Sanes,J.R.、Schneggenburger,R.ら(2004年)、「The synaptic vesicle protein CSP alpha prevents presynaptic degeneration」、Neuron、第42巻、237〜51ページ
19.Sagot,Y.、Vejsada,R.、およびKato,A.C.(1997年)、「Clinical and molecular aspects of motoneurone diseases:animal models,neurotrophic factors and Bcl−2 oncoprotein」、Trends Pharmacol Sci、第18巻、330〜7ページ
20.Mitsumoto,H.、Ikeda,K.、Klinkosz,B.、Cedarbaum,J.M.、Wong,V.、およびLindsay,R.M.(1994年)、「Arrest of motor neuron disease in wobbler mice cotreated with CNTF and BDNF」、Science、第265巻、1107〜10ページ
21.Miller,R.G.、Petajan,J.H.、Bryan,W.W.、Armon,C.、Barohn,R.J.、Goodpasture,J.C.、Hoagland,R.J.、Parry,G.J.、Ross,M.A.、およびStromatt,S.C.(1996年)、「A placebo−controlled trial of recombinant human ciliary neurotrophic(rhCNTF)factor in amyotrophic lateral sclerosis」、rhCNTF ALS Study Group.Ann Neurol、第39巻、256〜60ページ
22.Thoenen,H.およびSendtner,M.(2002年)、「Neurotrophins:from enthusiastic expectations through sobering experiences to rational therapeutic approaches」、Nat Neurosci、第5巻増刊、1046〜50ページ
23.Diener,P.S.およびBregman,B.S.(1994年)、「Neurotrophic factors prevent the death of CNS neurons after spinal cord lesions in newborn rats」、Neuroreport、第5巻、1913〜7ページ
24.Vejsada,R.、Sagot,Y.、およびKato,A.C.(1995年)、「Quantitative comparison of the transient rescue effects of neurotrophic factors on axotomized motoneurons in vivo」、Eur J Neurosci、第7巻、108〜15ページ
25.Sagot,Y.、Tan,S.A.、Baetge,E.、Schmalbruch,H.、Kato,A.C.、およびAebischer,P.(1995年)、「Polymer encapsulated cell lines genetically engineered to release ciliary neurotrophic factor can slow down progressive motor neuronopathy in the mouse」、Eur J Neurosci、第7巻、1313〜22ページ
26.Haase,G.、Kennel,P.、Pettmann,B.、Vigne,E.、Akli,S.、Revah,F.、Schmalbruch,H.、およびKahn,A.(1997年)、「Gene therapy of murine motor neuron disease using adenoviral vectors for neurotrophic factors」、Nat Med、第3巻、429〜36ページ
27.Bibel,M.およびBarde,Y.A.(2000年)、「Neurotrophins:key regulators of cell fate and cell shape in the vertebrate nervous system」、Genes Dev、第14巻、2919〜37ページ
28.Knusel,B.、Gao,H.、Okazaki,T.、Yoshida,T.、Mori,N.、Hefti,F.、およびKaplan,D.R.(1997年)、「Ligand−induced down−regulation of Trk messenger RNA、protein and tyrosine phosphorylation in rat cortical neurons」、Neuroscience、第78巻、851〜62ページ
29.Sendtner,M.(1997年)、「Gene therapy for motor neuron disease」、Nat Med、第3巻、380〜1ページ
30.Oppenheim,R.W.(1996年)、「Neurotrophic survival molecules for motoneurons:an embarrassment of riches」、Neuron、第17巻、195〜7ページ
31.Yan,Q.、Elliott,J.L.、Matheson,C.、Sun,J.、Zhang,L.、Mu,X.、Rex,K.L.、およびSnider,W.D.(1993年)、「Influences of neurotrophins on mammalian motoneurons in vivo」、J Neurobiol、第24巻、1555〜77ページ
32.Duberley,R.M.、Johnson,I.P.、Anand,P.、Leigh,P.N.、およびCairns,N.J.(1997年)、「Neurotrophin−3−like immunoreactivity and Trk C expression in human spinal motoneurones in amyotrophic lateral sclerosis」、J Neurol Sci、第148巻、33〜40ページ
33.Wang,T.、Xie,K.、およびLu,B.(1995年)、「Neurotrophins promote maturation of developing neuromuscular synapses」、J Neurosci、第15巻、4796〜805ページ
34.Liou,J.C.およびFu,W.M.(1997年)、「Regulation of quantal secretion from developing motoneurons by postsynaptic activity−dependent release of NT−3」、J Neurosci、第17巻、2459〜68ページ
35.Sendtner,M.(1998年)、「Neurotrophic factors:effects in modulating properties of the neuromuscular endplate」、Cytokine Growth Factor Rev、第9巻、1〜7ページ
36.Poo,M.M.(2001年)、「Neurotrophins as synaptic modulators」、Nat Rev Neurosci、第2巻、24〜32ページ
37.Lohof,A.M.、Ip,N.Y.、およびPoo,M.M.(1993年)、「Potentiation of developing neuromuscular synapses by the neurotrophins NT−3 and BDNF」、Nature、第363巻、350〜3ページ
38.Kang,H.およびSchuman,E.M.(1995年)、「Long−lasting neurotrophin−induced enhancement of synaptic transmission in the adult hippocampus」、Science、第267巻、1658〜62ページ
39.Loeb,J.A.およびFischbach,G.D.(1997年)、「Neurotrophic factors increase neuregulin expression in embryonic ventral spinal cord neurons」、J Neurosci、第17巻、1416〜24ページ
40.Griesbeck,O.、Parsadanian,A.S.、Sendtner,M.、およびThoenen,H.(1995年)、「Expression of neurotrophins in skeletal muscle:quantitative comparison and significance for motoneuron survival and maintenance of function」、J Neurosci Res、第42巻、21〜33ページ
41.Henderson,C.E.、Phillips,H.S.、Pollock,R.A.、Davies,A.M.、Lemeulle,C.、Armanini,M.、Simmons,L.、Moffet,B.、Vandlen,R.A.、Simpson,L.C.ら(1994年)、「GDNF:a potent survival factor for motoneurons present in peripheral nerve and muscle」、Science、第266巻、1062〜4ページ
42.Koliatsos,V.E.、Clatterbuck,R.E.、Winslow,J.W.、Cayouette,M.H.、およびPrice,D.L.(1993年)、「Evidence that brain−derived neurotrophic factor is a trophic factor for motor neurons in vivo」、Neuron、第10巻、359〜67ページ
43.Funakoshi,H.、Frisen,J.、Barbany,G.、Timmusk,T.、Zachrisson,O.、Verge,V.M.、およびPersson,H.(1993年)、「Differential expression of mRNAs for neurotrophins and their receptors after axotomy of the sciatic nerve」、J Cell Biol、第123巻、455〜65ページ
44.Loeb,J.A.、Hmadcha,A.、Fischbach,G.D.、Land,S.J.、およびZakarian,V.L.(2002年)、「Neuregulin expression at neuromuscular synapses is modulated by synaptic activity and neurotrophic factors」、J Neurosci、第22巻、2206〜14ページ
45.Bommel,H.、Xie,G.、Rossoll,W.、Wiese,S.、Jablonka,S.、Boehm,T.、およびSendtner,M.(2002年)、「Missense mutation in the tubulin−specific chaperone E(Tbce)gene in the mouse mutant progressive motor neuronopathy,a model of human motoneuron disease」、J Cell Biol、第159巻、563〜9ページ
46.Sagot,Y.、Rosse,T.、Vejsada,R.、Perrelet,D.、およびKato,A.C.(1998年)、「Differential effects of neurotrophic factors on motoneuron retrograde labeling in a murine model of motoneuron disease」、J Neurosci、第18巻、1132〜41ページ
47.DiStefano,P.S.、Friedman,B.、Radziejewski,C.、Alexander,C.、Boland,P.、Schick,C.M.、Lindsay,R.M.、およびWiegand,S.J.(1992年)、「The neurotrophins BDNF,NT−3,and NGF display distinct patterns of retrograde axonal transport in peripheral and central neurons」、Neuron、第8巻、983〜93ページ
48.Yan,Q.、Elliott,J.、およびSnider,W.D.(1992年)、「Brain−derived neurotrophic factor rescues spinal motor neurons from axotomy−induced cell death」、Nature、第360巻、753〜5ページ
49.Davies,A.M.、Horton,A.、Burton,L.E.、Schmelzer,C.、Vandlen,R.、およびRosenthal,A.(1993年)、「Neurotrophin−4/5 is a mammalian−specific survival factor for distinct populations of sensory neurons」、J Neurosci、第13巻、4961〜7ページ
50.Rogers,S.D.、Demaster,E.、Catton,M.、Ghilardi,J.R.、Levin,L.A.、Maggio,J.E.、およびMantyh,P.W.(1997年)、「Expression of endothelin−B receptors by glia in vivo is increased after CNS injury in rats,rabbits,and humans」、Exp Neurol、第145巻、180〜95ページ
51.McComas,A.J.(1991年)、「Motor unit estimation:methods,results,and present status」、Muscle Nerve、第14巻、585〜97ページ
52.Shefner,J.M.およびGooch,C.L.(2002年)、「Motor unit number estimation in neurologic disease」、Adv Neurol、第88巻、33〜52ページ
53.Ruiz,R.、Lin,J.、Forgie,A.、Foletti,D.、Shelton,D.、Rosenthal,A.、およびTabares,L.(2005年)、「Treatment with trkC agonist antibodies delays disease progression in neuromuscular degeneration(nmd)mice」、Human Molecular Genetics、第14巻、1825〜1837ページ
【0174】
略語
BDNF:脳由来神経栄養因子
bpm:1分あたりの呼吸数
CMAP:複合筋活動電位
CNTF:繊毛様神経栄養因子
GDNF:グリア細胞由来神経栄養因子
MG:内側腓腹筋
IGHMBP2:免疫グロブリンμ結合タンパク質2遺伝子
nmd:神経筋変性
MUNE:運動単位数推定
NGF:神経成長因子
NMJ:神経筋接合部
NT−3:ニューロトロフィン3
NT−4/5:ニューロトロフィン4/5
SMA:脊髄性筋萎縮症
SMARD1:脊髄性筋萎縮症1型
SMN:生存運動神経細胞遺伝子
SMUAP:単一運動単位活動電位
TI:吸息時間
trkA:チロシンキナーゼ受容体A
trkB:チロシンキナーゼ受容体B
trkC:チロシンキナーゼ受容体C
【図面の簡単な説明】
【0175】
【図1】モノクローナル抗体Mab2256がtrkC受容体を活性化し、培養物中の三叉神経ニューロンの生存を支え得ることを示すグラフである。(A)漸増濃度のNT3(左パネル)、すなわち内因性のtrkCリガンド、またはモノクローナル抗体Mab2256(右パネル)、すなわちtrkC抗体によって、増大し飽和状態になるレベルのtrkC受容体のリン酸化(OD450で示す、y軸)が誘発された。(B)培養物で48時間生存するラット胚三叉神経ニューロン数は、培地中の様々な濃度のNT3(左パネル)またはMab2256(右パネル)の存在によって支えられて増加し、飽和状態になった。
【図2】nmdマウスの疾患徴候および寿命を示すグラフおよび図である。(A)P20〜P70の野生型(+/+)(n=8)、nmd(−/−)(n=8)、およびヘテロ接合体(+/−)(n=8)マウスの平均体重。(B)モノクローナル抗体Mab2256を腹腔内注射した3〜11週の野生型(+/+Ab)(n=10)およびnmdマウス(−/−Ab)(n=16)の平均体重。(C〜D)PBSを注射した野生型(+/+、空の記号)(n=14)、ヘテロ接合体(+/−、灰色の記号)(n=8)、およびnmd(−/−、塗りつぶされた四角形)(n=5)、ならびにMab2256を注射したnmdマウス(−/−Ab、三角形)(n=9)の前肢握り時間(C)および棒上での平衡能(D)。(E〜F)棒上での野生型およびMab2256処理した突然変異体。突然変異体は、その尾を使って棒を掴むことができなかったが、それにもかかわらず、自身を回転している棒上に数秒間維持することができた。Mab2256で処置していないnmdマウスは、自身を1秒より長く棒上に維持することができなかった(表示なし)。(G)未処置(nmd)(n=30)およびMab2256処置したnmd(nmd Ab)(n=15)マウスのKaplan−Meier生存率分析。マンホイットニーの順位和検定で2つの群について有意差は得られなかった。
【図3】野生型およびnmdマウスの内側腓腹筋(MG)におけるCMAP振幅のEMG測定から、nmdマウスにおける神経伝達の効力の低下が明らかになることを示すグラフである。(A)未処置の野生型(+/+)(n=14)、PBSを注射したまたは注射していないnmd(−/−)(n=9)、Mab2256で処置した野生型(+/+Ab)(n=8)、Mab2256で処置したnmd(−/−Ab)(n=7)、およびNT−4/5で処置したnmdマウス(−/−NT4/5)(n=3)における、過最大の刺激に応答してのCMAPの絶対振幅(平均±SEM)。(B〜C)野生型(B)およびnmdマウス(C)における対パルスプロトコルに対する典型となるCMAP応答。刺激間の間隔:10ms。経時的なCMAPのピークからピークまでの振幅(A1およびA2)を矢印で表示する。(D)野生型(上方のトレース)ならびに3種のnmdマウス:未処置(2番目のトレース)、Mab2256処置(3番目のトレース)、およびNT−4/5処置(4番目のトレース)における、100Hzでの一続きの刺激の間の典型となる記録。(E)未処置の野生型マウス(+/+)(n=6)、Mab2256処置した野生型(+/+Ab)(n=8)、PBS注射したnmd(−/−)(n=6)、Mab2256処置したnmd(−/−Ab)マウス(上方のグラフ、n=6)、およびNT−4/5処置したマウス(−/−NT4/5)(下方のグラフ、n=3)における、100Hzで250msの一続きの刺激の間の(最初の応答を標準として比較した)CMAP振幅の縮小。(F)10〜100Hzの刺激周波数についての疑似定常状態レベルでのCMAP振幅の縮小パーセント。すべてのデータは、P69〜71で行ったEMG記録からのものである。
【図4】背側の足筋におけるCMAP振幅のEMG測定を示すグラフである。(A)野生型マウスにおいて単一のパルス刺激によって誘発したM波およびH波。(B)野生型(+/+)(n=6)およびnmdマウス(−/−)(n=6)における10〜100Hzの刺激周波数についての疑似定常状態レベルでのCMAP振幅の縮小パーセント。(C)野生型マウス(n=6)およびPBS注射したnmd(n=6)における、100Hzでの250msの一続きの刺激の間の(最初の応答を標準として比較した)CMAP振幅の縮小の時間経過。すべてのデータは、P69〜71のマウスからのものである。
【図5】P215〜230のマウスでのMGからの運動単位数推定(MUNE)を示すグラフである。(A〜B)野生型(A)およびnmd(B)マウスの運動単位トレース。(C〜F)対照(CおよびE)ならびにnmd同胞突然変異体(DおよびF)マウスにおける、段々と大きさの増す刺激に応答した単一運動単位活動電位(SMUAP)の振幅。X軸の各数字は、応答の振幅の増大を誘発した刺激を表す。対照動物はヘテロ接合体であった。
【図6】in vivoで記録した横隔膜の自発性の電気活性を示すグラフである。(AおよびB)対照(A)およびnmdマウス(B)の典型となる記録。下方のトレースは未加工の記録であり、上方のトレースは下方のトレースの積分である。円が心臓からの電気活性を表示する。(C)PBS注射した突然変異体(黒色のバー)、およびMab2256処置した突然変異体マウスにおける吸息バーストの平均持続時間(TI)をmsで示すヒストグラム。(D)対照およびnmd横隔神経の組織断面。(E)対照とnmd同腹仔との有意差を示していない4匹の38週齢マウスにおける有髄軸索数の定量化。バー:7μm。
【技術分野】
【0001】
本出願は、その全体が参照により本明細書に援用される2005年4月26日出願の米国仮特許出願第60/675393号の優先権の利益を主張するものである。
【0002】
本発明は、下位運動ニューロン疾患の治療および/または予防におけるアゴニスト抗trkC抗体の使用に関する。
【背景技術】
【0003】
運動ニューロン疾患は、運動ニューロンが変性し、死に至る障害である。上位運動ニューロンおよび下位運動ニューロンを含む運動ニューロンは、随意筋に影響を及ぼし、随意筋を刺激して収縮させる。上位運動ニューロンは、大脳皮質に始まり、脳幹および脊髄を経由して線維を送り、下位運動ニューロンの制御に関与する。下位運動ニューロンは、脳幹および脊髄に位置し、線維を筋肉に送る。下位運動ニューロン疾患は、下位運動ニューロンの変性を含む疾患である。下位運動ニューロンが変性すると、通常は活性化する筋線維が連絡切断され、収縮せず、筋力低下および反射の減弱が引き起こされる。どちらの型のニューロンの損失も、脱力、すなわち筋萎縮(消耗)をもたらし、無痛性の脱力は、運動ニューロン疾患の臨床的な特徴である。
【0004】
SMARD1は、第2の一般的な常染色体劣性遺伝疾患である脊髄性筋萎縮症(SMA)の臨床的な変形形態であり、小児期における最も一般的な遺伝的死因である。SMAおよびSMARD1は、進行性の筋肉運動麻痺を随伴する下位運動ニューロンの変性を特徴とする。SMA症例の大多数が、生存運動神経細胞遺伝子(SMN)の突然変異によるものである(1)のに対し、SMARD1は、異なる遺伝子、すなわち免疫グロブリンμ−結合タンパク質2遺伝子(IGHMBP2)の突然変異によって引き起こされる(2〜5)。SMARD1患者は、横隔膜の関与により呼吸器機能の早期の機能障害を患う(4)。一部の予備的な臨床試験で肯定的な結果が出ているにもかかわらず(7)、現在、一般にSMARD1またはSMAの有効な療法は存在しない(6)。
【0005】
nmdマウスは、DNA/RNAヘリカーゼ/ATPaseタンパク質ファミリーの一員であるマウスIghmbp2遺伝子の自然突然変異を有する(8〜10)。この遺伝的欠陥は、イントロン4における単一の突然変異(A→G)からなるため、転写物の80%が異常にスプライスされ、20%が全長である(9)。マウスではIGHMBP2の機能発現が完全に消滅しないので、nmdマウスは、より軽度の形のヒトSMARD1に類似した疾患表現型を示す(9)。筋力低下は、出生から2週間後に発現し始め、四肢の重度の神経性筋萎縮に進行する(8〜11)。
【0006】
神経栄養因子は、運動神経疾患の有望な治療薬であると考えられている。この期待は、培養中の動物胚の運動ニューロンにおけるそれら分子の生存促進特性、すなわち軸索切断後の神経に対するそのプラスの生物学的効果、および神経変性疾患動物モデルにおける病理学的症状の軽減(12〜15)に基づいている。in vitroおよびin vivo研究のほとんどで得られる有望な結果を踏まえて、外来のニューロトロフィンが、アルツハイマー病、筋萎縮性側索硬化症(ALS)、末梢ニューロパチー、パーキン病、およびハンチントン病患者のための臨床試験で使用されている。
【発明の開示】
【発明が解決しようとする課題】
【0007】
神経栄養因子を適用する際の1つの実際的な難点は、これらのタンパク質がすべて、比較的短い半減期を有する一方で、神経変性疾患は慢性的であり、長期の治療が必要であることである。ニューロトロフィン受容体を標的とする治療用のアゴニスト抗体は、特異性が高く半減期が長いために、神経変性疾患のための新規な手法となり得る。
【0008】
特許出願および特許公開を含む、本明細書で引用するすべての参考文献は、その全体が参照により援用される。
【課題を解決するための手段】
【0009】
本発明は、個体における(下位運動ニューロン変性を含む)下位運動ニューロン疾患の治療方法を提供する。下位運動ニューロン疾患の例は、脊髄性筋萎縮症(SMA)および呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)である。この方法は、個体への有効量のアゴニスト抗trkC抗体の投与を含む。
【0010】
一態様では、本発明は、個体における下位運動ニューロン疾患の治療方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。別の態様では、本発明は、個体において下位運動ニューロン疾患に関連する症状の出現を遅らせる方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。別の態様では、本発明は、個体において下位運動ニューロン疾患の症状を緩和させる方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。
【0011】
一部の実施形態では、アゴニスト抗trkC抗体の投与後に個体の筋肉強度が向上する。一部の実施形態では、アゴニスト抗trkC抗体の投与後に個体の筋肉強度の低下を遅らせる。
【0012】
一部の実施形態では、個体は、ヒトなどの哺乳動物である。
【0013】
アゴニスト抗trkC抗体は、当業界で知られている。一部の実施形態では、アゴニスト抗trkC抗体は、モノクローナル抗体である。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトtrkCを結合する。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトtrkCを特異的に結合する。アゴニスト抗trkC抗体は、ヒトおよびげっ歯類trkCを結合するものでよい。アゴニスト抗trkC抗体は、(抗体6.4.1(PCT公開WO01/98361)などの)ヒト抗体でもよいし、または(ヒト化モノクローナル抗体2256を含む)ヒト化抗体でもよい。別の実施形態では、アゴニスト抗trkC抗体は、PCT WO2004/058190および本明細書に記載のヒト化抗体A5である。さらに他の実施形態では、抗trkCアゴニスト抗体は、表1に示す重鎖可変部アミノ酸配列(配列番号1)と表2に示す軽鎖可変部アミノ酸配列(配列番号2)とを含む。他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。CDRの同定は、多分に当分野の技能の範囲内である。一部の実施形態では、CDRはKabatCDRを含む。他の実施形態では、CDRはChothiaCDRである。さらに他の実施形態では、CDRは、KabatおよびChothiaの両方のCDRを含む。一部の実施形態では、抗体は、受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる軽鎖を含む。一部の実施形態では、抗体は、受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖を含む。一部の実施形態では、抗体は、(a)受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる軽鎖と、(b)受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖とを含む。一部の実施形態では、抗体は、(a)受託番号がATCC No.PTA−5682であるベクター中のポリヌクレオチドによってコードされる1個または複数のCDR、および/または(b)受託番号がATCC No.PTA−5683であるベクター中のポリヌクレオチドによってコードされる重鎖を含む。
【0014】
抗体は、次のもの、すなわち6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256のいずれか1種または複数から選択される抗体と本質的に同じtrkCエピトープを結合するものでも、またはそうした抗体と結合をめぐって競合するものでもよい。PCT公開番号WO01/98361を参照されたい。抗体は、免疫学的に不活性である、例えば補体によって媒介される溶解のきっかけとならず、または抗体依存性の細胞によって媒介される細胞傷害性(ADCC)を刺激しない定常部などの、改変された定常部を含むものでもよい。他の実施形態では、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ、PCT出願番号PCT/GB99/01441、および/または英国特許出願第9809951.8号に記載の通りに定常部を改変する。
【0015】
抗体は、次のもの、すなわち、Fab、Fab’、F(ab’)2、Fv断片、ダイアボディ、抗体断片から生成される単鎖抗体分子および多特異性抗体、ならびに単鎖Fv(scFv)分子のうちの1種または複数から選択される抗体断片などの抗体断片でもよい。抗体はキメラでもよく、二重特異性でもよい。
【0016】
アゴニスト抗trkC抗体の投与は、次の手段、すなわち、静脈内、皮下、吸入、動脈内、筋肉内、心臓内、脳室内、くも膜下腔内、脊髄内、および腹腔内のうちの1種または複数を含めて、当業界で知られている適切な任意の方法でよい。投与は、全身(例えば静脈内)および/または局所でよい。投与は、短時間および/または長期間でよい。投与は、下位運動ニューロン疾患の発症前に行うことができる。
【0017】
別の態様では、本発明は、本発明の方法のいずれかで使用するための、アゴニスト抗trkC抗体を含む組成物およびキットを提供する。これらのキットは、アゴニスト抗trkC抗体を本明細書に記載の方法のいずれかで使用するための説明書をさらに含むこともある。本発明は、本明細書に記載の方法のいずれかで使用するための、アゴニスト抗trkC抗体と薬学的に許容できる担体とを含む医薬組成物も提供する。
【0018】
本発明はまた、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べた組成物およびキットのいずれかを提供する。
【発明を実施するための最良の形態】
【0019】
本発明は、個体における下位運動ニューロン疾患の治療方法であって、その個体への有効量のアゴニスト抗trkC抗体の投与を含む方法を提供する。下位運動ニューロン疾患の例には、SMA、およびSMAの臨床的な変形形態であるSMARD1が含まれる。
【0020】
一般技術
本発明の実践では、別段の指定がない限り、当分野の技能の範囲内にある(組換え技術を含む)分子生物学、微生物学、細胞生物学、生化学、および免疫学の従来の技術が使用される。このような技術は、「Molecular Cloning:A Laboratory Manual」、第2版(Sambrookら、1989年)、Cold Spring Harbor Press;「Oligonucleotide Synthesis」(M.J.Gait編、1984年);「Methods in Molecular Biology」、Humana Press;「Cell Biology:A Laboratory Notebook」(J.E.Cellis編、1998年)、Academic Press;「Animal Cell Culture」(R.I.Freshney編、1987年);「Introduction to Cell and Tissue Culture」(J.P.MatherおよびP.E.Roberts、1998年)、Plenum Press;「Cell and Tissue Culture:Laboratory Procedures」(A.Doyle、J.B.Griffiths、およびD.G.Newell編、1993〜8年)、J.Wiley and Sons;「Methods in Enzymology」(Academic Press,Inc.);「Handbook of Experimental Immunology」(D.M.WeirおよびC.C.Blackwell編);「Gene Transfer Vectors for Mammalian Cells」(J.M.MillerおよびM.P.Calos編、1987年);「Current Protocols in Molecular Biology」(F.M.Ausubelら編、1987年);「PCR:The Polymerase Chain Reaction」(Mullisら編、1994年);「Current Protocols in Immunology」(J.E.Coliganら編、1991年);「Short Protocols in Molecular Biology」(Wiley and Sons、1999年);「Immunobiology」(C.A.JanewayおよびP.Travers、1997年);「Antibodies」(P.Finch、1997);「Antibodies:a practical approach」(D.Catty編、IRL Press、1988〜1989年);「Monoclonal antibodies:a practical approach」(P.ShepherdおよびC.Dean編、Oxford University Press、2000年);「Using antibodies:a laboratory manual」(E.HarlowおよびD.Lane(Cold Spring Harbor Laboratory Press、1999年);「The Antibodies」(M.ZanettiおよびJ.D.Capra編、Harwood Academic Publishers、1995年);および「Cancer:Principles and Practice of Oncology」(V.T.DeVitaら編、J.B.Lippincott Company、1993年)などの文献で十分に説明されている。
【0021】
定義
(複数形でも区別なく使用される)「抗体」とは、炭水化物、ポリヌクレオチド、脂質、ポリペプチドなどの標的に、免疫グロブリン分子の可変部に位置する少なくとも1個の抗原認識部位を介して特異的に結合することのできる免疫グロブリン分子である。本明細書では、この用語は、無処置のポリクローナルまたはモノクローナル抗体だけでなく、(Fab、Fab’、F(ab’)2、Fvなどの)これらの断片、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ヒト化抗体、キメラ抗体、ダイアボディ、線形抗体、単鎖抗体、多特異性抗体(例えば二重特異性抗体)、および必要な特異性の抗原認識部位を含む他の任意の改変された形状の免疫グロブリン分子も含む。抗体には、IgG、IgA、IgMなどのどんなクラスの抗体も含まれ、抗体は、任意の特定のクラスのものである必要はない。その重鎖の定常ドメインの抗体アミノ酸配列に応じて、免疫グロブリンを異なるクラスに割り振ることができる。免疫グロブリンの5つの主要なクラスは、IgA、IgG、IgD、IgE、およびIgMであり、これらのいくつかは、さらにサブクラス(アイソタイプ)、例えば、IgG1、IgG2、IgG3、IgG4、IgA1、およびIgA2に分けられる場合もある。異なるクラスの免疫グロブリンに対応する重鎖定常ドメインは、それぞれα、γ、δ、ε、およびμと呼ばれる。軽鎖にも、κおよびλと呼ばれる2つのクラスがある。異なるクラスの免疫グロブリンのサブユニット構造および三次元立体配置はよく知られている。
【0022】
「モノクローナル抗体」とは、抗原の選択的な結合に関与するアミノ酸(自然に存在するものおよび自然に存在しないもの)からなる同種の抗体集団を指す。モノクローナル抗体の集団は、単一の抗原部位に向けられ、高度に特異的である。用語「モノクローナル抗体」は、無処置のモノクローナル抗体および全長モノクローナル抗体だけでなく、(Fab、Fab’、F(ab’)2、Fvなどの)これらの断片、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ヒト化モノクローナル抗体、キメラモノクローナル抗体、ならびに必要な特異性および抗原結合能の抗原認識部位を含む他の任意の改変された形状の免疫グロブリン分子も含む。抗体の供給源または(例えば、ハイブリドーマ、ファージ選択、組換え発現、トランスジェニック動物などによる)その製法については、限定されないものとする。
【0023】
「ヒト化」抗体とは、実質的にヒトでない種からの免疫グロブリンに由来する抗原結合部位を有し、分子の残りの免疫グロブリン構造がヒト免疫グロブリンの構造および/または配列に基づく分子を指す。抗原結合部位は、定常ドメインに融合した完全な可変ドメインを含んでもよいし、または可変ドメイン中の適切なフレームワーク領域に接ぎ合わされた相補性決定領域(CDR)のみを含んでもよい。抗原結合部位は、野生型でも、または1つまたは複数のアミノ酸置換によって改変されたもの、例えばヒト免疫グロブリンにより近似するように改変されたものでもよい。ヒト化抗体の一部の形態は、すべてのCDR配列を保存している(例えば、マウス抗体からの6個のCDRすべてを含むヒト化マウス抗体)。他の形態のヒト化抗体は、もとの抗体に関して変更されている1個または複数のCDR(1、2、3、4、5、6個)を有する。
【0024】
本明細書では、「ヒト抗体」とは、ヒトによって産生される抗体に相当するアミノ酸配列を有する抗体を意味し、かつ/または当業界で知られ、または本明細書で開示するヒト抗体の製造技術のいずれかを使用して製造されている。このヒト抗体の定義には、少なくとも1本のヒト重鎖ポリペプチドまたは少なくとも1本のヒト軽鎖ポリペプチドを含む抗体が含まれる。そのような1つの例は、ネズミ軽鎖ポリペプチドとヒト重鎖ポリペプチドとを含む抗体である。ヒト抗体は、当業界で知られている様々な技術を使用して生成することができる。一実施形態では、ヒト抗体は、ヒト抗体を発現させるファージライブラリーから選択される(Vaughanら、1996年、Nature Biotechnology、第14巻:309〜314ページ;Sheetsら、1998年、PNAS(米)、第95巻:6157〜6162ページ;HoogenboomおよびWinter、1991年、J.Mol.Biol.、227〜381ページ;Marksら、1991年、J.Mol.Biol.、第222巻:581ページ)。ヒト抗体は、ヒト免疫グロブリン座を、内在性の免疫グロブリン遺伝子が部分的または完全に不活化されているトランスジェニック動物、例えばマウスに導入して作ることもできる。この手法は、米国特許第5545807号、同第5545806号、同第5569825号、同第5625126号、同第5633425号、および同第5661016号に記載されている。別法として、ヒト抗体は、標的抗原に向けられる抗体を産生するヒトBリンパ球を不死化して調製することができる(そのようなBリンパ球は、個体から回収してもよいし、またはin vitroで免疫感作したものでもよい)。例えば、Coleら、「Monoclonal Antibodies and Cancer Therapy」、Alan R.Liss、77ページ(1985年);Boernerら、1991年、J.Immunol.、第147巻(1):86〜95ページ;および米国特許第5750373号を参照されたい。
【0025】
「キメラ抗体」とは、重鎖および軽鎖のアミノ酸配列それぞれの一部分が、ある特定の種に由来し、またはある特定のクラスに属する抗体中の対応する配列と相同であり、鎖の残りの区域が別の抗体中の対応する配列と相同である抗体を指す。通常、これらのキメラ抗体では、軽鎖および重鎖の両方の可変部が、哺乳動物のある種に由来する抗体の可変部を模倣し、定常部分は、別のものに由来する抗体中の配列と相同である。そのようなキメラ形態の1つの明白な利点は、例えば、非ヒト宿主生物から容易に入手可能なハイブリドーマまたはB細胞を使用して、現在知られている供給源から、例えばヒト細胞調製物由来の定常部と組み合わせて、可変部を好都合に得ることができる点である。可変部は、調製が容易であるという利点を有し、特異性はその供給源によって影響を受けないが、ヒトである定常部は、抗体が注射されたときに、ヒトでない供給源からの定常部よりもヒト対象からの免疫応答を誘発しにくい。しかし、この定義は、この特定の例に限定されない。
【0026】
抗体またはポリペプチドに「特異的に結合する」または「優先的に結合する」(本明細書では区別なく使用する)エピトープとは、当業界でよく理解されている用語であり、そのような特異的または優先的な結合を測定する方法も当業界でよく知られている。分子は、ある特定の細胞または物質と、別の細胞または物質より頻繁に、より迅速に、より長期間、かつ/またはより強い親和性で反応または結合するならば、「特異的結合」または「優先的な結合」を示すと言われる。抗体は、他の物質に結合するよりも強い親和性、すなわち結合力で、より容易に、かつ/またはより長い期間で結合するならば、標的に「特異的に結合する」または「優先的に結合する」。例えば、trkCエピトープに特異的または優先的に結合する抗体とは、他のtrkCエピトープまたは非trkCエピトープに結合するのより強い親和性、すなわち結合力で、より容易に、かつ/またはより長い期間、このtrkCエピトープを結合する抗体である。例えば、第1の標的に特異的または優先的に結合する抗体(または部分もしくはエピトープ)が、第2の標的に特異的または優先的に結合してもしなくてもよいことも、この定義を読むことで理解される。「特異的結合」または「優先的な結合」は、それ自体として、必ずしも独占的な結合を(含むことがあるとしても)必要としない。必ずしもそうでないが、一般に、結合への言及は優先的な結合を意味する。
【0027】
「機能Fc領域」は、未変性配列Fc領域の少なくとも1種のエフェクター機能を有する。好例となる「エフェクター機能」には、C1q結合、補体依存性細胞傷害性(CDC)、Fc受容体結合、抗体依存性細胞傷害(ADCC)、食作用、細胞表面受容体(例えばB細胞受容体、BCR)の下方調節などが含まれる。そのようなエフェクター機能には、一般に、Fc領域が結合ドメイン(例えば、抗体可変ドメイン)と合体することが必要であり、エフェクター機能は、そのような抗体エフェクター機能を評価するための当業界で知られている様々なアッセイを使用して評価することができる。
【0028】
「未変性配列Fc領域」は、自然界で見られるFc領域のアミノ酸配列と同一のアミノ酸配列を含む。「変異体Fc領域」は、少なくとも1個のアミノ酸の変更によって未変性配列Fc領域と異なるが、未変性配列Fc領域の少なくとも1種のエフェクター機能を保持するアミノ酸配列を含む。変異体Fc領域は、未変性配列Fc領域または親ポリペプチドFc領域と比べて少なくとも1つのアミノ酸置換を有することが好ましく、例えば、未変性配列Fc領域または親ポリペプチドFc領域中に約1〜約10個のアミノ酸置換、好ましくは約1〜約5個のアミノ酸置換を有する。変異体Fc領域は、本明細書では、未変性配列Fc領域および/または親ポリペプチドFc領域と、少なくとも約80%の配列同一性、最も好ましくは少なくとも約90%の配列同一性、より好ましくは少なくとも約95%の配列同一性を有することが好ましい。
【0029】
本明細書では、「抗体依存性細胞傷害」および「ADCC」とは、Fc受容体(FcR)を発現する非特異的な細胞傷害性細胞(例えば、ナチュラルキラー(NK)細胞、好中球、およびマクロファージ)が、標的細胞上の結合した抗体を認識し、その後標的細胞の溶解を引き起こす細胞性の反応を指す。対象とする分子のADCC活性は、米国特許第5500362号または同第5821337号に記載のものなどのin vitroADCCアッセイを使用して評価することができる。そのようなアッセイに有用なエフェクター細胞には、末梢血単核球(PBMC)およびナチュラルキラー細胞が含まれる。別法としてまたは追加として、対象とする分子のADCC活性は、in vivoで、例えばClynesら、1998年、PNAS(USA)、第95巻:652〜656で開示されているものなどの動物モデルで評価することもできる。
【0030】
「アゴニスト抗trkC抗体」(区別なく「抗trkCアゴニスト抗体」とも呼ぶ)とは、trkC受容体を結合し、および/またはtrkCシグナル伝達機能によって媒介される下流の経路を活性化することのできる抗体を指す。例えば、アゴニスト抗体は、trkC受容体の細胞外ドメインに結合し、それによって受容体の二量体化を引き起こし、細胞内の触媒キナーゼドメインの活性化をもたらすことができる。したがって、アゴニスト抗体は、in vitroおよび/またはin vivoで受容体を発現する細胞の増殖および/または分化を刺激し得る。一部の実施形態では、アゴニスト抗trkC抗体は、trkCに結合し、trkC生物活性を活性化する。一部の実施形態では、本発明の方法で有用なアゴニスト抗体は、trkCのドメインVおよび/またはドメインIVを認識する。Urferら、J.Biol.Chem.第273巻:5829〜5840ページ(1998年)を参照されたい。
【0031】
抗体の「可変部」とは、単独または複合語で、抗体軽鎖の可変部または抗体重鎖の可変部を指す。重鎖および軽鎖の可変部はそれぞれ、超可変領域としても知られている3つの相補性決定領域(CDR)によって連結された4つのフレームワーク領域(FR)からなる。それぞれの鎖のCDRは、FRによってきわめて接近してまとまり、他の鎖のCDRと共に、抗体の抗原結合性部位の形成に寄与する。CDRを決定する技術は、少なくとも2種存在する。すなわち(1)異種間の配列可変性に基づく手法(すなわち、Kabatら、「Sequences of Proteins of Immunological Interest」(第5版、1991年、National Institutes of Health、米メリーランド州ベセズダ))、および(2)抗原−抗体複合体の結晶学的研究に基づく手法(Al−lazikaniら(1997年)、J.Molec.Biol.第273巻:927〜948ページ))。本明細書では、CDRとは、どちらかの手法または両方の手法の組合せによって規定されるCDRを指す場合もある。
【0032】
抗体の「定常部」とは、単独または複合語で抗体軽鎖の定常部または抗体重鎖の定常部を指す。
【0033】
本明細書では、「Fc受容体」および「FcR」とは、抗体のFc領域に結合する受容体について述べるものである。好ましいFcRは、未変性配列ヒトFcRである。さらに、好ましいFcRは、IgG抗体(γ受容体)を結合するものであり、これには、そのアレル変異体および選択的にスプライスされた形態を含めてFcγRI、FcγRII、およびFcγRIIIサブクラスの受容体が含まれる。FcγRII受容体には、主にその細胞質ドメインが異なる同様のアミノ酸配列を有するFcγRIIA(「活性化する受容体」)およびFcγRIIB(「抑制する受容体」)が含まれる。FcRは、RavetchおよびKinet、1991年、Ann.Rev.Immunol.、第9巻:457〜92ページ;Capelら、1994年、Immunomethods、第4巻:25〜34ページ;およびde Haasら、1995年、J.Lab.Clin.Med.、第126巻:330〜41ページで総説されている。「FcR」には、母親のIgGの胎仔への移動を司る新生児受容体、すなわちFcRnも含まれる(Guyerら、1976年、J.Immunol.、第117巻:587ページ;およびKimら、1994年、J.Immunol.、第24巻:249ページ)。
【0034】
「補体依存性細胞傷害性」および「CDC」とは、補体の存在下で標的を溶解させることを指す。補体活性化経路は、補体系(C1q)の第1の構成成分が、同族抗原と複合体を形成した分子(例えば抗体)に結合することで開始される。補体活性化を評価するために、例えば、Gazzano−Santoroら、J.Immunol.Methods、第202巻:163ページ(1996年)に記載されているようにCDCアッセイを実施することができる。
【0035】
本明細書では、「親和性成熟」抗体とは、その1個または複数のCDR中に、抗体の抗原に対する親和性を、変更をもたない親抗体と比べて向上させる1つまたは複数の変更を有する抗体を意味する。一部の実施形態では、親和性成熟抗体は、標的抗原に対してナノモルまたはさらにはピコモルの親和性を有する。親和性成熟抗体は、当業界で知られている手順によって生成される(Marksら、1992年、Bio/Technology、第10巻:779〜783ページ;Barbasら、1994年、Proc Nat.Acad.Sci、USA第91巻:3809〜3813ページ;Schierら、1995年、Gene、第169巻:147〜155ページ;Yeltonら、1995年、J.Immunol.、第155巻:1994〜2004ページ;Jacksonら、1995年、J.Immunol.、第154巻(7):3310〜9ページ;Hawkinsら、1992年、J.Mol.Biol.、第226巻:889〜896ページ)。
【0036】
本明細書では、「trkC」とは、チロシンキナーゼスーパーファミリーの一員であるtrkC受容体ポリペプチドを指す。trkCは、その限りでないが、ヒト、イヌ、ネコ、ウシ、ウマ、霊長類、ならびに(マウスおよびラットを含む)げっ歯類を含めて、任意の哺乳類種の未変性trkC受容体を含む。全長未変性trkCの細胞外ドメインは、様々な他のタンパク質で同定されている相同またはさもなければ同様の構造に即して規定されている。そのドメインは、成熟trkC受容体のN末端から始めて、1)アミノ酸1からアミノ酸48まで伸びる第1のシステインリッチドメイン、2)アミノ酸49からアミノ酸120まで伸びるロイシンリッチドメイン、3)アミノ酸121からアミノ酸177まで伸びる第2のシステインリッチドメイン、4)約アミノ酸196からアミノ酸257まで伸びる第1の免疫グロブリン様ドメイン、および5)約アミノ酸288からアミノ酸351まで伸びる第2の免疫グロブリン様ドメインと呼ばれている。例えば、PCT公開第WO0198361号を参照されたい。ヒトtrkC受容体のドメイン構造は、結晶構造に即して次のようにも呼ばれている。すなわち、アミノ酸1〜アミノ酸47のドメイン1、アミノ酸48〜アミノ酸130のドメイン2、アミノ酸131〜アミノ酸177のドメイン3、アミノ酸178〜アミノ酸165のドメイン4、およびアミノ酸166〜アミノ酸381のドメイン5。例えば、PCT公開WO0198361;Urferら、J.Biol.Chem.第273巻:5829〜5840ページ(1998年)を参照されたい。trkCの変異体も含まれ、その例には、その限りでないが、キナーゼドメインを含まない変異体(Sheltonら.、J.Neurosci.第15巻(1):477〜491ページ(1995年))および改変されたキナーゼドメインを含む変異体(Sheltonら.、J.Neurosci.第15巻(1):477〜491ページ(1995年))が含まれる。
【0037】
「生物活性」とは、本発明のアゴニスト抗trkC抗体と共に使用するとき、一般にtrkC受容体チロシンキナーゼを結合し、および/またはtrkCシグナル伝達機能によって媒介される下流の経路を活性化する能力を有することを指す。本明細書では、「生物活性」は、trkCの未変性リガンドであるNT−3のtrkC発現細胞に対する作用によって誘発されるものと同じような、1種または複数のエフェクター機能を含む。trkCの「生物活性」は、下流のシグナル伝達経路、またはNT−3の作用によって誘発されるものと異なるエフェクター機能も含み得る。限定するものではないが、生物活性には、次のもの、すなわち、trkCを結合し活性化する能力;trkC受容体の二量体化を促進する能力;(損傷を受けた細胞を含む)細胞、特に、末梢(交感神経系、感覚、および腸管)ニューロンおよび中枢(脳および脊髄)ニューロンを含むin vitroまたはin vivoでのニューロン、ならびにニューロンでない細胞、例えば末梢血白血球の発生、生存、機能、維持、および/または再生を促進する能力のいずれか1種または複数が含まれる。特に好ましい生物活性は、下位運動ニューロン疾患の1つまたは複数の症状を(予防を含めて)治療し、および/または下位運動ニューロンの機能を修復および/もしくは改善する能力である。
【0038】
本明細書では、「治療」とは、有益なまたは所望の臨床的な成果を得るための手法である。本発明の目的では、有益なまたは所望の臨床的な成果には、その限りでないが、次のもの、すなわち、下位運動ニューロン疾患に関連する1種または複数の症状の軽減(例えば、下位運動ニューロンの変性、進行性の筋力低下または運動麻痺、呼吸不全、および低下する反射);下位運動ニューロン疾患の進行を遅らせまたは緩慢にすること;下位運動ニューロンの変性を安定化する(すなわち、悪化させない)こと;下位運動ニューロンの変性および/または筋肉の脱力の回復;車いす使用の必要を遅らせること;人工呼吸の必要を遅らせること;ならびに最終的に全体としての生存時間を延長することのうちの1種または複数が含まれる。
【0039】
下位運動ニューロン疾患または下位運動ニューロン疾患の1種または複数の症状を「軽減する」とは、本発明によるアゴニスト抗trkC抗体による治療を受ける個体または個体集団において、疾患の望ましくない臨床病態の程度および/または時間経過を減らすことを意味する。
【0040】
下位運動ニューロン疾患の「症状の重症度を軽減する」または下位運動ニューロン疾患の「症状を緩和させる」とは、下位運動ニューロン疾患の1つまたは複数の症状を和らげ、かつ/またはアゴニスト抗trkC抗体を投与しないものと比べて改善させることを意味する。「重症度を低減する」には、症状の期間の短縮または縮小も含まれる。
【0041】
本明細書では、下位運動ニューロン疾患の出現を「遅らせる」とは、この疾患の出現を先送りし、妨げ、緩慢にし、遅延させ、安定化し、かつ/または延期することを意味する。この遅れは、治療対象となる疾患および/または個体の経歴に応じて様々な長さの時間となり得る。当業者には明らかなように、十分または有意な遅れは、個体が下位運動ニューロンの変性ならびに筋力低下または運動麻痺にかからないという点で、事実上予防を含む。下位運動ニューロン疾患の出現を「遅らせる」方法は、その方法を使用しないのと比べて、所与の時間枠で下位運動ニューロンの変性が出現する確率を低下させ、かつ/または所与の時間枠で下位運動ニューロンの変性の程度を低減する方法である。そのような比較は、通常は、統計学的に有意な数の対象を使用する臨床的な調査に基づく。
【0042】
下位運動ニューロン疾患の「出現」とは、個体内での下位運動ニューロン疾患に関連する症状(例えば、下位運動ニューロン機能の低下ならびに筋力低下および運動麻痺)の発現および/または進行を意味する。出現は、本明細書に記載の標準の臨床的な技術を使用して検出可能となり得る。しかし、出現は、最初に検出不可能な場合もある疾患の進行をも指す。本発明の目的では、進行とは、この場合では、標準の神経学的検査または患者の問診によって決定され、あるいはより専門の量的試験によって決定される場合もある、疾患状態の生物学的な経過を指す。これらのより専門の量的試験には、その限りでないが、微小神経電図検査による冒されたニューロンの伝導速度の測定、専門の平衡性試験、反射試験、専門の固有受容および/または運動感覚の試験、力の試験(例えば、その限りでないが量的な筋力測定、筋電図検査、MRIを含む、筋力の臨床検査)、その限りでないが血圧コントロール試験、様々な生理的および薬理学的な刺激に対する心拍応答試験を含む自律神経機能の試験を含めることができる。例として、SMAでは、試験には、その限りでないが量的な筋力測定、筋電図検査、および/またはMRIを含む筋力の臨床検査などの、運動技能および/または運動力の試験を含めることができる。「出現」には、発生、再発、および発症が含まれる。本明細書では、下位運動ニューロン疾患の「発症」または「発生」には、最初の発症および/または再発が含まれる。
【0043】
本明細書では、「リスクのある」個体とは、下位運動ニューロン疾患が出現するリスクのある個体である。「リスクのある」個体は、検出可能な疾患にかかっていてもいなくてもよく、本明細書に記載の治療方法の前に検出可能な疾患を呈していてもいなくてもよい。「リスクのある」とは、個体が、生存運動神経細胞遺伝子(SMN)または免疫グロブリンμ結合タンパク質2遺伝子(IGHMBP2)の突然変異などの疾患の出現と相互に関係がある測定可能なパラメータである、1種または複数のいわゆるリスクファクターを有することを意味する。これらのリスクファクターの1つまたは複数を有する個体は、これらのリスクファクターなしの個体よりも下位運動ニューロン疾患にかかる確率が高い。
【0044】
「有効量」とは、疾患の発症を遅らせる臨床的な成果を含む、有益なまたは所望の臨床的な成果をもたらすのに十分な量である。有効量は、1回または複数の投与で投与することができる。本発明の目的では、本明細書に記載のアゴニスト抗trkC抗体の有効量は、下位運動ニューロン疾患を緩和させ、安定化し、逆戻りさせ、その進行を緩慢にしかつ/または遅らせ、あるいは予防するのに十分な量である。臨床的な状況の中で理解されるように、薬物、化合物、または医薬組成物の有効量は、別の薬物、化合物、または医薬組成物と共に実現されることもあれば、そうでないこともある。したがって、「有効量」は、1種または複数の治療薬を投与するという状況で考えることができ、1種または複数の他の薬剤と共に望ましい成果が実現される場合がありまたは実現されるならば、単剤が有効量で与えられたと考えることができる。
【0045】
本明細書では、「共に」投与は、同時の投与および/または異なる時点での投与を含む。共に投与は、合剤(co−formulation)としての投与または別個の組成物としての投与も含む。本明細書では、共投与は、アゴニスト抗trkC抗体および別の下位運動ニューロン疾患治療薬またはこれらの組合せが個体に投与され、それが同時および/または別々に起こり得る任意の状況を含む意味である。本明細書でさらに論述するように、アゴニスト抗trkC抗体と他の治療薬を異なる投薬回数または投薬間隔で投与できることは理解される。アゴニスト抗trkC抗体と他の治療薬を、同じ投与経路または異なる投与経路を使用して投与できることも理解される。
【0046】
「個体」は、哺乳動物、より好ましくはヒトである。哺乳動物には、その限りでないが、飼養動物、競技動物、愛玩動物、霊長類、ウマ、乳牛、ネコ、イヌ、および(マウスやラットなどの)げっ歯類も含まれる。
【0047】
本明細書では、単数形の「a」、「an」、および「the」の形は、別段の指摘がない限り、複数形の言及を含む。例えば、「an」antibodyは1種または複数の抗体を含み、「a symptom」は1種または複数の症状を意味する。
【0048】
本明細書では、「ベクター」とは、宿主細胞中で1種または複数の対象とする遺伝子または配列を放出する、好ましくは発現させることのできる構築物を意味する。ベクターの例には、その限りでないが、ウイルスベクター、裸DNAまたはRNA発現ベクター、プラスミド、コスミドまたはファージベクター、カチオン縮合剤と結合したDNAまたはRNA発現ベクター、リポソーム中にカプセル封入されたDNAまたはRNA発現ベクター、ならびに産生細胞などのある種の真核細胞が含まれる。
【0049】
本明細書では、「発現制御配列」とは、核酸の転写を指示する核酸配列を意味する。発現制御配列は、構成的もしくは誘導型プロモーターなどのプロモーター、またはエンハンサーでよい。発現制御配列は、転写される核酸配列に作動可能に連結される。
【0050】
本明細書では、「核酸」または「ポリヌクレオチド」とは、一本鎖または二本鎖の形のデオキシリボヌクレオチドまたはリボヌクレオチドポリマーを指し、別段の制限がない限り、自然に存在するヌクレオチドと同様にして核酸とハイブリッド形成する天然ヌクレオチドの既知の類似体を含む。
【0051】
本明細書では、「薬学的に許容できる担体」には、活性成分と組み合わせたとき、活性成分が生物活性を保持するのを可能にし、対象の免疫系と反応性でない任意の材料が含まれる。例には、その限りでないが、リン酸緩衝溶液、水、水中油型乳濁液などの乳濁液、および様々な種類の湿潤剤などの標準の医薬担体のいずれかが含まれる。エアロゾルまたは非経口投与用の好ましい希釈剤は、リン酸緩衝溶液または通常の(0.9%)生理食塩水である。
【0052】
そのような担体を含む組成物は、よく知られている従来の方法によって製剤される(例えば、「Remington’s Pharmaceutical Sciences」、第18版、A.Gennaro編、Mack Publishing Co.、米ペンシルヴェニア州イーストン、1990年;およびRemington、「The Science and Practice of Pharmacy」、第20版、Mack Publishing、2000年を参照されたい)。
【0053】
本発明の方法
本明細書に記載のすべての方法に関して、アゴニスト抗trkC抗体への言及は、これらの抗体の1種または複数を含む組成物も含む。これらの組成物は、緩衝剤を含めて薬学的に許容できる賦形剤などの、当業界でよく知られている適切な賦形剤をさらに含んでもよい。
【0054】
アゴニスト抗trkC抗体を使用して下位運動ニューロン疾患を治療する方法
本発明は、アゴニスト抗trkC抗体を使用して、下位運動ニューロン疾患を治療し、予防し、その症状の出現を遅らせ、かつ/または軽減する方法を含む。この方法は、その必要のある個体にこれらの抗体を有効量投与するものである(様々な適応症および態様は本明細書に記載する)。有効量のアゴニスト抗trkC抗体は、他の治療薬と共に投与しても、またはそれなしで投与してもよい。個体は、下位運動ニューロン疾患と診断されているものでもよいし、下位運動ニューロン疾患の出現にかかるリスクのあるものでもよい。一部の実施形態では、個体はヒトである。しかし、記載する方法は、獣医学の状況(例えば、イヌ、ネコ、ウシ、ウマなどのヒトでない哺乳動物)にも適用できる。
【0055】
SMAやSMARD1などの下位運動ニューロン疾患を評価する方法およびその治療は、当業界で知られており、本明細書に記載される。
【0056】
アゴニスト抗trkC抗体
本発明の方法は、trkCを活性化する方法でtrkCと相互作用する抗trkC抗体を使用するものである。抗trkCアゴニスト抗体は、以下の特性のいずれか1つまたは複数を示す。すなわち、(a)trkC受容体に結合する、(b)trkC受容体の1つまたは複数のエピトープに結合する、(c)trkC受容体に結合し、trkC生物活性、またはtrkCシグナル伝達機能によって媒介される1つまたは複数の下流経路を活性化する、(d)trkC受容体二量体化を促進し、(e)trkC受容体の活性化を増大させ、(f)好ましい薬物動態および生体利用度特性を示し、(g)細胞の発生、生存、機能、維持、および/または再生を促進する、(h)trkC受容体に結合し、下位運動ニューロン疾患の1つまたは複数の症状を治療し、予防し、逆戻りさせ、または緩和させる。
【0057】
アゴニスト抗trkC抗体は、当業界で知られている。PCT公開WO01/98361;Urferら、J.Biol.Chem.(1998年)第273巻:5829〜5840ページを参照されたい。一部の実施形態では、抗trkCアゴニスト抗体は、抗体「A5」と呼ばれるヒト化マウス抗trkCアゴニスト抗体であり、A330P331→S330S331の突然変異(アミノ酸の番号付けは野生型IgG2a配列に即したKabatの番号付けに基づく;Eur.J.Immunol.(1999年)第29巻:2613〜2624ページを参照されたい)を含むヒト重鎖IgG2a定常部;ヒト軽鎖κ定常部;ならびに表1および2に示す重鎖および軽鎖可変部を含む。
【0058】
表1:A5重鎖可変部。ChothiaCDRは下線を付したイタリック体として示し、KabatCDRは太字として示す。
QVQLVQSGAEVKKPGASVKVSCKASGYTFTSYRIHWVRQAPGQGLEWMGEIYPSNARTNYNEKFKSRVTMTRDTSTSTVYMELSSLRSEDTAVYYCARKYYYGNTRRSWYFDVWGQGTTVTVS(配列番号1)
【0059】
表2:A5軽鎖可変部。KabatCDRは下線を付したイタリック体として示し、ChothiaCDRは太字として示す。
【0060】
DIQMTQSPSSLSASVGDRVTITCRASESIDNYGISFLAWYQQKPGKAPKLLIYAASNRGSGVPSRFSGSGSGTDFTFTISSLQPEDIATYYCQQSKTVPRTFGQGTKLEIKRT(配列番号2)
【0061】
A5の重鎖可変部または軽鎖可変部をコードする以下のポリヌクレオチドは、アメリカ培養細胞系統保存機関、10801 University Boulevard、Manassas、Virginia、USA(ATCC)に寄託した。
【0062】
【表1】
【0063】
ベクターEb.pur.2256.A5は、A5軽鎖可変部をコードするポリヌクレオチドであり、ベクターDb.2256.A5は、A5重鎖可変部をコードするポリヌクレオチドである。
【0064】
他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。CDR領域の決定は、多分に当分野の技能の範囲内である。CDRを決定するいくつかの技術は次の通りである。(1)異種間の配列可変性に基づく手法(すなわち、Kabatら、「Sequences of Proteins of Immunological Interest」(第5版、1991年、National Institutes of Health、米メリーランド州ベセズダ))、(2)抗原−抗体複合体の結晶学的研究に基づく手法(Al−lazikaniら(1997年)、J.Molec.Biol.第273巻:927〜948ページ))。CDRの同定は、多分に当分野の技能の範囲内である。一部の実施形態では、CDRはKabatCDRを含む。他の実施形態では、CDRはChothiaCDRである。さらに他の実施形態では、CDRは、KabatおよびChothia両方のCDRを含む。
【0065】
抗体は、モノクローナル抗体、ポリクローナル抗体、抗体断片(例えば、Fab、Fab’、F(ab’)2、Fv、Fcなど)、キメラ抗体、単鎖(ScFv)、これらの突然変異体、抗体部分を含む融合タンパク質、ならびに必要な特異性の抗原認識部位を含む他の改変された形状の免疫グロブリン分子を含み得る。抗体は、(ヒト化抗体を含めて)ネズミ、ラット、ヒト、または他の任意の起源のものでよい。したがって、アゴニスト抗trkC抗体は、(抗体6.4.1(PCT公開WO01/98361)などの)ヒト抗体でもよいし、または(ヒト化モノクローナル抗体A5を含む)ヒト化抗体でもよい。
【0066】
アゴニスト抗trkC抗体は、ヒトtrkCを結合するものでよい。アゴニスト抗trkC抗体は、ヒトおよびげっ歯類のtrkCを結合するものでもよい。一部の実施形態では、アゴニスト抗trkC抗体は、ヒトおよびラットのtrkCを結合するものでよい。一部の実施形態では、抗trkC抗体は、ヒトおよびマウスのtrkCを結合するものでよい。一実施形態では、抗体は、ヒトtrkC細胞外ドメイン上の1つまたは複数のエピトープを認識する抗体である。別の実施形態では、抗体は、ヒトtrkC細胞外ドメイン上の1つまたは複数のエピトープを認識するマウスまたはラット抗体である。一部の実施形態では、抗体はヒトtrkCを結合し、別の哺乳類種(一部の実施形態では脊椎動物種)からのtrkCはほとんど結合しない。一部の実施形態では、抗体は、ヒトtrkCならびに別の哺乳類種(一部の実施形態では脊椎動物種)からの1つまたは複数のtrkCを結合する。別の実施形態では、抗体は、霊長類、イヌ、ネコ、ウマ、およびウシのもののうちの1種または複数から選択されるtrkC上の1つまたは複数のエピトープを認識する。一部の実施形態では、抗体は、trkCを結合し、(同類のニューロトロフィン受容体であるtrkAおよび/またはtrkBなどの)他のニューロトロフィン受容体とほとんど交差反応(結合)しない。一部の実施形態では、抗体はtrkCを結合し、さらにtrkAおよび/またはtrkBを結合する。
【0067】
trkCアゴニスト抗体によって認識されるエピトープは、連続的なものでも、連続的でないものでもよい。一部の実施形態では、抗体は、6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256のいずれか1種または複数から選択される抗体と本質的に同じtrkCエピトープを結合する。PCT公開WO01/98361を参照されたい。抗体が向かう先となり得るエピトープの例には、その限りでないが、trkCのドメインVおよび/またはドメインIVが含まれる。別の実施形態では、エピトープには、ヒトtrkCの残基L284、E287、およびN335のうちの1種または複数が含まれる。Urferら、J.Biol.Chem.(1998年)第273巻:5829〜5840ページ)を参照されたい。さらに他の実施形態では、抗体は、免疫学的に不活性な定常部、例えば、補体によって媒介される溶解のきっかけとならず、または抗体依存性細胞によって媒介される細胞傷害性(ADCC)を刺激しない定常部などの、改変された定常部を含む。他の実施形態では、定常部は、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ;PCT出願PCT/GB99/01441;および/または英国特許出願第9809951.8号に記載の通りに改変する。一部の実施形態では、定常部は、A330P331→S330S331の突然変異(野生型IgG2a配列に即したアミノ酸の番号付け;Eur.J.Immunol.(1999年)第29巻:2613〜2624ページを参照されたい)を含むヒト重鎖IgG2a定常部を含む。
【0068】
抗trkCアゴニスト抗体のtrkCに対する結合親和性は、約500nM、400nM、300nM、200nM、100nM、約50nM、約10nM、約1nM、約500pM、約100pM、または約50pMのいずれかから、約2pM、約5pM、約10pM、約15pM、約20pM、または約40pMのいずれかまでである場合がある。一部の実施形態では、結合親和性は、約100nM、約50nM、約10nM、約1nM、約500pM、約100pM、約50pM、または約50pM未満のいずれかである。一部の実施形態では、結合親和性は、約100nM、約50nM、約10nM、約1nM、約500pM、約100pM、または約50pM未満のいずれかである。さらに他の実施形態では、結合親和性は、約2pM、約5pM、約10pM、約15pM、約20pM、約40pM、または約40pM超である。当業界でよく知られているように、結合親和性は、KDまたは解離定数として表すことができ、結合親和性の増大は、KDの低下に一致する。BIAcore分析を使用して評価したマウス抗trkCアゴニストモノクローナル抗体2256のヒトtrkCに対する結合親和性は、約40nMであり、BIAcore分析を使用して評価した(本明細書に記載の)ヒト化抗trkCアゴニスト抗体A5のヒトtrkCに対する結合親和性は、約0.28nMである。
【0069】
抗体のtrkCに対する結合親和性を測定する1つの方法は、抗体の単機能Fab断片の結合親和性を測定することによるものである。単機能Fab断片を得るために、抗体(例えばIgG)をパパインで切断し、または組換えによって発現させることができる。抗体の抗trkC Fab断片の親和性は、表面プラズモン共鳴(BlAcore3000(商標)表面プラズモン共鳴(SPR)システム、BIAcore,INC、米ニュージャージー州Piscaway)によって測定することができる。CM5チップは、供給元の説明書に従って、N−エチル−N’−(3−ジメチルアミノプロピル)−カルボジイニド塩酸塩(EDC)およびN−ヒドロキシスクシンイミド(NHS)で活性化することができる。ヒトtrkC−Fc融合タンパク質(「htrkC」)(またはラットtrkCなどの他の任意のtrkC)を10mMの酢酸ナトリウムpH5.0に希釈し、0.0005mg/mLの濃度で活性化したチップ上に通液(inject)することができる。個々のチップチャネルでの変動する流動時間を使用して、詳細な動力学研究のための200〜400応答単位(RU)とスクリーニングアッセイのための500〜1000RUの2通りの範囲の抗原密度を実現することができる。チップは、エタノールアミンでブロックすることができる。再生研究では、Pierce溶離緩衝液(製品番号21004、Pierce Biotechnology、米イリノイ州ロックフォード)と4M NaCl(2:1)の混合物が、200回を超える通液の間チップ上のhtrkCの活性を保ちながら、結合したFabを効果的に除去することが示されている。HBS−EP緩衝液(0.01MのHEPES、pH7.4、0.15のNaCl、3mMのEDTA、0.005%のSurfactantP20)をBIAcoreアッセイ用の実施緩衝液として使用する。精製したFabサンプルの段階希釈物(推定されるKDの0.1〜10倍)を100μL/分で1分間かけて通液し、最長で2時間の解離時間を与える。Fabタンパク質の濃度は、(アミノ酸分析によって測定した)既知の濃度のFabを標準として使用するELISAおよび/またはSDS−PAGE電気泳動によって測定する。BIAevaluationプログラムを使用してデータを1:1のLangmuir結合モデルに当てはめることによって(Karlsson,R.、Roos,H.、Fagerstam,L.、Petersson,B.(1994年)、Methods Enzymology第6巻:99〜110ページ)、動力学的な結合速度(kon)と解離速度(koff)を同時に得る。平衡解離定数(KD)値をkoff/konとして算出する。
【0070】
別の態様では、ヒトtrkC受容体を活性化することのできる(例えば、ヒト、ヒト化、マウス、キメラの)抗体は、trkCの1つまたは複数の細胞外ドメインを発現する免疫原を使用して生成することもできる。免疫原の一例は、trkCを高度に発現する細胞であり、本明細書に記載の通りに得ることができる。使用することのできる免疫原の別の例は、trkC受容体の細胞外ドメインまたは細胞外ドメインの部分を含む(trkCイムノアドヘシンなどの)可溶性タンパク質である。
【0071】
宿主動物を免疫化する経路およびスケジュールは一般に、本明細書でさらに記載する、抗体の刺激および生成のための確立した技術および従来の技術に合わせることにある。ヒトおよびマウス抗体を生成するための一般技術は、当業界で知られており、本明細書に記載されている。
【0072】
ヒトを含む任意の哺乳類対象またはその抗体産生細胞は、ヒトを含む哺乳類のハイブリドーマ細胞系を生成する基盤として役立つように操作することができると予想される。通常は、宿主動物に、本明細書に記載のものを含むある量の免疫原を腹腔内接種する。
【0073】
ハイブリドーマは、Kohler,B.およびMilstein,C.(1975年)、Nature第256巻:495〜497ページの一般の体細胞ハイブリッド形成技術を使用して、またはBuck,D.W.ら(1982年)、In Vitro、第18巻:377〜381ページによって変更されている通りに、リンパ球および不死化した骨髄腫細胞から調製することができる。その限りでないが、X63−Ag8.653、およびSalk Institute、Cell Distribution Center、米カリフォルニア州サンディエゴからのものを含めて、入手可能な骨髄腫系統をハイブリッド形成で使用することができる。一般に、この技術は、ポリエチレングリコールなどの融合源(fusogen)を使用して、または当業界でよく知られている電気的な手段によって骨髄腫細胞とリンパ球を融合させるものである。融合させた後、細胞を融合培地から分離し、ヒポキサンチン−アミノプテリン−チミジン(HAT)培地などの選択的な増殖培地で増殖させて、ハイブリッド形成していない親細胞を排除する。モノクローナル抗体を分泌するハイブリドーマの培養には、血清を補充し、または補充しない本明細書に記載の培地のいずれかを使用することができる。細胞融合技術の別の選択肢として、EBVによって不死化したB細胞を使用して、本発明の抗trkCモノクローナル抗体を生成することができる。ハイブリドーマを拡張させ、所望ならばサブクローニングし、従来のイムノアッセイ手順(例えば、放射性免疫測定法、酵素免疫測定法、または蛍光免疫測定法)によって抗免疫原活性があるかどうか上清を検定する。
【0074】
抗体供給源として使用することのできるハイブリドーマとは、trkCまたはその部分に特異的なモノクローナル抗体を産生するすべての派生物、すなわち親ハイブリドーマの子孫細胞を含む。
【0075】
そのような抗体を産生するハイブリドーマは、知られている手順を使用して、in vitroまたはin vivoで増殖させることができる。モノクローナル抗体は、所望ならば、硫安塩析、ゲル電気泳動、透析、クロマトグラフィー、限外ろ過などの従来の免疫グロブリン精製手順によって培地または体液から単離してもよい。所望でない活性が存在するならば、例えば、固相に結合している免疫原でできた吸着剤上での調製を実施し、所望の抗体を免疫原から溶離または解放することによって除去できる。ヒトもしくは他の種のtrkC受容体またはヒトもしくは他の種のtrkC受容体の断片、あるいは免疫化する種において免疫原性であるタンパク質、例えば、キーホールリンペットヘモシアニン、血清アルブミン、ウシのサイログロブリン、または大豆トリプシンインヒビターに、二官能性または誘導体化の薬剤、例えば、マレイミドベンゾイルスルホスクシンイミドエステル(システイン残基を介して結合)、N−ヒドロキシスクシンイミド(リジン残基を介して)、グルタルアルデヒド、無水コハク酸、SOCl2、またはR1N=C=NR(RおよびR1は異なるアルキル基である)を使用して結合させたターゲットアミノ酸配列を含むヒトまたは他の種のtrkC受容体または断片で宿主動物を免疫化すると、抗体の集団(例えばモノクローナル抗体)を得ることができる。免疫原の別の例は、trkCを高度に発現する細胞であり、これは、組換えの手段によって、あるいは高レベルのtrkCを発現する天然供給源から細胞を単離または濃縮して得ることができる。これらの細胞は、ヒトまたは他の動物に由来するものでよく、直接に単離された免疫原として使用してもよいし、または免疫原性が増大し、または(trkC断片の)trkCの発現が増加しもしくは濃縮されるように処理してもよい。その処理には、その限りでないが、細胞またはその断片を、例えばホルムアルデヒド、グルタルアルデヒド、エタノール、アセトン、および/または様々な酸などの、その安定性または免疫原性を増大させるように設計された薬剤で処理することが含まれる。さらに、このような処理の前または後に、所望の免疫原、この場合ではtrkCまたはその断片を濃縮するため、細胞の処理を行ってもよい。これらの処理ステップには、当業界でよく知られている膜分画法が含まれる。
【0076】
所望ならば、目的の抗trkC抗体(モノクローナルまたはポリクローナル)の配列を決定してもよく、次いでポリヌクレオチド配列をベクターにクローン化して発現または増殖させてもよい。目的の抗体をコードする配列は、宿主細胞中のベクターの中で維持することができ、次いで将来の使用に向けて宿主細胞を拡張させ、凍結することができる。別法として、抗体を「ヒト化」し、または抗体の親和性もしくは他の特性を向上させるために、ポリヌクレオチド配列を遺伝子操作に使用してもよい。例えば、抗体をヒトの臨床試験および治療で使用するならば、定常部をヒト定常部により似るように操作して、免疫応答を回避する場合もある。抗体配列を遺伝子操作して、trkC受容体に対するより強い親和性およびtrkC受容体の活性化におけるより高い有効性を得ることが望ましい場合もある。抗trkC抗体のポリヌクレオチドに1つまたは複数の変更が加えられても、trkC細胞外ドメインまたはtrkCエピトープへのその結合能力が維持され得ることは、当業者には明白であろう。
【0077】
モノクローナル抗体をヒト化するには、大まかな4ステップを踏む。すなわち、(1)出発抗体の軽鎖および重鎖可変ドメインのヌクレオチドおよび推定されるアミノ酸配列を決定するステップ、(2)ヒト化抗体を設計する、すなわち、ヒト化処理の際にどちらの抗体フレームワーク領域を使用するかを決定するステップ、(3)実際のヒト化方法/技術のステップ、および(4)ヒト化抗体の形質移入および発現のステップ。例えば、米国特許第4816567号、同第5807715号、同第5866692号、同第6331415号、同第5530101号、同第5693761号、同第5693762号、同第5585089号、同第6180370号、および同第6548640号を参照されたい。例えば、抗体をヒトの臨床試験および治療で使用するならば、定常部をヒト定常部により似るように操作して、免疫応答を回避する場合もある。例えば、米国特許第5997867号および同第5866692号を参照されたい。
【0078】
組換えヒト化抗体では、Fcγ部分を変更して、Fcγ受容体および補体免疫系との相互作用を回避することができる。この種類の変更は、ケンブリッジ大学Department of PathologyのMike Clark博士によって設計されたものであり、その抗体を調製する技術は、1999年11月18日公開のPCT公開WO99/58572に記載されている。
【0079】
ヒト定常ドメインと融合させたげっ歯類可変部または改変されたげっ歯類可変部ならびにそれに伴う相補性決定領域(CDR)を有するキメラ抗体を含めて、非ヒト免疫グロブリンに由来する抗原結合部位を含むいくつかの「ヒト化」抗体分子が記載されている。例えば、Winterら、Nature第349巻:293〜299ページ(1991年)、Lobuglioら、Proc.Nat.Acad.Sci.USA第86巻:4220〜4224ページ(1989年)、Shawら、J Immunol.第138巻:4534〜4538ページ(1987年)、およびBrownら、Cancer Res.第47巻:3577〜3583ページ(1987年)を参照されたい。他の参考文献は、適切なヒト抗体定常ドメインと融合させる前にヒト支持フレームワーク領域(FR)に接ぎ合わせたげっ歯類CDRについて述べている。例えば、Riechmannら、Nature第332巻:323〜327ページ(1988年)、Verhoeyenら、Science第239巻:1534〜1536ページ(1988年)、およびJonesら、Nature第321巻:522〜525ページ(1986年)を参照されたい。別の参考文献は、組換えによって張り合わせたげっ歯類フレームワーク領域を支持体とするげっ歯類CDRについて述べている。例えば、欧州特許公開第519596号を参照されたい。これらの「ヒト化」分子は、ヒトレシピエントの治療にこれらの部分を適用する期間および有効性を制限する、げっ歯類抗ヒト抗体分子に対する不必要な免疫応答を最小限に抑えるように設計される。抗体定常部は、免疫学的に不活性になるように、例えば、補体によって媒介される溶解のきっかけとならず、抗体依存性の細胞によって媒介される細胞傷害性(ADCC)を刺激しないように操作することができる。他の実施形態では、定常部は、Eur.J.Immunol.(1999年)第29巻:2613〜2624ページ;PCT出願PCT/GB99/01441;および/または英国特許出願第9809951.8号に記載の通りに改変する。例えば、PCT/GB99/01441;英国特許出願第9809951.8号を参照されたい。利用することのできる、抗体をヒト化する他の方法は、Daughertyら、Nucl.Acids Res.第19巻:2471〜2476ページ(1991年)および米国特許第6180377号、同第6054297号、同第5997867号、同第5866692号、同第6210671号、同第6350861号、およびPCT公開WO01/27160号で開示されている。
【0080】
さらに別の選択肢では、特異的なヒト免疫グロブリンタンパク質を発現させるように操作された市販のマウスを使用して、完全なヒト抗体を得ることもできる。より望ましい(例えば、完全なヒト抗体)またはより強固な免疫応答を生み出すように設計されたトランスジェニック動物を、ヒト化抗体またはヒト抗体の産生に使用してもよい。そのような技術の例は、Abgenix,Inc.(米カリフォルニア州フリーモント)のXenomouse(商標)ならびにHuMAb−Mouse(登録商標)およびMedarex,Inc.(米ニュージャージー州プリンストン)のTC Mouse(商標)である。
【0081】
別法では、抗体を組換えによって生成し、当業界で知られている任意の方法を使用して発現させることができる。別の選択肢では、抗体をファージディスプレイ技術によって組換えで生成することができる。例えば、米国特許第5565332号、同第5580717号、同第5733743号、および同第6265150号、およびWinterら、Annu.Rev.Immunol.第12巻:433〜455ページ(1994年)を参照されたい。別法として、ファージディスプレイ技術(McCaffertyら、Nature第348巻:552〜553ページ(1990年))を使用すると、未免疫ドナーの免疫グロブリン可変(V)ドメイン遺伝子レパートリーからin vitroでヒト抗体および抗体断片を生成することができる。この技術によれば、抗体Vドメイン遺伝子を、M13やfdなどの糸状のバクテリオファージの主要または非主要外殻タンパク質遺伝子にインフレームでクローン化し、ファージ粒子表面上に機能性の抗体断片として提示する。糸状の粒子は、ファージゲノムの一本鎖DNAコピーを含むので、抗体の機能の特性に基づく選択は、これらの特性を示す抗体をコードする遺伝子の選択ともなる。したがって、ファージは、B細胞の特性の一部を模倣する。ファージディスプレイは、様々な形式で実施することができ、総説については、例えば、Johnson,Kevin S.およびChiswell,David J.、Current Opinion in Structural Biology第3巻、564〜571ページ(1993年)を参照されたい。いくつかのV遺伝子区域供給源をファージディスプレイに使用することができる。Clacksonら、Nature第352巻:624〜628ページ(1991年)は、免疫感作されたマウスの脾臓に由来するV遺伝子の小規模のランダムなコンビナトリアルライブラリーから、多様な並び方の抗オキサゾロン抗体を単離した。本質的にマークら、J.Mol.Biol.第222巻:581〜597ページ(1991年)またはGriffithら、EMBO J.第12巻:725〜734ページ(1993年)に記載の技術に従って、未免疫のヒトドナーからの一レパートリーのV遺伝子を構築することができ、多様なアレイの抗原に対する抗体(自己抗原を含む)を単離することができる。自然の免疫応答では、抗体遺伝子は、高い比率で突然変異を蓄積する(体細胞突然変異)。導入される変化の一部は、より高い親和性を付与することになり、高親和性の表面免疫グロブリンを示すB細胞は、その後の抗原による攻撃の際に優先的に複製され、分化する。この自然の過程は、「鎖シャフリング」として知られている技術を使用して模倣することができる。Marksら、Bio/Technol.第10巻:779〜783ページ(1992年)。この方法では、重鎖および軽鎖可変部遺伝子を、未免疫のドナーから得られたVドメイン遺伝子の(レパートリー)の自然に存在する変異体のレパートリーと順次交換することによって、ファージディスプレイによって得られた「一次」ヒト抗体の親和性を改良することができる。この技術は、pM〜nMの範囲で親和性を有する抗体および抗体断片の生成を可能にする。(「すべてのライブラリーの母なるもの」としても知られている)非常に大きなファージ抗体レパートリーを作製する戦略は、Waterhouseら、Nucl.Acids Res.第21巻:2265〜2266ページ(1993年)に記載されている。遺伝子シャフリングを使用して、げっ歯類抗体から、その出発げっ歯類抗体に対して同様の親和性および特異性を有するヒト抗体を得ることもできる。「エピトープインプリンティング」とも呼ばれるこの方法によれば、ファージディスプレイ技術によって得られたげっ歯類抗体の重鎖または軽鎖Vドメイン遺伝子が、ヒトVドメイン遺伝子のレパートリーと交換され、げっ歯類−ヒトキメラが作り出される。抗原上での選択により、機能性の抗原結合部位を回復させることのできるヒト可変部が単離される、すなわち、エピトープがパートナーの選択を支配(刷り込み)する。この過程を繰り返して残りのげっ歯類Vドメインを交換すると、ヒト抗体が得られる(1993年4月1日公開のPCT公開WO93/06213を参照されたい)。CDR移植によるげっ歯類抗体の伝統的なヒト化とは異なって、この技術は、げっ歯類由来のフレームワークまたはCDR残基をもたない完全にヒトの抗体を提供する。上記論述はヒト化抗体に関係するものであるが、論述した一般原理が、例えばイヌ、ネコ、霊長類、ウマ、およびウシで使用するための抗体のカスタマイズに適用できることは言うまでもない。
【0082】
抗体は、二重特異性抗体、すなわち、少なくとも2種の異なる抗原に対する結合特異性を有するモノクローナル抗体である場合もあり、本明細書で開示する抗体を使用して調製することができる。二重特異性抗体の生成方法は、当業界で知られている(例えば、Sureshら、1986年、Methods in Enzymology第121巻:210ページを参照されたい)。伝統的に、二重特異性抗体の組換え生成は、2種の重鎖が異なる特異性を有する2種の免疫グロブリン重鎖−軽鎖対の同時発現に基づくものであった(MillsteinおよびCuello、1983年、Nature第305巻、537〜539ページ)。
【0083】
二重特異性抗体を生成する1つの手法によれば、所望の結合特異性を有する抗体可変ドメイン(抗体−抗原合体部位)を、免疫グロブリン定常ドメイン配列と融合させる。ヒンジ、CH2、およびCH3領域の少なくとも一部を含む免疫グロブリン重鎖定常ドメインと融合させることが好ましい。第1の重鎖定常部(CH1)が、融合物の少なくとも1つに存在する軽鎖結合に必要な部位を含むことが好ましい。免疫グロブリン重鎖融合物、および所望ならば免疫グロブリン軽鎖をコードするDNAを、別個の発現ベクターに挿入し、適切な宿主生物に同時形質移入する。これは、その構築で使用される3種のポリペプチド鎖の不均等な割合が最適な収率をもたらすとき、実施形態の3種のポリペプチド断片の相互の割合の調整において高い柔軟性をもたらす。しかし、同等の割合の少なくとも2種のポリペプチド鎖の発現が高い収率をもたらすとき、またはその割合が特定の意味をもたないものであるとき、1つの発現ベクター中に2種または3種すべてのポリペプチド鎖のコード配列を挿入することは可能である。
【0084】
1つの手法では、二重特異性抗体は、一方の腕の第1の結合特異性を有するハイブリッド免疫グロブリン重鎖と、他方の腕のハイブリッド免疫グロブリン重鎖−軽鎖対(第2の結合特異性をもたらす)から構成される。二重特異性の半分にしか免疫グロブリン軽鎖がないこの非対称の構造は、望ましくない免疫グロブリン鎖の組合せからの所望の二重特異性化合物の分離を容易にする。この手法は、1994年3月3日公開のPCT公開WO94/04690に記載されている。
【0085】
共有結合によって結合した2種の抗体を含むヘテロコンジュゲート抗体も、本発明の範囲内である。そのような抗体は、免疫系細胞を望ましくない細胞に向けること(米国特許第4676980号)、およびHIV感染の治療(PCT公開WO91/00360およびWO92/200373、ならびにEP03089)に使用されている。ヘテロコンジュゲート抗体は、任意の好都合な架橋結合法を使用して作製することができる。適切な架橋剤および技術は、当業界でよく知られており、米国特許第4676980号に記載されている。
【0086】
抗体は、最初に宿主動物から作られた抗体を単離し、遺伝子配列を得、その遺伝子配列を使用して宿主細胞(例えばCHO細胞)中で組換えによって抗体を発現させることで、組換えで作製することができる。使用することのできる別の方法は、植物(例えばタバコ)、トランスジェニック乳汁、または他の生物において抗体配列を発現させることである。植物または乳汁において抗体を組換え発現させる方法は開示されている。例えば、Peetersら(2001年)、Vaccine第19巻:2756ページ;Lonberg,N.およびD.Huszar(1995年)、Int.Rev.Immunol第13巻:65ページ;およびPollockら(1999年)、J Immunol Methods第231巻:147ページを参照されたい。例えばヒト化、単鎖などの抗体派生物を生成する方法は、当業界で知られている。
【0087】
キメラまたはハイブリッド抗体も、架橋剤を使用するものを含む既知の合成タンパク質化学の方法を使用して、in vitroで調製することができる。例えば、ジスルフィド交換反応を使用して、またはチオエーテル結合を形成して、免疫毒素を構築することができる。この目的に適する試薬の例には、イミノチオラートおよびメチル−4−メルカプトブチルイミダートが含まれる。
【0088】
単鎖Fv断片も、Iliadesら、1997年、FEBS Letters、第409巻:437〜441ページに記載されているように生成することができる。様々なリンカーを使用するそのような単鎖断片の結合は、Korttら、1997年、Protein Engineering、第10巻:423〜433ページに記載されている。組換え産生および抗体の操作のための様々な技術は、当業界でよく知られている。
【0089】
抗体は、1999年11月18日公開のPCT公開WO99/58572号に記載の通りに改変することができる。これらの抗体は、標的分子に向けられる結合ドメインに加え、ヒト免疫グロブリン重鎖の定常ドメインの全部または一部と実質的に相同であるアミノ酸配列を有するエフェクタードメインを含む。これらの抗体は、著しい補体依存性の溶解または細胞性の標的の破壊を誘発することなく、標的分子を結合することができる。エフェクタードメインは、FcRnおよび/またはFcγRIIbを特異的に結合し得ることが好ましい。これらは通常、2種以上のヒト免疫グロブリン重鎖CH2ドメインから得られるキメラドメインに基づくものである。このようにして改変された抗体は、長期にわたる抗体療法で使用して、従来の抗体療法に対する炎症性および他の副作用を回避するのに好ましい。
【0090】
宿主動物の免疫化によってまたは組換えによって生成される抗体は、次の特性のいずれか1つまたは複数を示すべきである。すなわち、(a)trkC受容体に結合する、(b)trkC受容体の1つまたは複数のエピトープに結合する、(c)trkC受容体に結合し、trkCの生物活性、またはtrkCシグナル伝達機能に媒介される1つまたは複数の下流の経路を活性化する、(d)trkC受容体の二量体化を促進する、(e)trkC受容体の活性化を増大させる、(f)好ましい薬物動態および生体利用度特性を示す、(g)細胞の発生、生存、機能、維持、および/または再生を促進する、(h)trkC受容体に結合し、下位運動ニューロン疾患の1つまたは複数の症状を治療し、予防し、逆戻りさせ、または緩和させる。
【0091】
イムノアッセイ、およびfluorescence activated cell sorting(FACS)などのフローサイトメトリー選別技術を使用すると、trkCに特異的な抗体を単離することもできる。
【0092】
抗体は、多くの異なる担体に結合させることができる。担体は、活性のあるものおよび/または不活性なものでよい。よく知られた担体の例には、ポリプロピレン、ポリスチレン、ポリエチレン、デキストラン、ナイロン、アミラーゼ、ガラス、天然および改変型のセルロース、ポリアクリルアミド、アガロース、ならびに磁鉄鉱が含まれる。担体の性質は、本発明の目的では可溶性または不溶性のどちらでもよい。当業者ならば、抗体の結合に適する他の担体について知っており、または常法通りの実験を使用してそれを突き止めることができるであろう。
【0093】
アゴニスト抗trkC抗体をコードするDNAは、当業界で知られているように配列決定することができる。PCT公開WO01/98361を参照されたい。一般に、モノクローナル抗体は、従来の手順を使用して(例えば、モノクローナル抗体の重鎖および軽鎖をコードする遺伝子に特異的に結合することのできるオリゴヌクレオチドプローブを使用して)容易に単離および配列決定がなされる。ハイブリドーマ細胞は、そのようなcDNAの好ましい供給源として役立つ。単離されたなら、DNAは、(PCT公開WO87/04462で開示されている発現ベクターなどの)発現ベクター中に入れ、次いでそれを、別に免疫グロブリンタンパク質を産生しない大腸菌細胞、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、骨髄腫細胞などの宿主細胞に形質移入して、組換え宿主細胞中でモノクローナル抗体の合成を実現することができる。例えば、PCT公開WO87/04462を参照されたい。DNAは、例えば、相同のネズミ配列の代わりにヒト重鎖および軽鎖定常ドメインのコード配列を用いて(Morrisonら、Proc.Nat.Acad.Sci.第81巻:6851ページ(1984年))、または非免疫グロブリンポリペプチドのコード配列の全部もしくは一部に免疫グロブリンコード配列を共有結合によって連結して、改変することもできる。そのようにして、本明細書では、抗trkCモノクローナル抗体の結合特異性を有する「キメラ」または「ハイブリッド」抗体が調製される。(その抗原結合断片などの)アゴニスト抗trkC抗体をコードするDNAは、ここに記載するような、所望の細胞におけるアゴニスト抗trkC抗体の送達および発現に使用することもできる。DNA送達技術については、本明細書でさらに記載する。
【0094】
抗trkC抗体は、当業界でよく知られている方法を使用して特徴付けることができる。例えば、1つの方法は、それが結合するエピトープを同定することであり、例えば、HarlowおよびLane、「Using Antibodies,a Laboratory Manual」、第11章、Cold Spring Harbor Laboratory Press、米ニューヨーク州コールドスプリング港、1999年に記載されているように、抗体−抗原複合体の結晶構造の解明、競合アッセイ、遺伝子断片発現アッセイ、および合成ペプチドに基づくアッセイがこれに含まれる。追加の例では、エピトープマッピングを使用して、抗trkC抗体が結合する配列を決定することができる。エピトープマッピングは、様々な供給元、例えば、Pepscan Systems(Edelhertweg 15、8219 PH Lelystad、The Netherlands)からの市販品を利用することができる。エピトープは、線形のエピトープ、すなわち、一続きのアミノ酸中に含まれるものである場合もあり、または必ずしも一続きの中に含まれなくてもよい、アミノ酸の三次元の相互作用によって形成される高次構造的なエピトープである場合もある。様々な長さ(例えば少なくとも4〜6アミノ酸長)のペプチドを単離し、または(例えば組換えによって)合成し、抗trkC抗体との結合アッセイに使用することができる。別の例では、trkC細胞外配列由来のオーバーラップペプチドを使用し、抗trkC抗体による結合を判定する系統的なスクリーニングにおいて抗trkC抗体が結合するエピトープを決定することができる。遺伝子断片発現アッセイによれば、trkCをコードするオープンリーディングフレームを、無作為に、または特異的な遺伝子構築物によって分断し、発現されるtrkC断片と試験される抗体との反応性を測定する。遺伝子断片は、例えば、PCRによって生成し、次いで、放射性アミノ酸の存在下、in vitroでタンパク質へと転写、翻訳することができる。次いで、放射標識されたtrkC断片への抗体の結合を、免疫沈殿法およびゲル電気泳動によって測定する。ある種のエピトープは、ファージ粒子表面上に示されるランダムなペプチド配列の大規模なライブラリー(ファージライブラリー)を使用して同定することもできる。
【0095】
抗trkC抗体の特徴付けに使用することのできるさらに別の方法は、同じ抗原、すなわちtrkC細胞外ドメインに結合することがわかっている他の抗体との競合アッセイを使用して、抗trkC抗体が他の抗体と同じエピトープに結合するかどうかを判定することである。競合アッセイは、当業者によく知られている。競合アッセイにおいて有用な抗体の例には、抗体6.1.2、6.4.1、2345、2349、2.5.1、2344、2248、2250、2253、および2256が含まれる。PCT公開WO01/98361を参照されたい。
【0096】
ドメインスワップ突然変異体を使用して、PCT公開WO01/98361に記載の通りにエピトープマッピングを実施してもよい。一般に、この手法は、trkAまたはtrkBとほとんど交差反応しない抗trkC抗体に有用である。trkCのドメインスワップ突然変異体は、trkCの細胞外ドメインをtrkBまたはtrkAの対応するドメインと交換して作製することができる。ELISAまたは当業界で知られている他の方法を使用して、各アゴニスト抗trkC抗体の様々なドメインスワップ突然変異体への結合を評価し、その野生型(未変性)trkCへの結合と比較することができる。別の手法では、アラニン走査を実施することができる。抗原、すなわちtrkC受容体の個々の残基を、意図的に別のアミノ酸(通常はアラニン)に突然変異させ、ELISAまたは当業界で知られている他の方法を使用して、その改変trkCの抗体への結合能を試験することによって、その変化の影響を評価する。
【0097】
アゴニスト抗trkC抗体の同定
アゴニスト抗体は、以下の方法の1つまたは複数を含む業界で認められている方法を使用して同定することができる。例えば、米国特許第5766863号および同第5891650号に記載されているキナーゼ受容体活性化(KIRA)アッセイを使用することができる。このELISA型アッセイは、受容体タンパク質チロシンキナーゼ(rPTK、例えばtrk受容体)のキナーゼドメインの自己リン酸化を測定することによるキナーゼ活性化の定性的または定量的な測定、ならびに選択されたrPTKの有望なアゴニストまたはアンタゴニストの同定および特徴付けに適する。アッセイの第1期は、キナーゼ受容体、この場合では真核細胞の細胞膜に存在するtrkC受容体のキナーゼドメインのリン酸化を含む。受容体は、内因性の受容体または受容体をコードする核酸でもよいし、受容体構築物を細胞に入れて形質転換してもよい。通常、第1の固相(例えば、第1のアッセイプレートのウェル)を、そのような細胞(普通は哺乳動物細胞系)の実質的に均一な集団でコートして、細胞が固相に付着するようにする。細胞はしばしば接着性であり、そのために自然に第1の固相に付着する。「受容体構築物」を使用する場合、通常これは、キナーゼ受容体とflagポリペプチドの融合物を含む。flagポリペプチドは、アッセイのELISAの部分で捕捉剤、しばしば捕捉抗体によって認識される。次いで、候補アゴニストなどの分析物を、付着細胞を有するウェルに加えて、チロシンキナーゼ受容体(例えばtrkC受容体)が、分析物にさらされる(または分析物と接触する)ようにする。このアッセイは、目的のチロシンキナーゼ受容体(例えばtrkC)のアゴニストリガンドの同定を可能にするものである。分析物にさらした後、(その中に可溶化洗浄剤を含む)溶解緩衝液を使用して付着細胞を可溶化し、穏やかに攪拌し、それによって細胞可溶化物を放出させ、それを細胞可溶化物の濃縮または清澄の必要なしに直接にアッセイのELISA部分にかけることができる。
【0098】
このように細胞可溶化物を調製し、次いでアッセイのELISAステージにかける準備をする。ELISA段階の第1ステップとして、第2の固相(通常ELISAマイクロタイタープレートのウェル)を、チロシンキナーゼ受容体、または受容体構築物の場合ではflagポリペプチドに特異的に結合する捕捉剤(しばしば捕捉抗体)でコートする。第2の固相のコーティングは、捕捉剤が第2の固相に付着するように実施する。捕捉剤は、一般にモノクローナル抗体であるが、本明細書で実施例に記載するように、ポリクローナル抗体または他の薬剤を使用してもよい。次いで得られる細胞可溶化物を付着性の捕捉剤にさらし、またはそれと接触させて、受容体または受容体構築物が第2の固相に付着する(またはそれに捕捉される)ようにする。次いで、捕捉された受容体または受容体構築物を残して非結合の細胞可溶化物を除去するように洗浄ステップを実施する。次いで、付着しまたは捕捉された受容体または受容体構築物を、チロシンキナーゼ受容体中のリン酸化されたチロシン残基を同定する抗ホスホチロシン抗体にさらし、または接触させる。好ましい実施形態では、抗ホスホチロシン抗体を、非放射性呈色試薬の色調変化を触媒する酵素に(直接または間接的に)結合させる。したがって、受容体のリン酸化は、その後の試薬の色調変化によって測定することができる。酵素は、抗ホスホチロシン抗体に直接に結合することもでき、または結合性の分子(例えばビオチン)を抗ホスホチロシン抗体に結合し、その後酵素をその結合性の分子を介して抗ホスホチロシン抗体に結合することもできる。最後に、捕捉された受容体または受容体構築物への抗ホスホチロシン抗体の結合を、例えば呈色試薬の色調変化によって測定する。
【0099】
最初の同定の後、候補抗体のアゴニスト活性は、標的指向化された生物活性を試験することが知られているバイオアッセイによってさらに確認し、さらに正確にすることができる。例えば、抗trkCモノクローナル抗体がtrkCのアゴニストになる能力を、全長ヒトtrkCを形質移入したPC12細胞を使用するPC12神経突起伸長アッセイで試験することができる(Urferら、Biochem.第36巻:4775〜4781ページ(1997年);Tsoulfasら、Neuron第10巻:975〜990ページ(1993年))。このアッセイは、適切なリガンドによる刺激に応答したラット好チトクロム腫(pheocytochroma)細胞(PC12)による神経突起伸長の過程を測定するものである。これらの細胞は、内因性のtrkAを発現し、したがってNGFに応答する。しかし、これらの細胞は、内因性のtrkCを発現せず、したがって、trkCアゴニストに対する応答を誘発するためにtrkC発現構築物を形質移入する。形質移入された細胞を抗trkC抗体と共にインキュベートした後、神経突起の伸長を測定、例えば、細胞の直径の2倍を超える神経突起を有する細胞をカウントする。形質移入されたPC12細胞において神経突起の伸長を刺激する抗trkC抗体は、trkCアゴニスト活性を実証するものである。
【0100】
trkCの活性化は、胚発生の特定の段階にある様々な特定のニューロンを使用して判定することもできる。適切に選択したニューロンの生存は、trkCの活性化に応じて決まる場合があり、これらのニューロンのin vitroでの生存に従ってtrkCの活性化を測定することが可能である。候補抗体がtrkCを活性化するならば、適切なニューロンの初代培養物に候補抗体を加えると、これらのニューロンが少なくとも数日間生存するようになる。これによって、候補抗体がtrkCを活性化する能力の判定が可能になる。この種類のアッセイの一例では、E11マウス胎仔の三叉神経節を解剖し、分離し、得られるニューロンを低い密度で組織培養皿に播く。次いで、候補抗体を培地およびプレートに加え、24〜48時間インキュベートする。その後、ニューロンの生存を様々な方法のいずれかによって評価する。アゴニスト抗trkC抗体を加えたサンプルは通常、対照抗体を加えたサンプルよりも増大した生存率を示し、これによってアゴニスト抗trkC抗体の存在を判定することができる。例えば、Buchmanら(1993年)、Development第118巻(3):989〜1001ページを参照されたい。
【0101】
アゴニスト抗体は、自然にまたはtrkCをコードするDNAを形質移入した後にtrkCを発現する様々な細胞型において下流のシグナル伝達を活性化するその能力によって同定することができる。そのtrkCは、ヒトまたは(げっ歯類や霊長類などの)他の哺乳類のtrkCでよい。下流のシグナルカスケードは、タンパク質のタンパク質発現またはタンパク質リン酸化のレベルなどの、trkC発現細胞の様々な生化学的または生理学的パラメータへの変化、あるいは(本明細書に記載するようなニューロンの生存および/または神経突起の伸長を含む)細胞の代謝的状態または増殖状態への変化によって検出することができる。関連する生化学的または生理学的パラメータの検出方法は、当業界で知られている。
【0102】
アゴニスト抗trkC抗体の投与
アゴニスト抗trkC抗体の様々な製剤を投与に使用することができる。一部の実施形態では、アゴニスト抗trkC抗体を希釈せずに投与する場合もある。一部の実施形態では、アゴニスト抗trkC抗体は、薬学的に許容できる賦形剤を含む組成物にして投与する。薬学的に許容できる賦形剤は、当業界で知られており、薬理学的に有効な物質の投与を容易にする比較的不活性な物質である。例えば、賦形剤は、形または稠性を与えることができ、または希釈剤として働く。適切な賦形剤には、その限りでないが、安定剤、湿潤剤および乳化剤、様々なモル浸透圧濃度にするための塩、カプセル化剤、緩衝剤、ならびに皮膚浸透性改善剤が含まれる。非経口および非経口でない薬物送達のための賦形剤ならびに製剤は、Remington、「The Science and Practice of Pharmacy」、第20版、Mack Publishing(2000年)に記載されている。
【0103】
アゴニスト抗trkC抗体は、注射(例えば、腹腔内、静脈内、皮下、筋肉内など)による投与向けに製剤することができる。したがって、抗体は、生理食塩水、リンガー液、デキストロース溶液などの薬学的に許容できる媒体と組み合わせることができる。特定の投与計画、すなわち、投与量、タイミング、および繰返しは、特定の個体およびその個体の治療歴に応じて決まることになる。一般に、約1ug/kg体重未満、少なくとも約1ug/kg体重;少なくとも約2ug/kg体重、少なくとも約5ug/kg体重、少なくとも約10ug/kg体重、少なくとも約20ug/kg体重、少なくとも約50ug/kg体重、少なくとも約100ug/kg体重、少なくとも約200ug/kg体重、少なくとも約500ug/kg体重、少なくとも約1mg/kg体重、少なくとも約2mg/kg体重、少なくとも約5mg/kg体重、少なくとも約10mg/kg体重、少なくとも約30mg/kg体重、またはこれより多い量(約50mg/kg、約100mg/kg、約200mg/kg、約500mg/kgなど)の投与量が投与される。
【0104】
半減期などの経験に基づく考慮事項は、一般に投与量決定の一因となる。ヒト化抗体や完全ヒト抗体などのヒト免疫系と適合性のある抗体を使用すると、抗体の半減期を延長し、抗体が宿主の免疫系によって攻撃されないようにすることができる。投与の頻度は、治療の過程で決定し調整することができ、一般に、必ずしもそうでないが、患者におけるアゴニスト抗trkC抗体の有効濃度の維持、ならびに下位運動ニューロン疾患の1つまたは複数の症状の抑制および/または回復および/または遅延に基づくものである。別法として、アゴニスト抗trkC抗体の持続型の連続放出製剤が適切な場合もある。持続放出を実現するための様々な製剤および装置が当業界で知られている。本発明の方法に従うアゴニスト抗trkC抗体の投与は、例えば、投与の目的が治療であろうと予防であろうとレシピエントの生理的な状態、および熟練した従業者に知られている他の要素に応じて、継続的である場合もあれば断続的である場合もある。アゴニスト抗trkC抗体の投与は、予め選択された期間にわたる本質的に継続的なものでもよいし、あるいは例えば、下位運動ニューロン疾患の症状の出現の前、最中、もしくは後のいずれか;下位運動ニューロン疾患の症状の出現の前および最中;前および後;最中および後;および/または前、最中、および後の、間隙を挟んだ一連の投与にしてもよい。
【0105】
一般に、アゴニスト抗trkC抗体の投与では、初期の候補投与量は、約2mg/kgでよい。本発明の目的では、典型的な1日投与量は、上述の要素に応じて約30μg/kg〜100mg/kgまたはそれ以上の範囲となるであろう。数日またはそれ以上にわたって繰り返される投与では、状態に応じて、疾患症状の所望の抑制が起こるまで、または下位運動ニューロン疾患を治療もしくは予防するのに十分な治療レベルが実現されるまで治療を継続する。好例となる投与計画は、約2mg/kgの初回量の後、毎週約1mg/kgの維持量または隔週で約1mg/kgの維持量のtrkCアゴニスト抗体の投与を含む。
【0106】
一実施形態では、抗体の投与量は、trkC受容体を活性化して下位運動ニューロン疾患を治療するアゴニスト抗trkC抗体の1回または複数の投与を受けたことのある個体において経験に基づいて決定する場合もある。個体に、アゴニスト抗trkC抗体の投与量を徐々に増やして与える。アゴニスト抗trkC抗体の有効性を評価するには、本明細書に記載するような下位運動ニューロン疾患状態の指標に従うことができる。
【0107】
他の製剤は、その限りでないがリポソームなどの担体を含めて、当業界で知られている適切な送達形態を含む。例えば、Mahatoら(1997年)、Pharm.Res.第14巻:853〜859ページを参照されたい。リポソーム製剤には、その限りでないが、シトフェクチン(cytofectin)、多層ベシクル、および単層ベシクルが含まれる。
【0108】
in vivo投与に使用する製剤は、無菌でなければならない。これは、例えば無菌濾過膜でのろ過によって容易に実現される。治療用のアゴニスト抗trkC抗体組成物は、一般に、無菌の出入口を有する容器、例えば、静脈内溶液バッグ、または皮下投与針によって突き刺すことのできる栓を有するバイアルに入れられる。
【0109】
アゴニスト抗trkC抗体は、筋肉内、腹腔内、皮下、経口、くも膜下腔内、または局所的な経路によって、例えば大量瞬時投与としてまたは一定期間にわたる継続的な注入による静脈内投与などの既知の方法に従って個体に投与する。アゴニスト抗trkC抗体は、吸入によって投与することもできる。ジェットネブライザーおよび超音波ネブライザーを含む市販の液体製剤用ネブライザーは、投与に有用である。液体製剤をそのまま噴霧することもでき、凍結乾燥された粉末を再形成した後に噴霧することもできる。別法として、アゴニスト抗trkC抗体は、フルオロカーボン製剤および計量投与吸入器を使用してエアロゾル化してもよいし、または凍結乾燥および粉砕した粉末として吸入してもよい。
【0110】
一部の実施形態では、複数種の抗体が存在する場合もある。その抗体は、同じもので場合もあれば、互いに異なる場合もある。一部の実施形態では、少なくとも1種、少なくとも2種、少なくとも3種、少なくとも4種、少なくとも5種の異なるtrkCアゴニスト抗体が存在する。これらの抗体は、互いに不利に影響を及ぼさない相補的な活性を有することが好ましい。
【0111】
(その抗原結合断片などの)アゴニスト抗trkC抗体をコードするポリヌクレオチドは、所望の細胞におけるアゴニスト抗trkC抗体の送達および発現に使用することもできる。発現ベクターを使用してアゴニスト抗trkC抗体の発現を管理できることは言うまでもない。発現ベクターは、腹腔内、静脈内、筋肉内、皮下、くも膜下腔内、脳室内、経口的、経腸的、非経口的、鼻腔内、経皮的に、または吸入によって投与することができる。例えば、発現ベクターの投与には、注射、経口投与、粒子銃、またはカテーテルによる投与を含む局部または全身の投与、ならびに局所投与が含まれる。当業者は、外来タンパク質の発現をin vivoで実現するための発現ベクターの投与に精通している。例えば、米国特許第6436908号、同第6413942号、および同第6376471号を参照されたい。
【0112】
アゴニスト抗trkC抗体をコードするポリヌクレオチドを含む治療用組成物の標的指向化された送達を使用することもできる。受容体を媒介とするDNA送達技術は、例えば、Findeisら、Trends Biotechnol.(1993年)第11巻:202ページ;Chiouら、「Gene Therapeutics:Methods And Applications Of Direct Gene Transfer」(J.A.Wolff編)(1994年);Wuら、J.Biol.Chem.(1988年)第263巻:621ページ;Wuら、J.Biol.Chem.(1994年)第269巻:542ページ;Zenkeら、Proc.Natl.Acad.Sci.(USA)(1990年)第87巻:3655ページ;Wuら、J.Biol.Chem.(1991年)第266巻:338ページに記載されている。遺伝子治療プロトコルでは、ポリヌクレオチドを含む治療用組成物を、局所投与ではDNA約100ng〜約200mgの範囲で投与する。遺伝子治療プロトコルの際に、約500ng〜約50mg、約1μg〜約2mg、約5μg〜約500μg、および約20μg〜約100μgの濃度範囲のDNAを使用することもできる。本発明の治療用ポリヌクレオチドおよびポリペプチドは、遺伝子送達媒体を使用して送達することができる。遺伝子送達媒体は、ウイルス由来のものでも、ウイルス由来でないものでもよい(一般に、Jolly、Cancer Gene Therapy(1994年)第1巻:51ページ;Kimura、Human Gene Therapy(1994年)第5巻:845ページ;Connelly、Human Gene Therapy(1995年)第1巻:185ページ;およびKaplitt、Nature Genetics(1994年)第6巻:148ページ)を参照されたい。そのようなコード配列の発現は、内因性の哺乳類プロモーターまたは異種プロモーターを使用して誘発することができる。コード配列の発現は、構成性または調節性のどちらでもよい。
【0113】
所望のポリヌクレオチドの送達および所望の細胞での発現のためのウイルス系ベクターは、当業界でよく知られている。好例となるウイルス系媒体には、その限りでないが、組換えレトロウイルス(例えば、PCT公開WO90/07936、WO94/03622、WO93/25698、WO93/25234、WO93/11230、WO93/10218、WO91/02805、米国特許第5219740号、同第4777127号、英国特許第2200651号、およびEP0345242を参照されたい)、アルファウイルス系ベクター(例えば、シンドビスウイルスベクター、セムリキ森林熱ウイルス(ATCC VR−67、ATCC VR−1247)、ロスリバーウイルス(ATCC VR−373、ATCC VR−1246)、およびベネズエラウマ脳炎ウイルス(ATCC VR−923、ATCC VR−1250、ATCC VR 1249、ATCC VR−532))、ならびにアデノ随伴ウイルス(AAV)ベクター(例えば、PCT公開WO94/12649、WO93/03769、WO93/19191、WO94/28938、WO95/11984、およびWO95/00655を参照されたい)が含まれる。Curiel、Hum.Gene Ther.(1992年)第3巻:147ページに記載の、死滅させたアデノウイルスに連結されたDNAの投与を使用することもできる。
【0114】
その限りでないが、死滅させたアデノウイルスのみに連結されまたは連結されていないポリカチオン縮合DNA(例えば、Curiel、Hum.Gene Ther.(1992年)第3巻:147ページを参照されたい)、リガンドに連結されたDNA(例えば、Wu、J.Biol.Chem.(1989年)第264巻:16985ページを参照されたい)、真核細胞送達媒体細胞(例えば、米国特許第5814482号、PCT公開WO95/07994、WO96/17072、WO95/30763、およびWO97/42338を参照されたい)、ならびに核電荷中和(nucleic charge neutralization)または細胞膜との融合を含めて、非ウイルスの送達媒体および方法を使用することもできる。裸DNAを用いてもよい。好例となる裸DNA導入方法は、PCT公開WO90/11092および米国特許第5580859号に記載されている。遺伝子送達媒体として働き得るリポソームは、米国特許第5422120号、PCT公開WO95/13796、WO94/23697、WO91/14445、およびEP0524968に記載されている。追加の手法は、Philip、Mol.Cell Biol.(1994年)第14巻:2411ページ、およびWoffendin、Proc.Natl.Acad.Sci.(1994年)第91巻:1581ページに記載されている。
【0115】
アゴニスト抗trkC抗体は、下位運動ニューロン疾患を治療するための(1種または複数のニューロトロフィンなどの)1種または複数の他の薬剤と共に、すなわち、1種または複数の薬剤と組み合わせて、一斉に、または順次投与することができる。例えば、アゴニスト抗trkC抗体は、NT−4と共に投与することができる。共に投与とは、本明細書では、同時投与および/または異なる時点での投与を含む。共に投与は、合剤(すなわち、アゴニスト抗trkC抗体と他の薬剤とが同じ組成物中に存在するもの(組み合わされたもの))としての投与、および/または別個の組成物としての投与も含む。本明細書では、「共投与」は、アゴニスト抗trkC抗体と他の薬剤が有効量で個体に投与される任意の状況を含む意味である。本明細書でさらに論述するように、アゴニスト抗trkC抗体と他の薬剤を、異なる投薬頻度および/または投薬間隔で投与できることは理解される。例えば、アゴニスト抗trkC抗体は、毎週投与することができ、その間他の薬剤(例えばNT−4)をより頻繁に投与することができる。アゴニスト抗trkC抗体と他の薬剤は、同じ投与経路または異なる投与経路を使用して投与することができ、その異なる投与計画を投与の過程で変化させてよいことは理解される。投与は、下位運動ニューロン疾患の発症前でよい。
【0116】
アゴニスト抗trkC抗体をニューロトロフィンと共に投与するものでは、ニューロトロフィン(例えば、CNTF、NT−4)をコードするポリヌクレオチドを、本明細書に記載の発現ベクターを利用する所望の細胞(例えば骨格筋細胞)でのニューロトロフィンの送達および発現に使用してもよい。
【0117】
アゴニスト抗trkC抗体による治療の有効性を評価する方法
治療有効性の評価は、いくつかの異なるレベルで実施することができる。評価は、臨床徴候(例えば、体力試験、電気生理学的な応答、または分子的な変化)をモニターして行うことができる。これらには、標準の神経学的な検査または患者への問診によって決定されるパラメータを含めることができ、あるいは例えば本明細書に記載のより専門の定量的試験によって測定できるものでよい。これらのより専門の定量的試験には、微小神経電図検査などの手段による冒されたニューロンの伝導速度測定、筋電図検査(EMG)、握力などの随意筋力測定、音節の反復、15フィート(4.57メートル)歩くのにかかる時間によって測定される歩行速度、努力性肺活量(FVC)の測定を含む呼吸器の機能試験、呼吸器の能力障害の進行に関連する選択された事象の発生率、聴力、平衡性の試験、固有受容または運動感覚の専門試験、体力試験、筋電図検査;その限りでないが、血圧コントロール試験、様々な生理的および薬理学的な刺激に対する心拍応答試験を含む自律神経機能の試験を含めることができるが、この限りでない。これらの試験には、運動技能または体力の試験を含めることもできる。
【0118】
下位運動ニューロン疾患の治療で使用するための組成物
本発明は、本明細書に記載の方法のいずれかで使用するための組成物も提供する。本発明の方法で使用する組成物は、有効量のアゴニスト抗trkC抗体を含む。そのような組成物の例ならびに製剤方法は、より前の項および以下でも記載する。本発明は、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べた組成物のいずれかも提供する。
【0119】
本発明で使用する組成物は、凍結乾燥製剤または水溶液の形で、薬学的に許容できる担体、賦形剤、または安定剤をさらに含む場合もある(「Remington:The Science and practice of Pharmacy」、第20版(2000年)、Lippincott Williams and Wilkins、K.E.Hoover編)。許容される担体、賦形剤、または安定剤は、その投与量および濃度でレシピエントに対して非毒性であり、リン酸塩、クエン酸塩、他の有機酸などの緩衝剤;アスコルビン酸およびメチオニンを含む抗酸化剤;保存剤(塩化オクタデシルジメチルベンジルアンモニウム;塩化ヘキサメソニウム;塩化ベンザルコニウム;塩化ベンゼトニウム;フェノール;ブチルアルコールまたはベンジルアルコール;メチルパラベンやプロピルパラベンなどのアルキルパラベン;カテコール;レゾルシノール;シクロヘキサノール;3−ペンタノール;m−クレゾールなど);低分子量(約10残基未満)のポリペプチド;血清アルブミン、ゼラチン、免疫グロブリンなどのタンパク質;ポリビニルピロリドンなどの親水性ポリマー;グリシン、グルタミン、アスパラギン、ヒスチジン、アルギニン、リジンなどのアミノ酸;単糖、二糖、ならびにグルコース、マンノース、もしくはデキストランを含む他の炭水化物;EDTAなどのキレート化剤;スクロース、マンニトール、トレハロース、ソルビトールなどの糖;ナトリウムなどの、塩を生成する対イオン;金属複合体(例えばZn−タンパク質複合体);および/またはTWEEN(商標)、PLURONICS(商標)、ポリエチレングリコール(PEG)などの非イオン性界面活性剤を含む場合もある。薬学的に許容できる賦形剤については本明細書でさらに述べる。
【0120】
一態様では、本発明は、アゴニスト抗trkC抗体を含む組成物を提供する。他の実施形態では、アゴニスト抗trkC抗体はヒトtrkCを認識する。さらに他の実施形態では、アゴニスト抗trkC抗体は、(本明細書に記載の抗体A5などの)ヒト化されたものである。他の実施形態では、抗trkCアゴニスト抗体は、抗体A5の1個または複数のCDR(A5からの1、2、3、4、5個、または一部の実施形態では6個すべてのCDRなど)を含む。さらに他の実施形態では、抗trkCアゴニスト抗体は、表1に示す重鎖可変部のアミノ酸配列(配列番号1)と表2に示す軽鎖可変部アミノ酸配列(配列番号2)とを含む。さらに他の実施形態では、アゴニスト抗trkC抗体はヒト抗体である。
【0121】
組成物が、複数種のアゴニスト抗trkC抗体(例えば、異なるtrkCエピトープを認識するアゴニスト抗trkC抗体の混合物)を含む場合もあることは理解される。他の好例となる組成物は、同じエピトープを認識する複数種のアゴニスト抗trkC抗体、または異なるtrkCエピトープに結合する異なる種のアゴニスト抗trkC抗体を含む。
【0122】
アゴニスト抗trkC抗体およびその組成物は、アゴニスト抗trkC抗体の有効性を強化および/または補完するのに役立つ他の薬剤と共に使用することもできる。例えば、そのような追加の化合物には、下位運動ニューロン疾患の治療に有用であることがわかっている化合物、(CNTF、NT−3、NT−4、BDNF、およびGDNFを含む)1種または複数のニューロトロフィン、およびアゴニスト抗trkBアゴニストを含めることができる。このような分子は、意図した目的に有効な量で組み合わされて存在することが適切である。trkCアゴニスト抗体およびその組成物は、抗体の有効性を強化および/または補完するのに役立つ他の薬剤と共に使用することもできる。
【0123】
キット
本発明は、この方法で使用するためのキットも提供する。本発明のキットは、抗trkCアゴニスト抗体を含む1個または複数の容器を含み、一部の実施形態では、(下位運動ニューロン疾患の治療方法などの)本明細書に記載の本発明の方法のいずれかに従って使用するための説明書をさらに含む。一部の実施形態では、これらの説明書は、個体が下位運動ニューロン疾患を有しているかどうか、かつ/または下位運動ニューロン疾患にかかるリスクがあるかどうかの確認に基づき治療に適する個体を選択することについての記述を含み、この疾患の治療および/または予防のためのtrkCアゴニスト抗体の投与についてさらに述べる場合もある。本発明はまた、医薬としての使用および/または医薬の製造のための使用のいずれの状況にせよ、本明細書に記載の任意の使用について述べたキットのいずれかを提供する。
【0124】
したがって、一実施形態では、本発明は、アゴニスト抗trkC抗体を含むキットを提供する。一部の実施形態では、本発明は、本明細書に記載の方法で使用するための、アゴニスト抗trkC抗体を含むキットを提供する。本発明のキットは、適切な包装が施されている。適切な包装には、その限りでないが、バイアル、瓶、広口瓶、フレキシブルな包装(例えば、密閉されたMylar製またはプラスチック製バッグ)などが含まれる。一部の実施形態では、キットは、容器と、容器に貼られまたは添えられるラベルまたは添付文書とを含む。ラベルまたは添付文書は、組成物が下位運動ニューロン疾患を治療し、予防し、または緩和させるのに有用であることを表示する。説明書は、本明細書に記載の方法のいずれかの実施に向けて用意することができる。容器は、下位運動ニューロン疾患の治療に有効な組成物を保持し、無菌の出入口を有する場合もある(例えば、容器は、静脈内溶液バッグ、または皮下投与針によって突き刺すことのできる栓を有するバイアルでよい)。組成物中の少なくとも1種の活性薬剤は、trkCアゴニスト抗体である。容器は、第2の薬学的に活性のある薬剤をさらに含む場合もある。キットは、緩衝剤や説明的な情報などの追加の構成成分を場合により提供することもある。
【0125】
以下の実施例は、本発明を限定するのでなく例示するためのものである。
【実施例】
【0126】
SMARD1動物モデルに対するアゴニスト抗trkC抗体およびNT−4の効果
呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)は、小児の致命的な常染色体劣性遺伝疾患である。この疾患は、下位運動ニューロンの変性、進行性の筋肉麻痺、および呼吸不全を特徴とし、これらに対して有効な治療は存在しない。nmd(神経筋変性)マウスの表現型は、ヒトSMARD1に酷似している。nmdマウスにおける変異型のマウス遺伝子、すなわちIghmbp2の同定は、SMARD1に罹患したヒトにおける相同遺伝子の突然変異の発見をもたらした。発明者らは、in vivoの電気生理学的な技術を用いてnmdマウスモデルを研究し、Mab2256、すなわちチロシンキナーゼ受容体C(trkC)に対してアゴニスト効果を有するモノクローナル抗体の、nmdマウスにおける疾患の進行に対する有効性を評価した。Mab2256による治療は、nmdマウスにおいて、著しいが一過性の筋力向上をもたらすだけでなく、高頻度の神経刺激の際の神経筋の衰弱を正常化した。これらの結果は、SMARD1などの下位運動ニューロン疾患においてニューロトロフィン受容体に対するモノクローナルアゴニスト抗体を使用することの可能性を示唆するものである。
【0127】
この研究では、SMARD1のnmdマウスモデルの後肢筋および横隔膜筋のin vivoの電気生理学的な特徴付けを実施し、このマウスモデルにおいてtrkC受容体に対するアゴニストのモノクローナル抗体(Mab2256)の潜在的な有効性を評価した。Mab2256治療が筋力の初期の衰えを予防し、電気刺激を繰返し与えた際の衰弱レベルが正常に回復したことからなる筋肉機能の電気生理学的な改善をもたらしたことが示される。しかし、そのような初期の改善は、筋線維の保存または生存の利益には至らず、この治療戦略をさらに最適化することが強調された。
【0128】
結果
trkC抗体Mab2256のアゴニスト特性および薬物動態特性
ヒトtrkC細胞外ドメインに対するモノクローナル抗体を作製し、アゴニスト活性があるかどうか、細胞に基づく受容体チロシンリン酸化アッセイ(16)を使用してスクリーニングした。ネズミIgG1アイソタイプの1つのモノクローナル抗体であるMab2256は、trkAまたはtrkBとの交差反応性なしにtrkC受容体の特異的なバインダーとなることがわかった(データは示さず)。trkCを発現する安定なCHO細胞中で、Mab2256は、最大半有効濃度(EC50)0.87nMでtrkCのチロシンリン酸化を誘発し、一方NT3は、EC50 1.09nMでtrkCのチロシンリン酸化を誘発する(図1A)。そのtrkC受容体活性化能と一致して、Mab2256は、ラット胚三叉神経ニューロン培養物の生存もEC50 2.58nMで用量依存的に援助し、一方NT3のEC50は0.73pMである(図1B)。
【0129】
次に、Mab2256の薬物動態特性を検討した。発明者らは、マウスの腹腔内に投与したとき、Mab2256の血清からの排出相半減期(t1/2)が約199時間であることを見出した。これは、抗体に期待される長く循環する半減期であり、発表されている、内因性trkCアゴニストNT3の約1.28分の血漿半減期と比較すると有利である(17)。
【0130】
nmdマウスの運動能力および生存に対するMab2256の効果
変異マウスは、出生時には野生型と識別不能に見受けられたが、出生の2週間後には摂食および成長が不十分になった。したがって、ホモ変異体は、そのより少ない体重によって野生型およびヘテロ接合体の同腹仔と容易に識別可能であった(図2A)。さらに、変異マウスは、筋肉質量が進行性に損失し、前肢および後肢の握力が顕著に低下した(8〜11)。変異マウスは、重力に対してその身体を支えられず、ケージカバーを掴むことができず、肩および骨盤帯の筋肉質量を徐々に失った。
【0131】
本発明者らは、Mab2256による処置(5mg/体重kg、週2回、出生後20日目、すなわちP20に開始)が、nmdマウスで起こる筋肉機能の進行性の損失を予防し得るかどうかを検討した。Mab2256(n=10)またはPBS(n=5)を6〜8.5週間注射したマウスにおいて、ELISA分析によって血清中のモノクローナル抗体の存在を確認した。血清中のMab2256抗体の平均濃度は、10.6±3.4μg/mLであった(Mab2256注射したマウスn=10)。野生型およびnmdのMab2256処置マウスに副作用は認められなかった。処置したマウスと未処置のマウスとの体重に有意差はなく(図2A、B)、P60では、Mab2256処置した突然変異体の体重は14.5±0.2g(n=16)であり、未処置または媒体対照の体重は14.2±0.83g(n=8)であった。
【0132】
未処置およびMab2256処置した4〜10週齢のマウスにおいて、標準の神経学的検査を実施した。前肢握力を調べるために、マウスをその前肢によって床から10cm上に金属ワイヤーに吊るした。マウスが吊るされたままであった期間を記録した。未処置のnmdマウス(PBS注射したものn=5、注射しなかったものn=9)は、試験全体を通してその前肢のほとんど完全な握力の損失を示した(図2C、塗りつぶされた四角形)。Mab2256処置したnmdマウスは、著しく数週間握力を保持した(図2C、三角形)。PBS注射したnmdマウスが自身をワイヤー上に保つことができた平均期間は、4週齢で0.8±0.6秒であり(n=5)、Mab2256処置したnmdマウスでは同じ年齢で5.1±1.6秒(9匹のマウス、6匹の同腹仔)(p<0.03)、1週間後には4.9±1.4秒(p<0.034)であった。ヘテロ接合体の能力(図2C、灰色四角形、n=8)は、野生型の同胞と同一であった(図2C、白色の四角)。
【0133】
nmdマウスが自身を一定のスピードの回転棒上に保つ能力によって、運動協調性も測定した(試験時間:10秒)。未処置のnmdマウスは、野生型および異型接合性の同腹仔と比べて平衡性が非常に不十分であった(図2D、E)(p<0.001)が、Mab2256処置したnmdマウスは、処置してから1週間後に有意により良好な運動協調性を示した(図2D、F)(p<0.018)。これらの結果は、Mab2256処置がnmdマウスにおいて疾患の進行を緩和させたことを示唆するものであった。
【0134】
Mab2256が生存に影響するかどうかに解答するために、未処置およびMab2256処置したnmdマウスの寿命を離乳から成人期まで記録した。著しい寿命の延長は認められず(マンホイットニーの順位和検定)、Mab2256処置した変異マウスの中央値の寿命は69日間(n=15)であり、未処置nmdマウスでは62日間(n=30)(図2G)であった。
【0135】
nmdマウスにおける電気的神経筋活性
P70に内側腓腹筋(MG)でin vivoEMG測定を実施した。まず最大上振幅の単一の電流パルスによって坐骨神経を刺激し、複合筋活動電位(CMAP)を記録した。nmdマウスにおけるCMAPの平均振幅は、対照値の<50%に縮小した(図3A;それぞれ22.9±5.6mV、n=9、および48.8±4.8mV、n=14;p<0.002)。Mab2256の投与は、P70のnmd(24±3.1mV、n=7)または野生型マウス(61±9.2mV、n=8)においてCMAPの平均振幅に著しい効果を与えず(図3A)、Mab2256による治療プロトコルでは運動線維の損失を止められないことが示唆された。trkBアゴニストがtrkCアゴニストより良好となり得るかどうかを問うため、一群の変異マウスをtrkBアゴニストNT−4/5(5mg/体重kg、週2回、P20に開始)でも処置した。この場合では、CMAPの平均振幅は、NT−4/5処置によっていずれも変化しなかった(P70で26.7±7.1mV、n=3)。
【0136】
Mab2256およびNT−4/5は、高周波によって誘発される神経筋衰弱を正常レベルに回復させる
nmdマウスにおける神経筋の電気的特性ならびにMab2256およびNT−4/5の効果をさらに調査するために、対パルス、および周波数の異なる短い一続きの刺激を使用し、筋電計の応答を調査した。MGでは、nmdマウスにおける対パルス最大上刺激(10秒間隔)に対するCMAP応答の振幅(A1およびA2)は、野生型のものと大いに異なっていた。野生型マウスでは、第2のCMAP(A2)の振幅は、第1の応答(A1)よりかすかに大きいまたは同等であった(図3B)。変異マウスでは、第2の応答の振幅は、第1の応答よりも21.4±0.03%小さかった(n=7)(図3C)(p<0.005)。したがって、CMAP曲線下の面積はその振幅によって変化し、活性化した線維数が実際に変化したものであり、見せかけの低下ではなかったことが示された(データは示さず)。
【0137】
250msの一続きの100Hz刺激では、未処置の野生型MGにおけるCMAPの振幅は、一貫したパターンを示した。すなわち、CMAP振幅は、最初の3〜4回の刺激の間に徐々に増大し、次いで縮小の疑似定常状態値に達するまでの刺激期間にわたって段々に低下した(図3D、上部のトレース;図3E、空の四角形)。このパターンは、その刺激の第1と第2のCMAPの間で振幅が最大に落ち込んだ後、記録期間にわたってさらに縮小することを特徴とする、未処置のnmdマウスで見られるものと非常に異なっていた(図3D、2番目のトレース;図3E、塗りつぶされた四角形)。未処置の突然変異体では、縮小が急速であり、CMAPの標準化された振幅は、刺激の終わりには野生型よりも大いに小さくなり(それぞれ55±6.9%、n=6、および82.5±7.4%、n=6)(p<0.02)、伝達が活動電位発生の閾値を下回っている線維数の増加が示唆された。刺激しない間隔(2〜4分)を挟んだ後同じ刺激パターンを繰り返すと、刺激のそれぞれの実施に対してほとんど同一の応答パターンが得られた。
【0138】
神経筋機能に対するMab2256の有効性を評価するために、モノクローナル抗体で処置した動物の記録を行い、結果を未処置マウスと比較した。繰返しの神経刺激(100Hz)では、疑似定常状態での標準化されたCMAP振幅は、Mab2256処置nmdマウス(74±4.1%、n=6)(図3D、3番目のトレース)において、PBS注射した突然変異体(55±6.9%、n=6)(図3E、上部のグラフ)よりも有意に大きい(p<0.05)。しかし、刺激の終わりの応答の低下量は、Mab2256処置マウスと未処置の野生型マウスで有意には異なっておらず(それぞれ78±9%、n=8、および82.5±7.4%、n=6)(図3E、上部のグラフ)(p<0.73)、モノクローナル抗体による処置が正常な神経筋機能を変化させなかったことが示唆された。
【0139】
NT−4/5処置したnmdマウスでも、最後の定常状態CMAPの標準化された振幅が、100HzでPBS注射した突然変異体より有意に大きかった(79±2.5%、n=3)(図3D、下方のトレース;図3E、下方のグラフの塗りつぶされた円)(p<0.02)。野生型、Mab2256処置、および未処置突然変異体における異なる周波数(10、20、50、および100Hz)での縮小量を図3Fに示す。これらの結果は、Mab2256およびNT−4/5の両方が、高周波によって誘発される神経筋の衰弱をほぼ完全に正常レベルに回復し得たことを示唆している。
【0140】
nmdマウスにおける紡錘からの求心性線維の機能状態
筋紡錘からの求心性線維の機能状態を評価するために、未処置およびMab2256処置nmdマウスにおいて、H波が容易に検出可能である背側の足筋からの記録をとった。H波は、運動線維の活性化によって、単シナプスの固有受容性感覚求心回路(挿入図4A)を通して誘発され、神経運動線維の直接の刺激によって誘発されるM波の後に続く(図4A)。M/H波の比は、野生型(6.1±1.3、n=6)、未処置nmd(6.1±0.9、n=5)、およびMab2256処置したnmdマウス(7.6±2.6、n=4)で異なっておらず、背側の足筋の伸展反射を司る筋紡錘線維が突然変異体で優先的に失われないこと、およびこの処置がこの回路に影響を及ぼさなかったことが示唆された。
【0141】
背側の足筋では、nmdマウスおよび対照マウスにおける異なる刺激周波数でのEMG応答の低下量も調査した。10〜50Hzの周波数で野生型マウスとnmdマウスとにCMAPの平均疑似定常状態低下の有意差はなかった(図4B)。100Hz(図4C)でのみ、低下量がnmdマウス(40±1.6%、n=6)において野生型(32±2.85%、n=6)よりわずかに大きく(P<0.036)、この動物モデルでは背側の足筋がMGほど影響を受けないことが示唆された。
【0142】
MUNE分析
nmdマウスのMGで記録されたCMAP振幅の縮小は、機能運動単位数の減少を示唆するものである。運動単位数推定(MUNE)を使用して、nmdマウスのMGにおける運動ニューロン損失の程度を評価した。機能運動単位数を変異マウス(P215〜P230)の老齢期に測定し、対照同腹仔と比較した。その年齢では、突然変異体においては後肢の筋萎縮症が非常に重篤であったが、ヘテロ接合体では筋肉消耗または体力減退の徴候が認められなかった。連続的な漸増刺激は、対照では運動単位電位の規則的な増加を生じるが(図5A)、nmdマウスでは異常に大きい運動単位電位を誘発した(図5B)。強度が段々と増す10回の「成功した」刺激(すなわち、応答の振幅を増大させた刺激)の後の最終電位サイズは、突然変異体では大きな歩幅の増大(すなわち、巨大な運動単位)が存在するために、対照同胞マウスよりも突然変異体においてはるかに大きかった。2匹の対照(ヘテロ接合体)および2匹の突然変異体同腹仔の電位サイズの定量化を、図5C〜Fのグラフに示す。平均の単一運動単位活動電位(SMUAP)振幅は、nmdマウスでは1.21±0.62mV(n=3)であり、対照マウスでは0.178±0.06mV(n=3)であった。したがって、MUNEは、nmdマウス(平均値は37.5±11.4、n=3)において異型接合性の同腹仔(105.5±12.4、n=3、P<0.0015)と比べて50%超の減少があった。さらに、対照マウスに十分な弱い強度の刺激は、突然変異体では少しの応答も誘発せず、突然変異体の残りの運動単位の大部分で線維活性化の閾値が増大したことが示唆された(データは示さず)。巨大な運動単位の存在は、おそらくは神経筋伝達の重度の損失を少なくとも部分的に代償する、軸索の出芽および除神経された筋線維の神経再支配の徴候である。しかし、突然変異体の力が主として巨大な運動単位に依存するならば、これらの単位の損失が後に筋肉機能の突然の不全をもたらす場合もある。
【0143】
nmdマウスにおける横隔膜の機能
呼吸不全は、早くも生まれてから最初の年にヒトSMARD1に特徴的なものである(4)。nmdマウスにおける呼吸機能およびモノクローナル抗体の潜在的な有効性を評価するために、麻酔条件下でのマウス横隔膜の電気生理学的な特性を検討する。10週齢を超える12匹のマウス、すなわち、対照(野生型およびヘテロ接合体、n=6)、未処置nmdマウス(n=3)、およびMab2256処置したnmdマウス(n=3)がこの研究に含まれた。横隔膜の電気活性は、交互に起こる、自発性の活動電位のバースト(吸息バースト)と呼息と同時に起こる無変化の期間を特徴とする。対照および未処置nmdマウスの横隔膜からの典型となる記録を、図6A、Bの下方のトレースに示す。一般に、対照およびnmdマウスで吸息後の電気活性は、認められなかった。本発明者らは、持続時間および吸息バーストの活性(TI)が少ししか変動しなかったことを発見した。対照マウスの呼吸回数(140.2±15.7bpm、n=6)は、未処置のnmd(141.7±6.2bpm、n=3)およびMab2256処置したnmdマウス(132.9±22.6、n=3)と同様であった。しかし、平均吸息持続時間(TI)は、未処置のnmdマウス(131.6±4.1ms)において対照同腹仔(184.1±11.7ms)と比べて著しく短縮し(28%)(P<0.005)、老年期のnmdマウスが軽度の異常な吸息運動の消失を有することが示唆された。Mab2256処置したnmdマウスでは、TIは146.3±8.2msであり、未処置のnmdマウスよりわずかに長かった。この差は、おそらくはサンプルサイズが小さいために統計学的有意性には到達しなかった(P<0.08)(図6C)。非常に高週齢の突然変異体(38週齢)の横隔神経横断面(図6D)の組織学的検査では、nmd(軸索337±18本、n=2)と対照同腹仔(軸索364±11本、n=2)の有髄軸索数に有意差は示されなかった(図6E)。このことは、nmdマウスにおける呼吸機能不全の根底にあるものが、横隔神経の解剖学的な損失というよりも機能の欠陥であることを示唆している。
【0144】
考察
本発明者らは、nmdマウスの筋電図記録の特性を調査し、疾患の臨床的および電気生理学的な進行に対するtrkC受容体のアゴニストモノクローナル抗体(Mab2256)の有効性を試験した。Mab2256処置が筋力低下を数週間遅らせ、筋肉機能の電気生理学的な改善をもたらしたことを示す。
【0145】
nmdマウスにおける神経筋機能障害
nmd神経筋機能に若干の著しい変化が認められた。主な欠陥には、運動神経線維の重度の損失、およびnmdマウスが繰返しの神経刺激で正常な神経筋伝達を維持できないことが含まれる。通常、繰返しの神経刺激では、CMAPは、いくつかのパルスの後、安定した低下レベルに到達するまで振幅が徐々に縮小し、最大低下度は刺激回数に正比例する。本発明者らは、nmdと野生型のMGでこの生理的応答を比較し、nmdマウスが、その臨床的な欠点と一致した、同腹仔対照よりもはるかに重度のCMAP振幅の縮小を示したことを発見したが、このことは、他の運動ニューロン機能障害マウスモデルでも共通の知見である(18)。nmdマウスにおけるこの現象が、シナプス前終末での欠陥によるものか、シナプス後の効力の低下によるものか、または筋肉線維のより疲労性の表現型への移行によるものかを確認することが将来的に問題となるはずである。
【0146】
CMAP平均振幅は、P70のnmdマウスMGにおいて50%を超えて縮小した。これは、この突然変異体で以前に記載されている5週間での腰椎運動ニューロンの41%までの低下に一致している(11)。疾患のその後の段階(P150〜230)では、MG筋に残存する運動単位数は、対照値の35%に減少したが、12〜14週間の腰髄に28%の運動ニューロンが残存するという以前の推定と離れてはいない(11)。巨大な運動単位電位は、活性化の閾値が高いので、補充が不十分な場合もある。運動単位の重度の損失と共に、繰返しの刺激で有効な伝達を維持できないことは、これらの動物の筋力の減弱を説明するものである。
【0147】
神経栄養因子の治療的な可能性
神経栄養因子は、脊髄運動ニューロンの生存を支え、運動ニューロン疾患の動物モデルにおいて病理学的な症状の軽減にプラスの影響を及ぼすことが示されている(14、19、20)。これらの結果に基づき、組換え型の神経栄養因子は、十年以上の間運動ニューロン疾患有望な治療薬であると考えられてきた。しかし、臨床試験では、不適当な投与量、副作用などの問題に遭遇している(21、22)。ニューロトロフィン受容体のアゴニストモノクローナル抗体は、外来の投与された神経栄養因子に優るいくつかの理論上の利点、例えば、副作用を低減し得る、所与の栄養受容体に対するその特異性、ならびに薬物投与および治療濃度の維持を容易にするその長い循環半減期を有する。それにもかかわらず、これらの分子は、動物モデルで調査し確認する必要がある。例えば、一部のモデルでは、神経栄養因子による運動ニューロンの救出が、一過性の性質のものであることが判明した(23、24)。本発明者らのnmdマウスでの実験では、Mab2256は、疾患の進行を遅らせるが阻止しなかった。
【0148】
繊毛様神経栄養因子(CNTF)およびNT3は、マウス突然変異体pmn(進行性運動神経細胞障害)の寿命を延長し得ることが示されている(14、25、26)。nmdマウスの平均寿命は、54日であることが記載されている(10)。本発明者らの動物施設でも、未処置nmdマウスの生存中央値は同様であった(64日)。出生後第3週〜第11週にnmdマウスをMab2256で処置しても、生存確率は有意に増大しなかった(中央値:69日)。この結果は、nmdマウスでの全長IGHMBP2のニューロン発現もその生存状態を改善し得なかったという事実に従うものである(10)。
【0149】
Mab2256の一過性の効果について考えられる説明は、ニューロトロフィンを用いる他の研究で示されているように、一部の動物が到達する、血漿中で比較的高レベルとなった薬物が、trkC受容体の下方制御を引き起こしたのではないかということである(27、28)。これが事実であるならば、治療的な投与量および投薬回数は、この効果を回避するために調整すべきである。
【0150】
また、Mab2256は、ニューロン生存バイオアッセイにおいてNT−3ほど強力でなく(EC50が約3000倍の差)、有効でもなかった(最大効果が約3倍の差)。したがって、nmdマウスモデルにおいてより長くより高い有効性を実現するために、Mab2256抗体のより高い親和性/活性のものが必要であると思われる。
【0151】
その上、下位運動ニューロンは、trkBおよびtrkCの両方の受容体を発現する。これらおよびことによると他のニューロトロフィン受容体の活性化は、nmdマウスにおける臨床的に有益な成果に必要であるかもしれない。将来的には、この疾患モデルにおいてtrkBおよびtrkCの両方のアゴニストの同時の適用がより大きな治療利益を提供するかどうかを試験することが重要となる。実際に、本発明者らは、Mab2256および内因性のtrkBアゴニストであるNT−4/5が、繰返しの刺激プロトコルにおいてなど、nmdマウス神経筋接合部の電気生理学的特性の側面を回復させ得ることを見出した。NT−3と他の神経栄養因子の組合せが適切な場合もあり、例えば、CNTF遺伝子およびNT−3遺伝子をコードするアデノウイルスベクターのpmnマウス骨格筋細胞への同時注射は、どちらかのベクター単独でよりも軸索の生存時間を大きく増大させる(26、29)。
【0152】
nmdマウスにおいてMab2256でin vivoで見られるプラスの影響の原因である正確な細胞ターゲットはまだ不明であるが、運動ニューロン、筋細胞、グリア細胞、または上記の組合せである可能性がある。運動ニューロンは、NT−3受容体であるtrkCを発現し、NT−3に応答し、生存時間が段々と増す(15、30〜32)。最近の証拠は、NT−3が、運動ニューロンの生存に対するその効果のほかに、神経筋伝達の有効性に影響を及ぼす場合もあることを示唆している(33〜36)。外来のBNDFまたはNT−3は(神経成長因子NGFではないが)、培養物中で神経筋シナプスを発生させる、自発性およびインパルス誘発型のシナプス活性、すなわち、trkC受容体を媒介とし、因子が存在する限り存続するように思われる効果を増強する(37)。その上、単離したニューロンをNT−3で2日間処理すると、新たに形成された神経−筋肉シナプスでの素量的なAChパケットの平均サイズが増大するのに対し、NT−3に対する抗体、またはチロシンキナーゼ受容体の特異的な阻害剤であるK252aで処理すると、存在するシナプスでの素量サイズが縮小するが、これは、NT−3が素量的なパケットの発生および維持を司ることを示唆している(34)。NT−3によるシナプス伝達の増強も成体海馬スライスで報告されている(38)。筋肉によって合成されるNT−3およびNT−4/5は、シナプス前の運動ニューロンに対して逆方向に働き、それによって、例えばAChおよびニューレグリンの合成を増大させることで、途切れないニューロンの機能分化に影響を及ぼす場合がある(39)。これらのシナプス前の効果のほかに、筋肉によって分泌されるニューロトロフィン(BDNF、NT−3、NT−4/5、およびGNDF(15、40〜43)は、筋線維それ自体に対して自己分泌の方法で働く場合がある。NT−3の外からの投与は、クラーレ処理した筋肉において、その処理によって予め変更されたNMJ構造を回復させることが示されている(44)。筋細胞からのNT−3の放出は、NMJでのシナプスの活性によって次々に調節されるようである(36)。
【0153】
アデノウイルスを媒介とする、NT−3の遺伝子導入は、pmnマウスにおいて運動線維の終端の出芽を促進するが、このことは、NT−3が遠位の軸索構造の維持および再生にも関与することを示唆している(26、29)。さらに、神経栄養因子は、運動神経から脊髄への軸索輸送にも影響を及ぼし得る。チューブリンの重合が変更されたために運動の軸索が変性しているpmnマウス(45)において、腓腹筋に注射された、または切断された坐骨神経に直接に適用された蛍光トレーサーの逆方向性の輸送は、CNTF、BDNF、またはNT−3によって改善することができるが、GDNFまたはNGFによって改善することはできない(46)。
【0154】
Mab2256は分子サイズが大きいため、無処置の脳血液関門を通過できないと思われる。しかし、NT−3および他のニューロトロフィン(NT−4/5、BDNF)は、運動ニューロンの軸索に沿って逆行してその細胞体に輸送することができる(31、42、47、48)。一度trkCに結合したこのモノクローナル抗体が、逆行して輸送され、運動ニューロンの細胞体に直接の効果を与え得るかどうかを知ることは大きな関心の的となるはずである。正確な部位およびtrkC抗体によって発揮される作用様式は、疑いなくさらなる研究が待たれる。
【0155】
横隔膜電気活性
SMARD1患者では、横隔膜の麻痺が生後最初の13カ月の間に現れるが(2)、nmdマウスでは、比較的遅くに呼吸異常が現れる(10)。疾患の最終段階(P150〜230)の際の横隔膜の機能状態を評価するために、変異マウスの横隔膜の自発性の電気活性をin vivoで記録した。麻酔下のマウスでは、対照と突然変異体同腹仔との平均呼吸回数に差は見られなかったが、本発明者らは、吸息持続時間(TI)の26%の短縮と、それに付随する、横隔膜強度のある種の低下を生み出す場合もある各吸息バーストの際の活動電位数の減少を見出した。電気的な活性のない帯域と探査電極の位置決め誤りを識別するのが困難であったので、その記録から、横隔膜内に機能していない領域があるかどうかを見分けることはできない(方法を参照されたい)。最近では、nmdマウス横隔膜中の大量の筋障害性変化の存在が記載されている(11)。さらに、これらのマウスは、二次的に呼吸困難の一因となり得るうっ血性心不全および筋ジストロフィー様の表現型も患う(10)。しかし、有髄の軸索数は、非常に高週齢のnmdマウス(38週齢)では減少しなかったが、これは14週齢nmdマウスで見られたものに従うものであり(11)、ある量の全長機能性のIGHMBP2タンパク質の発現に一致するものである(9)。
【0156】
要するに、本発明者らは、nmdマウスへのMab2256処置が、著しいが一過性の筋力の改善、ならびに高頻度の神経刺激の際の神経筋の機能低下量の正常化を生じたことを発見した。この効果の作用機序を解明する別の研究、ならびに神経筋接合部での神経伝達の際のシナプス前およびシナプス後の事象のより完全な特徴付けは、我々がnmdマウスにおける神経筋の欠陥、運動ニューロン中のIGHMBP2の生理的役割を理解し、このひどい疾患の合理的な治療手法を探索する助けとなるはずである。
【0157】
材料および方法
マウスの繁殖および遺伝子型
B6.BLKS−nmd2JマウスをThe Jackson Laboratoryから入手した。nmd2Jについてヘテロ接合体のマウスを交雑し、野生型、ヘテロ接合体、および突然変異体マウスを実験に使用した。マウスを繁殖させ、突然変異体については食物および水をケージの床の高さで利用可能にしたことを除き、標準の条件で維持した。
【0158】
マウスの遺伝子型を記載されている通りに同定した(9)。簡潔に述べると、(その突然変異についてホモ接合性である)nmdマウスの表現型の原因である点突然変異は、野生型マウスには存在しない新しいDdeI制限部位を生じる。罹患していない子孫の中の保因動物(carrier)を特定するPCRアッセイは、突然変異が存在する694bpのPCR産物を増幅する、順方向プライマー:5’−GCTGGAAACGATCACATACCG−3’および逆方向プライマー:5’−AGCTCCTGATGATCCAATGG−3’の2種のオリゴヌクレオチドプライマーを用いて実施した。
【0159】
マウスの処置
両方の性別のコード化された同腹仔マウスのランダムな群に、モノクローナル抗体Mab2256(5mg/体重kg、週2回、20〜21日齢から;Rinat Neuroscience、米カリフォルニア州パロアルト)、ヒト組換え型NT−4/5(5mg/体重kg、週2回、21日齢から;Genentech、米カリフォルニア州サンフランシスコ)、またはPBSのいずれかを腹腔内注射した。すべての動物処置は、制度上の指針および認可に従って実施した。
【0160】
受容体チロシンリン酸化アッセイ
Mab2256のアゴニスト活性は、細胞に基づくtrkC受容体チロシンリン酸化アッセイで、以前に記載されている通りに評価した(16)。Prism Software(GraphPad、米サンディエゴ)を使用する非線形カーブフィッティングによって、最大半有効濃度を推定した。
【0161】
胚三叉神経ニューロン生存アッセイ
E12 Sprague Dawleyラットから、三叉神経ニューロンの分離した培養物を樹立した。解体した神経節をトリプシン処理し、摩砕によって分断した(49)。ニューロンを、96ウェル組織培養プレート中のポリオルニチン/ラミニン基層上の限定された無血清培地に低密度で播いた。プレーティング時に、様々な濃度のNT3およびMab2256抗体を、それぞれ3通りおよび4通りにして培養物に加えた。異なるそれぞれの条件下でのニューロンの生存を定量化するために、プレーティングから48時間後に生存していたニューロン数をカウントした。Prism Software(GraphPad、サンディエゴ)を使用する非線形カーブフィッティングによって、最大半量有効濃度を推定した。
【0162】
薬物動態研究
メスの成体CD−1マウス(n=3)にMab2256を2mg/kgで腹腔内注射した。次いで、注射してから24、48、136、および184時間後に動物の採血を行った。各時点でのMab2256の血清濃度を以下で述べる通りに測定した。
【0163】
モノクローナル抗体濃度の測定
マウスモノクローナル抗体Mab2256の血清レベルは、標準のサンドイッチELISAによって測定した。96ウェルMaxisorpプレート(Nunc)に、HEK293細胞への一過性の形質移入によって発現させた組換え型ヒトtrkC細胞外ドメイン−IgG Fc融合タンパク質(Rinat Neuroscience)をプロテインAカラムで精製したもの0.2mg/mLを一晩かけて4℃で予め吸収させた。次いで、trkCコートされたプレートを、0.5%のウシ血清アルブミンおよび0.05%のTween−20を含むリン酸緩衝溶液(PBS)で25℃で1時間かけてブロックした。プレートを、0.05%のTween−20を含むPBSで3回洗浄した。Mab2256の標準の希釈系列、ならびに適切に希釈した血清サンプルをプレートに入れて25℃で1時間インキュベートした後、3回洗浄した。西洋わさびペルオキシダーゼ(HRP)結合ウサギ抗マウスIgG(Jackson ImmunoResearch)を1:5000で適用し、25℃で1時間インキュベートして、結合したマウスモノクローナル抗体を検出した。最後に、HRPとTMB基質(KPL、デンマーク)の比色反応によってシグナルを検出した。
【0164】
神経学的試験
突然変異体および同腹仔対照マウスに標準の神経学的検査を行い、神経筋の欠陥の発生および拡大を定量化した(SHIRPAプロトコル;www.mgu.har.mrc.ac.uk/mutabase/shirpa_summary.html)(50)。平衡能測定では、マウスを尾から吊り下げて四角い薄い棒切れの上に降ろし、逆立ちさせた。次いで、棒切れを10秒間1秒毎に手で回転させ、マウスが棒切れに留まる能力を秒数で測定した。
【0165】
前肢握力については、マウスを水平な針金の上に保持し、降ろして前肢で針金を握らせるようにした。マウスが前肢によってつながっている能力を10秒間にわたり記録した。
【0166】
筋電図検査(EMG)
以前に記載されている通り(18)に、麻酔下(100mg/Kgケタミン+10mg/Kgキシラジン)のP70の野生型およびnmdマウスからの筋肉内複合活動電位(CMAP)を記録した。簡潔に述べると、記録針電極を、第5指骨の基部に不関電極を添えて背側の足筋中に置き、または腓腹筋の内側部分(MG)中に置いた。アース電極を尾の付け根に置いた。刺激針電極を坐骨の切込みおよび腓骨上部に置いた。過最大電流パルスの刺激プロトコル(持続時間0.05ms、振幅5mA)を単一パルスまたは10、20、50、および100Hzの短い持続時間のパルス列として適用した。刺激パルスは、孤立したパルス刺激装置(A−MSystems、2100モデル)によって作成した。記録された出力を示差的に増幅し(Brownlee Precision、210Aモデル)、20000サンプル/秒でデジタル方式で取得し(ADInstruments、PowerLab/4SP)、後の分析に向けてコンピュータに保存した。
【0167】
MGからの運動単位数推定(MUNE)は、全運動単位の和である最大CMAPを単一の運動単位電位の平均サイズで割ることによって算出した。単一の運動単位電位のサンプリングは、精密に制御された電流を、10回の応答が漸進的に補充されるまで閾値下のレベルから非常に小さな歩幅で適用するものである漸増法(51、52)によって行った。各電流振幅を3回適用し、安定であるとみなされたので、それらが同一であったなら受け入れた。個々の運動単位振幅は、各応答の振幅を以前の応答の振幅から差し引いて得た。個々の値の平均が単一の運動単位活動電位(SMUAP)サイズの推定となった。
【0168】
in vivo横隔膜記録は、針電極を、中線からわずかにそれた剣状突起の後ろでどちらかの部位に挿入して実施した。不関電極を胸の上、アース電極を尾の付け根に置いた。横隔膜は、呼吸と同時に起こるリズミカルなバーストの発生によって容易に確認された。ピーク振幅および記録の積分による面積によって吸息を定量化した。吸息持続時間(TI)も分析した。6連続の呼吸サイクルから平均値を算出した。
【0169】
研究は、コード化されたマウスを用い、どのマウスを試験しているかについて筋電図記録者は盲検化されるように実施した。
【0170】
すべてのデータは平均±SEMとして報告する。統計学的有意性は、スチューデントのt検定を使用して評価した。統計学的有意性判定のための判定基準レベルは、すべての実験についてP<0.05に設定した。
【0171】
組織像
マウスを過量のケタミン/キシラジンで屠殺し、右側横隔神経を、その横隔膜筋への入口近くで採取した。神経を、4%のパラホルムアルデヒドおよび2.5%のグルタルアルデヒドのPBS中固定液で固定し、次いで2%の四酸化オスミウムで固定した後、spurr樹脂中に包埋した(プラスチック包埋)。厚さ2μmの切片をトルイジンブルーで染色し、光学顕微鏡(Axiovert35、Zeiss)で調べた。各神経について有髄の軸索をカウントした。
【0172】
前述の発明は、理解を明確にする目的で図解および例として若干詳細に記載してきたが、ある種の変更および修正を実行してもよいことは当業者には明白となろう。したがって、その記述および例は、添付の請求項によって叙述される本発明の範囲を限定するものと解釈すべきでない。
【0173】
参考文献
1.Lefebvre,S.、Burglen,L.、Reboullet,S.、Clermont,O.、Burlet,P.、Viollet,L.、Benichou,B.、Cruaud,C.、Millasseau,P.、Zeviani,M.ら(1995年)、「Identification and characterization of a spinal muscular atrophy−determining gene」、Cell、第80巻、155〜65ページ
2.Grohmann,K.、Wienker,T.F.、Saar,K.、Rudnik−Schoneborn,S.、Stoltenburg−Didinger,G.、Rossi,R.、Novelli,G.、Nurnberg,G.、Pfeufer,A.、Wirth,B.ら(1999年)、「Diaphragmatic spinal muscular atrophy with respiratory distress is heterogeneous,and one form Is linked to chromosome 11q13−q21」、Am J Hum Genet、第65巻、1459〜62ページ
3.Grohmann,K.、Schuelke,M.、Diers,A.、Hoffmann,K.、Lucke,B.、Adams,C.、Bertini,E.、Leonhardt−Horti,H.、Muntoni,F.、Ouvrier,R.ら(2001年)、「Mutations in the gene encoding immunoglobulin mu−binding protein 2 cause spinal muscular atrophy with respiratory distress type 1」、Nat Genet、第29巻、75〜7ページ
4.Grohmann,K.、Varon,R.、Stolz,P.、Schuelke,M.、Janetzki,C.、Bertini,E.、Bushby,K.、Muntoni,F.、Ouvrier,R.、Van Maldergem,L.ら(2003年)、「Infantile spinal muscular atrophy with respiratory distress type 1(SMARD1)」、Ann Neurol、第54巻、719〜24ページ
5.Pitt,M.、Houlden,H.、Jacobs,J.、Mok,Q.、Harding,B.、Reilly,M.、およびSurtees,R.(2003年)、「Severe infantile neuropathy with diaphragmatic weakness and its relationship to SMARD1」、Brain、第126巻、2682〜92ページ
6.Wirth,B.(2000年)、「An update of the mutation spectrum of the survival motor neuron gene(SMN1)in autosomal recessive spinal muscular atrophy(SMA)」、Hum Mutat、第15巻、228〜37ページ
7.Mercuri,E.、Bertini,E.、Messina,S.、Pelliccioni,M.、D’Amico,A.、Colitto,F.、Mirabella,M.、Tiziano,F.D.、Vitali,T.、Angelozzi,C.ら(2004年)、「Pilot trial of phenylbutyrate in spinal muscular atrophy」、Neuromuscul Disord、第14巻、130〜5ページ
8.Cook,S.A.、Johnson,K.R.、Bronson,R.T.、およびDavisson,M.T.(1995年)、「Neuromuscular degeneration(nmd):a mutation on mouse chromosome 19 that causes motor neuron degeneration」、Mamm Genome、第6巻、187〜91ページ
9.Cox,G.A.、Mahaffey,C.L.、およびFrankel,W.N.(1998年)、「Identification of the mouse neuromuscular degeneration gene and mapping of a second site suppressor allele」、Neuron、第21巻、1327〜37ページ
10.Maddatu,T.P.、Garvey,S.M.、Schroeder,D.G.、Hampton,T.G.、およびCox,G.A.(2004年)、「Transgenic rescue of neurogenic atrophy in the nmd mouse reveals a role for Ighmbp2 in dilated cardiomyopathy」、Hum Mol Genet、第13巻、1105〜15ページ
11.Grohmann,K.、Rossoll,W.、Kobsar,I.、Holtmann,B.、Jablonka,S.、Wessig,C.、Stoltenburg−Didinger,G.、Fischer,U.、Hubner,C.、Martini,R.ら(2004年)、「Characterization of Ighmbp2 in motor neurons and implications for the pathomechanism in a mouse model of human spinal muscular atrophy with respiratory distress type1(SMARD1)」、Hum Mol Genet
12.Arakawa,Y.、Sendtner,M.、およびThoenen,H.(1990年)、「Survival effect of ciliary neurotrophic factor(CNTF)on chick embryonic motoneurons in culture:comparison with other neurotrophic factors and cytokines」、J Neurosci、第10巻、3507〜15ページ
13.Sendtner,M.、Kreutzberg,G.W.、およびThoenen,H.(1990年)、「Ciliary neurotrophic factor prevents the degeneration of motor neurons after axotomy」、Nature、第345巻、440〜1ページ
14.Sendtner,M.、Schmalbruch,H.、Stockli,K.A.、Carroll,P.、Kreutzberg,G.W.、およびThoenen,H.(1992年)、「Ciliary neurotrophic factor prevents degeneration of motor neurons in mouse mutant progressive motor neuronopathy」、Nature、第358巻、502〜4ページ
15.Henderson,C.E.、Camu,W.、Mettling,C.、Gouin,A.、Poulsen,K.、Karihaloo,M.、Rullamas,J.、Evans,T.、McMahon,S.B.、Armanini,M.P.ら(1993年)、「Neurotrophins promote motor neuron survival and are present in embryonic limb bud」、Nature、第363巻、266〜70ページ
16.Sadick,M.D.、Galloway,A.、Shelton,D.、Hale,V.、Weck,S.、Anicetti,V.、およびWong,W.L.(1997年)、「Analysis of neurotrophin/receptor interactions with a gD−flag−modified quantitative kinase receptor activation(gD.KIRA)enzyme−linked immunosorbent assay」、Exp Cell Res、第234巻、354〜61ページ
17.Poduslo,J.F.およびCurran,G.L.(1996年)、「Permeability at the blood−brain and blood−nerve barriers of the neurotrophic factors:NGF,CNTF,NT−3,BDNF」、Brain Res Mol Brain Res、第36巻、280〜6ページ
18.Fernandez−Chacon,R.、Wolfel,M.、Nishimune,H.、Tabares,L.、Schmitz,F.、Castellano−Munoz,M.、Rosenmund,C.、Montesinos,M.L.、Sanes,J.R.、Schneggenburger,R.ら(2004年)、「The synaptic vesicle protein CSP alpha prevents presynaptic degeneration」、Neuron、第42巻、237〜51ページ
19.Sagot,Y.、Vejsada,R.、およびKato,A.C.(1997年)、「Clinical and molecular aspects of motoneurone diseases:animal models,neurotrophic factors and Bcl−2 oncoprotein」、Trends Pharmacol Sci、第18巻、330〜7ページ
20.Mitsumoto,H.、Ikeda,K.、Klinkosz,B.、Cedarbaum,J.M.、Wong,V.、およびLindsay,R.M.(1994年)、「Arrest of motor neuron disease in wobbler mice cotreated with CNTF and BDNF」、Science、第265巻、1107〜10ページ
21.Miller,R.G.、Petajan,J.H.、Bryan,W.W.、Armon,C.、Barohn,R.J.、Goodpasture,J.C.、Hoagland,R.J.、Parry,G.J.、Ross,M.A.、およびStromatt,S.C.(1996年)、「A placebo−controlled trial of recombinant human ciliary neurotrophic(rhCNTF)factor in amyotrophic lateral sclerosis」、rhCNTF ALS Study Group.Ann Neurol、第39巻、256〜60ページ
22.Thoenen,H.およびSendtner,M.(2002年)、「Neurotrophins:from enthusiastic expectations through sobering experiences to rational therapeutic approaches」、Nat Neurosci、第5巻増刊、1046〜50ページ
23.Diener,P.S.およびBregman,B.S.(1994年)、「Neurotrophic factors prevent the death of CNS neurons after spinal cord lesions in newborn rats」、Neuroreport、第5巻、1913〜7ページ
24.Vejsada,R.、Sagot,Y.、およびKato,A.C.(1995年)、「Quantitative comparison of the transient rescue effects of neurotrophic factors on axotomized motoneurons in vivo」、Eur J Neurosci、第7巻、108〜15ページ
25.Sagot,Y.、Tan,S.A.、Baetge,E.、Schmalbruch,H.、Kato,A.C.、およびAebischer,P.(1995年)、「Polymer encapsulated cell lines genetically engineered to release ciliary neurotrophic factor can slow down progressive motor neuronopathy in the mouse」、Eur J Neurosci、第7巻、1313〜22ページ
26.Haase,G.、Kennel,P.、Pettmann,B.、Vigne,E.、Akli,S.、Revah,F.、Schmalbruch,H.、およびKahn,A.(1997年)、「Gene therapy of murine motor neuron disease using adenoviral vectors for neurotrophic factors」、Nat Med、第3巻、429〜36ページ
27.Bibel,M.およびBarde,Y.A.(2000年)、「Neurotrophins:key regulators of cell fate and cell shape in the vertebrate nervous system」、Genes Dev、第14巻、2919〜37ページ
28.Knusel,B.、Gao,H.、Okazaki,T.、Yoshida,T.、Mori,N.、Hefti,F.、およびKaplan,D.R.(1997年)、「Ligand−induced down−regulation of Trk messenger RNA、protein and tyrosine phosphorylation in rat cortical neurons」、Neuroscience、第78巻、851〜62ページ
29.Sendtner,M.(1997年)、「Gene therapy for motor neuron disease」、Nat Med、第3巻、380〜1ページ
30.Oppenheim,R.W.(1996年)、「Neurotrophic survival molecules for motoneurons:an embarrassment of riches」、Neuron、第17巻、195〜7ページ
31.Yan,Q.、Elliott,J.L.、Matheson,C.、Sun,J.、Zhang,L.、Mu,X.、Rex,K.L.、およびSnider,W.D.(1993年)、「Influences of neurotrophins on mammalian motoneurons in vivo」、J Neurobiol、第24巻、1555〜77ページ
32.Duberley,R.M.、Johnson,I.P.、Anand,P.、Leigh,P.N.、およびCairns,N.J.(1997年)、「Neurotrophin−3−like immunoreactivity and Trk C expression in human spinal motoneurones in amyotrophic lateral sclerosis」、J Neurol Sci、第148巻、33〜40ページ
33.Wang,T.、Xie,K.、およびLu,B.(1995年)、「Neurotrophins promote maturation of developing neuromuscular synapses」、J Neurosci、第15巻、4796〜805ページ
34.Liou,J.C.およびFu,W.M.(1997年)、「Regulation of quantal secretion from developing motoneurons by postsynaptic activity−dependent release of NT−3」、J Neurosci、第17巻、2459〜68ページ
35.Sendtner,M.(1998年)、「Neurotrophic factors:effects in modulating properties of the neuromuscular endplate」、Cytokine Growth Factor Rev、第9巻、1〜7ページ
36.Poo,M.M.(2001年)、「Neurotrophins as synaptic modulators」、Nat Rev Neurosci、第2巻、24〜32ページ
37.Lohof,A.M.、Ip,N.Y.、およびPoo,M.M.(1993年)、「Potentiation of developing neuromuscular synapses by the neurotrophins NT−3 and BDNF」、Nature、第363巻、350〜3ページ
38.Kang,H.およびSchuman,E.M.(1995年)、「Long−lasting neurotrophin−induced enhancement of synaptic transmission in the adult hippocampus」、Science、第267巻、1658〜62ページ
39.Loeb,J.A.およびFischbach,G.D.(1997年)、「Neurotrophic factors increase neuregulin expression in embryonic ventral spinal cord neurons」、J Neurosci、第17巻、1416〜24ページ
40.Griesbeck,O.、Parsadanian,A.S.、Sendtner,M.、およびThoenen,H.(1995年)、「Expression of neurotrophins in skeletal muscle:quantitative comparison and significance for motoneuron survival and maintenance of function」、J Neurosci Res、第42巻、21〜33ページ
41.Henderson,C.E.、Phillips,H.S.、Pollock,R.A.、Davies,A.M.、Lemeulle,C.、Armanini,M.、Simmons,L.、Moffet,B.、Vandlen,R.A.、Simpson,L.C.ら(1994年)、「GDNF:a potent survival factor for motoneurons present in peripheral nerve and muscle」、Science、第266巻、1062〜4ページ
42.Koliatsos,V.E.、Clatterbuck,R.E.、Winslow,J.W.、Cayouette,M.H.、およびPrice,D.L.(1993年)、「Evidence that brain−derived neurotrophic factor is a trophic factor for motor neurons in vivo」、Neuron、第10巻、359〜67ページ
43.Funakoshi,H.、Frisen,J.、Barbany,G.、Timmusk,T.、Zachrisson,O.、Verge,V.M.、およびPersson,H.(1993年)、「Differential expression of mRNAs for neurotrophins and their receptors after axotomy of the sciatic nerve」、J Cell Biol、第123巻、455〜65ページ
44.Loeb,J.A.、Hmadcha,A.、Fischbach,G.D.、Land,S.J.、およびZakarian,V.L.(2002年)、「Neuregulin expression at neuromuscular synapses is modulated by synaptic activity and neurotrophic factors」、J Neurosci、第22巻、2206〜14ページ
45.Bommel,H.、Xie,G.、Rossoll,W.、Wiese,S.、Jablonka,S.、Boehm,T.、およびSendtner,M.(2002年)、「Missense mutation in the tubulin−specific chaperone E(Tbce)gene in the mouse mutant progressive motor neuronopathy,a model of human motoneuron disease」、J Cell Biol、第159巻、563〜9ページ
46.Sagot,Y.、Rosse,T.、Vejsada,R.、Perrelet,D.、およびKato,A.C.(1998年)、「Differential effects of neurotrophic factors on motoneuron retrograde labeling in a murine model of motoneuron disease」、J Neurosci、第18巻、1132〜41ページ
47.DiStefano,P.S.、Friedman,B.、Radziejewski,C.、Alexander,C.、Boland,P.、Schick,C.M.、Lindsay,R.M.、およびWiegand,S.J.(1992年)、「The neurotrophins BDNF,NT−3,and NGF display distinct patterns of retrograde axonal transport in peripheral and central neurons」、Neuron、第8巻、983〜93ページ
48.Yan,Q.、Elliott,J.、およびSnider,W.D.(1992年)、「Brain−derived neurotrophic factor rescues spinal motor neurons from axotomy−induced cell death」、Nature、第360巻、753〜5ページ
49.Davies,A.M.、Horton,A.、Burton,L.E.、Schmelzer,C.、Vandlen,R.、およびRosenthal,A.(1993年)、「Neurotrophin−4/5 is a mammalian−specific survival factor for distinct populations of sensory neurons」、J Neurosci、第13巻、4961〜7ページ
50.Rogers,S.D.、Demaster,E.、Catton,M.、Ghilardi,J.R.、Levin,L.A.、Maggio,J.E.、およびMantyh,P.W.(1997年)、「Expression of endothelin−B receptors by glia in vivo is increased after CNS injury in rats,rabbits,and humans」、Exp Neurol、第145巻、180〜95ページ
51.McComas,A.J.(1991年)、「Motor unit estimation:methods,results,and present status」、Muscle Nerve、第14巻、585〜97ページ
52.Shefner,J.M.およびGooch,C.L.(2002年)、「Motor unit number estimation in neurologic disease」、Adv Neurol、第88巻、33〜52ページ
53.Ruiz,R.、Lin,J.、Forgie,A.、Foletti,D.、Shelton,D.、Rosenthal,A.、およびTabares,L.(2005年)、「Treatment with trkC agonist antibodies delays disease progression in neuromuscular degeneration(nmd)mice」、Human Molecular Genetics、第14巻、1825〜1837ページ
【0174】
略語
BDNF:脳由来神経栄養因子
bpm:1分あたりの呼吸数
CMAP:複合筋活動電位
CNTF:繊毛様神経栄養因子
GDNF:グリア細胞由来神経栄養因子
MG:内側腓腹筋
IGHMBP2:免疫グロブリンμ結合タンパク質2遺伝子
nmd:神経筋変性
MUNE:運動単位数推定
NGF:神経成長因子
NMJ:神経筋接合部
NT−3:ニューロトロフィン3
NT−4/5:ニューロトロフィン4/5
SMA:脊髄性筋萎縮症
SMARD1:脊髄性筋萎縮症1型
SMN:生存運動神経細胞遺伝子
SMUAP:単一運動単位活動電位
TI:吸息時間
trkA:チロシンキナーゼ受容体A
trkB:チロシンキナーゼ受容体B
trkC:チロシンキナーゼ受容体C
【図面の簡単な説明】
【0175】
【図1】モノクローナル抗体Mab2256がtrkC受容体を活性化し、培養物中の三叉神経ニューロンの生存を支え得ることを示すグラフである。(A)漸増濃度のNT3(左パネル)、すなわち内因性のtrkCリガンド、またはモノクローナル抗体Mab2256(右パネル)、すなわちtrkC抗体によって、増大し飽和状態になるレベルのtrkC受容体のリン酸化(OD450で示す、y軸)が誘発された。(B)培養物で48時間生存するラット胚三叉神経ニューロン数は、培地中の様々な濃度のNT3(左パネル)またはMab2256(右パネル)の存在によって支えられて増加し、飽和状態になった。
【図2】nmdマウスの疾患徴候および寿命を示すグラフおよび図である。(A)P20〜P70の野生型(+/+)(n=8)、nmd(−/−)(n=8)、およびヘテロ接合体(+/−)(n=8)マウスの平均体重。(B)モノクローナル抗体Mab2256を腹腔内注射した3〜11週の野生型(+/+Ab)(n=10)およびnmdマウス(−/−Ab)(n=16)の平均体重。(C〜D)PBSを注射した野生型(+/+、空の記号)(n=14)、ヘテロ接合体(+/−、灰色の記号)(n=8)、およびnmd(−/−、塗りつぶされた四角形)(n=5)、ならびにMab2256を注射したnmdマウス(−/−Ab、三角形)(n=9)の前肢握り時間(C)および棒上での平衡能(D)。(E〜F)棒上での野生型およびMab2256処理した突然変異体。突然変異体は、その尾を使って棒を掴むことができなかったが、それにもかかわらず、自身を回転している棒上に数秒間維持することができた。Mab2256で処置していないnmdマウスは、自身を1秒より長く棒上に維持することができなかった(表示なし)。(G)未処置(nmd)(n=30)およびMab2256処置したnmd(nmd Ab)(n=15)マウスのKaplan−Meier生存率分析。マンホイットニーの順位和検定で2つの群について有意差は得られなかった。
【図3】野生型およびnmdマウスの内側腓腹筋(MG)におけるCMAP振幅のEMG測定から、nmdマウスにおける神経伝達の効力の低下が明らかになることを示すグラフである。(A)未処置の野生型(+/+)(n=14)、PBSを注射したまたは注射していないnmd(−/−)(n=9)、Mab2256で処置した野生型(+/+Ab)(n=8)、Mab2256で処置したnmd(−/−Ab)(n=7)、およびNT−4/5で処置したnmdマウス(−/−NT4/5)(n=3)における、過最大の刺激に応答してのCMAPの絶対振幅(平均±SEM)。(B〜C)野生型(B)およびnmdマウス(C)における対パルスプロトコルに対する典型となるCMAP応答。刺激間の間隔:10ms。経時的なCMAPのピークからピークまでの振幅(A1およびA2)を矢印で表示する。(D)野生型(上方のトレース)ならびに3種のnmdマウス:未処置(2番目のトレース)、Mab2256処置(3番目のトレース)、およびNT−4/5処置(4番目のトレース)における、100Hzでの一続きの刺激の間の典型となる記録。(E)未処置の野生型マウス(+/+)(n=6)、Mab2256処置した野生型(+/+Ab)(n=8)、PBS注射したnmd(−/−)(n=6)、Mab2256処置したnmd(−/−Ab)マウス(上方のグラフ、n=6)、およびNT−4/5処置したマウス(−/−NT4/5)(下方のグラフ、n=3)における、100Hzで250msの一続きの刺激の間の(最初の応答を標準として比較した)CMAP振幅の縮小。(F)10〜100Hzの刺激周波数についての疑似定常状態レベルでのCMAP振幅の縮小パーセント。すべてのデータは、P69〜71で行ったEMG記録からのものである。
【図4】背側の足筋におけるCMAP振幅のEMG測定を示すグラフである。(A)野生型マウスにおいて単一のパルス刺激によって誘発したM波およびH波。(B)野生型(+/+)(n=6)およびnmdマウス(−/−)(n=6)における10〜100Hzの刺激周波数についての疑似定常状態レベルでのCMAP振幅の縮小パーセント。(C)野生型マウス(n=6)およびPBS注射したnmd(n=6)における、100Hzでの250msの一続きの刺激の間の(最初の応答を標準として比較した)CMAP振幅の縮小の時間経過。すべてのデータは、P69〜71のマウスからのものである。
【図5】P215〜230のマウスでのMGからの運動単位数推定(MUNE)を示すグラフである。(A〜B)野生型(A)およびnmd(B)マウスの運動単位トレース。(C〜F)対照(CおよびE)ならびにnmd同胞突然変異体(DおよびF)マウスにおける、段々と大きさの増す刺激に応答した単一運動単位活動電位(SMUAP)の振幅。X軸の各数字は、応答の振幅の増大を誘発した刺激を表す。対照動物はヘテロ接合体であった。
【図6】in vivoで記録した横隔膜の自発性の電気活性を示すグラフである。(AおよびB)対照(A)およびnmdマウス(B)の典型となる記録。下方のトレースは未加工の記録であり、上方のトレースは下方のトレースの積分である。円が心臓からの電気活性を表示する。(C)PBS注射した突然変異体(黒色のバー)、およびMab2256処置した突然変異体マウスにおける吸息バーストの平均持続時間(TI)をmsで示すヒストグラム。(D)対照およびnmd横隔神経の組織断面。(E)対照とnmd同腹仔との有意差を示していない4匹の38週齢マウスにおける有髄軸索数の定量化。バー:7μm。
【特許請求の範囲】
【請求項1】
個体に有効量のアゴニスト抗trkC抗体を投与することを含む、個体における下位運動ニューロン疾患の治療方法。
【請求項2】
下位運動ニューロン疾患が脊髄性筋萎縮症(SMA)である、請求項1に記載の方法。
【請求項3】
下位運動ニューロン疾患が、呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)である、請求項1に記載の方法。
【請求項4】
アゴニストtrkC抗体を投与した後に個体の筋力が改善される、請求項3に記載の方法。
【請求項5】
アゴニスト抗trkC抗体がヒトtrkCを結合する、請求項1に記載の方法。
【請求項6】
アゴニスト抗trkC抗体が、アゴニスト抗trkC抗体のFab断片によって評価したとき約1nM未満の親和性でヒトtrkCを結合する、請求項1に記載の方法。
【請求項7】
アゴニスト抗trkC抗体がヒトtrkCおよびげっ歯類trkCを結合する、請求項1に記載の方法。
【請求項8】
アゴニスト抗trkC抗体が、trkCのドメイン4にあるエピトープを結合する、請求項1に記載の方法。
【請求項9】
アゴニスト抗trkC抗体がヒト抗体である、請求項1に記載の方法。
【請求項10】
アゴニスト抗trkC抗体がヒト化抗体である、請求項1に記載の方法。
【請求項11】
アゴニスト抗trkC抗体がモノクローナル抗体である、請求項1に記載の方法。
【請求項12】
アゴニスト抗trkC抗体が、配列番号1からの3個のCDRを含む重鎖可変部と、配列番号2からの3個のCDRを含む軽鎖可変部とを含む、請求項1に記載の方法。
【請求項13】
アゴニスト抗trkC抗体が、配列番号1に示す重鎖可変部アミノ酸配列と、配列番号2に示す軽鎖可変部アミノ酸配列とを含む、請求項1に記載の方法。
【請求項14】
アゴニスト抗trkC抗体が抗体A5である、請求項1に記載の方法。
【請求項15】
アゴニスト抗trkC抗体と薬学的に許容できる担体とを含む、個体において下位運動ニューロン疾患を治療するための医薬組成物。
【請求項16】
a)アゴニスト抗trkC抗体および薬学的に許容できる担体を含む医薬組成物と、
b)有効量の前記医薬組成物を個体に投与するための説明書とを含む、下位運動ニューロン疾患を治療するためのキット。
【請求項1】
個体に有効量のアゴニスト抗trkC抗体を投与することを含む、個体における下位運動ニューロン疾患の治療方法。
【請求項2】
下位運動ニューロン疾患が脊髄性筋萎縮症(SMA)である、請求項1に記載の方法。
【請求項3】
下位運動ニューロン疾患が、呼吸困難を伴う脊髄性筋萎縮症1型(SMARD1)である、請求項1に記載の方法。
【請求項4】
アゴニストtrkC抗体を投与した後に個体の筋力が改善される、請求項3に記載の方法。
【請求項5】
アゴニスト抗trkC抗体がヒトtrkCを結合する、請求項1に記載の方法。
【請求項6】
アゴニスト抗trkC抗体が、アゴニスト抗trkC抗体のFab断片によって評価したとき約1nM未満の親和性でヒトtrkCを結合する、請求項1に記載の方法。
【請求項7】
アゴニスト抗trkC抗体がヒトtrkCおよびげっ歯類trkCを結合する、請求項1に記載の方法。
【請求項8】
アゴニスト抗trkC抗体が、trkCのドメイン4にあるエピトープを結合する、請求項1に記載の方法。
【請求項9】
アゴニスト抗trkC抗体がヒト抗体である、請求項1に記載の方法。
【請求項10】
アゴニスト抗trkC抗体がヒト化抗体である、請求項1に記載の方法。
【請求項11】
アゴニスト抗trkC抗体がモノクローナル抗体である、請求項1に記載の方法。
【請求項12】
アゴニスト抗trkC抗体が、配列番号1からの3個のCDRを含む重鎖可変部と、配列番号2からの3個のCDRを含む軽鎖可変部とを含む、請求項1に記載の方法。
【請求項13】
アゴニスト抗trkC抗体が、配列番号1に示す重鎖可変部アミノ酸配列と、配列番号2に示す軽鎖可変部アミノ酸配列とを含む、請求項1に記載の方法。
【請求項14】
アゴニスト抗trkC抗体が抗体A5である、請求項1に記載の方法。
【請求項15】
アゴニスト抗trkC抗体と薬学的に許容できる担体とを含む、個体において下位運動ニューロン疾患を治療するための医薬組成物。
【請求項16】
a)アゴニスト抗trkC抗体および薬学的に許容できる担体を含む医薬組成物と、
b)有効量の前記医薬組成物を個体に投与するための説明書とを含む、下位運動ニューロン疾患を治療するためのキット。
【図1】


【図2】

【図3】
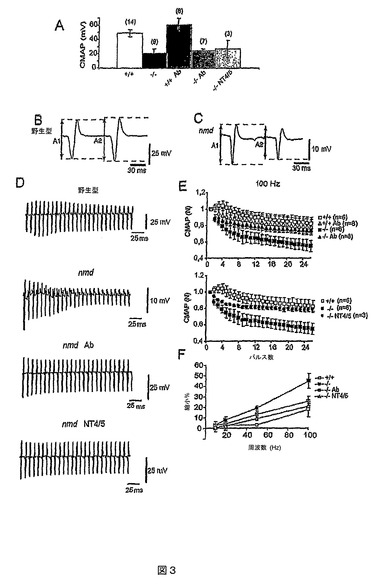
【図4】
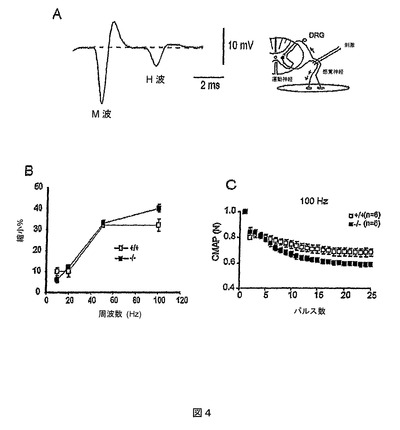
【図5】
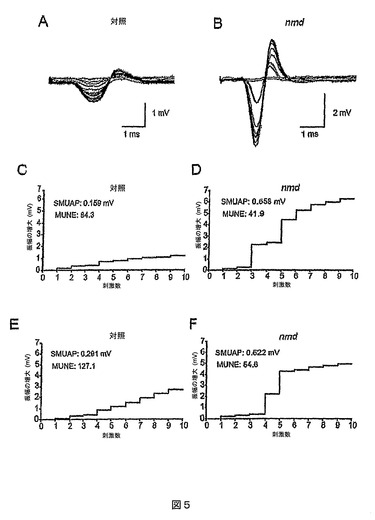
【図6】

【図2】
【図3】
【図4】
【図5】
【図6】
【公表番号】特表2008−539266(P2008−539266A)
【公表日】平成20年11月13日(2008.11.13)
【国際特許分類】
【出願番号】特願2008−509125(P2008−509125)
【出願日】平成18年4月26日(2006.4.26)
【国際出願番号】PCT/US2006/016046
【国際公開番号】WO2006/116609
【国際公開日】平成18年11月2日(2006.11.2)
【出願人】(505129415)ライナット ニューロサイエンス コーポレイション (33)
【Fターム(参考)】
【公表日】平成20年11月13日(2008.11.13)
【国際特許分類】
【出願日】平成18年4月26日(2006.4.26)
【国際出願番号】PCT/US2006/016046
【国際公開番号】WO2006/116609
【国際公開日】平成18年11月2日(2006.11.2)
【出願人】(505129415)ライナット ニューロサイエンス コーポレイション (33)
【Fターム(参考)】
[ Back to top ]