
キメラウイルス様粒子
gagポリペプチドを含むウイルス様キメラ粒子が記載されている。ウイルス様粒子は、gagポリペプチドと、抗原に連結された脂質ラフト会合ポリペプチドとから生成される。上記抗原は、脂質ラフトと天然には会合しないものである。好ましい生成方法は、昆虫細胞内で発現させる工程を含んでいる。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルス様粒子の分野に関する。本明細書には、特に(a)gagポリペプチド、および(b)脂質ラフトと天然には会合しない抗原に連結された脂質ラフト会合ポリペプチドを有する、キメラウイルス様粒子が開示される。
【背景技術】
【0002】
ウイルス様粒子(VLP)は、従来のワクチン技術を凌駕する利点をいくつか有している。ワクチン開発のためのVLPの重要な利点は、3次元構造の観点と、エピトープの一次構造および立体構造の両方に対する中和抗体応答を誘導できるという観点から、VLPが天然のウイルスと似ていることである。それ故、VLPは他のワクチン製剤よりも免疫原性が強いことが分かっているといえる。ウイルスベクターを使ったアプローチとは異なり、VLPは、既存の免疫に関する問題を孕んでいないために、繰り返し使用することができる。
【0003】
伝統的なワクチンの多くは、非経口投与され、それらの大部分は下気道を防御する全身性IgG応答を引き起こす(または増幅する)。粘膜応答を誘導するウイルスを使った新しいアプローチは、該アプローチが上気道および下気道でのウイルスの増殖を制限することができるので、より望ましいものであり、この新しいアプローチは、それぞれを防御したり、感染を減少させたりするための、ワクチンを使った最も優れたアプローチであると思われる(15)。鼻腔内ワクチンは、上気道および下気道を防御することに加えて、針を使った接種の複雑な作業を避けたり、微粒子状の抗原および/または可溶性の抗原と鼻咽頭会合リンパ性組織(NALT)との相互作用による、粘膜ならびに全身性の体液性応答および細胞性応答を誘導する手段を提供したりする(16−19)。VLPは一般的に、ロタウイルス、ノロウイルス、およびパピローマウイルスのVLPについて示されたような、鼻腔内投与の後に、粘膜免疫および全身免疫を誘導するのに十分適していると思われる(28−31)。
【0004】
VLPのこうした利点にも拘らず、ワクチンとしてのエンベロープVLP(内在性膜タンパク質((integral membrane protein)を含むエンベロープウイルスに由来するVLP)の開発は、現在のところ数種の問題によって制限されている。最も重要な問題の1つは、抗原の範囲が制限されていることであり、このためエンベロープVLPワクチンを開発できる病気も制限されている。VLPへの抗原の組み込みには、ウイルスのキャプシドタンパク質が、ウイルス粒子の集合を開始する脂質ラフトドメインと会合することが必要になると考えられる。VLPを製造するための現在の方法では、脂質ラフトと天然に会合する天然ウイルス抗原を含むVLPを形成するための、ウイルスキャップシドタンパク質の使用が制限されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、(脂質ラフトと天然に会合しないウイルス抗原、キャプシドタンパク質の源以外のウイルス由来の抗原、他の病原体(細菌、真菌、原虫、蠕虫、酵母など)由来の抗原、腫瘍抗原、ならびにアレルゲンを含む)あらゆる種類の抗原を含有するVLPを製造することができるVLPの基盤技術が必要とされている。
【0006】
既存のVLP技術が有するもう1つ別の重要な問題は、十分な収量のVLPを製造することができないことである。例えば、インフルエンザマトリックスに由来するVLPは免疫原性を有しているが、該VLPは、その収量が低いために、今までは、インフルエンザワクチンの従来のものに取って代わるものとして良好な選択ではなかった。したがって、ワクチンを製造するための十分な量のVLPを生成することができるVLPワクチンの基盤技術も必要とされている。
【課題を解決するための手段】
【0007】
本発明は、キメラVLPを製造および使用するための、本明細書に開示されているような種々の方法および組成物を提供することによって、これらの必要性を満たすものである。上記キメラVLPは、あらゆる種類の病原体の脂質ラフトと天然に会合しない抗原を含んでおり、ワクチンを製造するために十分な量で生成され得るものである。
【0008】
1態様では、本発明は、gagポリペプチドと、抗原に連結された脂質ラフト会合ポリペプチドとを有しているキメラウイルス様粒子であって、上記抗原は脂質ラフトと天然には会合しないものであるキメラウイルス様粒子を提供する。結合は、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であってもよい。共有結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合であってもよい。gagポリペプチドは、レトロウイルスに由来するものであることが好ましい。レトロウイルスには、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、デルタレトロウイルス、およびレンチウイルスが含まれてもよい。
【0009】
脂質ラフト会合ポリペプチドは、脂質ラフトと直接的にまたは間接的に会合する任意のポリペプチドであってもよく、例えば、内在性膜タンパク質であってもよい。好ましい実施形態では、脂質ラフト会合ポリペプチドは、ヘマグルチニンのポリペプチド、ノイラミニダーゼのポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、またはエンベロープタンパク質のポリペプチドである。
【0010】
抗原は、免疫応答を誘発することができる任意の物質であってもよい。そのような抗原としては、タンパク質、ポリペプチド、糖ポリペプチド(glycopolypeptide)、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドのペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、あるいは炭水化物などが挙げられる。抗原は、ウイルス抗原、細菌抗原、真核生物の病原体の抗原、腫瘍抗原、またはアレルゲンであることが好ましい。
【0011】
また別の態様では、本明細書に記載のウイルス様粒子は、ウイルス様粒子と会合したアジュバントを含んでいる。アジュバントは、VLPの内側に配置されてもよい。なお、該アジュバントは、gagポリペプチドに共有連結されていることによって、VLPの内側に配置されていることが好ましい。他の実施形態では、アジュバントは、VLPの外側に配置されている。なお、該アジュバントは、脂質ラフト会合ポリペプチドに共有連結されていることによって、VLPの外側に配置されていることが好ましい。ポリペプチドアジュバントの好ましい例には、フラジェリンおよびそのアジュバント活性断片、サイトカイン、コロニー刺激因子(例えばGM−CSFおよびCSFなど)、インターフェロン、腫瘍壊死因子、インターロイキン−2,−7,−12、ならびに他の成長因子が含まれる。
【0012】
さらに別の態様では、本発明は、gagポリペプチドをコードする第1ヌクレオチド配列と、抗原に連結される脂質ラフト会合ポリペプチドをコードする第2ヌクレオチド配列とを有しており、上記抗原は脂質ラフトと天然には会合しないものであり、上記ポリペプチドは、宿主細胞内で発現すると、ウイルス様粒子を形成するものである、ウイルス様粒子の発現ベクターシステムを提供する。1実施形態では、第1ヌクレオチド配列および第2ヌクレオチド配列は、1つの発現ベクター内に存在しており、別々のプロモーターに作動可能に連結されていることが好ましいが、1つのプロモーターに作動可能に連結されていてもよい。別の実施形態では、第1ヌクレオチド配列および第2ヌクレオチド配列は、複数の発現ベクター内に存在している。
【0013】
さらに別の態様では、本発明は、(a)gagポリペプチドと抗原に連結される脂質ラフト会合ポリペプチドとを一緒に発現する、1つ以上の発現ベクターを提供する工程、(b)上記1つ以上の発現ベクターを細胞に導入する工程、および、(c)上記gagポリペプチドと上記抗原に連結される脂質ラフト会合ポリペプチドとを発現させて、ウイルス様粒子を製造する工程とによって、ウイルス様粒子を製造する方法を提供する。ここで、上記抗原は、脂質ラフトと天然には会合しないものである。好ましい実施形態では、1つ以上の発現ベクターはウイルスベクターである。該ウイルスベクターは、バキュロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、またはレトロウイルスであってもよい。上記細胞は、昆虫細胞または哺乳類細胞であってもよい。ある実施形態では、上記方法は、上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる。
【0014】
別の態様では、本発明は、免疫原性量の本明細書に記載のキメラインフルエンザウイルス様粒子の何れかの投与によって、免疫系の症状あるいは病気を治療または予防する方法を提供する。ある実施形態では、上記投与は、感染防御免疫応答を被検体に誘導するものである。ある実施形態では、上記投与は、皮下投与、経皮投与、皮内投与、真皮下投与、筋肉内投与、経口投与(peroral delivery、oral delivery)、鼻腔内投与、バッカル投与、舌下投与、腹腔内投与、膣内投与、肛門投与、または頭蓋内投与である。
【0015】
本明細書に開示されているキメラインフルエンザウイルス様粒子の別の態様は、免疫原性量または治療量の本明細書に開示のキメラウイルス様粒子の何れかを含んでいてもよい薬学的組成物である。そのような薬学的組成物は、薬学的に許容可能な担体を含んでいることが好ましい。該担体は、好ましい投与方法用に製剤化されていることが好ましい。
【0016】
別の態様では、本発明は、本明細書に開示されているようなVLPを含んでいる薬学的組成物を提供する。好ましい実施形態では、該薬学的組成物は、薬学的に許容可能な担体を含んでいる。
【0017】
別の態様では、本発明は、gagポリペプチドおよび非ウイルス性脂質ラフト会合ポリペプチドを含んでいる、キメラウイルス様粒子を提供する。そのようなVLPは、本明細書に開示された他のVLPの種々の実施形態の全てを含んでいる。ある実施形態では、脂質ラフト会合ポリペプチドは、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、シグナル伝達のポリペプチド、または膜輸送のポリペプチド、あるいは好ましくはカベオリン(cavelin)のポリペプチド、フロチリンのポリペプチド、シンタキシン−1のポリペプチド、シンタキシン−4のポリペプチド、シナプシンIのポリペプチド、アデューシンのポリペプチド、VAMP2のポリペプチド、VAMP/シナプトブレビンのポリペプチド、シナプトブレビンIIのポリペプチド、SNAREのポリペプチド、SNAP−25のポリペプチド、SNAP−23のポリペプチド、シナプトタグミンIのポリペプチド、またはシナプトタグミンIIのポリペプチドであってもよい。また、上記gagポリペプチドおよび非ウイルス性脂質ラフト会合ポリペプチドを含んでいるキメラウイルス様粒子は、本明細書のあらゆる箇所に開示されている上記態様および実施形態の全て(特に限定されないが、発現ベクターシステム、製造の方法、治療および予防の方法、ならびに薬学的組成物を含む)を含んでいる。
【図面の簡単な説明】
【0018】
【図1】Gag、HAまたはコントロールベクターのそれぞれで、およびHA−gag−NAのトリプルベクターに感染された、Sf9細胞からの培養液のウエスタンブロットを示す図であり、(A)は、抗Gag抗体によるウエスタンブロットを示す図であり、(B)は抗HA抗体によるウエスタンブロットを示す図である。
【図2】サッカロースステップ勾配を用いて、ペレット状のHA−gag−NA VLPを再遠心分離することによって得られた画分のウエスタンブロットを示す図であり、(A)は、抗Gag抗体によるウエスタンブロットを示す図であり、(B)は抗HA抗体によるウエスタンブロットを示す図である。
【図3】以下の実施例1に関するトリプル発現ベクターのコード配列の配置を示す図である。
【図4】PA修飾されたHA、ならびにGagおよびNAをコードするトリプルバキュロウイルス発現ベクターのコード要素の配置を示す図である。
【図5】RSV Fタンパク質およびGagをコードするダブルバキュロウイルス発現ベクター内のコード要素の配置を示す図である。
【発明を実施するための形態】
【0019】
本発明は、キメラウイルス様粒子を形成するための基礎となるgagポリペプチドを含んでいる。VLPを生成する好ましい方法は、昆虫細胞内での発現によるものである。該昆虫細胞内での発現は、ポリペプチド抗原の共発現を含んでいることが好ましい。これは、バキュロウイルス発現システムの様々なレトロウイルスによって、gagVLPの収量を大幅に上げることができるからである(23、24、46、49、52−58)。gagポリペプチドは、天然のレトロウイルスの組立過程の際には、本質的にC末端延長領域を含んでいる。つまり、機能的なgagタンパク質は、レトロウイルスのプロテアーゼ、逆転写酵素、およびリボソームのフレームシフトによるインテグラーゼ活性を含んでいる巨大なC末端延長領域を天然に有している。人工の延長領域を有する機能的なgagタンパク質の製造は、RSVgag(59)およびMLVgag(60)に対して達成されている。gagC末端の操作におけるこの適応性は、他の抗原および免疫賦活タンパク質の配列などの他のポリペプチドを含有させるための重要な部位を提供する。
【0020】
あるウイルス由来のコア粒子と、それとは異なるウイルス由来の表面抗原を含んでいるキメラVLPの製造は、シュードタイピングと呼ばれる。gagポリペプチドは、インフルエンザHAおよびNAでもって効率よくシュードタイプされている。これは、おそらく、これらタンパク質が脂質ラフトドメイン(61、62)の内部に濃縮する一方、ミリストイル化されたgagタンパク質は、出芽過程の間に、脂質ラフトドメインの内表面に濃縮する(63)からである。
【0021】
本明細書に記載の本発明は、キメラVLPを形成するための基礎としての、脂質ラフトと天然には会合しない抗原に、連結された脂質ラフト会合ポリペプチドをさらに含んでいる。どのような理論にも束縛されることなく、該抗原が上記脂質ラフト会合ポリペプチドへ会合するという長所によって、該抗原を含んでいるキメラVLPが形成されると考えられる。
【0022】
本明細書に開示の方法およびプロトコールは、他に示されない限り、当業者が実施できる範囲内の従来の化学的手法、分子生物学的手法、微生物学的手法、組み換えDNAの手法、および免疫学的手法を使用するものである。そのような手法は、文献において説明されており、例えば、「J. Sambrook, E. F. Fritsch, and T. Maniatis, 1989, Molecular Cloning: A Laboratory Manual, Second Edition, Books 1-3, Cold Spring Harbor Laboratory Press」、「Ausubel, F. M.ら (1995 and periodic supplements; Current Protocols in Molecular Biology, ch. 9, 13, and 16, John Wiley & Sons, New York, N.Y.)」、「B. Roe, J. Crabtree, and A. Kahn, 1996, DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons」、「J. M. Polak and James O'D. McGee, 1990, In Situ Hybridization: Principles and Practice; Oxford University Press」、「M. J. Gait (Editor), 1984, Oligonucleotide Synthesis: A Practical Approach, Irl Press」、および「D. M. J. Lilley and J. E. Dahlberg, 1992, Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology, Academic Press」を参照のこと。これら一般的なテキストのそれぞれは、参照することによって本明細書に組み込まれる。
【0023】
〔定義〕
本明細書で使用されるような「Gagポリペプチド」は、本明細書に記載のウイルス様粒子の形成に関与するレトロウイルス由来の構造ポリペプチドである。ある実施形態では、RNAをパッケージングするための傾向(propensity)、または粒子の形成および出芽の効率などのある種の特徴に影響を及ぼすために、gagポリペプチドは、意図的に突然変異させられる。レトロウイルスのゲノムは、gag、pol、およびenvという3つの主要遺伝子産物をコードしている。gag遺伝子は、構造タンパク質をコードするものであり、pol遺伝子は、逆転者酵素、関連するタンパク質分解性のポリペプチド、ヌクレアーゼ、およびインテグラーゼに関連する機能をコードするものであり、envは、コードされた糖タンパク質の膜タンパク質が感染細胞の表面上で検出され、成熟し放出されたウイルス粒子の表面上で検出されるものである。全てのレトロウイルスのgag遺伝子の全体構造は類似しており、レトロウイルスの各グループ内では、gag遺伝子はアミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除く、コアタンパク質を生じさせるものである。
【0024】
MLVに関しては、gagの前躯体タンパク質は、Pr65Gagであり、前躯体での順番がNH2−p15−pp12−p30−p10−COOHとなっている4つのタンパク質に開裂される。これらの開裂は、ウイルスプロテアーゼに仲介され、ウイルスに依存するが、ウイルスの放出の前後に起こり得る。MLVのGagタンパク質は、グリコシル化された形態およびグリコシル化されていない形態で存在する。グリコシル化された形態は、gPr80Gagから開裂される。なおgPr80Gagは、グリコシル化されていないPr65GagのAUGコドンから上流に位置する、異なったインフレームの開始コドンから合成されるものである。グリコシル化されるGagを合成しないMLVの欠失変異体でも感染性を有し、グリコシル化されないGagでもウイルス様粒子を形成することができる。このことから、グリコシル化が起こることの重要性について疑問が生じる。HIV−1のGag前躯体であるpr55Gagの、ウイルスにコードされたプロテアーゼによる翻訳後開裂から、N−ミリストイル化され、内部にリン酸化されたp17マトリックスタンパク質(P17MA)、リン酸化されたp24キャップシドタンパク質(p24CA)、およびヌクレオキャップシドタンパク質P15(p15NC)が生じる。p15NCは、p9およびp6にさらに開裂される。
【0025】
構造的に原型のGagポリタンパク質は、マトリックスタンパク質(MA)、キャプシドタンパク質(CA)、およびヌクレオキャプシドタンパク質(NC)の3つの主要タンパク質に分けられる。これら主要タンパク質は、レトロウイルスのgag遺伝子に常に同じ順番で存在している。なお、マトリックスタンパク質(MA)は、インフルエンザのマトリックスタンパク質M1と混同されるものではない。マトリックスという名称を共有しているが、M1はMAと異なるタンパク質である。Gagポリタンパク質の成熟タンパク質へのプロセッシングは、レトロウイルスにコードされたプロテアーゼによって触媒され、新たに出芽するウイルス粒子として成熟させる。機能上、Gagポリタンパク質は、膜結合ドメイン、相互作用ドメイン、レイトドメインの3つのドメインに分けられる。膜結合ドメインは、Gagポリタンパク質に細胞膜を狙わせるものであり、相互作用ドメインは、Gagの重合化を促進するものであり、レイトドメインは、宿主細胞から発生するウイルス粒子の放出を促進するものである。組立を仲介するGagタンパク質の形態は、ポリタンパク質である。このため組立ドメインが、より後に形成する開裂産物の何れかの内にきちんと位置する必要は無い。このように、本明細書に包含されるようなGagポリペプチドは、VLPを形成および放出させるために重要な機能要素を含んでいる。これらの重要な機能要素に関して、最新技術はかなり進んでいる。例えば「Hansenら J. Virol 64, 5306-5316, 1990」、「Willら, AIDS 5, 639-654, 1991」、「Wangら J. Virol. 72, 7950-7959, 1998」、「McDonnellら, J. Mol. Biol. 279, 921-928, 1998」、「Schultz and Rein, J. Virol. 63, 2370-2372, 1989」、「Accolaら, J. Virol. 72, 2072-2078, 1998」、「Borsettiら, J. Virol., 72, 9313-9317, 1998」、「Bowzardら, J. Virol. 72, 9034-9044, 1998」、「Krishnaら, J. Virol. 72, 564-577, 1998」、「Willsら, J. Virol. 68, 6605-6618, 1994」、「Xiangら, J. Virol. 70, 5695-5700, 1996」、「Garnierら, J. Virol. 73, 2309-2320, 1999」を参照のこと。
【0026】
本発明のVLPにおいて使用されるようなgagポリペプチドは、最低でもVLPを形成するための機能要素を含んでいる。gagポリペプチドは、場合によっては1つ以上の別のポリペプチドを含んでいてもよい。該1つ以上の別のポリペプチドは、gagポリペプチドをコードする配列へ、該1つ以上の別のポリペプチドをコードする配列をスプラスすることによって生成されるものであってもよい。別のポリペプチドをgagポリペプチドに挿入するための部位は、C末端であることが好ましい。
【0027】
Gagポリペプチドのための好ましいレトロウイルスの源は、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス(avian leucosis virus)、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス,ヤーグジークテヒツジレトロウイルスおよびマソン・ファイザー・サルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトT細胞白血病ウイルスおよびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、あるいはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ伝染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)を包含する。
【0028】
本明細書で使用されているような「脂質ラフト」は、gagポリペプチドがウイルス粒子の組立過程の間に濃縮する細胞膜の微小ドメインのことをいう。
【0029】
本明細書で使用されているような「脂質ラフト会合ポリペプチド」は、脂質ラフトと直接的にまたは間接的に会合される任意のポリペプチドのことをいう。本発明において使用される具体的な脂質ラフト会合ポリペプチドは、キメラウイルス様粒子の望まれる使用に依存する。
【0030】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質、膜との会合を引き起こすタンパク質の修飾を介して脂質ラフトと直接的に会合されるタンパク質、または脂質ラフト会合ポリペプチドを介して脂質ラフトと間接的に会合されるポリペプチドであってもよい。
【0031】
脂質アンカーを有する多くのタンパク質は、脂質ラフトと会合する。ポリペプチドを脂質ラフトに連結させる脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、およびダブルアセチル化が含まれる。
【0032】
多種多様なポリペプチドが脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスの組立、および組立られた粒子の出芽を含む無数の生物学的活動の基盤として機能する。それ故、これらの過程に関連する様々なポリペプチドが脂質ラフトと会合する。
【0033】
シグナル伝達カスケードに関与する様々な種類のポリペプチドは、シグナル伝達の基盤として機能する脂質ラフトと会合する。シグナル伝達の基盤として機能する脂質ラフトの一つの形式は、カベオラと呼ばれる。カベオラは、形質膜のフラスコ型の陥入であり、カベオリンファミリー(例えばカベオリンおよび/またはフロチリン)に由来するポリペプチドを含んでいる。
【0034】
膜輸送ポリペプチドは、膜輸送の基盤として機能する脂質ラフトと会合する。膜輸送ポリペプチドの例には、エンドサイトーシスおよびエキソサイトーシスに関与するタンパク質、例えば、シンタキシン−1、シンタキシン−4、シナプシンI、アデューシン、VAMP2、VAMP/シナプトブレビン、シナプトブレビンII、SNAREタンパク質、SNAP−25、SNAP−23、シナプトタグミンI、およびシナプトタグミンIIなどが含まれる。
【0035】
ウイルス受容体、受容体−補助受容体の複合体、侵入過程を調節するのに役立つ他の任意の成分は、ウイルスの侵入のために特殊化した膜輸送の基盤として機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体には、例えば、分解促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー型膜糖タンパク質、グループAのロタウイルスの受容体、複数の成分(ガングリオシド、HSC70タンパク質、アルファ2−ベータ1インテグリン、およびアルファ5−ベータ2インテグリンを含む)をガ乳する複合体;数種のエンベロープウイルス(HIV、MLV、麻疹ウイルス、エボラウイルスなど)の糖タンパク質、およびHIVの侵入に関与するポリペプチド(CD5、CCR5およびnefなど)が含まれる。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. & Mol. Bio. Rev. 67(2):226-237」を参照のこと。
【0036】
ウイルス粒子の組立に関与するポリペプチドは、ウイルスの組立の基盤として機能する脂質ラフトと会合される。そのようなポリペプチドには、例えば、HAおよびNAというインフルエンザのエンベロープ糖タンパク質、麻疹ウイルスのHおよび成熟F1−F2融合タンパク質、ならびにHIVのgp160、gp41、およびPr55gagが含まれる。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. And Mol. Bio. Rev. 67(2):226-237」を参照のこと。
【0037】
組立られたウイルスの出芽に関与するポリペプチドは、ウイルスの出芽の基盤として機能する脂質ラフトと会合される。宿主細胞からのHIV−1の出芽が膜のラフトにおいて起こっていることを示唆するデータがある。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. And Mol. Bio. Rev. 67(2):226-237」を参照のこと。ウイルスの出芽に関与するポリペプチドに関する一般的な情報は、「Fields Virology (4th ed.) 2001」に見出される。
【0038】
好ましい脂質ラフト会合ポリペプチドには、ヘマグルチニンポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、およびエンベロープタンパク質のポリペプチドなどのウイルスポリペプチドが含まれる。これらポリペプチドのそれぞれは、あらゆる種類のウイルスに由来するものであってもよい。しかしながら、ある実施形態では、HIV−1ウイルスに由来するエンベロープタンパク質、呼吸器合胞体ウイルスまたは麻疹ウイルスに由来する融合タンパク質、呼吸器合胞体ウイルス、単純疱疹ウイルスまたはエボラウイルスに由来する糖タンパク質、および麻疹ウイルスに由来するヘマグルチニンタンパク質が含まれる。
【0039】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、以下の原虫、蠕虫、および他の真核微生物病原体から得られてもよい:特に限定されないが、マラリア原虫(Plasmodium)(熱帯熱マラリア原虫(Plasmodium falciparum)、四日熱マラリア原虫(Plasmodium malariae)、卵形マラリア原虫(Plasmodium ovale)、および三日熱マラリア原虫(Plasmodium vivax)など);トキソプラズマ・ゴンディ(Toxoplasma gondii);トリパノソーマ・ブルーセイ(Trypanosoma brucei)、トリパノソーマ・クルージ(Trypanosoma cruzi);ビルハルツ住血吸虫(Schistosoma haematobium)、マンソン住血吸虫(Schistosoma mansoni)、日本住血吸虫(Schistosoma japonicum);ドノバンリーシュマニア(Leishmania donovani);ランブル鞭毛虫(Giardia intestinalis);およびクリプトスポリジウム・パルバム(Cryptosporidium parvum)など。そのような非ウイルス性脂質ラフト会合ポリペプチドは、該脂質ラフト会合ポリペプチドそれ自体が抗原として作用するときは、脂質ラフトと天然に会合しない抗原と連結されることなく使用されてもよい。
【0040】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、ヘマグルチニンポリペプチドである。本明細書で使用されているような「ヘマグルチニンポリペプチド」は、インフルエンザウイルスのタンパク質に由来するものであり、感染される細胞へのウイルスの結合を仲介するものである。またヘマグルチニンポリペプチドは、類似する麻疹ウイルスのタンパク質に由来するものであってもよい。該タンパク質は、1つの膜貫通ドメインを介して、インフルエンザウイルスの表面に固定されることが発見された抗原性糖タンパク質である。インフルエンザのヘマグルチニンには、少なくとも16のサブタイプが同定されており、H1からH16と名づけられている。H1、H2およびH3は、ヒトのインフルエンザウイルスにおいて発見されているものである。H5またはH7のヘマグルチニンを有する病原性が高いトリインフルエンザウイルスは、割合が低いもののヒトに感染することが発見されている。ヒトの患者において、アミノ酸が1つ変更しているトリインフルエンザウイルス株のH5型のヘマグルチニンが発見されており、アミノ酸のこの変更によって、H5のヘマグルチニンがH5N1のトリインフルエンザウイルスの受容体特異性を顕著に変更することができるように、受容体特異性が変化し、該ウイルスがヒトの受容体に結合できるようになったと報告されている(109および110)。この発見は、普通はヒトに感染しないH5N1型ウイルスがどのようにして突然変異し、効率よくヒト細胞に感染できるようになったかを説明するものである。
【0041】
ヘマグルチニンは、ホモトリマーの内在性膜ポリペプチドである。膜貫通ドメインは、ラフト脂質ドメインと天然に会合しているため、ヘマグルチニンがVLPに組み込まれるためのgagポリペプチドと会合することができる。ヘマグルチニンはシリンダーのような形状をしており、約135Åの長さである。HAを構成する3つの同じモノマーは、中心のコイルドコイルと、VLPの表面に露出されるシアル酸結合部位を含む球状の頭部とを形成する。HAのモノマーは、1つのポリペプチド前躯体として合成され、該前躯体は、グリコシル化されて、そして2つのより小さいポリペプチド(HA1サブユニットおよびHA2サブユニット)に開裂される。HA2サブユニットは、膜に固定されるトリマーのコイルドコイルを形成し、HA1サブユニットは球状の頭部を形成する。
【0042】
本発明のVLPに使用されるようなヘマグルチニンポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。ヘマグルチニンポリペプチドは、任意のインフルエンザウイルスの型、亜型、株または亜株に由来するものであってもよく、好ましくはH1、H2、H3、H5、H7およびH9のヘマグルチニンに由来するものであってもよい。さらに、ヘマグルチニンポリペプチドは、様々なインフルエンザのへマグルチニンのキメラであってもよい。ヘマグルチニンポリペプチドは、脂質ラフトと天然に会合しない1つ以上の別の抗原を含んでいることが好ましい。該抗原は、ヘマグルチニンポリペプチドをコードする配列へ、上記1つ以上の別のポリペプチドをコードする配列をスプライスすることによって生成されるものであってもよい。ヘマグルチニンポリペプチドに別のポリペプチドを挿入する部位は、N末端であることが好ましい。
【0043】
ウイルス性脂質ラフト会合ポリペプチドの好ましいもう1つ別の例は、ノイラミニダーゼポリペプチドである。本明細書で使用されているような「ノイラミニダーゼポリペプチド」は、インフルエンザウイルスのタンパク質に由来し、糖タンパク質から末端のシアル酸残基を開裂することによって、インフルエンザウイルスの細胞からの放出を仲介するタンパク質である。ノイラミニダーゼ糖タンパク質は、ウイルスの表面に発現される。ノイラミニダーゼタンパク質は、四量体であり、ベータ−ピンホイール構造を有する球状頭部、細い茎領域、および小さい疎水性領域からなる共通の構造を有している。この疎水性領域は、1つの膜貫通ドメインにより、ウイルス膜にノイラミニダーゼタンパク質を固定させるものである。シアル酸残基の開裂のための活性部位は、全てのインフルエンザAウイルスに保存されている15個の荷電アミノ酸によって形成される各サブユニットの表面上のポケットを含んでいる。インフルエンザのノイラミニダーゼには、少なくとも9のサブタイプが同定はされており、N1からN9と名づけられている。
【0044】
本発明のVLPに使用されるようなノイラミニダーゼポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。機能領域についての最先端の技術水準は、極めて高い。例えば、「Vargheseら, Nature 303, 35-40, 1983」、「Colmanら, Nature 303, 41-44, 1983」、「Lentzら, Biochem, 26, 5321-5385, 1987」、「Websterら, Virol. 135, 30-42, 1984」を参照のこと。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルスの型、亜型、株または亜株に由来するものであってもよく、好ましくはN1およびN2のノイラミダーゼに由来するものであってもよい。さらに、ノイラミニダーゼポリペプチドは、様々なインフルエンザのノイラミニダーゼのキメラであってもよい。ノイラミニダーゼポリペプチドは、脂質ラフトと天然に会合しない1つ以上の別の抗原を1つ以上含んでいることが好ましい。該抗原は、ヘマグルチニンポリペプチドをコードする配列へ、上記1つ以上の別のポリペプチドをコードする配列をスプライスすることによって生成されるものであってもよい。ノイラミニダーゼポリペプチドをコードする配列に別のポリペプチドを挿入する部位は、C末端であることが好ましい。
【0045】
脂質ラフト会合ペプチドの好ましいもう1つ別の例は、ファシクリンI(FasI)と呼ばれる昆虫由来の接着タンパク質である。本明細書で使用されているような「ファシクリンIポリペプチド」は、胚発生に関与する昆虫タンパク質に由来するものである。この非ウイルス性タンパク質が、昆虫細胞バキュロウイルス発現系にて発現されると、FasIの脂質ラフト会合が引き起こされ得る(J. Virol. 77, 6265-6273, 2003))。それ故、異種抗原がファシクリンIポリペプチドに接着されることによって、gagと共発現されたとき、キメラ分子がVLPへ組み込まれるということになる。本発明のVLPに使用されるようなファシクリンIポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。
【0046】
脂質ラフト会合ペプチドのもう1つ別の好ましい例は、G糖タンパク質と名づけられたRSVからのウイルス由来の接着タンパク質である。本明細書で使用されているような「G糖タンパク質」は、RSVのG糖タンパク質に由来するものである。最近のデータによれば、脂質ラフトドメインは、該ドメインがインフルエンザの出芽に重要であるように、RSV粒子の出芽にも重要であることが実証されている(Virol 327, 175-185, 2004; Arch. Virol. 149, 199-210, 2004; Virol. 300, 244-254, 2002)。RSVのG糖タンパク質は、32.5kdの内在性膜タンパク質であり、ウイルスの接着タンパク質として機能するだけでなく、RSV感染のための感染防御抗原としても機能する。インフルエンザウイルスのヘマグルチニンと同じように、その抗原性は、それに接着される非脂質ラフト抗原の抗原性を増強し得る。RSVは、ノイラミニダーゼ活性を有するタンパク質を天然に発現しないために、gagおよびRSVのG糖タンパク質から構成されるVLPは、産生および放出を効率よくするためのNAが存在することを必要としない。それ故、MLVのgagおよびG糖ポリペプチドをコードする発現ベクターを開発することにより、膜に組み込まれるG糖ポリペプチドを含むVLPが産生されることになる。非脂質ラフト外来抗原接着をするように、該G糖ポリペプチドを任意に修飾することにより、該外来抗原に対する顕著な免疫応答を誘導することができるキメラVLPを得ることができる。
【0047】
用語「キメラウイルス様粒子」と「VLP」とは、VLPがその文脈から、本明細書に開示されているようなgagポリペプチドを用いて形成されないウイルス様粒子のことを言っている場合を除いて、本明細書を通して交換可能に使用される。
【0048】
〔抗原〕
本発明は、gagポリペプチドおよび脂質ラフト会合ポリペプチドを、脂質ラフトと天然に会合しない抗原を含んでいるVLPを形成するための、容易に改変できる基盤として、提供する。この項には、開示されたVLPと一緒に使用されるのが好ましい抗原が記載されている。
【0049】
<抗原と脂質ラフト会合ポリペプチドとの結合>
脂質ラフトと天然に会合しない抗原を含んでいるVLPを形成する手段として、抗原と脂質ラフト会合ポリペプチドとの間に結合が形成される。該脂質ラフト会合ポリペプチドは、1つの抗原に連結されてもよいし、複数の抗原に連結されてもよい。これにより、VLPの免疫原性が増加されたり、様々な病原体に対する免疫原性が付与されたり、または特定の病原体の様々な株に対する免疫原性が付与されたりする。
【0050】
抗原と脂質ラフト会合ポリペプチドとの結合は、抗原がVLPへ組み込まれるのに十分などのような種類の結合であってもよい。該結合は、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であってもよい。好ましい実施形態では、結合は、共有結合(ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合など)である。
【0051】
抗原は、組み換えによって、脂質ラフト会合ポリペプチドに既に結合した状態で製造されてもよいし、あるいは分離した物質として製造され、その後脂質ラフト会合ポリペプチドに連結されてもよい。
【0052】
<抗原の種類>
本明細書で使用されているような抗原は、免疫応答を誘発することができ、脂質ラフトと天然には会合しない任意の物質であってもよい。抗原には、特に限定されないが、タンパク質、ポリペプチド(活性タンパク質、およびタンパク質内の各ポリペプチドエピトープ)、糖ポリペプチド、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドおよび他の分子のペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、ならびに炭水化物が含まれる。抗原が脂質ラフトと直接的にまたは間接的に天然に会合しない場合、脂質ラフト会合ポリペプチドと結合することなく、VLPに組み込まれることは考えられない。抗原は、病気または疾患に関与する任意の抗原であってもよく、例えば微生物抗原(例えばウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、およびアレルゲンなどである。
【0053】
<抗原の源>
本明細書に記載の抗原は、化学的にまたは酵素を用いて合成されてもよいし、組み換えによって産生されてもよいし、天然源から単離されてもよいし、あるいはそれらの組み合わせによって得られてもよい。抗原は、精製されてもよいし、部分的に精製されてもよいし、粗抽出物であってもよい。
【0054】
ポリペプチド抗原は、技術的に知られているタンパク質精製の標準的な方法を使用して、天然源から単離されてもよい。そのような方法には、特に限定されないが、液体クロマトグラフィー(例えば高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ排除クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含む)、親和性クロマトグラフィー、または他の精製手法が包含される。多くの実施形態では、抗原は、例えば、約50%から約75%までの純度、約75%から約85%までの純度、約85%から約90%までの純度、約90%から約95%までの純度、約95%から約98%までの純度、約98%から約99%までの純度、あるいは99%よりも高い純度の精製抗原である。
【0055】
固相ペプチド合成手法を使用してもよい。そのような手法は、当業者に公知である。「Jones, The Chemical Synthesis of Peptides (Clarendon Press, Oxford) (1994)」参照。一般的に、そのような方法では、ペプチドは、活性化されたモノマーユニットが、固相に結合されている成長ペプチド鎖に連続的に付加されることによって産生される。
【0056】
十分に確立されている組み換えDNA手法を、脂質ラフト会合ポリペプチドと同じベクターにおいて、ポリペプチドを製造するために使用してもよい。例えば、ポリペプチドをコードするヌクレオチド配列を含んでいる発現コンストラクトが、適切な宿主細胞(例えば、インビトロの細胞培養において単細胞として成長する真核宿主細胞、例えば酵母細胞、昆虫細胞、哺乳類細胞など)、あるいは原核細胞(インビトロの細胞培養において成長するもの)に導入されて、遺伝的に修飾された宿主細胞が生成される場合、適切な培養条件下において、タンパク質は該遺伝的に修飾された宿主細胞によって製造される。
【0057】
<ウイルス抗原>
適したウイルス抗原には、以下のグループの1つ以上のウイルスと関連するもの(例えば以下のグループの1つ以上のウイルスによって合成されたもの)を包含する:レトロウイルス科(例えばHIV−1などのヒト免疫不全ウイルス(HTLV−III、LAVまたはHTLV−III/LAVまたはHIV−IIIとも呼ばれる);およびHIV−LPなどの他の単離種;ピコルナウイルス科(例えばポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス);カリシウイルス科(Calciviridae)(例えば、ノーウォークウイルスおよび関連ウイルスを含む、胃腸炎を引き起こす株);トガウイルス科(例えばウマ脳脊髄炎ウイルス、三日はしかウイルス);フラビウイルス科(Flaviridae)(例えばデング熱ウイルス、脳炎ウイルス、黄熱ウイルス);コロナウイルス科(例えばコロナウイルス);ラブドウイルス科(例えば水疱性口炎ウイルス、狂犬病ウイルス);フィロウイルス科(例えばエボラウイルス);パラミクソウイルス科(例えばパラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、呼吸器合胞体ウイルス、メタニューモウイルス科(例えばトリニューモウイルス、ヒトメタニューモウイルス);オルソミクソウイルス科(例えばインフルエンザウイルス);ブニヤウイルス科(Bungaviridae)(例えばハンタンウイルス、ブニヤウイルス(bungavirus)、フレボウイルス、およびナイロウイルス);アレナウイルス科(出血熱ウイルス);レオウイルス科(例えばレオウイルス、オルビウイルス、およびロタウイルス);ビルナウイルス科(Bimaviridae);ヘパドナウイルス科(B型肝炎ウイルス);パルボウイルス科(パルボウイルス);パポバウイルス科(乳頭腫ウイルス、ポリオームウイルス);アデノウイルス科(大部分のアデノビールス);ヘルペスウイルス科(単純疱疹ウイルス(HSV)1型(HHV−1)および2型(HHV−2)、水痘帯状疱疹ウイルス(HHV−3)、EBウイルス(エプスタインバーウイルス)(HHV−4)、サイトメガロウイルス(CMV)(HHV−5));ポックスウイルス(Poxyiridae)(天然痘ウイルス、ワクシニア・ウイルス、発疹ウイルス);およびイリドウイルス科(例えばアフリカ豚コレラウイルス);ならびに、分類されていないウイルス(例えば海綿状脳症の病原因子、デルタ肝炎の病原因子(B型肝炎ウイルスの欠陥サテライトウイルスであると考えられる)、非A非B型肝炎ウイルスの病原因子、(クラス1=体内伝染されるもの;クラス2=非経口的に伝染されるもの(すなわち肝炎C);ならびに、アストロウイルス。
【0058】
<非ウイルス性抗原>
本明細書に開示のVLPは、ノロウイルスファミリーに由来する様々な抗原を含んでいることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、カリシウイルス科のウイルスファミリーに含まれる4つの属のうちの1つを代表するものである。ノロウイルス属には、GenogroupIおよびGenogroupIIと命名された2つの主要な遺伝群が存在する。GenogroupIのノロウイルス株は、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスを包含している。GenogroupIIのノロウイルス株は、ヒューストンウイルス、ハワイウイルス、ローズデイルウイルス(Lordsdale virus)、グリムズビーウイルス(Grimsby virus)、メキシコウイルスおよびスノーマウンテンエージェント(Snow Mountain agent)を包含している(「Parker, T.D., ら J Virol. (2005) 79(12):7402-9」、「Hale, A.D., ら J Clin. Micro. (2000) 38(4):1656-1660」))。ノーウォークウイルス(NV)は、急性ウイルス性胃腸炎の世界規模の流行的発生の大部分に関与する、ヒトカリシウイルスの1つのグループの原型株である。ノーウォークウイルスのキャプシドタンパク質は、シェルドメイン(S)および突出ドメイン(P)という2つのドメインを有している。Pドメイン(ノーウォーク株の番号付けで、aa226−530)は、P1およびP2という2つのサブドメインに分けられる。P2ドメインは、P1ドメイン内の127aaの挿入部分(aa279−405)であり、折りたたまれたモノマーの最も末端の表面に位置する。P2ドメインは、ノロウイルス株の中で、VP1の最も保存されていない領域であり、P2内の超可変領域は、受容体結合と免疫反応性とに重要な役割を果たしていると考えられる。Pドメインが外側に位置していることを考慮すると、P2ドメインは、本明細書に開示のVLPワクチン用の抗原として使用するために、好ましい抗原またはポリペプチドエピトープの源であるといえる。P2ドメインは、GenogroupIまたはGenogroupIIのノロウイルス株に対する好ましい抗原である。各種ノロウイルス株を超えて保存されたP2ドメインの領域に位置していると、同定されたmAb 61.21エピトープ、およびmAb54.6エピトープが、さらに好ましい(「Lochridge, V.P., ら J Gen. Virol. (2005) 86:2799-2806)」)。
【0059】
<インフルエンザ抗原>
本明細書に開示のVLPは、ヘマグルチニンおよびノイラミニダーゼ以外の、またはそれらに加えて、インフルエンザに由来する様々な抗原を含んでいてもよい。追加のインフルエンザ抗原としては、M2ポリペプチドが好ましい。インフルエンザウイルスのM2ポリペプチドは、スプライシングが起こった後のRNAの断片7(マトリックス断片)にコードされているクラスIIIの小さい内在性膜タンパク質(97アミノ酸)である(80、81)。M2はウイルス粒子上にはほとんど存在しないが、感染細胞上にはより豊富に発見され得る。M2は、ウイルスの侵入に必要なプロトン選択的なイオンチャネルとして機能する(82、83)。感染または従来のワクチン接種の間、M2の免疫原性は最小であるが(このことは、その保存性を説明している)、別の形態で存在するときには、M2の免疫原性および防御性は、より強くなる(84−86)。このことは、M2のモノクローナル抗体をインビボにおいて受動的に移動させると、ウイルスの排除が促進され、その結果、防御がもたらされるという観察と一致する(87)。M2の外部ドメインエピトープがHBVのコア粒子に融合タンパク質として連結されるとき、該エピトープはマウスでは非経口で接種されたり、鼻腔内から接種されたりすることによって、防御をもたらし、さらに3つの直列に配置されたコピーが、コアタンパク質のN末端に融合されたとき、免疫原性が最も強くなる(88−90)。このことは、エピトープの密度が増加すると、免疫原性が増加するということを示す他の担体−ハプテンのデータと一致する(91)。
【0060】
M2ワクチンの鼻腔内投与に関して、良好な防御を得るためにアジュバントが必要であり、良好な結果は、LTR192G(88、90)およびCTA1−DD(89)を用いたときに得られている。また上記ペプチドは、KLHなどの担体、または髄膜炎菌(N.meningitides)の外膜タンパク質複合体、またはヒトパピローマウイルスのVLPと化学的に接合されてもよく、上記ペプチドは、ワクチンとしてマウスおよび他の動物に防御をもたらす(92、93)。
【0061】
M2タンパク質が高度に保存されている限りにおいて、M2タンパク質の配列は相違点を必ず有している。共通株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2の外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除く、他の配列決定された現代のヒト株の全てと、免疫学的に交差反応することが示された(92)。また、インフルエンザのデータベース上の配列を調査することにより、A/Vietnam/1203/04などの他のより最近見つかった病原性のヒト単離株(H5N1)のM2配列に、同様の相違点が存在することが示された。この発見によって明らかになることは、M2エピトープを組み込んだ、有効でかつH5に特異的なパンデミックワクチンは、ヒトのH1およびH3の単離株において現在流行しているM2配列というよりは、病原性のトリ株に独特なM2の配列を反映することが必要になるであろうということである。
【0062】
インフルエンザウイルス由来の別のタンパク質(HA、NAおよびM2以外のタンパク質)は、共発現されることによって、あるいは別の抗原の全てもしくは一部をgagポリペプチドまたはHAポリペプチドに結合されることによって、VLPワクチンに含まれてもよい。これらの別の抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、NS2が包含される。これらの後者の抗原は、一般的に中和抗体応答の標的ではないが、T細胞によって認識される重要なエピトープを含んでいる可能性がある。VLPワクチンによってそのようなエピトープに対して誘導されるT細胞応答が、感染防御免疫を高めることに有益であると判明する可能性がある。
【0063】
<他の病原性抗原>
適切な細菌抗原には、以下の様々な病原性細菌の何れかと関連する抗原(例えば、様々な病原性細菌の何れかによって合成される抗原、および様々な病原性細菌の何れかの内部に生じた抗原)が包含される:例えば、病原性グラム陽性菌(病原性パスツレラ種(pathogenic Pasteurella species)、スタフィロコッカス種(Staphylococci species)、およびストレプトコッカス種(Streptococcus species)など);ならびに、病原性グラム陰性菌(ナイセリア(Neisseria)属、エシェリキア(Escherichia)属、ボルデテラ(Bordetella)属、カンピロバクター(Campylobacter)属、レジオネラ(Legionella)属、シュードモナス(Pseudomonas)属、シゲラ(Shigella)属、ビブリオ(Vibrio)属、エルシニア(Yersinia)属、サルモネラ(Salmonella)属、ヘモフィルス(Haemophilus)属、ブルセラ(Brucella)属、フランシセラ(Francisella)属、およびバクレリオイデス(Bacterioides)属の病原性グラム陰性菌など)など。例えば「Schaechter, M, H. Medoff, D. Schlesinger, Mechanisms of Microbial Disease. Williams and Wilkins, Baltimore (1989))」を参照のこと。
【0064】
感染性病原性真菌と関連する適切な抗原(例えば、感染性病原性真菌によって合成される抗原、および感染性病原性真菌の内部に生じた抗原)には、以下の感染性真菌と関連する抗原が包含される:特に限定されないが、クリプトコックス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラスマ・カプスラーツム(Histoplasma capsulatum)、コクシジオイデス・イミチス(Coccidioides immitis)、ブラストミセス・デルマティティディス(Blastomyces dermatitidis)、およびカンジダ・アルビカンス(Candida albicans)、カンジダ・グラブラタ(Candida glabrata)、アスペルギルス・フリガータ(Aspergillus fumigata)、アスペルギルス・フラバス(Aspergillus flavus)、およびスポロトリックス・シェンキィ(Sporothrix schenckii)など。
【0065】
病原性の原虫、蠕虫および他の真核微生物病原体と関連する適切な抗原(例えばこれらによって合成される抗原、およびこれらの内部に生じた抗原)には、以下の原虫、蠕虫および他の真核微生物病原体と関連する抗原が包含される:特に限定されないが、マラリア原虫(Plasmodium)(熱帯熱マラリア原虫(Plasmodium falciparum)、四日熱マラリア原虫(Plasmodium malariae)、卵形マラリア原虫(Plasmodium ovale)、および三日熱マラリア原虫(Plasmodium vivax)など);トキソプラズマ・ゴンディ(Toxoplasma gondii);トリパノソーマ・ブルーセイ(Trypanosoma brucei)、トリパノソーマ・クルージ(Trypanosoma cruzi);ビルハルツ住血吸虫(Schistosoma haematobium)、マンソン住血吸虫(Schistosoma mansoni)、日本住血吸虫(Schistosoma japonicum);ドノバンリーシュマニア(Leishmania donovani);ランブル鞭毛虫(Giardia intestinalis);およびクリプトスポリジウム・パルバム(Cryptosporidium parvum)など。
【0066】
適切な抗原には、以下の病原性微生物と関連する抗原(例えば病原性微生物によって合成される抗原、および病原性微生物の内部に生じた抗原)が包含される:ヘリコバクター・ピロリ(Helicobacter pyloris)、ボレリア・ブルグドルフェリ(Borelia burgdorferi)、レジオネラ・ニューモフィラ(Legionella pneumophila)、マイコバクテリウム属(Mycobacteria sps)(例えばマイコバクテリウム・ツベルクローシス(M. tuberculosis)、マイコバクテリウム・アビウム(M. avium)、マイコバクテリウム・イントラセルラーレ(M. intracellulare)、マイコバクテリウム・カンサイ(M. kansaii)、マイコバクテリウム・ゴルドナエ(M. gordonae))、スタフィロコッカス・アウレウス(Staphylococcus aureus)、ナイセリア・ゴノレー(Neisseria gonorrhoeae)、ナイセリア・メニンギティディス(Neisseria meningitidis)、リステリア・モノサイトゲネス(Listeria monocytogenes)、クラミジア・トラコーマティス(Chlamydia trachomatis)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)(A群連鎖球菌(グループAストレプトコッカス(Group A Streptococcus))、ストレプトコッカス・アガラクチア(Streptococcus agalactiae)(B群連鎖球菌(グループAストレプトコッカス(Group B Streptococcus)))、ストレプトコッカス(ビリダンス群(viridans group))、ストレプトコッカス・フェカリス(Streptococcus faecalis)、ストレプトコッカス・ボビス(Streptococcus bovis)、ストレプトコッカス(嫌気性(anaerobic sps.))、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)、病原性のカンピロバクター(pathogenic Campylobacter sp.)、エンテロコッカス(Enterococcus sp.)、ヘモフィルス・インフルエンザエ(Haemophilus influenzae)、バチルス・アントラシス(Bacillus anthracis)、コリネバクテリウム・ジフセリエ(Corynebacterium diphtheriae)、コリネバクテリア(corynebacterium sp.)、エリジペロスリックス・ルジオパシエ(Erysipelothrix rhusiopathiae)、クロストリジウム・パーフリンジェンス(Clostridium perfringens)、クロストリジウム・テタニ(Clostridium tetani)、エンテロバクター・エロゲネス(Enterobacter aerogenes)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、パスツレラ・マルトシダ(Pasturella multocida)、バクテロイド(Bacteroides sp.)、フソバクテリウム・ヌクレアタム(Fusobacterium nucleatum)、ストレプトバシラス・モニリフォルミス(Streptobacillus moniliformis)、トレポネーマ・パリダム(Treponema pallidium)、トレポネーマ・ペルテニュ(Treponema pertenue)、レプトスピラ(Leptospira)、リケッチア(Rickettsia)、およびアクチノマイセス・イスラエリイ(Actinomyces israeli)など。病原性エシェリキア・コリ(E. coli)株としては、例えば、アメリカ培養細胞系統保存機関(ATCC)の整理番号31618、23505、43886、43892、35401、43896、33985、31619、および31617が挙げられるが、これらに限定されない。
【0067】
細胞内病原体と関連する様々なポリペプチドもしくは他の抗原の何れかは、VLPに含まれてもよい。細胞内病原体と関連するポリペプチドおよびペプチドのエピトープは、細胞内病原体と関連する(例えば細胞内病原体によってコードされている)ポリペプチドであって、該ポリペプチドの断片がMHCクラスI分子と一緒に感染細胞の表面上に提示され、該断片が、CD8.sup.+のリンパ球の表面上のT細胞抗原受容体に認識される(例えば該受容体に結合される)というポリペプチドであれば、どのようなものでもよい。細胞内病原体と関連するポリペプチドおよびペプチドのエピトープは、技術的に知られており、特に限定されないが、ヒト免疫不全ウイルス(例えばHIV gp120)と関連する抗原、またはその抗原断片;サイトメガロウイルス抗原;マイコバクテリウム抗原(例えばマイコバクテリウム・アビウム(Mycobacterium avium)、およびマイコバクテリウム・ツベルクローシス(Mycobacterium tuberculosis)など);ニューモシスチス・カリニ(Pneumocystic carinii)(PCP)抗原;マラリア抗原(特に限定されないが、熱帯熱マラリア原虫または他のあらゆるマラリア種と関連する抗原(41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど)を含む);真菌抗原;酵母抗原(例えばカンジダ(Candida spp.)の抗原);トキソプラズマ(toxoplasma)抗原(特に限定されないが、トキソプラズマ・ゴンディ(Toxoplasma gondii)、トキソプラズマ脳炎(Toxoplasma encephalitis)、または他のあらゆるトキソプラズマ種と関連する抗原を含む);EBウイルス(EBV)抗原;プラスモディウム(Plasmodium)抗原(例えばgp190/MSP1など);などを包含する。
【0068】
好ましいVLPワクチンは、バチルス・アントラシス(Bacillus anthracis)を対象とするものであってもよい。バチルス・アントラシスは、好気性または通性嫌気性でグラム陽性の、幅1.0μm、長さ3.0−5.0μmの非運動性桿菌である。悪条件下では、バチルス・アントラシスは、高耐性の内生胞子を形成する。この内生胞子は、感染動物が以前に死亡した場所の土壌で発見され得る。本明細書に開示されているようなVLPワクチンに使用するための好ましい抗原は、感染防御抗原(PA)である。該感染防御抗原は、83kDaのタンパク質であって、哺乳類細胞の受容体に結合し、バチルス・アントラシスが病気を引き起こすことができるのに極めて重要なものである。より好ましい抗原は、バチルス・アントラシスのPAにおけるC末端側の140アミノ酸の断片であり、該断片はバチルス・アントラシスに対する感染防御免疫を被検体に誘導するために使用され得るものである。炭疽菌に対するVLPワクチンに使用するための他の典型的な抗原は、炭疽菌の胞子に由来する抗原(例えばBclA)、この細菌の栄養生長期からの抗原(例えば、細胞壁抗原、莢膜抗原(例えばポリ−ガンマ−D−グルタミン酸、すなわちPGA)、分泌抗原(例えば感染防御抗原、致死因子、または浮腫因子などの外毒素)である。VLPワクチンに使用するためのもう1つ別の好ましい抗原は、バチルス・アントラシスに独特な、アントロース(anthrose)を含むテトラ−サッカリドである(Daubenspeck J.M., ら J. Biol. Chem. (2004), 279:30945)。該テトラ−サッカリドは、脂質ラフト会合ポリペプチドと共役されてもよい。これにより、抗原をVLPワクチンと会合させることができる。
【0069】
<腫瘍関連抗原>
公知の様々な腫瘍特異的抗原または腫瘍関連抗原(TAA)の何れかを、VLPに含めてもよい。完全なTAAが使用されてもよいが、使用されなくてもよい。その代わりに、TAAの一部(例えばエピトープ)を使用してもよい。VLPに使用されてもよい腫瘍関連抗原(またはエピトープを含むその断片)には、特に限定されないが、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量のメラノーマ関連抗原MAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原OV−TL3およびMOV18、TUAN、アルファ−フェトタンパク質(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原G250、EGP−40(EpCAMとしても知られている)、S100(悪性メラノーマ関連抗原)、p53、およびp21rasが包含される。(上述したTAAのいずれかを含む)任意のTAA(またはそのエピトープ)の合成類似体が、使用されてもよい。さらに、1つ以上のTAA(またはそのエピトープ)の組み合わせが、組成物に含まれてもよい。
【0070】
<アレルゲン>
1態様では、VLPワクチンの一部である抗原は、様々なアレルゲンの何れかであってもよい。アレルゲンを基本とするワクチンは、アレルゲンに対する寛容を被検体に誘導するために使用されてもよい。チロシンとの共沈に関与するアレルゲンワクチンの例は、米国特許第3,792,159号明細書、米国特許第4,070,455号明細書、および米国特許第6,440,426号明細書において見つけることができる。なお、特にアレルゲンワクチンの製剤化に関して、これら文献のそれぞれは参照することによって本明細書に組み込まれる。
【0071】
様々なアレルゲンの何れかは、VLPに含まれてもよい。アレルゲンには、特に限定されないが、環境エアロアレルゲン;ブタクサ/枯草熱などの植物の花粉;雑草の花粉のアレルゲン;草の花粉のアレルゲン;セイバンモロコシ(Johnson grass);木の花粉のアレルゲン;ライグラス;イエダニのアレルゲン(例えばDer p I、Der f Iなど)などのクモ類のアレルゲン;ストレージダニのアレルゲン;スギ花粉/枯草熱;カビ胞子のアレルゲン;動物のアレルゲン(例えばイヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどのアレルゲン);食物のアレルゲン(例えば甲殻類;ピーナッツなどのナット;柑橘系果物のアレルゲン);昆虫のアレルゲン;毒:(膜翅目、イエロー・ジャケット、ミツバチ、ワスプ、ホーネット、ハリアリ(fire ant));ゴキブリ、ノミ、蚊などからの他の環境昆虫のアレルゲン;ストレプトコッカスの抗原などの細菌のアレルゲン;アスカリス属抗原などの寄生虫のアレルゲン;ウイルス抗原;真菌の胞子;薬物のアレルゲン;抗生物質;ペニシリンおよび関連化合物;他の抗生物質;ホルモン(インシュリン)、酵素(ストレプトキナーゼ)などの総タンパク質;不完全な抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝産物;ハプテンとして作動することが可能な、およびアレルゲンとして機能することが可能な、工業用化学薬品および代謝産物(例えば酸無水物(無水トリメリット酸など)およびイソシアネート(トルエンジイソシアネートなど));小麦粉(例えばパン屋喘息(Baker’s asthma)を起こすアレルゲン)、トウゴマ、コーヒー豆、および上記工業用化学薬品などの職業性アレルゲン;ノミのアレルゲン;ならびにヒト以外の動物の中のヒトタンパク質が包含される。
【0072】
アレルゲンには、特に限定されないが、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドおよび他の分子のペプチド模擬体および非ペプチド模擬体、低分子、脂質、糖脂質、ならびに炭水化物が包含される。
【0073】
具体的な天然の動物および植物のアレルゲンには、例えば特に限定されないが、以下の属に特有のタンパク質が包含される:イヌ科(カニスファミリアリス(Canis familiaris));デルマトファゴイデス属(Dermatophagoides)(例えばデルマトファゴイデス・ファリナエ(コナヒョウヒダニ(Dermatophagoides farinae)));ネコ属(フェリス・ドメスチカス(Felis domesticus));アンプローシア属(Ambrosia)(アンプローシア・アルテミスフォリア(Ambrosia artemiisfolia);ロリウム属(Lolium)(例えばペレニアルライグラス(Lolium perenne)またはイタリアンライグラス(Lolium multiflorum));スギ属(Cryptomeria)(スギ(Cryptomeria japonica));アルテマリア属(Altemaria)(アルテマリア・アルテマタ(Altemaria altemata));アルダー属(Alder);ハンノキ属(アルヌス・グルチノアス(Alnus gultinoas));ベツラ属(Betula)(ベツラ・ベルコサ(Betula verrucosa));ケルクス属(Quercus)(ケルクス・アルバ(Quercus alba));オリーブ属(Olea)(オリーブ(Olea europa));アルテミシア属(Artemisia)(アルテミシア・バルガリス(Artemisia vulgaris));プランタゴ属(例えばヘラオオバコ(Plantago lanceolata));パリエタリア属(Parietaria)(例えばパリエタリア・オフィシナリス(Parietaria officinalis)またはパリエタリア・ジュダイカ(Parietaria judaica));チャバネゴキブリ属(Blattella)(例えばチャバネゴキブリ(Blattella germanica));アピス属(Apis)(例えばアピス・ムルチフロルム(Apis multiflorum));カプレサス属(Cupressus)(例えばカプレサス・センパーヴァイレンス(Cupressus sempervirens)、カプレサス・アリゾニカ(Cupressus arizonica)、およびカプレサス・マクロカルパ(Cupressus macrocarpa));ジュニペルス属(例えばジュニペルス・サビノイデス(Juniperus sabinoides)、ジュニペルス・ヴィルギニアナ(Juniperus virginiana)、ジュニペルス・コムニス(Juniperus communis)、およびジュニペルス・アシェイ(Juniperus ashei));ツヤ属(Thuya)(例えばツヤ・オリエンタリス(Thuya orientalis));ヒノキ属(Chamaecyparis)(例えばヒノキ(Chamaecyparis obtusa));ペリプラネタ属(Periplaneta)(例えばワモンゴキブリ(Periplaneta americana));アグロピロン属(例えばシバムギ(Agropyron repens));セカレ属(Secale)(例えばライムギ(Secale cereale));トリチカム属(Triticum)(例えばコムギ(Triticum aestivum));ダクチリス属(Dactylis)(例えばオーチャードグラス(Dactylis glomerata));フェスツカ属(Festuca)(例えばフェスツカ・エラチオル(Festuca elatior));ポア属(Poa)(例えばポアプラテンシス(Poapratensis)またはポア・コンプレッサ(Poa compressa));カラスムギ属(Avena)(例えばマカラスムギ(Avena sativa));ホルクス属(Holcus)(例えばホルクス・ラナタス(Holcus lanatus));アンソキサスム属(Anthoxanthum)(例えばハルガヤ(Anthoxanthum odoratum));アルヘナセラム属(Arrhenatherum)(例えばアルヘナセラム・エラチウス(Arrhenatherun elatius));アグロスチス属(例えばアグロスチス・アルバ(Agrostis alba));フレウム属(Phleum)(例えばチモシー(Phleum pratense));ファラリス(Phalaris)属(例えばファラリス・アルンジナセア(Phalaris arundinacea));スズメノヒエ属(Paspalum)(例えばバヒアグラス)(Paspalum notatum);ソルガム属(Sorghum)(例えばソルガム・ハレペンシス(Sorghum halepensis));そしてブロムス属(Bromus)(例えばブロムス・イネルミス(Bromus inermis))。
【0074】
<VLPを作成する好ましい方法>
VLPは、(a)脂質ラフトと天然に会合しない抗原に連結された脂質ラフト会合ポリペプチドおよび(b)gagポリペプチドを含んでいるVLPが、組立られることが好ましい技術分野の当業者の一人に利用可能な任意の方法によって、容易に組立られ得る。好ましい実施形態では、上記ポリペプチドは、あらゆる利用可能なタンパク質発現システムにおいて共発現されてもよい。また該タンパク質発現システムは、脂質内にラフト脂質ドメインを含む細胞を基本とするシステム(哺乳類細胞発現システム、および昆虫細胞発現システムなど)であることが好ましい。
【0075】
gagポリペプチドを用いて形成されたVLPの発現の多くの例は、公開されており、VLPを作製するのに利用可能な発現システムの範囲が実証されている。いくつかのレトロウイルスを用いた研究によれば、他のウイルス成分の非存在下において発現されたGagポリペプチドは、VLPの形成および細胞表面における出芽に十分であることが実証された(「Wills and Craven AIDS 5, 639-654, 1991」、「Zhouら, 3. Virol. 68, 2556-2569, 1994」、「Morikawaら, Virology 183, 288-297, 1991」、「Royerら, Virology 184, 417-422, 1991」、「Gheysenら, Cell 59, 103-112, 1989」、「Hughesら, Virology 193, 242-255, 1993」、「Yamshchikovら, Virology 214, 50-58, 1995」)。バキュロウイルスベクターを用いた、昆虫細胞内でのGag前躯体の発現によるVLPの形成は、いくつかのグループによって実証されている(「Delchambreら, EMBO J. 8, 2653-2660, 1989」、「Luoら, Virology 179, 874-880, 1990」、「Royerら, Virology 184, 417-422, 1991」、「Morikawaら, Virology 183, 288-297, 1991」、「Zhouら, J. Virol. 68, 2556-2569, 1994」、「Gheysenら, Cell 59, 103-112, 1989」、「Hughesら, Virology 193, 242-255, 1993」、「Yamshchikovら, Virology 214, 50-58, 1995」)。これらのVLPは、未成熟のレンチウイルス粒子に類似しており、効率的に組立られ、昆虫細胞の形質膜から出芽によって放出される。
【0076】
Gag前躯体のアミノ末端領域が、細胞表面への移送と、ウイルスの組立に必要な膜結合とのための、標的シグナルであることが報告されている(「Yuら, J. Virol. 66, 4966-4971, 1992」、「an, Xら, J. Virol. 67, 6387-6394, 1993」、「Zhouら, J. Virol. 68, 2556-2569, 1994」、「Lee and Linial J. Virol. 68, 6644-6654, 1994」、「Dorfmanら, J. Virol. 68, 1689-1696, 1994」、「Fackeら, J. Virol. 67, 4972-4980, 1993」)。ワクシニア・ウイルス発現システムを用いた、Gag構造タンパク質とEnv糖タンパク質であるgp120およびgp41とを含む、組み換えHIVを基本とするVLPの組立が、報告されている(「Haffarら, J. Virol. 66, 4279-4287, 1992」)。
【0077】
VLPのためのポリペプチドの組み換え発現には、上記ポリペプチドの1つ以上をコードするポリヌクレオチドを含んでいる発現ベクターのコンストラクションが必要である。上記ポリペプチドの1つ以上をコードするポリヌクレオチドが得られれば、該ポリペプチドを製造するためのベクターは、公知手法を用いる組み換えDNA技術によって製造され得る。したがって、本明細書には、VLPポリペプチドをコードするヌクレオチド配列の何れかを含んでいるポリヌクレオチドを発現させることによって、タンパク質を調製する方法が記載されている。当業者に公知の方法を用いることにより、VLPポリペプチドをコードする配列、適切な転写制御シグナル、および適切な翻訳制御シグナルを含んでいる発現ベクターがコンストラクトされることができる。これらの方法には、例えば、イントロでの組み換えDNA手法、合成の手法、およびインビボでの遺伝子組み換えが包含される。したがって、本発明は、gagポリペプチドと、抗原に連結される脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含んでおり、これらの全てが1つ以上のプロモーターに作動可能に連結されている、複製可能なベクターを提供する。
【0078】
上記発現ベクターは、従来手法によって宿主細胞に移動させられてもよく、トランスフェクションされた細胞は、それからVLPポリペプチドを製造するように従来手法によって培養される。したがって、本発明は、異種プロモーターに作動可能に連結されたVLPポリペプチドの1つ以上をコードするポリヌクレオチドを含んでいる宿主細胞を包含する。VLPを生成するための好ましい実施形態では、gagポリペプチド、および抗原に連結される脂質ラフト会合ポリペプチドの両方をコードするベクターは、VLPを生成するための宿主細胞内で共発現されてもよい。この点について以下で詳細に説明する。
【0079】
様々な宿主−発現ベクターシステムが、VLPポリペプチドを発現させるために利用されてもよい。そのような宿主−発現システムは、VLPを生成するために(好ましくは共発現によって)VLPポリペプチドが製造され得るための媒体を表す。多様な宿主が、適切な発現ベクターのコンストラクトに使用されてもよく、好ましい宿主−発現システムは、VLPの組立に適した脂質ラフトを有する宿主である。該宿主には、VLPポリペプチドをコードする配列(VLPポリペプチドコード配列)を含んでいる、組み換えバクテリオファージDNA、プラスミドDNA、またはコスミドDNAの発現ベクターで形質転換される、細菌などの微生物(例えばエシェリキア・コリ、バシラス・サブティリス(B. subtilis));VLPポリペプチドコード配列を含んでいる組み換え酵母発現ベクターで形質転換される、酵母(例えばサッカロマイセス(Saccharomyces)、ピキア(Pichia));VLPポリペプチドコード配列を含んでいる組み換えウイルス発現ベクター(例えばバキュロウイルス)で感染される昆虫細胞システム;VLPポリペプチドコード配列を含んでいる組み換えウイルス発現ベクター(例えばカリフラワーモザイクウイルス、CaMV、タバコモザイクウイルス、TMV)で感染されるか、VLPポリペプチドコード配列を含んでいる組み換えプラスミド発現ベクター(例えばTiプラスミド)で形質転換される、植物細胞システム;あるいは、哺乳類細胞のゲノム由来のプロモーター(例えばメタロチオネインのプロモーター)、または哺乳類ウイルス由来のプロモーター(例えばアデノウイルスの後期プロモーター;ワクシニア・ウイルスの7.5Kのプロモーター)由来のプロモーター、を含んでいる組み換え発現コンストラクトが導入される、哺乳類細胞システム(例えばCOS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)が包含されるが、これらに限定されない。好ましくは哺乳類細胞、より好ましくは昆虫細胞が、VLPの組立に適したラフト脂質を有しているので、VLPポリペプチドの発現のために使用される。例えば、ヒトサイトメガロウイルス由来の主要中間初期遺伝子プロモーターエレメントなどのベクターと組み合わせられる、チャイニーズハムスター卵巣(CHO)細胞などの哺乳類細胞は、VLPポリペプチドにとって効果的な発現システムであるといえる(「Foeckingら, Gene 45:101 (1986)」、「Cockettら, Bio/Technology 8:2 (1990)」)。
【0080】
昆虫システムでは、キンウワバ科(Autographa californica)の核多角体病ウイルス(AcNPV)が、外来遺伝子を発現させるベクターとして使用され得る。該ウイルスは、スポドプテラ・フルギペルダ(Spodoptera frugiperda)細胞において成長する。VLPポリペプチドコード配列は、ウイルスの非必須領域(例えばポリヘドリン遺伝子)にそれぞれクローニングされて、AcNPVプロモーター(例えばポリヘドリンプロモーター)の制御下に置かれてもよい。
【0081】
哺乳類宿主細胞では、ウイルスを基本とする発現システムの多くが利用され得る。アデノウイルスが発現ベクターとして使用される場合、関心のあるVLPポリペプチド配列が、アデノウイルスの転写/翻訳制御複合体(例えば後期プロモーターおよび3部分からなるリーダー配列)に結合されてもよい。このキメラ遺伝子は、その後、インビトロまたはインビボでの組み換えによってアデノウイルスのゲノムに挿入されてもよい。ウイルスゲノムの非必須領域(例えば領域E1またはE3)に挿入することによって、感染宿主内で生存することができ、VLPポリペプチドを発現することができる組み換えウイルスが得られる(例えば、「Logan & Shenk, Proc. Natl. Acad. Sci. USA 81:355-359 (1984)」参照)。また特定の開始シグナルが、挿入されたVLPポリペプチドコード配列を効率よく翻訳するために必要とされてもよい。該シグナルには、ATG開始コドンおよび隣接配列が包含される。さらに、開始コドンは、挿入物全体の翻訳を確実にするために、所望のコード配列のリーディングフレームと一致していなければならない。これら外来性の翻訳制御シグナルおよび開始コドンは、天然および合成の様々なものに由来してもよい。発現効率は、適切な転写エンハンサー要素、翻訳ターミネーターなどを含めることによって、上昇され得る(「Bittnerら, Methods in Enzymol. 153:51-544 (1987)」参照)。
【0082】
また、挿入された配列の発現を調節するか、遺伝子産物を特定の望まれる方法で修飾および加工する宿主細胞株が、選択されてもよい。タンパク質産物のそのような修飾(例えばグリコシル化)および加工(例えば開裂または膜への移送)は、VLPの生成、VLPポリペプチドの機能、または別のポリペプチド(アジュバントもしくは別の抗原)にとって重要である可能性がある。種々の宿主細胞は、タンパク質ならびに遺伝子産物の翻訳後加工および翻訳後修飾に対して、特徴的かつ特異的な機構を有している。適切な細胞または宿主システムは、発現された外来タンパク質の正しい修飾および加工が確実に行われるように選択されてもよい。これのためには、一次転写産物の適切なプロセッシングや、遺伝子産物のグリコシル化およびリン酸化に対する細胞機構を有する、真核宿主細胞が使用されてもよい。
【0083】
宿主細胞は、本発明の2つの発現ベクターで共トランスフェクションされてもよい(第1ベクターは、gagポリペプチドをコードするベクターであり、第2ベクターは、抗原に連結される脂質ラフト会合ポリペプチドをコードするベクターである)。2つのベクタ−は、各VLPポリペプチドを等しく発現させることができる、同じ選択マーカーを含んでいてもよい。また代わりに、gagポリペプチドと、脂質に連結される脂質ラフト会合ポリペプチドとの両方をコードし、これらを発現させることができる1つのベクターを使用してもよい。
【0084】
VLPが宿主細胞によって産生されれば、該VLPは、ポリペプチドの精製に関して技術的に知られている任意の方法によって、精製され得る。そのような方法としては、例えば、クロマトグラフィー(例えばイオン交換クロマトグラフィー、親和性クロマトグラフィー(特にポリペプチドに付加された任意の親和性精製タグに対する親和性を利用する、親和性クロマトグラフィー)、大きさを分けるクロマトグラフィー)、遠心、または溶解度差を用いる方法、あるいはタンパク質もしくは他の高分子を精製するための他の任意の標準的な手法を用いる方法が挙げられる。また、VLPポリペプチドは、VLPの精製を容易にするための、本明細書に記載されているか、さもなければ技術的に公知である異種のポリペプチド配列と融合されてもよい。精製後、別の抗原またはアジュバントなどの別の要素が、VLPポリペプチドに対する共有結合を介するか、または他の非共有結合の機構によってVLPに物理的に連結されてもよい。VLPポリペプチドが、ラフト脂質ドメインを有する宿主細胞(哺乳類細胞および昆虫細胞など)内で共発現される好ましい実施形態では、VLPは、自ら組立られて放出される。これにより、上記方法の何れかによってVLPが精製され得る。
【0085】
〔VLPを使用する好ましい方法〕
<製剤>
本明細書に記載のVLPは、ワクチン調製物として使用されることが好ましい。通常、そのようなワクチンは、注射剤として液体溶液または懸濁液として調製される。また該ワクチンは、注射前の時点では、液体溶液または懸濁液に適した固形剤として調製されてもよい。またそのような調製物は、乳化されてもよいし、乾燥粉末として製造されてもよい。しばしば、活性免疫原性成分が、該活性免疫原性成分と適合する薬学的に許容可能な賦形剤と混合される。適した賦形剤は、例えば、水、生理食塩水、右旋性グルコース、グリセロール、エタノール、または同類のもの、およびそれらの組み合わせである。さらに、希望に応じて、ワクチンは、補助物質(湿潤剤、乳化剤、pH緩衝剤、またはワクチンの有効性を向上させるアジュバントなど)を含んでもよい。
【0086】
ワクチンは、従来どおり、例えば皮下に、経皮的に、皮内に、真皮下に、または筋肉内に注入されることによって非経口的に投与されてもよい。他の投与様式に適した別の製剤は、坐薬を含んでおり、場合によっては経口製剤、鼻腔内製剤、バッカル製剤、舌下製剤、腹腔内製剤、膣内製剤、肛門製剤および頭蓋内製剤を含んでいる。坐薬については、在来の結合剤および担体は、例えばポリアルキレングリコールまたはトリグリセリドを含んでいてもよい。そのような坐薬は、上記活性免疫原性成分を、0.5%から10%、好ましくは1%から2%の範囲で含んでいる混合物から形成されてもよい。ある実施形態では、低融点のワックス(脂肪酸グリセリドまたはココアバターの混合物など)が初めに溶かされ、本明細書記載のVLPが攪拌などによって均質に分散させられる。溶解した均一混合物は、それから従来の大きさの型に注がれ、冷却されてから凝固される。
【0087】
鼻腔内投与に適した製剤には、液体および乾燥粉末が包含される。製剤には、普通に使用される賦形剤(例えば医薬品等級のマンニトール、ラクトース、サッカロース、トレハロースおよびキトサンなど)が含まれる。キトサンなどの粘膜接着剤(Mucosadhesive agent)は、鼻腔内投与された製剤の粘液線毛クリアランスを遅らせるために、液体製剤または乾燥粉末製剤に使用されることができる。マンニトールおよびサッカロースなどの糖は、液体製剤においては安定剤として、乾燥製剤においては安定剤および充填剤として使用されることができる。また、アジュバント(例えばモノホスホリルリピドA(MPL)、および単なる例であり特に限定されないが、二本鎖ポリ(I:C)、ポリイノシン酸、CpG含有オリゴヌクレオチド、イミキモド(imiquimod)、コレラ毒素およびその誘導体、易熱性エンテロトキシンおよびその誘導体、ならびに本明細書の全体に挙げられているアジュバントの多く)は、液体製剤および乾燥粉末製剤に、免疫賦活性アジュバントとして使用されることができる。
【0088】
経口投与に適した製剤には、液体、固体、半固体、ゲル、錠剤、カプセル、およびトローチ剤などが包含される。経口投与に適した製剤には、錠剤、トローチ剤、カプセル、ゲル、液体、食品、飲み物、および栄養補給食品などが包含される。製剤には、普通に使用される賦形剤(例えば医薬品等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、および炭酸マグネシウムなど)が含まれる。他のVLPワクチン組成物は、溶液、懸濁液、丸剤、徐放性製剤、または粉末の形態を取ってもよく、10%から95%、好ましくは25%から70%の活性免疫原性成分を含んでいる。経口製剤に関しては、コレラ毒素が製剤の相手として興味深い物質である(また接合の相手としても可能性がある)。
【0089】
VLPワクチンは、膣内投与用に製剤化されるときは、ペッサリー、タンポン、クリーム、ゲル、ペースト、泡、またはスプレーの形態になってもよい。上記製剤の何れかは、VLPに加えて、技術的に公知の適切な作用物質を含んでいてもよい。
【0090】
ある実施形態では、VLPワクチンは、全身投与用に、または局所投与用に製剤化されてもよい。そのような製剤は、技術的によく知られている。非経口投与用の媒体には、塩化ナトリウム溶液、リンガー右旋性グルコース、右旋性グルコースおよび塩化ナトリウムの溶液、乳酸加リンゲル溶液、または不揮発性油が包含される。静脈内投与用の媒体には、流動性で栄養性の補充液、および電解質補充液(リンガー右旋性グルコースに基づくものなど)などが包含される。全身投与および局所投与の経路には、経皮投与、皮内投与、局所適用、静脈内投与、筋肉内投与などが包含される。
【0091】
VLPは、中性製剤または塩に基づく製剤を包含するワクチンに製剤化されてもよい。薬学的に許容可能な塩には、酸付加塩(ペプチドの遊離アミノ基と一緒に形成された塩)が包含される。なお、該酸付加塩は、無機酸(例えば塩酸またはリン酸)、あるいは有機酸(酢酸、シュウ酸、酒石酸、およびマンデル酸など)と一緒に形成された塩である。また、遊離カルボキシル基と一緒に形成された塩は、無機塩基(例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化第二鉄など)、および有機塩基(イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、およびプロカインなど)から得られてもよい。
【0092】
ワクチンは、製剤の用量(dosage formulation)と適合するように、治療効果および免疫原性がもたらされるような量で、投与されてもよい。投与される量は、治療される被検体(例えば免疫応答を開始するための個々の免疫系の能力、および希望される防御の程度など)に依存する。適切な用量の範囲では、接種一回の当たりの活性成分は、数百マイクログラムの桁であり、(1から10mgの範囲というより多い量が考慮されるが)好ましくは約0.1μgから2000μgの範囲であり、例えば、約0.5μgから1000μgの範囲であり、好ましくは1μgから500μgの範囲であり、特に好ましくは約10μgから100μgの範囲である。また、初回投与および追加免疫に適した投薬計画は、変更可能であるが、初回投与が行われてから、その後の接種または他の投与が行われるというのが典型である。
【0093】
適用の様式は大きく変更されてもよい。ワクチンを投与するための従来の方法の何れかを適用することができる。これらには、生理学的に許容可能な固形の基質を利用して経口投与する方法、または生理学的に許容可能な分散形態で注射などによって投与する方法などが含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるものの年齢および抗原の剤形に基づいて変更される。
【0094】
ワクチン製剤には、それ自体ワクチンとして十分な免疫原性を有しているものがあるが、残りのワクチン製剤のいくつかには、ワクチンがアジュバント物質をさらに含んでいる場合に、免疫応答が増強されるものがある。
【0095】
また、粘膜接着を改善するための送達剤が使用されることにより、特に鼻腔内投与製剤、経口投与製剤、肺に基づく投与製剤に対する、送達および免疫原性が改善され得る。そのような化合物の1つであるキトサン(キチンのN−デアセチル化形態)が、多くの薬学的製剤に使用される(32)。キトサンは、粘液線毛クリアランスを遅らせることができ、粘膜での抗原の取り込みおよび加工により長く時間を費やすことができるため、鼻腔内ワクチンにとって魅力的な粘膜接着剤である(33、34)。さらに、キトサンは、一時的に密着結合を広げることができ、抗原のNALTへの経上皮輸送を向上させる可能性がある。最近のヒトでの試験において、他のアジュバントとは一緒に投与されず、キトサンと一緒に鼻腔内投与された3価の不活性化インフルエンザワクチンは、抗体陽転を引き起こすとともに、筋肉内接種の後に得られるHIの力価よりも僅かに低いHIの力価をもたらした(33)。
【0096】
またキトサンは、遺伝的に解毒されたE. coliの易熱性エンテロトキシンの突然変異体であるLTK63などの、鼻腔内で良好に機能するアジュバントと一緒に製剤化されることもできる。これにより、キトサンによって与えられる送達と接着との利点に加えて、免疫賦活効果が追加される。その結果、粘膜応答および全身応答が増強されることになる(35)。
【0097】
最後に、キトサンの製剤は、乾燥粉末の形式で製剤され得ることに注意されたい。この形式は、ワクチンの安定性を改善することが示されており、液体製剤よりもさらに粘液線毛クリアランスを遅らせることができる(42)。このことは、キトサンと一緒に製剤化された鼻腔内乾燥粉末であるジフテリアトキソイドワクチンを用いた、最近のヒトでの臨床試験において確かめられた。さらに、該臨床試験では、鼻腔内経路は、従来の筋肉内経路と同じくらい効果的であり、分泌型のIgA応答の利点が加えられることが確かめられた(43)。また上記ワクチンの耐性は、非常に強かった。キトサンおよびMPLを含んでいる炭疽菌用の鼻腔内乾燥粉末ワクチンは、筋肉内接種よりも強い応答をウサギに誘導し、またエアロゾルである胞子によるアレルギーの誘発に対する防御も誘導した(44)。
【0098】
鼻腔内ワクチンは、それが、非経口投与されるワクチンとは対照的に上気道および下気道に影響を与えることができるので、好ましい製剤である(なお、非経口投与されるワクチンは、下気道に影響を与える点においてより優れている)。このことは、アレルゲンを基本とするワクチンに対する寛容を誘導することや、また病原体を基本とするワクチンに対する免疫を誘導することにとって利点となり得る。
【0099】
鼻腔内ワクチンが使用されることにより、上気道および下気道に防御が与えられることに加えて、針を使った接種につきまとう厄介な問題が回避されるし、粒状および/または可溶性の抗原と鼻咽頭関連リンパ組織(NALT)との相互作用により、粘膜性ならびに全身性の体液性応答および細胞性応答の両方を引き起こすという手段が提供される(16−19)。鼻腔内経路の効果は、歴史的に、非経口接種の効果よりも弱かったが、新しい投与製剤、アジュバント、およびVLPの使用は、このパラダイムを変え始めている。実際のところ、機能的なヘマグルチニンポリペプチドを含んでいるVLPは、鼻腔内投与に特に適している可能性がある。なぜなら、鼻粘膜に豊富に存在するシアル酸含有受容体が、HA抗原の結合を向上させ、粘液線毛クリアランスを減少させることになり得るからである。
【0100】
<アジュバント>
ワクチンに対するアジュバント効果を得る方法としては、種々の方法が知られており、それらは本明細書に開示のVLPと併用して使用され得る。一般的な原理および方法の詳細は、「”The Theory and Practical Application of Adjuvants", 1995, Duncan E. S. Stewart-Tull (ed.), John Wiley & Sons Ltd, ISBN 0-471-95170-6」、および「”Vaccines: New Generation Immunological Adjuvants”, 1995, Gregoriadis Gら (eds.), Plenum Press, New York, ISBN 0-306-45283-9」に記載されている。これら文献は、参照することによって本明細書に組み込まれる。
【0101】
ある実施形態では、VLPワクチンは、少なくとも1つのアジュバントと混合されたVLPを含んでいる。上記VLPワクチンに含まれている上記VLPと上記アジュバントとの割合は、重量比で約10:1から約1010:1までであり、例えば、約10:1から約100:1まで、約100:1から約103:1まで、約103:1から約104:1まで、約104:1から約105:1まで、約105:1から約106:1まで、約106:1から約107:1まで、約107:1から約108:1まで、約108:1から約109:1まで、または約109:1から約1010:1までである。当業者の一人は、アジュバントおよび最適な比を決定するための慣例的な実験を通して、適切な比を容易に決定することができる。本明細書に記載されているようなVLPとアジュバントとの混合物は、当業者に利用可能なあらゆる組み合わせの形態を含んでいてもよい。そのような形態には、特に限定されないが、同じ溶液内においてVLPとアジュバントとが分離されているという混合物、VLPとアジュバントとが共有連結されているという混合物、VLPとアジュバントとがイオン的に連結されているという混合物、VLPとアジュバントとが疎水的に連結されているという混合物(VLP膜に部分的にまたは完全に埋め込まれている形態を含む)、VLPとアジュバントとが親水的に連結されているという混合物、ならびにそれらの任意の組み合わせが包含される。
【0102】
アジュバントの好ましい例はポリペプチドアジュバントである。該ポリペプチドアジュバントは、VLPポリペプチドと一緒に共発現されることによって、またはVLPポリペプチドと融合されてキメラポリペプチドが製造されることによって、本明細書に記載のVLPに容易に付加されるものであってもよい。鞭毛の主要タンパク質成分である細菌のフラジェリンは、Toll様受容体TLR5により先天性免疫系から認識されるために、アジュバントタンパク質として注目されている(65)。TLR5を介したフラジェリンのシグナルは、DCの成熟および遊走を誘導することによって、ならびに前炎症性メディエータの産生をもたらすマクロファージ、好中球および腸上皮細胞の活性化を誘導することによって、先天性免疫機能と獲得免疫機能との両方に効果を及ぼす(66−72)。
【0103】
TLR5は、フラジェリンモノマーに保存された、このタンパク質に独特で鞭毛の機能に必須の構造を認識するが、免疫学的圧力に応じた該構造の突然変異体は認識しない(73)。TLR5は、100fMの濃度に対して感受性があるが、完全なフィラメントを認識しない。鞭毛のモノマーへの分解には、結合と刺激とが要求される。
【0104】
アジュバントとしてのフラジェリンは、非経口でまたは鼻腔内に投与される異種抗原に対する防御応答を誘導するための強力な活性を有しているし(66、74−77)、DNAワクチンに対するアジュバント効果も報告されている(78)。フラジェリンが使用されたとき、Th2への偏りが観察され、このことはインフルエンザなどの呼吸器ウイルスに対して適しているであろうが、マウスまたはサルにおいてIgEが誘導されたという証拠は見つかっていない。また、サルに鼻腔内投与または全身投与した後に、局所的炎症反応または全身性炎症反応が起こらなかったということも報告されている(74)。MyD88に依存する様式でTLR5を経由するフラジェリンシグナル、およびTLRを経由する他の全てのMyD88依存性のシグナルは、Th1へ偏ることになることが示されていたために、フラジェリンを使用した後に誘発される応答に関するTh2の特徴には、ある程度驚かされる(67、79)。重要なことは、フラジェリンに対する予め存在していた抗体が、アジュバントの有効性に対して、あまり効果を及ぼさず(74)、このことが、フラジェリンを複数回使用されるアジュバント(multi-use adjuvant)として魅力あるものにしていることである。
【0105】
多くの最近の鼻腔内ワクチンの試験に共通するテーマは、ワクチンの有効性を改善するためのアジュバントおよび/または投与系の使用である。そのような研究によれば、遺伝的に解毒されたE. coliの易熱性エンテロトキシンアジュバント(LT R192G)を含んでいるインフルエンザのH3ワクチンを鼻腔内投与した後にだけ、H5の暴露に対するヘテロサブタイプな防御がもたらされている。防御は、中和抗体の交差の誘導に基づいており、新たなワクチンを開発する上で鼻腔内経路が重要であることを実証した(22)。
【0106】
またサイトカイン、コロニー刺激因子(例えばGM−CSFおよびCSFなど);腫瘍壊死因子;インターロイキン−2、−7、−12、インターフェロン、および他の成長因子もアジュバントとして使用されてもよく、それらはVLPポリペプチドと混合するか、融合することによってVLPワクチン内に容易に含まれ得るために、好ましいものでもある。
【0107】
ある実施形態では、本明細書に開示のVLPワクチンは、Toll様受容体を介して作用する他のアジュバントを含んでいてもよい。そのようなアジュバントとしては、TLR9のリガンドであるCpGオリゴヌクレオチドを含んでいる核酸;TLR7のリガンドであるイミダゾキノリン;TLR7/8のリガンドである置換グアニン;ロキソリビン(Loxoribine)、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、二本鎖ポリ(I:C)、ポリイノシン酸、イミキモド(Imiquimod)(R−837)およびレシキモド(Resiquimod)(R−848)などのTLR7受容体;あるいはMPL(登録商標)または合成誘導体などのTLR4のアゴニストなどが挙げられる。
【0108】
あるアジュバントは、樹状細胞などのAPCによって、ワクチン分子の取り込みを促進し、該ワクチン分子を活性化する。非限定的な例は、免疫標的アジュバント;免疫調節アジュバント(毒素、サイトカインおよび細菌の誘導体など);油製剤;ポリマー;ミセル形成アジュバント;サポニン;免疫賦活複合体マトリックス(ISCOMマトリックス(ISCOM matrix));粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;およびカプセル化アジュバント(encapsulating adjuvant)からなる群から選択される。
【0109】
アジュバントのさらに別の例には、水酸化アルミニウムまたはリン酸アルミニウムなどのアルミニウム塩(みょうばん)(通常、0.05%から0.1%のアルミニウム塩の緩衝生理食塩水溶液として使用される(例えばNicklas (1992) Res. Immunol. 143:489-493参照))などの作用物質、0.25%の溶液として使用される糖の合成ポリマーとの混合物(例えばカルボポル(Carbopol(登録商標)))、70℃から101℃の範囲の温度で、30秒から2分間熱処理することによって得られる、ワクチン中のタンパク質の凝集体が包含される。また、アジュバントとしては、架橋剤を使って得られた凝集体も可能である。さらに、(i)ペプシン処理されたアルブミンに対する抗体(Fab断片)を用いた再活性化による凝集体、(ii)クリプトスポリジウム・パルバム(C. parvum)などの細菌細胞、またはグラム陰性菌のエンドトキシンもしくはリポ多糖体成分との混合物、(iii)マンニドモノオレイン酸などの生理学的に許容可能な油媒体の乳濁液(アラセルA(Aracel A))、あるいは(iv)ブロック置換体として使用されるパーフルオロカーボンの20%溶液との乳濁液(フルオソル−DA(Fluosol-DA))、が使われてもよい。スクアレンおよびIFAなどの油との混合物も好ましい。
【0110】
DDA(ジメチルジオクタデシルアンモニウムブロマイド)は、アジュバントの興味深い候補であるし、フロイント完全アジュバントおよびフロイント不完全アジュバント、ならびにキラヤ属のサポニン(クイルA(QuilA)およびQS21など)も興味深い候補である。他の候補には、リポ多糖体のポリ[ジ(エアロボキシラトフェノキシ)ホスファゼン(PCPP)の誘導体(モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など)が包含される。リポ多糖類を基本とするアジュバントは、優勢なTh1型応答を産生するためのものであることが好ましい。そのようなアジュバントとしては例えば、モノホスホリルリピドA(好ましくは3−デ−O−アシル化モノホスホリルリピドA)とアルミニウム塩との組み合わせが挙げられる。MPL(登録商標)アジュバントは、グラクソスミスクラインから入手することができる(例えば米国特許第4,436,727号明細書、米国特許第4,877,611号明細書、米国特許第4,866,034号明細書、および米国特許第4,912,094号免疫原性参照。これら文献のそれぞれは、リポ多糖類に関連する教示に特に関して、参照することによって完全に組み込まれる)。
【0111】
またリポソーム製剤も、アジュバント効果を奏することが知られており、それ故、リポソームアジュバントがVLPと併用される好ましい例である。
【0112】
免疫賦活複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、特にこの形式のアジュバントがMHCクラスIIの発現をAPCによって上方調節できることが示されたために、本発明にとって好ましい選択肢である。ISCOMマトリックスは、キラヤ由来の(場合によっては分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質から成るものである。免疫原性タンパク質(例えばVPL中の免疫原性タンパク質)と混合されたときに生じる粒状製剤は、ISCOM粒子として知られているものである。サポニンは、該ISCOM粒子の60−70%w/wを構成してもよく、コレステロールおよびリン脂質は該ISCOM粒子の10−15%w/wを構成してもよく、上記タンパク質は該ISCOM粒子の10−15%w/wを構成してもよい。免疫賦活複合体の組成および使用に関する詳細は、例えば、アジュバントを扱っている上記テキストブックにおいて見つけることができるが、例えば、「Morein Bら, 1995, Clin. Immunother. 3: 461-475」および「Barr I G and Mitchell G F, 1996, Immunol. and Cell Biol. 74: 8-25」(これらは参照することによって本明細書に組み込まれる)にも、完全な免疫賦活複合体を調製するために役に立つ説明が記載されている。
【0113】
本明細書に開示のVLPワクチンを有するアジュバント組み合わせに使用されてもよいサポニンは、ISCOMの形態であってもなくても、クイルAと呼ばれるキラヤサポナリアモリナ(Quillaja Saponaria Molina)の皮から得られたもの、およびそれの画分を包含する。なお、これらは、米国特許第5,057,540号明細書(特にクイルAの画分ならびにその単離方法および使用に関して、参照されることによって本明細書に完全に組み込まれる)、「”Saponins as vaccine adjuvants”, Kensil, C. R., Crit Rev Ther Drug Carrier Syst, 1996, 12 (1-2):1-55」、および欧州特許第0362279号明細書に記載されている。クイルAの特に好ましい画分は、QS21、QS7、およびQS17である。
【0114】
β−エスチンは、本発明のアジュバント組成物に使用される、もう1つ別の好ましい溶血性サポニンである。エスチンは、メルクの索引(第12版:項目3737)に、トチノキの実に生じるサポニンの混合物として記載されており、ラテン語は、Aesculus hippocastanumである。それのクロマトグラフィーおよび精製による単離は、「Fiedler, Arzneimittel-Forsch. 4, 213 (1953)」に記載されており、それのイオン交換樹脂による単離は、「Erbringら, 米国特許第3,238,190号明細書」」に記載されている。エスチンの画分は、精製されており、生物活性を有することが示されている(「Yoshikawa M, ら (Chem Pharm Bull (Tokyo) 1996 August;44(8):1454-1464)」)。β−エスチンは、アセスチン(aescin)としても知られている。
【0115】
本発明に使用されるもう1つべつの好ましい溶血性サポニンは、ジギトニンである。ジギトニンは、メルクの索引(上記第12版:項目3204)に、ジギタリス・プルプレア(Digitalis purpurea)の実から得られ、「Gisvoldら, J.Am.Pharm.Assoc., 1934, 23, 664」および「Ruhenstroth-Bauer, Physiol.Chem., 1955, 301, 621」に記載の手法によって精製される、サポニンとして記載されている。その使用は、コレステロール測定のための臨床試薬として記載されている。
【0116】
アジュバント効果を得るもう1つ別の(そして好ましい)可能性は、「Gosselinら, 1992」(参照によって、本明細書に組み込まれる)に記載の手法を使うことである。要するに、本発明の抗原などの関連抗原の提示は、単球/マクロファージ上のFC受容体に対する抗体(または抗原結合性の抗体断片)へ、該抗原を接合することによって改善されることができる。特に抗原と抗−FCRIとの間の接合によって、ワクチン接種のための免疫原性が向上されることが実証された。上記抗体は、発現によって、VLPポリペプチドの何れか1つとの融合体として生成した後に、または該融合体の生成の一環として、VLPに接合されてもよい。
【0117】
他の候補には、標的物質および免疫調節物質(すなわちサイトカイン)の使用が含まれる。また、サイトカインの合成誘発物質(ポリI:Cなど)が使用されてもよい。
【0118】
適切な微生物誘導体は、ムラミルジペプチド、フロイント完全アジュバント、RIBI(Ribi ImmunoChem Research Inc., Hamilton, Mont.)、およびトレハロースのジエステル体(TDMおよびTDEなど)から成る群から選択されてもよい。
【0119】
適切な免疫標的アジュバントには、例えばCD40リガンドおよびCD40抗体、またはその特異的な結合性断片(上記の考察を参照)、マンノース、Fab断片、CTLA−4が包含される。
【0120】
適切なポリマーアジュバントには、例えばデキストラン、PEG、デンプン、マンナン、およびマンノースなどの炭水化物;プラスチックポリマー;およびラテックスビーズなどのラテックスが包含される。
【0121】
免疫応答を調節するさらにもう1つ別の興味深い方法は、免疫源を、「仮想リンパ節(VLN)」に含ませることである(場合によってはアジュバントならびに薬学的に許容可能な担体および溶媒と一緒に含ませることである)。VLNは、「ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017-6501」によって開発された専売医療機器である。VLN(細い管状の機器)は、リンパ節の構造および機能を模倣している。VLNが皮下へ挿入されることによって、サイトカインおよびケモカインの激増を伴う無菌の炎症部位が生まれる。T細胞、B細胞、およびAPCは、この危険シグナルに迅速に反応し、炎症部位に向かい、VLNの多孔性マトリックス中に蓄積する。VLNを使用すると、抗原に対する免疫応答を開始するために要求される必須抗原の用量が減少すること、およびVLNを使ったワクチン接種によって得られる免疫防御は、アジュバントとしてRibiを使った従来の免疫化を凌ぐことが示された。この技術の概要は、「Gelber Cら, 1998, ”Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node”」、および「”From the Laboratory to the Clinic, Book of Abstracts, Oct. 12-15, 1998, Seascape Resort, Aptos, Calif.”」に記載されている。
【0122】
オリゴヌクレオチドが、VLPワクチンと併用されるアジュバントとして使用されてもよい。該オリゴヌクレオチドは、少なくとも3つ、より好ましくは少なくとも6つ、またはそれ以上のヌクレオチドによって分離されたジヌクレオチドCpGモチーフを、2つ以上含んでいることが好ましい。(CpGジヌクレオチドがメチル化されていない)CpG含有オリゴヌクレオチドは、優勢なTh1応答を誘導する。そのようなオリゴヌクレオチドは、公知であり、例えば国際公開第96/02555号パンフレット、国際公開第99/33488号パンフレット、米国特許第6,008,200号明細書、および米国特許第5,856,462号明細書に記載されている(これら文献のそれぞれは、特にアジュバントとしてのCpGオリゴヌクレオチドを作製および使用する方法に関して、参照することによって完全に本明細書に組み込まれる)。
【0123】
そのようなオリゴヌクレオチドアジュバントは、デオキシヌクレオチドであってもよい。好ましい実施形態では、オリゴヌクレオチドのヌクレオチド骨格は、ホスホロジチオエート、より好ましくはホスホロチオエート結合であるが、ホスホジエステルおよび他のヌクレオチド骨格(PNAなど)も、(混合された骨格結合を有するオリゴヌクレオチドを含む)本発明の範囲に含まれる。ホスホロチオエートのオリゴヌクレオチドまたはホスホロジチオエートを製造するための方法は、米国特許第5,666,153号明細書、米国特許第5,278,302号明細書、および国際公開第95/26204号パンフレットに記載されており、これら文献のそれぞれは、特にホスホロチオエートおよびホスホロジチオエートの教示に関して、参照することによって完全に本明細書に組み込まれる。
【0124】
好ましいオリゴヌクレオチドは、例えば以下の配列を有している。これら配列は、ホスホロチオエートで修飾されたヌクレオチド骨格を含んでいることが好ましい。
【0125】
(配列番号1)オリゴ1:TCC ATG ACG TTC CTG ACG TT (CpG 1826);
(配列番号2)オリゴ2:TCT CCC AGC GTG CGC CAT (CpG 1758);
(配列番号3)オリゴ3:ACC GAT GAC GTC GCC GGT GAC GGC ACC ACG;
(配列番号4)オリゴ4:TCG TCG TTT TGT CGT TTT GTC GTT (CpG 2006);
(配列番号5)オリゴ5:TCC ATG ACG TTC CTG ATG CT (CpG 1668)。
【0126】
他の好ましいCpGオリゴヌクレオチドには、重要でない欠失または付加を有する上記配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、技術的に公知の任意の方法(例えば欧州特許公開第468520号明細書)によって合成されてもよい。好ましくは、そのようなオリゴヌクレオチドは、自動合成機を利用して合成されてもよい。そのようなオリゴヌクレオチドアジュバントの長さは、10−50塩基の間であってもよい。もう1つ別のアジュバントシステムは、CpG含有オリゴヌクレオチドと、サポニン誘導体との組み合わせを含んでいる。特にCpGとQS21との組み合わせは、国際公開第00/09159号パンフレットに開示されている。
【0127】
多くの一相または多相乳濁液システムが記載されている。当業者の一人は、乳濁液によってVLPの構造が破壊されないように、そのような乳濁液システムをVLPとの使用に容易に適応させることができる。水中油型乳濁液のアジュバントは、それ自体、アジュバント組成物として有用であると示唆されており(欧州特許第0399843号明細書)、水中油型乳濁液と他の活性剤との組み合わせもワクチン用のアジュバントとして記載されている(国際公開第95/17210号パンフレット、国際公開第98/56414号パンフレット、国際公開第99/12565号パンフレット、国際公開第99/11241号パンフレット)。油中水型乳濁液、および水中油中水型乳濁液(water in oil in water emulsion)などの他の油乳濁液のアジュバントは、それぞれ、米国特許第5,422,109号明細書および欧州特許第0480982号明細書、ならびに米国特許第5,424,067号明細書および欧州特許第0480981号明細書に記載されている。
【0128】
本明細書に記載されたVLPワクチンとともに使用するための油乳濁液アジュバントは、天然または合成されたものであってもよく、かつ無機的または有機的なものであってもよい。無機的および有機的な油の例は、当業者に容易に理解される。
【0129】
人への投与に好適な任意の水中油型組成物のために、乳濁系の上記油層は、好ましくは新陳代謝させることができる油を含む。新陳代謝させることができる油という用語の意味は、当業者によく知られている。新陳代謝させることができるという用語は「新陳代謝により変化される能力を有する」と定義できる(Dorland's Illustrated Medical Dictionary, W. B. Sanders Company, 25th edition (1974))。上記油は、受容者に有毒ではなく、かつ新陳代謝により変化する能力を有する任意の植物性油、魚油、動物油、または合成油であってよい。ナッツ(ピーナッツ油など)、種、および種子は、植物油の一般的な源である。合成油も本発明の一部であり、かつNEOBEE(登録商標)などのような市販されている油を包含できる。スクアレン(2,6,10,15, 19,23-Hexamethyl-2,6,10,14,18,22-tetracosahexaene)は、サメの肝臓油中に多量に見出され、かつオリーブ油、小麦の胚種油、ぬか油、および酵母に少量見出される不飽和油である。該スクアレンは、本発明の使用に特に好ましい油である。スクアレンは、新陳代謝させることができる油であり、事実、コレステロールの生合成における中間体であるという長所を有する(Merck index, 10th Edition, entry no.8619)。
【0130】
特に好ましい油乳濁液は、水乳濁液中の油であり、特にスクアレンの水乳濁液が好ましい。
【0131】
加えて、加工していない酸化防止剤において本発明の最も好ましい油乳濁液アジュバントは、好ましくはα−トコフェロール油(vitamin E, EP 0 382 271 Bl)である。
【0132】
国際公開第95/17210号パンフレットおよび国際公開第99/11241号パンフレットは、随意的に免疫賦活剤QS21および/または3D−MPLとともに製剤されたスクアレン、α−トコフェロール、およびTWEEN80に基づく乳濁液アジュバントを開示している。国際公開第99/12565号パンフレットは、油層にステロールを添加したこれらのスクアレン乳濁液の改良点について開示している。さらに、トリカプリリン(tricaprylin)(C27H50O6)のようなトリグリセリドは、乳濁液の安定化のために油層を添加してもよい。
【0133】
水乳濁液における安定した油内に見出される油滴の大きさは、好ましくは1ミクロンよりも小さく、実質的に30〜600nmの範囲であってよく、好ましくは、実質的に直径約30〜500nmであり、最も好ましくは実質的に直径150〜500nmであり、特に光相関分光法により測定した直径において約150nmである。この点で、油滴の80%は、型どおりに好ましい範囲内であるべきで、より好ましくは90%、最も好ましくは95%の油滴が定義された範囲の大きさ内であるべきである。本発明の油乳濁液に存在する構成要素の量は、スクアレンのように通常2〜10%の範囲が油である。また、構成要素が存在する場合、2から10%のアルファトコフェロール;およびポリオキシエチレンソルビタンモノオレエートのような0.3%〜3%の界面活性剤が存在する。好ましい油の比率は、アルファトコフェロールは、1に等しいか、または1よりも小さく、この条件では、より安定な乳濁液が提供される。スパン85(Span 85)は、約1%のレベルで存在してもよい。いくつかの場合では、本明細書に開示されたVLPワクチンは、さらに安定化剤を含むことが好都合であってよい。
【0134】
水乳濁液中に油を形成する方法は、当業者によく知られている。通常、上記方法は、PBS/TWEEN80(登録商標)のような界面活性剤と油層を混合する工程を含み、その後のホモジナイザーを用いるホモジナイゼーションにおいて、上記ミキサーにシリンジの針を通して2回混合することを含むことは、少量の液体のホモジナイゼーションにとって好適であることは当業者にとって明らかである。
【0135】
同様に、マイクロフルイダイザー(M110Sマイクロフルイディクス機、最大50回、6バールの入力最大圧において2分間(約850バールの出力圧力))における乳状化処理は、当業者によって少量または大量の乳濁液を形成するために採用できる。この採用は、調製物が所望の直径の油滴により達成されるまで、得られた乳濁液の測定を含む通常の実験により達成できる。
【0136】
本明細書において開示された上記VLPワクチンの調製物は、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮的投与、静脈内投与、または皮下投与によるワクチン投与によってウイルスに感染しやすい、またはウイルスの感染に苦しんでいる哺乳類または鳥を保護または処理するために用いられてよい。ワクチン調製物の全身性の投与方法は、従来型のシリンジおよび針、または固体のワクチンの弾道輸送(ballistic delivery)のために設計された装置(国際公開第99/27961号パンフレット)、無針圧液体噴出装置(米国特許出願公開第4,596,556号明細書;米国特許出願公開第5,993,412号明細書)、または経皮貼布(transdermal patches)(国際公開第97/48440号パンフレット;国際公開第98/28037号パンフレット)を含み得る。VLPワクチンは、皮膚にも適用され得る(経皮的(transdermal)または経皮投与(transcutaneous delivery)(国際公開第98/20734号パンフレット;国際公開98/28037号パンフレット)。本明細書において開示されている上記VLPワクチンは、それゆえ、全身投与のための輸送VLPワクチンまたはアジュバント組成物で前もって満たされた輸送装置を含む。従って、本明細書においては、好ましくは哺乳類または鳥といった個体において、本明細書に記載された任意のVLP組成物を含むワクチンの投与を含み、かつ任意に個体へのアジュバントおよび/またはキャリアを含む免疫応答を誘導する方法を提供する。ここで、ワクチンは、非経口または全身経路を介して投与される。
【0137】
好ましくは、本発明の上記ワクチン調製物は、経口(oral)/消化管(alimentary)または鼻(nasal)経路のような粘膜(mucosal)経路を介するワクチンの投与方法により、ウイルス感染に影響を受けやすい、また、ウイルス感染に苦しんでいる哺乳類または鳥の保護または処理のために用いられてよい。選択的な粘膜岐路は、膣内および直腸内(intra-rectal)である。投与の好ましい粘膜経路は鼻経路を介し、鼻腔ワクチン接種と称される。鼻腔ワクチン接種の方法は、当業者に公知であり、免疫性をあたえられた個体の鼻咽頭中のワクチンにおいて、小滴、噴霧、または乾燥粉末形態の投与を含む。霧状またはエアロゾル化されたワクチンの製剤は、それゆえ、本明細書に開示されたVLPワクチンの好ましい形状である。経口投与のための腹痛抵抗性カプセルおよび顆粒のような腸溶性の製剤、直腸または膣内投与のための座薬も、本明細書において開示されたVLPワクチンの製剤である。
【0138】
本明細書において開示された好ましいVLPワクチン組成物は、ワクチン接種による全身性ワクチン接種に換えて、ヒトにおいて好適に適用できる粘膜ワクチンの種類を表す。
【0139】
上記VLPワクチンは、経口経路を介して投与されてもよい。このような場合、薬学的に許容可能な賦形剤は、アルカリ緩衝液、または腸溶性カプセルもしくは顆粒を含んでもよい。上記VLPワクチンは、膣経路により投与されてもよい。このような場合、薬学的に許容可能な賦形剤は、乳化剤、CARBOPOL(登録商標)のようなポリマー、およびその他の公知の膣クリームおよび座薬の安定剤を含んでもよい。上記VLPワクチンは、直腸経路により投与されてもよい。この場合、賦形剤は、直腸にいれる座薬を形作るために当業者に知られたワックスおよびポリマーを含んでもよい。
【0140】
また、上記VLPワクチン製剤は、キトサン(上述したように)または他のポリカチオン性ポリマー、ポリラクチド粒子およびポリラクチド−コグリコライド(polylactide-coglycolide)粒子、ポリ−N−アセチルグリコサミンに基づくポリマーマトリックス、ポリサッカライドからなる粒子または化学的に修飾されたポリサッカライドからなる粒子、リポソームに基づく粒子および脂質に基づく粒子、ならびにグリセロールモノマーからなる粒子などからなるワクチン媒体と結合してもよい。上記サポニンは、リポソームまたはISCOMsのような粒子の構造を形作るためにコレステロールの存在下で製剤されてもよい。さらに、上記サポニンは、非粒子的な溶液または懸濁液のどちらか、もしくは、小ラメラリポソーム(paucilamelar liposome)またはISCOMのような粒子構造において、ポリオキシエチレンエーテルまたはエステルと共に製剤されてもよい。
【0141】
本明細書に記載されたような薬学的組成物およびワクチン組成物における使用のための追加の実例となるアジュバントは、SAF(Chiron, Calif., United States)、MF‐59(Chiron, 例えば、Granoffら(1997) Infect Immun. 65 (5):1710-1715を参照のこと)、アジュバンドのSBASシリーズ(例えば、SB‐AS2(SmithKline Beecham adjuvant system #2; an oil-in- water emulsion containing MPL and QS21));SmithKline Beechamから市販されているSBAS‐4(SmithKline Beecham adjuvant system #4; contains alum and MPL)、Detox(Enhanzyn)(登録商標)(GlaxoSmithKline)、RC‐512、RC‐522、RC‐527、RC‐529、RC‐544、およびRC‐560(GlaxoSmithKline)ならびにその全体が本明細書に組み込まれ開示される米国特許第08/853,826号明細書および米国特許第09/074,720号明細書に記載されたような他のアミノアルキルグルコサミド 4−ホスフェート(AGPs)を含む。
【0142】
アジュバントの他の実施例は、限定されないが、ハンターズタイターマックス(Hunter's TiterMax)(登録商標)アジュバント(CytRx Corp., Norcross, Ga.);ゲルブアジュバント(Gerbu adjuvants)(Gerbu Biotechnik GmbH, Gaiberg, Germany);ニトロセルロース(Nilsson and Larsson (1992) Res. Immunol. 143:553-557);モンタミドアジュバントセピックISAシリーズ(Seppic ISA series of Montamide adjuvants)(例えば、ISA‐51、ISA‐57、ISA‐720、ISA‐151など;Seppic, Paris, France)のような鉱物油、非鉱物油、油中水型乳剤、または水中油型乳剤を含む製剤に基づくミョウバン(alum)(例えば、アルミニウム水酸化物(aluminum hydroxide)、アルミニウムリン酸塩(aluminum phosphate)乳濁液;およびPROV AX(登録商標)(IDEC Pharmaceuticals);OM‐174(脂質Aに関連するグルコサミンニ糖類);リーシュマニア属伸長因子(Leishmania elongation factor);CRL1005のようなミセルを形作る非イオン化ブロック共重合体;およびシンテックスアジュバント製剤(Syntex Adjuvant Formulation)を含む。例えば、O'Haganら(2001) Biomol Eng. 18(3):69-85;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」D. O'Hagan, ed. (2000) Humana Pressを参照のこと。
【0143】
その他の好ましいアジュバントは、以下の製剤のアジュバント分子を含む。
【0144】
HO(CH2CH2O)n-A-R、(I)
ここで、nは1〜50であり、Aは結合または−C(O)−であり、RはC1〜50のアルキル基またはフェニルC1〜50のアルキル基である。
【0145】
本発明の実施形態の1つは一般式(I)のポリオキシエチレンエーテルを含む。ここで、nは1から50の間であり、好ましくは4から24であり、最も好ましくは9である;上記Rの構成要素は、C.sub.1−50であり、好ましくはC.sub.4‐C.sub.20アルキルおよび最も好ましくはC.sub.12アルキルであり、かつAは結合である。ポリオキシエチレンエーテルの濃度は、0.1〜20%であり、好ましくは0.1〜10%であり、最も好ましくは0.1〜1%の範囲である。好ましくは、ポリオキシエチレンエーテルは以下の群から選択される:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステオリルエーテル(polyoxyethylene-9-steoryl ether)、ポリオキシエチレン−8−ステオリルエーテル(polyoxyethylene-8-steoryl ether)、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテル。ポリオキシエチレンラウリルエーテルのようなポリオキシエチレンエーテルは、メルクインデックスに記載されている(12.sup.th edition: entry 7717)。これらのアジュバント分子は、国際公開第99/52549号パンフレットに記載されている。
【0146】
上記一般式(I)によるポリオキシエチレンエーテルは、もし望むのであれば、他のアジュバントに結合されてもよい。例えば、好ましいアジュバントの組み合わせは、好ましくは上記したようなCpGとの組み合わせである。
【0147】
本明細書に開示されたVLPワクチンと共に使用するための薬学的に許容可能な好ましい賦形剤の例は、水、リン酸緩衝液、および等浸透圧緩衝液を含む。
【0148】
本発明は、以下の限定されない参考文献により、さらに理解される。本明細書に記載したように、本発明は、天然に脂質ラフトと関連しない抗原に結合する脂質ラフト関連ポリペプチドの任意のものに組み込まれるキメラVLPを含む。以下の例は、インフルエンザ抗原とキメラVLPについての本発明の代表的な実施形態を記載する。
【実施例】
【0149】
〔実施例1−キメラインフルエンザVLPの生成〕
上記MLVgagをコードする配列は、全モロニームリー白血病(Moloney murin leukemia)ウイルス両宿主性のプロウイルス配列を含むプラスミドpAMS(ATCC)からPCRにより得られた。上記gagをコードする配列は、pFastBacI(インビトロジェン)のポリヘドリンプロモーター後に挿入された。また、得られたプラスミドは、バキュロウイルスゲノムに組み込むためにDH10Bacコンピテント細胞にトランスフォームされた。高分子量のBacmidDNAは、続いて、精製され、gagを発現する組み換えバキュロウイルスの創出のためにSf9細胞にトランスフェクトされた。A/PR/8/34(H1N1)において、それぞれ、ヘマグルチニンおよびノイラミニダーゼをコードする2つの他の組み換えバキュロウイルスは、ウイルスRNAに由来するHAおよびNAをコードする配列のRT‐PCRクローニング後、同じように(similar fashion)形成された。最後に、全ての3つの産生物(HA−gag−NA)をコードしている単一のバキュロウイルスベクターは、各pFastBacIプラスミドから個々のpFastBacIプラスミドに由来するHA、gag、およびNA発現ユニット(ポリへドリンプロモーターをコードする配列―ポリA部位)を単一のpFastBacIベクターに組み込むことで形成された。最初の分析について、gagまたはHAまたはgag‐HA‐NAをコードする組み換えバキュロウイルスは、MOI>1で6ウェルプレートにおいてSf9細胞に感染された。感染の3日後、培地上清は、夾雑物を取り除かれ、続いて、20%スクロース勾配により100,000×gで沈殿された。沈殿は、gagおよびH1N1−特異的抗血清ウエスタンブロット分析により分析された(図1AおよびBを参照のこと)。
【0150】
図1AおよびBにおいて、それぞれ左側の3つのレーンは、gagまたはHAまたはコントロール(EV=空ベクター)のそれぞれに感染したSf9細胞の結果を示す。予期したように、gagのみを感染させたバキュロウイルスの結果は、VLP出芽のために高分子量培地においてgag抗原に顕著な量をもたらす(図1A、レーン「Gag」)。対照的に、HAのみに感染したバキュロウイルスは、培地にHAをほとんど放出していない。しかし、HA−gag‐NAのトリプルベクターによるSf9細胞の感染は、gagの発現が細胞外のHAを引き付けられることを示す100,000×g画分(レーン1−9、図1AおよびB)において出現するgagおよびHAの両者に顕著な量をもたらす。
【0151】
図2AおよびBは、20−60%スクロースステップ勾配を用いて、ペレット状のHA−gag‐NA VLPを再遠心分離した後に、各勾配の画分をウエスタンブロットした結果を示す図である。gagおよびHAを誘導する同じ画分におけるgagおよびHAの両者のピークは、約1.16g/mlの密度において共結合していることを表し、このことはgagとHAはVLP中に存在することを示す。
【0152】
〔実施例2−HAに結合した炭疽菌PAエピトープを含んでいるHA-gag‐NA VLPの産生、特徴付け、および免疫原性試験〕
実施例1に記載したように、A/PR/8/34の生産物MLVgag、およびHA、およびNAを発現している個々のバキュロウイルスが作製された。加えて、全ての3つの生成物をコードしているトリプル発現組み換えバキュロウイルスもコンストラクトされ、感染した昆虫細胞由来の沈殿した培地上清のスクロース密度勾配遠心分離は、HAで形成されたVLPを誘導するウェスタンブロッティングにより検出されたため、gagおよびHAの同時結合を示す。トリプル発現ベクターにおけるコーディング配列の配置を、図3に示した。HA、gag、およびNAをコードしている要素は、頭−尾形態(head-to-tail fashion)において自身のプロモーター(Pr)およびポリアデニル化配列(pA)と共に並んでいる。単一のバキュロウイルスへの全てのコーディング配列の結合は、分離されたウイルスに共通感染する必要を回避し、3つの分離されたウイルスの感染の一貫的な多様性(consistent multiplicity)を達成することの困難さと関連する。
【0153】
実施例2における上記VLPワクチンは、HA遺伝子がB.anthracisの保護抗原(PA)によるHA免疫原性の決定の大部分を入れ替えることにより修飾されたHA遺伝子を除いて、同じように生成される。このことは、HA遺伝子(アミノ酸位置18−343)のHA1部位をコードしている配列(PAのC末端断片の140アミノ酸)を入れ替えることにより達成される。上記PAをコードしている配列は、HA1をコードしている配列(アミノ酸位置18−343)(図4)の直接入れ替えとして、HAシグナルペプチドをコードしている配列(アミノ酸位置1−17)とHA2をコードしている配列(アミノ酸位置344−565)の間のHA遺伝子に挿入される。修飾されたPA−HA遺伝子を含むトリプルバキュロウイルス発現ベクターは、MLVgagポリペプチド、NAポリペプチド、および入れ替え要素を含む修飾されたPA−HAポリペプチドを発現する。培養中の感染したSf9細胞に上記トリプル発現ベクターが使用された場合、炭疽菌PAエピトープを含むVLPが培地中に観察される。これは、修飾されたPA−HAポリペプチドが、脂質ラフトホーミング配列に残っており、脂質ラフトドメインから出芽している修飾されたPA粒子に組み込まれるからである。この証拠は、(PA)HA−gag−NAトリプル発現ベクターに感染した細胞由来の培地の回収、夾雑物の培地のクリーニング、および20%以上のスクロース勾配において100,000×gで遠心分離によりキメラVLPの回収により示される。VLPへのPAの組み込みの証拠は、PAに特異的な抗体を用いるウエスタンブロットにより得られる。定義により、これらの条件下において20%スクロース緩衝(sucrose cushion)による沈殿物は、天然には粒子であり、VLP形成の証拠を提供する。
【0154】
(サイズ排除クロマトグラフィー)
VLPを精製したスクロース勾配は、セファロースCL−4Bを用いたサイズ排除クロマトグラフィーにより実行される。また、画分はウエスタンブロットによりMLV gagおよび炭疽菌PAエピトープを観察される。VLPは、空隙容量で抽出され、MLV gag、NA、およびPAに修飾されたHAを含んでいる。
【0155】
(電子顕微鏡)
スクロース勾配に由来するVLP試料は、2%のグルタルアルデヒドで処理され、EMグリッド上で吸収され、ナトリウムホスホタングステート(sodium phosphotungstate)でネガティブ染色され、電子顕微鏡で観察される。
【0156】
(免疫原性)
キメラVLP(炭疽菌PA修飾HA含有VLP)の免疫原性は、参考文献(44)に記載のように、メスBalb/cマウスにおいて、炭疽菌感染防御抗原(PA)構造に類似する鼻腔内キトサン/MPL構造を用いて評価することができる。VLPは、Tris緩衝生理食塩水に再懸濁して、20〜60%スクロース密度勾配(sucrose density gradient)にバンドした後、20%スクロースクッションを介したVLP含有培養培地を、100,000×gで1時間、ペレット形成することによって精製することができる。VLP含有画分は、SDS−PAGEまたはウエスタンブロットによって同定して、保管することができる。VLPサンプルをPBS中において透析し、遠心マイクロ濃縮器(centrifuge microconcentrators)を用いて、または100,000×gで遠心分離することによって、濃縮させることができる。
【0157】
免疫処置のために、キトサン 40μg、VLP 20μg(gagに基づく)およびMPL 5μgを含む液体構成物(15μl)を、免疫処置一人分のための、2つの鼻孔用に分割することができる。0〜4週間の鼻腔内投与の前に、イソフルレンを用いて、動物を軽度に麻酔にかけることができる。また、ポジティブコントロールとしての腹腔内予防接種のために、VLPをMPLまたはコレラ毒素と一緒にPBS中で製剤化することができる。全身IgG反応を、PA特異的免疫反応のためのELISA法によって観察することができる。犠牲の時点において、ブロンコバスター洗浄(broncoaveolar lavage)サンプルもまた、PA特異的IgA反応の決定のために収集することができる。
【0158】
実施例2および以下の他の実施例における典型的な免疫処置実験は、スチューデントの対立t−検定(Student’s unpaired t-test)を用いて示されるように、統計的優位性に達する適切な可能性を提供し得る、1グループにつき最小限の8匹のマウスを使用することができる。免疫処置は、典型的に、各免疫処置の後10〜14日間行う血液サンプリングから4週間間隔をあけて、最初のおよび追加の免疫予防接種を伴うことができる。上述したように、ブロンコバスター洗浄サンプルは、IgA決定のための犠牲動物から収集することができる。
【0159】
〔実施例3:機能強化VLPの生成および免疫原性試験〕
本実施例3は、NAに取り付けられた炭疽菌PAエピトープに対する適応免疫反応の長さを促進するために、TLR5アゴニストフラジェリンを組み込むことによる、免疫原性および防御の向上のためのVLPの機能強化を証明することができる。
【0160】
(フラジェリン組み込みによるアジュバント効果)
フラジェリンコード配列は、最近、S.ティピムリアム(S. typimurium)のゲノムDNAからクローン化し、gagコード配列の3’末端に、ちょうど5’から終止コドンまで挿入した。フラジェリンコード配列はまた、A/PR/8/34 HAコード配列のN末端に、シグナルペプチドと成熟コード配列との間の境界に位置するPstI部位を利用して組み込むことも可能である。HAにおけるこの部位の組み込みは、SDS PAGEによって示されるように、予想される分子量増加を伴う、キメラHA分子の適切な発現を引き起こす。NAポリペプチドは、そのC末端に融合された140アミノ酸炭疽菌C末端PAドメインを有することができる。NAのC末端は、C末端の伸長部分が分子の外側に露出することが予想されるように、分子包囲の外側において見られる。フラジェリン修飾gagおよびHAコード配列は、三種類のバキュロウイルスの組換え体(HA−gag−NA(PA))の生成に用いることができる。フラジェリン修飾gagまたはフラジェリン修飾HAを有するVLPをコードするバキュロウイルス組換え体を生成し、基準VLP欠失フラジェリン配列に対する免疫原性試験のための、VLPを生成するために用いることができる(全てのVLPは、標的エピトープとしてPA修飾NAを含むことが可能であり、gagまたはHAに取り付けられたフラジェリン配列を含んでいても欠失していてもよい。実施例2において述べたように、全ての免疫処理実験は、最初のおよび追加の免疫予防接種を4週間間隔で採用することができる。免疫学的計測は、上述したように、全身IgGおよび粘膜IgA反応の両方を試験する、ELISA分析を介して行うことができる。
【0161】
フラジェリン組み込みのためのHAおよびgag挿入部位は、VLPの外側および内側のそれぞれにあるので、異なる程度のアジュバント効果が観察され得る。HAのN末端におけるフラジェリン挿入は、上皮粘膜の細胞におけるTLR5レセプターへのフラジェリンの接近を容易にするという結果をもたらし得る。一方、gag部位における挿入は、異なる接近をもたらす。細胞に対するVLP結合、および正常なインフルエンザウイルス侵入経路を介した内在化は、細胞内におけるgag−フラジェリン生成の堆積という結果をもたらし得る。これは、gag−フラジェリンのコンストラクトとHA−フラジェリンのコンストラクトとの間の異なるTLR5−介在アジュバント効果の結果をもたらし得る。自身の粘膜上皮細胞への結合および侵入のためのVLPの能力は、免疫原性において効果を有しているので、本発明者らは、TPCK−トリプシン処理を伴うおよび伴わない、フラジェリン修飾VLPおよび正常なVLPにおけるVLP免疫原性実験を行い得る。トリプシン処理したVLPのHA開裂は、VLP侵入の重要性を試験する免疫原性試験の開始前に、ウエスタンブロットによって確認することができる。さらに、細胞に融合および侵入するためのトリプシン処理したVLPの能力を、緑色蛍光タンパク質(GFP)修飾gag産物を含むVLPを使用して、in vitroにおける蛍光顕微鏡検査法によって試験することができる。この試験が、その出芽活性を破棄せずに、MLV gagがそのC末端においてGFPにより修飾され得ることが、すでに示されている(60)。
【0162】
フラジェリンの非TLR5結合領域の多くを除去することによりgag出芽活性を最大化するための、フラジェリンコード配列のサブフラグメントの使用もまた試験することができる。フラジェリンモノマー内におけるTLR5認識部位の最近の解析は、この努力によって促進することができる(73)。
【0163】
【表1】
【0164】
〔実施例4:gag−NA−(Norwalk−P2)VLPの生成、特徴付け、および免疫原性試験〕
本実施例4におけるVLPワクチンは、実施例3と同様のベクターにおいて生成することができる。ベクターは、MLV gagポリペプチドおよびNAポリペプチドのC末端に融合したノーウォーク(Norwalk)ウイルスカプシドタンパク質(VP1)のP2ドメインを有するNAポリペプチドを発現し得る。
【0165】
(初期分析)
本実施例において生成されたVLPは、ノーウォークウイルスキャプシドタンパク質特異的抗体を用いたウエスタンブロットによって特徴付けることが可能である。ノーウォークキャプシドタンパク質のP2ドメインは、優勢抗体結合部位であるので、ノーウォークキャプシド特異的抗体は、P2−修飾NAタンパク質を認識することが期待され得る。
【0166】
(サイズ排除クロマトグラフィー)
スクロース密度勾配で精製したVLPは、セファロースCL−4Bを用いたサイズ排除クロマトグラフィーの対象となり得、画分をウエスタンブロットすることによって、MLV gagおよびノーウォークP2ドメインを観察することができる。VLPは、空隙容量(void volume)において抽出され、MLV gag、P2修飾NAを含むことができる。
【0167】
(電子顕微鏡検査)
スクロース勾配からのVLPサンプルを、2% グルタルアルデヒドを用いて処理し、EMグリッド(EM grids)上に吸収させ、タングストリン酸ナトリウムを用いてネガティブ染色し、電子顕微鏡検査によって試験した。
【0168】
(免疫原性)
VLP免疫原性は、参考文献(44)に記載のように、メスBalb/cマウスにおいて、炭疽菌感染防御抗原(PA)構造に類似する鼻腔内キトサン/MPL構造を用いて評価することができる。VLPは、Tris緩衝生理食塩水中に再懸濁して、20〜60%スクロース密度勾配にバンドした後、20% スクロースクッションを介したVLP含有培養培地を、100,000×gで1時間、ペレット形成することによって精製することができる。VLP含有分画を、ウエスタンブロット分析によって同定し、保管することができる。VLPサンプルをPBS中において透析し、遠心マイクロ濃縮器を用いて、または100,000×gで遠心分離して、濃縮することができる。
【0169】
免疫処理のために、キトサン 40μg、VLP 20μg(gagに基づく)およびMPL 5μgを含む液体構成物(15μl)を、免疫処置一人分のための、2つの鼻孔用に分割することができる。0〜4週間の鼻腔内投与の前に、イソフルレンを用いて、動物を軽度に麻酔にかけることができる。また、ポジティブコントロールとしての腹腔内予防接種のために、VLPをMPLまたはコレラ毒素を有するPBS中に構築することができる。追加のポジティブコントロール動物は、ELISAによって観察され得るノーウォーク P2に特異的な全身IgG反応を伴う筋肉内予防接種を受けることができる。ELISAのための抗原の源は、バキュロウイルス生成ノーウォークVP1キャプシドタンパク質であることができる。ELISAによるノーウォークPA特異的IgA反応の測定のために、ブロンコバスター洗浄(broncoaveolar lavage)サンプルもまた、最終免疫処理後10〜14日間収集することができる。
【0170】
〔実施例5:機能強化VLPの生成および免疫原性試験〕
本実施例5は、適応免疫反応の長さを促進するために、TLR5アゴニストフラジェリンを組み込むことによる、免疫原性および防御を向上させるためのVLPの機能強化を証明することができる。
【0171】
(フラジェリン組み込みのためのアジュバントの効果)
フラジェリンコード配列は、最近、S.ティピムリアム(S. typimurium)のゲノムDNAからクローン化し、gagコード配列の3’末端に、ちょうど5’から終止コドンまで挿入した。フラジェリンコード配列はまた、A/PR/8/34 HAコード配列のN末端に、シグナルペプチドと成熟コード配列との間の境界に位置するPstI部位を利用して組み込むことも可能である。HAにおけるこの部位の組み込みは、SDS PAGEによって示されるように、予想される分子量増加を伴う、キメラHA分子の適切な発現を引き起こす。NAポリペプチドは、そのC末端に融合されたノーウォークウイルス(アミノ酸279〜405)のP2ドメインを有することができる。フラジェリン修飾gagおよびHAコード配列は、実施例5に示すように、三種類のバキュロウイルスの組換え体(HA−gag−NA(P2))を生成するために用いることができる。VLPをコードし、フラジェリン修飾gagまたはフラジェリン修飾HAを有する組換えバキュロウイルスを生成し、基準VLP欠失フラジェリン配列に対する免疫原性試験のためのVLPを生成するために用いることができる(全てのVLPは、標的エピトープとしてノーウォーク P2修飾NAを含むことが可能であり、gagまたはHAに取り付けられたフラジェリン配列を含んでいても欠失していてもよい。上述した実施例で述べたように、全ての免疫処理実験は、最初のおよび追加の免疫予防接種を4週間間隔で採用することができる。免疫学的計測は、上述したように、全身IgGおよび粘膜IgA反応の両方で試験する、ELISA分析を介して行うことができる。
【0172】
フラジェリン組み込みのためのHAおよびgag挿入部位は、VLPの外側および内側のそれぞれにあるので、異なる程度のアジュバント効果が観察され得る。HAのN末端におけるフラジェリン挿入は、上皮粘膜の細胞におけるTLR5レセプターへのフラジェリンの接近を容易にするという結果をもたらし得る。一方、gag部位における挿入は、異なる接近をもたらし得る。細胞に対するVLP結合および正常なインフルエンザウイルス侵入経路を介した内在化は、細胞内におけるgag−フラジェリン生成の堆積という結果をもたらし得る。これは、gag−フラジェリン構築物とHA−フラジェリン構築物との間の異なるTLR5−介在アジュバント効果の結果をもたらし得る。自身の粘膜上皮細胞への結合および侵入のためのVLPの能力は、免疫原性において効果を有しているので、本発明者らは、TPCK−トリプシン処理を伴うおよび伴わない、フラジェリン修飾VLPおよび正常なVLPにおけるVLP免疫原性実験を行い得る。トリプシン処理したVLPのHA開裂は、VLP侵入の重要性を試験する免疫原性試験の開始前に、ウエスタンブロットによって確認することができる。さらに、細胞に融合および侵入するためのトリプシン処理したVLPの能力を、緑色蛍光タンパク質(GFP)修飾gag産物を含むVLPを使用して、in vitroにおける蛍光顕微鏡検査法によって試験することができる。この試験がその出芽活性を破棄せずに、MLV gagがそのC末端においてGFPにより修飾され得ることはすでに示されている(60)。
【0173】
フラジェリンの非TLR5結合領域の多くを除去することによりgag出芽活性を最大化するための、フラジェリンコード配列のサブフラグメントの使用もまた試験することができる。フラジェリンモノマー内におけるTLR5認識部位の最近の解析は、この努力によって促進することができる(73)。
【0174】
【表2】
【0175】
〔実施例6:RSV−F−gag VLPの生成、特徴付けおよび免疫原性試験〕
本実施例における呼吸器合胞体ウイルス(RSV)のためのVLPワクチンは、インフルエンザHAおよびNAの脂質ラフト標的特性と同様である、RSV融合(F)タンパク質の脂質ラフト標的特性を利用することができる。これらの特性のお陰で、RSV Fタンパク質自身がポリペプチドに関連する脂質ラフトであり、それゆえに、キメラタンパク質の形成を必要とせずに、インフルエンザHAおよびNAと酷似のgagベースのVLPに直接組み込むことができる。
【0176】
RSV Fタンパク質は、最後まで、以下に示す5’プライマーおよび3’プライマーを用いて、標準RT−PCRクローン技術によってクローン化することができる(プライマー中に下線を付した配列は、RSV F 5’末端および3’末端コード配列のホモログであり、残りの配列は、pFastBac1ベクター中にクローニングするのに便利な制限酵素認識部位を含む)。
【0177】
(SEQ ID NO:6)
5’プライマー: ATATAGGCGCGCCACCATGGAGTTGCTAATCCTCAAAGC
(SEQ ID NO:7)
3’プライマー: ATATAGCGGCCGCTTAGTTACTAAATGCAATATTATTTATACCACTCAG
【0178】
上述したプライマーを用いたRT−PCRによるRSV F遺伝子の生成によって、pFB−Fと称するベクターとなるpFastBac1に挿入するための結合性のある端部を生成するために、AscIおよびNotIのいずれかの端部において切断され得るフラグメントを得ることができる。pFB−Fベクターの完成において、このベクターは、pFB−gagからのSnaBI−HpaIフラグメントの挿入のために、HpaIにおいて切断されることができる。これにより。MLV−gagおよびRSV−Fの両方をコードする二種類のバキュロウイルスを発現するベクターを得ることができる。このプラスミドのFおよびGag発現領域の主要な特徴の配置図を、図5に示す。
【0179】
ダブル発現(F−Gag)ベクターを、培地中においてSf9細胞の感染のために用いた場合、F遺伝子産物は、脂質ラフトホーミング配列(lipid raft homing sequence)を保持することが可能であり、脂質ラフトドメインから出芽した分子に組み込むことが可能であるので、RSV F産物を含むVLPを、培養培地中において観察することができる。F−Gagダブル発現ベクターに感染した細胞からの培養培地を収穫し、培地の残骸を取り除き、20% スクロースクッション上において100,000×gで遠心分離して収集することによって、この証拠を示すことができる。FのVLPへの組み込みの証拠は、特異的抗体を用いてウエスタンブロット解析によって得ることができる。当然、これらの条件下での20% スクロースクッションを介して堆積する材料は、本質的に粒子状物質であり、VLP構成物の証拠を提供する。
【0180】
同様のアプローチを、RSV G 糖タンパク質のような、追加のRSV抗原を組み込むために用いることができる。
【0181】
(免疫原性)
ヒトRSVストックA2のような、アレルギーの誘発に適切なRSVを用いること以外は、他のVLPの免疫原性を測定するために前述の実施例において用いた技術によって、RSV VLPの免疫原性を測定することができる。
【0182】
〔追加の参考文献〕
以下の参考文献は、それらが教示する全てのものに関して、参照することによって本明細書に組み込まれる。
【0183】
1.Katz, J. M., W. Lim, C. B. Bridges, T. Rowe, J. Hu-Primmer, X. Lu, R. A. Abernathy, M. Clarke, L. Conn, H. Kwong, M. Lee, G. Au, Y. Y. Ho, K. H. Mak, N. J. Cox, and K. Fukuda. 1999. Antibody response in individuals infected with avian influenza A (H5N1) viruses and detection of anti-H5 antibody among household and social contacts. J Infect Dis 180:1763.
2.Peiris, J. S., W. C. Yu, C. W. Leung, C. Y. Cheung, W. F. Ng, J. M. Nicholls, T. K. Ng, K. H. Chan, S. T. Lai, W. L. Lim, K. Y. Yuen, and Y. Guan. 2004. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet 363:617.
3.Horimoto, T., N. Fukuda, K. Iwatsuki-Horimoto, Y. Guan, W. Lim, M. Peiris, S. Sugii, T. Odagiri, M. Tashiro, and Y. Kawaoka. 2004. Antigenic differences between H5N1 human influenza viruses isolated in 1997 and 2003. J Vet Med Sci 66:303.
4.Tran, T. H., T. L. Nguyen, T. D. Nguyen, T. S. Luong, P. M. Pham, V. C. Nguyen, T. S. Pham, C. D. Vo, T. Q. Le, T. T. Ngo, B. K. Dao, P. P. Le, T. T. Nguyen, T. L. Hoang, V. T. Cao, T. G. Le, D. T. Nguyen, H. N. Le, K. T. Nguyen, H. S. Le, V. T. Le, D. Christiane, T. T. Tran, J. Menno de, C. Schultsz, P. Cheng, W. Lim, P. Horby, and J. Farrar. 2004. Avian influenza A (H5N1) in 10 patients in Vietnam. N Engl J Med 350:1179.
5.Li, K. S., Y. Guan, J. Wang, G. J. Smith, K. M. Xu, L. Duan, A. P. Rahardjo, P. Puthavathana, C. Buranathai, T. D. Nguyen, A. T. Estoepangestie, A. Chaisingh, P. Auewarakul, H. T. Long, N. T. Hanh, R. J. Webby, L. L. Poon, H. Chen, K. F. Shortridge, K. Y. Yuen, R. G. Webster, and J. S. Peiris. 2004. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature 430:209.
6.Lipatov, A. S., E. A. Govorkova, R. J. Webby, H. Ozaki, M. Peiris, Y. Guan, L. Poon, and R. G. Webster. 2004. Influenza: emergence and control. J Virol 78:8951.
7.Lipatov, A. S., R. J. Webby, E. A. Govorkova, S. Krauss, and R. G. Webster. 2005. Efficacy of H5 influenza vaccines produced by reverse genetics in a lethal mouse model. J Infect Dis 191:1216.
8.Stephenson, I., K. G. Nicholson, J. M. Wood, M. C. Zambon, and J. M. Katz. 2004. Confronting the avian influenza threat: vaccine development for a potential pandemic. Lancet Infect Dis 4:499.
9.Liu, M., J. M. Wood, T. Ellis, S. Krauss, P. Seiler, C. Johnson, E. Hoffmann, J. Humberd, D. Hulse, Y. Zhang, R. G. Webster, and D. R. Perez. 2003. Preparation of a standardized, efficacious agricultural H5N3 vaccine by reverse genetics. Virology 314:580.
10.Subbarao, K., H. Chen, D. Swayne, L. Mingay, E. Fodor, G. Brownlee, X. Xu, X. Lu, J. Katz, N. Cox, and Y. Matsuoka. 2003. Evaluation of a genetically modified reassortant H5N1 influenza A virus vaccine candidate generated by plasmid-based reverse genetics. Virology 305:192.
11.Webby, R. J., D. R. Perez, J. S. Coleman, Y. Guan, J. H. Knight, E. A. Govorkova, L. R. McClain-Moss, J. S. Peiris, J. E. Rehg, E. I. Tuomanen, and R. G. Webster. 2004. Responsiveness to a pandemic alert: use of reverse genetics for rapid development of influenza vaccines. Lancet 363:1099.
12.Treanor, J. J., B. E. Wilkinson, F. Masseoud, J. Hu-Primmer, R. Battaglia, D. O'Brien, M. Wolff, G. Rabinovich, W. Blackwelder, and J. M. Katz. 2001. Safety and immunogenicity of a recombinant hemagglutinin vaccine for H5 influenza in humans. Vaccine 19:1732.
13.Stephenson, I., R. Bugarini, K. G. Nicholson, A. Podda, J. M. Wood, M. C. Zambon, and J. M. Katz. 2005. Cross-reactivity to highly pathogenic avian influenza H5N1 viruses after vaccination with nonadjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a potential priming strategy. J Infect Dis 191:1210.
14.Nicholson, K. G., A. E. Colegate, A. Podda, I. Stephenson, J. Wood, E. Ypma, and M. C. Zambon. 2001. Safety and antigenicity of non-adjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a randomised trial of two potential vaccines against H5N1 influenza. Lancet 357:1937.
15.Subbarao, K., B. R. Murphy, and A. S. Fauci. 2006. Development of effective vaccines against pandemic influenza. Immunity 24:5.
16.Kuper, C. F., P. J. Koornstra, D. M. Hameleers, J. Biewenga, B. J. Spit, A. M. Duijvestijn, P. J. van Breda Vriesman, and T. Sminia. 1992. The role of nasopharyngeal lymphoid tissue. Immunol Today 13:219.
17.Liang, B., L. Hyland, and S. Hou. 2001. Nasal-associated lymphoid tissue is a site of long-term virus-specific antibody production following respiratory virus infection of mice. J Virol 75:5416.
18.Zuercher, A. W., S. E. Coffin, M. C. Thurnheer, P. Fundova, and J. J. Cebra. 2002. Nasal-associated lymphoid tissue is a mucosal inductive site for virus-specific humoral and cellular immune responses. J Immunol 168:1796.
19.Brandtzaeg, P. 1989. Overview of the mucosal immune system. Curr Top Microbiol Immunol 146:13.
20.Takada, A., S. Matsushita, A. Ninomiya, Y. Kawaoka, and H. Kida. 2003. Intranasal immunization with formalin-inactivated virus vaccine induces a broad spectrum of heterosubtypic immunity against influenza A virus infection in mice. Vaccine 21:3212.
21.Tamura, S. I., H. Asanuma, Y. Ito, Y. Hirabayashi, Y. Suzuki, T. Nagamine, C. Aizawa, T. Kurata, and A. Oya. 1992. Superior cross-protective effect of nasal vaccination to subcutaneous inoculation with influenza hemagglutinin vaccine. Eur J Immunol 22:477.
22.Tumpey, T. M., M. Renshaw, J. D. Clements, and J. M. Katz. 2001. Mucosal delivery of inactivated influenza vaccine induces B-cell-dependent heterosubtypic cross-protection against lethal influenza A H5N1 virus infection. J Virol 75:5141.
23.Kang, S. M., L. Guo, Q. Yao, I. Skountzou, and R. W. Compans. 2004. Intranasal immunization with inactivated influenza virus enhances immune responses to coadministered simian-human immunodeficiency virus-like particle antigens. J Virol 78:9624.
24.Guo, L., X. Lu, S. M. Kang, C. Chen, R. W. Compans, and Q. Yao. 2003. Enhancement of mucosal immune responses by chimeric influenza HA/SHIV virus-like particles. Virology 313:502.
25.Yao, Q., R. Zhang, L. Guo, M. Li, and C. Chen. 2004. Th cell-independent immune responses to chimeric hemagglutinin/simian human immunodeficiency virus-like particles vaccine. J Immunol 173:1951.
26.Latham, T., and J. M. Galarza. 2001. Formation of wild-type and chimeric influenza virus-like particles following simultaneous expression of only four structural proteins. J Virol 75:6154.
27.Galarza, J. M., T. Latham, and A. Cupo. 2005. Virus-like particle vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol 18:365.
28.Fromantin, C., B. Jamot, J. Cohen, L. Piroth, P. Pothier, and E. Kohli. 2001. Rotavirus 2/6 virus-like particles administered intranasally in mice, with or without the mucosal adjuvants cholera toxin and Escherichia coli heat-labile toxin, induce a Th1/Th2-like immune response. J Virol 75:11010.
29.Harrington, P. R., B. Yount, R. E. Johnston, N. Davis, C. Moe, and R. S. Baric. 2002. Systemic, mucosal, and heterotypic immune induction in mice inoculated with Venezuelan equine encephalitis replicons expressing Norwalk virus-like particles. J Virol 76:730.
30.Shi, W., J. Liu, Y. Huang, and L. Qiao. 2001. Papillomavirus pseudovirus: a novel vaccine to induce mucosal and systemic cytotoxic T-lymphocyte responses. J Virol 75:10139.
31.Han, M. G., S. Cheetham, M. Azevedo, C. Thomas, and L. J. Saif. 2006. Immune responses to bovine norovirus-like particles with various adjuvants and analysis of protection in gnotobiotic calves. Vaccine 24:317.
32.Illum, L. 1998. Chitosan and its use as a pharmaceutical excipient. Pharm Res 15:1326.
33.Illum, L., I. Jabbal-Gill, M. Hinchcliffe, A. N. Fisher, and S. S. Davis. 2001. Chitosan as a novel nasal delivery system for vaccines. Adv Drug Deliv Rev 51:81.
34.Soane, R. J., M. Hinchcliffe, S. S. Davis, and L. Illum. 2001. Clearance characteristics of chitosan based formulations in the sheep nasal cavity. Int J Pharm 217:183.
35.Baudner, B. C., M. M. Giuliani, J. C. Verhoef, R. Rappuoli, H. E. Junginger, and G. D. Giudice. 2003. The concomitant use of the LTK63 mucosal adjuvant and of chitosan-based delivery system enhances the immunogenicity and efficacy of intranasally administered vaccines. Vaccine 21:3837.
36.Fujihashi, K., T. Koga, F. W. van Ginkel, Y. Hagiwara, and J. R. McGhee. 2002. A dilemma for mucosal vaccination: efficacy versus toxicity using enterotoxin-based adjuvants. Vaccine 20:2431.
37.Mutsch, M., W. Zhou, P. Rhodes, M. Bopp, R. T. Chen, T. Linder, C. Spyr, and R. Steffen. 2004. Use of the inactivated intranasal influenza vaccine and the risk of Bell's palsy in Switzerland. N Engl J Med 350:896.
38.Baldridge, J. R., Y. Yorgensen, J. R. Ward, and J. T. Ulrich. 2000. Monophosphoryl lipid A enhances mucosal and systemic immunity to vaccine antigens following intranasal administration. Vaccine 18:2416.
39.Baldrick, P., D. Richardson, G. Elliott, and A. W. Wheeler. 2002. Safety evaluation of monophosphoryl lipid A (MPL): an immunostimulatory adjuvant. Regul Toxicol Pharmacol 35:398.
40.Baldridge, J. R., P. McGowan, J. T. Evans, C. Cluff, S. Mossman, D. Johnson, and D. Persing. 2004. Taking a Toll on human disease: Toll-like receptor 4 agonists as vaccine adjuvants and monotherapeutic agents. Expert Opin Biol Ther 4:1129.
41.Baldridge, J. GlaxoSmithKline, Personal Communication.
42.Huang, J., R. J. Garmise, T. M. Crowder, K. Mar, C. R. Hwang, A. J. Hickey, J. A. Mikszta, and V. J. Sullivan. 2004. A novel dry powder influenza vaccine and intranasal delivery technology: induction of systemic and mucosal immune responses in rats. Vaccine 23:794.
43.Mills, K. H., C. Cosgrove, E. A. McNeela, A. Sexton, R. Giemza, I. Jabbal-Gill, A. Church, W. Lin, L. Illum, A. Podda, R. Rappuoli, M. Pizza, G. E. Griffin, and D. J. Lewis. 2003. Protective levels of diphtheria-neutralizing antibody induced in healthy volunteers by unilateral priming-boosting intranasal immunization associated with restricted ipsilateral mucosal secretory immunoglobulin a. Infect Immun 71:726.
44.Wimer-Mackin, S., M. Hinchcliffe, C. R. Petrie, S. J. Warwood, W. T. Tino, M. S. Williams, J. P. Stenz, A. Cheff, and C. Richardson. 2006. An intranasal vaccine targeting both the Bacillus anthracis toxin and bacterium provides protection against aerosol spore challenge in rabbits. Vaccine in press.
45.Noad, R., and P. Roy. 2003. Virus-like particles as immunogens. Trends Microbiol 11:438.
46.Yao, Q., V. Vuong, M. Li, and R. W. Compans. 2002. Intranasal immunization with SIV virus-like particles (VLPs) elicits systemic and mucosal immunity. Vaccine 20:2537.
47.Pushko, P., T. M. Tumpey, F. Bu, J. Knell, R. Robinson, and G. Smith. 2005. Influenza virus-like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine 23:5751.
48.Baumert, T. F., S. Ito, D. T. Wong, and T. J. Liang. 1998. Hepatitis C virus structural proteins assemble into viruslike particles in insect cells. J Virol 72:3827.
49.Yao, Q., Z. Bu, A. Vzorov, C. Yang, and R. W. Compans. 2003. Virus-like particle and DNA-based candidate AIDS vaccines. Vaccine 21:638.
50.Tacket, C. O., M. B. Sztein, G. A. Losonsky, S. S. Wasserman, and M. K. Estes. 2003. Humoral, mucosal, and cellular immune responses to oral Norwalk virus-like particles in volunteers. Clin Immunol 108:241.
51.Gomez-Puertas, P., C. Albo, E. Perez-Pastrana, A. Vivo, and A. Portela. 2000. Influenza virus matrix protein is the major driving force in virus budding. J Virol 74:11538.
52.Gheysen, D., E. Jacobs, F. de Foresta, C. Thiriart, M. Francotte, D. Thines, and M. De Wilde. 1989. Assembly and release of HIV-1 precursor Pr55gag virus-like particles from recombinant baculovirus-infected insect cells. Cell 59:103.
53.Johnson, M. C., H. M. Scobie, and V. M. Vogt. 2001. PR domain of rous sarcoma virus Gag causes an assembly/budding defect in insect cells. J Virol 75:4407.
54.Kakker, N. K., M. V. Mikhailov, M. V. Nermut, A. Burny, and P. Roy. 1999. Bovine leukemia virus Gag particle assembly in insect cells: formation of chimeric particles by domain-switched leukemia/lentivirus Gag polyprotein. Virology 265:308.
55.Luo, L., Y. Li, and C. Y. Kang. 1990. Expression of gag precursor protein and secretion of virus-like gag particles of HIV-2 from recombinant baculovirus-infected insect cells. Virology 179:874.
56.Morikawa, S., T. F. Booth, and D. H. Bishop. 1991. Analyses of the requirements for the synthesis of virus-like particles by feline immunodeficiency virus gag using baculovirus vectors. Virology 183:288.
57.Takahashi, R. H., K. Nagashima, T. Kurata, and H. Takahashi. 1999. Analysis of human lymphotropic T-cell virus type II-like particle production by recombinant baculovirus-infected insect cells. Virology 256:371.
58.Yamshchikov, G. V., G. D. Ritter, M. Vey, and R. W. Compans. 1995. Assembly of SIV virus-like particles containing envelope proteins using a baculovirus expression system. Virology 214:50.
59.Weldon, R. A., Jr., C. R. Erdie, M. G. Oliver, and J. W. Wills. 1990. Incorporation of chimeric gag protein into retroviral particles. J Virol 64:4169.
60.Andrawiss, M., Y. Takeuchi, L. Hewlett, and M. Collins. 2003. Murine leukemia virus particle assembly quantitated by fluorescence microscopy: role of Gag-Gag interactions and membrane association. J Virol 77:11651.
61.Leser, G. P., and R. A. Lamb. 2005. Influenza virus assembly and budding in raft-derived microdomains: a quantitative analysis of the surface distribution of HA, NA and M2 proteins. Virology 342:215.
62.Takeda, M., G. P. Leser, C. J. Russell, and R. A. Lamb. 2003. Influenza virus hemagglutinin concentrates in lipid raft microdomains for efficient viral fusion. Proc Natl Acad Sci U S A 100:14610.
63.Campbell, S. M., S. M. Crowe, and J. Mak. 2001. Lipid rafts and HIV-1: from viral entry to assembly of progeny virions. J Clin Virol 22:217.
64.Sandrin, V., and F. L. Cosset. 2006. Intracellular versus cell surface assembly of retroviral pseudotypes is determined by the cellular localization of the viral glycoprotein, its capacity to interact with Gag, and the expression of the Nef protein. J Biol Chem 281:528.
65.Salazar-Gonzalez, R. M., and S. J. McSorley. 2005. Salmonella flagellin, a microbial target of the innate and adaptive immune system. Immunol Lett 101:117.
66.Cuadros, C., F. J. Lopez-Hernandez, A. L. Dominguez, M. McClelland, and J. Lustgarten. 2004. Flagellin fusion proteins as adjuvants or vaccines induce specific immune responses. Infect Immun 72:2810.
67.Didierlaurent, A., I. Ferrero, L. A. Otten, B. Dubois, M. Reinhardt, H. Carlsen, R. Blomhoff, S. Akira, J. P. Kraehenbuhl, and J. C. Sirard. 2004. Flagellin promotes myeloid differentiation factor 88-dependent development of Th2-type response. J Immunol 172:6922.
68.Tsujimoto, H., T. Uchida, P. A. Efron, P. O. Scumpia, A. Verma, T. Matsumoto, S. K. Tschoeke, R. F. Ungaro, S. Ono, S. Seki, M. J. Clare-Salzler, H. V. Baker, H. Mochizuki, R. Ramphal, and L. L. Moldawer. 2005. Flagellin enhances NK cell proliferation and activation directly and through dendritic cell-NK cell interactions. J Leukoc Biol 78:888.
69.Hayashi, F., T. K. Means, and A. D. Luster. 2003. Toll-like receptors stimulate human neutrophil function. Blood 102:2660.
70.Hayashi, F., K. D. Smith, A. Ozinsky, T. R. Hawn, E. C. Yi, D. R. Goodlett, J. K. Eng, S. Akira, D. M. Underhill, and A. Aderem. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 410:1099.
71.Gewirtz, A. T., P. O. Simon, Jr., C. K. Schmitt, L. J. Taylor, C. H. Hagedorn, A. D. O'Brien, A. S. Neish, and J. L. Madara. 2001. Salmonella typhimurium translocates flagellin across intestinal epithelia, inducing a proinflammatory response. J Clin Invest 107:99.
72.Means, T. K., F. Hayashi, K. D. Smith, A. Aderem, and A. D. Luster. 2003. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J Immunol 170:5165.
73.Smith, K. D., E. Andersen-Nissen, F. Hayashi, K. Strobe, M. A. Bergman, S. L. Barrett, B. T. Cookson, and A. Aderem. 2003. Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat Immunol 4:1247.
74.Honko, A. N., N. Sriranganathan, C. J. Lees, and S. B. Mizel. 2006. Flagellin is an effective adjuvant for immunization against lethal respiratory challenge with Yersinia pestis. Infect Immun 74:1113.
75.Jeon, S. H., T. Ben-Yedidia, and R. Arnon. 2002. Intranasal immunization with synthetic recombinant vaccine containing multiple epitopes of influenza virus. Vaccine 20:2772.
76.Levi, R., and R. Arnon. 1996. Synthetic recombinant influenza vaccine induces efficient long-term immunity and cross-strain protection. Vaccine 14:85.
77.Lee, S. E., S. Y. Kim, B. C. Jeong, Y. R. Kim, S. J. Bae, O. S. Ahn, J. J. Lee, H. C. Song, J. M. Kim, H. E. Choy, S. S. Chung, M. N. Kweon, and J. H. Rhee. 2006. A bacterial flagellin, Vibrio vulnificus FlaB, has a strong mucosal adjuvant activity to induce protective immunity. Infect Immun 74:694.
78.Applequist, S. E., E. Rollman, M. D. Wareing, M. Liden, B. Rozell, J. Hinkula, and H. G. Ljunggren. 2005. Activation of innate immunity, inflammation, and potentiation of DNA vaccination through mammalian expression of the TLR5 agonist flagellin. J Immunol 175:3882.
79.Ramos, H. C., M. Rumbo, and J. C. Sirard. 2004. Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol 12:509.
80.Holsinger, L. J., and R. A. Lamb. 1991. Influenza virus M2 integral membrane protein is a homotetramer stabilized by formation of disulfide bonds. Virology 183:32.
81.Lamb, R. A., S. L. Zebedee, and C. D. Richardson. 1985. Influenza virus M2 protein is an integral membrane protein expressed on the infected-cell surface. Cell 40:627.
82.Holsinger, L. J., D. Nichani, L. H. Pinto, and R. A. Lamb. 1994. Influenza A virus M2 ion channel protein: a structure-function analysis. J Virol 68:1551.
83.Takeda, M., A. Pekosz, K. Shuck, L. H. Pinto, and R. A. Lamb. 2002. Influenza a virus M2 ion channel activity is essential for efficient replication in tissue culture. J Virol 76:1391.
84.Frace, A. M., A. I. Klimov, T. Rowe, R. A. Black, and J. M. Katz. 1999. Modified M2 proteins produce heterotypic immunity against influenza A virus. Vaccine 17:2237.
85.Neirynck, S., T. Deroo, X. Saelens, P. Vanlandschoot, W. M. Jou, and W. Fiers. 1999. A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat Med 5:1157.
86.Slepushkin, V. A., J. M. Katz, R. A. Black, W. C. Gamble, P. A. Rota, and N. J. Cox. 1995. Protection of mice against influenza A virus challenge by vaccination with baculovirus-expressed M2 protein. Vaccine 13:1399.
87.Treanor, J. J., E. L. Tierney, S. L. Zebedee, R. A. Lamb, and B. R. Murphy. 1990. Passively transferred monoclonal antibody to the M2 protein inhibits influenza A virus replication in mice. J Virol 64:1375.
88.De Filette, M., W. Min Jou, A. Birkett, K. Lyons, B. Schultz, A. Tonkyro, S. Resch, and W. Fiers. 2005. Universal influenza A vaccine: optimization of M2-based constructs. Virology 337:149.
89.De Filette, M., A. Ramne, A. Birkett, N. Lycke, B. Lowenadler, W. Min Jou, X. Saelens, and W. Fiers. 2006. The universal influenza vaccine M2e-HBc administered intranasally in combination with the adjuvant CTA1-DD provides complete protection. Vaccine 24:544.
90.Fiers, W., M. De Filette, A. Birkett, S. Neirynck, and W. Min Jou. 2004. A "universal" human influenza A vaccine. Virus Res 103:173.
91.Liu, W., Z. Peng, Z. Liu, Y. Lu, J. Ding, and Y. H. Chen. 2004. High epitope density in a single recombinant protein molecule of the extracellular domain of influenza A virus M2 protein significantly enhances protective immunity. Vaccine 23:366.
92.Fan, J., X. Liang, M. S. Horton, H. C. Perry, M. P. Citron, G. J. Heidecker, T. M. Fu, J. Joyce, C. T. Przysiecki, P. M. Keller, V. M. Garsky, R. Ionescu, Y. Rippeon, L. Shi, M. A. Chastain, J. H. Condra, M. E. Davies, J. Liao, E. A. Emini, and J. W. Shiver. 2004. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine 22:2993.
93.Ionescu, R. M., C. T. Przysiecki, X. Liang, V. M. Garsky, J. Fan, B. Wang, R. Troutman, Y. Rippeon, E. Flanagan, J. Shiver, and L. Shi. 2006. Pharmaceutical and immunological evaluation of human papillomavirus viruslike particle as an antigen carrier. J Pharm Sci 95:70.
94.Hatziioannou, T., E. Delahaye, F. Martin, S. J. Russell, and F. L. Cosset. 1999. Retroviral display of functional binding domains fused to the amino terminus of influenza hemagglutinin. Hum Gene Ther 10:1533.
95.Li, Z. N., S. N. Mueller, L. Ye, Z. Bu, C. Yang, R. Ahmed, and D. A. Steinhauer. 2005. Chimeric influenza virus hemagglutinin proteins containing large domains of the Bacillus anthracis protective antigen: protein characterization, incorporation into infectious influenza viruses, and antigenicity. J Virol 79:10003.
96.Haynes, J. R., S. X. Cao, B. Rovinski, C. Sia, O. James, G. A. Dekaban, and M. H. Klein. 1991. Production of immunogenic HIV-1 viruslike particles in stably engineered monkey cell lines. AIDS Res Hum Retroviruses 7:17.
97.Rovinski, B., J. R. Haynes, S. X. Cao, O. James, C. Sia, S. Zolla-Pazner, T. J. Matthews, and M. H. Klein. 1992. Expression and characterization of genetically engineered human immunodeficiency virus-like particles containing modified envelope glycoproteins: implications for development of a cross-protective AIDS vaccine. J Virol 66:4003.
98.Fynan, E. F., R. G. Webster, D. H. Fuller, J. R. Haynes, J. C. Santoro, and H. L. Robinson. 1995. DNA vaccines: a novel approach to immunization. Int J Immunopharmacol 17:79.
99.Fynan, E. F., R. G. Webster, D. H. Fuller, J. R. Haynes, J. C. Santoro, and H. L. Robinson. 1993. DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc Natl Acad Sci U S A 90:11478.
100.Kodihalli, S., J. R. Haynes, H. L. Robinson, and R. G. Webster. 1997. Cross-protection among lethal H5N2 influenza viruses induced by DNA vaccine to the hemagglutinin. J Virol 71:3391.
101.Robinson, H. L., S. Lu, D. M. Feltquate, C. T. Torres, J. Richmond, C. M. Boyle, M. J. Morin, J. C. Santoro, R. G. Webster, D. Montefiori, Y. Yasutomi, N. L. Letvin, K. Manson, M. Wyand, and J. R. Haynes. 1996. DNA vaccines. AIDS Res Hum Retroviruses 12:455.
102.Drape, R. J., M. D. Macklin, L. J. Barr, S. Jones, J. R. Haynes, and H. J. Dean. 2006. Epidermal DNA vaccine for influenza is immunogenic in humans. Vaccine in press.
103.Kretzchmar, E., R. Geyer, and H. D. Klenk. 1994. Baculovirus infection does not alter N-glycosylation in Spodoptera frugiperda cells. Biol Chem Hoppe Seyler 375:23.
104.Lu, D., and A. J. Hickey. 2005. Liposomal dry powders as aerosols for pulmonary delivery of proteins. AAPS PharmSciTech 6:E641.
105.Cowdery, S., M. Frey, S. Orlowski, and A. Gray. 1976. Stability characteristics of freeze-dried human live virus vaccines. Dev Biol Stand 36:297.
106.Peetermans, J., G. Colinet, A. Bouillet, E. D'Hondt, and J. Stephenne. 1976. Stability of live, freeze-dried virus vaccines. Dev Biol Stand 36:291.
107.Yannarell, D. A., K. M. Goldberg, and R. N. Hjorth. 2002. Stabilizing cold-adapted influenza virus vaccine under various storage conditions. J Virol Methods 102:15.
108.Sampson, H. A., J. Bernhisel-Broadbent, E. Yang, and S. M. Scanlon. 1991. Safety of casein hydrolysate formula in children with cow milk allergy. J Pediatr 118:520.
109.Gambaryan A, A. Tuzikov, G. Pazynina, N. Bovin, A. Balish, and A. Klimov. 2005. Evolution of the receptor binding phenotype of influenza A (H5) viruses. Virology (electronic publication ahead of print version).
110.Suzuki, Y, 2005. Sialobiology of Influenza: Molecular Mechanism of Host Range Variation of Influenza Viruses. Biological and Pharmaceutical Bulletin 28:399-408.
【0184】
〔政府の支援〕
本発明は、米国政府の支援によって、米国陸軍医療研究および材料部隊(U.S. Army Medical Research and Material Command)からの助成金番号W81XWH-05-C-0135および助成金番号W81XWH-05-C-0150のもとでなされた。米国政府は、本発明の一定の権利を有する。
【技術分野】
【0001】
本発明は、ウイルス様粒子の分野に関する。本明細書には、特に(a)gagポリペプチド、および(b)脂質ラフトと天然には会合しない抗原に連結された脂質ラフト会合ポリペプチドを有する、キメラウイルス様粒子が開示される。
【背景技術】
【0002】
ウイルス様粒子(VLP)は、従来のワクチン技術を凌駕する利点をいくつか有している。ワクチン開発のためのVLPの重要な利点は、3次元構造の観点と、エピトープの一次構造および立体構造の両方に対する中和抗体応答を誘導できるという観点から、VLPが天然のウイルスと似ていることである。それ故、VLPは他のワクチン製剤よりも免疫原性が強いことが分かっているといえる。ウイルスベクターを使ったアプローチとは異なり、VLPは、既存の免疫に関する問題を孕んでいないために、繰り返し使用することができる。
【0003】
伝統的なワクチンの多くは、非経口投与され、それらの大部分は下気道を防御する全身性IgG応答を引き起こす(または増幅する)。粘膜応答を誘導するウイルスを使った新しいアプローチは、該アプローチが上気道および下気道でのウイルスの増殖を制限することができるので、より望ましいものであり、この新しいアプローチは、それぞれを防御したり、感染を減少させたりするための、ワクチンを使った最も優れたアプローチであると思われる(15)。鼻腔内ワクチンは、上気道および下気道を防御することに加えて、針を使った接種の複雑な作業を避けたり、微粒子状の抗原および/または可溶性の抗原と鼻咽頭会合リンパ性組織(NALT)との相互作用による、粘膜ならびに全身性の体液性応答および細胞性応答を誘導する手段を提供したりする(16−19)。VLPは一般的に、ロタウイルス、ノロウイルス、およびパピローマウイルスのVLPについて示されたような、鼻腔内投与の後に、粘膜免疫および全身免疫を誘導するのに十分適していると思われる(28−31)。
【0004】
VLPのこうした利点にも拘らず、ワクチンとしてのエンベロープVLP(内在性膜タンパク質((integral membrane protein)を含むエンベロープウイルスに由来するVLP)の開発は、現在のところ数種の問題によって制限されている。最も重要な問題の1つは、抗原の範囲が制限されていることであり、このためエンベロープVLPワクチンを開発できる病気も制限されている。VLPへの抗原の組み込みには、ウイルスのキャプシドタンパク質が、ウイルス粒子の集合を開始する脂質ラフトドメインと会合することが必要になると考えられる。VLPを製造するための現在の方法では、脂質ラフトと天然に会合する天然ウイルス抗原を含むVLPを形成するための、ウイルスキャップシドタンパク質の使用が制限されている。
【発明の概要】
【発明が解決しようとする課題】
【0005】
したがって、(脂質ラフトと天然に会合しないウイルス抗原、キャプシドタンパク質の源以外のウイルス由来の抗原、他の病原体(細菌、真菌、原虫、蠕虫、酵母など)由来の抗原、腫瘍抗原、ならびにアレルゲンを含む)あらゆる種類の抗原を含有するVLPを製造することができるVLPの基盤技術が必要とされている。
【0006】
既存のVLP技術が有するもう1つ別の重要な問題は、十分な収量のVLPを製造することができないことである。例えば、インフルエンザマトリックスに由来するVLPは免疫原性を有しているが、該VLPは、その収量が低いために、今までは、インフルエンザワクチンの従来のものに取って代わるものとして良好な選択ではなかった。したがって、ワクチンを製造するための十分な量のVLPを生成することができるVLPワクチンの基盤技術も必要とされている。
【課題を解決するための手段】
【0007】
本発明は、キメラVLPを製造および使用するための、本明細書に開示されているような種々の方法および組成物を提供することによって、これらの必要性を満たすものである。上記キメラVLPは、あらゆる種類の病原体の脂質ラフトと天然に会合しない抗原を含んでおり、ワクチンを製造するために十分な量で生成され得るものである。
【0008】
1態様では、本発明は、gagポリペプチドと、抗原に連結された脂質ラフト会合ポリペプチドとを有しているキメラウイルス様粒子であって、上記抗原は脂質ラフトと天然には会合しないものであるキメラウイルス様粒子を提供する。結合は、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であってもよい。共有結合は、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合であってもよい。gagポリペプチドは、レトロウイルスに由来するものであることが好ましい。レトロウイルスには、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス、ベータレトロウイルス、ガンマレトロウイルス、デルタレトロウイルス、デルタレトロウイルス、およびレンチウイルスが含まれてもよい。
【0009】
脂質ラフト会合ポリペプチドは、脂質ラフトと直接的にまたは間接的に会合する任意のポリペプチドであってもよく、例えば、内在性膜タンパク質であってもよい。好ましい実施形態では、脂質ラフト会合ポリペプチドは、ヘマグルチニンのポリペプチド、ノイラミニダーゼのポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、またはエンベロープタンパク質のポリペプチドである。
【0010】
抗原は、免疫応答を誘発することができる任意の物質であってもよい。そのような抗原としては、タンパク質、ポリペプチド、糖ポリペプチド(glycopolypeptide)、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドのペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、あるいは炭水化物などが挙げられる。抗原は、ウイルス抗原、細菌抗原、真核生物の病原体の抗原、腫瘍抗原、またはアレルゲンであることが好ましい。
【0011】
また別の態様では、本明細書に記載のウイルス様粒子は、ウイルス様粒子と会合したアジュバントを含んでいる。アジュバントは、VLPの内側に配置されてもよい。なお、該アジュバントは、gagポリペプチドに共有連結されていることによって、VLPの内側に配置されていることが好ましい。他の実施形態では、アジュバントは、VLPの外側に配置されている。なお、該アジュバントは、脂質ラフト会合ポリペプチドに共有連結されていることによって、VLPの外側に配置されていることが好ましい。ポリペプチドアジュバントの好ましい例には、フラジェリンおよびそのアジュバント活性断片、サイトカイン、コロニー刺激因子(例えばGM−CSFおよびCSFなど)、インターフェロン、腫瘍壊死因子、インターロイキン−2,−7,−12、ならびに他の成長因子が含まれる。
【0012】
さらに別の態様では、本発明は、gagポリペプチドをコードする第1ヌクレオチド配列と、抗原に連結される脂質ラフト会合ポリペプチドをコードする第2ヌクレオチド配列とを有しており、上記抗原は脂質ラフトと天然には会合しないものであり、上記ポリペプチドは、宿主細胞内で発現すると、ウイルス様粒子を形成するものである、ウイルス様粒子の発現ベクターシステムを提供する。1実施形態では、第1ヌクレオチド配列および第2ヌクレオチド配列は、1つの発現ベクター内に存在しており、別々のプロモーターに作動可能に連結されていることが好ましいが、1つのプロモーターに作動可能に連結されていてもよい。別の実施形態では、第1ヌクレオチド配列および第2ヌクレオチド配列は、複数の発現ベクター内に存在している。
【0013】
さらに別の態様では、本発明は、(a)gagポリペプチドと抗原に連結される脂質ラフト会合ポリペプチドとを一緒に発現する、1つ以上の発現ベクターを提供する工程、(b)上記1つ以上の発現ベクターを細胞に導入する工程、および、(c)上記gagポリペプチドと上記抗原に連結される脂質ラフト会合ポリペプチドとを発現させて、ウイルス様粒子を製造する工程とによって、ウイルス様粒子を製造する方法を提供する。ここで、上記抗原は、脂質ラフトと天然には会合しないものである。好ましい実施形態では、1つ以上の発現ベクターはウイルスベクターである。該ウイルスベクターは、バキュロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、またはレトロウイルスであってもよい。上記細胞は、昆虫細胞または哺乳類細胞であってもよい。ある実施形態では、上記方法は、上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる。
【0014】
別の態様では、本発明は、免疫原性量の本明細書に記載のキメラインフルエンザウイルス様粒子の何れかの投与によって、免疫系の症状あるいは病気を治療または予防する方法を提供する。ある実施形態では、上記投与は、感染防御免疫応答を被検体に誘導するものである。ある実施形態では、上記投与は、皮下投与、経皮投与、皮内投与、真皮下投与、筋肉内投与、経口投与(peroral delivery、oral delivery)、鼻腔内投与、バッカル投与、舌下投与、腹腔内投与、膣内投与、肛門投与、または頭蓋内投与である。
【0015】
本明細書に開示されているキメラインフルエンザウイルス様粒子の別の態様は、免疫原性量または治療量の本明細書に開示のキメラウイルス様粒子の何れかを含んでいてもよい薬学的組成物である。そのような薬学的組成物は、薬学的に許容可能な担体を含んでいることが好ましい。該担体は、好ましい投与方法用に製剤化されていることが好ましい。
【0016】
別の態様では、本発明は、本明細書に開示されているようなVLPを含んでいる薬学的組成物を提供する。好ましい実施形態では、該薬学的組成物は、薬学的に許容可能な担体を含んでいる。
【0017】
別の態様では、本発明は、gagポリペプチドおよび非ウイルス性脂質ラフト会合ポリペプチドを含んでいる、キメラウイルス様粒子を提供する。そのようなVLPは、本明細書に開示された他のVLPの種々の実施形態の全てを含んでいる。ある実施形態では、脂質ラフト会合ポリペプチドは、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、シグナル伝達のポリペプチド、または膜輸送のポリペプチド、あるいは好ましくはカベオリン(cavelin)のポリペプチド、フロチリンのポリペプチド、シンタキシン−1のポリペプチド、シンタキシン−4のポリペプチド、シナプシンIのポリペプチド、アデューシンのポリペプチド、VAMP2のポリペプチド、VAMP/シナプトブレビンのポリペプチド、シナプトブレビンIIのポリペプチド、SNAREのポリペプチド、SNAP−25のポリペプチド、SNAP−23のポリペプチド、シナプトタグミンIのポリペプチド、またはシナプトタグミンIIのポリペプチドであってもよい。また、上記gagポリペプチドおよび非ウイルス性脂質ラフト会合ポリペプチドを含んでいるキメラウイルス様粒子は、本明細書のあらゆる箇所に開示されている上記態様および実施形態の全て(特に限定されないが、発現ベクターシステム、製造の方法、治療および予防の方法、ならびに薬学的組成物を含む)を含んでいる。
【図面の簡単な説明】
【0018】
【図1】Gag、HAまたはコントロールベクターのそれぞれで、およびHA−gag−NAのトリプルベクターに感染された、Sf9細胞からの培養液のウエスタンブロットを示す図であり、(A)は、抗Gag抗体によるウエスタンブロットを示す図であり、(B)は抗HA抗体によるウエスタンブロットを示す図である。
【図2】サッカロースステップ勾配を用いて、ペレット状のHA−gag−NA VLPを再遠心分離することによって得られた画分のウエスタンブロットを示す図であり、(A)は、抗Gag抗体によるウエスタンブロットを示す図であり、(B)は抗HA抗体によるウエスタンブロットを示す図である。
【図3】以下の実施例1に関するトリプル発現ベクターのコード配列の配置を示す図である。
【図4】PA修飾されたHA、ならびにGagおよびNAをコードするトリプルバキュロウイルス発現ベクターのコード要素の配置を示す図である。
【図5】RSV Fタンパク質およびGagをコードするダブルバキュロウイルス発現ベクター内のコード要素の配置を示す図である。
【発明を実施するための形態】
【0019】
本発明は、キメラウイルス様粒子を形成するための基礎となるgagポリペプチドを含んでいる。VLPを生成する好ましい方法は、昆虫細胞内での発現によるものである。該昆虫細胞内での発現は、ポリペプチド抗原の共発現を含んでいることが好ましい。これは、バキュロウイルス発現システムの様々なレトロウイルスによって、gagVLPの収量を大幅に上げることができるからである(23、24、46、49、52−58)。gagポリペプチドは、天然のレトロウイルスの組立過程の際には、本質的にC末端延長領域を含んでいる。つまり、機能的なgagタンパク質は、レトロウイルスのプロテアーゼ、逆転写酵素、およびリボソームのフレームシフトによるインテグラーゼ活性を含んでいる巨大なC末端延長領域を天然に有している。人工の延長領域を有する機能的なgagタンパク質の製造は、RSVgag(59)およびMLVgag(60)に対して達成されている。gagC末端の操作におけるこの適応性は、他の抗原および免疫賦活タンパク質の配列などの他のポリペプチドを含有させるための重要な部位を提供する。
【0020】
あるウイルス由来のコア粒子と、それとは異なるウイルス由来の表面抗原を含んでいるキメラVLPの製造は、シュードタイピングと呼ばれる。gagポリペプチドは、インフルエンザHAおよびNAでもって効率よくシュードタイプされている。これは、おそらく、これらタンパク質が脂質ラフトドメイン(61、62)の内部に濃縮する一方、ミリストイル化されたgagタンパク質は、出芽過程の間に、脂質ラフトドメインの内表面に濃縮する(63)からである。
【0021】
本明細書に記載の本発明は、キメラVLPを形成するための基礎としての、脂質ラフトと天然には会合しない抗原に、連結された脂質ラフト会合ポリペプチドをさらに含んでいる。どのような理論にも束縛されることなく、該抗原が上記脂質ラフト会合ポリペプチドへ会合するという長所によって、該抗原を含んでいるキメラVLPが形成されると考えられる。
【0022】
本明細書に開示の方法およびプロトコールは、他に示されない限り、当業者が実施できる範囲内の従来の化学的手法、分子生物学的手法、微生物学的手法、組み換えDNAの手法、および免疫学的手法を使用するものである。そのような手法は、文献において説明されており、例えば、「J. Sambrook, E. F. Fritsch, and T. Maniatis, 1989, Molecular Cloning: A Laboratory Manual, Second Edition, Books 1-3, Cold Spring Harbor Laboratory Press」、「Ausubel, F. M.ら (1995 and periodic supplements; Current Protocols in Molecular Biology, ch. 9, 13, and 16, John Wiley & Sons, New York, N.Y.)」、「B. Roe, J. Crabtree, and A. Kahn, 1996, DNA Isolation and Sequencing: Essential Techniques, John Wiley & Sons」、「J. M. Polak and James O'D. McGee, 1990, In Situ Hybridization: Principles and Practice; Oxford University Press」、「M. J. Gait (Editor), 1984, Oligonucleotide Synthesis: A Practical Approach, Irl Press」、および「D. M. J. Lilley and J. E. Dahlberg, 1992, Methods of Enzymology: DNA Structure Part A: Synthesis and Physical Analysis of DNA Methods in Enzymology, Academic Press」を参照のこと。これら一般的なテキストのそれぞれは、参照することによって本明細書に組み込まれる。
【0023】
〔定義〕
本明細書で使用されるような「Gagポリペプチド」は、本明細書に記載のウイルス様粒子の形成に関与するレトロウイルス由来の構造ポリペプチドである。ある実施形態では、RNAをパッケージングするための傾向(propensity)、または粒子の形成および出芽の効率などのある種の特徴に影響を及ぼすために、gagポリペプチドは、意図的に突然変異させられる。レトロウイルスのゲノムは、gag、pol、およびenvという3つの主要遺伝子産物をコードしている。gag遺伝子は、構造タンパク質をコードするものであり、pol遺伝子は、逆転者酵素、関連するタンパク質分解性のポリペプチド、ヌクレアーゼ、およびインテグラーゼに関連する機能をコードするものであり、envは、コードされた糖タンパク質の膜タンパク質が感染細胞の表面上で検出され、成熟し放出されたウイルス粒子の表面上で検出されるものである。全てのレトロウイルスのgag遺伝子の全体構造は類似しており、レトロウイルスの各グループ内では、gag遺伝子はアミノ酸レベルで保存されている。gag遺伝子は、逆転写酵素を除く、コアタンパク質を生じさせるものである。
【0024】
MLVに関しては、gagの前躯体タンパク質は、Pr65Gagであり、前躯体での順番がNH2−p15−pp12−p30−p10−COOHとなっている4つのタンパク質に開裂される。これらの開裂は、ウイルスプロテアーゼに仲介され、ウイルスに依存するが、ウイルスの放出の前後に起こり得る。MLVのGagタンパク質は、グリコシル化された形態およびグリコシル化されていない形態で存在する。グリコシル化された形態は、gPr80Gagから開裂される。なおgPr80Gagは、グリコシル化されていないPr65GagのAUGコドンから上流に位置する、異なったインフレームの開始コドンから合成されるものである。グリコシル化されるGagを合成しないMLVの欠失変異体でも感染性を有し、グリコシル化されないGagでもウイルス様粒子を形成することができる。このことから、グリコシル化が起こることの重要性について疑問が生じる。HIV−1のGag前躯体であるpr55Gagの、ウイルスにコードされたプロテアーゼによる翻訳後開裂から、N−ミリストイル化され、内部にリン酸化されたp17マトリックスタンパク質(P17MA)、リン酸化されたp24キャップシドタンパク質(p24CA)、およびヌクレオキャップシドタンパク質P15(p15NC)が生じる。p15NCは、p9およびp6にさらに開裂される。
【0025】
構造的に原型のGagポリタンパク質は、マトリックスタンパク質(MA)、キャプシドタンパク質(CA)、およびヌクレオキャプシドタンパク質(NC)の3つの主要タンパク質に分けられる。これら主要タンパク質は、レトロウイルスのgag遺伝子に常に同じ順番で存在している。なお、マトリックスタンパク質(MA)は、インフルエンザのマトリックスタンパク質M1と混同されるものではない。マトリックスという名称を共有しているが、M1はMAと異なるタンパク質である。Gagポリタンパク質の成熟タンパク質へのプロセッシングは、レトロウイルスにコードされたプロテアーゼによって触媒され、新たに出芽するウイルス粒子として成熟させる。機能上、Gagポリタンパク質は、膜結合ドメイン、相互作用ドメイン、レイトドメインの3つのドメインに分けられる。膜結合ドメインは、Gagポリタンパク質に細胞膜を狙わせるものであり、相互作用ドメインは、Gagの重合化を促進するものであり、レイトドメインは、宿主細胞から発生するウイルス粒子の放出を促進するものである。組立を仲介するGagタンパク質の形態は、ポリタンパク質である。このため組立ドメインが、より後に形成する開裂産物の何れかの内にきちんと位置する必要は無い。このように、本明細書に包含されるようなGagポリペプチドは、VLPを形成および放出させるために重要な機能要素を含んでいる。これらの重要な機能要素に関して、最新技術はかなり進んでいる。例えば「Hansenら J. Virol 64, 5306-5316, 1990」、「Willら, AIDS 5, 639-654, 1991」、「Wangら J. Virol. 72, 7950-7959, 1998」、「McDonnellら, J. Mol. Biol. 279, 921-928, 1998」、「Schultz and Rein, J. Virol. 63, 2370-2372, 1989」、「Accolaら, J. Virol. 72, 2072-2078, 1998」、「Borsettiら, J. Virol., 72, 9313-9317, 1998」、「Bowzardら, J. Virol. 72, 9034-9044, 1998」、「Krishnaら, J. Virol. 72, 564-577, 1998」、「Willsら, J. Virol. 68, 6605-6618, 1994」、「Xiangら, J. Virol. 70, 5695-5700, 1996」、「Garnierら, J. Virol. 73, 2309-2320, 1999」を参照のこと。
【0026】
本発明のVLPにおいて使用されるようなgagポリペプチドは、最低でもVLPを形成するための機能要素を含んでいる。gagポリペプチドは、場合によっては1つ以上の別のポリペプチドを含んでいてもよい。該1つ以上の別のポリペプチドは、gagポリペプチドをコードする配列へ、該1つ以上の別のポリペプチドをコードする配列をスプラスすることによって生成されるものであってもよい。別のポリペプチドをgagポリペプチドに挿入するための部位は、C末端であることが好ましい。
【0027】
Gagポリペプチドのための好ましいレトロウイルスの源は、マウス白血病ウイルス、ヒト免疫不全ウイルス、アルファレトロウイルス(トリ白血病ウイルス(avian leucosis virus)、またはラウス肉腫ウイルスなど)、ベータレトロウイルス(マウス乳癌ウイルス,ヤーグジークテヒツジレトロウイルスおよびマソン・ファイザー・サルウイルスなど)、ガンマレトロウイルス(マウス白血病ウイルス、ネコ白血病ウイルス、細網内皮症ウイルス、およびテナガザル白血病ウイルスなど)、デルタレトロウイルス(ヒトT細胞白血病ウイルスおよびウシ白血病ウイルスなど)、イプシロンレトロウイルス(ウォールアイ皮膚肉腫ウイルスなど)、あるいはレンチウイルス(1型ヒト免疫不全ウイルス、HIV−2、サル免疫不全ウイルス、ネコ免疫不全ウイルス、ウマ伝染性貧血ウイルス、およびヤギ関節炎脳炎ウイルス)を包含する。
【0028】
本明細書で使用されているような「脂質ラフト」は、gagポリペプチドがウイルス粒子の組立過程の間に濃縮する細胞膜の微小ドメインのことをいう。
【0029】
本明細書で使用されているような「脂質ラフト会合ポリペプチド」は、脂質ラフトと直接的にまたは間接的に会合される任意のポリペプチドのことをいう。本発明において使用される具体的な脂質ラフト会合ポリペプチドは、キメラウイルス様粒子の望まれる使用に依存する。
【0030】
脂質ラフト会合ポリペプチドは、内在性膜タンパク質、膜との会合を引き起こすタンパク質の修飾を介して脂質ラフトと直接的に会合されるタンパク質、または脂質ラフト会合ポリペプチドを介して脂質ラフトと間接的に会合されるポリペプチドであってもよい。
【0031】
脂質アンカーを有する多くのタンパク質は、脂質ラフトと会合する。ポリペプチドを脂質ラフトに連結させる脂質アンカーには、GPIアンカー、ミリストイル化、パルミトイル化、およびダブルアセチル化が含まれる。
【0032】
多種多様なポリペプチドが脂質ラフトと会合する。脂質ラフトは、シグナル伝達、膜輸送、ウイルスの侵入、ウイルスの組立、および組立られた粒子の出芽を含む無数の生物学的活動の基盤として機能する。それ故、これらの過程に関連する様々なポリペプチドが脂質ラフトと会合する。
【0033】
シグナル伝達カスケードに関与する様々な種類のポリペプチドは、シグナル伝達の基盤として機能する脂質ラフトと会合する。シグナル伝達の基盤として機能する脂質ラフトの一つの形式は、カベオラと呼ばれる。カベオラは、形質膜のフラスコ型の陥入であり、カベオリンファミリー(例えばカベオリンおよび/またはフロチリン)に由来するポリペプチドを含んでいる。
【0034】
膜輸送ポリペプチドは、膜輸送の基盤として機能する脂質ラフトと会合する。膜輸送ポリペプチドの例には、エンドサイトーシスおよびエキソサイトーシスに関与するタンパク質、例えば、シンタキシン−1、シンタキシン−4、シナプシンI、アデューシン、VAMP2、VAMP/シナプトブレビン、シナプトブレビンII、SNAREタンパク質、SNAP−25、SNAP−23、シナプトタグミンI、およびシナプトタグミンIIなどが含まれる。
【0035】
ウイルス受容体、受容体−補助受容体の複合体、侵入過程を調節するのに役立つ他の任意の成分は、ウイルスの侵入のために特殊化した膜輸送の基盤として機能する脂質ラフトと会合する。脂質ラフトと会合するウイルス受容体には、例えば、分解促進因子(DAFまたはCD55)、多くのエンテロウイルスの受容体であるGPIアンカー型膜糖タンパク質、グループAのロタウイルスの受容体、複数の成分(ガングリオシド、HSC70タンパク質、アルファ2−ベータ1インテグリン、およびアルファ5−ベータ2インテグリンを含む)をガ乳する複合体;数種のエンベロープウイルス(HIV、MLV、麻疹ウイルス、エボラウイルスなど)の糖タンパク質、およびHIVの侵入に関与するポリペプチド(CD5、CCR5およびnefなど)が含まれる。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. & Mol. Bio. Rev. 67(2):226-237」を参照のこと。
【0036】
ウイルス粒子の組立に関与するポリペプチドは、ウイルスの組立の基盤として機能する脂質ラフトと会合される。そのようなポリペプチドには、例えば、HAおよびNAというインフルエンザのエンベロープ糖タンパク質、麻疹ウイルスのHおよび成熟F1−F2融合タンパク質、ならびにHIVのgp160、gp41、およびPr55gagが含まれる。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. And Mol. Bio. Rev. 67(2):226-237」を参照のこと。
【0037】
組立られたウイルスの出芽に関与するポリペプチドは、ウイルスの出芽の基盤として機能する脂質ラフトと会合される。宿主細胞からのHIV−1の出芽が膜のラフトにおいて起こっていることを示唆するデータがある。「Chazal and Gerlier, 2003, Virus Entry, Assembly, Budding, and Membrane Rafts, Microbiol. And Mol. Bio. Rev. 67(2):226-237」を参照のこと。ウイルスの出芽に関与するポリペプチドに関する一般的な情報は、「Fields Virology (4th ed.) 2001」に見出される。
【0038】
好ましい脂質ラフト会合ポリペプチドには、ヘマグルチニンポリペプチド、ノイラミニダーゼポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、およびエンベロープタンパク質のポリペプチドなどのウイルスポリペプチドが含まれる。これらポリペプチドのそれぞれは、あらゆる種類のウイルスに由来するものであってもよい。しかしながら、ある実施形態では、HIV−1ウイルスに由来するエンベロープタンパク質、呼吸器合胞体ウイルスまたは麻疹ウイルスに由来する融合タンパク質、呼吸器合胞体ウイルス、単純疱疹ウイルスまたはエボラウイルスに由来する糖タンパク質、および麻疹ウイルスに由来するヘマグルチニンタンパク質が含まれる。
【0039】
好ましい非ウイルス性病原体の脂質ラフト会合ポリペプチドは、以下の原虫、蠕虫、および他の真核微生物病原体から得られてもよい:特に限定されないが、マラリア原虫(Plasmodium)(熱帯熱マラリア原虫(Plasmodium falciparum)、四日熱マラリア原虫(Plasmodium malariae)、卵形マラリア原虫(Plasmodium ovale)、および三日熱マラリア原虫(Plasmodium vivax)など);トキソプラズマ・ゴンディ(Toxoplasma gondii);トリパノソーマ・ブルーセイ(Trypanosoma brucei)、トリパノソーマ・クルージ(Trypanosoma cruzi);ビルハルツ住血吸虫(Schistosoma haematobium)、マンソン住血吸虫(Schistosoma mansoni)、日本住血吸虫(Schistosoma japonicum);ドノバンリーシュマニア(Leishmania donovani);ランブル鞭毛虫(Giardia intestinalis);およびクリプトスポリジウム・パルバム(Cryptosporidium parvum)など。そのような非ウイルス性脂質ラフト会合ポリペプチドは、該脂質ラフト会合ポリペプチドそれ自体が抗原として作用するときは、脂質ラフトと天然に会合しない抗原と連結されることなく使用されてもよい。
【0040】
ウイルス性脂質ラフト会合ポリペプチドの好ましい例は、ヘマグルチニンポリペプチドである。本明細書で使用されているような「ヘマグルチニンポリペプチド」は、インフルエンザウイルスのタンパク質に由来するものであり、感染される細胞へのウイルスの結合を仲介するものである。またヘマグルチニンポリペプチドは、類似する麻疹ウイルスのタンパク質に由来するものであってもよい。該タンパク質は、1つの膜貫通ドメインを介して、インフルエンザウイルスの表面に固定されることが発見された抗原性糖タンパク質である。インフルエンザのヘマグルチニンには、少なくとも16のサブタイプが同定されており、H1からH16と名づけられている。H1、H2およびH3は、ヒトのインフルエンザウイルスにおいて発見されているものである。H5またはH7のヘマグルチニンを有する病原性が高いトリインフルエンザウイルスは、割合が低いもののヒトに感染することが発見されている。ヒトの患者において、アミノ酸が1つ変更しているトリインフルエンザウイルス株のH5型のヘマグルチニンが発見されており、アミノ酸のこの変更によって、H5のヘマグルチニンがH5N1のトリインフルエンザウイルスの受容体特異性を顕著に変更することができるように、受容体特異性が変化し、該ウイルスがヒトの受容体に結合できるようになったと報告されている(109および110)。この発見は、普通はヒトに感染しないH5N1型ウイルスがどのようにして突然変異し、効率よくヒト細胞に感染できるようになったかを説明するものである。
【0041】
ヘマグルチニンは、ホモトリマーの内在性膜ポリペプチドである。膜貫通ドメインは、ラフト脂質ドメインと天然に会合しているため、ヘマグルチニンがVLPに組み込まれるためのgagポリペプチドと会合することができる。ヘマグルチニンはシリンダーのような形状をしており、約135Åの長さである。HAを構成する3つの同じモノマーは、中心のコイルドコイルと、VLPの表面に露出されるシアル酸結合部位を含む球状の頭部とを形成する。HAのモノマーは、1つのポリペプチド前躯体として合成され、該前躯体は、グリコシル化されて、そして2つのより小さいポリペプチド(HA1サブユニットおよびHA2サブユニット)に開裂される。HA2サブユニットは、膜に固定されるトリマーのコイルドコイルを形成し、HA1サブユニットは球状の頭部を形成する。
【0042】
本発明のVLPに使用されるようなヘマグルチニンポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。ヘマグルチニンポリペプチドは、任意のインフルエンザウイルスの型、亜型、株または亜株に由来するものであってもよく、好ましくはH1、H2、H3、H5、H7およびH9のヘマグルチニンに由来するものであってもよい。さらに、ヘマグルチニンポリペプチドは、様々なインフルエンザのへマグルチニンのキメラであってもよい。ヘマグルチニンポリペプチドは、脂質ラフトと天然に会合しない1つ以上の別の抗原を含んでいることが好ましい。該抗原は、ヘマグルチニンポリペプチドをコードする配列へ、上記1つ以上の別のポリペプチドをコードする配列をスプライスすることによって生成されるものであってもよい。ヘマグルチニンポリペプチドに別のポリペプチドを挿入する部位は、N末端であることが好ましい。
【0043】
ウイルス性脂質ラフト会合ポリペプチドの好ましいもう1つ別の例は、ノイラミニダーゼポリペプチドである。本明細書で使用されているような「ノイラミニダーゼポリペプチド」は、インフルエンザウイルスのタンパク質に由来し、糖タンパク質から末端のシアル酸残基を開裂することによって、インフルエンザウイルスの細胞からの放出を仲介するタンパク質である。ノイラミニダーゼ糖タンパク質は、ウイルスの表面に発現される。ノイラミニダーゼタンパク質は、四量体であり、ベータ−ピンホイール構造を有する球状頭部、細い茎領域、および小さい疎水性領域からなる共通の構造を有している。この疎水性領域は、1つの膜貫通ドメインにより、ウイルス膜にノイラミニダーゼタンパク質を固定させるものである。シアル酸残基の開裂のための活性部位は、全てのインフルエンザAウイルスに保存されている15個の荷電アミノ酸によって形成される各サブユニットの表面上のポケットを含んでいる。インフルエンザのノイラミニダーゼには、少なくとも9のサブタイプが同定はされており、N1からN9と名づけられている。
【0044】
本発明のVLPに使用されるようなノイラミニダーゼポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。機能領域についての最先端の技術水準は、極めて高い。例えば、「Vargheseら, Nature 303, 35-40, 1983」、「Colmanら, Nature 303, 41-44, 1983」、「Lentzら, Biochem, 26, 5321-5385, 1987」、「Websterら, Virol. 135, 30-42, 1984」を参照のこと。ノイラミニダーゼポリペプチドは、任意のインフルエンザウイルスの型、亜型、株または亜株に由来するものであってもよく、好ましくはN1およびN2のノイラミダーゼに由来するものであってもよい。さらに、ノイラミニダーゼポリペプチドは、様々なインフルエンザのノイラミニダーゼのキメラであってもよい。ノイラミニダーゼポリペプチドは、脂質ラフトと天然に会合しない1つ以上の別の抗原を1つ以上含んでいることが好ましい。該抗原は、ヘマグルチニンポリペプチドをコードする配列へ、上記1つ以上の別のポリペプチドをコードする配列をスプライスすることによって生成されるものであってもよい。ノイラミニダーゼポリペプチドをコードする配列に別のポリペプチドを挿入する部位は、C末端であることが好ましい。
【0045】
脂質ラフト会合ペプチドの好ましいもう1つ別の例は、ファシクリンI(FasI)と呼ばれる昆虫由来の接着タンパク質である。本明細書で使用されているような「ファシクリンIポリペプチド」は、胚発生に関与する昆虫タンパク質に由来するものである。この非ウイルス性タンパク質が、昆虫細胞バキュロウイルス発現系にて発現されると、FasIの脂質ラフト会合が引き起こされ得る(J. Virol. 77, 6265-6273, 2003))。それ故、異種抗原がファシクリンIポリペプチドに接着されることによって、gagと共発現されたとき、キメラ分子がVLPへ組み込まれるということになる。本発明のVLPに使用されるようなファシクリンIポリペプチドは、最低でも膜アンカードメインを含んでいるべきである。
【0046】
脂質ラフト会合ペプチドのもう1つ別の好ましい例は、G糖タンパク質と名づけられたRSVからのウイルス由来の接着タンパク質である。本明細書で使用されているような「G糖タンパク質」は、RSVのG糖タンパク質に由来するものである。最近のデータによれば、脂質ラフトドメインは、該ドメインがインフルエンザの出芽に重要であるように、RSV粒子の出芽にも重要であることが実証されている(Virol 327, 175-185, 2004; Arch. Virol. 149, 199-210, 2004; Virol. 300, 244-254, 2002)。RSVのG糖タンパク質は、32.5kdの内在性膜タンパク質であり、ウイルスの接着タンパク質として機能するだけでなく、RSV感染のための感染防御抗原としても機能する。インフルエンザウイルスのヘマグルチニンと同じように、その抗原性は、それに接着される非脂質ラフト抗原の抗原性を増強し得る。RSVは、ノイラミニダーゼ活性を有するタンパク質を天然に発現しないために、gagおよびRSVのG糖タンパク質から構成されるVLPは、産生および放出を効率よくするためのNAが存在することを必要としない。それ故、MLVのgagおよびG糖ポリペプチドをコードする発現ベクターを開発することにより、膜に組み込まれるG糖ポリペプチドを含むVLPが産生されることになる。非脂質ラフト外来抗原接着をするように、該G糖ポリペプチドを任意に修飾することにより、該外来抗原に対する顕著な免疫応答を誘導することができるキメラVLPを得ることができる。
【0047】
用語「キメラウイルス様粒子」と「VLP」とは、VLPがその文脈から、本明細書に開示されているようなgagポリペプチドを用いて形成されないウイルス様粒子のことを言っている場合を除いて、本明細書を通して交換可能に使用される。
【0048】
〔抗原〕
本発明は、gagポリペプチドおよび脂質ラフト会合ポリペプチドを、脂質ラフトと天然に会合しない抗原を含んでいるVLPを形成するための、容易に改変できる基盤として、提供する。この項には、開示されたVLPと一緒に使用されるのが好ましい抗原が記載されている。
【0049】
<抗原と脂質ラフト会合ポリペプチドとの結合>
脂質ラフトと天然に会合しない抗原を含んでいるVLPを形成する手段として、抗原と脂質ラフト会合ポリペプチドとの間に結合が形成される。該脂質ラフト会合ポリペプチドは、1つの抗原に連結されてもよいし、複数の抗原に連結されてもよい。これにより、VLPの免疫原性が増加されたり、様々な病原体に対する免疫原性が付与されたり、または特定の病原体の様々な株に対する免疫原性が付与されたりする。
【0050】
抗原と脂質ラフト会合ポリペプチドとの結合は、抗原がVLPへ組み込まれるのに十分などのような種類の結合であってもよい。該結合は、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、または抗体−抗原相互作用であってもよい。好ましい実施形態では、結合は、共有結合(ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、またはジスルフィド結合など)である。
【0051】
抗原は、組み換えによって、脂質ラフト会合ポリペプチドに既に結合した状態で製造されてもよいし、あるいは分離した物質として製造され、その後脂質ラフト会合ポリペプチドに連結されてもよい。
【0052】
<抗原の種類>
本明細書で使用されているような抗原は、免疫応答を誘発することができ、脂質ラフトと天然には会合しない任意の物質であってもよい。抗原には、特に限定されないが、タンパク質、ポリペプチド(活性タンパク質、およびタンパク質内の各ポリペプチドエピトープ)、糖ポリペプチド、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドおよび他の分子のペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、ならびに炭水化物が含まれる。抗原が脂質ラフトと直接的にまたは間接的に天然に会合しない場合、脂質ラフト会合ポリペプチドと結合することなく、VLPに組み込まれることは考えられない。抗原は、病気または疾患に関与する任意の抗原であってもよく、例えば微生物抗原(例えばウイルス抗原、細菌抗原、真菌抗原、原虫抗原、蠕虫抗原、酵母抗原など)、腫瘍抗原、およびアレルゲンなどである。
【0053】
<抗原の源>
本明細書に記載の抗原は、化学的にまたは酵素を用いて合成されてもよいし、組み換えによって産生されてもよいし、天然源から単離されてもよいし、あるいはそれらの組み合わせによって得られてもよい。抗原は、精製されてもよいし、部分的に精製されてもよいし、粗抽出物であってもよい。
【0054】
ポリペプチド抗原は、技術的に知られているタンパク質精製の標準的な方法を使用して、天然源から単離されてもよい。そのような方法には、特に限定されないが、液体クロマトグラフィー(例えば高速液体クロマトグラフィー、高速タンパク質液体クロマトグラフィーなど)、サイズ排除クロマトグラフィー、ゲル電気泳動(一次元ゲル電気泳動、二次元ゲル電気泳動を含む)、親和性クロマトグラフィー、または他の精製手法が包含される。多くの実施形態では、抗原は、例えば、約50%から約75%までの純度、約75%から約85%までの純度、約85%から約90%までの純度、約90%から約95%までの純度、約95%から約98%までの純度、約98%から約99%までの純度、あるいは99%よりも高い純度の精製抗原である。
【0055】
固相ペプチド合成手法を使用してもよい。そのような手法は、当業者に公知である。「Jones, The Chemical Synthesis of Peptides (Clarendon Press, Oxford) (1994)」参照。一般的に、そのような方法では、ペプチドは、活性化されたモノマーユニットが、固相に結合されている成長ペプチド鎖に連続的に付加されることによって産生される。
【0056】
十分に確立されている組み換えDNA手法を、脂質ラフト会合ポリペプチドと同じベクターにおいて、ポリペプチドを製造するために使用してもよい。例えば、ポリペプチドをコードするヌクレオチド配列を含んでいる発現コンストラクトが、適切な宿主細胞(例えば、インビトロの細胞培養において単細胞として成長する真核宿主細胞、例えば酵母細胞、昆虫細胞、哺乳類細胞など)、あるいは原核細胞(インビトロの細胞培養において成長するもの)に導入されて、遺伝的に修飾された宿主細胞が生成される場合、適切な培養条件下において、タンパク質は該遺伝的に修飾された宿主細胞によって製造される。
【0057】
<ウイルス抗原>
適したウイルス抗原には、以下のグループの1つ以上のウイルスと関連するもの(例えば以下のグループの1つ以上のウイルスによって合成されたもの)を包含する:レトロウイルス科(例えばHIV−1などのヒト免疫不全ウイルス(HTLV−III、LAVまたはHTLV−III/LAVまたはHIV−IIIとも呼ばれる);およびHIV−LPなどの他の単離種;ピコルナウイルス科(例えばポリオウイルス、A型肝炎ウイルス;エンテロウイルス、ヒトコクサッキーウイルス、ライノウイルス、エコーウイルス);カリシウイルス科(Calciviridae)(例えば、ノーウォークウイルスおよび関連ウイルスを含む、胃腸炎を引き起こす株);トガウイルス科(例えばウマ脳脊髄炎ウイルス、三日はしかウイルス);フラビウイルス科(Flaviridae)(例えばデング熱ウイルス、脳炎ウイルス、黄熱ウイルス);コロナウイルス科(例えばコロナウイルス);ラブドウイルス科(例えば水疱性口炎ウイルス、狂犬病ウイルス);フィロウイルス科(例えばエボラウイルス);パラミクソウイルス科(例えばパラインフルエンザウイルス、ムンプスウイルス、麻疹ウイルス、呼吸器合胞体ウイルス、メタニューモウイルス科(例えばトリニューモウイルス、ヒトメタニューモウイルス);オルソミクソウイルス科(例えばインフルエンザウイルス);ブニヤウイルス科(Bungaviridae)(例えばハンタンウイルス、ブニヤウイルス(bungavirus)、フレボウイルス、およびナイロウイルス);アレナウイルス科(出血熱ウイルス);レオウイルス科(例えばレオウイルス、オルビウイルス、およびロタウイルス);ビルナウイルス科(Bimaviridae);ヘパドナウイルス科(B型肝炎ウイルス);パルボウイルス科(パルボウイルス);パポバウイルス科(乳頭腫ウイルス、ポリオームウイルス);アデノウイルス科(大部分のアデノビールス);ヘルペスウイルス科(単純疱疹ウイルス(HSV)1型(HHV−1)および2型(HHV−2)、水痘帯状疱疹ウイルス(HHV−3)、EBウイルス(エプスタインバーウイルス)(HHV−4)、サイトメガロウイルス(CMV)(HHV−5));ポックスウイルス(Poxyiridae)(天然痘ウイルス、ワクシニア・ウイルス、発疹ウイルス);およびイリドウイルス科(例えばアフリカ豚コレラウイルス);ならびに、分類されていないウイルス(例えば海綿状脳症の病原因子、デルタ肝炎の病原因子(B型肝炎ウイルスの欠陥サテライトウイルスであると考えられる)、非A非B型肝炎ウイルスの病原因子、(クラス1=体内伝染されるもの;クラス2=非経口的に伝染されるもの(すなわち肝炎C);ならびに、アストロウイルス。
【0058】
<非ウイルス性抗原>
本明細書に開示のVLPは、ノロウイルスファミリーに由来する様々な抗原を含んでいることが好ましい。「ノーウォーク様ウイルス」とも呼ばれるノロウイルスは、カリシウイルス科のウイルスファミリーに含まれる4つの属のうちの1つを代表するものである。ノロウイルス属には、GenogroupIおよびGenogroupIIと命名された2つの主要な遺伝群が存在する。GenogroupIのノロウイルス株は、ノーウォークウイルス、サウサンプトンウイルス、デザートシールドウイルス、およびチバウイルスを包含している。GenogroupIIのノロウイルス株は、ヒューストンウイルス、ハワイウイルス、ローズデイルウイルス(Lordsdale virus)、グリムズビーウイルス(Grimsby virus)、メキシコウイルスおよびスノーマウンテンエージェント(Snow Mountain agent)を包含している(「Parker, T.D., ら J Virol. (2005) 79(12):7402-9」、「Hale, A.D., ら J Clin. Micro. (2000) 38(4):1656-1660」))。ノーウォークウイルス(NV)は、急性ウイルス性胃腸炎の世界規模の流行的発生の大部分に関与する、ヒトカリシウイルスの1つのグループの原型株である。ノーウォークウイルスのキャプシドタンパク質は、シェルドメイン(S)および突出ドメイン(P)という2つのドメインを有している。Pドメイン(ノーウォーク株の番号付けで、aa226−530)は、P1およびP2という2つのサブドメインに分けられる。P2ドメインは、P1ドメイン内の127aaの挿入部分(aa279−405)であり、折りたたまれたモノマーの最も末端の表面に位置する。P2ドメインは、ノロウイルス株の中で、VP1の最も保存されていない領域であり、P2内の超可変領域は、受容体結合と免疫反応性とに重要な役割を果たしていると考えられる。Pドメインが外側に位置していることを考慮すると、P2ドメインは、本明細書に開示のVLPワクチン用の抗原として使用するために、好ましい抗原またはポリペプチドエピトープの源であるといえる。P2ドメインは、GenogroupIまたはGenogroupIIのノロウイルス株に対する好ましい抗原である。各種ノロウイルス株を超えて保存されたP2ドメインの領域に位置していると、同定されたmAb 61.21エピトープ、およびmAb54.6エピトープが、さらに好ましい(「Lochridge, V.P., ら J Gen. Virol. (2005) 86:2799-2806)」)。
【0059】
<インフルエンザ抗原>
本明細書に開示のVLPは、ヘマグルチニンおよびノイラミニダーゼ以外の、またはそれらに加えて、インフルエンザに由来する様々な抗原を含んでいてもよい。追加のインフルエンザ抗原としては、M2ポリペプチドが好ましい。インフルエンザウイルスのM2ポリペプチドは、スプライシングが起こった後のRNAの断片7(マトリックス断片)にコードされているクラスIIIの小さい内在性膜タンパク質(97アミノ酸)である(80、81)。M2はウイルス粒子上にはほとんど存在しないが、感染細胞上にはより豊富に発見され得る。M2は、ウイルスの侵入に必要なプロトン選択的なイオンチャネルとして機能する(82、83)。感染または従来のワクチン接種の間、M2の免疫原性は最小であるが(このことは、その保存性を説明している)、別の形態で存在するときには、M2の免疫原性および防御性は、より強くなる(84−86)。このことは、M2のモノクローナル抗体をインビボにおいて受動的に移動させると、ウイルスの排除が促進され、その結果、防御がもたらされるという観察と一致する(87)。M2の外部ドメインエピトープがHBVのコア粒子に融合タンパク質として連結されるとき、該エピトープはマウスでは非経口で接種されたり、鼻腔内から接種されたりすることによって、防御をもたらし、さらに3つの直列に配置されたコピーが、コアタンパク質のN末端に融合されたとき、免疫原性が最も強くなる(88−90)。このことは、エピトープの密度が増加すると、免疫原性が増加するということを示す他の担体−ハプテンのデータと一致する(91)。
【0060】
M2ワクチンの鼻腔内投与に関して、良好な防御を得るためにアジュバントが必要であり、良好な結果は、LTR192G(88、90)およびCTA1−DD(89)を用いたときに得られている。また上記ペプチドは、KLHなどの担体、または髄膜炎菌(N.meningitides)の外膜タンパク質複合体、またはヒトパピローマウイルスのVLPと化学的に接合されてもよく、上記ペプチドは、ワクチンとしてマウスおよび他の動物に防御をもたらす(92、93)。
【0061】
M2タンパク質が高度に保存されている限りにおいて、M2タンパク質の配列は相違点を必ず有している。共通株であるA/PR/8/34(H1N1)およびA/Aichi/68(H3N2)のM2の外部ドメインエピトープは、A/Hong Kong/156/97(H5N1)を除く、他の配列決定された現代のヒト株の全てと、免疫学的に交差反応することが示された(92)。また、インフルエンザのデータベース上の配列を調査することにより、A/Vietnam/1203/04などの他のより最近見つかった病原性のヒト単離株(H5N1)のM2配列に、同様の相違点が存在することが示された。この発見によって明らかになることは、M2エピトープを組み込んだ、有効でかつH5に特異的なパンデミックワクチンは、ヒトのH1およびH3の単離株において現在流行しているM2配列というよりは、病原性のトリ株に独特なM2の配列を反映することが必要になるであろうということである。
【0062】
インフルエンザウイルス由来の別のタンパク質(HA、NAおよびM2以外のタンパク質)は、共発現されることによって、あるいは別の抗原の全てもしくは一部をgagポリペプチドまたはHAポリペプチドに結合されることによって、VLPワクチンに含まれてもよい。これらの別の抗原には、PB2、PB1、PA、核タンパク質、マトリックス(M1)、NS1、NS2が包含される。これらの後者の抗原は、一般的に中和抗体応答の標的ではないが、T細胞によって認識される重要なエピトープを含んでいる可能性がある。VLPワクチンによってそのようなエピトープに対して誘導されるT細胞応答が、感染防御免疫を高めることに有益であると判明する可能性がある。
【0063】
<他の病原性抗原>
適切な細菌抗原には、以下の様々な病原性細菌の何れかと関連する抗原(例えば、様々な病原性細菌の何れかによって合成される抗原、および様々な病原性細菌の何れかの内部に生じた抗原)が包含される:例えば、病原性グラム陽性菌(病原性パスツレラ種(pathogenic Pasteurella species)、スタフィロコッカス種(Staphylococci species)、およびストレプトコッカス種(Streptococcus species)など);ならびに、病原性グラム陰性菌(ナイセリア(Neisseria)属、エシェリキア(Escherichia)属、ボルデテラ(Bordetella)属、カンピロバクター(Campylobacter)属、レジオネラ(Legionella)属、シュードモナス(Pseudomonas)属、シゲラ(Shigella)属、ビブリオ(Vibrio)属、エルシニア(Yersinia)属、サルモネラ(Salmonella)属、ヘモフィルス(Haemophilus)属、ブルセラ(Brucella)属、フランシセラ(Francisella)属、およびバクレリオイデス(Bacterioides)属の病原性グラム陰性菌など)など。例えば「Schaechter, M, H. Medoff, D. Schlesinger, Mechanisms of Microbial Disease. Williams and Wilkins, Baltimore (1989))」を参照のこと。
【0064】
感染性病原性真菌と関連する適切な抗原(例えば、感染性病原性真菌によって合成される抗原、および感染性病原性真菌の内部に生じた抗原)には、以下の感染性真菌と関連する抗原が包含される:特に限定されないが、クリプトコックス・ネオフォルマンス(Cryptococcus neoformans)、ヒストプラスマ・カプスラーツム(Histoplasma capsulatum)、コクシジオイデス・イミチス(Coccidioides immitis)、ブラストミセス・デルマティティディス(Blastomyces dermatitidis)、およびカンジダ・アルビカンス(Candida albicans)、カンジダ・グラブラタ(Candida glabrata)、アスペルギルス・フリガータ(Aspergillus fumigata)、アスペルギルス・フラバス(Aspergillus flavus)、およびスポロトリックス・シェンキィ(Sporothrix schenckii)など。
【0065】
病原性の原虫、蠕虫および他の真核微生物病原体と関連する適切な抗原(例えばこれらによって合成される抗原、およびこれらの内部に生じた抗原)には、以下の原虫、蠕虫および他の真核微生物病原体と関連する抗原が包含される:特に限定されないが、マラリア原虫(Plasmodium)(熱帯熱マラリア原虫(Plasmodium falciparum)、四日熱マラリア原虫(Plasmodium malariae)、卵形マラリア原虫(Plasmodium ovale)、および三日熱マラリア原虫(Plasmodium vivax)など);トキソプラズマ・ゴンディ(Toxoplasma gondii);トリパノソーマ・ブルーセイ(Trypanosoma brucei)、トリパノソーマ・クルージ(Trypanosoma cruzi);ビルハルツ住血吸虫(Schistosoma haematobium)、マンソン住血吸虫(Schistosoma mansoni)、日本住血吸虫(Schistosoma japonicum);ドノバンリーシュマニア(Leishmania donovani);ランブル鞭毛虫(Giardia intestinalis);およびクリプトスポリジウム・パルバム(Cryptosporidium parvum)など。
【0066】
適切な抗原には、以下の病原性微生物と関連する抗原(例えば病原性微生物によって合成される抗原、および病原性微生物の内部に生じた抗原)が包含される:ヘリコバクター・ピロリ(Helicobacter pyloris)、ボレリア・ブルグドルフェリ(Borelia burgdorferi)、レジオネラ・ニューモフィラ(Legionella pneumophila)、マイコバクテリウム属(Mycobacteria sps)(例えばマイコバクテリウム・ツベルクローシス(M. tuberculosis)、マイコバクテリウム・アビウム(M. avium)、マイコバクテリウム・イントラセルラーレ(M. intracellulare)、マイコバクテリウム・カンサイ(M. kansaii)、マイコバクテリウム・ゴルドナエ(M. gordonae))、スタフィロコッカス・アウレウス(Staphylococcus aureus)、ナイセリア・ゴノレー(Neisseria gonorrhoeae)、ナイセリア・メニンギティディス(Neisseria meningitidis)、リステリア・モノサイトゲネス(Listeria monocytogenes)、クラミジア・トラコーマティス(Chlamydia trachomatis)、ストレプトコッカス・ピオゲネス(Streptococcus pyogenes)(A群連鎖球菌(グループAストレプトコッカス(Group A Streptococcus))、ストレプトコッカス・アガラクチア(Streptococcus agalactiae)(B群連鎖球菌(グループAストレプトコッカス(Group B Streptococcus)))、ストレプトコッカス(ビリダンス群(viridans group))、ストレプトコッカス・フェカリス(Streptococcus faecalis)、ストレプトコッカス・ボビス(Streptococcus bovis)、ストレプトコッカス(嫌気性(anaerobic sps.))、ストレプトコッカス・ニューモニエ(Streptococcus pneumoniae)、病原性のカンピロバクター(pathogenic Campylobacter sp.)、エンテロコッカス(Enterococcus sp.)、ヘモフィルス・インフルエンザエ(Haemophilus influenzae)、バチルス・アントラシス(Bacillus anthracis)、コリネバクテリウム・ジフセリエ(Corynebacterium diphtheriae)、コリネバクテリア(corynebacterium sp.)、エリジペロスリックス・ルジオパシエ(Erysipelothrix rhusiopathiae)、クロストリジウム・パーフリンジェンス(Clostridium perfringens)、クロストリジウム・テタニ(Clostridium tetani)、エンテロバクター・エロゲネス(Enterobacter aerogenes)、クレブシエラ・ニューモニエ(Klebsiella pneumoniae)、パスツレラ・マルトシダ(Pasturella multocida)、バクテロイド(Bacteroides sp.)、フソバクテリウム・ヌクレアタム(Fusobacterium nucleatum)、ストレプトバシラス・モニリフォルミス(Streptobacillus moniliformis)、トレポネーマ・パリダム(Treponema pallidium)、トレポネーマ・ペルテニュ(Treponema pertenue)、レプトスピラ(Leptospira)、リケッチア(Rickettsia)、およびアクチノマイセス・イスラエリイ(Actinomyces israeli)など。病原性エシェリキア・コリ(E. coli)株としては、例えば、アメリカ培養細胞系統保存機関(ATCC)の整理番号31618、23505、43886、43892、35401、43896、33985、31619、および31617が挙げられるが、これらに限定されない。
【0067】
細胞内病原体と関連する様々なポリペプチドもしくは他の抗原の何れかは、VLPに含まれてもよい。細胞内病原体と関連するポリペプチドおよびペプチドのエピトープは、細胞内病原体と関連する(例えば細胞内病原体によってコードされている)ポリペプチドであって、該ポリペプチドの断片がMHCクラスI分子と一緒に感染細胞の表面上に提示され、該断片が、CD8.sup.+のリンパ球の表面上のT細胞抗原受容体に認識される(例えば該受容体に結合される)というポリペプチドであれば、どのようなものでもよい。細胞内病原体と関連するポリペプチドおよびペプチドのエピトープは、技術的に知られており、特に限定されないが、ヒト免疫不全ウイルス(例えばHIV gp120)と関連する抗原、またはその抗原断片;サイトメガロウイルス抗原;マイコバクテリウム抗原(例えばマイコバクテリウム・アビウム(Mycobacterium avium)、およびマイコバクテリウム・ツベルクローシス(Mycobacterium tuberculosis)など);ニューモシスチス・カリニ(Pneumocystic carinii)(PCP)抗原;マラリア抗原(特に限定されないが、熱帯熱マラリア原虫または他のあらゆるマラリア種と関連する抗原(41−3、AMA−1、CSP、PFEMP−1、GBP−130、MSP−1、PFS−16、SERPなど)を含む);真菌抗原;酵母抗原(例えばカンジダ(Candida spp.)の抗原);トキソプラズマ(toxoplasma)抗原(特に限定されないが、トキソプラズマ・ゴンディ(Toxoplasma gondii)、トキソプラズマ脳炎(Toxoplasma encephalitis)、または他のあらゆるトキソプラズマ種と関連する抗原を含む);EBウイルス(EBV)抗原;プラスモディウム(Plasmodium)抗原(例えばgp190/MSP1など);などを包含する。
【0068】
好ましいVLPワクチンは、バチルス・アントラシス(Bacillus anthracis)を対象とするものであってもよい。バチルス・アントラシスは、好気性または通性嫌気性でグラム陽性の、幅1.0μm、長さ3.0−5.0μmの非運動性桿菌である。悪条件下では、バチルス・アントラシスは、高耐性の内生胞子を形成する。この内生胞子は、感染動物が以前に死亡した場所の土壌で発見され得る。本明細書に開示されているようなVLPワクチンに使用するための好ましい抗原は、感染防御抗原(PA)である。該感染防御抗原は、83kDaのタンパク質であって、哺乳類細胞の受容体に結合し、バチルス・アントラシスが病気を引き起こすことができるのに極めて重要なものである。より好ましい抗原は、バチルス・アントラシスのPAにおけるC末端側の140アミノ酸の断片であり、該断片はバチルス・アントラシスに対する感染防御免疫を被検体に誘導するために使用され得るものである。炭疽菌に対するVLPワクチンに使用するための他の典型的な抗原は、炭疽菌の胞子に由来する抗原(例えばBclA)、この細菌の栄養生長期からの抗原(例えば、細胞壁抗原、莢膜抗原(例えばポリ−ガンマ−D−グルタミン酸、すなわちPGA)、分泌抗原(例えば感染防御抗原、致死因子、または浮腫因子などの外毒素)である。VLPワクチンに使用するためのもう1つ別の好ましい抗原は、バチルス・アントラシスに独特な、アントロース(anthrose)を含むテトラ−サッカリドである(Daubenspeck J.M., ら J. Biol. Chem. (2004), 279:30945)。該テトラ−サッカリドは、脂質ラフト会合ポリペプチドと共役されてもよい。これにより、抗原をVLPワクチンと会合させることができる。
【0069】
<腫瘍関連抗原>
公知の様々な腫瘍特異的抗原または腫瘍関連抗原(TAA)の何れかを、VLPに含めてもよい。完全なTAAが使用されてもよいが、使用されなくてもよい。その代わりに、TAAの一部(例えばエピトープ)を使用してもよい。VLPに使用されてもよい腫瘍関連抗原(またはエピトープを含むその断片)には、特に限定されないが、MAGE−2、MAGE−3、MUC−1、MUC−2、HER−2、高分子量のメラノーマ関連抗原MAA、GD2、癌胎児性抗原(CEA)、TAG−72、卵巣関連抗原OV−TL3およびMOV18、TUAN、アルファ−フェトタンパク質(AFP)、OFP、CA−125、CA−50、CA−19−9、腎腫瘍関連抗原G250、EGP−40(EpCAMとしても知られている)、S100(悪性メラノーマ関連抗原)、p53、およびp21rasが包含される。(上述したTAAのいずれかを含む)任意のTAA(またはそのエピトープ)の合成類似体が、使用されてもよい。さらに、1つ以上のTAA(またはそのエピトープ)の組み合わせが、組成物に含まれてもよい。
【0070】
<アレルゲン>
1態様では、VLPワクチンの一部である抗原は、様々なアレルゲンの何れかであってもよい。アレルゲンを基本とするワクチンは、アレルゲンに対する寛容を被検体に誘導するために使用されてもよい。チロシンとの共沈に関与するアレルゲンワクチンの例は、米国特許第3,792,159号明細書、米国特許第4,070,455号明細書、および米国特許第6,440,426号明細書において見つけることができる。なお、特にアレルゲンワクチンの製剤化に関して、これら文献のそれぞれは参照することによって本明細書に組み込まれる。
【0071】
様々なアレルゲンの何れかは、VLPに含まれてもよい。アレルゲンには、特に限定されないが、環境エアロアレルゲン;ブタクサ/枯草熱などの植物の花粉;雑草の花粉のアレルゲン;草の花粉のアレルゲン;セイバンモロコシ(Johnson grass);木の花粉のアレルゲン;ライグラス;イエダニのアレルゲン(例えばDer p I、Der f Iなど)などのクモ類のアレルゲン;ストレージダニのアレルゲン;スギ花粉/枯草熱;カビ胞子のアレルゲン;動物のアレルゲン(例えばイヌ、モルモット、ハムスター、アレチネズミ、ラット、マウスなどのアレルゲン);食物のアレルゲン(例えば甲殻類;ピーナッツなどのナット;柑橘系果物のアレルゲン);昆虫のアレルゲン;毒:(膜翅目、イエロー・ジャケット、ミツバチ、ワスプ、ホーネット、ハリアリ(fire ant));ゴキブリ、ノミ、蚊などからの他の環境昆虫のアレルゲン;ストレプトコッカスの抗原などの細菌のアレルゲン;アスカリス属抗原などの寄生虫のアレルゲン;ウイルス抗原;真菌の胞子;薬物のアレルゲン;抗生物質;ペニシリンおよび関連化合物;他の抗生物質;ホルモン(インシュリン)、酵素(ストレプトキナーゼ)などの総タンパク質;不完全な抗原またはハプテンとして作用することが可能なすべての薬物およびそれらの代謝産物;ハプテンとして作動することが可能な、およびアレルゲンとして機能することが可能な、工業用化学薬品および代謝産物(例えば酸無水物(無水トリメリット酸など)およびイソシアネート(トルエンジイソシアネートなど));小麦粉(例えばパン屋喘息(Baker’s asthma)を起こすアレルゲン)、トウゴマ、コーヒー豆、および上記工業用化学薬品などの職業性アレルゲン;ノミのアレルゲン;ならびにヒト以外の動物の中のヒトタンパク質が包含される。
【0072】
アレルゲンには、特に限定されないが、細胞、細胞抽出物、タンパク質、ポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドおよび他の分子のペプチド模擬体および非ペプチド模擬体、低分子、脂質、糖脂質、ならびに炭水化物が包含される。
【0073】
具体的な天然の動物および植物のアレルゲンには、例えば特に限定されないが、以下の属に特有のタンパク質が包含される:イヌ科(カニスファミリアリス(Canis familiaris));デルマトファゴイデス属(Dermatophagoides)(例えばデルマトファゴイデス・ファリナエ(コナヒョウヒダニ(Dermatophagoides farinae)));ネコ属(フェリス・ドメスチカス(Felis domesticus));アンプローシア属(Ambrosia)(アンプローシア・アルテミスフォリア(Ambrosia artemiisfolia);ロリウム属(Lolium)(例えばペレニアルライグラス(Lolium perenne)またはイタリアンライグラス(Lolium multiflorum));スギ属(Cryptomeria)(スギ(Cryptomeria japonica));アルテマリア属(Altemaria)(アルテマリア・アルテマタ(Altemaria altemata));アルダー属(Alder);ハンノキ属(アルヌス・グルチノアス(Alnus gultinoas));ベツラ属(Betula)(ベツラ・ベルコサ(Betula verrucosa));ケルクス属(Quercus)(ケルクス・アルバ(Quercus alba));オリーブ属(Olea)(オリーブ(Olea europa));アルテミシア属(Artemisia)(アルテミシア・バルガリス(Artemisia vulgaris));プランタゴ属(例えばヘラオオバコ(Plantago lanceolata));パリエタリア属(Parietaria)(例えばパリエタリア・オフィシナリス(Parietaria officinalis)またはパリエタリア・ジュダイカ(Parietaria judaica));チャバネゴキブリ属(Blattella)(例えばチャバネゴキブリ(Blattella germanica));アピス属(Apis)(例えばアピス・ムルチフロルム(Apis multiflorum));カプレサス属(Cupressus)(例えばカプレサス・センパーヴァイレンス(Cupressus sempervirens)、カプレサス・アリゾニカ(Cupressus arizonica)、およびカプレサス・マクロカルパ(Cupressus macrocarpa));ジュニペルス属(例えばジュニペルス・サビノイデス(Juniperus sabinoides)、ジュニペルス・ヴィルギニアナ(Juniperus virginiana)、ジュニペルス・コムニス(Juniperus communis)、およびジュニペルス・アシェイ(Juniperus ashei));ツヤ属(Thuya)(例えばツヤ・オリエンタリス(Thuya orientalis));ヒノキ属(Chamaecyparis)(例えばヒノキ(Chamaecyparis obtusa));ペリプラネタ属(Periplaneta)(例えばワモンゴキブリ(Periplaneta americana));アグロピロン属(例えばシバムギ(Agropyron repens));セカレ属(Secale)(例えばライムギ(Secale cereale));トリチカム属(Triticum)(例えばコムギ(Triticum aestivum));ダクチリス属(Dactylis)(例えばオーチャードグラス(Dactylis glomerata));フェスツカ属(Festuca)(例えばフェスツカ・エラチオル(Festuca elatior));ポア属(Poa)(例えばポアプラテンシス(Poapratensis)またはポア・コンプレッサ(Poa compressa));カラスムギ属(Avena)(例えばマカラスムギ(Avena sativa));ホルクス属(Holcus)(例えばホルクス・ラナタス(Holcus lanatus));アンソキサスム属(Anthoxanthum)(例えばハルガヤ(Anthoxanthum odoratum));アルヘナセラム属(Arrhenatherum)(例えばアルヘナセラム・エラチウス(Arrhenatherun elatius));アグロスチス属(例えばアグロスチス・アルバ(Agrostis alba));フレウム属(Phleum)(例えばチモシー(Phleum pratense));ファラリス(Phalaris)属(例えばファラリス・アルンジナセア(Phalaris arundinacea));スズメノヒエ属(Paspalum)(例えばバヒアグラス)(Paspalum notatum);ソルガム属(Sorghum)(例えばソルガム・ハレペンシス(Sorghum halepensis));そしてブロムス属(Bromus)(例えばブロムス・イネルミス(Bromus inermis))。
【0074】
<VLPを作成する好ましい方法>
VLPは、(a)脂質ラフトと天然に会合しない抗原に連結された脂質ラフト会合ポリペプチドおよび(b)gagポリペプチドを含んでいるVLPが、組立られることが好ましい技術分野の当業者の一人に利用可能な任意の方法によって、容易に組立られ得る。好ましい実施形態では、上記ポリペプチドは、あらゆる利用可能なタンパク質発現システムにおいて共発現されてもよい。また該タンパク質発現システムは、脂質内にラフト脂質ドメインを含む細胞を基本とするシステム(哺乳類細胞発現システム、および昆虫細胞発現システムなど)であることが好ましい。
【0075】
gagポリペプチドを用いて形成されたVLPの発現の多くの例は、公開されており、VLPを作製するのに利用可能な発現システムの範囲が実証されている。いくつかのレトロウイルスを用いた研究によれば、他のウイルス成分の非存在下において発現されたGagポリペプチドは、VLPの形成および細胞表面における出芽に十分であることが実証された(「Wills and Craven AIDS 5, 639-654, 1991」、「Zhouら, 3. Virol. 68, 2556-2569, 1994」、「Morikawaら, Virology 183, 288-297, 1991」、「Royerら, Virology 184, 417-422, 1991」、「Gheysenら, Cell 59, 103-112, 1989」、「Hughesら, Virology 193, 242-255, 1993」、「Yamshchikovら, Virology 214, 50-58, 1995」)。バキュロウイルスベクターを用いた、昆虫細胞内でのGag前躯体の発現によるVLPの形成は、いくつかのグループによって実証されている(「Delchambreら, EMBO J. 8, 2653-2660, 1989」、「Luoら, Virology 179, 874-880, 1990」、「Royerら, Virology 184, 417-422, 1991」、「Morikawaら, Virology 183, 288-297, 1991」、「Zhouら, J. Virol. 68, 2556-2569, 1994」、「Gheysenら, Cell 59, 103-112, 1989」、「Hughesら, Virology 193, 242-255, 1993」、「Yamshchikovら, Virology 214, 50-58, 1995」)。これらのVLPは、未成熟のレンチウイルス粒子に類似しており、効率的に組立られ、昆虫細胞の形質膜から出芽によって放出される。
【0076】
Gag前躯体のアミノ末端領域が、細胞表面への移送と、ウイルスの組立に必要な膜結合とのための、標的シグナルであることが報告されている(「Yuら, J. Virol. 66, 4966-4971, 1992」、「an, Xら, J. Virol. 67, 6387-6394, 1993」、「Zhouら, J. Virol. 68, 2556-2569, 1994」、「Lee and Linial J. Virol. 68, 6644-6654, 1994」、「Dorfmanら, J. Virol. 68, 1689-1696, 1994」、「Fackeら, J. Virol. 67, 4972-4980, 1993」)。ワクシニア・ウイルス発現システムを用いた、Gag構造タンパク質とEnv糖タンパク質であるgp120およびgp41とを含む、組み換えHIVを基本とするVLPの組立が、報告されている(「Haffarら, J. Virol. 66, 4279-4287, 1992」)。
【0077】
VLPのためのポリペプチドの組み換え発現には、上記ポリペプチドの1つ以上をコードするポリヌクレオチドを含んでいる発現ベクターのコンストラクションが必要である。上記ポリペプチドの1つ以上をコードするポリヌクレオチドが得られれば、該ポリペプチドを製造するためのベクターは、公知手法を用いる組み換えDNA技術によって製造され得る。したがって、本明細書には、VLPポリペプチドをコードするヌクレオチド配列の何れかを含んでいるポリヌクレオチドを発現させることによって、タンパク質を調製する方法が記載されている。当業者に公知の方法を用いることにより、VLPポリペプチドをコードする配列、適切な転写制御シグナル、および適切な翻訳制御シグナルを含んでいる発現ベクターがコンストラクトされることができる。これらの方法には、例えば、イントロでの組み換えDNA手法、合成の手法、およびインビボでの遺伝子組み換えが包含される。したがって、本発明は、gagポリペプチドと、抗原に連結される脂質ラフト会合ポリペプチドとをコードするヌクレオチド配列を含んでおり、これらの全てが1つ以上のプロモーターに作動可能に連結されている、複製可能なベクターを提供する。
【0078】
上記発現ベクターは、従来手法によって宿主細胞に移動させられてもよく、トランスフェクションされた細胞は、それからVLPポリペプチドを製造するように従来手法によって培養される。したがって、本発明は、異種プロモーターに作動可能に連結されたVLPポリペプチドの1つ以上をコードするポリヌクレオチドを含んでいる宿主細胞を包含する。VLPを生成するための好ましい実施形態では、gagポリペプチド、および抗原に連結される脂質ラフト会合ポリペプチドの両方をコードするベクターは、VLPを生成するための宿主細胞内で共発現されてもよい。この点について以下で詳細に説明する。
【0079】
様々な宿主−発現ベクターシステムが、VLPポリペプチドを発現させるために利用されてもよい。そのような宿主−発現システムは、VLPを生成するために(好ましくは共発現によって)VLPポリペプチドが製造され得るための媒体を表す。多様な宿主が、適切な発現ベクターのコンストラクトに使用されてもよく、好ましい宿主−発現システムは、VLPの組立に適した脂質ラフトを有する宿主である。該宿主には、VLPポリペプチドをコードする配列(VLPポリペプチドコード配列)を含んでいる、組み換えバクテリオファージDNA、プラスミドDNA、またはコスミドDNAの発現ベクターで形質転換される、細菌などの微生物(例えばエシェリキア・コリ、バシラス・サブティリス(B. subtilis));VLPポリペプチドコード配列を含んでいる組み換え酵母発現ベクターで形質転換される、酵母(例えばサッカロマイセス(Saccharomyces)、ピキア(Pichia));VLPポリペプチドコード配列を含んでいる組み換えウイルス発現ベクター(例えばバキュロウイルス)で感染される昆虫細胞システム;VLPポリペプチドコード配列を含んでいる組み換えウイルス発現ベクター(例えばカリフラワーモザイクウイルス、CaMV、タバコモザイクウイルス、TMV)で感染されるか、VLPポリペプチドコード配列を含んでいる組み換えプラスミド発現ベクター(例えばTiプラスミド)で形質転換される、植物細胞システム;あるいは、哺乳類細胞のゲノム由来のプロモーター(例えばメタロチオネインのプロモーター)、または哺乳類ウイルス由来のプロモーター(例えばアデノウイルスの後期プロモーター;ワクシニア・ウイルスの7.5Kのプロモーター)由来のプロモーター、を含んでいる組み換え発現コンストラクトが導入される、哺乳類細胞システム(例えばCOS細胞、CHO細胞、BHK細胞、293細胞、3T3細胞)が包含されるが、これらに限定されない。好ましくは哺乳類細胞、より好ましくは昆虫細胞が、VLPの組立に適したラフト脂質を有しているので、VLPポリペプチドの発現のために使用される。例えば、ヒトサイトメガロウイルス由来の主要中間初期遺伝子プロモーターエレメントなどのベクターと組み合わせられる、チャイニーズハムスター卵巣(CHO)細胞などの哺乳類細胞は、VLPポリペプチドにとって効果的な発現システムであるといえる(「Foeckingら, Gene 45:101 (1986)」、「Cockettら, Bio/Technology 8:2 (1990)」)。
【0080】
昆虫システムでは、キンウワバ科(Autographa californica)の核多角体病ウイルス(AcNPV)が、外来遺伝子を発現させるベクターとして使用され得る。該ウイルスは、スポドプテラ・フルギペルダ(Spodoptera frugiperda)細胞において成長する。VLPポリペプチドコード配列は、ウイルスの非必須領域(例えばポリヘドリン遺伝子)にそれぞれクローニングされて、AcNPVプロモーター(例えばポリヘドリンプロモーター)の制御下に置かれてもよい。
【0081】
哺乳類宿主細胞では、ウイルスを基本とする発現システムの多くが利用され得る。アデノウイルスが発現ベクターとして使用される場合、関心のあるVLPポリペプチド配列が、アデノウイルスの転写/翻訳制御複合体(例えば後期プロモーターおよび3部分からなるリーダー配列)に結合されてもよい。このキメラ遺伝子は、その後、インビトロまたはインビボでの組み換えによってアデノウイルスのゲノムに挿入されてもよい。ウイルスゲノムの非必須領域(例えば領域E1またはE3)に挿入することによって、感染宿主内で生存することができ、VLPポリペプチドを発現することができる組み換えウイルスが得られる(例えば、「Logan & Shenk, Proc. Natl. Acad. Sci. USA 81:355-359 (1984)」参照)。また特定の開始シグナルが、挿入されたVLPポリペプチドコード配列を効率よく翻訳するために必要とされてもよい。該シグナルには、ATG開始コドンおよび隣接配列が包含される。さらに、開始コドンは、挿入物全体の翻訳を確実にするために、所望のコード配列のリーディングフレームと一致していなければならない。これら外来性の翻訳制御シグナルおよび開始コドンは、天然および合成の様々なものに由来してもよい。発現効率は、適切な転写エンハンサー要素、翻訳ターミネーターなどを含めることによって、上昇され得る(「Bittnerら, Methods in Enzymol. 153:51-544 (1987)」参照)。
【0082】
また、挿入された配列の発現を調節するか、遺伝子産物を特定の望まれる方法で修飾および加工する宿主細胞株が、選択されてもよい。タンパク質産物のそのような修飾(例えばグリコシル化)および加工(例えば開裂または膜への移送)は、VLPの生成、VLPポリペプチドの機能、または別のポリペプチド(アジュバントもしくは別の抗原)にとって重要である可能性がある。種々の宿主細胞は、タンパク質ならびに遺伝子産物の翻訳後加工および翻訳後修飾に対して、特徴的かつ特異的な機構を有している。適切な細胞または宿主システムは、発現された外来タンパク質の正しい修飾および加工が確実に行われるように選択されてもよい。これのためには、一次転写産物の適切なプロセッシングや、遺伝子産物のグリコシル化およびリン酸化に対する細胞機構を有する、真核宿主細胞が使用されてもよい。
【0083】
宿主細胞は、本発明の2つの発現ベクターで共トランスフェクションされてもよい(第1ベクターは、gagポリペプチドをコードするベクターであり、第2ベクターは、抗原に連結される脂質ラフト会合ポリペプチドをコードするベクターである)。2つのベクタ−は、各VLPポリペプチドを等しく発現させることができる、同じ選択マーカーを含んでいてもよい。また代わりに、gagポリペプチドと、脂質に連結される脂質ラフト会合ポリペプチドとの両方をコードし、これらを発現させることができる1つのベクターを使用してもよい。
【0084】
VLPが宿主細胞によって産生されれば、該VLPは、ポリペプチドの精製に関して技術的に知られている任意の方法によって、精製され得る。そのような方法としては、例えば、クロマトグラフィー(例えばイオン交換クロマトグラフィー、親和性クロマトグラフィー(特にポリペプチドに付加された任意の親和性精製タグに対する親和性を利用する、親和性クロマトグラフィー)、大きさを分けるクロマトグラフィー)、遠心、または溶解度差を用いる方法、あるいはタンパク質もしくは他の高分子を精製するための他の任意の標準的な手法を用いる方法が挙げられる。また、VLPポリペプチドは、VLPの精製を容易にするための、本明細書に記載されているか、さもなければ技術的に公知である異種のポリペプチド配列と融合されてもよい。精製後、別の抗原またはアジュバントなどの別の要素が、VLPポリペプチドに対する共有結合を介するか、または他の非共有結合の機構によってVLPに物理的に連結されてもよい。VLPポリペプチドが、ラフト脂質ドメインを有する宿主細胞(哺乳類細胞および昆虫細胞など)内で共発現される好ましい実施形態では、VLPは、自ら組立られて放出される。これにより、上記方法の何れかによってVLPが精製され得る。
【0085】
〔VLPを使用する好ましい方法〕
<製剤>
本明細書に記載のVLPは、ワクチン調製物として使用されることが好ましい。通常、そのようなワクチンは、注射剤として液体溶液または懸濁液として調製される。また該ワクチンは、注射前の時点では、液体溶液または懸濁液に適した固形剤として調製されてもよい。またそのような調製物は、乳化されてもよいし、乾燥粉末として製造されてもよい。しばしば、活性免疫原性成分が、該活性免疫原性成分と適合する薬学的に許容可能な賦形剤と混合される。適した賦形剤は、例えば、水、生理食塩水、右旋性グルコース、グリセロール、エタノール、または同類のもの、およびそれらの組み合わせである。さらに、希望に応じて、ワクチンは、補助物質(湿潤剤、乳化剤、pH緩衝剤、またはワクチンの有効性を向上させるアジュバントなど)を含んでもよい。
【0086】
ワクチンは、従来どおり、例えば皮下に、経皮的に、皮内に、真皮下に、または筋肉内に注入されることによって非経口的に投与されてもよい。他の投与様式に適した別の製剤は、坐薬を含んでおり、場合によっては経口製剤、鼻腔内製剤、バッカル製剤、舌下製剤、腹腔内製剤、膣内製剤、肛門製剤および頭蓋内製剤を含んでいる。坐薬については、在来の結合剤および担体は、例えばポリアルキレングリコールまたはトリグリセリドを含んでいてもよい。そのような坐薬は、上記活性免疫原性成分を、0.5%から10%、好ましくは1%から2%の範囲で含んでいる混合物から形成されてもよい。ある実施形態では、低融点のワックス(脂肪酸グリセリドまたはココアバターの混合物など)が初めに溶かされ、本明細書記載のVLPが攪拌などによって均質に分散させられる。溶解した均一混合物は、それから従来の大きさの型に注がれ、冷却されてから凝固される。
【0087】
鼻腔内投与に適した製剤には、液体および乾燥粉末が包含される。製剤には、普通に使用される賦形剤(例えば医薬品等級のマンニトール、ラクトース、サッカロース、トレハロースおよびキトサンなど)が含まれる。キトサンなどの粘膜接着剤(Mucosadhesive agent)は、鼻腔内投与された製剤の粘液線毛クリアランスを遅らせるために、液体製剤または乾燥粉末製剤に使用されることができる。マンニトールおよびサッカロースなどの糖は、液体製剤においては安定剤として、乾燥製剤においては安定剤および充填剤として使用されることができる。また、アジュバント(例えばモノホスホリルリピドA(MPL)、および単なる例であり特に限定されないが、二本鎖ポリ(I:C)、ポリイノシン酸、CpG含有オリゴヌクレオチド、イミキモド(imiquimod)、コレラ毒素およびその誘導体、易熱性エンテロトキシンおよびその誘導体、ならびに本明細書の全体に挙げられているアジュバントの多く)は、液体製剤および乾燥粉末製剤に、免疫賦活性アジュバントとして使用されることができる。
【0088】
経口投与に適した製剤には、液体、固体、半固体、ゲル、錠剤、カプセル、およびトローチ剤などが包含される。経口投与に適した製剤には、錠剤、トローチ剤、カプセル、ゲル、液体、食品、飲み物、および栄養補給食品などが包含される。製剤には、普通に使用される賦形剤(例えば医薬品等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、および炭酸マグネシウムなど)が含まれる。他のVLPワクチン組成物は、溶液、懸濁液、丸剤、徐放性製剤、または粉末の形態を取ってもよく、10%から95%、好ましくは25%から70%の活性免疫原性成分を含んでいる。経口製剤に関しては、コレラ毒素が製剤の相手として興味深い物質である(また接合の相手としても可能性がある)。
【0089】
VLPワクチンは、膣内投与用に製剤化されるときは、ペッサリー、タンポン、クリーム、ゲル、ペースト、泡、またはスプレーの形態になってもよい。上記製剤の何れかは、VLPに加えて、技術的に公知の適切な作用物質を含んでいてもよい。
【0090】
ある実施形態では、VLPワクチンは、全身投与用に、または局所投与用に製剤化されてもよい。そのような製剤は、技術的によく知られている。非経口投与用の媒体には、塩化ナトリウム溶液、リンガー右旋性グルコース、右旋性グルコースおよび塩化ナトリウムの溶液、乳酸加リンゲル溶液、または不揮発性油が包含される。静脈内投与用の媒体には、流動性で栄養性の補充液、および電解質補充液(リンガー右旋性グルコースに基づくものなど)などが包含される。全身投与および局所投与の経路には、経皮投与、皮内投与、局所適用、静脈内投与、筋肉内投与などが包含される。
【0091】
VLPは、中性製剤または塩に基づく製剤を包含するワクチンに製剤化されてもよい。薬学的に許容可能な塩には、酸付加塩(ペプチドの遊離アミノ基と一緒に形成された塩)が包含される。なお、該酸付加塩は、無機酸(例えば塩酸またはリン酸)、あるいは有機酸(酢酸、シュウ酸、酒石酸、およびマンデル酸など)と一緒に形成された塩である。また、遊離カルボキシル基と一緒に形成された塩は、無機塩基(例えば、水酸化ナトリウム、水酸化カリウム、水酸化アンモニウム、水酸化カルシウム、または水酸化第二鉄など)、および有機塩基(イソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジン、およびプロカインなど)から得られてもよい。
【0092】
ワクチンは、製剤の用量(dosage formulation)と適合するように、治療効果および免疫原性がもたらされるような量で、投与されてもよい。投与される量は、治療される被検体(例えば免疫応答を開始するための個々の免疫系の能力、および希望される防御の程度など)に依存する。適切な用量の範囲では、接種一回の当たりの活性成分は、数百マイクログラムの桁であり、(1から10mgの範囲というより多い量が考慮されるが)好ましくは約0.1μgから2000μgの範囲であり、例えば、約0.5μgから1000μgの範囲であり、好ましくは1μgから500μgの範囲であり、特に好ましくは約10μgから100μgの範囲である。また、初回投与および追加免疫に適した投薬計画は、変更可能であるが、初回投与が行われてから、その後の接種または他の投与が行われるというのが典型である。
【0093】
適用の様式は大きく変更されてもよい。ワクチンを投与するための従来の方法の何れかを適用することができる。これらには、生理学的に許容可能な固形の基質を利用して経口投与する方法、または生理学的に許容可能な分散形態で注射などによって投与する方法などが含まれる。ワクチンの用量は、投与経路に依存し、ワクチン接種されるものの年齢および抗原の剤形に基づいて変更される。
【0094】
ワクチン製剤には、それ自体ワクチンとして十分な免疫原性を有しているものがあるが、残りのワクチン製剤のいくつかには、ワクチンがアジュバント物質をさらに含んでいる場合に、免疫応答が増強されるものがある。
【0095】
また、粘膜接着を改善するための送達剤が使用されることにより、特に鼻腔内投与製剤、経口投与製剤、肺に基づく投与製剤に対する、送達および免疫原性が改善され得る。そのような化合物の1つであるキトサン(キチンのN−デアセチル化形態)が、多くの薬学的製剤に使用される(32)。キトサンは、粘液線毛クリアランスを遅らせることができ、粘膜での抗原の取り込みおよび加工により長く時間を費やすことができるため、鼻腔内ワクチンにとって魅力的な粘膜接着剤である(33、34)。さらに、キトサンは、一時的に密着結合を広げることができ、抗原のNALTへの経上皮輸送を向上させる可能性がある。最近のヒトでの試験において、他のアジュバントとは一緒に投与されず、キトサンと一緒に鼻腔内投与された3価の不活性化インフルエンザワクチンは、抗体陽転を引き起こすとともに、筋肉内接種の後に得られるHIの力価よりも僅かに低いHIの力価をもたらした(33)。
【0096】
またキトサンは、遺伝的に解毒されたE. coliの易熱性エンテロトキシンの突然変異体であるLTK63などの、鼻腔内で良好に機能するアジュバントと一緒に製剤化されることもできる。これにより、キトサンによって与えられる送達と接着との利点に加えて、免疫賦活効果が追加される。その結果、粘膜応答および全身応答が増強されることになる(35)。
【0097】
最後に、キトサンの製剤は、乾燥粉末の形式で製剤され得ることに注意されたい。この形式は、ワクチンの安定性を改善することが示されており、液体製剤よりもさらに粘液線毛クリアランスを遅らせることができる(42)。このことは、キトサンと一緒に製剤化された鼻腔内乾燥粉末であるジフテリアトキソイドワクチンを用いた、最近のヒトでの臨床試験において確かめられた。さらに、該臨床試験では、鼻腔内経路は、従来の筋肉内経路と同じくらい効果的であり、分泌型のIgA応答の利点が加えられることが確かめられた(43)。また上記ワクチンの耐性は、非常に強かった。キトサンおよびMPLを含んでいる炭疽菌用の鼻腔内乾燥粉末ワクチンは、筋肉内接種よりも強い応答をウサギに誘導し、またエアロゾルである胞子によるアレルギーの誘発に対する防御も誘導した(44)。
【0098】
鼻腔内ワクチンは、それが、非経口投与されるワクチンとは対照的に上気道および下気道に影響を与えることができるので、好ましい製剤である(なお、非経口投与されるワクチンは、下気道に影響を与える点においてより優れている)。このことは、アレルゲンを基本とするワクチンに対する寛容を誘導することや、また病原体を基本とするワクチンに対する免疫を誘導することにとって利点となり得る。
【0099】
鼻腔内ワクチンが使用されることにより、上気道および下気道に防御が与えられることに加えて、針を使った接種につきまとう厄介な問題が回避されるし、粒状および/または可溶性の抗原と鼻咽頭関連リンパ組織(NALT)との相互作用により、粘膜性ならびに全身性の体液性応答および細胞性応答の両方を引き起こすという手段が提供される(16−19)。鼻腔内経路の効果は、歴史的に、非経口接種の効果よりも弱かったが、新しい投与製剤、アジュバント、およびVLPの使用は、このパラダイムを変え始めている。実際のところ、機能的なヘマグルチニンポリペプチドを含んでいるVLPは、鼻腔内投与に特に適している可能性がある。なぜなら、鼻粘膜に豊富に存在するシアル酸含有受容体が、HA抗原の結合を向上させ、粘液線毛クリアランスを減少させることになり得るからである。
【0100】
<アジュバント>
ワクチンに対するアジュバント効果を得る方法としては、種々の方法が知られており、それらは本明細書に開示のVLPと併用して使用され得る。一般的な原理および方法の詳細は、「”The Theory and Practical Application of Adjuvants", 1995, Duncan E. S. Stewart-Tull (ed.), John Wiley & Sons Ltd, ISBN 0-471-95170-6」、および「”Vaccines: New Generation Immunological Adjuvants”, 1995, Gregoriadis Gら (eds.), Plenum Press, New York, ISBN 0-306-45283-9」に記載されている。これら文献は、参照することによって本明細書に組み込まれる。
【0101】
ある実施形態では、VLPワクチンは、少なくとも1つのアジュバントと混合されたVLPを含んでいる。上記VLPワクチンに含まれている上記VLPと上記アジュバントとの割合は、重量比で約10:1から約1010:1までであり、例えば、約10:1から約100:1まで、約100:1から約103:1まで、約103:1から約104:1まで、約104:1から約105:1まで、約105:1から約106:1まで、約106:1から約107:1まで、約107:1から約108:1まで、約108:1から約109:1まで、または約109:1から約1010:1までである。当業者の一人は、アジュバントおよび最適な比を決定するための慣例的な実験を通して、適切な比を容易に決定することができる。本明細書に記載されているようなVLPとアジュバントとの混合物は、当業者に利用可能なあらゆる組み合わせの形態を含んでいてもよい。そのような形態には、特に限定されないが、同じ溶液内においてVLPとアジュバントとが分離されているという混合物、VLPとアジュバントとが共有連結されているという混合物、VLPとアジュバントとがイオン的に連結されているという混合物、VLPとアジュバントとが疎水的に連結されているという混合物(VLP膜に部分的にまたは完全に埋め込まれている形態を含む)、VLPとアジュバントとが親水的に連結されているという混合物、ならびにそれらの任意の組み合わせが包含される。
【0102】
アジュバントの好ましい例はポリペプチドアジュバントである。該ポリペプチドアジュバントは、VLPポリペプチドと一緒に共発現されることによって、またはVLPポリペプチドと融合されてキメラポリペプチドが製造されることによって、本明細書に記載のVLPに容易に付加されるものであってもよい。鞭毛の主要タンパク質成分である細菌のフラジェリンは、Toll様受容体TLR5により先天性免疫系から認識されるために、アジュバントタンパク質として注目されている(65)。TLR5を介したフラジェリンのシグナルは、DCの成熟および遊走を誘導することによって、ならびに前炎症性メディエータの産生をもたらすマクロファージ、好中球および腸上皮細胞の活性化を誘導することによって、先天性免疫機能と獲得免疫機能との両方に効果を及ぼす(66−72)。
【0103】
TLR5は、フラジェリンモノマーに保存された、このタンパク質に独特で鞭毛の機能に必須の構造を認識するが、免疫学的圧力に応じた該構造の突然変異体は認識しない(73)。TLR5は、100fMの濃度に対して感受性があるが、完全なフィラメントを認識しない。鞭毛のモノマーへの分解には、結合と刺激とが要求される。
【0104】
アジュバントとしてのフラジェリンは、非経口でまたは鼻腔内に投与される異種抗原に対する防御応答を誘導するための強力な活性を有しているし(66、74−77)、DNAワクチンに対するアジュバント効果も報告されている(78)。フラジェリンが使用されたとき、Th2への偏りが観察され、このことはインフルエンザなどの呼吸器ウイルスに対して適しているであろうが、マウスまたはサルにおいてIgEが誘導されたという証拠は見つかっていない。また、サルに鼻腔内投与または全身投与した後に、局所的炎症反応または全身性炎症反応が起こらなかったということも報告されている(74)。MyD88に依存する様式でTLR5を経由するフラジェリンシグナル、およびTLRを経由する他の全てのMyD88依存性のシグナルは、Th1へ偏ることになることが示されていたために、フラジェリンを使用した後に誘発される応答に関するTh2の特徴には、ある程度驚かされる(67、79)。重要なことは、フラジェリンに対する予め存在していた抗体が、アジュバントの有効性に対して、あまり効果を及ぼさず(74)、このことが、フラジェリンを複数回使用されるアジュバント(multi-use adjuvant)として魅力あるものにしていることである。
【0105】
多くの最近の鼻腔内ワクチンの試験に共通するテーマは、ワクチンの有効性を改善するためのアジュバントおよび/または投与系の使用である。そのような研究によれば、遺伝的に解毒されたE. coliの易熱性エンテロトキシンアジュバント(LT R192G)を含んでいるインフルエンザのH3ワクチンを鼻腔内投与した後にだけ、H5の暴露に対するヘテロサブタイプな防御がもたらされている。防御は、中和抗体の交差の誘導に基づいており、新たなワクチンを開発する上で鼻腔内経路が重要であることを実証した(22)。
【0106】
またサイトカイン、コロニー刺激因子(例えばGM−CSFおよびCSFなど);腫瘍壊死因子;インターロイキン−2、−7、−12、インターフェロン、および他の成長因子もアジュバントとして使用されてもよく、それらはVLPポリペプチドと混合するか、融合することによってVLPワクチン内に容易に含まれ得るために、好ましいものでもある。
【0107】
ある実施形態では、本明細書に開示のVLPワクチンは、Toll様受容体を介して作用する他のアジュバントを含んでいてもよい。そのようなアジュバントとしては、TLR9のリガンドであるCpGオリゴヌクレオチドを含んでいる核酸;TLR7のリガンドであるイミダゾキノリン;TLR7/8のリガンドである置換グアニン;ロキソリビン(Loxoribine)、7−デアザデオキシグアノシン、7−チア−8−オキソデオキシグアノシン、二本鎖ポリ(I:C)、ポリイノシン酸、イミキモド(Imiquimod)(R−837)およびレシキモド(Resiquimod)(R−848)などのTLR7受容体;あるいはMPL(登録商標)または合成誘導体などのTLR4のアゴニストなどが挙げられる。
【0108】
あるアジュバントは、樹状細胞などのAPCによって、ワクチン分子の取り込みを促進し、該ワクチン分子を活性化する。非限定的な例は、免疫標的アジュバント;免疫調節アジュバント(毒素、サイトカインおよび細菌の誘導体など);油製剤;ポリマー;ミセル形成アジュバント;サポニン;免疫賦活複合体マトリックス(ISCOMマトリックス(ISCOM matrix));粒子;DDA;アルミニウムアジュバント;DNAアジュバント;MPL;およびカプセル化アジュバント(encapsulating adjuvant)からなる群から選択される。
【0109】
アジュバントのさらに別の例には、水酸化アルミニウムまたはリン酸アルミニウムなどのアルミニウム塩(みょうばん)(通常、0.05%から0.1%のアルミニウム塩の緩衝生理食塩水溶液として使用される(例えばNicklas (1992) Res. Immunol. 143:489-493参照))などの作用物質、0.25%の溶液として使用される糖の合成ポリマーとの混合物(例えばカルボポル(Carbopol(登録商標)))、70℃から101℃の範囲の温度で、30秒から2分間熱処理することによって得られる、ワクチン中のタンパク質の凝集体が包含される。また、アジュバントとしては、架橋剤を使って得られた凝集体も可能である。さらに、(i)ペプシン処理されたアルブミンに対する抗体(Fab断片)を用いた再活性化による凝集体、(ii)クリプトスポリジウム・パルバム(C. parvum)などの細菌細胞、またはグラム陰性菌のエンドトキシンもしくはリポ多糖体成分との混合物、(iii)マンニドモノオレイン酸などの生理学的に許容可能な油媒体の乳濁液(アラセルA(Aracel A))、あるいは(iv)ブロック置換体として使用されるパーフルオロカーボンの20%溶液との乳濁液(フルオソル−DA(Fluosol-DA))、が使われてもよい。スクアレンおよびIFAなどの油との混合物も好ましい。
【0110】
DDA(ジメチルジオクタデシルアンモニウムブロマイド)は、アジュバントの興味深い候補であるし、フロイント完全アジュバントおよびフロイント不完全アジュバント、ならびにキラヤ属のサポニン(クイルA(QuilA)およびQS21など)も興味深い候補である。他の候補には、リポ多糖体のポリ[ジ(エアロボキシラトフェノキシ)ホスファゼン(PCPP)の誘導体(モノホスホリルリピドA(MPL(登録商標))、ムラミルジペプチド(MDP)、およびトレオニルムラミルジペプチド(tMDP)など)が包含される。リポ多糖類を基本とするアジュバントは、優勢なTh1型応答を産生するためのものであることが好ましい。そのようなアジュバントとしては例えば、モノホスホリルリピドA(好ましくは3−デ−O−アシル化モノホスホリルリピドA)とアルミニウム塩との組み合わせが挙げられる。MPL(登録商標)アジュバントは、グラクソスミスクラインから入手することができる(例えば米国特許第4,436,727号明細書、米国特許第4,877,611号明細書、米国特許第4,866,034号明細書、および米国特許第4,912,094号免疫原性参照。これら文献のそれぞれは、リポ多糖類に関連する教示に特に関して、参照することによって完全に組み込まれる)。
【0111】
またリポソーム製剤も、アジュバント効果を奏することが知られており、それ故、リポソームアジュバントがVLPと併用される好ましい例である。
【0112】
免疫賦活複合体マトリックス型(ISCOM(登録商標)マトリックス)アジュバントは、特にこの形式のアジュバントがMHCクラスIIの発現をAPCによって上方調節できることが示されたために、本発明にとって好ましい選択肢である。ISCOMマトリックスは、キラヤ由来の(場合によっては分画された)サポニン(トリテルペノイド)、コレステロール、およびリン脂質から成るものである。免疫原性タンパク質(例えばVPL中の免疫原性タンパク質)と混合されたときに生じる粒状製剤は、ISCOM粒子として知られているものである。サポニンは、該ISCOM粒子の60−70%w/wを構成してもよく、コレステロールおよびリン脂質は該ISCOM粒子の10−15%w/wを構成してもよく、上記タンパク質は該ISCOM粒子の10−15%w/wを構成してもよい。免疫賦活複合体の組成および使用に関する詳細は、例えば、アジュバントを扱っている上記テキストブックにおいて見つけることができるが、例えば、「Morein Bら, 1995, Clin. Immunother. 3: 461-475」および「Barr I G and Mitchell G F, 1996, Immunol. and Cell Biol. 74: 8-25」(これらは参照することによって本明細書に組み込まれる)にも、完全な免疫賦活複合体を調製するために役に立つ説明が記載されている。
【0113】
本明細書に開示のVLPワクチンを有するアジュバント組み合わせに使用されてもよいサポニンは、ISCOMの形態であってもなくても、クイルAと呼ばれるキラヤサポナリアモリナ(Quillaja Saponaria Molina)の皮から得られたもの、およびそれの画分を包含する。なお、これらは、米国特許第5,057,540号明細書(特にクイルAの画分ならびにその単離方法および使用に関して、参照されることによって本明細書に完全に組み込まれる)、「”Saponins as vaccine adjuvants”, Kensil, C. R., Crit Rev Ther Drug Carrier Syst, 1996, 12 (1-2):1-55」、および欧州特許第0362279号明細書に記載されている。クイルAの特に好ましい画分は、QS21、QS7、およびQS17である。
【0114】
β−エスチンは、本発明のアジュバント組成物に使用される、もう1つ別の好ましい溶血性サポニンである。エスチンは、メルクの索引(第12版:項目3737)に、トチノキの実に生じるサポニンの混合物として記載されており、ラテン語は、Aesculus hippocastanumである。それのクロマトグラフィーおよび精製による単離は、「Fiedler, Arzneimittel-Forsch. 4, 213 (1953)」に記載されており、それのイオン交換樹脂による単離は、「Erbringら, 米国特許第3,238,190号明細書」」に記載されている。エスチンの画分は、精製されており、生物活性を有することが示されている(「Yoshikawa M, ら (Chem Pharm Bull (Tokyo) 1996 August;44(8):1454-1464)」)。β−エスチンは、アセスチン(aescin)としても知られている。
【0115】
本発明に使用されるもう1つべつの好ましい溶血性サポニンは、ジギトニンである。ジギトニンは、メルクの索引(上記第12版:項目3204)に、ジギタリス・プルプレア(Digitalis purpurea)の実から得られ、「Gisvoldら, J.Am.Pharm.Assoc., 1934, 23, 664」および「Ruhenstroth-Bauer, Physiol.Chem., 1955, 301, 621」に記載の手法によって精製される、サポニンとして記載されている。その使用は、コレステロール測定のための臨床試薬として記載されている。
【0116】
アジュバント効果を得るもう1つ別の(そして好ましい)可能性は、「Gosselinら, 1992」(参照によって、本明細書に組み込まれる)に記載の手法を使うことである。要するに、本発明の抗原などの関連抗原の提示は、単球/マクロファージ上のFC受容体に対する抗体(または抗原結合性の抗体断片)へ、該抗原を接合することによって改善されることができる。特に抗原と抗−FCRIとの間の接合によって、ワクチン接種のための免疫原性が向上されることが実証された。上記抗体は、発現によって、VLPポリペプチドの何れか1つとの融合体として生成した後に、または該融合体の生成の一環として、VLPに接合されてもよい。
【0117】
他の候補には、標的物質および免疫調節物質(すなわちサイトカイン)の使用が含まれる。また、サイトカインの合成誘発物質(ポリI:Cなど)が使用されてもよい。
【0118】
適切な微生物誘導体は、ムラミルジペプチド、フロイント完全アジュバント、RIBI(Ribi ImmunoChem Research Inc., Hamilton, Mont.)、およびトレハロースのジエステル体(TDMおよびTDEなど)から成る群から選択されてもよい。
【0119】
適切な免疫標的アジュバントには、例えばCD40リガンドおよびCD40抗体、またはその特異的な結合性断片(上記の考察を参照)、マンノース、Fab断片、CTLA−4が包含される。
【0120】
適切なポリマーアジュバントには、例えばデキストラン、PEG、デンプン、マンナン、およびマンノースなどの炭水化物;プラスチックポリマー;およびラテックスビーズなどのラテックスが包含される。
【0121】
免疫応答を調節するさらにもう1つ別の興味深い方法は、免疫源を、「仮想リンパ節(VLN)」に含ませることである(場合によってはアジュバントならびに薬学的に許容可能な担体および溶媒と一緒に含ませることである)。VLNは、「ImmunoTherapy, Inc., 360 Lexington Avenue, New York, N.Y. 10017-6501」によって開発された専売医療機器である。VLN(細い管状の機器)は、リンパ節の構造および機能を模倣している。VLNが皮下へ挿入されることによって、サイトカインおよびケモカインの激増を伴う無菌の炎症部位が生まれる。T細胞、B細胞、およびAPCは、この危険シグナルに迅速に反応し、炎症部位に向かい、VLNの多孔性マトリックス中に蓄積する。VLNを使用すると、抗原に対する免疫応答を開始するために要求される必須抗原の用量が減少すること、およびVLNを使ったワクチン接種によって得られる免疫防御は、アジュバントとしてRibiを使った従来の免疫化を凌ぐことが示された。この技術の概要は、「Gelber Cら, 1998, ”Elicitation of Robust Cellular and Humoral Immune Responses to Small Amounts of Immunogens Using a Novel Medical Device Designated the Virtual Lymph Node”」、および「”From the Laboratory to the Clinic, Book of Abstracts, Oct. 12-15, 1998, Seascape Resort, Aptos, Calif.”」に記載されている。
【0122】
オリゴヌクレオチドが、VLPワクチンと併用されるアジュバントとして使用されてもよい。該オリゴヌクレオチドは、少なくとも3つ、より好ましくは少なくとも6つ、またはそれ以上のヌクレオチドによって分離されたジヌクレオチドCpGモチーフを、2つ以上含んでいることが好ましい。(CpGジヌクレオチドがメチル化されていない)CpG含有オリゴヌクレオチドは、優勢なTh1応答を誘導する。そのようなオリゴヌクレオチドは、公知であり、例えば国際公開第96/02555号パンフレット、国際公開第99/33488号パンフレット、米国特許第6,008,200号明細書、および米国特許第5,856,462号明細書に記載されている(これら文献のそれぞれは、特にアジュバントとしてのCpGオリゴヌクレオチドを作製および使用する方法に関して、参照することによって完全に本明細書に組み込まれる)。
【0123】
そのようなオリゴヌクレオチドアジュバントは、デオキシヌクレオチドであってもよい。好ましい実施形態では、オリゴヌクレオチドのヌクレオチド骨格は、ホスホロジチオエート、より好ましくはホスホロチオエート結合であるが、ホスホジエステルおよび他のヌクレオチド骨格(PNAなど)も、(混合された骨格結合を有するオリゴヌクレオチドを含む)本発明の範囲に含まれる。ホスホロチオエートのオリゴヌクレオチドまたはホスホロジチオエートを製造するための方法は、米国特許第5,666,153号明細書、米国特許第5,278,302号明細書、および国際公開第95/26204号パンフレットに記載されており、これら文献のそれぞれは、特にホスホロチオエートおよびホスホロジチオエートの教示に関して、参照することによって完全に本明細書に組み込まれる。
【0124】
好ましいオリゴヌクレオチドは、例えば以下の配列を有している。これら配列は、ホスホロチオエートで修飾されたヌクレオチド骨格を含んでいることが好ましい。
【0125】
(配列番号1)オリゴ1:TCC ATG ACG TTC CTG ACG TT (CpG 1826);
(配列番号2)オリゴ2:TCT CCC AGC GTG CGC CAT (CpG 1758);
(配列番号3)オリゴ3:ACC GAT GAC GTC GCC GGT GAC GGC ACC ACG;
(配列番号4)オリゴ4:TCG TCG TTT TGT CGT TTT GTC GTT (CpG 2006);
(配列番号5)オリゴ5:TCC ATG ACG TTC CTG ATG CT (CpG 1668)。
【0126】
他の好ましいCpGオリゴヌクレオチドには、重要でない欠失または付加を有する上記配列が含まれる。アジュバントとしてのCpGオリゴヌクレオチドは、技術的に公知の任意の方法(例えば欧州特許公開第468520号明細書)によって合成されてもよい。好ましくは、そのようなオリゴヌクレオチドは、自動合成機を利用して合成されてもよい。そのようなオリゴヌクレオチドアジュバントの長さは、10−50塩基の間であってもよい。もう1つ別のアジュバントシステムは、CpG含有オリゴヌクレオチドと、サポニン誘導体との組み合わせを含んでいる。特にCpGとQS21との組み合わせは、国際公開第00/09159号パンフレットに開示されている。
【0127】
多くの一相または多相乳濁液システムが記載されている。当業者の一人は、乳濁液によってVLPの構造が破壊されないように、そのような乳濁液システムをVLPとの使用に容易に適応させることができる。水中油型乳濁液のアジュバントは、それ自体、アジュバント組成物として有用であると示唆されており(欧州特許第0399843号明細書)、水中油型乳濁液と他の活性剤との組み合わせもワクチン用のアジュバントとして記載されている(国際公開第95/17210号パンフレット、国際公開第98/56414号パンフレット、国際公開第99/12565号パンフレット、国際公開第99/11241号パンフレット)。油中水型乳濁液、および水中油中水型乳濁液(water in oil in water emulsion)などの他の油乳濁液のアジュバントは、それぞれ、米国特許第5,422,109号明細書および欧州特許第0480982号明細書、ならびに米国特許第5,424,067号明細書および欧州特許第0480981号明細書に記載されている。
【0128】
本明細書に記載されたVLPワクチンとともに使用するための油乳濁液アジュバントは、天然または合成されたものであってもよく、かつ無機的または有機的なものであってもよい。無機的および有機的な油の例は、当業者に容易に理解される。
【0129】
人への投与に好適な任意の水中油型組成物のために、乳濁系の上記油層は、好ましくは新陳代謝させることができる油を含む。新陳代謝させることができる油という用語の意味は、当業者によく知られている。新陳代謝させることができるという用語は「新陳代謝により変化される能力を有する」と定義できる(Dorland's Illustrated Medical Dictionary, W. B. Sanders Company, 25th edition (1974))。上記油は、受容者に有毒ではなく、かつ新陳代謝により変化する能力を有する任意の植物性油、魚油、動物油、または合成油であってよい。ナッツ(ピーナッツ油など)、種、および種子は、植物油の一般的な源である。合成油も本発明の一部であり、かつNEOBEE(登録商標)などのような市販されている油を包含できる。スクアレン(2,6,10,15, 19,23-Hexamethyl-2,6,10,14,18,22-tetracosahexaene)は、サメの肝臓油中に多量に見出され、かつオリーブ油、小麦の胚種油、ぬか油、および酵母に少量見出される不飽和油である。該スクアレンは、本発明の使用に特に好ましい油である。スクアレンは、新陳代謝させることができる油であり、事実、コレステロールの生合成における中間体であるという長所を有する(Merck index, 10th Edition, entry no.8619)。
【0130】
特に好ましい油乳濁液は、水乳濁液中の油であり、特にスクアレンの水乳濁液が好ましい。
【0131】
加えて、加工していない酸化防止剤において本発明の最も好ましい油乳濁液アジュバントは、好ましくはα−トコフェロール油(vitamin E, EP 0 382 271 Bl)である。
【0132】
国際公開第95/17210号パンフレットおよび国際公開第99/11241号パンフレットは、随意的に免疫賦活剤QS21および/または3D−MPLとともに製剤されたスクアレン、α−トコフェロール、およびTWEEN80に基づく乳濁液アジュバントを開示している。国際公開第99/12565号パンフレットは、油層にステロールを添加したこれらのスクアレン乳濁液の改良点について開示している。さらに、トリカプリリン(tricaprylin)(C27H50O6)のようなトリグリセリドは、乳濁液の安定化のために油層を添加してもよい。
【0133】
水乳濁液における安定した油内に見出される油滴の大きさは、好ましくは1ミクロンよりも小さく、実質的に30〜600nmの範囲であってよく、好ましくは、実質的に直径約30〜500nmであり、最も好ましくは実質的に直径150〜500nmであり、特に光相関分光法により測定した直径において約150nmである。この点で、油滴の80%は、型どおりに好ましい範囲内であるべきで、より好ましくは90%、最も好ましくは95%の油滴が定義された範囲の大きさ内であるべきである。本発明の油乳濁液に存在する構成要素の量は、スクアレンのように通常2〜10%の範囲が油である。また、構成要素が存在する場合、2から10%のアルファトコフェロール;およびポリオキシエチレンソルビタンモノオレエートのような0.3%〜3%の界面活性剤が存在する。好ましい油の比率は、アルファトコフェロールは、1に等しいか、または1よりも小さく、この条件では、より安定な乳濁液が提供される。スパン85(Span 85)は、約1%のレベルで存在してもよい。いくつかの場合では、本明細書に開示されたVLPワクチンは、さらに安定化剤を含むことが好都合であってよい。
【0134】
水乳濁液中に油を形成する方法は、当業者によく知られている。通常、上記方法は、PBS/TWEEN80(登録商標)のような界面活性剤と油層を混合する工程を含み、その後のホモジナイザーを用いるホモジナイゼーションにおいて、上記ミキサーにシリンジの針を通して2回混合することを含むことは、少量の液体のホモジナイゼーションにとって好適であることは当業者にとって明らかである。
【0135】
同様に、マイクロフルイダイザー(M110Sマイクロフルイディクス機、最大50回、6バールの入力最大圧において2分間(約850バールの出力圧力))における乳状化処理は、当業者によって少量または大量の乳濁液を形成するために採用できる。この採用は、調製物が所望の直径の油滴により達成されるまで、得られた乳濁液の測定を含む通常の実験により達成できる。
【0136】
本明細書において開示された上記VLPワクチンの調製物は、鼻腔内投与、筋肉内投与、腹腔内投与、皮内投与、経皮的投与、静脈内投与、または皮下投与によるワクチン投与によってウイルスに感染しやすい、またはウイルスの感染に苦しんでいる哺乳類または鳥を保護または処理するために用いられてよい。ワクチン調製物の全身性の投与方法は、従来型のシリンジおよび針、または固体のワクチンの弾道輸送(ballistic delivery)のために設計された装置(国際公開第99/27961号パンフレット)、無針圧液体噴出装置(米国特許出願公開第4,596,556号明細書;米国特許出願公開第5,993,412号明細書)、または経皮貼布(transdermal patches)(国際公開第97/48440号パンフレット;国際公開第98/28037号パンフレット)を含み得る。VLPワクチンは、皮膚にも適用され得る(経皮的(transdermal)または経皮投与(transcutaneous delivery)(国際公開第98/20734号パンフレット;国際公開98/28037号パンフレット)。本明細書において開示されている上記VLPワクチンは、それゆえ、全身投与のための輸送VLPワクチンまたはアジュバント組成物で前もって満たされた輸送装置を含む。従って、本明細書においては、好ましくは哺乳類または鳥といった個体において、本明細書に記載された任意のVLP組成物を含むワクチンの投与を含み、かつ任意に個体へのアジュバントおよび/またはキャリアを含む免疫応答を誘導する方法を提供する。ここで、ワクチンは、非経口または全身経路を介して投与される。
【0137】
好ましくは、本発明の上記ワクチン調製物は、経口(oral)/消化管(alimentary)または鼻(nasal)経路のような粘膜(mucosal)経路を介するワクチンの投与方法により、ウイルス感染に影響を受けやすい、また、ウイルス感染に苦しんでいる哺乳類または鳥の保護または処理のために用いられてよい。選択的な粘膜岐路は、膣内および直腸内(intra-rectal)である。投与の好ましい粘膜経路は鼻経路を介し、鼻腔ワクチン接種と称される。鼻腔ワクチン接種の方法は、当業者に公知であり、免疫性をあたえられた個体の鼻咽頭中のワクチンにおいて、小滴、噴霧、または乾燥粉末形態の投与を含む。霧状またはエアロゾル化されたワクチンの製剤は、それゆえ、本明細書に開示されたVLPワクチンの好ましい形状である。経口投与のための腹痛抵抗性カプセルおよび顆粒のような腸溶性の製剤、直腸または膣内投与のための座薬も、本明細書において開示されたVLPワクチンの製剤である。
【0138】
本明細書において開示された好ましいVLPワクチン組成物は、ワクチン接種による全身性ワクチン接種に換えて、ヒトにおいて好適に適用できる粘膜ワクチンの種類を表す。
【0139】
上記VLPワクチンは、経口経路を介して投与されてもよい。このような場合、薬学的に許容可能な賦形剤は、アルカリ緩衝液、または腸溶性カプセルもしくは顆粒を含んでもよい。上記VLPワクチンは、膣経路により投与されてもよい。このような場合、薬学的に許容可能な賦形剤は、乳化剤、CARBOPOL(登録商標)のようなポリマー、およびその他の公知の膣クリームおよび座薬の安定剤を含んでもよい。上記VLPワクチンは、直腸経路により投与されてもよい。この場合、賦形剤は、直腸にいれる座薬を形作るために当業者に知られたワックスおよびポリマーを含んでもよい。
【0140】
また、上記VLPワクチン製剤は、キトサン(上述したように)または他のポリカチオン性ポリマー、ポリラクチド粒子およびポリラクチド−コグリコライド(polylactide-coglycolide)粒子、ポリ−N−アセチルグリコサミンに基づくポリマーマトリックス、ポリサッカライドからなる粒子または化学的に修飾されたポリサッカライドからなる粒子、リポソームに基づく粒子および脂質に基づく粒子、ならびにグリセロールモノマーからなる粒子などからなるワクチン媒体と結合してもよい。上記サポニンは、リポソームまたはISCOMsのような粒子の構造を形作るためにコレステロールの存在下で製剤されてもよい。さらに、上記サポニンは、非粒子的な溶液または懸濁液のどちらか、もしくは、小ラメラリポソーム(paucilamelar liposome)またはISCOMのような粒子構造において、ポリオキシエチレンエーテルまたはエステルと共に製剤されてもよい。
【0141】
本明細書に記載されたような薬学的組成物およびワクチン組成物における使用のための追加の実例となるアジュバントは、SAF(Chiron, Calif., United States)、MF‐59(Chiron, 例えば、Granoffら(1997) Infect Immun. 65 (5):1710-1715を参照のこと)、アジュバンドのSBASシリーズ(例えば、SB‐AS2(SmithKline Beecham adjuvant system #2; an oil-in- water emulsion containing MPL and QS21));SmithKline Beechamから市販されているSBAS‐4(SmithKline Beecham adjuvant system #4; contains alum and MPL)、Detox(Enhanzyn)(登録商標)(GlaxoSmithKline)、RC‐512、RC‐522、RC‐527、RC‐529、RC‐544、およびRC‐560(GlaxoSmithKline)ならびにその全体が本明細書に組み込まれ開示される米国特許第08/853,826号明細書および米国特許第09/074,720号明細書に記載されたような他のアミノアルキルグルコサミド 4−ホスフェート(AGPs)を含む。
【0142】
アジュバントの他の実施例は、限定されないが、ハンターズタイターマックス(Hunter's TiterMax)(登録商標)アジュバント(CytRx Corp., Norcross, Ga.);ゲルブアジュバント(Gerbu adjuvants)(Gerbu Biotechnik GmbH, Gaiberg, Germany);ニトロセルロース(Nilsson and Larsson (1992) Res. Immunol. 143:553-557);モンタミドアジュバントセピックISAシリーズ(Seppic ISA series of Montamide adjuvants)(例えば、ISA‐51、ISA‐57、ISA‐720、ISA‐151など;Seppic, Paris, France)のような鉱物油、非鉱物油、油中水型乳剤、または水中油型乳剤を含む製剤に基づくミョウバン(alum)(例えば、アルミニウム水酸化物(aluminum hydroxide)、アルミニウムリン酸塩(aluminum phosphate)乳濁液;およびPROV AX(登録商標)(IDEC Pharmaceuticals);OM‐174(脂質Aに関連するグルコサミンニ糖類);リーシュマニア属伸長因子(Leishmania elongation factor);CRL1005のようなミセルを形作る非イオン化ブロック共重合体;およびシンテックスアジュバント製剤(Syntex Adjuvant Formulation)を含む。例えば、O'Haganら(2001) Biomol Eng. 18(3):69-85;および「Vaccine Adjuvants: Preparation Methods and Research Protocols」D. O'Hagan, ed. (2000) Humana Pressを参照のこと。
【0143】
その他の好ましいアジュバントは、以下の製剤のアジュバント分子を含む。
【0144】
HO(CH2CH2O)n-A-R、(I)
ここで、nは1〜50であり、Aは結合または−C(O)−であり、RはC1〜50のアルキル基またはフェニルC1〜50のアルキル基である。
【0145】
本発明の実施形態の1つは一般式(I)のポリオキシエチレンエーテルを含む。ここで、nは1から50の間であり、好ましくは4から24であり、最も好ましくは9である;上記Rの構成要素は、C.sub.1−50であり、好ましくはC.sub.4‐C.sub.20アルキルおよび最も好ましくはC.sub.12アルキルであり、かつAは結合である。ポリオキシエチレンエーテルの濃度は、0.1〜20%であり、好ましくは0.1〜10%であり、最も好ましくは0.1〜1%の範囲である。好ましくは、ポリオキシエチレンエーテルは以下の群から選択される:ポリオキシエチレン−9−ラウリルエーテル、ポリオキシエチレン−9−ステオリルエーテル(polyoxyethylene-9-steoryl ether)、ポリオキシエチレン−8−ステオリルエーテル(polyoxyethylene-8-steoryl ether)、ポリオキシエチレン−4−ラウリルエーテル、ポリオキシエチレン−35−ラウリルエーテル、およびポリオキシエチレン−23−ラウリルエーテル。ポリオキシエチレンラウリルエーテルのようなポリオキシエチレンエーテルは、メルクインデックスに記載されている(12.sup.th edition: entry 7717)。これらのアジュバント分子は、国際公開第99/52549号パンフレットに記載されている。
【0146】
上記一般式(I)によるポリオキシエチレンエーテルは、もし望むのであれば、他のアジュバントに結合されてもよい。例えば、好ましいアジュバントの組み合わせは、好ましくは上記したようなCpGとの組み合わせである。
【0147】
本明細書に開示されたVLPワクチンと共に使用するための薬学的に許容可能な好ましい賦形剤の例は、水、リン酸緩衝液、および等浸透圧緩衝液を含む。
【0148】
本発明は、以下の限定されない参考文献により、さらに理解される。本明細書に記載したように、本発明は、天然に脂質ラフトと関連しない抗原に結合する脂質ラフト関連ポリペプチドの任意のものに組み込まれるキメラVLPを含む。以下の例は、インフルエンザ抗原とキメラVLPについての本発明の代表的な実施形態を記載する。
【実施例】
【0149】
〔実施例1−キメラインフルエンザVLPの生成〕
上記MLVgagをコードする配列は、全モロニームリー白血病(Moloney murin leukemia)ウイルス両宿主性のプロウイルス配列を含むプラスミドpAMS(ATCC)からPCRにより得られた。上記gagをコードする配列は、pFastBacI(インビトロジェン)のポリヘドリンプロモーター後に挿入された。また、得られたプラスミドは、バキュロウイルスゲノムに組み込むためにDH10Bacコンピテント細胞にトランスフォームされた。高分子量のBacmidDNAは、続いて、精製され、gagを発現する組み換えバキュロウイルスの創出のためにSf9細胞にトランスフェクトされた。A/PR/8/34(H1N1)において、それぞれ、ヘマグルチニンおよびノイラミニダーゼをコードする2つの他の組み換えバキュロウイルスは、ウイルスRNAに由来するHAおよびNAをコードする配列のRT‐PCRクローニング後、同じように(similar fashion)形成された。最後に、全ての3つの産生物(HA−gag−NA)をコードしている単一のバキュロウイルスベクターは、各pFastBacIプラスミドから個々のpFastBacIプラスミドに由来するHA、gag、およびNA発現ユニット(ポリへドリンプロモーターをコードする配列―ポリA部位)を単一のpFastBacIベクターに組み込むことで形成された。最初の分析について、gagまたはHAまたはgag‐HA‐NAをコードする組み換えバキュロウイルスは、MOI>1で6ウェルプレートにおいてSf9細胞に感染された。感染の3日後、培地上清は、夾雑物を取り除かれ、続いて、20%スクロース勾配により100,000×gで沈殿された。沈殿は、gagおよびH1N1−特異的抗血清ウエスタンブロット分析により分析された(図1AおよびBを参照のこと)。
【0150】
図1AおよびBにおいて、それぞれ左側の3つのレーンは、gagまたはHAまたはコントロール(EV=空ベクター)のそれぞれに感染したSf9細胞の結果を示す。予期したように、gagのみを感染させたバキュロウイルスの結果は、VLP出芽のために高分子量培地においてgag抗原に顕著な量をもたらす(図1A、レーン「Gag」)。対照的に、HAのみに感染したバキュロウイルスは、培地にHAをほとんど放出していない。しかし、HA−gag‐NAのトリプルベクターによるSf9細胞の感染は、gagの発現が細胞外のHAを引き付けられることを示す100,000×g画分(レーン1−9、図1AおよびB)において出現するgagおよびHAの両者に顕著な量をもたらす。
【0151】
図2AおよびBは、20−60%スクロースステップ勾配を用いて、ペレット状のHA−gag‐NA VLPを再遠心分離した後に、各勾配の画分をウエスタンブロットした結果を示す図である。gagおよびHAを誘導する同じ画分におけるgagおよびHAの両者のピークは、約1.16g/mlの密度において共結合していることを表し、このことはgagとHAはVLP中に存在することを示す。
【0152】
〔実施例2−HAに結合した炭疽菌PAエピトープを含んでいるHA-gag‐NA VLPの産生、特徴付け、および免疫原性試験〕
実施例1に記載したように、A/PR/8/34の生産物MLVgag、およびHA、およびNAを発現している個々のバキュロウイルスが作製された。加えて、全ての3つの生成物をコードしているトリプル発現組み換えバキュロウイルスもコンストラクトされ、感染した昆虫細胞由来の沈殿した培地上清のスクロース密度勾配遠心分離は、HAで形成されたVLPを誘導するウェスタンブロッティングにより検出されたため、gagおよびHAの同時結合を示す。トリプル発現ベクターにおけるコーディング配列の配置を、図3に示した。HA、gag、およびNAをコードしている要素は、頭−尾形態(head-to-tail fashion)において自身のプロモーター(Pr)およびポリアデニル化配列(pA)と共に並んでいる。単一のバキュロウイルスへの全てのコーディング配列の結合は、分離されたウイルスに共通感染する必要を回避し、3つの分離されたウイルスの感染の一貫的な多様性(consistent multiplicity)を達成することの困難さと関連する。
【0153】
実施例2における上記VLPワクチンは、HA遺伝子がB.anthracisの保護抗原(PA)によるHA免疫原性の決定の大部分を入れ替えることにより修飾されたHA遺伝子を除いて、同じように生成される。このことは、HA遺伝子(アミノ酸位置18−343)のHA1部位をコードしている配列(PAのC末端断片の140アミノ酸)を入れ替えることにより達成される。上記PAをコードしている配列は、HA1をコードしている配列(アミノ酸位置18−343)(図4)の直接入れ替えとして、HAシグナルペプチドをコードしている配列(アミノ酸位置1−17)とHA2をコードしている配列(アミノ酸位置344−565)の間のHA遺伝子に挿入される。修飾されたPA−HA遺伝子を含むトリプルバキュロウイルス発現ベクターは、MLVgagポリペプチド、NAポリペプチド、および入れ替え要素を含む修飾されたPA−HAポリペプチドを発現する。培養中の感染したSf9細胞に上記トリプル発現ベクターが使用された場合、炭疽菌PAエピトープを含むVLPが培地中に観察される。これは、修飾されたPA−HAポリペプチドが、脂質ラフトホーミング配列に残っており、脂質ラフトドメインから出芽している修飾されたPA粒子に組み込まれるからである。この証拠は、(PA)HA−gag−NAトリプル発現ベクターに感染した細胞由来の培地の回収、夾雑物の培地のクリーニング、および20%以上のスクロース勾配において100,000×gで遠心分離によりキメラVLPの回収により示される。VLPへのPAの組み込みの証拠は、PAに特異的な抗体を用いるウエスタンブロットにより得られる。定義により、これらの条件下において20%スクロース緩衝(sucrose cushion)による沈殿物は、天然には粒子であり、VLP形成の証拠を提供する。
【0154】
(サイズ排除クロマトグラフィー)
VLPを精製したスクロース勾配は、セファロースCL−4Bを用いたサイズ排除クロマトグラフィーにより実行される。また、画分はウエスタンブロットによりMLV gagおよび炭疽菌PAエピトープを観察される。VLPは、空隙容量で抽出され、MLV gag、NA、およびPAに修飾されたHAを含んでいる。
【0155】
(電子顕微鏡)
スクロース勾配に由来するVLP試料は、2%のグルタルアルデヒドで処理され、EMグリッド上で吸収され、ナトリウムホスホタングステート(sodium phosphotungstate)でネガティブ染色され、電子顕微鏡で観察される。
【0156】
(免疫原性)
キメラVLP(炭疽菌PA修飾HA含有VLP)の免疫原性は、参考文献(44)に記載のように、メスBalb/cマウスにおいて、炭疽菌感染防御抗原(PA)構造に類似する鼻腔内キトサン/MPL構造を用いて評価することができる。VLPは、Tris緩衝生理食塩水に再懸濁して、20〜60%スクロース密度勾配(sucrose density gradient)にバンドした後、20%スクロースクッションを介したVLP含有培養培地を、100,000×gで1時間、ペレット形成することによって精製することができる。VLP含有画分は、SDS−PAGEまたはウエスタンブロットによって同定して、保管することができる。VLPサンプルをPBS中において透析し、遠心マイクロ濃縮器(centrifuge microconcentrators)を用いて、または100,000×gで遠心分離することによって、濃縮させることができる。
【0157】
免疫処置のために、キトサン 40μg、VLP 20μg(gagに基づく)およびMPL 5μgを含む液体構成物(15μl)を、免疫処置一人分のための、2つの鼻孔用に分割することができる。0〜4週間の鼻腔内投与の前に、イソフルレンを用いて、動物を軽度に麻酔にかけることができる。また、ポジティブコントロールとしての腹腔内予防接種のために、VLPをMPLまたはコレラ毒素と一緒にPBS中で製剤化することができる。全身IgG反応を、PA特異的免疫反応のためのELISA法によって観察することができる。犠牲の時点において、ブロンコバスター洗浄(broncoaveolar lavage)サンプルもまた、PA特異的IgA反応の決定のために収集することができる。
【0158】
実施例2および以下の他の実施例における典型的な免疫処置実験は、スチューデントの対立t−検定(Student’s unpaired t-test)を用いて示されるように、統計的優位性に達する適切な可能性を提供し得る、1グループにつき最小限の8匹のマウスを使用することができる。免疫処置は、典型的に、各免疫処置の後10〜14日間行う血液サンプリングから4週間間隔をあけて、最初のおよび追加の免疫予防接種を伴うことができる。上述したように、ブロンコバスター洗浄サンプルは、IgA決定のための犠牲動物から収集することができる。
【0159】
〔実施例3:機能強化VLPの生成および免疫原性試験〕
本実施例3は、NAに取り付けられた炭疽菌PAエピトープに対する適応免疫反応の長さを促進するために、TLR5アゴニストフラジェリンを組み込むことによる、免疫原性および防御の向上のためのVLPの機能強化を証明することができる。
【0160】
(フラジェリン組み込みによるアジュバント効果)
フラジェリンコード配列は、最近、S.ティピムリアム(S. typimurium)のゲノムDNAからクローン化し、gagコード配列の3’末端に、ちょうど5’から終止コドンまで挿入した。フラジェリンコード配列はまた、A/PR/8/34 HAコード配列のN末端に、シグナルペプチドと成熟コード配列との間の境界に位置するPstI部位を利用して組み込むことも可能である。HAにおけるこの部位の組み込みは、SDS PAGEによって示されるように、予想される分子量増加を伴う、キメラHA分子の適切な発現を引き起こす。NAポリペプチドは、そのC末端に融合された140アミノ酸炭疽菌C末端PAドメインを有することができる。NAのC末端は、C末端の伸長部分が分子の外側に露出することが予想されるように、分子包囲の外側において見られる。フラジェリン修飾gagおよびHAコード配列は、三種類のバキュロウイルスの組換え体(HA−gag−NA(PA))の生成に用いることができる。フラジェリン修飾gagまたはフラジェリン修飾HAを有するVLPをコードするバキュロウイルス組換え体を生成し、基準VLP欠失フラジェリン配列に対する免疫原性試験のための、VLPを生成するために用いることができる(全てのVLPは、標的エピトープとしてPA修飾NAを含むことが可能であり、gagまたはHAに取り付けられたフラジェリン配列を含んでいても欠失していてもよい。実施例2において述べたように、全ての免疫処理実験は、最初のおよび追加の免疫予防接種を4週間間隔で採用することができる。免疫学的計測は、上述したように、全身IgGおよび粘膜IgA反応の両方を試験する、ELISA分析を介して行うことができる。
【0161】
フラジェリン組み込みのためのHAおよびgag挿入部位は、VLPの外側および内側のそれぞれにあるので、異なる程度のアジュバント効果が観察され得る。HAのN末端におけるフラジェリン挿入は、上皮粘膜の細胞におけるTLR5レセプターへのフラジェリンの接近を容易にするという結果をもたらし得る。一方、gag部位における挿入は、異なる接近をもたらす。細胞に対するVLP結合、および正常なインフルエンザウイルス侵入経路を介した内在化は、細胞内におけるgag−フラジェリン生成の堆積という結果をもたらし得る。これは、gag−フラジェリンのコンストラクトとHA−フラジェリンのコンストラクトとの間の異なるTLR5−介在アジュバント効果の結果をもたらし得る。自身の粘膜上皮細胞への結合および侵入のためのVLPの能力は、免疫原性において効果を有しているので、本発明者らは、TPCK−トリプシン処理を伴うおよび伴わない、フラジェリン修飾VLPおよび正常なVLPにおけるVLP免疫原性実験を行い得る。トリプシン処理したVLPのHA開裂は、VLP侵入の重要性を試験する免疫原性試験の開始前に、ウエスタンブロットによって確認することができる。さらに、細胞に融合および侵入するためのトリプシン処理したVLPの能力を、緑色蛍光タンパク質(GFP)修飾gag産物を含むVLPを使用して、in vitroにおける蛍光顕微鏡検査法によって試験することができる。この試験が、その出芽活性を破棄せずに、MLV gagがそのC末端においてGFPにより修飾され得ることが、すでに示されている(60)。
【0162】
フラジェリンの非TLR5結合領域の多くを除去することによりgag出芽活性を最大化するための、フラジェリンコード配列のサブフラグメントの使用もまた試験することができる。フラジェリンモノマー内におけるTLR5認識部位の最近の解析は、この努力によって促進することができる(73)。
【0163】
【表1】
【0164】
〔実施例4:gag−NA−(Norwalk−P2)VLPの生成、特徴付け、および免疫原性試験〕
本実施例4におけるVLPワクチンは、実施例3と同様のベクターにおいて生成することができる。ベクターは、MLV gagポリペプチドおよびNAポリペプチドのC末端に融合したノーウォーク(Norwalk)ウイルスカプシドタンパク質(VP1)のP2ドメインを有するNAポリペプチドを発現し得る。
【0165】
(初期分析)
本実施例において生成されたVLPは、ノーウォークウイルスキャプシドタンパク質特異的抗体を用いたウエスタンブロットによって特徴付けることが可能である。ノーウォークキャプシドタンパク質のP2ドメインは、優勢抗体結合部位であるので、ノーウォークキャプシド特異的抗体は、P2−修飾NAタンパク質を認識することが期待され得る。
【0166】
(サイズ排除クロマトグラフィー)
スクロース密度勾配で精製したVLPは、セファロースCL−4Bを用いたサイズ排除クロマトグラフィーの対象となり得、画分をウエスタンブロットすることによって、MLV gagおよびノーウォークP2ドメインを観察することができる。VLPは、空隙容量(void volume)において抽出され、MLV gag、P2修飾NAを含むことができる。
【0167】
(電子顕微鏡検査)
スクロース勾配からのVLPサンプルを、2% グルタルアルデヒドを用いて処理し、EMグリッド(EM grids)上に吸収させ、タングストリン酸ナトリウムを用いてネガティブ染色し、電子顕微鏡検査によって試験した。
【0168】
(免疫原性)
VLP免疫原性は、参考文献(44)に記載のように、メスBalb/cマウスにおいて、炭疽菌感染防御抗原(PA)構造に類似する鼻腔内キトサン/MPL構造を用いて評価することができる。VLPは、Tris緩衝生理食塩水中に再懸濁して、20〜60%スクロース密度勾配にバンドした後、20% スクロースクッションを介したVLP含有培養培地を、100,000×gで1時間、ペレット形成することによって精製することができる。VLP含有分画を、ウエスタンブロット分析によって同定し、保管することができる。VLPサンプルをPBS中において透析し、遠心マイクロ濃縮器を用いて、または100,000×gで遠心分離して、濃縮することができる。
【0169】
免疫処理のために、キトサン 40μg、VLP 20μg(gagに基づく)およびMPL 5μgを含む液体構成物(15μl)を、免疫処置一人分のための、2つの鼻孔用に分割することができる。0〜4週間の鼻腔内投与の前に、イソフルレンを用いて、動物を軽度に麻酔にかけることができる。また、ポジティブコントロールとしての腹腔内予防接種のために、VLPをMPLまたはコレラ毒素を有するPBS中に構築することができる。追加のポジティブコントロール動物は、ELISAによって観察され得るノーウォーク P2に特異的な全身IgG反応を伴う筋肉内予防接種を受けることができる。ELISAのための抗原の源は、バキュロウイルス生成ノーウォークVP1キャプシドタンパク質であることができる。ELISAによるノーウォークPA特異的IgA反応の測定のために、ブロンコバスター洗浄(broncoaveolar lavage)サンプルもまた、最終免疫処理後10〜14日間収集することができる。
【0170】
〔実施例5:機能強化VLPの生成および免疫原性試験〕
本実施例5は、適応免疫反応の長さを促進するために、TLR5アゴニストフラジェリンを組み込むことによる、免疫原性および防御を向上させるためのVLPの機能強化を証明することができる。
【0171】
(フラジェリン組み込みのためのアジュバントの効果)
フラジェリンコード配列は、最近、S.ティピムリアム(S. typimurium)のゲノムDNAからクローン化し、gagコード配列の3’末端に、ちょうど5’から終止コドンまで挿入した。フラジェリンコード配列はまた、A/PR/8/34 HAコード配列のN末端に、シグナルペプチドと成熟コード配列との間の境界に位置するPstI部位を利用して組み込むことも可能である。HAにおけるこの部位の組み込みは、SDS PAGEによって示されるように、予想される分子量増加を伴う、キメラHA分子の適切な発現を引き起こす。NAポリペプチドは、そのC末端に融合されたノーウォークウイルス(アミノ酸279〜405)のP2ドメインを有することができる。フラジェリン修飾gagおよびHAコード配列は、実施例5に示すように、三種類のバキュロウイルスの組換え体(HA−gag−NA(P2))を生成するために用いることができる。VLPをコードし、フラジェリン修飾gagまたはフラジェリン修飾HAを有する組換えバキュロウイルスを生成し、基準VLP欠失フラジェリン配列に対する免疫原性試験のためのVLPを生成するために用いることができる(全てのVLPは、標的エピトープとしてノーウォーク P2修飾NAを含むことが可能であり、gagまたはHAに取り付けられたフラジェリン配列を含んでいても欠失していてもよい。上述した実施例で述べたように、全ての免疫処理実験は、最初のおよび追加の免疫予防接種を4週間間隔で採用することができる。免疫学的計測は、上述したように、全身IgGおよび粘膜IgA反応の両方で試験する、ELISA分析を介して行うことができる。
【0172】
フラジェリン組み込みのためのHAおよびgag挿入部位は、VLPの外側および内側のそれぞれにあるので、異なる程度のアジュバント効果が観察され得る。HAのN末端におけるフラジェリン挿入は、上皮粘膜の細胞におけるTLR5レセプターへのフラジェリンの接近を容易にするという結果をもたらし得る。一方、gag部位における挿入は、異なる接近をもたらし得る。細胞に対するVLP結合および正常なインフルエンザウイルス侵入経路を介した内在化は、細胞内におけるgag−フラジェリン生成の堆積という結果をもたらし得る。これは、gag−フラジェリン構築物とHA−フラジェリン構築物との間の異なるTLR5−介在アジュバント効果の結果をもたらし得る。自身の粘膜上皮細胞への結合および侵入のためのVLPの能力は、免疫原性において効果を有しているので、本発明者らは、TPCK−トリプシン処理を伴うおよび伴わない、フラジェリン修飾VLPおよび正常なVLPにおけるVLP免疫原性実験を行い得る。トリプシン処理したVLPのHA開裂は、VLP侵入の重要性を試験する免疫原性試験の開始前に、ウエスタンブロットによって確認することができる。さらに、細胞に融合および侵入するためのトリプシン処理したVLPの能力を、緑色蛍光タンパク質(GFP)修飾gag産物を含むVLPを使用して、in vitroにおける蛍光顕微鏡検査法によって試験することができる。この試験がその出芽活性を破棄せずに、MLV gagがそのC末端においてGFPにより修飾され得ることはすでに示されている(60)。
【0173】
フラジェリンの非TLR5結合領域の多くを除去することによりgag出芽活性を最大化するための、フラジェリンコード配列のサブフラグメントの使用もまた試験することができる。フラジェリンモノマー内におけるTLR5認識部位の最近の解析は、この努力によって促進することができる(73)。
【0174】
【表2】
【0175】
〔実施例6:RSV−F−gag VLPの生成、特徴付けおよび免疫原性試験〕
本実施例における呼吸器合胞体ウイルス(RSV)のためのVLPワクチンは、インフルエンザHAおよびNAの脂質ラフト標的特性と同様である、RSV融合(F)タンパク質の脂質ラフト標的特性を利用することができる。これらの特性のお陰で、RSV Fタンパク質自身がポリペプチドに関連する脂質ラフトであり、それゆえに、キメラタンパク質の形成を必要とせずに、インフルエンザHAおよびNAと酷似のgagベースのVLPに直接組み込むことができる。
【0176】
RSV Fタンパク質は、最後まで、以下に示す5’プライマーおよび3’プライマーを用いて、標準RT−PCRクローン技術によってクローン化することができる(プライマー中に下線を付した配列は、RSV F 5’末端および3’末端コード配列のホモログであり、残りの配列は、pFastBac1ベクター中にクローニングするのに便利な制限酵素認識部位を含む)。
【0177】
(SEQ ID NO:6)
5’プライマー: ATATAGGCGCGCCACCATGGAGTTGCTAATCCTCAAAGC
(SEQ ID NO:7)
3’プライマー: ATATAGCGGCCGCTTAGTTACTAAATGCAATATTATTTATACCACTCAG
【0178】
上述したプライマーを用いたRT−PCRによるRSV F遺伝子の生成によって、pFB−Fと称するベクターとなるpFastBac1に挿入するための結合性のある端部を生成するために、AscIおよびNotIのいずれかの端部において切断され得るフラグメントを得ることができる。pFB−Fベクターの完成において、このベクターは、pFB−gagからのSnaBI−HpaIフラグメントの挿入のために、HpaIにおいて切断されることができる。これにより。MLV−gagおよびRSV−Fの両方をコードする二種類のバキュロウイルスを発現するベクターを得ることができる。このプラスミドのFおよびGag発現領域の主要な特徴の配置図を、図5に示す。
【0179】
ダブル発現(F−Gag)ベクターを、培地中においてSf9細胞の感染のために用いた場合、F遺伝子産物は、脂質ラフトホーミング配列(lipid raft homing sequence)を保持することが可能であり、脂質ラフトドメインから出芽した分子に組み込むことが可能であるので、RSV F産物を含むVLPを、培養培地中において観察することができる。F−Gagダブル発現ベクターに感染した細胞からの培養培地を収穫し、培地の残骸を取り除き、20% スクロースクッション上において100,000×gで遠心分離して収集することによって、この証拠を示すことができる。FのVLPへの組み込みの証拠は、特異的抗体を用いてウエスタンブロット解析によって得ることができる。当然、これらの条件下での20% スクロースクッションを介して堆積する材料は、本質的に粒子状物質であり、VLP構成物の証拠を提供する。
【0180】
同様のアプローチを、RSV G 糖タンパク質のような、追加のRSV抗原を組み込むために用いることができる。
【0181】
(免疫原性)
ヒトRSVストックA2のような、アレルギーの誘発に適切なRSVを用いること以外は、他のVLPの免疫原性を測定するために前述の実施例において用いた技術によって、RSV VLPの免疫原性を測定することができる。
【0182】
〔追加の参考文献〕
以下の参考文献は、それらが教示する全てのものに関して、参照することによって本明細書に組み込まれる。
【0183】
1.Katz, J. M., W. Lim, C. B. Bridges, T. Rowe, J. Hu-Primmer, X. Lu, R. A. Abernathy, M. Clarke, L. Conn, H. Kwong, M. Lee, G. Au, Y. Y. Ho, K. H. Mak, N. J. Cox, and K. Fukuda. 1999. Antibody response in individuals infected with avian influenza A (H5N1) viruses and detection of anti-H5 antibody among household and social contacts. J Infect Dis 180:1763.
2.Peiris, J. S., W. C. Yu, C. W. Leung, C. Y. Cheung, W. F. Ng, J. M. Nicholls, T. K. Ng, K. H. Chan, S. T. Lai, W. L. Lim, K. Y. Yuen, and Y. Guan. 2004. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet 363:617.
3.Horimoto, T., N. Fukuda, K. Iwatsuki-Horimoto, Y. Guan, W. Lim, M. Peiris, S. Sugii, T. Odagiri, M. Tashiro, and Y. Kawaoka. 2004. Antigenic differences between H5N1 human influenza viruses isolated in 1997 and 2003. J Vet Med Sci 66:303.
4.Tran, T. H., T. L. Nguyen, T. D. Nguyen, T. S. Luong, P. M. Pham, V. C. Nguyen, T. S. Pham, C. D. Vo, T. Q. Le, T. T. Ngo, B. K. Dao, P. P. Le, T. T. Nguyen, T. L. Hoang, V. T. Cao, T. G. Le, D. T. Nguyen, H. N. Le, K. T. Nguyen, H. S. Le, V. T. Le, D. Christiane, T. T. Tran, J. Menno de, C. Schultsz, P. Cheng, W. Lim, P. Horby, and J. Farrar. 2004. Avian influenza A (H5N1) in 10 patients in Vietnam. N Engl J Med 350:1179.
5.Li, K. S., Y. Guan, J. Wang, G. J. Smith, K. M. Xu, L. Duan, A. P. Rahardjo, P. Puthavathana, C. Buranathai, T. D. Nguyen, A. T. Estoepangestie, A. Chaisingh, P. Auewarakul, H. T. Long, N. T. Hanh, R. J. Webby, L. L. Poon, H. Chen, K. F. Shortridge, K. Y. Yuen, R. G. Webster, and J. S. Peiris. 2004. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature 430:209.
6.Lipatov, A. S., E. A. Govorkova, R. J. Webby, H. Ozaki, M. Peiris, Y. Guan, L. Poon, and R. G. Webster. 2004. Influenza: emergence and control. J Virol 78:8951.
7.Lipatov, A. S., R. J. Webby, E. A. Govorkova, S. Krauss, and R. G. Webster. 2005. Efficacy of H5 influenza vaccines produced by reverse genetics in a lethal mouse model. J Infect Dis 191:1216.
8.Stephenson, I., K. G. Nicholson, J. M. Wood, M. C. Zambon, and J. M. Katz. 2004. Confronting the avian influenza threat: vaccine development for a potential pandemic. Lancet Infect Dis 4:499.
9.Liu, M., J. M. Wood, T. Ellis, S. Krauss, P. Seiler, C. Johnson, E. Hoffmann, J. Humberd, D. Hulse, Y. Zhang, R. G. Webster, and D. R. Perez. 2003. Preparation of a standardized, efficacious agricultural H5N3 vaccine by reverse genetics. Virology 314:580.
10.Subbarao, K., H. Chen, D. Swayne, L. Mingay, E. Fodor, G. Brownlee, X. Xu, X. Lu, J. Katz, N. Cox, and Y. Matsuoka. 2003. Evaluation of a genetically modified reassortant H5N1 influenza A virus vaccine candidate generated by plasmid-based reverse genetics. Virology 305:192.
11.Webby, R. J., D. R. Perez, J. S. Coleman, Y. Guan, J. H. Knight, E. A. Govorkova, L. R. McClain-Moss, J. S. Peiris, J. E. Rehg, E. I. Tuomanen, and R. G. Webster. 2004. Responsiveness to a pandemic alert: use of reverse genetics for rapid development of influenza vaccines. Lancet 363:1099.
12.Treanor, J. J., B. E. Wilkinson, F. Masseoud, J. Hu-Primmer, R. Battaglia, D. O'Brien, M. Wolff, G. Rabinovich, W. Blackwelder, and J. M. Katz. 2001. Safety and immunogenicity of a recombinant hemagglutinin vaccine for H5 influenza in humans. Vaccine 19:1732.
13.Stephenson, I., R. Bugarini, K. G. Nicholson, A. Podda, J. M. Wood, M. C. Zambon, and J. M. Katz. 2005. Cross-reactivity to highly pathogenic avian influenza H5N1 viruses after vaccination with nonadjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a potential priming strategy. J Infect Dis 191:1210.
14.Nicholson, K. G., A. E. Colegate, A. Podda, I. Stephenson, J. Wood, E. Ypma, and M. C. Zambon. 2001. Safety and antigenicity of non-adjuvanted and MF59-adjuvanted influenza A/Duck/Singapore/97 (H5N3) vaccine: a randomised trial of two potential vaccines against H5N1 influenza. Lancet 357:1937.
15.Subbarao, K., B. R. Murphy, and A. S. Fauci. 2006. Development of effective vaccines against pandemic influenza. Immunity 24:5.
16.Kuper, C. F., P. J. Koornstra, D. M. Hameleers, J. Biewenga, B. J. Spit, A. M. Duijvestijn, P. J. van Breda Vriesman, and T. Sminia. 1992. The role of nasopharyngeal lymphoid tissue. Immunol Today 13:219.
17.Liang, B., L. Hyland, and S. Hou. 2001. Nasal-associated lymphoid tissue is a site of long-term virus-specific antibody production following respiratory virus infection of mice. J Virol 75:5416.
18.Zuercher, A. W., S. E. Coffin, M. C. Thurnheer, P. Fundova, and J. J. Cebra. 2002. Nasal-associated lymphoid tissue is a mucosal inductive site for virus-specific humoral and cellular immune responses. J Immunol 168:1796.
19.Brandtzaeg, P. 1989. Overview of the mucosal immune system. Curr Top Microbiol Immunol 146:13.
20.Takada, A., S. Matsushita, A. Ninomiya, Y. Kawaoka, and H. Kida. 2003. Intranasal immunization with formalin-inactivated virus vaccine induces a broad spectrum of heterosubtypic immunity against influenza A virus infection in mice. Vaccine 21:3212.
21.Tamura, S. I., H. Asanuma, Y. Ito, Y. Hirabayashi, Y. Suzuki, T. Nagamine, C. Aizawa, T. Kurata, and A. Oya. 1992. Superior cross-protective effect of nasal vaccination to subcutaneous inoculation with influenza hemagglutinin vaccine. Eur J Immunol 22:477.
22.Tumpey, T. M., M. Renshaw, J. D. Clements, and J. M. Katz. 2001. Mucosal delivery of inactivated influenza vaccine induces B-cell-dependent heterosubtypic cross-protection against lethal influenza A H5N1 virus infection. J Virol 75:5141.
23.Kang, S. M., L. Guo, Q. Yao, I. Skountzou, and R. W. Compans. 2004. Intranasal immunization with inactivated influenza virus enhances immune responses to coadministered simian-human immunodeficiency virus-like particle antigens. J Virol 78:9624.
24.Guo, L., X. Lu, S. M. Kang, C. Chen, R. W. Compans, and Q. Yao. 2003. Enhancement of mucosal immune responses by chimeric influenza HA/SHIV virus-like particles. Virology 313:502.
25.Yao, Q., R. Zhang, L. Guo, M. Li, and C. Chen. 2004. Th cell-independent immune responses to chimeric hemagglutinin/simian human immunodeficiency virus-like particles vaccine. J Immunol 173:1951.
26.Latham, T., and J. M. Galarza. 2001. Formation of wild-type and chimeric influenza virus-like particles following simultaneous expression of only four structural proteins. J Virol 75:6154.
27.Galarza, J. M., T. Latham, and A. Cupo. 2005. Virus-like particle vaccine conferred complete protection against a lethal influenza virus challenge. Viral Immunol 18:365.
28.Fromantin, C., B. Jamot, J. Cohen, L. Piroth, P. Pothier, and E. Kohli. 2001. Rotavirus 2/6 virus-like particles administered intranasally in mice, with or without the mucosal adjuvants cholera toxin and Escherichia coli heat-labile toxin, induce a Th1/Th2-like immune response. J Virol 75:11010.
29.Harrington, P. R., B. Yount, R. E. Johnston, N. Davis, C. Moe, and R. S. Baric. 2002. Systemic, mucosal, and heterotypic immune induction in mice inoculated with Venezuelan equine encephalitis replicons expressing Norwalk virus-like particles. J Virol 76:730.
30.Shi, W., J. Liu, Y. Huang, and L. Qiao. 2001. Papillomavirus pseudovirus: a novel vaccine to induce mucosal and systemic cytotoxic T-lymphocyte responses. J Virol 75:10139.
31.Han, M. G., S. Cheetham, M. Azevedo, C. Thomas, and L. J. Saif. 2006. Immune responses to bovine norovirus-like particles with various adjuvants and analysis of protection in gnotobiotic calves. Vaccine 24:317.
32.Illum, L. 1998. Chitosan and its use as a pharmaceutical excipient. Pharm Res 15:1326.
33.Illum, L., I. Jabbal-Gill, M. Hinchcliffe, A. N. Fisher, and S. S. Davis. 2001. Chitosan as a novel nasal delivery system for vaccines. Adv Drug Deliv Rev 51:81.
34.Soane, R. J., M. Hinchcliffe, S. S. Davis, and L. Illum. 2001. Clearance characteristics of chitosan based formulations in the sheep nasal cavity. Int J Pharm 217:183.
35.Baudner, B. C., M. M. Giuliani, J. C. Verhoef, R. Rappuoli, H. E. Junginger, and G. D. Giudice. 2003. The concomitant use of the LTK63 mucosal adjuvant and of chitosan-based delivery system enhances the immunogenicity and efficacy of intranasally administered vaccines. Vaccine 21:3837.
36.Fujihashi, K., T. Koga, F. W. van Ginkel, Y. Hagiwara, and J. R. McGhee. 2002. A dilemma for mucosal vaccination: efficacy versus toxicity using enterotoxin-based adjuvants. Vaccine 20:2431.
37.Mutsch, M., W. Zhou, P. Rhodes, M. Bopp, R. T. Chen, T. Linder, C. Spyr, and R. Steffen. 2004. Use of the inactivated intranasal influenza vaccine and the risk of Bell's palsy in Switzerland. N Engl J Med 350:896.
38.Baldridge, J. R., Y. Yorgensen, J. R. Ward, and J. T. Ulrich. 2000. Monophosphoryl lipid A enhances mucosal and systemic immunity to vaccine antigens following intranasal administration. Vaccine 18:2416.
39.Baldrick, P., D. Richardson, G. Elliott, and A. W. Wheeler. 2002. Safety evaluation of monophosphoryl lipid A (MPL): an immunostimulatory adjuvant. Regul Toxicol Pharmacol 35:398.
40.Baldridge, J. R., P. McGowan, J. T. Evans, C. Cluff, S. Mossman, D. Johnson, and D. Persing. 2004. Taking a Toll on human disease: Toll-like receptor 4 agonists as vaccine adjuvants and monotherapeutic agents. Expert Opin Biol Ther 4:1129.
41.Baldridge, J. GlaxoSmithKline, Personal Communication.
42.Huang, J., R. J. Garmise, T. M. Crowder, K. Mar, C. R. Hwang, A. J. Hickey, J. A. Mikszta, and V. J. Sullivan. 2004. A novel dry powder influenza vaccine and intranasal delivery technology: induction of systemic and mucosal immune responses in rats. Vaccine 23:794.
43.Mills, K. H., C. Cosgrove, E. A. McNeela, A. Sexton, R. Giemza, I. Jabbal-Gill, A. Church, W. Lin, L. Illum, A. Podda, R. Rappuoli, M. Pizza, G. E. Griffin, and D. J. Lewis. 2003. Protective levels of diphtheria-neutralizing antibody induced in healthy volunteers by unilateral priming-boosting intranasal immunization associated with restricted ipsilateral mucosal secretory immunoglobulin a. Infect Immun 71:726.
44.Wimer-Mackin, S., M. Hinchcliffe, C. R. Petrie, S. J. Warwood, W. T. Tino, M. S. Williams, J. P. Stenz, A. Cheff, and C. Richardson. 2006. An intranasal vaccine targeting both the Bacillus anthracis toxin and bacterium provides protection against aerosol spore challenge in rabbits. Vaccine in press.
45.Noad, R., and P. Roy. 2003. Virus-like particles as immunogens. Trends Microbiol 11:438.
46.Yao, Q., V. Vuong, M. Li, and R. W. Compans. 2002. Intranasal immunization with SIV virus-like particles (VLPs) elicits systemic and mucosal immunity. Vaccine 20:2537.
47.Pushko, P., T. M. Tumpey, F. Bu, J. Knell, R. Robinson, and G. Smith. 2005. Influenza virus-like particles comprised of the HA, NA, and M1 proteins of H9N2 influenza virus induce protective immune responses in BALB/c mice. Vaccine 23:5751.
48.Baumert, T. F., S. Ito, D. T. Wong, and T. J. Liang. 1998. Hepatitis C virus structural proteins assemble into viruslike particles in insect cells. J Virol 72:3827.
49.Yao, Q., Z. Bu, A. Vzorov, C. Yang, and R. W. Compans. 2003. Virus-like particle and DNA-based candidate AIDS vaccines. Vaccine 21:638.
50.Tacket, C. O., M. B. Sztein, G. A. Losonsky, S. S. Wasserman, and M. K. Estes. 2003. Humoral, mucosal, and cellular immune responses to oral Norwalk virus-like particles in volunteers. Clin Immunol 108:241.
51.Gomez-Puertas, P., C. Albo, E. Perez-Pastrana, A. Vivo, and A. Portela. 2000. Influenza virus matrix protein is the major driving force in virus budding. J Virol 74:11538.
52.Gheysen, D., E. Jacobs, F. de Foresta, C. Thiriart, M. Francotte, D. Thines, and M. De Wilde. 1989. Assembly and release of HIV-1 precursor Pr55gag virus-like particles from recombinant baculovirus-infected insect cells. Cell 59:103.
53.Johnson, M. C., H. M. Scobie, and V. M. Vogt. 2001. PR domain of rous sarcoma virus Gag causes an assembly/budding defect in insect cells. J Virol 75:4407.
54.Kakker, N. K., M. V. Mikhailov, M. V. Nermut, A. Burny, and P. Roy. 1999. Bovine leukemia virus Gag particle assembly in insect cells: formation of chimeric particles by domain-switched leukemia/lentivirus Gag polyprotein. Virology 265:308.
55.Luo, L., Y. Li, and C. Y. Kang. 1990. Expression of gag precursor protein and secretion of virus-like gag particles of HIV-2 from recombinant baculovirus-infected insect cells. Virology 179:874.
56.Morikawa, S., T. F. Booth, and D. H. Bishop. 1991. Analyses of the requirements for the synthesis of virus-like particles by feline immunodeficiency virus gag using baculovirus vectors. Virology 183:288.
57.Takahashi, R. H., K. Nagashima, T. Kurata, and H. Takahashi. 1999. Analysis of human lymphotropic T-cell virus type II-like particle production by recombinant baculovirus-infected insect cells. Virology 256:371.
58.Yamshchikov, G. V., G. D. Ritter, M. Vey, and R. W. Compans. 1995. Assembly of SIV virus-like particles containing envelope proteins using a baculovirus expression system. Virology 214:50.
59.Weldon, R. A., Jr., C. R. Erdie, M. G. Oliver, and J. W. Wills. 1990. Incorporation of chimeric gag protein into retroviral particles. J Virol 64:4169.
60.Andrawiss, M., Y. Takeuchi, L. Hewlett, and M. Collins. 2003. Murine leukemia virus particle assembly quantitated by fluorescence microscopy: role of Gag-Gag interactions and membrane association. J Virol 77:11651.
61.Leser, G. P., and R. A. Lamb. 2005. Influenza virus assembly and budding in raft-derived microdomains: a quantitative analysis of the surface distribution of HA, NA and M2 proteins. Virology 342:215.
62.Takeda, M., G. P. Leser, C. J. Russell, and R. A. Lamb. 2003. Influenza virus hemagglutinin concentrates in lipid raft microdomains for efficient viral fusion. Proc Natl Acad Sci U S A 100:14610.
63.Campbell, S. M., S. M. Crowe, and J. Mak. 2001. Lipid rafts and HIV-1: from viral entry to assembly of progeny virions. J Clin Virol 22:217.
64.Sandrin, V., and F. L. Cosset. 2006. Intracellular versus cell surface assembly of retroviral pseudotypes is determined by the cellular localization of the viral glycoprotein, its capacity to interact with Gag, and the expression of the Nef protein. J Biol Chem 281:528.
65.Salazar-Gonzalez, R. M., and S. J. McSorley. 2005. Salmonella flagellin, a microbial target of the innate and adaptive immune system. Immunol Lett 101:117.
66.Cuadros, C., F. J. Lopez-Hernandez, A. L. Dominguez, M. McClelland, and J. Lustgarten. 2004. Flagellin fusion proteins as adjuvants or vaccines induce specific immune responses. Infect Immun 72:2810.
67.Didierlaurent, A., I. Ferrero, L. A. Otten, B. Dubois, M. Reinhardt, H. Carlsen, R. Blomhoff, S. Akira, J. P. Kraehenbuhl, and J. C. Sirard. 2004. Flagellin promotes myeloid differentiation factor 88-dependent development of Th2-type response. J Immunol 172:6922.
68.Tsujimoto, H., T. Uchida, P. A. Efron, P. O. Scumpia, A. Verma, T. Matsumoto, S. K. Tschoeke, R. F. Ungaro, S. Ono, S. Seki, M. J. Clare-Salzler, H. V. Baker, H. Mochizuki, R. Ramphal, and L. L. Moldawer. 2005. Flagellin enhances NK cell proliferation and activation directly and through dendritic cell-NK cell interactions. J Leukoc Biol 78:888.
69.Hayashi, F., T. K. Means, and A. D. Luster. 2003. Toll-like receptors stimulate human neutrophil function. Blood 102:2660.
70.Hayashi, F., K. D. Smith, A. Ozinsky, T. R. Hawn, E. C. Yi, D. R. Goodlett, J. K. Eng, S. Akira, D. M. Underhill, and A. Aderem. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 410:1099.
71.Gewirtz, A. T., P. O. Simon, Jr., C. K. Schmitt, L. J. Taylor, C. H. Hagedorn, A. D. O'Brien, A. S. Neish, and J. L. Madara. 2001. Salmonella typhimurium translocates flagellin across intestinal epithelia, inducing a proinflammatory response. J Clin Invest 107:99.
72.Means, T. K., F. Hayashi, K. D. Smith, A. Aderem, and A. D. Luster. 2003. The Toll-like receptor 5 stimulus bacterial flagellin induces maturation and chemokine production in human dendritic cells. J Immunol 170:5165.
73.Smith, K. D., E. Andersen-Nissen, F. Hayashi, K. Strobe, M. A. Bergman, S. L. Barrett, B. T. Cookson, and A. Aderem. 2003. Toll-like receptor 5 recognizes a conserved site on flagellin required for protofilament formation and bacterial motility. Nat Immunol 4:1247.
74.Honko, A. N., N. Sriranganathan, C. J. Lees, and S. B. Mizel. 2006. Flagellin is an effective adjuvant for immunization against lethal respiratory challenge with Yersinia pestis. Infect Immun 74:1113.
75.Jeon, S. H., T. Ben-Yedidia, and R. Arnon. 2002. Intranasal immunization with synthetic recombinant vaccine containing multiple epitopes of influenza virus. Vaccine 20:2772.
76.Levi, R., and R. Arnon. 1996. Synthetic recombinant influenza vaccine induces efficient long-term immunity and cross-strain protection. Vaccine 14:85.
77.Lee, S. E., S. Y. Kim, B. C. Jeong, Y. R. Kim, S. J. Bae, O. S. Ahn, J. J. Lee, H. C. Song, J. M. Kim, H. E. Choy, S. S. Chung, M. N. Kweon, and J. H. Rhee. 2006. A bacterial flagellin, Vibrio vulnificus FlaB, has a strong mucosal adjuvant activity to induce protective immunity. Infect Immun 74:694.
78.Applequist, S. E., E. Rollman, M. D. Wareing, M. Liden, B. Rozell, J. Hinkula, and H. G. Ljunggren. 2005. Activation of innate immunity, inflammation, and potentiation of DNA vaccination through mammalian expression of the TLR5 agonist flagellin. J Immunol 175:3882.
79.Ramos, H. C., M. Rumbo, and J. C. Sirard. 2004. Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol 12:509.
80.Holsinger, L. J., and R. A. Lamb. 1991. Influenza virus M2 integral membrane protein is a homotetramer stabilized by formation of disulfide bonds. Virology 183:32.
81.Lamb, R. A., S. L. Zebedee, and C. D. Richardson. 1985. Influenza virus M2 protein is an integral membrane protein expressed on the infected-cell surface. Cell 40:627.
82.Holsinger, L. J., D. Nichani, L. H. Pinto, and R. A. Lamb. 1994. Influenza A virus M2 ion channel protein: a structure-function analysis. J Virol 68:1551.
83.Takeda, M., A. Pekosz, K. Shuck, L. H. Pinto, and R. A. Lamb. 2002. Influenza a virus M2 ion channel activity is essential for efficient replication in tissue culture. J Virol 76:1391.
84.Frace, A. M., A. I. Klimov, T. Rowe, R. A. Black, and J. M. Katz. 1999. Modified M2 proteins produce heterotypic immunity against influenza A virus. Vaccine 17:2237.
85.Neirynck, S., T. Deroo, X. Saelens, P. Vanlandschoot, W. M. Jou, and W. Fiers. 1999. A universal influenza A vaccine based on the extracellular domain of the M2 protein. Nat Med 5:1157.
86.Slepushkin, V. A., J. M. Katz, R. A. Black, W. C. Gamble, P. A. Rota, and N. J. Cox. 1995. Protection of mice against influenza A virus challenge by vaccination with baculovirus-expressed M2 protein. Vaccine 13:1399.
87.Treanor, J. J., E. L. Tierney, S. L. Zebedee, R. A. Lamb, and B. R. Murphy. 1990. Passively transferred monoclonal antibody to the M2 protein inhibits influenza A virus replication in mice. J Virol 64:1375.
88.De Filette, M., W. Min Jou, A. Birkett, K. Lyons, B. Schultz, A. Tonkyro, S. Resch, and W. Fiers. 2005. Universal influenza A vaccine: optimization of M2-based constructs. Virology 337:149.
89.De Filette, M., A. Ramne, A. Birkett, N. Lycke, B. Lowenadler, W. Min Jou, X. Saelens, and W. Fiers. 2006. The universal influenza vaccine M2e-HBc administered intranasally in combination with the adjuvant CTA1-DD provides complete protection. Vaccine 24:544.
90.Fiers, W., M. De Filette, A. Birkett, S. Neirynck, and W. Min Jou. 2004. A "universal" human influenza A vaccine. Virus Res 103:173.
91.Liu, W., Z. Peng, Z. Liu, Y. Lu, J. Ding, and Y. H. Chen. 2004. High epitope density in a single recombinant protein molecule of the extracellular domain of influenza A virus M2 protein significantly enhances protective immunity. Vaccine 23:366.
92.Fan, J., X. Liang, M. S. Horton, H. C. Perry, M. P. Citron, G. J. Heidecker, T. M. Fu, J. Joyce, C. T. Przysiecki, P. M. Keller, V. M. Garsky, R. Ionescu, Y. Rippeon, L. Shi, M. A. Chastain, J. H. Condra, M. E. Davies, J. Liao, E. A. Emini, and J. W. Shiver. 2004. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine 22:2993.
93.Ionescu, R. M., C. T. Przysiecki, X. Liang, V. M. Garsky, J. Fan, B. Wang, R. Troutman, Y. Rippeon, E. Flanagan, J. Shiver, and L. Shi. 2006. Pharmaceutical and immunological evaluation of human papillomavirus viruslike particle as an antigen carrier. J Pharm Sci 95:70.
94.Hatziioannou, T., E. Delahaye, F. Martin, S. J. Russell, and F. L. Cosset. 1999. Retroviral display of functional binding domains fused to the amino terminus of influenza hemagglutinin. Hum Gene Ther 10:1533.
95.Li, Z. N., S. N. Mueller, L. Ye, Z. Bu, C. Yang, R. Ahmed, and D. A. Steinhauer. 2005. Chimeric influenza virus hemagglutinin proteins containing large domains of the Bacillus anthracis protective antigen: protein characterization, incorporation into infectious influenza viruses, and antigenicity. J Virol 79:10003.
96.Haynes, J. R., S. X. Cao, B. Rovinski, C. Sia, O. James, G. A. Dekaban, and M. H. Klein. 1991. Production of immunogenic HIV-1 viruslike particles in stably engineered monkey cell lines. AIDS Res Hum Retroviruses 7:17.
97.Rovinski, B., J. R. Haynes, S. X. Cao, O. James, C. Sia, S. Zolla-Pazner, T. J. Matthews, and M. H. Klein. 1992. Expression and characterization of genetically engineered human immunodeficiency virus-like particles containing modified envelope glycoproteins: implications for development of a cross-protective AIDS vaccine. J Virol 66:4003.
98.Fynan, E. F., R. G. Webster, D. H. Fuller, J. R. Haynes, J. C. Santoro, and H. L. Robinson. 1995. DNA vaccines: a novel approach to immunization. Int J Immunopharmacol 17:79.
99.Fynan, E. F., R. G. Webster, D. H. Fuller, J. R. Haynes, J. C. Santoro, and H. L. Robinson. 1993. DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc Natl Acad Sci U S A 90:11478.
100.Kodihalli, S., J. R. Haynes, H. L. Robinson, and R. G. Webster. 1997. Cross-protection among lethal H5N2 influenza viruses induced by DNA vaccine to the hemagglutinin. J Virol 71:3391.
101.Robinson, H. L., S. Lu, D. M. Feltquate, C. T. Torres, J. Richmond, C. M. Boyle, M. J. Morin, J. C. Santoro, R. G. Webster, D. Montefiori, Y. Yasutomi, N. L. Letvin, K. Manson, M. Wyand, and J. R. Haynes. 1996. DNA vaccines. AIDS Res Hum Retroviruses 12:455.
102.Drape, R. J., M. D. Macklin, L. J. Barr, S. Jones, J. R. Haynes, and H. J. Dean. 2006. Epidermal DNA vaccine for influenza is immunogenic in humans. Vaccine in press.
103.Kretzchmar, E., R. Geyer, and H. D. Klenk. 1994. Baculovirus infection does not alter N-glycosylation in Spodoptera frugiperda cells. Biol Chem Hoppe Seyler 375:23.
104.Lu, D., and A. J. Hickey. 2005. Liposomal dry powders as aerosols for pulmonary delivery of proteins. AAPS PharmSciTech 6:E641.
105.Cowdery, S., M. Frey, S. Orlowski, and A. Gray. 1976. Stability characteristics of freeze-dried human live virus vaccines. Dev Biol Stand 36:297.
106.Peetermans, J., G. Colinet, A. Bouillet, E. D'Hondt, and J. Stephenne. 1976. Stability of live, freeze-dried virus vaccines. Dev Biol Stand 36:291.
107.Yannarell, D. A., K. M. Goldberg, and R. N. Hjorth. 2002. Stabilizing cold-adapted influenza virus vaccine under various storage conditions. J Virol Methods 102:15.
108.Sampson, H. A., J. Bernhisel-Broadbent, E. Yang, and S. M. Scanlon. 1991. Safety of casein hydrolysate formula in children with cow milk allergy. J Pediatr 118:520.
109.Gambaryan A, A. Tuzikov, G. Pazynina, N. Bovin, A. Balish, and A. Klimov. 2005. Evolution of the receptor binding phenotype of influenza A (H5) viruses. Virology (electronic publication ahead of print version).
110.Suzuki, Y, 2005. Sialobiology of Influenza: Molecular Mechanism of Host Range Variation of Influenza Viruses. Biological and Pharmaceutical Bulletin 28:399-408.
【0184】
〔政府の支援〕
本発明は、米国政府の支援によって、米国陸軍医療研究および材料部隊(U.S. Army Medical Research and Material Command)からの助成金番号W81XWH-05-C-0135および助成金番号W81XWH-05-C-0150のもとでなされた。米国政府は、本発明の一定の権利を有する。
【特許請求の範囲】
【請求項1】
(a)gagポリペプチドと、
(b)非ウイルス性脂質ラフト会合ポリペプチドとを含んでいる、キメラウイルス様粒子。
【請求項2】
上記脂質ラフト会合ポリペプチドが、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、シグナル伝達のポリペプチド、および膜輸送のポリペプチドから成る群から選択される、請求項1に記載のウイルス様粒子。
【請求項3】
上記脂質ラフト会合ポリペプチドが、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、カベオリンのポリペプチド、フロチリンのポリペプチド、シンタキシン−1のポリペプチド、シンタキシン−4のポリペプチド、シナプシンIのポリペプチド、アデューシンのポリペプチド、VAMP2のポリペプチド、VAMP/シナプトブレビンのポリペプチド、シナプトブレビンIIのポリペプチド、SNAREのポリペプチド、SNAP−25のポリペプチド、SNAP−23のポリペプチド、シナプトタグミンIのポリペプチド、およびシナプトタグミンIIのポリペプチドから成る群から選択される、請求項1に記載のウイルス様粒子。
【請求項4】
ヘマグルチニンのポリペプチドをさらに含んでいる、請求項1に記載のウイルス様粒子。
【請求項5】
ノイラミニダーゼのポリペプチドをさらに含んでいる、請求項4に記載のウイルス様粒子。
【請求項6】
(a)gagポリペプチドと、
(b)抗原に連結されて結合を形成している脂質ラフト会合ポリペプチドとを含んでおり、
上記抗原が、脂質ラフトと天然に会合しないものである、キメラウイルス様粒子。
【請求項7】
上記結合が、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用から成る群から選択される、請求項6に記載のウイルス様粒子。
【請求項8】
上記結合が共有結合である、請求項6に記載のウイルス様粒子。
【請求項9】
上記共有結合が、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合から成る群から選択される、請求項8に記載のウイルス様粒子。
【請求項10】
上記脂質ラフト会合ポリペプチドが内在性膜タンパク質である、請求項6に記載のウイルス様粒子。
【請求項11】
上記脂質ラフト会合ポリペプチドが、ヘマグルチニンのポリペプチド、ノイラミニダーゼのポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、およびエンベロープタンパクのポリペプチドから選択される、請求項6に記載のウイルス様粒子。
【請求項12】
上記脂質ラフト会合ポリペプチドがヘマグルチニンのポリペプチドである、請求項11に記載のウイルス様粒子。
【請求項13】
ノイラミニダーゼのポリペプチドをさらに含んでいる、請求項12に記載のウイルス様粒子。
【請求項14】
上記脂質ラフト会合ポリペプチドがノイラミニダーゼのポリペプチドである、請求項11に記載のウイルス様粒子。
【請求項15】
ヘマグルチニンポリペプチドをさらに含んでいる、請求項14に記載のウイルス様粒子。
【請求項16】
上記抗原が、タンパク質、ポリペプチド、糖ポリペプチド、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドのペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、および炭水化物から成る群から選択される、請求項6に記載のウイルス様粒子。
【請求項17】
上記ウイルス様粒子が昆虫細胞のグリコシル化を含んでいる、請求項6に記載のウイルス様粒子。
【請求項18】
上記ウイルス様粒子が哺乳類細胞のグリコシル化を含んでいる、請求項6に記載のウイルス様粒子。
【請求項19】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項6に記載のウイルス様粒子。
【請求項20】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項6に記載のウイルス様粒子。
【請求項21】
上記アジュバントが、上記ウイルス様粒子の内側に配置されている、請求項20に記載のウイルス様粒子
【請求項22】
上記アジュバントが、上記gagポリペプチドに共有的に連結されて共有結合を形成している、請求項21に記載のウイルス様粒子。
【請求項23】
上記アジュバントが、上記ウイルス様粒子の外側に配置されている、請求項20に記載のウイルス様粒子。
【請求項24】
上記アジュバントが、上記脂質ラフト会合ポリペプチドに共有的に連結されて共有結合を形成している、請求項23に記載のウイルス様粒子。
【請求項25】
上記アジュバントがフラジェリンのアジュバント活性断片を含んでいる、請求項20に記載のウイルス様粒子。
【請求項26】
(a)gagポリペプチドと、
(b)RSVの脂質ラフト会合ポリペプチドとを含んでいるキメラウイルス様粒子。
【請求項27】
上記ウイルス様粒子が昆虫細胞のグリコシル化を含んでいる、請求項26に記載のウイルス様粒子。
【請求項28】
上記ウイルス様粒子が哺乳類細胞のグリコシル化を含んでいる、請求項26に記載のウイルス様粒子。
【請求項29】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項30】
RSV脂質ラフト会合ポリペプチドに連結されて結合を形成している抗原をさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項31】
上記第2脂質ラフト会合ポリペプチドが、第2RSV抗原であるか、または第2RSV抗原に連結されている、請求項29に記載のウイルス様粒子。
【請求項32】
上記第2脂質ラフト会合ポリペプチドが、RSV抗原に連結されている、請求項29に記載のウイルス様粒子。
【請求項33】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項34】
(a)gagポリペプチドと、
(b)エンベロープウイルスの脂質ラフト会合ポリペプチドとを含んでおり、
昆虫細胞のグリコシル化を含んでいる、キメラウイルス様粒子。
【請求項35】
上記エンベロープウイルスの脂質ラフト会合ポリペプチドが、パラミクソウイルス科の脂質ラフト会合ポリペプチド、およびヘルペスウイルス科の脂質ラフト会合ポリペプチドから成る群から選択される、請求項34に記載のウイルス様粒子。
【請求項36】
上記脂質ラフト会合ポリペプチドが、パラインフルエンザウイルスの脂質ラフト会合ポリペプチド、ムンプスウイルスの脂質ラフト会合ポリペプチド、麻疹ウイルスの脂質ラフト会合ポリペプチド、呼吸器合胞体ウイルスの脂質ラフト会合ポリペプチド、トリニューモウイルスの脂質ラフト会合ポリペプチド、ヒトメタニューモウイルスの脂質ラフト会合ポリペプチド、単純疱疹ウイルス1型(HHV−I)の脂質ラフト会合ポリペプチド、単純疱疹ウイルス2型(HHV−2)の脂質ラフト会合ポリペプチド、水痘帯状疱疹ウイルス(HHV−3)の脂質ラフト会合ポリペプチド、EBウイルス(エプスタインバーウイルス)(HHV−4)の脂質ラフト会合ポリペプチド、およびサイトメガロウイルス(CMV)(HHV−5)の脂質ラフト会合ポリペプチドから成る群から選択される、請求項35に記載のウイルス様粒子。
【請求項37】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項34に記載のウイルス様粒子。
【請求項38】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項34に記載のウイルス様粒子。
【請求項39】
(a)gagポリペプチドをコードする第1ヌクレオチド配列と、
(b)抗原に連結される脂質ラフト会合ポリペプチドをコードする第2ヌクレオチド配列とを含んでおり、
上記抗原は脂質ラフトと天然には会合しないものであり、
上記ポリペプチドは、宿主細胞内で発現すると、ウイルス様粒子を形成するものである、キメラウイルス様粒子の発現ベクターシステム。
【請求項40】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、1つの発現ベクター内に存在している、請求項39に記載のウイルス様粒子の発現ベクターシステム。
【請求項41】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、1つのプロモーターに作動可能に連結されている、請求項40に記載のウイルス様粒子の発現ベクターシステム。
【請求項42】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、複数の発現ベクター内に存在している、請求項39に記載のウイルス様粒子の発現ベクターシステム。
【請求項43】
(a)gagポリペプチドと抗原に連結される脂質ラフト会合ポリペプチドとを一緒に発現する、1つ以上の発現ベクターを提供する工程、
(b)上記1つ以上の発現ベクターを細胞に導入する工程、および、
(c)上記gagポリペプチドと上記抗原に連結される脂質ラフト会合ポリペプチドとを発現させて、キメラウイルス様粒子を製造する工程を含んでおり、
上記抗原は、脂質ラフトと天然には会合しないものである、キメラウイルス様粒子を製造する方法。
【請求項44】
上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる、請求項43に記載の方法。
【請求項45】
上記1つ以上の発現ベクターがウイルスベクターである、請求項43に記載の方法。
【請求項46】
上記ウイルスベクターが、バキュロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスから成る群から選択される、請求項45に記載の方法。
【請求項47】
上記ウイルスベクターがバキュロウイルスである、請求項46に記載の方法。
【請求項48】
上記細胞が昆虫細胞および哺乳類細胞から成る群から選択される、請求項43に記載の方法。
【請求項49】
上記細胞が昆虫細胞である、請求項48に記載の方法。
【請求項50】
上記1つ以上の発現ベクターがバキュロウイルスであり、上記細胞が昆虫細胞である、請求項43に記載の方法。
【請求項51】
上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる、請求項43に記載の方法。
【請求項52】
免疫系の症状あるいは病気を治療または予防する方法であって、
免疫原性量のキメラウイルス様粒子の投与を含んでおり、
上記粒子は、gagポリペプチドと、抗原に連結された脂質ラフト会合ポリペプチドとを含んでおり、
上記抗原は、脂質ラフトと天然には会合しないものである、方法。
【請求項53】
上記投与は、感染防御免疫応答を被検体に誘導するものである、請求項52に記載の方法。
【請求項54】
上記投与は、皮下投与、経皮投与、皮内投与、真皮下投与、筋肉内投与、経口投与、鼻腔内投与、バッカル投与、舌下投与、腹腔内投与、膣内投与、肛門投与、および頭蓋内投与から成る群から選択される、請求項52に記載の方法。
【請求項55】
免疫原性量のキメラウイルス様粒子を含んでいる薬学的組成物であって、上記キメラウイルス様粒子は、gagポリペプチド、および抗原に連結される脂質ラフト会合ポリペプチドを含んでおり、上記抗原は脂質ラフトと天然には会合しないものである、薬学的組成物。
【請求項56】
薬学的に許容可能な担体をさらに含んでいる、請求項55に記載の薬学的組成物。
【請求項1】
(a)gagポリペプチドと、
(b)非ウイルス性脂質ラフト会合ポリペプチドとを含んでいる、キメラウイルス様粒子。
【請求項2】
上記脂質ラフト会合ポリペプチドが、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、シグナル伝達のポリペプチド、および膜輸送のポリペプチドから成る群から選択される、請求項1に記載のウイルス様粒子。
【請求項3】
上記脂質ラフト会合ポリペプチドが、GPIアンカーのポリペプチド、ミリストイル化配列のポリペプチド、パルミトイル化配列のポリペプチド、ダブルアセチル化配列のポリペプチド、カベオリンのポリペプチド、フロチリンのポリペプチド、シンタキシン−1のポリペプチド、シンタキシン−4のポリペプチド、シナプシンIのポリペプチド、アデューシンのポリペプチド、VAMP2のポリペプチド、VAMP/シナプトブレビンのポリペプチド、シナプトブレビンIIのポリペプチド、SNAREのポリペプチド、SNAP−25のポリペプチド、SNAP−23のポリペプチド、シナプトタグミンIのポリペプチド、およびシナプトタグミンIIのポリペプチドから成る群から選択される、請求項1に記載のウイルス様粒子。
【請求項4】
ヘマグルチニンのポリペプチドをさらに含んでいる、請求項1に記載のウイルス様粒子。
【請求項5】
ノイラミニダーゼのポリペプチドをさらに含んでいる、請求項4に記載のウイルス様粒子。
【請求項6】
(a)gagポリペプチドと、
(b)抗原に連結されて結合を形成している脂質ラフト会合ポリペプチドとを含んでおり、
上記抗原が、脂質ラフトと天然に会合しないものである、キメラウイルス様粒子。
【請求項7】
上記結合が、共有結合、イオン相互作用、水素結合、イオン結合、ファンデルワールス力、金属−配位子相互作用、および抗体−抗原相互作用から成る群から選択される、請求項6に記載のウイルス様粒子。
【請求項8】
上記結合が共有結合である、請求項6に記載のウイルス様粒子。
【請求項9】
上記共有結合が、ペプチド結合、炭素−酸素結合、炭素−硫黄結合、炭素−窒素結合、炭素−炭素結合、およびジスルフィド結合から成る群から選択される、請求項8に記載のウイルス様粒子。
【請求項10】
上記脂質ラフト会合ポリペプチドが内在性膜タンパク質である、請求項6に記載のウイルス様粒子。
【請求項11】
上記脂質ラフト会合ポリペプチドが、ヘマグルチニンのポリペプチド、ノイラミニダーゼのポリペプチド、融合タンパク質のポリペプチド、糖タンパク質のポリペプチド、およびエンベロープタンパクのポリペプチドから選択される、請求項6に記載のウイルス様粒子。
【請求項12】
上記脂質ラフト会合ポリペプチドがヘマグルチニンのポリペプチドである、請求項11に記載のウイルス様粒子。
【請求項13】
ノイラミニダーゼのポリペプチドをさらに含んでいる、請求項12に記載のウイルス様粒子。
【請求項14】
上記脂質ラフト会合ポリペプチドがノイラミニダーゼのポリペプチドである、請求項11に記載のウイルス様粒子。
【請求項15】
ヘマグルチニンポリペプチドをさらに含んでいる、請求項14に記載のウイルス様粒子。
【請求項16】
上記抗原が、タンパク質、ポリペプチド、糖ポリペプチド、リポポリペプチド、ペプチド、ポリサッカリド、ポリサッカリドの接合体、ポリサッカリドのペプチド模擬体または非ペプチド模擬体、低分子、脂質、糖脂質、および炭水化物から成る群から選択される、請求項6に記載のウイルス様粒子。
【請求項17】
上記ウイルス様粒子が昆虫細胞のグリコシル化を含んでいる、請求項6に記載のウイルス様粒子。
【請求項18】
上記ウイルス様粒子が哺乳類細胞のグリコシル化を含んでいる、請求項6に記載のウイルス様粒子。
【請求項19】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項6に記載のウイルス様粒子。
【請求項20】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項6に記載のウイルス様粒子。
【請求項21】
上記アジュバントが、上記ウイルス様粒子の内側に配置されている、請求項20に記載のウイルス様粒子
【請求項22】
上記アジュバントが、上記gagポリペプチドに共有的に連結されて共有結合を形成している、請求項21に記載のウイルス様粒子。
【請求項23】
上記アジュバントが、上記ウイルス様粒子の外側に配置されている、請求項20に記載のウイルス様粒子。
【請求項24】
上記アジュバントが、上記脂質ラフト会合ポリペプチドに共有的に連結されて共有結合を形成している、請求項23に記載のウイルス様粒子。
【請求項25】
上記アジュバントがフラジェリンのアジュバント活性断片を含んでいる、請求項20に記載のウイルス様粒子。
【請求項26】
(a)gagポリペプチドと、
(b)RSVの脂質ラフト会合ポリペプチドとを含んでいるキメラウイルス様粒子。
【請求項27】
上記ウイルス様粒子が昆虫細胞のグリコシル化を含んでいる、請求項26に記載のウイルス様粒子。
【請求項28】
上記ウイルス様粒子が哺乳類細胞のグリコシル化を含んでいる、請求項26に記載のウイルス様粒子。
【請求項29】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項30】
RSV脂質ラフト会合ポリペプチドに連結されて結合を形成している抗原をさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項31】
上記第2脂質ラフト会合ポリペプチドが、第2RSV抗原であるか、または第2RSV抗原に連結されている、請求項29に記載のウイルス様粒子。
【請求項32】
上記第2脂質ラフト会合ポリペプチドが、RSV抗原に連結されている、請求項29に記載のウイルス様粒子。
【請求項33】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項26に記載のウイルス様粒子。
【請求項34】
(a)gagポリペプチドと、
(b)エンベロープウイルスの脂質ラフト会合ポリペプチドとを含んでおり、
昆虫細胞のグリコシル化を含んでいる、キメラウイルス様粒子。
【請求項35】
上記エンベロープウイルスの脂質ラフト会合ポリペプチドが、パラミクソウイルス科の脂質ラフト会合ポリペプチド、およびヘルペスウイルス科の脂質ラフト会合ポリペプチドから成る群から選択される、請求項34に記載のウイルス様粒子。
【請求項36】
上記脂質ラフト会合ポリペプチドが、パラインフルエンザウイルスの脂質ラフト会合ポリペプチド、ムンプスウイルスの脂質ラフト会合ポリペプチド、麻疹ウイルスの脂質ラフト会合ポリペプチド、呼吸器合胞体ウイルスの脂質ラフト会合ポリペプチド、トリニューモウイルスの脂質ラフト会合ポリペプチド、ヒトメタニューモウイルスの脂質ラフト会合ポリペプチド、単純疱疹ウイルス1型(HHV−I)の脂質ラフト会合ポリペプチド、単純疱疹ウイルス2型(HHV−2)の脂質ラフト会合ポリペプチド、水痘帯状疱疹ウイルス(HHV−3)の脂質ラフト会合ポリペプチド、EBウイルス(エプスタインバーウイルス)(HHV−4)の脂質ラフト会合ポリペプチド、およびサイトメガロウイルス(CMV)(HHV−5)の脂質ラフト会合ポリペプチドから成る群から選択される、請求項35に記載のウイルス様粒子。
【請求項37】
第2脂質ラフト会合ポリペプチドをさらに含んでいる、請求項34に記載のウイルス様粒子。
【請求項38】
上記ウイルス様粒子と混合されたアジュバントをさらに含んでいる、請求項34に記載のウイルス様粒子。
【請求項39】
(a)gagポリペプチドをコードする第1ヌクレオチド配列と、
(b)抗原に連結される脂質ラフト会合ポリペプチドをコードする第2ヌクレオチド配列とを含んでおり、
上記抗原は脂質ラフトと天然には会合しないものであり、
上記ポリペプチドは、宿主細胞内で発現すると、ウイルス様粒子を形成するものである、キメラウイルス様粒子の発現ベクターシステム。
【請求項40】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、1つの発現ベクター内に存在している、請求項39に記載のウイルス様粒子の発現ベクターシステム。
【請求項41】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、1つのプロモーターに作動可能に連結されている、請求項40に記載のウイルス様粒子の発現ベクターシステム。
【請求項42】
上記第1ヌクレオチド配列および第2ヌクレオチド配列が、複数の発現ベクター内に存在している、請求項39に記載のウイルス様粒子の発現ベクターシステム。
【請求項43】
(a)gagポリペプチドと抗原に連結される脂質ラフト会合ポリペプチドとを一緒に発現する、1つ以上の発現ベクターを提供する工程、
(b)上記1つ以上の発現ベクターを細胞に導入する工程、および、
(c)上記gagポリペプチドと上記抗原に連結される脂質ラフト会合ポリペプチドとを発現させて、キメラウイルス様粒子を製造する工程を含んでおり、
上記抗原は、脂質ラフトと天然には会合しないものである、キメラウイルス様粒子を製造する方法。
【請求項44】
上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる、請求項43に記載の方法。
【請求項45】
上記1つ以上の発現ベクターがウイルスベクターである、請求項43に記載の方法。
【請求項46】
上記ウイルスベクターが、バキュロウイルス、アデノウイルス、ヘルペスウイルス、ポックスウイルス、およびレトロウイルスから成る群から選択される、請求項45に記載の方法。
【請求項47】
上記ウイルスベクターがバキュロウイルスである、請求項46に記載の方法。
【請求項48】
上記細胞が昆虫細胞および哺乳類細胞から成る群から選択される、請求項43に記載の方法。
【請求項49】
上記細胞が昆虫細胞である、請求項48に記載の方法。
【請求項50】
上記1つ以上の発現ベクターがバキュロウイルスであり、上記細胞が昆虫細胞である、請求項43に記載の方法。
【請求項51】
上記細胞が培養されている培地から上記ウイルス様粒子を回収する工程をさらに含んでいる、請求項43に記載の方法。
【請求項52】
免疫系の症状あるいは病気を治療または予防する方法であって、
免疫原性量のキメラウイルス様粒子の投与を含んでおり、
上記粒子は、gagポリペプチドと、抗原に連結された脂質ラフト会合ポリペプチドとを含んでおり、
上記抗原は、脂質ラフトと天然には会合しないものである、方法。
【請求項53】
上記投与は、感染防御免疫応答を被検体に誘導するものである、請求項52に記載の方法。
【請求項54】
上記投与は、皮下投与、経皮投与、皮内投与、真皮下投与、筋肉内投与、経口投与、鼻腔内投与、バッカル投与、舌下投与、腹腔内投与、膣内投与、肛門投与、および頭蓋内投与から成る群から選択される、請求項52に記載の方法。
【請求項55】
免疫原性量のキメラウイルス様粒子を含んでいる薬学的組成物であって、上記キメラウイルス様粒子は、gagポリペプチド、および抗原に連結される脂質ラフト会合ポリペプチドを含んでおり、上記抗原は脂質ラフトと天然には会合しないものである、薬学的組成物。
【請求項56】
薬学的に許容可能な担体をさらに含んでいる、請求項55に記載の薬学的組成物。
【図1】

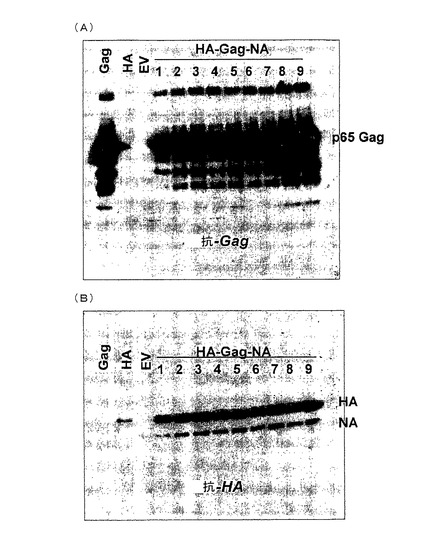
【図2】
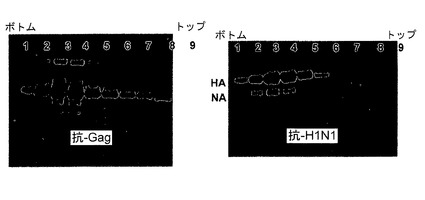
【図3】
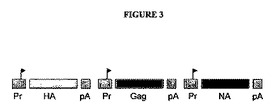
【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公表番号】特表2009−544322(P2009−544322A)
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願番号】特願2009−521858(P2009−521858)
【出願日】平成19年7月27日(2007.7.27)
【国際出願番号】PCT/US2007/016938
【国際公開番号】WO2008/094200
【国際公開日】平成20年8月7日(2008.8.7)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
【公表日】平成21年12月17日(2009.12.17)
【国際特許分類】
【出願日】平成19年7月27日(2007.7.27)
【国際出願番号】PCT/US2007/016938
【国際公開番号】WO2008/094200
【国際公開日】平成20年8月7日(2008.8.7)
【出願人】(509024190)リゴサイト ファーマシューティカルズ インコーポレイテッド (6)
【氏名又は名称原語表記】LIGOCYTE PHARMACEUTICALS,INC.
【住所又は居所原語表記】2155 Analysis Drive,Bozeman,MT 59718−6831,United States of America
【Fターム(参考)】
[ Back to top ]