
目的の化合物を産生するための改善された宿主細胞
本発明は、目的の化合物を産生するための組換え宿主細胞に関する。本発明はさらに、そのような宿主細胞の産生方法に関する。本発明はさらに、目的の化合物の産生に関する。本発明はさらに、単離されたポリヌクレオチドならびに前記ポリヌクレオチドを含むベクターおよび宿主細胞に関する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、目的の化合物を産生するための組換え宿主細胞に関する。本発明はさらに、そのような宿主細胞の産生方法に関する。本発明はさらに、目的の化合物の産生に関する。本発明はさらに、単離されたポリヌクレオチドならびに前記ポリヌクレオチドを含むベクターおよび宿主細胞に関する。
【0002】
[背景技術]
本発明は、目的の化合物を産生するための組換え宿主細胞に関する。
【0003】
そのような宿主細胞は、とりわけ、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットから知られており、ここで、宿主細胞は、前記宿主細胞の染色体の少なくとも2つの実質的に相同なDNAドメインの1つに目的のポリヌクレオチドを含み、かつ目的のポリヌクレオチドのコピー数は、遺伝子変換によって増加している。
【0004】
しかしながら、高コピーの遺伝子変換株(例えば、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットによる方法によって得られる株)を得ることは、多くの場合、長い時間と労力を要すると考えられることが示されている。それゆえ、長い時間と労力をあまり要さない高コピーの遺伝子変換株の構築を可能にするために方法を改善することができれば有利であろう。
【図面の簡単な説明】
【0005】
【図1】置換ベクターpGBDELの物理的地図を示す。フランキング領域をクローニングするための多重クローニング部位がamdSマーカーに対して示されている。
【図2】アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))のhdfA遺伝子を不活化するために用いられる置換ベクターpDEL−HDFAの物理的地図を示す。この置換ベクターは、hdfAフランキング領域、amdSマーカーおよび大腸菌(E.coli)DNAを含む。大腸菌(E.coli)DNAは、A.ニガー(A.niger)株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図3】宿主株から遺伝子または遺伝子断片を欠失させるために用いられる戦略を示す。使用されるDNAコンストラクトは、欠失させようとする遺伝子の相同領域(5’および3’)が両隣にあるamdS選択マーカーを含む(1)。このコンストラクトは、対応するゲノム遺伝子座での二重相同組換え(X)によって組み込まれ(2)、ゲノム遺伝子コピーに置き換わる(3)。その後、直接反復上での組換え(U)によって、amdSマーカーが除去され、欠失させようとする遺伝子または遺伝子断片の正確な切除が生じる(4)。
【図4】親A.ニガー(A.niger)株CBS124.903におけるglaA(グルコアミラーゼ)遺伝子座および組換え株GBA300における3つの切断された「Xの印がある」ΔglaA遺伝子座(「X」は、BamHI、SalIまたはBglII制限部位を表す)の物理的地図を示す。
【図5】A.ニガー(A.niger)CBS124.903由来の組換え株GBA300のglaA DNAアンプリコンにおける3つのΔglaA遺伝子座の概略図を示す。各々の遺伝子座は、異なる制限部位(BamHI、SalIまたはBglII)でマークが付けられている。glaA遺伝子座は各々、約20〜60bpだけ長さが異なっており、PCRに基づくDNAフラッグテストによって、切断された異なるglaA遺伝子座を可視化することができる。
【図6】実質的に相同なDNAドメインを適合させるために用いられるベクターpGBGLA−65の物理的地図を示す。基本的に、このベクターは、2つのPglaA断片と3’glaAおよび3”glaA領域の間にamdS選択マーカーを含む。3’glaAおよび3”glaA領域は、ΔglaA遺伝子座の対応するゲノム領域への標的化および組込みに用いられる。2つのPglaA断片は、図3に見られるような対抗選択によるamdS選択マーカーの切除に用いられる。1つのPglaA断片は、(MluI部位の3’にあるPglaAプロモーターの最後の600bpを欠く)切断されたPglaAプロモーター断片であり、これは、amdS対抗選択後にゲノム中に存在したまま残る。
【図7】適合されたBamHIアンプリコンを有する株のglaA DNAアンプリコンにおける3つのΔglaA遺伝子座の物理的地図を示す。BamHIアンプリコンは、ゲノムglaA遺伝子座の相同な3’glaA遺伝子座と3”glaA遺伝子座の間に切断された相同なグルコアミラーゼプロモーター断片を導入することによって適合された。
【図8】新規のアセトアミダーゼ選択マーカーおよび適合されたBamHIアンプリコン標的化ベクターpGBAAS−3の物理的地図を示す。glaAプロモーターは、標的化領域としておよびamdS遺伝子のプロモーターとして機能する。
【図9】新規のアセトアミダーゼ選択マーカーおよび適合されたBamHIアンプリコン標的化ベクターpGBAAS−4の物理的地図を示す。(切断された)glaAプロモーターは標的化領域として機能し、gpdAプロモーターは、amdS遺伝子の発現を駆動する。
【図10】新規の発現および適合されたBamHIアンプリコン標的化ベクターpGBTOP−11の物理的地図を示す。HindIII制限酵素を用いて、ベクターを線状化し、大腸菌(E.coli)部分を除去することができる。
【図11】新規の発現および適合されたBamHIアンプリコン標的化ベクターpGBTOP−12の物理的地図を示す。NotI制限酵素を用いて、ベクターを線状化し、大腸菌(E.coli)部分を除去することができる。
【図12】置換ベクターpDEL−AMYBIIの物理的地図を示す。5’amyBIIフランキング領域、3’amyBIIフランキング領域がamdSマーカーに対して示されている。amyBIIの3’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図13】置換ベクターpGBDEL−FUM3の物理的地図を示す。fumBコード配列の一部を含む5’fumB領域と3’fumBフランキング領域とがamdSマーカーに対して示されている。fumBの5’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図14】置換ベクターpGBDEL−OCH2の物理的地図を示す。フランキング領域としてのAn15g07860遺伝子配列の一部とAn15g07930遺伝子配列の一部がamdSマーカーに対して示されている。An15g07930遺伝子の配列は少なくとも2〜300bp重複し、相同組換えによる対抗選択を可能にしている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図15】置換ベクターpGBDEL−PRT2の物理的地図を示す。5’prtTフランキング領域、3’prtTフランキング領域がamdSマーカーに対して示されている。prtT3’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素BstBIおよびXmaIによる消化によって除去することができる。
【図16】置換ベクターpGBDEL−Sec61*の物理的地図を示す。Sec61*突然変異体遺伝子および3’Sec61*突然変異体遺伝子断片がamdSマーカーに対して示されている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図17】PLA2発現ベクターpGBTOPPLA−2aの物理的地図を示す。glaAフランキング領域が、glaAプロモーター、切断されたglaA遺伝子およびpro−pla2コード配列に対して示されている。大腸菌(E.coli)DNAは、宿主株に形質転換する前に、制限酵素NotIによる消化によって除去することができる。
【図18】PLA2発現ベクターpGBTOPPLA−2bの物理的地図を示す。glaAプロモーター、切断されたglaA遺伝子、pro−pla2コード配列および3’glaAフランキング配列が示されている。大腸菌(E.coli)DNAは、宿主株への形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図19】発現ベクターpGBFINFUA−2の物理的地図を示す。glaAプロモーターおよびA.ニガー(A.niger)amyBコドン最適化配列に対してglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図20】FUA発現ベクターpGBTOPFUA−3の物理的地図を示す。glaAプロモーター、切断されたglaA遺伝子、A.ニガー(A.niger)amyBコドン最適化配列および3’glaAフランキング配列が示されている。大腸菌(E.coli)DNAは、宿主株への形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図21】(例えば、実施例中のGBA303に基づく株のような)適合されたBamHIアンプリコンを有する株における単一の相同組換えによる組込みの概略図を示す。発現ベクターは、特定の標的化配列を有する選択可能なamdSマーカーと目的の遺伝子とを含む。標的化配列は、最も高い頻度の遺伝子変換を有する適合されたアンプリコンへの特異的なゲノム標的化(「賢明な組込み戦略」)を可能にする。
【0006】
[発明の詳細な説明]
驚くことに、宿主細胞(例えば、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットに記載されているような宿主細胞)が少なくとも2つの実質的に相同なDNAドメインを含む場合、これらのDNAドメインの遺伝子変換頻度は必ずしも同じというわけではないことが解析から明らかになった。これらのDNAドメインのうちのいくつかは、他の実質的に相同なDNAドメインの遺伝子変換頻度よりも実質的に低い遺伝子変換頻度を有することが明らかになった。そのような場合、目的のポリヌクレオチドが、高い遺伝子変換頻度を有するDNAドメインではなく、低い遺伝子変換頻度を有するDNAドメインに組み込まれるのは不利である。より高い遺伝子変換頻度を有するDNAドメインは、大方の場合、より低い遺伝子変換頻度を有する(目的のポリヌクレオチドを含む)DNAドメインに優る。その結果として、より低い遺伝子変換頻度を有するDNAドメインに組み込まれた目的のポリヌクレオチドを有する細胞から高コピーの遺伝子変換宿主細胞を得るには、より多くの時間と労力を要する。
【0007】
それゆえ、目的のポリヌクレオチドを実質的により高い遺伝子変換頻度を有する実質的に相同なDNAドメインに含めることができれば有利であろう。というのは、これによって、あまり時間と労力を要さないで目的のポリヌクレオチドの複数コピーを含む宿主株(高コピーの遺伝子変換株)を構築することが可能になるからである。
【0008】
その結果として、目的のポリヌクレオチドが実質的により高い遺伝子変換頻度を有する相同なDNAドメインに優先的に組み込まれる宿主細胞が必要とされている。それゆえ、本発明の目的はそのような宿主細胞を提供することである。
【0009】
したがって、本発明は、目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている、組換え宿主細胞を提供する。前記組換え宿主細胞は、本明細書中、本発明による組換え宿主細胞と称される。
【0010】
本発明はまた、目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることを含む方法を提供する。
【0011】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で本発明による宿主細胞を培養することと、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0012】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で組換え宿主細胞を培養すること、前記宿主細胞は目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドの少なくとも1コピー以上を有する、と、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0013】
したがって、本発明による組換え宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている。前記目的のポリヌクレオチドの1コピー以上の組込みに好適な前記実質的に相同なDNAドメインは、好ましくは、互いの実質的な複製物であり、より好ましくは、前記DNAドメインはアンプリコンである。
【0014】
本発明との関連で、「組換え」という用語は、天然のプロセスおよび/または宿主細胞をランダムな突然変異誘発に供することによって誘導される遺伝子改変を含む任意の遺伝子改変だけでなく、例えば、遺伝子の破壊および/または欠失および/または特異的な突然変異誘発も指す。その結果として、組換えによるプロセスと天然のプロセスの組合せおよび/または宿主細胞をランダムな突然変異誘発に供することによって誘導される遺伝子改変は、組換えであると解釈される。
【0015】
本発明による宿主細胞は、そのゲノム中に目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含む。野生型株では、これらのDNAドメインは、単一コピーとしてまたは複数コピーとして前記細胞のゲノム中に生じ得る。天然の複数コピーDNAドメインの例は、rDNAドメインである。したがって、rDNAドメインを本発明で好ましく用いてもよいし、または用いなくてもよい。本発明によって提供されるような、これらの実質的に相同なDNAドメインの複数のコピーを含む株は、例えば、前記DNAドメイン中の遺伝子によってコードされる遺伝子産物の産生が改善された株を選択することによる古典的な株改良によって、DNAドメインの単一コピーを含む株から得ることができる。多くの場合、そのような産生改善は、選択された株におけるDNAドメインの増幅の結果である。そのような増幅されたDNAドメインは、これ以降アンプリコンとも称される。そのようなアンプリコンを含む宿主細胞の例は、例えば、van Dijckら、2003,Regulatory Toxicology and Pharmacology 28;27−35:On the safety of a new generation of DSM Aspergillus niger enzyme production strainsに記載されている。van Dijckらには、7つの増幅されたグルコアミラーゼ遺伝子座(すなわち、アンプリコン)を含むアスペルギルス・ニガー(Aspergillus niger)株が記載されている。本発明は、そのようなアンプリコンを組換えDNA分子の組込み用のDNAドメインとして用いることが好ましいが、本発明は、決してそれに限定されない。実際、1)DNAドメインは、目的のポリヌクレオチドの組込みを受け入れるのに好適であるべきである;2)DNAドメインは、遺伝子変換による組み込まれた組換えDNA分子の増加を達成するために、真菌ゲノム中のDNAドメインの他の実質的に相同なバージョンとの組換えが可能であるべきであるという2つの機能的基準が満たされる限り、2つ以上の実質的に相同なバージョンが宿主細胞のゲノム中に生じる任意のDNAドメインを用いることができる。
【0016】
第1の基準を満たすには、DNAドメインは、相同組換えによって目的のポリヌクレオチドをDNAドメインに標的化するのを可能にするために十分な長さでなければならない。この目的のために、DNAドメインは、少なくとも100bp、好ましくは少なくとも500bp、より好ましくは少なくとも1kbおよびより好ましくは少なくとも2kbを含むべきである。好ましくは、目的のポリヌクレオチドのその中への組込みに対するDNAドメインの好適性は、DNAドメインへの組込みが、問題になっている宿主細胞の生存に必須の機能を破壊するべきではないという要件によってさらに決定される。
【0017】
第2の機能的基準、すなわち、宿主細胞のゲノム中の他の実質的に相同なバージョンのDNAドメインとの組換えの能力は、異なるバージョンのDNAドメイン間の遺伝子変換を可能にするために必要とされる。この目的のための最低限の必要条件は、フランキング配列間の相同組換えを可能にするよう、各々のバージョンのDNAドメインが、DNAドメインの両端で、他のバージョンのDNAドメインの対応するフランキング配列に十分相同なDNA配列に隣接していることである。この相同組換えの結果が遺伝子変換であり、その場合、これらのバージョンのDNAドメインの1つは、組み込まれた組換えDNA分子を含む複製されたバージョンの他のDNAドメインに置き換えられる。遺伝子変換をなおも可能にするフランキング配列の長さおよび相同性の程度に関する最低限の必要条件は、問題となっている生物によって異なり得る。おそらく、全体的な相同性が少なくとも60%の最低100bpの長さであれば、遺伝子変換がなおも可能となるであろう。明らかに、遺伝子変換の頻度は、フランキング配列の長さと相同性が増大するとともに増加する。好ましくは、異なるDNAドメインは、少なくとも80%の相同性を共有する少なくとも1kbのフランキング配列を含む。さらにより好ましくは、異なるDNAドメインは、少なくとも95%の相同性を共有する少なくとも5kbのフランキング配列を含む。より好ましい実施形態では、異なるDNAドメインは、少なくとも99%の相同性を共有する少なくとも10kbのフランキング配列を含む。最も好ましい実施形態では、異なるDNAドメインは、少なくとも99%の相同性を共有する少なくとも50kbのフランキング配列を含む。本明細書で実質的な複製物として示される、互いの完全なコピーではない実質的に相同なDNAドメインの例は、アレル変異体、遺伝子ファミリーおよび/またはアイソザイムをコードする遺伝子である。最も好ましいのは、組み込まれた目的のポリヌクレオチドが存在するときにせいぜい異なる互いの正確なコピーである実質的に相同なDNAドメインである。そのような同一なDNAドメインの例は、アンプリコンである。あるいは、人工DNAドメインをDNA合成および/または組換えDNA技術を用いて作り出してもよく、また次に、そのような人工DNAドメインを所望の真菌に導入し、増幅させてもよい。したがって、そのような人工DNAドメインを本発明で好ましく用いてもよいし、または用いなくてもよい。
【0018】
実質的に相同なDNAドメインの全体的な長さ、すなわち、フランキング配列を含む全体的な長さは、1kb未満から数百kbまで様々であってもよく、例えば、DNAドメインの長さは、アスペルギルス・オリザエ(Aspergillus oryzae)の親株IF04177(国際公開第9812300号パンフレットおよび米国特許第5,766,912号明細書でA1560として開示されている)中に存在する3つのTAKAアミラーゼ遺伝子の1ユニット当たり2〜3kbの範囲であることが以前に示されており、これらの遺伝子は、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)中の増幅されたペニシリンクラスターの1ユニット当たり約57kbおよびアスペルギルス・ニガー(Aspergillus niger)の増幅されたglaA遺伝子座の1ユニット当たり80kb超までA.オリザエ(A.oryzae)BECh2(国際公開第0039322号パンフレットに開示されている)において不活化されている(国際公開第9846772号パンフレットを参照されたい)。
【0019】
遺伝子変換は、目的のポリヌクレオチドを含むDNAドメインが増加している宿主細胞を同定することによりモニタリングすることができる。例えば、目的のポリヌクレオチドによってコードされる産物の産生レベルがより高い宿主細胞を単にスクリーニングすることなどの、当業者に公知の任意の方法を用いることができる。また、例えば、DNAコピー数を決定するための定量的方法(例えば、本明細書で以下に概説するような「DNAフラッグ」テスト、定量的PCR、サザン解析もしくは比較ゲノムハイブリダイゼーションによる宿主細胞の遺伝子型の解析など)、またはRNAレベルを決定するための定量的方法(例えば、RT−PCR、ノーザン解析もしくはGeneChipもしくはマイクロアレイ解析など)を用いることもできる。
【0020】
本発明によれば、各々のバージョンの実質的に相同なDNAドメインは、好ましくは、同定目的のための独特の配列タグによって糸状菌中の他のバージョンのDNAドメインと区別される。そのような配列タグは、異なるDNAドメイン間の遺伝子変換の検出およびモニタリングを可能にし、それにより、所望の遺伝子型を有する遺伝子変換体のスクリーニングおよび/または選択を容易にする。異なるバージョンのDNAドメインの検出を可能にする限り、例えば、サザンブロット上で検出される制限部位から簡単にアッセイ可能な表現型を提供する完全な選択可能マーカー遺伝子にまで及ぶ任意の形態の配列タグを用いることができる。特に有用な配列タグ法は、国際公開第9846772号パンフレット、国際公開第9846774号パンフレットおよびvan Dijckら(上記)に記載されている。この方法は、単一のオリゴヌクレオチドPCRプライマー対を用いる単一のPCRにおいて各々のDNAドメインを検出することを可能にする。DNAドメインは、PCRにおいて、各々のバージョンのDNAドメインが独特の長さのPCR断片を生じるように改変される。得られるPCR断片の長さおよび強度は、それぞれ、各々のDNAドメインの存在およびコピー数を示す。「DNAフラッグ」と称されるこの形態の配列タグは、所望の遺伝子型を有する変換体を得るために、多数の変換体コロニーの遺伝子型の迅速な解析を可能にする。
【0021】
本発明によれば、本発明による宿主細胞の少なくとも2つの実質的に相同なDNAドメインの1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている。好ましくは、宿主細胞の少なくとも2つの実質的に相同なDNAドメインの1つの標的化DNAドメインは、相同組換えによって目的の組換えDNA分子/ポリヌクレオチドを適合された標的化DNAドメインにより特異的に標的化することを可能にするように適合されている。標的化DNA配列またはDNAドメインが宿主細胞中に2回以上現れる場合、目的のポリヌクレオチドの相同組換えは、宿主細胞の複数の標的化DNAドメインについて見出される。標的化DNAドメインを実質的に相同なDNAドメインの1つに適合させて、宿主中の他の実質的に相同なバージョンのDNAドメインとの組換えに影響を及ぼすことなく、その適合された標的化DNAドメインへのより特異的な標的化を支持することが可能である。最適な標的化選好のために、標的化に用いられるDNA配列の適合は、目的の組換えDNA分子/ポリヌクレオチド中および宿主中で行なわれる。したがって、マッチした宿主と組込みベクターの組が得られる。
【0022】
好ましくは、標的化DNAドメインの適合は、増強された組込み選好を有する配列を実質的に相同なDNAドメインに導入することによって実施される。好ましくは、増強された組込み選好を有する前記配列は、遺伝子置換によって導入される。したがって、本発明はまた、目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、ここで、適合された実質的に相同なDNAドメインは標的化DNAドメインを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、組換え宿主細胞を提供する。より好ましくは、標的化ドメインの適合は、本明細書中の実施例に従って実施される。
【0023】
本発明によれば、標的化に用いられる適合されたDNAドメインの標的化配列は、好ましくは、宿主のゲノム内で独特であるように適合される。好ましくは、宿主のゲノム内で独特であるように適合される標的化配列は、異種DNAポリヌクレオチド配列、宿主から事前に除去され、再導入されるDNAポリヌクレオチド配列、宿主内の野生型状況と比較して異なる順序で組み合わされた相同なDNAポリヌクレオチド断片、および人工DNAポリヌクレオチドの群から選択される配列を含む。
【0024】
本発明によれば、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、好ましくは宿主のゲノム内で独特である。好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、異種DNAポリヌクレオチド配列、宿主から事前に除去され、再導入されるDNAポリヌクレオチド配列、宿主内の野生型状況と比較して異なる順序で組み合わされた相同なDNAポリヌクレオチド断片、および人工DNAポリヌクレオチドの群から選択される配列を含む。
【0025】
好ましくは、適合されたDNAドメインの標的化配列が宿主のゲノム内で独特であるように適合されているとともに、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列も宿主のゲノム内で独特である。
【0026】
本発明によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現に用いられる遺伝子エレメントである。遺伝子エレメントの例は、遺伝子、プロモーター、ターミネーター、cDNA、イントロン、遺伝子間領域、またはそれらの一部および/もしくは組合せである。より好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上の異種遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上の異種遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上のA.ニガー(A.niger)遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上のA.ニガー(A.niger)遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼ遺伝子エレメントおよび/またはフランキング領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼ遺伝子エレメントおよび/またはフランキング領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼプロモーターおよび/またはグルコアミラーゼターミネーター領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼプロモーターおよび/またはグルコアミラーゼターミネーター領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の、野生型状況と比較して逆の順序のA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の、野生型状況と比較して逆の順序のA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。最も好ましくは、適合されたDNAドメインへの標的化に用いられる配列および適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの両方のフランキング配列は同じであり、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。
【0027】
したがって、本発明は、配列番号:2によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドならびに配列番号:2によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドを含むベクターおよび宿主細胞を提供する。機能性は、プロモーター活性を有することおよび増強された組込み選好を有する標的化配列であることと本明細書では解釈されるべきである。好ましくは、配列番号:2によるポリヌクレオチドの機能性断片は、少なくとも100bp、より好ましくは少なくとも200bp、より好ましくは少なくとも300bp、より好ましくは少なくとも500bp、より好ましくは少なくとも600bp、より好ましくは少なくとも700bp、より好ましくは少なくとも800bp、より好ましくは少なくとも900bp、より好ましくは少なくとも1000bp、より好ましくは少なくとも1200bp、より好ましくは少なくとも1500bp、より好ましくは少なくとも2000bp、より好ましくは少なくとも3000bp、およびさらにより好ましくは少なくとも4000bpを含む。好ましくは、機能性断片は、少なくとも約50%同一な、より好ましくは少なくとも約60%同一な、さらにより好ましくは少なくとも約70%同一な、さらにより好ましくは少なくとも約80%同一な、さらにより好ましくは少なくとも約85%同一な、さらにより好ましくは少なくとも約90%同一な、さらにより好ましくは少なくとも約95%同一な、さらにより好ましくは少なくとも約97%同一な、さらにより好ましくは少なくとも約98%同一な、さらにより好ましくは少なくとも約99%同一な一致率(すなわち、位置同一性)を有し、かつさらに好ましくは、一致率(すなわち、同一性)は100%に等しい。
【0028】
最も好ましくは、配列番号:2によるポリヌクレオチドを含むポリヌクレオチドは、配列番号:3またはその機能性断片を含む。機能性は、プロモーター活性を有することおよび増強された組込み選好を有する標的化配列であることと本明細書では解釈されるべきである。
【0029】
したがって、本発明は、配列番号:3によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドならびに配列番号:3によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドを含むベクターおよび宿主細胞を提供する。好ましくは、配列番号:3による機能性断片は、少なくとも100bp、より好ましくは少なくとも200bp、より好ましくは少なくとも300bp、より好ましくは少なくとも500bp、より好ましくは少なくとも600bp、より好ましくは少なくとも700bp、より好ましくは少なくとも800bp、より好ましくは少なくとも900bp、より好ましくは少なくとも1000bp、より好ましくは少なくとも1200bp、より好ましくは少なくとも1300bp、およびさらにより好ましくは少なくとも1400bpを含む。好ましくは、機能性断片は、少なくとも約50%同一な、より好ましくは少なくとも約60%同一な、さらにより好ましくは少なくとも約70%同一な、さらにより好ましくは少なくとも約80%同一な、さらにより好ましくは少なくとも約85%同一な、さらにより好ましくは少なくとも約90%同一な、さらにより好ましくは少なくとも約95%同一な、さらにより好ましくは少なくとも約97%同一な、さらにより好ましくは少なくとも約98%同一な、さらにより好ましくは少なくとも約99%同一な一致率(すなわち、位置同一性)を有し、かつ最も好ましくは、一致率(すなわち、同一性)は100%に等しい。
【0030】
ポリヌクレオチドの例は、一本鎖または二本鎖のいずれかの、リボ核酸(RNA)分子およびデオキシリボ核酸(DNA)分子であるが、これらに限定されない。当業者であれば、本明細書で特許請求されるポリヌクレオチドの相補体および逆相補体が本発明の範囲内に含まれることが意図されることを理解するであろう。
【0031】
本発明の実施形態によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現用の遺伝子エレメントであり、ここで、適合されたDNAドメイン中の遺伝子エレメントの機能性はコンストラクトの組込みによって破壊される。本発明の別の実施形態によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現用の遺伝子エレメントであり、ここで、適合されたDNAドメイン中の遺伝子エレメントの機能性はコンストラクトの組込みによって回復される。
【0032】
実質的に相同なDNAドメインは遺伝子変換によっておよび/または遺伝子増幅によって増加することができることが以前に証明されている。本明細書における使用では、異なるものであるが、遺伝子変換および遺伝子増幅という用語は両方とも互換的に用いられる。実質的に相同な異なるDNAドメインは、異なる増加頻度(すなわち、遺伝子変換頻度)を有することが明らかになった。最も高い遺伝子変換頻度を有する実質的に相同なDNAドメインの標的化DNAドメインを適合させる利点は、とりわけ、遺伝子変換および/または増幅によって目的のポリヌクレオチドのコピー数が増加した宿主のスクリーニングおよび選択がはるかにより効率的であることである。
【0033】
したがって、適合された実質的に相同なDNAドメインが由来する実質的に相同なDNAドメインは、好ましくは、他の少なくとも2つの実質的に相同なDNAドメインの1つよりも少なくとも10%高い遺伝子変換頻度を有する。より好ましくは、遺伝子変換頻度は、少なくとも15%高く、さらにより好ましくは少なくとも20%高く、さらにより好ましくは少なくとも30%高く、さらにより好ましくは少なくとも40%高く、さらにより好ましくは少なくとも50%高く、さらにより好ましくは少なくとも60%高く、さらにより好ましくは少なくとも70%高く、さらにより好ましくは少なくとも80%高く、さらにより好ましくは少なくとも90%高く、さらにより好ましくは少なくとも100%高く、さらにより好ましくは少なくとも150%高く、さらにより好ましくは少なくとも200%高く、さらにより好ましくは少なくとも300%高く、さらにより好ましくは少なくとも400%高く、さらにより好ましくは少なくとも500%より高く、およびさらにより好ましくは少なくとも1000%高い。
【0034】
本発明によれば、より高い遺伝子変換頻度を有する実質的に相同なDNAドメインは、宿主細胞の実質的に相同なDNAドメインの遺伝子変換頻度を決定することによって同定することができる。遺伝子変換頻度は、当業者に公知の任意の手段によって決定することができる。好ましい方法は、本明細書中、実施例3.1に記載されている方法である。この方法では、3つの実質的に相同なDNAドメインを含む宿主細胞を目的のポリヌクレオチドで形質転換する。各々、単一の実質的に相同なDNAドメイン中に目的のポリヌクレオチドの単一コピーを含み、各々の形質転換体がそのポリヌクレオチドを異なる実質的に相同なDNAドメインに組み込んでいる、3つの形質転換体を選択する。形質転換体を子孫クローンへと増殖させた後、同定目的のための独特の配列タグによって、すなわち、本明細書で上に記載した「フラッグDNA」解析を用いて、子孫クローンの遺伝子型を決定する。次に、前記実質的に相同なDNAドメインを含む形質転換体の遺伝子変換体の量を前記特定の形質転換体のクローンの総数で割ることによって、実質的に相同なDNAドメインの遺伝子変換頻度を算出する。
【0035】
好ましくは、本発明による宿主細胞は、実質的に相同なDNAドメインのうちの少なくとも2つが各々、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する細胞である。より好ましくは、各々の前記ドメインは、組み込まれた目的のポリヌクレオチドを少なくとも2コピー、さらにより好ましくは少なくとも3コピー、さらにより好ましくは4コピー、さらにより好ましくは4〜6コピー有する。好ましくは、本発明による宿主細胞の各々の実質的に相同なDNAドメインは、同じ数の組み込まれた目的のポリヌクレオチドを有する。
【0036】
目的のポリヌクレオチドのコピー数は、当業者に利用可能な任意の方法、例えば、サザン解析または(半)定量的PCRによる遺伝子型の解析、ノーザン解析、GeneChipまたはマイクロアレイ解析または(半)定量的RT−PCRによる発現解析、ウェスタンブロット、ELISA、酵素活性アッセイによる目的のポリヌクレオチドによってコードされる産物の解析によって決定することができる。目的のポリヌクレオチドのコピー数を決定する方法は国際公開第9846772号パンフレットに広範に記載されており、目的のポリヌクレオチドのコピー数を決定する好ましい方法は本発明によるものである。
【0037】
好ましくは、本発明による宿主細胞は、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合された少なくとも2つの実質的に相同なDNAドメイン、より好ましくは3つの適合されたDNAドメイン、さらにより好ましくは4つの適合されたDNAドメイン、さらにより好ましくは5つの適合されたDNAドメイン、さらにより好ましくは6つの適合されたDNAドメインを含む。最も好ましくは、本発明による前記宿主細胞の各々の実質的に相同なDNAドメインは、適合されたDNAドメインである。好ましくは、適合されたDNAドメインは、少なくとも2コピーの組み込まれた目的のポリヌクレオチドを少なくとも2コピー有し、さらにより好ましくは少なくとも3コピー、さらにより好ましくは少なくとも4コピーの組み込まれた目的のポリヌクレオチドを有し、さらにより好ましくは適合されたDNAドメインは、少なくとも5コピー、さらにより好ましくは少なくとも6コピー、少なくとも7コピー、少なくとも8コピー、少なくとも9コピー、少なくとも10コピーの組み込まれた目的のポリヌクレオチドを有する。最も好ましくは、適合されたDNAドメインは、組み込まれた目的のポリヌクレオチドの5または6コピーを有する。好ましくは、各々の前記適合されたDNAドメインは、同じ数の組み込まれた目的のポリヌクレオチドを有する。
【0038】
好ましくは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、ここで、好ましくは、各々の適合されたDNAドメインは複数コピーの組み込まれた目的のポリヌクレオチドを有する、複数の実質的に相同なDNAドメインを含む前記宿主細胞は、複数コピーの組み込まれた目的のポリヌクレオチドを有する単一の適合されたDNAドメインを含む宿主細胞から得られる;すなわち、複数コピーの組み込まれた目的のポリヌクレオチドを有するさらなるコピーの適合されたDNAドメインは、遺伝子変換によって生成される。
【0039】
好ましくは、本発明による宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率は、組込み経路をHRの方向に導くことによって増大する。そのような誘導は、HR経路の効率を高めることによって、および/またはNHR経路の効率を下げる(低下させるという意味)ことによっておよび/またはNHR/HR比を減少させることによって達成することができる。
【0040】
真核細胞は、少なくとも2つの別々の経路(相同組換え(HR)による経路および非相同組換え(NHR)による経路)を有しており、これらの経路によって、ポリヌクレオチドを宿主ゲノム中に組み込むことができる。本発明との関連で、HR経路は、宿主のゲノム中へのポリヌクレオチドの標的化組込みの制御に関与する全ての遺伝子およびエレメントとして定義され、前記ポリヌクレオチドは、組込みが標的化される宿主のゲノムのある所定の部位との一定の相同性を有する。NHR経路は、前記ポリヌクレオチドと宿主のゲノム配列との相同性の程度とは無関係な、宿主のゲノム中へのポリヌクレオチドの標的化組込みの制御に関与する全ての遺伝子およびエレメントとして定義される。
【0041】
非相同組換えと相同組換えの比(NHR/HR)は、宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率を決定する。NHR/HR比が高い場合、標的化組込みの効率は比較的低い。NHR/HR比が低い場合、標的化組込みの効率は比較的高い。NHR/HRの比は、NHRとHRの頻度を評価し、その後、それぞれの頻度を割り算することにより、当業者に公知の方法によって決定することができ、方法の一例は、国際公開第02/052026号パンフレットに記載されている方法である。好ましくは、本発明による宿主細胞は、改変を含むNHR構成要素をコードするポリヌクレオチドを含み、ここで、前記宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記NHR構成要素の産生を欠損している。前記改変は、NHR/HR比の減少をもたらし、それに伴って、宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率を増大させる。前記NHR構成要素の欠損をもたらす、NHR構成要素をコードするポリヌクレオチドの改変によって、宿主細胞のNHR/HR比がHRの方向に導かれ、その結果として、標的化組込みの効率が増大することになる。
【0042】
好ましくは、本発明による宿主細胞は、前記NHR構成要素の少なくとも5%欠損、より好ましくは少なくとも10%欠損、より好ましくは少なくとも20%欠損、より好ましくは少なくとも30%欠損、より好ましくは少なくとも40%欠損、より好ましくは少なくとも50%欠損、より好ましくは少なくとも60%欠損、より好ましくは少なくとも70%欠損、より好ましくは少なくとも80%欠損、より好ましくは少なくとも90%欠損、より好ましくは少なくとも95%欠損、より好ましくは少なくとも97%欠損、より好ましくは少なくとも99%欠損を示す。最も好ましくは、宿主細胞は、前記NHR構成要素の100%欠損を示す。
【0043】
改変すべきNHR構成要素は、当業者に公知の任意のNHR構成要素であることができる。好ましい改変すべきNHR構成要素は、KU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4、またはそれらのホモログの群から選択される。より好ましい改変すべきNHR構成要素は、KU70およびKU80、hdfAおよびhdfBまたはそれらのホモログである。最も好ましい改変すべきNHR構成要素は、KU70もしくはhdfA、またはそれらのホモログである。好ましくは、本発明による宿主細胞は、国際公開第2005/095624号パンフレットに記載されているような破壊または欠失を含むNHR構成要素をコードするポリヌクレオチドを含む。そのような宿主細胞を得る方法は当業者に公知であり、かつ国際公開第2005/095624号パンフレットに広範に記載されている。好ましくは、本発明によるNHR欠損宿主細胞におけるNHR/HR比は、同一の条件下でアッセイしたとき、非欠損親宿主細胞と比較して、少なくとも10%、より好ましくは少なくとも20%、より好ましくは少なくとも30%、より好ましくは少なくとも40%、より好ましくは少なくとも50%、より好ましくは少なくとも100%、より好ましくは少なくとも200%、およびさらにより好ましくは少なくとも1000%減少している。
【0044】
その代わりに、またはNHR構成要素の欠損と組み合わせて、本発明による宿主細胞は、宿主細胞が由来している細胞内の同じ遺伝子の発現レベルと比較して、HRに関与する少なくとも1つ遺伝子の発現レベルが上方調節されている。これは、HRに関与する構成要素をコードする遺伝子の発現レベルを増大させることによっておよび/またはHRに関与する構成要素の発現レベルを増大させることによっておよび/またはHRに関与する構成要素の活性を(一時的に)増大させることによって達成することができる。
【0045】
その代わりに、または本明細書で上に記載したようなNHR/HR比の改変と組み合わせて、国際公開2007/115886号パンフレットおよび国際公開2007/115887号パンフレットに記載されている方法を用いて、目的のポリヌクレオチドが相同組換えによって組み込まれている宿主細胞を前向きに選択し、かつ/または組込みが非相同組換えによって生じた細胞を除外することができる。国際公開2007/115886号パンフレットおよび国際公開2007/115887号パンフレットには、そのような選択または除外用の(さらなる)選択マーカーの使用が記載されている。そのような選択の例は、組込みが非相同組換えによって生じた細胞をジフテリア毒素Aによって選択的に死滅させることである(国際公開2007/115887号パンフレット)。
【0046】
ポリヌクレオチドの改変は、ポリヌクレオチドの配列の変化をもたらす任意の事象として本明細書で定義される。改変は1つ以上の改変と解釈される。改変は、ヌクレオチド配列中の1つ以上のヌクレオチドまたはポリヌクレオチドの転写もしくは翻訳に必要とされる調節エレメントの導入(挿入)、置換または除去(欠失)によって遂行することができる。例えば、ヌクレオチドは、終止コドンの導入、開始コドンの除去またはポリヌクレオチドのオープンリーディングフレームの変化もしくはフレームシフトをもたらすように挿入または除去することができる。配列またはその調節エレメントの改変は、当技術分野で公知の方法に従って、部位特異的もしくはランダムな突然変異誘発、DNAシャッフリング法、DNA再構築法、遺伝子合成(例えば、YoungおよびDong(2004),Nucleic Acids Research 32(7)電子アクセスhttp://nar.oupjournals.org/cgi/reprint/32/7/e59もしくはGuptaら(1968),Proc.Natl.Acad.Sci USA,60:1338−1344;Scarpullaら(1982),Anal.Biochem.121:356−365;Stemmerら(1995),Gene 164:49−53を参照されたい)、またはPCRによる突然変異誘発によって遂行することができる。ランダムな突然変異誘発手順の例は当技術分野で周知であり、例えば、化学的(例えば、NTG)突然変異誘発または物理的(例えば、UV)突然変異誘発などがある。特異的突然変異誘発手順の例は、クイックチェンジ(QuickChange)(商標)部位特異的突然変異誘発キット(Stratagene Cloning Systems,La Jolla,CA)、The Altered Sites(登録商標)II インビトロ突然変異誘発システム(Promega Corporation)またはGene.1989 Apr 15;77(1):51−9(Ho SN,Hunt HD,Horton RM,Pullen JK,Pease LR “Site−directed mutagenesis by overlap extension using the polymerase chain reaction”)に記載されているようなPCRを用いるかもしくはMolecular Biology:Current Innovations and Future Trends.(A.M.GriffinおよびH.G.Griffin編.ISBN 1−898486−01−8;1995 Horizon Scientific Press,PO Box 1,Wymondham,Norfolk,U.K.)に記載されているようなPCRを用いる重複伸長によるものである。
【0047】
好ましい改変の方法は、遺伝子置換、遺伝子欠失、または遺伝子破壊の手法を基にしている。例えば、遺伝子破壊法では、内在性ポリヌクレオチドに対応するポリヌクレオチドにインビトロで突然変異を誘発して、欠損のあるポリヌクレオチドを生じさせる。次に、これを親細胞に形質転換して、欠損のあるポリヌクレオチドを生じさせる。相同組換えによって、欠損のあるポリヌクレオチドは内在性ポリヌクレオチドに置き換わる。欠損のあるポリヌクレオチドは、核酸配列が改変された形質転換体の選択に使用し得るマーカーもコードすることが望ましい場合がある。内在性ポリヌクレオチドの欠失または置換の場合、適切なDNA配列は、欠失または置換しようとする標的遺伝子座に導入しなければならない。適切なDNA配列は、好ましくは、クローニングベクター上に存在する。好ましい組込みクローニングベクターは、この所定の遺伝子座へのクローニングベクターの組込みを標的化するために欠失または置換しようとするポリヌクレオチドに相同なDNA断片を含む。標的化組込みを促進するために、宿主細胞の形質転換前に、クローニングベクターを線状化することが好ましい。線状化は、クローニングベクターの少なくとも一方の末端、しかし、好ましくは両方の末端に、欠失または置換しようとするDNA配列に相同な配列が隣接するように行なわれることが好ましい。
【0048】
その代わりにまたは他の言及された手法と組み合わせて、A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans(2000) Chaveroche,M−K.,Ghico,J−M.およびd’Enfert C;Nucleic acids Research,第28巻,第22号に記載されているような、大腸菌(E.coli)でのコスミドのインビボ組換えに基づく手法を用いることができる。
【0049】
あるいは、前記宿主細胞がポリヌクレオチドの産生を欠損する改変を、ポリヌクレオチドの核酸配列に相補的なヌクレオチド配列を用いる確立されたアンチセンス技術によって実施することもできる。より具体的には、宿主細胞によるポリヌクレオチドの発現は、細胞内で転写されることができ、かつ細胞内で産生されるmRNAにハイブリダイズすることが可能な、ポリヌクレオチドの核酸配列に相補的なヌクレオチド配列を導入することによって低下させるかまたは削減することができる。相補的なアンチセンスヌクレオチド配列がmRNAにハイブリダイズするのを可能にする条件下で、翻訳されるタンパク質の量をこのように低下させるかまたは削減する。アンチセンスRNAを発現する例は、Appl Environ Microbiol.2000 Feb;66(2):775−82.(Characterization of a foldase,protein disulfide isomerase A,in the protein secretory pathway of Aspergillus niger.Ngiam C,Jeenes DJ,Punt PJ,Van Den Hondel CA,Archer DB)または(Zrenner R,Willmitzer L,Sonnewald U.Analysis of the expression of potato uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993);190(2):247−52.)に示されている。
【0050】
さらに、遺伝子の改変、下方調節または不活化は、RNA干渉(RNAi)技術によって得ることができる(FEMS Microb.Lett.237(2004):317−324)。この方法では、その発現に影響を及ぼそうとするヌクレオチド配列の同一のセンス部分およびアンチセンス部分を、ヌクレオチドスペーサーを間に入れて、互いの背後にクローニングし、発現ベクターに挿入する。そのような分子が転写された後で、小さいヌクレオチド断片の形成により、影響を及ぼそうとするmRNAの標的化分解がもたらされる。特定のmRNAの削減は様々な程度までであることができる。国際公開第2008/053019号パンフレット、国際公開第2005/05672A1号パンフレット、国際公開第2005/026356A1号パンフレット、Oliveiraら,“Efficient cloning system for construction of gene silencing vectors in Aspergillus niger”(2008) Appl Microbiol and Biotechnol 80(5):917−924および/またはBarnesら,“siRNA as a molecular tool for use in Aspergillus niger”(2008) Biotechnology Letters 30(5):885−890に記載されているRNA干渉技術を、遺伝子の下方調製、改変または不活化に用いることができる。
【0051】
ポリヌクレオチドの改変後、得られた株を、ポリヌクレオチドによってコードされる産物の欠損についてスクリーニングする。ポリヌクレオチドの発現レベルの下方調節および/または上方調節は、例えば、ノーザン解析(SambrookおよびRussell(2001) “Molecular Cloning:A Laboratory Manual(第3版),Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,New York.)によって細胞内に存在する対応するmRNAの量を定量化することによっておよび/または例えば、ウェスタンブロッティングもしくはELISAによって細胞内に存在する対応するタンパク質の量を定量化することによってモニタリングすることができる。mRNA量の違いは、DNAアレイ解析によって定量化することもできる(Eisen,M.B.およびBrown,P.O.DNA arrays for analysis of gene expression.Methods Enzymol.1999:303:179−205)。
【0052】
宿主細胞の欠損は、細胞が、未改変のポリヌクレオチドを含む親細胞と比較したときに、改変されたポリヌクレオチドによってコードされる産物をそれほど産生せず、かつ/または改変されたポリヌクレオチドの発現レベルが低下し(すなわち、mRNAレベルが低下し)、かつ/または改変されたポリヌクレオチドによってコードされる産物の特定の(タンパク質)活性が減少する表現型特徴、およびこれらの可能性の組合せとして本明細書で定義される。
【0053】
本発明によれば、実質的に相同なDNAドメインは、そのネイティブ(native)の状態で高レベルの発現が可能な内在性遺伝子を含むDNAドメインまたは遺伝子座(これらの用語は、本明細書で互換的に用いられる)である。「内在性」遺伝子という用語は、問題になっている生物のゲノム中の天然の遺伝子コピーとして本明細書で定義される。
【0054】
組み込まれた組換え遺伝子の発現レベルは、その遺伝子が組み込まれるゲノム遺伝子座によって大きく異なり得ることが一般に知られている。発現させようとする組換え遺伝子の組込みのために、高発現されるDNAドメインを用いる利点は、これらのDNAドメインが、少なくとも内在性遺伝子の高レベル発現を支持することができるということである。それゆえ、そのようなDNAドメインは、組み込まれた組換え遺伝子の高レベル発現も支持する可能性が高い。これまでに、A.ニガー(A.niger)のglaA遺伝子座における組込みおよびP.クリソゲヌム(P.chrysogenum)のペニシリンクラスターにおける組込みは、いくつかの他のゲノム遺伝子座における組込みと比較して遺伝子1コピー当たりでより高い発現レベルを提供することが明らかにされた(国際公開第9846772号パンフレットを参照されたい)。この関連で、高レベル発現が可能な遺伝子は、最大レベルで発現されたときに、全mRNA集団の少なくとも0.1%、好ましくは全mRNAの少なくとも0.5%および最も好ましくは全mRNAの少なくとも1%を占めるmRNAを産生する遺伝子と定義されることが理解されるであろう。そのような高発現可能な内在性遺伝子(これらがその中に含まれるDNAドメインは、本発明による実質的に相同なDNAドメインとして特に好適である)の例は、糖分解酵素、デンプン分解酵素、セルロース分解酵素および/または抗生物質生合成酵素をコードする遺伝子である。さらにより好ましいのは、グルコアミラーゼ遺伝子、TAKAアミラーゼ遺伝子、セロビオヒドロラーゼ遺伝子およびペニシリン生合成遺伝子などの、工業プロセスに関与し、かつ高レベルで発現されることが知られている遺伝子を含む遺伝子座である。
【0055】
好ましくは、本発明による宿主細胞の少なくとも2つの実質的に相同なDNAドメインは、glaAの遺伝子座であるかまたはamyAの遺伝子座であるかまたはamyBの遺伝子座であるかまたはこれらの遺伝子座の断片もしくはホモログである。より好ましい遺伝子座は、アスペルギルス(Aspergillus)のglaA、amyAおよびamyB遺伝子座である。さらにより好ましい遺伝子座は、アスペルギルス・ニガー(Aspergillus niger)のglaA、およびamyB遺伝子座である。さらにより好ましい遺伝子座は、アスペルギルス・ニガー(Aspergillus niger)CBS513.88のglaA、amyBIおよびamyBII遺伝子座である(遺伝子は、国際公開第2005/095624号パンフレットと同じように命名されている)。さらにより好ましい遺伝子座は、glaA、amyBIおよびamyBII遺伝子座(glaA遺伝子のヌクレオチド配列(An03g06550)およびそのゲノムコンテクストa.o.はhttp://www.ncbi.nlm.nih.gov/から得ることができる;amyBI遺伝子配列(An12g06930)およびamyBII遺伝子配列(An05g02100)についても同上かまたはこれらの遺伝子座の断片もしくはホモログである。
【0056】
本発明の目的のために、「相同性」および「同一性」という用語は、互換的に用いられる。2つの核酸配列間の相同性(同一性)の程度は、本明細書ではBLASTプログラムによって決定されることが好ましい。BLAST解析を実施するためのソフトウェアは、全米バイオテクノロジー情報センター(National Center for Biotechnology Information)(http://www.ncbi.nlm.nih.gov/)から公に入手可能である。BLASTアルゴリズムパラメータのW、T、およびXは、アラインメントの感度と速度を決定する。BLASTプログラムでは、ワード長(W)11、BLOSUM62スコアリングマトリックス(Henikoff & Henikoff,Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照されたい)アラインメント(B)50、期待値(E)10、M=5、N=−4、および両方の鎖の比較がデフォルトとして用いられる。
【0057】
本発明によれば、内在性遺伝子の発現が必要ない場合、高度に発現される内在性遺伝子は、本発明による宿主細胞において各々のコピーのDNAドメインで不活化されていることが好ましい。そのような場合、内在性遺伝子の高レベル発現の不活化によって、目的のポリヌクレオチドの発現にさらに利用することができるエネルギーおよび資源が放出される。さらに、目的のポリヌクレオチドの組込みによって産生されるべき所望の化合物と内在性遺伝子によってコードされる酵素が両方とも分泌型酵素である場合、内在性酵素の不活化によって、所望の酵素のより純粋な調製がもたらされる。不活化の逆転を排除するために、内在性遺伝子は、内在性遺伝子の少なくとも一部の不可逆的な欠失によって不活化されていることが好ましい。内在性遺伝子の不活化は、プロモーターおよび上流の活性化配列の少なくとも一部を含む不可逆的な欠失によってもたらされることがより好ましい。これは、組換えDNA分子の組込みによって産生されるべき酵素をコードする所望の遺伝子の発現が内在性遺伝子に由来するプロモーターから駆動される場合に、特に有利であるが、それは、所望の遺伝子の発現に必要な転写因子を潜在的に制限する競合を排除するからである。
【0058】
目的のポリヌクレオチドは任意のポリヌクレオチドであってもよい。目的のポリヌクレオチドは、任意の原核生物、真核生物、または他の供給源から得ることができる。目的のポリヌクレオチドおよびそれと関連するプロモーターは、宿主細胞にとって相同であり、自己クローンとなる組換え宿主細胞を生じさせることが好ましい。
【0059】
本発明によれば、目的のポリヌクレオチドは、例えば、国際公開第2006077258号パンフレットに記載されているような、最適化されたターミネーター配列を含む、変異型の最適化されたポリヌクレオチドであってもよい。目的のポリヌクレオチドは、部分的合成ポリヌクレオチドまたは完全合成核酸配列であってもよい。目的のポリヌクレオチドは、好ましくは、参照により本明細書に組み込まれる国際公開第2006/077258号パンフレットおよび/または国際公開第2008/000632号パンフレットに記載の方法に従って、そのコドン使用について最適化されてもよい。国際公開第2008/000632号パンフレットは、コドン対最適化に対処している。コドン対最適化は、ポリペプチドをコードするヌクレオチド配列の改善された発現および/またはコードされたポリペプチドの改善された産生を得るために、ポリペプチドをコードするヌクレオチド配列がそのコドン使用頻度(特に、使用されるコドン対)に関して改変されている方法である。コドン対は、コード配列中の2つの引き続くトリプレット(コドン)の組として定義される。
【0060】
したがって、本発明によれば、目的のポリヌクレオチドは、コドン最適化されたポリヌクレオチドであることが好ましい。
【0061】
ポリヌクレオチドはコード配列を含み、かつ目的のポリヌクレオチドによってコードされる化合物のクローニング、形質転換、発現および/または産生を促進するプロモーターおよび制御配列をさらに含む、核酸コンストラクト中および/またはベクター中に含めることができることが好ましい。
【0062】
「核酸コンストラクト」は、天然の遺伝子から単離されるかまたはさもなければ自然には存在しないような形で組み合わされ、並置された核酸のセグメントを含むように改変されている一本鎖または二本鎖のいずれかの核酸分子として本明細書で定義される。核酸コンストラクトという用語は、核酸コンストラクトがコード配列の発現に必要な全ての制御配列を含む場合、「発現カセット」という用語と同義である。本明細書で定義するような「コード配列」という用語は、mRNAに転写され、ポリペプチドに翻訳される配列である。コード配列の境界は、通常、mRNAの5’末端のATG開始コドンおよびmRNAの3’末端でオープンリーディングフレームを終結させる翻訳停止コドン配列によって決定される。コード配列は、DNA、cDNA、および組換え核酸配列を含むことができるが、これらに限定されない。
【0063】
「制御配列」という用語は、ポリペプチドの発現に必要または有利な全ての構成要素を含むよう、本明細書で定義される。各々の制御配列は、ポリペプチドをコードする核酸配列にとってネイティブまたは外来性であってもよい。そのような制御配列には、リーダー、最適な翻訳開始配列(Kozak,1991,J.Biol.Chem.266:19867−19870に記載されているもの)、ポリアデニル化配列、プロ−ペプチド配列、プレ−プロ−ペプチド配列、プロモーター、シグナル配列、および転写ターミネーターが含まれるが、これらに限定されない。最低でも、制御配列は、プロモーター、ならびに転写および翻訳停止シグナルを含む。
【0064】
制御配列には、制御配列とポリペプチドをコードする核酸配列のコード領域とのライゲーションを容易にする特定の制限部位を導入する目的のためのリンカーが備えられていてもよい。「機能的に連結された」という用語は、制御配列がポリペプチドの産生を導くよう、制御配列がDNA配列のコード配列に対して適切な位置に置かれている配置として本明細書で定義される。
【0065】
制御配列は、核酸配列の発現用の宿主細胞によって認識される適切なプロモーター配列、核酸配列であってもよい。プロモーター配列は、ポリペプチドの発現を仲介する転写制御配列を含む。プロモーターは、突然変異体プロモーター、切断型プロモーター、およびハイブリッドプロモーターを含む、細胞で転写活性を示す任意の核酸配列であってもよく、また、宿主細胞にとって相同または異種のいずれかの細胞外または細胞内ポリペプチドをコードする遺伝子から得られてもよい。
【0066】
プロモーターはまた、mRNAへの転写後の翻訳のための(プロモーターと翻訳開始点の間にある)5’非コード領域、シス作用転写制御エレメント(例えば、エンハンサー)、および転写因子と相互作用することができる他のヌクレオチド配列と理解されるので。
【0067】
プロモーターは、発現させようとするコード配列とネイティブに関連するプロモーターであってもよい。プロモーターはまた、発現させようとするコード配列にとって外来性の構成的または誘導性プロモーターであってもよい。哺乳動物細胞での使用に好適なプロモーターの例は、例えば、SambrookおよびRussell(上記)に記載されている。酵母での使用に好適なプロモーターの例には、例えば、糖分解プロモーターが含まれる。
【0068】
使用可能な好ましい誘導性プロモーターの例は、デンプン、銅、オレイン酸誘導性プロモーターである。
【0069】
プロモーターは、A.オリザエ(A.oryzae)のTAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)のアスパラギン酸プロテイナーゼ、A.ニガー(A.niger)の中性α−アミラーゼ、A.ニガー(A.niger)の酸安定性α−アミラーゼ、A.ニガー(A.niger)またはA.アワモリ(A.awamori)のグルコアミラーゼ(glaA)、R.ミエヘイ(R.miehei)のリパーゼ、A.オリザエ(A.oryzae)のアルカリプロテアーゼ、A.オリザエ(A.oryzae)のトリオースリン酸イソメラーゼ、A.ニデュランス(A.nidulans)のアセトアミダーゼをコードする遺伝子から得られるプロモーター、NA2−tpiプロモーター(A.ニガー(A.niger)の中性α−アミラーゼをコードする遺伝子由来のプロモーターとA.オリザエ(A.oryzae)のトリオースリン酸イソメラーゼをコードする遺伝子由来のプロモーターのハイブリッド、国際公開第03/008575号パンフレット)、ならびにそれらの突然変異体プロモーター、切断型プロモーター、およびハイブリッドプロモーターを含むが、これらに限定されない群から選択されることが好ましい。糸状菌細胞での使用に特に好ましいプロモーターは、プロテアーゼ遺伝子由来(例えば、F.オキシスポルム(F.oxysporum)のトリプシン様プロテアーゼ遺伝子(米国特許第4,288,627号明細書)、A.オリザエ(A.oryzae)のアルカリプロテアーゼ遺伝子(alp)、A.ニガー(A.niger)のpacA遺伝子、A.オリザエ(A.oryzae)のアルカリプロテアーゼ遺伝子、A.オリザエ(A.oryzae)の中性メタロプロテアーゼ遺伝子、A.ニガー(A.niger)のアスペルギロペプシンプロテアーゼpepA遺伝子、もしくはF.ベネナツム(F.venenatum)のトリプシン遺伝子、A.ニガー(A.niger)のアスパラギン酸プロテアーゼpepB遺伝子由来)のプロモーター、またはその機能性部分である。
【0070】
単一の適切なプロモーター配列を目的のポリヌクレオチドの発現用の制御配列として用いてもよいし、または複数の別個のプロモーター配列を目的のポリヌクレオチドの発現用の制御配列として用いてもよい。複数の別個のプロモーター配列を用いる場合、少なくとも2つの別個のプロモーター配列が各々、国際公開第2008/098933号パンフレットに従って、目的のポリヌクレオチドに機能的に連結されていることが好ましい。
【0071】
制御配列はまた、好適な転写ターミネーター配列、すなわち、糸状菌細胞によって認識されて、転写を終結させる配列であってもよい。ターミネーター配列は、ポリペプチドをコードする核酸配列の3’末端に機能的に連結されている。細胞内で機能的な任意のターミネーターを本発明で用いてもよい。
【0072】
糸状菌細胞のための好適なターミネーターは、A.オリザエ(A.oryzae)のTAKAアミラーゼ、A.ニガー(A.niger)のグルコアミラーゼ(glaA)、A.ニデュランス(A.nidulans)のアントラニル酸シンターゼ、A.ニガー(A.niger)のα−グルコシダーゼ、trpC遺伝子およびフサリウム・オキシスポルム(Fusarium oxysporum)のトリプシン様プロテアーゼをコードする遺伝子から得られる。
【0073】
制御配列はまた、好適なリーダー配列、糸状菌細胞による翻訳に重要なmRNAの非翻訳領域であってもよい。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端に機能的に連結されている。細胞内で機能的な任意のリーダー配列を本発明で使用してもよい。
【0074】
糸状菌細胞のための好適なリーダーは、A.オリザエ(A.oryzae)のTAKAアミラーゼおよびA.ニデュランス(A.nidulans)のトリオースリン酸イソメラーゼおよびA.ニガー(A.niger)のglaAをコードする遺伝子から得られる。
【0075】
他の制御配列は、ペニシリウムIPNS遺伝子、またはpcbC遺伝子、βチューブリン遺伝子から単離されてもよい。国際公開第01/21779号パンフレットに引用されている全ての制御配列は、本発明での使用が想定されることが企図される。
【0076】
制御配列はまた、ポリアデニル化配列、すなわち、核酸配列の3’末端に機能的に連結され、かつ転写されたときに、糸状菌細胞によって、転写されたmRNAにポリアデノシン残基を付加するシグナルとして認識される配列であってもよい。細胞内で機能的な任意のポリアデニル化配列を本発明で用いてもよい。
【0077】
糸状菌細胞のための好ましいポリアデニル化配列は、A.オリザエ(A.oryzae)のTAKAアミラーゼ、A.ニガー(A.niger)のグルコアミラーゼ、A.ニデュランス(A.nidulans)のアントラニル酸シンターゼ、フサリウム・オキシスポルム(Fusarium oxysporum)のトリプシン様プロテアーゼおよびA.ニガー(A.niger)のα−グルコシダーゼをコードする遺伝子から得られる。
【0078】
好ましくは、DNAコンストラクトは、目的の化合物をコードする目的のポリヌクレオチドと、本明細書で上に記載したようなプロモーターDNA配列と、好ましい制御配列、例えば、
− 以下の配列リスト:TAAG、TAGAおよびTAAA、好ましくはTAAAから選択される5’から3’の方向に配向された1つの翻訳終結配列、ならびに/または
− 以下の配列リスト:GCTACCCCC;GCTACCTCC;GCTACCCTC;GCTACCTTC;GCTCCCCCC;GCTCCCTCC;GCTCCCCTC;GCTCCCTTC;GCTGCCCCC;GCTGCCTCC;GCTGCCCTC;GCTGCCTTC;GCTTCCCCC;GCTTCCTCC;GCTTCCCTC;およびGCTTCCTTC、好ましくはGCT TCC TTCから選択される5’から3’の方向に配向された1つの翻訳イニシエーターコード配列、ならびに/または
− 以下の配列リスト:ヌクレオチドの多義性暗号:m(A/C);w(ATT);y(C/T);k(GTT);h(A/C/T)を用いた、5’−mwChkyCAAA−3’;5’−mwChkyCACA−3’もしくは5’−mwChkyCAAG−3’、好ましくは5’−CACCGTCAAA−3’もしくは5’−CGCAGTCAAG−3’から選択される1つの翻訳イニシエーター配列と
を含む。
【0079】
本発明との関連で、「翻訳イニシエーターコード配列」という用語は、DNAコード配列のオープンリーディングフレームのイニシエーターまたは開始コドンのすぐ下流にある9個のヌクレオチドとして定義される。イニシエーターまたは開始コドンは、AAメチオニンをコードする。イニシエーターコドンは、典型的にはATGであるが、GTGなどの任意の機能的な開始コドンであってもよい。
【0080】
本発明との関連で、「翻訳終結配列」という用語は、オープンリーディングフレームまたはヌクレオチドコード配列の3’末端にある翻訳停止コドンから始まり、5’から3’の方向に配向されている4個のヌクレオチドとして定義される。
【0081】
本発明との関連で、「翻訳イニシエーター配列」という用語は、ポリペプチドをコードするDNA配列のオープンリーディングフレームのイニシエーターまたは開始コドンのすぐ上流にある10個のヌクレオチドとして定義される。イニシエーターまたは開始コドンは、AAメチオニンをコードする。イニシエーターコドンは、典型的にはATGであるが、GTGなどの任意の機能的な開始コドンであってもよい。RNAでは、ウラシル(U)がデオキシヌクレオチドのチミン(T)に置き換わることが当技術分野で周知である。
【0082】
目的の化合物をコードする目的のポリヌクレオチド、または目的のポリヌクレオチドおよび上記の制御配列を含むDNAコンストラクトを連結させて、最適なポリヌクレオチド配列の挿入または置換を可能にする1つ以上の好都合な制限部位を含み得る組換え発現ベクターを産生することができる。
【0083】
あるいは、目的のポリヌクレオチドは、配列または目的のポリヌクレオチドを含む核酸コンストラクトを発現用の適切なベクターに挿入することによって発現させることができる。発現ベクターを作製する際、コード配列が発現、および場合によっては分泌用の適切な制御配列と機能的に連結されるようにコード配列をベクター内に位置させる。
【0084】
組換え発現ベクターは、組換えDNA手順に好都合に供することができ、かつ目的のポリヌクレオチドの発現をもたらすことができる任意のベクター(例えば、プラスミドまたはウイルス)であってもよい。ベクターの選択は、通常、ベクターとベクターを導入しようとする宿主細胞との適合性によって決まる。ベクターは、線状または閉環状プラスミドであってもよい。ベクターは、自律複製ベクター、すなわち、その複製が染色体複製に依存しない染色体外実体として存在するベクター(例えば、プラスミド、染色体外エレメント、ミニ染色体、または人工染色体)であってもよい。自律的に維持されるクローニングベクターは、AMA1配列を含んでいてもよい(例えば、AleksenkoおよびClutterbuck(1997),Fungal Genet.Biol.21:373−397を参照されたい)。
【0085】
あるいは、ベクターは、宿主細胞に導入したときに、ゲノム中に組み込まれ、それが組み込まれた染色体とともに複製するベクターであってもよい。組込み型クローニングベクターは、この所定の遺伝子座へのクローニングベクターの組込みを標的化するために、宿主細胞のゲノム中の所定の標的遺伝子座のDNA配列と相同なDNA断片を含むことが好ましい。標的化組込みを促進するために、宿主細胞の形質転換前に、クローニングベクターを線状化することが好ましい。線状化は、クローニングベクターの少なくとも一方の、しかし、好ましくは両方の末端に、標的遺伝子座と相同な配列が隣接するように行なわれることが好ましい。標的遺伝子座に隣接する相同配列の長さは、好ましくは少なくとも30bp、好ましくは少なくとも50bp、好ましくは少なくとも0.1kb、さらに好ましくは少なくとも0.2kb、より好ましくは少なくとも0.5kb、さらにより好ましくは少なくとも1kb、最も好ましくは少なくとも2kbである。
【0086】
ベクター系は、単一のベクターもしくはプラスミドまたは糸状菌細胞のゲノム中に導入しようとする全DNAを一緒に含有する、2つ以上のベクターもしくはプラスミド、またはトランスポゾンであってもよい。
【0087】
ベクターは、形質転換された細胞の容易な選択を可能にする1つ以上の選択可能マーカーを含むことが好ましい。共形質転換の方法を用いると、1つのベクターは選択可能マーカーを含むことができ、一方、別のベクターは、目的のポリヌクレオチドまたは目的の核酸コンストラクトを含むことができる。これらのベクターは、宿主細胞の形質転換に同時に用いられる。本発明による宿主細胞は、共形質転換効率が非常に高く、最大で100%となることができる能力を有する(実施例12.2を参照されたい)。
【0088】
選択可能マーカーは、その産物が殺生物剤またはウイルスに対する耐性、重金属に対する耐性、栄養要求株に対する原栄養性などを提供する遺伝子である。糸状菌宿主細胞で用いられる選択可能マーカーは、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、bleA(フレオマイシン結合)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、およびtrpC(アントラニル酸シンターゼ)、ならびに他の種由来の等価物を含むが、これらに限定されない群から選択することができる。アスペルギルス(Aspergillus)およびペニシリウム(Penicillium)細胞での使用に好ましいのは、amdS(欧州特許第635574B1号明細書、国際公開第97/06261号パンフレット)およびA.ニデュランス(A.nidulans)またはA.オリザエ(A.oryzae)のpyrG遺伝子およびストレプトミセス・ハイグロスコピクス(Streptomyces hygroscopicus)のbar遺伝子である。より好ましくはamdS遺伝子、さらにより好ましくはA.ニデュランス(A.nidulans)またはA.ニガー(A.niger)由来のamdS遺伝子を用いる。最も好ましい選択マーカー遺伝子は、A.ニデュランス(A.nidulans)のgpdAプロモーターに融合されたA.ニデュランス(A.nidulans)のamdSコード配列である(欧州特許第635574B1号明細書を参照されたい)。他の糸状菌由来のAmdS遺伝子も用いることができる(国際公開第97/06261号パンフレット)。
【0089】
上記のエレメントを連結して本発明の組換え発現ベクターを構築するために用いられる手順は、当業者に周知である(例えば、SambrookおよびRussell(上記);ならびにAusubelら,Current Protocols in Molecular Biology,Wiley InterScience,NY,1995を参照されたい)。
【0090】
好ましくは、本発明による宿主細胞は、適合されたDNAドメインおよび場合により標的化組込みの増大した効率に加えて、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドおよびprtTの群から選択されるポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、改変を含むポリヌクレオチドによってコードされる産物を欠損している。改変は本明細書で先に定義されている。
【0091】
好ましくは、本発明による宿主細胞はSec61の改変をさらに含む。宿主細胞の欠損は、先に本明細書にあるように、細胞が、未改変のポリヌクレオチドを含む親細胞と比較したとき、改変されたポリヌクレオチドによってコードされる産物をそれほど産生せず、かつ/または改変されたポリヌクレオチドの発現レベルが低下し(すなわち、mRNAレベルが低下し)、かつ/または改変されたポリヌクレオチドによってコードされる産物の特定の(タンパク質)活性が減少する表現型特徴、およびこれらの特徴の組合せとして本明細書で定義される。
【0092】
好ましくは、本発明による宿主細胞は、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドまたはprtTのうちの少なくとも1つの少なくとも5%の欠損、より好ましくは少なくとも10%の欠損、より好ましくは少なくとも20%の欠損、より好ましくは少なくとも30%の欠損、より好ましくは少なくとも40%の欠損、より好ましくは少なくとも50%の欠損、より好ましくは少なくとも60%の欠損、より好ましくは少なくとも70%の欠損、より好ましくは少なくとも80%の欠損、より好ましくは少なくとも90%の欠損、より好ましくは少なくとも95%の欠損、より好ましくは少なくとも97%の欠損、より好ましくは少なくとも99%の欠損を示す。最も好ましくは、宿主細胞は、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドまたはprtTのうちの少なくとも1つの100%の欠損を示す。
【0093】
目的の化合物の産生におけるグルコアミラーゼ(glaA)、酸安定性α−アミラーゼ(amyA)、中性α−アミラーゼ(amyBIおよびamyBII)ならびにシュウ酸ヒドロラーゼ(oahA)の群から選択される1つ以上のポリペプチドの欠損の利点は、エネルギーおよび資源がこれらの副産物に利用されないとうことである。このエネルギーおよび資源は、目的のポリヌクレオチドによってコードされる化合物の産生に用いることができる。さらに、副産物がほとんど存在しないので、下流での目的の化合物の処理が簡単になる。
【0094】
シュウ酸ヒドロラーゼ(oahA)は、多くの宿主細胞におけるシュウ酸の合成経路の構成要素である。oahAを欠損している宿主細胞は、シュウ酸を欠損している。シュウ酸は、食品用途などの多くの用途において望ましくない副産物である。さらに、シュウ酸は、この構成要素を産生する宿主細胞の培地培養のpHを低下させ、収率の低下をもたらす。すなわち、シュウ酸欠損宿主細胞では収率が増加する。それゆえ、本発明による宿主細胞がoahAを欠損していれば有利である。OahA欠損宿主細胞および前記宿主細胞を産生する好ましい方法は、国際公開第2000/50576号パンフレットおよび国際公開第2004/070022号パンフレットに広範に記載されている。oahA欠損宿主細胞を産生する好ましい方法は、国際公開第2000/50576号パンフレットに記載されている組換えによる破壊法である。好ましくは、本発明による宿主細胞は、oahAを欠損している。好ましくは、oahAは、真菌のoahAである。より好ましくは、oahAは、アスペルギルス(Aspergillus)由来のoahAである。さらにより好ましくは、oahAは、アスペルギルス・ニガー(Aspergillus niger)由来のoahAである。さらにより好ましくは、oahAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のoahAである。最も好ましくは、oahAは、An10g00820の配列を含む。
【0095】
prtTは、真核細胞におけるプロテアーゼの転写アクチベーターである。最近、プロテアーゼのいくつかの真菌転写アクチベーターが、国際公開第00/20596号パンフレット、国際公開第01/68864号パンフレット、国際公開第2006/040312号パンフレットおよび国際公開第2007/062936号パンフレットに記載されている。これらの転写アクチベーターは、アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))、アスペルギルス・フミガーツス(Aspergillus fumigatus)(A.フミガーツス(A.fumigatus))、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)(P.クリソゲヌム(P.chrysogenum))およびアスペルギルス・オリザエ(Aspergillus oryzae)(A.オリザエ(A.oryzae))から単離された。プロテアーゼ遺伝子のこれらの転写アクチベーターは、真菌細胞でポリペプチドを産生するための方法を改善するために用いることができ、その場合、ポリペプチドは、プロテアーゼ分解に感受性がある。宿主細胞がprtTを欠損しているとき、宿主細胞は、prtTの転写制御下にあるプロテアーゼをあまり産生しない。それゆえ、本発明による宿主細胞がprtTを欠損していれば有利である。prtT欠損宿主およびこれらの宿主を産生する好ましい方法は、国際公開第01/68864号パンフレット、国際公開第2006/040312号パンフレットに広範に記載されている。国際公開第01/68864号パンフレットおよび国際公開第2006/040312号パンフレットには、prtTコード配列を破壊するための組換えによる、古典的な方法が記載されている。国際公開第2007/062936号パンフレットには、プロテアーゼプロモーター内のprtT結合部位の破壊が記載されている。結合部位の破壊は、prtTが結合部位に結合するのを妨げる。その結果として、プロテアーゼの転写はprtTによって活性化されず、プロテアーゼはあまり産生されない。
【0096】
好ましくは、本発明による宿主細胞は、prtTをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、prtTの産生を欠損している。好ましくは、prtTは、真菌のprtTである。より好ましくは、prtTは、アスペルギルス(Aspergillus)由来のprtTである。さらにより好ましくは、prtTは、アスペルギルス・ニガー(Aspergillus niger)由来のprtTである。さらにより好ましくは、prtTは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のprtTである。最も好ましくは、prtTは、An04g06940の配列を含む。
【0097】
「グルコアミラーゼ」(glaA)という用語は、「アミログルコシダーゼ」という用語と同一であり、1,4結合α−D−グルコース残基および末端1,4結合α−D−グルコース残基の鎖中の分岐点における1,6−α−D−グルコシド結合のエンド加水分解を触媒するデキストリン6−α−D−グルカノヒドロラーゼ活性を有する酵素として本明細書で定義される。グルコアミラーゼ活性は、基質のp−ニトロフェニル−a−D−グルコピラノシド(シグマ(Sigma))からのパラニトロフェノールの遊離を測定することによって、AGIU/mlとして測定することができる。これにより黄色い色が生じ、その吸光度は、分光光度計を用いて405nmで測定することができる。1AGIUは、可溶性のデンプン基質からpH4.3および60℃で1分間に1μモルのグルコースを生じさせる酵素の分量である。国際公開第98/46772号パンフレットに、アッセイのさらなる詳細を見出すことができる。
【0098】
好ましくは、本発明による宿主細胞は、glaAをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、glaAの産生を欠損している。好ましくは、glaAは、真菌のglaAである。より好ましくは、glaAは、アスペルギルス(Aspergillus)由来のglaAである。さらにより好ましくは、glaAは、アスペルギルス・ニガー(Aspergillus niger)由来のglaAである。さらにより好ましくは、glaAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のglaAである。最も好ましくは、glaAは、An03g06550の配列を含む。
【0099】
「α−アミラーゼ」という用語は、水の存在下で3つ以上のα−1,4結合グルコースユニットを有する多糖のマルトオリゴ糖へのエンド加水分解を触媒する1,4−α−D−グルカングルカノヒドロラーゼ活性として本明細書で定義される。(中性)α−アミラーゼ活性を測定するために、供給元のプロトコルに従って、メガザイム(Megazyme)の穀物α−アミラーゼキットを用いる(メガザイム、CERALPHAαアミラーゼアッセイキット、カタログ参照番号K−CERA、2000−2001年)。測定される活性は、過剰のグルコアミラーゼおよびα−グルコシダーゼの存在下、pH7.0での、非還元末端がブロックされたp−ニトロフェニルマルトヘプタオシドの加水分解に基づく。形成されたp−ニトロフェノールの量は、試料中に存在するα−アミラーゼ活性の尺度である。
【0100】
「酸安定性α−アミラーゼ」(amyA)という用語は、酸性pH範囲で最適な活性を有するα−アミラーゼ活性を有する酵素として本明細書で定義される。酸安定性α−アミラーゼ活性を測定するために、これも、供給元のプロトコルに従うが、酸性pHでメガザイムの穀物α−アミラーゼキットを用いる(メガザイム、CERALPHAαアミラーゼアッセイキット、カタログ参照番号K−CERA、2000−2001年)。測定される活性は、過剰のグルコアミラーゼおよびα−グルコシダーゼの存在下、pH4.5での、非還元末端がブロックされたp−ニトロフェニルマルトヘプタオシドの加水分解に基づく。形成されたp−ニトロフェノールの量は、試料中に存在する酸安定性α−アミラーゼ活性の尺度である。
【0101】
好ましくは、本発明による宿主細胞は、AmyAをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、amyAを欠損している。好ましくは、amyAは、真菌のamyAである。より好ましくは、amyAは、アスペルギルス(Aspergillus)由来のamyAである。さらにより好ましくは、amyAは、アスペルギルス・ニガー(Aspergillus niger)由来のamyAである。さらにより好ましくは、amyAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyAである。最も好ましくは、amyAは、An11g03340の配列を含む。
【0102】
「中性α−アミラーゼ活性」(amy)という用語は、中性pH範囲で最適な活性を有するα−アミラーゼ活性を有する酵素として本明細書で定義される。
【0103】
好ましくは、本発明による宿主細胞は、AmyBをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、amyBIおよび/またはamyBIIの産生を欠損している。より好ましくは、宿主細胞は、amyBIおよびamyBIIを欠損している。好ましくは、amyBは、真菌のamyBである。より好ましくは、amyBは、アスペルギルス(Aspergillus)由来のamyBである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)由来のamyBIである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyBIである。最も好ましくは、amyBIは、An12g06930の配列を含む。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)由来のamyBIIである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyBIIである。最も好ましくは、amyBIIは、An05g02100の配列を含む。
【0104】
毒素関連ポリヌクレオチドという用語は、化合物をコードする遺伝子クラスター、多数の遺伝子、遺伝子もしくはその部分、または少なくとも1つの毒素もしくは毒素中間体化合物の生合成もしくは分泌に関与する生化学的経路として本明細書で定義される。前記化合物は、例えば、ポリペプチドであり、これは、酵素であってもよい。
【0105】
目的のポリペプチドの産生において宿主細胞として用いられるいくつかの宿主細胞(とりわけ、真菌)は、様々な毒素の生合成に関与する酵素をコードする遺伝子を有する。例えば、シクロピアゾン酸、コウジ酸、3−ニトロプロピオン酸およびアフラトキシンは、例えば、アスペルギルス・フラブス(Aspergillus flavus)で形成される既知の毒素である。同様に、トリコテセンは、いくつかの真菌、例えば、フサリウム(Fusarium)属の種(例えば、フサリウム・ベネナツム(Fusarium venenatum)およびトリコデルマ(Trichoderma)で形成され、オクラトキシンは、アスペルギルス(Aspergillus)によって産生され得る。最近、工業用アスペルギルス・ニガー(Aspergillus niger)宿主株のゲノムの配列決定により、フモニシン遺伝子クラスターが明らかになった(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88”.Nat Biotechnol.2007 Feb;25(2):221−231)。これらの毒素は、操作者、消費者および環境に健康被害を提示する可能性があるので、目的の化合物の発酵時のそのような毒素の形成は極めて望ましくない。その結果として、毒素欠損宿主細胞は目的の化合物を、毒素を含まずに産生するのを可能にする。産物から毒素を取り除く必要がないので、毒素を含まない化合物は産生するのがより簡単である。さらに、化合物の規制認可手続きは、より簡単である。
【0106】
好ましくは、本発明による宿主細胞は、化合物(例えば、酵素であり得るポリペプチドであってもよい)または生化学的経路をコードする毒素関連ポリヌクレオチドを含み、前記毒素関連ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記毒素または毒素中間体化合物の産生を欠損している。好ましくは、毒素または毒素中間体化合物は、真菌の毒素または毒素中間体化合物である。より好ましくは、毒素または毒素中間体化合物は、アスペルギルス(Aspergillus)由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、アスペルギルス・ニガー(Aspergillus niger)由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、フモニシンまたはフモニシン中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、オクラトキシンまたはオクラトキシン中間体化合物である。最も好ましくは、毒素または毒素中間体化合物は、オクラトキシンまたはフモニシンまたはオクラトキシン中間体またはフモニシン中間体化合物である。
【0107】
好ましくは、毒素関連ポリヌクレオチドは、化合物(例えば、酵素であり得るポリペプチドであってもよい)または真菌の毒素もしくは毒素中間体化合物の産生に関与する生化学的経路をコードする。より好ましくは、アスペルギルス(Aspergillus)由来の毒素または毒素中間体化合物。さらにより好ましくは、アスペルギルス・ニガー(Aspergillus niger)由来の毒素または毒素中間体化合物。さらにより好ましくは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来の毒素または毒素中間体化合物。さらにより好ましくは、フモニシンまたはフモニシン中間体化合物。さらにより好ましくは、フモニシン−Bまたはフモニシン−B中間体化合物。さらにより好ましくは、フモニシン−B2またはフモニシン−B2中間体化合物。さらにより好ましくは、毒素関連ポリヌクレオチドは、An01g06820からAn01g06930までのフモニシンクラスターの配列を含む。最も好ましくは、毒素関連ポリヌクレオチドは、An01g06930の配列を含む。
【0108】
別の好ましい実施形態では、毒素関連ポリヌクレオチドは、化合物(例えば、酵素であり得るポリペプチドであってもよい)またはオクラトキシンもしくはオクラトキシン中間体化合物の産生に関与する生化学的経路をコードする。より好ましくは、オクラトキシンAまたはオクラトキシンA中間体化合物。より好ましくは、毒素関連ポリヌクレオチドは、An15g07880からAn15g07930までのクラスターの配列を含む。最も好ましくは、毒素関連ポリヌクレオチドは、An15g07910の配列および/またはAn15g07920の配列を含む。
【0109】
好ましくは、本発明による宿主細胞は、化合物(例えば、酵素であり得るポリペプチドであってもよい)または生化学的経路をコードする少なくとも1つの毒素関連ポリヌクレオチドを含み、前記毒素関連ポリヌクレオチドは少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、毒素または毒素中間体化合物の産生を欠損している。
【0110】
より好ましくは、本発明による宿主細胞は、2つの毒素関連ポリヌクレオチドを含み、前記2つの毒素関連ポリヌクレオチドは各々、少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、フモニシンおよびオクラトキシンの産生を欠損していることが好ましい。
【0111】
さらにより好ましくは、本発明による宿主細胞は、3つ以上の毒素関連ポリヌクレオチドを含み、前記3つ以上の毒素関連ポリヌクレオチドは各々、少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、フモニシン、オクラトキシンおよび少なくとも1つのさらなる毒素または毒素中間体化合物の産生を欠損していることが好ましい。
【0112】
好ましくは、本発明による宿主細胞は、glaAおよび前記構成要素をコードするポリヌクレオチドに改変を有するために、glaAならびにamyA、amyBI、amyBII、oahA、毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。より好ましくは、本発明による宿主細胞は、glaA、oahAならびにamyA、amyBI、amyBII、毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、毒素関連化合物ならびにamyA、amyBI、amyBII、およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBIIならびに毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBII、prtTおよび毒素関連化合物を欠損している。最も好ましくは、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBII、prtTおよび毒素関連化合物を欠損しており、かつNHR構成要素、好ましくはku70またはそのホモログをさらに欠損している。
【0113】
本明細書で上に記載した構成要素の欠損または改変の他に、本発明による宿主細胞は、他の構成要素(例えば、pepAのような主要なプロテアーゼ)を欠損していてもよい。好ましくは、pepAは、真菌のpepAである。より好ましくは、pepAは、アスペルギルス(Aspergillus)由来のpepAである。さらにより好ましくは、pepAは、アスペルギルス・ニガー(Aspergillus niger)由来のpepAである。さらにより好ましくは、pepAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のpepAである。最も好ましくは、pepAは、An14g04710の配列を含む。好ましくは、本発明による宿主細胞は、pepAの少なくとも5%の欠損、より好ましくは少なくとも10%の欠損、より好ましくは少なくとも20%の欠損、より好ましくは少なくとも30%の欠損、より好ましくは少なくとも40%の欠損、より好ましくは少なくとも50%の欠損、より好ましくは少なくとも60%の欠損、より好ましくは少なくとも70%の欠損、より好ましくは少なくとも80%の欠損、より好ましくは少なくとも90%の欠損、より好ましくは少なくとも95%の欠損、より好ましくは少なくとも97%の欠損、より好ましくは少なくとも99%の欠損を示す。最も好ましくは、宿主細胞は、pepAの100%の欠損を示す。
【0114】
その代わりに、または本明細書で上に記載した欠損と組み合わせて、宿主細胞は、目的のポリペプチドの産生能力を高めるために、野生型細胞と比較して上昇した小胞体ストレス応答(UPR)を含む。UPRは、米国特許出願公開第2004/0186070A1号明細書および/または米国特許出願公開第2001/0034045A1号明細書および/または国際公開第01/72783A2号明細書および/または国際公開第2005/123763号パンフレットに記載の手法によって増大させることができる。より具体的には、UPRが上昇した宿主細胞を得るために、HAC1および/もしくはIRE1および/もしくはPTC2のタンパク質レベルが調節され、かつ/またはSEC61タンパク質が操作されている。好ましいSEC61改変は、SEC61の一方向の突然変異体、すなわち、デノボ合成タンパク質がSEC61を介してERに侵入することができるが、タンパク質がSEC61を介してERを離れることができない突然変異体を生じさせる改変である。そのような改変は、国際公開第2005/123763号パンフレットに広範に記載されている。最も好ましくは、SEC61改変は、セリン376がトリプトファンに置き換えられているS376W突然変異である。
【0115】
本発明による好ましい宿主細胞は、目的の化合物を産生するための組換えアスペルギルス・ニガー(Aspergillus niger)組換え宿主細胞であり、前記宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ適合された実質的に相同なDNAドメインが由来する実質的に相同なDNAドメインは、少なくとも2つの実質的に相同なDNAドメインの他の1つよりも少なくとも10%高い遺伝子変換頻度を有し、ここで、適合された実質的に相同なDNAドメインは、独特の配列タグによって他のバージョンの実質的に相同なDNAドメインと区別され、かつ適合された実質的に相同なDNAドメインは、標的化DNAドメインを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含み、かつ前記宿主細胞は、glaA、pepA、hdfA、amyBII、amyBI、amyA、oahA、fumB、ochを欠損している。好ましくは、前記宿主細胞は、sec61 S376W突然変異、すなわち、セリン376がトリプトファンに置き換えられているS376W突然変異を有する。より好ましくは、前記宿主細胞はさらに、prtTを欠損している。そのような宿主細胞は、実施例9、10および11に記載されている。
【0116】
好ましくは、本発明による宿主細胞は、選択マーカーを含まない宿主細胞である。
【0117】
この状況は、アスペルギルス・ニデュランス(Aspergillus nidulans)由来のアセトアミーゼ遺伝子(amdS)などの、二方向性の優性選択マーカーの対抗選択によって得てもよい。このマーカーを用いて、アセトアミドを唯一の炭素および/または窒素源として用いて選択することによって、この遺伝子を有する形質転換体を選択することができる一方、例えば、フルオロアセトアミドで、対抗選択(この用語は、以下、マーカー遺伝子の不在についての選択に用いられる)を行なうことができる。特定の宿主細胞で作用する他の二方向性の優性選択マーカーを類似のやり方で用いることができる。二方向性マーカー遺伝子の除去組換え(out−recombination)は、導入しようとするプラスミド上に直接反復が両側にある遺伝子を挿入することによって容易になる。
【0118】
欧州特許第0635574B1号明細書に開示されているように、amdS二方向性マーカーは両方向に優性であり、これは、任意の遺伝子バックグラウンドの形質転換された細胞を(アセトアミドを唯一の炭素源および/または唯一の窒素源として用いて)マーカーの存在について選択することができることを意味する。他の二方向性マーカーは、URA3、LYS2、pyrG、facAなどである。
【0119】
マーカーを含まない細胞の使用はより簡単に規制当局に認可され、同時に、工業用の発酵条件下における酵母のエネルギー収支にあまり負担をかけない。
【0120】
本発明による宿主細胞は、任意の宿主細胞であってもよい。本発明による宿主細胞で産生される化合物の特定の使用について、宿主細胞の選択は、そのような使用に従って行なうことができる。例えば、本発明による宿主細胞で産生される化合物を食品用途で用いることになっている場合、宿主細胞は、出芽酵母(Saccharomyces cerevisiae)などの食品等級の生物から選択することができる。特定の使用には、食品用途、(動物)飼料用途、医薬品用途、農業用途(例えば、作物保護)、および/またはパーソナルケア用途が含まれるが、これらに限定されない。
【0121】
一実施形態によれば、本発明による宿主細胞は、真核宿主細胞である。好ましくは、真核細胞は、哺乳動物細胞、昆虫細胞、植物細胞、真菌細胞、または藻類細胞である。好ましい哺乳動物細胞には、例えば、チャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマが含まれる。好ましい昆虫細胞には、例えば、Sf9およびSf21細胞ならびにその派生物が含まれる。より好ましくは、真核細胞は、真菌細胞、すなわち、カンジダ(Candida)、ハンセヌラ(Hansenula)、クルイベロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、スキゾサッカロミセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)株などの酵母細胞である。より好ましくは、クルイベロミセス・ラクチス(Kluyveromyces lactis)、出芽酵母(S.cerevisiae)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)およびピキア・パストリス(Pichia pastoris)、または糸状菌細胞由来のものである。最も好ましくは、真核細胞は糸状菌細胞である。
【0122】
「糸状菌」には、(AinsworthおよびBisbyのDictionary of The Fungi,第8版,1995,CAB International,University Press,Cambridge,UKでHawksworthらによって定義されているような)全ての糸状形態の真菌類および卵菌類という亜門が含まれる。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖から構成される菌糸壁を特徴とする。栄養増殖は菌糸伸長によるものであり、炭素代謝は偏性好気性である。糸状菌株には、アクレモニウム(Acremonium)、アガリクス(Agaricus)、アスペルギルス(Aspergillus)、アウレオバシジウム(Aureobasidium)、クリソスポリウム(Chrysosporium)、コプリヌス(Coprinus)、クリプトコッカス(Cryptococcus)、フィリバシジウム(Filibasidium)、フサリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、パエシロミセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、パネロカエテ(Panerochaete)、ヒラタケ(Pleurotus)、スエヒロタケ(Schizophyllum)、タラロミセス(Talaromyces)、テルモアスクス(Thermoascus)、ティエラビア(Thielavia)、トリポクラジウム(Tolypocladium)、およびトリコデルマ(Trichoderma)の株が含まれるが、これらに限定されない。
【0123】
好ましい糸状菌細胞は、アスペルギルス(Aspergillus)属、クリロスポリウム(Chrysosporium)属、ペニシリウム(Penicillium)属、タラロミセス(Talaromyces)属、フサリウム(Fusarium)属またはトリコデルマ(Trichoderma)属の種、および最も好ましくはアスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティヅス(Aspergillus foetidus)、アスペルギルス・ソヤエ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニイ(Talaromyces emersonii)、アスペルギルス・オリザエ(Aspergillus oryzae)、クリソスポリウム・ルクノウェンス(Chrysosporium lucknowense)、フサリウム・オキシスポルム(Fusarium oxysporum)、トリコデルマ・リーゼイ(Trichoderma reesei)またはペニシリウム・クリソゲヌム(Penicillium chrysogenum)の種に属する。より好ましい宿主細胞は、アスペルギルス・ニガー(Aspergillus niger)である。本発明による宿主細胞がアスペルギルス・ニガー(Aspergillus niger)宿主細胞である場合、宿主細胞は、好ましくは、CBS513.88、CBS124.903またはそれらの派生物である。最も好ましくは、本発明による組換え宿主細胞は、本明細書中の実施例に記載されているような宿主細胞、好ましくは実施例4〜11に見られるような宿主細胞である。
【0124】
糸状菌のいくつかの株は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM)、Centraalbureau Voor Schimmelcultures(CBS)、およびAgricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL)アスペルギルス・ニガー(Aspergillus niger)CBS513.88、CBS124.903、アスペルギルス・オリザエ(Aspergillus oryzae)ATCC20423、IFO4177、ATCC1011、CBS205.89、ATCC9576、ATCC14488−14491、ATCC11601、ATCC12892、P.クリソゲヌム(P.chrysogenum)CBS455.95、ペニシリウム・シトリヌム(Penicillium citrinum)ATCC38065、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)P2、タラロミセス・エメルソニイ(Talaromyces emersonii)CBS124.902、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・リーゼイ(Trichoderma reesei)ATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソヤエ(Aspergillus sojae)ATCC11906、クリソスポリウム・ルクノウェンス(Chrysosporium lucknowense)ATCC44006およびそれらの派生物などのいくつかの培養コレクションにおいて一般に容易に入手可能なものがある。
【0125】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核宿主細胞は細菌細胞である。「細菌細胞」という用語には、グラム陰性微生物とグラム陽性微生物の両方が含まれる。好適な細菌は、例えば、エシェリキア(Escherichia)、アナベナ(Anabaena)、カウロバクター(Caulobacter)、グルコノバクター(Gluconobacter)、ロドバクター(Rhodobacter)、シュードモナス(Pseudomonas)、パラコッカス(Paracoccus)、バシルス(Bacillus)、ブレビバクテリウム(Brevibacterium)、コリネバクテリウム(Corynebacterium)、リゾビウム(Rhizobium)(シノリゾビウム(Sinorhizobium))、フラボバクテリウム(Flavobacterium)、クレブシエルラ(Klebsiella)、エンテロバクター(Enterobacter)、乳酸桿菌(Lactobacillus)、ラクトコッカス(Lactococcus)、メチロバクテリウム(Methylobacterium)ブドウ球菌(Staphylococcus)またはストレプトミセス(Streptomyces)から選択してもよい。好ましくは細菌細胞は、枯草菌(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニホルミス(B.licheniformis)、B.プンティス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロヅランス(B.halodurans)、B.プミルス(B.pumilus)、G.オキシダンス(G.oxydans)、カウロバクター・クレスケンツス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スファエロイデス(Rhodobacter sphaeroides、シュードモナス・ゼアキサンティニファシエンス(Pseudomonas zeaxanthinifaciens)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(glutamicum)、スタフィロコッカス・カルノースス(Staphylococcus carnosus)、ストレプトミセス・リウィダンス(Streptomyces lividans)、シノリゾビウム・メリオティ(Sinorhizobium melioti)およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0126】
本発明による宿主細胞は、様々な目的、例えば、目的の化合物の産生ならびに国際公開第1999/032617号パンフレットおよび国際公開第2008/138835号パンフレットに記載されているような発現クローニングに好都合に用いることができる。これらの特許出願では、糸状菌細胞を宿主細胞として用いる発現クローニングの便宜性が記載されている。
【0127】
本発明はまた、目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、好ましくはより高い遺伝子変換頻度を有する実質的に相同なDNAドメインに標的化DNAドメインを提供することによって、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、方法を提供する。
【0128】
別の実質的に相同なDNAドメインと比較してより高い遺伝子変換比率を有する実質的に相同なDNAドメインを、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることは、当業者に公知の任意の手段によって実施することができる。好ましくは、適合された実質的に相同なDNAドメインは、宿主のゲノム内で部分的にまたは完全に独特である。実施例3に示すように、A.ニガー(A.niger)におけるアンプリコンは、事前にゲノムから除去されたグルコアミラーゼプロモーター配列を用いて適合させる。これにより、独特の標的化配列を有する実質的に相同なDNAドメインが得られた。目的のポリヌクレオチドのプロモーターおよびターミネーター部分は、実施例11に示すような形質転換の後、適合された実質的に相同なDNAドメインでの組込みに好適なDNA配列として機能する。当業者のための、これらの実験を実施するための全ての手段は、本明細書ならびにSambrookおよびRussel(上記)に記載されている。
【0129】
好ましくは、目的の化合物を産生するための組換え宿主細胞の産生方法は、
a.得られた細胞を、目的のポリヌクレオチドを含む、より高い遺伝子変換比率を有する適合された実質的に相同なDNAドメインで形質転換する工程と、
b.実質的に相同なDNAドメインの少なくとも1つに組み込まれた目的のポリヌクレオチドを有する細胞を選択またはスクリーニングする工程と、
c.(b)で得られた細胞を繁殖させ、さらなるコピーの実質的に相同なDNAドメインに組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングする工程と
をさらに含む。
【0130】
より好ましくは、目的の化合物を産生するための組換え宿主細胞の産生方法は、本明細書中の実施例で記載されているような方法である。
【0131】
細胞の形質転換のための方法は当業者に周知である。任意のそのような方法を本発明の目的のために用いてもよい。形質転換は、それ自体公知の方法でのプロトプラスト形成、プロトプラストの形質転換、および細胞壁の再生からなるプロセスを含んでいてもよい。アスペルギルス(Aspergillus)細胞の形質転換の好適な手順は、欧州特許第238023号明細書およびYeltonら,1984,Proceedings of the National Academy of Sciences USA 81:1470−1474に記載されている。アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を用いるアスペルギルス(Aspergillus)および他の糸状菌宿主細胞の形質転換の好適な手順は、例えば、Nat Biotechnol.1998 Sep;16(9):839−42.(誤記あり(Erratum in:))Nat Biotechnol 1998 Nov;16(11):1074.Agrobacterium tumefaciens−mediated transformation of filamentous fungi.de Groot MJ,Bundock P,Hooykaas PJ,Beijersbergen AG.Unilever Research Laboratory Vlaardingen,The Netherlands中の訂正に記載されている。フサリウム(Fusarium)種を形質転換する好適な方法は、Malardierら,1989,Gene 78:147156によってまたは国際公開第96/00787号パンフレットに記載されている。Biolistic transformation of the obligate plant pathogenic fungus,Erysiphe graminis f.sp.hordei.Christiansen SK,Knudsen S,Giese H.Curr Genet.1995 Dec;29(1):100−2に記載されているような微粒子銃による形質転換を用いる方法などの他の方法を適用することができる。酵母は、Abelson,J.N.およびSimon,M.I.編,Guide to Yeast Genetics and Molecular Biology,Methods in Enzymology,第194巻,182−187頁,Academic Press,Inc.,New York中のBeckerおよびGuarente;ltoら,1983,Journal of Bacteriology 153:163;ならびにHinnenら,1978,Proceedings of the National Academy of Sciences USA 75:1920によって記載されている手順を用いて形質転換してもよい。
【0132】
得られた形質転換体のうちのどれが、そのDNAドメインの1つに組換えDNA分子の組込みを有するのかを明らかにするために、いくつかの日常的な手法が当業者に利用可能である。
【0133】
さらなる工程では、選択された形質転換体を繁殖させ、その子孫から、DNAドメインのうちの少なくとも2つが組み込まれた目的のポリヌクレオチドを含む株を選択する。これは、組み込まれた目的のポリヌクレオチドを含むDNAドメインが、「空の」DNAドメインによる遺伝子変換を通じてかまたは増幅を通じてかのいずれかで増加している株を選択することを意味する。そのような遺伝子変換および/または増幅事象は、低い頻度で自然に起こる。これらの事象が起こる正確な頻度は、問題となっている宿主細胞ならびにDNAドメインの数、長さおよび相同性の程度を含むいくつかの変数によって決まり得る。しかしながら、これらの頻度は、これらの事象が起こった株を当業者に公知の解析手法を用いてスクリーニングおよび選択するのを可能にするほど十分に高い。組み込まれた目的のポリヌクレオチドを含むDNAドメインが増加している株は、例えば、目的のポリヌクレオチドによってコードされる産物の産生レベルがより高い株を単にスクリーニングすることによって、またはあるいはその遺伝子型を、例えば、上で概説したような「DNAフラッグ」テストで解析することによって同定することができる。
【0134】
本発明による組換え宿主細胞の産生方法は、組み込まれた目的のポリヌクレオチドを含むDNAドメインの増加が起こった株の1つを繁殖させ、その子孫から、さらなるコピーのDNAドメインが、組み込まれた目的のポリヌクレオチドを含む株を選択する、さらなる工程を含んでいてもよい。その後、各々のDNAドメインが、組み込まれた目的のポリヌクレオチドを含む株が得られるまで、これらの株に再びこの手順を受けさせてもよい。
【0135】
好ましくは、上記の方法の工程(c)を実質的に相同なDNAドメインのうちの少なくとも3つが組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有するまで繰り返す。より好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも4つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。さらにより好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも5つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。さらにより好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも6つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。最も好ましくは、工程(c)を実質的に相同なDNAドメインの各々が少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。
【0136】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で本発明による宿主細胞を培養することと、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0137】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で組換え宿主細胞を培養することであって、前記宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドを少なくとも1コピー有する、と、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0138】
本発明による産生方法では、当技術分野で公知の方法を用いて、目的の化合物(例えば、ポリペプチドまたは代謝産物)の産生に好適な栄養培地中で宿主細胞を培養する。本発明を限定するものと解釈されるべきでない培養方法の例は、深部発酵、固体状態での表面発酵および液体基質上での表面発酵である。例えば、細胞は、好適な培地中ならびにコード配列の発現および/またはポリペプチドの単離を可能にする条件下での実験用または工業用発酵槽における振盪フラスコ培養、小規模または大規模発酵(連続発酵、回分発酵、流加発酵、または固体状発酵を含む)によって培養することができる。培養は、当技術分野で公知の手順を用いて、炭素および窒素源ならびに無機塩を含む好適な栄養培地中で起こる。好適な培地は、商業的供給業者から入手可能であるかまたは(例えば、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)のカタログで)公表されている組成に従って調製することができる。ポリペプチドまたは代謝産物が栄養培地中に分泌される場合、ポリペプチドまたは代謝産物を培地から直接回収することができる。ポリペプチドまたは代謝産物が分泌されない場合、細胞ライセートから回収することができる。
【0139】
ポリペプチドは、ポリペプチドに特異的な当技術分野で公知の方法を用いて検出することができる。これらの検出方法は、特異的抗体の使用、酵素産物の形成、または酵素基質の消失が含むことができる。
【0140】
得られる目的の化合物(例えば、ポリペプチドまたは代謝産物)は、当技術分野で公知の方法によって回収することができる。例えば、ポリペプチドまたは代謝産物は、遠心分離、濾過、抽出、噴霧乾燥、蒸発、または沈殿を含むが、これらに限定されない、従来の手順によって栄養培地から回収することができる。
【0141】
ポリペプチドは、クロマトグラフィー(例えば、イオン交換、親和性、疎水性、クロマトフォーカシング、およびサイズ排除)、電気泳動手順(例えば、分取等電点電気泳動法)、示差溶解度(例えば、硫酸アンモニウム沈殿)、SDS−PAGE、または抽出を含むが、これらに限定されない、当技術分野で公知の種々の手順によって精製することができる(例えば、Protein Purification,J.−C.JansonおよびLars Ryden編,VCH Publishers,New York,1989を参照されたい)。
【0142】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような選択マーカーを含まない宿主細胞である。
【0143】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような真菌宿主細胞である。
【0144】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような糸状菌宿主細胞である。
【0145】
本発明の目的の化合物は、任意の生体化合物であることができる。生体化合物は、任意の生体ポリマーまたは代謝産物であってもよい。生体化合物は、生合成経路もしくは代謝経路を構成する単一のポリヌクレオチドもしくは一連のポリヌクレオチドによってコードされていてもよいし、または単一のポリヌクレオチドの産物もしくは一連のポリヌクレオチドの産物の直接の結果であってもよい。生体化合物は、宿主細胞にとってネイティブであっても、または異種であってもよい。
【0146】
「異種の生体化合物」という用語は、細胞にとってネイティブでない生体化合物;またはネイティブな生体化合物を変化させるために構造的な改変がなされたネイティブな生体化合物として本明細書で定義される。
【0147】
「生体ポリマー」という用語は、同一の、同様の、または異なるサブユニット(モノマー)の鎖(またはポリマー)として本明細書で定義される。生体ポリマーは、任意の生体ポリマーであってもよい。生体ポリマーは、例えば、核酸、ポリアミン、ポリオール、ポリペプチド(もしくはポリアミド)、または多糖であってもよいが、これらに限定されない。
【0148】
生体ポリマーはポリペプチドであってもよい。ポリペプチドは、目的の生体活性を有する任意のポリペプチドであってもよい。「ポリペプチド」という用語は、本明細書では、特定の長さのコードされた産物を指すことを意味するものではなく、それゆえ、ペプチド、オリゴペプチド、およびタンパク質を包含する。ポリペプチドには、上述のポリペプチドおよびハイブリッドポリペプチドの天然のアレルバリエーションおよび操作されたバリエーションがさらに含まれる。ポリペプチドは、宿主細胞にとってネイティブであっても、または異種であってもよい。ポリペプチドは、コラーゲンもしくはゼラチン、またはそれらの変異体もしくはハイブリッドであってもよい。ポリペプチドは、抗体またはその部分、抗原、凝固因子、酵素、ホルモンまたはホルモン変異体、受容体またはその部分、調節タンパク質、構造タンパク質、受容体、または輸送体タンパク質、分泌過程に関与するタンパク質、フォールディング過程に関与するタンパク質、シャペロン、ペプチドアミノ酸輸送体、グリコシル化因子、転写因子、合成ペプチドまたはオリゴペプチド、細胞内タンパク質であってもよい。細胞内タンパク質は、プロテアーゼ、セラミダーゼ、エポキシドヒドロラーゼ、アミノペプチダーゼ、アシラーゼ、アルドラーゼ、ヒドロキシラーゼ、アミノペプチダーゼ、リパーゼなどの酵素であってもよい。ポリペプチドは、細胞外に分泌される酵素であってもよい。そのような酵素は、オキシドレダクターゼ、トランスフェラーゼ、ヒドロラーゼ、リアーゼ、イソメラーゼ、リガーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼの群に属してもよい。酵素は、カルボヒドラーゼ、例えば、セルラーゼ(例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドロラーゼ、もしくはβ−グルコシダーゼ)、ヘミセルラーゼもしくはペクチン分解酵素(例えば、キシラナーゼ、キシロシダーゼ、マンナナーゼ、ガラクタナーゼ、ガラクトシダーゼ、ペクチンメチルエステラーゼ、ペクチンリアーゼ、ペクチン酸リアーゼ、エンドポリガラクツロナーゼ、エキソポリガラクツロナーゼ、ラムノーガラクツロナーゼ、アラバナーゼ、アラビノフラノシダーゼ、アラビノキシランヒドロラーゼ、ガラクツロナーゼ、リアーゼ、もしくはデンプン分解酵素);ヒドロラーゼ、イソメラーゼ、もしくはリガーゼ、ホスファターゼ(例えば、フィターゼ)、エステラーゼ(例えば、リパーゼ)、タンパク質分解酵素、オキシドレダクターゼ(例えば、オキシダーゼ)、トランスフェラーゼ、またはイソメラーゼであってもよい。酵素はフィターゼであってもよい。酵素は、アミノペプチダーゼ、アスパラギナーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシペプチダーゼ、エンドプロテアーゼ、メタロプロテアーゼ、セリンプロテアーゼカタラーゼ、キチナーゼ、クチナーゼ、シクロデキストリングリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α−ガラクトシダーゼ、β−ガラクトシダーゼ、グルコアミラーゼ、α−グルコシダーゼ、β−グルコシダーゼ、ハロペルオキシダーゼ、タンパク質デアミナーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチン分解酵素、ペルオキシダーゼ、ホスホリパーゼ、ポリフェノールオキシダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはグルコースオキシダーゼ、ヘキソースオキシダーゼ、モノオキシゲナーゼであってもよい。
【0149】
本発明の方法では、ポリペプチドは、別のポリペプチドがポリペプチドまたはその断片のN末端もしくはC末端で融合された融合ポリペプチドまたはハイブリッドポリペプチドであることもできる。融合されたポリペプチドは、あるポリペプチドをコードする核酸配列(またはその一部)を別のポリペプチドをコードする核酸配列(またはその一部)に融合することによって産生される。
【0150】
融合ポリペプチドを産生するための技術は当技術分野で公知であり、かつそれらがインフレームとなり、融合されたポリペプチドの発現が同じプロモーターおよびターミネーターの制御下となるようにポリペプチドをコードするコード配列を連結させることを含む。ハイブリッドポリペプチドは、1つまたは複数が宿主細胞にとって異種であってもよい少なくとも2つの異なるポリペプチドから得られる部分的なまたは完全なポリペプチド配列の組合せを含んでいてもよい。
【0151】
生体ポリマーは多糖であってもよい。多糖は、ムコ多糖(例えば、ヘパリンおよびヒアルロン酸)ならびに窒素含有多糖(例えば、キチン)を含むが、これらに限定されない、任意の多糖であってもよい。より好ましい選択肢では、多糖はヒアルロン酸である。
【0152】
本発明による目的のポリヌクレオチドは、有機酸、カロテノイド、(β−ラクタム)抗生物質、およびビタミンなどの、一次または二次代謝産物の合成に関与する酵素をコードしていてもよい。そのような代謝産物は、本発明による生体化合物とみなしてもよい。
【0153】
「代謝産物」という用語は、一次代謝産物と二次代謝産物の両方を包含し、代謝産物は、任意の代謝産物であってもよい。好ましい代謝産物は、クエン酸、グルコン酸およびコハク酸である。
【0154】
代謝産物は、例えば、生合成経路または代謝経路内の1つ以上の遺伝子によってコードされていてもよい。一次代謝産物は、エネルギー代謝、増殖、および構造に関する細胞の一次代謝または一般的な代謝の産物である。二次代謝産物は、二次代謝の産物である(例えば、R.B.Herbert,The Biosynthesis of Secondary Metabolites,Chapman and Hall,New York,1981を参照されたい)。
【0155】
一次代謝産物は、アミノ酸、脂肪酸、ヌクレオシド、ヌクレオチド、糖、トリグリセリド、またはビタミンであってもよいが、これらに限定されない。
【0156】
二次代謝産物は、アルカロイド、クマリン、フラボノイド、ポリケチド、キニーネ、ステロイド、ペプチド、またはテルペンであってもよいが、これらに限定されない。二次代謝産物は、抗生物質、摂食阻害物質、誘引物質、殺細菌物質、殺真菌物質、ホルモン、殺虫物質、または殺鼠物質であってもよい。好ましい抗生物質は、セファロスポリンおよびβ−ラクタムである。
【0157】
生体化合物はまた、選択可能マーカーの産物であってもよい。選択可能マーカーは、目的のポリヌクレオチドの産物であり、この産物は、殺生物剤またはウイルスに対する耐性、重金属に対する耐性、栄養要求株に対する原栄養性などを提供する。選択可能マーカーには、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、trpC(アントラニル酸シンターゼ)、ble(フレオマイシン耐性タンパク質)、およびそれらの等価物が含まれるが、これらに限定されない。
【0158】
本発明によれば、本発明による方法における目的の化合物は、本明細書に記載したようなポリペプチドであることが好ましい。
【0159】
本発明による方法におけるポリペプチドは、本明細書に記載したような酵素であることが好ましい。
【0160】
本発明によれば、本発明による方法における目的の化合物は、代謝産物であることが好ましい。
【0161】
本発明の別の態様によれば、宿主細胞の少なくとも2つの実質的に相同なDNAドメインの適合されたDNAドメインは、遺伝子変換および/または増幅によって、最も低い増加頻度を有する。異なる実質的に相同なDNAドメインは、異なる増加頻度、すなわち、遺伝子変換頻度を有することが明らかになった。最も低い増加頻度を有する実質的に相同なDNAドメインの標的化DNAドメインを適合させる利点は、実質的に相同なDNAドメインにおける組換えの頻度が低い遺伝的に安定な宿主が得られることである。
【0162】
誤って同定された塩基を含む必要があるため、本明細書で提供されている配列情報は、それほど狭義に解釈されるべきではない。本明細書に開示されている特定の配列を容易に用いて、それぞれの宿主細胞から完全な遺伝子を単離することができ、次に、この完全な遺伝子をさらなる配列解析に容易にかけ、それにより、シークエンシングのエラーを同定することができる。
【0163】
別途示されない限り、本明細書中のDNA分子をシークエンシングすることによって決定される全てのヌクレオチド配列は、自動化DNAシークエンサーを用いて決定され、本明細書で決定されるDNA分子によってコードされるポリペプチドの全てのアミノ酸配列は、上記のように決定された核酸配列の翻訳によって予想された。それゆえ、この自動化された手法によって決定される任意のDNA配列について当技術分野で知られているように、本明細書で決定される任意のヌクレオチド配列は、いくつかのエラーを含んでいてもよい。自動化によって決定されるヌクレオチド配列は、典型的には、シークエンシングされたDNA分子の実際のヌクレオチド配列と少なくとも約90%同一、より典型的には、少なくとも約95%から少なくとも約99.9%同一である。実際の配列は、当技術分野で周知の手動のDNAシークエンシング法を含む他の手法によってより正確に決定することができる。これも当技術分野で知られているように、実際の配列と比較した、決定されたヌクレオチド配列における単一の挿入または欠失は、決定されたヌクレオチド配列によってコードされる予想アミノ酸配列が、そのような挿入点または欠失点から、シークエンシングされたDNA分子によって実際にコードされるアミノ酸配列と完全に異なるように、ヌクレオチド配列の翻訳においてフレームシフトを生じさせる。
【0164】
当業者は、そのような誤って同定された塩基を同定することができ、そのようなエラーを訂正する方法を知っている。
【0165】
本明細書に含まれる特定の実施形態は、本発明のいくつかの態様の実例であることが意図されるので、本明細書に記載され、特許請求された本発明は、これらの実施形態によって範囲が限定されるべきではない。等価な実施形態はいずれも、本発明の範囲内であることが意図される。実際、本明細書に示され、記載された変更に加えた、本発明の様々な変更が、前述の記載から当業者に明らかになるであろう。そのような変更も、添付の特許請求の範囲の範囲内に含まれることが意図される。矛盾がある場合、定義を含む本開示が指針と考えられる。
【0166】
[実施例]
[株]
WT1:このアスペルギルス・ニガー(Aspergillus niger)株を野生型株として用いる。この株は、寄託番号CBS513.88でCBS研究所(CBS Institute)に寄託されている。
【0167】
WT2:このA.ニガー(A.niger)突然変異体株は、国際公開第98/46772号パンフレットに本質的に記載されているような方法による古典的な株改良によってA.ニガー(A.niger)CBS513.88から得られる。NTGまたはUVによるWT1の胞子の突然変異誘発後、振盪フラスコにおける突然変異体株の改善されたグルコアミラーゼ産生について選択を行なった。いくつかの改善されたA.ニガー(A.niger)株が同定され、そのうち、優良な株は、3〜4倍増加したグルコアミラーゼ活性レベルを生じさせた。この突然変異体株は、寄託番号CBS124.903でCBS研究所に寄託されている。振盪フラスコ解析および遺伝子解析から、A.ニガー(A.niger)CBS124.903株は、glaA遺伝子座(アンプリコン)の増幅に起因する(3)glaA遺伝子コピー数の増加を伴って、グルコアミラーゼ産生が3〜4倍増加しているという結論が得られた。
【0168】
GBA300:このA.ニガー(A.niger)株は、3つのglaAアンプリコンの改変を含むWT2株である。GBA300株の構築は、国際公開第98/46772号パンフレットに記載の方法に従って行なわれた。この特許出願では、A.ニガー(A.niger)ゲノムを含むアンプリコンから3つのglaA特異的DNA配列を欠失させて、3つの切断されたglaAアンプリコンを有するamdS陰性株を生じさせる方法が広範に記載されている。この手順により、最終的に外来性のDNA配列を全く有さない、マーカー遺伝子を含まない(MARKER−GENE FREE)ΔglaA組換えA.ニガー(A.niger)WT2株が得られた。この3つの切断されたglaAアンプリコンをBamHI切断アンプリコン、SalI切断アンプリコンおよびBglII切断アンプリコンと命名する。
【0169】
[A.ニガー(A.niger)振盪フラスコ発酵]
A.ニガー(A.niger)株を、国際公開第99/32617号パンフレットの実施例「アスペルギルス・ニガー(Aspergillus niger)振盪フラスコ発酵」の節に記載されているように、20mlの前培養培地中で前培養した。一晩増殖させた後、この培養物10mlを、国際公開第99/32617号パンフレットに記載されているような7%グルコースを含む発酵培地1(FM1)に移した。このFM1は、1リットル当たりに、以下のものを含有する:カゼイン加水分解物25g、酵母抽出物12.5g、KH2PO4 1g、K2SO4 2g、MgSO4.7H2O 0.5g、ZnCl2 0.03g、CaCl2 0.02g、MnSO4.4H2O 0.01g、FeSO4.7H2O 0.3g、Pen−Strep(5000IU/mlのPen−5mg/mlのStrep) 10ml、4NのH2SO4でpH5.6に調整。発酵は、表示された日数の間、34℃および170rpmで、100mlの発酵ブロスを含むバッフル付きの500mlフラスコ中で行なわれる。
【0170】
発酵培地2(FM2)はPLA2発酵に用いられ、1リットル当たりに、以下のものを含有する:グルコース.1H2O 82.5g、Maldex15(ブーム・メッペル(Boom Meppel)、オランダ) 25g、クエン酸2g、NaH2PO4.1H2O 4.5g、KH2PO4 9g、(NH4)2SO4 15g、ZnCl2 0.02g、MnSO4.1H2O 0.1g、CuSO4.5H2O 0.015g、CoCl2.6H2O 0.015g、MgSO4.7H2O 1g、CaCl2.2H2O 0.1g、FeSO4.7H2O 0.3g、MES(2−[N−モルホリノ]エタンスルホン酸) 30g、pH=6。
【0171】
[毒素測定用のA.ニガー(A.niger)寒天培地]
以下の培地上で株を培養し、オクラトキシンA(OTA)およびフモニシンB2(FB2)の産生について試験した:FrisvadおよびFiltenborg,1989(Terverticillate Penicillia:chemotaxonomy and mycotoxin production,Mycologia 81:837−861)ならびにFrisvadおよびThrane,1993(Liquid column chromatography of mycotoxinsm In:Betina(編):Chromatography of mycotoxins.Techniques and applications.J.of Chromatography Library 54:253−372 Elsevier,Amsterdam)によって本質的に記載されているようなツァペック酵母自己分解物寒天(CYA)および酵母抽出物ショ糖寒天(YES)。ペトリ皿を暗所24℃で7日間インキュベートし、その後、抽出用に単一コロニーから寒天プラグを採取した。84%CH3CN水を用い、超音波浴に1時間置いて抽出を行ない、その後、0.45μmのPTFEフィルターに通して溶媒を濾過した。
【0172】
規定量のFB2およびOTAになるよう内部標準溶液をサンプルに添加して、定量を行なった。Mogensenら,2010(Production of fumonisin B2 and B4 by Aspergillus niger in raisins and grapes,J of Agricultural and Food Chemistry 58:954−958)によって記載され、Nielsenら、2009(Review of secondary metabolites and mycotoxins from the Aspergillus niger group,Anal Bioanal Chem 395:1225−1242)に概説されているようにLC−MS/MSを行なった。
【0173】
[実施例1.アスペルギルス・ニガー(Aspergillus niger)GBA301(ΔglaA、ΔpepA)の構築]
このA.ニガー(A.niger)株は、主要な細胞外アスパラギン酸プロテアーゼPepAをコードするpepA遺伝子の欠失を含むGBA300株である。GBA301株は、欧州特許第0635574号明細書に記載されているような「マーカー遺伝子を含まない(MARKER−GENE FREE)」アプローチを用いて構築される。この特許に記載されている方法を用いて、van den Homberghら(van den Hombergh JP,Sollewijn Gelpke MD,van de Vondervoort PJ,Buxton FP,Visser J.(1997)−Disruption of three acid proteases in Aspergillus niger−effects on protease spectrum,intracellular proteolysis,and degradation of target proteins−Eur J Biochem.247(2):605−13)によって記載されているように、GBA300のゲノム中のpepA特異的DNA配列を欠失させる。この手順により、GBA300株バックグラウンドでpepA遺伝子が不活化されているマーカー遺伝子を含まないGBA301株が得られた。
【0174】
[実施例2.アスペルギルス・ニガー(Aspergillus niger)GBA302(ΔglaA、ΔpepA、ΔhdfA)の構築]
既知の原理に従ってhdfA用の遺伝子置換ベクターを設計し、日常的なクローニング手順に従って構築した。基本的には、これらのベクターは、所定のゲノム遺伝子座での相同組換え用に、hdfA ORFの約1〜2kbのフランキング領域を含む。さらに、これらは、直接反復と直接反復の間にA.ニデュランス(A.nidulans)二方向性amdS選択マーカーを含む。欠失ベクターの一般的な設計は、以前に欧州特許出願公開第635574B号明細書および国際公開第98/46772号パンフレットに記載されており、欠失ベクターを構築するための一般的なクローニングベクターpGBDEL(図1)の使用は、国際公開第06/040312号パンフレットに記載されている。
【0175】
ベクターpDEL−HDFA(図2)は、相同組換え用にhdfA ORFの約1kbフランキング領域を含む。A.ニガー(A.niger)遺伝子の全ヌクレオチド配列およびそのゲノムコンテクストは、例えば、NCBI(http://www.ncbi.nlm.nih.gov/)またはEMBL(http://www.ebi.ac.uk/embl/)から得ることができる。欠失ベクターpDEL−HDFAの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に詳細に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA301を形質転換し、hdfA遺伝子を欠失させた。本明細書中の全ての実施例における遺伝子欠失に適用された方法では、線状DNAが用いられた。この線状DNAは、二重交差によってフランキング配列の相同な遺伝子座でゲノム中に組み込まれ、その結果、欠失しようとする遺伝子をamdS遺伝子に置き換える。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。遺伝子破壊の一般的手順は、図3に示されている。GBA301株バックグラウンドでhdfA遺伝子が不活化されている代表的な株として株GBA302を選択した。
【0176】
[実施例3.アスペルギルス・ニガー(Aspergillus niger)GBA303(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン)の構築]
この実施例では、最も高い頻度の遺伝子変換を有するΔglaAアンプリコンを適合させて、標的化組込み用の独特の遺伝子座を生じさせ、賢明な組込み戦略を利用できるようにした。適合されたアンプリコンは、この実施例中のグルコアミラーゼプロモーターおよびターミネーターなどのフランキング制御配列を標的化領域として利用できるようにもする。
【0177】
[実施例3.1 最も高い頻度の遺伝子変換を有するアンプリコンの同定]
株GBA301は、ゲノム中に3つの改変されたΔglaA遺伝子座を含む。3つ全ての遺伝子座について、約4.3kbのglaA配列(2kbのglaAプロモーターおよび全glaAコード配列)が欠失している(国際公開第98/46772号パンフレットの図4に見られる)。3つの異なる欠失ベクターを用いたので、各々のΔglaA遺伝子座はわずかに異なっており、その特徴を用いて、PCRテストで各々の切断されたΔglaA遺伝子座を可視化することができる(国際公開第98/46772号パンフレットの図5)。組み込まれたこの違いを用いて、切断されたglaAアンプリコンの中に遺伝子変換事象を追跡することもできる。3つの切断されたglaAアンプリコンはBamHI切断アンプリコン、SalI切断アンプリコンおよびBglII切断アンプリコンと命名されており、いわゆる「DNAフラッグ」テストにおいて、それぞれ、240、260および300bpのサイズのバンドを示す(国際公開第98/46772号パンフレットの図5)。
【0178】
株GBA301から、GBA301の3つの切断されたglaAアンプリコンの1つに全て標的化される複数のフィターゼ発現カセットを含む、複数のフィターゼ産生株を(pGBAAS−1とpGBTOPFYT−1を共形質転換することによって)作製した(国際公開第98/46772号パンフレットの実施例1.4に本質的に記載されている)。得られる株は各々、3つの切断されたglaAアンプリコンの1つにフィターゼカセットを有し、フィターゼカセット組込み部位に従って、BAM−PHY、SAL−PHYおよびBGL−PHYと呼ばれる。
【0179】
3つの異なる株、BAM−PHY、SAL−PHYおよびBGL−PHYについてフィターゼカセットのコピー数が増加した変換体を同定した(国際公開第98/46772号パンフレットの実施例1.5に本質的に記載されている)。フィターゼコピー数が増加した変換体について、フィターゼ含有glaAアンプリコンの特定のアンプリコンの頻度および遺伝子型を「DNAフラッグ」テストを用いて決定した。「DNAフラッグ」テストによってそれぞれの株の何千もの個々の子孫を解析することにより、特定の切断されたglaAアンプリコン(例えば、SalI)の欠失と同時に、別の特定の切断されたglaAアンプリコン(例えば、BamHI)の増幅によって遺伝子変換を検出することができる。
【0180】
3つの異なる株の何百もの個々の子孫のPCRスコアリングおよびタイピングの結果を表1に見ることができる。さらに、いくつかの単離された株について単一の遺伝子変換事象後に同様の手法を取った。これらの株は、同じ種類の2つのアンプリコンを含んでいた。3種類の株の何百もの個々の子孫をPCRでスコアリングしたこれらの結果を表2に見ることができる。この3つの異なる株に関しては、第1の遺伝子変換事象と第2の遺伝子変換事象の両方について、BamHIアンプリコン中にあるフィターゼカセットを遺伝子変換によって最も高い頻度で増加させることができる。
【0181】
【表1】
【0182】
【表2】
【0183】
[実施例3.2 最も高い頻度の遺伝子変換を有するBamHIアンプリコンの適合]
この実施例のこの部分では、(株GBA300を生じさせるCBS124.903中のglaA遺伝子座の3倍の改変によって)事前に除去されたグルコアミラーゼプロモーターPglaAを、ゲノム中の単一の遺伝子座に、しかし、適合させようとするアンプリコン中の異なる位置に再導入し、標的化組込み用の独特のかつ改善された遺伝子座を生じさせる方法が記載されている。
【0184】
アンプリコンを適合させることができるように、ΔglaA遺伝子座用の遺伝子置換型のベクターを既知の原理に従って設計し、日常的なクローニング手順に従って構築した。この目的のために、ベクターpGBGLA−65を構築した(図6)。基本的に、このベクターは、2つのPglaA断片と3’glaAおよび3”glaA領域の間にamdS選択マーカーを含む。3’glaAおよび3”glaA領域は、ΔglaA遺伝子座の同一のゲノム領域中への標的化および組込みに用いられる。2つのPglaA断片は、対抗選択によるamdS選択マーカーのループアウトに用いられる(図3)。1つのPglaA断片は、切断されたPglaAプロモーター断片(MluI部位の3’にあるPglaAプロモーターの最後の600bpを欠く)であり、これは、amdS対抗選択後にゲノム中に存在したまま残る。
【0185】
ゲノム中にBamHIアンプリコンを適合させることができるように、ベクターpDEL−GLA65(図6)をHindIIIで消化し、ゲル精製して、大腸菌(E.coli)骨格を除去した。欠失ベクターpDEL−GLA65の線状DNAを用い、国際公開第05/095624号パンフレットに以前に詳細に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA302を形質転換した。pDEL−GLA65の線状DNAは、二重交差によって3’glaA配列および3”glaA配列でアンプリコンに組み込まれ得る。いくつかのコロニーを純化し、3つのglaA遺伝子座の存在およびBamHIアンプリコンでのカセットの組込みについてPCRで解析した(標的PCR)。BamHIアンプリコンの標的化頻度は、20〜30%と推定された。合計7つの株をフルオルアセトアミド対抗選択について選択した。フルオルアセトアミド耐性コロニーをPDAに移し、「フラッグ」−PCRでテストした。適合されたBamHI遺伝子座を有し、組み込まれたpDEL−GLA65カセットのPglaA反復上での組換えによってamdSマーカーが正確に欠失している代表的な株としてある株を選択した(図7)。この選択された株をGBA303と命名した。これは、pepAおよびhdfA遺伝子が不活化され、BamHIアンプリコンが適合された、3つ全てのΔglaA遺伝子座を有するGBA300株バックグラウンドを表す。glaAアンプリコン全体はサイズが大きい(>80kb)ので、BamHIアンプリコンが適合されている(2〜6kb)としても、かなりの実質的に相同なDNAドメインを有する断片が依然としてゲノム中に存在したまま残る。
【0186】
ΔglaAアンプリコンの3’断片および3”断片を含むゲノム領域の配列(図7)は、配列番号:1の下に見出すことができる。3’断片、切断されたPglaAプロモーター断片および適合されたBamHIアンプリコンの3”断片を含むゲノム領域の配列は、配列番号:2の下に見出すことができる。
【0187】
[実施例3.3 新規の組込みベクターの構築]
欧州特許出願公開第0635574A1号明細書、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットには、標的化された株構築、とりわけ、単一の交差を用いる標的化株構築で用いられる組込みおよび発現ベクターpGBAAS−1(欧州特許第0635574号明細書中のpGBGLA−50)およびpGBTOP−8が記載されている。これらの組込みベクターの調整は、GBA302型の株の適合されたBamHIアンプリコンへの特異的標的化を可能にするために必要である。
【0188】
amdSマーカーを含み、かつ共形質転換で用いられるpGBAAS−1ベクターのうち、2つの異なる変異体が、適合されたBamHIアンプリコンへの標的化組込み用に構築されている。グルコアミラーゼプロモーターPglaAの制御下で発現されるamdSマーカー遺伝子を有するpGBAAS−3(図8)と呼ばれるベクターを構築した。PglaAプロモーターは、amdSマーカー遺伝子の発現を駆動する際に機能を有する。さらに、PglaAプロモーターおよび3’glaA断片は、特に、適合されたBamHI ΔglaAアンプリコンの標的化配列として機能する(図21)。pGBAAS−1のamdSマーカーカセット(gpdAプロモーターの制御下のamdS)を含むpGBAAS−4と呼ばれる第2のベクター(図9)を構築した。ここで、切断されたPglaAプロモーター断片および3’glaA断片は、適合されたBamHI ΔglaAアンプリコンに標的化する際に機能を有する(図21)。
【0189】
pGBTOP型のベクターについては、適合されたBamHI ΔglaAアンプリコンへの標的化を可能にするために、2つの新規の組込みおよび発現ベクターを構築した。pGBTOP−11(図10)およびpGBTOP−12(図11)ベクターは、基本的には、それぞれ、3”glaA断片が除去されたpGBTOP−8ベクターおよび大腸菌(E.coli)ベクター除去用の制限部位のさらなる改変を含むpGBTOP−8ベクターである。ここで、PglaAプロモーターおよび3’glaAターミネーター断片は二重の機能を有する。すなわち、目的の遺伝子を発現させるためのプロモーターおよびターミネーターとしてのその役割の他に、適合されたBamHI ΔglaAアンプリコンに標的化する役割を有する(図21)。
【0190】
[実施例4.アスペルギルス・ニガー(Aspergillus niger)GBA304(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII)の構築]
α−アミラーゼをコードするamyBII ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYBIIを得た(図12)。欠失ベクターpDEL−AMYBIIの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA303を形質転換し、amyBII遺伝子を欠失させた。二重交差事象による相同なamyBII遺伝子座でのゲノム中へのpDEL−AMYBIIの組込みによってamyBII ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA303株バックグラウンドでamyBII遺伝子が不活化された代表的な株として株GBA304を選択した。
【0191】
[実施例5.アスペルギルス・ニガー(Aspergillus niger)GBA305(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI)の構築]
α−アミラーゼをコードするamyBI ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYBIを得た(pDEL−AMYBIのレイアウトの代表的な図を図12に見ることができる)。欠失ベクターpDEL−AMYBIの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA304を形質転換し、amyBI遺伝子を欠失させた。二重交差事象による相同なamyBI遺伝子座でのゲノム中へのpDEL−AMYBIの組込みによってamyBI ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA304株バックグラウンドでamyBI遺伝子が不活化された代表的な株として株GBA305を選択した。
【0192】
[実施例6.アスペルギルス・ニガー(Aspergillus niger)GBA306(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA)の構築]
酸安定性α−アミラーゼをコードするamyA ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYAを得た(pDEL−AMYAのレイアウトの代表的な図を図12に見ることができる)。欠失ベクターpDEL−AMYAの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA305を形質転換し、amyA遺伝子を欠失させた。二重交差事象による相同なamyA遺伝子座でのゲノム中へのpDEL−AMYBIの組込みによってamyA ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA305株バックグラウンドでamyA遺伝子が不活化された代表的な株として株GBA306を選択した。
【0193】
[実施例7.アスペルギルス・ニガー(Aspergillus niger)GBA307(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA)の構築]
このA.ニガー(A.niger)のオキサラート欠損株は、オキサロ酢酸ヒドロラーゼをコードするoahA遺伝子の欠失によって得ることができる。これは、欧州特許第1157100号明細書および米国特許第6,936,438号明細書に詳細に記載されている。GBA306株バックグラウンドでoahA遺伝子が不活化された代表的な株として株GBA307を選択した。
【0194】
あるいは、国際公開第04/070022号パンフレットおよび欧州特許第1590444号明細書に記載されているような古典的な株改良によってA.ニガー(A.niger)GBA306から突然変異体株を得ることができる。これらの文献では、オキサラート欠損A.ニガー(A.niger)株についてスクリーニングする方法が広範に記載されている。これらを欧州特許第1590444号明細書の実施例1および2の方法に従って単離した。
【0195】
[実施例8.アスペルギルス・ニガー(Aspergillus niger)GBA308(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB)の構築]
CBS513.88のゲノム配列において、マイコトキシンのフモニシンをコードする、ギベレラ・モニリフォルミス(Gibberella moniliformis)中の遺伝子クラスターとの相同性に基づいて、推定上のフモニシン遺伝子クラスターを同定した(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88”.Nat Biotechnol.2007 Feb;25(2):221−231)。
【0196】
【表3】
【0197】
遺伝子An01g06930は、フモニシン産生に関与する可能性があるポリケチド合成酵素(PKS)をコードする。プロモーター側に推定上のfumB遺伝子(An01g06930)のコード配列のごく一部分も含む、遺伝子フランキング領域を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているように、ベクターpGBDEL(図1)にクローニングし、ベクターpGBDEL−FUM3(図13)を得た。欠失ベクターpGBDEL−FUM3の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA307を形質転換し、fumB遺伝子を欠失させた。二重交差事象による相同なAn01g06930遺伝子座でのゲノム中へのpGBDEL−FUM3の組込みによってfumB ORFが除去され、結果として約8kbのゲノム配列が除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA307株バックグラウンドでfumB遺伝子が不活化された代表的な株として株GBA308を選択した。
【0198】
株WT1およびGBA308をCYAおよびYES寒天培地上で増殖させ、フモニシン産生を測定した。FB2測定の結果を以下の表に示す。
【0199】
【表4】
【0200】
これらの結果から、PKSをコードするfumB遺伝子の破壊によってフモニシン産生が障害された株が得られることは明らかである。フモニシン陰性株バックグラウンドは、商業的なタンパク質産生にとって明らかな利点がある。
【0201】
さらに、遺伝子An01g06820を含む巨大な遺伝子クラスターが(約38kbのゲノム配列が除去された)An01g06930まで破壊されている株は、同一の表現型を示す、すなわち、上で詳述したような試験でフモニシン産生が陰性であることが示された(データは示さない)。
【0202】
[実施例9.アスペルギルス・ニガー(Aspergillus niger)GBA309(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch)の構築]
CBS513.88のゲノム配列において、オクラトキシン産生に関与するA.オクラセウス(A.ochraceus)のPKS断片に基づいて、推定上のオクラトキシン遺伝子クラスターを同定した(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88.” Nat Biotechnol.2007 Feb;25(2):221−231)。
【0203】
【表5】
【0204】
このPKS様遺伝子は、An15g07920(ochAと呼ばれる)と注釈付けられている。さらに、遺伝子An15g07910(ochBと呼ばれる)は、オクラトキシン産生に関与する潜在的なペプチド合成酵素に相当し得る。ペプチド合成酵素および/またはPKSをコードする遺伝子の破壊は、オクラトキシン産生が障害された株をもたらし得る。
【0205】
二重交差用の標的化配列を用いて完全なオクラトキシン遺伝子クラスターを破壊することができるように、一方の側のAn15g07860 ORFの遺伝子配列断片およびもう一方の側のAn15g07930 ORFの遺伝子配列断片を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているようにクローニングして、ベクターpGBDEL−OCH2(図14)を得た。欠失ベクターpGBDEL−OCH2の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA308を形質転換し、推定上のオクラトキシン遺伝子クラスターを欠失させた。二重交差事象による相同な遺伝子座でのゲノム中へのpGBDEL−OCH2の組込みによってoch ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA308株バックグラウンドでオクラトキシンゲノム遺伝子クラスター(遺伝子An15g07860と遺伝子An15g07930の間にある;遺伝子型はΔochと表示される)が除去された代表的な株として株GBA309を選択した。
【0206】
株WT1およびGBA309をCYAおよびYES寒天培地上で増殖させ、オクラトキシンAを測定した。OTA測定の結果を以下の表に示す。
【0207】
【表6】
【0208】
これらの結果から、OTA生合成に関与する推定上のペプチド合成酵素およびPKSを含むオクラトキシン遺伝子クラスターの破壊によって、オクラトキシンA産生が低下した株が得られることは明らかである。オクラトキシンA陰性および場合によってはフモニシン陰性との組合せの株バックグラウンドは、商業的なタンパク質産生にとって明らかな利点がある。
【0209】
[実施例10.アスペルギルス・ニガー(Aspergillus niger)GBA310(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch、ΔprtT)の構築]
プロテアーゼ調節因子をコードするprtT ORFの遺伝子フランキング領域を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているようにクローニングし、ベクターpGBDEL−PRT2(図15)を得た。欠失ベクターpGBDEL−PRT2の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA309を形質転換し、prtT遺伝子を欠失させた。二重交差事象による相同なprtT遺伝子座でのゲノム中へのpGBDEL−PRT2の組込みによってprtT ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA309株バックグラウンドでprtT遺伝子が不活化された代表的な株として株GBA310を選択した。
【0210】
[実施例11.アスペルギルス・ニガー(Aspergillus niger)GBA311(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch、ΔprtT、sec61 S376W突然変異)の構築]
この実施例では、セリン376がトリプトファンに置き換えられているSec61*タンパク質をコードする改変されたsec61遺伝子(sec61*と呼ぶ)の導入を記載する。そうするために、遺伝子フランキング配列および改変されたSec61輸送チャネルをコードするsec61*コード配列を、国際公開第2005123763号パンフレットに本質的に詳細に記載されているようにクローニングし、ベクターpGBDEL−SEC61*(図16)を得た。欠失ベクターpGBDEL−SEC61*の線状DNAを単離し、これを用いて、国際公開第2005123763号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA310を形質転換し、Sec61遺伝子を改変した。二重交差事象による相同なSec61遺伝子座でのゲノム中へのpGBDEL−SEC61*の組込みによってSec61*ORFが改変された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA310株バックグラウンドでSec61遺伝子が改変された代表的な株として株GBA311を選択した。
【0211】
[実施例12.適合されたアスペルギルス・ニガー(Aspergillus niger)株およびプラスミドを用いる改善された酵素産生および産生株構築]
ブタホスホリパーゼA2(PLA2)タンパク質(Roberts I.N.,Jeenes D.J.,MacKenzie D.A.,Wilkinson A.P.,Sumner I.G.およびArcher D.B.(1992) “Heterologous gene expression in Aspergillus niger.a glucoamylase−porcine pancreatic phospholipase A2 fusion protein is secreted and processed to yield mature enzyme” Gene 122:155−161)を、A.ニガー(A.niger)のCBS513.88&GBA株系統における酵素発現のモデルタンパク質として選択した。PLA2の過剰発現用の断片をproPLA2とA.ニガー(A.niger)のネイティブなグルコアミラーゼA遺伝子との融合体として作製し、原則的に、Robertsら(1992)によって記載されているように調製した。このglaA−pla2融合遺伝子をpGBTOP−8およびpGBTOP−12にクローニングし、それぞれ、pGBTOPPLA−2a(図17)およびpGBTOPPLA−2b(図18)を得た。
【0212】
さらに、A.ニガー(A.niger)のα−アミラーゼを、A.ニガー(A.niger)のCBS513.88&GBA株系統における酵素発現の第2のモデルタンパク質として用いた。過剰発現に用いたamyBコード配列は、(国際公開第2008/000632号パンフレットに詳細に記載されているように)amyB遺伝子をコードするα−アミラーゼの単一コドンおよびコドン対が最適化されたコード配列を含んでいた。グルコアミラーゼglaAプロモーターの翻訳開始配列は、(これも国際公開第2006/077258号パンフレットに詳述されているように)作製されたamyB発現コンストラクトにおいて5’−CACCGTCAAA ATG−3’へと改変されている。さらに、最適な翻訳終結配列が用いられたため、野生型5’−TGA−3’翻訳終結配列は、全ての発現コンストラクトにおいて、(国際公開第2006/077258号パンフレットに詳述されているように)5’−TAAA−3’に置き換えられた。最適化されたamyB cDNA配列を含むEcoRI−SnaBI断片を完全に合成し、サブクローニングし、配列解析によって配列を確認した。amyB断片をpGBTOP−8およびpGBTOP−12にクローニングし、それぞれ、pGBTOPFUA−2(図19)およびpGBTOPFUA−3(図20)を得た。
【0213】
[実施例12.1 適合された発現ベクターのA.ニガー(A.niger)株のΔglaA遺伝子座への改善された標的化]
構築された組込みベクター(実施例3.3および12)は2つのタイプ、すなわち、適合されていないΔglaA遺伝子座に対して3’glaA配列および3”glaA配列に標的化するために用いることができる組込みベクターと、ベクターpGBTOPPLA−2a、pGBTOPFUA−2およびpGBAAS−1である組込みベクターとに分けることができる。さらに、切断されたPglaAプロモーター断片および3’glaA断片に適合されたBamHI ΔglaAアンプリコン(適合されたΔglaA遺伝子座)に特異的に標的化するために用いることができる新規の組込みベクターを構築した。適合されたBamHIアンプリコンに標的化するために用いることができるベクターは、pGBTOPPLA−2b、pGBTOPFUA−3、pGBAAS−3およびpGBAAS−4である。
【0214】
以下のスキーム(表3)に従って、2つのPLA2発現ベクター(pGBTOPPLA−2aおよびpGBTOPPLA−2b)を、pGBBAAS−1およびpGBAAS−4との共形質転換によって、A.ニガー(A.niger)のGBA301(ΔglaA、ΔpepA)、GBA302(ΔglaA、ΔpepA、ΔhdfA)およびGBA303(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン)に導入した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、4つ全てのベクターの線状DNAを単離し、これを用いて、3つのA.ニガー(A.niger)株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によってΔglaA遺伝子座の1つで起こる。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。3つ全ての株について、amdSマーカー遺伝子とPLA2発現カセットの両方を有する数百個の形質転換体を、amdSおよびPLA2特異的プライマーを用いるPCRにより同定した。同定された共形質転換体に対して(国際公開第98/46772号パンフレットの実施例1.4に詳述されているように)標的化PCRテストを実施し、amdSおよび/またはPLA2カセットのいずれかがΔglaA遺伝子座に組み込まれている株のパーセンテージを決定した。
【0215】
さらに、フルオロアセトアミド培地上にプレーティングすることにより、同定されたΔglaA標的化共形質転換体からamdSマーカー遺伝子を含まない株を選択した。PCRに基づくDNA「フラッグテスト」を実施して、amdSを含まない子孫におけるアンプリコンの潜在的な喪失を検出した。国際公開第98/46772号パンフレットの実施例1.4.fで説明されているように、これは、組換えによるamdSマーカーと全glaAアンプリコンの同時喪失を示すものであり、そのようなものとして元のカセットの位置を示すものである。この試験を用いて、3つ全ての株におけるBamHIアンプリコンへの標的化頻度を決定した。データを表3に示す。
【0216】
【表7】
【0217】
表3から結論付けることができるように、適合された発現ベクターを適合された標的化DNAドメイン(適合されたBamHIアンプリコン)と組み合わせて用いると、標的化された株構築における明らかな利点が与えられ、より集中的でかつ効率的な株構築がもたらされる。
【0218】
[実施例12.2 適合されたコンストラクトおよびA.ニガー(A.niger)株を用いる改善された共形質転換]
以下のスキーム(表4)に従って、FUA発現ベクターpGBTOPFUA−3を、参照セットとしてのpGBTOPFUA−2およびpGBAASとともに、pGBAAS−3およびpGBAAS−4の共形質転換によって、A.ニガー(A.niger)GBA307(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA)に導入した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、5つのベクターの線状DNAを単離し、これを用いて、A.ニガー(A.niger)GBA307株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によって、それぞれ、適合されたBamHI ΔglaAアンプリコンまたはΔglaAアンプリコンで起こる。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。amdSおよびamyB特異的(コドン対が最適化されたamyBコード配列)プライマーを用いるPCRにより、各々のプラスミド組合せの50個の形質転換体について(国際公開第98/46772号パンフレットの実施例1.4に原理的に詳述されているような)共形質転換パーセンテージを決定した。データを表4に示す。
【0219】
【表8】
【0220】
これまでに知られているpGBAAS−1および改善された標的化のためのその変異体pGBAAS−4の場合、典型的な共形質転換頻度が得られる(通常25〜75%)。驚くことに、表4から結論付けることができるように、pGBBAAS−3変異体(glaAプロモーターの制御下のamdS遺伝子)を適合された標的化コンストラクト(例えば、pGBTOPFUA−3)と組み合わせて用いると、非常に高い共形質転換頻度が得られる。pGBAAS−3を用いると、共形質転換頻度が劇的に増加し、標的化された株構築において株をさらに操作する前に共形質転換体を選択する必要が回避され、そのため、より効率的な株構築がもたらされる。
【0221】
これらの形質転換体のFM1における振盪フラスコ解析は、GBA307で発現された異なるコンストラクトについて、遺伝子1コピー当たりに同様のFUA発現レベルを示した。
【0222】
[実施例12.3 適合されたA.ニガー(A.niger)株における改善された株構築]
工業用の組換えA.ニガー(A.niger)産生株を構築する1つの方法は、ΔglaA遺伝子座の1つに発現カセットを組み込み、その後、組み込まれた発現カセットを有するΔglaA遺伝子座を遺伝子変換または増幅によって増加させることである。
【0223】
発現カセットは、例えば、国際公開第2005/100573号パンフレット、国際公開第2006/077258号パンフレット、国際公開第2006/092396号パンフレットまたは国際公開第2008/000632号パンフレットに詳述されているようないくつかの方法によって改善することができる。株構築での遺伝子変換または増幅の使用は、例えば、国際公開第1998/46772号パンフレットにより詳細に記載されている。先の実施例では、株およびプラスミド適合によって、どのようにして共形質転換および/または特定の遺伝子座への標的化を改善することができるかということが示されている。この実施例では、新規のGBAタイプの株を用いる株構築での遺伝子変換または増幅の使用が示されている。
【0224】
A.ニガー(A.niger)GBA311(実施例11)株に、発現ベクターの2つの組合せ(すなわち、pGBTOPPLA−2aとpGBBAAS−1の混合物;pGBTOPPLA−2bとpGBBAAS−3の混合物)を共形質転換した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、使用した全てのベクターの線状DNAを単離し、これを用いて、GBA311 A.ニガー(A.niger)株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によって、ΔglaA遺伝子座で起こる(標的化配列の詳細については、実施例12.1を参照されたい)。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。amdSマーカー遺伝子とPLA2発現カセットの両方を有する数百個の形質転換体を、amdSおよびPLA2特異的プライマーを用いるPCRにより同定した。同定された共形質転換体に対して、(国際公開第98/46772号パンフレットの実施例1.4に詳述されているように)標的化PCRテストを実施し、amdSおよび/またはPLA2カセットのいずれかがΔglaA遺伝子座に組み込まれている株のパーセンテージを決定した。さらに、PCRを用いて、コロニーをPLA2コピー数について診断した。サザン解析(国際公開第98/46772号パンフレット)を用いて、高コピーと考えられる株のコピー数をより正確に決定した。pGBTOPPLA−2とpGBBAAS−1の混合物については、さらなる形質転換体をスクリーニングし、純化して、より高いコピー数のPLA2発現カセットを有する形質転換体を単離した。600個の個々の形質転換体をスクリーニングした後、6コピーのBglII標的化形質転換体を単離した。得られたデータを表5に示す。
【0225】
【表9】
【0226】
GBA300バックグラウンドの2つの高コピー株(すなわち、6コピーのBglII標的化株および7コピーのBamHI標的化株)について、PLA2コピー数が増加している変換体を単離した(実験の詳細については、本明細書中の実施例3.1および国際公開第98/46772号パンフレットを参照されたい)。両方の株の500個の個々の子孫をスクリーニングすることによって、BamHI標的化株については、遺伝子変換の結果としてPLA2コピー数が増加した2つの株が同定されたが、BglII標的化株については全く同定されなかった。この2つの変換体は、それぞれ、13コピーおよび14コピーのPLA−2を含んでいた。この実施例およびデータは、(例えば、増強された遺伝子変換または増幅頻度を有する)ゲノム標的化DNAドメインの適合は、明確でかつ制御された多コピー(産生)株の構築の過程を促進および加速することによって、株構築に大きな影響を及ぼすことを明らかに示している。これらの形質転換体のFM2における振盪フラスコ解析は、GBA311で発現された異なるコンストラクトについて、遺伝子1コピー当たりに同様のPLA2発現レベルを示した。
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、目的の化合物を産生するための組換え宿主細胞に関する。本発明はさらに、そのような宿主細胞の産生方法に関する。本発明はさらに、目的の化合物の産生に関する。本発明はさらに、単離されたポリヌクレオチドならびに前記ポリヌクレオチドを含むベクターおよび宿主細胞に関する。
【0002】
[背景技術]
本発明は、目的の化合物を産生するための組換え宿主細胞に関する。
【0003】
そのような宿主細胞は、とりわけ、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットから知られており、ここで、宿主細胞は、前記宿主細胞の染色体の少なくとも2つの実質的に相同なDNAドメインの1つに目的のポリヌクレオチドを含み、かつ目的のポリヌクレオチドのコピー数は、遺伝子変換によって増加している。
【0004】
しかしながら、高コピーの遺伝子変換株(例えば、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットによる方法によって得られる株)を得ることは、多くの場合、長い時間と労力を要すると考えられることが示されている。それゆえ、長い時間と労力をあまり要さない高コピーの遺伝子変換株の構築を可能にするために方法を改善することができれば有利であろう。
【図面の簡単な説明】
【0005】
【図1】置換ベクターpGBDELの物理的地図を示す。フランキング領域をクローニングするための多重クローニング部位がamdSマーカーに対して示されている。
【図2】アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))のhdfA遺伝子を不活化するために用いられる置換ベクターpDEL−HDFAの物理的地図を示す。この置換ベクターは、hdfAフランキング領域、amdSマーカーおよび大腸菌(E.coli)DNAを含む。大腸菌(E.coli)DNAは、A.ニガー(A.niger)株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図3】宿主株から遺伝子または遺伝子断片を欠失させるために用いられる戦略を示す。使用されるDNAコンストラクトは、欠失させようとする遺伝子の相同領域(5’および3’)が両隣にあるamdS選択マーカーを含む(1)。このコンストラクトは、対応するゲノム遺伝子座での二重相同組換え(X)によって組み込まれ(2)、ゲノム遺伝子コピーに置き換わる(3)。その後、直接反復上での組換え(U)によって、amdSマーカーが除去され、欠失させようとする遺伝子または遺伝子断片の正確な切除が生じる(4)。
【図4】親A.ニガー(A.niger)株CBS124.903におけるglaA(グルコアミラーゼ)遺伝子座および組換え株GBA300における3つの切断された「Xの印がある」ΔglaA遺伝子座(「X」は、BamHI、SalIまたはBglII制限部位を表す)の物理的地図を示す。
【図5】A.ニガー(A.niger)CBS124.903由来の組換え株GBA300のglaA DNAアンプリコンにおける3つのΔglaA遺伝子座の概略図を示す。各々の遺伝子座は、異なる制限部位(BamHI、SalIまたはBglII)でマークが付けられている。glaA遺伝子座は各々、約20〜60bpだけ長さが異なっており、PCRに基づくDNAフラッグテストによって、切断された異なるglaA遺伝子座を可視化することができる。
【図6】実質的に相同なDNAドメインを適合させるために用いられるベクターpGBGLA−65の物理的地図を示す。基本的に、このベクターは、2つのPglaA断片と3’glaAおよび3”glaA領域の間にamdS選択マーカーを含む。3’glaAおよび3”glaA領域は、ΔglaA遺伝子座の対応するゲノム領域への標的化および組込みに用いられる。2つのPglaA断片は、図3に見られるような対抗選択によるamdS選択マーカーの切除に用いられる。1つのPglaA断片は、(MluI部位の3’にあるPglaAプロモーターの最後の600bpを欠く)切断されたPglaAプロモーター断片であり、これは、amdS対抗選択後にゲノム中に存在したまま残る。
【図7】適合されたBamHIアンプリコンを有する株のglaA DNAアンプリコンにおける3つのΔglaA遺伝子座の物理的地図を示す。BamHIアンプリコンは、ゲノムglaA遺伝子座の相同な3’glaA遺伝子座と3”glaA遺伝子座の間に切断された相同なグルコアミラーゼプロモーター断片を導入することによって適合された。
【図8】新規のアセトアミダーゼ選択マーカーおよび適合されたBamHIアンプリコン標的化ベクターpGBAAS−3の物理的地図を示す。glaAプロモーターは、標的化領域としておよびamdS遺伝子のプロモーターとして機能する。
【図9】新規のアセトアミダーゼ選択マーカーおよび適合されたBamHIアンプリコン標的化ベクターpGBAAS−4の物理的地図を示す。(切断された)glaAプロモーターは標的化領域として機能し、gpdAプロモーターは、amdS遺伝子の発現を駆動する。
【図10】新規の発現および適合されたBamHIアンプリコン標的化ベクターpGBTOP−11の物理的地図を示す。HindIII制限酵素を用いて、ベクターを線状化し、大腸菌(E.coli)部分を除去することができる。
【図11】新規の発現および適合されたBamHIアンプリコン標的化ベクターpGBTOP−12の物理的地図を示す。NotI制限酵素を用いて、ベクターを線状化し、大腸菌(E.coli)部分を除去することができる。
【図12】置換ベクターpDEL−AMYBIIの物理的地図を示す。5’amyBIIフランキング領域、3’amyBIIフランキング領域がamdSマーカーに対して示されている。amyBIIの3’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図13】置換ベクターpGBDEL−FUM3の物理的地図を示す。fumBコード配列の一部を含む5’fumB領域と3’fumBフランキング領域とがamdSマーカーに対して示されている。fumBの5’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図14】置換ベクターpGBDEL−OCH2の物理的地図を示す。フランキング領域としてのAn15g07860遺伝子配列の一部とAn15g07930遺伝子配列の一部がamdSマーカーに対して示されている。An15g07930遺伝子の配列は少なくとも2〜300bp重複し、相同組換えによる対抗選択を可能にしている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図15】置換ベクターpGBDEL−PRT2の物理的地図を示す。5’prtTフランキング領域、3’prtTフランキング領域がamdSマーカーに対して示されている。prtT3’配列の配列は少なくとも2〜300bp重複している。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素BstBIおよびXmaIによる消化によって除去することができる。
【図16】置換ベクターpGBDEL−Sec61*の物理的地図を示す。Sec61*突然変異体遺伝子および3’Sec61*突然変異体遺伝子断片がamdSマーカーに対して示されている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素AscIおよびNotIによる消化によって除去することができる。
【図17】PLA2発現ベクターpGBTOPPLA−2aの物理的地図を示す。glaAフランキング領域が、glaAプロモーター、切断されたglaA遺伝子およびpro−pla2コード配列に対して示されている。大腸菌(E.coli)DNAは、宿主株に形質転換する前に、制限酵素NotIによる消化によって除去することができる。
【図18】PLA2発現ベクターpGBTOPPLA−2bの物理的地図を示す。glaAプロモーター、切断されたglaA遺伝子、pro−pla2コード配列および3’glaAフランキング配列が示されている。大腸菌(E.coli)DNAは、宿主株への形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図19】発現ベクターpGBFINFUA−2の物理的地図を示す。glaAプロモーターおよびA.ニガー(A.niger)amyBコドン最適化配列に対してglaAフランキング領域が示されている。大腸菌(E.coli)DNAは、宿主株の形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図20】FUA発現ベクターpGBTOPFUA−3の物理的地図を示す。glaAプロモーター、切断されたglaA遺伝子、A.ニガー(A.niger)amyBコドン最適化配列および3’glaAフランキング配列が示されている。大腸菌(E.coli)DNAは、宿主株への形質転換前に、制限酵素NotIによる消化によって除去することができる。
【図21】(例えば、実施例中のGBA303に基づく株のような)適合されたBamHIアンプリコンを有する株における単一の相同組換えによる組込みの概略図を示す。発現ベクターは、特定の標的化配列を有する選択可能なamdSマーカーと目的の遺伝子とを含む。標的化配列は、最も高い頻度の遺伝子変換を有する適合されたアンプリコンへの特異的なゲノム標的化(「賢明な組込み戦略」)を可能にする。
【0006】
[発明の詳細な説明]
驚くことに、宿主細胞(例えば、国際公開第1998/046774号パンフレットおよび国際公開第1998/46772号パンフレットに記載されているような宿主細胞)が少なくとも2つの実質的に相同なDNAドメインを含む場合、これらのDNAドメインの遺伝子変換頻度は必ずしも同じというわけではないことが解析から明らかになった。これらのDNAドメインのうちのいくつかは、他の実質的に相同なDNAドメインの遺伝子変換頻度よりも実質的に低い遺伝子変換頻度を有することが明らかになった。そのような場合、目的のポリヌクレオチドが、高い遺伝子変換頻度を有するDNAドメインではなく、低い遺伝子変換頻度を有するDNAドメインに組み込まれるのは不利である。より高い遺伝子変換頻度を有するDNAドメインは、大方の場合、より低い遺伝子変換頻度を有する(目的のポリヌクレオチドを含む)DNAドメインに優る。その結果として、より低い遺伝子変換頻度を有するDNAドメインに組み込まれた目的のポリヌクレオチドを有する細胞から高コピーの遺伝子変換宿主細胞を得るには、より多くの時間と労力を要する。
【0007】
それゆえ、目的のポリヌクレオチドを実質的により高い遺伝子変換頻度を有する実質的に相同なDNAドメインに含めることができれば有利であろう。というのは、これによって、あまり時間と労力を要さないで目的のポリヌクレオチドの複数コピーを含む宿主株(高コピーの遺伝子変換株)を構築することが可能になるからである。
【0008】
その結果として、目的のポリヌクレオチドが実質的により高い遺伝子変換頻度を有する相同なDNAドメインに優先的に組み込まれる宿主細胞が必要とされている。それゆえ、本発明の目的はそのような宿主細胞を提供することである。
【0009】
したがって、本発明は、目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている、組換え宿主細胞を提供する。前記組換え宿主細胞は、本明細書中、本発明による組換え宿主細胞と称される。
【0010】
本発明はまた、目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることを含む方法を提供する。
【0011】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で本発明による宿主細胞を培養することと、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0012】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で組換え宿主細胞を培養すること、前記宿主細胞は目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドの少なくとも1コピー以上を有する、と、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0013】
したがって、本発明による組換え宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている。前記目的のポリヌクレオチドの1コピー以上の組込みに好適な前記実質的に相同なDNAドメインは、好ましくは、互いの実質的な複製物であり、より好ましくは、前記DNAドメインはアンプリコンである。
【0014】
本発明との関連で、「組換え」という用語は、天然のプロセスおよび/または宿主細胞をランダムな突然変異誘発に供することによって誘導される遺伝子改変を含む任意の遺伝子改変だけでなく、例えば、遺伝子の破壊および/または欠失および/または特異的な突然変異誘発も指す。その結果として、組換えによるプロセスと天然のプロセスの組合せおよび/または宿主細胞をランダムな突然変異誘発に供することによって誘導される遺伝子改変は、組換えであると解釈される。
【0015】
本発明による宿主細胞は、そのゲノム中に目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含む。野生型株では、これらのDNAドメインは、単一コピーとしてまたは複数コピーとして前記細胞のゲノム中に生じ得る。天然の複数コピーDNAドメインの例は、rDNAドメインである。したがって、rDNAドメインを本発明で好ましく用いてもよいし、または用いなくてもよい。本発明によって提供されるような、これらの実質的に相同なDNAドメインの複数のコピーを含む株は、例えば、前記DNAドメイン中の遺伝子によってコードされる遺伝子産物の産生が改善された株を選択することによる古典的な株改良によって、DNAドメインの単一コピーを含む株から得ることができる。多くの場合、そのような産生改善は、選択された株におけるDNAドメインの増幅の結果である。そのような増幅されたDNAドメインは、これ以降アンプリコンとも称される。そのようなアンプリコンを含む宿主細胞の例は、例えば、van Dijckら、2003,Regulatory Toxicology and Pharmacology 28;27−35:On the safety of a new generation of DSM Aspergillus niger enzyme production strainsに記載されている。van Dijckらには、7つの増幅されたグルコアミラーゼ遺伝子座(すなわち、アンプリコン)を含むアスペルギルス・ニガー(Aspergillus niger)株が記載されている。本発明は、そのようなアンプリコンを組換えDNA分子の組込み用のDNAドメインとして用いることが好ましいが、本発明は、決してそれに限定されない。実際、1)DNAドメインは、目的のポリヌクレオチドの組込みを受け入れるのに好適であるべきである;2)DNAドメインは、遺伝子変換による組み込まれた組換えDNA分子の増加を達成するために、真菌ゲノム中のDNAドメインの他の実質的に相同なバージョンとの組換えが可能であるべきであるという2つの機能的基準が満たされる限り、2つ以上の実質的に相同なバージョンが宿主細胞のゲノム中に生じる任意のDNAドメインを用いることができる。
【0016】
第1の基準を満たすには、DNAドメインは、相同組換えによって目的のポリヌクレオチドをDNAドメインに標的化するのを可能にするために十分な長さでなければならない。この目的のために、DNAドメインは、少なくとも100bp、好ましくは少なくとも500bp、より好ましくは少なくとも1kbおよびより好ましくは少なくとも2kbを含むべきである。好ましくは、目的のポリヌクレオチドのその中への組込みに対するDNAドメインの好適性は、DNAドメインへの組込みが、問題になっている宿主細胞の生存に必須の機能を破壊するべきではないという要件によってさらに決定される。
【0017】
第2の機能的基準、すなわち、宿主細胞のゲノム中の他の実質的に相同なバージョンのDNAドメインとの組換えの能力は、異なるバージョンのDNAドメイン間の遺伝子変換を可能にするために必要とされる。この目的のための最低限の必要条件は、フランキング配列間の相同組換えを可能にするよう、各々のバージョンのDNAドメインが、DNAドメインの両端で、他のバージョンのDNAドメインの対応するフランキング配列に十分相同なDNA配列に隣接していることである。この相同組換えの結果が遺伝子変換であり、その場合、これらのバージョンのDNAドメインの1つは、組み込まれた組換えDNA分子を含む複製されたバージョンの他のDNAドメインに置き換えられる。遺伝子変換をなおも可能にするフランキング配列の長さおよび相同性の程度に関する最低限の必要条件は、問題となっている生物によって異なり得る。おそらく、全体的な相同性が少なくとも60%の最低100bpの長さであれば、遺伝子変換がなおも可能となるであろう。明らかに、遺伝子変換の頻度は、フランキング配列の長さと相同性が増大するとともに増加する。好ましくは、異なるDNAドメインは、少なくとも80%の相同性を共有する少なくとも1kbのフランキング配列を含む。さらにより好ましくは、異なるDNAドメインは、少なくとも95%の相同性を共有する少なくとも5kbのフランキング配列を含む。より好ましい実施形態では、異なるDNAドメインは、少なくとも99%の相同性を共有する少なくとも10kbのフランキング配列を含む。最も好ましい実施形態では、異なるDNAドメインは、少なくとも99%の相同性を共有する少なくとも50kbのフランキング配列を含む。本明細書で実質的な複製物として示される、互いの完全なコピーではない実質的に相同なDNAドメインの例は、アレル変異体、遺伝子ファミリーおよび/またはアイソザイムをコードする遺伝子である。最も好ましいのは、組み込まれた目的のポリヌクレオチドが存在するときにせいぜい異なる互いの正確なコピーである実質的に相同なDNAドメインである。そのような同一なDNAドメインの例は、アンプリコンである。あるいは、人工DNAドメインをDNA合成および/または組換えDNA技術を用いて作り出してもよく、また次に、そのような人工DNAドメインを所望の真菌に導入し、増幅させてもよい。したがって、そのような人工DNAドメインを本発明で好ましく用いてもよいし、または用いなくてもよい。
【0018】
実質的に相同なDNAドメインの全体的な長さ、すなわち、フランキング配列を含む全体的な長さは、1kb未満から数百kbまで様々であってもよく、例えば、DNAドメインの長さは、アスペルギルス・オリザエ(Aspergillus oryzae)の親株IF04177(国際公開第9812300号パンフレットおよび米国特許第5,766,912号明細書でA1560として開示されている)中に存在する3つのTAKAアミラーゼ遺伝子の1ユニット当たり2〜3kbの範囲であることが以前に示されており、これらの遺伝子は、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)中の増幅されたペニシリンクラスターの1ユニット当たり約57kbおよびアスペルギルス・ニガー(Aspergillus niger)の増幅されたglaA遺伝子座の1ユニット当たり80kb超までA.オリザエ(A.oryzae)BECh2(国際公開第0039322号パンフレットに開示されている)において不活化されている(国際公開第9846772号パンフレットを参照されたい)。
【0019】
遺伝子変換は、目的のポリヌクレオチドを含むDNAドメインが増加している宿主細胞を同定することによりモニタリングすることができる。例えば、目的のポリヌクレオチドによってコードされる産物の産生レベルがより高い宿主細胞を単にスクリーニングすることなどの、当業者に公知の任意の方法を用いることができる。また、例えば、DNAコピー数を決定するための定量的方法(例えば、本明細書で以下に概説するような「DNAフラッグ」テスト、定量的PCR、サザン解析もしくは比較ゲノムハイブリダイゼーションによる宿主細胞の遺伝子型の解析など)、またはRNAレベルを決定するための定量的方法(例えば、RT−PCR、ノーザン解析もしくはGeneChipもしくはマイクロアレイ解析など)を用いることもできる。
【0020】
本発明によれば、各々のバージョンの実質的に相同なDNAドメインは、好ましくは、同定目的のための独特の配列タグによって糸状菌中の他のバージョンのDNAドメインと区別される。そのような配列タグは、異なるDNAドメイン間の遺伝子変換の検出およびモニタリングを可能にし、それにより、所望の遺伝子型を有する遺伝子変換体のスクリーニングおよび/または選択を容易にする。異なるバージョンのDNAドメインの検出を可能にする限り、例えば、サザンブロット上で検出される制限部位から簡単にアッセイ可能な表現型を提供する完全な選択可能マーカー遺伝子にまで及ぶ任意の形態の配列タグを用いることができる。特に有用な配列タグ法は、国際公開第9846772号パンフレット、国際公開第9846774号パンフレットおよびvan Dijckら(上記)に記載されている。この方法は、単一のオリゴヌクレオチドPCRプライマー対を用いる単一のPCRにおいて各々のDNAドメインを検出することを可能にする。DNAドメインは、PCRにおいて、各々のバージョンのDNAドメインが独特の長さのPCR断片を生じるように改変される。得られるPCR断片の長さおよび強度は、それぞれ、各々のDNAドメインの存在およびコピー数を示す。「DNAフラッグ」と称されるこの形態の配列タグは、所望の遺伝子型を有する変換体を得るために、多数の変換体コロニーの遺伝子型の迅速な解析を可能にする。
【0021】
本発明によれば、本発明による宿主細胞の少なくとも2つの実質的に相同なDNAドメインの1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている。好ましくは、宿主細胞の少なくとも2つの実質的に相同なDNAドメインの1つの標的化DNAドメインは、相同組換えによって目的の組換えDNA分子/ポリヌクレオチドを適合された標的化DNAドメインにより特異的に標的化することを可能にするように適合されている。標的化DNA配列またはDNAドメインが宿主細胞中に2回以上現れる場合、目的のポリヌクレオチドの相同組換えは、宿主細胞の複数の標的化DNAドメインについて見出される。標的化DNAドメインを実質的に相同なDNAドメインの1つに適合させて、宿主中の他の実質的に相同なバージョンのDNAドメインとの組換えに影響を及ぼすことなく、その適合された標的化DNAドメインへのより特異的な標的化を支持することが可能である。最適な標的化選好のために、標的化に用いられるDNA配列の適合は、目的の組換えDNA分子/ポリヌクレオチド中および宿主中で行なわれる。したがって、マッチした宿主と組込みベクターの組が得られる。
【0022】
好ましくは、標的化DNAドメインの適合は、増強された組込み選好を有する配列を実質的に相同なDNAドメインに導入することによって実施される。好ましくは、増強された組込み選好を有する前記配列は、遺伝子置換によって導入される。したがって、本発明はまた、目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、ここで、適合された実質的に相同なDNAドメインは標的化DNAドメインを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、組換え宿主細胞を提供する。より好ましくは、標的化ドメインの適合は、本明細書中の実施例に従って実施される。
【0023】
本発明によれば、標的化に用いられる適合されたDNAドメインの標的化配列は、好ましくは、宿主のゲノム内で独特であるように適合される。好ましくは、宿主のゲノム内で独特であるように適合される標的化配列は、異種DNAポリヌクレオチド配列、宿主から事前に除去され、再導入されるDNAポリヌクレオチド配列、宿主内の野生型状況と比較して異なる順序で組み合わされた相同なDNAポリヌクレオチド断片、および人工DNAポリヌクレオチドの群から選択される配列を含む。
【0024】
本発明によれば、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、好ましくは宿主のゲノム内で独特である。好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、異種DNAポリヌクレオチド配列、宿主から事前に除去され、再導入されるDNAポリヌクレオチド配列、宿主内の野生型状況と比較して異なる順序で組み合わされた相同なDNAポリヌクレオチド断片、および人工DNAポリヌクレオチドの群から選択される配列を含む。
【0025】
好ましくは、適合されたDNAドメインの標的化配列が宿主のゲノム内で独特であるように適合されているとともに、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列も宿主のゲノム内で独特である。
【0026】
本発明によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現に用いられる遺伝子エレメントである。遺伝子エレメントの例は、遺伝子、プロモーター、ターミネーター、cDNA、イントロン、遺伝子間領域、またはそれらの一部および/もしくは組合せである。より好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上の異種遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上の異種遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上のA.ニガー(A.niger)遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の1つ以上のA.ニガー(A.niger)遺伝子エレメントを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼ遺伝子エレメントおよび/またはフランキング領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼ遺伝子エレメントおよび/またはフランキング領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼプロモーターおよび/またはグルコアミラーゼターミネーター領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用のA.ニガー(A.niger)glaAグルコアミラーゼプロモーターおよび/またはグルコアミラーゼターミネーター領域を含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および/または発現用の、野生型状況と比較して逆の順序のA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および/または発現用の、野生型状況と比較して逆の順序のA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる配列は、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。さらにより好ましくは、適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの一方または両方のフランキング配列は、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。最も好ましくは、適合されたDNAドメインへの標的化に用いられる配列および適合されたDNAドメインへの標的化に用いられる目的のポリヌクレオチドの両方のフランキング配列は同じであり、目的のポリヌクレオチドの導入および発現用の、配列番号:2で特定されるようなA.ニガー(A.niger)glaAグルコアミラーゼターミネーター領域およびglaAグルコアミラーゼプロモーターを含む。
【0027】
したがって、本発明は、配列番号:2によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドならびに配列番号:2によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドを含むベクターおよび宿主細胞を提供する。機能性は、プロモーター活性を有することおよび増強された組込み選好を有する標的化配列であることと本明細書では解釈されるべきである。好ましくは、配列番号:2によるポリヌクレオチドの機能性断片は、少なくとも100bp、より好ましくは少なくとも200bp、より好ましくは少なくとも300bp、より好ましくは少なくとも500bp、より好ましくは少なくとも600bp、より好ましくは少なくとも700bp、より好ましくは少なくとも800bp、より好ましくは少なくとも900bp、より好ましくは少なくとも1000bp、より好ましくは少なくとも1200bp、より好ましくは少なくとも1500bp、より好ましくは少なくとも2000bp、より好ましくは少なくとも3000bp、およびさらにより好ましくは少なくとも4000bpを含む。好ましくは、機能性断片は、少なくとも約50%同一な、より好ましくは少なくとも約60%同一な、さらにより好ましくは少なくとも約70%同一な、さらにより好ましくは少なくとも約80%同一な、さらにより好ましくは少なくとも約85%同一な、さらにより好ましくは少なくとも約90%同一な、さらにより好ましくは少なくとも約95%同一な、さらにより好ましくは少なくとも約97%同一な、さらにより好ましくは少なくとも約98%同一な、さらにより好ましくは少なくとも約99%同一な一致率(すなわち、位置同一性)を有し、かつさらに好ましくは、一致率(すなわち、同一性)は100%に等しい。
【0028】
最も好ましくは、配列番号:2によるポリヌクレオチドを含むポリヌクレオチドは、配列番号:3またはその機能性断片を含む。機能性は、プロモーター活性を有することおよび増強された組込み選好を有する標的化配列であることと本明細書では解釈されるべきである。
【0029】
したがって、本発明は、配列番号:3によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドならびに配列番号:3によるポリヌクレオチドまたはその機能性断片を含むポリヌクレオチドを含むベクターおよび宿主細胞を提供する。好ましくは、配列番号:3による機能性断片は、少なくとも100bp、より好ましくは少なくとも200bp、より好ましくは少なくとも300bp、より好ましくは少なくとも500bp、より好ましくは少なくとも600bp、より好ましくは少なくとも700bp、より好ましくは少なくとも800bp、より好ましくは少なくとも900bp、より好ましくは少なくとも1000bp、より好ましくは少なくとも1200bp、より好ましくは少なくとも1300bp、およびさらにより好ましくは少なくとも1400bpを含む。好ましくは、機能性断片は、少なくとも約50%同一な、より好ましくは少なくとも約60%同一な、さらにより好ましくは少なくとも約70%同一な、さらにより好ましくは少なくとも約80%同一な、さらにより好ましくは少なくとも約85%同一な、さらにより好ましくは少なくとも約90%同一な、さらにより好ましくは少なくとも約95%同一な、さらにより好ましくは少なくとも約97%同一な、さらにより好ましくは少なくとも約98%同一な、さらにより好ましくは少なくとも約99%同一な一致率(すなわち、位置同一性)を有し、かつ最も好ましくは、一致率(すなわち、同一性)は100%に等しい。
【0030】
ポリヌクレオチドの例は、一本鎖または二本鎖のいずれかの、リボ核酸(RNA)分子およびデオキシリボ核酸(DNA)分子であるが、これらに限定されない。当業者であれば、本明細書で特許請求されるポリヌクレオチドの相補体および逆相補体が本発明の範囲内に含まれることが意図されることを理解するであろう。
【0031】
本発明の実施形態によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現用の遺伝子エレメントであり、ここで、適合されたDNAドメイン中の遺伝子エレメントの機能性はコンストラクトの組込みによって破壊される。本発明の別の実施形態によれば、適合されたDNAドメインへの標的化に用いられる配列は、好ましくは、目的のポリヌクレオチドの導入および/または発現用の遺伝子エレメントであり、ここで、適合されたDNAドメイン中の遺伝子エレメントの機能性はコンストラクトの組込みによって回復される。
【0032】
実質的に相同なDNAドメインは遺伝子変換によっておよび/または遺伝子増幅によって増加することができることが以前に証明されている。本明細書における使用では、異なるものであるが、遺伝子変換および遺伝子増幅という用語は両方とも互換的に用いられる。実質的に相同な異なるDNAドメインは、異なる増加頻度(すなわち、遺伝子変換頻度)を有することが明らかになった。最も高い遺伝子変換頻度を有する実質的に相同なDNAドメインの標的化DNAドメインを適合させる利点は、とりわけ、遺伝子変換および/または増幅によって目的のポリヌクレオチドのコピー数が増加した宿主のスクリーニングおよび選択がはるかにより効率的であることである。
【0033】
したがって、適合された実質的に相同なDNAドメインが由来する実質的に相同なDNAドメインは、好ましくは、他の少なくとも2つの実質的に相同なDNAドメインの1つよりも少なくとも10%高い遺伝子変換頻度を有する。より好ましくは、遺伝子変換頻度は、少なくとも15%高く、さらにより好ましくは少なくとも20%高く、さらにより好ましくは少なくとも30%高く、さらにより好ましくは少なくとも40%高く、さらにより好ましくは少なくとも50%高く、さらにより好ましくは少なくとも60%高く、さらにより好ましくは少なくとも70%高く、さらにより好ましくは少なくとも80%高く、さらにより好ましくは少なくとも90%高く、さらにより好ましくは少なくとも100%高く、さらにより好ましくは少なくとも150%高く、さらにより好ましくは少なくとも200%高く、さらにより好ましくは少なくとも300%高く、さらにより好ましくは少なくとも400%高く、さらにより好ましくは少なくとも500%より高く、およびさらにより好ましくは少なくとも1000%高い。
【0034】
本発明によれば、より高い遺伝子変換頻度を有する実質的に相同なDNAドメインは、宿主細胞の実質的に相同なDNAドメインの遺伝子変換頻度を決定することによって同定することができる。遺伝子変換頻度は、当業者に公知の任意の手段によって決定することができる。好ましい方法は、本明細書中、実施例3.1に記載されている方法である。この方法では、3つの実質的に相同なDNAドメインを含む宿主細胞を目的のポリヌクレオチドで形質転換する。各々、単一の実質的に相同なDNAドメイン中に目的のポリヌクレオチドの単一コピーを含み、各々の形質転換体がそのポリヌクレオチドを異なる実質的に相同なDNAドメインに組み込んでいる、3つの形質転換体を選択する。形質転換体を子孫クローンへと増殖させた後、同定目的のための独特の配列タグによって、すなわち、本明細書で上に記載した「フラッグDNA」解析を用いて、子孫クローンの遺伝子型を決定する。次に、前記実質的に相同なDNAドメインを含む形質転換体の遺伝子変換体の量を前記特定の形質転換体のクローンの総数で割ることによって、実質的に相同なDNAドメインの遺伝子変換頻度を算出する。
【0035】
好ましくは、本発明による宿主細胞は、実質的に相同なDNAドメインのうちの少なくとも2つが各々、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する細胞である。より好ましくは、各々の前記ドメインは、組み込まれた目的のポリヌクレオチドを少なくとも2コピー、さらにより好ましくは少なくとも3コピー、さらにより好ましくは4コピー、さらにより好ましくは4〜6コピー有する。好ましくは、本発明による宿主細胞の各々の実質的に相同なDNAドメインは、同じ数の組み込まれた目的のポリヌクレオチドを有する。
【0036】
目的のポリヌクレオチドのコピー数は、当業者に利用可能な任意の方法、例えば、サザン解析または(半)定量的PCRによる遺伝子型の解析、ノーザン解析、GeneChipまたはマイクロアレイ解析または(半)定量的RT−PCRによる発現解析、ウェスタンブロット、ELISA、酵素活性アッセイによる目的のポリヌクレオチドによってコードされる産物の解析によって決定することができる。目的のポリヌクレオチドのコピー数を決定する方法は国際公開第9846772号パンフレットに広範に記載されており、目的のポリヌクレオチドのコピー数を決定する好ましい方法は本発明によるものである。
【0037】
好ましくは、本発明による宿主細胞は、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合された少なくとも2つの実質的に相同なDNAドメイン、より好ましくは3つの適合されたDNAドメイン、さらにより好ましくは4つの適合されたDNAドメイン、さらにより好ましくは5つの適合されたDNAドメイン、さらにより好ましくは6つの適合されたDNAドメインを含む。最も好ましくは、本発明による前記宿主細胞の各々の実質的に相同なDNAドメインは、適合されたDNAドメインである。好ましくは、適合されたDNAドメインは、少なくとも2コピーの組み込まれた目的のポリヌクレオチドを少なくとも2コピー有し、さらにより好ましくは少なくとも3コピー、さらにより好ましくは少なくとも4コピーの組み込まれた目的のポリヌクレオチドを有し、さらにより好ましくは適合されたDNAドメインは、少なくとも5コピー、さらにより好ましくは少なくとも6コピー、少なくとも7コピー、少なくとも8コピー、少なくとも9コピー、少なくとも10コピーの組み込まれた目的のポリヌクレオチドを有する。最も好ましくは、適合されたDNAドメインは、組み込まれた目的のポリヌクレオチドの5または6コピーを有する。好ましくは、各々の前記適合されたDNAドメインは、同じ数の組み込まれた目的のポリヌクレオチドを有する。
【0038】
好ましくは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、ここで、好ましくは、各々の適合されたDNAドメインは複数コピーの組み込まれた目的のポリヌクレオチドを有する、複数の実質的に相同なDNAドメインを含む前記宿主細胞は、複数コピーの組み込まれた目的のポリヌクレオチドを有する単一の適合されたDNAドメインを含む宿主細胞から得られる;すなわち、複数コピーの組み込まれた目的のポリヌクレオチドを有するさらなるコピーの適合されたDNAドメインは、遺伝子変換によって生成される。
【0039】
好ましくは、本発明による宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率は、組込み経路をHRの方向に導くことによって増大する。そのような誘導は、HR経路の効率を高めることによって、および/またはNHR経路の効率を下げる(低下させるという意味)ことによっておよび/またはNHR/HR比を減少させることによって達成することができる。
【0040】
真核細胞は、少なくとも2つの別々の経路(相同組換え(HR)による経路および非相同組換え(NHR)による経路)を有しており、これらの経路によって、ポリヌクレオチドを宿主ゲノム中に組み込むことができる。本発明との関連で、HR経路は、宿主のゲノム中へのポリヌクレオチドの標的化組込みの制御に関与する全ての遺伝子およびエレメントとして定義され、前記ポリヌクレオチドは、組込みが標的化される宿主のゲノムのある所定の部位との一定の相同性を有する。NHR経路は、前記ポリヌクレオチドと宿主のゲノム配列との相同性の程度とは無関係な、宿主のゲノム中へのポリヌクレオチドの標的化組込みの制御に関与する全ての遺伝子およびエレメントとして定義される。
【0041】
非相同組換えと相同組換えの比(NHR/HR)は、宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率を決定する。NHR/HR比が高い場合、標的化組込みの効率は比較的低い。NHR/HR比が低い場合、標的化組込みの効率は比較的高い。NHR/HRの比は、NHRとHRの頻度を評価し、その後、それぞれの頻度を割り算することにより、当業者に公知の方法によって決定することができ、方法の一例は、国際公開第02/052026号パンフレットに記載されている方法である。好ましくは、本発明による宿主細胞は、改変を含むNHR構成要素をコードするポリヌクレオチドを含み、ここで、前記宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記NHR構成要素の産生を欠損している。前記改変は、NHR/HR比の減少をもたらし、それに伴って、宿主細胞のゲノム中の所定の部位へのポリヌクレオチドの標的化組込みの効率を増大させる。前記NHR構成要素の欠損をもたらす、NHR構成要素をコードするポリヌクレオチドの改変によって、宿主細胞のNHR/HR比がHRの方向に導かれ、その結果として、標的化組込みの効率が増大することになる。
【0042】
好ましくは、本発明による宿主細胞は、前記NHR構成要素の少なくとも5%欠損、より好ましくは少なくとも10%欠損、より好ましくは少なくとも20%欠損、より好ましくは少なくとも30%欠損、より好ましくは少なくとも40%欠損、より好ましくは少なくとも50%欠損、より好ましくは少なくとも60%欠損、より好ましくは少なくとも70%欠損、より好ましくは少なくとも80%欠損、より好ましくは少なくとも90%欠損、より好ましくは少なくとも95%欠損、より好ましくは少なくとも97%欠損、より好ましくは少なくとも99%欠損を示す。最も好ましくは、宿主細胞は、前記NHR構成要素の100%欠損を示す。
【0043】
改変すべきNHR構成要素は、当業者に公知の任意のNHR構成要素であることができる。好ましい改変すべきNHR構成要素は、KU70、KU80、MRE11、RAD50、RAD51、RAD52、XRS2、SIR4、LIG4、またはそれらのホモログの群から選択される。より好ましい改変すべきNHR構成要素は、KU70およびKU80、hdfAおよびhdfBまたはそれらのホモログである。最も好ましい改変すべきNHR構成要素は、KU70もしくはhdfA、またはそれらのホモログである。好ましくは、本発明による宿主細胞は、国際公開第2005/095624号パンフレットに記載されているような破壊または欠失を含むNHR構成要素をコードするポリヌクレオチドを含む。そのような宿主細胞を得る方法は当業者に公知であり、かつ国際公開第2005/095624号パンフレットに広範に記載されている。好ましくは、本発明によるNHR欠損宿主細胞におけるNHR/HR比は、同一の条件下でアッセイしたとき、非欠損親宿主細胞と比較して、少なくとも10%、より好ましくは少なくとも20%、より好ましくは少なくとも30%、より好ましくは少なくとも40%、より好ましくは少なくとも50%、より好ましくは少なくとも100%、より好ましくは少なくとも200%、およびさらにより好ましくは少なくとも1000%減少している。
【0044】
その代わりに、またはNHR構成要素の欠損と組み合わせて、本発明による宿主細胞は、宿主細胞が由来している細胞内の同じ遺伝子の発現レベルと比較して、HRに関与する少なくとも1つ遺伝子の発現レベルが上方調節されている。これは、HRに関与する構成要素をコードする遺伝子の発現レベルを増大させることによっておよび/またはHRに関与する構成要素の発現レベルを増大させることによっておよび/またはHRに関与する構成要素の活性を(一時的に)増大させることによって達成することができる。
【0045】
その代わりに、または本明細書で上に記載したようなNHR/HR比の改変と組み合わせて、国際公開2007/115886号パンフレットおよび国際公開2007/115887号パンフレットに記載されている方法を用いて、目的のポリヌクレオチドが相同組換えによって組み込まれている宿主細胞を前向きに選択し、かつ/または組込みが非相同組換えによって生じた細胞を除外することができる。国際公開2007/115886号パンフレットおよび国際公開2007/115887号パンフレットには、そのような選択または除外用の(さらなる)選択マーカーの使用が記載されている。そのような選択の例は、組込みが非相同組換えによって生じた細胞をジフテリア毒素Aによって選択的に死滅させることである(国際公開2007/115887号パンフレット)。
【0046】
ポリヌクレオチドの改変は、ポリヌクレオチドの配列の変化をもたらす任意の事象として本明細書で定義される。改変は1つ以上の改変と解釈される。改変は、ヌクレオチド配列中の1つ以上のヌクレオチドまたはポリヌクレオチドの転写もしくは翻訳に必要とされる調節エレメントの導入(挿入)、置換または除去(欠失)によって遂行することができる。例えば、ヌクレオチドは、終止コドンの導入、開始コドンの除去またはポリヌクレオチドのオープンリーディングフレームの変化もしくはフレームシフトをもたらすように挿入または除去することができる。配列またはその調節エレメントの改変は、当技術分野で公知の方法に従って、部位特異的もしくはランダムな突然変異誘発、DNAシャッフリング法、DNA再構築法、遺伝子合成(例えば、YoungおよびDong(2004),Nucleic Acids Research 32(7)電子アクセスhttp://nar.oupjournals.org/cgi/reprint/32/7/e59もしくはGuptaら(1968),Proc.Natl.Acad.Sci USA,60:1338−1344;Scarpullaら(1982),Anal.Biochem.121:356−365;Stemmerら(1995),Gene 164:49−53を参照されたい)、またはPCRによる突然変異誘発によって遂行することができる。ランダムな突然変異誘発手順の例は当技術分野で周知であり、例えば、化学的(例えば、NTG)突然変異誘発または物理的(例えば、UV)突然変異誘発などがある。特異的突然変異誘発手順の例は、クイックチェンジ(QuickChange)(商標)部位特異的突然変異誘発キット(Stratagene Cloning Systems,La Jolla,CA)、The Altered Sites(登録商標)II インビトロ突然変異誘発システム(Promega Corporation)またはGene.1989 Apr 15;77(1):51−9(Ho SN,Hunt HD,Horton RM,Pullen JK,Pease LR “Site−directed mutagenesis by overlap extension using the polymerase chain reaction”)に記載されているようなPCRを用いるかもしくはMolecular Biology:Current Innovations and Future Trends.(A.M.GriffinおよびH.G.Griffin編.ISBN 1−898486−01−8;1995 Horizon Scientific Press,PO Box 1,Wymondham,Norfolk,U.K.)に記載されているようなPCRを用いる重複伸長によるものである。
【0047】
好ましい改変の方法は、遺伝子置換、遺伝子欠失、または遺伝子破壊の手法を基にしている。例えば、遺伝子破壊法では、内在性ポリヌクレオチドに対応するポリヌクレオチドにインビトロで突然変異を誘発して、欠損のあるポリヌクレオチドを生じさせる。次に、これを親細胞に形質転換して、欠損のあるポリヌクレオチドを生じさせる。相同組換えによって、欠損のあるポリヌクレオチドは内在性ポリヌクレオチドに置き換わる。欠損のあるポリヌクレオチドは、核酸配列が改変された形質転換体の選択に使用し得るマーカーもコードすることが望ましい場合がある。内在性ポリヌクレオチドの欠失または置換の場合、適切なDNA配列は、欠失または置換しようとする標的遺伝子座に導入しなければならない。適切なDNA配列は、好ましくは、クローニングベクター上に存在する。好ましい組込みクローニングベクターは、この所定の遺伝子座へのクローニングベクターの組込みを標的化するために欠失または置換しようとするポリヌクレオチドに相同なDNA断片を含む。標的化組込みを促進するために、宿主細胞の形質転換前に、クローニングベクターを線状化することが好ましい。線状化は、クローニングベクターの少なくとも一方の末端、しかし、好ましくは両方の末端に、欠失または置換しようとするDNA配列に相同な配列が隣接するように行なわれることが好ましい。
【0048】
その代わりにまたは他の言及された手法と組み合わせて、A rapid method for efficient gene replacement in the filamentous fungus Aspergillus nidulans(2000) Chaveroche,M−K.,Ghico,J−M.およびd’Enfert C;Nucleic acids Research,第28巻,第22号に記載されているような、大腸菌(E.coli)でのコスミドのインビボ組換えに基づく手法を用いることができる。
【0049】
あるいは、前記宿主細胞がポリヌクレオチドの産生を欠損する改変を、ポリヌクレオチドの核酸配列に相補的なヌクレオチド配列を用いる確立されたアンチセンス技術によって実施することもできる。より具体的には、宿主細胞によるポリヌクレオチドの発現は、細胞内で転写されることができ、かつ細胞内で産生されるmRNAにハイブリダイズすることが可能な、ポリヌクレオチドの核酸配列に相補的なヌクレオチド配列を導入することによって低下させるかまたは削減することができる。相補的なアンチセンスヌクレオチド配列がmRNAにハイブリダイズするのを可能にする条件下で、翻訳されるタンパク質の量をこのように低下させるかまたは削減する。アンチセンスRNAを発現する例は、Appl Environ Microbiol.2000 Feb;66(2):775−82.(Characterization of a foldase,protein disulfide isomerase A,in the protein secretory pathway of Aspergillus niger.Ngiam C,Jeenes DJ,Punt PJ,Van Den Hondel CA,Archer DB)または(Zrenner R,Willmitzer L,Sonnewald U.Analysis of the expression of potato uridinediphosphate−glucose pyrophosphorylase and its inhibition by antisense RNA.Planta.(1993);190(2):247−52.)に示されている。
【0050】
さらに、遺伝子の改変、下方調節または不活化は、RNA干渉(RNAi)技術によって得ることができる(FEMS Microb.Lett.237(2004):317−324)。この方法では、その発現に影響を及ぼそうとするヌクレオチド配列の同一のセンス部分およびアンチセンス部分を、ヌクレオチドスペーサーを間に入れて、互いの背後にクローニングし、発現ベクターに挿入する。そのような分子が転写された後で、小さいヌクレオチド断片の形成により、影響を及ぼそうとするmRNAの標的化分解がもたらされる。特定のmRNAの削減は様々な程度までであることができる。国際公開第2008/053019号パンフレット、国際公開第2005/05672A1号パンフレット、国際公開第2005/026356A1号パンフレット、Oliveiraら,“Efficient cloning system for construction of gene silencing vectors in Aspergillus niger”(2008) Appl Microbiol and Biotechnol 80(5):917−924および/またはBarnesら,“siRNA as a molecular tool for use in Aspergillus niger”(2008) Biotechnology Letters 30(5):885−890に記載されているRNA干渉技術を、遺伝子の下方調製、改変または不活化に用いることができる。
【0051】
ポリヌクレオチドの改変後、得られた株を、ポリヌクレオチドによってコードされる産物の欠損についてスクリーニングする。ポリヌクレオチドの発現レベルの下方調節および/または上方調節は、例えば、ノーザン解析(SambrookおよびRussell(2001) “Molecular Cloning:A Laboratory Manual(第3版),Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,New York.)によって細胞内に存在する対応するmRNAの量を定量化することによっておよび/または例えば、ウェスタンブロッティングもしくはELISAによって細胞内に存在する対応するタンパク質の量を定量化することによってモニタリングすることができる。mRNA量の違いは、DNAアレイ解析によって定量化することもできる(Eisen,M.B.およびBrown,P.O.DNA arrays for analysis of gene expression.Methods Enzymol.1999:303:179−205)。
【0052】
宿主細胞の欠損は、細胞が、未改変のポリヌクレオチドを含む親細胞と比較したときに、改変されたポリヌクレオチドによってコードされる産物をそれほど産生せず、かつ/または改変されたポリヌクレオチドの発現レベルが低下し(すなわち、mRNAレベルが低下し)、かつ/または改変されたポリヌクレオチドによってコードされる産物の特定の(タンパク質)活性が減少する表現型特徴、およびこれらの可能性の組合せとして本明細書で定義される。
【0053】
本発明によれば、実質的に相同なDNAドメインは、そのネイティブ(native)の状態で高レベルの発現が可能な内在性遺伝子を含むDNAドメインまたは遺伝子座(これらの用語は、本明細書で互換的に用いられる)である。「内在性」遺伝子という用語は、問題になっている生物のゲノム中の天然の遺伝子コピーとして本明細書で定義される。
【0054】
組み込まれた組換え遺伝子の発現レベルは、その遺伝子が組み込まれるゲノム遺伝子座によって大きく異なり得ることが一般に知られている。発現させようとする組換え遺伝子の組込みのために、高発現されるDNAドメインを用いる利点は、これらのDNAドメインが、少なくとも内在性遺伝子の高レベル発現を支持することができるということである。それゆえ、そのようなDNAドメインは、組み込まれた組換え遺伝子の高レベル発現も支持する可能性が高い。これまでに、A.ニガー(A.niger)のglaA遺伝子座における組込みおよびP.クリソゲヌム(P.chrysogenum)のペニシリンクラスターにおける組込みは、いくつかの他のゲノム遺伝子座における組込みと比較して遺伝子1コピー当たりでより高い発現レベルを提供することが明らかにされた(国際公開第9846772号パンフレットを参照されたい)。この関連で、高レベル発現が可能な遺伝子は、最大レベルで発現されたときに、全mRNA集団の少なくとも0.1%、好ましくは全mRNAの少なくとも0.5%および最も好ましくは全mRNAの少なくとも1%を占めるmRNAを産生する遺伝子と定義されることが理解されるであろう。そのような高発現可能な内在性遺伝子(これらがその中に含まれるDNAドメインは、本発明による実質的に相同なDNAドメインとして特に好適である)の例は、糖分解酵素、デンプン分解酵素、セルロース分解酵素および/または抗生物質生合成酵素をコードする遺伝子である。さらにより好ましいのは、グルコアミラーゼ遺伝子、TAKAアミラーゼ遺伝子、セロビオヒドロラーゼ遺伝子およびペニシリン生合成遺伝子などの、工業プロセスに関与し、かつ高レベルで発現されることが知られている遺伝子を含む遺伝子座である。
【0055】
好ましくは、本発明による宿主細胞の少なくとも2つの実質的に相同なDNAドメインは、glaAの遺伝子座であるかまたはamyAの遺伝子座であるかまたはamyBの遺伝子座であるかまたはこれらの遺伝子座の断片もしくはホモログである。より好ましい遺伝子座は、アスペルギルス(Aspergillus)のglaA、amyAおよびamyB遺伝子座である。さらにより好ましい遺伝子座は、アスペルギルス・ニガー(Aspergillus niger)のglaA、およびamyB遺伝子座である。さらにより好ましい遺伝子座は、アスペルギルス・ニガー(Aspergillus niger)CBS513.88のglaA、amyBIおよびamyBII遺伝子座である(遺伝子は、国際公開第2005/095624号パンフレットと同じように命名されている)。さらにより好ましい遺伝子座は、glaA、amyBIおよびamyBII遺伝子座(glaA遺伝子のヌクレオチド配列(An03g06550)およびそのゲノムコンテクストa.o.はhttp://www.ncbi.nlm.nih.gov/から得ることができる;amyBI遺伝子配列(An12g06930)およびamyBII遺伝子配列(An05g02100)についても同上かまたはこれらの遺伝子座の断片もしくはホモログである。
【0056】
本発明の目的のために、「相同性」および「同一性」という用語は、互換的に用いられる。2つの核酸配列間の相同性(同一性)の程度は、本明細書ではBLASTプログラムによって決定されることが好ましい。BLAST解析を実施するためのソフトウェアは、全米バイオテクノロジー情報センター(National Center for Biotechnology Information)(http://www.ncbi.nlm.nih.gov/)から公に入手可能である。BLASTアルゴリズムパラメータのW、T、およびXは、アラインメントの感度と速度を決定する。BLASTプログラムでは、ワード長(W)11、BLOSUM62スコアリングマトリックス(Henikoff & Henikoff,Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照されたい)アラインメント(B)50、期待値(E)10、M=5、N=−4、および両方の鎖の比較がデフォルトとして用いられる。
【0057】
本発明によれば、内在性遺伝子の発現が必要ない場合、高度に発現される内在性遺伝子は、本発明による宿主細胞において各々のコピーのDNAドメインで不活化されていることが好ましい。そのような場合、内在性遺伝子の高レベル発現の不活化によって、目的のポリヌクレオチドの発現にさらに利用することができるエネルギーおよび資源が放出される。さらに、目的のポリヌクレオチドの組込みによって産生されるべき所望の化合物と内在性遺伝子によってコードされる酵素が両方とも分泌型酵素である場合、内在性酵素の不活化によって、所望の酵素のより純粋な調製がもたらされる。不活化の逆転を排除するために、内在性遺伝子は、内在性遺伝子の少なくとも一部の不可逆的な欠失によって不活化されていることが好ましい。内在性遺伝子の不活化は、プロモーターおよび上流の活性化配列の少なくとも一部を含む不可逆的な欠失によってもたらされることがより好ましい。これは、組換えDNA分子の組込みによって産生されるべき酵素をコードする所望の遺伝子の発現が内在性遺伝子に由来するプロモーターから駆動される場合に、特に有利であるが、それは、所望の遺伝子の発現に必要な転写因子を潜在的に制限する競合を排除するからである。
【0058】
目的のポリヌクレオチドは任意のポリヌクレオチドであってもよい。目的のポリヌクレオチドは、任意の原核生物、真核生物、または他の供給源から得ることができる。目的のポリヌクレオチドおよびそれと関連するプロモーターは、宿主細胞にとって相同であり、自己クローンとなる組換え宿主細胞を生じさせることが好ましい。
【0059】
本発明によれば、目的のポリヌクレオチドは、例えば、国際公開第2006077258号パンフレットに記載されているような、最適化されたターミネーター配列を含む、変異型の最適化されたポリヌクレオチドであってもよい。目的のポリヌクレオチドは、部分的合成ポリヌクレオチドまたは完全合成核酸配列であってもよい。目的のポリヌクレオチドは、好ましくは、参照により本明細書に組み込まれる国際公開第2006/077258号パンフレットおよび/または国際公開第2008/000632号パンフレットに記載の方法に従って、そのコドン使用について最適化されてもよい。国際公開第2008/000632号パンフレットは、コドン対最適化に対処している。コドン対最適化は、ポリペプチドをコードするヌクレオチド配列の改善された発現および/またはコードされたポリペプチドの改善された産生を得るために、ポリペプチドをコードするヌクレオチド配列がそのコドン使用頻度(特に、使用されるコドン対)に関して改変されている方法である。コドン対は、コード配列中の2つの引き続くトリプレット(コドン)の組として定義される。
【0060】
したがって、本発明によれば、目的のポリヌクレオチドは、コドン最適化されたポリヌクレオチドであることが好ましい。
【0061】
ポリヌクレオチドはコード配列を含み、かつ目的のポリヌクレオチドによってコードされる化合物のクローニング、形質転換、発現および/または産生を促進するプロモーターおよび制御配列をさらに含む、核酸コンストラクト中および/またはベクター中に含めることができることが好ましい。
【0062】
「核酸コンストラクト」は、天然の遺伝子から単離されるかまたはさもなければ自然には存在しないような形で組み合わされ、並置された核酸のセグメントを含むように改変されている一本鎖または二本鎖のいずれかの核酸分子として本明細書で定義される。核酸コンストラクトという用語は、核酸コンストラクトがコード配列の発現に必要な全ての制御配列を含む場合、「発現カセット」という用語と同義である。本明細書で定義するような「コード配列」という用語は、mRNAに転写され、ポリペプチドに翻訳される配列である。コード配列の境界は、通常、mRNAの5’末端のATG開始コドンおよびmRNAの3’末端でオープンリーディングフレームを終結させる翻訳停止コドン配列によって決定される。コード配列は、DNA、cDNA、および組換え核酸配列を含むことができるが、これらに限定されない。
【0063】
「制御配列」という用語は、ポリペプチドの発現に必要または有利な全ての構成要素を含むよう、本明細書で定義される。各々の制御配列は、ポリペプチドをコードする核酸配列にとってネイティブまたは外来性であってもよい。そのような制御配列には、リーダー、最適な翻訳開始配列(Kozak,1991,J.Biol.Chem.266:19867−19870に記載されているもの)、ポリアデニル化配列、プロ−ペプチド配列、プレ−プロ−ペプチド配列、プロモーター、シグナル配列、および転写ターミネーターが含まれるが、これらに限定されない。最低でも、制御配列は、プロモーター、ならびに転写および翻訳停止シグナルを含む。
【0064】
制御配列には、制御配列とポリペプチドをコードする核酸配列のコード領域とのライゲーションを容易にする特定の制限部位を導入する目的のためのリンカーが備えられていてもよい。「機能的に連結された」という用語は、制御配列がポリペプチドの産生を導くよう、制御配列がDNA配列のコード配列に対して適切な位置に置かれている配置として本明細書で定義される。
【0065】
制御配列は、核酸配列の発現用の宿主細胞によって認識される適切なプロモーター配列、核酸配列であってもよい。プロモーター配列は、ポリペプチドの発現を仲介する転写制御配列を含む。プロモーターは、突然変異体プロモーター、切断型プロモーター、およびハイブリッドプロモーターを含む、細胞で転写活性を示す任意の核酸配列であってもよく、また、宿主細胞にとって相同または異種のいずれかの細胞外または細胞内ポリペプチドをコードする遺伝子から得られてもよい。
【0066】
プロモーターはまた、mRNAへの転写後の翻訳のための(プロモーターと翻訳開始点の間にある)5’非コード領域、シス作用転写制御エレメント(例えば、エンハンサー)、および転写因子と相互作用することができる他のヌクレオチド配列と理解されるので。
【0067】
プロモーターは、発現させようとするコード配列とネイティブに関連するプロモーターであってもよい。プロモーターはまた、発現させようとするコード配列にとって外来性の構成的または誘導性プロモーターであってもよい。哺乳動物細胞での使用に好適なプロモーターの例は、例えば、SambrookおよびRussell(上記)に記載されている。酵母での使用に好適なプロモーターの例には、例えば、糖分解プロモーターが含まれる。
【0068】
使用可能な好ましい誘導性プロモーターの例は、デンプン、銅、オレイン酸誘導性プロモーターである。
【0069】
プロモーターは、A.オリザエ(A.oryzae)のTAKAアミラーゼ、リゾムコール・ミエヘイ(Rhizomucor miehei)のアスパラギン酸プロテイナーゼ、A.ニガー(A.niger)の中性α−アミラーゼ、A.ニガー(A.niger)の酸安定性α−アミラーゼ、A.ニガー(A.niger)またはA.アワモリ(A.awamori)のグルコアミラーゼ(glaA)、R.ミエヘイ(R.miehei)のリパーゼ、A.オリザエ(A.oryzae)のアルカリプロテアーゼ、A.オリザエ(A.oryzae)のトリオースリン酸イソメラーゼ、A.ニデュランス(A.nidulans)のアセトアミダーゼをコードする遺伝子から得られるプロモーター、NA2−tpiプロモーター(A.ニガー(A.niger)の中性α−アミラーゼをコードする遺伝子由来のプロモーターとA.オリザエ(A.oryzae)のトリオースリン酸イソメラーゼをコードする遺伝子由来のプロモーターのハイブリッド、国際公開第03/008575号パンフレット)、ならびにそれらの突然変異体プロモーター、切断型プロモーター、およびハイブリッドプロモーターを含むが、これらに限定されない群から選択されることが好ましい。糸状菌細胞での使用に特に好ましいプロモーターは、プロテアーゼ遺伝子由来(例えば、F.オキシスポルム(F.oxysporum)のトリプシン様プロテアーゼ遺伝子(米国特許第4,288,627号明細書)、A.オリザエ(A.oryzae)のアルカリプロテアーゼ遺伝子(alp)、A.ニガー(A.niger)のpacA遺伝子、A.オリザエ(A.oryzae)のアルカリプロテアーゼ遺伝子、A.オリザエ(A.oryzae)の中性メタロプロテアーゼ遺伝子、A.ニガー(A.niger)のアスペルギロペプシンプロテアーゼpepA遺伝子、もしくはF.ベネナツム(F.venenatum)のトリプシン遺伝子、A.ニガー(A.niger)のアスパラギン酸プロテアーゼpepB遺伝子由来)のプロモーター、またはその機能性部分である。
【0070】
単一の適切なプロモーター配列を目的のポリヌクレオチドの発現用の制御配列として用いてもよいし、または複数の別個のプロモーター配列を目的のポリヌクレオチドの発現用の制御配列として用いてもよい。複数の別個のプロモーター配列を用いる場合、少なくとも2つの別個のプロモーター配列が各々、国際公開第2008/098933号パンフレットに従って、目的のポリヌクレオチドに機能的に連結されていることが好ましい。
【0071】
制御配列はまた、好適な転写ターミネーター配列、すなわち、糸状菌細胞によって認識されて、転写を終結させる配列であってもよい。ターミネーター配列は、ポリペプチドをコードする核酸配列の3’末端に機能的に連結されている。細胞内で機能的な任意のターミネーターを本発明で用いてもよい。
【0072】
糸状菌細胞のための好適なターミネーターは、A.オリザエ(A.oryzae)のTAKAアミラーゼ、A.ニガー(A.niger)のグルコアミラーゼ(glaA)、A.ニデュランス(A.nidulans)のアントラニル酸シンターゼ、A.ニガー(A.niger)のα−グルコシダーゼ、trpC遺伝子およびフサリウム・オキシスポルム(Fusarium oxysporum)のトリプシン様プロテアーゼをコードする遺伝子から得られる。
【0073】
制御配列はまた、好適なリーダー配列、糸状菌細胞による翻訳に重要なmRNAの非翻訳領域であってもよい。リーダー配列は、ポリペプチドをコードする核酸配列の5’末端に機能的に連結されている。細胞内で機能的な任意のリーダー配列を本発明で使用してもよい。
【0074】
糸状菌細胞のための好適なリーダーは、A.オリザエ(A.oryzae)のTAKAアミラーゼおよびA.ニデュランス(A.nidulans)のトリオースリン酸イソメラーゼおよびA.ニガー(A.niger)のglaAをコードする遺伝子から得られる。
【0075】
他の制御配列は、ペニシリウムIPNS遺伝子、またはpcbC遺伝子、βチューブリン遺伝子から単離されてもよい。国際公開第01/21779号パンフレットに引用されている全ての制御配列は、本発明での使用が想定されることが企図される。
【0076】
制御配列はまた、ポリアデニル化配列、すなわち、核酸配列の3’末端に機能的に連結され、かつ転写されたときに、糸状菌細胞によって、転写されたmRNAにポリアデノシン残基を付加するシグナルとして認識される配列であってもよい。細胞内で機能的な任意のポリアデニル化配列を本発明で用いてもよい。
【0077】
糸状菌細胞のための好ましいポリアデニル化配列は、A.オリザエ(A.oryzae)のTAKAアミラーゼ、A.ニガー(A.niger)のグルコアミラーゼ、A.ニデュランス(A.nidulans)のアントラニル酸シンターゼ、フサリウム・オキシスポルム(Fusarium oxysporum)のトリプシン様プロテアーゼおよびA.ニガー(A.niger)のα−グルコシダーゼをコードする遺伝子から得られる。
【0078】
好ましくは、DNAコンストラクトは、目的の化合物をコードする目的のポリヌクレオチドと、本明細書で上に記載したようなプロモーターDNA配列と、好ましい制御配列、例えば、
− 以下の配列リスト:TAAG、TAGAおよびTAAA、好ましくはTAAAから選択される5’から3’の方向に配向された1つの翻訳終結配列、ならびに/または
− 以下の配列リスト:GCTACCCCC;GCTACCTCC;GCTACCCTC;GCTACCTTC;GCTCCCCCC;GCTCCCTCC;GCTCCCCTC;GCTCCCTTC;GCTGCCCCC;GCTGCCTCC;GCTGCCCTC;GCTGCCTTC;GCTTCCCCC;GCTTCCTCC;GCTTCCCTC;およびGCTTCCTTC、好ましくはGCT TCC TTCから選択される5’から3’の方向に配向された1つの翻訳イニシエーターコード配列、ならびに/または
− 以下の配列リスト:ヌクレオチドの多義性暗号:m(A/C);w(ATT);y(C/T);k(GTT);h(A/C/T)を用いた、5’−mwChkyCAAA−3’;5’−mwChkyCACA−3’もしくは5’−mwChkyCAAG−3’、好ましくは5’−CACCGTCAAA−3’もしくは5’−CGCAGTCAAG−3’から選択される1つの翻訳イニシエーター配列と
を含む。
【0079】
本発明との関連で、「翻訳イニシエーターコード配列」という用語は、DNAコード配列のオープンリーディングフレームのイニシエーターまたは開始コドンのすぐ下流にある9個のヌクレオチドとして定義される。イニシエーターまたは開始コドンは、AAメチオニンをコードする。イニシエーターコドンは、典型的にはATGであるが、GTGなどの任意の機能的な開始コドンであってもよい。
【0080】
本発明との関連で、「翻訳終結配列」という用語は、オープンリーディングフレームまたはヌクレオチドコード配列の3’末端にある翻訳停止コドンから始まり、5’から3’の方向に配向されている4個のヌクレオチドとして定義される。
【0081】
本発明との関連で、「翻訳イニシエーター配列」という用語は、ポリペプチドをコードするDNA配列のオープンリーディングフレームのイニシエーターまたは開始コドンのすぐ上流にある10個のヌクレオチドとして定義される。イニシエーターまたは開始コドンは、AAメチオニンをコードする。イニシエーターコドンは、典型的にはATGであるが、GTGなどの任意の機能的な開始コドンであってもよい。RNAでは、ウラシル(U)がデオキシヌクレオチドのチミン(T)に置き換わることが当技術分野で周知である。
【0082】
目的の化合物をコードする目的のポリヌクレオチド、または目的のポリヌクレオチドおよび上記の制御配列を含むDNAコンストラクトを連結させて、最適なポリヌクレオチド配列の挿入または置換を可能にする1つ以上の好都合な制限部位を含み得る組換え発現ベクターを産生することができる。
【0083】
あるいは、目的のポリヌクレオチドは、配列または目的のポリヌクレオチドを含む核酸コンストラクトを発現用の適切なベクターに挿入することによって発現させることができる。発現ベクターを作製する際、コード配列が発現、および場合によっては分泌用の適切な制御配列と機能的に連結されるようにコード配列をベクター内に位置させる。
【0084】
組換え発現ベクターは、組換えDNA手順に好都合に供することができ、かつ目的のポリヌクレオチドの発現をもたらすことができる任意のベクター(例えば、プラスミドまたはウイルス)であってもよい。ベクターの選択は、通常、ベクターとベクターを導入しようとする宿主細胞との適合性によって決まる。ベクターは、線状または閉環状プラスミドであってもよい。ベクターは、自律複製ベクター、すなわち、その複製が染色体複製に依存しない染色体外実体として存在するベクター(例えば、プラスミド、染色体外エレメント、ミニ染色体、または人工染色体)であってもよい。自律的に維持されるクローニングベクターは、AMA1配列を含んでいてもよい(例えば、AleksenkoおよびClutterbuck(1997),Fungal Genet.Biol.21:373−397を参照されたい)。
【0085】
あるいは、ベクターは、宿主細胞に導入したときに、ゲノム中に組み込まれ、それが組み込まれた染色体とともに複製するベクターであってもよい。組込み型クローニングベクターは、この所定の遺伝子座へのクローニングベクターの組込みを標的化するために、宿主細胞のゲノム中の所定の標的遺伝子座のDNA配列と相同なDNA断片を含むことが好ましい。標的化組込みを促進するために、宿主細胞の形質転換前に、クローニングベクターを線状化することが好ましい。線状化は、クローニングベクターの少なくとも一方の、しかし、好ましくは両方の末端に、標的遺伝子座と相同な配列が隣接するように行なわれることが好ましい。標的遺伝子座に隣接する相同配列の長さは、好ましくは少なくとも30bp、好ましくは少なくとも50bp、好ましくは少なくとも0.1kb、さらに好ましくは少なくとも0.2kb、より好ましくは少なくとも0.5kb、さらにより好ましくは少なくとも1kb、最も好ましくは少なくとも2kbである。
【0086】
ベクター系は、単一のベクターもしくはプラスミドまたは糸状菌細胞のゲノム中に導入しようとする全DNAを一緒に含有する、2つ以上のベクターもしくはプラスミド、またはトランスポゾンであってもよい。
【0087】
ベクターは、形質転換された細胞の容易な選択を可能にする1つ以上の選択可能マーカーを含むことが好ましい。共形質転換の方法を用いると、1つのベクターは選択可能マーカーを含むことができ、一方、別のベクターは、目的のポリヌクレオチドまたは目的の核酸コンストラクトを含むことができる。これらのベクターは、宿主細胞の形質転換に同時に用いられる。本発明による宿主細胞は、共形質転換効率が非常に高く、最大で100%となることができる能力を有する(実施例12.2を参照されたい)。
【0088】
選択可能マーカーは、その産物が殺生物剤またはウイルスに対する耐性、重金属に対する耐性、栄養要求株に対する原栄養性などを提供する遺伝子である。糸状菌宿主細胞で用いられる選択可能マーカーは、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、bleA(フレオマイシン結合)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、およびtrpC(アントラニル酸シンターゼ)、ならびに他の種由来の等価物を含むが、これらに限定されない群から選択することができる。アスペルギルス(Aspergillus)およびペニシリウム(Penicillium)細胞での使用に好ましいのは、amdS(欧州特許第635574B1号明細書、国際公開第97/06261号パンフレット)およびA.ニデュランス(A.nidulans)またはA.オリザエ(A.oryzae)のpyrG遺伝子およびストレプトミセス・ハイグロスコピクス(Streptomyces hygroscopicus)のbar遺伝子である。より好ましくはamdS遺伝子、さらにより好ましくはA.ニデュランス(A.nidulans)またはA.ニガー(A.niger)由来のamdS遺伝子を用いる。最も好ましい選択マーカー遺伝子は、A.ニデュランス(A.nidulans)のgpdAプロモーターに融合されたA.ニデュランス(A.nidulans)のamdSコード配列である(欧州特許第635574B1号明細書を参照されたい)。他の糸状菌由来のAmdS遺伝子も用いることができる(国際公開第97/06261号パンフレット)。
【0089】
上記のエレメントを連結して本発明の組換え発現ベクターを構築するために用いられる手順は、当業者に周知である(例えば、SambrookおよびRussell(上記);ならびにAusubelら,Current Protocols in Molecular Biology,Wiley InterScience,NY,1995を参照されたい)。
【0090】
好ましくは、本発明による宿主細胞は、適合されたDNAドメインおよび場合により標的化組込みの増大した効率に加えて、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドおよびprtTの群から選択されるポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、改変を含むポリヌクレオチドによってコードされる産物を欠損している。改変は本明細書で先に定義されている。
【0091】
好ましくは、本発明による宿主細胞はSec61の改変をさらに含む。宿主細胞の欠損は、先に本明細書にあるように、細胞が、未改変のポリヌクレオチドを含む親細胞と比較したとき、改変されたポリヌクレオチドによってコードされる産物をそれほど産生せず、かつ/または改変されたポリヌクレオチドの発現レベルが低下し(すなわち、mRNAレベルが低下し)、かつ/または改変されたポリヌクレオチドによってコードされる産物の特定の(タンパク質)活性が減少する表現型特徴、およびこれらの特徴の組合せとして本明細書で定義される。
【0092】
好ましくは、本発明による宿主細胞は、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドまたはprtTのうちの少なくとも1つの少なくとも5%の欠損、より好ましくは少なくとも10%の欠損、より好ましくは少なくとも20%の欠損、より好ましくは少なくとも30%の欠損、より好ましくは少なくとも40%の欠損、より好ましくは少なくとも50%の欠損、より好ましくは少なくとも60%の欠損、より好ましくは少なくとも70%の欠損、より好ましくは少なくとも80%の欠損、より好ましくは少なくとも90%の欠損、より好ましくは少なくとも95%の欠損、より好ましくは少なくとも97%の欠損、より好ましくは少なくとも99%の欠損を示す。最も好ましくは、宿主細胞は、glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドまたはprtTのうちの少なくとも1つの100%の欠損を示す。
【0093】
目的の化合物の産生におけるグルコアミラーゼ(glaA)、酸安定性α−アミラーゼ(amyA)、中性α−アミラーゼ(amyBIおよびamyBII)ならびにシュウ酸ヒドロラーゼ(oahA)の群から選択される1つ以上のポリペプチドの欠損の利点は、エネルギーおよび資源がこれらの副産物に利用されないとうことである。このエネルギーおよび資源は、目的のポリヌクレオチドによってコードされる化合物の産生に用いることができる。さらに、副産物がほとんど存在しないので、下流での目的の化合物の処理が簡単になる。
【0094】
シュウ酸ヒドロラーゼ(oahA)は、多くの宿主細胞におけるシュウ酸の合成経路の構成要素である。oahAを欠損している宿主細胞は、シュウ酸を欠損している。シュウ酸は、食品用途などの多くの用途において望ましくない副産物である。さらに、シュウ酸は、この構成要素を産生する宿主細胞の培地培養のpHを低下させ、収率の低下をもたらす。すなわち、シュウ酸欠損宿主細胞では収率が増加する。それゆえ、本発明による宿主細胞がoahAを欠損していれば有利である。OahA欠損宿主細胞および前記宿主細胞を産生する好ましい方法は、国際公開第2000/50576号パンフレットおよび国際公開第2004/070022号パンフレットに広範に記載されている。oahA欠損宿主細胞を産生する好ましい方法は、国際公開第2000/50576号パンフレットに記載されている組換えによる破壊法である。好ましくは、本発明による宿主細胞は、oahAを欠損している。好ましくは、oahAは、真菌のoahAである。より好ましくは、oahAは、アスペルギルス(Aspergillus)由来のoahAである。さらにより好ましくは、oahAは、アスペルギルス・ニガー(Aspergillus niger)由来のoahAである。さらにより好ましくは、oahAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のoahAである。最も好ましくは、oahAは、An10g00820の配列を含む。
【0095】
prtTは、真核細胞におけるプロテアーゼの転写アクチベーターである。最近、プロテアーゼのいくつかの真菌転写アクチベーターが、国際公開第00/20596号パンフレット、国際公開第01/68864号パンフレット、国際公開第2006/040312号パンフレットおよび国際公開第2007/062936号パンフレットに記載されている。これらの転写アクチベーターは、アスペルギルス・ニガー(Aspergillus niger)(A.ニガー(A.niger))、アスペルギルス・フミガーツス(Aspergillus fumigatus)(A.フミガーツス(A.fumigatus))、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)(P.クリソゲヌム(P.chrysogenum))およびアスペルギルス・オリザエ(Aspergillus oryzae)(A.オリザエ(A.oryzae))から単離された。プロテアーゼ遺伝子のこれらの転写アクチベーターは、真菌細胞でポリペプチドを産生するための方法を改善するために用いることができ、その場合、ポリペプチドは、プロテアーゼ分解に感受性がある。宿主細胞がprtTを欠損しているとき、宿主細胞は、prtTの転写制御下にあるプロテアーゼをあまり産生しない。それゆえ、本発明による宿主細胞がprtTを欠損していれば有利である。prtT欠損宿主およびこれらの宿主を産生する好ましい方法は、国際公開第01/68864号パンフレット、国際公開第2006/040312号パンフレットに広範に記載されている。国際公開第01/68864号パンフレットおよび国際公開第2006/040312号パンフレットには、prtTコード配列を破壊するための組換えによる、古典的な方法が記載されている。国際公開第2007/062936号パンフレットには、プロテアーゼプロモーター内のprtT結合部位の破壊が記載されている。結合部位の破壊は、prtTが結合部位に結合するのを妨げる。その結果として、プロテアーゼの転写はprtTによって活性化されず、プロテアーゼはあまり産生されない。
【0096】
好ましくは、本発明による宿主細胞は、prtTをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、prtTの産生を欠損している。好ましくは、prtTは、真菌のprtTである。より好ましくは、prtTは、アスペルギルス(Aspergillus)由来のprtTである。さらにより好ましくは、prtTは、アスペルギルス・ニガー(Aspergillus niger)由来のprtTである。さらにより好ましくは、prtTは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のprtTである。最も好ましくは、prtTは、An04g06940の配列を含む。
【0097】
「グルコアミラーゼ」(glaA)という用語は、「アミログルコシダーゼ」という用語と同一であり、1,4結合α−D−グルコース残基および末端1,4結合α−D−グルコース残基の鎖中の分岐点における1,6−α−D−グルコシド結合のエンド加水分解を触媒するデキストリン6−α−D−グルカノヒドロラーゼ活性を有する酵素として本明細書で定義される。グルコアミラーゼ活性は、基質のp−ニトロフェニル−a−D−グルコピラノシド(シグマ(Sigma))からのパラニトロフェノールの遊離を測定することによって、AGIU/mlとして測定することができる。これにより黄色い色が生じ、その吸光度は、分光光度計を用いて405nmで測定することができる。1AGIUは、可溶性のデンプン基質からpH4.3および60℃で1分間に1μモルのグルコースを生じさせる酵素の分量である。国際公開第98/46772号パンフレットに、アッセイのさらなる詳細を見出すことができる。
【0098】
好ましくは、本発明による宿主細胞は、glaAをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、glaAの産生を欠損している。好ましくは、glaAは、真菌のglaAである。より好ましくは、glaAは、アスペルギルス(Aspergillus)由来のglaAである。さらにより好ましくは、glaAは、アスペルギルス・ニガー(Aspergillus niger)由来のglaAである。さらにより好ましくは、glaAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のglaAである。最も好ましくは、glaAは、An03g06550の配列を含む。
【0099】
「α−アミラーゼ」という用語は、水の存在下で3つ以上のα−1,4結合グルコースユニットを有する多糖のマルトオリゴ糖へのエンド加水分解を触媒する1,4−α−D−グルカングルカノヒドロラーゼ活性として本明細書で定義される。(中性)α−アミラーゼ活性を測定するために、供給元のプロトコルに従って、メガザイム(Megazyme)の穀物α−アミラーゼキットを用いる(メガザイム、CERALPHAαアミラーゼアッセイキット、カタログ参照番号K−CERA、2000−2001年)。測定される活性は、過剰のグルコアミラーゼおよびα−グルコシダーゼの存在下、pH7.0での、非還元末端がブロックされたp−ニトロフェニルマルトヘプタオシドの加水分解に基づく。形成されたp−ニトロフェノールの量は、試料中に存在するα−アミラーゼ活性の尺度である。
【0100】
「酸安定性α−アミラーゼ」(amyA)という用語は、酸性pH範囲で最適な活性を有するα−アミラーゼ活性を有する酵素として本明細書で定義される。酸安定性α−アミラーゼ活性を測定するために、これも、供給元のプロトコルに従うが、酸性pHでメガザイムの穀物α−アミラーゼキットを用いる(メガザイム、CERALPHAαアミラーゼアッセイキット、カタログ参照番号K−CERA、2000−2001年)。測定される活性は、過剰のグルコアミラーゼおよびα−グルコシダーゼの存在下、pH4.5での、非還元末端がブロックされたp−ニトロフェニルマルトヘプタオシドの加水分解に基づく。形成されたp−ニトロフェノールの量は、試料中に存在する酸安定性α−アミラーゼ活性の尺度である。
【0101】
好ましくは、本発明による宿主細胞は、AmyAをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、amyAを欠損している。好ましくは、amyAは、真菌のamyAである。より好ましくは、amyAは、アスペルギルス(Aspergillus)由来のamyAである。さらにより好ましくは、amyAは、アスペルギルス・ニガー(Aspergillus niger)由来のamyAである。さらにより好ましくは、amyAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyAである。最も好ましくは、amyAは、An11g03340の配列を含む。
【0102】
「中性α−アミラーゼ活性」(amy)という用語は、中性pH範囲で最適な活性を有するα−アミラーゼ活性を有する酵素として本明細書で定義される。
【0103】
好ましくは、本発明による宿主細胞は、AmyBをコードするポリヌクレオチドを含み、前記ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、amyBIおよび/またはamyBIIの産生を欠損している。より好ましくは、宿主細胞は、amyBIおよびamyBIIを欠損している。好ましくは、amyBは、真菌のamyBである。より好ましくは、amyBは、アスペルギルス(Aspergillus)由来のamyBである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)由来のamyBIである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyBIである。最も好ましくは、amyBIは、An12g06930の配列を含む。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)由来のamyBIIである。さらにより好ましくは、amyBは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のamyBIIである。最も好ましくは、amyBIIは、An05g02100の配列を含む。
【0104】
毒素関連ポリヌクレオチドという用語は、化合物をコードする遺伝子クラスター、多数の遺伝子、遺伝子もしくはその部分、または少なくとも1つの毒素もしくは毒素中間体化合物の生合成もしくは分泌に関与する生化学的経路として本明細書で定義される。前記化合物は、例えば、ポリペプチドであり、これは、酵素であってもよい。
【0105】
目的のポリペプチドの産生において宿主細胞として用いられるいくつかの宿主細胞(とりわけ、真菌)は、様々な毒素の生合成に関与する酵素をコードする遺伝子を有する。例えば、シクロピアゾン酸、コウジ酸、3−ニトロプロピオン酸およびアフラトキシンは、例えば、アスペルギルス・フラブス(Aspergillus flavus)で形成される既知の毒素である。同様に、トリコテセンは、いくつかの真菌、例えば、フサリウム(Fusarium)属の種(例えば、フサリウム・ベネナツム(Fusarium venenatum)およびトリコデルマ(Trichoderma)で形成され、オクラトキシンは、アスペルギルス(Aspergillus)によって産生され得る。最近、工業用アスペルギルス・ニガー(Aspergillus niger)宿主株のゲノムの配列決定により、フモニシン遺伝子クラスターが明らかになった(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88”.Nat Biotechnol.2007 Feb;25(2):221−231)。これらの毒素は、操作者、消費者および環境に健康被害を提示する可能性があるので、目的の化合物の発酵時のそのような毒素の形成は極めて望ましくない。その結果として、毒素欠損宿主細胞は目的の化合物を、毒素を含まずに産生するのを可能にする。産物から毒素を取り除く必要がないので、毒素を含まない化合物は産生するのがより簡単である。さらに、化合物の規制認可手続きは、より簡単である。
【0106】
好ましくは、本発明による宿主細胞は、化合物(例えば、酵素であり得るポリペプチドであってもよい)または生化学的経路をコードする毒素関連ポリヌクレオチドを含み、前記毒素関連ポリヌクレオチドは改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記毒素または毒素中間体化合物の産生を欠損している。好ましくは、毒素または毒素中間体化合物は、真菌の毒素または毒素中間体化合物である。より好ましくは、毒素または毒素中間体化合物は、アスペルギルス(Aspergillus)由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、アスペルギルス・ニガー(Aspergillus niger)由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来の毒素または毒素中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、フモニシンまたはフモニシン中間体化合物である。さらにより好ましくは、毒素または毒素中間体化合物は、オクラトキシンまたはオクラトキシン中間体化合物である。最も好ましくは、毒素または毒素中間体化合物は、オクラトキシンまたはフモニシンまたはオクラトキシン中間体またはフモニシン中間体化合物である。
【0107】
好ましくは、毒素関連ポリヌクレオチドは、化合物(例えば、酵素であり得るポリペプチドであってもよい)または真菌の毒素もしくは毒素中間体化合物の産生に関与する生化学的経路をコードする。より好ましくは、アスペルギルス(Aspergillus)由来の毒素または毒素中間体化合物。さらにより好ましくは、アスペルギルス・ニガー(Aspergillus niger)由来の毒素または毒素中間体化合物。さらにより好ましくは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来の毒素または毒素中間体化合物。さらにより好ましくは、フモニシンまたはフモニシン中間体化合物。さらにより好ましくは、フモニシン−Bまたはフモニシン−B中間体化合物。さらにより好ましくは、フモニシン−B2またはフモニシン−B2中間体化合物。さらにより好ましくは、毒素関連ポリヌクレオチドは、An01g06820からAn01g06930までのフモニシンクラスターの配列を含む。最も好ましくは、毒素関連ポリヌクレオチドは、An01g06930の配列を含む。
【0108】
別の好ましい実施形態では、毒素関連ポリヌクレオチドは、化合物(例えば、酵素であり得るポリペプチドであってもよい)またはオクラトキシンもしくはオクラトキシン中間体化合物の産生に関与する生化学的経路をコードする。より好ましくは、オクラトキシンAまたはオクラトキシンA中間体化合物。より好ましくは、毒素関連ポリヌクレオチドは、An15g07880からAn15g07930までのクラスターの配列を含む。最も好ましくは、毒素関連ポリヌクレオチドは、An15g07910の配列および/またはAn15g07920の配列を含む。
【0109】
好ましくは、本発明による宿主細胞は、化合物(例えば、酵素であり得るポリペプチドであってもよい)または生化学的経路をコードする少なくとも1つの毒素関連ポリヌクレオチドを含み、前記毒素関連ポリヌクレオチドは少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、毒素または毒素中間体化合物の産生を欠損している。
【0110】
より好ましくは、本発明による宿主細胞は、2つの毒素関連ポリヌクレオチドを含み、前記2つの毒素関連ポリヌクレオチドは各々、少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、フモニシンおよびオクラトキシンの産生を欠損していることが好ましい。
【0111】
さらにより好ましくは、本発明による宿主細胞は、3つ以上の毒素関連ポリヌクレオチドを含み、前記3つ以上の毒素関連ポリヌクレオチドは各々、少なくとも1つの改変を含み、ここで、宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、フモニシン、オクラトキシンおよび少なくとも1つのさらなる毒素または毒素中間体化合物の産生を欠損していることが好ましい。
【0112】
好ましくは、本発明による宿主細胞は、glaAおよび前記構成要素をコードするポリヌクレオチドに改変を有するために、glaAならびにamyA、amyBI、amyBII、oahA、毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。より好ましくは、本発明による宿主細胞は、glaA、oahAならびにamyA、amyBI、amyBII、毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、毒素関連化合物ならびにamyA、amyBI、amyBII、およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBIIならびに毒素関連化合物およびprtTの群から選択される構成要素のうちの少なくとも1つを欠損している。さらにより好ましくは、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBII、prtTおよび毒素関連化合物を欠損している。最も好ましくは、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合されている、本発明による宿主細胞は、glaA、oahA、amyA、amyBI、amyBII、prtTおよび毒素関連化合物を欠損しており、かつNHR構成要素、好ましくはku70またはそのホモログをさらに欠損している。
【0113】
本明細書で上に記載した構成要素の欠損または改変の他に、本発明による宿主細胞は、他の構成要素(例えば、pepAのような主要なプロテアーゼ)を欠損していてもよい。好ましくは、pepAは、真菌のpepAである。より好ましくは、pepAは、アスペルギルス(Aspergillus)由来のpepAである。さらにより好ましくは、pepAは、アスペルギルス・ニガー(Aspergillus niger)由来のpepAである。さらにより好ましくは、pepAは、アスペルギルス・ニガー(Aspergillus niger)CBS513.88由来のpepAである。最も好ましくは、pepAは、An14g04710の配列を含む。好ましくは、本発明による宿主細胞は、pepAの少なくとも5%の欠損、より好ましくは少なくとも10%の欠損、より好ましくは少なくとも20%の欠損、より好ましくは少なくとも30%の欠損、より好ましくは少なくとも40%の欠損、より好ましくは少なくとも50%の欠損、より好ましくは少なくとも60%の欠損、より好ましくは少なくとも70%の欠損、より好ましくは少なくとも80%の欠損、より好ましくは少なくとも90%の欠損、より好ましくは少なくとも95%の欠損、より好ましくは少なくとも97%の欠損、より好ましくは少なくとも99%の欠損を示す。最も好ましくは、宿主細胞は、pepAの100%の欠損を示す。
【0114】
その代わりに、または本明細書で上に記載した欠損と組み合わせて、宿主細胞は、目的のポリペプチドの産生能力を高めるために、野生型細胞と比較して上昇した小胞体ストレス応答(UPR)を含む。UPRは、米国特許出願公開第2004/0186070A1号明細書および/または米国特許出願公開第2001/0034045A1号明細書および/または国際公開第01/72783A2号明細書および/または国際公開第2005/123763号パンフレットに記載の手法によって増大させることができる。より具体的には、UPRが上昇した宿主細胞を得るために、HAC1および/もしくはIRE1および/もしくはPTC2のタンパク質レベルが調節され、かつ/またはSEC61タンパク質が操作されている。好ましいSEC61改変は、SEC61の一方向の突然変異体、すなわち、デノボ合成タンパク質がSEC61を介してERに侵入することができるが、タンパク質がSEC61を介してERを離れることができない突然変異体を生じさせる改変である。そのような改変は、国際公開第2005/123763号パンフレットに広範に記載されている。最も好ましくは、SEC61改変は、セリン376がトリプトファンに置き換えられているS376W突然変異である。
【0115】
本発明による好ましい宿主細胞は、目的の化合物を産生するための組換えアスペルギルス・ニガー(Aspergillus niger)組換え宿主細胞であり、前記宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ適合された実質的に相同なDNAドメインが由来する実質的に相同なDNAドメインは、少なくとも2つの実質的に相同なDNAドメインの他の1つよりも少なくとも10%高い遺伝子変換頻度を有し、ここで、適合された実質的に相同なDNAドメインは、独特の配列タグによって他のバージョンの実質的に相同なDNAドメインと区別され、かつ適合された実質的に相同なDNAドメインは、標的化DNAドメインを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含み、かつ前記宿主細胞は、glaA、pepA、hdfA、amyBII、amyBI、amyA、oahA、fumB、ochを欠損している。好ましくは、前記宿主細胞は、sec61 S376W突然変異、すなわち、セリン376がトリプトファンに置き換えられているS376W突然変異を有する。より好ましくは、前記宿主細胞はさらに、prtTを欠損している。そのような宿主細胞は、実施例9、10および11に記載されている。
【0116】
好ましくは、本発明による宿主細胞は、選択マーカーを含まない宿主細胞である。
【0117】
この状況は、アスペルギルス・ニデュランス(Aspergillus nidulans)由来のアセトアミーゼ遺伝子(amdS)などの、二方向性の優性選択マーカーの対抗選択によって得てもよい。このマーカーを用いて、アセトアミドを唯一の炭素および/または窒素源として用いて選択することによって、この遺伝子を有する形質転換体を選択することができる一方、例えば、フルオロアセトアミドで、対抗選択(この用語は、以下、マーカー遺伝子の不在についての選択に用いられる)を行なうことができる。特定の宿主細胞で作用する他の二方向性の優性選択マーカーを類似のやり方で用いることができる。二方向性マーカー遺伝子の除去組換え(out−recombination)は、導入しようとするプラスミド上に直接反復が両側にある遺伝子を挿入することによって容易になる。
【0118】
欧州特許第0635574B1号明細書に開示されているように、amdS二方向性マーカーは両方向に優性であり、これは、任意の遺伝子バックグラウンドの形質転換された細胞を(アセトアミドを唯一の炭素源および/または唯一の窒素源として用いて)マーカーの存在について選択することができることを意味する。他の二方向性マーカーは、URA3、LYS2、pyrG、facAなどである。
【0119】
マーカーを含まない細胞の使用はより簡単に規制当局に認可され、同時に、工業用の発酵条件下における酵母のエネルギー収支にあまり負担をかけない。
【0120】
本発明による宿主細胞は、任意の宿主細胞であってもよい。本発明による宿主細胞で産生される化合物の特定の使用について、宿主細胞の選択は、そのような使用に従って行なうことができる。例えば、本発明による宿主細胞で産生される化合物を食品用途で用いることになっている場合、宿主細胞は、出芽酵母(Saccharomyces cerevisiae)などの食品等級の生物から選択することができる。特定の使用には、食品用途、(動物)飼料用途、医薬品用途、農業用途(例えば、作物保護)、および/またはパーソナルケア用途が含まれるが、これらに限定されない。
【0121】
一実施形態によれば、本発明による宿主細胞は、真核宿主細胞である。好ましくは、真核細胞は、哺乳動物細胞、昆虫細胞、植物細胞、真菌細胞、または藻類細胞である。好ましい哺乳動物細胞には、例えば、チャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマが含まれる。好ましい昆虫細胞には、例えば、Sf9およびSf21細胞ならびにその派生物が含まれる。より好ましくは、真核細胞は、真菌細胞、すなわち、カンジダ(Candida)、ハンセヌラ(Hansenula)、クルイベロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、スキゾサッカロミセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)株などの酵母細胞である。より好ましくは、クルイベロミセス・ラクチス(Kluyveromyces lactis)、出芽酵母(S.cerevisiae)、ハンセヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)およびピキア・パストリス(Pichia pastoris)、または糸状菌細胞由来のものである。最も好ましくは、真核細胞は糸状菌細胞である。
【0122】
「糸状菌」には、(AinsworthおよびBisbyのDictionary of The Fungi,第8版,1995,CAB International,University Press,Cambridge,UKでHawksworthらによって定義されているような)全ての糸状形態の真菌類および卵菌類という亜門が含まれる。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖から構成される菌糸壁を特徴とする。栄養増殖は菌糸伸長によるものであり、炭素代謝は偏性好気性である。糸状菌株には、アクレモニウム(Acremonium)、アガリクス(Agaricus)、アスペルギルス(Aspergillus)、アウレオバシジウム(Aureobasidium)、クリソスポリウム(Chrysosporium)、コプリヌス(Coprinus)、クリプトコッカス(Cryptococcus)、フィリバシジウム(Filibasidium)、フサリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、パエシロミセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、パネロカエテ(Panerochaete)、ヒラタケ(Pleurotus)、スエヒロタケ(Schizophyllum)、タラロミセス(Talaromyces)、テルモアスクス(Thermoascus)、ティエラビア(Thielavia)、トリポクラジウム(Tolypocladium)、およびトリコデルマ(Trichoderma)の株が含まれるが、これらに限定されない。
【0123】
好ましい糸状菌細胞は、アスペルギルス(Aspergillus)属、クリロスポリウム(Chrysosporium)属、ペニシリウム(Penicillium)属、タラロミセス(Talaromyces)属、フサリウム(Fusarium)属またはトリコデルマ(Trichoderma)属の種、および最も好ましくはアスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティヅス(Aspergillus foetidus)、アスペルギルス・ソヤエ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニイ(Talaromyces emersonii)、アスペルギルス・オリザエ(Aspergillus oryzae)、クリソスポリウム・ルクノウェンス(Chrysosporium lucknowense)、フサリウム・オキシスポルム(Fusarium oxysporum)、トリコデルマ・リーゼイ(Trichoderma reesei)またはペニシリウム・クリソゲヌム(Penicillium chrysogenum)の種に属する。より好ましい宿主細胞は、アスペルギルス・ニガー(Aspergillus niger)である。本発明による宿主細胞がアスペルギルス・ニガー(Aspergillus niger)宿主細胞である場合、宿主細胞は、好ましくは、CBS513.88、CBS124.903またはそれらの派生物である。最も好ましくは、本発明による組換え宿主細胞は、本明細書中の実施例に記載されているような宿主細胞、好ましくは実施例4〜11に見られるような宿主細胞である。
【0124】
糸状菌のいくつかの株は、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)、Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM)、Centraalbureau Voor Schimmelcultures(CBS)、およびAgricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL)アスペルギルス・ニガー(Aspergillus niger)CBS513.88、CBS124.903、アスペルギルス・オリザエ(Aspergillus oryzae)ATCC20423、IFO4177、ATCC1011、CBS205.89、ATCC9576、ATCC14488−14491、ATCC11601、ATCC12892、P.クリソゲヌム(P.chrysogenum)CBS455.95、ペニシリウム・シトリヌム(Penicillium citrinum)ATCC38065、アクレモニウム・クリソゲヌム(Acremonium chrysogenum)P2、タラロミセス・エメルソニイ(Talaromyces emersonii)CBS124.902、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)ATCC36225またはATCC48272、トリコデルマ・リーゼイ(Trichoderma reesei)ATCC26921またはATCC56765またはATCC26921、アスペルギルス・ソヤエ(Aspergillus sojae)ATCC11906、クリソスポリウム・ルクノウェンス(Chrysosporium lucknowense)ATCC44006およびそれらの派生物などのいくつかの培養コレクションにおいて一般に容易に入手可能なものがある。
【0125】
別の実施形態によれば、本発明による宿主細胞は原核細胞である。好ましくは、原核宿主細胞は細菌細胞である。「細菌細胞」という用語には、グラム陰性微生物とグラム陽性微生物の両方が含まれる。好適な細菌は、例えば、エシェリキア(Escherichia)、アナベナ(Anabaena)、カウロバクター(Caulobacter)、グルコノバクター(Gluconobacter)、ロドバクター(Rhodobacter)、シュードモナス(Pseudomonas)、パラコッカス(Paracoccus)、バシルス(Bacillus)、ブレビバクテリウム(Brevibacterium)、コリネバクテリウム(Corynebacterium)、リゾビウム(Rhizobium)(シノリゾビウム(Sinorhizobium))、フラボバクテリウム(Flavobacterium)、クレブシエルラ(Klebsiella)、エンテロバクター(Enterobacter)、乳酸桿菌(Lactobacillus)、ラクトコッカス(Lactococcus)、メチロバクテリウム(Methylobacterium)ブドウ球菌(Staphylococcus)またはストレプトミセス(Streptomyces)から選択してもよい。好ましくは細菌細胞は、枯草菌(B.subtilis)、B.アミロリケファシエンス(B.amyloliquefaciens)、B.リケニホルミス(B.licheniformis)、B.プンティス(B.puntis)、B.メガテリウム(B.megaterium)、B.ハロヅランス(B.halodurans)、B.プミルス(B.pumilus)、G.オキシダンス(G.oxydans)、カウロバクター・クレスケンツス(Caulobacter crescentus)CB15、メチロバクテリウム・エキストロクエンス(Methylobacterium extorquens)、ロドバクター・スファエロイデス(Rhodobacter sphaeroides、シュードモナス・ゼアキサンティニファシエンス(Pseudomonas zeaxanthinifaciens)、パラコッカス・デニトリフィカンス(Paracoccus denitrificans)、大腸菌(E.coli)、C.グルタミクム(glutamicum)、スタフィロコッカス・カルノースス(Staphylococcus carnosus)、ストレプトミセス・リウィダンス(Streptomyces lividans)、シノリゾビウム・メリオティ(Sinorhizobium melioti)およびリゾビウム・ラジオバクター(Rhizobium radiobacter)からなる群から選択される。
【0126】
本発明による宿主細胞は、様々な目的、例えば、目的の化合物の産生ならびに国際公開第1999/032617号パンフレットおよび国際公開第2008/138835号パンフレットに記載されているような発現クローニングに好都合に用いることができる。これらの特許出願では、糸状菌細胞を宿主細胞として用いる発現クローニングの便宜性が記載されている。
【0127】
本発明はまた、目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、好ましくはより高い遺伝子変換頻度を有する実質的に相同なDNAドメインに標的化DNAドメインを提供することによって、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、方法を提供する。
【0128】
別の実質的に相同なDNAドメインと比較してより高い遺伝子変換比率を有する実質的に相同なDNAドメインを、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させることは、当業者に公知の任意の手段によって実施することができる。好ましくは、適合された実質的に相同なDNAドメインは、宿主のゲノム内で部分的にまたは完全に独特である。実施例3に示すように、A.ニガー(A.niger)におけるアンプリコンは、事前にゲノムから除去されたグルコアミラーゼプロモーター配列を用いて適合させる。これにより、独特の標的化配列を有する実質的に相同なDNAドメインが得られた。目的のポリヌクレオチドのプロモーターおよびターミネーター部分は、実施例11に示すような形質転換の後、適合された実質的に相同なDNAドメインでの組込みに好適なDNA配列として機能する。当業者のための、これらの実験を実施するための全ての手段は、本明細書ならびにSambrookおよびRussel(上記)に記載されている。
【0129】
好ましくは、目的の化合物を産生するための組換え宿主細胞の産生方法は、
a.得られた細胞を、目的のポリヌクレオチドを含む、より高い遺伝子変換比率を有する適合された実質的に相同なDNAドメインで形質転換する工程と、
b.実質的に相同なDNAドメインの少なくとも1つに組み込まれた目的のポリヌクレオチドを有する細胞を選択またはスクリーニングする工程と、
c.(b)で得られた細胞を繁殖させ、さらなるコピーの実質的に相同なDNAドメインに組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングする工程と
をさらに含む。
【0130】
より好ましくは、目的の化合物を産生するための組換え宿主細胞の産生方法は、本明細書中の実施例で記載されているような方法である。
【0131】
細胞の形質転換のための方法は当業者に周知である。任意のそのような方法を本発明の目的のために用いてもよい。形質転換は、それ自体公知の方法でのプロトプラスト形成、プロトプラストの形質転換、および細胞壁の再生からなるプロセスを含んでいてもよい。アスペルギルス(Aspergillus)細胞の形質転換の好適な手順は、欧州特許第238023号明細書およびYeltonら,1984,Proceedings of the National Academy of Sciences USA 81:1470−1474に記載されている。アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を用いるアスペルギルス(Aspergillus)および他の糸状菌宿主細胞の形質転換の好適な手順は、例えば、Nat Biotechnol.1998 Sep;16(9):839−42.(誤記あり(Erratum in:))Nat Biotechnol 1998 Nov;16(11):1074.Agrobacterium tumefaciens−mediated transformation of filamentous fungi.de Groot MJ,Bundock P,Hooykaas PJ,Beijersbergen AG.Unilever Research Laboratory Vlaardingen,The Netherlands中の訂正に記載されている。フサリウム(Fusarium)種を形質転換する好適な方法は、Malardierら,1989,Gene 78:147156によってまたは国際公開第96/00787号パンフレットに記載されている。Biolistic transformation of the obligate plant pathogenic fungus,Erysiphe graminis f.sp.hordei.Christiansen SK,Knudsen S,Giese H.Curr Genet.1995 Dec;29(1):100−2に記載されているような微粒子銃による形質転換を用いる方法などの他の方法を適用することができる。酵母は、Abelson,J.N.およびSimon,M.I.編,Guide to Yeast Genetics and Molecular Biology,Methods in Enzymology,第194巻,182−187頁,Academic Press,Inc.,New York中のBeckerおよびGuarente;ltoら,1983,Journal of Bacteriology 153:163;ならびにHinnenら,1978,Proceedings of the National Academy of Sciences USA 75:1920によって記載されている手順を用いて形質転換してもよい。
【0132】
得られた形質転換体のうちのどれが、そのDNAドメインの1つに組換えDNA分子の組込みを有するのかを明らかにするために、いくつかの日常的な手法が当業者に利用可能である。
【0133】
さらなる工程では、選択された形質転換体を繁殖させ、その子孫から、DNAドメインのうちの少なくとも2つが組み込まれた目的のポリヌクレオチドを含む株を選択する。これは、組み込まれた目的のポリヌクレオチドを含むDNAドメインが、「空の」DNAドメインによる遺伝子変換を通じてかまたは増幅を通じてかのいずれかで増加している株を選択することを意味する。そのような遺伝子変換および/または増幅事象は、低い頻度で自然に起こる。これらの事象が起こる正確な頻度は、問題となっている宿主細胞ならびにDNAドメインの数、長さおよび相同性の程度を含むいくつかの変数によって決まり得る。しかしながら、これらの頻度は、これらの事象が起こった株を当業者に公知の解析手法を用いてスクリーニングおよび選択するのを可能にするほど十分に高い。組み込まれた目的のポリヌクレオチドを含むDNAドメインが増加している株は、例えば、目的のポリヌクレオチドによってコードされる産物の産生レベルがより高い株を単にスクリーニングすることによって、またはあるいはその遺伝子型を、例えば、上で概説したような「DNAフラッグ」テストで解析することによって同定することができる。
【0134】
本発明による組換え宿主細胞の産生方法は、組み込まれた目的のポリヌクレオチドを含むDNAドメインの増加が起こった株の1つを繁殖させ、その子孫から、さらなるコピーのDNAドメインが、組み込まれた目的のポリヌクレオチドを含む株を選択する、さらなる工程を含んでいてもよい。その後、各々のDNAドメインが、組み込まれた目的のポリヌクレオチドを含む株が得られるまで、これらの株に再びこの手順を受けさせてもよい。
【0135】
好ましくは、上記の方法の工程(c)を実質的に相同なDNAドメインのうちの少なくとも3つが組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有するまで繰り返す。より好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも4つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。さらにより好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも5つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。さらにより好ましくは、工程(c)を実質的に相同なDNAドメインのうちの少なくとも6つが少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。最も好ましくは、工程(c)を実質的に相同なDNAドメインの各々が少なくとも1コピーの組み込まれた目的のポリヌクレオチドを有するまで繰り返す。
【0136】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で本発明による宿主細胞を培養することと、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0137】
本発明はまた、目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で組換え宿主細胞を培養することであって、前記宿主細胞は、目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する実質的に相同なDNAドメインと比較して目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドを少なくとも1コピー有する、と、
b.培養培地から目的の化合物を回収することと
を含む方法を提供する。
【0138】
本発明による産生方法では、当技術分野で公知の方法を用いて、目的の化合物(例えば、ポリペプチドまたは代謝産物)の産生に好適な栄養培地中で宿主細胞を培養する。本発明を限定するものと解釈されるべきでない培養方法の例は、深部発酵、固体状態での表面発酵および液体基質上での表面発酵である。例えば、細胞は、好適な培地中ならびにコード配列の発現および/またはポリペプチドの単離を可能にする条件下での実験用または工業用発酵槽における振盪フラスコ培養、小規模または大規模発酵(連続発酵、回分発酵、流加発酵、または固体状発酵を含む)によって培養することができる。培養は、当技術分野で公知の手順を用いて、炭素および窒素源ならびに無機塩を含む好適な栄養培地中で起こる。好適な培地は、商業的供給業者から入手可能であるかまたは(例えば、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)のカタログで)公表されている組成に従って調製することができる。ポリペプチドまたは代謝産物が栄養培地中に分泌される場合、ポリペプチドまたは代謝産物を培地から直接回収することができる。ポリペプチドまたは代謝産物が分泌されない場合、細胞ライセートから回収することができる。
【0139】
ポリペプチドは、ポリペプチドに特異的な当技術分野で公知の方法を用いて検出することができる。これらの検出方法は、特異的抗体の使用、酵素産物の形成、または酵素基質の消失が含むことができる。
【0140】
得られる目的の化合物(例えば、ポリペプチドまたは代謝産物)は、当技術分野で公知の方法によって回収することができる。例えば、ポリペプチドまたは代謝産物は、遠心分離、濾過、抽出、噴霧乾燥、蒸発、または沈殿を含むが、これらに限定されない、従来の手順によって栄養培地から回収することができる。
【0141】
ポリペプチドは、クロマトグラフィー(例えば、イオン交換、親和性、疎水性、クロマトフォーカシング、およびサイズ排除)、電気泳動手順(例えば、分取等電点電気泳動法)、示差溶解度(例えば、硫酸アンモニウム沈殿)、SDS−PAGE、または抽出を含むが、これらに限定されない、当技術分野で公知の種々の手順によって精製することができる(例えば、Protein Purification,J.−C.JansonおよびLars Ryden編,VCH Publishers,New York,1989を参照されたい)。
【0142】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような選択マーカーを含まない宿主細胞である。
【0143】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような真菌宿主細胞である。
【0144】
好ましくは、本発明による方法における宿主細胞は、本発明で先に記載したような糸状菌宿主細胞である。
【0145】
本発明の目的の化合物は、任意の生体化合物であることができる。生体化合物は、任意の生体ポリマーまたは代謝産物であってもよい。生体化合物は、生合成経路もしくは代謝経路を構成する単一のポリヌクレオチドもしくは一連のポリヌクレオチドによってコードされていてもよいし、または単一のポリヌクレオチドの産物もしくは一連のポリヌクレオチドの産物の直接の結果であってもよい。生体化合物は、宿主細胞にとってネイティブであっても、または異種であってもよい。
【0146】
「異種の生体化合物」という用語は、細胞にとってネイティブでない生体化合物;またはネイティブな生体化合物を変化させるために構造的な改変がなされたネイティブな生体化合物として本明細書で定義される。
【0147】
「生体ポリマー」という用語は、同一の、同様の、または異なるサブユニット(モノマー)の鎖(またはポリマー)として本明細書で定義される。生体ポリマーは、任意の生体ポリマーであってもよい。生体ポリマーは、例えば、核酸、ポリアミン、ポリオール、ポリペプチド(もしくはポリアミド)、または多糖であってもよいが、これらに限定されない。
【0148】
生体ポリマーはポリペプチドであってもよい。ポリペプチドは、目的の生体活性を有する任意のポリペプチドであってもよい。「ポリペプチド」という用語は、本明細書では、特定の長さのコードされた産物を指すことを意味するものではなく、それゆえ、ペプチド、オリゴペプチド、およびタンパク質を包含する。ポリペプチドには、上述のポリペプチドおよびハイブリッドポリペプチドの天然のアレルバリエーションおよび操作されたバリエーションがさらに含まれる。ポリペプチドは、宿主細胞にとってネイティブであっても、または異種であってもよい。ポリペプチドは、コラーゲンもしくはゼラチン、またはそれらの変異体もしくはハイブリッドであってもよい。ポリペプチドは、抗体またはその部分、抗原、凝固因子、酵素、ホルモンまたはホルモン変異体、受容体またはその部分、調節タンパク質、構造タンパク質、受容体、または輸送体タンパク質、分泌過程に関与するタンパク質、フォールディング過程に関与するタンパク質、シャペロン、ペプチドアミノ酸輸送体、グリコシル化因子、転写因子、合成ペプチドまたはオリゴペプチド、細胞内タンパク質であってもよい。細胞内タンパク質は、プロテアーゼ、セラミダーゼ、エポキシドヒドロラーゼ、アミノペプチダーゼ、アシラーゼ、アルドラーゼ、ヒドロキシラーゼ、アミノペプチダーゼ、リパーゼなどの酵素であってもよい。ポリペプチドは、細胞外に分泌される酵素であってもよい。そのような酵素は、オキシドレダクターゼ、トランスフェラーゼ、ヒドロラーゼ、リアーゼ、イソメラーゼ、リガーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼの群に属してもよい。酵素は、カルボヒドラーゼ、例えば、セルラーゼ(例えば、エンドグルカナーゼ、β−グルカナーゼ、セロビオヒドロラーゼ、もしくはβ−グルコシダーゼ)、ヘミセルラーゼもしくはペクチン分解酵素(例えば、キシラナーゼ、キシロシダーゼ、マンナナーゼ、ガラクタナーゼ、ガラクトシダーゼ、ペクチンメチルエステラーゼ、ペクチンリアーゼ、ペクチン酸リアーゼ、エンドポリガラクツロナーゼ、エキソポリガラクツロナーゼ、ラムノーガラクツロナーゼ、アラバナーゼ、アラビノフラノシダーゼ、アラビノキシランヒドロラーゼ、ガラクツロナーゼ、リアーゼ、もしくはデンプン分解酵素);ヒドロラーゼ、イソメラーゼ、もしくはリガーゼ、ホスファターゼ(例えば、フィターゼ)、エステラーゼ(例えば、リパーゼ)、タンパク質分解酵素、オキシドレダクターゼ(例えば、オキシダーゼ)、トランスフェラーゼ、またはイソメラーゼであってもよい。酵素はフィターゼであってもよい。酵素は、アミノペプチダーゼ、アスパラギナーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシペプチダーゼ、エンドプロテアーゼ、メタロプロテアーゼ、セリンプロテアーゼカタラーゼ、キチナーゼ、クチナーゼ、シクロデキストリングリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α−ガラクトシダーゼ、β−ガラクトシダーゼ、グルコアミラーゼ、α−グルコシダーゼ、β−グルコシダーゼ、ハロペルオキシダーゼ、タンパク質デアミナーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ムタナーゼ、オキシダーゼ、ペクチン分解酵素、ペルオキシダーゼ、ホスホリパーゼ、ポリフェノールオキシダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはグルコースオキシダーゼ、ヘキソースオキシダーゼ、モノオキシゲナーゼであってもよい。
【0149】
本発明の方法では、ポリペプチドは、別のポリペプチドがポリペプチドまたはその断片のN末端もしくはC末端で融合された融合ポリペプチドまたはハイブリッドポリペプチドであることもできる。融合されたポリペプチドは、あるポリペプチドをコードする核酸配列(またはその一部)を別のポリペプチドをコードする核酸配列(またはその一部)に融合することによって産生される。
【0150】
融合ポリペプチドを産生するための技術は当技術分野で公知であり、かつそれらがインフレームとなり、融合されたポリペプチドの発現が同じプロモーターおよびターミネーターの制御下となるようにポリペプチドをコードするコード配列を連結させることを含む。ハイブリッドポリペプチドは、1つまたは複数が宿主細胞にとって異種であってもよい少なくとも2つの異なるポリペプチドから得られる部分的なまたは完全なポリペプチド配列の組合せを含んでいてもよい。
【0151】
生体ポリマーは多糖であってもよい。多糖は、ムコ多糖(例えば、ヘパリンおよびヒアルロン酸)ならびに窒素含有多糖(例えば、キチン)を含むが、これらに限定されない、任意の多糖であってもよい。より好ましい選択肢では、多糖はヒアルロン酸である。
【0152】
本発明による目的のポリヌクレオチドは、有機酸、カロテノイド、(β−ラクタム)抗生物質、およびビタミンなどの、一次または二次代謝産物の合成に関与する酵素をコードしていてもよい。そのような代謝産物は、本発明による生体化合物とみなしてもよい。
【0153】
「代謝産物」という用語は、一次代謝産物と二次代謝産物の両方を包含し、代謝産物は、任意の代謝産物であってもよい。好ましい代謝産物は、クエン酸、グルコン酸およびコハク酸である。
【0154】
代謝産物は、例えば、生合成経路または代謝経路内の1つ以上の遺伝子によってコードされていてもよい。一次代謝産物は、エネルギー代謝、増殖、および構造に関する細胞の一次代謝または一般的な代謝の産物である。二次代謝産物は、二次代謝の産物である(例えば、R.B.Herbert,The Biosynthesis of Secondary Metabolites,Chapman and Hall,New York,1981を参照されたい)。
【0155】
一次代謝産物は、アミノ酸、脂肪酸、ヌクレオシド、ヌクレオチド、糖、トリグリセリド、またはビタミンであってもよいが、これらに限定されない。
【0156】
二次代謝産物は、アルカロイド、クマリン、フラボノイド、ポリケチド、キニーネ、ステロイド、ペプチド、またはテルペンであってもよいが、これらに限定されない。二次代謝産物は、抗生物質、摂食阻害物質、誘引物質、殺細菌物質、殺真菌物質、ホルモン、殺虫物質、または殺鼠物質であってもよい。好ましい抗生物質は、セファロスポリンおよびβ−ラクタムである。
【0157】
生体化合物はまた、選択可能マーカーの産物であってもよい。選択可能マーカーは、目的のポリヌクレオチドの産物であり、この産物は、殺生物剤またはウイルスに対する耐性、重金属に対する耐性、栄養要求株に対する原栄養性などを提供する。選択可能マーカーには、amdS(アセトアミダーゼ)、argB(オルニチンカルバモイルトランスフェラーゼ)、bar(ホスフィノトリシンアセチルトランスフェラーゼ)、hygB(ハイグロマイシンホスホトランスフェラーゼ)、niaD(硝酸還元酵素)、pyrG(オロチジン−5’−リン酸デカルボキシラーゼ)、sC(硫酸アデニルトランスフェラーゼ)、trpC(アントラニル酸シンターゼ)、ble(フレオマイシン耐性タンパク質)、およびそれらの等価物が含まれるが、これらに限定されない。
【0158】
本発明によれば、本発明による方法における目的の化合物は、本明細書に記載したようなポリペプチドであることが好ましい。
【0159】
本発明による方法におけるポリペプチドは、本明細書に記載したような酵素であることが好ましい。
【0160】
本発明によれば、本発明による方法における目的の化合物は、代謝産物であることが好ましい。
【0161】
本発明の別の態様によれば、宿主細胞の少なくとも2つの実質的に相同なDNAドメインの適合されたDNAドメインは、遺伝子変換および/または増幅によって、最も低い増加頻度を有する。異なる実質的に相同なDNAドメインは、異なる増加頻度、すなわち、遺伝子変換頻度を有することが明らかになった。最も低い増加頻度を有する実質的に相同なDNAドメインの標的化DNAドメインを適合させる利点は、実質的に相同なDNAドメインにおける組換えの頻度が低い遺伝的に安定な宿主が得られることである。
【0162】
誤って同定された塩基を含む必要があるため、本明細書で提供されている配列情報は、それほど狭義に解釈されるべきではない。本明細書に開示されている特定の配列を容易に用いて、それぞれの宿主細胞から完全な遺伝子を単離することができ、次に、この完全な遺伝子をさらなる配列解析に容易にかけ、それにより、シークエンシングのエラーを同定することができる。
【0163】
別途示されない限り、本明細書中のDNA分子をシークエンシングすることによって決定される全てのヌクレオチド配列は、自動化DNAシークエンサーを用いて決定され、本明細書で決定されるDNA分子によってコードされるポリペプチドの全てのアミノ酸配列は、上記のように決定された核酸配列の翻訳によって予想された。それゆえ、この自動化された手法によって決定される任意のDNA配列について当技術分野で知られているように、本明細書で決定される任意のヌクレオチド配列は、いくつかのエラーを含んでいてもよい。自動化によって決定されるヌクレオチド配列は、典型的には、シークエンシングされたDNA分子の実際のヌクレオチド配列と少なくとも約90%同一、より典型的には、少なくとも約95%から少なくとも約99.9%同一である。実際の配列は、当技術分野で周知の手動のDNAシークエンシング法を含む他の手法によってより正確に決定することができる。これも当技術分野で知られているように、実際の配列と比較した、決定されたヌクレオチド配列における単一の挿入または欠失は、決定されたヌクレオチド配列によってコードされる予想アミノ酸配列が、そのような挿入点または欠失点から、シークエンシングされたDNA分子によって実際にコードされるアミノ酸配列と完全に異なるように、ヌクレオチド配列の翻訳においてフレームシフトを生じさせる。
【0164】
当業者は、そのような誤って同定された塩基を同定することができ、そのようなエラーを訂正する方法を知っている。
【0165】
本明細書に含まれる特定の実施形態は、本発明のいくつかの態様の実例であることが意図されるので、本明細書に記載され、特許請求された本発明は、これらの実施形態によって範囲が限定されるべきではない。等価な実施形態はいずれも、本発明の範囲内であることが意図される。実際、本明細書に示され、記載された変更に加えた、本発明の様々な変更が、前述の記載から当業者に明らかになるであろう。そのような変更も、添付の特許請求の範囲の範囲内に含まれることが意図される。矛盾がある場合、定義を含む本開示が指針と考えられる。
【0166】
[実施例]
[株]
WT1:このアスペルギルス・ニガー(Aspergillus niger)株を野生型株として用いる。この株は、寄託番号CBS513.88でCBS研究所(CBS Institute)に寄託されている。
【0167】
WT2:このA.ニガー(A.niger)突然変異体株は、国際公開第98/46772号パンフレットに本質的に記載されているような方法による古典的な株改良によってA.ニガー(A.niger)CBS513.88から得られる。NTGまたはUVによるWT1の胞子の突然変異誘発後、振盪フラスコにおける突然変異体株の改善されたグルコアミラーゼ産生について選択を行なった。いくつかの改善されたA.ニガー(A.niger)株が同定され、そのうち、優良な株は、3〜4倍増加したグルコアミラーゼ活性レベルを生じさせた。この突然変異体株は、寄託番号CBS124.903でCBS研究所に寄託されている。振盪フラスコ解析および遺伝子解析から、A.ニガー(A.niger)CBS124.903株は、glaA遺伝子座(アンプリコン)の増幅に起因する(3)glaA遺伝子コピー数の増加を伴って、グルコアミラーゼ産生が3〜4倍増加しているという結論が得られた。
【0168】
GBA300:このA.ニガー(A.niger)株は、3つのglaAアンプリコンの改変を含むWT2株である。GBA300株の構築は、国際公開第98/46772号パンフレットに記載の方法に従って行なわれた。この特許出願では、A.ニガー(A.niger)ゲノムを含むアンプリコンから3つのglaA特異的DNA配列を欠失させて、3つの切断されたglaAアンプリコンを有するamdS陰性株を生じさせる方法が広範に記載されている。この手順により、最終的に外来性のDNA配列を全く有さない、マーカー遺伝子を含まない(MARKER−GENE FREE)ΔglaA組換えA.ニガー(A.niger)WT2株が得られた。この3つの切断されたglaAアンプリコンをBamHI切断アンプリコン、SalI切断アンプリコンおよびBglII切断アンプリコンと命名する。
【0169】
[A.ニガー(A.niger)振盪フラスコ発酵]
A.ニガー(A.niger)株を、国際公開第99/32617号パンフレットの実施例「アスペルギルス・ニガー(Aspergillus niger)振盪フラスコ発酵」の節に記載されているように、20mlの前培養培地中で前培養した。一晩増殖させた後、この培養物10mlを、国際公開第99/32617号パンフレットに記載されているような7%グルコースを含む発酵培地1(FM1)に移した。このFM1は、1リットル当たりに、以下のものを含有する:カゼイン加水分解物25g、酵母抽出物12.5g、KH2PO4 1g、K2SO4 2g、MgSO4.7H2O 0.5g、ZnCl2 0.03g、CaCl2 0.02g、MnSO4.4H2O 0.01g、FeSO4.7H2O 0.3g、Pen−Strep(5000IU/mlのPen−5mg/mlのStrep) 10ml、4NのH2SO4でpH5.6に調整。発酵は、表示された日数の間、34℃および170rpmで、100mlの発酵ブロスを含むバッフル付きの500mlフラスコ中で行なわれる。
【0170】
発酵培地2(FM2)はPLA2発酵に用いられ、1リットル当たりに、以下のものを含有する:グルコース.1H2O 82.5g、Maldex15(ブーム・メッペル(Boom Meppel)、オランダ) 25g、クエン酸2g、NaH2PO4.1H2O 4.5g、KH2PO4 9g、(NH4)2SO4 15g、ZnCl2 0.02g、MnSO4.1H2O 0.1g、CuSO4.5H2O 0.015g、CoCl2.6H2O 0.015g、MgSO4.7H2O 1g、CaCl2.2H2O 0.1g、FeSO4.7H2O 0.3g、MES(2−[N−モルホリノ]エタンスルホン酸) 30g、pH=6。
【0171】
[毒素測定用のA.ニガー(A.niger)寒天培地]
以下の培地上で株を培養し、オクラトキシンA(OTA)およびフモニシンB2(FB2)の産生について試験した:FrisvadおよびFiltenborg,1989(Terverticillate Penicillia:chemotaxonomy and mycotoxin production,Mycologia 81:837−861)ならびにFrisvadおよびThrane,1993(Liquid column chromatography of mycotoxinsm In:Betina(編):Chromatography of mycotoxins.Techniques and applications.J.of Chromatography Library 54:253−372 Elsevier,Amsterdam)によって本質的に記載されているようなツァペック酵母自己分解物寒天(CYA)および酵母抽出物ショ糖寒天(YES)。ペトリ皿を暗所24℃で7日間インキュベートし、その後、抽出用に単一コロニーから寒天プラグを採取した。84%CH3CN水を用い、超音波浴に1時間置いて抽出を行ない、その後、0.45μmのPTFEフィルターに通して溶媒を濾過した。
【0172】
規定量のFB2およびOTAになるよう内部標準溶液をサンプルに添加して、定量を行なった。Mogensenら,2010(Production of fumonisin B2 and B4 by Aspergillus niger in raisins and grapes,J of Agricultural and Food Chemistry 58:954−958)によって記載され、Nielsenら、2009(Review of secondary metabolites and mycotoxins from the Aspergillus niger group,Anal Bioanal Chem 395:1225−1242)に概説されているようにLC−MS/MSを行なった。
【0173】
[実施例1.アスペルギルス・ニガー(Aspergillus niger)GBA301(ΔglaA、ΔpepA)の構築]
このA.ニガー(A.niger)株は、主要な細胞外アスパラギン酸プロテアーゼPepAをコードするpepA遺伝子の欠失を含むGBA300株である。GBA301株は、欧州特許第0635574号明細書に記載されているような「マーカー遺伝子を含まない(MARKER−GENE FREE)」アプローチを用いて構築される。この特許に記載されている方法を用いて、van den Homberghら(van den Hombergh JP,Sollewijn Gelpke MD,van de Vondervoort PJ,Buxton FP,Visser J.(1997)−Disruption of three acid proteases in Aspergillus niger−effects on protease spectrum,intracellular proteolysis,and degradation of target proteins−Eur J Biochem.247(2):605−13)によって記載されているように、GBA300のゲノム中のpepA特異的DNA配列を欠失させる。この手順により、GBA300株バックグラウンドでpepA遺伝子が不活化されているマーカー遺伝子を含まないGBA301株が得られた。
【0174】
[実施例2.アスペルギルス・ニガー(Aspergillus niger)GBA302(ΔglaA、ΔpepA、ΔhdfA)の構築]
既知の原理に従ってhdfA用の遺伝子置換ベクターを設計し、日常的なクローニング手順に従って構築した。基本的には、これらのベクターは、所定のゲノム遺伝子座での相同組換え用に、hdfA ORFの約1〜2kbのフランキング領域を含む。さらに、これらは、直接反復と直接反復の間にA.ニデュランス(A.nidulans)二方向性amdS選択マーカーを含む。欠失ベクターの一般的な設計は、以前に欧州特許出願公開第635574B号明細書および国際公開第98/46772号パンフレットに記載されており、欠失ベクターを構築するための一般的なクローニングベクターpGBDEL(図1)の使用は、国際公開第06/040312号パンフレットに記載されている。
【0175】
ベクターpDEL−HDFA(図2)は、相同組換え用にhdfA ORFの約1kbフランキング領域を含む。A.ニガー(A.niger)遺伝子の全ヌクレオチド配列およびそのゲノムコンテクストは、例えば、NCBI(http://www.ncbi.nlm.nih.gov/)またはEMBL(http://www.ebi.ac.uk/embl/)から得ることができる。欠失ベクターpDEL−HDFAの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に詳細に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA301を形質転換し、hdfA遺伝子を欠失させた。本明細書中の全ての実施例における遺伝子欠失に適用された方法では、線状DNAが用いられた。この線状DNAは、二重交差によってフランキング配列の相同な遺伝子座でゲノム中に組み込まれ、その結果、欠失しようとする遺伝子をamdS遺伝子に置き換える。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。遺伝子破壊の一般的手順は、図3に示されている。GBA301株バックグラウンドでhdfA遺伝子が不活化されている代表的な株として株GBA302を選択した。
【0176】
[実施例3.アスペルギルス・ニガー(Aspergillus niger)GBA303(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン)の構築]
この実施例では、最も高い頻度の遺伝子変換を有するΔglaAアンプリコンを適合させて、標的化組込み用の独特の遺伝子座を生じさせ、賢明な組込み戦略を利用できるようにした。適合されたアンプリコンは、この実施例中のグルコアミラーゼプロモーターおよびターミネーターなどのフランキング制御配列を標的化領域として利用できるようにもする。
【0177】
[実施例3.1 最も高い頻度の遺伝子変換を有するアンプリコンの同定]
株GBA301は、ゲノム中に3つの改変されたΔglaA遺伝子座を含む。3つ全ての遺伝子座について、約4.3kbのglaA配列(2kbのglaAプロモーターおよび全glaAコード配列)が欠失している(国際公開第98/46772号パンフレットの図4に見られる)。3つの異なる欠失ベクターを用いたので、各々のΔglaA遺伝子座はわずかに異なっており、その特徴を用いて、PCRテストで各々の切断されたΔglaA遺伝子座を可視化することができる(国際公開第98/46772号パンフレットの図5)。組み込まれたこの違いを用いて、切断されたglaAアンプリコンの中に遺伝子変換事象を追跡することもできる。3つの切断されたglaAアンプリコンはBamHI切断アンプリコン、SalI切断アンプリコンおよびBglII切断アンプリコンと命名されており、いわゆる「DNAフラッグ」テストにおいて、それぞれ、240、260および300bpのサイズのバンドを示す(国際公開第98/46772号パンフレットの図5)。
【0178】
株GBA301から、GBA301の3つの切断されたglaAアンプリコンの1つに全て標的化される複数のフィターゼ発現カセットを含む、複数のフィターゼ産生株を(pGBAAS−1とpGBTOPFYT−1を共形質転換することによって)作製した(国際公開第98/46772号パンフレットの実施例1.4に本質的に記載されている)。得られる株は各々、3つの切断されたglaAアンプリコンの1つにフィターゼカセットを有し、フィターゼカセット組込み部位に従って、BAM−PHY、SAL−PHYおよびBGL−PHYと呼ばれる。
【0179】
3つの異なる株、BAM−PHY、SAL−PHYおよびBGL−PHYについてフィターゼカセットのコピー数が増加した変換体を同定した(国際公開第98/46772号パンフレットの実施例1.5に本質的に記載されている)。フィターゼコピー数が増加した変換体について、フィターゼ含有glaAアンプリコンの特定のアンプリコンの頻度および遺伝子型を「DNAフラッグ」テストを用いて決定した。「DNAフラッグ」テストによってそれぞれの株の何千もの個々の子孫を解析することにより、特定の切断されたglaAアンプリコン(例えば、SalI)の欠失と同時に、別の特定の切断されたglaAアンプリコン(例えば、BamHI)の増幅によって遺伝子変換を検出することができる。
【0180】
3つの異なる株の何百もの個々の子孫のPCRスコアリングおよびタイピングの結果を表1に見ることができる。さらに、いくつかの単離された株について単一の遺伝子変換事象後に同様の手法を取った。これらの株は、同じ種類の2つのアンプリコンを含んでいた。3種類の株の何百もの個々の子孫をPCRでスコアリングしたこれらの結果を表2に見ることができる。この3つの異なる株に関しては、第1の遺伝子変換事象と第2の遺伝子変換事象の両方について、BamHIアンプリコン中にあるフィターゼカセットを遺伝子変換によって最も高い頻度で増加させることができる。
【0181】
【表1】
【0182】
【表2】
【0183】
[実施例3.2 最も高い頻度の遺伝子変換を有するBamHIアンプリコンの適合]
この実施例のこの部分では、(株GBA300を生じさせるCBS124.903中のglaA遺伝子座の3倍の改変によって)事前に除去されたグルコアミラーゼプロモーターPglaAを、ゲノム中の単一の遺伝子座に、しかし、適合させようとするアンプリコン中の異なる位置に再導入し、標的化組込み用の独特のかつ改善された遺伝子座を生じさせる方法が記載されている。
【0184】
アンプリコンを適合させることができるように、ΔglaA遺伝子座用の遺伝子置換型のベクターを既知の原理に従って設計し、日常的なクローニング手順に従って構築した。この目的のために、ベクターpGBGLA−65を構築した(図6)。基本的に、このベクターは、2つのPglaA断片と3’glaAおよび3”glaA領域の間にamdS選択マーカーを含む。3’glaAおよび3”glaA領域は、ΔglaA遺伝子座の同一のゲノム領域中への標的化および組込みに用いられる。2つのPglaA断片は、対抗選択によるamdS選択マーカーのループアウトに用いられる(図3)。1つのPglaA断片は、切断されたPglaAプロモーター断片(MluI部位の3’にあるPglaAプロモーターの最後の600bpを欠く)であり、これは、amdS対抗選択後にゲノム中に存在したまま残る。
【0185】
ゲノム中にBamHIアンプリコンを適合させることができるように、ベクターpDEL−GLA65(図6)をHindIIIで消化し、ゲル精製して、大腸菌(E.coli)骨格を除去した。欠失ベクターpDEL−GLA65の線状DNAを用い、国際公開第05/095624号パンフレットに以前に詳細に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA302を形質転換した。pDEL−GLA65の線状DNAは、二重交差によって3’glaA配列および3”glaA配列でアンプリコンに組み込まれ得る。いくつかのコロニーを純化し、3つのglaA遺伝子座の存在およびBamHIアンプリコンでのカセットの組込みについてPCRで解析した(標的PCR)。BamHIアンプリコンの標的化頻度は、20〜30%と推定された。合計7つの株をフルオルアセトアミド対抗選択について選択した。フルオルアセトアミド耐性コロニーをPDAに移し、「フラッグ」−PCRでテストした。適合されたBamHI遺伝子座を有し、組み込まれたpDEL−GLA65カセットのPglaA反復上での組換えによってamdSマーカーが正確に欠失している代表的な株としてある株を選択した(図7)。この選択された株をGBA303と命名した。これは、pepAおよびhdfA遺伝子が不活化され、BamHIアンプリコンが適合された、3つ全てのΔglaA遺伝子座を有するGBA300株バックグラウンドを表す。glaAアンプリコン全体はサイズが大きい(>80kb)ので、BamHIアンプリコンが適合されている(2〜6kb)としても、かなりの実質的に相同なDNAドメインを有する断片が依然としてゲノム中に存在したまま残る。
【0186】
ΔglaAアンプリコンの3’断片および3”断片を含むゲノム領域の配列(図7)は、配列番号:1の下に見出すことができる。3’断片、切断されたPglaAプロモーター断片および適合されたBamHIアンプリコンの3”断片を含むゲノム領域の配列は、配列番号:2の下に見出すことができる。
【0187】
[実施例3.3 新規の組込みベクターの構築]
欧州特許出願公開第0635574A1号明細書、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットには、標的化された株構築、とりわけ、単一の交差を用いる標的化株構築で用いられる組込みおよび発現ベクターpGBAAS−1(欧州特許第0635574号明細書中のpGBGLA−50)およびpGBTOP−8が記載されている。これらの組込みベクターの調整は、GBA302型の株の適合されたBamHIアンプリコンへの特異的標的化を可能にするために必要である。
【0188】
amdSマーカーを含み、かつ共形質転換で用いられるpGBAAS−1ベクターのうち、2つの異なる変異体が、適合されたBamHIアンプリコンへの標的化組込み用に構築されている。グルコアミラーゼプロモーターPglaAの制御下で発現されるamdSマーカー遺伝子を有するpGBAAS−3(図8)と呼ばれるベクターを構築した。PglaAプロモーターは、amdSマーカー遺伝子の発現を駆動する際に機能を有する。さらに、PglaAプロモーターおよび3’glaA断片は、特に、適合されたBamHI ΔglaAアンプリコンの標的化配列として機能する(図21)。pGBAAS−1のamdSマーカーカセット(gpdAプロモーターの制御下のamdS)を含むpGBAAS−4と呼ばれる第2のベクター(図9)を構築した。ここで、切断されたPglaAプロモーター断片および3’glaA断片は、適合されたBamHI ΔglaAアンプリコンに標的化する際に機能を有する(図21)。
【0189】
pGBTOP型のベクターについては、適合されたBamHI ΔglaAアンプリコンへの標的化を可能にするために、2つの新規の組込みおよび発現ベクターを構築した。pGBTOP−11(図10)およびpGBTOP−12(図11)ベクターは、基本的には、それぞれ、3”glaA断片が除去されたpGBTOP−8ベクターおよび大腸菌(E.coli)ベクター除去用の制限部位のさらなる改変を含むpGBTOP−8ベクターである。ここで、PglaAプロモーターおよび3’glaAターミネーター断片は二重の機能を有する。すなわち、目的の遺伝子を発現させるためのプロモーターおよびターミネーターとしてのその役割の他に、適合されたBamHI ΔglaAアンプリコンに標的化する役割を有する(図21)。
【0190】
[実施例4.アスペルギルス・ニガー(Aspergillus niger)GBA304(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII)の構築]
α−アミラーゼをコードするamyBII ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYBIIを得た(図12)。欠失ベクターpDEL−AMYBIIの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA303を形質転換し、amyBII遺伝子を欠失させた。二重交差事象による相同なamyBII遺伝子座でのゲノム中へのpDEL−AMYBIIの組込みによってamyBII ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA303株バックグラウンドでamyBII遺伝子が不活化された代表的な株として株GBA304を選択した。
【0191】
[実施例5.アスペルギルス・ニガー(Aspergillus niger)GBA305(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI)の構築]
α−アミラーゼをコードするamyBI ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYBIを得た(pDEL−AMYBIのレイアウトの代表的な図を図12に見ることができる)。欠失ベクターpDEL−AMYBIの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA304を形質転換し、amyBI遺伝子を欠失させた。二重交差事象による相同なamyBI遺伝子座でのゲノム中へのpDEL−AMYBIの組込みによってamyBI ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA304株バックグラウンドでamyBI遺伝子が不活化された代表的な株として株GBA305を選択した。
【0192】
[実施例6.アスペルギルス・ニガー(Aspergillus niger)GBA306(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA)の構築]
酸安定性α−アミラーゼをコードするamyA ORFの遺伝子フランキング領域を実施例2で本質的に記載したようにクローニングし、ベクターpDEL−AMYAを得た(pDEL−AMYAのレイアウトの代表的な図を図12に見ることができる)。欠失ベクターpDEL−AMYAの線状DNAを単離し、これを用い、国際公開第05/095624号パンフレットに以前に記載されているような方法を用いて、アスペルギルス・ニガー(Aspergillus niger)GBA305を形質転換し、amyA遺伝子を欠失させた。二重交差事象による相同なamyA遺伝子座でのゲノム中へのpDEL−AMYBIの組込みによってamyA ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA305株バックグラウンドでamyA遺伝子が不活化された代表的な株として株GBA306を選択した。
【0193】
[実施例7.アスペルギルス・ニガー(Aspergillus niger)GBA307(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA)の構築]
このA.ニガー(A.niger)のオキサラート欠損株は、オキサロ酢酸ヒドロラーゼをコードするoahA遺伝子の欠失によって得ることができる。これは、欧州特許第1157100号明細書および米国特許第6,936,438号明細書に詳細に記載されている。GBA306株バックグラウンドでoahA遺伝子が不活化された代表的な株として株GBA307を選択した。
【0194】
あるいは、国際公開第04/070022号パンフレットおよび欧州特許第1590444号明細書に記載されているような古典的な株改良によってA.ニガー(A.niger)GBA306から突然変異体株を得ることができる。これらの文献では、オキサラート欠損A.ニガー(A.niger)株についてスクリーニングする方法が広範に記載されている。これらを欧州特許第1590444号明細書の実施例1および2の方法に従って単離した。
【0195】
[実施例8.アスペルギルス・ニガー(Aspergillus niger)GBA308(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB)の構築]
CBS513.88のゲノム配列において、マイコトキシンのフモニシンをコードする、ギベレラ・モニリフォルミス(Gibberella moniliformis)中の遺伝子クラスターとの相同性に基づいて、推定上のフモニシン遺伝子クラスターを同定した(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88”.Nat Biotechnol.2007 Feb;25(2):221−231)。
【0196】
【表3】
【0197】
遺伝子An01g06930は、フモニシン産生に関与する可能性があるポリケチド合成酵素(PKS)をコードする。プロモーター側に推定上のfumB遺伝子(An01g06930)のコード配列のごく一部分も含む、遺伝子フランキング領域を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているように、ベクターpGBDEL(図1)にクローニングし、ベクターpGBDEL−FUM3(図13)を得た。欠失ベクターpGBDEL−FUM3の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA307を形質転換し、fumB遺伝子を欠失させた。二重交差事象による相同なAn01g06930遺伝子座でのゲノム中へのpGBDEL−FUM3の組込みによってfumB ORFが除去され、結果として約8kbのゲノム配列が除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA307株バックグラウンドでfumB遺伝子が不活化された代表的な株として株GBA308を選択した。
【0198】
株WT1およびGBA308をCYAおよびYES寒天培地上で増殖させ、フモニシン産生を測定した。FB2測定の結果を以下の表に示す。
【0199】
【表4】
【0200】
これらの結果から、PKSをコードするfumB遺伝子の破壊によってフモニシン産生が障害された株が得られることは明らかである。フモニシン陰性株バックグラウンドは、商業的なタンパク質産生にとって明らかな利点がある。
【0201】
さらに、遺伝子An01g06820を含む巨大な遺伝子クラスターが(約38kbのゲノム配列が除去された)An01g06930まで破壊されている株は、同一の表現型を示す、すなわち、上で詳述したような試験でフモニシン産生が陰性であることが示された(データは示さない)。
【0202】
[実施例9.アスペルギルス・ニガー(Aspergillus niger)GBA309(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch)の構築]
CBS513.88のゲノム配列において、オクラトキシン産生に関与するA.オクラセウス(A.ochraceus)のPKS断片に基づいて、推定上のオクラトキシン遺伝子クラスターを同定した(Pelら,“Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88.” Nat Biotechnol.2007 Feb;25(2):221−231)。
【0203】
【表5】
【0204】
このPKS様遺伝子は、An15g07920(ochAと呼ばれる)と注釈付けられている。さらに、遺伝子An15g07910(ochBと呼ばれる)は、オクラトキシン産生に関与する潜在的なペプチド合成酵素に相当し得る。ペプチド合成酵素および/またはPKSをコードする遺伝子の破壊は、オクラトキシン産生が障害された株をもたらし得る。
【0205】
二重交差用の標的化配列を用いて完全なオクラトキシン遺伝子クラスターを破壊することができるように、一方の側のAn15g07860 ORFの遺伝子配列断片およびもう一方の側のAn15g07930 ORFの遺伝子配列断片を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているようにクローニングして、ベクターpGBDEL−OCH2(図14)を得た。欠失ベクターpGBDEL−OCH2の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA308を形質転換し、推定上のオクラトキシン遺伝子クラスターを欠失させた。二重交差事象による相同な遺伝子座でのゲノム中へのpGBDEL−OCH2の組込みによってoch ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA308株バックグラウンドでオクラトキシンゲノム遺伝子クラスター(遺伝子An15g07860と遺伝子An15g07930の間にある;遺伝子型はΔochと表示される)が除去された代表的な株として株GBA309を選択した。
【0206】
株WT1およびGBA309をCYAおよびYES寒天培地上で増殖させ、オクラトキシンAを測定した。OTA測定の結果を以下の表に示す。
【0207】
【表6】
【0208】
これらの結果から、OTA生合成に関与する推定上のペプチド合成酵素およびPKSを含むオクラトキシン遺伝子クラスターの破壊によって、オクラトキシンA産生が低下した株が得られることは明らかである。オクラトキシンA陰性および場合によってはフモニシン陰性との組合せの株バックグラウンドは、商業的なタンパク質産生にとって明らかな利点がある。
【0209】
[実施例10.アスペルギルス・ニガー(Aspergillus niger)GBA310(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch、ΔprtT)の構築]
プロテアーゼ調節因子をコードするprtT ORFの遺伝子フランキング領域を、実施例2で本質的に記載したように、かつ国際公開第06/040312号パンフレットにさらに詳述されているようにクローニングし、ベクターpGBDEL−PRT2(図15)を得た。欠失ベクターpGBDEL−PRT2の線状DNAを単離し、これを用いて、国際公開第06/040312号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA309を形質転換し、prtT遺伝子を欠失させた。二重交差事象による相同なprtT遺伝子座でのゲノム中へのpGBDEL−PRT2の組込みによってprtT ORFが除去された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA309株バックグラウンドでprtT遺伝子が不活化された代表的な株として株GBA310を選択した。
【0210】
[実施例11.アスペルギルス・ニガー(Aspergillus niger)GBA311(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA、ΔfumB、Δoch、ΔprtT、sec61 S376W突然変異)の構築]
この実施例では、セリン376がトリプトファンに置き換えられているSec61*タンパク質をコードする改変されたsec61遺伝子(sec61*と呼ぶ)の導入を記載する。そうするために、遺伝子フランキング配列および改変されたSec61輸送チャネルをコードするsec61*コード配列を、国際公開第2005123763号パンフレットに本質的に詳細に記載されているようにクローニングし、ベクターpGBDEL−SEC61*(図16)を得た。欠失ベクターpGBDEL−SEC61*の線状DNAを単離し、これを用いて、国際公開第2005123763号パンフレットに以前に記載されているように、アスペルギルス・ニガー(Aspergillus niger)GBA310を形質転換し、Sec61遺伝子を改変した。二重交差事象による相同なSec61遺伝子座でのゲノム中へのpGBDEL−SEC61*の組込みによってSec61*ORFが改変された形質転換体を選択した。その後、フルオロアセトアミド培地上にプレーティングすることによって、amdSマーカーを除去し、マーカー遺伝子を含まない株を選択した。GBA310株バックグラウンドでSec61遺伝子が改変された代表的な株として株GBA311を選択した。
【0211】
[実施例12.適合されたアスペルギルス・ニガー(Aspergillus niger)株およびプラスミドを用いる改善された酵素産生および産生株構築]
ブタホスホリパーゼA2(PLA2)タンパク質(Roberts I.N.,Jeenes D.J.,MacKenzie D.A.,Wilkinson A.P.,Sumner I.G.およびArcher D.B.(1992) “Heterologous gene expression in Aspergillus niger.a glucoamylase−porcine pancreatic phospholipase A2 fusion protein is secreted and processed to yield mature enzyme” Gene 122:155−161)を、A.ニガー(A.niger)のCBS513.88&GBA株系統における酵素発現のモデルタンパク質として選択した。PLA2の過剰発現用の断片をproPLA2とA.ニガー(A.niger)のネイティブなグルコアミラーゼA遺伝子との融合体として作製し、原則的に、Robertsら(1992)によって記載されているように調製した。このglaA−pla2融合遺伝子をpGBTOP−8およびpGBTOP−12にクローニングし、それぞれ、pGBTOPPLA−2a(図17)およびpGBTOPPLA−2b(図18)を得た。
【0212】
さらに、A.ニガー(A.niger)のα−アミラーゼを、A.ニガー(A.niger)のCBS513.88&GBA株系統における酵素発現の第2のモデルタンパク質として用いた。過剰発現に用いたamyBコード配列は、(国際公開第2008/000632号パンフレットに詳細に記載されているように)amyB遺伝子をコードするα−アミラーゼの単一コドンおよびコドン対が最適化されたコード配列を含んでいた。グルコアミラーゼglaAプロモーターの翻訳開始配列は、(これも国際公開第2006/077258号パンフレットに詳述されているように)作製されたamyB発現コンストラクトにおいて5’−CACCGTCAAA ATG−3’へと改変されている。さらに、最適な翻訳終結配列が用いられたため、野生型5’−TGA−3’翻訳終結配列は、全ての発現コンストラクトにおいて、(国際公開第2006/077258号パンフレットに詳述されているように)5’−TAAA−3’に置き換えられた。最適化されたamyB cDNA配列を含むEcoRI−SnaBI断片を完全に合成し、サブクローニングし、配列解析によって配列を確認した。amyB断片をpGBTOP−8およびpGBTOP−12にクローニングし、それぞれ、pGBTOPFUA−2(図19)およびpGBTOPFUA−3(図20)を得た。
【0213】
[実施例12.1 適合された発現ベクターのA.ニガー(A.niger)株のΔglaA遺伝子座への改善された標的化]
構築された組込みベクター(実施例3.3および12)は2つのタイプ、すなわち、適合されていないΔglaA遺伝子座に対して3’glaA配列および3”glaA配列に標的化するために用いることができる組込みベクターと、ベクターpGBTOPPLA−2a、pGBTOPFUA−2およびpGBAAS−1である組込みベクターとに分けることができる。さらに、切断されたPglaAプロモーター断片および3’glaA断片に適合されたBamHI ΔglaAアンプリコン(適合されたΔglaA遺伝子座)に特異的に標的化するために用いることができる新規の組込みベクターを構築した。適合されたBamHIアンプリコンに標的化するために用いることができるベクターは、pGBTOPPLA−2b、pGBTOPFUA−3、pGBAAS−3およびpGBAAS−4である。
【0214】
以下のスキーム(表3)に従って、2つのPLA2発現ベクター(pGBTOPPLA−2aおよびpGBTOPPLA−2b)を、pGBBAAS−1およびpGBAAS−4との共形質転換によって、A.ニガー(A.niger)のGBA301(ΔglaA、ΔpepA)、GBA302(ΔglaA、ΔpepA、ΔhdfA)およびGBA303(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン)に導入した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、4つ全てのベクターの線状DNAを単離し、これを用いて、3つのA.ニガー(A.niger)株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によってΔglaA遺伝子座の1つで起こる。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。3つ全ての株について、amdSマーカー遺伝子とPLA2発現カセットの両方を有する数百個の形質転換体を、amdSおよびPLA2特異的プライマーを用いるPCRにより同定した。同定された共形質転換体に対して(国際公開第98/46772号パンフレットの実施例1.4に詳述されているように)標的化PCRテストを実施し、amdSおよび/またはPLA2カセットのいずれかがΔglaA遺伝子座に組み込まれている株のパーセンテージを決定した。
【0215】
さらに、フルオロアセトアミド培地上にプレーティングすることにより、同定されたΔglaA標的化共形質転換体からamdSマーカー遺伝子を含まない株を選択した。PCRに基づくDNA「フラッグテスト」を実施して、amdSを含まない子孫におけるアンプリコンの潜在的な喪失を検出した。国際公開第98/46772号パンフレットの実施例1.4.fで説明されているように、これは、組換えによるamdSマーカーと全glaAアンプリコンの同時喪失を示すものであり、そのようなものとして元のカセットの位置を示すものである。この試験を用いて、3つ全ての株におけるBamHIアンプリコンへの標的化頻度を決定した。データを表3に示す。
【0216】
【表7】
【0217】
表3から結論付けることができるように、適合された発現ベクターを適合された標的化DNAドメイン(適合されたBamHIアンプリコン)と組み合わせて用いると、標的化された株構築における明らかな利点が与えられ、より集中的でかつ効率的な株構築がもたらされる。
【0218】
[実施例12.2 適合されたコンストラクトおよびA.ニガー(A.niger)株を用いる改善された共形質転換]
以下のスキーム(表4)に従って、FUA発現ベクターpGBTOPFUA−3を、参照セットとしてのpGBTOPFUA−2およびpGBAASとともに、pGBAAS−3およびpGBAAS−4の共形質転換によって、A.ニガー(A.niger)GBA307(ΔglaA、ΔpepA、ΔhdfA、適合されたBamHIアンプリコン、ΔamyBII、ΔamyBI、ΔamyA、ΔoahA)に導入した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、5つのベクターの線状DNAを単離し、これを用いて、A.ニガー(A.niger)GBA307株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によって、それぞれ、適合されたBamHI ΔglaAアンプリコンまたはΔglaAアンプリコンで起こる。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。amdSおよびamyB特異的(コドン対が最適化されたamyBコード配列)プライマーを用いるPCRにより、各々のプラスミド組合せの50個の形質転換体について(国際公開第98/46772号パンフレットの実施例1.4に原理的に詳述されているような)共形質転換パーセンテージを決定した。データを表4に示す。
【0219】
【表8】
【0220】
これまでに知られているpGBAAS−1および改善された標的化のためのその変異体pGBAAS−4の場合、典型的な共形質転換頻度が得られる(通常25〜75%)。驚くことに、表4から結論付けることができるように、pGBBAAS−3変異体(glaAプロモーターの制御下のamdS遺伝子)を適合された標的化コンストラクト(例えば、pGBTOPFUA−3)と組み合わせて用いると、非常に高い共形質転換頻度が得られる。pGBAAS−3を用いると、共形質転換頻度が劇的に増加し、標的化された株構築において株をさらに操作する前に共形質転換体を選択する必要が回避され、そのため、より効率的な株構築がもたらされる。
【0221】
これらの形質転換体のFM1における振盪フラスコ解析は、GBA307で発現された異なるコンストラクトについて、遺伝子1コピー当たりに同様のFUA発現レベルを示した。
【0222】
[実施例12.3 適合されたA.ニガー(A.niger)株における改善された株構築]
工業用の組換えA.ニガー(A.niger)産生株を構築する1つの方法は、ΔglaA遺伝子座の1つに発現カセットを組み込み、その後、組み込まれた発現カセットを有するΔglaA遺伝子座を遺伝子変換または増幅によって増加させることである。
【0223】
発現カセットは、例えば、国際公開第2005/100573号パンフレット、国際公開第2006/077258号パンフレット、国際公開第2006/092396号パンフレットまたは国際公開第2008/000632号パンフレットに詳述されているようないくつかの方法によって改善することができる。株構築での遺伝子変換または増幅の使用は、例えば、国際公開第1998/46772号パンフレットにより詳細に記載されている。先の実施例では、株およびプラスミド適合によって、どのようにして共形質転換および/または特定の遺伝子座への標的化を改善することができるかということが示されている。この実施例では、新規のGBAタイプの株を用いる株構築での遺伝子変換または増幅の使用が示されている。
【0224】
A.ニガー(A.niger)GBA311(実施例11)株に、発現ベクターの2つの組合せ(すなわち、pGBTOPPLA−2aとpGBBAAS−1の混合物;pGBTOPPLA−2bとpGBBAAS−3の混合物)を共形質転換した。形質転換およびその後の形質転換体の選択は、国際公開第98/46772号パンフレットおよび国際公開第99/32617号パンフレットに記載されているように実施した。原則的に、使用した全てのベクターの線状DNAを単離し、これを用いて、GBA311 A.ニガー(A.niger)株を共形質転換した。両方の線状化プラスミドの組込みは、単一の交差事象によって、ΔglaA遺伝子座で起こる(標的化配列の詳細については、実施例12.1を参照されたい)。形質転換体をアセトアミド培地上で選択し、標準的な手順に従ってコロニーを純化した。amdSマーカー遺伝子とPLA2発現カセットの両方を有する数百個の形質転換体を、amdSおよびPLA2特異的プライマーを用いるPCRにより同定した。同定された共形質転換体に対して、(国際公開第98/46772号パンフレットの実施例1.4に詳述されているように)標的化PCRテストを実施し、amdSおよび/またはPLA2カセットのいずれかがΔglaA遺伝子座に組み込まれている株のパーセンテージを決定した。さらに、PCRを用いて、コロニーをPLA2コピー数について診断した。サザン解析(国際公開第98/46772号パンフレット)を用いて、高コピーと考えられる株のコピー数をより正確に決定した。pGBTOPPLA−2とpGBBAAS−1の混合物については、さらなる形質転換体をスクリーニングし、純化して、より高いコピー数のPLA2発現カセットを有する形質転換体を単離した。600個の個々の形質転換体をスクリーニングした後、6コピーのBglII標的化形質転換体を単離した。得られたデータを表5に示す。
【0225】
【表9】
【0226】
GBA300バックグラウンドの2つの高コピー株(すなわち、6コピーのBglII標的化株および7コピーのBamHI標的化株)について、PLA2コピー数が増加している変換体を単離した(実験の詳細については、本明細書中の実施例3.1および国際公開第98/46772号パンフレットを参照されたい)。両方の株の500個の個々の子孫をスクリーニングすることによって、BamHI標的化株については、遺伝子変換の結果としてPLA2コピー数が増加した2つの株が同定されたが、BglII標的化株については全く同定されなかった。この2つの変換体は、それぞれ、13コピーおよび14コピーのPLA−2を含んでいた。この実施例およびデータは、(例えば、増強された遺伝子変換または増幅頻度を有する)ゲノム標的化DNAドメインの適合は、明確でかつ制御された多コピー(産生)株の構築の過程を促進および加速することによって、株構築に大きな影響を及ぼすことを明らかに示している。これらの形質転換体のFM2における振盪フラスコ解析は、GBA311で発現された異なるコンストラクトについて、遺伝子1コピー当たりに同様のPLA2発現レベルを示した。
【特許請求の範囲】
【請求項1】
目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、前記少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ前記適合された実質的に相同なDNAドメインが由来する前記実質的に相同なDNAドメインは、前記少なくとも2つの実質的に相同なDNAドメインの他の1つよりも少なくとも10%高い遺伝子変換頻度を有する、組換え宿主細胞。
【請求項2】
前記適合された実質的に相同なDNAドメインが、独特の配列タグによって前記実質的に相同なDNAドメインの他のバージョンと区別される、請求項1に記載の宿主細胞。
【請求項3】
前記適合された実質的に相同なDNAドメインが、標的化DNAドメインを含み、前記標的化DNAドメインが、増強された組込み選好を有する配列を含む、請求項1または2のいずれか一項に記載の宿主細胞。
【請求項4】
前記実質的に相同なDNAドメインのうちの少なくとも2つが各々、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する、請求項1〜3のいずれか一項に記載の宿主細胞。
【請求項5】
組込み経路をHRの方向に導くことによる、前記宿主細胞のゲノム中の所定の部位への標的化されたポリヌクレオチドの組込み効率の増大をさらに含む、請求項1〜4のいずれか一項に記載の宿主細胞。
【請求項6】
前記少なくとも2つの実質的に相同なDNAドメインが、glaAの遺伝子座であるかまたはamyBI、もしくはamyBII)の遺伝子座である、請求項1〜5のいずれか一項に記載の宿主細胞。
【請求項7】
前記目的のポリヌクレオチドがコドン最適化されたポリヌクレオチドである、請求項1〜6のいずれか一項に記載の宿主細胞。
【請求項8】
glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドおよびprtTの群から選択されるポリヌクレオチドをさらに含み、前記ポリヌクレオチドが改変を含み、ここで、前記宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記改変を含むポリヌクレオチドによってコードされる産物を欠損している、請求項1〜7のいずれか一項に記載の宿主細胞。
【請求項9】
pepAの欠損をさらに含む、請求項8に記載の宿主細胞。
【請求項10】
SEC61の改変をさらに含む、請求項8または9のいずれか一項に記載の宿主細胞。
【請求項11】
前記宿主細胞が選択マーカーを含まない宿主細胞である、請求項1〜10のいずれか一項に記載の宿主細胞。
【請求項12】
前記宿主細胞が真菌宿主細胞である、請求項1〜11のいずれか一項に記載の宿主細胞。
【請求項13】
前記宿主細胞が糸状菌宿主細胞である、請求項12に記載の宿主細胞。
【請求項14】
目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させ、前記より高い遺伝子変換頻度を有する実質的に相同なDNAドメインに標的化DNAドメインを提供することを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、方法。
【請求項15】
a.請求項14で得られた前記細胞を目的のポリヌクレオチドで形質転換することと、
b.前記実質的に相同なDNAドメインのうちの少なくとも1つに組み込まれた少なくとも1コピーの前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングすることと、
c.(b)で得られた前記細胞を繁殖させ、実質的に相同なDNAドメインのさらなるコピーに組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングすることと
をさらに含む、請求項14に記載の方法。
【請求項16】
前記実質的に相同なDNAドメインの各々のコピーが、組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有するまで、工程(c)を繰り返す工程をさらに含む、請求項15に記載の方法。
【請求項17】
目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で請求項1〜13のいずれか一項に記載の宿主細胞を培養することと、
b.前記培養培地から前記目的の化合物を回収することと
を含む方法。
【請求項18】
目的の化合物を産生する方法であって、
a.組換え宿主細胞を前記化合物の産生をもたらす条件下で培養することであって、前記宿主細胞は目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、前記少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ前記実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する、と、
b.前記培養培地から前記目的の化合物を回収することと
を含む方法。
【請求項19】
前記宿主細胞が選択マーカーを含まない宿主細胞である、請求項17または18に記載の方法。
【請求項20】
前記宿主細胞が真菌宿主細胞である、請求項17〜19のいずれか一項に記載の方法。
【請求項21】
前記宿主細胞が糸状菌宿主細胞である、請求項20に記載の方法。
【請求項22】
前記目的の化合物がポリペプチドである、請求項17〜21のいずれか一項に記載の方法。
【請求項23】
前記ポリペプチドが酵素である、請求項22に記載の方法。
【請求項24】
前記目的の化合物が代謝産物である、請求項17〜21のいずれか一項に記載の方法。
【請求項25】
配列番号:2によるポリヌクレオチドを含む、単離されたポリヌクレオチド、またはその機能性断片。
【請求項26】
配列番号:3によるポリヌクレオチドを含む、単離されたポリヌクレオチド、またはその機能性断片。
【請求項27】
請求項25または26に記載のポリヌクレオチドを含むベクター。
【請求項28】
請求項25もしくは26に記載のポリヌクレオチドを含むか、または請求項27に記載のベクターを含む宿主細胞。
【請求項1】
目的の化合物を産生するための組換え宿主細胞であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、前記少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ前記適合された実質的に相同なDNAドメインが由来する前記実質的に相同なDNAドメインは、前記少なくとも2つの実質的に相同なDNAドメインの他の1つよりも少なくとも10%高い遺伝子変換頻度を有する、組換え宿主細胞。
【請求項2】
前記適合された実質的に相同なDNAドメインが、独特の配列タグによって前記実質的に相同なDNAドメインの他のバージョンと区別される、請求項1に記載の宿主細胞。
【請求項3】
前記適合された実質的に相同なDNAドメインが、標的化DNAドメインを含み、前記標的化DNAドメインが、増強された組込み選好を有する配列を含む、請求項1または2のいずれか一項に記載の宿主細胞。
【請求項4】
前記実質的に相同なDNAドメインのうちの少なくとも2つが各々、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する、請求項1〜3のいずれか一項に記載の宿主細胞。
【請求項5】
組込み経路をHRの方向に導くことによる、前記宿主細胞のゲノム中の所定の部位への標的化されたポリヌクレオチドの組込み効率の増大をさらに含む、請求項1〜4のいずれか一項に記載の宿主細胞。
【請求項6】
前記少なくとも2つの実質的に相同なDNAドメインが、glaAの遺伝子座であるかまたはamyBI、もしくはamyBII)の遺伝子座である、請求項1〜5のいずれか一項に記載の宿主細胞。
【請求項7】
前記目的のポリヌクレオチドがコドン最適化されたポリヌクレオチドである、請求項1〜6のいずれか一項に記載の宿主細胞。
【請求項8】
glaA、amyA、amyBI、amyBII、oahA、毒素関連ポリヌクレオチドおよびprtTの群から選択されるポリヌクレオチドをさらに含み、前記ポリヌクレオチドが改変を含み、ここで、前記宿主細胞は、同等の条件下で培養したとき、それが由来する親細胞と比較して、前記改変を含むポリヌクレオチドによってコードされる産物を欠損している、請求項1〜7のいずれか一項に記載の宿主細胞。
【請求項9】
pepAの欠損をさらに含む、請求項8に記載の宿主細胞。
【請求項10】
SEC61の改変をさらに含む、請求項8または9のいずれか一項に記載の宿主細胞。
【請求項11】
前記宿主細胞が選択マーカーを含まない宿主細胞である、請求項1〜10のいずれか一項に記載の宿主細胞。
【請求項12】
前記宿主細胞が真菌宿主細胞である、請求項1〜11のいずれか一項に記載の宿主細胞。
【請求項13】
前記宿主細胞が糸状菌宿主細胞である、請求項12に記載の宿主細胞。
【請求項14】
目的の化合物を産生するための組換え宿主細胞の産生方法であって、前記宿主細胞が目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、別の実質的に相同なDNAドメインと比較してより高い遺伝子変換頻度を有する実質的に相同なDNAドメインを、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合させ、前記より高い遺伝子変換頻度を有する実質的に相同なDNAドメインに標的化DNAドメインを提供することを含み、ここで、前記標的化DNAドメインは、増強された組込み選好を有する配列を含む、方法。
【請求項15】
a.請求項14で得られた前記細胞を目的のポリヌクレオチドで形質転換することと、
b.前記実質的に相同なDNAドメインのうちの少なくとも1つに組み込まれた少なくとも1コピーの前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングすることと、
c.(b)で得られた前記細胞を繁殖させ、実質的に相同なDNAドメインのさらなるコピーに組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有する細胞を選択またはスクリーニングすることと
をさらに含む、請求項14に記載の方法。
【請求項16】
前記実質的に相同なDNAドメインの各々のコピーが、組み込まれた前記目的のポリヌクレオチドの少なくとも1コピーを有するまで、工程(c)を繰り返す工程をさらに含む、請求項15に記載の方法。
【請求項17】
目的の化合物を産生する方法であって、
a.前記化合物の産生をもたらす条件下で請求項1〜13のいずれか一項に記載の宿主細胞を培養することと、
b.前記培養培地から前記目的の化合物を回収することと
を含む方法。
【請求項18】
目的の化合物を産生する方法であって、
a.組換え宿主細胞を前記化合物の産生をもたらす条件下で培養することであって、前記宿主細胞は目的のポリヌクレオチドの1コピー以上の組込みに好適な少なくとも2つの実質的に相同なDNAドメインを含み、ここで、前記少なくとも2つの実質的に相同なDNAドメインのうちの少なくとも1つは、それが由来する前記実質的に相同なDNAドメインと比較して前記目的のポリヌクレオチドに対する増強された組込み選好を有するように適合され、かつ前記実質的に相同なDNAドメインのうちの少なくとも2つは、組み込まれた目的のポリヌクレオチドの少なくとも1コピーを有する、と、
b.前記培養培地から前記目的の化合物を回収することと
を含む方法。
【請求項19】
前記宿主細胞が選択マーカーを含まない宿主細胞である、請求項17または18に記載の方法。
【請求項20】
前記宿主細胞が真菌宿主細胞である、請求項17〜19のいずれか一項に記載の方法。
【請求項21】
前記宿主細胞が糸状菌宿主細胞である、請求項20に記載の方法。
【請求項22】
前記目的の化合物がポリペプチドである、請求項17〜21のいずれか一項に記載の方法。
【請求項23】
前記ポリペプチドが酵素である、請求項22に記載の方法。
【請求項24】
前記目的の化合物が代謝産物である、請求項17〜21のいずれか一項に記載の方法。
【請求項25】
配列番号:2によるポリヌクレオチドを含む、単離されたポリヌクレオチド、またはその機能性断片。
【請求項26】
配列番号:3によるポリヌクレオチドを含む、単離されたポリヌクレオチド、またはその機能性断片。
【請求項27】
請求項25または26に記載のポリヌクレオチドを含むベクター。
【請求項28】
請求項25もしくは26に記載のポリヌクレオチドを含むか、または請求項27に記載のベクターを含む宿主細胞。
【図1】


【図2】
【図3】

【図7】

【図8】
【図9】

【図10】
【図11】
【図12】
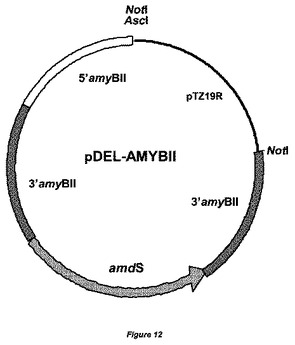
【図13】
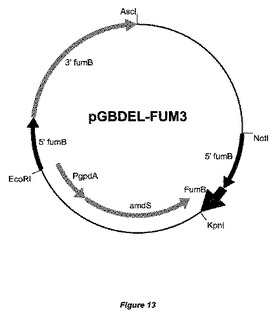
【図15】
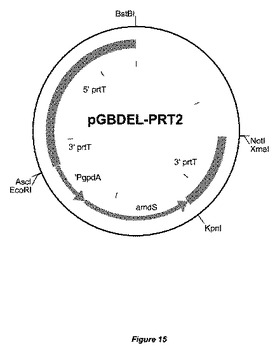
【図16】
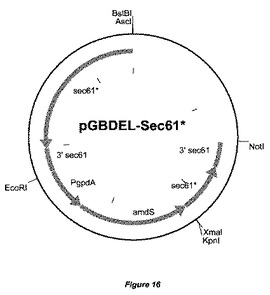
【図17】

【図18】

【図19】
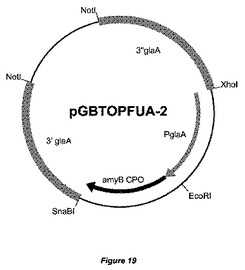
【図20】

【図21】
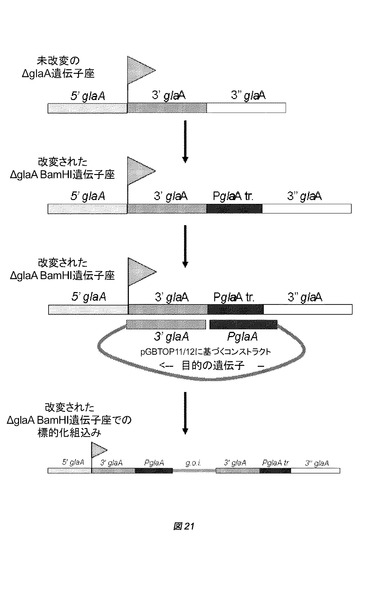
【図4】
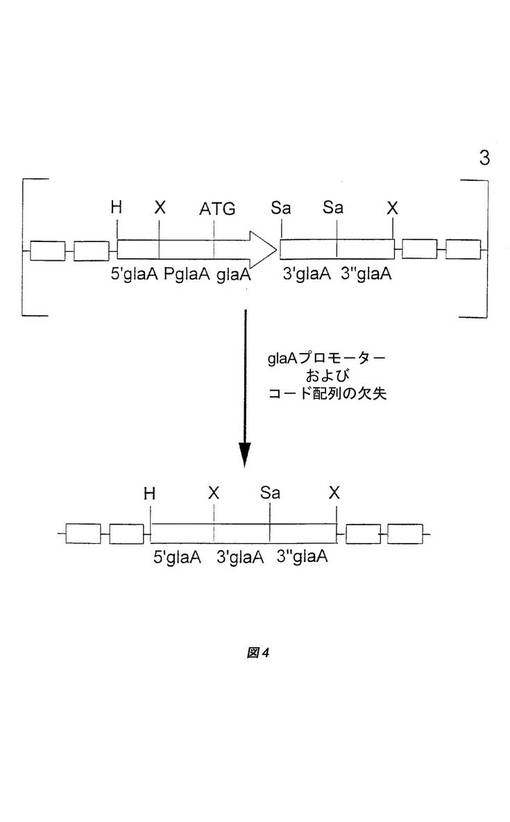
【図5】
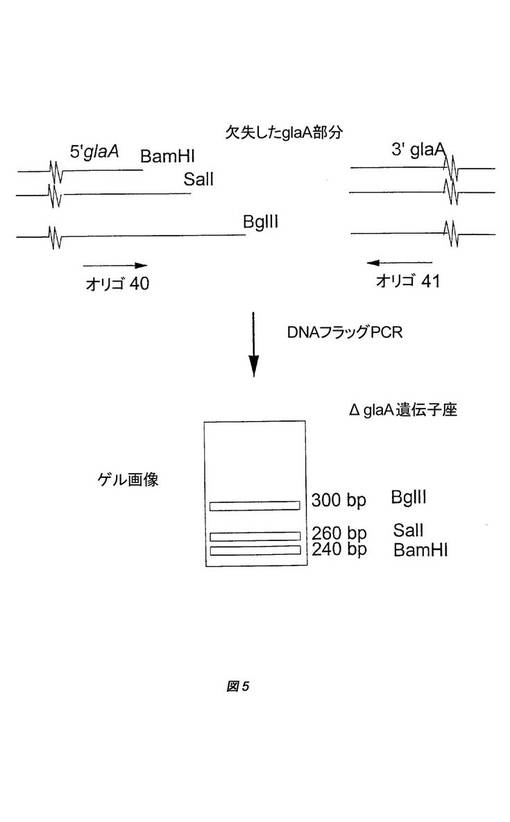
【図6】
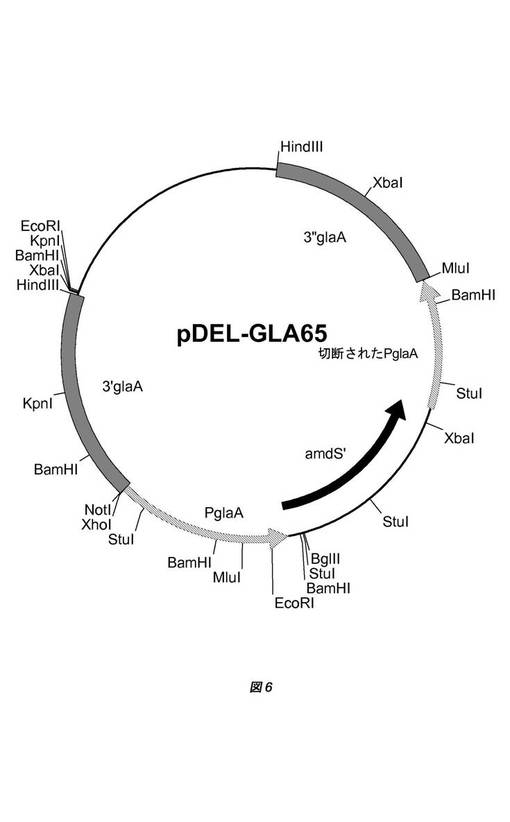
【図14】
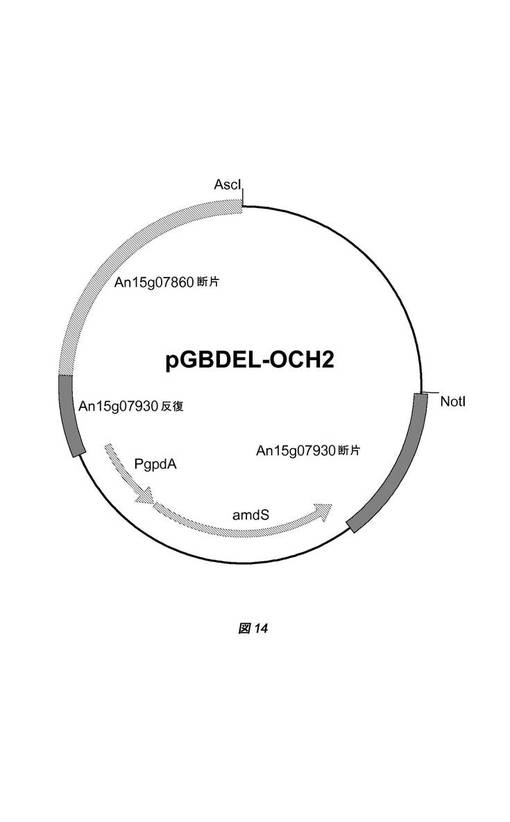
【図2】
【図3】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図4】
【図5】
【図6】
【図14】
【公表番号】特表2012−533312(P2012−533312A)
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2012−520984(P2012−520984)
【出願日】平成22年7月1日(2010.7.1)
【国際出願番号】PCT/EP2010/059390
【国際公開番号】WO2011/009700
【国際公開日】平成23年1月27日(2011.1.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願日】平成22年7月1日(2010.7.1)
【国際出願番号】PCT/EP2010/059390
【国際公開番号】WO2011/009700
【国際公開日】平成23年1月27日(2011.1.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]