
プロトプラスト形質転換効率の向上方法
【課題】形質転換効率の高いプロトプラスト形質転換方法の提供。
【解決手段】宿主微生物の細胞膜脂質の生合成に係るugtP遺伝子を欠失又は不活性化すること。
【解決手段】宿主微生物の細胞膜脂質の生合成に係るugtP遺伝子を欠失又は不活性化すること。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、プロトプラスト形質転換効率の向上方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、あるいは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。更に、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。ゲノム情報の公開されている産業的に有用な宿主微生物としては、枯草菌Bacillus subtilis Marburg No.168(非特許文献1)、大腸菌Escherichia coli K−12 MG1655(非特許文献2)、コリネバクテリウムCorynebacterium glutamicum ATCC132032などが挙げられ、これらのゲノム情報を利用し、改良を加えた菌株が開発されている。
【0004】
また、特に最近では、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われている。DNAなどの核酸分子を外部から枯草菌に導入して形質転換する場合、自然形質転換法(コンピテントセル法)(非特許文献3)、プロトプラスト法(非特許文献4)、エレクトロポレーション法(非特許文献5)が主に用いられる。これらの方法のうち、自然形質転換法は、コンピテント培養により調製した、DNA取り込み能を有する細胞(コンピテント細胞)を用いた形質転換である。この自然形質転換は、枯草菌マーブルグ(Marburg)系の菌株(168、166、160株)でのみ生じるが、納豆菌等では自然形質転換の頻度は極めて低く、これらは形質転換能(コンピテンシー)を有しないともいわれる。一方で、プロトプラスト法及びエレクトロポレーション法は、多くの菌種に適用可能である。ここで、プロトプラスト法とは、等張液中で細胞壁をリゾチームによって分解してプロトプラスト細胞を調製し、DNAとプロトプラスト細胞を高濃度ポリエチレングリコールにより凝集させ、更にポリエチレングリコールの濃度を急激に低下させることでDNAを取り込ませる手法である。またエレクトロポレーション法とは、細胞懸濁液に電気パルスを負荷して細胞膜に微小な孔を形成させ、そこからDNAを細胞内に導入する手法である(非特許文献6)。
【0005】
しかしながら枯草菌を含む多くの細菌では、導入された外来DNAが修飾(メチル化)されている自己のDNAと区別され、更に当該外来DNAが、そのDNA中の特定の塩基配列を認識して切断する制限酵素により分解されるため、形質転換は一般には起こりにくい。形質転換においてはこれらの制限修飾系が少ないほど好ましいと考えられ、例えば魚住らは、枯草菌マーブルグ系の株を基に、制限修飾能を欠失させた株RM125株を作製し、形質転換能を10〜100倍に向上させている(非特許文献7)。
【0006】
ugtP遺伝子は1,2−ジアシルグリセロールにグルコース残基を付加するUDP−グリコシルトランスフェラーゼをコードする遺伝子である。In vitroの実験から、本酵素は1,2−ジアシルグリセロールにグルコースを4個まで連続して付加することが可能であることがわかっているが、生体内ではジグリコシルアシルグリセロールが主要な糖脂質である(非特許文献8)。ugtP遺伝子に関しては、その欠損により細胞の膜脂質の脂肪酸組成に影響を与えることが報告されている(非特許文献9)が、ugtP遺伝子と形質転換効率の向上との関係についてはこれまで報告されていない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Nature,390(6657),p249−256(1997)
【非特許文献2】Science,277(5331),p1453−1462(1997)
【非特許文献3】J.Bacteriol.81,p741(1961)
【非特許文献4】Mol.Gen.Genet.168,p111(1979)
【非特許文献5】FEMS Microbiol.Lett.55,p135(1990)
【非特許文献6】Molecular Biological Method for Bacillus,Edited by Harwood,C.R.ら,John Wiley&Sons,p98−103
【非特許文献7】Mol.Gen.Genne.,152,p65−9(1977)
【非特許文献8】Mol.Microbiol.,1998,29(2),p.419−430
【非特許文献9】J.Bacteriol.,2008,190,7797−7807
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、形質転換効率の高いプロトプラスト形質転換方法に関する。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題に鑑み検討した結果、宿主微生物の細胞膜脂質の生合成に係るugtP遺伝子を欠失又は不活性化することにより、プロトプラスト形質転換効率が向上することを見出した。
【0010】
すなわち本発明は、以下に関する。
(1) ugtP遺伝子が欠失又は不活性化された宿主微生物のプロトプラストを用いることを特徴とする、形質転換方法。
(2) 前記宿主微生物がグラム陽性細菌である、前記(1)記載の形質転換方法。
(3) 前記宿主微生物がバチルス属細菌である、前記(2)記載の形質転換方法。
(4) 前記宿主微生物が枯草菌である、前記(3)記載の形質転換方法。
【発明の効果】
【0011】
本発明の形質転換方法を用いることにより、目的のDNA、例えば目的タンパク質又はポリペプチドをコードする遺伝子を宿主微生物に効率的に導入することが可能となる。その結果、例えばタンパク質又はポリペプチドを高生産する優良株の選抜が容易になり、更に当該タンパク質又はポリペプチドの生産性向上が期待できる。
【図面の簡単な説明】
【0012】
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
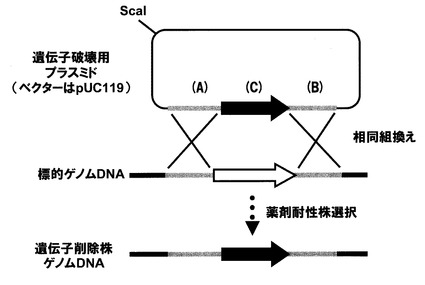
【図2】遺伝子欠失用プラスミドを用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
【図3】形質転換効率を示すグラフ。(a)はpHY300PLK、(b)はpC194を用いて形質転換した場合の、168株(野生株)及びΔugtP株における結果である。結果はN=3の平均値を示し、エラーバーは標準偏差を示す。
【発明を実施するための形態】
【0013】
本発明の方法においては、宿主微生物のugtP遺伝子が、欠失又は不活性化されている。このugtP遺伝子は、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)において、遺伝子番号 BG11611として登録・公開されている(http://bacillus.genome.ad.jp/、2006年1月18日更新)。
【0014】
ugtP遺伝子(配列番号1)と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、更に好ましくは95%以上、更により好ましくは98%以上の同一性を有し、1,2−ジアシルグリセロールにグルコース残基を付加するUDP−グリコシルトランスフェラーゼ活性を有するタンパク質をコードする遺伝子も、当該ugtP遺伝子に相当する遺伝子と考えられ、本発明において欠失又は不活性化されるugtP遺伝子に含まれるものとする。
【0015】
アミノ酸配列及び塩基配列の同一性はLipman−Pearson法 (Science,227,1435,(1985))によって計算することができる。具体的には、遺伝情報処理ソフトウェアGenetyx−Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0016】
また、枯草菌UgtPのホモログをKEGG Sequence Similarity DataBase(SSDB)にて検索を行った結果、Staphylococcus carnosusのSca_0619(Soc.Appl.Bacteriol.Symp.Ser.(1990)19,49S−53S)、Desulfitobacterium hafnienseのDhaf_3178、及びDSY3206(FEMS Microbiol.Rev.,(2006)30,706−733)、Desulfotomaculum reducensのDred_1966(Environ Microbiol.(2009)12,3007−3017)、Clostridium cellulolyticumのCcel_0034(FEMS Microbiol.Rev.(2005)29,741−764)、Clostridium thermocellumのCthe_2414(Microbiol Mol.Biol.Rev.,(2005)69,124−154)、Clostridium cellulovoransのClocel_3086(Annu.Rev.Microbiol.,(2004)58,521−554)、Clostridium ljungdahliiのCLJU_c02160(Bioprocess Biosyst.Eng.,(2009)32,369−380)、Brevibacillus brevisのBBR47_41080(Proc.Natl.Acad.Sci.USA.(1988)86:3589−93)、Variovorax paradoxusのVapar_4986及びVarpa_5682(Arch.Microbiol.,(2000),174,111−119)、及びDeinococcus radioduransのDR_1076(Curr.Opin.Biotechnol.(2000)11,280−285)は枯草菌UgtPのホモログと推察されることより、これらをコードする遺伝子は本発明において欠失又は不活性化されるugtP遺伝子に含まれるものとする。
【0017】
本発明で使用される宿主微生物としては、バチルス(Bacillus)属細菌や、上述のスタフィロコッカス(Staphylococcus)属細菌、ブレビバシラス(Brevibacillus)属細菌、クロストリジウム(Clostridium)属細菌等のグラム陽性細菌が挙げられ、中でもバチルス属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質を菌体外に分泌生産させる能力を有する点から、枯草菌(Bacillus subtilis)がより好ましい。
【0018】
上記宿主微生物からのugtP遺伝子の欠失又は不活性化は、標的遺伝子の一部若しくは全部をゲノム中から除去するか又は他の遺伝子と置き換える、当該遺伝子中に他のDNA断片を挿入する、あるいは当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって行い、好適には当該遺伝子を物理的に欠失させる。より具体的には、ugtP遺伝子を計画的に欠失又は不活性化する方法、及びランダムな遺伝子の欠失又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行って所望の変異を選択する方法が挙げられる。
【0019】
ugtP遺伝子を欠失又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、ugtP遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、ugtP遺伝子の一部領域における相同組換えによって宿主微生物ゲノム上のugtP遺伝子を分断して不活性化することが可能である。あるいは、塩基置換や塩基挿入等の変異によって不活性化したugtP遺伝子、又は図1のようにugtP遺伝子の上流、下流領域を含むがugtP遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを宿主微生物細胞内に取り込ませて宿主微生物ゲノムのugtP遺伝子内の変異箇所の外側の2ヶ所、又はugtP遺伝子上流側、下流側で2回交差の相同組換えを起こさせることにより、ゲノム上のugtP遺伝子を欠失あるいは他の遺伝子断片と置換させることによって不活性化させることが可能である。
【0020】
例えば、本発明の方法に使用する微生物を構築するための宿主微生物として枯草菌を用いる場合、相同組換えによりugtP遺伝子を欠失又は不活性化する方法については、既に幾つかの報告例があり(Mol.Gen.Genet.,223,268,1990等)、かかる方法を繰り返すことにより、本発明で用いられる微生物を得ることができる。
【0021】
また、ランダムな遺伝子の欠失又は不活性化については、ランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、宿主微生物にγ線等を照射する方法等が実施可能である。
【0022】
以下に、具体例として、SOE(splicing by overlap extension)−PCR法(Gene,77,61,1989)によって調製される欠失用DNA断片を用いた二重交差法による欠失方法について説明するが、本発明における遺伝子欠失又は不活性化方法は下記に限定されるものではない。
【0023】
欠失用DNA断片は、例えば、欠失対象遺伝子の上流に隣接する約1.0kb断片と、同じく下流に隣接する約1.0kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、欠失対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0024】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列において、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子を挿入したDNA断片を得ることができる(図2)。以上のPCRは、市販のPCR用酵素キット等を用いて、成書(PCR Protocols.Current Methods and Applications,Edited by B.A.White,Humana Press pp251,1993、Gene,77,61,1989)等に示される通常の条件により行うことができる。
【0025】
かくして得られた遺伝子欠失用DNA断片を、コンピテント法あるいは下記のプロトプラスト法等の定法によって微生物細胞内に導入すると、対応する欠失対象遺伝子の上流及び下流の相同領域において、細胞内での遺伝子組換えが生じ、ugtP遺伝子が薬剤耐性遺伝子と置換された細胞、あるいはugtP遺伝子内に薬剤耐性遺伝子が挿入された細胞が生じ得る(図2)。これを薬剤耐性マーカーによる選択によって分離する。即ち、遺伝子導入した微生物を、上記薬剤を含む寒天培地上で培養し、生育するコロニーを分離した後、ゲノムを鋳型としたPCR法などによってゲノム上のugtP遺伝子が薬剤耐性遺伝子と置換されていることを確認すれば良い。
【0026】
上記により得られた、ugtP遺伝子が欠失又は不活性化された微生物は、プロトプラストとして調製することにより、外部からの核酸分子の導入を効率的に行うことができる。ここで、微生物のプロトプラストの調製は、公知の方法に従い実施でき、例えば細菌のプロトプラストを調製する場合には、まず一般的に用いられる細菌培養用培地(LB培地等)に該細菌を接種して培養し、菌体を得、次いでこの菌体を、浸透圧を調整する物質(スクロース、ソルビトール等)を含有する高張緩衝液に懸濁し、更にリゾチームなどの細胞壁分解酵素を作用させて細胞壁を除去することにより、プロトプラストを調製することができる。
【0027】
上記で得られたプロトプラストを、例えば、ポリエチレングリコール及び薬剤耐性マーカーを有する核酸分子(プラスミドDNA等)と混合することにより、プラスミドが当該プロトプラスト内に取り込まれ、宿主微生物が形質転換される。
【0028】
下記の実施例に示すように、ugtP遺伝子を欠失させた宿主微生物のプロトプラストを用いてプラスミドにより形質転換することにより、その形質転換効率は、当該ugtP遺伝子が欠失又は不活性化されていない微生物を用いた場合と比較し高い。
【0029】
上記により宿主微生物を形質転換した後、形質転換された宿主微生物のクローンを得る場合、例えば一定時間培養した後、クロラムフェニコール等の薬剤を含む再生培地で培養し、導入された核酸分子上に存在する薬剤耐性マーカーによる選抜を行えばよい(Mol.Gen.Genet.168,111(1979))。
【0030】
また、本発明のプロトプラスト形質転換に用いる核酸分子は、形質転換された微生物の用途に応じて適宜選択でき、特に限定されない。例えば、宿主微生物に導入される核酸分子は、目的タンパク質又は目的ポリペプチドをコードする遺伝子を有するプラスミドであってもよく、当該目的タンパク質又は目的ポリペプチドには、例えば洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが挙げられる。産業用酵素としては、機能別に、酸化還元酵素(オキシドレダクターゼ)、転移酵素(トランスフェラーゼ)、加水分解酵素(ヒドロラーゼ)、脱離酵素(リアーゼ)、異性化酵素(イソメラーゼ)、合成酵素(リガーゼ/シンセターゼ)等が含まれる。好適には、セルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素やクロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられ、より好適には、セルラーゼが挙げられる。
【実施例】
【0031】
以下に、本発明の組換え微生物の構築方法及び当該組換え微生物を用いたタンパク質の製造方法について具体的に説明する。
【0032】
以下の実施例におけるDNA断片増幅のためのポリメラーゼ連鎖反応(PCR)には、GeneAmp PCR System(アプライドバイオシステムズ)を使用し、Pyrobest DNA Polymerase(タカラバイオ)と付属の試薬類を用いてDNA増幅を行った。PCRの反応液組成は、適宜希釈した鋳型DNAを1μL、センスプライマー及びアンチセンスプライマーを各々20pmol、及びPyrobest DNA Polymeraseを2.5U添加して、反応液総量を50μLとした。PCR反応は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30回繰り返した後、72℃で5分間反応させる条件で行った。
【0033】
また、以下の実施例において、遺伝子の上流・下流とは、複製開始点からの位置ではなく、上流とは各操作・工程において対象となる遺伝子の開始コドンの5’側に続く領域のことを示し、一方、下流とは各操作・工程において対象となる遺伝子の終始コドンの3’側に続く領域のことを示す。
【0034】
更に、以下の実施例における各遺伝子及び遺伝子領域の名称は、Nature,390,249−256(1997)で報告され、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2004年3月10日更新)された枯草菌ゲノムデータの記載に基づくものである。
【0035】
枯草菌の形質転換はコンピテントセル法(J.Bacteriol.,93,1925(1967))にて行った。すなわち、枯草菌をSPI培地(0.20%(w/v)硫酸アンモニウム、1.40%(w/v)リン酸水素二カリウム、0.60%(w/v)リン酸二水素カリウム、0.10%(w/v)クエン酸三ナトリウム二水和物、0.50%(w/v)グルコース、0.02%(w/v)カザミノ酸(Difco)、5mM硫酸マグネシウム、0.25μM塩化マンガン、50μg/mlトリプトファン)において、37℃で、生育度(OD600)の値が1程度になるまで振盪培養し、振盪培養後、培養液の一部を9倍量のSPII培地(0.20%(w/v)硫酸アンモニウム、1.40%(w/v)リン酸水素二カリウム、0.60%(w/v)リン酸二水素カリウム、0.10%(w/v)クエン酸三ナトリウム二水和物、0.50%(w/v)グルコース、0.01%(w/v)カザミノ酸(Difco)、5mM硫酸マグネシウム、0.40μM塩化マンガン、5μg/mLトリプトファン)に接種し、更に生育度(OD600)の値が0.4程度になるまで振盪培養して枯草菌のコンピテントセルを調製し、次いで得られたコンピテントセルをSPII培地で懸濁し、当該懸濁液100μLに各種DNA断片を含む溶液(SOE−PCRの反応液等)5μLを添加し、37℃で1時間振盪培養後、適切な薬剤を含むLB寒天培地(1%(w/v)トリプトン、0.5%(w/v)酵母エキス、1%(w/v)NaCl、1.5%(w/v)寒天)に全量を塗沫し、37℃で静置培養した後、生育したコロニーを形質転換体として分離した。更に得られた形質転換体のゲノムを抽出し、これを鋳型とするPCRによって、目的とするゲノム構造の改変がなされたことを確認した。
【0036】
目的のタンパク質を発現するプラスミドの宿主微生物への導入は、プロトプラスト形質転換法(Mol.Gen.Genet.,168,111(1979))により行った。
【0037】
実施例1(ugtP欠失株の構築)
表1に示したypfP−337FとypfP+201R及びypfP+827FとypfP+529Rの各プライマーセットを用いて、配列番号1に示すugtP遺伝子の上流を含む5’末端側の504bp断片(A)、及びugtP遺伝子の下流を含む3’末端側の823bp断片(B)をそれぞれ調製した。得られた断片(A)はSphI及びSalI、(B)はBamHI及びScaI処理した。一方、プラスミドpDG780(Gene,167,335,(1995))のSalI及びSmaI制限酵素切断点よりカナマイシン耐性遺伝子領域を切り出した(C)。次に、3断片を(A)(C)(B)の順になる様に、pUC119(TAKARA)に(A)はSphI及びSalI、(C)はSalI及びBamHI、(B)はBamHI及びSmaI制限酵素切断点にそれぞれ挿入した。この結果得られた組換えプラスミドDNAを制限酵素ScaIで処理して直鎖状DNAにし、形質転換用の供与体DNAとした(図2参照)。このDNA断片を用いてコンピテント法による枯草菌168株の形質転換を行い、カナマイシン(10μg/mL)を含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってugtP遺伝子が欠失してカナマイシン耐性遺伝子に置換していることを確認した。また相同組換えに利用した領域の(A)及び(B)のDNA配列についてシーケンスを行った。(A)領域でugtP遺伝子上流−9bpのTがCに置換されていたが、ugtP遺伝子の上流metA遺伝子及び下流cspD遺伝子の発現に影響を及ぼさないことを確認した。以上の様にして、枯草菌のugtP遺伝子が欠失した菌株を構築し、ΔugtP株と命名した。
【0038】
【表1】

【0039】
実施例2(形質転換効率の算出)
プラスミドpHY300PLK(Jpn.J.Genet.,60,485(1985))及びpC194(J.Bacteriol.,150,815(1982))を用いて、実施例1にて構築したΔugtP株及び親株である168株の形質転換をプロトプラスト法により行った。pHY300PLKはStreptococcus faecalis strain DS−5由来のプラスミドpAMα1(J.Bacteriol.,117,283(1974))由来であり、pC194はStaphylococcus aureus由来のプラスミドである。pHY300PLKは大腸菌HB101株(タカラバイオ)を用いて調製し、pC194は枯草菌168株を用いて調製した。プロトプラスト法は以下の手順で行った。まず、各菌をLB液体培地(1%(w/v)トリプトン、0.5%(w/v)酵母エキス、1%(w/v)塩化ナトリウム)にて、30℃、150rpmで10時間培養した培養液を、再度LB液体培地に1%植菌し、3時間培養した。培養液を12000rpmで5分間遠心し、上清を除去した後、4mg/mL リゾチームを含むSMMP溶液(35g/L Antibiotic Medium 3、171.5g/L スクロース、3.2g/L マレイン酸2ナトリウム、4.06g/L 塩化マグネシウム6水和物)500μLに懸濁し、37℃で1時間静置した。3500rpmで10分間遠心した後、SMMP溶液400μLに懸濁し、そのうち13μLを50ngのプラスミドDNAと混合した。更にPEGを100μL添加し、ボルテックスした後、SMMPを350μL添加して転倒混和した。その後、30℃、150rpmで1時間培養した後100μLを抗生物質(pHY300PLKの場合50mg/L テトラサイクリン、pC194の場合20mg/L クロラムフェニコール)を含むDM3再生培地(81g/L コハク酸ナトリウム6水和物、5g/L 酵母エキス、5g/L カザミノ酸、10g/L 寒天、10g/L カルボキシメチルセルロース、1.5g/L リン酸2水素カリウム、3.5g/L リン酸水素2カリウム、0.5g/L グルコース、0.01g/L BSA、0.005g/L トリパンブルー、20mM 塩化マグネシウム)に塗布した。また、生菌数を測定する目的で、抗生物質を含まないDM3再生培地にも等量塗布した。30℃で3日間静置培養し、コロニー数をカウントした。形質転換効率を図2に示す。抗生物質を含むプレート上のコロニー数を、抗生物質を含まないプレート上のコロニー数で割った値を、形質転換効率とした。図2より、ΔugtP株は親株である168株よりも有意に形質転換効率が向上していることが判明した(P<0.05、t−検定)。
【技術分野】
【0001】
本発明は、プロトプラスト形質転換効率の向上方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、アルコール飲料や味噌、醤油等の食品類をはじめとし、アミノ酸、有機酸、核酸関連物質、抗生物質、糖質、脂質、タンパク質等、その種類は多岐に渡っており、またその用途についても食品、医薬や、洗剤、化粧品等の日用品、あるいは各種化成品原料に至るまで幅広い分野に広がっている。
【0003】
こうした微生物による有用物質の工業生産においては、その生産性の向上が重要な課題の一つであり、その手法として、突然変異等の遺伝学的手法による生産菌の育種が行われてきた。更に、近年のゲノム解析技術の急速な発展を受けて、対象とする微生物のゲノム情報を解読し、これらを積極的に産業に応用しようとする試みもなされている。ゲノム情報の公開されている産業的に有用な宿主微生物としては、枯草菌Bacillus subtilis Marburg No.168(非特許文献1)、大腸菌Escherichia coli K−12 MG1655(非特許文献2)、コリネバクテリウムCorynebacterium glutamicum ATCC132032などが挙げられ、これらのゲノム情報を利用し、改良を加えた菌株が開発されている。
【0004】
また、特に最近では、バイオテクノロジーの発展により、遺伝子組換え技術等を用いたより効率的な生産菌の育種が行われている。DNAなどの核酸分子を外部から枯草菌に導入して形質転換する場合、自然形質転換法(コンピテントセル法)(非特許文献3)、プロトプラスト法(非特許文献4)、エレクトロポレーション法(非特許文献5)が主に用いられる。これらの方法のうち、自然形質転換法は、コンピテント培養により調製した、DNA取り込み能を有する細胞(コンピテント細胞)を用いた形質転換である。この自然形質転換は、枯草菌マーブルグ(Marburg)系の菌株(168、166、160株)でのみ生じるが、納豆菌等では自然形質転換の頻度は極めて低く、これらは形質転換能(コンピテンシー)を有しないともいわれる。一方で、プロトプラスト法及びエレクトロポレーション法は、多くの菌種に適用可能である。ここで、プロトプラスト法とは、等張液中で細胞壁をリゾチームによって分解してプロトプラスト細胞を調製し、DNAとプロトプラスト細胞を高濃度ポリエチレングリコールにより凝集させ、更にポリエチレングリコールの濃度を急激に低下させることでDNAを取り込ませる手法である。またエレクトロポレーション法とは、細胞懸濁液に電気パルスを負荷して細胞膜に微小な孔を形成させ、そこからDNAを細胞内に導入する手法である(非特許文献6)。
【0005】
しかしながら枯草菌を含む多くの細菌では、導入された外来DNAが修飾(メチル化)されている自己のDNAと区別され、更に当該外来DNAが、そのDNA中の特定の塩基配列を認識して切断する制限酵素により分解されるため、形質転換は一般には起こりにくい。形質転換においてはこれらの制限修飾系が少ないほど好ましいと考えられ、例えば魚住らは、枯草菌マーブルグ系の株を基に、制限修飾能を欠失させた株RM125株を作製し、形質転換能を10〜100倍に向上させている(非特許文献7)。
【0006】
ugtP遺伝子は1,2−ジアシルグリセロールにグルコース残基を付加するUDP−グリコシルトランスフェラーゼをコードする遺伝子である。In vitroの実験から、本酵素は1,2−ジアシルグリセロールにグルコースを4個まで連続して付加することが可能であることがわかっているが、生体内ではジグリコシルアシルグリセロールが主要な糖脂質である(非特許文献8)。ugtP遺伝子に関しては、その欠損により細胞の膜脂質の脂肪酸組成に影響を与えることが報告されている(非特許文献9)が、ugtP遺伝子と形質転換効率の向上との関係についてはこれまで報告されていない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Nature,390(6657),p249−256(1997)
【非特許文献2】Science,277(5331),p1453−1462(1997)
【非特許文献3】J.Bacteriol.81,p741(1961)
【非特許文献4】Mol.Gen.Genet.168,p111(1979)
【非特許文献5】FEMS Microbiol.Lett.55,p135(1990)
【非特許文献6】Molecular Biological Method for Bacillus,Edited by Harwood,C.R.ら,John Wiley&Sons,p98−103
【非特許文献7】Mol.Gen.Genne.,152,p65−9(1977)
【非特許文献8】Mol.Microbiol.,1998,29(2),p.419−430
【非特許文献9】J.Bacteriol.,2008,190,7797−7807
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明は、形質転換効率の高いプロトプラスト形質転換方法に関する。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題に鑑み検討した結果、宿主微生物の細胞膜脂質の生合成に係るugtP遺伝子を欠失又は不活性化することにより、プロトプラスト形質転換効率が向上することを見出した。
【0010】
すなわち本発明は、以下に関する。
(1) ugtP遺伝子が欠失又は不活性化された宿主微生物のプロトプラストを用いることを特徴とする、形質転換方法。
(2) 前記宿主微生物がグラム陽性細菌である、前記(1)記載の形質転換方法。
(3) 前記宿主微生物がバチルス属細菌である、前記(2)記載の形質転換方法。
(4) 前記宿主微生物が枯草菌である、前記(3)記載の形質転換方法。
【発明の効果】
【0011】
本発明の形質転換方法を用いることにより、目的のDNA、例えば目的タンパク質又はポリペプチドをコードする遺伝子を宿主微生物に効率的に導入することが可能となる。その結果、例えばタンパク質又はポリペプチドを高生産する優良株の選抜が容易になり、更に当該タンパク質又はポリペプチドの生産性向上が期待できる。
【図面の簡単な説明】
【0012】
【図1】SOE−PCRによる遺伝子欠失用DNA断片の調製、及び当該DNA断片を用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
【図2】遺伝子欠失用プラスミドを用いて標的遺伝子を欠失する(薬剤耐性遺伝子と置換)方法を模式的に示した図。
【図3】形質転換効率を示すグラフ。(a)はpHY300PLK、(b)はpC194を用いて形質転換した場合の、168株(野生株)及びΔugtP株における結果である。結果はN=3の平均値を示し、エラーバーは標準偏差を示す。
【発明を実施するための形態】
【0013】
本発明の方法においては、宿主微生物のugtP遺伝子が、欠失又は不活性化されている。このugtP遺伝子は、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)において、遺伝子番号 BG11611として登録・公開されている(http://bacillus.genome.ad.jp/、2006年1月18日更新)。
【0014】
ugtP遺伝子(配列番号1)と塩基配列において70%以上、好ましくは80%以上、より好ましくは90%以上、更に好ましくは95%以上、更により好ましくは98%以上の同一性を有し、1,2−ジアシルグリセロールにグルコース残基を付加するUDP−グリコシルトランスフェラーゼ活性を有するタンパク質をコードする遺伝子も、当該ugtP遺伝子に相当する遺伝子と考えられ、本発明において欠失又は不活性化されるugtP遺伝子に含まれるものとする。
【0015】
アミノ酸配列及び塩基配列の同一性はLipman−Pearson法 (Science,227,1435,(1985))によって計算することができる。具体的には、遺伝情報処理ソフトウェアGenetyx−Win(ソフトウェア開発)のホモロジー解析(Search homology)プログラムを用いて、Unit size to compare(ktup)を2として解析を行うことにより算出される。
【0016】
また、枯草菌UgtPのホモログをKEGG Sequence Similarity DataBase(SSDB)にて検索を行った結果、Staphylococcus carnosusのSca_0619(Soc.Appl.Bacteriol.Symp.Ser.(1990)19,49S−53S)、Desulfitobacterium hafnienseのDhaf_3178、及びDSY3206(FEMS Microbiol.Rev.,(2006)30,706−733)、Desulfotomaculum reducensのDred_1966(Environ Microbiol.(2009)12,3007−3017)、Clostridium cellulolyticumのCcel_0034(FEMS Microbiol.Rev.(2005)29,741−764)、Clostridium thermocellumのCthe_2414(Microbiol Mol.Biol.Rev.,(2005)69,124−154)、Clostridium cellulovoransのClocel_3086(Annu.Rev.Microbiol.,(2004)58,521−554)、Clostridium ljungdahliiのCLJU_c02160(Bioprocess Biosyst.Eng.,(2009)32,369−380)、Brevibacillus brevisのBBR47_41080(Proc.Natl.Acad.Sci.USA.(1988)86:3589−93)、Variovorax paradoxusのVapar_4986及びVarpa_5682(Arch.Microbiol.,(2000),174,111−119)、及びDeinococcus radioduransのDR_1076(Curr.Opin.Biotechnol.(2000)11,280−285)は枯草菌UgtPのホモログと推察されることより、これらをコードする遺伝子は本発明において欠失又は不活性化されるugtP遺伝子に含まれるものとする。
【0017】
本発明で使用される宿主微生物としては、バチルス(Bacillus)属細菌や、上述のスタフィロコッカス(Staphylococcus)属細菌、ブレビバシラス(Brevibacillus)属細菌、クロストリジウム(Clostridium)属細菌等のグラム陽性細菌が挙げられ、中でもバチルス属細菌が好ましい。更に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点、またタンパク質を菌体外に分泌生産させる能力を有する点から、枯草菌(Bacillus subtilis)がより好ましい。
【0018】
上記宿主微生物からのugtP遺伝子の欠失又は不活性化は、標的遺伝子の一部若しくは全部をゲノム中から除去するか又は他の遺伝子と置き換える、当該遺伝子中に他のDNA断片を挿入する、あるいは当該遺伝子の転写・翻訳開始領域に変異を与える等の方法によって行い、好適には当該遺伝子を物理的に欠失させる。より具体的には、ugtP遺伝子を計画的に欠失又は不活性化する方法、及びランダムな遺伝子の欠失又は不活性化変異を与えた後、適当な方法によりタンパク質生産性の評価及び遺伝子解析を行って所望の変異を選択する方法が挙げられる。
【0019】
ugtP遺伝子を欠失又は不活性化するには、例えば相同組換えによる方法を用いればよい。すなわち、ugtP遺伝子の一部を含むDNA断片を適当なプラスミドベクターにクローニングして得られる環状の組換えプラスミドを親微生物細胞内に取り込ませ、ugtP遺伝子の一部領域における相同組換えによって宿主微生物ゲノム上のugtP遺伝子を分断して不活性化することが可能である。あるいは、塩基置換や塩基挿入等の変異によって不活性化したugtP遺伝子、又は図1のようにugtP遺伝子の上流、下流領域を含むがugtP遺伝子を含まない直鎖状のDNA断片等をPCR等の方法によって構築し、これを宿主微生物細胞内に取り込ませて宿主微生物ゲノムのugtP遺伝子内の変異箇所の外側の2ヶ所、又はugtP遺伝子上流側、下流側で2回交差の相同組換えを起こさせることにより、ゲノム上のugtP遺伝子を欠失あるいは他の遺伝子断片と置換させることによって不活性化させることが可能である。
【0020】
例えば、本発明の方法に使用する微生物を構築するための宿主微生物として枯草菌を用いる場合、相同組換えによりugtP遺伝子を欠失又は不活性化する方法については、既に幾つかの報告例があり(Mol.Gen.Genet.,223,268,1990等)、かかる方法を繰り返すことにより、本発明で用いられる微生物を得ることができる。
【0021】
また、ランダムな遺伝子の欠失又は不活性化については、ランダムにクローニングしたDNA断片を用いて上述の方法と同様な相同組換えを起こさせる方法や、宿主微生物にγ線等を照射する方法等が実施可能である。
【0022】
以下に、具体例として、SOE(splicing by overlap extension)−PCR法(Gene,77,61,1989)によって調製される欠失用DNA断片を用いた二重交差法による欠失方法について説明するが、本発明における遺伝子欠失又は不活性化方法は下記に限定されるものではない。
【0023】
欠失用DNA断片は、例えば、欠失対象遺伝子の上流に隣接する約1.0kb断片と、同じく下流に隣接する約1.0kb断片の間に、薬剤耐性マーカー遺伝子断片を挿入した断片である。まず、1回目のPCRによって、欠失対象遺伝子の上流断片及び下流断片、並びに薬剤耐性マーカー遺伝子断片の3断片を調製するが、この際、例えば、上流断片の下流末端に薬剤耐性マーカー遺伝子の上流側10〜30塩基対配列、逆に下流断片の上流末端には薬剤耐性マーカー遺伝子の下流側10〜30塩基対配列が付加される様にデザインしたプライマーを用いる(図1)。
【0024】
次いで、1回目に調製した3種類のPCR断片を鋳型とし、上流断片の上流側プライマーと下流断片の下流側プライマーを用いて2回目のPCRを行うことによって、上流断片の下流末端及び下流断片の上流末端に付加した薬剤耐性マーカー遺伝子配列において、薬剤耐性マーカー遺伝子断片とのアニールが生じ、PCR増幅の結果、上流側断片と下流側断片の間に、薬剤耐性マーカー遺伝子を挿入したDNA断片を得ることができる(図2)。以上のPCRは、市販のPCR用酵素キット等を用いて、成書(PCR Protocols.Current Methods and Applications,Edited by B.A.White,Humana Press pp251,1993、Gene,77,61,1989)等に示される通常の条件により行うことができる。
【0025】
かくして得られた遺伝子欠失用DNA断片を、コンピテント法あるいは下記のプロトプラスト法等の定法によって微生物細胞内に導入すると、対応する欠失対象遺伝子の上流及び下流の相同領域において、細胞内での遺伝子組換えが生じ、ugtP遺伝子が薬剤耐性遺伝子と置換された細胞、あるいはugtP遺伝子内に薬剤耐性遺伝子が挿入された細胞が生じ得る(図2)。これを薬剤耐性マーカーによる選択によって分離する。即ち、遺伝子導入した微生物を、上記薬剤を含む寒天培地上で培養し、生育するコロニーを分離した後、ゲノムを鋳型としたPCR法などによってゲノム上のugtP遺伝子が薬剤耐性遺伝子と置換されていることを確認すれば良い。
【0026】
上記により得られた、ugtP遺伝子が欠失又は不活性化された微生物は、プロトプラストとして調製することにより、外部からの核酸分子の導入を効率的に行うことができる。ここで、微生物のプロトプラストの調製は、公知の方法に従い実施でき、例えば細菌のプロトプラストを調製する場合には、まず一般的に用いられる細菌培養用培地(LB培地等)に該細菌を接種して培養し、菌体を得、次いでこの菌体を、浸透圧を調整する物質(スクロース、ソルビトール等)を含有する高張緩衝液に懸濁し、更にリゾチームなどの細胞壁分解酵素を作用させて細胞壁を除去することにより、プロトプラストを調製することができる。
【0027】
上記で得られたプロトプラストを、例えば、ポリエチレングリコール及び薬剤耐性マーカーを有する核酸分子(プラスミドDNA等)と混合することにより、プラスミドが当該プロトプラスト内に取り込まれ、宿主微生物が形質転換される。
【0028】
下記の実施例に示すように、ugtP遺伝子を欠失させた宿主微生物のプロトプラストを用いてプラスミドにより形質転換することにより、その形質転換効率は、当該ugtP遺伝子が欠失又は不活性化されていない微生物を用いた場合と比較し高い。
【0029】
上記により宿主微生物を形質転換した後、形質転換された宿主微生物のクローンを得る場合、例えば一定時間培養した後、クロラムフェニコール等の薬剤を含む再生培地で培養し、導入された核酸分子上に存在する薬剤耐性マーカーによる選抜を行えばよい(Mol.Gen.Genet.168,111(1979))。
【0030】
また、本発明のプロトプラスト形質転換に用いる核酸分子は、形質転換された微生物の用途に応じて適宜選択でき、特に限定されない。例えば、宿主微生物に導入される核酸分子は、目的タンパク質又は目的ポリペプチドをコードする遺伝子を有するプラスミドであってもよく、当該目的タンパク質又は目的ポリペプチドには、例えば洗剤、食品、繊維、飼料、化学品、医療、診断など各種産業用酵素や、生理活性ペプチドなどが挙げられる。産業用酵素としては、機能別に、酸化還元酵素(オキシドレダクターゼ)、転移酵素(トランスフェラーゼ)、加水分解酵素(ヒドロラーゼ)、脱離酵素(リアーゼ)、異性化酵素(イソメラーゼ)、合成酵素(リガーゼ/シンセターゼ)等が含まれる。好適には、セルラーゼ、α−アミラーゼ、プロテアーゼ等の加水分解酵素やクロラムフェニコールアセチルトランスフェラーゼ(CAT)等の転移酵素が挙げられ、より好適には、セルラーゼが挙げられる。
【実施例】
【0031】
以下に、本発明の組換え微生物の構築方法及び当該組換え微生物を用いたタンパク質の製造方法について具体的に説明する。
【0032】
以下の実施例におけるDNA断片増幅のためのポリメラーゼ連鎖反応(PCR)には、GeneAmp PCR System(アプライドバイオシステムズ)を使用し、Pyrobest DNA Polymerase(タカラバイオ)と付属の試薬類を用いてDNA増幅を行った。PCRの反応液組成は、適宜希釈した鋳型DNAを1μL、センスプライマー及びアンチセンスプライマーを各々20pmol、及びPyrobest DNA Polymeraseを2.5U添加して、反応液総量を50μLとした。PCR反応は、98℃で10秒間、55℃で30秒間及び72℃で1〜5分間(目的増幅産物に応じて調整。目安は1kbあたり1分間)の3段階の温度変化を30回繰り返した後、72℃で5分間反応させる条件で行った。
【0033】
また、以下の実施例において、遺伝子の上流・下流とは、複製開始点からの位置ではなく、上流とは各操作・工程において対象となる遺伝子の開始コドンの5’側に続く領域のことを示し、一方、下流とは各操作・工程において対象となる遺伝子の終始コドンの3’側に続く領域のことを示す。
【0034】
更に、以下の実施例における各遺伝子及び遺伝子領域の名称は、Nature,390,249−256(1997)で報告され、JAFAN:Japan Functional Analysis Network for Bacillus subtilis(BSORF DB)でインターネット公開(http://bacillus.genome.ad.jp/、2004年3月10日更新)された枯草菌ゲノムデータの記載に基づくものである。
【0035】
枯草菌の形質転換はコンピテントセル法(J.Bacteriol.,93,1925(1967))にて行った。すなわち、枯草菌をSPI培地(0.20%(w/v)硫酸アンモニウム、1.40%(w/v)リン酸水素二カリウム、0.60%(w/v)リン酸二水素カリウム、0.10%(w/v)クエン酸三ナトリウム二水和物、0.50%(w/v)グルコース、0.02%(w/v)カザミノ酸(Difco)、5mM硫酸マグネシウム、0.25μM塩化マンガン、50μg/mlトリプトファン)において、37℃で、生育度(OD600)の値が1程度になるまで振盪培養し、振盪培養後、培養液の一部を9倍量のSPII培地(0.20%(w/v)硫酸アンモニウム、1.40%(w/v)リン酸水素二カリウム、0.60%(w/v)リン酸二水素カリウム、0.10%(w/v)クエン酸三ナトリウム二水和物、0.50%(w/v)グルコース、0.01%(w/v)カザミノ酸(Difco)、5mM硫酸マグネシウム、0.40μM塩化マンガン、5μg/mLトリプトファン)に接種し、更に生育度(OD600)の値が0.4程度になるまで振盪培養して枯草菌のコンピテントセルを調製し、次いで得られたコンピテントセルをSPII培地で懸濁し、当該懸濁液100μLに各種DNA断片を含む溶液(SOE−PCRの反応液等)5μLを添加し、37℃で1時間振盪培養後、適切な薬剤を含むLB寒天培地(1%(w/v)トリプトン、0.5%(w/v)酵母エキス、1%(w/v)NaCl、1.5%(w/v)寒天)に全量を塗沫し、37℃で静置培養した後、生育したコロニーを形質転換体として分離した。更に得られた形質転換体のゲノムを抽出し、これを鋳型とするPCRによって、目的とするゲノム構造の改変がなされたことを確認した。
【0036】
目的のタンパク質を発現するプラスミドの宿主微生物への導入は、プロトプラスト形質転換法(Mol.Gen.Genet.,168,111(1979))により行った。
【0037】
実施例1(ugtP欠失株の構築)
表1に示したypfP−337FとypfP+201R及びypfP+827FとypfP+529Rの各プライマーセットを用いて、配列番号1に示すugtP遺伝子の上流を含む5’末端側の504bp断片(A)、及びugtP遺伝子の下流を含む3’末端側の823bp断片(B)をそれぞれ調製した。得られた断片(A)はSphI及びSalI、(B)はBamHI及びScaI処理した。一方、プラスミドpDG780(Gene,167,335,(1995))のSalI及びSmaI制限酵素切断点よりカナマイシン耐性遺伝子領域を切り出した(C)。次に、3断片を(A)(C)(B)の順になる様に、pUC119(TAKARA)に(A)はSphI及びSalI、(C)はSalI及びBamHI、(B)はBamHI及びSmaI制限酵素切断点にそれぞれ挿入した。この結果得られた組換えプラスミドDNAを制限酵素ScaIで処理して直鎖状DNAにし、形質転換用の供与体DNAとした(図2参照)。このDNA断片を用いてコンピテント法による枯草菌168株の形質転換を行い、カナマイシン(10μg/mL)を含むLB寒天培地上に生育したコロニーを形質転換体として分離した。得られた形質転換体のゲノムを抽出し、PCRによってugtP遺伝子が欠失してカナマイシン耐性遺伝子に置換していることを確認した。また相同組換えに利用した領域の(A)及び(B)のDNA配列についてシーケンスを行った。(A)領域でugtP遺伝子上流−9bpのTがCに置換されていたが、ugtP遺伝子の上流metA遺伝子及び下流cspD遺伝子の発現に影響を及ぼさないことを確認した。以上の様にして、枯草菌のugtP遺伝子が欠失した菌株を構築し、ΔugtP株と命名した。
【0038】
【表1】
【0039】
実施例2(形質転換効率の算出)
プラスミドpHY300PLK(Jpn.J.Genet.,60,485(1985))及びpC194(J.Bacteriol.,150,815(1982))を用いて、実施例1にて構築したΔugtP株及び親株である168株の形質転換をプロトプラスト法により行った。pHY300PLKはStreptococcus faecalis strain DS−5由来のプラスミドpAMα1(J.Bacteriol.,117,283(1974))由来であり、pC194はStaphylococcus aureus由来のプラスミドである。pHY300PLKは大腸菌HB101株(タカラバイオ)を用いて調製し、pC194は枯草菌168株を用いて調製した。プロトプラスト法は以下の手順で行った。まず、各菌をLB液体培地(1%(w/v)トリプトン、0.5%(w/v)酵母エキス、1%(w/v)塩化ナトリウム)にて、30℃、150rpmで10時間培養した培養液を、再度LB液体培地に1%植菌し、3時間培養した。培養液を12000rpmで5分間遠心し、上清を除去した後、4mg/mL リゾチームを含むSMMP溶液(35g/L Antibiotic Medium 3、171.5g/L スクロース、3.2g/L マレイン酸2ナトリウム、4.06g/L 塩化マグネシウム6水和物)500μLに懸濁し、37℃で1時間静置した。3500rpmで10分間遠心した後、SMMP溶液400μLに懸濁し、そのうち13μLを50ngのプラスミドDNAと混合した。更にPEGを100μL添加し、ボルテックスした後、SMMPを350μL添加して転倒混和した。その後、30℃、150rpmで1時間培養した後100μLを抗生物質(pHY300PLKの場合50mg/L テトラサイクリン、pC194の場合20mg/L クロラムフェニコール)を含むDM3再生培地(81g/L コハク酸ナトリウム6水和物、5g/L 酵母エキス、5g/L カザミノ酸、10g/L 寒天、10g/L カルボキシメチルセルロース、1.5g/L リン酸2水素カリウム、3.5g/L リン酸水素2カリウム、0.5g/L グルコース、0.01g/L BSA、0.005g/L トリパンブルー、20mM 塩化マグネシウム)に塗布した。また、生菌数を測定する目的で、抗生物質を含まないDM3再生培地にも等量塗布した。30℃で3日間静置培養し、コロニー数をカウントした。形質転換効率を図2に示す。抗生物質を含むプレート上のコロニー数を、抗生物質を含まないプレート上のコロニー数で割った値を、形質転換効率とした。図2より、ΔugtP株は親株である168株よりも有意に形質転換効率が向上していることが判明した(P<0.05、t−検定)。
【特許請求の範囲】
【請求項1】
ugtP遺伝子が欠失又は不活性化された宿主微生物のプロトプラストを用いることを特徴とする、形質転換方法。
【請求項2】
前記宿主微生物がグラム陽性細菌である、請求項1記載の形質転換方法。
【請求項3】
前記宿主微生物がバチルス属細菌である、請求項2記載の形質転換方法。
【請求項4】
前記宿主微生物が枯草菌である、請求項3記載の形質転換方法。
【請求項1】
ugtP遺伝子が欠失又は不活性化された宿主微生物のプロトプラストを用いることを特徴とする、形質転換方法。
【請求項2】
前記宿主微生物がグラム陽性細菌である、請求項1記載の形質転換方法。
【請求項3】
前記宿主微生物がバチルス属細菌である、請求項2記載の形質転換方法。
【請求項4】
前記宿主微生物が枯草菌である、請求項3記載の形質転換方法。
【図1】


【図2】
【図3】

【図2】
【図3】
【公開番号】特開2012−254041(P2012−254041A)
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願番号】特願2011−129180(P2011−129180)
【出願日】平成23年6月9日(2011.6.9)
【出願人】(000000918)花王株式会社 (8,290)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
【公開日】平成24年12月27日(2012.12.27)
【国際特許分類】
【出願日】平成23年6月9日(2011.6.9)
【出願人】(000000918)花王株式会社 (8,290)
【出願人】(504180239)国立大学法人信州大学 (759)
【Fターム(参考)】
[ Back to top ]