
生体内でヒト肝細胞を増大させる方法
フマリルアセト酢酸ヒドラーゼ(Fah)がさらに欠損している免疫不全マウスを用いて、生体内でヒト肝細胞を増大させる方法を、本明細書に記載する。該方法は、該免疫不全およびFah欠損マウスにヒト肝細胞を移植するステップと、該肝細胞を増大させるステップと、増大させた該ヒト肝細胞を採取するステップと、を含む。該方法はまた、二次、または三次、または四次、または追加次のマウスへの、該ヒト肝細胞の連続移植も可能にする。また、Fah、Rag2、およびIl2rg遺伝子において、ホモ接合体欠失または点変異を含む、変異体マウスも提供する。

【発明の詳細な説明】
【技術分野】
【0001】
関連特許の相互参照
本願は、2007年6月5日に出願された米国暫定特許出願第60/933,432号の利益を主張するものであり、参照することによりその全体として本明細書に組み込まれる。
【0002】
政府支援の声明
本発明は、米国立衛生研究所の国立糖尿病・消化器病・腎臓病研究所からの授与番号DK051592に基づいて、米国政府の支援でもってなされた。米国政府は、本発明における一定の権利を有する。
【0003】
本開示は、ヒト肝細胞を増大させる方法、具体的には、免疫不全マウスを利用して、ヒト肝細胞を増大させる方法を目的とする。
【背景技術】
【0004】
肝臓は、医薬を含む生体異物化合物を代謝する主要な部位である。多くの肝酵素が種特異的であるため、培養された一次ヒト肝細胞またはそれらのミクロソーム分画を用いて、候補医薬の代謝を評価することが必要である(非特許文献1、非特許文献2)。ミクロソーム肝細胞分画を用いて、一部の代謝機能を解明することができる一方、他の試験は、生きている肝細胞に依存する。例えば、一部の化合物は、肝酵素を誘導し、ひいては、それらの代謝は、経時的に変化する。酵素誘導を分析するために、肝細胞は、生存能力があるだけでなく、十分に分化し、かつ機能的でなければならない。
【0005】
薬物代謝および他の研究に関しては、肝細胞は、一般に、死体臓器ドナーから単離し、試験が行われる場所に輸送される。死体源からの肝細胞の状態(生存能力および分化の状態)は、極めて変わりやすく、多くの細胞調製物の品質は、あまり良くない。高品質のヒト肝細胞の利用可能性は、組織培養において有意に増大することができないという事実によりさらに妨げられる(非特許文献3、非特許文献4)。播種した後、該細胞は、生存するが、分裂しない。マウス等のすぐに利用できる哺乳類種からの肝細胞は、代謝酵素の異なる相補体を有し、誘導研究において異なって反応するため、薬物試験には適さない。不死化ヒト肝細胞(肝細胞腫)または胎児肝芽細胞も、十分に分化した成熟細胞の適切な代用物ではない。ヒト肝細胞はまた、微生物学の分野における研究に必要である。肝炎を生じるウイルス等の多くのヒトウイルスは、任意の他の細胞型で複製することができない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Brandon et al.Toxicol.Appl.Pharmacol.189:233−246,2003
【非特許文献2】Gomez−Lechon et al.Curr.Drug Metab.4:292−312,2003
【非特許文献3】Runge et al.Biochem.Biophys.Res.Commun.274:1−3,2000
【非特許文献4】Cascio S.M.,Artif.Organs 25:529−538,2001
【発明の概要】
【発明が解決しようとする課題】
【0007】
これらの限界を考えると、一次ヒト肝細胞を増大させる方法が、非常に望ましい。よって、ヒト肝細胞を増大させるための堅牢なシステムを本明細書に提供する。
【課題を解決するための手段】
【0008】
生体内でヒト肝細胞を増大させる方法を本明細書に提供する。該方法は、免疫不全マウスに単離したヒト肝細胞を注入するステップと、該肝細胞を増大させるステップと、該ヒト肝細胞を採取するステップと、を含む。
【0009】
いくつかの実施形態において、ヒト肝細胞は、免疫不全受容マウスに投与され、該マウスは、フマリルアセト酢酸ヒドラーゼ(Fah)がさらに欠損している。一部の実施形態において、該マウスは、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである。他の実施形態において、該マウスは、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである。該方法の一部の実施形態において、ヒトウロキナーゼをコード化するベクターが、ヒト肝細胞の注入前、マウスに投与される。他の実施形態において、マクロファージは、肝細胞注入前、マウスから減損させる。
【0010】
一部の実施例において、該ヒト肝細胞は、FRGマウスまたはFpmRGマウスのような受容マウスにおいて、少なくとも約2週間、増大する。その増大させたヒト肝細胞は、該受容マウスから採取される。一実施形態において、その単離されたヒト肝細胞は、該受容マウスの脾臓に注入され、その増大させたヒト肝細胞は、該マウスの肝臓から採取される。その採取された肝細胞は、さらなる増大のために、FRGマウスまたはFpmRGマウスのような別の受容マウスに導入することができる。よって、該方法は、ヒト肝細胞の追加の増大のために繰り返して使用することができる。
【0011】
本明細書に提供される方法の一実施形態において、Fah欠損マウスに、肝細胞注入前、肝臓疾患の発症を阻害、または予防、または遅延する薬剤が投与される。このような一薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。NTBC、または他の適した薬剤は、飲料水内投与、食物内投与、または注入が挙げられるが、これらに限定されない、任意の適した手段により投与することができる。NTBCは、任意に、肝細胞注入後、少なくとも約3日間から少なくとも約6日間投与される。
【0012】
いくつかの実施形態において、ヒト肝細胞は、臓器ドナーから、または外科的切除術の肝臓から得られる。一部の実施形態において、該ヒト肝細胞は、幹細胞に由来する。追加の実施形態において、ヒト肝細胞は、注入前、凍結保存される。さらなる実施形態において、該ヒト肝細胞は、肝細胞の細胞株からのものである。
【0013】
また、遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、該欠失または点変異がホモ抱合体であり、該マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞を、該マウスに増大させることができる、遺伝子組み換えマウスを、本明細書に提供する。一実施形態において、該欠失は、B細胞、T細胞、およびNK細胞の完全喪失をもたらす。別の実施形態において、該マウスは、ヒトウロキナーゼを発現する。
【0014】
前述ならびに他の特性および利点は、いくつかの以下の実施形態の発明を実施するための形態からさらに明らかになり、添付の図を参照して進める。
【図面の簡単な説明】
【0015】
【図1a】ヒト肝細胞により生着および再増殖した後、三重変異体FRGマウス(#471)および2匹のヘテロ接合同腹子(#469、#470)の相対的重量を示すグラフ。該FRGマウスは、移植後6週間、その体重を維持したが、Il2rg遺伝子のヘテロ接合同腹子は、継続的に減量した。NTBCは、第1週目および第4週目にのみ投与され、陰影で示す。
【図1b】肝細胞受容肝臓からのゲノムDNAにおけるヒトAlu配列のPCR増幅産物を示すゲルのデジタル画像。FRGマウスのみが陽性であった。
【図1c】野生型(図1c)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1d】Fah(−/−)(図1d)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1e】ヒト化マウス肝臓(図1e)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1f】80%を超える肝細胞がFAHに対して陽性であることを示す(暗染および大きな矢印により示される)、再増殖したマウス肝臓におけるFAH免疫染色のデジタル画像。小さい矢印は、FAH陰性細胞を画定する。
【図1g】ヒト肝細胞は、好酸性が低いことを示す(矢印により示される)、同一の肝切片のH&E染色のデジタル画像。原拡大率×200
【図2a】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。FAH陽性ヒト肝細胞は、マウス肝臓組織に統合したが、受容者の肝臓微細構造を阻害しなかったことを示すデジタル画像。
【図2b】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。高度に再増殖したキメラ肝臓はまた、正常構造を持つことを示すデジタル画像。
【図2c】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。ヒト肝細胞クラスターは、好酸性が低いことを示すH&E染色のデジタル画像。
【図2d】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。ヒト肝細胞クラスターは、好酸性が低いことを示すH&E染色のデジタル画像。
【図2e】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。FAHの染色した連続切片を示すデジタル画像。
【図2f】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。HepParの染色した連続切片を示すデジタル画像。
【図2g】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。移植してから4ヶ月後でさえ、管状または糸球体の破壊がないことを示す、高度に再増殖したマウスからの腎臓切片のデジタル画像。
【図2h】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。脾臓におけるFAH陽性ヒト肝細胞を示すデジタル画像。原拡大率×100(図2c)、×200(図1a、2b、2e、2f、および2g)、×400(図2dおよび2h)。
【図3a】キメラマウス肝臓からのRT−PCR産物を示す、一連のゲル切片。ヒトALB、FAH、TAT、TF、TTR、およびUGT1A1遺伝子を、キメラマウス肝臓(#697および#785)において発現させた。ヒト肝細胞およびマウス肝細胞はそれぞれ、陽性および陰性対照として用いた。
【図3b】図3b(正常プロット)は、ELISAを用いて、一次肝細胞受容者のヒトアルブミン血中濃度を示すグラフである。システムの閾値濃度は、約0.005μg/mlである。
【図3c】図3c(対数プロット)は、ELISAを用いて、一次肝細胞受容者のヒトアルブミン血中濃度を示すグラフである。システムの閾値濃度は、約0.005μg/mlである。
【図3d】図3d(正常プロット)は、二次受容者のヒトアルブミン濃度を示すグラフである。対数プロットは、アルブミン濃度の倍加時間が約1週間であることを示す。
【図3e】図3e(対数プロット)は、二次受容者のヒトアルブミン濃度を示すグラフである。対数プロットは、アルブミン濃度の倍加時間が約1週間であることを示す。
【図4a】一次細胞(極左の暗ボックス)を始めとして連続移植スキームを示す概略図。暗ボックスは、再増殖した連続受容者を示し、白ボックスは、非生着マウスを示す。一次受容者の1/4のみが、再増殖されたが、6匹すべての二次受容者が、生着された。
【図4b】連続的に移植された受容者の肝臓からのAlu配列のPCR増幅産物を示すゲルのデジタル画像。
【図4c】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4d】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4e】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4f】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。一次(図4f)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図4g】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。二次(図4g)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図4h】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。三次(図4h)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図5a】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5b】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5c】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5d】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5e】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5f】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5g】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。HLA−A、B、C(図5g)に対する対照ヒト肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5h】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。H−2Kb(図5h)に対する対照ヒト肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5i】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。HLA−A、B、C(図5i)に対する対照マウス肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5j】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。H−2Kb(図5j)に対する対照マウス肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5k】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。高キメラマウス(図5k)からの肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5l】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。高キメラマウス(図5l)からの肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図6a】エトキシレソルフィン−O−デエチラーゼ(CYP1A1依存)の代謝(図6a)を示すグラフ。ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスから培養された肝細胞を分析した。
【図6b】6−β−ヒドロキシルテストステロン(CYP3A4媒介)へのテストステロンの転換(図6b)を示すグラフ。ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスから培養された肝細胞を分析した。
【図6c】定量的RT−PCRにより判定される、薬物代謝、薬物移送、および薬物抱合に関する、ヒト特異遺伝子のmRNAレベルを示すグラフ。ヒト薬物代謝遺伝子の比率は、成熟ヒト肝細胞に特有である。
【図7a】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。マウスアクチンmRNAに対して正規化した、3つの試料における肝臓特異的遺伝子の基底発現の棒グラフ。
【図7b】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CYP3A4(図7b)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7c】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CYP2B6(図7c)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7d】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CAR(核ホルモン受容体)(図7d)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7e】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。MDR1(輸送体)(図7e)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7f】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。MRP(図7f)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7g】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。BSEP(輸送体)(図7g)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7h】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。PXR(核ホルモン受容体)(図7h)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【発明を実施するための形態】
【0016】
I. 略語
AAV アデノ随伴ウイルス
ALB アルブミン
ALT アラニン・アミノトランスフェラーゼ
AST アスパラギン酸アミノトランスフェラーゼ
BNF ベータナフトフラボン
DAB ジアミノベンジジン
ELISA 酵素免疫測定法
EROD エトキシレソルフィン−O−デエチラーゼ
FACS 蛍光活性化細胞分類
FAH フマリルアセト酢酸ヒドラーゼ
FISH 蛍光原位置ハイブリダイゼーション
FITC フルオレセインイソチオシアネート
FRG Fah−/−/Rag2−/−/Il2rg−/−三重変異体マウス
HLA ヒト白血球抗原
IL−2Rγ インターロイキン2受容体γ
MHC 主要組織適合性複合体
NTBC 2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン
PB フェノバルビタール
PBS リン酸緩衝生理食塩水
PCR ポリメラーゼ連鎖反応
PE フィコエリトリン
PFU プラーク形成単位
RAG リコンビナーゼ活性化遺伝子
Rif リファンピシン
RT−PCR 逆転写ポリメラーゼ連鎖反応
TAT チロシンアミノ転移酵素
TF トランスフェリン
TTR トランスサイレチン
UGT1A1 UDPグルクロノシルトランスフェラーゼ1ファミリー、ポリペプチドA1
uPA ウロキナーゼプラミノーゲン活性化因子
II. 用語
別途記載のない限り、技術用語は、従来の用法に準拠して使用される。分子生物学における一般用語の定義は、Benjamin Lewin,Genes V,Oxford University Pressより出版、1994(ISBN0−19−854287−9)、Kendrew et al.(eds.),The Encyclopedia of Molecular Biology,published by Blackwell Science Ltd.,1994(ISBN0−632−02182−9)、およびRobert A.Meyers(ed.),Molecular Biology and Biotechnology:a Comprehensive Desk Reference,VCH Publishers,Inc.より出版、1995(ISBN1−56081−569−8)において見出すことができる。
【0017】
本発明の種々の実施形態の概説を容易にするために、具体的な用語の以下の説明を提供する。
【0018】
肝臓疾患の発症を阻害するまたは予防する薬剤:FRGマウス, またはFpmRGマウス、または他の型のFah欠損マウスに投与される場合、マウスにおける肝臓疾患の発症を予防、または遅延、または阻害する化合物または組成物。肝臓疾患または肝機能障害は、肝組織像の変化(壊死、炎症、異形成、または肝臓癌等)、肝臓特異的酵素および他のタンパク質(アスパラギン酸アミノトランスフェラーゼ、アラニン・アミノトランスフェラーゼ、ビリルビン、アルカリフォスファターゼ、およびアルブミン等)のレベルの変化、または全身性肝不全が挙げられるが、これらに限定されない、兆候または症状のうちのいずれか1つにより特徴付けられる。一実施形態において、肝臓疾患を阻害する薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。
【0019】
羊膜細胞:胎児を取り囲む羊水中に見られる細胞。
【0020】
抗体:免疫グロブリン遺伝子または免疫グロブリン遺伝子のフラグメントにより実質的にコード化された1つもしくは複数のポリペプチドを含むタンパク質(またはタンパク質複合体)。認識された免疫グロブリン遺伝子は、カッパ、ラムダ、アルファ、ガンマ、デルタ、イプシロンおよびミュー定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖はカッパまたはラムダのいずれかに分類される。重鎖はガンマ、またはミュー、またはアルファ、またはデルタ、またはイプシロンに分類され、同様に、各々IgG、IgM、IgA、IgD、およびIgEの免疫グロブリンを表す。
【0021】
基本的な免疫グロブリン(抗体)の構造単位は、通常、テトラマーである。各々のテトラマーは、2つの同一のペアのポリペプチド鎖から成り、各々のペアは、1つの「軽い」鎖(約25kD)と1つの「重い」鎖(約50〜70kD)を有する。各々の鎖のN末端は、抗原認識に主に関与する約100〜110個またはそれより多いアミノ酸の可変領域を定義する。「可変軽鎖」(VL)および「可変重鎖」(VH)という用語はそれぞれ、これらの軽鎖および重鎖を指す。
【0022】
本明細書に使用される、「抗体」という用語は、完全な免疫グロブリン、ならびによく特徴付けられたフラグメントをいくつか含む。例えば、標的タンパク質(またはタンパク質または融合タンパク質内のエピトープ)に結合するFabs、Fvs、および一本鎖Fvs(SCFvs)は、そのタンパク質(またはエピトープ)のための特異的結合剤でもある。これらの抗体フラグメントは、以下のように定義される。(1)Fab、完全な軽鎖および1つの重鎖の一部分を産生するために、酵素パパインによる全抗体の消化により生成された抗体分子の一価抗原結合フラグメントを含むフラグメント、(2)Fab’、完全な軽鎖および1つの重鎖の一部分を産生するために、ペプチンで全抗体を処理し、次いで、還元することにより得られる、抗体分子のフラグメント、(3)(Fab’)2、後続の還元を伴わずに酵素ペプチンで全抗体を処理することにより得られる、抗体のフラグメント、(4)F(ab’)2、2個のジスルフィド結合により2つのFab’フラグメントが一緒にされた二量体、(5)Fv、2本の鎖として発現された軽鎖の可変域および重鎖の可変域を含む遺伝子組み換えフラグメント、および(6)1本鎖抗体、遺伝子融合1本鎖分子として適切なポリペプチドリンカーにより連結された、軽鎖の可変域、重鎖の可変域を含む遺伝子組み換え分子, として定義される。これらのフラグメントを作製する方法は、日常的である(例えば、Harlow and Lane,Using Antibodies:A Laboratory Manual,CSHL,New York,1999を参照)。
【0023】
本開示の方法に用いられる抗体は、モノクローナルまたはポリクローナルであり得る。単に一例として、モノクローナル担体は、Kohler and Milsteinの古典的方法(Nature 256:495−97,1975)またはその誘導的な方法に従って、ネズミハイブリドーマから調製することができる。モノクローナル抗体生成のための詳細な手順は、Harlow and Lane,Using Antibodies:A Laboratory Manual,CSHL,New York,1999に記載されている。
【0024】
B細胞:体液性免疫反応に大きな役割を果たすリンパ球の一型。B細胞の主要機能は、可溶性抗原に対して抗体を作製することである。B細胞は、適応免疫系の不可欠な要素である。
【0025】
採取:本明細書に使用される増大されたヒト肝細胞を「採取する(collecting)」とは、単離されたヒト肝細胞を注入されているマウス(または受容マウスとも称される)から増大された肝細胞を除去するプロセスをさす。採取は、任意に、肝細胞を他の細胞型から分離することを含む。一実施形態において、その増大されたヒト肝細胞は、Fah欠損マウスの肝臓から採取される。一部の例において、その増大されたヒト肝細胞は、FRGマウスまたはFpmRGマウスの肝臓から採取される。
【0026】
インターロイキン受容体の一般的なγ鎖(Il2rg):インターロイキン受容体の一般的なガンマ鎖をコード化する遺伝子。Il2rgは、IL−2、IL−4、IL−7、およびIL−15を含む、いくつかのインターロイキンに対する受容体の構成要素である(Di Santo et al.Proc.Natl.Acad.Sci.U.S.A.92:377−381,1995)。Il2rgの欠損した動物は、B細胞およびT細胞の減少を示し、ナチュラルキラー細胞が欠如する。
【0027】
凍結保存:本明細書に使用される「凍結保存」とは、77Kまたは−196℃(液体窒素の沸点)等の低い氷点下の温度まで冷却することにより保存または維持されている細胞または組織を指す。これらの低温度で、細胞死をもたらす生化学的反応を含む、いかなる生物学的活性も、効果的に停止する。
【0028】
肝機能の低下:肝臓の健康または機能を測定する多くのパラメータのうちのいずれか1つの異常変化。肝機能の低下はまた、本明細書では、「肝機能障害」と称される。肝機能は、肝組織像の検査および肝臓酵素または他のタンパク質の測定等に限定されない、当技術分野で周知のような多くの手段のうちのいずれか1つにより評価することができる。例えば、肝機能障害は、肝臓の壊死、または炎症、または酸化的損傷、または異形成で示され得る。一部の例において、肝機能障害は、肝細胞癌等の肝臓癌で示される。肝機能障害を評価するために試験され得る肝臓酵素およびタンパク質の例としては、アラニン・アミノトランスフェラーゼ(ALT)、アスパラギン酸アミノトランスフェラーゼ(AST)、ビリルビン、アルカリフォスファターゼ、およびアルブミンが挙げられるが、これらに限定されない。肝機能障害はまた、全身性肝不全をもたらし得る。肝機能を試験するための手順は、Grompeら(Genes Dev.7:2298−2307,1993)およびManningら(Proc.Natl.Acad.Sci.U.S.A.96:11928−11933,1999)により教示されるもの等、当技術分野で周知であり、それらは参照することにより本明細書に組み込まれる。
【0029】
欠損:本明細書に使用される「Fah欠損」または「Fahの欠損」とは、Fahの突然変異を含む、マウス等の動物を指し、それは、Fah mRNA発現および/または機能的FAHタンパク質の実質的減少、またはその欠損をもたらす。一実施形態において、該Fah欠損動物は、Fah遺伝子のホモ接合体欠失を含む。一例として、該ホモ接合体欠失は、Fahのエクソン5にある。別の実施形態において、該Fah欠損動物は、Fah遺伝子の1つもしくは複数の点変異を含む。適切なFah点変異の例は、当技術分野で知られている(例えば、Aponte et al.Proc.Natl.Acad.Sci.U.S.A.98(2):641−645,2001参照、参照することにより本明細書に組み込まれる)。
【0030】
減損する:減じるまたは除去すること。本明細書に使用される「マクロファージ減損」とは、動物におけるマクロファージを消去する、または除去する、または減じる、または殺滅するプロセスを指す。マクロファージが減損している動物は、必ずしもマクロファージが完全に欠如しているとは限らないが、少なくともマクロファージの数または活性の減少を示す。一実施形態において、マクロファージ減損は、機能的マクロファージの少なくとも10%、少なくとも25%、少なくとも50%、少なくとも75%、少なくとも90%、または100%の減少をもたらす。
【0031】
生着する:動物に細胞または組織を移植すること。本明細書に使用される、受容マウスにおいて、ヒト肝細胞の生着とは、注入後、ヒト肝細胞が受容マウスに移植されるプロセスを指す。生着されたヒト肝細胞は、受容マウスにおいて、増大することができる。本明細書に記載される、「有意な生着」とは、肝臓において、少なくとも約1%の肝細胞がヒト肝細胞である、受容マウスを指す。「高度に生着された」マウスは、肝細胞の少なくとも約30%がヒト肝細胞である、肝臓を有するマウスである。しかしながら、生着効率はさらに高くなり得、マウス肝臓において、例えば、肝細胞の少なくとも約40%、または少なくとも約50%、または少なくとも約60%、または少なくとも約70%、または少なくとも約80%、または少なくとも約90%、または少なくとも約95%がヒト肝細胞である。
【0032】
胚幹(ES)細胞:発達する胚盤胞の内細胞塊から単離された多能性細胞。ES細胞は、多能性細胞であり、それらが、体内(骨、筋肉、脳細胞等)に存在するすべての細胞を作製することができることを意味する。ネズミES細胞を産生するための方法は、米国特許第5,670,372号に見出され得、参照することにより本明細書に組み込まれる。ヒトES細胞を産生するための方法は、米国特許第6,090,622号、PCT公開第WO 00/70021号、およびPCT公開第WO 00/27995号に見出され得、これらのそれぞれが、参照することにより本明細書に組み込まれる。
【0033】
増大させる:量を増加させること。本明細書に使用される、ヒト肝細胞を「増大させる(expanding)」とは、ヒト肝細胞の数が増加するような、細胞分裂を生じることが可能なプロセスを指す。本明細書に記載される、ヒト肝細胞は、受容マウスにおいて、少なくとも約4週間、または少なくとも約6週間、または少なくとも約8週間、または少なくとも約12週間、または少なくとも約16週間、または少なくとも約20週間、または少なくとも約24週間、または少なくとも約28週間、増大可能である。一実施形態において、該ヒト肝細胞は、最長約6ヶ月まで増大可能である。増大から結果として生じるヒト肝細胞の数は、変化し得る。一実施形態において、増大は、少なくとも1,000万、または少なくとも2,000万、または少なくとも3,000万、または少なくとも4,000万、または少なくとも5,000万の肝細胞をもたらす。100万の肝細胞が最初に注入され、約10%生着されると仮定すると、肝細胞の増大は、約10倍〜約500倍の範囲であり得る。一部の実施形態において、受容マウスにおいて、ヒト肝細胞の増大は、少なくとも10倍、または少なくとも50倍、または少なくとも100倍、または少なくとも150倍、または少なくとも200倍、または少なくとも250倍、または少なくとも300倍、または少なくとも400倍、または少なくとも500倍、または少なくとも1,000倍の増加をもたらす。
【0034】
FRGマウス:フマリルアセト酢酸ヒドラーゼ(Fah)、リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)遺伝子において、ホモ接合体欠失を有する、変異体マウス。また、本明細書では、Fah−/−/Rag2−/−/Il2rg−/−と称される。本明細書に使用される、Fah、Rag2、およびIl2rg遺伝子においてホモ接合体欠失は、機能的FAH、RAG−2、およびIL−2Rγタンパク質が、突然変異を含むマウスに発現しないことを示す。
【0035】
FpmRGマウス:リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)遺伝子のホモ接合体欠失、ならびにフマリルアセト酢酸ヒドラーゼ(Fah)のホモ接合体点変異を有する、変異体マウス。FpmRGマウスのFah遺伝子の点変異は、mRNAにおいて、エクソン7のミススプライシングおよび喪失をもたらす(Aponte et al.,Proc.Natl.Acad.Sci.USA 98:641−645,2001)。また、本明細書では、Fahpm/Rag2−/−/Il2rg−/−と称される。本明細書に使用される、Rag2およびIl2rg遺伝子のホモ接合体欠失は、機能的RAG−2およびIL−2Rγタンパク質が、該突然変異を含むマウスに発現しないことを示す。さらに、Fah遺伝子のホモ接合体の点変異を有するマウスは、機能的FAHタンパク質を発現しない。
【0036】
フマリルアセト酢酸ヒドラーゼ(FAH):チロシン異化の最終ステップに触媒作用を及ぼす代謝酵素。Fah遺伝子のホモ接合体欠失を有するマウスは、肝mRNA発現の変化および重度の肝機能障害を示す(Grompe et al. Genes Dev.7:2298−2307,1993、参照することにより本明細書に組み込まれる)。該Fah遺伝子の点変異はまた、肝不全および出生後の致死を生じることも示している(Aponte et al.Proc.Natl.Acad.Sci.U.S.A.98(2):641−645,2001、参照することにより本明細書に組み込まれる)。
【0037】
漸減させる:本明細書に使用される、NTBCの用量を「漸減する」とは、数日にわたるような時間とともに、Fah欠損マウスに投与されるNTBCの用量を漸減するプロセスを指す。一実施形態において、該NTBC用量は、約6日間にわたり、漸減し、該用量は、約1週間後、NTBCがもはや投与されなくなるように、約1または2日間間隔で減少する。NTBCの漸減は、短期間または長期間にわたり行うことができ、用量の減少の間の時間間隔もまた、短くする、または長くすることができる。
【0038】
肝細胞:肝臓の細胞質塊のうちの70〜80%まで作製する細胞型。肝細胞は、タンパク質合成、タンパク質保存、および炭水化物の形質転換、コレステロール、胆汁塩、およびホスホリン脂質の合成、ならびに外因性および内因性物質の解毒、修飾、および排出に関与する。該肝細胞はまた、胆汁の形成および分泌も引き起こす。肝細胞は、血清アルブミン、フィブリノゲン、および凝固因子のプロトロンビン基を製造し、リポタンパク質、セルロプラスミン、トランスフェリン、相補体、および糖タンパク質の合成のための主部位である。さらに、肝細胞は、薬物および殺虫剤のような外因性化合物、およびステロイドのような内因性化合物を代謝する、解毒する、および不活性化する能力を有する。
【0039】
ホモ接合:1つもしくは複数の遺伝子座で同一の対立遺伝子を有すること。本明細書に使用される、「ホモ接合体欠失」とは、1つの遺伝子の両方の対立遺伝子が同一の欠失を有する生物を指す。
【0040】
免疫不全:免疫系のうちの少なくとも1つの不可欠な機能を欠如すること。本明細書に使用される、「免疫不全」マウスとは、免疫系の特異成分を欠如する、または免疫系の特異成分の機能を欠如するマウスである。一実施形態において、免疫不全マウスは、機能的B細胞、T細胞、および/またはNK細胞を欠如する。別の実施形態において、免疫不全マウスは、さらに、マクロファージを欠如する。
【0041】
単離された:「単離された」肝細胞とは、臓器ドナー等の特定の源から得られている、および他の細胞型から実質的に分離または精製された、肝細胞を指す。
【0042】
マクロファージ:単球と呼ばれる特定の白血球から由来する組織内の細胞。単球およびマクロファージは、食細胞であり、脊椎動物の非特異的防御(または先天性免疫)、ならびに特異的防御(または細胞性免疫)の両方において作用する。これらの役割は、細胞残屑、および固定細胞あるいは移動細胞としての病原体を食菌する(飲み込んだ後、消化する)こと、および病原体に対して反応するために、リンパ球および他の免疫細胞を刺激することである。
【0043】
ナチュラルキラー(NK)細胞:内因性免疫系の主成分を構成する細胞傷害性リンパ球型。NK細胞は、腫瘍およびウイルス感染した細胞の両方の宿主の拒絶反応において大きな役割を果たす。
【0044】
受容:本明細書に使用される、「受容マウス」とは、本明細書に記載される単離されたヒト肝細胞を注入されているマウスである。一般に、ヒト肝細胞の一部(割合は異なり得る)は、受容マウスに生着する。一実施形態において、該受容マウスは、Fahがさらに欠損している免疫不全マウスである。別の実施形態において、該受容マウスは、Fahがさらに欠損しているRag2−/−/Il2rg−/−マウスである。別の実施形態において、該受容マウスはFRGマウスである。別の実施形態において、該受容マウスは、FpmRGマウスである。
【0045】
リコンビナーゼ活性化遺伝子2(Rag2):免疫グロブリン遺伝子座とT細胞受容体遺伝子座の組み換えに関与する遺伝子。Rag2遺伝子が欠損する動物は、V(D)J組み換えを行うことができず、機能的T細胞およびB細胞の完全喪失をもたらす(Shinkai et al.Cell 68:855−867,1992)。
【0046】
連続移植:一次マウスにおいて増大された肝細胞が、採取され、さらなる増大のために二次マウスに注入等により移植される、生体内でヒト肝細胞を増大させるためのプロセス。連続移植は、三次、または四次、または追加次のマウスをさらに含むことができる。
【0047】
幹細胞:不変の娘細胞を生成する(自己再増殖;親細胞と同一である少なくとも1つの娘細胞を生成する細胞分裂)、および特殊化細胞型を形成する(潜在力)ための特異な能力を有する細胞。幹細胞としては、胚幹(ES)細胞、胚性生殖(EG)細胞、生殖系列幹(GS)細胞、ヒト間充織幹細胞(hMSC)、脂肪組織由来幹細胞(ADSC)、多能性成体幹細胞(MAPC)、多能性成体生殖系列幹細胞(maGSC)、および非制限体幹細胞(USSC)が挙げられるが、これらに限定されない。生体内の幹細胞の役割は、動物の正常な生活中、破壊される細胞を置換することである。通常、幹細胞は、無制限に分裂することができる。分裂後、該幹細胞は、幹細胞として残る、または前駆細胞になる、または末端分化に移り得る。前駆細胞は、少なくとも1つの所与の細胞型の十分に分化した機能細胞を作製することができる細胞である。通常、前駆細胞は、分裂することができる。分裂後、前駆細胞は、前駆細胞のままであり得る、または末端分化に移り得る。一実施形態において、該幹細胞は、肝細胞を形成する。
【0048】
T細胞:細胞性免疫において中心的役割を果たす白血球型、またはリンパ球型。T細胞は、T細胞受容体(TCR)と呼ばれるB細胞およびNK細胞の細胞表面上にある特別受容体の存在により、B細胞およびNK細胞のようなリンパ球の他型と区別される。胸腺は、通常、T細胞発達のための主要臓器であると考えられる。
【0049】
導入遺伝子:細胞または生命体のゲノムに導入される外因性核酸配列。
【0050】
トランスジェニック動物:動物の細胞の一部において染色体外要素として存在するか、または動物の生殖細胞系DNAに(すなわち、その細胞のほとんどまたはすべてのゲノム配列に)安定的に組み込まれている、非内因性(異種)核酸配列を有する、通常、哺乳類である、ヒト以外の動物。異種核酸は、当技術分野で周知のような方法に従って、例えば、宿主動物の胚または胚幹細胞の遺伝子操作により、このようなトランスジェニック動物の生殖細胞系に導入される。「導入遺伝子」は、このような異種核酸、例えば、(例えば「ノックイン」トランスジェニック動物の作製用の)発現コンストラクトの形状をした異種核酸、または標的遺伝子内またはその近傍に挿入されて、(例えば「ノックアウト」トランスジェニック動物の作製用の)標的遺伝子の発現低下を生じる異種核酸を意味する。遺伝子の「ノックアウト」とは、標的遺伝子の機能低下を生じる遺伝子の配列の変化を意味し、好ましくは、このような標的遺伝子の発現は検出されないか、または有意ではない。トランスジェニックのノックアウト動物は、標的遺伝子についてヘテロ接合のノックアウト、または標的遺伝子についてホモ接合のノックアウトを含むことができる。「ノックアウト」はまた、条件的なノックアウトも含み、標的遺伝子の変化が、例えば、標的遺伝子の変化を促進する物質に動物を曝露した際、標的遺伝子部位における組換えを促進する酵素(例えばCre−lox系のCre)を導入した際、または標的遺伝子の変化を出生後に指令する他の方法で生じる場合がある。
【0051】
ベクター:宿主細胞において複製する、および/または挿入するための、ベクターの能力を崩壊することなく、外部の核酸の挿入を可能にする核酸分子。ベクターは、複製の複製起点のような、宿主細胞において、核酸配列の複製を可能にする核酸配列を含むことができる。ベクターはまた、1つもしくは複数の選択可能なマーカー遺伝子および他の遺伝要素を含むこともできる。挿入ベクターは、それ自体、宿主核酸に挿入することができる。発現ベクターは、一つまたは複数の挿入遺伝子の転写および翻訳を可能にするのに必要な調節塩基配列を含むベクターである。本明細書に記載される一実施形態において、該ベクターは、ヒトウロキナーゼのような、ウロキナーゼをコード化する配列を含有する。一実施形態において、該ベクターは、プラスミドベクターである。別の実施形態において、該ベクターは、アデノウイルスベクターまたはアデノ随伴ウイルス(AAV)ベクターのような、ウイルスベクターである。
【0052】
ウロキナーゼ:また、ウロキナーゼ型プラミノーゲン活性化因子(uPA)と呼ばれる、ウロキナーゼは、セリンプロテアーゼである。ウロキナーゼは、本来、人尿から分離されたが、血流および細胞外マトリックスのような、いくつかの生理学的位置に存在する。主要な生理学的基質は、プラスミノーゲンであり、これはセリンプロテアーゼプラスミンの不活性な酵素前駆体である。プラスミンの活性化は、生理学的環境によって、血栓溶解または細胞外マトリックスの分解に関与するタンパク質分解カスケードを誘発する。本明細書に提供される方法の一実施形態において、ウロキナーゼは、肝細胞注入前、受容マウスに投与される。一部の実施形態において、ウロキナーゼは、ヒトウロキナーゼである。一部の実施形態において、該ヒトウロキナーゼは、分泌型のウロキナーゼである。一部の実施形態において、該ヒトウロキナーゼは、修飾された、非分泌型のウロキナーゼである(米国特許第5,980,886号参照、参照することにより本明細書に組み込まれる)。
【0053】
別途説明されない限り、本明細書に使用されるすべての技術および科学用語は、本発明が属する技術分野の当業者によって一般に理解される用語と同じ意味を持つ。文脈で特に指定しない限り、単数形、「一つの(a)」「一つの(an)」、および「その(the)」は複数形の指示物を含む。同様に、文脈で特に指定しない限り、「または(or)」という語は、「および(and)」を含むことを意図する。したがって、「AまたはBを含む(comprising A or B)」とは、AまたはB、あるいはAおよびBの両方を含むことを意味する。核酸またはポリペプチドに対して規定したすべての塩基サイズまたはアミノ酸サイズ、およびすべての分子量または分子量値は概算値であり、説明のために提供するものであることがさらに理解されるべきである。本発明の実施または検討のために本明細書に記載したものと同様または同等の方法および材料を用いることができるが、適した方法および材料は以下に説明する。本明細書で言及するすべての刊行物、特許出願、特許、および他の参考文献は、その全体を参照することにより本明細書に組み込まれる。矛盾する場合には、用語の説明を含め、本明細書が優先するものとする。さらに、材料、方法、および例は、例示的なものに過ぎず、限定を意図したものではない。
【0054】
III. いくつかの実施形態の説明
生体内でヒト肝細胞を増大させる着実な方法を本明細書に提供する。該方法は、チロシン異化酵素、フマリルアセト酢酸ヒドラーゼ(Fah)が欠損している免疫不全マウスに単離されたヒト肝細胞を移植するステップを含む。一実施形態において、該免疫不全マウスは、Rag2−/−/Il2rg−/−マウスである。一実施形態において、該Fah欠損マウスは、Fahのホモ接合体欠失を含む。別の実施形態において、該Fah欠損マウスは、タンパク質の機能、および/または作製が実質的に低下するように、Fahにおいて、1つもしくは複数の点変異を含む。本明細書に記載されるように、Fah、リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)が欠損した三重変異体マウスは、生体内でヒト肝細胞を増大させるための十分な生体内系を提供する。一部の実施形態において、該マウスは、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである。一部の実施形態において、該マウスは、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである。
【0055】
生体内でヒト肝細胞を増大させる方法を、本明細書に開示し、単離されたヒト肝細胞を注入等により、免疫不全およびFah欠損マウス(また、受容マウスと称される)に移植するステップと、少なくとも約2週間ヒト肝細胞を増大させるステップと、増大させたヒト肝細胞をマウスから採取するステップと、を含む。該肝細胞は、当技術分野で知られている任意の適した手段を用いて移植することができる。一実施形態において、該単離されたヒト肝細胞は、注入等により、該受容マウスの脾臓に移植される。別の実施形態において、該増大されたヒト肝細胞は、該受容マウスの肝臓から採取される。該ヒト肝細胞は、受容マウスにおいて、ヒト肝細胞の増大を可能にするのに十分な期間、増大可能である。増大のための正確な期間は、日常の実験を用いて実験的に判定することができる。一実施形態において、該ヒト肝細胞は、最長6ヶ月まで増大可能である。別の実施形態において、該ヒト肝細胞は、少なくとも約4週間、または少なくとも約6週間、または少なくとも約8週間、または少なくとも約12週間、または少なくとも約16週間、または少なくとも約20週間、または少なくとも約24週間、または少なくとも約28週間、増大可能である。肝細胞増大の程度は、異なり得る。一部の実施形態において、受容マウスにおいて、ヒト肝細胞の増大は、少なくとも約10倍、または少なくとも約50倍、または少なくとも約100倍、または少なくとも約150倍、または少なくとも約200倍、または少なくとも約250倍、または少なくとも約300倍、または少なくとも約400倍、または少なくとも約500倍、または少なくとも約1,000倍の増加をもたらす。
【0056】
また、生体内でヒト肝細胞を増大させる方法を提供し、受容マウスは、該ヒト肝細胞の注入前、ウロキナーゼ遺伝子をコード化するベクターが投与される。一実施形態において、該ウロキナーゼ遺伝子は、ヒトウロキナーゼである。野生型ウロキナーゼは、分泌タンパク質である。したがって、一部の実施形態において、該ヒトウロキナーゼは、分泌型のウロキナーゼである(Nagai et al.,Gene 36:183−188,1985、参照することにより本明細書に組み込まれる)。ヒトウロキナーゼ(分泌型)の配列は、当技術分野で知られており、GenBank受入番号AH007073(1993年8月3日受入)、D11143(1996年5月9日受入)、A18397(1994年7月21日受入)、BC002788(2003年8月19日受入)、X02760(1993年4月21日受入)、BT007391(2003年5月13日受入)、NM_002658(2004年10月1日受入)、X74039(1994年2月20日受入)に限定されず、参照することにより本明細書に組み込まれる。
【0057】
一部の実施形態において、該ヒトウロキナーゼは、非分泌型のウロキナーゼである。例えば、Lieberら(Proc.Natl.Acad.Sci.92:6210−6214,1995、参照することにより本明細書に組み込まれる)は、ウロキナーゼのカルボキシル末端で小胞体保留シグナルをコード化する配列を挿入することにより、またはpre−uPAシグナルペプチド(Strubin et al.,Cell 47:619−625,1986、Schutze et al.,EMBO J.13:1696−1705,1994、これらの双方は、参照することにより本明細書に組み込まれる)をアミノ末端RR保留シグナルと置換することにより作製された非分泌型のウロキナーゼ、ならびに細胞膜IIタンパク質Iip33(Strubin et al.,Cell 47:619−625,1986)からスペーサーペプチドにより引き離された膜貫通アンカーを記載する。非分泌型のウロキナーゼはまた、米国特許第5,980,886号に記載され、参照することにより本明細書に組み込まれる。
【0058】
ウロキナーゼをコード化するベクターは、マウスへの送達に適している任意の型のベクターであり得、ウロキナーゼ遺伝子を発現することができる。このようなベクターは、ウイルスベクターまたはプラスミドベクターを含む。一実施形態において、該ベクターは、アデノウイルスベクターである。別の実施形態において、該ベクターは、AAVベクターである。ウロキナーゼをコード化するベクターは、当技術分野で知られている任意の適した手段により投与され得る。一実施形態において、該ベクターは、静脈内投与される。一態様において、該ベクターは、後眼窩注射により投与される。ウロキナーゼをコード化するベクターは、該ヒト肝細胞の注入前、いつでも投与され得る。一般に、該ベクターは、ウロキナーゼが十分な時間で発現できるように、投与される。一実施形態において、該ベクターは、肝細胞注入の24〜48時間前に投与される。
【0059】
生体内でヒト肝細胞を増大させる方法を、本明細書にさらに提供し、該受容マウスは、該ヒト肝細胞の注入前、マクロファージが減損している。一実施形態において、該受容マウスに、マクロファージ減損前、ウロキナーゼをコード化するベクターが投与される。別の実施形態において、該受容マウスに、マクロファージ減損後、ウロキナーゼをコード化するベクターが投与される。別の実施形態において、該マクロファージ減損受容マウスに、ウロキナーゼをコード化するベクターが投与されない。マクロファージを、化学物質または抗体を用いるような、当技術分野で周知のようないくつかの方法のうちのいずれか1つを用いて、該受容マウスから減損することができる。例えば、マクロファージを、C12MDP、またはマクロファージ発達、機能、および/または生存力を変える抗体を含む、有害物質のようなアンタゴニストの投与により減損することができる。アンタゴニストの投与は、周知の技法により行われ、欧州特許第1552740号に記載されるようなリポソームの使用を含み、参照することにより本明細書に組み込まれる。クロドロネート含有リポソームはまた、van Rijnら(Blood 102:2522−2531,2003)により記載されるように、マクロファージを減損するために使用することができ、これは参照することにより本明細書に組み込まれる。
【0060】
本明細書に記載される方法の一実施形態において、肝細胞注入前、該Fah欠損マウスに、該マウスにおける肝臓疾患の発症を阻害、予防、または遅延する薬剤が投与される。該薬剤は、肝臓疾患を阻害するための当技術分野で知られている任意の化合物または組成物であり得る。このような一薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。NTBCは、該Fah欠損マウスにおける肝臓疾患の発症を調節するために投与される。用量、投与スケジュール、および投与方法は、該Fah欠損マウスにおける肝機能障害を予防するために必要に応じ、調整することができる。一実施形態において、該NTBCは、約0.01mg/kg/日から約0.50mg/kg/日の用量で投与される。別の実施形態において、該NTBCは、約0.05mg/kg/日、または約0.06mg/kg/日、または約0.07mg/kg/日、または約0.08mg/kg/日、または約0.09mg/kg/日、または約0.10mg/kg/日のような、約0.05mg/kg/日から約0.10mg/kg/日の用量で投与される。NTBCは、ヒト肝細胞の注入前、および/または肝細胞注入後、選択された期間投与することができる。NTBCは、肝細胞の増大の期間中、必要に応じ、中止または再投与することができる。一実施形態において、該Fah欠損マウスに、肝細胞注入前、および肝細胞注入後、少なくとも約3日間、NTBCが投与される。別の実施形態において、該Fah欠損マウスに、肝細胞注入前、および肝細胞注入後、少なくとも約6日間、NTBCが投与される。一態様において、NTBCの用量は、肝細胞注入後、6日間の過程にわたり漸減される。NTBCは、飲料水内投与、食物内投与、または注入が挙げられるが、これらに限定されない、任意の適した手段により投与することができる。一実施形態において、肝細胞注入前、飲料水内投与されるNTBCの濃度は、約1mg/L、または約2mg/L、または約3mg/L、または約4mg/L、または約5mg/L、または約6mg/L、または約7mg/L、または約8mg/Lのような、約1mg/Lから約8mg/Lである。別の実施形態において、肝細胞注入前、飲料水内投与されるNTBCの濃度は、約1.0mg/L、または約1.2mg/L、または約1.4mg/L、または約1.6mg/L、または約1.8mg/L、または約2.0mg/Lのような、約1mg/Lから約2mg/Lである。
【0061】
単離されたヒト肝細胞は、多くの異なる源のうちのいずれか1つから得ることができる。一実施形態において、該ヒト肝細胞は、臓器ドナーの肝臓から単離された。別の実施形態において、該ヒト肝細胞は、外科的切除から単離された。別の実施形態において、該ヒト肝細胞は、胚幹細胞、または間葉由来幹細胞、または脂肪組織由来幹細胞、または多能性成体幹細胞、または非制限体幹細胞のような、幹細胞に由来した。別の実施形態において、該ヒト肝細胞は、単球または羊膜細胞に由来し、ひいては、幹細胞または前駆細胞は、肝細胞を産生するために、体外で得られる。別の実施形態において、該ヒト肝細胞は、注入前、凍結保存された。
【0062】
該Fah欠損受容マウスにおけるヒト肝細胞の連続移植の方法を、本明細書にさらに提供する。該方法は、第1の受容マウスからの増大されたヒト肝細胞を採取するステップと、二次、または三次、または四次、または追加次の受容マウスにおいて、肝細胞をさらに増大させるステップを含む。ヒト肝細胞は、いくつかの技法のうちのいずれか1つを用いて、マウスから採取することができる。例えば、該肝細胞は、下述の実施例に記載されるように、マウス肝臓を潅流し、次いで、徐々に細分化することにより採取することができる。さらに、該肝細胞は、ヒト細胞またはヒト肝細胞を特異的に認識する抗体を用いるような、周知の方法を用いて、他の細胞型、組織、および/または、破片から分離することができる。このような抗体としては、抗ヒトHLA−A、B、Cのような、クラスI主要組織適合性抗原に特異的に結合する抗体が挙げられるが、これらに限定されない(Markus et al.Cell Transplantation 6:455−462,1997)。抗体結合肝細胞は、その後、パンニング(固体マトリックスに付着したモノクローナル抗体を利用する)、蛍光活性化細胞分類(FACS)、磁気ビーズ分離等により、分離することができる。肝細胞を採取する代替的な方法は、当技術分野で周知である。
【0063】
また、遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、該欠失または点変異がホモ抱合体であり、該マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞を、該マウスに増大させることができる、遺伝子組み換えマウスを、本明細書に提供する。一実施形態において、該欠失は、B細胞、T細胞、およびN細胞の完全喪失をもたらす。別の実施形態において、該マウスは、ウロキナーゼを発現する。一実施形態において、ウロキナーゼは、ヒトウロキナーゼである。一態様において、ウロキナーゼの発現は、ウロキナーゼをコード化する導入遺伝子の取り込みに起因する。別の態様において、ウロキナーゼの発現は、分泌型または非分泌型のウロキナーゼのような、ウロキナーゼをコード化するベクターの投与に起因する。ウロキナーゼをコード化するベクターは、マウスへの送達に適している任意の型のベクターであり得、ウロキナーゼ遺伝子を発現することができる。一実施形態において、該ベクターは、アデノウイルスベクターである。別の実施形態において、該ベクターは、AAVベクターである。一部の実施形態において、該マウスは、FRGマウスである。一部の実施形態において、該マウスは、FpmRGマウスである。
【0064】
IV. ヒト肝細胞の増大のための遺伝子組み換えマウス株
いくつかのグループは、齧歯動物において、一次ヒト肝細胞を生着および増大しようとした(米国特許第6,509,514号、PCT公開第WO 01/07338号、米国公開第2005−0255591号)。Dandriら(Hepatology 33:981−988,2001)が、ヒト肝細胞を用いてマウス肝臓の再増殖の成功を初めて報告した。その後、他のグループは、マウスにおいて、成功を収めたヒト肝臓細胞の生着を報告している。これらの研究のすべてにおいて、使用された動物は、アルブミンプロモータの転写制御下で、ウロキナーゼプラミノーゲン活性化因子(uPA)を発現する、トランスジェニック動物であった(Sandgren et al.Cell 66:245−256,1991)。uPAの過剰発現は、代謝破壊を引き起こし、移植されたヒト肝細胞に影響を及ぼすことなく、マウス肝細胞の細胞死をもたらし、それは導入遺伝子を発現しない。該alb−uPA導入遺伝子は、様々な免疫不全のバックグラウンドと交錯し、ヒト細胞の拒絶を阻止した(Tateno et al.Am.J.Pathol.165:901−912,2004、Katoh et al.J.Pharm.Sci.96:428−437,2007、Turrini et al.Transplant.Proc.38:1181−1184,2006)。
【0065】
最大70%の生着レベルが、これらのモデルにおいて報告されているが、該系は、広範な使用を阻止しているいくつかの主要な不利点がある。第1に、該alb−uPA導入遺伝子は、若年期に、不活性化または喪失する。この理由のため、非常に初期(14日齢)にヒト細胞を移植し、導入遺伝子がホモ接合体であるマウスを使用することが必須である。この狭い移植の時間窓が、モデルの柔軟性を著しく制限する。第2に、該導入遺伝子の自発的不活性化は、導入遺伝子陰性の健康なマウス肝細胞のプールを作成する。これらの復帰突然変異体マウス肝細胞は、再増殖中、ヒト細胞と効率的に競合する。したがって、該ヒト細胞を連続移植すると同時に二次受容者を再増殖することは不可能である。第3に、肝臓疾患は、このモデルにおいて、非常に早発型であり、ひいては、トランスジェニックマウスの生存力を低下させる。したがって、十分な数の実験動物を繁殖することが困難である。さらに、該トランスジェニックマウスには、手術中、死亡率を増加する出血傾向がある。最後に、alb−uPAトランスジェニック動物は、一旦ヒト細胞を用いる再増殖が50%を超えると、腎疾患を発症する。これは、腎臓上皮上のヒト補体によるためであると考えられる。ヒト生着の非常に高いレベルを得るために、抗相補性プロテアーゼ阻害剤で移植されたマウスを処理することが必要である(Tateno et al.Am.J.Pathol.165:901−912,2004)。これらの多くの限定のため、ヒト肝細胞を増大させるためのさらに堅牢なシステムが非常に望ましい。遺伝子欠失の独自の組み合わせを有する遺伝子組み換えマウスを用いて、生体内でヒト肝細胞を増大させるための高効率な方法を、本明細書に記載する。マウス肝臓においてヒト肝細胞の成功を収めた生着および増大は、ある程度の肝機能障害がある免疫不全マウスを必要とする。マウス肝臓は、様々な異なる型の免疫不全マウスにおいて、ヒト肝細胞を用いて再増殖し、RAG−2ノックアウトまたはSCIDマウスを含み、これらの両方は、B細胞およびT細胞を欠如する(米国特許第6,509,514号、PCT公開第WO 01/07338号、米国公開第2005−0255591号)。肝機能障害に達するために、免疫不全マウスを、ウロキナーゼプラスミノーゲン活性化因子(uPA)トランスジェニックマウスと交配させた。マウス肝臓において、uPAの発現は、マウス肝細胞の成長の不利点を生じ、移植したヒト肝細胞の増大を促進する(PCT公開第WO 01/07338号)。該uPA導入遺伝子の限界を避けるために、Fah欠損マウスは、ヒト肝細胞の増大を可能にさせる能力について分析された。FAHは、チロシン異化の最後のステップを触媒する代謝酵素である。Fah遺伝子のホモ接合体欠失を有するマウスは、肝mRNA発現の変化および重度の肝機能障害を示す(Grompe et al.Genes Dev.7:2298−2307,1993)。該Fah突然変異体が、ヌードまたはRag1−/−(Mombaerts et al.Cell 68:869−877,1992)バックグラウンドと交錯した場合、ヒト肝細胞を有するマウス肝臓の再増殖は、免疫拒絶による可能性が最も高いため、成功しなかった。NOD/SCIDマウスとのFah欠損マウスの交配(Dick et al.Stem Cell 15:199−203,1997)は、ヒト肝細胞の副次的な生着が観察されるマウスを産生したが、これらの動物は、SCIDマウスに存在する二本鎖切断DNA修復の欠陥の可能性があるため、急速な肝不全を発症した。Fah−/−/Rag2−/−/Il2rg−/−(FRG)三重変異体マウスは、T細胞、B細胞、およびNK細胞を欠如することを、本明細書に開示する。Rag2−/−/Il2rg−/−マウスは、当技術分野で知られている(Traggiai et al.Science 304:104−107,2004、Gorantla et al.J.Virol.81:2700−2712,2007)。
【0066】
以下の実施例に記載されるように、ヒト肝細胞の生着および増大は、驚くべきことに、FRGマウスにおいて、高効率である。例えば、FRGマウスに、100万の単離されたヒト肝細胞を注入できる。10%の効率を仮定すると、100,000のヒト肝細胞が、受容マウスに生着される。増大後のFRGマウスからの平均収量は、その後、約3,000万から約4,500万のヒト肝細胞であり、ヒト肝細胞の300倍から450倍の増加と同じである。FRGマウスはまた、ヒト肝細胞の連続移植にも使用することができる。連続移植は、多重マウスを含み得、マウスあたりヒト肝細胞の少なくとも約150倍の増大をもたらし得る。
【0067】
Fah欠損を含む任意の免疫不全マウスは、本明細書に記載される諸方法に適している。一実施形態において、該マウスは、Fahが欠損しているRag2−/−/Il2rg−/−マウスでもある。該Fah欠損マウスは、例えば、Fahのホモ接合体欠失、またはFahの1つもしくは複数の点変異を含むことができる。Fah欠損(点変異またはホモ接合体欠失等による)は、Fah mRNA発現および/または機能的FAHタンパク質の実質的減少、またはその欠損をもたらす。該FRGマウスに加えて、Fah遺伝子において、点変異がホモ抱合体である免疫不全マウス(Rag2−/−/Il2rg−/−)(本明細書では、FpmRGマウスと称される)はまた、生体内でヒト肝細胞の生着および増大に適したマウスでもあることを、本明細書に記載する。
【0068】
V. ヒト肝細胞の単離および送達
ヒト肝細胞の生体内の増大のためにFah欠損マウスを用いることの有意な利点は、様々な源から採取したヒト肝細胞を該マウスに生着する能力である。下述の実施例に記載されるように、ヒト肝細胞は、死体ドナーまたは肝臓切除術から採取し得る、または市販の源から得ることができる。さらに、本明細書に示されるように、FRGマウスに、すべての年齢のドナーからのヒト肝細胞、または凍結保存された肝細胞を、うまく移植することができる。多くの場合、ヒト肝細胞の単離と移植との間の遅延(一般に、1〜2日)があり、これは該肝細胞の生存能力の低下をもたらし得る。しかしながら、該FRGマウス系は、限定された生存能力がある肝細胞を生着した場合でさえ、ヒト肝細胞を増大させることができる。
【0069】
ヒト肝細胞を単離する方法は、当技術分野で周知である。例えば、臓器ドナーまたは肝臓切除術からのヒト肝細胞を単離する諸方法が、PCT公開第WO 2004/009766号および第WO 2005/028640号、ならびに米国特許第6,995,299号および第6,509,514号に記載され、それらのすべては、参照することにより本明細書に組み込まれる。肝細胞は、経皮的に、または開腹手術を介して採取された肝生検から得ることができる。FRGマウスのような、受容動物への移植手術のためのヒト肝細胞は、当技術分野で知られている任意の従来の方法によりヒト肝臓組織から単離される。肝臓組織は、機械的に、または酵素的に分離し、単一細胞の懸濁液に供給することができる、さもなければ完全なヒト肝組織のフラグメントを使用してもよい。例えば、該肝細胞は、所定のコラゲナーゼ潅流によりドナー組織から摘出し(Ryan et al.Meth.Cell Biol.13:29,1976)、次いで、低速遠心分離を行う。肝細胞は、その後、ステンレス製のメッシュで濾過することにより精製し、次いで、密度勾配遠心分離法を行うことができる。あるいは、肝細胞を濃縮するための他の諸方法、例えば、蛍光活性化細胞分類、またはパンニング、または磁気ビーズ分離、または遠心力場内の水簸、または当技術分野で周知のような任意の他の方法を使用することができる。類似の肝細胞単離法が、受容マウスの肝臓から増大されたヒト肝細胞を採取するために使用することができる。
【0070】
あるいは、ヒト肝細胞は、Guguen−Guillouzoら(Cell Biol.Int.Rep.6:625−628,1982)により記載される技法を用いて調製され得、これは参照することにより組み込まれる。簡潔に述べると、肝臓またはその一部は、単離され、カニューレは、門脈または門脈管に導入される。該肝臓組織は、その後、37℃で、1分間あたり30から70ミリリットルの流量で、無カルシウム緩衝液、次いで、HEPES緩衝液中で、塩化カルシウム溶液(約0.075%塩化カルシウム等)のコラゲナーゼ(約0.025%コラゲナーゼ)を含有する酵素溶液で、該カニューレを介して潅流される。潅流された該肝臓組織は、小部分(約1立法ミリメートル等)に細分化される。該酵素消化を、上記に記載されるように、同一の緩衝液で、37℃で約10〜20分間、徐々に撹拌しながら行い、細胞懸濁液を生成する。放出した肝細胞は、該細胞懸濁液を60〜80マイクロメーターのナイロン製のメッシュで濾過することにより採取する。採取した肝細胞は、その後、pH7.0で、低温HEPES緩衝液中で洗浄し、低速遠心分離を用いて、コラゲナーゼおよび細胞残屑を除去することができる。非実質細胞は、メトリザミド勾配遠心法により除去してもよい(米国特許第6,995,299号参照)。
【0071】
ヒト肝細胞は、新鮮組織(死亡してから数時間内に得られた組織等)、または新鮮冷凍された組織(約0℃以下で、冷凍保存された新鮮組織等)から得ることができる。好ましくは、該ヒト組織は、検出可能な病原体がなく、形態学および組織学において正常であり、原則的に無病である。生着に使用される肝細胞は、最近、例えば、数時間以内に単離され得る、または細胞が適切な保存培地で保存される場合、より長期間後でも移植することができる。下述の実施例に記載されるこのような一培地は、VIASPAN(登録商標)(腹腔内臓器の保存用の汎用大動脈洗浄および保冷液、ウィスコンシン大学液、またはUWとも称される)である。肝細胞はまた、移植手術前に、凍結保存することができる。肝細胞を凍結保存する諸方法は、当技術分野で周知であり、米国特許第6,136,525号に記載され、これは参照することにより本明細書に組み込まれる。
【0072】
臓器提供者または肝臓切除術からヒト肝細胞を得るステップに加えて、生着のために使用される該細胞は、ヒト幹細胞または、受容動物への移植手術後、増大可能なヒト肝細胞に発達または分化する肝細胞前駆細胞であり得る。ES細胞の性質を有するヒト細胞は、内胚盤胞細胞塊(Thomson et al.,Science 282:1145−1147,1998)、および成長中の生殖細胞(Shamblott et al.,Proc.Natl.Acad.Sci.USA 95:13726−13731,1998)から単離され、ヒト胚幹細胞を産生している(米国特許第6,200,806号参照、参照することにより本明細書に組み込まれる)。米国特許第6,200,806号に開示されるように、ES細胞は、ヒトおよび非ヒト霊長類から産生することができる。通常、霊長類ES細胞は、ES細胞培地の存在下で、マウス胎児線維芽細胞の融合層上に単離される。ES培地は、通常、20%ウシ胎仔血清(FBS;Hyclone)、0.1mM β−メルカプトエタノール(Sigma)、1%非必須アミノ酸液(Gibco BRL)を有する、80%ダルベッコ改変イーグル培地(DMEM;ピルビン酸塩なし、高グルコース、Gibco BRL)からなる。ES細胞の際立った特徴は、成人に存在する前駆「多分化能性」幹細胞と比較して、培養中に無制限に分化されない状態を維持するES細胞の能力、およびES細胞があらゆる異なる細胞型に発達する必要がある潜在能力を含む。ヒトES(hES)は、特異的なモノクローナル抗体により認識される糖脂質細胞表面抗原であるSSEA−4、を発現する(例えば、Amit et al.,Devel.Biol.227:271−278,2000を参照)。
【0073】
ヒト間葉幹細胞(hMSC)に由来したヒト肝細胞もまた、本明細書に記載される諸方法に使用することができる。骨髄由来hMSCの肝性因子への順次暴露は、肝細胞の性質を有する細胞への、幹細胞の分化をもたらす(Snykers et al.BMC Dev Biol.7:24,2007、Aurich et al.Gut.56(3):405−15,2007、これらのそれぞれは、参照することにより本明細書に組み込まれる)。骨髄由来間充織幹細胞と脂肪組織由来幹細胞(ADSC)の肝細胞の分化もまた、記載されている(Talens−Visconti et al.World J Gastroenterol.12(36):5834−45,2006参照、参照することにより本明細書に組み込まれる)。ヒト肝細胞はまた、単球から作製することができる。Ruhnkeら(Transplantation 79(9):1097−103,2005、参照することにより本明細書に組み込まれる)は、最終分化した末梢血単球からの肝細胞様(NeoHep)細胞の生成を記載する。該NeoHep細胞は、形態、肝細胞マーカーの発現、様々な分泌および代謝機能、ならびに薬物解毒活性に関して主要なヒト肝細胞と類似している。さらに、羊膜細胞に由来したヒト肝細胞もまた、本明細書に記載される諸方法に使用することができる。
【0074】
ヒトES細胞株は、存在し、本明細書に開示される諸方法に使用することができる。ヒトES細胞はまた、体外受精した(IVF)胚からの着床前の胚から採取し得る。未使用のヒトIVFから産生した胚は、該胚が14日齢未満である場合、シンガポールおよび英国のような多くの国で許可される。高品質の胚のみが、ES単離に適している。増大された胚盤胞への1つの細胞ヒト胚を培養するための現在の明確な培地条件が、記載されている(Bongso et al.,Hum Reprod.4:706−713,1989参照)。ヒト卵管細胞とのヒト胚の共培養は、高品質の胚盤胞の産生をもたらす。細胞共培養、または改善された合成培地において成長したIVF由来の増大されたヒト胚盤胞は、ヒトES細胞の単離を可能にする(米国特許第6,200,806号参照)。
【0075】
一実施形態において、ヒト肝細胞は、脾臓への注入等の、移植手術により受容マウスに送達される。肝細胞は、肝臓柔組織または門脈への注入等の、他の手段により送達され得る。受容マウスに注入されたヒト肝細胞の数は、異なり得る。一実施形態において、約105〜約107のヒト肝細胞が、注入される。別の実施形態において、約5×105〜約5×106のヒト肝細胞が、注入される。例示的な一実施形態において、約106のヒト肝細胞が、注入される。
【0076】
VI. Fah欠損マウスにおいて増大されたヒト肝細胞の使用
ヒト肝細胞は、当技術分野で知られているいくつかの技法のうちのいずれかを用いて、受容マウスから採取することができる。例えば、マウスは、麻酔することができ、門脈または下大静脈に、カテーテルを用いてカニューレを挿入した。該肝臓は、その後、適切な緩衝液(0.5mM EGTAおよび10mM HEPESが追加された無カルシウムおよび無マグネシウムEBSS)で潅流し、次いで、該肝臓にコラゲナーゼ処理(例えば、0.1mg/ml コラゲナーゼXIおよび0.05mg/ml DNase Iが追加されたEBSSからなる溶液を用いて)を行うことができる。該肝臓は、徐々に細分化し、ナイロン製のメッシュで濾過(70μmと40μmのナイロン製のメッシュでの連続等)し、次いで、遠心分離し、該細胞の洗浄を行った。受容マウスから採取したヒト肝細胞は、当技術分野で周知の任意の技法を用いて、非ヒト細胞または他の混入物質(組織または細胞残屑等)から分離することができる。例えば、このような方法は、ヒト肝細胞に選択的に結合する抗体を用いるステップを含む。このような抗体としては、抗ヒトHLA−A、B、Cのような、クラスI主要組織適合性抗原に特異的に結合する抗体が挙げられるが、これらに限定されない(Markus et al.Cell Transplantation 6:455−462,1997)。ヒト細胞またはヒト肝細胞に対して特異的な抗体は、FACS、またはパンニング、または磁気ビーズ分離を含む、様々な異なる技法に使用することができる。FACSは、他のさらに高度な検出レベルのうちで、複数の色チャネル、ローアングルかつ鈍角の光散乱検出チャネル、およびインピーダンスチャネルを利用して、抗体が結合した細胞を分離または分類する(米国特許第5,061,620号参照)。磁気分離は、1)ヒト特異抗体と接合する、または2)ヒト特異抗体と結合することができる検出抗体と接合する、または3)ビオチン化抗体と結合することができるアビジンと接合する、常磁性粒子の使用を含む。パンニングは、アガロースビーズ、またはポリスチレンビーズ、または中空糸膜、またはプラスチック製のペトリ皿のような、固体マトリックスに結合したモノクローナル抗体を含む。該抗体で結合した細胞は、試料から固体支持体を単に物理的に分離することにより試料から単離させることができる。
【0077】
下述の実施例に記載されるように、いくつかの肝細胞輸送タンパク質を含む、薬物抱合および解毒に関与する遺伝子の発現を、受容マウスから採取した増大されたヒト肝細胞において検出した。最近の研究では、薬物毒性を予測する際に、これらの抱合経路(Kostrubsky et al.Drug.Metab.Dispos.28:1192−1197,2000)、および肝細胞輸送タンパク質(Kostrubsky et al.Toxicol.Sci.90:451−459,2006)が、重要な役割を果たすことを示している。リファンピシンもしくはPB、またはBNFのような外因性薬物によるCYP誘導に対する正常ヒト反応に加えて、核ホルモン受容体転写因子の発現、抱合経路、およびFRGマウスにおいて増大されたヒト肝細胞による主要な輸送タンパク質は、生体内で、ヒト薬物代謝および毒性において、これらの遺伝子産物の役割の評価を可能にする。単離された肝細胞において、化合物の毒性を試験する諸方法は、当技術分野で周知であり、例えば、PCT公開第WO 2007/022419号に記載され、これは参照することにより本明細書に組み込まれる。
【0078】
本開示は、肝臓再構成の治療を必要とする対象体において、肝臓再構成のためにヒト肝細胞の源として、受容マウスにおいて増大され、採取したヒト肝細胞の使用をさらに考慮する。肝細胞の導入による患者における肝臓組織の再構成は、肝臓移植の想定における応急処置として、あるいは単離された代謝低下に罹患する患者に対する最終的治療として、急性肝不全に罹患する患者に対する潜在的な治療法の選択肢である(Bumgardner et al.Transplantation 65:53−61,1998)。肝細胞の再構成は、例えば、遺伝子治療のために遺伝子組み換え肝細胞を導入するために、または疾患、物理的損傷もしくは化学的損傷、または悪性腫瘍によって喪失した肝細胞を交換するために、使用してもよい(米国特許第6,995,299号、参照することにより本明細書に組み込まれる)。例えば、家族性高コレステロール血症を患っている患者の遺伝子治療において、導入された肝細胞の使用が、報告されている(Grossman et al.Nat.Genet.6:335,1994)。さらに、増大されたヒト肝細胞は、人工肝臓補助装置を装着するために使用することができる。
【0079】
FRGマウスにおいて増大され、採取したヒト肝細胞はまた、様々な微生物学的研究のためにも有用である。C型肝炎ウイルスおよびB型肝炎ウイルスを含む、いくつかの病原性ウイルスは、ヒト宿主、またはヒトの初代肝細胞において、唯一複製することができる。したがって、ヒトの初代肝細胞の十分な源を有することは、これらの病原体の研究にとって重要である。増大されたヒト肝細胞は、ウイルス感染および複製の研究および肝臓のウイルスの感染を調整する化合物を同定するための研究に使用することができる。肝臓のウイルスの研究のためにヒトの初代肝細胞を使用する諸方法が、欧州特許第1552740号、米国特許第6,509,514号、およびPCT公開第WO 00/17338号に記載され、これらのそれぞれは、参照することにより本明細書に組み込まれる。
【0080】
VII. ヒト肝臓疾患に対するモデル系としてのFah欠損マウスの使用
免疫不全、かつFah欠損マウスはまた、ヒト肝臓疾患に対するモデル系として使用することもできる。ヒト肝細胞を生着したFah欠損をさらに含む、Rag2−/−/Il2rg−/−マウスを含む、これらのマウス(例えば、FRGまたはFpmRGマウス等)は、例えば、毒素への暴露、感染症、遺伝病、または悪性腫瘍が挙げられるが、これらに限定されない、様々な異なる病因に起因する肝臓疾患のモデルを作製するために使用することができる。ヒト肝細胞を生着し、再構成したFah欠損マウスは、これらの疾患の理解をより深め、疾患経過を予防、または遅延、または逆行し得る薬剤を同定するために、使用することができる。例えば、該Fah欠損マウスは、遺伝子治療ベクターを試験するために使用することができる。着目遺伝子治療用ベクターは、Fah欠損マウスに投与することができ、生着されたヒト肝細胞におけるベクターの効果を評価することができる。
【0081】
同様に、ヒト肝細胞を含むFah欠損マウスは、生体内の設定において、ヒト肝細胞におけるそれらの効果に対して、毒素または医薬品のような、化合物をスクリーニングするために使用することができる。肝疾患を引き起こす、または寄与する疑いがある薬剤は、Fah欠損マウスに対する薬剤の有効量を投与する、および生着されたヒト細胞の機能における薬剤効果を評価することにより、スクリーニングすることができる。一実施例として、Fah欠損マウスは、ヒト向肝性(hepatotrophic)病原体により感染を阻害する、または予防する薬剤を同定するために使用することができる。薬剤等を同定するために、試験薬剤を投与後、または投与前に、Fah欠損マウスを病原体に暴露させる、あるいは該マウスに病原体を接種することができる。該マウスは、その後、感染の兆候に対して評価することができ、任意に、該薬剤で処理されていない、および/または、該病原体により感染させられていない、対照マウスと比較することができる。
【0082】
Fah欠損マウスが、毒素により引き起こされる肝臓疾患のモデルとして使用される場合には、注入されたヒト肝細胞を生着し、該毒剤への暴露前に、好適な期間、増大させる必要がある。肝細胞の増大のために必要とされる時間量は、経験的に判定され得、当業者の能力範囲内である。対応するヒト条件を最も厳密に模倣する成果物を産生するために必要とされる毒剤量は、毒剤の増分量に対して暴露されるいくつかのFah欠損マウスを用いることにより判定されてもよい。毒剤の例としては、アルコール、アセトアミノフェン、フェニトイン、メチルドパ、イソニアジド、四塩化炭素、黄リン、およびファロイジンが挙げられるが、これらに限定されない。
【0083】
Fah欠損マウスが悪性肝臓疾患のモデルとして使用される場合の実施形態において、該悪性腫瘍は、形質転換剤への暴露により、または、悪性細胞の導入により生成されてもよい。形質転換剤または悪性細胞は、ヒト肝細胞の初期のコロナイジング導入とともに、または好ましくは、ヒト肝細胞が宿主動物において増殖開始後、導入されてもよい。形質転換剤の場合、ヒト肝細胞が活発に増殖する時に、該薬剤を投与することが好ましい場合がある。このような形質転換剤は、動物に全身的に、あるいは、肝臓自体に局所的に、投与されてもよい。悪性細胞は、肝臓へ直接接種することができる。一実施例として、Fah欠損マウスに、ヒト肝細胞が移植される。該ヒト肝細胞の生着後、該マウスは、形質転換剤が投与される、または悪性細胞が接種される。あるいは、該形質転換剤または該悪性細胞は、ヒト肝細胞とともに投与することができる。悪性腫瘍が該マウスに発症した後、当技術分野で知られているいくつの方法のうちのいずれか1つにより投与することができ、Fah欠損マウスは、ヒト肝臓癌のモデルとして使用することができる。
【0084】
VIII. ウロキナーゼをコード化するベクター
本明細書に記載される方法の一部の実施形態において、Fah欠損マウスに、ヒト肝細胞の移植手術前、ウロキナーゼをコード化するベクターが投与される。一実施形態において、該ウロキナーゼ(ウロキナーゼプラスミノーゲン活性化因子(uPA)としても知られる)は、分泌型のヒトウロキナーゼである。別の実施形態において、該ウロキナーゼは、修飾された、非分泌型のウロキナーゼである(米国特許第5,980,886号参照、参照することにより本明細書に組み込まれる)。マウスにおけるウロキナーゼの発現に適した任意の型のベクターが企図される。このようなベクターは、プラスミドベクターまたはウイルスベクターを含む。適したベクターとしては、DNAベクター、アデノウイルスベクター、レトロウイルスベクター、偽型レトロウイルスベクター、AAVベクター、テナガザル白血病のベクター、VSV−G、VL30ベクター、リポソーム媒介ベクター等が挙げられるが、これらに限定されない。一実施形態において、該ウイルスベクターはアデノウイルスベクターである。該アデノウイルスベクターは、任意のアデノウイルス血清型を含む、任意の適したアデノウイルスに由来し得る(Ad2およびAd5等であるが、これらに限定されない)。アデノウイルスベクターは、第1、第2、第3、および/または第4世代のアデノウイルスベクター、またはアデノウイルスゲノムをほぼ完全に欠失させた(gutless)アデノウイルスベクターであり得る。非ウイルスベクターは、プラスミド、ホスホリン脂質、異なる構造のリポソーム(陽イオンおよび陰イオン)により構成することができる。別の実施形態において、該ウイルスベクターは、AAVベクターである。該AAVベクターは、当技術分野で知られている任意の適したAAVベクターであり得る。
【0085】
ウロキナーゼをコード化するウイルスおよび非ウイルスベクターは、当技術分野で周知である。例えば、ヒトウロキナーゼをコード化するアデノウイルスベクターは、米国特許第5,980,886号、およびLieberら(Proc.Natl.Acad.Sci.U.S.A.92(13):6210−4,1995)に記載される。米国特許出願公開第2005−176129号および米国特許第5,585,362号は、組み換えアデノウイルスベクターを記載し、米国特許第6,025,195号は、肝臓特的な発現のアデノウイルスベクターを開示する。米国特許出願公開第2003−0166284号は、ウロキナーゼを含む、着目遺伝子の肝臓特異的な発現のアデノ随伴ウイルス(AAV)ベクターを記載する。米国特許第6,521,225号および第5,589,377号は、組み換えAAVベクターを記載する。PCT公開第WO 0244393号は、ヒトウロキナーゼプラスミノーゲン活性化因子遺伝子を含む、ウイルスおよび非ウイルスベクターを記載する。該ヒトウロキナーゼ遺伝子の高レベルの発現が可能である発現ベクターは、PCT公開第WO 03087393号に開示される。前述の特許出願および刊行物のそれぞれは、参照することにより本明細書に組み込まれる。
【0086】
ウロキナーゼをコード化するベクターは、任意に、適切なプロモータ、エンハンサ、転写ターミネータ、タンパク質をコード化する遺伝子の前にある開始コドン(すなわち、ATG)、イントロンのスプライシングシグナル、mRNAの適切な翻訳を可能にするためのその遺伝子の正しいリーディングフレームの維持、および終止コドンを含む、発現調節配列を含むことができる。通常、発現調節配列は、転写を指示するのに十分な最小配列である、プロモータを含む。
【0087】
該発現ベクターは、複製起点、プロモータ、ならびに形質転換細胞の表現型選抜を可能にする特異的遺伝子(抗生物質耐性カセット等)を含むことができる。通常、該発現ベクターは、プロモータを含み得る。該プロモータは、誘導性または構成的であり得る。該プロモータは、組織特異的であり得る。適したプロモータとしては、チミジンキナーゼプロモータ(TK)プロモータ、またはメタロチオネインIプロモータ、またはポリヒドロンプロモータ、またはニューロン特異的エノラーゼプロモータ、またはチロシンヒドロキシラーゼプロモータ、またはベータアクチンプロモータ、または他のプロモータが挙げられる。一実施形態において、該プロモータは、異種プロモータである。
【0088】
一実施例において、ウロキナーゼをコード化する配列は、所望のプロモータの下流にある。任意に、エンハンサ要素もまた、含まれ、通常、該ベクター上のどこかにあり得、増強効果も有する。しかしながら、増大した活性量は、通常、距離とともに減少する。
【0089】
ウロキナーゼをコード化する該ベクターは、静脈内投与、または腹腔内投与、または門脈を介して血管内注入が挙げられるが、これらに限定されない、様々な経路により投与することができる。投与されたベクター量は、異なり、日常の実験を用いて決定され得る。一実施形態において、FRGマウスに、約1×108〜約1×1010のプラーク形成単位の用量で、アデノウイルスウロキナーゼをコード化するベクターが投与される。好ましい一実施形態において、該用量は、約5×109のプラーク形成単位である。
【0090】
例示的な一実施形態において、FRGマウスに、ヒトウロキナーゼをコード化するアデノウイルスベクターが投与される。アデノウイルスベクターは、非常に高い力価のある感染粒子に作製され得る、多種多様の細胞に感染する、分裂してない細胞へ遺伝子を効率的に転送する、およびゲストゲノムにほとんど組み込まれない(これは挿入突然変異により細胞形質転換のリスクを避ける)等の、他の型のウイルスベクターに優るいくつかの利点がある(Douglas and Curiel,Science and Medicine, March/April 1997,pages 44−53、Zern and Kresinam,Hepatology:25(2),484−491,1997)。ウロキナーゼをコード化するために使用することができる代表的なアデノウイルスベクターは、Stratford−Perricaudetら(J.Clin.Invest.90:626−630,1992)、Graham and Prevec(In Methods in Molecular Biology:Gene Transfer and Expression Protocols 7:109−128,1991)、およびBarrら(Gene Therapy,2:151−155,1995)に記載され、これらは参照することにより本明細書に組み込まれる。
【0091】
以下の実施例を提供し、ある特定の特性、および/または実施形態を説明する。これらの実施例は、本発明を、記載される特定の特性または実施形態に制限すると解釈されるべきではない。
【実施例】
【0092】
以下の実施例に記載されるように、ヒト肝細胞の生着および増大は、FRGマウスにおいて、高効率である。FRGマウスの肝臓は、大きさおよび形において、肉眼的に正常であり、組織学的検査は、免疫補体Fah−/−マウスとの間で有意な差異を示さなかった。さらに、FRGマウスは、NTBCを投与した場合、よく生育し、十分に繁殖力があった。
【0093】
一実施形態において、FRGマウスにおける肝臓疾患および選択圧の限度は、NTBCを投与および中止することにより制御することができる(Grompe et al.Nat.Genet.10:453−460,1995)。NTBCの中止は、移植したヒト肝細胞に対する選択的な利点を与える。肝臓疾患の程度を容易に制御することができることは、畜産および手術をさらに容易にする。さらに、該Fah欠損突然変異体は、欠失であり、導入遺伝子の不活性化により野生型に戻すことができない。したがって、内因性復帰マウス細胞からの競合は、Fahノックアウト肝臓には存在しない。これは、FRGマウスは、年齢を問わず、ヒト肝細胞を生着することができ、連続移植は、実現可能であることを意味する。また、重要なことに、FRGマウスは、ヒト細胞を用いた再増殖が50%を超える場合、腎疾患を予防するために必要とされることが前述の研究によって示されている(Tateno et al.Am.J.Pathol.165:901−912,2004)、補体阻害剤の投与なく、高度に再増殖することができる。これは、畜産において実質的に重要だけでなく、薬理学的干渉の潜在源を除去する。
【0094】
FRGマウスにおいて、ヒト肝細胞の再増殖率は、約70%を超えることができる(マウス肝臓において、70%以上の肝細胞がヒト肝細胞である)。単一の再増殖したFRGマウスからの平均収量は、約3,000万から約4,500万のヒト肝細胞である。単一FRGマウスから増大された肝細胞を、その後、連続移植と称されるプロセスにおいて、最高100匹の二次FRG受容マウスで再増殖するために使用することができる。
【0095】
連続移植は、本明細書に記載される新規のシステムにおいて達成することができる。連続移植は、ヒト肝細胞を大規模に増大できるだけでなく、受容マウスのいくつかの世代を通して同一の遺伝子型のヒト細胞を増大させるための手段を提供する。また、さらなる移植手術のために高品質の源のヒト肝細胞を所有することは、重要である。少なくとも4回の肝細胞移植手術が実現可能であることを本明細書に実証している。さらに、連続移植から単離されたヒト肝細胞の生存能力は、約80%を超えることができ、該肝細胞は、コラーゲンで被覆したプレートに容易に結合する。10%の生着効率(100,000の細胞)、および再増殖後の約1,500万の増大されたヒト肝細胞の最終採取の概算に基づいて、少なくとも150倍の生体内での増大を毎回達成することができる。したがって、FRGマウスにおいて、ヒト肝細胞の全増大は、5億倍を超え得る。
【0096】
ヒト肝細胞の生着および増大は、様々な源および可変品質からの単離されたヒト肝細胞を用いて、FRGマウスにおいて、可能である。ヒト肝細胞の源としては、死体ドナー、肝臓切除術、および商業的に入手可能な源が挙げられるが、これらに限定されない。本明細書に示されるように、ヒト肝細胞の生着および増大は、あらゆる年齢のドナーを用いて、成功した。さらに、ヒト肝細胞を、該肝細胞があらかじめ凍結保存された場合でさえ、FRGマウスにおいてうまく増大させることが、本明細書に実証される。これは、肝細胞単離後、直ちに移植手術を必要とするシステムに優る重要な利点である。
【0097】
実施例1:Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスの作製
免疫不全Fahノックアウトマウスのいくつかの株を作製した。Fah突然変異体をヌード、nod/scidまたはRag1−/−バックグラウンドと交錯することは、不成功であった。T細胞、B細胞、およびNK細胞が完全に欠如するが、DNA修復欠損のない、免疫不全Fah−/−マウス株を作製するために、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスを作製した。オスFah−/−129S4マウス(Grompe et al.Genes Dev.7:2298−2307,1993)を、メスRag2−/−/Il2rg−/−マウス(Taconic)と交配させた。すべての動物に、1.6mg/Lの濃度で、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン(NTBC)を含有する飲料水を与えてすべての動物を維持した(Grompe et al.Nat.Genet.10:453−460,1995)。各動物の遺伝子型を確認するために、PCRベースの遺伝子タイピングを、足指組織から単離した200ngのゲノムDNAに実行した(Grompe et al.Genes Dev7:2298−2307,1993、Traggiai et al.Science 304:104−107,2004)。
【0098】
飲料水内にNTBCを継続して与えた場合、FRGマウスはよく生育し、十分な繁殖力があった。FRGマウス肝臓は、大きさおよび形において、肉眼的に正常であり、組織学的検査は、従来のFah−/−マウスと該FRGマウスとの間で差異を示さなかった。従来のFah−/−マウスの場合のように、NTBCの中止は、FRGマウスにおいて、漸次の肝臓細胞損傷をもたらし、4〜8週間後、FRGマウスは最終的に死亡した(Overturf et al.Nat.Genet.12:266−273,1996)。
【0099】
実施例2:組織学および生着検出アッセイ
組織学および免疫細胞化学
FAH免疫組織化学を、前述のように行った(Wang et al.Am.J.Pathol.161:565−574,2002)。簡潔に述べると、pH7.4で、10%リン酸緩衝ホルマリン中で固定された肝臓組織および腎臓組織は、100%エタノール中で脱水され、58℃で、パラフィンワックスに包埋された。脱パラフィン処理した4μmの切片を、ヘマトキシリンおよびエオシンで染色した。免疫組織化学のために、切片は、3%H2O2を含むメタノールで、内因性ペルオキシダーゼのブロッキングのために処理した。また、アビジンおよびビオチンブロッキングは、一次抗体でインキュベーションする前に、行なった。切片は、室温で2時間、抗FAHウサギ抗体またはHepPar抗体(DAKO)でインキュベートし、次いで、HRPに接合した二次抗体でインキュベートした。シグナルは、ジアミノベンジジン(DAB)により検出した。
【0100】
FAH酵素アッセイ
フマリルアセト酢酸は、受容者の肝臓からの細胞質の肝臓分画とともにインキュベートし、消失速度を、330nmで分光学的に測定した。野生型およびFah−/−肝臓はそれぞれ、陽性対照および陰性対照として使用した。フマリルアセト酢酸は、ホモゲンチジン酸から酵素的に調製した(Knox et al.Methods Enzymol.2:287−300,1955)。
【0101】
Alu配列のためのゲノムPCR
ゲノムDNAは、DNeasy組織キット(Qiagen)を用いて、肝臓から単離した。ヒトAlu配列は、以下のプライマー5’−GGCGCGGTGGCTCACG−3’(配列番号1)および5’−TTTTTTGAGACGGAGTCTCGCTC−3’(配列番号2)を用いた標準手順に従って、PCRにより増幅した。
【0102】
肝細胞特異遺伝子発現のためのRT−PCR
総RNAは、Rneasyミニキット(Qiagen)を用いて、肝臓から単離した。相補的DNAは、オリゴdTプライマーを用いて逆転写酵素により合成した。表1に示されるプライマーは、ヒトまたはマウス特異的cDNA増幅のために使用した。
【0103】
【表1】
【0104】
ヒトアルブミンの測定
少量の血液を、ヘパリン化された毛細血管を有する左側の伏在静脈から1週間に1回採取した。トリス緩衝食塩水で1,000倍または10,000倍希釈した後、ヒトアルブミン濃度は、製造業者のプロトコルに従って、ヒトアルブミンELISA Quantitation Kit(Bethyl)を用いて測定した。
【0105】
蛍光免疫細胞化学
ヒト化マウス肝臓からの肝細胞を、ダルベッコ改変イーグル培地(DMEM)中で懸濁し、コラーゲン型1被覆6ウェルプレート上に播種した。付着した細胞を、4%パラホルムアルデヒドで、15分間固定し、5%スキムミルクでブロックした。ウサギ抗FAH、ヤギ抗ヒトアルブミン(Bethyl)、ヤギ抗マウスアルブミン(Bethyl)を、1/200の希釈で、一次抗体として使用した。ALEXA(登録商標)フルオロ488抗ヤギIgG(Invitrogen)またはALEXA(登録商標)フルオロ555抗ウサギIgG(Invitrogen)を、二次抗体として使用した。該画像は、Nikonのデジタルカメラを用いることによりAXIOVERT(登録商標)200顕微鏡で捕捉した。
【0106】
FACS解析
受容者の肝臓の解離後、実質細胞を、フルオレセインイソチオシアネート(FITC)接合した抗ヒトヒト白血球抗原(HLA)−A、B、C(BD Pharmingen)、およびフィコエリトリン(PE)を接合した抗マウスH2−K(b)(BD Pharmingen)抗体と共に、4℃で30分間インキュベートした。その後、これらをPBSで2回洗浄し、FACS CALIBUR(登録商標)(Becton Dickinson)フローサイトメーターで解析した。FITCを接合したIgGおよびPEを接合したIgGを、負の対照として使用した。
【0107】
蛍光原位置ハイブリダイゼーション
全ゲノムDNAプローブは、全マウスゲノムDNAおよび全ヒトゲノムDNAのニックトランスレーション法により作製した。Cy3−dUTPの取り込みは、製造業者の推奨(Invitrogen)に従って実行した。最終プローブ濃度は、200ng/μlであった。付着した細胞のあるスライドを、37℃で1時間、100mg/mlのRnaseで処理し、2X SSC中で3回(1回3分間)洗浄した。洗浄ステップ後、スライドを、70%、90%、および100%エタノール中で、3分間、それぞれ脱水した。染色体を、75℃で3分間、70%ホルムアミド/2X SSC中で変性し、次いで、冷70%、90%、および100%エタノールで3分間、それぞれ脱水した。プローブカクテルを、75℃で10分間変性し、37℃で30分間あらかじめハイブリダイズした。プローブを、スライドに適用し、湿潤チャンバ中で、37℃で一晩インキュベートした。ハイブリダイゼーション後の洗浄は、50%ホルムアミド/2X SSC中で3回の3分洗浄、PN緩衝液(0.1M Na2HPO4、0.1M NaH2PO4、pH8.0、2.5% NONIDET(登録商標)P−40)中で3回の3分洗浄において、すべて45℃で行われた。スライドは、その後、紫外線蛍光(Zeiss)下で、ヘキスト(0.2ug/ml)で対比染色し、カバースリップをかぶせ、観察した。
【0108】
実施例3:ヒト肝細胞の単離および凍結保存
ヒト肝細胞は、前述の手順に従って、肝臓移植手術のために使用されないドナー肝臓から単離した(Strom et al. Cell Transplant.15:S105−110,2006)。簡潔に述べると、肝臓組織を、0.5mM EGTA(Sigma)およびHEPES(Cellgro)が追加された無カルシウムおよび無マグネシウムのハンクス液(Cambrex)で潅流し、次いで、既存の脈管構造を通してイーグル最小必須培地(Cambrex)中で、100mg/LのコラゲナーゼXI(Sigma)および50mg/LのデオキシリボヌクレアーゼI(Sigma)を用いて消化した。該細胞を、イーグル最小必須培地と7%小ウシ血清(Hyclone)で、50xgで2分間、3回洗浄した。ペレット化された肝細胞は、冷VIASPAN(登録商標)(腹腔内臓器の保存用の汎用大動脈洗浄および保冷液、ウィスコンシン大学液、またはUWとも称される)に移した。
【0109】
輸送された肝細胞は、1mlあたり5×106の肝細胞で、10%ウシ胎仔血清および10%ジメチルスルホキシドが追加されたVIASPAN(登録商標)溶液に移した。該クリオチューブ(cryotube)を、紙タオルで厚く覆い、−80℃で1日間保存し、液体窒素に最終的に移した。解凍のために、細胞を、37℃の水浴中で急速に再加熱し、DMEMを徐々に加え、DMSO濃度の変化速度を最小化した。
【0110】
実施例4:ヒト肝細胞を用いたFRGマウス肝臓の再増殖
ウロキナーゼの過剰発現は、いくつかの系において、肝細胞生着を強化することが示されている(Lieber et al.Hum.Gene Ther.6:1029−1037,1995)。したがって、実験を行い、ヒト肝細胞の移植手術前、ウロキナーゼを発現するアデノウイルスの投与が有益であるかどうかを判定した。分泌型のヒトウロキナーゼ(ウロキナーゼプラスミノーゲン活性化因子;uPA)を発現するアデノウイルスベクターは、先に説明されている(Lieber et al.Proc.Natl.Acad.Sci.U.S.A.92:6210−6214,1995および米国特許第5,980,886号参照、参照することによって本明細書に組み込まれる)。
【0111】
ドナー肝細胞は、単離し、単離してから24〜36時間後に、移植した。大抵の場合、細胞は、VIASPAN(登録商標)溶液中に保存し、輸送中、4℃に保った。しかしながら、2つの実験において、凍結保存した肝細胞を移植した。ドナー肝細胞の生存能力および品質は、10%〜60%の範囲の平板効率を伴い、大いに変化した。
【0112】
移植手術のために、以下の汎用プロトコルを使用した。成熟(6〜15週齢)オスまたはメスFRGマウスに、移植手術する24〜48時間前に、uPAアデノウイルス(マウスあたり5×109のプラーク形成単位(PFU))の静脈注射(後眼窩注射)をした。100μlのダルベッコ改変イーグル培地中で、100万の生存能力のあるヒト肝細胞(トリパンブルー色素排除試験法により判定された)を、27ゲージ針で脾臓内に注入した。NTBCの投与を、次の6日間(1.6mg/L 0〜2日目、0.8mg/L 3〜4日目、0.4mg/L 5〜6日目)にわたり徐々に中止し、移植手術から1週間後、完全に中止した。NTBCの投与を停止してから2週間後、動物は、5日間薬物の投与を再開した後、再度中止した。
【0113】
3つの別個の移植手術において、ヒト肝細胞の一次生着は、最初にuPAアデノウイルスを受けた受容者であるFRGマウスにおいて観察した。したがって、uPAの前処理レジメンを、ほとんどのその後の移植手術実験において使用した。
【0114】
全体で、9つの異なるドナーからのヒト肝細胞をうまく使用して、uPAアデノウイルスレジメンの導入後、生着の失敗は生じなかった。これらのうち、7つは、脳死ドナーの肝臓から単離し、2つは、外科的肝臓切除術から単離した。ドナーの年齢は、1.2歳から64歳の範囲で異なった(表2)。
【0115】
【表2】
【0116】
すべての実験において、少なくとも1つの受容者に、細胞バッチの使用にかかわらず、本プロトコルを用いて、ヒト肝細胞が有意に生着された(>1%ヒト細胞)。生着は、組織学、DNA解析、酵素アッセイ、および後の実験である、ヒト血清アルブミンを含む、異なる方法により実証された。アルブミンレベルによりモニターされた移植手術において、43のうちの17(39.5%;12〜67%)の一次受容者は再増殖した(表2および図3)。これらのうち、7つは、高度に再増殖し(30〜90%)、アルブミンレベル>1mg/mlに達した。死体肝臓からの肝細胞だけでなく、肝臓切除術からの肝細胞も生着した。さらに、凍結保存細胞も、無事に生着された。
【0117】
高度に生着されたマウス(>30%再増殖)において、移植されたFRGマウスの体重は、第2のNTBC中止中に、安定したのに対して、同一の細胞を与えたIl2rg(Fah−/−/Rag2−/−/Il2rg+/−)のヘテロ接合型である免疫不全同腹仔のうち減量し続けたものは少なかった(図1a)。三重変異体マウスにおいて、この体重の安定は、移植されたヒト肝細胞が、罹患Fah−/−受容肝細胞の機能を置換していることを示唆した。完全に体重が安定化するや否や(初期移植手術から2〜3ヶ月後)、受容者の肝臓は、その後、採取された。肉眼的に、FRG肝臓は、大きさおよび形において、正常であり、肉眼的結節はなかった。ヒト特異的Alu DNA配列のゲノムPCRは、FRG受容者の肝臓において陽性であったのに対して、Il2rgヘテロ接合体は、すべて陰性であった(図1b)。再増殖細胞の肝細胞機能を直接確認するために、FAH酵素活性を、アッセイした(Knox et al.Methods Enzymol.2:287−300,1955)。
【0118】
受容マウスの肝臓は、正常マウス肝臓と同等またはそれを超える、多量のFAH酵素活性があった(図1c〜e)。FAHは、十分に分化した肝細胞において、単独に発現するため、移植されたヒト肝細胞は、マウス肝臓に生着された場合、脱分化または異常がないことを示唆した。FAH免疫染色により、70%以上の肝実質が、FAH陽性ヒト肝細胞によって再増殖されたことが確認された(図1fおよび図1g)。
【0119】
組織学的および免疫組織化学的試験は、追加の受容者の肝臓を用いて行った(図2aおよび図2b)。FAH陽性ヒト肝細胞は、受容者の肝臓の構造に完全に統合されたようであった。いくつかの受容者において、生着された肝細胞は、受容者の肝臓組織を阻害することなく、80%以上の柔組織を占めた(図2b、2e、および2f)。クローン的に増加しているヒト肝細胞は、形態学的に、大きさにより、およびそれらの蒼白の細胞質により、マウス肝細胞とは明らかに区別することができた(図2cおよび図2d)。ヒト肝細胞のその大きさは、恐らく、上述に報告されるようにグリコーゲン蓄積のため、比較的大きく、それらの細胞質は、鮮明であった(Meuleman et al.Hepatology 41:847−856,2005)。FAH陽性肝細胞はまた、HepPar抗体に対しても陽性であり、この抗体はヒト肝細胞を特異的に標識化するが、マウス肝細胞は、標識化しない(図2eおよび図2f)。対照的に、FAH陰性領域は、NTBC投与の中止後、従来のFah−/−マウスにおいて所見と一致する、壊死炎症(necroinflammation)を示し、異形成肝細胞を含んだ。
【0120】
再増殖したヒト肝細胞が、成熟肝細胞特異遺伝子を発現するかどうかを検査するために、RT−PCRを、受容者の肝臓から抽出されたメッセンジャーRNAに行った。ヒトアルブミン(ALB)遺伝子、FAH遺伝子、トランスフェリン(TF)遺伝子、トランスサイレチン(TTR)遺伝子、チロシンアミノ転移酵素(TAT)遺伝子、およびUGT1A1遺伝子が、受容者の肝臓において、多量に発現した(図3a、図6c、および図7)。肝細胞の機能性をまた、ヒトアルブミンの血中濃度を測定することにより評価した。ヒトアルブミンに対して特異的なELISAキットを使用し、検出の閾値は、1:100に希釈した試料を用いて、0.05μg/mlであった。ヒトアルブミンは、まず、一次受容者において、移植手術から4〜10週間後に検出された。当初は、ヒトアルブミンレベルに変動があったが、その後、ヒトアルブミンの濃度は、さらに数週間、相対的に漸増した(図3bおよび図3c)。
【0121】
薬理作用のアルプロティナーゼ阻害剤は、高度に再増殖したマウスの生存能力を長期間維持するために必要であり得、該ドナー肝細胞により産生されたヒト補体は、受容者の腎臓に損害を与える可能性がある(Tateno et al. Am.J.Pathol.165:901−912,2004参照)。したがって、いくつかの(n=3)高度に再増殖したマウスを、長期間観察した。これらのマウスは、NTBCの投与を中止した4ヶ月間減量しなくて、マウスのヒトアルブミン濃度は、安定していた。さらに、該マウスの腎臓は、採取時に、肉眼的かつ組織学的に正常であった(図2g)。
【0122】
実施例5:ヒト肝細胞の連続移植
前述の肝臓異種再増殖モデルの限界のうちの1つは、生着されたヒト肝細胞をさらに増大させる能力の低下である。該FRGマウス系において、連続移植の実行可能性を試験するために、高度に再増殖した一次受容者の肝臓(約70%ヒト細胞)を潅流し、実質肝細胞を、標準コラゲナーゼ消化プロトコルを用いて採取した。
【0123】
ヒト細胞で再増殖したマウスに麻酔をかけ、門脈または下大静脈に、24ゲージのカテーテルを用いてカニューレを挿入した。該肝臓を、0.5mM EGTAおよび10mM HEPESが追加された無カルシウムおよび無マグネシウムのEBSSで5分間潅流した。該溶液は、0.1mg/mlのコラゲナーゼXI(sigma)および0.05mg/mlのDNase I(sigma)が追加されたEBSSに10分間変更した。該肝臓は、第2の溶液中で徐々に細分化し、70μmおよび40μmのナイロン製のメッシュで連続して濾過した。5分間、150xgで遠心分離した後、該ペレットを、50xgで2分間、さらに2回洗浄した。細胞の数および生存能力は、トリパンブルー色素排除試験法により評価した。
【0124】
100μlのDMEM中に懸濁された100万の生存能力のある細胞を、27ゲージ針で受容者の脾臓に注入した。Fah陽性ヒト肝細胞とFah陰性マウス肝細胞に分離することなく、二次FRG受容者への肝細胞の移植手術を行った。一次生着のために使用した該細胞と対照的に、本方法において採取されたヒト肝細胞の生存能力は、>80%であり、それらは、コラーゲンで被覆した培養プレートに容易に付着した(図4c〜e)。二次受容者の生着後、該連続移植は、類似の方法において、三次および四次受容者に対して継続された。各世代において、すべてではないが、一部の受容マウスのヒト血清アルブミンは、高度に陽性になった(図4a)。高度に再増殖したマウスの割合は、順次移植受容者においてさらに高く(7/43と比較して17/28)、アルブミンの増加率は、さらに一貫していた(図3e)。これは、連続継代の肝細胞が、最も移植可能なヒト肝細胞の質を高めることを表す可能性がある、あるいは、ドナーマウスから新たに採取した細胞のさらなる高品質および生存能力を単に反映する可能性がある。アルブミン陽性マウスからの肝臓試料のゲノムPCRは、各世代において、ヒトDNAの存在を示した(図4b)。ヒト肝細胞による肝臓再増殖はまた、FAHに対する蛍光免疫染色により確認された(図4c〜e)。組織学的検査は、生着されたヒト肝細胞が、各世代において、形態学的に類似し、明らかにFAH陽性であることを示した(図4f〜h)。
【0125】
実施例6:肝細胞再増殖は、細胞融合の結果ではない。
【0126】
ウロキナーゼトランスジェニックマウスにおいて、一次細胞を用いた肝臓再増殖の最新報告は、細胞融合が、明らかな「肝細胞再増殖」を潜在的に明らかにすることを実証した(Okamura et al.Histochem.Cell Biol.125:247−257,2006)。uPAトランスジェニックマウスが、その研究に使用されたため、これらの所見は、細胞融合がまた、他の報告において、マウス肝臓ヒト化のメカニズムでもあることの可能性を高めた。造血細胞と肝細胞との間の細胞融合はまた、Fah欠損マウスにおいても観察されている(Wang et al.Nature 422:897−901,2003)。マウス細胞とヒト細胞との間の細胞融合は、薬学的応用のために、ヒト化マウス肝臓の価値を大幅に減退し得る。該再増殖した肝細胞が、元来は真にヒト肝細胞であったことを確認するために、ヒトまたはマウス特異的アルブミン、およびFAHに対して二重免疫染色を行った。ほとんど(>95%)のマウスアルブミン陽性肝細胞は、実際には、FAH陰性であり、ほとんどのFAH陽性肝細胞は、マウスアルブミン陰性であった(図5a〜c)。他方では、ほとんどすべて(>90%)のヒトアルブミン陽性肝細胞はまた、FAH陽性であるが、一方残りの肝細胞は、二重陰性であった(図5d〜f)。
【0127】
アルブミンは、分泌タンパク質であり、故に、細胞は、他の細胞から異種アルブミンを得ることにより抗体陽性を示すことができた。細胞融合の欠如をさらに確認するために、ヒトおよびマウス抗主要組織適合性複合体(MHC)抗体を、フローサイトメトリーのために使用した。各抗体は、種特異であることが確認された(図5g〜j)。両方の種の表面マーカーとも肝細胞陽性でないことが、高度に再増殖した肝臓に認められた(図5kおよび図5l)。
【0128】
最終的に、蛍光原位置ハイブリダイゼーション(FISH)を、高度に再増殖した移植受容者から得た肝細胞に、ヒトおよびマウス全ゲノムプローブを用いて行った。高度に再増殖した一次(キメラマウス#531)および三次(キメラマウス#631)マウスから得た肝細胞を、ヒト全ゲノムDNAあるいはマウス全ゲノムDNAを用いてハイブリダイズした。該ヒトプローブまたは該マウスプローブに対して陽性である細胞の割合を記録した(表3)。対照は、純ヒトおよびマウス肝細胞、またはヒト肝細胞とマウス肝細胞の等混合であった。キメラ肝臓において見出されたヒト細胞が、細胞融合の産物であった場合、多くの肝細胞は、ヒトプローブとマウスプローブの両方に対して二重陽性であり、故に、マウスDNAおよびヒトDNAが陽性である細胞の割合は、100%を超え得る。それどころか、割合合計は、ヒト肝細胞とマウス肝細胞の混合の場合と同じように、ほぼ100%であると概算された。さらに、ヒト肝細胞は、脾臓が、ヒト細胞の融合パートナーとしての役割をし得るマウス肝細胞を欠如するという事実にもかかわらず、高度に再増殖したマウスの脾臓に検出された(図2h)。したがって、二重陽性細胞(融合産物)は、ヒト細胞の大部分を占めることができなかった。
【0129】
【表3】
総合すれば、これらの結果は、融合事象が生じる場合、融合事象は、まれであり、再増殖する細胞の大部分は、連続移植を行う場合でさえ、純ヒト由来からなることを示す。したがって、FRGマウスにおいて増大されるヒト肝細胞は、ヒトの遺伝的および生化学的性質のみ有する。
【0130】
実施例7:ヒト化マウスにおける薬物代謝の機能的特性
キメラマウスにおける、ヒト肝臓特異的遺伝子の基底の発現および誘導を、検査した。培養された肝細胞におけるテストステロン代謝およびエトキシレソルフィン−O−デエチラーゼ(EROD)活性の評価を、それぞれ、Kostrubskyら(Drug Metab. Dispos.27:887−894,1999)、およびWenら(Drug Metab.Dispos.30:977−984,2002)に記載されるように実行した。RNA単離、cDNA合成、およびリアルタイムPCRを、Komoroskiら(Drug Metab.Dispos.32:512−518,2004)に記載されるように実行した。Applied Biosystemsから得られたプライマーは、ヒトCYP1A1(Hs00153120_m1)、CYP1A2(Hs00167927_m1)、CYP3A4(Hs00430021_m1)、CYP3A7(Hs00426361_a1)、CYP2B6(Hs00167937_g1)、CYP2D6(Hs00164385_a1)、多剤体制関連タンパク質MRP2(Hs00166123_m1)、胆汁塩輸出ポンプBSEP、(Hs00184829_m1)、CAR(Hs00231959_m1)アルブミン(Hs00609411_m1)、HNF4α(Hs00230853_m1)、シクロフィリン(Hs99999904_m1)、およびマウスアクチン(Ma00607939_s1)に対して特異的であった。
単離された肝細胞の培養物を、構築し、シトクロムP450遺伝子の原型誘導物質に暴露した。該結果は、シトクロム(CYP1A1、CYP1A2、CYP2B6、CYP3A4、CYP3A7)、輸送(BSEP、MRP2)、および薬物抱合酵素(UGT1A1)の基底遺伝子発現レベルは、培養された正常成熟ヒト肝細胞に見られる、まさにそのものであった(図6c、図7)。さらに、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)のような化合物により誘発された遺伝子型は、正常ヒト細胞から予期されたとおりであった。ヒト薬物代謝遺伝子のmRNA発現レベルに加えて、ヒトCYP1Aおよび3Aファミリーメンバーの酵素活性を測定した。エトキシレソルフィン−O−デエチラーゼ活性(EROD)は、ヒト肝臓において、CYP1A1および1A2により媒介されることが知られている。該結果は、EROD活性が、ヒト化マウス肝臓細胞において、BNFへの事前暴露により特異的かつ堅牢的に誘導されたことを示す(図6a)。逆に、PBまたはリファンピシンへの事前暴露は、CYP3A4を介した代謝の特異的測定である6−ベータ−ヒドロキシルテストステロンへのテストステロンの転換を特異的に誘導した(図6b)。したがって、再増殖したFRG肝臓からの肝細胞は、これらの標準薬物代謝アッセイにおいて、正常成熟ヒト肝細胞との区別がつかなかった。
【0131】
実施例8:肝細胞再増殖前のマクロファージの減損
一次生着は、ウロキナーゼアデノウイルスの事前投与を行っても、100%のFRG受容マウスで生じなかった。FRGマウスにおいて、正常数存在する肝臓マクロファージは、先天性免疫反応を促進することにより、ヒト細胞生着を限定することが可能である。
【0132】
潜在的なマクロファージにより惹起される免疫反応を中止するために、FRGマウスから、ヒト肝細胞の移植手術前に、マクロファージを減損させる。マクロファージ減損は、化学的減損(Schiedner et al. Mol.Ther.7:35−43,2003)、または抗体(McKenzie et al.Blood 106:1259−1261,2005)を用いることを含む、当技術分野で周知のいくつかの方法のうちのいずれか1つを用いて達成され得る。マクロファージはまた、クロドロネート含有リポソームを用いて減損され得る(van Rijn et al.Blood 102:2522−2531,2003)。マクロファージを減損するための追加の化合物および組成物は、米国特許公開第2004−0141967号およびPCT公開第WO 02/087424号に記載され、参照することにより本明細書に組み込まれる。マクロファージ減損後、FRGマウスに、本明細書の前述の実施例に記載される方法に従って、ヒト肝細胞を移植する、または連続移植する。
【0133】
実施例9:FpmRGマウスにおけるヒト肝細胞の生着
FRGマウスは、Fah遺伝子のエクソン5(FahΔexon5)に欠失を含む。ヒト肝細胞を、Fah欠損の他のモデルにおいて、生着および増大し得ることを確認するために、Fah遺伝子に点変異を含むマウス株を作製した。Fah点変異(Fahpm)マウスと呼ばれる、これらのマウスは、ミススプライシングを起させFah mRNAにおいて、エクソン7の喪失を生じさせる該Fah遺伝子に点変異を有する(Aponte et al.,Proc.Natl.Acad.Sci.USA 98:641−645,2001、参照することにより本明細書に組み込まれる)。表現型における差異は、FahpmマウスとFahΔexon5マウスとの間で検出されなかった。
【0134】
FahpmマウスをRag2−/−/Il2rg−/−マウス(実施例1に記載されるような)と交配させて、ホモ接合性Fahpm/Rag2−/−/Il2rg−/−(FpmRG)三重変異体マウスを作製した。FpmRGマウスの2つのコホートに、実施例4に記載される方法に従って、ヒト肝細胞を移植した。肝細胞移植手術の約24〜48時間前、マウスは、uPAアデノウイルスの静脈注射(後眼窩注射)を受けた。比較するために、同時に、FRGマウスに、ヒト肝細胞を移植した。ヒト血清アルブミンを、移植手術から2ヶ月および3ヶ月後に、極めて有意なレベル(23μg/ml)で、FpmRGマウスの末梢血に検出した。ヒト血清アルブミンのこれらの血中濃度は、同時に移植されたFRGマウスに見られる濃度と似ていた。
【0135】
これらの結果は、FpmRGマウスが、FRGマウスと同程度に、ヒト肝細胞で再増殖することができることを示す。したがって、ヒト化肝臓再増殖は、FahΔexon5変異を有するFRGマウスに特有ではないが、Fah欠損マウスの任意の株で達成することができる。
【0136】
本開示は、生体内でヒト肝細胞を増大させるための方法を提供する。本開示は、生体内でヒト肝細胞を増大させるのに有用な遺伝子組み換え型免疫不全マウスをさらに提供する。記載される方法の正確な詳細は、本発明の趣旨から逸脱することなく、変更または修正することができることは明らかであろう。下記の特許請求の範囲および趣旨に入る範囲内ですべてのこのような修正および変更を請求する。
【0137】
配列表
添付の配列表に記載される核酸およびアミノ酸配列は、37C.F.R.1.822に定義されるように、ヌクレオチド塩基の標準的な文字略語、およびアミノ酸の三文字コードを示す。各核酸配列の1本鎖のみを示すが、該相補鎖は、表示された該鎖に言及するときは常にそれに含まれるものとして理解する。添付の配列表において、
配列番号1および2は、ヒトAlu配列を増幅するためのPCRプライマーの核酸配列である。
【0138】
配列番号3は、ヒトALB順方向RT−PCRプライマーの核酸配列である。
【0139】
配列番号4は、ヒトALB逆方向RT−PCRプライマーの核酸配列である。
【0140】
配列番号5は、マウスAlb順方向RT−PCRプライマーの核酸配列である。
【0141】
配列番号6は、マウスAlb逆方向RT−PCRプライマーの核酸配列である。
【0142】
配列番号7は、ヒトTAT順方向RT−PCRプライマーの核酸配列である。
【0143】
配列番号8は、ヒトTAT逆方向RT−PCRプライマーの核酸配列である
配列番号9は、ヒトTF順方向RT−PCRプライマーの核酸配列である。
【0144】
配列番号10は、ヒトTF逆方向RT−PCRプライマーの核酸配列である。
【0145】
配列番号11は、ヒトFAH順方向RT−PCRプライマーの核酸配列である。
【0146】
配列番号12は、ヒトFAH逆方向RT−PCRプライマーの核酸配列である。
【0147】
配列番号13は、ヒトTTR順方向RT−PCRプライマーの核酸配列である。
【0148】
配列番号14は、ヒトTTR逆方向RT−PCRプライマーの核酸配列である。
【0149】
配列番号15は、ヒトUGT1A1順方向RT−PCRプライマーの核酸配列である。
【0150】
配列番号16は、ヒトUGT1A1逆方向RT−PCRプライマーの核酸配列である。
【技術分野】
【0001】
関連特許の相互参照
本願は、2007年6月5日に出願された米国暫定特許出願第60/933,432号の利益を主張するものであり、参照することによりその全体として本明細書に組み込まれる。
【0002】
政府支援の声明
本発明は、米国立衛生研究所の国立糖尿病・消化器病・腎臓病研究所からの授与番号DK051592に基づいて、米国政府の支援でもってなされた。米国政府は、本発明における一定の権利を有する。
【0003】
本開示は、ヒト肝細胞を増大させる方法、具体的には、免疫不全マウスを利用して、ヒト肝細胞を増大させる方法を目的とする。
【背景技術】
【0004】
肝臓は、医薬を含む生体異物化合物を代謝する主要な部位である。多くの肝酵素が種特異的であるため、培養された一次ヒト肝細胞またはそれらのミクロソーム分画を用いて、候補医薬の代謝を評価することが必要である(非特許文献1、非特許文献2)。ミクロソーム肝細胞分画を用いて、一部の代謝機能を解明することができる一方、他の試験は、生きている肝細胞に依存する。例えば、一部の化合物は、肝酵素を誘導し、ひいては、それらの代謝は、経時的に変化する。酵素誘導を分析するために、肝細胞は、生存能力があるだけでなく、十分に分化し、かつ機能的でなければならない。
【0005】
薬物代謝および他の研究に関しては、肝細胞は、一般に、死体臓器ドナーから単離し、試験が行われる場所に輸送される。死体源からの肝細胞の状態(生存能力および分化の状態)は、極めて変わりやすく、多くの細胞調製物の品質は、あまり良くない。高品質のヒト肝細胞の利用可能性は、組織培養において有意に増大することができないという事実によりさらに妨げられる(非特許文献3、非特許文献4)。播種した後、該細胞は、生存するが、分裂しない。マウス等のすぐに利用できる哺乳類種からの肝細胞は、代謝酵素の異なる相補体を有し、誘導研究において異なって反応するため、薬物試験には適さない。不死化ヒト肝細胞(肝細胞腫)または胎児肝芽細胞も、十分に分化した成熟細胞の適切な代用物ではない。ヒト肝細胞はまた、微生物学の分野における研究に必要である。肝炎を生じるウイルス等の多くのヒトウイルスは、任意の他の細胞型で複製することができない。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Brandon et al.Toxicol.Appl.Pharmacol.189:233−246,2003
【非特許文献2】Gomez−Lechon et al.Curr.Drug Metab.4:292−312,2003
【非特許文献3】Runge et al.Biochem.Biophys.Res.Commun.274:1−3,2000
【非特許文献4】Cascio S.M.,Artif.Organs 25:529−538,2001
【発明の概要】
【発明が解決しようとする課題】
【0007】
これらの限界を考えると、一次ヒト肝細胞を増大させる方法が、非常に望ましい。よって、ヒト肝細胞を増大させるための堅牢なシステムを本明細書に提供する。
【課題を解決するための手段】
【0008】
生体内でヒト肝細胞を増大させる方法を本明細書に提供する。該方法は、免疫不全マウスに単離したヒト肝細胞を注入するステップと、該肝細胞を増大させるステップと、該ヒト肝細胞を採取するステップと、を含む。
【0009】
いくつかの実施形態において、ヒト肝細胞は、免疫不全受容マウスに投与され、該マウスは、フマリルアセト酢酸ヒドラーゼ(Fah)がさらに欠損している。一部の実施形態において、該マウスは、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである。他の実施形態において、該マウスは、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである。該方法の一部の実施形態において、ヒトウロキナーゼをコード化するベクターが、ヒト肝細胞の注入前、マウスに投与される。他の実施形態において、マクロファージは、肝細胞注入前、マウスから減損させる。
【0010】
一部の実施例において、該ヒト肝細胞は、FRGマウスまたはFpmRGマウスのような受容マウスにおいて、少なくとも約2週間、増大する。その増大させたヒト肝細胞は、該受容マウスから採取される。一実施形態において、その単離されたヒト肝細胞は、該受容マウスの脾臓に注入され、その増大させたヒト肝細胞は、該マウスの肝臓から採取される。その採取された肝細胞は、さらなる増大のために、FRGマウスまたはFpmRGマウスのような別の受容マウスに導入することができる。よって、該方法は、ヒト肝細胞の追加の増大のために繰り返して使用することができる。
【0011】
本明細書に提供される方法の一実施形態において、Fah欠損マウスに、肝細胞注入前、肝臓疾患の発症を阻害、または予防、または遅延する薬剤が投与される。このような一薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。NTBC、または他の適した薬剤は、飲料水内投与、食物内投与、または注入が挙げられるが、これらに限定されない、任意の適した手段により投与することができる。NTBCは、任意に、肝細胞注入後、少なくとも約3日間から少なくとも約6日間投与される。
【0012】
いくつかの実施形態において、ヒト肝細胞は、臓器ドナーから、または外科的切除術の肝臓から得られる。一部の実施形態において、該ヒト肝細胞は、幹細胞に由来する。追加の実施形態において、ヒト肝細胞は、注入前、凍結保存される。さらなる実施形態において、該ヒト肝細胞は、肝細胞の細胞株からのものである。
【0013】
また、遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、該欠失または点変異がホモ抱合体であり、該マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞を、該マウスに増大させることができる、遺伝子組み換えマウスを、本明細書に提供する。一実施形態において、該欠失は、B細胞、T細胞、およびNK細胞の完全喪失をもたらす。別の実施形態において、該マウスは、ヒトウロキナーゼを発現する。
【0014】
前述ならびに他の特性および利点は、いくつかの以下の実施形態の発明を実施するための形態からさらに明らかになり、添付の図を参照して進める。
【図面の簡単な説明】
【0015】
【図1a】ヒト肝細胞により生着および再増殖した後、三重変異体FRGマウス(#471)および2匹のヘテロ接合同腹子(#469、#470)の相対的重量を示すグラフ。該FRGマウスは、移植後6週間、その体重を維持したが、Il2rg遺伝子のヘテロ接合同腹子は、継続的に減量した。NTBCは、第1週目および第4週目にのみ投与され、陰影で示す。
【図1b】肝細胞受容肝臓からのゲノムDNAにおけるヒトAlu配列のPCR増幅産物を示すゲルのデジタル画像。FRGマウスのみが陽性であった。
【図1c】野生型(図1c)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1d】Fah(−/−)(図1d)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1e】ヒト化マウス肝臓(図1e)のFAH酵素活性を示すグラフ。FAH基質濃度は、野生型マウス肝臓において低下したが、Fah−/−マウス肝臓では変化しなかった。ヒト化マウス肝臓は、十分な酵素活性を示した。
【図1f】80%を超える肝細胞がFAHに対して陽性であることを示す(暗染および大きな矢印により示される)、再増殖したマウス肝臓におけるFAH免疫染色のデジタル画像。小さい矢印は、FAH陰性細胞を画定する。
【図1g】ヒト肝細胞は、好酸性が低いことを示す(矢印により示される)、同一の肝切片のH&E染色のデジタル画像。原拡大率×200
【図2a】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。FAH陽性ヒト肝細胞は、マウス肝臓組織に統合したが、受容者の肝臓微細構造を阻害しなかったことを示すデジタル画像。
【図2b】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。高度に再増殖したキメラ肝臓はまた、正常構造を持つことを示すデジタル画像。
【図2c】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。ヒト肝細胞クラスターは、好酸性が低いことを示すH&E染色のデジタル画像。
【図2d】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。ヒト肝細胞クラスターは、好酸性が低いことを示すH&E染色のデジタル画像。
【図2e】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。FAHの染色した連続切片を示すデジタル画像。
【図2f】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。HepParの染色した連続切片を示すデジタル画像。
【図2g】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。移植してから4ヶ月後でさえ、管状または糸球体の破壊がないことを示す、高度に再増殖したマウスからの腎臓切片のデジタル画像。
【図2h】キメラマウスからの組織学的および免疫組織化学的組織切片のデジタル画像。脾臓におけるFAH陽性ヒト肝細胞を示すデジタル画像。原拡大率×100(図2c)、×200(図1a、2b、2e、2f、および2g)、×400(図2dおよび2h)。
【図3a】キメラマウス肝臓からのRT−PCR産物を示す、一連のゲル切片。ヒトALB、FAH、TAT、TF、TTR、およびUGT1A1遺伝子を、キメラマウス肝臓(#697および#785)において発現させた。ヒト肝細胞およびマウス肝細胞はそれぞれ、陽性および陰性対照として用いた。
【図3b】図3b(正常プロット)は、ELISAを用いて、一次肝細胞受容者のヒトアルブミン血中濃度を示すグラフである。システムの閾値濃度は、約0.005μg/mlである。
【図3c】図3c(対数プロット)は、ELISAを用いて、一次肝細胞受容者のヒトアルブミン血中濃度を示すグラフである。システムの閾値濃度は、約0.005μg/mlである。
【図3d】図3d(正常プロット)は、二次受容者のヒトアルブミン濃度を示すグラフである。対数プロットは、アルブミン濃度の倍加時間が約1週間であることを示す。
【図3e】図3e(対数プロット)は、二次受容者のヒトアルブミン濃度を示すグラフである。対数プロットは、アルブミン濃度の倍加時間が約1週間であることを示す。
【図4a】一次細胞(極左の暗ボックス)を始めとして連続移植スキームを示す概略図。暗ボックスは、再増殖した連続受容者を示し、白ボックスは、非生着マウスを示す。一次受容者の1/4のみが、再増殖されたが、6匹すべての二次受容者が、生着された。
【図4b】連続的に移植された受容者の肝臓からのAlu配列のPCR増幅産物を示すゲルのデジタル画像。
【図4c】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4d】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4e】三次マウスから培養された70%を超える肝細胞が、FAHに対して陽性であったことを実証するFAH免疫細胞化学により解析された肝細胞のデジタル画像。
【図4f】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。一次(図4f)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図4g】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。二次(図4g)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図4h】連続的に移植されたマウス肝臓のFAH免疫組織化学を示す、組織切片のデジタル画像。三次(図4h)受容者の肝臓が、ヒト肝細胞により再増殖された。
【図5a】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5b】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5c】キメラマウス肝細胞の抗マウスアルブミンおよび抗FAH免疫細胞化学のデジタル画像。キメラ肝臓からのほとんどの肝細胞は、マウスアルブミンまたはFAH単一陽性であった。
【図5d】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5e】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5f】キメラマウス肝細胞の抗ヒトアルブミンおよび抗FAH免疫細胞化学のデジタル画像。ほとんどの肝細胞は、ヒトアルブミンおよびFAH二重陽性であった。原拡大倍率×100。
【図5g】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。HLA−A、B、C(図5g)に対する対照ヒト肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5h】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。H−2Kb(図5h)に対する対照ヒト肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5i】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。HLA−A、B、C(図5i)に対する対照マウス肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5j】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。H−2Kb(図5j)に対する対照マウス肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5k】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。高キメラマウス(図5k)からの肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図5l】キメラマウス肝細胞のフローサイトメトリー解析を示すグラフ。FITC結合抗HLA A、B、CおよびPE結合抗H−2Kbを用いた。高キメラマウス(図5l)からの肝細胞を示し、それらは、HLAまたはH−2Kbのいずれか一つに対して単一に陽性であった。
【図6a】エトキシレソルフィン−O−デエチラーゼ(CYP1A1依存)の代謝(図6a)を示すグラフ。ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスから培養された肝細胞を分析した。
【図6b】6−β−ヒドロキシルテストステロン(CYP3A4媒介)へのテストステロンの転換(図6b)を示すグラフ。ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスから培養された肝細胞を分析した。
【図6c】定量的RT−PCRにより判定される、薬物代謝、薬物移送、および薬物抱合に関する、ヒト特異遺伝子のmRNAレベルを示すグラフ。ヒト薬物代謝遺伝子の比率は、成熟ヒト肝細胞に特有である。
【図7a】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。マウスアクチンmRNAに対して正規化した、3つの試料における肝臓特異的遺伝子の基底発現の棒グラフ。
【図7b】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CYP3A4(図7b)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7c】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CYP2B6(図7c)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7d】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。CAR(核ホルモン受容体)(図7d)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7e】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。MDR1(輸送体)(図7e)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7f】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。MRP(図7f)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7g】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。BSEP(輸送体)(図7g)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【図7h】ヒト肝細胞ヒト肝細胞の再増殖の異なるレベル(M790 10%、M697 30%、およびM785 60%)を有する3匹のマウスからの肝細胞における、肝臓特異的遺伝子および薬物代謝と関連する遺伝子の遺伝子発現の基底レベルを示すグラフ。非誘導培養物中の誘導と比較した、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)に反応する薬物代謝に関連するmRNAの誘導の棒グラフ。PXR(核ホルモン受容体)(図7h)を示す。フェノバルビタールによるCYP3A4の誘導は、酵素量における誘導よりもさらになお著しかった。
【発明を実施するための形態】
【0016】
I. 略語
AAV アデノ随伴ウイルス
ALB アルブミン
ALT アラニン・アミノトランスフェラーゼ
AST アスパラギン酸アミノトランスフェラーゼ
BNF ベータナフトフラボン
DAB ジアミノベンジジン
ELISA 酵素免疫測定法
EROD エトキシレソルフィン−O−デエチラーゼ
FACS 蛍光活性化細胞分類
FAH フマリルアセト酢酸ヒドラーゼ
FISH 蛍光原位置ハイブリダイゼーション
FITC フルオレセインイソチオシアネート
FRG Fah−/−/Rag2−/−/Il2rg−/−三重変異体マウス
HLA ヒト白血球抗原
IL−2Rγ インターロイキン2受容体γ
MHC 主要組織適合性複合体
NTBC 2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン
PB フェノバルビタール
PBS リン酸緩衝生理食塩水
PCR ポリメラーゼ連鎖反応
PE フィコエリトリン
PFU プラーク形成単位
RAG リコンビナーゼ活性化遺伝子
Rif リファンピシン
RT−PCR 逆転写ポリメラーゼ連鎖反応
TAT チロシンアミノ転移酵素
TF トランスフェリン
TTR トランスサイレチン
UGT1A1 UDPグルクロノシルトランスフェラーゼ1ファミリー、ポリペプチドA1
uPA ウロキナーゼプラミノーゲン活性化因子
II. 用語
別途記載のない限り、技術用語は、従来の用法に準拠して使用される。分子生物学における一般用語の定義は、Benjamin Lewin,Genes V,Oxford University Pressより出版、1994(ISBN0−19−854287−9)、Kendrew et al.(eds.),The Encyclopedia of Molecular Biology,published by Blackwell Science Ltd.,1994(ISBN0−632−02182−9)、およびRobert A.Meyers(ed.),Molecular Biology and Biotechnology:a Comprehensive Desk Reference,VCH Publishers,Inc.より出版、1995(ISBN1−56081−569−8)において見出すことができる。
【0017】
本発明の種々の実施形態の概説を容易にするために、具体的な用語の以下の説明を提供する。
【0018】
肝臓疾患の発症を阻害するまたは予防する薬剤:FRGマウス, またはFpmRGマウス、または他の型のFah欠損マウスに投与される場合、マウスにおける肝臓疾患の発症を予防、または遅延、または阻害する化合物または組成物。肝臓疾患または肝機能障害は、肝組織像の変化(壊死、炎症、異形成、または肝臓癌等)、肝臓特異的酵素および他のタンパク質(アスパラギン酸アミノトランスフェラーゼ、アラニン・アミノトランスフェラーゼ、ビリルビン、アルカリフォスファターゼ、およびアルブミン等)のレベルの変化、または全身性肝不全が挙げられるが、これらに限定されない、兆候または症状のうちのいずれか1つにより特徴付けられる。一実施形態において、肝臓疾患を阻害する薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。
【0019】
羊膜細胞:胎児を取り囲む羊水中に見られる細胞。
【0020】
抗体:免疫グロブリン遺伝子または免疫グロブリン遺伝子のフラグメントにより実質的にコード化された1つもしくは複数のポリペプチドを含むタンパク質(またはタンパク質複合体)。認識された免疫グロブリン遺伝子は、カッパ、ラムダ、アルファ、ガンマ、デルタ、イプシロンおよびミュー定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子を含む。軽鎖はカッパまたはラムダのいずれかに分類される。重鎖はガンマ、またはミュー、またはアルファ、またはデルタ、またはイプシロンに分類され、同様に、各々IgG、IgM、IgA、IgD、およびIgEの免疫グロブリンを表す。
【0021】
基本的な免疫グロブリン(抗体)の構造単位は、通常、テトラマーである。各々のテトラマーは、2つの同一のペアのポリペプチド鎖から成り、各々のペアは、1つの「軽い」鎖(約25kD)と1つの「重い」鎖(約50〜70kD)を有する。各々の鎖のN末端は、抗原認識に主に関与する約100〜110個またはそれより多いアミノ酸の可変領域を定義する。「可変軽鎖」(VL)および「可変重鎖」(VH)という用語はそれぞれ、これらの軽鎖および重鎖を指す。
【0022】
本明細書に使用される、「抗体」という用語は、完全な免疫グロブリン、ならびによく特徴付けられたフラグメントをいくつか含む。例えば、標的タンパク質(またはタンパク質または融合タンパク質内のエピトープ)に結合するFabs、Fvs、および一本鎖Fvs(SCFvs)は、そのタンパク質(またはエピトープ)のための特異的結合剤でもある。これらの抗体フラグメントは、以下のように定義される。(1)Fab、完全な軽鎖および1つの重鎖の一部分を産生するために、酵素パパインによる全抗体の消化により生成された抗体分子の一価抗原結合フラグメントを含むフラグメント、(2)Fab’、完全な軽鎖および1つの重鎖の一部分を産生するために、ペプチンで全抗体を処理し、次いで、還元することにより得られる、抗体分子のフラグメント、(3)(Fab’)2、後続の還元を伴わずに酵素ペプチンで全抗体を処理することにより得られる、抗体のフラグメント、(4)F(ab’)2、2個のジスルフィド結合により2つのFab’フラグメントが一緒にされた二量体、(5)Fv、2本の鎖として発現された軽鎖の可変域および重鎖の可変域を含む遺伝子組み換えフラグメント、および(6)1本鎖抗体、遺伝子融合1本鎖分子として適切なポリペプチドリンカーにより連結された、軽鎖の可変域、重鎖の可変域を含む遺伝子組み換え分子, として定義される。これらのフラグメントを作製する方法は、日常的である(例えば、Harlow and Lane,Using Antibodies:A Laboratory Manual,CSHL,New York,1999を参照)。
【0023】
本開示の方法に用いられる抗体は、モノクローナルまたはポリクローナルであり得る。単に一例として、モノクローナル担体は、Kohler and Milsteinの古典的方法(Nature 256:495−97,1975)またはその誘導的な方法に従って、ネズミハイブリドーマから調製することができる。モノクローナル抗体生成のための詳細な手順は、Harlow and Lane,Using Antibodies:A Laboratory Manual,CSHL,New York,1999に記載されている。
【0024】
B細胞:体液性免疫反応に大きな役割を果たすリンパ球の一型。B細胞の主要機能は、可溶性抗原に対して抗体を作製することである。B細胞は、適応免疫系の不可欠な要素である。
【0025】
採取:本明細書に使用される増大されたヒト肝細胞を「採取する(collecting)」とは、単離されたヒト肝細胞を注入されているマウス(または受容マウスとも称される)から増大された肝細胞を除去するプロセスをさす。採取は、任意に、肝細胞を他の細胞型から分離することを含む。一実施形態において、その増大されたヒト肝細胞は、Fah欠損マウスの肝臓から採取される。一部の例において、その増大されたヒト肝細胞は、FRGマウスまたはFpmRGマウスの肝臓から採取される。
【0026】
インターロイキン受容体の一般的なγ鎖(Il2rg):インターロイキン受容体の一般的なガンマ鎖をコード化する遺伝子。Il2rgは、IL−2、IL−4、IL−7、およびIL−15を含む、いくつかのインターロイキンに対する受容体の構成要素である(Di Santo et al.Proc.Natl.Acad.Sci.U.S.A.92:377−381,1995)。Il2rgの欠損した動物は、B細胞およびT細胞の減少を示し、ナチュラルキラー細胞が欠如する。
【0027】
凍結保存:本明細書に使用される「凍結保存」とは、77Kまたは−196℃(液体窒素の沸点)等の低い氷点下の温度まで冷却することにより保存または維持されている細胞または組織を指す。これらの低温度で、細胞死をもたらす生化学的反応を含む、いかなる生物学的活性も、効果的に停止する。
【0028】
肝機能の低下:肝臓の健康または機能を測定する多くのパラメータのうちのいずれか1つの異常変化。肝機能の低下はまた、本明細書では、「肝機能障害」と称される。肝機能は、肝組織像の検査および肝臓酵素または他のタンパク質の測定等に限定されない、当技術分野で周知のような多くの手段のうちのいずれか1つにより評価することができる。例えば、肝機能障害は、肝臓の壊死、または炎症、または酸化的損傷、または異形成で示され得る。一部の例において、肝機能障害は、肝細胞癌等の肝臓癌で示される。肝機能障害を評価するために試験され得る肝臓酵素およびタンパク質の例としては、アラニン・アミノトランスフェラーゼ(ALT)、アスパラギン酸アミノトランスフェラーゼ(AST)、ビリルビン、アルカリフォスファターゼ、およびアルブミンが挙げられるが、これらに限定されない。肝機能障害はまた、全身性肝不全をもたらし得る。肝機能を試験するための手順は、Grompeら(Genes Dev.7:2298−2307,1993)およびManningら(Proc.Natl.Acad.Sci.U.S.A.96:11928−11933,1999)により教示されるもの等、当技術分野で周知であり、それらは参照することにより本明細書に組み込まれる。
【0029】
欠損:本明細書に使用される「Fah欠損」または「Fahの欠損」とは、Fahの突然変異を含む、マウス等の動物を指し、それは、Fah mRNA発現および/または機能的FAHタンパク質の実質的減少、またはその欠損をもたらす。一実施形態において、該Fah欠損動物は、Fah遺伝子のホモ接合体欠失を含む。一例として、該ホモ接合体欠失は、Fahのエクソン5にある。別の実施形態において、該Fah欠損動物は、Fah遺伝子の1つもしくは複数の点変異を含む。適切なFah点変異の例は、当技術分野で知られている(例えば、Aponte et al.Proc.Natl.Acad.Sci.U.S.A.98(2):641−645,2001参照、参照することにより本明細書に組み込まれる)。
【0030】
減損する:減じるまたは除去すること。本明細書に使用される「マクロファージ減損」とは、動物におけるマクロファージを消去する、または除去する、または減じる、または殺滅するプロセスを指す。マクロファージが減損している動物は、必ずしもマクロファージが完全に欠如しているとは限らないが、少なくともマクロファージの数または活性の減少を示す。一実施形態において、マクロファージ減損は、機能的マクロファージの少なくとも10%、少なくとも25%、少なくとも50%、少なくとも75%、少なくとも90%、または100%の減少をもたらす。
【0031】
生着する:動物に細胞または組織を移植すること。本明細書に使用される、受容マウスにおいて、ヒト肝細胞の生着とは、注入後、ヒト肝細胞が受容マウスに移植されるプロセスを指す。生着されたヒト肝細胞は、受容マウスにおいて、増大することができる。本明細書に記載される、「有意な生着」とは、肝臓において、少なくとも約1%の肝細胞がヒト肝細胞である、受容マウスを指す。「高度に生着された」マウスは、肝細胞の少なくとも約30%がヒト肝細胞である、肝臓を有するマウスである。しかしながら、生着効率はさらに高くなり得、マウス肝臓において、例えば、肝細胞の少なくとも約40%、または少なくとも約50%、または少なくとも約60%、または少なくとも約70%、または少なくとも約80%、または少なくとも約90%、または少なくとも約95%がヒト肝細胞である。
【0032】
胚幹(ES)細胞:発達する胚盤胞の内細胞塊から単離された多能性細胞。ES細胞は、多能性細胞であり、それらが、体内(骨、筋肉、脳細胞等)に存在するすべての細胞を作製することができることを意味する。ネズミES細胞を産生するための方法は、米国特許第5,670,372号に見出され得、参照することにより本明細書に組み込まれる。ヒトES細胞を産生するための方法は、米国特許第6,090,622号、PCT公開第WO 00/70021号、およびPCT公開第WO 00/27995号に見出され得、これらのそれぞれが、参照することにより本明細書に組み込まれる。
【0033】
増大させる:量を増加させること。本明細書に使用される、ヒト肝細胞を「増大させる(expanding)」とは、ヒト肝細胞の数が増加するような、細胞分裂を生じることが可能なプロセスを指す。本明細書に記載される、ヒト肝細胞は、受容マウスにおいて、少なくとも約4週間、または少なくとも約6週間、または少なくとも約8週間、または少なくとも約12週間、または少なくとも約16週間、または少なくとも約20週間、または少なくとも約24週間、または少なくとも約28週間、増大可能である。一実施形態において、該ヒト肝細胞は、最長約6ヶ月まで増大可能である。増大から結果として生じるヒト肝細胞の数は、変化し得る。一実施形態において、増大は、少なくとも1,000万、または少なくとも2,000万、または少なくとも3,000万、または少なくとも4,000万、または少なくとも5,000万の肝細胞をもたらす。100万の肝細胞が最初に注入され、約10%生着されると仮定すると、肝細胞の増大は、約10倍〜約500倍の範囲であり得る。一部の実施形態において、受容マウスにおいて、ヒト肝細胞の増大は、少なくとも10倍、または少なくとも50倍、または少なくとも100倍、または少なくとも150倍、または少なくとも200倍、または少なくとも250倍、または少なくとも300倍、または少なくとも400倍、または少なくとも500倍、または少なくとも1,000倍の増加をもたらす。
【0034】
FRGマウス:フマリルアセト酢酸ヒドラーゼ(Fah)、リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)遺伝子において、ホモ接合体欠失を有する、変異体マウス。また、本明細書では、Fah−/−/Rag2−/−/Il2rg−/−と称される。本明細書に使用される、Fah、Rag2、およびIl2rg遺伝子においてホモ接合体欠失は、機能的FAH、RAG−2、およびIL−2Rγタンパク質が、突然変異を含むマウスに発現しないことを示す。
【0035】
FpmRGマウス:リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)遺伝子のホモ接合体欠失、ならびにフマリルアセト酢酸ヒドラーゼ(Fah)のホモ接合体点変異を有する、変異体マウス。FpmRGマウスのFah遺伝子の点変異は、mRNAにおいて、エクソン7のミススプライシングおよび喪失をもたらす(Aponte et al.,Proc.Natl.Acad.Sci.USA 98:641−645,2001)。また、本明細書では、Fahpm/Rag2−/−/Il2rg−/−と称される。本明細書に使用される、Rag2およびIl2rg遺伝子のホモ接合体欠失は、機能的RAG−2およびIL−2Rγタンパク質が、該突然変異を含むマウスに発現しないことを示す。さらに、Fah遺伝子のホモ接合体の点変異を有するマウスは、機能的FAHタンパク質を発現しない。
【0036】
フマリルアセト酢酸ヒドラーゼ(FAH):チロシン異化の最終ステップに触媒作用を及ぼす代謝酵素。Fah遺伝子のホモ接合体欠失を有するマウスは、肝mRNA発現の変化および重度の肝機能障害を示す(Grompe et al. Genes Dev.7:2298−2307,1993、参照することにより本明細書に組み込まれる)。該Fah遺伝子の点変異はまた、肝不全および出生後の致死を生じることも示している(Aponte et al.Proc.Natl.Acad.Sci.U.S.A.98(2):641−645,2001、参照することにより本明細書に組み込まれる)。
【0037】
漸減させる:本明細書に使用される、NTBCの用量を「漸減する」とは、数日にわたるような時間とともに、Fah欠損マウスに投与されるNTBCの用量を漸減するプロセスを指す。一実施形態において、該NTBC用量は、約6日間にわたり、漸減し、該用量は、約1週間後、NTBCがもはや投与されなくなるように、約1または2日間間隔で減少する。NTBCの漸減は、短期間または長期間にわたり行うことができ、用量の減少の間の時間間隔もまた、短くする、または長くすることができる。
【0038】
肝細胞:肝臓の細胞質塊のうちの70〜80%まで作製する細胞型。肝細胞は、タンパク質合成、タンパク質保存、および炭水化物の形質転換、コレステロール、胆汁塩、およびホスホリン脂質の合成、ならびに外因性および内因性物質の解毒、修飾、および排出に関与する。該肝細胞はまた、胆汁の形成および分泌も引き起こす。肝細胞は、血清アルブミン、フィブリノゲン、および凝固因子のプロトロンビン基を製造し、リポタンパク質、セルロプラスミン、トランスフェリン、相補体、および糖タンパク質の合成のための主部位である。さらに、肝細胞は、薬物および殺虫剤のような外因性化合物、およびステロイドのような内因性化合物を代謝する、解毒する、および不活性化する能力を有する。
【0039】
ホモ接合:1つもしくは複数の遺伝子座で同一の対立遺伝子を有すること。本明細書に使用される、「ホモ接合体欠失」とは、1つの遺伝子の両方の対立遺伝子が同一の欠失を有する生物を指す。
【0040】
免疫不全:免疫系のうちの少なくとも1つの不可欠な機能を欠如すること。本明細書に使用される、「免疫不全」マウスとは、免疫系の特異成分を欠如する、または免疫系の特異成分の機能を欠如するマウスである。一実施形態において、免疫不全マウスは、機能的B細胞、T細胞、および/またはNK細胞を欠如する。別の実施形態において、免疫不全マウスは、さらに、マクロファージを欠如する。
【0041】
単離された:「単離された」肝細胞とは、臓器ドナー等の特定の源から得られている、および他の細胞型から実質的に分離または精製された、肝細胞を指す。
【0042】
マクロファージ:単球と呼ばれる特定の白血球から由来する組織内の細胞。単球およびマクロファージは、食細胞であり、脊椎動物の非特異的防御(または先天性免疫)、ならびに特異的防御(または細胞性免疫)の両方において作用する。これらの役割は、細胞残屑、および固定細胞あるいは移動細胞としての病原体を食菌する(飲み込んだ後、消化する)こと、および病原体に対して反応するために、リンパ球および他の免疫細胞を刺激することである。
【0043】
ナチュラルキラー(NK)細胞:内因性免疫系の主成分を構成する細胞傷害性リンパ球型。NK細胞は、腫瘍およびウイルス感染した細胞の両方の宿主の拒絶反応において大きな役割を果たす。
【0044】
受容:本明細書に使用される、「受容マウス」とは、本明細書に記載される単離されたヒト肝細胞を注入されているマウスである。一般に、ヒト肝細胞の一部(割合は異なり得る)は、受容マウスに生着する。一実施形態において、該受容マウスは、Fahがさらに欠損している免疫不全マウスである。別の実施形態において、該受容マウスは、Fahがさらに欠損しているRag2−/−/Il2rg−/−マウスである。別の実施形態において、該受容マウスはFRGマウスである。別の実施形態において、該受容マウスは、FpmRGマウスである。
【0045】
リコンビナーゼ活性化遺伝子2(Rag2):免疫グロブリン遺伝子座とT細胞受容体遺伝子座の組み換えに関与する遺伝子。Rag2遺伝子が欠損する動物は、V(D)J組み換えを行うことができず、機能的T細胞およびB細胞の完全喪失をもたらす(Shinkai et al.Cell 68:855−867,1992)。
【0046】
連続移植:一次マウスにおいて増大された肝細胞が、採取され、さらなる増大のために二次マウスに注入等により移植される、生体内でヒト肝細胞を増大させるためのプロセス。連続移植は、三次、または四次、または追加次のマウスをさらに含むことができる。
【0047】
幹細胞:不変の娘細胞を生成する(自己再増殖;親細胞と同一である少なくとも1つの娘細胞を生成する細胞分裂)、および特殊化細胞型を形成する(潜在力)ための特異な能力を有する細胞。幹細胞としては、胚幹(ES)細胞、胚性生殖(EG)細胞、生殖系列幹(GS)細胞、ヒト間充織幹細胞(hMSC)、脂肪組織由来幹細胞(ADSC)、多能性成体幹細胞(MAPC)、多能性成体生殖系列幹細胞(maGSC)、および非制限体幹細胞(USSC)が挙げられるが、これらに限定されない。生体内の幹細胞の役割は、動物の正常な生活中、破壊される細胞を置換することである。通常、幹細胞は、無制限に分裂することができる。分裂後、該幹細胞は、幹細胞として残る、または前駆細胞になる、または末端分化に移り得る。前駆細胞は、少なくとも1つの所与の細胞型の十分に分化した機能細胞を作製することができる細胞である。通常、前駆細胞は、分裂することができる。分裂後、前駆細胞は、前駆細胞のままであり得る、または末端分化に移り得る。一実施形態において、該幹細胞は、肝細胞を形成する。
【0048】
T細胞:細胞性免疫において中心的役割を果たす白血球型、またはリンパ球型。T細胞は、T細胞受容体(TCR)と呼ばれるB細胞およびNK細胞の細胞表面上にある特別受容体の存在により、B細胞およびNK細胞のようなリンパ球の他型と区別される。胸腺は、通常、T細胞発達のための主要臓器であると考えられる。
【0049】
導入遺伝子:細胞または生命体のゲノムに導入される外因性核酸配列。
【0050】
トランスジェニック動物:動物の細胞の一部において染色体外要素として存在するか、または動物の生殖細胞系DNAに(すなわち、その細胞のほとんどまたはすべてのゲノム配列に)安定的に組み込まれている、非内因性(異種)核酸配列を有する、通常、哺乳類である、ヒト以外の動物。異種核酸は、当技術分野で周知のような方法に従って、例えば、宿主動物の胚または胚幹細胞の遺伝子操作により、このようなトランスジェニック動物の生殖細胞系に導入される。「導入遺伝子」は、このような異種核酸、例えば、(例えば「ノックイン」トランスジェニック動物の作製用の)発現コンストラクトの形状をした異種核酸、または標的遺伝子内またはその近傍に挿入されて、(例えば「ノックアウト」トランスジェニック動物の作製用の)標的遺伝子の発現低下を生じる異種核酸を意味する。遺伝子の「ノックアウト」とは、標的遺伝子の機能低下を生じる遺伝子の配列の変化を意味し、好ましくは、このような標的遺伝子の発現は検出されないか、または有意ではない。トランスジェニックのノックアウト動物は、標的遺伝子についてヘテロ接合のノックアウト、または標的遺伝子についてホモ接合のノックアウトを含むことができる。「ノックアウト」はまた、条件的なノックアウトも含み、標的遺伝子の変化が、例えば、標的遺伝子の変化を促進する物質に動物を曝露した際、標的遺伝子部位における組換えを促進する酵素(例えばCre−lox系のCre)を導入した際、または標的遺伝子の変化を出生後に指令する他の方法で生じる場合がある。
【0051】
ベクター:宿主細胞において複製する、および/または挿入するための、ベクターの能力を崩壊することなく、外部の核酸の挿入を可能にする核酸分子。ベクターは、複製の複製起点のような、宿主細胞において、核酸配列の複製を可能にする核酸配列を含むことができる。ベクターはまた、1つもしくは複数の選択可能なマーカー遺伝子および他の遺伝要素を含むこともできる。挿入ベクターは、それ自体、宿主核酸に挿入することができる。発現ベクターは、一つまたは複数の挿入遺伝子の転写および翻訳を可能にするのに必要な調節塩基配列を含むベクターである。本明細書に記載される一実施形態において、該ベクターは、ヒトウロキナーゼのような、ウロキナーゼをコード化する配列を含有する。一実施形態において、該ベクターは、プラスミドベクターである。別の実施形態において、該ベクターは、アデノウイルスベクターまたはアデノ随伴ウイルス(AAV)ベクターのような、ウイルスベクターである。
【0052】
ウロキナーゼ:また、ウロキナーゼ型プラミノーゲン活性化因子(uPA)と呼ばれる、ウロキナーゼは、セリンプロテアーゼである。ウロキナーゼは、本来、人尿から分離されたが、血流および細胞外マトリックスのような、いくつかの生理学的位置に存在する。主要な生理学的基質は、プラスミノーゲンであり、これはセリンプロテアーゼプラスミンの不活性な酵素前駆体である。プラスミンの活性化は、生理学的環境によって、血栓溶解または細胞外マトリックスの分解に関与するタンパク質分解カスケードを誘発する。本明細書に提供される方法の一実施形態において、ウロキナーゼは、肝細胞注入前、受容マウスに投与される。一部の実施形態において、ウロキナーゼは、ヒトウロキナーゼである。一部の実施形態において、該ヒトウロキナーゼは、分泌型のウロキナーゼである。一部の実施形態において、該ヒトウロキナーゼは、修飾された、非分泌型のウロキナーゼである(米国特許第5,980,886号参照、参照することにより本明細書に組み込まれる)。
【0053】
別途説明されない限り、本明細書に使用されるすべての技術および科学用語は、本発明が属する技術分野の当業者によって一般に理解される用語と同じ意味を持つ。文脈で特に指定しない限り、単数形、「一つの(a)」「一つの(an)」、および「その(the)」は複数形の指示物を含む。同様に、文脈で特に指定しない限り、「または(or)」という語は、「および(and)」を含むことを意図する。したがって、「AまたはBを含む(comprising A or B)」とは、AまたはB、あるいはAおよびBの両方を含むことを意味する。核酸またはポリペプチドに対して規定したすべての塩基サイズまたはアミノ酸サイズ、およびすべての分子量または分子量値は概算値であり、説明のために提供するものであることがさらに理解されるべきである。本発明の実施または検討のために本明細書に記載したものと同様または同等の方法および材料を用いることができるが、適した方法および材料は以下に説明する。本明細書で言及するすべての刊行物、特許出願、特許、および他の参考文献は、その全体を参照することにより本明細書に組み込まれる。矛盾する場合には、用語の説明を含め、本明細書が優先するものとする。さらに、材料、方法、および例は、例示的なものに過ぎず、限定を意図したものではない。
【0054】
III. いくつかの実施形態の説明
生体内でヒト肝細胞を増大させる着実な方法を本明細書に提供する。該方法は、チロシン異化酵素、フマリルアセト酢酸ヒドラーゼ(Fah)が欠損している免疫不全マウスに単離されたヒト肝細胞を移植するステップを含む。一実施形態において、該免疫不全マウスは、Rag2−/−/Il2rg−/−マウスである。一実施形態において、該Fah欠損マウスは、Fahのホモ接合体欠失を含む。別の実施形態において、該Fah欠損マウスは、タンパク質の機能、および/または作製が実質的に低下するように、Fahにおいて、1つもしくは複数の点変異を含む。本明細書に記載されるように、Fah、リコンビナーゼ活性化遺伝子2(Rag2)、およびインターロイキン受容体の共通γ鎖(Il2rg)が欠損した三重変異体マウスは、生体内でヒト肝細胞を増大させるための十分な生体内系を提供する。一部の実施形態において、該マウスは、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである。一部の実施形態において、該マウスは、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである。
【0055】
生体内でヒト肝細胞を増大させる方法を、本明細書に開示し、単離されたヒト肝細胞を注入等により、免疫不全およびFah欠損マウス(また、受容マウスと称される)に移植するステップと、少なくとも約2週間ヒト肝細胞を増大させるステップと、増大させたヒト肝細胞をマウスから採取するステップと、を含む。該肝細胞は、当技術分野で知られている任意の適した手段を用いて移植することができる。一実施形態において、該単離されたヒト肝細胞は、注入等により、該受容マウスの脾臓に移植される。別の実施形態において、該増大されたヒト肝細胞は、該受容マウスの肝臓から採取される。該ヒト肝細胞は、受容マウスにおいて、ヒト肝細胞の増大を可能にするのに十分な期間、増大可能である。増大のための正確な期間は、日常の実験を用いて実験的に判定することができる。一実施形態において、該ヒト肝細胞は、最長6ヶ月まで増大可能である。別の実施形態において、該ヒト肝細胞は、少なくとも約4週間、または少なくとも約6週間、または少なくとも約8週間、または少なくとも約12週間、または少なくとも約16週間、または少なくとも約20週間、または少なくとも約24週間、または少なくとも約28週間、増大可能である。肝細胞増大の程度は、異なり得る。一部の実施形態において、受容マウスにおいて、ヒト肝細胞の増大は、少なくとも約10倍、または少なくとも約50倍、または少なくとも約100倍、または少なくとも約150倍、または少なくとも約200倍、または少なくとも約250倍、または少なくとも約300倍、または少なくとも約400倍、または少なくとも約500倍、または少なくとも約1,000倍の増加をもたらす。
【0056】
また、生体内でヒト肝細胞を増大させる方法を提供し、受容マウスは、該ヒト肝細胞の注入前、ウロキナーゼ遺伝子をコード化するベクターが投与される。一実施形態において、該ウロキナーゼ遺伝子は、ヒトウロキナーゼである。野生型ウロキナーゼは、分泌タンパク質である。したがって、一部の実施形態において、該ヒトウロキナーゼは、分泌型のウロキナーゼである(Nagai et al.,Gene 36:183−188,1985、参照することにより本明細書に組み込まれる)。ヒトウロキナーゼ(分泌型)の配列は、当技術分野で知られており、GenBank受入番号AH007073(1993年8月3日受入)、D11143(1996年5月9日受入)、A18397(1994年7月21日受入)、BC002788(2003年8月19日受入)、X02760(1993年4月21日受入)、BT007391(2003年5月13日受入)、NM_002658(2004年10月1日受入)、X74039(1994年2月20日受入)に限定されず、参照することにより本明細書に組み込まれる。
【0057】
一部の実施形態において、該ヒトウロキナーゼは、非分泌型のウロキナーゼである。例えば、Lieberら(Proc.Natl.Acad.Sci.92:6210−6214,1995、参照することにより本明細書に組み込まれる)は、ウロキナーゼのカルボキシル末端で小胞体保留シグナルをコード化する配列を挿入することにより、またはpre−uPAシグナルペプチド(Strubin et al.,Cell 47:619−625,1986、Schutze et al.,EMBO J.13:1696−1705,1994、これらの双方は、参照することにより本明細書に組み込まれる)をアミノ末端RR保留シグナルと置換することにより作製された非分泌型のウロキナーゼ、ならびに細胞膜IIタンパク質Iip33(Strubin et al.,Cell 47:619−625,1986)からスペーサーペプチドにより引き離された膜貫通アンカーを記載する。非分泌型のウロキナーゼはまた、米国特許第5,980,886号に記載され、参照することにより本明細書に組み込まれる。
【0058】
ウロキナーゼをコード化するベクターは、マウスへの送達に適している任意の型のベクターであり得、ウロキナーゼ遺伝子を発現することができる。このようなベクターは、ウイルスベクターまたはプラスミドベクターを含む。一実施形態において、該ベクターは、アデノウイルスベクターである。別の実施形態において、該ベクターは、AAVベクターである。ウロキナーゼをコード化するベクターは、当技術分野で知られている任意の適した手段により投与され得る。一実施形態において、該ベクターは、静脈内投与される。一態様において、該ベクターは、後眼窩注射により投与される。ウロキナーゼをコード化するベクターは、該ヒト肝細胞の注入前、いつでも投与され得る。一般に、該ベクターは、ウロキナーゼが十分な時間で発現できるように、投与される。一実施形態において、該ベクターは、肝細胞注入の24〜48時間前に投与される。
【0059】
生体内でヒト肝細胞を増大させる方法を、本明細書にさらに提供し、該受容マウスは、該ヒト肝細胞の注入前、マクロファージが減損している。一実施形態において、該受容マウスに、マクロファージ減損前、ウロキナーゼをコード化するベクターが投与される。別の実施形態において、該受容マウスに、マクロファージ減損後、ウロキナーゼをコード化するベクターが投与される。別の実施形態において、該マクロファージ減損受容マウスに、ウロキナーゼをコード化するベクターが投与されない。マクロファージを、化学物質または抗体を用いるような、当技術分野で周知のようないくつかの方法のうちのいずれか1つを用いて、該受容マウスから減損することができる。例えば、マクロファージを、C12MDP、またはマクロファージ発達、機能、および/または生存力を変える抗体を含む、有害物質のようなアンタゴニストの投与により減損することができる。アンタゴニストの投与は、周知の技法により行われ、欧州特許第1552740号に記載されるようなリポソームの使用を含み、参照することにより本明細書に組み込まれる。クロドロネート含有リポソームはまた、van Rijnら(Blood 102:2522−2531,2003)により記載されるように、マクロファージを減損するために使用することができ、これは参照することにより本明細書に組み込まれる。
【0060】
本明細書に記載される方法の一実施形態において、肝細胞注入前、該Fah欠損マウスに、該マウスにおける肝臓疾患の発症を阻害、予防、または遅延する薬剤が投与される。該薬剤は、肝臓疾患を阻害するための当技術分野で知られている任意の化合物または組成物であり得る。このような一薬剤は、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3シクロヘキサンジオン(NTBC)である。NTBCは、該Fah欠損マウスにおける肝臓疾患の発症を調節するために投与される。用量、投与スケジュール、および投与方法は、該Fah欠損マウスにおける肝機能障害を予防するために必要に応じ、調整することができる。一実施形態において、該NTBCは、約0.01mg/kg/日から約0.50mg/kg/日の用量で投与される。別の実施形態において、該NTBCは、約0.05mg/kg/日、または約0.06mg/kg/日、または約0.07mg/kg/日、または約0.08mg/kg/日、または約0.09mg/kg/日、または約0.10mg/kg/日のような、約0.05mg/kg/日から約0.10mg/kg/日の用量で投与される。NTBCは、ヒト肝細胞の注入前、および/または肝細胞注入後、選択された期間投与することができる。NTBCは、肝細胞の増大の期間中、必要に応じ、中止または再投与することができる。一実施形態において、該Fah欠損マウスに、肝細胞注入前、および肝細胞注入後、少なくとも約3日間、NTBCが投与される。別の実施形態において、該Fah欠損マウスに、肝細胞注入前、および肝細胞注入後、少なくとも約6日間、NTBCが投与される。一態様において、NTBCの用量は、肝細胞注入後、6日間の過程にわたり漸減される。NTBCは、飲料水内投与、食物内投与、または注入が挙げられるが、これらに限定されない、任意の適した手段により投与することができる。一実施形態において、肝細胞注入前、飲料水内投与されるNTBCの濃度は、約1mg/L、または約2mg/L、または約3mg/L、または約4mg/L、または約5mg/L、または約6mg/L、または約7mg/L、または約8mg/Lのような、約1mg/Lから約8mg/Lである。別の実施形態において、肝細胞注入前、飲料水内投与されるNTBCの濃度は、約1.0mg/L、または約1.2mg/L、または約1.4mg/L、または約1.6mg/L、または約1.8mg/L、または約2.0mg/Lのような、約1mg/Lから約2mg/Lである。
【0061】
単離されたヒト肝細胞は、多くの異なる源のうちのいずれか1つから得ることができる。一実施形態において、該ヒト肝細胞は、臓器ドナーの肝臓から単離された。別の実施形態において、該ヒト肝細胞は、外科的切除から単離された。別の実施形態において、該ヒト肝細胞は、胚幹細胞、または間葉由来幹細胞、または脂肪組織由来幹細胞、または多能性成体幹細胞、または非制限体幹細胞のような、幹細胞に由来した。別の実施形態において、該ヒト肝細胞は、単球または羊膜細胞に由来し、ひいては、幹細胞または前駆細胞は、肝細胞を産生するために、体外で得られる。別の実施形態において、該ヒト肝細胞は、注入前、凍結保存された。
【0062】
該Fah欠損受容マウスにおけるヒト肝細胞の連続移植の方法を、本明細書にさらに提供する。該方法は、第1の受容マウスからの増大されたヒト肝細胞を採取するステップと、二次、または三次、または四次、または追加次の受容マウスにおいて、肝細胞をさらに増大させるステップを含む。ヒト肝細胞は、いくつかの技法のうちのいずれか1つを用いて、マウスから採取することができる。例えば、該肝細胞は、下述の実施例に記載されるように、マウス肝臓を潅流し、次いで、徐々に細分化することにより採取することができる。さらに、該肝細胞は、ヒト細胞またはヒト肝細胞を特異的に認識する抗体を用いるような、周知の方法を用いて、他の細胞型、組織、および/または、破片から分離することができる。このような抗体としては、抗ヒトHLA−A、B、Cのような、クラスI主要組織適合性抗原に特異的に結合する抗体が挙げられるが、これらに限定されない(Markus et al.Cell Transplantation 6:455−462,1997)。抗体結合肝細胞は、その後、パンニング(固体マトリックスに付着したモノクローナル抗体を利用する)、蛍光活性化細胞分類(FACS)、磁気ビーズ分離等により、分離することができる。肝細胞を採取する代替的な方法は、当技術分野で周知である。
【0063】
また、遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、該欠失または点変異がホモ抱合体であり、該マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞を、該マウスに増大させることができる、遺伝子組み換えマウスを、本明細書に提供する。一実施形態において、該欠失は、B細胞、T細胞、およびN細胞の完全喪失をもたらす。別の実施形態において、該マウスは、ウロキナーゼを発現する。一実施形態において、ウロキナーゼは、ヒトウロキナーゼである。一態様において、ウロキナーゼの発現は、ウロキナーゼをコード化する導入遺伝子の取り込みに起因する。別の態様において、ウロキナーゼの発現は、分泌型または非分泌型のウロキナーゼのような、ウロキナーゼをコード化するベクターの投与に起因する。ウロキナーゼをコード化するベクターは、マウスへの送達に適している任意の型のベクターであり得、ウロキナーゼ遺伝子を発現することができる。一実施形態において、該ベクターは、アデノウイルスベクターである。別の実施形態において、該ベクターは、AAVベクターである。一部の実施形態において、該マウスは、FRGマウスである。一部の実施形態において、該マウスは、FpmRGマウスである。
【0064】
IV. ヒト肝細胞の増大のための遺伝子組み換えマウス株
いくつかのグループは、齧歯動物において、一次ヒト肝細胞を生着および増大しようとした(米国特許第6,509,514号、PCT公開第WO 01/07338号、米国公開第2005−0255591号)。Dandriら(Hepatology 33:981−988,2001)が、ヒト肝細胞を用いてマウス肝臓の再増殖の成功を初めて報告した。その後、他のグループは、マウスにおいて、成功を収めたヒト肝臓細胞の生着を報告している。これらの研究のすべてにおいて、使用された動物は、アルブミンプロモータの転写制御下で、ウロキナーゼプラミノーゲン活性化因子(uPA)を発現する、トランスジェニック動物であった(Sandgren et al.Cell 66:245−256,1991)。uPAの過剰発現は、代謝破壊を引き起こし、移植されたヒト肝細胞に影響を及ぼすことなく、マウス肝細胞の細胞死をもたらし、それは導入遺伝子を発現しない。該alb−uPA導入遺伝子は、様々な免疫不全のバックグラウンドと交錯し、ヒト細胞の拒絶を阻止した(Tateno et al.Am.J.Pathol.165:901−912,2004、Katoh et al.J.Pharm.Sci.96:428−437,2007、Turrini et al.Transplant.Proc.38:1181−1184,2006)。
【0065】
最大70%の生着レベルが、これらのモデルにおいて報告されているが、該系は、広範な使用を阻止しているいくつかの主要な不利点がある。第1に、該alb−uPA導入遺伝子は、若年期に、不活性化または喪失する。この理由のため、非常に初期(14日齢)にヒト細胞を移植し、導入遺伝子がホモ接合体であるマウスを使用することが必須である。この狭い移植の時間窓が、モデルの柔軟性を著しく制限する。第2に、該導入遺伝子の自発的不活性化は、導入遺伝子陰性の健康なマウス肝細胞のプールを作成する。これらの復帰突然変異体マウス肝細胞は、再増殖中、ヒト細胞と効率的に競合する。したがって、該ヒト細胞を連続移植すると同時に二次受容者を再増殖することは不可能である。第3に、肝臓疾患は、このモデルにおいて、非常に早発型であり、ひいては、トランスジェニックマウスの生存力を低下させる。したがって、十分な数の実験動物を繁殖することが困難である。さらに、該トランスジェニックマウスには、手術中、死亡率を増加する出血傾向がある。最後に、alb−uPAトランスジェニック動物は、一旦ヒト細胞を用いる再増殖が50%を超えると、腎疾患を発症する。これは、腎臓上皮上のヒト補体によるためであると考えられる。ヒト生着の非常に高いレベルを得るために、抗相補性プロテアーゼ阻害剤で移植されたマウスを処理することが必要である(Tateno et al.Am.J.Pathol.165:901−912,2004)。これらの多くの限定のため、ヒト肝細胞を増大させるためのさらに堅牢なシステムが非常に望ましい。遺伝子欠失の独自の組み合わせを有する遺伝子組み換えマウスを用いて、生体内でヒト肝細胞を増大させるための高効率な方法を、本明細書に記載する。マウス肝臓においてヒト肝細胞の成功を収めた生着および増大は、ある程度の肝機能障害がある免疫不全マウスを必要とする。マウス肝臓は、様々な異なる型の免疫不全マウスにおいて、ヒト肝細胞を用いて再増殖し、RAG−2ノックアウトまたはSCIDマウスを含み、これらの両方は、B細胞およびT細胞を欠如する(米国特許第6,509,514号、PCT公開第WO 01/07338号、米国公開第2005−0255591号)。肝機能障害に達するために、免疫不全マウスを、ウロキナーゼプラスミノーゲン活性化因子(uPA)トランスジェニックマウスと交配させた。マウス肝臓において、uPAの発現は、マウス肝細胞の成長の不利点を生じ、移植したヒト肝細胞の増大を促進する(PCT公開第WO 01/07338号)。該uPA導入遺伝子の限界を避けるために、Fah欠損マウスは、ヒト肝細胞の増大を可能にさせる能力について分析された。FAHは、チロシン異化の最後のステップを触媒する代謝酵素である。Fah遺伝子のホモ接合体欠失を有するマウスは、肝mRNA発現の変化および重度の肝機能障害を示す(Grompe et al.Genes Dev.7:2298−2307,1993)。該Fah突然変異体が、ヌードまたはRag1−/−(Mombaerts et al.Cell 68:869−877,1992)バックグラウンドと交錯した場合、ヒト肝細胞を有するマウス肝臓の再増殖は、免疫拒絶による可能性が最も高いため、成功しなかった。NOD/SCIDマウスとのFah欠損マウスの交配(Dick et al.Stem Cell 15:199−203,1997)は、ヒト肝細胞の副次的な生着が観察されるマウスを産生したが、これらの動物は、SCIDマウスに存在する二本鎖切断DNA修復の欠陥の可能性があるため、急速な肝不全を発症した。Fah−/−/Rag2−/−/Il2rg−/−(FRG)三重変異体マウスは、T細胞、B細胞、およびNK細胞を欠如することを、本明細書に開示する。Rag2−/−/Il2rg−/−マウスは、当技術分野で知られている(Traggiai et al.Science 304:104−107,2004、Gorantla et al.J.Virol.81:2700−2712,2007)。
【0066】
以下の実施例に記載されるように、ヒト肝細胞の生着および増大は、驚くべきことに、FRGマウスにおいて、高効率である。例えば、FRGマウスに、100万の単離されたヒト肝細胞を注入できる。10%の効率を仮定すると、100,000のヒト肝細胞が、受容マウスに生着される。増大後のFRGマウスからの平均収量は、その後、約3,000万から約4,500万のヒト肝細胞であり、ヒト肝細胞の300倍から450倍の増加と同じである。FRGマウスはまた、ヒト肝細胞の連続移植にも使用することができる。連続移植は、多重マウスを含み得、マウスあたりヒト肝細胞の少なくとも約150倍の増大をもたらし得る。
【0067】
Fah欠損を含む任意の免疫不全マウスは、本明細書に記載される諸方法に適している。一実施形態において、該マウスは、Fahが欠損しているRag2−/−/Il2rg−/−マウスでもある。該Fah欠損マウスは、例えば、Fahのホモ接合体欠失、またはFahの1つもしくは複数の点変異を含むことができる。Fah欠損(点変異またはホモ接合体欠失等による)は、Fah mRNA発現および/または機能的FAHタンパク質の実質的減少、またはその欠損をもたらす。該FRGマウスに加えて、Fah遺伝子において、点変異がホモ抱合体である免疫不全マウス(Rag2−/−/Il2rg−/−)(本明細書では、FpmRGマウスと称される)はまた、生体内でヒト肝細胞の生着および増大に適したマウスでもあることを、本明細書に記載する。
【0068】
V. ヒト肝細胞の単離および送達
ヒト肝細胞の生体内の増大のためにFah欠損マウスを用いることの有意な利点は、様々な源から採取したヒト肝細胞を該マウスに生着する能力である。下述の実施例に記載されるように、ヒト肝細胞は、死体ドナーまたは肝臓切除術から採取し得る、または市販の源から得ることができる。さらに、本明細書に示されるように、FRGマウスに、すべての年齢のドナーからのヒト肝細胞、または凍結保存された肝細胞を、うまく移植することができる。多くの場合、ヒト肝細胞の単離と移植との間の遅延(一般に、1〜2日)があり、これは該肝細胞の生存能力の低下をもたらし得る。しかしながら、該FRGマウス系は、限定された生存能力がある肝細胞を生着した場合でさえ、ヒト肝細胞を増大させることができる。
【0069】
ヒト肝細胞を単離する方法は、当技術分野で周知である。例えば、臓器ドナーまたは肝臓切除術からのヒト肝細胞を単離する諸方法が、PCT公開第WO 2004/009766号および第WO 2005/028640号、ならびに米国特許第6,995,299号および第6,509,514号に記載され、それらのすべては、参照することにより本明細書に組み込まれる。肝細胞は、経皮的に、または開腹手術を介して採取された肝生検から得ることができる。FRGマウスのような、受容動物への移植手術のためのヒト肝細胞は、当技術分野で知られている任意の従来の方法によりヒト肝臓組織から単離される。肝臓組織は、機械的に、または酵素的に分離し、単一細胞の懸濁液に供給することができる、さもなければ完全なヒト肝組織のフラグメントを使用してもよい。例えば、該肝細胞は、所定のコラゲナーゼ潅流によりドナー組織から摘出し(Ryan et al.Meth.Cell Biol.13:29,1976)、次いで、低速遠心分離を行う。肝細胞は、その後、ステンレス製のメッシュで濾過することにより精製し、次いで、密度勾配遠心分離法を行うことができる。あるいは、肝細胞を濃縮するための他の諸方法、例えば、蛍光活性化細胞分類、またはパンニング、または磁気ビーズ分離、または遠心力場内の水簸、または当技術分野で周知のような任意の他の方法を使用することができる。類似の肝細胞単離法が、受容マウスの肝臓から増大されたヒト肝細胞を採取するために使用することができる。
【0070】
あるいは、ヒト肝細胞は、Guguen−Guillouzoら(Cell Biol.Int.Rep.6:625−628,1982)により記載される技法を用いて調製され得、これは参照することにより組み込まれる。簡潔に述べると、肝臓またはその一部は、単離され、カニューレは、門脈または門脈管に導入される。該肝臓組織は、その後、37℃で、1分間あたり30から70ミリリットルの流量で、無カルシウム緩衝液、次いで、HEPES緩衝液中で、塩化カルシウム溶液(約0.075%塩化カルシウム等)のコラゲナーゼ(約0.025%コラゲナーゼ)を含有する酵素溶液で、該カニューレを介して潅流される。潅流された該肝臓組織は、小部分(約1立法ミリメートル等)に細分化される。該酵素消化を、上記に記載されるように、同一の緩衝液で、37℃で約10〜20分間、徐々に撹拌しながら行い、細胞懸濁液を生成する。放出した肝細胞は、該細胞懸濁液を60〜80マイクロメーターのナイロン製のメッシュで濾過することにより採取する。採取した肝細胞は、その後、pH7.0で、低温HEPES緩衝液中で洗浄し、低速遠心分離を用いて、コラゲナーゼおよび細胞残屑を除去することができる。非実質細胞は、メトリザミド勾配遠心法により除去してもよい(米国特許第6,995,299号参照)。
【0071】
ヒト肝細胞は、新鮮組織(死亡してから数時間内に得られた組織等)、または新鮮冷凍された組織(約0℃以下で、冷凍保存された新鮮組織等)から得ることができる。好ましくは、該ヒト組織は、検出可能な病原体がなく、形態学および組織学において正常であり、原則的に無病である。生着に使用される肝細胞は、最近、例えば、数時間以内に単離され得る、または細胞が適切な保存培地で保存される場合、より長期間後でも移植することができる。下述の実施例に記載されるこのような一培地は、VIASPAN(登録商標)(腹腔内臓器の保存用の汎用大動脈洗浄および保冷液、ウィスコンシン大学液、またはUWとも称される)である。肝細胞はまた、移植手術前に、凍結保存することができる。肝細胞を凍結保存する諸方法は、当技術分野で周知であり、米国特許第6,136,525号に記載され、これは参照することにより本明細書に組み込まれる。
【0072】
臓器提供者または肝臓切除術からヒト肝細胞を得るステップに加えて、生着のために使用される該細胞は、ヒト幹細胞または、受容動物への移植手術後、増大可能なヒト肝細胞に発達または分化する肝細胞前駆細胞であり得る。ES細胞の性質を有するヒト細胞は、内胚盤胞細胞塊(Thomson et al.,Science 282:1145−1147,1998)、および成長中の生殖細胞(Shamblott et al.,Proc.Natl.Acad.Sci.USA 95:13726−13731,1998)から単離され、ヒト胚幹細胞を産生している(米国特許第6,200,806号参照、参照することにより本明細書に組み込まれる)。米国特許第6,200,806号に開示されるように、ES細胞は、ヒトおよび非ヒト霊長類から産生することができる。通常、霊長類ES細胞は、ES細胞培地の存在下で、マウス胎児線維芽細胞の融合層上に単離される。ES培地は、通常、20%ウシ胎仔血清(FBS;Hyclone)、0.1mM β−メルカプトエタノール(Sigma)、1%非必須アミノ酸液(Gibco BRL)を有する、80%ダルベッコ改変イーグル培地(DMEM;ピルビン酸塩なし、高グルコース、Gibco BRL)からなる。ES細胞の際立った特徴は、成人に存在する前駆「多分化能性」幹細胞と比較して、培養中に無制限に分化されない状態を維持するES細胞の能力、およびES細胞があらゆる異なる細胞型に発達する必要がある潜在能力を含む。ヒトES(hES)は、特異的なモノクローナル抗体により認識される糖脂質細胞表面抗原であるSSEA−4、を発現する(例えば、Amit et al.,Devel.Biol.227:271−278,2000を参照)。
【0073】
ヒト間葉幹細胞(hMSC)に由来したヒト肝細胞もまた、本明細書に記載される諸方法に使用することができる。骨髄由来hMSCの肝性因子への順次暴露は、肝細胞の性質を有する細胞への、幹細胞の分化をもたらす(Snykers et al.BMC Dev Biol.7:24,2007、Aurich et al.Gut.56(3):405−15,2007、これらのそれぞれは、参照することにより本明細書に組み込まれる)。骨髄由来間充織幹細胞と脂肪組織由来幹細胞(ADSC)の肝細胞の分化もまた、記載されている(Talens−Visconti et al.World J Gastroenterol.12(36):5834−45,2006参照、参照することにより本明細書に組み込まれる)。ヒト肝細胞はまた、単球から作製することができる。Ruhnkeら(Transplantation 79(9):1097−103,2005、参照することにより本明細書に組み込まれる)は、最終分化した末梢血単球からの肝細胞様(NeoHep)細胞の生成を記載する。該NeoHep細胞は、形態、肝細胞マーカーの発現、様々な分泌および代謝機能、ならびに薬物解毒活性に関して主要なヒト肝細胞と類似している。さらに、羊膜細胞に由来したヒト肝細胞もまた、本明細書に記載される諸方法に使用することができる。
【0074】
ヒトES細胞株は、存在し、本明細書に開示される諸方法に使用することができる。ヒトES細胞はまた、体外受精した(IVF)胚からの着床前の胚から採取し得る。未使用のヒトIVFから産生した胚は、該胚が14日齢未満である場合、シンガポールおよび英国のような多くの国で許可される。高品質の胚のみが、ES単離に適している。増大された胚盤胞への1つの細胞ヒト胚を培養するための現在の明確な培地条件が、記載されている(Bongso et al.,Hum Reprod.4:706−713,1989参照)。ヒト卵管細胞とのヒト胚の共培養は、高品質の胚盤胞の産生をもたらす。細胞共培養、または改善された合成培地において成長したIVF由来の増大されたヒト胚盤胞は、ヒトES細胞の単離を可能にする(米国特許第6,200,806号参照)。
【0075】
一実施形態において、ヒト肝細胞は、脾臓への注入等の、移植手術により受容マウスに送達される。肝細胞は、肝臓柔組織または門脈への注入等の、他の手段により送達され得る。受容マウスに注入されたヒト肝細胞の数は、異なり得る。一実施形態において、約105〜約107のヒト肝細胞が、注入される。別の実施形態において、約5×105〜約5×106のヒト肝細胞が、注入される。例示的な一実施形態において、約106のヒト肝細胞が、注入される。
【0076】
VI. Fah欠損マウスにおいて増大されたヒト肝細胞の使用
ヒト肝細胞は、当技術分野で知られているいくつかの技法のうちのいずれかを用いて、受容マウスから採取することができる。例えば、マウスは、麻酔することができ、門脈または下大静脈に、カテーテルを用いてカニューレを挿入した。該肝臓は、その後、適切な緩衝液(0.5mM EGTAおよび10mM HEPESが追加された無カルシウムおよび無マグネシウムEBSS)で潅流し、次いで、該肝臓にコラゲナーゼ処理(例えば、0.1mg/ml コラゲナーゼXIおよび0.05mg/ml DNase Iが追加されたEBSSからなる溶液を用いて)を行うことができる。該肝臓は、徐々に細分化し、ナイロン製のメッシュで濾過(70μmと40μmのナイロン製のメッシュでの連続等)し、次いで、遠心分離し、該細胞の洗浄を行った。受容マウスから採取したヒト肝細胞は、当技術分野で周知の任意の技法を用いて、非ヒト細胞または他の混入物質(組織または細胞残屑等)から分離することができる。例えば、このような方法は、ヒト肝細胞に選択的に結合する抗体を用いるステップを含む。このような抗体としては、抗ヒトHLA−A、B、Cのような、クラスI主要組織適合性抗原に特異的に結合する抗体が挙げられるが、これらに限定されない(Markus et al.Cell Transplantation 6:455−462,1997)。ヒト細胞またはヒト肝細胞に対して特異的な抗体は、FACS、またはパンニング、または磁気ビーズ分離を含む、様々な異なる技法に使用することができる。FACSは、他のさらに高度な検出レベルのうちで、複数の色チャネル、ローアングルかつ鈍角の光散乱検出チャネル、およびインピーダンスチャネルを利用して、抗体が結合した細胞を分離または分類する(米国特許第5,061,620号参照)。磁気分離は、1)ヒト特異抗体と接合する、または2)ヒト特異抗体と結合することができる検出抗体と接合する、または3)ビオチン化抗体と結合することができるアビジンと接合する、常磁性粒子の使用を含む。パンニングは、アガロースビーズ、またはポリスチレンビーズ、または中空糸膜、またはプラスチック製のペトリ皿のような、固体マトリックスに結合したモノクローナル抗体を含む。該抗体で結合した細胞は、試料から固体支持体を単に物理的に分離することにより試料から単離させることができる。
【0077】
下述の実施例に記載されるように、いくつかの肝細胞輸送タンパク質を含む、薬物抱合および解毒に関与する遺伝子の発現を、受容マウスから採取した増大されたヒト肝細胞において検出した。最近の研究では、薬物毒性を予測する際に、これらの抱合経路(Kostrubsky et al.Drug.Metab.Dispos.28:1192−1197,2000)、および肝細胞輸送タンパク質(Kostrubsky et al.Toxicol.Sci.90:451−459,2006)が、重要な役割を果たすことを示している。リファンピシンもしくはPB、またはBNFのような外因性薬物によるCYP誘導に対する正常ヒト反応に加えて、核ホルモン受容体転写因子の発現、抱合経路、およびFRGマウスにおいて増大されたヒト肝細胞による主要な輸送タンパク質は、生体内で、ヒト薬物代謝および毒性において、これらの遺伝子産物の役割の評価を可能にする。単離された肝細胞において、化合物の毒性を試験する諸方法は、当技術分野で周知であり、例えば、PCT公開第WO 2007/022419号に記載され、これは参照することにより本明細書に組み込まれる。
【0078】
本開示は、肝臓再構成の治療を必要とする対象体において、肝臓再構成のためにヒト肝細胞の源として、受容マウスにおいて増大され、採取したヒト肝細胞の使用をさらに考慮する。肝細胞の導入による患者における肝臓組織の再構成は、肝臓移植の想定における応急処置として、あるいは単離された代謝低下に罹患する患者に対する最終的治療として、急性肝不全に罹患する患者に対する潜在的な治療法の選択肢である(Bumgardner et al.Transplantation 65:53−61,1998)。肝細胞の再構成は、例えば、遺伝子治療のために遺伝子組み換え肝細胞を導入するために、または疾患、物理的損傷もしくは化学的損傷、または悪性腫瘍によって喪失した肝細胞を交換するために、使用してもよい(米国特許第6,995,299号、参照することにより本明細書に組み込まれる)。例えば、家族性高コレステロール血症を患っている患者の遺伝子治療において、導入された肝細胞の使用が、報告されている(Grossman et al.Nat.Genet.6:335,1994)。さらに、増大されたヒト肝細胞は、人工肝臓補助装置を装着するために使用することができる。
【0079】
FRGマウスにおいて増大され、採取したヒト肝細胞はまた、様々な微生物学的研究のためにも有用である。C型肝炎ウイルスおよびB型肝炎ウイルスを含む、いくつかの病原性ウイルスは、ヒト宿主、またはヒトの初代肝細胞において、唯一複製することができる。したがって、ヒトの初代肝細胞の十分な源を有することは、これらの病原体の研究にとって重要である。増大されたヒト肝細胞は、ウイルス感染および複製の研究および肝臓のウイルスの感染を調整する化合物を同定するための研究に使用することができる。肝臓のウイルスの研究のためにヒトの初代肝細胞を使用する諸方法が、欧州特許第1552740号、米国特許第6,509,514号、およびPCT公開第WO 00/17338号に記載され、これらのそれぞれは、参照することにより本明細書に組み込まれる。
【0080】
VII. ヒト肝臓疾患に対するモデル系としてのFah欠損マウスの使用
免疫不全、かつFah欠損マウスはまた、ヒト肝臓疾患に対するモデル系として使用することもできる。ヒト肝細胞を生着したFah欠損をさらに含む、Rag2−/−/Il2rg−/−マウスを含む、これらのマウス(例えば、FRGまたはFpmRGマウス等)は、例えば、毒素への暴露、感染症、遺伝病、または悪性腫瘍が挙げられるが、これらに限定されない、様々な異なる病因に起因する肝臓疾患のモデルを作製するために使用することができる。ヒト肝細胞を生着し、再構成したFah欠損マウスは、これらの疾患の理解をより深め、疾患経過を予防、または遅延、または逆行し得る薬剤を同定するために、使用することができる。例えば、該Fah欠損マウスは、遺伝子治療ベクターを試験するために使用することができる。着目遺伝子治療用ベクターは、Fah欠損マウスに投与することができ、生着されたヒト肝細胞におけるベクターの効果を評価することができる。
【0081】
同様に、ヒト肝細胞を含むFah欠損マウスは、生体内の設定において、ヒト肝細胞におけるそれらの効果に対して、毒素または医薬品のような、化合物をスクリーニングするために使用することができる。肝疾患を引き起こす、または寄与する疑いがある薬剤は、Fah欠損マウスに対する薬剤の有効量を投与する、および生着されたヒト細胞の機能における薬剤効果を評価することにより、スクリーニングすることができる。一実施例として、Fah欠損マウスは、ヒト向肝性(hepatotrophic)病原体により感染を阻害する、または予防する薬剤を同定するために使用することができる。薬剤等を同定するために、試験薬剤を投与後、または投与前に、Fah欠損マウスを病原体に暴露させる、あるいは該マウスに病原体を接種することができる。該マウスは、その後、感染の兆候に対して評価することができ、任意に、該薬剤で処理されていない、および/または、該病原体により感染させられていない、対照マウスと比較することができる。
【0082】
Fah欠損マウスが、毒素により引き起こされる肝臓疾患のモデルとして使用される場合には、注入されたヒト肝細胞を生着し、該毒剤への暴露前に、好適な期間、増大させる必要がある。肝細胞の増大のために必要とされる時間量は、経験的に判定され得、当業者の能力範囲内である。対応するヒト条件を最も厳密に模倣する成果物を産生するために必要とされる毒剤量は、毒剤の増分量に対して暴露されるいくつかのFah欠損マウスを用いることにより判定されてもよい。毒剤の例としては、アルコール、アセトアミノフェン、フェニトイン、メチルドパ、イソニアジド、四塩化炭素、黄リン、およびファロイジンが挙げられるが、これらに限定されない。
【0083】
Fah欠損マウスが悪性肝臓疾患のモデルとして使用される場合の実施形態において、該悪性腫瘍は、形質転換剤への暴露により、または、悪性細胞の導入により生成されてもよい。形質転換剤または悪性細胞は、ヒト肝細胞の初期のコロナイジング導入とともに、または好ましくは、ヒト肝細胞が宿主動物において増殖開始後、導入されてもよい。形質転換剤の場合、ヒト肝細胞が活発に増殖する時に、該薬剤を投与することが好ましい場合がある。このような形質転換剤は、動物に全身的に、あるいは、肝臓自体に局所的に、投与されてもよい。悪性細胞は、肝臓へ直接接種することができる。一実施例として、Fah欠損マウスに、ヒト肝細胞が移植される。該ヒト肝細胞の生着後、該マウスは、形質転換剤が投与される、または悪性細胞が接種される。あるいは、該形質転換剤または該悪性細胞は、ヒト肝細胞とともに投与することができる。悪性腫瘍が該マウスに発症した後、当技術分野で知られているいくつの方法のうちのいずれか1つにより投与することができ、Fah欠損マウスは、ヒト肝臓癌のモデルとして使用することができる。
【0084】
VIII. ウロキナーゼをコード化するベクター
本明細書に記載される方法の一部の実施形態において、Fah欠損マウスに、ヒト肝細胞の移植手術前、ウロキナーゼをコード化するベクターが投与される。一実施形態において、該ウロキナーゼ(ウロキナーゼプラスミノーゲン活性化因子(uPA)としても知られる)は、分泌型のヒトウロキナーゼである。別の実施形態において、該ウロキナーゼは、修飾された、非分泌型のウロキナーゼである(米国特許第5,980,886号参照、参照することにより本明細書に組み込まれる)。マウスにおけるウロキナーゼの発現に適した任意の型のベクターが企図される。このようなベクターは、プラスミドベクターまたはウイルスベクターを含む。適したベクターとしては、DNAベクター、アデノウイルスベクター、レトロウイルスベクター、偽型レトロウイルスベクター、AAVベクター、テナガザル白血病のベクター、VSV−G、VL30ベクター、リポソーム媒介ベクター等が挙げられるが、これらに限定されない。一実施形態において、該ウイルスベクターはアデノウイルスベクターである。該アデノウイルスベクターは、任意のアデノウイルス血清型を含む、任意の適したアデノウイルスに由来し得る(Ad2およびAd5等であるが、これらに限定されない)。アデノウイルスベクターは、第1、第2、第3、および/または第4世代のアデノウイルスベクター、またはアデノウイルスゲノムをほぼ完全に欠失させた(gutless)アデノウイルスベクターであり得る。非ウイルスベクターは、プラスミド、ホスホリン脂質、異なる構造のリポソーム(陽イオンおよび陰イオン)により構成することができる。別の実施形態において、該ウイルスベクターは、AAVベクターである。該AAVベクターは、当技術分野で知られている任意の適したAAVベクターであり得る。
【0085】
ウロキナーゼをコード化するウイルスおよび非ウイルスベクターは、当技術分野で周知である。例えば、ヒトウロキナーゼをコード化するアデノウイルスベクターは、米国特許第5,980,886号、およびLieberら(Proc.Natl.Acad.Sci.U.S.A.92(13):6210−4,1995)に記載される。米国特許出願公開第2005−176129号および米国特許第5,585,362号は、組み換えアデノウイルスベクターを記載し、米国特許第6,025,195号は、肝臓特的な発現のアデノウイルスベクターを開示する。米国特許出願公開第2003−0166284号は、ウロキナーゼを含む、着目遺伝子の肝臓特異的な発現のアデノ随伴ウイルス(AAV)ベクターを記載する。米国特許第6,521,225号および第5,589,377号は、組み換えAAVベクターを記載する。PCT公開第WO 0244393号は、ヒトウロキナーゼプラスミノーゲン活性化因子遺伝子を含む、ウイルスおよび非ウイルスベクターを記載する。該ヒトウロキナーゼ遺伝子の高レベルの発現が可能である発現ベクターは、PCT公開第WO 03087393号に開示される。前述の特許出願および刊行物のそれぞれは、参照することにより本明細書に組み込まれる。
【0086】
ウロキナーゼをコード化するベクターは、任意に、適切なプロモータ、エンハンサ、転写ターミネータ、タンパク質をコード化する遺伝子の前にある開始コドン(すなわち、ATG)、イントロンのスプライシングシグナル、mRNAの適切な翻訳を可能にするためのその遺伝子の正しいリーディングフレームの維持、および終止コドンを含む、発現調節配列を含むことができる。通常、発現調節配列は、転写を指示するのに十分な最小配列である、プロモータを含む。
【0087】
該発現ベクターは、複製起点、プロモータ、ならびに形質転換細胞の表現型選抜を可能にする特異的遺伝子(抗生物質耐性カセット等)を含むことができる。通常、該発現ベクターは、プロモータを含み得る。該プロモータは、誘導性または構成的であり得る。該プロモータは、組織特異的であり得る。適したプロモータとしては、チミジンキナーゼプロモータ(TK)プロモータ、またはメタロチオネインIプロモータ、またはポリヒドロンプロモータ、またはニューロン特異的エノラーゼプロモータ、またはチロシンヒドロキシラーゼプロモータ、またはベータアクチンプロモータ、または他のプロモータが挙げられる。一実施形態において、該プロモータは、異種プロモータである。
【0088】
一実施例において、ウロキナーゼをコード化する配列は、所望のプロモータの下流にある。任意に、エンハンサ要素もまた、含まれ、通常、該ベクター上のどこかにあり得、増強効果も有する。しかしながら、増大した活性量は、通常、距離とともに減少する。
【0089】
ウロキナーゼをコード化する該ベクターは、静脈内投与、または腹腔内投与、または門脈を介して血管内注入が挙げられるが、これらに限定されない、様々な経路により投与することができる。投与されたベクター量は、異なり、日常の実験を用いて決定され得る。一実施形態において、FRGマウスに、約1×108〜約1×1010のプラーク形成単位の用量で、アデノウイルスウロキナーゼをコード化するベクターが投与される。好ましい一実施形態において、該用量は、約5×109のプラーク形成単位である。
【0090】
例示的な一実施形態において、FRGマウスに、ヒトウロキナーゼをコード化するアデノウイルスベクターが投与される。アデノウイルスベクターは、非常に高い力価のある感染粒子に作製され得る、多種多様の細胞に感染する、分裂してない細胞へ遺伝子を効率的に転送する、およびゲストゲノムにほとんど組み込まれない(これは挿入突然変異により細胞形質転換のリスクを避ける)等の、他の型のウイルスベクターに優るいくつかの利点がある(Douglas and Curiel,Science and Medicine, March/April 1997,pages 44−53、Zern and Kresinam,Hepatology:25(2),484−491,1997)。ウロキナーゼをコード化するために使用することができる代表的なアデノウイルスベクターは、Stratford−Perricaudetら(J.Clin.Invest.90:626−630,1992)、Graham and Prevec(In Methods in Molecular Biology:Gene Transfer and Expression Protocols 7:109−128,1991)、およびBarrら(Gene Therapy,2:151−155,1995)に記載され、これらは参照することにより本明細書に組み込まれる。
【0091】
以下の実施例を提供し、ある特定の特性、および/または実施形態を説明する。これらの実施例は、本発明を、記載される特定の特性または実施形態に制限すると解釈されるべきではない。
【実施例】
【0092】
以下の実施例に記載されるように、ヒト肝細胞の生着および増大は、FRGマウスにおいて、高効率である。FRGマウスの肝臓は、大きさおよび形において、肉眼的に正常であり、組織学的検査は、免疫補体Fah−/−マウスとの間で有意な差異を示さなかった。さらに、FRGマウスは、NTBCを投与した場合、よく生育し、十分に繁殖力があった。
【0093】
一実施形態において、FRGマウスにおける肝臓疾患および選択圧の限度は、NTBCを投与および中止することにより制御することができる(Grompe et al.Nat.Genet.10:453−460,1995)。NTBCの中止は、移植したヒト肝細胞に対する選択的な利点を与える。肝臓疾患の程度を容易に制御することができることは、畜産および手術をさらに容易にする。さらに、該Fah欠損突然変異体は、欠失であり、導入遺伝子の不活性化により野生型に戻すことができない。したがって、内因性復帰マウス細胞からの競合は、Fahノックアウト肝臓には存在しない。これは、FRGマウスは、年齢を問わず、ヒト肝細胞を生着することができ、連続移植は、実現可能であることを意味する。また、重要なことに、FRGマウスは、ヒト細胞を用いた再増殖が50%を超える場合、腎疾患を予防するために必要とされることが前述の研究によって示されている(Tateno et al.Am.J.Pathol.165:901−912,2004)、補体阻害剤の投与なく、高度に再増殖することができる。これは、畜産において実質的に重要だけでなく、薬理学的干渉の潜在源を除去する。
【0094】
FRGマウスにおいて、ヒト肝細胞の再増殖率は、約70%を超えることができる(マウス肝臓において、70%以上の肝細胞がヒト肝細胞である)。単一の再増殖したFRGマウスからの平均収量は、約3,000万から約4,500万のヒト肝細胞である。単一FRGマウスから増大された肝細胞を、その後、連続移植と称されるプロセスにおいて、最高100匹の二次FRG受容マウスで再増殖するために使用することができる。
【0095】
連続移植は、本明細書に記載される新規のシステムにおいて達成することができる。連続移植は、ヒト肝細胞を大規模に増大できるだけでなく、受容マウスのいくつかの世代を通して同一の遺伝子型のヒト細胞を増大させるための手段を提供する。また、さらなる移植手術のために高品質の源のヒト肝細胞を所有することは、重要である。少なくとも4回の肝細胞移植手術が実現可能であることを本明細書に実証している。さらに、連続移植から単離されたヒト肝細胞の生存能力は、約80%を超えることができ、該肝細胞は、コラーゲンで被覆したプレートに容易に結合する。10%の生着効率(100,000の細胞)、および再増殖後の約1,500万の増大されたヒト肝細胞の最終採取の概算に基づいて、少なくとも150倍の生体内での増大を毎回達成することができる。したがって、FRGマウスにおいて、ヒト肝細胞の全増大は、5億倍を超え得る。
【0096】
ヒト肝細胞の生着および増大は、様々な源および可変品質からの単離されたヒト肝細胞を用いて、FRGマウスにおいて、可能である。ヒト肝細胞の源としては、死体ドナー、肝臓切除術、および商業的に入手可能な源が挙げられるが、これらに限定されない。本明細書に示されるように、ヒト肝細胞の生着および増大は、あらゆる年齢のドナーを用いて、成功した。さらに、ヒト肝細胞を、該肝細胞があらかじめ凍結保存された場合でさえ、FRGマウスにおいてうまく増大させることが、本明細書に実証される。これは、肝細胞単離後、直ちに移植手術を必要とするシステムに優る重要な利点である。
【0097】
実施例1:Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスの作製
免疫不全Fahノックアウトマウスのいくつかの株を作製した。Fah突然変異体をヌード、nod/scidまたはRag1−/−バックグラウンドと交錯することは、不成功であった。T細胞、B細胞、およびNK細胞が完全に欠如するが、DNA修復欠損のない、免疫不全Fah−/−マウス株を作製するために、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスを作製した。オスFah−/−129S4マウス(Grompe et al.Genes Dev.7:2298−2307,1993)を、メスRag2−/−/Il2rg−/−マウス(Taconic)と交配させた。すべての動物に、1.6mg/Lの濃度で、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン(NTBC)を含有する飲料水を与えてすべての動物を維持した(Grompe et al.Nat.Genet.10:453−460,1995)。各動物の遺伝子型を確認するために、PCRベースの遺伝子タイピングを、足指組織から単離した200ngのゲノムDNAに実行した(Grompe et al.Genes Dev7:2298−2307,1993、Traggiai et al.Science 304:104−107,2004)。
【0098】
飲料水内にNTBCを継続して与えた場合、FRGマウスはよく生育し、十分な繁殖力があった。FRGマウス肝臓は、大きさおよび形において、肉眼的に正常であり、組織学的検査は、従来のFah−/−マウスと該FRGマウスとの間で差異を示さなかった。従来のFah−/−マウスの場合のように、NTBCの中止は、FRGマウスにおいて、漸次の肝臓細胞損傷をもたらし、4〜8週間後、FRGマウスは最終的に死亡した(Overturf et al.Nat.Genet.12:266−273,1996)。
【0099】
実施例2:組織学および生着検出アッセイ
組織学および免疫細胞化学
FAH免疫組織化学を、前述のように行った(Wang et al.Am.J.Pathol.161:565−574,2002)。簡潔に述べると、pH7.4で、10%リン酸緩衝ホルマリン中で固定された肝臓組織および腎臓組織は、100%エタノール中で脱水され、58℃で、パラフィンワックスに包埋された。脱パラフィン処理した4μmの切片を、ヘマトキシリンおよびエオシンで染色した。免疫組織化学のために、切片は、3%H2O2を含むメタノールで、内因性ペルオキシダーゼのブロッキングのために処理した。また、アビジンおよびビオチンブロッキングは、一次抗体でインキュベーションする前に、行なった。切片は、室温で2時間、抗FAHウサギ抗体またはHepPar抗体(DAKO)でインキュベートし、次いで、HRPに接合した二次抗体でインキュベートした。シグナルは、ジアミノベンジジン(DAB)により検出した。
【0100】
FAH酵素アッセイ
フマリルアセト酢酸は、受容者の肝臓からの細胞質の肝臓分画とともにインキュベートし、消失速度を、330nmで分光学的に測定した。野生型およびFah−/−肝臓はそれぞれ、陽性対照および陰性対照として使用した。フマリルアセト酢酸は、ホモゲンチジン酸から酵素的に調製した(Knox et al.Methods Enzymol.2:287−300,1955)。
【0101】
Alu配列のためのゲノムPCR
ゲノムDNAは、DNeasy組織キット(Qiagen)を用いて、肝臓から単離した。ヒトAlu配列は、以下のプライマー5’−GGCGCGGTGGCTCACG−3’(配列番号1)および5’−TTTTTTGAGACGGAGTCTCGCTC−3’(配列番号2)を用いた標準手順に従って、PCRにより増幅した。
【0102】
肝細胞特異遺伝子発現のためのRT−PCR
総RNAは、Rneasyミニキット(Qiagen)を用いて、肝臓から単離した。相補的DNAは、オリゴdTプライマーを用いて逆転写酵素により合成した。表1に示されるプライマーは、ヒトまたはマウス特異的cDNA増幅のために使用した。
【0103】
【表1】
【0104】
ヒトアルブミンの測定
少量の血液を、ヘパリン化された毛細血管を有する左側の伏在静脈から1週間に1回採取した。トリス緩衝食塩水で1,000倍または10,000倍希釈した後、ヒトアルブミン濃度は、製造業者のプロトコルに従って、ヒトアルブミンELISA Quantitation Kit(Bethyl)を用いて測定した。
【0105】
蛍光免疫細胞化学
ヒト化マウス肝臓からの肝細胞を、ダルベッコ改変イーグル培地(DMEM)中で懸濁し、コラーゲン型1被覆6ウェルプレート上に播種した。付着した細胞を、4%パラホルムアルデヒドで、15分間固定し、5%スキムミルクでブロックした。ウサギ抗FAH、ヤギ抗ヒトアルブミン(Bethyl)、ヤギ抗マウスアルブミン(Bethyl)を、1/200の希釈で、一次抗体として使用した。ALEXA(登録商標)フルオロ488抗ヤギIgG(Invitrogen)またはALEXA(登録商標)フルオロ555抗ウサギIgG(Invitrogen)を、二次抗体として使用した。該画像は、Nikonのデジタルカメラを用いることによりAXIOVERT(登録商標)200顕微鏡で捕捉した。
【0106】
FACS解析
受容者の肝臓の解離後、実質細胞を、フルオレセインイソチオシアネート(FITC)接合した抗ヒトヒト白血球抗原(HLA)−A、B、C(BD Pharmingen)、およびフィコエリトリン(PE)を接合した抗マウスH2−K(b)(BD Pharmingen)抗体と共に、4℃で30分間インキュベートした。その後、これらをPBSで2回洗浄し、FACS CALIBUR(登録商標)(Becton Dickinson)フローサイトメーターで解析した。FITCを接合したIgGおよびPEを接合したIgGを、負の対照として使用した。
【0107】
蛍光原位置ハイブリダイゼーション
全ゲノムDNAプローブは、全マウスゲノムDNAおよび全ヒトゲノムDNAのニックトランスレーション法により作製した。Cy3−dUTPの取り込みは、製造業者の推奨(Invitrogen)に従って実行した。最終プローブ濃度は、200ng/μlであった。付着した細胞のあるスライドを、37℃で1時間、100mg/mlのRnaseで処理し、2X SSC中で3回(1回3分間)洗浄した。洗浄ステップ後、スライドを、70%、90%、および100%エタノール中で、3分間、それぞれ脱水した。染色体を、75℃で3分間、70%ホルムアミド/2X SSC中で変性し、次いで、冷70%、90%、および100%エタノールで3分間、それぞれ脱水した。プローブカクテルを、75℃で10分間変性し、37℃で30分間あらかじめハイブリダイズした。プローブを、スライドに適用し、湿潤チャンバ中で、37℃で一晩インキュベートした。ハイブリダイゼーション後の洗浄は、50%ホルムアミド/2X SSC中で3回の3分洗浄、PN緩衝液(0.1M Na2HPO4、0.1M NaH2PO4、pH8.0、2.5% NONIDET(登録商標)P−40)中で3回の3分洗浄において、すべて45℃で行われた。スライドは、その後、紫外線蛍光(Zeiss)下で、ヘキスト(0.2ug/ml)で対比染色し、カバースリップをかぶせ、観察した。
【0108】
実施例3:ヒト肝細胞の単離および凍結保存
ヒト肝細胞は、前述の手順に従って、肝臓移植手術のために使用されないドナー肝臓から単離した(Strom et al. Cell Transplant.15:S105−110,2006)。簡潔に述べると、肝臓組織を、0.5mM EGTA(Sigma)およびHEPES(Cellgro)が追加された無カルシウムおよび無マグネシウムのハンクス液(Cambrex)で潅流し、次いで、既存の脈管構造を通してイーグル最小必須培地(Cambrex)中で、100mg/LのコラゲナーゼXI(Sigma)および50mg/LのデオキシリボヌクレアーゼI(Sigma)を用いて消化した。該細胞を、イーグル最小必須培地と7%小ウシ血清(Hyclone)で、50xgで2分間、3回洗浄した。ペレット化された肝細胞は、冷VIASPAN(登録商標)(腹腔内臓器の保存用の汎用大動脈洗浄および保冷液、ウィスコンシン大学液、またはUWとも称される)に移した。
【0109】
輸送された肝細胞は、1mlあたり5×106の肝細胞で、10%ウシ胎仔血清および10%ジメチルスルホキシドが追加されたVIASPAN(登録商標)溶液に移した。該クリオチューブ(cryotube)を、紙タオルで厚く覆い、−80℃で1日間保存し、液体窒素に最終的に移した。解凍のために、細胞を、37℃の水浴中で急速に再加熱し、DMEMを徐々に加え、DMSO濃度の変化速度を最小化した。
【0110】
実施例4:ヒト肝細胞を用いたFRGマウス肝臓の再増殖
ウロキナーゼの過剰発現は、いくつかの系において、肝細胞生着を強化することが示されている(Lieber et al.Hum.Gene Ther.6:1029−1037,1995)。したがって、実験を行い、ヒト肝細胞の移植手術前、ウロキナーゼを発現するアデノウイルスの投与が有益であるかどうかを判定した。分泌型のヒトウロキナーゼ(ウロキナーゼプラスミノーゲン活性化因子;uPA)を発現するアデノウイルスベクターは、先に説明されている(Lieber et al.Proc.Natl.Acad.Sci.U.S.A.92:6210−6214,1995および米国特許第5,980,886号参照、参照することによって本明細書に組み込まれる)。
【0111】
ドナー肝細胞は、単離し、単離してから24〜36時間後に、移植した。大抵の場合、細胞は、VIASPAN(登録商標)溶液中に保存し、輸送中、4℃に保った。しかしながら、2つの実験において、凍結保存した肝細胞を移植した。ドナー肝細胞の生存能力および品質は、10%〜60%の範囲の平板効率を伴い、大いに変化した。
【0112】
移植手術のために、以下の汎用プロトコルを使用した。成熟(6〜15週齢)オスまたはメスFRGマウスに、移植手術する24〜48時間前に、uPAアデノウイルス(マウスあたり5×109のプラーク形成単位(PFU))の静脈注射(後眼窩注射)をした。100μlのダルベッコ改変イーグル培地中で、100万の生存能力のあるヒト肝細胞(トリパンブルー色素排除試験法により判定された)を、27ゲージ針で脾臓内に注入した。NTBCの投与を、次の6日間(1.6mg/L 0〜2日目、0.8mg/L 3〜4日目、0.4mg/L 5〜6日目)にわたり徐々に中止し、移植手術から1週間後、完全に中止した。NTBCの投与を停止してから2週間後、動物は、5日間薬物の投与を再開した後、再度中止した。
【0113】
3つの別個の移植手術において、ヒト肝細胞の一次生着は、最初にuPAアデノウイルスを受けた受容者であるFRGマウスにおいて観察した。したがって、uPAの前処理レジメンを、ほとんどのその後の移植手術実験において使用した。
【0114】
全体で、9つの異なるドナーからのヒト肝細胞をうまく使用して、uPAアデノウイルスレジメンの導入後、生着の失敗は生じなかった。これらのうち、7つは、脳死ドナーの肝臓から単離し、2つは、外科的肝臓切除術から単離した。ドナーの年齢は、1.2歳から64歳の範囲で異なった(表2)。
【0115】
【表2】
【0116】
すべての実験において、少なくとも1つの受容者に、細胞バッチの使用にかかわらず、本プロトコルを用いて、ヒト肝細胞が有意に生着された(>1%ヒト細胞)。生着は、組織学、DNA解析、酵素アッセイ、および後の実験である、ヒト血清アルブミンを含む、異なる方法により実証された。アルブミンレベルによりモニターされた移植手術において、43のうちの17(39.5%;12〜67%)の一次受容者は再増殖した(表2および図3)。これらのうち、7つは、高度に再増殖し(30〜90%)、アルブミンレベル>1mg/mlに達した。死体肝臓からの肝細胞だけでなく、肝臓切除術からの肝細胞も生着した。さらに、凍結保存細胞も、無事に生着された。
【0117】
高度に生着されたマウス(>30%再増殖)において、移植されたFRGマウスの体重は、第2のNTBC中止中に、安定したのに対して、同一の細胞を与えたIl2rg(Fah−/−/Rag2−/−/Il2rg+/−)のヘテロ接合型である免疫不全同腹仔のうち減量し続けたものは少なかった(図1a)。三重変異体マウスにおいて、この体重の安定は、移植されたヒト肝細胞が、罹患Fah−/−受容肝細胞の機能を置換していることを示唆した。完全に体重が安定化するや否や(初期移植手術から2〜3ヶ月後)、受容者の肝臓は、その後、採取された。肉眼的に、FRG肝臓は、大きさおよび形において、正常であり、肉眼的結節はなかった。ヒト特異的Alu DNA配列のゲノムPCRは、FRG受容者の肝臓において陽性であったのに対して、Il2rgヘテロ接合体は、すべて陰性であった(図1b)。再増殖細胞の肝細胞機能を直接確認するために、FAH酵素活性を、アッセイした(Knox et al.Methods Enzymol.2:287−300,1955)。
【0118】
受容マウスの肝臓は、正常マウス肝臓と同等またはそれを超える、多量のFAH酵素活性があった(図1c〜e)。FAHは、十分に分化した肝細胞において、単独に発現するため、移植されたヒト肝細胞は、マウス肝臓に生着された場合、脱分化または異常がないことを示唆した。FAH免疫染色により、70%以上の肝実質が、FAH陽性ヒト肝細胞によって再増殖されたことが確認された(図1fおよび図1g)。
【0119】
組織学的および免疫組織化学的試験は、追加の受容者の肝臓を用いて行った(図2aおよび図2b)。FAH陽性ヒト肝細胞は、受容者の肝臓の構造に完全に統合されたようであった。いくつかの受容者において、生着された肝細胞は、受容者の肝臓組織を阻害することなく、80%以上の柔組織を占めた(図2b、2e、および2f)。クローン的に増加しているヒト肝細胞は、形態学的に、大きさにより、およびそれらの蒼白の細胞質により、マウス肝細胞とは明らかに区別することができた(図2cおよび図2d)。ヒト肝細胞のその大きさは、恐らく、上述に報告されるようにグリコーゲン蓄積のため、比較的大きく、それらの細胞質は、鮮明であった(Meuleman et al.Hepatology 41:847−856,2005)。FAH陽性肝細胞はまた、HepPar抗体に対しても陽性であり、この抗体はヒト肝細胞を特異的に標識化するが、マウス肝細胞は、標識化しない(図2eおよび図2f)。対照的に、FAH陰性領域は、NTBC投与の中止後、従来のFah−/−マウスにおいて所見と一致する、壊死炎症(necroinflammation)を示し、異形成肝細胞を含んだ。
【0120】
再増殖したヒト肝細胞が、成熟肝細胞特異遺伝子を発現するかどうかを検査するために、RT−PCRを、受容者の肝臓から抽出されたメッセンジャーRNAに行った。ヒトアルブミン(ALB)遺伝子、FAH遺伝子、トランスフェリン(TF)遺伝子、トランスサイレチン(TTR)遺伝子、チロシンアミノ転移酵素(TAT)遺伝子、およびUGT1A1遺伝子が、受容者の肝臓において、多量に発現した(図3a、図6c、および図7)。肝細胞の機能性をまた、ヒトアルブミンの血中濃度を測定することにより評価した。ヒトアルブミンに対して特異的なELISAキットを使用し、検出の閾値は、1:100に希釈した試料を用いて、0.05μg/mlであった。ヒトアルブミンは、まず、一次受容者において、移植手術から4〜10週間後に検出された。当初は、ヒトアルブミンレベルに変動があったが、その後、ヒトアルブミンの濃度は、さらに数週間、相対的に漸増した(図3bおよび図3c)。
【0121】
薬理作用のアルプロティナーゼ阻害剤は、高度に再増殖したマウスの生存能力を長期間維持するために必要であり得、該ドナー肝細胞により産生されたヒト補体は、受容者の腎臓に損害を与える可能性がある(Tateno et al. Am.J.Pathol.165:901−912,2004参照)。したがって、いくつかの(n=3)高度に再増殖したマウスを、長期間観察した。これらのマウスは、NTBCの投与を中止した4ヶ月間減量しなくて、マウスのヒトアルブミン濃度は、安定していた。さらに、該マウスの腎臓は、採取時に、肉眼的かつ組織学的に正常であった(図2g)。
【0122】
実施例5:ヒト肝細胞の連続移植
前述の肝臓異種再増殖モデルの限界のうちの1つは、生着されたヒト肝細胞をさらに増大させる能力の低下である。該FRGマウス系において、連続移植の実行可能性を試験するために、高度に再増殖した一次受容者の肝臓(約70%ヒト細胞)を潅流し、実質肝細胞を、標準コラゲナーゼ消化プロトコルを用いて採取した。
【0123】
ヒト細胞で再増殖したマウスに麻酔をかけ、門脈または下大静脈に、24ゲージのカテーテルを用いてカニューレを挿入した。該肝臓を、0.5mM EGTAおよび10mM HEPESが追加された無カルシウムおよび無マグネシウムのEBSSで5分間潅流した。該溶液は、0.1mg/mlのコラゲナーゼXI(sigma)および0.05mg/mlのDNase I(sigma)が追加されたEBSSに10分間変更した。該肝臓は、第2の溶液中で徐々に細分化し、70μmおよび40μmのナイロン製のメッシュで連続して濾過した。5分間、150xgで遠心分離した後、該ペレットを、50xgで2分間、さらに2回洗浄した。細胞の数および生存能力は、トリパンブルー色素排除試験法により評価した。
【0124】
100μlのDMEM中に懸濁された100万の生存能力のある細胞を、27ゲージ針で受容者の脾臓に注入した。Fah陽性ヒト肝細胞とFah陰性マウス肝細胞に分離することなく、二次FRG受容者への肝細胞の移植手術を行った。一次生着のために使用した該細胞と対照的に、本方法において採取されたヒト肝細胞の生存能力は、>80%であり、それらは、コラーゲンで被覆した培養プレートに容易に付着した(図4c〜e)。二次受容者の生着後、該連続移植は、類似の方法において、三次および四次受容者に対して継続された。各世代において、すべてではないが、一部の受容マウスのヒト血清アルブミンは、高度に陽性になった(図4a)。高度に再増殖したマウスの割合は、順次移植受容者においてさらに高く(7/43と比較して17/28)、アルブミンの増加率は、さらに一貫していた(図3e)。これは、連続継代の肝細胞が、最も移植可能なヒト肝細胞の質を高めることを表す可能性がある、あるいは、ドナーマウスから新たに採取した細胞のさらなる高品質および生存能力を単に反映する可能性がある。アルブミン陽性マウスからの肝臓試料のゲノムPCRは、各世代において、ヒトDNAの存在を示した(図4b)。ヒト肝細胞による肝臓再増殖はまた、FAHに対する蛍光免疫染色により確認された(図4c〜e)。組織学的検査は、生着されたヒト肝細胞が、各世代において、形態学的に類似し、明らかにFAH陽性であることを示した(図4f〜h)。
【0125】
実施例6:肝細胞再増殖は、細胞融合の結果ではない。
【0126】
ウロキナーゼトランスジェニックマウスにおいて、一次細胞を用いた肝臓再増殖の最新報告は、細胞融合が、明らかな「肝細胞再増殖」を潜在的に明らかにすることを実証した(Okamura et al.Histochem.Cell Biol.125:247−257,2006)。uPAトランスジェニックマウスが、その研究に使用されたため、これらの所見は、細胞融合がまた、他の報告において、マウス肝臓ヒト化のメカニズムでもあることの可能性を高めた。造血細胞と肝細胞との間の細胞融合はまた、Fah欠損マウスにおいても観察されている(Wang et al.Nature 422:897−901,2003)。マウス細胞とヒト細胞との間の細胞融合は、薬学的応用のために、ヒト化マウス肝臓の価値を大幅に減退し得る。該再増殖した肝細胞が、元来は真にヒト肝細胞であったことを確認するために、ヒトまたはマウス特異的アルブミン、およびFAHに対して二重免疫染色を行った。ほとんど(>95%)のマウスアルブミン陽性肝細胞は、実際には、FAH陰性であり、ほとんどのFAH陽性肝細胞は、マウスアルブミン陰性であった(図5a〜c)。他方では、ほとんどすべて(>90%)のヒトアルブミン陽性肝細胞はまた、FAH陽性であるが、一方残りの肝細胞は、二重陰性であった(図5d〜f)。
【0127】
アルブミンは、分泌タンパク質であり、故に、細胞は、他の細胞から異種アルブミンを得ることにより抗体陽性を示すことができた。細胞融合の欠如をさらに確認するために、ヒトおよびマウス抗主要組織適合性複合体(MHC)抗体を、フローサイトメトリーのために使用した。各抗体は、種特異であることが確認された(図5g〜j)。両方の種の表面マーカーとも肝細胞陽性でないことが、高度に再増殖した肝臓に認められた(図5kおよび図5l)。
【0128】
最終的に、蛍光原位置ハイブリダイゼーション(FISH)を、高度に再増殖した移植受容者から得た肝細胞に、ヒトおよびマウス全ゲノムプローブを用いて行った。高度に再増殖した一次(キメラマウス#531)および三次(キメラマウス#631)マウスから得た肝細胞を、ヒト全ゲノムDNAあるいはマウス全ゲノムDNAを用いてハイブリダイズした。該ヒトプローブまたは該マウスプローブに対して陽性である細胞の割合を記録した(表3)。対照は、純ヒトおよびマウス肝細胞、またはヒト肝細胞とマウス肝細胞の等混合であった。キメラ肝臓において見出されたヒト細胞が、細胞融合の産物であった場合、多くの肝細胞は、ヒトプローブとマウスプローブの両方に対して二重陽性であり、故に、マウスDNAおよびヒトDNAが陽性である細胞の割合は、100%を超え得る。それどころか、割合合計は、ヒト肝細胞とマウス肝細胞の混合の場合と同じように、ほぼ100%であると概算された。さらに、ヒト肝細胞は、脾臓が、ヒト細胞の融合パートナーとしての役割をし得るマウス肝細胞を欠如するという事実にもかかわらず、高度に再増殖したマウスの脾臓に検出された(図2h)。したがって、二重陽性細胞(融合産物)は、ヒト細胞の大部分を占めることができなかった。
【0129】
【表3】
総合すれば、これらの結果は、融合事象が生じる場合、融合事象は、まれであり、再増殖する細胞の大部分は、連続移植を行う場合でさえ、純ヒト由来からなることを示す。したがって、FRGマウスにおいて増大されるヒト肝細胞は、ヒトの遺伝的および生化学的性質のみ有する。
【0130】
実施例7:ヒト化マウスにおける薬物代謝の機能的特性
キメラマウスにおける、ヒト肝臓特異的遺伝子の基底の発現および誘導を、検査した。培養された肝細胞におけるテストステロン代謝およびエトキシレソルフィン−O−デエチラーゼ(EROD)活性の評価を、それぞれ、Kostrubskyら(Drug Metab. Dispos.27:887−894,1999)、およびWenら(Drug Metab.Dispos.30:977−984,2002)に記載されるように実行した。RNA単離、cDNA合成、およびリアルタイムPCRを、Komoroskiら(Drug Metab.Dispos.32:512−518,2004)に記載されるように実行した。Applied Biosystemsから得られたプライマーは、ヒトCYP1A1(Hs00153120_m1)、CYP1A2(Hs00167927_m1)、CYP3A4(Hs00430021_m1)、CYP3A7(Hs00426361_a1)、CYP2B6(Hs00167937_g1)、CYP2D6(Hs00164385_a1)、多剤体制関連タンパク質MRP2(Hs00166123_m1)、胆汁塩輸出ポンプBSEP、(Hs00184829_m1)、CAR(Hs00231959_m1)アルブミン(Hs00609411_m1)、HNF4α(Hs00230853_m1)、シクロフィリン(Hs99999904_m1)、およびマウスアクチン(Ma00607939_s1)に対して特異的であった。
単離された肝細胞の培養物を、構築し、シトクロムP450遺伝子の原型誘導物質に暴露した。該結果は、シトクロム(CYP1A1、CYP1A2、CYP2B6、CYP3A4、CYP3A7)、輸送(BSEP、MRP2)、および薬物抱合酵素(UGT1A1)の基底遺伝子発現レベルは、培養された正常成熟ヒト肝細胞に見られる、まさにそのものであった(図6c、図7)。さらに、ベータナフトフラボン(BNF)、フェノバルビタール(PB)、およびリファンピシン(Rif)のような化合物により誘発された遺伝子型は、正常ヒト細胞から予期されたとおりであった。ヒト薬物代謝遺伝子のmRNA発現レベルに加えて、ヒトCYP1Aおよび3Aファミリーメンバーの酵素活性を測定した。エトキシレソルフィン−O−デエチラーゼ活性(EROD)は、ヒト肝臓において、CYP1A1および1A2により媒介されることが知られている。該結果は、EROD活性が、ヒト化マウス肝臓細胞において、BNFへの事前暴露により特異的かつ堅牢的に誘導されたことを示す(図6a)。逆に、PBまたはリファンピシンへの事前暴露は、CYP3A4を介した代謝の特異的測定である6−ベータ−ヒドロキシルテストステロンへのテストステロンの転換を特異的に誘導した(図6b)。したがって、再増殖したFRG肝臓からの肝細胞は、これらの標準薬物代謝アッセイにおいて、正常成熟ヒト肝細胞との区別がつかなかった。
【0131】
実施例8:肝細胞再増殖前のマクロファージの減損
一次生着は、ウロキナーゼアデノウイルスの事前投与を行っても、100%のFRG受容マウスで生じなかった。FRGマウスにおいて、正常数存在する肝臓マクロファージは、先天性免疫反応を促進することにより、ヒト細胞生着を限定することが可能である。
【0132】
潜在的なマクロファージにより惹起される免疫反応を中止するために、FRGマウスから、ヒト肝細胞の移植手術前に、マクロファージを減損させる。マクロファージ減損は、化学的減損(Schiedner et al. Mol.Ther.7:35−43,2003)、または抗体(McKenzie et al.Blood 106:1259−1261,2005)を用いることを含む、当技術分野で周知のいくつかの方法のうちのいずれか1つを用いて達成され得る。マクロファージはまた、クロドロネート含有リポソームを用いて減損され得る(van Rijn et al.Blood 102:2522−2531,2003)。マクロファージを減損するための追加の化合物および組成物は、米国特許公開第2004−0141967号およびPCT公開第WO 02/087424号に記載され、参照することにより本明細書に組み込まれる。マクロファージ減損後、FRGマウスに、本明細書の前述の実施例に記載される方法に従って、ヒト肝細胞を移植する、または連続移植する。
【0133】
実施例9:FpmRGマウスにおけるヒト肝細胞の生着
FRGマウスは、Fah遺伝子のエクソン5(FahΔexon5)に欠失を含む。ヒト肝細胞を、Fah欠損の他のモデルにおいて、生着および増大し得ることを確認するために、Fah遺伝子に点変異を含むマウス株を作製した。Fah点変異(Fahpm)マウスと呼ばれる、これらのマウスは、ミススプライシングを起させFah mRNAにおいて、エクソン7の喪失を生じさせる該Fah遺伝子に点変異を有する(Aponte et al.,Proc.Natl.Acad.Sci.USA 98:641−645,2001、参照することにより本明細書に組み込まれる)。表現型における差異は、FahpmマウスとFahΔexon5マウスとの間で検出されなかった。
【0134】
FahpmマウスをRag2−/−/Il2rg−/−マウス(実施例1に記載されるような)と交配させて、ホモ接合性Fahpm/Rag2−/−/Il2rg−/−(FpmRG)三重変異体マウスを作製した。FpmRGマウスの2つのコホートに、実施例4に記載される方法に従って、ヒト肝細胞を移植した。肝細胞移植手術の約24〜48時間前、マウスは、uPAアデノウイルスの静脈注射(後眼窩注射)を受けた。比較するために、同時に、FRGマウスに、ヒト肝細胞を移植した。ヒト血清アルブミンを、移植手術から2ヶ月および3ヶ月後に、極めて有意なレベル(23μg/ml)で、FpmRGマウスの末梢血に検出した。ヒト血清アルブミンのこれらの血中濃度は、同時に移植されたFRGマウスに見られる濃度と似ていた。
【0135】
これらの結果は、FpmRGマウスが、FRGマウスと同程度に、ヒト肝細胞で再増殖することができることを示す。したがって、ヒト化肝臓再増殖は、FahΔexon5変異を有するFRGマウスに特有ではないが、Fah欠損マウスの任意の株で達成することができる。
【0136】
本開示は、生体内でヒト肝細胞を増大させるための方法を提供する。本開示は、生体内でヒト肝細胞を増大させるのに有用な遺伝子組み換え型免疫不全マウスをさらに提供する。記載される方法の正確な詳細は、本発明の趣旨から逸脱することなく、変更または修正することができることは明らかであろう。下記の特許請求の範囲および趣旨に入る範囲内ですべてのこのような修正および変更を請求する。
【0137】
配列表
添付の配列表に記載される核酸およびアミノ酸配列は、37C.F.R.1.822に定義されるように、ヌクレオチド塩基の標準的な文字略語、およびアミノ酸の三文字コードを示す。各核酸配列の1本鎖のみを示すが、該相補鎖は、表示された該鎖に言及するときは常にそれに含まれるものとして理解する。添付の配列表において、
配列番号1および2は、ヒトAlu配列を増幅するためのPCRプライマーの核酸配列である。
【0138】
配列番号3は、ヒトALB順方向RT−PCRプライマーの核酸配列である。
【0139】
配列番号4は、ヒトALB逆方向RT−PCRプライマーの核酸配列である。
【0140】
配列番号5は、マウスAlb順方向RT−PCRプライマーの核酸配列である。
【0141】
配列番号6は、マウスAlb逆方向RT−PCRプライマーの核酸配列である。
【0142】
配列番号7は、ヒトTAT順方向RT−PCRプライマーの核酸配列である。
【0143】
配列番号8は、ヒトTAT逆方向RT−PCRプライマーの核酸配列である
配列番号9は、ヒトTF順方向RT−PCRプライマーの核酸配列である。
【0144】
配列番号10は、ヒトTF逆方向RT−PCRプライマーの核酸配列である。
【0145】
配列番号11は、ヒトFAH順方向RT−PCRプライマーの核酸配列である。
【0146】
配列番号12は、ヒトFAH逆方向RT−PCRプライマーの核酸配列である。
【0147】
配列番号13は、ヒトTTR順方向RT−PCRプライマーの核酸配列である。
【0148】
配列番号14は、ヒトTTR逆方向RT−PCRプライマーの核酸配列である。
【0149】
配列番号15は、ヒトUGT1A1順方向RT−PCRプライマーの核酸配列である。
【0150】
配列番号16は、ヒトUGT1A1逆方向RT−PCRプライマーの核酸配列である。
【特許請求の範囲】
【請求項1】
生体内でヒト肝細胞を増大させる方法であって、
i)単離されたヒト肝細胞をRag2−/−/Il2rg−/−マウスに移植するステップであって、前記マウスが、Fahの発現が欠損している、ステップと、
ii)前記ヒト肝細胞を、少なくとも約2週間増大させるステップと、
iii)前記マウスからヒト肝細胞を採取するステップと、を含む、方法。
【請求項2】
前記マウスが、前記Fah遺伝子において、欠失に対してホモ接合体である、請求項1に記載の方法。
【請求項3】
前記マウスが、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである、請求項2に記載の方法。
【請求項4】
前記マウスが、前記Fah遺伝子において、点変異に対してホモ接合体である、請求項1に記載の方法。
【請求項5】
前記マウスが、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである、請求項4に記載の方法。
【請求項6】
ヒトウロキナーゼをコード化するベクターが、前記ヒト肝細胞を注入する前に、前記マウスに投与される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記ウロキナーゼが、分泌型のウロキナーゼである、請求項6に記載の方法。
【請求項8】
前記ウロキナーゼが、非分泌型のウロキナーゼである、請求項6に記載の方法。
【請求項9】
前記ベクターが、アデノウイルスベクターである、請求項6〜8のいずれか1項に記載の方法。
【請求項10】
前記ベクターが、アデノ随伴ウイルスベクターである、請求項6〜8のいずれか1項に記載の方法。
【請求項11】
前記ベクターが、静脈内投与される、請求項6〜10のいずれか1項に記載の方法。
【請求項12】
前記ベクターが、肝細胞注入の約24〜48時間前に投与される、請求項6〜11のいずれか1項に記載の方法。
【請求項13】
前記マウスに、肝細胞注入前に、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン(NTBC)が投与される、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記NTBCが、約0.05mg/kg/日から約0.10mg/kg/日の用量で投与される、請求項13に記載の方法。
【請求項15】
前記マウスに、肝細胞注入後、少なくとも約3日間、NTBCがさらに投与される、請求項13または請求項14に記載の方法。
【請求項16】
前記マウスに、肝細胞注入後、少なくとも約6日間、NTBCがさらに投与される、請求項13または請求項14に記載の方法。
【請求項17】
NTBCの前記用量が、肝細胞注入後、6日間の過程にわたり漸減される、請求項16に記載の方法。
【請求項18】
前記NTBCが、飲料水内投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項19】
肝細胞注入前、飲料水内投与されるNTBCの濃度が、約1mg/Lから約8mg/Lである、請求項18に記載の方法。
【請求項20】
前記NTBCが、食物内投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項21】
前記NTBCが、注入することにより投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項22】
前記ヒト肝細胞が、少なくとも約6週間、増大可能である、請求項1〜21のいずれか1項に記載の方法。
【請求項23】
前記ヒト肝細胞が、最長約6ヶ月間、増大可能である、請求項1〜21のいずれか1項に記載の方法。
【請求項24】
前記単離されたヒト肝細胞が、前記マウスの脾臓または門脈に注入される、請求項1〜23のいずれか1項に記載の方法。
【請求項25】
前記増大されたヒト肝細胞は、前記マウスの肝臓から採取される、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
前記ヒト肝細胞が、臓器ドナーの肝臓から単離された、または外科的切除術から単離された、または幹細胞もしくは単球もしくは羊膜細胞に由来した、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
前記肝細胞は、注入前、凍結保存された、請求項26に記載の方法。
【請求項28】
肝細胞注入前、前記FRGマウスからマクロファージを減損させるステップをさらに含む、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
マクロファージを減損させるステップが、化学的減損を含む、請求項28に記載の方法。
【請求項30】
マクロファージを減損させるステップが、マクロファージに特異的に結合する抗体の使用を含む、請求項28に記載の方法。
【請求項31】
連続移植により採取されたヒト肝細胞を増大させるステップをさらに含む、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、前記欠失または1つもしくは複数の点変異に対して、ホモ接合体であり、前記マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞は、前記マウスにおいて、増大させることができる、遺伝子組み換えマウス。
【請求項33】
前記欠失または点変異が、B細胞、T細胞、およびNK細胞の完全喪失をもたらす、請求項32に記載のマウス。
【請求項34】
前記マウスが、FRGマウスである、請求項32または33に記載のマウス。
【請求項35】
前記マウスが、FpmRGマウスである、請求項32または33に記載のマウス。
【請求項36】
前記マウスが、ヒトウロキナーゼを発現する、請求項32〜35のいずれか1項に記載のマウス。
【請求項37】
前記ウロキナーゼが、分泌型のウロキナーゼである、請求項36に記載のマウス。
【請求項38】
前記ウロキナーゼが、非分泌型のウロキナーゼである、請求項36に記載のマウス。
【請求項39】
ヒトウロキナーゼの発現が、ヒトウロキナーゼをコード化する導入遺伝子の取り込みに起因する、請求項36〜38のいずれか1項に記載のマウス。
【請求項40】
ヒトウロキナーゼの発現が、ヒトウロキナーゼをコード化するベクターの投与に起因する、請求項36〜38のいずれか1項に記載のマウス。
【請求項41】
前記ベクターが、アデノウイルスベクターである、請求項40に記載のマウス。
【請求項42】
前記ベクターが、アデノ随伴ウイルスベクターである、請求項40に記載のマウス。
【請求項1】
生体内でヒト肝細胞を増大させる方法であって、
i)単離されたヒト肝細胞をRag2−/−/Il2rg−/−マウスに移植するステップであって、前記マウスが、Fahの発現が欠損している、ステップと、
ii)前記ヒト肝細胞を、少なくとも約2週間増大させるステップと、
iii)前記マウスからヒト肝細胞を採取するステップと、を含む、方法。
【請求項2】
前記マウスが、前記Fah遺伝子において、欠失に対してホモ接合体である、請求項1に記載の方法。
【請求項3】
前記マウスが、Fah−/−/Rag2−/−/Il2rg−/−(FRG)マウスである、請求項2に記載の方法。
【請求項4】
前記マウスが、前記Fah遺伝子において、点変異に対してホモ接合体である、請求項1に記載の方法。
【請求項5】
前記マウスが、Fahpm/Rag2−/−/Il2rg−/−(FpmRG)マウスである、請求項4に記載の方法。
【請求項6】
ヒトウロキナーゼをコード化するベクターが、前記ヒト肝細胞を注入する前に、前記マウスに投与される、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
前記ウロキナーゼが、分泌型のウロキナーゼである、請求項6に記載の方法。
【請求項8】
前記ウロキナーゼが、非分泌型のウロキナーゼである、請求項6に記載の方法。
【請求項9】
前記ベクターが、アデノウイルスベクターである、請求項6〜8のいずれか1項に記載の方法。
【請求項10】
前記ベクターが、アデノ随伴ウイルスベクターである、請求項6〜8のいずれか1項に記載の方法。
【請求項11】
前記ベクターが、静脈内投与される、請求項6〜10のいずれか1項に記載の方法。
【請求項12】
前記ベクターが、肝細胞注入の約24〜48時間前に投与される、請求項6〜11のいずれか1項に記載の方法。
【請求項13】
前記マウスに、肝細胞注入前に、2−(2−ニトロ−4−トリフルオロ−メチル−ベンゾイル)−1,3 シクロヘキサンジオン(NTBC)が投与される、請求項1〜12のいずれか1項に記載の方法。
【請求項14】
前記NTBCが、約0.05mg/kg/日から約0.10mg/kg/日の用量で投与される、請求項13に記載の方法。
【請求項15】
前記マウスに、肝細胞注入後、少なくとも約3日間、NTBCがさらに投与される、請求項13または請求項14に記載の方法。
【請求項16】
前記マウスに、肝細胞注入後、少なくとも約6日間、NTBCがさらに投与される、請求項13または請求項14に記載の方法。
【請求項17】
NTBCの前記用量が、肝細胞注入後、6日間の過程にわたり漸減される、請求項16に記載の方法。
【請求項18】
前記NTBCが、飲料水内投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項19】
肝細胞注入前、飲料水内投与されるNTBCの濃度が、約1mg/Lから約8mg/Lである、請求項18に記載の方法。
【請求項20】
前記NTBCが、食物内投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項21】
前記NTBCが、注入することにより投与される、請求項13〜17のいずれか1項に記載の方法。
【請求項22】
前記ヒト肝細胞が、少なくとも約6週間、増大可能である、請求項1〜21のいずれか1項に記載の方法。
【請求項23】
前記ヒト肝細胞が、最長約6ヶ月間、増大可能である、請求項1〜21のいずれか1項に記載の方法。
【請求項24】
前記単離されたヒト肝細胞が、前記マウスの脾臓または門脈に注入される、請求項1〜23のいずれか1項に記載の方法。
【請求項25】
前記増大されたヒト肝細胞は、前記マウスの肝臓から採取される、請求項1〜24のいずれか1項に記載の方法。
【請求項26】
前記ヒト肝細胞が、臓器ドナーの肝臓から単離された、または外科的切除術から単離された、または幹細胞もしくは単球もしくは羊膜細胞に由来した、請求項1〜25のいずれか1項に記載の方法。
【請求項27】
前記肝細胞は、注入前、凍結保存された、請求項26に記載の方法。
【請求項28】
肝細胞注入前、前記FRGマウスからマクロファージを減損させるステップをさらに含む、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
マクロファージを減損させるステップが、化学的減損を含む、請求項28に記載の方法。
【請求項30】
マクロファージを減損させるステップが、マクロファージに特異的に結合する抗体の使用を含む、請求項28に記載の方法。
【請求項31】
連続移植により採取されたヒト肝細胞を増大させるステップをさらに含む、請求項1〜30のいずれか1項に記載の方法。
【請求項32】
遺伝子組み換えマウスであって、そのゲノムは、欠失または点変異が、機能的FAH、RAG−2、およびIL−2Rγタンパク質の発現の喪失をもたらすような、Fah、Rag2、およびIl2rg遺伝子において、前記欠失または1つもしくは複数の点変異に対して、ホモ接合体であり、前記マウスは、免疫不全であり、肝機能の低下を示し、ヒト肝細胞は、前記マウスにおいて、増大させることができる、遺伝子組み換えマウス。
【請求項33】
前記欠失または点変異が、B細胞、T細胞、およびNK細胞の完全喪失をもたらす、請求項32に記載のマウス。
【請求項34】
前記マウスが、FRGマウスである、請求項32または33に記載のマウス。
【請求項35】
前記マウスが、FpmRGマウスである、請求項32または33に記載のマウス。
【請求項36】
前記マウスが、ヒトウロキナーゼを発現する、請求項32〜35のいずれか1項に記載のマウス。
【請求項37】
前記ウロキナーゼが、分泌型のウロキナーゼである、請求項36に記載のマウス。
【請求項38】
前記ウロキナーゼが、非分泌型のウロキナーゼである、請求項36に記載のマウス。
【請求項39】
ヒトウロキナーゼの発現が、ヒトウロキナーゼをコード化する導入遺伝子の取り込みに起因する、請求項36〜38のいずれか1項に記載のマウス。
【請求項40】
ヒトウロキナーゼの発現が、ヒトウロキナーゼをコード化するベクターの投与に起因する、請求項36〜38のいずれか1項に記載のマウス。
【請求項41】
前記ベクターが、アデノウイルスベクターである、請求項40に記載のマウス。
【請求項42】
前記ベクターが、アデノ随伴ウイルスベクターである、請求項40に記載のマウス。
【図1a】

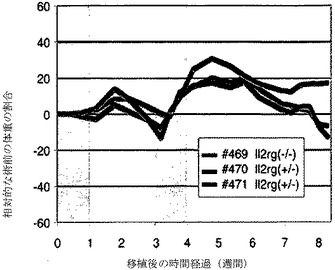
【図1b】
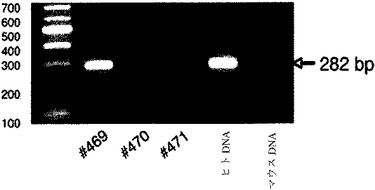
【図1c】

【図1d】
【図1e】
【図1f】
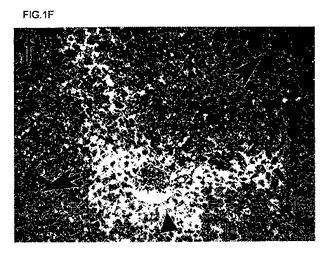
【図1g】
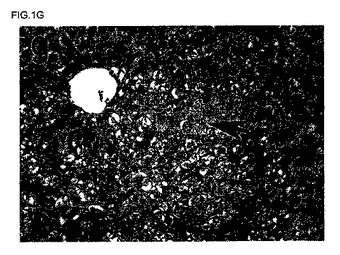
【図2A】

【図2B】

【図2C】

【図2D】

【図2E】

【図2F】

【図2G】

【図2H】

【図3a】
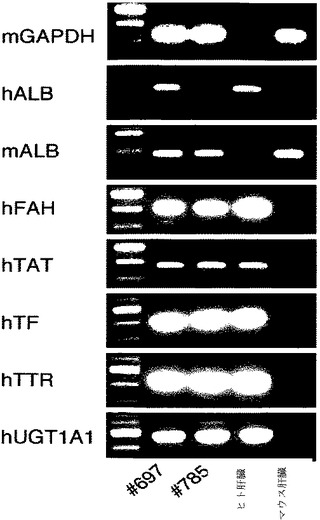
【図3b】

【図3c】

【図3d】

【図3e】
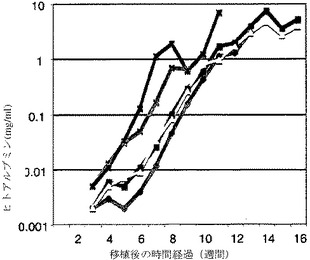
【図4a】
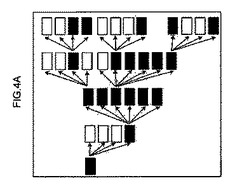
【図4b】

【図4c】
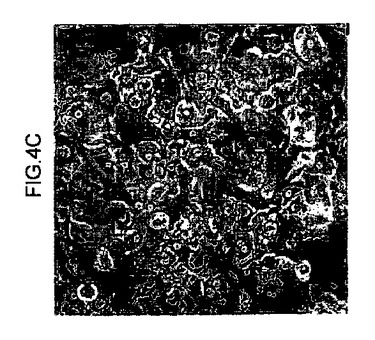
【図4d】

【図4e】

【図4f】

【図4g】
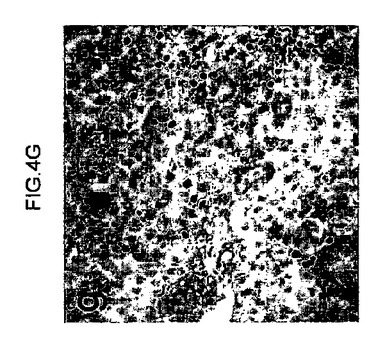
【図4h】

【図5a】

【図5b】

【図5c】

【図5d】

【図5e】

【図5f】
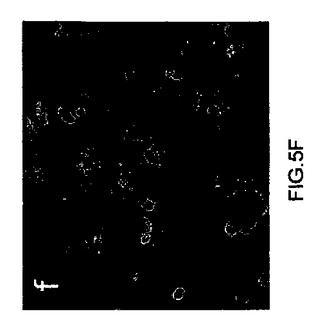
【図5g】
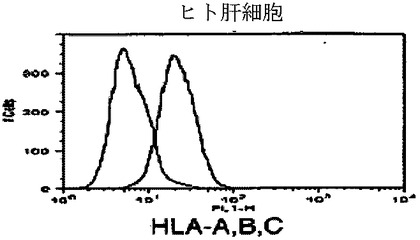
【図5h】
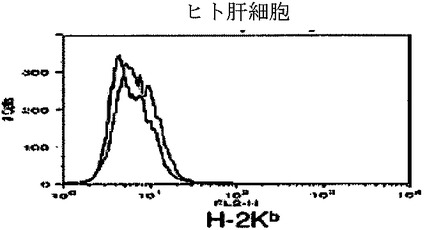
【図5i】
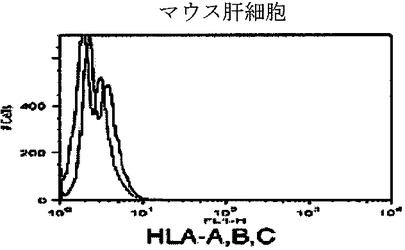
【図5j】

【図5k】

【図5l】
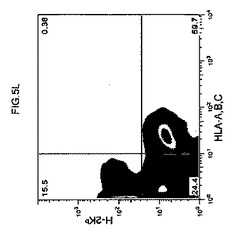
【図6a】
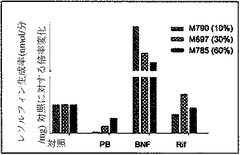
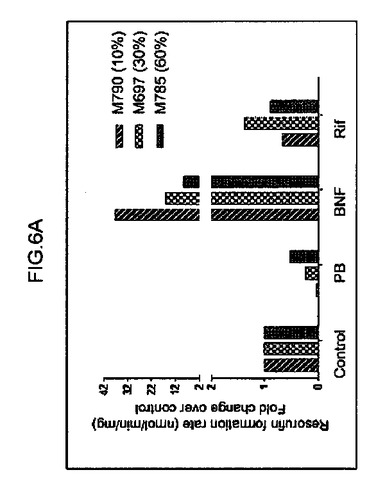
【図6b】

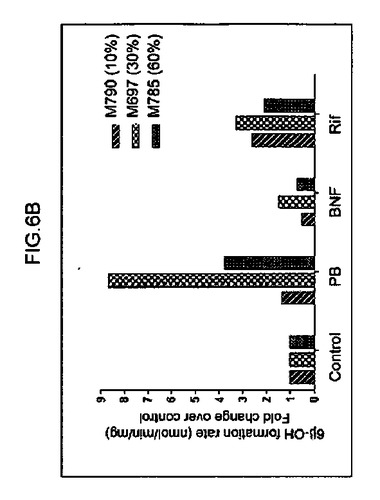
【図6c】
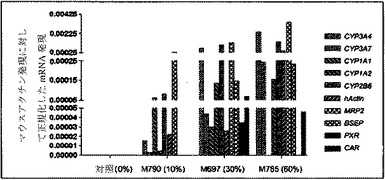
【図7a】

【図7b】
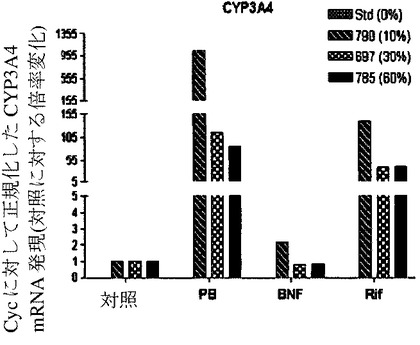
【図7c】

【図7d】
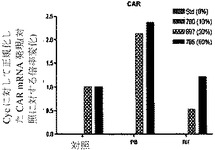
【図7e】
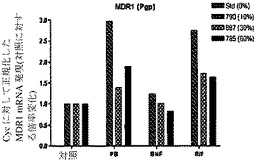
【図7f】

【図7g】
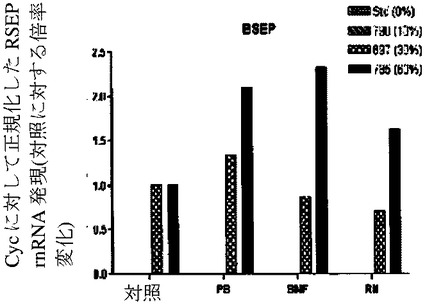
【図7h】
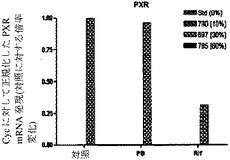
【図1b】
【図1c】
【図1d】
【図1e】
【図1f】
【図1g】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図2H】
【図3a】
【図3b】
【図3c】
【図3d】
【図3e】
【図4a】
【図4b】
【図4c】
【図4d】
【図4e】
【図4f】
【図4g】
【図4h】
【図5a】
【図5b】
【図5c】
【図5d】
【図5e】
【図5f】
【図5g】
【図5h】
【図5i】
【図5j】
【図5k】
【図5l】
【図6a】
【図6b】
【図6c】
【図7a】
【図7b】
【図7c】
【図7d】
【図7e】
【図7f】
【図7g】
【図7h】
【公表番号】特表2010−528661(P2010−528661A)
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願番号】特願2010−511327(P2010−511327)
【出願日】平成20年6月5日(2008.6.5)
【国際出願番号】PCT/US2008/065937
【国際公開番号】WO2008/151283
【国際公開日】平成20年12月11日(2008.12.11)
【出願人】(506124239)オレゴン ヘルス アンド サイエンス ユニバーシティ (8)
【出願人】(503115205)ザ ボード オブ トラスティーズ オブ ザ リランド スタンフォード ジュニア ユニヴァーシティ (69)
【Fターム(参考)】
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願日】平成20年6月5日(2008.6.5)
【国際出願番号】PCT/US2008/065937
【国際公開番号】WO2008/151283
【国際公開日】平成20年12月11日(2008.12.11)
【出願人】(506124239)オレゴン ヘルス アンド サイエンス ユニバーシティ (8)
【出願人】(503115205)ザ ボード オブ トラスティーズ オブ ザ リランド スタンフォード ジュニア ユニヴァーシティ (69)
【Fターム(参考)】
[ Back to top ]